
ASPECTOS ECOLÓGICOS DE LA COMUNIDAD DE PECES EN …biblio.uabcs.mx/tesis/te2888.pdf ·...
74
UNIVERSIDAD AUTÓNOMA DE BAJA CALIFORNIA SUR ÁREA DE CONOCIMIENTO DE CIENCIAS DEL MAR DEPARTAMENTO ACADÉMICO DE BIOLOGÍA MARINA ASPECTOS ECOLÓGICOS DE LA COMUNIDAD DE PECES EN HÁBITATS DE MANGLAR EN LA BAHÍA DE LA PAZ, B. C. S. MÉXICO TESIS Que para obtener el título de: BIÓLOGO MARINO PRESENTA: Juan Ángel Payan Alcacio DIRECTOR: M.C. Gustavo De la Cruz Agüero La Paz, Baja California Sur, México 14/02/2013
Transcript of ASPECTOS ECOLÓGICOS DE LA COMUNIDAD DE PECES EN …biblio.uabcs.mx/tesis/te2888.pdf ·...

UNIVERSIDAD AUTÓNOMA DE BAJA CALIFORNIA SUR
ÁREA DE CONOCIMIENTO DE CIENCIAS DEL MAR
DEPARTAMENTO ACADÉMICO DE BIOLOGÍA MARINA
ASPECTOS ECOLÓGICOS DE LA COMUNIDAD DE PECES EN
HÁBITATS DE MANGLAR EN LA BAHÍA DE LA PAZ, B. C. S.
MÉXICO
TESIS
Que para obtener el título de:
BIÓLOGO MARINO
PRESENTA:
Juan Ángel Payan Alcacio
DIRECTOR:
M.C. Gustavo De la Cruz Agüero
La Paz, Baja California Sur, México 14/02/2013

I
Dedicatoria
A mi Madre: Por todo el amor recibido, la paciencia, la fe y las dificultades que sufrimos a
través de este camino, gracias por enseñarme que el trabajo duro y constante es
recompensado pero que nunca debemos de trabajar esperando que se nos recompense.
A mi Padre: Por introducirme en este bello camino que es la biología marina, por sus
consejos y amor, gracias por enseñarme a vivir y pensar como humano antes que como un
patriota, y así como humanos es nuestra obligación mejorar el mundo en el que vivimos.
A mi hermana: Por la fe que depósito en mí, el amor y todas sus atenciones, siempre me ha
mostrado que la tenacidad nos impulsa hacia adelante no importa lo escabroso del camino.
Así y no por menos el resto de mi familia: Mi hermano Salvador, Primos: Sergio, Alicia,
Cristina, Lolita, Gaby, Víctor, Memo, Ocky y Pollo, Tías: Rosa, Norma, Lupe, Rosalba,
Lolis, Juani, Tíos: Guillermo y Paco, abuelo Juan, y Roni e Inu, gracias por todo su cariño
y amor.
Los humanos somos una pequeña galaxia de células que interactúa en el
vasto universo que llamamos sociedad (Payan-Alcacio, 2009)

II
Agradecimientos
Al Lector por tomarse el tiempo de hojear el resultado de este esfuerzo conjunto.
M. en C. Gustavo De la Cruz: director, gracias por su dirección, dedicación, consejos,
tiempo, comentarios, apoyo, paciencia, siempre correctas correcciones, amistad, así como
la gran calidez y calidad humana que me ha y sigue brindado, pero principalmente por ser
un gran mentor tanto dentro como fuera del aula.
Dr. José De La Cruz: asesor, por su apoyo, consejos, tiempo, paciencia, orientación, por su
gran calidez y calidad humana que me ha y sigue brindando, correcciones, pero
principalmente por romper su regla y aceptarme en su equipo de trabajo, lo que originó mi
amor por lo peces.
Dra. Liliana Olande: asesora, por todo el tiempo, orientación actual y previa, consejos,
paciencia, apoyo, correcciones y amistad, así como su gran calidez y calidad humana que
me ha brindado desde el día que la conocí.
Tec. Víctor Cota: por formar parte de mi temprano desarrollo en el campo de la ictiología,
por su paciencia (con y sin comillas), sus comentarios, sugerencias y principalmente por su
amistad.

III
Al Dr. Carlos Salomón y al Dr. Oscar Trujillo, por aceptar pertenecer a mi comité de tesis.
A todos los maestros nuevos y viejos que me inspiraron y me motivaron a exigirme más,
así como me encaminaron a seguir los pasos que ahora doy.
Al honorable gremio y cofradía de la Colección Ictiológica CICIMAR-IPN por su apoyo,
consejos y amistad brindada desde mi inclusión al equipo hasta la fecha, Jorge Chollet,
Francisco Vergara, Adriana Martínez, Armando Medina, Víctor Tapia, Daniel Galindo y
Dr. Francisco García.
Y a mis amigos por su apoyo y cariño en esos momentos felices y difíciles: Héctor, Gengis,
Cecy, Mario, Tai-Coh, Jorge, Caro, Vaio, Javi, Charlangas, Alfredo, Cuyo, Jan Carlo,
Diana, Ember, Iván, Lu, Vicky, Ara, Miguel, Belén, Ilse, Ana, René, Coché, Jario, Yekka,
Eliot, Ricky, Andrés, Sergio, Diego, Luna, Nubs, Artemio, Berto, Fer (L.A.B.) y a todos
aquellos que escapan de mi mente, NO por OLVIDO sino por premura.

IV
Índice de contenido
Resumen……………………………………………………………………………….…….1
Introducción…………………………………………………………………………………2
Antecedentes………………………………………………………………………………...5
Objetivos…………………………………………………………………………………….8
Justificación………………………………………………………………………………….9
Área de estudio………………………………………………………………………………9
Metodología………………………………………………………………………………..13
Resultados………………………………………………………………………………….17
Discusión………………………………………………………………………………...…39
Conclusiones……………………………………………………………………………….52
Recomendaciones…………………………………………………………………………..53
Bibliografía………………………………………………………………………………...55
Anexo I. Mezcla frigorífera………………………………………………………………...66
Anexo II. Tabla de temperaturas…………………………………………………………...67
Índice de figuras y tablas
Figura 1. Área de estudio y localización de sitios de muestreo.……………………….......12
Figura 2. Intervalos de temperatura promedio por época del año………………………….18
Figura 3. Abundancia relativa por familias por épocas de muestreo…………………..…..23
Figura 4. Abundancia relativa por familias por épocas climáticas en Balandra…………...24
Figura 5. Abundancia relativa por familias por épocas climáticas en Zacatecas.………….25

V
Figura 6. Porcentaje del I.V.I. general……………………………………………………..30
Figura 7. Porcentaje del I.V.I. en Balandra.………………………………………………..31
Figura 8. Porcentaje del I.V.I. en Zacatecas ..……………………………………………..32
Figura 9. Análisis de similitud general por épocas………………………………………...38
Figura 10. Análisis de similitud entre épocas de Balandra y Zacatecas………...…………38
Tabla I. Listado taxonómico……………………………………………………………….20
Tabla II. Estructura taxonómica general, por localidad y época del año…………………..22
Tabla III. Abundancia de clases y ordenes de peces en el muestreo……………………….26
Tabla IV. Abundancia de las familias y especies de peces en el muestreo………………...27
Tabla V. Dominancia de las especies en el muestreo……………………………………...33
Tabla VI. Tabla de Riqueza específica, Diversidad ecológica y Equidad…………………37
Tabla VII. Listado taxonómico de Balandra……………………………………………….42
Tabla VIII. Listado taxonómico de Zacatecas.…………………………………………….45

1
Resumen
Los manglares son sitios de reclutamiento importante para los juveniles de diferentes
especies de animales; un grupo que sobresale en particular en este ambiente es el de los
peces. Aunque existen numerosos trabajos sobre los peces en el área, los estudios
ecológicos son escasos. Se realizaron recolectas mensuales durante un ciclo anual (2010-
2011) en dos localidades con manglar, una comunicada directamente (Balandra) y la otra
indirectamente (Zacatecas) con la Bahía de La Paz, B.C.S. Los peces fueron capturados
mediante lances con chinchorro playero por duplicado en cada ocasión. Se caracterizaron
tres épocas climáticas: Cálida (julio - noviembre), Transición (abril - junio) y Templada
(diciembre - marzo). Para el análisis de datos obtenidos se aplicaron estadísticos básicos así
como índices ecológicos de dominancia, diversidad, equidad y pruebas de similitud entre
épocas y entre localidades. Se capturaron 4055 peces de los cuales se determinaron 54
especies pertenecientes a 21 familias. Para Balandra se identificaron 37 especies mientras
que en Zacatecas se identificaron 35; en ambas localidades la época cálida presentó un
mayor número de especies que en las otras temporadas. Seis especies de cinco familias se
capturaron durante todo el año en ambas localidades. Las familias Haemulidae y Gerreidae
fueron las más diversas presentando siete especies cada una, mientras que la más abundante
fue la familia Gerreidae. Mugil curema (Mugilidae) fue la especie más abundante y
dominante en Zacatecas mientras que Eucinostomus dowii (Gerreidae) lo fue en Balandra.
La mayor diversidad ecológica y equidad se presentaron en Balandra. De la misma manera,
las mayores diversidades ocurrieron en las épocas cálidas, mientras que las mayores
equidades en las épocas templadas. El análisis de similitud entre localidades corroboró que
la estructura comunitaria es diferente entre sitios y que las épocas cálidas y templadas
presentan asociaciones ictiofaunísticas diferentes.
Palabras clave: Épocas, Índices ecológicos, Balandra y Zacatecas

2
Introducción
Podemos definir a un pez como un organismo vertebrado acuático ectotérmico,
generalmente con apéndices modificados en forma de aleta, que respiran mediante
branquias en algún momento de su desarrollo y cuyo cuerpo, en su mayoría, está cubierto
por escamas (Stein y Lavenberg, 1997; Nelson, 2006). Existen aproximadamente de
25,000 a 28,000 especies de peces en el planeta distribuidas en dos ambientes, el marino
con aproximadamente un 58% de la especies y el dulceacuícola con un 42% de las especies
(Nelson, 2006; Helfman et al., 2009).
De manera general, los organismos acuáticos como los peces presentan múltiples
estrategias ecológicas, por lo que para su estudio se tienen tres tipos de enfoques: el
primero es a nivel geográfico, el segundo a nivel local y el tercero a nivel de los
microambientes (Salazar-Vallejo y González, 1993). En lo que respecta al nivel local,
existen divisiones basadas en la estratificación vertical u horizontal de las especies en la
columna de agua. Desde la perspectiva de la distribución horizontal en el ambiente marino
se suelen diferenciar los ambientes litorales, neríticos y oceánicos (Bone y Moore, 2008;
Helfman et al., 2009).
Los manglares son ecosistemas litorales con alta productividad que funcionan como
sitio de crianza, alimentación y reclutamiento para diversas especies de aves, moluscos,
crustáceos, peces en diferentes estadios de desarrollo (larvas, juveniles y adultos)

3
(Kathiresan y Bingham, 2001; González-Acosta et al., 2005; Ellison, 2008) y hasta de
algunos mamíferos (Moreno-Bejarano y Álvarez-León, 2003; Marcín-Medina, 2010).
Dichos ecosistemas constituyen un lugar físico protegido y una fuente de
nutrimentos para esa variada fauna (Parrish, 1989; Laegdsgaard y Craig, 1995, 2001; Beck
et al., 2001; Hindell y Jenkins, 2004). Los manglares son típicamente tropicales, aunque se
pueden encontrar en latitudes intermedias (González-Zamorano, 2002); los árboles llegan a
medir más de 30 metros de altura y llegan a abarcar extensiones de miles de hectáreas
(Lara-Lara et al., 2008), tienden a ser favorecidos por condiciones de bajo oleaje (Carter,
1988) y generalmente son sitios con alta productividad, por lo que comúnmente son
considerados hábitats de crianza (Laegdsgaard y Craig, 1995, 2001; Blaber, 1980; 2000;
Nagelkerken et al., 2000; Beck et al., 2001; Kathiresan y Bingham, 2001; Ellison, 2008).
Es así que, juveniles y adultos de peces de áreas costeras explotan estos sitios como
áreas de alimentación, refugio y reproducción, lo que propicia un gran dinamismo gracias a
los intercambios de energía y materia entre los sistemas de manglar y los ecosistemas
marinos adyacentes. Desde un punto de vista funcional, se reconoce que uno de los roles
ecológicos principales que desempeñan los peces en estos ecosistemas es el control
específico y numérico de los consumidores a través de la competencia y la depredación, a
la vez que contribuyen al transporte y flujo energético entre el manglar y las aéreas
circundantes (Begon et al., 2006; Carbajal-Fajardo et al., 2009; Yagi et al., 2011) debido al
proceso de migración.

4
Siendo las áreas costeras rodeadas de manglar uno de los ambientes más vulnerables
al cambio climático (Yañez-Arancibia et al., 1998), las presiones antropogénicas (e.g. uso
recreativo, acuacultura, desarrollo costero) (Linares et al., 2004) así como su ictiofauna
unos de los componentes más conspicuos e importantes (Unsworth et al., 2009), es que se
plantea un estudio ecológico de la ictiofauna de los esteros de la bahía con el ánimo de
contribuir al conocimiento de este recurso.
La Bahía de La Paz se sitúa en la porción suroccidental del Golfo de California; por
su geografía física e historia geológica presenta una gran variedad de ambientes litorales
entre los cuales sobresalen las playas arenosas, las costas rocosas, los fondos coralinos, así
como los esteros y áreas de manglar (Urbán–Ramírez y Ramírez–Rodríguez, 1997).
La bahía sobresale a nivel nacional como una de las más diversas ya que se conocen
522 especies de peces (Balart et al., 1995) del total de las 875 reconocidas para todo el
Golfo de California (Thomson et al., 2000)); sin embargo, la mayoría de los estudios sobre
los peces de la región han sido más bien faunísticos mientras que aquellos con enfoque
ecológico han sido menos frecuentes tanto por los hábitats considerados como por el hecho
de que no se conocen trabajos sobre aspectos ecológicos de la ictiofauna de la bahía en los
últimos ocho años.

5
Antecedentes
Los estudios de la ictiofauna en la Bahía de la Paz han estado enfocados
principalmente a inventarios taxonómicos, mientras que los estudios ecológicos han sido
más bien escasos.
Chávez (1985) realizó un trabajo de compilación bibliográfica registrando 188
artículos entre 1881 y 1983, los cuales presentaban información ictiológica sobre el
Pacífico y el Golfo de California incluyendo a la Bahía de la Paz, entre los que sobresalen
34 trabajos taxonómicos y 19 trabajos ecológicos de los cuales, solo cuatro pertenecen a la
Bahía de la Paz (Chávez, 1985).
En la perspectiva taxonómica. Abitia-Cárdenas et al. (1994) presentan el primer
elenco sistemático de la ictiofauna de la Bahía de La Paz, B.C.S; en la lista se encuentran
390 especies agrupadas en 250 géneros y 106 familias. El inventario se generó con base en
especímenes provenientes de muestreos realizados en 30 localidades entre 1981 y 1986 de
los que se obtuvieron 83 familias, 161 géneros y 227 especies, además de las especies
registradas en la zona por otros autores.
Balart et al., (1995) presentaron un listado taxonómico actualizado de la ictiofauna
de la bahía en el que se adicionan 132 registros nuevos, resultando en 522 el número total
de especies nominales conocidas. Dicha cifra integra registros previos, muestreos con

6
redes, observaciones sistemáticas en campos pesqueros e identificación de otolitos
proveniente de copros de lobo marino.
De La Cruz-Agüero et al. (1997) realizaron un catálogo de peces de la Bahía de la
Paz, B.C.S., registrando un total de 40 órdenes que se dividen en 205 familias y
comprenden 281 especies. Todos los organismos mencionados en el trabajo se encuentran
preservados en la Colección Ictiológica (CI) del Centro Interdisciplinario de Ciencias del
Mar - Instituto Politécnico Nacional (CICIMAR-IPN) (http://coleccion.cicimar.ipn.mx)
En la perspectiva bioecológica, Maeda-Martínez (1981) estudió la composición,
abundancia, diversidad y alimentación de la ictiofauna en tres zonas de manglar en la bahía,
Enfermería (E) con 23 especies, Puerto Balandra (B) con 49 especies y Estero Zacatecas
(Z) con 28 especies, describiendo cuatro categorías ecológicas de la ictiofauna: residentes
[E 2 sp; B 6 sp; Z 1 sp], peces que los utilizan como aéreas de crianza [E, 8 sp; B, 12 sp; Z,
7 sp], visitantes ocasionales [E, 6 sp; B, 8 sp; Z, 9 sp] y visitantes excepcionales [E, 7 sp;
B, 21 sp; Z, 11 sp].
Balart et al. (1997) realizaron un estudio en nueve sitios con fondos blandos en la
Bahía de La Paz (Pichilingue, Bahía Falsa, Enfermería, Coromuel, El Esterito, frente a
CICIMAR, frente a la planta de tratamiento de aguas residuales, entre Centenario y
Comitán, y en el estero Zacatecas), el estudio fue de dos años no consecutivos y se
encontraron 103 especies en 21,383 especímenes, donde las familias Gerreidae, Mugilidae,

7
Engraulidae y Clupeidae dominaron numéricamente ambos años en la mayoría de las
localidades. Se observó una variación de diversidad de 2.95 a 3.37 bits/ind a lo largo del
estudio.
Galván-Piña (1998) por su parte presenta información básica acerca de la estructura
de las comunidades de peces de diversos ambientes costeros de la Bahía de La Paz
determinando 57 especies en tres grupos (permanentes, temporales y ocasionales) basado
en cuatro muestreos estacionales durante un ciclo anual. Especies pertenecientes a las
familias Gerreidae, Ariidae, Lutjanidae, Haemulidae, Ephippidae y Kypsodidae fueron las
dominantes en abundancia, mientras que según la temporada del año, diferentes
asociaciones de peces utilizan las diferentes zonas estudiadas (e.g. San Juan de la Costa, la
isla La Partida, Zacatecas y El Mogote).
Malpica-Maury (1999) realizó un estudio espacio-temporal de la composición,
riqueza específica y abundancia de la ictiofauna de la Ensenada de La Paz. El estudio
consideró muestras con red de arrastre de prueba en cuatro sitios de la laguna frente a
Zacatecas, Comitán, Chametla y CICIMAR en cinco campañas realizadas en julio,
septiembre y noviembre de 1995 y enero y mayo de 1996; se capturaron 1224 individuos
pertenecientes a 42 especies; de éstas el 12% se clasificaron residentes y 50% temporales.
Las mayores riquezas, abundancias y biomasa aparecieron en noviembre y enero, por lo
que se consideró que los factores que determinaron una mayor influencia para la
composición de la comunidad fueron la temperatura y la marea.

8
González-Acosta et al. (2005) evaluaron cambios estacionales durante la temporada
de Octubre de 1996 a Agosto de 1997 y como afectan éstos la estructura poblacional de los
peces asociados al manglar de El Conchalito, B. C. S., encontrando que se pueden
distinguir dos temporadas en la estructura de la comunidad íctica, una cálida (verano-otoño)
y una templada (primavera-invierno). Las mayores abundancias en la comunidad se
presentan durante el verano, mientras que las especies dominantes para este sitio son la
mojarra bandera Eucinostomus currani (Zahuranec en Yáñez 1980) en otoño, el machete
Albula nemoptera (Fowler 1911) en invierno y la anchoa chicotera Anchoa ischana (Jordan
y Gilbert 1882) en primavera-verano.
Objetivos
General
Analizar la variación estacional de la comunidad ictiofaunística de los
esteros Balandra y Zacatecas de la Bahía de La Paz.
Particulares
Conocer la composición y abundancia de la ictiofauna de los esteros
Balandra y Zacatecas en tres épocas del año (templada, transición y cálida,).
Comparar y analizar las diferencias en composición y abundancia de la
ictiofauna en los dos esteros y entre tres épocas del año.

9
Justificación
Según Faunce y Serafy (2006), en los últimos 50 años de estudios ictiológicos
asociados al manglar, la mitad de las investigaciones realizadas se han centrado en la
variación espacio-temporal de las comunidades que utilizan estos ambientes y en menor
proporción a los inventarios taxonómicos de las especie que las integran. En lo que respecta
a la Bahía de la Paz ocurre lo contrario, los trabajos sobre listados e inventarios
taxonómicos presentan una mayor proporción que los trabajos ecológicos; incluso, entre
estos mismos encontramos un intervalo carente de información o de estudios en los
últimos ocho años.
El conocimiento de los cambios estacionales de la ictiofauna en las áreas de crianza
como son los manglares es importante para coadyuvar a un manejo sustentable del recurso
pesquero y para la conservación de las especies residentes (Unsworth et al., 2009), así
como para la elaboración de mapas de distribución de las mismas. De acuerdo con ello, se
espera contribuir al conocimiento científico de la composición y dinámica de la abundancia
y distribución de las especies ícticas en dos aéreas de crianza seleccionadas como lo son los
esteros Balandra y Zacatecas en la Bahía de la Paz, B.C.S.
Área de estudio
La Bahía de La Paz es la bahía más grande que existe en la costa occidental del
Golfo de California. Se localiza entre los 24° 07´ y 24° 21´ Longitud Norte y entre los
meridianos 110° 17´y 110° 40´ Longitud Oeste, cerca del extremo sur de la península de

10
Baja California en el estado de Baja California Sur. La bahía está protegida por la Isla San
José al Norte y al Este por las islas del archipiélago Espíritu Santo. No obstante presenta
una comunicación continua con el Golfo de California a través del canal San Lorenzo y la
propia boca de la bahía en su porción Noreste (Flores, 1998).
La Ensenada de La Paz es el accidente geográfico más notable de la bahía; se
localiza en su extremo Sur-Suroeste entre los paralelos 24° 10´ y 24° 06´ Latitud Norte y
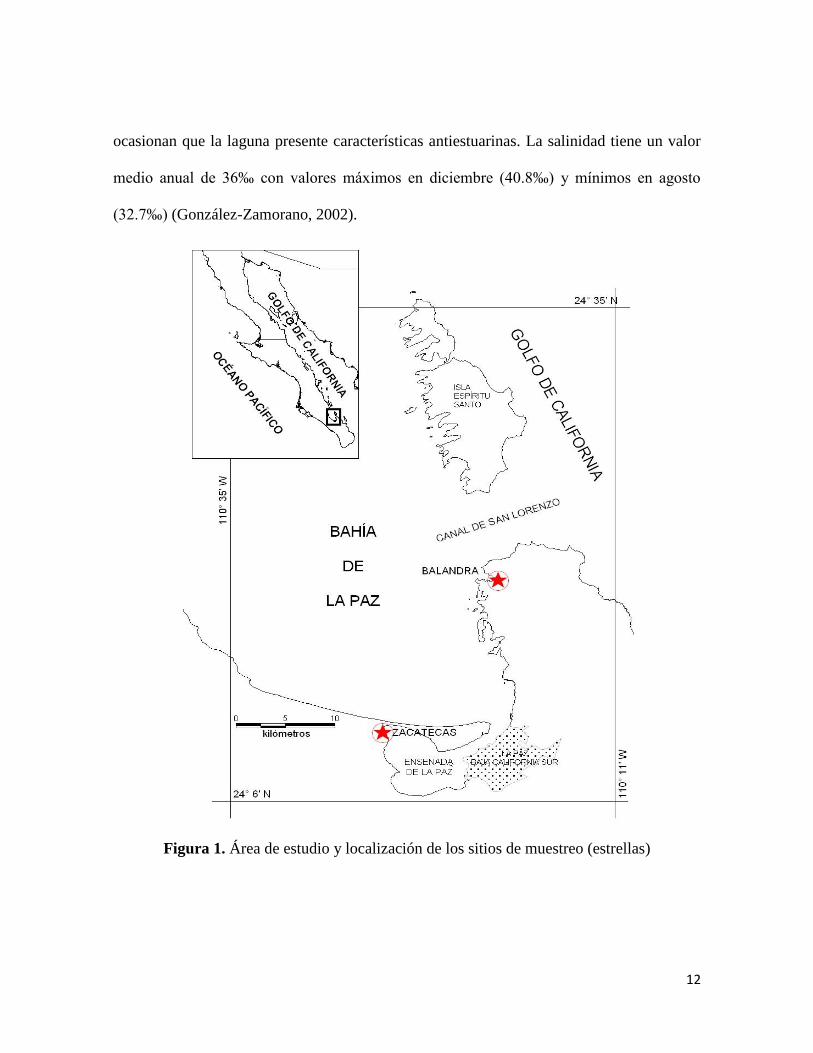
entre los meridianos 110° 19´ y 110° 26´ Longitud Oeste (Fig. 1) . Este cuerpo de agua se
comunica con la bahía a través de un canal de mareas que conforma la boca del sistema,
con una profundidad mediana entre los 7 - 10m y una anchura entre 0.6 – 1 Km. En su parte
central, la recorre un canal principal que disminuye de profundidad hasta los cuatro metros
en su porción más interna. Asimismo, la ensenada representa una cuenca con una superficie
aproximada de 45 km2 al nivel medio del mar (González-Acosta et al., 2005)
El clima de la región es seco árido. La temperatura oscila alrededor de 18.7°C. La
precipitación anual es de 166.5 mm en promedio; la evaporación excede a la precipitación.
El oleaje es mínimo y la marea, que es semidiurna mixta y con una amplitud de alrededor
de un metro ejerce una gran influencia en sus playas, debido principalmente a la suave
pendiente (Suárez-Altamirano, 2005).
En la Bahía de La Paz podemos encontrar numerosos esteros con manglares como
sitio de reclutamiento de la ictiofauna como lo son Enfermería, Zacatecas, Puerto Balandra,

11
El Conchalito, Pichilingue, El Mogote, Eréndira y Zacatal (Mendoza et al., 1984;
González-Zamorano, 2002).
En este trabajo nos centraremos en dos de estos manglares, uno dentro de la
Ensenada de La Paz y otro fuera de ella. Dentro de la ensenada se seleccionó al Estero
Zacatecas. Éste se encuentra en los 24° 11´ 15´´ de Latitud Norte y 110° 26´ 00´´ de
Longitud Oeste en el extremo occidental de la ensenada (Fig. 1). Su superficie total es de
57.19 hectáreas, el manglar cubre 22.44 ha, el cuerpo de agua es una canal de mareas
sinuoso bordeado de manglar, cuenta con tres zonas de inundación situadas hacia el oeste,
su profundidad máxima es de 2.5 m al nivel medio del mar, su sustrato es principalmente
fangoso y está rodeado por una comunidad de mangles que está compuesta por Rhizophora
mangle, Laguncularia racemosa y Avicennia germinans (Mendoza, 1983).
La segunda localidad se encuentra fuera de la ensenada y es conocida como Puerto o
Bahía Balandra; es una cuenca lagunar de sustrato arenoso/rocoso rodeada por vegetación
de manglar compuesta por Avicennia germinans, Laguncularia racemosa y Rhizophora
mangle. Dicha laguna se encuentra comunicada a la bahía hacia el oeste a través de una
boca de 140 metros de anchura aproximadamente; su superficie comprende alrededor de
52.5 h de los cuales 30 h son área lagunar y el resto vegetación. La salinidad se incrementa
gradualmente hacia el interior de la laguna principalmente por lo somero de esta zona lo
cual aunado a factores tales como la elevada tasa de evaporación, escasa precipitación
pluvial y a la casi nulidad de los escurrimientos provenientes de sistemas adyacentes,
12
ocasionan que la laguna presente características antiestuarinas. La salinidad tiene un valor
medio anual de 36‰ con valores máximos en diciembre (40.8‰) y mínimos en agosto
(32.7‰) (González-Zamorano, 2002).
Figura 1. Área de estudio y localización de los sitios de muestreo (estrellas)

13
Metodología
Trabajo de campo
Se realizaron lances mensuales durante el periodo julio 2010 – agosto 2011 con una
red chinchorro playero con un centímetro de luz de malla, 50 metros de largo y una caída
de tres metros y medio. Por cada lance se realizó una réplica. Las recolectas se llevaron a
cabo aproximadamente entre las 7:30 am y las 12:00 m en presencia de marea alta dado que
las características topográficas de los sitios (pendiente muy suave) dificultaban el trabajo
durante la marea baja.
Los individuos capturados fueron colocados en una bolsa de plástico, la cual
contenía una etiqueta con los datos correspondientes a la fecha y el número de lance así
como la localidad de la cual fueron extraídos. Los organismos se trasladaron en una hielera
con una mezcla frigorífica (Anexo 1) a la Colección Ictiológica (CI) del CICIMAR-IPN.
Una vez en el laboratorio los especímenes se almacenaron en un congelador para evitar el
proceso de descomposición y para su procesamiento posterior.
Trabajo de laboratorio
La identificación de los organismos se realizó hasta el menor taxón posible con
ayuda de claves especializadas; para los niveles superiores se recurrió a las “Claves de
Familias de peces marinos del noroeste de México” (De La Cruz Agüero, 1994) y para la
identificación a nivel específico la “Guía de FAO para los fines de pesca, Pacífico Centro-
Oriental” (Fisher et al., 1995). Posterior a la identificación se tomaron medidas de longitud

14
(Ictiómetro, precisión= 1 mm) y peso (balanza Ohaus Navigator modelo N1D110,
precisión = 0.1 g). Finalmente los organismos muestreados se catalogaron en la base de
datos de la CI del CICIMAR.
Análisis de datos
Para definir las temporadas climáticas del muestreo se realizó una compilación de
datos de temperatura en la Bahía de La Paz. Con base en los datos mensuales, se realizaron
promedios de las temperaturas para cada mes y de acuerdo con el patrón de variación, se
distinguieron tres temporadas climáticas, seleccionando de julio a noviembre para la
temporada cálida, de abril a junio para la temporada de transición y de diciembre a marzo
para la temporada templada; se realizó un análisis de varianza de una vía para corroborar
que las temporadas climáticas muestran diferencias significativas en este factor (Anexo 1).
Los datos obtenidos de la captura se ordenaron en una hoja de cálculo de MsExcel
donde la información se organizó por medio de tablas dinámicas. Para el análisis de los
datos se utilizó el programa ANACOM versión 3.0 (De La Cruz-Agüero, 1994) donde se
realizaron los siguientes análisis:
Se utilizaron estadísticas básicas para representar los valores de composición y
abundancia ictiofaunística; se sumó el número de individuos mensuales por épocas
climáticas y se dividió entre el número de meses encontrados en la época. Para la
composición específica se cuantificó el número de especies por época climática.

15
Índice del Valor de Importancia (IVI) es usado para determinar la importancia total
de cada especie en la estructura comunitaria; el índice calcula el porcentaje de la frecuencia
y de la abundancia numérica para un valor máximo de 200% (Mueller-Dombois y
Ellenberg, 1974)
Donde:
Rni= número de individuos de la especie con respecto a número total de individuos de
todas las especies
Rfi= número de veces en la que apareció la especie con respecto a la frecuencia de todas las
especies
Diversidad ecológica medida con el índice de Shannon-Wiener, refleja la
complejidad de la comunidad en la medida en que el índice mide la incertidumbre de la
identidad (especie) a la que puede pertenecer un individuo tomando al azar de la muestra
(Ludwig y Reynolds. 1988).
Donde:
H´= índice de Shannon Wiener (bits/individuos)
S= número de especies

16
pi= proporción de la abundancia con respecto al total de la muestra perteneciente a la i-
ésima especie
Índice de equidad de Pielou, se refiere a la distribución de la abundancia en la
comunidad, es un derivado del índice de diversidad de Shannon y presenta resultados entre
0 y 1; mientras más cercano a 1 existe una distribución de la abundancia más uniforme
(Pielou, 1984; Brower et al., 1998).
Donde:
H´: Índice de diversidad observado
H´max: valor máximo posible del índice de diversidad, dado por S especies y N individuos y
se alcanza cuando la abundancia es la misma para todas las especies (H´max=log2(S)).
Se hizo un análisis de similitud basando en el índice de Morisita para estimar la
afinidad entre las localidades y entre las épocas del año analizadas. Los grupos se formaron
utilizando la unión media no ponderada de acuerdo con la implementación disponible en el
programa PAST versión 2.15 (Hammer et al., 2001)

17
Donde:
Cj,k= índice de similitud de Morisita entre las muestras j y k
pi,j= abundancia relativa de la especie i en la muestra j
pi,k= abundancia relativa de la especie i en la muestra k
S= número de especies
Se utilizó el programa FigTree versión 1.3.1 (Rambaut, 2009) para darle el formato
final al dendograma obtenido con el programa Past.
Resultados
Temporadas climáticas
El análisis de varianza realizado arrojó diferencias significativas entre los intervalos
de temperaturas para las épocas climáticas, definiendo a la época templada con 20.8±0.7
°C, la de transición 23.6±0.7 °C y la época cálida 27.9±0.8 °C (Fig. 2). Así pues al 95% de
confianza se obtuvo una p< 0.001 que indica que los intervalos de temperatura y las
combinaciones entre ellos son diferentes entre sí (Fig. 2 y Anexo 2).

18
Figura 2. Temperatura media e intervalo de variación por época del año en la Bahía de La
Paz B.C.S.(2010-2011). Se considera la temperatura superficial del mar
Composición específica
En el muestreo general se encontraron dos clases de peces Actinopterygii y
Chondrichthyes. La clase Actinopterygii es mucho más diversa en todos los niveles
taxonómicos (orden, familia, género y especie) presentes, que la clase Chondrichthyes, lo
mismo ocurre por localidad (Balandra y Zacatecas). A nivel de orden Perciformes es el más
diverso con 11 familias, mientras que por familia, las más diversas fueron Gerreidae y
Haemilidae con cuatro géneros y siete especies respectivamente. Por su parte Balandra fue
ligeramente más diversa que Zacatecas superando de uno a cinco componentes en cada
categoría taxonómica. De las 54 especies encontradas en el muestreo, seis especies
(Ctenogobius sagittula, Eucinostomus currani, Eucinostomus dowii, Mugil curema,
Paralabrax maculatofasciatus y Sphoeroides annulatus) tuvieron presencia en todos los
muestreos. En el estero Balandra las seis especies que aparecieron en todo el muestreo

19
fueron Ctenogobius sagittula, Eucinostomus currani, Eucinostomus dowii, Mugil curema,
Paralabrax maculatofasciatus y Sphoeroides annulatus, mientras que en Zacatecas las
especies encontradas fueron 11; Ctenogobius sagittula, Eucinostomus currani,
Eucinostomus dowii, Mugil curema, Paralabrax maculatofasciatus y Sphoeroides
annulatus, Diapterus brevirostris, Diodon hystrix, Eucinostomus entomelas,
Hyporhamphus naos y Pomadasys branickii) más a las registradas para el muestreo
general que se capturaron el Zacatecas en todo el año. Por épocas climáticas la temporada
cálida fue la más diversa en todos los niveles taxonómicos presentados, llegando a
presentar un 79% de las especies totales en el muestreo (Tabla I y II).
En Balandra encontramos un incremento en la diversidad de las diferentes
categorías taxonómicas conforme incrementaba la temperatura del agua. El orden
Perciformes fue el más diverso en todas las épocas climáticas, mientras que a nivel familia,
las más diversas fueron Gerreidae y Gobiidae en la época templada, Gerreidae y
Lutjanidae en la época de transición y Gerreidae en la época cálida. En Zacatecas se
presenta un decremento en la diversidad en la categoría de clase, caso opuesto a nivel de
orden, familia, género y especie cuyos valores aumentaron conforme se incrementa la
temperatura del agua. El orden Perciformes y la familia Gerreidae fueron las categorías
taxonómicas más diversas en todas las temporadas climáticas (Tabla I y II).

20
Tabla I. Listado taxonómico de las especies de peces encontradas en los esteros de
Balandra y Zacatecas, Bahía de la Paz, B.C.S.
Phylum Chordata
Subphylum Vertebrata
Superclase Gnathostomata
Clase Chondrichthyes
Orden Myliobatiformes
Familia Urolophidae
Urolophus halleri Cooper, 1863
Urolophus maculatus (Garman, 1913)
Clase Actinopterygii
División Teleostei
Orden Elopiformes
Familia Elopidae
Elops affinis Regan, 1909
Orden Albuliformes
Familia Albulidae
Albula nemoptera (Fowler, 1911)
Orden Clupeiformes
Familia Clupeidae
Harengula thrissina (Jordan & Gilbert, 1882)
Opisthonema libertate (Günther, 1867)
Sardinops sagax (Jenyns, 1842)
Orden Mugiliformes
Familia Mugilidae
Mugil cephalus Linnaeus, 1758
Mugil curema Valenciennes, 1836
Orden Beloniformes
Familia Hemiramphidae
Hyporhamphus naos Banford & Collette, 2001
Orden Gasterosteiformes
Familia Fistulariidae
Fistularia commersonii Rüppell, 1838
Orden Perciformes
Familia Serranidae
Paralabrax maculatofasciatus (Steindachner, 1868)
Familia Carangidae
Caranx caninus Günther, 1867
Caranx vinctus Jordan & Gilbert, 1882
Hemicaranx zelotes Gilbert, 1898
Oligoplites saurus (Bloch & Schneider, 1801)

21
Oligoplites saurus (Bloch & Schneider, 1801)
Familia Lutjanidae
Hoplopagrus guentherii Gill 1862
Lutjanus aratus Günther, 1864
Lutjanus argentiventris Peters, 1869
Lutjanus colorado Jordan & Gilbert, 1882
Lutjanus novemfasciatus Gill, 1862
Familia Haemulidae
Haemulon sexfasciatum Gill, 1862
Haemulon steindachneri (Jordan & Gilbert, 1882)
Haemulopsis leuciscus (Günther, 1864)
Orthopristis reddingi Jordan & Richardson, 1895
Pomadasys branickii (Steindachner, 1879)
Pomadasys elongatus (Steindachner, 1879)
Pomadasys macracanthus (Günther, 1864)
Familia Gerreidae
Eucinostomus currani Zahuranec en Yañez-Aranciba, 1980
Eucinostomus dowii (Gill, 1863)
Eucinostomus entomelas Zahuranec en Yañez-Aranciba, 1980
Eucinostomus gracilis (Gill, 1862)
Eugerres lineatus (Humboldt, 1821)
Diapterus brevirostris (Sauvage, 1879)
Gerres cinereus (Walbaum, 1792)
Familia Sciaenidae
Cynoscion stolzmanni Steindachner, 1879
Cynoscion xanthulus Jordan & Gilbert, 1882
Familia Blenniidae
Hypsoblennius gentilis Girard, 1854
Familia Labrisomidae
Exerpes asper (Jenkins & Evermann, 1889)
Paraclinus mexicanus (Gilbert, 1904)
Paraclinus sini Hubbs, 1952
Familia Gobiidae
Ctenogobius manglicola (Jordan & Starks, 1895)
Ctenogobius sagittula (Günther, 1862)
Quietula y-cauda (Jenkins & Evermann, 1889)
Familia Ephippidae
Chaetodipterus zonatus (Girard, 1858)
Familia Scombridae
Auxis thazard (Lacepède, 1800)
Orden Pleuronectiformes
Paralichthyidae
Cyclopsetta panamensis (Steindachner, 1876)

22
Etropus crossotus Jordan & Gilbert, 1882
Paralichthys woolmani Jordan & Williams, 1897
Achiridae
Achirus mazatlanus (Steindachner, 1869)
Cynoglossidae
Symphurus chabanaudi Mahadeva & Munroe, 1990
Symphurus leei Jordan & Bollman, 1890
Orden Tetraodontiformes
Familia Tetraodontidae
Sphoeroides annulatus Jenyns, 1842
Tabla II. Estructura taxonómica general, por localidad y época del año. Los valores
representan el número de entidades de cada nivel. Época templada (Tem), transición (Tra),
cálida (Cal) y total (T).
Balandra Zacatecas General
Tem Tra Cal T Tem Tra Cal T Tem Tra Cal T
Clase 1 2 2 2 2 1 1 2 2 2 2 2
Orden 6 8 8 10 5 5 6 7 8 8 9 10
Familia 8 13 15 17 9 12 14 17 12 16 18 21
Género 9 16 22 27 11 15 21 25 15 22 30 36
Especie 13 18 31 37 10 21 29 35 17 28 43 54
Abundancia
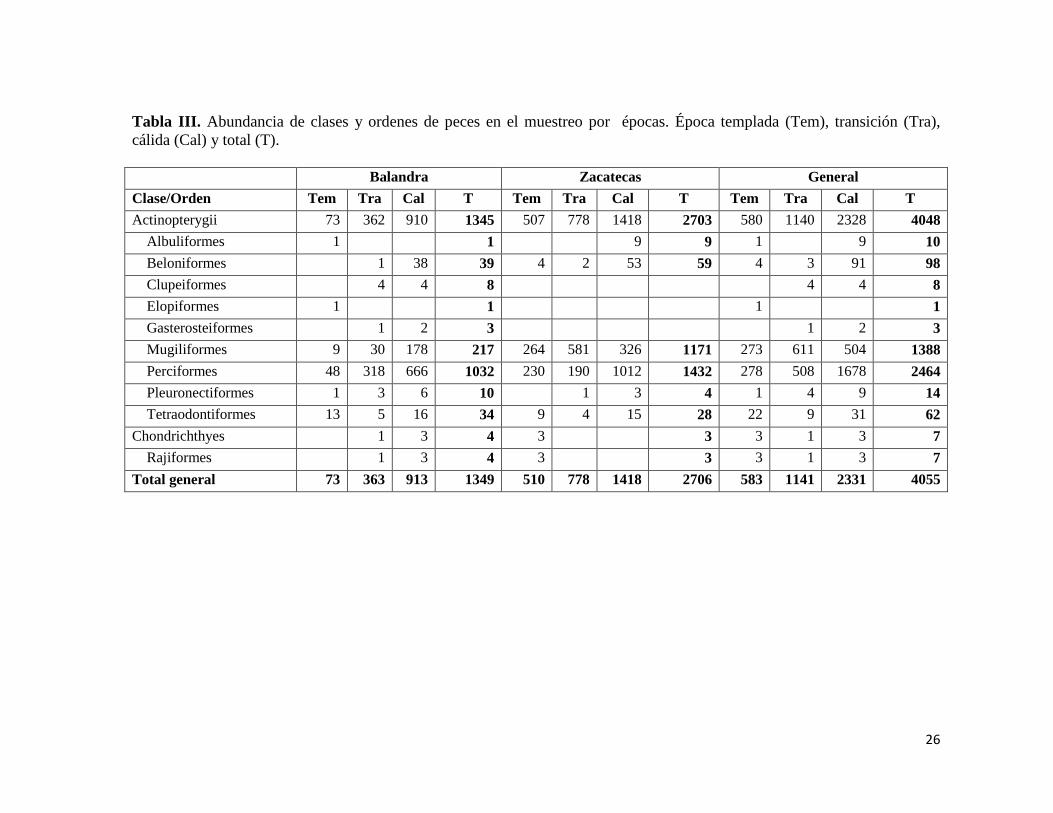
Se capturaron un total de 4055 especímenes; la clase Actinopterygii fue la más
abundante con 99.9% de la abundancia; a nivel de orden, Perciformes presentó la mayor
abundancia con 2464 organismos (Tabla III), por familia, los gerréidos presentaron mayor
abundancia con 1556 organismos, a nivel de especie, Mugil curema presentó mayor
abundancia con 1380 individuos mientras que en el otro extremo encontramos 11 especies
con solo un individuo (Tabla IV, Fig. 3). En todas las épocas la clase Actinopterygii fue la
más abundante, mientras que la abundancia por orden se ve modificada por la temperatura,

23
durante las temperaturas bajas en la época templada presenta un empate de abundancia con
los órdenes Perciformes y Mugiliformes, al incrementar la temperatura en la época de
transición el orden más abundante es Mugiliformes, y al presentarse los mayores valores de
temperatura en la época cálida el orden de mayor abundancia es Perciformes. En la
categoría de familia y especie, Mugilidae y Mugil curema presentaron los mayores valores
de dominancia para la época templada y de transición, mientras que para la época cálida fue
la familia Gerreidae y la especie Eucinostomus dowii (Tabla IV) (Fig. 3).
Figura 3. Abundancia relativa de familias por épocas en el muestreo. Época templada
(Tem), transición (Tra), cálida (Cal).
0 10 20 30 40 50 60
Albulidae
Blenniidae
Carangidae
Clupeidae
Diodontidae
Elopidae
Ephippidae
Fistulariidae
Gerreidae
Gobiidae
Haemulidae
Hemiramphidae
Labrisomidae
Lutjanidae
Mugilidae
Paralichthyidae
Sciaenidae
Scombridae
Serranidae
Tetraodontidae
Urolophidae
Número de organismos
Tem
Tra
Cal

24
En Balandra la clase Actinopterygii fue la más abundante, mientras que el orden
más abundante fue Perciformes con 1032, por su parte la familia Gerreidae presentó 756
organismos siendo así la categoría más abundante, mientras que Eucinostomus dowii fue la
especie con mayor número de individuos capturados. En todas las épocas la clase
Actinopterygii presentó los mayores valores de abundancia al igual que el orden
Perciformes; lo mismo ocurre a nivel de familia con Gerreidae y a nivel de especie, donde
Eucinostomus dowii presenta los mayores valores de abundancia con la excepción de la
temporada templada se empatan los valores de abundancia con Eucinostomus currani (Fig.
4).
Figura 4. Abundancia relativa de familias en las épocas climáticas en la localidad de
Balandra. Época templada (Tem), transición (Tra), cálida (Cal).
0 10 20 30 40 50 60 70
Albulidae
Carangidae
Diodontidae
Ephippidae
Gerreidae
Haemulidae
Labrisomidae
Mugilidae
Sciaenidae
Serranidae
Urolophidae
Número de organismos
Tem
Tra
Cal

25
En Zacatecas la clase Actinopterygii fue la más abundante con 2703 organismos. El
orden Perciformes fue el más abundante con 1432 individuos, mientras que Rajiformes
presentó solo tres organismos. La familia Mugilidae y la especie Mugil curema presentaron
la mayor abundancia con 1171 organismos. Con excepción de la temporada cálida, el
orden Perciformes es el más abundante, el resto del muestreo es dominado en abundancia
por el orden Mugiliforme y sus categorías taxonómicas inferiores hasta llegar a nivel de
especie (familia Mugilidae y especie Mugil curema).
Figura 5. Abundancia relativa de las épocas climáticas en la localidad de Zacatecas. Época
templada (Tem), transición (Tra), cálida (Cal).
0 10 20 30 40 50 60 70 80
Albulidae
Blenniidae
Carangidae
Diodontidae
Ephippidae
Gerreidae
Gobiidae
Haemulidae
Hemiramphidae
Lutjanidae
Mugilidae
Paralichthyidae
Sciaenidae
Scombridae
Serranidae
Tetraodontidae
Urolophidae
Número de organismos
Tem
Tra
Cal
26
Tabla III. Abundancia de clases y ordenes de peces en el muestreo por épocas. Época templada (Tem), transición (Tra),
cálida (Cal) y total (T).
Balandra Zacatecas General
Clase/Orden Tem Tra Cal T Tem Tra Cal T Tem Tra Cal T
Actinopterygii 73 362 910 1345 507 778 1418 2703 580 1140 2328 4048
Albuliformes 1 1 9 9 1 9 10
Beloniformes 1 38 39 4 2 53 59 4 3 91 98
Clupeiformes 4 4 8 4 4 8
Elopiformes 1 1 1 1
Gasterosteiformes 1 2 3 1 2 3
Mugiliformes 9 30 178 217 264 581 326 1171 273 611 504 1388
Perciformes 48 318 666 1032 230 190 1012 1432 278 508 1678 2464
Pleuronectiformes 1 3 6 10 1 3 4 1 4 9 14
Tetraodontiformes 13 5 16 34 9 4 15 28 22 9 31 62
Chondrichthyes 1 3 4 3 3 3 1 3 7
Rajiformes 1 3 4 3 3 3 1 3 7
Total general 73 363 913 1349 510 778 1418 2706 583 1141 2331 4055

27
Tabla IV. Abundancia de familias y especies de peces en el muestreo, Época templada (Tem), transición (Tra), cálida
(Cal) y total (T).
Balandra Zacatecas General
Tem Tra Cal T Tem Tra Cal T Tem Tra Cal T
Albulidae 1 1 9 9 1 9 10
Albula nemoptera 1 1 9 9 1 9 10
Blenniidae 1 1 1 1
Hypsoblennius gentilis 1 1 1 1
Carangidae 3 3 15 15 18 18
Caranx caninus 1 1 1 1
Caranx vinctus 1 1 1 1 2 2
Hemicaranx zelotes 1 1 1 1
Oligoplites altus 3 3 3 3
Oligoplites saurus 11 11 11 11
Clupeidae 4 4 8 4 4 8
Harengula thrissina 1 1 1 1
Opisthonema libertate 3 3 3 3
Sardinops sagax 4 4 4 4
Diodontidae 1 1 2 2 1 4 7 2 2 5 9
Diodon hystrix 1 1 2 2 1 4 7 2 2 5 9
Elopidae 1 1 1 1
Elops affinis 1 1 1 1
Ephippidae 14 14 14 14
Chaetodipterus zonatus 14 14 14 14
Fistulariidae 1 2 3 1 2 3
Fistularia commersonii 1 2 3 1 2 3

28
Gerreidae 28 227 510 765 166 110 515 791 194 337 1025 1556
Diapterus brevirostris 65 65 44 13 104 161 44 13 169 226
Eucinostomus currani 14 22 57 93 34 22 22 78 48 44 79 171
Eucinostomus dowii 14 150 321 485 62 54 258 374 76 204 579 859
Eucinostomus entomelas 42 64 106 26 16 119 161 26 58 183 267
Eucinostomus gracilis 13 1 14 8 8 13 9 22
Eugerres lineatus 5 3 8 5 3 8
Gerres cinereus 2 2 1 1 3 3
Gobiidae 13 38 1 52 5 1 7 13 18 39 8 65
Ctenogobius manglicola 1 1 1 1
Ctenogobius sagittula 7 16 1 24 5 1 6 12 12 17 7 36
Quietula y-cauda 6 22 28 6 22 28
Haemulidae 17 17 6 58 445 509 6 58 462 526
Haemulon sexfasciatum 1 1 1 1
Haemulon steindachneri 12 12 12 12
Haemulopsis leuciscus 7 1 8 7 1 8
Orthopristis reddingi 2 2 2 2
Pomadasys branickii 5 5 5 45 437 487 5 45 442 493
Pomadasys elongatus 1 1 1 1
Pomadasys macracanthus 6 4 10 6 4 10
Hemiramphidae 1 38 39 4 2 53 59 4 3 91 98
Hyporhamphus naos 1 38 39 4 2 53 59 4 3 91 98
Lisomidae 3 5 8 3 5 8
Exerpes asper 3 3 3 3
Paraclinus mexicanus 3 3 3 3
Paraclinus sini 2 2 2 2
Lutjanidae 7 60 67 1 3 4 8 63 71

29
Hoplopagrus guentherii 1 1 2 1 1 2
Lutjanus aratus 1 1 1 1
Lutjanus argentiventris 4 40 44 1 2 3 5 42 47
Lutjanus colorado 9 9 9 9
Lutjanus novemfasciatus 1 10 11 1 1 1 11 12
Mugilidae 9 30 178 217 264 581 326 1171 273 611 504 1388
Mugil cephalus 8 8 8 8
Mugil curema 9 30 178 217 264 581 318 1163 273 611 496 1380
Paralichthyidae 1 3 6 10 1 3 4 1 4 9 14
Citharichthys gilberti 1 1 1 1
Citharichthys platophrys 2 2 2 2
Cyclopsetta panamensis 2 2 2 2
Etropus crossotus 3 4 7 1 1 2 4 5 9
Sciaenidae 6 6 6 6
Cynoscion stolzmanni 4 4 4 4
Cynoscion xanthulus 2 2 2 2
Scombridae 2 2 2 2
Auxis thazard 2 2 2 2
Serranidae 7 43 70 120 53 17 7 77 60 60 77 197
Paralabrax maculatofasciatus 7 43 70 120 53 17 7 77 60 60 77 197
Tetraodontidae 13 4 15 32 7 3 11 21 20 7 26 53
Sphoeroides annulatus 13 4 13 30 7 3 11 21 20 7 24 51
Sphoeroides lobatus 2 2 2 2
Urolophidae 1 3 4 3 3 3 1 3 7
Urolophus halleri 3 3 3 3
Urolophus maculatus 1 3 4 1 3 4
Total general 73 363 913 1349 510 778 1418 2706 583 1141 2331 4055

30
Dominancia
De manera general en el muestreo encontramos a Mugil curema como la especie
dominante seguida por Eucinostomus dowii y Pomadasys branickii acumulando entre las
tres el 81.5% en la dominancia total (Fig. 6). En ambos esteros estudiados, encontramos de
igual manera a Mugil curema y Eucinostomus dowii en las primeras dos posiciones de
dominancia, con la excepción de que para Balandra, encontramos a Paralabrax
maculatofaciatus como la tercera especie dominante, mientras que en Zacatecas la tercera
es Pomadasys branickii.
Figura 6. Porcentaje de dominancia (I.V.I) de las especies en el muestreo
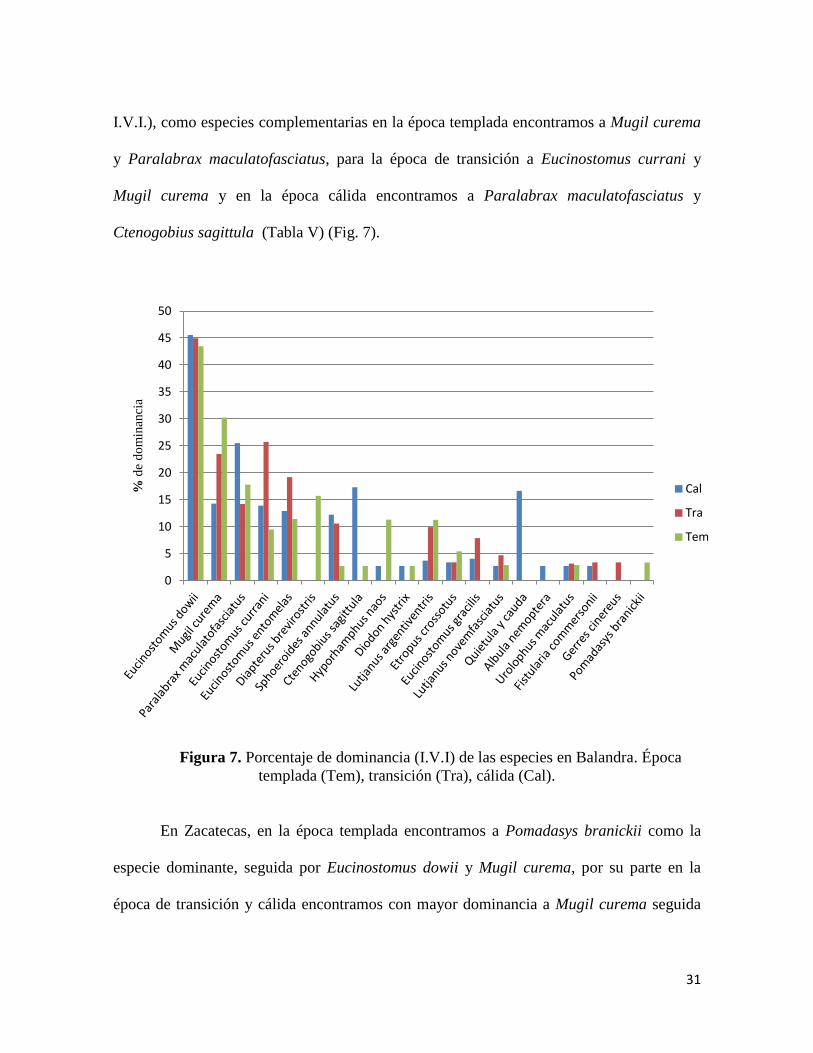
En Balandra Eucinostomus dowii se encuentra como la especie dominante en todas
las épocas climáticas y con el mismo valor aproximado de dominancia (43.5 - 45.6%
0
5
10
15
20
25
30
35
40
45
% d
e d
om
inan
cia
31
I.V.I.), como especies complementarias en la época templada encontramos a Mugil curema
y Paralabrax maculatofasciatus, para la época de transición a Eucinostomus currani y
Mugil curema y en la época cálida encontramos a Paralabrax maculatofasciatus y
Ctenogobius sagittula (Tabla V) (Fig. 7).
Figura 7. Porcentaje de dominancia (I.V.I) de las especies en Balandra. Época
templada (Tem), transición (Tra), cálida (Cal).
En Zacatecas, en la época templada encontramos a Pomadasys branickii como la
especie dominante, seguida por Eucinostomus dowii y Mugil curema, por su parte en la
época de transición y cálida encontramos con mayor dominancia a Mugil curema seguida
0
5
10
15
20
25
30
35
40
45
50
% d
e d
om
inan
cia
Cal
Tra
Tem

32
por Eucinostomus dowii, mientras que para la tercera especie dominante, en la época de
transición fue Diapterus brevirostris y en la época cálida fue Paralabrax
maculatofasciatus. A diferencia de Balandra, Zacatecas no presentó la especie de mayor
dominancia de manera continua, sino que la segunda especie fue Eucinostomus dowii.
(Tabla V) (Fig. 8).
Figura 8. Porcentaje de dominancia (I.V.I) de las especies en Zacatecas. Época
templada (Tem), transición (Tra), cálida (Cal).
0
10
20
30
40
50
60
70
80
90
% d
e d
om
inan
cia
Cal
Tra
Tem

33
Tabla V. Dominancia de especies en el muestreo I.V.I%. Época templada (Tem), transición (Tra), cálida (Cal) y total
(T).
Balandra Zacatecas General
Tem Tra Cal T Tem Tra Cal T Tem Tra Cal T
Mugil curema 30.2 23.5 14.3 19.9 24.8 48.5 85.1 53.1 52.7 57.1 23.6 42.3
Eucinostomus dowii 43.5 44.9 45.6 41.6 26.7 25 18.4 17.5 18.9 21.5 27.2 25.4
Pomadasys branickii 3.4 2 44.8 9.5 7.6 20.3 6.7 7.5 21.3 13.8
Eucinostomus entomelas 11.5 19.2 12.9 11.7 17.6 14.3 10.8 10.2 10.3 8.7 10.2 10.4
Paralabrax maculatofasciatus 17.8 14.2 25.5 14.4 5.7 10.1 16.7 7.9 16.2 8.8 5.6 10
Eucinostomus currani 9.5 25.8 14 11.9 3.2 15.6 11.7 7.9 14.1 7.4 5.7 9.2
Diapterus brevirostris 15.7 5.7 13.8 22 10.2 10.2 13.4 4.7 9.6 8.2
Sphoeroides annulatus 2.7 10.6 12.3 7.2 3.7 9.3 5.6 9.3 4.2 3.4 6.2
Hyporhamphus naos 11.3 2.7 5.8 9.6 6.8 6.1 6.7 6.6 3.8 6.2 6.1
Ctenogobius sagittula 2.7 17.4 7.3 5.7 2 6.2 5.3 7.9 5.1 2.6 5.9
Diodon hystrix 2.7 2.7 3.5 5.6 4.1 2.9 5.1 6.2 3.7 2.5 4.3
Lutjanus argentiventris 11.3 9.9 3.7 6.2 2.7 4.1 3.3 4 4.1 4.3
Etropus crossotus 5.5 3.4 3.4 3.9 2.7 2.9 3.3 3.9 2.5 3.5
Eucinostomus gracilis 7.9 4.1 4.7 3.2 1.9 4.7 2.7 3.1
Lutjanus novemfasciatus 2.9 4.7 2.7 4.1 2.7 1.6 3.7 2.8 2.7
Quietula y cauda 16.7 6.1 6.9 5.5 2.5
Pomadasys macracanthus 3.4 3.6 4.1 2.5 1.9
Haemulopsis leuciscus 2.7 3 3.6 4.2 2.4 1.9
Eugerres lineatus 2.9 2.7 3.6 4 2.5 1.9
Albula nemoptera 2.7 1.8 3 2.7 1.9 6.1 2.7 1.9
Urolophus maculatus 2.9 3.2 2.7 3.6 3.7 2.5 1.7
Fistularia commersonii 3.4 2.7 3.6 3.7 2.4 1.7
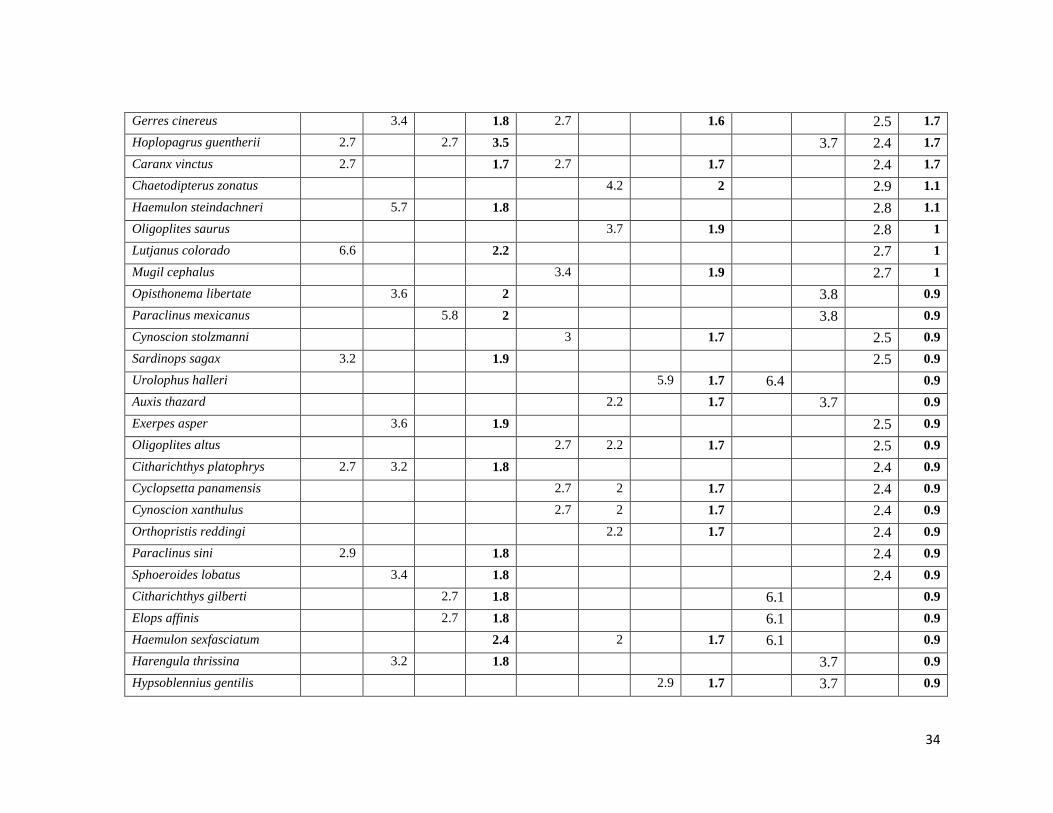
34
Gerres cinereus 3.4 1.8 2.7 1.6 2.5 1.7
Hoplopagrus guentherii 2.7 2.7 3.5 3.7 2.4 1.7
Caranx vinctus 2.7 1.7 2.7 1.7 2.4 1.7
Chaetodipterus zonatus 4.2 2 2.9 1.1
Haemulon steindachneri 5.7 1.8 2.8 1.1
Oligoplites saurus 3.7 1.9 2.8 1
Lutjanus colorado 6.6 2.2 2.7 1
Mugil cephalus 3.4 1.9 2.7 1
Opisthonema libertate 3.6 2 3.8 0.9
Paraclinus mexicanus 5.8 2 3.8 0.9
Cynoscion stolzmanni 3 1.7 2.5 0.9
Sardinops sagax 3.2 1.9 2.5 0.9
Urolophus halleri 5.9 1.7 6.4 0.9
Auxis thazard 2.2 1.7 3.7 0.9
Exerpes asper 3.6 1.9 2.5 0.9
Oligoplites altus 2.7 2.2 1.7 2.5 0.9
Citharichthys platophrys 2.7 3.2 1.8 2.4 0.9
Cyclopsetta panamensis 2.7 2 1.7 2.4 0.9
Cynoscion xanthulus 2.7 2 1.7 2.4 0.9
Orthopristis reddingi 2.2 1.7 2.4 0.9
Paraclinus sini 2.9 1.8 2.4 0.9
Sphoeroides lobatus 3.4 1.8 2.4 0.9
Citharichthys gilberti 2.7 1.8 6.1 0.9
Elops affinis 2.7 1.8 6.1 0.9
Haemulon sexfasciatum 2.4 2 1.7 6.1 0.9
Harengula thrissina 3.2 1.8 3.7 0.9
Hypsoblennius gentilis 2.9 1.7 3.7 0.9

35
Lutjanus aratus 3.2 1.8 3.7 0.9
Caranx caninus 2.7 1.7 2.4 0.8
Ctenogobius manglicola 2.7 1.6 2.4 0.8
Hemicaranx zelotes 2.7 1.7 2.4 0.8
Pomadasys elongatus 3 2 1.6 2.4 0.8

36
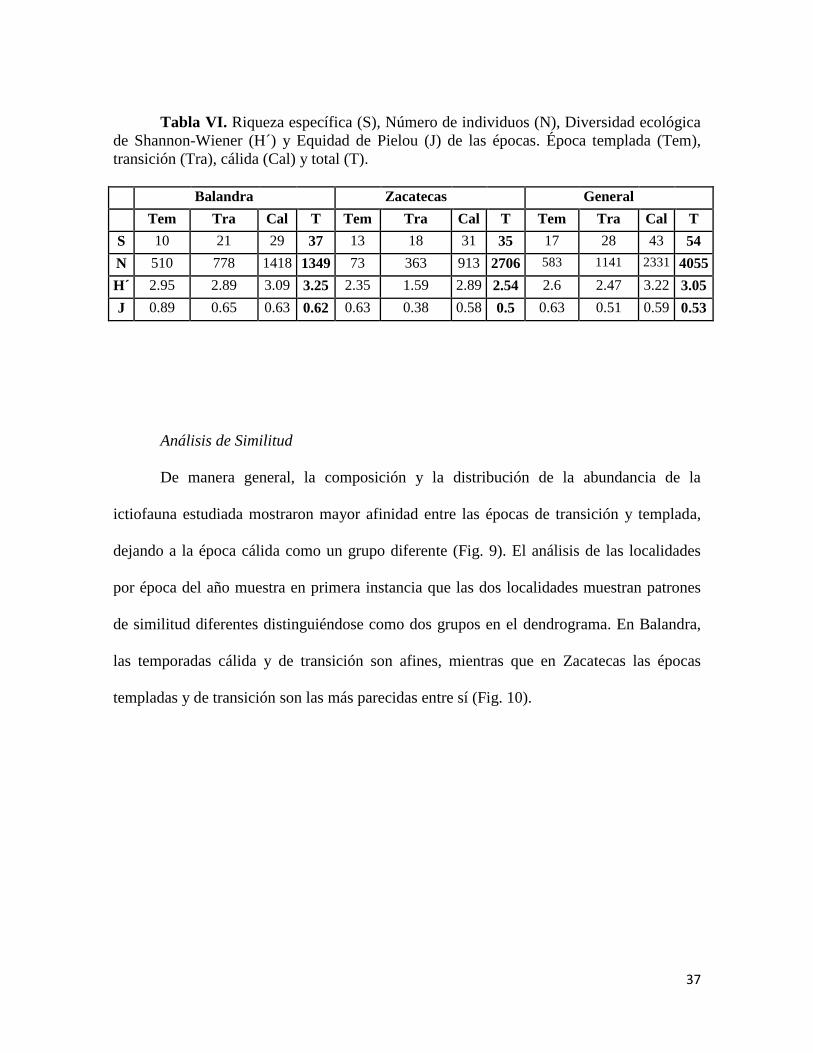
Diversidad
Encontramos una diversidad general de 3.05 bits/ind, la época cálida presentó el
mayor valor de diversidad con 3.22 bits/ind, mientras que el menor valor lo presentó la
época de transición con 2.47 bits/ind. En Balandra se presentó una diversidad de 3.25
bits/ind, por época encontramos el mayor valor en la temporada cálida (3.09 bits/ind) y la
menor en la época de transición (2.89 bits/ind). En Zacatecas el valor de diversidad fue 2.59
bits/ind; la época cálida presentó el mayor valor de diversidad (2.89 bits/ind) mientras que
en la época de transición fue el menor (1.59 bits/ind). Por épocas climáticas los valores más
altos los presenta Balandra, mientras que para las localidades los valores más altos se
presentan en las épocas cálidas (Tabla VI).
Equidad
La equidad del muestreo general fue 0.53, mientras que por épocas el mayor valor
lo presenta la temporada templada (0.63) y el menor la temporada cálida (0.51). Para
Balandra el valor en general fue de 0.62, siendo la época templada la temporada de mayor
equidad (0.89) y la temporada cálida como la menor (0.63). Zacatecas presentó una equidad
de 0.50, y por temporadas la época templada presentó el mayor valor (0.63) mientras que la
época de transición presentó el menor (0.38). Entre localidades Balandra presentó valores
más equitativos que en Zacatecas, mientras que por temporadas climáticas, los valores más
altos de equidad los presenta las épocas templadas (Tabla VI).
37
Tabla VI. Riqueza específica (S), Número de individuos (N), Diversidad ecológica
de Shannon-Wiener (H´) y Equidad de Pielou (J) de las épocas. Época templada (Tem),
transición (Tra), cálida (Cal) y total (T).
Balandra Zacatecas General
Tem Tra Cal T Tem Tra Cal T Tem Tra Cal T
S 10 21 29 37 13 18 31 35 17 28 43 54
N 510 778 1418 1349 73 363 913 2706 583 1141 2331 4055
H´ 2.95 2.89 3.09 3.25 2.35 1.59 2.89 2.54 2.6 2.47 3.22 3.05
J 0.89 0.65 0.63 0.62 0.63 0.38 0.58 0.5 0.63 0.51 0.59 0.53
Análisis de Similitud
De manera general, la composición y la distribución de la abundancia de la
ictiofauna estudiada mostraron mayor afinidad entre las épocas de transición y templada,
dejando a la época cálida como un grupo diferente (Fig. 9). El análisis de las localidades
por época del año muestra en primera instancia que las dos localidades muestran patrones
de similitud diferentes distinguiéndose como dos grupos en el dendrograma. En Balandra,
las temporadas cálida y de transición son afines, mientras que en Zacatecas las épocas
templadas y de transición son las más parecidas entre sí (Fig. 10).

38
Figura 9. Análisis de similitud general de Balandra-Zacatecas. Época templada (Tem),
transición (Tra), cálida (Cal).
Figura 10. Análisis de similitud para las épocas climáticas de Balandra y Zacatecas.
Localidades Balandra (Bal) y Zacatecas (Zac), época templada (Tem), transición (Tra),
cálida (Cal).

39
Discusión
Temperatura
Jiménez-Illescas et al. (1997) utilizando 20 años de registros de precipitación (sin
considerar los de la temperatura) definen tres temporadas climáticas en La Bahía de La Paz.
Época I: de Marzo a Junio; época II: de Julio a Octubre; y la época III: de Noviembre a
Febrero. Suárez-Altamirano (2005) realizó un estudio evaluando diferentes factores
oceanográficos (mareas vivas, mareas muertas y clorofilas) y fisicoquímicos (amonios,
silicatos, fosfatos, pH, nitratos y temperatura) en el manglar El Conchalito. En su trabajo
considera tres temporadas climáticas de las que define un mes representativo; la época
cálida en junio, transición en noviembre y templada en febrero; sin embargo, no definió
intervalos para cada época. En nuestro análisis, considerando valores de temperaturas
compiladas de diferentes fuentes (Anexo 2), se definieron tres temporadas climáticas: época
templada (diciembre-marzo), de transición (abril-junio) y cálida (julio-noviembre) según la
significancia estadística de la diferencia en la temperatura para cada temporada (Fig. 2). En
otras regiones de México se conocen las variaciones de abundancia de los organismos
basados en las épocas de lluvias, nortes y secas; otras veces se consideran las estaciones
convencionales de primavera, verano, otoño e invierno sin corroborar si es adecuado o no a
la región de estudio; en el estado de B.C.S. estos eventos son menos típicos por lo que se
debe de explorar otro tipo de criterios para estandarizar épocas que nos permitan explicar
de manera más eficiente el cambio en la distribución de la abundancia de las especies.
Siendo la temperatura un factor determinante en la distribución de los organismos así como
el factor climático preponderante en la región, este criterio resultó con una mayor bondad

40
en nuestro estudio. Sin bien, el trabajo de Suárez-Altamirano (2005) es más extenso en este
ámbito, no presenta un intervalo definido con respecto a los meses a considerar en cada una
de las épocas propuestas.
Composición específica
Las 54 especies encontradas en este trabajo representan alrededor del 10% de las
especies reportadas por Balart et al. (1995) y 18% de las familias reportadas por Abitia-
Cárdenas et al. (1994) para la Bahía de La Paz. De manera particular, para localidades de
fondos blandos Balart et al. (1997) reportan 103 especies en nueve localidades en toda la
Bahía de La Paz. En Zacatecas, con sustrato fangoso encontramos 35 especies que
representa un 33% de los peces conocidos de fondos blandos en la bahía; el número de
localidades en este trabajo es menor al considerado en el trabajo previamente mencionado,
por lo que pensamos que en un estudio con una mayor cantidad de sitios ó recolectas, el
porcentaje de especies representadas sin duda sería mayor. Considerando las especies
reportada previamente para Balandra por Maeda-Martínez (1981) (Tabla VII) y para
Zacatecas por Maeda-Martínez (1981) y Malpica-Maury (1999) (Tabla VIII); las listas
faunísticas alcanzan 62 especies para Balandra y 61 especies para Zacatecas, haciendo un
total de 92 para ambas localidades bajo estudio.
Para la Bahía de La Paz, la familia Serranidae es la más diversificada con 24
especies (Abitia-Cárdenas et al., 1994; Balart et al., 1995). Para Balandra y Zacatecas, las
familias Gerreidae y Haemulidae fueron las más diversas con siete especies

41
respectivamente. La diferencia en diversidad se puede deber a dos factores, la edad de los
organismos y la preferencia de hábitats. Los serránidos habitan zonas rocosas de la bahía
(Avilés-Quevedo, 2005), mientras que los hemúlidos y gerréidos se reportan con mayor
frecuencias para áreas costeras (Fisher et al., 1995). Por otro lado los serránidos reportados
son adultos en sus hábitats preferenciales (Abitia-Cárdenas et al., 1994) mientras que los
encontrados en este trabajo de acuerdo con su talla son considerados en su mayoría
juveniles que se encuentran estrechamente relacionados a zonas de manglar (Laegdsgaard y
Craig, 1995; 2001); resultados semejantes a lo obtenido en el presente trabajo lo obtuvieron
González–Acosta et al., (2005) en El Conchalito B.C.S. donde reportan las familias
Gerreidae y Haemulidae como las más diversas con seis especies; cabe aclarar que el
número de especies es semejante aunque su esfuerzo de captura fue mayor que el realizado
en nuestro trabajo. De manera semejante, Rodríguez-Romero et al. (2011) encuentran 10
especies de la familia Haemulidae en el área de manglar de Rancho Bueno, B.C.S. Tal
riqueza puede relacionarse con el número de especies de hemúlidos registradas para el
Pacífico (40 sp) (Tavera, 2012) que las registradas para el Golfo de California (25 sp)
(Thomson et al., 2000), por lo que el potencial de especies de esta familia para estos
ambientes de manglar en el Océano Pacífico es mayor que la encontrada en los manglares
de el Golfo de California.
Para Balandra se registraron 37 especies y 17 familias, mientras que para Zacatecas
se encontraron 35 especies y 17 familias, números cercanos a los de González-Acosta et al.
(2005) con 42 especies y 21 familias en el manglar de El Conchalito. Por su parte, Malpica-

42
Maury (1999) reporta 14 especies para Zacatecas y 19 especies para Chametla, mientras
que Maeda-Martínez (1981) habiendo estudiado tres manglares de la bahía, reporta 49
especies para Balandra y 28 para Zacatecas. Con esto podemos encontrar un intervalo muy
amplio en el número de especies encontradas para Balandra y Zacatecas, probablemente
relacionado con el esfuerzo de muestreo, la periodicidad, así como las artes de captura
utilizadas.
Tabla VII. Listado taxonómico de Balandra, la tabla comprende las especies del presente
trabajo [1]
más las reportados por Maeda-Martínez (1981) [2]
Phylum Chordata
Subphylum Vertebrata
Superclase Gnathostomata
Clase Chondrichthyes
Orden Rajiformes
Familia Rhinobatidae
Rhinobatos sp. [2]
Orden Myliobatiformes
Familia Urolophidae
Urolophus maculatus (Garman, 1913) [1, 2]
Clase Actinopterygii
División Teleostei
Orden Elopiformes
Familia Elopidae
Elops affinis Regan, 1909 [1, 2]
Orden Albuliformes
Familia Albulidae
Albula nemoptera (Fowler, 1911) [1]
Albula vulpes (Linnaeus, 1758) [2]
Orden Anguiliformes
Familia Ophistognathidae
Myrichthys xysturus (Jordan & Gilbert, 1882) [2]
Myrophis vafer Jordan & Gilbert, 1883 [2]
Orden Clupeiformes
Familia Clupeidae
Harengula thrissina (Jordan & Gilbert, 1882) [1, 2]

43
Opisthonema libertate (Günther, 1867) [1, 2]
Sardinops sagax (Jenyns, 1842) [1]
Familia Engraulidae
Anchoa ischana (Jordan & Gilbert, 1882) [2]
Orden Mugiliformes
Familia Mugilidae
Mugil cephalus Linnaeus, 1758 [2]
Mugil curema Valenciennes, 1836 [1, 2]
Orden Beloniformes
Familia Hemiramphidae
Hyporhamphus naos Banford & Collette, 2001 [1]
Hyporhamphus unifasciatus (Ranzani, 1841) [2]
Orden Gasterosteiformes
Familia Fistulariidae
Fistularia commersonii Rüppell, 1838 [1]
Fistularia petimba Lacepède, 1803 [2]
Familia Syngnathidae
Syngnathus sp. [2]
Orden Perciformes
Familia Serranidae
Paralabrax maculatofasciatus (Steindachner, 1868) [1, 2]
Familia Carangidae
Caranx caninus Günther, 1867 [1]
Caranx vinctus Jordan & Gilbert, 1882 [1]
Hemicaranx zelotes Gilbert, 1898 [1]
Oligoplites saurus (Bloch & Schneider, 1801) [2]
Familia Lutjanidae
Hoplopagrus guentherii Gill 1862 [1, 2]
Lutjanus aratus Günther, 1864 [1, 2]
Lutjanus argentiventris Peters, 1869 [1, 2]
Lutjanus colorado Jordan & Gilbert, 1882 [1]
Lutjanus novemfasciatus Gill, 1862 [1, 2]
Familia Gerreidae
Eucinostomus currani Zahuranec en Yañez-Aranciba, 1980 [1,
2]
Eucinostomus dowii (Gill, 1863) [1, 2]
Eucinostomus entomelas Zahuranec en Yañez-Aranciba, 1980 [1]
Eucinostomus gracilis (Gill, 1862) [1]
Diapterus brevirostris (Sauvage, 1879) [1, 2]
Gerres cinereus (Walbaum, 1792) [1, 2]
Familia Haemulidae
Conodon serrifer Jordan y Gilbert, 1882 [2]
Haemulon flaviguttatum Gill, 1862 [2]

44
Haemulon maculicauda (Gill, 1862) [2]
Haemulon steindachneri (Jordan & Gilbert, 1882) [1]
Microlepidotus inornatus Gill, 1862 [2]
Orthopristis chalceus (Günther, 1864) [2]
Orthopristis reddingi Jordan & Richardson, 1895 [2]
Pomadasys branickii (Steindachner, 1879) [1, 2]
Familia Sciaenidae
Bairdiella icistia (Jordan & Gilbert, 1882) [2]
Cynoscion xanthulus Jordan & Gilbert, 1882 [2]
Familia Mullidae
Psedupeneus grandisquamis (Gill, 1863) [2]
Familia Kyphosidae
Kyphosus analogus (Gill, 1862) [2]
Familia Scaridae
Scarus sp. [2]
Familia Labrisomidae
Exerpes asper (Jenkins & Evermann, 1889) [1, 2]
Paraclinus mexicanus (Gilbert, 1904) [1]
Paraclinus sini Hubbs, 1952 [1]
Familia Gobiidae
Ctenogobius sagittula (Günther, 1862) [1, 2]
Eucyclogobius newberryi (Girard, 1856) [2]
Gobiosoma chiquita (Jenkins y Evermman, 1889) [2]
Quietula y-cauda (Jenkins & Evermann, 1889) [1, 2]
Orden Pleuronectiformes
Familia Paralichthyidae
Citharichthys gilberti Jenkins & Evermann, 1889 [1]
Citharichthys platophrys Gilbert, 1891 [1]
Etropus crossotus Jordan & Gilbert, 1882 [1, 2]
Paralichthys woolmani Jordan & Williams, 1897 [2]
Orden Tetraodontiformes
Familia Diodontidae
Diodon holocanthus (Linnaeus, 1758) [2]
Diodon hystrix Linnaeus, 1758 [1]
Familia Tetraodontidae
Sphoeroides annulatus (Jenyns, 1842) [1, 2]
Sphoeroides lobatus (Steindachner, 1870) [1, 2]

45
Tabla VIII. Listado taxonómico de Zacatecas, la tabla comprende las especies del presente
trabajo [1]
más las reportados por Maeda-Martínez (1981) [2]
y por Malpica-Maury (2000) [3]
.
Phylum Chordata
Subphylum Vertebrata
Superclase Gnathostomata
Clase Chondrichthyes
Orden Rajiformes
Familia Rhinobatidae
Rhinobatos glaucostigma Jordan & Gilbert, 1883 [3]
Orden Myliobatiformes
Familia Urolophidae
Urolophus halleri Cooper, 1863 [1, 3]
Urolophus maculatus (Garman, 1913) [3]
Familia Gymnuridae
Gymnura marmorata (Cooper, 1894) [3]
Clase Actinopterygii
División Teleostei
Orden Elopiformes
Familia Elopidae
Elops affinis Regan, 1909 [2]
Orden Albuliformes
Familia Albulidae
Albula nemoptera (Fowler, 1911) [1]
Albula vulpes (Linnaeus, 1758) [2]
Orden Clupeiformes
Familia Engraulidae
Anchoa ischana (Jordan & Gilbert, 1882) [2]
Orden Aulopiformes
Familia Synodontidae
Synodus scituliceps Jordan & Gilbert 1882 [3]
Orden Mugiliformes
Familia Mugilidae
Mugil cephalus Linnaeus, 1758 [1, 2]
Mugil curema Valenciennes, 1836 [1, 2]
Orden Beloniformes
Familia Hemiramphidae
Hyporhamphus naos Banford & Collette, 2001 [1]
Hyporhamphus unifasciatus (Ranzani, 1841) [2]
Orden Gasterosteiformes
Familia Syngnathidae
Hippocampus ingens Girard, 1858 [3]

46
Orden Scorpaeniformes
Familia Scorpaenidae
Sebastes cortezi (Beebe & Tee Van, 1938) [3]
Familia Triglidae
Prionotus horrens Richardson, 1844 [3]
Orden Perciformes
Familia Serranidae
Diplectrum pacificum Meek & Hildebrand, 1925 [3]
Paralabrax maculatofasciatus (Steindachner, 1868) [1, 2, 3]
Familia Carangidae
Caranx vinctus Jordan & Gilbert, 1882 [1]
Caranx sexfasciatus (Quoy & Gaimard, 1825) [2]
Oligoplites altus (Günther, 1868) [1]
Oligoplites saurus (Bloch & Schneider, 1801) [1, 2]
Familia Lutjanidae
Lutjanus aratus Günther, 1864 [2]
Lutjanus argentiventris Peters, 1869 [1, 2]
Lutjanus novemfasciatus Gill, 1862 [1, 2]
Familia Gerreidae
Diapterus brevirostris (Sauvage, 1879) [1, 2, 3]
Eucinostomus currani Zahuranec en Yañez-Aranciba, 1980 [1,
2]
Eucinostomus dowii (Gill, 1863) [2]
Eucinostomus entomelas Zahuranec en Yañez-Aranciba, 1980 [1]
Eucinostomus gracilis (Gill, 1862) [1, 3]
Eugerres axilaris (Günter, 1864) [2]
Eugerres lineatus (Humboldt, 1821) [1]
Gerres cinereus (Walbaum, 1792) [1, 2]
Familia Haemulidae
Conodon serrifer Jordan y Gilbert, 1882 [2]
Haemulon sexfasciatum Gill, 1862 [1]
Haemulopsis leuciscus (Günther, 1864) [1]
Orthopristis chalceus (Günther, 1864) [2, 3]
Orthopristis reddingi Jordan & Richardson, 1895 [1, 2]
Pomadasys branickii (Steindachner, 1879) [1, 2]
Pomadasys elongatus (Steindachner, 1879) [1]
Pomadasys macracanthus (Günther, 1864) [1]
Pomadasys panamensis (Steindachner, 1879) [2, 3]
Familia Sparidae
Calamus brachysomus (Lockington, 1980) [3]
Familia Sciaenidae
Cynoscion stolzmanni Steindachner, 1879 [1]
Cynoscion xanthulus Jordan & Gilbert, 1882 [1, 2]

47
Familia Blenniidae
Hypsoblennius gentilis Girard, 1854 [1]
Familia Labrisomidae
Exerpes asper (Jenkins & Evermann, 1889) [2]
Familia Gobiidae
Ctenogobius manglicola (Jordan & Starks, 1895) [1]
Ctenogobius sagittula (Günther, 1862) [1, 2]
Quietula y-cauda (Jenkins & Evermann, 1889) [2]
Familia Ephippidae
Chaetodipterus zonatus (Girard, 1858) [1, 3]
Familia Scombridae
Auxis thazard (Lacepède, 1800) [1]
Orden Pleuronectiformes
Paralichthyidae
Cyclopsetta panamensis (Steindachner, 1876) [1]
Etropus crossotus Jordan & Gilbert, 1882 [1, 2, 3]
Paralichthys woolmani Jordan & Williams, 1897 [3]
Achiridae
Achirus mazatlanus (Steindachner, 1869) [3]
Cynoglossidae
Symphurus chabanaudi Mahadeva & Munroe, 1990 [3]
Symphurus leei Jordan & Bollman, 1890 [3]
Orden Tetraodontiformes
Familia Balistidae
Balistes polylepis Steindachner, 1876 [3]
Familia Diodontidae
Diodon hystrix Linnaeus, 1758 [1]
Familia Tetraodontidae
Sphoeroides annulatus Jenyns, 1842 [1, 2]
Abundancia
La mayoría de los estudios en manglares en la región tropical del país reportan un
mayor número de individuos capturados respecto a este trabajo; 6,474 en Río Lagartos,
Yucatán (Arceo-Carranza y Vega-Cedeja, 2008), 7,386 en Laguna Chacahua, Oaxaca
(Mendoza et al., 2010) y 9,108 en Pueblo Viejo, Veracruz (Castillo-Rivera y Zarate-
Hernández, 2001), mientras que para la parte subtropical, los datos de este trabajo son
mayores a los reportados: 1,168 en Laguna Teacapan-Agua Brava, Nayarit (Flores-

48
Verdugo et al., 1990), 1,224 en la Ensenada de La Paz (Malpica-Maury, 1999) con la
excepción para González-Acosta et al., (2005) donde capturaron 35,138 ejemplares en El
Conchalito B.C.S. con un esfuerzo mayor, con recolectas durante ciclos de 24h cada mes
en 13 meses.
La abundancia total de peces reportados en otros estudios alrededor del mundo son
muy variables: para América: 1,591 (Feutry et al., 2010), 3,296 (Sedberry y Carter, 1993);
Australia: 23,630 en dos años de estudio [13,516 en el primer año y 10,114 en el segundo
año] (Laegdsgaard y Johnson, 1995), 3,320 (Hallidy y Young, 1996); Asia: 85,427 (Nip y
Chong, 2010); Africa: 1,316 (Okyere et al., 2011). Con respecto a Asia y Australia nuestros
números son menores, mientras que para los demás continentes existe una mayor
semejanza a los números de capturas realizados en un periodo semejante de tiempo.
Las especies con mayor abundancia en diferentes sitios de fondos blandos en la
Bahía de La Paz también varía ampliamente; Balart et al., (1997) reportan a Diapterus
aureolus, Eucinostomus currani y Mugil cephalus como las más abundantes; Malpica-
Maury (1999) para El Comitán a Achirus mazatlanus, para Chametla a Paralabrax
maculatofasciatus y para Zacatecas a Diapterus brevirostris, Paralabrax
maculatofasciatus, Orthopristis chalceus y Achirus mazatlanus; González-Acosta et al.
(2005) encuentran en el estero El Conchalito a Eucinostomus currani, Diapterus
brevirostris y Albula vulpes; Mendonza et al. (2010) en Oaxaca encuentran a Diapterus
brevirostris, Lutjanus novemfascitus, Lile stolifera y Harengula thrissina; Vega-Cedeja y

49
Hernandez de Santillana (2004) encuentran en Río Lagartos, Yucatán a Floridichthys
polyommus, Eucinostomus argenteus y Sphoeroides testudineus. En este trabajo las
especies más abundantes fueron: Mugil curema, Eucinostomus dowii, Eucinostomus
entomelas, Diapterus brevirostris y Pomadasys branickii. En todos los casos se demuestra
que en este tipo de sistemas, las especies más abundantes son principalmente de las familias
Gerreidae, Serranidae, Haemulidae y Tetraodontidae.
Dominancia
Cuatro especies (Eucinostomus dowii, Mugil curema, Paralabrax maculatofasciatus
y Pomadasys branickii) resaltaron con los valores obtenidos así como se presentaron en
todas las épocas climáticas; en Zacatecas, Mugil curema, Eucinostomus dowii y Pomadasys
branickii fueron las especies con mayores valores de dominancia, mientras que en Balandra
fueron Eucinostomus dowii, Mugil curema y Paralabrax maculatofasciatus. Desde un
punto de vista trófico, en ambos sitios encontramos una especie omnívora, una iliófaga y un
depredador, sugiriendo que la estructura comunitaria de los peces asociados a los
manglares está bien definida. Esto probablemente se deba a las características morfológicas
y funcionales que presentan las mojarras y las lisas. Por un lado, parte de los gerréidos son
poco accesibles a los depredadores por sus grandes espinas en las aletas dorsal y anal
(Fisher et al., 1995), mientras que los mugílidos poseen gran velocidad de natación y
pueden saltar fuera del agua (González-Sansón et al., 1978). En lo que respecta a
Pomadasys y Paralabrax sus diferencias biológicas se reflejan en el tipo de hábitat
preferencial, ya que los miembros de la familia Serranidae prefieren sustratos arenosos con

50
hábitats rocosos (Aviles-Quevedo, 2005) como los observados en Balandra (Torres-Alfaro,
2010) a diferencia de los Pomadasys spp., que prefieren sustratos blandos y lodosos
(Fisher et al., 1995), como los observados en Zacatecas.
De manera particular, la variación en la dominancia entre Mugil curema y
Eucinostomus dowii, seguramente depende del éxito en la apropiación de recursos
alimenticios siendo un factor determinante para la presencia y abundancia de las especies
ícticas (Bone y Moore, 2008; Helfmann et al. 2009). Trabajos sobre la ecología trófica de la
familia Mugilidae han demostrado que sus hábitos alimenticios son iliófagos (Osorio-
Dualiby, 1988), lo que explica los valores obtenidos en el área de manglar fangoso, y de
alguna manera justifica su posición en la tabla de valores de Balandra (Tabla VI).
Diversidad
La diversidad ecológica obtenida en este trabajo se encuentra por encima de lo
reportado para muchas zonas del territorio nacional: 3.11 bits/ind en El Conchalito, B.C.S.
(González-Acosta et al., 2005), 3.37 bits/ind para la ensenada de La Paz. (Balart et al.,
1997), 3.6 bits/ind en Bahía Magdalena (Rodríguez-Romero et al., 2011), 0.9 bits/ind en
Oaxaca (Mendoza et al., 2010), 1.58 bits/ind en Veracruz (Castillo-Rivera et al., 2001),
2.16 bits/ind en Nayarit (Benítez-Valles et al., 2007), 2.9 bits/ind en Yucatán (Arceo-
Carrera y Vega-Cedejas, 2009).

51
La ictiofauna asociada a manglares con fondos arenosos en Estados Unidos reporta
2.46 bits/ind (Philiphs, 1981) mientras que para zonas similares en manglares de África,
Okyere et al., (2011) arrojan 2.7 bits/ind, mientras que en Balandra, los valores son
mayores independientemente de la época del año (3.09, 2.95 y 2.89 bits/ind para las épocas
cálida, templada y transición, respectivamente).
Para trabajos en manglares con fondo fangoso encontramos a Lakra et al., (2010)
con valores de 3.51 bits/ind en India, Jalal et al., (2012) con 2.09 bits/ind en Malasia y
Shih-Rong et al., (2001) con 4.1 bits/ind, en Taiwán; en Zacatecas los valores para este
tipo de sustrato fueron menores (2.89, 2.35 y 1.59 bits/ind para las épocas cálida, templada
y transición, respectivamente). Si bien los manglares con aporte de agua dulce tienden a
presentar una mayor diversidad que los manglares sin este aporte (Barletta et al., 2003),
cabe resaltar que las localidades muestreadas presentan un patrón contrario, ya que
muestran valores superiores de diversidad a los que normalmente se reportarían en otro
caso.
Equidad
La equidad general fue mayor que la observada para Zacatecas, mientras que fue
menor que la obtenida para Balandra, lo que sugiere que cuando las dos matrices se
unieron, los datos de Balandra homogeneizaron a los de Zacatecas y por ende,
incrementaron el valor de la prueba de manera general.

52
Por lo mismo, Balandra presenta mayores valores de diversidad, puesto que la
distribución de abundancia entre sus especies es más equitativa, esto ocurre porque los
manglares costeros tienen una mayor dinámica y flujo de organismos, por lo que presentan
un alto número de especies temporales lo que homogeniza el número de organismos/
especies presentes en el hábitat, esta homogenización reduce la dominancia de algunas
especies (Laegdsgaard y Craig, 2001; Unsworth et al., 2009).
Análisis de Similitud
En el dendrograma no se muestran patrones de similitud entre localidades pero si
entre épocas climáticas (Fig. 9, 10), esto sugiere que cada sitio presenta una composición y
una abundancia propia. Según Toepfer et al., (1998) unas de las razones por la cual estos
grupos faunísticos podrían estar bien definidos, serían las propiedades del sustrato y los
patrones de corrientes, sin embargo los patrones de corrientes de la Bahía de La Paz son
similares a los de la ensenada (Gómez-Valdés et al., 2003; Guevara-Guillen, 2009) lo que
sugiere que el sustrato es realmente el que define la composición y abundancia encontrada
(Mirera et al., 2010), limoso/fangoso en Zacatecas y arenoso/rocoso en Balandra (Maeda-
Martínez, 1981), así como es la temperatura la que define la composición faunística entre
las épocas climáticas (Bone & Moore, 2008).
Conclusiones
Las familias de peces características de los manglares estudiados son:
Gerreidae, Mugilidae, Haemuliade, Serranidae, Tetraodontidae y Gobiidae. Las

53
especie típicas de este tipo de hábitat resultaron ser Eucinostomus dowii, Mugil
curema, Pomadasys branickii, Paralabrax maculatofasciatus, Eucinostomus
currani, Sphoeroides annulatus y Ctenogobius sagittula.
La composición de la comunidad íctica en los dos manglares estudiados,
debe de considerarse diferente como consecuencia de procesos de reclutamiento
derivados de la diferencia entre los hábitats.
El estero Balandra es más diverso y presenta una distribución de la
abundancia entre las especies más homogénea que las observadas en el estero
Zacatecas.
Para ambas localidades, la época cálida presenta la mayor diversidad de
especies, mientras que la época templada presenta la mayor homogeneidad en la
abundancia de las especies presentes.
En ambas localidades, la comunidad de peces se encuentra dominada por
una especie depredadora (Paralabrax maculatofasciatus en Balandra y Pomadasys
branickiin en Zacatecas), así como por una iliófaga (Mugil curema) y una omnívora
(Eucinostomus dowii) comunes.
Recomendaciones
La mezcla frigorífera resultó ser una herramienta adecuada para el manejo y
conservación en el campo de los especímenes capturados, permitiendo realizar el muestreo

54
durante una jornada completa y el regreso al laboratorio en excelentes condiciones para el
almacenamiento temporal o definitivo de las muestras (en nuestro caso, en congelador hasta
su procesamiento). De esta manera se evita el manejo y los riegos de emplear en el campo
sustancias peligrosas como el formol. Por lo anterior, se considera ampliamente
recomendable considerar este procedimiento para conservar muestras de material biológico
en trabajos de campo de corto plazo.
Las épocas climáticas establecidas para el análisis de nuestros datos (templada,
transición y cálida) permiten superar la convención climática de dividir el año en cuatro
estaciones (primavera, verano, otoño e invierno), representando de mejor manera la marcha
de la temperatura en la Bahía de La Paz y las diferencias significativas entre las épocas.
Con base en ello, para la bahía se recomienda hacer el análisis de patrones de variación
temporal de procesos biológicos con respecto a este factor, considerando las tres épocas
climáticas de este trabajo.
La comprensión de la estructura comunitaria, así como de los factores involucrados
en la determinación de los patrones observados dejan abiertos una multitud de preguntas
relacionadas con el reclutamiento de las poblaciones involucradas, la competencia por
recursos alimenticios, las adaptaciones de las especies a las peculiaridades del hábitat y
demás. De acuerdo con ello, se recomienda ampliar los estudios sobre las comunidades
ícticas en hábitats de manglar y considerar en las investigaciones aspectos sobre la biología
y reproducción, los hábitos alimenticios así como los atributos morfo-funcionales que

55
permitan una mejor explicación de la distribución, abundancia y diversidad de estas
comunidades.
Bibliografía
Abitia-Cárdenas L. A, Rodríguez-Romero J., Galván-Magaña F., De La Cruz-Agüero J. &
H. Chávez-Ramos. 1994. Lista sistemática de la ictiofauna de Bahía de La Paz,
Baja California Sur, México. Ciencias Marinas. 20(2): 159-181.
Arceo-Carranza D. & M. E. Vega-Cedejas. 2009. Spatial and temporal characterization
of fish assemblages in a tropical costal system influenced by freshwater inputs,
northeastern Yucatan península. Revista de Biología Tropical. 57(1-2): 89-103
Avilés-Quevedo M. A. 2005. Calidad de huevos y larvas según el manejo de los
reproductores de la cilla (Paralabrax maculatofasciatus, Pisces:Serranidae). Tesis
de doctorado. Universidad de Barcelona. 199p.
Balart E. F., Castro-Aguirre J. L., Aurioles-Gamboa D., García-Rodríguez F. & C.
Villavicencio-Garayzar. 1995. Adiciones a la ictiofauna de Bahía de La Paz, Baja
California Sur, México. Hidrobiológica. 5(1-2): 79-85.
Balart E. F., Castro-Aguirre J. L. & F. de Lachica-Bonilla. 1997. Análisis comparativo de
las comunidades ícticas de fondos blandos y someros de la Bahía de La Paz, p.
20-35 en Urbán-Ramírez J. & M. Ramírez-Rodríguez (Eds). La Bahía de La Paz
Investigación y Conservación. Universidad Autónoma de Baja California Sur.
México.

56
Barletta M., Barletta-Bergan A., Saint-Paul U., & G. Hubold. 2003. Seasonal changes in
density, biomass, and diversity of estuarine fishes in tidal mangrove creeks of the
lower Caeté Estuary (northern Brazilian coast, east Amazon). Marine Ecology
Progress Series. 256: 217-228
Beck M., Heck K. Jr., Kenneth A., Childers D., Eggleston D., Gillanders B., Halpern B.,
Hays C., Hoshino K., Minello T., Orth T., Sheridan P. & M. P. Weinstein. 2001. The
identification, conservation, and management of estuarine and marine nurseries
for fish and invertebrates. BioScience. 51(8): 633-641
Begon M., Colin R., & J. Harper. 2006. Ecology. From individual to ecosystems. 4th
Ed.
Blackwell. E.U.A. 736p.
Benítez-Valles C., Ruiz-Velazco-Arce J. M. J., Peña-Messina E., Blanco y Correa M.,
López-Rivas C., López-Lugo P. & A. Castañeda Martínez. 2007. Diversidad y
abundancia de la comunidad de peces del estero “El Custodio” Municipio de
Compostela, Nayarit, México. Revista Electrónica de Veterinaria. 8(5): 1-13
Blaber S. J. M. & G. Blaber T. 1980. Factors affecting the distribution of juvenile
estuarine and inshore fish. Journal of Fish Biology. 17: 143-162
Blaber S. J. M. 2000. Tropical Estuarine Fishes: Ecology, exploitation and
conservation. Blackwell Science. Australia. 357p
Bone Q., & R. H. Moore. 2008. Biology of fishes. 3rd
Ed. Blackie Academic and
Professional. E.U.A. 332p.
Brower J. E., Zar J. H. & C. N. von Ende. 1998. Field and laboratory methods for
general Ecology. 4th
Ed. McGraw-Hill. E.U.A. 273p.

57
Carbajal-Fajardo Z. S., Franco-López J., Barrera-Escorcia H., Abarca-Arenas L. G., Bedia-
Sánchez C., Moran-Silva A.. & H. Vázquez-López. 2009. Trophic seasonal
behavior of the icthyofauna of Camaronera Lagoon, Veracruz. Journal of
Fisheries and Aquatic Science 4(2): 75-89
Carter R. W. G. 1988. Coastal Environments, An introduction to the physical,
ecological and cultural systems of costalines. Academic Press. San Diego,
California. 617p.
Castillo-Rivera M. & R. Zarate-Hernández. 2001. Patrones espacio-temporales de la
abundancia de los peces en la Laguna de Pueblo Viejo, Veracruz. Hidrobiológica.
11(1): 75-84
Chávez, H., 1985. Bibliografía sobre peces de la Bahía de La Paz, Baja California Sur,
México. Investigaciones Marinas CICIMAR. 2(No. Esp. 2):1-75.
De la Cruz-Agüero G., 1994. ANACOM: Sistema para el Análisis de Comunidades.
Versión 3.0. Manual del Usuario. CINVESTAV-IPN, Unidad Mérida, Yucatán,
México, 99
De La Cruz Agüero J. 1994. Peces marinos del noroeste de México, identificación de
familias. Centro Interdisciplinario de Ciencias Marinas-Instituto Politécnico
Nacional. Baja California Sur. Manuscrito sin publicar. 100p.
De La Cruz-Agüero J., Arellano-Martínez M., Cota-Gómez V., & G. De la Cruz-Agüero.
1997. Catálogo de los peces marinos de Baja California Sur. CONABIO. México.
340p.

58
Ellison A. 2008. Managing mangroves with benthic biodiversity in mind: Moving
beyond roving banditry. Journal of Sea Research. 29: 2-15.
Faunce C. H. & J. E. Serafy 2006. Mangroves as fish habitat: 50 years of field studies.
Marine Ecology Progress Series. 318: 1–18
Feutry P., Hartmann H. J., Casaconnet H. & G. Umaña. 2010. Preliminary analysis of the
fish species of the Pacific Central American mangrove of Zancudo, Golfo Dulce,
Costa Rica. Westlands Ecology Management. 18: 637-650.
Fischer W., Krupp F., Schneider W., Sommer C., Carpenter K.E. & V. H. Niem.
1995. Guía FAO para identificación de especies para los fines de la pesca.
Pacífico centro-oriental. Volumenes II y III. Vertebrados. Roma. 647-1813
Flores Z. E. 1998. Geosudcalifornia. Geografía, agua y ciclones. Universidad Autónoma
de Baja California Sur. México 277p.
Flores-Verdugo F., González-Farías F., Amenzcua-Linares F., Yañez-Ararcibia A.,
Alvarez-Rubio M., Ramírez-Flores O. M. & J. W. Day. 1990. Mangrove ecology,
aquatic primary production and fish community dynamic in the Teacan-Agua
Brava lagoon estuary system (Mexican Pacific). Estuaries. 13:213-230
Galván-Piña V. H. 1998. Estructura de la comunidad de peces capturadas con redes
agallera y charalera en Bahía de La Paz. B.C.S. Tesis de Maestría. Centro
Interdisciplinario de Ciencias Marinas - Instituto Politécnico Nacional. La Paz,
B.C.S. México. 117p.

59
Gómez-Valdés J., A. Delgado J., & J. A. Dwaorak. 2003. Overtides, compound tides, and
tidal-residual current in Ensenada de La Paz lagoon, Baja California Sur,
México. Geofísica Internacional 42(4): 623-634
González-Acosta A. F., De la Cruz-Agüero G., De La Cruz-Agüero J. & G. Ruiz-Campos.
2005. Seasonal pattern of the fish assemblage of El Conchalito, mangrove
swamp, La Paz Bay, Baja California Sur, México. Hidrobiológica. 15(2): 204-214
González-Sansón G., Álvarez-Lajonchere L. S. & M. Baez. 1978. Lista preliminar de
peces presentes en las lagunas costeras de Tunas de Zaza, Cuba. Revista de
Investigación Marina. 32: 1-15
González-Zamorano P. 2002. Estructura y análisis espacial de la cobertura del manglar
“El Conchalito”, B.C.S., México. Tesis de Maestría. Centro Interdisciplinario de
Ciencias Marinas Centro Interdisciplinario de Ciencias Marinas - Instituto
Politécnico Nacional. 88p
Guevara-Guillén C. 2009. Hidrografía en la Bahía de La Paz y zona oceánica
adyacente. Tesis de Maestría. Centro Interdisciplinario de Ciencias Marinas -
Instituto Politécnico Nacional. La Paz, B.C.S. México. 170p.
Hammer O., Harper, D. A. T., & P.D. Ryan. 2001. PAST: Paleontological statistics
software package for education and data analysis. Palaeontologia Electronica 4(1):
9
Helfman G., Collette B., Facey D. & B. Bowen. 2009. The diversity of fishes. Biology,
evolution and ecology. 1st Ed. Blackwell Science. E.U.A. 737p.

60
Hindell J. S. & G. P. Jenkins. 2004. Spatial and temporal variability in the assemblage
structure of fishes associated with mangroves (Avicennia marina) and intertidal
mudflats in temperate Australian embayments. Marine Biology. 144: 385–395p.
Jalal K. C. A., Ahmad A. M. Akbar J. B., Kamaruzzaman Y.B. & S. Shahbudin. 2012.
Diversity and community composition of fishes in tropical estuaries Pahang
Malaysia. Pakistan Journal of Zoology. 44(1) 181-187
Jiménez-Illescas A., Obeso-Nieblas M., & D. Salas de León. Oceanografía física de la
Bahía de La Paz, B.C.S. en: Urbán-Ramírez J. & M. Ramírez-Rodríguez. La Bahía
de La Paz Investigación Conservación. Universidad Autónoma de Baja California
Sur. México. 188p
Kathiresan K. & B. I. Bingham. 2001. Biology of mangroves and mangrove ecosystems.
Advances in Marine Biology. 40: 81-251.
Laegdsgaard P. & J. R. Craig. 1995. Mangrove habitats as nurseries: unique
assemblages of juvenile fish in subtropical mangroves in eastern Australia.
Marine Ecology Progress Series. 126: 67 – 81.
Laegdsgaard P. & J. R. Craig. 2001. Why do juvenile fish utilize mangrove habitats?
Journal of Experimental Marine Biology and Ecology. 257: 229–253.
Lakra, W. S., Sarkar U. K., Kumar R. S., Pandey A., Dubey V. K. & O. P. Gusain, 2010.
Fish diversity, habitat ecology and their conservation and management issues of
a tropical River in Ganga basin, India. Environmentalist. 30: 306-319
Lara-Lara J. R., Arreola-Lizárraga J. A., Calderón-Aguilera L. E., Camacho-Ibar V. F., De
la Lanza-Espino G., Escofet-Giansone A., Espejel-Carbajal M. I., Guzmán-Arroyo

61
M., Ladah L., López-Hernández M., Meling-López E. A., Moreno-Casasola B. P.,
Reyes-Bonilla H., Ríos-Jara E. & J. A. Zertuche-González. 2008. Los ecosistemas
costeros, insulares y epicontinentales, en CONABIO (Eds). Capital natural de
México, Vol. I: Conocimiento actual de la biodiversidad. CONABIO, México. 109-
134p.
Linares Mazariegos. R. M., Tovilla Hernández. C. & J. C. De la Presa-Pérez. 2004.
Educación ambiental: una alternativa para la conservación del manglar. Madera
y Bosques. Número especial 2: 105-114.
Ludwig J. & J. Reynolds. 1988. Statistical ecology: a primer on methods and
computing. John Wiley & Sons, Inc. E.U.A. 337p.
Maeda-Martínez. A. N. 1981. Composición abundancia diversidad y alimentación de la
ictiofauna en tres lagunas costeras del Golfo de California. Tesis de licenciatura.
Universidad Autónoma de Nuevo León. Nuevo León, México. 140p.
Malpica-Maury O. M. 1999. Ictiofauna de la Ensenada de La Paz B.C.S. México. Tesis
de Maestría. . Centro Interdisciplinario de Ciencias Marinas - Instituto Politécnico
Nacional. La Paz, B.C.S. México. 85p.
Marcín-Medina R. 2010. Uso del hábitat del tursion (Tursiops truncatus) en la relación
a factores ambientales y antropogenicos en la ensenada y sur de la bahía de La
Paz. B.C.S. México. Tesis de doctorado. Universidad Autónoma de Baja California
Sur. La Paz, B.C.S. México 133p.
Mendoza E., Castillo-Rivera M., Zárate-Hernández R. & S. Ortiz-Burgos. 2010. Seasonal
variations in the diversity, abundance, and composition of species in an

62
estuarine fish community in the Tropical Eastern Pacific, Mexico. Icthyological
Research. 56: 330-339
Mendoza R. 1983. Identificación, distribución y densidad de la avifauna marina en los
manglares: Puerto Balandra, Enfermería y Zacatecas en la Bahía de La Paz,
Baja California Sur, México. Tesis de Licenciatura. Universidad Autónoma de Baja
California Sur.
Mendoza R., Fuentes H. & M. Espinosa. 1984. Avances de estudios de avifauna marina
de los manglares: Balandra, Enfermerias y Zacatecas, de la Bahía de la Paz, B.
C. S., México. Informe General de Labores CIB La Paz, B.C.S. 40-51p.
Mirera D., Kairo J.G., Kimani E. N. & F. K. Waweru. 2010. A comparison between fish
assemblages in mangrove forest and on interdial flats at Ungwana Bay Kenya.
African Journal of Aquatic Science. 35(2): 165-171
Moreno-Bejarano L. M. & R. Álvarez-León. 2003. Fauna asociada a los manglares y
otros humedales en el delta-estuario del Río Magdalena, Colombia. Revista
Académica Colombiana de Ciencia. 27 (105): 518-534
Mueller-Dombois D. & H. Ellemberg. 1974. Aims and methods of vegetation ecology. 1st
Ed. John Wiley & Sons, New York. 547p.
Nagelkerken I., Van Der Velde G, Gorissen M. W., Meijer G. J., Vna´t Hof T. & Den
Hartog. 2000. Importance of mangrove, seagrass beds and the shallow coral reef
as a nursery for important coral reef fishes, using visual census technique.
Estuarine, Coastal and Shelf Science. 51: 31-44.
Nelson J. 2006. Fishes of the world. 4th
ed. John Wiley & Sons, Inc. E.U.A. 601p

63
Nip T. H. & K. Chong-Wong. 2010. Juvenile fish assemblages in mangrove and non-
mangrove soft-shore habitats in eastern Hong Kong. Zoological Studies, 49(6):
760-778
Okyere I., Blay J., Aggrey-Fynn J. & D. Worlanyo-Aheto. 2011. Composition, diversity
and food habitas of the fish community of a costal wetland in Ghana. Journal of
Environmental and Ecology. 2(1): 1-18
Osorio-Dualiby, D. 1988. Ecología trófica de Mugil curema, M. incilis y M. liza. (Pisces:
Mugilidae) en la Ciénaga Grande de Santa Marta, Caribe colombiano. I. análisis
cualitativo y cuantitativo. Anales Instituto investigación Marina de Punta de Betín.
18: 113-126
Parrish J. 1989. Fish communities interacting shallow-water habitats in tropical
oceanic regions. Marine Ecology Progress Series. 58: 143-160.
Philips P. C. 1981. Diversity and fish community structure in a Central America
mangrove embayment. Revista de Biología Tropical. 29(2): 227-236
Pielou E.C. 1984. The interpretation of ecological data. A primer on classification and
ordination. John Wiley & Sons, New York. 263p.
Rambaut A. 2009. FigTree ver.1.3.1. [Online].Disponible:
http://tree.bio.ed.ac.uk/software/figtree/
Rodríguez-Romero J., López-González L. C., Galván-Magaña F., Sánchez-Gutiérrez F. J.,
Inohuye-Rivera R. B. & J. C. Pérez-Urbiola. 2011. Seasonal changes in a fish
assemblage associated with mangrove in a costal lagoon of Baja California Sur,
Mexico. Latin American Journal of Aquatic Research. 39(2): 250-260

64
Salazar-Vallejo S. & N. E. González (Eds). 1993. Biodiversidad marina y costera de
México. CONABIO. México. 865p.
Sedberry R. G. & J. Carter. 1993. The fish community of a shallow tropical lagoon in
Belize, Central America. Estuaries. 16(2): 198-215
Shih-Rong K., Hsing-Juh L. & S. Kwang-Tsao. 2001. Seasonal changes in abundance
and composition of the fish assemblage in Chiku lagoon southwestern Taiwan.
Bulletin of Marine Science. 68(1): 85-99.
Stein G. J. & R. Lavenberg J. 1997. The fishes of the Galápagos Islands. Stanford Press.
E.U.A. 863p.
Suárez-Altamirano J. M. 2005. Flujos de material suspendido y disuelto entre el estero
“El Conchalito” y la laguna “Ensenada de La Paz”, B.C.S., México. Tesis de
Maestría. Centro Interdisciplinario Ciencias Marinas – Instituto Politécnico Nacional.
La Paz, B.C.S. México. 119p.
Tavera J. J. 2012. Evolución, filogenia y biogeografía de la subfamilia Haemulinae
(PERCOIDEI:HAEMULIDAE) en el nuevo mundo. Tesis de doctorado. Centro de
Investigaciones Biológicas del Noroeste. La Paz, B.C.S. México. 343p.
Thomson D., Findley L. & A. Kerstitch. 2000. Reef fishes of the Sea of Cortez.
University of Texas Press. E.U.A. 353p.
Toepfer C. S., Williams L. R., Martínez A. D. & W. L. Fisher. 1998. Fish and habitat
heterogenecity in four streams in the central Oklahoma/Texas plains ecoregion.
Procceding of the Oklahoma Academic of Science. 78: 41-48

65
Torres-Alfaro G. M. 2010. Ecología de la taxocenosis de peracáridos macrobentónicos
en playas arenosas protegidas: análisis y consideraciones para el manejo
sustentable. Tesis de doctorado. Centro de Investigaciones Biológicas del Noroeste.
La Paz, B.C.S. México. 176p.
Unsworth R. K. F., Garrad I. S., Salinas L. P., Cullen L. C., Smith D. J., Sloman K. A. & J.
J. Bell. 2009. Structuring of Indo Pacific fish assemblages along the mangrove-
seagrass continuum. Aquatic Biology. 5: 85-95
Urbán–Ramírez J. & M. Ramírez–Rodríguez (Eds.). 1997. La Bahía de La Paz,
Investigación y Conservación. Universidad Autónoma de Baja California Sur,
Centro Interdisciplinario de Ciencias Marinas, IPN, SCRIPPS Institution of
Oceanography. La Paz, B.C.S. México. 345p
Vega-Cedejas M. E. & M. Hernández de Santillana. 2004. Fish community structure and
dynamics in a costal hypersaline lagoon: Rio Lagartos, Yucatan, Mexico.
Estuarine Costal and Shelf Science. 60: 285-299
Yagi Y., Kinoshita I., Fujita S., Aoyama D. & Y. Kawamura. 2011. Importance of the
upper estuary as a nursery ground for fishes in Ariake Bay, Japan.
Environmental Biology of Fishes. 91(3): 337-352
Yáñez-Arancibia A., R. Twilley R. & A. L. Lara-Domínguez. 1998. Los ecosistemas de
manglar frente al cambio climático global. Madera y Bosques 4(2): 3-19

66
Anexo 1
Receta Mezcla Frigorífica
1 bolsa de hielo (4-5 kilos)
2 litros de agua (puede ser marina)
1 kilo de Sal
1 litro de alcohol etílico
Se agrega el hielo desagregado y se vierte el agua a lo largo del hielo, se agrega la sal
tratando de cubrir toda la parte superior del hielo y luego se adiciona el alcohol en la parte
de las esquinas y el centro. La mezcla puede alcanzar temperaturas menores a 0°C.

67
Anexo 2
Tabla I. Datos de temperatura utilizados para definir las épocas climáticas.
Autor\Mes Ene Feb Mar Abr May Jun Jul Ago Sep Oct Nov Dic
Vázquez-Hurtado, 2011 21.0 20.0 21.0 22.0 24.0 24.0 27.0 29.0 29.0 28.0 26.0 22.0
Salinas-Zacarías, 2000 19.4 19.8 22.2 23.5 22.5 26.5 23.8 30.3 27.9 27.1
Hernández-Rivas, 2007 (con per) 24.2 27.0 28.0 29.0 26.0 21.5
Hernández-Rivas, 2008 (con per) 19.5 18.0 21.0 22.5 23.5 21.5
Verdugo-Díaz, 2000 23.0 28.0 29.0 27.0 23.0
Verdugo-Díaz, 2001 20.0 19.0 20.0 21.0 24.0 23.5
Martínez-López et al., 1993 23.9 24.5 25.9 30.1 30.0 30.1 28.0 25.1
Martínez-López et al., 1994 20.4 20.3 23.2 24.0 25.7 28.6 32.5 29.2 29.0 27.5 23.0
Martínez-López et al., 1995 21.1 21.1 22.1
20.2 19.7 21.1 22.7 23.7 24.3 27.5 28.4 29.3 28.2 25.9 22.9
Referencias:
Hernández Rivas M. (Com. Per.) 2007; Hernández Rivas M. (Com. Per.) 2008
Martínez- López et al., 1993; 1994; 1995; Martínez-López A., Cervantes-Duarte R., Reyes-Salinas A. & J. E. Valdez-Holguín. 2001.
Cambios estacional de la clorofila a en la Bahía de La Paz B.C.S. México. Hidrobiológica. 11(1): 45-52
Salinas-Zacarías M. A. 2000. Ecología de tursiones, Tursiops truncatus en la Bahía de la Paz B. C.S. Tesis de doctorado. Centro
Interdisciplinario de Ciencias Marinas-Instituto Politécnico Nacional. 104p
Verdugo-Díaz, 2000; 2001; Verdugo-Díaz G. 2003. Respuesta ecofisiológica del fitoplancton ante la variabilidad ambiental de
una laguna costera sub-tropical de Baja California Sur, México. Tesis de doctorado. Centro Interdisciplinario de Ciencias
Marinas-Instituto Politécnico Nacional. 139p

68
Tabla II. Tabla de estadísticos por época climáticadistribución de frecuencias
Factor N Media DesvEst ErrorEst Temp-
95%
Temp+95%
67 24.4 3.5 0.4 23.6 25.3
Templada 21 20.8 1.5 0.3 20.1 21.5
Transición 19 23.6 1.4 0.3 22.9 24.3
Cálida 27 27.9 2.1 0.4 27.1 28.7
Tabla III. Análisis de varianza por épocas climáticas
Efecto SumCuadr g.l. Cuad.
Medios
F p Sig
Intercepto 38166.26 1 38166.26 12349.36 0.00 ***
Época 616.13 2 308.07 99.68 0.00 ***
Error 197.79 64 3.09


















