mecanismos de evolucion en celula eucariota
18
MECANISMOS EVOLUCIONARIOS PARA EL ESTABLECIMIENTO DE LA COMPLEJIDAD EN CELULAS EUCARIOTAS Gracias a un enfoque comparativo, la biología celular evolutiva hace el uso de la genómica, bioinformática, y la biología celular de eucariontes modelos para proporcionar nuevas vías para la comprensión de los procesos básicos de la célula. Este enfoque ha dado lugar a proponer mecanismos que sustentan la evolución de la organización de las células eucariotas incluyendo endosimbiosis y procesos autógenos y procesos neutros y adaptativos. Junto con estos mecanismos se contribuyó al origen y complejidad de las organelas, complejos moleculares y la arquitectura del genoma. Nosotros revisamos estos mecanismos y sugieren que una mayor apreciación de la diversidad de formas eucariotas llevo a una comprensión más completa de las conexiones evolutivas entre las organelas y las rutas inesperadas por las cuales se ha llegado a tal diversidad. REUNIENDO A LA BIOLOGIA CELULAR Y LA BIOLOGIA EVOLUTIVA La aparición de los eucariontes fue hace casi 2 billones de años transformando la vida en la Tierra. Los esfuerzos para desentrañar los mecanismos evolutivos que dieron forma y siguen dando forma, las células eucariotas están comenzando a abortar este monumental cambio evolutivo. La comprensión de estos mecanismos nos ayudara a hacer conexiones conceptuales entre la biología celular de eucariontes modernos taxonómicamente diferentes, portar conocimiento derivado de sistemas modelos sobre los organismos menos estudiados de agricultura (e.j. cultivos y fitopatogenos), el medio ambiental (e.j productores primarios acuáticos como las haptofitas y diatomeas) o relevancia médica (e.j paracitos como Plasmodium falciparum, agente causal de la malaria. Este amplio enfoque comparativo conocido como biología celular evolutiva facilita la generación de hipótesis que intentan explicar las funciones biológicas de las células entre toda la gama de eucariontes. Este enfoque se ha aplicado con éxito a muchos aspectos de la célula eucariota. La combinación de la ultraestructura y la biología celular molecular con datos genómicos de un muestreo de organismos que abarcan la amplitud taxonómica de eucariontes ha proporcionado una gran cantidad de conocimiento sobre la evolución de las células eucariotas y su diversidad. Desde la perspectiva de un biólogo celular, esta riqueza de datos permite la integración de la teoría de la evolución establecida con el estudio de los mecanismos celulares. Glosario Complejidad: Una medida de la cantidad de componentes e interacciones de un sistema relativo a otro sistema equivalente.
-
Upload
rosa-amelie -
Category
Documents
-
view
47 -
download
5
description
traduccion de review
Transcript of mecanismos de evolucion en celula eucariota

MECANISMOS EVOLUCIONARIOS PARA EL ESTABLECIMIENTO DE LA COMPLEJIDAD EN CELULAS EUCARIOTASGracias a un enfoque comparativo, la biología celular evolutiva hace el uso de la genómica, bioinformática, y la biología celular de eucariontes modelos para proporcionar nuevas vías para la comprensión de los procesos básicos de la célula. Este enfoque ha dado lugar a proponer mecanismos que sustentan la evolución de la organización de las células eucariotas incluyendo endosimbiosis y procesos autógenos y procesos neutros y adaptativos. Junto con estos mecanismos se contribuyó al origen y complejidad de las organelas, complejos moleculares y la arquitectura del genoma. Nosotros revisamos estos mecanismos y sugieren que una mayor apreciación de la diversidad de formas eucariotas llevo a una comprensión más completa de las conexiones evolutivas entre las organelas y las rutas inesperadas por las cuales se ha llegado a tal diversidad.
REUNIENDO A LA BIOLOGIA CELULAR Y LA BIOLOGIA EVOLUTIVA
La aparición de los eucariontes fue hace casi 2 billones de años transformando la vida en la Tierra. Los esfuerzos para desentrañar los mecanismos evolutivos que dieron forma y siguen dando forma, las células eucariotas están comenzando a abortar este monumental cambio evolutivo. La comprensión de estos mecanismos nos ayudara a hacer conexiones conceptuales entre la biología celular de eucariontes modernos taxonómicamente diferentes, portar conocimiento derivado de sistemas modelos sobre los organismos menos estudiados de agricultura (e.j. cultivos y fitopatogenos), el medio ambiental (e.j productores primarios acuáticos como las haptofitas y diatomeas) o relevancia médica (e.j paracitos como Plasmodium falciparum, agente causal de la malaria. Este amplio enfoque comparativo conocido como biología celular evolutiva facilita la generación de hipótesis que intentan explicar las funciones biológicas de las células entre toda la gama de eucariontes.
Este enfoque se ha aplicado con éxito a muchos aspectos de la célula eucariota. La combinación de la ultraestructura y la biología celular molecular con datos genómicos de un muestreo de organismos que abarcan la amplitud taxonómica de eucariontes ha proporcionado una gran cantidad de conocimiento sobre la evolución de las células eucariotas y su diversidad. Desde la perspectiva de un biólogo celular, esta riqueza de datos permite la integración de la teoría de la evolución establecida con el estudio de los mecanismos celulares.
Glosario
Complejidad: Una medida de la cantidad de componentes e interacciones de un sistema relativo a otro sistema equivalente.
Endosimbiosis (primaria):El proceso por el cual una célula procariota (endosymbionte) se incorpora en el citoplasma de una célula eucariota (host), con una relación establecida a través de la integración metabólica y EGT tal que ninguno de los dos puede sobrevivir por sí mismo.
Endosimbiosis (secundaria): el mismo proceso que la endosimbiosis primaria excepto que el endosimbionte es una célula eucariota que posee un plastidio primario.El proceso se puede extender a endosimbiosis terciaria (el endosimbionte es una célula que posee un plastidio secundaria) y endosimbiosis secundario de serie (un linaje que posee un tipo de plástidio secundario reemplaza su plásmido secundario con un plastidio secundario de un linaje diferente).
La transferencia de genes endosimbiótica (EGT): un caso especial de transferencia horizontal de genes (ver más abajo), mediante el cual el gen en cuestión es adquirida por el linaje del hospedero desde el genoma del endosimbionte.
Evolutiva biología celular: Una disciplina emergente que incorpora perspectivas y técnicas comparativas de la biología celular, protozoologia, evolución molecular, y la teoría evolutiva matemática para abordar las cuestiones de los orígenes y la diversidad de las células.

En primer ancestro común eucariota (FECA):La célula (o población de células), perteneciente al linaje que dio origen a la línea moderna de los eucariotas en el punto más temprano en la que poseía características biológicas de células distintas de las de células procariotas similares. Aunque se deduce que este organismo ha existido, una manera útil para el tratamiento del FECA es con una reconstrucción teórica con los rasgos que la definen y con una emocionante pregunta de investigación abierta.
Horizontal transferencia de genes:La adquisición de un gen por un genoma a partir de una fuente que no sea del linaje parental inmediata.
Último ancestro común eucariota (LECA): La célula (o población de células), perteneciente al linaje que dio origen a la línea moderna de los eucariotas en el punto más reciente en la que los distintos linajes descendientes divergieron dejar los linajes eucariotas existentes. Una vez más, este concepto es más útil como un punto de la reconstrucción teórica o de referencia para evaluar la antigüedad de varias características biológicas celulares.
Monofilético: un grupo se considera "monofilético 'cuando abarca todos los descendientes de un único ancestro.
Parafilético: un grupo se considera 'parafilético "cuando se incluye a algunos, pero no todos, los descendientes de un único ancestro.
Paralog: genes que son el resultado de un proceso de duplicación de genes.
Selección:El proceso por el que un factor (incluyendo la presencia de otro organismo) presenta una circunstancia que resulta en la muerte preferencial de algunos organismos en el medio ambiente sobre otros.
Se han hecho notables progresos en revelar los mecanismos que sustentan el surgimiento de la complejidad de la célula eucariota. El número y la sofisticación de los compartimentos internos en eucariotas los distinguen de los procariotas y es esta sofisticación que se traduce en mucho mayor "complejidad" en la configuración de células eucariotas, que se mide por el número de componentes en un sistema y el número de interacciones entre ellos [4,5] en comparación con otro, sistema equivalente (ej, célula vs célula). Existe un acuerdo general de entre los análisis moleculares filogenéticos [6,7] y la evidencia del registro fósil ([8,9], pero ver [10]) que los procariotas eran anteriores a los eucariotas. Esta evidencia sugiere que los eucariotas tienen que haber surgido de un estado que se asemeja a un procariota; es decir, que la adquisición de orgánulos celulares y máquinas complejas en eucariotas debe explicarse desde un estado celular que carecen de la amplia presencia de estas características.
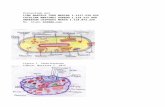
Evolución de menor complejidad (estado procariota) a un aumento de la complejidad de la arquitectura celular (estado eucariota) se puede dividir en al menos tres etapas evolutivas sucesivas (Figura 1). En primer lugar debe ocurrir, una transición de un organismo que carece de algunas o de todas las membranas internas a un organismo que posee algunas de las características celulares que definen a un eucariota [un primer ancestro común eucariota (FECA)]. A continuación, el organismo debe pasar del FECA a un último ancestro común eucariota (LECA). Por último, la evolución y divergencia después de la LECA deben ocurrir para formar los principales linajes de eucariotas existentes [2,11].
La investigación sobre la transición de procariotas a eucariotas (por ejemplo, [1]) ha reconstruido una sorprendentemente sofisticada LECA que posee un citoesqueleto bien establecido de actina / tubulina, un sistema endomembranoso elaborado, un núcleo, mitocondrias y maquinaria para procesos tales como la eliminación de intrones y meiosis (Figura 1).Además, al menos una innovación celular adicional importante de influencia posterior al LECA que aumentaron la complejidad: la adquisición de plastidios.
Por lo tanto, una cuestión importante para la biología celular evolutiva son los mecanismos de accionamiento que aumentaron la complejidad celular a nivel de máquinas moleculares y la formación de orgánulos. En este artículo revisamos el progreso que se ha logrado en abordar esta cuestión.

Figura 1. Volviendo sobre la aparición de los eucariotas modernos. (A) La aparición de
los eucariotas se puede dividir en al menos tres etapas evolutivas sucesivas: (i) la transición desde un punto de partida procariota como a la primera ancestro común eucariota (FECA); (ii) transición de el FECA a la última eucariota ancestro común (LECA); y (iii) la evolución y divergencia después de la LECA para formar los principales linajes de eucariotas como los conocemos hoy en día [2,11]. (B) Después de la LECA, seis linajes eucariotas existentes (Recuadro 1) se han caracterizado [11]. Estos son los Opisthokonta, Amoebozoa, Excavata y Archaeplastida, el stramenopiles, alveolates y rhizarians (SAR), y la agrupación contenciosa de cryptomonads, Centrohelida, telonemids y haptophytes (CCTH).
Mecanismos para la adquisición de orgánulos
Antes de 1974, la hipótesis para la evolución de los compartimentos de membrana interna (orgánulos) dentro de la célula eucariota fue un proceso autógeno; es decir, las células eucariotas se construyen de una manera escalonada a partir de bloques de construcción individuales presentes en el ancestro pre-eucariota [12,13]. Sin embargo, la evidencia demuestra que tanto las mitocondrias y cloroplastos son de origen bacteriano (endosimbiosis) [14,15] esto cambio la base teórica sobre la evolución eucariótica de tal manera que tanto la endosimbiótica y explicaciones autógenas son alternativas viables ahora para considerar a la hora de abordar los orígenes de una organela eucariota. Nuestra comprensión científica de ambos mecanismos está madurando, con la endosimbiosis siendo mucho mejor entendida. A continuación, se discute la situación actual en el surgimiento de la complejidad de las células eucariotas a través de la endosimbiosis y procesos autógenos.
Endosimbiosis
Endosimbiosis es la incorporación y permanencia de un organismo, el endosimbionte, dentro de otro, el anfitrión. Más allá de los ejemplos familiares de endosimbiosis (es decir, el cloroplasto de las plantas y las mitocondrias casi omnipresente) también existe una amplia gama de organismos con orgánulos derivados de eventos endosimbióticos adicionales. Estudios biológicos y genéticos celulares recientes de estos organismos han revelado mucho sobre los mecanismo de la endosimbiosis. De hecho, una razón por que la endosimbiosis se entiende mejor que los mecanismos autógenos de adquisición de orgánelas es la riqueza de los productos intermedios endosimbióticos disponibles para el estudio (por ejemplo, [16]). Acontecimientos recientes e independientes de la endosimbiosis han revelado las primeras etapas del proceso, incluyendo varios ejemplos de orgánulos primarias (por ejemplo, Paulinella) y secundarias (por ejemplo, Hatena) organelas plastidios derivados así como plástidos transitoriamente adquiridos denominados cleptoplastia. Otros ejemplos de donde el hospedero y el simbionte están en el comienzo de su integración incluyen dinotoms, algas en el que el huésped es un linaje dinoflagellate que posee un endosimbionte diatomea mínimamente reducido. Trabajos recientes han comenzado a descubrir el alcance y la naturaleza de sus organelas y la integración metabólica [17]. La secuenciación del genoma de organismos tales como el cryptophyte y algas chlorarachniophyte, cuyo orgánulo fotosintético contiene

tanto un genoma de plástidios secundarios y el resto del genoma nuclear de algas de color rojo y verde (nucleomorph) y el citoplasma [18], también han permitido el examen de la reducción del genoma y la integración celular en endosimbiosis (Recuadro 2). Debido a que la fotosíntesis se ha ganado, robado y co-optado a lo largo de la historia de los eucariotas, la adquisición de plastidios ha sido un modelo particularmente útil para la comprensión de las primeras etapas de la endosimbiosis. Sin embargo, un raro ejemplo de una potencial endosimbiosis mitocondrial secundaria recientemente ha sido descrito por métodos genómicos [19]. El fishpathogen Neoparamoeba contiene lo que parece ser un simbionte intracelular relacionado con Ichthyobodo Necator, un kinetoplástido. Aunque la mitocondria atípica de este simbionte ocupa casi la mitad de su volumen citoplasmático, el grado en que la endosimbionte ha progresado hasta convertirse en un orgánulo no está claro. El genoma nuclear del simbionte no parece que hayan sido objeto de una amplia reducción en comparación con la de otros kinetoplastids. Todos estos ejemplos ayudan a enfocar la cuestión de que tan reducido un endosimbionte tiene que ser para que se le considere un orgánulo y no un organismo.
En el otro extremo de la integración endosimbiótica existen organelas aparentemente reducidas del estado eucariota canónico, como los apicoplasts no fotosintéticos de apicomplexans y los hidrogenosomas y mitosomes, algunos de los cuales ya no poseen genomas organellar. Inicialmente estas últimas organelas fueron vistas como clases distintas; Sin embargo, estudios recientes ellos han establecido claramente como derivados de las mitocondrias y encontrado diversos intermedios que poseen metabolismos aeróbicos o anaeróbicos y diferentes organizaciones genómicas.La gama de minimización genómica y citoplasmática encontrados para endosimbiosis aliado a organelas derivadas plantea la cuestión de qué mecanismo determina y limita el alcance de esta tendencia reductora de cualquier linaje dado. El paso del tiempo no puede explicar esta reducción debido a una amplia gama de reducción que se observó en organelas claramente derivados del mismo evento fundación (por ejemplo, la mitocondria). Sin embargo, se propuso que debido a que el principal mecanismo de la transferencia de ADN al núcleo de acogida proviene de orgánulos lisaron, la velocidad de transferencia es proporcional al número de copia de la endosimbionte en la célula (Figura 2B).Esta idea se conoce como la " transfer-window hipótesis" [21], lo que implica que la transferencia no puede continuar una vez que el número de orgánulos ha llegado a un solo ejemplar. De hecho, los análisis genómicos experimentales y comparativos revelaron muchos menos transferencias de plastidios a genomas nucleares en los organismos que poseen un número de copias de plástidos baja.Además, los genomas nucleares de un cryptophyte y de un alga chlorarachniophyte, cada una con un único nucleomorph, se han secuenciado y se informaron en 2012 Estos hallazgos revelaron una completa falta de transferencia de ADN reciente, ya sea del plastidio o el genoma nucleomorph pesar de la evidencia de la transferencia de las mitocondrias, lo cual es consistente con una reducción a un solo orgánulo que es responsable de detener la transferencia de genes endosimbiótica (EGT) y por lo tanto la orgánulo reducción .
Endosimbiosis ha permitido en varias ocasiones para aumentar la complejidad general en las células eucariotas en comparación con su estado pre-fusionada. Irónicamente, porque la célula se encuentra en su etapa más compleja inmediatamente después de que comience la endosimbiosis, con la integración avanza principalmente a través de GET o geneloss, el proceso de endosimbiosis realmente implican disminuciones en complejidad.
1. La Diversidad Eucariótica
Diversidad eucariota (Figura 1B en el texto principal) está dividido en seis grandes grupos taxonómicos o supergrupos [2]. El Opisthokonta abarca los linajes de animales y hongos, así como sus parientes unicelulares. El Amoebozoa alberga una diversidad de linajes ameboides con, y sin, etapas flageladas. Incluye los patógenos Balamuthia, Acanthamoeba, y, lo más famoso, Entamoeba histolytica, el agente causante de la disentería amebiana.
El Opisthokonta y Amoebozoa se unen en los análisis filogenéticos grandes a escala molecular y se cree que representan un grupo monofilético, nombrado el Amorphea.El Archae-plastida incorpora los linajes de algas rojas, algas verdes (incluyendo plantas de la tierra), y los glaucophytes, que se derivan de un único evento primario de endosimbiósis fundador.El SARclade une los linajes aparentemente dispares de stramenopiles (diatomeas, algas pardas, y el agente causal de la gran hambruna irlandesa, Phytophthora)

y alveolates (ciliados como Paramecium, los dinoflagelados causantes de mareas rojas y apicomplexans como Plasmodium, que causa la malaria ). El supergrupo Excavata Incluye agentes causantes de enfermedades importantes, como Trypanosoma, Leishmania, Giardia y Trichomonas, así como de vida libre, o parientes no patógenos. Por último, el supergrupo CCTH contiene actualmente los linajes de criptofitas, Centrohelida, telonemids y haptophytes; Sin embargo, los análisis de evolución molecular a gran escala más recientes han puesto en duda la unidad de estos en un solo grupo y el CCTH debe ser tratado como provisional en el mejor de los casos.
Recuadro 2. Endosimbiosis
Los tipos de endosimbiosis se clasifican en función de la naturaleza del host y del endosimbionte. La forma más simple, de la endosimbiosis primaria, implica un huésped eucariota y una endosimbionte bacteriana (Figura 2A en el texto principal). Dos de esos eventos primarios han sido transformadores en la historia de los eucariotas y contó con la incorporación de un a-proteobacteria y una cianobacteria para dar lugar a la mitocondria y orgánulos plastidios derivados, respectivamente. Ambos eventos se sabe que han ocurrido en la historia temprana eucariota, con el evento mitocondrial ahora se muestra de manera convincente que han precedido el LECA [20].Una endosimbiosis primaria de plastidios es muy probable que haya ocurrido en la base del Linaje Archaeplastida, que confirió la capacidad fotosintética y dando lugar a todas las algas y plantas terrestres rojo y verde. La capacidad fotosintética fue claramente ventajosa, ya que dio lugar a la posterior evolución de plastidios complejos [16] a través de la endosimbiosis secundaria y terciaria (Figura 2A en el texto principal).
Como mecanismo, el proceso de endosimbiosis se puede dividir en la iniciación y la integración. Iniciación puede provenir de varias posibles asociaciones microbianas, incluyendo el intercambio mutualista de metabolitos, la invasión intracelular del "host" por un parásito, o ingestión depredadora de un endosimbionte eventual por un phagotroph. Después de la iniciación, el éxito del organismo resultante depende de la capacidad de sincronizar el crecimiento celular y la división de los ciclos endosimbionte hospedero. En todos los casos, la transferencia gradual de material genético del genoma endosimbionte para el genoma del hospedero promueve esta sincronización (Figura 2B en el texto principal). Este mecanismo de trinquete de EGT impulsa el establecimiento de una relación obligada entre el endosimbionte y su hospedero. Después de la adquisición de un endosimbionte, el organismo tiene dos genomas, uno en el núcleo y una en el endosimbionte (Figura 2B, i en el texto principal). Ya sea por lisis o por eventos incorrectos de fisión y fusión del endosimbionte durante la replicación, el ADN endosimbionte liberado en el citoplasma se puede integrar en el genoma del huésped (Figura 2B, ii en el texto principal) .Con un gen endosimbionte ahora codificado y expresado por el hospedero, debe ser dirigido correctamente al endosimbionte (Figura 2B, iii en el texto principal). Cuando esto ocurre, la copia endosimbionte es redundante y sostiene la decadencia mutacional, y el genoma endosimbionte se reduce (Figura 2B, iv en el texto principal).La direccionalidad impuesta por esta transferencia se traduce en un mecanismo reiterativo. La ventana de oportunidad que permite EGG permanece abierta hasta que sólo queda un único genoma endosimbionte (Figura 2 B, V y VI en el texto principal) [16,20]. La pérdida de genes, coincidente con pérdida de la función, en la transición-organismo-a orgánulo es también una fuente principal de la reducción del genoma [16,20].

Figura 2.
Mecanismos de la evolución celular. (A) La variedad de plastidios derivan de adquisición reiterativa de endosimbiontes fotosintéticos. (i) la endosimbiosis primaria se estableció tras inmersión de una cianobacteria (verde) por una célula huésped eucariota. Un proceso de endosimbiosis primaria similar habría producido a la mitocondria de una proteobacteria. (ii) En la endosimbiosis secundaria de plástidos, una célula de alga verde o rojo está envuelta por una nueva célula huésped. (iii) Este proceso se repite en la endosimbiosis terciaria. Aunque endosimbiosis primaria, secundaria y terciaria son conceptualmente interconectados, no son "pasos consecutivos de un solo colonización. (B) Las etapas de la transferencia de genes endosimbiótica (EGT) de un endosimbionte recién adquirido: la lisis de la endosimbionte y (ii) la transferencia del gen al núcleo del hospedero; (iii) reorientación y (iv) la pérdida de genes codificados de un endosimbionte c; y (v) la repetición hasta que(vi) un único endosimbionte permanece. (C) La hipótesis organelle-paralogy (OF). (a) Las diferentes familias de proteínas interactúan cooperativamente para especificar las propiedades de orgánulos que define como la inmovilización, de acoplamiento, la fisión o fusión. (ii) las familias de proteínas Specificity-encoding evolucionan por duplicación de genes y la divergencia, como se representa por esta hipótesis filogenetica. (iii) Aumento en la complejidad de la especificidad de codificación de las familias de proteínas se refleja en incrementos en la complejidad del sistema de membranas. Los parálogos de la familia de proteínas de codificación especifica residen en y tienen su efecto en compartimentos distintos.Modificado a partir de [59]. (D) Un esquema generalizado de evolución neutral constructiva (CNE). (i) factor de proteína "A" posee una actividad determinada. (ii) A través de colisiones estéricas aleatorias, interactua con un separado factor de 'B' ; que tiene poco o ningún efecto sobre la actividad del factor A. (iii) una mutación (representado por una estrella amarilla) se produce en factor que reduce su actividad , pero debido a la interacción del factor A

con factor B, la mutación se suprime y la actividad de factor A se mantiene a niveles casi originales. Esto podría ser debido a la estabilización de la estructura del factor A, enmascaramiento de su carga o residuos hidrofóbicos expuestos, o la localización alterada del factor A lo que permite un mejor acceso a su sustrato. (iv) las mutaciones posteriores del factor A originales y mutaciones compensatorias en el factor de interactuar B, integrar aún más el factor B en la actividad del factor de A via un mecanismo de trinquete que también puede dar lugar a la contratación de factores adicionales.
Organelas autógenas (no endosimbióticas)Aunque la endosimbiosis ha sido, sin duda, una fuerza poderosa en la construcción de algunos aspectos de la complejidad celular eucariótica, no explica todos ellos. Una explicación alternativa más simple para el origen de los orgánulos delimitados por una sola bicapa lipídica y desprovisto de material genético es que son autógenas. Los orgánulos más comúnmente propuestos a tener un origen autógeno son las del sistema de transporte de membrana, incluyendo el retículo endoplasmático (ER), aparato de Golgi, endosomas, y membrana plasmática [25]. Aunque estos orgánulos de endomembrana están conectados dinámicamente entre sí, están contenidos en distintos compartimentos a través de la acción de los mecanismos de transporte de membrana, tales como los complejos Rabs, SNAREs, coatomer, y Adaptin (AP) [26]. Estas familias proteicas de codificación específica tienen diferentes miembros que realizan la misma función (por ejemplo, la inducción de la curvatura de la membrana o facilitar la fusión de membranas) en lugares distintos dentro del sistema de transporte de membrana [26]. Aunque cada familia de proteínas podría desempeñar un papel individual, parte de la información de codificación especificada en el transporte de membrana parece ser el resultado de las interacciones combinatorias proteína-proteína entre los miembros de las diferentes familias [27]. Comparativos análisis genómicos y filogenéticos de estas diferentes familias de proteínas han revelado detalles de su diversificación primaria por la duplicación de genes (por ejemplo, [1]). Sorprendentemente, las duplicaciones que dieron lugar a parálogos de las diversas proteínas de codificación específica asociados con cada localización celular se produjeron antes del LECA. Sin embargo, una examinación de los parálogos endocíticos de las familias SNARE, Rab, y AP reveló un patrón por el cual algunos parálogos específicos de una organela no habían duplicado antes del LECA, con duplicaciones paralelas que ocurren en vez de linajes después del LECA [28]. Estos patrones proporcionan un entendimiento que data del tiempo de estos eventos y sugieren un

Tendencias en Biología Celular de julio de 2014, vol. 24, N ° 7
posible mecanismo que sustenta ellos, que es formalizado en la hipótesis de la paralogía organular (OPH) [28,29]. El OPH (Figura 2C) propone que las propiedades de orgánulos finas de-un conjunto de proteínas especificidad codifica con funciones complementarias que producen juegos de interacción para-troncos al someterse a duplicaciones. A través de la coevolución, estos conjuntos de proteínas especificidad codifica acumulan mutaciones que fijan sus orgánulos separados de unión funcionales, definiendo así específicas [30]. Por lo tanto, las iteraciones de este proceso podría ser responsable de la serie de orgánulos en los sistemas endomembrane de eucariotas existentes que surgieron a través de la diferenciación de un compartimento interno prototipo original en la FECA.
Recientemente, la OPH (Hipótesis Organo-Paralógica) ha sido probada en simulaciones computarizadas. El modelamiento matemático de genes codificadores de especifidad en poblaciones de vesículas mostró que la duplicación de genes y las interacciones diferenciales entre paralogos producen nuevos compartimentos vesiculares [31]. Aún más, la OPH predice que el orden de emergencia evolucionaria para cada miembro de una familia de proteínas codificadoras de especifidad debe corresponder al orden de aparición de las diferentes organelas que definen y en las cuales tienen efecto. Dos estudios recientes reportaron resoluciones filogenéticas para familias de proteínas codificadores de especifidad importantes, permitiendo de este modo a las hipótesis ser propuestas basándose en evidencia empírica en cuanto a un orden de aparición evolutiva después del establecimiento de la complejidad extensiva en el tráfico intracelular de membranas en el último ancestro común eucariótico. Los complejos AP ayudan a clasificar el tráfico vesicular entre las organelas encontradas entre e incluyendo la membrana plasmática y la red trans-Golgi [32,33]. La genómica comparativa y el análisis filogenético concluyo el orden de aparición de los miembros de la familia compleja AP, siendo AP3 y AP5 las primeros en divergir de los otros complejos AP, seguidos por AP4 y AP1/2 [32]. Basados en sus ubicaciones de acción conocidas, este orden sugiere que las adaptinas actuaron primeramente en una interfaz organular entre los sistemas secretor y fagocítico, antes de establecerse la red trans-Golgi. En adición, evidencias recientes proveen pistas de la conservación mediante la familia Rab de GTPasas, que son interruptores moleculares involucrados en la especificación de la identidad organular en el sistema de tráfico intracelular de membrana [34]. Aunque está bien establecido que las Rab GTPasas son ancestrales y que el LECA poseía un gran complemento de tales proteínas [35], el grado de qué familias Rab están conservados permanece desconocido. La búsqueda rigorosa de homólogos resultó en el aumento de complementos Rab en el LECA a 15 subfamilias [36]. Sin embargo, grandes resoluciones filogenéticas entre los paralogos de las familias genéticas Rab incrementaron el número estimado de subfamilias Rab en el Leca a entre 19 y 23 [37]. Sorprendentemente, este análisis revelo también dos conjuntos de Rabs, uno inferido a estar involucrado en la exocitosis y otro predominantemente en endocitosis, reflejando potencialmente el establecimiento más temprano de estas vías. Como las mejoras de los métodos comparativos y philogenéticos están aplicadas a otras familias de tráfico, será importante comparar los patrones evolutivos que emergen y profundizar en los eventos pre-LECA.
Aunque el OPH es un mecanismo para la evolución del incrementado número y especialización de compartimientos dentro de un sistema de organelas, se limita actualmente al
439

sistema de transporte en membranas. Sin embargo, una idea que complementa la OPH es la hipótesis del protocoatomer, que propone, basado en la evidencia estructural de proteínas, que existe homología entre los componentes de la deformación de la membrana del transporte vesicular y el poro nuclear [38]. En concreto, las proteínas integradas en la COP I, COP II, clatrina, y los complejos del poro nuclear comparten una estructura de hélice enrollada seguido de un dominio α-solenoid. Estas proteínas están sugeridas a ser homólogos y por lo tanto derivadas de una sola proteína protocoatomer ancestral [39]. Análisis recientes también han establecido firmemente las relaciones entre las proteínas derivadas de protocoatomer del complejo intraflagelar [40]. Estas proteínas, que se dispersan por toda la célula y son esenciales para las funciones de una específica organela, parecen haberse expandido junto con sus orgánulos a través del proceso descrito en la OPH. Por lo tanto, la superposición entre las dos hipótesis extiende el mecanismo de la evolución organular autógena potencialmente a todos las organelas para las que parece probable un origen no endosimbiótica.
Existen ejemplos de organelas cuyos orígenes enturbian las divisiones entre la autogénesis y la evolución organular endosimbiótica. El origen del peroxisoma se ha explicado contenciosamente por ambos mecanismos. Aunque la evidencia, tanto funcional [41] como evolutivamente [42], favorece fuertemente un origen autógeno para los peroxisomas, hay sin duda han sido, y siguen siendo, interacciones moleculares y funcionales entre los peroxisomas y organelas de origen endosimbiótico, en particular la mitocondria [43]. Muchas proteínas que se localizan en el peroxisoma son codificadas por genes de origen bacteriano y funcionan en los procesos metabólicos compartidos con mitocondrias (por ejemplo, la oxidación de ácidos grasos). Determinar cómo las organelas endosimbióticas se han integrado dentro de la célula e interactúan con los sistemas derivados no endosimbióticamente es un área emergente de investigación para la biología celular y biología celular de la evolución. El trabajo en los últimos años ha descubierto varios complejos de proteínas que involucran proteínas, lípidos, y transportes iónicos entre el RE y las mitocondrias [44] y se ha demostrado recientemente que los complejos de proteínas que recorren el RE y las mitocondrias en los hongos están más ampliamente presentes en las eucariotas que se sospechaba [45,46].
Evolución neutral Constructiva (CNE)Los procesos evolutivos no se limitan al nivel organular. La maquinaria celular individual en la eucariota (por ejemplo, ribosomas, proteasomas) también muestran una mayor complejidad sobre sus homólogos procariotas. En algunos casos, este aumento de la complejidad podría dar lugar a nuevas funciones, proveyendo una ventaja selectiva a la célula eucariota. Sin embargo, está cada vez más en tela de juicio el papel de la selección como el único piloto en la evolución de la complejidad.
La teoría de la CNE [47] postula que pueden surgir muchos fenómenos biológicos, o ser elaborados, por procesos evolutivos neutrales que promuevan una mayor complejidad sin funcionalidad adicional [48]. CNE se basa en una idea de presupresión (Figura 2D); es decir, las interacciones entre los factores que son el resultado inicial de colisiones aleatorias o hacinamiento citosólica y esa función mínimamente afectada [49] puede estabilizarse debido a una mutación aleatoria en medida de un factor o en ambos factores. En cada una,

Tendencias en Biología Celular de julio de 2014, vol. 24, N ° 7estas mutaciones pueden ser ligeramente deletéreo para la función original, pero si la unión de la pareja restaura la funcionalidad, la interacción se vuelve estable. Por lo tanto, la mutación no es selectiva en el sentido tradicional, pero tiene que ser suficientemente compensatoria para evitar la selección negativa y para permitir que el organismo pueda sobrevivir. Estas mutaciones pueden ser extremadamente raras; sin embargo, una vez establecidos resultan ser un trinquete que promueve una unión más estrecha y, potencialmente, recluta a otros factores. Estas interacciones podrían implicar interacciones de proteínas con casi cualquier molecula o superficie de la célula (por ejemplo, pequeñas moléculas difusibles, membranas celulares).
Entre estos fenómenos biológicos, el origen de la spliceosome ha sido propuesto para requerir CNE [47,48]. Estudios genómicos comparativos de componentes del spliceosoma han demostrado que la spliceosome es una innovación eucariota que estaba presente en su estado altamente elaborada antes del LECA [50]. Consta de más de 100 proteínas diferentes y componentes de ARN, el spliceosome es un candidato para una de las más complejas maquinarias celulares de la existencia. Sin embargo, desde hace tiempo se ha apreciado que el proceso esencial subyacente podría haber evolucionado a partir de un simple grupo de auto-splicing intrón de II-clase. En lugar de ser el resultado de fuerzas selectivas, el spliceosome se explica mejor como un producto de la CNE mediante el cual las mutaciones en la molécula ARN de auto- splicing fueron suprimidos a través de una pre-existente interacción con un ARN o proteína compleja ARN [47,48]. Como componentes de proteína y ARN acumulados a lo largo tiempo, la función básica del splicing se mantuvo inalterada.
Un ejemplo reciente bien elaborado que involucra pruebas experimentales de los procesos CNE hipotéticos es el anillo V0-ATPasa vacuolar de la levadura [49,51]. Aunque el antepasado del anillo V0-ATPasa de levadura comprende dos subunidades, algunas levaduras requieren tres subunidades, con la tercera subunidad resultante de una antigua duplicación de genes que era seguida por una supresión de genes. Para verificar esta secuencia de eventos, los investigadores reconstruyeron el gen ancestral común de los existentes anillos de dos subunidades y de tres subunidades y revelaron interacciones supresoras específicas necesarias para cumplir la adopción del sistema de tres subunidades [51]. Cuando la supresión tiene éxito, el sistema, debido a esta dependencia, es más compleja; sin embargo, el efecto neto de la mayor complejidad se mantiene neutral en que ninguna alteración en la capacidad de la célula para producir el fenotipo ha ocurrido. Por lo tanto, CNE permite la acumulación de una mayor complejidad combinado con una dilución de responsabilidad para el mantenimiento de un fenotipo entre múltiples factores. Irónicamente, esta dilución, a través de la funcionalidad redundante de los componentes, podría reducir el riesgo de selección negativa en un solo objetivo mutacional y, como tal, el mecanismo de CNE en sí puede estar bajo presión selectiva positiva [48].
Observaciones finalesLo anterior fue organizado en los procesos que actúan a nivel de la organela o en el nivel del complejo molecular subyacente, pero esas divisiones no son de ninguna manera absolutas. Las maquinarias moleculares claramente cooperan para construir y definir las organelas. Al mismo tiempo, la compartimentación de específicos mecanismos moleculares por el cual un orgánulo dado limita el rango de proteínas con las que estas
440

Reseñamaquinarias moleculares frecuentemente interactuarán y por lo tanto aumenta las oportunidades para los distintos ambientes que conduzcan a la complejidad a través de los mecanismos del CNE.
En la actualidad, existen grandes oportunidades para el avance de la biología celular evolutiva como disciplina. Mientras que el campo trae a la biología evolutiva de la población y los grandes organismos a la escala de la célula, también trae un enfoque comparativo sobre las especies y el espacio a los biólogos celulares centrados en organismos específicos u organelas. Sin embargo, hay también un potencial para conceptos erróneos. En muchos aspectos, el estudio biología celular brinda similitudes conceptuales con la disciplina de la ingeniería inversa [52]. Biología celular suele entenderse desde un enfoque reduccionista por el que se desmonta la célula, tanto conceptual como físicamente, en sus componentes (proteínas, orgánulos y complejos) y luego establecidos, manipulados, y comprendidos. Por lo tanto, no es sorprendente que las cuestiones relativas a los mecanismos evolutivos que dan origen a las células a veces se malinterpreten como un problema de la ingeniería avanzada; es decir, '¿Cómo encontró la célula la forma más eficiente de llevar a cabo el proceso' 'x' '?' Sin embargo, hay una diferencia fundamental entre la evolución e ingeniería. La evolución no siempre procede a lo largo de una ruta optimizada al estado moderno observado.
Viendo cada rasgo como el resultado de un proceso iterativo y mecanicista, en lugar de teleológica, que lleva a estas soluciones cambia la forma en que las investigaciones son entendidas y los datos son interpretados. Aunque puede seguir siendo útil preguntar '¿Cuál es la ventaja selectiva de un rasgo dado?', sabiendo que el camino evolutivo no siempre es directo y constante permite al investigador considerar múltiples ventajas y posiblemente considerar explicaciones alternativas más allá de la selección. Por lo tanto, puede ser más productivo responder el "cómo" detrás de interrogantes biológicas evolutivas celulares y reconstruir los pasos y detalles evolucionarios para la aparición de un rasgo dado, proceso que deriva tanto de los patrones observados a través de varios ejemplos.
Aunque se han logrado avances significativos en el desarrollo de los sistemas biológicos celulares de eucariotas (por ejemplo,Dictyostelium, Toxoplasma, Trypanosoma, Arabidopsis) y el análisis de la evolución molecular de deducir el origen de complejos proteicos y sus organelas residentes, produciendo algunos de los descubrimientos descritos anteriormente, muchas áreas aún se mantienen inexplorados. Por ejemplo, las consecuencias de la genética de poblaciones no se han explorado completamente en el contexto de la evolución celular [53]. Del mismo modo, aunque ha habido intentos para correlacionar la geología con la evolución celular [54], particularmente en relación con el origen de la vida [55], este aspecto es a menudo pasado por alto por los biólogos celulares. Por otra parte, los mecanismos de la aparición de innovaciones evolutivas, como herencia de organelas, que combinan múltiples, componentes celulares bien adaptados quedan por ser mejor aclarados [56]. Por último, como nuestra comprensión de los sistemas biológicos crece y los datos e información sobre ómica son cada

vez más accesibles, vamos a ser capaces de integrar la información sobre el tiempo y el contexto de genes y proteínas en varios modelos de evolución celular [57].
Con este progreso tratable en progreso con preguntas mecanicistas concretas, es realmente un momento emocionante como la biología de la célula ahora se puede analizar 'a la luz de la evolución " [58].


![CELULA EUCARIOTA COMPLETOP[1][1]](https://static.fdocuments.mx/doc/165x107/5571fd3f497959916998b16b/celula-eucariota-completop11.jpg)














