Informe Rev Src
23
PONTIFICIA UNIVERSIDAD CATÓLICA DE VALPARAÍSO FACULTAD DE INGENIERÍA ESCUELA DE INGENIERÍA BIOQUÍMICA Tópicos de Cultivo en Continuo (EIB7032) “Análisis de crecimiento celular en quimiostato bajo simple y doble limitación por sustrato” Paulina Aguirre. Silvia Rojas. Luciana Santibáñez.
-
Upload
silvia-rojas-caro -
Category
Documents
-
view
36 -
download
0
Transcript of Informe Rev Src

PONTIFICIA UNIVERSIDAD CATÓLICA DE VALPARAÍSO
FACULTAD DE INGENIERÍA
ESCUELA DE INGENIERÍA BIOQUÍMICA
Tópicos de Cultivo en Continuo (EIB7032)
“Análisis de crecimiento celular en quimiostato bajo simple y doble limitación por sustrato”
Paulina Aguirre.
Silvia Rojas.
Luciana Santibáñez.
Profesor: Fernando Acevedo
Mayo, 2013.

RESUMEN
Las condiciones de cultivo influyen notablemente en la composición celular y en el
funcionamiento metabólico de las células microbianas. Uno de los parámetros más importantes en el
cultivo microbiano es la disponibilidad de nutrientes, por lo que se han planteado diversos modelos
para predecir el comportamiento bajo simple y doble limitación de nutrientes. El objetivo de este
trabajo fue realizar un análisis comparativo de las predicciones de dos modelos de limitación doble
por sustrato: modelos aditivo y ponderado, tomando como referencia el modelo de Monod para
limitación simple y los para doble limitación, en un caso de estudio determinado que consiste en un
cultivo continuo bajo limitación por fuente de carbono y nitrógeno, donde se analizó el
comportamiento de la productividad volumétrica, concentración de biomasa y sustratos. Para lo
anterior fue necesario desarrollar ecuaciones que describieran el comportamiento de cada variable
en función de la tasa de dilución a partir de balances de masa y la relación entre la velocidad de
crecimiento y las concentraciones de sustrato residual presentadas por cada modelo. Los resultados
indican que el comportamiento de las variables analizadas en base a los distintos modelos muestra
similitudes entre sí, lo que estaría dado por la estrecha relación entre el modelo básico de limitación
simple de Monod y los modelos de limitación doble utilizados en este caso. El modelo ponderado
sería el más consistente dado que se obtiene un perfil similar al obtenido por limitación simple según
Monod en función de la tasa de dilución.

ÍNDICE
1. INTRODUCCIÓN................................................................................................................................2
2.1 PLANTEAMIENTO DEL PROBLEMA..............................................................................................2
2.2 DESARROLLO...............................................................................................................................2
2.2.1 DISEÑO DE MEDIO DE CULTIVO..................................................................................................3
2.2.2 BALANCES DE CULTIVO CONTINUO........................................................................................3
2.2.3 LIMITACIÓN SIMPLE...............................................................................................................5
2.2.4 DOBLE LIMITACIÓN POR CARBONO Y NITRÓGENO, SEGÚN MODELO ADITIVO.....................5
2.2.5 MODELO PONDERADO DE MANKAD Y BUNGAY....................................................................7
3.1 LIMITACIÓN SIMPLE POR LA FUENTE DE CARBONO......................................................................8
3.2 COMPORTAMIENTO DEL CULTIVO CONTINUO POR LIMITACION DOBLE DE SUSTRATO SEGÚN MODELO ADITIVO...............................................................................................................................9
3.3 Comportamiento del cultivo continuo por limitación doble de sustrato de acuerdo al modelo ponderado...........................................................................................................................................9
4. DISCUSIÓN.....................................................................................................................................10
5. CONCLUSIONES..............................................................................................................................12
REFERENCIAS.........................................................................................................................................13
ÍNDICE DE FIGURAS
Figura 3.1: Comportamiento de un cultivo continuo bajo limitación de la fuente de carbono...............7
Figura 3.2. Comportamiento de un cultivo continuo bajo limitación de la fuente de nitrógeno............8
Figura 3.3. Comportamiento de un cultivo continuo bajo limitación doble de la fuente de carbono y nitrógeno según el modelo aditivo..........................................................................................................9
Figura 3.4. Comportamiento de un cultivo continuo bajo limitación doble de la fuente de carbono y nitrógeno según el modelo ponderado.................................................................................................10
ÍNDICE DE TABLAS
Tabla 3.1: Resumen de los datos de Productividad volumétrica de Biomasa máxima (Qx máx), Tasa de Dilución de Inflexión (Dinflexión) y Tasa de Dilución crítica (Dc).................................................................10
Silvia, 28/05/13,
REVISAR INDICE

1. INTRODUCCIÓN
Existen varias características de los microorganismos que están directamente influenciadas por el
ambiente y su composición química. Esto se cumple de tal forma que no podemos hablar de la
composición química de un microorganismo sin al mismo tiempo especificar las condiciones medio
ambientales en las que éste se produce. En este contexto, la disponibilidad de nutrientes es un
parámetro clave en el cultivo de células microbianas. En efecto, la limitación de nutrientes
específicos es utilizada en gran cantidad de procesos biotecnológicos para inducir u optimizar la
producción de ciertos productos microbianos (Egli and Zinn, 2003). Frecuentemente, los medios de
cultivo para microorganismos han sido diseñados bajo el principio básico de nutrición biológica,
formulado por Justus von Liebig (1840), la “ley del mínimo”. Este principio planteaba que en
microbiología siempre existía un nutriente en particular que determina la máxima cantidad de
biomasa que puede ser producida en un medio de cultivo definido (Egli and Zinn, 2003; Gottschal,
1993). El modelo más utilizado en la limitación simple por sustrato es el de Monod, que indica una
relación algebraica simple y continua que permite realizar fácilmente un análisis teórico del
crecimiento celular. Plantea que la dependencia del substrato de la velocidad de crecimiento
específica (µ) es de forma que si la cantidad de substrato es muy grande la tasa específica se
aproxima al valor máximo y si la concentración del substrato tiende a cero, se aproxima a cero (Bader,
1978; Monod, 1949).
Sin embargo, existen evidencias experimentales en la literatura que contradicen el principio de
von Liebig, ya que muestran que el crecimiento de cultivos microbianos puede verse limitado por dos
o más nutrientes simultáneamente (Bader, 1978; Zinn et al., 2004). A la par con lo anterior, se
desarrollaron modelos matemáticos capaces de describir el crecimiento microbiano bajo distintos
tipos de doble limitación por sustrato. Tomando en cuenta el crecimiento de un microorganismo en
presencia de varios sustratos y asumiendo que cada uno de ellos tiene un efecto de captación
competitiva sobre la captación de otros sustratos, se planteó el modelo aditivo (Blanch and Clark,
1997). Otro modelo empleado en la limitación por más de un nutriente es el modelo aditivo
ponderado (Mankad and Bungay, 1988). En este caso, la tasa de crecimiento bajo doble limitación de
sustrato es expresada en términos del promedio ponderado de estas tasas bajo limitación simple por
sustrato.
1

El objetivo de este trabajo fue comparar los comportamientos predichos por el modelo aditivo y
el modelo ponderado de Mankad y Bungay. Además lo anterior fue comparado también con el
comportamiento bajo limitación simple por glucosa.
2. METODOLOGÍA
2.1. PLANTEAMIENTO DEL PROBLEMA
Analizar el comportamiento de un quimiostato en función de D (X, S i, Qx, µ) sometido a doble
limitación por carbono (glucosa) y nitrógeno (sulfato de amonio) bajo las siguientes condiciones:
Tabla 2.1. Condiciones de operación conocidas
X a D = 0,30 h-1 7,5 g L-1
µmax 0,56 h-1
Ks glucosa 4,5 mg·L-1
Ks NH4+ 0,13 mg·L-1
C en célula 49%
N en célula 13%
Cultivo Continuo aerobio
Rendimientos Constantes con µ
2.2. DESARROLLO
Bajo las condiciones mencionadas, se comparó el comportamiento de un cultivo continuo de
acuerdo a lo siguiente:
i) Limitación simple por fuente de carbono (glucosa).
2

ii) Limitación doble por carbono y nitrógeno, según Modelo Aditivo (suponiendo un solo
µmax).
iii) Limitación doble de acuerdo al Modelo Ponderado de Mankad y Bungay (Mankad and
Bungay, 1988).
A partir de las ecuaciones de balance de masa propias del sistema de cultivo continuo y la
relación entre la velocidad de crecimiento y la concentración de sustrato en el reactor, de cada
modelo, se obtuvo las ecuaciones que describen el comportamiento de la productividad, la
concentración de biomasa (X) y sustrato (Si), en función la tasa de dilución (D).
2.2.1. DISEÑO DE MEDIO DE CULTIVO
Para desarrollar el medio de cultivo es necesario conocer algunos factores como: concentración
celular, composición elemental de biomasa además los rendimientos de los nutrientes en la células,
así como la fuente de cada elemento utilizado como nutriente. Si tomamos como referencia la base
indicada en la tabla 2.1 sobre la tasa de dilución (D) igual 0,3 h -1 con limitación de nitrógeno y carbono
se puede recurrir al cálculo de los rendimientos (YX/C) e (YX/N), considerando el contenido del nutriente
en la célula y en el sustrato, planteando las siguientes ecuaciones:
Para la fuente de carbono:
(ec. 1)
Para la fuente de nitrógeno:
(ec.2)
Para un microorganismo aerobio, se debe considerar un factor de corrección (f ) igual a 0,6
en la ecuación (ec.1), debido a que no todo el carbono de la fuente de carbono y energía, es
3
Silvia, 28/05/13,
Verificar que se vean bien las ecuaciones.

metabolizado hacia la generación de biomasa. A partir de este valor, se puede obtener la
concentración de los sustratos en la alimentación del quimiostato.
El porcentaje de carbono en glucosa es de un 40% y el de nitrógeno en amonio de un 76%.
Por lo tanto los valores de rendimientos para glucosa y nitrógeno son YX/c= 49, YX/N= 1,63,
respectivamente.
2.2.2. BALANCES DE CULTIVO CONTINUO
Se consideró un sistema de quimiostato de mezcla completa, con alimentación estéril,
volumen de fermentación constante, ausencia de inhibidores y en el cual el único producto obtenido
es biomasa (Fig. 1). En base a lo anterior, se plantearon los siguientes balances:
Figura 2.1. Esquema de un quimiostato simple.
Balance general de biomasa
(ec 3)
Considerando volumen constante
(ec 4)
4
Silvia, 28/05/13,
Verificar que todos queden en la misma posición

Y sea:
(ec 5)
Se obtiene:
(ec 6)
En estado estacionario, donde , se cumple que:
(ec 7)
Balance sustratos:
(ec 8)
Suponiendo estado estacionario
(ec 9)
Dado que :
(ec 10)
Despejando la concentración de sustrato en el reactor, se obtiene que
(ec 11)
5

Productividad volumétrica:
(ec 12)
Se realizaron los cálculos correspondientes con la finalidad de determinar la relación SC0/SN0
sin omitir las concentraciones Sc y SN de nuestro sistema planteado bajo los modelos aditivo y
ponderado de Mankad y Bungay.
2.2.3. LIMITACIÓN SIMPLE
De acuerdo a lo descrito en la sección 2.2.2 y utilizando la ecuación de Monod, que describe la
velocidad de crecimiento en función de la concentración de sustrato en un cultivo con limitación
simple, tenemos que:
; para carbono (ec 13)
; para nitrógeno (ec 14)
Reemplazando la ecuación 11 en la ecuación 13 y 14, para carbono y nitrógeno respectivamente, y
despejando la variable X tenemos que:
(ec 15)
(ec 16)
6

A partir de la ecuación 15 y 16 se obtienen los perfiles de biomasa (X), Concentración de carbono (S c)
y productividad volumétrica (Qx) en función de la tasa de dilución (D).
2.2.4. DOBLE LIMITACIÓN POR CARBONO Y NITRÓGENO, SEGÚN MODELO ADITIVO.
Considerando el modelo aditivo para doble limitación por carbono y nitrógeno tenemos:
(ec 17)
En base a la ecuación 17 y reemplazando la ecuación 11 para concentración de carbono y nitrógeno
obtenemos la siguiente ecuación:
(ec 18)
De la ecuación 18, obtenemos SCO, que posteriormente sirve para determinar la relación SCo/ SNo
(ec 19)
Reemplazando por los valores numéricos de μ, μmax, ksc, ksn, X, y simplificando la ecuación 19 se tiene:
Luego
7

Así mismo, resolviendo el sistema compuesto por las ecuaciones 13 a la 17, se obtiene la
relación el perfil de comportamiento de la biomasa (X) en función de Sco, Sno, bajo
limitación doble. Biomasa (X) estará definida por:
(ec 20)
Donde:
2.2.5 MODELO PONDERADO DE MANKAD Y BUNGAY
En el modelo ponderado (Mankad and Bungay, 1988) se define la siguiente relación:
(ec 21)
Donde W1 y W2 son los factores de ponderación en función de Ki y Si, y están definidos de la siguiente
manera:
Reemplazando W1 y W2, la ecuación 21 se puede reescribir de la siguiente manera:
(ec 22)
8

Utilizando el mismo procedimiento de las secciones 2.2.3 y 2.2.4, se obtuvo las ecuaciones con la
finalidad de obtener Sco y posteriormente la relación SCO/Sno, y concentración de biomasa. Para este
casos en particular, debido la dificultad para visualizar las ecuaciones por su complejidad, se las
presenta por secciones.
(ec 23)
Donde:
Reemplazando por los valores numéricos de μ, μmax, ksc, ksn, X, se tiene:
Para el cálculo de la concentración de biomasa (X):
Donde:
9

Todos los cálculos de este trabajo fueron realizados con la aplicación Mathematica 7.0 (Wolfram,
2010).
3. RESULTADOS
Como es de esperar según lo reportado en la literatura, los resultados muestran diferencias en el
comportamiento obtenido bajo la predicción de los modelos aditivo y ponderado de limitación doble.
Para esquematizar los comportamientos en los distintos casos, se resolvió los sistemas de ecuaciones
planteados anteriormente de modo de representar la concentración celular (X), concentración del
nutriente limitante (Si) y productividad volumétrica de biomasa (Qx) en función de la velocidad de
dilución (D).
3.1. LIMITACIÓN SIMPLE POR LA FUENTE DE CARBONO
En el caso de limitación simple de la fuente de carbono, se utilizó una concentración de
alimentación de glucosa (SC0) de 15,31 g·L-1, valor que se obtuvo a partir de los cálculos de diseño del
medio de cultivo. En la Figura 3.1 podemos observar que la concentración celular comienza a
disminuir aproximadamente a una D = 0,5 h-1, alcanzando las condiciones de lavado a una tasa de
dilución crítica (Dc)= 0,55 h-1. La concetración de glucosa residual (Sc) se mantiene constante con
valores cercanos a cero, y aumenta exponencialmente a partir de D= 0,5 h -1. La productividad
volumétrica (Qx) alcanza un valor máximo igual a 3,5 L-1h-1 a un valor de D= a 0,5 h-1, donde
posteriormente disminuye drásticamente.
10

Figura 3.1: Comportamiento de un cultivo continuo bajo limitación de la fuente de carbono.
3.2. COMPORTAMIENTO DEL CULTIVO CONTINUO POR LIMITACION DOBLE DE SUSTRATO SEGÚN MODELO ADITIVO
Con respecto a la limitación doble de sustrato de acuerdo al modelo aditivo, se utilizó una
concentración de alimentación de glucosa (SC0) de 15,31 g·L-1 y de nitrógeno (Sno) de 4,60 g·L-1, que se
obtuvieron a partir del diseño del medio de cultivo. En la Figura 3.2 se representa X, S c, Sn y QX en
función de la tasa de dilución (D). Podemos observar que los valores de la concentración de biomasa
disminuyen a partir de un valor de Dc = 1,1 h-1, alcanzando las condiciones de lavado; a medida que
aumenta la velocidad de crecimiento, aumenta la productividad volumétrica hasta un Q x máximo de
8,23g·L-1 ·h-1, a la misma tasa de dilución, la concentración de Sc y Sn comienzan a incrementarse.
11

Figura 3.2. Comportamiento de un cultivo continuo bajo limitación doble de la fuente de carbono y
nitrógeno según el modelo aditivo.
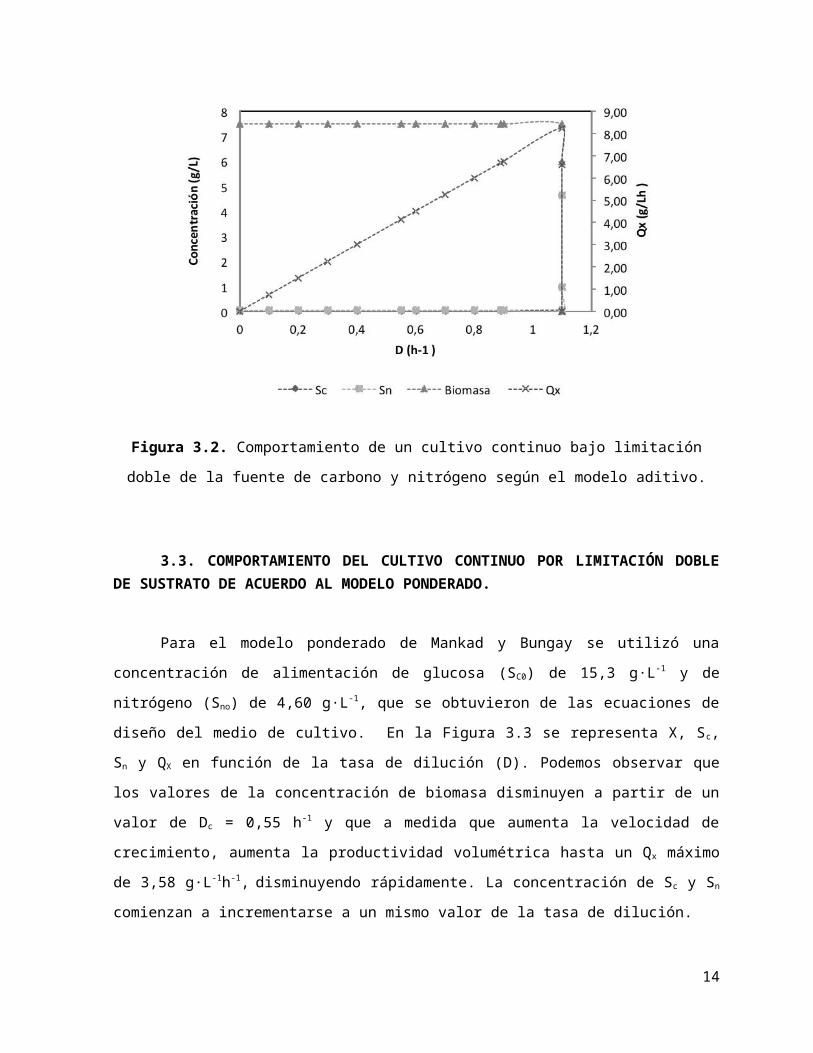
3.3. COMPORTAMIENTO DEL CULTIVO CONTINUO POR LIMITACIÓN DOBLE DE SUSTRATO DE ACUERDO AL MODELO PONDERADO.
Para el modelo ponderado de Mankad y Bungay se utilizó una concentración de alimentación
de glucosa (SC0) de 15,3 g·L-1 y de nitrógeno (Sno) de 4,60 g·L-1, que se obtuvieron de las ecuaciones de
diseño del medio de cultivo. En la Figura 3.3 se representa X, Sc, Sn y QX en función de la tasa de
dilución (D). Podemos observar que los valores de la concentración de biomasa disminuyen a partir
de un valor de Dc = 0,55 h-1 y que a medida que aumenta la velocidad de crecimiento, aumenta la
productividad volumétrica hasta un Qx máximo de 3,58 g·L-1h-1, disminuyendo rápidamente. La
concentración de Sc y Sn comienzan a incrementarse a un mismo valor de la tasa de dilución.
12

Figura 3.3. Comportamiento de un cultivo continuo bajo limitación doble de la fuente de carbono y nitrógeno según el modelo ponderado.
4. DISCUSIÓN
Realizando un análisis comparativo de los modelos antes mencionados podemos decir lo
siguiente:
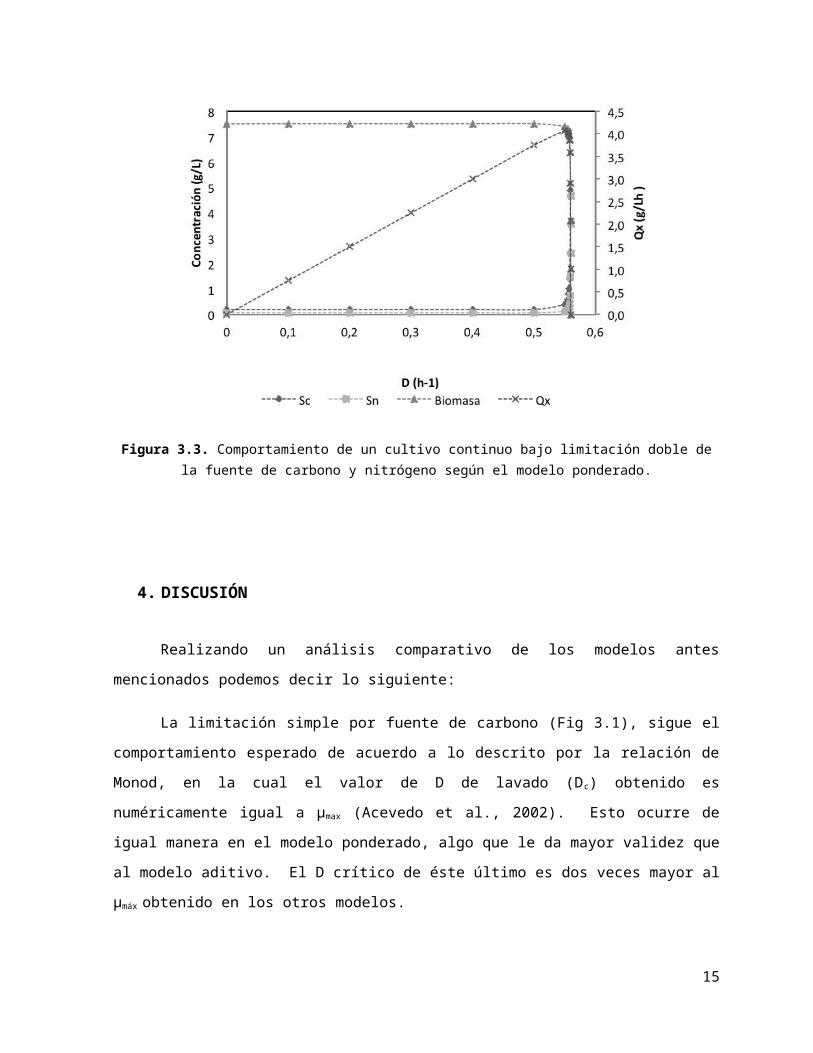
La limitación simple por fuente de carbono (Fig 3.1), sigue el comportamiento esperado de
acuerdo a lo descrito por la relación de Monod, en la cual el valor de D de lavado (D c) obtenido es
numéricamente igual a µmax (Acevedo et al., 2002). Esto ocurre de igual manera en el modelo
ponderado, algo que le da mayor validez que al modelo aditivo. El D crítico de éste último es dos
veces mayor al µmáx obtenido en los otros modelos.
Por otro lado visualizando la Figura 3.2, podemos observar como la suma de las velocidades
de crecimiento (µmáx) en el modelo aditivo, sobredimensionan el sistema provocando un alto valor de
D crítico, lo que le resta validez al modelo, puesto que en sistemas con alimentación estéril no es
13

posible alcanzar D críticos mayores a la velocidad de crecimiento máximo del microorganismo.
Lamentablemente no se encontró referencia donde el modelo aditivo sea válido experimentalmente.
De igual manera se puede señalar que los valores entregados por el modelo aditivo, a tasas
de dilución menores a µmáx concuerdan con los obtenidos por el modelo ponderado y la ecuación de
Monod, lo que indica que hasta este rango el modelo se ajusta. Matemáticamente, en sistemas
donde KS2 posee valores muy elevados, el modelo se ajusta a la ecuación de Monod debido que el
segundo término de la ecuación es despreciable.
Analizando la ecuación 22, se puede ver que el modelo ponderado guarda similitud con el
modelo multiplicativo, donde el factor , siempre mayor que 1, corrige los bajos
valores de μ que suele entregar el modelo multiplicativo cuando las constantes de afinidad (Ks) son
mayores a uno; sin embargo, dado los bajos valores de Ks para este caso, este factor no causa un
efecto significativo sobre μ.
Se debe indicar además, que los sistemas de ecuaciones obtenidos para cada caso (modelo
aditivo y ponderado) poseen solución analítica, sin embargo la complejidad de las ecuaciones
resultantes hacen difícil su evaluación, por lo que es necesario el uso de software matemáticos
avanzados.
5. CONCLUSIÓN
El comportamiento de las variables obtenidas mediante el modelo ponderado es similar a los
perfiles de limitación simple por Monod en función de la tasa de dilución. Sin embargo se requieren
datos experimentales para comprobar la representatividad de este modelo. En cuanto al modelo
aditivo, en este caso carece de validez, dado que sobredimensiona el sistema.
14
REFERENCIAS
Acevedo F, Gentina JC, Illanes A. 2002. Fundamentos de Ingeniería Bioquímica 3rd ed. Valparaíso: Ediciones Universitarias de Valparaíso. Pontifica Universidad Católica de Valparaíso.
Bader FG. 1978. Analysis of Double-Substrate Limited Growth. Biotechnology and bioengineering XX:183–202.
Blanch HW, Clark DS. 1997. Biochemical Engineering 2nd ed. New York: Marcel Dekker, Inc.
Egli T, Zinn M. 2003. The concept of multiple-nutrient-limited growth of microorganisms and its application in biotechnological processes. Biotechnology Advances 22:35–43.
Gottschal JC. 1993. Growth kinetics and competition - some contemporary comments. Antonie van Leeuwenhoek 63:299–313.
Mankad T, Bungay HR. 1988. Model for microbial growth with more than one limiting nutrient. Journal of Biotechnology 7:161–166.
Monod J. 1949. The Growth of Bacterial Cultures. Annual review of microbiology 3:371–394.
Wolfram. 2010. Mathematica. Champaign, Illinois: Wolfram Research, Inc.
Zinn M, Witholt B, Egli T. 2004. Dual nutried limited growth: models, experimental observations, and applications. Journal of Biotechnology 113:263–279.
15
NOMENCLATURA
D = Tasa de dilución (h-1)
X = Concentración de biomasa en el quimiostato (g·L-1)
Si = Concentración de sustrato i en la alimentación (g·L-1)
Qx = Productividad volumétrica de biomasa (g·L-1·h-1).
µ = Velocidad especifica de crecimiento (h-1)
µmax = Velocidad máxima de crecimiento (h-1)
ksc = Constante de saturación con respecto a la fuente de carbono (g·L-1)
ksn = Constante de saturación con respecto a la fuente de nitrógeno (g·L-1)
Y(X⁄C) = Rendimiento de producción de biomasa con respecto a la fuente de carbono (g biomasa/g glucosa)
Y(X⁄n)= Rendimiento de producción de biomasa con respecto a la fuente de nitrógeno (g biomasa/g sulfato amonio.)
F = Flujo de alimentación (L· h-1)
V = Volumen del quimiostato (L)
SC = Concentración de la fuente de carbono en el quimiostato (g·L-1)
Sn = Concentración de la fuente de nitrógeno en el quimiostato (g·L-1)
Sco = Concentración de la fuente de carbono en la alimentación del quimiostato (g·L-1)
Sno= Concentración de la fuente de nitrógeno en la alimentación del quimiostato (g·L-1)
16