
METABOLITOS SEC LA PATAGONIA AR BIOLÓGICA ...
226
METABO LA PATA BIOLÓG Tesista: Directora: Codirector: OLITOS SECUNDARIOS DE AGONIA ARGENTINA. SU A GICA Lic. José Miguel Gilabert Valero Dra. Alicia Bardón Dr. Mario E. Arena Cas E HEPÁTICAS DE ACTIVIDAD Foto: Guillermo Palavecino. scada del lago Steffen, Rio Negro, Argentina.
Transcript of METABOLITOS SEC LA PATAGONIA AR BIOLÓGICA ...

METABOLITOS SECUNDARIOS DE HEPÁTICAS DE
LA PATAGONIA ARGENTINA. SU ACTIVIDAD
BIOLÓGICA
Tesista:
Directora :
Codirector:
METABOLITOS SECUNDARIOS DE HEPÁTICAS DE
PATAGONIA ARGENTINA. SU ACTIVIDAD
BIOLÓGICA
Lic. José Miguel Gilabert Valero
Dra. Alicia Bardón
Dr. Mario E. Arena
Cascada del lago Steffen, Rio Negro, Argentina.
METABOLITOS SECUNDARIOS DE HEPÁTICAS DE
PATAGONIA ARGENTINA. SU ACTIVIDAD
Foto: Guillermo Palavecino.
Cascada del lago Steffen, Rio Negro, Argentina.

UNIVERSIDAD NACIONAL DE TUCUMÁN
FACULTAD DE BIOQUÍMICA, QUÍMICA Y FARMACIA HONORABLE CONSEJO DIRECTIVO Dra. Silvia Fernández de Brunetti Dra. María Graciela Agüero Lic. María Cristina Torres Mag. Adriana Correa Zeballos Master María Regina Rintoul Dra. María Ángela Jure Bioq. Farm. David José Merep Sr. Mario Luis Rodríguez Sr. Joaquín Di Pasquale Srta. Cecilia Lludgar Sr. Maximiliano Gastal
DECANA Dra. Silvia Nelina González
VICE-DECANA Dra. Aida Ben Altabef
SECRETARIA DE ASUNTOS ACADEMICOS Dra. Marta Elena Cecilia
JEFA DEL DEPARTAMENTO POSGRADO Lic. Marta Quinteros

DEPARTAMENTO DE POSGRADO
AUTORIDADES
DIRECTORA
Dra. Florencia Fagalde
REPRESENTANTE TITULAR EN EL DEPARTAMENTO DE POSGRADO DE LA UNT
Dra. Rosana Chehín
CONSEJO TITULAR
Dra. Florencia Fagalde Dra. María Inés Ybarra Dra. Adriana María Sales Dra. Nancy Roxana Vera Dra. María Eugenia Bibas Bonet Dra. Rosana Chehín Farm. Mónica Eugenia Nacir
CONSEJO SUPLENTE
Dra. Sonia Beatriz Díaz Dra. María Cristina Rubio de Recúpero

TRABAJO DE POSGRADO PARA LA OBTENCIÓN DEL GRADO ACADÉMICO SUPERIOR DE DOCTOR EN CIENCIAS
QUÍMICAS
PROGRAMA DE POSGRADO EN CIENCIAS QUÍMICAS
Acreditado y Categorizado A-Doctorado según Resolución nº: 115/03 y B-Maestría según Resolución nº: 114/03 ante la
Comisión Nacional de Acreditación Universitaria (CONEAU)
Directora
Dra. Aída Ben Altabef
Comité Académico
Dra. Aída Ben Altabef
Dr. Eleuterio Luis Arancibia Dra. Carola Schuff

TRABAJO DE POSGRADO TITULADO
METABOLITOS SECUNDARIOS DE HEPÁTICAS DE LA PATAGONIA ARGENTINA. SU ACTIVIDAD BIOLÓGICA
TESISTA
Lic. José Miguel Gilabert Valero
DIRECTORA
Dra. Alicia Bardón
CODIRECTOR
Dr. Mario E. Arena
COMISIÓN DE SUPERVISIÓN
Dra. Adriana Neske Dr. Juan Carlos Valdéz

A mi madre y hermanos.
A mi padre quien me incitó a recorrer este camino y a quien extraño
profundamente.
A Marina.

AGRADECIMIENTOS
A la directora de este trabajo de tesis, la Dra. Alicia Bardón, por sus enseñanzas y
sus precisos consejos no sólo en lo relacionado al trabajo científico.
Al codirector de este trabajo de tesis, el Dr. Mario Arena, por su ayuda, confianza y
amistad.
Al Dr. Manuel González Sierra por sus enseñanzas sobre espectroscopia, su
amabilidad y predisposición para registrar los espectros de RMN y por su amistad.
A la Dra. María Schiavone por su invalorable labor en la determinación taxonómica
de las especies estudiadas.
Al Dr. Juan Carlos Valdés y Dra. Adriana Neske, quienes formaron parte de la
comisión de supervisión, por el interés demostrado y sus oportunas sugerencias.
A las personas que colaboraron en el trabajo experimental durante las pasantías
que realizaron en la cátedra, Lic. Karenina Marcinkevicius, Lic. Rodrigo Gibilisco y Lic.
Álvaro Guzmán.
Al Lic. Livio Corzo por la realización del estudio histológico en los ensayos con
insectos.
Al Dr. Alberto Ramos por su ayuda en los ensayos biológicos con bacterias.
A mis amigos y compañeros de trabajo Lic. Federico Arrighi, Lic. Lilian Di Toto
Blessing, Dr. Marcos Derita, Lic. Lucrecia Arias, Lic. Abel Arrollo Aguilar, Lic. Mariana
Díaz y Lic. Eduardo Parellada por su ayuda, compañía y afecto.
A los docentes del Instituto de Química Orgánica y el laboratorio de cromatografía,
Dra. Elena Cartagena, Dra. Nancy Vera, Lic. Marta Ramírez, Dra. María Inés Ibarra y
Dra. Susana Kousal por su buena predisposición para responder a mis preguntas.
A la Dra. Cecilia Socolsky por registrar los espectros de RMN de 500 MHz de algunos
de los compuestos.
Al Consejo de Investigaciones Científicas y Técnicas (CONICET) por las becas
otorgadas para la realización de esta tesis.

“I believe the simplest explanation is, there is no God. No one created the universe
and no one directs our fate. This leads me to a profound realization that there
probably is no heaven and no afterlife either. We have this one life to appreciate the
grand design of the universe and for that, I am extremely grateful.”
“Creo que la explicación más simple es, no hay Dios. Nadie creó el universo y nadie
dirige nuestro destino. Esto me lleva a la profunda reflexión de que probablemente no
hay cielo ni tampoco vida después de la muerte. Tenemos sólo esta vida para apreciar
el magnífico diseño del universo y por eso, estoy extremadamente agradecido.”
― Stephen Hawking

INDICE
Resumen…………….……………………………………………..…………………………………….. 13
Capítulo I: Introducción
1.1 Generalidades.…………..……………………………………………………………......... 15
1.2 Antecedentes químicos………………………………………………………………... 19
1.2.1 Terpenoides………………………………………………………………………… 20
1.2.1.1 Monoterpenoides……………………....………………………………………… 20
1.2.1.2 Sesquiterpenoides…………………..………..………………………………….. 20
1.2.1.3 Diterpenoides……………………….……………………………………………... 24
1.2.1.4 Esteroides y triterpenoides…………………………………………………... 25
1.2.2 Compuestos Aromáticos………………………………………………………. 26
1.2.2.1 Derivados del ácido benzoico y del ácido cinámico…………………26
1.2.2.2 Bibencilos……………………………………………………...……………………. 26
1.2.2.3 Bis-bibencilos……………………………………………………………………… 27
1.2.2.4 Dímeros de bis-bibencilos…………………………………………………….. 27
1.2.2.5 Fenoles de cadena alquílica larga………………………………………….. 28
1.2.2.6 Naftalenos e isocumarinas………………………………………………........ 28
1.2.2.7 Neolignanos, fenantrenos, ftálidos y otros compuestos
aromáticos…………………………………………………………………………... 28
1.2.2.8 Flavonoides…………………………………………………………………………. 29
1.2.3 Lipidos………………………………………………………………………………… 30
1.2.3.1 n-Alcanos………………….…………………………………………………………. 30
1.2.3.2 Ácidos grasos………………………………………………………………………. 30
1.2.4 Carbohidratos……………………………………………………………………… 30
1.2.5 Compuestos con azufre………………………………………………………… 30
1.3 Antecedentes biológicos…..………………………………………………………….. 30
1.3.1 Sabores picantes, amargos y dulces.………..…………………………….. 33
1.3.2 Dermatitis alérgica por contacto.………………………………………….. 33

1.3.3 Actividad antitumoral…..………………………………………………………. 34
1.3.4 Actividad molusquicida…..……………………………………………………. 35
1.3.5 Actividad antialimentaria de insectos………….…..……………………. 35
1.3.6 Actividad inhibidora de la liberación de anión superóxido.......... 35
1.3.7 Actividades antimicrobianas y antifúngicas..….…….………………... 36
1.3.8 Actividad piscicida……………..……………….……………………………….. 36
1.4 Referencias………………………………………………………………….……..………… 38
Capítulo II: Materiales y métodos
2.1 Recolección, clasificación y preparación del material
Vegetal………………………………………………………………………………………….. 42
2.2 Extracción….…………………………………………………………………………………. 42
2.3 Purificación y aislamiento..………………………………………………………….. 43
2.4 Identificación de los compuestos..……………………………………………….. 45
2.4.1 Espectroscopía infrarroja (IR)………………………………………………. 45
2.4.2 Espectroscopía ultravioleta-visible (UV)……………………………….. 45
2.4.3 Espectrometría de masas (EM)……………………………………………... 46
2.4.4 Resonancia magnética nuclear (RMN)…………………………………... 47
2.4.5 Dicroísmo circular (DC)……………………………………………………….. 47
2.5 Actividad biológica……………………………………………………………………… 49
2.5.1 Actividad anti-patogénica sobre P. aeruginosa y S. aureus……….49
2.5.1.1 Determinación del crecimiento bacteriano…………………………… 50
2.5.1.2 Determinación de la formación de biofilm……………………….......... 50
2.5.1.3 Evaluación de los efectos sobre el proceso de QS……………………51
2.5.1.4 Ensayo sobre la actividad elastasa………………………………………… 52
2.5.2 Actividad insecticida sobre S. frugiperda……………………………….. 53
2.5.2.1 Insectos y dieta……………………………………………………………………. 53
2.5.2.2 Bioensayo de toxicidad………………………………………………………… 53
2.5.2.3 Análisis histológico………………………………………………………………. 54
2.6 Referencias…………………………………………………………………………………… 55

Capítulo III: Estudio químico de Porella chilensis
3.1 Antecedentes químicos de la familia Porellaceae….……………………. 58
3.2 Clasificación botánica de P. chilensis……………..………………..…………… 58
3.3 Experimental…….………....……………...……………………………………...………... 59
3.3.1 Material vegetal……...………….………………………………......................... 59
3.3.2 Extracción y aislamiento…………………….…………….………………….. 59
3.4 Resultados……………………………………………………………….…...……..……….. 60
3.4.1 Elucidación estructural de los nuevos compuestos 1 y 2………... 60
3.4.2 Determinación de la configuración absoluta de 2………..…............ 64
3.4.3 Determinación de la configuración absoluta mediante cálculos
computacionales y DC………………………………………………………….. 67
3.4.4 Estudio quimiotaxonómico de P. chilensis……………..……….……… 69
3.4.5 Datos espectrales de 1 y 2………………………….…………………..…….. 70
3.5 Referencias…………….…………………………………………..…………..…………….. 72
Capítulo IV: Actividad anti-patogénica de Porella
chilensis
4.1 Generalidades……………………………………………..………………….…………….. 76
4.2 Experimental………………...………………………………………………….…………... 76
4.2.1 Crecimiento bacteriano……………………………………….……………….. 77
4.2.2 Ensayos sobre la formación de biofilm……………..…………….……... 77
4.2.3 Ensayo de efectos sobre el QS…………………………………….…………. 77
4.2.4 Análisis estadístico…..………………………………………………….……….. 77
4.3 Resultados………………………………………………………………………..…….…….. 77
4.3.1 Fusicoccanos……………………………………………………………………….. 78
4.3.2 Pinguisanos…………………………………………………………………………. 78
4.3.3 Aromadendranos…………………………………………………………………. 78
4.4 Referencias….……………………………………………………………………………….. 82

Capítulo V: Actividad insecticida de Porella
chilensis
5.1 Generalidades………………………………………………………………………………. 85
5.2 Experimental…………….…………………………………………………………………. 86
5.2.1 Análisis estadístico......................................................................................... 86
5.3 Resultados y discusión…………………………………………………………………. 87
5.4 Referencias…………………………………………………………………………………… 92
Capítulo VI: Estudio químico de Lepidozia
chordulifera
6.1 Consideraciones botánicas…………………………………………………………... 95
6.2 Clasificación botánica de L. chordulifera……………………………………… 97
6.3 Antecedentes químicos de la familia Lepidoziaceae…………………… 97
6.4 Experimetal………………………………………………………………………………….. 98
6.4.1 Material vegetal..………………………………………………………………….. 98
6.4.2 Extracción y aislamiento………………………………………………………. 98
6.5 Resultados……………………………………………………………………………………. 99
6.6 Estudio quimiotaxonómico de L. chordulifera…….……………..………....104
6.7 Referencias…………………………………………………………………………………… 105
Capítulo VII: Actividad anti-patogénica de
Lepidozia chordulifera
7.1 Generalidades………………………………………………………………………………. 107
7.2 Antecedentes………………………………………………………………………………... 108
7.3 Experimetnal………………………………………………………………………………... 108
7.3.1 Crecimiento bacteriano…….…………………………………….……………. 108

7.3.2 Ensayos sobre la formación de biofilm………………………….………. 109
7.3.3 Ensayo de efectos sobre el QS……………………………………….………. 109
7.3.4 Ensayos sobre la actividad elastasa………………………………………. 109
7.3.5 Análisis estadístico…..…………………………………………………….…….. 109
7.4 Resultados………………………………………………………………………..…….…….. 110
7.4.1 L. chorduliera contra P. aeruginosa……………………………………….. 110
7.4.1.1 Inhibición del crecimiento bacteriano……………………………….….. 111
7.4.1.2 Inhibición de la formación de biofilm……………………………………. 111
7.4.1.3 Inhibición del proceso de QS (inhibición de la producción de
autoinductores)…………………………………………………………………… 111
7.4.1.4 Actividad elastasa…………………………………………………….………….. 111
7.4.1.5 Relación estructura – actividad…………………………………………….. 112
7.4.2 L. chorduliera contra S. aureus……….……………………………………… 114
7.4.2.1 Inhibición del crecimiento bacteriano…………………………………… 114
7.4.2.2 Inhibición de la formación de biofilm……………….…………………… 114
7.4.2.3 Relación estructura – actividad…………………………………………….. 115
7.5 Referencias…………………………………………………………………………………… 116
Capítulo VIII: Resumen de resultados y
conclusiones
8.1 Porella chilensis............................................................................................. ………... 119
8.2 Lepidozia chordulifera…………………………………………………………………. 121
8.3 Actividad biológica………………………………………………………………………. 123
8.4 Referencias…………………………………………………………………………………… 125
Apéndice
Espectros………………………………………………………………………………………………... 130

Resumen
Nuestro laboratorio es el pionero en el estudio químico de especies nativas de hepáticas del norte
argentino y en la descripción estructural de numerosos nuevos compuestos así como de sus actividades
biológicas. Esos resultados unidos a los aportes del presente trabajo de tesis constituyen los únicos
abordajes a la química y actividad biológica de hepáticas en la República Argentina.
Se purificaron e identificaron los metabolitos secundarios de dos especies foliosas de la clase
Hepaticae que crecen en la Patagonia Argentina, y se evaluaron algunos efectos biológicos de los
compuestos mayoritarios aislados de dichas especies, en lo que constituye el primer aporte a la química
de hepáticas de la Patagonia.
Las especies estudiadas fueron Porella chilensis y Lepidozia chordulifera. De la primera se aislaron
cuatro diterpenoides de tipo fusicoccano, dos de ellos nuevos productos naturales, como así también
cuatro sesquiterpenoides de tipo pinguisano y dos de tipo aromadendrano. De Lepidozia chordulifera se
aislaron dos aromadendranos y diez triterpenoides, todos ellos conocidos. Entre los triterpenoides se
encuentran dos taraxanos, tres ácidos triterpénicos frecuentemente encontrados en plantas superiores
y cinco dammaranos.
El aislamiento de los metabolitos se logró mediante el empleo exhaustivo de MPLC y HPLC. La
identificación de los productos aislados se realizó por espectrometría de masas, espectroscopía de
infrarrojo, ultravioleta, RMN protónico y de 13
C con técnicas mono y bidimensionales. Para la
determinación de la estereoquímica absoluta de los diterpenoides nuevos se empleó la técnica de
dicroísmo circular electrónico (medido en el rango del UV).
Se evaluaron las propiedades antipatogénicas de los constituyentes mayoritarios sobre bacterias
patógenas oportunistas de humanos, una cepa de Pseudomonas aeruginosa (Gram -) y otra de
Staphylococcus aureus (Gram +). Estos ensayos se llevaron a cabo sobre bacterias en estado fenotípico
plantónico (vida libre) y en estado fenotípico biofilm. Siendo estos los primeros estudios sobre
compuestos de hepáticas actuando sobre biofilms bacterianos, el sistema de comunicación bacteriana o
quorum sensing (QS), y factores de virulencia. Uno de los compuestos del tipo fusicoccano y los dos del
tipo aromadendrano de P. chilensis inhiben significativamente la formación de biofilm, el crecimiento
bacteriano y el QS. Entre los terpenoides de L. chorudilifera, los cinco compuestos dammaranos y un
aromadendrano inhiben la formación de biofilm de P. aeruginos., Sólo tres de los dammaranos y el
compuesto aromadendrano, previamente mencionado, inhiben también la formación de biofilm de S.
aureus. El crecimiento plantónico no es inhibido por ninguno de estos compuestos. La enzima elastasa,
factor de virulencia, es inhibida por todos los terpenoides de L. chorudilifera, siendo los más activos dos
de los ácidos triterpénicos.
También se determinaron los efectos tóxicos e insecticidas sobre el lepidóptero Spodoptera
frugiperda (insecto plaga del maíz en el norte Argentino), siendo los compuestos del tipo pinguisano los
más activos.

Capítulo I
Introducción
I

METABOLITOS SECUNDARIOS DE HEPÁTICAS DE LA PATAGONIA
ARGENTINA. SU ACTIVIDAD BIOLÓGICA
15 | P á g i n a
I
Capítulo I
Introducción
1.1 Generalidades
Las Briofitas, que comprenden más de 25.000 especies en todo el mundo,
taxonómicamente se encuentran ubicadas entre las algas y las pteridofitas. Las
briofitas representan varias líneas evolutivas muy distintas y se encuentran clasificadas
en tres clases: Musci (musgos, 14.000 especies), Hepaticae (hepáticas, 6.000 especies)
y Athocerotae (antoceros, 300 especies) (Asakawa y col., 2012) (Organigrama 1.1).
Organigrama 1.1. Ubicación de las hepáticas dentro de la división Briophyta.
Las hepáticas son plantas pequeñas y verdes que carecen de un verdadero tejido
vascular. En su cuerpo se distinguen 3 partes: rizoide, caulidios y filidios, análogos
Hierba con cuerno, Antocero
megaceros sp.
Hepática
Bazzania peruviana (Nees) Trev
Musgo
Zygodon pentastichus (Mont.) Müll. Hal.
Musci (musgos) Hepaticae (hepáticas) Athocerotae (antoceros)
División Briophyta

METABOLITOS SECUNDARIOS DE HEPÁTICAS DE LA PATAGONIA
ARGENTINA. SU ACTIVIDAD BIOLÓGICA
16 | P á g i n a
I
(pero no homólogos) a la raíz, tallo y hojas de los vegetales superiores,
respectivamente. Además, las hepáticas no poseen semillas.
Según presenten o no caulidio y filidios, las hepáticas pueden separarse en dos
grandes grupos, hepáticas foliosas y hepáticas talosas (Figura 1.1).
Figura 1.1. (A) Hepática talosa Conocephalum cunicum (B) Hepática foliosa Plagiochila asplenoides.
El término “hepática” (del latín hepaticus, relativo al hígado) surgió en el siglo IX. La
doctrina “Signatura Rerum” (“Las señales de las cosas”) era un concepto medieval
según el cual la apariencia externa de un cuerpo y su semejanza con otros revelaban su
propósito. El parecido con los lóbulos del hígado de algunas hepáticas talosas llevó a la
gente de la época a creer que sería útil contra dolencias hepáticas. En inglés, se usa el
término “liverwort” que resulta de “liver”, hígado, y la terminación “wort”,
proveniente de “wyrt”, que en inglés antiguo significa “hierba”. Actualmente, no hay
evidencias de que las hepáticas tengan uso alguno en el tratamiento de enfermedades
de hígado.
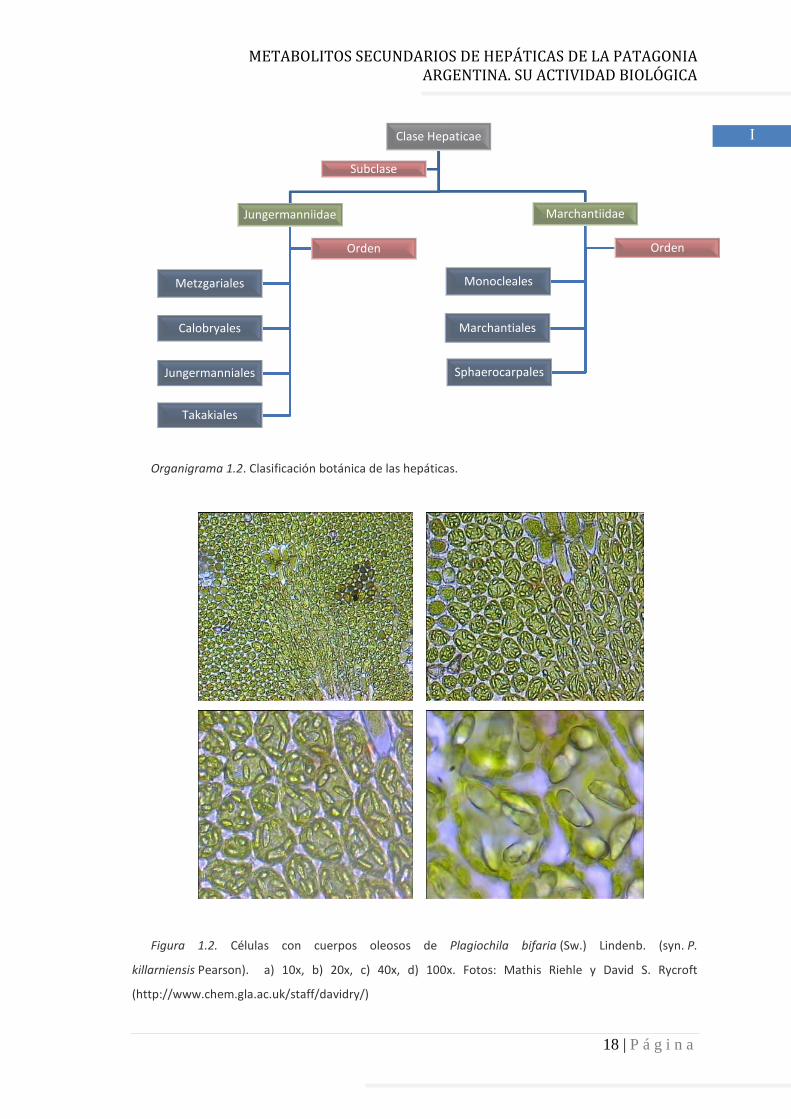
En cuanto a la clasificación taxonómica, la clase Hepaticae se divide en dos
subclases, Jungermanniidae y Marchantiidae. La primera está constituida por cuatro
órdenes y la segunda por tres (Organigrama 1.2). La clasificación morfológica de las
hepáticas es extremadamente difícil, por lo que un estudio de sus metabolitos
secundarios es invaluable para la asignación de especies.
Respecto a la fitoquímica de las briofitas, cabe destacar que hasta mediados de la
década de mil novecientos setenta era casi completamente ignorada. Esto se debía,

METABOLITOS SECUNDARIOS DE HEPÁTICAS DE LA PATAGONIA
ARGENTINA. SU ACTIVIDAD BIOLÓGICA
17 | P á g i n a
I
quizás en primer término, a que eran consideradas plantas pequeñas de uso
insignificante en la dieta humana. Por otro lado, la dificultad para colectar una
cantidad suficiente de material puro de una determinada especie desalentaba su
estudio. En la mayoría de los casos sucede que las matas de la especie que se desea
investigar están entremezcladas con varias otras especies, ramas, corteza y raíces de
plantas superiores, y con parte del suelo de donde fueron extraídas. Separar tales
mezclas bajo la lupa o bien a simple vista consume mucho tiempo y convierte esta
tarea en algo tedioso. En otros casos la determinación taxonómica a nivel de especie
es difícil, sino imposible, debido a la falta de estudios botánicos suficientes que
permitan confirmar dicha clasificación.
Sin embargo, la extraordinaria riqueza química por la diversidad estructural de sus
metabolitos secundarios, así como el uso de las hepáticas en la medicina popular en
China y Japón y el interés acerca de su actividad biológica, dieron origen en los últimos
veinte años, al estudio de numerosas especies de hepáticas de todo el mundo. Los
primeros estudios de metabolitos secundarios de hepáticas argentinas fueron
realizados en nuestro laboratorio, con el estudio de los constituyentes químicos de
Dumortiera hirsuta, Plagiochasma rupestre, Marchantia plicata, (Bardón y col., 1999a;
Bardón y col., 1999b; Kamiya, 2000) Porella swartziana, Frulania brasilensis (Bovi Mitre
y col., 2004; Bardón y col., 2002; Asakawa y Bardón, 2001) y Plagiochila bursata
(Ramírez y col., 2010). En el presente trabajo de tesis continuamos con el estudio de
hepáticas argentinas, y en particular, de la Patagonia argentina.
Por las razones antes mencionadas es que estudiamos los productos naturales
presentes en hepáticas, presentes particularmente en los cuerpos oleosos (figura 1.2).
Los cuerpos oleosos producen además de una variedad de terpenoides lipofílicos con
diferentes esqueletos carbonados, una variedad de compuestos aromáticos,
especialmente fenólicos. Muchos de estos constituyentes son particulares de
hepáticas y muestran interesantes actividades biológicas, como antimicrobiana,
antifúngica, citotóxica, antialimentaria de insectos, insecticida, inhibitoria de algunas
enzimas y apoptósis (Asakawa, 2004).
METABOLITOS SECUNDARIOS DE HEPÁTICAS DE LA PATAGONIA
Organigrama 1.2. Clasificación
Figura 1.2. Células con cuerpos oleosos de
killarniensis Pearson). a) 10x, b) 20x, c) 40x, d) 100x
(http://www.chem.gla.ac.uk/staff/davidry/)
Jungermanniidae
Calobryales
Jungermanniales
Takakiales
Metzgariales
METABOLITOS SECUNDARIOS DE HEPÁTICAS DE LA PATAGONIA
ARGENTINA. SU ACTIVIDAD BIOLÓGICA
Clasificación botánica de las hepáticas.
Células con cuerpos oleosos de Plagiochila bifaria (Sw.)
Pearson). a) 10x, b) 20x, c) 40x, d) 100x. Fotos: Mathis Riehle y David S. Rycroft
(http://www.chem.gla.ac.uk/staff/davidry/)
Clase Hepaticae
Jungermanniidae
Orden
Marchantiidae
Sphaerocarpales
Monocleales
Marchantiales
Subclase
METABOLITOS SECUNDARIOS DE HEPÁTICAS DE LA PATAGONIA
ARGENTINA. SU ACTIVIDAD BIOLÓGICA
18 | P á g i n a
I
(Sw.) Lindenb. (syn. P.
Fotos: Mathis Riehle y David S. Rycroft
Marchantiidae
Orden

METABOLITOS SECUNDARIOS DE HEPÁTICAS DE LA PATAGONIA
ARGENTINA. SU ACTIVIDAD BIOLÓGICA
19 | P á g i n a
I
1.2 Antecedentes químicos
Los productos naturales aislados de plantas y microorganismos se han dividido, con
cierta arbitrariedad, en dos grandes grupos: metabolitos primarios y metabolitos
secundarios. Esta división se basa en las características detalladas en la Tabla 1.1.
En esta clasificación, los metabolitos primarios comprenden aquellas sustancias que
se encuentran en todos los organismos vivos, siendo por lo tanto, metabólicamente
esenciales. Un grupo reducido de estos metabolitos primarios, sirve como precursor de
los metabolitos secundarios.
Para el químico el término “producto natural” es sinónimo de metabolito
secundario y una definición más pragmática de metabolito secundario es: “Toda
sustancia que aparenta no tener un papel explicito en la economía interna del
organismo que lo produce”.
Metabolito primario Metabolito secundario
Producto del metabolismo general Producto del metabolismo especial.
Biosintetizado a partir de metabolitos
primarios.
Ampliamente distribuidos en plantas y
microorganismos.
Con distribución restringida a ciertas
plantas y microorganismos (a veces es
característico de un género dado o una
especie en particular).
Ejemplos: Aminoácidos de proteínas
comunes, monosacáridos comunes,
nucleótidos, ácidos carboxílicos del ciclo del
ácido cítrico, lípidos, glicéridos, entre otros.
Ejemplos: Alcaloides, terpenoides,
flavonoides, oligosacáridos, entre otros.
Tabla 1.1. Metabolitos primarios vs metabolitos secundarios.
Se detallan a continuación los metabolitos secundarios de hepáticas aislados hasta
el presente.

METABOLITOS SECUNDARIOS DE HEPÁTICAS DE LA PATAGONIA
ARGENTINA. SU ACTIVIDAD BIOLÓGICA
20 | P á g i n a
I
1.2.1 Terpenoides
Bajo este nombre se agrupan un gran número de compuestos cuyos esqueletos
carbonados presentan como factor común la unión de unidades de isopreno (2-metil-
1,3-butadieno)
1.2.1.1 Monoterpenoides
Se agrupan bajo este nombre los compuestos formados por la unión de dos
unidades de isopreno, es decir, poseen diez átomos de carbono.
La mayoría de los monoterpenoides fueron separados e identificados por
cromatografía gaseosa acoplada a espectrometría de masas, razón por la cual las
configuraciones de los centros asimétricos no fueron establecidas.
Se detectaron monoterpenoides tanto en la subclase Jungermanniidae como en la
sub clase Marchantiidae. Los más frecuentemente hallados son α-pineno, β-pineno y
limoneno.
1.2.1.2 Sesquiterpenoides
Las hepáticas son ricas fuentes de sesquiterpenoides. No obstante, una
característica importante de la clase Hepaticae es que muchos de los
sesquiterpenoides aislados de ella, difieren de aquellos encontrados en plantas
superiores en la configuración de uno o más centros quirales. Un claro ejemplo de esta
situación es la que se presenta para los sesquiterpenoides del tipo aromadendranos.

METABOLITOS SECUNDARIOS DE HEPÁTICAS DE LA PATAGONIA
ARGENTINA. SU ACTIVIDAD BIOLÓGICA
21 | P á g i n a
I
Todos los aromadendranos aislados de hepáticas poseen configuración ent, es decir,
son enantiómeros de los que se encuentran en plantas superiores (Gijsen, 1993).
En general, los organismos vivos biosintetizan estéreo-específicamente uno de los
compuestos del par enantiomérico mediante reacciones enzimáticas estero-
específicas. Por ejemplo, los aminoácidos constituyentes de proteínas y los azucares
poseen normalmente la forma D y L, respectivamente, y sus contrapartes antipodales
no son sintetizadas en absoluto. Sin embargo, salvo raras excepciones, las hepáticas
fabrican sesquiterpenoides que son imágenes especulares de los sintetizados por otros
tipos de plantas. Esto indica que el complejo enzima-sustrato para la síntesis de
sesquiterpenoides en hepáticas puede poseer una conformación inversa a la que
posee en plantas superiores. Este es un hecho importante no solo desde la
quimiotaxonomía, sino también desde la filogenia o historia evolutiva de las hepáticas
(Matsuo, 1982).
Se han aislado gran cantidad y variedad de sesquiterpenoides, en su mayoría de la
subclase Jungermanniidae. A continuación se exponen ejemplos de sesquiterpenoides
aislados de hepáticas que presentan distintos tipos de esqueletos.
METABOLITOS SECUNDARIOS DE HEPÁTICAS DE LA PATAGONIA
ARGENTINA. SU ACTIVIDAD BIOLÓGICA
22 | P á g i n a
I
OHC
Isobiciclogermacrano
OHC
H H
Lepidozanos
H
Bisabolanos
H
H
Bourbonanos
Calamenanos Calacoranos Cadalenos
O
Cariofilenos
METABOLITOS SECUNDARIOS DE HEPÁTICAS DE LA PATAGONIA
ARGENTINA. SU ACTIVIDAD BIOLÓGICA
23 | P á g i n a
I
METABOLITOS SECUNDARIOS DE HEPÁTICAS DE LA PATAGONIA
ARGENTINA. SU ACTIVIDAD BIOLÓGICA
24 | P á g i n a
I
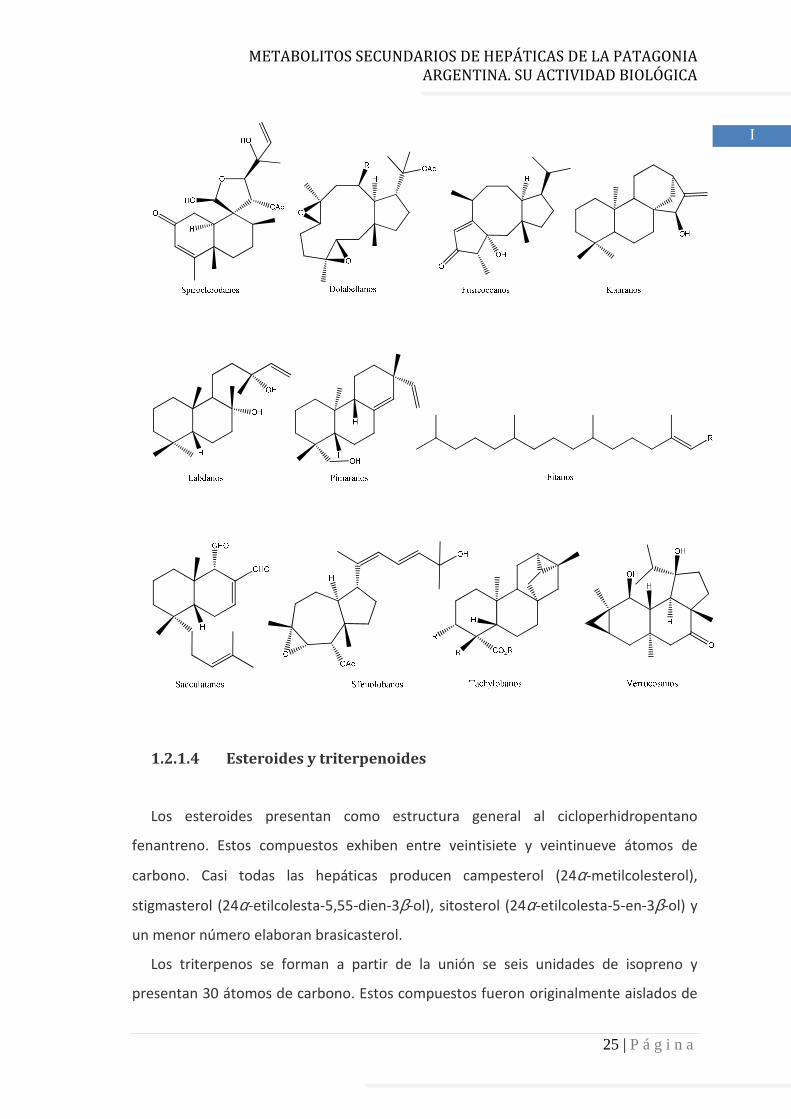
1.2.1.3 Diterpenoides
Estos compuestos se forman por la unión de cuatro unidades de isopreno, es decir,
poseen veinte átomos de carbono.
Las hepáticas también son ricas en diterpenoides. Al igual que en el caso de los
sesquiterpenoides, la mayoría de los diterpenoides de hepáticas poseen la
configuración absoluta opuesta a la encontrada en plantas superiores.
La mayoría de los diterpenoides de hepáticas se aislaron de la subclase
Jungermanniidae, solo se encontraron unos pocos en la subclase Marchantiidae.
A continuación se exponen algunos diterpenoides aislados de hepáticas que
representan los esqueletos más frecuentes.
METABOLITOS SECUNDARIOS DE HEPÁTICAS DE LA PATAGONIA
ARGENTINA. SU ACTIVIDAD BIOLÓGICA
25 | P á g i n a
I
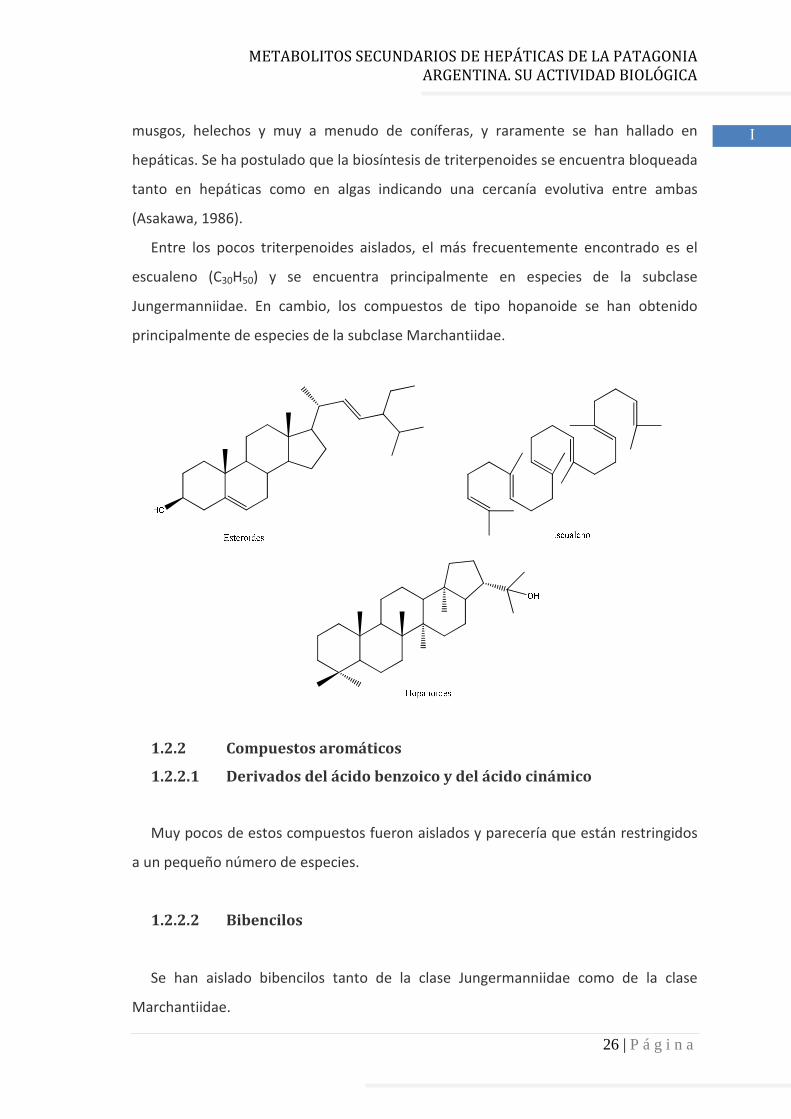
1.2.1.4 Esteroides y triterpenoides
Los esteroides presentan como estructura general al cicloperhidropentano
fenantreno. Estos compuestos exhiben entre veintisiete y veintinueve átomos de
carbono. Casi todas las hepáticas producen campesterol (24α-metilcolesterol),
stigmasterol (24α-etilcolesta-5,55-dien-3β-ol), sitosterol (24α-etilcolesta-5-en-3β-ol) y
un menor número elaboran brasicasterol.
Los triterpenos se forman a partir de la unión se seis unidades de isopreno y
presentan 30 átomos de carbono. Estos compuestos fueron originalmente aislados de
METABOLITOS SECUNDARIOS DE HEPÁTICAS DE LA PATAGONIA
ARGENTINA. SU ACTIVIDAD BIOLÓGICA
26 | P á g i n a
I
musgos, helechos y muy a menudo de coníferas, y raramente se han hallado en
hepáticas. Se ha postulado que la biosíntesis de triterpenoides se encuentra bloqueada
tanto en hepáticas como en algas indicando una cercanía evolutiva entre ambas
(Asakawa, 1986).
Entre los pocos triterpenoides aislados, el más frecuentemente encontrado es el
escualeno (C30H50) y se encuentra principalmente en especies de la subclase
Jungermanniidae. En cambio, los compuestos de tipo hopanoide se han obtenido
principalmente de especies de la subclase Marchantiidae.
1.2.2 Compuestos aromáticos
1.2.2.1 Derivados del ácido benzoico y del ácido cinámico
Muy pocos de estos compuestos fueron aislados y parecería que están restringidos
a un pequeño número de especies.
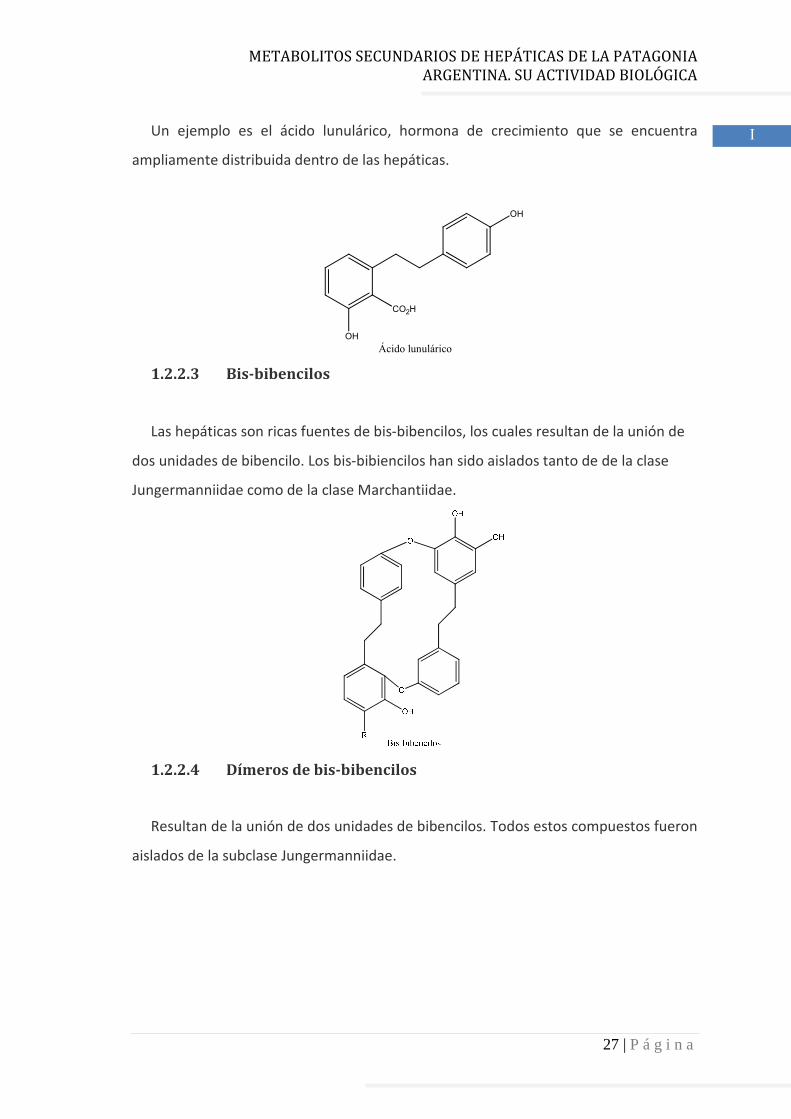
1.2.2.2 Bibencilos
Se han aislado bibencilos tanto de la clase Jungermanniidae como de la clase
Marchantiidae.
METABOLITOS SECUNDARIOS DE HEPÁTICAS DE LA PATAGONIA
ARGENTINA. SU ACTIVIDAD BIOLÓGICA
27 | P á g i n a
I
Un ejemplo es el ácido lunulárico, hormona de crecimiento que se encuentra
ampliamente distribuida dentro de las hepáticas.
OH
CO2H
OH
Ácido lunulárico
1.2.2.3 Bis-bibencilos
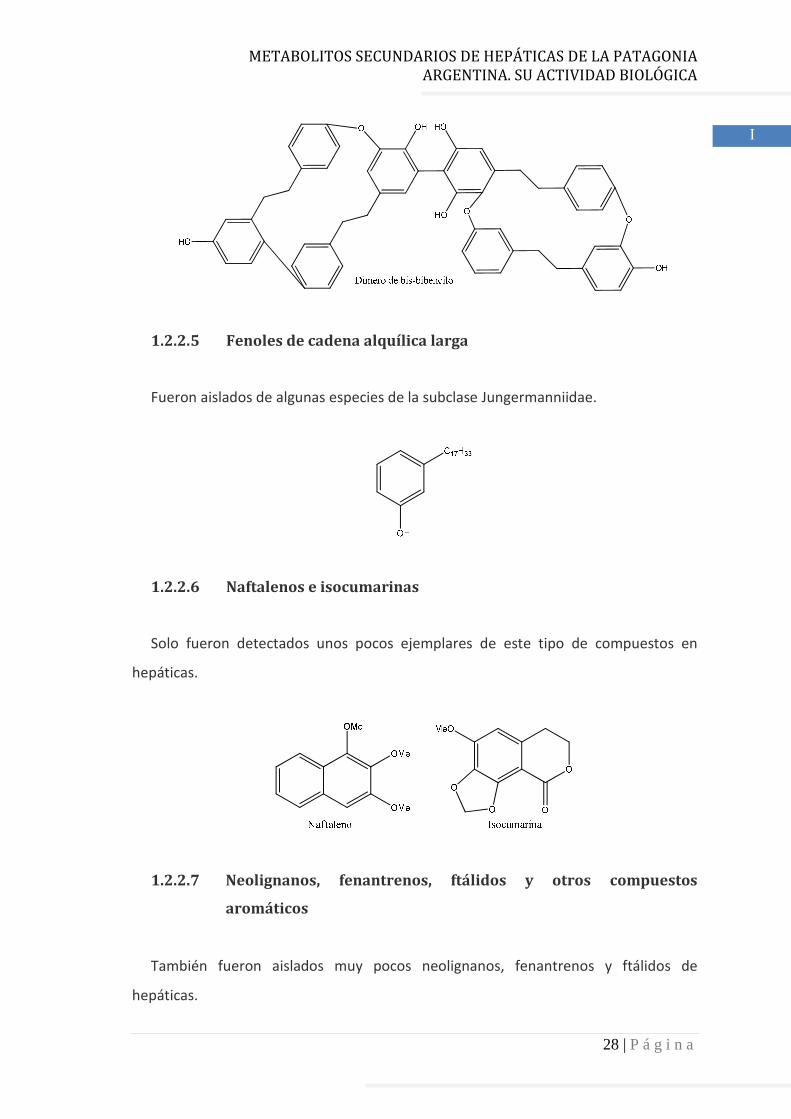
Las hepáticas son ricas fuentes de bis-bibencilos, los cuales resultan de la unión de
dos unidades de bibencilo. Los bis-bibiencilos han sido aislados tanto de de la clase
Jungermanniidae como de la clase Marchantiidae.
1.2.2.4 Dímeros de bis-bibencilos
Resultan de la unión de dos unidades de bibencilos. Todos estos compuestos fueron
aislados de la subclase Jungermanniidae.
METABOLITOS SECUNDARIOS DE HEPÁTICAS DE LA PATAGONIA
ARGENTINA. SU ACTIVIDAD BIOLÓGICA
28 | P á g i n a
I
1.2.2.5 Fenoles de cadena alquílica larga
Fueron aislados de algunas especies de la subclase Jungermanniidae.
1.2.2.6 Naftalenos e isocumarinas
Solo fueron detectados unos pocos ejemplares de este tipo de compuestos en
hepáticas.
1.2.2.7 Neolignanos, fenantrenos, ftálidos y otros compuestos
aromáticos
También fueron aislados muy pocos neolignanos, fenantrenos y ftálidos de
hepáticas.

METABOLITOS SECUNDARIOS DE HEPÁTICAS DE LA PATAGONIA
ARGENTINA. SU ACTIVIDAD BIOLÓGICA
29 | P á g i n a
I
1.2.2.8 Flavonoides
Los flavonoides están ampliamente distribuidos en hepáticas, reportes previos
listan la presencia de flavonoides en más de 100 especies pertenecientes a 60 géneros
diferentes de hepáticas.
Entre el 70 y 100% de las Marchantiidae y el 50% de las Jungermanniidae presentan
flavonoides.
Los principales tipos de flavonoides aislados de Marchantiidae son flavonas O-
glucurónidos y C-glicósidos, mientras que los flavonoles son muy escasos.
Dentro de la subclase Jungermanniidae, las Metzgariales son caracterizadas por
flavonas C-glicósidos, las Jungermanniales pro flavonas C y O-glicósidos, y dentro de las
Marchantiidae, Marchantiales, Sphaerocarpales y Monocleales, por flavonas O-
glucurónidos. Los dos últimos órdenes se pueden diferenciar de Marchantiales por la
completa ausencia de flavonas C-glicósidos.

METABOLITOS SECUNDARIOS DE HEPÁTICAS DE LA PATAGONIA
ARGENTINA. SU ACTIVIDAD BIOLÓGICA
30 | P á g i n a
I
1.2.3 Lípidos
1.2.3.1 n-Alcanos
En hepáticas, la proporción entre n-alcanos con número par e impar de átomos de
carbono es la misma, a diferencia de lo que sucede en plantas superiores en las que
son predominantes los n-alcanos con número par de átomos de carbono.
1.2.3.2 Ácidos grasos
La mayoría de las hepáticas poseen varios ácidos grasos libres. Solo en especies de
los géneros Riccia y Monoclea se encontraron ácidos grasos acetilénicos. En especies
del género Monoclea, se encontraron además ácidos grasos en-ino que están
ampliamente distribuidos en musgos.
1.2.4 Carbohidratos
Se aislaron diferentes carbohidratos de hepáticas, como (-)-D-manitol, (+)-glucosa,
(+)-sacarosa y (+)-trealosa, cuya configuración absoluta coincide con la encontrada en
plantas superiores.
1.2.5 Compuestos con azufre
Las hepáticas presentan un fuerte olor a algas cuando son secadas. Uno de los
responsables de este aroma es el sulfuro de dimetilo.
También se han encontrado ésteres que contienen azufre en sus moléculas.
1.3 Antecedentes biológicos
Los estudios realizados en busca de productos naturales bioactivos, se han
incrementado en los últimos años debido a causas diversas. Una de ellas es la
necesidad de conseguir más alimentos y medicamentos para una población humana

METABOLITOS SECUNDARIOS DE HEPÁTICAS DE LA PATAGONIA
ARGENTINA. SU ACTIVIDAD BIOLÓGICA
31 | P á g i n a
I
que crece en forma progresiva y para la cual se calcula que, en unos años, será
insuficiente la producción proyectada. Otra causa es el problema de la extinción de
especies vegetales y animales que ocurre día a día debido al exterminio de
ecosistemas realizado a mano del hombre. Esto produce pérdidas irreparables ya que
con ellas se extingue toda la información de los productos naturales que las especies
son capaces de biosintetizar. Otra razón importante se origina en la creciente pérdida
del conocimiento etnobotánico empírico, adquirido por los pueblos a través de miles
de años, debido a la influencia de culturas extrañas a las autóctonas. Todas estas
causas generaron como consecuencia que los investigadores formaran equipos de
trabajo multidisciplinarios para poder afrontar la búsqueda de información sobre
especies bioactivas, la recolección del material y su posterior estudio (Molyneux y
Colegate, 2007).
Por otra parte, el interés en aislar productos naturales biológicamente activos para
su aplicación y no emplear la planta entera u otras preparaciones crudas con motivos
experimentales o terapéuticos, tiene las siguientes justificaciones: las variaciones en
las cantidades de constituyentes activos según el lugar geográfico, las estaciones del
año, condiciones climáticas y ecológicas, las diferentes partes de la planta que se
empleen, la existencia de compuestos indeseables que causen modulaciones
impredecibles de la bioactividad y también la pérdida de la misma debido a
variabilidad en la recolección, almacenado y preparación del material crudo. El empleo
de compuestos puros, además, trae aparejado una serie de ventajas. Entre ellas, la
posibilidad de producir sintéticamente los productos naturales luego de su
determinación estructural, modificarlos estructuralmente y comprender su mecanismo
de acción. Estas investigaciones pueden también complementarse con estudios acerca
de las relaciones estructura-actividad, facilitando el desarrollo de nuevos compuestos
con bioactividad similares o más deseables. También admite la posibilidad de ser
administrado en dosis exactas, reproducibles, con los consiguientes beneficios desde el
punto de vista experimental o terapéutico.
Generalmente, las briofitas no son dañadas por bacterias, hongos, insectos,
caracoles o babosas. Es un hecho conocido que las briofitas contienen aleloquímicos y
hay registros de su uso como plantas medicinales en Norte América, China y Europa.

METABOLITOS SECUNDARIOS DE HEPÁTICAS DE LA PATAGONIA
ARGENTINA. SU ACTIVIDAD BIOLÓGICA
32 | P á g i n a
I
Hace más de 400 años es conocido el hecho de que algunas especies de Fissidens y
Poytrichum poseen actividad diurética y sus cenizas promueven el crecimiento del
cabello humano. La mayoría de las briofitas con usos medicinales fueron usadas como
decocciones, o bien, trituradas y el polvo resultante mezclado con aceite para hacer un
ungüento con el cual curar cortes, quemaduras y/o heridas externas. Los aborígenes
nativos de América del Norte usaron especies de Bryum, Mnium y Philotonis, y
Politrychum juniperinum como musgos medicinales para curar quemaduras,
contusiones y heridas. Marchantia polymorpha ha sido usada en Europa como
diurético (Asakawa y col., 2012).
En tabla 1.2 se listan las hepáticas usadas en medicina popular, así como los efectos
y actividades fisiológicas atribuidas.
Especie Actividades fisiológicas y efectos
Conocephalum cunicum Antimicrobiana, antifúngica y antipirética. Se emplea
sobre heridas, quemaduras, escaldaduras, fracturas,
tejidos inflamados, picaduras de serpientes venenosas y
tratamiento de cálculos biliares.
Frulania tramarisci Antiséptica
Marchantia polymorpha Antipirética y diurética. Se emplea en heridas,
fracturas, picaduras de serpientes venenosas,
quemaduras, escaldaduras y heridas abiertas.
Reboulia hemisphaerica Se emplea sobre ronchas, heridas abiertas y
contusiones
Tabla 1.2. Hepáticas utilizadas en medicina popular y propiedades atribuidas.
Las actividades biológicas atribuidas a las hepáticas, se deben principalmente a los
terpenoides y compuestos aromáticos presentes en los cuerpos oleosos (figura 1.1).
Además de las actividades biológicas descriptas arriba, algunas briofitas emiten
fragancias características y, dependiendo de la fuente, sabores intensamente picantes,
amargos o dulces.

METABOLITOS SECUNDARIOS DE HEPÁTICAS DE LA PATAGONIA
ARGENTINA. SU ACTIVIDAD BIOLÓGICA
33 | P á g i n a
I
1.3.1 Sabores picantes, amargos y dulces
Algunos géneros de briofitas producen sustancias que provocan intensos sabores
picantes, amargos o dulces. En ciertos casos estos sabores se deben a la presencia de
sesquiterpenos (Toyota y col., 1991) y diterpenos (Asakawa, 1990). Los más intensos y
picantes se deben a dialdehidos como poygodial y sacculatal (Asakawa y col., 2012).
1.3.2 Dermatitis alérgica por contacto
Muchas hepáticas producen dermatitis alérgica por contacto, especialmente
algunas especies de Frullania. Esta reacción se ha atribuido a la presencia de α-
metilen-γ-lactonas sesquiterpénicas ((+)-frullanolide y (-)-frullanolide) (Molyneux y
Colegate, 2007). En Schistochila appendiculata, se ha atribuido a la presencia de alquil
fenoles, ácidos salicílicos de cadena alquílica larga y sus sales de potasio, como así
también a catecol de cadena larga (Asakawa y col., 1987). Poligodial, antes citado,
también produce este tipo de irritación.

METABOLITOS SECUNDARIOS DE HEPÁTICAS DE LA PATAGONIA
ARGENTINA. SU ACTIVIDAD BIOLÓGICA
34 | P á g i n a
I
1.3.3 Actividad antitumoral
A través del Instituto Nacional de Cáncer, USA, se investigaron 184 especies de 97
géneros de musgos, 23 especies de 16 géneros de hepáticas y 1 especie de
antocerotales, buscando encontrar posibles agentes antitumorales (Asakawa, 1990).
Se realizaron ensayos sobre la línea celular P388 correspondiente a leucemia linfocítica
de ratón.
El nivel de actividad antitumoral entre las especies de hepáticas fue relativamente
bajo. Entre las 23 especies de hepáticas investigadas solo cuatro presentaron actividad
P388. Ellas fueron las Jungermanniales Bazzania trilobata y Porella bolanderi y las
Marchantiales Conocephalum conicum y Dumortiera hirsuta. De las especies de
musgos estudiadas, 43 de ellas resultaron activas y la única especie de antoceros
estudiada resultó inactiva.
Los constituyentes químicos de hepáticas, musgos y antoceros difieren
considerablemente. En particular, el procedimiento de extracción en los ensayos
mencionados arriba (EtOH caliente durante 8 hs), pudo haber sido demasiado vigoroso
para los constituyentes de hepáticas entre los que se incluyen bibencilos, bis-
bibencilos cíclicos, mono-, sesqui- y diterpenoides que contienen funciones
hemiacetales, dialdehidos, etc, propensos a descomponerse bajo estas condiciones
(Asakawa y col., 2012).
Algunos guaianolidos de Conocephalum conicum también demostraron poseer
actividad anti cáncer contra la línea celular de leucemia linfocítica P388.
Las sustancias ricardin A y B, y el pinguisano dehidropinguisenol aisladas de
Riccardia multifida fueron activas contra las células KB de carcinoma humano. También
las sustancias plagiochilina A de diferentes especies de Plagiochila; eremofrulanolide y
oxifrulanolide de Frullania dilatata; y marchantin A, B y C de diferentes especies de
Marchantia mostraron actividad citotóxica contra células KB (Asakawa, 1982).

METABOLITOS SECUNDARIOS DE HEPÁTICAS DE LA PATAGONIA
ARGENTINA. SU ACTIVIDAD BIOLÓGICA
35 | P á g i n a
I
1.3.4 Actividad molusquicida
Algunos sesquiterpenoides de tipo cuprano y monociclofarnesano, aislados de
Ricciocarpos natans, el ácido lunulárico y dos bibencilos relacionados con este último,
mostraron tener actividad molusquicida contra Biomphalaria glabrata, vector de la
schistosomiasis (Wurzel y Becker, 1990).
1.3.5 Actividad antialimentaria de insectos
Algunas especies demostraron tener actividad antialimentaria de insectos debido a
ciertos terpenoides presentes en ellas. El más activo resultó ser plagiochilina A, un
sesquiterpeno del tipo secoaromadendrano, aislado de varias especies del genero
Plagiochila, sobre el lepidóptero Spodoptera exempta. Polygodial también mostró
poseer actividad antialimentaria de áfidos (Asakawa, 1990).
1.3.6 Actividad inhibidora de la liberación de anión superóxido
El exceso de anión radical superóxido (O2-) en los organismos es la causa de varias
angiopatías. Se ha demostrado que norpinguisona de Porella vernicosa, el éster
metílico de norpinguisona aislado de P. elegantula, y Perrottetianal A de P.
perrottetiana, inhiben la liberación de superóxido en experimentos realizados con
animales de laboratorio.

METABOLITOS SECUNDARIOS DE HEPÁTICAS DE LA PATAGONIA
ARGENTINA. SU ACTIVIDAD BIOLÓGICA
36 | P á g i n a
I
1.3.7 Actividades antimicrobianas y antifúngicas
Varias hepáticas mostraron actividad antimicrobiana y otras tantas exhibieron
actividad antifúngica (Molyneux y Colegate, 2007). Ciertos bibencilos prenilados fueron
activos contra Staphylococcus aureus (Asakawa y col., 1991).
Los diterpenoides α-santonina, 8-hidroxi-9-hidroperrotetianal, perrotetianal A y
perrotetianal B también mostraron actividad antimicrobiana contra S. aureus y Bacillus
subtilis. Marchantin A fue activo contra S. aureus (Asakawa, 1990) y Streptococcus
pyogenes (Kamory y col., 1995).
Los diterpenoides del tipo dolabelano, de Odontoshisma denudatum (Matsuo y col.,
1988) y algunos de sus derivados, como así también 3-metoxi-4’-hidroxibibencilo
aislado de Plagiochila stephensoniana (Lorimer y col., 1993), y algunos sesquiterpenos
fenólicos de Herbertus aduncus (Matsuo y col., 1981), demostraron poseer actividad
fungicida. Además se observó que las concentraciones de acido lunulárico en muchas
hepáticas llegan a ser suficientes para prevenir casi completamente el ataque de
hongos (Schwabe, 1990) y que ciertas hidroxi-acetofenonas aisladas de Plagiochila
fasciculata, también mostraron actividad antifúngica (Lorimer y Perry, 1994).
1.3.8 Actividad piscicida
Se observó que los extractos crudos que contenían sustancias picantes poseían
fuerte actividad hemolítica (Asakawa, 1990).

METABOLITOS SECUNDARIOS DE HEPÁTICAS DE LA PATAGONIA
ARGENTINA. SU ACTIVIDAD BIOLÓGICA
37 | P á g i n a
I
Tanto polygodial como sacculatal, exhiben la mayor actividad piscicida (Asakawa y
col., 2012). Otros sesquiterpenoides como diplophyllin, (+)-frullanolide y plagiochilina
A también mostraron actividad piscicida (Molyneux y Colegate, 2007).
Los estudios antimicrobianos reportados en bibliografía fueron llevados a cabo
sobre bacterias en estado fenotípico plantónico (vida libre)(Ríos y Recio, 2005). En la
actualidad, es un hecho conocido que el 99% de las bacterias viven en comunidades,
utilizando señales químicas para su comunicación y que son capaces de formar biofilm
como mecanismo de resistencia. No parece haber estudios realizados sobre
compuestos de hepáticas actuando sobre biofilms bacterianos o comunicación
bacteriana.
Dado el amplio espectro de actividades biológicas reportados para los productos
aislados de hepáticas, su diversidad estructural, y los estudios previos llevados a cabo
en nuestro laboratorio, consideramos más que interesante la realización de estudios
químicos sobre las especies argentinas de hepáticas, y consecuentemente estudios
biológicos sobre los metabolitos aislados de ellas.

METABOLITOS SECUNDARIOS DE HEPÁTICAS DE LA PATAGONIA
ARGENTINA. SU ACTIVIDAD BIOLÓGICA
38 | P á g i n a
I
Referencias
Asakawa Y. Chemical constituents of Hepaticae. Springer, Vienna. 1982.
Asakawa Y. Chemical relationships between algae, bryophytes, and pteridophytes. Journal
of Bryology 1986, 14, 59-70.
Asakawa Y. Terpenoids and aromatic compounds with pharmaceutical activity from
bryophytes. in Bryophytes, their chemistry and chemical taxonomy; Clarendon Press; 1990,
pp. 369-410.
Asakawa Y. Chemosystematics of the Hepaticae. Phytochemistry 2004, 65, 623-669.
Asakawa Y. and Bardón A. Constituyentes bioactivos de hepáticas. Importancia de las
especies americanas. Revista Latinoamericana de química 2001, 29, 5-22.
Asakawa Y., Kondo K., Takikawa N. K., Tori M., Hashimoto T. and Ogawa S. Prenyl
bibenzyls from the liverwort Radula kojana. Phytochemistry 1991, 30, 219-234.
Asakawa Y., Ludwiczuk A., Nagashima F., Kinghorn A. D., Falk H. and Kobayashi J.
Chemical Constituents of Bryophytes: Bio- And Chemical Diversity, Biological Activity,
and Chemosystematics. Springer. 2012.
Asakawa Y., Masuya T., Tori M. and Campbell E. O. Long chain alkyl phenols from the
liverwort Schistochila appendiculata. Phytochemistry 1987, 26, 735-738.
Bardón A., Bovi Mitre G., Kamiya N., Toyota M. and Asakawa Y. Eremophilanolides and
other constituents from the Argentine liverwort Frullania brasiliensis. Phytochemistry
2002, 59, 205-213.

METABOLITOS SECUNDARIOS DE HEPÁTICAS DE LA PATAGONIA
ARGENTINA. SU ACTIVIDAD BIOLÓGICA
39 | P á g i n a
I
Bardón A., Kamiya N., Toyota M. and Asakawa Y. A 7-nordumortenone and other
dumortane derivatives from the Argentine liverwort Dumortiera hirsuta. Phytochemistry
1999a, 51, 281-287.
Bardón A., Kamiya N., Toyota M., Takaoka S. and Asakawa Y. Sesquiterpenoids,
hopanoids and bis(bibenzyls) from the argentine liverwort Plagiochasma rupestre.
Phytochemistry 1999b, 52, 1323-1329.
Bovi Mitre G., Kamiya N., Bardón A. and Asakawa Y. Africane-type sesquiterpenoids from
the Argentine liverwort Porella swartziana and their antibacterial activity. J. Nat. Prod.
2004, 67, 31-36.
Gijsen, H. J. M. (+)-Aromadendrene as chiral starting material for thesynthesis of
sesquiterpenes. Ph D Thesis, 1993, Landbouwuniversiteit, pp 2-3.
Kamiya, N. Metabolitos secundarios de hepáticas del NOA y su actividad biológica. Ph. D
Thesis, 2000, Universidad Nacional de Tucumán, pp 61-152.
Kamory E., Keseru G. M. and Papp B. Isolation and antibacterial activity of marchantin A,
a cyclic bis(bibenzyl) constituent of Hungarian Marchantia polymorpha. Planta Med. 1995,
61, 387-388.
Lorimer S. D. and Perry N. B. Antifungal hydroxy-acetophenones from the New Zealand
liverwort, Plagiochila fasciculata. Planta Med. 1994, 60, 386-387.
Lorimer S. D., Perry N. B. and Tangney R. S. An antifungal bibenzyl from the New
Zealand liverwort, Plagiochila stephensoniana. Bioactivity-directed isolation, synthesis,
and analysis. J. Nat. Prod. 1993, 56, 1444-1450.

METABOLITOS SECUNDARIOS DE HEPÁTICAS DE LA PATAGONIA
ARGENTINA. SU ACTIVIDAD BIOLÓGICA
40 | P á g i n a
I
Matsuo A. Selected chemotaxonomic characteristics of liverwort sesquiterpenoids. The
Journal of the Hattori Botanical Laboratory = Hattori Shokubutsu Kenkyujo hokoku 1982,
295-304.
Matsuo A., Kamio K., Uohama K., Yoshida K., Connolly J. D. and Sim G. A. Dolabellane
diterpenoids from the liverwort Odontoschisma denudatum. Phytochemistry 1988, 27,
1153-1159.
Matsuo A., Yuki S., Nakayama M. and Hayashi S. (-)-Herbertene, an aromatic
sesquiterpene with a novel carbon skeleton from the liverwort Herberta adunca. J. Chem.
Soc. , Chem. Commun. 1981, 864-865.
Molyneux R. J. and Colegate S. M. An Introduction and Overview. in Bioactive Natural
Products; CRC Press; 2007, pp. 1-9.
Ramírez M., Kamiya N., Popich S., Asakawa Y. and Bardón A. Insecticidal constituents
from the Argentine Liverwort Plagiochila bursata. Chemistry and Biodiversity 2010, 7,
1855-1861.
Ríos J. L. and Recio M. C. Medicinal plants and antimicrobial activity. J. Ethnopharmacol.
2005, 100, 80-84.
Schwabe W. W. Lunularic acid in growth and dormancy of liverworts. in Physiology and
Biochemistry; Chopra R. N. and Bhatla S. C., eds; CRC Press; Boca Raton; 1990, pp. 1-
245.
Toyota M., Ueda A. and Asakawa Y. Sesquiterpenoids from the liverwort Porella acutifolia
subsp. Tosana. Phytochemistry 1991, 30, 567-573.
Wurzel G. and Becker H. Sesquiterpenoids from the liverwort Ricciocarpos natans.
Phytochemistry 1990, 29, 2565-2568.

Capítulo II
Materiales y Métodos
II

METABOLITOS SECUNDARIOS DE HEPÁTICAS DE LA PATAGONIA
ARGENTINA. SU ACTIVIDAD BIOLÓGICA
42 | P á g i n a
II
Capítulo II
Materiales y métodos
La estrategia planificada para el estudio químico de las especies vegetales plantea el
seguimiento de una serie de pasos para llevar a cabo el aislamiento e identificación de
los metabolitos secundarios, como así también la posterior determinación de sus
actividades biológicas. Dichos pasos se describen a continuación.
2.1 Recolección, clasificación y preparación del material vegetal
La búsqueda y recolección del material vegetal se realizó en Febrero de 2005 en
lago Steffen, Provincia de Río Negro, Argentina. La búsqueda se realizó en zonas
principalmente húmedas y propicias para el crecimiento de las hepáticas.
Para facilitar la clasificación taxonómica del material vegetal, es conveniente que las
plantas presenten sus estructuras reproductoras. La reproducción de las briofitas se
realiza recién cuando empieza la época lluviosa. Esto se debe a que la fecundación se
lleva a cabo cuando las gotas de agua transportan los espermatozoides desde los
anteridios hasta los arquegonios, donde se encuentran los óvulos.
Una etapa imprescindible previa a la extracción, es la limpieza del material vegetal
que implica quitar las demás plantas (musgos, líquenes, helechos y otras plantas) que
por lo general se encuentran mezclados con la especie que se desea estudiar. Como así
también se debe retirar la tierra adherida a los rizoides.
2.2 Extracción
El material vegetal fue secado al aire y triturado con el fin de aumentar la superficie
de contacto con el solvente y asegurar un buen rendimiento en el proceso de
extracción. Las extracciones fueron llevadas a cabo por maceración a temperatura
ambiente. El material así preparado fue colocado en un erlenmeyer, se cubrió con

METABOLITOS SECUNDARIOS DE HEPÁTICAS DE LA PATAGONIA
ARGENTINA. SU ACTIVIDAD BIOLÓGICA
43 | P á g i n a
II
solvente y se dejó en reposo por 10 días con agitación ocasional. Luego, el extracto
obtenido se filtró y el solvente se eliminó en evaporador rotatorio. Se realizaron dos
extracciones con éter etílico, obteniéndose un extracto de baja polaridad (extracto
etéreo) y luego, una vez seco el material, se realizaron dos extracciones con metanol
obteniéndose un extracto polar (extracto metanólico).
2.3 Purificación y aislamiento
En primer lugar, el extracto obtenido se somete a un desclorofilado mediante
cromatografía por exclusión con Sephadex LH-20. La cromatografía por exclusión de
tamaño, también denominada cromatografía en gel, es eficiente para separar grupos
de solutos en base a su tamaño efectivo en solución. Las fases estacionarias utilizadas
para lograr estas separaciones son polímeros que han sido entrecruzados de modo que
formen una red abierta con numerosos poros de tamaño uniforme. El grado de
entrecruzamiento en una serie de geles define los diferentes tamaños de poro y rango
de fraccionamiento. Cuando se hace pasar una fase móvil que contiene disuelta una
serie de solutos de distintos tamaños a través de una columna que contiene este tipo
de materiales, las moléculas demasiado grandes para entrar en los poros son
“excluidas” y permanecen en la fase móvil sin interactuar con la fase estacionaria, por
lo tanto, estas son eluidas con rapidez. Las moléculas de menor tamaño pueden
difundir libremente dentro y fuera de los poros del gel, de modo que, en efecto su
trayectoria a través de la columna es más larga y eluirán mas tarde.
Este efecto selectivo de tamaño es utilizado para separar moléculas de gran tamaño
presentes en un extracto, como son las clorofilas, de los metabolitos secundarios de
menor tamaño. La fase estacionaria utilizada para este procedimiento es Sephadex LH-
20, y la fase móvil una mezcla 1:1 CH2Cl2:MeOH.
Una vez que se posee el extracto desclorofilado, se procede a someterlo a una
cratografía en columna de mesada (CC) con el fin de realizar una primera separación
de los componentes químicos agrupados en “paquetes” de compuestos de polaridad
similar. Esta se realiza con Sílica gel de 70-230 Mesh como fase estacionaria y mezcla
de n-hexano – acetato de etilo (100:0 � 0:100) como fase móvil para el extracto

METABOLITOS SECUNDARIOS DE HEPÁTICAS DE LA PATAGONIA
ARGENTINA. SU ACTIVIDAD BIOLÓGICA
44 | P á g i n a
II
etéreo y acetato de etilo - metanol (100:0 � 0:100) para el extracto metanólico. La
evolución de cada columna de mesada fue monitoreada por cromatografía en capa
fina. Para ello se utilizaron cromatofolios Merck F254 (con indicador fluorescente a 254
nm). El revelado de las placas se realizó por métodos físicos y químicos. El revelado
físico fue logrado por la observación de la placa bajo luz ultravioleta de 254 nm y 350
nm. Este método no destructivo de revelado permite la detección de sustancias que
absorben radiación ultravioleta a la longitud de onda mencionada. El revelador
químico elegido fue el reactivo de Godin (Godin, 1954), seguido de asperjado con una
solución de H2SO4 al 30% y calentamiento en estufa a 110 ºC. Este revelador es útil en
la detección de sustancias oxidables como alquenos, alcoholes y aldehídos y según el
color que presentan las manchas permite tener una idea de qué tipo de compuesto se
trata.
Las fracciones de columna obtenidas fueron procesadas en algunos casos por
cromatografía líquida de mediana presión (MPLC) y por cromatografía de líquida de
alta presión (HPLC) en otros casos. Estas técnicas presentan una mayor resolución que
la cromatografía en columna y difieren entre sí en el tamaño y empaquetamiento de
las columnas, siendo las de HPLC más pequeñas y empaquetadas. Estas técnicas
permiten la resolución de mezclas y la obtención de compuestos puros. Tanto la
técnica de MPLC como la de HPLC, pueden ser llevadas a cabo ya sea en fase normal o
en fase reversa. En fase normal se emplea como adsorbente sílica gel y como fase
móvil mezclas de solventes de baja polaridad (generalmente hexano – acetato de
etilo). En fase reversa, la fase estacionaria es no polar (octilsilano C8 u octadecilsilano
C18) y la fase móvil es polar, usándose con frecuencia mezclas MeOH – H2O o CH3CN –
H2O.
Las cromatografías de MPLC y HPLC se realizaron en un cromatógrafo Gilson con un
refractómetro diferencial como detector con las siguientes columnas: MPLC (A) Merck
LiChroprep Si 60 (40-60 μm, fase normal), HPLC (B) Chemo Pack Develosil 60 (5 μm, 10
mm i.d. x 250 mm, fase normal), (C) Phenomenex Luna C8 (5 μm, 10 mm i.d. x 250 mm,
fase reversa), and (D) Phenomenex Luna C18 (5 μm, 10 mm i.d. x 250 mm, fase
reversa)

METABOLITOS SECUNDARIOS DE HEPÁTICAS DE LA PATAGONIA
ARGENTINA. SU ACTIVIDAD BIOLÓGICA
45 | P á g i n a
II
2.4 Identificación de los compuestos
La identificación de los compuestos puros obtenidos fue lograda mediante métodos
espectroscópicos. Las técnicas empleadas fueron las siguientes: espectroscopía
infrarroja (IR) y ultravioleta-visible (UV), espectrometría de masas (EM), resonancia
magnética nuclear (RMN) y dicroísmo circular (DC).
2.4.1 Espectroscopía infrarroja (IR)
La espectroscopía infrarroja (IR) permite obtener información sobre los grupos
funcionales presentes en la molécula analizada, siendo una técnica complementaria en
la elucidación estructural. Los espectros de IR fueron obtenidos en un
espectrofotómetro Perkin Elmer GX1.
2.4.2 Espectroscopía ultravioleta-visible (UV)
La espectroscopía ultravioleta-visible es una técnica que brinda información sobre
los grupos cromóforos presentes en la molécula de interés. Para que una molécula
absorba radiación ultravioleta a longitudes de onda de entre 200 y 400 nm, debe
presentar por lo menos uno de los siguientes grupos:
- Dos o más dobles enlaces conjugados
- Un anillo aromático
- Un grupo carbonilo α,β-insaturado
- Un doble enlace adyacente a un átomo adyacente a un átomo con un
par de electrones no compartidos (O, N, S, etc.)
El espectro ultravioleta-visible de una sustancia es sencillo y consta de una o unas
pocas bandas anchas de absorción. Sin embargo la posición exacta de máximos de
absorción puede brindar información sobre el grupo cromóforo presente en la
molécula. Los espectros de UV fueron obtenidos en un espectrofotómetro Shimadzu
UV-160.

METABOLITOS SECUNDARIOS DE HEPÁTICAS DE LA PATAGONIA
ARGENTINA. SU ACTIVIDAD BIOLÓGICA
46 | P á g i n a
II
2.4.3 Espectrometría de masas (EM)
Esta técnica a baja resolución brinda información estructural por análisis de
fragmentaciones sufridas por la molécula, y en algunos casos permite obtener su peso
molecular redondeado al número entero más cercano. La comparación de los
espectros de masas de baja resolución con espectros de sustancias de referencia
depositados en una biblioteca informática, permite en algunos casos determinar o
confirmar una estructura.
Para analizar una muestra por espectrometría de masas, esta debe ser ionizada.
Con este fin pueden emplearse diferentes métodos, entre las cuales utilizamos las
ionizaciones por electrones (EI) y química (CI).
Los iones formados por ionización por electrones presentan un gran exceso de
energía, que tienden a perder por fragmentaciones del ion molecular formado. Este
hecho hace que el pico correspondiente al ion molecular sea de baja intensidad en
algunos casos e incluso esté ausente en otros.
En la técnica de ionización química se utiliza un gas auxiliar, generalmente metano o
isobutano. Este gas auxiliar es ionizado por impacto electrónico, los iones así formados
reaccionan con la muestra transfiriéndole un protón. Esta reacción da lugar a iones
cuasimoleculares [M+1]+ (m/z = peso molecular + 1), esta ionización es más suave que
la lograda mediante ionización por electrones y los iones cuasimoleculares formados,
de menor energía, sufren menos fragmentaciones dando picos de mayor intensidad.
Los espectros de masas de baja resolución fueron obtenidos en un espectrómetro
Thermo Polaris Q. Este espectrómetro de masas utiliza como detector una trampa de
iones y no un detector de cuadrupolo como la mayoría de los espectrómetros. El
detector de trampa de iones es capaz de producir en algunas situaciones iones
cuasimoleculares cuando se está utilizando ionización por electrones (EI). Este es un
fenómeno observado frecuentemente y está descripto en bibliografía como auto-
ionización química en detectores de trampa de iones (Eichelberger y col., 1987).
Cuando se realiza a alta resolución (HR), la espectrometría de masas permite
obtener el peso molecular exacto del compuesto, y a partir de este valor la fórmula
molecular del compuesto de interés. Esto se debe a que los pesos atómicos no son

METABOLITOS SECUNDARIOS DE HEPÁTICAS DE LA PATAGONIA
ARGENTINA. SU ACTIVIDAD BIOLÓGICA
47 | P á g i n a
II
números enteros como resultado de la presencia de isótopos. Sólo unas pocas
combinaciones de átomos son capaces de presentar un determinado peso molecular
expresado con cuatro decimales, y es así como se determina la formula molecular.
Los espectros de masas de alta resolución fueron obtenidos en un espectrómetro
JEOL JMS AX 500.
2.4.4 Resonancia magnética nuclear (RMN)
Esta técnica es la más importante en la elucidación estructural de compuestos
orgánicos ya que brinda información sobre la conectividad de los átomos en la
molécula y sobre la disposición espacial de los átomos.
Existen técnicas de RMN unidimensionales y bidimensionales. Dentro de las técnicas
unidimensionales encontramos la resonancia magnética nuclear protónica (1H-RMN) y
de 13
C (13
C-RMN). Dentro de las bidimensionales encontramos las técnicas COSY (en
inglés: “Correlated Spectroscopy” o ”Espectroscopía correlacionada”), HSQC (en inglés:
“Heteronuclear Simple Quantum Coherence” o “Coherencia Cuántica Simple
Heteronuclear”), HMBC (en inglés: “Hetronuclear Multiple Bond Coherence” o
“Coherencia Cuántica Heteronuclear múltiple”) y NOESY (en inglés: “Nuclear
Overhauser Enhanced Spectroscopy” o “Espectroscopía por efecto Nuclear
Overhauser”).
Los espectros de RMN de este trabajo fueron registrados con los siguientes equipos:
- Varian Unity 500 MHz para 1H y 125 MHz para
13C
- Bruker 300 MHz para 1H y 75 MHz para
13C
- Bruker 200 MHz para 1H y 50 MHz para
13C
2.4.5 Dicroísmo circular (DC)
Un rayo de luz polarizado en un plano puede considerarse formado por dos
componentes polarizados circularmente, uno a la derecha y el otro a la izquierda. Estos
componentes están en fase y son de la misma amplitud. Al pasar por un medio
ópticamente activo, cada componente interactúa de manera diferente con los centros
quirales de las moléculas presentes. La interacción de la radiación con la muestra

METABOLITOS SECUNDARIOS DE HEPÁTICAS DE LA PATAGONIA
ARGENTINA. SU ACTIVIDAD BIOLÓGICA
48 | P á g i n a
II
induce un desfasamiento y un cambio de magnitud diferenciales en ambos
componentes circularmente polarizados de la luz, y estos fenómenos provocan una
rotación del plano de polarización en un ángulo α y la distorsión de este plano genera
una elipse. La rotación del plano y la diferente absorción de las componentes
circularmente polarizadas varían de acuerdo con la longitud de onda, pudiéndose
obtener espectros de estos fenómenos, esto es, gráficas de la rotación o elipticidad
versus la longitud de onda.
Los espectros de dicroísmo circular (DC) permiten obtener configuraciones
absolutas de los compuestos quirales por comparación, ya que la misma configuración
absoluta de dos compuestos con configuraciones electrónicas semejantes, darán
espectros DC del mismo signo. Además, los enantiómeros tendrán espectros de DC que
serán imágenes especulares uno del otro. Existen además reglas empíricas, como la
regla del octante, las cuales permiten predecir el signo de ciertas bandas del espectro
de CD para algunos sistemas específicos (Djerassi, 1960).
Avances producidos en química computacional ofrecen nuevas alternativas en lo
referido a la determinación de configuraciones absolutas de moléculas orgánicas. Es
posible mediante cálculos computacionales obtener los espectros de DC teóricos para
ambos enantiómeros y por comparación con el espectro de CD experimental
determinar la configuración absoluta de una determinada sustancia (Wu y col., 2010).
Los cálculos de los espectros de CD teóricos realizados en este trabajo fueron
realizados con Gaussian 09. Para los cálculos se utilizó la teoría del funcional densidad
(DFT) al nivel B3LYP/6-31G(d) de modo de optimizar la geometría de la molécula.
Luego, se hicieron los cálculos para los correspondientes estados excitados para las
geometrías optimizadas del estado fundamental. Se utilizó DFT dependiente del
tiempo combinado con el modelo PCM (TD-DFT/PCM) con la misma base para calcular
la energía de excitación y la fuerza rotatoria para los 50 primeros estados excitados. El
espectro de ECD (Dicroísmo Circular Electrónico) final fue calculado a partir de las
siguientes ecuaciones:

METABOLITOS SECUNDARIOS DE HEPÁTICAS DE LA PATAGONIA
2.5 Actividad biológica
Los bioensayos ejecutados
o Actividad
aeruginosa
el crecimiento
inductores
o Actividad
esta actividad se realizaron ensayos sobre el crecimiento
y la formación de
o Actividad insecticida contra el lepidóptero
2.5.1 Actividad
Las cepas utilizadas en los ensayos microbiológicos fueron
biofilm (figura 2.1) provenientes de la colección internacional
Collection” (ATCC): P. aeruginosa
aureus ATCC 6538 P, coco Gram positivo
Figura 2.1. Microorganismos formadores de biofilm. A)
aureus
A
METABOLITOS SECUNDARIOS DE HEPÁTICAS DE LA PATAGONIA
ARGENTINA. SU ACTIVIDAD BIOLÓGICA
Actividad biológica
ejecutados en este trabajo de tesis permitieron evaluar
Actividad anti-patogénica contra el patógeno oportunista
aeruginosa. Para determinar esta actividad se realizaron
el crecimiento bacteriano, formación de biofilm, producción de auto
inductores y actividad elastasa.
Actividad anti-patogénica contra Staphylococcus aureus
esta actividad se realizaron ensayos sobre el crecimiento
la formación de biofilm.
Actividad insecticida contra el lepidóptero Spodoptera frugiperda.
Actividad anti-patogénica sobre P. aeruginosa y
Las cepas utilizadas en los ensayos microbiológicos fueron dos cepas formadoras de
provenientes de la colección internacional “American Type Culture
P. aeruginosa ATCC 27853, bacilo Gram negativo
6538 P, coco Gram positivo.
Microorganismos formadores de biofilm. A) Pseudomonas aeruginosa
B
METABOLITOS SECUNDARIOS DE HEPÁTICAS DE LA PATAGONIA
ARGENTINA. SU ACTIVIDAD BIOLÓGICA
49 | P á g i n a
II
permitieron evaluar:
contra el patógeno oportunista Pseudomonas
ara determinar esta actividad se realizaron ensayos sobre
producción de auto-
Staphylococcus aureus. Para evaluar
esta actividad se realizaron ensayos sobre el crecimiento antibacteriano
Spodoptera frugiperda.
y S. aureus
dos cepas formadoras de
“American Type Culture
negativo y Staphylococcus
Pseudomonas aeruginosa B) Staphylococcus

METABOLITOS SECUNDARIOS DE HEPÁTICAS DE LA PATAGONIA
ARGENTINA. SU ACTIVIDAD BIOLÓGICA
50 | P á g i n a
II
Los medios de cultivos empleados fueron:
- Medio Luria-Bertani (LB) para la cepa de P. aeruginosa. (NaCl 1% p/v, extracto de
levadura 0,5% p/v, peptona 0,5% p/v, pH 6,5).
- Medio Müller-Hinton (MH) para la cepa de S. aureus. (Almidon 0,15% p/v, infusión
de carne 0,2% p/v, peptona de caseína hidrolizada 1,75% p/v, pH 7,4).
2.5.1.1 Determinación del crecimiento bacteriano
Un cultivo de 12 h del microorganismo (DO ≥ 1) fue diluido al 10% (v/v) en medio de
cultivo adecuado. El cultivo diluido (190 µl) fue vertido en cada uno de los 96 pocillos
de una microplaca. Además, se prepararon soluciones conteniendo 1 y 0.1 mg/ml de
los compuestos a testear en DMSO – H2O destilada (1:1) y 10 µl de cada una fueron
agregadas individualmente a cada pocillo de la microplaca (8 repeticiones) de modo de
obtener concentraciones finales de 5 y 50 µg/ml. A los pocillos para el control negativo
(8 repeticiones) conteniendo 190 µl de cultivo diluido se les adicionó 10 µl de DMSO –
H2O destilada (1:1). Las placas fueron incubadas a 37 ºC durante 24 h y se determinó el
crecimiento bacteriano espectrofotométricamente a 600 nm en lector de microplaca
(PowerWave XS2, Biotek, VT, USA).
2.5.1.2 Determinación de la formación de biofilm
Para la determinación de la concentración de biofilm se empleó una micro técnica
basada en un protocolo previamente reportado (O'Toole y Kolter, 1998). Luego de
medir el crecimiento bacteriano, el líquido de los pocillos fue descartado y el material
que se mantuvo adherido al plástico (biofilm) fue lavado 3 veces con PBS. El biofilm
que permaneció adherido después del lavado fue teñido con una solución acuosa de
cristal violeta (0.1% p/v) durante 20 min. El exceso de colorante fue eliminado
mediante lavados con H2O y el cristal violeta unido al biofilm fue posteriormente
disuelto en 200 µl de EtOH absoluto. Se determinó la absorbancia a 540 nm usando un
lector de microplaca (PowerWave XS2, Biotek, VT, USA). Ciprofloxacina, un conocido
inhibidor de biofilm (Sandasi y col., 2011), fue incorporado al bioensayo a 5 µg/ml

METABOLITOS SECUNDARIOS DE HEPÁTICAS DE LA PATAGONIA
ARGENTINA. SU ACTIVIDAD BIOLÓGICA
51 | P á g i n a
II
como control positivo en las mismas condiciones experimentales que las utilizadas
para evaluar los compuestos.
2.5.1.3 Evaluación de los efectos sobre el proceso de QS (sólo P.
aeruginosa)
Un cultivo de 12 h a 37 ºC de la cepa reportera P. aeruginosa qsc 119 (DO ≥ 1)
(Whiteley y col., 1999) en LB fue diluido 10 veces en el mismo medio. Una porción de
25 µl de esta suspensión fue mezclada en cada pocillo de una microplaca con 25 µl de
un sobrenadante libre de células obtenido de un cultivo de P. aeruginosa ATCC 27853
incubado en LB conteniendo 5 y 50 µg/ml de los compuestos a testear durante 24 h.
Azitromicina, capaz de intervenir en los procesos de QS (Tateda y col., 2001), fue usado
a 5 µl/ml como control positivo de inhibición de QS bajo las mismas condiciones que
los compuestos. Los pocillos de control negativos (8 repeticiones) conteniendo 50 µl de
sobrenadante libre de células obtenido de un cultivo de P. aeruginosa ATCC 27853
incubado en LB conteniendo 10 µl de DMSO – H2O (1:1).
P. aeruginosa qsc 119 es una cepa mutante que no produce AHL (moléculas señal
del QS), pero responde a las AHL exógenas con la producción de β-galactosidasa. En
consecuencia, la actividad de β-galactosidasa está bajo el control del QS y en relación
directa con la actividad de autoinductores. La actividad β-galactosidasa fue medida
espectrofotométricamente por el test de Miller (O'Toole y Kolter, 1998).
Luego de una incubación de entre 4 y 5 h de la cepa P. aeruginosa qsc 119 en
presencia del sobrenadante, a cada pocillo se le agregaron 50 µl de buffer Z, 10 µl de
CHCl3 y 5 µl del detergente dodecilsulfato de sodio (SDS) 0,1%. Se incubó a 28 ºC
durante 15 min con agitación suave. A continuación se agregaron 20 µl de solución de
orto-nitrofenilgalactopiranósido (ONPG) (4 mg/ml) a cada pocillo, este compuesto es
un sustrato de la β-galactosidasa y la solución debe ser preparada en el mismo día de
su uso debido a que se descompone en presencia de la luz. Esta se puede guardar en la
heladera por unos pocos días protegida de la luz con papel aluminio. Una vez que se
desarrolló color amarillo claro, entre 3 y 5 h, se registró el tiempo t y se detuvo la
reacción con el agregado de 65 µl de Na2CO3 1M. Finalmente se midió la DO a 420, 550

METABOLITOS SECUNDARIOS DE HEPÁTICAS DE LA PATAGONIA
ARGENTINA. SU ACTIVIDAD BIOLÓGICA
52 | P á g i n a
II
y 600 nm y se calcularon las unidades de β-galactosidasa mediante la siguiente
fórmula:
�������� �� ���� �������� �1000 � ���420�� 1,75 � ��550���
�� � � � ��600���
Donde:
DO420nm = Absorbancia del producto de reacción entre la enzima β-galactosidasa y el
ONPG
DO550nm = Indica la dispersión de luz (partículas)
DO600nm= Indica la densidad celular antes del ensayo
t = Tiempo de desarrollo de color en horas
V = Volumen de cultivo qsc en mililitros
Composición del Buffer Z pH 7,0. Para 1000 ml de agua destilada
Nombre Cantidad (g)
Fosfato dibásico de sodio heptahidratado 16,1
Fosfato monobásico de sodio monohidrato 5,5
Cloruro de potasio 0,75
Sulfato de magnesio heptahidratado 0,246
β-mercaptoetanol 2,7 ml
El compuesto β-mercaptoetanol debe ser agregado en el momento de usar. Se debe
llevar a pH 7 con HCl.
2.5.1.4 Ensayo sobre la actividad elastasa (solo P. aeruginosa)
Las producción de las distintas elastasas de P. aeruginosa está regulada por el
proceso de QS. Para evaluar el efecto de los compuestos aislados sobre la actividad
elastasa de P. aeruginosa se hizo crecer la misma en caldo LB durante 24 horas a 37ºC
en presencia de los compuestos a testear a una concentración de 50 µg/ml.
Posteriormente se separaron las células mediante centrifugación (15 minutos a 11000
rpm). Se midió la actividad elastasa en los sobrenadantes obtenidos luego de la
incubación con los compuestos.

METABOLITOS SECUNDARIOS DE HEPÁTICAS DE LA PATAGONIA
ARGENTINA. SU ACTIVIDAD BIOLÓGICA
53 | P á g i n a
II
La enzima elastasa degrada la elastina, en presencia del sustrato insoluble rojo
congo-elastina (RC-E), libera el colorante unido a la elastina y este se solubiliza. La
concentración de rojo congo liberado al medio se determina
espectrofotométricamente a 495 nm (Gambello y Iglewski, 1991).
2.5.2 Actividad insecticida sobre S. frugiperda
2.5.2.1 Insectos y dieta
Las larvas de S. frugiperda fueron obtenidas de nuestras colonias de laboratorio,
originalmente colectadas de plantas de maíz (Zea mays) y mantenidas en dieta
artificial durante dos generaciones. Las colonias no fueron previamente expuestas a
insecticidas. Los individuos fueron mantenidos en cámara de cría a 25±1 ºC, 60-70% de
humedad relativa, y un fotoperiodo de 12 h Luz/Oscuridad. La dieta larval consiste de
una mezcla de levadura (3g), poroto hervido y molido (224g), germen de trigo (52g),
agar agar (60g), acido ascórbico (9g), metil-p-hidroxibenzoato (3g), formaldehido (3 ml
de una solución al 40% en agua), y agua (1.5 l).
2.5.2.2 Bioensayo de toxicidad
Una porción de dieta artificial fue impregnada con solvente (acetona) y, después de
remover el solvente, esta porción fue empleada como dieta control. Otras porciones
fueron impregnadas y cuidadosamente mezcladas con una solución en acetona de los
compuestos puros de forma de obtener 100 µg y 200 µg de cada uno por gramo de
dieta. Luego de evaporar el solvente, las dietas control y tratadas fueron puestas en
tubos de ensayos (15 x 1.5 cm, 20 para el control y 20 para tratado). Larvas
cuidadosamente pesadas y de tamaño homogéneo (2do
estadío) fueron puestas
individualmente en cada tubo y mantenidas a 25 ± 1 ºC y 60-70% HR en cámara de cría
(Electro Lab). Los tubos de ensayo fueron cubiertos con un pequeño trozo de algodón
húmedo para prevenir la desecación de la dieta. A las larvas se les permitió
alimentarse con cantidades pesadas ( 50 mg/d) de las dietas control y tratadas hasta el
estado de pupa. Luego de 14 días del inicio del experimento, se pesaron nuevamente

METABOLITOS SECUNDARIOS DE HEPÁTICAS DE LA PATAGONIA
ARGENTINA. SU ACTIVIDAD BIOLÓGICA
54 | P á g i n a
II
las larvas para obtener el índice de crecimiento (GR) durante ese periodo. El peso de
dieta larval fue también calculado para el mismo período (14 días) de modo de
determinar el índice de consumo (CR). Además, fueron evaluadas las malformaciones
en adultos.
GR = (A-B)/t, es el promedio del incremento del peso larval por día, donde A = peso
larval final, B = peso larval inicial, y t = 14 días (periodo de evaluación) (Ramírez y col.,
2010).
CR = D/t, es el promedio de dieta larval consumida por día, donde D es el peso total
de dieta consumida durante los 14 días del experimento (Ramírez y col., 2010).
2.5.2.3 Análisis histológico
Las larvas (tratadas y control) fueron fijadas con el fijador de Bouin (Carson, 1997)
durante 24 h. Después de la deshidratación en soluciones acuosas de etanol al 70 y
90%, y finalmente en etanol puro, las larvas fueron incorporadas en n-butanol puro
durante 24 h para ablandar la cutícula. Las larvas fueron llevadas a una mezcla de n-
butanol-histowax (1/1) a 60 ºC durante 24 h. Luego, se cortaron secciones de 5-7 µm
de espesor en un micrótomo minot (LEICA RM 2035) y teñidas con hematoxilina-eosina
(H-E). EL análisis histológico fue realizado en un microscópio LEICA DM 1000 y se
tomaron fotografías con una microcámara SONY DSC-W 100.

METABOLITOS SECUNDARIOS DE HEPÁTICAS DE LA PATAGONIA
ARGENTINA. SU ACTIVIDAD BIOLÓGICA
55 | P á g i n a
II
Referencias
Carson F. L. Histotechnology: A self-instructional text. ASCP Press, Chicago. 1997.
Djerassi C. Optical Rotary Dispersion: Applications to Organic Chemistry. McGraw-
Hill Book Co, New York. 1960.
Eichelberger J. W., Budde W. L. and Slivon L. E. Existence of self chemical ionization
in the ion trap detector. Anal. Chem. 1987, 59, 2730-2732.
Gambello M. J. and Iglewski B. H. Cloning and characterization of the Pseudomonas
aeruginosa lasR gene, a transcriptional activator of elastase expression. Journal of
Bacteriology 1991, 173, 3000-3009.
Godin P. A new spray reagent for paper chromatography of polyols and cetoses. Nature
1954, 174, 134.
O'Toole G. A. and Kolter R. Flagellar and twitching motility are necessary for
Pseudomonas aeruginosa biofilm development. Molecular Microbiology 1998, 30, 295-
304.
Ramírez M., Kamiya N., Popich S., Asakawa Y. and Bardón A. Insecticidal constituents
from the Argentine Liverwort Plagiochila bursata. Chemistry and Biodiversity 2010, 7,
1855-1861.
Sandasi M., Leonard C. M., van Vuuren S. F. and Viljoen A. M. Peppermint (Mentha
piperita) inhibits microbial biofilms in vitro. South African Journal of Botany 2011, 77,
80-85.

METABOLITOS SECUNDARIOS DE HEPÁTICAS DE LA PATAGONIA
ARGENTINA. SU ACTIVIDAD BIOLÓGICA
56 | P á g i n a
II
Tateda K., Comte R., Pechere J. C., Köhler T., Yamaguchi K. and van Delden C.
Azithromycin inhibits quorum sensing in Pseudomonas aeruginosa. Antimicrobial
Agents and Chemotherapy 2001, 45, 1930-1933.
Whiteley M., Lee K. M. and Greenberg E. P. Identification of genes controlled by
quorum sensing in Pseudomonas aeruginosa. Proceedings Of The National Academy Of
Sciences Of The United States Of America 1999, 96, 13904-13909.
Wu X. F., Hu Y. C., Yu S. S., Jiang N., Ma J., Tan R. X., Li Y., Lv H. N., Liu J. and Ma
S. G. Lysidicins F-H, three new phloroglucinols from Lysidice rhodostegia. Organic
Letters 2010, 12, 2390-2393.

Capítulo III
Estudio químico de
Porella chilensis
III

METABOLITOS SECUNDARIOS DE HEPÁTICAS DE LA PATAGONIA
ARGENTINA. SU ACTIVIDAD BIOLÓGICA
58 | P á g i n a
III
Capitulo III:
Estudio químico de Porella chilensis
3.1 Antecedentes químicos de la familia Porellaceae
Particularmente, las especies de Porella producen sesquiterpenoides de los tipos
africano, pinguisano, germacrano, guaiano, drimano y aromadendrano, como así
también diterpenoides del tipo sacculatano. (Asakawa y col., 2012; Asakawa, 2004;
Van Klink y col., 2002; Fukuyama y col., 1988; Toyota y col., 1989b; Toyota y col.,
1991a; Quang y Asakawa, 2010; Asakawa y col., 1987; Tori y col., 1993; Tori y col.,
1994; Bovi Mitre y col., 2004; Toyota y col., 1989a). Todos los aromadendranos
encontrados en hepáticas poseen configuracion ent y a algunos de ellos se les conoce
su contraparte antipodal en plantas superiores. (Gijsen, 1993)
3.2 Clasificación botánica de P. chilensis.
Jungermanniidae Marchantiidae
Metzgeriales Takakiales Calobryales Jungermanniales
Suborden
Jungermanniineae Lepidoziineae Ptilidiineae Porellineae Radulineae Pleuroziineae
Familia
Porellaceae
Genero
Porella
Especie
Porella chilensis
Subclase
Clase Hepaticae
Orden
Jubulineae

METABOLITOS SECUNDARIOS DE HEPÁTICAS DE LA PATAGONIA
ARGENTINA. SU ACTIVIDAD BIOLÓGICA
59 | P á g i n a
III
3.3 Experimental
3.3.1 Material vegetal
El material vegetal de Porella chilensis (Lehm. & Lindenb.) Trevis. (Figura 3.1), fue
recolectado en Febrero de 2005 en lago Steffen, Provincia de Río Negro, Argentina.
Una muestra fue depositada en el herbario Miguel Lillo (LIL Nº 3411), Tucumán,
Argentina.
Figura 3.1. Porella chilensis (Lehm. & Lindenb.) Trevis
3.3.2 Extracción y aislamiento
El material vegetal (573 g) fue extraído a temperatura ambiente por 7 días con éter
etílico para dar 4,8 g de extracto después de la remoción del solvente con rotavapor
(rendimiento 0,84 %). El extracto fue sometido a cromatografía en columna (CC) con
Sephadex LH-20 (MeOH-CH2Cl2, 1:1) y luego a CC con silica gel (70 – 230 mesh) con
hexano y cantidades crecientes de acetato de etilo (AcOEt) y finalmente MeOH como
eluyentes para dar 5 fracciones.

METABOLITOS SECUNDARIOS DE HEPÁTICAS DE LA PATAGONIA
ARGENTINA. SU ACTIVIDAD BIOLÓGICA
60 | P á g i n a
III
El compuesto 6 (250 mg) fue obtenido como material cristalino a partir de la
fracción I. Después de la separación de 6 por filtración, el material sobrenadante fue
sometido a MPLC con columna A, con hexano y cantidades crecientes de AcOEt (0 –
100%) como eluyente, para dar el compuesto 5 (19 mg) y una nueva cantidad de 6 (12
mg).
La fracción II (98 mg), fue sometida a HPLC con columna B (hexano-AcOEt, 23:2),
para dar los compuestos 8 (6,5 mg) y 9 (6mg).
La fracción III (296 mg) fue procesada por HPLC con columna A con hexano y
cantidades crecientes de AcOEt (5 – 100%) como eluyente, para dar los compuestos 3
(5 mg) y 7 (39 mg).
Por recromatografía en columna de la fr IV con silica gel, usando hexano y
cantidades crecientes de AcOEt (10 – 100%) se obtuvieron 2 subfracciones. El
compuesto 3 (39 mg) fue obtenido de la fracción 1. La fracción 2 fue procesada por
HPLC con columna C (MeOH-H2O 17:3), para dar los compuestos 1 (1,6 mg), 2 (4,3 mg),
4 (7 mg) y una nueva cantidad del compuesto 3 (7 mg).
La fracción V fue sometida a CC con Sephadex LH20 y luego a HPLC con columna D
(MeOH-H2O 3:1) para dar el compuesto 10 (9 mg).
3.4 Resultados
Del estudio del extracto etéreo se obtuvieron dos nuevos diterpenoides del tipo
fusicoccano (1 y 2) junto con dos fusicoccanos conocidos (3 y 4), cuatro
sesquiterpenoides conocidos del tipo pinguisano (5 - 8), y dos aromadendranos
conocidos (9 y 10), siendo norpinguisona (6) el compuesto mayoritario.
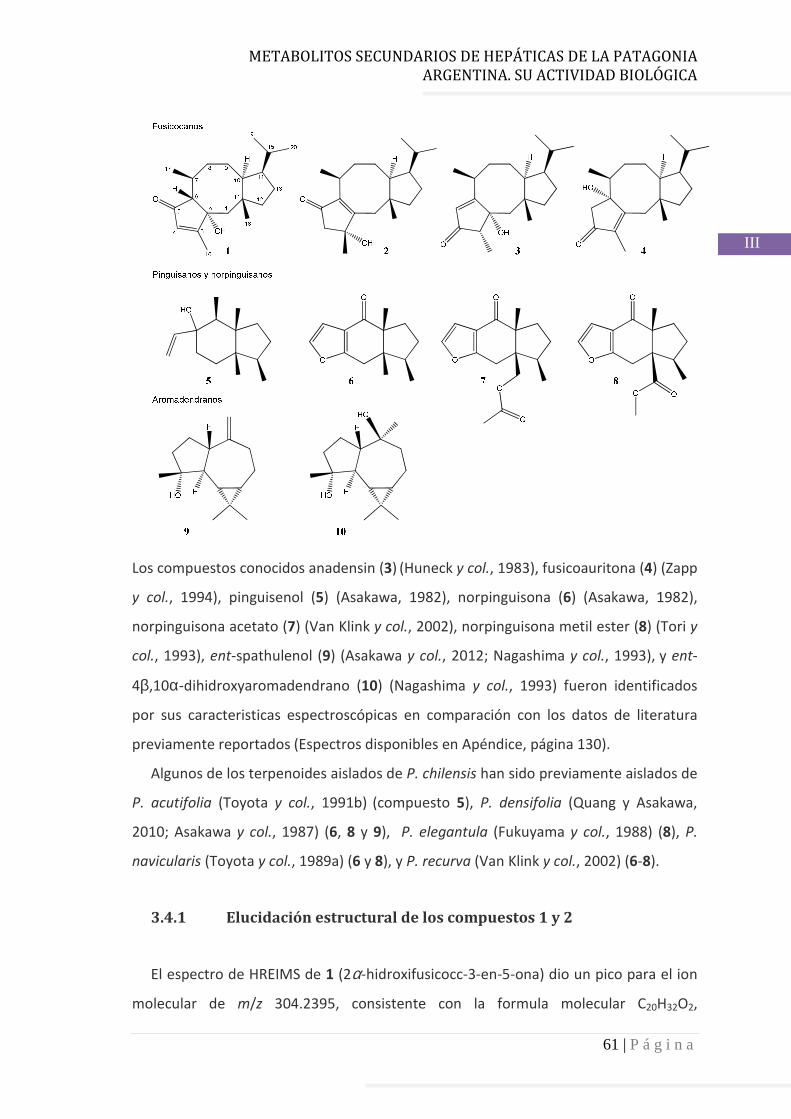
METABOLITOS SECUNDARIOS DE HEPÁTICAS DE LA PATAGONIA
ARGENTINA. SU ACTIVIDAD BIOLÓGICA
61 | P á g i n a
III
Los compuestos conocidos anadensin (3) (Huneck y col., 1983), fusicoauritona (4) (Zapp
y col., 1994), pinguisenol (5) (Asakawa, 1982), norpinguisona (6) (Asakawa, 1982),
norpinguisona acetato (7) (Van Klink y col., 2002), norpinguisona metil ester (8) (Tori y
col., 1993), ent-spathulenol (9) (Asakawa y col., 2012; Nagashima y col., 1993), y ent-
4β,10α-dihidroxyaromadendrano (10) (Nagashima y col., 1993) fueron identificados
por sus caracteristicas espectroscópicas en comparación con los datos de literatura
previamente reportados (Espectros disponibles en Apéndice, página 130).
Algunos de los terpenoides aislados de P. chilensis han sido previamente aislados de
P. acutifolia (Toyota y col., 1991b) (compuesto 5), P. densifolia (Quang y Asakawa,
2010; Asakawa y col., 1987) (6, 8 y 9), P. elegantula (Fukuyama y col., 1988) (8), P.
navicularis (Toyota y col., 1989a) (6 y 8), y P. recurva (Van Klink y col., 2002) (6-8).
3.4.1 Elucidación estructural de los compuestos 1 y 2
El espectro de HREIMS de 1 (2α-hidroxifusicocc-3-en-5-ona) dio un pico para el ion
molecular de m/z 304.2395, consistente con la formula molecular C20H32O2,

METABOLITOS SECUNDARIOS DE HEPÁTICAS DE LA PATAGONIA
ARGENTINA. SU ACTIVIDAD BIOLÓGICA
62 | P á g i n a
III
correspondiente a cinco grados de instauración. En el espectro 1H RMN (tabla 3.1) se
encontraron señales para cinco metilos, tres de ellos secundarios (δ 0.80, d, J=6.7Hz,
H-19; δ 0.90, d, J=6.6Hz, H-20; δ 1.34, d, J=6.8Hz, H-17) y dos terciarios (δ 0.95, s, H-18;
δ 2.07, s, J=1.0Hz, H-16). El espectro 1H
1H COSY reveló un acoplamiento alílico entre
H-16 y el protón a δ 5.77 (q, J=1.0Hz), indicando su ubicación y su naturaleza olefínica.
En adición, el espectro HMBC (tabla 3.2) mostró picos cruzados entre el sistema AB
metilénico correspondiente a H-1 (δ 1.85, d, J=15.3Hz; δ 1.95, d, J=15.3) y C-2, C-3, C-6,
C-10, C-11, C-12 y C-18, indicando su ubicación en C-1. Las señales a δ 128.5 y 178.1 en
el espectro 13
C RMN (tabla 3.1) corresponden al único doble enlace presente en la
molécula y su ubicación fue deducida por la correlación en el espectro de HMBC entre
el protón olefínico (δ 5.77) y el C-16 (δ 13.3). La única señal carbonílica en el espectro
13C RMN (δ 209.0), fue asignada al C-5 a través de su relación en el espectro HMBC con
H-4 y el protón del metino H-6 (δ 2.50, d, J=10.2), mientras que la única señal de
carbinol en el espectro de 13
C RMN (δ 82.6) fue asignada al C-2 basado en su
correlación HMBC con H-1, H-4, H-6 y H-16. La estereoquímica relativa del compuesto
1, como se representa en la figura 3.2, fue establecida por las correlaciones observadas
en el espectro NOESY entre H-18 y (i) H-1β, (ii) H-6 (con orientación β), (iii) H-9β, (iv)
H-12β, (v) H-13β, (vi) H-17 y (vii) H-19, indicando que todos ellos están ubicados en el
lado β de la molécula. En adición, la estructura conformacional de menor energía
(Figura 3.3) calculada con el programa hyperchem (v8.0.6) (54.73 kcal/mol) es
coincidente con lo propuesto en la figura 3.2. La estereoquímica relativa para C-6 y C-7
(Figura 3.2) es consistente con la constante de acoplamiento observada en el espectro
1H RMN (J=10.2 Hz) entre H-6 (con orientación β) y H-7 (con orientación α) como
muestra la tabla 3.1. Cálculos adicionales con el programa Pcmodel (v6.0) para el
confórmero de minima energía indican un ángulo diedro de 147.3º para HβC(6)-
HαC(7), con un acoplamiento calculado de J=9.1 Hz.
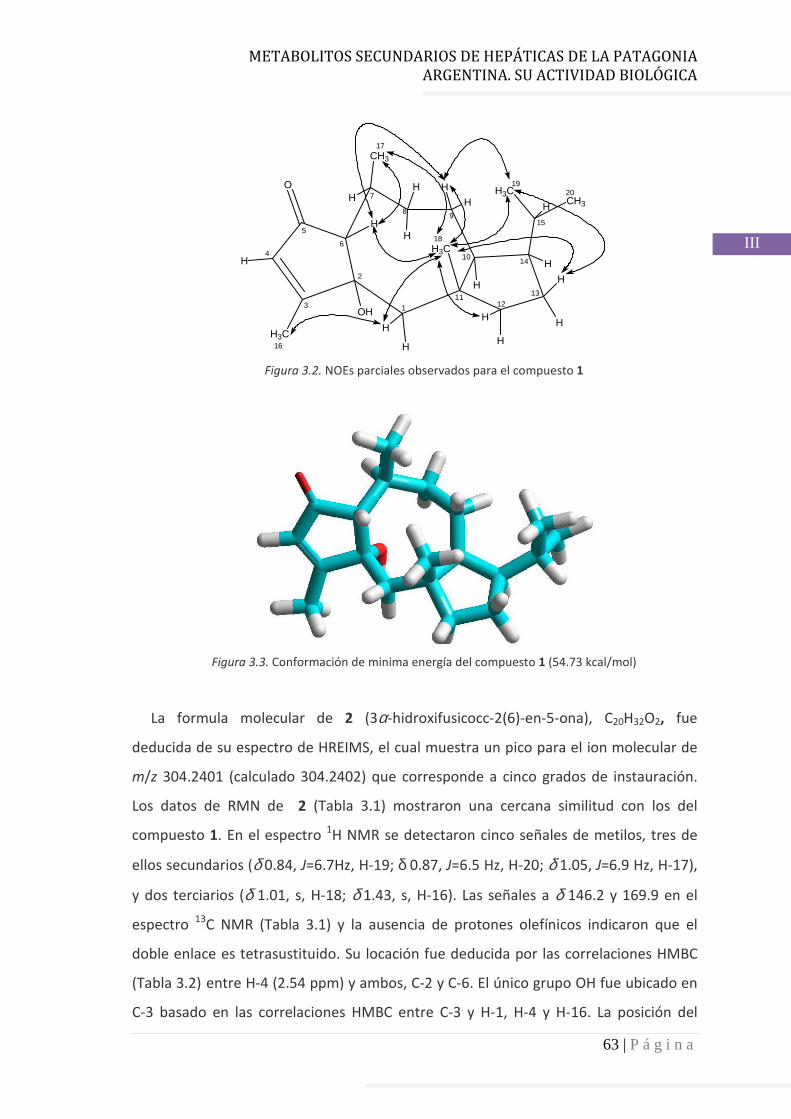
METABOLITOS SECUNDARIOS DE HEPÁTICAS DE LA PATAGONIA
ARGENTINA. SU ACTIVIDAD BIOLÓGICA
63 | P á g i n a
III
2
111
1213
1410
15
H3C19
CH3
20
6
7
89
5
4
3
O
H3C16
H
H
H
H3C18
H
HH
H
H
H
H
H
H
H
H
CH3
17
H
H
OH
Figura 3.2. NOEs parciales observados para el compuesto 1
Figura 3.3. Conformación de minima energía del compuesto 1 (54.73 kcal/mol)
La formula molecular de 2 (3α-hidroxifusicocc-2(6)-en-5-ona), C20H32O2, fue
deducida de su espectro de HREIMS, el cual muestra un pico para el ion molecular de
m/z 304.2401 (calculado 304.2402) que corresponde a cinco grados de instauración.
Los datos de RMN de 2 (Tabla 3.1) mostraron una cercana similitud con los del
compuesto 1. En el espectro 1H NMR se detectaron cinco señales de metilos, tres de
ellos secundarios (δ 0.84, J=6.7Hz, H-19; δ 0.87, J=6.5 Hz, H-20; δ 1.05, J=6.9 Hz, H-17),
y dos terciarios (δ 1.01, s, H-18; δ 1.43, s, H-16). Las señales a δ 146.2 y 169.9 en el
espectro 13
C NMR (Tabla 3.1) y la ausencia de protones olefínicos indicaron que el
doble enlace es tetrasustituido. Su locación fue deducida por las correlaciones HMBC
(Tabla 3.2) entre H-4 (2.54 ppm) y ambos, C-2 y C-6. El único grupo OH fue ubicado en
C-3 basado en las correlaciones HMBC entre C-3 y H-1, H-4 y H-16. La posición del

METABOLITOS SECUNDARIOS DE HEPÁTICAS DE LA PATAGONIA
ARGENTINA. SU ACTIVIDAD BIOLÓGICA
64 | P á g i n a
III
único carbonilo (δ 205.04) en el espectro 13
C RMN pudo ser inferida por la correlación
HMBC entre C-5 y H-1. La estereoquímica relativa del compuesto 2 fue establecida por
correlaciones NOESY (Figura 3.4) observadas entre H-18 y (i) H-1β, (ii) H-8β, (iii) H-
12β, (iv) H-16 y (v) H-19, indicando que CH3-16, CH3-18 y el grupo isopropilo en C-14,
están todos en posición β. Correlaciones entre H-14 y (i) H-10 y (ii) H-15 indican que
todos ellos permanecen en el lado α de la molécula.
Figura 3.4. NOEs parciales observados para el compuesto 2
3.4.2 Determinación de la configuración absoluta de 2
La configuración absoluta de 2 fue determinada en base a su espectro de CD el cual
mostró una traza similar a la de las ciclopentenonas sesquiterpénicas A y B, (Bovi Mitre
y col., 2004) las cuales comparten caracteristicas estructurales con el compuesto 2. En
ciclopentenonas α,β-insaturadas, la transición n→π* del grupo carbonilo produce una
banda de absorción débil entre 310 y 350 nm. Para este tipo de compuestos se
observan efectos Cotton a las longitudes de onda mencionadas (Stöcklin y col., 1970;
Djerassi y col., 1957). El espectro de CD de 2 muestra un efecto Cotton negativo (Δε317
−5.11) como puede ser predicho por la regla del octante (Djerassi y col., 1960; Moffitt y
col., 1961). De acuerdo al espectro de NOESY, el CH3-16 cae en el cuadrante superior
derecho (−) dada su orientación pseudoecuatorial, mientras que el resto de los
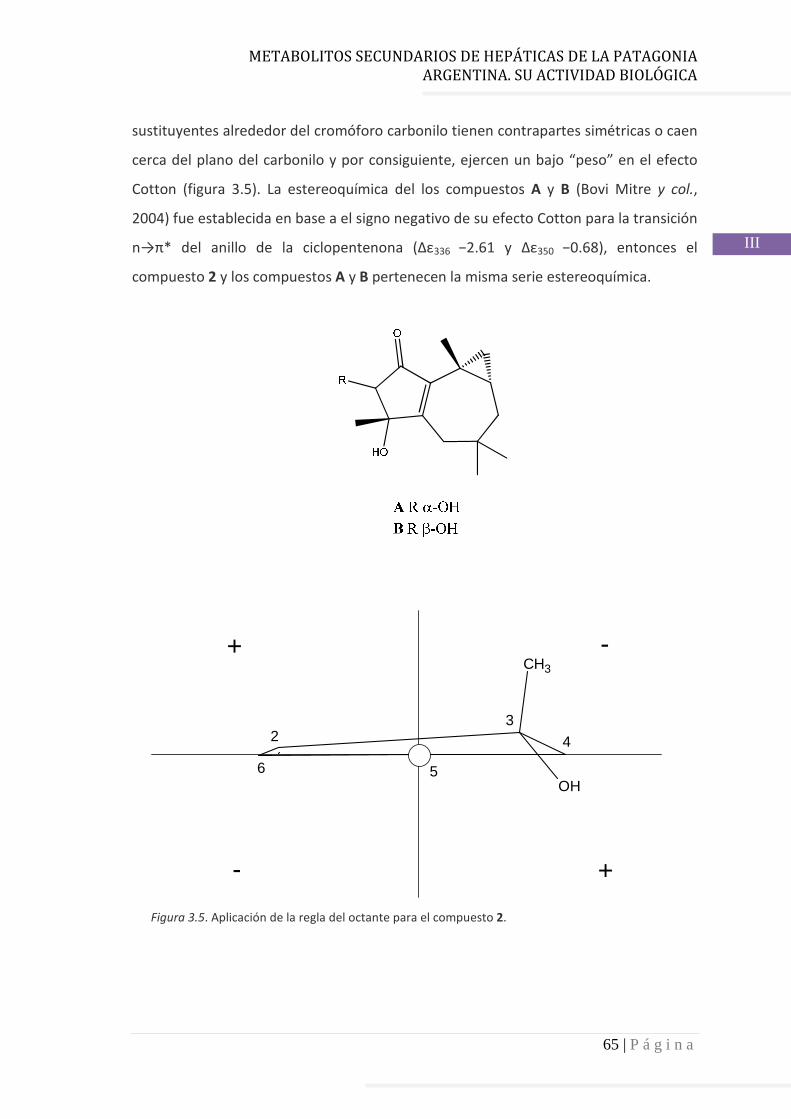
METABOLITOS SECUNDARIOS DE HEPÁTICAS DE LA PATAGONIA
ARGENTINA. SU ACTIVIDAD BIOLÓGICA
65 | P á g i n a
III
sustituyentes alrededor del cromóforo carbonilo tienen contrapartes simétricas o caen
cerca del plano del carbonilo y por consiguiente, ejercen un bajo “peso” en el efecto
Cotton (figura 3.5). La estereoquímica del los compuestos A y B (Bovi Mitre y col.,
2004) fue establecida en base a el signo negativo de su efecto Cotton para la transición
n→π* del anillo de la ciclopentenona (Δε336 −2.61 y Δε350 −0.68), entonces el
compuesto 2 y los compuestos A y B pertenecen la misma serie estereoquímica.
OH
CH3
+
+
-
-
5
4
32
6
Figura 3.5. Aplicación de la regla del octante para el compuesto 2.
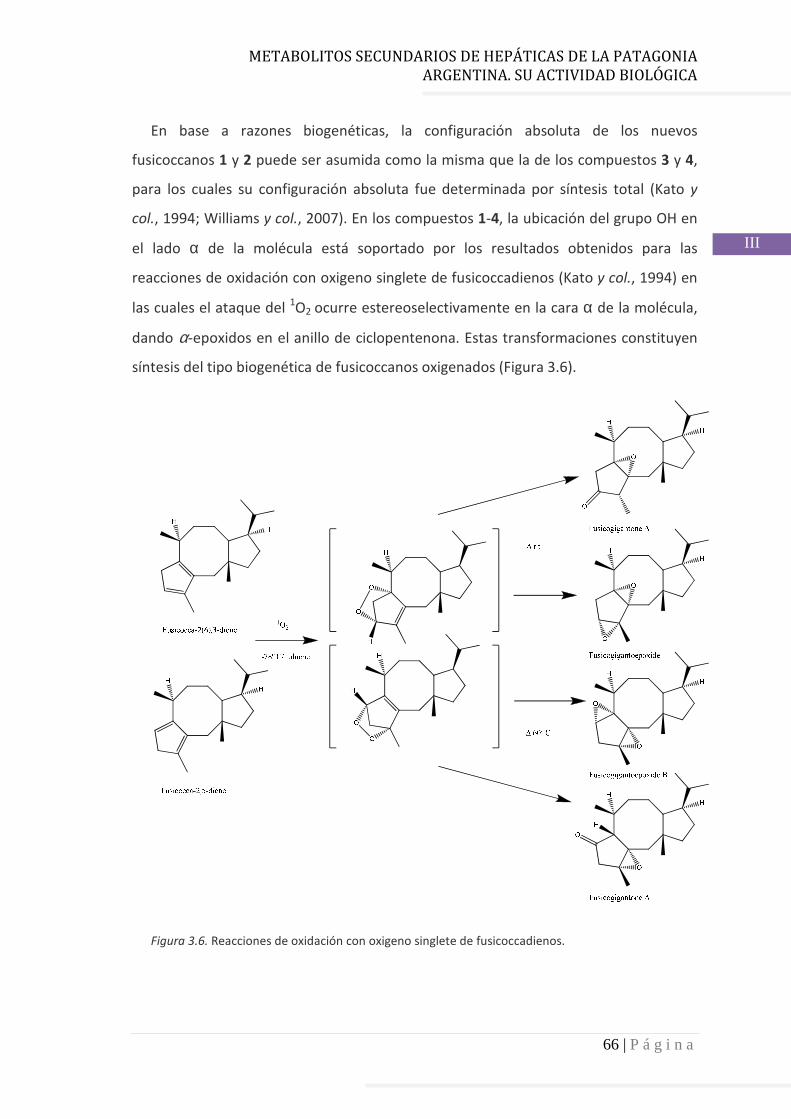
METABOLITOS SECUNDARIOS DE HEPÁTICAS DE LA PATAGONIA
ARGENTINA. SU ACTIVIDAD BIOLÓGICA
66 | P á g i n a
III
En base a razones biogenéticas, la configuración absoluta de los nuevos
fusicoccanos 1 y 2 puede ser asumida como la misma que la de los compuestos 3 y 4,
para los cuales su configuración absoluta fue determinada por síntesis total (Kato y
col., 1994; Williams y col., 2007). En los compuestos 1-4, la ubicación del grupo OH en
el lado α de la molécula está soportado por los resultados obtenidos para las
reacciones de oxidación con oxigeno singlete de fusicoccadienos (Kato y col., 1994) en
las cuales el ataque del 1O2 ocurre estereoselectivamente en la cara α de la molécula,
dando α-epoxidos en el anillo de ciclopentenona. Estas transformaciones constituyen
síntesis del tipo biogenética de fusicoccanos oxigenados (Figura 3.6).
Figura 3.6. Reacciones de oxidación con oxigeno singlete de fusicoccadienos.
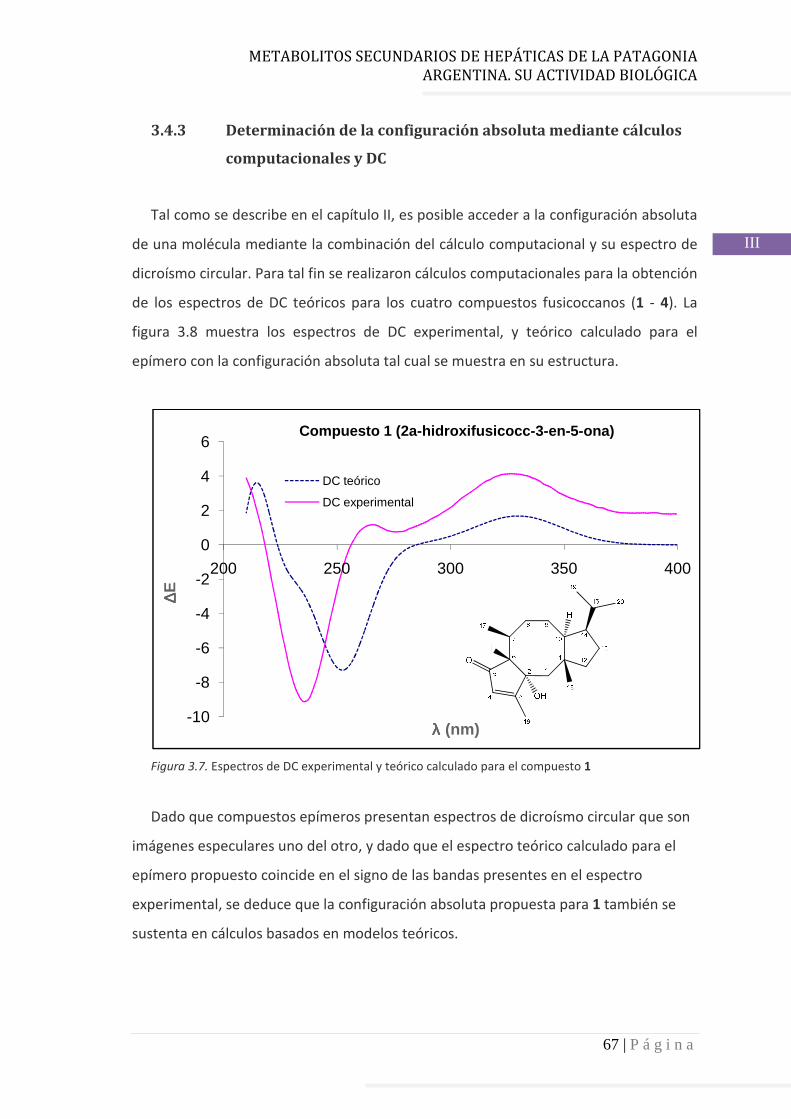
METABOLITOS SECUNDARIOS DE HEPÁTICAS DE LA PATAGONIA
ARGENTINA. SU ACTIVIDAD BIOLÓGICA
67 | P á g i n a
III
3.4.3 Determinación de la configuración absoluta mediante cálculos
computacionales y DC
Tal como se describe en el capítulo II, es posible acceder a la configuración absoluta
de una molécula mediante la combinación del cálculo computacional y su espectro de
dicroísmo circular. Para tal fin se realizaron cálculos computacionales para la obtención
de los espectros de DC teóricos para los cuatro compuestos fusicoccanos (1 - 4). La
figura 3.8 muestra los espectros de DC experimental, y teórico calculado para el
epímero con la configuración absoluta tal cual se muestra en su estructura.
Figura 3.7. Espectros de DC experimental y teórico calculado para el compuesto 1
Dado que compuestos epímeros presentan espectros de dicroísmo circular que son
imágenes especulares uno del otro, y dado que el espectro teórico calculado para el
epímero propuesto coincide en el signo de las bandas presentes en el espectro
experimental, se deduce que la configuración absoluta propuesta para 1 también se
sustenta en cálculos basados en modelos teóricos.
-10
-8
-6
-4
-2
0
2
4
6
200 250 300 350 400
∆∆ ∆∆E
λλλλ (nm)
Compuesto 1 (2a-hidroxifusicocc-3-en-5-ona)
DC teórico
DC experimental
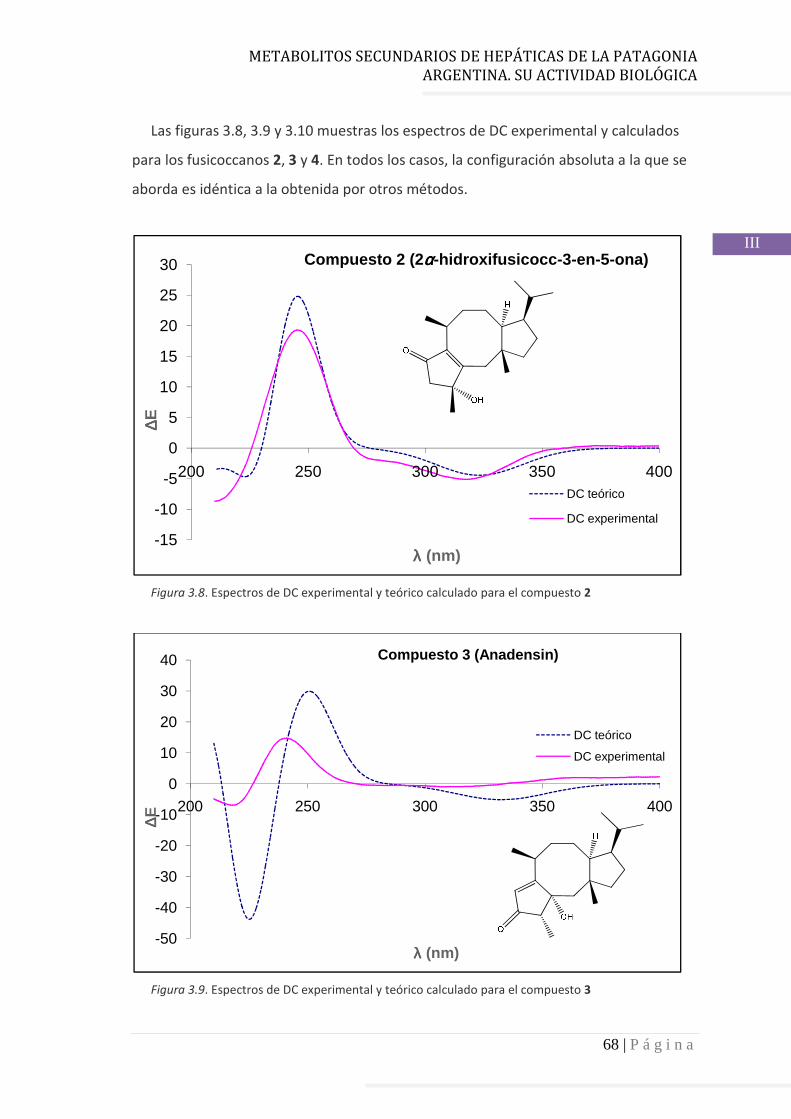
METABOLITOS SECUNDARIOS DE HEPÁTICAS DE LA PATAGONIA
ARGENTINA. SU ACTIVIDAD BIOLÓGICA
68 | P á g i n a
III
Las figuras 3.8, 3.9 y 3.10 muestras los espectros de DC experimental y calculados
para los fusicoccanos 2, 3 y 4. En todos los casos, la configuración absoluta a la que se
aborda es idéntica a la obtenida por otros métodos.
Figura 3.8. Espectros de DC experimental y teórico calculado para el compuesto 2
Figura 3.9. Espectros de DC experimental y teórico calculado para el compuesto 3
-15
-10
-5
0
5
10
15
20
25
30
200 250 300 350 400
∆∆ ∆∆E
λλλλ (nm)
Compuesto 2 (2αααα-hidroxifusicocc-3-en-5-ona)
DC teórico
DC experimental
-50
-40
-30
-20
-10
0
10
20
30
40
200 250 300 350 400
∆∆ ∆∆E
λλλλ (nm)
Compuesto 3 (Anadensin)
DC teórico
DC experimental
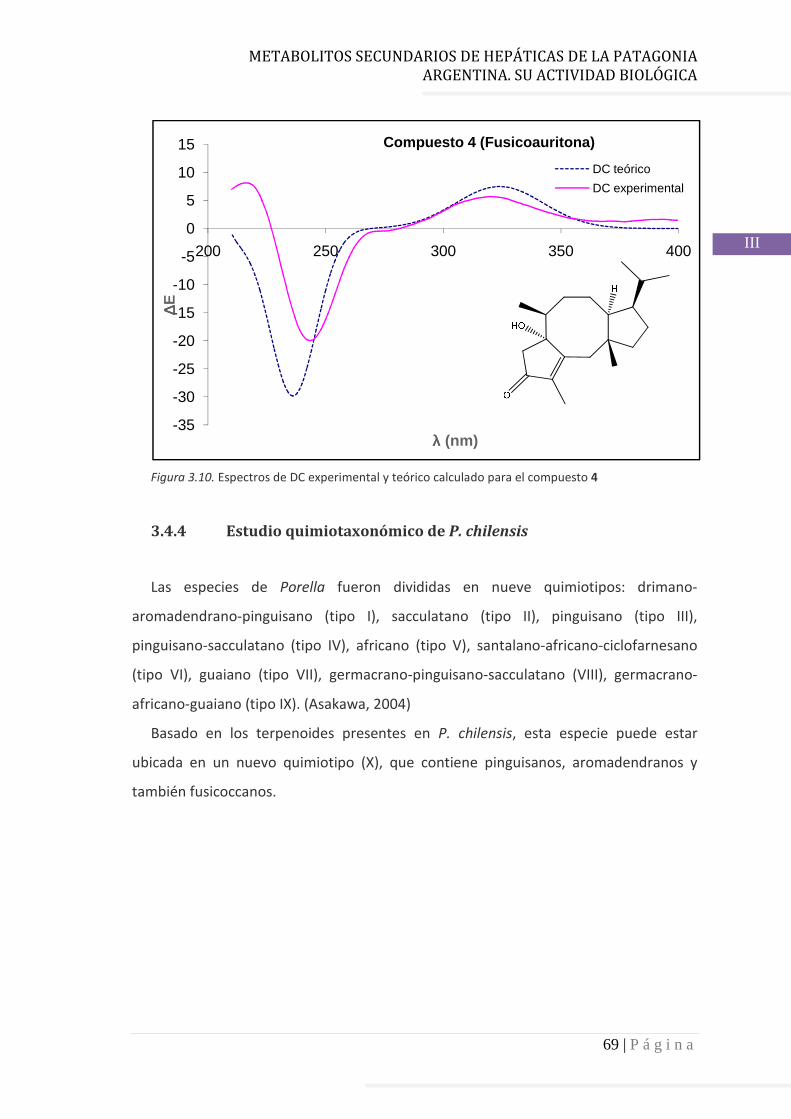
METABOLITOS SECUNDARIOS DE HEPÁTICAS DE LA PATAGONIA
ARGENTINA. SU ACTIVIDAD BIOLÓGICA
69 | P á g i n a
III
Figura 3.10. Espectros de DC experimental y teórico calculado para el compuesto 4
3.4.4 Estudio quimiotaxonómico de P. chilensis
Las especies de Porella fueron divididas en nueve quimiotipos: drimano-
aromadendrano-pinguisano (tipo I), sacculatano (tipo II), pinguisano (tipo III),
pinguisano-sacculatano (tipo IV), africano (tipo V), santalano-africano-ciclofarnesano
(tipo VI), guaiano (tipo VII), germacrano-pinguisano-sacculatano (VIII), germacrano-
africano-guaiano (tipo IX). (Asakawa, 2004)
Basado en los terpenoides presentes en P. chilensis, esta especie puede estar
ubicada en un nuevo quimiotipo (X), que contiene pinguisanos, aromadendranos y
también fusicoccanos.
-35
-30
-25
-20
-15
-10
-5
0
5
10
15
200 250 300 350 400
∆∆ ∆∆E
λλλλ (nm)
Compuesto 4 (Fusicoauritona)
DC teórico
DC experimental
METABOLITOS SECUNDARIOS DE HEPÁTICAS DE LA PATAGONIA
ARGENTINA. SU ACTIVIDAD BIOLÓGICA
70 | P á g i n a
III
3.4.5 Datos espectrales de 1 y 2
Compuesto 1 Compuesto 2
H δδδδH C δδδδC H δδδδH C δδδδC
1a 1.85 d (15.3) 1 51.1 1a 2.49 d (14.1) 1 37.2
1b 1.95 d (15.3) 2 82.6 1b 2.73 d (14.0) 2 169.9
4 5.77 q (1.0) 3 178.1 4a,b 2.54 s 3 76.1
6 2.50 d (10.2) 4 128.5 7 2.60 m 4 50.8
7 2.01 m 5 209.0 8a 1.59 m 5 205.0
8a 1.64 m 6 54.2 8b 2.00 m 6 146.2
8b 2.28 m 7 30.5 9a,b 1.68 m 7 31.3
9a 1.44 m 8 36.3 10 2.01 m 8 31.9
9b 1.72 m 9 19.4 12a 1.24 dd (11.3, 6.5) 9 26.3
10 2.23 m 10 48.6 12b 1.69 m 10 43.2
12a 1.38 m 11 43.9 13a 1.46 m 11 44.4
12b 1.56 m 12 45.9 13b 1.57 m 12 40.3
13a,b 1.37 m 13 23.4 14 1.94 m 13 24.9
14 1.50 m 14 49.3 15 1.74 m 14 46.3
15 1.80 m 15 28.3 16 1.43 s 15 28.6
16 2.07 d (1.0) 16 13.3 17 1.05 d (6.9) 16 28.4
17 1.34 d (6.8) 17 20.7 18 1.01 s 17 21.5
18 0.95 s 18 24.4 19 0.84 d (6.7) 18 22.9
19 0.80 d (6.7) 19 19.4 20 0.87 d (6.5) 19 20.3
20 0.90 d (6.6) 20 23.9 - - 20 24.4
Tabla 3.1. Datos espectrales 1H NMR (500 MHz) y
13C NMR (125 MHz) de los compuestos 1 y 2
(CDCl3).
Constantes de acoplamiento (J en Hz) son dadas en paréntesis.
Señales indicadas con m fueron multipletes no resueltos o superpuestos.
Las asignaciones para 13
C fueron confirmadas por mediciones HSQC y DEPT.

METABOLITOS SECUNDARIOS DE HEPÁTICAS DE LA PATAGONIA
ARGENTINA. SU ACTIVIDAD BIOLÓGICA
71 | P á g i n a
III
Compuesto 1 Compuesto 2
H C H C
1a 2, 3, 10, 11, 18 1a 2, 3, 6, 10, 11, 12
1b 2, 6, 11, 12, 18 1b 2, 3, 6, 11, 12, 18
4 2, 3, 5, 6 ,16 4a,b 2, 3, 5, 6, 16
6 1, 2, 5, 7, 8, 17 9a,b 11
8a 10 10 8, 9, 11,14, 15, 18
8b 7, 9, 10, 17 12a 10, 11, 14, 18
9b 7, 8, 10, 11 12b 1, 11, 13, 18
10 8, 9, 11, 14, 15, 18 13a 14, 15
12a 13 13b 10, 11, 12, 14, 15
12b 10, 18 15 10,13,14, 19, 20
15 14, 19, 20 16 2, 3, 4
16 2, 3, 4 17 7
17 6, 7, 8 18 1, 10, 11, 12
18 1, 10, 11, 12 19 14, 15, 20
19 14, 15, 20 20 14, 15, 19
20 14, 15, 19 - -
Tabla 3.2. Correlaciones HMBC para los compuestos 1 y 2
Compuesto 1 (2α-hidroxifusicocc-3-en-5-ona): Solido blanco amorfo; [α]D -10º (c
0.42, CHCl3)27D; UV (MeOH) λmax (log ε) 221 (4.0) nm; CD (EtOH) ∆ε326 +4.14,
∆ε234 −8.96, ∆ε209 +3.96 (c 2.88x10-4
M); IR νmax CHCl3 cm-1
: 3416, 2920, 1690, 1633,
1452, 1378; 1H y
13C NMR (tabla 1); HRMS: encontrado 304.2395; C20H36O2 requiere
304.2402; EIMS: m/z (rel. int.): 304 [M]+ (11), 286 [M - H2O]
+ (69), 271 [M - H2O - Me]
+
(8), 261 [M - C3H7]+ (8), 243 [M - H2O - C3H7]
+ (62), 176 [M - 128]
+ (97), 135 [M - 169]
+
(100).
Compuesto 2 (3α-hidroxifusicocc-2(6)-en-5-ona): Aceite incoloro; [α]D -4º (c 0.56,
CHCl3)29D; UV (MeOH) λmax (log ε) 241 (4.0) nm; CD (EtOH) ∆ε317 −5.11, ∆ε245 +19.29,
∆ε210 −8.69 (c 4.11x10-4
M); IR νmax CHCl3 cm-1
: 3407, 2920, 1685, 1624, 1462, 1372; 1H y
13C NMR (table 1); HRMS: encontrado 304.2401; C20H36O2 requiere 304.2402; EIMS:
m/z (rel. int.): 304 [M]+ (47), 286 [M – H2O]
+ (27), 271 [M - H2O - Me]
+ (5), 261 [M –
C3H7]+ (80), 243 [M - H2O - C3H7]
+ (100), 225 [M - 79]
+ (95), 201 [M - 103]
+ (37).

METABOLITOS SECUNDARIOS DE HEPÁTICAS DE LA PATAGONIA
ARGENTINA. SU ACTIVIDAD BIOLÓGICA
72 | P á g i n a
III
Referencias
Asakawa Y. Chemical constituents of Hepaticae. Vienna. 1982.
Asakawa Y. Chemosystematics of the Hepaticae. Phytochemistry 2004, 65, 623-669.
Asakawa Y., Ludwiczuk A., Nagashima F., Kinghorn A. D., Falk H. and Kobayashi J.
Chemical constituents of bryophytes: Bio- and chemical diversity, biological activity,
and chemosystematics. Springer. 2012.
Asakawa Y., Takikawa K., Toyota M., Ueda A., Tori M. and Kumar S. S. Sesqui- and
diterpenoids from the liverworts Porella densifolia subsp. appendiculata and Porella
densifolia var. fallax. Phytochemistry 1987, 26, 1019-1022.
Bovi Mitre G., Kamiya N., Bardón A. and Asakawa Y. Africane-type sesquiterpenoids
from the Argentine liverwort Porella swartziana and their antibacterial activity. Journal
of Natural Products 2004, 67, 31-36.
Djerassi C., Geller L. E. and Eisenbraun E. J. Optical rotatory dispersion studies. XXVI.
α-haloketones (part 4). Demonstration of conformational mobility in α-
halocyclohexanones. Journal of Organic Chemistry 1960, 25, 1-6.
Djerassi C., Osiecki J. and Herz W. Optical rotatory dispersion studies. XIII.
Assignment of absolute configuration to certain members of the guaianolide series of
sesquiterpenes. Journal of Organic Chemistry 1957, 22, 1366-1369.
Fukuyama Y., Tori M., Wakamatsu M. and Asakawa Y. Norpinguisone methyl ester
and norpinguisanolide, pinguisane-type norsesquiterpenoids from Porella elegantula.
Phytochemistry 1988, 27, 3557-3561.

METABOLITOS SECUNDARIOS DE HEPÁTICAS DE LA PATAGONIA
ARGENTINA. SU ACTIVIDAD BIOLÓGICA
73 | P á g i n a
III
Gijsen, H. J. M. (+)-Aromadendrene as chiral starting material for thesynthesis of
sesquiterpenes. Ph D Thesis, 1993, Landbouwuniversiteit, pp 2-3.
Huneck S., Baxter G., Cameron A. F., Connolly J. D. and Rycroft D. S. Anadensin, a
new fusicoccane diterpenoid from the liverwort Anastrepta orcadensis. Crystal structure
analysis. Tetrahedron Letters 1983, 24, 3787-3788.
Kato N., Nakanishi K., Wu X., Nishikawa H. and Takeshita H. Total synthesis of
fusicogigantones A and B and fusicogigantepoxide via the singlet oxygen-oxidation of
fusicoccadienes. Fusicogigantepoxide B', a missing congener metabolite. Tetrahedron
Letters 1994, 35, 8205-8208.
Moffitt W., Woodward R. B., Moscowitz A., Klyne W. and Djerassi C. Structure and
the optical rotatory dispersion of saturated ketones. Journal of the American Chemical
Society 1961, 83, 4013-4018.
Nagashima F., Sari Y., Tori M., Asakawa Y. and Huneck S. Sesquiterpenoids from
some European liverworts. Phytochemistry 1993, 34, 1341-1343.
Quang D. N. and Asakawa Y. Chemical constituents of the Vietnamese liverwort
Porella densifolia. Fitoterapia 2010, 81, 659-661.
Stöcklin W., Waddell T. G. and Geissman T. A. Circular dichroism and optical rotatory
dispersion of sesquiterpene lactones. Tetrahedron 1970, 26, 2397-2409.
Tori M., Nakashima K., Takaoka S. and Asakawa Y. Fusicorrugatol from the
Venezuelan liverwort Plagiochila corrugata. Chemical and Pharmaceutical Bulletin
1994, 42, 2650-2652.

METABOLITOS SECUNDARIOS DE HEPÁTICAS DE LA PATAGONIA
ARGENTINA. SU ACTIVIDAD BIOLÓGICA
74 | P á g i n a
III
Tori M., Nakashima K., Takeda T. and Asakawa Y. Structure of secoswartzianins A and
B isolated from the liverwort Porella swartziana. Tetrahedron Letters 1993, 34, 3753-
3754.
Toyota M., Nagashima F. and Asakawa Y. Striatane- and pinguisane-type
sesquiterpenoids and phytane-type diterpenoid from the liverwort Porella navicularis.
Phytochemistry 1989a, 28, 1661-1665.
Toyota M., Nagashima F. and Asakawa Y. Volatile components and pinguisane-type
sesquiterpenoids from the liverwort Porella cordaeana. Phytochemistry 1989b, 28,
3383-3387.
Toyota M., Nagashima F., Shima K. and Asakawa Y. Africane-and santalane-type
sesquiterpenoids from the liverwort Porella caespitans var. setigera. Phytochemistry
1991a, 31, 183-189.
Toyota M., Ueda A. and Asakawa Y. Sesquiterpenoids from the liverwort Porella
acutifolia subsp. Tosana. Phytochemistry 1991b, 30, 567-573.
Van Klink J. W., Zapp J. and Becker H. Pinguisane-type sesquiterpenes from the South
American liverwort Porella recurva (Taylor) Kuhnemann. Zeitschrift fur
Naturforschung - Section C Journal of Biosciences 2002, 57, 413-417.
Williams D. R., Robinson L. A., Nevill C. R. and Reddy J. P. Strategies for the
synthesis of fusicoccanes by nazarov reactions of dolabelladienones: Total synthesis of
(+)-fusicoauritone. Angewandte Chemie - International Edition 2007, 46, 915-918.
Zapp J., Burkhardt G. and Becker H. Sphenolobane and fusicoccane diterpenoids from
the liverwort Anastrophyllum auritum. Phytochemistry 1994, 37, 787-793.

Capítulo IV
Actividad anti-patogénica
de Porella chilensis
IV

METABOLITOS SECUNDARIOS DE HEPÁTICAS DE LA PATAGONIA
ARGENTINA. SU ACTIVIDAD BIOLÓGICA
76 | P á g i n a
IV
Capítulo IV
Actividad anti-patogénica de Porella
chilensis
4.1 Generalidades
Los biofilms bacterianos son complejas comunidades de bacterias embebidas en
una matriz de exopolisacaridos. Esta matriz es producida por las bacterias incluidas en
ella y la misma se encuentra adherida a superficies inertes o vivas. Los
microorganismos en estado de biofilm son más resistentes a antibióticos y al sistema
inmunológico del huésped que las células planctónicas. En Pseudomonas aeruginosa la
formación de biofilm, la producción de factores de virulencia y la resistencia a
antimicrobianos son regulados por el proceso de quórum sensing (QS) (Costerton y
col., 1999). A través de este proceso, las bacterias son capaces de interactuar unas con
otras y coordinar la expresión de genes de acuerdo a la densidad de población
empleando comunicación química. En bacterias Gram negativas, el proceso de QS
toma lugar vía la emisión y recepción de señales moleculares difusibles,
particularmente N-acilhomoserin lactonas (AHL), conocidas como autoinductores de
QS (Greenberg, 1997). Controlar la producción de biofilm y/o el proceso de QS de P.
aeruginosa empleando compuestos naturales puede ser una buena estrategia en la
búsquedas de nuevos antimicrobianos.
Productos naturales, como lactonas sesquiterpénicas y acetogeninas anonáceas,
son capaces de alterar la formación de biofilm de P. aeruginosa (Cartagena y col.,
2007).
Los 10 terpenoides aislados de P. chilensis fueron evaluados en su habilidad para
inhibir el crecimiento bacteriano, la formación de biofilm y la producción de
autoinductores en una cepa de P. aeruginosa.

METABOLITOS SECUNDARIOS DE HEPÁTICAS DE LA PATAGONIA
ARGENTINA. SU ACTIVIDAD BIOLÓGICA
77 | P á g i n a
IV
4.2 Experimental
4.2.1 Crecimiento bacteriano
El crecimiento bacteriano fue determinado espectrofotométricamente a 600 nm en
lector de microplaca (ver capítulo II). A los pocillos para el control negativo se les
adicionó un volumen de una mezcla DMSO – H2O destilada (1:1) en vez de los
compuestos a evaluar. El valor de la absorbancia del control negativo fue 1.34.
4.2.2 Ensayo de formación de biofilm
La formación de biofilm fue determinada por la técnica de cristal violeta en
microplaca (ver capítulo II). La absorbancia del control negativo fue 2.75.
Ciprofloxacina, un conocido inhibidor de biofilm (Sandasi y col., 2011), fue
incorporado al bioensayo a 5 µg/ml como control positivo en las mismas condiciones
experimentales que las utilizadas para evaluar los compuestos.
4.2.3 Ensayo de efectos sobre el QS
La cuantificación de autoinductores para determinar los efectos sobre el QS fue
determinada mediante el uso de la cepa reportera P. aeruginosa qsc 119 (ver capítulo
II). La actividad del control negativo fue 15.2 expresado como unidades de miller.
Azitromicina, capaz de intervenir en los procesos de QS (Tateda y col., 2001), fue
usado a 5 µl/ml como control positivo de inhibición de QS bajo las mismas condiciones
que los compuestos testeados.
4.2.4 Análisis estadístico
Los datos son presentados como media ± desviación estándar (DS). Las diferencias
significativas entre valores se establecieron con el test de Student. Valores de p ≤ 0.05
fueron considerados significativos.

METABOLITOS SECUNDARIOS DE HEPÁTICAS DE LA PATAGONIA
ARGENTINA. SU ACTIVIDAD BIOLÓGICA
78 | P á g i n a
IV
4.3 Resultados
4.3.1 Fusicoccanos
Como se muestra en la figura 4.1a (página 80), los compuestos del tipo fuscicoccano
alteraron el crecimiento bacteriano en formas diferentes. El compuesto 1 incrementó
el crecimiento en un 15 % a 50 µg/ml, mientras que los compuestos 3 y 4 inhibieron
significativamente el crecimiento bacteriano a la misma dosis (16 y 25 %) en relación al
control negativo. A 50 µg/ml, los fusicoccanos 1, 2 y 4 produjeron un fuerte aumento
en la producción de biofilm (95, 93 y 105 %, respectivamente) comparado con el
control negativo. Estos resultados indican que la formación de biofilm depende del
incremento en la producción de autoinductores más que del crecimiento bacteriano
(figura 4.1a). El compuesto 3 es el único fusicoccano que fue capaz de inhibir el
crecimiento bacteriano y la formación de autoinductores. Estos efectos conllevan a
una inhibición en la producción de biofilm.
4.3.2 Pinguisanos
El crecimiento bacteriano fue levemente acrecentado por todos los pinguisanos,
tanto a 50 como a 5 µg/ml (figura 4.1b, página 80). Para los compuestos 5 y 6 se
observó una buena correlación entre crecimiento bacteriano, formación de biofilm y
producción de autoinductores. El compuesto 7 fue capaz de inhibir en 27 y 20 % la
producción de biofilm a 50 y 5 µg/ml, respectivamente. Es importante resaltar que el
compuesto 8 produjo incrementos en el crecimiento bacteriano y la formación de
autoinductores que no resultaron en un aumento en el porcentaje de biofilm.
4.3.3 Aromadendranos
Los compuestos 9 y 10 inhibieron significativamente no sólo el crecimiento
bacteriano, sino también la producción de biofilm y autoinductores incluso a 5 µg/ml
(figura 4.1c). La inhibición de autoiductores producida por 10 (80 %) no está sólo

METABOLITOS SECUNDARIOS DE HEPÁTICAS DE LA PATAGONIA
ARGENTINA. SU ACTIVIDAD BIOLÓGICA
79 | P á g i n a
IV
relacionada con la inhibición sobre el crecimiento bacteriano (24 %), sugiriendo que el
proceso de QS también fue alterado.
Sólo el fusicoccano 3 es tan potente como ciprofloxacina en la inhibición de
formación de biofilm (figura 4.1a), mientras que los aromadendranos 9 y 10 son menos
potentes que el antibiótico a la misma concentración (figura 4.1c).
El antibiótico ciprofloxacina, utilizado como control positivo de inhibición de
formación de biofilm, produjo una inhibición del crecimiento de 73 % e impidió en un
51 % la formación de biofilm a 5 µg/ml, mientras que el antibiótico azitromicina,
utilizado como control positivo de inhibición de QS, no alteró significativamente el
crecimiento bacteriano pero inhibió en un 76 % el proceso de QS a la misma dosis.
Los compuestos fusicoccanos y pinguisanos comparten algunas características
estructurales con algunos productos naturales capaces de inhibir la producción de
autoinductores de P. aeruginosa. Algunas furanonas halogenadas (Givskov y col., 1996;
Manefield y col., 2002), como así también el ácido penicilico y patulina (Rasmussen y
col., 2005) portan anillos de furanona, los cuales aparentemente juegan un papel
importante en la inhibición de la formación de biofilm. Desde nuestros resultados, es
difícil evaluar los requerimientos estructurales para la actividad. En la serie de
fusicoccanos (todos contienen un anillo de ciclopentanona), pequeñas diferencias en la
estructura molecular alteran el proceso de QS, inhibiéndolo o estimulándolo. Estos
efectos diferenciales (inhibición o estimulación) fueron también previamente
reportados para una serie de análogos de N-acilhomoserinlactonas estructuralmente
relacionados (Smith y col., 2003). Es importante destacar que los aromadendranos 9 y
10, que inhibieron ambos parámetros (formación de biofilm y autoinductores), no
están estructuralmente relacionados a ningún compuesto activo previamente
reportado. Mientras que los inhibidores de QS más potentes (dos análogos de AHLs
sintéticos) (Devienne y Raddi, 2002) producen una inhibición del 50 % a 2 y 6 µM. El
efecto producido por los terpenoides de P. chilensis puede ser considerado moderado
(9 produce 33% de inhibición a 16.4 µM).
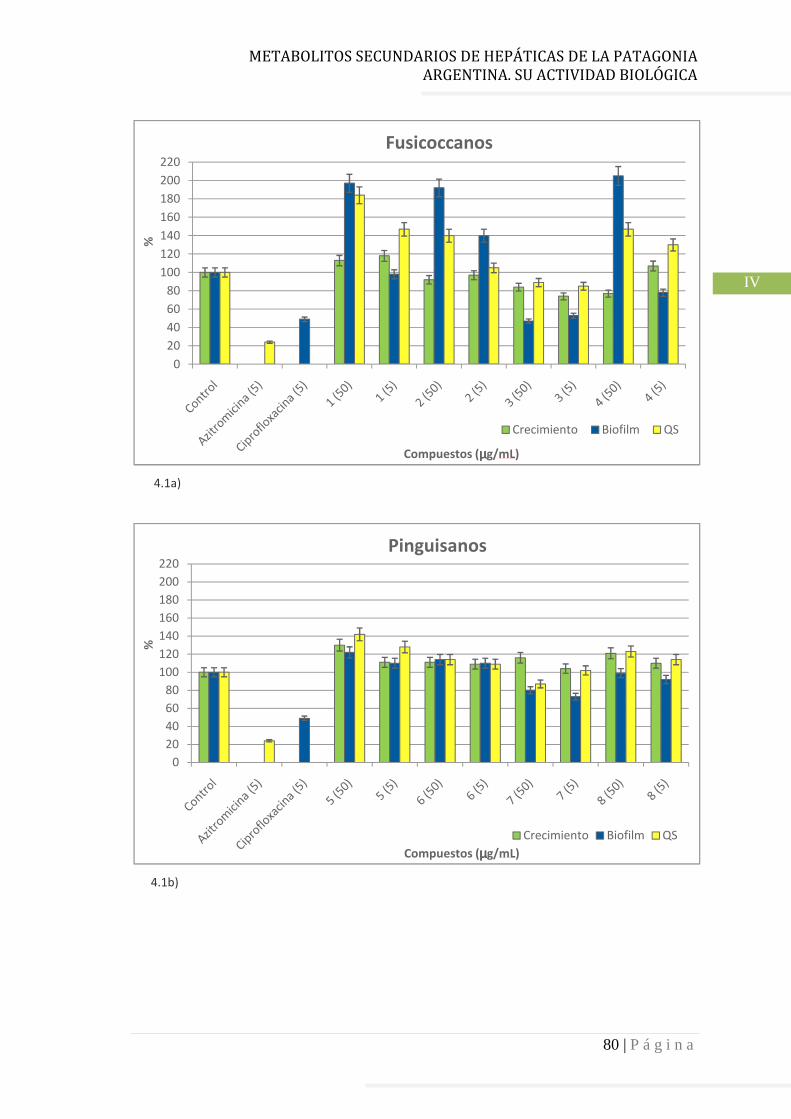
METABOLITOS SECUNDARIOS DE HEPÁTICAS DE LA PATAGONIA
ARGENTINA. SU ACTIVIDAD BIOLÓGICA
80 | P á g i n a
IV
4.1a)
4.1b)
0
20
40
60
80
100
120
140
160
180
200
220%
Compuestos (µµµµg/mL)
Fusicoccanos
Crecimiento Biofilm QS
0
20
40
60
80
100
120
140
160
180
200
220
%
Compuestos (µµµµg/mL)
Pinguisanos
Crecimiento Biofilm QS
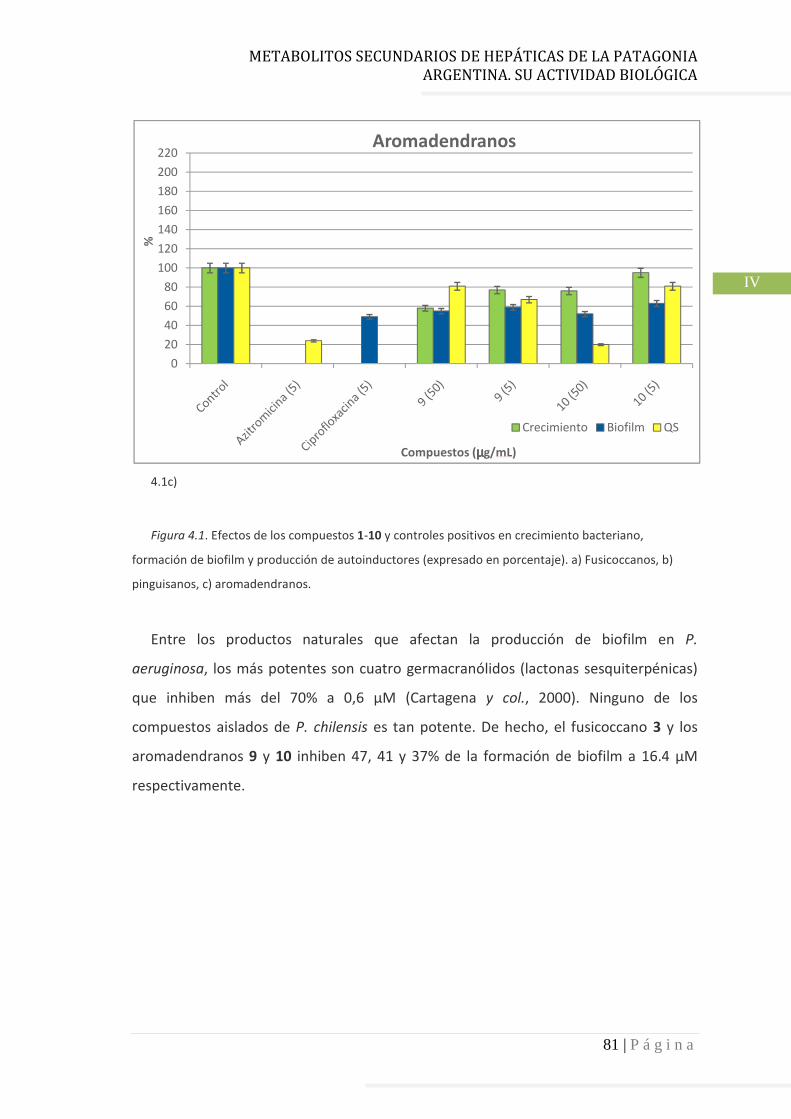
METABOLITOS SECUNDARIOS DE HEPÁTICAS DE LA PATAGONIA
ARGENTINA. SU ACTIVIDAD BIOLÓGICA
81 | P á g i n a
IV
4.1c)
Figura 4.1. Efectos de los compuestos 1-10 y controles positivos en crecimiento bacteriano,
formación de biofilm y producción de autoinductores (expresado en porcentaje). a) Fusicoccanos, b)
pinguisanos, c) aromadendranos.
Entre los productos naturales que afectan la producción de biofilm en P.
aeruginosa, los más potentes son cuatro germacranólidos (lactonas sesquiterpénicas)
que inhiben más del 70% a 0,6 µM (Cartagena y col., 2000). Ninguno de los
compuestos aislados de P. chilensis es tan potente. De hecho, el fusicoccano 3 y los
aromadendranos 9 y 10 inhiben 47, 41 y 37% de la formación de biofilm a 16.4 µM
respectivamente.
0
20
40
60
80
100
120
140
160
180
200
220%
Compuestos (µµµµg/mL)
Aromadendranos
Crecimiento Biofilm QS

METABOLITOS SECUNDARIOS DE HEPÁTICAS DE LA PATAGONIA
ARGENTINA. SU ACTIVIDAD BIOLÓGICA
82 | P á g i n a
IV
Referencias
Cartagena E., Bardon A., Catalan C. A. N., De Hernadez Z. N. J., Hernandez L. R. and
Joseph-Nathan P. Germacranolides and a new type of guaianolide from
Acanthospermum hispidum. Journal of Natural Products 2000, 63, 1323-1328.
Cartagena E., Colom O. A., Neske A., Valdez J. C. and Bardón A. Effects of plant
lactones on the production of biofilm of Pseudomonas aeruginosa. Chemical and
Pharmaceutical Bulletin 2007, 55, 22-25.
Costerton J. W., Stewart P. S. and Greenberg E. P. Bacterial biofilms: A common cause
of persistent infections. Science 1999, 284, 1318-1322.
Devienne K. F. and Raddi M. S. G. Screening for antimicrobial activity of natural
products using a microplate photometer. Brazilian Journal of Microbiology 2002, 33,
166-168.
Givskov M., De Nys R., Manefield M., Gram L., Maximilien R., Eberl L., Molin S.,
Steinberg P. D. and Kjelleberg S. Eukaryotic interference with homoserine lactone-
mediated prokaryotic signalling. Journal of Bacteriology 1996, 178, 6618-6622.
Greenberg E. P. Quorum Sensing in Gram-Negative Bacteria. ASM News 1997, 63, 371-
377.
Manefield M., Rasmussen T. B., Henzter M., Andersen J. B., Steinberg P., Kjelleberg S.
and Givskov M. Halogenated furanones inhibit quorum sensing through accelerated
LuxR turnover. Microbiology 2002, 148, 1119-1127.
Rasmussen T. B., Skindersoe M. E., Bjarnsholt T., Phipps R. K., Christensen K. B.,
Jensen P. O., Andersen J. B., Koch B., Larsen T. O., Hentzer M., Eberl L., Hoiby N.

METABOLITOS SECUNDARIOS DE HEPÁTICAS DE LA PATAGONIA
ARGENTINA. SU ACTIVIDAD BIOLÓGICA
83 | P á g i n a
IV
and Givskov M. Identity and effects of quorum-sensing inhibitors produced by
Penicillium species. Microbiology 2005, 151, 1325-1340.
Sandasi M., Leonard C. M., van Vuuren S. F. and Viljoen A. M. Peppermint (Mentha
piperita) inhibits microbial biofilms in vitro. South African Journal of Botany 2011, 77,
80-85.
Smith K. M., Bu Y. and Suga H. Library screening for synthetic agonists and
antagonists of a Pseudomonas aeruginosa autoinducer. Chemistry and Biology 2003,
10, 563-571.
Tateda K., Comte R., Pechere J. C., Köhler T., Yamaguchi K. and van Delden C.
Azithromycin inhibits quorum sensing in Pseudomonas aeruginosa. Antimicrobial
Agents and Chemotherapy 2001, 45, 1930-1933.

Capítulo V
Actividad insecticida de
Porella chilensis
V

METABOLITOS SECUNDARIOS DE HEPÁTICAS DE LA PATAGONIA
ARGENTINA. SU ACTIVIDAD BIOLÓGICA
85 | P á g i n a
V
Capítulo V
Actividad insecticida de Porella chilensis
5.1 Generalidades
Los estudios relacionados a las interacciones insectos-plantas son relevantes para el
desarrollo de nuevos pesticidas basados en plantas. Estas interacciones involucran
metabolitos secundarios de plantas que actúan como reguladores de crecimiento y
antialimentarios. Estos metabolitos secundarios actúan afectando los sistemas
hormonal y nervioso, como así también el estomago y tejidos musculares (Senthil-
Nathan y col., 2008).
El canal alimentario de insectos está constituido por 3 zonas, el intestino anterior,
medio y posterior. En Lepidóptera, el intestino medio está constituido por células
columnares, caliciformes, endócrinas y regenerativas. Estas células se encuentran
involucradas en procesos de absorción y secreción de enzimas, homeostasis iónica y
renovación del epitelio (de Sousa y col., 2009). Las células epiteliales del intestino
medio están protegidas del contenido del intestino por una delicada película de quitina
llamada membrana peritrófica (MP). La MP es secretada por las células columnares del
epitelio digestivo envolviendo los alimentos consumidos por el insecto y puede actuar
como una barrera física y química para la entrada de patógenos. Esta membrana
compartimenta la digestión y protege las microvellosidades de las células epiteliales
del contacto directo con las partículas de alimento, permitiendo así el pasaje de
enzimas digestivas hacia el lumen, y consecuentemente, la absorción de los productos
de digestión (Lehane, 1997; Terra, 2001). La MP actúa como una potencial línea de
defensa reduciendo o eliminando la absorción de ciertos aleloquímicos (Barbehenn,
2001) y controlando los procesos de detoxificación.
Previamente fue reportado (de Sousa y col., 2009) que las alteraciones en el
intestino medio afectan el crecimiento y desarrollo de insectos como resultado de
cambios fisiológicos en los cuales están involucrados la transformación y absorción de

METABOLITOS SECUNDARIOS DE HEPÁTICAS DE LA PATAGONIA
ARGENTINA. SU ACTIVIDAD BIOLÓGICA
86 | P á g i n a
V
alimento. Muchos reportes indican que los productos naturales de plantas pueden
producir alteraciones histológicas en el canal alimentario de insectos cuando son
incorporados a la dieta larval. Las larvas del lepidóptero Plodia interpunctella,
alimentadas con el producto natural harmalina a 250 µg/g y 20-hidroxiecdisona a 50
µg/g, exhibieron severos daños en las células epiteliales del intestino medio (Rharrabe
y col., 2007; Rharrabe y col., 2009), mientras que larvas del S. frugiperda alimentadas
con 60 µg/g de aceite de neem (Azadirachta indica) mostraron necrosis de células
columnares, regenerativas y caliciformes (Roel y col., 2010). Las larvas de S. exigua
mostraron daño en la MP después del tratamiento con el producto natural
goniothalamin a 10 µg/g (Senthil-Nathan y col., 2008). En base a los resultados
mencionados anteriormente y dado que las larvas de S. frugiperda causan daño por
consumo de follaje a mas de 80 especies de plantas, muchas de las cuales tienen
importancia agrícola como algodón, sorgo y diferentes pastos y forrajes (Murúa y Virla,
2004), decidimos evaluar la toxicidad y efectos en el canal alimentario producido por
los terpenoides 2-8 y 10, en larvas de S. frugiperda. El objetivo de este estudio fue
evaluar los efectos de los terpenoides aislados de P. chilensis en el desarrollo y
supervivencia de larvas de S. frugiperda evaluando las alteraciones producidas en el
intestino medio.
5.2 Experimental
Los procedimientos experimentales para las determinaciones se describen en
capítulo II.
5.2.1 Análisis estadístico
Los resultados son reportados como Media ± DS. Las diferencias en los valores
medios fueron evaluadas por análisis de varianza (ANOVA). El test de Dunnett fue
utilizado para la comparación de los diferentes grupos con el grupo control. En todos
los análisis estadísticos, un valor de P > 0,05 fue considerado no significante. Los
valores fueron calculados con el software Sigmaplot 12.0.

METABOLITOS SECUNDARIOS DE HEPÁTICAS DE LA PATAGONIA
ARGENTINA. SU ACTIVIDAD BIOLÓGICA
87 | P á g i n a
V
5.3 Resultados y discusión
Los compuestos 2-8, y 10 fueron incorporados a la dieta larval de S. frugiperda a
100 µg por g de dieta mientras que 3 y 5-8 fueron agregados a 200 µg/g.
No se detectaron diferencias significativas (P > 0,05) en los valores del índice de
consumición (CR) en las dosis testeadas. Sin embargo, la determinación del índice de
crecimiento (GR) durante 14 días, mostró que en el mencionado periodo, los
compuestos 3 y 5-8, a 200 µg/g, produjeron una disminución del 42, 70, 55, 51 y 63 %
en el crecimiento larval respectivamente (figura 5.1), en relación al control. Además, a
200 µg/g, el tratamiento con los compuestos 3 y 5-8 produjo mortalidades del 10, 40,
50, 30 y 45% respectivamente (tabla 5.1). A 100 µg/g, solo los compuestos 2, 6 y 8
redujeron significativamente el crecimiento larval en 26, 19 y 35% respectivamente,
comparado con el control, indicando un efecto dosis dependiente. A la misma dosis,
los compuestos 2-8 y 10 mataron 20, 5, 15, 25, 30, 20, 20 y 30% de la población larval,
respectivamente (tabla 5.1). Los valores de mortalidad mostrados no fueron ajustados
a la mortalidad natural observada en el grupo de control (5%). Los tratamientos no
produjeron mortalidad pupal ni mortalidad o malformaciones de adultos.
Mortalidad larval (%)
Compuesto 100 µg/g 200 µg/g
Control 5 5
2 20 ND1 3 5 10
4 15 ND1 5 25 40
6 30 50
7 20 30
8 20 45
10 30 ND1
1No Determinado
Tabla 5.1. Índices de mortalidad producidos por los compuestos 2-8 y 10 en larvas de Spodoptera
frugiperda a 100 y 200 µg/g, calculados a los 14 días de exposición (n=20).
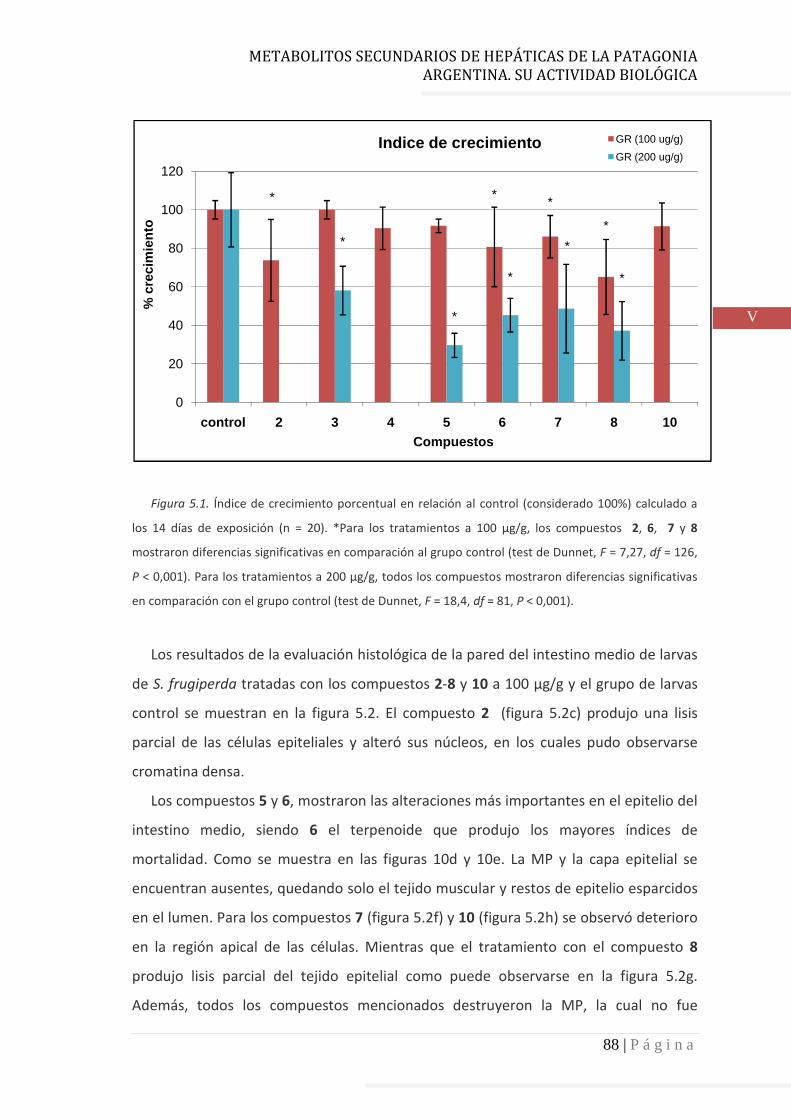
METABOLITOS SECUNDARIOS DE HEPÁTICAS DE LA PATAGONIA
ARGENTINA. SU ACTIVIDAD BIOLÓGICA
88 | P á g i n a
V
Figura 5.1. Índice de crecimiento porcentual en relación al control (considerado 100%) calculado a
los 14 días de exposición (n = 20). *Para los tratamientos a 100 µg/g, los compuestos 2, 6, 7 y 8
mostraron diferencias significativas en comparación al grupo control (test de Dunnet, F = 7,27, df = 126,
P < 0,001). Para los tratamientos a 200 µg/g, todos los compuestos mostraron diferencias significativas
en comparación con el grupo control (test de Dunnet, F = 18,4, df = 81, P < 0,001).
Los resultados de la evaluación histológica de la pared del intestino medio de larvas
de S. frugiperda tratadas con los compuestos 2-8 y 10 a 100 µg/g y el grupo de larvas
control se muestran en la figura 5.2. El compuesto 2 (figura 5.2c) produjo una lisis
parcial de las células epiteliales y alteró sus núcleos, en los cuales pudo observarse
cromatina densa.
Los compuestos 5 y 6, mostraron las alteraciones más importantes en el epitelio del
intestino medio, siendo 6 el terpenoide que produjo los mayores índices de
mortalidad. Como se muestra en las figuras 10d y 10e. La MP y la capa epitelial se
encuentran ausentes, quedando solo el tejido muscular y restos de epitelio esparcidos
en el lumen. Para los compuestos 7 (figura 5.2f) y 10 (figura 5.2h) se observó deterioro
en la región apical de las células. Mientras que el tratamiento con el compuesto 8
produjo lisis parcial del tejido epitelial como puede observarse en la figura 5.2g.
Además, todos los compuestos mencionados destruyeron la MP, la cual no fue
* **
**
*
*
*
*
0
20
40
60
80
100
120
control 2 3 4 5 6 7 8 10
% c
reci
mie
nto
Compuestos
Indice de crecimiento GR (100 ug/g)
GR (200 ug/g)

METABOLITOS SECUNDARIOS DE HEPÁTICAS DE LA PATAGONIA
ARGENTINA. SU ACTIVIDAD BIOLÓGICA
89 | P á g i n a
V
detectada en el analisis histológico de las larvas tratadas. Los tratamientos con los
compuestos 3 y 4 a 100 µg/g no produjeron alteraiones histológicas en el intestino
medio. Como se mencionó en el parrafo anterior, los compuestos 3 y 4 no inhibieron el
crecimiento larval a la dosis mencionada.
Las alteraciones histológicas son en parte responsables de la disminución en el
crecimiento bacteriano de las larvas. Los compuestos testeados produjeron
alteraciones histológicas similares a las reportadas para el aceite de neem (A. indica)
(Roel y col., 2010) incoporado a la dieta de larvas de S. frugiperda a 400 ppm. Es
importante resaltar que el compuesto activo del aceite de neemes el
nortetraterpenoide azadirachtin. Comparado con los productos naturales gonithalamin
(Senthil-Nathan y col., 2008) y 20-hidroxiecdisona (Rharrabe y col., 2009), previamenta
evaluados en su toxicidad contra larvas de lepidopteros, los terpenoides reportados
aquí prodecen el mismo efecto a concentraciones mayores. Sin embargo, los
terpenoides testeados son mas toxicos (producen el mismo efecto a dosis menores)
que harmalina (Rharrabe y col., 2007), un compuesto natural reportado capaz de
generar daños histológicos en larvas del lepidoptero P. intepunctella.
Este es el primer reporte sobre los efectos y daños histológicos producidos por los
terpenoides de P. chilensis en el epiteliodel intestino medio de S. frugiperda. Los
terpenoides de hepáticas, que en muchos casos son enantiomeros de los encontrados
en plantas superiores, han sido raramente investigados en su potencial para controlar
insectos, a pesar de que algunos pocos mostraron actividades promisorias.
La exploración del potencial insecticida de los terpenoides de P. chilensis indica que
se requieren mas estudios para caracterizar el espectro completo de los efectos letales
y subletales que estos pueden producir. Aunque los terpenoides de P. chilensis no son
tan toxicos como otros productos naturales como azadirachtin (Martinez y Van Emden
2001) o los esteroles de Myrtillocactus geometrizans (Cespedes y col. 2005), sus
estructuras son bastantes mas simples, y en consecuencia, más faciles de obtener por
procedimientos de sintesis.
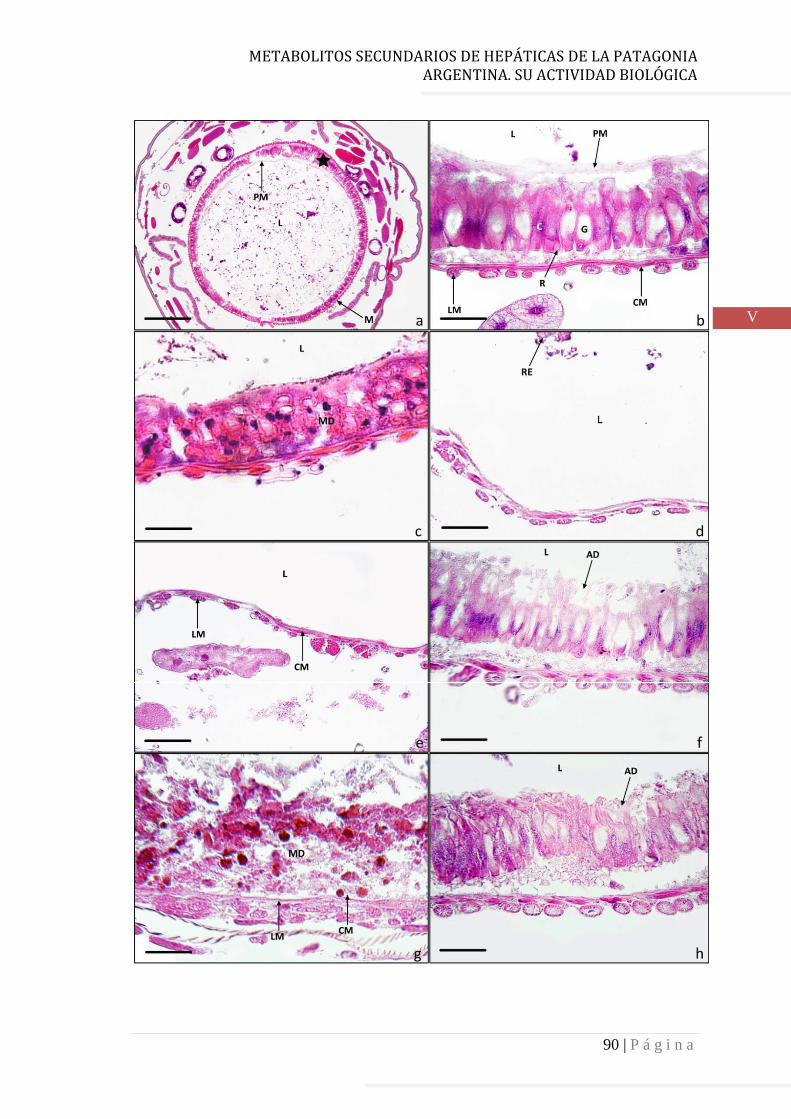
METABOLITOS SECUNDARIOS DE HEPÁTICAS DE LA PATAGONIA
METABOLITOS SECUNDARIOS DE HEPÁTICAS DE LA PATAGONIA
ARGENTINA. SU ACTIVIDAD BIOLÓGICA
METABOLITOS SECUNDARIOS DE HEPÁTICAS DE LA PATAGONIA
ARGENTINA. SU ACTIVIDAD BIOLÓGICA
90 | P á g i n a
V

METABOLITOS SECUNDARIOS DE HEPÁTICAS DE LA PATAGONIA
ARGENTINA. SU ACTIVIDAD BIOLÓGICA
91 | P á g i n a
V
Figura 5.2. Microscopia óptica del intestino medio de Spodoptera frugiperda (teñidas con
hematoxilina-eosina). a: Sección de larva donde el intestino medio es claramente observado (10X, barra
de escala: 400 µm). b: Fragmento de intestino medio de larva control donde la capa epitelial es
observada en detalle (100X, barra de escala: 40 µm). c: Fragmento de intestino medio de larva tratada
con 100 µg/g de compuesto 2 donde se observa lisis parcial de las células epiteliales (100X, barra de
escala: 40 µm). d: Fragmento de intestino medio de larva tratada con 100 µg/g de compuesto 5 donde la
capa epitelial está ausente (100X, barra de escala: 40 µm). e: Fragmento de intestino medio de larva
tratada con 100 µg/g de compuesto 6 donde la capa epitelial está ausente (100X, barra de escala: 40
µm). f: Fragmento de intestino medio de larva tratada con 100 µg/g de compuesto 7 donde se observan
células epiteliales con destrucción apical (100X, barra de escala: 40 µm). g: Fragmento de intestino
medio de larva tratada con 100 µg/g de compuesto 8 donde se observa lisis parcial del tejido epitelial
(100X, barra de escala: 40 µm). h: Fragmento de intestino medio de larva tratada con 100 µg/g de
compuesto 10 donde las células epiteliales muestran destrucción apical (100X, barra de escala: 40 µm).
(PM) membrana peritrófica; (M) tejido muscular; (�) capa epitelial; (L) lumen; (C) células columnares,
(G) células caliciformes; (R) células regenerativas; (CM) musculo circular; (LM) musculo longitudinal; (AD)
destrucción apical de las células; (RE) epitelio remanente; (DM) destrucción de la membrana celular.

METABOLITOS SECUNDARIOS DE HEPÁTICAS DE LA PATAGONIA
ARGENTINA. SU ACTIVIDAD BIOLÓGICA
92 | P á g i n a
V
Referencias
Barbehenn R. V. Roles of peritrophic membranes in protecting herbivorous insects from
ingested plant allelochemicals. Archives of Insect Biochemistry and Physiology 2001,
47, 86-99.
de Sousa M. E. C., Wanderley-Teixeira V., Teixeira A. A. C., de Siqueira H. A. A.,
Santos F. A. B. and Alves L. C. Ultrastructure of the Alabama argillacea (Hübner)
(Lepidoptera: Noctuidae) midgut. Micron 2009, 40, 743-749.
Lehane M. J. Peritrophic matrix structure and function. Annual Review of Entomology
1997, 42, 525-550.
Murúa G. and Virla E. Population parameters of Spodoptera frugiperda (J E Smith)
(Lepidoptera: Noctuidae) fed on corn and two predominant grasses in Tucumán
(Argentina). Acta Zoológica Mexicana (n. s. ) 2004, 20, 199-210.
Rharrabe K., Bakrim A., Ghailani N. and Sayah F. Bioinsecticidal effect of harmaline
on Plodia interpunctella development (Lepidoptera: Pyralidae). Pesticide Biochemistry
and Physiology 2007, 89, 137-145.
Rharrabe K., Bouayad N. and Sayah F. Effects of ingested 20-hydroxyecdysone on
development and midgut epithelial cells of Plodia interpunctella (Lepidoptera,
Pyralidae). Pesticide Biochemistry and Physiology 2009, 93, 112-119.
Roel A. R., Dourado D. M., Matias R., Porto K. R. A., Bednaski A. V. and da Costa R.
B. The effect of sub-lethal doses of Azadirachta indica (Meliaceae) oil on the midgut of
Spodoptera frugiperda (Lepidoptera, Noctuidae). Revista Brasileira de Entomologia
2010, 54, 505-510.

METABOLITOS SECUNDARIOS DE HEPÁTICAS DE LA PATAGONIA
ARGENTINA. SU ACTIVIDAD BIOLÓGICA
93 | P á g i n a
V
Senthil-Nathan S., Choi M. Y., Paik C. H. and Kalaivani K. The toxicity and
physiological effect of goniothalamin, a styryl-pyrone, on the generalist herbivore,
Spodoptera exigua Hübner. Chemosphere 2008, 72, 1393-1400.
Terra W. R. The origin and functions of the insect peritrophic membrane and
peritrophic gel. Archives of Insect Biochemistry and Physiology 2001, 47, 47-61.

Capítulo VI
Estudio químico de
Lepidozia chordulifera
VI

METABOLITOS SECUNDARIOS DE HEPÁTICAS DE LA PATAGONIA
Capítulo VI
Estudio químico de
6.1 Consideraciones botánicas
La especie Lepidozia chordulifera
constituida por plantas de color verde claro a verde
longitud y hojas pinnadas o bipinnadas,
raramente flageliformes, curvadas hacia abajo en la punta
inserción de la hoja es oblicua. Hojas del tallo distantes a imbricadas, asimétricas,
aovado–truncadas, 0,4 mm de longitud, 0,3 a 0,4 mm de ancho en la base
dorsal fuertemente convexo,
ventrales enteros o dentados, divididas en 4 segmentos desiguales desde la mitad de
su longitud, triangulares
sobre troncos, base de árboles o suelo
Figura 6.1. Lepidozia chordulifera
METABOLITOS SECUNDARIOS DE HEPÁTICAS DE LA PATAGONIA
ARGENTINA. SU ACTIVIDAD BIOLÓGICA
Estudio químico de Lepidozia chordulifera
Consideraciones botánicas
Lepidozia chordulifera pertenceciente a la familia Lepidoziaceae
constituida por plantas de color verde claro a verde–marrón con
longitud y hojas pinnadas o bipinnadas, las ramas laterales de 5 a 10 mm de longitud,
raramente flageliformes, curvadas hacia abajo en la punta (figura 6.1)
inserción de la hoja es oblicua. Hojas del tallo distantes a imbricadas, asimétricas,
truncadas, 0,4 mm de longitud, 0,3 a 0,4 mm de ancho en la base
dorsal fuertemente convexo, ya sea entero o con 1 a 2 dientes conspicuos
ntrales enteros o dentados, divididas en 4 segmentos desiguales desde la mitad de
su longitud, triangulares. Esta especie crece formando tapices sueltos o almohadillas
sobre troncos, base de árboles o suelo (Ardiles y col., 2008).
Lepidozia chordulifera (Ardiles y col., 2008).
METABOLITOS SECUNDARIOS DE HEPÁTICAS DE LA PATAGONIA
ARGENTINA. SU ACTIVIDAD BIOLÓGICA
95 | P á g i n a
VI
Lepidozia chordulifera
pertenceciente a la familia Lepidoziaceae está
tallos de 5 cm de
las ramas laterales de 5 a 10 mm de longitud,
(figura 6.1). La línea de
inserción de la hoja es oblicua. Hojas del tallo distantes a imbricadas, asimétricas,
truncadas, 0,4 mm de longitud, 0,3 a 0,4 mm de ancho en la base con margen
entero o con 1 a 2 dientes conspicuos y márgenes
ntrales enteros o dentados, divididas en 4 segmentos desiguales desde la mitad de
formando tapices sueltos o almohadillas

METABOLITOS SECUNDARIOS DE HEPÁTICAS DE LA PATAGONIA
ARGENTINA. SU ACTIVIDAD BIOLÓGICA
96 | P á g i n a
VI
6.2 Clasificación botánica de L. chordulifera
6.3 Antecedentes químicos de la familia Lepidoziaceae
Las especies de Lepidozia producen como constituyente mayoritario un
sesquiterpeno del tipo bazzanano llamado β-bazzaneno. Este sesquiterpeno es
también un marcador químico de las especies de Bazzania pertenecientes a la misma
familia (Asakawa y col., 2012). Sin embargo, una investigación previa de Lepidozia
chordulifera (Zapp y col., 2008) no arrojó como resultado el aislamiento de bazzaneno,
en cambio, esta especie de Lepidozia demostró ser capaz de sintetizar una variada
gama de triterpenos, entre ellos taraxerol y los ácidos carboxílicos triterpénicos
shoreico, betulínico, betulónico y ursólico.
De otras especies del género Lepidozia se aislaron compuestos de los tipos
isobiciclogermacrano, lepidozano, cadinano, eudesmano, maaliano, vitrano y bibencilo
(Asakawa y col., 2012).
Jungermanniidae Marchantiidae
Metzgeriales Takakiales Calobryales Jungermanniales
Suborden
Jungermanniineae Lepidoziineae Ptilidiineae Porellineae Radulineae Pleuroziineae
Familia
Lepidoziaceae
Genero
Lepidozia
Especie
Lepidozia chordulifera
Subclase
Clase Hepaticae
Orden
Jubulineae

METABOLITOS SECUNDARIOS DE HEPÁTICAS DE LA PATAGONIA
ARGENTINA. SU ACTIVIDAD BIOLÓGICA
97 | P á g i n a
VI
β-bazzaneno
6.4 Experimental
6.4.1 Material vegetal
El material vegetal de Lepidozia chordulifera T. Taylor (Figura 5.1), fue recolectado
en Febrero de 2005 en lago Steffen, Provincia de Río Negro, Argentina. Una muestra
fue depositada en el herbario de la Fundación Miguel Lillo (LIL Nº 3414), Tucumán,
Argentina.
6.4.2 Extracción y aislamiento
El material vegetal (240 g) fue extraído a temperatura ambiente por 7 días con éter
etílico obteniéndose 5,9 g de extracto después de la remoción del solvente con
rotavapor (rendimiento 2,46 %). El extracto fue sometido a cromatografía en columna
(CC) con Sephadex LH-20 (MeOH-CH2Cl2, 1:1) y luego a CC con silica gel (70 – 230
mesh) con hexano y cantidades crecientes de AcOEt y finalmente MeOH como
eluyentes para dar 5 fracciones.
La fracción I (370 mg), fue sometida a HPLC de fase normal con columna B (hexano-
AcOEt, 49:1) para dar el compuesto 11 (17 mg).
A partir de la fracción II (103 mg), la cual fue sometida a HPLC con columna B
(hexano-AcOEt, 26:1) se obtuvieron los compuestos 12 (34 mg), 21 (10 mg) y 9 (15
mg).
Por tratamiento de la fracción III (80 mg) mediante HPLC de fase reversa y columna
C (MeOH:H2O, 9:1) se obtuvo 18 (7 mg).
La fracción IV (150 mg), fue sometida a HPLC de fase reversa con columna C
(MeOH:H2O, 17:1) para dar 19 (3 mg), 20 (4 mg) y una sub-fracción, la cual fue
recromatografiada con columna D (MeOH:H2O, 17:1 + ácido acético 1%) obteniéndose
13 (11 mg), 14 (3 mg) y 15 (6 mg).

METABOLITOS SECUNDARIOS DE HEPÁTICAS DE LA PATAGONIA
ARGENTINA. SU ACTIVIDAD BIOLÓGICA
98 | P á g i n a
VI
La fracción V (120 mg) fue sometida a CC con Sephadex LH20 y luego a
cromatografía de fase reversa con columna C (MeOH:H2O, 22:3 + HAc 1%) para dar 16
(14 mg) y una sub-fracción. De esta última, luego de ser reprocesada por
cromatografía con columna D (MeOH:H2O, 21:4 + HAc 1%), se obtuvo 17 (3 mg) y una
nueva cantidad de 16 (3 mg).
6.5 Resultados
Los compuestos 11 y 12 fueron identificados como taraxerona y taraxerol en base a
sus características espectroscópicas y datos previamente reportados en bibliografía
(Hernandez-Chavez y col., 2012). El espectro de RMN protónico de taraxerona (11)
mostró ocho señales de metilos singletes a 0,83, 0,91, 0,92, 0,96, 1,07, 1,08, 1,09 y
1,14 para H-26, H-30, H-28, H-29, H-24, H-25, H-23 y H-27, respectivamente y una
señal doble doblete para a un protón olefínico a δ 5.56 asignado a H-15, con
constantes de acoplamiento J1=8.1 y J2=3.2 Hz, que corresponden a los acoplamientos
con cada uno de los protones H-16.
El espectro de RMN protónico de taraxerol (12) mostró características similares a las
de 11, ocho señales para metilos terciarios a 0.80, 0.82, 0.90 (6H), 0.92, 0.95, 0.98 y
1.09 para H-23, H-29, H-27, H-30, H-25, H-28, H-24 y H-26, respectivamente y también
una señal doble doblete para a un protón olefínico a δ 5.53 correspondiente a H-15
(J=8.2, 3.2 Hz) debido a los acoplamientos con cada uno de los H-16. A diferencia del
compuesto 11, taraxerol (12) muestra una señal doble doblete para un protón unido a
carbinol (H-3) a δ 3.19 (J=10.5, 5.0 Hz) acoplado con los protones 2β y 2α.
Los compuestos 13, 14 y 15 fueron identificados como los ácidos oleanólico,
ursólico y betulínico, respectivamente, en base a sus características espectroscópicas y
por comparación con datos de bibliografía (Gohari y col., 2009; Ayatollahi y col., 2011;
Muhit y col., 2010).
El espectro de RMN protónico de 13 presentó características similares a 12, salvo
por la presencia de siete y no ocho metilos singletes a δ 0.75, 0.77, 0.90, 0.91, 0.93,
0.98 y 1.13 para los hidrógenos H-26, H-23, H-29, H-25, H-30, H-24 y H-27
respectivamente y una señal triplete para un protón olefínico a δ 5.28 (J=3.5 Hz). El

METABOLITOS SECUNDARIOS DE HEPÁTICAS DE LA PATAGONIA
ARGENTINA. SU ACTIVIDAD BIOLÓGICA
99 | P á g i n a
VI
espectro también presentó una señal doble doblete para un protón unido a carbinol
(H-3) a δ 3.22 (J=5.0 Hz) debido a los acoplamientos con los protones en C-2. Además,
el espectro muestra una señal doble doblete para un hidrógeno a δ 2.82
correspondiente a H-18 (J1=13.7, 4.3 Hz) acoplado con cada uno de los protones H19.
El espectro de RMN protónico de 14 (ácido ursólico), mostró 5 señales singletes a δ
0.78, 0.79, 0.93, 0.98 y 1.08 correspondientes a los metilos H-23, H-26, H-25, H-24 y H-
27, respectivamente, y dos señales dobletes a δ 0.86 (J=6.8 Hz) y 0.94 (J=6.4 Hz)
correspondientes a los metilos CH3-29 y CH3-30, respectivamente. Al igual que 13, el
compuesto 14 también presentó una señal triplete para un protón olefínico a δ 5.25

METABOLITOS SECUNDARIOS DE HEPÁTICAS DE LA PATAGONIA
ARGENTINA. SU ACTIVIDAD BIOLÓGICA
100 | P á g i n a
VI
con una constante de acoplamiento de 3,4 Hz. También, una señal doble doblete para
un protón unido a carbinol (H-3) a δ 3.22 con constantes de acoplamiento J1=10.9 y
J2=4.8 Hz que corresponden a los acoplamientos con los protones en C-2.
El espectro de RMN protónico de 15 (ácido betulínico) mostró seis señales singletes
para metilos a δ 0.75, 0.82, 0.93, 0.96, 0.97 y 1.68 para los protones H-23, H-25, H-26,
H-24, H-27 y H-30. Dos señales singletes ensanchadas para protones olefínicos a δ 4.61
y 4.73 correspondientes a los protones H-29a y H-29b. Además, una señal doble
doblete para un protón unido a carbinol (H-3) a δ 3.19 (J=10.8, 5.2 Hz) acoplado con los
protones en C-2, como así también una señal triple doblete a δ 2.98 para el protón H-
19 con constantes de acoplamiento J1=10.9 y J2=4.9 Hz debido al acoplamiento con los
protones H-21 y el protón H-18, respectivamente.
La información obtenida de una revisión bibliográfica sobre los datos
espectroscópicos de RMN de 1H y
13C de los ácidos 13, 14 y 15 en CDCl3, muestra datos
muy disimiles y en otros casos se informan desplazamientos químicos para mezclas de
solventes, quizás debido a la baja solubilidad de estos compuestos en CHCl3. En la tabla
6.1 se muestran valores comparativos para las señales diagnóstico de los compuestos
13, 14 y 15 disueltos en CDCl3.
Los compuestos 16, 17, 18 y 19 fueron identificados en base a sus características
espectroscópicas y por comparación con datos de bibliografía como ácido shoreico
(Govindachari y col., 1994), eichlerialactona (Rao y col., 1975), cabraleona y
cabraleadiol (Hisham y col., 1996) respectivamente. El espectro de RMN protónico del
ácido shoreico (16) mostró 7 señales para metilos terciarios a δ 0.85, 0.88, 1.01, 1.11,
1.14, 1.19 y 1.73 que corresponden a H-18, H-19, H-30, H-21, H-26, H-27 y H-29
respectivamente, como así también dos señales singletes para hidrógenos vinílicos a δ
4.65 y 4.84 correspondientes a los hidrógenos en C-28, y una señal para un hidrógeno
unido a un carbono base de oxigeno a δ 3.64 correspondiente a H-24. Mientras que el
espectro de 1H RMN de eichlerialactona (17) sólo mostró 5 señales para metilos
terciarios a 0.86, 0.90, 1.01, 1.34 y 1.73 correspondientes a los hidrógenos H-18, H-19,
H-30, H-21 y H-29, respectivamente, y dos señales singletes para hidrógenos olefínicos
a δ 4.66 y 4.86 que corresponden a los hidrógenos en C-28.
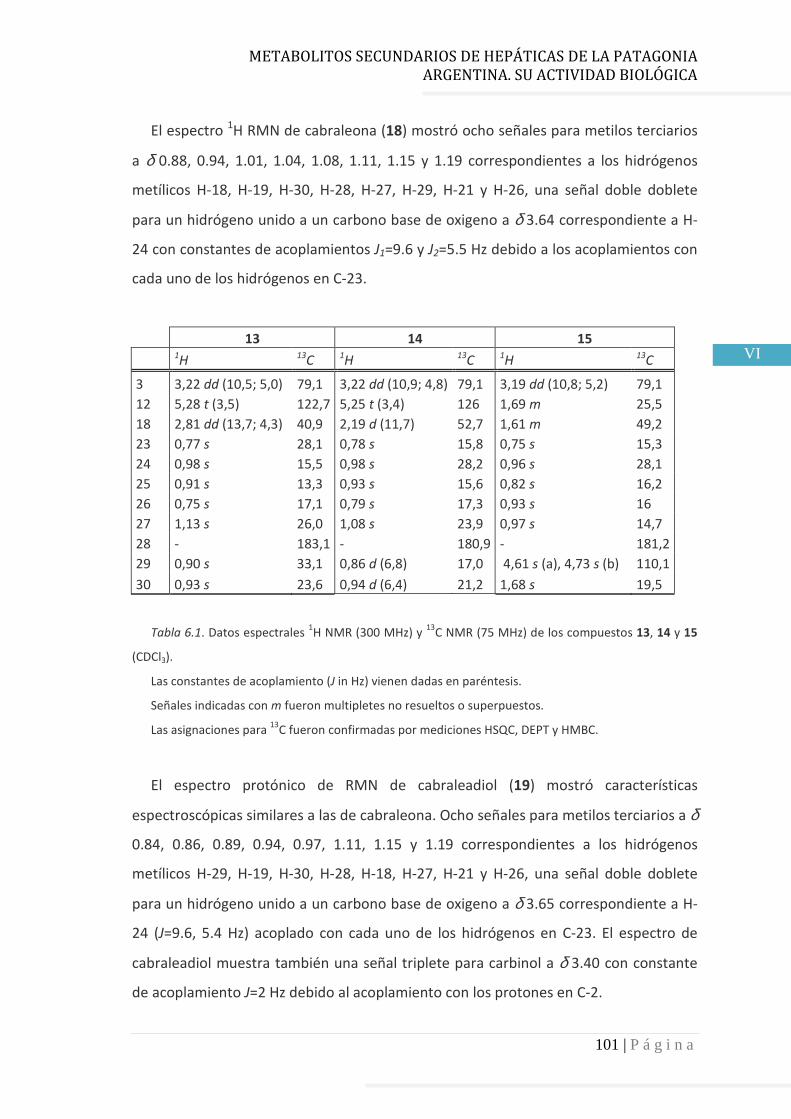
METABOLITOS SECUNDARIOS DE HEPÁTICAS DE LA PATAGONIA
ARGENTINA. SU ACTIVIDAD BIOLÓGICA
101 | P á g i n a
VI
El espectro 1H RMN de cabraleona (18) mostró ocho señales para metilos terciarios
a δ 0.88, 0.94, 1.01, 1.04, 1.08, 1.11, 1.15 y 1.19 correspondientes a los hidrógenos
metílicos H-18, H-19, H-30, H-28, H-27, H-29, H-21 y H-26, una señal doble doblete
para un hidrógeno unido a un carbono base de oxigeno a δ 3.64 correspondiente a H-
24 con constantes de acoplamientos J1=9.6 y J2=5.5 Hz debido a los acoplamientos con
cada uno de los hidrógenos en C-23.
13 14 15
1H 13
C 1H 13
C 1H 13
C
3 3,22 dd (10,5; 5,0) 79,1 3,22 dd (10,9; 4,8) 79,1 3,19 dd (10,8; 5,2) 79,1
12 5,28 t (3,5) 122,7 5,25 t (3,4) 126 1,69 m 25,5
18 2,81 dd (13,7; 4,3) 40,9 2,19 d (11,7) 52,7 1,61 m 49,2
23 0,77 s 28,1 0,78 s 15,8 0,75 s 15,3
24 0,98 s 15,5 0,98 s 28,2 0,96 s 28,1
25 0,91 s 13,3 0,93 s 15,6 0,82 s 16,2
26 0,75 s 17,1 0,79 s 17,3 0,93 s 16
27 1,13 s 26,0 1,08 s 23,9 0,97 s 14,7
28 - 183,1 - 180,9 - 181,2
29 0,90 s 33,1 0,86 d (6,8) 17,0 4,61 s (a), 4,73 s (b) 110,1
30 0,93 s 23,6 0,94 d (6,4) 21,2 1,68 s 19,5
Tabla 6.1. Datos espectrales 1H NMR (300 MHz) y
13C NMR (75 MHz) de los compuestos 13, 14 y 15
(CDCl3).
Las constantes de acoplamiento (J in Hz) vienen dadas en paréntesis.
Señales indicadas con m fueron multipletes no resueltos o superpuestos.
Las asignaciones para 13
C fueron confirmadas por mediciones HSQC, DEPT y HMBC.
El espectro protónico de RMN de cabraleadiol (19) mostró características
espectroscópicas similares a las de cabraleona. Ocho señales para metilos terciarios a δ
0.84, 0.86, 0.89, 0.94, 0.97, 1.11, 1.15 y 1.19 correspondientes a los hidrógenos
metílicos H-29, H-19, H-30, H-28, H-18, H-27, H-21 y H-26, una señal doble doblete
para un hidrógeno unido a un carbono base de oxigeno a δ 3.65 correspondiente a H-
24 (J=9.6, 5.4 Hz) acoplado con cada uno de los hidrógenos en C-23. El espectro de
cabraleadiol muestra también una señal triplete para carbinol a δ 3.40 con constante
de acoplamiento J=2 Hz debido al acoplamiento con los protones en C-2.

METABOLITOS SECUNDARIOS DE HEPÁTICAS DE LA PATAGONIA
ARGENTINA. SU ACTIVIDAD BIOLÓGICA
102 | P á g i n a
VI
La estereoquímica relativa para la cadena lateral que contiene el grupo
tetrahidrofurano de los compuestos 16, 18 y 19, fue establecida como CH3-21β y el
grupo isopropilol en C-24 también en posición β en base al desplazamiento químico de
H-24 en el espectro de RMN protónico, como así también en base al desplazamiento
químico de C-24 en el espectro de RMN de 13
C en CDCl3. Para los compuestos 16, 18 y
19, la señal correspondiente a C-24 aparece a 86.2, mientras que en los epímeros con
el grupo isopropilol en C-24 en posición α (ácido eichleriánico, ocotillona y ocotillol)
esta señal es encontrada a 83.2. Además, 16, 18 y 19 presentan la señal de H-24 como
un doble doblete a δ 3.65, mientras que sus epímeros muestran esta señal como un
triplete a δ 3.71 (Hisham y col., 1996).
El compuesto 20 fue identificado como 3β-hidroxihexanordammaran-20-ona
(Tanaka y col., 1987) en base a sus características espectroscópicas y por comparación
con datos de bibliografía. El espectro de 1H RMN mostró cinco señales de metilos
terciarios a δ 0.77, 0.84, 0.87, 0.97 y 0.98 correspondientes a H-28, H-19, H-30, H-29 y
H-18, una señal singlete a δ 2.13 para una metilcetona (H-21), una señal triple doblete
a δ 2.58 (H-17) con constantes de acoplamiento J=10.9, 6.1 Hz, y una señal doble
doblete para un carbinol a δ 3.20 (H-3) (J=10.9, 5.4 Hz).
El compuesto 21 fue identificado en base a sus características espectroscópicas y
por comparación con datos de bibliografía como viridiflorol (Bombarda y col., 2001). El
espectro de 1H RMN mostró señales para tres metilos terciarios a δ 0.99, 1.02 y 1.15
que corresponden a H-13, H-12 y H-14 respectivamente, un metilo secundario a δ 0.92
(H-15), un triplete para el metino H-6 con constante de acoplamiento J=9.1 Hz y un
multiplete para el metino H-7 a δ = 0.62.
6.6 Estudio quimiotaxonómico de Lepidozia chordulifera
Como se mencionó anteriormente, los compuestos taraxerol (12) y los ácidos
carboxílicos triterpénicos ursólico (14), betulinico (15) y shoreico (16), y 3β-
hidroxihexanordammaran-20-ona (20) fueron aislados en una investigación previa de
L. chordulifera (Zapp y col., 2008). Taraxerona (11), Taraxerol (12), ácido ursólico (14) y

METABOLITOS SECUNDARIOS DE HEPÁTICAS DE LA PATAGONIA
ARGENTINA. SU ACTIVIDAD BIOLÓGICA
103 | P á g i n a
VI
ácido betulínico (15) fueron previamente aislados de L. raptans (Zhang, 2011).
Cabraleadiol (19) fue previamente reportado en Blepharidophyllum densifolium (Flegel
y Becker, 1999) y viridiflorol (21) fue previamente aislado de Bazzania trilobata,
Locopholea bidentata y L. heterophylla (Asakawa y col., 2012). Sin embargo, parece ser
la primera vez que los compuestos ácido oleanólico (13), eichleralactona (17), y
cabraleona (18) son reportados en una especie de hepáticas.
También, es importante remarcar que ni la actual investigación de L. chordulifera, ni
la realizada previamente por Zapp y col. (2008) arrojó como resultado el aislamiento
de bazzaneno, considerado un marcador químico del género. Teniendo en cuenta que
la presencia de bazzaneno en las especies de Lepidozia fue registrada para especies
asiáticas, sugerimos reconsiderar la presencia de este como marcador químico del
género, o por lo menos considerarlo sólo para las especies asiáticas y no así para las
americanas, dado que ambos estudios, el actual y el realizado por Zapp y col. (2008),
fueron realizados sobre material vegetal recolectado en sud-América.

METABOLITOS SECUNDARIOS DE HEPÁTICAS DE LA PATAGONIA
ARGENTINA. SU ACTIVIDAD BIOLÓGICA
104 | P á g i n a
VI
Referencias
Ardiles V., Cuvertino J. and Osorio F. Briófitas de los bosques templados de Chile.
Chile. 2008.
Asakawa Y., Ludwiczuk A., Nagashima F., Kinghorn A. D., Falk H. and Kobayashi J.
Chemical constituents of bryophytes: Bio- and chemical diversity, biological activity,
and chemosystematics. Springer. 2012.
Ayatollahi A. M., Ghanadian M., Afsharypour S., Abdella O. M., Mirzai M. and Askari
G. Pentacyclic triterpenes in Euphorbia microsciadia with their t-cell proliferation
activity. Iranian Journal of Pharmaceutical Research 2011, 10, 287-294.
Bombarda I., Raharivelomanana P., Ramanoelina P. A. R., Faure R., Bianchini J. P. and
Gaydou E. M. Spectrometric identifications of sesquiterpene alcohols from niaouli
(Melaleuca quinquenervia) essential oil. Analytica Chimica Acta 2001, 447, 113-123.
Flegel M. and Becker H. Di- and triterpenoids from the liverwort Blepharidophyllum
densifolium. Zeitschrift fur Naturforschung - Section C Journal of Biosciences 1999, 54,
481-487.
Gohari A. R., Saeidnia S., Hadjiakhoondi A., Abdoullahi M. and Nezafati M. Isolation
and quantificative analysis of oleanolic acid from Satureja mutica Fisch. & C. A. Mey.
Journal of Medicinal Plants 2009, 8, 65-69.
Govindachari T. R., Suresh G. and Kumari G. N. K. Triterpenoids from Dysoxylum
malabaricum. Phytochemistry 1994, 37, 1127-1129.

METABOLITOS SECUNDARIOS DE HEPÁTICAS DE LA PATAGONIA
ARGENTINA. SU ACTIVIDAD BIOLÓGICA
105 | P á g i n a
VI
Hernandez-Chavez I., Torrez-Tapia L. W., Sima-Polanco P., Cedillo-Rivera R., Moo-
Puc R. and Peraza-Sanchez S. R. Antigiardial activity of Cupania dentata bark and its
constituents. Journal of the Mexican Chemical Society 2012, 56, 105-108.
Hisham A., Ajitha Bai M. D., Fujimoto Y., Hara N. and Shimada H. Complete 1H and
13C NMR spectral assignment of cabraleadiol, a dammarane triterpene from Dysoxylum
malabaricum bedd. Magn. Reson. Chem. 1996, 34, 146-150.
Muhit M. A., Tareq S. M., Apu A. S., Basak D. and Islam M. S. Isolation and
identification of compounds from the leaf extract of Dillenia Indica Linn. Bangladesh
Pharmaceutical Journal 2010, 13, 49-53.
Rao M. M., Meshulam H., Zelnik R. and Lavie D. Cabralea eichleriana dc.
(meliaceae)−I structure and stereochemistry of wood extractives. Tetrahedron 1975, 31,
333-339.
Tanaka R., atsuda M. and atsunaga S. 3β-Hydroxyhexanordammaran-20-one from
Euphorbia supina. Phytochemistry 1987, 26, 3365-3366.
Zapp H., Orth K., Zapp J., Connolly J. D. and Becker H. 21,28-Epoxy-18β,21β-
dihydroxbaccharan-3-one and other terpenoids from the liverwort Lepidozia
chordulifera T. Taylor. Natural Product Communications 2008, 3, 1755-1758.
Zhang, Y. L. Chemical Constituents Of Liverworts Lepidozia Reptans And Asterella
Multiflora. Master, 2011, China

Capítulo VII
Actividad anti-patogénica
de Lepidozia chordulifera
VII

METABOLITOS SECUNDARIOS DE HEPÁTICAS DE LA PATAGONIA
ARGENTINA. SU ACTIVIDAD BIOLÓGICA
107 | P á g i n a
VII
Capítulo VII
Actividad antibiofilm de Lepidozia
chordulifera
7.1 Generalidades
La patogenicidad bacteriana está asociada a factores de virulencia. Estos, o bien
pueden estar unidos a las células bacterianas o ser sintetizados por ella y excretados al
tejido circundante. En bibliografía pueden encontrarse numerosos reportes
relacionados a las proteasas o elastasas elaboradas por las bacterias como
componentes de sus toxinas. Inyecciones de elastasas producidas por Pseudomonas
aeruginosa en la piel de animales induce en pocos minutos a lesiones hemorrágicas, y
en 24 horas estas lesiones se vuelven necróticas (Liu, 1974). En infecciones sistémicas
provocadas por P. aeruginosa se producen comunmente hemorragias de órganos
internos, particularmente en los pulmones, y se cree que las elastasas y otras enzimas
proteolíticas son responsables de la patogénesis de estas lesiones (Morihara y col.,
1979; Morihara y Tsuzuki, 1977; Liu, 1974). Por los descripto arriba, es que la
producción de elastasas es considerado uno de los factores de virulencia clave en P.
aeruginosa (Shaver y Hauser, 2004).
Como se mencionó en el capítulo IV, la formación de biofilm, la producción de
factores de virulencia y la resistencia a antimicrobianos son regulados por el proceso
de quórum sensing (QS). La posibilidad de inhibir el proceso de QS o algunos de los
factores de virulencia relacionados a la patogenicidad de bacterias, aparece como una
estrategia para hacerlas más susceptibles al sistema inmune del hospedador. Es
importante encontrar productos naturales capaces de producir dichos efectos sin
afectar directamente el crecimiento bacteriano puesto que al no producir una presión
selectiva sobre el microorganismo el producto no produciría generación de resistencia
por parte del microorganismo. Por lo expuesto, controlar la producción de biofilm y/o

METABOLITOS SECUNDARIOS DE HEPÁTICAS DE LA PATAGONIA
ARGENTINA. SU ACTIVIDAD BIOLÓGICA
108 | P á g i n a
VII
el proceso de QS de patógenos mediante compuestos naturales puede ser una buena
estrategia en la búsqueda de nuevos antimicrobianos.
Diez de los doce terpenoides de L. chordulifera fueron evaluados en su habilidad
para inhibir el crecimiento bacteriano, la formación de biofilm, la producción de
autoinductores y la actividad elastasa en una cepa de Pseudomonas aeruginosa.
Otros estudios se realizaron para determinar la capacidad de estos compuestos de
inhibir el crecimiento bacteriano y la formación de biofilm en una cepa de
Staphylococcus aureus.
El compuesto 11 (taraxerona) mostró un cierto grado de descomposición por lo que
no pudo ser incluido en el bioensayo, mientras que el compuesto 9 (ent-spatulenol) no
fue incluido debido a que ya había sido testeado con los demás constituyentes de P.
chilensis (ver capítulo III).
7.2 Antecedentes
Algunos de los compuestos aislados de L. chordulifera poseen interesantes
actividades biológicas previamente reportadas. Taraxerol y taraxerona (11 -12)
producen efectos alelopáticas, antifúngicas y antiparasitarios (Macias-Rubalcava y col.,
2007; Magadula y Erasto, 2009; Hernandez-Chavez y col., 2012). Taraxerol produce
efectos analgésicos y antiinflamatorios (Ghosh y col., 2009). Los ácidos oleanólico y
ursólico (13 - 14) son bactericidas (Horiuchi y col., 2007) y anticancerígenos (Yan y col.,
2010; Li y col., 2002). El compuesto 15 (ácido betulínico) presenta una gama de
actividades, entre ellas anti HIV, antibacteriana, antinflamatoria, antihelmíntica y
antioxidante (Yogeeswari y Sriram, 2005). El compuesto 17 produce efectos anti
micobacterianos (Phongmaykin y col., 2008).
7.3 Experimental
7.3.1 Crecimiento bacteriano
El crecimiento bacteriano fue determinado espectrofotométricamente a 600 nm en
lector de microplaca (ver capítulo II). A los pocillos para el control negativo se les

METABOLITOS SECUNDARIOS DE HEPÁTICAS DE LA PATAGONIA
ARGENTINA. SU ACTIVIDAD BIOLÓGICA
109 | P á g i n a
VII
adicionó un volumen de una mezcla DMSO – H2O destilada (1:1) en vez de los
compuestos a evaluar. El valor de la absorbancia control fue 1.59 para P. aeruginosa y
1.35 para S. aureus.
7.3.2 Ensayo de formación de biofilm
La formación de biofilm fue determinada por la técnica de cristal violeta en
microplaca (ver capítulo II). La absorbancia del control negativo fue 2.09 para P.
aeruginosa y 0.35 para S. aureus.
Ciprofloxacina, un conocido inhibidor de biofilm (Sandasi y col., 2011), fue
incorporado al bioensayo a 5 µg/ml como control positivo en las mismas condiciones
experimentales que las utilizadas para evaluar los compuestos.
7.3.3 Ensayo de efectos sobre el QS
La cuantificación de autoinductores para determinar los efectos sobre el QS fue
determinada mediante el uso de la cepa reportera P. aeruginosa qsc 119 (ver capítulo
II). La actividad del control negativo fue 13.8 expresado como unidades de miller.
Azitromicina, capaz de intervenir en los procesos de QS (Tateda y col., 2001), fue
usado a 5 µl/ml como control positivo de inhibición de QS bajo las mismas condiciones
que los compuestos evaluados.
7.3.4 Ensayo sobre la actividad elastasa
Los ensayos para determinar la actividad elastasa fueron descriptos en capítulo II.
Dicha actividad se determinó espectrofotométricamente a 495 nm. A esta longitud de
onda absorbe el colorante rojo congo, el cual es liberado al medio luego de que la
enzima elastasa degrada el sustrato insoluble rojo congo-elastina (RC-E). La
absorbancia para el control negativo fue 6.11.
7.3.5 Análisis estadístico
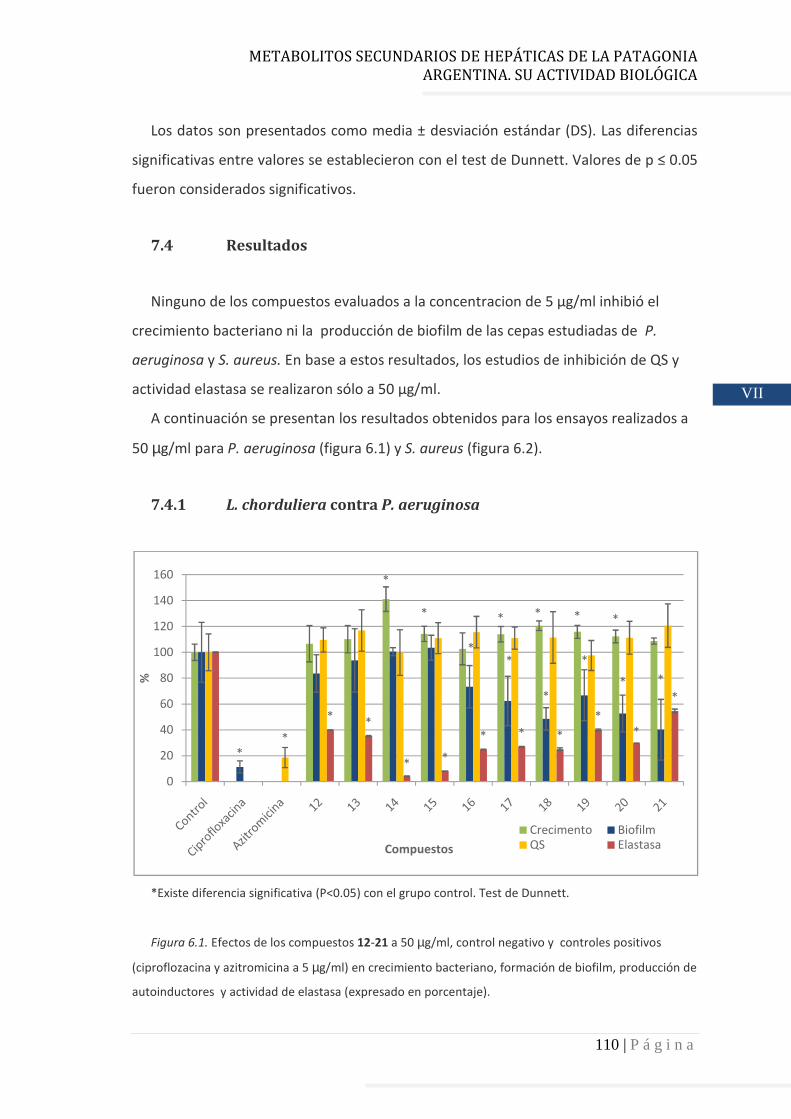
METABOLITOS SECUNDARIOS DE HEPÁTICAS DE LA PATAGONIA
ARGENTINA. SU ACTIVIDAD BIOLÓGICA
110 | P á g i n a
VII
Los datos son presentados como media ± desviación estándar (DS). Las diferencias
significativas entre valores se establecieron con el test de Dunnett. Valores de p ≤ 0.05
fueron considerados significativos.
7.4 Resultados
Ninguno de los compuestos evaluados a la concentracion de 5 µg/ml inhibió el
crecimiento bacteriano ni la producción de biofilm de las cepas estudiadas de P.
aeruginosa y S. aureus. En base a estos resultados, los estudios de inhibición de QS y
actividad elastasa se realizaron sólo a 50 µg/ml.
A continuación se presentan los resultados obtenidos para los ensayos realizados a
50 µg/ml para P. aeruginosa (figura 6.1) y S. aureus (figura 6.2).
7.4.1 L. chorduliera contra P. aeruginosa
*Existe diferencia significativa (P<0.05) con el grupo control. Test de Dunnett.
Figura 6.1. Efectos de los compuestos 12-21 a 50 µg/ml, control negativo y controles positivos
(ciproflozacina y azitromicina a 5 µg/ml) en crecimiento bacteriano, formación de biofilm, producción de
autoinductores y actividad de elastasa (expresado en porcentaje).
*
* * * * *
*
**
*
*
* *
*
**
**
* * *
*
*
*
0
20
40
60
80
100
120
140
160
%
Compuestos
Crecimento Biofilm
QS Elastasa

METABOLITOS SECUNDARIOS DE HEPÁTICAS DE LA PATAGONIA
ARGENTINA. SU ACTIVIDAD BIOLÓGICA
111 | P á g i n a
VII
7.4.1.1 Inhibición del crecimiento bactetriano
Ningún compuesto mostró actividad antimicrobiana a la concentración de 50 µg/ml,
de hecho los compuestos 14, 15, 17, 18, 19 y 20 produjeron estimulaciones de 41, 14,
14, 20, 16 y 12 % en el crecimiento bacteriano, respectivamente. En presencia de los
compuestos 12, 13, 16 y 21 no se registraron diferencias significativas con respecto al
control.
7.4.1.2 Inhibición de la formación de biofilm
Si bien, a la concentración testeada, los compuestos 16-21 no inhibieron el
crecimiento bacteriano. Estos inhibieron significativamente la producción de biofilm en
27, 38, 51, 33, 47 y 60 %, respectivamente. Los demás compuestos no presentaron
diferencias significativas con el control. El antimicrobiano ciprofloxacina a 5 µg/ml,
utilizado como control positivo, produjo una inhibición del 89% en la producción de
biofilm.
7.4.1.3 Inhibición del proceso de QS (inhibición de la producción de
autoinductores)
Ninguno de los compuestos testeados produjo modificaciones significativas en la
producción de autoinductores. Azitromicina, utilizado a 5 µg/ml como control positivo
de inhibición de QS, redujo la producción de autoinductores en 81 %.
7.4.1.4 Inhibición de la actividad elastasa
Todos los compuestos fueron capaces de producir disminuciones significativas en la
elastasa, siendo los ácidos triterpénicos 14 y 15 los más activos. Los compuestos 12 y
13 y 16 - 21 produjeron disminuciones de 60, 65, 75, 73, 75, 60, 70 y 55 %,
respectivamente, mientras que los compuestos 14 y 15 produjeron una disminución en
la actividad elastasa en 96 y 92 %, respectivamente.

METABOLITOS SECUNDARIOS DE HEPÁTICAS DE LA PATAGONIA
ARGENTINA. SU ACTIVIDAD BIOLÓGICA
112 | P á g i n a
VII
Los tritepenoides taraxerol y acido oleanólico 12 y 13 produjeron una disminución
en la actividad elastasa alrededor de un 60 %.
El compuesto 21, el único compuesto aromadendrano de la serie, fue capaz de
inhibir un 60 % la producción de biofilm y un 45 % la producción de elastasa sin
producir una modificación significativa en el crecimiento bacteriano. Una situación
similar se observa para el acido shoerico (16), el cual produjo una disminución de 27 %
en la formación de biofilm y 75 % en la actividad elastasa.
La situación mencionada en el párrafo anterior, puede ser deseable en la
postulación de compuestos antipatogénicos, ya que al no afectar el crecimiento
bacteriano estos compuestos no producen una presión selectiva sobre el
microorganismo que conduzca a la generación de resistencia, sin embargo, al tener
disminuido su principal mecanismo de defensa (biofilm) y estar atenuado uno de sus
principales factores de virulencia (actividad elastasa) el microorganismo puede ser
susceptible al ataque por parte del sistema inmune del hospedador
Los ácidos treiterpénicos ursólico y betulínico (14 y 15) a pesar de no mostrar
diferencias significativas en la formación total de biofilm, inhiben casi totalmente la
actividad elastasa (> 90 %).
Los compuestos 17 - 20 estimularon levemente el crecimiento bacteriano (entre 10
y 20 %) pero fueron capaces de inhibir tanto la formación de biofilm (entre 30 y 60%)
como la actividad elastasa (entre 60 y 70 %). Es decir, se produce un incremento en la
población bacteriana (mayor crecimiento) pero con individuos menos patógenos
(menor actividad elastasa) y menos resistentes (menor formación de biofilm).
7.4.1.5 Relación estructura – actividad
Viridiflorol (21), el único sesquiterpeno evaluado, fue menos activo en su actividad
elastasa que los triterpenoides 12-20, sin embargo, 21 fue más potente que los
triterpenoides mencionados en su capacidad para inhibir el biofilm. Esta capacidad fue
previamente observada en otros aromadendranos aislados de Porella chilensis.
Entre los triterpenoides analizados, aquellos que poseen esqueleto dammarano (16
– 20) fueron más potentes en la inhibición de la formación de biofilm. La presencia o

METABOLITOS SECUNDARIOS DE HEPÁTICAS DE LA PATAGONIA
ARGENTINA. SU ACTIVIDAD BIOLÓGICA
113 | P á g i n a
VII
ausencia del anillo tetrahidrofurano (THF) sobre el carbono 17 o la presencia de un OH
o carbonilo en carbono 3 no es importante para la actividad citada.
Con respecto a la actividad elastasa, los compuestos 14 y 15 resultaron los más
activos. Ambos portan grupos carboxilos en carbono 17, y aunque el triterpeno 13
también posee un grupo carboxilo en carbono 17 este no resultó tan activo, por lo
tanto, es difícil establecer una certera relación estructura-actividad para los ácidos 13,
14 y 15.
2
3
4
5
10
1
6
7
8
9 14
13
12
11
15
16
17
18
22
21
20
19
O
23
25 26
24
27
3029
28
HO HO
OH
O
HO
OH
O
HO
OH
O2
29
4
5
10
1
6
7
8
9 14
13
12
11
19 18
30
3
HO
O
28
15
16
17
20
O
24
23
22
21
25
26
27
OH
HO
O
O
O
O
O
OH
HO
O
OH
O
HO
3
4
5
1
2
6
7
8
9
10
15
H
11
12
13
14
OH
H
11 12 13
14 15 16
17 18 19
20 21
H H
H H H H H H
H
H
H H H H
H
H
H H
H
H
H H
H
H
H H
H
H
H
H
H
H
HOH
H
9
H
H
H
H
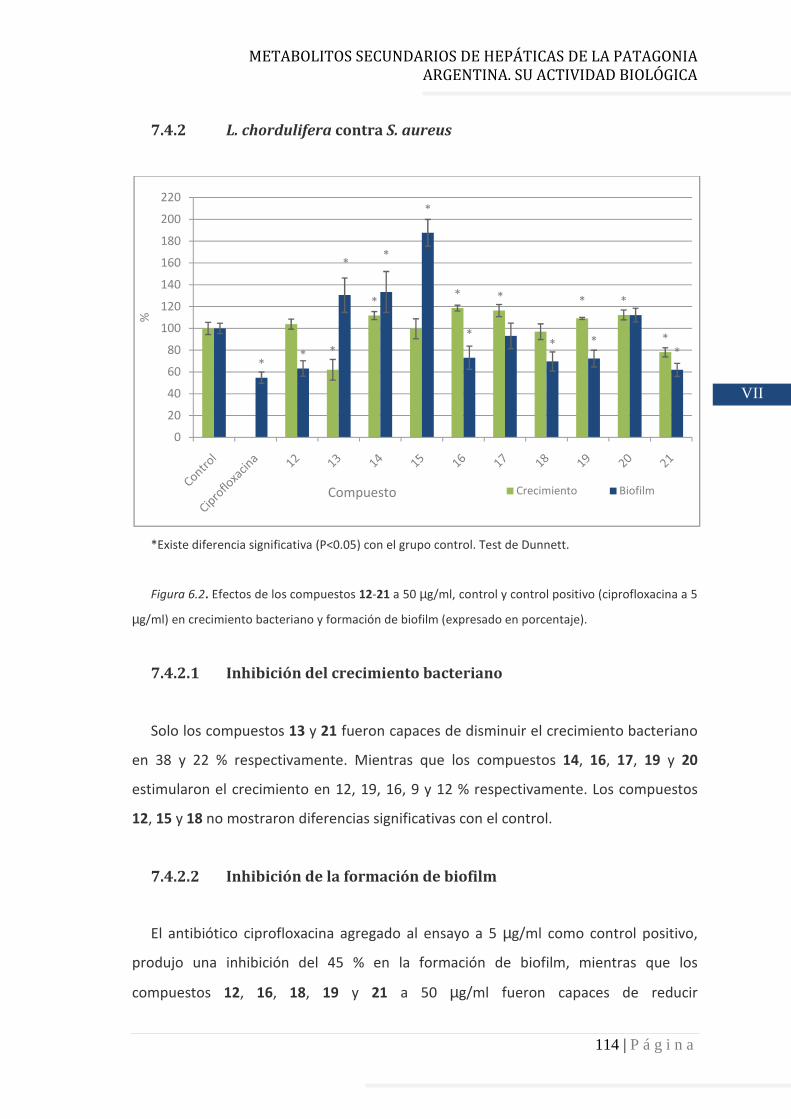
METABOLITOS SECUNDARIOS DE HEPÁTICAS DE LA PATAGONIA
ARGENTINA. SU ACTIVIDAD BIOLÓGICA
114 | P á g i n a
VII
7.4.2 L. chordulifera contra S. aureus
*Existe diferencia significativa (P<0.05) con el grupo control. Test de Dunnett.
Figura 6.2. Efectos de los compuestos 12-21 a 50 µg/ml, control y control positivo (ciprofloxacina a 5
µg/ml) en crecimiento bacteriano y formación de biofilm (expresado en porcentaje).
7.4.2.1 Inhibición del crecimiento bacteriano
Solo los compuestos 13 y 21 fueron capaces de disminuir el crecimiento bacteriano
en 38 y 22 % respectivamente. Mientras que los compuestos 14, 16, 17, 19 y 20
estimularon el crecimiento en 12, 19, 16, 9 y 12 % respectivamente. Los compuestos
12, 15 y 18 no mostraron diferencias significativas con el control.
7.4.2.2 Inhibición de la formación de biofilm
El antibiótico ciprofloxacina agregado al ensayo a 5 µg/ml como control positivo,
produjo una inhibición del 45 % en la formación de biofilm, mientras que los
compuestos 12, 16, 18, 19 y 21 a 50 µg/ml fueron capaces de reducir
*
** * * *
*
**
**
*
** *
*
0
20
40
60
80
100
120
140
160
180
200
220
%
Compuesto Crecimiento Biofilm

METABOLITOS SECUNDARIOS DE HEPÁTICAS DE LA PATAGONIA
ARGENTINA. SU ACTIVIDAD BIOLÓGICA
115 | P á g i n a
VII
significativamente la formación de biofilm por parte de S. aureus en 37, 27, 30, 28 y 38
% respectivamente. Mientras que 13, 14 y 15 estimularon significativamente la
formación de biofilm en 30, 33 y 88 % a la misma concentración. Los compuestos 17 y
20 no produjeron modificaciones en relación al control.
El único de los compuestos testeados capaz de modificar tanto el crecimiento
bacteriano como la formación de biofilm, fue el aromadendrano viridiflorol (21),
mientras que 12 y 18 produjeron una disminución en la formación de biofilm (37 y 30
%) sin alterar el crecimiento bacteriano.
7.4.2.3 Relación estructura – actividad
Los ácidos oleanólico, ursólico y betulínico (13 – 15) produjeron estimulaciones en
la formación de biofilm y no así taraxerol (12), indicando que la presencia del grupo
carboxílico es importante para dicha estimulación.
Con respecto a los dammaranos (16 - 20), sólo tres de ellos (16, 18 y 19) fueron
capaces de inhibir la formación de biofilm de S. aureus. Esto indica que el anillo THF
sobre C-17 unido a un grupo isopropilol en C-24 son requeridos para la actividad.
Tanto los aromadendranos de P. chilensis como de L. chordulifera inhiben la
formación de biofilm y el crecimiento bacteriano de P. aeruginosa.
Vistos los resultados expuestos sobre los efectos antipatogénicos de los terpenoides
evaluados, consideramos que se deben ampliar los estudios con un grupo de
terpenoides con mayor diversidad estructural a fin de establecer relaciones más
precisas entre la estructura y la actividad.

METABOLITOS SECUNDARIOS DE HEPÁTICAS DE LA PATAGONIA
ARGENTINA. SU ACTIVIDAD BIOLÓGICA
116 | P á g i n a
VII
Referencias
Ghosh A., Biswas K., Biswas M. and Haldar P. A pentacyclic triterpenoid possessing
analgesic activity from the fruits of Dregea volubilis. Phcog Mag 2009, 5, 90-92.
Hernandez-Chavez I., Torrez-Tapia L. W., Sima-Polanco P., Cedillo-Rivera R., Moo-
Puc R. and Peraza-Sanchez S. R. Antigiardial activity of Cupania dentata bark and its
constituents. Journal of the Mexican Chemical Society 2012, 56, 105-108.
Horiuchi K., Hiota S., Hatano T., Yoshida T., Turoda T. and Tsuchiya T. Antimicrobial
activity of oleanolic acid from Salvia officinalis and related compounds on vancomycin-
resistant enterococci (VRE). Biological and Pharmaceutical Bulletin 2007, 30, 1147-
1149.
Li J., Guo W. J. and Yang Q. Y. Effects of ursolic acid and oleanolic acid on human
colon carcinoma cell line HCT15. World Journal of Gastroenterology 2002, 8, 493-495.
Liu P. V. Extracellular Toxins of Pseudomonas aeruginosa. Journal of Infectious
Diseases 1974, 130, S94-S99.
Macias-Rubalcava M., Hernandez-Bautista B., Jimenez-Estrada M., Cruz-Ortega R. and
Anaya A. Pentacyclic triterpenes with selective bioactivity from Sebastiania
adenophora; leaves, Euphorbiaceae. Journal of Chemical Ecology 2007, 33, 147-156.
Magadula J. and Erasto P. Bioactive natural products derived from the East African
flora. Nat. Prod. Rep. 2009, 26, 1535-1554.
Morihara K. and Tsuzuki H. Production of protease and elastase by Pseudomonas
aeruginosa strains isolated from patients. Infection and Immunity 1977, 15, 679-685.

METABOLITOS SECUNDARIOS DE HEPÁTICAS DE LA PATAGONIA
ARGENTINA. SU ACTIVIDAD BIOLÓGICA
117 | P á g i n a
VII
Morihara K., Tsuzuki H. and Oda K. Protease and elastase of Pseudomonas aeruginosa:
inactivation of human plasma alpha 1-proteinase inhibitor. Infection and Immunity
1979, 24, 188-193.
Phongmaykin J., Kumamoto T., Ishikawa T., Suttisri R. and Saifah E. A new
sesquiterpene and other terpenoid constituents of Chisocheton penduliflorus. Archives of
Pharmacal Research 2008, 31, 21-27.
Sandasi M., Leonard C. M., van Vuuren S. F. and Viljoen A. M. Peppermint (Mentha
piperita) inhibits microbial biofilms in vitro. South African Journal of Botany 2011, 77,
80-85.
Shaver C. M. and Hauser A. R. Relative contributions of Pseudomonas aeruginosa
exoU, exoS, and exoT to virulence in the lung. Infection and Immunity 2004, 72, 6969-
6977.
Tateda K., Comte R., Pechere J. C., Köhler T., Yamaguchi K. and van Delden C.
Azithromycin inhibits quorum sensing in Pseudomonas aeruginosa. Antimicrobial
Agents and Chemotherapy 2001, 45, 1930-1933.
Yan S., Huang C., Wu S. and Yin M. Oleanolic acid and ursolic acid induce apoptosis
in four human liver cancer cell lines. Toxicology in Vitro 2010, 24, 842-848.
Yogeeswari P. and Sriram D. Betulinic acid and its derivatives: A review on their
biological properties. Current Medicinal Chemistry 2005, 12, 657-666.

Capítulo VIII
Resumen de resultados y
conclusiones
VII
VIII

METABOLITOS SECUNDARIOS DE HEPÁTICAS DE LA PATAGONIA
ARGENTINA. SU ACTIVIDAD BIOLÓGICA
119 | P á g i n a
VIII
Capítulo VIII
Resumen de resultados y conclusiones
Este trabajo de tesis nos planteó el desafío de continuar con el trabajo pionero, en
nuestro país, en las investigaciones químicas y de actividad biológica de especies de la
clase Hepaticae.
Las investigaciones de hepáticas en Tucumán se iniciaron con el estudio de las
especies talosas endémicas en el Norte Argentino, pertenecientes al orden
Marchantiales, Dumortiera hirsuta, Plagiochasma rupestre y Marchantia plicata. A
continuación se estudiaron las hepáticas foliosas pertenecientes al orden
Jungermaniales Frullania brasiliensis y Porella swartziana también endémicas del
Norte Argentino.
Esta tesis doctoral tuvo como objetivo la investigación de dos especies foliosas,
también pertenecientes al orden Jungermaniales: Porella chilensis y Lepidozia
chordulifera, que crecen en la Patagonia Argentina.
Se aislaron e identificaron 21 metabolitos secundarios. Dichos productos constituían
mezclas complejas resueltas mediante el empleo exhaustivo de cromatografía líquida
de alta performance en fase normal y reversa.
8.1 Porella chilensis
Del estudio del extracto etéreo de P. chilensis resultó el aislamiento e identificación
de los diterpenos de tipo fusicoccano 1-4, de los cuales 1 y 2 fueron descriptos por
primera vez. Se aislaron e identificaron seis sesquiterpenoides conocidos, cuatro de
tipo pinguisano (5 - 8) y dos aromadendranos (9 y 10), siendo norpinguisona (6) el
compuesto mayoritario.
Anadensina (3)
(Huneck y col., 1983), fusicoauritona (4) (Zapp y col., 1994),
pinguisenol (5) (Asakawa, 1982), norpinguisona (6) (Asakawa, 1982), norpinguisona
acetato (7) (Van Klink y col., 2002), norpinguisona metil éster (8) (Tori y col., 1993), ent-

METABOLITOS SECUNDARIOS DE HEPÁTICAS DE LA PATAGONIA
ARGENTINA. SU ACTIVIDAD BIOLÓGICA
120 | P á g i n a
VIII
spathulenol (9) (Asakawa y col., 2012; Nagashima y col., 1993),
y ent-4β,10α-
dihidroxiaromadendrano (10) (Nagashima y col., 1993) fueron identificados por sus
características espectroscópicas en comparación con los datos de literatura
previamente reportados.
Figura 8.1. Terpenoides aislados de Porella chilensis
La configuración absoluta de los nuevos fusicoccanos 1 y 2 se obtuvo mediante el
análisis de dicroísmo circular (DC) en la zona del espectro de UV y posterior
comparación con espectros de dicroísmo circular de moléculas estructuralmente
relacionadas con 1 y 2, de configuración absoluta conocida. La estereoquímica
absoluta de 1 y 2 resultó ser la misma que la previamente establecida para los
compuestos 3 y 4, determinada por síntesis total (Kato y col., 1994; Williams y col.,
2007). Adicionalmente, y para apuntalar la propuesta estereoquímica, se realizaron
cálculos computacionales que permitieron deducir los espectros de DC teóricos que
coincidieron con los datos experimentales de DC (figura 8.2), confirmando así la
configuración absoluta de los nuevos compuestos.
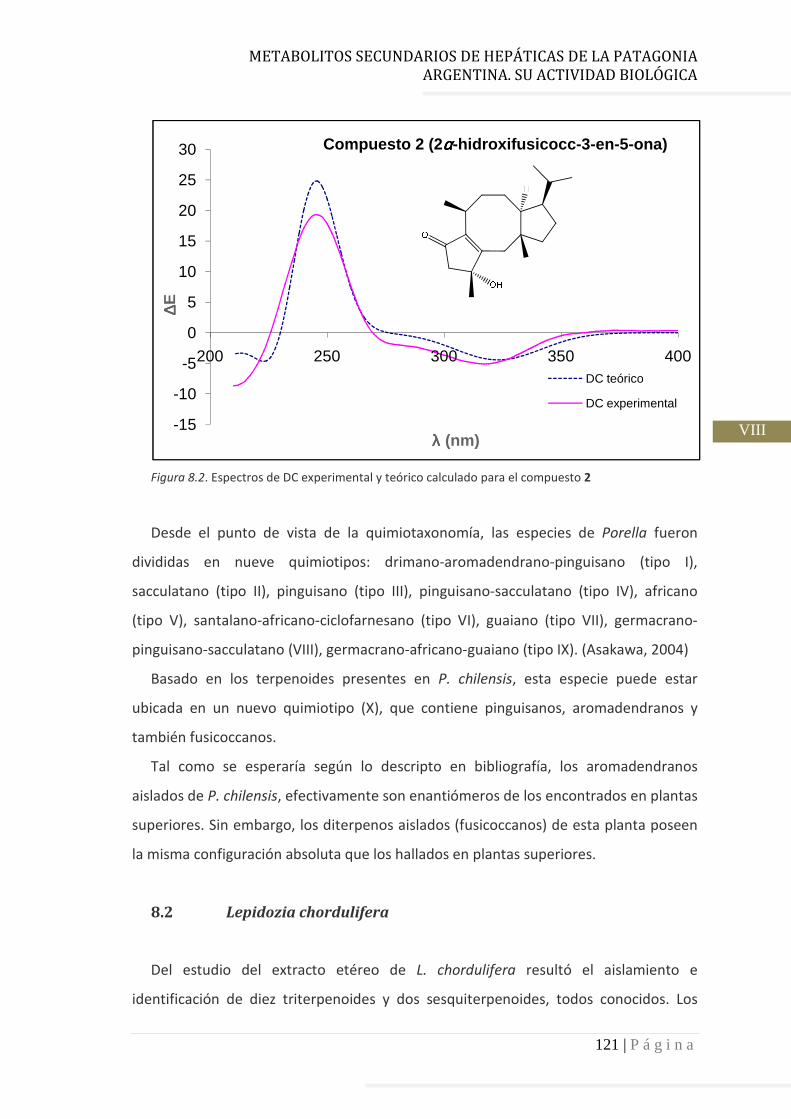
METABOLITOS SECUNDARIOS DE HEPÁTICAS DE LA PATAGONIA
ARGENTINA. SU ACTIVIDAD BIOLÓGICA
121 | P á g i n a
VIII
Figura 8.2. Espectros de DC experimental y teórico calculado para el compuesto 2
Desde el punto de vista de la quimiotaxonomía, las especies de Porella fueron
divididas en nueve quimiotipos: drimano-aromadendrano-pinguisano (tipo I),
sacculatano (tipo II), pinguisano (tipo III), pinguisano-sacculatano (tipo IV), africano
(tipo V), santalano-africano-ciclofarnesano (tipo VI), guaiano (tipo VII), germacrano-
pinguisano-sacculatano (VIII), germacrano-africano-guaiano (tipo IX). (Asakawa, 2004)
Basado en los terpenoides presentes en P. chilensis, esta especie puede estar
ubicada en un nuevo quimiotipo (X), que contiene pinguisanos, aromadendranos y
también fusicoccanos.
Tal como se esperaría según lo descripto en bibliografía, los aromadendranos
aislados de P. chilensis, efectivamente son enantiómeros de los encontrados en plantas
superiores. Sin embargo, los diterpenos aislados (fusicoccanos) de esta planta poseen
la misma configuración absoluta que los hallados en plantas superiores.
8.2 Lepidozia chordulifera
Del estudio del extracto etéreo de L. chordulifera resultó el aislamiento e
identificación de diez triterpenoides y dos sesquiterpenoides, todos conocidos. Los
-15
-10
-5
0
5
10
15
20
25
30
200 250 300 350 400
∆∆ ∆∆E
λλλλ (nm)
Compuesto 2 (2αααα-hidroxifusicocc-3-en-5-ona)
DC teórico
DC experimental

METABOLITOS SECUNDARIOS DE HEPÁTICAS DE LA PATAGONIA
ARGENTINA. SU ACTIVIDAD BIOLÓGICA
122 | P á g i n a
VIII
triterpenoides aislados fueron dos taraxanos, taraxerona (11) y taraxerol (12)
(Hernandez-Chavez y col., 2012); los ácidos oleanólico (13) (Gohari y col., 2009),
ursólico (14) (Ayatollahi y col., 2011) y betulínico (15) (Muhit y col., 2010); y los
dammaranos ácido shoerico (16) (Govindachari y col., 1994), eichlerialactona (17) (Rao
y col., 1975), cabraleona (18), cabraleadiol (19) (Hisham y col., 1996) y 3β-
hidroxihexanordammaran-20-ona (20) (Tanaka y col., 1987). Los sesquiterpenos
aislados fueron los aromadendranos viridiflorol (21) (Bombarda y col., 2001) y ent-
spathulenol aislado también de P. chilensis.
Figura 8.3. Terpenoides aislados de Lepidozia chordulifera

METABOLITOS SECUNDARIOS DE HEPÁTICAS DE LA PATAGONIA
ARGENTINA. SU ACTIVIDAD BIOLÓGICA
123 | P á g i n a
VIII
Es la primera vez que los compuestos ácido oleanólico (13), eichleralactona (17), y
cabraleona (18) se aíslan de una especie de la clase Hepaticae.
También es importante remarcar que ni la actual investigación de L. chordulifera, ni
la realizada previamente por Zapp y col. (2008) arrojó como resultado el aislamiento
de β-bazzaneno, considerado un marcador químico del género. Teniendo en cuenta
que la presencia de β-bazzaneno en las especies de Lepidozia fue registrada para
especies asiáticas, sugerimos reconsiderar la presencia de este como marcador
químico del género, o por lo menos considerarlo solo para las especies asiáticas y no
así para las americanas, dado que ambos estudios, el actual y el realizado por Zapp y
col. (2008), fueron realizados sobre material vegetal recolectado en sud-América.
Otro aspecto a resaltar es que la presencia de triterpenoides en hepáticas es
bastante poco habitual, por lo que se ha sugerido que la biosíntesis de triterpenoides
se encuentra bloqueada tanto en hepáticas como en algas (Asakawa, 1986). Sin
embargo, la cantidad de trierpenoides aislados de esta especie pone a prueba esta
hipótesis. Este tipo de triterpenos son más comunes en musgos y evidencia la cercanía
evolutiva entre ambas clases.
8.3 Actividad biológica
Cuando se estudiaron los efectos de los compuestos aislados de P. chilensis sobre la
formación de biofilm de Pseudomonas aeruginosa, sólo el fusicoccano 3 a 5 µg/ml
resultó tan potente como el antibiótico ciprofloxacina a la misma concentración,
mientras que los aromadendranos 9 y 10 son menos potentes que el antibiótico a la
misma concentración. Los tres compuestos fueron capaces de inhibir
significativamente, además de la formación de biofilm, el crecimiento bacteriano y la
producción de autoinductores de QS.
Es importante destacar que los aromadendranos 9 y 10, no están estructuralmente
relacionados a ningún compuesto previamente reportado como capaz de inhibir la
formación de biofilm o el proceso de QS de P. aeruginosa. Mientras que los
fusicoccanos comparten algunas características estructurales con algunos productos

METABOLITOS SECUNDARIOS DE HEPÁTICAS DE LA PATAGONIA
ARGENTINA. SU ACTIVIDAD BIOLÓGICA
124 | P á g i n a
VIII
naturales capaces de inhibir la producción de autoinductores de P. aeruginosa (Givskov
y col., 1996; Manefield y col., 2002; Rasmussen y col., 2005).
En cuanto a la capacidad de los compuestos de L. chordulifera para inhibir la
formación de biofilm de P. aeruginosa, el compuesto más potente fue el
aromadendrano viridiflorol (21) a 50 µg/ml, aunque no resultó tan activo como el
antibiótico ciprofloxacina utilizado como control positivo a 5 µg/ml. Todos los
compuestos dammaranos (16 - 20) a 50 µg/ml también fueron capaces de inhibir la
formación de biofilm indicando que tanto el esqueleto dammarano como el
aroadendrano son importantes para esta actividad.
Con respecto a la actividad elastasa, actividad relacionada a la patogenicidad de
Pseudomonas aeruginosa, todos los compuestos fueron capaces de inhibirla
significativamente, siendo los ácidos ursólico y betulínico (14 - 15) los más potentes
produciendo inhibiciones superiores al 90 %. a 50 µg/ml.
El aromadendrano viridiflorol (21) inhibió el desarrollo bacteriano y la formación de
biofilm de Staphylococcus aureus. Los aromadendranos de Porella chilensis también
produjeron los mismos efectos sobre P. aeruginosa, indicando que los compuestos de
esta familia son promisorios como antibacterianos de cepas patógenas humanas.
El pinguisano 6, aislado de P. chilensis, produjo el mayor porcentaje de mortalidad
de larvas de Spodoptera frugiperda entre los compuestos evaluados. Esta importante
actividad se relaciona con la capacidad de alterar fuertemente el epitelio del intestino
medio de las larvas.
Los compuestos que fueron más activos contra los microorganismos no provocaron
efectos tóxicos importantes a las células eucariotas del epitelio intestinal de S.
frugiperda. Estos resultados pueden ser alentadores en la postulación de sustancias
anti-patogénicas bacterianas que, para ser usadas como fármacos, no deberían afectar
células eucariotas.
Si bien nuestros resultados mostraron que los terpenoides evaluados producen
efectos moderados sobre los sistemas biológicos estudiados, los logros obtenidos
plantean un futuro promisorio para la aplicación de los constituyentes químicos de
hepáticas argentinas en formulaciones farmacéuticas e insecticidas.

METABOLITOS SECUNDARIOS DE HEPÁTICAS DE LA PATAGONIA
ARGENTINA. SU ACTIVIDAD BIOLÓGICA
125 | P á g i n a
VIII
Referencias
Asakawa Y. Chemical constituents of hepaticae. Springer, Vienna. 1982.
Asakawa Y. Chemical Relationships Between Algae, Bryophytes, and Pteridophytes.
Journal of Bryology 1986, 14, 59-70.
Asakawa Y. Chemosystematics of the Hepaticae. Phytochemistry 2004, 65, 623-669.
Asakawa Y., Ludwiczuk A., Nagashima F., Kinghorn A. D., Falk H. and Kobayashi J.
Chemical Constituents of Bryophytes: Bio- And Chemical Diversity, Biological Activity,
and Chemosystematics. Springer. 2012.
Ayatollahi A. M., Ghanadian M., Afsharypour S., Abdella O. M., Mirzai M. and Askari
G. Pentacyclic triterpenes in Euphorbia microsciadia with their t-cell proliferation
activity. Iranian Journal of Pharmaceutical Research 2011, 10, 287-294.
Bombarda I., Raharivelomanana P., Ramanoelina P. A. R., Faure R., Bianchini J. P. and
Gaydou E. M. Spectrometric identifications of sesquiterpene alcohols from niaouli
(Melaleuca quinquenervia) essential oil. Analytica Chimica Acta 2001, 447, 113-123.
Givskov M., De Nys R., Manefield M., Gram L., Maximilien R., Eberl L., Molin S.,
Steinberg P. D. and Kjelleberg S. Eukaryotic interference with homoserine lactone-
mediated prokaryotic signalling. Journal of Bacteriology 1996, 178, 6618-6622.
Gohari A. R., Saeidnia S., Hadjiakhoondi A., Abdoullahi M. and Nezafati M. Isolation
and quantificative analysis of oleanolic acid from Satureja mutica Fisch. & C. A. Mey.
Journal of Medicinal Plants 2009, 8, 65-69.

METABOLITOS SECUNDARIOS DE HEPÁTICAS DE LA PATAGONIA
ARGENTINA. SU ACTIVIDAD BIOLÓGICA
126 | P á g i n a
VIII
Govindachari T. R., Suresh G. and Kumari G. N. K. Triterpenoids from Dysoxylum
malabaricum. Phytochemistry 1994, 37, 1127-1129.
Hernandez-Chavez I., Torrez-Tapia L. W., Sima-Polanco P., Cedillo-Rivera R., Moo-
Puc R. and Peraza-Sanchez S. R. Antigiardial Activity of Cupania dentata Bark and its
Constituents. Journal of the Mexican Chemical Society 2012, 56, 105-108.
Hisham A., Ajitha Bai M. D., Fujimoto Y., Hara N. and Shimada H. Complete 1H and
13C NMR Spectral Assignment of Cabraleadiol, a Dammarane Triterpene from
Dysoxylum malabaricum Bedd. Magn. Reson. Chem. 1996, 34, 146-150.
Huneck S., Baxter G., Cameron A. F., Connolly J. D. and Rycroft D. S. Anadensin, a
new fusicoccane diterpenoid from the liverwort Anastrepta orcadensis. Crystal structure
analysis. Tetrahedron Letters 1983, 24, 3787-3788.
Kato N., Nakanishi K., Wu X., Nishikawa H. and Takeshita H. Total synthesis of
fusicogigantones A and B and fusicogigantepoxide via the singlet oxygen-oxidation of
fusicoccadienes. Fusicogigantepoxide B', a missing congener metabolite. Tetrahedron
Letters 1994, 35, 8205-8208.
Manefield M., Rasmussen T. B., Henzter M., Andersen J. B., Steinberg P., Kjelleberg S.
and Givskov M. Halogenated furanones inhibit quorum sensing through accelerated
LuxR turnover. Microbiology 2002, 148, 1119-1127.
Muhit M. A., Tareq S. M., Apu A. S., Basak D. and Islam M. S. Isolation and
Identification of Compounds from the leaf extract of Dillenia Indica Linn. Bangladesh
Pharmaceutical Journal 2010, 13, 49-53.

METABOLITOS SECUNDARIOS DE HEPÁTICAS DE LA PATAGONIA
ARGENTINA. SU ACTIVIDAD BIOLÓGICA
127 | P á g i n a
VIII
Nagashima F., Sari Y., Tori M., Asakawa Y. and Huneck S. Sesquiterpenoids from
some European liverworts. Phytochemistry 1993, 34, 1341-1343.
Rao M. M., Meshulam H., Zelnik R. and Lavie D. Cabralea eichleriana dc.
(meliaceae)−I structure and stereochemistry of wood extractives. Tetrahedron 1975, 31,
333-339.
Rasmussen T. B., Skindersoe M. E., Bjarnsholt T., Phipps R. K., Christensen K. B.,
Jensen P. O., Andersen J. B., Koch B., Larsen T. O., Hentzer M., Eberl L., Hoiby N.
and Givskov M. Identity and effects of quorum-sensing inhibitors produced by
Penicillium species. Microbiology 2005, 151, 1325-1340.
Tanaka R., atsuda M. and atsunaga S. 3β-Hydroxyhexanordammaran-20-one from
Euphorbia supina. Phytochemistry 1987, 26, 3365-3366.
Tori M., Nakashima K., Takeda T. and Asakawa Y. Structure of secoswartzianins A and
B isolated from the liverwort Porella swartziana. Tetrahedron Letters 1993, 34, 3753-
3754.
Van Klink J. W., Zapp J. and Becker H. Pinguisane-type sesquiterpenes from the South
American liverwort Porella recurva (Taylor) Kuhnemann. Zeitschrift fur
Naturforschung - Section C Journal of Biosciences 2002, 57, 413-417.
Williams D. R., Robinson L. A., Nevill C. R. and Reddy J. P. Strategies for the
synthesis of fusicoccanes by nazarov reactions of dolabelladienones: Total synthesis of
(+)-fusicoauritone. Angewandte Chemie - International Edition 2007, 46, 915-918.

METABOLITOS SECUNDARIOS DE HEPÁTICAS DE LA PATAGONIA
ARGENTINA. SU ACTIVIDAD BIOLÓGICA
128 | P á g i n a
VIII
Zapp J., Burkhardt G. and Becker H. Sphenolobane and fusicoccane diterpenoids from
the liverwort Anastrophyllum auritum. Phytochemistry 1994, 37, 787-793.

Apéndice
Espectros
VII
A
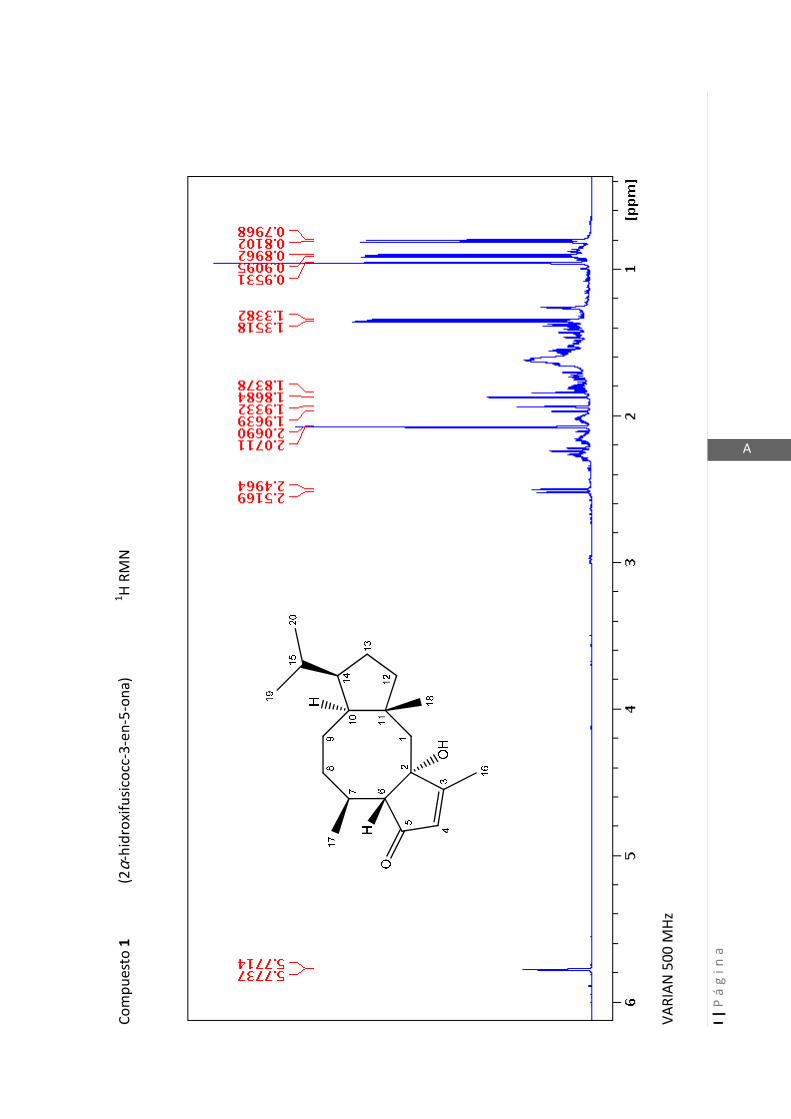
I |
Pá
gi
na
A
Co
mp
ue
sto
1
(2
α-h
idro
xifu
sico
cc-3
-en
-5-o
na
)
1H
RM
N
VA
RIA
N 5
00
MH
z
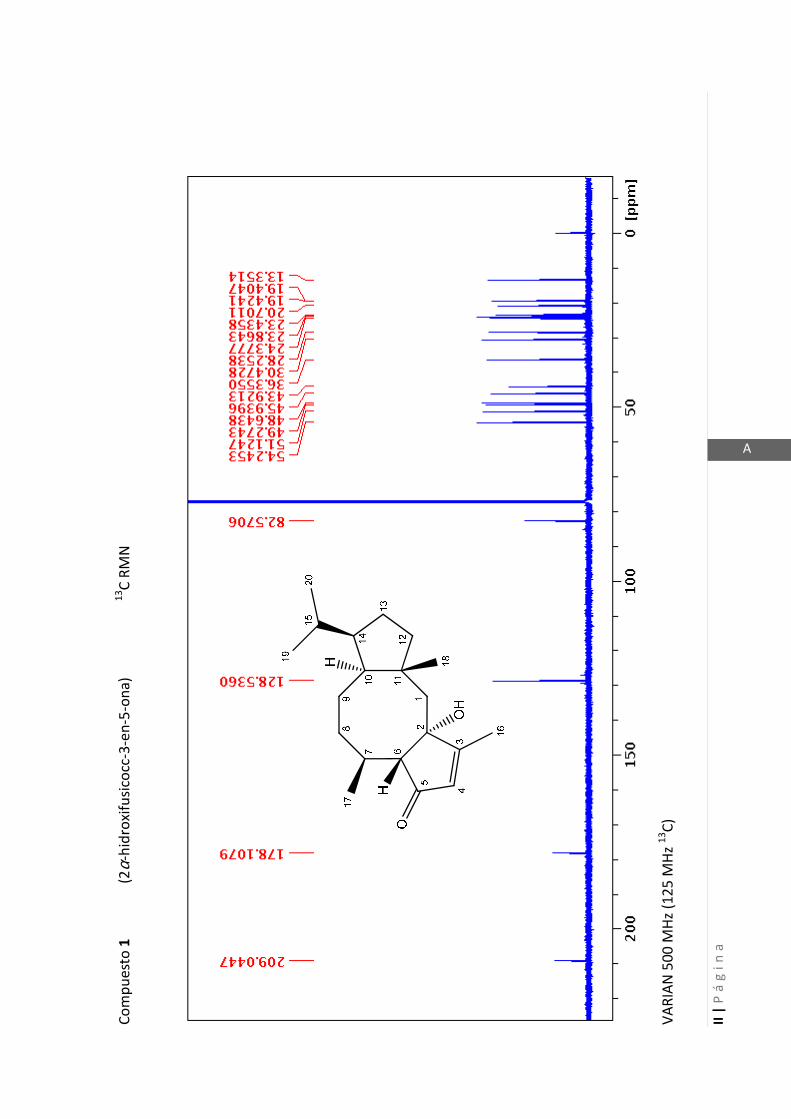
II |
Pá
gi
na
A
Co
mp
ue
sto
1
(2
α-h
idro
xifu
sico
cc-3
-en
-5-o
na
)
13C
RM
N
VA
RIA
N 5
00
MH
z (1
25
MH
z 1
3C
)
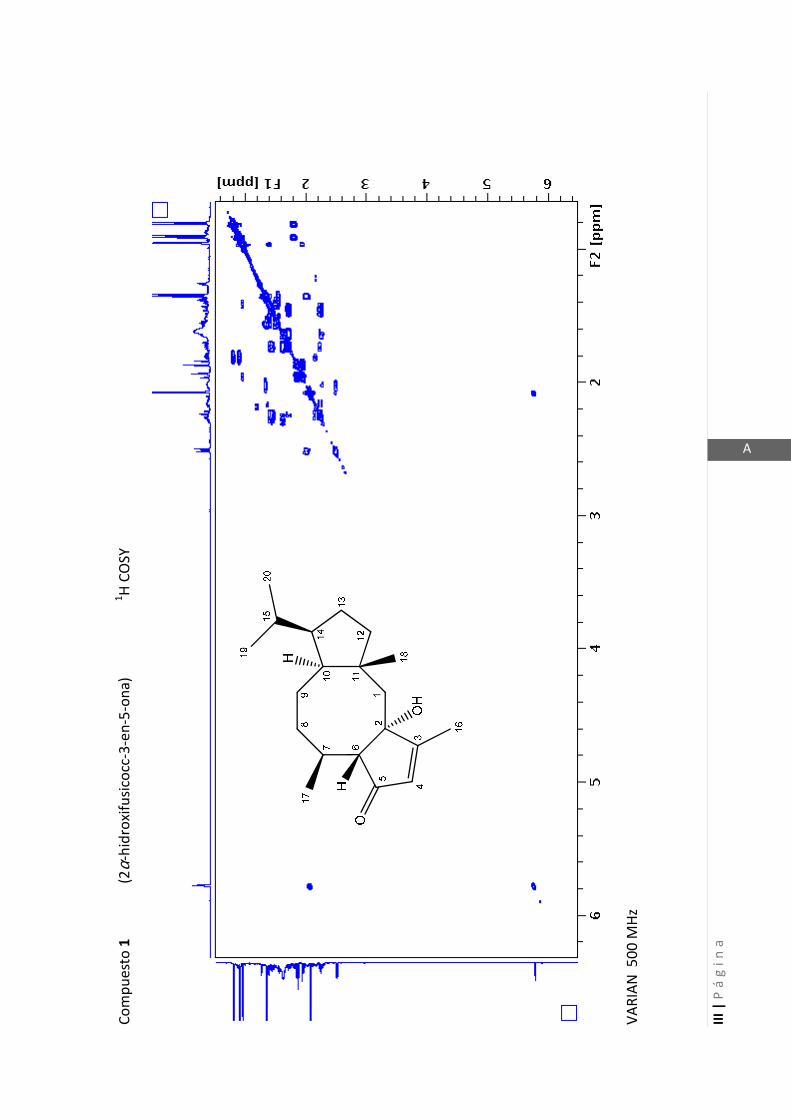
III
| P
ág
in
a
A
Co
mp
ue
sto
1
(2
α-h
idro
xifu
sico
cc-3
-en
-5-o
na
)
1H
CO
SY
VA
RIA
N 5
00
MH
z
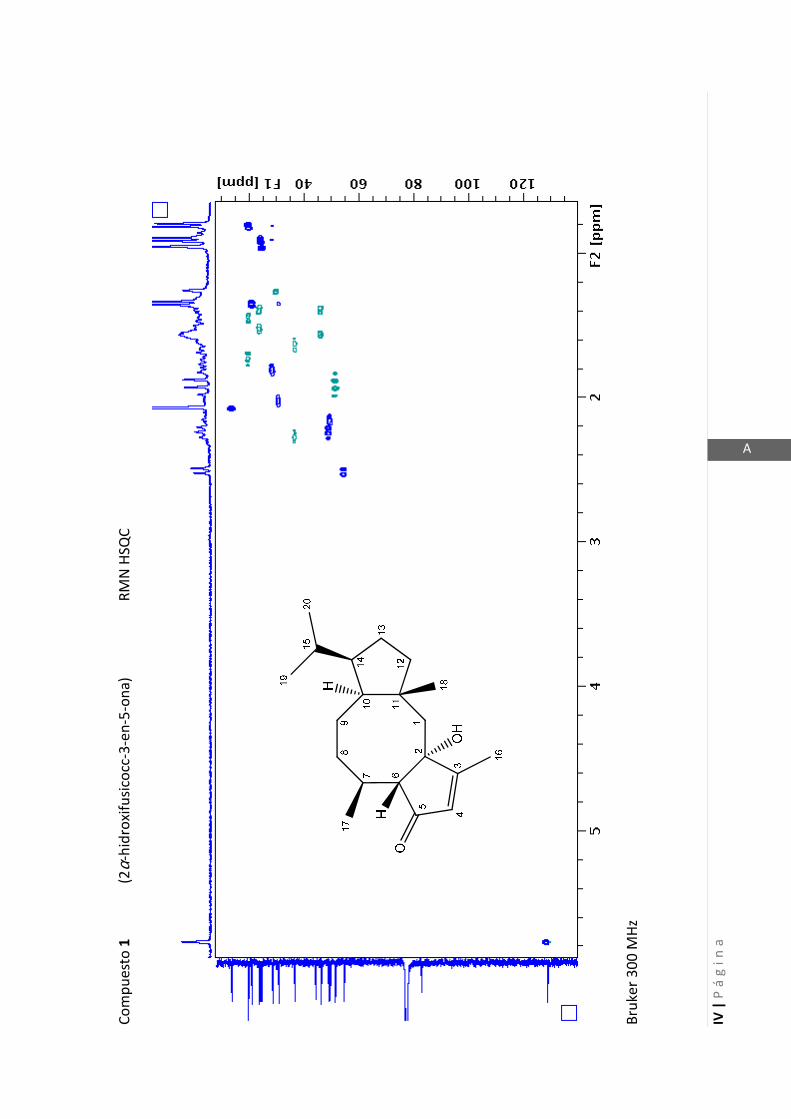
IV |
Pá
gi
na
A
Co
mp
ue
sto
1
(2
α-h
idro
xifu
sico
cc-3
-en
-5-o
na
)
RM
N H
SQ
C
Bru
ke
r 3
00
MH
z
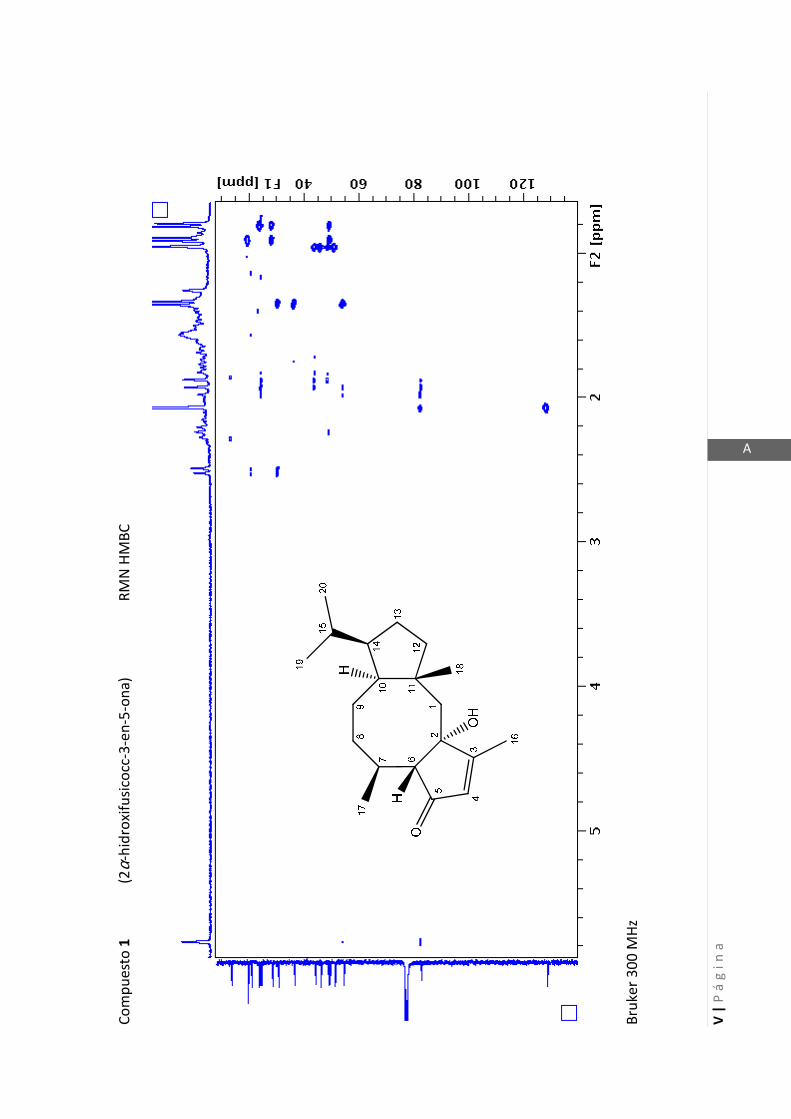
V |
Pá
gi
na
A
Co
mp
ue
sto
1
(2
α-h
idro
xifu
sico
cc-3
-en
-5-o
na
)
RM
N H
MB
C
Bru
ke
r 3
00
MH
z
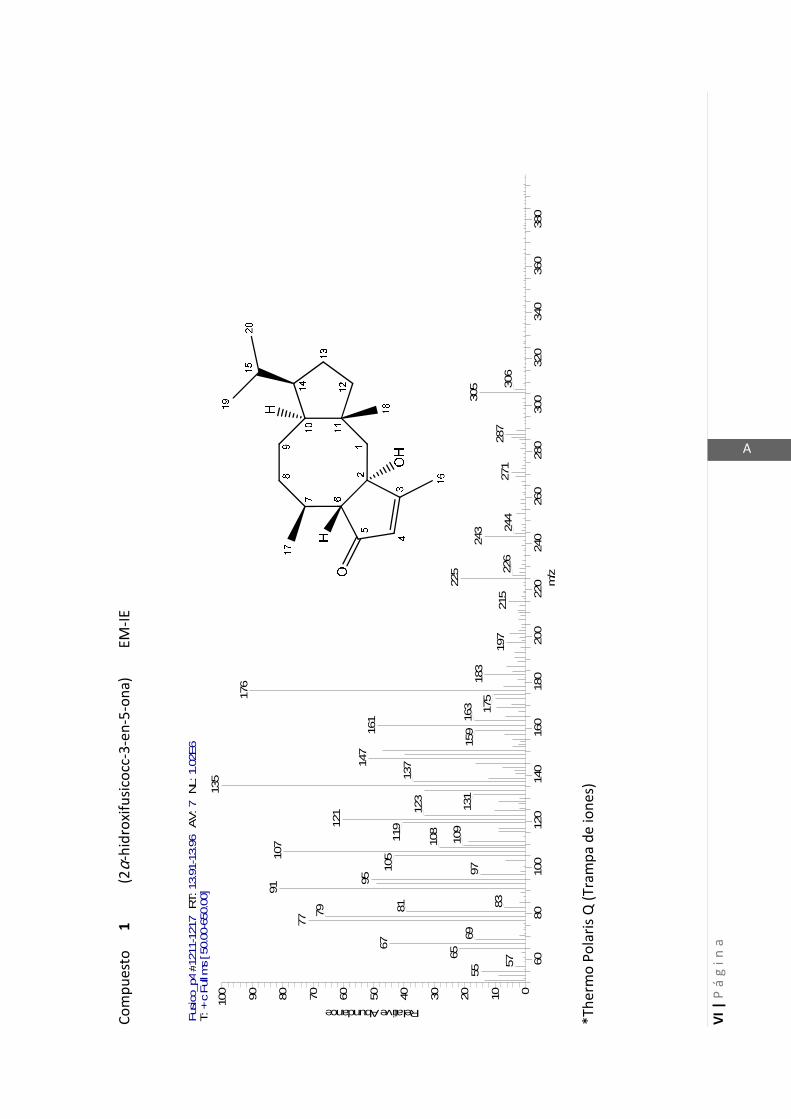
VI
| P
ág
in
a
A
Co
mp
ue
sto
1
(2
α-h
idro
xifu
sico
cc-3
-en
-5-o
na
) E
M-I
E
*T
he
rmo
Po
lari
s Q
(T
ram
pa
de
io
ne
s)
Fus
ico_
p4 #
1211
-121
7RT:
13.9
1-13
.96
AV:7
NL:
1.02
E6
T:+
c Full m
s [ 5
0.00
-650
.00]
6080
100
120
140
160
180
200
220
240
260
280
300
320
340
360
380
m/z
0102030405060708090100
Relative Abundance
135
176
9110
7
77
79
121
147
9516
167
105
119
8113
712
3
108
6522
510
913
116
369
159
305
9755
243
183
175
8328
719
721
527
122
630
624
457
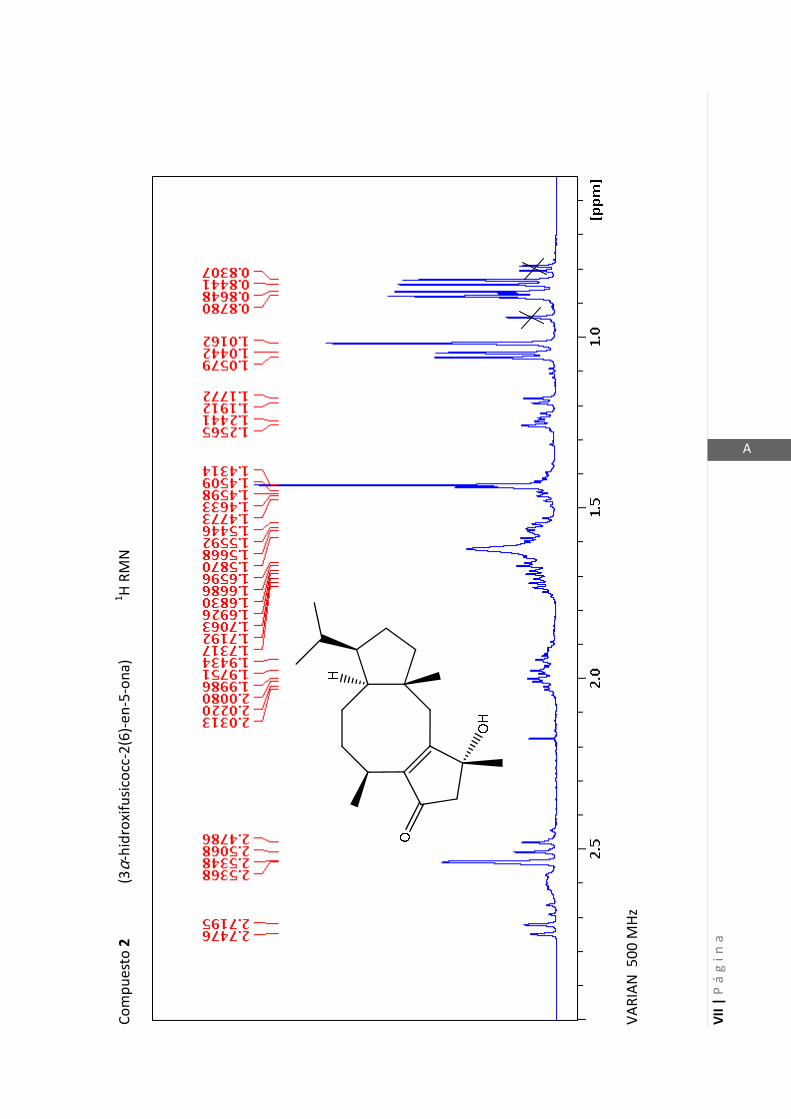
VII
| P
ág
in
a
A
Co
mp
ue
sto
2
(3
α-h
idro
xifu
sico
cc-2
(6)-
en
-5-o
na
)
1H
RM
N
VA
RIA
N 5
00
MH
z
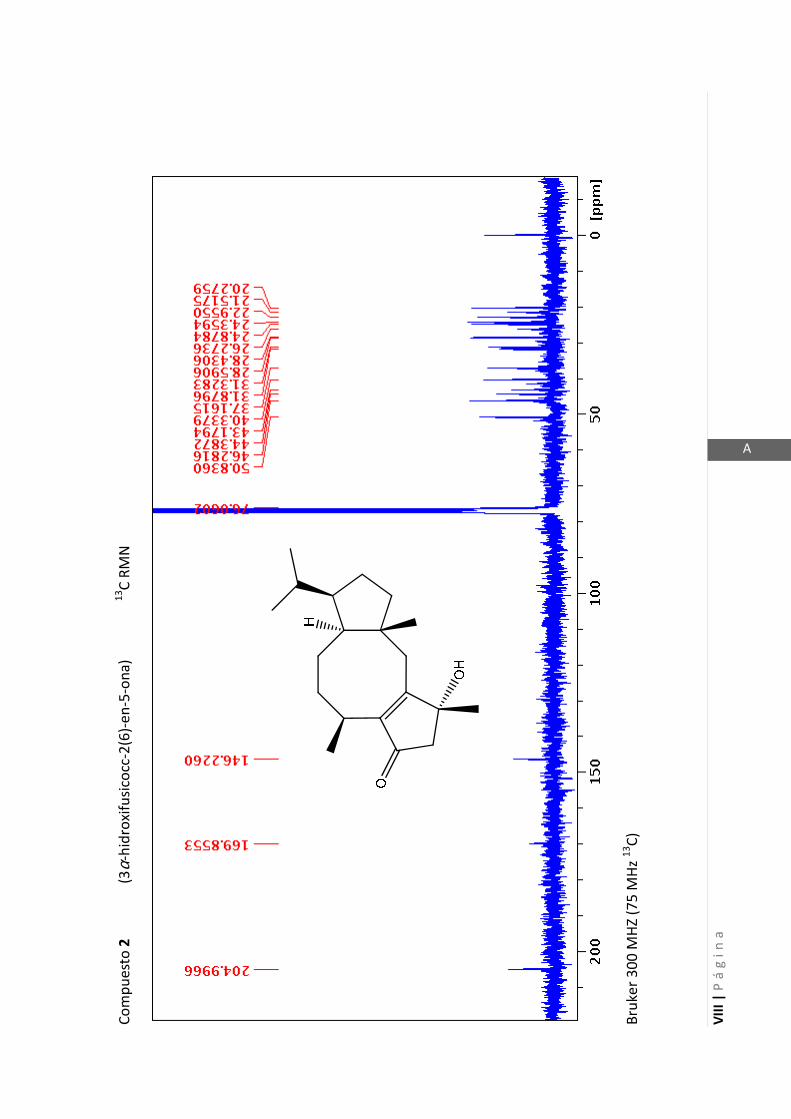
VII
I |
Pá
gi
na
A
Co
mp
ue
sto
2
(3
α-h
idro
xifu
sico
cc-2
(6)-
en
-5-o
na
)
13C
RM
N
Bru
ke
r 3
00
MH
Z (
75
MH
z 1
3C
)
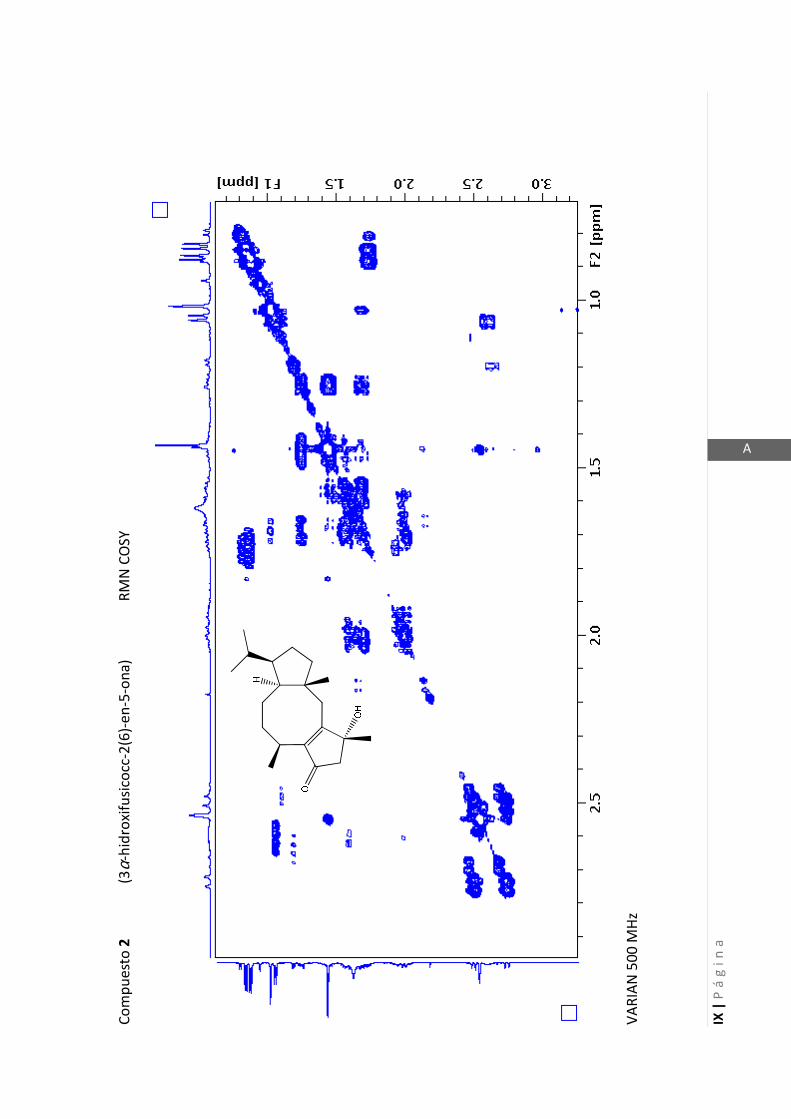
IX |
Pá
gi
na
A
Co
mp
ue
sto
2
(3
α-h
idro
xifu
sico
cc-2
(6)-
en
-5-o
na
)
RM
N C
OS
Y
VA
RIA
N 5
00
MH
z
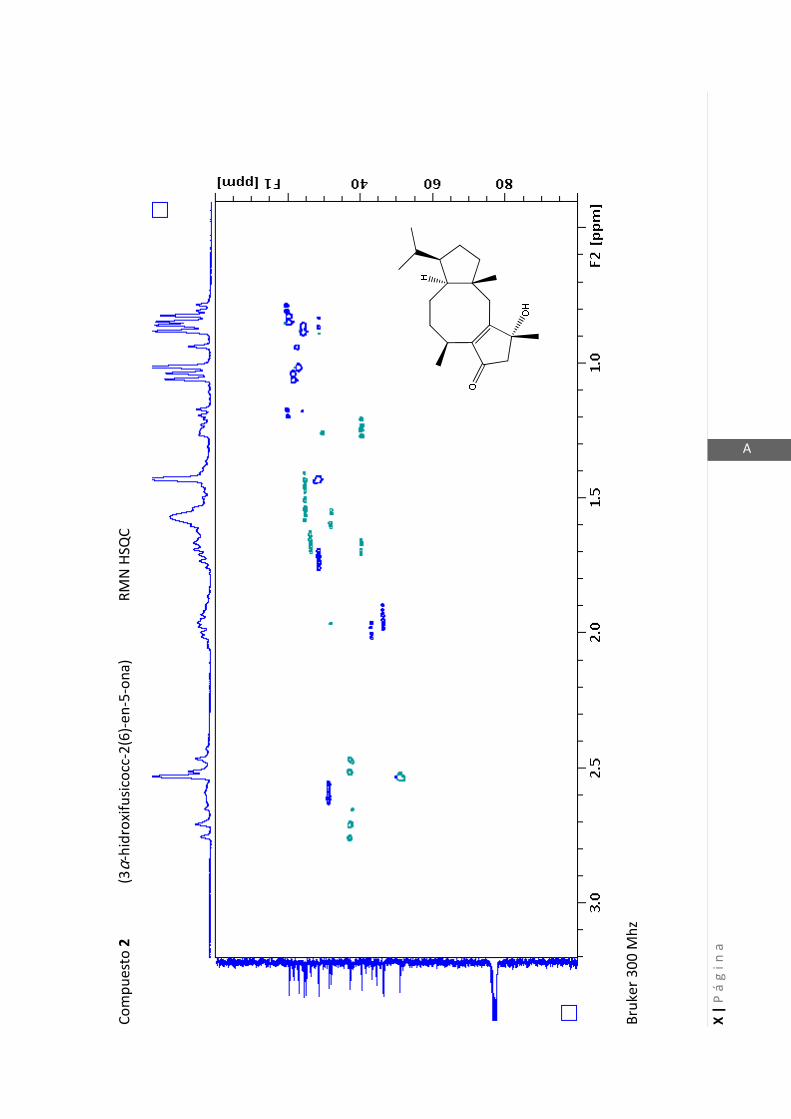
X |
Pá
gi
na
A
Co
mp
ue
sto
2
(3
α-h
idro
xifu
sico
cc-2
(6)-
en
-5-o
na
)
RM
N H
SQ
C
Bru
ke
r 3
00
Mh
z
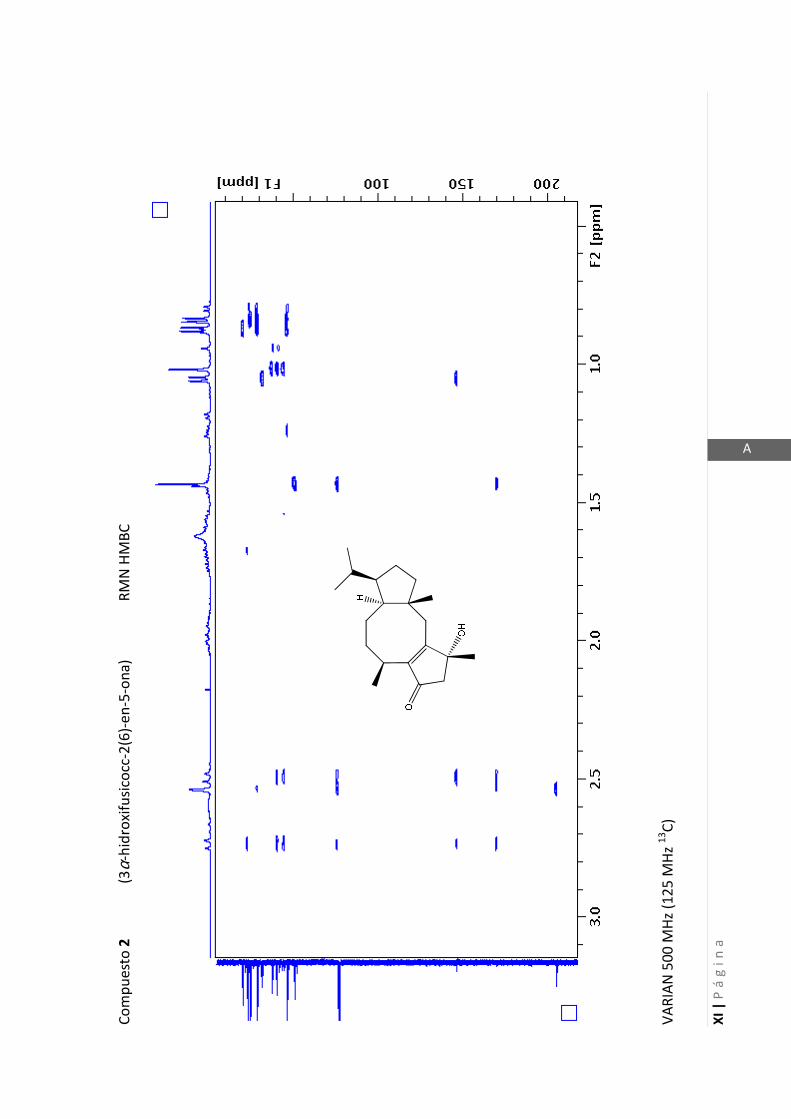
XI
| P
ág
in
a
A
Co
mp
ue
sto
2
(3
α-h
idro
xifu
sico
cc-2
(6)-
en
-5-o
na
)
RM
N H
MB
C
VA
RIA
N 5
00
MH
z (1
25
MH
z 1
3C
)
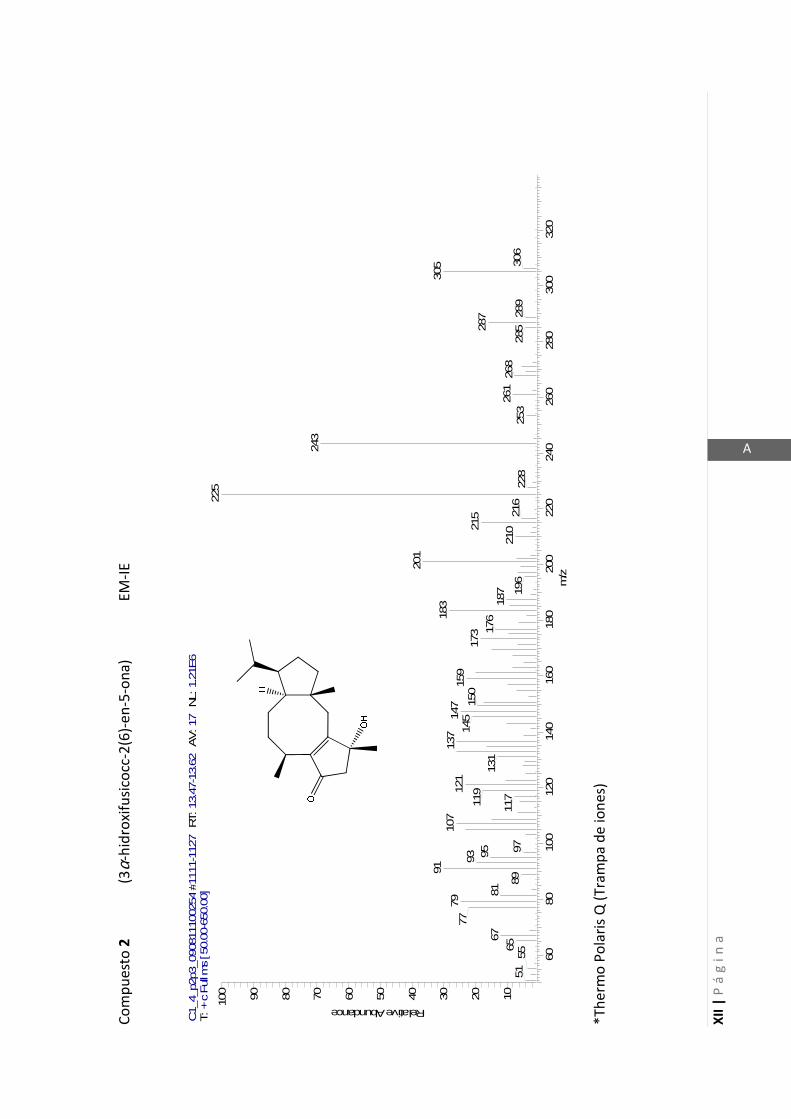
XII
| P
ág
in
a
A
Co
mp
ue
sto
2
(3
α-h
idro
xifu
sico
cc-2
(6)-
en
-5-o
na
)
EM
-IE
*T
he
rmo
Po
lari
s Q
(T
ram
pa
de
io
ne
s)
C1_
4_p2
p3_0
9081
1100
254
#111
1-11
27RT:
13.4
7-13
.62
AV:17
NL:
1.21
E6
T:+
c Full m
s [ 5
0.00
-650
.00]
6080
100
120
140
160
180
200
220
240
260
280
300
320
m/z
102030405060708090100
Relative Abundance
225
243
201
305
9118
310
713
714
779
121
159
7714
593
150
173
215
119
287
9517
613
181
6718
726
111
726
821
065
8921
697
306
196
5128
928
525
322
855
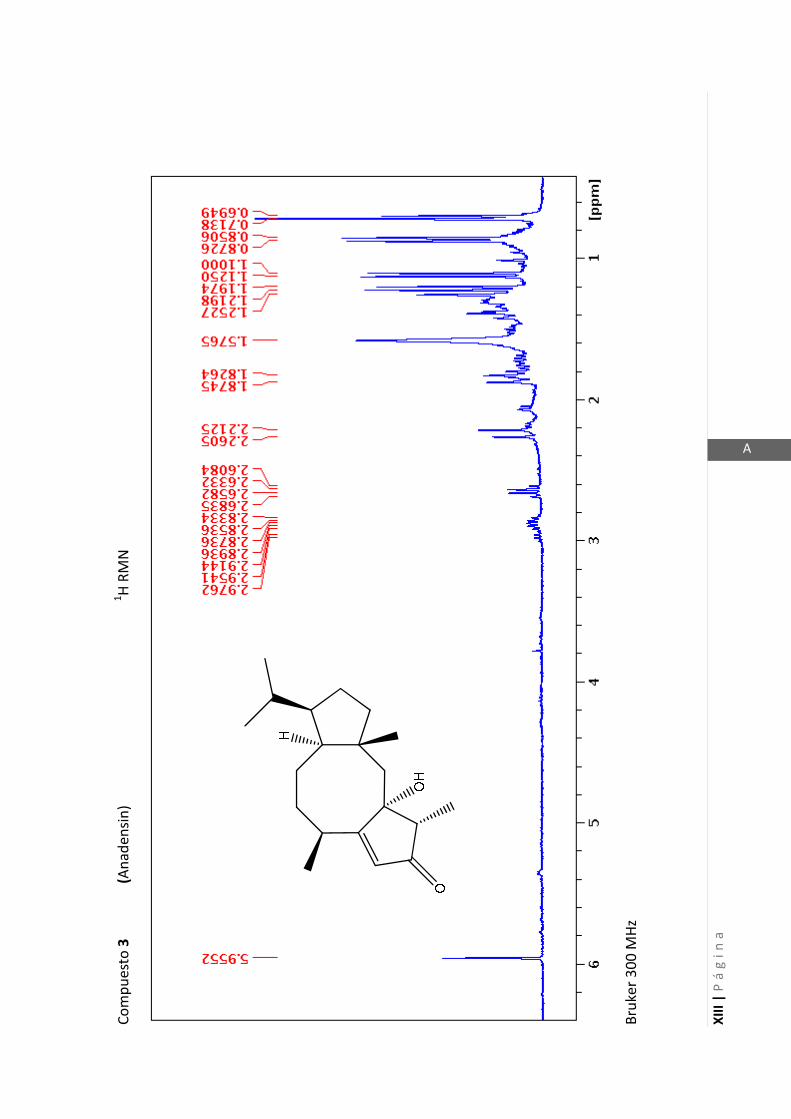
XII
I |
Pá
gi
na
A
Co
mp
ue
sto
3
(A
na
de
nsi
n)
1H
RM
N
Bru
ke
r 3
00
MH
z
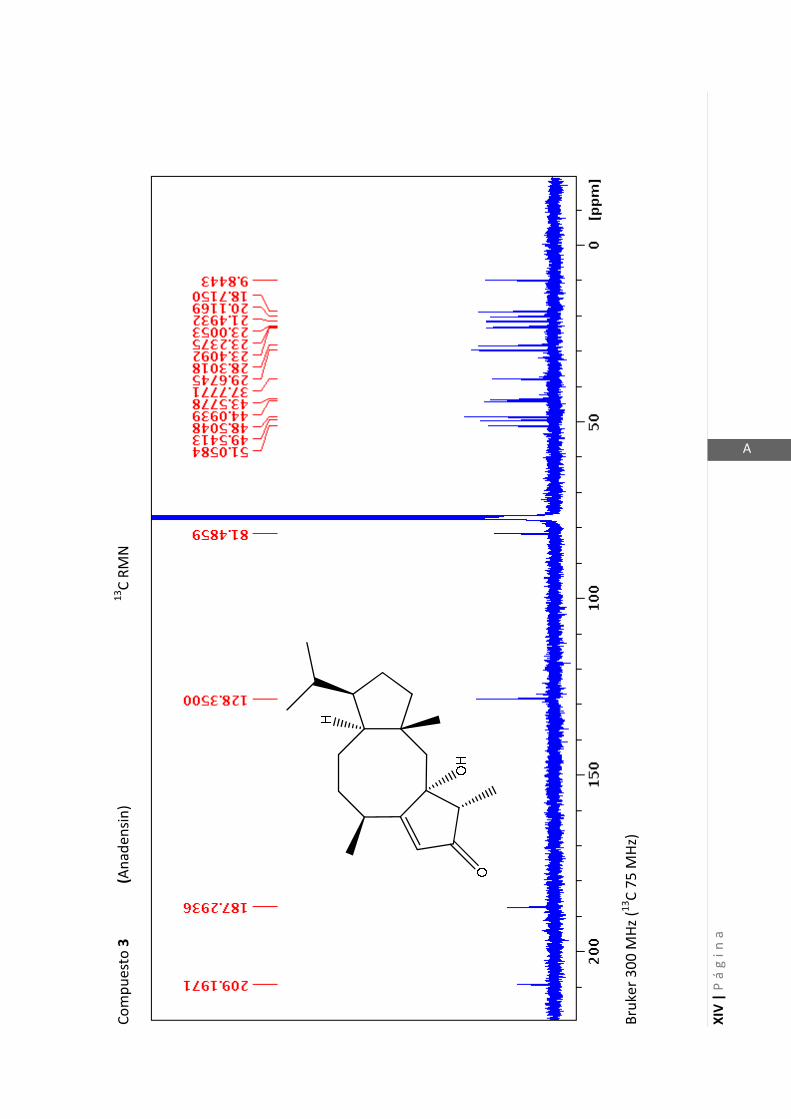
XIV
| P
ág
in
a
A
Co
mp
ue
sto
3
(A
na
de
nsi
n)
13C
RM
N
Bru
ke
r 3
00
MH
z (1
3C
75
MH
z)
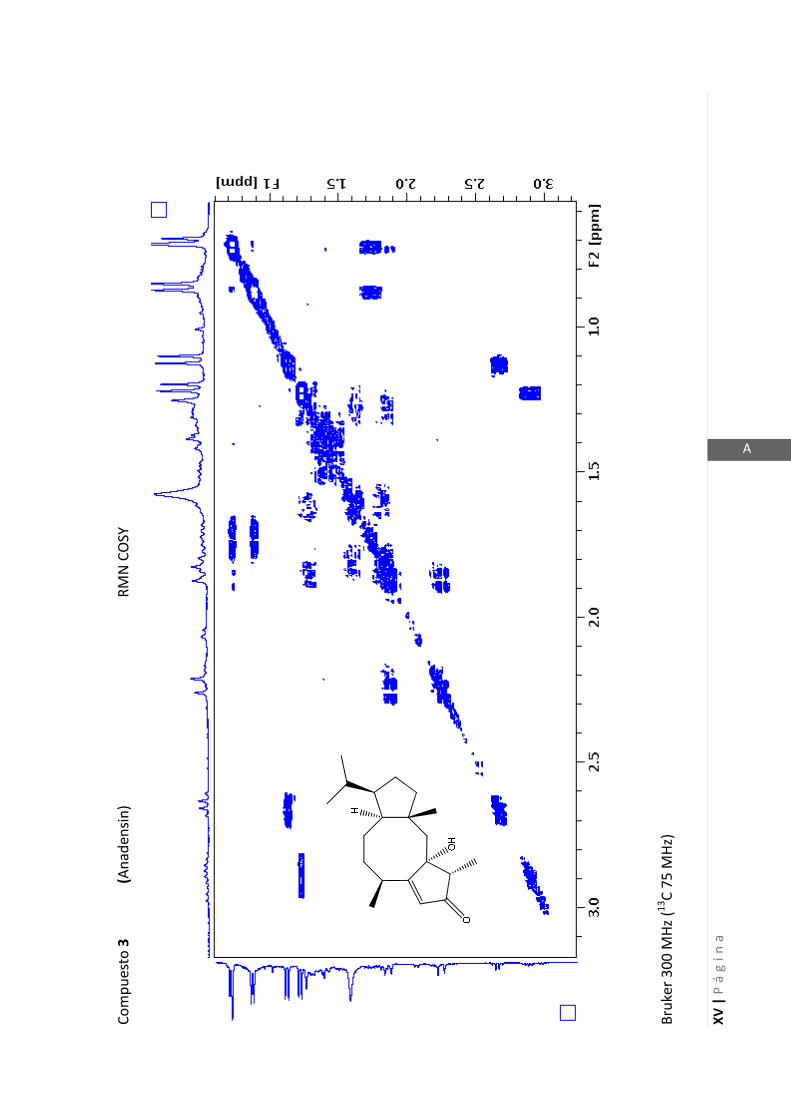
XV
| P
ág
in
a
A
Co
mp
ue
sto
3
(A
na
de
nsi
n)
RM
N C
OS
Y
Bru
ke
r 3
00
MH
z (1
3C
75
MH
z)
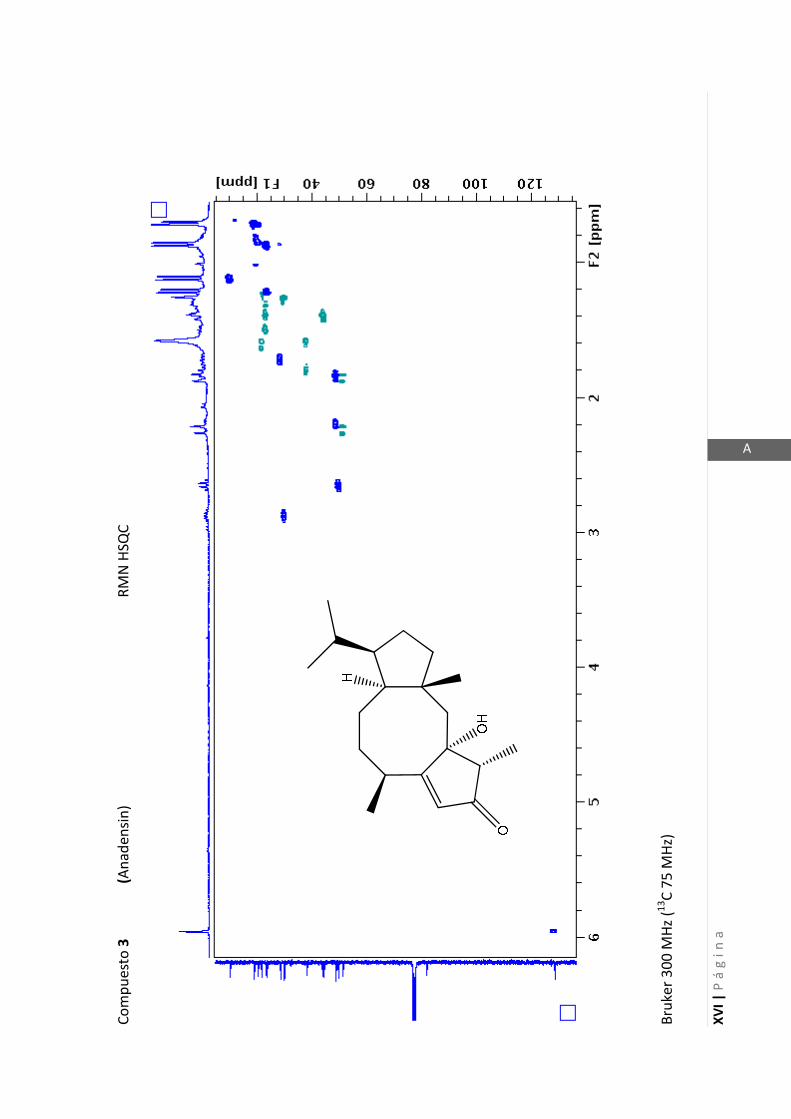
XV
I |
Pá
gi
na
A
Co
mp
ue
sto
3
(A
na
de
nsi
n)
RM
N H
SQ
C
Bru
ke
r 3
00
MH
z (1
3C
75
MH
z)
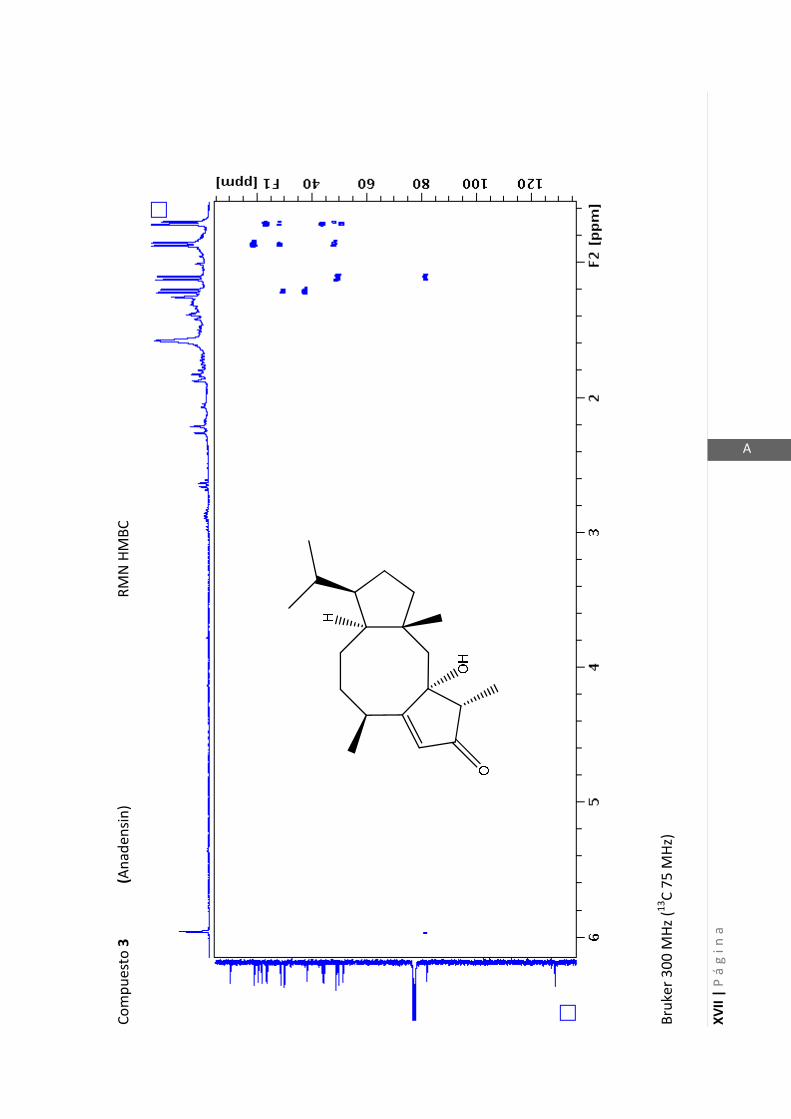
XV
II |
Pá
gi
na
A
Co
mp
ue
sto
3
(A
na
de
nsi
n)
RM
N H
MB
C
Bru
ke
r 3
00
MH
z (1
3C
75
MH
z)
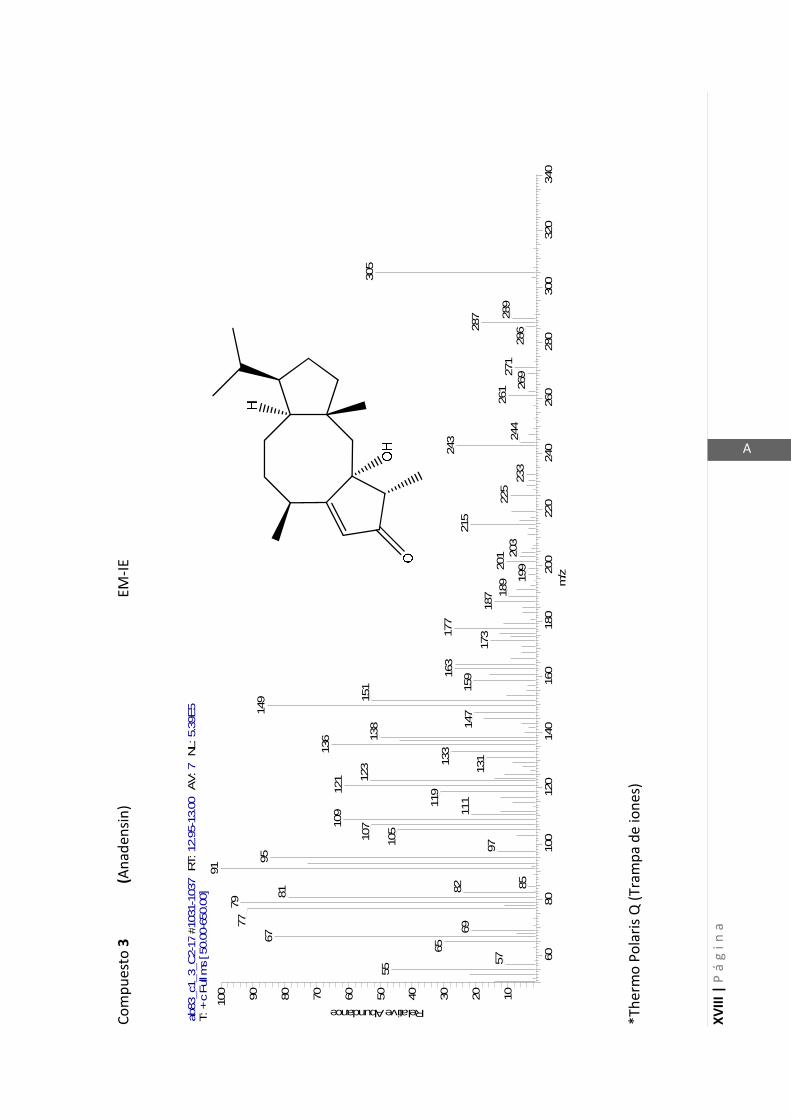
XV
III
| P
ág
in
a
A
Co
mp
ue
sto
3
(A
na
de
nsi
n)
EM
-IE
*T
he
rmo
Po
lari
s Q
(T
ram
pa
de
io
ne
s)
ab83
_c1_
3_C
2-17
#10
31-1
037
RT:
12.9
5-13
.00
AV:7
NL:
5.39
E5
T:+
c Full m
s [ 5
0.00
-650
.00]
6080
100
120
140
160
180
200
220
240
260
280
300
320
340
m/z
102030405060708090100
Relative Abundance
91
7977
149
9567
81
136
109
121 12
315
110
730
513
855
105
119
6513
317
716
324
382
215
111
6915
914
728
713
117
318
797
5720
126
118
922
528
927
120
324
423
328
626
919
985
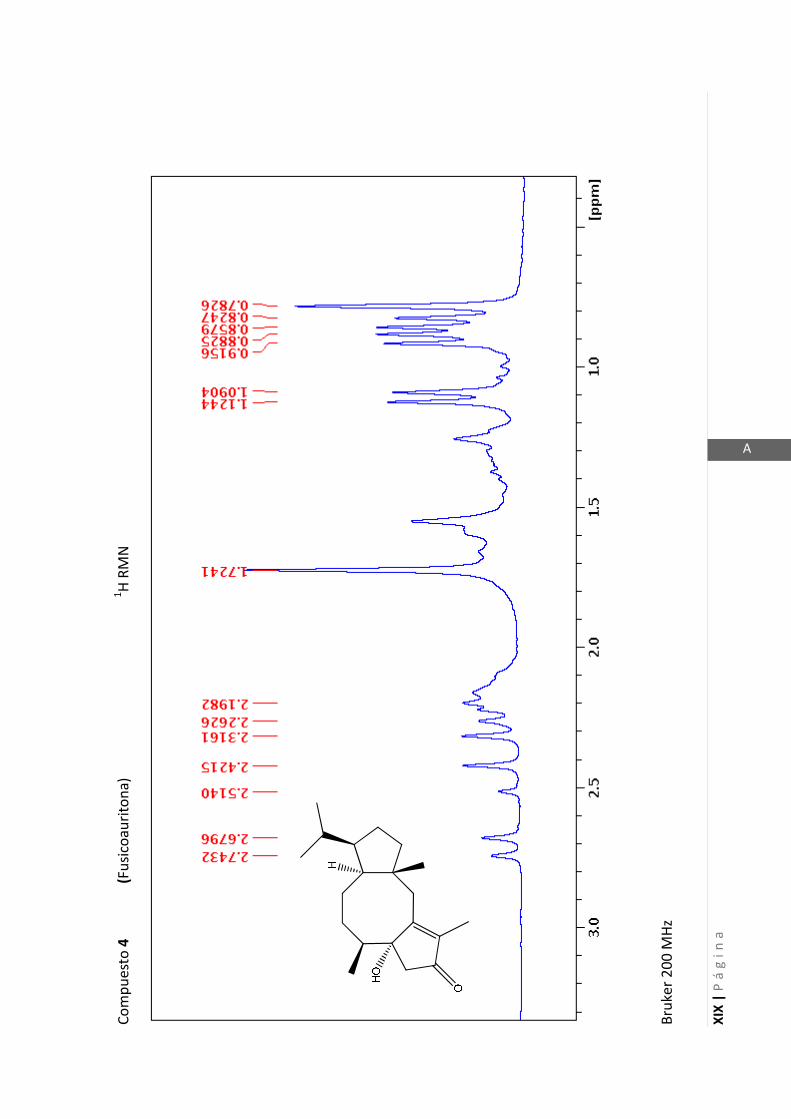
XIX
| P
ág
in
a
A
Co
mp
ue
sto
4
(F
usi
coa
uri
ton
a)
1
H R
MN
Bru
ke
r 2
00
MH
z
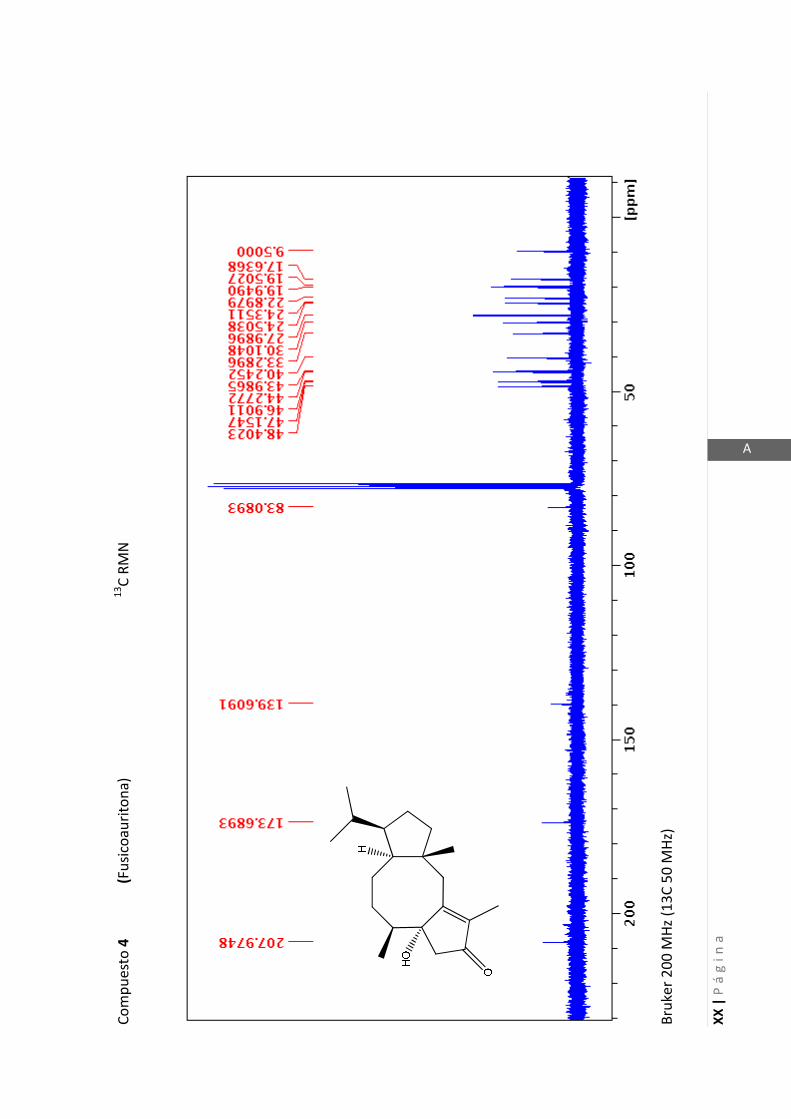
XX
| P
ág
in
a
A
Co
mp
ue
sto
4
(F
usi
coa
uri
ton
a)
1
3C
RM
N
Bru
ke
r 2
00
MH
z (1
3C
50
MH
z)
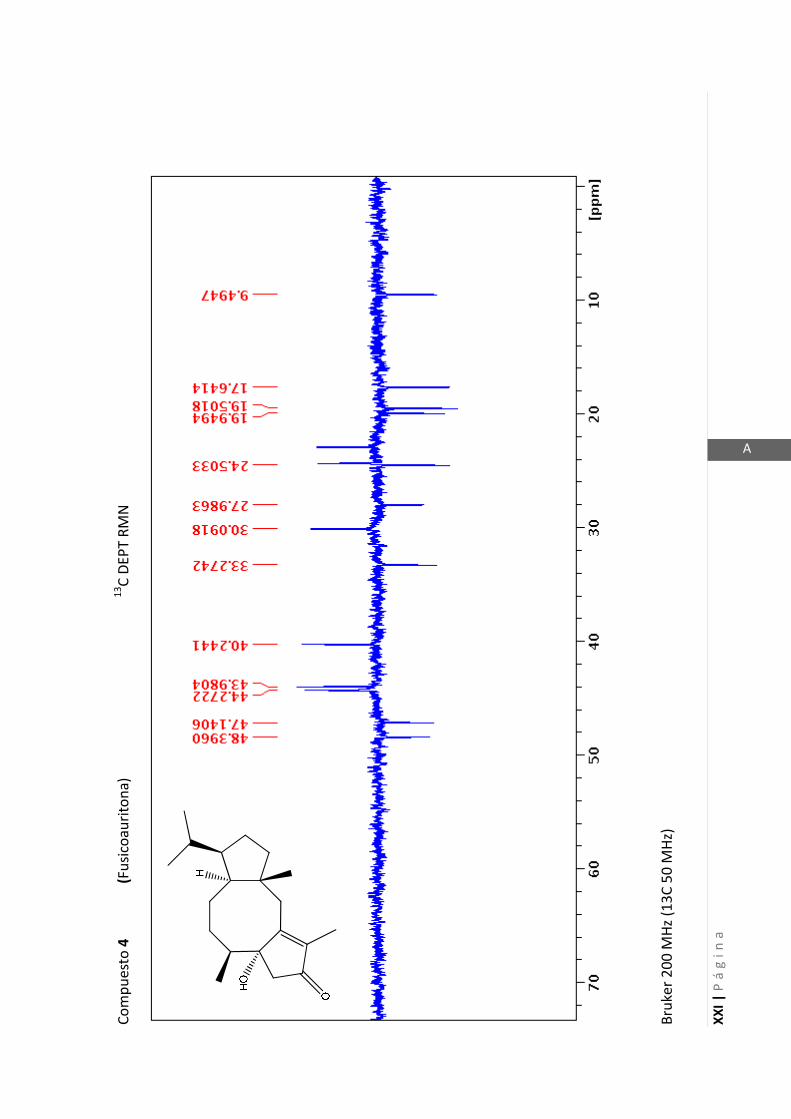
XX
I |
Pá
gi
na
A
Co
mp
ue
sto
4
(F
usi
coa
uri
ton
a)
1
3C
DE
PT
RM
N
Bru
ke
r 2
00
MH
z (1
3C
50
MH
z)
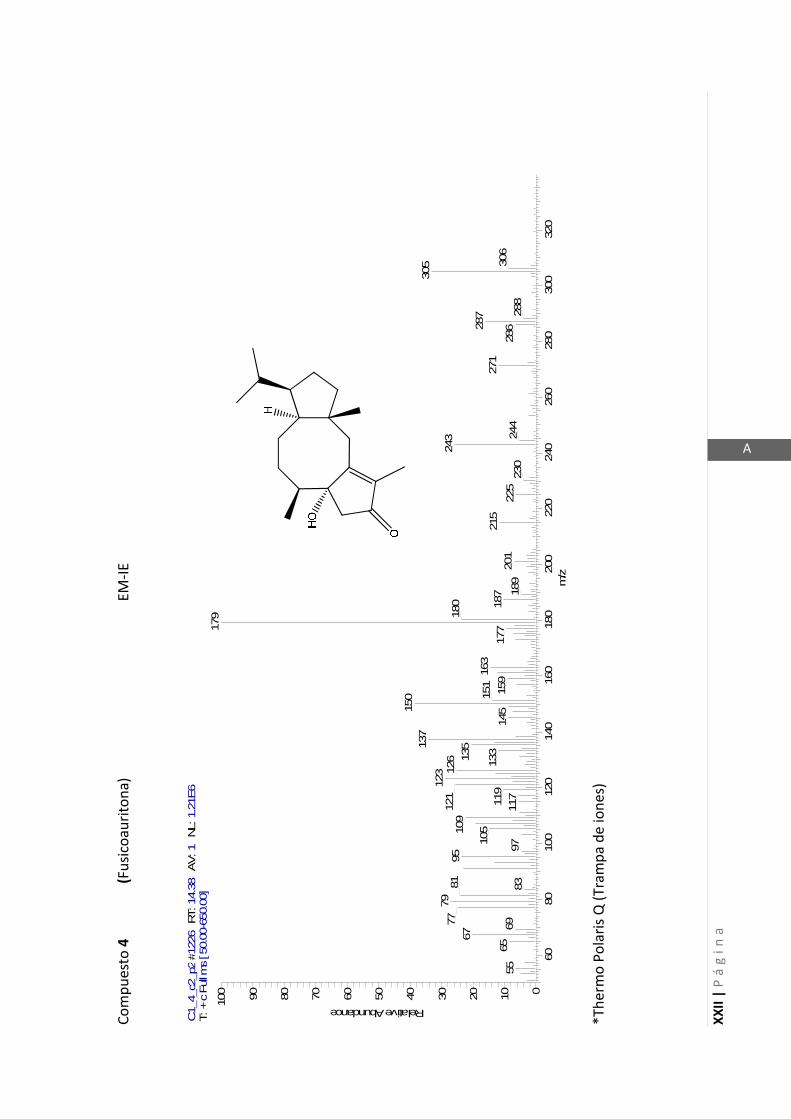
XX
II |
Pá
gi
na
A
Co
mp
ue
sto
4
(F
usi
coa
uri
ton
a)
E
M-I
E
*T
he
rmo
Po
lari
s Q
(T
ram
pa
de
io
ne
s)
C1_
4_c2
_p2
#122
6RT:
14.3
8AV:1
NL:
1.21
E6
T:+
c Full m
s [ 5
0.00
-650
.00]
6080
100
120
140
160
180
200
220
240
260
280
300
320
m/z
0102030405060708090100
Relative Abundance
179
150
137
305
123
7924
312
112
677
8118
095
109
135
6728
710
516
315
127
113
321
511
918
717
715
914
530
665
201
6928
622
555
117
244
9718
928
823
083
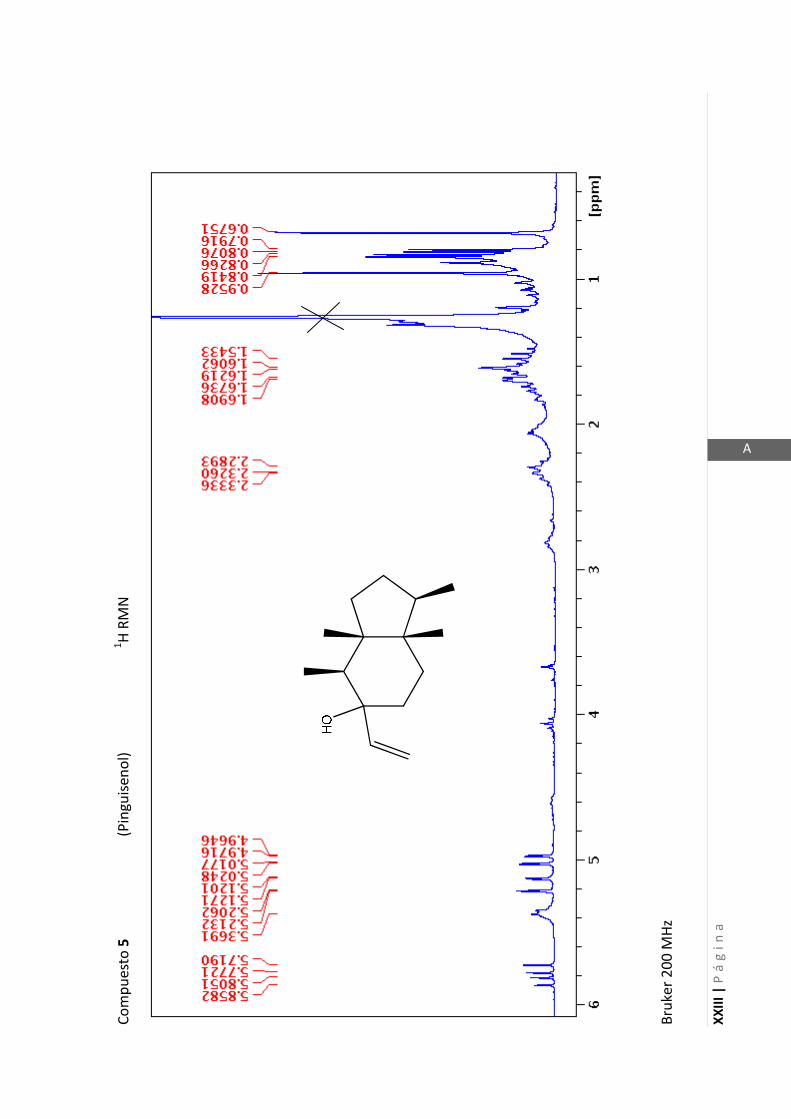
XX
III
| P
ág
in
a
A
Co
mp
ue
sto
5
(Pin
gu
ise
no
l)
1H
RM
N
Bru
ke
r 2
00
MH
z
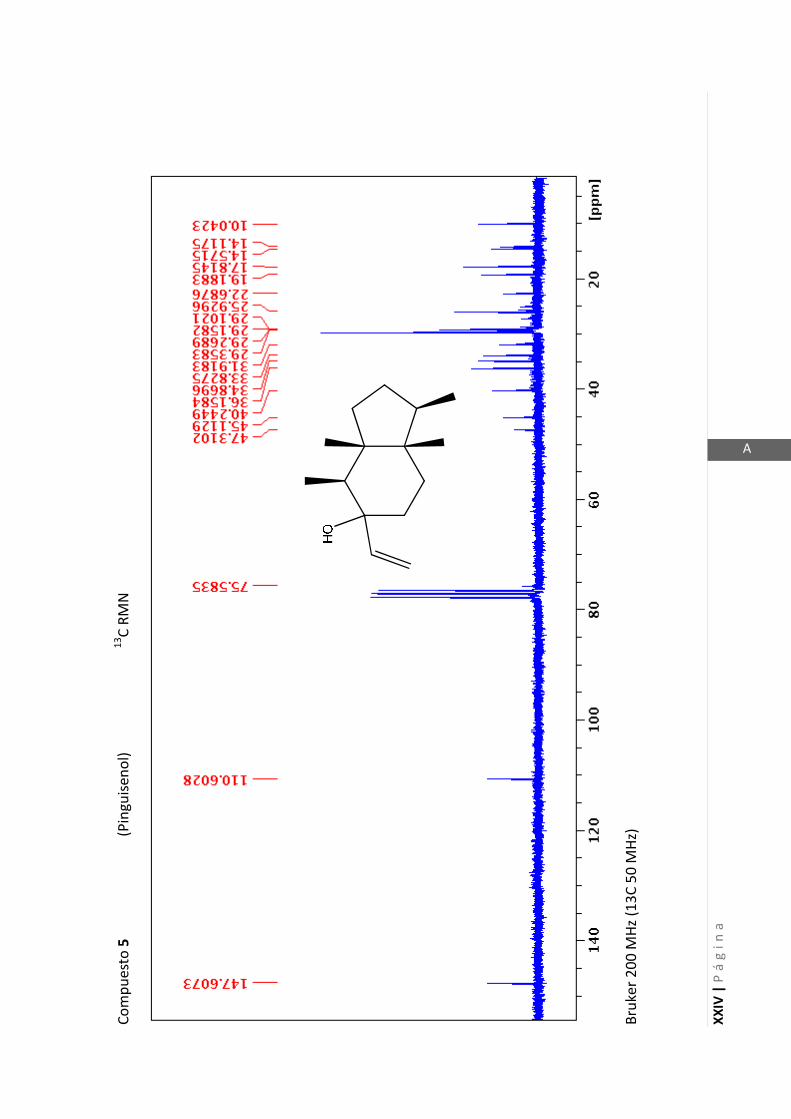
XX
IV |
Pá
gi
na
A
Co
mp
ue
sto
5
(Pin
gu
ise
no
l)
13C
RM
N
Bru
ke
r 2
00
MH
z (1
3C
50
MH
z)
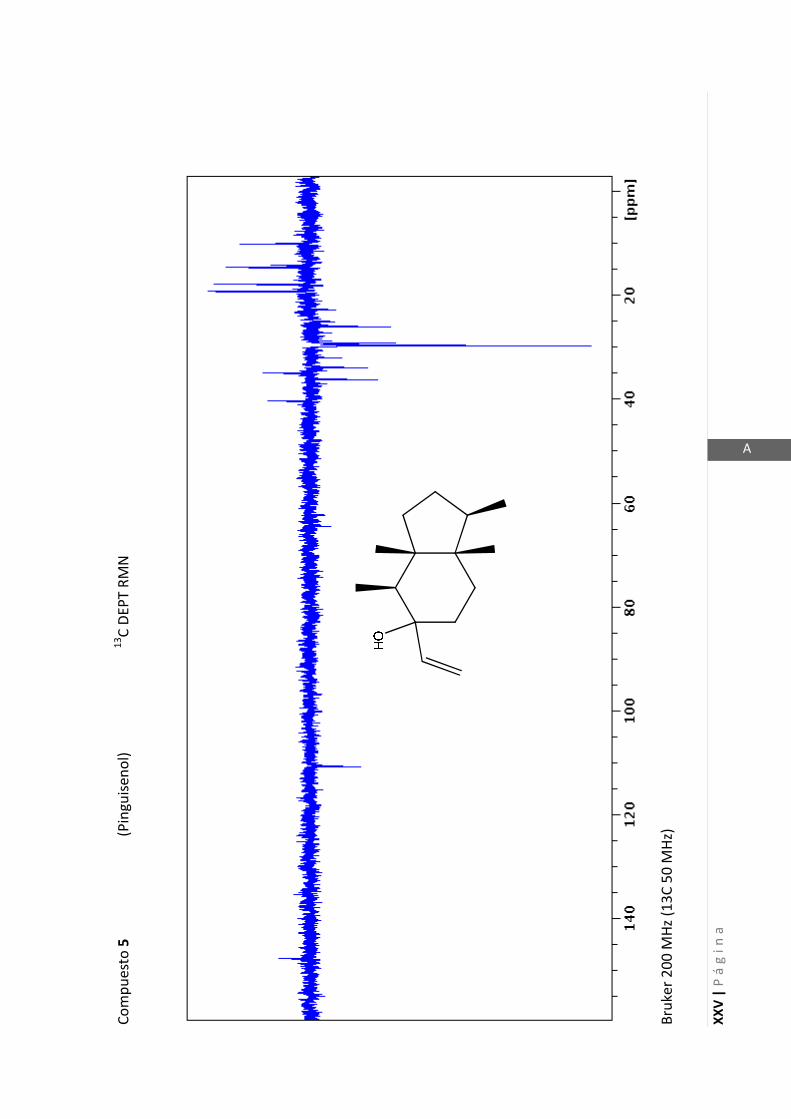
XX
V |
Pá
gi
na
A
Co
mp
ue
sto
5
(Pin
gu
ise
no
l)
13C
DE
PT
RM
N
Bru
ke
r 2
00
MH
z (1
3C
50
MH
z)
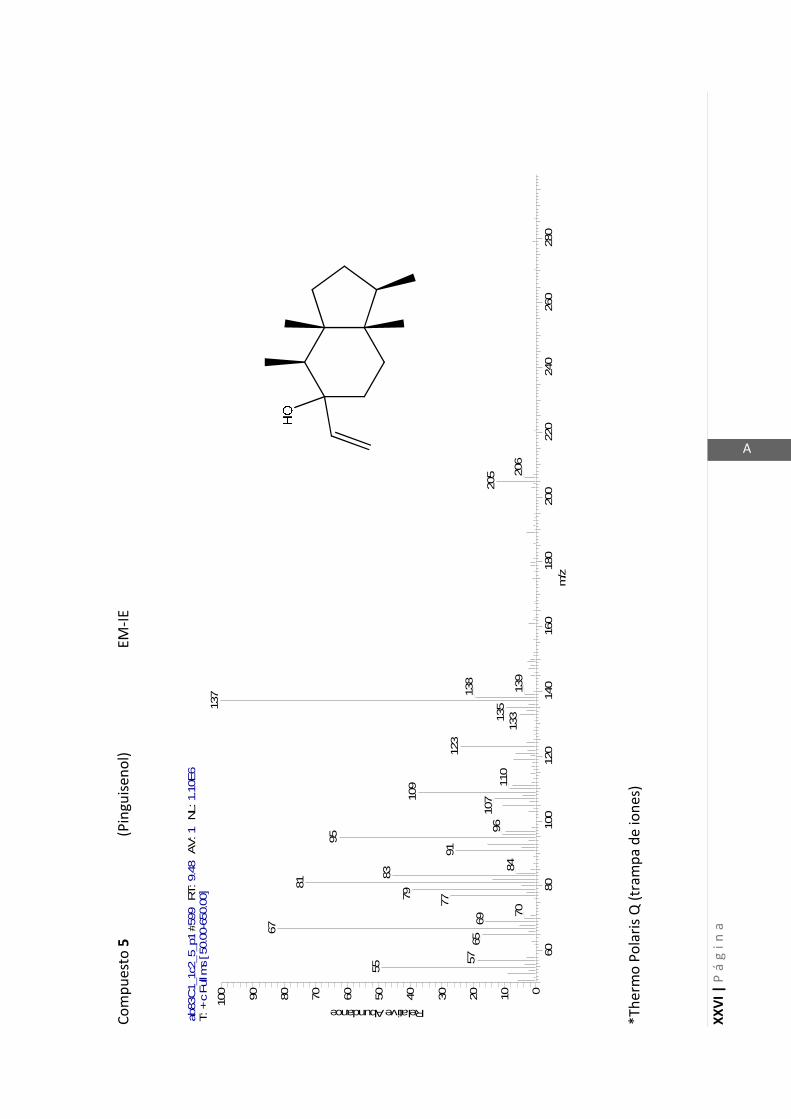
XX
VI
| P
ág
in
a
A
Co
mp
ue
sto
5
(Pin
gu
ise
no
l)
EM
-IE
*T
he
rmo
Po
lari
s Q
(tr
am
pa
de
io
ne
s)
ab83
C1_
1c2_
5_p1
#59
9RT:
9.48
AV:1
NL:
1.10
E6
T:+
c Full m
s [ 5
0.00
-650
.00]
6080
100
120
140
160
180
200
220
240
260
280
m/z
0102030405060708090100
Relative Abundance
137
67
81
95
5583
7910
9
7791
123
138
5765
6910
720
596
135
110
8413
320
670
139
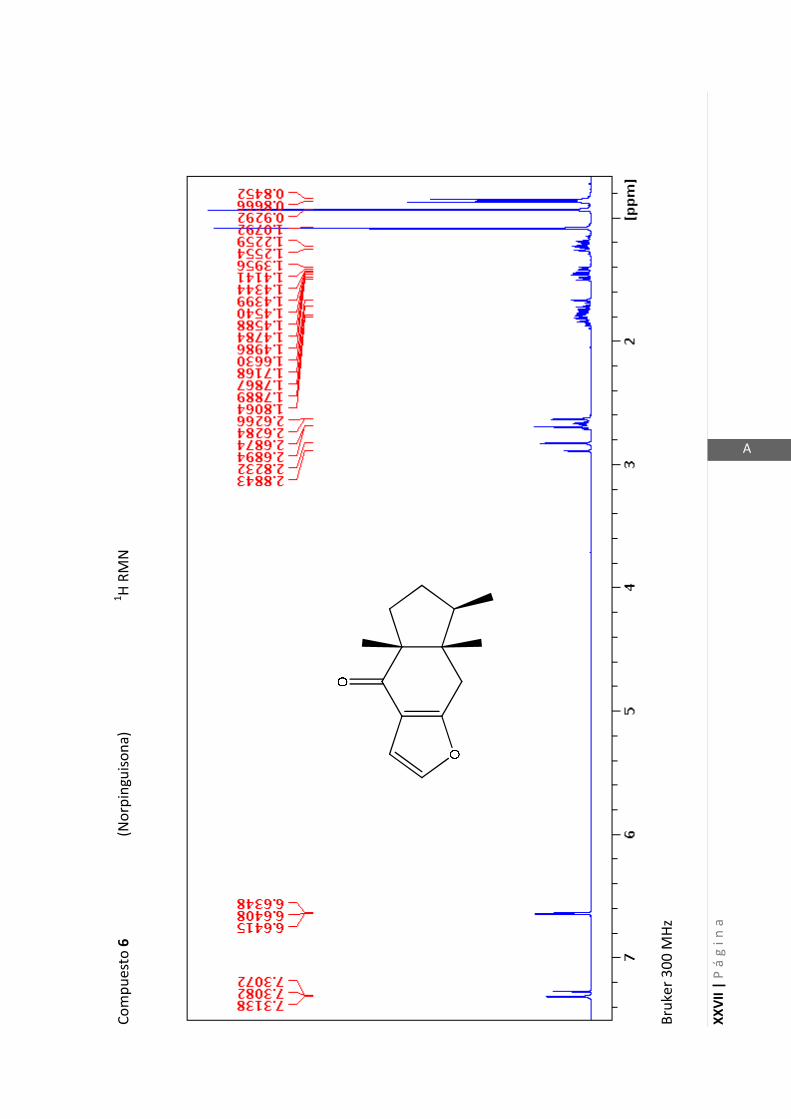
XX
VII
| P
ág
in
a
A
Co
mp
ue
sto
6
(No
rpin
gu
iso
na
)
1H
RM
N
Bru
ke
r 3
00
MH
z
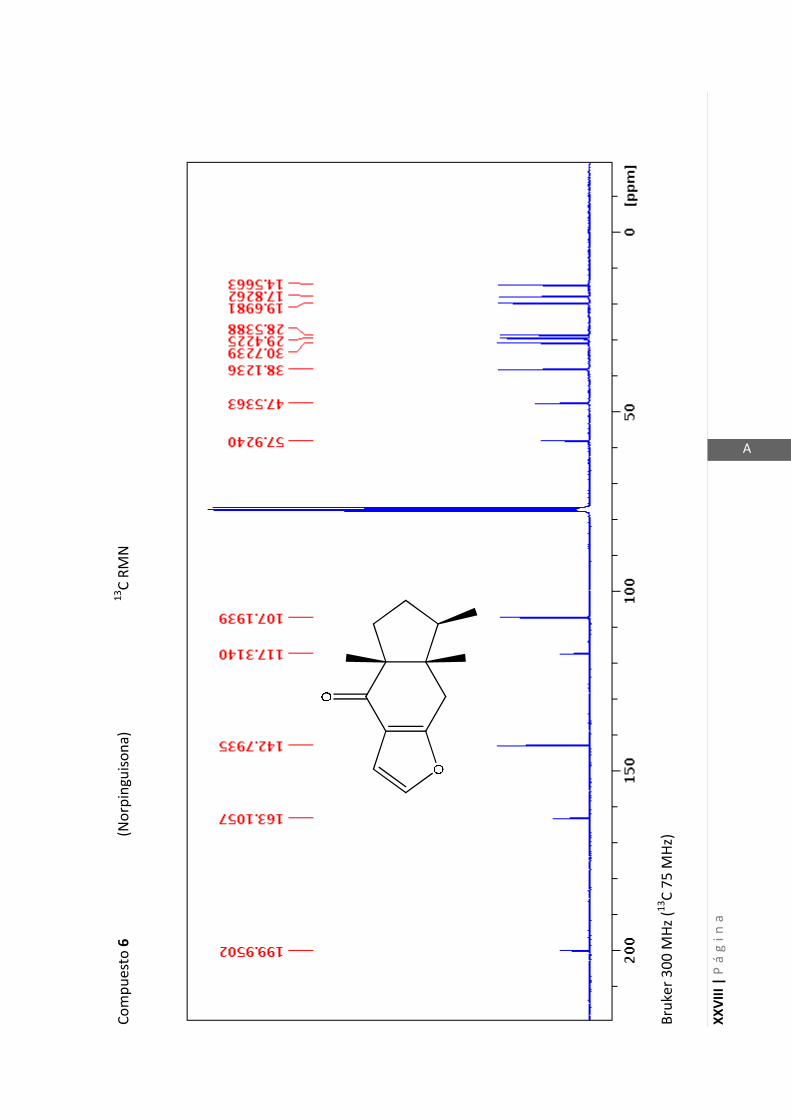
XX
VII
I |
Pá
gi
na
A
Co
mp
ue
sto
6
(No
rpin
gu
iso
na
)
1
3C
RM
N
Bru
ke
r 3
00
MH
z (1
3C
75
MH
z)
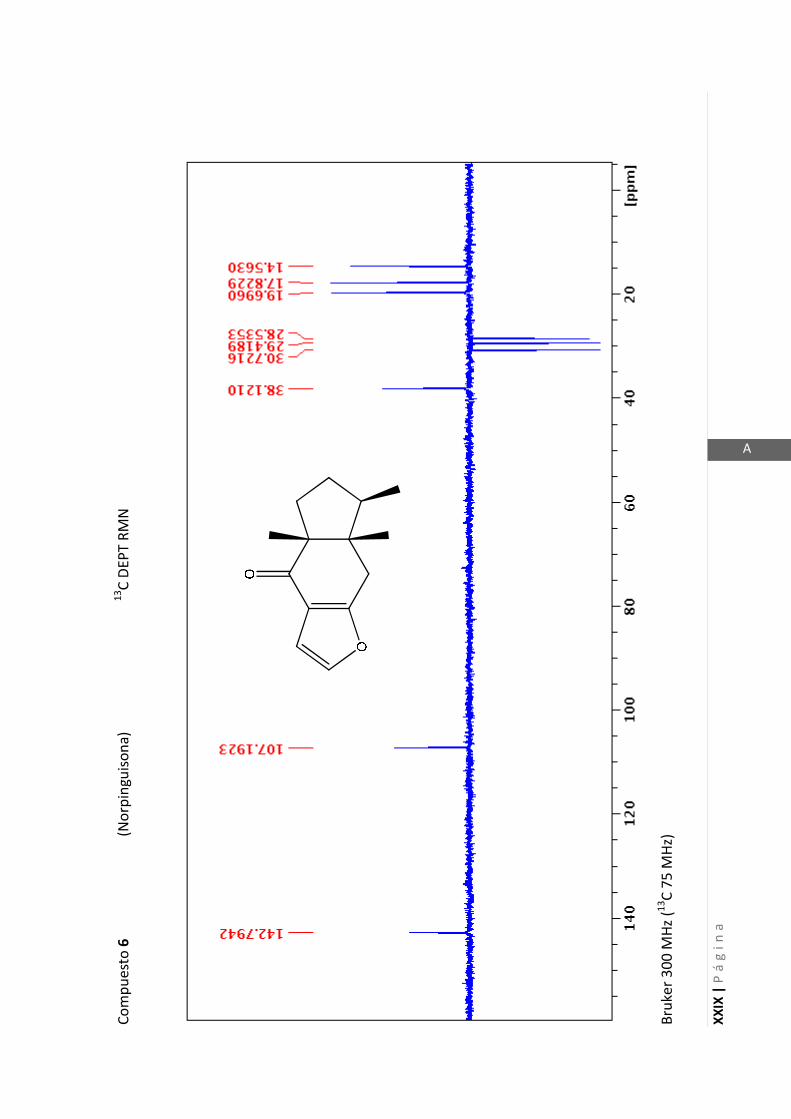
XX
IX |
Pá
gi
na
A
Co
mp
ue
sto
6
(No
rpin
gu
iso
na
)
1
3C
DE
PT
RM
N
Bru
ke
r 3
00
MH
z (1
3C
75
MH
z)
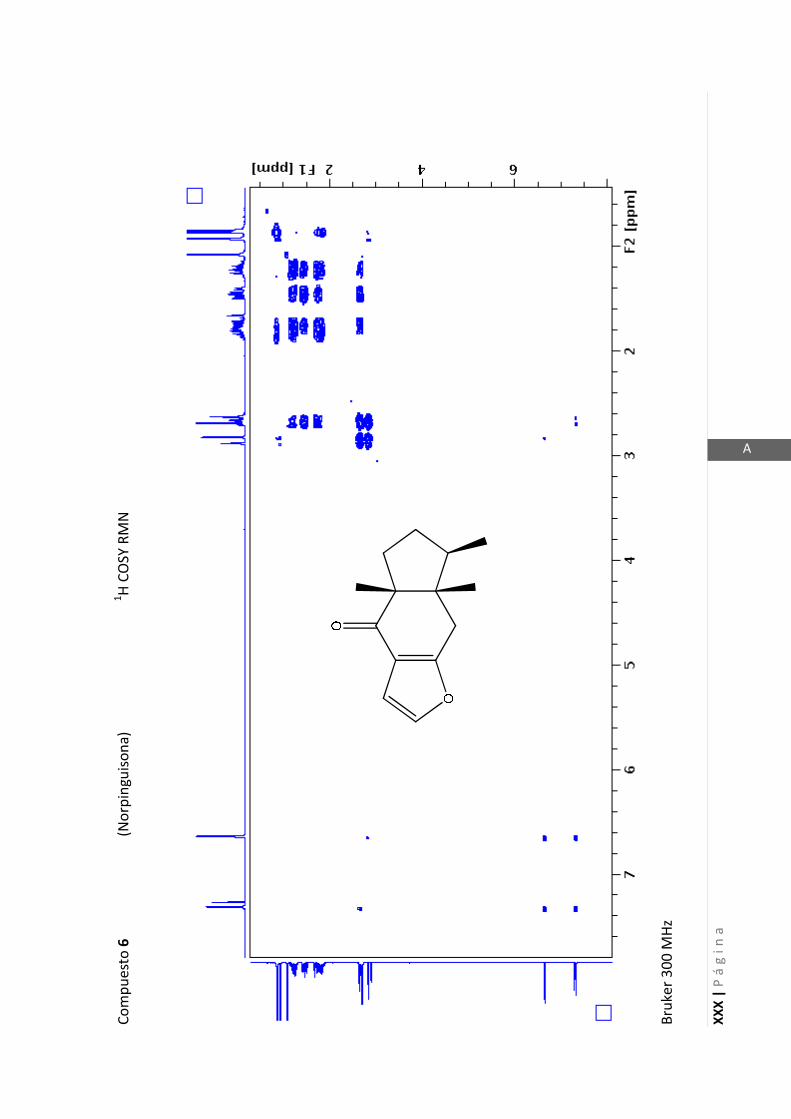
XX
X |
Pá
gi
na
A
Co
mp
ue
sto
6
(No
rpin
gu
iso
na
)
1H
CO
SY
RM
N
Bru
ke
r 3
00
MH
z
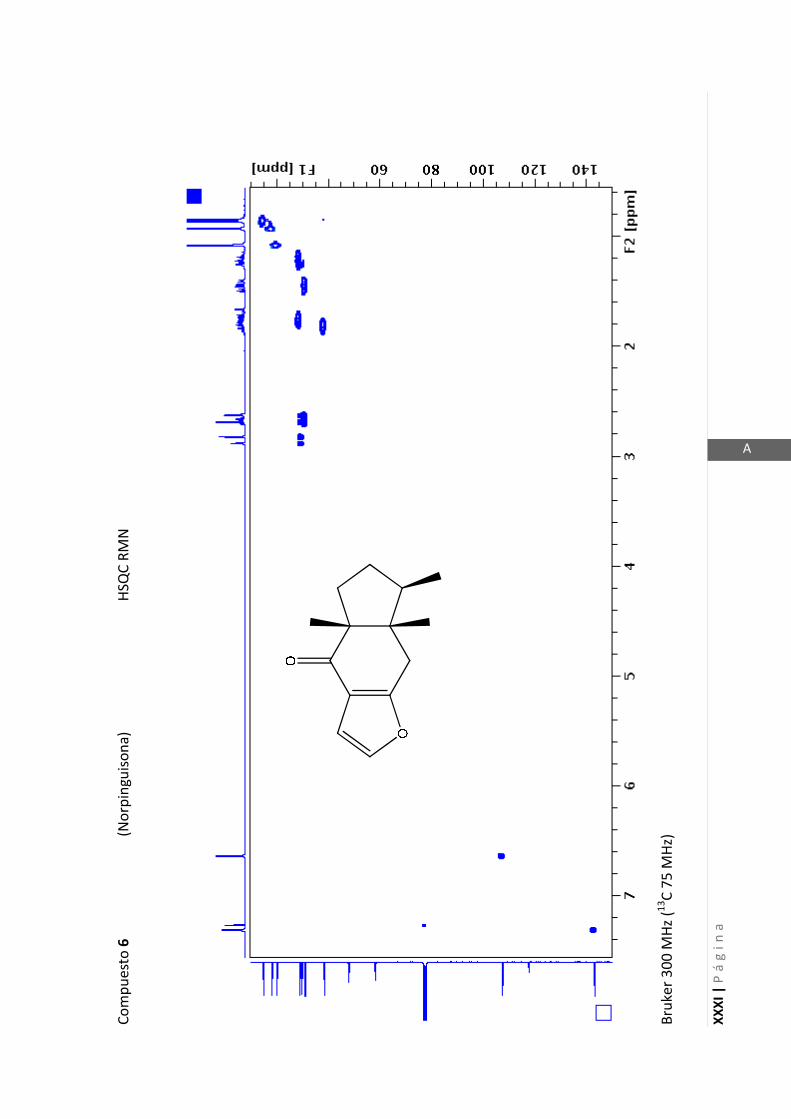
XX
XI
| P
ág
in
a
A
Co
mp
ue
sto
6
(No
rpin
gu
iso
na
)
H
SQ
C R
MN
Bru
ke
r 3
00
MH
z (1
3C
75
MH
z)
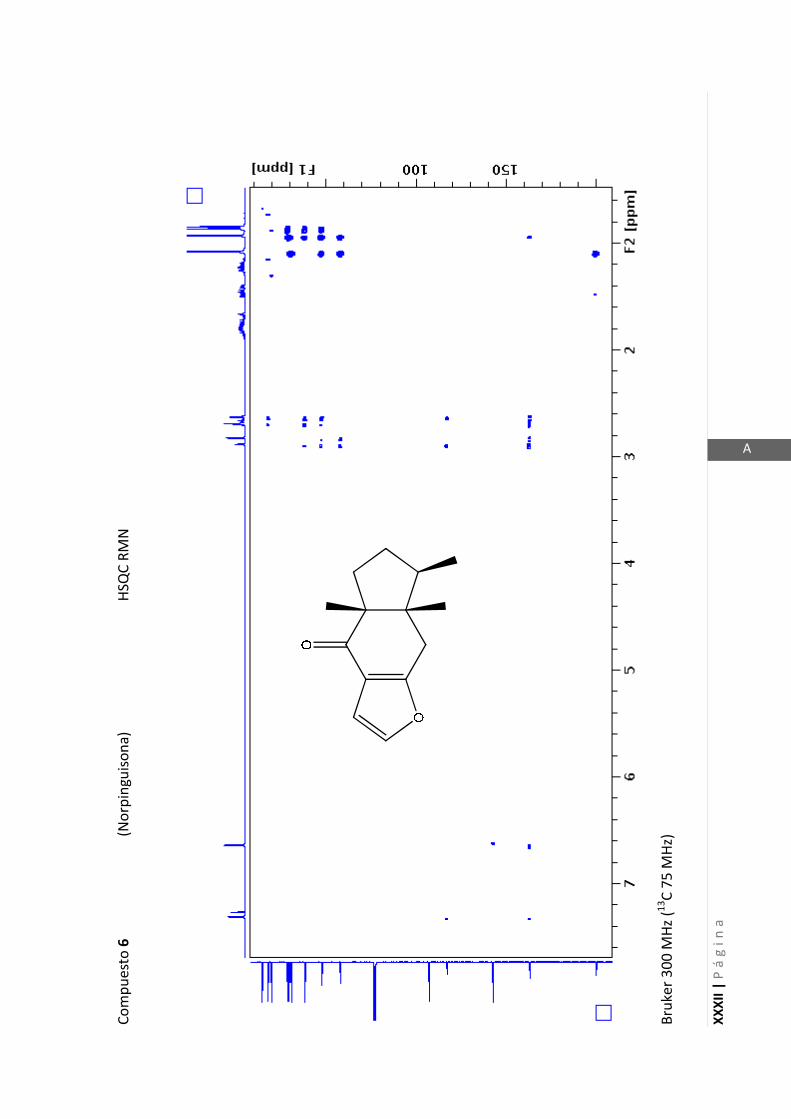
XX
XII
| P
ág
in
a
A
Co
mp
ue
sto
6
(No
rpin
gu
iso
na
)
H
SQ
C R
MN
Bru
ke
r 3
00
MH
z (1
3C
75
MH
z)
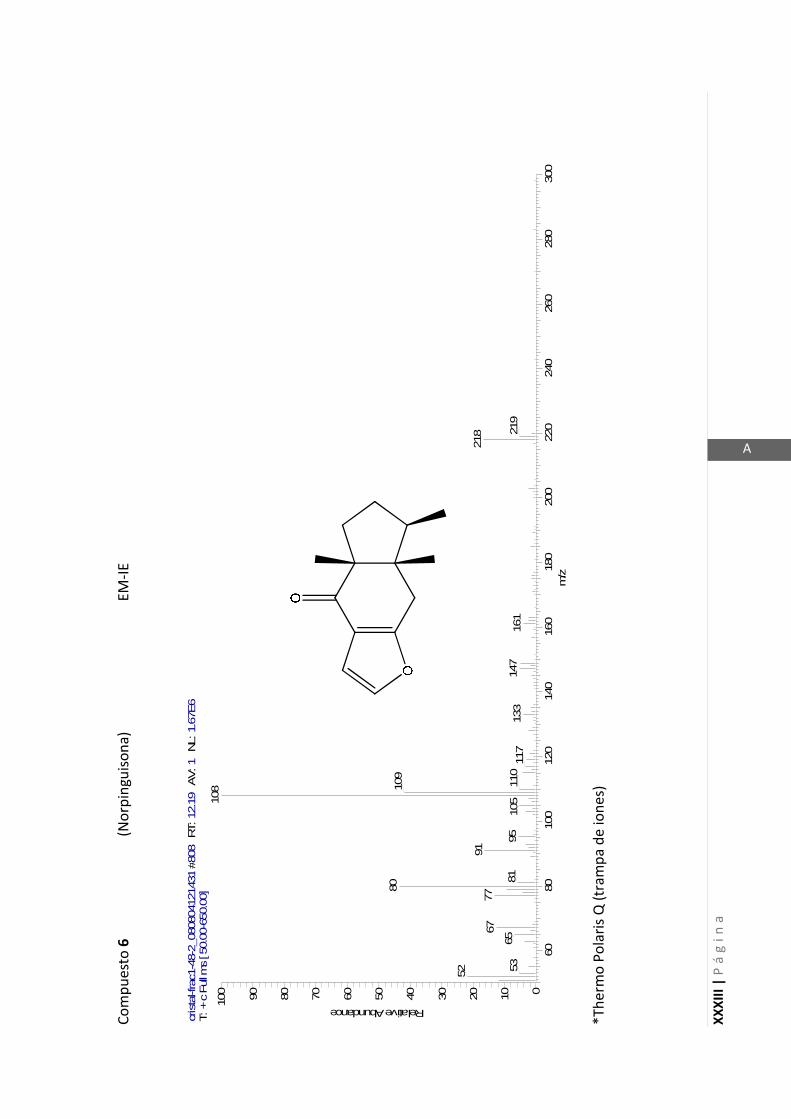
XX
XII
I |
Pá
gi
na
A
Co
mp
ue
sto
6
(No
rpin
gu
iso
na
)
E
M-I
E
*T
he
rmo
Po
lari
s Q
(tr
am
pa
de
io
ne
s)
cris
tal-f
rac1
-48-
2_08
0804
12143
1 #8
08RT:
12.1
9AV:1
NL:
1.67
E6
T:+
c Full m
s [ 5
0.00
-650
.00]
6080
100
120
140
160
180
200
220
240
260
280
300
m/z
0102030405060708090100
Relative Abundance
108
8010
9
52
218
9177
67
6581
9514
711
021
953
105
133
161
117
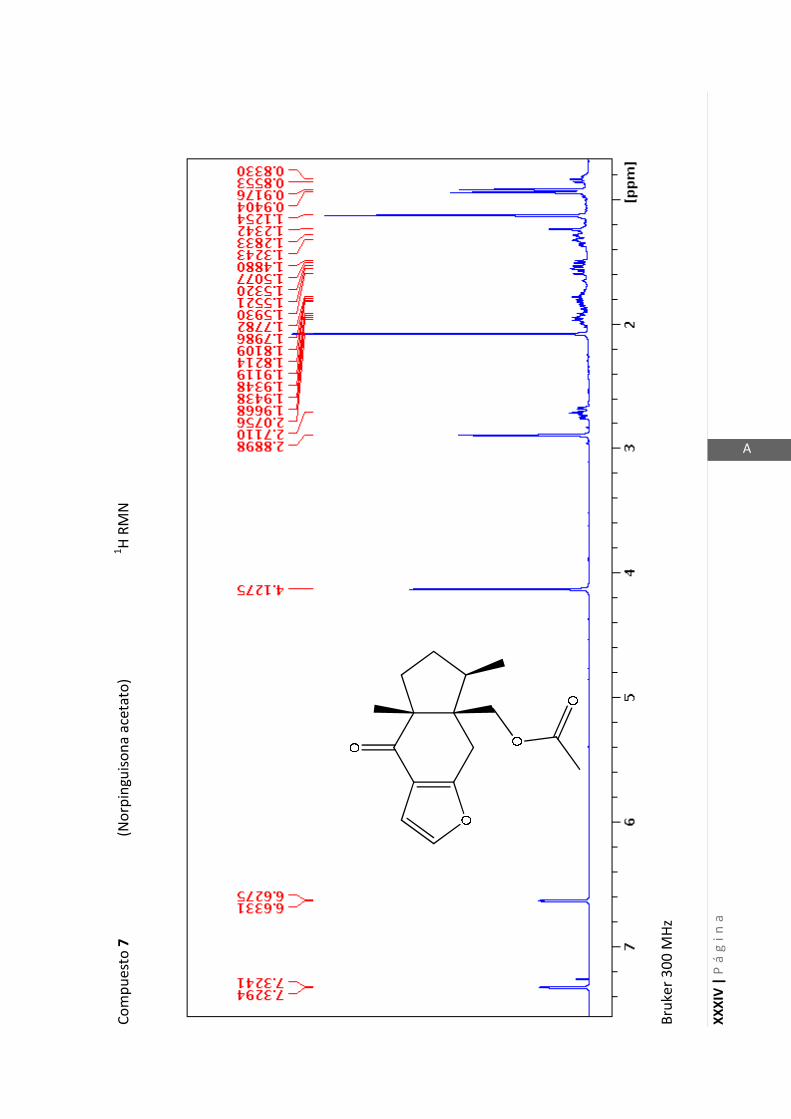
XX
XIV
| P
ág
in
a
A
Co
mp
ue
sto
7
(No
rpin
gu
iso
na
ace
tato
)
1H
RM
N
Bru
ke
r 3
00
MH
z
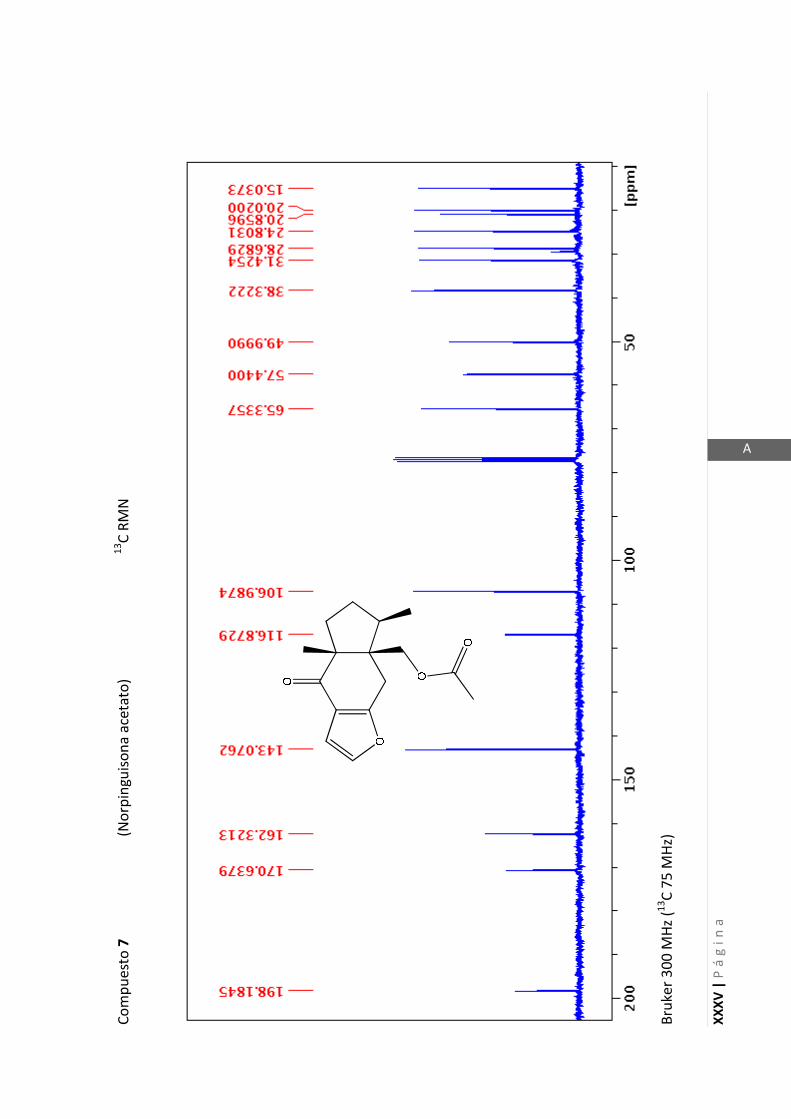
XX
XV
| P
ág
in
a
A
Co
mp
ue
sto
7
(No
rpin
gu
iso
na
ace
tato
)
1
3C
RM
N
Bru
ke
r 3
00
MH
z (1
3C
75
MH
z)
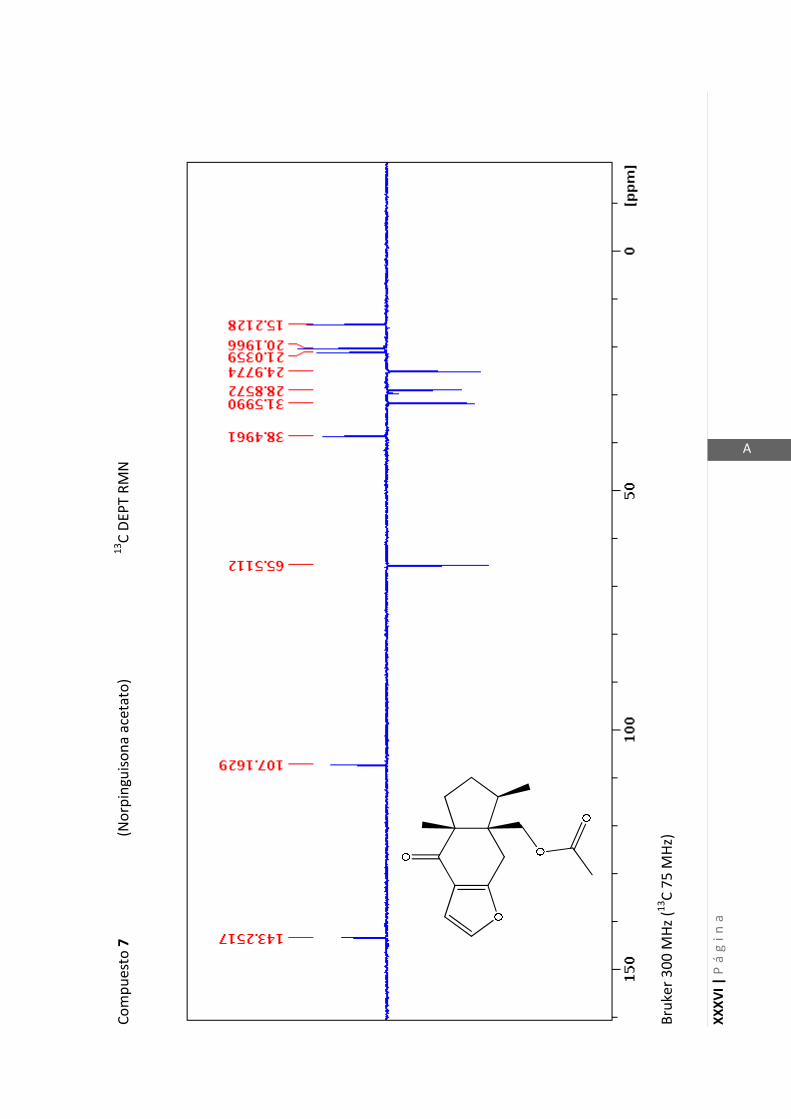
XX
XV
I |
Pá
gi
na
A
Co
mp
ue
sto
7
(No
rpin
gu
iso
na
ace
tato
)
1
3C
DE
PT
RM
N
Bru
ke
r 3
00
MH
z (1
3C
75
MH
z)
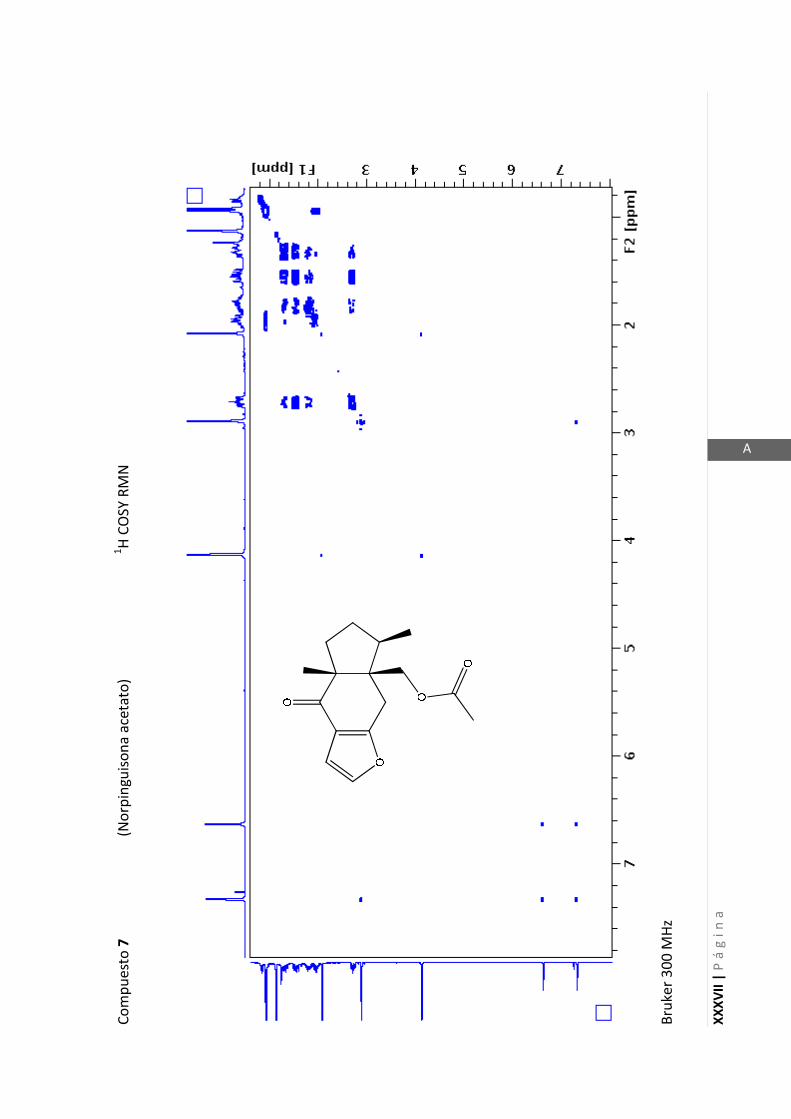
XX
XV
II |
Pá
gi
na
A
Co
mp
ue
sto
7
(No
rpin
gu
iso
na
ace
tato
)
1H
CO
SY
RM
N
Bru
ke
r 3
00
MH
z
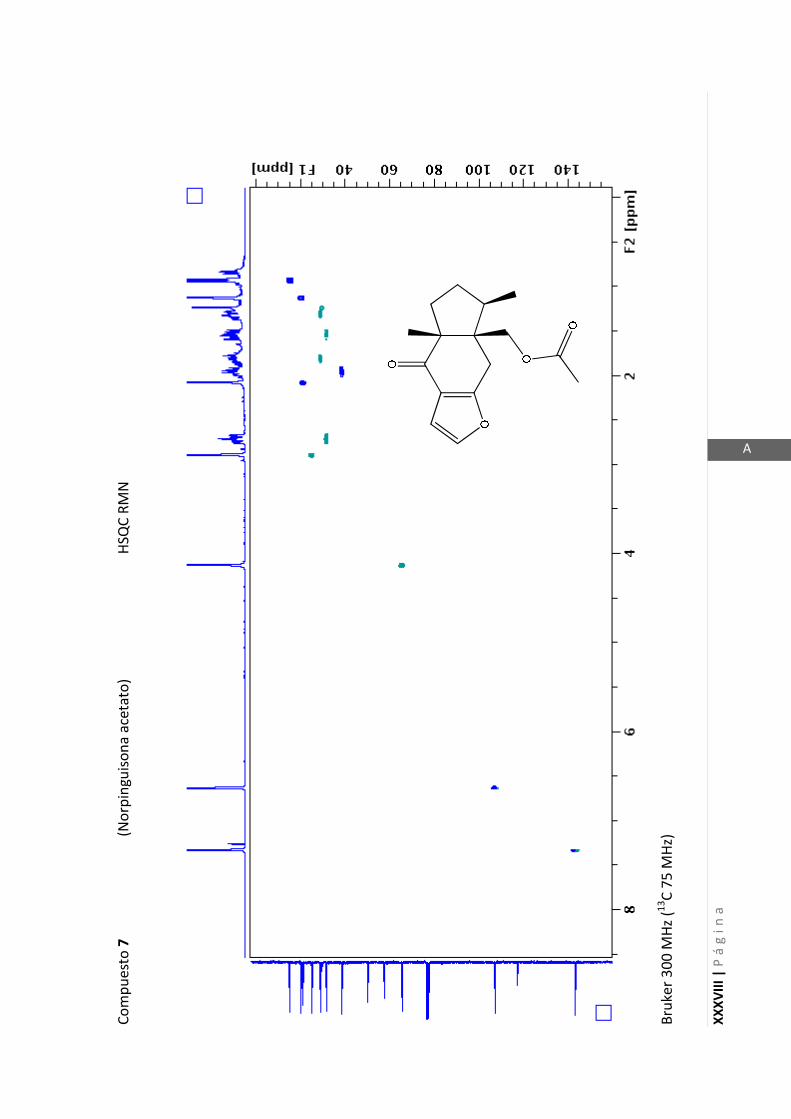
XX
XV
III
| P
ág
in
a
A
Co
mp
ue
sto
7
(No
rpin
gu
iso
na
ace
tato
)
H
SQ
C R
MN
Bru
ke
r 3
00
MH
z (1
3C
75
MH
z)
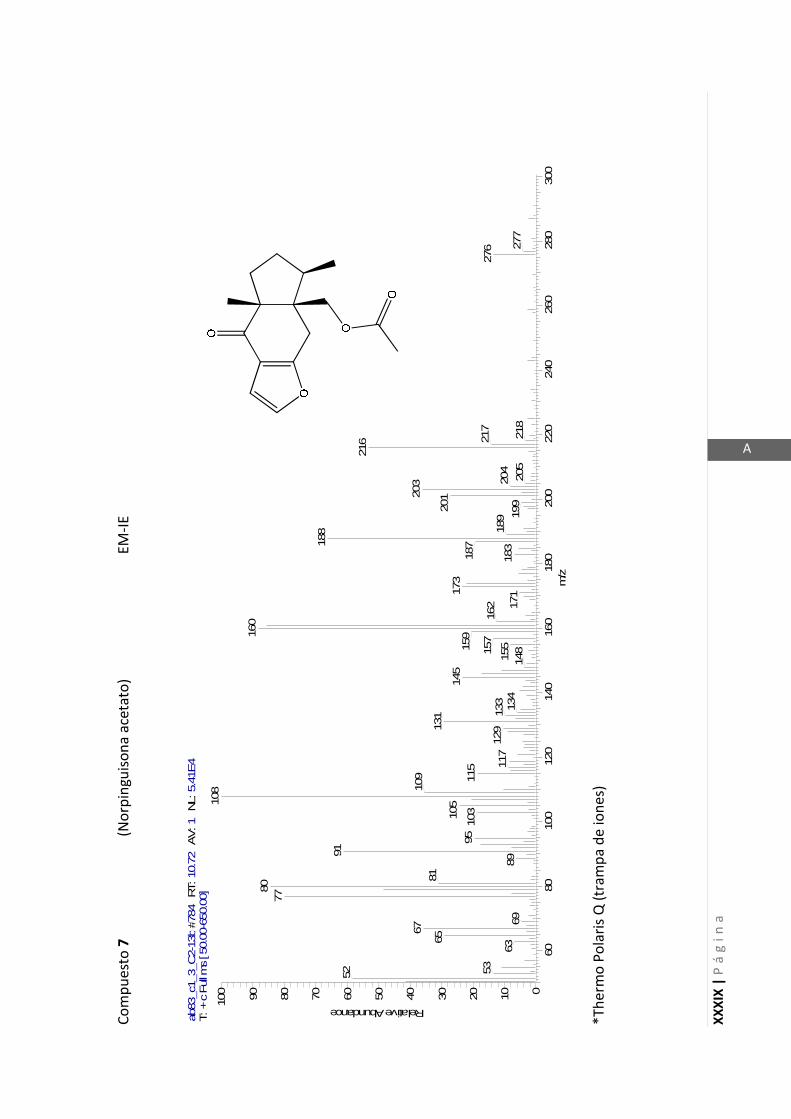
XX
XIX
| P
ág
in
a
A
Co
mp
ue
sto
7
(No
rpin
gu
iso
na
ace
tato
)
E
M-I
E
*T
he
rmo
Po
lari
s Q
(tr
am
pa
de
io
ne
s)
ab83
_c1_
3_C
2-13
b #7
84RT:
10.7
2AV:1
NL:
5.41
E4
T:+
c Full m
s [ 5
0.00
-650
.00]
6080
100
120
140
160
180
200
220
240
260
280
300
m/z
0102030405060708090100
Relative Abundance
108
160
8077
188
9152
216
203
6710
981
131
6520
110
514
517
315
995
187
115
103
217
5327
615
716
212
913
318
911
720
415
518
363
8913
417
169
199
277
148
218
205
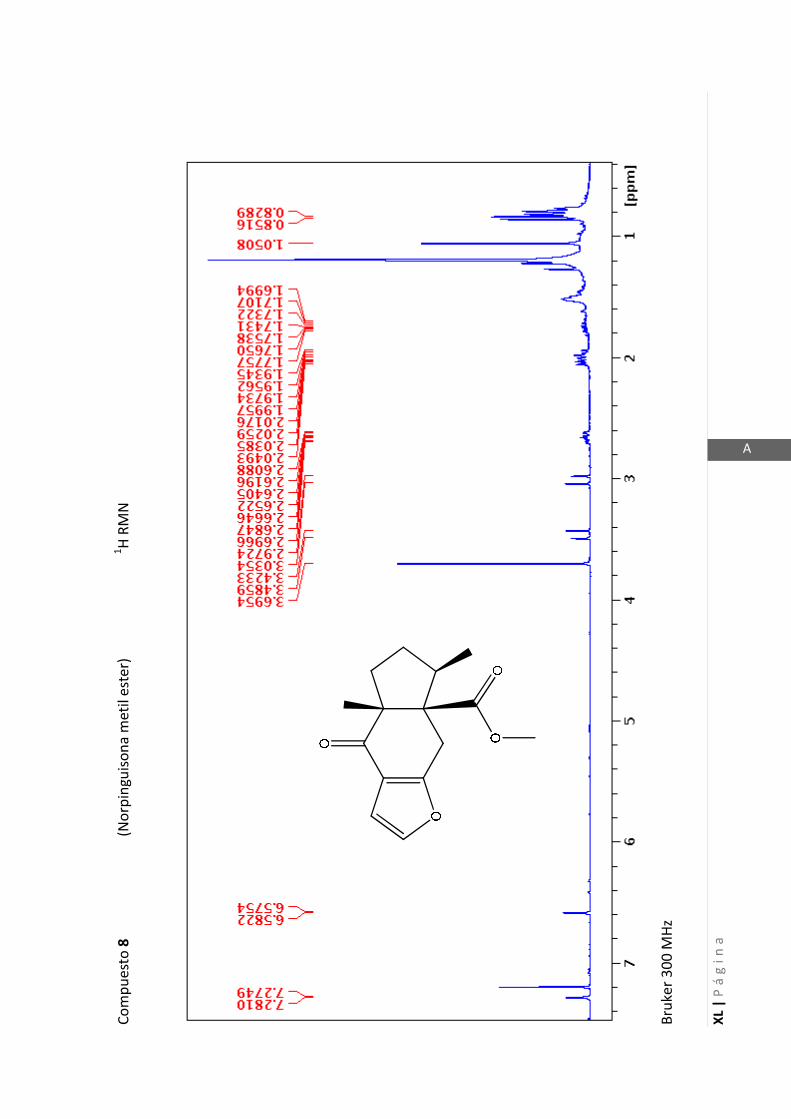
XL
| P
ág
in
a
A
Co
mp
ue
sto
8
(No
rpin
gu
iso
na
me
til e
ste
r)
1H
RM
N
Bru
ke
r 3
00
MH
z
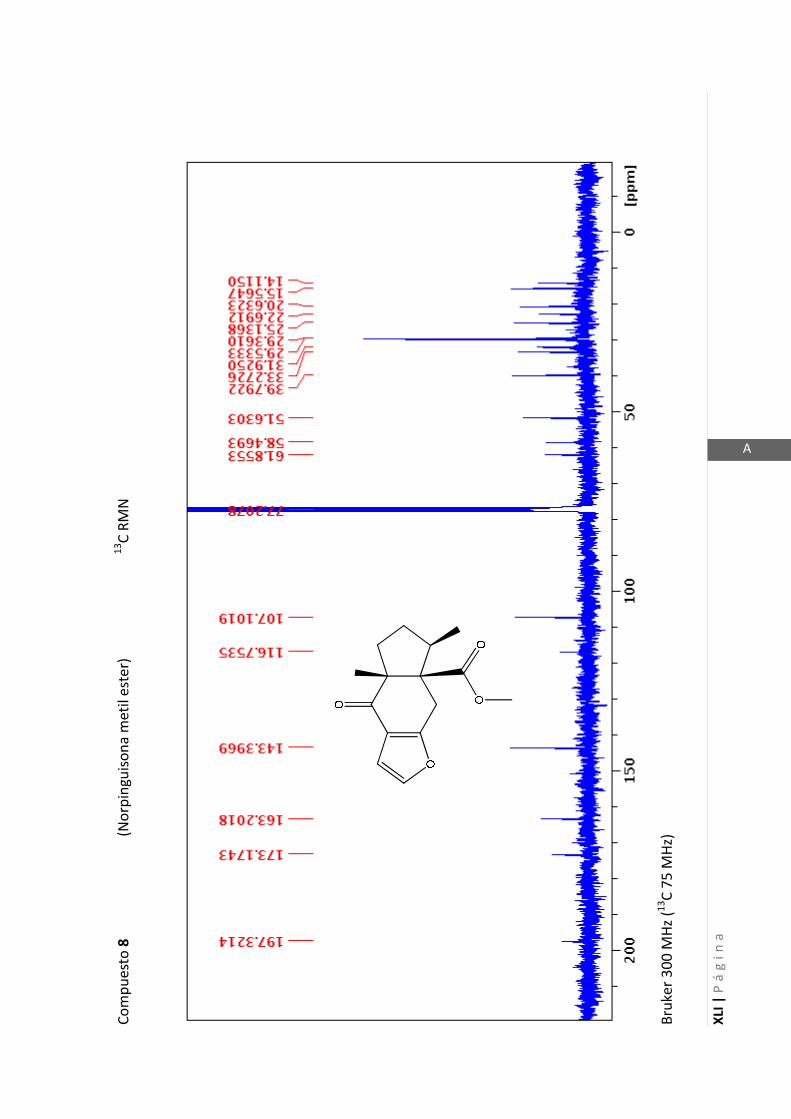
XLI
| P
ág
in
a
A
Co
mp
ue
sto
8
(No
rpin
gu
iso
na
me
til e
ste
r)
13C
RM
N
Bru
ke
r 3
00
MH
z (1
3C
75
MH
z)
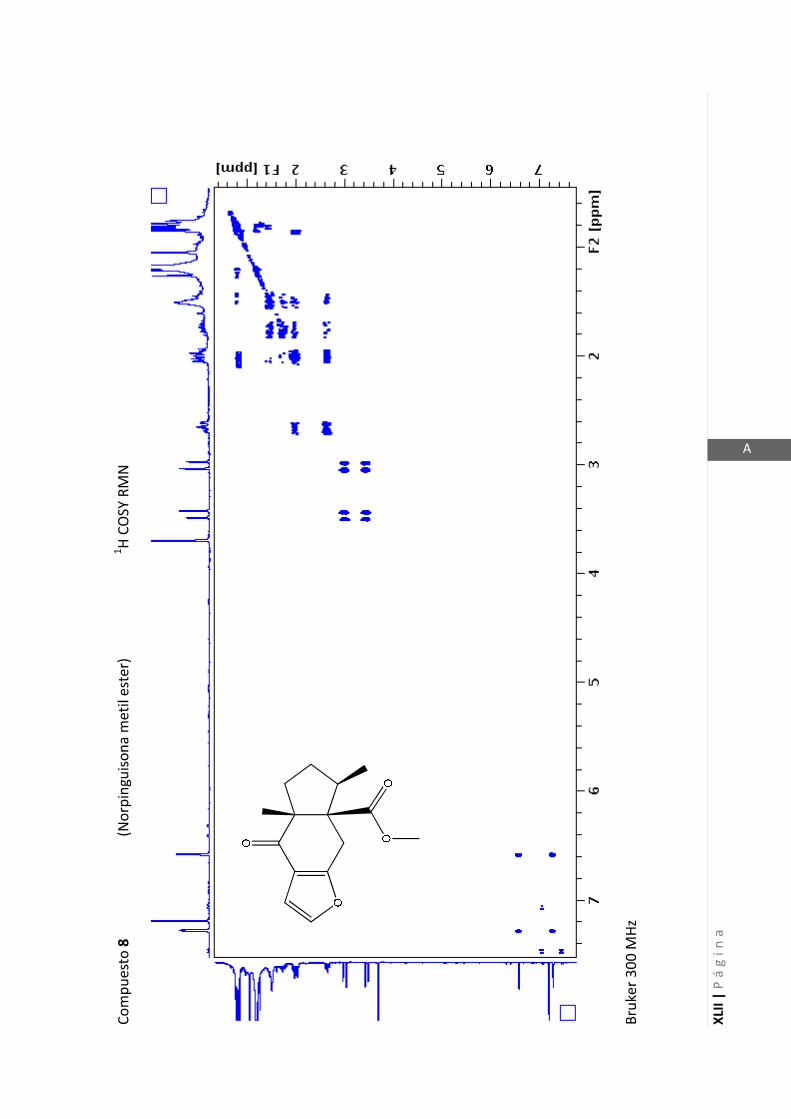
XLI
I |
Pá
gi
na
A
Co
mp
ue
sto
8
(No
rpin
gu
iso
na
me
til e
ste
r)
1H
CO
SY
RM
N
Bru
ke
r 3
00
MH
z
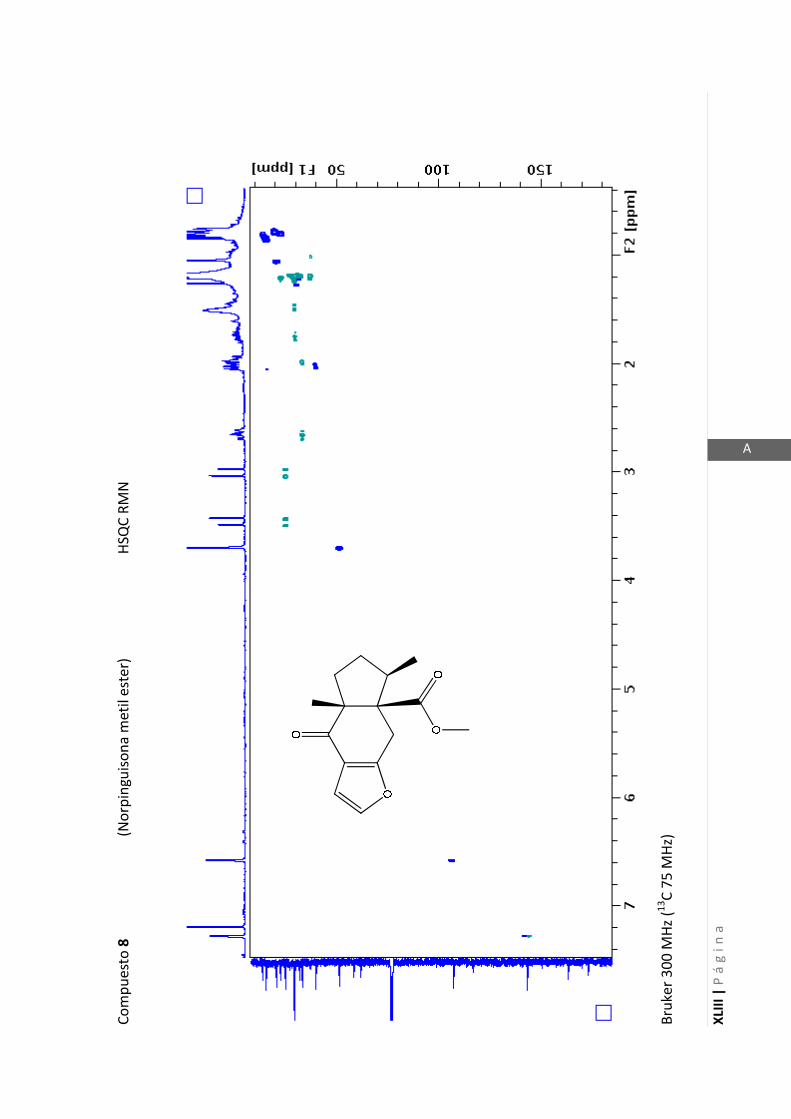
XLI
II |
Pá
gi
na
A
Co
mp
ue
sto
8
(No
rpin
gu
iso
na
me
til e
ste
r)
HS
QC
RM
N
Bru
ke
r 3
00
MH
z (1
3C
75
MH
z)
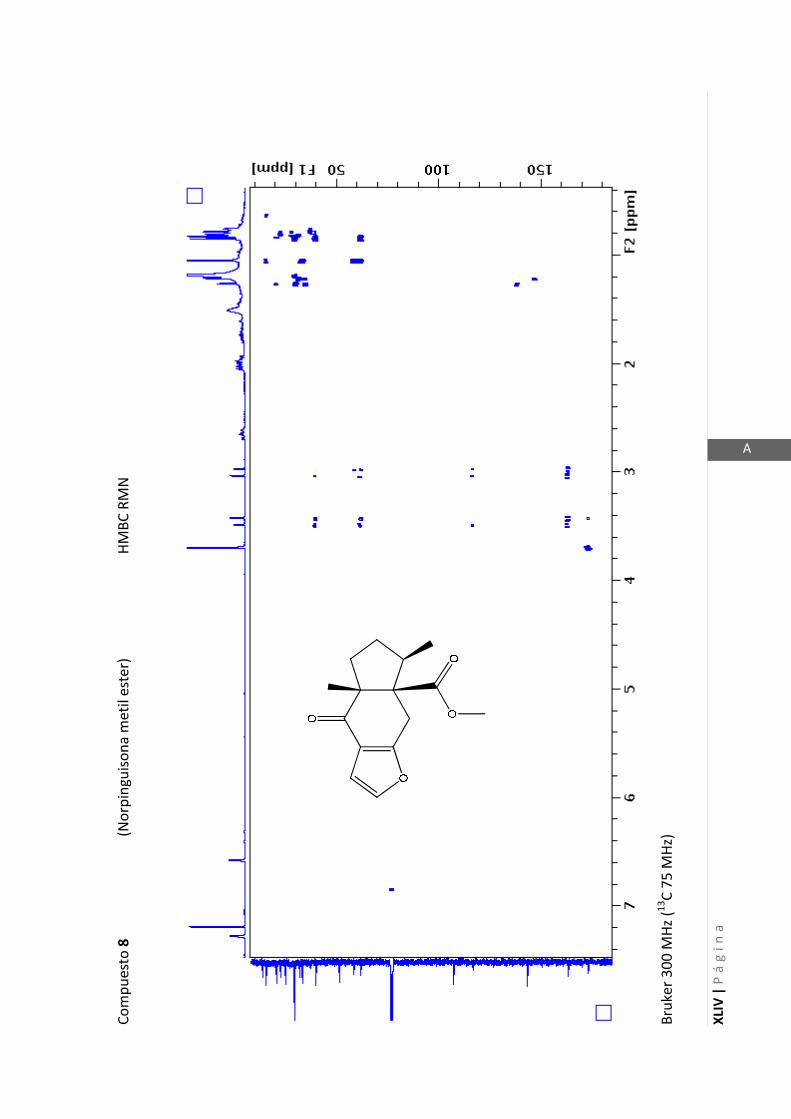
XLI
V |
Pá
gi
na
A
Co
mp
ue
sto
8
(No
rpin
gu
iso
na
me
til e
ste
r)
HM
BC
RM
N
Bru
ke
r 3
00
MH
z (1
3C
75
MH
z)
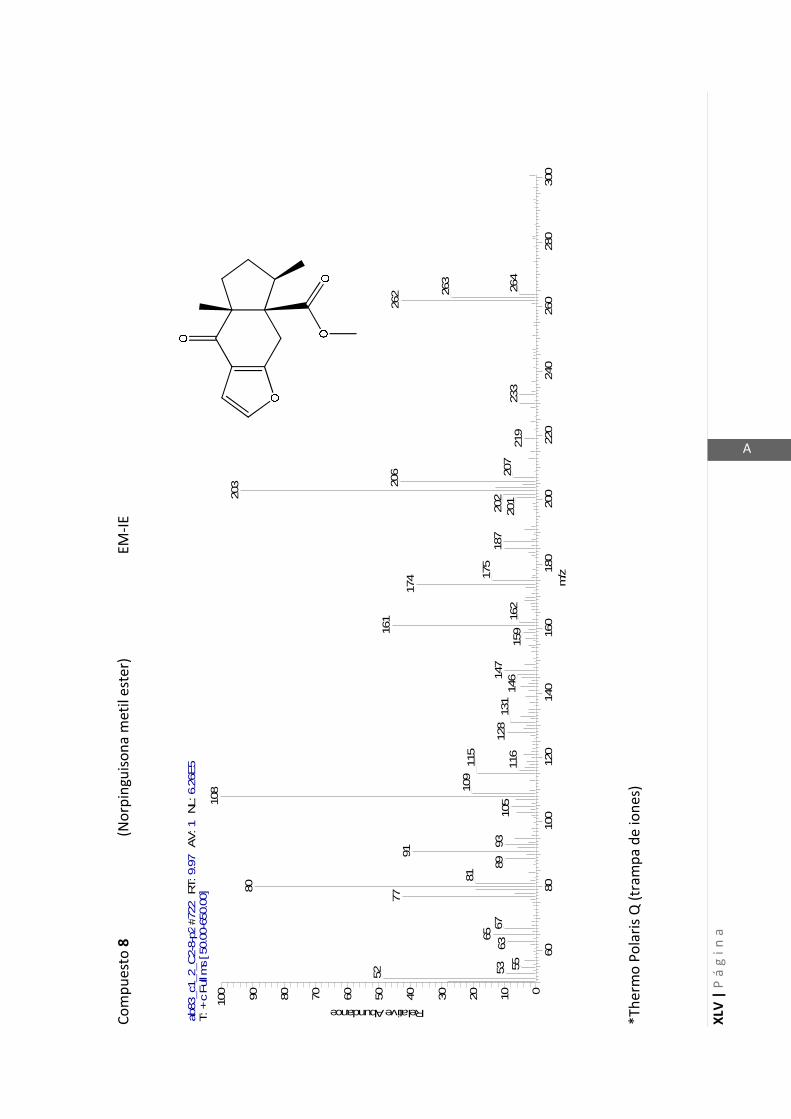
XLV
| P
ág
in
a
A
Co
mp
ue
sto
8
(No
rpin
gu
iso
na
me
til e
ste
r)
EM
-IE
*T
he
rmo
Po
lari
s Q
(tr
am
pa
de
io
ne
s)
ab83
_c1_
2_C
2-8-
p2 #
722
RT:
9.97
AV:1
NL:
6.26
E5
T:+
c Full m
s [ 5
0.00
-650
.00]
6080
100
120
140
160
180
200
220
240
260
280
300
m/z
0102030405060708090100
Relative Abundance
108
203
80
5216
120
626
277
9117
4
263
109
8111
5
175
6520
218
714
767
9389
5312
863
131
105
207
201
146
116
264
233
162
5515
921
9
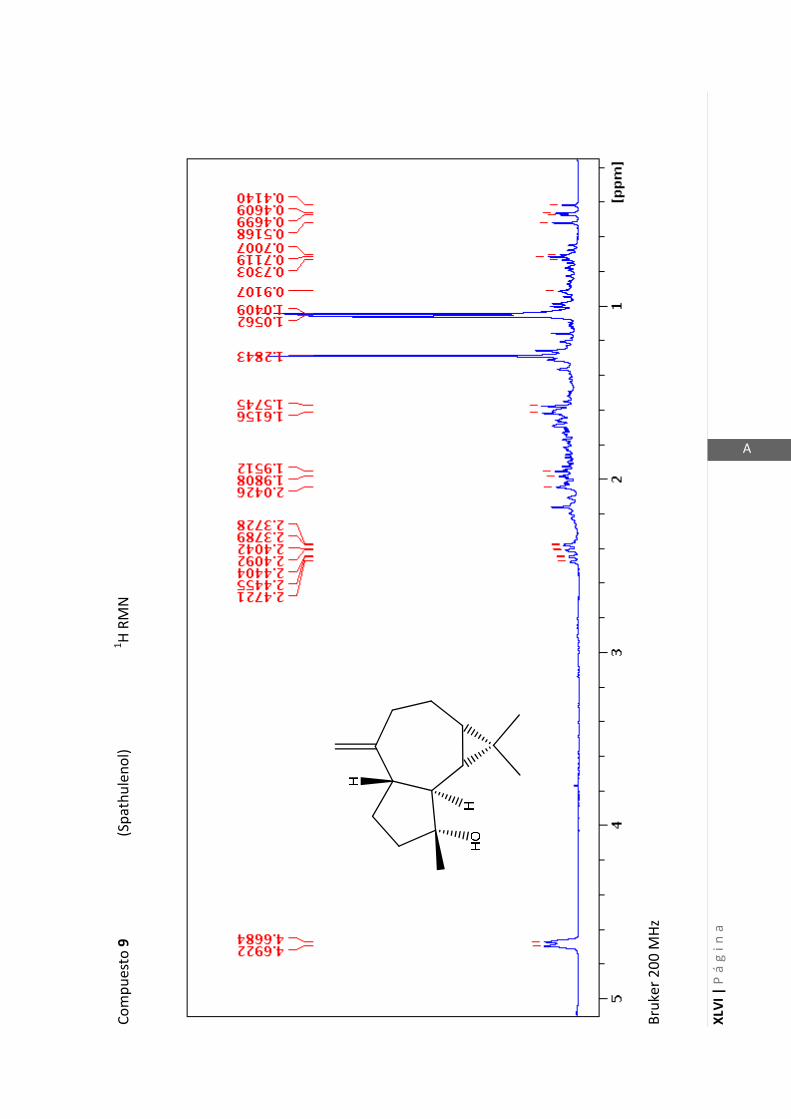
XLV
I |
Pá
gi
na
A
Co
mp
ue
sto
9
(Sp
ath
ule
no
l)
1H
RM
N
Bru
ke
r 2
00
MH
z
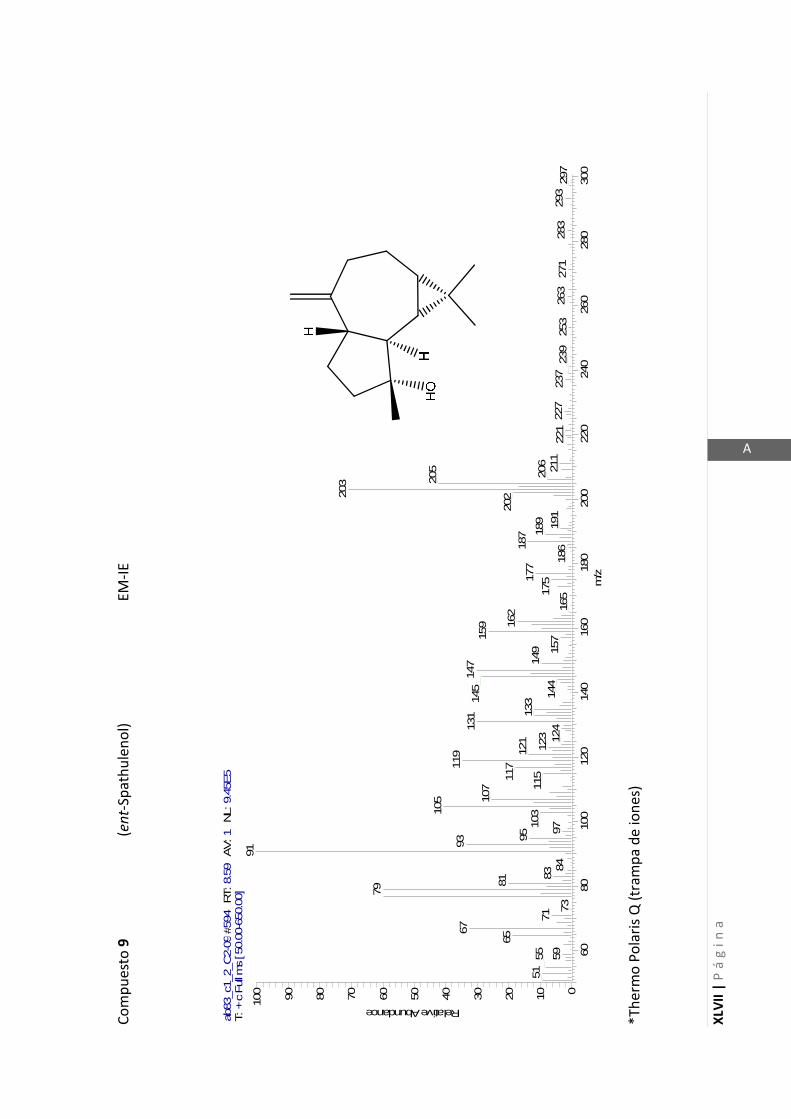
XLV
II |
Pá
gi
na
A
Co
mp
ue
sto
9
(ent-S
pa
thu
len
ol)
E
M-I
E
*T
he
rmo
Po
lari
s Q
(tr
am
pa
de
io
ne
s)
ab83
_c1_
2_C
2-09
#59
4RT:
8.59
AV:1
NL:
9.45
E5
T:+
c Full m
s [ 5
0.00
-650
.00]
6080
100
120
140
160
180
200
220
240
260
280
300
m/z
0102030405060708090100
Relative Abundance
91
203
79
205
105
119
9367
147
131
145
159
107
8165
202
117
162
187
121
9513
317
710
314
951
115
5518
920
612
317
571
8314
421
119
115
712
459
9722
723
729
322
184
263
283
186
271
253
239
165
297
73
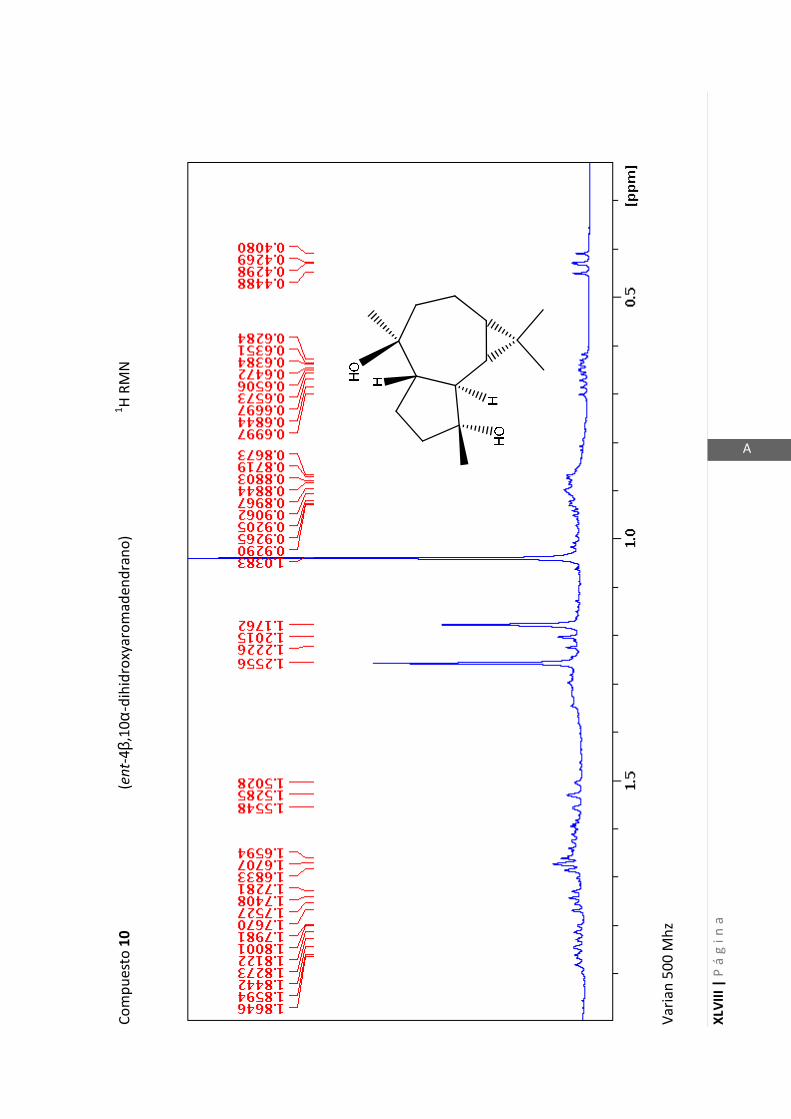
XLV
III
| P
ág
in
a
A
Co
mp
ue
sto
10
(ent-4
β,1
0α-
dih
idro
xya
rom
ad
en
dra
no
)
1H
RM
N
Va
ria
n 5
00
Mh
z
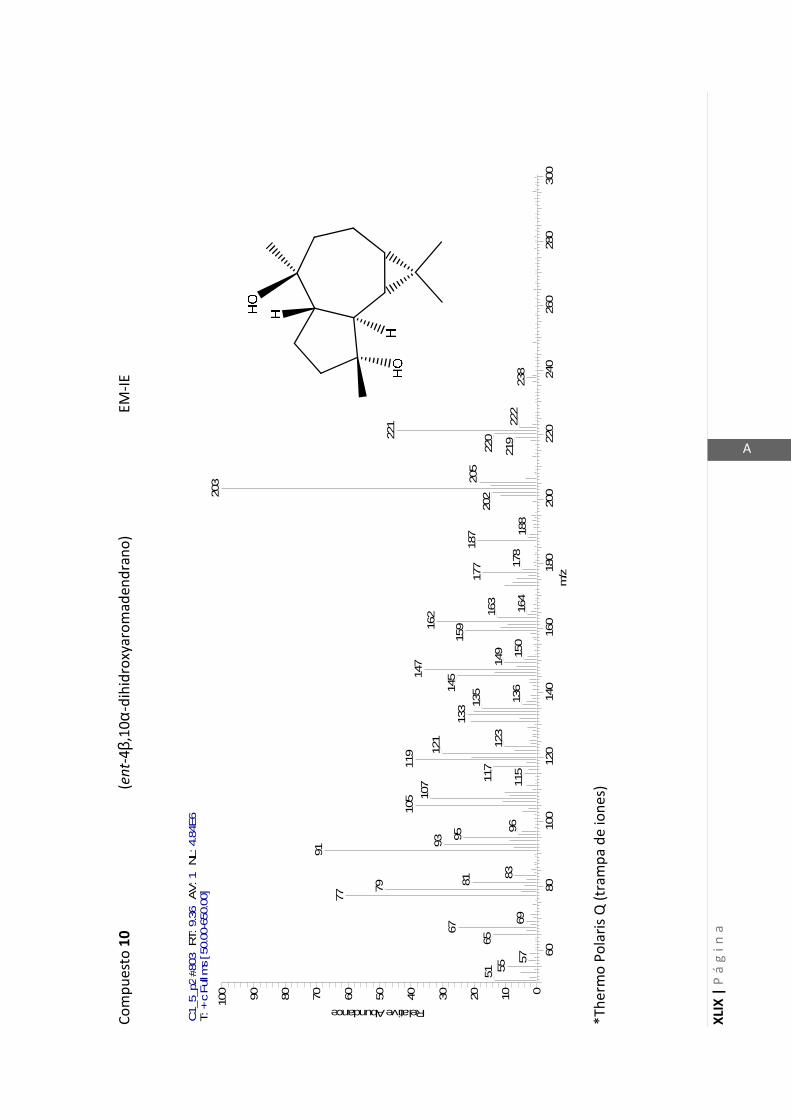
XLI
X |
Pá
gi
na
A
Co
mp
ue
sto
10
(ent-4
β,1
0α-
dih
idro
xya
rom
ad
en
dra
no
)
E
M-I
E
*T
he
rmo
Po
lari
s Q
(tr
am
pa
de
io
ne
s)
C1_
5_p2
#80
3RT:
9.36
AV:1
NL:
4.84
E6
T:+
c Full m
s [ 5
0.00
-650
.00]
6080
100
120
140
160
180
200
220
240
260
280
300
m/z
0102030405060708090100
Relative Abundance
203
91
77
7922
1
119
105
147
107
162
121
9314
567
9515
913
381
187
205
135
177
202
6511
722
051
163
123
149
5583
219
9622
217
813
611
515
069
238
164
188
57
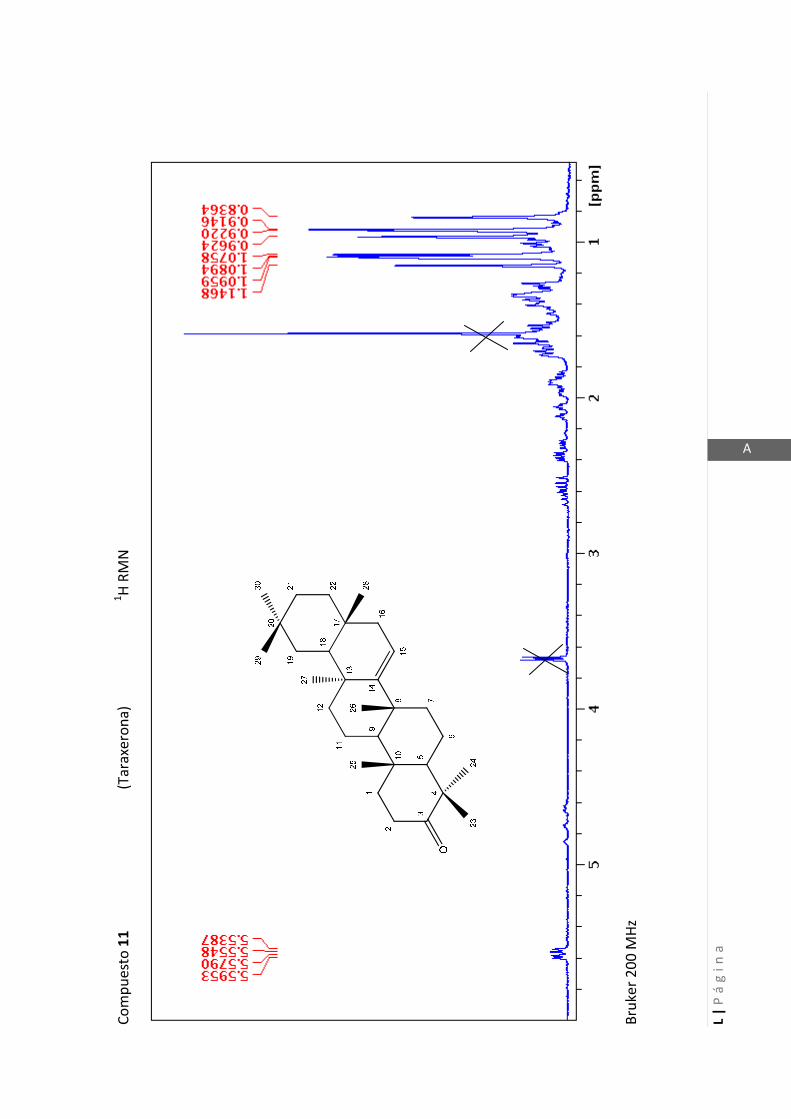
L |
Pá
gi
na
A
Co
mp
ue
sto
11
(T
ara
xero
na
)
1H
RM
N
Bru
ke
r 2
00
MH
z
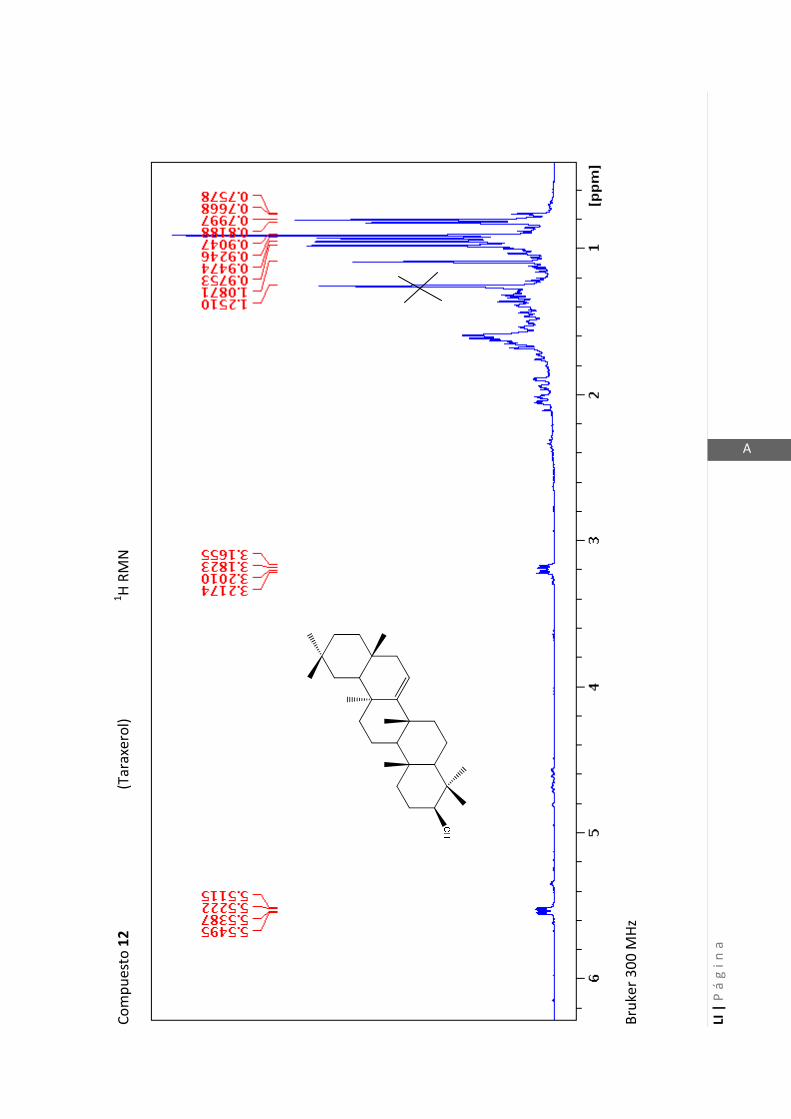
LI |
Pá
gi
na
A
Co
mp
ue
sto
12
(T
ara
xero
l)
1H
RM
N
Bru
ke
r 3
00
MH
z
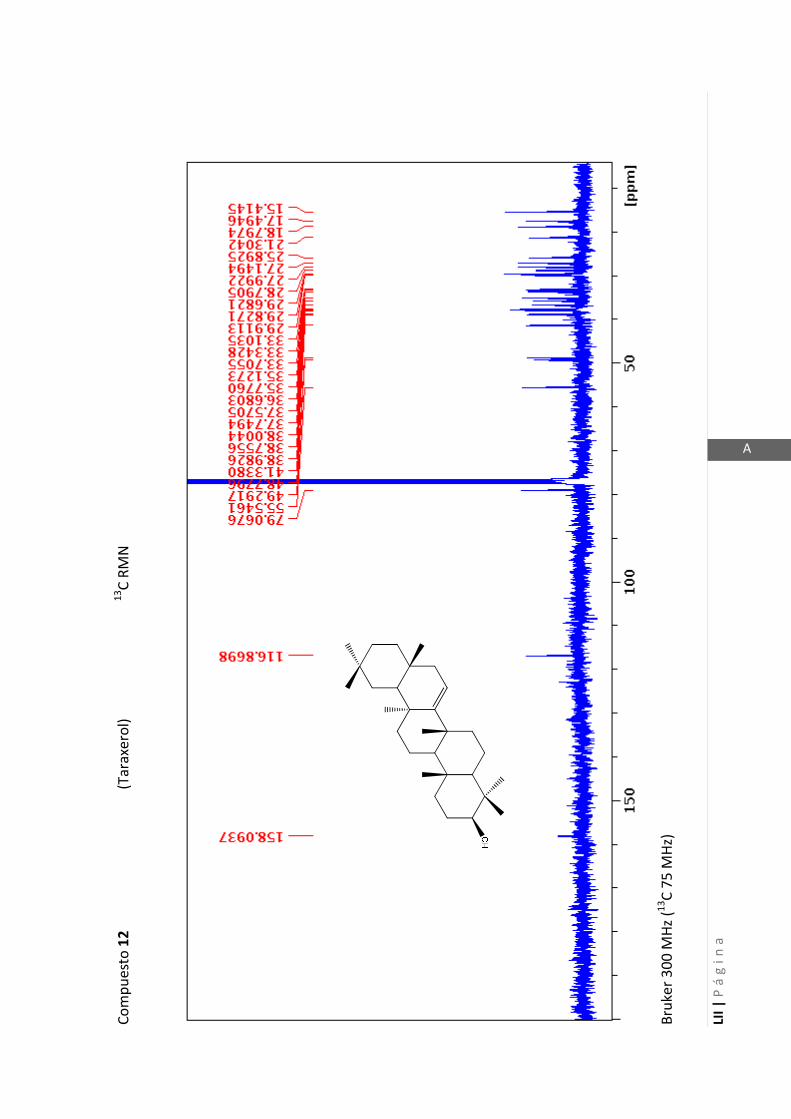
LII
| P
ág
in
a
A
Co
mp
ue
sto
12
(T
ara
xero
l)
13C
RM
N
Bru
ke
r 3
00
MH
z (1
3C
75
MH
z)
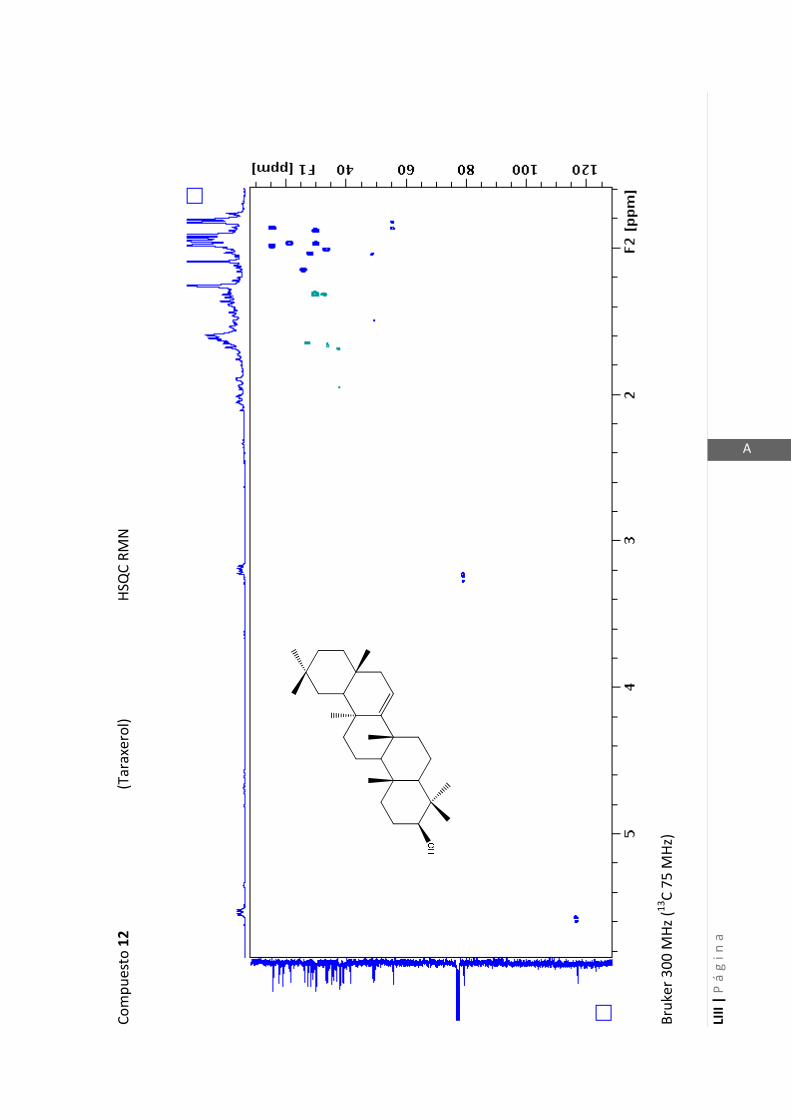
LIII
| P
ág
in
a
A
Co
mp
ue
sto
12
(T
ara
xero
l)
HS
QC
RM
N
Bru
ke
r 3
00
MH
z (1
3C
75
MH
z)
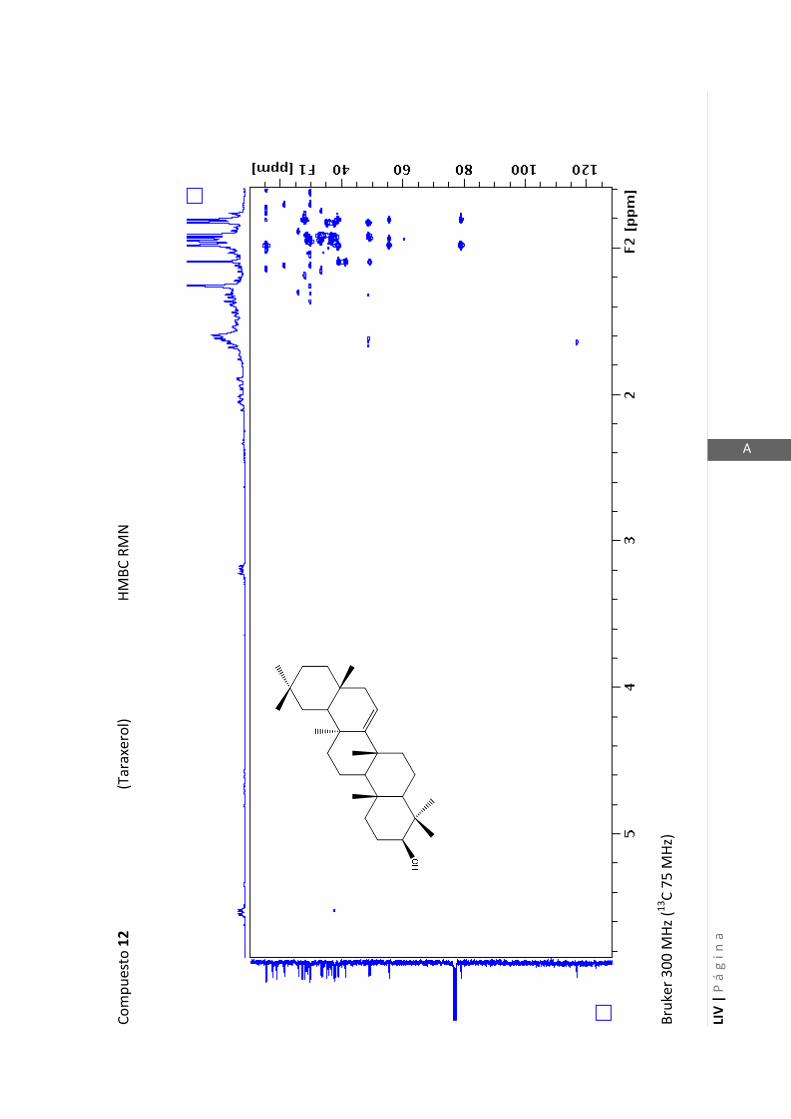
LIV
| P
ág
in
a
A
Co
mp
ue
sto
12
(T
ara
xero
l)
HM
BC
RM
N
Bru
ke
r 3
00
MH
z (1
3C
75
MH
z)
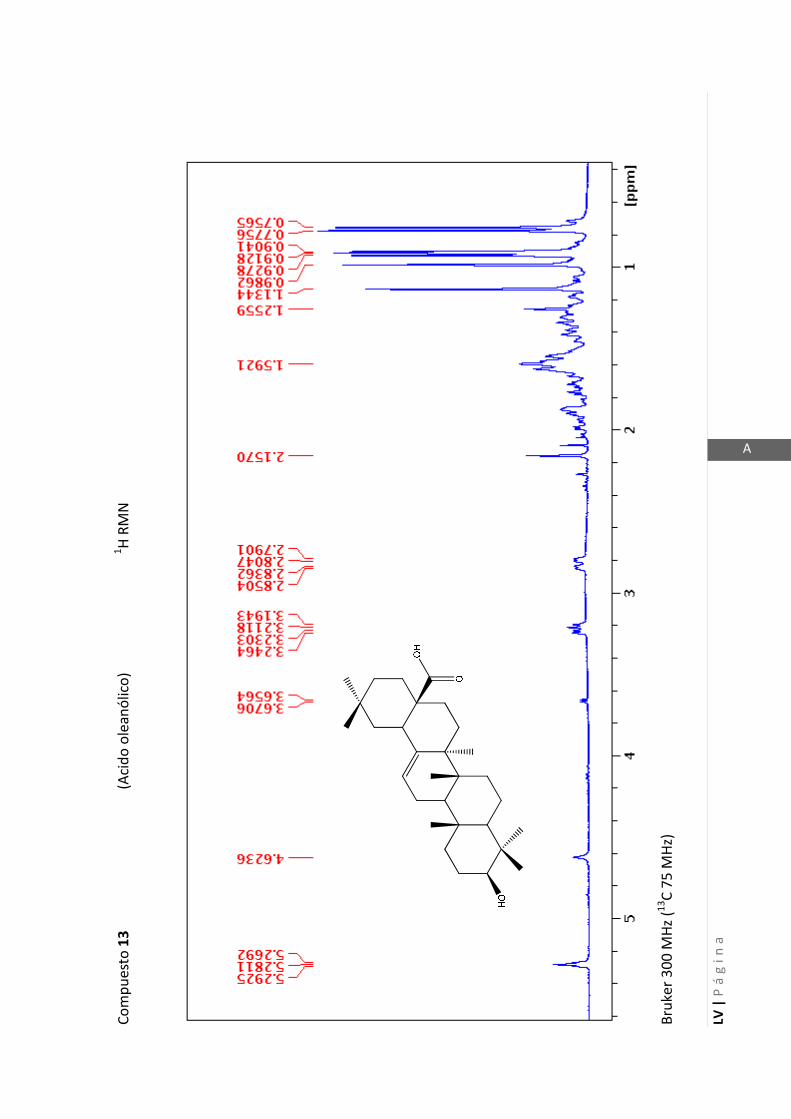
LV |
Pá
gi
na
A
Co
mp
ue
sto
13
(A
cid
o o
lea
nó
lico
)
1H
RM
N
Bru
ke
r 3
00
MH
z (1
3C
75
MH
z)
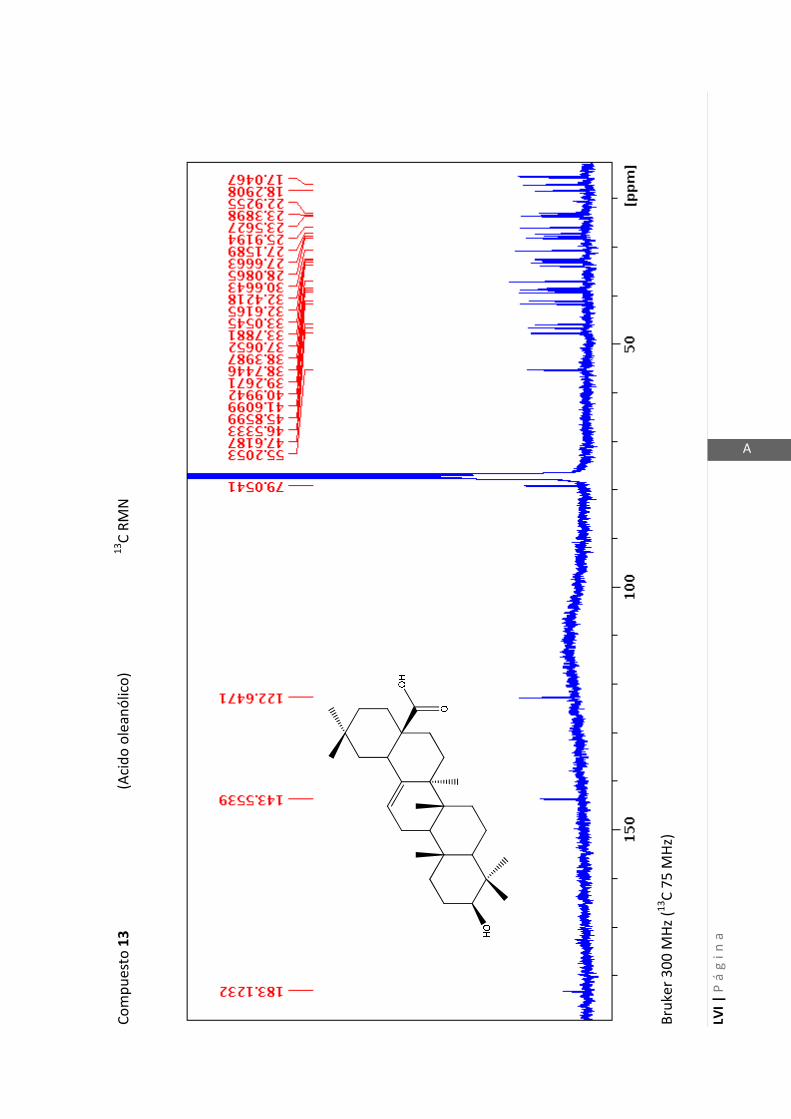
LVI
| P
ág
in
a
A
Co
mp
ue
sto
13
(A
cid
o o
lea
nó
lico
)
1
3C
RM
N
Bru
ke
r 3
00
MH
z (1
3C
75
MH
z)
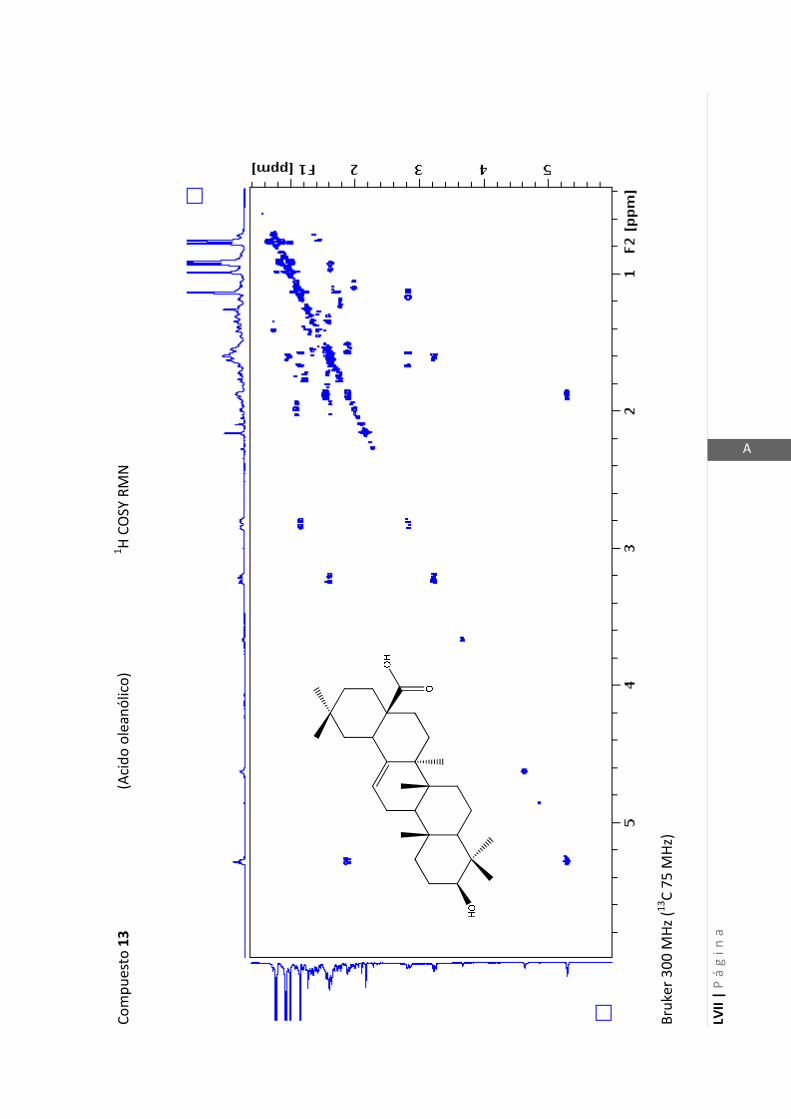
LVII
| P
ág
in
a
A
Co
mp
ue
sto
13
(A
cid
o o
lea
nó
lico
)
1H
CO
SY
RM
N
Bru
ke
r 3
00
MH
z (1
3C
75
MH
z)
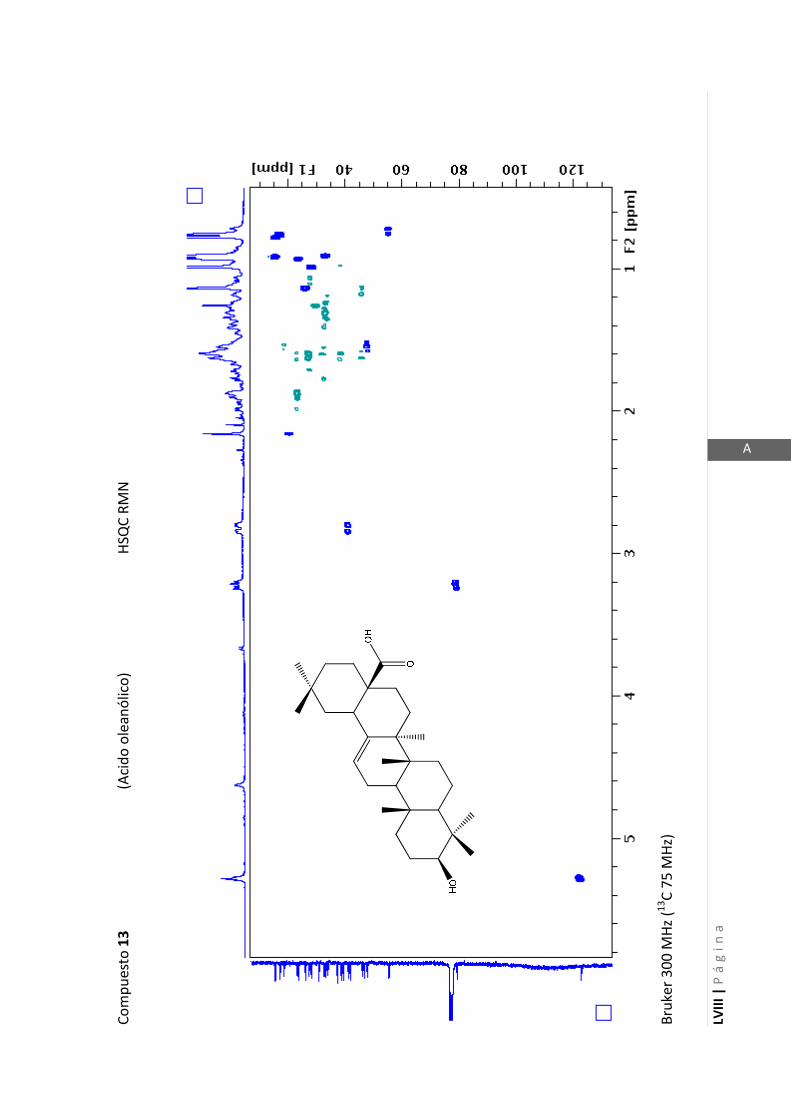
LVII
I |
Pá
gi
na
A
Co
mp
ue
sto
13
(A
cid
o o
lea
nó
lico
)
H
SQ
C R
MN
Bru
ke
r 3
00
MH
z (1
3C
75
MH
z)
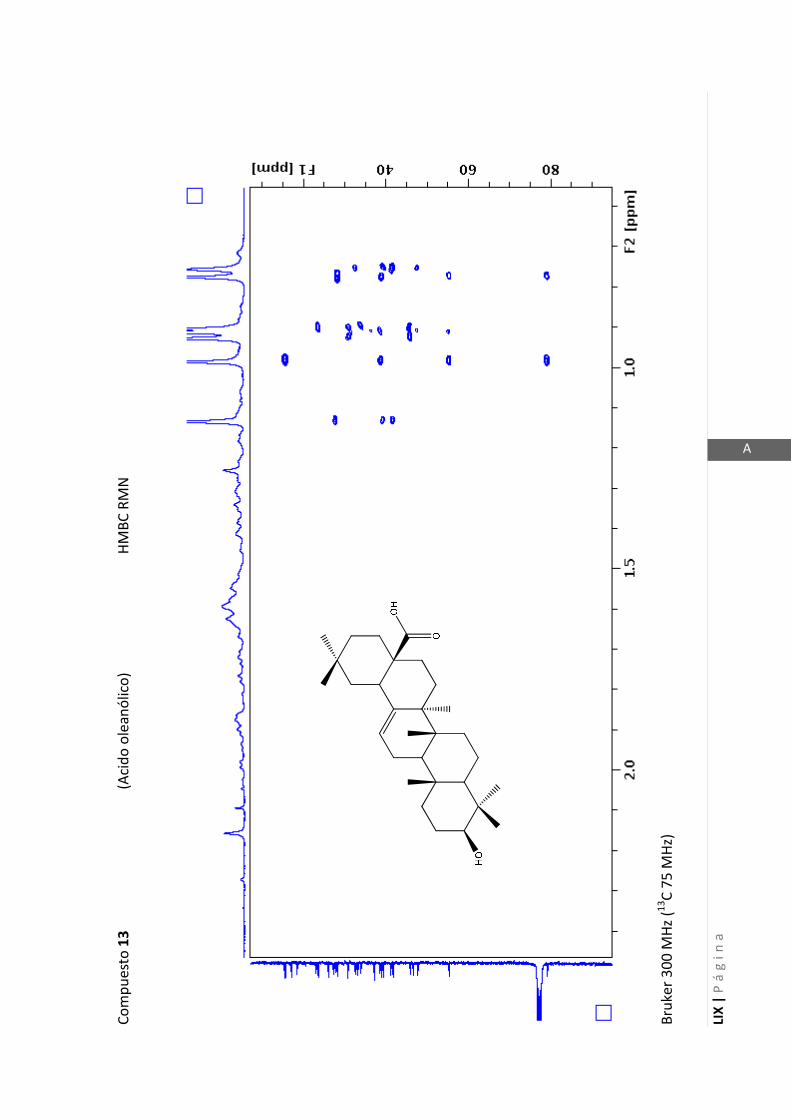
LIX
| P
ág
in
a
A
Co
mp
ue
sto
13
(A
cid
o o
lea
nó
lico
)
H
MB
C R
MN
Bru
ke
r 3
00
MH
z (1
3C
75
MH
z)
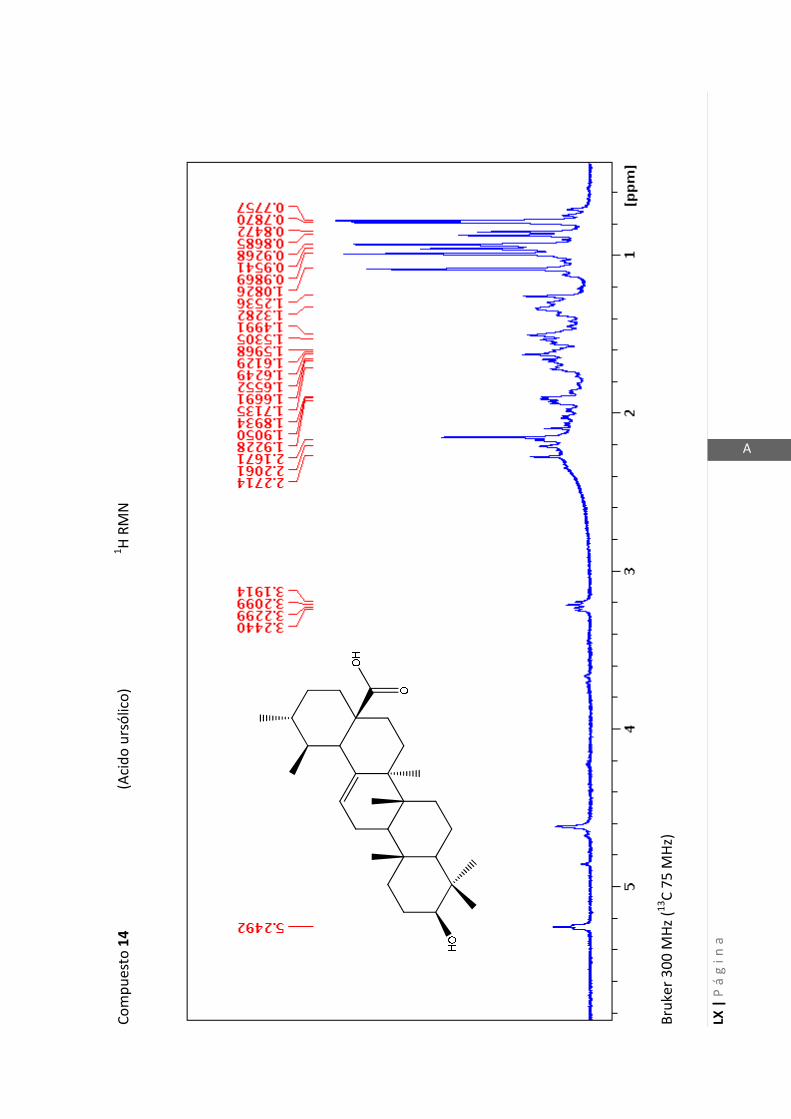
LX |
Pá
gi
na
A
Co
mp
ue
sto
14
(A
cid
o u
rsó
lico
)
1H
RM
N
Bru
ke
r 3
00
MH
z (1
3C
75
MH
z)
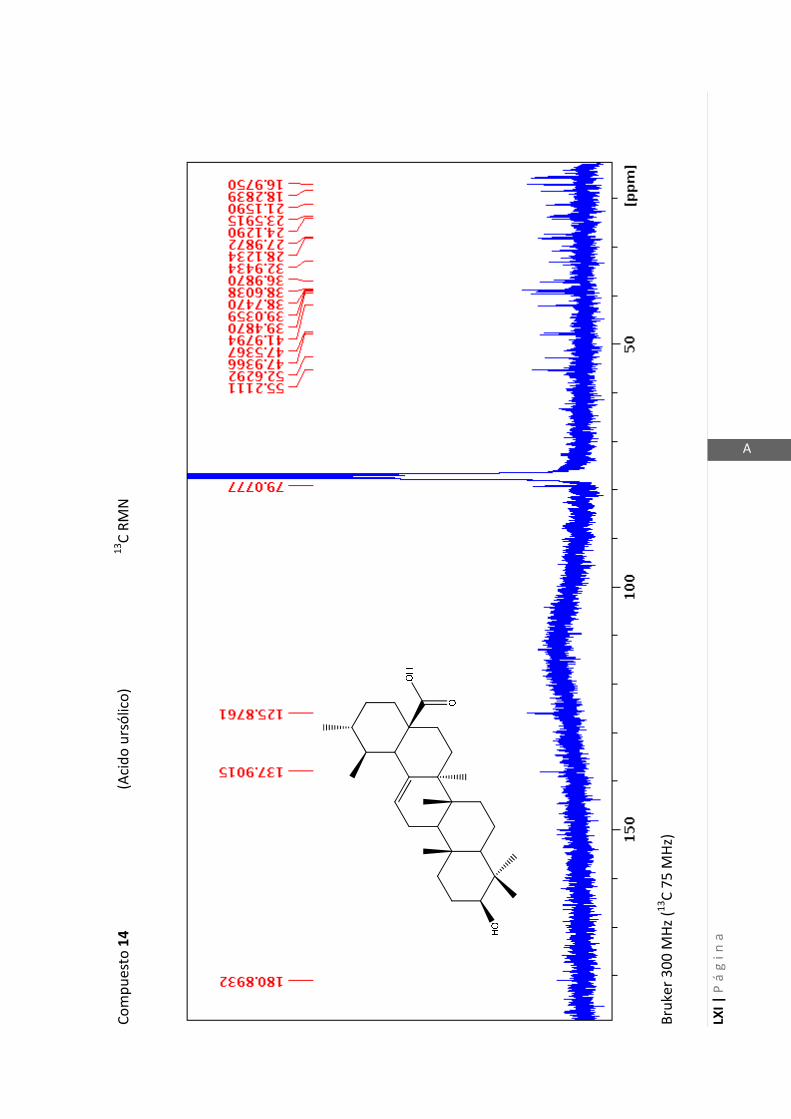
LXI
| P
ág
in
a
A
Co
mp
ue
sto
14
(A
cid
o u
rsó
lico
)
1
3C
RM
N
Bru
ke
r 3
00
MH
z (1
3C
75
MH
z)
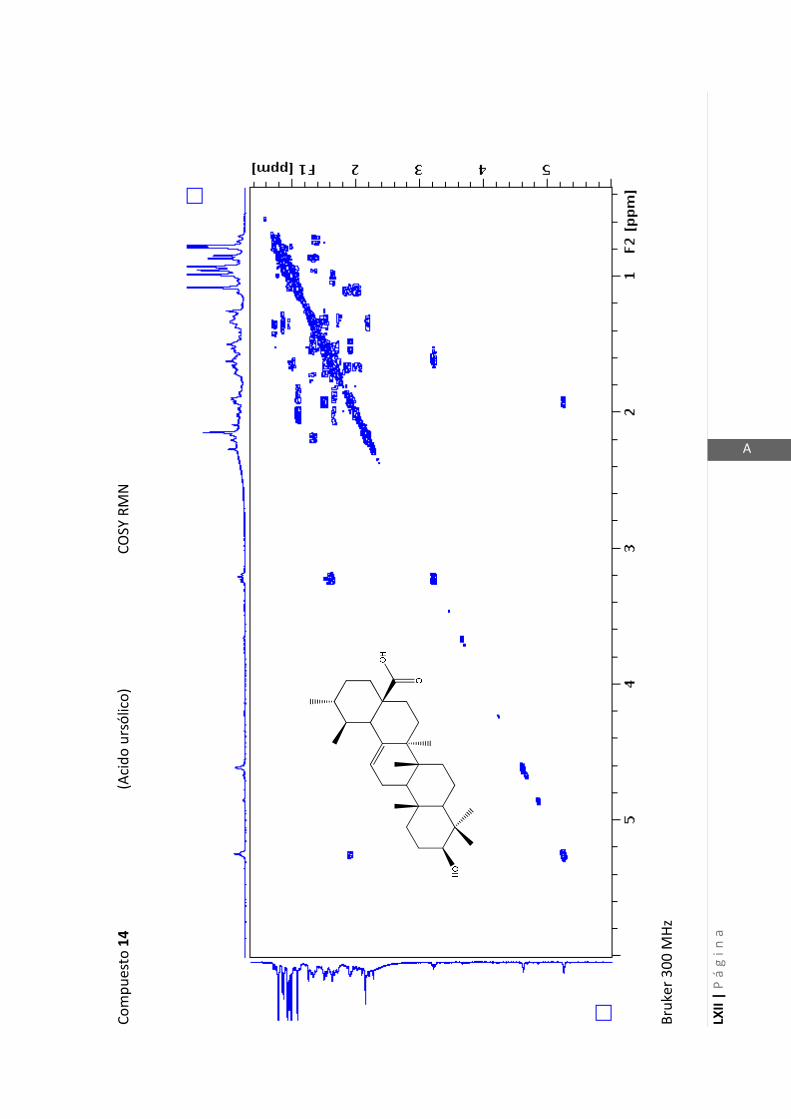
LXII
| P
ág
in
a
A
Co
mp
ue
sto
14
(A
cid
o u
rsó
lico
)
C
OS
Y R
MN
Bru
ke
r 3
00
MH
z
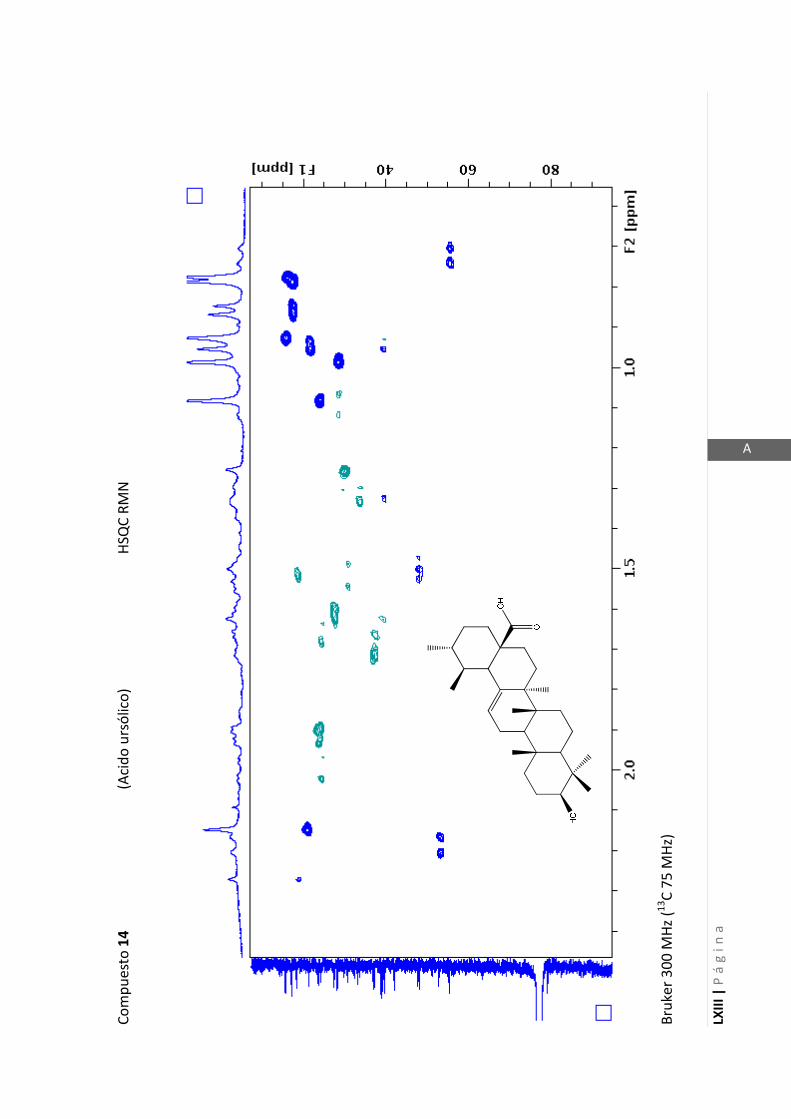
LXII
I |
Pá
gi
na
A
Co
mp
ue
sto
14
(A
cid
o u
rsó
lico
)
H
SQ
C R
MN
Bru
ke
r 3
00
MH
z (1
3C
75
MH
z)
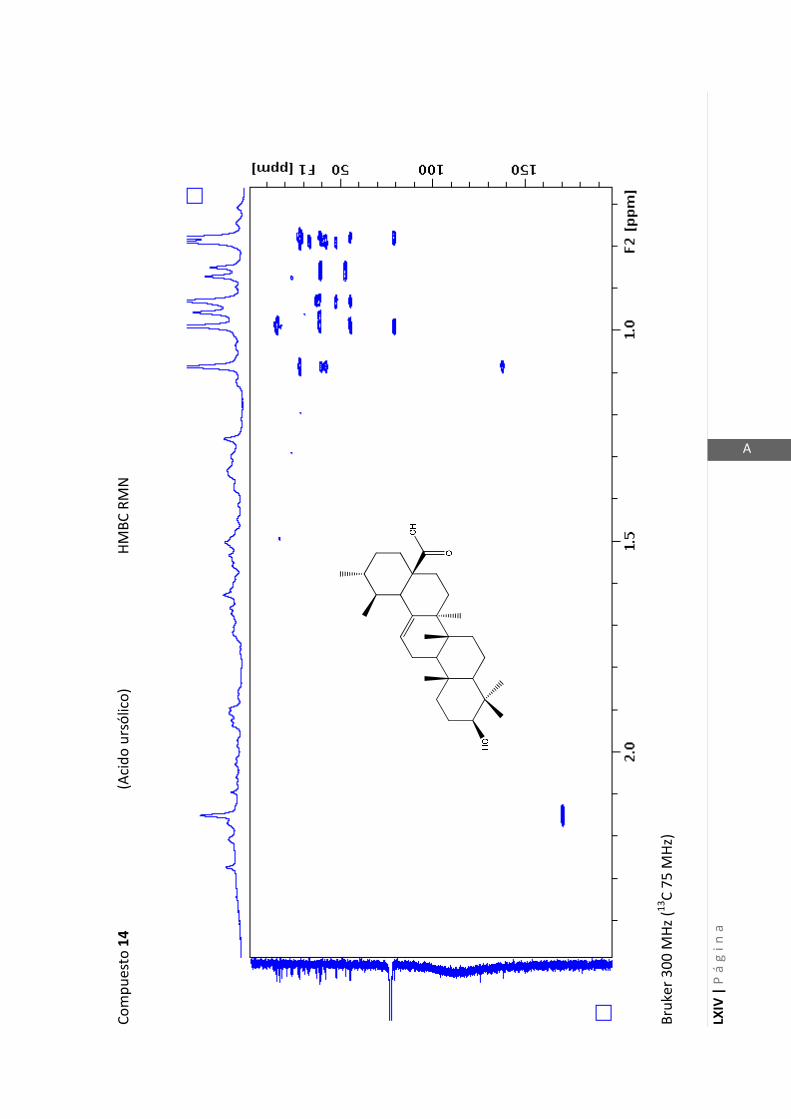
LXIV
| P
ág
in
a
A
Co
mp
ue
sto
14
(A
cid
o u
rsó
lico
)
H
MB
C R
MN
Bru
ke
r 3
00
MH
z (1
3C
75
MH
z)
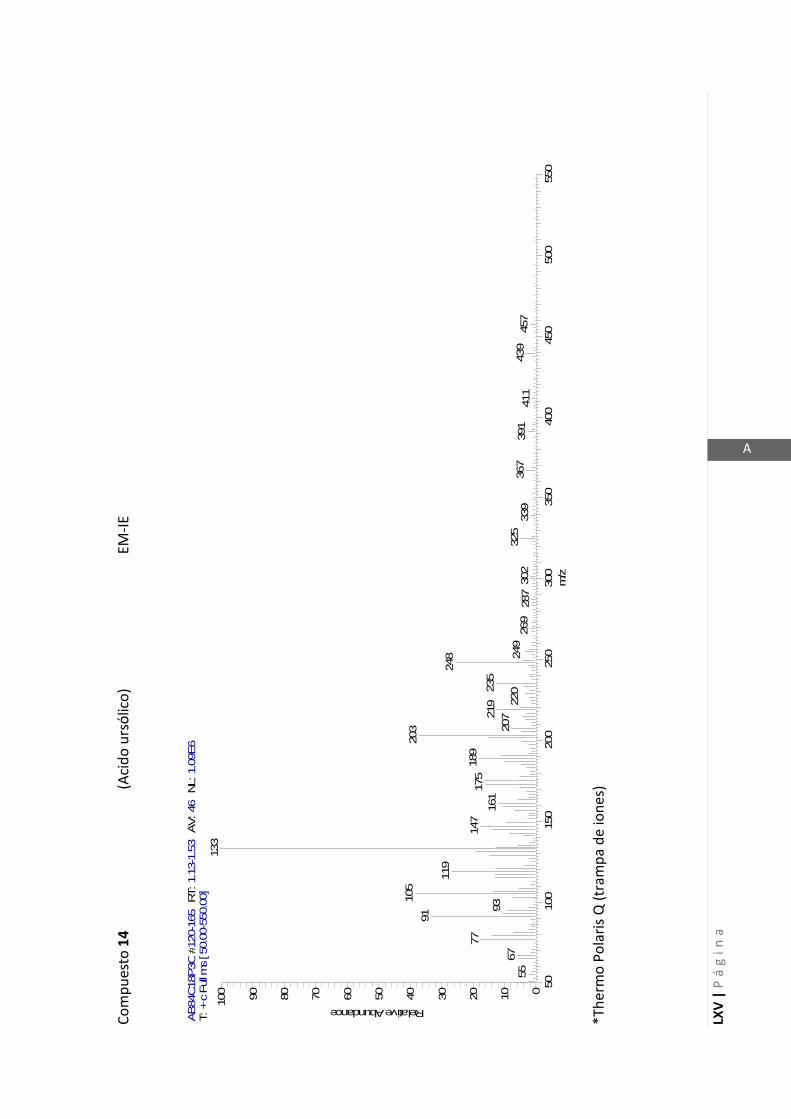
LXV
| P
ág
in
a
A
Co
mp
ue
sto
14
(A
cid
o u
rsó
lico
)
E
M-I
E
*T
he
rmo
Po
lari
s Q
(tr
am
pa
de
io
ne
s)
AB84
C18
P3C
#12
0-16
5RT:
1.13
-1.5
3AV:46
NL:
1.09
E6
T:+
c Full m
s [ 5
0.00
-550
.00]
5010
015
020
025
030
035
040
045
050
055
0m
/z
0102030405060708090100
Relative Abundance
133
105
203
91
119
248
189
147
7717
523
521
916
193
207
6722
032
524
943
936
739
155
457
269
302
339
287
411
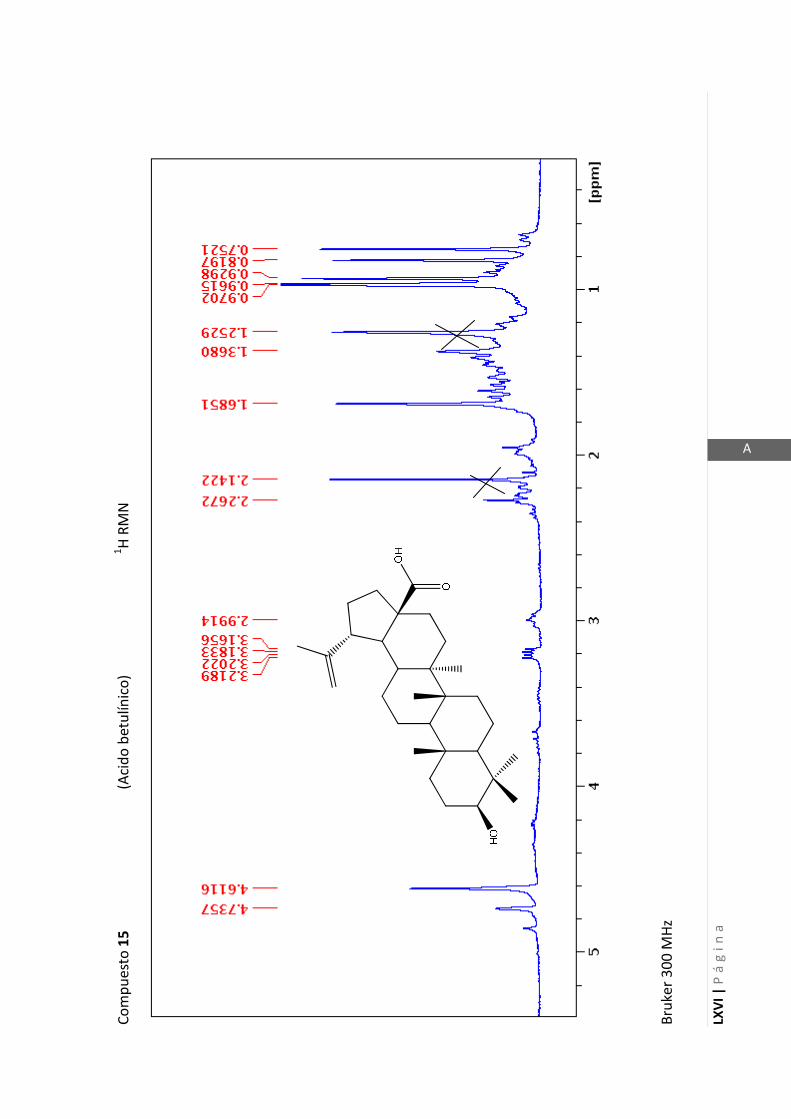
LXV
I |
Pá
gi
na
A
Co
mp
ue
sto
15
(A
cid
o b
etu
lín
ico
)
1H
RM
N
Bru
ke
r 3
00
MH
z
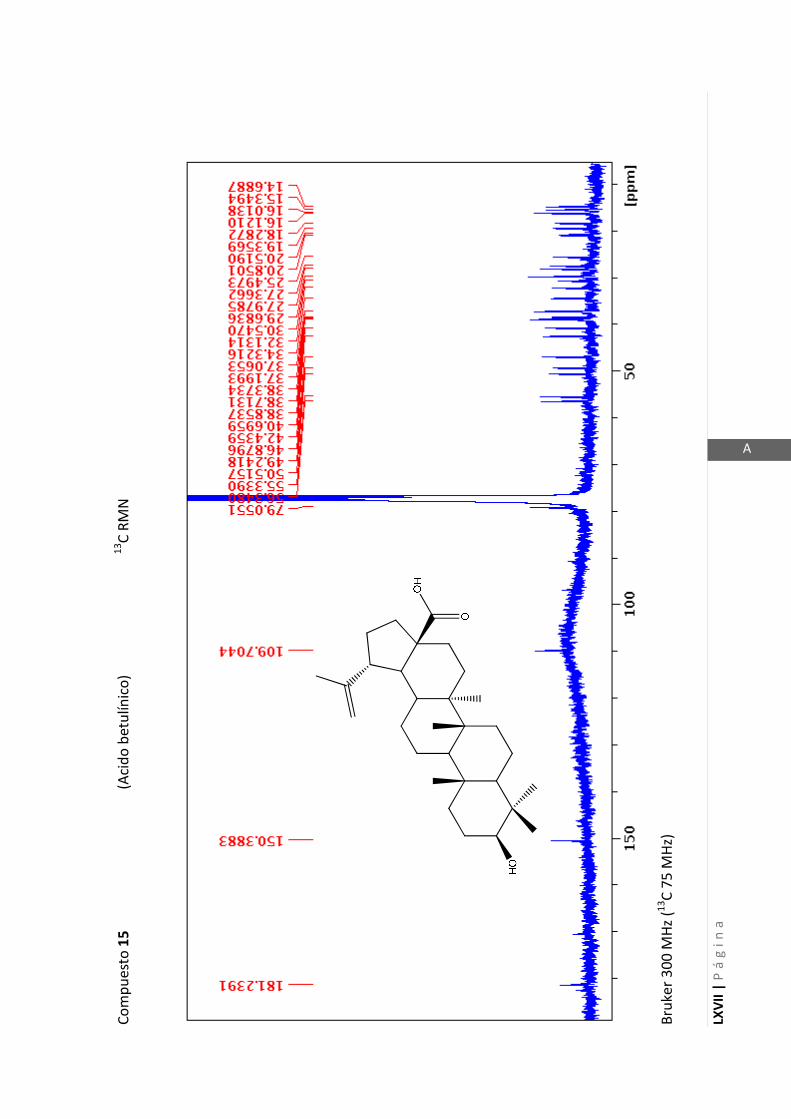
LXV
II |
Pá
gi
na
A
Co
mp
ue
sto
15
(A
cid
o b
etu
lín
ico
)
1
3C
RM
N
Bru
ke
r 3
00
MH
z (1
3C
75
MH
z)
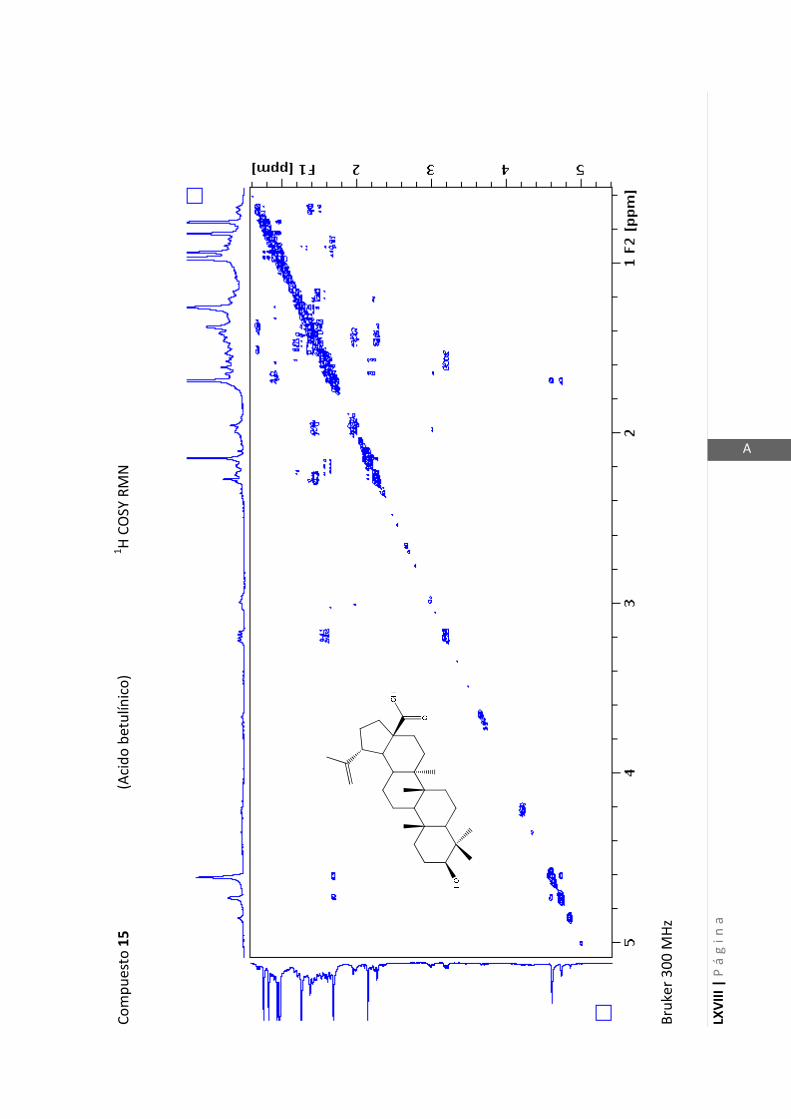
LXV
III
| P
ág
in
a
A
Co
mp
ue
sto
15
(A
cid
o b
etu
lín
ico
)
1H
CO
SY
RM
N
Bru
ke
r 3
00
MH
z
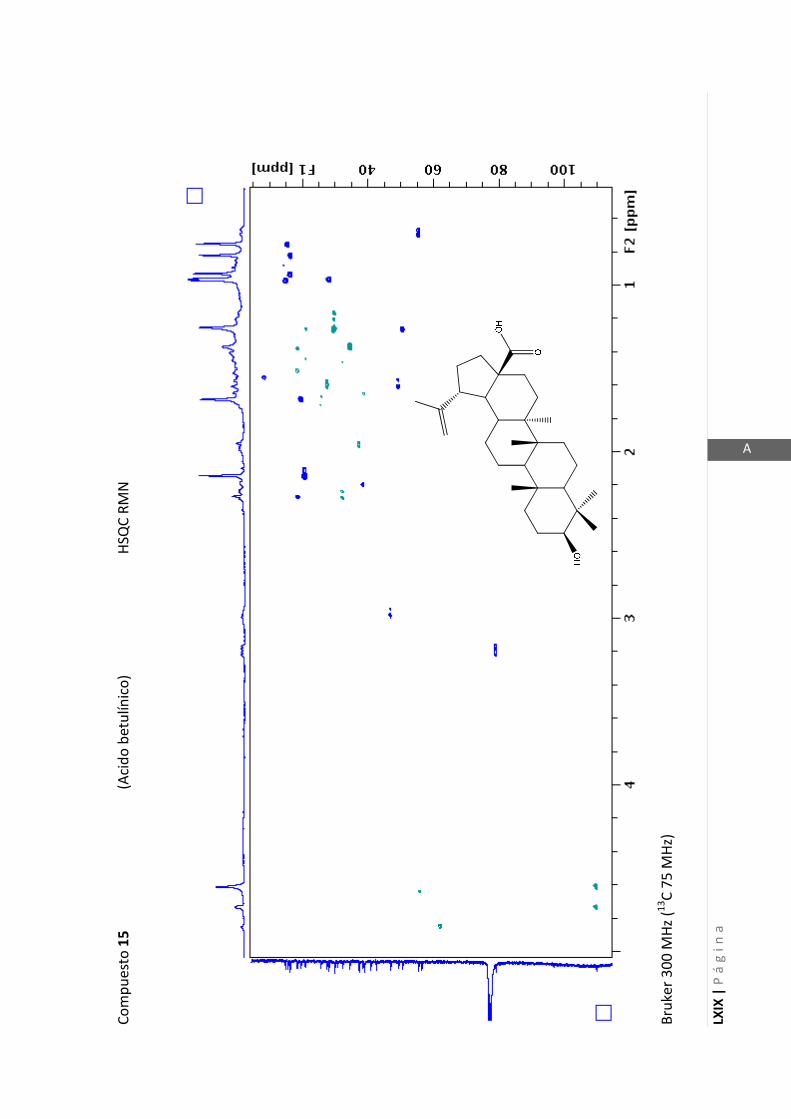
LXIX
| P
ág
in
a
A
Co
mp
ue
sto
15
(A
cid
o b
etu
lín
ico
)
H
SQ
C R
MN
Bru
ke
r 3
00
MH
z (1
3C
75
MH
z)
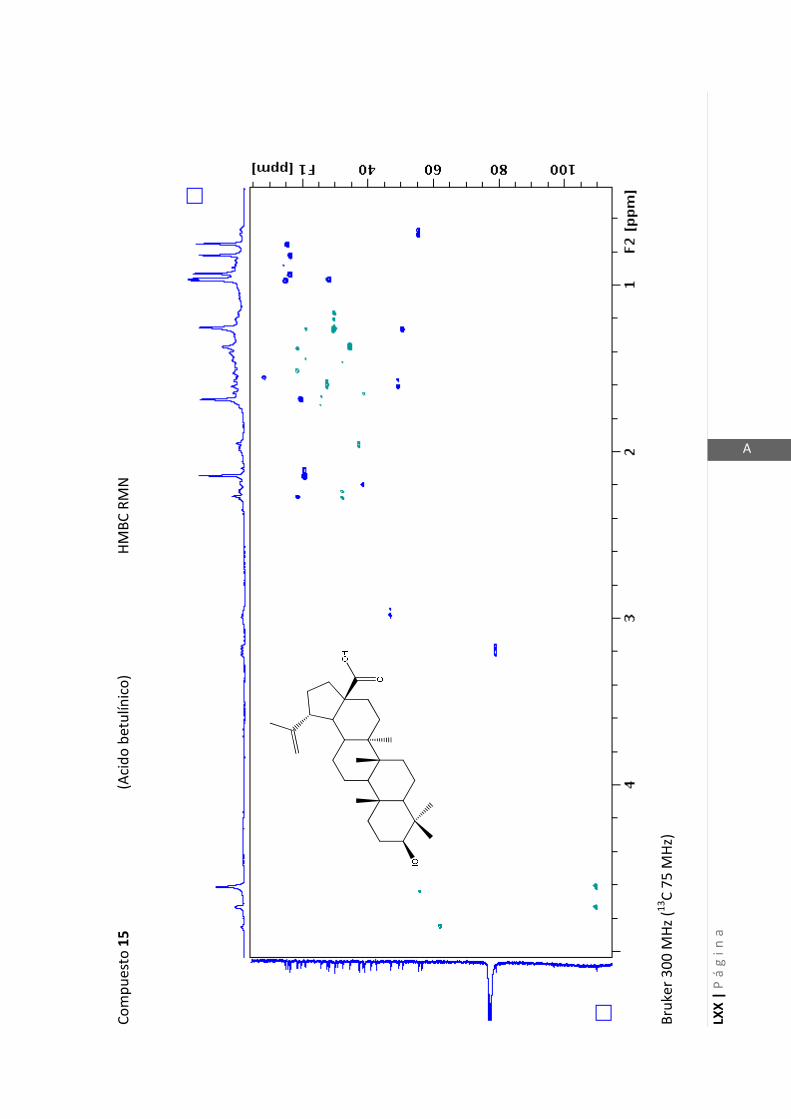
LXX
| P
ág
in
a
A
Co
mp
ue
sto
15
(A
cid
o b
etu
lín
ico
)
H
MB
C R
MN
Bru
ke
r 3
00
MH
z (1
3C
75
MH
z)
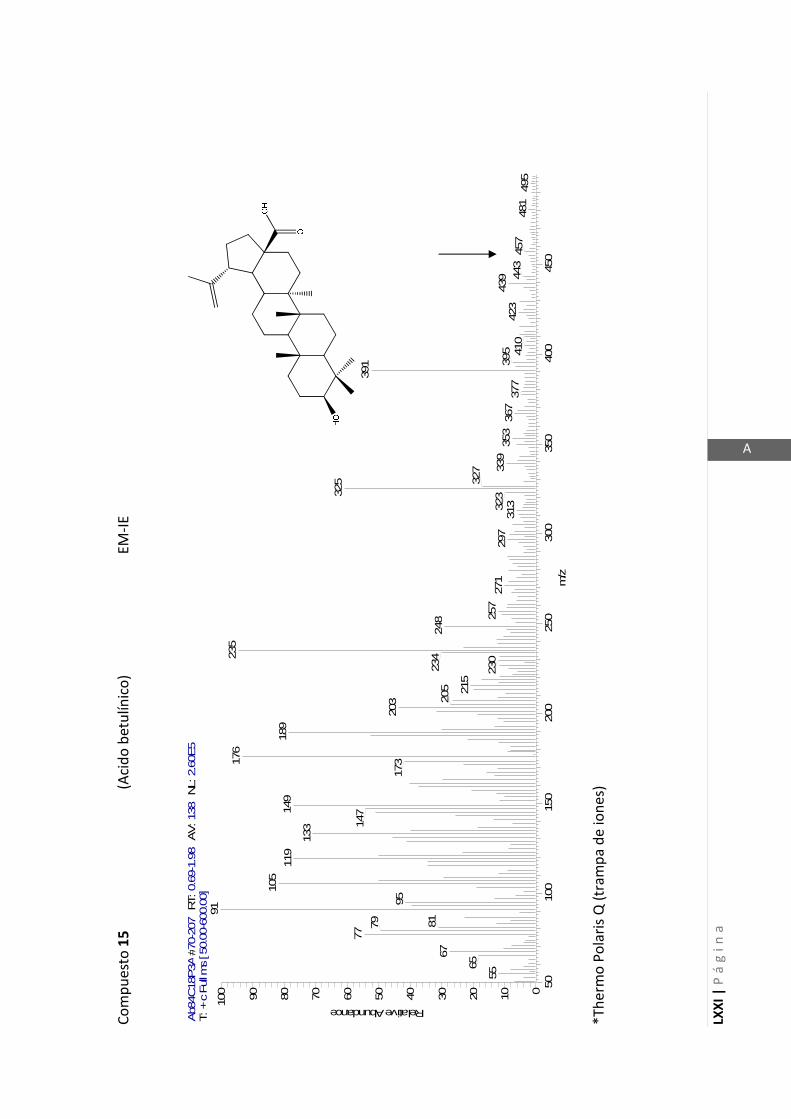
LXX
I |
Pá
gi
na
A
Co
mp
ue
sto
15
(A
cid
o b
etu
lín
ico
)
E
M-I
E
*T
he
rmo
Po
lari
s Q
(tr
am
pa
de
io
ne
s)
Ab8
4C18
P3A
#70
-207
RT:
0.69
-1.9
8AV:13
8NL:
2.60
E5
T:+
c Full m
s [ 5
0.00
-600
.00]
5010
015
020
025
030
035
040
045
0m
/z
0102030405060708090100
Relative Abundance
91
235
176
105
189
149
119
133
325
7714
739
179
203
173
95
8123
424
867
205 21
565
327
230
257
5527
132
333
929
743
935
339
536
731
342
337
744
341
045
748
149
5
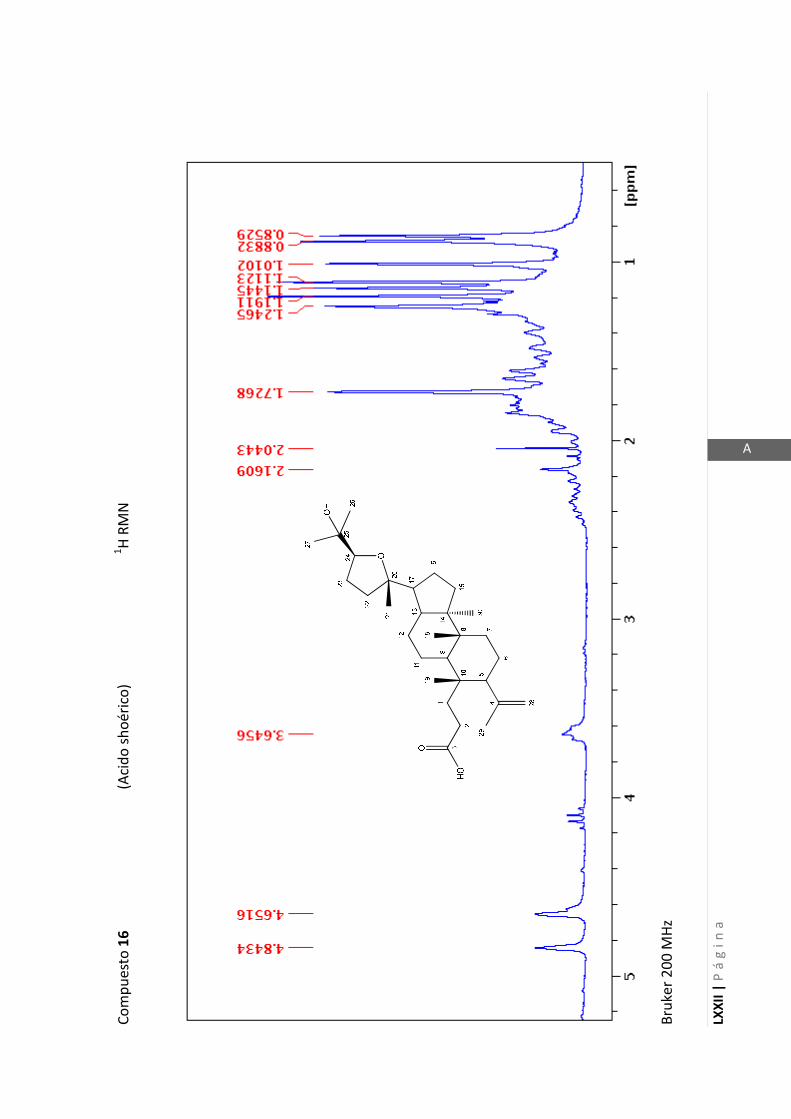
LXX
II |
Pá
gi
na
A
Co
mp
ue
sto
16
(A
cid
o s
ho
éri
co)
1H
RM
N
Bru
ke
r 2
00
MH
z
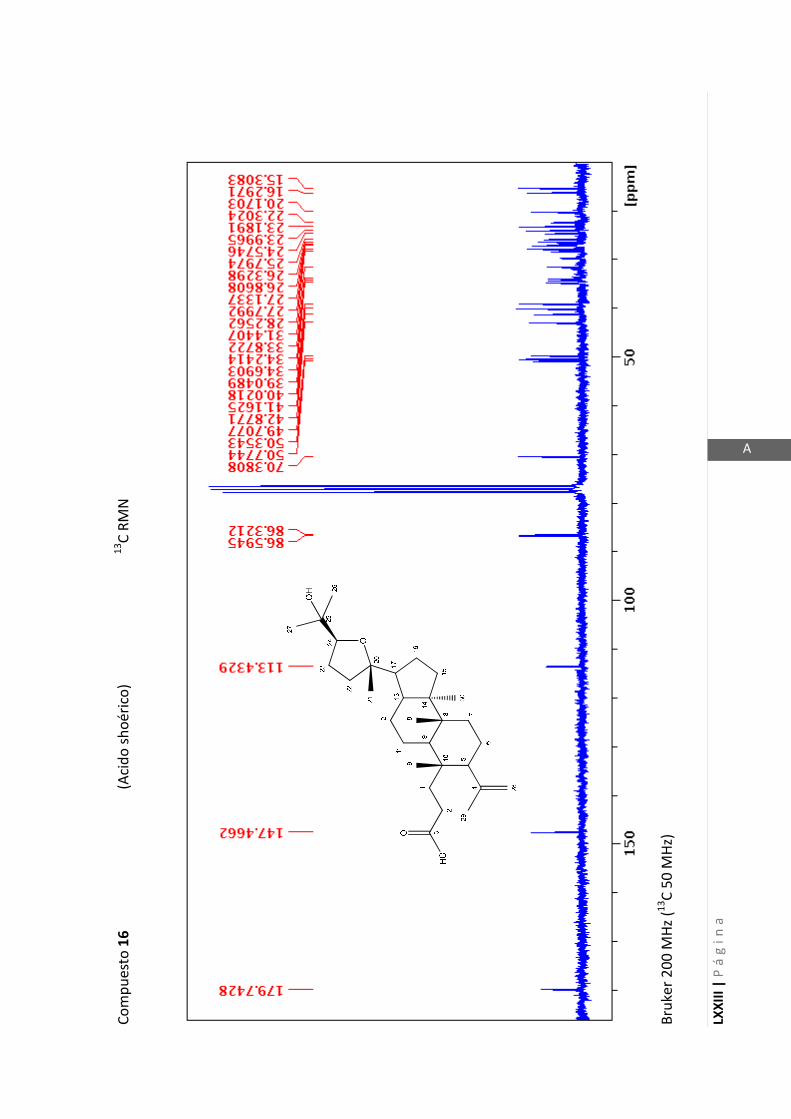
LXX
III
| P
ág
in
a
A
Co
mp
ue
sto
16
(A
cid
o s
ho
éri
co)
13C
RM
N
Bru
ke
r 2
00
MH
z (1
3C
50
MH
z)
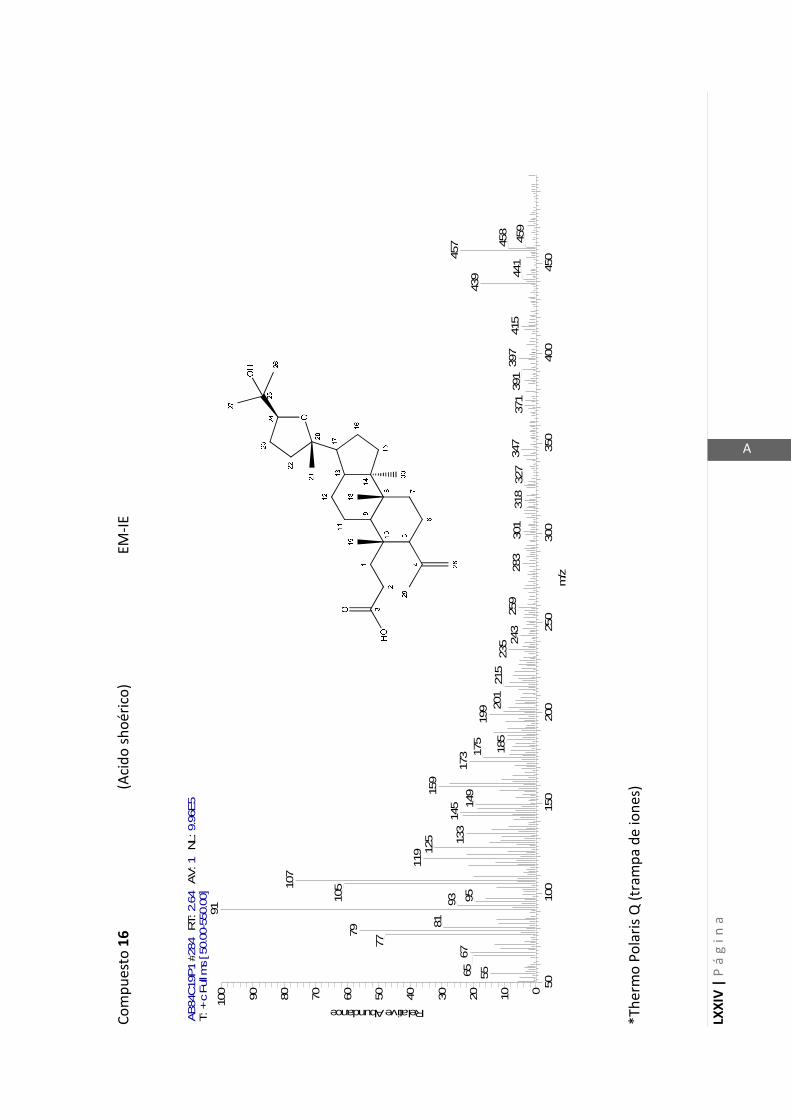
LXX
IV |
Pá
gi
na
A
Co
mp
ue
sto
16
(A
cid
o s
ho
éri
co)
EM
-IE
*T
he
rmo
Po
lari
s Q
(tr
am
pa
de
io
ne
s)
AB84
C19
P1
#284
RT:
2.64
AV:1
NL:
9.96
E5
T:+
c Full m
s [ 5
0.00
-550
.00]
5010
015
020
025
030
035
040
045
0m
/z
0102030405060708090100
Relative Abundance
91
107
105
79
77
119 12
515
981
9345
714
513
317
367
6595
149
439
175
199
5520
121
518
523
545
825
939
724
341
534
728
339
130
144
131
832
737
145
9
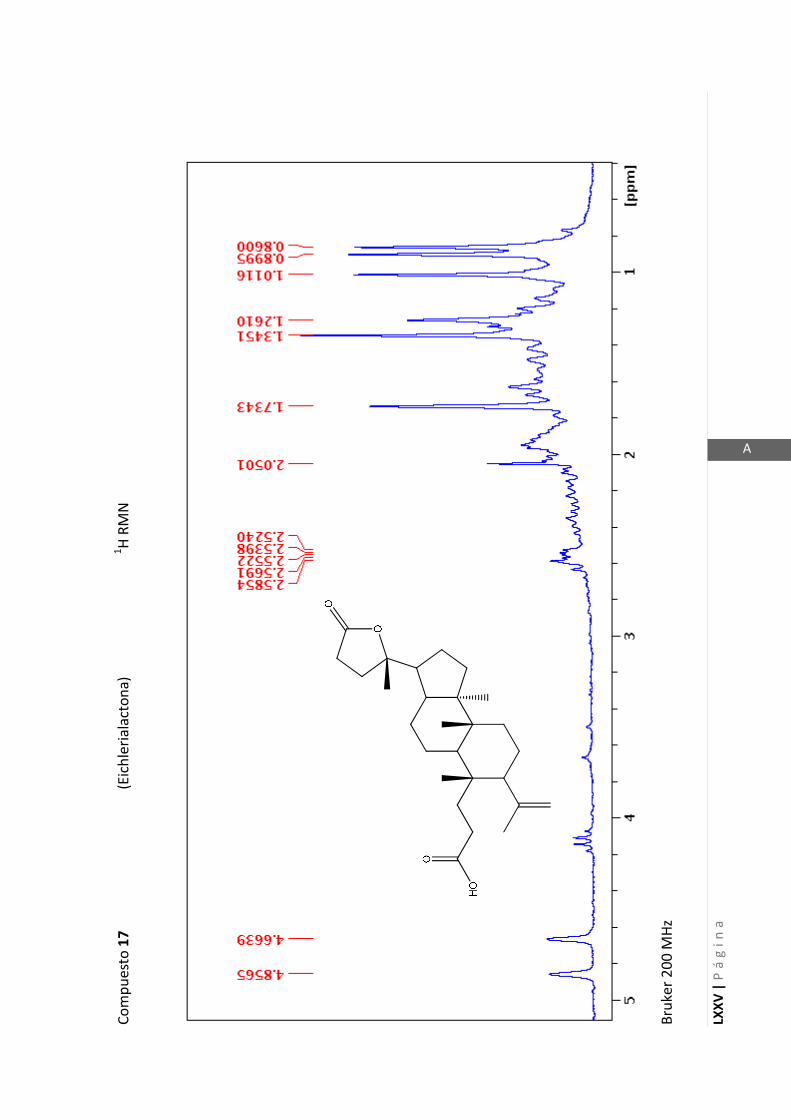
LXX
V |
Pá
gi
na
A
Co
mp
ue
sto
17
(E
ich
leri
ala
cto
na
)
1H
RM
N
Bru
ke
r 2
00
MH
z
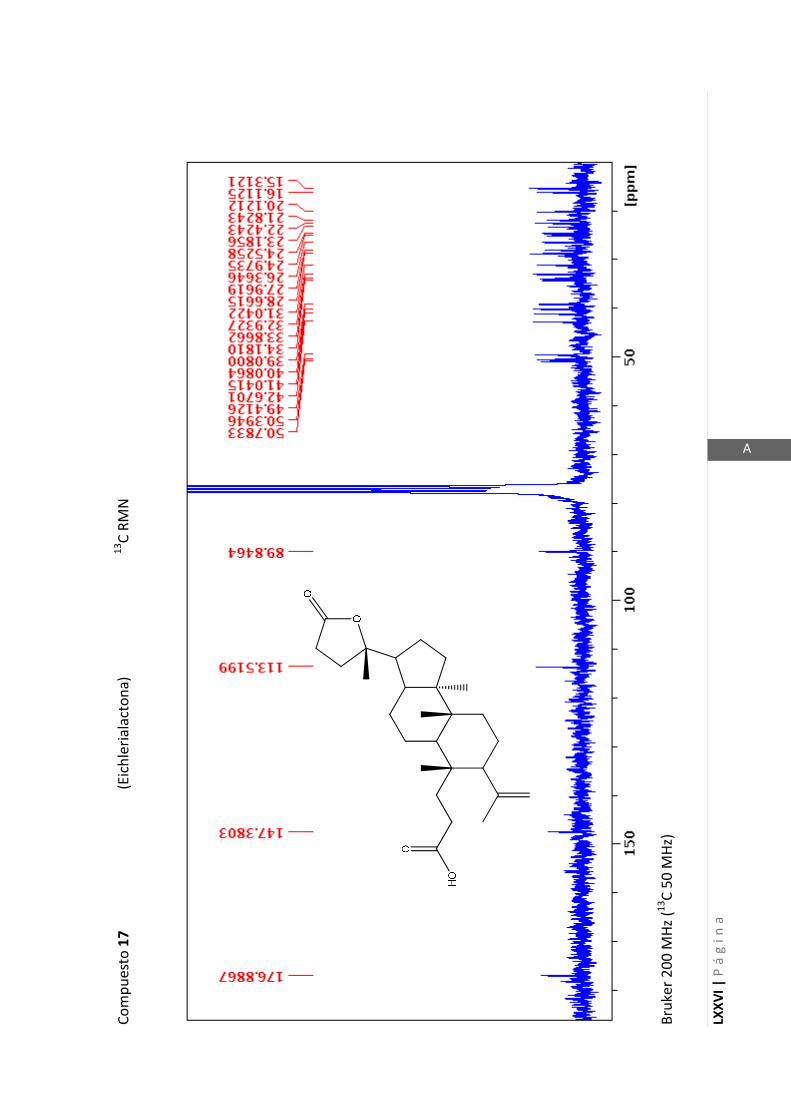
LXX
VI
| P
ág
in
a
A
Co
mp
ue
sto
17
(E
ich
leri
ala
cto
na
)
1
3C
RM
N
Bru
ke
r 2
00
MH
z (1
3C
50
MH
z)
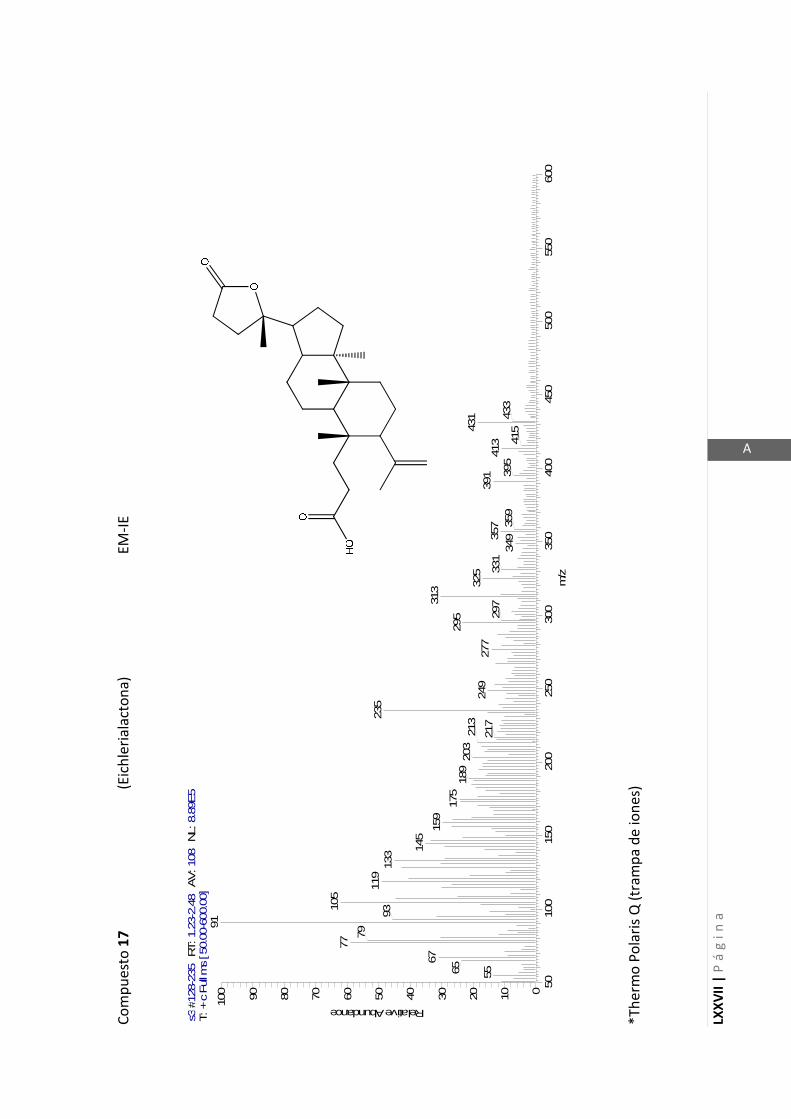
LXX
VII
| P
ág
in
a
A
Co
mp
ue
sto
17
(E
ich
leri
ala
cto
na
)
E
M-I
E
*T
he
rmo
Po
lari
s Q
(tr
am
pa
de
io
ne
s)
s3 #
128-
235
RT:
1.23
-2.4
8AV:10
8NL:
8.89
E5
T:+
c Full m
s [ 5
0.00
-600
.00]
5010
015
020
025
030
035
040
045
050
055
060
0m
/z
0102030405060708090100
Relative Abundance
91
105
77
7911
923
593
133 14
567
313
159
175
6529
518
920
343
121
332
524
927
739
155
217
357
413
331
297
433
395
359
349
415
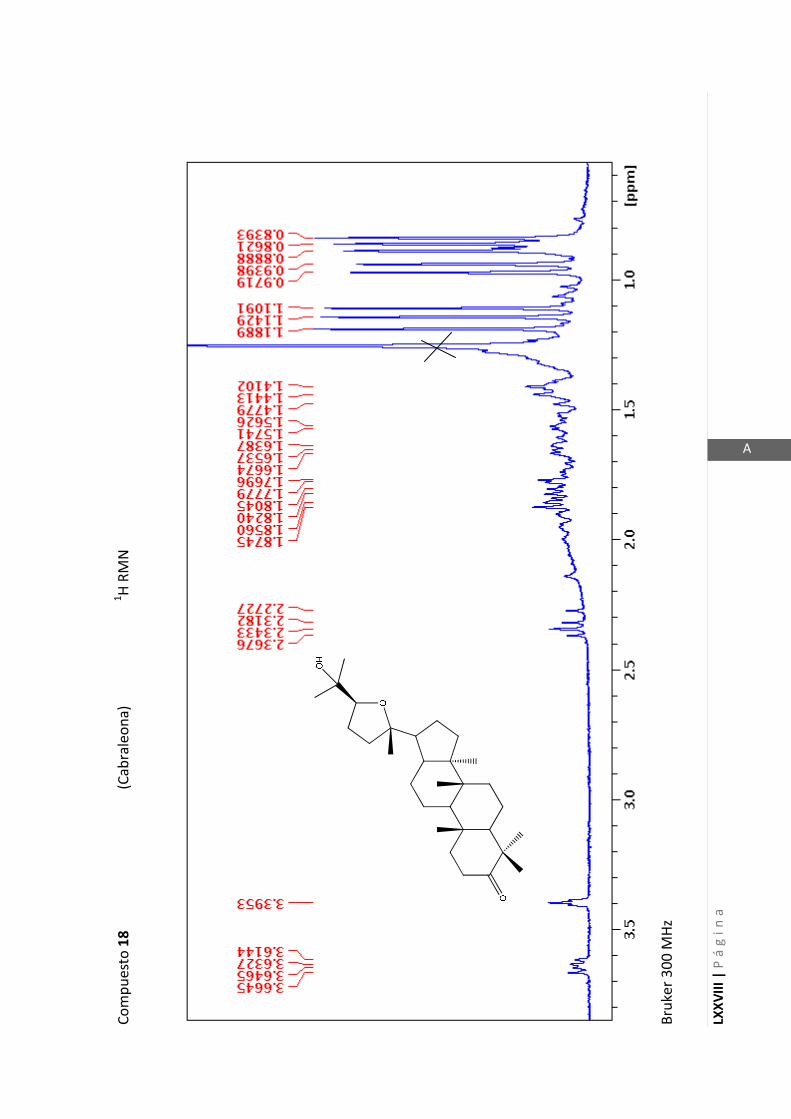
LXX
VII
I |
Pá
gi
na
A
Co
mp
ue
sto
18
(C
ab
rale
on
a)
1H
RM
N
Bru
ke
r 3
00
MH
z
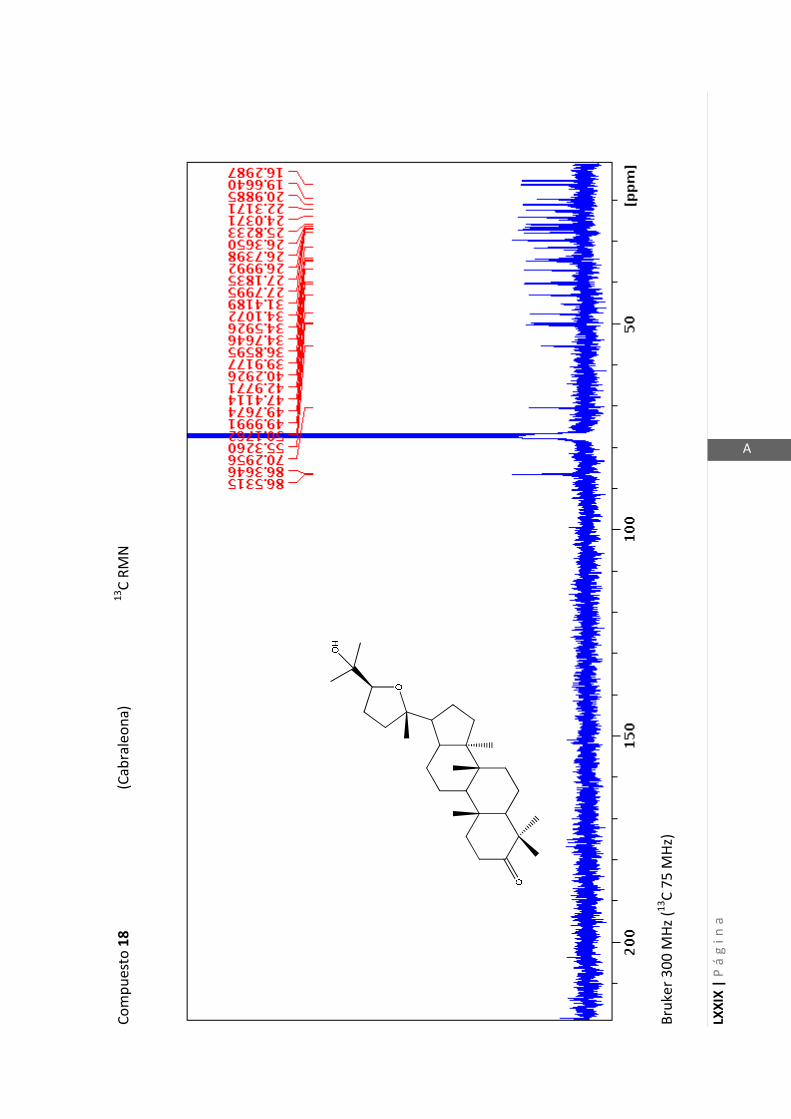
LXX
IX |
Pá
gi
na
A
Co
mp
ue
sto
18
(C
ab
rale
on
a)
13C
RM
N
Bru
ke
r 3
00
MH
z (1
3C
75
MH
z)
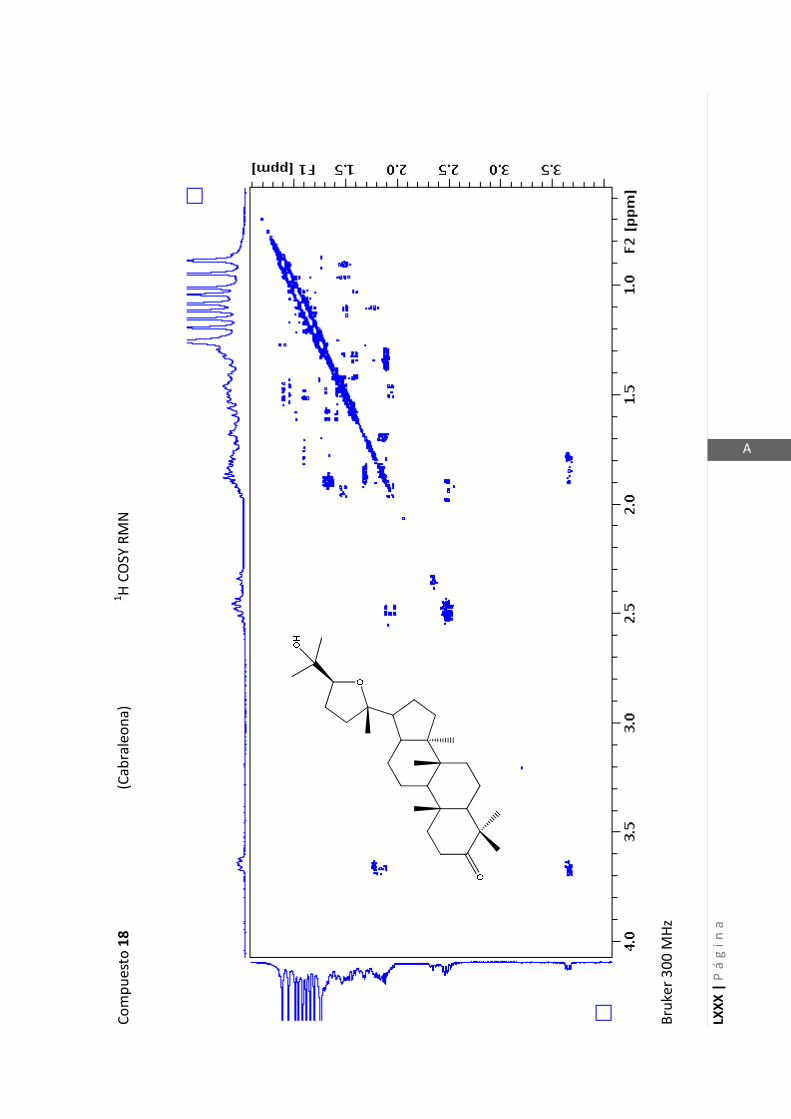
LXX
X |
Pá
gi
na
A
Co
mp
ue
sto
18
(C
ab
rale
on
a)
1H
CO
SY
RM
N
Bru
ke
r 3
00
MH
z
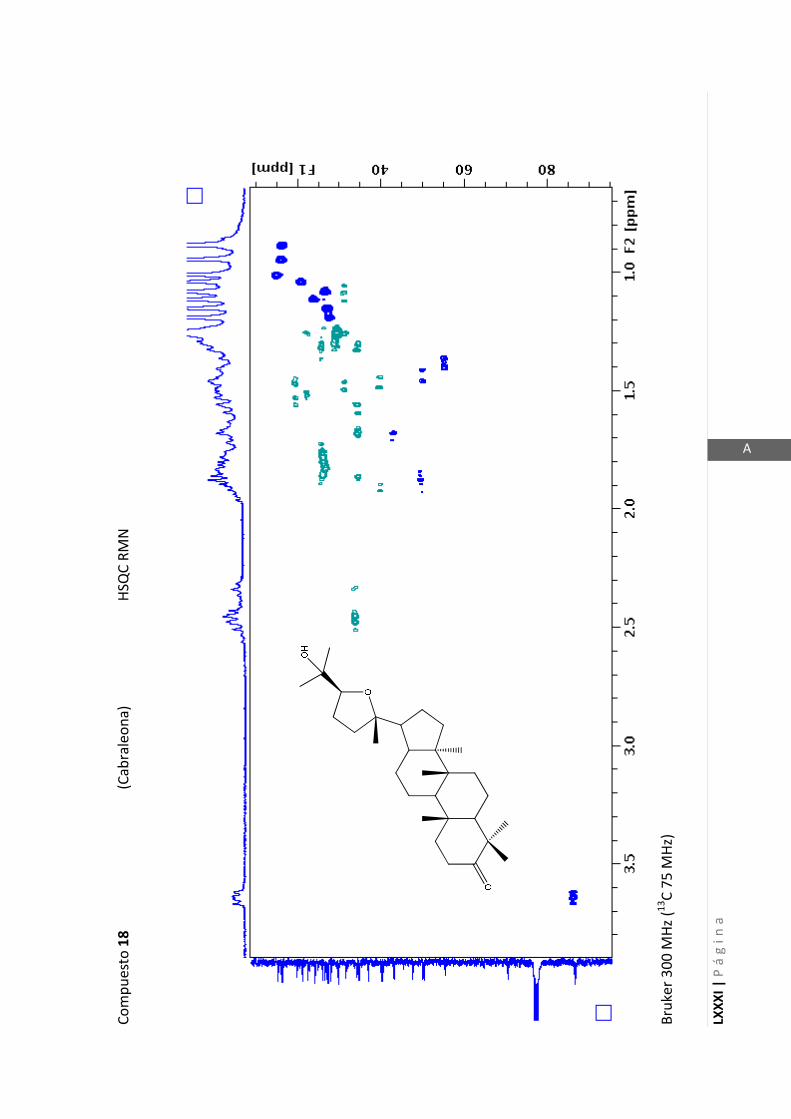
LXX
XI
| P
ág
in
a
A
Co
mp
ue
sto
18
(C
ab
rale
on
a)
HS
QC
RM
N
Bru
ke
r 3
00
MH
z (1
3C
75
MH
z)
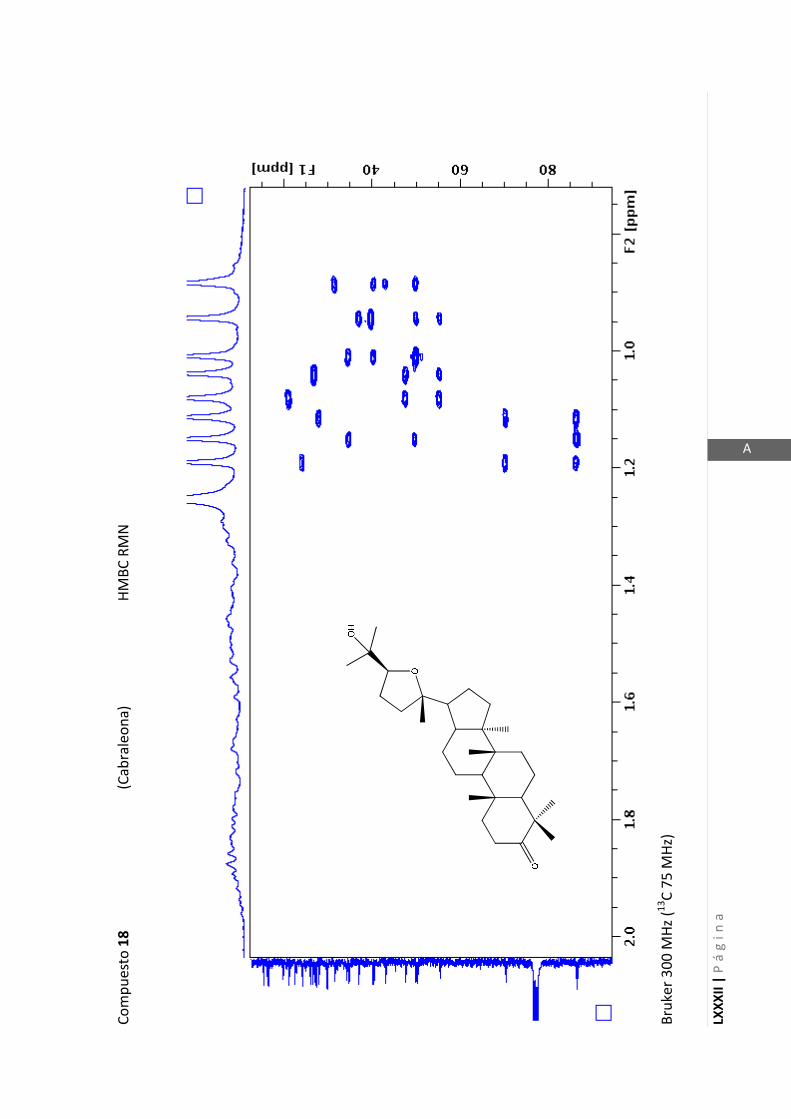
LXX
XII
| P
ág
in
a
A
Co
mp
ue
sto
18
(C
ab
rale
on
a)
HM
BC
RM
N
Bru
ke
r 3
00
MH
z (1
3C
75
MH
z)

LXX
XII
I |
Pá
gi
na
A
Co
mp
ue
sto
18
(C
ab
rale
on
a)
EM
-IE
*T
he
rmo
Po
lari
s Q
(tr
am
pa
de
io
ne
s)
AB84
C16
P5
#105
-131
RT:
1.03
-1.3
5AV:27
NL:
1.74
E6
T:+
c Full m
s [ 5
0.00
-600
.00]
5010
015
020
025
030
035
040
045
050
0m
/z
0102030405060708090100
Relative Abundance
441
143
107
126
205
442
145
109
423
175
9296
120
161
177
207
7956
203
279
6713
514
938
139
921
759
459
424
7030
140
031
322
142
524
525
528
527
3
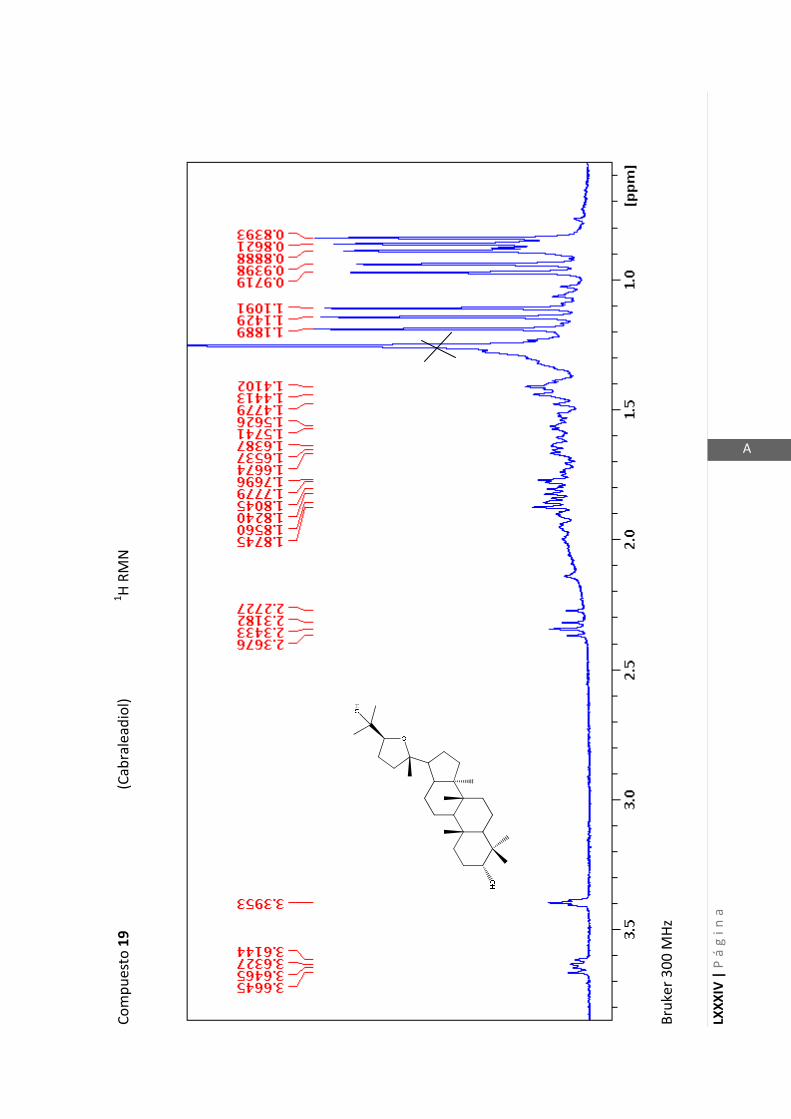
LXX
XIV
| P
ág
in
a
A
Co
mp
ue
sto
19
(C
ab
rale
ad
iol)
1H
RM
N
Bru
ke
r 3
00
MH
z
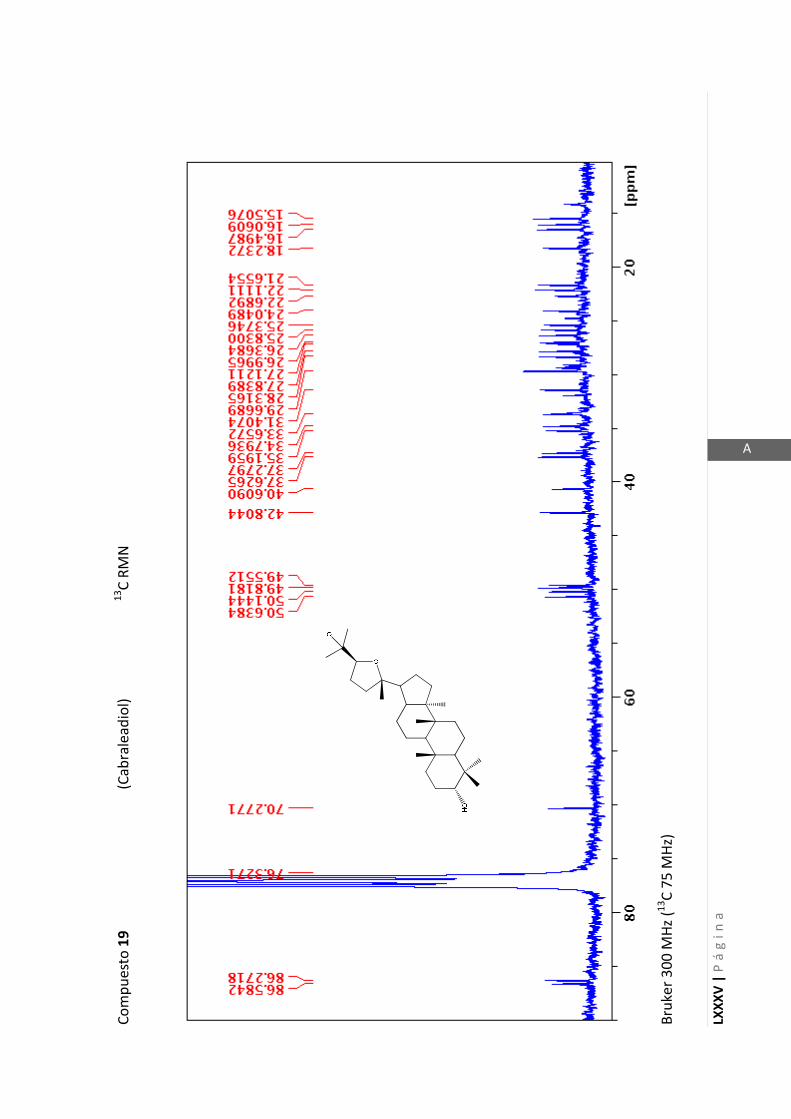
LXX
XV
| P
ág
in
a
A
Co
mp
ue
sto
19
(C
ab
rale
ad
iol)
13C
RM
N
Bru
ke
r 3
00
MH
z (1
3C
75
MH
z)
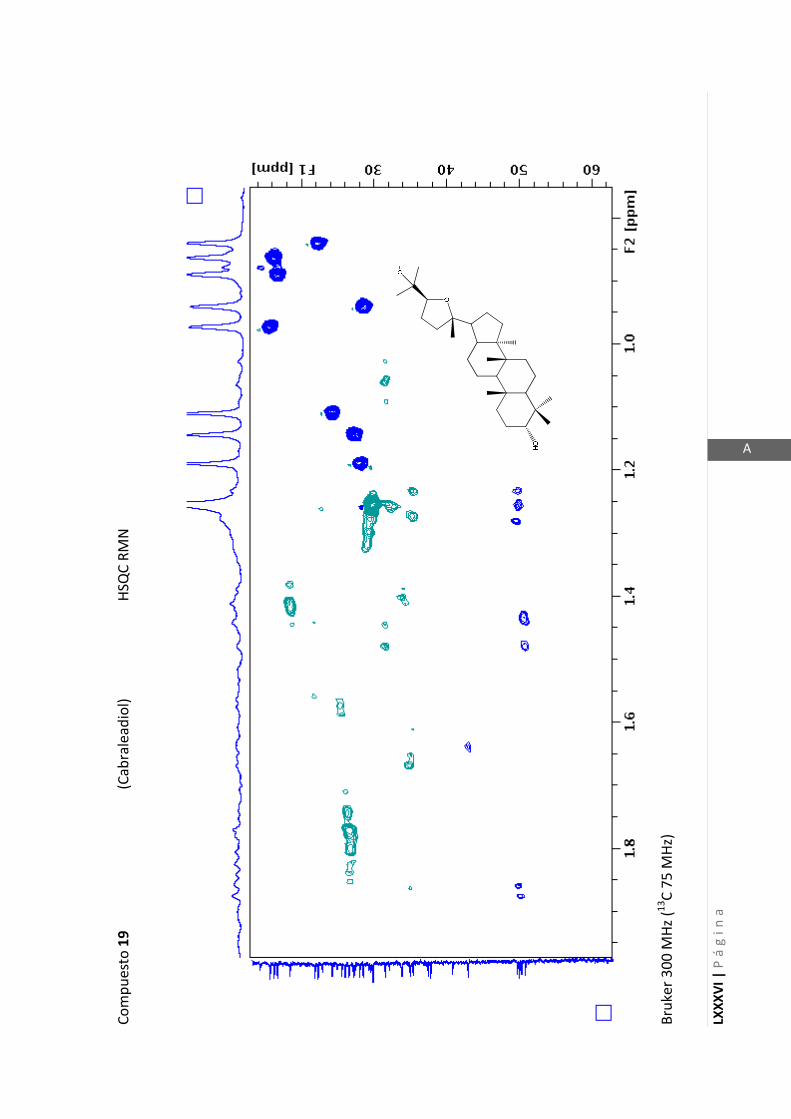
LXX
XV
I |
Pá
gi
na
A
Co
mp
ue
sto
19
(C
ab
rale
ad
iol)
HS
QC
RM
N
Bru
ke
r 3
00
MH
z (1
3C
75
MH
z)
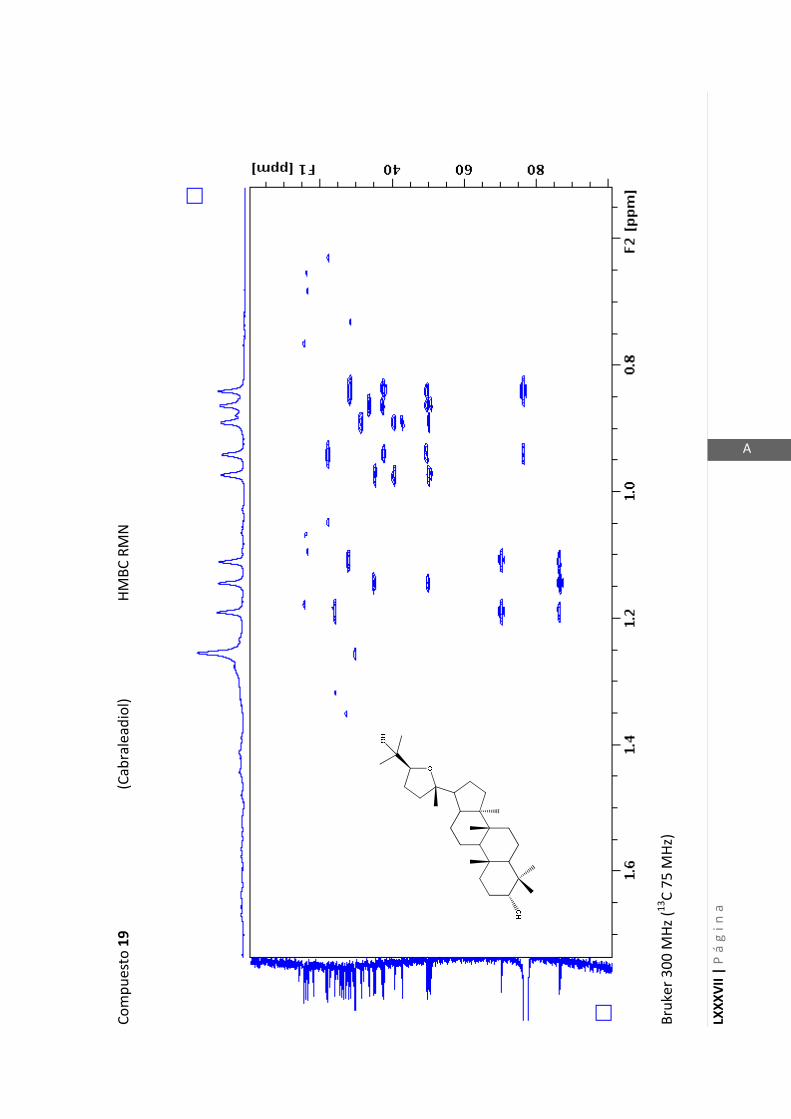
LXX
XV
II |
Pá
gi
na
A
Co
mp
ue
sto
19
(C
ab
rale
ad
iol)
HM
BC
RM
N
Bru
ke
r 3
00
MH
z (1
3C
75
MH
z)
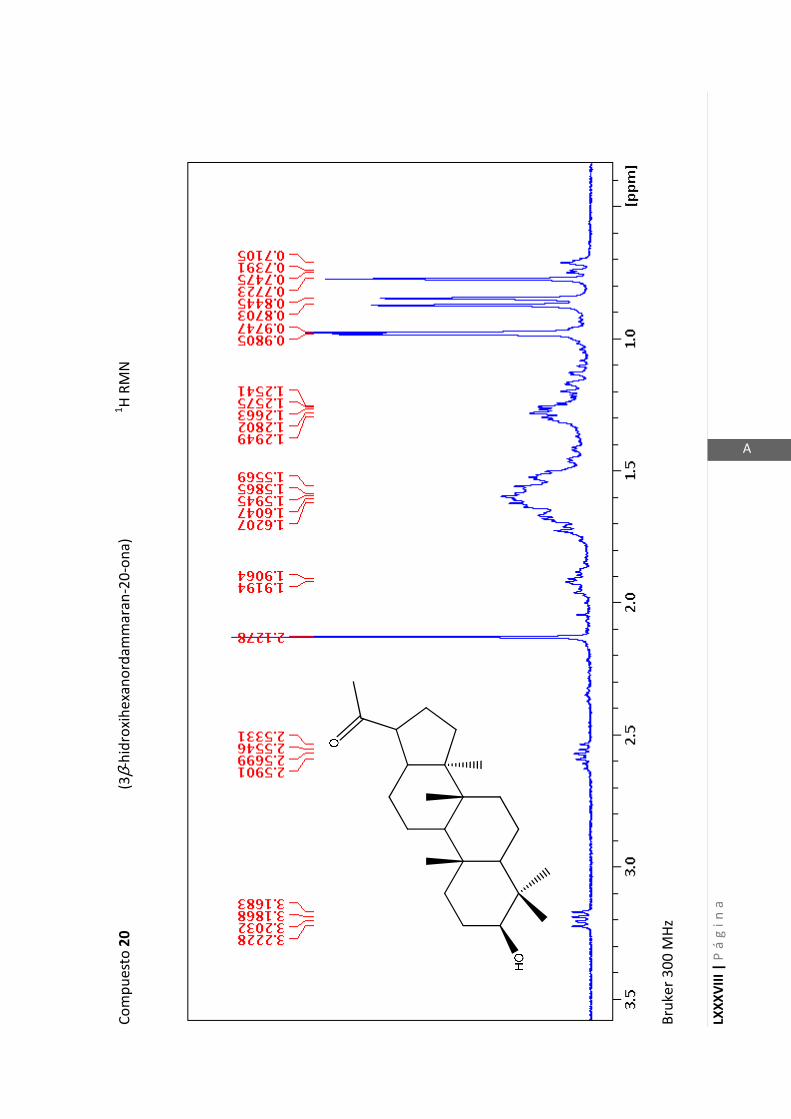
LXX
XV
III
| P
ág
in
a
A
Co
mp
ue
sto
20
(3
β-h
idro
xih
exa
no
rda
mm
ara
n-2
0-o
na
)
1H
RM
N
Bru
ke
r 3
00
MH
z
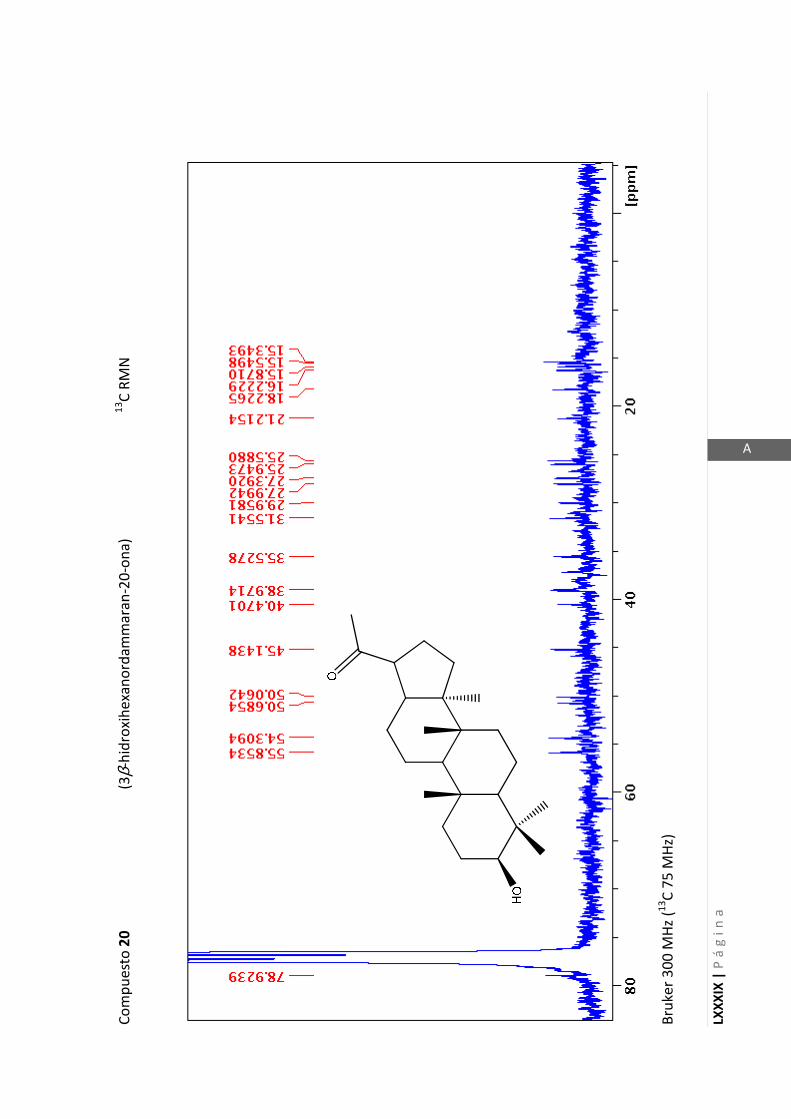
LXX
XIX
| P
ág
in
a
A
Co
mp
ue
sto
20
(3
β-h
idro
xih
exa
no
rda
mm
ara
n-2
0-o
na
)
1
3C
RM
N
Bru
ke
r 3
00
MH
z (1
3C
75
MH
z)
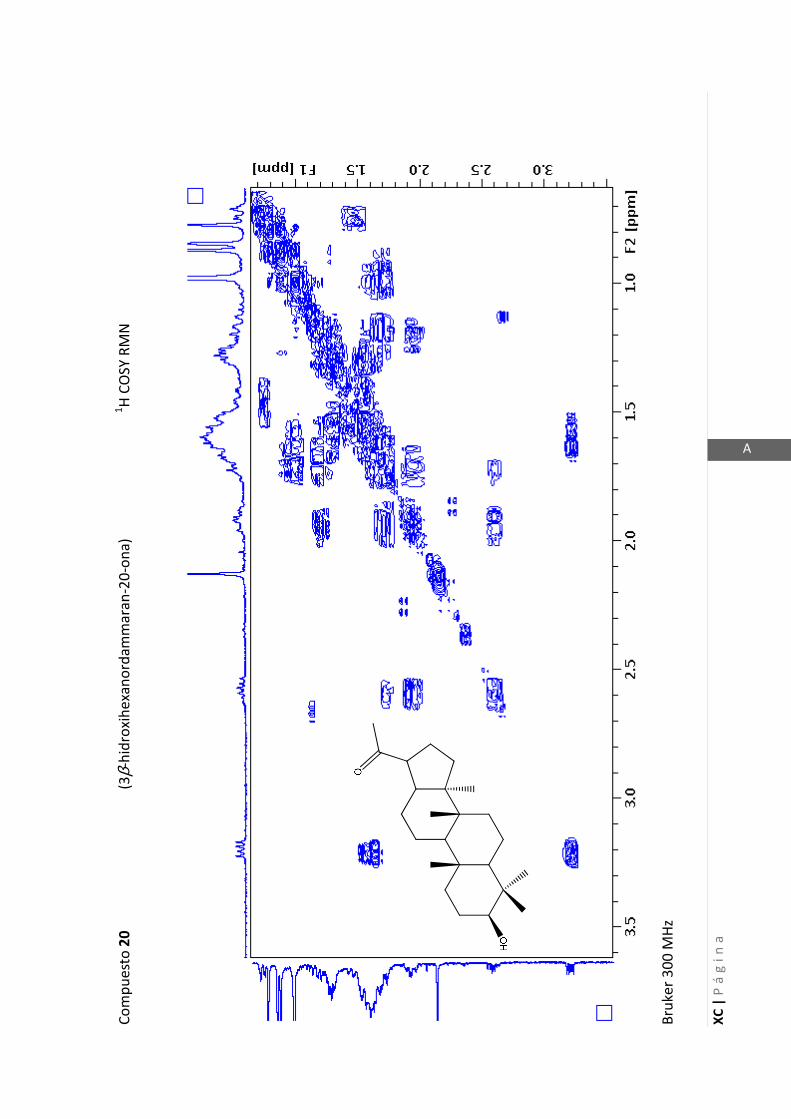
XC
| P
ág
in
a
A
Co
mp
ue
sto
20
(3
β-h
idro
xih
exa
no
rda
mm
ara
n-2
0-o
na
)
1H
CO
SY
RM
N
Bru
ke
r 3
00
MH
z
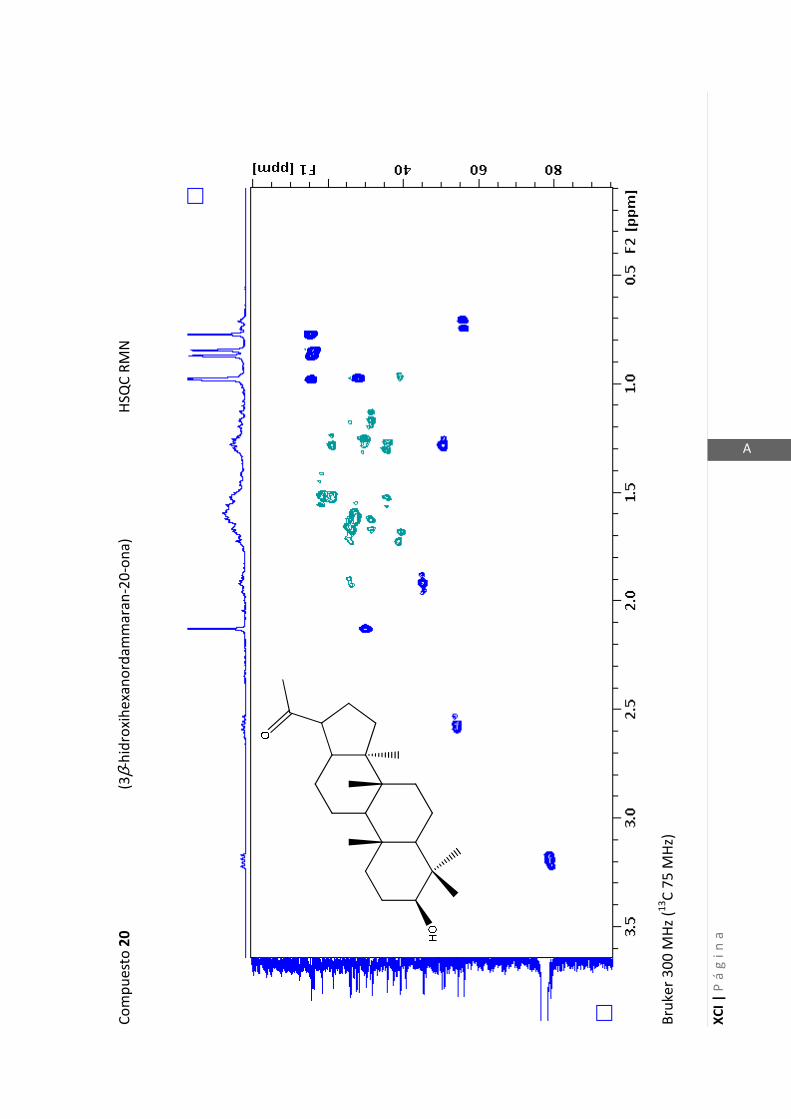
XC
I |
Pá
gi
na
A
Co
mp
ue
sto
20
(3
β-h
idro
xih
exa
no
rda
mm
ara
n-2
0-o
na
)
H
SQ
C R
MN
Bru
ke
r 3
00
MH
z (1
3C
75
MH
z)
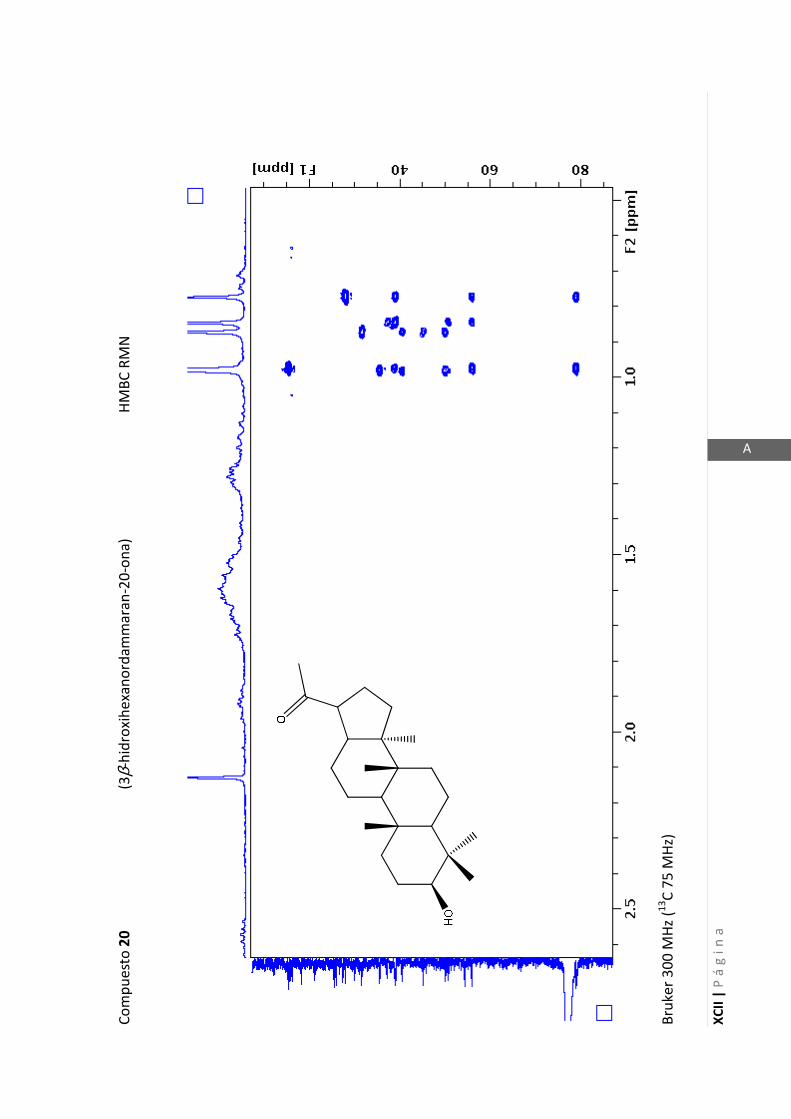
XC
II |
Pá
gi
na
A
Co
mp
ue
sto
20
(3
β-h
idro
xih
exa
no
rda
mm
ara
n-2
0-o
na
)
H
MB
C R
MN
Bru
ke
r 3
00
MH
z (1
3C
75
MH
z)
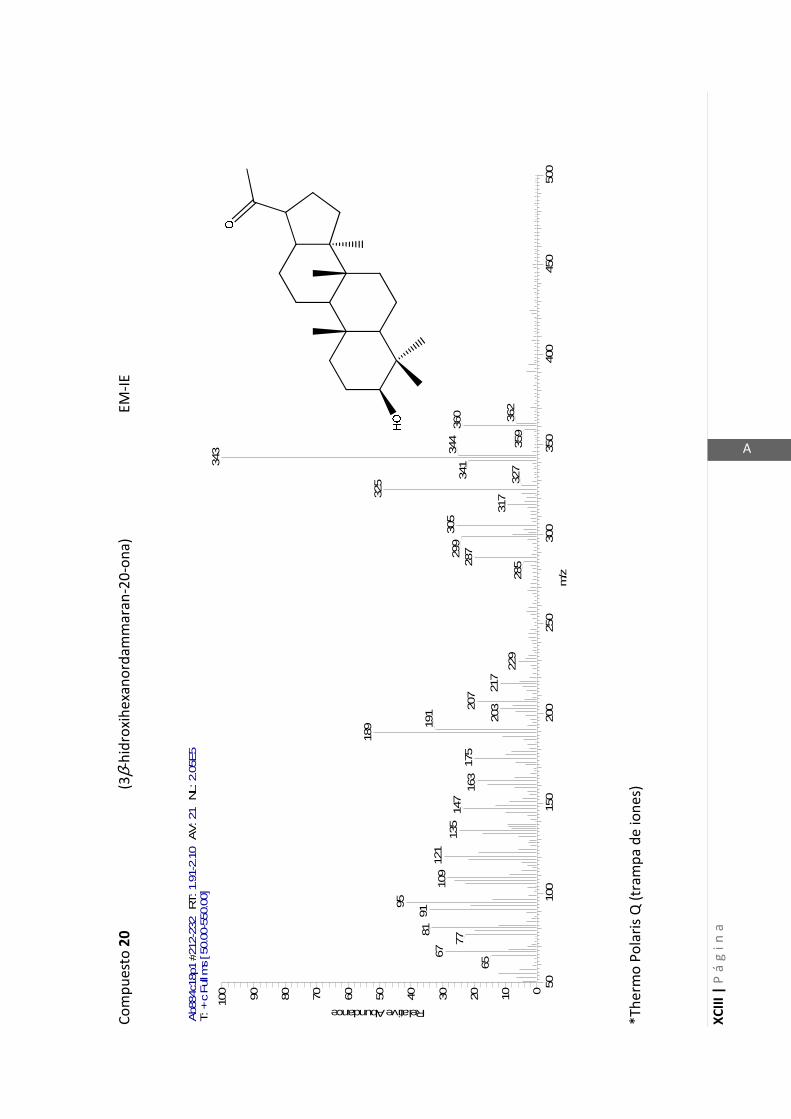
XC
III
| P
ág
in
a
A
Co
mp
ue
sto
20
(3
β-h
idro
xih
exa
no
rda
mm
ara
n-2
0-o
na
)
E
M-I
E
*T
he
rmo
Po
lari
s Q
(tr
am
pa
de
io
ne
s)
Ab8
84c1
8p1
#212
-232
RT:
1.91
-2.1
0AV:21
NL:
2.05
E5
T:+
c Full m
s [ 5
0.00
-550
.00]
5010
015
020
025
030
035
040
045
050
0m
/z
0102030405060708090100
Relative Abundance
343
189
325
95
9181
191
121
6710
930
534
413
529
936
014
777
341
175
287
163
207
6521
720
331
736
222
932
728
535
9
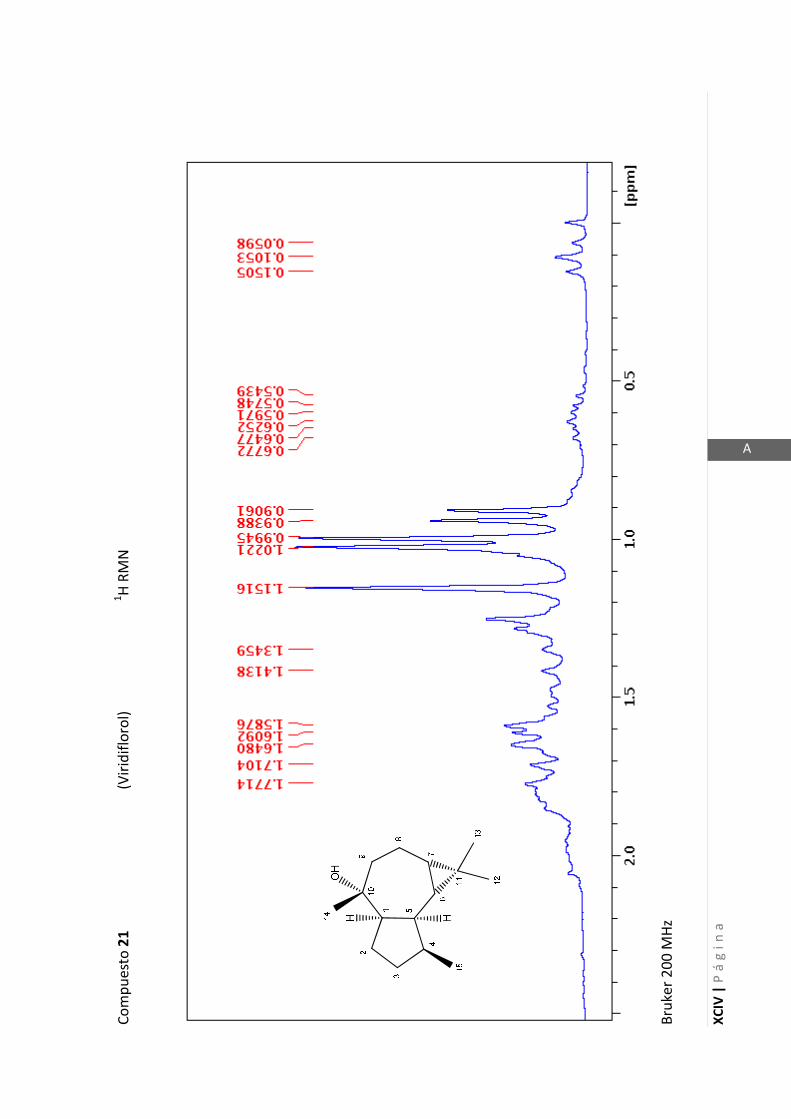
XC
IV |
Pá
gi
na
A
Co
mp
ue
sto
21
(V
irid
iflo
rol)
1H
RM
N
Bru
ke
r 2
00
MH
z
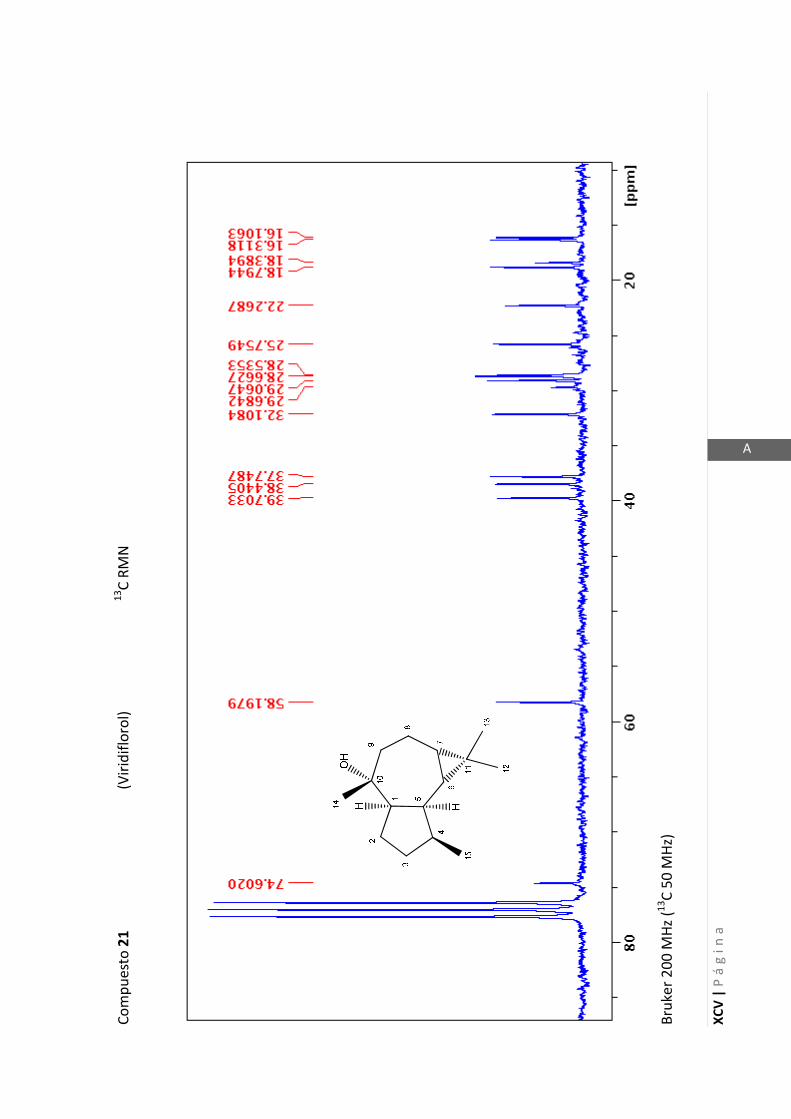
XC
V |
Pá
gi
na
A
Co
mp
ue
sto
21
(V
irid
iflo
rol)
1
3C
RM
N
Bru
ke
r 2
00
MH
z (1
3C
50
MH
z)

XC
VI
| P
ág
in
a
A
*Lo
s e
spe
ctro
s d
e m
asa
s o
bte
nid
os
con
el e
spe
ctró
me
tro
Th
erm
o P
ola
ris
Q (
tra
mp
a d
e i
on
es)
pu
ed
en
pre
sen
tar
pic
os
corr
esp
on
die
nte
s a
l io
n
cua
sim
ole
cula
r [M
+ H
]+ d
eb
ido
al fe
nó
me
no
de
au
to i
on
iza
ció
n q
uím
ica
(V
er
cap
ítu
lo I
I).














![Metabolitos Prim y Sec[1].](https://static.fdocuments.mx/doc/165x107/55cf8e2c550346703b8f4ff0/metabolitos-prim-y-sec1.jpg)


