Fisiologia de La Membrana Celular
33
FISIOLOGIA DE LA MEMBRANA CELULAR Contenido INTRODUCCIÓN...................................................3 MEMBRANAS CELULARES Y TRANSPORTE TRASNMEMBRANA DE SOLUTOS Y AGUA ...............................................................4 EQUILIBRIOS IÓNICOS Y POTENCIALES DE MEMBRANA EN REPOSO.......14 GENERACIÓN Y CONDUCCIÓN DEL POTENCIAL DE ACCIÓN...............18 RECEPTORES DE MEMBRANA, SEGUNDOS MENSAJEROS Y VÍAS DE TRANSDUCCIÓN DE SEÑALES.......................................22 CONCLUSIONES................................................. 25 BIBLIOGRAFÍA................................................. 26 ANEXOS.........................................................27 [Escribir texto] Página 1
Transcript of Fisiologia de La Membrana Celular

FISIOLOGIA DE LA MEMBRANA CELULAR
Contenido
INTRODUCCIÓN............................................................................................................................3
MEMBRANAS CELULARES Y TRANSPORTE TRASNMEMBRANA DE SOLUTOS Y AGUA..................4
EQUILIBRIOS IÓNICOS Y POTENCIALES DE MEMBRANA EN REPOSO.........................................14
GENERACIÓN Y CONDUCCIÓN DEL POTENCIAL DE ACCIÓN.......................................................18
RECEPTORES DE MEMBRANA, SEGUNDOS MENSAJEROS Y VÍAS DE TRANSDUCCIÓN DE SEÑALES.....................................................................................................................................22
CONCLUSIONES.........................................................................................................................25
BIBLIOGRAFÍA............................................................................................................................26
ANEXOS........................................................................................................................................27
[Escribir texto] Página 1

Seminario de Histología
INTRODUCCIÓN
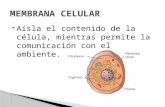
La membrana plasmática o citoplasmática es una estructura laminar que engloba a las células, define sus límites y contribuye a mantener el equilibrio entre el interior y el exterior de éstas.
La composición de la membrana celular le confiere la propiedad de selectividad con la cual algunas sustancias van a pasar con mucha facilidad hacia el citoplasma, como otras se quedarán fuera por el tamaño de su molécula.
Es importante también describir el potencial de acción que pueden desencadenar por la gradiente de sus iones intracelulares y extracelulares.
La membrana plasmática cumple diferente e importantes funciones que le permiten mantener la homeostasis de la misma célula.
En el presente trabajo estudiaremos la fisiología de la membrana celular y la importancia que tiene esta parta llevar a cabo las funciones del cuerpo humano.
Página 2

Seminario de Histología
MEMBRANAS CELULARES Y TRANSPORTE TRASNMEMBRANA DE SOLUTOS Y AGUA
LAS MEMBRANAS DIVIDEN LA CÉLULA EN COMPARTIMIENTOS CON FUNCIONES BIOQUÍMICAS ESPECÍFICAS.
Las membranas son una barrera de permeabilidad de la célula permitiéndole mantener su composición diferente a la del liquido extracelular; contiene enzimas, receptores y antígenos para interactuar con otras células, hormonas y otros agentes reguladores del liquido extracelular. Las proteínas de la membrana interactúan con las del citoesqueleto y la matriz extracelular interviniendo en la traducción de señales.
Las membranas que engloban organelas dividen su célula en compartimientos independientes en el que se llevan a cabo procesos bioquímicos particulares en organelas especificas. Ejemplo: procesos de transporte de electrones y fosforilación oxidativa que se da sobre, dentro y a través de la membrana mitocondrial interna.
Las membranas celulares presentan características comunes, sin embargo de acuerdo a la función que realice su composición y estructura difiere una célula de otra.
La matriz de las membranas, constituida por una bicapa lipídica, es una barrera para la permeabilidad de la mayoría de las sustancias.
Las proteínas y los fosfolípidos son los constituyentes más abundantes de las membranas celulares. La bicapa fosfolipídica es responsable de las propiedades de permeabilidad pasiva de las membranas. Las sustancias que son muy hidrosolubles suelen atravesar la membrana muy lentamente; en cambio los compuestos no polares hidrófobos, la cruzan con mayor rapidez.
La mayoría de las membranas consisten en un “mosaico fluido” de fosfolípidos y proteínas.
El modelo del mosaico fluido explica la mayoría de las propiedades de las membranas biológicas. Los fosfolípidos se encuentran formando la bicapa y también encontramos proteínas de 2 clases: integrales o intrínsecas incrustadas en la bicapa de fosfolípidos y extrínsecas o periféricas, situadas en su superficie. La interacción de la proteína con la membrana se estabiliza mediante “anclajes” hidrofóbicos asociados covalentemente y se insertan en la bicapa lipídica (un ácido graso unido a la proteína que se intercala entre las cadenas de ácidos grasos). Las proteínas integrales presentan interacciones hidrófobas aun más extensas con el interior de la membrana.
Las membranas celulares son estructuras fluidas, los lípidos y proteínas pueden moverse sobre el plano de la membrana, el intercambio de estas moléculas de una monocapa a otra es infrecuente. Existe algunos casos en el que los componentes de la membrana no pueden difundirse libremente en el plano de esta, ejemplo de ello es el secuestro de los receptores de
Página 3

Seminario de Histología
acetilcolina (proteína integral de membrana) en la placa motora terminal del musculo esquelético.
LAS MEMBRANAS ESTÁN COMPUESTAS DE LÍPIDOS Y PROTEÍNAS
Los fosfolípidos y el colesterol constituyen los principales componentes lipídicos de las membranas.
Los fosfolípidos más abundantes son los que contienen colina: lecitinas (fosfatidilcolinas), y las esfingomielinas. Los siguientes en abundancia son los aminofosfolípidos: fosfatidilserina y fosfatidiletanolamina. Otros fosfolípidos importantes presentes son el fosfatidilglicerol, el fostatidilinositol y la cardiolipina.
En el proceso de transducción de señales a nivel celular participan determinados fosfolípidos como el bifosfato de fosfatidilinositol (IP3) y diacilglicerol. El IP3 se libera en el citosol, donde actúa sobre los receptores del retículo endoplasmático para provocar la liberación del Ca+2
almacenado, lo que afecta una gran diversidad de procesos celulares. El diacilglicerol permanece en la membrana plasmática, donde interviene, junto con el Ca+2, en la activación de la proteincinasa C, una proteína muy importante en la transducción de señales.
El colesterol es un constituyente fundamental de las membranas celulares, su núcleo esteroideo se encuentra paralelo a las cadenas de ácidos grasos de los fosfolípidos de membrana. Actúa como amortiguador de la fluidez de la membrana, su presencia mantiene un rango intermedio de fluidez de la región de cadenas de ácidos grasos de la bicapa de
Página 4

Seminario de Histología
fosfolípidos en presencia de ciertos agentes, como los alcoholes y los anestésicos generales, que tienden a fluidificar las membranas biológicas.
Las porciones hidrocarbonadas orientadas hacia el exterior de los glucolípidos y las glucoproteinas actúan como receptores y antígenos.
A pesar de no ser abundantes los glucolípidos desempeñan importantes funciones. La parte de hidratos de carbono se encuentra en la cara externa de la membrana y suelen actuar como receptor o antígeno.
Los fosfolípidos se distribuyen asimétricamente entre las monocapas lipídicas interna y externa de la membrana.
Los componentes lipídicos en la bicapa no se distribuyen uniformemente. Los glucolípidos se ubican casi de forma exclusiva en la monocapa externa de la membrana. Los fosfolípidos también se distribuyen asimétricamente en las monocapas interna y externa, en la membrana de los eritrocitos por ejemplo la monocapa externa posee una mayor proporción de fosfolípidos que contienen colina por el contrario la monocapa interna alberga una mayoría de aminofosfolípidos.
Las proteínas de membrana son enzimas, transportadores y receptores
La composición proteica de las membranas puede ser simple o compleja. Las membranas funcionalmente especializadas del retículo sarcoplásmico del musculo esquelético y los discos del segmento externo de la capa de bastones de la retina contienen solamente unas pocas proteínas diferentes. Por el contrario, las membranas celulares, que realizan muchas funciones, pueden tener más de 100 constituyentes proteicos distintos. Las proteínas de membrana incluyen enzimas, proteínas transportadoras y receptoras para hormonas y neurotransmisoras.
Las membranas constituyen barreras de permeabilidad
Las membranas son impermeables para la mayoría de las sustancias hidrosolubles
Las membranas biológicas actúan como barreras de permeabilidad. La mayoría de moléculas en los sistemas vivos son muy hidrosolubles y muy poco solubles en disolventes no polares entonces estas moléculas no atraviesan la bicapa debido a su entorno no polar de su interior. Es por ello que la membrana celular proporciona una barrera de permeabilidad de la mayoría de moléculas hidrosolubles manteniendo la diferencia de concentración entre el citoplasma y el líquido extracelular para muchas sustancias.
Las sustancias pueden atravesar las membranas sin pasar entre las moléculas que las componen
Página 5

Seminario de Histología
Algunas sustancias ingresan a la célula sin pasar a través de la membrana mediante el proceso de endocitosis que incluye la fagocitosis (captación de partículas) y la pinocitosis (captación de partículas solubles).
Las moléculas son expulsadas de la célula mediante un proceso que se parece a una endocitosis inversa: exocitosis. Ejemplo de ello es la liberación de neurotransmisores. La exocitosis es responsable de la salida de las proteínas segregadas por muchas células, la liberación de enzimas pancreáticas desde las células acinares del páncreas es un ejemplo de ello.
El transporte de moléculas a través de las membranas se produce por difusión, osmosis y procesos mediados por proteínas.
El paso de moléculas se da por difusión entre las moléculas que constituyen la membrana, en otros casos por medio de proteínas transportadoras especificas de la membrana.
El O2 y CO2 son moléculas pequeñas solubles en disolventes no polares, atravesando la membrana biológica por difusión entre sus moléculas lipídicas. En cambio la glucosa que es una molécula mucho mayor no es muy soluble en los lípidos de la membrana sin embargo la atraviesa por medio de proteínas específicas transportadoras de glucosa.
Las membranas son más permeables a las sustancias liposolubles que a las hidrosolubles.
La permeabilidad de las membranas a una molécula concreta es proporcional a su solubilidad en el interior de la bicapa lipídica.
Página 6
Proceso de endocitosis

Seminario de Histología
Mientras más soluble sea el compuesto en disolventes no polares, mejor atravesará la membrana biológica. Ej.: Las vitaminas liposolubles se absorben en el intestino delgado por sus células epiteliales mediante difusión simple (membrana plasmática luminal). Las vitaminas hidrosolubles no difunden fácilmente por lo que requieren en mayoría proteínas especializadas trasportadoras de membrana.
Sólo las moléculas hidrosolubles bastante pequeñas pueden difundir con rapidez a través de la membrana.
Por ejemplo el agua penetra las membranas unas 100 veces más deprisa que lo que podría predecirse a partir de su radio molecular y liposolubilidad, esto debido a 2 motivos. Al igual que otras sustancias muy pequeñas hidrosolubles puede pasar a través de las moléculas de fosfolípidos adyacentes sin disolverse en la región ocupada por las cadenas laterales de ácidos grasos. La mayoría de membranas plasmáticas de las células contienen proteínas denominadas acuaporinas.
La permeabilidad de las membranas a las moléculas hidrosolubles no cargadas eléctricamente desciende a medida que aumenta su tamaño, generalmente se permite el paso con un PM inferior a 200. Los iones por su carga neta son relativamente insolubles en los lípidos de la membrana, su difusión se da por medio de canales iónicos proteicos que cruzan la membrana (algunos dejan pasar cationes otros aniones).
Algunos canales iónicos son muy específicos respecto a los iones que dejan pasar, mientras que otros permiten el paso a todos los iones inferiores a cierto tamaño o con una carga del mismo signo. Algunos canales son controlados por la diferencia de voltaje a través de la membrana, y otros lo son por neurotransmisores u otros tipos de moléculas reguladoras.
El agua fluye por osmosis cuando existe una diferencia en la concentración de solutos a través de la membrana
La ósmosis se define como el flujo de agua a través de una membrana semipermeable desde un compartimento en el que la concentración de solutos es más baja hacia otro compartimiento de mayor concentración. El proceso de ósmosis tiene lugar porque la presencia de soluto reduce el potencial químico del agua. El agua tiende a fluir desde la zona de mayor potencial químico hacia el área donde éste es menor. Otros efectos originados por el descenso del potencial químico del agua son la reducción de la presión de vapor, un descenso del punto de congelación y una elevación del punto de ebullición de la solución en comparación con el agua pura. Todas estas propiedades dependen fundamentalmente de la concentración de los solutos presentes por eso se denominan propiedades coligativas.
Página 7

Seminario de Histología
Y entendemos por presión osmótica, a aquella que sería necesaria para detener el flujo de agua a través de la membrana semipermeable. Al considerar como semipermeable a la membrana plasmática, las células de los organismos pluricelulares deben permanecer en equilibrio osmótico con los líquidos tisulares que los bañan
Las células se hinchan o se retraen en respuesta a los cambios en el contenido de solutos del líquido extracelular
Las membranas plasmáticas de la mayoría de las células corporales son relativamente impermeables a muchos de los solutos del líquido extracelular, pero muy permeables al agua. Por lo tanto cuando la presión osmótica del líquido extracelular aumenta, el agua sale de la célula por ósmosis y la célula se retrae hasta que las presiones osmóticas celular y extracelular se igualan.
En un determinado rango de concentraciones de solutos externos, los eritrocitos se comportan como un osmómetro, ya que su volumen se relaciona inversamente con la concentración de solutos en el medio extracelular.
El comportamiento osmótico de los eritrocitos humanos en soluciones de NaCl, en soluciones más concentradas (hipotónicas), se retrae y en soluciones más diluidas (hipertónicas) se hinchan.
La solución isotónica de NaCl también denominada solución salina isotónica se utiliza para la rehidratación intravenosa o para la administración de medicamentos. La solución salina isotónica se administra a los pacientes porque al ser isotónica, no modifica el volumen celular.
Página 8

Seminario de Histología
Cuanto más permeable sea una membrana a un soluto determinado, menor será el flujo osmótico que ese soluto puede ocasionar. El flujo osmótico del agua a través de una membrana es directamente proporcional a la diferencia de presión osmótica entre las soluciones de ambos lados de la membrana. Cuanto mayor sea la permeabilidad de un soluto, menor será el flujo osmótico que provoca y cuanto mayor sea la molécula se soluto, más impermeable resultara la membrana para él y mayor el flujo osmótico de agua que origina.
Las proteínas transportadoras son las responsables del desplazamiento de las sustancias relevantes a través de las membranas
Determinadas sustancia entran y salen de la células por medio de transportadores o canales específicos que corresponden a proteínas intrínsecas de la membrana plasmática. A esto se denomina transporte mediado por proteínas o transporte mediado. Estos sistemas incluyen procesos de transporte activo y transporte facilitado, que tienen diversas propiedades en común, la diferencia fundamental es que el transporte activo requiere de energía y es capaz de bombear una sustancia contra un gradiente químico o electroquímico, mientras que el transporte facilitado sólo participa en la mediación del transporte de la sustancia a favor de su gradiente químico o electroquímico.
Página 9

Seminario de Histología
El transporte mediado por proteínas tiene algunas de las propiedades de la catálisis enzimática.
Una sustancia que pasa por transporte mediado lo hace mucho más rápidamente que las moléculas con un peso molecular y una solubilidad lipídica semejantes, pero que atraviesan la membrana por difusión simple.
La velocidad de transporte se comporta según una cinetica de saturación: cuando la concentración del compuesto transportado aumenta, la velocidad de transporte se incrementa al principio, pero finalmente se alcanza una concentración a partir de la cual dicha velocidad ya no aumenta más. Llegado a ese punto se dice que el sistema de transporte se ha saturado con el compuesto transportado.
La proteína transportadora tiene especificidad química: solo se transportan las moléculas con la estructura química requerida.
La moléculas relacionadas estructuralmente pueden competir por su transporte. En general el tranporte de un sustrato reducirá la tasa de transporte de un segundo sustrato, debido a la competición por la unión a una proteína transportadora. Esta competición es análoga a la inhibición competitiva de una enzima.
El transporte puede quedar inhibido por compuestos sin una relación estructural con los sustratos transportados. Un inhibidor puede unirse a la proteína transportadora de forma que reduzca su afinidad por el sustrato transportado normal. El compuesto florentina no se parece a la molécula de glucosa y, sin embargo, inhibe fuertemente su transporte en los eritrocitos.
El transporte facilitado aumenta su velocidad de flujo de una sustancia a favor de su gradiente de concentración o electroquímico.
Denominado en ocasiones difusión facilitada, el transporte facilitado se produce por medio de una proteína transportadora que no depende de la energía metabólica. Los inhibidores metabólicos no suelen disminuirlo, estos sistemas actúan para igualar concentraciones de las sustancias transportada a ambos lados de la membrana.
Una de las acciones principales de la insulina es estimular el transporte facilitado de glucosa a través de la membrana plasmática de las células musculares y los adipocitos. Las personas con diabetes tipo 1 segregan insulina en cantidades anormalmente bajas, la velocidad de captación es tan lenta que la capacidad de estos tejidos para utilizar glucosa como combustible metabólico está muy limitado.
Una proteína de transporte activo puede desplazar una sustancia de las zonas de menor a las de mayor concentración, este proceso necesita energía.
Estos sistemas son capaces de concentrar sus sustratos en contra del gradiente de concentración o de potenciales electroquímicos. Los procesos de transporte activo deben ir ligados de alguna forma de metabolismo energético, pueden utilizar el ATP directamente o
Página 10

Seminario de Histología
estar relacionados con el metabolismo en forma indirecta, por esta dependencia sus procesos pueden ser inhibidos por sustancias que interfieran con el metabolismo energético.
Un transporte activo, ligado de forma directa al metabolismo celular se denomina transporte activo primario, en el citoplasma de la mayoría de las células animales, la concentración se Na+ es mucho menor y la de K+ es mucho mayor que sus niveles extracelulares. Estos gradientes de concentración se originan por la acción de una Na+, K+- ATPasa, una proteína integral de la membrana plasmática. La Na+, K+- ATPasa utiliza la energía del ATP para bombear Na+ hasta fuera de la célula y K+ hacia su interior. Esta ATPasa transporta tres iones de sodio fuera de la célula y dos de potasio a través de la misma por cada molécula de ATP hidrolizada, ya que utiliza la energía del enlace del fosfato terminal del ATP para activar el ciclo de transporte.
Una proteína de transporte activo secundario obtiene su energía del gradiente de concentración de otra sustancia que se transporte de forma activa, una vez creado el gradiente de concentración representa un almacén de energía potencial química que se puede aprovechar para generar trabajo. En la mayoría de los tipos de células, el gradiente de concentración de Na+ generado por Na+, K+- ATPasa se utiliza para transportar activamente al interior de la célula otros solutos. Muchas de las células captan aminoácidos neutros hidrófilos mediante proteínas transportadoras de membrana, que vinculan el transporte de Na+ hacia el interior de la célula a favor de su gradiente de potencial electroquímico con el transporte hacia el interior de la célula de los aminoácidos en contra de sus gradientes de concentración. La energía para el transporte de los aminoácidos no esta proporcionada directamente por el ATP ni por ningún otro compuesto de alta energía, sino de forma indirecta por el gradiente de Na+ que sí se transporta activamente. Algunos otros procesos de transporte activo secundario, como la absorción de pequeños péptidos en el intestino delgado están activados por un gradiente transmembrana de potencial electroquímico de H+.
Las proteínas transportadoras de membrana pueden clasificarse según el mecanismo por el que intervienen en el transporte a través de la membrana
La acuaporina seria unos de los mecanismos de transporte más sencillos, el canal de agua parece estar siempre abierto y disponible para las moléculas de agua de ambos lados de la membrana. Por el contrario, las proteínas transportadoras, como los transportadores de glucosa o la Na+, K+- ATPasa, se unen al sustrato transportado situado a uno de los lados de la membrana y a continuación deben sufrir un cambio de conformación antes de liberarlo en el otro lado de la misma. Estos cambios de conformación son relativamente lentos es por eso que los transportadores presentan las tasas más lentas de transporte de sustratos. Los iones de transportan a través de las membranas por canales iónicos de naturaleza proteica, suelen estar regulados por un mecanismo de compuerta, según el cual alterna entre un estado abierto (de conducción rápida) y cerrado (de conducción lenta).
Las células dependen de algunas proteínas de transporte de membrana, el Ca+2 se transporta a través de las membranas por Ca+2- ATPasas y por proteínas antiportadoras de 3Na+-1Ca2+.
Página 11

Seminario de Histología
La glucosa se transporta al interior del músculo y a los adipocitos mediante transportadores facilitados, se difunde a través de la membrana plasmática muy lentamente. Las membranas plasmáticas de muchos tipos celulares contienen varias proteínas diferentes transportadoras de glúcidos que median el trasporte facilitado de glucosa y otros monosacáridos relacionados. Los eritrocitos, los adipocitos y las células musculares poseen todas ellas transportadores de glucosa de la familia GLUT, no dependen ni de la diferencia de potencial electroquímico del Na+ a través de la membrana plasmática ni de ninguna via directa del metabolismo celular. En los adipocitos y en las células musculares la insulina aumenta el transporte de glucosa a través de la membrana plasmática, lo que hace que se inserten más proteínas transportadoras en ella.
La síntesis proteica es necesaria para el recambio de las células y los tejidos en procesos como la cicatrización de las heridas. Los transportes de aminoácidos incluyen tres clases distintas de transportadores: para loa aminoácidos neutros, para los básicos y para los ácidos.
En el estudio de la fisiología se encuentra en numerosos transportadores:
H+,K+-ATPasas que expulsan H+ de las células parietales gástricas y lo intercambian con K+.
Antiportadores aniónicos, intercambian CL- con HCO3- a través de la membrana eritrocitaria.
Antiportadores de Na+-H+ que segregan H+ que absorbe pequeños péptidos del intestino delgado
Simportadores de Na+-HCO3- Simportadores de Na+- Cl- Simportadores de K+- CL-
Las concentraciones citosólicas de iones en equilibrio estacionario representan el resultado neto de las acciones de las bombas iónicas y los flujos transmembrana de iones a través de simportadores y antiportadores.
Las células epiteliales están polarizadas: las membranas plamáticas apical y basolateral contienen diferentes proteínas transportadoras. Los transportadores de la membrana plasmática de un lado de la capa de células epiteliales son diferentes a los del otro lado, buenos ejemplos son las células epiteliales del intestino delgado y del túbulo proximal del riñón.
Las uniones hermeticas que conectan los lados de las células epiteliales impiden que se mezclen las proteínas de las membranas plasmáticas luminal y basolateral.
Las uniones hermeticas intercelulares son algo permeables al agua y a los iones y pequeñas moléculas hidrosolubles. La oclusión de las uniones herméticas varía entre los epitelios. Por tanto hay dos tipos de rutas para el transporte a través de los epitelios:
1) Rutas transcelulares (a través de las células)2) Rutas paracelulares (entre las células)
Página 12

Seminario de Histología
Las neuronas también son células polarizadas. Los axones neuronales contienen proteínas que no aparecen en las dentritas y viceversa. La polaridad de la neurona favorece las diferentes funciones muy especializadas de los axones y las dentritas.
EQUILIBRIOS IÓNICOS Y POTENCIALES DE MEMBRANA EN REPOSO
Es un tipo especial de equilibrio químico, caracterizado por la presencia de especies químicas en solución acuosa, las cuales producen iones
Las especies que producen en solución cargas son denominadas electrolitos. Un electrolito es cualquier especie que permite la conducción de la corriente eléctrica.
En base a esto, se clasifica a los electrolitos en base a dos criterios:
Comportamiento en solución: electrolitos ácidos, básicos, y neutros Capacidad conductora: electrolitos fuertes y débiles
La ecuación de Nernst
Se aplica para un ion que está en equilibrio con una membrana, ∆µ viene a ser la fuerza neta sobre el ion, mientras que el logaritmo es la fuerza provocada por la diferencia de concentración, y ZF ( EA - EB ) es la fuerza originada por la diferencia de potencial eléctrico, cuando las dos fuerzas son iguales y opuestas, ∆µ es igual a cera y no hay una fuerza neta sobre el ion. Si este es el caso, n ose producirá un desplazamiento neto del mismo, y se dice que el ion esta en equilibrio electroquímico a través de la membrana.
DESPEJANDO EA – EB, SE OBTIENE
Esta es la ecuación de Nernst, una vez derivada asume la condición de equilibrio, y la ecuación de Nernst solo se cumple para los iones en equilibrio. Esta fórmula permite calcular la
Página 13
∆µ = RT ln [X]A + ZF ( EA - EB ) = 0
[X]B
EA - EB = -RT ln [X]A
ZF [X]B

Seminario de Histología
diferencia de potencial eléctrico, necesaria para producir una fuerza eléctrica que es justo igual y opuesta a la fuerza de concentración. Dicha fuerza eléctrica es igual a:
En resumen la ecuación de Nernst se puede utilizar para predecir el sentido en el que tienen a fluir los iones:
Si la diferencia de potencial medida a través de una membrana es igual a la diferencia de potencial calculada mediante la ecuación de Nernst para un ion concreto, entonces este ion esta en equilibrio electroquímico a través de la membrana y no tiene un flujo neto a través de la misma.
Si el potencial eléctrico medido es del mismo signo que el calculado mediante la ecuación de Nernst para un ion determinado, pero tiene una magnitud mayor que este valor, entonces la fuerza eléctrica es mayor que la fuerza de concentración y el flujo neto de dicho ion tenderá a producirse en el sentido determinado por la fuerza eléctrica.
Cuando la diferencia medida de potencial eléctrico es de mismo signo pero numéricamente menor que la calculada mediante la ecuación de Nernst para un ion en particular, entonces la fuerza de concentración es mayor que la fuerza eléctrica y el flujo neto de dicho ion tenderá a producirse en el sentido determinado por la diferencia de concentración.
Si la diferencia de potencial eléctrico medida a través de la membrana es de signo opuesto a la pronosticada mediante la ecuación de Nernst para un ion en particular, entonces las fuerzas eléctricas y de concentración actúan en el mismo sentido. Por tanto este ion nop odra estar en equilibrio y tenderá a desplazarse en el sentido determinado por ambas fuerzas, la eléctrica y la concentración
Equilibrio de Gibbs-Donnan
Se da cuando una especie iónica no puede atravesar la membrana.
En general el citoplasma contiene proteínas, polifosfatos orgánicos, ac. Nucleicos y otras sustancias ionizadas que no pueden atravesar la membrana plasmática. La presencia intercelular de aniones no permeantes afecta a la distribución de cationes e iones permeantes a través de la membrana plasmática y, como resultado, hace lo propio sobre las concentraciones de iones permeantes en el citosol e influye en el potencial de membrana en reposo. Las propiedades de equilibrio estacionario de esta mezcla de iones permeantes y no permeantes se describen según el Equilibrio de Gibbs-Donnan
Página 14
-RT ln [X]A = RT ln [X]B
ZF [X]B ZF [X]A
[K+]A [Cl-]A = [K+]B [Cl-]B

Seminario de Histología
Este índice de Gibbs-Donnan o ecuación de Gibbs-Donnan, se cumple para cualquier par de anión y catión monovalentes en equilibrio entre las dos cámaras. Si estuvieran presentes otros iones monovalentes que pudieran alcanzar una distribución equilibrada, se podría aplicar el mismo razonamiento y una ecuación similar para cada par catión-anión.
Potencial de membrana en reposo
El citosol de una célula en reposo es electronegativo respecto al líquido extracelular
La comunicación entre las células nerviosas depende de una modificación eléctrica, denominada potencial de acción, que se propaga por la membrana plasmática de dichas células. En el músculo estriado, el potencial de acción se propaga rápidamente sobre toda la superficie celular y permite que la célula se contraiga de manera sincrónica.
Todas las células que pueden producir potenciales de acción tienen potenciales de membrana en reposo considerables (citoplasma negativo) a través de sus membranas plasmáticas.
Las diferencias de potencial de membrana se expanden como el voltaje en el citoplasma menos el voltaje en el líquido extracelular. Un valor negativo indica que el citoplasma es eléctricamente negativo con respecto al líquido extracelular.
El potencial de membrana en reposo es necesario para que la célula desencadene un potencial de acción.
El gradiente de concentración de un tipo de ion permeante a través de una membrana produce una diferencia de potencial eléctrico en la misma
Mediante una grafica podemos esquematizar dicha diferencia de potencial eléctrico.
Tenemos que la membrana se separa en dos cámaras una A y la otra B esta es permeable a los cationes pero no a los aniones. En un principio no existe ninguna diferencia de potencial eléctrico a ambos lados de la membrana el K+ pasará desde A hacia B por la fuerza de concentración que actúa sobre él. El Cl- está sometido a la misma fuerza pero no puede pasar porque la membrana es impermeable a los aniones.
El flujo de K+ desde A hacia B transferirá una carga neta positiva al lado B y dejara un exceso muy ligero de cargas negativas en el lado A, por lo que el lado A se volverá eléctricamente negativo con respecto al B. esta fuerza tiene el sentido opuesto a la fuerza de concentración sobre el K+. Cuando más K+ pase a las membranas, mayor será la fuerza eléctrica en sentido opuesto. El flujo neto de K+ se detendrá cuando la fuerza eléctrica equilibre exactamente la fuerza de concentración, lo que sucede cuando la diferencia de potencial eléctrico sea igual al potencial de equilibrio (Nernst) para el K+. Es decir:
Página 15
EA – EB = -60mV log K + A = - (60mV) log 0.1
+1 K+ B 0.01

Seminario de Histología
Solo una cantidad muy pequeña de K+ influye desde A hacia B antes de que alcance el equilibrio. Esto se debe a que la separación de cargas negativas y positivas necesita una gran cantidad de energía. La diferencia de potencial eléctrico que se genera para oponerse a los nuevos desplazamientos del K+ es una manifestación de dicha energía.
Esto se debe a que cuando la célula no estimulada por corrientes despolarizantes supra umbrales, se dice que se encuentra en un potencial de membrana en reposo.
Los cambios en la permeabilidad de la membrana y el establecimiento y cese de corrientes iónicas durante el potencial de acción refleja la apertura y cierre de los canales iónicos que forman zonas de paso a través de membrana para los iones.
Diversos factores que contribuyen a la generacion de potencial de membrana en reposo
Los gradientes de concentraciones iónicas a través de la membrana plasmática
En la mayoría de tejidos, los iones no están en equilibrio entre el líquido extracelular y el citoplasma. El potencial de membrana en reposo de una célula esquelética es de unos -90 mV. El Cl- está casi en equilibrio a través de la membrana plasmática de este tipo de células. El K +
tiene una fuerza de concentración que tiene a hacerle salir de la célula. La fuerza eléctrica sobre el K+ tiene un sentido opuesto a la fuerza de concentración. Po tanto la fuerza de concentración como la fuerza eléctrica sobre el Na+ es el más alejado de una distribución de equilibrio.
El Na+, K+ y ATPasa (contribuyen directamente)
La bomba de Na+ y K- es responsable de la elevada concentración de K+ y la baja concentración de Na+ dentro de la célula, debido a que saca mas Na del K que entra (3-2), existe una transferencia neta de cargas positivas al exterior que contribuye al potencial de membrana en
Página 16

Seminario de Histología
reposo por lo que se le denomina electrógena. En algunas células, el Na+, K+ y ATPasa son responsables de una gran parte del potencial de membrana en reposo.
La difusión de iones a favor de sus gradientes de potencial electroquímico
Cuando varios iones están distribuidos a través de una membrana, habiendo suprimido su equilibrio electroquímico, cada ion tendera a aproximar el potencial transmembrana a su propio potencial de equilibrio, calculado mediante la ecuación de Nernst, cuanto más permeable sea la membrana para un ion concreto, mayor será la fuerza con la que éste tratara de acercar el potencial de membrana a su potencial de equilibrio.
La ecuación de conductancia armonizada describe la aportación de iones permeantes al potencial de membrana en reposo. La forma en que la interacción de los gradientes iónicos genera el potencial de membrana en reposo (Em) se explica mediante un modelo matemático sencillo. Si se considera la distribución de Na+, K+, Cl- a través de la membrana plasmática de una célula, la siguiente ecuación predice la diferencia de potencial transmembrana a través de dicha membrana:
La conductancia es la inversa de la resistencia ( g= 1/R), por lo que; cuando más permeable sea la membrana para un ion concreto, mayor será su conducta
GENERACIÓN Y CONDUCCIÓN DEL POTENCIAL DE ACCIÓN
Un potencial de ACCION ES UN CAMBIO rápido en el potencial de membrana que se propaga a lo largo de toda la longitud de la célula, seguido de un retorno al potencial de reposo. Los potenciales de acción tienen formas diferentes en cada tejido.
Potencial de membrana
El potencial de membrana es el voltaje de la diferencia de potencial eléctrico a un lado y al otro de la membrana plasmática de una célula. La membrana de las células está polarizada, debido a que hay un reparto desigual de cargas eléctricas entre el interior y el exterior de la célula. Esto crea una diferencia de potencial, siendo el exterior positivo respecto al interior.
Página 17
Em = gK EK + gNa ENa + gCl ECl
∑g ∑g ∑g
g= conductancia de la membrana del ion indicado en el subíndice
∑g= ( gK + gNa + gCl )
E= potenciales de equilibrio de los iones señalados en los subíndices

Seminario de Histología
El potencial de membrana de una célula puede medirse al atravesar su membrana plasmática con un micro electrodo. Al penetrar un microelectrodo por la membrana se observa una diferencia de potencial entre el microelectrodo con su punta introducida en la célula y el electrodo extracelular.
Si a través de la membrana plasmática de una célula fluye un pulso de corriente, el potencial de membrana cambia.
Los pulsos de corriente son:
Despolarización
Proceso químico mediante el cual una célula neuronal cambia su potencial eléctrico, normalmente negativo, a positivo mediante el intercambio de iones con ayuda de canales de cloruro y canales de sodio. Este proceso forma parte de la transmisión sináptica.
Consiste en el aumento de la permeabilidad para el Na+ (sodio), el cual ingresa a la célula cambiando la polaridad de la membrana: interior positivo y exterior negativo.
La despolarización significa un descenso de la diferencia de potencial a través de la membrana.
Hiperpolarización
es cualquier cambio en el potencial de membrana de la célula, que hace que esté más polarizada. Es decir, la hiperpolarización es un incremento en el valor absoluto del potencial de membrana de la célula. Así pues, los cambios en el voltaje de la membrana en los que el potencial de membrana es más netamente positivo o negativo, son hiperpolarizaciones.
Página 18

Seminario de Histología
Potencial de membrana en reposo de los nervios
El potencial de membrana en reposo de las fibras nerviosas grandes cuando no transmiten señales nerviosas es de aproximadamente -90mV.
Propiedades de transporte de la membrana para sodio y potasio.
Transporte activo de los iones de sodio y potasio a través de la membrana: Bomba de sodio y potasio.
Todas las membranas celulares tienen una potente bomba de sodio y potasio, que bombea continuamente iones sodio hacia el exterior de la célula e iones potasio hacia el interior.
Fuga de potasio y de sodio a través de la membrana nerviosa.
Factores que determinan de potencial de membrana en reposo.
Contribución del potencial de difusión de potasio.
Se pone énfasis en la fuga de potasio porque, en promedio, los canales son mucho más permeables al potasio que al sodio, normalmente aproximadamente 100 veces más permeables.
Debido al elevado cociente de los iones potasio entre el interior y el exterior, 35:1, el potencial de reposo en el interior de la fibra seria igual a -94mV.
Contribución de la difusión de sodio a través de la membrana nerviosa.
Contribución de la bomba Na+ - K-
El hecho de que se bombeen más iones sodio hacia el exterior que iones potasio hacia el interior da lugar a una perdida continua de cargas positivas desde el interior de la membrana; esto genera un grado adicional de negatividad (aproximadamente de -4 mV mas) en el interior además del que se puede explicar por la difusión de manera aislada.
Los potenciales de difusión aislados que produce la difusión del sodio y potasio darían un potencial de membrana de aproximadamente -86mV, casi todo determinado por la difusión de potasio. Además se generan – 4mV adicionales al potencial de membrana por la acción continua de la bomba de Na+ - K- electrógena, generándose un potencial neto de membrana de -90mV.
Potencial de acción nervioso
Página 19

Seminario de Histología
Las señales nerviosas se transmiten mediante potenciales de acción que son cambios rápidos del potencial de membrana que se extienden rápidamente a lo largo de la membrana de la fibra nerviosa.
Cada potencial de acción comienza con un cambio súbito desde el potencial de membrana negativo en reposo normal hasta un potencial positivo y después termina con un cambio casi igual de rápido de nuevo hacia el potencial negativo.
Las sucesivas fases del potencial de acción son las siguientes:
Fase de reposo
Potencial de membrana en reposo antes del comienzo del potencial de acción.se dice que la membrana esta polarizada durante esta fase debido al potencial de membrana negativo de -90mV que esta presente.
Fase de despolarización
En este momento la membrana se hace súbitamente permeable a los iones sodio. El estado polarizado normal de -90mV se neutraliza inmediatamente por la entrada de iones sodio cargados positivamente, y el potencial aumenta rápidamente en dirección positiva. Esto se denomina despolarización.
Fase de repolarización
Después que la membrana se haya hecho muy permeable a los iones sodio, los canales de sodio comienzan a cerrarse y los canales de potasio se abren mas de lo normal. De tal manera que la rápida difusión de los iones potasio hacia el exterior restablece el potencial de membrana en reposo negativo normal. Esto se denomina re polarización de la membrana.
RECEPTORES DE MEMBRANA, SEGUNDOS MENSAJEROS Y VÍAS DE TRANSDUCCIÓN DE SEÑALES
Los procesos celulares básicos están regulados por numerosas sustancias. Algunas de ellas, como las hormonas en las células y se unen a receptores que son factores de transcripción activados por la unión a la hormona. Otras sustancias reguladoras (agonistas) ejercen influencia desde el exterior de la célula. Sus acciones se inician cuando los agonistas se unen a proteínas receptoras en la membrana plasmática, lo que inicia una secuencia de fenómenos denominada via de transducción de señales que provoca las acciones finales de los agonistas.
Página 20

Seminario de Histología
La proteincinasa dependiente de segundo mensajero es modulada por el contenido celular de un segundo mensajero
La proteincinasa dependiente de AMPc participa en la regulación de importantes vías metabólicas. En ausencia de AMPc, la proteincinasa dependiente de AMPc esta compuesta por cuatro subunidades: dos reguladoras y dos catalíticas. La presencia de subunidades reguladoras inhibe reguladoras inhibe fuertemente la actividad enzimática del complejo. En presencia de concentraciones micromoleculares de AMPc, cada subunidad reguladora une dos moléculas de AMPc. La unión de AMPc provoca que las subunidades reguladoras se disocien de las subunidades catalíticas y de esta forma se activan estas ultimas. La subunidad catalítica activa fosforila las proteínas objetivo en determinados residuos de serina y tronina.
Las proteincinasas dependientes de la calmodulina se activan por el complejo de Ca+2 con calmodulinaUna de las formas por las que el Ca+2 ejerce su control s uniéndose a la calmodulina.
Las proteincinasas dependientes de calmodulina exclusivas, tales como la cinasas de la cadena ligera de miosina y la fosforilasa-cinasa, tienen un único sustrato celular. Las proteincinasas dependientes de calmodulina multifuncionales fosforilan más de un sustrato proteico.
La cinasa de la cadena ligera de miosina desempeña un papel fundamental para regular la contracción del musculo liso. La elevación de la concentración citosólica de Ca+2 en una celula muscular lisa estimula la actividad cinasa de la cadena ligera de miosina; la fosforilación resultante de las cadenas ligeras de miosina reguladoras permite que se mantenga la contracción de las células del musculo liso.
La proteincinasa C se activa por el Ca+2 y los lípidos de la membrana.
La principal acción de determinadas subunidades lipófilas estimuladoras de tumores, sobre todo de los esteres de forbol, es activar directamente la proteincinasa C. Esto favorece intensamente la división celular en numerosos tipos celulares y convierte las células normales con propiedades de crecimiento controlado en células transformadas, como las células tumorales, que crecen de modo incontrolado.
Página 21

Seminario de Histología
En una célula no estimulada, la mayor parte de la proteincinasa C se encuentra en el citosol y está inactivada. Cuando los niveles citosólicos de Ca+2 se elevan, el Ca+2 se une a la proteincinasaC. Esto hace que la proteincinasa C se adhiera a la superficie interna de la membrana plásmatica, donde puede ser activada pro el diacilglicerol producido por hidrólisis del fosfatidilinositol 4,5-difosfato.
Las tirosincinasas desempeñan funciones clave en el control de la proliferación celular
Los receptores de algunos factores de crecimiento son tirosincinasas Los receptores tirosina.proteincinasa conocidos hasta ahora se agrupan en ocho subfamilias. La unión del ligando al receptor determina la dimerización de los complejos receptor-ligando. La dimerización mejora la afinidad de unión y activa la actividad tirosina- proteincinasa.
Las familias monoméricas que ligan GTP de la familia Ras están implicadas en el acoplamiento de la unión de los ligandos mitógenos a sus receptores tirosina-proteincinasa para dar lugar a los efectos intracelulares resultantes sobre la proliferación celular. Cuando la proteína Ras esta inactivada, las células o pueden responder a los factores de crecimiento que actúan a través de los receptores tirosincinasas.
La activación de la proteína Ras por un receptor tirosincinasa activado desencadena a su vez una vía de transducción de señales que finalmente pone en marcha la transcripción de ciertos genes claves que favorecen el crecimiento celular. La cascada e proteincinasa activada pro un mitógeno (MAP) participa en respuestas a la proteína Ras activada. La proteincinasa C también activa la cascada cinasa de la MAp. Esta cascada es un punto destacado de convergencia de múltiples efectos que fomentan la proliferación celular.
Otros receptores de factores de crecimiento forman complejos con tirosincinasas intracelulares Los receptores de la hormona de crecimiento prolactina y eritropoyetina no son en sí mismo poteincinasa. Sin embargo, al activarse, estos receptores forman complejos de señales con las tirosincinasas intracelulares que originan sus efectos intracelulares.
Las proteinfosfatasas deshacen el trabajo de las proteincinasas.
El grado de folforilacion de una proteína regulada es el resultado de las actividades de la proteincinasa que fosforila esa proteína y la proteinfosfatasa que la desfosforila. Además de los diferentes tipos de proteincinasas comentados, todas las células también contienen proteinfosfatasa cuya tarea es revertir los efectos de la fosforilacion de las proteínas. Las proteincinaas se clasifican en serina-tronina-proteinfosfatasas y tirosina-proteinfosfatasas.
Página 22

Seminario de Histología
Los receptores del péptido natriurético auricular tienen actividad guanilciclasa.
El péptido natriurético auricular (PNA) se libera por las células auriculares cardiacas en respuesta a una elevación de la presión auricular. Esta hormona aumenta la excreción renal del NaCl y agua y disminuye la constriccion de determinados vasos sanguíneos. Los receptores de la membrana para el PNA poseen en sí mismos actividad guanililciclasa que se estimula cuando el PNA se une al receptor. No se necesita un segundo mensajero para activar la ganililciclasa. Los receptores del PNA tienen un dominio de unión extracelular, una única hélice transmembrana y un dominio guanililciclasa intracelular. La unión del PNA estimula la actividad guanililciclasa y eleva las concentraciones intracelulares del segundo mensajero GMPc.
El oxido nítrico es un mediador paracrino de vida corta
El oxido nítrico (NO) es un mediador pancreático que se libera en las células endoteliales y algunas neuronas. El NO se oxida con rapidez, por lo que su vida biológica solo es de varios segundos. Por esta razón, el No sólo afecta a las células en inmediata proximidad a la célula que lo produce. El NO estimula la guanililciclasa soluble en la célula objetivo, lo que eleva la concentración intracelular de GMPc en dicha célula, estimulando por tanto la proteincinasa dependiente de GMPc.
La producción de NO es catalizada por la NO sintasa, una enzima dependiente de Ca+2- calmodulina que acelera la conversión de la arginina en cicutrina y No. El aumento de la concentración citosólica de Ca+2 suele ser el estímulo para el incremento de la formación y liberación de NO.
Los componentes de la transducción de señales se localizan en el interior celular.
Si las cinasas, fosfatasa y todas las proteínas que las regulan pudieran difundir con libertad en el citoplasma, sería difícil lograr la especificidad temporal y espacial de las respuestas iniciadas por un agonista concreto. Las proteincinasas estimuladas por AMPc, por ejemplo, se unen a proteínas intracelularesespecificas por miembros de la familia de proteínas de anclaje de cinasa A (AKAP). Las serina-treonina-proteinfosfatasa suelen anclarse a estructuras subcelulares concretas mediante subunidades objetivo que se asocian a la subunidad fosfatasa catalítica.
Página 23

Seminario de Histología
Página 24