
BIOENERGETICA DE LAS flBAAS MUSCULARES Y EIERCICIO E
61
Transcript of BIOENERGETICA DE LAS flBAAS MUSCULARES Y EIERCICIO E


En nuestra experiencia docente relacionada con el ejercicio y aplicada a diferentes areas de conocimiento hemos aprendido a distinguir entre lo importante y lo imprescindible, y esta obra, concisa pero rigurosa, nace con la vocaci6n de ser de utilidad para estudiantes, profesionales del ejercicio e incluso para profesores vinculados directa o indirectamente a las ciencias de la actividad fisica y el ejercicio.
Este libro trata de abordar desde un punto de vista conceptual un area tan compleja como es la Bioquimica del Ejercicio, apostando por ofrecer un enfoque a la vez cientifico y practice, renunciando conscientemente a la complejidad de la materia.
E 1 estudio de las respuestas y adaptaciones de 6rganos y sistemas del cuerpo humano al ejercicio es el objetivo mas relevante de la Fisiologia del Ejercicio. Gran parte de esas respuestas y de las
adaptaciones obtenidas con el entrenamiento tienen en la bioenergetica muscular la base en la que se asientan tanto la capacidad de respuesta como los cambios morfofuncionales de las grandes sistemas provocados par el ejercicio repetido. Par consiguiente, conocer c6mo obtienen las fibras musculares la energia para la contracci6n en funci6n de las caracteristicas del ejercicio desarrollado es esencial para entender las respuestas fisiol6gicas.
BIOENERGETICA DE LAS flBAAS MUSCULARES Y EIERCICIO

117 ---- BIBLIOGRAFiA
1 lnteracci6n de las diferentes
sistemas energeticos durante el ejercicia
L
80 • Metabalisma de las_protefnas_ CAPITULO 4
68 CAPITULO 3 • Metabalisma de las lfpidas
27 , Metabalisma de las hidratas de carbono CA TULO 2
11
7
CONTENIDOS INT£l0DUCCION ---~---~-
CAPITULO 1 • Metabalis!T1_()_9e l_ci~ fosfagenos

Durante el ejercicio, el rmisculo esqueletico satisface sus demandas energeticas utilizando sustratos que proceden de las reservas del organismo gracias a la ingestion diaria de nutrientes.
Los sustratos energeticos de las que el musculo esqueletico obtiene la energia quimica para transformarla en energia mecanica o estatica son, fundamentalmente, las grasas y las hidratos de carbono. Las proteinas actuan tambien coma sustratos energeticos en condiciones especiales (ejercicios muy prolongados, ejercicio en ayuno, etc.), si bien son otras muy diferentes sus funciones fundamentales en el organismo (sintesis de tejidos, sintesis de hormonas, sintesis de enzimas, etc).
Los sustratos que hemos mencionado no son utilizados directamente par la celula muscular. Todos ellos deben ceder la energfa contenida en sus enlaces quimicos para la fosforilacion del adenos{n trifosfato (ATP), ya que la celula muscular solo es capaz de obtener directamente la energia quimica de este compuesto de alta energia y transformarla en energia mecanica, De manera que el metabolismo energetico de nuestras celulas musculares va a consistir esencialmente en una serie de transferencias de energia para conseguir que la celula disponga de las cantidades de ATP necesarias para satisfacer las demandas energeticas en cada momenta. El ATP es un nucleotido presente en todas las celulas de nuestro organismo, con diferentes funciones, coma vamos aver a continuacion.
11 Metabolismo de los f osf agenos

Cada uno de los tres grupos fosfatos del ATP se identifican con una letra griega (a,~,Y), teniendo cargas negativas y estando unidos por enlaces anhidros. Estos enlaces cuando se rompen al hidrolizarse generan una importante cantidad de energia para la celula.
Ademas del ATP, ADP y AMP, existen en la celula otros tres tipos de nucleotidos: GTP o guanosin trifosfato, que contiene como base purina la guanina, UTP o uridin trifosfato, que contiene la base pirimidina, uracilo, y CTP o citidina trifosfato que incluye la base pirimidina citosina. Estos compuestos aportan energia para procesos concretos del funcionamiento celular diferentes del proceso contractil. Asi, el GTP esta implicado en la sintesis de proteinas, y el UTP se utiliza para la sintesis de gluc6geno.
Los acldos nucleicos, ARN y ADN, estan compuestos por nucleotides. Los nucleotides del ADN contienen el monosacarido desoxirribosa, y los del ARN, ribosa. Ademas las bases nitrogenadas que contienen pueden ser puricas (citosina, timina o uracilo) o pirimidinicas (adenina, guanina). En el ADN los nucleotides se disponen formando una doble helice, mientras que en el ARN la molecula consta solo de una hebra. Los nucleotides son por tanto la base de las macrornoleculas que contienen toda la informacion genetica de los seres vivos.
Los nucleotidos pueden presentar uno, dos o tres grupos fosfato. Los enlaces entre el fosfato 1 y 2 y entre el fosfato 2 y 3 son considerados en laces de alta enerqia, pues almacenan gran cantidad de energia quimica "utilizable" por la celula, En los procesos en los que se utilizan los nucleotides, tanto estructurales como metabolicos, la transferencia de energia se produce constantemente, produciendose la perdida de uno o dos de estos grupos fosfatos. Para incorporarlos de nuevo a la molecula, es necesario aportar energia (anabolismo). Por tanto, la celula necesita mantener el balance de
o- o- 0 I I II
o--p- 0-P- 0-P- 0-Adenosina II II I 0 0 o-
Mg2+ -ATP
Figura 2. Figura 2 . ATP unido al Mg2+.
Mg2+ .. , '•. . ·- .. .. En la celula el ATP se esta cargado negativamente y encuentra asociado a un ion magnesio formando Mg2+-ATP (figura 2).
1IMetabolismo de los fosfagenos
Cuando la adenina se combina con la ribosa, tenemos una molecula (nucleosido) denominada adenosina. Si se afiade un grupo fosfato a la posicion 5' de la ribosa en la adenosina, tendremos un nucleotide denominado adenosin-mono-fosfato (AMP). Si afiadimos otro grupo fosfato conseguiremos adenosin-difosfato (ADP), y si afiadimos otro mas, se formara adenosin-trifosfato (ATP) (figura 1).
Figura 1. Esquema de la estructura qufmica del ATP.
1, = en laces de alta energfa
ATP Adenosina
Ribosa
Grupos fosfato
Adenina Desde el punto de vista
energetico, el ATP, ADP ( adenos{n difosfato) y el AMP (adenos{n monofosfato) son los maximos representantes de los nucleotides. Estas moleculas tienen en cornun poseer tres componentes: una base de pirimidina o purina ( adenina en el caso del ATP), un monosacarido ribosa o desoxirribosa, y al menos un grupo fosfato.
Los nucleotides son moleculas omnipresentes en nuestras celulas. Adernas de ser los sillares estructurales del ADN y ARN, los nucleotides desempefian en las celulas otras funciones no menos importantes. Algunos contienen enlaces de alta energia, lo que les permite actuar como transportadores de energia. En concreto, el trifosfato de adenosina (ATP) actua universalmente en todas las celulas como intermediario energetico. En algunas reacciones del metabolismo, otros nucleotidos trifosfato como el GTP, CTP y UTP, pueden sustituir al ATP en este papel. Por otra parte, algunos nucleotides o sus derivados pueden actuar como coenzimas, como NAD, NADP, FAD o FMN, que actuan como transportadores de electrones en reacciones metabolicas de oxidacion-reduccion. Otros nucleotides como el cAMP, actuan como mediadores en multiples procesos de sefializacion celular, transmitiendo al citoplasma sefiales quimicas procedentes del exterior.
LOS NUCLE6TIDOS DE LA CELULA MUSCULAR l

ENERGIA Fosfato inorganico ADP
+ 7,3kcal/ molATP
0 u
HO-P-o- 1
OH
+
ATP I I~ H.O agua
+ H.O----• agua
Figura 3. Hidr61isis del ATP. Proceso exerg6nico que libera 7,3 kilocalorias por cada mol.
El ATP es la fuente de energfa mas rapida o inmediata en la celula. Todas las celulas de nuestro organismo disponen de una determinada concentracion de ATP imprescindible para la vida celular. Estas concentraciones de ATP deben mantenerse constantes para permitir la funcion muscular. Como ya hemos visto, el desprendimiento por hidrolisis del fosfato terminal del ATP da lugar a ADP.
ADENOSiN TAIFOSFATO (ATP)
.....
La direccion de la reaccion dependera de las concentraciones relativas de ATP, ADP y otros NTPs y NDPs. La nucle6sido difosfato cinasa es inespecifca. Para convertir NMP a NDP, enzimas nucleotide especificas transfieren un grupo fosfato del ATP al nucleotide monofosfato, obteniendo ADP y un nucleotide difosfato. Asi, la adenilato cinasa cataliza la reaccion:
Nudeotidos Tnfosfato
Nucleotides Oifosfato La hidrolisis del ATP para convertirse en ADP es un proceso exergonico
por el cual se libera parte de la energia quimica contenida en el ATP, en concreto, la energia contenida en el enlace del tercer grupo fosfato. La hidrolisis del ATP puede acoplarse a cualquier proceso celular que precise energia, no es exclusivo de la contraccion muscular (transporte activo a traves de membranas, sintesis proteica, etc). La energia liberada en la celula muscular por la hidrolisis del ATP varia con las condiciones de temperatura y pH, pero se ha calculado que durante la contraccion muscular en condiciones normales es de 7 ,3 kcal/mol. El musculo esqueletico utiliza esa energia para provocar cambios conformacionales en su ultraestructura (interaccion actina- miosina) que se manifiestan internamente por acortamiento del sarcomero o aumento de tension, y externamente por generar fuerza aplicada, y en su caso movimiento. Una vez experimentada la hidrolisis, el ADP debera volver a reincorporar un grupo fosfato para transformarse de nuevo en ATP. En este caso, la reaccion necesita un aporte energetico (es una reaccion endergonica). La energia contenida en los distintos sustratos energeticos que utiliza la celula muscular por distintas vias metabolicas sera utilizada para tal fin.
ADP
nucleotides trifosfato de forma adecuada, mediante la fosforilacion de sus precursores y la aplicacion de energia. El objetivo es mantener un balance en la relacion entre NTP y NDP.
El proceso inmediato para la conversion de energfa quimica en energia estatica y/o dinamica esta catalizado por la enzima ATPasa y acopla la hidrolisis del ATP a la contraccion muscular (figura 3).
1IMetabolismo de los fosfagen~ ~~~~~~~~~~~~~~~~~~~~

Las reservas de ATP en la celula muscular son pequefias. La energia liberada en la celula muscular por la hidrolisis del ATP varia con las condiciones de temperatura y pH, pero en condiciones de reposo se ha calculado que es de unos 70 kJ /mol pudiendo descender durante el ejercicio intenso hasta 52.1
kJ/mol 6 7,3 kcal/mol en condiciones estandar que pueden llegar a ser 10
kcal/mol o mas en la celula. El musculo esqueletico contiene unicamente 5 x 10-6 mol-g', de manera que si durante un ejercicio ponemos en marcha
Figura 5. La utilizaci6n de ATP durante el ejercicio se lleva a cabo emdiante la acci6n de tres enzimas ATPasas durante el proceso de la contracci6n: la actomiosfn ATPasa que permite el movimiento de las molecules de miosina modificando su conformaci6n, la SERCA ATPasa que introduce de nuevo el calcio al interior del retlculo sarcoplasrnico para permitir la relajaci6n y la ATPasa Na•/K• que reestablece el equilibria i6nico tras la despolarizaci6n para recuperar el potencial de membrana de reposo. La fosforilaci6n oxidativa, la glucolisis y la transferencia de grupos fosfato de alta energfa de determinados compuestos como la fosfocreatina al ADP, son los procesos que permiten recuperar los niveles celulares de ATP.
Utilizaci6n Producci6n
Bombeo Na•/K• Na•/K' ATPasa (<10%) GLUC6LISIS
SERCA ATPasa 20 - 30 %
HOMEOSTASIS DEL ATP
Cicio de los puentes )ru~dos Cicio del calcio
ACTOMIOSfN ATPasa 60 - 70 %
FOSFORILACl6N OXIDATIVA
relajaci6n muscular. Por otra parte, la enzima ATPasa de la bomba sodio- potasio tambien hidroliza ATP para restaurar el potencial de membrana durante la relajaci6n muscular, aunque su contribuci6n en la hidrolisis total de ATP durante la actividad muscular es de menos del 10% de los ATP
consumidos (figura 5).
menor concentraci6n ( citosol) hacia otras de mayor concentraci6n (reticulo) se requiere energia para conseguir este transporte. Se ha sugerido que entre un 20-30% del ATP utilizado durante la contracci6n muscular puede ser hidrolizado por la ATPasa del reticulo sarcoplasmico durante la fase de
,,
1IMetabolismo de los fosfagenos [
La enzima ATPasa SERCA (sarcoplasmic-endoplasmic reticulum calcium
ATPase) del reticulo sarcoplarnico bombea iones calcio desde el citoplasma al interior de las vesiculas del reticulosarcoplasmico durante la fase de relajaci6n muscular. Ya que el calcio ha de ser transportado desde areas de
Figura 4. Resumen de los pasos de la contracci6n muscular en la que se destaca el papel de las ATPasas del sarcolema (ATPasa Na•/K•) y de la membrana del retfculo sarcoplasrnico (SERCA ATPas?) e~,cargad~s de reestablecer el equilibrip del Na, Ky Ca respe~tivamente tras el proceso de activacion nervioso que desencadena el fenomeno de la contraccion.
'- Union de CaH a troponina. Contracci6n. I. Liberaci6n de Ca" de troponina. I. Entrada de Ca" ill RS (SERCA). Relajaci6n 7. Recuperaci6n de potencial de membrana.
,. Liberaci6n de ach en placa motora. 2. Apertura de canales de Na·• en Sy TI:
entrada de Na•. I.Apertura de canales de Ca", salida de Ca+de RS.
TT TObulosT Relajaci6n Contracci6n
RS Retfculo sarcoplasrruco
S Sarcolema •·
0 ATPasa Serca Na' I K•
ecaH •K·
_.........._ ATPasa Serca .T;
.+j---!+J O OAch
, )Na+
e-. 0 •• 0 s o: -..:,O
118:J
:~ :J\oD
• 00
~· o 00 •
Ii·.,+-
i !' • - ... ... ..
La mayoria de la hidr61isis del ATP durante el ejercicio muscular ( 60- 70%) es debido a la actividad ATPasa de la miosina. La miosina, proteina contractil de la celula muscular, tiene capacidad para hidrolizar ATP en ADP y Pi. Dicha actividad ATPasa es el acontecimiento principal en la utilizaci6n del ATP durante la contracci6n muscular. Hay otras dos enzimas ATPasa (ATPasa Na+/K+ y ATPasa SERCA) que contribuyen a la hidr61isis total del ATP durante el ejercicio, y son las que controlan los movimientos de calcio, sodio y potasio a traves de las membranas (figura 4).
lacTIVIDAD ATPasa EN LA CELULA MUSCULAR

Tabla 1. Maxima tasa de producci6n de Pde alta energfa a partir de diferentes sustratos y cantidad disponible en un hombre de 70 kg de eso (masa muscular estimada 28 kg). Modificada de Greenhaff y col, 1993.
I TASA, mrncl-mln" CANTIDAO DlSPONtBLE, mol
ATP, PCr ~ ADP, Cr 4,4 0,67
GLUCOGENO ~ LACTATO 2,35 1,67 ENTOTAL6,7
GLUCOGENO ~ C02 0,85-1, 14 84
GLUCOGENO HEPATIC~~ co, 0,37 19
Acmos GRAS OS~ (02 0,40 4000
La necesidad de mantener en las celulas una elevada relaci6n ATP/ADP justifica la existencia de diferentes rutas metabolicas para la sintesis de ATP. La selecci6n de estas vias energeticas o rutas metabolicas va a depender especialmente de la tasa de utilizaci6n de ATP o intensidad del ejercicio, es decir, de la necesidad de energia por unidad de tiempo, que podemos denominar "potencia energetica".
En la tabla 1 podemos observar la tasa a la que se regenera el ATP a partir del metabolismo de cada una de las diferentes fuentes energia (expresada en moles.min:') de la celula muscular, asi coma la cantidad disponible de cada una de ellas (expresada en moles) en un sujeto de 70 kg de peso.
,,
macronutrientes antes mencionados. En este proceso es imprescindible la presencia de oxigeno y se le denomina metabolismo "aerobico". Implica el acoplamiento entre las reacciones de oxidacion ( en la que se donan electrones) y las de reducci6n (en la que se aceptan electrones), ya que cada oxidacion coincide con una reducci6n. Este proceso proporciona de forma continuada atomos de hidr6geno procedentes del catabolismo de los hidratos de carbono, grasas y proteinas almacenados.
consideramos que esta denominaci6n puede inducir al error. No obstante, la mayor parte de la reposici6n de ATP en la celula muscular tiene lugar mediante un proceso complejo que acontece en las mitocondrias denominado fosforilaci6n oxidativa (u oxidaci6n celular) a partir de la combustion de los
1IMetabolismo de los fosfagenos I ~~~~~~~~~~~~~~~~~~~~
En determinadas circunstancias, estos procesos quimicos pueden llevarse a cabo sin la participacion del oxigeno molecular procedente del aire atmosferico. En estos casos todas las reacciones quimicas que en ellos acontecen tienen lugar en el citoplasma celular. Clasicamente a estas rutas metabolicas se les ha denominado "anaerobicas", aunque nosotros
ADP
Al igual que el ATP, la fosfocreatina pertenece a un grupo de moleculas denominadas fosfagenos celulares, ya que contienen enlaces fosfato con un elevado nivel energetico,
Pero la mayoria de los ATP de la celula se recuperan gracias a la energia liberada en el catabolismo de macronutrientes (grasas, hidratos de carbono, proteinas), Esas moleculas energeticas se oxidan y forman compuestos simples coma C02 y H20. Durante el proceso liberan energia quimica que se aplica en la fosforilaci6n del ADP para generar ATP.
c, • i''~· n . 18
I··.,.,
En funcion del trabajo muscular, la reposicion de ATP durante el ejercicio puede llegar a realizarse a tasas varios cientos de veces superiores a lo que acontece en reposo sin que se modifiquen las concentraciones intracelulares de ATP. Para que esto ocurra, debe obtenerse un equilibria perfecta entre la hidrolisis del ATP y su resintesis. La forma mas inmediata de reponer el ATP es a partir de la energia liberada en la hidrolisis de la fosfocreatina (PCr), de la cual existe cierta cantidad en todas nuestras celulas, en la siguiente reacci6n muy rapida catalizada por la creatin cinasa:
RUTAS METABOLICAS PARA LA AESiNTESIS DE ATP DURANTE EL EIEACICIO
j 20 kg de rnusculo, las reservas totales no superan 10-• moles, que permite obtener una energia de 1 kcal. Esta reserva se agotara en teoria despues de 0,5 segundos de un esfuerzo maxima.

4. Metabolismo de las proteinas ( oxidacion)
3. Metabolismo de las grasas ( oxidacion)
HIDRATOS DE CARBO NO
FOSFOCREATINA
CITOSOL MITOCONDRIA
Metabolismo glucofftico
2. Metabolismo de los hidratos de carbono (glucolisis citosolica y oxidacion)
En la Tabla se agrupan las sistemas de producci6n de ATP en el rnusculo
esqueletico,
1. Metabolismo de los fosfagenos o de la fosfocreatina
Desde un punto de vista didactico y conceptual, se puede establecer una clasificaci6n de los sistemas energeticos en funcion del tipo de sustrato utilizado, dejando claro en cualquier caso, que in vivo no se produce la participacion unitaria de sistemas, y mucho menos durante el ejercicio.
Figura 7. Participaci6n de las diferentes sistemas energeticos en la formaci6n de ATP en funci6n del tiempo que dura la actividad.
Tiempo de ejercicio
O Sistema aer6bico ] Sistema anaer6bico lactico ~ Sistema de fosfagenos (ATP-PC) "E ~~~~~~~~~~--::::_..100 .E 100.., :I "' c. ~ (U
"O (U
"iii' .... c: (U v ... 0 c.
Es importante sefialar que en condiciones fisiologicas es practicamente imposible la participaci6n unica de uno solo de estos sistemas (figura 7).
En todo momenta la celula dispone de un metabolismo mixto en el que predominara un tipo d: sistema energetico sobre el resto en funcion de las caracteristicas del ejercicio (ej. intensidad), disponibilidad de sustratos energeticos, condiciones ambientales, aspectos nutricionales, etc.
1IMetabolismo de las fosfagenos I
Figura 6. Sistemas de producci6n de ATP en la fibra muscular esqueletica
[
-creatinaf--JI 1~ Lactato
Fosfu"eat;n,~ ~,.L ~
... H20+C02
O o, - Glucosa
A. grasos ADP----
ATP·----
Como la concentraci6n de ATP en las celulas es muy baja (3-8 mmol/litro de agua celular 6 2-6 mmol/kg de tejido ), la produccion de ATP debera adaptarse al consumo del mismo. Si la necesidad energetica de una celula muscular en reposo es de -1 mmol/kg/min de ATP, durante un sprint, un atleta de elite es capaz de hidrolizar hasta 240 mmol/kg/min de ATP. Si la celula no fuera capaz de ir reponiendo el ATP que hidroliza, toda la energia disponible en forma de ATP en la celula se consumiria en uno o dos segundos durante un ejercicio de alta intensidad. Esto nos lleva a concluir que dependiendo de la necesidad energetica en cada momenta, uno u otro sistema energetico contribuiran en mayor o menor medida en la produccion de ATP, siendo este un concepto fundamental para entender la bioenergetica de las celulas musculares durante el ejercicio. En la figura 6, se resumen las diferentes rutas metabolicas que utiliza la celula muscular para producir ATP.
,·.~;.~ · 20 I· -
11.
Tabla 2. Tasa de utilizaci6n y cantidad total de fosfatos de alta energfa durante diferentes pruebas deportivas.
~TIVIDAD TASA, mmo~min-1 CANTIDAD DISPONIBLE, mol
DESCANSO O,Q7
SPRINT100M 2,6 0,43
SPRIN'T 400 M 2,3 1,72 ---- CORRERBOOM 2,0 3,43
CORRER ·1500 M 1,7 6,00
MARATON 42,200 M 1.0 L iso.oo ----- ----
La tabla 2 expresa la tasa de utilizacion de ATP y el gasto total de moles de ATP estimado en reposo y durante diferentes pruebas de atletismo.
•

La reaccion es reversible, de manera que durante la recuperacion cambia el sentido de la reaccion favoreciendo la regeneracion de la PCr. Dado que la energia liberada de la hidrolisis de la PCr es pequefia en comparacion con la liberada por el ATP, las concentraciones de PCr caeran rapidamente si la necesidad energetica de la celula, y por consiguiente la hidrolisis de ATP, es muy alta. En la figura s se muestra como la concentracion intracelular de fosfocreatina disminuye mas rapidamente que la de ATP a medida que va aumentando la intensidad del ejercicio expresada en consumo de oxigeno (VOJ
La concentraci6n celular de fosfocreatina es de 3 a 5 veces superior a la de ATP, por lo que este compuesto esta considerado como una verdadera reserva energetica celular. Su agotamiento seria completo tras unos 2 segundos de esfuerzo maxima si no se produjera simultaneamente su reposicion. En las fibras musculares tipo I, la concentracion de fosfocreatina es ligeramente mas baja queen las fibras tipo II (entre un 15% y un 20%).
-,
celular, si bien existe tambien una isoforma mitocondrial. En los mamiferos cuatro genes codifican 4 monomeros de proteinas relacionados con la CK. Dos de esos monomeros son designados B (cerebra) y M (rnusculo), y otros dos se sinian en la mitocondria. De las subunidades mitocondriales, una se expresa en musculo esqueletico y cardiaco, y la otra en el cerebra y otros tejidos. La subunidad mitocondrial de la CK facilita la fosforilaclon oxidativa, mientras que la forma no mitocondrial de la CK en el musculo esqueletico es activa coma un dimero, conteniendo dos subunidades M, y expresandose como MM.
Como se observa en la ecuacion, la ruptura de la PCr produce una reaccion alcalinizante al consumir un proton (H•) en la formacion de ATP. El proton se requiere para reemplazar el grupo fosfato de la fosfocreatina, completando al segundo grupo funcional amino (NH2) de la creatina.
La creat(ncinasa que hidroliza la fosfocreatina y transfiere el fosfato de alta energia al ATP, se activa cuando aumentan las concentraciones citoplasmaticas de ADP, mientras que las concentraciones elevadas de ATP ejercen un efecto inhibitorio. Es una enzima muy abundante en el citoplasma
1IMetabolismo de las fosfagenos J ~~~~~~~~-
La utilizacion de los termlnos "aerobico" y "anaer6bico" en fisiologfa del ejercicio es mas una cuestion didactica que realmente fisiol6gica. El denominado metabolismo "anaer6bico" nose refiere a una ruta metabolica que funciona en ausencia de oxigeno, sino que no utiliza el oxfgeno. Adernas, la provision de energfa para diferentes modalidades de ejercicio (ej. intensidad) se basa realmente en la participacion slrnultanea de las tres rutas rnetabolicas principales (fosfocreatina o de fosfagenos, glucolisis anaer6bica y fosforilaci6n oxidativa) con mayor o menor protagonismo de una sob re otra. Los ejercicios de muy corta duraci6n ( 1-6 s) y maxima intensidad no son dependientes solo de la via de los fosfagenos, sino que tarnbien interviene la glucolisis (19,20-- t-Ilbroz.ed). Por otro lado, en esfuerzos de maxima intensidad de 30 s (ej. Test de Wingate), vinculados a la glucolisis anaerobica, la contribuci6n
I energetics de esta ruta metab61ica oscila entre un 28% y un 45%, dependiendo del perfil del atleta (7--11ibro4ed). En pruebas atleticas como los 400 m realizados en alrededor de 52 s, los ultimas 20 s del esfuerzo se realizan en Vozrnax mostrando
I asf una activaci6n maxima de la fosforilaci6n oxidativa o metabolismo "aerobico" en tiempos mucho mas breve de lo que clasicarnente se pensaba (21-1 libroz.ed). Ho; en
I dfa es aceptado que la provision de energfa para cada intensidad de ejercicio implica la participaci6n simultanea de las tres rutas principales de energfa con predominio de
\ unas sobre otras (21-1 libro4ed).(Chaman y Padulo, 2015) ', ,~
CUESTIONANDO LOS TERMINOS "AEROBICO" Y ANAEROBICO" EN FISIOLOG(A DEL EJERCICIO
I',,, - - - - - - - - - - - ...... ~ .. - - - - - - - - ......... - - - - ... - - - - - - - - - - - .....
I
Cr ;f- H++creatfncinasa-+
Como hemos visto, el ATP tiene que ser resintetizado constantemente en las celulas, ya que se encuentra en concentraciones muy pequefias y se esta gastando continuamente para mantener las funciones celulares. Parte de este ATP se resintetiza gracias a la energia contenida en otro fosfageno: la fosfocreatina. La transferencia de energia desde la fosfocreatina al ATP es un proceso rapido ( una (mica reaccion catalizada por la creati ncinasa [ CK]) que no requiere la participacion del oxigeno. Consiste en la transf erencia de energia desde la fosfocreatina al ADP para que este pueda reincorporar un grupo fosfato a su molecula y transformarse en ATP. Por su parte, la fosfocreatina pierde su grupo fosfato (de cuyo enlace procede la energia liberada) y queda transformada en creatina. Se produce de la siguiente manera, de forma que la energia que se ha liberado en la hidrolisis de la fosfocreatina es directamente utilizada para resintetizar ATP a partir de ADP.
METABOUSMO DE LOS FOSFiGENOS O DE LA FOSFOCREATINA
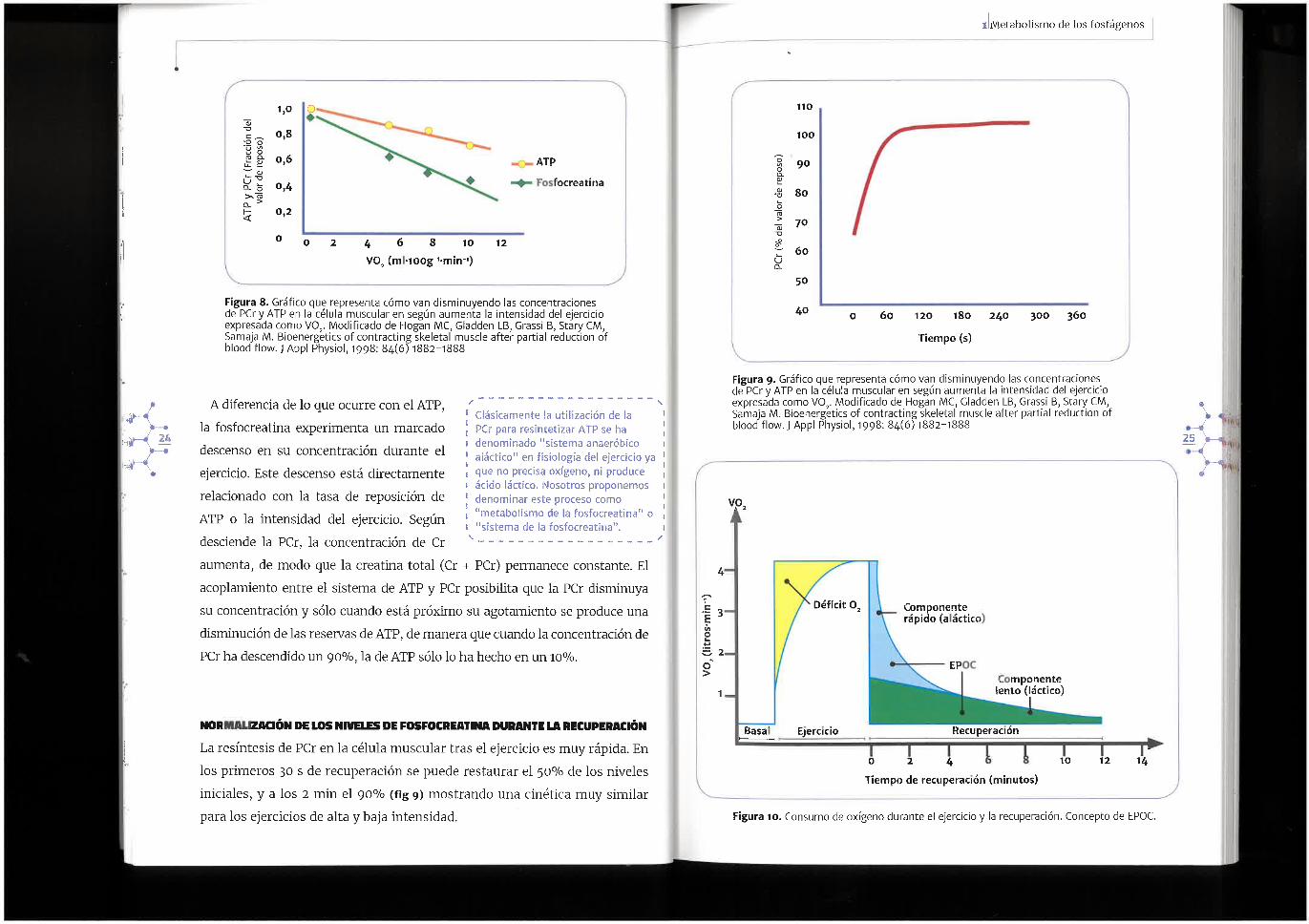
Figura 10. Consumo de oxigeno durante el ejercicio y la recuperaci6n. Concepto de EPOC.
14 12 0 10 4 2
Ejercicio Basal
Componente rapido (alactico)
....... c: ·e 3 .;, 0 .. ..... 52 ~N
4
vo2
Tiempo de recuperaci6n (minutos)
Figura 9. Grafico que representa c6mo van disminuyendo las concentraciones de PCr y ATP en la celula muscular en segun aumenta la intensidad del ejercicio expresada coma V02• Modificado de Hogan MC, Gladden LB, Grassi B, Stary CM, Samaja M. Bioenergetics of contracting skeletal muscle after partial reduction of blood flow. J Appl Physiol, 1998: 84(6) 1882-1888
110
100
0 90 Vl 0 <.l. ~ Qj So ~o 0
-;;; > 70 "'iii
'1!l
t. 60 u C'..
50
40 0 60 120 180 240 300 360
Tiempo (s)
1IMetabolismo de los fosfagenos
NOAMALIZACION DE LOS NIVELES DE FOSFOCREATINA DURANTE LA RECUPERACION
La resintesis de PCr en la celula muscular tras el ejercicio es muy rapida. En los primeros 30 s de recuperacion se puede restaurar el 50% de los niveles iniciales, ya los 2 min el 90% (fig 9) mostrando una cinetica muy similar para los ejercicios de alta y baja intensidad.
desciende la PCr, la concentracion de Cr ' - - - - - - - - - - - - - - - - - - / aumenta, de modo que la creatina total (Cr + PCr) permanece constante. El acoplamiento entre el sistema de ATP y PCr posibilita que la PCr disminuya su concentracion y solo cuando esta proximo su agotamiento se produce una disminucion de las reservas de ATP, de manera que cuando la concentracion de PCr ha descendido un 90%, la de ATP solo lo ha hecho en un 10%.
/ ' I Clasicamente la utilizaci6n de la I
I PCr para resintetizar ATP se ha I denominado "sistema anaer6bico I a lactico" en fisiologfa del ejercicio ya I
que no precisa oxfgeno, ni produce acido lactico. Nosotros proponemos
I denominar este proceso como "metabolismo de la fosfocreatina" o "sistema de la fosfocreatina".
A diferencia de lo que ocurre con el ATP, la fosfocreatina experimenta un marcado descenso en su concentracion durante el ejercicio. Este descenso esta directamente relacionado con la tasa de reposicion de ATP o la intensidad del ejercicio. Segtin
Figura 8. Grafico que representa c6mo van disminuyendo las concentraciones de PCr y ATP en la celula muscular en segun aumenta la intensidad del ejercicio expresada coma V02• Modificado de Hogan MC, Gladden LB, Grassi B, Stary CM, Samaja M. Bioenergetics of contracting skeletal muscle after partial reduction of blood flow. J Appl Physiol, 1998: 84(6) 1882-1888
10 8 0 12 6 4 2 0
...,._ Fosfocreatina
ATP
0,2
0,4
o,6
o,8 1,0
l

Figura 11. Procesos citos61icos (anaer6bicos) y mitocondriales (aer6bicos) par las que las cetulas abtienen energfa a partir de la glucosa. En las celulas de nuestro arganismo, no se produce la fermentaci6n alcoh61ica .
-, -~-------------------------------- l ATP
Perdida de lr- hidr6genos - oxidaci6n .J.~ Compuestos de +_O..::.•-- - 3atomosdeC , ~~ • t
Fermentaci6n / \ Fermentaci6n alcoh61ica ,I.. 2 ATP ~ lactica 3
--
Glucosa Compuestos de 6 atomos de C
~ 2ATP
EN LA MITOCONDRIA (AEROBICO)
EN EL CITOSOL (ANAEROBICO)
-: I
Los hidratos de carbono (concretamente la glucosa), suponen el unico sustrato que nuestras celulas son capaces de utilizar para obtener energia con o sin la presencia de oxigeno. El proceso par
el cual las celulas obtienen energia de la glucosa se denomina glucolisis, Dependiendo, par un lado, de la tasa de activaci6n de la gluc61isis (par necesidad energetica), y par otro, de la capacidad oxidativa rnitocondrial muscular, el metabolismo de la glucosa finalizara en la transformaci6n de piruvato en lactato (lo que clasicamente se denomin6 glucolisis anaer6bica, y que podemos referir coma glucolisis citosolica), o en la introducci6n del piruvato en la mitocondria (glucolisis mitocondrial u oxidativa) para despues de transforrnarse en acetil-CoA incorporarse al ciclo de las acidos tricarboxflicos o "ciclo de Krebs", que finaliza con la fosforilaci6n oxidativa (figura 11)
2IMetabolismo de las hidratos de carbono
..... -- - -- --- - - ----- -- - - - - - -- --- -- --- -- - -
., ' SUPLEMENTACION CON CREATINA
lES NECESARIA? lMEJORA EL RENDIMIENTO? La creatina se sintetiza en el hfgado a partir de los arninoacldos glicina y arginina, y se absorbe de los alimentos, especialmente de la came. Las necesidades de creatina en el ser humano son de aproximadamente 2 g/dfa. La creatina pasa al interior de la celula muscular mediante un transportador espedfico, formando fosfocreatina. La perdida de creatina en las celulas musculares se hace en forma de creatinina (proceso de transformaci6n no enzirnatico), siendo esta rnolecula aclarada de la sangre en el rifi6n y excretada en la orina. Normalmente se pierden unos 2 g/dfa de creatina por la orina (-115% del total), cantidad que necesitamos reponer. La excreci6n de creatinina refleja indirectamente la masa muscular del individuo (aproximadamente 1 g de creatinina eliminada por cada 20 kg de rnusculo) y se utiliza coma pararnetro clfnico para valorar la funci6n renal. Diversos estudios han demostrado que las concentraciones basales de PCr en la celula pueden elevarse gracias a la suplementaci6n con creatina, y que esto puede mejorar el rendimiento durante actividades de intensidad elevada (Casey, 2000). En concreto, se ha comprobado que en los corredores de resistencia aer6bica el tiempo de recuperaci6n de la fosfocreatina tras el ejercicio es menor si han recibido suplementos de creatina, lo que refleja una mejor capacidad oxidativa de sus celulas musculares. Parece que existe una correlaci6n significativa entre el V02 max y el tiempo de resfntesis de la PCr (Takahashi, 1995). Adernas, este efecto es dependiente de la edad, de forma que la suplementaci6n con creatina tiene un mayor efecto sabre la disponibilidad y la tasa de resfntensis en las I
personas de mediana edad que en las mas j6venes, cuyos niveles de PCr basales son mas elevados. No obstante, si bien la suplementaci6n con creatina mejora el rendimiento durante ejercicios dinamicos, especialmente los intervalados, 1
I no parece que acelere la recuperaci6n de PCr tras la realizaci6n de ejercicios I isometrlcos intermitentes. (Vanderberghe y cols. 1999) I ' .,
~------------------------------------,
Parad6jicamente la sintesis de PCr a partir de Pi y creatina precisa la energia procedente de la hidr61isis delATP, que debe ser generado par las rutas aer6bicas de obtenci6n de energia, lo que elevara el consumo de oxigeno de la celula. Este exceso de consumo de oxigeno de la celula despues del ejercicio se denomina exceso de consumo de oxigeno post-ejercicio o EPOC (fig 10.). Este proceso de EPOC, no solo representa el consumo de oxigeno necesario para reponer la PCr, sino que tambien esta influenciado (largo plaza) par otros factores, coma el aumento de temperatura, influencia hormonal o el aclaramiento de lactato.
1IMetabolismo de los fosfagenos I
·~i ;,' ,., 26 hr· - -::

Figura 13. Esquema general del proceso de almacenamiento de glucosa en forma de gluc6geno (glucogenogenesis) por acci6n de la gluc6geno sintetasa
j ~co::J-+ Glucosa 6P -+ lL_ -~ ~) Glua,,a 1P ? GIU<6geno JI
gluc6geno sintetasa .
Posteriormente, si la celula no necesita en estos momentos utilizar la glucosa que le llega, la glucosa 6P se isomeriza a glucosa 1P, la cual se va almacenando en la celula en forma del polimero gluc6geno por accion de la gluc6geno sintetasa. A este proceso se le denomina glucogenogenesis (Figura 13). De manera que en el hepatocito yen la celula muscular, existe un almacen de hidratos de carbono en forma de glucogeno, cuya capacidad de almacenaje es limitada. Tambien los rifiones son capaces de almacenar cierta cantidad de glucogeno.
Glucosa _. Glucosa &P ATP-----+ ADP+ Pi
si bien por sus caracteristicas metabolicas las celulas hepaticas y las celulas musculares son cuantitativamente el destino mas importante.
Para que la glucosa circulante entre en el interior de las celulas, es necesaria la presencia de la hormona pancreatica insulina, salvo en determinadas circunstancias. Las celulas musculares esqueleticas activas y las neuronas no precisan la presencia de insulina para captar glucosa. Mas adelante explicaremos por que. Cuando tras la ingestion de hidratos de carbono se produce una elevacion de la glucemia, el pancreas aumenta la produccion y liberacion de insulina a la sangre de manera que la glucosa tendera a entrar en las celulas volviendose a normalizar los niveles glucemicos, Los hepatocitos y las celulas musculares ( en reposo) cap tan gran parte de esta glucosa. Cuando la glucosa atraviesa la membrana celular, inmediatamente experimenta una fosforilacion, transformandose en glucosa-6-fosfato. Este compuesto, adquiere carga negativa por lo cual queda "atrapado" dentro de la celula, y tiene un nivel energetico superior al de la glucosa. Para fosforilar una molecula de glucosa, la celula tiene que aportar la energia procedente de la hidrolisis de un ATP. A este proceso inicial de aumento del nivel energetico de un compuesto para posteriormente ser catabolizado se denomina enerqia de activaci6n.
2IMetabolismo de los hidratos de carbono
ABSORCION Y DISTRIBUD6N DE LOS HIDRATOS DE CARBONO
Los hidratos de carbono que ingerimos con la dieta se absorben en forma de monosacaridos en las celulas intestinales tras someterse a las enzimas de la digestion (amilasas y disacaridasas principalmente). La mayor parte de estos monosacaridos son transformados fundamentalmente en glucosa y, en menor cantidad en fructosa y galactosa. Una vez que alcanza el torrente sanguineo, la glucosa es puesta a disposicion de la totalidad de las celulas del organismo,
Figura 12. Esquema general de la utilizaci6n de los nutrientes por la celula muscular para la obtenci6n de energfa.
Beta - oxidaclon
Citosol
( Gl~,,,.a ·1 G•;na_J _._ )
o, I
.: I Sa1~_:_ __
I Addo, grasos
Estos procesos mitocondriales ( ciclo de Krebs y fosforilacion oxidativa) no son exclusivos del metabolismo de los hidratos de carbono, sino que, como iremos viendo a continuacion, las grasas e incluso las proteinas cuando se utilizan como fuente energetica confluyen para experimentar la actividad enzimatica de estos procesos que caracterizan a lo que hemos denominado metabolismo oxidativo (Figura 12). En este contexto, los hidratos de carbono, las grasas y las proteinas son combustibles que experimentan realmente una combustion en el interior de nuestras celulas ( obviamente en presencia del oxigeno que acnia como comburente) de la cual se obtienen como productos finales C02 y H20 ademas de una gran cantidad de energia libre.
•
Mitocondria J I '--------)
•

EL EJERCICIO REALIZADO CON BAJOS NIVELES DE GLUCOGE O MEJORA LA BIOGENESIS MITOCONDRIAL Se comparan los efectos del ejercicio con bajo nivel de gluc6geno (LG) y gluc6geno normal (NG) sobre diferentes factores moleculares implicados en la regulacion de la biogenesis rnitocondrlal. Diez ciclistas entrenados (V02max: 65±1 ml/kg/min; Wmax: 387±8) se ejercitaron durante 6omin a! 61~0/o V02max con NG (478 rnrnol/kg) o LG (166 mmol/kg) obtenido por medio de un ejercicio previo. El estudio de la biopsia muscular obtenida mostr6 que el mRNA del PGC-10. aument6 mas (8,1 veces vs. 2,5 veces) cuando el ejercicio se realiz6 con LG frente a NG. El mRNA de la subunidad I de la citocromo c oxidasa y la lsoenzima piruvato deshidrogenasa kinasa aumentaron despues de LG, pero no despues de NG. Ei analisis de los resultados de este estudio invita a pensar que el ejercicio realizado con LG amplifica la expresi6n de los principales marcadores geneticos de la biogenesis mitocondriai en ciclistas entrenados, es decir, que esta intervenci6n puede ser beneficiosa para mejorar la capacidad oxidativa muscular. Psilander N, y cols. Eur J Appl Physiol, 2013
' ,, '--------------------------------------
,, I
TRANSPORTADORES CELULARES DE GLUCOSA: GLUT-le. Para que la glucosa atraviese las membranas celulares, existen una serie de transportadores denominados de forma global coma GLUT. Estos transportadores no son iguales en todas las celulas del organismo. En las celulas musculares, encontramos el tipo GLUT-4, transportador que se activa en dos condiciones: 1) en presencia de insulina y 2) cuando se eleva la concentracion de calcio intracelular. Esto explica par que la celula muscular activa no precisa la presencia de insulina para captar glucosa, ya que el proceso de contraccion por la interaccion actina-miosina se acompafia de una elevacion importante de la concentracion intracelular de calcio que par si misma activa a las GLUT-4. En otras celulas, el transportador es diferente y, coma es el caso de las neuronas, no precisa la presencia de insulina (GLUT-1). Esto explica par que las neuronas captan glucosa independientemente de la concentracion de insulina plasmatica (ilas neuronas solo utilizan glucosa coma fuente energetical), Segun un estudio realizado par Kraniou y cols en el 2000 el entrenamiento de resistencia aerobica aumenta la concentracion de GLUT- 4 en las celulas musculares, par lo que la capacidad oxidativa se correlaciona directamente con la concentracion de GLUT-4.
--- 2_I_M_etabolismo de los hidratos de carbono j
GLUCOGENO MUSCULAR: El gluc6geno muscular, sin embargo, tiene coma mision suministrar glucosa a la propia celula muscular en la que se encuentra. Tradicionalmente se asume que el proceso de defosforilaci6n de la glucosa (glucosa 6P a glucosa) no parece posible en la celula muscular, ya que esta carece de la enzima responsable, la fosfatasa. Par tan to, la glucosa que entra en la celula muscular ya no la abandona y queda a su disposicion coma sustrato energetico. No obstante, Shieh y col en 2004 propusieron la presencia de una fosfatasa (G6PC) similar a la expresada en higado, rifion e intestino, en el tejido muscular esqueletico (Glucosa-6-fosfatasa ~) que podria permitir la salida de glucosa de la celula muscular, contribuyendo con ello a la homeostasis de la glucosa.
La cantidad de gluc6geno almacenado en el tejido muscular es variable, ya que dependera de la mas a muscular de cada sujeto, pero en terminos generales podemos hablar de unos 350-400 g en total. El musculo cardiaco tambien almacena glucosa en forma de gluc6geno. Otras celulas del organismo, coma las del cerebra y las globules rojos, dependen del aporte continuado de glucosa sanguinea para funcionar correctamente, ya que no almacenan gluc6geno.
GLUCOGENO HEPATICO: La finalidad del gluc6geno hepatico es mantener las niveles de glucemia. Es decir, cuando finaliza la absorcion de glucosa y la captacion de glucosa par las celulas del organismo provoca un descenso de la glucemia, en el hepatocito se produce un proceso inverso al que acabamos de describir gracias al cual la glucosa se desprende del polimero gluc6geno (glucogenolisis) en forma de glucosa lP. La glucosa 1P posteriormente se defosforila y finalmente sale de la celula para dirigirse a la sangre circulante. Se estima que la cantidad maxima de hidratos de carbono almacenados en forma de gluc6geno en el higado es de aproximadamente 100 g para un sujeto con talla y peso media. El limite de almacenaje se debe a la necesidad de almacenar agua junta con la glucosa para mantener la osmolaridad celular. El limite indefinido de un soluto en el interior del citoplasma celular conduciria a la muerte celular par balonizacion, Cuando la glucosa absorbida supera la capacidad de almacenaje de gluc6geno hepatico, el higado la metaboliza transforrnandola en acidos grasos que son enviados al tejido adiposo para almacenarse en forma de trigliceridos.

Uno de los efectos mas caracteristicos del entrenamiento de resistencia aerobica es la disminucion de la oxidaci6n de los hidratos de carbono durante el ejercicio debida a un descenso de la glucogen61isis, a favor de la utilizaci6n de acidos grasos libres como fuente energetica. Por otra parte, numerosos estudios sugieren que la tasa de glucogen61isis depende de la cantidad inicial de gluc6geno almacenado en la celula muscular, de manera que cuanto mayor sea esta mayor sera la utilizaci6n de gluc6geno para la obtenci6n de ATP.
Figura 14. Esquema general de la glucogen61isis en la celula muscular. -----
Glucogen61isis
Glucosa - - _ _. I
ll .1__ No en el rmisculo - .,~,~
Glucosa 1P
l ATP
Gluc6geno
Adrenalina
io Ca•• AMPco
fosforilasa A •4-...:..!._0 __ _..:,:_j._ fosforilasa B (activa) (inactiva) ,,.... i
0 // ,,
r- -----~
respuesta lenta y que, por tanto, no justifica la rapida actividad glucolitica al inicio del ejercicio. El otro mecanismo esta mediado por la liberaci6n de calcio desde el reticulo sarcoplasmico al citoplasma. Se ha sugerido que el efecto de la .. adrenalina es mas importante sobre la glucogen61isis hepatica que sobre la muscular. La fosforilasa se inactiva en presencia de concentraciones elevadas de ATP y de glucosa-6-P.
2 IMetabolismo de los hidratos de carbono
GWCOGENOLISIS MUSCULAR. La glucosa-6-fosfato procedente del gluc6geno muscular como consecuencia de la acci6n de la fosforilasa entra en la via metabolica de la gluc61isis. La carencia de la enzima fosfatasa hace que la glucosa no sea capaz de abandonar la celula muscular, por lo cual se utiliza directamente coma fuente energetica propia en el proceso catabolico de la gluc6lisis (figura 14). Este proceso dependiente de la actividad de la enzima fosforilasa, esta regulada por varios mecanismos. Un mecanismo hormonal por la concentraci6n sanguinea de adrenalina y la accion del AMPc, de
GLUCOGENOLISIS HEPATICA. En el caso del higado, la glucosa-6P resultante de la hidrolisis del gluc6geno se transforma de nuevo en glucosa gracias a la acci6n de la glucosa-6-fosfatasa, y sale de los hepatocitos a la sangre circulante. El objetivo fundamental de la glucogenolisis hepatica es mantener la glucemia en cifras normales. Parte de esta glucosa hepatica que sale a la sangre sera captada por la celula muscular, ya que el musculo esqueletico es el principal tejido que capta y utiliza la glucosa circulante durante el ejercicio. Si la tasa de utilizacion de la glucosa es mayor que la tasa de liberaci6n de glucosa hepatica, la glucemia disminuye (hipoglucemia) lo que se manifiesta con diversos sintomas como consecuencia, fundamentalmente, de reducir el aporte energetico al sistema nervioso. Este fen6meno es mas frecuente durante la realizaci6n de ejercicio de mas de 2-3 horas de duracion.
GLUCOGENOLISIS. Cuando pasamos de una situacion de reposo a ejercicio, hay que movilizar la glucosa almacenada en la celula como glucogeno para poder resintetizar el ATP que se va utilizando en la contraccion muscular gracias a la energia liberada en su catabolismo. El proceso por el cual el polimero gluc6geno va desprendiendo moleculas de glucosa para ser utilizadas (bien en la propia celula, como es el caso del musculo, bien por otras celulas como ocurre con el gluc6geno hepatico) es la glucogenolisis. Esta regulado por la accion de la fosforilasa, enzima que se activa fundamentalmente cuando la adrenalina se une a su receptor en la celula muscular, o cuando esta
experimenta un descenso de la concentraci6n de ATP.
MOVILIZACION Y UTILIZACION DE LOS HIDRATOS DE CAABONO

Figura 15. Represen!a~i6n ~sguematica de \os procesos de transformaci6n de la glucosa en acido lactico en la glucolisis citos6\ica (anaer6bica).
2 NADH -+ 2 NAD
2X piruvato
2 ADP-+ 2 ATP
2X fosfoenolpiruvato
2X 3 fosfoglicerato
2X 1-3 difosfoglicerato
2X lactato
2 ADP-+ 2 ATP
2 NAD -+ 2 NADH
{§j~ --~ Fructosa 1-6 diP
@~~ ~rr(i) Gliceraldehfdo 3P
~ ~
(F@@ 2X gliceraldehfdo 3P
(I)"
Fosfato de di OH acetona
ATP-+ ADP
Glucosa 6P
Glucosa
Fructosa 6P
ATP-+ ADP
GLUC6USIS ANAER6BICA
2IMetabolismo de los hidratos de carbono
Por lo que el rendimiento neto de la gluc61isis es de 4 - 2 moleculas de ATP.
A partir de cada molecula de glucosa ( 6 atomos de carbono) estas reacciones se producen por duplicado, ya que tanto el difosfoglicerato como el fosfoenolpiruvato son triosas. De manera que con la gluc61isis la obtenci6n de energia de forma directa equivaldria a la resintesis de 4 moleculas de ATP. Sin embargo, la activaci6n previa de la molecula de glucosa exige a la
celula la aportaci6n de energia en dos de sus reacciones previas:
Conversion de glucosa en piruvato: Vfa de Embden-Meverhof Las fuentes de glucosa para la celula muscular son dos: la que procede del gluc6geno almacenado (en este caso esta ya fosforilada) y la que entra directamente de la glucosa circulante. En el caso en que la glucosa proceda de la glucosa circulante, es inicialmente activada y transformada en glucosa-6- P para continuar su catabolismo en la via de Embden- Meyerhof. Si la glucosa procede del gluc6geno, la activaci6n ya no es necesaria. La primera parte del catabolismo de la glucosa tiene lugar en el citoplasma de la celula muscular por acci6n de una serie de enzimas que transforman la glucosa (procedente de la sangre o del gluc6geno almacenado en la propia celula) en acido piruvico y finalmente en acido lactico (Figura 15). De forma muy resumida, en este proceso la glucosa (que es una hexosa) es modificada por la acci6n de diferentes enzimas y fracturada en dos moleculas de 3 atomos de carbono cada una ( triosas) que finalmente se transforman en acido pinivico. La transformaci6n de glucosa en acido piruvico permite la fosforilaci6n directa del ADP en ATP (y por tanto, obtenci6n de energia
celular util) en dos de sus reacciones:
GLUCOLISIS CITOSOLICA

•
La produccion celular de acido lactico depende del equilibria resultante de la competencia por el piruvato y el NADH entre la lactato deshidrogenasa (LDH), Y los transportadores de piruvato y las lanzaderas de protones al interior de la mitocondria. Segun aumenta la intensidad de trabajo, la tasa glucolftica aumenta debido a la activacion de enzimas reguladoras de la glucolisis como la fosfofructocinasa (PFK) y la fosforilasa. Desde una perspectiva global, la produccion de lactato depende del balance entre la actividad de la fosforilasa Y la PFK frente a la actividad de la piruvato deshidrogenasa (PDH, complejo enzimatico responsable de introducir el piruvato a la mitocondria). Segun aumenta la intensidad de ejercicio, dorninara la actividad de la PFK/ fosforilasa sobre la de PDH.
CONTROL DE LA GLUCOLISIS
La cantidad de ATP que se puede resintetizar mediante la formacion de lactato es de unos 60-75 mmol por kg de musculo en caso de que el lactato quedara retenido dentro de la celula. De ser asi, los H· disociados ejercerfan un ef ecto de retroalimentacion negativa o de inhibicion de la glucolisis para impedir la formacion de mas lactato. Sin embargo, si la celula es capaz de eliminar los H+, en teorfa todo el glucogeno almacenado podria transformarse en lactato y seria suficiente para mantener la produccion de ATP a una tasa maxima durante 2,5 minutos.
En resumen, durante la glucolisis citosolica una molecula de glucosa procedente de la sangre o del glucogeno almacenado es transformada en dos moleculas de acido lactico o lactato. Esto tiene lugar gracias a una serie de reacciones enzimaticas citoplasmaticas, aportando un balance energetico neto para la resintesis de 2 ATP (4-2) ( en el caso de que la glucosa proceda del glucogeno almacenado, el balance energetico neto corresponde a 3 ATP, ya que la glucosa ya se encuentra fosforilada) y dejando intacto el equilibria redox citoplasmatico, ya que inicialmente se reduce un NAD, pero finalmente se oxida cuando el acido pinivico se transforma en acido lactico. Esta forma de obtencion de energfa no requiere la participacion de oxfgeno Y generalmente responde a una demanda de tasas elevadas de reposicion de ATP.
---------------- 2IMetabolismo de los hidratos de carbono J
Figura 16. cuando la mitocondria es ~apaz de oxidar los ~ producidos en el citoplasma, el piruvato se atraviesa la membrana mttocondnat Y se transforma en acetil CoA por acci6n de la p1~~vato des_h1drogenasa, desprendrendose co2 para continuar su degradac1on en el ciclo de Krebs
Acetil CoA
piruvato
----·-----·---------------
En este proceso citoplasmatico denominado glicolisis, se produce en
definitiva una reaccion de oxidacion controlada, ya que en diversos puntos
del proceso se extrae un proton (H•) con dos electrones (2e) durante el desacoplamiento de la hexosa. El proton y los electrones se unen al dinucleotido de adenina nicotinamida (NAD) para reducirlo formando NADH. Durante el catabolismo de una molecula de glucosa y su transformacion a piruvato se forman 2 NADH, que deben ser nuevamente oxidados para mantener el equilibria redox (equilibria quimico del conjunto de reacciones de reduccion-oxidacion) de la celula y que las reacciones citoplasmaticas puedan continuar. En este contexto, si la mitocondria mediante sus lanzaderas de protones es capaz de aceptar el par de electrones con el H • transportado por el NADH, quedaran disponibles para la cadena respiratoria mitocondrial permitiendo conseguir mas ATP. Si es asi, el piruvato entrara en la mitocondria para continuar aqui su degradacion; todo este proceso se denomina gluc6lisis mitocondrial u oxidativa (Figura 16). Si por el contrario la actividad mitocondrial no es capaz de aceptar estos complejos reductores, el NADH se vuelve a oxidar en el citoplasma mediante una reaccion catalizada por la lactato deshidrogenasa y por la cual el piruvato se reduce y se transforma en lactato. simultaneamente, el NADH se oxida transformandose en NAD y dejando asi en equilibria el redox citosolico. Esto es lo que ocurre en el proceso de glucolisis citosolica o de obtencion de energfa a partir de glucosa
y sin participacion del metabolismo oxidativo mitocondrial.

Cicio de Krebs v fosforilaci6n oxidativa
Cuando el NADH formado en el citoplasma por efecto del proceso glucolitico puede ser reoxidado por la lanzadera de protones de la membrana mitocondrial, que acepta H+ y los transfiere a los coenzimas mitocondriales
' el piruvato, en lugar de transformarse en lactato coma ocurre enla glucolisis citosolica, abandona el citosol y entra en la mitocondria para completar aqui su metabolismo hasta transformarse finalmente en H o y co con 2 2
la presencia del oxigeno. De esta manera, se conserva tarnbien el redox citosolico y se completa la obtencion de energia en condiciones aerobicas a partir de la glucosa. A este tipo de gluc61isis que finaliza en la introduccion de acido pinivico en la mitocondria en lugar de llegar a la transformacion de acido lactico, la hemos denominado glucolisis mitocondrial u oxidativa.
Las fases de este metabolismo podemos separarlas virtualmente en cuatro '
teniendo siempre presente que los tres ultimas procesos metabolicos son mitocondriales:
t. Glucolisis ( transforrnacion de glucosa en piruvato) 2. Transformacion del piruvato en acetil coenzima A (acetil CoA)
GLUCOLISIS ITOCONDAIAL
de hexosas-fosfato (glucosa-6-P, fructosa-6-P, fructosa-1,6-diP) regula la gluc61isis de manera que al elevarse activan a la PFK. Por el contrario, las triosas (3-fosfoglicerato, 2-fosfoglicerato, 2,3-difosfoglicerato, fosfoenolpiruvato) tienen probablemente un efecto inhibidor sabre la PFK. La concentraci6n de calcio tambien se asocia a la activaci6n de la PFK.
Aparte de este control enzimatico (Figura 17), la disponibilidad de sustratos condiciona obviamente la tasa de gluc61isis. Para que la glucolisis pueda desarrollarse, la celula debe disponer de glucosa ode gluc6geno almacenado. Durante el ejercicio, el descenso de la concentraci6n de insulina plasmatica favorece la entrada de glucosa en las celulas musculares, ya que dificulta que otras celulas del organismo la capten, dejandola a disposicion del rmisculo activo y del sistema nervioso. Sin embargo, cuando se produce un descenso de la glucemia durante el ejercicio la glucolisis muscular se detendra por falta de sustrato, lo que inevitablemente se asociara con fatiga.
2 IMetabolismo de los hidratos de carbo~
Lactato deshidrogenasa
•••••••4~· , Acetil CoA 11-----
( Fosfoen~lpiruvato
Piruvato deshidrogenasa
~fosfoglicerato
Figura 17. Principa!es enzimas que parncrpan en la regulaci6n de la gluc61isis.
Lafosforilasa es la enzima que permite "extraer" las moleculas de glucosa contenidas en el polimero gluc6geno. Existen dos formas, fosforilasa a Y fosforilasa b. La fosforilasa a es la forma activa y solo constituye de un 5% a un 20% en el musculo en reposo. Los principales factores que activan
la fosforilasa b transformandola en fosforilasa a son el aumento de la concentraci6n de calcio (que se produce simultaneamente al inicio de la contracci6n muscular), la adrenalina mediante la activaci6n de receptores ~ -adrenergicos y el aumento de la concentraci6n de AMP. La activaci6n de la fosforilasa permite la glucogenolisis. Sin embargo, es la fosfofructocinasa (PFK) la enzima clave que cataliza la reacci6n que determina la tasa glucolitica. La disminucion en la concentraci6n de ATP activa de forma sensible esta enzima de manera que potencia la utilizacion de la glucosa al principio del ejercicio. La elevaci6n de la concentraci6n de H+ tiene un potente efecto inhibidor sabre la fosforilasa y sabre la PFK. La concentraci6n

de acciones enzimaticas que finalmente lo transformaran de nuevo a oxalacetato (Figura 18). Este es un proceso ciclico a lo largo del cual se produce la degradaci6n completa del acetil y que consiste fundamentalmente en extraer los H+ (reduciendo coenzimas como el NAD y el FAD) y los atomos de carbono en forma de C02. Ademas, durante el ciclo de Krebs se produce una reacci6n en la que hay una transferencia directa de energfa, en la que se fosforila un GDP para transforrnarse en GTP.
Para que este ciclo se mantenga en funcionamiento es necesario aportarle grupos acetilo y Hp. Como podemos observar, aun no hemos recurrido al oxigeno molecular procedente de la respiraci6n ni tampoco hemos conseguido la transferencia de energfa al ATP. Estos dos eventos se produciran en el complejo proceso de la fosforilacion oxidativa que describiremos mas adelante.
Resumiendo, la principal funci6n del ciclo de Krebs es la de extraer los hidr6genos contenidos inicialmente en las moleculas de combustible (en este caso de la glucosa ya transformada en acetilo) para poder oxidarlos en el proceso de la fosforilaci6n oxidativa. Pero de forma sorprendente, y para rentabilizar el proceso, en el ciclo de Krebs no solo se extraen los hidr6genos procedentes de las combustibles, sino que se utiliza el H20 celular para extraer hidr6genos y volver a ser oxidados para la obtenci6n de energfa. Es decir, finalmente la energfa procede de la oxidaci6n de los hidr6genos que estan contenidos en los combustibles, pero tambien del agua que ocupa el media intemo celular.
En un ciclo de Krebs completo en el que se introduce un acetilo se reducen 3 NAD, 1 FAD, se genera un GTP a partir de GDP, se obtienen 2 moleculas de C02 (que se eliminan a traves de la respiraci6n) y se gastan 3 moleculas de H20. No olvidemos que par cada molecula de glucosa que comienza en el citoplasma su catabolismo ( 6 atomos de carbono) se obtienen 2 de piruvato (3 atomos de carbono), y por tanto, el ciclo de Krebs se alimenta dos veces, de manera que para calcular el rendimiento energetico obtenido a partir de una molecula de glucosa hemos de multiplicar por 2 la energfa obtenida a partir de la entrada del piruvato en la mitocondria.
2IMetabolismo de los hidratos de carbono
CICLO DE KREBS (Cicio de las acidos tricarboxnicos) . ,
Una vez obtenido el acetil CoA, en este caso a partir del piruvato, la porcion acetil se incorpora a un compuesto mitocondrial denominado oxalaceta~o para formar un nuevo compuesto ( citrato) que sera sometido a una sene
NADH
TRANSFORMACION DEL PIRUVATO EN ACOIL·CoA 1 · la acci6n del El piruvato atraviesa la membrana mitocondria gracias a
complejo enzimattco piruvato deshidrogenasa (PDH) situado en la _membr~na intema mitocondrial que lo transforma de forma esencialmente irreversible en acetil-CoA (vease Figura 16). La accion de este complejo compuesto par tres
. 1 piruvato· a) produce una enzimas provoca diversas transformac10nes en e . . . , , . NAD· b) produce una decarbox1lac1on deshidrogenacion, reduciendo a su vez un ' , .
un atomo de carbono de la molecula de piruvato en por lo que se separa . , . enzima A La incorporacicn forma de C02 y c) finalmente le mcorpora un co .
del coenzima A supone la activaci6n energetics del piruvato. Como produ~to . , . 't' de la PDH se obtiene una molecula de tan solo final de la accton enzima tea
dos atomos de carbono, unida a un coenzima A (queen su conjunto for~_an acetil CoA) y un NADH. La elevaci6n de los niveles de adrenalina.plasmat1ca durante el ejercicio provoca la activaci6n de la piruvato deshidrogenasa, lo que da como resultado una mayor utilizaci6n del metabolismo de los hidratos de carbono. Aun no se conoce el mecanismo por el cual se produce esta activaci6n, aunque probablemente esta relacionado con un aumento del
calcio intracitoplasmatico inducido por la adrenalina (Watt, 2001).
1 f 3 y 4 son comunes para Es importante destacar en este punto que as ases . , . d 1 t ' as ya que el acetil el metabolismo energetico de los lipidos y e as pro em '
CoA y los elementos intermedios del ciclo de Krebs pueden proceder de otras
fuentes energeticas diferentes del piruvato.
3. Entrada del acetil CoA en el ciclo de Krebs para su ~egr~~aci6n 4. Fosforilacion oxidativa (fosforilaci6n del ADP y oxidacion de los
H+ extraidos a lo largo de todo el proceso glucolitico
•

GXIDACION
Los hidr6genos liberados en la glucolisis, en la transformaci6n de piruvato en acetil CoA y en el ciclo de Krebs ( asi como en otros procesos metab61icos que participan en el metabolismo de las grasas como la J3-oxidaci6n) son transportados por coenzimas celulares coma el NAD y el FAD has ta el interior de la matriz mitocondrial. Alli los hidr6genos se liberan, descomponiendoss a su vez en H+ y un e. Los electrones liberados van pasando de un compuesto a otro, entre los compuestos que componen la cadena respiratoria o de transporte de electrones. Segun el electron va pasando de uno a otro de estos compuestos, se libera energfa. El ultimo compuesto de la cadena respiratoria es el citocromo A3 el cual transfiere los electrones al oxigeno. Finalmente, los electrones los acepta el oxigeno molecular obtenido del aire atmosferico, transformandose en oxigeno ionico, el cual se une al H+ para formar en Hp.
• Utilizacion de la energfa Figura 19.
I ATP ADP+ Pi Acoplamiento de las procesos de oxidaci6n-
l1 fosforilaci6n en la mitocondria de la celula
Fosforltaclon --+ UNION DE Pi A LA MOLECULA DE ADP -- muscular. esqueletica
Acoplamiento de los procesos de cxidacion
y fosforilacion
(:idacion--. TRANSFERENCIA DE H+ Y ELECTRONES Al OXfGENO
I Compuestos que
I NAO NADH + w +-- contiene.n ,w de /\ ,,.• /\ ... /\ e" /\ e"j alta energia y , / V V V V electrones
Ii 0 Aceptor final de Cadena transportadora de electrones +----- las electrones
I H20 • I 0:1 en la cadena .... respiratoria
i I I +-- I I I
La cadena o transporte de electrones se realiza en la membrana intema mitocondrial y gracias a ella se lleva a cabo el proceso de la oxidacion.
2IMetabolismo de los hidratos de carbono J
CADENA DE ELECTRONES. Fosforilaci6n oxidativa. . d bi imico que se refiere a os La fosforilaci6n oxidativa es un concepto Ioqui . . .
1 n en el interior de la mitocondna (Figura 19). procesos que se acop a , , . o en el cual el oxigeno es el t. Un proceso de oxidacion espontaneo, exergomc ' , d hidrogeno
lib de los atomos e aceptor final de los electrones que se 1 eran h obtenidos de los nutrientes a lo largo de los diferentes procesos que emos
descrito hasta ahora, Y . u O
fosfato al ADP de fosforilaci6n por el cual se mcorpora un gr p
2. un proceso , . e para poderse . t. el ATP Este es un proceso endergomco qu para resmte 1zar · . ,
llevar a cabo utiliza la energia liberada en la oxtdacion.
. de\ ciclo de Krebs o de \os acidos , · d \as reacc1ones ti en Figura 18. Representc:ci6n esq~emat1ca r ciclo uniendose al oxa\acetato para conver irse tricarboxflicos. El acetil CoA se mcorpora ay libera H (reduciendo FAD y NAO) y C02
citrato. El cido consume agua
Cetoglutarato H20
Citrato
H20
Piruvato
+l .. e Acetil CoA NAO.+ NADH

Gluconeogenesis v glucogenesls
La gluconeogenesis y la glucogenesis son procesos de sintesis de glucosa a partir de diferentes sustratos. La gluconeogenesis es la sintesis de glucosa utilizando aminoacidos (fundamentalmente alanina, Figura 20) y lactato (ciclo de Cori, Figura 21) mientras que si obtenemos la glucosa a partir de piruvato, se denomina glucogenesis (Figura 21).
SiNTESIS DE HIDRATOS DE CAABONO
Los hidr6genos transportados por el FADH hasta la fosforilaci6n oxidativa en la cadena de electrones proporcionan la energia equivalente a la fosforilaci6n de 1, 5 ADP:
· (2 x 1,5) = 3 ATP obtenidos del FADH mitocondrial • 2 ATP de la gluc61isis • 2 ATP de la fosforilaci6n del GDP
En total, conseguimos la energia necesaria para fosforilar 34 moleculas de ADP y transformarlas en 34 ATP con la oxidaci6n completa de una molecula de glucosa. Si la glucosa procede del gluc6geno almacenado en la celula en lugar de proceder de la sangre circulante, tendremos 1 ATP mas, puesto que la glucosa almacenada en forma de gluc6geno ya se encuentra fosforilada (recordemos que se polimeriza a partir de glucosa-6-P).
Asi que:
· (2 x 1,5) = 3 ATP obtenidos del NADH citoplasmatico si los hidr6genos pasan a la fosforilaci6n oxidativa
· (8 x 3) = 24 ATP obtenidos del NADH mitocondrial
La lanzadera que predomina en el musculo de los mamiferos (y en el cerebra) es la bomba del glicerol 3 fosfato. Mediante esta lanzadera, los electrones del NADH producidos en el citsol son transferidos al FAD, lo cual resulta en la producci6n de 1,5 ATP. Por tanto, por cada molecula de glucosa, la cantidad de ATP mitocondrial que se obtiene de los hidr6genos contenidos en el NADH producidos en el citosol e introducidos en la mitocondria sera de 3 (1.5 x 2).
-------- 2IMetabolismo de los hidratos de carbonj
b . d en el citoplasma a la fosforilaci6n Para introducir los H+ o tern os . . que actue una lanzadera de protones a mvel de la oxidativa, es necesano
membrana mitocondrial.
•(4 - 2) = 2 ATP citoplasmaticos •8 NADH
mitocondriales
•2 FADH mitocondriales •2 NADH
citoplasmaticos
•2 x (Succinil CoA + GDP~ succinato + GTP)
En el ciclo de Krebs
•Glucosa +ATP~ Glucosa-6-P + ADP
•Fructosa-6-P +ATP~ Fructosa-1,6-diP + ADP
•2 x (,,3-difosfoglicerato + ADP ~ 3-tosfoglicerato + ATP)
•2 x (Fosfoenolpiruvato + ADP ~ Piruvato + ATP)
En la lucolisis
•2 x (Succinato + FAD ~ Fumarato + FADH2)
z DEFOSFORILACIONES Y FOSFORILACIONES DIRECTAS
•2 x (lsocitrato + NAO ~ oxoglutarato + NADH + H+)
•2 x (a-cetoglutarato + NAO~ Succinil CoA + NADH + H+)
•2 x (Malato + NAO~
EN LA GLICOLISIS ANAEROB/CA (NADH citoplasmatico)
•2 x (Gliceraldehfdo-3-fosfato + NAO ~ 1,3-difosfoglicerato + NADH + H+)
EN LOS PROCESOS AEROBICOS (NADH y FADH mitocondria/)
•2 x (Piruvato + NAO+ CoA ~ Acetil CoA + (02 + NADH + H+)
1. OBTE~CION DE iTOMOS DE HIDROGENO i•ti,hGU:HY\U@•»tl•t1•i
DE MANERA QUE EN TOTAL SE OBTIENEN •2 GTP = 2 ATP
mitocondriales
BALANCE ENEAGETICO OBTENIDO DE LA OXIDACl6N COMPLETA DE UNA OLECULA DE GLUCOSA
oxalacetato + NADH + H+)
FOSFORILACION · 1 tili da para refosfonlar e , liberada en este proceso es u 1 iza Toda la energia 1 ·1 · ,
. 1 /\'TP Por tanto los productos finales de la fosfon acion ADP y convertir o en rt ·
oxidativa son Hp Y C02, Y ATP.

La enzima que cataliza esta reacci6n reversible es la lactato deshidrogenasa. Esta transformaci6n de glucosa en dos moleculas de lactato es lo que denominamos la glucolisis citos6lica ( denominada glucolisis anaer6bica de forma desafortunada a nuestro entender, pues si bien es cierto que no precisa la presencia de oxigeno, la celula muscular si dispone de oxigeno cuando se produce esta deriva del metabolismo de la glucosa). Podemos afirmar en lineas generales que es la saturacion del proceso de oxidacion mitocondrial- y no la anaerobiosis en si misma- la responsable de que se oxide el NADH + H· en el citoplasma gracias a la accion de la lactato deshidrogenasa lo que implica la formacion de acido lactico. Como resultado de esta "deriva", la celula no obtiene toda la energia que contiene una molecula de glucosa, ya que su metabolismo no pasa por la fase mitocondrial. Si la concentraci6n de lactato producido es elevada, este comienza a salir de la celula a la sangre. No olvidemos que este lactato posteriormente puede ser utilizado como sustrato energetico en diferentes celulas en los que se mantiene el equilibria redox, siendo convertido a piruvato y posteriormente introducido en la mitocondria a traves del complejo de la piruvato deshidrogenasa para finalizar su metabolismo en el ciclo de Krebs junto con la fosforilacion oxidativa.
De manera que cuando la celula muscular esta consumiendo ATP a una tasa cuya reposici6n no es capaz de satisfacer la mitocondria (o lo que es lo mismo, esta reduciendo mas NAD del que es capaz de reoxidar la
t,G'0= -25.1 kJ/mol
/actato deshidrogenasa
O o- \ I
c I ~~~~~~~~~~~~~• HO-C- H I CH3
L-Lactato
NADH + W NAD• v O o- \ /
c I C=O I CH3
Piruvato
el redox citoplasmatico comenzaria a disminuir por un desequilibrio entre NADH + H+ y NAD (a favor del primero). En estas circunstancias, es el propio piruvato el que reoxida el NADH + H+, quedando este reducido en forma de lactato:
2IMetabolismo de las hidratos de carbono I
Fi ura 21. Esqu~~a ~,eneral ?~\ ci~o de Cori. Utilizacion h~pat1ca de\ \actato producido en el musculo para transformarlo en g\ucosa
--------------- CICLO DE CORI
METABOLISMO DE LACTATO EN LAS CELULAS MUSCULARES DURANTE EL EIEACICIO
la celula muscular, el Durante la oxidaci6n completa de la glucosa en d 1 NADH + w que se re uce estado redox del citosol se mantiene ya que e , . . , .
mo consecuencia de la transformaci6n de la glucosa hasta acido piruvico co t nes de la membrana en el citosol se reoxida porque la lanzadera de pr~ o . 1 ( decir mitocondrial transfiere los H+ a los coenzimas m1tocon:nal es a::ptore~ 1 s introduce en el mitocondria). Sin embargo, cuan o os 0
· d la lanzadera de protones, . d . 1 s (FAD) no son capaces de reoxi ar mitocon nae
I .. Piruvato ..--- Glucosa
ti tJ Lactato Glucogeno
Glicerol~Glurrno
Aminoacidos ~ Glucosa
-> La<tato? Piruvato
Fi ura 20. Esquema general de\ c)clo . d:la alanina-glucosa. G\uc_oneogenes1s hepatica a partir de la alanine procedente de la celula muscular
I \
i CICLO DE ALANINA - GLUCOSA ~)
I I . Glucosa ~ Piruvato +-A anma il 1 Gluc6geoo Urea +- ~NHi)
Alanina plasmatlca
Glucosa plasmatica
Gluc6geno Aminoacidos .,t: (NH,)L~ .
Glucosa _., Piruvato-~ Alan ma

Po~ co~siguiente, la capacidad mitocondrial es fundamental para entender la acidosis generada por el ejercicio. De manera que la acumulacion de lactato durante el ejercicio de alta intensidad no seria la causa de la acumulacion de :rotones y el descenso resultante del pH. De hecho si se inhibe la produccion
~ ~actato durante el ejercicio, el pH desciende incluso de manera mas rapida, ya que la produccion de lactato actua coma un sistema amortiguador O buff er consumiend H • o , y como transportador de protones a traves del sarcolema pa t 1 , ' ra pro eger a a celula contra la acidosis metabolica
/---------------- I Como se puede observar la '1 I ., ' I
reaccion catalizada por la lactato I deshidrogenasa es alcalinizante I en la celula, no acidificante lo I ' I que se opone al concepto clasico I de la acidosis lactlca . Segtin
algunos autores, (Robergs y col) I no hay evidencia bioqufmica I que sostenga que la producci6n : de lactato libera un proton. La I interpretaci6n fisiol6gica correcta I
I de estos hechos bioqufmicos : es que la producci6n de lactato I
I retarda el desarrollo metab61ico de I
I la acidosis. ; ' ________________ /
que exceden la capacidad mitocondrial, la tasa de liberacion de protones puede exceder el amortiguamiento de los mismos por parte del Pi, aminoacidos y proteinas. Si a esto se une una saturacion de la capacidad de expulsar protones por el sistema Iactato/Hr, se producira inevitablemente un aumento de la concentracion de protones ( descenso del pH) 0
acidosis metabolica.
2IMetabolismo de los hidratos de carbono
C) Los protones producidos en exceso, pueden ser eliminados de la celula por diversos mecanismos (transporte al interior de la mitocondria por los transportadores MCTl - transportadores de monocarboxilatos intracelulares-, salida de la celula a traves del sarcolema
D) En las condiciones metabolicas en las que hay una participacion pref erente de la glucolisis citosolica.distintas ATPasas rompen las moleculas de ATP para generar energia Y posibilitar la contraccion muscular rapida, Si el NADH y los protones de las reacciones citosolicas se producen a tasas
buffer celulares como (C03H- , residuos de histidina, fosfocreatina, fosfatos .... ). Estos compuestos son capaces de mantener el pH celular sin cambios a pesar de que aumente la produccion de protones (independientemente
de cual sea su origen).
B) Al activarse la glucolisis citosolica la cantidad protones que se disocian de los acidos que se van formando como intermediarios de la glucolisis va aumentando. Estos protones ( que se suman a los resultantes de la hidrolisis del ATP) seran amortiguados o tamponados por diferentes elementos
A) La conversion piruvato a lactato implica la ((desaparicion" de un proton, ya que la reaccion consiste en la rcduccion del piruvato para oxidar el NADH + W. En este sentido, la produccion de lactato reduciria la concentracion de protones.
LA FORMACION DE LACTATO lES RESPONSABLE DE LA ACIDOSIS CELULAR?
En los ultimas afios, algunos autores han polemizado acerca de cual es la causa de la acidosis metabolica que se asocia al ejercicio de elevada intensidad, cuando este se mantiene en el tiempo. Podemos extraer las
siguientes conclusiones:
mitocondria), se acelera la glicolisis citosouca para proporcionar ATP y los
NAD reducidos se oxidan gracias a la conversion del acido piruvico en acido
lactico. El acido lactico formado coma consecuencia de este proceso, presenta una
pKa de 3,86 lo que significa que al pH intracelular (de 7,0 aproximadamente) practtcamente se encuentra disociado en un anion lactate- y un proton (H•). Este es el motivo por el cual nos referiremos a este compuesto coma
"Iactato" y no como "acido lactico".
por el intercambiador Na/H
... ). Sin embargo, al aumentar
la intensidad del ejercicio y
la demanda de ATP, habra
un momenta en el cual la
produccion de H· excedera a
la capacidad de amortiguacion
de la celula, e incluso a la
capacidad que tiene la celula
muscular para eliminar
estos protones a traves del
sarcolema. En esta situacion es
cuando se instaura la acidosis
metabolica.

La respuesta general de la concentraci6n de lactato sanguineo en individuos entrenados y no entrenados a intensidades de ejercicio progresivamente crecientes sigue un patron similar al que se muestra en la figura 22.
Si analizamos la grafica, observamos estabilidad en las concentraciones de lactato manteniendose la concentraci6n que hay en reposo en las cargas de trabajo de baja y moderada intensidad. A partir de una intensidad determinada comienza a observarse un aumento progresivo de la concentraci6n de lactato en sangre. La intensidad de ejercicio asociada al inicio de aumento progresivo de lactato en sangre se denomina umbral lactico (flecha en el grafico ).
El concepto de umbral lcictico se ha relacionado con el momenta metab6lico en el que se compromete la formaci6n inmediata de ATP a traves de la fosforilaci6n oxidativa, lo que obligaria a una activaci6n mayor de la gluc61isis
CONCEPTO DE UMBRAL LACTICO
METABOLISMO DEL LACTATO DURANTE EL EJERCICIO
La valoraci6n e interpretaci6n de las variaciones en la concentraci6n sanguinea de lactato durante el ejercicio es uno de los focos principales de interes para los fisi6logos del ejercicio en relaci6n con el entrenamiento de resistencia aer6bica.
La participaci6n de estos sistemas para la regulaci6n del equilibria acido- base en la celula, de manera que cuando la producci6n de H+ excede la tasa de amortiguaci6n o buffering y eliminaci6n o aclaramiento de protones en el musculo esqueletico, se instaura una acidosis metab61ica. Es importante resaltar que la producci6n de lactato actua tanto como un sistema amortiguador (buffer) al consumir un proton, como eliminando protones mediante el transporte de H+ a traves del sarcolema, protegiendo asi a la celula de la acidosis metab61ica.
Las conclusiones anteriormente expuestas no impiden que, desde un pun to de vista practice, sigamos utilizando la valoraci6n de la concentraci6n de lactato en sangre durante el ejercicio para obtener informaci6n indirecta de la actividad glucolitica citos6lica y la acidosis generada, lo cual supone una gran ayuda en el control del entrenamiento.
2IMetabolismo de los hidratos de carbono j
Tabla 4. Causas de acidosis y tamponamiento de protones en el musculo esqueletico Modificado de Robergs RA y cols, 2004.
Hidr61isis del ATP
Protefnas mitocondrial H++ HC03-
Aminoacidos Cotransporte
Hidr61isis de PCr Lactate I H+ HiC03 lntercambio de
Producci6n de lactato membrana Na+ I H+ Formaci6n de IMP lntercambio
H20 + C02 HC03- HC03-1C1-
Glicolisis
La causa de la acidosis metab6lica asociada al ejercicio de alta intensidad
mantenida no es debido en sentido estricto a la liberaci6n de protones,
sino mas bien a un desequilibrio entre la tasa de producci6n de protones,
frente a la tasa de amortiguaci6n y eliminaci6n de los mismos. Como hemos
t . id d la liberaci6n de H+ se produce esencialmente comentado con an enon a , desde la glucolisis citos6lica e hidr6lisis del ATP; sin embargo,, e~ pH
celular no desciende inmediatamente debido a la capacidad de multiples
componentes celulares de amortiguaci6n y eliminaci6n de protones (Tabla
...... ). Estos ultimas deben ser tamponados por los sistemas buffer en las
celulas musculares (C03H-, residuos de histidina, fosfocreatina, fosf~to~, . . . . , d 1 CO 8 _ que se ha estimado puede contnbuu carnosma). La participacion e 3 ,
en un 16% con la capacidad buffer total de la celula muscular en ejercicio,
se asociara a un aumento de la producci6n de C02 (C03H- + H+ ~ C03H2 ~ CO +HO) durante el ejercicio intenso (>UL), en una tasa equilibrada con el
2 2 . lta del CO H- El aumento del lactato junta con el descenso descenso simu aneo 3 · del CO H- intracelular se va equilibrando por intercambio de estos iones a traves 3del sarcolema y, como consecuencia, disminuira la concentraci6n de
co H- y aumentara la de lactato sanguineo. ;especto a la eliminaci6n de los protones, estos son expulsados del citosol
via transporte mitocondrial, transporte del sarcolema (lactatorl+; Na+/H+)
e intercambiador dependiente del bicarbonato (HC03-/Cl-).

'
, Parece que la formaci6n de lactato I a partir del gluc6geno muscular
o desde la glucosa sangufnea no I es causa de formaci6n neta de I protones, Y por tanto no serfa
responsable de la acidosis inducida , por el ejercicio
Como ya hemos mencionado el acido lactico tiene una constante de disociaci6n que expresa un pK de 3 8 lo q . ·t. . , , ' ' ue sigru ica que a pH fisiologico celular la for~ac1on de acido lactico proporciona hidrogeniones ( que se utilizan en gran medida para la reduccion de los nucleotides celulares).
. Com~ hemos mencionado anteriormente, durante el ejercicio de alta intensidad se produce una acidosis sistemica que se ha atribuido a la acumul . , de , · d
1, . acion
aci o actico y su disociacion a lactato + H•· sin emba 1 • rgo actua mente se asume que la mayor contribucion a la acidosis - - - - - - / ---------- muscular durante el ejercicio de alta intensidad , La asociaci6n entre la acumulaci6n 'i corre d 1 de lactato con la acidificaci6n ,
span e a a hidrolisis del ATP a ADP, la I muscular durante el ejercicio de I
cual genera la liberacion de un proton (H•). alta intensidad es mas coincidente que causal.
El hecho de que la glucolisis citosolica no consume protones y que la micotondria satura sus mecanismos de oxidacion de los protones producidos en el citosol, favorece el aumento intracelular de la concentracion de protones.
-\ 8 Lactate
vo2 \ 4 I
7 • - - .. vo, • I I
6 I I
* :::::::. I
I 0 I 3
E 5 • < E
/ ~o 0
; ,......
I ... 4 ; ... ,
"' ,. ... 3
u
"' ; ~
-' 3 ; 2 I /
; • ' 2 ;
I ; • - - .. - - . - - ..... - - . ;
I 0 0 10 20 30 40 50 60 70 So 90 120j 100 110
lntensidad (watios)
~igura 23. Respue~!a del lactato sangufneo y del VO urante la realizacion de un ejercicio incremental. 2
ETABOLISMO DEL LACTATO
.----~-------- -·---------·-----~---
------- 2IMetabolismo de los hidratos de carbono [
;
I Aunque en condiciones de deficit de aporte de oxigeno (isquemia muscular) se facilita la acti'laci6n de la gluc61isis cltosclica y por tanto la formaci6n de lactate, las cineticas del V02 y del lactate sangu[neo durante un ejerclclo de inter,sidad creciente (figura 2::1:), demuestran que sunque el V02 aumenta !ir.ealmente respecto a la intensidad de ejerciclo, la concentrnci6n de tactato rompe el comportamiento lineal incrementando exponencialmente sus valores a partir de una dete,minada irrtensldad. Esta cinetica del comportamiento de! \/02 dernuestra que el aumento en la concentraci6n de lactate no es directarnente atribuib!e a un deficit de oxfgeno en la celula muscular. No obstante, el urnbral !actico es considerado tambien corno umbral am1er6bico, denominaci6n claslca que expresa que a partir de una determinada intensidad de ejercicio el aporte de las vias citos6iicas a la obtencion de energfa se hace mucho mas
\ relevante, a pesar de mantener su actividad el metabolismo oxidative. J / '----------------------------------------~
~---- - ------------- - - ---- --- --
citos61ica (piruvato ~ lactato) lo que provoca un aumento en la concentraci6n
celular -y secundariamente- sanguinea de lactato. Hay que dejar claro en este punto que la participaci6n metabolica de la
glucolisis citos61ica o anaer6bica no implica necesariamente el cese del metabolismo oxidativo, sino que existe un solapamiento de ambos sistemas con predominio de uno u otro segun la actividad fisica desarrollada (figura 23).
Por otra parte, el aumento de la concentraci6n de lactato en sangre no implica
necesariamente condiciones anaer6bicas. Por otra parte, el aumento de actividad simpatico-adrenal que se produce
a partir de una intensidad de ejercicio determinada favorece la tormacion de lactato en el musculo mediante la activaci6n de la glucogenolisis y la gluc61isis
muscular por efecto de las catecolaminas.
Figura 22. Comportamiento de la concentraci6n sangu[nea de lactato (mmol/1) durante la realizaci6n de un ejercicio de intensidad creciente.
~~-~--R 6 7 8_v_e_i_:_ci_d_a_d_::_m_Jh_:_1 __ 1_2--13--1-4 _ __,
3
::::::. -c5 E E 0 . ., "' ..... u 11:l -'
4
5
2
6
--------------------------------------- i

[ LACTATO I ~IRUVATOI
LDH H+ -. +
o~ .,...o- c I C= 0 + I
H-C-H I H
o~ .,...o- f r
HQ-C- H + ./ I
H-C-H t H
H H ~~o ll ::JJ C....._NH2
N
H --"'\
?' o I (J('NH2 N
Glucosa + 2 ADP + 2 Pi -+ 2 lactato + 2 ATP + 2 H O 2
Glucogenon + I ATP + 3 Pi + 1 H+ -+glucogeno + 2 I 9
"'TP n-, actato +;aN +2HO 2
2 IMetabolismo de los hidratos de carbono J
*La lanzadera aspartato-malato en la membrana . . de NADH que introduce en la mitocondria para su ox~1toc_ondnal consume un ATP por cada molecula por 1 NADH producido en el citosol e introducid 1 acion, par lo que el ,rendimiento neto es de 2 ATP o en a mitocondria a traves de esta lanzad era.
Sin embargo, esto solo ocurrira si el FAD mitocond . 1 1 na es capaz de reoxidar la anzadera de protones de la membrana mitocondrial· si no es , ,
t , asi, no se podra
man ener una relacion NADH + H + I NAD . + normal, con lo que el estado redox del citosol se reducira y la glu T . , . . . . co isis se vena inhibida si no hubiera una via alternativa capaz de reoxidar el NADH , . + H+. Esto se consigue en el citosol a traves del piruvato, que es capaz de reoxidar el NADH H d . , , id r ti h + +, re uciendose a aci o ac ico, echo que ocurre sin la participacion del oxigeno At , d 1 T · · , . · raves e esta g uco isis citosolica solo se forman 2 l ' 1 , mo ecu as ATP en lugar de 36 ATP obteman por oxidaci6n. que se
A partir de una molecula de glucosa intracelular, la produccion de . lactato (Figura
25) , prruvato a , n~ supone una produccion neta de protones; si la glucosa es
procedente del glucogeno intracelular incluso existe un des d 1
. censo e un proton y a ganancia de 1 ATP adicional.
ifii.fk.i,i,IBil ____. G ATP + 2 NADH(C) + 8 NADH + 2 FAD + 2 GTP J ., .. .,, EQUIVALENTES ENERGETICOS
NADH+3 ATP II FADH+2 ATP II GTP+1 ATP II Figura 24 NADH (C)+2 ATP (citosol) Bomba aspartato malat~
De Gluc6geno almacenado
UU4ift!I --t~ & ij\jf.fi,j.j.rffi• ___.. ( 2 ATP + 2 NADH(C) + 8 NADH + 2 FAD + 2 GTP}
Gii-Siii
De glucosa circulante
1) PRODUCCION DE Acmo LA.cTICO (LACTATO) Durante el catabolismo de glucosa a piruvato en el citosol (glucolisis citosolica), el rendimiento energetico neto equivale a la resintesis de 3 moleculas de ATP, si el NADH + H+ que se forma, puede ser reoxidado por la lanzadera de protones de la membrana mitocondrial. Esta manera de regenerar NAD+ en el citoplasma celular mantiene el estado redox del citosol y permite que la glucolisis pueda continuar. Por tanto, de las 6 moleculas de ATP formadas con la energia procedente de la glucosa hasta su descomposicion en piruvato, 2 ATP se forman en el citosol y 4 ATP en la
mitocondria por la reoxidacion del NADH. El catabolismo de la glucosa en estas condiciones de actividad mitocondrial y
mantenimiento del redox citosouco continua en la mitocondria. La formacion de acetil-CoA desde piruvato y su posterior entrada al ciclo de Krebs aporta un rendimiento neto de 4 NADH + H+, 1 FADH2 y 1 GTP. Ya que el proceso de reoxidacion de cada NADH + H+ por la cadena de electrones de la mitocondria rinde 3 moleculas de ATP y el de FADH2, 2 moleculas de ATP, el total de moleculas de ATP generadas sera de 15; en realidad seran 30 ATP ya que por cada molecula de glucosa ( 6 atomos de carbono) se forman dos moleculas de piruvato (3 atomos de carbono ). Si a esto le anadimos las 2 moleculas de ATP formadas en la glucolisis (a partir de glucosa) y las 4 obtenidos de la reoxidacion del NADH + H+ citosolico*, la ganancia total de ATP de la oxidacion completa de una molecula de glucosa sera de 36 ATP (Figura 24).

Figura 2:7.· Diag:ramas para representar el metabolis , . , ~In ejercrcio a diferentes intensidades. (A): estado e~ob~nerget1~0 de la celula muscul~r durante pir~~ato, NADH y W producidos en la licoli . a 60 Vo de VO,max. A esta intensidad
Tarnbien los pro9uc~~s de la hidr61isis d~I ATi1(;~pcons~men pnnc1~alm~~te en la mitocondria. mant1e~e. (B): ejercicro breve al 110% d I VO y P1). En esta _s1tuac1on, el pH muscular se escenar!o, la hidr61isis de! ATP es ta! qu! la ~~ax q~e provoca fatiga en 2-3 minutos. En este necesana mediante la oxidaci6n or lo I ocon _na n.~ es capaz de proporcionar la ener fa PCr ~ la glic61isis citos61ica. Por ~fda ADiu;u:\nec~f·ano utilizar de forma significativa la via degla pro~~n en el citosol, generando una producci6n ~e~a1~za ent estas reacciones, se produce un Pi y un
OXI ar O amortiguar. Esto se traduce en un descenso ed~ro ~n~s qdu•e sdupera a la que la celula puede P · o 111ca ode Robergs RAycols, 2004.
~----- 1
MITOCONDRIA ,--~I--- ~~~~~---i---..1'!'
..
No hay acurnulacldn de H" y Pi
(B) al 110% del V02max respectivamente. En el caso de intensidad mas elevada (figura 27 B), el metabolismo que utiliza la celula para obt , . , ener energ1a provoca una produccion neta de protones y fosfato inorganico.
2IMetabolismo de los hidratos de carbono I
Figura 26. La capacidad de la lanzadera de protones a traves de la membrana mitocondrial para reoxidar el NADH+H+ condiciona la conversion de piruvico a iactico
BOMBA i,MHI
-, \
I
I
I 2) AUMENTO DE LA CONCENTRACION DE LACTATO EN SANGRE
Por tanto, el lactato se acumulara en sangre durante el ejercicio si: 1) el desarrollo de la gluc6lisis es mas rapido que la utilizacion del piruvato por la mitocondria, aumentando su concentraci6n en el citoplasma celular, dando lugar a un aumento del lactato por la ley de acci6n de masas. Este proceso se vincula a una baja (absoluta o relativa) actividad oxidativa mitocondrial. En este caso no se alteraria la relaci6n lactato/piruvato; y 2) si el NADH + H+ no puede ser reoxidado rapidamente por la lanzadera de protones de la membrana mitocondrial. La lanzadera de protones de la membrane mitocondrial, que normalmente oxida el NADH + H citoplasmatico al lanzar protones y electrones al 02 mitocondrial, es demasiado lenta para reoxidar el NADH citoplasmatico reducido. Este mecanismo si alteraria la relaci6n
lactato/piruvato. En cualquier caso, la celula muscular esqueletica produce lactato por dos
razones: (1) porque la gluc6lisis citos6lica al inicio de la actividad muscular es mas rapida al aumentar la velocidad de sus reacciones que la via oxidativa mitocondrial; y (2) porque la capacidad glucolitica maxima excede a la capacidad oxidativa maxima. Como ya hemos explicado, la producci6n de lactato en musculo normalmente no se debe a una disponibilidad insuficiente de oxigeno. En la figura 26 se representa el metabolismo cnergenco en el musculo esqueletico a dos intensidades de ejercicio diferentes: (A) al 60% y

, ffi•IE••M
Fibras oxidativas (Tipo I)
Fibras glicolfticas (Tipo II) H+
, iiili·M
iigura 2_8: Acoplamiento metab61ico en el mu I , . . a~igluc9l1s_1s y expresan niveles elevados de GL~~ o esqueletico. Las fibras tipo II estan muy adaptadas ara Po~otlact1co, el cu~I es eliminado a traves de Mc\fa~a pot~ captar la glucose circulante y convertirl~ en
o ra tf rte, las fibras tipo I son altamente oxidati~a: an I rasa carbonica (AC) facilita este transporte exceso de lactato pasa a los vasos sangufne~i e(:r~?fan Md CTd1 par_a captar el lactato y oxidarlo:
· 0 1 rca o e Adijanto y Philp, 2012)
3) DISTRIBUCION DEL LACTATO
Cuando la produccion de lactato en las celulas musculares activas aumenta, parte se almacena en la celula muscular para ser utilizado, y parte difunde a traves del sarcolema, para posteriormente ser aclarado de la sangre por distintos mecanismos (eliminacion, transforrnacion ... ).
Durante mucho tiempo, se estudiaron tan solo las cantidades y localizaciones de la produccion de lactato y no se dedico la atencion suficiente al estudio de los mecanismos de transporte del mismo. Se considero al lactato coma una sustancia "Iibre y rapidamente difusible", que podia ser transportada inmediatamente desde las celulas donde se producia a todos los compartimentos del organismo. Sin embargo, despues de observar las diferentes concentraciones de lactato que aparecen tras el ejercicio en sangre arterial, sangre venosa y tejido muscular, se postulo una distribucion espacial y temporalmente hornogenea. El lactato generado por la glucolisis citosolica puede transportarse a fibras musculares adyacentes con menores concentraciones de lactato o pasar a la sangre de los capilares contiguos, desde donde se distribuira al resto de la circulacion sangufnea. Por consiguiente parece importante para un adecuado funcionamiento muscular un optima mecanismo de transporte de lactato a traves del sarcolema. En este sentido, conocemos desde hace tiempo que el lactato no puede moverse libremente desde el musculo a la sangre, sino que el transporte a traves del sarcolema esta mediado por un sistema que es saturable y estereoespedfico que implica un acoplamiento obligatorio 1:1 entre el lactato- y el H•. Diferentes isoformas del transportador MCT (monocarboxilate transporter) catalizan el transporte de monocarboxilatos coma el L- lactato ligado a un proton a traves de las membranas celulares. Ademas existe un proceso de difusion simple del lactato no disociado. En el musculo esqueletico estas isoformas son MCT1 y MCT 4.
La funcion fisiologica principal de MCTl es facilitar la entrada y/o salida de L-lactato de las celulas, dependiendo de su situacion metabolica, Por ejernplo, en el higado y en los tubules contorneados de rifion, MCTl mete lactato especialmente despues del ejercicio para que se realice gluconeogenesis. En el musculo esqueletico, al igual que en el corazon, se necesita MCT1 para que el lactato y los cuerpos cetonicos entren a los miocitos para ser oxidados
[ 2 IMetabolismo de los hidratos de carbono [
en aquellas circunstancias en las que sus concentr . , aciones estan elevadas De ~echo, en el miisculo esqueletico existe una fuerte correlacion entre I~ cantidad de MCT1 expresada en la fib . . . s 1 ras y su capacidad oxidativa (masa m1tocondna~). MCT1 tarnbien facilita el transporte de estos combustibles monocarbox1latos a traves de la barrera hematoencefalica para que las neuronas puedan utilizarlos coma combustible Dentro d 1 . , . · · e rrusrno musculo las isoformas de MCT coordinan el . . ' , mov1m1ento de lactato entre diferentes celulas ya que el transporte de lactato a traves del sarcolem d . . . a pue e ocurnr en ambas direcciones dependiendo del gradiente establecido para el lactato y los protones. De manera que las fibras tipo I con baja concentracion de lactato pueden capta~-lactato y su proton para oxidarlo en sus mitocondrias cuando la concentrac10n en el exterior es mas elevada (figura 28).
·~ ,,
l~.it>'.:' ,, .// ' . ,,
.l,.i.' l.',.._ SS ~ ... ;,-,,,... - I
1~· •• .r ' ( ..

c1cLO DE CORI. Una parte importante del lactato procedente de la celula muscular que alcanza el torrente sanguineo es captado por el higado para ser transformado en glucosa en el interior del hepatocito durante la recuperaci6n. Adernas, este proceso gluconeogenico contribuye a la recuperaci6n del gluc6geno hepatico. Aunque en la literatura encontramos una gran variabilidad en las cantidades, se calcula que aproximadamente un 50-60% del lactato producido es metabolizado en el higado, donde difunde libremente a traves de la membrana celular del hepatocito y se transforma de inmediato en piruvato a traves de la reacci6n lactato-deshidrogenasa NAD dependiente. Esta reacci6n representa la entrada del acido lactico en la via gluconeogenica. El piruvato entra en la mitocondria del hepatocito para convertirse en oxalacetato gracias a la acci6n de la piruvato-carboxilasa. La piruvato carboxilasa es un enzima mitocondrial, mientras que el resto de enzimas de la gluconogenesis son citos61icos. Por tanto se debe transportar el oxalacetato producido fuera de la mitocondria para lo cual se reduce a malato el cual es transportado al citosol par el sistema lanzadera aspartatc- malato. Una vez en el citosol, el malato es reoxidado a oxalacetato par una malato deshidrogenasa citos6lica y a partir de aqui todas las reacciones de la gluconeogenesis tienen lugar en el citosol. La gluconeogenesis no es exactamente el proceso inverso de la glicolisis par motivos termodinamicos ya que hay tres reacciones de la glicolisis que estan muy desplazadas del equilibria y son practicarnente irreversibles. En la gluconeogenesis estas reacciones son sustituidas par reacciones nuevas (Figura 29).
El consumo hepatico de lactato aumenta con la intensidad del ejercicio alcanzando su pico durante tasas subrnaximas de trabajo. Se piensa incluso que el entrenamiento elevaria la capacidad del higado de aclarar lactato par las adaptaciones que se generan coma consecuencia de la realizaci6n de ejercicio repetido (aumento del gasto cardiaco, mantenimiento del
ELIMINACION O ACLA.RAMIENTO DEL LACTATO
Durante el ejercicio el aumento de la liberaci6n muscular de lactato se acompafia de un incremento de su eliminaci6n plasmatica, de manera que la tasa de aclaramiento del lactato plasmatico aumenta en intensidades submaximas de trabajo en comparaci6n con el reposo.
2IMetabolismo de los hidratos de carbono
La capacidad de transporte lactate /W en las fibras lenta~ oxid.ativas es practicamente el doble queen las fibras rapidas glucoliticas. S~ c~ns1deramos
t d de lactato-/H+ como un sistema espec1ahzado en la el transpor a or . liberaci6n rapida de lactato durante las contracciones musculares mtensas, parece sorprendente que la mayor capacidad de transporte se encuentre en las fibras musculares menos glucoliticas. Sin embargo, hay que tener
n cuenta que las fibras lentas oxidativas rambien producen lactato a :ntensidades altas de trabajo y que son reclutadas durante mas tiempo que las fibras rapidas, por lo que parece importante para estas fibras poder
liberarse rapidamente del lactato producido. . . expresa especialmente en los tejidos cuyo Por su parte, MCT4 se
. b la glicolisis como el musculo esqueletico, los metabohsmo se asa en ' , . , . hematies y otras celulas. Su principal funci6n es sacar acido lactico de las
celulas, en este caso las fibras musculares. La relaci6n MCT4 /M~T~ es fib d f ma que las fibras rapidas diferente en los distintos tipos de 1 ras, e or ,
, MTC4 mientras que las mas oxidativas expresan mas MCTl expresan mas , 1
. , MCT4 Sin embargo si bien la isoforma MCT 4 se expresa mas en re acion a · ' . en las fibras rapidas, los estudios realizados demuestran que la cant1da~ ~e MCTl es muy similar en las fibras tipo I y tipo II por unidad de superf1c1e. Con respecto al efecto del entrenamiento, podemos afirmar que las personas
Id d MTCl (y una tendencia a presentar entrenadas tienen mayor contem o e cifras mas elevadas de MCT4) que las sedentarias, lo mismo que ocurre con la capacidad de transporte de lactato que es mayor en las entrenadas que en las no entrenados, y dentro del grupo de entrenados, es mayor cuanto
1 Vo max Por tanto parece confirmada la hip6tesis de que la mayor es e 2 • • , .
actividad contractil mantenida de forma cr6nica es un est1mulo i~po.rtante
1 tenido de MCT. Si nos cuestionamos que tipo de para aumentar e con . entrenamiento y cuanto (intensidad-duraci6n) entrenamiento es necesano para aumentar al maxima el contenido en MCT' todavia no podemos dar una respuesta clara. Incluso hay algunos estudios (pocos) ,en los que no se han observado cambios en la expresi6n de estas protemas. Podemoe~ concluir que el tipo de entrenamiento parece ser poco importante, y que entrenamiento con intensidades elevadas parece ser el mas ef ectivo.

El musculo esqueletico desempefia un importante papel en el metabolismo del lactato producido, estimandose que aproximadamente un 75% del lactato presente en los musculos activos es metabolizado en ellos durante la recuperaci6n. No se conoce bien el papel cuantitativo de los musculos inactivos en este proceso de aclaramiento.
Messonnier y col (2013) estudiaron la cinetica del lactato y su tasa de aclaramiento a intensidades +/-10% del umbral lactico (UL) en sujetos entrenados y no entrenados. Los autores observaron que el flujo de lactato en el UL es mayor en los entrenados que en desentrenados, comprobando que a estas intensidades de ejercicio asociadas al UL se produce un descenso de la tasa de aclaramiento. Los autores sugieren que aunque el entrenamiento de resistencia aerobica aumenta la capacidad de producci6n de lactato y tambien de su eliminaci6n o aclaramiento, el UL representa la intensidad en la que el aclaramiento del lactato comienza a estar limitado, independientemente del estado de entrenamiento. Adernas concluyen que el UL quedaria determinado, precisamente, por una limitaci6n en la tasa de aclaramiento.
En relaci6n a los procesos de aclaramiento, cuando el lactato circulante entra en las celulas para ser oxidado directamente en las mismas, se denomina oxidacion directa del lactato circulante. Cuando el lactato es convertido a glucosa en los procesos de gluconeogenesis hepatica y renal
ORGANO moles·dia·1.70 .-,
Hfgado 0.72 Rifi6n 0.12
Corazon o.os
Existen otros 6rganos que tambien participan en el aclaramiento del lactato:
El consumo hepatico de lactato aumenta con la intensidad del ejercicio alcanzando su pico durante tasas submaximas de trabajo. Se piensa incluso que el entrenamiento elevaria la capacidad del higado de aclarar lactato por las adaptaciones que se generan coma consecuencia de la realizaci6n de ejercicio repetido (aumento del gasto cardiaco, mantenimiento del flujo hepatico durante el ejercicio, induccion de enzimas hepaticas gluconeogenicas y amortiguacion de la respuesta aut6noma).
2IMetabolismo de los hidratos de carbono J
. . lucolisis la gluconeogenesis. Solo se han indicado algunos Figura 29. Esquema de las reacciones de la g on re~ersibles y son sustituidas por otras.
detalles en las reacc1ones que no s
Ill - Ill LLI z LLI c, 0 u ::, ~ c, 0 LI.I z
En la . mitocondna
2 fosfoglicerato
+t 3 fosfoglicerato
fosfato de diOHacetona
( ATP ) G~ct~~ 6PJ /0 . ~I t ( ffuctosa 1,6- bifosfatasa
fosfofrutoqwna~'t ::::--...._
( ADP ) Fru::-sa 1,6 diP @
(ATP) ( Glucosa ]~0 h . ~I t glucosa 6-fosfasa
exoqwnasa /,'
( ADP ) G~~:~~i.J @ t
Ill - Ill - _. 0 u ::,
1 . . . inducci6n de enzimas hepaticas flujo hepatico durante e e1ere1c10, , gluconeogenicas y amortiguaci6n de la respuesta autonoma).

RECUPERACION TRAS EL EJERCICIO
Como una parte importante del lactato producido es oxidado en 1 , 1 esq l 'f . os muscu os
due ~ icos activados a baja intensidad, y la redistribucion del lactato
pro ucido se establece ' , . . . via sangumea, la recuperacion activa despues de ejercicios que pr 1 ovocan a tas concentraciones de lactato se muestra ma fi que la recu . , . as e 1caz 1 . peracion pasiva en las procesos de aclaramiento o elirninacion de actato Sm embarg h . o, no ay una estrategia clara acerca de la intensidad idonea
Figura 30. Esquema que muestra la relaci6n en I . . La glucogenolisis muscular es la princi al fuent tree m<?"'.l'.111ento de glucosa y lactato en la san re
Ygl~co~a/a~gdufnea. E! aclaramiento del~actato s! i~o~~~~cp1~nlde l~dcta~9, se_guida de la glicolisis d! I~
sin es1s. e glucogeno (esto ultimo en menor . r a oxt a.c1.9n directa, la gluconeogenssj, gl~co~eno!1s1s y gluconeogenesls hepatica mientra~ed1d~). Lia apar1c1on de la glucosa se debe a la y a stntesis de gluc6geno. Una e uefia ' . , que e ac ~ram1ento se produce por la oxidaci6
ge: pool del lactate. La oxidaci6npin~irect!~;f1~~t~~~ a~laram1e~tp dde la glucosa vuelve a entra e~
ucosa rnediante la gluconeo , . . s a porc1on e lactato que se · Messonnier, L. A.1 Homing, M.A., F~~;o~~tlra~lfiitto~mente oxidarse. Adaptado de Em~~?t~.~~ ~n
glycogenolys1s dunng exercise at the lactate thre~h. ~d J Broo~s, fG. A. (2013). Gluconeogenesis and hepati~ o . ouma o Applied Physiology, 114(3), 297-306.
Oxidacion directa
Sfntesis glucogeno MS
0 ..... e s
Glucogenolisis hepatlca
~ Glucosa sangufnea
0 U S(ntesis de glucogeno ms
:, .... \:J Oxidacion
Glucogenolisis muscular
__________________ 2_IM_et_a_b_ol_is_m_o_de los hidratos de carbono j
de la sangre se produce par la oxidacion de la . , . , misma Y par la sintesis de glucogeno muscular O hepatico,
para posteriormente ser oxidado coma glucosa, se denomina oxidacion
indirecta. El entrenamiento de resistencia aerobica aumenta la capacidad
de oxidacion (aclaramiento), especialmente la contribucion de la oxidaclon
directa. Emhoff y col, 2013a comprobaron que este proceso de oxidacion
directa suponia aproximadamente el 75% de la oxidacion total del lactato
en sujetos no entrenados, llegando al 90% en deportistas entrenados.
Tambien son relevantes las adaptaciones que llevan a un aumento de la
oxidacion indirecta de lactato, mediante la conversion del mismo a glucosa
y la oxidacion de la misma par las celulas musculares especialmente.
Recientemente, Emhoff y col, 2013b comprobaron que la oxidacion
indirecta (neoglucogenesis) aumentaba durante el ejercicio cuando las
niveles de lactato se incrementaban, sugiriendo que la contribucion de
la neoglucogenesis a la produccion total de glucosa puede estar limitada
par la disponibilidad de precursores gluconeogenicos, especialmente en el
tejido hepatico y alga menos en corteza renal. Asi, debido a que el aporte
sangufneo al higado y rifion disminuyen proporcionalmente al aumento de
intensidad del ejercicio, se producira un descenso del aporte de sustratos
gluconeogenicos especialmente a la celula hepatica y con ello un descenso
de la capacidad gluconeogenica en ejercicios de alta intensidad. Los
autores tambien comprobaron que, aunque el entrenamiento de resistencia
aerobica aumenta la intensidad absoluta y relativa asociada al umbral
lactico, las sujetos entrenados no manifiestan descensos en la capacidad
gluconeogenica a intensidad umbral. Se ha sugerido que un aumento en
la capacidad de transporte de lactato en el sarcolema pueda contribuir al
mayor aclaramiento de lactato despues del entrenamiento.
En la figura 29 se muestra la relacion entre el movimiento de lactato y glucosa en la sangre. La aparicion de lactato se debe principalmente a la glugogenolisis muscular, yen menor grado, a la glucolisis de la glucosa de la sangre. La eliminacion del lactato sanguineo se debe a la gluconeogenesis y a la oxidacion directa, contribuyendo solo discretamente la sintesis de glucogeno muscular. El lactato que entra en la gluconeogenesis se destina a la sintesis de glucogeno hepatico o a loa produccion de glucosa, lo que tambien depende de la glucogenolisis hepatica. El aclaramiento de la glucosa

LACTATO ARTERIAL y LACTATO VENOSO
Se ha ~bservado q~e el umbral lactico ocurre en intensidades de trabajo mas altas si se determma en sangre ven . osa que si se valora en sangre arterial ~alcausa probable de este hecho es el consumo de lactato por los rmisculos
e antebrazo antes de que la sangre llegue al lugar destinado para la toma venosa (en general, vena antecubital).
Dado que el umbral ventilatorio ha demostrado una elevada correlacion con el umbral · lactico cuando se utiliza sa art
. 1· d ngre venosa, sangre capilar ena iza a o sangre a t . 1
1 r ena ' es probable que las posibles diferencias entre
ta es tomas sean pequ - enas, y por tanto pueda utilizarse indistintamente sangre venosa, arterial o arterializada si lo que se t d . . pre en e es analizar el comportarruento del lactato con el ejercicio Es ne . . · cesano precisar el lugar exacto de l~ toma de muestra sanguinea si el objetivo es comparar o analizar
tconcdentrac10n~~ absolutas de lactato sanguineo, obtenidas en diferentes est e valoracion,
' I /------------------- -----
' El lactato no es un "producto de - - - - - - - - - - - - - - - : des~e~ho'.': En 1984, Brooks expuso I~ coordinaci6n del metabolismo en
su hipotesis acerca de la lanzadera dlferentes tejidos. La lanzadera actiia d_e lactato (Brooks, 1985), la cual ha • en todas las situaciones metab61icas sido posteriormente confirmada por Lo mas destacado de esta hip6tesis es numerosos estudios. La hip6tesis expone : que el lactato deja de ser un producto
i q~e l_a fo~~aci6n de lactato y su posterior : me.tabolico de deshecho causante de distribucion por todo el cuerpo es un fatrga ~~ra pasar a ser un intermediario mecanismo fundamental que permite rr,ie~abolrco que puede intercambiarse
- - - _ _ _ _ _ _ _ _ _ _ _ _ _ rapidarnente entre los tejidos. ------ --------
________________ 2_IM_etabolismo de los hidratos de carbono I
LACTATO MUSCULAR Y LACTATO SANGUINEO Existen algunas evidencias que sugieren que la acumulacton de lactato en el interior de la celula muscular podria ocurrir antes de observar el aumento de la concentracion de lactato en sangre periferica. Los procesos de aclaramiento de lactato por el propio musculo y por tejidos perifericos podrian explicar este acontecimiento. Chwalbinska-Moneta y col, 1989, examinaron las concentraciones de lactato en sangre y musculo, llegando a la conclusion de que se produce un umbral lactico muscular durante el ejercicio incremental. Estos autores observaron que a pesar de que el aumento de la concentracion de lactato en el musculo precede al que se observa en la sangre venosa,
existe una elevada correlacion entre ambos. Por otra parte, los lugares utilizados habitualmente para obtener las
muestras de sangre arterial (arteria braquial) estan muy lejos del punto de liberacion del lactato por el musculo (generalmente en los musculos de las piernas). En estas situaciones, el lactato debe atravesar organos conocidos coma «metabolizadores'' antes de llegar al territorio de obtencion de la muestra. Por ultimo hay que tener en cuenta, que el lactato puede ser parcialmente oxidado en el musculo antes de que aparezca en sangre.
que optimice estos procesos de aclaramiento. Estudios previos sugirieron el rango idoneo en el aclaramiento del lactato a una intensidad correspondiente al 25-63% vomax. Sin embargo, en estos estudios los autores relacionaban
la intensidad de recuperacion a un determinado %V02max, pero dado que el acumulo del lactato no mantiene una relacion lineal con el %V02max, esta referenda puede no ser la [donea. En este sentido, determinar la intensidad optima de recuperacion en base al UL puede ser mas apropiado y preciso. Menzies y col, (2010) mostraron que el descenso de las concentraciones de lactato en sangre despues de un ejercicio en tapiz rodante al 90% vomax (2,1-6,7 mM/1) fue mayor con recuperacion activa que con recuperacion pasiva. La intensidad correspondiente al 80-100% UL fue mas eficaz que la recuperacion activa a menor intensidad (40-60% UL). La intensidad del 60%
UL fue tambien mas eficaz que la correspondiente al 40% UL, de rnanera que el aclaramiento del lactato durante la recuperacion activa mantiene una
relacion dosis- respuesta.
I

i, .~ i I
1 ~:s aat~::sg~:s~:·r~:~:e ~~~g~on:p:~~~t~s de las gras~s, esta~ compuestos de cadenas , d , • - I rogeno Y se dlferencian unos de otros I
~um~ro.,. e atomos de.carbono y el grade de saturaci6n de atomos de hidrog~~: ;i os c asmcarnos atendiendo a la presencia o no de dobl 1- , . de carbon I es en aces entre los atomos
O que os componen tenemos una crime di · ·, , . saturados (no presentan dobles 'enlaces' ; . '" ". era rvrsion e~ acidos grasos doble en lace. Los acidos grasos saturado~' c::;~:~~~aai°s, ,~~e pre~enli,m al ;11enos un hidr6geno posib!e y tienen todos sus enlace , .. e rnaxrrno numero de atomos de
~n~aturados, 1si~i
emb~r~o, pueden absorber :t~~~:t~: ~~:~~~~s~ ~~~qauc~d~~ ;i~a~;~ o os sus en aces qurrrucos ocupados L , id ·· rnonolnsaturado 1i . os ac'. os grasos insaturados se clasifican en
• , s, aque os que presentan solo un doble enlace nudiendo ab rb por tanto dos atornos de hidro 1.. - . · so, er enlace~ pudlendo ~bs~r~..,; , . vtg,eno, Y_ !~ nnsaturad~s, ~ue contienen varios dobles
-· cua ro o mas atornos de hidrdzerro.
~:b!::::nr!~~:d~ns6~i~::\;~:sos saturados. normal.m:nte. estan a temperature
Las grasas ricas en acid;s gras::ui::r~;:.i~:e ~~e~:i:e:s~:,'.r:~ (:.!~;;anteq~~lla). temperature ambiente v son mas abundantes en 1- - .d " ~~ o iqurco a aceites de pal ma y de coco su onen una exc - . , as .grasas e ongen vegetal (los concentraclon de acidos grasos .saturadcs). xcepcron, ya que presentan una elevada
Dentro_de los acidos grasos poliinsaturados encontramos los ome - . denornmados asf debido a la localizaci6n del doble enlace en 1- cga. ~·e3 y 1;1s ?mega-d6, carbono. a u na ae atornos e
I Los tres tipos principales de ornez 3 14 .. , linolenico (ALA) ~ id ·. nega . en a nutrtcion hurnana son el acido alfa El ALO. se con. s•~ a~1 ~ e1so~dapentaeno1co (.~- P_A) y el acido docosahexaeonoico (Dl-!A).
• - , ,..ie, a un ac1 o graso esenclal · d funciones fisiolozicas. el org-ni~mo n ~ ' -· ';or~ue, _s1en o necesario para clertas en la dieta El ALA · _ "' ' - ~ '0 '0 pue. e sintetizar por lo que debe obtenerse coma fruto~}~cos, ::e~~~::~;:a 1~J::~7i,:~~~::es~~~
;~:~:~~to~ed;e ~~i~.en. v:getal ~~~A a pa:L!r de ALA o se_ pueden obtener en alimeritos como pe~cado d~n;;~1;;~~::
' sco o, en menor rnedida, la yema de huevo. . ' I L~ in?esta de acidos ornega-j, especialmente EPA v DH -t · disrninucion del rles
O rl 'I "" , • , ~ pod, 1a estar asoc1ada a una
ben f . . g '."e paaec~, eme, medades card1ovasculares por SU" efoc•o
Ta:b;~~:~s~:oab::~i:~~1i~e/n;:stC:~e:t:~o~g:~3s;~;r~n: ;;ti~:c~~~~:dlode 1: sa;,;r~~ en la smtes1s de e1cosanoides antiin£1amat . E . s precursores • • 1 • onos n "St<> 5°r,t1rlo re I que la ingesta de acidos omega-3 d ,_ _ . · · ~ ·- ~ ' ~ ·, _, - 1a propLJesto inflamatoria muscular asociad~ al ~~ n~ ,aso;-1arse a. un d~scenso de la respuesta recupe'.aci6n. _Los posibles efectos be~:;~~~o;~ea;:: ~::~:i:~de,ga~~oradn~o asf la potenc1ados si 5· It, - · ::, "JO nan verse ' imu aneamente, se disrninuye la ingest:;i de acidos o . - ~;:~s i~::aa~~e~~a ~!s'.11a ruta rnetab61ica. De esta forma, un exce:eegna !a6i~;e~~:
I e 't 1
' pe ma que se metabolicen los AG omega-3 en EPA y DHA p _ I v1 are exceso de AG omega-6 . d . . . a, a \, ~ritos (incluyendo frutos secos fr~t::;,c;:~e~' ~~~l~~~ro!:i'i::~~~sd:r~~::~~~~ como
- - - - - --------- ----------- - - - - - - - - - - - - - -- -:;....._--
,.-------------- / ACIDOS GRAS~; ~~-E~~-; (L~~ - .. -B-U-E~~~~,) - - - - ..
V OMEGA 6 (LOS "MALOS")
1IMetabolismo de Ios lipidos
A diferencia de los hidratos de carbono, cuya principal funcion y casi unica es actuar como fuente energetica, las grasas son fundamentales para diferentes funciones biol6gicas, entre las que se encuentra actuar como combustible en el metabolismo aer6bico celular. El espectro de tipos de grasas y lipidos que ingerimos en la dieta es mas amplio que el de los hidratos de carbono (fosfolipidos, colesterol, trigliceridos, acidos grasos ... ) estando destinado cada uno de ellos a una funci6n principal. Son los acidos grasos ( componentes esenciales de los mgltcenoos jun to con el glicerol) los
que van a actuar como fuente energetica. Una vez ingeridos, y tras su complejo proceso digestivo con ayuda de
las sales biliares y la lipasa, los acidos grasos se absorben en el intestino delgado y alcanzan la linfa recubiertos de una capa proteica proporcionada por los enterocitos dando lugar a los quilomicrones. Los quilomicrones estan compuestos por triglicendos y proteinas. Desde la linfa, estos quilomicrones alcanzan la circulaci6n sistemica. De manera que los acidos grasos procedentes de la dieta Hegan al torrente sanguineo en forma de
trigliceridos contenidos en quilomicrones.
ABSOACION, DISTRIBUCION Y ALMACENAMIENTO DE LAS GAASAS EN EL ORGANISMO
Los lipidos almacenados en el organismo en forma de trigliceridos (TG) representan la principal reserva energetica y constituyen una fuente casi inagotable de energia durante el ejercicio flsico.
La utilizaci6n de los lipidos como fuente energetica tiene consecuencias metab61icas determinantes, como el ahorro de gluc6geno muscular y hepatico, que inciden en la capacidad de resistencia del organismo.
La celula muscular utiliza los acidos grasos (AG) contenidos en los TG como combustible para obtener energia. Estos AG se obtienen de los trigliceridos almacenados en el tejido adiposo o en el propio musculo, y de
las lipoproteinas circulantes.
3\Metabolismo de las Ifpidos

DEL TEHDO ADIPOSO A LA CELULA MUSCULAR. Los principales estimulos lipoliticos al inicio del ejercicio en el hombre son el aumento de la concentraci6n de adrenalina y la disminucion de la de insulina circulantes. El cortisol tarnbien es un agente lipolitoco. La tasa lipolitica depende en parte de la capacidad de la sangre para transportar acidos grasos, 0 lo que es lo mismo, de la concentraci6n serica de albumina, ya que los AG en plasma van transportados por esta proteina plasmatica (y se denominan entonces AG libres, AGL). Por otra parte, depende del flujo sanguineo en el tejido adiposo Y del proceso de reesterificaci6n en los adipocitos. El aumento del flujo sanguineo al tejido adiposo durante el ejercicio favorece enormemente la movilizaci6n de los acidos grasos. Al mismo tiempo, parece que esta regulaci6n vascular esta mediada por la adenosina y que es secundaria a eventos conectados con la lip61isis.
La Iipolisis de los trigliceridos almacenados se lleva a cabo por accion de la enzima denominada lipasa hormono sensible (LHS) la cual, como acabamos de mencionar, se activa por la presencia de concentraciones elevadas de cortisol, adrenalina y noradrenalina y concentraciones bajas de insulina (Figura 31). La LHS cataliza la reacci6n de triacilglicerol a diacilglicerol y monoacilglicerol. La hidr61isis del tercer acido graso esta catalizada por la LHS asi coma por una monoacilglicero/ lipasa, lo que garantiza que la lipolisis sea completa. Como resultado de la lipolisis, se liberan al torrente sanguineo dcidos grasos fibres y glicerol. Los primeros (AGL) se transportan hasta el tejido muscular unidos a la albumina plasmatica y el glicerol lo encontramos disuelto en el plasma. El glicerol derivado de la hidrolisis de los trigllcendos puede dirigirse al higado para constituir un sustrato gluconeogenico o entrar en la celula muscular e incorporarse a la glucolisis anaer6bica.
La captaci6n de AGL por el musculo es un proceso dependiente de la concentraci6n en sangre circulante que no requiere un gasto energetico. No obstante, existen mecanismos reguladores que impiden que la celula muscular capte mas AGL de los que necesita (solo entra en la celula el AG
'
LIPOLISIS Y MOVILIZACION DE LOS iCIDOS GAASOS DURANTE EL EIEACICIO
Por otra parte, el higado sintetiza otro complejo lipoproteico, las lipoproteinas, que se encargan de distribuir las grasas por el organismo. El higado supone la otra fuente de trigliceridos para el tejido adiposo Y muscular. La proporci6n y el tipo de grasas que va transportado en las diferentes lipoproteinas varian, siendo las VLDL o proteinas de muy baja densidad las que transportan mayor cantidad de acidos grasos ya en forma
de trigliceridos. Los quilomicrones y VLDL circulantes son los responsables de transportar
la mayor parte de los acidos grasos (o trigliceridos) al tejido adiposo Ya las celulas musculares para su almacenamiento. Este proceso se lleva a cabo gracias a la accion de la lipoprotein lipasa (LPL) que se encarga de romper los complejos lipoproteicos permitiendo la entrada de la porcion lipidica en el interior de los adipocitos o de las celulas musculares. La LPL esta ampliamente distribuida por el organismo, localizada en el endotelio vascular, aunque presenta concentraciones mas elevadas en el tejido adiposo, en los musculos en los que hay predominio de fibras tipo I u oxidativas y en el corazon. Entre otros facto res, la adrenalina y la contracci6n muscular per se provocan la activaci6n de la LPL en el tejido muscular. En el tejido adiposo, el control hormonal se ejerce sabre la sintesis intracelular de LPL, ya que una vez localizada en el exterior de la celula, la activaci6n de la enzima no depende de las variaciones hormonales. Por otra parte, la actividad de la LPL esta regulada por el estado metabolico del organismo, de manera que LPL es activa en el tejido adiposo en los periodos postprandiales, mientras queen etapas de ayuno, es mas activa en el tejido muscular. De esta forma, el almacen de grasas esta favorecido en un tipo u otro de tejido en funcion de si hay necesidad
de oxidarlas o no. Los acidos grasos, por tanto, se almacenan en el tejido adiposo Y en la
celula muscular en forma de trigliceridos, es decir, tras la esterificaci6n de 3 acidos grasos y una molecula de glicerol (triacilgliclerol). En realidad, los quilomicrones y las VLDL transportan TG, los cuales son hidrolizados
a acidos grasos y glicerol por accion de la LPL.
3f Metabolismo de los lipidos ---------------- --------------------..:, ------- ----------------- __,

TRIGLICIERIDOS INTRAMUSCULARES. Se ha estimado que en ejercicios de mas de 1 h de duracion aproximadamente el 90% de los AG oxidados en el musculo durante el ejercicio procede de los AG circulantes, mientras que
LIPOPROTEiNAS CIRCULANTES. El papel de los trigliceridos circulantes contenidos en las VLDL y los quilomicrones como fuente de energia durante el ejercicio se ha considerado como relativamente poco importante. El ejercicio intenso y prolongado produce un descenso en las concentraciones sanguineas de estos dos complejos lipoproteicos. Sin embargo, durante la actividad fisica de intensidad moderada, las concentraciones plasmaticas de las lipoproteinas transportadoras de trigliceridos no se modifican.
quedando la proteina albumina en la sangre). En general, la captacion de AGL desde la sangre circulante a la celula consiste en: 1) disociacion del AGL de la albiimina, 2) union del AGL a proteinas de la membrana celular o integracion en la bicapa lipidica, 3) transporte del AGL a traves de la membrana. El mecanismo por el cual los AGL atraviesan la membrana en la celula muscular (y otras celulas en las que la captacion de AGL es elevada como el hepatocito y adipocito) no se reduce exclusivamente a su difusion pasiva a traves de la misma, sino que existen procesos mediados por proteinas especificos para cada AG, saturables y regulados hormonalmente. Estos mecanismos son los responsables de la captacion de mas del 90% de los AGL que capta la celula muscular cuando las concentraciones de albumina y AGL se encuentran en rango fisiologico. El complejo proteico mas importante en el musculo esqueletico es F ATPl.
Los acidos grasos se oxidan principalmente en las fibras oxidativas o tipo I que fundamentalmente se activan durante los ejercicios de baja y moderada intensidad. Por tanto, la contribucion de la oxidacion de lipidos al metabolismo oxidativo total depende de la carga de trabajo relativa. Durante las actividades intensas, los hidratos de carbono contribuyen con casi el 80% de la energia mientras que durante el ejercicio moderado de duracion prolongada la combustion de lipidos puede cubrir hasta un 90% de los sustratos utilizados. Cuanto mas entrenado esta un musculo, mayor capacidad tiene para oxidar grasas.
31Metabolismo de los lipidos
--------··- ------- ,------ / PERDIDA DE GRASA CORPORAL:
EJERCICIO coNTfNUO vs INTERMITENTE . . . . • ltente de alta intensidad (HIIT) con el
Si compararnos el e1erc1c10 l~c:rrn1_ ' d de forma que se realice la misma . . . tl uo de menos m,ens1aa u , dld e1erc1c10 con m . HIIT orovoca una mavor per I a
cantidad de traba]o total, se observa qu~ e1 Se investigaron las' respuestas I e el ejercicio contmuo. . de grasa corpora q~. . • rotocolos de ejercicio, semepntes
fisio16gicas y metabohcas a ~osdd1fCerenLeS ~amana do intervalo, sujetos sanos I b . total realiza o on una .,_ - d respecto a tra ajo L a • . 1 00/. VO max O intervalos de 20 s e
pedalearon en bicicleta durante 30 mm a 5 o d~s p~~ 40 s de recuperaci6n. , imo al 150% VO max separa di ejercicio supramax1 dif . 2 . 'en la frecuencia cardiaca y V02 me ro,
Los resultados no mostraron ,_,erenc1as bos protocolos. Con el protocolo ni en glicerol o acidos gra~os hbres, en~;:;;de lactato e hipoxantina y hubo de HIIT se elevaron los nive~es plahs.ma tl a v acldo urico. Estos resultados
,. . ion urlnarta de ipoxan m , . , d una myor e.1mmac1 a • 1 , 1 en HllT La restauracon e , did neta de ATP aesde e rnuscu o · f reflejan una per I a L • • , d balance energetico negativo justi ica los niveles de ATP en esta situacron e I b ervada con HIIT vs el ejercicio
i on arte la mayor perdida de grasa corpor~ o s \ ~o!inuo. Gerber y col, 2014; (Eur J Appl P~y~,~ ~1~: ::_~2~)- _
' ------- ..... _
-------- - - - - -
. uerna de la activaci6n de la lip61isis f~y~~~a3~~;f.; acci6n de la lipasa horrnonosensible.
Musculo
Acidos grasos
9. · · · Catecolaminas ~
Lipasa - hormosensible •· GH
0
Tejido Adiposo
Trigliceridos

acilo de cadena corta, media y larga. Estas acil-carnitin transferasas estan localizadas en la membrana mitocondrial en dos formas: en la membrana externa, una forma (acil-carnit{n transferasa 1) convierte el acil-CoA en acil- carnitina, uniendo el grupo acil a la carnitina, y en la membrana interna, el complejo acil-carnitina vuelve a transformarse en acil-CoA por otra forma del enzima que libera la carnitina y une un CoA de nuevo al acil (acil- carnitin transferasa 2) (Figura 33). Es decir, las acil-carnitin transferasas 1 y 2 y la carnitina permiten el paso exclusivo del radical acil, pero sin que pierda su nivel energetico, ya que al atravesar la membrana interna mitocondrial se unen de nuevo a un CoA Este mecanismo de transporte, si bien no es directamente dependiente del oxigeno, si depende de la masa mitocondrial
' de manera que la concentraci6n de acil-carnitin transferasas sera mayor cuanto mayor sea el numero y tamafio de las mitocondrias, por lo que los acidos grasos son oxidados preferentemente en las fibras tipo I.
Una vez en el interior de la mitocondria, los acil-CoA estan preparados para someterse a la f3-oxidaci6n. La f3-oxidaci6n es un proceso metab6lico por el cual se oxida el radical acilo en el carbono f3, es decir, el que es ta en
Figura 32. Activaci?~ de los grupo~ ac!I, una v.ez introducidos en la celula muscular. El gasto energetico de esta actrvadon equivale a la fosforilaci6n de dos ATP.
IAcil O
D
AG-Alb
____________________ 3_1M_e_ta_b_o_lismo de los lipidos j
ACTIVACION Y OXIDACION DE LOS LiPIDOS EN LA CiLULA MUSCULAR Independientemente de la procedencia de los AG, una vez en el interior del miocito y antes de ser oxidados experimentan un proceso de ccnvccion por el cual elevan su nivel energetico para posteriormente ceder su energia. Este proceso tiene lugar en el sarcoplasma y consiste en la union de una coenzima A ( Co A) al AG, dando lugar al complejo acil-CoA El acil-CoA puede, segun las necesidades celulares, reesterificarse y almacenarse en forma de mgliceridos en la propia celula muscular, o bien oxidarse. La activaci6n del grupo acil, por ser un proceso enderg6nico, tiene un coste energetico para la celula, equivalente a la defosforilaci6n de 2 ATP (en realidad, un ATP se transforma en AMP, perdiendo sus dos enlaces de alta energia) (Figura 32).
El acil-CoA debe entrar en el interior de la mitocondria para oxidarse. En la membrana mitocondrial existe un sistema transportador espedfico de los grupos acilo, dependiente absolutamente de la carnitina Y situado en el espacio intermembrana, cuya funci6n es transportar los grupos acilo a traves de la membrana mitocondrial. La carnitina se acompafia de diversas formas de acil-carnit{n transferasas con afinidad por los grupos
solo el 10% procede de los trigliceridos almacenados en la propia celula muscular. Los sujetos entrenados en resistencia aerobica poseen mayores dep6sitos de TG intramusculares y una mayor capacidad de activaci6n de la lipasa hormono-sensible y, aunque los resultados de las investigaciones pueden tener discrepancias, parece que los sujetos entrenados son capaces de oxidar mas triglicerldos musculares durante el ejercicio especialmente cuando el ejercicio se realiza en ayunas. Este efecto parece que queda enmascarado por el papel inhibitorio de la insulina sobre la lipolisis incluso hasta 6 horas despues de haber ingerido hidratos de carbono. Las dietas ricas en hidratos de carbono anulan esta adaptaci6n (Horowitz Y cols. 1997). Sin embargo, la utilizaci6n muscular de los trigliceridos almacenados en el tejido adiposo, no parece mejorar con el entrenamiento de resistencia,
especialmente a corto plazo (Camera y cols, 2010).

90 150 180 120 60
Figura 35. Comportamiento de la concentraci6n de acidos grasos libres en sangre durante la realizaci6n de un ejercicio de intensidad submaxima. (Modificado de Horowitz y col, 1997)
Veamos que es lo que ocurre con los AGL circulantes durante la realizacion de un ejercicio prolongado en el que se mantiene la intensidad constante (moderada). En general, al inicio de un ejercicio se observa una ligera disminucion de la concentracion de AGL ya que la mayor captacion de AGL por el musculo supera inicialmente la capacidad de liberacion de AGL por los adipocitos. Segun avanza el ejercicio la Iiberacion de AGL desde los adipocitos supera a la captacion, y
ACIDOS GRASOS LIBRES Y EIERCICIO
0
_...._ ' 2000 0 E 3 ra E 1500 Ill ra 0.. c: (1J
...J 1000 (.'.) <i: (1J ~ c: '0 "iJ 500 ra .... ... c:
(1J u c: 0 u 0
segunda posicion, dando lugar de nuevo a un acil CoA ( esta vez con n- 2
atomos de carbono) mas un acetil-CoA. Consta de 4 reacciones enzimaticas, y se transforma en un proceso ciclico hasta que todo el acil o acido graso haya quedado dividido en fragmentos de 2 atornos de carbono en forma de acetil-CoA (Figura 34). Durante el proceso de la ~-oxidaci6n, se produce la extracci6n de atomos de hidr6geno uni dos a sus electrones, de man era que en cada ~-oxidaci6n se reduce un NAD y un FAD, transtormandose en NADH y FADH respectivamente. Asi, un acido graso que tenga, por ejemplo, 18 atomos de carbono en su radical acilo, experimenta 8 veces la p-oxtdaoon, permitiendo la reduccion de 8 NAD y 8 FAD y dando lugar a 9 moleculas de
acetil CoA. A continuacion, el acetil CoA entra en el ciclo de Krebs, que ya hemos
comentado, de la misma manera que el procedente del piruvato. A partir de aqui, el metabolismo oxidativo es cornun con el de los hidratos de carbono, teniendo en cuenta que a la fosforilaci6n oxidativa iran los hidr6genos obtenidos en el ciclo de Krebs mas los transportados por el NADH y F ADH
reducidos en el proceso de la B -oxidacion.
Figura 33. Utilizaci6n de los acidos grasos P?r la cel~I? muscular. Papel de la carnitina en el mstabolismo l1p1d1co
Figu~a 34,; Esquem~ ~el proceso de la p-ox1dac1on de los acidos grasos en la mitocondria de la celula muscular
~~) .. ·---- rMatriz mitocondria~ ~oxi~] L'..:.----~ _l
..,....._A..,. .... r;cil-CoA+Acil-CoA J--· ~~ l~--------·
~ ;.,.t---------~( Carnitina J-··-- + Acil-Carnitina Espacio intermembrana
H,0- - ·~ Hidratasa
~-~~d;;xiacil-CoA
NAO···:· NADH..,,..
Giiribi,fi ... · - · Li 2-Enoil-CoA
r--;cil-C~ . x.; ~_) : FAD·--- . :
FADH .a4· •• Desnidroqenasa :
Duraci6n del ejercicio (min)
30
coma resultado la concentracion de AGL en sangre arterial aumenta (figura 35). Si el ejercicio se realiza despues de ingerir alimentos (especialmente ricos en hidratos de carbono) el aumento asociado de la insulina "frenara" parcialmente el estimulo de activaci6n de la lipolisis por lo que los niveles de AGL arteriales seran menores durante el periodo de ejercicio.
,------------------------------------.I-- 1_I_M_e_ta_bolismo de los lfpidos I

El musculo puede utilizar como combustible estos cuerpos cetonicos (especialmente cuando la disponibilidad de glucosa es baja), ya que se transforman en acetil-CoA, pudiendose incorporar de esta manera al ciclo de Krebs. No obstante, la obtencion de energia a partir del aceto-acetato Y el 3-hidroxibutirato constituye una proporcion pequefia con respecto a la energia total. El entrenamiento de resistencia aerobica mejora la capacidad de oxidar cuerpos cetonicos mediante un aumento de la sintesis de las enzimas responsables de su metabolismo. Durante el ejercicio de baja intensidad, la tasa de cuerpos cetonicos utilizados es baja, no superando la de la situacion de reposo. Por otra parte, la tasa de utilizacion de los cuerpos cetonicos durante ejercicios de intensidad moderada depende de la cetonemia. Sin embargo, hay que tener en cuenta que incluso con concentraciones sanguineas altas de cuerpos cetonicos la oxidacion en el rnusculo esqueletico no puede contribuir mas que con un 7% como maxima a la produccion de energia en la celula muscular.
En situaciones de hipoglucemia los cuerpos cetonicos pueden ser utilizados corno sustrato energetico por otras celulas del organismo, coma es el caso de las neuronas.
Figu~a 36. Formaci6n de cuerpos cet6nicos en el higado y su utilizaci6n en la celula muscular.
(3- Hidroxibutirato
____________________ 1_IM_et_abolismo de los lipidosj
METABOLISMO DE LOS CUERPOS CETONICOS DURANTE EL EIERCICIO Parte de los AGL movilizados desde el tejido adiposo se dirigen al higado donde son degradados hasta formar una serie de sustancias denominadas genericamente cuerpos cet6nicos (aceto-acetato y 3-hidroxibutirato). Estos cuerpos cetonicos, son vertidos de nuevo a la circulacion sangufnea (Figura 36).
La cantidad de cuerpos cetonicos vertidos a la sangre depende de la cantidad de acidos grasos libres que lleguen al higado, asi como de la cetonemia.
Cuando los depositos hepaticos ymusculares de hidratos de carbono disminuyen aumenta la formacion de cuerpos cetonicos, como ocurre en situacion de ayuno prolongado, en dietas con muy bajo aporte de hidratos de carbono, o durante el ejercicio de duracion prolongada (sin aporte de hidratos de carbono). Los bajos niveles de insulina plasmatica asociados a los estados referidos anteriormente producen un desequilibrio entre la forrnacion de trigliceridos y la liberacion de acidos grasos a la sangre a favor de estos ultimas, lo que provoca un aumento importante de los niveles sericos de AGL. El higado extrae mas del 30% de los AGL que le Hegan por la circulacion, utilizandolos para formar acil-CoA, que servira bien para formar trigliceridos o fosfolipidos, bien para introducirlo en la matriz mitocondrial. La beta-oxidacion del acil-CoA aumentara, formando acetil-CoA a una tasa que puede exceder la capacidad mitocondrial hepatica de su oxidacion en el ciclo de Krebs. Si esto sucede en el hepatocito, la parte excedente del acetil-CoA se transformara en acetoacetato, reduciendose este en parte a hidroxibutirato. Como resultado, el higado comienza a liberar al plasma cuerpos cetonicos (ya que el higado no utiliza los cuerpos cetonicos como combustible).
En ejercicios realizados por encima del umbral lactico (intensidad->65%
VO max) la liberacion de AGL desde el tejido adiposo es menor, y ello a pesar 2
de un aumento de la activacion simpatico-adrenal y por tanto del estimulo lipolf tico de las catecolaminas. Se han propuesto dos mecanismos para explicar este hecho: por una parte, el aumento de los niveles de lactato en sangre (el lactato inhibe la lipolisis aumentando la reesterlficacion de los acidos grasos a triglicerldos en el tejido adiposo, gliceroneogenesis) y por otro el descenso del flujo sangufneo al tejido adiposo asociado a la intensidad relativa de ejercicio.

,.
En la degradaci6n de los arninoacidos, el grupo amino se libera quedando un esqueleto de atomos de carbono que se convierte en un intermediario metab61ico. La mayoria de los aminoacidos se convierten en piruvato, acetil CoA o en uno de los intermediarios del ciclo de Krebs (Figura 37). La perdida del grupo amino se produce por desaminaci6n o transaminaci6n.
Se han descrito al menos seis aminoacidos que pueden utilizarse como combustible: alanina, aspartato, glutamato, y los tres arninoacidos de cadena ramificada (valina, leucina e isoleucina). Parece ser que este ultimo grupo de aminoacidos es el que preferentemente oxida la celula muscular esqueletica Tanto la valina como la isoleucina producen succinil CoA como producto. Los productos del catabolismo de la leucina, aceto-acetato y acetil-CoA, no pueden ser convertidos en glucosa, considerandose un aminoacido cetogenico. Por otra parte, algunos arninoacidos musculares actuan como precursores gluconeogenicos durante el ejercicio (hasta 3,8 gramos/hora de glucosa), como es el caso de la alanina.
Es importante destacar que la aportaci6n de las proteinas como sustrato energetico varia en funci6n de las reservas de otros sustratos energeticos: asi, la oxidaci6n de los aminoacidos es mayor cuando los niveles previos de gluc6geno muscular son bajos. La temperatura parece ser tarnbien un factor
' I La glutamina es un arninoacldo , : especial no esencial cuyo I contenido es muy elevado en : muchas celulas yen la sangre, I representando alrededor , del 60% def pool total de , : aminoacidos, Aunque se obtiene I alga de glutamina en la dieta, : la mayorfa de la glutamina I se sintetiza en el organismo I a partir del glutamato par fa : acci6n de la enzima glutamina , sintetasa. La glutamina : constituye un importante • combustible para las celulas def ' est6mago y def sistema inmune : (macr6fagos y linfocitos). , Adernas, la glutamina muscular I , parece tener un papel relevante , en las procesos de regulaci6n de ' sfntesis de protefnas. ...
/------------- proteinas pueden ser utilizadas durante la actividad muscular, especialmente durante ejercicios muy prolongados. Las proteinas que se utilizan en el miisculo como combustible parecen ser las proteinas no contractiles sin , que se produzca la degradaci6n importante de los elementos proteicos que participan en la contracci6n muscular. El aporte energetico procedente de los aminoacidos se situa entre el 3% Y el 10% de la energia total generada. En general, podemos considerar que en un ejercicio cuya duraci6n sea inferior a 60
minutos, el aporte energetico por parte de las proteinas no resulta significativo.
41Metabolismo de las proteinas coma sustrato energetico durante el ejercicio
aminoacidos, del catabolismo de las proteinas celulares y de la sintesis de amtnoacidos por el
higado. Durante el ejercicio, se produce una elevaci6n
de la producci6n de urea sanguinea y de la excreci6n de nitr6geno. Esto indica que las
El 80% de los aminoacidos libres que hay en el cuerpo se encuentra en el musculo esqueletico. El musculo puede utilizar estos aminoacidos como sustrato energetico mediante su oxidaci6n (principalmente los amlnoactdos de cadena ramificada), o bien liberarlos al plasma (fundamentalmente alanina y glutamina), desde donde pueden dirigirse al higado para constituir precursores de glucosa. Asi, el pool de aminoacidos de la sangre Y los liquidos
- - - - - - - - - - - - - - - , extracelulares es el resultado de la ingesta de ,. , Aminoacidos no esenciales: I alanina, asparagina, aspartato, 1 I , I . I cistema, glutamato, g utamma, I glicina, prolina, serina y tirosina I
I Aminoacidos esenciales: I arginina, histidina, isoleucina, I leucina, lisina, metionina, I . , fenilalanina, treonma, ', tript6fano y valina ,.
"'--·---·----------
Las proteinas pueden utilizarse para obtener energia en algunas
circunstancias, aunque. pa,r~ ello primero es necesario s~parar el grupo amino de los aminoacidos que las componen. Postenormente
a su desaminaci6n casi todos ellos (18 de los 20) pueden convertirse en glucosa en el higado en el proceso de la gluconeogenesis. Se les denomina por ello amtnoacidos glucogenicos. Los otros dos son cetogenicos. Sin embargo, para la obtenci6n de energia la mayor parte de los armnoacidos tras ser desaminados se incorporan al ciclo de Krebs. En estados de ayuno prolongado, las proteinas incluso pueden generar acidos grasos libres con fines energeticos. El proceso de conversion de proteinas a acidos grasos se
denomina lipogenesis. El higado es el principal regulador de la composici6n del pool de
aminoacidos del organismo, siendo capaz de sintetizar algunos de los 20 aminoacidos necesarios para formar las proteinas. Esos aminoacidos se denominan aminocicidos no esenciales. Otro grupo de aminoacidos, denominados aminocicidos esenciales, no pueden ser sintetizados por lo que
debemos obtenerlos a traves de la dieta.
4\Metabolismo de las proteinas coma sustrato energetico durante el ejercicio
Figura 38. Cicio de las purinas.
C'\,. /H Fumarato C = C
/ " C H
Ribosa 5-P Adenilo
succinato
Ribosa 5-P Adenosfn
monofosfato (AMP)
H-C-C-C I
N
Aspartato
I c I NH3-~ -H
I f c
Ribosa 5-P lnosfn
monofosfato (IMP)
NP'z I
-<=O c
0 II w----
La celula no es capaz de provocar la combustion completa de los arninoacidos (no oxida los componentes nitrogenados) por lo que los grupos amino son previamente liberados para oxidar solo el resto oxoacido. Estos grupos amino se liberan al plasma para su posterior metabolismo.
Durante el ejercicio, la fuente inmediata de amonio es la desaminacion de adenosina monofosfato (AMP) en el musculo dando lugar a la formacion de inosina monofosfato (IMP), proceso que tiene lugar esencialmente en las fibras musculares tipo II. El musculo esqueletico puede producir amonio tambien a partir de aspartato. La conversion de aspartato en fumarato y amonio ocurre a traves de un proceso ciclico denominado ciclo de las purinas,
(Figura 38) el cual se ha observado activado en situaciones de aumento de la tasa glucolitica. De hecho, la (mica fuente de amonio durante el ejercicio intenso de corta duracion es el ciclo de las purinas.
PRODUCCION DE AMONIO
41Metabolismo de las proteinas coma sustrato energetico durante el ejercicio --- :.:J
Figura 37. Oxidaci6n de arninoacldos mediante su conversion en piruvato o en intermediarios del ciclo de Krebs. (Poortmans JR. Protein Metabolism).
Alanina Glicina Serina
( Glucosa ) Cisteina Acidos grasos
I /. Pirutato Lt 1 PDH lsoleu~ina , Leucma
Fosfoenolpiruvato Acetil-CoA Fenialanina
~ l T~;;;!:~o
PEPC ~ - ( Citratosintetasa r ::::.1
ir .~::r· :r:J / Fuma,ato 2-oxoglutararo +-+l'.:utamato J
p:ri::~a l:H :KGDHJ ~ldlna Tirosina Succinato - Succinil-CoA Prolina l Succ. CoA S ) t Arginina
~:~~~n-cia \ etionina reonina --
a tener en cuenta en el catabolismo de las proteinas durante el ejercicio,
siendo mayor a temperaturas bajas que altas. Ademas de su contribucion al aporte energetico, la oxidacion de los
aminoacidos en el musculo durante el ejercicio tiene una funcion anapletorica
importante reponiendo los intermediarios del ciclo de Krebs, los cuales van
reduciendo sus concentraciones en la mitocondria segun avanza el ejercicio.
El descenso en las concentraciones de estos intermediarios se postula como
una causa de fatiga.

/
320
INFLUENCIA DEL ENTRENAMIENTO
Las adaptaciones de la capacidad oxidativa muscular al entrenamiento aerobico permiten una mayor capacidad del musculo esqueletico para fosforilar el ADP lib t , d 1 . . , . . . , re a raves e a respirac10n mitocondrial, con lo que se disminuiria la deplecion de ATP la acumulacion de IMP 1 ' Ye aumento s , d angumeo e NH3 que se observa durante el ejercicio intenso.
El entrenamiento anaerobico conlleva una menor deplecion muscular de ATP, junta con un descenso de la acumulacion de sus productos de deg~adaci6n (IMP y NH) y una menor actividad de la AMP desaminasa con el ejercicio, indicando un mejor equilibria entre la hid '1· . 1 , . ro ISIS Y a resintesis de ATP con este tipo de entrenamiento.
DISTRIBUCION DEL TIPO DE FIBRA
Se ha suger~do ~~e _las fibras rapidas (IIx) son la fuente del NH3 producido durante ~l ejercicio mtenso, mientras que durante el ejercicio prolongado, el reclutamiento predominante de fibras I hace pensar que estas pudieran ser una fuente importante de NH3 en este tipo de ejercicio.
las de cadena ramificada, parece ser la fuente potencial de NH durante el • • • 3
ejercicio de larga duracion.
Potencia (W)
Figura 39. Respuesta del amonio sangufne · · · · un aumento exponencial a partir de 200 va~i~sun ejercicro ,ncredmental ~n la que se observa de la potencia maxima. ' que correspon e aprox,madamente al 60%
300 250 200 150 100 so o---r--r----,-----i----r---r-------- Reposo
20
40
60
So
100
Amonio (µmoJ-1-1)
120
140
_______ 4_fMetabolismo de las proteinas coma sustrato energetico durante el ejercicio
DURACION DEL EJERCICIO
En general, las niveles plasmaticos de NH3 aumentan segun lo hace la duracion del ejercicio. El NH3 que aparece en la sangre durante el ejercicio de corta duracion proviene de la desaminacion del AMP a IMP en el ciclo de las purinas, mientras que la degradaci6n de las aminoacidos, sabre todo
INTENSIDAD DE EJERCICIO
Los niveles sanguineos de NH3 varian poco durante intensidades de ejercicio ligeras a moderadas ( <50% V02max), y aumentan exponencialmente con cargas de trabajo > 60-70% V02max, siendo el aumento desproporcionado en intensidades 2'.: 100% V02max (figura 39). No es infrecuente observar aumentos en la concentracion sanguinea de amonio de 200-300% despues de un ejercicio de corta duracion y elevada intensidad (>100% V02max).
Durante el ejercicio de larga duracion e intensidad moderada.(60-70% del
V02max) la produccion de amonio (50 veces superior a las niveles de reposo) excede con mucho la tasa de desaminacion de AMP. Parece claro, par lo tanto, que durante este tipo de ejercicio parte del amonio producido se debe a la desaminacion de aminoacidos en el musculo esqueletico.
Dado que la desaminacion del AMP no es reversible en condiciones fisiologicas, la recuperaci6n del contenido de las nucleotides de adenina requiere el proceso de reaminacion de IMP a AMP, catalizado par las enzimas adenilosuccinato sintetasa y adenilosuccinato liasa. En el proceso de reaminacion, el grupo amino lo aporta el aspartato consumiendose energia a partir del GTP. Par lo que, un ciclo completo conlleva la desaminacion de una molecula de aspartato y la produccion de amonio y fumarato. Estos procesos de desaminacion y reaminacion no son simultaneos durante la contraccion muscular, ya que la desaminacion del AMP se produce durante el ejercicio y la reaminacion durante el periodo de recuperaci6n.
La acumulacion sangufnea de NH3 en el ejercicio varia ampliamente entre sujetos, dependiendo de la distribucicn del tipo de fibra muscular, el estado de entrenamiento o la disponibilidad del sustrato energetico (gluc6geno muscular). El aumento de las niveles sanguineos de NH3 con el ejercicio tambien depende esencialmente de la intensidad y duracion del esfuerzo.

, •
La concentraci6n de urea en sangre durante el ejercicio de corta duraci6n se mantiene estable, siendo a partir de los 60-70 minutos de duraci6n de la actividad cuando se empieza a observar un aumento de dicha concentraci6n. A partir de ese momenta se observa una correlaci6n significativa entre la duraci6n del ejercicio y la concentraci6n de urea en sangre. Se ha observado tarnbien una correlaci6n significativa entre el aumento en la concentraci6n de urea en sangre y el descenso de nitr6geno a-amino, que parece indicar un aumento en la oxidaci6n de proteinas durante el ejercicio prolongado. Sin embargo, debemos tener en cuenta que el aporte energetico procedente de la oxidaci6n de las proteinas es en cualquier caso moderado. Parte de los arninoacidos procedentes del catabolismo de las proteinas durante el ejercicio prolongado pueden servir de precursores para la formaci6n de glucosa.
Figura ~o. Esquema ~e~eral de/ cicl? de la urea que acontece en el hepatocito con el fin de poder elirninar el arnoruo (NH4-) de la sangre a traves de la orina.
Fumarato
Cicio de la urea 0 II
H2N-C-NH2
Urea
Ornitina
I I 1 _N~~+ ~ _c~,-, -r-----1~ Foslato de carbamllo ====u
Citrulina llr:~·· Argi n i nosucci nato
Con el fin de eliminar el amonio (y otros compuestos aminados) de la sangre, el higado transforma en urea en el proceso metab6lico denominado ciclo de la urea (figura 40).
PRODUCDON DE UREA
4[Metabolismo de las proteinas coma sustrato energetico durante el ejercicio
FUNCIONES DE LA DESAMINACION DE AMP
•l\'lantenimiento de la relacion ATP/ADP durante la contracci6n. Atenuando el aumento de AMP por acci6n de la AMP-deaminasa se permite la activaci6n de la adenilasa-cinasa para la formaci6n de ATP a partir de 2 ADP. Esta actividad por tanto disminuye la cantidad de ADP procedente de la gran actividad de la ATPasa, y por tanto facilita el mantenimiento de la relaci6n ATP/ADP. Se ha demostrado que las fibras I mantienen mejor la relaci6n ATP/ADP y por tanto producen menos IMP que las fibras tipo IL
-Prevencion de descenso de nucleotides de adenina. La producci6n de IMP via AMP-deaminasa durante contracciones intensas permite conservar los nucle6tidos celulares (no-Hargreaves). El IMP permanece en la celula Y puede ser reaminado cuando las condiciones celulares lo permitan.
·Amonio como buffer de protones. El NH3 esta presente en el musculo en forma de NH y de NH - (NH + H+-+ NH-). La formaci6n de amonio consume H+ Y
3 4 3 4
esto ha llevado a algunos investigadores a pensar que la formaci6n de NH4-
sirve como amortiguador (buffer) de la elevada formaci6n de H+ durante el ejercicio intenso, si bien parece que esta funci6n no es muy importante.
, Regulaci6n del metabolismo de los hidratos de carbono. El amonio ( NH 4 - ) activa la PFK in vitro, y el IMP parece estimular lafosforilasa b, con lo que aumenta la glucogenolisis, aunque el estimulo no es muy significativo.
ACLARAMIENTO DEL NH, PAODUDDO DURANTE EL ao El higado metaboliza NH3 a traves del ciclo de la urea, aunque durante el ejercicio no parece el lugar principal de aclaramiento. Otros posibles mecanismos de eliminaci6n los constituyen el aclaramiento renal, la excreci6n por el sudor, o el musculo esqueletico. Tambien el pulm6n puede ser un sistema clave para el aclaramiento de NH3 durante el ejercicio.
DISPONIBILIDAD DE SUSTRATO ENERGETICO
El metabolismo del NH se ve modificado por la disponibilidad de gluc6geno 3
muscular, de forma que cuando existen niveles bajos de gluc6geno muscular, la liberaci6n muscular de NH3 puede ser hasta 5 veces mayor, lo que puede ocasionar unos niveles plasmaticos de NH3 tres veces mayores despues de
ejercicios de duraci6n similar.
I • tJ;r:,.,~. ' .. tJJ;/~~,,. •, 86 j 1 ••
iN·JA,• • ,,·.· ..

•
EJERCICIO DE ALTA INTENSIDAD
Durante el ejercicio de alta intensidad, se produce una menor liberaci6n de glutamina y alanina desde el miisculo. La entrada de glutamato al musculo no parece aumentar significativamente durante el ejercicio de alta intensidad, Y dado que la sintesis de glutamina consume ATP y su liberaci6n par el musculo representa una perdida de glutamato en el rmisculo, teniendo en cuenta que este ultimo es una fuente de formaci6n del intermediario del ciclo de Krebs, el a-cetoglutarato, no parece que la sintesis y liberaci6n de glutamina sean procesos productivos cuando la necesidad de ATPs es elevada.
La mayoria de las aminoacidos resultantes del catabolismo de las protefnas son liberados desde el musculo a la sangre. La principal excepci6n a este comportamiento es el glutamato, ya que se observa una elevada captaci6n del mismo durante el ejercicio. El glutamato se utiliza coma precursor de la glutamina, que junta con la alanina, son liberados del musculo en funci6n de la intensidad y duraci6n del ejercicio, habiendose propuesto que durante el ejercicio el musculo aumenta la tasa neta de sfntesis y liberaci6n de las mismos (Figura 41). Del piruvato obtenido en la glicolisis, aproxiamdamente el 1% se convierte en alanina. Durante el ejercicio en ayunas, con bajos niveles de gluc6geno, se genera menos piruvato desde la glucolisis, y par tanto se formara menos alanina. En este estado el catabolismo proteico es mayor, con un aumento de liberaci6n de la mayoria de las aminoacidos desde el musculo. La alanina que se libera desde la celula muscular coma resultado de la transaminaci6n a partir de las aminoacidos de cadena ramificada (AACR) puede ser un sustrato glucogenico en el higado, para formar glucosa que posteriormente puede de nuevo ir al musculo para volver a entrar en la glicolisis. Este proceso se conoce coma el ciclo de la alanina-glucosa. Adernas de contribuir a mantener las niveles de glucemia, este ciclo sirve para transportar desde el musculo hasta el higado las grupos amino de las AACR y que alli se transformen en urea para su eliminaci6n. Cuando el ejercicio tiene una duraci6n muy prolongada, la producci6n de alanina es menor ya que las dep6sitos de gluc6geno se van agotando.
41Metabolismo de las protefnas coma sustrato energetico durante el ejercicio
i
Figura 41. Durante el ejercicio se observa una liberaci6n neta de los ~n:iinoacidos desde la. ~elula musuclar hacia la sangre, exceptuando el glutamato (GLU) y l_os a~~noac1dos _de cadena rarnificada (AACR) que experimentan una captaci6n neta, provocando una liberacion ~~ a)anina (ALA) Y glutam1na). La conversion de AMP en IMP es muy activa en el musculo durante ejercicro 1n~enso (AMP deaminasa ,
por lo que el musculo en estas circunstancias libera NH/. (Mod1f1cado de T11/dus, 2012, P 258)
EJERCICIO DE INTENSIDAD MODERADA
Durante el ejercicio en estado post-absortivo, el musculo esqueletico adopta un perfil catabolico, donde la ruptura de proteinas excede la tasa de sfntesis.
OXIDACION DE AMINOACIDOS El estudio del metabolismo de las aminoacidos durante el ejercicio es complejo, determinandose habitualmente mediante la diferencia de concentraci6n arterio-venosa de aminoacidos y el flujo de sangre al tejido, y asi poder conocer la liberaci6n y captaci6n de aminoacidos par el tejido muscular activo. Adernas, se pueden obtener biopsias musculares, Y medir concentraciones de aminoacidos, antes, durante y despues del ejercicio. La utilizaci6n de la infusion de is6topos estables de aminoacidos espedficos esta permitiendo en la actualidad determinar con mas exactitud las tasas de
captaci6n y oxidaci6n de aminoacidos durante el ejercicio.

• • ejercicio a la maxima intensidad posible, desde las velocistas de distancias
mas cortas (actividades que duran menos de 10 segundos a la maxima intensidad) hasta las atletas que realizan eventos de resistencia (mas de 30 minutos), cada uno de las tres sistemas celulares que nos proporcionan energia esta contribuyendo a las necesidades energeticas totales del organismo. Lo que ocurre es que en las diferentes tipos de ejercicio, y sabre todo en funci6n de la intensidad, predomina un sistema energetico sabre las otros, excepto en las circunstancias en las que se producen transiciones de la predominancia de un sistema energetico a otro (Figura
42). Par ejemplo, en un evento muy intenso y breve, coma puede ser una carrera de 100 m que se realiza en 10 segundos, el mayor aporte de energia procede del sistema de la fosfocreatina (PCr-ATP) o sistema anaer6bico alactico, pero tanto las sistemas anaer6bico lactico (gluc61isis anaer6bica) coma el oxidativo o aer6bico estan proporcionando parte de la energia necesaria. En el otro extrema del rango, en una carrera de 30 minutos (10.000 m) predomina el sistema oxidativo, si bien contribuyen tambien las dos sistemas anaer6bicos.
n las celulas musculares, las diferentes sistemas energeticos (anaer6bico alactico, anaer6bico lactico y aer6bico) no actuan • de forma independiente. Cuando un individuo esta realizando
I Interaccion de las diferentes sistemas energeticos durante el ejercicio
I •
1-+1:i~V·--· , ; ·. ·--·
,li \Ju >-- e 90 •'l)',r:•J!,• - ; ... - ..
l,J~A,{ .. ·-. 'i '. ' •
La oxidaci6n de leucina (utilizado coma indicador del catabolismo . · 1 · · · decir a la proteico total) es proporcional a la intensidad de eJerc1c10, es . . .
demanda energetica del mismo. En las intensidades mas altas de e1erc1c10 la
tasa de oxidaci6n de este aminoacido puede llegar a ser 30 veces superior a
la de reposo. Ademas de la intensidad y duraci6n del ejercicio, la ing~sta de
hidratos de carbono, grasas y proteinas tambien parece afectar el mvel de
oxidaci6n de aminoacidos. Asi, la ingesta de hidratos de carbono disminuye
la oxidaci6n de aminoacldos de cadena ramificada, mientras que el ayuno
y las dietas ricas en grasas y en proteinas parecen aumentar el nivel de
oxidaci6n de arninoacidos. Adernas del musculo esqueletico, hay otros 6rganos coma el higado y
el intestino delgado que suponen las fuentes mas importantes de estos
aminoacidos oxidados.

• • •
•
El agotamiento de las dep6sitos de gluc6geno muscular se produce tras la realizaci6n de ejercicios prolongados ( de aproximadamente unas 2 horas de duraci6n) a una intensidad relativamente alta (alrededor del 85% del V02max). A intensidades alga menores, el gluc6geno muscular dura, 16gicamente, mas tiempo (Figura 44). Si la intensidad es mas elevada, las dep6sitos de gluc6geno no se agotan, ya que dicha intensidad no es posible mantenerla durante el tiempo suficiente para que esto ocurra. En estos casos, las causas de fatiga no se relacionan con el vaciamiento de las dep6sitos de gluc6geno. Sin embargo, si puede llegar a agotarse el gluc6geno cuando se realizan ejercicios de alta intensidad intervalados, en las que existen pausas de recuperaci6n que permiten al rnusculo recuperar su situaci6n metab61ica basal.
Comparados con las grasas y las proteinas, las hidratos de carbono siguen siendo el combustible favorito durante ejercicio aer6bico de alta intensidad ya que es el que proporciona energia de forma mas rapida mediante las procesos oxidativos.
Figur: 43, Descenso del glucogeno muscular en el gemelo durante tres horas de carrera al 7.0 Yodel VC\max. ~Mod1f1cada de CDL Cost1II, _1986, Inside ~unning: Basics of sports
physiology. lnd1anapol1_s, Benchmark Press. _En: _Wilmore y Costili, Physiology of Sport and Exercise, 3.a ed. Human Kinetics, 2004; 149, fig. 4.19.)
Musculo gastronemio
175
,...._ 150 b.o 7 0 125 E s 100 ..... ~ :::, u Vl ::I 75 E 0 c:: QI 50 b.O '0 u :::,
"6 25
0 0 2 3
Tiempo de ejercicio (h)
linteracci6n. de los diferentes sistemas energeticos durante el ejercic~
Tiempo de la prueba Segundos ------------!~ minutos
0
Sistema del acido lactlco
1. DINiMICA DE LOS HIDRATOS DE CARBONO DURANTE EL EIERCICIO
Los resultados obtenidos a partir de la realizaci6n de biopsias musculares indican que las principales factores que determinan la mezcla de sustratos energeticos durante el ejercicio son, par un lado, la intensidad y la duraci6n del ejercicio, y par otro, la forma fisica y el estado nutricional del individuo.
El hfgado aumenta significativamente la liberaci6n de glucosa a las musculos activos segun progresa la intensidad del ejercicio. Simultaneamente, el gluc6geno muscular constituye la principal fuente de energfa durante las fases iniciales del ejercicio y segun va aumentando la intensidad. La tasa de utilizaci6n del gluc6geno es mas elevada en las primeros 15 a 20 minutos de
ejercicio. (Figura 43).
0
Figura 42 Relacion entre el porcentaje de ATP aportado por las diferentes sistemas energeticos en relaci6n con el tiempo de la prueba.
~
~
100 100
• • • • •
92
J '• • . ~itt,f._ • ' ', ~ ., ,,,
,\j\r I':!'?'
•
GWCOGENO HEPATICO. La utilizacion de la glucosa circulante durante el ejercicio prolongado depende fundamentalmente de la liberacion de glucosa hepatica a la sangre. La tasa de liberacion esta relacionada con la duracion y la intensidad del ejercicio. La mayoria de esta glucosa procedente del higado procede a su vez de la degradaci6n del gluc6geno hepatico, siendo la otra fuente menos importante desde el punto de vista cuantitativo los precursores gluconeogenicos como el lactato, el glicerol o la alanina captadas por el higado. El gluc6geno hepatico se reduce aproximadamente a la mitad al cabo de una hora de ejercicio intenso; al cabo de unas dos horas de ejercicio intenso casi todo el gluc6geno almacenado en el hfgado Y en los rmisculos activos se ha consumido. En la (figura 45) se muestra la relacion entre la producci6n de glucosa hepatica y la duracion e intensidad del ejercicio.
EIERCICIO IWODEAADO Y PROLONGADO. La mezcla de sustratos energeticos depende fundamentalmente de la intensidad del ejercicio. Cuanto mas baja es la intensidad, mayor protagonismo adquieren las grasas. Por otra parte, los hidratos de carbono que se utilizan proceden al principio del gluc6geno almacenado, pero segun este se va consumiendo, va ganando mayor protagonismo la glucosa circulante, a la vez que las grasas aportan cada vez mayor parte de los requerimientos energeticos.
EIERCICIO INTENSO. La realizacion de ejercicio de alta intensidad desencadena una respuesta neurohormonal caracterizada por un aumento de la liberaci6n de adrenalina, noradrenalina y glucagon, y por un descenso de insulina. Esta respuesta hormonal activa a la gluc6geno fosforilasa muscular de manera que el gluc6geno muscular es el principal sustrato energetico en los minutos iniciales del ejercicio, ya que se puede utilizar en condiciones anaerobicas cuando el organismo aun no ha sido capaz de adaptar la oferta de 02 a la demanda existente. Segiin va pasando el tiempo, la glucosa sangumea va aumentando su contribucion al aporte energetico, llegando a proporcionar aproximadamente un 30% de los requerimientos energeticos
LA DISPONIBILIDAD DE HIDAATOS DE CARBONO REGULA SU UTILIZACION PARA APORTAR
ENERGiA. Cuando los niveles de glucosa en sangre se elevan durante el ejercicio, debido, por ejemplo, a la ingesta de alimentos con alto Indice glucernico antes de la realizacion del mismo, se inhibe la movilizacion y la oxidacion de los acidos grasos por el musculo esqueletico,
·' ' · d I ' en hfgado y rnusculo en funci6n Figura 44. Tasas de util_i~ac1on ~e l~s_depos1tof~ edgducoJen~ TD Lore of Running 3.a ed. de la intensidad y duracion del_ ejerocio. (Modi ica a e oa es · , Leisure Press. Illinois: Cahrnpaign, 1991; 68.)
Duracl6n de! ejercicio (horas)
5 4 3 2 0 0
85% del V02max 50
70% del V02max
50% del V02mc1x --- --- --- - ...... - --- --- --- --- ---- ---
250
Durad6n del ejercicio (horas)
4 3 2
Ritmo de marat6n o de triatl6n ( «A» o «B»)
(85% del vo.rnax)
/
Ritmo de ultramarat6n, ironman o etapa
ciclista (70-75% dei V02max)
________ f r_nt_e_ra_cci6n de los diferentes sistemas energeticos durante el ejercicio J -----~- -----
o 100 c: OJ co '0 u :::, rs
0 S:i a., E E 200 U!l -" -.... ~ s ,_ 150 ~
... 94 , .
• I
160
140 :§ 0 120 u ·.;::;
't'\'.l o, (!J .c 100 0 c: a., eo Bo '0 u ::, To a., 60 -0 v, 0 .....
"iii 40 '0 Cl. a., 0 20
0 0
i--

•
•
I. UTILIZACION DE AMINOiCIDOS COMO FUENTE ENERGETICA
La oxidaci6n de aminoacidos durante el ejercicio es minima (>1% de la energia necesaria). Incluso en situaciones extremas (es decir, ejercicio muy intenso y prolongado o situaciones de ayuno prolongado) la oxidaci6n de aminoacidos representa unicarnente una fracci6n relativamente pequefia de la utilizaci6n total de sustratos ( <10% ).
pueden conseguir mayor utilizaci6n (uno de los efectos del entrenamiento de resistencia es el aumento de los dep6sitos intramusculares de trigliceridos).
La activaci6n simpatica y la disminuci6n de los niveles de insulina son los factores que favorecen la movilizaci6n de los acidos grasos desde el tejido adiposo. En ejercicios de intensidad moderada, los trigliceridos y los hidratos de carbono aportan energia aproximadamente a partes iguales, pero segun se va prolongando la duraci6n de la actividad (una hora o mas), la utilizaci6n de los acidos grasos va siendo mayor, de forma paralela a c6mo se van vaciando los dep6sitos de gluc6geno. El descenso de la glucosa sangumea, y por tanto de la insulina, son estimulos potentes para la lip61isis .
A intensidades inferiores al 40% del maxima, las grasas constituyen la principal fuente energetica, mientras que a una intensidad del 85%, la cantidad de grasas oxidada es la misma que al 25%, lo que significa que su participaci6n relativa es mucho menor.
El entrenamiento favorece la utilizaci6n de grasas y el 11 ahorro" de hidratos de carbono, de manera que para una misma intensidad relativa, un sujeto entrenado obtiene un mayor porcentaje del aporte energetico de las grasas que un individuo sedentario.
--
Los hidratos de carbono se almacenan en forma de glucogeno fundamentalmente en hfgado (pequefias cantidades en el rifion) y en el rmisculo estriado. Los trigllcerldos en el tejido adiposo yen el rnusculo. El hfgado contiene entre 80 y 100 gramos de glucogeno en los periodos postprandiales, mientras que el glucogeno muscular puede variar desde 50 gramos despues de un ejercicio extenuante hasta 900 gramos en un sujeto bien musculado, bien entrenado y bien nutrido. Los almacenes de grasa son mucho mayores, y presentan muchas mas cliferencias individuales, constituyendo la grasa corporal entre un 8% y un 35% de la rnasa corporal total. Aunque la mayor parte se almacena en el tejido adiposo, podemos encontrar aproximadamente 300 gramos de triglicerldos intramuscu!ares.
---------- ---------- ----------- linteracci6n de los diferentes sistemas energeticos durante el ejercicio I
• z. DINiMJCA DE LAS GRASAS DURANTE EL EIERCICIO
La oxidaci6n de los acidos grasos en el musculo esqueletlco depende de
2 factores fundamentales para cualquier tasa metab61ica. Estos son la disponibilidad de acido grasos libres (AGL)- determinada a su vez por la movilizaci6n de AG-, y la capacidad de los tejidos para oxidar dichos AG.
La grasa intracelular y extracelular proporciona entre un 30% Y un 80% de la energia necesaria para la actividad fisica, dependiendo del estado nutricional, del grado de entrenamiento y de la intensidad Y duraci6n del
ejercicio. Durante los ejercicios de baja intensidad, el flujo sanguineo al tejido adiposo
aumenta, lo que permite una mayor movilizaci6n de acidos grasos ~ara ~~e puedan ser utilizados por el musculo. Sin embargo, esta vasodilatacion no se observa durante los ejercicios de intensidades elevadas por lo que la concentraci6n de AGL en sangre en estas situaciones no es mayor que la de reposo. Esto favorece la utilizaci6n de los trigliceridos intramusculares Y del gluc6geno. Los trigliceridos intramusculares pueden llegar a aportar entre un 15% y un 350;0
de la energia, siendo los deportistas de resistencia los que
• • ~ . ,.
10 20 30 30 Tiempo de ejercicio (min)
Figura 45. Liberaci6n de glucosa he~atica dur~~~fb~r~~~~o d~ c~~~is~n~~ti:j~e:ed~t;;:~~s (30, 50-60, 75 y 85%iel V~2~~xh~~! f!~~tensidad. (Modificada de Greenhaff! Hultman,
H:~r1t~1riih~~~~~t~t~bs~[~. En: Poortmans JR led.I. Principles of Ex~rcis)e B1ochem1stry, · 3.a rev ed. Med Sport Sci. Basel: Karger, 2004, vol. 46:133, f1g.5 · •
120 0
6,o
~ c: 5,0 .E 0 E 5 4,0 ro u ·.;:::;
-rc 0. 3,0 a, ..c ro VI 0 u :::J
bl) 2,0 a, ·o c: '0 ·.::; 1,0 u :::J
'U 0 a:
0
•
lntensidad de ejercicio (%FC max)
90 70 80 50 60 40
-+- Activos
.....,_ Sedentarios
lntensidad de ejercicio (%V02 max) 90 70 80 50 40 60 30 20 10
-+- Activos
.....,_ Sedentarios
Figura 47. Tasa de oxidaci6n de grasas en funci6n de la intensidad expresada como % VO max o %FC max. Cornparacioh entre sujetos activos y sedentarios. (modificado de Rami y cols, 2014)
0,30 ,....._ \:: .E 0,25 bD '-' VI ro 0,20 VI ~ b.O Qj
"C 0,15 c: '0 'c ro 0,10 "C ·x 0 Qj
"C 0,05 ro VI ro I-
0 30
0,30 c- 'c: .E 0,25 bD '-' VI ro 0,20 VI ro ... b.O Qj
"C 0,15 c: ,o 'c ro 0,10 "C ·x 0 Qj
"C 0,05 ro VI ro I-
0 0
Segun aumenta la intensidad del ejercicio, la contribuci6n relativa de los hidratos de carbono coma combustible tambien aumenta; al mismo tiempo, se reduce la contribuci6n relativa de las grasas. Sin embargo, en terminos absolutos, la oxidaci6n de los hidratos de carbono aumentara proporcionalmente a la intensidad del ejercicio, mientras que la tasa de oxidaci6n de las grasas aumentara inicialmente, pero a partir de cierta intensidad, comienza a disminuir. En el afio 2001, Jaukendrup defini6 coma fat-max a la intensidad de ejercicio (expresada como % de VO max ode la
2
FC max) a la cual la tasa de oxidaci6n de grasas (g/mirr-) es maxima, (Figura 46).
lrnteracci6n de los diferentes sistemas energeticos durante el ejercicio
45 50 55 60 65 70 75 80 85 90 95 lntensidad de ejercicio (% FC max)
•
Fat Max: intensidad de ejercicio a
la que la oxidaci6n de grasas es maxima
Figura 46. Tasa de oxidaci6n de grasas durante la realizaci6n de un ejercicio incremental. Concepto de "Fat max".
0.1
0.2
0.3
0.4
0.5 VI ("IJ VI ("IJ L.. b.() Cl)
""C c: ,o
·c; ("IJ
""C ·x 0 Cl)
""C ("IJ VI ("IJ f-
o.6 c: E - b.() .__..
0.7
•
• • 98 '•--;-
•
En reposo, el cociente de intercambio respiratorio (RER=VC02/VO) es de 0,75-0,82, lo que indica que las lipidos constituyen el sustrato oxidado prioritario par el organismo. Durante el ejercicio diferentes factores modifican esa relaci6n de oxidaci6n de sustratos con fines energeticos. Mas adelante en este capitulo analizaremos cuales son las interacciones de las diferentes sistemas energeticos durante el ejercicio.
La variable que mas condiciona el tipo de sustrato energetico utilizado par el miisculo activo es, sin d uda, la intensidad. A in tensidades baj as de ej ercicio, la utilizaci6n absoluta de grupos acil par la mitocondria va aumentando segun lo hace la intensidad hasta llegar a una intensidad aproximada del 50% del vo.max. Par encima de dicho valor, la participaci6n de las grasas coma combustible comienza a descender a favor de las hidratos de carbono. Esto quiere decir que la aportaci6n relativa de grasas frente a hidratos de carbono comienza a ser menor a partir de cierta intensidad.

•
ELECCION DEL SUSTRATO EN FUNCION DE LA INTENSIDAD DEL EIERCICIO
En general, las AGL constituyen el principal sustrato energetico en reposo y durante el ejercicio de baja intensidad. No obstante, el retardo en la movilizacion de las reservas de grasas y en el transporte de las AGL
La regulaci6n de la elecci6n del combustible durante el ejercicio no esta totalmente aclarada. Los mecanismos par las cuales las celulas musculares seleccionan las sustratos que van a utilizar en cada situacion son complejos. Si bien se pensaba que el principal mecanismo regulador era el ciclo glucosa- AGL ( el aumento de acidos grasos libres disponibles en plasma permite un aumento de su oxidacion, de manera que aumenta la concentraci6n de acetil CoA sin tener que utilizar glucosa-) en la actualidad se piensa que es mas bien la disponibilidad de glucosa la que regula la oxidaci6n de las acidos grasos actuando sabre la enzima carnitin pa/mi toil transferasa 1 que regula la entrada de las grupos acilo en la mitocondria.
A continuacion destacaremos las principales factores que condicionan la elecci6n de las sustratos disponibles durante la realizaci6n de ejercicio.
• La intensidad del ejercicio • La duraci6n del ejercicio • La condicion fisica del sujeto ( entrenamiento) • La composicion fibrilar del rmisculo • La dieta (disponibilidad de sustratos) • El sexo • Determinados factores ambientales
Adernas de estos factores, existe una gran variabilidad interindividual en la capacidad de oxidacion de las grasas. En un estudio realizado par Venables y col. con 300 sujetos se concluy6 que variables coma el nivel de actividad fisica, el V02max y el genera solo explicaron el 12% de la variacion interindividual en la tasa maxima de oxidacion de grasas durante el ejercicio, mientras que el contenido de grasa corporal no fue un predictor significativo. La variaci6n interindividual en la oxidacion de las grasas permanece en gran parte sin explicar.
lrnteraccion de los diferentes sistemas energeticos durante el ejercicio I
lntensidad de ejercicio (% V02max)
I ib ·' de los hidratos de carbono y de las grasas Figura 48. Grafica que representa a_ ~ontri UCIOfa durante un eiercicio incremental. El punto como compbustlbles para la producdc1f~n de energunto de Cross-dver (Brooks, 1985)esqueletica donde ambas graficas se cruzan, se e me como P
-,R. 0 0. lb
100 lb :::J lb -,
90 O'Q iii' - 80 n DJ .::::;
70 "C -, 0
60 n lb c.. lb
50 :::J ,..+ lb
40 0. lb ::r
30 0: -, DJ
i ,..+ 20 0 Ill 0. 10 lb n DJ 0 -, O" 0 :::J 0
80 60 40 20
Cross over: intensida~ de ejercicio a la que la energia
aportada por HC supera a la aportada por las grasas
Ill 100 ro Ill ro 90 .... b.O 111 80 "C ~,
70 c 111
"C CII 60 u 0 .... 0. 50 i1 40 -~ 30 b.O .... 111 c 20 111
\._! 10 0 Basal
• p •
•
---- ----------------------------.........---- Esta intensidad de ejercicio varia en funcion de diversos parametros, sabre todo
con la duraci6n. Es diffcil calcularla a nivel individual, aunque la mayoria de las estudios realizados la describen entre el 50%-75% del V02max. Algunos autores la describen coma la intensidad previa a la aparicion del umbral lactico. Obviamente factores coma el grado de entrenamiento del sujeto y el estado nutricional son condicionamientos muy importantes para el calculo de este parametro. Rami Y cols c2014) demostraron que el fat-max no varia demasiado entre sujetos sedentarios y entrenados si se expresa en valores relativos al V02 max o FC maxima, a ~esar de que en ambos casos la tasa de oxidaci6n de grasas es mas elevada en sujetos
activos para todas las intensidades de ejercicio (figura 47).
Anteriormente, en 1985 Brooks habia propuesto el concepto "crossover" coma la intensidad relativa de ejercicio en la que la Jormaci6n de ATP procedente de los hidratos de carbono excede a la derivada de las grasas (figura 48); a partir de esa intensidad relativa las hidratos de carbono aumentaran su aporte energetico, mientras que
' (( t disminuira el de las grasas. La intensidad metab6lica de este crossover o pun o de cruce" se situaria par encima de la correspondiente al fat max.
•

•
• • 103 • . ...
•
• •
F!gura 50. Esta figura ~epr~senta ,la utilizaci6n de_ la glucosa y de acidos grasos libres (AGL) rnculantes durante el ejercrcio, ~s1 coma del .glucogeno intramuscular. Elaborada por Brooks
(1997) con datos de diversos estudios prevros publicados en la literatura.
Tiempo de marcha (min)
120 100 So 40 60 20 0
Hidratos de carbono --- -- ----
(J) :a ·.;:::; Ill ::,
..0 E 0 u (lJ
-0 (J) i: 0 0.
<t:
Lf pidos 1 ----------
100% _.__
aer6bica del gluc6geno hepatico y muscular se va hacienda progresivamente mas importante, quedando la utilizaci6n anaer6bica de la glucosa y de la PCr para intensidades de trabajo mas elevadas. El gluc6geno muscular es el sustrato mas importante cuando la intensidad del ejercicio esta por encima del 70% del V02max. A intensidades superiores al 95% del VO max solo se
2
oxidarian hidratos de carbono (Figura 49).
El motivo por el cual a intensidades mas elevadas predomina la energia procedente de los hidratos de carbono se debe a la velocidad de los procesos catabolicos de los diferentes sustratos, o potencia energetica de los sustratos. La maxima tasa de recuperaci6n de ATP a partir de la oxidaci6n de grasas se ha calculado que es de 0,24 mmol·seg-1·kg-1, mientras que la oxidaci6n de la glucosa permite resintetizar ATP a una tasa maxima de o 51-0 68
' ' mmol · seg-'- kg',
Asi, el hecho es que realizar ejercicio por encima de una intensidad relativa aumenta la contribuci6n energetica (ATPs) de los hidratos de carbono y desciende la procedente de las grasas (figura 50). Paralelamente, la liberaci6n de glucosa a la sangre desde el tejido hepatico aumenta con la intensidad del ejercicio, aumentando exponencialmente la utilizacion de gluc6geno con fines energeticos.
51Interacci6n de los diferentes sistemas energeticos durante el ejercicio I
Figura 49. Al disminuir la intensidad ~el. ejercicio (y aume1;t?r la duraci6n), las grasas constituyen el principal sustrato metabollco
20 So
Baja lntensidad Alta
100 0 Larga Duraci6n Corta
40 60
60
80 (J)
~ ·,;::; "' :::J
.0 E 0 u 0 E 0 u 0 .:: 0 .c ,_ ro u (J) ·o Ill 0 ...... ~
TJ :c: (J)
-0 -J:.
(lJ
15 ·.;:::; V1 :::J
.0 E 0 u 0 E 0 u V1 ro V1
~ co (lJ
-0 -J:.
100
limplica cierta utilizacion de hidratos de carbono incluso a intensidades de ejercicio muy bajas, al menos en las fases iniciales. Segun aumenta la intensidad de ejercicio, aumenta la oxidaci6n de grasas hasta intensidades correspondientes al 50%-75% del V02max aproximadamente (figura 35).
A pesar de que la intensidad a la que se produce la maxima tasa de oxidaci6n de grasas es muy similar entre los sujetos entrenados y no entrenados, estudios realizados con atletas de resistencia como el de Knechtle en 2004
han concluido que estos deportistas (varones y mujeres) presentan la maxima tasa de oxidaci6n de grasas a intensidades significativamente mas altas (-::75% del V02max). En poblaciones sedentarias de hombres y mujeres, la maxima tasa de oxidaci6n de las grasas se ha observado a intensidades pr6ximas al 50% del V02max. Si la intensidad continua creciendo, se observa un descenso en la tasa de oxidacion de las grasas. A diferencia de lo que ocurre con el metabolismo de los hidratos de carbono, el cual aumenta en funci6n de la tasa de trabajo, la oxidaci6n de las grasas esta disminuida a intensidades elevadas. A intensidades de trabajo mas elevadas, la utilizacion
• • •• • •
• 102 • •

•
• • 105 • •
Figura 52. Cambios en la contribuci6n relativa de los diferentes sustratos (grasas, glucosa sangufnea y gluc6geno muscular) durante un ejercicio de cuatro horas de duraci6n a una intensidad del 30% del V02max. (Modificada de Ahlborg G, Felig P, Hagenfeldt L, Hendler R, Wahren J. Splanchnic and
leg metabolism of glucose, free fatty acids and aminoacids. J Clin Invest, 1974; 53:1086
Duracion del ejerciclo (horas) 4 3 2 0
Gluc6geno muscular 0
30°/o 20
40
27°/o
.. - - - --- - .. - - - - - ... - ... - - - .. .... - ...... .. - .... - ..... Glucosa sanguf nea ....... " .. -
60
Grasas So 37°/o
% del gasto energetico 100
aporte energetico proporcionado por las grasas aumento progresivamente hasta llegar a constituir el 62% del total, mientras que al principio de dicho ejercicio, las grasas constituyen solo el 37%. Por el contrario, el glucogeno muscular paso de contribuir con un 36% a un 8%. El resto, y con una proporcion casi constante, lo proporciona la glucosa sanguinea procedente del glucogeno hepatico y de la neoglucogenesis hepatica, siendo esta ultima via mas importante segun e prolonga la duracion del ejercicio.
-~
El "rnuro" para los corredores de marat6n se refiere a un conjunto de sensaciones negativas que percibe el corredor alrededor del kil6metro 30 de carrera, que se ateruian pasacios urros ki16metros, pero que obligan generalmente a disminuir el ritmo de carrera hasta la llegada a meta. La sensaci6n de falta de energfa y de fuerza muscular es la consecuencia de un problerna energetlco, que se debe fundamentalments al vaciamiento de los depositos de gluc6geno muscular y hepatico, En estas circunstancias rnetab61icas, los acidos grasos tornaran casi en exclusiva la responsabilidad de proporcionar energfa a las celulas musculares activas para proseguir la carrera. Dada ia menor potencia energetica de las grasas en reiaci6n a ios hidratos de carbono, el ritmo de carrera se vera inevitablemente afectado.
II
----------------------------------------, ,. I
-f rnteracci6n de los diferentes sistemas energeticos durante el ejercicio j
20 40 0
0 0
100 60 80
10
. . • . •• • .. · •• •• . •• .. ••• ·····
20 20
30 30
40 40
50 50 • •
10
...J \..!) <( QJ
"CJ >, ro VI 0 u :J ~
b.O ~ ~~ c: bo -o ..::£ ·c;..:.. ro o
.!::! E = :::1. ~- QJ
"CJ ro VI ro I-
• • ·~ • •
,-.... c: :J c -·l"'t" o..= QJ -· 0..N (I) QJ VI Q.
OQ o- - :J c: 0.. 8 (I)
!!!.OQ . c:
7'n OQ o-
. .:.OQ 3 (I) -· :J ~o
60------------r-----------,~:-----,60 • . . • Gluc6geno:
• . • . • . .
• •
l~ ELECCION DEL SUSTAATO EN FUNCION DE LA DURACION DEL EIEACICIO
Como ya hemos mencionado anteriormente, en los ejercicios de intensidad baja O moderada, las grasas iran ganando protagonismo coma principal sustrato energetico segun va avanzando el ejercicio, y la oxidacion de los
hidratos de carbono va siendo menor (Figura 51).
La contribucion de las grasas al aporte energetico puede llegar a ser hasta del 90% en ejercicios de muy larga duracion. La tasa de oxidacion de las grasas se situa normalmente en valores de entre 0.2 Y 0.5 g-rnin', pero se han descrito valores de hasta 1.0-1.5 g-min' despues de 6 horas de carrera. La disponibilidad de glucogeno muscular es uno de los principales determinantes para que, segun progresa la duracion del ejercicio, aun manteniendose una misma intensidad submaxima, las grasas, junta con la glucosa circulante, vayan aumentando su aportacion al gasto energetico total. En la figura 52 se muestran las conclusiones obtenidas en un estudio clasico que demuestra coma al cabo de 2 horas de ejercicio de baja intensidad (30% del vo.max) el

• • • • • •
107 •
•
2,2 2,4 0,2 0,4 o,6 o.s 1,2 1,4 1,6 1,8 2
Concentracion de sustratos (mM·l-1)
Figu_ra 53, Co~ el entrenarniento aumenta la tasa de producci6n de ener fa a ~=r~~lfn~~~ g~1dici't ~e ~ustr~tos_. especialrnents de las grasas. (Modifi~ada h a in · y~ot esis: significance of skeletal muscle oxidative
enzyme en ancement with endurance training. Clin Physiol, 1982; 4.)
0
Preentrenamiento
' #
' ' + ' I
I
~-----+--+--+--+-----+ - +- .. ""T"
;+f .... K Postentrenamiento 0.
~ a.,
"O c: '0 'o u :::,
-0 e c, a.,
"'Cl re V\
~
explicana las diferencias observadas en la oxidaci6n de las AGL en el rmisculo coma consecuencia de una adaptaci6n local al entrenamiento.
En definitiva, en las rmisculos entrenados en actividades de resistencia se produce una adaptaci6n a una utilizacion mas econ6mica de las sustratos energeticos, La capacidad del ciclo de Krebs y de la cadena transportadora de electrones aumenta, lo que perrnite una mayor tasa de resintesis de ATP (Figura
53). Esto se debe fundamentalmente a un aumento de la masa mitocondrial - ' acompanado de un aumento importante de su actividad enzimatica. El efecto es un descenso de la tasa glucogenolitica y glucolitica para una determinada carga de trabajo con un aumento simultaneo de la oxidaci6n de las grasas. Parece que la mayor capacidad de un musculo para oxidar las acidos grasos es la consecuencia de un una mayor utilizaci6n de las trigliceridos intramusculares.
Al aumentar la capacidad de resintesis del ATP, se produce un efecto directo sabre uno de las principales factores reguladores de la glucolisis: el cociente ATP/(ADP + Pi). A medida que va disminuyendo este Indice, se va produciendo una mayor activaci6n de la gluc61isis. Cuando la masa mitocondrial tiene gran capacidad de resintentizar ATP, este cociente disminuye de forma menos
,J1ntera~ci6n de los diferentes sistemas energeticos durante el ejercicio J
EFECTOS DEL ENTRENIIMIENTO El entrenamiento de resistencia aerobica aumenta la capacidad muscular de oxidar grasas durante el ejercicio de intensidad submaxima mejorando la eficiencia energetics de las procesos metabolicos, incluyendo la potencia energetica. Asi, despues de unas semanas de entrenamiento, a una misma velocidad submaxima de carrera (ej.12 km/h) el consumo de grasas aumentara y el de hidratos de carbono descendera, lo que se traduce coma un aumento potencial de rendimiento al retrasar la fatiga asociada a la depleci6n de las niveles de gluc6geno. Sin embargo, durante intensidades de ejercicio realizadas a la misma intensidad relativa (ej. 60% vo.max), existen ciertas discrepancias entre las investigadores a la hara de verificar un mayor consumo de grasas post- entrenamiento. Seguramente gran parte de esas diferencias en las resultados esten motivadas par considerar al %V02max coma un marcador fiable de intensidad relativa, cuando puede reflejar estados metabolicos diferentes.
En cualquier caso, las deportistas de resistencia aerobica poseen mayor capacidad para almacenar trigliceridos en sus rnusculos, mayor actividad de la enzima lipoprotein- lipasa y una capacidad mas elevada para oxidar las lipoproteinas (VLDL) circulantes par la sangre, lo que les permite una mayor utilizacion de las grasas con fines energeticos (aporte de ATP a la celula muscular) a mayores intensidades de ejercicio lo que supone un ahorro de las
dep6sitos de gluc6geno hepatico y muscular. La movilizaci6n de las AGL desde el tejido adiposo va aumentando con la
duraci6n del ejercicio, lo que se refleja en un aumento de la concentraci6n de AGL en plasma. En ejercicios de larga duracion (3h), las sujetos entrenados mantienen e incluso elevan las tasas de captaci6n de AGL par las musculos ejercitante, mientras que las sujetos desentrenados no son capaces de mantener dicha tasa a partir de la tercera hara de ejercicio, a pesar de aumentar la disponibilidad de las mismos. Simultaneamente se produce en este grupo de sujetos una mayor utilizacion de la glucosa coma sustrato energetico. Podemos concluir par el resultado de estos estudios que en las sujetos no entrenados el sistema de captaci6n y utilizacion de las grasas se satura con el tiempo, lo que
l
• • •
•

•
• 3
27%
DIETAS RICAS EN GRASA: En el otro sentido, las dietas ricas en grasas y mas pobres en hidratos de carbono permiten una mayor tasa de utilizacion de acidos grasos desde el principio del ejercicio si bien la aparicion de la fatiga es mas precoz.
Por otra parte, diferentes estudios han mostrado que una dieta rica en grasas mantenida durante mas de 7 dias se asocia a un incremento de la oxidacion de las grasas en intensidades submaximas de ejercicio en detrimento de los hidratos de carbono. Este hecho parece ligado a un descenso de activacion de la piruvato-deshidrogenasa (PDH) quinasa, lo que reduce la produccion de acetil a partir de hidratos de carbono en la mitocondria.
La administraci6n de diferentes dietas ricas en grasas antes del ejercicio tiene distintos efectos metabolicos dependiendo de si estas son saturadas (S) o poliinsaturadas (PI). Las grasas PI ( en concreto AGPI omega 6) son las que
H~s H~s
Figura ~4· Efectos de la dieta preyi~ (tres dfas antes del ejercicio) en la oxidacion de los diferentes sustratos ener_g_et1cos durante un ejercicio (ciclismo al 70% del V02max) de tres horas de duracion. (Modificada de Bosch AN Dennis SC Noakes TD. lnfluenc~ of carbohydrate loading on fuel substrate tur~over and o'xidation
during prolonged exercise. J Appl Physiol, 1993; 74 [4]):1993.
2 3 O 2 0
27% Gluc6geno muscular
46%
76%
- - ... - - - - .. - - 27°/o 7% -----·-·- ... - - .
ill - - .. - ... - - .... 1111 ...
43% Grasas 17% Grasas 18%
Gluc6geno muscular
3 horas de ciciisrno al 70% del VO max con sobrecarga de hidratos de carbono (3 dfas)
3 horns de ciclisrno al 70% del VO max sin sobrecarga de hidratos de car~ono
las reservas musculares de gluc6geno en reposo se asocia con concentraciones sanguineas de AGL mas bajas, lo que indica que hay una menor movilizaci6n de las grasas.
)1nteracci6n de los diferentes sistemas energeticos durante el ejercicio
AUIIIENTO EN LA DISPONIBILIDAD DE GLUCOSA: Cuanto mayor es la cantidad de gluc6geno almacenada, mas tiempo somos capaces de realizar ejercicio sin que aparezca la fatiga, si bien es verdad que al mismo tiempo la tasa de utilizacion de hidratos de carbono es mas elevada desde el principio del ejercicio.
La sobrecarga de hidratos de carbono permite la utilizacion de mayor cantidad de gluc6geno y glucosa (Figura 54). Al elevar las reservas musculares de gluc6geno, se utiliza mas glucosa y se produce mas lactato en comparaci6n con los valores observados a la misma intensidad de ejercicio pero con depositos bajos de gluc6geno muscular. Ademas, se ha observado que una elevaci6n de
CONCENTRACIONES DE SUSTRATOS PREVIAS AL EIERCICIO: EFECTO DE LA DIETA.
Indudablemente, las concentraciones de sustratos disponibles a la hora de realizar ejercicio condicionaran el empleo predominante de alguno de ellos. En lineas generales, las manipulaciones dieteticas van destinadas fundamentalmente a intentar controlar los niveles de gluc6geno muscular y
hepatico disponibles al empezar un ejercicio.
COMPOSICION FIBRILAR DEL MOSCULO. Las fibras rapidas O fibras tipo Ilx presentan unas caracteristicas metabolicas dirigidas a la produccion de energia preferentemente citosolica (anaerobica). Por tanto, en los musculos en los que predominan las fibras tipo II, sera mayor la utilizacion de hidratos de carbono queen los que predominan las fibras oxidativas o tipo I. Ya hemos mencionado que son estas fibras las principales responsables del metabolismo oxidativo, Y
por tanto de la oxidaci6n de las grasas y la glucosa.
acusada, incluso durante ejercicios de intensidad aumentada, por lo que los
estimulos glucoliticos son menores.
Como hemos comentado anteriormente, los dos factores que condicionan la
utilizacion de acidos grasos son la disponibilidad de los mismos y la capacidad
de la celula muscular para oxidarlos. Como la disponibilidad de acidos grasos
supera siempre a las necesidades del organismo de oxidarlos, la mejora en
la combustion de los mismos con el entrenamiento se debe probablemente
a la mayor capacidad de la celula muscular para oxidar los trigliceridos
intramusculares y a una mayor expresion de los transportadores de acidos
grasos al interior de la celula muscular.

•
• 4( .!.!!. , . '
.. •
El "entrenamiento en ayunas" se ha propuesto corno una estrategia mas en la planificaci6n del entrenamiento de resistencia aer6bica. Los principales objetivos de esta actuac!6n _s,erfan la optimizaci6n del metabolismo de los acidos grasos, y una mayor activacion de la proliferaci6n mitocondrial. En la practica se denomina er~trenamiento en ayunas (fasted) a entrenar por la maiiana sin haber ingerido allmentos desde la noche anterior. Ouizas la mayor ventaja observada de entrenar en ayunas sea la mejora de la capacidad oxidativa muscular, aunque no esta nada claro que se mejore la oxidaci6n de los acidos grasos con fines energeticos. Esta ventaja bioqufmica parece darse mas en hombres que en mujeres. Parece que hay una adaptaci6n asociada a esta modalidad de entrenamiento consistente en la mayor utilizaci6n de los trigliceridos intramusculares, asi coma un desplazamiento significativo del FATmax. Adernas, al entrenar con baja disponibilidad de gluc6geno muscular parece que se potencian las adaptaciones mticocondriales a traves de una activaci6n de la sefializacion molecular (PGC-1a) para la biogenesis
· NIVELES BAJOS DE GLUC6GE O ------------- ,------ ------
Continua siendo una controversia la existencia de diferencias significativas en la utilizacion relativa de hidratos de carbono y grasas durante el ejercicio entre sexos. Cada vez hay mas estudios que demuestran que la contribucion de la energia procedente de la oxidacion de las grasas es mayor en las mujeres, si bien no todos los autores estan de acuerdo. En un estudio realizado por Venables Y col. en 2005, en 300 hombres y mujeres, entrenados y no entrenados, se
Mientras los depositos de glucogeno hepatico permitan por un lado reponer la glucemia y por otro, abastecer al musculo con glucosa, nuestro musculo podra seguir utilizando hidratos de carbono, pero cuando el gluc6geno hepatico se agota, la posibilidad de mantener la glucemia sin aporte extrinseco de glucosa desaparece y se produce la situacion de hipoglucemia. Esta situacion clinica se caracteriza por un cuadro sintomatico bastante tipico (malestar general, incoordinacion, perdida de la capacidad de concentracion, disminucion del rendimiento muy acusada, diplopia o vision borrosa, perdida de conciencia y, si progresa, situacion de coma). Silos depositos de gluc6geno hepatico y muscular eran adecuados antes de comenzar el ejercicio, este cuadro puede aparecer al cabo de 3-4 horas de ejercicio intenso (si nose produce la adrninistracion de glucosa durante el ejercicio ).
,IInteraccion de los diferentes sistemas energeticos durante el ejercicio J
Cuando se realiza ejercicio de intensidad submaxima durante un tiempo prolongado (> 90 min) sin ingerir hidratos de carbono, los niveles de glucemia pueden disminuir como reflejo de un descenso en la disponibilidad de gluc6geno de los depositos hepatico y muscular. En ese escenario, el ejercicio solo podra continuar si se produce un aumento de oxidacion de los acidos grasos libres que supla la necesidad energetica de las celulas musculares para una intensidad de ejercicio determinada; sin embargo, la capacidad de generar energia por media de los acidos grasos siempre va a ser menor en terminos de potencia energetica ( energia por unidad de tiempo) que la obtenida por los hidratos de carbono, lo que se traducira inevitablemente con un descenso de la intensidad de ejercicio,
es decir, en fatiga muscular.
DI PONI ILID D INNlliDIAT u
... 110
•
permiten un mayor ahorro de los hidratos de carbono, debido a la inhibicion
de la PDH. Esto se acompafia de un descenso de los AGL circulantes tanto en
reposo como durante el ejercicio, asi como de una mayor tasa de oxidacion de
las grasas. Estos resultados indican que el musculo esqueletico oxida mas grasas
cuando previamente se han administrado AGPI que si se administran AGS,
debido en parte al papel regulador de la PDH cuya actividad se ve claramente
atenuada en esta circunstancia.
Por el contrario, si la manipulacion dietetica consiste en la administracion
de una dieta escasa en grasas con abundantes hidratos de carbono, se observa
una reduccion de la oxidacion de grasa durante el ejercicio gracias a una
disminucion de los procesos lipoliticos que se asocia con una reduccion de los
depositos de TG intracelulares.
La manipulacion cronica de la cornposicion de la dieta (hidratos de carbono/
grasas) parece ten er su ef ecto mas importante sob re la oxidacion de sustratos
en el ejercicio mediante la modificacion de los depositos intramusculares
(trigliceridos y gluc6geno). Segun Coyle y cols. (2001) las dietas bajas en grasa a largo plaza pueden estimular la lipogenesis en deportistas entrenados, lo cual podria compensar la reduccion del a po rte de grasas de la dieta y restaurar, al menos parcialmente, los trigliceridos intramusculares.

•
• • 113 ••
•
La figura 56 representa de forma grafica la aportaci6n de los diferentes sistemas energeticos en funci6n de la duracion de diferentes tipos de esfuerzos maxirnos. Podemos observar en un extrema que los fosfagenos intramusculares de alta energia aportan casi toda la energia necesaria para el ejercicio. La fosfocreatina puede llegar a aportar un 15% de la energia durante la realizacion de series de corta duracion (30 segundos) y elevada intensidad
Figura 55, Contribuci6n relativa de l?s.fuentes de energfa aer6bicas y anaer6bicas durante ejercrcros maxrmos de duraci6n diferente.
2 4 10 30 60 120 •••••• ••••••
10 30 60 2 4 10 30 60 120 Segundos Minutos
Tiempo de ejercicio rnaxirno
Segundos Minutos
100 re -,:; re .µ
0 So a. re ,!!! e..o Q) c: 60 <lJ
..<:!1 <lJ -,:; <lJ
"iij' 40 .... c: <lJ u .... 0
Cl.
20
120 -,---i--------i---,---,-----.--.----r----..--~
En la figura 55 se ilustra la contribucion relativa de las fuentes energeticas citos61icas -anaerootcas- (alactica y lactica) y aerobicas en relacion con la duracion de ejercicios maximos.
________ lr_n_teracci6n de los diferentes sistemas energeticos durante el ejercicioJ - --........--
Destacaremos de todos ellos la temperatura. Se ha demostrado que la utilizacion de gluc6geno aumenta cuando se realiza ejercicio en ambientes muy calurosos. El aumento de la temperatura central del organismo produce un aumento de la oxidacion de hidratos de carbono con un descenso concomitante de la oxidacion de grasas. Los facto res a los que se atribuye este ef ecto metabolico son el aumento de la temperatura muscular y de la concentracion plasmatica de adrenalina, factores que activan la glic61isis. Por otra parte, la utilizacion de sustratos durante la exposici6n al frio produce un aumento de la contribucion de los hidratos de carbono al aporte energetico, especialmente si se tirita. Tiritar provoca la salida de glucosa hepatica y un aumento de la tasa de utilizacion de glucosa por la celula muscular.
•
• • • ill • •
•
observ6 que la utilizaci6n de sustratos energeticos durante un ejercicio de
intensidad creciente hasta el agotamiento, las mujeres presentaron mayores
tasas de oxidaci6n de las grasas, y que el aumento de la oxidacion de los
hidratos de carbono se producia a una mayor intensidad relativa. Otros estudios
confirman que la oxidacion total de grasas es la misma en las mujeres que en
los varones en ejercicios de resistencia de intensidad moderada a pesar de que
las mujeres oxidan mas acidos grasos procedentes del plasma que los varones,
lo que indica que probablemente la lipolisis del tejido adiposo sea mayor. Sin
embargo, hay estudios en los que se concluye que las mujeres tienen mayores
dep6sitos de trigliceridos intramusculares asi como una mayor actividad de las
enzimas implicadas en el metabolismo de las grasas.
Los estr6genos parecen predisponer a las mujeres a utilizar las grasas de
forma mas eficiente con fines energeticos mediante la activaci6n de la AMPK.
Algunos autores consideran que esto supone una ventaja metabolica para la
mujer durante actividades de intensidad submaxima y duracion prolongada, en
los que la fuerza muscular no esta tan ligada al rendimiento.
El papel otorgado a los estr6genos ha llevado a proponer posibles dif erencias
en relacion al rendimiento en la mujer dependiendo de la fase del ciclo menstrual en la que compita. El rendimiento al final de la fase folicular (niveles elevados de estr6genos y basales de progesterona) podria mejorar potencialmente por los ef ectos metabolicos asociados.

Figura 56. Contribuci6n de los diferentes sistemas energeticos.
Duraclon del ejercicio maxlrno
30 s 10 s 5 min 1 min 2 min •
• • •, 114 • •
Sistema aer6bico
~ -:f2. 0
VI~ O VI -o a, u "'O ·.p c: ,a,
'0 i:_Jl ·o w ::::, c: .0 a, 'i:: V1 .... "' CE 8~
VI 'vi
•
100%
Sistema de fosfagenos
en las que otro 15% es proporcionado por la glucolisis citosolica y el resto por el metabolismo aerobico, Los sistemas anaerobicos (ATP- PCr y glucolisis anaerobica, aportan aproximadamente el 50% de la energia en eventos que duran aproximadamente 2 minutos; en el resto, predominan las reacciones aerobicas con una participacion mayor segun va disminuyendo la intensidad del ejercicio realizado.

Bosch AN, Dennis SC, Noakes TD. (1993)
Influence of carbohydrate loading on
Bonen, A. (2001). The expression of
lactate transporters (MCT1 and MCT4)
in heart and muscle. Eur J Appl Physiol,
86, 6-11.
Bonen A, Belcastro AN. (1976 ).
Comparison of self-selected recovery
methods on lactic acid removal rates.
Med Sci Sports Exerc, 8, 176-178.
Boileau RA, Misner JE, Dykstra JE y col.
(1983). Blood lactic acid removal during
treadmill and bicycle exercise at various
intensities. J Sports Med Phys Fitness, 2,
159-167.
Blomstrand E, Saltin B. (1999). Effect
of muscle glycogen on glucose, lactate
and amino acid metabolism during
exercise and recovery in human subjects.
J Physiol, 514, 293-302.
Barclay CJ, Woledge RC, Curin NA.
(2007). Energy turnover for Ca2+ cycling
in skeletal muscle. J Muscle Res Cell Motil,
28, 259-274.
I Bibliografia
Bangsbo J, Gollnick PD, Graham TE,
Saltin B. (1991). Substrates for muscle
glycogen sintesis in recovery from
intense exercise in man. J Physiol,
434,423-440.
Astrand PO, Hultman E, Juhlin-Dannfelt
A, Reynolds GG. (1986). Disposal of
lactate during and after strenuous
exercise in humans. J Appl Physiol, 61,
338-343.
Ahlborg G, Felig P, Hagenfeldt L,
Hendler R, Wahren J. (1974). Sustrate
turnover during prolonged exercise in
man: Splachnic and leg metabolism of
glucose, free fatty acids and amino acids.
J Clin Invest, 53, 1080-1090.
Adijanto J, & Philp NJ. (2012). The
SLC16A Family of Monocarboxylate
Transporters. Co-Transport Systems, 70,
275.
Achten J, Gleeson M, Jeukendrup AE.
(2002). Determination of the exercise
intensity that elicits maximal fat
oxidation. Med Sci Sports Exerc, 34, 92-9.
_____________________________ I Bibliografia I r---

Greenhaff PL, Hultman E, Harris RC.
(1993) Carbohydrate metabolism. En:
Poortmans JR (ed): Principles of Exercise
Gladden LB. (1996 ). Lactate transport
and exchange during exercise. En: Rowell
LB, Shepherd JT, editors. Handbook
of Physiology, Exercise: Regulation and
Integration of Multiple Systems. pp.
614-648. section 12.NewYork: Oxford
University Press.
Gerber T, Borg ML, Hayes A, Stathis
CG. (2014). High-intensity intermittent
cycling increases purine loss compared
with workload-matched continuous
moderate intensity cycling. Eur J Appl
Physiol, 114(7),1513-20.
Gaesser GA, Brooks GA.(1984). Metabolic
bases of excess post exercise oxygen
consumption: A review. Med Sci Sports
Exerc, 16,29-43.
Freund H, Zouloumian P. (1981a). Lactate
after exercise in man: Evolution kinetics
in arterial blood. Eur J Appl Physiol Occup
Physiol. 46, 121-133.
TF, Meyer RA. (2008). Phosphocreatine
recovery kinetics following low- and
high-intensity exercise in human triceps
surae and rat posterior hindlimb muscles.
Am] Physiol, Regul Integr Compr Physiol
296 (1), R161-R170.
Forbes SC, Paganini AT, Slade JM, Towse
Decombaz J, Reindhardt P, Anantharaman
K, Glutz GV, Poortmans JR. (1979).
Biochemical changes in a 100 km run:
Free amino acids, urea and creatinine. Eur
J Appl Physiol, 41, 61-72.
Emhoff, CAW, Messonnier, LA, Horning
MA, Fattor JA, Carlson TJ, Brooks GA.
(2013). Gluconeogenesis and hepatic
glycogenolysis during exercise at the
lactate threshold. J Appl Physiol, 114(3),
297-306.
Coyle EF, Jeukendrup AE, Oseto
MC, Hodgkinson BJ, Zderic TW. (2001).
Low-fat diet alters intramuscular
substrates and reduces lipolysis and fat
oxidation during exercise. Am] Physiol -
Endocrinol Me tab. 280(3), E391- E398
Emhoff CAW, Messonnier LA, Horning
MA, Fattor JA, Carlson TJ, Brooks GA.
(2013a). Direct and indirect lactate
oxidation in trained and untrained men. J
Appl Physiol, 115, 829-838.
Dood S, Powers SK, Callender T, Brooks
E. (1984). Blood lactate disappearance at
various intensities of recovery exercise. J
Appl Physiol, 57, 1462-1465.
Chwalbinska-Moneta J, Robergs
RA, Costill DL, FinkWJ. (1989) Threshold
for muscle lactate accumulation during
progressive exercise. J Appl Physiol,
66(6),2710-6.
Dolny DG, Lemon PWR. (1988). Effect
of ambient temperature on protein
breakdown during prolonged exercise. J
Appl Physiol, 64, 550-555.
Chamari K, Padula J. (2015). "Aerobic"
and "anaerobic" term used in exercise
physiology: a critical terminology
reflection. Sports Medicine Open, 1, 9.
Doege H, Stahl A. Protein-Mediated Fatty
Acid Uptake: Novel Insights from In Vivo
Models. (2006). Physiology, 21(4), 259-
268.
Caset A, Greenhaff PL. (2000). Does
dietary creatine supplementation play a
role in skeletal muscle metabolism and
performance? Am] Clin Nutr, 2000, 72,
S607-S617-
Djurhuus CB, Gravholt CH, Nielsen S,
mengel A, Christiansen JS, Schmitz OE,
Moller N. (2002). Effects of cortisol on
lipolisis and regional interstitial glycerol
levels in humans. Am] Physiol Endocrinol
Metab, 283:E, 172-177.
JA,Carey AL. (2010). Short-term
endurance training does not alter the
oxidative capacity of human subcutaneous
adipose tissue. Eur J Appl Physiol, 109(2),
307-316.
I Bibliografia
Camera DM, Anderson MJ, Hawley
Brooks, GA. (1985). Lactate: glycolytic
product and oxidative substrate during
sustained exercise in mammals -the
'lactate shuttle'; en Gilles R (ed):
Comparative Physiology and biochemistry;
Current Topics and Trends. Volume A: p
459-460. Respiration. Metabolism-
Circulation. Berlin, Springer, 1985,
Brooks GA. (2002). Lactate shuttles in
nature. Biochemical Society Transactions,
30, 258-263.
Brooks GA, Brown MA, Butz CE, Sicurella
JP, Dubouchaud H. (1999). Cardiac and
skeletal muscle mitochondria have a
monocarboxylate transporter MCTl. J Appl
Physiol, 87,1713-1718.
Brooks GA. (1997). Importance of
the 'crossover' concept in exercise
metabolism. Clinical and Exp Pharmacol
physiol, 24, 889-895.
Bradley NS, Heigenhauser GJ, Roy
BD, Staples EM, Inglis JG, LeBlanc
PJ, Peters SJ. (2008). The acute effects of
differential dietary fatty acids on human
skeletal muscle pyruvate dehydrogenase
activity. J Appl Physiol.104, 1-9.
fuel substrate turnover and oxidation
during prolonged exercise. J Appl Physiol,
74(4),1921-7.

Pagliassotti MJ, Donovan CM. (1990)
Glycogenesis from lactate in rabbit
Mittendorfer B, Horowitz JF, Klein S.
(2002). Effect of gender on lipid kinetics
during endurance exercise of moderate
intensity in untrained subjects. Am J
Physiol Endocrinol Metab, 283, E58-E65.
Messonnier LA, Emhoff Ch W, Fattor
JA, Horning MA, Carlson TJ, Brooks GA.
(2013) Lactate kinetics at the lactate
threshold in trained and untrained men. J
Appl Physiol, 114: 1593-1602.
Menzies P, Menzies C, Mcintyre Ly col.
(2010) Blood lactate clearance during
active recovery after an intense running
bout depends on the intensity of the
active recovery. J Sports Sci, 28: 975-982.
Melanson EL, MacLean PS, Hill JO. (2009)
Exercise improves fat metabolism in
muscle but does not increase 24-h fat
oxidation. Exerc Sport Sci Review 37, 93-101.
McLane JA, Holloszy JO. (1979). Glycogen
synthesis from lactate in the three types
of skeletal muscle.] Biol Chem, 254, 6548-
6553.
Tipton KF, O'Brien M, Bonen A. (1997).
Chronic electrical stimulation increases
MCTl and lactate uptake in red and
white skeletal muscle. A] Physiol.273,
E239-E246.
I Bibliografia
Mccullagh KJA, Poole RC, Halestrap AP, Knechtle B, Muller G, Willmann F, Kotteck
Lemon, PWR, Mullin JP. (1980) Effect of
initial muscle glycogen levels on protein
catabolism during exercise. J Appl Physiol,
48,624-629.
Juel C. Halestrap AP. (1999) Lactate
transport in skeletal muscle-role and
regulation of the monocarboxylate
transporter. J Physiol. 517, 633-642.
Lemon, PWR. (1987) Protein and exercise:
update. Med Sci Sports Exerc, 19,S179-190.
Juel C. (1997) Lactate-proton cotransport
in skeletal muscle. Physiol Rev, 77 (2),
321-358.
Laurent D, Hundal RS, Dresner A, Price
TB, Vogel SM, Petersen KF, Shulman
GI.(2000) Mechanism of muscle glycogen
autoregulation in humans. Am J Physiol
Endocrinol Metab, 278(4) E663-8.
Juel C. (2001) Current aspects of lactate
exchange: Lactate/H+ transport in human
skeletal muscle. Eur J Appl Physiol, 86,
12-16.
Jeukendrup AE. (2003). Modulation of
carbohydrate and fat utilization by diet,
exercise and environment. Biochemical
Society Transactions, 31: 1270-1273.
Lamont LS, McCullough AJ, Kalhan SC.
(2001). Relationship between leucine
oxidation and oxygen consumption during
steady-state exercise. Med Sci Sports Exerc,
33: 237-241.
Jeukendrup AE, Achten J. Fatmax : A new
concept to optimize fat oxidation during
exercise? (2001) Eur J Sport Sci, (5), 1-5.
Krebs HA. (1963) Glyconeogenesis.
Croonian lecture. Proc R Soc Lond [Bl
159, 545-560.
Jeppesen J, Kiens B. Regulation and
limitations to fatty acid oxidation during
exercise. (2012) J Physiol, 590(5), 1059-
1068.
Kraniou Y, Cameron-Smith D, Misso
M, Collier G, and Hargreaves M. (2000)
Effects of exercise on GLUT-4 and
glycogenin gene expression in human
skeletal muscle. Appl Physiol, 88, 794-796.
Horton TJ, Pagliassotti MJ, Hobbs K, Hill
JO. (1998) Fuel metabolism in men and
women during and after long-duration
exercise. J Appl Physiol, 85,1823-1832.
K, Eser P, Knecht H. (2004) Fat oxidation
in men and women endurance athletes in
running and cycling. Int] Sports Med, 25,
38-44.
fat oxidation during exercise. Am J Physiol
Endocrinol Metab. 273(4), E768-E775.
Horowitz JF, Mora-Rodriguez R, Byerley
LO, Coyle EF. (1997) Lipolytic suppression
following carbohydrate ingestion limits
Hogan MC, Gladden LB, Grassi B, Stary
CM, Samaja M. (1988). Bioenergetics
of contracting skeletal muscle after
partial reduction of blood flow. J Appl
Physiol, 84( 6 ), 1882-1888.
Hermansen L, Vaage 0. (1977) Lactate
disappearance and glycogen synthesis in
human muscle after maximal exercise. Am
J Physiol, 233,E422-E429.
Hermansen L, Stensvold I. (1972)
Production and removal of lactate during
exercise in man. Acta Physiol Scand, 86,
191-201.
Haralambie G, Berg A. (1976) Serum urea
and amino nitrogen changes with exercise
duration. Eur J Appl Physiol, 36,39-48.
Hamann JJ, Kelley KM, Gladden LB.
(2001) Effect of epinephrine on net lactate
uptake by contracting skeletal muscle. J
Appl Physiol, 91, 635-2641.
Halestrap AP, Wilson MC. (2012) The
Monocarboxylate Transporter Family-
Role and Regulation. IUBMB Life, 64(2),
109-119.
Biochemistry, zved. Vol 38 pp 89-136. Med
Sport Sci. Basel, Karger.
l

Thomas C, Bishop DJ, Lambert K, Mercier
J, Brooks GA (2012). Effects of acute and
J Appl Physiol and occupational
physiology, 71(5), 396-404.
human quadriceps muscles. European
exercise in trained and untrained
of phosphocreatine resynthesis after
& Itai, Y. (1995). Control of the rate
K., Katsuta, S., Anno, I., Nutsu, M.,
Takahashi, H., Inaki, M., Fujimoto,
Steffensen CH, Roepstorff C, Madsen
M, Kiens B. (2002). Myocellular
triacylglycerol breakdown in females but
not in males during exercise. Am J Physiol
Endocrinol Metab, 282, E634-E642.
.44 (Sl), 87-96.
'',.--, ~
123 • • -, ---,
Spriet LL. (2041) New insights into the
interaction of carbohydrate and fat
metabolism during exercise. Sports Med
exercise. J Appl Physiol, 85(4), 1349-1356.
Smith SA, Montain SJ, Ralph P, Zientara
MGP, Jolesz FA, Fielding RA. (1998)
Creatine supplementation and age
influence muscle metabolism during
Chem,279 (25), 26215-26219.
Shieh JJ, Pan CJ, Mansfield BC, Chou JY.
(2004) A potential new role for muscle
in blood glucose homeostasis. J Biol
exercising human skeletal muscle. J Appl
Physiol, 95(6), 2328-32.
content is increased after exercise in non-
Kooi ME. (2003) Intramyocellular lipid urine during and after prolonged heavy
exercise. ScandJ Clin Lab Invest, 35,775-780.
LJ, Koopman R, Nicolay K, Saris WH, creatinine production and excretion in
Schrauwen-Hinderling VB, Van Loon Refsum HE, Str6tarne SB. (1974) Urea and
muscle. J Physiol, 541, 569-574. 541, 575-580.
mouse vastus lateralis muscle. J Physiol,
Tonkonogi M. (2002) No evidence of an
intracellular lactate shuttle in rat skeletal
a mitochrondial enzyme in human and
Sahlin K, Fernstrom M. Svensson M. UF. (2002) Lactate dehydrogenase is not
adults. Eur J Appl Physiol, 111, 2063-8. Rasmussen HN, van Hall G, Rasmussen
maximal fat oxidation rate in untrained 386-392.
resistance exercise. J Appl Physiol, 88,
enhances muscle protein anabolism after
Rynders CA, Angadi SS, Weltman
NY, Gaesser GA, Weltman A (2011)
Oxygen uptake and ratings of perceived
exertion at the lactate threshold and
amino acid-carbohydrate supplement
E435-E447. Rasmussen BB, Tipton KD, Miller SL, Wolf
SE, Wolfe RR. (2000) An oral essential
Am J Physiol Endocrinol Me tab, 282, 5(2),53-64.
sedentary males. Jentashapir J Health Res,
substrate utilization during submaximal
exercise in endurance-trained subjects.
Comparison between FATmax and
maximal fat oxidation in the active and
Rami M, Habibi AH, Shakerian S. (2014)
Roepstorff C, Steffensen CH, Madsen M,
Stallknecht B, Kanstrup IL, Richter EA,
Kiens B. (2002) Gender differences in
European J Appl Physiol, 113(4), 951-963.
M., Sahlin, K. (2013) Exercise with
low glycogen increases PGC-la gene
expression in human skeletal muscle.
Ro bergs RA, Ghiasvand F, Parker D.
(2004) Biochemistry of exercise-induced
metabolic acidosis. Am J Physiol Regul
Integr Comp Physiol 287, R502- R516.
Psilander N, Frank, P., Flockhart,
639. Miami, Symposia Specialists,
protein turnover in man. Clin Sci, 61,627-
W (eds): En Regulatory Mechanisms in
Metabolism during Exercise. pp 159 184.
Krywawych S, Halliday D, Waterlow JC,
Millward DJ. (1981) Effect of exercise on
during exercise; in Landry F, Orban
Rennie MJ, Edwards RHT, Davies M, Poortmans JR. (1978) Protein turnover
I Bibliograffa j
Birkhliuser.
lasting exercise and training on protein
metabolism; in Howald IL Poortmans JR
(eds): Metabolic Adaptation to Prolonged
Physical Excreise. pp 212-228. Basel,
Poortmans JR. (1975) Effects of long-
Renal implications of exogenous creatine
monohydrate supplementation. Am] Med
Sports, 4,212-216.
Poortmans JR, Francaux M. (2002).
pp 87-107. Med Sport Sci. Basel, Karger,
exercise biochemistry, 3rd, ed rev. Vol 46,
Poortmans JR (ed): (2004) Principles of
Physiol, 113(4), 951-963.
human skeletal muscle. European J Appl
increases PGC-la gene expression in
Psilander N, Frank P, Flockhart M, Sahlin
K. (2013) Exercise with low glycogen
Pilegaard H, Domino K, Noland T, Juel
C, Hellsten Y, Halestrap AP, Bangsbo J.
(1999) Effect of high-intensity exercise
training on lactate/H+ transport capacity
in human skeletal muscle. Am. J. Physiol. -
Endocrinol. Metab. 276, E255-E261.
glycogen resynthesis after short term,
high intensity exercise and resistance
exercise. Sports Med, 21, 98-118.
Pascoe DD, Gladden LB. (1996) Muscle
258, R903-R911.
skeletal muscle fiber types. Am J Physiol,

elevates resting intramuscular triglyceride
concentration and whole body lipolysis
during exercise. Am J Physiol Endocrinol
Metab. 286, E217-25.
Zderic TW, Davidson CJ, Schenk S, Byerley
LO, Coyle EF. (2004) High-fat diet
Biol. 441:249-261.
mechanisms regulating fatty acid
oxidation during exercise. Adv Exp Med
Winder WW. (1998) Intramuscular
Watt MJ, Howlett KF, Febbraio MA, Spriet
LL, Hargreaves M. (2001) Adrenaline
Am] Physiol, 260, E883-890.
supplementation, glycogen depletion, and
amino acid metabolism during exercise.
GJ, Saris WHM. (1991) Carbohydrate
Wagenmakers AJM, Beckers EJ, Brouns
F, Kuipers H, Soeters PB, Van der Vusse
performance in athletes. Med Sci Sports
Exerc. 2003 Jun, 35(6),952-60.
fat on muscle substrates, metabolism, and
Vogt M, Puntschart A, Howald H, Mueller
B, Mannhart C, Gfeller-Tuescher L, Mullis
P, Hoppeler H. (2003) Effects of dietary
Physiol, 98, 160-7.
Venables MC, Achten J, Jeukendrup AE.
(2005) Determinants of fat oxidation
during exercise in healthy men and
women: a cross-sectional study. J Appl
Vandenberghe K, Van Hecke P, Van
Leemputte M, Vanstapel F, Hespel P.
(1999) Phosphocreatine resynthesis is not
affected by creatine loading. Med Sci Sports
Exerc. 31(2),236-42.
Wilkerson JE, Batterton DL, Horvath SM.
(1997) Exercise induced changes in blood
ammonia levels in humans. Eur J Appl
Physiol, 37, 255-263.
The Netherlands.
278. Thesis, University of Limburg, Maastricht,
ammonia and exercise in man. PhD
Van Hall, G. (1996). Amino acids,
Acta Physiologica, 199: 499-508.
increases skeletal muscle glycogenolysis
pyruvate dehydrogenase activation and
carbohydrate oxidation during moderate
exercise in humans. J Physiol, 534, 269-
human tissues at rest and during exercise.
Van Hall G. (2010) Lactate kinetics in
Physiol. 262(6 Pt 1):E791-9.
Turcotte LP, Richter EA, Kiens B. (1992)
Increased plasma FF A uptake and
oxidation during prolonged exercise
in trained vs. untrained humans. Am J
Trump ME, Heigenhauser GJ, Putman
CT, Spriet LL. (1996). Importance
of muscle phosphocreatine during
intermittent maximal cycling. J Appl
Physiol, 80 (5), 1574-1580.
Am] Physiol, 281,E197-E206.
response of muscle to resistance exercise.
Tipton KD, Rasmussen BB, Miller SL,
Wolf SE, Owens-Stovall SK, Petrini BE,
Wolfe RR. (2001) Timing of amino acid-
carbohydrate ingestion alters anabolic
J Physiol, 276, E628-E634.
net protein synthesis in human muscle
from orally administered amino acids. Am
Tipton KD, Ferrando AA, Philips SM,
Doyle DJ, Wolfe RR. (1999). Postexercise
Champaign.
science 4a ed. Ed. Human Kinetics,
Tiidus PM, Tupling AR, Houston ME.
(2012) Biochemical primer for exercise
Int Comp Physiol, 302(1), Rt - R14.
and MCT 4 contents in human skeletal
muscles: current status. Am J Physiol Reg
chronic exercise on sarcolemmal MCTl I
J Bibliografia


















