
APORTACIONES AL CONOCIMIENTO DE LA NUTRICIÓN · 2015-03-24 · Archivos de Zootecnia ISSN:...
33
Archivos de Zootecnia ISSN: 0004-0592 [email protected] Universidad de Córdoba España Aguilera, J.F. Aportaciones al conocimiento de la nutrición energética de pequeños rumiantes, con particular referencia al ganado caprino Archivos de Zootecnia, vol. 50, núm. 192, 2001, pp. 565-596 Universidad de Córdoba Córdoba, España Disponible en: http://www.redalyc.org/articulo.oa?id=49519208 Cómo citar el artículo Número completo Más información del artículo Página de la revista en redalyc.org Sistema de Información Científica Red de Revistas Científicas de América Latina, el Caribe, España y Portugal Proyecto académico sin fines de lucro, desarrollado bajo la iniciativa de acceso abierto
Transcript of APORTACIONES AL CONOCIMIENTO DE LA NUTRICIÓN · 2015-03-24 · Archivos de Zootecnia ISSN:...

Archivos de Zootecnia
ISSN: 0004-0592
Universidad de Córdoba
España
Aguilera, J.F.
Aportaciones al conocimiento de la nutrición energética de pequeños rumiantes, con particular
referencia al ganado caprino
Archivos de Zootecnia, vol. 50, núm. 192, 2001, pp. 565-596
Universidad de Córdoba
Córdoba, España
Disponible en: http://www.redalyc.org/articulo.oa?id=49519208
Cómo citar el artículo
Número completo
Más información del artículo
Página de la revista en redalyc.org
Sistema de Información Científica
Red de Revistas Científicas de América Latina, el Caribe, España y Portugal
Proyecto académico sin fines de lucro, desarrollado bajo la iniciativa de acceso abierto

Arch. Zootec. 50: 565-596. 2001.
APORTACIONES AL CONOCIMIENTO DE LA NUTRICIÓNENERGÉTICA DE PEQUEÑOS RUMIANTES, CON PARTICULAR
REFERENCIA AL GANADO CAPRINO
CONTRIBUTIONS TO THE KNOWLEDGE OF THE ENERGY NUTRITION OF SMALLRUMINANTS, WITH PARTICULAR REFERENCE TO THE GOAT
Aguilera, J.F.
Unidad de Nutrición Animal. Estación Experimental del Zaidín (CSIC). Camino del Jueves s/n. 18100
Armilla. Granada. España. E-mail: [email protected]
PALABRAS CLAVE ADICIONALES
Valoración energética. Metano. Necesidadesenergéticas. Mantenimiento. Producción. Activi-dad física. Técnica CER.
ADDITIONAL KEYWORDS
Energy evaluation. Methane. Energy requi-rements. Maintenance. Production. Physicalactivity. CER technique.
RESUMEN
El artículo describe aportaciones notablesdel departamento de Nutrición Animal de la Esta-ción Experimental del Zaidín del Consejo Supe-rior de Investigaciones Científicas a la valoraciónenergética de los alimentos en pequeños rumian-tes y al conocimiento de las necesidades energé-ticas del ganado caprino.
Distintos trabajos sugieren la inexistencia dediferencias interespecíficas entre ovino y caprinoen la digestibilidad de nutrientes de forrajes ydietas mixtas de calidad nutritiva media o alta.Cuando se les alimenta con dietas de composi-ción equilibrada, ovino y caprino son mutuamen-te reemplazables como donadores de inóculopara estudios de digestibilidad o degradabilidadin vitro. Nuestras observaciones permiten afirmarque es correcto asignar a un mismo alimento omezcla de alimentos un único valor energéticoaplicable indistintamente a ovino o caprino. No esposible aún establecer conclusión alguna res-pecto a dietas desequilibradas o de muy bajacalidad.
Se exponen los resultados de pruebasmetabólicas realizadas para determinar las nece-sidades energéticas del ganado caprino en situa-ciones fisiológicas diversas. Mantenimiento y pro-ducción son objeto de consideración especial. Seestablecen los costes energéticos de la actividadmuscular relacionada con la locomoción y laingestión de alimento y se describen los ensayosllevados a cabo para estimar el gasto energéticototal del ganado caprino en pastoreo. Se analizanlos resultados alcanzados con la aplicación de latécnica CER mediante la infusión de bicarbonatomarcado con 13C y su validación para la estima-ción de la producción de calor del animal enpastoreo.
SUMMARY
This paper describes significant contributionsof the department of Animal Nutrition of EstaciónExperimental del Zaidín of the Spanish Council of

Archivos de zootecnia vol. 50, núm. 192, p. 566.
AGUILERA
Scientific Research (CSIC) to the energy evaluationof feeds for small ruminants and to the assessmentof the energy requirements of the goat.
Several observations suggest that there is noevidence of differences between sheep and goatsin nutrient digestibility for good- or medium quality-forages or feed mixtures. When fed on balanceddiets these two species are mutually exchangeableas inoculum donors for in vitro digestibility ordegradability studies. Our observations allow usto indicate the correctness of assigning a singleenergy value to a particular, given feed or feedmixture to be applied either in sheep or goat.
Experimental results are given from metabolictrials which were designed to assess the energyrequirement of the goat under several physio-logical situations. Special emphasis is given tomaintenance and production. The energy costs ofmuscular activity involved in locomotion and foodingestion are stated and a account is given oftrials carried out to estimate total energy expensesin the grazing goat. An analysis is made of dataobtained by using the so-called CER techniqueand 13C-bicarbonate and its validation to estimatethe total heat production for the unrestricted goat.
INTRODUCCIÓN
Cincuenta años de edición de unarevista científica, cualquiera que sea elcampo de la Ciencia y Tecnología quele corresponda, es un periodo extraor-dinariamente amplio, en el que el rit-mo vertiginoso de adquisición de nue-vos conocimientos que ha caracteriza-do a la época ha tenido que producircambios conceptuales necesariamenteimportantes. Qué duda cabe que Ar-chivos de Zootecnia puede dar testi-monio riguroso de la extraordinariaevolución que se ha producido en elámbito de las especialidades que leconciernen, adscritas al campo cientí-fico de la Producción Animal. En lo
que respecta a la Nutrición Animal, losconocimientos sobre digestión y meta-bolismo de los animales de interészootécnico y, en particular, de los ru-miantes han evolucionado notablemen-te, lo que ha obligado a modificarmuchos de los conceptos preexistentes.En tanto la mayoría de los países de laUnión Europea han incorporado estosavances a la metodología en uso parala valoración nutritiva de sus alimen-tos y materias primas y actualizado sussistemas de alimentación, España (jun-to a Portugal y Grecia) no ha consegui-do extraer de su importante produc-ción científica la información integra-da que pueda incidir favorablementeen el sistema productivo, con propues-tas útiles al sector sobre sistemas devaloración y recomendaciones nutriti-vas con las que mejorar la práctica dela alimentación en nuestras razasautóctonas y favorecer la utilizaciónde recursos propios. Almacenar laamplia información generada para quepueda ser analizada y ayude a la tomade decisiones es, pues, una tarea im-prescindible sobre la que existen algu-nas iniciativas que es necesario impul-sar. Mucha información relevante so-bre pruebas de alimentación o valora-ción nutritiva procedente de ensayosllevados a cabo en centros públicos deinvestigación nacionales ha sido pu-blicada en Archivos de Zootecnia que,consiguientemente, puede contribuirnotablemente a esta tarea.
Soy consciente de las dificultadesque Archivos de Zootecnia ha encon-trado en el desempeño de su actividady de la decisión encomiable de la Di-rección y el Consejo de redacción encuanto a la exigencia de la calidadcientífica como requisito ineludible

Archivos de zootecnia vol. 50, núm. 192, p. 567.
NUTRICIÓN ENERGÉTICA DE PEQUEÑOS RUMIANTES
para la publicación en sus páginas delos resultados de la investigación. Con-cedo especial importancia a la labor dedifusión de la actividad científica querealiza tanto en el ámbito nacionalcomo en el internacional, especialmen-te valiosa la relacionada con los paísescon quienes compartimos un idiomacomún. Finalmente, quiero subrayar laimportantísima contribución que Ar-chivos de Zootecnia presta a mantenerviva la literatura científica relaciona-da con el área de la Producción Animalen el idioma español, seriamente ame-nazada por la supremacía del ingléscomo idioma de uso común por lacomunidad científica.
Sólo unos años después de que vie-se la luz el primer número de Archivosde Zootecnia se creaba, en 1957, en laEstación Experimental del Zaidín delConsejo Superior de InvestigacionesCientíficas, una unidad departamentalactualmente denominada Unidad deNutrición Animal. Las líneas de inves-tigación tradicionales de este Departa-mento han sido:
-la valoración nutritiva de alimen-tos y materias primas.
-la determinación de necesidadesde nutrientes y energía de animales deinterés zootécnico, particularmente, derazas autóctonas.
La importancia de una y otra líneade investigación en el contexto de laNutrición Animal es extraordinaria,dado que la alimentación animal sesustenta básicamente en los conoci-mientos desarrollados en ambas líneas.Efectivamente, de su interacción sur-ge el diseño de los llamados sistemasde alimentación, que permiten prede-cir la respuesta del animal a tratamien-tos alimentarios específicos o formu-
lar una ración para obtener del animaluna respuesta concreta.
Las publicaciones más tempranasdel departamento corresponden a laprimera de las líneas de investigaciónmencionadas y contienen informaciónsobre el valor nutritivo de alimentospara especies monogástricas y pararumiantes (Boza y Varela, 1960; Bozay Varela, 1961; Varela y Boza, 1961;Varela et al., 1961). A ellas han segui-do numerosos trabajos que, en gene-ral, se han orientado a determinar elvalor nutritivo de dietas completas. Enalgunos trabajos el diseño experimen-tal ha permitido realizar el cálculo o ladeterminación directa del valor nutri-tivo de un determinado ingrediente. Seha puesto énfasis en la evaluación derecursos propios, principalmente en lavaloración nutritiva de leguminosasgrano de producción nacional y desubproductos, generalmente con elpropósito de facilitar su inclusión enraciones para pequeños rumiantes(Boza et al., 1966; Boza et al., 1970;Pérez Cuesta y Boza, 1977; Boza etal., 1981a y b; Molina et al., 1983;Fonollá et al., 1984; Aguilera y Molina,1986; Boza et al., 1987; Fonollá et al.,1988; Molina y Aguilera, 1988;Aguilera, 1989; Aguilera et al., 1992;Fonollá y Boza, 1993; Isac et al., 1994;Sanz Sampelayo et al., 1999; MolinaAlcaide et al., 2000). En el futuropróximo es probable que los resulta-dos que nuestro departamento puedaaportar en esta línea de investigaciónprocedan de estudios no dirigidosespecíficamente a este propósito.
La segunda línea de investigaciónes de aparición más tardía y está rela-cionada con los estudios realizadossobre la utilización de la proteína y de

Archivos de zootecnia vol. 50, núm. 192, p. 568.
AGUILERA
la energía de los alimentos en el orga-nismo animal (Prieto y Aguilera, 1986;Aguilera et al., 1986) y fuertementevinculada a la disponibilidad de cáma-ras de respirometría para animales dediverso tamaño (Aguilera y Prieto,1985 y 1986). En los ensayos de balan-ce energético la retención de energíaen el animal se ha obtenido general-mente por diferencia entre la ingestade energía metabolizable y la produc-ción de calor del animal, calculadaésta mediante la medida del consumode oxígeno y de la producción deanhídrido carbónico y de metano(Brouwer, 1965). No obstante, se hanllevado a cabo estudios metabólicosen los que la retención energética seobtuvo directamente en ensayos desacrificio comparado (Sanz Sampelayoet al., 1987, 1990, 1994).
En este artículo se sintetiza la in-formación generada a lo largo de va-rios años en la Unidad de NutriciónAnimal como contribución al desarro-llo de un sistema de alimentación parael ganado caprino. En un primer apar-tado se aborda la valoración energéti-ca de los alimentos en los rumiantes,en tanto que un segundo apartado sededica a exponer los resultados obte-nidos que definen las necesidades delganado caprino en situaciones fisioló-gicas diversas.
VALORACIÓN NUTRITIVA DE LOSALIMENTOS EN RUMIANTES.ESTIMACIÓN DE SU VALOR
ENERGÉTICO
Como es sabido, la cuantificaciónde los contenidos en energía digestibley, especialmente, en energía metabo-
lizable constituye el criterio univer-salmente aceptado de expresión delvalor energético de un alimento. Ladeterminación directa de dichos con-tenidos mediante ensayos de balancerequiere disponer de una bombacalorimétrica y presenta dificultadesque derivan de la complejidad y eleva-do costo del equipo necesario paramedir la producción de metano. Afor-tunadamente, se puede estimar el con-tenido en energía metabolizable (EM,MS) a partir de la fracción digestiblede la materia seca (MSD, g/kg MS) omateria orgánica (MOD, g/kg MS) dela dieta asumiendo que su valor ener-gético es, respectivamente, 18,4 o 19,0MJ/kg y que la suma de las pérdidasenergéticas relacionadas con la forma-ción de metano y excreción de produc-tos en la orina supone un 18 p.100 deaquel valor:
EM = MSD/1000 x 18,4 x 0,82
EM = MOD/1000 x 19,0 x 0,82
Se considera que las pérdidasenergéticas en metano y orina varían,respectivamente, entre el 5 y 12 p.100y el 3 y 7 p.100 de la energía brutaingerida.
Sólo disponemos de una pequeñabase de datos con la que establecerrelaciones entre parámetros nutritivostales como materia seca digestible(MSD) o materia orgánica digestible(MOD) y contenido energético (tablaI) . Dos son las limitaciones que cabeatribuir a la validez de los resultadosde este estudio: (a) el reducido númerode datos experimentales disponibles y(b) su procedencia, limitada a mezclasde cebada grano y heno de alfalfa. Por

Archivos de zootecnia vol. 50, núm. 192, p. 569.
NUTRICIÓN ENERGÉTICA DE PEQUEÑOS RUMIANTES
otro lado, dado que los correspondien-tes ensayos se llevaron a cabo en gana-do caprino, conviene validar suextrapolación a otras especies rumian-tes. En este sentido, el Departamentode Nutrición Animal de la EstaciónExperimental del Zaidín ha realizadovarios trabajos para poder discernir sies correcto asignar a un mismo ali-mento o mezcla de alimentos un únicovalor energético, aplicable indistinta-mente a ganado ovino o caprino, ycuales son los límites de tal aplicación.El subapartado siguiente contiene in-formación a este respecto.
ESTUDIOS COMPARATIVOS REALIZADOSEN EL ÁREA DE LA NUTRICIÓN DEL GANA-DO OVINO Y CAPRINO. PREDICCIÓN DELVALOR ENERGÉTICO DE LOS ALIMENTOSPARA SU EMPLEO EN OVINO Y CAPRINO
El Agricultural and Food ResearchCouncil's Technical Committee onResponses to Nutrients publicó en 1997una revisión sobre el estado de conoci-mientos existente con respecto a lanutrición del ganado caprino (AFRC,1997). La revisión comprende publi-caciones existentes hasta 1991, inclu-sive. En ella se señalaba la falta detrabajos de investigación y de exten-
Tabla I. Valores medios de ingesta de nutrientes, de producción de metano y pérdidasenergéticas urinarias de ganado caprino en mantenimiento y producción láctea1. (Mean values
of nutrient intake, methane production and urinary energy losses of goats fed at maintenance or at
production levels).
Nivel de alimentación Mantenimiento Mantenimiento ProducciónExperimento núm. 1 (n=6) 2 - 5 (n=32) 6 - 11 (n=70)
MSD, g/kg MS 208,4 (169,9 - 240,6) 352,9 (272,3 - 419,3) -
MOD, g/kg MS 196,6 (159,7 - 225,5) 331,4 (252,5 - 395,7) 22211 (14933 - 36443)
EB, KJ/día 5270 (4583 - 6084) 11461 (10046 - 13555)
ED, KJ/día 3806 (3119 - 4414) 6198 (4991 - 7543) 14880 (9950 - 24765)
EM, KJ/día 3192 (2646 - 3753) 5185 (4155 - 6523) 12459 (8065 - 20779)
EO, KJ/día 232,7 (174,2 - 324,4) 373,1 (117,5 - 648,4) 911,3 (134,6 - 2008)
ECH4, KJ/día 380,5 (294,9 - 456,6) 637,9 (471,7 - 854,4) 1534 (722,4 - 2651)
CC, d/día - 269,5 (196,8 - 297,4) 690,6 (432,8 - 1140)
FND, g/día - 298,3 (224,6 - 398,4) 452,0 (298,4 - 747,9)
FAD, g/día - 204,8 (133,7 - 294,9) 228,6 (124,4 - 385,2)
HC, g/día - 98,1 (72,4 - 127,7) 223,5 (154,2 - 362,7)
Cel, g/día - 136,1 (90,3 - 194,3) 157,4 (88,5 - 265,3)
1MSD: Materia seca digestible; MOD: Materia orgánica digestible; EB: Energía bruta; ED: Energíadigestible; EM: Energía metabolizable; EO: Energía en orina; ECH4: Energía en metano; CC:Contenidos celulares; FND: Fibra neutro detergente; FAD: Fibra ácido detergente; HC: Hemicelulosas;Cel: Celulosa.

Archivos de zootecnia vol. 50, núm. 192, p. 570.
AGUILERA
sión realizados en caprino en el ReinoUnido, en contraste con otros paísesdesarrollados (Francia, Estados Uni-dos de Norteamérica, Países escandi-navos, etc.). El ganado ovino se hautilizado con preferencia al caprinocomo modelo en investigación especí-fica del área de Nutrición y a ello cabeatribuir la escasez de información cien-tífica y técnica existente en este áreadel conocimiento. Algunos países hanelaborado recomendaciones nutritivasque el AFRC no duda en calificar deoscuras en su base científica. En suopinión, son poco numerosos los tra-bajos científicos que se han publicadoy muchos de estos estudios se hanrealizado en países tropicales osubtropicales, cuyos resultados no sonaplicables a países de clima templado.Nosotros entendimos que este análisisera objetivo y que, por tanto, compar-tía nuestro punto de vista sobre la ne-cesidad, patente al inicio de la décadade los noventa, de obtener mayor in-formación con la que formar una basecientífica suficientemente amplia paraabordar rigurosamente la nutrición delganado caprino y su aplicación a laproducción caprina.
El AFRC dedica un apartado a ana-lizar aspectos de la fisiología digestivade esta especie animal in view of thefrequent suggestions in the literaturethat in goats, digestive processes maydiffer in important aspects from thoseof cattle and sheep y otro, a estudiarcomparativamente frente a ovino ovacuno datos de capacidad de inges-tión, con el propósito de discernir so-bre la validez de extrapolaciones quepudieran realizarse entre especies.Algo más tarde de 1991 iniciamos ennuestro departamento una serie de tra-
bajos en los que establecíamos compa-raciones entre ovino y caprino respec-to a aspectos diversos de su fisiologíadigestiva: digestibilidad de nutrientes(in vivo e in vitro), ritmos de degrada-ción y de paso de la digesta, capacidadde ingestión y pautas de fermentaciónruminal (Isac et al., 1994; García etal., 1994 y 1995; Molina Alcaide etal., 1997 y 2000). Los estudios sellevaron a cabo con alimentos de cali-dad nutritiva diversa. Existía un doblepropósito para realizar tales trabajos:(a) evaluar desde un punto de vistateórico los mecanismos que pudieranexplicar en su caso la existencia dediferencias interespecíficas en capaci-dad de ingesta de nutrientes digestibles;y (b) obtener una base científica ytécnica suficiente que facilitara el di-seño de un modelo de producción inte-grada en sistemas áridos y semiáridos.Parcialmente esta información se re-fiere a animales en situación no pro-ductiva, sujetos a planos de alimenta-ción próximos a mantenimiento y con-finados en células metabólicas (Isac etal., 1994; Molina Alcaide et al., 2000);por lo tanto, bajo condiciones experi-mentales que minimizan posibles dife-rencias entre especies animales en com-portamiento selectivo ante el alimentoque pudieran explicar diferencias eneficiencia digestiva. En estos trabajosno observamos diferencias significa-tivas entre ovino y caprino en ladigestibilidad aparente de nutrientescuando los animales consumieron fo-rrajes o dietas mixtas de calidad nutri-tiva media o alta (henos de alfalfa,paja de veza, dietas mixtas, con coefi-cientes de digestibilidad de la materiaorgánica (DMO) situados entre 0,58 y0,76). Tampoco se detectaron diferen-

Archivos de zootecnia vol. 50, núm. 192, p. 571.
NUTRICIÓN ENERGÉTICA DE PEQUEÑOS RUMIANTES
cias interespecíficas en ritmos de fer-mentación (0,126-0,023, en caprino;0,120-0,035, en ovino) o de tránsito departículas a través del rumen (0,034-0,024, en caprino; 0,032-0,020, enovino). Por el contrario, los parámetrosde fermentación ruminal arrojaron re-sultados contradictorios.
Sin embargo, la mayor parte de losdatos que hemos publicado procedende ensayos que se llevaron a cabo enanimales no productivos en co-pasto-reo (un único rebaño) en tierras áridaso semiáridas y, consecuentemente, enganado ovino y caprino que consumiópastos naturales de media o baja cali-dad nutricional (García et al., 1994 y1995; Molina Alcaide et al., 1997). Enestos ensayos obtuvimos resultados quearrojaron diferencias, en algún casosignificativas, en digestibilidad in vitrode los pastos seleccionados, que fue-ron favorables al ganado caprino cuan-do el pasto consumido fue de bajacalidad. Se observaron también pe-queñas diferencias en algunos pará-metros de la fermentación ruminal,dependientes del estado fenológico delpasto disponible. Los resultados de lostrabajos que llevamos a cabo con ani-males en pastoreo demuestran clara-mente que el ganado caprino tienemayor capacidad de ingesta que elovino, la cual sería resultado del ritmode paso de partículas a través del rumensignificativamente mayor (0,035, encaprino; 0,027, en ovino), así como dela tendencia manifiesta a degradar amayor velocidad el material colocadoen bolsas de nylon suspendidas en elrumen. El tamaño del compartimentoruminal también fue significativamentesuperior en el caprino en comparacióncon el ovino (649 vs 470 g de materia
seca). Por lo tanto y en concordanciacon datos analizados por el AFRC(1997), concluimos que, aunque nocabe descartar que las diferencias endigestibilidad observadas (calculadasa partir de datos in vitro) pudieran sóloser el resultado de pequeñas diferen-cias en composición nutritiva del ma-terial consumido por una y otra espe-cie animal, no detectadas por el méto-do de muestreo utilizado, el ganadocaprino es capaz de mantener un ma-yor tamaño del compartimento ruminalsin que aumente el grado de distensiónde la pared del rumen. Ello le capaci-taría notablemente para ingerir ma-yores cantidades de alimento, en com-paración con el ganado ovino. Tam-poco hay que olvidar que en el animalno productivo la ingesta voluntaria estálimitada por la baja demanda denutrientes que requiere la atención asus necesidades de mantenimiento.Bajo estas condiciones las mayoresnecesidades energéticas para el man-tenimiento observadas en el ganadocaprino en relación con el ovino(Aguilera et al., 1986 y 1991; Prieto etal., 1990) contribuirían a explicar unaingestión de alimento mayor en aque-lla especie animal. No cabe duda deque sería necesario disponer de mayornúmero de datos de digestibilidad invivo y valor energético de forrajes ydietas mixtas, que habrían de obtenersemediante ensayos paralelos llevados acabo en ovino y caprino bajo condicio-nes idénticas en el laboratorio. Hastatanto no se disponga de tales datos debeaceptarse la información disponible, fa-vorable a utilizar en ovino y caprino unúnico valor energético aplicable a am-bas especies animales para alimentos decalidad media o alta (DMO>0,58).

Archivos de zootecnia vol. 50, núm. 192, p. 572.
AGUILERA
Pruebas de digestibilidad in vitro(Tilley y Terry, 1963) llevadas a caboen nuestro laboratorio con ovino ycaprino alimentados con heno de alfal-fa de buena calidad a nivel aproxima-do de mantenimiento (Aguilera et al.,1986 y 1991; Prieto et al., 1990), dise-ñadas para discernir si la procedenciadel contenido ruminal utilizado comoinóculo en el medio de incubaciónpudiera afectar al valor de digestibi-lidad, demostraron la ausencia de dife-rencias significativas en la digestibi-lidad de la materia seca (DMSvitro),materia orgánica (DMOvitro) y frac-ción de constituyentes de la pared ce-lular (DFNDvitro) (García, 1992). Seincubaron 16 muestras de pastospolifitos o de especies simples herbá-ceas o arbustivas, cuya DMOvitro va-rió entre 0,44 y 0,70. Por lo tanto, estosresultados indican que ovino y caprinopudieran ser mutuamente reemplaza-bles como donadores de inóculo cuan-do se les alimente con dietas equili-bradas de composición próxima.
ESTIMACIÓN DEL CONTENIDO EN ENER-GÍA DIGESTIBLE Y EN ENERGÍA METABO-LIZABLE
La energía digestible ingerida estáestrechamente correlacionada con elconsumo de materia seca o de materiaorgánica digestibles, prefiriéndose estaúltima ya que de la primera formanparte sustancias minerales (5-10 p.100)de nulo valor energético para el ani-mal, aunque los valores obtenidos conla segunda tiendan a ser algo más va-riables.
A partir de datos proporcionadospor 38 balances individuales obteni-dos en ganado caprino de peso vivovariable y alimentado a nivel próximo
a mantenimiento (ver tabla I ), hemosobtenido las siguientes ecuaciones quepermiten calcular el contenido en ener-gía digestible (ED) de la dieta conoci-do su contenido en materia seca o ma-teria orgánica digestibles (MSD yMOD, respectivamente):
ED, KJ = 17,56 (±0,13) MSD, g; p<0,001 (1)
ED, KJ = 18,71 (±0,11) MOD, g; p<0,001 (2)
cuyos errores absolutos respecto alvalor medio de la variable dependienteson 3,92 p.100 y 2,76 p.100, respecti-vamente.
Numerosos ensayos realizados enganado ovino y en vacuno, utilizandoraciones de composición ampliamentevariable, han servido de base a distin-tos investigadores para establecer re-laciones con las que obtener el conte-nido en energía metabolizable (EM)de los alimentos conocida su riquezaen MO digestible o en distintosnutrientes digestibles. Nosotros hemosintentado obtener una ecuación de estetipo utilizando los datos de los 38 ba-lances individuales en caprino, cita-dos anteriormente:
EM, KJ = 16,0 MOD, g - 9,1 ND, g
donde ND indica contenido en nitró-geno digestible. Sin embargo el se-gundo término no es significativo. Laecuación lineal correspondiente queobtenemos eliminando dicho términoes la siguiente:
EM, KJ = 15,66 (±0,12) MOD, g; p<0,001 (3)
cuyo error absoluto respecto al valormedio de la variable dependiente es3,72 p.100. El coeficiente de la varia-

Archivos de zootecnia vol. 50, núm. 192, p. 573.
NUTRICIÓN ENERGÉTICA DE PEQUEÑOS RUMIANTES
ble independiente no difiere signifi-cativamente del valor 15,58 (19 x 0,82)aceptado por el Agricultural ResearchCouncil (1980 y 1984).
El estudio estadístico realizado so-bre 102 balances energéticos indivi-duales en ganado caprino de raza gra-nadina, 32 de ellos con machos adultoscastrados y 70 con hembras en lactación(ver tabla I ), nos ha permitido relacio-nar mediante regresión lineal lasingestas de energía digestible (EDI) ymetabolizable (EMI) de la dieta. Enestos ensayos los animales se mantu-vieron en jaulas individuales y se lesofreció en una sola comida la dietacorrespondiente, formada por heno dealfalfa granulada (Proteína bruta, 187g/kg MS; MO digestible, 456 g/kgMS) y cebada grano (Proteína bruta,108 g/kg MS; MO digestible, 835 g/kgMS), en proporciones relativas am-pliamente variables. El balance ener-gético individual se determinó en cá-maras de respirometría en circuitoabierto. Los procedimientos experi-mentales y analíticos aparecen publica-dos en detalle en Aguilera et al. (1990)y Prieto et al. (1990). El nivel de ingestaenergética osciló entre 0,80 y 3,02 vecesel correspondiente a mantenimiento.
La ecuación de regresión
EMI (KJ/kg0,75 y día) = 7,87+ 0,837 (±0,0039)
EDI (KJ/kg0,75 y día); r = 0,999; p<0,001
tiene un término independiente quecarece de significación estadística, porlo que hemos construido una nuevaregresión en la que se fuerza que estetérmino sea igual a cero:
EMI (KJ/Kg0,75 y día) = 0,836 (±0,0016) EDI (KJ/
kg0,75 y día); p<0,001 (4)
Esta ecuación permite predecir elcontenido en energía metabolizable dela dieta cuando se conoce el de energíadigestible; indica que las pérdidas ener-géticas en orina y metano suponen el16,4 p.100 de la energía digestibleingerida o, lo que es igual, la ingesta deenergía metabolizable alcanza 0,836KJ/KJ de energía digestible ingerida.El coeficiente de la variable indepen-diente no difiere significativamente delfactor 0,82 y podría utilizarse indis-tintamente en el cálculo de EM en lavaloración energética de dietas pararumiantes.
ESTIMACIÓN DE LA PRODUCCIÓN DEMETANO A PARTIR DE LA INGESTA DEMATERIA ORGÁNICA O ENERGÍA DIGES-TIBLE
La formación de metano, como re-sultado de la fermentación microbianaruminal, constituye una vía de pérdidaenergética cuya cuantificación es ne-cesario hacer si se desea expresar elvalor nutritivo del alimento en energíametabolizable. La cuantificación pre-cisa de la producción de metano re-quiere la utilización de equipos com-plejos, generalmente cámaras derespirometría, y de técnicas calorimé-tricas adecuadas para la medida delintercambio de gases. De aquí que sehayan realizado esfuerzos para esti-mar aquellas pérdidas energéticas enfunción de la naturaleza de la dieta yde su ingesta. Blaxter y Clapperton(1965), en ovino y vacuno de carne;Neergaard (1973), en terneros, y Moey Tyrrell (1979), en vacuno lechero,realizaron estudios de esta naturalezay demostraron la existencia de relacio-nes entre la formación de metano y lacomposición nutritiva de la ración. Más

Archivos de zootecnia vol. 50, núm. 192, p. 574.
AGUILERA
tarde Lindgren (1980) sometió a igualtipo de análisis las más de 2500 deter-minaciones publicadas sobre la pro-ducción de metano en ovino y vacuno.Nosotros analizamos la producción demetano de ganado caprino alimentadocon dietas de naturaleza conocida, paraobtener ecuaciones con las que esti-mar dicha producción a partir de lacomposición nutritiva de la ración con-sumida (Aguilera y Prieto, 1991). Elestudio se realizó sobre los 102 balan-ces energéticos individuales, mencio-nados anteriormente, llevados a caboen ganado caprino de raza granadina,32 de ellos con machos adultos castra-dos y 70 con hembras en lactación. Losanimales se mantuvieron en jaulas in-dividuales y se les ofreció en una solacomida la dieta correspondiente, for-mada por heno de alfalfa granulada(Proteína bruta, 187 g/kg MS; MOdigestible, 456 g/kg MS) y cebada gra-no (Proteína bruta, 108 g/kg MS; MOdigestible, 835 g/kg MS), en propor-ciones relativas ampliamente variables.El balance energético individual sedeterminó en cámaras de respirometríaen circuito abierto. Como valor calorí-fico del metano se tomó la cifra de39,539 KJ/litro (Brouwer, 1965).Como ya se ha dicho, los procedimien-tos experimentales y las técnicas ana-líticas usadas han sido publicados porAguilera et al. (1990) y Prieto et al.(1990). El nivel de ingesta energéticaosciló entre 0,80 y 3,02 veces el co-rrespondiente a mantenimiento y laproducción de metano se situó entre11,63 y 67,05 litros/día, en función delnivel de alimentación y la composi-ción de la dieta, con un valor medio de33,50 litros/día. La producción demetano se registró a lo largo de 24
horas a intervalos de 4,8 minutos.Dentro de estas pruebas, concreta-
mente a lo largo de dos series de medi-das de balance, todos los animales re-cibieron la misma cantidad de una die-ta única basada en heno de alfalfagranulado y cebada grano en propor-ción relativa constante. Estos ensayosnos permiten cuantificar la variabili-dad individual en la formación de me-tano, juzgada a través del coeficientede variación. Éste es del ±8 p.100,igual al observado en ovino por Blaxtery Clapperton (1965).
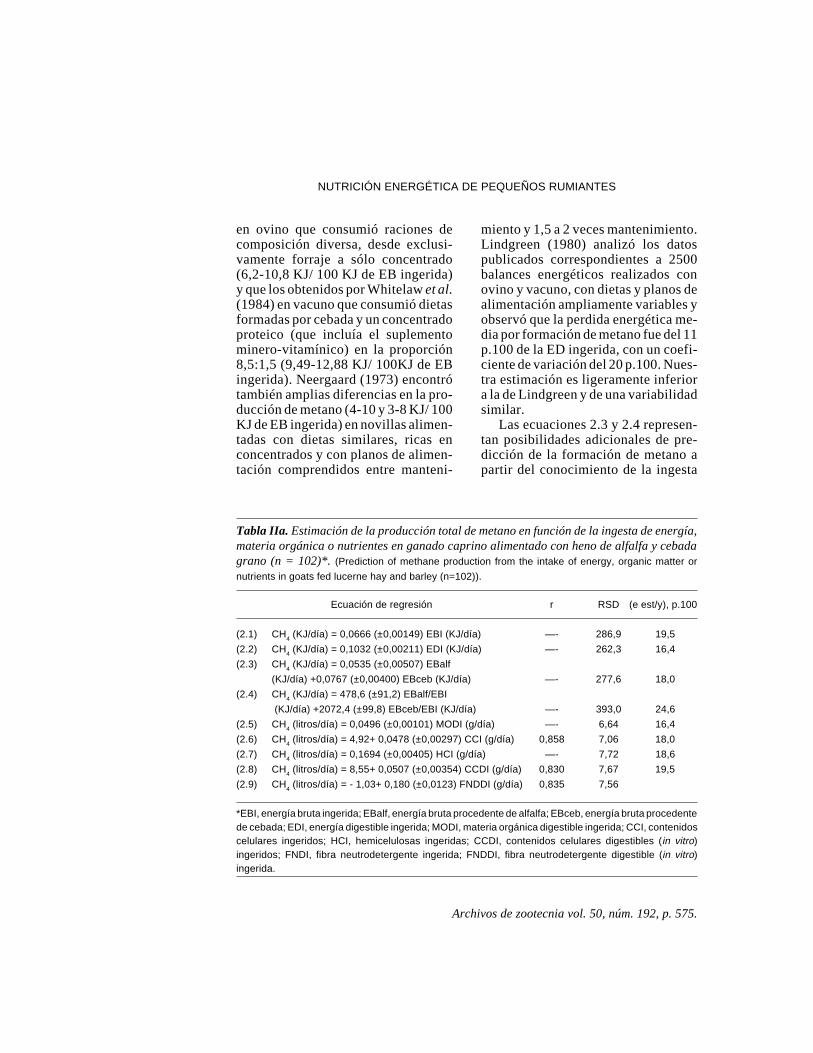
Recientemente hemos revisadoecuaciones previamente publicadas pornosotros (Aguilera y Prieto, 1991) querelacionan la producción de metanocon parámetros de composición nutri-tiva de la ración consumida. Hemosobtenido así las ecuaciones de regre-sión lineal que figuran en la tabla IIa ,con las que estimar la producción demetano en función de la ingesta ener-gética o de la ingestión de MOdigestible (ecuaciones 2.1 – 2.5) o dela composición química (Goering yvan Soest, 1970) y digestibilidad invitro (Tilley y Terry, 1963) de loscarbohidratos ingeridos (ecuaciones2.6 –2.9).
Las ecuaciones 2.1 y 2.2 indicanque la producción de metano alcanza6,66 KJ/100 KJ de energía bruta (EB)ingerida o 10,32 KJ/100 KJ de energíadigestible (ED) ingerida. Las pérdidasenergéticas debidas a la formación demetano se situan entre 3,58 y 9,57 KJ/100 KJ de EB ingerida y entre 5,94 y14,45 KJ/ 100 KJ de ED ingerida. Losintervalos de variación son más am-plios que los observados por Blaxter yClapperton (1965) al analizar los re-sultados de 391 balances energéticos
Archivos de zootecnia vol. 50, núm. 192, p. 575.
NUTRICIÓN ENERGÉTICA DE PEQUEÑOS RUMIANTES
en ovino que consumió raciones decomposición diversa, desde exclusi-vamente forraje a sólo concentrado(6,2-10,8 KJ/ 100 KJ de EB ingerida)y que los obtenidos por Whitelaw et al.(1984) en vacuno que consumió dietasformadas por cebada y un concentradoproteico (que incluía el suplementominero-vitamínico) en la proporción8,5:1,5 (9,49-12,88 KJ/ 100KJ de EBingerida). Neergaard (1973) encontrótambién amplias diferencias en la pro-ducción de metano (4-10 y 3-8 KJ/ 100KJ de EB ingerida) en novillas alimen-tadas con dietas similares, ricas enconcentrados y con planos de alimen-tación comprendidos entre manteni-
miento y 1,5 a 2 veces mantenimiento.Lindgreen (1980) analizó los datospublicados correspondientes a 2500balances energéticos realizados conovino y vacuno, con dietas y planos dealimentación ampliamente variables yobservó que la perdida energética me-dia por formación de metano fue del 11p.100 de la ED ingerida, con un coefi-ciente de variación del 20 p.100. Nues-tra estimación es ligeramente inferiora la de Lindgreen y de una variabilidadsimilar.
Las ecuaciones 2.3 y 2.4 represen-tan posibilidades adicionales de pre-dicción de la formación de metano apartir del conocimiento de la ingesta
Tabla IIa. Estimación de la producción total de metano en función de la ingesta de energía,materia orgánica o nutrientes en ganado caprino alimentado con heno de alfalfa y cebadagrano (n = 102)*. (Prediction of methane production from the intake of energy, organic matter or
nutrients in goats fed lucerne hay and barley (n=102)).
Ecuación de regresión r RSD (e est/y), p.100
(2.1) CH4 (KJ/día) = 0,0666 (±0,00149) EBI (KJ/día) —- 286,9 19,5
(2.2) CH4 (KJ/día) = 0,1032 (±0,00211) EDI (KJ/día) —- 262,3 16,4
(2.3) CH4 (KJ/día) = 0,0535 (±0,00507) EBalf
(KJ/día) +0,0767 (±0,00400) EBceb (KJ/día) —- 277,6 18,0
(2.4) CH4 (KJ/día) = 478,6 (±91,2) EBalf/EBI
(KJ/día) +2072,4 (±99,8) EBceb/EBI (KJ/día) —- 393,0 24,6
(2.5) CH4 (litros/día) = 0,0496 (±0,00101) MODI (g/día) —- 6,64 16,4
(2.6) CH4 (litros/día) = 4,92+ 0,0478 (±0,00297) CCI (g/día) 0,858 7,06 18,0
(2.7) CH4 (litros/día) = 0,1694 (±0,00405) HCI (g/día) —- 7,72 18,6
(2.8) CH4 (litros/día) = 8,55+ 0,0507 (±0,00354) CCDI (g/día) 0,830 7,67 19,5
(2.9) CH4 (litros/día) = - 1,03+ 0,180 (±0,0123) FNDDI (g/día) 0,835 7,56
*EBI, energía bruta ingerida; EBalf, energía bruta procedente de alfalfa; EBceb, energía bruta procedentede cebada; EDI, energía digestible ingerida; MODI, materia orgánica digestible ingerida; CCI, contenidoscelulares ingeridos; HCI, hemicelulosas ingeridas; CCDI, contenidos celulares digestibles (in vitro)ingeridos; FNDI, fibra neutrodetergente ingerida; FNDDI, fibra neutrodetergente digestible (in vitro)ingerida.

Archivos de zootecnia vol. 50, núm. 192, p. 576.
AGUILERA
de energía bruta procedente del forrajey grano utilizados o de su contribucióna la ingesta energética total. Estasecuaciones no mejoran en precisión alas anteriores. La ecuación 2.5, queestima la formación de metano a partirdel consumo de materia orgánica dige-rida, es similar a la obtenida por Murrayet al. (1978) en ovino alimentado conalfalfa:
CH4 (litros/día)= 2,81 + 0,042 (±0,003) MO
digestible ingerida (g/día)
aunque en nuestros ensayos se observauna mayor formación de metano porunidad de materia orgánica digerida.
Del resto de las ecuaciones que apa-recen en la tabla IIa la mejor estima-ción de la producción de metano laproporcionan las ecuaciones 2.6 y 2.7,que predicen la formación de metano apartir de las cantidades ingeridas delos contenidos celulares (CC) yhemicelulosas (HC), respectivamente.Las fracciones digestibles in vitro delos contenidos celulares o de los com-ponentes de la pared celular no mejo-ran dicha estimación. Wilkerson et al.(1994) analizaron la bondad de predic-ción en la producción de metano desiete ecuaciones publicadas para ga-nado vacuno utilizando como base decomparación datos de su propio labo-ratorio: 660 observaciones con pro-ducciones de metano situadas en tér-minos energéticos entre 3,72 y 30,17MJ/día. Observaron que los erroresmenores derivaban de las ecuacionesque relacionaban la producción demetano con la ingesta de celulosa,hemicelulosa y contenidos celulares osus fracciones digestibles y concluye-ron que la ecuación de Moe y Tyrrell
(1979) basada en la ingesta total decarbohidratos es la más exacta y preci-sa (error absoluto de predicción: 11,5p.100 respecto al valor medio de pro-ducción de metano).
Con el objeto de determinar el efec-to del nivel de alimentación (L), ex-presado como múltiplo del correspon-diente a mantenimiento, sobre la pro-ducción de metano, analizamos losdatos correspondientes a los 70 balan-ces energéticos realizados en caprinoen lactación, comprendiendo planosde ingestión situados entre 1,28 y 3,02veces mantenimiento. La digestibilidadaparente media de la energía de ladieta osciló en el estrecho margen de65,7±0,37 y 67,9±0,45. Las ecuacionescorrespondientes (2.10 y 2.11) figuranen la tabla IIb e indican que un au-mento del plano de alimentación en unmúltiplo del mantenimiento conduce auna reducción de las pérdidas energé-ticas por formación de metano de 1,51KJ/ 100KJ de EB ingerida o de 2,19KJ/ 100KJ de ED ingerida. Estos valo-res son superiores a los observados porBlaxter y Clapperton (1965) en ovino(1,32 KJ/ 100 KJ de EB ingerida, paradietas mixtas) y por Lindgreen (1980)en ovino (1,70 KJ/ 100 KJ de EDingerida) y vacuno (1,44 KJ/ 100 KJ deED ingerida).
También en la tabla IIb figura laecuación de regresión (2.12) que rela-ciona la producción de metano con ladigestibilidad aparente de la energíade la dieta (D, p.100). Se ha calculadoa partir de los datos proporcionadospor los 32 balances energéticos indivi-duales obtenidos en machos castradosalimentados con heno de alfalfa o biencon mezclas de heno de alfalfa y ceba-da grano. El plano de alimentación se

Archivos de zootecnia vol. 50, núm. 192, p. 577.
NUTRICIÓN ENERGÉTICA DE PEQUEÑOS RUMIANTES
mantuvo en la proximidad del mante-nimiento. Esta ecuación indica que seproduce un incremento en la produc-ción de metano de 0,151 KJ/ 100 KJ deEB ingerida por cada unidad porcen-tual de aumento en la digestibilidadaparente de la energía de la dieta. Elcoeficiente de la variable independientede la ecuación es superior a los deecuaciones similares publicadas pararaciones mixtas por Blaxter y Clapper-ton (1965) ensayadas en ovino (0,074KJ/ 100 KJ de EB ingerida) y porLindgreen (1980) en vacuno (0,030KJ/ 100 KJ de EB ingerida). Sin em-bargo, las pérdidas energéticas debi-das a la formación de metano de dietascon coeficiente de digestibilidad apa-rente de la energía de 50 y 70 p.100 seestimarían en 6,73 y 8,21 KJ/ 100 KJde EB ingerida, de acuerdo con Blaxtery Clapperton (1965) y 4,97 y 7,99 KJ/100 KJ de EB ingerida, mediante laecuación (2.12).
En conclusión, los resultados pre-sentados aquí sugieren que no existendiferencias importantes en formaciónde metano durante la fermentación
ruminal de alimentos entre ovino, ca-prino y vacuno. Ello daría validez a lasextrapolaciones entre especies anima-les que pudieran llevarse a cabo, porejemplo, con fines de evaluación ener-gética. No obstante, una vez más, cree-mos necesario puntualizar que los da-tos de nuestro laboratorio proceden dedietas formuladas exclusivamente conheno de alfalfa y cebada, por lo quehabrá que incorporar datos derivadosde dietas de origen ampliamente di-verso para alcanzar conclusiones defi-nitivas.
ESTIMACIÓN DE LAS PÉRDIDAS ENERGÉ-TICAS CORRESPONDIENTES A LA EXCRE-CIÓN URINARIA A PARTIR DE LA INGESTADE ENERGÍA DIGESTIBLE
El análisis de los datos procedentesde los 102 balances energéticos indivi-duales obtenidos en ganado caprino enmantenimiento y producción láctea nospermite estimar las pérdidas de ener-gía a través de la excreción de orina(EO), conocida la ingesta de energíadigestible (EDI). La ecuación de re-gresión correspondiente:
Tabla IIb. Efecto del nivel de alimentación (L; múltiplo de mantenimiento) y de la digestibilidadde la energía de la dieta (D, p.100) sobre la producción de metano en ganado caprinoalimentado con heno de alfalfa y cebada grano*. (Effect of the feeding level (L; times maintenance)
and the digestibility of dietary energy (D, p.100) on methane production in goats fed on lucerne hay and
barley).
Ecuación de regresión n r RSD
(2.10) CH4 (KJ/ 100 KJ EBI) = 10,1 – 1,51 (±0,381) L 70 0,451 1,43(2.11) CH4 (KJ/ 100 KJ EDI) = 15,1 – 2,19 (±0,573) L 70 0,440 2,14(2.12) CH4 (KJ/ 100 KJ EBI) = - 2,58 + 0,151 (±0,0147) D 32 0,883 0,635
*EBI, energía bruta ingerida; EDI, energía digestible ingerida.

Archivos de zootecnia vol. 50, núm. 192, p. 578.
AGUILERA
EO (KJ/día) = 0,0625 (±0,00292) EDI (KJ/día);
p<0,001 (5)
estima las pérdidas de energía en orinaen 0,0625 KJ/ KJ de incremento pro-ducido en la ingesta de energíadigestible. La ecuación anterior carecede ordenada en el origen, ya que en laecuación de regresión previamentecalculada este término no alcanzó sig-nificación estadística. Como puedecomprobarse, la suma de los coefi-cientes del término independiente deesta ecuación (0,0625) y de la ecua-ción (2.2), que estima las pérdidas deenergía a causa de la formación demetano (0,1032), explica sensiblemen-te el valor del coeficiente de la ecua-ción (4) que relaciona los contenidosde energía digestible y metabolizablede la dieta (0,836).
DETERMINACIÓN DE LASNECESIDADES ENERGÉTICAS DEL
GANADO CAPRINO
Conocer los mecanismos de trans-ferencia de energía en el organismoanimal y cómo éste utiliza la energíaen los procesos fisiológicos relaciona-dos con el mantenimiento de lahomeostasis interna (por ejemplo,mantenimiento del tono muscular, ac-tividad cardiaca, actividad renal, trans-porte iónico a través de membranas,transmisión del impulso nervioso, etc.)o con la producción (por ejemplo, sín-tesis de proteína y grasa orgánicas,secreción de leche, producción de lana,etc.) es de enorme importancia si sequiere establecer las necesidades nu-tritivas de los animales bajo situacio-nes fisiológicas y ambientales diver-
sas y la capacidad de los alimentospara satisfacerlas. Como ya se ha di-cho, tal conocimiento es fundamentalpara diseñar sistemas de alimentaciónprecisos, con los que predecir riguro-samente el balance energético del ani-mal, esto es, en términos energéticos,la cantidad de producto animal quederiva de la ingesta de cantidades con-cretas de alimento. No es fácil, porqueun análisis cuantitativo del metabolis-mo del animal requiere ineludible-mente conocer en términos cualita-tivos y cuantitativos los nutrientes ab-sorbidos. Los nutriólogos están deacuerdo en que esto es relativamentesimple de obtener en animales mono-gástricos, pero presenta dificultadesextraordinarias en el animal rumiante,a causa de la existencia de los procesosfermentativos ruminales. No existe unmétodo único que proporcione unaestimación cuantitativa de los produc-tos de la digestión ruminal, debidofundamentalmente a la incompatibili-dad del empleo simultáneo en un mis-mo animal de ciertas técnicas experi-mentales y a las propias limitacionesde éstas para estimar las velocidadesde formación y absorción de los pro-ductos de la digestión ruminal. La si-tuación se complica más aún, porquela cantidad de nutrientes disponiblesen el animal rumiante para su utiliza-ción como fuente energética y comosubstratos en procesos de síntesis yrenovación no es necesariamente iguala la cantidad absorbida en el tractodigestivo, a causa de la existencia detransformaciones metabólicas en elanimal. Esto dificulta extraordinaria-mente el análisis desde un punto devista bioquímico del metabolismo ener-gético del animal, la constatación de

Archivos de zootecnia vol. 50, núm. 192, p. 579.
NUTRICIÓN ENERGÉTICA DE PEQUEÑOS RUMIANTES
los mecanismos causales y la elabora-ción de modelos metabólicos que pro-porcionen explicaciones satisfactoriasde las transformaciones que observa-mos y cuantificamos en los trabajoscalorimétricos. A pesar de todo elloresulta relativamente simple explicardesde un punto de vista meramentetermodinámico una de las causas de laconocida baja eficiencia de utilizaciónde la energía y de la mayor producciónde calor asociada al metabolismo delos alimentos en los rumiantes, com-parativamente frente a los animalesmonogástricos: las grandes pérdidasenergéticas inherentes a la fermenta-ción ruminal, que suponen por términomedio un 22 p.100 de la energía delsustrato digerido (15 p.100 del calorde combustión del CH
4 y 7 p.100 del
calor de fermentación), a las que sesuma el residuo indigestible del mate-rial microbiano (el material microbianosupone en términos energéticos un 8p.100 de la energía digerida). En elapartado 1 hemos realizado un análisisde las pérdidas energéticas debidas ala formación de CH
4 en el ganado ca-
prino.Cuando en 1983, terminada la cons-
trucción de una planta de respirometríapara pequeños rumiantes, iniciábamosen nuestro departamento los trabajosde ajuste y calibrado de sus elementosde medida, éramos perfectamente cons-cientes de que habían transcurrido nadamenos que veinte años desde queBlaxter y su equipo del RowettResearch Institute de Aberdeen (Esco-cia) presentara sus propuestas que ha-brían de desembocar, tres años des-pués, en la adopción por el Reino Uni-do de un nuevo sistema de evaluaciónenergética, que ha constituido la base
científica para el desarrollo de poste-riores sistemas. Una gran cantidad detrabajos calorimétricos realizados enel curso de esos veinte años ha elimi-nado numerosas lagunas e incertidum-bres. Ha quedado bien establecido quela eficiencia de utilización de la ener-gía metabolizable para el mantenimien-to y para el crecimiento-cebo; que laparticipación de la energía metabo-lizable hacia la secreción de leche ohacia formación de proteína y grasatisular; y que la composición de laproducción láctea son determinadasfundamentalmente por la naturalezade la mezcla de los productos de ladigestión que son absorbidos en eltracto digestivo.
La evidencia experimental muestra(tabla III) que para el mantenimientola mezcla de AGV absorbidos en elrumen se utiliza con una eficiencia del83 p.100, mientras que la glucosa y los
Tabla III. Eficiencia con que la energía delos distintos nutrientes se utiliza en los pro-cesos de mantenimiento y de producción enel animal rumiante. (Energetic efficiency of
nutrients for maintenance and production in the
ruminant animal).
Producto final Estado fisiológicode la digestión Mantenimiento Crecimiento-
cebo
AGV 0,83 0,57-0,64Glucosa 1,00 0,70Aminoácidos 0,81 0,65Lípidos 0,97 0,80Acetato 0,59 0,30Propionato 0,87 0,52Butirato 0,76 0,62

Archivos de zootecnia vol. 50, núm. 192, p. 580.
AGUILERA
aminoácidos absorbidos en el intesti-no lo hacen con eficiencias del 100 y81p.100, respectivamente. La eficien-cia de utilización de los lípidos pareceestar situada en la vecindad del 97p.100. En el animal en crecimiento-cebo las eficiencias de utilización de laglucosa, aminoácidos y ácidos grasosde cadena larga absorbidos en el intes-tino delgado son, respectivamente, del70, 65 y 80 p.100, mientras que para elacetato, propionato y butirato taleseficiencias son 30, 52 y 62 p.100. Labaja eficiencia de utilización delacetato se produce cuando es insufi-ciente el aporte de glucosa o de precur-sores de glucosa. Las mezclas de AGVse utilizan con eficiencias que oscilanentre 57 y 64 p.100 en función de susproporciones molares relativas. Ade-más, la infusión de nutrientes indivi-duales en rumiantes en lactación con-duce a respuestas específicas, de modoque se han propuesto mecanismosmetabólicos que explican los efectosobservados. Por ejemplo, el efecto es-timulante del acetato sobre la síntesisde grasa, proteína y lactosa en leche seexplica a través del aumento que pro-duce en precursores para la síntesis deácidos grasos en la glándula mamaria
y, al propio tiempo, en el aporte desubstratos para la formación de ATPen dicha glándula.
La técnica más usada en el estudiodel metabolismo energético es la debalance calorimétrico: el cambio encontenido energético del organismoen un período de tiempo, debido acambios en su contenido en grasa yproteína, es estimado directamente,deduciendo las pérdidas energéticas(heces, orina, gases combustibles yproducción de calor del animal) de laingesta energética. La producción decalor del animal la determinamos ennuestro caso por calorimetría indirecta(Brouwer, 1965). En algún momentodel ensayo de digestibilidad coloca-mos al animal en una cámara respiro-métrica y, tras un período de adapta-ción de 24 o 48 horas, medimos duran-te otras 24 o 48 horas su consumo de 0
2y su producción de CO
2 y CH
4 Para
determinar la producción de calor basalprivamos al animal de alimento duran-te 72 horas y, previo control del co-ciente respiratorio y de la producciónde CH
4, con el propósito de verificar
que los animales han alcanzado lascondiciones basales, cuantificamos elconsumo de 0
2 y la producción de CO
2
Tabla IV. Ensayos calorimétricos realizados en ganado caprino con anterioridad a 1985.(Calorimetric trials carried out in goats before 1985).
Estado fisiológico Producción de calor basal, kl Autoresdel animal KJ/kg0.75y día
Hembras no lactantes 386 - Brody (1945)Hembras en lactación 236 0,691 Armtrong y Blaxter (1965)Hembras no lactantes 357 - Fujihara et al. (1973)Machos castrados 331 Roy-Smith (1980)

Archivos de zootecnia vol. 50, núm. 192, p. 581.
NUTRICIÓN ENERGÉTICA DE PEQUEÑOS RUMIANTES
y a partir de ellos la producción decalor. La planta de respirometría exis-tente en nuestro departamento cuentacon cámaras de circuito abierto(Aguilera y Prieto, 1986) y de confina-miento (Lachica et al., 1995) adecua-das para l levar a cabo ensayosmetabólicos de larga duración en pe-queños rumiantes y en animalesmonogástricos. Como resultado de laactividad metabólica del animal se pro-ducen alteraciones en la composicióndel aire existente en la cámara. Sumedida junto a la del volumen de aireque circula por ella o está contenido enella permite conocer el consumo de O2y la producción de CO
2 y CH
4. La
concentración de CH4, CO
2, y 0
2 se
obtiene mediante métodos físicos: ab-sorción en el infrarrojo para CO
2 y CH
4y paramagnetismo para el O
2. El siste-
ma está totalmente automatizado einformatizado.
NECESIDADES ENERGÉTICAS DE MANTE-NIMIENTO Y PRODUCCIÓN DEL GANADO
CAPRINO
Probablemente como resultado dela menor importancia económica de laespecie caprina en los países del mun-do occidental se le ha prestado unaescasa atención. No es esto lo quesucede en nuestro entorno ni en paísesque comprendan zonas geográficas enlas que prevalezca la aridez. Es más,las actuales tendencias en producciónanimal que dan preferencia a la calidadde los productos sobre la eficiencia dela producción; al control del impactode la producción animal sobre el me-dio y al bienestar animal, concedenespecial interés a la extensificación.Qué duda cabe de que en los ecosis-
temas mediterráneos la produccióncaprina es de elección por su fáciladecuación a estos requisitos en com-patibilidad con niveles aceptables derentabilidad de las producciones. Ha-cia 1985 las recomendaciones energé-ticas para la producción de caprinodescansaban en pruebas de alimenta-ción (Mohammed y Owen, 1980;Morand-Fher, 1981; NRC, 1981).Frente a la enorme cantidad de estu-dios calorimétricos realizados en otrasespecies, la tabla IV recoge los únicostrabajos hasta entonces realizados encaprino. En todos ellos se determinó laactividad metabólica basal en anima-les en situación no productiva, conresultados muy variables. Sólo uno deestos estudios se llevó a cabo, aunquecon objetivos muy distintos a los per-seguidos por nosotros, en hembras enlactación.
Ha existido tradicionalmente la ten-dencia a considerar que el ganado ca-prino tiene iguales requerimientosenergéticos que el ovino con respectoal mantenimiento y a asumir que secomporta como el vacuno en lo concer-niente a sus necesidades energéticas parala producción de leche. Sin embargo, labibliografía que sustenta tales afirma-ciones es escasísima. De aquí que nues-tro departamento decidiera desarrollarun programa de investigación que tu-viera por objeto aportar informacióncon la que establecer las necesidadesenergéticas del ganado caprino. Esteprograma ha prestado mucha atencióna la evaluación del gasto energéticorelacionado con la actividad física paraasí obtener estimaciones precisas delas necesidades globales del animal enproducción extensiva.

Archivos de zootecnia vol. 50, núm. 192, p. 582.
AGUILERA
La tabla V muestra los resultadosde medidas del metabolismo basal deganado caprino y ovino adulto castra-do sometido a protocolos experimen-tales idénticos (Aguilera et al., 1986;Prieto et al., 1990). Los resultadossugieren que la actividad metabólicadel ganado caprino es alrededor de un20 p.100 superior a la del ovino.
En nuestros trabajos con ganadocaprino siempre hemos dedicado es-pecial atención al mantenimiento. Yello porque parece demostrado que lasnecesidades energéticas para el man-tenimiento varían considerablementecon el plano de nutrición, estado fisio-lógico, edad, nivel de productividad,etc. Nosotros hemos pretendido esti-mar en el ganado caprino la magnitudde esta variación. Ciertamente no sehan realizado suficientes trabajos deinvestigación dirigidos a definir loscambios fisiológicos y metabólicosresponsables de la variación del man-tenimiento, tanto en el animal mono-gástrico como en el rumiante. En cam-bio sí que existe bastante controversia.
En primer lugar son muchos los cien-tíficos que argumentan que el concep-to de mantenimiento, que está incor-porado en todos los sistemas de eva-luación energética actuales, es inade-cuado y, consecuentemente, debesuprimirse. Argumentan, y es absolu-tamente cierto, que en un animal cuyaingesta le proporciona una retenciónenergética igual a cero puede tenerlugar síntesis neta de proteína a costade movilización de grasa corporal. Elmantenimiento, considerado como unaconstante, sólo tendría valor académi-co. Para otros autores no hay problemaen tener en cuenta las variaciones delmantenimiento, una vez que sean co-nocidas y cuantificadas. Finalmente,otros consideran que el aumento de lasnecesidades de mantenimiento en losanimales en producción es un costoenergético de la producción y debe re-flejarse en las estimaciones de utiliza-ción de la energía metabolizable para laproducción. Para estos autores las nece-sidades energéticas de mantenimientohan de considerarse constantes.
Tres son los métodos comúnmenteutilizados para estimar por balancecalorimétrico las necesidades de man-tenimiento: a) Un método consiste encorregir las ingestas de energíametabolizable que conducen a reten-ciones ligeramente positivas o negati-vas, empleando valores teóricos paralas eficiencias de utilización de la ener-gía metabolizable por debajo y porencima de mantenimiento. Siempre quelas retenciones energéticas obtenidasen los ensayos estén próximas a cero,el error que implica este procedimien-to es realmente despreciable; b) Unsegundo método consiste en emplearecuaciones de regresión del tipo ER =
Tabla V. Estimación del metabolismo basalmediante calorimetría indirecta en ganadoovino de raza Segureña y caprino adulto deraza Granadina. (Estimation of fasting
metabolism by indirect calorimetry in sheep of the
Segureña breed and in adult Granadina goats).
Especie Edad Peso vivo Producción deanimal (años) (kg) calor basal**
Ovino* 1,5 40 272Caprino* 2-3 26-33 324
*macho castrado; **(KJ/kg0,75 y día).

Archivos de zootecnia vol. 50, núm. 192, p. 583.
NUTRICIÓN ENERGÉTICA DE PEQUEÑOS RUMIANTES
a IEM - b, obtenidas a partir de ingestasenergéticas variables que promuevenretenciones positivas y calcular laingesta de energía metabolizable (IEM)necesaria para obtener una retenciónenergética (ER) igual a cero. Este es elmétodo que hemos usado con hembrasen lactación; y c) Con hembras encrecimiento y con machos castradoshemos empleado un tercer método con-sistente en determinar la producciónde calor de los animales en condicio-nes basales y aquella también con ni-veles de ingesta ligeramente inferioresal valor esperado para mantenimiento.Este procedimiento ofrece la ventajade que nos permite conocer, al propiotiempo, la eficiencia con que la ener-gía metabolizable de la dieta es utiliza-da por el animal para atender a losprocesos de mantenimiento.
La tabla VI resume los resultadosde los ensayos realizados (Prieto et al.,1990; Aguilera et al., 1990; Aguileraet al., 1991). Nuestros datos indicanque las necesidades de mantenimientode ganado caprino, independientemen-te de su situación fisiológica, son su-periores a las correspondientes a gana-do ovino de peso similar (318 KJ EM/kg0,75 y día, ARC, 1980; 374 KJ EM/
kg0.75 y día, Aguilera et al., 1986) yestán en el rango de valores publica-dos para vacuno de engorde y lecheroseco (418-469 KJ EM/kg0,75 y día;ARC, 1980). El NRC (1981) da unacifra media de 424 KJ EM/kg0,75 y díaobtenida mediante ensayos calorimé-tricos. A partir de los datos que figuranen la tabla VI hemos propuesto fijaren 422 KJ/kg0,75 y día la ingesta deenegía metabolizable que cubre lasnecesidades de mantenimiento del ga-nado caprino cualquiera que sea susituación fisiológica.
La estimación de la eficiencia conque la hembra en lactación utiliza laenergía metabolizable de la dieta parasus procesos productivos puede hacer-se según tres procedimientos: a) Cuan-tificar las diferencias entre determina-ciones de balance realizadas a dos ni-veles de producción; b) Cuantificar lasdiferencias entre un punto medido yotro estimado, correspondiente a man-tenimiento; y c) Realizar un análisis deregresión con datos correspondientesa determinaciones llevadas a cabo adistintos niveles de producción. Estaúltima técnica ha sido la utilizada ennuestros ensayos. Los planos deingesta se fijaron en función del ni-
Tabla VI. Estimación de las necesidades energéticas de mantenimiento (EMm) y del metabo-
lismo basal (producción de calor en ayuno, PCA) en ganado caprino (KJ/kg0,75 y día).(Estimation of maintenance energy requirements (EMm) and fasting metabolism (fasting heat production,
PCA) in the goat (KJ/kg0.75 day)).
Animal Ecuación de regresión PCA km EMm
Macho adulto castrado ER = 0,732 IEM - 324 234 0,732 443Hembra en crecimiento ER = 0,760 IEM - 320 320 0.760 421Hembra en lactación ER + EL = 0,669 IEM - 268 - - 401

Archivos de zootecnia vol. 50, núm. 192, p. 584.
AGUILERA
vel de producción láctea esperado.La determinación de la eficiencia
con que la energía metabolizable seutiliza para la producción de lechepresenta complicaciones especiales,porque en el animal en lactación, juntoa la producción de leche, pueden tenerlugar procesos concomitantes de de-posición de grasa corporal o de movi-lización de reservas para atender a lalactación, cuyas eficiencias difierende la que corresponde a la producciónláctea.
En nuestros ensayos sobre la utili-zación de la energía de la dieta paraatender a las necesidades de la lactaciónen el ganado caprino de raza Granadi-na (Aguilera et al., 1990) relaciona-mos la retención energética total, estoes, la energía excretada en leche (EL)más la retención corporal aparente(ER), con la ingesta de energíametabolizable (IEM) mediante regre-sión (tabla VII) . La ecuación (7.1)proporciona una estimación de la efi-ciencia con que la energía metabo-lizable se utiliza en los procesos pro-ductivos (0.669) y nos permite calcu-
lar la ingesta de energía metabolizablepara el mantenimiento cuando el tér-mino (EL + ER) es igual a cero. IEM
m= 268/0,669 = 401 KJ/kg0,75 y día. Estevalor es inferior a los valores mediospublicados para vacuno en producciónláctea (510 KJ EM/kg0,75 y día; Moe etal., 1970; 523 KJ EM/kg0,75 y día; vander Honing y van Es, 1974).
La eficiencia de utilización de laenergía metabolizable para la lactacióncuando no existe deposición ni movi-lización de reservas corporales (k
l) se
calculó de acuerdo con el ARC (1980).Para ello asumimos que la energía per-dida por el organismo, indicadora demovilización de grasa corporal en apo-yo a la secreción de leche, se utilizapara la síntesis de leche con una efi-ciencia del 84 p.100 y que la deposi-ción de energía en el organismo, con-comitante con la lactación, tiene lugarcon una eficiencia que es 0,95 veces lade la propia lactación: ELc = EL +(0,84 x ER(-)) + (1,05 x ER(+)). Cuan-do construimos la regresión usandocomo función este valor corregido deexcreción energética en leche y como
Tabla VII. Estimación de las necesidades energéticas de mantenimiento (IEMm) y producción
en la cabra en lactación (KJ/kg0,75 y día). (Estimation of energy requirements for maintenance (IEMm)
and production in the lactating goat (KJ/kg0.75 day)).
Ecuación de regresión n EMm kl
(7.1) EL + ER = 0,669 IEM - 268 70 401 -(7.2) ELc = 0,667 IEM - 258 70 - 0.667(7.3) ELc = 0,657 IEM - 237 46 0.657(7.4) ER/100 Kj IEMp* = - 0,907EL/100 Kj IEMp +67,6 46 -(7.5) ELc = EL + 0,84 ER + 1,05 ER
*IEMp = IEM - EMm

Archivos de zootecnia vol. 50, núm. 192, p. 585.
NUTRICIÓN ENERGÉTICA DE PEQUEÑOS RUMIANTES
variable independiente la ingesta deenergía metabolizable, se obtiene laecuación (7.2) de la tabla VII , queindica que en el ganado caprino laeficiencia de uso de la energíametabolizable para la lactación es del66,7 p.100. La eficiencia obtenida esligeramente superior al rango de valo-res teóricos basados en consideracio-nes bioquímicas (60-65 p.100) y nosupone, por tanto, diferenciación al-guna con los valores de eficiencia deuso de la energía metabolizable para lalactación en otras especies rumiantes:vacuno y ovino. Como el contenidoenergético medio de la leche de cabragranadina (con el 5,88 p.100 de gra-sa) es de 3,59 MJ/kg, se necesitan3,59/0,667 = 5,38 MJ EM/kg de lechedel 5,88 p.100 de grasa (4,20 MJ EM/kg de leche con un contenido en grasadel 4 p.100).
Cuando para construir la regresiónutilizamos exclusivamente los datoscorrespondientes a animales en los quejunto a la producción de leche hubouna deposición energética tisular con-comitante, la eficiencia de uso de laenergía metabolizable para la lactaciónes muy semejante: 65,7 p.100 (ecua-ción (7.3) de la tabla VII ). En estecaso, si la producción energética enforma de leche y la retención energéti-ca corporal se expresan como porcen-taje de la ingesta de energía metabo-lizable dada por encima de manteni-miento, utilizando para EM
m el valor
de 401 KJ/kg0,75 y día, y se estableceuna regresión con ambas series de da-tos, con RE/l00 KJ EM
p como función
y ER/100 KJ EMp como variable inde-
pendiente, se obtiene la ecuación (7.4)que indica que en el animal en lactaciónuna caída en la producción de leche de
1 KJ está asociada a un incremento enla deposición de grasa corporal de 0,907KJ. Ello indica que la cabra en lactacióndeposita grasa con una eficiencia quesería el 90,7 p.100 de la eficiencia parala lactación, es decir, con una eficien-cia del 0,907 x 0,667= 0,605. Estosresultados demuestran, en concordan-cia con otras investigaciones, que en elanimal en lactación la eficiencia conque la energía metabolizable se utilizapara la deposición de grasa corporal essuperior a la que presenta el animal nolactante. Las cifras bibliográficas os-cilan entre 0,78 y 1,13. El ARC (1980)ha adoptado un valor de 0,95 veces laeficiencia de uso de la energíametabolizable para la lactación. Encabras en lactación Armstrong yBlaxter (1965) obtuvieron una cifra de0,964 veces k
l, siendo k
l 69,1 p.100.
LA ACTIVIDAD FÍSICADe gran interés son los datos que se
ofrecen a continuación procedentes deensayos llevados a cabo con el propó-sito de cuantificar los gastos energéti-cos (transformación de energía quími-ca en trabajo mecánico) relacionadoscon la locomoción (Lachica et al.,1997a) y con la ingestión de alimento(Lachica et al., 1997b), actividades deespecial significación para el animalen pastoreo. Se recomienda su incor-poración, a través de un sistemafactorial, a las necesidades energéti-cas de mantenimiento y producciónestimadas con ganado caprino en con-finamiento. Respecto a la locomoción,apenas existen datos obtenidos en es-pecies de interés ganadero acerca delcosto energético del desplazamientosobre plano descendente. En cuanto ala ingestión, es poco lo que se conoce

Archivos de zootecnia vol. 50, núm. 192, p. 586.
AGUILERA
sobre el costo energético de la activi-dad muscular relacionada con la apre-hensión, masticación, deglución yrumia y la posible influencia que lanaturaleza físico-química del alimen-to tiene sobre dicho costo.
El gasto energético de la locomo-ción contribuye significativamente alos requerimientos energéticos de losanimales que viven en libertad y hande tenerse en cuenta si se desean eva-luar de modo preciso las necesidadesenergéticas de los animales en pasto-reo. Este gasto energético está relati-vamente bien definido en el vacuno yovino y, sin embargo, no se habíanrealizado hasta nuestros ensayos estu-dios sistemáticos con ganado caprino,con una sola excepción. La existenciade diferencias interespecíficas en efi-ciencia locomotora está demostrada yson consecuencia de adaptacionesmorfológicas, fisiológicas y de com-portamiento. Nuestros resultados, re-sumidos en la tabla VIII , indican queen la cabra de raza Granadina el costoenergético de la locomoción aumentadesde 1,91 a 6,44 J/kg peso vivo ymetro de desplazamiento al elevarse lapendiente desde -10 a +10 p.100. Di-
cho costo (ECw) puede ser calculado
en función de la pendiente (Sl) según la
ecuación exponencial:
ECw = 3,39 e 0,063 Sl r = 0,996; RSD = 0,049;
n = 5 (6)
La ecuación (7) permite obtener losvalores medios de dicho coste, calcu-lados para cada animal por separaciónde sus componentes horizontal y verti-cal y mediante regresión múltiple de laproducción de calor (PC, J/kg pesovivo y hora sobre las distancias reco-rridas horizontal (D
h) y verticalmente
(Du y D
d, para ascenso y descenso,
respectivamente), dadas en metros:
PC = 6724 + 3,31 Dh + 31,7 Du - 13,2 Dd (7)
Los coeficientes de regresión deD
h, D
u y D
d indican los valores del
coste energético neto (J/kg peso vivo ymetro) para el desplazamiento en hori-zontal (EC
w), en vertical con sentido
ascendente (ECu) o descendente (EC
d).
De acuerdo con nuestros ensayos,el coste energético neto del desplaza-miento en horizontal es en la cabra deraza Granadina de 3,35 J/kg peso vivo
Tabla VIII . Valores medios (n=6) del gasto energético de la locomoción (ECw, J/kg peso vivo
y metro de desplazamiento) obtenidos en ganado caprino mediante regresión de la produc-ción total de calor (PC, J/kg peso vivo/hora) sobre la distancia recorrida (m). (Mean values
(n=6) of the energy cost of walking (ECw, J/kg body weight per meter) for goats estimated by regression
of heat production (HP, kJ/kg body weight per hour) vs. distance travelled (m)).
Pendiente, p.100- 10 - 5 0 + 5 +10
ECw 1,91±0,129 2,33±0,130 3,35±0,127 4,68±0,148 6,44±0,245PC de pie, sin desplazamiento 6960±365 6910±366 6883±342 6841±333 6838±329

Archivos de zootecnia vol. 50, núm. 192, p. 587.
NUTRICIÓN ENERGÉTICA DE PEQUEÑOS RUMIANTES
y metro, cifra algo inferior a la de 3,63J/kg peso vivo y metro que puede cal-cularse a partir de los datos de Tayloret al. (1974) obtenidos en otra razacaprina. Los datos que ofrece la bi-bliografía sobre estos costes energéti-cos (J/kg peso vivo y metro) son muyinferiores, tanto en ovino (2,47,Clapperton, 1964; 2,83, Farrell et al.,1972; 2,30, Brockway y Boyne, 1980)como en vacuno (2,09, Ribeiro etal.,1977; 1,54, Shibata et al., 1981;1,91, Lawrence y Stibbards, 1990).
Nuestras investigaciones indicaronque la eficiencia energética media deldesplazamiento en sentido vertical as-cendente, calculada como la relaciónentre el trabajo realizado y el costeenergético de llevarle a cabo y expre-sada como porcentaje, es en la cabra deraza Granadina del 30,9 p.100. El ARC(1980) asume tanto para ovino comovacuno el valor de 28 J/kg peso vivo ymetro de desplazamiento en vertical,equivalente a una eficiencia energéti-ca del 35 p.100.
La mayor parte del incremento delgasto energético relacionado con laactividad física procede de la activi-dad muscular ligada a la locomoción eingestión de alimento. Los procesoscontinuos de contracción y relajaciónmuscular que tienen lugar durante laaprehensión, masticación y propulsióndel alimento en el tracto digestivo con-tribuyen notablemente a aumentar estecomponente. El coste energético de laingestión del alimento está bastantedocumentado en ovino (Osuji et al.,1975) y vacuno (Adam et al., 1984),sin que existiesen datos publicadossobre observaciones llevadas a caboen caprino con anterioridad a nuestrosensayos.
La medida del coste energético dela ingestión de alimento se llevó a cabocon cuatro cabras adultas dotadas defístula ruminal en ensayos individua-les en los que se determinó su produc-ción de calor durante periodos deaproximadamente 15 minutos mien-tras permanecían alojadas en la cáma-ra de confinamiento.
Los animales consumieron alimen-tos de naturaleza y composición dis-tintas, desde arbustos a concentrados.El coste energético de la ingestión dealimento se calculó a partir del incre-mento en producción de calor obser-vado respecto al periodo previo al su-ministro de alimento. Dicho costo serelacionó con el tipo y cantidad delalimento consumido y con el tiempoempleado en ingerir el alimento (tablaIX) .
En un ensayo paralelo se colocaronen el rumen a través de una fístularuminal cantidades similares a las con-sumidas por los animales en las prue-bas anteriormente citadas. Los incre-mentos en producción de calor obser-vados respecto al periodo inmediata-mente anterior en que los animalespermanecieron en ayunas, fueron des-preciables, lo que demostraba que elaumento observado en la producciónde calor de los animales durante laingestión de los alimentos ensayadosestaba relacionado exclusivamente conla ingestión per se, esto es, con laaprehensión, masticación y propulsióndel alimento hacia el rumen. El ritmode ingestión (g de materia seca/minu-to) osciló entre 6,3 para alfalfa frescay 46-99 para los concentrados. El cos-te energético medio de la ingestión dealimento (J/kg peso vivo/g MS) fue7,08 para la alfalfa fresca; 9,02 para
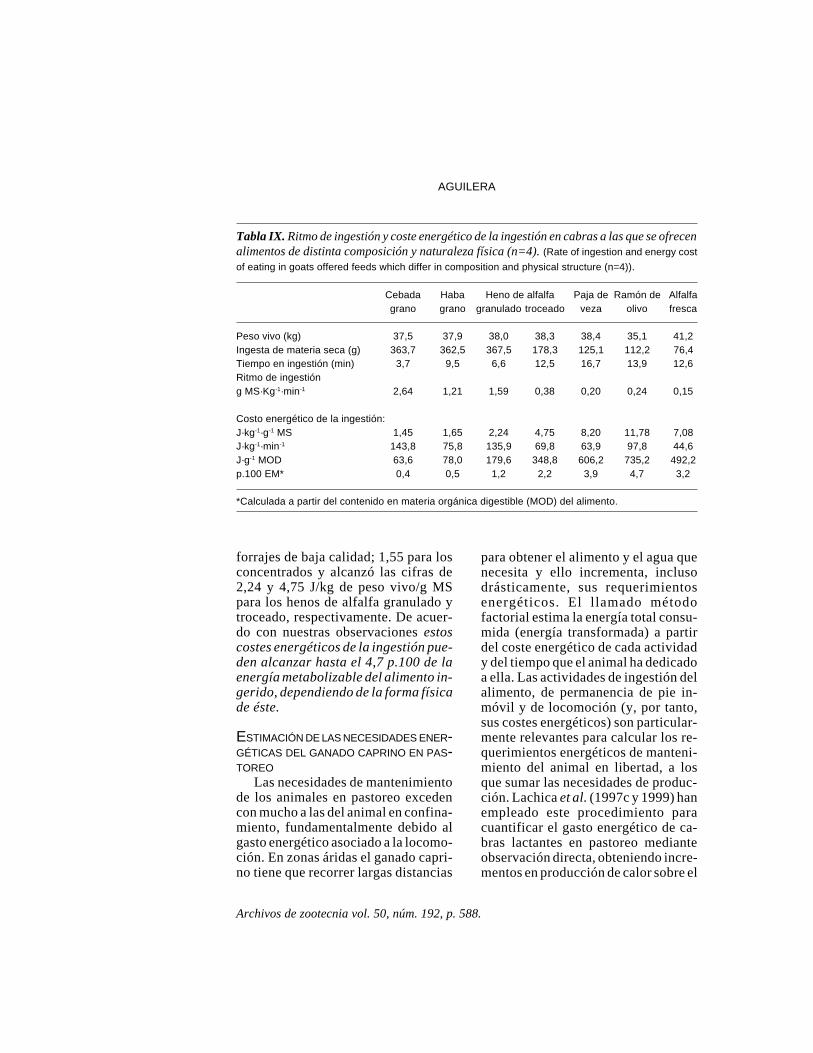
Archivos de zootecnia vol. 50, núm. 192, p. 588.
AGUILERA
forrajes de baja calidad; 1,55 para losconcentrados y alcanzó las cifras de2,24 y 4,75 J/kg de peso vivo/g MSpara los henos de alfalfa granulado ytroceado, respectivamente. De acuer-do con nuestras observaciones estoscostes energéticos de la ingestión pue-den alcanzar hasta el 4,7 p.100 de laenergía metabolizable del alimento in-gerido, dependiendo de la forma físicade éste.
ESTIMACIÓN DE LAS NECESIDADES ENER-GÉTICAS DEL GANADO CAPRINO EN PAS-TOREO
Las necesidades de mantenimientode los animales en pastoreo excedencon mucho a las del animal en confina-miento, fundamentalmente debido algasto energético asociado a la locomo-ción. En zonas áridas el ganado capri-no tiene que recorrer largas distancias
para obtener el alimento y el agua quenecesita y ello incrementa, inclusodrásticamente, sus requerimientosenergéticos. El l lamado métodofactorial estima la energía total consu-mida (energía transformada) a partirdel coste energético de cada actividady del tiempo que el animal ha dedicadoa ella. Las actividades de ingestión delalimento, de permanencia de pie in-móvil y de locomoción (y, por tanto,sus costes energéticos) son particular-mente relevantes para calcular los re-querimientos energéticos de manteni-miento del animal en libertad, a losque sumar las necesidades de produc-ción. Lachica et al. (1997c y 1999) hanempleado este procedimiento paracuantificar el gasto energético de ca-bras lactantes en pastoreo medianteobservación directa, obteniendo incre-mentos en producción de calor sobre el
Tabla IX. Ritmo de ingestión y coste energético de la ingestión en cabras a las que se ofrecenalimentos de distinta composición y naturaleza física (n=4). (Rate of ingestion and energy cost
of eating in goats offered feeds which differ in composition and physical structure (n=4)).
Cebada Haba Heno de alfalfa Paja de Ramón de Alfalfagrano grano granulado troceado veza olivo fresca
Peso vivo (kg) 37,5 37,9 38,0 38,3 38,4 35,1 41,2Ingesta de materia seca (g) 363,7 362,5 367,5 178,3 125,1 112,2 76,4Tiempo en ingestión (min) 3,7 9,5 6,6 12,5 16,7 13,9 12,6Ritmo de ingestióng MS·Kg-1·min-1 2,64 1,21 1,59 0,38 0,20 0,24 0,15
Costo energético de la ingestión:J·kg-1·g-1 MS 1,45 1,65 2,24 4,75 8,20 11,78 7,08J·kg-1·min-1 143,8 75,8 135,9 69,8 63,9 97,8 44,6J·g-1 MOD 63,6 78,0 179,6 348,8 606,2 735,2 492,2p.100 EM* 0,4 0,5 1,2 2,2 3,9 4,7 3,2
*Calculada a partir del contenido en materia orgánica digestible (MOD) del alimento.

Archivos de zootecnia vol. 50, núm. 192, p. 589.
NUTRICIÓN ENERGÉTICA DE PEQUEÑOS RUMIANTES
valor aceptado de mantenimiento (401KJ EM/kg0,75 y día; Aguilera et al.,1990) variables entre 10,8 y 26,3 p.100.Factores tales como composición bo-tánica, disponibilidad del sustrato ve-getal, manejo, etc. afectan a esteparámetro, de gran importancia para elcálculo de la carga animal. El métodofactorial parece ser adecuado para si-mular y cuantificar las actividades delganado caprino en pastoreo por obser-vación directa. Es simple y fácil deaplicar en campo, siendo suficiente unúnico observador.
Por su importancia e innovación hede mencionar los trabajos llevados acabo recientemente en nuestro depar-tamento para facilitar la aplicación dela técnica CER (iniciales tomadas delinglés carbon dioxide entry rate) a laestimación del gasto energético totalde la cabra en pastoreo. El método sebasa en la medida de la tasa aparentede ingreso de CO
2 en los compar-
timentos corporales utilizando –y aquíreside el carácter innovador- H13CO
3-
como marcador. Este procedimientoisotópico requiere la infusión constan-te del bicarbonato marcado y elmuestreo frecuente o continuo de unfluido fisiológico para determinar ladilución del isótopo cuando se alcanzael equilibrio. Se estima así la produc-ción de CO
2 del animal que ha de
relacionarse con su producción de ca-lor asumiendo un determinado valorpara el cociente respiratorio.
Los primeros intentos de utilizar latécnica CER para obtener informaciónsobre el gasto energético de rumiantesen pastoreo se deben a Corbett et al.(1971) usando H14CO
3- en ganado ovi-
no. Otros autores le han usado tanto enovino como en vacuno (Plaza, 1989;
Sahlu et al., 1989 y Sánchez y Morris,1984; Di Marco et al., 1996, respecti-vamente, por citar los trabajos másrecientes). El uso de un isótopo establedel carbono en lugar del 14C ofrecegran ventaja desde el punto de vista dela seguridad del operador y del entor-no, si bien presenta dos tipos de difi-cultades:
-El carbono 13 alcanza en la natu-raleza aproximadamente el 1,1 p.100del carbono total. Las plantas C
3, que
fijan el CO2 según el ciclo de Calvin,
difieren en abundancia natural de esteisótopo de las plantas C
4 que fijan el
CO2 a través del ciclo dicarboxílico.
Este distinto fraccionamiento isotópicopuede ocurrir también en componen-tes de los tejidos animales y, así, elenriquecimiento natural en carbono 13de grasas y carbohidratos puede dife-rir significativamente, lo que es im-portante en relación con la magnituddel cambio que se desea cuantificar.Este problema se soluciona midiendola abundancia isotópica en muestrastomadas inmediatamente antes de ini-ciar la infusión del marcador.
-El problema más serio que presen-ta la técnica CER se debe al secuestrodel carbono marcado en ciertosmetabolitos corporales, que impide larecuperación cuantitativa del isótopoy que, si no es tenido en cuenta, condu-ce a una sobreestimación de la produc-ción de CO
2. Esta fijación del CO
2puede ocurrir en distintas vías metabó-licas; por ejemplo, síntesis de urea,reacciones del ciclo tricarboxílico, es-pecialmente las catalizadas por lapiruvato carboxilasa y la malatodeshidrogenasa, o en compartimentosque se renuevan muy lentamente; porejemplo, el carbonato óseo (Elia et al.,

Archivos de zootecnia vol. 50, núm. 192, p. 590.
AGUILERA
1988; Rocha et al., 1994).La validación del método ha reque-
rido que realicemos observacionesacerca de los efectos de factores diver-sos sobre la abundancia natural; paradiscernir cual es el tiempo necesariopara alcanzar el equilibrio isotópico y,finalmente, para determinar la recupe-ración de 13C. Y así, Prieto et al. (2001)han estudiado el efecto del ejerciciofísico sobre la abundancia natural de13C. Las muestras analizadas eran desaliva recogidas mediante un catéterimplantado en la glándula parótida decabras sometidas o no a un programade locomoción intermitente, observán-dose que la actividad física provocauna caída significativa en abundancianatural de 13C. En colaboración coninvestigadores del Instituto (Kika) dela Garza, de la Universidad de Langston(Oklahoma) uno de nosotros ha estu-diado en caprino los efectos de loscambios en el plano de alimentación(Lachica et al., 2001a) y de la exposi-ción al frío (Lachica et al., 2001b)sobre la abundancia natural de carbo-no 13. Tras tres días de ayuno se pro-duce un descenso en abundancia natu-ral frente al animal alimentado adlibitum. Un efecto similar es observa-do por la exposición al frío, que pareceestar asociado con cambios en los rit-mos relativos de oxidación de sustratosmetabólicos ligados a la termogénesis.En ambos casos se produce un aumen-to en la oxidación de grasas, que pre-sentan menor abundancia natural encarbono 13. Por otro lado, cuando seutiliza saliva como sustrato en el quedeterminar los cambios en enriqueci-miento isotópico el tiempo necesariopara alcanzar el equilibrio en loscompartimentos corporales tras la in-
fusión del isótopo es de 8-16 horas(Prieto et al., 1991; Sahlu et al., 1992;Lachica, 1993; Herselman et al.,1998).
Prieto et al. (1997) determinaron laproducción de CO
2 en cabras que per-
manecían de pie inmóviles o camina-ban sobre la cinta de un ergómetrocolocado en el interior de una de lascámaras de respirometría de circuitoabierto existentes en nuestro departa-mento. Los animales recibían una in-fusión continua de solución deNaH13CO
3 en suero fisiológico a tra-
vés de un catéter insertado en la yugu-lar, iniciada 15 horas antes de comen-zar el muestreo de saliva de la glándulaparótida. Al propio tiempo se registra-ba la producción de CO
2. Se observó la
existencia de una relación significati-va entre el CO
2 producido, cuantifica-
do por respirometría, y la producciónestimada según la técnica CER. Larecuperación aparente media del isóto-po, calculada mediante la relación CO
2producido / CER, es la que establecenlas ecuaciones (8), obtenida cuandolos animales permanecieron inmóvi-les, y (9), calculada cuando realizaronun ejercicio físico intermitente:
CO2r = 0,667 CO2CER (8)
CO2r = 0,744 CO2CER (9)
donde CO2r indica el volumen de CO
2expirado (ml/kg peso vivo y hora),determinado por respirometría, yCO
2CER es la producción de CO
2 (ml/kg
peso vivo y hora) estimada por la téc-nica CER. En estos ensayos los valo-res estimados por la técnica CER exce-den a los respirométricos, indicando elsecuestro de carbono marcado enmetabolitos corporales, lo que impide

Archivos de zootecnia vol. 50, núm. 192, p. 591.
NUTRICIÓN ENERGÉTICA DE PEQUEÑOS RUMIANTES
su excreción cuantitativa. En un poste-rior ensayo (Prieto et al., 2001) corro-boramos la existencia del efecto de laactividad física sobre la fijación delcarbono marcado: se obtuvo una recu-peración menor (0,72) de 13C cuandolas cabras permanecieron inmóvilesque cuando desarrollaron un programade locomoción intermitente (0,78). Larecuperación del isótopo varía amplia-mente. En caprino Prieto et al. (1991)y Junghans et al. (1997) han obtenidocoeficientes de 0,87 y 0,92. Sin embar-go, es improbable que estas diferen-cias introduzcan desviaciones importan-tes en las estimaciones de la producciónde calor del animal en pastoreo si larelación CO
2 producido/CER ha sido
establecida en animales alimentados.Basados en nuestras propias obser-
vaciones (Aguilera et al., 1990; Lachi-ca, 1993) hemos propuesto utilizar lassiguientes ecuaciones de regresión paracalcular la producción total de calor(PC, J/kg peso vivo y hora) del animalen pastoreo a partir de la estimacióndel CO
2 producido (CO
2 = coeficiente
de recuperación aparente de 13CO2 x
valor CER, ml/kg peso vivo y hora):
PC = 19,82 CO2; RQ = 1,0 (10)
PC = 24,07 CO2; RQ = 0,87 (11)
Muy recientemente se han llevadoa cabo con total satisfacción variosensayos (Lachica et al., 2001a y b)usando la técnica CER y 13C para esti-mar la producción de calor de caprinoen pastoreo. Se han empleado machoscastrados a los que se les ha adaptadoun arnés en el que se ha colocado unabomba de infusión y bolsas depolietileno que contenían la soluciónde NaH13CO
3 y para la recogida conti-
nua de saliva. Los animales estabanprovistos de dos catéteres: uno enparótida y otro en yugular. Los anima-les pastaban en una parcela de unahectárea. La producción de calor al-canzó 401 y 574 KJ/kg0,75 y día, en elanimal estabulado y en pastoreo, res-pectivamente. La distancia total reco-rrida por los animales en pastoreo nosuperó los 2 km, lo que explicó sólo un9 p.100 del incremento en producciónde calor, por lo que el resto ha de seratribuido a las demás actividades inhe-rentes al pastoreo. Como conclusiónpodemos indicar que el empleo de unainfusión intravenosa de NaH13CO
3 y
muestreo continuo de saliva pareceser, por tanto, una técnica adecuadapara determinar la producción de CO
2y, consecuentemente, la producción decalor de ganado caprino en pastoreo.
BIBLIOGRAFÍA
Adam, I., B.A. Young, A.M. Nicol and A.A. Degen.1984. Energy cost of eating in cattle givendiets of different form. Anim. Prod., 38: 53-56.
AFRC Technical Committee on Responses toNutrients. Report No. 10, 1997. En: Thenutrition of Goats. Nutrition Abstracts andReviews (Series B), 67: 765-830.
Agricultural Research Council. 1980. The NutrientRequirements of Ruminant Livestock.Technical review by an Agricultural ResearchCouncil Working Party. CommonwealthAgricultural Bureaux, Farnham Royal, UK.
Agricultural Research Council. 1984. The NutrientRequirements of Ruminant Livestock.

Archivos de zootecnia vol. 50, núm. 192, p. 592.
AGUILERA
Supplement No 1. Slough, UK; Common-wealth Agricultural Bureaux.
Aguilera, J.F. 1989. Aprovechamiento desubproductos agroindustriales en la alimen-tación de rumiantes. Rev. Arg. Nutr. Anim., 9:253-267.
Aguilera, J.F. and C. Prieto. 1985. An openrespiration unit for calorimetric studies withsmall animals. Arch. Anim. Nutr., 35: 825-833.
Aguilera, J.F. and C. Prieto. 1986. Descriptionand function of an open-circuit respirationplant for pigs and small ruminants and thetechniques used to measure energy metabo-lism. Arch. Anim. Nutr., 36: 1009-1018.
Aguilera, J.F. and C. Prieto. 1991. Methaneproduction in goats given diets based onlucerne hay and barley. Arch. Anim. Nutr., 41:77-82.
Aguilera, J.F. et E. Molina. 1986. Valorisationnutritive d’un grignon d’olive traité à la soude.Ann. Zootech., 35: 205-218.
Aguilera, J.F., C. Prieto and J. Fonollá. 1990.Protein and energy metabolism of lactatingGranadina goats. Br. J. Nutr., 63: 165-175.
Aguilera, J.F., E. Molina, C. Prieto y J. Boza.1986. Estimación de las necesidades ener-géticas de mantenimiento en ganado ovinode raza Segureña. Arch. Zootec., 35: 89-96.
Aguilera, J.F., L. Lara, E. Molina and C. Prieto.1991. Energy balance studies with growingGranadina goats during fasting and mainte-nance. Small Rum. Res., 5: 109-115.
Aguilera, J.F., M. Bustos and E. Molina. 1992.The degradability of legume seed meals inthe rumen: effect of heat treatment. Anim.Feed Sci. Technol., 36: 101-112.
Armstrong, D.G. and K.L. Blaxter. 1965. Effects ofacetic and propionic acids on energy retentionand milk secretion in goats. Proc. 3rd Symp.on Energy Metabolism of Farm Animals. EAAPPubl. No. 11, p. 59-70 (KL Blaxter, ed). London:Academic Press.
Blaxter, K.L. and J.L. Clapperton. 1965. Predictionof the amount of methane produced byruminants. Br. J. Nutr., 19: 511-522.
Boza, J., G. Varela, J. Fonollá y C. Rodríguez.1966. Influencia de las pajas de trigo y habassobre la digestibilidad de la harina de algo-dón. Rev. Nutr. Anim., 4: 191-196.
Boza, J. y G. Varela. 1960. Experiencias dedigestibilidad en cerdos retintos de tipo ibéri-co. Ars Pharm., 1: 181-195.
Boza, J. y G. Varela. 1961. Digestibilidad y valornutritivo en cerdos ibéricos del orujo bruto deaceituna. Rev. Granja, 9: 31-49.
Boza, J., F.J. Muñoz, J.F. Aguilera y E. Molina.1987. Valoración nutritiva del cañote de gira-sol (Helianthus annus L.) tratado con hidróxi-do sódico en ganado cabrío. Arch. Zootec.,36: 253-259.
Boza, J., G. Varela y J. Fonollá. 1967. Estudio delvalor nutritivo de la harina de cártamo enóvidos. Rev. Nutr. Anim., 5: 228-234.
Boza, J., J. Fonollá y J. F. Aguilera. 1970. Aprove-chamiento de subproductos agrícolas- indus-triales en la alimentación del ganado ovino. I.Estudio de la digestibilidad de dietas a basede orujo de aceituna y melazas de remola-cha. Rev. Nutr. Anim., 8: 13-22.
Boza, J., J.E. Guerrero, J.F. Aguilera y V.Escandón. 1981a. Estudio de la cascarilla dela semilla de girasol (Helianthus annus) en laalimentación del ganado cabrío. AYMA, 22:305-307.
Boza, J., J.E. Guerrero, J.F. Aguilera, R. Sanz yE. Molina. 1981b. Utilización de dietas conbiuret como fuente de nitrógeno no proteicoen la alimentación del ganado cabrío. Arch.Zootec., 30: 157-169.
Brockway, J.M. and A.W. Boyne. 1980. The energycost for sheep of walking on gradients. Proc.8th Symp. Energy Metabolism, EAAP Publ.No. 26. (L.E. Mount ed.), p.449-453. London:Butterworths.
Brody, S. 1945. Bioenergetics and growth, 1974ed.. New York: Hafner Press.
Brouwer, E. 1965. Report of Sub-Committee onConstants and Factors. Proc. 3rd Symp. onenergy metabolism, EAAP Publ. No. 11, p.441-443 (K.L. Blaxter, ed.). London: AcademicPress.

Archivos de zootecnia vol. 50, núm. 192, p. 593.
NUTRICIÓN ENERGÉTICA DE PEQUEÑOS RUMIANTES
Clapperton, J.L. 1964. The energy metabolism ofsheep walking on the level and on gradients.Br. J. Nutr., 18: 47-54.
Corbett, J.L., D.J. Farrell, R.A. Leng, G.L.McClymont and B.A. Young. 1971. Determi-nation of the energy expenditure of pennedand grazing sheep from estimates of carbondioxide entry rate. Br. J. Nutr., 26: 277-291.
Di Marco, O.N., M.S. Aello and D.G. Mendez.1996. Energy expenditure of cattle grazing onpastures of low and high availability. Anim.Sci., 63: 45-50.
Elia, M., N. Fuller and O. Murgatroid. 1988. Thepotential use of the labelled bicarbonatemethod for estimating energy expenditure inman. Proc. Nutr. Soc., 47: 247-258.
Farrell, D.J., R.A. Leng and J.L. Corbett. 1972.Undernutrition in grazing sheep. II. Calori-metric measurements on sheep taken frompasture. Aust. J. Agric. Res., 23: 466-509.
Fonollá J. y J. Boza. 1993. Utilización de losresiduos del espárrago, procedentes de laindustria conservera, en la alimentación derumiantes. AYMA, 33: 163-165.
Fonollá, J., J. Silva y J. Boza. 1988. Estudio delvalor nutritivo de algunos subproductos agrí-colas procedentes de cultivos intensivos.AYMA, 28: 107-110.
Fonollá, J., R. Sanz Sampelayo y V. Escandón.1984. Utilización de subproductos agrícolas-industriales en la alimentación de animalesherbívoros. II. Bagazo de cerveza. AYMA, 25:471-477.
Fujihara, T., I. Tasaki and T. Furuhashi. 1973.Energetic utilization of starch introduced intoabomasum of goats. Proc. 6th Symp. on EnergyMetabolism of Farm AnimaIs. EAAP Publ.No. 14, p. 67-70. (KH. Menke, H.J. Lantzschy J.R. Reichl, ed.). Stuttgart: Univ. Hohenheim.
García, M.A. 1992. Estudio de la ingestión volun-taria y de la fermentación ruminal de pastosnaturales de zonas semiáridas en ganadocaprino y ovino. Tesis doctoral. Universidadde Granada.
García, M. A., J.F. Aguilera and E. Molina Alcai-de. 1995. Voluntary intake and kinetics of
degradation and passage of unsupplementedand supplemented pastures from semiaridlands in grazing goats and sheep. Livest.Product. Sci., 44: 245-255.
García, M.A., M.D. Isac, J.F. Aguilera and E.Molina Alcaide. 1994. Rumen fermentationpattern in goats and sheep grazing pasturesfrom semiarid Spanish lands unsupplementedor supplemented with barley grain or barleygrain-urea. Livest. Prod. Sci., 39: 81-84.
Goering, H.H. and P.J. van Soest. 1970. Foragefiber analyses (Apparatus, Reagents,Procedure and some applications). AgricultureHandbook no. 379. ARS, USDA, 20 pp.
Herselman, M.J., T. Sahlu, S.P. Hart and A.L.Goetsch. 1998. Energy expenditure by dryand lactating Alpine does estimated by entryrate of carbon dioxide. J. Dairy Sci., 81: 2469-2474.
Isac, M.D., M.A. García, J.F. Aguilera and E.Molina Alcaide. 1994. A comparative study ofnutrient digestibility, kinetics of digestión andpasaje and rumen fermentation pattern ingoats and sheep offered médium qualityforages at the maintenance level of feeding.Arch. Anim. Nutr., 46: 37-50.
Junghans, P., M. Derno, M. Gehre, Kowski, P.,Hofhing, G. Stranch, W. Jentsch, J. Voigt andV. Henning. 1997. Calorimetric validation of13C bicarbonate and doubly labeled watermethod for determining the energy expenditu-re in goats. Z. Ernahrungswiss., 36: 268-272.
Lachica, M. 1993. Estimación del gasto energéti-co de la actividad física en ganado caprino.Comparación de técnicas calorimétricas y nocalorimétricas. Tesis doctoral, Universidadde Granada.
Lachica, M., A.L. Goetsch y T. Sahlu. 2001a y b.Comunicación personal.
Lachica, M., C. Prieto and J.F. Aguilera. 1997a.The energy cost of walking on the level and onnegative and positive slopes in the Granadinagoat (Capra hircus). Br. J. Nutr., 77: 73-81.
Lachica, M., F.G. Barroso and C. Prieto. 1997c.Seasonal variation of locomotion and energyexpenditure in goats under range grazing

Archivos de zootecnia vol. 50, núm. 192, p. 594.
AGUILERA
conditions. J. Range Manage., 50: 234-238.Lachica, M., J.F. Aguilera and C. Prieto. 1995. A
confinement respiration chamber for shortgaseous exchange measurements. Arch.Anim. Nutr., 48: 329-336.
Lachica, M., J.F. Aguilera and C. Prieto. 1997b.Energy expenditure related to the act of eatingin Granadina goats given diets of differentphysical form. Br. J. Nutr., 77: 417-426.
Lachica, M., R. Somlo, F.G. Barroso, J. Boza andC. Prieto. 1999. Goats locomotion energyexpenditure under range grazing conditions:Seasonal variation. J. Range Manage., 52:431-435.
Lawrence, P.R. and R.J. Stibbards. 1990. Theenergy cost of walking, carrying and pullingloads on flat surfaces by Brahman cattle andswamp buffalo. Anim. Prod., 50: 29-39.
Lindgreen, E. 1980. Skattning av energiförlusteri metan och urin hos idisslare. SwedishUniversity of Agricultural Sciences. Depart-ment of Animal Nutrition. Report 47.
Moe, P.W. and H.F. Tyrrell. 1979. Methaneproduction in dairy cows. J. Dairy Sci., 62:1583-1586.
Moe, P.W., H.F. Tyrrell, and W.P. Flatt. 1970.Partial efficiency of energy use for mainte-nance, lactation, body gain and gestation inthe dairy cow. 5th Symp. on EnergyMetabolism of Farm Animals. EAAP Publ.No. 13, p. 65-68 (A. Schurch y C. Wenk, ed.).Zurich: Juris Druck & Verlag.
Mohammed, H.H. and E. Owen. 1980. Comparisonof the maintenance energy requirements ofsheep and goat. Anim. Prod., 30: 479.
Molina Alcaide, E., A.I. Martín García and J.F.Aguilera. 2000. A comparative study of nutrientdigestibility, kinetics of degradation andpassage and rumen fermentation pattern ingoats and sheep offered good-quality diets.Livest. Prod. Sci., 64: 215-223.
Molina Alcaide, E., M.A. García and J.F. Aguilera.1997. The voluntary intake and rumen diges-tión by grazing goats and sheep of a low-quality pasture from a semi-arid land. Livest.Prod. Sci., 52: 39-47.
Molina, E. et J.F. Aguilera. 1988. Valorisationnutritive d’un grignon d’olive traité à la soude.Utilisation digestive des constituents desparois cellulaires. Ann. Zootech., 37: 63-72.
Molina, E., J. Boza and J.F. Aguilera. 1983.Nutritive value for ruminants of sugar canevagase ensiled after spray treatment withdifferent levels of NaOH. Anim. Feed Sci.Technol., 9: 1-17.
Morand-Fehr, P. 1981. Nutrition and Feeding ofgoats: Application to temperate climaticconditions. En: Goat Nutrition (C. Gall, ed.),pp. 193-232. London: Academic Press.
Murray, R.M., A.M. Bryant and R.A. Leng. 1978.Methane production in the rumen and lowergut of sheep given Lucerne chaff: effect oflevel of intake. Br. J. Nutr., 39: 337-345.
National Rescarch Council. 1981. Nutrientrequirements of domestic animals, Publ. No.15. Washington: National Academy Press.
Neergaard, L. 1973. Methane production ingrowing calves in relation to the compositionof the food. Proc. 6th Symp. on energymetabolism, EAAP Publ. No. 14, p. 103-107(K.H. Menke, H.J. Lantzsch and J.R. Reichl,ed.). Stuttgart: Univ. Hohenheim.
Osuji, P.O., J.G. Gordon and A.J.F. Webster.1975. Energy exchanges associated witheating and rumination in sheep given grassdiets of different physical form. Br. J. Nutr.,34: 59-71.
Pérez Cuesta, M. y J. Boza. 1977. Valoraciónnutritiva del Lupinus albus var. Neuland enóvidos. Pastos, 7: 307-310.
Plaza, J. 1989. Necesidades energéticas de ove-jas gestantes en pastoreo. Tesis doctoral.Universidad de León.
Prieto, C. and J.F. Aguilera. 1986. The effect ofthe supplementation with methionine andlysine of diets based on lupin seed (Lupinusalbus var. multolupa) on protein and energyutilization in growing rats. J. Anim. Physiol. A.Anim. Nutr., 55: 239-246.
Prieto, C., E. Reyes, M. Lachica, E. Caballero andJ.F. Aguilera. 1991. Applications of the labelled13C-bicarbonate method for estimating energy

Archivos de zootecnia vol. 50, núm. 192, p. 595.
NUTRICIÓN ENERGÉTICA DE PEQUEÑOS RUMIANTES
expenditure of the goat under field conditions.En: International Symp. on Nuclear andRelated Techniques in Animal, InternationalAtomic Energy Agency, Food and AgricultureOrganization of the United Nations (FAO),Vienna, pp. 150.
Prieto, C., J.F. Aguilera, L. Lara and J. Fonollá.1990. Protein and energy requirements formaintenance of indigenous Granadina goats.Br. J. Nutr., 63: 155-163.
Prieto, C., M. Lachica, F. Bermúdez y J.F. Aguilera.1997. Aplicación del 13C para la estimacióndel gasto energético en cabras de raza Gra-nadina. 1997. En: Asociación Interprofesionalpara el Desarrollo Agrario (A.I.D.A.), XXIXjornadas de estudio, ITEA-VII jornadas sobreproducción animal, p. 16-18. Zaragoza.
Prieto, C., M. Lachica, R. Nieto and J.F. Aguilera.2001. The 13C-bicarbonate method: itssuitability for estimating the energy expen-diture in grazing goats. Livest. Prod. Sci., 69:207-215.
Ribeiro, J.M. de C.R., J.M. Brockway and A.J.F.Webster. 1977. A note on the energy cost ofwalking in cattle. Anim. Prod., 25: 107-110.
Rocha, H.J.G., A. Connell and G.E. Lobley. 1994.Whole body and trans-organ bicarbonatekinetics in sheep: effects of nutritional statusProc. 13th Symp. on energy metabolism, EAAPPubl. No. 76, p. 27-30 (J.F. Aguilera, ed.).Madrid: CSIC Publishing Service.
Roy-Smith, F. 1980. The fasting metabolism andrelative energy intake of goats compared withsheep. Anim. Prod., 30: 491.
Sahlu, T., H.G. Jung and J.G. Morris. 1989.Influence of grazing pressure on energy costof grazing by sheep on smooth bromegrass.J. Anim. Sci., 67: 2098-2105.
Sahlu, T., S.P. Hart, P.D. Klein, N. Jacquementand H. Carneiro. 1992. Aplication of 13CO2
measurements to the estimation of energyexpenditure in goats. Can. J. Anim. Sci., 72:245-252.
Sánchez, M.D. and J.G. Morris. 1984. Energyexpenditure of beef cattle grazing annualgrassland. Can. J. Anim. Sci., 64: 332-334.
Sanz Sampelayo, M.R., L. Pérez, F. Gil Extremera,J.J. Boza and J. Boza. 1999. Use of differentdietary sources for lactating goats: Milkproduction and composition as functions ofprotein degradability and amino acidcomposition. J. Dairy Sci., 82: 555-565.
Sanz Sampelayo, M.R., F.J. Muñoz, L. Lara, F.Gil Extremera and J. Boza. 1987. Factorsaffecting pre- and post-weaning growth andbody composition in kid goats of the Granadi-na breed. Anim. Prod., 45: 233-238.
Sanz Sampelayo, M.R., I. Prieto, L. Lara, F. GilExtremera and J. Boza. 1994. Granadina kidgoats v. Segureña lambs: food intake andperformance during milk feeding from birth to60 days. Anim. Prod., 58: 257-261.
Sanz Sampelayo, M.R., O.D. Hernández-Clua,J.A. Naranjo, F. Gil y J. Boza. 1990. Utilizationof goat milk vs. milk replacer for Granadinagoat kids. Small Rum. Res., 3: 37-46.
Shibata, M., A. Mukai and S. Kume. 1981.Estimation of energy expenditure in dairyheifers walking on the level and on gradients.Bull. Kyushu Natl. Agric. Exp. Stn., 21: 589-609.
Taylor, C.R., A. Shkolnik, R. Dmi'el, D. Baharavand A. Borut. 1974. Running in cheetahs,gazelles and goats: energy cost and limbconfiguration. Am. J. Physiol., 227: 848-850.
Tilley, J.M.A. and R.A. Terry. 1963. A two-stagetechnique for in vitro digestion of forage crops.J. Br. Grassl. Soc., 18: 104-111.
Van der Honing, Y. and A.J.H. van Es. 1974.Utilization of energy from pelleted roughagesin dairy cattle rations. Proc. 6th Symp. onEnergy Metabolism of Farm Animals. EAAPPubl. No. 14, p. 209-212. (KH. Menke, H.J.Lantzsch y J.R. Reichl, ed.). Stuttgart: Univ.Hohenheim.
Varela, G. y J. Boza. 1961. Digestibilidad y valornutritivo de las cabezas de remolacha azuca-rera en óvidos. AYMA, 2: 645-650.
Varela, G., J. Ferrer y J. Boza. 1961. Experien-cias de digestibilidad en cabras de raza gra-nadina. Zootecnia, 10: 3-10.
Whitelaw, F.G., J. Margaret Eadie, L.A. Bruce

Archivos de zootecnia vol. 50, núm. 192, p. 596.
AGUILERA
and W.J. Shand. 1984. Methane formation infaunated and ciliate-free cattle and itsrelationship with rumen volatile fatty acidproportion. Br. J. Nutr., 52: 261-275.
Wilkerson, V.A., D.P. Casper, D.R. Mertens and
H.F. Tyrrell. 1994. Evaluation of severalmethane prediction equations for dairy cows.Proc. 13th Symp. on energy metabolism, EAAPPubl. No. 76, p. 395-398 (J.F. Aguilera, ed.).Madrid: CSIC Publishing Service.


















