
ALCANCES DE LOS EFECTOS DEL ESTRÉS AGUDO SOBRE LA ...
159
1 Universidad de los Andes Ciencias sociales Departamento de psicología ALCANCES DE LOS EFECTOS DEL ESTRÉS AGUDO SOBRE LA INFLAMACIÓN DEPENDIENTE DE LA MICROGLÍA EN ESTRUCTURA HIPOCAMPALES Y SU RELACIÓN CON LA CONSOLIDACIÓN DE LA MEMORIA EMOCIONAL Y ESPACIAL EN UN BIOMODELO DE ANSIEDAD Trabajo de grado para optar al título de PSICÓLOGA Maria Alejandra Tangarife Cabrera Bajo la dirección de Fernando Cárdenas P. (Universidad de los Andes, Bogotá, Colombia) Bajo la asesoría de J. Landeira-Fernández (Pontifícia Uuniversidade Católica de Rio de Janeiro, Brasil - PUC) Bogotá, D.C., enero de 2018.
Transcript of ALCANCES DE LOS EFECTOS DEL ESTRÉS AGUDO SOBRE LA ...

1
Universidad de los Andes
Ciencias sociales
Departamento de psicología
ALCANCES DE LOS EFECTOS DEL ESTRÉS AGUDO SOBRE LA INFLAMACIÓN
DEPENDIENTE DE LA MICROGLÍA EN ESTRUCTURA HIPOCAMPALES Y SU
RELACIÓN CON LA CONSOLIDACIÓN DE LA MEMORIA EMOCIONAL Y
ESPACIAL EN UN BIOMODELO DE ANSIEDAD
Trabajo de grado para optar al título de
PSICÓLOGA
Maria Alejandra Tangarife Cabrera
Bajo la dirección de Fernando Cárdenas P. (Universidad de los Andes, Bogotá, Colombia)
Bajo la asesoría de J. Landeira-Fernández (Pontifícia Uuniversidade Católica de Rio de Janeiro,
Brasil - PUC)
Bogotá, D.C., enero de 2018.

2

3
A Gumball, del Increíble Mundo de Gumball.

4
Agradecimientos
Considero que no me alcanzan las palabras para expresar lo profundamente agradecida que estoy
con todas las personas que me apoyaron durante la realización de mi tesis, pero procederé con los
puntos que considero más importantes. En primer lugar, quisiera agradecerle al profesor J.
Landeira-Fernández, director del departamento de psicología y del laboratorio de Neurociencia
de la PUC en Rio de Janeiro y a todo el equipo de su laboratorio (Silvia, Flavia y a todos sus
estudiantes) por haberme apoyado en el 100% del financiamiento del presente trabajo y por haber
sido tan amables conmigo durante mi estadía en Brasil. En segundo lugar, quiero agradecer al
equipo del laboratorio de Neurociencia y Comportamiento de la Universidad de los Andes, pues
la culminación del trabajo no hubiera podido ser posible sin ellos. Así pues, quiero agradecerle al
profe Fernando por su admirable paciencia, dedicación, liderazgo y por supuesto, muchas
correcciones. Junto a él, Karencita, Diana y Martita (de manera paralela) resolvimos preguntas y
aclaramos dudas como si fuésemos un equipo; gracias por su apoyo. Por otro lado, quisiera
agradecerles a mis papás por haberme dado mucho amor y obligarme a ir al médico. Finalmente,
quisiera agradecerle a mi amiga Catalina por acompañarme durante noches enteras a mirarme
escribir, a mi amigo Sergio por no hacer nada, pero ser cool, a mi amigo Mateo por ser muy
paciente, nuevamente a Martita por darme muchos ánimos y especialmente a mi novio David por
darme tranquilidad. Muchas gracias, a todos los quiero.

5
Índice
Lista de Figuras………………………………………………………………………6
Resumen……………………………………………………………………………...7
Introducción………………………………………………………………………….8
Aproximaciones a la microglía.………………………………………...…………...10
Aproximaciones al hipocampo……………………………………………………...14
Aproximaciones al estrés…………………………………………………………....24
Aproximaciones a los efectos del estrés sobre el hipocampo……………………….33
Aproximaciones a los efectos del estrés sobre la microglía…………………...……43
Aportes a la inhibición de la microglía de fenotipo M1……………….…...……….53
Introducción a las cepas Carioca High Freezing y Carioca Low Freezing…………57
Método………………………………………………………….…………….…….64
Resultados…………………………………………………….………………….…71
Discusión……………………………………………………………………............95
Referencias……………………………………………………………………..….107

6
Lista de figuras
Figura 1. División de grupos experimentales……………………………………65
Figura 2. Ilustración de una caja de condicionamiento aversivo…………....…..67
Figura 3. Esquema general de Xplorat………………………………………..…67
Figura 4. Test de reconocimiento de objetos……………………………………69
Figura 5. Morfología de la microglía……………..………………………….….71
Figura 6. Media del tiempo de congelamiento durante el protocolo de miedo
condicionado…………………………………………………………….………73
Figura 7. Motricidad por el campo abierto……… ………………………….….76
Figura 8. Promedios del tiempo de congelamiento y frecuencia del congelamiento en los grupos
experimentales……………………………………………………….….… …..80
Figura 9. Promedios de tiempo y frecuencia de acicalamiento………..…..….. 82
Figura 10. Promedios de tiempo y frecuencia de estar parado en las patas
traseras………………………………………………………………………….84
Figura 11. Porcentaje de exploración de los objetos durante la fase de
familiarización………………………………………………………………….86
Figura 12. Porcentajes de exploración en el objeto familiar y novedoso………87
Figura 13. Microglía activada en el giro dentado…………………………..…..89
Figura 14. Microglía activada en el área CA3 del hipocampo estresado…….....91
Figura 15. Microglía activada en el área CA3 de los animales Nv……………..92
Figura 16. Microglía activada en el área CA1 de los grupos
experimentales…………………………………………………………………..94
Figura 17. Microglía activada en el CPU de los grupos
experimentales………………………………………………………………......96

7
Resumen
La microglía, principal célula reguladora de los procesos inmunes en el ambiente cerebral (Streit
et al., 2004), se destaca durante el estrés por su colaboración en procesos de proinflamación y
fagocitosis (Sugama et al., 2007). Se encuentra en grandes concentraciones en el hipocampo
(Lawson et al., 1990) y se ha observado que en condiciones de estrés fuertes puede responder de
manera poco adaptativa, al favorecer procesos de muerte neuronal e impedir la correcta
consolidación de memorias espaciales, en contraposición con emocionales (Milior et al., 2016;
Tanaka et al., 2006). El presente experimento evaluó los efectos de la respuesta inmune,
dependiendo del grado de vulnerabilidad ante el estrés usando un biomodelo de ansiedad
desarrollado en el laboratorio de neurociencia de la PUC-Rio (de Castro Gómes et al., 2011). El
modelo, compuesto por las cepas CHF (más congelamiento, “vulnerable”) y CLF (menos
congelamiento, “no vulnerable”) fue dividido en dos grupos (Si Estrés vs. No estrés); la mitad
recibió un estímulo aversivo (choque 0.7mA). Posteriormente, fue evaluado en protocolos
comportamentales relacionados con la consolidación de la memoria emocional y espacial, y
finalmente se midieron los niveles de inflamación del hipocampo y el CPU. Se identificó que el
estrés afectó los patrones de pro-inflamación el hipocampo de CHF, quien se caracterizó por
consolidar y mantener las respuestas relacionadas con el estímulo aversivo. En CLF, por el
contrario se observó que la pro-inflamación estuvo presente en los animales estresados y no
estresados sin diferencia. En cuanto al comportamiento, se observaron impedimentos en la
consolidación de las memorias, además del aumento de la motricidad, lo que permite sugerir
problemas cognitivos. Se cree que los patrones de inflamación pueden tener bases genéticas
derivadas del proceso de fenotipaje, pero no se considera que la información sea concluyente; se
recomiendan posteriores estudios para aclarar los interrogantes del experimento.

8
Introducción
El uso de modelos animales para la investigación en neurociencia del comportamiento ha
facilitado el entendimiento sobre los principios anatómicos y fisiológicos de la conducta humana
en la actualidad (Bovenkerk & Kaldewaij, 2015; Heeger, 2011; Panksepp, 2011).
Uno de los temas relacionados con el comportamiento que se ha pretendido explorar a
través del tiempo es el estrés y sus consecuencias cuando no es regulado correctamente por el
sistema. El estrés es una condición que ha cobrado una relevancia representativa en la sociedad,
pues es reconocido como un “gatillo” para el desarrollo de otras condiciones, tales como el
aumento de la propensión a los trastornos psicológicos y enfermedades como la depresión, la
ansiedad, la demencia, el cáncer, la diabetes, (Barnay, 2016; McEwen, 2000; Waugh, Hamilton,
Chen, Joormann, & Gotlib, 2012); y otros problemas cognitivos como la pérdida de la
concentración, la dificultad frente a la toma de decisiones y la pérdida de la memoria a corto
plazo (McEwen & Sapolsky, 1995).
Esencialmente, el estrés es una característica compartida a través de las especies, pues
permite al organismo movilizar sus recursos energéticos para adaptarse al ambiente (McEwen &
Gianaros, 2010) y regresar a un estado de homeostasis (Juster, McEwen, & Lupien, 2010). En
otras palabras, se reconoce como una reacción de supervivencia potencialmente adaptativa ante
condiciones normales y de cambio medioambiental (McEwen, 2007; de, V & Landeira-
Fernandez, 2008a; de Castro Gómes, Barroso Silva, & Landeira-Fernandez, 2011). Sin embargo,
los efectos sobre el sistema se encuentran en un dialogo constante con la intensidad, la duración,
la frecuencia y el grado de controlabilidad del estímulo (McEwen, 2000; Lucassen et al., 2014).
Se sabe que su mala regulación puede conducir al desarrollo de enfermedades (Rosch, 1979) y es
por esta razón existe la necesidad de explorar sus características biológicas y fisiológicas en
diferentes contextos experimentales y de vulnerabilidad.
El estrés es una condición que tiene una complicada fisiología que ha ser explorada con
determinación. Hoy en día se considera como un problema representativo que prevalece en
nuestra sociedad, hasta el punto de ser considerado uno de los principales causantes de
enfermedad en la población trabajadora (Chrousos, 2009). Su incidencia se ha reportado a través
del incremento de los costos de mantenimiento de las pensiones y los subsidios para ciudadanos

9
inhabilitados por causas de enfermedades relacionadas. Por ejemplo, en EEUU se ha superado
una suma media de 225,8 billones de dólares anuales para el tratamiento de enfermedades
(Stewart, Ricci, Chee, & Morganstein, 2003). De este dinero, aquel que sólo es dedicado al
tratamiento de la depresión o la ansiedad relacionadas al estrés laboral es de aproximadamente
42, 3 billones de dólares, de los cuales tan sólo para 1990, trataron a una tercera parte de los
diagnosticados (Lepine, 2002). En Europa se han reportado gastos que pueden oscilar entre 500€
y 1600€ anuales por persona (Andlin-Sobocki & Wittchen, 2005). Al mismo tiempo, su
prevalencia y la de otros trastornos relacionados aumenta alrededor del mundo tanto en las zonas
urbanas como en las zonas rurales (Bandelow & Michaelis, 2015; Kessler & Bromet, 2013).
En respuesta a esta situación, los gobiernos de EEUU y Europa, por ejemplo, han sugerido
nuevos programas de intervención que promueven la sostenibilidad laboral y el aumento del
bienestar de los trabajadores, por medio del mejoramiento del ambiente de trabajo, la adaptación
al trabajo y la prevención de accidentes y enfermedades (Barnay, 2016). Estos programas
proponen estrategias de relajación, asistencia psicológica e intervenciones relacionadas con los
individuos y las organizaciones (van der Klink, Blonk, Schene, & van Dijk, 2001; Murphy,
1996), pues la mejoría del trabajador consecuentemente generaría efectos positivos sobre la
eficiencia de las empresas (Royer Ghiglione, 2011). No obstante, los resultados no son los
mismos para todos los intervenidos, dado que las diferencias individuales establecen otras
estrategias de afrontamiento que cambian de persona a persona. Por lo tanto, es necesario seguir
explorando este problema desde otras perspectivas y campos de conocimiento, pues de esta
manera se podrían ofrecer soluciones efectivas, duraderas y por supuesto baratas. Desde la
neurociencia, se planea realizar un acercamiento a los efectos del estrés y sus relaciones con la
inflamación y los posteriores problemas cognitivos partiendo de un biomodelo de ansiedad. Todo
esto con el propósito de agregar nueva información que pueda ser utilizada en el futuro para
estudios o tratamientos relacionados.
A continuación, en los siguientes capítulos se realizará una aclaración general de los conceptos
más importantes que se deben tener en cuenta para entender los propósitos y objetivos de la presente
investigación. Si se tiene alguna duda sobre la organización de estos, por favor remítase al índice y escoja
el título que más le interese. Sin embargo, se recomienda fuertemente que se haga una revisión de todos
los capítulos para hacer más enriquecedor el aprendizaje después de la lectura.

10
Aproximaciones a la microglía
El cerebro es un órgano que va cambiando de acuerdo a las contingencias del ambiente
por medio de un mecanismo de reorganización, muerte y eventual nacimiento de células
nerviosas que modulan su balance inhibitorio y excitatorio (Pascual, Achour, Rostaing, Triller, &
Bessis, 2012). Estos cambios ayudan a modificar, en condiciones normales, la función de los
organismos, al abarcar el trabajo, tanto de funciones fisiológicas como de mecanismos de
aprendizaje y comportamiento a lo largo de la vida (Tay, Savage, Hui, Bisht, & Tremblay, 2016).
Durante la trasmisión sináptica, más de un grupo de células nerviosas se encuentra participando,
mediando los procesos de plasticidad; se ha observado que las células de la glía, como los
astrocitos y la microglía, medían la actividad de las neuronas (Pascual et al., 2012). En
circunstancias particulares como es el caso de proteger al cerebro de un peligro potencial (Cherry,
Olschowka, & O'Banion, 2014), los cambios estructurales y de funcionamiento se ven afectados
por los efectos inflamatorios, neurotóxicos y neuroprotectores de la microglía (Streit, Mrak, &
Griffin, 2004).
La microglía es, por excelencia, la célula inmune del sistema nervioso central (SNC) y es
una de las mediadoras principales de los efectos proinflamatorios y antiinflamatorios (Streit et al.,
2004). Esta célula, de origen hematopoyético, comparte su nacimiento en el saco vitelino con el
sistema de células llamadas fagocitos mononucleares (Hanisch & Kettenmann, 2007) sin
embargo, antes de la vascularización se diferencia y migra al SNC (Kissa & Herbomel, 2010;
Arnold & Betsholtz, 2013). Este tipo de células se diferencian de los otros macrófagos, pues
poseen una expresión de genes que sólo se encuentran dentro del SNC (Geissmann et al., 2010;
Gautier et al., 2012; Wolf, Yona, Kim, & Jung, 2013). Se ha evidenciado que se encuentra
ubicada en la sustancia gris, correspondiendo a un 5-20% de la población glial del cerebro
(Dalmau, Finsen, Zimmer, Gonzalez, & Castellano, 1998; Kim, Son, & Mook-Jung, 2013) y se
divide en fenotipos con funciones diferentes, de los cuales se han encontrado en mayor
concentración, en el hipocampo (Dalmau et al., 1998), en el cuerpo calloso y en la substancia
nigra (Kierdorf & Prinz, 2013). Dos fenotipos allí encontrados son el ameboideo y el ramificado
(Dalmau et al., 1998), los cuales median su activación de acuerdo a cambios ambientales, dentro
y fuera del cerebro (Meireles et al., 2016), como por ejemplo con las respuestas de estrés (Milior
et al., 2016). También se pueden identificar otros estados de que se encargan de establecer

11
funciones diferentes y restringidas de acuerdo a la naturaleza de la situación (Boche, Perry, &
Nicoll, 2013). De ahí se estimulan cambios en la morfología, la expresión de los genes y la
composición de las proteínas (Pascual, Ben, Rostaing, Triller, & Bessis, 2012; Boche et al.,
2013).
La microglía, como macrófago, se encarga de mantener la homeostasis del sistema por
medio de la comunicación con las neuronas (Wolf et al., 2013). Todo esto es desencadenado por
medio de señales endógenas y exógenas de cambio (Doens & Fernandez, 2014), detectadas
gracias al efecto de las moléculas liberadas por las células nerviosas sobre varios tipos de
receptores que se encuentran en la membrana de la célula microglial (Kierdorf & Prinz, 2013).
Entre estos, Kierdorf y Pritz (2013) han reportado los siguientes: los receptores CD200,
los cuales al estar activados ayudan a mantener el estado de reposo de la microglía; los receptores
inhibitorios CD47, cuyos ligandos transmiten el mensaje a la microglía de no fagocitar; la
glicoproteína TREM-2, que es asociada con la proteína DAP12 y estimula la fagocitosis y la
reparación del tejido en la microglía; el factor de estimulación de colonias 1 (Csfr1), activado por
la Interleucina 34 (IL-34); el factor estimulador de colonias (Csf1), que es recibido como una
señal de sobrevivencia para la célula; y el receptor de quimosina (CX3C), que también es
encontrado en otro tipo de células inmunes en el cuerpo. Este se encarga de ejercer una respuesta
inhibitoria que ayuda a mantener el fenotipo de reposo de la microglía y ha sido estudiado in
vitro, puesto que la deficiencia de su eje (CX3CL1-CX3CR1) es asociada con el aumento de la
neuroinflamación y el desarrollo de enfermedades neurodegenerativas (Cho et al., 2011; Sellner
et al., 2016). No obstante, estudios en organismos vivientes demuestran la presencia de receptores
purinérgicos como el receptor P2X7, los cuales se encuentran expresados en presencia de ATP
extracelular liberado por células heridas. También hace parte del tejido excitable de la microglía,
en el que su canal de iones ligandodependientes estimula la liberación de citosinas
proinflamatorias como la famosa IL-1β (Vonend et al., 2004) y otros receptores purinérgicos
ionotrópicos como P2X, o metabotrópicos como P2Y1, P2Y2/4 y P2Y6, los cuales responden de
maneras diferentes ante la presencia de distintas concentraciones de ATP en el espacio
extracelular (Boucsein et al., 2003).
Ahora bien, se han registrado cuatro estados morfológicos principales en la microglía, que
son mediados gracias a sus mecanismos de comunicación autocrinos y paracrinos (Cherry et al.,

12
2014). Todos estos se encargan de cumplir cuatro funciones principales: la de vigilancia, la de
fagocitosis, la de citotoxicidad y la de neuroprotección (Maysinger & Zhang, 2016), que han sido
definidas gracias a la observación del efecto de las sustancias liberadas por cada una (Martinez &
Gordon, 2014).
El primer estado morfológico se conoce como inflamatorio, M1, en estado activo o en
forma de ameba, pero también se ha observado un fenotipo hiperramificado que también se
considera como “activo” (Mantovani et al., 2004; Reus et al., 2015). Dicho estado se interpreta
como una respuesta inmune impulsada debido a un posible daño en el tejido del SNC o a causa
de la presencia de microbios (Aloisi, 2001; Cherry et al., 2014). El “estado activo” puede ser
precipitado por la inducción del compuesto lipopolisacarido (LPS), el ATP extracelular
(Boucsein et al., 2003; Kierdorf & Prinz, 2013), por células de tipo th1, activadas por el
interferón-y (Gordon & Martinez, 2010; Cherry et al., 2014; Lisi, Stigliano, Lauriola, Navarra, &
Dello, 2014), por la respuesta de receptores de tipo Toll (Gurley et al., 2008), receptores tipo nod
(NLR) (Martinon, Burns, & Tschopp, 2002) etc.
A grandes rasgos, la respuesta ante los patógenos o el daño del tejido, es mediada a través
de los también llamados receptores reconocedores de patrones (PRR) (receptores tipo toll, NRL),
cuya labor principal es reconocer los patrones moleculares asociados a patógenos (PAMPs) y a
daños (DAMPs), para luego establecer la respuesta inmune e inflamatoria (Walker, Kavelaars,
Heijnen, & Dantzer, 2014; Kigerl, de Rivero Vaccari, Dietrich, Popovich, & Keane, 2014). En
consecuencia libera TNF-α, IL1-β, IL-6, iNOS (Mantovani, Sozzani, Locati, Allavena, & Sica,
2002; Meireles et al., 2016), IL-17, IL-23 (Kawanokuchi et al., 2008), IL-12 (Aloisi, Penna,
Cerase, Menendez, & Adorini, 1997; Gurley et al., 2008), IL-8 (Fernandez, Britton, & Rao, 2013;
Doens & Fernandez, 2014), respuestas desde los marcadores superficiales de las células, CD86,
CD68, CD40 (Sun et al., 2008; Kobayashi et al., 2013; Yang et al., 2016) y otras quimiocinas
(Fernandez et al., 2013) que ejercen un efecto citotóxico, inflamatorio y fagocítico sobre las
neuronas (Mantovani et al., 2004; Doens & Fernandez, 2014). Estando en este estado, la célula
tiende a liberar citocinas pro-inflamatorias, especies reactivas de oxígeno y especies reactivas a
nitrógeno para combatir con el peligro que afecta al tejido. Posteriormente, se activa el factor
nuclear κB (NF-κB) y las MAPks, cuya activación favorece la transcripción de genes, entre los
cuales se destacan aquellos asociados al inflamasoma (Martinon et al., 2002; Walker et al., 2014),

13
que quiere decir el complejo de proteínas responsable de la activación de la capatasa 1,
promotora de la posterior liberación de citosinas proinflamatorias (Petrilli, Papin, & Tschopp,
2005; Franchi, Eigenbrod, Munoz-Planillo, & Nunez, 2009).
La respuesta de neuroinflamación del estado M1 de la microglía no solo se ha observado
ante la presencia de agentes patógenos o lesiones, sino también durante la exposición a
situaciones de potencial peligro, en donde se aumenta la activación y cantidad de su fenotipo
proinflamatorio M1 (Reus et al., 2015). Aunque el proceso de neuroinflamación sea considerado
como adaptativo para sistema inmune en el cerebro (Streit et al., 2004), cuando se presenta de
manera crónica o muy fuerte, la inflamación puede causar problemas que se han visto
relacionados con el padecimiento de trastornos psiquiátricos, neurodegenerativos y del desarrollo
(Streit et al., 2004; Reus et al., 2015), que aumentan su riesgo con la edad (Cho et al., 2011;
Vukovic, Colditz, Blackmore, Ruitenberg, & Bartlett, 2012).
Ahora bien, el fenotipo M2, en forma ramificada, o en “estado de alternativo” (Kitamura,
Tsuchihashi, & Fujita, 1978; Morris, Clark, Zinn, & Vissel, 2013; Cherry et al., 2014), se encarga
de producir IL-10 para disminuir la inflamación y reproducir matrices de proteínas protectoras
extracelulares YM1, que son poliaminas para la reparación de las heridas y activación de
receptores asociados con la inhibición de la fagocitosis (Cherry et al., 2014; Yang et al., 2016).
Este fenotipo se divide en tres fenotipos complementarios llamados M2a, M2b y M2c, los
cuales actúan de manera agregada a las funciones del fenotipo principal (Meireles et al., 2016).
La respuesta de la microglía M2a, se caracteriza por ser activada después de la liberación de la
citosina anti-inflamatoria IL-4, la elevación de AMPc (Ghosh, Xu, & Pearse, 2016; Meireles et
al., 2016) y la IL-13, secretadas por las células de tipo Th2 (Doyle et al., 1994; Cherry et al.,
2014). Tanto la IL-4 como la IL-13 son los inductores prototípicos del estado alternativo de la
microglía (M2) (Gordon & Martinez, 2010). Después, se encuentra el M2b, que es el fenotipo
inmunoregulatorio, en el que se evidencia el incremento de la concentración de IL-10 y factor de
crecimiento tumoral beta (TGF-β) (Lisi et al., 2014). El estado morfológico el M2c es activado
por IL-10, cuyo fenotipo es impulsado por complejos inmunes, para desactivar la microglía
“activada” (Boche et al., 2013; Meireles et al., 2016). Con la activación de M2c se ha observado
la liberación de otras moléculas que también ejercen un rol neuroprotector sobre las neuronas y
estos son arignasa-1 (arg-1), CD206 y SOCS3 (Chhor et al., 2013; Kobayashi et al., 2013).

14
Se ha observado que el fenotipo M2 se encarga de cicatrizar y reparar el tejido, después
de desactivar la respuesta inmune e inflamatoria (Cherry et al., 2014), no obstante a lo largo del
desarrollo también se ha encargado de otros procesos esenciales para el mantenimiento del
sistema nervioso, como lo son la poda neuronal, la plasticidad sináptica (Kim et al., 2013;
Meireles et al., 2016) y la neurogénesis (Vukovic et al., 2012; Kierdorf & Prinz, 2013; Sellner et
al., 2016). Durante el desarrollo, el mantener el estado ramificado de la microglía es esencial para
promover la plasticidad sináptica y estructural, pues por medio de la liberación de las citosinas
anti-inflamatorias y los factores de crecimiento (Vukovic et al., 2012) se define el soporte los
procesos cognitivos, como lo son el aprendizaje y la memoria (Morris et al., 2013).
En el siguiente capítulo se va a exponer una introducción sobre el hipocampo y su
funcionamiento. Tener un conocimiento sobre esta estructura permite entender de manera más
profunda la función y los efectos de la microglía sobre esta estructura.
Aproximaciones al hipocampo
El hipocampo es una estructura que juega un papel importante en una gran variedad de
funciones heterogéneas alrededor del SNC (Fanselow & Dong, 2010); entre estas, se puede
observar aquellas que involucran particularmente, la codificación y el procesamiento de las
memorias episódicas a corto plazo (Squire, 1992; Spiers, Maguire, & Burgess, 2001) que son
dependientes de la potenciación a largo plazo (LTP) (Squire, 1992; Izquierdo et al., 2008; Kim,
Mahmoud, & Grover, 2005), el reconocimiento y representación espacial y contextual (Fanselow,
2000; Pothuizen, Zhang, Jongen-Relo, Feldon, & Yee, 2004; Solstad, Yousif, & Sejnowski,
2014), la neurogénesis (Alvarez-Buylla & Lim, 2004; Ehninger & Kempermann, 2006), la
plasticidad neuronal (Zhao, Deng, & Gage, 2008; Perederiy & Westbrook, 2013), algunos
procesos cognitivos (Eichenbaum, Yonelinas, & Ranganath, 2007), la regulación de la activación
fisiológica durante respuestas de estrés (Jacobson & Sapolsky, 1991; Dedovic, Duchesne,
Andrews, Engert, & Pruessner, 2009), el control emocional (Blanchard & Blanchard, 1972), entre
otros.
A continuación, se va a elaborar un viaje por el hipocampo, sus estructuras y sus
funciones más importantes.

15
Como se dijo anteriormente las funciones del hipocampo han sido ampliamente
estudiadas en cuanto a su papel en la respuesta de aprendizaje contextual, el condicionamiento
aversivo y el control emocional. Varios estudios han observado estas funciones por medio de
lesiones en dicha estructura. De esta manera, lesiones en el hipocampo reducen el tiempo de
congelamiento frente a estímulos potencialmente peligrosos en ratas, como por ejemplo el olor de
los gatos o la presencia de estos mismos; no obstante también se aumentan los comportamientos
de evitación y la disminución del congelamiento, dependiendo de la región (Blanchard &
Blanchard, 1972). Otros estudios corroboran estas observaciones por medio de aumentos en la
agresividad y la evitación después de lesiones en el hipocampo en las pruebas de evitación activa
y pasiva (Blanchard, Blanchard, & Fial, 1970). Todo esto permitiría sugerir una disminución en
el control emocional.
Ahora bien, el estudio de esta región se ha abarcado de maneras cada vez más específicas
a lo largo del tiempo. De esta forma, al hipocampo se le ha dividido en dos estructuras que
funcionan de manera diferencial; estas son el hipocampo dorsal (HD) (hipocampo posterior en
primates (Fanselow & Dong, 2010)), que mantiene conexiones con el neo-córtexy el hipocampo
ventral (HV) (hipocampo anterior en primates (Fanselow & Dong, 2010)), que mantiene
conexiones con la amígdala y el hipotálamo (Fanselow & Dong, 2010; Dorey, Pierard, Chauveau,
David, & Beracochea, 2012). Se tiene conocimiento de que el HD regula las funciones
cognitivas, mientras que el HV se asocia con el control emocional y la inhibición del
comportamiento (Segal, Richter-Levin, & Maggio, 2010; Kouvaros & Papatheodoropoulos,
2016).
Estudios previos han identificado mecanismos divergentes de ambas estructuras por
medio de lesiones selectivas. Si la lesión es en HD, se provoca un deterioro de la formación de la
memoria espacial, la flexibilidad cognitiva y provoca alteraciones en la motivación y problemas
en la extinción de comportamientos aprendidos; pero una mejoría en el aprendizaje instrumental
(Moser, Moser, Forrest, Andersen, & Morris, 1995; Pothuizen et al., 2004; Segal et al., 2010;
Busse & Schwarting, 2016). Otros estudios identifican que HV regula y procesa información
sobre el estado motivacional y homeostático, asociado con el aprendizaje emocional y el
comportamiento defensivo (Pentkowski, Blanchard, Lever, Litvin, & Blanchard, 2006; Fanselow
& Dong, 2010; Segal et al., 2010).

16
En la literatura se han aclarado los roles de estas dos estructuras. En primer lugar, se ha
demostrado que las lesiones del HV y HD en el contexto del aprendizaje aversivo, después de una
estimulación en la sustancia gris periacueductal dorsal (SGP), en un test de miedo condicionado
por medio de un choque en las patas, disminuye el tiempo de congelamiento, el cual, en HD es
interrumpido después de 24 horas desde el entrenamiento; y en HV inmediatamente después del
entrenamiento (Ballesteros, de Oliveira, Maisonette, & Landeira-Fernandez, 2014). Otros
estudios corroboran dicho hallazgo, como por ejemplo, (Kjelstrup et al., 2002), quienes
observaron que las lesiones en el HV no causaban deterioros sustanciales en el desempeño en el
laberinto acuático de Morris (pues sugerían que este aprendizaje era más dependiente de HD),
pero aumentaban significativamente el tiempo en los brazos abiertos de un laberinto en cruz
elevado. Lo anterior, permite sugerir que las manipulaciones en HV disminuyen las respuestas el
miedo, la ansiedad en las ratas y esto se debe gracias a sus conexiones funcionales (Fanselow &
Dong, 2010).
De la misma manera, se han se han reportado estudios en los que las lesiones de HV
hacen de los animales experimentales un blanco para los efectos nocivos del estrés y se ha
observado por medio del agravamiento de las úlceras gástricas en un test de restricción y frío
(Henke, 1990). Por otro lado, las lesiones selectivas de HD con NMDA en ésta área (Maren,
Aharonov, & Fanselow, 1997), provocan déficits en la memoria anterógrada y aumentan
sustancialmente el tiempo de entrenamiento para la expresión de miedo condicionado, muestran
impedimentos en el congelamiento después de la presentación de un tono, muestran
impedimentos en el reconocimiento de los contextos, muestran aumentos en la actividad
locomotora y disminuyen el comportamiento exploratorio en comparación con un grupo control
(Maren et al., 1997). A partir de la evidencia, puede quedar claro que el HD se encarga de
procesos cognitivos asociados con la locomoción, la exploración y la navegación y HV se
encarga de funciones emocionales y motivacionales (Fanselow & Dong, 2010).
Una serie de estructuras funcionales que conforman el hipocampo también han sido
ampliamente estudiadas por su conectividad y sus funciones. Así, dicha conectividad categoriza
al hipocampo desde las áreas corticales que lo rodean, que son la corteza perirrinal, la corteza
postrinal y las dos subdivisiones de la corteza entorrinal (lateral y medial) (Burwell & Amaral,
1998). Estas estructuras se encargan de recibir aferencias, con intensidades desiguales, desde

17
distintas áreas corticales, especialmente de las modalidades sensoriales y otras áreas como la
corteza visual y el neo-cortex (Burwell & Amaral, 1998; Kesner, Lee, & Gilbert, 2004; Dolorfo
& Amaral, 1998; Dong, Swanson, Chen, Fanselow, & Toga, 2009; Fanselow & Dong, 2010). De
manera recíproca, proyectan hacia distintas áreas corticales y desde la corteza entorrinal hacia
estructuras hipocampales (Agster & Burwell, 2009). La evidencia ha demostrado el rol de estas
estructuras en varios aspectos del aprendizaje y la memoria. Se ha sugerido que la corteza
entorrinal contribuye a procesos de representación espacial (Fyhn, Molden, Witter, Moser, &
Moser, 2004; Lipton, White, & Eichenbaum, 2007) y de memoria de trabajo (Fransen,
Tahvildari, Egorov, Hasselmo, & Alonso, 2006); que la corteza perirrinal contribuye a los
procesos de percepción (Bussey, Saksida, & Murray, 2003), de reconocimiento de objetos, en
compañía del fornix (Ennaceur & Aggleton, 1997; Bussey, Dias, Amin, Muir, & Aggleton, 2001)
y (junto con las otras cortezas) al aprendizaje y al reconocimiento contextual (Bucci, Phillips, &
Burwell, 2000; Burwell, Saddoris, Bucci, & Wiig, 2004); y la corteza postrrinal contribuye al
aprendizaje contextual y el reconocimiento de objetos (Burwell et al., 2004; Eacott & Gaffan,
2005).
El camino de comunicación del hipocampo comienza con la corteza entorrinal, que juega
un papel importante en la interconexión entre este y las estructuras corticales de asociación
(Dong et al., 2009). Esta corteza recibe la información cortical y desde las otras cortezas
parahipocampales y envía proyecciones hacia el hipocampo desde sus diferentes capas (Ino,
Kaneko, & Mizuno, 1998). De ahí se han observado inervaciones excitatorias unidireccionales
desde su capa II, por medio de la trayectoria perforante hacia la capa molecular de GD, (Spiller &
Racine, 1994; Amaral, Scharfman, & Lavenex, 2007) y en menos abundancia, a otras estructuras
como CA1, CA3 y el complejo subicular (Guzowski, Knierim, & Moser, 2004; Ino et al., 1998).
Desde la corteza entorrinal se promueve la excitación de GD, el cual por medio de la
estimulación de las fibras musgosas dirige sus inputs excitatorios hacia las neuronas piramidales
del área dorsal de CA3 (Wenzel, Buckmaster, Anderson, Wenzel, & Schwartzkroin, 1997;
Kesner et al., 2004; Henze & Buzsaki, 2007; Jonas & Lisman, 2014). En esta región se observan
respuestas de la actividad de la estructura al responder a cambios sensoriales y motivacionales
(Fyhn, Hafting, Treves, Moser, & Moser, 2007). Finalmente la proyección de CA3 hacia CA1
por medio de las colaterales de shaffer (Huang & Kandel, 2005) se promueve a través de las

18
neuronas piramidales dependientes de NMDA. Su excitación prolongada puede inducir la
potenciación a largo plazo (LTP) (de la cual se va a hablar posteriormente) (Stepan et al., 2012).
Después de haber sido procesada la información, CA1 envía proyecciones hacia el
subículo (Cenquizca & Swanson, 2007), el cual proyecta sus porciones septales hacia las áreas
más caudales y laterales de la corteza entorrinal y sus porciones temporales hacia las áreas
rostrales y mediales de esta misma (Ino, Kaneko, & Mizuno, 2001). El área CA1 se encarga de
extender sus axones por diferentes vías de asociación alrededor del cerebro. En primer lugar,
envía proyecciones de realimentación hacia las áreas más rostrales de la corteza entorrinal, hasta
las capas II y III (Ino et al., 2001). No obstante, también se han reportado outputs hacia
hipotálamo, el tálamo y otras regiones corticales, como el área perirrinal, postrrinal y la corteza
cingulada, además de las áreas visuales, auditivas, somatosensoriales, gustativas, viscerales, e
inclusive, la amígdala basolateral, la zona agranular de la ínsula y las áreas orbitales (Kesner et
al., 2004; Cenquizca & Swanson, 2007; Stepan et al., 2012). Se ha observado la actividad de
estas conexiones, especialmente por parte de CA1, excitada por CA3y es importante para los
procesos de consolidación de la memoria episódica en la corteza (Malerba, Krishnan, Fellous, &
Bazhenov, 2016).
Ahora bien, es pertinente para los propósitos de este trabajo anotar algunos respetos de
otras estructuras hipocampales. En primer lugar, se ha considerado al GD una región importante
debido a su participación en la neurogénesis (Zhao et al., 2008; Mongiat & Schinder, 2011). La
neurogénesis se ha observado en las células granulares de la zona subgranular y subventricular,
siendo la zona subventricular, la que se dedica a la proliferación de células que viajan al bulbo
olfatorio y la zona subgranular, la que se dedica a la proliferación de neuronas que permanecen
en el hipocampo (Zhao et al., 2008; Jessberger & Gage, 2014; Chesnokova, Pechnick, &
Wawrowsky, 2016). Se ha reportado que estas células inmaduras carecen de sinapsis funcional y
poseen una alta resistencia a inputs de otras células (Ambrogini et al., 2004). También se ha
observado que pueden durar varias semanas antes de poder ejecutar contactos sinápticos
correctamente, no obstante son altamente receptivas a los inputs GABAérgicos que facilitan su
despolarización y su diferenciación (Ambrogini et al., 2004; Tozuka, Fukuda, Namba, Seki, &
Hisatsune, 2005; Zhao et al., 2008; Mongiat & Schinder, 2011; Sibbe & Kulik, 2016). Con el
paso del tiempo, estas células pueden responder a pequeños contactos sinápticos glutamatérgicos

19
de la corteza entorrinal (Mongiat, Esposito, Lombardi, & Schinder, 2009). Otro de sus
reguladores principales es el BDNF, un factor de crecimiento que ayuda a modular la
neurogénesis y la sinaptogénesis (Lee & Son, 2009).
Una vez las neuronas han madurado lo suficiente, ofrecen un aumento de la plasticidad y
la capacidad de generar LTP (Ge, Yang, Hsu, Ming, & Song, 2007), además de proyecciones
hacia CA3. La proliferación y las posteriores conexiones de estas células es relevante para
perdurar los procesos de memoria y aprendizaje espacial y contextual a lo largo del tiempo (Zhao
et al., 2008; Vivar & van, 2013; Jessberger & Gage, 2014).
Teniendo en cuenta que las células inmaduras son vulnerables a los cambios ambientales
que facilitan o interrumpen los procesos de maduración y sobrevivencia (Ehninger &
Kempermann, 2006), se ha registrado que su funcionamiento es dependiente de mecanismos de
aprendizaje, como por ejemplo el aprendizaje espacial (Dupret et al., 2007). En experimentos que
involucran el aprendizaje y la neurogénesis, se ha observado que la sobrevivencia de las neuronas
que maduran, perdura en el tiempo, mientras que la sobrevivencia de las más jóvenes no, pues
aún no han generado conexiones efectivas (Dupret et al., 2007). Otros estudios han demostrado
los mismos resultados (Dobrossy et al., 2003). Sin embargo, a largo plazo, las actividades que
fomentan el aprendizaje tienden a aumentar la sobrevivencia de estas neuronas (Sisti, Glass, &
Shors, 2007).
Siguiendo esta misma línea, la neurogénesis en el área granular del GD es mayor en
roedores expuestos a ambientes enriquecidos, de los cuales se ha observado una mejora en
diversas pruebas aprendizaje, como el test de reconocimiento de objetos (Kempermann, Brandon,
& Gage, 1998; Bruel-Jungerman, Laroche, & Rampon, 2005). No obstante, otro tipo de estímulos
ambientales, como lo son el estrés físico y psicológico regulan el nacimiento de neuronas y
promueven el retraimiento de las conexiones sinápticas (Ehninger & Kempermann, 2006). En
estudios preliminares se ha evidenciado que una sola exposición a estímulos de estrés agudo
promueve la reducción de la proliferación de la neurogénesis en el GD del hipocampo adulto
(Gould, Tanapat, McEwen, Flugge, & Fuchs, 1998). Por el contrario, se considera que
actividades como el ejercicio voluntario es un factor protector e inductor de neurogénesis y
plasticidad en presencia de estrés (Lee et al., 2013; Lee, Yau, Lee, Lau, & So, 2016).

20
Se ha concretado en párrafos anteriores que el GD es proyectado hacia el área dorsal de
CA3 por medio de las fibras musgosas. Estudios anteriores han documentado que estas fibras
producen transmisiones GABAérgicas y glutamatérgicas, pero su sinapsis GABAérgica sólo se
presenta durante periodos de hiperexitabilidad, estimulación repetida o epilepsia (Gutierrez et al.,
2003). La importancia de este sistema de comunicación es esencial para los procesos de memoria
y aprendizaje contextual, puesto que GD procesa y releva sus señales, por medio de estas fibras,
hacia CA3, influenciando la cantidad de disparos excitatorios en CA1 (Urban, Henze, &
Barrionuevo, 2001; Henze, Wittner, & Buzsaki, 2002; Gruart, Munoz, & Delgado-Garcia, 2006).
Se ha sugerido que la modulación de la sinapsis de CA3-CA1 podría verse impedida con el daño
de GD u otra estructura que haga parte del sistema hipocampal; y estudios que corroboran esta
observación, demuestran que el daño de GD impide la consolidación de la memoria contextual en
tareas de adicción a la cocaína y en procesos de condicionamiento, después de la exposición a un
estímulo aversivo (Hernandez-Rabaza et al., 2008). Al mismo tiempo, la lesión de esta estructura
puede impedir la consolidación de memorias y bloquear el aprendizaje que depende de la
plasticidad neuronal (Madronal et al., 2016).
Otras características sobre la funcionalidad de GD se evidencian en tareas de modalidad
espacial y contextual al funcionar de la mano de CA1 y CA3. Durante la adquisición y
consolidación de las memorias, GD prima codificando aquellas de tipo contextual; de este modo
sus lesiones impedirían una consolidación correcta, además de problemas en otros procesos de
aprendizaje que funcionan de la mano de CA3 y CA1 (Solstad et al., 2014; Lana, Iovino, Nosi,
Wenk, & Giovannini, 2016; Madronal et al., 2016). Al mismo tiempo, las neuronas de GD y CA3
son receptivas a características espaciales en donde se observan disparos relacionados con
cambios específicos en el ambiente. Es por eso que (Leutgeb & Moser, 2007) identificaron que el
número de células activas en GD son más sensitivas a los cambios pequeños de las características
del ambiente, mientras que CA3 y CA1 no asemejan dichos cambios (Hasselmo, 2005a), pero si
establecen re-mapeos. (Leutgeb et al., 2005) mostraron que estos re-mapeos se encargan de
cambiar las intensidades de los disparos postsinápticos dependiendo de modificaciones en la
ubicación espacial, o cambios en la frecuencia de los disparos sobre una misma locación, en la
que se han alterado características del entorno. También son observados cuando en un mismo
ambiente las tareas del animal se ven alteradas (Guzowski et al., 2004). De ahí definieron que

21
CA3 mudaba sus patrones de disparo cuando las distribuciones de los objetos se ubicaban de
manera diferente en un mismo espacio. El rol de CA1 en esta misma situación demuestra que,
con la colaboración de la corteza entorrinal, procesa detalles selectivos de localizaciones para
codificar representaciones específicas de la memoria espacial (Lipton et al., 2007).
Ahora bien, en cuanto a CA3, se ha establecido que posee una red celular que se encarga
de representar y reorganizar el espacio y la distribución del ambiente en el que el animal se
encuentra, puesto que genera representaciones de dichos espacios en mapas espaciales y
contextuales (Leutgeb et al., 2005; Johnson & Redish, 2007; Kesner, 2007). Al mismo tiempo,
también es posible identificar un aumento en la actividad de sus células en pruebas de toma de
decisiones, como el laberinto en T, en donde se observa que son capaces de representar
posiciones de otros espacios y otras temporalidades (Johnson & Redish, 2007). Los procesos de
toma de decisiones de índole espacial pueden ser representados por medio de los cambios
dinámicos en la representación del animal en un espacio futuro (Johnson & Redish, 2007). De
manera interesante, en otros estudios se ha observado que esta área codifica información que
asocia los objetos con el espacio y también con sus olores (Ji & Maren, 2008; Solstad et al.,
2014). Así es como la lesión de esta área impediría la codificación de las memorias espaciales,
además del reconocimiento del espacio en el contexto (Ji & Maren, 2008).
Hasselmo (2009), comentando los modelos propuestos por Lee, Hunsaker y Kesner
(2005), quienes han trabajado la memoria espacial de ratas experimentales en un test de
reconocimiento de objetos después de lesiones en CA1 y CA3, ilustró la labor de estos autores, al
destacar sus resultados. Ellos observaron que el tiempo de exploración de los objetos novedosos
disminuía en un mismo espacio con lesiones en CA1, pero el efecto era aún mayor desde CA3.
Al mismo tiempo, explicó que el tiempo de habituación a los espacios puede disminuir con
lesiones en esta estructura, por lo cual esto impide que los animales evoquen una recuperación de
memorias contextuales incluso en el espacio de experimentación.
Las consideraciones sobre el área CA1 se enfocan en que posee funciones diferenciales
que dependen del agrupamiento y las conexiones que establecen sus neuronas piramidales
glutamatérgicas (Cenquizca & Swanson, 2006; Dong et al., 2009). Dividida esta estructura en
varias regiones, se le ha dado importancia a la porción dorsal (CA1d) y ventral (CA1v). Se ha

22
definido que las conexiones desde CA1v, como las que se ubican de manera bidireccional con la
amígdala (von Bohlen und & Albrecht, 2002) y otras subcorticales, están asociadas con procesos
autónomos, neuroendocrinos, emocionales y afectivos, además de proyectar hacia las regiones
ventrales y laterales del núcleo septal, la stria terminalis, las regiones preópticas, el tálamo, varios
núcleos del hipotálamoy áreas mamilares y premamilares incluyendo el paraventricular (NPV) y
el lateral (Cenquizca & Swanson, 2006; Dong et al., 2009; Dong et al., 2009). Al mismo tiempo,
CA1d mantiene relaciones con las estructuras corticales, por medio de la corteza entrorrinal y el
subículo (Cenquizca & Swanson, 2007). Lo anterior ha sugerido su papel en la modulación de los
ritmos theta y la navegación espacial, pues concentra neuronas de lugar que están envueltas en
procesos cognitivos que se encargan del procesamiento de la información visual (Dong et al.,
2009). Todo esto permite sugerir que las conexiones de CA1 soportan de manera independiente
respuestas emocionales y cognitivas relacionadas con el procesamiento la memoria dependiente
del contexto (Ji & Maren, 2008).
Finalmente pasando de la funcionalidad celular a los procesos moleculares, la
potenciación a largo plazo (LTP), es uno de los mecanismos más importantes que representa el
aprendizaje y la memoria en el hipocampo, puesto que altera los procesos de plasticidad
sináptica, aumenta la liberación de neurotransmisores de la neurona presináptica y magnifica las
respuestas de la postsináptica, para el establecimiento de nuevas conexiones (Barria, Muller,
Derkach, Griffith, & Soderling, 1997; Bernabeu et al., 1997; Martin & Morris, 2002). El LTP es
presentado como una estimulación neuronal que manifiesta respuestas postsinápticas excitatorias
que se encargan de extender la liberación de glutamato por varias horas, días e incluso meses
(Martin & Morris, 2002; Izquierdo et al., 2008). Al mismo tiempo, la expresión de LTP juega un
papel en el incremento de la sensibilidad de la neurona postsináptica (Manabe & Nicoll, 1994).
Los mecanismos de memoria y aprendizaje en el hipocampo dependientes de LTP, han sido
ampliamente estudiados en las aferencias desde CA3, por medio de los colaterales de Shaffer
hasta CA1 3(Izquierdo et al., 2008; Stepan et al., 2012).
Un resumen sobre los estudios de las dinámicas moleculares del LTP en el hipocampo
puede ser observado de la siguiente manera. El LTP requiere de la estimulación de los receptores
glutamatérgicos ionotrópicos AMPA (Dingledine, Borges, Bowie, & Traynelis, 1999), los cuales
ejercen una acción de despolarización de la célula postsináptica y colaboran con la

23
sensibilización de los receptores de NMDA (Dingledine et al., 1999). El influjo de Ca2+ cambia la
polaridad dentro de la célula y promueve la liberación del Mg+2 dependiente de voltaje que
bloquea la entrada de NMDA (Dingledine et al., 1999; Bloodgood, Giessel, & Sabatini, 2009;
Luscher & Malenka, 2012). Dentro de la célula, dicho influjo de Ca2+ también promueve la
estimulación de cascadas enzimáticas, entre las que se observa la autofosforilación de la
calcio/calmodulina-dependiente de proteína kinasa II (CaMKII) (Lisman, Schulman, & Cline,
2002). La CaMKII, a su vez, promueve la fosforilación de los receptores AMPA (Hayashi et al.,
1997). En estudios previos, se ha evidenciado que el bloqueo de CaMKII no afecta la transmisión
sináptica, pero si detiene la expresión de LTP y la fosforilación de AMPA (Barria et al., 1997).
En cierto modo los componentes rápidos de la entrada de Ca2+ son afectados por la modulación
de los receptores AMPA que igualmente son sensibilizados por medio del aumento de la
transmisión sináptica y la activación de CaMKII (Bloodgood et al., 2009; Luscher & Malenka,
2012).
Más adelante este efecto lento de la apertura de NMDA también permite la entrada de
Ca2+ a la neurona (Hayashi, Zviman, Brand, Teeter, & Restrepo, 1996) y coopera como inductor
de LTP y plasticidad (Bloodgood et al., 2009; Luscher & Malenka, 2012). Con el incremento de
la concentración Ca2+, se disminuye el potencial de membrana y aumenta el chance, tanto de una
despolarización, como de la entrada de más Ca2+ en la neurona postsináptica (Hayashi et al.,
1996). Esto mismo promueve la expresión de la proteína quinasa C (PKC), la cual por medio de
su autofosforilación promueve otras cascadas enzimáticas como la fosforilación de su substrato
GAP43 (proteína B50), que se expresa durante inducción de LTP (Gianotti, Nunzi, Gispen, &
Corradetti, 1992) y ayuda a promover la la transmisión glutamatérgica (Gianotti et al., 1992;
Roberson, English, & Sweatt, 1996; Izquierdo et al., 2008). De igual manera, se activan las
proteínas quinasas activadas por mitogeno (MAPKs) y la kinasa extracelular relacionada con la
señal (ERK) que juegan un papel importante en la regulación de la plasticidad dependiente de la
inducción de LTP (English & Sweatt, 1996; Vanhoutte et al., 1999). Las cascadas enzimáticas de
MAPK (p44 y p42) y ERK se translocan al núcleo para promover la fosforilación de los factores
de transcripción ELK-1, CREB y la aparición de la proteína fos (Vanhoutte et al., 1999). La
señalización de cascadas enzimáticas provocadas por MAPK juega un rol importante en la
regulación de los genes controlados por CRE, vía la fosforlación de CREB (Davis, Vanhoutte,

24
Pages, Caboche, & Laroche, 2000; Kanterewicz et al., 2000). Se ha sugerido de CREB es
necesario para la inducción de LTP, la formación de nuevas memorias y el mantenimiento de la
plasticidad (Bernabeu et al., 1997; Martin & Morris, 2002; Izquierdo et al., 2008).
Paralelamente, la estimulación de NMDA de la cual devienen cascadas enzimáticas y la
inducción de factores de crecimiento (English & Sweatt, 1996), también promueve la actividad
de la adenilil ciclasa, que impulsa al segundo mensajero, adenosín monofosfato cíclico (AMPc),
para aumentar el influjo de Ca2+ hacia la célula (Chetkovich, Gray, Johnston, & Sweatt, 1991).
AMPc promueve la activación de la proteína quinasa dependiente de AMPc (PKA), requerida
para la inducción de LTP y la formación de memorias (Bernabeu et al., 1997; Yang et al., 2004).
Inmediatamente después de que los niveles de AMPc aumentan, en la fase tardía del LTP, la
actividad de PKA empieza a ser evidente (Bernabeu et al., 1997), no obstante existen estudios
que argumentan que PKA también hace parte de la fase inicial (Yang et al., 2004). PKA se
encarga de mantener la autofosforilación de CaMKII por medio de la inhibición del inhibidor de
fosforilación 1 (I-1) (Yang et al., 2004), pero también se ha observado fosforilando otros
compuestos como el receptor AMPA y CREB, el cual se encargará de sintetizar las proteínas
requeridas para el mantenimiento y la plasticidad estructural que sigue después de la inducción de
LTP (Luscher & Malenka, 2012).
Para los propósitos de este estudio, es fundamental entender al estrés y los efectos que
este produce sobre el organismo y el hipocampo, pues esta es una estructura que influencia y se
ve influenciada por este proceso.
Aproximaciones al estrés
Todos los organismos vivos invierten sus recursos energéticos para adaptarse a las
circunstancias ambientales que aseguran su sobrevivencia (Habib, Gold, & Chrousos, 2001).
Cuando los recursos ambientales son menos óptimos, las respuestas de autorregulación
adaptativas comprometen mayores cantidades de energía para regresar a la línea de base, ya sea
en una situación verdadera o percibida (Morilak et al., 2005; Dijkstra & Homan, 2016). Todos
estos procesos se ubican bajo los conceptos que muchos autores, entre ellos McEwen (McEwen
& Wingfield, 2003) llaman homeostasis, alostasis, carga alostática y sobrecarga alostática, de las
cuales el estrés es inherente. En primer lugar, la homeostasis es definida como los procesos de

25
balance interno de un organismo para mantener la vida (McEwen & Wingfield, 2003; McEwen,
2007). En segundo lugar, el mantenimiento de esta homeostasis por medio de la generación de
respuestas adaptativas a cambios medioambientales, es denominado alostasis (Goymann &
Wingfield, 2004; McEwen & Wingfield, 2003; McEwen, 2007). El concepto incluye los procesos
que se llevan a cabo para alcanzar la estabilidad (McEwen & Wingfield, 2003). Otras
definiciones refieren a la alostasis como los niveles de actividad requeridos para que se mantenga
la estabilidad durante situaciones de cambio (Goldstein & McEwen, 2002). De la alostasis,
también se deriva el concepto de carga alostática, el cual se refiere a los ajustes fisiológicos y
morfológicos y la inversión de energía del cuerpo para adaptarse a cambios transitorios del
ambiente como las estaciones, el estatus social, las migraciones, los ciclos circadianos, la
búsqueda de comida, etc. (Juster et al., 2010). En otras palabras, la carga alostática se refiere a los
efectos prolongados de manera continua, o la intermitente activación de la alostasis (Goldstein &
McEwen, 2002).
En situaciones de cambios medioambientales, la percepción y la evaluación del entorno
sirven como punto de partida para que se pongan en marcha ciertos mecanismos de
autoregulación que movilizan la energía con el fin de regresar al estado de homeostasis (Juster et
al., 2010). Durante el incremento de la carga alostática, se han observado dos mecanismos
importantes, cuya función es servir como mediadores de estos procesos. De ahí se derivan el eje
simpático-adrenal-medular (SAM), que se encarga de liberar catecolaminas y el eje hipotálamo-
pituitario-adrenal (HPA), cuya labor principal es la liberación de glucocorticoides (Nicolaides,
Charmandari, Chrousos, & Kino, 2014; Goymann & Wingfield, 2004). De estos se va a hablar en
párrafos posteriores.
En condiciones en las que la carga alostática aumenta de manera impredecible y las
demandas de energía exceden los suplementos disponibles para regresar a la homeostasis, se
produce un estado de emergencia, cuyo propósito es producir cambios comportamentales y
fisiológicos rápidos en respuesta a la perturbación; a este se le denomina sobrecarga alostática
tipo I (Goymann & Wingfield, 2004; Goldstein & McEwen, 2002; McEwen & Wingfield, 2003).
Una vez la emergencia ha terminado, los niveles de energía disminuyen nuevamente a la línea de
base o incluso menos, aumentando la probabilidad de consecuencias dañinas o patológicas para el
sistema (Goymann & Wingfield, 2004). En general, la sobrecarga alostática tipo I ayuda al

26
organismo a sobrellevar las situaciones con el fin de obtener, nuevamente, los recursos necesarios
para la sobrevivencia (McEwen & Wingfield, 2003). Por el contario, otro tipo de sobrecarga
alostática, la tipo II, se encuentra situada en un contexto en el que los suplementos de energía se
mantienen constantes, pero las situaciones percibidas, que pueden ser derivadas, por ejemplo, de
una disfunción social, no desencadenan una respuesta adaptativa de lucha o huida, sino que
requieren de otros mecanismos de aprendizaje para que el organismo se adapte a la naturaleza de
la situación (McEwen & Wingfield, 2003).
Ahora bien, el concepto de estrés ha sido formulado desde diferentes disciplinas y
contextos (Selye, 1976). En primer lugar (McEwen & Wingfield, 2003) lo han definido como una
situación de amenaza real o implícita a la homeostasis, en la que un evento o sucesión de eventos
causan respuestas comportamentales y fisiológicas específicas para mediar dicha situación. No
obstante, en otros contextos no se ha precisado como una situación, sino como la respuesta hacia
una situación (McEwen & Wingfield, 2003). También se ha ubicado como un proceso que ayuda
al organismo a ajustarse a los cambios medioambientales diarios por medio de modificaciones en
el metabolismo (McEwen & Wingfield, 2003), funciones fisiológicas y funciones
neuroendocrinas (Nicolaides et al., 2014). De igual forma se ha explicado como la percepción de
un organismo hacia una interrupción o discrepancia en la homeostasis, cuyo trabajo es conducir
una acción fisiológica de regulación y una realimentación negativa (Goldstein & McEwen, 2002).
Normalmente, los mecanismos después del estrés, permiten que el cuerpo retome sus funciones
biológicas normales, sin embargo cuando este es severo o crónico, los costos biológicos de las
respuestas interrumpen estas funciones (Carstens & Moberg, 2000).
Por la misma línea (Selye, 1976) expuso una de las definiciones del estrés con el nombre
de Síndrome de Adaptación General (GAS), el cual era precisado como una respuesta no
específica del cuerpo ante una demanda apetitiva o aversiva. De ahí, al estrés positivo, lo
denominó eustrés y al negativo lo denominó distrés. En todo caso, todas las respuestas de estrés,
ya sean apetitivas o aversivas exhiben cuatro mecanismos biológicos de defensa, denominados
como comportamentales, autónomos, neuroendocrinos e inmunes (Carstens & Moberg, 2000). El
distrés, al contrario del eustrés involucra la percepción o el reconocimiento, consciente o
inconsciente, de que una situación es peligrosa para el mantenimiento de la homeostasis
(Carstens & Moberg, 2000). En animales de laboratorio, los indicadores del distrés usualmente se

27
encuentran en la presencia de patologías, automutilación o muerte. Es más, el distrés suele
acontecer cuando los costos biológicos que se derivan del estrés son negativos para las funciones
del cuerpo (Carstens & Moberg, 2000), o cuando se evocan respuestas con base a experiencias
aversivas, en las que la alostasis no es suficiente para mantener o restaurar la homeostasis
(Goldstein & McEwen, 2002).
Todas estas definiciones del concepto de estrés van ser abarcadas en este texto con el
mismo nombre, estrés.
Ahora bien, se va explicar la manera en la que estos cambios homeostáticos producen
respuestas a estímulos internos y externos, amenazantes e impredecibles, cuyas funciones van de
la mano de cambios fisiológicos, afectivos y cognitivos (Gagnon & Wagner, 2016). Cuando un
organismo se ve expuesto a una situación que provoca estrés, se puede asegurar la identificación
de ciertas reacciones fisiológicas que van a comprometer las respuestas del eje SAM y la función
del eje HPA (McEwen & Wingfield, 2003; Carstens & Moberg, 2000). La primera reacción
(SAM) ha sido considerada como una respuesta “rápida”, mientras que la segunda (HPA), es más
“lenta” (Sutanto & de Kloet, 1994) y predominante cuando el estímulo estresor se mantiene en el
tiempo (Zárate, Cárdenas, Acevedo-Triana, Sarmiento-Bolaños, & Leon, 2014).
El estrés de respuesta "rápida" o SAM, se destaca por la activación del sistema nervioso
simpático, en el que se observa una liberación de catecolaminas como epinefrina (Epi)
norepinefrina (NE) y por otra parte, acetilcolina (ACh) (Schwabe, Joels, Roozendaal, Wolf, &
Oitzl, 2012). Su respuesta “rápida” se encarga de movilizar los recursos metabólicos, es decir la
presión arterial, la respiración y el envío de sangre hacia las extremidades para que se produzca
una respuesta aguda de "lucha o huida" (Cacioppo et al., 1998; Lucassen et al., 2014). Bien es
sabido que muchas de estas reacciones son adaptativas para la sobrevivencia de las especies, no
obstante, es necesario tener en cuenta que estas situaciones ya no hacen parte del contexto en el
que se desarrolla el estilo de vida de las poblaciones humanas. Por esto, es interesante revisar las
reacciones fisiológicas que normalmente se dan durante situaciones potencialmente peligrosas,
pues ciertamente se expresan a través de los eventos que vivimos en la actualidad, pero no
solemos interpretamos de manera correcta (Cacioppo et al., 1998).

28
Ahora bien, al iniciar la respuesta de estrés, Epi es liberada desde la medula adrenal y el
tallo cerebral y comunica la información sensorial con el sistema nervioso central (SNC) por
medio de la estimulación de los β-adrenoreceptores del nervio vago (Wong et al., 2012). Una de
las labores principales de Epi, abarca la modulación de la energía durante la respuesta de estrés,
puesto que actúa en el hígado para liberar glucógeno e incrementar los niveles de glucosa en la
sangre (Osborne, Pearson-Leary, & McNay, 2015; Gold, 2014). Desde la periferia, Epi no sólo
modula la energía del cuerpo sino que también establece relaciones con los circuitos que
cooperan con el procesamiento de la memoria y aprendizaje en el hipocampo (Gold, 2014). De
manera paralela, habiendo sido estimulado por Epi, el nervio vago excita al núcleo del tracto
solitario (NTS) el cual dirige sus proyecciones al locus coeruleus (LC) (Roosevelt, Smith,
Clough, Jensen, & Browning, 2006; Wong et al., 2012) y de ahí, ambos desplazan sus
conexiones a estructuras tales como la amígdala (McGaugh & Roozendaal, 2002), en particular
al núcleo basolateral (BLA), que se encarga de ejercer una actividad excitatoria sobre el
hipotálamo, el neocortex, el hipocampo, el núcleo caudado y otras regiones que ayudan a
modular, por ejemplo, la consolidación de las memorias (McGaugh & Roozendaal, 2002;
Wong et al., 2012; Gagnon & Wagner, 2016); a la amígdala medial (MeA) (Morilak et al.,
2005); a la amígdala central (Cea) (Cecchi, Khoshbouei, & Morilak, 2002); a LC (McGaugh &
Roozendaal, 2002); al núcleo paraventricular del hipotálamo (NPV) en donde se estimula la
producción de CRF (Francis, Caldji, Champagne, Plotsky, & Meaney, 1999; Nicolaides et al.,
2014; Charmandari, Tsigos, & Chrousos, 2005); al hipocampo, SGP y a otras áreas en la corteza
(Belujon & Grace, 2015) que van a permitir la modulación de las respuestas adaptativas de lucha
o huida (McEwen & Wingfield, 2003; Gagnon & Wagner, 2016).
Es importante resaltar el papel de LC durante la respuesta simpática del estrés, pues sus
funciones son imperantes para las posteriores respuestas de activación. Esta estructura también es
activada por medio del factor liberador de corticotropina (CRF) que es un péptido que, por una
parte, se encarga de iniciar la respuesta neuroendocrina del estrés y por la otra, funciona como un
neurotransmisor excitatorio que activa las respuestas noradrenérgicas (Van Bockstaele, Colago,
& Valentino, 1996; Curtis, Leiser, Snyder, & Valentino, 2012; Snyder, Wang, Han, McFadden,
& Valentino, 2012; Sun, Hunt, & Sah, 2015). Estudios recientes demuestran que los
comportamientos de ansiedad y aversión inducidos a través del estrés por restricción son

29
producto de la liberación de CRF en LC desde el núcleo central de la amígdala (CeA) (Sah,
Faber, Lopez De, & Power, 2003a; McCall et al., 2015). El incremento de la actividad de las vías
noradrenérgicas de LC, por medio de la activación de sus β-adrenoreceptores y sus α-
adrenoreceptores, aumentan los comportamientos de ansiedad y de aversión respectivamente
(McCall et al., 2015). Otros estudios han identificado que la administración
intracerebroventricular de CRF aumenta las concentraciones norepinefrina (NE) y dopamina
(DA) en el hipotálamo medial y la corteza prefrontal (CPF) (Lavicky & Dunn, 1993). Por otro
lado, si LC llega a ser inhibido, llevaría consigo, como consecuencia, una disminución
significativa de la hormona corticotrópica (ACTH) y CRF en NPV y provocarían una
desregulación en la respuesta de estrés (Francis et al., 1999).
Está claro que los principales impactos emocionales que se efectúan a raíz de estímulos
estresantes, son producto de las estructuras amigdalinas como CeA, pues de esta región se envían
eferencias de CRF hacia LC y al núcleo del tracto solitario (NTS) (Sah, Faber, Lopez De, &
Power, 2003b). El 34% de las proyecciones de CeA hacia LC son de neuronas que contienen
CRF, no obstante, esta estructura también dirige sus inputs excitatorios hacia otras regiones del
cerebro (Kravets, Reyes, Unterwald, & Van Bockstaele, 2015; Sah et al., 2003b). Esta estructura
juega un papel importante en el procesamiento de los estímulos emocionales derivados del
estrés y es la responsable de mediar las respuestas autónomas y endocrinas con estímulos
emocionales salientes (Kravets et al., 2015). Desde LC, NE también ejerce acciones sobre BLA,
que envía sus proyecciones hacia el hipocampo y la corteza perirrinal (CPeri) y sus neuronas
glutamatérgicas también cooperan con la regulación de la respuesta de estrés al cooperar en la
consolidación las memorias emocionalmente significativas (McGaugh, 2000; Roozendaal,
Brunson, Holloway, McGaugh, & Baram, 2002; Sah et al., 2003a).
La liberación de NE es adaptativa, por ejemplo, en situaciones como el estar ante la
presencia de un depredador, eso quiere decir todas aquellas condiciones que representen un
peligro potencial para la sobrevivencia de un individuo (Curtis et al., 2012; Snyder et al., 2012).
Ciertamente estudios han indicado que, frente a una exposición a estrés agudo por restricción en
ratas, la liberación de NE es aumentada en el espacio extracelular y este es captado por la
amígdala, que ejecuta una acción activadora en el eje HPA (Morilak et al., 2005).

30
Relacionada con estresores psicológicos agudos o muy fuertes, también se evidencia la
proliferación de respuestas inmunes. Los detalles de estos efectos van a ser detallados más
adelante en el capítulo sobre los efectos inmunes durante las respuestas de estrés (para más
información vea capítulo de acercamientos a los efectos del estrés sobre la microglía).
Aunque NE es una de las sustancias más características del eje SAM ante las situaciones
de estrés agudo (Belujon & Grace, 2015), en los párrafos anteriores se mencionó que el eje HPA
también se encarga de mediar las respuestas estresoras por medio de otra sustancia. Los
protagonistas de este proceso son los glucocorticoides (o corticosteroides) (GC) (Cecchi et al.,
2002).
A partir de este punto, se describe la respuesta “lenta” del estrés, que es mediada por el
eje HPA (Lucassen et al., 2014). Este mismo desencadena una secreción de hormonas en el
sistema y por un mecanismo de realimentación negativa, regula su actividad (Herman &
Cullinan, 1997; Nicolaides et al., 2014). Ante la percepción de un estresor, se ha identificado que
NPV se encarga de regular la acción del eje (Herman, Cullinan, Ziegler, & Tasker, 2002). Las
neuronas de NPV se responsabilizan por excretar y transmitir CRF (Heinrichs, Menzaghi, Merlo,
Britton, & Koob, 1995; Herman & Cullinan, 1997) y vasopresina arginina (VPA) (Scott & Dinan,
1998) para estimular la liberación y el aumento significativo de la concentración de ACTH desde
la glándula pituitaria anterior (Ono, Bedran de Castro, & McCann, 1985; Herman & Cullinan,
1997; Gallo-Payet, 2016). La liberación de ACTH compromete la generación del estímulo más
potente para la corteza adrenal (Gallo-Payet, 2016), puesto que viaja por la sangre hasta la zona
Fasiculata (ZF) de las glándulas adrenales, en donde promueve la liberación de GC (Szalay, De,
Stark, & Folly, 1985; Payne et al., 2007).
De manera interesante, en condiciones normales la liberación de GC, circadiana y
ultradiana (liberación aproximadamente cada hora) es esencial para mantener la homeostasis del
sistema (Sarabdjitsingh et al., 2010). Los GC viajan por el torrente sanguíneo, organizando la
movilización y el metabolismo de sustancias importantes, como la glucosa en la sangre, además
de controlar los ritmos energéticos por todo el cuerpo (Herman et al., 2003; Myers, McKlveen, &
Herman, 2014; Oster et al., 2016). La glucosa es la principal fuente de energía para el buen
funcionamiento del sistema y ésta sólo puede ser modulada a través de la insulina (Kuo,
McQueen, Chen, & Wang, 2015). En ese caso los GC estimulan la glucogénesis a través del

31
antagonismo de la producción de insulina en el páncreas (Kuo et al., 2015). Los GC actúan de
manera genómica como reguladores de la transcripción de los genes que responden a ellos
mismos (Lucassen et al., 2014).
No obstante, ante una respuesta urgente o de estrés, los GC viajan por el torrente
sanguíneo hasta traspasar la barrera hematoencefálica y ejercen un efecto de realimentación
negativa (de Kloet, 2013) en el hipotálamo, la corteza prefrontal medial (CPFm), la hipófisis y el
hipocampo (de Kloet, Joels, & Holsboer, 2005; de Kloet, 2013). La realimentación negativa
ocurre por medio de la adhesión de GC a los receptores de glucocorticoides (GR), de poca
afinidad y de mineralocorticoides (MR), de alta afinidad, en los núcleos de las células de estas
áreas (Zhu et al., 2014; de Kloet et al., 2005). No obstante, ha y que tener en cuenta que los GC
no sólo tienen un efecto inhibidor de la actividad, sino que esto depende de la estructura a la que
se adhieren, por ejemplo, si estimulan la conexión amígdala-CRF-NE, llevarán a un aumento de
la actividad, que induce ansiedad (de Kloet, 2013).
En condiciones estresantes MR se encarga de responder a las estimulaciones iniciales y de
manera prolongada GR permite la finalización de la realimentación negativa (Oster et al., 2016).
Es de resaltar que los receptores MR se encuentran situados especialmente en las áreas límbicas,
particularmente el hipocampo, mientras que los receptores para GR se ubican por todo el cerebro,
incluyendose, en menos concentración, en las áreas límbicas (Herman, Patel, Akil, & Watson,
1989; Sutanto & de Kloet, 1994; Zhe, Fang, & Yuxiu, 2008; Oster et al., 2016). En el hipocampo
se ha considerado que la actividad de los MR ayuda a mantener la excitabilidad de las neuronas,
pero GR envuelve la supresión o la normalización de esta actividad (Liu, Wang, Ni, & Sun,
2007). En condiciones normales existe mayor afinidad hacia los receptores GR, que sólo se
activan a partir de grandes concentraciones de GC, como las observadas durante los picos de
liberación circadianos y ultradianos (Sarabdjitsingh et al., 2010; de Kloet, 2013).
En cuanto a las acciones no genómicas al nivel de la membrana, las cuales son evidentes
en presencia de estrés, se ha evidenciado que los MR promueven una neurotransmisión
excitatoria al aumentar la liberación presináptica de glutamato y que los GR promueven una
transmisión inhibitoria al influenciar la neurotransmisión GABAérgica y de endocanabinoides
(de Kloet, 2013). De esta manera, el rol de MR estaría modulando la adquisición y el
procesamiento de la información y la respuesta comportamental ante el estresor, mientras que GR

32
suprimiría la actividad hasta llegar a la condición de base, además de permitir la consolidación de
las experiencias traumáticas y eliminar las memorias de poca importancia en ese momento (de
Kloet, 2013).
La funcionalidad de la relación entre estos receptores es de importancia para la regulación
de muchos estados que se encargan de organizar los cambios homeostáticos del sistema; es por
eso ha sido interesante estudiar la interrupción de esta dinámica. Por ejemplo, estudios han
indicado que la desregulación de la función de los receptores MR y GR, durante la respuesta del
eje HPA tiene varias consecuencias en cuanto a los comportamientos poco adaptativos después
de la exposición a un estímulo estresor (Feldman & Weidenfeld, 1999). Harris, Holmes, de Kloet,
Chapman y Seckl (2013) observaron impactos en la memoria y la actividad cognitiva debido al
aumento de la expresión de MR y la disminución de expresión de GR después de la exposición
de ratas a un modelo de estrés por restricción. Ellos observaron que el aumento de la expresión de
MR facilitaba la consolidación de las memorias, pero disminuía la flexibilidad cognitiva,
mientras que la disminución de GR incrementaba los comportamientos depresivos y también
disminuía la flexibilidad cognitiva. Todo esto impediría una realimentación negativa efectiva.
Durante la realimentación negativa, una de las estructuras principales es NPV. Aunque los
inputs sinápticos de NPV dependen de transmisiones de glutamato, la realimentación negativa de
las respuestas del eje HPA consiste en la liberación de GABA desde sus neuronas GABAérgicas
(Herman et al., 2002). Se ha demostrado que la regulación del eje HPA en NPV se efectúa por
medio del aumento de la concentración de glucocorticoides en el área, los cuales también
promueven la síntesis de anandamida, 2-araquidonilglicerol (2-AG) (endocanabinoides) y la
activación de los receptores CB1 (cannabinoides), que se encuentran en las neuronas
presinápticas (Di, Malcher-Lopes, Marcheselli, Bazan, & Tasker, 2005). En efecto, CB1 se
encarga de disminuir la secreción de neurotransmisores hacia el espacio sináptico, por lo tanto,
inhibe el flujo de glutamato de las neuronas glutamatérgicas en NPV y disminuye su actividad
(Evanson, Tasker, Hill, Hillard, & Herman, 2010), pero a su vez facilita la liberación de GABA
(Di et al., 2005). Se ha observado que la liberación de GABA es bloqueada si se inhibe de manera
selectiva CB1, no obstante, los agonistas de endocanabinoides no aumentan la actividad (Di et
al., 2005). Es así como NPV, por medio del receptor CB1, facilita una rápida realimentación

33
negativa por medio de la inhibición del eje HPA durante la exposición a estrés (Evanson et al.,
2010).
Ahora bien, para los propósitos de esta investigación, es interesante tener en cuenta las
respuestas de un organismo cuando se ve expuesto a una situación de estrés severa, puesto que
esta condición lleva consigo una serie de respuestas que no son adaptativas para el sistema. Este
tipo de situaciones se caracteriza por la desregulación en el eje HPA y provocan un aumento en la
velocidad de la realimentación negativa desde los GC (Belda et al., 2008; Perrine et al., 2016).
Todo esto es debido a una disminución en los niveles basales de GC, una baja respuesta
adrenocortical (Kanter et al., 2001), una modesta supresión de ACTH, el cual no es suprimido de
manera correcta por causa de la falta de GC en los linfocitos (Kanter et al., 2001; Belda et al.,
2008) y un aumento de la concentración de GR, específicamente en el hipocampo (Belda et al.,
2008). Esto mismo trae como consecuencia una hiperresponsividad de ACTH a la inducción de
estresores no severos, como la exposición a ambientes novedosos (Belda et al., 2008; Yehuda,
Lowy, Southwick, Shaffer, & Giller, Jr., 1991).
A continuación, se van a explicar los efectos del estrés sobre el hipocampo y la microglía.
Aproximaciones a los efectos del estrés sobre el hipocampo
Se han estudiado los efectos del estrés sobre las actividades del hipocampo (Feldman &
Weidenfeld, 1999; Kim, Song, & Kosten, 2006) y se ha identificado que este mismo provoca
cambios adaptativos y no adaptativos en el sistema dependiendo de la intensidad del estímulo, su
anticipación y la percepción sobre su controlabilidad (McEwen, 2000; Lucassen et al., 2014).
Existe evidencia de que el hipocampo es una estructura dependiente de la actividad de los GC y
es indispensable para la regulación de la realimentación negativa (Joels, Sarabdjitsingh, & Karst,
2012). Las acciones del estrés sobre el hipocampo son muy fuertes, a tal punto que promueven
cambios morfológicos en las neuronas del área CA3, en GD y en CA1 (no muy fuertemente)
(Joels, 2008; Alfarez et al., 2009). En estado de desarrollo también se ha observado la reducción
de la complejidad dendrítica en las neuronas piramidales de CA1 y una disminución en las
conexiones con CA3 (Alfarez et al., 2009). Otros ejemplos sobre dichos efectos se han observado
por medio de algunos modelos animales que se encargan de estudiar acciones las acciones del
estrés sobre la plasticidad sináptica (LTP/LTD), con la cual depende el aprendizaje y la memoria

34
(Kim & Yoon, 1998; Ryan, Vollmayr, Klyubin, Gass, & Rowan, 2010; Tse, Bagot, Hutter,
Wong, & Wong, 2011); la neurogénesis en GD (Bain, Dwyer, & Rusak, 2004); la consolidación
de memorias emocionalmente significativas (McEwen & Gianaros, 2010; Okuda, Roozendaal, &
McGaugh, 2004); los procesos cognitivos; los procesos afectivos (Sapolsky, 1985) y otros
cambios morfológicos y funcionales en la región (Fuchs & Flugge, 1998).
En efecto, el estrés afecta las funciones básicas que comprometen la actividad del
hipocampo no obstante, esta estructura también ejecuta una acción importante en el proceso de
realimentación negativa de GC, que es mediado a través de su gran concentración de MR y GR
(Kim & Yoon, 1998; Feldman & Weidenfeld, 1999; Sapolsky, Krey, & McEwen, 1984). Existen
resultados que soportan la hipótesis de que el hipocampo juega un rol importante en la inhibición
del eje HPA (Sapolsky et al., 1984) y se ha puesto en evidencia a través del daño en esta
estructura. Daños en el hipocampo producen el aumento de la expresión del ARNm que codifica
ACTH en la pituitaria; del ARNm que codifica AVP en la subdivisión medial del NPV; del
ARNm que codifica CRF en NPV; además de un aumento de la concentración de GC (Herman et
al., 1989). También pueden resultar en una hipersecresión de GC, epecialmente en cerebros de
mayor edad (Lucassen et al., 2014; Sapolsky et al., 1984; Sapolsky, 1985). Todo esto se debe a
una disminución en los receptores GR y MR en las neuronas piramidales del hipocampo; no
obstante, estos efectos son reversibles (Sapolsky et al., 1984). También se ha investigado que el
impedimento de la secreción de GC durante el estrés está relacionado con la pérdida de neuronas
hipocampales. Estudios demuestran que la muerte de células granulares del hipocampo después
de una adrenalectomía corroboran la desaparición de receptores GR en la capa granular del
hipocampo junto con la posterior muerte de células granulares (Hu et al., 1997). Sapolsky y
colegas (1984), demostraron que la ablación completa del hipocampo provoca efectos similares al
síndrome de hipersecreción de corticosteroides, pues al ser asociados con la disminución de
receptores GR y MR, se aumenta la concentración de GC en la zona y se disminuye el chance de
realizar una realimentación negativa efectiva. Así mismo, mostraron que el impedimento de la
terminación de la respuesta del eje HPA prolonga la percepción del estresor en el tiempo.
Finalmente, los autores comentan sobre un aumento en la concentración de CRF después de las
lesiones del hipocampo, que también desinhiben la supresión de la realimentación negativa.

35
Los GR y MR en NPV son regulados por la liberación de GC y sin la acción inhibitoria
del hipocampo sobre esta estructura, el eje HPA no funcionaría correctamente (Feldman &
Weidenfeld, 1999). Aunque es necesario tener presente que los efectos inhibitorios del eje HPA
no sólo dependen de la actividad del hipocampo, sino también de la glándula pituitaria, el
hipotálamo, la corteza prefrontal y las proyecciones indirectas desde la amígdala (Sapolsky et al.,
1984). Como ya se ha dicho, daños en la función inhibitoria del hipocampo y la disminución de
su actividad pueden resultar en una hipersecreción del eje HPA (Herman et al., 1989; Kim &
Yoon, 1998) y evidencia suficiente corrobora esta observación (Feldman & Weidenfeld, 1999;
Herman et al., 1989).
En cuanto a los mecanismos de los GC que han sido examinados durante situaciones
estresantes, también se identifica que los cambios de su concentración en el hipocampo pueden
provocar la muerte de las neuronas de la región, pero la adrenalectomía atenúa estos daños
(Sapolsky, 1985). Al mismo tiempo, lesiones con neurotoxinas son exacerbadas por medio del
aumento de la concentración de GC, los cuales reducen el número de neuronas hipocampales
(Sapolsky, 1985). Se ha documentado que los GC son importantes en los cambios metabólicos de
las células y su incremento está asociado con un efecto catabólico en el tejido al que se adhiere
(May, Bailey, Mitch, Masud, & England, 1996). Por consiguiente, participarían en la degradación
de las células hipocampales, las cuales van a tender a oxidarse y a degradar sus proteínas y
aminoácidos más velozmente (Sapolsky, 1985; May et al., 1996; McEwen & Sapolsky, 1995).
Ahora bien, se sabe que el estrés facilita la formación de memorias cuando el estresor se
encuentra directamente relacionado con el contexto de aprendizaje, no obstante la consolidación
de otras memorias (de valencia emocional neutra) se ve impedida (Wiegert, Joels, & Krugers,
2006; Payne et al., 2007). El aumento de la concentración de GC incrementa la potenciación
sináptica en el área CA1 cuando los niveles altos de la hormona coinciden con la frecuencia de
estimulación (Wiegert et al., 2006). En otros estudios se ha observado que un estímulo estresor
después de una habituación a un ambiente disminuye la tendencia de recuperar memorias no
relacionadas con el estresor, que ya han sido consolidadas con anterioridad (Okuda et al., 2004).
Por otro lado, si el estímulo estresor se encuentra presente durante la presentación de un ambiente
novedoso, la consolidación en ese ambiente se va a ver aumentada, pero una reconsolidación a
nuevos estímulos no (Okuda et al., 2004). Las diferencias que compelen la consolidación y

36
reconsolidación de la memoria de los animales no habituados también depende de la actividad de
BLA, mediada por sus receptores adrenérgicos (Okuda et al., 2004).
El estrés afecta todos los procesos que convienen a la consolidación, recuperación,
reconsolidación y extinción de las memorias en un ambiente dado (Schwabe et al., 2012). De
hecho, la fuerza de la consolidación de este tipo de memorias no solo es trabajo del hipocampo,
sino también de BLA, la cual influencia la consolidación de las memorias a largo plazo después
de estímulos estresantes o emocionalmente significativos (McGaugh, 2000; Roozendaal et al.,
2002). Un aumento en la consolidación de las memorias estresantes se ve mediada por esta
estructura cuando es inducida por hormonas del estrés, glucosa y neurotransmisores como NE
(Lee, Graham, & Gold, 1988; Roozendaal et al., 2002; McGaugh, 2000). Varios estudios han
indicado que la relación entre los efectos del estrés sobre la consolidación de las memorias toma
rumbos diferentes sobre dicha consolidación. Por un lado, las memorias dependientes de la
liberación de NE, se observan mediante sus efectos sobre BLA (Roozendaal et al., 2002) la cual
recibe inputs de NE para incrementar la señalización de endocanabinoides desde receptor CB1 y
finalmente fomentar la consolidación de la memoria (Roozendaal et al., 2009; Roozendaal,
McEwen, & Chattarji, 2009). En específico, la activación de los receptores α y β-adrenérgicos de
BLA estan directamente relacionados con la consolidación de las memorias en el hipocampo
(Roozendaal & McGaugh, 1996; McGaugh & Roozendaal, 2002; Roozendaal et al., 2002). Así
pues, la actividad de la amígdala se enfoca en la consolidación de la memoria emocionalmente
significativa a través de sus conexiones con el hipocampo y se encarga de aparear rápidamente
estímulos neutros con eventos aversivos para que estos se reconozcan con propiedades aversivas
(Phelps, 2004).
Por esta misma línea, se ha reportado que los GC también se adhieren a los receptores GR
de la BLA y nuevamente aumentan la consolidación de las memorias de valencia emocional
(Roozendaal et al., 2009). Por ejemplo, si en la estructura son administradas infusiones de
antagonistas de CRF, su función se inhibe antes de la exposición a estresores fuertes, lo que
conyeva a que no se pueda consolidar la memoria estresante, pero si otras memorias que se
encuentren en el ambiente a través de la actividad del hipocampo (Roozendaal et al., 2002). Otro
ejemplo se observa cuando después de un entrenamiento aversivo, las infusiones de NE en la
amígdala aumentan significativamente la consolidación de las memorias y el LTP en

37
comparación con los grupos control (McGaugh & Roozendaal, 2002; Kim et al., 2005), pero sus
lesiones inhiben dichos impedimentos (Kim et al., 2005).
Todo esto nos hace reflexionar sobre el papel de la amígdala como una de las estructuras
implicadas en los comportamientos y la consolidación de la memoria estresante gracias a sus
influencias excitatorias en el hipocampo (Eagle et al., 2013; Walker & Davis, 2002). Así pues, al
observar que estas estructuras se encuentran directamente relacionadas, sus conexiones
anatómicas permiten identificar la influencia de la una sobre la otra (Canning & Leung, 1997;
Henke, 1990; Pelletier & Pare, 2004), en otras palabras, la amígdala envía proyecciones directas
(desde las neuronas parvocelulares y magnocelulares del BLA hacia CA1 y el súbiculo) e
indirectas (hacia la corteza entorrinal) hacia el hipocampo (Canning & Leung, 1997).
Ahora bien, como se ha expuesto en la sección anterior, la respuesta de estrés SAM tiene
como predominio la liberación de Epi y NE en el sistema y sus efectos regulan de manera
indirecta la función del hipocampo. Comenzando por el nervio vago, se ha demostrado que su
estimulación afecta los caminos noradrenérgicos de NTS y LC, los cuales aumentan la
concentración de NE la corteza y el hipocampo (Morilak et al., 2005; Habib et al., 2001). Los
efectos específicos de NE también se han constatado por medio de sus acciones sobre la
neurogénesis, en las que se ha demostrado, por medio de la activación de los receptores β3-
adrenérgicos de la ZSG del GD, que aumentan la posibilidad de sobrevivencia de las nuevas
neuronas, pero su inhibición impide por completo los efectos de NE sobre esa región (Jhaveri et
al., 2010).
Paralelamente, la liberación de Epi también participa en las influencias sobre la
consolidación de la memoria, tanto de estímulos emocionalmente significativos como de
estímulos neutros, gracias al funcionamiento de los receptores β-adrenérgicos de las estructuras
que estimula y su posterior acción sobre el hipocampo (Dornelles et al., 2007). Como ya se sabe,
los receptores β-adrenérgicos NTS se proyectan hacia la amígdala, la cual dirige sus proyecciones
hacia LC (Nordby, Torras-Garcia, Portell-Cortes, & Costa-Miserachs, 2006; Dornelles et al.,
2007). La administración de Epi poco tiempo después del entrenamiento aumenta aún más la
consolidación durante eventos emocionalmente significativos (Dornelles et al., 2007; Nordby et
al., 2006). Nordby y colegas (2006) argumentaron que es común observar que consolidación de

38
las memorias es mayor en situaciones en las que el cuerpo requiere una gran demanda de energía
para sobrevivir. Este es el caso de la liberación de Epi permite que se mantenga el rendimiento
por más tiempo y se acelere el proceso de adquisición de las memorias emocionales por más
tiempo.
Por otro lado, el papel de la glucosa en el hipocampo también es importante y esta ha sido
evaluada por medio del aumento de la consolidación después de un condicionamiento apetitivo y
un condicionamiento aversivo (Lee et al., 1988; Bueno, de Paiva, Correa, Tiba, & Fornari, 2017;
Dickinson, 2012). Como ya se ha dicho, Epi no puede cruzar la barrera hematoencefálica, pero
sus efectos sobre la memoria dependen de los mecanismos periféricos de respuesta a su
incremento en el plasma (McGaugh, 2000). En este caso, inyecciones de glucosa después de un
entrenamiento aversivo aumentan de manera significativa la consolidación de la memoria (Lee et
al., 1988) y estas repuestas son consistentes con otros resultados que indican que el aumento de la
concentración de Epi en la sangre aumenta los niveles de glucosa y la posterior consolidación de
la memoria (Lee et al., 1988; McGaugh, 2000; Wiegert et al., 2006).
A continuación, se van a abordar los efectos directos del estrés sobre el hipocampo. En
primer lugar, el estrés influencia las funciones neuronales (Keralapurath, Clark, Hammond, &
Wagner, 2014) que conceden la posibilidad de aumentar o disminuir los procesos de aprendizaje,
memoria y mediar la regulación emocional y la posterior adaptación comportamental en un
contexto dado (McEwen & Sapolsky, 1995; de Kloet, Oitzl, & Joels, 1999). Precisamente, la
regulación comportamental depende de la función de GC sobre MR y GRy se ha observado que
su activación es indispensable para la consolidación de las memorias (de Kloet et al., 1999; Oitzl
& de Kloet, 1992). En ratas, el bloqueo selectivo de MR y GR puede interferir con el aprendizaje
espacial. Particularmente el bloqueo de MR altera las estrategias de escape en el laberinto
acuático de Morris, mientras que el bloqueo de GR interrumpe la consolidación de información
espacialy la adrenalectomía provoca los mismos efectos (Oitzl & de Kloet, 1992). En cuanto al
aprendizaje emocional (Zhou et al., 2010) han detallado que, durante situaciones estresantes, GR
se bloquea durante y después de la adquisición de la tarea de estrés, lo que conlleva
impedimentos en la memoria contextual, pero después de unas horas, el rendimiento aumenta.
Dicho aumento del rendimiento, según los autores, es debido a que la liberación de GC,
predominante para la activación de MR, aumenta la formación de memoria espacial y emocional.

39
Por otra parte, examinaron que el bloqueo de MR no tiene efectos inmediatos en la memoria
contextual, pero si disminuye el rendimiento en una tarea de estrés. Esto permite sugerir que las
respuestas de adaptación ante situaciones estresantes se ejercen por medio de GR y MR que
facilitan la formación de memorias emocionales de corto y largo plazo.
Partiendo de lo anterior, no se puede dejar de lado la noción que el estrés afecta esta
estructura en cuanto a todas funciones. Si se examinan los efectos del estrés sobre LTP, es
interesante analizar como un único estímulo de estrés agudo puede provocar cambios en el LTP
que son diferenciados en HD (disminución de LTP) y en HV (aumento de LTP), incluso 2
semanas después de la exposición. En condiciones normales, el LTP de HV es de menor
magnitud que HD, pero se observa un incremento significativo de este después de la exposición a
un estímulo estresante (Maggio & Segal, 2007; Keralapurath et al., 2014). Con esto, desde otras
perspectivas, los efectos comportamentales del estrés, como por ejemplo la consolidación de las
memorias emocionales se hace más claro, en tanto que los circuitos límbicos dependientes de HV
son fortificados, pero los de HD que se encargan de la consolidación de memorias que no son
emocionales, como la navegación espacial se ven disminuidos (Pothuizen et al., 2004;
Keralapurath et al., 2014). Cabe resaltar que específicamente en HV, el aumento del LTP es
mediado por medio de la activación de MR, pero la supresión del LTP es mediada por la
activación de GR (Maggio & Segal, 2007). Desde HV se envían eferencias hacia NPV para
inhibir la respuesta de estrés (Maggio & Segal, 2007). Todo esto compromete cambios en el
rendimiento cognitivo y los comportamientos relacionados con el estrés (McEwen & Sapolsky,
1995; Maggio & Segal, 2007; Keralapurath et al., 2014).
Nuevamente, durante el estrés, GR y MR van a impedir y facilitar LTP en el hipocampo
de manera respectiva. Específicamente la activación de MR facilita el LTP y la de GR lo inhibe
(Korz & Frey, 2003; Maggio & Segal, 2007). Estudios antecedentes han exhibido que una prueba
de nado bloquea GR y mantiene el proceso del LTP, pero la inactivación de MR obstaculiza este
último (Korz & Frey, 2003). Todo esto permite sugerir el papel que juegan estos receptores sobre
la modulación de LTP durante la respuesta de estrés y la regulación de los comportamientos
ocasionados a partir de estas reacciones. En cambio, si el estímulo estresor es muy fuerte, la
plasticidad en el hipocampo se tornará más difícil, sin importar el receptor por lo tanto, se alterará
el aprendizaje (Ryan et al., 2010).

40
Diferentes tipos de estrés agudo pueden alterar los procesos LTP por algunos minutos e
incluso varias semanas y de manera más específica sus efectos se observarán en las corrientes
sinápticas sobre los canales de calcio dependientes de voltaje (VDCC) y los receptores NMDA
(Ryan et al., 2010). En todo caso el estrés, desde moderado hasta incontrolable, causa una
modulación en la plasticidad sináptica del hipocampo, la cual es relacionada con la inducción de
comportamientos de depresión o ansiedad (Joels, 2008; Lucassen et al., 2014) o la falta de
rendimiento durante tareas cognitivas (Cho et al., 2011; Korz & Frey, 2003; McEwen &
Sapolsky, 1995).
En cuanto a los efectos sobre la regulación de los receptores glutamatérgicos NMDA, se
ha observado que la liberación de GC inhibe su actividad en el hipocampo (Hardingham, 2009;
Ryan et al., 2010; Feldman & Weidenfeld, 1999). Estas acciones inhibitorias de los GC afectan
las corrientes de NMDA y protegen las neuronas que están expuestas al incremento de su
concentración durante el estrés (Liu et al., 2007). Por otro lado, la concentración de GC puede
modular de manera aguda la inhibición de NMDA por medio de sus efectos sobre el aumento de
la concentración de Ca2+ intracelular (Liu et al., 2007).
Otros estudios que corroboran esta observación sugieren que la potenciación de NMDA
se ve inhibida después de la administración de GC en el área CA1 (Krugers et al., 2005). Se debe
aclarar que la potenciación por medio de NMDA es diferente a la que se efectúa en VDCC, pues
la inducción de GC efectivamente aumenta la potenciación de VDCC, más no precisamente de
NMDA (Krugers et al., 2005). La activación de los receptores MR y GR cooperan con la
modulación de los niveles de Ca2+ intracelular, al aumentar sus corrientes de entrada, pero en
estos casos NMDA se ve inhibida, pues el aumento de la concentración de glucocorticoides vía
GR aumenta fuertemente la inducción de Ca2+ intracelular y suprime su función (Krugers et al.,
2005). Grandes concentraciones de Ca2+, atribuye a una mayor inhibición de las corrientes de
NMDA inducidas por inhibidores de PKA (Liu et al., 2007).
Por otro lado, también se ha reportado que la actividad de NMDA después de la
exposición un estímulo estresor incrementa su función sináptica por medio de la actividad de sus
subunidades, GluN2A y GluN2B, incluso una hora después de la exposición inicial (Tse et al.,
2011). Se ha observado que los antagonistas de dichas subunidades inhiben de manera selectiva
el proceso de LTP o LTD respectivamente, haciendo de estos factores importantes para

41
determinar la polaridad y plasticidad sináptica (Liu et al., 2004). Adicionalmente, los efectos de
PKA en la actividad funcional de NMDA incluso depende de estas subunidades (Krugers et al.,
2005).
Es preciso entender que los efectos del estrés agudo y el estrés crónico divergen en las
diferentes estructuras hipocampales y se han evidenciado diferencias significativas en el
desarrollo y actividad de las neuronas en las áreas de GD y CA3. Observaciones sobre los
cambios en la proliferación y la sobrevivencia de las neuronas en ratas experimentales que fueron
sometidas a una sola sesión de estrés agudo y a sesiones repetidas de estrés crónico por tres
semanas, permiten argumentar que muchos factores que regulan la neurogénesis en el hipocampo
participan en la inducción de plasticidad estructural (Pham, Nacher, Hof, & McEwen, 2003),
como la remodelación dendrítica de las neuronas piramidales de CA3, que con el exceso de
glucocorticoides o bajo el efecto de los mecanismos excitatorios de NMDA promueve cambios
(Kole, Czeh, & Fuchs, 2004; Pham et al., 2003). Otros estudios han indicado que los efectos del
estrés agudo sobre la neurogénesis de GD evidencian una disminución de la sobrevivencia y
proliferación de las nuevas neuronas (Schoenfeld & Gould, 2012).
Ahora bien, es importante recalcar la importancia de la sensación de controlabilidad que
se tiene sobre la situación estresante. (Kavushansky, Vouimba, Cohen, & Richter-Levin, 2006),
demostraron en humanos que los efectos del estrés incontrolable efectuaron una supresión en la
plasticidad CA1 y disminuyeron LTP, pero aumentaron la plasticidad en GD. Además,
observaron que los niveles de corticoides en el plasma eran mayores que el grupo control. En
estos casos, el aumento de la reactividad de la amígdala media los efectos de LTP en el
hipocampo. Es posible que bajo estas condiciones la memoria dependa más de GD que de CA1.
Pero el estrés crónico, provoca efectos diferentes, este se caracteriza por liberar una
cantidad sustanciosa de GC en el cerebro y vía la estimulación de GR, inhibe la actividad de las
células (Lau et al., 2016). Dichos efectos generan, a largo plazo, déficits cognitivos y problemas
en la memoria (Magarinos, Verdugo, & McEwen, 1997). En CA3, después de la exposición a
estrés crónico, se observa que la resistencia de la membrana disminuye, se incrementa la
amplitud del estado de hiperpolarización, se disminuye el tamaño de las espinas dendríticas, se
aumenta la amplitud de la hiperpolarización, se aumenta la respuesta a la liberación de GC
dependiente la actividad a través de MR y GR, se interrumpe el proceso de LTP y se aumenta la

42
actividad del eje HPA (Kole et al., 2004), no obstante la excitabilidad somática se mantiene y no
se observan cambios sustanciales en las propiedades de la membrana, incluyendo el patrón de
despolarización, los voltajes de los picos y los voltajes del estado de base en las neuronas del
hipocampo (Kole et al., 2004).
Por otro lado, para los propósitos de esta investigación es importante tener claros los
efectos del estrés muy fuerte y prolongado sobre el hipocampo, debido a que a través de
estímulos estresores muy fuertes, promoverían la disminución del volumen del hipocampo
(Bonne et al., 2008). Estos efectos se pueden vincular con las consecuencias derivadas del
trastorno de estrés postraumático (PTSD). De ahí, (Zhe et al., 2008), observaron que la
disminución de la realimentación negativa que se encuentra aumentada en pacientes con PTSD y
una pérdida en el volumen del hipocampo, se deben a una disminución en los niveles de
glucocorticodes. Con esto, argumentaron que una sola exposición prolongada a un estímulo
estresante induce una disminución de la concentración de MR en el hipocampo (particularmente
en CA1) y un cambio de distribución de GR, los cuales se observan no solo en el núcleo, sino
también en el citoplasma. Lo anterior sugiere que la realimentación negativa ofrecida por el
hipocampo puede aumentar su velocidad en este tipo de situaciones y estos datos se relacionaron
con el aumento de los comportamientos de ansiedad y miedo. Los receptores de MR son aquellos
que controlan el eje HPA, por lo que su pérdida figuraría en una menor responsividad durante el
control de la realimentación negativa de dicho eje. También se ha observado que los receptores
GR ejercen un rol neuroprotector sobre el tejido hipocampal y su desregulación también
provocaría efectos nocivos en la estructura (Hu et al., 1997).
A diferencia de la observación anterior, otros estudios han demostrado que estímulos de
estrés muy fuerte aumentan la expresión de GR, que es regulado a través de un aumento en la
fosforilación de proteina quinasa B (PKB) (Eagle et al., 2013), una serina/taurina proteína kinasa
que se encuentra expresada en los circuitos emocionales y su fosforilación es observada durante
eventos como el estrés y en la regulación de las proteínas celulares involucradas en el
metabolismo, la sobrevivencia/apoptosis, la diferenciación y la proliferación (Fayard, Tintignac,
Baudry, & Hemmings, 2005). A lo largo del estímulo de estrés agudo, el aumento de la
concentración de estos dos contribuye al robustecimiento de comportamientos relacionados con

43
ansiedad después del estímulo estresor en el hipocampo, más no en la amígdala (Eagle et al.,
2013).
Por otro lado, también el mRNA de GR y MR en esta estructura también han sido objeto
de estudio durante el estrés (Liberzon, Lopez, Flagel, Vazquez, & Young, 1999). Existe evidencia
sobre los efectos en GR y MR después de la exposición prolongada a un estresor. Se reportó que
después de una semana de la exposición al estresor, se evidenció un aumento en el mRNA de GR,
pero una disminución del mRNA de MR. Esto permite sugerir que la estructura se torna más
sensible a responder a la realimentación negativa de los GC. Después de dos semanas, se observó
un aumento de GR específicamente en CA1 y GD y una disminución de MR en CA2 y GD
(Liberzon et al., 1999).
Finalmente, las hormonas del estrés, también actúan en conjunto con respuestas inmunes que
también responden de manera adaptativa a la interrupción en la homeostasis (Hass & Barnstable,
2016). En el siguiente capítulo se van a tratar los efectos de la respuesta inmune, específicamente
de la microglía sobre el estrés y sus efectos en el hipocampo.
Aproximaciones a los efectos del estrés sobre la microglía
En capítulos anteriores se hizo un breve comentario acerca de los mecanismos inmunes
que se encuentran involucrados en la regulación de las respuestas de estrés. Siendo cada vez
más aceptada la idea de que el estrés viene acompañado de una respuesta inmune, se ha
considerado que esta actividad también funciona como una mediadora de la regulación
homeostática en el sistema (McEwen & Wingfield, 2003). En el cerebro, el protagonismo de la
respuesta inmune en cuanto al estrés recae en la microglía y sus acciones proinflamatorias y
antiinflamatorias. No obstante, hace más de una década, estas no eran consideradas como
actores sustanciales debido a que la evidencia vinculaba a GC como los principales agentes
antiinflamatorios del sistema y resultaba ser una inconsistencia pensar que iban a permitir
efectos de inflamación (Johnson, O'Connor, Watkins, & Maier, 2004; Smoak & Cidlowski,
2004). Hoy en día, estas consideraciones han sido reevaluadas y van a ser expuestas en el
presente capítulo, cuyo tema principal aborda los efectos del estrés sobre las acciones de la
microglía.

44
Las células de la glía están comprometidas en la interacción con las neuronas y el
mantenimiento de la homeostasis neuronal por medio de la modulación de los electrolitos, los
neurotransmisores, las citosinas y los factores neurotróficos (Maletic et al., 2007). Durante el
estrés, la hiperactividad del eje HPA en conjunción con el eje SAM promueven el aumento en el
tono simpatético del sistema y posteriormente la liberación de citosinas proinflamatorias
asociadas con la pérdida de insulina y la sensibilidad de los MR y GR (Maletic et al., 2007;
Payne et al., 2007).
Se ha observado que, dependiendo de la naturaleza y la intensidad del estresor, se
aumentará o se disminuirá la acción inmune en el cuerpo. Estudios previos han resaltado que el
estrés psicológico aumenta la proliferación de citosinas proinfalamatorias e inmunoregulatorias
(Maes et al., 1998) como IL-6 (Maes et al., 1999), IL-1β (Spivak et al., 1997), TNF-α, INF-γ, e
inmunoregulatorias como IL-4 e IL-10 (Maes et al., 1998), además de las catecolaminas y
neurohormonas usualmente encontradas durante las respuestas de estrés.
La evidencia de que la regulación del eje HPA es mediada por medio de la liberación de
citocinas proinflamatorias (Besedovsky et al., 1991), se ha observado a través de las
consecuencias específicas sobre los niveles de IL-1y en menor intensidad IL-6 y TNF-α en el
cerebro, pues su estimulación induce una regulación positiva de CRF y ACTH que conllevan la
posterior liberación de glucocorticoides desde las glándulas adrenales (Besedovsky et al.,
1991; Spivak et al., 1997; Sweep, Rijnkels, & Hermus, 1991; Smoak & Cidlowski, 2004). Se
ha observado el interesante efecto indirecto del polipéptido activador de adenil-ciclasa de la
pituitaria (PACAP)-el cual se encarga de incrementar los niveles del ARNm productor de CRF
en la región parvocelular del núcleo paraventricular del hipotálamo (Kageyama & Suda, 2009)-
en la producción de IL-6 (Nezi, Mastorakos, & Mouslech, 2000).
Se parte de que GC es una neurohormona que causa un efecto de inmunosupresión en el
sistema. En efecto, estas sustancias se han considerado como los immunosupresores más
importantes del cuerpo y sus mecanismos han sido descritos por medio de sus acciones
genómicas en las que, vía GR, inhiben la síntesis de genes proinflamatorios como el factor
nuclear-κB (NF-κB) o la proteína activadora-1 (AP-1) cuyas acciones también se encuentran
presentes en la microglía (Smoak & Cidlowski, 2004; Rhen & Cidlowski, 2005). Los primeros
estudios que mostraron el interés en este tema comenzaron a presentar modestas aproximaciones

45
hacia los efectos inmunes que podrían estar implicados en el estrés, pero con bastante precaución
por causa de la inconsistencia que representaba el rol antiinflamatorio de GC ante las respuestas
proinflamatorias del estrés.
Así pues, en estudios precedentes se observa que la administración de IL-1 aumenta la
respuesta de ACTH y GC por varias semanas y provoca efectos directos en el aumento de la
secreción de AVP y CRF en NPV (Schmidt et al., 2001). Estos planteamientos demuestran su rol
en la sensibilización del eje HPA.
Otras líneas de estudio han dirigido sus investigaciones hacia la observación de los
efectos del aumento de la proliferación de ciertas citosinas proinflamatorias después de la
inducción a un estímulo de estrés agudo y la administración de LPS. En la mayoría se ha
observado que, por ejemplo un choque eléctrico inescapable aumenta la concentración de IL-1β,
la cual es exacerbada con la administración de LPS o una adrenalectomía (Nguyen et al., 1998).
Pero, nuevamente, estas observaciones fueron discutidas por causa de las ya presentes
inconsistencias con los efectos antiinflamatorios de GC.
En relación con el apartado anterior, es importante recalcar que el estrés efectivamente
promueve respuestas proinflamatorias y sin la necesidad de administrar LPS. Por ejemplo,
después de un estímulo de estrés severo, como choque eléctrico inescapable, se ha evidenciado
un incremento del mARN de IL-1β en las regiones del hipotálamo, el hipocampo, la glándula
pituitaria y el esplenio hasta por 8 horas después del estímulo; pero no aumenta
significativamente la expresión del ARNm de TNF-α e IL-6 en Cortex parietal frontal en la
glándula pituitaria y en el hígado (O'Connor et al., 2003). Por otro lado, otros estudios si han
observado el incremento de la concentración en el plasma de IL-6 la cual, incluso al lado de la
acción de los GC (Zhou, Kusnecov, Shurin, DePaoli, & Rabin, 1993). En todo caso, se ha
considerado que IL-1β es una citosina que medía las consecuencias comportamentales y
fisiológicas de los estímulos de estrés agudo. No obstante, también se debe tener en cuenta que
las moléculas clásicas que reaccionan frente al estrés, GC y NE, también regulan dicha expresión,
por medio de otros mecanismos que alteran la glía (O'Connor et al., 2003).
Ciertamente los resultados no se alejaban de esta tendencia, pero sus aportes eran valiosos
para el entendimiento del papel de la inflamación en el estrés. Observaciones como el aumento de

46
la concentración de citosinas proinflamatorias (IL-1β y TNF-α o IL-6) posteriores a estímulos de
estrés incontrolable (como el choque eléctrico en la cola inescapable), el interesante aporte sobre
el aumento en el plasma de estas sustancias (Johnson et al., 2002) e incluso estudios con la
administración de antagonistas de IL-1β, que provocan un bloqueo completo en la liberación de
otras citosinas proinflamatoiras después de la exposición a un estímulo de choque eléctrico
inescapable en la cola (Johnson et al., 2004), demuestran cambios en la investigación sobre el
estrés junto al rol de las citosinas proinflamatorias, pero sin considerar la microglía como un
actor importante.
Ahora bien, los primeros estudios que tuvieron en cuenta el papel de la microglía
propendieron por observar los efectos de su bloqueo en el sistema. Antes de ser indagada con
profundidad se tenía conocimiento de que este tipo de células inducían la proliferación de
metabolitos de NO, de IL-1β y de P38 MAPK cuando era estimulada con glutamato o kainatoy
que su acción proinflamatoria era inhibida a través de la administración de la tetraciclina
antiinflamatoria, minocicina (Tikka, Fiebich, Goldsteins, Keinanen, & Koistinaho, 2001).
El papel de la microglía como la principal fuente de IL-1β también ha sido observada a
través de la modulación de NE (Blandino, Jr., Barnum, & Deak, 2006). El tratamiento con
antagonistas de receptores β-adrenergicos, como el propanolol, bloquea completamente la
liberación de IL-1β, por otro lado el bloqueo de la recaptación de NE (con dispramina)
incrementa las concentraciones de IL-1β (Blandino, Jr. et al., 2006). En efecto, se ha observado
que la producción de citosinas proinflamatorias durante el estrés es aumentada por medio de la
estimulación de los receptores β-adrenérgicos. (Johnson, Zimomra, & Stewart, 2013), observaron
las maneras en las que la microglía podía incrementar la liberación de estas citosinas después de
la estimulación de receptores β-adrenérgicos antes a una administración de LPS en el hipocampo.
Estas mismas conclusiones se han comprobado después de examinar la estimulación y la
inhibición de la microglía después de la inducción de agonistas y antagonistas de receptores β-
adrenérgicos y α1-adrenérgicos (Wang et al., 2010). Los resultados concluyeron que el
antagonismo de los receptores α1-adrenérgicos no causaron efectos significativos en la acción de
la microglía, pero los agonistas de los receptores β-adrenérgicos, incluso en ausencia de una
estimulación proinflamatoria incrementaron el ARNm de IL-1β, TNF-α e IL-6 en un cultivo de
microglía (Wang et al., 2010). La expresión de citosinas proinflamatorias durante un trauma

47
cerebral aumenta después de la administración de agonistas β-adrenérgicos, pero se atenúan por
antagonistas de estos mismos. La estimulación de la microglía por medio de los agonistas de
receptores β-adrenérgicos también aumenta la concentración de ERK y p38 MAPK (Wang et al.,
2010). Ciertamente, todo esto demuestra que los receptores β-adrenérgicos medían la función
proinflamatoria de la microglía.
Estos efectos de la microglía se ven expresados en comportamientos de ansiedad
inducidos por estrés social. Nuevamente es observado un aumento en la expresión de citosinas
proinflamatorias liberadas desde la microglía M1 en el cerebro, que es dependiente de la
actividad de las vías noradrenérgicas (Wohleb et al., 2011). Un estudio sobre estrés social
evidenció que la estimulación repetida del estresor incrementaba la concentración de microglía de
fenotipo M1 en la amígdala, la CPF y el hipocampo, además de mostrar un aumento del mARN
de IL-1β y una disminución en genes que responden a GC (Wohleb et al., 2011).
Por consiguiente, la microglía ejerce un papel activo durante las respuestas de estrés, por
ejemplo, estímulos de estrés agudo pueden provocar un aumento robusto en su fenotipo M1 tanto
en el tálamo, como en el hipotálamo, el hipocampo, la substancia nigra y la sustancia gris central,
aunque no precisamente se ha observado acompañado por marcadores de inflamación, es decir
IL-1β, IL-6 e iNOS (Sugama, Fujita, Hashimoto, & Conti, 2007). De manera adicional, es
importante tener en cuenta que durante dicha respuesta no se deben ignorar las otras funciones de
la microglía, pues más allá de la liberación de citosinas proinflamatorias, también se ejecuta su
acción de fagocitar otras células afectadas por la excitotoxicidad producida por el estrés (Sugama
et al., 2007). Entre las funciones de la microglía, el fenotipo proinflamatorio también se encarga
de fagocitar las células que han muerto y esto mismo se observa por medio del marcador CD68
(Croisier, Moran, Dexter, Pearce, & Graeber, 2005). La microglía que expresa la presencia de
este marcador regularmente se encuentra alrededor de neuronas necróticas o que han pasado por
un curso de enfermedad (Croisier et al., 2005) ejerciendo efectos de limpieza sobre el tejido
celular.
Por otro lado, el hipocampo es una estructura vulnerable, no solo ante los efectos del
estrés, sino también ante la actividad de la microglía, pues es en esta región donde se encuentra
en grandes concentraciones (Lawson, Perry, Dri, & Gordon, 1990). En el hipocampo, la
microglía se distribuye principalmente en las áreas CA1 y CA3 (Dalmau et al., 1998) y se ha

48
reportado una fuerte proliferación en las neuronas granulares del giro dentado, Stratum
Pyramidale (SP), Stratum Lucidum (SL) and Stratum Radiatum (SR) (Vukovic et al., 2012; Lana
et al., 2016). Siendo considerada la microglía como un substrato de las respuestas de estrés, no
cabe duda de que las funciones hipocampales se van a ver afectadas de manera significativa. Por
ejemplo, hay estudios que tratan los efectos del estrés en la expresión de la microglía y
demuestran que un solo choque incontrolable promueve en las regiones CA3 y GD, más no muy
fuertemente en CA1, la activación microglial (MHC II, CD11b, Iba-1y GFAP) y la disminución
de sus marcadores regulatorios (CD200) (Frank, Baratta, Sprunger, Watkins, & Maier, 2007).
Las acciones del LPS sobre la activación de la microglía en el hipocampo cuando es
inyectado en el área CA1, demuestran aumentos en la expresión de IL-1 y TNF-α, incluso 6 horas
después de la inducción (Tanaka et al., 2006). Ciertamente la proliferación de la microglía M1 es
observada a la par con el incremento de especies reactivas a oxígeno, pero también se observa el
aumento del gen de oxigenasa-1 (HO-1), un marcador del estrés oxidativo (Block, Zecca, &
Hong, 2007). Es importante tener en cuenta que HO-1 no se encarga precisamente de efectuar
acciones proinflamatorias, sino que juega un rol en los efectos de neuroprotección durante
actividades que promueven daños oxidativos y exitotóxicos (Satoh et al., 2003).
El estrés, a largo plazo, provoca la activación prolongada de la microglía M1 y puede
resultar en una desregulación de la transmisión glutamatérgica en el hipocampo, promoviendo
problemas de memoria y aprendizaje (Tanaka et al., 2006). De manera interesante, la morfología
dinámica y la motilidad de estas células también son reguladas a través de la actividad excitatoria
glutamatérgica e inhibitoria GABAérgica (Fontainhas et al., 2011). De hecho, la microglía
expresa receptores de glutamato (GluR) con los cuales se puede aumentar su actividad
proinflamatoria y neurotóxica (Murugan, Ling, & Kaur, 2013). Por ejemplo, se ha observado que
después de una inducción de isquemia en el área CA1 del hipocampo, la expresión de GluR desde
las células microgliales reactivas (M1) incrementa en las áreas afectadas por el daño isquémico
(Gottlieb & Matute, 1997).
Otros estudios han evaluado los efectos proinflamatorios de la microglía en el proceso de
neurogénesis. Así pues, estudios que evalúan los efectos del estrés agudo sobre calitrícidos
adultos, evidencian una disminución en la proliferación de neuronas subgranulares de GD y
cambios en la morfología de células inmunes agrupadas alrededor de la estructura (Gould et al.,

49
1998). Conforme a este tipo de estudios, se ha evidenciado que la microglía ejerce un rol
importante en la neurogénesis del hipocampo adulto (Sierra et al., 2010).
En cuanto al LTP, se debe tener presente que los procesos de comunicación entre las
neuronas del hipocampo y la microglía han destacado el rol del receptor (CX3CR1) y su respetivo
ligando (CX3CL1), también conocidos por fractalkina (Meireles et al., 2016). Se ha reportado que
la señalización por medio del eje CX3CR1-CX3CL1 resulta ser neuroprotectora y tiene efectos
moduladores positivos en la neurogénesis y el LTP del hipocampo (Vukovic et al., 2012).
(Vukovic et al., 2012) observaron que el aumento de CX3CL1 es el responsable de mediar la
activación de los precursores de células (NCP) de la microglía en ratones después de 14 días de
ejercicio, especialmente en el giro dentado. La deficiencia de CX3CR1 impide la función de la
microglía, provocando la liberación de citosinas proinflamatorias. Otros estudios han reportado
que la deficiencia de este receptor impide la neurogénesis y el LTP en el hipocampo adulto,
cuando se elevan los niveles de citosinas proinflamatorias como IL-1β (Kierdorf & Prinz, 2013).
Retomando la idea anterior, este eje se encarga de mediar los cambios morfológicos de la
microglía y en presencia de estrés crónico y se ha observado que reduce la plasticidad en el
hipocampo (Milior et al., 2016). Su deficiencia previene que la microglía pueda ejercer una
respuesta morfológica ante el estrés, pues no permite su mantenimiento en estado de reposo
después de que dicho estrés ha terminado (Milior et al., 2016). En efecto, la exposición a
estímulos de estrés disminuye las arborizaciones microgliales y mantienen la morfología
ameboidea por más tiempo (Milior et al., 2016). A largo plazo se han observado problemas
cognitivos que son dependientes de las acciones neurotóxicas de la microglía cuando no se tiene
regulada la actividad de este receptor (Milior et al., 2016).
En cuanto al estrés crónico, también se ha distinguido que este provoca el aumento
dramático de la proliferación de microglía M1, que trae como consecuencia la pérdida de
conexiones sinápticas, el retraimiento de los axones, la sobreexpresión de citosinas
proinflamatorias IL-1 y TNF-α, la sobreexpresión de especies reactivas a oxígeno y la pérdida de
neuronas piramidales (Espinosa-Oliva et al., 2011). Ciertamente, cuando la inflamación es
crónica se inhibe la neurogénesis en el hipocampo (Monje, Toda, & Palmer, 2003). Ya se ha
mencionado que grandes concentraciones de microglía se encuentran ubicadas alrededor de las
zonas subgranular y granular de GD, en la proximidad de la zona de nacimiento de las neuronas

50
nuevas (Ekdahl, Claasen, Bonde, Kokaia, & Lindvall, 2003). Si a esta estructura se le administra
una dosis de LPS, muy posiblemente aumentará la proliferación de microglía de fenotipo M1,
seguida de la disminución de hasta un 85% de la neurogenesis (Ekdahl et al., 2003). No hay que
ignorar que estos efectos también son evidentes en otros cambios de la sinaptogénesis en
neuroblastos (Ekdahl et al., 2003; Chugh, Nilsson, Afjei, Bakochi, & Ekdahl, 2013). Impactos de
estos procesos, se pueden observar sobre la sinapsis GABAérgica e incrementos en las espinas
dendríticas de las nuevas neuronas, cuyos cambios pueden afectar de manera significativa la
transmisión sináptica en GD (Chugh et al., 2013).
Es cierto que la inflamación crónica previene el desarrollo los mecanismos en los que se
desencadena la neurogénesis y fomenta el progreso de deficiencias cognitivas, emocionales y de
memoria (Chesnokova et al., 2016). Asociado con la neuroinflamación crónica, una serie de
desórdenes cognitivos y problemas en la memoria y el aprendizaje, también son producto de
problemas inmediatos en la plasticidad, los cuales han sido relacionados con el gen Arc (proteína
regulada y asociada con la actividad del citoesqueleto) (Rosi et al., 2009; Belarbi, Arellano,
Ferguson, Jopson, & Rosi, 2012). Su actividad es asociada con la exploración espacial y los
procesos de plasticidad necesarios para la consolidación y el mantenimiento de la memoria
dependientes del contexto (Vazdarjanova, McNaughton, Barnes, Worley, & Guzowski, 2002). La
integración funcional de las nuevas neuronas de GD durante los procesos de inflamación, suelen
provocar cambios en la plasticidad y el procesamiento contextual y espacial, especialmente en
CA1, CA3 y GD (Rosi et al., 2009; Belarbi et al., 2012).
En párrafos anteriores se expuesto que la proliferación de microglía es producto de la
liberación de glutamato en el espacio sináptico al estimular NMDA. El estrés agudo que induce la
actividad de NMDA provoca un aumento del influjo de Ca2+ en las neuronas y un posterior
aumento en la exitotoxicidad, además de la disfunción en la mitocondria (Hardingham, 2009).
Como el NMDA funciona bajo una curva de toxicidad, muy poca actividad o mucha actividad
puede promover la muerte neuronal (Hardingham, 2009). Se sugeriría que este efecto induce la
posterior sobreactividad de la microglía. Otros estudios demuestran que la neuroinflamación
crónica afecta la expresión de la plasticidad en CA3 y GD, a través de un incremento
pronunciado de la actividad de la microglía, no obstante la memantina, un agonista NMDA,

51
reduce la neuroinflamación y retorna la expresión de Arc y sus posteriores actividades sobre la
cognición y la memoria (Rosi et al., 2009).
Al mismo tiempo, esta actividad inmune se asocia con la desregulación de varios aspectos
del eje HPA después de estímulos de estrés muy fuertes, como por ejemplo los causantes del
PTSD (Gill, Vythilingam, & Page, 2008). Investigaciones en humanos con dicho trastorno
también han demostrado que el aumento de la sensibilidad de los GC en los tejidos inmunes
(Rohleder, Joksimovic, Wolf, & Kirschbaum, 2004) revelan bajos niveles de IL-4, los cuales son
asociados con comportamientos de ansiedad excesiva; con una mayor circulación de IL-1β (von
et al., 2007); un aumento en la concentración de IL-6; de TNF-α y una disminución significativa
de la concentración de GC (von et al., 2007; Gill et al., 2008). Partiendo de esto, la
realimentación negativa es más veloz, por lo tanto, la supresión del eje HPA es reemplazada por
el incremento de la sensibilidad de las citosinas proinflamatorias (Rohleder et al., 2004). La
afinidad de los GC se encuentra distribuida alrededor de todo el cuerpo, no obstante, su supresión
conlleva un mecanismo de sobre reactividad en el sistema inmune (Rohleder et al., 2004). En
efecto el PTSD en el hipocampo aumenta la exacerbación de células microgliales M1 y provocan
degeneración celular y reducción en la neurogénesis de GD (Acosta et al., 2013).
De manera consistente con estas observaciones, los estímulos de estrés agudo en ratas
adrenalectomizadas y no adrenalectomizadas demuestran que hay incrementos en la liberación de
IL-1β y otras citosinas proinflamatorias, cuando el GC no se encuentra presente o se libera en
pocas intensidades (Nguyen et al., 2000; Rohleder et al., 2004). La posterior administración de
GC atenúa este efecto de manera significativa (Nguyen et al., 2000).
Hoy en día son cada vez son más estudiados los procesos de inflamación de la microglía
M1 durante situaciones estresantes, en las cuales también se observa el aumento de GC. (Nair &
Bonneau, 2006) hicieron parte de las primeras investigaciones que pretendían determinar la
interacción entre la microglía y los GC después de un protocolo de restricción de movimientos en
ratones, por 15 horas al día, durante 6 días y sin la posibilidad de recibir comida o beber agua. De
este protocolo se registró un aumento en la proliferación de microglía M1, GCy la función de los
receptores de NMDA. Entre los días 1 y 4, los niveles de microglía aumentaron de manera
dramática en el hipocampo, pero después del cuarto día el pico comenzó a disminuir. Los
resultados evidenciaron que efectivamente, durante una situación estresante los GC si

52
estimulaban la proliferación de microglía in vivo, es más también se ha registrado que la
actividad de GC sobre la microglía es directa, sin la necesidad de ningún intermediario, debido a
que posee GR y MR en su membrana (Tanaka et al., 2006; Barrientos et al., 2015; Sierra,
Gottfried-Blackmore, Milner, McEwen, & Bulloch, 2008; Tanaka et al., 1997). A partir de este
punto, los cuestionamientos que giraban en torno a las inconsistencias entre las actividades
antiinflamatorias de GC y la presencia de microglía M1 durante situaciones estresantes se pueden
responder si se tienen en cuenta los efectos de estos receptores en esta célula.
Así pues, se ha evidenciado que GR y MR se encuentran presentes en la microglía, más
no en el núcleo, sino en la membrana y el citosol (Sierra et al., 2008; Tanaka et al., 1997). Las
funciones específicas de GR en la microglía han sido estudiadas en ratones que carecen de GR en
los macrófagos. Con una dosis de LPS, se activa el receptor de tipo toll 4 (TLR4) que promueve
lesiones en las células y daños en los axones. La inhibición global de GR demuestra que,
específicamente en la microglía, se previene la degeneración neuronal por todo el cerebro y otras
células (Carrillo-de Sauvage et al., 2013). In vivo e in vitro GR tiene funciones en cuanto a la
diferenciación microglial, su proliferación y su motilidad. Se ha observado que después de
lesiones isquémicas, los MR y GR que se encuentran en la microglía inician su actividad
inmunoreactiva en el área afectada, aumentando la muerte neuronal incluso 7 días después de la
lesión. Precisamente en el experimento de (Carrillo-de Sauvage et al., 2013), los resultados
indicaron que la degeneración neuronal en las ratas después de una inyección de LPS es dañina
para las neuronas después de un estímulo estresante, pero GR disminuyó los efectos provocados
por la inducción de LPS y controló la proliferación de radicales de oxígeno, es decir que se
encargó de detener la acción de la microglía M1. Por el contrario, se ha sugerido que MR ejecuta
efectos proinflamatorios cuando la microglía se encuentra en estado de activación M1 (Tanaka et
al., 1997).
De manera más específica, MR se encarga de promover la acción inmune de la microglía
M1, en respuesta a daños en el cerebro. Por ejemplo, (Frieler et al., 2011) revisaron la activación
de la microglía, vía MR durante situaciones de accidente cerebrovascular y comprobaron que este
receptor permite el incremento del daño vascular y la isquemia. Partiendo de esta observación,
sugirieron que MR funciona como un regulador de la polarización de los macrófagos, pero en
algunas ocasiones también promueve la actividad de la microglía M2. Los autores, identificaron

53
que la eliminación de MR disminuía de manera significativa el volumen de daño del accidente
cerebrovascular. Partiendo de lo anterior, se puede apreciar que los GC están implicados en la
regulación de la función de MR y su liberación durante situaciones potencialmente peligrosas
incrementa el daño de la zona afectada (Frieler et al., 2011). En efecto la señalización de GC
representa un equilibrio entre la actividad inmune y la actividad neuroendocrina, en tanto que
posibilita el control de los efectos proinflamatorios y antiinflamatorios de la microglía (Sorrells,
Caso, Munhoz, & Sapolsky, 2009).
Como se ha observado a lo largo de este capítulo, la actividad de la microglía afecta de
manera significativa todas las funciones del hipocampo. En situaciones de estrés, tanto agudo
como crónico, el fenotipo proinflamatorio de la microglía puede afectar de manera significativa
la sobrevivencia de las células hipocampales. En el siguiente capítulo, se va a abordar el tema de
las futuras aproximaciones desde las cuales se puede llegar a inhibir la microglía M1 por medio
de la administración del fármaco Rosiglitazona.
Aportes a la inhibición de la microglía de fenotipo M1.
Una de las moléculas que se encuentran expresadas durante el estado activado de la
microglía proinflamatoria es CD40 (Sun et al., 2016; Sun et al., 2008; Fernandez et al., 2013). El
CD40 es un miembro de la superfamilia de los factores de necrosis tumoral, cuya expresión se
ubica en las células presentadoras de antígeno, las células dendríticas, los monocitos y otras
células no inmunes como las epiteliales, las endoteliales, las mesenquimales y en las plaquetas y
tumores (Kawabe, Matsushima, Hashimoto, Imaizumi, & Hasegawa, 2011). Estas moléculas
actúan como un transductor de las señales de la membrana celular, puesto que, una vez activadas
permiten la estimulación de quinasas y factores de transcripción encargados de iniciar las
actividades humorales e inmunes de las células (Kawabe et al., 2011). En el SNC, la interacción
entre este compuesto y su respectivo ligando (CD40-CD40L) ejerce un papel importante en las
respuestas de inflamación, pues dirige a la microglía hacia el aumento de la secreción de TNF-α,
IL-6, IL-1β. Posteriormente, esto provocaría un aumento de las heridas celulares (Sun et al.,
2008; Fernandez et al., 2013).
En contraposición, los efectos antiinflamatorios y neuroprotectores de la microglía son
mediados a través del aumento de la expresión de la enzima arginasa-1 (arg-1), considerada como

54
un sello de respuesta a la IL-4 (Pourcet & Pineda-Torra, 2013). Recientemente se ha definido que
la arg-1 es expresada mediante la actividad de otro compuesto: el receptor proliferador-activado
de peroxisoma gamma (PPAR-γ), que también se considera como un regulador de la polarización
de la microglía M2 (Song et al., 2016).
El PPAR-γ es un factor de transcripción nuclear activado por ligando, asociado con un
tipo de receptores nucleares en las células (Wang et al., 2014). Entre sus funciones principales, se
ha observado la regulación de la transcripción de genes proinflamatorios y lípidos microgliales
(GlaxoSmithKline, 2007), o en la utilización y transporte de glucosa por medio de la facilitación
de los genes que responden a la insulina y el metabolismo de los ácidos grasos
(GlaxoSmithKline, 2007). Últimamente se ha observado que su actividad inhibe la producción de
citosinas proinflamatorias en de la micoglía M1 (Wang et al., 2015). Es por eso, que se ha
afirmado que ejerce efectos neuroprotectores sobre el SNC (Luo, He, Kuang, Jiang, & Yang,
2014). No obstante también se encuentran presentes en los tejidos adiposos, los músculos
esqueléticos, el hígado y otros macrófagos.
Retomando a la arg-1, esta es una enzima producto del mismo substrato con que se
sintetizan los iNOS1, esta es la L-arginina. Las dos sustancias (arg-1 e iNOS), son los marcadores
más representativos de los fenotipos M2 y M1 respectivamente (Cherry et al., 2014). La
regulación de la liberación y transcripción de la arg-1 no se podría realizar si no fuese por la
activación del PPAR-γ (Gallardo-Soler et al., 2008) en el núcleo de la célula, o por la actividad
que ejerce la IL-4, puesto que activan el gen de unión (3kb) que le permite comenzar con la
transcripción y proliferación de la enzima (Pourcet & Pineda-Torra, 2013). Se ha observado que
la una (IL-4) activa a la otra (PPAR-γ) en el contexto de la inflamación. En estudios previos, se
ha reportado que la actividad de la IL-4 sobre la arg-1 potencia los efectos antiinflamatorios que
se observan durante el aumento de la expresión de arg-1 después de la administración de un
agonista de PPAR-γ (Gallardo-Soler et al., 2008).
Recientemente se ha observado que la administración de agonistas de PPAR-γ ejercen un
efecto antiinflamatoro y neuroprotector en modelos animales de Parkinson, daño cerebral
1 Es interesante destacar que arg-1 es un poderoso inhibidor de iNOs (Cherry et al., 2014).

55
isquémico e inflamación relacionada con la edad (Pisanu et al., 2014; Song et al., 2016), así como
efectos antiarerogénicos en modelos murinos de arteroesclerosis (Gallardo-Soler et al., 2008).
Uno de los fármacos que funciona como agonista selectivo de PPAR-γ es la Rosiglitazona
Maleate, un fármaco miembro de la familia de las tiazolidinelionas, que se caracterizan por ser
agentes antidiabéticos para el tratamiento de la diabetes tipo II (Park, Kim, Kang, Kim, & Shin,
2004; GlaxoSmithKline, 2007). En un estudio sobre los efectos neuroprotectores de la
Rosiglitazona en la enfermedad de epilepsia, se observó que el incremento de la exitotoxicidad
durante los episodios epilépticos intensificaba la muerte neuronal en el giro dentado y las
regiones CA1 y CA3 pero la Rosiglitazona atenuaba dicho efecto (Sun et al., 2008).
Al mismo tiempo, los efectos más importantes de la Rosiglitazona se observan en la
inhibición significativa de CD40 en el área CA1 del hipocampo y la inhibición de la expresión de
TNFα, 24h después de la inducción de epilepsia con inyecciones de hidrocloruro de policarpina
administradas intraperitonialmente (Sun et al., 2008). Lo anterior permite que la Rosiglitazona
sea considerada como un potente neuroprotector, pues demostró una disminución significativa de
la muerte neuronal después de los ataques epilépticos (Sun et al., 2008). Se considera a este
fármaco (al igual que otros agonistas de PPARγ) como un protector efectivo de las enfermedades
del SNC (Luo et al., 2014). No obstante, aún no se tienen bien definidos sus efectos después de la
neuroinflamación inducida por estrés agudo.
Ahora bien, se ha relacionado la activación de GR con la actividad de PPAR-γ, en tanto
que este receptor ejecuta una activación parcial sobre el primero, pero no al contrario
(Yamamoto, Kakuta, & Sugimoto, 2014). Tanto la Pioglitazona como la Rosiglitazona efectúan
una acción antiinflamatoria al inducir una traslocación nuclear de GR hacia el núcleo de otras
células, es más se han observado actividades antiinflamatorias desde los agonistas de PPAR-γ al
reversar las acciones de los antagonistas de GR (RU486) (Ialenti et al., 2005). Partiendo de esta
observación, se puede evidenciar sus posibles efectos sobre la regulación del eje HPA, que
también depende de la correcta actividad de GR.
Siguiendo esta misma línea, otros efectos de la Rosiglitazona han sido revisados para
tartar la diabetes y se cree que sus mecanismos de acción también pueden afectar la actividad del
eje HPA. Torres, Magalhaes, Martinsy Carvalho (2016) observaron que en ratas diabéticas el
tratamiento con este fármaco inhibe la hipertrofia adrenal debido a la enfermedad. También

56
definieron que la Rosiglitazona bloquea los efectos de la diabetes al aumentar la presencia de los
receptores de ACTH, los receptores de melanocortina-2 (MC2R) en la corteza adrenal (Roy et al.,
2012), disminuir la expresión de los péptidos derivados de la pro-opiomelanocortina (POMC) en
la pituitaria, que se relacionan con la ingesta de comida y la utilización de la energía por todo el
sistema y homeostasis de la energía (Mountjoy, 2010; Mountjoy, 2015) y todo esto asociado con
el aumento de la concentración de PI3K, un regulador del crecimiento y supervivencia celular
(Pinzón, Serrano, & Sanabria, 2017). Pero el tratamiento no tuvo efectos significativos sobre la
disminución de GR y MR. Sus resultados indican que la activación de PPAR-γ con Rosiglitazona
normaliza el eje HPA de las ratas diabéticas, al regular la producción de ACTH y GC en la
glándula pituitaria y las adrenales, no obstante, no se observó que las ratas pudieran realizar una
realimentación negativa correctamente.
Las anteriores observaciones fueron evaluadas de manera más específica por Luo, He,
Quang, Jiang y Yang de la universidad de Chongquing, en China (Luo et al., 2014). Estos
investigadores observaron que la Rosiglitazona juega un papel protector en el daño hipocampal
después de un daño de isquemia cerebral, puesto que inhiben la respuesta de estrés oxidativo y la
inflamación. En su experimento, administraron diferentes volúmenes de Rosiglitazona y
observaron que 7.2mg/kg de Rosiglitazona significaban una dosis efectiva para la inhibición de la
microglía proinflamatoria después de la inducción de isquemia.
Con la información procedente de los estudios anteriores, el observar los efectos de este
fármaco como un antiinflamatorio que actúa de manera específica sobre la actividad de la
microglía activada M1, sería interesante para proceder a futuros estudios que caractericen sus
acciones neuroprotectoras durante o después de las respuestas de estrés. Se creería que por medio
de su efecto agonista sobre los receptores PPAR-γ, podría cambiar la polarización de la microglía
desde el fenotipo citotóxico y proinflamatorio M1, hacia el fenotipo antiinflamatorio y
neuroprotector M2 (Pisanu et al., 2014; Song et al., 2016). Por el lado de la regulación del eje
HPA, se ha observado que los agonistas de PPAR-γ ejercen una activación parcial de los
receptores GR y se encargan de aumentar la presencia de ACTH y la actividad de MC2R (Roy et
al., 2012; Yamamoto et al., 2014). Lo anterior permite sugerir que la administración sistémica de
Rosiglitazona ejecutará una respuesta moduladora de la realimentación negativa del eje HPA y

57
durante la inducción robusta del estrés, todos estos mecanismos que se ven desregulados van a
ser atenuados.
Se sabe que esta estructura es fuertemente afectada por los efectos proinflamatorios de la
microglía M1 (Lana et al., 2016) durante el estrés agudo y puede afectar la funciones cognitivas e
impedir la neurogénesis (Ekdahl et al., 2003; Monje et al., 2003). Es por eso que para identificar
los efectos neuroprotectores de esta sustancia, va a ser importante observar su acción en durante
la consolidación de memorias afectivas y espaciales. En estudios previos se ha evidenciado que la
consolidación de las memorias espaciales y de objetos novedosos se ve impedida después de
lesiones en el GD y en CA3, mas no en CA1, la cual se ve más involucrada en la retención de las
memorias de tipo temporal (Kesner et al., 2004; Hasselmo, 2005b). La evaluación de estas áreas
en el laberinto acuático de Morris puede facilitar la evaluación de los efectos de la Rosiglitazona,
después del protocolo de estrés agudo. En cuanto a las memorias episódicas de corte emocional,
se esperaría que después de la administración intraperitonial de Rosiglitazona no afecte
significativamente la consolidación de memorias emocionales.
Se ha reportado que una sola exposición a estímulos de estrés agudo puede provocar
cambios a nivel cognitivo, comportamental, además de alteraciones en la respuesta del eje HPA
que duran por un plazo de tiempo más largo, por lo menos 28 días (Sutanto & de Kloet, 1994), no
obstante otros estudios consideran que después de la inducción a estrés agudo, una recuperación
de 7 días puede ser suficiente para que el animal vuelva a su estado basal (Yohe, Suzuki, &
Lucas, 2012). Se esperaría que, en experimentos futuros, se revisen los efectos que podría generar
el fármaco en animales experimentales que han recibido estrés, con el fin de observar si es
posible proteger los patrones de consolidación de la memoria espacial.
Introducción a las cepas Carioca High Freezing y Carioca Low Freezing
El estudio de la bioquímica de las reacciones comportamentales hace necesario que se
exploren y evalúen las diferencias entre las condiciones que definen las características innatas y
adquiridas de los sistemas vivientes (Lerner, 1978). Se ha observado que las discrepancias adaptativas de
los seres vivos ante condiciones estresantes pueden dirigir al sistema hacia arreglos funcionales que lo
protegen del estrés, o por el contrario, que facilitan el desarrollo de patologías o comportamientos
derivados del estímulo (Macri, Zoratto, & Laviola, 2011; McEwen, 2000; Lucassen et al., 2014). Lo

58
anterior es posible gracias un dialogo constante entre la intensidad del estímulo y su grado de
controlabilidad (Lucassen et al., 2014), los cuales vía elementos epigenéticos (Ebner & Singewald, 2017)
y gracias a diferencias innatas en la expresión de las moléculas relacionadas con el funcionamiento del
eje HPA, desencadenan procesos adaptativos o no adaptativos después de la percepción del cambio
medioambiental (Scharf & Schmidt, 2012; Ebner & Singewald, 2017; Kabbaj, Devine, Savage, & Akil,
2000).
Algunas aproximaciones que evalúan el desarrollo del comportamiento de las especies
desde perspectivas evolucionistas, han demostrado que la activación neuronal en diferentes
contextos es similar en humanos y otros mamíferos (de Castro Gómes et al., 2011). Entre los
comportamientos, está el estrés (ver capítulo de introducción al estrés), una característica
compartida a través de las especies que se destaca por ser una reacción de supervivencia que
genera conductas potencialmente adaptativas ante situaciones peligrosas (McEwen, 2007). La
expresión de los genes reactivos al estrés dependen, por un lado de los niveles de plasticidad neuronaly
por el otro, de los tipos de estímulos y de la favorabilidad del contexto (Belsky & Pluess, 2009).
Adicionalmente, los factores genéticos dependientes de los cambios producidos por esta condición
pueden ser heredados (Sullivan, Neale, & Kendler, 2000), aumentando la probabilidad de que se
produzca una predisposición a cambios en el sistema en generaciones futuras (Sullivan et al., 2000).
Aproximaciones acerca del estudio de las conductas ansiosas y otras patologías del
comportamiento, suelen usar como base el modelo de diátesis estrés (Belsky & Pluess, 2009),
conocido como la vulnerabilidad genética de algunos individuos a verse mayormente afectados por un
estresor ambiental (Belsky & Pluess, 2009). Por medio de esta teoría y otros conocimientos sobre la
bioquímica vinculada con las diferencias individuales en las reacciones comportamentales del estrés y la
ansiedad (Sweis, Veverka, Dhillon, Urban, & Lucas, 2013), se han elaborado diseños experimentales,
desde los cuales se ha podido responder a la necesidad de entender ciertas condiciones vividas en las
poblaciones humanas (Bovenkerk & Kaldewaij, 2015; Ramos, 2008).
Entre las herramientas de análisis para estos propósitos, los modelos animales con roedores han
realizado aproximaciones a la explicación y el entendimiento del comportamiento normal y
patológico desde una perspectiva que antes no se había podido estudiar con detalle en humanos
(Bovenkerk & Kaldewaij, 2015). La rata, como modelo animal, ha resultado ser efectivo para el
estudio del estrés (Ramos, 2008), pues ha aportado información valiosa sobre sus características

59
comportamentales y fisiológicas en diferentes contextos experimentales (ver Introducción al
estrés). Así mismo, sirven como punto de partida para explorar otros mecanismos del estrés, los
cuales aún no han sido suficientemente estudiados, podrían ser gatillos para conducir a los
sistemas a una mala regulación y posteriormente al desarrollo de enfermedades (Rosch, 1979).
Un modelo animal que ha pretendido realizar un acercamiento a estos temas, se ha desarrollado
en el departamento de psicología la Pontificia Universidad Católica de Rio de Janeiro (PUC), en
Brasil. Dos linajes de ratas que difieren en características asociadas a los comportamientos de
miedo y de ansiedad, Carioca Hight Freezing (CHF) y Carioca Low Freezing (CLF), fueron
desarrolladas desde el 2006 a través de la separación de ciertos rasgos fenotípicos en las ratas
Wistar, hasta tener comportamientos de miedo definidos. Las cepas, fueron desarrolladas con
base a la selección de distintivos emocionales descritos en la literatura y la evaluación del
congelamiento condicionado. El congelamiento2 determinó la separación de los individuos
durante el proceso de fenotipaje. Las ratas que contenían mayor cantidad y menor cantidad de
congelamiento fueron cruzadas por varias generaciones hasta desarrollar los fenotipos de miedo y
de ansiedad (de Castro Gómes et al., 2011).
Se ha afirmado que las cepas Carioca High Freezing y Carioca Low Freezing representan
un modelo novedoso para el estudio de ciertas afecciones vinculadas con la ansiedad, el pánico,
el miedo, etc. Debido a que sus características comportamentales, en estado natural, se mantienen
con comportamientos relacionados con la presencia o ausencia de congelamiento, un predictor
de ansiedad (de Castro Gómes et al., 2011; de Oliveira Galvão, de Castro Gómes, Masionette, &
Landeira-Fernandez, 2011). En estudios que se han realizado a través de sus generaciones, se
observa de manera consistente que el tiempo de congelamiento en las pruebas de miedo
condicionado es más prolongado para CHF que para CLF, cuya actividad locomotora se
encuentra presente con mayor intensidad que CHF antes y después de los protocolos
experimentales (de Castro Gómes et al., 2011). Estos comportamientos se han relacionado con la
capacidad de desarrollar, o bien conductas de pánico y respuestas de miedo exageradas y
2 Definido como un comportamiento defensivo en respuesta a un peligro caracterizado por la ausencia de
movimiento del animal con excepción de la respiración (Fanselow, 1984).

60
espontaneas, o bien ningún tipo de respuesta después de la inducción de los estímulos estresantes
(de Oliveira Galvão et al., 2011).
Se considera que a través del estudio de las cepas, se podrían entender las razones por las cuales
los sistemas biológicos reaccionan de maneras diferentes ante estímulos aversivos. De la literatura, se ha
destacado la necesidad por explorar su etología a través de pruebas comportamentales (Dias,
Bevilaqua, Silveira, Landeira-Fernandez, & Gardino, 2009). No obstante, los resultados aún
dejan cuestiones abiertas sobre su desarrollo, incluso en estado natural.
Así pues, se resaltan algunos detalles en que las diferencian las cepas de las ratas Wistar
comunes. Por ejemplo, se ha observado que la interacción social con animales desconocidos es
significativamente menor en CHF que en un grupo control. O también que los comportamientos
relacionados con la depresión no difieren en las características de la patología entre CHF y el
grupo control. En cuanto a la cognición espacial durante el reconocimiento de objetos, tampoco
han evidenciado diferencias significativas durante la exploración, a pesar de que CHF si mantiene
una menor exploración que la contraparte de control durante la fase de familiarización (Dias et
al., 2009). De ahí, se concluiría que la etología del modelo animal sólo cuenta con diferencias
significativas en variables emocionales, más no cognitivas. Sin embargo, la información
recolectada no se ha adentrado en el estudio de CLF o de todas las cepas después de protocolos
relacionados al estrés.
Es interesante resaltar que en otros estudios se han aclarado algunas diferencias
funcionales en el cerebro de estas ratas que refuerzan las observaciones preliminares del
comportamiento relacionado con el miedo y la defensa. Se destaca una diferencia de función y
conectividad en SGP (de Oliveira Galvão et al., 2011), los núcleos de la amígdala como Cea y BLA
(de, V & Landeira-Fernandez, 2008b) y los niveles de algunas especies reactivas a oxígeno y la
corticosteriona plasmática (Hassan, Gomes, Pinton, Batista Teixeira da, & Landeira-Fernandez, 2013).
Nuevas aproximaciones hacia las raíces del comportamiento de las cepas han resultado
interesantes para la presente investigación, puesto que, al aprovechar las ventajas
comportamentales, se podrían evaluar nuevos efectos sobre las interacciones entre la genética y el
ambiente cuando hay un estímulo de estrés de por medio. Teniendo en cuenta que la inflamación
juega un papel importante en estos procesos, se pretende evaluar la relación del estrés y la

61
expresión de la microglía activada, dado el estado de vulnerabilidad de los animales
experimentales, sobre la consolidación de la memoria espacial y emocional en las ratas CHF y
CLF.
Ahora bien, en el presente experimento se espera ver diferencias en cuanto a las
características comportamentales y neuroquímicas relacionadas con el estrés y la consolidación
de las memorias emocionales y espaciales, pues se conoce que este representa un impedimento para
la consolidación dichas memorias, en comparación con aquellas que si son emocionales (Bodnoff et al.,
1995). Además de esto, también se espera realizar acercamientos a las características
neurofisiológicas referentes a la expresión de microglía activada, pues en el hipocampo estresado
provoca efectos en detrimento de la consolidación de las memorias neutras y el LTP (ver capítulo
de microglía y estrés). Aunque en las cepas no ha sido suficientemente estudiado, el presente
estudio pretende agregar más información a su caracterización, explorando las respuestas
comportamentales y su relación con los niveles de inflamación en el hipocampo de los animales
estresados y no estresados.
Del presente estudio, se tiene presente que la información es limitada en cuanto a la
alteración o deterioro del aprendizaje después de la inducción de estrés tanto en CHF como en
CLF. También se tiene en cuenta que no se han establecido relaciones específicas de acuerdo con
los niveles de inflamación observados en cada cepa. Lo anterior, podría ser una limitación para la
explicación de su biología y sus comportamientos, hasta que no se hagan más estudios
relacionados al tema en cuestión. Es por eso que se tiene la intención de aclarar algunos vacíos de
conocimiento relacionados con las cepas por medio de la realización de un estudio con propósitos
exploratorios.
Se postula una hipótesis que va a guiar los objetivos del presente estudio. Esta es, la
inducción de estrés agudo estimula los efectos proinflamatorios y citotóxicos de la microglía M1
en el hipocampo de la cepa vulnerable (CHF) y actúa en detrimento de la consolidación de la
memoria espacial, en contraposición con el robustecimiento de la emocional, gracias a la muerte
neuronal por excitotoxicidad en las regiones GD, CA3 y CA1. Lo anterior también se va a ver
expresado en el aumento de los comportamientos ansiosos y la disminución de la locomoción.

62
Para evaluar estas condiciones se realizará un protocolo de estrés agudo en la mitad de los
sujetos experimentales de cada grupo CHF, CLF y control (CON) en una caja de
condicionamiento aversivo. Posteriormente, todos los animales serán pasados por diferentes
protocolos de aprendizaje para medir la consolidación de la memoria emocional. La memoria
espacial, va a ser evaluada en un protocolo de reconocimiento de objetos. Y los niveles de
ansiedad y de locomoción en un test de campo abierto. Finalmente, los animales serán
perfundidos y su tejido cerebral se va a analizar para contabilizar el número de microglía activada
por cada 0.1mm en varias regiones del hipocampo (y el cuerpo estriado). Todos los estudios
fueron avalados por el organismo ético de la universidad Pontifica Universidad Católica de Rio
de Janeiro (PUC) en Brasil.
De los resultados, se esperan observar diferencias entre todos los grupos en los
protocolos. En primer lugar, de manera consistente a la literatura (de Castro Gómes et al., 2011),
se espera un aumento significativo en el congelamiento de las ratas CHF, pero no en las CLF
después del condicionamiento aversivo. Luego, durante la evaluación de la consolidación la
memoria a largo plazo, se espera que CHF y el grupo control (CON) evoquen el recuerdo, pero
CLF no.
En el campo abierto, se espera observar que el aumento del congelamiento después del
condicionamiento aversivo disminuirá tanto los niveles de exploración como de locomoción a
través de los cuadrantes en CHF en comparación con los grupos CLF y CON estresados. Esto
mismo podría compararse con los resultados del congelamiento y la exploración en las CHF no
estresadas las cuales, al no haber sido expuestas a una situación de estrés significativo, no van a
exponer el miedo condicionado.
En comparación a CHF, se cree que CLF va a contar con un nivel de exploración intenso
en el campo abiertoy que este resultado no se va a diferenciar de manera significativa en
comparación con su contraparte no estresada. De CLF se puede destacar que no responde de
manera consistente al miedo condicionado como los animales de los otros grupos, pues
disminuye su tiempo de congelamiento cuando es reexpuesto al contexto después del
condicionamiento (de Castro Gómes et al., 2011; de Oliveira Galvão et al., 2011). Es por esa
razón que se sugieren dos posibles explicaciones que se van a tener en cuenta durante la

63
realización del experimento, la primera, sugiere que CLF puede tener problemas para consolidar
las memorias aversivas, por lo que se espera observar inflamación en estructuras hipocampales
ventrales, sin embargo, debido al comportamiento de locomoción incrementado que se observa
durante el condicionamiento aversivo y otras pruebas, cuyos resultados también afirman el
aumento de la locomoción en protocolos de catatonía en modelos de Parkinson (estudio en
proceso), también se podría considerar que el comportamiento de la cepa se relacione con
problemas alteraciones en los circuitos relacionados con la automatización de los movimientos
(Cromwell & Berridge, 1996).
Posteriormente, en la prueba de memoria declarativa, no se espera que los grupos
estresados generen una buena evocación del recuerdo de los objetos, aun habiéndolos explorado
durante la fase de familiarización. Este detalle se debería a los efectos que tiene el estrés en
detrimento de la consolidación de las memorias neutras (diríjase al capítulo de efectos del estrés
sobre el hipocampo). Sin embargo, no se considera que el déficit de aprendizaje en CLF se derivé
directamente del estrés, sino más bien, de otros elementos cognitivos que pueden estar jugando
un papel importante en sus procesos de consolidación. Esto se podría concluir debido a que el
déficit de aprendizaje que tiene CLF, por ejemplo, a la hora de evocar recuerdo aversivo durante
el condicionamiento podría extrapolarse a otros contextos de diferentes naturalezas. Es por eso
que tampoco se espera identificar diferencias con las ratas de su mismo grupo que no han sido
estresadas, cuyo déficit de aprendizaje, puede ser el mismo.
Es también de interés observar los efectos que puede ejercer la microglía, principal célula
nerviosa encargada de las respuestas inmunes y el establecimiento de la homeostasis durante
cambios en el microambiente (Hanisch & Kettenmann, 2007) después del estrés en la región
hipocampal. Finalmente, se esperan observar cambios en la concentración de la microglía
activada por región, dependiendo de la naturaleza comportamental de cada cepa.
En este caso, se espera identificar distintas concentraciones de microglía dependiendo, por
un lado de la cepa (CHF, CLF y CON) y por el otro, del tratamiento (Estrés vs. No estrés). Se
aclara que la principal estructura para analizar en el experimento es el hipocampo, por lo que se
esperó hacer el conteo de microglía activada en porciones hipocampales, anteriores, mediales en
los animales de los grupos experimentales.

64
Se esperan observar las siguientes características: primero, que los hipocampos de los
animales estresados tengan una mayor cantidad de células activadas por área que en los animales
no estresados. En los estresados, se cree que las porciones dorsales del hipocampo posterior se
van a ver más inflamadas que las ventrales. Todo esto se deduciría, debido a que el aumento de la
exitotoxicidad sobre el hipocampo, el aumento de la excitabilidad de la amígdala basolateral
sobre consolidación de memorias de valencia neutra y la activación de la respuesta inmune en
CA3, CA1 y GD impedirían directamente la consolidación de las memorias de valencia neutra
correctamente.
en segundo lugar, en los animales estresados, se cree que en CHF se va a observar una
mayor cantidad de microglía activada en las estructuras dependientes de la consolidación de los
recuerdos espaciales, en comparación con CLF. En este caso, se espera observar microglía
activada en el giro dentado, CA3y en el hipocampo dorsal posterior. En cuanto a CLF, aunque se
cree que sus niveles de inflamación son menores que CHF, se considera que dados los problemas
de aprendizaje demostrados en las pruebas emocionales y dependiendo del aprendizaje en la
prueba de reconocimiento de objetos, tenga microglía activada por todo el hipocampo, pero en
menor concentración. Es interesante explorar sus niveles de inflamación, puesto que la cepa, aún
no ha sido completamente caracterizada como para llegar a especulaciones sobre su cognición y
aprendizaje emocional y espacial. Por último, no se esperan diferencias en cuanto a los niveles de
inflamación de los hipocampos de los grupos no estresados.
A continuación, se va a exponer el método y los resultados obtenidos la presente
investigación.
Método
Animales
Se utilizaron 60 ratas macho adultas, de edades entre 2 y 3 meses y pesos oscilando entre
250 y 350 gramos, del bioterio del laboratorio de Neurociencia de la Pontificia Universidad
Católica de Rio de Janeiro, Brasil. Los animales experimentales fueron divididos en 3 grupos,
que a su vez fueron subdivididos en otros dos (ver figura 1). Cada grupo fue sometido a los
mismos procedimientos experimentales para evaluar la consolidación de la memoria emocional y

65
espacial, sin embargo de cada grupo, una mitad fue sometida a un protocolo de estrés en una caja
de condicionamiento aversivo (de Oliveira Galvão et al., 2011). El número de animales
seleccionado por grupo corresponde al n óptimo requerido para el cumplimiento de los objetivos
(Leon et al., 2013; Hassan et al., 2013). Los sujetos serán alojados bajo condiciones controladas
de temperatura (22±2°C) y humedad (57%), mantenidas en un ciclo de luz de 12:12 h, con inicio
de luz a las 5:00am.
Figura 1. División de grupos experimentales
Procedimiento
El trabajo constó de varias fases experimentales que se enfocaron en evaluaciones
comportamentales y procedimientos de análisis histológicos en el hipocampo. Las fases
exploraron el efecto del estrés agudo en una tarea de consolidación de la memoria emocional, la
memoria espacial y finalmente se realizaron análisis en el tejido que permitieron explorar sus
efectos sobre las células hipocampales (explicados más abajo). En cada uno de los experimentos,
los animales fueron divididos en dos grupos de los cuales uno fue expuesto a un protocolo de
estrés en una caja de condicionamiento aversivo (de Oliveira Galvão et al., 2011). 24 horas
después, fueron puestos en las tareas de memoria. Finalizado el último día de tarea, se revisó de
nuevo el tiempo de congelamiento en el contexto aversivo, para luego ser perfundidos. Las
perfusiones fueron realizadas o inmediatamente después de la exposición al contexto aversivo o
24 horas después de este mismo, pues se pretendía hacer comparaciones que tuviesen.

66
Antes de los registros, las ratas fueron manipuladas por 5 minutos después de realizar la
toma de peso todos los días para que se acostumbraran a las manos del experimentador. Al
mismo tiempo, se procuró que los animales se aclimataran a la sala experimental, por lo que
fueron sacados del bioterio y colocados en las salas durante 1 hora antes de los registros. Todos
los procedimientos fueron avalados por el comité de ética de la universidad.
Inducción del estrés y evaluación de la memoria emocional
Ambos, la inducción del estrés y el condicionamiento aversivo fueron evaluados por
medio de un protocolo de miedo condicionado. Este procedimiento es generalmente utilizado
para medir los recuerdos asociados al miedo a un contexto. Dihco miedo fue medido a través de
la cantidad de tiempo de congelamiento de la rata después de la inducción de un choque eléctrico
(0.7-0.8mA) que causó estrés.
Ahora bien, el procedimiento consistió en ubicar a los animales experimentales en una
caja de 30x 25x 30 con un piso electrificable de 15 tubos de metal de 4mm de diámetro,
separados a 1,5 el uno del otro (ver figura 2). Los tubos estaban conectados a un generador de
choques eléctricos y a un computador. La caja se encontraba ubicada entre otras cajas
atenuadoras de ruido para eliminar estímulos ambientales no relacionados al contexto. Cada
animal fue colocado en las cajas, uno por uno por un tiempo de 8 minutos. Después del quinto
minuto, se indujeron 3 choques eléctricos de 1 segundo de duración, con intervalos de 20
segundos por un minuto y se midió el tiempo de congelamiento después del último choque
durante los siguientes tres minutos. Posteriormente, cada rata fue colocada en su caja de
alojamiento permanente y se limpió el espacio.
Figura 2. Ilustración de una caja de condicionamiento aversivo.

67
24 horas después, se colocó a los animales en el contexto por los 8 minutos y se midió el
tiempo de congelamiento, esta vez sin inducción de choque. Tres días después, las ratas fueron
puestas en el contexto para evaluar el reconocimiento al contexto a largo plazo.
Es importante agregar que todos los procesos fueron realizados a la misma hora del día
durante la mañana.
Campo abierto
Los resultados comportamentales fueron medidos a través de distintos métodos ajustados
para cada prueba. Para la prueba de reconocimiento de objetos, se consideró necesario realizar
una medición tanto de la ansiedad como de la motricidad durante la fase de habituación al
contexto, por lo que se ejecutó un protocolo de campo abierto. Específicamente, se evaluó la
distancia recorrida en el campo y los comportamientos de congelamiento, acicalamiento y estar
parado en las patas traseras, pues brindaron información adicional sobre los niveles de ansiedad
de cada animal durante el experimento (Seibenhener & Wooten, 2015). Para esta, se hizo uso del
programa XploRat 3.3.0, creado por el profesor Fernando Cárdenas de la Universidad de los
Andes. Este consistió en dividir el campo en 9 cuadrantes que me permitieron medir el tiempo de
permanencia total en cada cuadrante, en el centro, la periferia y contar los cruces de la rata a
través de estos (ver figura 3).
Figura 3. Esquema general de X-plo-rat (versión 2.2).

68
Test de reconocimiento de objetos
Para evaluar la memoria espacial se van a someter a los animales experimentales a un
protocolo modificado de reconocimiento de objetos. Las modificaciones consistieron en utilizar
un test de campo abierto para evaluar el reconocimiento dependiente de la porción dorsal
hipocampo (Moser et al., 1995), cuya activación está relacionada a factores de reconocimiento
espacial (Broadbent, Gaskin, Squire, & Clark, 2010; Murray & Mishkin, 1986). Este proceso se
realizó para evitar que la consolidación dependiera de las regiones parahipocampales, las cuales
son usualmente activadas durante el reconocimiento de los detalles de los objetos dentro del
contexto (Winters, Saksida, & Bussey, 2008; Meunier, Hadfield, Bachevalier, & Murray, 1996).
Por esa razón, se utilizó una caja de campo abierto, cuyas dimensiones son suficientes para
reconocer el desplazamiento de los objetos en el espacio (Winters et al., 2008) y uno de los
objetos, además de ser cambiado por otro novedoso, también fue desplazado a 90° de la posición
inicial.
Para evaluar la efectividad de la prueba, se realizó una serie de estudios piloto que
consistieron en cambiar la posición de los objetos, moverlos por diferentes cuadrantes y aumentar
el tiempo de habituación al campo, con el propósito de asegurarse de que el aprendizaje, o su
ausencia, se debiera a la inducción de estrés y no a la falta de reconocimiento del contexto.
Al finalizar la prueba piloto, el experimento consistió en evaluar la memoria espacial en
tres fases. En primera, o fase de habituación, se colocó a las ratas, una por una, en el campo
abierto para que exploraran el espacio por cinco minutos. Durante el procedimiento, se evaluaron
los comportamientos relacionados con la ansiedad y la motricidad. Inmediatamente después, se
colocó a las ratas en sus cajas de alojamiento permanente. La efectividad del procedimiento
estuvo sujeta a la variable de habituación, por lo que se repitió el procedimiento veces antes
comenzar con la familiarización a los objetos- una en el primer día de condicionamiento aversivo,
la otra en el segundo día y la tercera y la cuarta 12 y 24 horas después respectivamente-. 24 horas
después se realizó la fase de familiarización. En esta, se volvió a ubicar a cada rata en el campo
abierto, solo que esta vez en la caja se situaron dos objetos de manera paralela en dos cuadrantes
exteriores. Se dejó a las ratas explorar los objetos durante 5 minutos. Una vez terminado el
tiempo, se limpió el contexto y fueron de nuevo colocadas en las cajas de alojamiento
permanente. 24 horas después, se realizó la fase de prueba, que consistió en cambiar uno de los

69
objetos y moverlo a 90° de la posición original (ver figura 4). Durante los 5 minutos se observó el
tiempo de exploración en el objeto familiar y el objeto novedoso. Finalmente se acomodó cada
rata en las cajas de alojamiento permanente.
Figura 4. Test de reconocimiento de objetos.
Histología
Después de finalizados los procedimientos experimentales, los animales fueron anestesiados con
una dosis letal de ketamina y Xilazine (5mg/kg y 20mg/kg respectivamente) para luego ser
perfundidos transcardialmente con 250ml de solución PBS (0,1M) seguida de 250ml de
paraformaldehido (4%), de acuerdo con protocolo operativo utilizado en el Laboratorio de
Neurociencia y Aprendizaje de la Universidad de los Andes. Los cerebros fueron entonces
extraídos y almacenados en una solución de sacarosa (30%) de una a dos noches para asegurar la
crioprotección y la consistencia de los tejidos durante el proceso de congelamiento y la
realización de los cortes en el criostato (Bondonna, Jacquet, & Wolf, 1977). Posteriormente
fueron ubicados en paraformaldehído (4%) cuatro días.
Para el congelamiento, unos por uno fueron los cerebros colocados en isopentano con hielo
seco y fueron ubicados en una nevera a -80°C por una semana. Luego, fueron cortados con un
criostato a -25°C en tajadas de 16µm. Se buscaron cortes que incluyeran CA1, CA3 y giro
dentado. A cada treinta cortes se tomó uno para realizar una coloración de Nissl, necesaria para la
localización estructural. Los demás cortes fueron almacenados en tubos eppendorf en solución
anticongelante, para su almacenamiento a -20°C.

70
Para la inmunohistoquímica, los cortes fueron procesados en baños de diferentes reactivos,
seguidos de PBS 0,1M por 5 minutos para limpiar residuos. Se realizó un pretratamiento del
tejido por medio de la incubación los cortes en una solución de H2O2 al 0,01% diluido al 30% en
PBS 0,1M de 5 a 7 minutos, seguida de 3 baños de PBS 0,1M. Posteriormente, los cortes fueron
colocados en suero de bloqueo UltraCruz® Blocking Reagent (sc-516214) por una hora e
inmediatamente después, se incubaron en el anticuerpo Iba1 (sc-32725; 200µg/ml; 1:100 en
UltraCruz® Blocking Reagent (sc-516214)) por una noche a temperatura ambiente. Al día
siguiente se bañaron de nuevo en PBS 0.1M y se incubaron en el anticuerpo secundario, la
proteína de unión a IgG kappa de ratón (m-IgGκ BP) conjugada a Horseradish Peroxidasa (HRP)
(sc-516102); 200µg/0,5ml; 1:36 en UltraCruz® Blocking Reagent (sc-516214)) por 120 minutos.
Al finalizar el tiempo fueron nuevamente bañados en PBS 0,1M. 15 minutos antes de acabar la
incubación en el anticuerpo secundario, se preparó la solución ABC, avidina y horseradish
peroxidasa biotinilado (ImmunoCruz® ABC Kit (sc-516216)) en PBS, por media hora a
temperatura ambiente. Se incubaron los cortes en la solución por media hora. Luego del
procedimiento y repetitivos lavados en PBS 0,1M se inició el proceso de visualización, en el que
se incubaron los cortes en solución de X1 de DAB (DAB, 50X (CAS 7411-49-6)) y peróxido de
hidrógeno al 0,3% en 100mM de Tris-HCl (pH 7.6) por 15 minutos. Finalmente, se hicieron
nuevos lavados con PBS 0,1M, se montaron los cortes en láminas y cuando estuvieron secas
fueron deshidratadas en baños de alcoholes a diferentes concentraciones (50%, 70% y 90%) por 1
minunto. Finalmente, los cortes se bañaron dos veces por 10 segundos en xilol.
El análisis morfológico y la cuantificación de la microglía se realizaron por medio de la
utilización de microscopía óptica con objetivo de 40X. Las imágenes se digitalizaron en formato
.tif. para observaciones posteriores. Para el conteo de las células inmunopositivas con Iba1, se
utilizó el software Image J.
Para el conteo de la microglía, se usó un marco de conteo de 220 x 220 µm (Diz-Chaves,
Astiz, Bellini, Garcia-Segura, 2013) (ilustración de la morfología de la microglía se encuentra en
la figura 5).

71
Figura 5. Morfología de la microglía.
Análisis estadístico
Los datos de comportamiento, fueron analizados por medio del uso de ANOVAs de uno y
dos factores que evaluaron los resultados (condicionamiento aversivo, campo abierto y
reconocimiento de objetos) dependiendo del grupo experimental (CHF, CLF y CON) y la
condición (estrés vs no estrés). En caso de ser necesario, usando la prueba t de Student Newman-
Keuls. En todos los casos, el alfa establecido fue de 0.05. Se destaca que algunas pruebas,
específicamente aquellas relacionadas con el condicionamiento aversivo, no presentaron
normalidad (Shapiro-Wilk <0,05), por lo que se procedió a realizar análisis estadísticos no
paramétricos, pruebas de Kruskal-Wallis y de Mann-Whitney.
Para el análisis de la cantidad de células inmunopositivas para Iba-1, se realizaron
ANOVAS con medidas repetidas comparando el número de células activadas de acuerdo con la
condición de estrés (estrés vs no estrés) en los respectivos grupos en distintas áreas
hipocampales. Finalmente, se hicieron regresiones lineales simples para observar el efecto del
estrés sobre la activación de la microglía y la consolidación de las memorias espaciales y
emocionales.
Resultados
Miedo condicionado
En la figura 6 se encuentran los promedios y las desviaciones estándar del tiempo de
congelamiento en cada uno de los grupos experimentales inmediatamente después de la
inducción del choque, 24 horas y 6 días después. Esta sección del diseño experimental posee más

72
de una variable independiente a tener en cuenta para los resultados sobre el tiempo de
congelamiento en los grupos experimentales.
Se observó que las distribuciones de los datos correspondientes a las mediciones del
congelamiento tanto en los momentos post, como 24 horas y a 6 días y tanto para los grupos
naïve como para los estresados, no presentaron normalidad (Shapiro-Wilk <0,05), razón por la
cual se optó por realizar análisis estadísticos no paramétricos. Para el caso de los animales naïve,
sólo fue posible realizar la comparación de los animales control con los animales CLF, pues no se
tuvieron datos de los animales CHF.
Sujetos estresados
El análisis Kruskal-Wallis para el congelamiento en la condición pre mostró que hubo
diferencias significativas entre los grupos (H[2]=26,40; p<0,001). La comparación post-hoc (test
de Dunn) mostró que los animales del grupo CLF presentaron un tiempo de congelamiento
significativamente inferior a los animales del grupo control (P<0,05).
En la condición post se hallaron diferencias significativas entre los grupos (H[2]=19,43;
p<0,001). La comparación post-hoc mostró que los animales del grupo CLF presentaron un
tiempo de congelamiento significativamente inferior a los animales del grupo control (P<0,05).
Para la condición 24 horas, fueron halladas diferencias significativas entre los grupos
(H[2]=23,38; p<0,001). La comparación post-hoc mostró que los animales del grupo CLF
presentaron un tiempo de congelamiento significativamente inferior a los animales del grupo
control (P<0,05).
En la condición 6 días, se encontraron diferencias significativas entre los grupos
(H[2]=22,24; p<0,001). La comparación post-hoc mostró que los animales del grupo CLF
presentaron un tiempo de congelamiento significativamente inferior a los animales del grupo
control (P<0,05).
Finalmente se compararon las condiciones 24 horas y 6 días. ANOVAs con medidas
repetidas determinaron diferencias significativas en el tiempo de congelamiento a lo largo del
tiempo (F[4,25]=40.313, p<0.001). Posteriores análisis de post-hoc revelaron que solamente los
animales del grupo CLF tuvieron la tendencia de disminuir su tiempo de congelamiento (p<0.05).
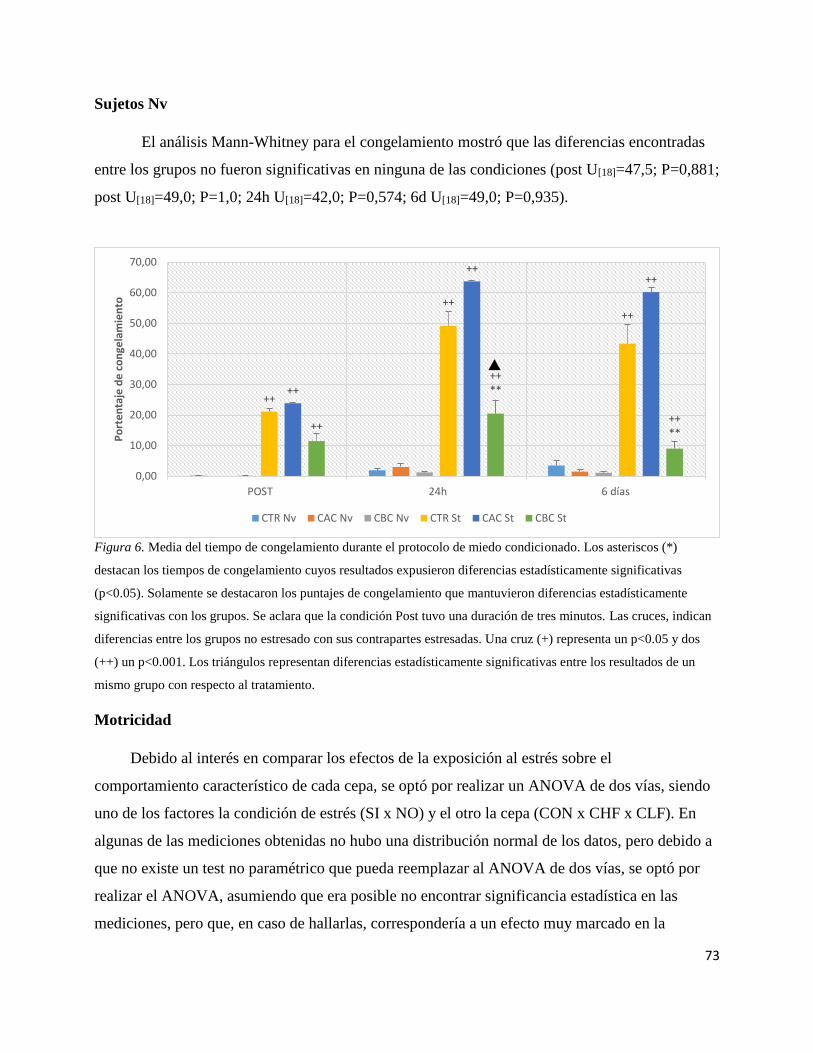
73
Sujetos Nv
El análisis Mann-Whitney para el congelamiento mostró que las diferencias encontradas
entre los grupos no fueron significativas en ninguna de las condiciones (post U[18]=47,5; P=0,881;
post U[18]=49,0; P=1,0; 24h U[18]=42,0; P=0,574; 6d U[18]=49,0; P=0,935).
Figura 6. Media del tiempo de congelamiento durante el protocolo de miedo condicionado. Los asteriscos (*)
destacan los tiempos de congelamiento cuyos resultados expusieron diferencias estadísticamente significativas
(p<0.05). Solamente se destacaron los puntajes de congelamiento que mantuvieron diferencias estadísticamente
significativas con los grupos. Se aclara que la condición Post tuvo una duración de tres minutos. Las cruces, indican
diferencias entre los grupos no estresado con sus contrapartes estresadas. Una cruz (+) representa un p<0.05 y dos
(++) un p<0.001. Los triángulos representan diferencias estadísticamente significativas entre los resultados de un
mismo grupo con respecto al tratamiento.
Motricidad
Debido al interés en comparar los efectos de la exposición al estrés sobre el
comportamiento característico de cada cepa, se optó por realizar un ANOVA de dos vías, siendo
uno de los factores la condición de estrés (SI x NO) y el otro la cepa (CON x CHF x CLF). En
algunas de las mediciones obtenidas no hubo una distribución normal de los datos, pero debido a
que no existe un test no paramétrico que pueda reemplazar al ANOVA de dos vías, se optó por
realizar el ANOVA, asumiendo que era posible no encontrar significancia estadística en las
mediciones, pero que, en caso de hallarlas, correspondería a un efecto muy marcado en la
++
++++
++
++++
++
++**
++**
0,00
10,00
20,00
30,00
40,00
50,00
60,00
70,00
POST 24h 6 días
Po
rte
nta
je d
e c
on
gela
mie
nto
CTR Nv CAC Nv CBC Nv CTR St CAC St CBC St

74
población. En este caso, las diferencias halladas han de ser tomadas con cautela, pero debe
recordarse que, posiblemente, si se tuvieran grupos de mayor tamaño y con igual número de
sujetos, con gran probabilidad el análisis paramétrico acusaría las mismas significancias.
Motricidad en el campo abierto
Primera exposición
El ANOVA de dos vías mostró que no hubo diferencias significativas entre las medias
obtenidas entre el factor de cepa y la condición de estrés (F[2,54]=1.824; P=0.171) para la distancia
recorrida en el campo abierto. Sin embargo, para los dos factores por separado, las diferencias
halladas si alcanzaron el valor de significancia. Para el factor cepa (F[2,54]=14.721, P<0.001), se
identificaron diferencias significativas entre los grupos experimentales. La comparación post-hoc
(test de Student Newman-Keuls) mostró que el grupo CLF recorrió una distancia
significativamente mayor que los demás grupos experimentales (CHF p<0.001; CON p=0.005).
CHF se destacó por haber recorrido la menor distancia en comparación con su respectivo control
(p=0.019).
En cuanto a la condición de estrés (F[1, 54]=4.075, p<0.05), las diferencias entre los grupos
también sugirieron un efecto del tratamiento en la distancia recorrida por el campo. La
comparación post-hoc mostró que los animales pertenecientes al grupo Nv recorrieron una
distancia significativamente mayor a los del grupo estresado.
Vale la pena destacar las interacciones obtenidas entre los factores. En grupos sometidos a
estrés, el análisis de post-hoc mostró diferencias estadísticamente significativas que constataron
que los animales de CLF recorrieron distancias mayores en el campo abierto en comparación con
sus respectivos CHF (p<0.001) y sus respectivos controles (P=0.004); y que los animales de CHF
recorrieron distancias significativamente menores que sus respectivos controles (p=0.036). En el
caso de los animales Nv, no se identificaron diferencias estadísticamente significativas entre los
grupos (figura 7). Posteriores análisis para los efectos del estrés entre los grupos con sus
contrapartes estresadas mostraron que CHF Nv recorrió una distancia significativamente mayor
que su CLF (p<0.05). En los otros grupos, no hubo diferencias que permitiesen afirmar un efecto
del estrés sobre la locomoción de los animales durante su tiempo en el campo (figura 7).

75
Segunda exposición
El ANOVA de dos vías para la segunda exposición al campo abierto tampoco mostró
diferencias estadísticamente significativas para el factor de condición de estrés entre las cepas
(F[2,54]=1.047, p=0.358). Por el contrario, para el factor cepa y condición de estrés por separado,
las diferencias si alcanzaron un valor de significancia. Para el factor cepa (F[2, 54]=14.754,
P<0.001), se observaron diferencias estadísticamente significativas entre los grupos
experimentales. La comparación post-hoc (test de Student Newman-Keuls) destacó que,
nuevamente, CLF recorrió una distancia significativamente mayor que CHF (p<0.001) y su
respectivo control (p=0.042). Se agrega que CLF fue el grupo cuya distancia recorrida fue la
menor en comparación con el grupo control (p=0.002).
En cuanto a la condición de estrés, (F[1, 54]=4.435, p<0.05), se resalta que los grupos Nv
recorrieron distancias significativamente mayores por el campo que los sujetos estresados.
Las interacciones entre los resultados obtenidos destacan que, para los animales sometidos
a estrés hubo diferencias estadísticamente significativas, las cuales destacaron que los animales
de CHF recorrieron distancias menores en el campo en comparación con CLF (p<0.001) y sus
respectivos controles (p=0.039). Se rescata que el grupo CLF obtuvo la mayor distancia recorrida
en el campo en comparación con su control (p=0.042). En los animales Nv, el grupo CHF Nv
recorrió distancias menores en el campo que CLF Nv (p=0.019) y su respectivo control
(p=0.017). Finalmente, los análisis sobre las diferencias entre los grupos Nv con sus contrapartes
estresadas no mostraron diferencias estadísticamente significativas (figura 7).
Tercera exposición
El ANOVA de dos vías para la tercera exposición al campo mostró que no hubo
diferencias estadísticamente significativas para el factor de condición de estrés entre las cepas
(F[2,54]=1.905, p=0.159) durante esta sesión. Sin embargo, al igual que en las exposiciones
pasadas, para el factor cepa y condición de estrés por separado, las diferencias si alcanzaron el
factor de significancia. Así pues, para el factor cepa (F[2, 54]=18.417, P<0.001), se observaron
diferencias estadísticamente significativas que destacaron que CLF recorrió distancias mayores
en comparación con las demás cepas experimentales (CHF p<0.001; CON P=0.001). El grupo
CHF obtuvo la menor distancia recorrida en comparación con su respectivo control (p=0.014).

76
Para las comparaciones referentes al factor de condición de estrés mostraron diferencias
estadísticamente significativas (F[1, 54]=7.844, p=0.007). La comparación post-hoc rescató que las
cepas que no recibieron inducción de estrés recorrieron distancias significativamente mayores a
sus contrapartes estresadas.
Las interacciones referentes a los resultados obtenidos ofrecieron información adicional
que permitió afirmar un efecto del tratamiento sobre la distancia recorrida por el campo.
Comparaciones de post-hoc aclararon que, en los animales sometidos a estrés, CHF recorrió
distancias significativamente menores que CLF (p=0.002) y sus respectivos controles (p=0.009)
(figura 7). En los animales del grupo CHF Nv también se mostró que las distancias recorridas
fueron menores que CLF Nv (p<0.001) y el grupo control (p<0.001). De manera interesante,
comparaciones entre los grupos y sus respectivas contrapartes estresadas solamente rescató
diferencias estadísticamente significativas en el grupo control, puesto que los animales estresados
recorrieron una distancia significativamente menor que su contraparte no estresada (p=0.003)
(figura 7).
Figura 7. Motricidad por el campo abierto. Los asteriscos representan diferencias estadísticamente significativas
entre los grupos de la misma naturaleza (Estrés vs. Estrés y No Estrés vs. No estrés). Un asterisco (*) representa un
p<0.05 y dos (**) representan un p<0.001. Las cruces, indican diferencias entre los grupos no estresado con sus
contrapartes estresadas. Una cruz (+) representa un p<0.05 y dos (++) un p<0.001.
Habituación al campo
**
+
+
+ ++
**
****
0,00
10,00
20,00
30,00
40,00
50,00
60,00
70,00
Primera Exposición Segunda Exposición Tercera Exposición
Dis
tan
cia
reco
rrid
a (m
s*0
.2) CON
NvCHF nV
CLF Nv
CON St
CHF St
CLF St

77
Con el fin de identificar las diferencias en cuanto a la distancia recorrida en la primera,
segunda y tercera sesión de habituación al campo, se realizaron análisis de varianza de una vía.
En primer lugar, en el grupo CHF estresado no se identificaron diferencias entre la
distancia recorrida en el campo durante las tres sesiones experimentales (F[2, 33]=0.416, p=0.633).
Para el grupo CLF estresado tampoco se identificaron diferencias en cuanto a la distancia
recorrida en ninguna de las sesiones (F[2, 36]=1.107, p=0.342). Para el grupo control, se
identificaron resultados similares (F[2, 21]=0.870, p=0.433). Por lo tanto, se concluye que los
animales experimentales pertenecientes a los grupos sometidos a estrés mantuvieron tendencias
de exploración similares durante las tres exposiciones al campo abierto.
En cuanto a los animales no estresados, tampoco se identificaron diferencias entre las
distancias recorridas en los grupos durante las tres exposiciones al campo abierto. Los grupos
también mostraron tendencias similares de exploración por el campo (CHFNv F[2, 18]=0.325,
p=0.727; CLFNv, F[2, 24]=0.270, p=0.766; CONNv, F[2, 30]=0.0264, p=0.974).
Comportamientos observados en el campo abierto
Se evaluaron los comportamientos correspondientes al congelamiento, el acicalamiento y
levantarse en las patas posteriores durante la primera exposición al protocolo del campo abierto.
Los promedios de tiempo de los comportamientos y sus respectivas frecuencias durante la sesión
experimental se encuentran ordenados de la siguiente manera: en la figura 8, los promedios del
tiempo de congelamiento y la frecuencia de ese comportamiento durante la sesión experimental;
en la figura 9, los promedios del tiempo de acicalamiento y la frecuencia de ese comportamiento
durante la sesión experimental; y en la figura 10, el promedio y las desviaciones estándar del
tiempo de estar de pie y la frecuencia de ese comportamiento durante la sesión experimental.
Debido al interés en comparar los efectos de la exposición al estrés sobre el
comportamiento característico de cada cepa, se optó por realizar un ANOVA de dos vías, siendo
uno de los factores la condición de estrés (SI x NO) y el otro la cepa (CON x CHF x CLF). En
algunas de las mediciones obtenidas, no hubo una distribución normal de los datos, pero debido a
que no existe un test no paramétrico que pueda reemplazar al ANOVA de dos vías, se optó por
realizar el ANOVA, asumiendo que era posible no encontrar significancia estadística en las
mediciones, pero que, en caso de hallarlas, correspondería a un efecto muy marcado en la

78
población. En este caso, las diferencias halladas han de ser tomadas con cautela, pero debe
recordarse que, posiblemente, si se tuvieran grupos de mayor tamaño y con igual número de
sujetos, con gran probabilidad el análisis paramétrico acusaría las mismas significancias.
Congelamiento
El ANOVA de dos vías para el tiempo de congelamiento mostró que hubo diferencias
estadísticamente significativa para los factores de condición de estrés y cepa (F[2, 54]=8.470,
p<0.001). Tanto en el factor cepa (F[2,54]=35.174, p<0.001) como condición de estrés (F[1,
54]=30.082, p<0.001) se observaron diferencias estadísticamente significativas entre los grupos
experimentales. El análisis post-hoc (test de Student Newman-Keuls) mostró los siguientes
resultados: En cuanto al factor cepa, la comparación mostró diferencias estadísticamente
significativas en todos los grupos. Para este caso, CHF mantuvo un tiempo de congelamiento
significativamente mayor que CLF (p<0.001) y el grupo control (p<0.001). CLF, por el contrario,
obtuvo un tiempo de congelamiento menor que su respectivo control (p=0.001). En segundo
lugar, para la condición de estrés, las diferencias resaltaron que los animales estresados
congelaron más que aquellos que no recibieron el estímulo de estrés (p<0.001).
Las comparaciones entre los factores exhibieron que las medias de tiempo de
congelamiento entre los grupos estresados destacaron a CHF por congelar por un tiempo
significativamente mayor que CLF (p<0.001) y su respectivo grupo control (p<0.001). Se aclara
que el grupo CLF congeló por un tiempo significativamente menor que los demás animales
experimentales (p<0.001). Con respecto a los animales Nv, nuevamente el grupo CHF obtuvo un
tiempo de congelamiento significativamente mayor que CLF (p<0.05) y su respectivo control
(p<0.05). Los efectos del tratamiento entre las cepas experimentales con sus contrapartes
estresadas mostraron que los grupos que fueron sometidos a estrés, con excepción de CLF
(p=0.888), tuvieron tiempos de congelamiento significativamente mayores (p<0.01 para CHF;
p=0.01 para CON).
Ahora bien, el análisis de la frecuencia del comportamiento, mostró que hubo diferencias
estadísticamente significativas para el factor de condición de estrés entre las cepas
experimentales (F[2,54]=4.711, p<0.05). Asimismo, las comparaciones de cada factor mostraron
grados de significancia. Se identificaron diferencias estadísticamente significativas para el factor
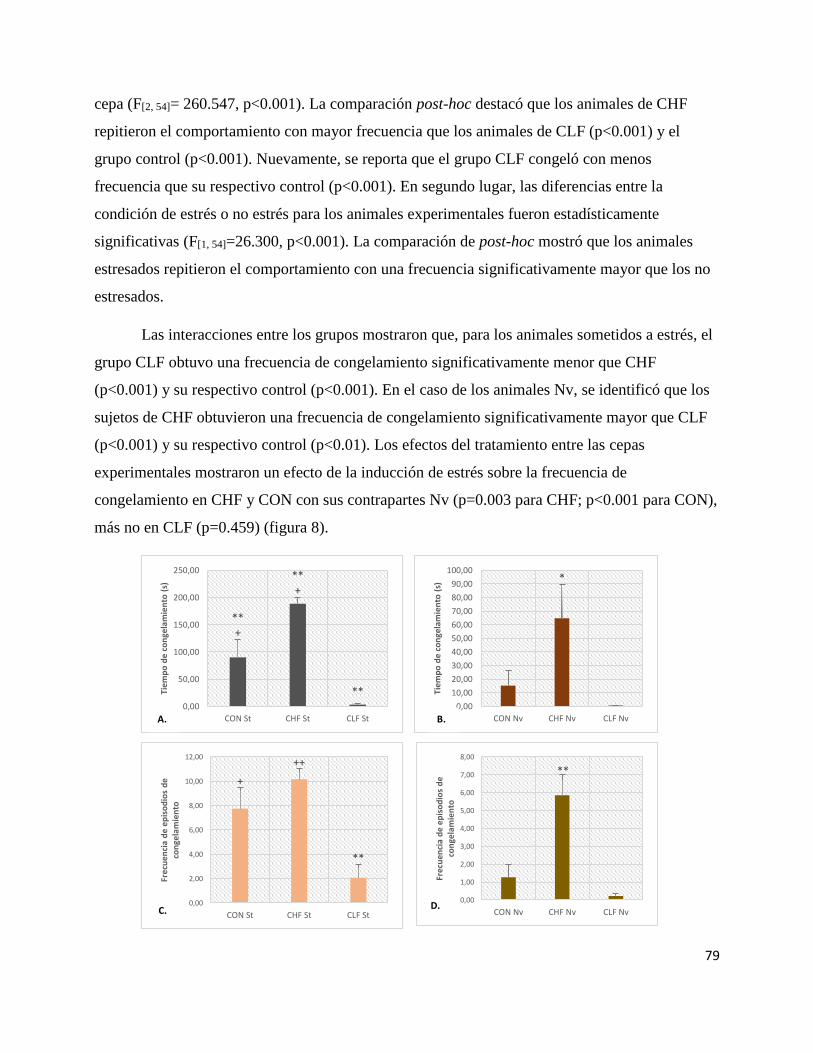
79
cepa (F[2, 54]= 260.547, p<0.001). La comparación post-hoc destacó que los animales de CHF
repitieron el comportamiento con mayor frecuencia que los animales de CLF (p<0.001) y el
grupo control (p<0.001). Nuevamente, se reporta que el grupo CLF congeló con menos
frecuencia que su respectivo control (p<0.001). En segundo lugar, las diferencias entre la
condición de estrés o no estrés para los animales experimentales fueron estadísticamente
significativas (F[1, 54]=26.300, p<0.001). La comparación de post-hoc mostró que los animales
estresados repitieron el comportamiento con una frecuencia significativamente mayor que los no
estresados.
Las interacciones entre los grupos mostraron que, para los animales sometidos a estrés, el
grupo CLF obtuvo una frecuencia de congelamiento significativamente menor que CHF
(p<0.001) y su respectivo control (p<0.001). En el caso de los animales Nv, se identificó que los
sujetos de CHF obtuvieron una frecuencia de congelamiento significativamente mayor que CLF
(p<0.001) y su respectivo control (p<0.01). Los efectos del tratamiento entre las cepas
experimentales mostraron un efecto de la inducción de estrés sobre la frecuencia de
congelamiento en CHF y CON con sus contrapartes Nv (p=0.003 para CHF; p<0.001 para CON),
más no en CLF (p=0.459) (figura 8).
**+
**+
**0,00
50,00
100,00
150,00
200,00
250,00
CON St CHF St CLF St
Tiem
po
de
con
gela
mie
nto
(s)
A.
*
0,00
10,00
20,00
30,00
40,00
50,00
60,00
70,00
80,00
90,00
100,00
CON Nv CHF Nv CLF Nv
Tiem
po
de
con
gela
mie
nto
(s)
B.
+
++
**
0,00
2,00
4,00
6,00
8,00
10,00
12,00
CON St CHF St CLF St
Fre
cuen
cia
de
epis
od
ios
de
con
gela
mie
nto
C.
**
0,00
1,00
2,00
3,00
4,00
5,00
6,00
7,00
8,00
CON Nv CHF Nv CLF Nv
Fre
cuen
cia
de
epis
od
ios
de
con
gela
mie
nto
D.

80
Figura 8. Promedios del tiempo de congelamiento y frecuencia del congelamiento en los grupos experimentales.
Los asteriscos representan diferencias estadísticamente significativas entre los grupos. Los dos asteriscos (**) hacen
referencia a aquel que obtuvo un valor p<0.001 en comparación con los otros grupos y un asterisco (*) hace
referencia a un p<0.05. Las cruces (+) representan diferencias estadísticamente significativas entre los grupos con
sus respectivas contrapartes no estresadas (nv) A. Promedio del tiempo de congelamiento en los grupos estresados y
Nv. En este se pueden identificar diferencias significativas en todos los grupos. Entre estos se destaca a CLF por
tener el menor tiempo de congelamiento durante toda la sesión y a CHF por tener el mayor tiempo de congelamiento
B. Promedio del tiempo de congelamiento de los grupos no estresados. En este se destaca que el tiempo de
congelamiento de CHF fue el mayor. C. Frecuencia de los episodios, de congelamiento en los grupos estresados. En
este se destaca que CLF tuvo la menor frecuencia en comparación con CHF y CON, cuya frecuencia fue similar
durante el protocolo experimental. D. Frecuencia de los episodios de congelamiento en los grupos no estresados. De
estos, nuevamente se destaca que CHF tuvo una frecuencia mayor que los otros dos grupos durante el protocolo
experimental.
Acicalamiento
Un ANOVA de dos vías para el tiempo de acicalamiento mostró que no hubo diferencias
estadísticamente significativas entre el factor de condición de estrés y las cepas experimentales
(F[2,54]=0.108, p=0.897). Sin embargo para cada factor, las diferencias si alcanzaron el grado de
significancia (cepa, F[2,54]=15.179, p<0.001;condición de estrés, F[1,54]=4.882, p<0.05). La
comparación post-hoc (test de Student Newman-Keuls) para el factor cepa, mostró que el grupo
CLF, en general, se acicaló por un tiempo significativamente mayor que los demás grupos
experimentales (CHF p<0.001; CON p<0.001). En cuanto a la condición de estrés, se rescató que
los animales Nv también se acicalaron por un tiempo significativamente mayor que los
estresados (p=0.032).
Procediendo a las interacciones entre los factores, se rescata lo siguiente. Entre los
animales sometidos a estrés, el grupo CLF tuvo un tiempo de acicalamiento significativamente
mayor que CHF (p<0.001) y su respectivo control (p<0.01). Para el caso de los Nv, se identificó
que los sujetos de CLF también obtuvieron un tiempo de acicalamiento significativamente mayor
que CHF (p<0.01) y su respectivo control (p<0.01). Los efectos del tratamiento entre las cepas
experimentales no mostraron diferencias significativas que permitiesen afirmar un efecto de la
inducción del estrés sobre el tiempo de acicalamiento en los grupos (CHF, p=138; CLF, p=0.157;
CON, p=0.375) (figura 9).
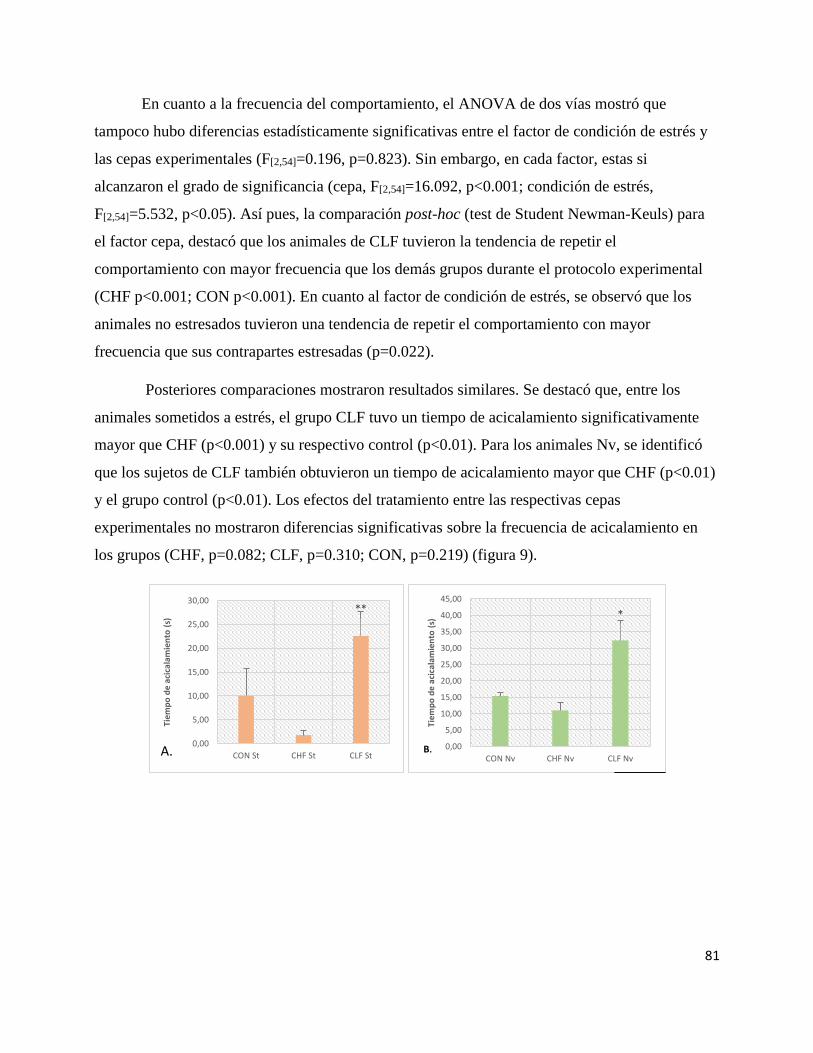
81
En cuanto a la frecuencia del comportamiento, el ANOVA de dos vías mostró que
tampoco hubo diferencias estadísticamente significativas entre el factor de condición de estrés y
las cepas experimentales (F[2,54]=0.196, p=0.823). Sin embargo, en cada factor, estas si
alcanzaron el grado de significancia (cepa, F[2,54]=16.092, p<0.001; condición de estrés,
F[2,54]=5.532, p<0.05). Así pues, la comparación post-hoc (test de Student Newman-Keuls) para
el factor cepa, destacó que los animales de CLF tuvieron la tendencia de repetir el
comportamiento con mayor frecuencia que los demás grupos durante el protocolo experimental
(CHF p<0.001; CON p<0.001). En cuanto al factor de condición de estrés, se observó que los
animales no estresados tuvieron una tendencia de repetir el comportamiento con mayor
frecuencia que sus contrapartes estresadas (p=0.022).
Posteriores comparaciones mostraron resultados similares. Se destacó que, entre los
animales sometidos a estrés, el grupo CLF tuvo un tiempo de acicalamiento significativamente
mayor que CHF (p<0.001) y su respectivo control (p<0.01). Para los animales Nv, se identificó
que los sujetos de CLF también obtuvieron un tiempo de acicalamiento mayor que CHF (p<0.01)
y el grupo control (p<0.01). Los efectos del tratamiento entre las respectivas cepas
experimentales no mostraron diferencias significativas sobre la frecuencia de acicalamiento en
los grupos (CHF, p=0.082; CLF, p=0.310; CON, p=0.219) (figura 9).
**
0,00
5,00
10,00
15,00
20,00
25,00
30,00
CON St CHF St CLF St
Tiem
po
de
acic
alam
ien
to (
s)
A.
*
0,00
5,00
10,00
15,00
20,00
25,00
30,00
35,00
40,00
45,00
CON Nv CHF Nv CLF Nv
Tie
mp
o d
e ac
ical
amie
nto
(s)
B.

82
Figura 9. Promedios de tiempo y frecuencia de acicalamiento. Los asteriscos hacen referencia a diferencias
estadísticamente significativas que vale la pena resaltar entre los grupos. Un asterisco (*) representa un p<0.05 y dos
(**) un p<0.001. Las cruces (+) representan diferencias estadísticamente significativas entre los grupos con sus
respectivas contrapartes no estresadas (Nv) A. Medias del tiempo de acicalamiento en las ratas estresadas. Se
destacan al grupo CLF haber presentado el comportamiento por un periodo de tiempo más prolongado que los
demás. B. Medias de tiempo de acicalamiento de los grupos no estresados. No se observaron diferencias entre las
medias de tiempo del comportamiento entre los grupos. Se destaca que CLFNv obtuvo un tiempo de acicalamiento
significativamente mayor que CHF y su respectivo control C. Frecuencia del comportamiento de acicalamiento de
los grupos estresados durante el protocolo experimental. Se destaca que CLF repitió el comportamiento con más
frecuencia de los otros grupos. D. Frecuencia del acicalamiento en los grupos no estresados durante la sesión
experimental. Para este caso, se rescata a CLF pues la frecuencia repitió el comportamiento con más frecuencia que
los otros grupos.
Levantarse en las patas traseras
Un ANOVA de dos vías para el tiempo de estar parado en las patas traseras mostró que no
hubo diferencias estadísticamente significativas entre el factor de condición de estrés y las cepas
experimentales (F[2,54]=0.00469, p=0.995). Por el contrario, cuando se evaluaron los factores por
separado, las diferencias si alcanzaron el grado de significancia (cepa, F[2,54]=28.565, p<0.001;
condición de estrés, F[1,54]=5.717, p<0.05). La comparación post-hoc (test de Student Newman-
Keuls) mostró que en el factor cepa si hubo diferencias estadísticamente significativas entre los
grupos experimentales. En este caso, se aclara que los animales del grupo CLF se pararon en las
patas traseras por un tiempo significativamente mayor que los demás grupos durante el protocolo
experimental (CHF p<0.001; CON, p<0.001). Por el contrario, los animales de CHF
permanecieron parados en las patas traseras por un tiempo significativamente menor que el grupo
control (p=0.002). En cuanto al factor de condición de estrés, se rescata que los animales
**
0,00
2,00
4,00
6,00
8,00
10,00
12,00
CON St CHF St CLF St
Frec
uen
cia
de
acic
alam
ien
to
C.
*
0,00
2,00
4,00
6,00
8,00
10,00
12,00
14,00
CON Nv CHF Nv CLF Nv
Frec
uen
cia
de
acic
alam
ien
to
D.

83
estresados mantuvieron este comportamiento por tiempo significativamente menor que los no
estresados (p=0.020).
Posteriores comparaciones entre los factores mostraron que, para los animales sometidos a
estrés hubo diferencias estadísticamente significativas en las medias de todos los grupos. Se
observó que el grupo CLF tuvo un tiempo de acicalamiento mayor que CHF (p<0.001) y su
respectivo control (p<0.01). En contraposición, el grupo CHF obtuvo el menor tiempo de estar
levantado en las patas durante la fase experimental. De manera interesante, en los animales Nv
también se identificaron diferencias estadísticamente significativas en el tiempo de estar parado
en las patas traseras. Nuevamente, los sujetos de CLF obtuvieron un tiempo de acicalamiento
significativamente mayor que CHF (p<0.01) y su respectivo control (p<0.01). Se agrega que CHF
Nv obtuvo el menor tiempo de estar de pie (figura 11). Los efectos del tratamiento entre las
respectivas cepas experimentales no mostraron diferencias significativas que permitiesen afirmar
un efecto de la inducción del estrés sobre la frecuencia de pararse en los grupos (CHF, p=0.188;
CLF, p=0.132; CON=0.201).
En cuanto a la frecuencia del comportamiento, el ANOVA de dos vías tampoco destacó
diferencias estadísticamente significativas entre la condición de estrés y las cepas experimentales
(F[2, 54]=0.855, p<0.431), pero cada factor alcanzó un nivel de significancia. El factor cepa
demostró diferencias estadísticamente significativas (F[2,54]=13.043, p<0.001). Este demostró,
durante la comparación post-hoc, que los animales del grupo CLF repitieron el comportamiento
con más frecuencia que las demás cepas experimentales (CHF p<0.001; CON p=0.010). CHF y
control no se diferenciaron. Ahora bien, para el factor de condición de estrés (F[1,54]=0.533,
p=0.469), no se identificaron diferencias que permitiesen afirmar que el estrés causó un efecto
sobre el tiempo de estar parado en las patas traseras entre los grupos (CHF, p>0.05; CLF, p>0.05;
CON>0.05).
Finalmente, las comparaciones entre los factores (test de Student Newman-Keuls)
mostraron que los animales del grupo CLF sometido a estrés obtuvieron una frecuencia del
comportamiento estadísticamente mayor que CHF (p<0.001) y su contraparte control (p<0.01).
En cuanto a los animales Nv, solamente se identificó que CLF repitió el comportamiento con
mayor frecuencia que los animales del grupo CHF (p<0.05). Los efectos del tratamiento entre las
respectivas cepas experimentales no mostraron diferencias significativas que permitiesen afirmar
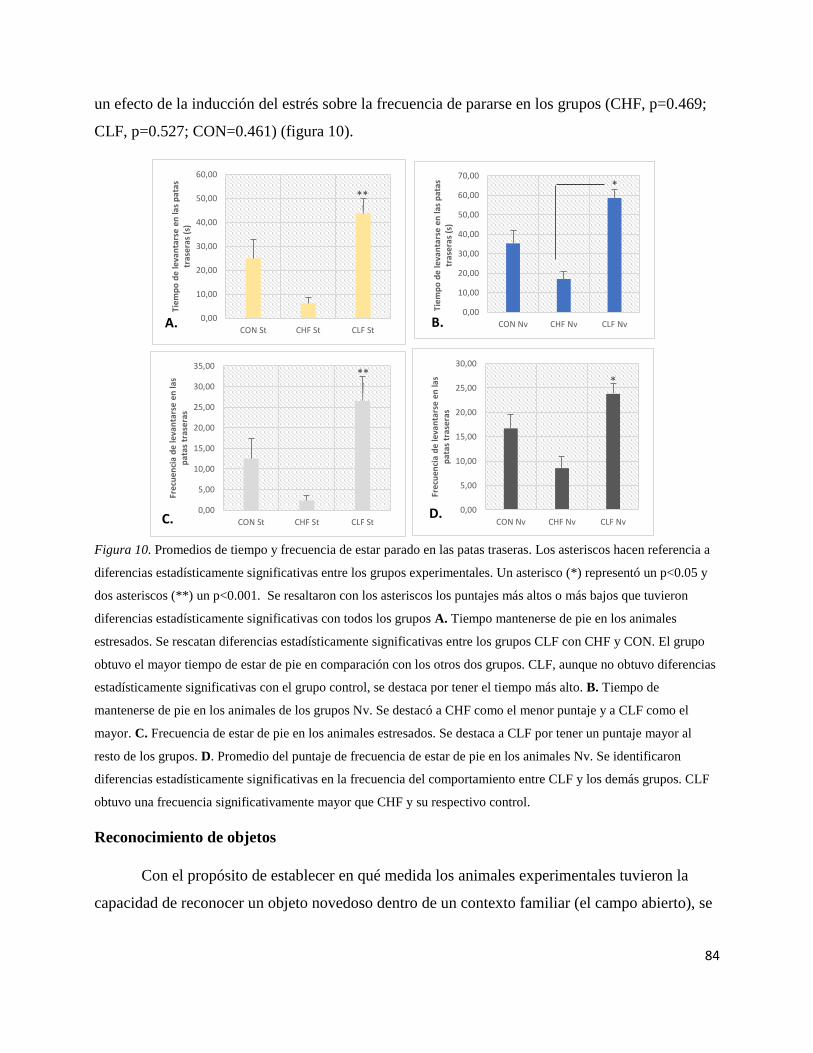
84
un efecto de la inducción del estrés sobre la frecuencia de pararse en los grupos (CHF, p=0.469;
CLF, p=0.527; CON=0.461) (figura 10).
Figura 10. Promedios de tiempo y frecuencia de estar parado en las patas traseras. Los asteriscos hacen referencia a
diferencias estadísticamente significativas entre los grupos experimentales. Un asterisco (*) representó un p<0.05 y
dos asteriscos (**) un p<0.001. Se resaltaron con los asteriscos los puntajes más altos o más bajos que tuvieron
diferencias estadísticamente significativas con todos los grupos A. Tiempo mantenerse de pie en los animales
estresados. Se rescatan diferencias estadísticamente significativas entre los grupos CLF con CHF y CON. El grupo
obtuvo el mayor tiempo de estar de pie en comparación con los otros dos grupos. CLF, aunque no obtuvo diferencias
estadísticamente significativas con el grupo control, se destaca por tener el tiempo más alto. B. Tiempo de
mantenerse de pie en los animales de los grupos Nv. Se destacó a CHF como el menor puntaje y a CLF como el
mayor. C. Frecuencia de estar de pie en los animales estresados. Se destaca a CLF por tener un puntaje mayor al
resto de los grupos. D. Promedio del puntaje de frecuencia de estar de pie en los animales Nv. Se identificaron
diferencias estadísticamente significativas en la frecuencia del comportamiento entre CLF y los demás grupos. CLF
obtuvo una frecuencia significativamente mayor que CHF y su respectivo control.
Reconocimiento de objetos
Con el propósito de establecer en qué medida los animales experimentales tuvieron la
capacidad de reconocer un objeto novedoso dentro de un contexto familiar (el campo abierto), se
*
0,00
5,00
10,00
15,00
20,00
25,00
30,00
CON Nv CHF Nv CLF Nv
Frec
uen
cia
de
leva
nta
rse
en la
s p
atas
tra
sera
s
D.
**
0,00
5,00
10,00
15,00
20,00
25,00
30,00
35,00
CON St CHF St CLF St
Frec
uen
cia
de
leva
nta
rse
en la
s p
atas
tra
sera
s
C.
*
0,00
10,00
20,00
30,00
40,00
50,00
60,00
70,00
CON Nv CHF Nv CLF Nv
Tiem
po
de
leva
nta
rse
en la
s p
atas
tr
aser
as (
s)
B.
**
0,00
10,00
20,00
30,00
40,00
50,00
60,00
CON St CHF St CLF St
Tiem
po
de
leva
nta
rse
en la
s p
atas
tr
aser
as (
s)
A.

85
analizaron distintas variables que permitieron establecer que tanto exploraron los objetos en la
fase de familiarización y exploraron el objeto novedoso en la fase de prueba.
Familiarización
Se realizaron comparaciones sobre las medias de porcentaje de exploración de los objetos
durante la fase de familiarización. Se realizó este proceso con el propósito identificar si hubo
exploración de los objetos por parte de los grupos experimentales. Para esto, se realizaron
ANOVAs con medidas repetidas, las cuales compararon las medias de los tiempos de exploración
de los objetos en los grupos experimentales estresados y no estresados. En la figura 11 se
encuentran las medias y desviaciones estándar del tiempo de exploración durante la sesión
experimental.
Sujetos estresados
Aunque se puede evidenciar que los animales exploraron los objetos, se observaron
diferencias significativas en cuanto a los tiempos de exploración (F[12, 5]=5.796, p<0.001).
Posteriores comparaciones de post-hoc mostraron que los animales del grupo CHF exploraron los
objetos significativamente menos que los otros grupos, tanto para el objeto P (p<0.05) como para
el objeto B (p<0.05).
Sujetos no estresados
En cuanto a los grupos no estresados, el análisis para la exploración de los objetos
muestra que no hubo diferencias estadísticamente significativas en los grupos (F[10, 5]=2.724,
p=0.488).

86
Figura 11. Porcentaje de exploración de los objetos durante la fase de familiarización. No se identificaron
diferencias estadísticamente significativas en las medias de exploración en los grupos Nv. En los grupos estresados
se puede evidenciar un tiempo de exploración de los objetos significativamente menor en CHF.
Fase de prueba
Una ANOVA de dos vías que comparó las medias de tiempo de exploración del objeto
novedoso entre los grupos experimentales, mostró que las distribuciones de los datos
correspondientes no obtuvo diferencias estadísticamente significativas (F[2, 54]=0.264, p=0.769).
Las diferencias no alcanzaron el grado de significancia, ni para el factor cepa (F[2,54]=3.000,
p=0.058), ni para el factor de condición de estrés (F[1, 54]=0.512, p=0.769). A partir de este punto,
se procede a afirmar que no hubo un efecto del estrés sobre el tiempo de exploración del objeto
novedoso entre las cepas.
Se procedió a realizar un ANOVA de dos vías para identificar las diferencias entre las
medias de tiempo de exploración del objeto familiar, sin embargo estas no se observaron entre el
factor cepa y condición de estrés (F[2, 54]=1.958, p=0.151). Cuando se evaluaron los factores por
separado, las diferencias sólo alcanzaron el grado de significancia para el factor cepa
(F[2,54]=5.015, p<0.01), más no para el factor de condición de estrés (F[1, 54]=0.0926, p=0.762). La
comparación post-hoc para este primer factor (test de Student Newman-Keuls) mostró que los
animales sometidos a estrés CLF tuvieron un tiempo de exploración del objeto familiar
significativamente mayor que CHF (p=0.004) y su respectivo control (p=0.012). En el caso de los
grupos Nv, no se observaron diferencias estadísticamente significativas entre los grupos
**
0,00
10,00
20,00
30,00
40,00
50,00
60,00
70,00
80,00
90,00
CON Nv CHF Nv CLF Nv CON St CHF St CLF St
Exp
lora
ció
n o
bje
tos
fam
iliar
izac
ión
(s)
objeto B Objeto P

87
experimentales. Comparaciones múltiples entre las cepas con sus respectivas contrapartes no
estresadas tampoco mostraron diferencias estadísticamente significativas (figura 12).
Comparaciones intra-grupo
se procedió a analizar los porcentajes de las medias de tiempo de exploración entre el
objeto familiar y el objeto novedoso intra-grupo por medio del uso de pruebas t pareadas.
En primer lugar, la prueba t pareada para el tiempo de exploración del objeto familiar y el
objeto novedoso en CHF estresado mostró que las diferencias entre las medias de exploración
fueron estadísticamente significativas en el grupo (t([1]=-2.830, p=0.016). Se identificó que los
animales tuvieron la tendencia de explorar el objeto novedoso en comparación con el familiar. En
cuanto al grupo Nv, el análisis mostró que las diferencias entre los tiempos de exploración del
objeto familiar y novedoso no fueron estadísticamente significativas (t[6]=-2.242, p=0.066).
Para CLF estresado, las diferencias entre el tiempo de exploración tampoco destacaron
diferencias estadísticamente significativas entre el objeto familiar y el objeto novedoso
(t[12]=1.582, p=0.234). En CLF Nv tampoco se identificaron diferencias entre las medias de
tiempo de exploración de los objetos familiar y novedoso (t[8]=-0.154, p=0.882).
Finalmente, las diferencias entre las medias de tiempo de exploración de los objetos en el
grupo control sometido a estrés fueron estadísticamente significativas (t[7]=-4.049, p<0.05),
permitiendo afirmar que se discriminó el objeto familiar del novedoso. De la misma manera, el
análisis rescató que para control Nv también hubo diferencias estadísticamente significativas
entre la exploración de los objetos (t[10]=3.125, p<0.01).

88
Figura 12. Porcentajes de exploración en el objeto familiar y novedoso. Los asteriscos representan diferencias
estadísticamente significativas entre las medias del grupo señalado con las de los demás grupos. Un asterisco
representa un p<0.05 Nota: Se identificaron diferencias estadísticamente significativas entre CLF St y sus respectivas
contrapartes no estresadas. En cuento a los análisis intra-grupo, se observan diferencias estadísticamente
significativas entre los grupos experimentales entre el tiempo de exploración entre la exploración del objeto familiar
y novedoso, las cuales destacan una discriminación entre estos dos. No se observaron diferencias entre los tiempos
de exploración en los animales del grupo CLF Nv y CLF St.
Histología
Ahora bien, se procedió a efectuar el análisis de las muestras de tejido cerebral en los
grupos experimentales. Debido al interés en comparar los efectos de la exposición al estrés sobre
la proliferación de microglía activada por área (0.1mm) en cada cepa, se optó por realizar un
ANOVA de dos vías, siendo uno de los factores la condición de estrés (SI x NO) y el otro la cepa
(CTR x CAC x CBC). En algunas de las mediciones obtenidas, no hubo una distribución normal
de los datos, pero debido a que no existe un test no paramétrico que pueda reemplazar al
ANOVA de dos vías, se optó por realizar el ANOVA, asumiendo que era posible no encontrar
significancia estadística en las mediciones, pero que, en caso de hallarlas, correspondería a un
efecto muy marcado en la población. En este caso, las diferencias halladas han de ser tomadas
con cautela, pero debe recordarse que, posiblemente, si se tuvieran grupos de mayor tamaño y
con igual número de sujetos, con gran probabilidad el análisis paramétrico acusaría las mismas
significancias.
Hipocampo
*
*
**
*
0,00
10,00
20,00
30,00
40,00
50,00
60,00
70,00
CON Nv CHF Nv CLF Nv CON St CHF St CLF St
Tie
mp
o d
e e
xplo
raci
ón
ob
jeto
s p
rue
ba
(s)
Objeto familiar
Objeto novedoso

89
Giro dentado
Un ANOVA de dos vías para el tiempo de pararse en las patas traseras mostró que hubo
interacciones estadísticamente significativas entre el factor de condición de estrés y la
proliferación de microglía activada en las cepas experimentales (F[2,50]=4.590, p=0.015). La
comparación post-hoc (test de Student Newman-Keuls) mostró que, entre los animales sometidos
a estrés, no se mostraron diferencias estadísticamente significativas en las medias de microglía de
fenotipo M1. En cuanto a los animales Nv, si se identificaron diferencias estadísticamente
significativas en la proliferación de microglía activada. Los sujetos de CLF Nv obtuvieron una
cantidad de microglía M1 significativamente mayor que acicalamiento significativamente mayor
que CHF Nv (p<0.04) y su respectivo control (p<0.01).
En cuando a los análisis entre los grupos con sus respectivas contrapartes estresadas, se
observaron diferencias estadísticamente significativas entre CHF sometido a estrés y su
contraparte no estresada (p=0.009). CHF St obtuvo una cantidad de microglía M1
significativamente mayor que CHF Nv. No se observaron diferencias estadísticamente
significativas entre CLF y CON. (figura 13).
*
*
0
2
4
6
8
10
12
14
16
18
CHF St CLF St CON St CHF Nv CLF Nv CON Nv
Mic
rogl
ía a
ctiv
ada
po
r ár
ea
(x0.
1mm
)
D.
A.
C.
B.

90
Figura 13. Microglía activada en el giro dentado. Se muestran las imágenes para aquellos grupos que obtuvieron
diferencias estadísticamente significativas durante el análisis. A. Microglía activada en una porción del giro dentado
ventral de CHF estresado. B. microglía activada de una porción de un giro dentado de un animal de CHF Nv. C.
Microglía activada para una porción de un giro dentado de un animal del grupo CLF Nv. D. Promedio de microglía
activada por área en los grupos experimentales. Se identificaron diferencias estadísticamente significativas entre los
grupos CHF estresado y Nv y el grupo CLF Nv con CHF Nv. Nota: el asterisco (*) hace referencia a un p<0.05.
Todas las imágenes fueron tomadas y procesadas en el Laboratorio de Neurociencia de la Universidad de los Andes.
CA3
En vista de que en CHF Nv no se contaron células de microglía M1, no fue posible el
ANOVA de dos vías, razón por la cual se optó por realizar análisis estadísticos por separado.
Para el caso de los animales naïve, sólo fue posible realizar la comparación de los animales
control con los animales CLF, pues no se tuvieron datos de los animales CHF.
Sujetos estresados
Ahora bien, una ANOVA de una vía comparó las cantidades de microglía por área en la
región anterior de CA3 en los animales estresados. Se identificaron diferencias estadísticamente
significativas entre los grupos experimentales (F[2, 12]=9.006, p<0.01). Las comparaciones de
post-hoc mostraron que el grupo CON inflamó significativamente menos que los otros dos
grupos (ver figura 14).
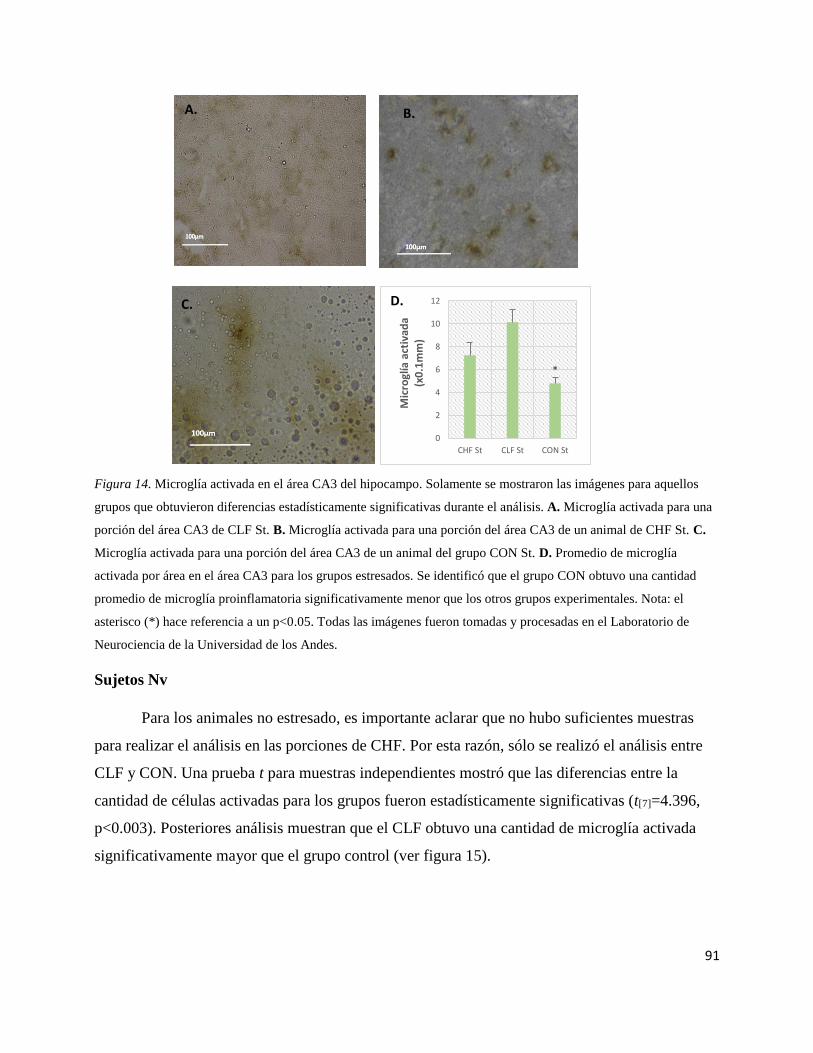
91
Figura 14. Microglía activada en el área CA3 del hipocampo. Solamente se mostraron las imágenes para aquellos
grupos que obtuvieron diferencias estadísticamente significativas durante el análisis. A. Microglía activada para una
porción del área CA3 de CLF St. B. Microglía activada para una porción del área CA3 de un animal de CHF St. C.
Microglía activada para una porción del área CA3 de un animal del grupo CON St. D. Promedio de microglía
activada por área en el área CA3 para los grupos estresados. Se identificó que el grupo CON obtuvo una cantidad
promedio de microglía proinflamatoria significativamente menor que los otros grupos experimentales. Nota: el
asterisco (*) hace referencia a un p<0.05. Todas las imágenes fueron tomadas y procesadas en el Laboratorio de
Neurociencia de la Universidad de los Andes.
Sujetos Nv
Para los animales no estresado, es importante aclarar que no hubo suficientes muestras
para realizar el análisis en las porciones de CHF. Por esta razón, sólo se realizó el análisis entre
CLF y CON. Una prueba t para muestras independientes mostró que las diferencias entre la
cantidad de células activadas para los grupos fueron estadísticamente significativas (t[7]=4.396,
p<0.003). Posteriores análisis muestran que el CLF obtuvo una cantidad de microglía activada
significativamente mayor que el grupo control (ver figura 15).
A. B.
C.
*
0
2
4
6
8
10
12
CHF St CLF St CON St
Mic
rogl
ía a
ctiv
ada
(x0
.1m
m)
D.

92
Figura 15. Microglía activada en el área CA3 de los animales Nv. Solamente se mostraron las imágenes para
aquellos grupos que obtuvieron diferencias estadísticamente significativas durante el análisis A. Microglía activada
en una porción del área CA3 de un animal perteneciente a CLF Nv. B. Microglía activada en una porción del área
CA3 de un animal perteneciente a CON Nv. C. Gráfico de promedio de microglía activada por área en el área CA3
para los grupos no estresados. Se identificó que el grupo CLF obtuvo una cantidad promedio de microglía
proinflamatoria significativamente mayor que el grupo control. Nota: el asterisco (*) hace referencia a un p<0.05.
Todas las imágenes fueron tomadas y procesadas en el Laboratorio de Neurociencia de la Universidad de los Andes.
Efectos del estrés sobre la inflamación
En cuanto a CHF, no se procedió a comparar los resultados entre el grupo estresado y no
estresado, pues el Nv no obtuvo un n suficiente de muestras para ser comparado con su
contraparte estresada.
En los grupos no estresados, la prueba t para muestras independientes mostró que las
medias de los grupos experimentales no fueron lo suficientemente diferentes como para afirmar
un efecto del estrés sobre la cantidad de microglía activada en esta área (t[4]=-0.198, p=0.853).
B.
*
0
2
4
6
8
10
12
14
CLF Nv CON Nv
Mic
rogl
ía a
ctiv
ada
(x0
.01
mm
)C.
A.

93
Finalmente, para el grupo control, una prueba t para muestras independientes mostró que
las diferencias entre la cantidad de microglía activada entre los grupos no fue estadísticamente
significativa (t[8]=0.081, p=0.311). Por lo tanto, no se puede afirmar un efecto del estrés sobre la
cantidad de microglía activada en esta área.
CA1
Al igual que en CA3, en CHF Nv no se contaron células de microglía activada, no fue posible el
ANOVA de dos vías, razón por la cual se optó por realizar análisis estadísticos por separado.
Para el caso de los animales naïve, sólo fue posible realizar la comparación de los animales
control con los animales CLF, pues no se tuvieron datos de los animales CHF.
Sujetos estresados
Para comparar la cantidad de microglía activada en el área CA1 del hipocampo de los
animales experimentales, se realizó una ANOVA de una vía. Los resultados indicaron que no
existen diferencias estadísticamente significativas entre los grupos experimentales (F[2, 5]=3.759,
p=0.101) (ver figura 18).
Sujetos Nv
Como no se contaron células de microglía activada en el grupo CHF Nv, se procedió a
hacer la comparación entre los grupos CLF y CON. Se compararon las medias de los grupos por
medio de una prueba t para muestras independientes. Del análisis se rescataron diferencias
estadísticamente significativas entre los grupos (t(5)=5.393, p<0.01). Comparaciones de post-hoc
mostraron que CLF obtuvo niveles de inflamación significativamente mayores que el grupo
control (ver figura 16).
Efectos del estrés sobre la inflamación
Al igual que CA3, no se realizó un análisis entre CHF estresada con su contraparte no
estresada, pues CHF Nv no contó con el n de muestras suficiente.
100µm
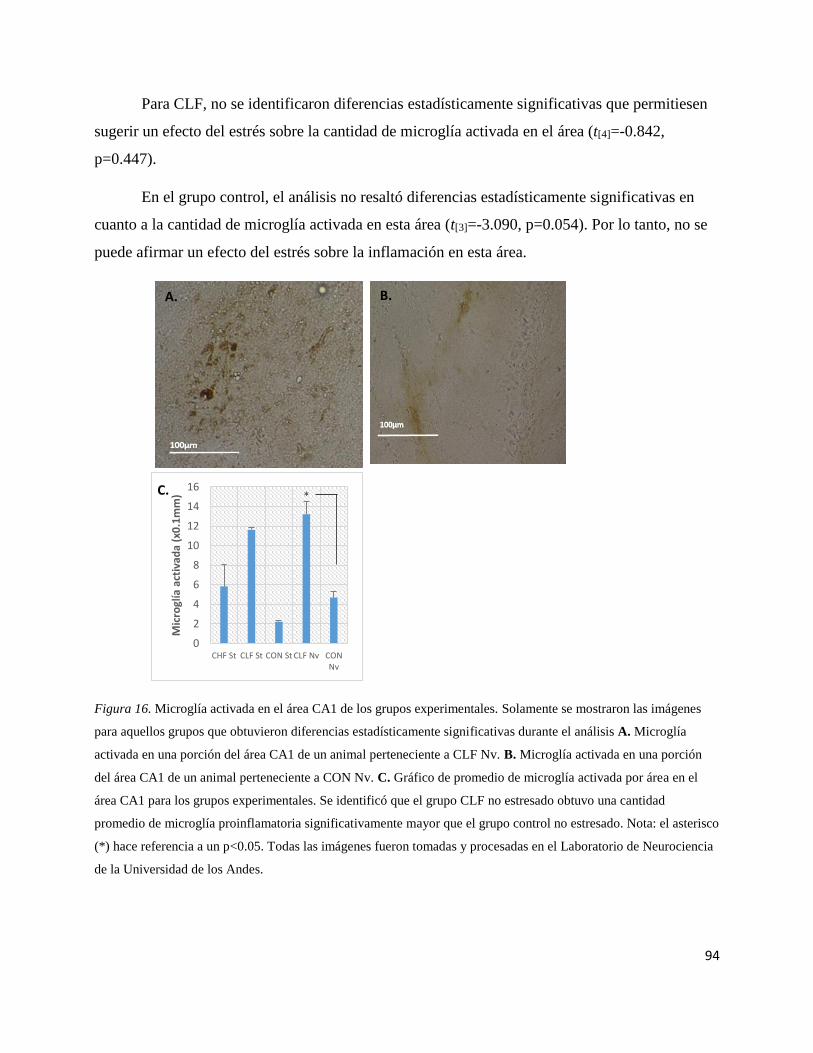
94
Para CLF, no se identificaron diferencias estadísticamente significativas que permitiesen
sugerir un efecto del estrés sobre la cantidad de microglía activada en el área (t[4]=-0.842,
p=0.447).
En el grupo control, el análisis no resaltó diferencias estadísticamente significativas en
cuanto a la cantidad de microglía activada en esta área (t[3]=-3.090, p=0.054). Por lo tanto, no se
puede afirmar un efecto del estrés sobre la inflamación en esta área.
Figura 16. Microglía activada en el área CA1 de los grupos experimentales. Solamente se mostraron las imágenes
para aquellos grupos que obtuvieron diferencias estadísticamente significativas durante el análisis A. Microglía
activada en una porción del área CA1 de un animal perteneciente a CLF Nv. B. Microglía activada en una porción
del área CA1 de un animal perteneciente a CON Nv. C. Gráfico de promedio de microglía activada por área en el
área CA1 para los grupos experimentales. Se identificó que el grupo CLF no estresado obtuvo una cantidad
promedio de microglía proinflamatoria significativamente mayor que el grupo control no estresado. Nota: el asterisco
(*) hace referencia a un p<0.05. Todas las imágenes fueron tomadas y procesadas en el Laboratorio de Neurociencia
de la Universidad de los Andes.
*
0
2
4
6
8
10
12
14
16
CHF St CLF St CON St CLF Nv CONNv
Mic
rogl
ía a
ctiv
ada
(x0
.1m
m)
A. B.
C.

95
CPU
Para los propósitos del experimento, fue de interés observar los niveles de inflamación de
algunos núcleos motores. En este caso, se destacó el cuerpo estriado.
Sujetos estresados
Se realizó una prueba de una vía que evaluó los niveles de inflamación en los grupos
estresados para esta estructura. Los resultados destacaron diferencias estadísticamente
significativas entre los grupos experimentales (F[2, 18]=3.721, p<0.05). Comparaciones de post-
hoc mostraron que el grupo CLF tuvo un nivel de inflamación significativamente mayor que CHF
(p<0.05) (ver figura 17).
Sujetos Nv
En los animales no estresados, no se observaron diferencias estadísticamente
significativas para las medias de inflamación en esta estructura (F[2, 8]=0.267, p=0.772).
Efectos del estrés sobre la inflamación
Para identificar las diferencias entre la inflamación de CHF estresado con su contraparte
no estresada, se realizó una prueba t para muestras independientes. De los resultados no se
rescataron diferencias estadísticamente significativas entre los niveles de inflamación entre los
grupos experimentales (t(8)=-0.975, p=0.354).
En CLF estresado y no estresado, tampoco se identificaron diferencias estadísticamente
significativas que destacaran efectos de la inducción del estrés sobre los niveles de inflamación
en esta área (t(14)=1.220, p=0.243).
Por último, en el grupo control no se observaron diferencias estadísticamente
significativas en cuanto a las medias de cantidad de microglía activada por área (t(4)=0.351,
p=0.743). Por lo tanto, no se puede afirmar que el estrés ejerza un efecto sobre la inflamación en
esta región.
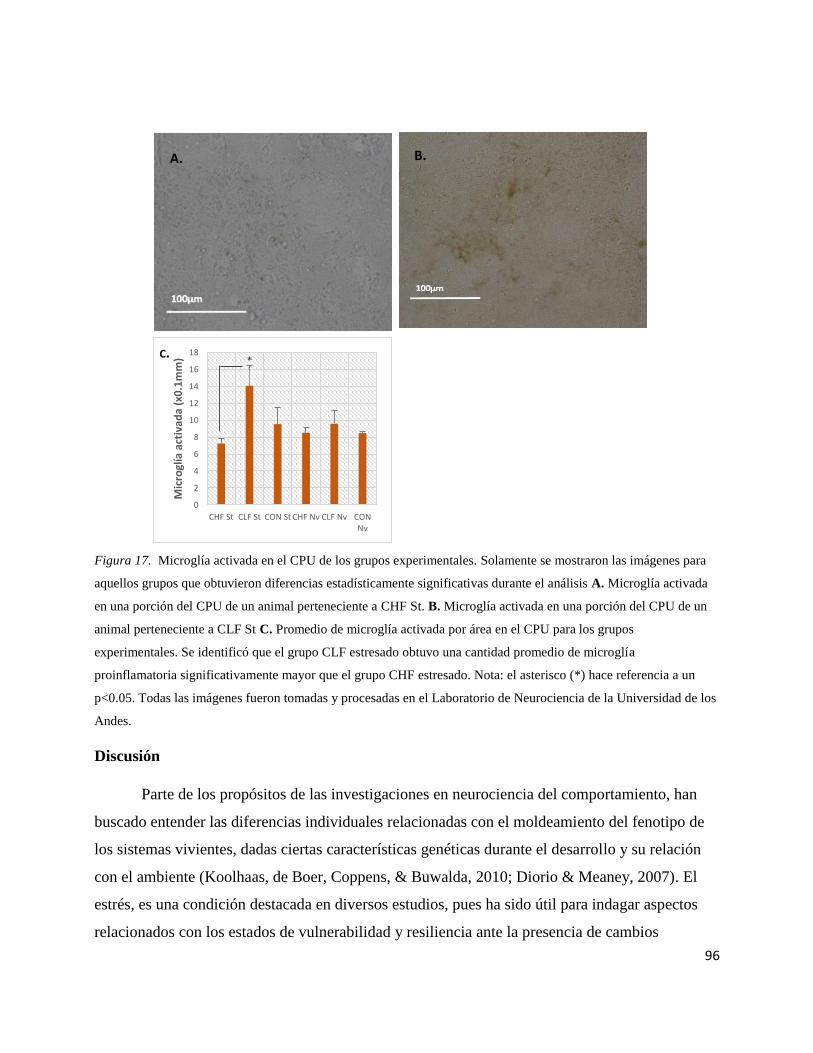
96
Figura 17. Microglía activada en el CPU de los grupos experimentales. Solamente se mostraron las imágenes para
aquellos grupos que obtuvieron diferencias estadísticamente significativas durante el análisis A. Microglía activada
en una porción del CPU de un animal perteneciente a CHF St. B. Microglía activada en una porción del CPU de un
animal perteneciente a CLF St C. Promedio de microglía activada por área en el CPU para los grupos
experimentales. Se identificó que el grupo CLF estresado obtuvo una cantidad promedio de microglía
proinflamatoria significativamente mayor que el grupo CHF estresado. Nota: el asterisco (*) hace referencia a un
p<0.05. Todas las imágenes fueron tomadas y procesadas en el Laboratorio de Neurociencia de la Universidad de los
Andes.
Discusión
Parte de los propósitos de las investigaciones en neurociencia del comportamiento, han
buscado entender las diferencias individuales relacionadas con el moldeamiento del fenotipo de
los sistemas vivientes, dadas ciertas características genéticas durante el desarrollo y su relación
con el ambiente (Koolhaas, de Boer, Coppens, & Buwalda, 2010; Diorio & Meaney, 2007). El
estrés, es una condición destacada en diversos estudios, pues ha sido útil para indagar aspectos
relacionados con los estados de vulnerabilidad y resiliencia ante la presencia de cambios
*
0
2
4
6
8
10
12
14
16
18
CHF St CLF St CON St CHF Nv CLF Nv CONNv
Mic
rogl
ía a
ctiv
ada
(x0
.1m
m)
A. B.
C.

97
medioambientales (Uchida et al., 2011; Campos, Fogaca, Aguiar, & Guimaraes, 2013). Hoy en
día, el uso de modelos animales ha tratado de refinar los procedimientos experimentales, con el
fin de ofrecer protocolos con mayor validez ecológica y facilitar el estudio de las conductas de
interés (Whitaker, Gilpin, & Edwards, 2014; Koolhaas, de Boer, Buwalda, & van, 2007).
Existen modelos animales que facilitan el entendimiento de la neurobiología del estrés.
Por ejemplo, estudios acerca del comportamiento de los ratones C57BL/J, los cuales incrementan
sus niveles de ansiedad y disminuyen su tiempo exploración después de la inducción de estrés
social cuando son vulnerables al estímulo (Krishnan et al., 2007); u otras investigaciones que han
aclarado diferencias individuales relacionadas con los estilos de afrontamiento en ratas durante la
adolescencia3 (Vidal, Buwalda, & Koolhaas, 2011); e incluso algunas delegan la caracterización
de ciertos rasgos entre linajes de ratas, sobre las diferencias en la exploración, el comportamiento
de escape y la desesperanza aprendida (Padilla, Barrett, Shumake & Gonzalez-Lima, 2009); o
que exploran las diferencias en cuanto al comportamiento ansioso en ratas (Rex, Voigt, Gustedt,
Beckett, & Fink, 2004), etc. Sirven como buenos puntos de partida para proponer nuevas
aproximaciones que aporten al conocimiento de temas relacionados.
Del linaje Wistar, fueron desarrolladas las cepas CHF y CLF (de Castro Gómes, Barroso
Silva, & Landeira-Fernandez, 2011), las cuales se caracterizan por poseer fenotipos distintos
relacionados con la respuesta de congelamiento. A lo largo de varias generaciones de cruzamiento,
demostraron diferencias de conectividad e integración de la información aversiva en el cerebro (de
Oliveira Galvão, de Castro Gómes, Masionette, & Landeira-Fernandez, 2011; de Castro Gómes et al.,
2011). En el presente estudio, se tuvo el propósito de determinar, de manera exploratoria, el
posible efecto de la inducción de estrés agudo sobre la inflamación y la generación de
comportamientos relacionados con la consolidación de las memorias emocionales y neutras y la
generación de otras conductas ansiosas que diferenciaron una cepa de la otra. Las suposiciones
sobre la posible relación entre la inflamación y la predisposición a comportamientos ansiosos en
los animales experimentales, fue evaluada a través de diversos protocolos experimentales, con los
3 Que han afirmado al final, menos estrategias por parte de las ratas Wistar en comparación con salvajes en tareas de
estrés social (Vidal, Buwalda, & Koolhaas, 2011).

98
cuales se relacionaron los patrones inflamatorios en el hipocampo con los comportamientos
subyacentes en los animales estresados y no estresados.
Se destacaron varias observaciones que vale la pena considerar; estas serán expuestas en
las secciones siguientes.
Consideraciones generales del experimento
Durante el protocolo de miedo condicionado, se observó que los comportamientos
provocados por el choque, efectivamente afectaron de manera desigual la habilidad de los
animales para establecer mecanismos de afrontamiento ante la situación estresora (Campos et al.,
2013). En concordancia con estudios pasados (de, V & Landeira-Fernandez, 2008a; Hassan,
Gomes, Pinton, Batista Teixeira da, & Landeira-Fernandez, 2013; Leon et al., 2013; de Oliveira
Galvão et al., 2011), se observó que los animales de CHF mantuvieron el congelamiento con la
misma intensidad durante todas las sesiones de la prueba (post, 24 horas y 6 días), permitiendo
sugerir que el recuerdo aversivo se consolidó fuertemente y se retuvo con la misma intensidad
cada vez que los animales fueron reexpuestos a la caja de condicionamiento. Por otro lado, el
grupo CLF tuvo la tendencia de disminuir su congelamiento durante las sesiones experimentales,
demostrando una dificultad en, o bien consolidar, o bien evocar (Genzel, Rossato, Jacobse,
Grieves, Spooner, Battaglia, Férnandez, & Morris, 2017) el recuerdo aversivo a mediano y largo
plazo.
El congelamiento es una respuesta de defensa producida por el animal una vez percibe
que no le es posible escapar de una amenaza subyacente (Steimer, 2002; Fanselow, 2000). No
obstante, cuando es evocado de manera exagerada, resulta poco adaptativo para la sobrevivencia
del sistema a largo plazo (McEwen & Gianaros, 2010). Después del protocolo, el congelamiento
fue el comportamiento más prevalente en CHF, hasta el punto de presentarse, incluso, en otros
contextos como el campo abierto. Cuando se compararon los resultados con los animales que no
recibieron el choque, se reafirmó su capacidad para evocar el recuerdo aversivo. En cuanto a
CLF, aunque también se observaron diferencias con su contraparte no estresada, se recalca que el
comportamiento fue disminuyendo de manera significativa después de la reexposición al
contexto.

99
Ahora bien, el campo abierto es un buen predictor de respuestas emocionales relacionadas
con la ansiedad y el estrés y también es utilizado para explorar los efectos de agentes ansiolíticos
y ansiogénicos (Prut & Belzung, 2003). Dentro del campo, usualmente se evalúan los niveles de
exploración de los animales dentro de un contexto abierto y desconocido, del cual no pueden
escapar (Choleris, Thomas, Kavaliers, & Prato, 2001; Seibenhener & Wooten, 2015).
En el experimento, se valoraron los niveles de deambulación y el tiempo y frecuencia de
los comportamientos de congelamiento, acicalamiento y estar parado en las patas traseras. Se
considera que estas conductas agregaron información importante sobre los niveles de ansiedad de
en los animales experimentales en el campo (Walsh & Cummins, 1976). Al finalizar la prueba, se
observaron resultados consistentes con aquellos reportados en el condicionamiento aversivo,
estos van a ser expuestos a continuación.
En resultados no reportados en el presente experimento, se observó que los animales
tuvieron una tendencia de deambular por los cuadrantes exteriores del campo abierto. El anterior
comportamiento, puede servir como un indicio de ansiedady también se puede considerar como
una conducta de supervivencia de la especie ante depredadores naturales (Choleris et al., 2001;
Seibenhener & Wooten, 2015). Aun así, dentro de las periferias, los animales se movilizaron de
manera diferentey los niveles de exploración revelaron diferencias estadísticamente significativas
en cuanto a la locomoción por el campo.
En primer lugar, CHF estresado exploró menos en comparación con el resto de los grupos
experimentales. Partiendo de este punto, se procede a considerar que después del estrés, los
animales del grupo no contaron con las habilidades para discriminar de manera adaptativa los
contextos parecidos a aquel del miedo condicionado (McEwen, 2007; Lissek, Rabin, McDowell,
Dvir, Bradford, Geraci, Pine & Grillon, 2009; Lissek et al., 2014)4 y por lo tanto se mantuvieron
congelados.
4 Se afirmó que también tuvieron una tendencia a sobregeneralizar el contexto condicionado a otros después del
entrenamiento. La generalización, es alcanzada cuando una respuesta de miedo adquirida a un estímulo particular es
transferida a otros estímulos (McLaren & Mackintosh, 2002). También se relaciona con la incapacidad para
discriminar el contexto peligroso del que no lo es después del aprendizaje (Lissek et al., 2009).

100
Por el contrario, los animales del grupo CLF no congelaron, pero evocaron otros
comportamientos, además de mostrar un aumento significativo en la locomoción por el campo. A
partir de este punto, se consideró que de acuerdo con lo observado tanto en la duración, como en
la frecuencia del acicalamiento y el pararse en dos patas, los animales de la cepa pudieron haber
tenido una tendencia al aumento de la ansiedad durante su tiempo en el campo. Sin embargo,
dicha ansiedad no se relacionaría con la inducción del choque, pues de los resultados no se
revelaron diferencias con su contraparte no estresada. Probablemente otros mecanismos de
afrontamiento diferentes del congelamiento característico de CHF, exactamente durante la
presencia de situaciones potencialmente amenazantes, podrían estar jugando un papel importante
en la conducta de la cepa (Wood & Bhatnagar, 2015). En párrafos posteriores se va a tratar el
tema con más detalle.
Durante la última fase del experimento, se tuvo la intención de evaluar la consolidación
de la memoria contextual y comparar los efectos del estrés entre los niveles de aprendizaje de los
grupos experimentales, haciendo uso de un protocolo de reconocimiento de objetos. Al finalizar
el análisis se observó que, para la familiarización, el grupo CHF estresado y no estresado obtuvo
el menor tiempo de exploración, en comparación con sus respectivos controles, para ambos
objetos. En CLF, por el contrario, se reportó un tiempo de exploración significativamente mayor,
el cual, nuevamente, no se diferenció del de su contraparte no estresada.
En cuanto a la fase de prueba, es importante resaltar que los animales estresados y no
estresados del grupo CHF presentaron una discriminación de los objetos en el campo, a pesar del
bajo tiempo de exploración. Es relevante destacar que en los animales de CLF (estresado y su
contraparte no estresada) se observó una incapacidad para consolidar el recuerdo, pues los
animales demostraron una tendencia por explorar los objetos familiar y novedoso de manera
indiscriminada. Por eso, se afirma que la incapacidad para consolidar de manera el recuerdo
relacionado con el objeto novedoso no depende de la inducción de estrés.
A continuación, se van a exponer las consideraciones complementarias sobre las dos
cepas, CHF y CLFy los posibles acercamientos desde la biología para la explicación del
comportamiento observado durante el experimento.
Consideraciones sobre CHF

101
De acuerdo con las últimas observaciones de la cepa, se procede a sugerir que las
alteraciones vinculadas con los comportamientos de congelamiento y el aumento de la
consolidación de la memoria emocional podrían verse mayormente evocados después de la inducción
del choque. Se propone que la cepa, a pesar de mostrar estados de vulnerabilidad frente al estímulo
estresor, podría no expresar patologías relacionas durante su vida si se encuentra en presencia de
ambientes no hostiles. Sin embargo, después de verse afectadas, se comienzan a destacar
comportamientos ansiosos, los cuales son evidentes en las sesiones del experimento. De los
resultados, se concluye que los animales comienzan a padecer estados relacionados con una
desregulación en la respuesta del eje HPA (Belda et al., 2008), un robustecimiento del
funcionamiento del circuito hipotálamo-amígdala-SGP para la iniciación de la respuesta de miedo
(de, V & Landeira-Fernandez, 2008a; Shvil, Rusch, Sullivan, & Neira, 2013; Patel, Spreng, Shin, &
Girard, 2012; McGaugh, 2000; Kim et al., 2013) y una sobreactividad de los núcleos hipocampales que
también la consolidación del recuerdo aversivo (Ballesteros, de Oliveira, Maisonette, & Landeira-
Fernandez, 2014; Fanselow & Dong, 2010; Belda et al., 2008), más no contextualy una consecuente
proinflamación derivada de la actividad de la microglía M1- la cual va a ser tratada en párrafos
posteriores-.
En primer lugar, se afirma que la respuesta de congelamiento y la reactividad frente al estrés en la
cepa, se relaciona con una estimulación paralela de SGP, una estructura relacionada con la integración de
la información aversiva para evocar respuestas inmediatas de miedo (congelamiento) y defensa (escape)
(Kincheski, Mota-Ortiz, Pavesi, Canteras, & Carobrez, 2012; Benarroch, 2012). SGP posee proyecciones
bidireccionales con el hipotálamo, la corteza y la amígdala desde su porción dorsal (Fendt & Fanselow,
1999; Graeff et al., 1993; Vianna & Brandao, 2003); y con el hipocampo en su porción ventral, que se
encarga de establecer un aprendizaje aversivo (Kim, Rison & Fanselow, 1993; Vianna, Graeff, Landeira-
Fernandez, & Brandao, 2001). Se ha reportado que la estimulación de SGPd, efectivamente aumenta el
tiempo de congelamiento, junto con otros comportamientos anticipatorios de ansiedad en CHF, mas no
en CLF (de Oliveira Galvão et al., 2011).
La propensión al miedo observada en el experimento ha sido corroborada a través de otras
evaluaciones en tareas de condicionamiento aversivo. Cuando esta región es estimulada eléctrica
o químicamente, se afirma que el aprendizaje asociativo se adquiere rápidamente y no es sencillo

102
realizar un proceso de extinción (Kincheski et al., 2012; Motta et al., 2017; Di, Mana, Jacobs, &
Phillips, 1987). Al mismo tiempo, se han registrado comportamientos relacionados con la
propensión al estrés, el aumento del aprendizaje emocional (Motta et al., 2017; Kincheski et al.,
2012), la disminución de la actividad locomotora (de Oliveira Galvão et al., 2011) y otros, como
el aumento de las presión arterial, la disnea, la piloerección, la hiperreactividad y la presencia de
un comportamiento de alerta (de Oliveira Galvão et al., 2011; Vianna et al., 2001). Las lesiones,
por el contrario, tienden a disminuir el congelamiento (Vianna et al., 2001), provocando problemas de
aprendizaje asociativo relacionado con el miedo, el cual fue un comportamiento recurrente en las ratas
CLF.
Desde SGP, también se destaca la liberación de sustancia P (SP)5 (Berton et al., 2007),
reconocida por ser uno de los precursores de procesos fisiológicos y patofisiológicos involucrados en la
regulación y el incremento del estrés y la ansiedad (Ebner & Singewald, 2006). Otros estudios afirman
que la estimulación de SGP puede iniciar de un proceso de inflamación neurogénica después de liberar
SP en el sistema (Corrigan, Vink, & Turner, 2016). La inmunoreactividad de esta sustancia se observa de
5 horas hasta incluso 7 días después accidentes cerebrovasculares inducidos (Corrigan et al., 2016). Este
proceso podría verse sobrelapado con la presencia de la microglía activada (ver capítulo de microglía),
pues en previos estudios se ha corroborado que la expresión del ARNm que codifica para el receptor
NK-1 (receptor de SP) en células microgliales (Rasley, Bost, Olson, Miller, & Marriott, 2002) tiende a
aumentar después del incremento de IL-4 e IFN-γ (Marriott & Bost, 2000). Otros, han observado que la
inducción del péptido (SP) es capaz promover los efectos proinflamatorios y citotóxicos de la microglía
M1 (explicada en el capítulo de introducción a la microglía), hasta por 24 horas después de su inducción
in vitro (Zhu, Qu, Lu, & Zhang, 2014).
Ahora bien, con relación a la respuesta de estrés, nuevamente se afirma que hubo una reactividad
más fuerte de los animales experimentales frente al estímulo, en comparación con sus respectivos
controles. En estudios antecedentes se ha reportado que la concentración corticosterona plasmática basal
y post choque son más altas en CHF que en CLF (Leon et al., 2013). El aumento de la corticosterona
suele afectar la respuesta del sistema frente al estrés, pues dicho aumento ha demostrado causar efectos
5 Un péptido de orígenes neuronales, cuyo receptor se encuentra de manera abundante en todo el sistema ya que es secretado tanto
por neuronas como de células gliales (O'Connor et al., 2004; Marriott & Bost, 2001).

103
de saturación sobre los receptores de glucocorticoides- y posteriormente una disminución de estos
mismos (Sapolsky et al., 1984)-, que los incapacita para realizar una realimentación negativa efectiva del
eje HPA (Kanter et al., 2001; Belda et al., 2008; Harris et al., 2013). Por lo tanto, se observa un
consecuente incremento significativo de liberación glutamatérgica derivada de la estimulación,
que causa impedimentos en el LTP dependiente de GD y CA36 (Alfarez, Joels & Krugers, 2003;
McEwen & Sapolsky, 1995), muerte por excitotoxicidad (Hu et al., 1997; Rosso, Crowley, Silveri,
Rauch & Jensen, 2017), reducción de la neurogénesis (Brummelte & Gales, 2010; Cameron &
Gould, 1994; Schoenfeld & Gould, 2012) -relacionada con el aumento de la ansiedad (Bergami,
Berninger & Canossa, 2009; Revest, Dupret, Koehl, Funk-Reiter, Grosjean & Abrous, 2009)- y
disminución en plasticidad sináptica de GD, CA3 y sus conexiones funcionales con CA1 (Alfarez et al.,
2009; Joels, 2008). Otros estudios sobre la administración con agonistas de esta sustancia, como la
dexametasona, igualmente han demostrado una mejora del recuerdo aversivo después del
condicionamiento de miedo (Roozendaal & McGaugh, 1996).
Directamente relacionado al tema anterior, se ha observado que la inducción de estrés
muy fuerte o crónico, también produce un incremento de la microglía proinflamatoria y
citotóxica, la cual comienza a acumularse sobre las áreas de concentración de glucocorticoides y
actúa en detrimento de la función normal del cerebro (Monje, Toda, & Palmer, 2003; Ekdahl,
Claasen, Bonde, Kokaia, & Lindvall, 2003). Aunque no se esperaba encontrar niveles mayores de
proinflamación en CLF, en CHF se identificó un número incrementado de microglía de fenotipo
M1 en el hipocampo que difirió significativamente con su contraparte no estresada y su
respectivo control. De acuerdo a la hipótesis propuesta, se observó una consistencia entre los resultados
reportados y la literatura correspondiente a los efectos del aumento de la inflamación hipocampal
después de la inducción de estrés (Jinno, Fleischer, Eckel, Schmidt, & Kosaka, 2007; Munhoz,
García-Bueno, Madrigal, Lepsh, Scavone & Leza, 2008).
6 dos estructuras altamente plásticas, cruciales para la adquisición y consolidación de la memoria relacionada con el
espacio y el contexto (Fellini, Florian, Courtney & Roullet, 2009; Florian & Roullet, 2004) y para la diferenciación de patrones
y detección de novedad (Kesner et al., 2004).

104
Se considera que la microglía, efectivamente fue estimulada a través del aumento de la
liberación glutamatérgica, noradrenérgica, de corticosterona y de otros factores derivados del
aumento de la degradación de proteínas y aminoácidos en las células de estructura (Hardingham, 2009;
Hassan et al., 2013; (Sapolsky, 1985; May et al., 1996; McEwen & Sapolsky, 1995; Murugan et
al., 2013; Tanaka et al., 2006; Wohleb et al., 2011), para ejercer sus efectos proinflamatorios y
citotóxicos. La anterior información se relaciona con los resultados obtenidos de otros experimentos,
que sugieren que los animales de CHF mantienen una mayor concentración de especies reactivas a
oxigeno (catalaza y glutaiona peroxidasa), indicadoras de aumento de radicales libres, en la corteza, el
hipocampo y el cerebelo (Hassan et al., 2013). El aumento de estas sustancias ha demostrado ser un
producto del mecanismo de defensa efectuado por la microglía en contra de patrones de muerte neuronal
o aparición de microorganismos potencialmente amenazantes (Hu, Sheng, Peterson & Chao, 1995;
Rock, Gekker, Hu, Sheng, Cheeran, Lokesgard & Peterson, 2004). En general, se afirma que la
reactividad frente al estrés causó en el hipocampo un ciclo de proinflamación derivado del mismo
estímulo, que aumentó las interferencias en los procesos de consolidación del recuerdo aversivo y
contextual (Marchand, Barbelivien, Seillier, Herbeaux, Sarrieau & Majchrzak, 2007).
Este problema ha sido de interés, debido a las aproximaciones que se han reportado acerca
de la presencia de neuroinflamación y aumento de radicales libres durante y después de estímulos
emocionalmente significativos (para más información vaya a capítulo de efectos del estrés sobre
la microglía); además de sus efectos sobre el desarrollo de patologías relacionadas, como la
depresión (Walker, Kavelaars, Heijnen, & Dantzer, 2014; Shibata & Suzuki, 2017; Hashioka et
al., 2007), la bipolaridad (Jakobsson et al., 2015; Knijff et al., 2007), la esquizofrenia (Monji,
Kato, & Kanba, 2009; Monji et al., 2013), los trastornos de ansiedad (Shigemori, Sakai, Takumi,
Itoh, & Suzuki, 2015), el trastorno de estrés posttraumático (PSTD) (Acosta et al., 2013; Sun et
al., 2016), las enfermedades neurodegenerativas (Cunningham, 2013; von, Eugenin-von, &
Eugenin, 2015; Gleichgerrcht, Ibanez, Roca, Torralva, & Manes, 2010), entre otros.
Consideraciones sobre CLF
La interesante ausencia del efecto del choque y la similaridad de los resultados con y sin
inducción de estrés en CLF, sugieren que el comportamiento podría estar relacionado con
impedimentos funcionales de otra naturaleza. Partiendo de lo anterior, se procede a sugerir que
las dificultades en el aprendizaje emocional y contextual durante el experimento se derivan de un

105
incremento en los niveles de neuroinflamación y se cree que la desregulación de los procesos
inflamatorios presentes en la cepa (Mantovani et al., 2002; Hoogman, Aarts, Zwiers, Slaats-
Willemse, Onnik, Cools, Kan, Buitelaar & Franke, 2011), son probables indicativos de otros
tipos de vulnerabilidad, los cuales, en concordancia con el proceso de fenotipaje, podrían ser de
naturaleza genética (Vazdarjanova et al., 2002; Rosi et al., 2009; Belarbi et al., 2012). Sin
embargo, este tema no ha sido suficientemente estudiado y se recomienda explorarlo en estudios
futuros. Aun así, se va a realizar un breve un acercamiento a los posibles procesos que podrían
estar jugando un papel en el desarrollo de estos animales.
Motricidad
Ahora bien, resulta conveniente comenzar por abordar el tema relacionado con el aumento
de la motricidad en la cepa. Como se observó en el presente experimento, los animales se
caracterizaron por mantenerse en constante movimiento a lo largo de las pruebas
comportamentales. Se identificó un incremento significativamente mayor que su respectivo
control en el acicalamiento, en el pararse en las patas traseras y en la distancia recorrida por el
campo abierto. Cuando se relacionó este patrón comportamental con los resultados obtenidos en
el condicionamiento aversivo y la prueba de reconocimiento de objetos, se consideraron dos
puntos: primero, que el aumento de la motricidad si puede ser considerado como un indicio de
aumento de ansiedad; y en segundo lugar, que el comportamiento ansioso es de corto plazo y solo
se expresa en presencia o poco después del estrés.
Entre los comportamientos que se evaluaron, se encuentra en primer lugar, el pararse en
las patas traseras, o mejor conocido como “rearing”. Este es un comportamiento exploratorio
natural de la especie, inducido en contextos novedosos (Lever, Burton, & O'Keefe, 2006). Su
aparición, juega un rol importante en el aprendizaje contextual dependiente del hipocampo (Lever
et al., 2006), sin embargo en contextos experimentales afirma que o bien, el aumento de su
tiempo y frecuencia es indicio de estrés (Borta & Schwarting, 2005), o bien su disminución
significativa es indicio de estrés (Costall, Jones, Kelly, Naylor, & Tomkins, 1989). Aunque aún
no es muy consistente su caracterización en contextos relacionados en la literatura (Seibenhener
& Wooten, 2015), en el presente experimento se sugiere que es consecuencia de un aumento

106
general de la actividad del sistema cuando el animal se encuentra en presencia contextos
potencialmente peligrosos.
Lo anterior se afirma en concordancia con el aumento significativo del acicalamiento.
Aunque en condiciones normales se encuentra involucrado en el mantenimiento de la higiene y
otros procesos que incluyen la termorregulación, la interacción social y también estados de
activación fisiológica, se ha reportado que, en condiciones estresantes, puede llegar a ser
patológico (Kalueff, Stewart, Song, Berridge, Graybiel & Fentress , 2016). En estudios previos,
se ha observado que la inducción de estrés, muestra incrementos en su frecuencia y duración,
además de evidenciar patrones erráticos de limpieza (Pires, Tufik, & Andersen, 2013). En CLF,
el acicalamiento se destacó, tanto en duración como en frecuencia durante toda la sesión
experimental. Por lo tanto, nuevamente se procede a relacionar su intensidad con un aumento en
la activación del sistema.
Relacionado con el tema anterior, en la literatura se ha destacado que las estructuras
pertenecientes a los núcleos que medían el control motor y su secuenciación7, mantienen
conexiones funcionales con algunas regiones encargadas del procesamiento emocional, como la
amígdala y el hipotálamo (Kalueff et al., 2016). De la amígdala, se ha reportado que la
estimulación glutamatérgica en MeA promueve el acicalamiento repetitivo, mientras que la
GABAérgica impide lo anterior (Hong et al., 2014). Resultados similares se han observado
durante estimulación de CeA con agonistas orexinérgicos (ORX-A), agonistas de NMDA y
antagonistas del receptor GABAa (Alo, Avolio, Mele, Di, & Canonaco, 2015).
Se sabe que la amígdala mantiene conexiones excitatorias con el hipotálamo, en especial
el NPV, uno de los núcleos encargados de la producción de CRF (Heinrichs, Menzaghi, Merlo,
Britton, & Koob, 1995; Herman & Cullinan, 1997). Se ha observado que ambas estructuras se
encuentran en estado de activación durante la secuenciación locomotora- junto con el cuerpo
estriado y los ganglios basales- en momentos de activación emocional (Kalueff et al., 2016).
Existe la posibilidad de que el aumento del funcionamiento de estas estructuras esté modulando
7 su circuito incorpora el funcionamiento los ganglios basales, el cuerpo estriado, los globos pálidos, la substancia nigra, el núcleo
acumbes y el núcleo subtalámico (Kalueff et al., 2016; Hong, Kim, & Anderson, 2014).

107
los patrones aumentados de motricidad en la cepa. Por lo que es recomendable explorar los
mecanismos relacionados a estos circuitos en contextos relacionados.
Consolidación de la memoria
En relación a los comportamientos observados en la cepa durante el condicionamiento
aversivo, se quiso destacar el papel de la amígdala, pues se considera que la de pérdida
progresiva del recuerdo condicionado por parte de la cepa, podría derivarse de que esta estructura
no está estableciendo conexiones efectivas sobre el hipocampo (McGaugh & Roozendaal, 2002;
Kim et al., 2005). En cuanto al reconocimiento de objetos, se considera que la inflamación
también actúa en detrimento del funcionamiento de la estructura.
En estudios antecedentes sobre el funcionamiento de la amígdala en CHF y CLF, se reportó
que las lesiones en los núcleos central (CeA) y basolateral (BLA) disminuyeron el congelamiento de
manera diferenciada en cada grupo. Si bien CHF tuvo la tendencia de reducir significativamente este
comportamiento después del condicionamiento, CLF no reveló una disminución significativa en el grupo
lesionado en comparación con el no lesionado (de, V & Landeira-Fernandez, 2008a). Podría resultar
interesante establecer que la disminución del congelamiento de CLF en comparación con CHF se
relaciona con el aumento en los niveles de proinflamación en la estructura, pues la microglía ejerce
acciones inhibitorias sobre el establecimiento del LTP y la plasticidad (Kelly, Lynch, Vereker,
Nolan, Queenlan, Whittaker, O’Neil & Lynch, 2001; Vereker, O’Donnell & Lynch, 2000). De
todos modos, se espera que a futuro se resuelvan los cuestionamientos relacionados con este
punto.
Adicionalmente, los altos niveles de inflamación también podrían estar afectando la
consolidación del recuerdo durante el sueño, pues el input excitatorio de la amígdala hacia las
cortezas hipocampales después del estrés (McGaugh, 2000), no facilitaría la producción de LTP y
la consolidación del recuerdo emocionales en las zonas ventrales de la estructura (hipocampo)
(McGaugh, 2000; Sapolsky, Krey, & McEwen, 1984). El robustecimiento de los recuerdos
aversivos se han observado durante el sueño por medio del incremento de las oscilaciones theta
desde BLA durante la activación emocional (Pelletier & Pare, 2004), que, al sincronizarse con el
hipocampo y la región infralímbica de corteza prefrontal durante el sueño REM (Hutchison &
Rathore, 2015)- a la vez que disminuyen el tiempo total de ondas de sueño lento (SOL) (Datta &

108
O'Malley, 2013)-. Información adicional muestra que, incluso 24 horas después de un
condicionamiento aversivo en ratas Wistar, existe un mejoramiento en la sincronía de frecuencias
theta entre dichas estructuras, especialmente entre BLA y el hipocampo (Narayanan et al., 2007), pero
una menor entre BLA y la corteza prefrontal medial (CPFm) (Lesting et al., 2011; Popa, Duvarci,
Popescu, Lena, & Pare, 2010).
De igual importancia, en vista de que la microglía se encuentra distribuida por todo el
hipocampo, incluso sin la necesidad de la presencia de estrés - con una mayor concentración en el
borde subgranular y granular del giro dentado (GD), CA1 y la porción dorsal de CA3- (Jinno,
Fleischer, Eckel, Schmidt, & Kosaka, 2007) y que los procesos de inflamación agudos tienden a
disminuir los procesos de plasticidad y LTP (Rodriguez, Davies, Errington, Verkhrasky, Bliss &
Stewart, 2007; Vazdarjanova, McNaughton, Barnes, Worley, & Guzowski, 2002), se sugiere que
las ratas de la cepa CLF tienen deficiencias cognitivas que afectan su desempeño a largo plazo
(Chesnokova et al., 2016; Pearson-Leary et al., 2017).
Acercamientos a la epigenética
Aunque se expresa que los resultados obtenidos en CLF aún no son conclusivos y
tampoco son suficientes para explicar la biología y la caracterización de los efectos observados
en el presente experimento, pues ni las variables comportamentales ni la inflamación pudieron
establecer un acercamiento preciso a la naturaleza de su comportamiento, se procede a establecer
otras observaciones, desde la evidencia, sobre los posibles mecanismos que podrían verse
involucrados en el aumento de la proinflamación a lo largo de las generaciones.
En primer lugar, se parte de que las secuencias genéticas relacionadas con el
funcionamiento de los sistemas pueden permanecer inalteradas durante toda la vida, sin embargo,
es muy probable que varios patrones de cambio regulen su expresión para adaptarse los cambios
del ambiente (McGowan, Sasaki, D’Alessio, Dymov, Labonté, Szyf, Turecki & Meaney, 2009;
Roth, Lubin, Funk, Sweatt, 2009). Los genes son modificados por medio de varios procesos,
conocidos como metilación de la citocina, modificación de las histonas, remodelación de la
cromatina y otros mecanismos relacionados con el funcionamiento del ARN (Jaenisch & Bird,
2003).

109
La adición del grupo metil a la citosina por medio de la ADN-metiltransferaza (Houvert,
Vallette & Cartron, 2009), facilita los procesos de inhibición del reconocimiento del ADN por
algunas proteínas, o facilitación de las conexiones entre estos (Prokhortchouk & Defossez, 2008).
Se ha observado que varias regiones de los dinucleotidos CpG (citosina-unión de fosfodiester-
guanina) a lo largo del genoma de los mamíferos son metilados (Lister et al., 2009), sin embargo,
se ha destacado que las células cerebrales también tienen otros complementos de citosinas
metiladas observadas en su contexto (Ladd-Acosta, Pevsner, Sabunciyan, Yolken, Webster,
Dinkins, Callinan, Fan Potash & Feinberg, 2007).
Específicamente en el cerebro, los mecanismos de metilación y modificación de histonas
se encuentran ligados con la expresión y regulación dinámica de los genes dependientes de
cambios medioambientales (Tsnakova, Renthal, Kumar & Nestler, 2007). Estos procesos, están
sujetos a cambios funcionales que, consecuentemente pueden formar cambios adaptativos,
necesarios para la sobrevivencia, incluso por generaciones (Lim & Brunet, 2013; Uchida et al.,
2011). Estudios que observan cambios en la conducta alimentaria y el metabolismo de los
alimentos (Wu, Suzuki, 2006; Dunn & Bale, 2009), la acción frente a la presencia de virus
(Rechavi, Minevich & Hobert, 2011) y la presencia de estrés ambiental (Franklin, Russig, Weiss,
Graff, Linder, Michalon, Vizi & Mansuy, 2010; Weiss, Franklin & Mansuy, 2011), corroboran
esta observación.
Específicamente en la microglía, es interesante observar que algunos tipos de ARN no
codificante, ARNmi (microARN), han sido identificados como promotores de procesos
proinflamatorios y antiinflamatorios a largo plazo (Garden, 2013). En estudios se ha destacado
que, por ejemplo, el aumento de los patrones de expresión de miARN-155 -conocido por su papel
en la estimulación de la secreción de IL-12 en las células CD, para la expresión de IFN- γ en
células NK durante la respuesta inmune y otros procesos de apoptosis (Lu, Huang, Zhang,
Roensch, Cao, Nakayama, Blazar, Zeng & Zhou, 2012)-, por medio de la estimulación de los
recetores de tipo toll 4 o mediante la administración con LPS, suprimen en la microglía al
supresor de la proteína supresora de IL-1 (SOCS-1) y aumenta los niveles de iNOs (Cardoso,

110
Guerdes, Pereira de Almeida & Pedroso de Lima, 2012)8. Otros estudios también han relacionado
la regulación de la expresión de iNOS, con la acción de miARN-155 después de la
administración de LPS (Wang et al., 2009).
Otros ejemplos relacionados han revelado que cambios en el ambiente celular pueden
afectar la expresión de genes antiinflamatorios (Garden, 2013). Se ha destacado que cuando las
células microgliales son expuestas a deprivación de oxígeno y glucosa, existen mayores niveles
de concentración de IL-1, iNOS y en especial TNF-α, en contraposición con la disminución de la
expresión de miR-181c en el hipocampo (Zhang, Dong, Li, Hong & Wei, 2012). También se ha
destacado que la sobreexpresión de el miR-424 que se encarga de detener la respuesta
proinflamatoria y citotóxica en áreas afectadas por isquemia cerebral, tiende a suprimir del Iba-1
y la disminución de la producción de TNF- α (Zhao et al., 2013).
La regulación molecular de los patrones de neuroinflamación es dependiente de
mecanismos epigenéticos afectan el desarrollo funcional y anatómico a largo plazo (Garden,
2013) y los cambios de dichos patrones han sugerido influencias hereditables de los sistemas
afectados (Kendler, Gardner & Presscott, 2006; Smith et al., 2011). Lo anterior se ha relacionado
con el proceso de selección artificial de las cepas efectuado en el Laboratorio de Neurociencia de
la PUC-Rio. De manera efectiva, se ha encargado de caracterizar y conservar a lo largo del
tiempo los fenotipos comportamentales característicos de CHF y CLF, sin embargo, aún existen
vacíos en el conocimiento sobre la naturaleza de estos animales y sus aportes a la ciencia. Para
los animales de CHF, se concluye que efectivamente representa un modelo comportamental
valido para el estudio de patologías relacionadas con la ansiedad el pánico y el estrés, ya que se
considera evidente su fenotipo vulnerable. En CLF, por el contrario, se observó que las ratas,
además de tener problemas cognitivos relacionados con la consolidación de la memoria, también
demostraron tener un aumento significativo de los niveles de inflamación, los cuales también
fueron relacionados con la ausencia de consolidación. Aunque se cree que las diferencias
8 Las iNOS son conocidas como mediadoras del control intrínseco de la excitabilidad neuronal y la plasticidad
(Steinert, Chernova & Forsythe, 2010; Steinert, Robinson, Tong, Haustein, Kopp-Scheinpflug & Forsythe, 2011). Se
ha reportado que en bajas concentraciones puede ejercer papeles neuroprotectores por medio de la mediación de la
señalización fisiológica (Brew & Forsythe, 1995) sin embargo, en altas medía otras acciones inflamatorias y
neurotóxicas durante la respuesta inmune (Wang et al., 2009).

111
obtenidas a partir del proceso de fenotipaje son un producto “funcional” de los cambios en la
expresión de algunos genes relacionados a los defectos desde las primeras generaciones, la
información no es suficiente para como afirmar de manera fehaciente las conjeturas relacionadas
durante el experimento con relación a CLF. Se espera que en el futuro se aclaren muchos de los
cuestionamientos obtenidos a partir del fenotipo tan particular de estos animales.
Reference List
Acosta, S. A., Diamond, D. M., Wolfe, S., Tajiri, N., Shinozuka, K., Ishikawa, H. et al. (2013). Influence
of post-traumatic stress disorder on neuroinflammation and cell proliferation in a rat model of
traumatic brain injury. PLoS.One., 8, e81585.
Agster, K. L. & Burwell, R. D. (2009). Cortical efferents of the perirhinal, postrhinal, and entorhinal
cortices of the rat. Hippocampus, 19, 1159-1186.
Alfarez, D. N., Joels, M., & Krugers, H. J. (2003). Chronic umpredictable stress impairs long-term potentiation
in rat hipocampal CA1 area and dentate gyrus in vitro. European Journal of Nouroscience., 17, 1928-34.
Alfarez, D. N., De, S. A., Velzing, E. H., Bracey, E., Joels, M., Edwards, F. A. et al. (2009).
Corticosterone reduces dendritic complexity in developing hippocampal CA1 neurons.
Hippocampus, 19, 828-836.
Alo, R., Avolio, E., Mele, M., Di Vito, A., Canonaco, M. (2015). Central amygdalar nucleus treated with
orexin neuropeptides evoke differing feeding and grooming responses in the hamster. J Neurol
Sci., 351, 46-51.
Aloisi, F. (2001). Immune function of microglia. Glia, 36, 165-179.
Aloisi, F., Penna, G., Cerase, J., Menendez, I. B., & Adorini, L. (1997). IL-12 production by central
nervous system microglia is inhibited by astrocytes. J.Immunol., 159, 1604-1612.
Alvarez-Buylla, A. & Lim, D. A. (2004). For the long run: maintaining germinal niches in the adult
brain. Neuron, 41, 683-686.

112
Amaral, D. G., Scharfman, H. E., & Lavenex, P. (2007). The dentate gyrus: fundamental
neuroanatomical organization (dentate gyrus for dummies). Prog.Brain Res., 163, 3-22.
Ambrogini, P., Lattanzi, D., Ciuffoli, S., Agostini, D., Bertini, L., Stocchi, V. et al. (2004). Morpho-
functional characterization of neuronal cells at different stages of maturation in granule cell layer
of adult rat dentate gyrus. Brain Res., 1017, 21-31.
Andlin-Sobocki, P. & Wittchen, H. U. (2005). Cost of anxiety disorders in Europe. Eur.J.Neurol., 12
Suppl 1, 39-44.
Bain, M. J., Dwyer, S. M., & Rusak, B. (2004). Restraint stress affects hippocampal cell proliferation
differently in rats and mice. Neurosci.Lett., 368, 7-10.
Ballesteros, C. I., de Oliveira, G. B., Maisonette, S., & Landeira-Fernandez, J. (2014). Effect of dorsal
and ventral hippocampal lesions on contextual fear conditioning and unconditioned defensive
behavior induced by electrical stimulation of the dorsal periaqueductal gray. PLoS.One., 9,
e83342.
Bandelow, B. & Michaelis, S. (2015). Epidemiology of anxiety disorders in the 21st century.
Dialogues.Clin.Neurosci., 17, 327-335.
Barnay, T. (2016). Health, work and working conditions: a review of the European economic literature.
Eur.J.Health Econ., 17, 693-709.
Barria, A., Muller, D., Derkach, V., Griffith, L. C., & Soderling, T. R. (1997). Regulatory
phosphorylation of AMPA-type glutamate receptors by CaM-KII during long-term potentiation.
Science, 276, 2042-2045.
Barrientos, R. M., Thompson, V. M., Kitt, M. M., Amat, J., Hale, M. W., Frank, M. G. et al. (2015).
Greater glucocorticoid receptor activation in hippocampus of aged rats sensitizes microglia.
Neurobiol.Aging, 36, 1483-1495.
Behbehani, M. M. (1995). Functional characteristics of the midbrain periaqueductal gray.
Prog.Neurobiol., 46, 575-605.

113
Belarbi, K., Arellano, C., Ferguson, R., Jopson, T., & Rosi, S. (2012). Chronic neuroinflammation
impacts the recruitment of adult-born neurons into behaviorally relevant hippocampal networks.
Brain Behav.Immun., 26, 18-23.
Belda, X., Rotllant, D., Fuentes, S., Delgado, R., Nadal, R., & Armario, A. (2008). Exposure to severe
stressors causes long-lasting dysregulation of resting and stress-induced activation of the
hypothalamic-pituitary-adrenal axis. Ann.N.Y.Acad.Sci., 1148, 165-173.
Belsky, J. & Pluess, M. (2009). Beyond diathesis stress: differential susceptibility to environmental
influences. Psychol.Bull., 135, 885-908.
Belujon, P. & Grace, A. A. (2015). Regulation of dopamine system responsivity and its adaptive and
pathological response to stress. Proc.Biol.Sci., 282.
Benarroch, E. E. (2012). Periaqueductal gray: an interface for behavioral control. Neurology, 78, 210-
217.
Bergami, M., Berninger, B., & Canossa, M., (2009). Conditional deletion of TrkB alters adult
hipocampal neurogénesis and axiety-related behavior. Communicative & Integrative Biology., 2,
14-16.
Bernabeu, R., Bevilaqua, L., Ardenghi, P., Bromberg, E., Schmitz, P., Bianchin, M. et al. (1997).
Involvement of hippocampal cAMP/cAMP-dependent protein kinase signaling pathways in a late
memory consolidation phase of aversively motivated learning in rats. Proc.Natl.Acad.Sci.U.S.A,
94, 7041-7046.
Berton, O., Covington, H. E., III, Ebner, K., Tsankova, N. M., Carle, T. L., Ulery, P. et al. (2007).
Induction of deltaFosB in the periaqueductal gray by stress promotes active coping responses.
Neuron, 55, 289-300.
Besedovsky, H. O., del, R. A., Klusman, I., Furukawa, H., Monge, A. G., & Kabiersch, A. (1991).
Cytokines as modulators of the hypothalamus-pituitary-adrenal axis. J.Steroid
Biochem.Mol.Biol., 40, 613-618.

114
Blanchard, R. J. & Blanchard, D. C. (1972). Effects of hippocampal lesions on the rat's reaction to a cat.
J.Comp Physiol Psychol., 78, 77-82.
Blanchard, R. J., Blanchard, D. C., & Fial, R. A. (1970). Hippocampal lesions in rats and their effect on
activity, avoidance, and aggression. J.Comp Physiol Psychol., 71, 92-101.
Blandino, P., Jr., Barnum, C. J., & Deak, T. (2006). The involvement of norepinephrine and microglia in
hypothalamic and splenic IL-1beta responses to stress. J.Neuroimmunol., 173, 87-95.
Block, M. L., Zecca, L., & Hong, J. S. (2007). Microglia-mediated neurotoxicity: uncovering the
molecular mechanisms. Nat.Rev.Neurosci., 8, 57-69.
Bloodgood, B. L., Giessel, A. J., & Sabatini, B. L. (2009). Biphasic synaptic Ca influx arising from
compartmentalized electrical signals in dendritic spines. PLoS.Biol., 7, e1000190.
Boche, D., Perry, V. H., & Nicoll, J. A. (2013). Review: activation patterns of microglia and their
identification in the human brain. Neuropathol.Appl.Neurobiol., 39, 3-18.
Bodnoff, S. R., Humphreys, A. G., Lehman, J. C., Diamond, D. M., Rose, G. M., & Meaney, M. J.
(1995). Enduring effects of chronic corticosterone treatment on spatial learning, synaptic
plasticity, and hippocampal neuropathology in young and mid-aged rats. J.Neurosci., 15, 61-69.
Bondonna, T. J., Jacquet, Y., & Wolf, G. (1977). Perfusion-fixation procedure for immediate histologic
processing of brain tissue. Physiol Behav., 19, 345-347.
Bonne, O., Vythilingam, M., Inagaki, M., Wood, S., Neumeister, A., Nugent, A. C. et al. (2008).
Reduced posterior hippocampal volume in posttraumatic stress disorder. J.Clin.Psychiatry, 69,
1087-1091.
Borta, A., & Scharting, R. K. (2005). Inhibitory avoidance, pain reactivity, and plus-maze behavior in
wistar rats with high versus rearing activiry. Physiol Behav., 84, 187-96.
Boucsein, C., Zacharias, R., Farber, K., Pavlovic, S., Hanisch, U. K., & Kettenmann, H. (2003).
Purinergic receptors on microglial cells: functional expression in acute brain slices and
modulation of microglial activation in vitro. Eur.J.Neurosci., 17, 2267-2276.

115
Bovenkerk, B. & Kaldewaij, F. (2015). The use of animal models in behavioural neuroscience research.
Curr.Top.Behav.Neurosci., 19, 17-46.
Brew, H.M., & Forsythe, I.D. (1995). Two voltaje-dependent K+ conductances with complementary
functions in postsynaptic integration at a central auditory synapse. J. Neurosci., 15, 8011-22.
Broadbent, N. J., Gaskin, S., Squire, L. R., & Clark, R. E. (2010). Object recognition memory and the
rodent hippocampus. Learn.Mem., 17, 5-11.
Bruel-Jungerman, E., Laroche, S., & Rampon, C. (2005). New neurons in the dentate gyrus are involved
in the expression of enhanced long-term memory following environmental enrichment.
Eur.J.Neurosci., 21, 513-521.
Brummelte, S., & Galea, L. A. (2010). Chronic high corticosterone reduces neurogénesis in the dentate
gyrus of adult male and female rats. Neuroscience., 168, 680-90.
Bucci, D. J., Phillips, R. G., & Burwell, R. D. (2000). Contributions of postrhinal and perirhinal cortex to
contextual information processing. Behav.Neurosci., 114, 882-894.
Bueno, A. P., de Paiva, J. P., Correa, M. D., Tiba, P. A., & Fornari, R. V. (2017). Corticosterone
administration after a single-trial contextual fear conditioning does not influence the strength and
specificity of recent and remote memory in rats. Physiol Behav., 171, 175-180.
Burwell, R. D. & Amaral, D. G. (1998). Cortical afferents of the perirhinal, postrhinal, and entorhinal
cortices of the rat. J.Comp Neurol., 398, 179-205.
Burwell, R. D., Saddoris, M. P., Bucci, D. J., & Wiig, K. A. (2004). Corticohippocampal contributions to
spatial and contextual learning. J.Neurosci., 24, 3826-3836.
Busse, S. & Schwarting, R. K. (2016). Decoupling Actions from Consequences: Dorsal Hippocampal
Lesions Facilitate Instrumental Performance, but Impair Behavioral Flexibility in Rats. Front
Behav.Neurosci., 10, 118.
Bussey, T. J., Dias, R., Amin, E., Muir, J. L., & Aggleton, J. P. (2001). Perirhinal cortex and place-
object conditional learning in the rat. Behav.Neurosci., 115, 776-785.

116
Bussey, T. J., Saksida, L. M., & Murray, E. A. (2003). Impairments in visual discrimination after
perirhinal cortex lesions: testing 'declarative' vs. 'perceptual-mnemonic' views of perirhinal cortex
function. Eur.J.Neurosci., 17, 649-660.
Cacioppo, J. T., Berntson, G. G., Malarkey, W. B., Kiecolt-Glaser, J. K., Sheridan, J. F., Poehlmann, K.
M. et al. (1998). Autonomic, neuroendocrine, and immune responses to psychological stress: the
reactivity hypothesis. Ann.N.Y.Acad.Sci., 840, 664-673.
Cameron, H. A., & Gould, E. (1994). Adult neurogénesis is regulated by adrenal steroids in the dentate
gyrus. Neuroscience., 61, 203-209.
Campos, A. C., Fogaca, M. V., Aguiar, D. C., & Guimaraes, F. S. (2013). Animal models of anxiety
disorders and stress. Rev.Bras.Psiquiatr., 35 Suppl 2, S101-S111.
Canning, K. J. & Leung, L. S. (1997). Lateral entorhinal, perirhinal, and amygdala-entorhinal transition
projections to hippocampal CA1 and dentate gyrus in the rat: a current source density study.
Hippocampus, 7, 643-655.
Cardoso, A. L., Guerdes, J. R., Pereira de Almeida, L., & Pedroso de Lima, M.C. (2012). miR-155
modulates microglía-mediated immune response by down-regulating SOCS-1 and promoting
cytokine and nitric oxide production. Immunology., 135, 73-88.
Carrillo-de Sauvage, M. A., Maatouk, L., Arnoux, I., Pasco, M., Sanz, D. A., Delahaye, M. et al. (2013).
Potent and multiple regulatory actions of microglial glucocorticoid receptors during CNS
inflammation. Cell Death.Differ., 20, 1546-1557.
Carstens, E. & Moberg, G. P. (2000). Recognizing pain and distress in laboratory animals. ILAR.J., 41,
62-71.
Cecchi, M., Khoshbouei, H., & Morilak, D. A. (2002). Modulatory effects of norepinephrine, acting on
alpha 1 receptors in the central nucleus of the amygdala, on behavioral and neuroendocrine
responses to acute immobilization stress. Neuropharmacology, 43, 1139-1147.
Cenquizca, L. A. & Swanson, L. W. (2006). Analysis of direct hippocampal cortical field CA1 axonal
projections to diencephalon in the rat. J.Comp Neurol., 497, 101-114.

117
Cenquizca, L. A. & Swanson, L. W. (2007). Spatial organization of direct hippocampal field CA1 axonal
projections to the rest of the cerebral cortex. Brain Res.Rev., 56, 1-26.
Charmandari, E., Tsigos, C., & Chrousos, G. (2005). Endocrinology of the stress response.
Annu.Rev.Physiol, 67, 259-284.
Cherry, J. D., Olschowka, J. A., & O'Banion, M. K. (2014). Neuroinflammation and M2 microglia: the
good, the bad, and the inflamed. J.Neuroinflammation., 11, 98.
Chesnokova, V., Pechnick, R. N., & Wawrowsky, K. (2016). Chronic peripheral inflammation,
hippocampal neurogenesis, and behavior. Brain Behav.Immun., 58, 1-8.
Chetkovich, D. M., Gray, R., Johnston, D., & Sweatt, J. D. (1991). N-methyl-D-aspartate receptor
activation increases cAMP levels and voltage-gated Ca2+ channel activity in area CA1 of
hippocampus. Proc.Natl.Acad.Sci.U.S.A, 88, 6467-6471.
Chhor, V., Le, C. T., Lebon, S., Ore, M. V., Celador, I. L., Josserand, J. et al. (2013). Characterization of
phenotype markers and neuronotoxic potential of polarised primary microglia in vitro. Brain
Behav.Immun., 32, 70-85.
Cho, S. H., Sun, B., Zhou, Y., Kauppinen, T. M., Halabisky, B., Wes, P. et al. (2011). CX3CR1 protein
signaling modulates microglial activation and protects against plaque-independent cognitive
deficits in a mouse model of Alzheimer disease. J.Biol.Chem., 286, 32713-32722.
Choleris, E., Thimas, A. W., Kavaliers, M., Prato, F. S. (2001). A detailed ethological analysis of the
mouse open field test: effects of diazepam, chlordiazepoxide and an extremely low frecuency
pulsed magnetic field. Neurosci Behav Rev., 25, 235-60.
Chrousos, G. P. (2009). Stress and disorders of the stress system. Nat.Rev.Endocrinol., 5, 374-381.
Chugh, D., Nilsson, P., Afjei, S. A., Bakochi, A., & Ekdahl, C. T. (2013). Brain inflammation induces
post-synaptic changes during early synapse formation in adult-born hippocampal neurons.
Exp.Neurol., 250, 176-188.

118
Corrigan, F., Vink, R., & Turner, R. J. (2016). Inflammation in acute CNS injury: a focus on the role of
substance P. Br.J.Pharmacol., 173, 703-715.
Costall, B., Jones, B. J., Kelly, M. E., Naylor, R. J., Tomkins, D. M. (1989). Exploration of mice in a
black and white test box: validation as a model of anxiety. Pharmacol Biochem Behav., 32, 777-
85.
Croisier, E., Moran, L. B., Dexter, D. T., Pearce, R. K., & Graeber, M. B. (2005). Microglial
inflammation in the parkinsonian substantia nigra: relationship to alpha-synuclein deposition.
J.Neuroinflammation., 2, 14.
Cromwell, H. C. & Berridge, K. C. (1996). Implementation of action sequences by a neostriatal site: a
lesion mapping study of grooming syntax. J.Neurosci., 16, 3444-3458.
Cunningham, C. (2013). Microglia and neurodegeneration: the role of systemic inflammation. Glia, 61,
71-90.
Curtis, A. L., Leiser, S. C., Snyder, K., & Valentino, R. J. (2012). Predator stress engages corticotropin-
releasing factor and opioid systems to alter the operating mode of locus coeruleus norepinephrine
neurons. Neuropharmacology, 62, 1737-1745.
Dalmau, I., Finsen, B., Zimmer, J., Gonzalez, B., & Castellano, B. (1998). Development of microglia in
the postnatal rat hippocampus. Hippocampus, 8, 458-474.
Datta, S. & O'Malley, M. W. (2013). Fear extinction memory consolidation requires potentiation of
pontine-wave activity during REM sleep. J.Neurosci., 33, 4561-4569.
Davis, S., Vanhoutte, P., Pages, C., Caboche, J., & Laroche, S. (2000). The MAPK/ERK cascade targets
both Elk-1 and cAMP response element-binding protein to control long-term potentiation-
dependent gene expression in the dentate gyrus in vivo. J.Neurosci., 20, 4563-4572.
de Castro Gómes, V., Barroso Silva, C., & Landeira-Fernandez, J. (2011). The Carioca High and Low
Conditioned Freezing Lines: A New Animal Model of Generalized Anxiety Disorder. In Anxiety
Disorders ( InTech.

119
de Kloet, E. R. (2013). Functional profile of the binary brain corticosteroid receptor system: mediating,
multitasking, coordinating, integrating. Eur.J.Pharmacol., 719, 53-62.
de Kloet, E. R., Joels, M., & Holsboer, F. (2005). Stress and the brain: from adaptation to disease.
Nat.Rev.Neurosci., 6, 463-475.
de Kloet, E. R., Oitzl, M. S., & Joels, M. (1999). Stress and cognition: are corticosteroids good or bad
guys? Trends Neurosci., 22, 422-426.
De Oca, B. M., DeCola, J. P., Maren, S., & Fanselow, M. S. (1998). Distinct regions of the
periaqueductal gray are involved in the acquisition and expression of defensive responses.
J.Neurosci., 18, 3426-3432.
de Oliveira Galvão, B., de Castro Gómes, V., Masionette, S., & Landeira-Fernandez, J. (2011). Panic-
like behaviors in Carioca High-and Low-conditioned
Freezing rats. Psychology & Neuroscience, 4, 205-210.
de, C. G., V & Landeira-Fernandez, J. (2008a). Amygdaloid lesions produced similar contextual fear
conditioning disruption in the Carioca high- and low-conditioned freezing rats. Brain Res., 1233,
137-145.
de, C. G., V & Landeira-Fernandez, J. (2008b). Amygdaloid lesions produced similar contextual fear
conditioning disruption in the Carioca high- and low-conditioned freezing rats. Brain Res., 1233,
137-145.
Dedovic, K., Duchesne, A., Andrews, J., Engert, V., & Pruessner, J. C. (2009). The brain and the stress
axis: the neural correlates of cortisol regulation in response to stress. Neuroimage., 47, 864-871.
Di, S., Malcher-Lopes, R., Marcheselli, V. L., Bazan, N. G., & Tasker, J. G. (2005). Rapid
glucocorticoid-mediated endocannabinoid release and opposing regulation of glutamate and
gamma-aminobutyric acid inputs to hypothalamic magnocellular neurons. Endocrinology, 146,
4292-4301.

120
Di, S. G., Mana, M. J., Jacobs, W. J., & Phillips, A. G. (1987). Evidence of Pavlovian conditioned fear
following electrical stimulation of the periaqueductal grey in the rat. Physiol Behav., 40, 55-63.
Dias, G. P., Bevilaqua, M. C., Silveira, A. C., Landeira-Fernandez, J., & Gardino, P. F. (2009).
Behavioral profile and dorsal hippocampal cells in carioca high-conditioned freezing rats. Behav
Brain Res., 205, 342-348.
Dickinson, A. (2012). Associative learning and animal cognition. Philos.Trans.R.Soc.Lond B Biol.Sci.,
367, 2733-2742.
Dijkstra, M. T. & Homan, A. C. (2016). Engaging in Rather than Disengaging from Stress: Effective
Coping and Perceived Control. Front Psychol., 7, 1415.
Dingledine, R., Borges, K., Bowie, D., & Traynelis, S. F. (1999). The glutamate receptor ion channels.
Pharmacol.Rev., 51, 7-61.
Diorio, J., & Meaney, M. J. (2007). Maternal programing of defensive responses through sustained
effects on gene expression. J Psychiatry Neurosci., 32, 275-84.
Dobrossy, M. D., Drapeau, E., Aurousseau, C., Le, M. M., Piazza, P. V., & Abrous, D. N. (2003).
Differential effects of learning on neurogenesis: learning increases or decreases the number of
newly born cells depending on their birth date. Mol.Psychiatry, 8, 974-982.
Doens, D. & Fernandez, P. L. (2014). Microglia receptors and their implications in the response to
amyloid beta for Alzheimer's disease pathogenesis. J.Neuroinflammation., 11, 48.
Dolorfo, C. L. & Amaral, D. G. (1998). Entorhinal cortex of the rat: organization of intrinsic
connections. J.Comp Neurol., 398, 49-82.
Dong, H. W., Swanson, L. W., Chen, L., Fanselow, M. S., & Toga, A. W. (2009). Genomic-anatomic
evidence for distinct functional domains in hippocampal field CA1. Proc.Natl.Acad.Sci.U.S.A,
106, 11794-11799.

121
Dorey, R., Pierard, C., Chauveau, F., David, V., & Beracochea, D. (2012). Stress-induced memory
retrieval impairments: different time-course involvement of corticosterone and glucocorticoid
receptors in dorsal and ventral hippocampus. Neuropsychopharmacology, 37, 2870-2880.
Dornelles, A., de Lima, M. N., Grazziotin, M., Presti-Torres, J., Garcia, V. A., Scalco, F. S. et al. (2007).
Adrenergic enhancement of consolidation of object recognition memory. Neurobiol.Learn.Mem.,
88, 137-142.
Doyle, A. G., Herbein, G., Montaner, L. J., Minty, A. J., Caput, D., Ferrara, P. et al. (1994). Interleukin-
13 alters the activation state of murine macrophages in vitro: comparison with interleukin-4 and
interferon-gamma. Eur.J.Immunol., 24, 1441-1445.
Dunn, G.A., Bale, T.L. (2009). Maternal high-fat diet promotes body length increases and insulin
insensitivity in second-generation mice. Endocrinology., 150, 4999-5009.
Dupret, D., Fabre, A., Dobrossy, M. D., Panatier, A., Rodriguez, J. J., Lamarque, S. et al. (2007). Spatial
learning depends on both the addition and removal of new hippocampal neurons. PLoS.Biol., 5,
e214.
Eacott, M. J. & Gaffan, E. A. (2005). The roles of perirhinal cortex, postrhinal cortex, and the fornix in
memory for objects, contexts, and events in the rat. Q.J.Exp.Psychol.B, 58, 202-217.
Eagle, A. L., Knox, D., Roberts, M. M., Mulo, K., Liberzon, I., Galloway, M. P. et al. (2013). Single
prolonged stress enhances hippocampal glucocorticoid receptor and phosphorylated protein
kinase B levels. Neurosci.Res., 75, 130-137.
Ebner, K. & Singewald, N. (2017). Individual differences in stress susceptibility and stress inhibitory
mechanisms. Current Opinion in Behavioral Sciences, 14, 54-64.
Ebner, K. & Singewald, N. (2006). The role of substance P in stress and anxiety responses. Amino.Acids,
31, 251-272.

122
Ehninger, D. & Kempermann, G. (2006). Paradoxical effects of learning the Morris water maze on adult
hippocampal neurogenesis in mice may be explained by a combination of stress and physical
activity. Genes Brain Behav., 5, 29-39.
Eichenbaum, H., Yonelinas, A. P., & Ranganath, C. (2007). The medial temporal lobe and recognition
memory. Annu.Rev.Neurosci., 30, 123-152.
Ekdahl, C. T., Claasen, J. H., Bonde, S., Kokaia, Z., & Lindvall, O. (2003). Inflammation is detrimental
for neurogenesis in adult brain. Proc.Natl.Acad.Sci.U.S.A, 100, 13632-13637.
English, J. D. & Sweatt, J. D. (1996). Activation of p42 mitogen-activated protein kinase in hippocampal
long term potentiation. J.Biol.Chem., 271, 24329-24332.
Ennaceur, A. & Aggleton, J. P. (1997). The effects of neurotoxic lesions of the perirhinal cortex
combined to fornix transection on object recognition memory in the rat. Behav.Brain Res., 88,
181-193.
Espinosa-Oliva, A. M., de Pablos, R. M., Villaran, R. F., Arguelles, S., Venero, J. L., Machado, A. et al.
(2011). Stress is critical for LPS-induced activation of microglia and damage in the rat
hippocampus. Neurobiol.Aging, 32, 85-102.
Evanson, N. K., Tasker, J. G., Hill, M. N., Hillard, C. J., & Herman, J. P. (2010). Fast feedback
inhibition of the HPA axis by glucocorticoids is mediated by endocannabinoid signaling.
Endocrinology, 151, 4811-4819.
Fanselow, M. S. (1984). Opiate modulation of the active and inactive components of the postshock
reaction: parallels between naloxone pretreatment and shock intensity. Behav.Neurosci., 98, 269-
277.
Fanselow, M. S. (2000). Contextual fear, gestalt memories, and the hippocampus. Behav.Brain Res.,
110, 73-81.
Fanselow, M. S. & Dong, H. W. (2010). Are the dorsal and ventral hippocampus functionally distinct
structures? Neuron, 65, 7-19.

123
Fayard, E., Tintignac, L. A., Baudry, A., & Hemmings, B. A. (2005). Protein kinase B/Akt at a glance.
J.Cell Sci., 118, 5675-5678.
Feldman, S. & Weidenfeld, J. (1999). Glucocorticoid receptor antagonists in the hippocampus modify
the negative feedback following neural stimuli. Brain Res., 821, 33-37.
Fellini, L., Florian, C., Courtney, J., & Roullet, P. (2009). Pharmacological intervention of hipocampal
CA3 NMDA receptors impairs acquisition and long-term memory retrieval of spatial pattern
completion task. Learn Mem., 16, 387-94.
Fendt, M. & Fanselow, M. S. (1999). The neuroanatomical and neurochemical basis of conditioned fear.
Neurosci.Biobehav.Rev., 23, 743-760.
Fernandez, P. L., Britton, G. B., & Rao, K. S. (2013). Potential immunotargets for Alzheimer's disease
treatment strategies. J.Alzheimers.Dis., 33, 297-312.
Florian, C., & Roullet, P. (2004). Hippocampal CA3-region is crucial for acquisition and memory
consolidation in Morris water maze task in mice. Behavioral Brain Research., 154, 365-374.
Fontainhas, A. M., Wang, M., Liang, K. J., Chen, S., Mettu, P., Damani, M. et al. (2011). Microglial
morphology and dynamic behavior is regulated by ionotropic glutamatergic and GABAergic
neurotransmission. PLoS.One., 6, e15973.
Franchi, L., Eigenbrod, T., Munoz-Planillo, R., & Nunez, G. (2009). The inflammasome: a caspase-1-
activation platform that regulates immune responses and disease pathogenesis. Nat.Immunol., 10,
241-247.
Francis, D. D., Caldji, C., Champagne, F., Plotsky, P. M., & Meaney, M. J. (1999). The role of
corticotropin-releasing factor--norepinephrine systems in mediating the effects of early
experience on the development of behavioral and endocrine responses to stress. Biol.Psychiatry,
46, 1153-1166.
Frank, M. G., Baratta, M. V., Sprunger, D. B., Watkins, L. R., & Maier, S. F. (2007). Microglia serve as
a neuroimmune substrate for stress-induced potentiation of CNS pro-inflammatory cytokine
responses. Brain Behav.Immun., 21, 47-59.

124
Franklin, T.B., Russig, H., Weiss, I.C., Graff, J., Linder, N., Michalon, A., Vizi, S., & Mansuy, I.M.
(2010). Epigenetic transmission of the impact of early stress across generations. Biol Psychiatry.,
68, 408-15.
Fransen, E., Tahvildari, B., Egorov, A. V., Hasselmo, M. E., & Alonso, A. A. (2006). Mechanism of
graded persistent cellular activity of entorhinal cortex layer v neurons. Neuron, 49, 735-746.
Frieler, R. A., Meng, H., Duan, S. Z., Berger, S., Schutz, G., He, Y. et al. (2011). Myeloid-specific
deletion of the mineralocorticoid receptor reduces infarct volume and alters inflammation during
cerebral ischemia. Stroke, 42, 179-185.
Fuchs, E. & Flugge, G. (1998). Stress, glucocorticoids and structural plasticity of the hippocampus.
Neurosci.Biobehav.Rev., 23, 295-300.
Fyhn, M., Hafting, T., Treves, A., Moser, M. B., & Moser, E. I. (2007). Hippocampal remapping and
grid realignment in entorhinal cortex. Nature, 446, 190-194.
Fyhn, M., Molden, S., Witter, M. P., Moser, E. I., & Moser, M. B. (2004). Spatial representation in the
entorhinal cortex. Science, 305, 1258-1264.
Gagnon, S. A. & Wagner, A. D. (2016). Acute stress and episodic memory retrieval: neurobiological
mechanisms and behavioral consequences. Ann.N.Y.Acad.Sci., 1369, 55-75.
Gallardo-Soler, A., Gomez-Nieto, C., Campo, M. L., Marathe, C., Tontonoz, P., Castrillo, A. et al.
(2008). Arginase I induction by modified lipoproteins in macrophages: a peroxisome proliferator-
activated receptor-gamma/delta-mediated effect that links lipid metabolism and immunity.
Mol.Endocrinol., 22, 1394-1402.
Gallo-Payet, N. (2016). 60 YEARS OF POMC: Adrenal and extra-adrenal functions of ACTH.
J.Mol.Endocrinol., 56, T135-T156.
Gautier, E. L., Shay, T., Miller, J., Greter, M., Jakubzick, C., Ivanov, S. et al. (2012). Gene-expression
profiles and transcriptional regulatory pathways that underlie the identity and diversity of mouse
tissue macrophages. Nat.Immunol., 13, 1118-1128.

125
Garden, G.A. (2013). Epigenetics and the modulation of neuroinflamation. Neurotherapeutics., 10, 782-
788.
Ge, S., Yang, C. H., Hsu, K. S., Ming, G. L., & Song, H. (2007). A critical period for enhanced synaptic
plasticity in newly generated neurons of the adult brain. Neuron, 54, 559-566.
Geissmann, F., Manz, M. G., Jung, S., Sieweke, M. H., Merad, M., & Ley, K. (2010). Development of
monocytes, macrophages, and dendritic cells. Science, 327, 656-661.
Genzel, L., Rossato, J. I., Jacobse, J., Grieves, R. M., Spooner, P. A., Battaglia, F. P., Fernandez, G., &
Morris, R. G. M. (2017). The Yin and Yang of memory consolidation: hippocampal and
neocortical. PLoS Biology.
Ghosh, M., Xu, Y., & Pearse, D. D. (2016). Cyclic AMP is a key regulator of M1 to M2a phenotypic
conversion of microglia in the presence of Th2 cytokines. J.Neuroinflammation., 13, 9.
Gianotti, C., Nunzi, M. G., Gispen, W. H., & Corradetti, R. (1992). Phosphorylation of the presynaptic
protein B-50 (GAP-43) is increased during electrically induced long-term potentiation. Neuron,
8, 843-848.
Gill, J., Vythilingam, M., & Page, G. G. (2008). Low cortisol, high DHEA, and high levels of stimulated
TNF-alpha, and IL-6 in women with PTSD. J.Trauma Stress., 21, 530-539.
GlaxoSmithKline (2007). PRESCRIBING INFORMATION AVANDIA® (rosiglitazone maleate).
http://www.fda.gov/downloads/Drugs/DrugSafety/PostmarketDrugSafetyInformationforPatientsa
ndProviders/UCM143413.pdf;%20Bob,%20Kremmerer,%20Brune%20&%20Namgaladze,%202
015
Gleichgerrcht, E., Ibanez, A., Roca, M., Torralva, T., & Manes, F. (2010). Decision-making cognition in
neurodegenerative diseases. Nat.Rev.Neurol., 6, 611-623.
Gold, P. E. (2014). Regulation of memory - from the adrenal medulla to liver to astrocytes to neurons.
Brain Res.Bull., 105, 25-35.
Goldstein, D. S. & McEwen, B. (2002). Allostasis, homeostats, and the nature of stress. Stress., 5, 55-58.

126
Gordon, S. & Martinez, F. O. (2010). Alternative activation of macrophages: mechanism and functions.
Immunity., 32, 593-604.
Gottlieb, M. & Matute, C. (1997). Expression of ionotropic glutamate receptor subunits in glial cells of
the hippocampal CA1 area following transient forebrain ischemia. J.Cereb.Blood Flow Metab,
17, 290-300.
Gould, E., Tanapat, P., McEwen, B. S., Flugge, G., & Fuchs, E. (1998). Proliferation of granule cell
precursors in the dentate gyrus of adult monkeys is diminished by stress.
Proc.Natl.Acad.Sci.U.S.A, 95, 3168-3171.
Goymann, W. & Wingfield, J. C. (2004). Allostatic load, social status and stress hormones: The costs of
social status matter. Anim Behav, 67, 591-602.
Graeff, F. G., Silveira, M. C., Nogueira, R. L., Audi, E. A., & Oliveira, R. M. (1993). Role of the
amygdala and periaqueductal gray in anxiety and panic. Behav.Brain Res., 58, 123-131.
Gruart, A., Munoz, M. D., & Delgado-Garcia, J. M. (2006). Involvement of the CA3-CA1 synapse in the
acquisition of associative learning in behaving mice. J.Neurosci., 26, 1077-1087.
Gurley, C., Nichols, J., Liu, S., Phulwani, N. K., Esen, N., & Kielian, T. (2008). Microglia and Astrocyte
Activation by Toll-Like Receptor Ligands: Modulation by PPAR-gamma Agonists. PPAR.Res.,
2008, 453120.
Gutierrez, R., Romo-Parra, H., Maqueda, J., Vivar, C., Ramirez, M., Morales, M. A. et al. (2003).
Plasticity of the GABAergic phenotype of the "glutamatergic" granule cells of the rat dentate
gyrus. J.Neurosci., 23, 5594-5598.
Guzowski, J. F., Knierim, J. J., & Moser, E. I. (2004). Ensemble dynamics of hippocampal regions CA3
and CA1. Neuron, 44, 581-584.
Habib, K. E., Gold, P. W., & Chrousos, G. P. (2001). Neuroendocrinology of stress. Endocrinol.Metab
Clin.North Am., 30, 695-728.

127
Hanisch, U. K. & Kettenmann, H. (2007). Microglia: active sensor and versatile effector cells in the
normal and pathologic brain. Nat.Neurosci., 10, 1387-1394.
Hardingham, G. E. (2009). Coupling of the NMDA receptor to neuroprotective and neurodestructive
events. Biochem.Soc.Trans., 37, 1147-1160.
Harris, A. P., Holmes, M. C., de Kloet, E. R., Chapman, K. E., & Seckl, J. R. (2013). Mineralocorticoid
and glucocorticoid receptor balance in control of HPA axis and behaviour.
Psychoneuroendocrinology, 38, 648-658.
Hashioka, S., Klegeris, A., Monji, A., Kato, T., Sawada, M., McGeer, P. L. et al. (2007). Antidepressants
inhibit interferon-gamma-induced microglial production of IL-6 and nitric oxide. Exp.Neurol.,
206, 33-42.
Hass, D. T. & Barnstable, C. J. (2016). Uncoupling protein 2 in the glial response to stress: implications
for neuroprotection. Neural Regen.Res., 11, 1197-1200.
Hassan, W., Gomes, V. C., Pinton, S., Batista Teixeira da, R. J., & Landeira-Fernandez, J. (2013).
Association between oxidative stress and contextual fear conditioning in Carioca high- and low-
conditioned freezing rats. Brain Res., 1512, 60-67.
Hasselmo, M. E. (2005a). The role of hippocampal regions CA3 and CA1 in matching entorhinal input
with retrieval of associations between objects and context: theoretical comment on Lee et al.
(2005). Behav.Neurosci., 119, 342-345.
Hasselmo, M. E. (2005b). The role of hippocampal regions CA3 and CA1 in matching entorhinal input
with retrieval of associations between objects and context: theoretical comment on Lee et al.
(2005). Behav.Neurosci., 119, 342-345.
Hayashi, Y., Ishida, A., Katagiri, H., Mishina, M., Fujisawa, H., Manabe, T. et al. (1997). Calcium- and
calmodulin-dependent phosphorylation of AMPA type glutamate receptor subunits by
endogenous protein kinases in the post-synaptic density. Brain Res.Mol.Brain Res., 46, 338-342.

128
Hayashi, Y., Zviman, M. M., Brand, J. G., Teeter, J. H., & Restrepo, D. (1996). Measurement of
membrane potential and [Ca2+]i in cell ensembles: application to the study of glutamate taste in
mice. Biophys.J., 71, 1057-1070.
Heeger, R. (2011). Experimenting on animals: does their size matter morally. In Large animals as
biological models: ethical, societal, legal, and biological aspects (pp. 13-23). Europaische
Academie.
Heinrichs, S. C., Menzaghi, F., Merlo, P. E., Britton, K. T., & Koob, G. F. (1995). The role of CRF in
behavioral aspects of stress. Ann.N.Y.Acad.Sci., 771, 92-104.
Henke, P. G. (1990). Hippocampal pathway to the amygdala and stress ulcer development. Brain
Res.Bull., 25, 691-695.
Henze, D. A. & Buzsaki, G. (2007). Hilar mossy cells: functional identification and activity in vivo.
Prog.Brain Res., 163, 199-216.
Henze, D. A., Wittner, L., & Buzsaki, G. (2002). Single granule cells reliably discharge targets in the
hippocampal CA3 network in vivo. Nat.Neurosci., 5, 790-795.
Herman, J. P. & Cullinan, W. E. (1997). Neurocircuitry of stress: central control of the hypothalamo-
pituitary-adrenocortical axis. Trends Neurosci., 20, 78-84.
Herman, J. P., Cullinan, W. E., Ziegler, D. R., & Tasker, J. G. (2002). Role of the paraventricular
nucleus microenvironment in stress integration. Eur.J.Neurosci., 16, 381-385.
Herman, J. P., Figueiredo, H., Mueller, N. K., Ulrich-Lai, Y., Ostrander, M. M., Choi, D. C. et al.
(2003). Central mechanisms of stress integration: hierarchical circuitry controlling hypothalamo-
pituitary-adrenocortical responsiveness. Front Neuroendocrinol., 24, 151-180.
Herman, J. P., Patel, P. D., Akil, H., & Watson, S. J. (1989). Localization and regulation of
glucocorticoid and mineralocorticoid receptor messenger RNAs in the hippocampal formation of
the rat. Mol.Endocrinol., 3, 1886-1894.

129
Herman, J. P., Schafer, M. K., Young, E. A., Thompson, R., Douglass, J., Akil, H. et al. (1989).
Evidence for hippocampal regulation of neuroendocrine neurons of the hypothalamo-pituitary-
adrenocortical axis. J.Neurosci., 9, 3072-3082.
Hernandez-Rabaza, V., Hontecillas-Prieto, L., Velazquez-Sanchez, C., Ferragud, A., Perez-Villaba, A.,
Arcusa, A. et al. (2008). The hippocampal dentate gyrus is essential for generating contextual
memories of fear and drug-induced reward. Neurobiol.Learn.Mem., 90, 553-559.
Hong, W., Kim, D. W., Anderson, D. J. (2014). ntagonistic control of social versus repetitive self-
grooming behaviors by separable amygdala neuronal subsets. Cell., 158, 1348-61.
Hoogman, M., Aarts, E., Slaats-Willemse, D., Naber, M., Onnink, M., Cools, R., Kan, C., Buitelaar, J.,
& Franke, B. (2011).Nitric oxide synthase genotype modulation of impulsivity and ventral
striatal activity in adult ADHD patients and healthy comparison subjects. Am J Psychiatry., 168,
1099-106.
Houvert, E., Vallette, F.M., & Cartron, P.F. (2009). Dnmt3/transcription factor interactions as crucial
layers in targeted DNA methylation. Epigenetics., 4, 487-99.
Hu, Z., Yuri, K., Ozawa, H., Lu, H., Yang, Y., Ito, T. et al. (1997). Adrenalectomy-induced granule cell
death is predicated on the disappearance of glucocorticoid receptor immunoreactivity in the rat
hippocampal granule cell layer. Brain Res., 778, 293-301.
Huang, Y. Y. & Kandel, E. R. (2005). Theta frequency stimulation up-regulates the synaptic strength of
the pathway from CA1 to subiculum region of hippocampus. Proc.Natl.Acad.Sci.U.S.A, 102,
232-237.
Hutchison, I. C. & Rathore, S. (2015). The role of REM sleep theta activity in emotional memory. Front
Psychol., 6, 1439.
Ialenti, A., Grassia, G., Di, M. P., Maffia, P., Di, R. M., & Ianaro, A. (2005). Mechanism of the anti-
inflammatory effect of thiazolidinediones: relationship with the glucocorticoid pathway.
Mol.Pharmacol., 67, 1620-1628.

130
Ino, T., Kaneko, T., & Mizuno, N. (1998). Direct projections from the entorhinal cortical layers to the
dentate gyrus, hippocampus, and subicular complex in the cat. Neurosci.Res., 32, 241-265.
Ino, T., Kaneko, T., & Mizuno, N. (2001). Projections from the hippocampal and parahippocampal
regions to the entorhinal cortex. An anterograde and retrograde tract-tracing study in the cat.
Neurosci.Res., 39, 51-69.
Izquierdo, I., Cammarota, M., Da Silva, W. C., Bevilaqua, L. R., Rossato, J. I., Bonini, J. S. et al. (2008).
The evidence for hippocampal long-term potentiation as a basis of memory for simple tasks.
An.Acad.Bras.Cienc., 80, 115-127.
Jacobson, L. & Sapolsky, R. (1991). The role of the hippocampus in feedback regulation of the
hypothalamic-pituitary-adrenocortical axis. Endocr.Rev., 12, 118-134.
Jaenisch, R., & Bird, A. (2003). Epigenetic regulation of gene expression: how the genome integrates
intrinsic and enviromental signals. Nature Genetics., 33, 245-254.
Jakobsson, J., Bjerke, M., Sahebi, S., Isgren, A., Ekman, C. J., Sellgren, C. et al. (2015). Monocyte and
microglial activation in patients with mood-stabilized bipolar disorder. J.Psychiatry Neurosci.,
40, 250-258.
Jessberger, S. & Gage, F. H. (2014). Adult neurogenesis: bridging the gap between mice and humans.
Trends Cell Biol., 24, 558-563.
Jhaveri, D. J., Mackay, E. W., Hamlin, A. S., Marathe, S. V., Nandam, L. S., Vaidya, V. A. et al. (2010).
Norepinephrine directly activates adult hippocampal precursors via beta3-adrenergic receptors.
J.Neurosci., 30, 2795-2806.
Ji, J. & Maren, S. (2008). Differential roles for hippocampal areas CA1 and CA3 in the contextual
encoding and retrieval of extinguished fear. Learn.Mem., 15, 244-251.
Jinno, S., Fleischer, F., Eckel, S., Schmidt, V., & Kosaka, T. (2007). Spatial arrangement of microglia in
the mouse hippocampus: a stereological study in comparison with astrocytes. Glia, 55, 1334-
1347.

131
Joels, M. (2008). Functional actions of corticosteroids in the hippocampus. Eur.J.Pharmacol., 583, 312-
321.
Joels, M., Sarabdjitsingh, R. A., & Karst, H. (2012). Unraveling the time domains of corticosteroid
hormone influences on brain activity: rapid, slow, and chronic modes. Pharmacol.Rev., 64, 901-
938.
Johnson, A. & Redish, A. D. (2007). Neural ensembles in CA3 transiently encode paths forward of the
animal at a decision point. J.Neurosci., 27, 12176-12189.
Johnson, J. D., O'Connor, K. A., Deak, T., Stark, M., Watkins, L. R., & Maier, S. F. (2002). Prior
stressor exposure sensitizes LPS-induced cytokine production. Brain Behav.Immun., 16, 461-476.
Johnson, J. D., O'Connor, K. A., Watkins, L. R., & Maier, S. F. (2004). The role of IL-1beta in stress-
induced sensitization of proinflammatory cytokine and corticosterone responses. Neuroscience,
127, 569-577.
Johnson, J. D., Zimomra, Z. R., & Stewart, L. T. (2013). Beta-adrenergic receptor activation primes
microglia cytokine production. J.Neuroimmunol., 254, 161-164.
Jonas, P. & Lisman, J. (2014). Structure, function, and plasticity of hippocampal dentate gyrus
microcircuits. Front Neural Circuits., 8, 107.
Juster, R. P., McEwen, B. S., & Lupien, S. J. (2010). Allostatic load biomarkers of chronic stress and
impact on health and cognition. Neurosci.Biobehav.Rev., 35, 2-16.
Kabbaj, M., Devine, D. P., Savage, V. R., & Akil, H. (2000). Neurobiological correlates of individual
differences in novelty-seeking behavior in the rat: differential expression of stress-related
molecules. J.Neurosci., 20, 6983-6988.
Kageyama, K. & Suda, T. (2009). Regulatory mechanisms underlying corticotropin-releasing factor gene
expression in the hypothalamus. Endocr.J., 56, 335-344.

132
Kalueff, A. V., Stewart, A. M., Song, C., Berridge, K. C., Graybiel, A. M., & Fentress, J. C. (2016).
Neurobiology of rodents self.-grooming and its value for translational neuroscience. Nature
Reviews., 17, 45-59.
Kanter, E. D., Wilkinson, C. W., Radant, A. D., Petrie, E. C., Dobie, D. J., McFall, M. E. et al. (2001).
Glucocorticoid feedback sensitivity and adrenocortical responsiveness in posttraumatic stress
disorder. Biol.Psychiatry, 50, 238-245.
Kanterewicz, B. I., Urban, N. N., McMahon, D. B., Norman, E. D., Giffen, L. J., Favata, M. F. et al.
(2000). The extracellular signal-regulated kinase cascade is required for NMDA receptor-
independent LTP in area CA1 but not area CA3 of the hippocampus. J.Neurosci., 20, 3057-3066.
Kavushansky, A., Vouimba, R. M., Cohen, H., & Richter-Levin, G. (2006). Activity and plasticity in the
CA1, the dentate gyrus, and the amygdala following controllable vs. uncontrollable water stress.
Hippocampus, 16, 35-42.
Kawabe, T., Matsushima, M., Hashimoto, N., Imaizumi, K., & Hasegawa, Y. (2011). CD40/CD40
ligand interactions in immune responses and pulmonary immunity. Nagoya J.Med.Sci., 73, 69-78.
Kawanokuchi, J., Shimizu, K., Nitta, A., Yamada, K., Mizuno, T., Takeuchi, H. et al. (2008). Production
and functions of IL-17 in microglia. J.Neuroimmunol., 194, 54-61.
Keegan, A. P., Paris, D., Luis, C. A., Abdullah, L., Ait-Ghezala, G., Beaulieu-Abdelahad, D. et al.
(2017). Plasma cytokine IL-6 levels and subjective cognitive decline: preliminary findings.
Int.J.Geriatr.Psychiatry.
Kelly, A., Lynch, A., Vereker, W., Nolan, Y., Queenan, P., Whittaker, E., O’Neill, L.A., & Lynch, M.A. (2001).
The anti-inflammtory cytokine, interleukin (IL)-10, bloks the inhibitory effect of IL-1beta on long term
potentiation. A role for JNK. J Biol Chem., 276, 45562-72.
Kempermann, G., Brandon, E. P., & Gage, F. H. (1998). Environmental stimulation of 129/SvJ mice
causes increased cell proliferation and neurogenesis in the adult dentate gyrus. Curr.Biol., 8, 939-
942.

133
Kendler, K.S., Gardner, C.O., & Prescott. C.A. (2006). Toward a comprehensive developmental model
for major depression in men. Am J Psychiatry., 163, 115-24.
Keralapurath, M. M., Clark, J. K., Hammond, S., & Wagner, J. J. (2014). Cocaine- or stress-induced
metaplasticity of LTP in the dorsal and ventral hippocampus. Hippocampus, 24, 577-590.
Kesner, R. P. (2007). Behavioral functions of the CA3 subregion of the hippocampus. Learn.Mem., 14,
771-781.
Kesner, R. P., Lee, I., & Gilbert, P. (2004). A behavioral assessment of hippocampal function based on a
subregional analysis. Rev.Neurosci., 15, 333-351.
Kessler, R. C. & Bromet, E. J. (2013). The epidemiology of depression across cultures. Annu.Rev.Public
Health, 34, 119-138.
Kierdorf, K. & Prinz, M. (2013). Factors regulating microglia activation. Front Cell Neurosci., 7, 44.
Kim, J. J., Rison, R. A., & Fanselow, M. S. (1993). Effects of amígdala, hippocampus, and
periaqueductal gray lesions on short- and long- term contextual fear. Behav Neurosci., 107, 1093-
8.
Kigerl, K. A., de Rivero Vaccari, J. P., Dietrich, W. D., Popovich, P. G., & Keane, R. W. (2014). Pattern
recognition receptors and central nervous system repair. Exp.Neurol., 258, 5-16.
Kim, E. J., Horovitz, O., Pellman, B. A., Tan, L. M., Li, Q., Richter-Levin, G. et al. (2013). Dorsal
periaqueductal gray-amygdala pathway conveys both innate and learned fear responses in rats.
Proc.Natl.Acad.Sci.U.S.A, 110, 14795-14800.
Kim, E. Y., Mahmoud, G. S., & Grover, L. M. (2005). REM sleep deprivation inhibits LTP in vivo in
area CA1 of rat hippocampus. Neurosci.Lett., 388, 163-167.
Kim, J. J., Song, E. Y., & Kosten, T. A. (2006). Stress effects in the hippocampus: synaptic plasticity
and memory. Stress., 9, 1-11.
Kim, J. J. & Yoon, K. S. (1998). Stress: metaplastic effects in the hippocampus. Trends Neurosci., 21,
505-509.

134
Kim, K. H., Son, S. M., & Mook-Jung, I. (2013). Contributions of microglia to structural synaptic
plasticity. J.Exp.Neurosci., 7, 85-91.
Kincheski, G. C., Mota-Ortiz, S. R., Pavesi, E., Canteras, N. S., & Carobrez, A. P. (2012). The
dorsolateral periaqueductal gray and its role in mediating fear learning to life threatening events.
PLoS.One., 7, e50361.
Kitamura, T., Tsuchihashi, Y., & Fujita, S. (1978). Initial response of silver-impregnated "resting
microglia" to stab wounding in rabbit hippocampus. Acta Neuropathol., 44, 31-39.
Kjelstrup, K. G., Tuvnes, F. A., Steffenach, H. A., Murison, R., Moser, E. I., & Moser, M. B. (2002).
Reduced fear expression after lesions of the ventral hippocampus. Proc.Natl.Acad.Sci.U.S.A, 99,
10825-10830.
Knijff, E. M., Breunis, M. N., Kupka, R. W., de Wit, H. J., Ruwhof, C., Akkerhuis, G. W. et al. (2007).
An imbalance in the production of IL-1beta and IL-6 by monocytes of bipolar patients:
restoration by lithium treatment. Bipolar.Disord., 9, 743-753.
Kobayashi, K., Imagama, S., Ohgomori, T., Hirano, K., Uchimura, K., Sakamoto, K. et al. (2013).
Minocycline selectively inhibits M1 polarization of microglia. Cell Death.Dis., 4, e525.
Kole, M. H., Czeh, B., & Fuchs, E. (2004). Homeostatic maintenance in excitability of tree shrew
hippocampal CA3 pyramidal neurons after chronic stress. Hippocampus, 14, 742-751.
Koolhaas, J. M., de Boer, S. F., Buwalda, B., van Reenen, K. (2007). Individual variation in coping with
stress: a multidimetional approach of ultimate and proximate mechanisms. Brain Behav Evol., 70,
218-26.
Koolhaas, J. M., de Boer, S. F., Coppens, C. M., & Buwalda. B. (2010). Neruroendocrinology of coping
styles: towards understanding the biology og individual variation. Front Neuroendocrinol., 31,
307-21.
Korz, V. & Frey, J. U. (2003). Stress-related modulation of hippocampal long-term potentiation in rats:
Involvement of adrenal steroid receptors. J.Neurosci., 23, 7281-7287.

135
Kouvaros, S. & Papatheodoropoulos, C. (2016). Theta burst stimulation-induced LTP: Differences and
similarities between the dorsal and ventral CA1 hippocampal synapses. Hippocampus, 26, 1542-
1559.
Kravets, J. L., Reyes, B. A., Unterwald, E. M., & Van Bockstaele, E. J. (2015). Direct targeting of
peptidergic amygdalar neurons by noradrenergic afferents: linking stress-integrative circuitry.
Brain Struct.Funct., 220, 541-558.
Krishnan, V., Han, M. H., Graham, D. L., Berton, O., Renthal, W., Laplant, Q., Graham, A., Lutter, M.,
Lagance, D. C., Ghose, S., Reister, R., Tannous, P., Green, T. A., Neve, R .L., Chakravarty, S.,
Kumar, A., Eish, A. J., Self, D. W.,Lee, F. S., Tamminga, C. A., Cooper, D. C., Gershenfeld, H.
K., & Nestler, E. J. (2007). Molecular adaptations underlying susceptibility and resistance to
social defeat in brain reward regions. Cell., 131, 391-404.
Krugers, H. J., Alfarez, D. N., Karst, H., Parashkouhi, K., van, G. N., & Joels, M. (2005). Corticosterone
shifts different forms of synaptic potentiation in opposite directions. Hippocampus, 15, 697-703.
Kuo, T., McQueen, A., Chen, T. C., & Wang, J. C. (2015). Regulation of Glucose Homeostasis by
Glucocorticoids. Adv.Exp.Med.Biol., 872, 99-126.
Ladd-Acosta, C., Pevsner, J., Sabunciyan, S, Yolken, R.H., Webster, M.J., Dinkins, T., Callinan, P.A., Fan, J.B.,
Potash, J.B., & Feinberg, A.P. (2007). DNA methylation signatures within the human brain. Am. J.
Hum. Genet., 81, 1304-15.
Lana, D., Iovino, L., Nosi, D., Wenk, G. L., & Giovannini, M. G. (2016). The neuron-astrocyte-
microglia triad involvement in neuroinflammaging mechanisms in the CA3 hippocampus of
memory-impaired aged rats. Exp.Gerontol., 83, 71-88.
Lau, C., Hebert, M., Vani, M. A., Walling, S., Hayley, S., Lagace, D. C. et al. (2016). Absence of
neurogenic response following robust predator-induced stress response. Neuroscience, 339, 276-
286.

136
Lavicky, J. & Dunn, A. J. (1993). Corticotropin-releasing factor stimulates catecholamine release in
hypothalamus and prefrontal cortex in freely moving rats as assessed by microdialysis.
J.Neurochem., 60, 602-612.
Lawson, L. J., Perry, V. H., Dri, P., & Gordon, S. (1990). Heterogeneity in the distribution and
morphology of microglia in the normal adult mouse brain. Neuroscience, 39, 151-170.
Lee, E. & Son, H. (2009). Adult hippocampal neurogenesis and related neurotrophic factors. BMB.Rep.,
42, 239-244.
Lee, J. C., Yau, S. Y., Lee, T. M., Lau, B. W., & So, K. F. (2016). Voluntary wheel running reverses the
decrease in subventricular zone neurogenesis caused by corticosterone. Cell Transplant..
Lee, M. C., Inoue, K., Okamoto, M., Liu, Y. F., Matsui, T., Yook, J. S. et al. (2013). Voluntary
resistance running induces increased hippocampal neurogenesis in rats comparable to load-free
running. Neurosci.Lett., 537, 6-10.
Lee, M. K., Graham, S. N., & Gold, P. E. (1988). Memory enhancement with posttraining
intraventricular glucose injections in rats. Behav.Neurosci., 102, 591-595.
Leon, L. A., Castro Gomez, V., Lira Brandao, M., Rodrigues, C., Cardenas, L. F., & Landeira-
Fernandez, J. (2013). Corticosterone plasma concentrations in Carioca Highand Low-conditioned
freezing rats after a fear conditioned task. Avances en Psicología Latinoamericana, 31, 279-287.
Lepine, J. P. (2002). The epidemiology of anxiety disorders: prevalence and societal costs.
J.Clin.Psychiatry, 63 Suppl 14, 4-8.
Lerner, R. M. (1978). Nature, Nurture, and Dynamic Interactionism. Human Development, 21, 20.
Lesting, J., Narayanan, R. T., Kluge, C., Sangha, S., Seidenbecher, T., & Pape, H. C. (2011). Patterns of
coupled theta activity in amygdala-hippocampal-prefrontal cortical circuits during fear extinction.
PLoS.One., 6, e21714.
Leutgeb, J. K. & Moser, E. I. (2007). Enigmas of the dentate gyrus. Neuron, 55, 176-178.

137
Leutgeb, S., Leutgeb, J. K., Barnes, C. A., Moser, E. I., McNaughton, B. L., & Moser, M. B. (2005).
Independent codes for spatial and episodic memory in hippocampal neuronal ensembles. Science,
309, 619-623.
Liberzon, I., Lopez, J. F., Flagel, S. B., Vazquez, D. M., & Young, E. A. (1999). Differential regulation
of hippocampal glucocorticoid receptors mRNA and fast feedback: relevance to post-traumatic
stress disorder. J.Neuroendocrinol., 11, 11-17.
Lim, J.P., & Brunet, A. (2013). Bridging the transgenerational gap with epigenetic memory. Trends
Genet., 29, 176-186.
Lipton, P. A., White, J. A., & Eichenbaum, H. (2007). Disambiguation of overlapping experiences by
neurons in the medial entorhinal cortex. J.Neurosci., 27, 5787-5795.
Lisi, L., Stigliano, E., Lauriola, L., Navarra, P., & Dello, R. C. (2014). Proinflammatory-activated
glioma cells induce a switch in microglial polarization and activation status, from a predominant
M2b phenotype to a mixture of M1 and M2a/B polarized cells. ASN.Neuro., 6, 171-183.
Lisman, J., Schulman, H., & Cline, H. (2002). The molecular basis of CaMKII function in synaptic and
behavioural memory. Nat.Rev.Neurosci., 3, 175-190.
Lissek, S., Rabin, S. T., McDowell, D. J., Dvir, S., Bradford, D. E., Graci, M., Pine, D. S., & GRillon, C.
(2009). Impaired discriminative fear-conditioning resultinf from elevated fear responding to
learned safesty cues among individuals with panic disorder. Behavior Research and Therapy., 47,
111-118.
Lissek, S., Kaczkurkin, A. N., Rabin, S., Geraci, M., Pine, D. S., & Grillon, C. (2014). Generalized
anxiety disorder is associated with overgeneralization of classically conditioned fear.
Biol.Psychiatry, 75, 909-915.
Lister R, Pelizzola M, Dowen RH, Hawkins RD, Hon G, et al. (2009). Human DNA methylomes at base
resolution show widespread epigenomic differences. Nature., 462, 315–322.
Liu, L., Wang, C., Ni, X., & Sun, J. (2007). A rapid inhibition of NMDA receptor current by
corticosterone in cultured hippocampal neurons. Neurosci.Lett., 420, 245-250.

138
Liu, L., Wong, T. P., Pozza, M. F., Lingenhoehl, K., Wang, Y., Sheng, M. et al. (2004). Role of NMDA
receptor subtypes in governing the direction of hippocampal synaptic plasticity. Science, 304,
1021-1024.
Lu, C., Huang, X., Zhang, X., Roensch, K., Cao, Q., Nakayama, K.I., Blazar, B.R., Zeng, Y., & Zhou, X.
(2011). miR-221 and miR-155 regulate human dendritic cell development, apoptosis, and IL-12
production through targeting of p27kip1, KPC1, and SOCS-1. Blood., 117, 4293-4303.
Lucassen, P. J., Pruessner, J., Sousa, N., Almeida, O. F., Van Dam, A. M., Rajkowska, G. et al. (2014).
Neuropathology of stress. Acta Neuropathol., 127, 109-135.
Luo, Y., He, Q., Kuang, G., Jiang, Q., & Yang, J. (2014). PPAR-alpha and PPAR-beta expression
changes in the hippocampus of rats undergoing global cerebral ischemia/reperfusion due to
PPAR-gamma status. Behav.Brain Funct., 10, 21.
Luscher, C. & Malenka, R. C. (2012). NMDA receptor-dependent long-term potentiation and long-term
depression (LTP/LTD). Cold Spring Harb.Perspect.Biol., 4.
Macri, S., Zoratto, F., & Laviola, G. (2011). Early-stress regulates resilience, vulnerability and
experimental validity in laboratory rodents through mother-offspring hormonal transfer.
Neurosci.Biobehav.Rev., 35, 1534-1543.
Madronal, N., Delgado-Garcia, J. M., Fernandez-Guizan, A., Chatterjee, J., Kohn, M., Mattucci, C. et al.
(2016). Rapid erasure of hippocampal memory following inhibition of dentate gyrus granule
cells. Nat.Commun., 7, 10923.
Maes, M., Lin, A. H., Delmeire, L., Van, G. A., Kenis, G., De, J. R. et al. (1999). Elevated serum
interleukin-6 (IL-6) and IL-6 receptor concentrations in posttraumatic stress disorder following
accidental man-made traumatic events. Biol.Psychiatry, 45, 833-839.
Maes, M., Song, C., Lin, A., De, J. R., Van, G. A., Kenis, G. et al. (1998). The effects of psychological
stress on humans: increased production of pro-inflammatory cytokines and a Th1-like response in
stress-induced anxiety. Cytokine, 10, 313-318.

139
Magarinos, A. M., Verdugo, J. M., & McEwen, B. S. (1997). Chronic stress alters synaptic terminal
structure in hippocampus. Proc.Natl.Acad.Sci.U.S.A, 94, 14002-14008.
Maggio, N. & Segal, M. (2007). Striking variations in corticosteroid modulation of long-term
potentiation along the septotemporal axis of the hippocampus. J.Neurosci., 27, 5757-5765.
Malerba, P., Krishnan, G. P., Fellous, J. M., & Bazhenov, M. (2016). Hippocampal CA1 Ripples as
Inhibitory Transients. PLoS.Comput.Biol., 12, e1004880.
Maletic, V., Robinson, M., Oakes, T., Iyengar, S., Ball, S. G., & Russell, J. (2007). Neurobiology of
depression: an integrated view of key findings. Int.J.Clin.Pract., 61, 2030-2040.
Manabe, T. & Nicoll, R. A. (1994). Long-term potentiation: evidence against an increase in transmitter
release probability in the CA1 region of the hippocampus. Science, 265, 1888-1892.
Mantovani, A., Sica, A., Sozzani, S., Allavena, P., Vecchi, A., & Locati, M. (2004). The chemokine
system in diverse forms of macrophage activation and polarization. Trends Immunol., 25, 677-
686.
Mantovani, A., Sozzani, S., Locati, M., Allavena, P., & Sica, A. (2002). Macrophage polarization:
tumor-associated macrophages as a paradigm for polarized M2 mononuclear phagocytes. Trends
Immunol., 23, 549-555.
Marchand, A. R., Barbelivien, A., Seillier, A., Herbeaux, K., Sarrieau, A., & Majchzak, M. (2007).
Contribution of corticosterone to cued versus contextual fear in rats. Behav Brain Res., 183, 101-
10.
Maren, S., Aharonov, G., & Fanselow, M. S. (1997). Neurotoxic lesions of the dorsal hippocampus and
Pavlovian fear conditioning in rats. Behav.Brain Res., 88, 261-274.
Marriott, I. & Bost, K. L. (2000). IL-4 and IFN-gamma up-regulate substance P receptor expression in
murine peritoneal macrophages. J.Immunol., 165, 182-191.
Marriott, I. & Bost, K. L. (2001). Expression of authentic substance P receptors in murine and human
dendritic cells. J.Neuroimmunol., 114, 131-141.

140
Martin, S. J. & Morris, R. G. (2002). New life in an old idea: the synaptic plasticity and memory
hypothesis revisited. Hippocampus, 12, 609-636.
Martinez, F. O. & Gordon, S. (2014). The M1 and M2 paradigm of macrophage activation: time for
reassessment. F1000Prime.Rep., 6, 13.
Martinon, F., Burns, K., & Tschopp, J. (2002). The inflammasome: a molecular platform triggering
activation of inflammatory caspases and processing of proIL-beta. Mol.Cell, 10, 417-426.
May, R. C., Bailey, J. L., Mitch, W. E., Masud, T., & England, B. K. (1996). Glucocorticoids and
acidosis stimulate protein and amino acid catabolism in vivo. Kidney Int., 49, 679-683.
Maysinger, D. & Zhang, I. (2016). Nutritional and Nanotechnological Modulators of Microglia. Front
Immunol., 7, 270.
McCall, J. G., Al-Hasani, R., Siuda, E. R., Hong, D. Y., Norris, A. J., Ford, C. P. et al. (2015). CRH
Engagement of the Locus Coeruleus Noradrenergic System Mediates Stress-Induced Anxiety.
Neuron, 87, 605-620.
McEwen, B. S. (2000). The neurobiology of stress: from serendipity to clinical relevance. Brain Res.,
886, 172-189.
McEwen, B. S. (2007). Physiology and neurobiology of stress and adaptation: central role of the brain.
Physiol Rev., 87, 873-904.
McEwen, B. S. & Gianaros, P. J. (2010). Central role of the brain in stress and adaptation: links to
socioeconomic status, health, and disease. Ann.N.Y.Acad.Sci., 1186, 190-222.
McEwen, B. S. & Sapolsky, R. M. (1995). Stress and cognitive function. Curr.Opin.Neurobiol., 5, 205-
216.
McEwen, B. S. & Wingfield, J. C. (2003). The concept of allostasis in biology and biomedicine.
Horm.Behav., 43, 2-15.
McGaugh, J. L. (2000). Memory--a century of consolidation. Science, 287, 248-251.

141
McGaugh, J. L. & Roozendaal, B. (2002). Role of adrenal stress hormones in forming lasting memories
in the brain. Curr.Opin.Neurobiol., 12, 205-210.
McGowan, P. O., Sasaki, A., D'Alessio, A. C., Dymov, S., Labonte, B., Szyf, M., Turecki, G., &
Meaney, M. J. (2009). Epigenetic regulation of the glucocorticoid receptor in human brain
associates with childhood abuse. Nat Neurosci., 12, 342-8.
McLaren, I. P. & Mackintosh, N. J. (2002). Associative learning and elemental representation: II.
Generalization and discrimination. Anim Learn.Behav, 30, 177-200.
Meireles, M., Marques, C., Norberto, S., Santos, P., Fernandes, I., Mateus, N. et al. (2016). Anthocyanin
effects on microglia M1/M2 phenotype: Consequence on neuronal fractalkine expression.
Behav.Brain Res., 305, 223-228.
Meunier, M., Hadfield, W., Bachevalier, J., & Murray, E. A. (1996). Effects of rhinal cortex lesions
combined with hippocampectomy on visual recognition memory in rhesus monkeys.
J.Neurophysiol., 75, 1190-1205.
Milior, G., Lecours, C., Samson, L., Bisht, K., Poggini, S., Pagani, F. et al. (2016). Fractalkine receptor
deficiency impairs microglial and neuronal responsiveness to chronic stress. Brain
Behav.Immun., 55, 114-125.
Mongiat, L. A., Esposito, M. S., Lombardi, G., & Schinder, A. F. (2009). Reliable activation of
immature neurons in the adult hippocampus. PLoS.One., 4, e5320.
Mongiat, L. A. & Schinder, A. F. (2011). Adult neurogenesis and the plasticity of the dentate gyrus
network. Eur.J.Neurosci., 33, 1055-1061.
Monje, M. L., Toda, H., & Palmer, T. D. (2003). Inflammatory blockade restores adult hippocampal
neurogenesis. Science, 302, 1760-1765.
Monji, A., Kato, T., & Kanba, S. (2009). Cytokines and schizophrenia: Microglia hypothesis of
schizophrenia. Psychiatry Clin.Neurosci., 63, 257-265.

142
Monji, A., Kato, T. A., Mizoguchi, Y., Horikawa, H., Seki, Y., Kasai, M. et al. (2013).
Neuroinflammation in schizophrenia especially focused on the role of microglia.
Prog.Neuropsychopharmacol.Biol.Psychiatry, 42, 115-121.
Morilak, D. A., Barrera, G., Echevarria, D. J., Garcia, A. S., Hernandez, A., Ma, S. et al. (2005). Role of
brain norepinephrine in the behavioral response to stress.
Prog.Neuropsychopharmacol.Biol.Psychiatry, 29, 1214-1224.
Morris, G. P., Clark, I. A., Zinn, R., & Vissel, B. (2013). Microglia: a new frontier for synaptic
plasticity, learning and memory, and neurodegenerative disease research. Neurobiol.Learn.Mem.,
105, 40-53.
Moser, M. B., Moser, E. I., Forrest, E., Andersen, P., & Morris, R. G. (1995). Spatial learning with a
minislab in the dorsal hippocampus. Proc.Natl.Acad.Sci.U.S.A, 92, 9697-9701.
Motta, S. C., Carobrez, A. P., & Canteras, N. S. (2017). The periaqueductal gray and primal emotional
processing critical to influence complex defensive responses, fear learning and reward seeking.
Neurosci.Biobehav.Rev., 76, 39-47.
Mountjoy, K. G. (2010). Functions for pro-opiomelanocortin-derived peptides in obesity and diabetes.
Biochem.J., 428, 305-324.
Mountjoy, K. G. (2015). Pro-Opiomelanocortin (POMC) Neurones, POMC-Derived Peptides,
Melanocortin Receptors and Obesity: How Understanding of this System has Changed Over the
Last Decade. J.Neuroendocrinol., 27, 406-418.
Munhoz, C. D., Garcia-Bueno, B., Madrigal, J. L., Lepsch, L. B., Scavone, C., & Leza, J.C. (2008).
Stress-induces neuroinflammation:mechanisms and new pharmacological targets. Braz J Med
Biol Res., 41, 1037-46.
Murphy, L. R. (1996). Stress management in work settings: a critical review of the health effects.
Am.J.Health Promot., 11, 112-135.
Murray, E. A. & Mishkin, M. (1986). Visual recognition in monkeys following rhinal cortical ablations
combined with either amygdalectomy or hippocampectomy. J.Neurosci., 6, 1991-2003.

143
Murugan, M., Ling, E. A., & Kaur, C. (2013). Glutamate receptors in microglia.
CNS.Neurol.Disord.Drug Targets., 12, 773-784.
Myers, B., McKlveen, J. M., & Herman, J. P. (2014). Glucocorticoid actions on synapses, circuits, and
behavior: implications for the energetics of stress. Front Neuroendocrinol., 35, 180-196.
Nair, A. & Bonneau, R. H. (2006). Stress-induced elevation of glucocorticoids increases microglia
proliferation through NMDA receptor activation. J.Neuroimmunol., 171, 72-85.
Narayanan, R. T., Seidenbecher, T., Kluge, C., Bergado, J., Stork, O., & Pape, H. C. (2007). Dissociated
theta phase synchronization in amygdalo- hippocampal circuits during various stages of fear
memory. Eur.J.Neurosci., 25, 1823-1831.
Nashold, B. S., Jr., Wilson, W. P., & Slaughter, D. G. (1969). Sensations evoked by stimulation in the
midbrain of man. J.Neurosurg., 30, 14-24.
Nezi, M., Mastorakos, G., & Mouslech, Z. (2000). Corticotropin Releasing Hormone And The
Immune/Inflammatory Response.
Nguyen, K. T., Deak, T., Owens, S. M., Kohno, T., Fleshner, M., Watkins, L. R. et al. (1998). Exposure
to acute stress induces brain interleukin-1beta protein in the rat. J.Neurosci., 18, 2239-2246.
Nguyen, K. T., Deak, T., Will, M. J., Hansen, M. K., Hunsaker, B. N., Fleshner, M. et al. (2000).
Timecourse and corticosterone sensitivity of the brain, pituitary, and serum interleukin-1beta
protein response to acute stress. Brain Res., 859, 193-201.
Nicolaides, N. C., Charmandari, E., Chrousos, G. P., & Kino, T. (2014). Circadian endocrine rhythms:
the hypothalamic-pituitary-adrenal axis and its actions. Ann.N.Y.Acad.Sci., 1318, 71-80.
Nordby, T., Torras-Garcia, M., Portell-Cortes, I., & Costa-Miserachs, D. (2006). Posttraining
epinephrine treatment reduces the need for extensive training. Physiol Behav, 89, 718-723.
O'Connor, K. A., Johnson, J. D., Hansen, M. K., Wieseler Frank, J. L., Maksimova, E., Watkins, L. R. et
al. (2003). Peripheral and central proinflammatory cytokine response to a severe acute stressor.
Brain Res., 991, 123-132.

144
O'Connor, T. M., O'Connell, J., O'Brien, D. I., Goode, T., Bredin, C. P., & Shanahan, F. (2004). The role
of substance P in inflammatory disease. J.Cell Physiol, 201, 167-180.
Oitzl, M. S. & de Kloet, E. R. (1992). Selective corticosteroid antagonists modulate specific aspects of
spatial orientation learning. Behav.Neurosci., 106, 62-71.
Okuda, S., Roozendaal, B., & McGaugh, J. L. (2004). Glucocorticoid effects on object recognition
memory require training-associated emotional arousal. Proc.Natl.Acad.Sci.U.S.A, 101, 853-858.
Ono, N., Bedran de Castro, J. C., & McCann, S. M. (1985). Ultrashort-loop positive feedback of
corticotropin (ACTH)-releasing factor to enhance ACTH release in stress.
Proc.Natl.Acad.Sci.U.S.A, 82, 3528-3531.
Osborne, D. M., Pearson-Leary, J., & McNay, E. C. (2015). The neuroenergetics of stress hormones in
the hippocampus and implications for memory. Front Neurosci., 9, 164.
Oster, H., Challet, E., Ott, V., Arvat, E., de Kloet, E. R., Dijk, D. J. et al. (2016). The functional and
clinical significance of the 24-h rhythm of circulating glucocorticoids. Endocr.Rev., er20151080.
Padilla, E., Barret, D., Shumake, J., & Gonzalez-Lima, F. (2009). Strain, sex, and open-field behavior:
factors underlying the genetic susceptibility to helplessness. Behav Brain Res., 201, 257-64.
Panksepp, J. (2011). Cross-species affective neuroscience decoding of the primal affective experiences
of humans and related animals. PLoS.One., 6, e21236.
Park, J. Y., Kim, K. A., Kang, M. H., Kim, S. L., & Shin, J. G. (2004). Effect of rifampin on the
pharmacokinetics of rosiglitazone in healthy subjects. Clin.Pharmacol.Ther., 75, 157-162.
Pascual, O., Achour, S., Rostaing, P., Triller, A., & Bessis, A. (2012). Microglia activation triggers
astrocyte-mediated modulation of excitatory neurotransmission. PNAS, 109, 1009-1010.
Pascual, O., Ben, A. S., Rostaing, P., Triller, A., & Bessis, A. (2012). Microglia activation triggers
astrocyte-mediated modulation of excitatory neurotransmission. Proc.Natl.Acad.Sci.U.S.A, 109,
E197-E205.

145
Payne, J. D., Jackson, E. D., Hoscheidt, S., Ryan, L., Jacobs, W. J., & Nadel, L. (2007). Stress
administered prior to encoding impairs neutral but enhances emotional long-term episodic
memories. Learn.Mem., 14, 861-868.
Pelletier, J. G. & Pare, D. (2004). Role of amygdala oscillations in the consolidation of emotional
memories. Biol.Psychiatry, 55, 559-562.
Pentkowski, N. S., Blanchard, D. C., Lever, C., Litvin, Y., & Blanchard, R. J. (2006). Effects of lesions
to the dorsal and ventral hippocampus on defensive behaviors in rats. Eur.J.Neurosci., 23, 2185-
2196.
Perederiy, J. V. & Westbrook, G. L. (2013). Structural plasticity in the dentate gyrus- revisiting a classic
injury model. Front Neural Circuits., 7, 17.
Perrine, S. A., Eagle, A. L., George, S. A., Mulo, K., Kohler, R. J., Gerard, J. et al. (2016). Severe,
multimodal stress exposure induces PTSD-like characteristics in a mouse model of single
prolonged stress. Behav.Brain Res., 303, 228-237.
Petrilli, V., Papin, S., & Tschopp, J. (2005). The inflammasome. Curr.Biol., 15, R581.
Pham, K., Nacher, J., Hof, P. R., & McEwen, B. S. (2003). Repeated restraint stress suppresses
neurogenesis and induces biphasic PSA-NCAM expression in the adult rat dentate gyrus.
Eur.J.Neurosci., 17, 879-886.
Phelps, E. A. (2004). Human emotion and memory: interactions of the amygdala and hippocampal
complex. Curr.Opin.Neurobiol., 14, 198-202.
Pinzón, C. E., Serrano, M. D., & Sanabria, M. D. (2017). Papel de la vía fosfatidilinositol 3 kinasa
(PI3K/Akt) en humanos. Rev.Cienc.Salud, 7.
Pires, G. N., Tufik, S., & Andersen, M.L. (2012). Grooming analysis algorithm: use in the relationship
between sleep deprivation and anxiety-like behavior.Progress in Neuro-psychopharmacology &
Biological Psychiatry., 41, 6-10.

146
Pisanu, A., Lecca, D., Mulas, G., Wardas, J., Simbula, G., Spiga, S. et al. (2014). Dynamic changes in
pro- and anti-inflammatory cytokines in microglia after PPAR-gamma agonist neuroprotective
treatment in the MPTPp mouse model of progressive Parkinson's disease. Neurobiol.Dis., 71,
280-291.
Popa, D., Duvarci, S., Popescu, A. T., Lena, C., & Pare, D. (2010). Coherent amygdalocortical theta
promotes fear memory consolidation during paradoxical sleep. Proc.Natl.Acad.Sci.U.S.A, 107,
6516-6519.
Pothuizen, H. H., Zhang, W. N., Jongen-Relo, A. L., Feldon, J., & Yee, B. K. (2004). Dissociation of
function between the dorsal and the ventral hippocampus in spatial learning abilities of the rat: a
within-subject, within-task comparison of reference and working spatial memory.
Eur.J.Neurosci., 19, 705-712.
Pourcet, B. & Pineda-Torra, I. (2013). Transcriptional regulation of macrophage arginase 1 expression
and its role in atherosclerosis. Trends Cardiovasc.Med., 23, 143-152.
Prokhortchouk, E., & Defossez, P. (2008). The cell biology of DNA methylation in mammals.
Biochimica et biophysica Acta (BBA)- Molecular Cell Research., 1783, 2167-2173.
Prut, L., & Belzung, C. (22003). The open field as a paradigm to measure the effects of drugs on axiety-
like behaviors: a review. Eur J Pharmacol., 28, 3-33.
Ramos, A. (2008). Animal models of anxiety: do I need multiple tests? Trends Pharmacol.Sci., 29, 493-
498.
Rasley, A., Bost, K. L., Olson, J. K., Miller, S. D., & Marriott, I. (2002). Expression of functional NK-1
receptors in murine microglia. Glia, 37, 258-267.
Rechavi, O., Minevich, G., & Hobert, O. (2011). Transgenerational inheritance o fan acquired small
RNA-based antiviral response in C. elegans. Cell., 147, 1248-56.
Reus, G. Z., Fries, G. R., Stertz, L., Badawy, M., Passos, I. C., Barichello, T. et al. (2015). The role of
inflammation and microglial activation in the pathophysiology of psychiatric disorders.
Neuroscience, 300, 141-154.

147
Revest, J. M., Dupret, M., Koehl, C., Funk-Reiter, N., Grosjean, P. V., & Abrous, D. N. (2009). Adulta
hipocampal neurogénesis is involver in axiety-related behaviors. Molecular Psychiatry., 14, 959-
967.
Rex, A., Voigt, J. P. Gustedt, C., Beckett, S., & Fink, H. (2004). Axiolytic-like profile in wistar, but not
Sprague-Dawley rats in the social interaction test. Psychopharmacology (Berl)., 177, 23-34.
Rhen, T. & Cidlowski, J. A. (2005). Antiinflammatory action of glucocorticoids--new mechanisms for
old drugs. N.Engl.J.Med., 353, 1711-1723.
Richter-Levin, G. & Akirav, I. (2000). Amygdala-hippocampus dynamic interaction in relation to
memory. Mol.Neurobiol., 22, 11-20.
Rizvi, T. A., Ennis, M., Behbehani, M. M., & Shipley, M. T. (1991). Connections between the central
nucleus of the amygdala and the midbrain periaqueductal gray: topography and reciprocity.
J.Comp Neurol., 303, 121-131.
Roberson, E. D., English, J. D., & Sweatt, J. D. (1996). A biochemist's view of long-term potentiation.
Learn.Mem., 3, 1-24.
Rodriguez, J.J., Davies, H.A., Errington, M.L., Verkhrasky, A., Bliss, T.V.P., & Stewart. M.G. (2007).
ARG3.1/ARC expression in hippocampal dentate gyrus astrocytes: ultrastructural evidence and
co-localization with glial fibrillary acidic protein. J Cell Mol Med., 12, 671-678.
Rohleder, N., Joksimovic, L., Wolf, J. M., & Kirschbaum, C. (2004). Hypocortisolism and increased
glucocorticoid sensitivity of pro-Inflammatory cytokine production in Bosnian war refugees with
posttraumatic stress disorder. Biol.Psychiatry, 55, 745-751.
Roosevelt, R. W., Smith, D. C., Clough, R. W., Jensen, R. A., & Browning, R. A. (2006). Increased
extracellular concentrations of norepinephrine in cortex and hippocampus following vagus nerve
stimulation in the rat. Brain Res., 1119, 124-132.
Roozendaal, B., Brunson, K. L., Holloway, B. L., McGaugh, J. L., & Baram, T. Z. (2002). Involvement
of stress-released corticotropin-releasing hormone in the basolateral amygdala in regulating
memory consolidation. Proc.Natl.Acad.Sci.U.S.A, 99, 13908-13913.

148
Roozendaal, B., McEwen, B. S., & Chattarji, S. (2009). Stress, memory and the amygdala.
Nat.Rev.Neurosci., 10, 423-433.
Roozendaal, B. & McGaugh, J. L. (1996). The memory-modulatory effects of glucocorticoids depend on
an intact stria terminalis. Brain Res., 709, 243-250.
Roozendaal, B., McReynolds, J. R., Van der Zee, E. A., Lee, S., McGaugh, J. L., & McIntyre, C. K.
(2009). Glucocorticoid effects on memory consolidation depend on functional interactions
between the medial prefrontal cortex and basolateral amygdala. J.Neurosci., 29, 14299-14308.
Rosch, P. (1979). Stress and Illness. JAMA, 242, 427-428.
Rosi, S., Ramirez-Amaya, V., Vazdarjanova, A., Esparza, E. E., Larkin, P. B., Fike, J. R. et al. (2009).
Accuracy of hippocampal network activity is disrupted by neuroinflammation: rescue by
memantine. Brain, 132, 2464-2477.
Rosso, I. M., Crowley, D. J., Silveri, M M., Rauch, S. L., & Jensen J.E. (2017). Hippocampus glutamae
and N-acetyl aspartate markers of excitotoxic neural compromise in posttraumatic stress disorder.
Neuropsychopharmacology., 42, 1698-1705.
Roth, T. L., Lubin, F. D., Funk, A. J., Sweatt, J. D. (2009). Lasting epigenetic influence of early-life
adversity on the BDNF gene. Biol Psychiatry., 65, 760-9.
Roy, S., Roy, S. J., Pinard, S., Agulleiro, M. J., Cerda-Reverter, J. M., Parent, J. L. et al. (2012). The C-
terminal domains of melanocortin-2 receptor (MC2R) accessory proteins (MRAP1) influence
their localization and ACTH-induced cAMP production. Gen.Comp Endocrinol., 176, 265-274.
Royer Ghiglione, C. (2011). Recensión de tesis: El mejoramiento de la calidad de vida laboral como
estrategia para vigorizar la capacidad de gestión municipal. DAAPGE, 157-162.
Ryan, B. K., Vollmayr, B., Klyubin, I., Gass, P., & Rowan, M. J. (2010). Persistent inhibition of
hippocampal long-term potentiation in vivo by learned helplessness stress. Hippocampus, 20,
758-767.

149
Sadowski, R. N., Jackson, G. R., Wieczorek, L., & Gold, P. E. (2009). Effects of stress, corticosterone,
and epinephrine administration on learning in place and response tasks. Behav.Brain Res., 205,
19-25.
Sah, P., Faber, E. S., Lopez De, A. M., & Power, J. (2003a). The amygdaloid complex: anatomy and
physiology. Physiol Rev., 83, 803-834.
Sah, P., Faber, E. S., Lopez De, A. M., & Power, J. (2003b). The amygdaloid complex: anatomy and
physiology. Physiol Rev., 83, 803-834.
Sapolsky, R. M. (1985). A mechanism for glucocorticoid toxicity in the hippocampus: increased
neuronal vulnerability to metabolic insults. J.Neurosci., 5, 1228-1232.
Sapolsky, R. M., Krey, L. C., & McEwen, B. S. (1984). Glucocorticoid-sensitive hippocampal neurons
are involved in terminating the adrenocortical stress response. Proc.Natl.Acad.Sci.U.S.A, 81,
6174-6177.
Sarabdjitsingh, R. A., Spiga, F., Oitzl, M. S., Kershaw, Y., Meijer, O. C., Lightman, S. L. et al. (2010).
Recovery from disrupted ultradian glucocorticoid rhythmicity reveals a dissociation between
hormonal and behavioural stress responsiveness. J.Neuroendocrinol., 22, 862-871.
Satoh, T., Baba, M., Nakatsuka, D., Ishikawa, Y., Aburatani, H., Furuta, K. et al. (2003). Role of heme
oxygenase-1 protein in the neuroprotective effects of cyclopentenone prostaglandin derivatives
under oxidative stress. Eur.J.Neurosci., 17, 2249-2255.
Scharf, S. H. & Schmidt, M. V. (2012). Animal models of stress vulnerability and resilience in
translational research. Curr.Psychiatry Rep., 14, 159-165.
Schmidt, E. D., Schoffelmeer, A. N., De Vries, T. J., Wardeh, G., Dogterom, G., Bol, J. G. et al. (2001).
A single administration of interleukin-1 or amphetamine induces long-lasting increases in evoked
noradrenaline release in the hypothalamus and sensitization of ACTH and corticosterone
responses in rats. Eur.J.Neurosci., 13, 1923-1930.
Schoenfeld, T. J. & Gould, E. (2012). Stress, stress hormones, and adult neurogenesis. Exp.Neurol., 233,
12-21.

150
Schwabe, L., Joels, M., Roozendaal, B., Wolf, O. T., & Oitzl, M. S. (2012). Stress effects on memory: an
update and integration. Neurosci.Biobehav.Rev., 36, 1740-1749.
Scott, L. V. & Dinan, T. G. (1998). Vasopressin and the regulation of hypothalamic-pituitary-adrenal
axis function: implications for the pathophysiology of depression. Life Sci., 62, 1985-1998.
Segal, M., Richter-Levin, G., & Maggio, N. (2010). Stress-induced dynamic routing of hippocampal
connectivity: a hypothesis. Hippocampus, 20, 1332-1338.
Seibenhener, M. L. & Wooten, M. C. (2015). Use of the Open Field Maze to measure locomotor and
anxiety-like behavior in mice. J.Vis.Exp., e52434.
Sellner, S., Paricio-Montesinos, R., Spiess, A., Masuch, A., Erny, D., Harsan, L. A. et al. (2016).
Microglial CX3CR1 promotes adult neurogenesis by inhibiting Sirt 1/p65 signaling independent
of CX3CL1. Acta Neuropathol.Commun., 4, 102.
Selye, H. (1976). Forty years of stress research: principal remaining problems and misconceptions.
Can.Med.Assoc.J., 115, 53-56.
Shibata, M. & Suzuki, N. (2017). Exploring the role of microglia in cortical spreading depression in
neurological disease. J.Cereb.Blood Flow Metab, 37, 1182-1191.
Shigemori, T., Sakai, A., Takumi, T., Itoh, Y., & Suzuki, H. (2015). Altered Microglia in the Amygdala
Are Involved in Anxiety-related Behaviors of a Copy Number Variation Mouse Model of
Autism. J.Nippon Med.Sch, 82, 92-99.
Sibbe, M. & Kulik, A. (2016). GABAergic Regulation of Adult Hippocampal Neurogenesis.
Mol.Neurobiol..
Sierra, A., Encinas, J. M., Deudero, J. J., Chancey, J. H., Enikolopov, G., Overstreet-Wadiche, L. S. et
al. (2010). Microglia shape adult hippocampal neurogenesis through apoptosis-coupled
phagocytosis. Cell Stem Cell, 7, 483-495.
Sierra, A., Gottfried-Blackmore, A., Milner, T. A., McEwen, B. S., & Bulloch, K. (2008). Steroid
hormone receptor expression and function in microglia. Glia, 56, 659-674.

151
Sisti, H. M., Glass, A. L., & Shors, T. J. (2007). Neurogenesis and the spacing effect: learning over time
enhances memory and the survival of new neurons. Learn.Mem., 14, 368-375.
Smith, A.K., Conneely, K.N., Kilaru, V., Mercer, K.B., Weiss, T.E., Bradley, B., Tang, Y., Gillespie,
C.F., Cubells, J.F., & Ressel, K.J. (2011). Differential immune system DNA methylation an
cytokine regulation in post-traumatic stress disorder. Am J Med Genet B Neurospsychiatry
Genet., 156B, 700-8.
Smoak, K. A. & Cidlowski, J. A. (2004). Mechanisms of glucocorticoid receptor signaling during
inflammation. Mech.Ageing Dev., 125, 697-706.
Snyder, K., Wang, W. W., Han, R., McFadden, K., & Valentino, R. J. (2012). Corticotropin-releasing
factor in the norepinephrine nucleus, locus coeruleus, facilitates behavioral flexibility.
Neuropsychopharmacology, 37, 520-530.
Solstad, T., Yousif, H. N., & Sejnowski, T. J. (2014). Place cell rate remapping by CA3 recurrent
collaterals. PLoS.Comput.Biol., 10, e1003648.
Song, G. J., Nam, Y., Jo, M., Jung, M., Koo, J. Y., Cho, W. et al. (2016). A novel small-molecule
agonist of PPAR-gamma potentiates an anti-inflammatory M2 glial phenotype.
Neuropharmacology, 109, 159-169.
Sorrells, S. F., Caso, J. R., Munhoz, C. D., & Sapolsky, R. M. (2009). The stressed CNS: when
glucocorticoids aggravate inflammation. Neuron, 64, 33-39.
Spencer, T. J., Biederman, J., & Mick, E. (2007). Attention-deficit/hyperactivity disorder: diagnosis,
lifespan, comorbidities, and neurobiology. J.Pediatr.Psychol., 32, 631-642.
Spiers, H. J., Maguire, E. A., & Burgess, N. (2001). Hippocampal amnesia. Neurocase., 7, 357-382.
Spiller, A. E. & Racine, R. J. (1994). Transfer kindling between sites in the entorhinal cortex-perforant
path-dentate gyrus system. Brain Res., 635, 130-138.

152
Spivak, B., Shohat, B., Mester, R., Avraham, S., Gil-Ad, I., Bleich, A. et al. (1997). Elevated levels of
serum interleukin-1 beta in combat-related posttraumatic stress disorder. Biol.Psychiatry, 42,
345-348.
Squire, L. R. (1992). Memory and the hippocampus: a synthesis from findings with rats, monkeys, and
humans. Psychol.Rev., 99, 195-231.
Steimer, T. (2002). The biology of fear-and anxiety-related behaviors. Dialogues Clin Neurosci., 4, 231-
249.
Steinert, J.R., Chernova, T., & Forsythe, I.D. (2010). Nitric oxide signaling in brain fuction, dysfunction,
and dementia. Neuroscientist., 16, 435-52.
Steinert, J.R., Robinson, S.W., Tong, H., Haustein, M.D., Kopp-Scheinpflug, C., & Forsythe, I.D.
(2011). Nitric Oxide Is an Activity-Dependent Regulator of Targer Neuron Intrinsic Excitability.
Neuron., 71, 291-305.
Stepan, J., Dine, J., Fenzl, T., Polta, S. A., von, W. G., Wotjak, C. T. et al. (2012). Entorhinal theta-
frequency input to the dentate gyrus trisynaptically evokes hippocampal CA1 LTP. Front Neural
Circuits., 6, 64.
Stewart, W. F., Ricci, J. A., Chee, E., & Morganstein, D. (2003). Lost productive work time costs from
health conditions in the United States: results from the American Productivity Audit.
J.Occup.Environ.Med., 45, 1234-1246.
Streit, W. J., Mrak, R. E., & Griffin, W. S. (2004). Microglia and neuroinflammation: a pathological
perspective. J.Neuroinflammation., 1, 14.
Sugama, S., Fujita, M., Hashimoto, M., & Conti, B. (2007). Stress induced morphological microglial
activation in the rodent brain: involvement of interleukin-18. Neuroscience, 146, 1388-1399.
Sullivan, P. F., Neale, M. C., & Kendler, K. S. (2000). Genetic epidemiology of major depression:
review and meta-analysis. Am.J.Psychiatry, 157, 1552-1562.

153
Sun, H., Huang, Y., Yu, X., Li, Y., Yang, J., Li, R. et al. (2008). Peroxisome proliferator-activated
receptor gamma agonist, rosiglitazone, suppresses CD40 expression and attenuates inflammatory
responses after lithium pilocarpine-induced status epilepticus in rats. Int.J.Dev.Neurosci., 26,
505-515.
Sun, R., Zhang, Z., Lei, Y., Liu, Y., Lu, C., Rong, H. et al. (2016). Hippocampal activation of microglia
may underlie the shared neurobiology of comorbid posttraumatic stress disorder and chronic
pain. Mol.Pain, 12.
Sun, Y., Hunt, S., & Sah, P. (2015). Norepinephrine and Corticotropin-Releasing Hormone: Partners in
the Neural Circuits that Underpin Stress and Anxiety. Neuron, 87, 468-470.
Sutanto, W. & de Kloet, E. R. (1994). The use of various animal models in the study of stress and stress-
related phenomena. Lab Anim, 28, 293-306.
Shvil, E., Rusch, H. L., Sullivan, G. M., Neira, Y. (2013). Neural, psychophysiological, and behavioral
markers of fear precessing in PTSD: a review of the literature. Curr Psychiatry Rep., 15.
Sweep, F., Rijnkels, C., & Hermus, A. (1991). Activation of the hypothalamus-pituitary-adrenal axis by
cytokines. Acta Endocrinol.(Copenh), 125 Suppl 1, 84-91.
Sweis, B. M., Veverka, K. K., Dhillon, E. S., Urban, J. H., & Lucas, L. R. (2013). Individual differences
in the effects of chronic stress on memory: behavioral and neurochemical correlates of resiliency.
Neuroscience, 246, 142-159.
Szalay, K. S., De, W. D., Stark, E., & Folly, G. (1985). Structure-activity studies with ACTH/alpha-
MSH fragments on corticosteroid secretion of isolated zona glomerulosa and fasciculata cells.
Regul.Pept., 11, 187-192.
Tanaka, J., Fujita, H., Matsuda, S., Toku, K., Sakanaka, M., & Maeda, N. (1997). Glucocorticoid- and
mineralocorticoid receptors in microglial cells: the two receptors mediate differential effects of
corticosteroids. Glia, 20, 23-37.

154
Tanaka, S., Ide, M., Shibutani, T., Ohtaki, H., Numazawa, S., Shioda, S. et al. (2006).
Lipopolysaccharide-induced microglial activation induces learning and memory deficits without
neuronal cell death in rats. J.Neurosci.Res., 83, 557-566.
Tay, T. L., Savage, J., Hui, C. W., Bisht, K., & Tremblay, M. E. (2016). Microglia across the lifespan:
from origin to function in brain development, plasticity and cognition. J.Physiol.
Tikka, T., Fiebich, B. L., Goldsteins, G., Keinanen, R., & Koistinaho, J. (2001). Minocycline, a
tetracycline derivative, is neuroprotective against excitotoxicity by inhibiting activation and
proliferation of microglia. J.Neurosci., 21, 2580-2588.
Torres, R. C., Magalhaes, N. S., PM, E. S., Martins, M. A., & Carvalho, V. F. (2016). Activation of
PPAR-gamma reduces HPA axis activity in diabetic rats by up-regulating PI3K expression.
Exp.Mol.Pathol., 101, 290-301.
Tozuka, Y., Fukuda, S., Namba, T., Seki, T., & Hisatsune, T. (2005). GABAergic excitation promotes
neuronal differentiation in adult hippocampal progenitor cells. Neuron, 47, 803-815.
Tse, Y. C., Bagot, R. C., Hutter, J. A., Wong, A. S., & Wong, T. P. (2011). Modulation of synaptic
plasticity by stress hormone associates with plastic alteration of synaptic NMDA receptor in the
adult hippocampus. PLoS.One., 6, e27215.
Tsnakova, N., Renthal, W., Kumar, A., & Nestler, E.J. (2007). Epigenetic regulation in psychiatric
disorders. Nat Rev Neurosci., 8, 355-67.
Urban, N. N., Henze, D. A., & Barrionuevo, G. (2001). Revisiting the role of the hippocampal mossy
fiber synapse. Hippocampus, 11, 408-417.
Uchida, S., Hara, K., Kobayashi, A., Otsuki, K., Yamagata, H., Hobara, T., Suzuki, T., Miyata, N., &
Watanabe, T. (2011). Epihenetic status of GDNF in the ventral striatum determines susceptibility
and adaptation to daily stressful events. Neuron., 69, 359-72.
Van Bockstaele, E. J., Colago, E. E., & Valentino, R. J. (1996). Corticotropin-releasing factor-containing
axon terminals synapse onto catecholamine dendrites and may presynaptically modulate other

155
afferents in the rostral pole of the nucleus locus coeruleus in the rat brain. J.Comp Neurol., 364,
523-534.
van der Klink, J. J., Blonk, R. W., Schene, A. H., & van Dijk, F. J. (2001). The benefits of interventions
for work-related stress. Am.J.Public Health, 91, 270-276.
Vanhoutte, P., Barnier, J. V., Guibert, B., Pages, C., Besson, M. J., Hipskind, R. A. et al. (1999).
Glutamate induces phosphorylation of Elk-1 and CREB, along with c-fos activation, via an
extracellular signal-regulated kinase-dependent pathway in brain slices. Mol.Cell Biol., 19, 136-
146.
Vazdarjanova, A., McNaughton, B. L., Barnes, C. A., Worley, P. F., & Guzowski, J. F. (2002).
Experience-dependent coincident expression of the effector immediate-early genes arc and
Homer 1a in hippocampal and neocortical neuronal networks. J.Neurosci., 22, 10067-10071.
Vereker, E., O’Donnell, W., & Lynch, M.A. (2000). The inhibitory effect of interleukin-1 beta o long-term
potentiation is couples with increased activity of stress-activated protein kinases. J Neurosci., 20, 6811-9.
Vianna, D. M. & Brandao, M. L. (2003). Anatomical connections of the periaqueductal gray: specific
neural substrates for different kinds of fear. Braz.J.Med.Biol.Res., 36, 557-566.
Vianna, D. M., Graeff, F. G., Landeira-Fernandez, J., & Brandao, M. L. (2001). Lesion of the ventral
periaqueductal gray reduces conditioned fear but does not change freezing induced by stimulation
of the dorsal periaqueductal gray. Learn.Mem., 8, 164-169.
Vidal, J., Buwalda, B., Koolhaas, J. M. (2011). Differential long-term effects of social stress during
adolescence on aiety in wistar and wild-type rats. Behav processes., 87, 176-82.
Vivar, C. & van, P. H. (2013). Functional circuits of new neurons in the dentate gyrus. Front Neural
Circuits., 7, 15.
von Bohlen und, H. O. & Albrecht, D. (2002). Reciprocal connections of the hippocampal area CA1, the
lateral nucleus of the amygdala and cortical areas in a combined horizontal slice preparation.
Neurosci.Res., 44, 91-100.

156
von, B. R., Eugenin-von, B. L., & Eugenin, J. (2015). Microglial cell dysregulation in brain aging and
neurodegeneration. Front Aging Neurosci., 7, 124.
von, K. R., Hepp, U., Kraemer, B., Traber, R., Keel, M., Mica, L. et al. (2007). Evidence for low-grade
systemic proinflammatory activity in patients with posttraumatic stress disorder. J.Psychiatr.Res.,
41, 744-752.
Vonend, O., Turner, C. M., Chan, C. M., Loesch, A., Dell'Anna, G. C., Srai, K. S. et al. (2004).
Glomerular expression of the ATP-sensitive P2X receptor in diabetic and hypertensive rat
models. Kidney Int., 66, 157-166.
Vukovic, J., Colditz, M. J., Blackmore, D. G., Ruitenberg, M. J., & Bartlett, P. F. (2012). Microglia
modulate hippocampal neural precursor activity in response to exercise and aging. J.Neurosci.,
32, 6435-6443.
Walker, A. K., Kavelaars, A., Heijnen, C. J., & Dantzer, R. (2014). Neuroinflammation and comorbidity
of pain and depression. Pharmacol.Rev., 66, 80-101.
Walker, D. L. & Davis, M. (2002). The role of amygdala glutamate receptors in fear learning, fear-
potentiated startle, and extinction. Pharmacol.Biochem.Behav., 71, 379-392.
Walsh, R. N., Cummins, R. A. (1976). The open-field test: a critical review. Psuchol Bull., 83, 482-504.
Wang, X., Zaho, Q., Matta, R., Meng, X., Liu, X., Liu, C., Nelin, L.D., & Liu, Y. (2009). Inductible
Nitric-oxide Synthase Expression Is Regulared by Mitogen-activated Protein Kinase
Phosphatase-1. J Biol Chem., 284, 27123-34.
Wang, J., Li, J., Sheng, X., Zhao, H., Cao, X. D., Wang, Y. Q. et al. (2010). Beta-adrenoceptor mediated
surgery-induced production of pro-inflammatory cytokines in rat microglia cells.
J.Neuroimmunol., 223, 77-83.
Wang, L., Waltenberger, B., Pferschy-Wenzig, E. M., Blunder, M., Liu, X., Malainer, C. et al. (2014).
Natural product agonists of peroxisome proliferator-activated receptor gamma (PPARgamma): a
review. Biochem.Pharmacol., 92, 73-89.

157
Wang, Y., Cella, M., Mallinson, K., Ulrich, J. D., Young, K. L., Robinette, M. L. et al. (2015). TREM2
lipid sensing sustains the microglial response in an Alzheimer's disease model. Cell, 160, 1061-
1071.
Waugh, C. E., Hamilton, J. P., Chen, M. C., Joormann, J., & Gotlib, I. H. (2012). Neural temporal
dynamics of stress in comorbid major depressive disorder and social anxiety disorder.
Biol.Mood.Anxiety.Disord., 2, 11.
Wenzel, H. J., Buckmaster, P. S., Anderson, N. L., Wenzel, M. E., & Schwartzkroin, P. A. (1997).
Ultrastructural localization of neurotransmitter immunoreactivity in mossy cell axons and their
synaptic targets in the rat dentate gyrus. Hippocampus, 7, 559-570.
Weiss, I.C., Franklin, T.B., & Mansuy, I.M. (2011). Inheritable effect of unpredictable maternal
separation on behavioral responses in mice. Front Behav Neurosci., 5:3.
Whitaker, A. M., Glipin, N. W., Edwards, S. (2014). Animal models of post-traumatic stress disorder
and recent neurobiological insights. Behav Pharmacol., 25, 398-409.
Wiegert, O., Joels, M., & Krugers, H. (2006). Timing is essential for rapid effects of corticosterone on
synaptic potentiation in the mouse hippocampus. Learn.Mem., 13, 110-113.
Winters, B. D., Saksida, L. M., & Bussey, T. J. (2008). Object recognition memory: neurobiological
mechanisms of encoding, consolidation and retrieval. Neurosci.Biobehav.Rev., 32, 1055-1070.
Wohleb, E. S., Hanke, M. L., Corona, A. W., Powell, N. D., Stiner, L. M., Bailey, M. T. et al. (2011).
beta-Adrenergic receptor antagonism prevents anxiety-like behavior and microglial reactivity
induced by repeated social defeat. J.Neurosci., 31, 6277-6288.
Wolf, Y., Yona, S., Kim, K. W., & Jung, S. (2013). Microglia, seen from the CX3CR1 angle. Front Cell
Neurosci., 7, 26.
Wong, D. L., Tai, T. C., Wong-Faull, D. C., Claycomb, R., Meloni, E. G., Myers, K. M. et al. (2012).
Epinephrine: a short- and long-term regulator of stress and development of illness : a potential
new role for epinephrine in stress. Cell Mol.Neurobiol., 32, 737-748.

158
Wood, S. K., & Bhatnagar, S. (2015). Resilience to the effect of social stress: Evidence fron clinical and
preclinical studies on the role of coping strategies. Neurobiol Stress., 1, 164-173.
Wright, B. J., & Jackson, M. B. Long-term potentiation in hilar circuitry modulates gating by the dentate gyrus. J
Neurosci., 34, 9743-53.
Wu, Q., & Suzuki, M. (2006). Parental obesity and overweight affect the body-fat accumulation in the
offspring: the posible effect of a high fat diet through epigenetic inheritance. Obes. Rev., 7, 201-
8.
Yamamoto, A., Kakuta, H., & Sugimoto, Y. (2014). Involvement of glucocorticoid receptor activation
on anti-inflammatory effect induced by peroxisome proliferator-activated receptor gamma
agonist in mice. Int.Immunopharmacol., 22, 204-208.
Yang, H.Y., & Iadarola, M.J. (2003). Activation of spinal neuropeptide FF and the neuropeptide FF
receptor 2 during inflammatory hyperalgesia in rats. Neuroscience., 118, 179-87.
Yang, H. W., Hu, X. D., Zhang, H. M., Xin, W. J., Li, M. T., Zhang, T. et al. (2004). Roles of CaMKII,
PKA, and PKC in the induction and maintenance of LTP of C-fiber-evoked field potentials in rat
spinal dorsal horn. J.Neurophysiol., 91, 1122-1133.
Yang, S. S., Lin, L., Liu, Y., Wang, J., Chu, J., Zhang, T. et al. (2016). High Morphologic Plasticity of
Microglia/Macrophages Following Experimental Intracerebral Hemorrhage in Rats.
Int.J.Mol.Sci., 17.
Yehuda, R., Lowy, M. T., Southwick, S. M., Shaffer, D., & Giller, E. L., Jr. (1991). Lymphocyte
glucocorticoid receptor number in posttraumatic stress disorder. Am.J.Psychiatry, 148, 499-504.
Yohe, L. R., Suzuki, H., & Lucas, L. R. (2012). Aggression is suppressed by acute stress but induced by
chronic stress: immobilization effects on aggression, hormones, and cortical 5-HT(1B)/ striatal
dopamine D(2) receptor density. Cogn Affect.Behav.Neurosci., 12, 446-459.
Zárate, S., Cárdenas, F. P., Acevedo-Triana, C., Sarmiento-Bolaños, M. J., & Leon, L. A. (2014).
Efectos del estrés sobre los procesos de plasticidad y neurogénesis: una revisión. Universitas
psicologica, 13, 1181-1214.

159
Zhang, L., Dong, L., Li, Y., Hong, Z., & Wei, W. (2012). The microRNA miR-181c controls microglía-
mediated neuronal apoptosis by suppressing tumor necrosis factor. J Neuroinflammation., 9.
Zhao, C., Deng, W., & Gage, F. H. (2008). Mechanisms and functional implications of adult
neurogenesis. Cell, 132, 645-660.
Zhao, H., Wang, J., Gao, L., Wang, R., Liu, X., Gao, Z., Tao, Z., Xu, C., Song, J., Ji, X., & Luo, Y.
(2013). MiRNA-424 Protects Against Permanent Focal Cerebral Ischemia Injury in Mice
Involving Suppressing Microglia Activation. Stroke., 44, 1706-13.
Zhe, D., Fang, H., & Yuxiu, S. (2008). Expressions of hippocampal mineralocorticoid receptor (MR) and
glucocorticoid receptor (GR) in the single-prolonged stress-rats. Acta Histochem.Cytochem., 41,
89-95.
Zhou, D., Kusnecov, A. W., Shurin, M. R., DePaoli, M., & Rabin, B. S. (1993). Exposure to physical
and psychological stressors elevates plasma interleukin 6: relationship to the activation of
hypothalamic-pituitary-adrenal axis. Endocrinology, 133, 2523-2530.
Zhou, M., Bakker, E. H., Velzing, E. H., Berger, S., Oitzl, M., Joels, M. et al. (2010). Both
mineralocorticoid and glucocorticoid receptors regulate emotional memory in mice.
Neurobiol.Learn.Mem., 94, 530-537.
Zhu, J., Qu, C., Lu, X., & Zhang, S. (2014). Activation of microglia by histamine and substance P. Cell
Physiol Biochem., 34, 768-780.
Zhu, L. J., Liu, M. Y., Li, H., Liu, X., Chen, C., Han, Z. et al. (2014). The different roles of
glucocorticoids in the hippocampus and hypothalamus in chronic stress-induced HPA axis
hyperactivity. PLoS.One., 9, e97689.


















