







Idiomas
Páginas
Jurídico


Matriz extracelular y uniones celulares IBQ. Alma K Tzec NahuatBioquímica Avanzada

Matriz extracelular
La matriz extracelular (MEC) es una entidad estructuralmente compleja que rodea y soporta las células que se encuentran en los tejidos de los mamíferos. La MEC también es comúnmente conocida como tejido conectivo.

Estructura :1. Proteínas Estructurales: Colágeno y elastina.
2. Proteínas Especializadas: Fibrilina, fibronectina y laminina.
3. Proteoglicanos: Estos están compuestos de una proteína central a la cual se unen cadenas largas de unidades de disacáridos repetitivos llamados glicosaminoglicanos (GAGs) formando así compuestos complejos de alto peso molecular que conforman la MEC

Colágeno Estos 30 genes generan una serie de
proteínas que se combinan de varias formas para crear 20 diferentes tipos de fibrillas de colágeno.
Los tipos de colágeno I, II y III son los más abundantes y forman fibrillas de estructura similares.

Colágeno: Los colágenos son las proteínas más
abundantes.


Fibronectina (especializada)
La fibronectina une las células a todas las matrices excepto la del tipo IV que involucra a la laminina como la molécula de adhesión.
Las fibronectinas son dímeros de 2 péptidos similares. Cada cadena mide 60–70nm de longitud y 2-3nm de grosor.

contienen por lo menos 6 dominios doblados estrechamente cada uno con una alta afinidad por un diferente sustrato.

o Establecen uniones entre moléculas de la matriz extracelular y entre moléculas de las células con la matriz extracelular.
o Forman fibras insolubles en los tejidos conectivos o solubles en el plasma de los fluidos corporales, como la sangre.
o Tienen un papel muy importante durante el desarrollo embrionario creando sendas por las que pueden migrar las células de un lugar a otro del embrión

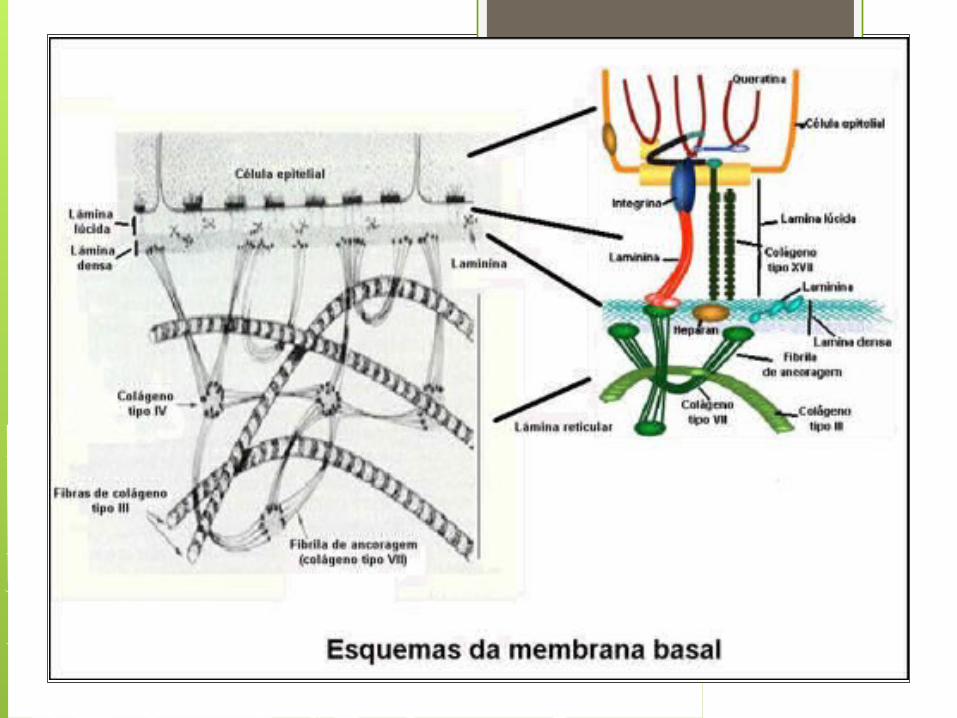
Laminina Todas las láminas basales contienen un mismo
grupo de proteínas determinadas y GAGs. Estas son el colágeno tipo IV, proteoglicanos de heparan sulfato, entactina y laminina
La lámina basal se la llama frecuentemente matriz tipo IV.
La laminina une las superficies celulares a la lámina basal.
La laminina es sintetizada por células epiteliales, musculares, neuronas y células de la médula ósea. La mayoría de estas células depositan la laminina principalmente en las láminas basales que las separan del tejido conectivo.

Glucosoaminoglicanos Los GAGs son moléculas negativamente
cargadas con una conformación extendida que brinda alta viscosidad a una solución.
Los GAGs están principalmente ubicados en la superficie de las células o en la matriz extracelular (MEC).
La viscosidad que brindan los GAGs también se observa una compresibilidad baja la cual hace que estas moléculas sean ideales como líquido lubricante de las articulaciones.
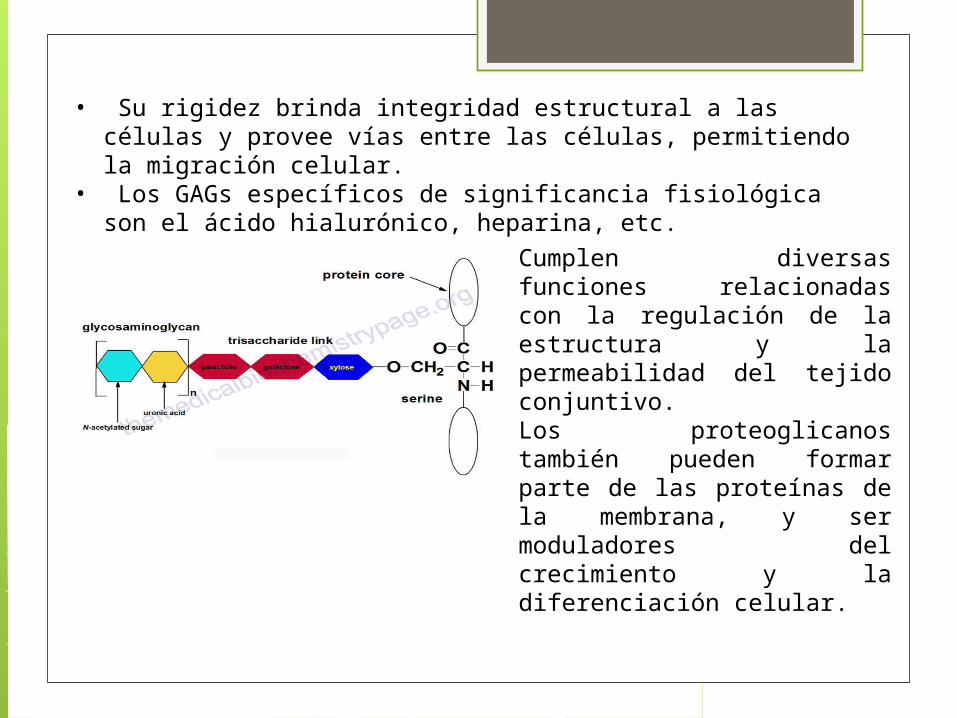
• Su rigidez brinda integridad estructural a las células y provee vías entre las células, permitiendo la migración celular.
• Los GAGs específicos de significancia fisiológica son el ácido hialurónico, heparina, etc.
Cumplen diversas funciones relacionadas con la regulación de la estructura y la permeabilidad del tejido conjuntivo. Los proteoglicanos también pueden formar parte de las proteínas de la membrana, y ser moduladores del crecimiento y la diferenciación celular.

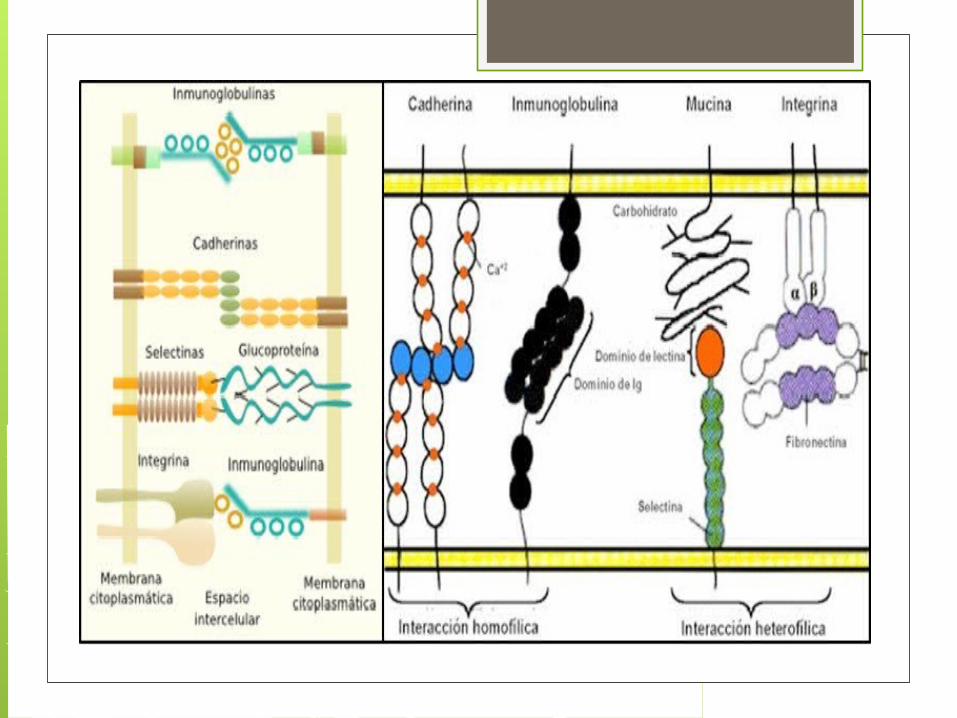
Familia de Receptores Receptores de la familia de integrinas, Receptores de la superfamilia de
inmunoglobulinas, Receptores de la familia de las
selectinas Receptores de la familia de las
cadherinas.

Integrinas
Las integrinas son la principal familia de receptores de la superficie celular que actúan mediando la unión de las células a la MEC.
Las integrinas son glucoproteínas formadas por cadenas a y b que atraviesan la membrana.
Las porciones extracelulares de las integrinas se unen a componentes de la MEC (fibronectina, laminina y algunas clases de colágeno) por reconocimiento.

El enganche entre los componentes de la MEC y el receptor de la integrina produce agrupamiento de los receptores y la formación de adhesiones locales, en donde las integrinas se conectan con los complejos del citoesqueleto intracelular.
Las proteínas que se fijan con las integrinas en esas adhesiones locales son la talina, vinculina, a-actinina, tensina y paxilina. Una vez ensambladas, los complejos formados por integrina-citoesqueleto funcionan igual que los receptores activados, y reclutan a los componentes de los sistemas de señalización intracelular.



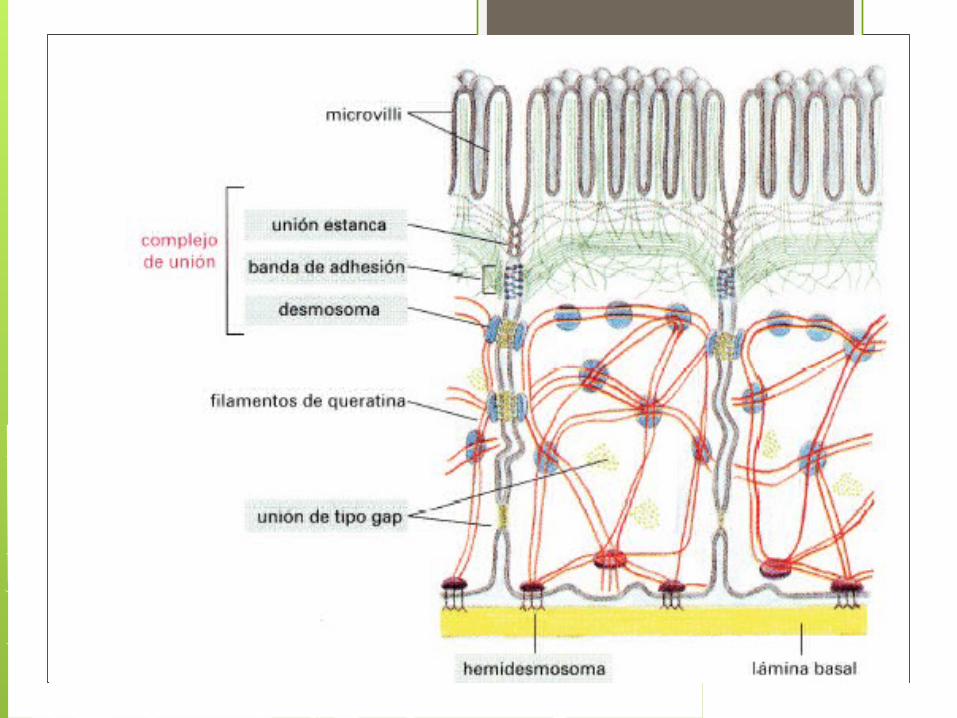
Uniones celularesSe pueden dividir en tres :
-uniones de oclusión -uniones de anclaje
-uniones de comunicación


Uniones de oclusión Funcionan como barreras selectivas
contra la libre difusión de las proteínas.

El transporte transcelular se da por proteínas en : Dominio apical (región membranosa en
contacto con la luz intestinal) bombea nutrientes hacia el interior de la célula epitelial.
Dominio basolateral (basal y lateral )trasporte de moléculas mediante difusión facilitada hacia el plasma


Ocludinas y claudinas Las uniones estrechas están formadas por la ocludina y
por una familia de moléculas denominadas claudinas, que son las proteínas transmembranal encargadas de establecer los contactos célula-célula.
Las claudinas parecen ser las más importantes en el establecimiento de la unión y en estas uniones forman unos poros que dejan pasar ciertos iones por el espacio extracelular, no más de 1 nm de diámetro.
Hay 20 tipos de claudinas, cada una de las cuales forma uno poro extracelular distinto y así los epitelios pueden modificar la selectividad de su permeabilidad intercelular según el tipo de claudina que expresen.

El dominio intracelular de estas moléculas interactúa con otras moléculas denominadas ZO, las cuales forman un entramado que interacciona con los filamentos de actina del citoesqueleto y con otras proteínas citosólicas que desencadenan cascadas de señalización.

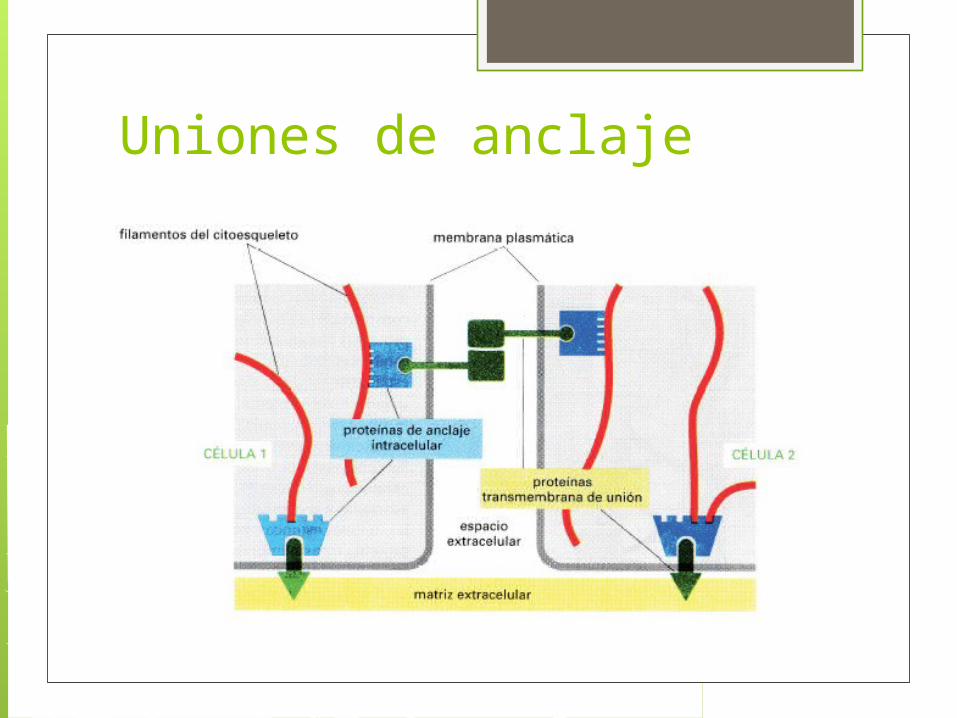
Uniones de anclaje Se dan entre célula- célula y célula- matriz, se presentan en tejidos con alta tensión mecánica.
Formada por: Proteínas de adhesión intracelular.(forman una placa
en la cara citoplasma conectan el complejo de unión a filamentos de actina o intermedios)
Glucoproteínas transmembranales de unión. Son dominios citoplasmáticos se unen a una o mas proteínas de adhesión y los dominios extracelulares interaccionan con la matriz o con otras glucoproteinas transmembranales.
Uniones de anclaje

Adhesión intracelular (caderinas) Bandas de adhesión (zonula adherens)
situadas cerca de la membrana luminal y por debajo de las uniones estanca. Construye un anillo contráctil de filamentos de actina, situado en la cara citoplasmática en la región membranosa implicada en la unión)

Caderinas Los filamentos de actina se unen célula-
célula mediante proteínas de unión (caderinas), cuyos dominios extracelulares se unen a otra molécula de caderina idéntica en la célula adyacente. Se cree que la contracción de esta red depende de proteínas motoras (miosina).
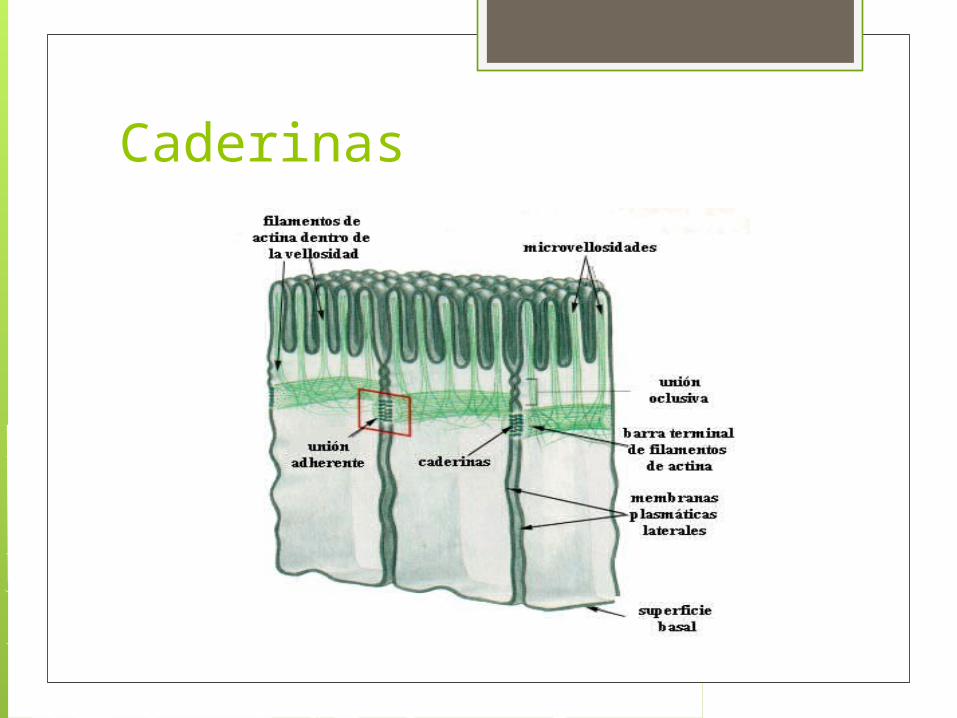
Caderinas

Los desmosomas o macula adherens conexiones puntuales en forma de disco entre
células vecinas, como si fuesen remaches. Son muy abundantes entre las células
epiteliales y entre las musculares, pero también en otros tejidos como el nervioso.
tipo caderinas denominadas desmogleínas y desmocolinas.
El dominio intracelular de estas caderinas contacta con los filamentos intermedios como las queratinas, gracias a proteínas intermediarias.

Los hemidesmosomas y las uniones focales establecen uniones fuertes entre las células y la
matriz extracelular. Las uniones se establecen por integrinas. Unen las células epiteliales a la lámina basal
gracias al dominio extracelular de la integrina, mientras que el dominio intracelular contacta con los filamentos intermedios citosólicos.
Las uniones focales unen a las células con diversos tipos de matrices extracelulares gracias a otro tipo de integrinas que en su dominio intracelular contacta con los filamentos de actina.


Uniones de comunicación Median la comunicación intercelular al
permitir el paso de iones inorgánicos,y otras pequeñas moléculas hidrosolubles (-1000 D), azucares, aa, nucleotidos y vitaminas entre los respectivos citoplasmas

Las uniones GAP, están organizadas en base a proteínas transmembrana forman estructuras = conexones.
Cada conexon esta compuesto por un anillo de 6 subunidades proteicas idénticas = conexinas.
Cada conexina tiene 4 hélices alfa que atraviesan la membrana formando un canal mayor y mas permeable que los canales neurotransmisores
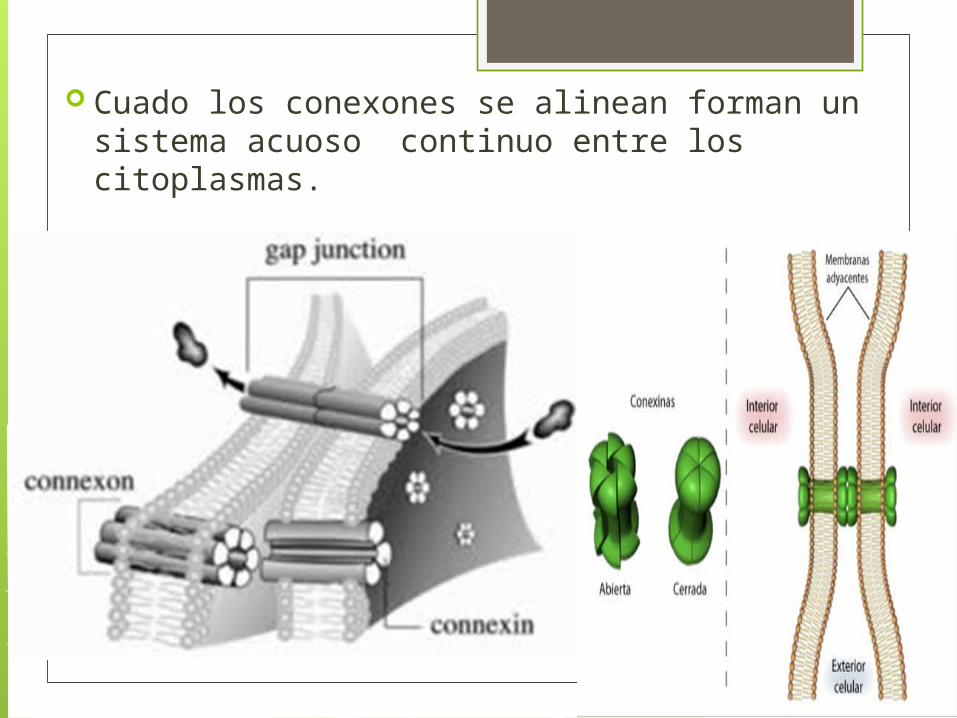
Cuado los conexones se alinean forman un sistema acuoso continuo entre los citoplasmas.
Top Related