







Idiomas
Páginas
Jurídico


Research ArticleMolecular Cloning and Optimization forHigh Level Expression of Cold-Adapted Serine Protease fromAntarctic Yeast Glaciozyma antarctica PI12
Norsyuhada Alias1 Mursquoadz Ahmad Mazian1 Abu Bakar Salleh1
Mahiran Basri2 and Raja Noor Zaliha Raja Abd Rahman1
1 Enzyme and Microbial Technology Research Centre Faculty of Biotechnology and Biomolecular SciencesUniversiti Putra Malaysia (UPM) 43400 Serdang Selangor Malaysia
2 Enzyme and Microbial Technology Research Centre Faculty of Science Universiti Putra Malaysia (UPM) 43400 SerdangSelangor Malaysia
Correspondence should be addressed to Raja Noor Zaliha Raja Abd Rahman rnzalihaupmedumy
Received 4 March 2014 Revised 6 May 2014 Accepted 14 May 2014 Published 30 June 2014
Academic Editor Raffaele Porta
Copyright copy 2014 Norsyuhada Alias et al This is an open access article distributed under the Creative Commons AttributionLicense which permits unrestricted use distribution and reproduction in any medium provided the original work is properlycited
Psychrophilic basidiomycete yeast Glaciozyma antarctica strain PI12 was shown to be a protease-producer Isolation of the PI12protease gene from genomic and mRNA sequences allowed determination of 19 exons and 18 introns Full-length cDNA of PI12protease gene was amplified by rapid amplification of cDNA ends (RACE) strategy with an open reading frame (ORF) of 2892 bpcoded for 963 amino acids PI12 protease showed low homology with the subtilisin-like protease from fungus Rhodosporidiumtoruloides (42 identity) and no homology to other psychrophilic proteases The gene encoding mature PI12 protease was clonedinto Pichia pastoris expression vector pPIC9 and positioned under the induction of methanol-alcohol oxidase (AOX) promoterThe recombinant PI12 protease was efficiently secreted into the culture medium driven by the Saccharomyces cerevisiae 120572-factorsignal sequence The highest protease production (283Uml) was obtained from P pastoris GS115 host (GpPro2) at 20∘C after 72hours of postinduction time with 05 (vv) of methanol inducer The expressed protein was detected by SDS-PAGE and activitystaining with a molecular weight of 99 kDa
1 Introduction
Eukaryotic as well as prokaryotic organisms can produceenzymes adapted to cold The majority of the cold-adaptedenzymes that have been characterized were originated frombacteria living in the Polar Regions inAntarctic andAntarcticseawaters [1 2] Other possible sources of cold enzymesare offered by the psychrophiles that inhabit at sim5∘C inother permanently cold environments such as the deepsea glaciers and mountain regions in soils and fresh orsaline waters related with cold-blooded animals such as fishor crustaceans and artificial sources such as refrigerationappliances and equipments [2ndash6] Although the studies ofcold adapted enzymes are focusing more on psychrophilicand psychrotrophic bacteria yeast fungi and unicellular
green algae that lived in cold environment are also listedas impending sources of cold enzymes The dominant taxaare ascomycetous and basidiomycetous yeasts and melanizedfungi [2 7]
The properties that characterized and distinguished coldadapted enzymes from enzymes of higher temperature originare their increased turnover number (119896cat) and inherentcatalytic efficiency (119896cat119870119898) at low temperatures which isconsidered to be an adaptive strategy of the psychrophiles tocompensate for low metabolic fluxes and slow reaction ratesat their physiological temperatures [8] Cold-active enzymesare more thermolabile and sensitive to other denaturantsrelative to their thermophilic and mesophilic counterpartsthat confer the conformational flexibility to the active site [9]As temperature decreases enzymes demonstrate a declining
Hindawi Publishing CorporationEnzyme ResearchVolume 2014 Article ID 197938 20 pageshttpdxdoiorg1011552014197938
2 Enzyme Research
catalytic rate owing to reduction of structural flexibility andundergo cold denaturation [7]
Psychrophilic enzymes possess remarkable prospectivein a wide variety of industrial applications Our interestis focused on cold-adapted protease as a useful biocata-lyst in industry such as low temperature clothes washingand enzyme-added detergents to hydrolyze macromolecularstains [10] An example of commercially available cold-active protease is from Novozyme (trade name Savinase)which is sold as an encapsulated detergent [11] Proteasesare degradative enzymes which catalyze the hydrolysis ofproteins and are commonly found among animal tissuesplant and microorganisms [12] Proteases execute a largevariety of functions and take part in numerous biochemicalreactions in living organisms including formation of sporeand germination coagulation cascade reactions posttransla-tion reactions modulation of gene expression enzymemodi-fication and secretion of various protein enzymes biocatalystOver one-third of all known proteolytic enzymes are serinepeptidases which constitute the largest group of peptidases[13]
Glaciozyma antarctica (formerly known as Leucosporid-ium antarcticum) is an obligate pyschrophilic yeast thatinhabit cold marin and terrestrial Antarctic ecosystemsGlaciozyma antarctica strain PI12 has an optimum growthtemperature of 12∘C [14] and can grow up to 18∘CThe reclas-sification of this yeast from L antarcticum to G antarcticawas proposed by Turchetti et al 2011 [15] Previous studyon this unique yeast has isolated several cold-active proteinsnamely antifreeze protein 120572-amylase and chitinase [16ndash18]In this work we described the morphological characteristicsofG antarctica through observation under scanning electronmicroscope (SEM) and transmission electron microscope(TEM) cloning of genomic DNA and cDNA sequencesencoding the PI12 protease gene phylogenetic study andexpression and optimization of the recombinant PI12 pro-tease expression in Pichia pastoris expression system
2 Materials and Methods
21 Culture and Isolation of Microorganism Glaciozymaantarctica strain PI12 was isolated from the Antarctic marinewater near Casey Station (66∘2110158402510158401015840S 110∘3710158400910158401015840E) Thestock culture was kept in 20 (vv) glycerol and stored atminus80∘C prior to experimentation
22 Microorganism Identification The isolated strain wasgrown on different types of solid media (nutrient agarsabouroud dextrose agar and potato dextrose agar) for 10days at 4∘C and the culture characteristics (colony colourshape and texture) were determined Simple and negativestaining were performed to identify the cell morphologyarrangement and size of the psychrophilic yeast SEM andTEM were conducted to study the surface features and theinternal ultrastructure in the thin sections of theG antarcticaPI12 cells Ribosomal RNA identification was performedthrough ITS1ITS2 region amplification The sequence has
been deposited in the GenBank database under accessionnumber JX896956 [14]
23 Nucleic Acid Isolation A single colony of the yeast wasinoculated into 50mL yeast peptone dextrose broth (YPD)and incubated for 10 days at 4∘C without shaking The cellpellets were frozen in liquid nitrogen and ground to a powderin a ceramic mortar Genomic DNA was extracted usinga phenol-chloroform method as described elsewhere [19]Removal of RNA from genomic DNA was performed by theaddition of 15120583L RNase (10mgmL) and incubated at 37∘Cfor 45min Total RNA was extracted by the guanidium isoth-iocyanate method using Trizol reagent (Invitrogen USA)according to the manufacturerrsquos instructions Approximately4 120583g of total RNA was digested with 05 120583g of RNAse-freeDNase in a 10 120583L reaction mixture containing 1x reactionbuffer (20mM Tris-HCl pH 84 50mM KCl and 2mMMgCl
2) prior to removal of DNAThemixture was incubated
at 37∘C for 15min DNaseI was later heat-activated at 65∘Cfor 15min DNA and RNA were quantified using Ultraspec2100 prospectrophotometer (Amersham Biosciences USA)The integrity of total RNAwas assessed by running an aliquotof the RNA sample on a denaturing agarose gel stainedwith ethidium bromide (EtBr) and by spectrophotometricanalysis (Ultraspec 2100 pro Amersham Biosciences USA)Total RNA was reverse-transcribed with an oligo(dT)
20and
SuperScript III Moloney Murine Leukemia Virus ReverseTranscriptase (InvitrogenUSA) following themanufacturerrsquosinstructions
24 Cloning of G antarctica PI12 Protease Gene Puta-tive partial protease gene of 1321 bp was derived througha genome survey sequence library of G antarctica Thegene has 27 similarity to subtilisin-like protease fromRhodosporidium toruloides (EMS20811) DNA walking ofpartial putative protease gene was conducted using DNAWalking SpeedUp Premix Kit (Seegene Korea) accord-ing to the manufacturerrsquos instructions Three target spe-cific primers (TSP) were designed from the upstreamregion of known sequences with the following condi-tions 18ndash23 nucleotides long with 40 lt GC content lt60 for TSP1 (51015840-AGGGTCAAGACGTTGCAGT-31015840) 55∘Cle Tm le 60∘C for TSP2 (51015840-ATGCGAAGTCAGAAG-CAGGATC-31015840) while 60∘C le Tm le 65∘C for TSP3 (51015840-GCAGCCAAATACCTGGAAGCAC-31015840) RACE was per-formed using the SMART RACE cDNA Amplification Kit(Clontech USA) Two gene-specific primers (GSPs) weresynthesized for the 51015840- and 31015840-RACE reactions based onthe sequence of the RT-PCR products as follows GSPI 51015840-ACCAGTGTCCAGCACCCCAATCTTAATCC-31015840 andGSP II 51015840-TCATCAGTGGGACGAGCATGTCGT-31015840 Bothprimers were paired with universal primers provided inthe kit to amplify the upstream and downstream regionof the gene of interest The PCR products were subclonedinto pGEM-T vector (Promega USA) and sequenced Rapidamplification of cDNA ends (RACE) produced two PCRproducts (51015840- RACE and 31015840-RACE) which were cloned andsequenced to capture the end-to-end sequence of both
Enzyme Research 3
products One set of primers was designed from the extreme51015840 and 31015840 ends of cDNA as follows forward 51015840 CAP 5-GCGGGGGCCGACAATAAAAAC-3 and reverse 31015840 A-Tail5-TTTTTTTTTTTTTTTTTTTTTTTTTTGAGGTGGC-31015840the 51015840-RACE-Ready cDNA served as a template to generatefull-length cDNA through long distance PCR (LDPCR)Amplification process was carried out in a reaction mixture(50120583L) containing 5 120583L of 50ndash100 ng cDNA template 1 120583L(10 pmole120583L) of each forward and reverse primers 1 120583L of50x dNTP mix 1 120583L of 1 U120583L 50x Advantage 2 polymerasemix 5 120583L of 10x Advantage 2 PCR buffer (Clontech USA)and 36 120583L of distilled water Predenaturation was performedat 95∘C for 1min followed by 30 PCR cycles of denaturation(95∘C 30 sec) annealing (61∘C 3min) and extension (68∘C3min) Final extension was conducted at 68∘C for 3minand held at 4∘C The gene was amplified using thermocycler(CG1-96 Corbett Research Australia) The amplification ofputative open reading frame (ORF) was conducted usingboth genomicDNAand cDNA to locate the introns and exonslocations The amplicons were examined by electrophoresiscloned into pJET12 blunt vector (Fermentas USA) and sentfor sequencing The sequence obtained was deposited inGenBank under accession number FM178559
25 Assembly and Analysis of PI12 Protease SequencesSequences derived from DNA walking and RACE strat-egy were assembled by conducting sequence alignmentin the Biology Workbench (httpworkbenchsdscedu)Analysis and interpretation of nucleic acid sequences wereconducted using several publicly available web serversand integrated software as listed in Table 1 The phylo-genetic tree was constructed based on a comparison ofthe PI12 protease protein sequence with the closest pro-tease sequence that was extracted from GeneBank database(httpwwwncbinlmnihgov) The protein sequences usedfor phylogenetic analysis were aligned using ClustalW Phy-logenetic tree was constructed using Molecular EvolutionaryGenetics Analysis 6 (MEGA 6) package by neighbor-joiningmethod
26 Cloning of PI12 Protease Gene in Pichia pastorisExpression System The gene encoding mature PI12 pro-tease was amplified by PCR using recombinant pJET12FLcDNA of PI12 protease as a template Sense primer51015840-CAAGCCCTAGGCTACGCACGGAACGAGAA-31015840 andantisense primer 51015840-CTGCCGAATTCTTCCTCAACCCA-GTTACCAAC-31015840 were designed based on mature PI12 pro-tease gene sequence (FM178559) for flanking the PCR prod-uct at the 51015840- and 31015840- terminus respectively Amplificationprocess was carried out in a reactionmixture (50 120583L) contain-ing 5 120583L of 50ndash100 ng DNA template 15 120583L (10 pmole120583L) ofeach sense and antisense primer 1 120583L of 10mM dNTP mix4 120583L of 25mM MgCl
2 1 120583L of 1 U120583L Taq DNA polymerase
5 120583L of 10x PCR buffer (MBI Fermentas USA) and 31120583L ofdH2O PCRwas commenced by predenaturation step at 94∘C
for 4min followed by 30 PCR cycles of denaturation (94∘C1min) annealing (65∘C 1min) and extension (72∘C 2min)
Final extension was conducted at 72∘C for 7min and held at4∘C
ThePCRproduct of themature PI12 protease gene and thevector pPIC9 (05ndash1 120583g) were separately added to restrictionenzyme digestion mixtures containing 3 120583L EcoRI (10U120583L)6 120583L AvrII (10U120583L) 6 120583L Y+ Tango buffer (10x) and dH
2O
to a final volume of 30 120583L The reactions were incubatedat 37∘C for 1 hour The digestion products were observedthrough agarose gel electrophoresis and gel purified usingthe QIAquick Gel Extraction Kit (Qiagen Germany) Theconstruct was added to a ligation mixture with a ratio of1 5 to pPIC9 vector The ligation mixture was comprised of1 120583L of 10x ligation buffer (MBI Fermentas USA) 1120583L T4DNA ligase (5U120583L MBI Fermentas USA) and dH
2O to a
final volume of 10 120583L The reaction was incubated overnight(14ndash16 h) at 16∘C Ligation mixture was transformed into Ecoli Top10 competent cells The procedures for preparationof competent cells and heat-shock transformation were per-formed according to the method described by Sambrook etal 1989 [19] The transformation mixture was spread on LBagar plates containing 100 120583gmL ampicillin and incubatedovernight at 37∘C The positive transformants were selectedfor transformation into Pichia pastoris electrocompetent cell
Prior to transformation into P pastoris cell the emptypPIC9 and pPIC9 recombinant plasmids harbouring PI12protease gene were linearized using PmeI (10U120583L) restric-tion enzyme digestion The digestion mixtures consisted of5 120583L buffer B (10x) 3 120583L of PmeI and 6 120583L plasmids whiledH2O was added to a final volume of 60 120583L The linearized
plasmids were purified using the QIAquick PCR purificationkit The preparation of electrocompetent cells was doneaccording to Pichia expression kit manual (Invitrogen USA)Later about 80ndash100 120583L of the electrocompetent cells weretransferred into an overnight prechilled (kept in minus20∘C)02 cm electroporation cuvette (BioRad USA) and mixedwith 10 120583L (1ndash5 120583g) recombinant DNA The transformationmixtures (mixture of cells and plasmids) were incubated onice for 5min in the electroporation cuvette and pulsedwith anelectroporator (Gene Pulser BioRad USA) at the followingparameters charging voltage of 1500V capacitance of 25120583Fand resistance of 400Ω which generated a pulse length ofsim5ndash10ms with a field strength of sim7500Vcm Iced-cold 1Msorbitol (1mL) was added immediately after the pulse intothe cuvettes The contents were transferred into sterile bijoubottles and incubated at 30∘C for 2 hours without shakingLater 10ndash200120583L of the cells were spread on the MD agar[Minimal dextrose medium 134 (wv) yeast nitrogen base(YNB) 4 times 10minus5 (wv) biotin 2 (wv) dextrose 2 (wv)agar]The plates were incubated at 30∘C for 3ndash5 days until thetransformant colonies were formed
27 Expression and Optimization of Recombinant pPIC9Mature PI12 Protease Single colonies of the recom-binantPichia pastoris carrying the pPIC9PI12 protease were grownin 3mL of BMGY [buffered glycerol-complex medium 1(wv yeast extract) 2 (wv) peptone 134 (wv) yeastnitrogen base (YNB) 4 times 10minus5 (wv) biotin 1 (wv)glycerol 100mM potassium phosphate buffer pH 60] at
4 Enzyme Research
Table 1 List of web servers applied for full-length cDNA of PI12 protease analysis
Analysis Web serverSoftware Website(1) Open reading frame (ORF)(2) Amino acid translation(3) Intron-exon boundaries
Augustus Web server httpaugustusgobicsde
(1) Amino acid composition(2) Molecular mass(3) Isoelectric point
ProtParam httpwwwexpasyorgtool
Homology search (1) NCBI BLASTX httpwwwncbinlmnihgov(2) NCBI BLASTP
Phylogenetic tree (1) Molecular Evolutionary Genetics Analysis (MEGA 6) httpwwwmegasoftwarenet(2) CLUSTALW httpseqtoolsdsceduCGIBWcgi
Propeptide ProP httpwwwcbsdtudkservicesProP-10Open reading frame ORF Finder httpwwwncbinlmnihgovprojectsgorfSignal peptide SignalP httpwwwcbsdtudkservicesSignalPHydrophobic profile ProtScale httpauexpasyorgcgi-binMultiple sequence alignment Biology Workbench httpworkbenchsdsceduProtein motifs ScanProsite httpprositeexpasyorgscanprositeN-glycosylation NetNGlyc httpwwwcbsdtudkservicesNetNGlyc
30∘C in a shake incubator (250 rpm) overnight The nextday 1mL of the starter cultures was used as inoculumand inoculated into 10mL of BMGY in a 50mL flask andincubated in a condition as described to generate cell biomassbefore induction Cells were harvested by centrifugation at1500timesg for 10min
The cells were resuspended in 50mL BMMY medium[bufferedmethanol-complexmedium 1 (wv) yeast extract2 (wv) peptone 134 (wv) yeast nitrogen base (YNB) 4times10minus5 (wv) biotin methanol 100mM potassium phosphatebuffer pH 60] and incubated with vigorous shaking at30∘C to induce expression The expression control was doneby applying the same protein expression condition to therecombinant Pichia bearing the empty vector (pPIC9) Thegrowth of Pichia pastoris harboring recombinant plasmidGpPro2 was optimized in four different parameters in orderto enhance the protease enzyme yield The yeast was grownup to 5 days for postinduction study and the concentrationof methanol was varied from 0 to 1 Assay temperature ofactive enzyme was optimized ranging from 10∘C until 35∘Cwith 5∘C intervals The last parameter was to find out thecapability of cell density in protease production at A
600 nmfrom 1 to 10
28 Protease Assay The proteolytic activity was measured bythemodifiedmethod of Brock et al 1982 [20] using azocaseinas substrate Azocasein (05) was dissolved in 01M Tris-HCl-2mM CaCl
2 pH 7 The reaction was initiated by the
addition of 100 120583L of enzyme solution for 30min at 20∘C Anequal volume of 10 (wv) trichloroacetic acid (TCA) wasadded to terminate the reaction The absorbance was read at450 nm using UVVisible spectrophotometer Ultraspec 2100pro (Amersham Biosciences USA) One unit of azocaseinaseactivity was defined as the rate of absorbance change of 0001per min at 20∘C under the standard assay condition
29 SDS-PAGE Sodium dodecyl sulphate polyacrylamidegel electrophoresis (SDS-PAGE) was prepared according tothe Laemmli [21] method using 12 polyacrylamide gel inthe Mini PROTEAN 3 Cell apparatus (BioRad USA) Afterelectrophoresis gels were stained with Coomassie BrilliantBlue R-250 (BioRad USA)
210 Activity Staining and Cup Plate Assay Activity stainingwas performed as described elsewhere [22] The supernatantfrom recombinant Pichia clones were applied in this methodand loaded into SDS-PAGE [12 (wv)] After electrophore-sis the gel was immersed in 20 isopropanol to remove SDSand washed with distilled water (2-3 times) and the gel wastransferred into 2 skim milk yeast peptone dextrose (YPD)agar plate Protease activity onPetri dishwas also tested by theactivity ring staining or cup plate assay using 2 skim milkYPD agar plate adapted from Poza et al 2001 [23] The platewas incubated at 20∘C for 5-6 hours
3 Results and Discussion
31 Microorganism and Culture Condition After 10 daysat 4∘C the growth of G antarctica PI12 on nutrient agarsabourouddextrose agar and potato dextrose agarwas cream-coloured smooth mucous runny and glisteningThe borderwas entire circular and convex while on skim milk agar theyeast produced hydrolytic colonies (Figure 1(a)) The cellsof G antarctica PI12 occurred in oval yeast-like shape withthe size of about 2ndash4 120583m and present singly when observedvia simple and negative staining (Figures 1(b) and 1(c)) SEMphotograph showed budding yeast form of the G antarcticastrain PI12 (Figure 1(d)) All cells clumped together andpossessed smooth and oval shape appearance
Figure 2 shows TEM photographs of the thin-sectionedof the basidiomycete yeast G antarctica strain PI12 which
Enzyme Research 5
(a)
2120583m
(b)
(c) (d)
Figure 1 (a) Casein hydrolysis on skim milk agar plate (b) Negative staining with enlargement of 10000x bar = 2120583m (c) Simple stainingphotograph (d) Scanning electron microscopy (SEM) photograph with magnification 2500x Bar = 10 120583m
reveals the intracellular organelles Figures 2(a) and 2(b)show budding yeast appearance of membrane-boundednucleus and easily recognized bud scarwhile Figure 2(c) illus-trates ultrastructure comprising of a dark and thick cell walla cell membrane an irregular shape nucleus mitochondriavacuoles golgi and bud scars
32 Ribosomal RNA Identification The ITS158S rDNAITS2 region of PI12 was successfully amplified by Booet al 2013 [14] with the predicted size of 646 bp TheITS158SITS2 sequence of Glaciozyma antarctica strainPI12 had been deposited in GenBank with the accessionnumber JX896956 Amplification of ITS region showed100 identity to ITS158SITS2 sequence of Antarctic yeastCBS 8938 (AY040659) and Glaciozyma antarctica strainCBS 8943 (AY033637) National Collection of Yeast Cul-tures (NCYC) identified PI12 as an isolate of Glaciozymaantarctica based on 26S rDNA D1D2 sequencing resultwhere it displayed 99 sequence identity to Glaciozymaantarctica strain CBS 5942 [formerly known as Leucosporid-ium antarcticum strain CBS 5942 (AF444529)] Phyloge-netic tree analysis was created based on comparison of ITSsequence of this strain with the closest strains that areavailable in the database with reference to BLAST result
(Figure 3) Thirty-nine of ITS sequences from Glaciozymaclade were comparedThe phylogenetic tree constructed con-sists of three main clusters cluster I (JX896956 Glaciozymaantarctica strain PI12 AY040659 Antarctic yeast CBS 8938AY033637 Glaciozyma antarctica strain CBS 8943 EU149809Glaciozyma antarctica strain CBS 10640 AB774460 Leu-cosporidium sp KGK-2 JQ857017 Glaciozyma antarcticaisolate T21Ga EU149808 Glaciozyma antarctica strain CBS10636 AY040663 Glaciozyma antarctica strain CBS 8939EU149806Glaciozyma antarctica strainCBS 10639 EU149805Glaciozyma antarctica strain CBS 10638 AY040657 Antarcticyeast CBS 8927 AF444529 Glaciozyma antarctica strainCBS 5942 and KF934487 Glaciozyma sp KP7-5-2) isthe sequences from strains originally considered as Lantarcticum which was reclassified as Glaciozyma antarc-tica [15] Cluster II (GQ336996 Leucosporidium sp AY30EU149804 Glaciozyma watsonii strain CBS 10641 AY040661Antarctic yeast ML 4515 AY040660 Glaciozyma watsoniistrain CBS 8940 AY033638 Glaciozyma watsonii strain CBS8944 AB774462 Leucosporidium sp BSS-2 KC785578Uncul-tured Glaciozyma clone 180 AB774461 Leucosporidium spBSS-1 EU149803 Glaciozyma watsonii strain CBS 10684HQ432823 Glaciozyma watsonii DBVPG4802 HQ432818Glaciozyma watsonii DBVPG4726 HQ432820 Glaciozyma
6 Enzyme Research
2120583m
(a)
2120583m
(b)
BS
V
N
CW
CM
GM
Length 314866nm
Length 226635nm
1120583m
(c)
Figure 2 Transmission electronmicroscopy (TEM) photographs of AntarcticG antarctica strain PI12 (a) 12500xmagnification (b) 10000xmagnification (c) 27500x magnificationThe scale bars represent 2 120583m for (a) and (b) and 1 120583m for (c)
watsonii DBVPG4760 JX171176 Glaciozyma watsonii iso-late LKF08-112 DQ402535 Leucosporidium sp LC-03-120isolate 03-120 JF900360 Glaciozyma watsonii strain CBS7009 KC333171 Glaciozyma watsonii isolate AU CryS06HQ432821 Glaciozyma watsoniiDBVPG4799) is categorizedas G watsonii while cluster III (AY040664 Glaciozyma mar-tinii strain CBS 8929 AY033641 Glaciozyma martinii strain
CBS 8928 HQ432816 Glaciozyma martinii DBVPG4841KF934486 Glaciozyma sp KP7-5-1 FR682434 UnculturedBasidiomycota HF934010 Glaciozyma sp K81b JF900362Glaciozyma martinii strain CBS 9639 JF900359 Glaciozymamartinii strain CBS 6581 JF900361 Glaciozyma martiniistrain CBS 7054) is categorized as G martini Recognitionof the Glaciozyma species has been done by comparing
Enzyme Research 7
JX896956 Glaciozyma antarctica strain PI12
AY040659 Antarctic yeast CBS 8938
AY033637 Glaciozyma antarctica strain CBS 8943
EU149809 Glaciozyma antarctica strain CBS 10640
AB774460 Leucosporidium sp KGK-2
JQ857017 Glaciozyma antarctica isolate T21Ga
EU149808 Glaciozyma antarctica strain CBS 10636
AY040663 Glaciozyma antarctica strain CBS 8939
EU149806 Glaciozyma antarctica strain CBS 10639
EU149805 Glaciozyma antarctica strain CBS 10638
AY040657 Antarctic yeast CBS 8927
AF444529 Glaciozyma antarctica strain CBS 5942
KF934487 Glaciozyma sp KP7-5-2
GQ336996 Leucosporidium sp AY30
EU149804 Glaciozyma watsonii strain CBS 10641
AY040661 Antarctic yeast ML 4515
AY040660 Glaciozyma watsonii strain CBS 8940
AY033638 Glaciozyma watsonii strain CBS 8944
EU149803 Glaciozyma watsonii strain CBS 10684
JX171176 Glaciozyma watsonii isolate LKF08-112
DQ402535 Leucosporidium sp LC-03-120 isolate 03-120
AB774462 Leucosporidium sp BSS-2
KC785578 uncultured Glaciozyma clone 180
HQ432818 Glaciozyma watsonii DBVPG4726
HQ432823 Glaciozyma watsonii DBVPG4802
AB774461 Leucosporidium sp BSS-1
HQ432820 Glaciozyma watsonii DBVPG4760
KC333171 Glaciozyma watsonii isolate AU CryS06
HQ432821 Glaciozyma watsonii DBVPG4799
JF900360 Glaciozyma watsonii strain CBS 7009
AY040664 Glaciozyma martinii strain CBS 8929
AY033641 Glaciozyma martinii strain CBS 8928
HQ432816 Glaciozyma martinii DBVPG4841
KF934486 Glaciozyma sp KP7-5-1
FR682434 uncultured Basidiomycota
HF934010 Glaciozyma sp K81b
JF900362 Glaciozyma martinii strain CBS 9639
JF900359 Glaciozyma martinii strain CBS 6581
JF900361 Glaciozyma martinii strain CBS 7054
001
Clus
ter 1
Glac
iozy
ma
anta
rctic
aCl
uste
r 3
Glac
iozy
ma
mar
tinii
Clus
ter 2
Glac
iozy
ma
watso
nii
Figure 3 Neighbor-joining phylogenetic analysis of internal transcribed spacer 1 (ITS1)58S rRNA geneinternal transcribed spacer 2 (ITS2)of Glaciozyma clade Numbers indicate percentage bootstrap values calculated on 1000 repeats of the alignment The tree was constructedusingMEGA6 software [39]The scale bar represents 001 substitutions per nucleotide position GenBank accession numbers of the sequencesare indicated before strain name ITS158SITS2 sequence of Glaciozyma antarctica strain PI12 had been deposited in GenBank with theaccession number JX896956 [14]
8 Enzyme Research
their molecular phylogenetic data and physiological andmorphological characteristics [15] Phylogenetic analysis ofITS158SITS2 sequence of strain PI12 showed that thesequence was closely related to ITS158SITS2 sequenceof Glaciozyma antarctica with high homology similarity toCluster I (99-100 identity) The ITS158SITS2 sequenceof G antarctica strain PI12 was differed from G watsonii(Cluster II) by 2-3 gap in the sequence (93 identity)and G martinii by 4-5 gap in the sequence (90ndash92identity)
33 Sequence Analysis of PI12 Protease cDNA In this studyamplification of the PI12 protease gene sequence was con-ducted using genomicDNAand cDNAas a template Putativepartial PI12 protease gene with the size of 1321 bp was initiallyacquired from the G antarctica genome survey sequencelibrary (Supplementary Figure see Supplementary Mate-rial available online at httpdxdoiorg1011552014197938)However the gene was discovered as an incomplete pro-tease gene sequence DNA walking strategy successfullyamplified 1500 bp of the upstream region of partial PI12protease Both sequences of about 1500 bp (derived fromthe DNA walking reaction) and 1321 bp (partial PI12 pro-tease) were assembled and made up a total length of2687 bp protease sequence (Supplementary figure)The puta-tive PI12 protease was predicted by Augustus webserver(httpaugustusgobicsde) having intervening sequencesintronsThus reverse-transcriptase PCR (RT-PCR) and rapidamplification of cDNA ends (RACE) were accomplished toobtain the full-length cDNA As a result the ORF comprising2892 bp encoding 963 amino acid residues were successfullydeduced
Nucleotide and deduced amino acid sequences ofgenomic DNA and cDNA fragment encoding PI12 proteasewere shown in Figure 4 The size of the ORF of putative PI12protease was 3867 bp with genomic DNA as the templatewhile the ORF comprised 2892 bp encoding 963 amino acidresidues were successfully deduced through RACE strategywith cDNA as the template The ORF was predicted usingORF Finder (httpwwwncbinlmnihgovprojectsgorf)while prediction using SignalP (httpwwwcbsdtudkser-vicesSignalP) webserver allowed the identification of theputative PI12 protease prepeptide where it predicted thepresence of signal peptidase I cleavage sites [24]The putativeN-terminal signal peptide (1 to 16 the underlined amino acid)was identified as a secretion signal There is no propeptidepredicted by ProP Webserver (httpwwwcbsdtudkser-vicesProP-10) Comparison of cDNA and gene sequencesallowed determination of gene structureTherefore sequencealignment was performed between the sequence of these twotemplates and it was confirmed that the coding sequence(consisted of 19 exons) was interrupted by 18 introns withtheir length varying from 48 to 60 nucleotides However theGT and AG rule for donor-acceptor sites was not obeyedOf 18 introns 10 introns contained canonical GT and AGdinucleotides at their 51015840 and 31015840 ends while 8 introns holdnoncanonical splice site pairs with TT-GG (2 introns)AG-TG GC-GT TG-GG TC-GG TA-GG and TG-AG
dinucleotides at the splice junction Upper case letters inblue colour represent introns while lower case letters inblack colour represent exons Splice donor and acceptor werehighlighted in grey
The amino acid composition (upper case letters) withinthe ORF amino acid residues was determined by theProtParam tool of the Expasy molecular biology server(httpwwwexpasyorgtool) Translation starts at a nucle-otide position 1 and translation stop is marked with anasterisk The nucleotide sequences ATG (1 to 3) and TAG(3865 to 3867) indicate the initiation codon and termi-nal codon respectively The presumed putative sequencesof promoter CAAT (minus101 to minus98) and TATA-box (minus92to minus88) were identified in the 51015840-untranslated (UTR)region The search was achieved via Neural Network Pro-moter Prediction (Berkeley Drosophila Genome Project)(httpwwwfruitflyorgseq toolspromoterhtml) In the 31015840-untranslated (UTR) region there was a putative polyadeny-lation signal attaa (3889 to 3893) detected from the TAGstop codon to the poly (A) tail The predicted molecularmass and pI of PI12 serine protease were 10099 kDa and641 respectively Out of 963 amino acid residues in thededuced sequence of the PI12 protease 70 are negativelycharged residues (Asp + Glu) while 66 are positively chargedresidues (Arg + Lys) at neutral pH (Table 2)The total chargedresidues (Arg Asp Glu His and Lys) are 150 amino acidsin the open reading frame of PI12 protease corresponded to156 while hydrophobic residues (Ala Phe Ile Leu MetPro Val and Trp) are 441 amino acids which correspondedto 458 of the total amino acids discovered The totaluncharged residues (Asp Cys Gln Gly SerThr and Tyr) areconsisted of 372 amino acids which corresponded to 386of the total amino acids of the open reading frame of PI12protease The aliphatic index is 9290 and the grand averageof hydropathicity (GRAVY) is 0141 The instability index (II)of PI12 protease was computed to be 3414 which classify theprotein as stable
N-glycosylation sites were predicted using NetNGlyc10 Server (httpwwwcbsdtudkservicesNetNGlyc)Therewere five putative N-glycosylation sites for the PI12 proteasegene predicted using the server For N-glycosylation thesequence motif Asn-Xaa-SerThr (Xaa any amino acid butnot Pro) has been defined as a prerequisite for glycosylationThe sequencemotifs in PI12 protease occurred in the position170 NETH 365 NATV 576 NLTP 634 NDTA and 833 NSSPAmong five predicted sites only three (except positions 576and 833) showed a high potential score (averaged output ofnine neural networks) which support the prediction Theputative N-glycosylation sites were demonstrated in boxesin Figure 4 The comparison of amino acid composition ofthis cold-adapted protease fromG antarctica strain PI12 withnine other subtilisin serine proteases fromdifferent families isshown in Table 2 Analysis of this comparison depicts that thepercentage of each amino acids in PI12 protease is almost thesamewith other proteaseswhileGly (G) is themost abundantaccounting for up to 104 of the total amino acid in PI12 pro-tease PI12 protease also contained the highest percentage ofLeu (L) with 99 of the total amino acids compared to other
Enzyme Research 9
Table2Aminoacid
compo
sitionandmolecular
weighto
fproteases
acrossdifferent
families
Amino
CAQ76821
CAG25549
EDR11679
CAG38357
CAD118
98CA
L25578
EDU50747
AAW
40780
ACB3
0121
ACB3
0119
acid
Qty
Qty
Qty
Qty
Qty
Qty
Qty
Qty
Qty
Qty
Ala
(A)
97101
8191
7887
7590
8299
101
115
8189
9196
8092
105
116
Arg
(R)
4042
2730
2224
1619
1721
1821
2932
2223
2124
3134
Asn
(N)
4446
3843
4348
4149
4251
4046
5257
5154
4552
4347
Asp
(D)
4951
6269
5460
6578
5567
5158
5358
6669
5260
6370
Cys
(C)
808
506
404
405
911
809
910
909
708
1011
Gln
(Q)
2324
3438
2326
2834
3036
3742
3539
2931
1922
2629
Glu
(E)
2122
1618
1921
2024
1518
2023
2528
2627
2832
2426
Gly
(G)
100
104
89100
8493
85102
85103
7788
8493
8084
8598
8695
His
(H)
1415
1416
1314
1619
1316
1113
1618
1415
1618
1213
Ile(I)
5355
4955
5157
5364
4150
3843
5156
4446
3540
4449
Leu
(L)
9599
6876
7179
6274
6073
6675
6774
8084
7485
6875
Lys
(K)
2627
2932
4449
2935
4959
5158
4044
3032
5058
5662
Met
(M)
1212
202
202
911
1113
809
1820
808
1113
1517
Phe
(F)
4143
5966
4752
5566
2935
3641
4449
3941
3439
3438
Pro
(P)
6264
6573
6774
5870
5870
5967
5055
5154
5564
5763
ser
(S)
7780
6977
91101
6072
6680
7080
7279
111
117
6373
6167
Thr
(T)
8790
89100
7583
83100
6579
7383
7178
7276
7182
6875
Trp
(W)
505
303
607
202
1215
1214
1213
909
1214
1415
Tyr
(Y)
3334
1213
2629
708
2935
2832
3943
5053
2630
2730
val
(V)
7679
8292
8089
6679
5769
7484
5864
6872
8295
6268
Total
963
1000
893
1000
900
1000
834
1000
825
1000
878
1000
906
1000
950
1000
866
1000
906
1000
MW
(Da)
1009898
932669
944365
869455
869549
924905
979343
1009629
917209
961988
Thea
ccessio
nnu
mbersof
proteasespu
tativ
eserinep
roteaseo
fGlacio
zymaantarctica
PI12
(CAQ
76821)sub
tilisin-lik
eproteaseP
leurotuso
streatus(CA
G25549)pyrolysin
LaccariabicolorS
238N
-H82
(EDR11679)
subtilisin
-like
proteasePh
anerochaetechrysosporiu
m(C
AG38357)sub
tilisin-lik
eserinep
roteaseP
R1CMetarhiziu
maniso
pliaev
aranisopliae(CA
D11898)serinee
ndop
eptid
aseH
ypocrealixii(C
AL2
5578)peptidase
Pyrenophoratritici-repentis
(EDU50747)p
eptid
aseCr
yptococcus
neoforman
svarn
eoform
ansJEC
21(A
AW40
780)sub
tilisin-lik
eprotease
Epich
loefestucae(A
CB30121)sub
tilisin-lik
eprotease
Epich
loefestucae
(ACB
30119
)
10 Enzyme Research
atgctcttcctccccgtcctcctcctcctccttcccggcgtcactgccttcctcaaccca 60
M L F L P V L L L L L P G V T A F L N Pgttaccaaccgcgcgaccaacgccatctcctcgactcaatacctctctaacgcctatatc 120
V T N R A T N A I S S T Q Y L S N A Y IctcgaattggacctctccacccccggcctcgtcaaacgggatagcacgcccgactctGTG 180
L E L D L S T P G L V K R D S T P D SCGTCCCGCGTTCCTCCCTCGCTGTCGAAGACGAGCTAACAATGACGTGGTAGatcctaga 240
I L Eggacgtactcacgtccgtaggccgcaacggtatcaagtaccaactccgccaccgctttat 300
D V L T S V G R N G I K Y Q L R H R F Ictccccgactctgttccacggcgcttcgatcactgtcccccctggaatctcccgctccca 360
S P T L F H G A S I T V P P G I S R S QaatcgcctctctccgcggtatcaaaGTCCGTCCCTCTCTCTCCCCTACTCCCTCCGTGCT 420
I A S L R G I KAACCCTGCACGCAACAGcgcgtctggcccgttcgaaagttctcccgacccagcgcagtag 480
R V W P V R K F S R P S A Vtggacgccgatggaggaggaagcgggttctcagggtcgcctatcaaggcggcgctcatgg 540
V D A D G G G S G F S G S P I K A A L Mgggtgaaagagctcgggaagcgcgcgaacgcttatgctggagatacgtttggaccgcatg 600
G V K E L G K R A N A Y A G D T F G P HtcatgacgggggttaatgagacgcatgaggcggggttgttgggagctgggattaagGTGT 660
V M T G V N E T H E A G L L G A G I KGTTTTCGTCTTTGTTTGGGGGAGGGGGGAGGGAGGAGCTGATTAATGGGTGCAGattggg 720
I GgtgctggacactggtgAGAAGCGGTTTGGAGGGAGGAGAGGATGGAGCTGATGAGTGTGC 780
V L D T GAGGTGttgattatttgaacccgattctgggaggctgctttggacctgggtgccatatgtc 840
V D Y L N P I L G G C F G P G C H M Sgtttgggtacgacttggttggcgatgattacgatggagataacgctcctgtgccggatgt 900
F G Y D L V G D D Y D G D N A P V P D VggatccttGTGCGTCCTTCCCTCCGCGATGGGCTGGAATCTGAGCTGACGGCGTTGTTGA 960
D PTCAGttgcgagctgcgatcctcatggTTCGTCTCGCTTTCCGCTCCTCTGATGCGCTCGC 1020
F A S C D P H GTGATGCACTTGGCAATAGGaactcacgttacgggaatcattggagcgctcccgaatgcgt 1080
T H V T G I I G A L P N AttggatttactggcgtcgcacccgccgctactctgggccactaccgagtGCGCTCCCTCT 1140
F G F T G V A P A A T L G H Y R VATCCGTTCTTCCTCGCAGCTCCCACTCACTCGTGCTTCCAGGTatttggctgcactggct 1200
F G C T GtcgtcggagaagatatcGTTCGTCCTCTCTCCAGTCCCGGCTAGCGATCCTGCTTCTGAC 1260
F V G E D ITTCGCATCTCCCTCTGAGattctcgctggactcatgcgaggagtcgaggacaactgcaac 1320
I L A G L M R G V E D N C Ngtcttgaccctctctctcggaggtccaggagggtgggtcaagggcacgccggcgtccatc 1380
V L T L S L G G P G G W V K G T P A S Icttatcgaccagatcgaagcgcaaggcattctcgtcaccgtcgccactggcaactcggga 1440
L I D Q I E A Q G I L V T V A T G N S GgctgagggTGAGCTCCTCGCTACTTGCGCGTCGAGACCAGTACTAACAGGGGCGTGGACG 1500
A E GCAGGcatgttcttctccgagtctcccgcctcgacgatcaacggcctttccatcgcatcca 1560
M F F S E S P A S T I N G L S I A ScgTCCGTCTCTCCCTTCCTTCGCACGCCACTCGCTCACACTCTCTCGCAGGgacgttacc 1620
T D V Tgacctcatcgcctacaacgccaccgtctcaggccaacctgcgatcccttacctctccgcc 1680
D L I A Y N A T V S G Q P A I P Y L S A
ggggggggggtctg minus194
ctagcgcgaatcgagaaaacacaaaagaacctcccgacttggcgttgcttctgctcttct minus180
ccttctcccaaccgccgacaataaaaactataacacacacacaccccatctcgacgcctg minus120
cagacttcacgactctcctcacgctctttaccgccccacgggtcccacccacgagccacg minus60
(a)
Figure 4 Continued
Enzyme Research 11
acgcccctcaacgtcgtcgccaacagcttccgcgtccacttcacctctaccgaccccaac 1740
T P L N V V A N S F R V H F T S T D P Naaccccgtcgacgcctgctctcctcttccggctggagcgcccgacttcgccaactatgtt 1800
N P V D A C S P L P A G A P D F A N Y Vacggtcgttcagcgtgggacttgtacgttcgttaccaagtaccagaacgttctcaatgct 1860
T V V Q R G T C T F V T K Y Q N V L N AggaggGTGCGTCGGCTTCTTCTCTCCCGACTGGATTGGGCGGAGACTGACCTTCTGAGCA 1920
G GGaaagatcgtattgttgtacaactcggagggagctgggaacctcccttacctcacgccca 1980
K I V L L Y N S E G A G N L P Y L T PacggtgtcggcatcgacgccgttgcaggtcttcgtcgttccgacggactcaagGTTCGTC 2040
N G V G I D A V A G L R R S D G L KTACGCTCTTCGGTGCTGAGGCCCTTCTCTGATCTCGCTCCGACAGcttctctcgtactat 2100
L L S Y Ycagaatgccaacaagcgtctcactctgcgcttccccaagggcaagatcgtcgcaggcttg 2160
Q N A N K R L T L R F P K G K I V A G LaccgataccatcaccggcggactcatctcgTTCGTTCCTCTCCGCCTCCCTCGCTGGCCG 2220
T D T I T G G L I SATGAGCTGACGCGGTCTTTGCAGGggttactcgacgtttggtccgacgaatgacctctac 2280
G Y S T F G P T N D L Yggtcagcctaccctctctgcccctggtggcaacatcctttcgaccttccctctctccgag 2340
G Q P T L S A P G G N I L S T F P L S EggaggagtggcggtcatcagtgggacgagcatgtcgtgcccctttgtcgGTTCGTCCTTC 2400
G G V A V I S G T S M S C P F VTCCTCGTTCGTTCGCTGAGCGTCGACTGATCTTGCACGATCGCAGctggatctgcggcgg 2460
A G S A Atcctcatggccgctcgcgcttcggagaacctcacgccgcttgagatcaggagtctcctta 2520
V L M A A R A S E N L T P L E I R S L Lctaccactgcgaagcttacgccggtctcgctcttgggatcgacgcctttggtgagcgtga 2580
T T T A K L T P V S L L G S T P L V S VttcgtcaaggaggaggTACGTTCCTTTCTCGCTGCTTTGAGGATCGAGGCTGATTAGCGA 2640
I R Q G G GGCTTCGCAGGactcgttcaggttgccaaggcgctcgcggccaagacgctaatctctcctc 2700
L V Q V A K A L A A K T L I S Pacgagctcctgctcaacgacactgcgaacgcgaactacgtccagactatcaagatcaaga 2760
H E L L L N D T A N A N Y V Q T I K I Kacaccaactcgtgggcgatgaagtacaccttctcctcggccgtcgcccaaggactcggaa 2820
N T N S W A M K Y T F S S A V A Q G L Gctttcgacgcttcgggcgatatcctccctaccctcgacccagtcgccgtctctggcgcac 2880
T F D A S G D I L P T L D P V A V S G Aaggctaccgtcgcgttcaacactcggatcctcagcgtcgcgcccggcgcgacggggtccg 2940
Q A T V A F N T R I L S V A P G A T G Stcgtggcgactatcacgccgccggttcttcccgtagcggacgctgcgaggttccctatct 3000
V V A T I T P P V L P V A D A A R F P Itctctgggtggatcagggtgaatgggcaaggcgcgagggatagcagtaggaacgaggcgt 3060
F S G W I R V N G Q G A R D S S R N E AacactgGTGCGTTTGGCTCGCTTTTTGGGAGGTGCCGAGCTAATCGTTTCTCGTAGtccc 3120
Y T V PgtactttgggcttgcggcgaagatgatcgatatgcaagGTGCGTCGATGGTTCTCTCGCT 3180
Y F G L A A K M I D M QTCTCTCTCGGTTGCTCACTTCAAAGTGCAGtcctcgacaccaccgagaccatttacggtc 3240
V L D T T E T I Y Gcgggctacgcctaccccttcgtgatcgacgacgcgattggagacatccaatccaccacaa 3300
P G Y A Y P F V I D D A I G D I Q S T TcgtcgtactccaggaacctcgggcccaccgtcttcgctcgctttgccaTGCGTCGACCGT 3360
T S Y S R N L G P T V F A R F ACCTCATTTCCTGCGGCAGAGCGAGAGCTGATTCCGACCTCGAACAGctggaacccttcac 3420
T G T L Htacagcctcgatctcgtcctagccgacatcgcctttacccccacctaccccaactcctcc 3480
Y S L D L V L A D I A F T P T Y P N S S
(b)
Figure 4 Continued
12 Enzyme Research
cccgccactcgtttcgtcaagcgctccctcacgcagcacacctctgccgcttcgcacctc 3540
P A T R F V K R S L T Q H T S A A S H Lgccaagcgccgcgtctccatcgccactatcaaccccaaagccaccctcgtcgccgatcga 3600
A K R R V S I A T I N P K A T L V A D Rcagctccactcggacgtccctatcgagggcaacatcttcacccaaccctttactggaagg 3660
Q L H S D V P I E G N I F T Q P F T G Rgattacctcgtcgacgcagccccgacgggatccaccgatcgtaccgtcacttttaacggg 3720
D Y L V D A A P T G S T D R T V T F N Gcagtacgccgagaacggcctcgtgaggacggctgtgacggggacttcgtaccgcttcctc 3780
Q Y A E N G L V R T A V T G T S Y R F Lcttcgggcgttgaagatctcgggagacgcgatgtacgaggatcagtatgagagctggctc 3840
L R A L K I S G D A M Y E D Q Y E S W L3900
S L P F S F R Attttgccacctcaaaaaaaaaaaaaaaaaaaaaaaaaa 3938
lowast
tcgctaccgttctcgttccgtgcgtaggcttgttgtggacaccactgcattaagttatga
(c)
Figure 4 Nucleotide and deduced amino acid sequences of genomic DNA and cDNA encoding protease from G antarctica strain PI12 Thenucleotides are numbered from the 51015840 end of the cDNA A putative CAAT TATA box and polyadenylation signal are bold italicThe putativeN-terminal signal peptide is underlined The initiation codon is boldfaced and termination codon is represented by an asterisk (lowast) boxedpeptide sequences showed N-glycosylated asparagines residues The amino acid residues that form the catalytic triad are double underlinedand bold while introns are represented in blue font Highlighted in grey are the intron donor-acceptor splice sites
proteases Hydrophobic profile of PI12 amino acid proteasewas performed by ProtScale tools with Kyte and Doolittlemethod at httpauexpasyorgcgi-bin The hydrophobicityprofile elucidation of PI12 protease amino acid was carriedout from amino acid 1 to 963 The amino acid residuesfrom 1 to 16 (M-L-F-L-P-V-L-L-L-L-L-P-G-V-T-A) and 548 to562 (V-A-V-I-S) were hydrophobic grooves The first regionwhich consisted of 16 amino acids was very hydrophobic andwas predicted to be the signal peptide by SignalP and ProPwebserver
The bold and double-underlined amino acids showedresidues of catalytic triad aspartic acid (D) histidine (H) andserine (S) active site respectively NCBI Conserved DomainsDatabase (CDD) identified the presence of subtilase proteaseand subtilisin-like serine protease domains in the amino acidsequence The predicted PI12 protease amino acid sequencewas found sharing significant homology to the subtilisinfamily of protease The highest similarity of PI12 proteasewas observed with 42 identity to subtilisin-like proteaseof Rhodosporidium toruloides (EMS20793) 37 identity tosubtilisin-like protease of Pleurotus ostreatus (CAG25549)and 35 identity to subtilisin-like protease of Gloeophyllumtrabeum ATCC 11539 (EPQ55564) The nucleotide sequenceof G antarctica strain PI12 showed low homology to thefungus subtilisin protease family and none to other psy-chrophilic proteases The multiple sequence alignment wasillustrated in Figure 5 The consensus sequence around thecatalytic triad aspartic acid (D) histidine (H) and serine(S) of PI12 protease showed high conservation with othersubtilisin (showed in black boxes) Glycine residues G142G173 G191 G262 and G327 that were highly conserved inmost of subtilases were conserved in PI12 protease [25]Therefore PI12 protease belongs to the subtilisin subgroup ofthe subtilase serine protease superfamilyThis serine protease
of G antarctica PI12 has been deposited into GenBank withaccession number CAQ76821
34 Phylogenetic Relationship of PI12 Protease The phylo-genetic tree was constructed to demonstrate the similaritiesand relationship of protein sequence between differentspecies or organisms Figure 6 illustrates the phylogeneticdendrogram indicating the relationship of PI12 proteasewith another 25 translated amino acid sequences ofserine protease The bar scale signifies the branch lengthto approximate variance along each branch Basicallythis tree comprises two main clusters cluster I was sub-divided into subcluster a (CAG25549 EPQ55564 ETW-80907 XP 007381302 XP 001877576 CAG38357 XP007341385 CCA68627 EJU04161 XP 007369624 EIW-62675 CCO37249 XP 001830835 XP 007342189) and sub-cluster b (XP 007589863 XP 003711175 EFQ32840 XP006694395 ENH74885 EON99722) cluster II was sub-divided into subcluster c (CBQ73425 GAC97617 CCF52619)and subcluster d [CAQ76821 (PI12 protease) EMS20793]
Serine protease constructed by the phylogenetic treecomprised large size of protein with about 801 to 1032 aminoacids The majority of the translated protease sequenceswere obtained from fungus All serine proteases from cluster1 subcluster a are derived from Agaricomycetes except forEJU04161 which belongs to Dacrymycetes whereby all serineproteases from cluster 1 subcluster b are from Sordari-omycetes Cluster II subcluster c consisted of serine proteasefrom fungus belonging to Ustilaginomycetes PI12 proteasewas categorized under cluster II subcluster d It possessedhomology similarity with the subtilisin-like protease fromRhodosporidium toruloides (EMS20793) (formerly known asRhodotorula glutinis or Rhodotorula gracilis) which is a
Enzyme Research 13
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
MKG-YSWSLLQLAFTGAALATVPLSDVKRISSAS--VVPNKFVVEVESASEIPGKR--SL
MKGVLVWSILS---AGCALAVTPLSSVKRVTNLP--IVANKFIVEVDSTSDIPTKR--S-
MKLPLLSLALT---AVSVGATVPLSSVQRTTTSPNNALAGEYIVEMNNVQGLGGKR--AY
---MAHGSLASLLLLLAASAQGAAAFGGKSATGGRSVVPNSYILQVNASAPALTKRGVSV
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease ---MLFLPVLLLLLPGVTAFLNPVTNRATNAISSTQYLSNAYILELDLSTPGLVKRDSTP
lowastlowast
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
YTPHELFYRSLSARGVSYEVNKEYNS-EGLFVGAVVTVSDANDVKALAETPGVKAIRPVR
-NPHDALYESLRKRAIGFKVDREFNT-QGLFVGAALTLSDAQDVGQIQSTTGVKSIRPVR
ASTHDTLYRSLRKRGISFDVKKEYNQ-PGIMVGAALKVASPQDVQQIATMAEVKAIRPVI
FEALDHTLAAVKQNGVKYTVRQRFDAIPEAFQAVSIQVEDGASMAELAQIPGVQRVWPVS
DSILEDVLTSVGRNGIKYQLRHRFIS-PTLFHGASITVPPGISRSQIASLRGIKRVWPVR
lowastlowast
TIKSPKPVKVHVVTGKDDPEIPA-------------------------DTESTHIMTGVN
AFDRPKPVKVQVVKGPDDPALPP-------------------------DSESTHVITGVD
AVPAPERINQFTVSSPNDSRITA-------------------------DSETTHIMTGVD
LIPRPVEPTVSDFTPVSSSSSSTKSKRD-THTLEKRGTNFPPASAYLNDTFYPHVQTGID
KFSRPSAVVDADGGGSGFSGSPIKAALMGVKELGKR------ANAYAGDTFGPHVMTGVN
lowast lowast lowast lowastlowast
KLHAQGIFGAGIKIGIIDTGIDYTHPALGGALGAGHKVIGGYDFVGDAYDGTNTPVPDND
KLHAQGITGAGIKIGILDTGIDYTHPTLGGAFGPGNKVIGGFDLVGDDYDGTNTPVPDPD
KVHAAGITGEGIKIGIIDTGIDYNHPLLGAGFGPGFKVIGGYDFVGDDYTGSNNPAPDDD
VLHNKGILGQGVKIVVVDEGVDYTNPILGGCFGPGCQISFGYDFVGDNYTGTNAPVPDSD
ETHEAGLLGAGIKIGVLDTGVDYLNPILGGCFGPGCHMSFGYDLVGDDYDGDNAPVPDVD
lowast lowast lowast lowastlowastlowast lowast lowast lowastlowast lowast lowastlowast lowast lowast lowast lowast lowastlowastlowast lowast lowast lowast lowast lowastlowast lowast
PLDQCNGHGTHVAGIIGANPDNQFNISGVSYGASLSSYRVFGCSGSVSDDIIIEALLRGV
PLDQCAGHGTHVAGIIGANPDNAFNISGVAFQASLSAYRVFGCVGFVTDDVLVDALLLGF
PLDNCNGHGTHVAGIIGANPGNDFGISGVAYGSSIASYRVFGCSGFVSDDIIIDALLRGY
PFSSCSNHGTHTTGTVGALAN-EYGFSGAAPMATIGHYRVISCSGATTDEILVAALTRAL
PFASCDPHGTHVTGIIGALPN-AFGFTGVAPAATLGHYRVFGCTGFVGEDIILAGLMRGV
lowast lowastlowastlowast lowastlowast lowast lowastlowast lowast lowastlowastlowast lowast lowast lowast
QDGQDILTLSLGGTDGWTE-SSSSVVASRIAASGKIVTIAAGNEGAYGSWYSSSPGNGID
NEGQDILTLSLGGADGWTE-SVSAVVASRIAATGKVVTIAAGNDGASGAWYTSSPGNGID
SDGMDILTLSLGGVEGWTE-SSSSVVASRIADQGRVVTIAAGNDGAYGSWYASSPGTGVD
QDNVNIVSMSLGSSVGWLDDSPVQIMASYLGSKGIHVVASVGNERNEGLFAADQPAASRI
EDNCNVLTLSLGGPGGWVKGTPASILIDQIEAQGILVTVATGNSGAEGMFFSESPASTIN
lowastlowastlowast lowastlowast lowast lowast lowastlowast lowast lowast
VISVASIDNTVDNIQNATVNGVEHGPIPYFKTLPLPFTNA-LPLYVISNDTTIANDACNA
VISVASLDNTVVPLQTAQVHGVTHDPIIYQDTFPLPINDT-RPIFATSTDVTVVDDACNP
VISVASVNNIAIPVQNATVQGVEHAPIPYLEALPLNVTGS-RPIYATSTDTTVADDACNA
GTGVGGVDPTYLAAYYAYLI--KRDPVPYISPTPFNLPNN-YLLYFTSTNTSVTNDACNP
GLSIASTDVTDLIAYNATVS--GQPAIPYLSATPLNVVANSFRVHFTSTDPNNPVDACSP
lowast lowast lowast lowast lowastlowastlowast
LPDNTPDLSGFLVLVRRGSCTFVQKLTNIAAKGGNAVFIYDNGNG-----FAGIDTGKFT
LPDSTPDLSGFVVIVRRGTCTFVQKLGNVAAKGAKVTLIYDNGSG-----FGAIDTGAFV
LPNDTPDLSGYVVVIKRGTCTFVTKLQNAAAKGANTFLIYNNGGS-----FSAISVGDYT
LPSTTPNLANRVVVVQRGTCGFTVKQQNVAAAGGKIVLIYNSKGSGLIPQLDVGSTGLTA
LPAGAPDFANYVTVVQRGTCTFVTKYQNVLNAGGKIVLLYNSEGAGNLPYLTPNGVGIDA
lowastlowast lowast lowastlowast lowast lowast lowast lowast lowast lowast lowast
(a)
Figure 5 Continued
14 Enzyme Research
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
NATLIQAADGVFLAQQVAAGKSVTITFPQTGGSAQITN-PSGGLVSSFTTYGPTNDFYFK
-ASFIQAADGEFLVQQFASGQNVSLSFPQSGGLIQFPD-PAGGLISSFTSYGPSNDFFFK
-AALISETDGTFLVNQFAAKANITISFPQSGSAWNYPDDVSGGLVSDFSSYGPSYDMYFK
VGSLRYEDGLKLLSYYQSSPRGQSISFPNGPLAPYVSNTVSGGVVASYSNFGPTNELFIY
VAGLRRSDGLKLLSYYQNANKRLTLRFPKGKIVAGLTDTITGGLISGYSTFGPTNDLYGQ
lowast lowastlowast lowastlowast lowastlowast
PAFGAPGGNILSTLPVPLGSYGVESGTSMATPYVAGSAALLLSVKGKSAAVALGARTLFQ
PAVAAPGGNILSTLPVNLGLFGIESGTSMATPFVAGSAALLFQAKGTSAAVGRSARTLFE
PAVAAPGGNILSTLPIALGSYGVESGTSMATPFTAGSAALILQAKGK--SVAKSIRNLFQ
PTLAAPGTDILSTV---TGGVALMRGTSMAAPLVAGAYALLLSVGNNLKLSPEEARTILM
PTLSAPGGNILSTFPLSEGGVAVISGTSMSCPFVAGSAAVLMAARASENLTPLEIRSLLT
lowast lowastlowastlowast lowastlowastlowastlowast lowast lowastlowastlowastlowast lowast lowastlowast lowast lowast
STAQRVPSSHTDGDPLQTLSQQGAGLIDVFKALYTETIVWPTELILNDTAHYKEVHTFTV
TTAQRVPSTHTDGDPLQTLTQQGAGLINVFNAIHTTTIVSPGELILNDTAHFKGTQQFTV
TTASAVASSKTDGDPLQTVSQQGAGLIQVDRAIGTTTVVSPGQLTLNDTAHFEGFKTFNI
STSQPSPVAY-GSETLDTVVSQGAGVINMTAAIAAHTVISPAQFNLNDTQYSNNTQTLTL
TTAKLTPVSLLGSTPLVSVIRQGGGLVQVAKALAAKTLISPHELLLNDTANANYVQTIKI
VNTGKTSKQYKLTHVPAGTAQTIT---SGTIFAADGPVPLSTDYATVKLSSSSFTLRPGQ
RNAGSSAKTYTLRHIPAGTAVTVT---PGTIFPADGPVPLSTDFASVSLSTSKFTLAPGK
QNTGSKAVTYKISHVPAGTAASVQ---SGSIFAADGPVPLTADYAQVTFSLNKVTVLPGV
RNNNRFPVSYQLSWVDSTGIVTYNDGATTDIIPSTTPNYVSTSVLRVAFSQRTVTVPGGQ
KNTNSWAMKYTFSSAVAQGLGTFD--ASGDILPTLDPVAVSGAQATVAFNTRILSVAPGA
lowast lowast lowast lowast lowast lowast
SKQVTTHINAP--SGVDASTYPVFSGFIEIATAT--------ESVHVTYLGLAASLKDKQ
TQTVTARFTPP--AAADPSTFPVFSGFIQIESGT--------EQVQVSYLGLKASLKDKQ
SVPILVWFKPP--QGVDASTYPVYSGFIEIAGLG--------ETLRVAYLGVAASLKDKA
TVKVTAQFIPPNLTAQQRNQFPIYSGFVTITGQGQGAGAEQMEKYNLPFFGLGARMVDMP
TGSVVATITPPVLPVADAARFPIFSGWIRVNGQG-ARDSSRNEAYTVPYFGLAAKMIDMQ
lowast lowast lowastlowast lowast lowast lowast lowast
IVDNTNEFFGITIPAVLNNTGDAQ-LSPTNYTFSGSDFPTLLLRLAFGTPLLRVDLVDAT
VIDNTDFFFGVPTPVLTDPNGEVQ-TSPRNYSFLASDFPTLIFRLAFGSPKVVVDLVSPT
VVDNTDEYFGVKIPALLDASGDVQ-GNSTNYTLVGDDVPSLLYRFAFGSAHVRVDLVEKA
VLDTTNVALGP--NLPFIAVGQN--IQTGPTTFSESNPAVVYFRLAAGTRRLRVDLVDAS
VLDTTETIYGPGYAYPFVIDDAIGDIQSTTTSYSRNLGPTVFARFATGTLHYSLDLVLAD
lowast lowast lowast lowast lowastlowastlowastlowast lowast
IDLKPTLNSREIDDKRHVFERESFSFPHPGPKGS-------------------TFAQVKI
INFKPTLNTRALGDTHGSF----FSFPHRVKTG--------------------SFAQVPT
STVSATLSKRNWWN---------WWWPQKPSN---------------------SYDKVPI
TSYNATVPAVTNPASRLVKRSNTELTPRAAFPT--------------------LYSQVRT
IAFTPTYPNSS-PATRFVKRSLTQHTSAASHLAKRRVSIATINPKATLVADRQLHSDVPI
lowast lowast
VGTLSEQSHIPRNSDASTVDDGGYSTFAFNKP------SFANGTTIPNGTYRWLLRALKV
LGTVFELDFTSRNNDDPTDN--GFNTVAIDPP------TFANGTTIPNGQYRLLLRALRV
VGTLAEYDYVPRNSDASTIDDNGYNVFQFT-D------SFVNGTKIAAGEYKVLLRALKV
VGSLFAPSYWPPRDYLFNNGGT-YSDYEIQLDGSFTFPNGTKGMAELGRSYKVLLRALKI
EGNIFTQPFTG-RDYLVDAAPTGSTDRTVTFNGQY-AENGLVRTAVTGTSYRFLLRALKI
lowast lowast lowastlowast lowast lowast lowast lowast lowastlowastlowastlowast
lowast lowast lowastlowastlowastlowastlowast
(b)
Figure 5 Continued
Enzyme Research 15
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
TGDPTKEADFETWLSPIVGITVA
TGDPTNEADFESFLSPIIGVNAP
TGNPAEEGDYETWLSPSFSVVPA
TADPTYTSSYESWLSPSFTFTA-
SGDAMYEDQYESWLSLPFSFRA-
lowast lowastlowast
ConservedSimilar
lowast All match
(c)
Figure 5 Multiple sequence alignment between amino acid sequence of PI12 protease with other subtilisin The serine protease sequenceswere obtained from the GenBank database Their accession numbers are EMS20793 (subtilisin-like protease of Rhodosporidium toruloidesNP11) CAG25549 (subtilisin-like protease of Pleurotus ostreatus) EPQ55564 (subtilisin-like protease of Gloeophyllum trabeumATCC 11539)and ETW80907 (serine protease S8 ofHeterobasidion irregulare TC 32-1) Putative ORF of serine protease fromG antarctica PI12 was labeledas PI12 protease
CAG38357 Phanerochaete chrysosporium
EIW62675 Trametes versicolor FP-101664 SS1
XP 007369624 Dichomitus squalens LYAD-421 SS1
XP 007381302 Punctularia strigosozonata HHB-11173 SS5
CAG25549 Pleurotus ostreatus
ETW80907 Heterobasidion irregulare TC 32-1
XP 001877576 Laccaria bicolor S238N-H82
EPQ55564 Gloeophyllum trabeum ATCC 11539
XP 001830835 Coprinopsis cinerea okayama7130
XP 007341385 Auricularia delicata TFB-10046 SS5
CCA68627 Piriformospora indica DSM 11827
CCO37249 Rhizoctonia solani AG-1 IB
XP 007342189 Auricularia delicata TFB-10046 SS5
EJU04161 Dacryopinax sp DJM-731 SS1
XP 007589863 Colletotrichum fioriniae PJ7
EFQ32840 Colletotrichum graminicola M1001
ENH74885 Fusarium oxysporum f sp cubense race 1
XP 003711175 Magnaporthe oryzae 70-15
XP 006694395 Chaetomium thermophilum DSM 1495
EON99722 Togninia minima UCRPA7
CBQ73425 Sporisorium reilianum SRZ2
GAC97617 Pseudozyma hubeiensis SY62
CCF52619 Ustilago hordei
EMS20793 Rhodosporidium toruloides NP11
CAQ76821 Glaciozyma antarctica PI12
01
Clus
ter 1
Clus
ter 2
a
b
c
d
Figure 6 Phylogenetic dendrogram among 25 translated amino acid sequences of serine proteaseThe tree was constructed using a neighbor-joining algorithm with 1000 bootstrappings by MEGA 6 software [39]The scale bar represents 01 substitutions per amino acid positionTheserine protease sequences were obtained from the GenBank database GenBank accession numbers of the sequences were indicated beforethe strain name The putative serine protease of G antarctica strain PI12 has been deposited in GenBank with accession number CAQ76821
16 Enzyme Research
0
2
4
6
8
10
12
0 24 48 72 96 110
Prot
ease
activ
ity (U
mL)
Post induction time (hours)
Figure 7 Effect of postinduction time towards protease activity The highest activity was detected at 72 hours after 110 hours of incubationwith 9UmL activity followed by 48 24 and 96 hoursThe substrate (azocasein) was dissolved in Tris HCL buffer and incubated for 30min at20∘CThe absorbance was measured at 450 nm after 15min incubation A one-way ANOVAwas used to test for preference differences amongsix different postinduction times The statistical value (119875 value lt005) between hours is 0003
pink-coloured nonpathogenic oleaginous yeast (42 iden-tity) Both G antarctica PI12 and Rhodosporidium toruloidesNP11 belong to Microbotryomycetes class PI12 protease didnot show any homology similarity to other psychrophilicproteases however it showed 37 identity to hypotheticalprotein from Pseudozyma antarctica T-34 which was isolatedfrom Japan (GAC76005) (not listed in the phylogenetic tree)
35 Cloning and Expression of Cold-Adapted PI12 Protease inthe Yeast Expression System The PCR product harbouringthe mature PI12 protease with two flanking restriction sites(EcoRI and AvrII) was cloned into pPIC9 expression vectorThe recombinant plasmid harbouring PI12 protease was usedto transform both GS115 and KM71 intended to produce bothHis+ Mut+ and His+ MutS Two recombinant clones fromGS115 (His+Mut+) strain (GpPro1 andGpPro2) and one fromKM71 (His+MutS) strain (KpPro1) were successfully obtainedwhich were further analyzed for protein expression study
36 Expression and Enzyme Assay of PI12 Protease in Pichiapastoris All positive recombinants were grown in small scalecultures and assayed for the cold-adapted protease activityin the culture supernatants The clones were first grown inBMGY medium with glycerol as the sole carbon sourceFor induction of AOX1 promoter and protein productionthe growth medium was transferred to the same mediumexcept with 05 methanol in place with glycerol as thecarbon source [26] The supernatant of the recombinant Ppastoris expression cultures after 48 h of incubation time wasanalyzed to determine the recombinant protease activity bythe modified method of Brock et al (1982) [20] All testedclones secreted protease into the supernatant in differentexpression levels This might be due to the variability ofthe strength of the promoter and copy number in each cellto secrete and express the recombinant product The clonewith the highest yield was GpPro2 with 63UmL activityfollowed by KpPro1 and GpPro1 with 25UmL and 37UmLactivities respectively (data not shown) It is perceptible thatexpression systemof Pichia pastoris is the ideal system for thisprotein The preliminary assay has fruitfully produced high
protease expression at 20∘C Yet further optimization of therecombinant PI12 protease production need to be conductedin order to examine the optimum condition of this proteinReported cold-adapted proteases showed optimum activity ata range of 15∘Cndash60∘C [27ndash30] This difference was possiblyrelated to the respective genetic and physiological adaptationof the strains There are several factors that influenced thedifferent rate of expression It includes different site of inte-gration between interest gene and Pichia genome variabilityof different expression host cell to secrete and express therecombinant enzyme and different sites in the polylinker forinserting foreign genes [31]
This is the second extracellular protease reported to besynthesized by this obligate psychrophilic yeastG antarcticaThe first cold-adapted extracellular subtilase from this yeasthas been purified and characterized previously [32 33] Theextracellular serine proteinase (LAP2) from this Antarcticyeast formerly known as Leucosporidium antarcticum strain171 is specific towards synthetic substrates of chymotrypsinand subtilisin The subtilase of L antarcticum strain 171showed specific activity of 4043Umg when utilized N-SucAAPFpNA as a substrate [33]
37 Optimization of Protease Production GpPro2 was chosenfor further optimization due to its highest protease activityat the preliminary expression stage High protease activitywas detected after 72 hours of incubation (Figure 7) byusing the standard protocol method as recommended forPichia pastoris expression system (Pichia Expression KitInvitrogen USA) The expression decreased after 72 hoursand no activity was detected at 110 hours The prolongedincubation may result in the decrease of biomass productionand will subsequently reduce the expression of recombinantprotein In addition oxygen and nutrient depletion duringprolonged incubationwill trigger the stress responses in orderto survive in such harsh environment [34]
Methanol was used as a sole carbon by P pastoris Themethanol was oxidized to formaldehyde with the aid of oxy-gen as electron acceptor by an alcohol oxidase enzyme (AOXgene) [35] The optimal methanol concentration is at 05(vv) with 14UmL activity The addition of more than 05
Enzyme Research 17
02468
10121416
0 01 03 05 07 09Pr
otea
se ac
tivity
(Um
L)Methanol concentration ()
Figure 8 Effect ofmethanol concentration towards protease activityThe recombinant was cultivated for 72 hours at 30∘C in BMMYmedium05 methanol gives the highest activity with 14UmL The substrate (azocasein) was dissolved in Tris HCL buffer and incubated for 30minat 20∘CThe mixture was left for 15min and absorbance was taken at 450 nm A one-way ANOVA was used to test for preference differencesamong six different concentrations The statistical value (119875 value lt005) between concentration is 002
0
5
10
15
20
25
10 15 20 25 30 35
Prot
eas a
ctiv
ity (
Um
L)
Temperature (∘C)
Figure 9 Effect of temperature towards protease activity The reaction was carried out in temperature ranging from 10∘C to 35∘C at pH 7 for30min A one-way ANOVA was used to test for preference differences among six different temperatures The statistical value (119875 value lt005)between temperature is 0067
(vv) methanol resulted in tremendous decrease of proteaseactivity (Figure 8) Production of PI12 protease was decreasedwhen the methanol concentration increased This might bedue to the toxic effect of methanol concentration on DNAreplication and membrane synthesis Basically methanol isused to increase the permeability of the cell membrane tohelp the secretion of foreign protein [26] However if thepermeability of cell membrane is too high lowmolecular sizemetabolites would leak from the cell and such leakage willseriously damage cell activities Such condition would abusethe cellular system and might reduce the yield of proteinexpression [36]
Psychrophilic microorganism dominated in low temper-ature habitat which most other microorganism are inca-pable of withstanding due to their inability to produceenzyme that can tolerate the extreme environment con-dition Psychrophilic enzymes are always associated withhigh catalytic efficiency at low and moderate temperature(0∘C to 30∘C) compared to the mesophilic counterpart thatare generally unable to catalyze efficiently under extremely
low temperature [2] The recombinant protein assay wasdone at various temperatures ranging from 10∘C to 35∘Cusing 05 azocasein as a substrate dissolved in 50mMTris HCl in presence of 2mM CaCl
2 pH 7 In the activity
assays expressed protease showed a highest activity at 20∘C(20UmL) and decreased tremendously once it exceededmaximum temperature due to the denatured state of theenzyme (Figure 9) The active site of an enzyme is easilyaltered when the high temperature is introduced duringincubation or activity assay resulting in a low reaction rate[37]
The recombinant construct was transferred into BMMYmedia at different A
600 nm ranging from 1 to 10 The high-est activity of active enzyme was detected at A
600 nm 60(283UmL) and above (Figure 10) The slope during expo-nential phase was much higher compared to other phasesindicating an increase in the doubling time At exponentialphase the cells are adapted to the growth condition andother processes like cell growth cycle and synthesis of RNAThe yield of active enzyme during expression of P pastoris
18 Enzyme Research
05
1015202530
1 2 3 4 5 6 7 8 9 10Prot
ease
activ
ity (U
mL)
Initial inoculum A600
Figure 10 Effect of inoculums size towards protease activity The recombinant was cultivated for 72 hours at 30∘C in BMMY mediumwith 05 methanol concentration A one-way ANOVA was used to test for preference differences among six different optical densitiesThe statistical value (119875 value lt005) between optical density value is 003
(kD
a)
1160
662
450
350
250
184
144
M 1 2 3
(a) (b) (c)
Figure 11 (a) Coomassie-stained SDS-PAGE of recombinant P pastoris clone 1mL supernatants collected from the third-day culturewere concentrated 10-fold using trichloroacetic acid and 10120583L resultant products were applied to SDS-PAGE analysis Lane 1 molecularweight marker (Fermentas) Lane 2 GS115 (Control) Lane 3 GS115pPIC9 empty (Control) Lane 4 GS115pPIC9PI12prot (GpPro2)The arrow indicates the secreted recombinant PI12 protease (b) Plate activity staining of different recombinant P pastoris clones GGS115 GP GS115pPIC9 GRI GS115pPIC9PI12prot1 (GpPro1) GRII GS115pPIC9PI12prot2 (GpPro2) K KM71 KP KM71pPIC9 KRKM71pPIC9PI12prot (KpPro) (c)Gel activity staining of recombinant PI12 protease inP pastorisGpPro2 clone Lane 1 prestainedmolecularweightmarker (Fermentas) Lane 2 GS115pPIC9 empty (Control) Lane 3 GS115pPIC9PI12prot1 (GpPro2)The arrow indicates the secretedrecombinant PI12 protease
dropped after 28 hours of incubation which is at stationaryor death phase [38]
38 Detection of Recombinant Protein by SDS-PAGE andActivity Staining TheSDS-PAGE gel of the GpPro2 recombi-nant clonesrsquo supernatant showed a putative expression bandat around 993 kDa Plate staining and gel activity stainingwere executed to examine if the recombinant PI12 proteaseexpressed in P pastoris was functionally active In the platestaining the transformants showed strong proteolytic activity(Figure 11(a)) In gel staining proteolytic activity was alsodetected around 993 kDa after incubation at 20∘C This is
evidence of protease secretion into the surrounding mediawhereby it catalyzed the breakdown of casein thus formedclearing zones around the activity ring and band (Figures11(b) and 11(c)) As a conclusion the cold-adapted PI12protease was functionally expressed in P pastoris GS115(GpPro2 clone) with the size of 993 kDa
4 Conclusions
Psychrophilic yeast which was originated frommarine waterin Casey Station Antarctica was identified as Glaciozymaantarctica strain PI12 A serine protease with an open reading
Enzyme Research 19
frame of 2892 bp encoded for 963 amino acids from thisAntarctic yeast has been cloned and expressed in Pichiaexpression system This is the second extracellular pro-tease reported being synthesized by this psychrophilic yeastSequence analysis through bioinformatics studies revealedfew homology identities of this protein with other ser-ine proteases The mature PI12 protease was successfullyexpressed and secreted into the culture medium driven bythe Saccharomyces cerevisiae 120572-factor signal sequence in Ppastoris The protease production in P pastoris was bestobtained from strain GS115 (GpPro2) with 283UmL after 72hours of induction time with 05 (vv) of methanol inducerThe starting optical density at 600 nm (OD600) was 60 andthe optimum assay temperature was 20∘C The expressedprotein was detected by SDS-PAGE and activity staining witha molecular weight of 99 kDaThis cold-active enzymemightoffer an opportunity for biotechnological exploitation basedon its high catalytic activity at low temperatures and lowthermostability Structural and functional study of this cold-adapted enzyme would give insight into its adaptation to anextreme cold environment
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgment
This research was supported financially by the InitiativeBiotechnology-GMBI-MOSTI (Project number 07-05-16-MG-GMB02) andRUGS-Initiative 6 (Project number 05-02-11-1414RU)
References
[1] J W Deming ldquoPsychrophiles and polar regionsrdquo CurrentOpinion in Microbiology vol 5 no 3 pp 301ndash309 2002
[2] C Gerday M Aittaleb M Bentahir et al ldquoCold-adaptedenzymes from fundamentals to biotechnologyrdquo Trends inBiotechnology vol 18 no 3 pp 103ndash107 2000
[3] A Svingor J Kardos I Hajdu A Nemeth and P Zavodszky ldquoAbetter enzyme to cope with cold comparative flexibility studieson psychrotrophic mesophilic and thermophilic IPMDHSrdquoThe Journal of Biological Chemistry vol 276 no 30 pp 28121ndash28125 2001
[4] G Antranikian C E Vorgias and C Bertoldo ldquoExtremeenvironments as a resource for microorganisms and novel bio-catalystsrdquo Advances in Biochemical EngineeringBiotechnologyvol 96 pp 219ndash262 2005
[5] T Lonhienne C Gerday andG Feller ldquoPsychrophilic enzymesrevisiting the thermodynamic parameters of activation mayexplain local flexibilityrdquoBiochimica et Biophysica Acta vol 1543no 1 pp 1ndash10 2000
[6] R Cavicchioli and TThomas ldquoExtremophilesrdquo in Encyclopediaof Microbiology pp 317ndash337 Academic Press San Diego CalifUSA 2000
[7] G Feller E Narinx J L Arpigny et al ldquoEnzymes frompsychrophilic organismsrdquo FEMS Microbiology Reviews vol 18no 2-3 pp 189ndash202 1996
[8] J Arnorsdottir R B Smaradottir T O Magnusson S HThorbjarnardottir G Eggertsson and M M KristjanssonldquoCharacterization of a cloned subtilisin-like serine proteinasefrom a psychrotrophic Vibrio speciesrdquo European Journal ofBiochemistry vol 269 no 22 pp 5536ndash5546 2002
[9] N J Russell ldquoPsychrophilic bacteriamdashmolecular adaptations ofmembrane lipidsrdquo Comparative Biochemistry and Physiology APhysiology vol 118 no 3 pp 489ndash493 1997
[10] G Feller and C Gerday ldquoPsychrophilic enzymes hot topics incold adaptationrdquo Nature Reviews Microbiology vol 1 no 3 pp200ndash208 2003
[11] P Raspor and J Zupan ldquoYeasts in extreme environmentsrdquo inBiodiversity and Ecophysiology of Yeasts The Yeast Handbookpp 371ndash417 Springer Berlin Germany 2006
[12] M B Rao A M Tanksale M S Ghatge and V V Desh-pande ldquoMolecular and biotechnological aspects of microbialproteasesrdquoMicrobiology andMolecular Biology Reviews vol 62no 3 pp 597ndash635 1998
[13] L Polgar ldquoThe catalytic triad of serine peptidasesrdquo Cellular andMolecular Life Sciences vol 62 no 19-20 pp 2161ndash2172 2005
[14] S Y Boo C M V L Wong K F Rodrigues N NajimudinA M A Murad and N M Mahadi ldquoThermal stress responsesin Antarctic yeastGlaciozyma antarctica PI12 characterized byreal-time quantitative PCRrdquoPolar Biology vol 36 no 3 pp 381ndash389 2013
[15] B Turchetti S R T Hall L B Connell et al ldquoPsychrophilicyeasts from Antarctica and European glaciers descriptionof Glaciozyma gen nov Glaciozyma martinii sp nov andGlaciozyma watsonii sp novrdquo Extremophiles vol 15 no 5 pp573ndash586 2011
[16] N H F Hashim I Bharudin D L S Nguong et al ldquoCharac-terization of Afp1 an antifreeze protein from the psychrophilicyeast Glaciozyma antarctica PI12rdquo Extremophiles vol 17 no 1pp 63ndash73 2013
[17] A N M Ramli M A Azhar M S Shamsir et al ldquoSequenceand structural investigation of a novel psychrophilic 120572-amylasefrom Glaciozyma antarctica PI12 for cold-adaptation analysisrdquoJournal ofMolecularModeling vol 19 no 8 pp 3369ndash3383 2013
[18] A N M Ramli N M Mahadi A Rabu A M A Murad F DA Bakar and R M Illias ldquoMolecular cloning expression andbiochemical characterisation of a cold-adapted novel recombi-nant chitinase from Glaciozyma antarctica PI12rdquoMicrobial CellFactories vol 10 no 1 article 94 2011
[19] J Sambrook E Fritsch and T Maniatis Molecular Cloning ALaboratoryManual Cold SpringHarbor Laboratory Press NewYork NY USA 1989
[20] F M Brock C W Forsberg and J G Buchanan-SmithldquoProteolytic activity of rumen microorganisms and effects ofproteinase inhibitorsrdquoApplied and EnvironmentalMicrobiologyvol 44 no 3 pp 561ndash569 1982
[21] U K Laemmli ldquoCleavage of structural proteins during theassembly of the head of bacteriophage T4rdquo Nature vol 227 no5259 pp 680ndash685 1970
[22] A Henne R A Schmitz M Bomeke G Gottschalk and RDaniel ldquoScreening of environmental DNA libraries for thepresence of genes conferring lipolytic activity on Escherichiacolirdquo Applied and Environmental Microbiology vol 66 no 7 pp3113ndash3116 2000
[23] M Poza T de Miguel C Sieiro and T G Villa ldquoCharacteri-zation of a broad pH range protease of Candida caseinolyticardquoJournal of Applied Microbiology vol 91 no 5 pp 916ndash921 2001
20 Enzyme Research
[24] J D Bendtsen H Nielsen G von Heijne and S BrunakldquoImproved prediction of signal peptides SignalP 30rdquo Journalof Molecular Biology vol 340 no 4 pp 783ndash795 2004
[25] R J Siezen and J A M Leunissen ldquoSubtilases the superfamilyof subtilisin-like serine proteasesrdquo Protein Science vol 6 no 3pp 501ndash523 1997
[26] Y Katakura W Zhang G Zhuang et al ldquoEffect of methanolconcentration on the production of human 1205732- glycoprotein Idomain V by a recombinant Pichia pastoris a simple system forthe control of methanol concentration using a semiconductorgas sensorrdquo Journal of Fermentation and Bioengineering vol 86no 5 pp 482ndash487 1998
[27] Q-FWang Y-HHou Z Xu J-LMiao andG-Y Li ldquoPurifica-tion and properties of an extracellular cold-active protease fromthe psychrophilic bacterium Pseudoalteromonas sp NJ276rdquoBiochemical Engineering Journal vol 38 no 3 pp 362ndash3682008
[28] S Vazquez L Ruberto and W Mac Cormack ldquoProper-ties of extracellular proteases from three psychrotolerantStenotrophomonas maltophilia isolated from Antarctic soilrdquoPolar Biology vol 28 no 4 pp 319ndash325 2005
[29] R Zeng R Zhang J Zhao and N Lin ldquoCold-active ser-ine alkaline protease from the psychrophilic bacterium Pseu-domonas strain DY-A enzyme purification and characteriza-tionrdquo Extremophiles vol 7 no 4 pp 335ndash337 2003
[30] M Turkiewicz E Gromek H Kalinowska and M ZielinskaldquoBiosynthesis and properties of an extracellularmetalloproteasefrom the Antarctic marine bacterium Sphingomonas paucimo-bilisrdquo Journal of Biotechnology vol 70 no 1ndash3 pp 53ndash60 1999
[31] A Vassileva D A Chugh S Swaminathan and N KhannaldquoEffect of copy number on the expression levels of hepatitisB surface antigen in the methylotrophic yeast Pichia pastorisrdquoProtein Expression andPurification vol 21 no 1 pp 71ndash80 2001
[32] M Pazgier M Turkiewicz H Kalinowska and S Bielecki ldquoTheunique cold-adapted extracellular subtilase from psychrophilicyeast Leucosporidium antarcticumrdquo Journal ofMolecular Catal-ysis B Enzymatic vol 21 no 1-2 pp 39ndash42 2003
[33] M Turkiewicz M Pazgier H Kalinowska and S BieleckildquoA cold-adapted extracellular serine proteinase of the yeastLeucosporidium antarcticumrdquo Extremophiles vol 7 no 6 pp435ndash442 2003
[34] D Mattanovich B Gasser H Hohenblum and M SauerldquoStress in recombinant protein producing yeastsrdquo Journal ofBiotechnology vol 113 no 1ndash3 pp 121ndash135 2004
[35] P Secades BAlvarez and JAGuijarro ldquoPurification andprop-erties of a new psychrophilic metalloprotease (Fpp2) in the fishpathogen Flavobacterium psychrophilumrdquo FEMS MicrobiologyLetters vol 226 no 2 pp 273ndash279 2003
[36] L Y Ling I Ithoi and F M Yik ldquoOptimization for high-level expression in Pichia pastoris and purification of truncatedand full length recombinant sag2 of toxoplasma gondii fordiagnostic userdquo Southeast Asian Journal of Tropical Medicineand Public Health vol 41 no 3 pp 507ndash513 2010
[37] A Illanes ldquoStability of biocatalystsrdquo Electronic Journal ofBiotechnology vol 2 no 1 pp 1ndash9 1999
[38] B A Plantz J Sinha L Villarete K W Nickerson and VL Schlegel ldquoPichia pastoris fermentation optimization energystate and testing a growth-associated modelrdquo Applied Microbi-ology and Biotechnology vol 72 no 2 pp 297ndash305 2006
[39] K Tamura G Stecher D Peterson A Filipski and S KumarldquoMEGA6molecular evolutionary genetics analysis version 60rdquo
Molecular Biology and Evolution vol 30 no 12 pp 2725ndash27292013
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

2 Enzyme Research
catalytic rate owing to reduction of structural flexibility andundergo cold denaturation [7]
Psychrophilic enzymes possess remarkable prospectivein a wide variety of industrial applications Our interestis focused on cold-adapted protease as a useful biocata-lyst in industry such as low temperature clothes washingand enzyme-added detergents to hydrolyze macromolecularstains [10] An example of commercially available cold-active protease is from Novozyme (trade name Savinase)which is sold as an encapsulated detergent [11] Proteasesare degradative enzymes which catalyze the hydrolysis ofproteins and are commonly found among animal tissuesplant and microorganisms [12] Proteases execute a largevariety of functions and take part in numerous biochemicalreactions in living organisms including formation of sporeand germination coagulation cascade reactions posttransla-tion reactions modulation of gene expression enzymemodi-fication and secretion of various protein enzymes biocatalystOver one-third of all known proteolytic enzymes are serinepeptidases which constitute the largest group of peptidases[13]
Glaciozyma antarctica (formerly known as Leucosporid-ium antarcticum) is an obligate pyschrophilic yeast thatinhabit cold marin and terrestrial Antarctic ecosystemsGlaciozyma antarctica strain PI12 has an optimum growthtemperature of 12∘C [14] and can grow up to 18∘CThe reclas-sification of this yeast from L antarcticum to G antarcticawas proposed by Turchetti et al 2011 [15] Previous studyon this unique yeast has isolated several cold-active proteinsnamely antifreeze protein 120572-amylase and chitinase [16ndash18]In this work we described the morphological characteristicsofG antarctica through observation under scanning electronmicroscope (SEM) and transmission electron microscope(TEM) cloning of genomic DNA and cDNA sequencesencoding the PI12 protease gene phylogenetic study andexpression and optimization of the recombinant PI12 pro-tease expression in Pichia pastoris expression system
2 Materials and Methods
21 Culture and Isolation of Microorganism Glaciozymaantarctica strain PI12 was isolated from the Antarctic marinewater near Casey Station (66∘2110158402510158401015840S 110∘3710158400910158401015840E) Thestock culture was kept in 20 (vv) glycerol and stored atminus80∘C prior to experimentation
22 Microorganism Identification The isolated strain wasgrown on different types of solid media (nutrient agarsabouroud dextrose agar and potato dextrose agar) for 10days at 4∘C and the culture characteristics (colony colourshape and texture) were determined Simple and negativestaining were performed to identify the cell morphologyarrangement and size of the psychrophilic yeast SEM andTEM were conducted to study the surface features and theinternal ultrastructure in the thin sections of theG antarcticaPI12 cells Ribosomal RNA identification was performedthrough ITS1ITS2 region amplification The sequence has
been deposited in the GenBank database under accessionnumber JX896956 [14]
23 Nucleic Acid Isolation A single colony of the yeast wasinoculated into 50mL yeast peptone dextrose broth (YPD)and incubated for 10 days at 4∘C without shaking The cellpellets were frozen in liquid nitrogen and ground to a powderin a ceramic mortar Genomic DNA was extracted usinga phenol-chloroform method as described elsewhere [19]Removal of RNA from genomic DNA was performed by theaddition of 15120583L RNase (10mgmL) and incubated at 37∘Cfor 45min Total RNA was extracted by the guanidium isoth-iocyanate method using Trizol reagent (Invitrogen USA)according to the manufacturerrsquos instructions Approximately4 120583g of total RNA was digested with 05 120583g of RNAse-freeDNase in a 10 120583L reaction mixture containing 1x reactionbuffer (20mM Tris-HCl pH 84 50mM KCl and 2mMMgCl
2) prior to removal of DNAThemixture was incubated
at 37∘C for 15min DNaseI was later heat-activated at 65∘Cfor 15min DNA and RNA were quantified using Ultraspec2100 prospectrophotometer (Amersham Biosciences USA)The integrity of total RNAwas assessed by running an aliquotof the RNA sample on a denaturing agarose gel stainedwith ethidium bromide (EtBr) and by spectrophotometricanalysis (Ultraspec 2100 pro Amersham Biosciences USA)Total RNA was reverse-transcribed with an oligo(dT)
20and
SuperScript III Moloney Murine Leukemia Virus ReverseTranscriptase (InvitrogenUSA) following themanufacturerrsquosinstructions
24 Cloning of G antarctica PI12 Protease Gene Puta-tive partial protease gene of 1321 bp was derived througha genome survey sequence library of G antarctica Thegene has 27 similarity to subtilisin-like protease fromRhodosporidium toruloides (EMS20811) DNA walking ofpartial putative protease gene was conducted using DNAWalking SpeedUp Premix Kit (Seegene Korea) accord-ing to the manufacturerrsquos instructions Three target spe-cific primers (TSP) were designed from the upstreamregion of known sequences with the following condi-tions 18ndash23 nucleotides long with 40 lt GC content lt60 for TSP1 (51015840-AGGGTCAAGACGTTGCAGT-31015840) 55∘Cle Tm le 60∘C for TSP2 (51015840-ATGCGAAGTCAGAAG-CAGGATC-31015840) while 60∘C le Tm le 65∘C for TSP3 (51015840-GCAGCCAAATACCTGGAAGCAC-31015840) RACE was per-formed using the SMART RACE cDNA Amplification Kit(Clontech USA) Two gene-specific primers (GSPs) weresynthesized for the 51015840- and 31015840-RACE reactions based onthe sequence of the RT-PCR products as follows GSPI 51015840-ACCAGTGTCCAGCACCCCAATCTTAATCC-31015840 andGSP II 51015840-TCATCAGTGGGACGAGCATGTCGT-31015840 Bothprimers were paired with universal primers provided inthe kit to amplify the upstream and downstream regionof the gene of interest The PCR products were subclonedinto pGEM-T vector (Promega USA) and sequenced Rapidamplification of cDNA ends (RACE) produced two PCRproducts (51015840- RACE and 31015840-RACE) which were cloned andsequenced to capture the end-to-end sequence of both
Enzyme Research 3
products One set of primers was designed from the extreme51015840 and 31015840 ends of cDNA as follows forward 51015840 CAP 5-GCGGGGGCCGACAATAAAAAC-3 and reverse 31015840 A-Tail5-TTTTTTTTTTTTTTTTTTTTTTTTTTGAGGTGGC-31015840the 51015840-RACE-Ready cDNA served as a template to generatefull-length cDNA through long distance PCR (LDPCR)Amplification process was carried out in a reaction mixture(50120583L) containing 5 120583L of 50ndash100 ng cDNA template 1 120583L(10 pmole120583L) of each forward and reverse primers 1 120583L of50x dNTP mix 1 120583L of 1 U120583L 50x Advantage 2 polymerasemix 5 120583L of 10x Advantage 2 PCR buffer (Clontech USA)and 36 120583L of distilled water Predenaturation was performedat 95∘C for 1min followed by 30 PCR cycles of denaturation(95∘C 30 sec) annealing (61∘C 3min) and extension (68∘C3min) Final extension was conducted at 68∘C for 3minand held at 4∘C The gene was amplified using thermocycler(CG1-96 Corbett Research Australia) The amplification ofputative open reading frame (ORF) was conducted usingboth genomicDNAand cDNA to locate the introns and exonslocations The amplicons were examined by electrophoresiscloned into pJET12 blunt vector (Fermentas USA) and sentfor sequencing The sequence obtained was deposited inGenBank under accession number FM178559
25 Assembly and Analysis of PI12 Protease SequencesSequences derived from DNA walking and RACE strat-egy were assembled by conducting sequence alignmentin the Biology Workbench (httpworkbenchsdscedu)Analysis and interpretation of nucleic acid sequences wereconducted using several publicly available web serversand integrated software as listed in Table 1 The phylo-genetic tree was constructed based on a comparison ofthe PI12 protease protein sequence with the closest pro-tease sequence that was extracted from GeneBank database(httpwwwncbinlmnihgov) The protein sequences usedfor phylogenetic analysis were aligned using ClustalW Phy-logenetic tree was constructed using Molecular EvolutionaryGenetics Analysis 6 (MEGA 6) package by neighbor-joiningmethod
26 Cloning of PI12 Protease Gene in Pichia pastorisExpression System The gene encoding mature PI12 pro-tease was amplified by PCR using recombinant pJET12FLcDNA of PI12 protease as a template Sense primer51015840-CAAGCCCTAGGCTACGCACGGAACGAGAA-31015840 andantisense primer 51015840-CTGCCGAATTCTTCCTCAACCCA-GTTACCAAC-31015840 were designed based on mature PI12 pro-tease gene sequence (FM178559) for flanking the PCR prod-uct at the 51015840- and 31015840- terminus respectively Amplificationprocess was carried out in a reactionmixture (50 120583L) contain-ing 5 120583L of 50ndash100 ng DNA template 15 120583L (10 pmole120583L) ofeach sense and antisense primer 1 120583L of 10mM dNTP mix4 120583L of 25mM MgCl
2 1 120583L of 1 U120583L Taq DNA polymerase
5 120583L of 10x PCR buffer (MBI Fermentas USA) and 31120583L ofdH2O PCRwas commenced by predenaturation step at 94∘C
for 4min followed by 30 PCR cycles of denaturation (94∘C1min) annealing (65∘C 1min) and extension (72∘C 2min)
Final extension was conducted at 72∘C for 7min and held at4∘C
ThePCRproduct of themature PI12 protease gene and thevector pPIC9 (05ndash1 120583g) were separately added to restrictionenzyme digestion mixtures containing 3 120583L EcoRI (10U120583L)6 120583L AvrII (10U120583L) 6 120583L Y+ Tango buffer (10x) and dH
2O
to a final volume of 30 120583L The reactions were incubatedat 37∘C for 1 hour The digestion products were observedthrough agarose gel electrophoresis and gel purified usingthe QIAquick Gel Extraction Kit (Qiagen Germany) Theconstruct was added to a ligation mixture with a ratio of1 5 to pPIC9 vector The ligation mixture was comprised of1 120583L of 10x ligation buffer (MBI Fermentas USA) 1120583L T4DNA ligase (5U120583L MBI Fermentas USA) and dH
2O to a
final volume of 10 120583L The reaction was incubated overnight(14ndash16 h) at 16∘C Ligation mixture was transformed into Ecoli Top10 competent cells The procedures for preparationof competent cells and heat-shock transformation were per-formed according to the method described by Sambrook etal 1989 [19] The transformation mixture was spread on LBagar plates containing 100 120583gmL ampicillin and incubatedovernight at 37∘C The positive transformants were selectedfor transformation into Pichia pastoris electrocompetent cell
Prior to transformation into P pastoris cell the emptypPIC9 and pPIC9 recombinant plasmids harbouring PI12protease gene were linearized using PmeI (10U120583L) restric-tion enzyme digestion The digestion mixtures consisted of5 120583L buffer B (10x) 3 120583L of PmeI and 6 120583L plasmids whiledH2O was added to a final volume of 60 120583L The linearized
plasmids were purified using the QIAquick PCR purificationkit The preparation of electrocompetent cells was doneaccording to Pichia expression kit manual (Invitrogen USA)Later about 80ndash100 120583L of the electrocompetent cells weretransferred into an overnight prechilled (kept in minus20∘C)02 cm electroporation cuvette (BioRad USA) and mixedwith 10 120583L (1ndash5 120583g) recombinant DNA The transformationmixtures (mixture of cells and plasmids) were incubated onice for 5min in the electroporation cuvette and pulsedwith anelectroporator (Gene Pulser BioRad USA) at the followingparameters charging voltage of 1500V capacitance of 25120583Fand resistance of 400Ω which generated a pulse length ofsim5ndash10ms with a field strength of sim7500Vcm Iced-cold 1Msorbitol (1mL) was added immediately after the pulse intothe cuvettes The contents were transferred into sterile bijoubottles and incubated at 30∘C for 2 hours without shakingLater 10ndash200120583L of the cells were spread on the MD agar[Minimal dextrose medium 134 (wv) yeast nitrogen base(YNB) 4 times 10minus5 (wv) biotin 2 (wv) dextrose 2 (wv)agar]The plates were incubated at 30∘C for 3ndash5 days until thetransformant colonies were formed
27 Expression and Optimization of Recombinant pPIC9Mature PI12 Protease Single colonies of the recom-binantPichia pastoris carrying the pPIC9PI12 protease were grownin 3mL of BMGY [buffered glycerol-complex medium 1(wv yeast extract) 2 (wv) peptone 134 (wv) yeastnitrogen base (YNB) 4 times 10minus5 (wv) biotin 1 (wv)glycerol 100mM potassium phosphate buffer pH 60] at
4 Enzyme Research
Table 1 List of web servers applied for full-length cDNA of PI12 protease analysis
Analysis Web serverSoftware Website(1) Open reading frame (ORF)(2) Amino acid translation(3) Intron-exon boundaries
Augustus Web server httpaugustusgobicsde
(1) Amino acid composition(2) Molecular mass(3) Isoelectric point
ProtParam httpwwwexpasyorgtool
Homology search (1) NCBI BLASTX httpwwwncbinlmnihgov(2) NCBI BLASTP
Phylogenetic tree (1) Molecular Evolutionary Genetics Analysis (MEGA 6) httpwwwmegasoftwarenet(2) CLUSTALW httpseqtoolsdsceduCGIBWcgi
Propeptide ProP httpwwwcbsdtudkservicesProP-10Open reading frame ORF Finder httpwwwncbinlmnihgovprojectsgorfSignal peptide SignalP httpwwwcbsdtudkservicesSignalPHydrophobic profile ProtScale httpauexpasyorgcgi-binMultiple sequence alignment Biology Workbench httpworkbenchsdsceduProtein motifs ScanProsite httpprositeexpasyorgscanprositeN-glycosylation NetNGlyc httpwwwcbsdtudkservicesNetNGlyc
30∘C in a shake incubator (250 rpm) overnight The nextday 1mL of the starter cultures was used as inoculumand inoculated into 10mL of BMGY in a 50mL flask andincubated in a condition as described to generate cell biomassbefore induction Cells were harvested by centrifugation at1500timesg for 10min
The cells were resuspended in 50mL BMMY medium[bufferedmethanol-complexmedium 1 (wv) yeast extract2 (wv) peptone 134 (wv) yeast nitrogen base (YNB) 4times10minus5 (wv) biotin methanol 100mM potassium phosphatebuffer pH 60] and incubated with vigorous shaking at30∘C to induce expression The expression control was doneby applying the same protein expression condition to therecombinant Pichia bearing the empty vector (pPIC9) Thegrowth of Pichia pastoris harboring recombinant plasmidGpPro2 was optimized in four different parameters in orderto enhance the protease enzyme yield The yeast was grownup to 5 days for postinduction study and the concentrationof methanol was varied from 0 to 1 Assay temperature ofactive enzyme was optimized ranging from 10∘C until 35∘Cwith 5∘C intervals The last parameter was to find out thecapability of cell density in protease production at A
600 nmfrom 1 to 10
28 Protease Assay The proteolytic activity was measured bythemodifiedmethod of Brock et al 1982 [20] using azocaseinas substrate Azocasein (05) was dissolved in 01M Tris-HCl-2mM CaCl
2 pH 7 The reaction was initiated by the
addition of 100 120583L of enzyme solution for 30min at 20∘C Anequal volume of 10 (wv) trichloroacetic acid (TCA) wasadded to terminate the reaction The absorbance was read at450 nm using UVVisible spectrophotometer Ultraspec 2100pro (Amersham Biosciences USA) One unit of azocaseinaseactivity was defined as the rate of absorbance change of 0001per min at 20∘C under the standard assay condition
29 SDS-PAGE Sodium dodecyl sulphate polyacrylamidegel electrophoresis (SDS-PAGE) was prepared according tothe Laemmli [21] method using 12 polyacrylamide gel inthe Mini PROTEAN 3 Cell apparatus (BioRad USA) Afterelectrophoresis gels were stained with Coomassie BrilliantBlue R-250 (BioRad USA)
210 Activity Staining and Cup Plate Assay Activity stainingwas performed as described elsewhere [22] The supernatantfrom recombinant Pichia clones were applied in this methodand loaded into SDS-PAGE [12 (wv)] After electrophore-sis the gel was immersed in 20 isopropanol to remove SDSand washed with distilled water (2-3 times) and the gel wastransferred into 2 skim milk yeast peptone dextrose (YPD)agar plate Protease activity onPetri dishwas also tested by theactivity ring staining or cup plate assay using 2 skim milkYPD agar plate adapted from Poza et al 2001 [23] The platewas incubated at 20∘C for 5-6 hours
3 Results and Discussion
31 Microorganism and Culture Condition After 10 daysat 4∘C the growth of G antarctica PI12 on nutrient agarsabourouddextrose agar and potato dextrose agarwas cream-coloured smooth mucous runny and glisteningThe borderwas entire circular and convex while on skim milk agar theyeast produced hydrolytic colonies (Figure 1(a)) The cellsof G antarctica PI12 occurred in oval yeast-like shape withthe size of about 2ndash4 120583m and present singly when observedvia simple and negative staining (Figures 1(b) and 1(c)) SEMphotograph showed budding yeast form of the G antarcticastrain PI12 (Figure 1(d)) All cells clumped together andpossessed smooth and oval shape appearance
Figure 2 shows TEM photographs of the thin-sectionedof the basidiomycete yeast G antarctica strain PI12 which
Enzyme Research 5
(a)
2120583m
(b)
(c) (d)
Figure 1 (a) Casein hydrolysis on skim milk agar plate (b) Negative staining with enlargement of 10000x bar = 2120583m (c) Simple stainingphotograph (d) Scanning electron microscopy (SEM) photograph with magnification 2500x Bar = 10 120583m
reveals the intracellular organelles Figures 2(a) and 2(b)show budding yeast appearance of membrane-boundednucleus and easily recognized bud scarwhile Figure 2(c) illus-trates ultrastructure comprising of a dark and thick cell walla cell membrane an irregular shape nucleus mitochondriavacuoles golgi and bud scars
32 Ribosomal RNA Identification The ITS158S rDNAITS2 region of PI12 was successfully amplified by Booet al 2013 [14] with the predicted size of 646 bp TheITS158SITS2 sequence of Glaciozyma antarctica strainPI12 had been deposited in GenBank with the accessionnumber JX896956 Amplification of ITS region showed100 identity to ITS158SITS2 sequence of Antarctic yeastCBS 8938 (AY040659) and Glaciozyma antarctica strainCBS 8943 (AY033637) National Collection of Yeast Cul-tures (NCYC) identified PI12 as an isolate of Glaciozymaantarctica based on 26S rDNA D1D2 sequencing resultwhere it displayed 99 sequence identity to Glaciozymaantarctica strain CBS 5942 [formerly known as Leucosporid-ium antarcticum strain CBS 5942 (AF444529)] Phyloge-netic tree analysis was created based on comparison of ITSsequence of this strain with the closest strains that areavailable in the database with reference to BLAST result
(Figure 3) Thirty-nine of ITS sequences from Glaciozymaclade were comparedThe phylogenetic tree constructed con-sists of three main clusters cluster I (JX896956 Glaciozymaantarctica strain PI12 AY040659 Antarctic yeast CBS 8938AY033637 Glaciozyma antarctica strain CBS 8943 EU149809Glaciozyma antarctica strain CBS 10640 AB774460 Leu-cosporidium sp KGK-2 JQ857017 Glaciozyma antarcticaisolate T21Ga EU149808 Glaciozyma antarctica strain CBS10636 AY040663 Glaciozyma antarctica strain CBS 8939EU149806Glaciozyma antarctica strainCBS 10639 EU149805Glaciozyma antarctica strain CBS 10638 AY040657 Antarcticyeast CBS 8927 AF444529 Glaciozyma antarctica strainCBS 5942 and KF934487 Glaciozyma sp KP7-5-2) isthe sequences from strains originally considered as Lantarcticum which was reclassified as Glaciozyma antarc-tica [15] Cluster II (GQ336996 Leucosporidium sp AY30EU149804 Glaciozyma watsonii strain CBS 10641 AY040661Antarctic yeast ML 4515 AY040660 Glaciozyma watsoniistrain CBS 8940 AY033638 Glaciozyma watsonii strain CBS8944 AB774462 Leucosporidium sp BSS-2 KC785578Uncul-tured Glaciozyma clone 180 AB774461 Leucosporidium spBSS-1 EU149803 Glaciozyma watsonii strain CBS 10684HQ432823 Glaciozyma watsonii DBVPG4802 HQ432818Glaciozyma watsonii DBVPG4726 HQ432820 Glaciozyma
6 Enzyme Research
2120583m
(a)
2120583m
(b)
BS
V
N
CW
CM
GM
Length 314866nm
Length 226635nm
1120583m
(c)
Figure 2 Transmission electronmicroscopy (TEM) photographs of AntarcticG antarctica strain PI12 (a) 12500xmagnification (b) 10000xmagnification (c) 27500x magnificationThe scale bars represent 2 120583m for (a) and (b) and 1 120583m for (c)
watsonii DBVPG4760 JX171176 Glaciozyma watsonii iso-late LKF08-112 DQ402535 Leucosporidium sp LC-03-120isolate 03-120 JF900360 Glaciozyma watsonii strain CBS7009 KC333171 Glaciozyma watsonii isolate AU CryS06HQ432821 Glaciozyma watsoniiDBVPG4799) is categorizedas G watsonii while cluster III (AY040664 Glaciozyma mar-tinii strain CBS 8929 AY033641 Glaciozyma martinii strain
CBS 8928 HQ432816 Glaciozyma martinii DBVPG4841KF934486 Glaciozyma sp KP7-5-1 FR682434 UnculturedBasidiomycota HF934010 Glaciozyma sp K81b JF900362Glaciozyma martinii strain CBS 9639 JF900359 Glaciozymamartinii strain CBS 6581 JF900361 Glaciozyma martiniistrain CBS 7054) is categorized as G martini Recognitionof the Glaciozyma species has been done by comparing
Enzyme Research 7
JX896956 Glaciozyma antarctica strain PI12
AY040659 Antarctic yeast CBS 8938
AY033637 Glaciozyma antarctica strain CBS 8943
EU149809 Glaciozyma antarctica strain CBS 10640
AB774460 Leucosporidium sp KGK-2
JQ857017 Glaciozyma antarctica isolate T21Ga
EU149808 Glaciozyma antarctica strain CBS 10636
AY040663 Glaciozyma antarctica strain CBS 8939
EU149806 Glaciozyma antarctica strain CBS 10639
EU149805 Glaciozyma antarctica strain CBS 10638
AY040657 Antarctic yeast CBS 8927
AF444529 Glaciozyma antarctica strain CBS 5942
KF934487 Glaciozyma sp KP7-5-2
GQ336996 Leucosporidium sp AY30
EU149804 Glaciozyma watsonii strain CBS 10641
AY040661 Antarctic yeast ML 4515
AY040660 Glaciozyma watsonii strain CBS 8940
AY033638 Glaciozyma watsonii strain CBS 8944
EU149803 Glaciozyma watsonii strain CBS 10684
JX171176 Glaciozyma watsonii isolate LKF08-112
DQ402535 Leucosporidium sp LC-03-120 isolate 03-120
AB774462 Leucosporidium sp BSS-2
KC785578 uncultured Glaciozyma clone 180
HQ432818 Glaciozyma watsonii DBVPG4726
HQ432823 Glaciozyma watsonii DBVPG4802
AB774461 Leucosporidium sp BSS-1
HQ432820 Glaciozyma watsonii DBVPG4760
KC333171 Glaciozyma watsonii isolate AU CryS06
HQ432821 Glaciozyma watsonii DBVPG4799
JF900360 Glaciozyma watsonii strain CBS 7009
AY040664 Glaciozyma martinii strain CBS 8929
AY033641 Glaciozyma martinii strain CBS 8928
HQ432816 Glaciozyma martinii DBVPG4841
KF934486 Glaciozyma sp KP7-5-1
FR682434 uncultured Basidiomycota
HF934010 Glaciozyma sp K81b
JF900362 Glaciozyma martinii strain CBS 9639
JF900359 Glaciozyma martinii strain CBS 6581
JF900361 Glaciozyma martinii strain CBS 7054
001
Clus
ter 1
Glac
iozy
ma
anta
rctic
aCl
uste
r 3
Glac
iozy
ma
mar
tinii
Clus
ter 2
Glac
iozy
ma
watso
nii
Figure 3 Neighbor-joining phylogenetic analysis of internal transcribed spacer 1 (ITS1)58S rRNA geneinternal transcribed spacer 2 (ITS2)of Glaciozyma clade Numbers indicate percentage bootstrap values calculated on 1000 repeats of the alignment The tree was constructedusingMEGA6 software [39]The scale bar represents 001 substitutions per nucleotide position GenBank accession numbers of the sequencesare indicated before strain name ITS158SITS2 sequence of Glaciozyma antarctica strain PI12 had been deposited in GenBank with theaccession number JX896956 [14]
8 Enzyme Research
their molecular phylogenetic data and physiological andmorphological characteristics [15] Phylogenetic analysis ofITS158SITS2 sequence of strain PI12 showed that thesequence was closely related to ITS158SITS2 sequenceof Glaciozyma antarctica with high homology similarity toCluster I (99-100 identity) The ITS158SITS2 sequenceof G antarctica strain PI12 was differed from G watsonii(Cluster II) by 2-3 gap in the sequence (93 identity)and G martinii by 4-5 gap in the sequence (90ndash92identity)
33 Sequence Analysis of PI12 Protease cDNA In this studyamplification of the PI12 protease gene sequence was con-ducted using genomicDNAand cDNAas a template Putativepartial PI12 protease gene with the size of 1321 bp was initiallyacquired from the G antarctica genome survey sequencelibrary (Supplementary Figure see Supplementary Mate-rial available online at httpdxdoiorg1011552014197938)However the gene was discovered as an incomplete pro-tease gene sequence DNA walking strategy successfullyamplified 1500 bp of the upstream region of partial PI12protease Both sequences of about 1500 bp (derived fromthe DNA walking reaction) and 1321 bp (partial PI12 pro-tease) were assembled and made up a total length of2687 bp protease sequence (Supplementary figure)The puta-tive PI12 protease was predicted by Augustus webserver(httpaugustusgobicsde) having intervening sequencesintronsThus reverse-transcriptase PCR (RT-PCR) and rapidamplification of cDNA ends (RACE) were accomplished toobtain the full-length cDNA As a result the ORF comprising2892 bp encoding 963 amino acid residues were successfullydeduced
Nucleotide and deduced amino acid sequences ofgenomic DNA and cDNA fragment encoding PI12 proteasewere shown in Figure 4 The size of the ORF of putative PI12protease was 3867 bp with genomic DNA as the templatewhile the ORF comprised 2892 bp encoding 963 amino acidresidues were successfully deduced through RACE strategywith cDNA as the template The ORF was predicted usingORF Finder (httpwwwncbinlmnihgovprojectsgorf)while prediction using SignalP (httpwwwcbsdtudkser-vicesSignalP) webserver allowed the identification of theputative PI12 protease prepeptide where it predicted thepresence of signal peptidase I cleavage sites [24]The putativeN-terminal signal peptide (1 to 16 the underlined amino acid)was identified as a secretion signal There is no propeptidepredicted by ProP Webserver (httpwwwcbsdtudkser-vicesProP-10) Comparison of cDNA and gene sequencesallowed determination of gene structureTherefore sequencealignment was performed between the sequence of these twotemplates and it was confirmed that the coding sequence(consisted of 19 exons) was interrupted by 18 introns withtheir length varying from 48 to 60 nucleotides However theGT and AG rule for donor-acceptor sites was not obeyedOf 18 introns 10 introns contained canonical GT and AGdinucleotides at their 51015840 and 31015840 ends while 8 introns holdnoncanonical splice site pairs with TT-GG (2 introns)AG-TG GC-GT TG-GG TC-GG TA-GG and TG-AG
dinucleotides at the splice junction Upper case letters inblue colour represent introns while lower case letters inblack colour represent exons Splice donor and acceptor werehighlighted in grey
The amino acid composition (upper case letters) withinthe ORF amino acid residues was determined by theProtParam tool of the Expasy molecular biology server(httpwwwexpasyorgtool) Translation starts at a nucle-otide position 1 and translation stop is marked with anasterisk The nucleotide sequences ATG (1 to 3) and TAG(3865 to 3867) indicate the initiation codon and termi-nal codon respectively The presumed putative sequencesof promoter CAAT (minus101 to minus98) and TATA-box (minus92to minus88) were identified in the 51015840-untranslated (UTR)region The search was achieved via Neural Network Pro-moter Prediction (Berkeley Drosophila Genome Project)(httpwwwfruitflyorgseq toolspromoterhtml) In the 31015840-untranslated (UTR) region there was a putative polyadeny-lation signal attaa (3889 to 3893) detected from the TAGstop codon to the poly (A) tail The predicted molecularmass and pI of PI12 serine protease were 10099 kDa and641 respectively Out of 963 amino acid residues in thededuced sequence of the PI12 protease 70 are negativelycharged residues (Asp + Glu) while 66 are positively chargedresidues (Arg + Lys) at neutral pH (Table 2)The total chargedresidues (Arg Asp Glu His and Lys) are 150 amino acidsin the open reading frame of PI12 protease corresponded to156 while hydrophobic residues (Ala Phe Ile Leu MetPro Val and Trp) are 441 amino acids which correspondedto 458 of the total amino acids discovered The totaluncharged residues (Asp Cys Gln Gly SerThr and Tyr) areconsisted of 372 amino acids which corresponded to 386of the total amino acids of the open reading frame of PI12protease The aliphatic index is 9290 and the grand averageof hydropathicity (GRAVY) is 0141 The instability index (II)of PI12 protease was computed to be 3414 which classify theprotein as stable
N-glycosylation sites were predicted using NetNGlyc10 Server (httpwwwcbsdtudkservicesNetNGlyc)Therewere five putative N-glycosylation sites for the PI12 proteasegene predicted using the server For N-glycosylation thesequence motif Asn-Xaa-SerThr (Xaa any amino acid butnot Pro) has been defined as a prerequisite for glycosylationThe sequencemotifs in PI12 protease occurred in the position170 NETH 365 NATV 576 NLTP 634 NDTA and 833 NSSPAmong five predicted sites only three (except positions 576and 833) showed a high potential score (averaged output ofnine neural networks) which support the prediction Theputative N-glycosylation sites were demonstrated in boxesin Figure 4 The comparison of amino acid composition ofthis cold-adapted protease fromG antarctica strain PI12 withnine other subtilisin serine proteases fromdifferent families isshown in Table 2 Analysis of this comparison depicts that thepercentage of each amino acids in PI12 protease is almost thesamewith other proteaseswhileGly (G) is themost abundantaccounting for up to 104 of the total amino acid in PI12 pro-tease PI12 protease also contained the highest percentage ofLeu (L) with 99 of the total amino acids compared to other
Enzyme Research 9
Table2Aminoacid
compo
sitionandmolecular
weighto
fproteases
acrossdifferent
families
Amino
CAQ76821
CAG25549
EDR11679
CAG38357
CAD118
98CA
L25578
EDU50747
AAW
40780
ACB3
0121
ACB3
0119
acid
Qty
Qty
Qty
Qty
Qty
Qty
Qty
Qty
Qty
Qty
Ala
(A)
97101
8191
7887
7590
8299
101
115
8189
9196
8092
105
116
Arg
(R)
4042
2730
2224
1619
1721
1821
2932
2223
2124
3134
Asn
(N)
4446
3843
4348
4149
4251
4046
5257
5154
4552
4347
Asp
(D)
4951
6269
5460
6578
5567
5158
5358
6669
5260
6370
Cys
(C)
808
506
404
405
911
809
910
909
708
1011
Gln
(Q)
2324
3438
2326
2834
3036
3742
3539
2931
1922
2629
Glu
(E)
2122
1618
1921
2024
1518
2023
2528
2627
2832
2426
Gly
(G)
100
104
89100
8493
85102
85103
7788
8493
8084
8598
8695
His
(H)
1415
1416
1314
1619
1316
1113
1618
1415
1618
1213
Ile(I)
5355
4955
5157
5364
4150
3843
5156
4446
3540
4449
Leu
(L)
9599
6876
7179
6274
6073
6675
6774
8084
7485
6875
Lys
(K)
2627
2932
4449
2935
4959
5158
4044
3032
5058
5662
Met
(M)
1212
202
202
911
1113
809
1820
808
1113
1517
Phe
(F)
4143
5966
4752
5566
2935
3641
4449
3941
3439
3438
Pro
(P)
6264
6573
6774
5870
5870
5967
5055
5154
5564
5763
ser
(S)
7780
6977
91101
6072
6680
7080
7279
111
117
6373
6167
Thr
(T)
8790
89100
7583
83100
6579
7383
7178
7276
7182
6875
Trp
(W)
505
303
607
202
1215
1214
1213
909
1214
1415
Tyr
(Y)
3334
1213
2629
708
2935
2832
3943
5053
2630
2730
val
(V)
7679
8292
8089
6679
5769
7484
5864
6872
8295
6268
Total
963
1000
893
1000
900
1000
834
1000
825
1000
878
1000
906
1000
950
1000
866
1000
906
1000
MW
(Da)
1009898
932669
944365
869455
869549
924905
979343
1009629
917209
961988
Thea
ccessio
nnu
mbersof
proteasespu
tativ
eserinep
roteaseo
fGlacio
zymaantarctica
PI12
(CAQ
76821)sub
tilisin-lik
eproteaseP
leurotuso
streatus(CA
G25549)pyrolysin
LaccariabicolorS
238N
-H82
(EDR11679)
subtilisin
-like
proteasePh
anerochaetechrysosporiu
m(C
AG38357)sub
tilisin-lik
eserinep
roteaseP
R1CMetarhiziu
maniso
pliaev
aranisopliae(CA
D11898)serinee
ndop
eptid
aseH
ypocrealixii(C
AL2
5578)peptidase
Pyrenophoratritici-repentis
(EDU50747)p
eptid
aseCr
yptococcus
neoforman
svarn
eoform
ansJEC
21(A
AW40
780)sub
tilisin-lik
eprotease
Epich
loefestucae(A
CB30121)sub
tilisin-lik
eprotease
Epich
loefestucae
(ACB
30119
)
10 Enzyme Research
atgctcttcctccccgtcctcctcctcctccttcccggcgtcactgccttcctcaaccca 60
M L F L P V L L L L L P G V T A F L N Pgttaccaaccgcgcgaccaacgccatctcctcgactcaatacctctctaacgcctatatc 120
V T N R A T N A I S S T Q Y L S N A Y IctcgaattggacctctccacccccggcctcgtcaaacgggatagcacgcccgactctGTG 180
L E L D L S T P G L V K R D S T P D SCGTCCCGCGTTCCTCCCTCGCTGTCGAAGACGAGCTAACAATGACGTGGTAGatcctaga 240
I L Eggacgtactcacgtccgtaggccgcaacggtatcaagtaccaactccgccaccgctttat 300
D V L T S V G R N G I K Y Q L R H R F Ictccccgactctgttccacggcgcttcgatcactgtcccccctggaatctcccgctccca 360
S P T L F H G A S I T V P P G I S R S QaatcgcctctctccgcggtatcaaaGTCCGTCCCTCTCTCTCCCCTACTCCCTCCGTGCT 420
I A S L R G I KAACCCTGCACGCAACAGcgcgtctggcccgttcgaaagttctcccgacccagcgcagtag 480
R V W P V R K F S R P S A Vtggacgccgatggaggaggaagcgggttctcagggtcgcctatcaaggcggcgctcatgg 540
V D A D G G G S G F S G S P I K A A L Mgggtgaaagagctcgggaagcgcgcgaacgcttatgctggagatacgtttggaccgcatg 600
G V K E L G K R A N A Y A G D T F G P HtcatgacgggggttaatgagacgcatgaggcggggttgttgggagctgggattaagGTGT 660
V M T G V N E T H E A G L L G A G I KGTTTTCGTCTTTGTTTGGGGGAGGGGGGAGGGAGGAGCTGATTAATGGGTGCAGattggg 720
I GgtgctggacactggtgAGAAGCGGTTTGGAGGGAGGAGAGGATGGAGCTGATGAGTGTGC 780
V L D T GAGGTGttgattatttgaacccgattctgggaggctgctttggacctgggtgccatatgtc 840
V D Y L N P I L G G C F G P G C H M Sgtttgggtacgacttggttggcgatgattacgatggagataacgctcctgtgccggatgt 900
F G Y D L V G D D Y D G D N A P V P D VggatccttGTGCGTCCTTCCCTCCGCGATGGGCTGGAATCTGAGCTGACGGCGTTGTTGA 960
D PTCAGttgcgagctgcgatcctcatggTTCGTCTCGCTTTCCGCTCCTCTGATGCGCTCGC 1020
F A S C D P H GTGATGCACTTGGCAATAGGaactcacgttacgggaatcattggagcgctcccgaatgcgt 1080
T H V T G I I G A L P N AttggatttactggcgtcgcacccgccgctactctgggccactaccgagtGCGCTCCCTCT 1140
F G F T G V A P A A T L G H Y R VATCCGTTCTTCCTCGCAGCTCCCACTCACTCGTGCTTCCAGGTatttggctgcactggct 1200
F G C T GtcgtcggagaagatatcGTTCGTCCTCTCTCCAGTCCCGGCTAGCGATCCTGCTTCTGAC 1260
F V G E D ITTCGCATCTCCCTCTGAGattctcgctggactcatgcgaggagtcgaggacaactgcaac 1320
I L A G L M R G V E D N C Ngtcttgaccctctctctcggaggtccaggagggtgggtcaagggcacgccggcgtccatc 1380
V L T L S L G G P G G W V K G T P A S Icttatcgaccagatcgaagcgcaaggcattctcgtcaccgtcgccactggcaactcggga 1440
L I D Q I E A Q G I L V T V A T G N S GgctgagggTGAGCTCCTCGCTACTTGCGCGTCGAGACCAGTACTAACAGGGGCGTGGACG 1500
A E GCAGGcatgttcttctccgagtctcccgcctcgacgatcaacggcctttccatcgcatcca 1560
M F F S E S P A S T I N G L S I A ScgTCCGTCTCTCCCTTCCTTCGCACGCCACTCGCTCACACTCTCTCGCAGGgacgttacc 1620
T D V Tgacctcatcgcctacaacgccaccgtctcaggccaacctgcgatcccttacctctccgcc 1680
D L I A Y N A T V S G Q P A I P Y L S A
ggggggggggtctg minus194
ctagcgcgaatcgagaaaacacaaaagaacctcccgacttggcgttgcttctgctcttct minus180
ccttctcccaaccgccgacaataaaaactataacacacacacaccccatctcgacgcctg minus120
cagacttcacgactctcctcacgctctttaccgccccacgggtcccacccacgagccacg minus60
(a)
Figure 4 Continued
Enzyme Research 11
acgcccctcaacgtcgtcgccaacagcttccgcgtccacttcacctctaccgaccccaac 1740
T P L N V V A N S F R V H F T S T D P Naaccccgtcgacgcctgctctcctcttccggctggagcgcccgacttcgccaactatgtt 1800
N P V D A C S P L P A G A P D F A N Y Vacggtcgttcagcgtgggacttgtacgttcgttaccaagtaccagaacgttctcaatgct 1860
T V V Q R G T C T F V T K Y Q N V L N AggaggGTGCGTCGGCTTCTTCTCTCCCGACTGGATTGGGCGGAGACTGACCTTCTGAGCA 1920
G GGaaagatcgtattgttgtacaactcggagggagctgggaacctcccttacctcacgccca 1980
K I V L L Y N S E G A G N L P Y L T PacggtgtcggcatcgacgccgttgcaggtcttcgtcgttccgacggactcaagGTTCGTC 2040
N G V G I D A V A G L R R S D G L KTACGCTCTTCGGTGCTGAGGCCCTTCTCTGATCTCGCTCCGACAGcttctctcgtactat 2100
L L S Y Ycagaatgccaacaagcgtctcactctgcgcttccccaagggcaagatcgtcgcaggcttg 2160
Q N A N K R L T L R F P K G K I V A G LaccgataccatcaccggcggactcatctcgTTCGTTCCTCTCCGCCTCCCTCGCTGGCCG 2220
T D T I T G G L I SATGAGCTGACGCGGTCTTTGCAGGggttactcgacgtttggtccgacgaatgacctctac 2280
G Y S T F G P T N D L Yggtcagcctaccctctctgcccctggtggcaacatcctttcgaccttccctctctccgag 2340
G Q P T L S A P G G N I L S T F P L S EggaggagtggcggtcatcagtgggacgagcatgtcgtgcccctttgtcgGTTCGTCCTTC 2400
G G V A V I S G T S M S C P F VTCCTCGTTCGTTCGCTGAGCGTCGACTGATCTTGCACGATCGCAGctggatctgcggcgg 2460
A G S A Atcctcatggccgctcgcgcttcggagaacctcacgccgcttgagatcaggagtctcctta 2520
V L M A A R A S E N L T P L E I R S L Lctaccactgcgaagcttacgccggtctcgctcttgggatcgacgcctttggtgagcgtga 2580
T T T A K L T P V S L L G S T P L V S VttcgtcaaggaggaggTACGTTCCTTTCTCGCTGCTTTGAGGATCGAGGCTGATTAGCGA 2640
I R Q G G GGCTTCGCAGGactcgttcaggttgccaaggcgctcgcggccaagacgctaatctctcctc 2700
L V Q V A K A L A A K T L I S Pacgagctcctgctcaacgacactgcgaacgcgaactacgtccagactatcaagatcaaga 2760
H E L L L N D T A N A N Y V Q T I K I Kacaccaactcgtgggcgatgaagtacaccttctcctcggccgtcgcccaaggactcggaa 2820
N T N S W A M K Y T F S S A V A Q G L Gctttcgacgcttcgggcgatatcctccctaccctcgacccagtcgccgtctctggcgcac 2880
T F D A S G D I L P T L D P V A V S G Aaggctaccgtcgcgttcaacactcggatcctcagcgtcgcgcccggcgcgacggggtccg 2940
Q A T V A F N T R I L S V A P G A T G Stcgtggcgactatcacgccgccggttcttcccgtagcggacgctgcgaggttccctatct 3000
V V A T I T P P V L P V A D A A R F P Itctctgggtggatcagggtgaatgggcaaggcgcgagggatagcagtaggaacgaggcgt 3060
F S G W I R V N G Q G A R D S S R N E AacactgGTGCGTTTGGCTCGCTTTTTGGGAGGTGCCGAGCTAATCGTTTCTCGTAGtccc 3120
Y T V PgtactttgggcttgcggcgaagatgatcgatatgcaagGTGCGTCGATGGTTCTCTCGCT 3180
Y F G L A A K M I D M QTCTCTCTCGGTTGCTCACTTCAAAGTGCAGtcctcgacaccaccgagaccatttacggtc 3240
V L D T T E T I Y Gcgggctacgcctaccccttcgtgatcgacgacgcgattggagacatccaatccaccacaa 3300
P G Y A Y P F V I D D A I G D I Q S T TcgtcgtactccaggaacctcgggcccaccgtcttcgctcgctttgccaTGCGTCGACCGT 3360
T S Y S R N L G P T V F A R F ACCTCATTTCCTGCGGCAGAGCGAGAGCTGATTCCGACCTCGAACAGctggaacccttcac 3420
T G T L Htacagcctcgatctcgtcctagccgacatcgcctttacccccacctaccccaactcctcc 3480
Y S L D L V L A D I A F T P T Y P N S S
(b)
Figure 4 Continued
12 Enzyme Research
cccgccactcgtttcgtcaagcgctccctcacgcagcacacctctgccgcttcgcacctc 3540
P A T R F V K R S L T Q H T S A A S H Lgccaagcgccgcgtctccatcgccactatcaaccccaaagccaccctcgtcgccgatcga 3600
A K R R V S I A T I N P K A T L V A D Rcagctccactcggacgtccctatcgagggcaacatcttcacccaaccctttactggaagg 3660
Q L H S D V P I E G N I F T Q P F T G Rgattacctcgtcgacgcagccccgacgggatccaccgatcgtaccgtcacttttaacggg 3720
D Y L V D A A P T G S T D R T V T F N Gcagtacgccgagaacggcctcgtgaggacggctgtgacggggacttcgtaccgcttcctc 3780
Q Y A E N G L V R T A V T G T S Y R F Lcttcgggcgttgaagatctcgggagacgcgatgtacgaggatcagtatgagagctggctc 3840
L R A L K I S G D A M Y E D Q Y E S W L3900
S L P F S F R Attttgccacctcaaaaaaaaaaaaaaaaaaaaaaaaaa 3938
lowast
tcgctaccgttctcgttccgtgcgtaggcttgttgtggacaccactgcattaagttatga
(c)
Figure 4 Nucleotide and deduced amino acid sequences of genomic DNA and cDNA encoding protease from G antarctica strain PI12 Thenucleotides are numbered from the 51015840 end of the cDNA A putative CAAT TATA box and polyadenylation signal are bold italicThe putativeN-terminal signal peptide is underlined The initiation codon is boldfaced and termination codon is represented by an asterisk (lowast) boxedpeptide sequences showed N-glycosylated asparagines residues The amino acid residues that form the catalytic triad are double underlinedand bold while introns are represented in blue font Highlighted in grey are the intron donor-acceptor splice sites
proteases Hydrophobic profile of PI12 amino acid proteasewas performed by ProtScale tools with Kyte and Doolittlemethod at httpauexpasyorgcgi-bin The hydrophobicityprofile elucidation of PI12 protease amino acid was carriedout from amino acid 1 to 963 The amino acid residuesfrom 1 to 16 (M-L-F-L-P-V-L-L-L-L-L-P-G-V-T-A) and 548 to562 (V-A-V-I-S) were hydrophobic grooves The first regionwhich consisted of 16 amino acids was very hydrophobic andwas predicted to be the signal peptide by SignalP and ProPwebserver
The bold and double-underlined amino acids showedresidues of catalytic triad aspartic acid (D) histidine (H) andserine (S) active site respectively NCBI Conserved DomainsDatabase (CDD) identified the presence of subtilase proteaseand subtilisin-like serine protease domains in the amino acidsequence The predicted PI12 protease amino acid sequencewas found sharing significant homology to the subtilisinfamily of protease The highest similarity of PI12 proteasewas observed with 42 identity to subtilisin-like proteaseof Rhodosporidium toruloides (EMS20793) 37 identity tosubtilisin-like protease of Pleurotus ostreatus (CAG25549)and 35 identity to subtilisin-like protease of Gloeophyllumtrabeum ATCC 11539 (EPQ55564) The nucleotide sequenceof G antarctica strain PI12 showed low homology to thefungus subtilisin protease family and none to other psy-chrophilic proteases The multiple sequence alignment wasillustrated in Figure 5 The consensus sequence around thecatalytic triad aspartic acid (D) histidine (H) and serine(S) of PI12 protease showed high conservation with othersubtilisin (showed in black boxes) Glycine residues G142G173 G191 G262 and G327 that were highly conserved inmost of subtilases were conserved in PI12 protease [25]Therefore PI12 protease belongs to the subtilisin subgroup ofthe subtilase serine protease superfamilyThis serine protease
of G antarctica PI12 has been deposited into GenBank withaccession number CAQ76821
34 Phylogenetic Relationship of PI12 Protease The phylo-genetic tree was constructed to demonstrate the similaritiesand relationship of protein sequence between differentspecies or organisms Figure 6 illustrates the phylogeneticdendrogram indicating the relationship of PI12 proteasewith another 25 translated amino acid sequences ofserine protease The bar scale signifies the branch lengthto approximate variance along each branch Basicallythis tree comprises two main clusters cluster I was sub-divided into subcluster a (CAG25549 EPQ55564 ETW-80907 XP 007381302 XP 001877576 CAG38357 XP007341385 CCA68627 EJU04161 XP 007369624 EIW-62675 CCO37249 XP 001830835 XP 007342189) and sub-cluster b (XP 007589863 XP 003711175 EFQ32840 XP006694395 ENH74885 EON99722) cluster II was sub-divided into subcluster c (CBQ73425 GAC97617 CCF52619)and subcluster d [CAQ76821 (PI12 protease) EMS20793]
Serine protease constructed by the phylogenetic treecomprised large size of protein with about 801 to 1032 aminoacids The majority of the translated protease sequenceswere obtained from fungus All serine proteases from cluster1 subcluster a are derived from Agaricomycetes except forEJU04161 which belongs to Dacrymycetes whereby all serineproteases from cluster 1 subcluster b are from Sordari-omycetes Cluster II subcluster c consisted of serine proteasefrom fungus belonging to Ustilaginomycetes PI12 proteasewas categorized under cluster II subcluster d It possessedhomology similarity with the subtilisin-like protease fromRhodosporidium toruloides (EMS20793) (formerly known asRhodotorula glutinis or Rhodotorula gracilis) which is a
Enzyme Research 13
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
MKG-YSWSLLQLAFTGAALATVPLSDVKRISSAS--VVPNKFVVEVESASEIPGKR--SL
MKGVLVWSILS---AGCALAVTPLSSVKRVTNLP--IVANKFIVEVDSTSDIPTKR--S-
MKLPLLSLALT---AVSVGATVPLSSVQRTTTSPNNALAGEYIVEMNNVQGLGGKR--AY
---MAHGSLASLLLLLAASAQGAAAFGGKSATGGRSVVPNSYILQVNASAPALTKRGVSV
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease ---MLFLPVLLLLLPGVTAFLNPVTNRATNAISSTQYLSNAYILELDLSTPGLVKRDSTP
lowastlowast
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
YTPHELFYRSLSARGVSYEVNKEYNS-EGLFVGAVVTVSDANDVKALAETPGVKAIRPVR
-NPHDALYESLRKRAIGFKVDREFNT-QGLFVGAALTLSDAQDVGQIQSTTGVKSIRPVR
ASTHDTLYRSLRKRGISFDVKKEYNQ-PGIMVGAALKVASPQDVQQIATMAEVKAIRPVI
FEALDHTLAAVKQNGVKYTVRQRFDAIPEAFQAVSIQVEDGASMAELAQIPGVQRVWPVS
DSILEDVLTSVGRNGIKYQLRHRFIS-PTLFHGASITVPPGISRSQIASLRGIKRVWPVR
lowastlowast
TIKSPKPVKVHVVTGKDDPEIPA-------------------------DTESTHIMTGVN
AFDRPKPVKVQVVKGPDDPALPP-------------------------DSESTHVITGVD
AVPAPERINQFTVSSPNDSRITA-------------------------DSETTHIMTGVD
LIPRPVEPTVSDFTPVSSSSSSTKSKRD-THTLEKRGTNFPPASAYLNDTFYPHVQTGID
KFSRPSAVVDADGGGSGFSGSPIKAALMGVKELGKR------ANAYAGDTFGPHVMTGVN
lowast lowast lowast lowastlowast
KLHAQGIFGAGIKIGIIDTGIDYTHPALGGALGAGHKVIGGYDFVGDAYDGTNTPVPDND
KLHAQGITGAGIKIGILDTGIDYTHPTLGGAFGPGNKVIGGFDLVGDDYDGTNTPVPDPD
KVHAAGITGEGIKIGIIDTGIDYNHPLLGAGFGPGFKVIGGYDFVGDDYTGSNNPAPDDD
VLHNKGILGQGVKIVVVDEGVDYTNPILGGCFGPGCQISFGYDFVGDNYTGTNAPVPDSD
ETHEAGLLGAGIKIGVLDTGVDYLNPILGGCFGPGCHMSFGYDLVGDDYDGDNAPVPDVD
lowast lowast lowast lowastlowastlowast lowast lowast lowastlowast lowast lowastlowast lowast lowast lowast lowast lowastlowastlowast lowast lowast lowast lowast lowastlowast lowast
PLDQCNGHGTHVAGIIGANPDNQFNISGVSYGASLSSYRVFGCSGSVSDDIIIEALLRGV
PLDQCAGHGTHVAGIIGANPDNAFNISGVAFQASLSAYRVFGCVGFVTDDVLVDALLLGF
PLDNCNGHGTHVAGIIGANPGNDFGISGVAYGSSIASYRVFGCSGFVSDDIIIDALLRGY
PFSSCSNHGTHTTGTVGALAN-EYGFSGAAPMATIGHYRVISCSGATTDEILVAALTRAL
PFASCDPHGTHVTGIIGALPN-AFGFTGVAPAATLGHYRVFGCTGFVGEDIILAGLMRGV
lowast lowastlowastlowast lowastlowast lowast lowastlowast lowast lowastlowastlowast lowast lowast lowast
QDGQDILTLSLGGTDGWTE-SSSSVVASRIAASGKIVTIAAGNEGAYGSWYSSSPGNGID
NEGQDILTLSLGGADGWTE-SVSAVVASRIAATGKVVTIAAGNDGASGAWYTSSPGNGID
SDGMDILTLSLGGVEGWTE-SSSSVVASRIADQGRVVTIAAGNDGAYGSWYASSPGTGVD
QDNVNIVSMSLGSSVGWLDDSPVQIMASYLGSKGIHVVASVGNERNEGLFAADQPAASRI
EDNCNVLTLSLGGPGGWVKGTPASILIDQIEAQGILVTVATGNSGAEGMFFSESPASTIN
lowastlowastlowast lowastlowast lowast lowast lowastlowast lowast lowast
VISVASIDNTVDNIQNATVNGVEHGPIPYFKTLPLPFTNA-LPLYVISNDTTIANDACNA
VISVASLDNTVVPLQTAQVHGVTHDPIIYQDTFPLPINDT-RPIFATSTDVTVVDDACNP
VISVASVNNIAIPVQNATVQGVEHAPIPYLEALPLNVTGS-RPIYATSTDTTVADDACNA
GTGVGGVDPTYLAAYYAYLI--KRDPVPYISPTPFNLPNN-YLLYFTSTNTSVTNDACNP
GLSIASTDVTDLIAYNATVS--GQPAIPYLSATPLNVVANSFRVHFTSTDPNNPVDACSP
lowast lowast lowast lowast lowastlowastlowast
LPDNTPDLSGFLVLVRRGSCTFVQKLTNIAAKGGNAVFIYDNGNG-----FAGIDTGKFT
LPDSTPDLSGFVVIVRRGTCTFVQKLGNVAAKGAKVTLIYDNGSG-----FGAIDTGAFV
LPNDTPDLSGYVVVIKRGTCTFVTKLQNAAAKGANTFLIYNNGGS-----FSAISVGDYT
LPSTTPNLANRVVVVQRGTCGFTVKQQNVAAAGGKIVLIYNSKGSGLIPQLDVGSTGLTA
LPAGAPDFANYVTVVQRGTCTFVTKYQNVLNAGGKIVLLYNSEGAGNLPYLTPNGVGIDA
lowastlowast lowast lowastlowast lowast lowast lowast lowast lowast lowast lowast
(a)
Figure 5 Continued
14 Enzyme Research
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
NATLIQAADGVFLAQQVAAGKSVTITFPQTGGSAQITN-PSGGLVSSFTTYGPTNDFYFK
-ASFIQAADGEFLVQQFASGQNVSLSFPQSGGLIQFPD-PAGGLISSFTSYGPSNDFFFK
-AALISETDGTFLVNQFAAKANITISFPQSGSAWNYPDDVSGGLVSDFSSYGPSYDMYFK
VGSLRYEDGLKLLSYYQSSPRGQSISFPNGPLAPYVSNTVSGGVVASYSNFGPTNELFIY
VAGLRRSDGLKLLSYYQNANKRLTLRFPKGKIVAGLTDTITGGLISGYSTFGPTNDLYGQ
lowast lowastlowast lowastlowast lowastlowast
PAFGAPGGNILSTLPVPLGSYGVESGTSMATPYVAGSAALLLSVKGKSAAVALGARTLFQ
PAVAAPGGNILSTLPVNLGLFGIESGTSMATPFVAGSAALLFQAKGTSAAVGRSARTLFE
PAVAAPGGNILSTLPIALGSYGVESGTSMATPFTAGSAALILQAKGK--SVAKSIRNLFQ
PTLAAPGTDILSTV---TGGVALMRGTSMAAPLVAGAYALLLSVGNNLKLSPEEARTILM
PTLSAPGGNILSTFPLSEGGVAVISGTSMSCPFVAGSAAVLMAARASENLTPLEIRSLLT
lowast lowastlowastlowast lowastlowastlowastlowast lowast lowastlowastlowastlowast lowast lowastlowast lowast lowast
STAQRVPSSHTDGDPLQTLSQQGAGLIDVFKALYTETIVWPTELILNDTAHYKEVHTFTV
TTAQRVPSTHTDGDPLQTLTQQGAGLINVFNAIHTTTIVSPGELILNDTAHFKGTQQFTV
TTASAVASSKTDGDPLQTVSQQGAGLIQVDRAIGTTTVVSPGQLTLNDTAHFEGFKTFNI
STSQPSPVAY-GSETLDTVVSQGAGVINMTAAIAAHTVISPAQFNLNDTQYSNNTQTLTL
TTAKLTPVSLLGSTPLVSVIRQGGGLVQVAKALAAKTLISPHELLLNDTANANYVQTIKI
VNTGKTSKQYKLTHVPAGTAQTIT---SGTIFAADGPVPLSTDYATVKLSSSSFTLRPGQ
RNAGSSAKTYTLRHIPAGTAVTVT---PGTIFPADGPVPLSTDFASVSLSTSKFTLAPGK
QNTGSKAVTYKISHVPAGTAASVQ---SGSIFAADGPVPLTADYAQVTFSLNKVTVLPGV
RNNNRFPVSYQLSWVDSTGIVTYNDGATTDIIPSTTPNYVSTSVLRVAFSQRTVTVPGGQ
KNTNSWAMKYTFSSAVAQGLGTFD--ASGDILPTLDPVAVSGAQATVAFNTRILSVAPGA
lowast lowast lowast lowast lowast lowast
SKQVTTHINAP--SGVDASTYPVFSGFIEIATAT--------ESVHVTYLGLAASLKDKQ
TQTVTARFTPP--AAADPSTFPVFSGFIQIESGT--------EQVQVSYLGLKASLKDKQ
SVPILVWFKPP--QGVDASTYPVYSGFIEIAGLG--------ETLRVAYLGVAASLKDKA
TVKVTAQFIPPNLTAQQRNQFPIYSGFVTITGQGQGAGAEQMEKYNLPFFGLGARMVDMP
TGSVVATITPPVLPVADAARFPIFSGWIRVNGQG-ARDSSRNEAYTVPYFGLAAKMIDMQ
lowast lowast lowastlowast lowast lowast lowast lowast
IVDNTNEFFGITIPAVLNNTGDAQ-LSPTNYTFSGSDFPTLLLRLAFGTPLLRVDLVDAT
VIDNTDFFFGVPTPVLTDPNGEVQ-TSPRNYSFLASDFPTLIFRLAFGSPKVVVDLVSPT
VVDNTDEYFGVKIPALLDASGDVQ-GNSTNYTLVGDDVPSLLYRFAFGSAHVRVDLVEKA
VLDTTNVALGP--NLPFIAVGQN--IQTGPTTFSESNPAVVYFRLAAGTRRLRVDLVDAS
VLDTTETIYGPGYAYPFVIDDAIGDIQSTTTSYSRNLGPTVFARFATGTLHYSLDLVLAD
lowast lowast lowast lowast lowastlowastlowastlowast lowast
IDLKPTLNSREIDDKRHVFERESFSFPHPGPKGS-------------------TFAQVKI
INFKPTLNTRALGDTHGSF----FSFPHRVKTG--------------------SFAQVPT
STVSATLSKRNWWN---------WWWPQKPSN---------------------SYDKVPI
TSYNATVPAVTNPASRLVKRSNTELTPRAAFPT--------------------LYSQVRT
IAFTPTYPNSS-PATRFVKRSLTQHTSAASHLAKRRVSIATINPKATLVADRQLHSDVPI
lowast lowast
VGTLSEQSHIPRNSDASTVDDGGYSTFAFNKP------SFANGTTIPNGTYRWLLRALKV
LGTVFELDFTSRNNDDPTDN--GFNTVAIDPP------TFANGTTIPNGQYRLLLRALRV
VGTLAEYDYVPRNSDASTIDDNGYNVFQFT-D------SFVNGTKIAAGEYKVLLRALKV
VGSLFAPSYWPPRDYLFNNGGT-YSDYEIQLDGSFTFPNGTKGMAELGRSYKVLLRALKI
EGNIFTQPFTG-RDYLVDAAPTGSTDRTVTFNGQY-AENGLVRTAVTGTSYRFLLRALKI
lowast lowast lowastlowast lowast lowast lowast lowast lowastlowastlowastlowast
lowast lowast lowastlowastlowastlowastlowast
(b)
Figure 5 Continued
Enzyme Research 15
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
TGDPTKEADFETWLSPIVGITVA
TGDPTNEADFESFLSPIIGVNAP
TGNPAEEGDYETWLSPSFSVVPA
TADPTYTSSYESWLSPSFTFTA-
SGDAMYEDQYESWLSLPFSFRA-
lowast lowastlowast
ConservedSimilar
lowast All match
(c)
Figure 5 Multiple sequence alignment between amino acid sequence of PI12 protease with other subtilisin The serine protease sequenceswere obtained from the GenBank database Their accession numbers are EMS20793 (subtilisin-like protease of Rhodosporidium toruloidesNP11) CAG25549 (subtilisin-like protease of Pleurotus ostreatus) EPQ55564 (subtilisin-like protease of Gloeophyllum trabeumATCC 11539)and ETW80907 (serine protease S8 ofHeterobasidion irregulare TC 32-1) Putative ORF of serine protease fromG antarctica PI12 was labeledas PI12 protease
CAG38357 Phanerochaete chrysosporium
EIW62675 Trametes versicolor FP-101664 SS1
XP 007369624 Dichomitus squalens LYAD-421 SS1
XP 007381302 Punctularia strigosozonata HHB-11173 SS5
CAG25549 Pleurotus ostreatus
ETW80907 Heterobasidion irregulare TC 32-1
XP 001877576 Laccaria bicolor S238N-H82
EPQ55564 Gloeophyllum trabeum ATCC 11539
XP 001830835 Coprinopsis cinerea okayama7130
XP 007341385 Auricularia delicata TFB-10046 SS5
CCA68627 Piriformospora indica DSM 11827
CCO37249 Rhizoctonia solani AG-1 IB
XP 007342189 Auricularia delicata TFB-10046 SS5
EJU04161 Dacryopinax sp DJM-731 SS1
XP 007589863 Colletotrichum fioriniae PJ7
EFQ32840 Colletotrichum graminicola M1001
ENH74885 Fusarium oxysporum f sp cubense race 1
XP 003711175 Magnaporthe oryzae 70-15
XP 006694395 Chaetomium thermophilum DSM 1495
EON99722 Togninia minima UCRPA7
CBQ73425 Sporisorium reilianum SRZ2
GAC97617 Pseudozyma hubeiensis SY62
CCF52619 Ustilago hordei
EMS20793 Rhodosporidium toruloides NP11
CAQ76821 Glaciozyma antarctica PI12
01
Clus
ter 1
Clus
ter 2
a
b
c
d
Figure 6 Phylogenetic dendrogram among 25 translated amino acid sequences of serine proteaseThe tree was constructed using a neighbor-joining algorithm with 1000 bootstrappings by MEGA 6 software [39]The scale bar represents 01 substitutions per amino acid positionTheserine protease sequences were obtained from the GenBank database GenBank accession numbers of the sequences were indicated beforethe strain name The putative serine protease of G antarctica strain PI12 has been deposited in GenBank with accession number CAQ76821
16 Enzyme Research
0
2
4
6
8
10
12
0 24 48 72 96 110
Prot
ease
activ
ity (U
mL)
Post induction time (hours)
Figure 7 Effect of postinduction time towards protease activity The highest activity was detected at 72 hours after 110 hours of incubationwith 9UmL activity followed by 48 24 and 96 hoursThe substrate (azocasein) was dissolved in Tris HCL buffer and incubated for 30min at20∘CThe absorbance was measured at 450 nm after 15min incubation A one-way ANOVAwas used to test for preference differences amongsix different postinduction times The statistical value (119875 value lt005) between hours is 0003
pink-coloured nonpathogenic oleaginous yeast (42 iden-tity) Both G antarctica PI12 and Rhodosporidium toruloidesNP11 belong to Microbotryomycetes class PI12 protease didnot show any homology similarity to other psychrophilicproteases however it showed 37 identity to hypotheticalprotein from Pseudozyma antarctica T-34 which was isolatedfrom Japan (GAC76005) (not listed in the phylogenetic tree)
35 Cloning and Expression of Cold-Adapted PI12 Protease inthe Yeast Expression System The PCR product harbouringthe mature PI12 protease with two flanking restriction sites(EcoRI and AvrII) was cloned into pPIC9 expression vectorThe recombinant plasmid harbouring PI12 protease was usedto transform both GS115 and KM71 intended to produce bothHis+ Mut+ and His+ MutS Two recombinant clones fromGS115 (His+Mut+) strain (GpPro1 andGpPro2) and one fromKM71 (His+MutS) strain (KpPro1) were successfully obtainedwhich were further analyzed for protein expression study
36 Expression and Enzyme Assay of PI12 Protease in Pichiapastoris All positive recombinants were grown in small scalecultures and assayed for the cold-adapted protease activityin the culture supernatants The clones were first grown inBMGY medium with glycerol as the sole carbon sourceFor induction of AOX1 promoter and protein productionthe growth medium was transferred to the same mediumexcept with 05 methanol in place with glycerol as thecarbon source [26] The supernatant of the recombinant Ppastoris expression cultures after 48 h of incubation time wasanalyzed to determine the recombinant protease activity bythe modified method of Brock et al (1982) [20] All testedclones secreted protease into the supernatant in differentexpression levels This might be due to the variability ofthe strength of the promoter and copy number in each cellto secrete and express the recombinant product The clonewith the highest yield was GpPro2 with 63UmL activityfollowed by KpPro1 and GpPro1 with 25UmL and 37UmLactivities respectively (data not shown) It is perceptible thatexpression systemof Pichia pastoris is the ideal system for thisprotein The preliminary assay has fruitfully produced high
protease expression at 20∘C Yet further optimization of therecombinant PI12 protease production need to be conductedin order to examine the optimum condition of this proteinReported cold-adapted proteases showed optimum activity ata range of 15∘Cndash60∘C [27ndash30] This difference was possiblyrelated to the respective genetic and physiological adaptationof the strains There are several factors that influenced thedifferent rate of expression It includes different site of inte-gration between interest gene and Pichia genome variabilityof different expression host cell to secrete and express therecombinant enzyme and different sites in the polylinker forinserting foreign genes [31]
This is the second extracellular protease reported to besynthesized by this obligate psychrophilic yeastG antarcticaThe first cold-adapted extracellular subtilase from this yeasthas been purified and characterized previously [32 33] Theextracellular serine proteinase (LAP2) from this Antarcticyeast formerly known as Leucosporidium antarcticum strain171 is specific towards synthetic substrates of chymotrypsinand subtilisin The subtilase of L antarcticum strain 171showed specific activity of 4043Umg when utilized N-SucAAPFpNA as a substrate [33]
37 Optimization of Protease Production GpPro2 was chosenfor further optimization due to its highest protease activityat the preliminary expression stage High protease activitywas detected after 72 hours of incubation (Figure 7) byusing the standard protocol method as recommended forPichia pastoris expression system (Pichia Expression KitInvitrogen USA) The expression decreased after 72 hoursand no activity was detected at 110 hours The prolongedincubation may result in the decrease of biomass productionand will subsequently reduce the expression of recombinantprotein In addition oxygen and nutrient depletion duringprolonged incubationwill trigger the stress responses in orderto survive in such harsh environment [34]
Methanol was used as a sole carbon by P pastoris Themethanol was oxidized to formaldehyde with the aid of oxy-gen as electron acceptor by an alcohol oxidase enzyme (AOXgene) [35] The optimal methanol concentration is at 05(vv) with 14UmL activity The addition of more than 05
Enzyme Research 17
02468
10121416
0 01 03 05 07 09Pr
otea
se ac
tivity
(Um
L)Methanol concentration ()
Figure 8 Effect ofmethanol concentration towards protease activityThe recombinant was cultivated for 72 hours at 30∘C in BMMYmedium05 methanol gives the highest activity with 14UmL The substrate (azocasein) was dissolved in Tris HCL buffer and incubated for 30minat 20∘CThe mixture was left for 15min and absorbance was taken at 450 nm A one-way ANOVA was used to test for preference differencesamong six different concentrations The statistical value (119875 value lt005) between concentration is 002
0
5
10
15
20
25
10 15 20 25 30 35
Prot
eas a
ctiv
ity (
Um
L)
Temperature (∘C)
Figure 9 Effect of temperature towards protease activity The reaction was carried out in temperature ranging from 10∘C to 35∘C at pH 7 for30min A one-way ANOVA was used to test for preference differences among six different temperatures The statistical value (119875 value lt005)between temperature is 0067
(vv) methanol resulted in tremendous decrease of proteaseactivity (Figure 8) Production of PI12 protease was decreasedwhen the methanol concentration increased This might bedue to the toxic effect of methanol concentration on DNAreplication and membrane synthesis Basically methanol isused to increase the permeability of the cell membrane tohelp the secretion of foreign protein [26] However if thepermeability of cell membrane is too high lowmolecular sizemetabolites would leak from the cell and such leakage willseriously damage cell activities Such condition would abusethe cellular system and might reduce the yield of proteinexpression [36]
Psychrophilic microorganism dominated in low temper-ature habitat which most other microorganism are inca-pable of withstanding due to their inability to produceenzyme that can tolerate the extreme environment con-dition Psychrophilic enzymes are always associated withhigh catalytic efficiency at low and moderate temperature(0∘C to 30∘C) compared to the mesophilic counterpart thatare generally unable to catalyze efficiently under extremely
low temperature [2] The recombinant protein assay wasdone at various temperatures ranging from 10∘C to 35∘Cusing 05 azocasein as a substrate dissolved in 50mMTris HCl in presence of 2mM CaCl
2 pH 7 In the activity
assays expressed protease showed a highest activity at 20∘C(20UmL) and decreased tremendously once it exceededmaximum temperature due to the denatured state of theenzyme (Figure 9) The active site of an enzyme is easilyaltered when the high temperature is introduced duringincubation or activity assay resulting in a low reaction rate[37]
The recombinant construct was transferred into BMMYmedia at different A
600 nm ranging from 1 to 10 The high-est activity of active enzyme was detected at A
600 nm 60(283UmL) and above (Figure 10) The slope during expo-nential phase was much higher compared to other phasesindicating an increase in the doubling time At exponentialphase the cells are adapted to the growth condition andother processes like cell growth cycle and synthesis of RNAThe yield of active enzyme during expression of P pastoris
18 Enzyme Research
05
1015202530
1 2 3 4 5 6 7 8 9 10Prot
ease
activ
ity (U
mL)
Initial inoculum A600
Figure 10 Effect of inoculums size towards protease activity The recombinant was cultivated for 72 hours at 30∘C in BMMY mediumwith 05 methanol concentration A one-way ANOVA was used to test for preference differences among six different optical densitiesThe statistical value (119875 value lt005) between optical density value is 003
(kD
a)
1160
662
450
350
250
184
144
M 1 2 3
(a) (b) (c)
Figure 11 (a) Coomassie-stained SDS-PAGE of recombinant P pastoris clone 1mL supernatants collected from the third-day culturewere concentrated 10-fold using trichloroacetic acid and 10120583L resultant products were applied to SDS-PAGE analysis Lane 1 molecularweight marker (Fermentas) Lane 2 GS115 (Control) Lane 3 GS115pPIC9 empty (Control) Lane 4 GS115pPIC9PI12prot (GpPro2)The arrow indicates the secreted recombinant PI12 protease (b) Plate activity staining of different recombinant P pastoris clones GGS115 GP GS115pPIC9 GRI GS115pPIC9PI12prot1 (GpPro1) GRII GS115pPIC9PI12prot2 (GpPro2) K KM71 KP KM71pPIC9 KRKM71pPIC9PI12prot (KpPro) (c)Gel activity staining of recombinant PI12 protease inP pastorisGpPro2 clone Lane 1 prestainedmolecularweightmarker (Fermentas) Lane 2 GS115pPIC9 empty (Control) Lane 3 GS115pPIC9PI12prot1 (GpPro2)The arrow indicates the secretedrecombinant PI12 protease
dropped after 28 hours of incubation which is at stationaryor death phase [38]
38 Detection of Recombinant Protein by SDS-PAGE andActivity Staining TheSDS-PAGE gel of the GpPro2 recombi-nant clonesrsquo supernatant showed a putative expression bandat around 993 kDa Plate staining and gel activity stainingwere executed to examine if the recombinant PI12 proteaseexpressed in P pastoris was functionally active In the platestaining the transformants showed strong proteolytic activity(Figure 11(a)) In gel staining proteolytic activity was alsodetected around 993 kDa after incubation at 20∘C This is
evidence of protease secretion into the surrounding mediawhereby it catalyzed the breakdown of casein thus formedclearing zones around the activity ring and band (Figures11(b) and 11(c)) As a conclusion the cold-adapted PI12protease was functionally expressed in P pastoris GS115(GpPro2 clone) with the size of 993 kDa
4 Conclusions
Psychrophilic yeast which was originated frommarine waterin Casey Station Antarctica was identified as Glaciozymaantarctica strain PI12 A serine protease with an open reading
Enzyme Research 19
frame of 2892 bp encoded for 963 amino acids from thisAntarctic yeast has been cloned and expressed in Pichiaexpression system This is the second extracellular pro-tease reported being synthesized by this psychrophilic yeastSequence analysis through bioinformatics studies revealedfew homology identities of this protein with other ser-ine proteases The mature PI12 protease was successfullyexpressed and secreted into the culture medium driven bythe Saccharomyces cerevisiae 120572-factor signal sequence in Ppastoris The protease production in P pastoris was bestobtained from strain GS115 (GpPro2) with 283UmL after 72hours of induction time with 05 (vv) of methanol inducerThe starting optical density at 600 nm (OD600) was 60 andthe optimum assay temperature was 20∘C The expressedprotein was detected by SDS-PAGE and activity staining witha molecular weight of 99 kDaThis cold-active enzymemightoffer an opportunity for biotechnological exploitation basedon its high catalytic activity at low temperatures and lowthermostability Structural and functional study of this cold-adapted enzyme would give insight into its adaptation to anextreme cold environment
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgment
This research was supported financially by the InitiativeBiotechnology-GMBI-MOSTI (Project number 07-05-16-MG-GMB02) andRUGS-Initiative 6 (Project number 05-02-11-1414RU)
References
[1] J W Deming ldquoPsychrophiles and polar regionsrdquo CurrentOpinion in Microbiology vol 5 no 3 pp 301ndash309 2002
[2] C Gerday M Aittaleb M Bentahir et al ldquoCold-adaptedenzymes from fundamentals to biotechnologyrdquo Trends inBiotechnology vol 18 no 3 pp 103ndash107 2000
[3] A Svingor J Kardos I Hajdu A Nemeth and P Zavodszky ldquoAbetter enzyme to cope with cold comparative flexibility studieson psychrotrophic mesophilic and thermophilic IPMDHSrdquoThe Journal of Biological Chemistry vol 276 no 30 pp 28121ndash28125 2001
[4] G Antranikian C E Vorgias and C Bertoldo ldquoExtremeenvironments as a resource for microorganisms and novel bio-catalystsrdquo Advances in Biochemical EngineeringBiotechnologyvol 96 pp 219ndash262 2005
[5] T Lonhienne C Gerday andG Feller ldquoPsychrophilic enzymesrevisiting the thermodynamic parameters of activation mayexplain local flexibilityrdquoBiochimica et Biophysica Acta vol 1543no 1 pp 1ndash10 2000
[6] R Cavicchioli and TThomas ldquoExtremophilesrdquo in Encyclopediaof Microbiology pp 317ndash337 Academic Press San Diego CalifUSA 2000
[7] G Feller E Narinx J L Arpigny et al ldquoEnzymes frompsychrophilic organismsrdquo FEMS Microbiology Reviews vol 18no 2-3 pp 189ndash202 1996
[8] J Arnorsdottir R B Smaradottir T O Magnusson S HThorbjarnardottir G Eggertsson and M M KristjanssonldquoCharacterization of a cloned subtilisin-like serine proteinasefrom a psychrotrophic Vibrio speciesrdquo European Journal ofBiochemistry vol 269 no 22 pp 5536ndash5546 2002
[9] N J Russell ldquoPsychrophilic bacteriamdashmolecular adaptations ofmembrane lipidsrdquo Comparative Biochemistry and Physiology APhysiology vol 118 no 3 pp 489ndash493 1997
[10] G Feller and C Gerday ldquoPsychrophilic enzymes hot topics incold adaptationrdquo Nature Reviews Microbiology vol 1 no 3 pp200ndash208 2003
[11] P Raspor and J Zupan ldquoYeasts in extreme environmentsrdquo inBiodiversity and Ecophysiology of Yeasts The Yeast Handbookpp 371ndash417 Springer Berlin Germany 2006
[12] M B Rao A M Tanksale M S Ghatge and V V Desh-pande ldquoMolecular and biotechnological aspects of microbialproteasesrdquoMicrobiology andMolecular Biology Reviews vol 62no 3 pp 597ndash635 1998
[13] L Polgar ldquoThe catalytic triad of serine peptidasesrdquo Cellular andMolecular Life Sciences vol 62 no 19-20 pp 2161ndash2172 2005
[14] S Y Boo C M V L Wong K F Rodrigues N NajimudinA M A Murad and N M Mahadi ldquoThermal stress responsesin Antarctic yeastGlaciozyma antarctica PI12 characterized byreal-time quantitative PCRrdquoPolar Biology vol 36 no 3 pp 381ndash389 2013
[15] B Turchetti S R T Hall L B Connell et al ldquoPsychrophilicyeasts from Antarctica and European glaciers descriptionof Glaciozyma gen nov Glaciozyma martinii sp nov andGlaciozyma watsonii sp novrdquo Extremophiles vol 15 no 5 pp573ndash586 2011
[16] N H F Hashim I Bharudin D L S Nguong et al ldquoCharac-terization of Afp1 an antifreeze protein from the psychrophilicyeast Glaciozyma antarctica PI12rdquo Extremophiles vol 17 no 1pp 63ndash73 2013
[17] A N M Ramli M A Azhar M S Shamsir et al ldquoSequenceand structural investigation of a novel psychrophilic 120572-amylasefrom Glaciozyma antarctica PI12 for cold-adaptation analysisrdquoJournal ofMolecularModeling vol 19 no 8 pp 3369ndash3383 2013
[18] A N M Ramli N M Mahadi A Rabu A M A Murad F DA Bakar and R M Illias ldquoMolecular cloning expression andbiochemical characterisation of a cold-adapted novel recombi-nant chitinase from Glaciozyma antarctica PI12rdquoMicrobial CellFactories vol 10 no 1 article 94 2011
[19] J Sambrook E Fritsch and T Maniatis Molecular Cloning ALaboratoryManual Cold SpringHarbor Laboratory Press NewYork NY USA 1989
[20] F M Brock C W Forsberg and J G Buchanan-SmithldquoProteolytic activity of rumen microorganisms and effects ofproteinase inhibitorsrdquoApplied and EnvironmentalMicrobiologyvol 44 no 3 pp 561ndash569 1982
[21] U K Laemmli ldquoCleavage of structural proteins during theassembly of the head of bacteriophage T4rdquo Nature vol 227 no5259 pp 680ndash685 1970
[22] A Henne R A Schmitz M Bomeke G Gottschalk and RDaniel ldquoScreening of environmental DNA libraries for thepresence of genes conferring lipolytic activity on Escherichiacolirdquo Applied and Environmental Microbiology vol 66 no 7 pp3113ndash3116 2000
[23] M Poza T de Miguel C Sieiro and T G Villa ldquoCharacteri-zation of a broad pH range protease of Candida caseinolyticardquoJournal of Applied Microbiology vol 91 no 5 pp 916ndash921 2001
20 Enzyme Research
[24] J D Bendtsen H Nielsen G von Heijne and S BrunakldquoImproved prediction of signal peptides SignalP 30rdquo Journalof Molecular Biology vol 340 no 4 pp 783ndash795 2004
[25] R J Siezen and J A M Leunissen ldquoSubtilases the superfamilyof subtilisin-like serine proteasesrdquo Protein Science vol 6 no 3pp 501ndash523 1997
[26] Y Katakura W Zhang G Zhuang et al ldquoEffect of methanolconcentration on the production of human 1205732- glycoprotein Idomain V by a recombinant Pichia pastoris a simple system forthe control of methanol concentration using a semiconductorgas sensorrdquo Journal of Fermentation and Bioengineering vol 86no 5 pp 482ndash487 1998
[27] Q-FWang Y-HHou Z Xu J-LMiao andG-Y Li ldquoPurifica-tion and properties of an extracellular cold-active protease fromthe psychrophilic bacterium Pseudoalteromonas sp NJ276rdquoBiochemical Engineering Journal vol 38 no 3 pp 362ndash3682008
[28] S Vazquez L Ruberto and W Mac Cormack ldquoProper-ties of extracellular proteases from three psychrotolerantStenotrophomonas maltophilia isolated from Antarctic soilrdquoPolar Biology vol 28 no 4 pp 319ndash325 2005
[29] R Zeng R Zhang J Zhao and N Lin ldquoCold-active ser-ine alkaline protease from the psychrophilic bacterium Pseu-domonas strain DY-A enzyme purification and characteriza-tionrdquo Extremophiles vol 7 no 4 pp 335ndash337 2003
[30] M Turkiewicz E Gromek H Kalinowska and M ZielinskaldquoBiosynthesis and properties of an extracellularmetalloproteasefrom the Antarctic marine bacterium Sphingomonas paucimo-bilisrdquo Journal of Biotechnology vol 70 no 1ndash3 pp 53ndash60 1999
[31] A Vassileva D A Chugh S Swaminathan and N KhannaldquoEffect of copy number on the expression levels of hepatitisB surface antigen in the methylotrophic yeast Pichia pastorisrdquoProtein Expression andPurification vol 21 no 1 pp 71ndash80 2001
[32] M Pazgier M Turkiewicz H Kalinowska and S Bielecki ldquoTheunique cold-adapted extracellular subtilase from psychrophilicyeast Leucosporidium antarcticumrdquo Journal ofMolecular Catal-ysis B Enzymatic vol 21 no 1-2 pp 39ndash42 2003
[33] M Turkiewicz M Pazgier H Kalinowska and S BieleckildquoA cold-adapted extracellular serine proteinase of the yeastLeucosporidium antarcticumrdquo Extremophiles vol 7 no 6 pp435ndash442 2003
[34] D Mattanovich B Gasser H Hohenblum and M SauerldquoStress in recombinant protein producing yeastsrdquo Journal ofBiotechnology vol 113 no 1ndash3 pp 121ndash135 2004
[35] P Secades BAlvarez and JAGuijarro ldquoPurification andprop-erties of a new psychrophilic metalloprotease (Fpp2) in the fishpathogen Flavobacterium psychrophilumrdquo FEMS MicrobiologyLetters vol 226 no 2 pp 273ndash279 2003
[36] L Y Ling I Ithoi and F M Yik ldquoOptimization for high-level expression in Pichia pastoris and purification of truncatedand full length recombinant sag2 of toxoplasma gondii fordiagnostic userdquo Southeast Asian Journal of Tropical Medicineand Public Health vol 41 no 3 pp 507ndash513 2010
[37] A Illanes ldquoStability of biocatalystsrdquo Electronic Journal ofBiotechnology vol 2 no 1 pp 1ndash9 1999
[38] B A Plantz J Sinha L Villarete K W Nickerson and VL Schlegel ldquoPichia pastoris fermentation optimization energystate and testing a growth-associated modelrdquo Applied Microbi-ology and Biotechnology vol 72 no 2 pp 297ndash305 2006
[39] K Tamura G Stecher D Peterson A Filipski and S KumarldquoMEGA6molecular evolutionary genetics analysis version 60rdquo
Molecular Biology and Evolution vol 30 no 12 pp 2725ndash27292013
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

Enzyme Research 3
products One set of primers was designed from the extreme51015840 and 31015840 ends of cDNA as follows forward 51015840 CAP 5-GCGGGGGCCGACAATAAAAAC-3 and reverse 31015840 A-Tail5-TTTTTTTTTTTTTTTTTTTTTTTTTTGAGGTGGC-31015840the 51015840-RACE-Ready cDNA served as a template to generatefull-length cDNA through long distance PCR (LDPCR)Amplification process was carried out in a reaction mixture(50120583L) containing 5 120583L of 50ndash100 ng cDNA template 1 120583L(10 pmole120583L) of each forward and reverse primers 1 120583L of50x dNTP mix 1 120583L of 1 U120583L 50x Advantage 2 polymerasemix 5 120583L of 10x Advantage 2 PCR buffer (Clontech USA)and 36 120583L of distilled water Predenaturation was performedat 95∘C for 1min followed by 30 PCR cycles of denaturation(95∘C 30 sec) annealing (61∘C 3min) and extension (68∘C3min) Final extension was conducted at 68∘C for 3minand held at 4∘C The gene was amplified using thermocycler(CG1-96 Corbett Research Australia) The amplification ofputative open reading frame (ORF) was conducted usingboth genomicDNAand cDNA to locate the introns and exonslocations The amplicons were examined by electrophoresiscloned into pJET12 blunt vector (Fermentas USA) and sentfor sequencing The sequence obtained was deposited inGenBank under accession number FM178559
25 Assembly and Analysis of PI12 Protease SequencesSequences derived from DNA walking and RACE strat-egy were assembled by conducting sequence alignmentin the Biology Workbench (httpworkbenchsdscedu)Analysis and interpretation of nucleic acid sequences wereconducted using several publicly available web serversand integrated software as listed in Table 1 The phylo-genetic tree was constructed based on a comparison ofthe PI12 protease protein sequence with the closest pro-tease sequence that was extracted from GeneBank database(httpwwwncbinlmnihgov) The protein sequences usedfor phylogenetic analysis were aligned using ClustalW Phy-logenetic tree was constructed using Molecular EvolutionaryGenetics Analysis 6 (MEGA 6) package by neighbor-joiningmethod
26 Cloning of PI12 Protease Gene in Pichia pastorisExpression System The gene encoding mature PI12 pro-tease was amplified by PCR using recombinant pJET12FLcDNA of PI12 protease as a template Sense primer51015840-CAAGCCCTAGGCTACGCACGGAACGAGAA-31015840 andantisense primer 51015840-CTGCCGAATTCTTCCTCAACCCA-GTTACCAAC-31015840 were designed based on mature PI12 pro-tease gene sequence (FM178559) for flanking the PCR prod-uct at the 51015840- and 31015840- terminus respectively Amplificationprocess was carried out in a reactionmixture (50 120583L) contain-ing 5 120583L of 50ndash100 ng DNA template 15 120583L (10 pmole120583L) ofeach sense and antisense primer 1 120583L of 10mM dNTP mix4 120583L of 25mM MgCl
2 1 120583L of 1 U120583L Taq DNA polymerase
5 120583L of 10x PCR buffer (MBI Fermentas USA) and 31120583L ofdH2O PCRwas commenced by predenaturation step at 94∘C
for 4min followed by 30 PCR cycles of denaturation (94∘C1min) annealing (65∘C 1min) and extension (72∘C 2min)
Final extension was conducted at 72∘C for 7min and held at4∘C
ThePCRproduct of themature PI12 protease gene and thevector pPIC9 (05ndash1 120583g) were separately added to restrictionenzyme digestion mixtures containing 3 120583L EcoRI (10U120583L)6 120583L AvrII (10U120583L) 6 120583L Y+ Tango buffer (10x) and dH
2O
to a final volume of 30 120583L The reactions were incubatedat 37∘C for 1 hour The digestion products were observedthrough agarose gel electrophoresis and gel purified usingthe QIAquick Gel Extraction Kit (Qiagen Germany) Theconstruct was added to a ligation mixture with a ratio of1 5 to pPIC9 vector The ligation mixture was comprised of1 120583L of 10x ligation buffer (MBI Fermentas USA) 1120583L T4DNA ligase (5U120583L MBI Fermentas USA) and dH
2O to a
final volume of 10 120583L The reaction was incubated overnight(14ndash16 h) at 16∘C Ligation mixture was transformed into Ecoli Top10 competent cells The procedures for preparationof competent cells and heat-shock transformation were per-formed according to the method described by Sambrook etal 1989 [19] The transformation mixture was spread on LBagar plates containing 100 120583gmL ampicillin and incubatedovernight at 37∘C The positive transformants were selectedfor transformation into Pichia pastoris electrocompetent cell
Prior to transformation into P pastoris cell the emptypPIC9 and pPIC9 recombinant plasmids harbouring PI12protease gene were linearized using PmeI (10U120583L) restric-tion enzyme digestion The digestion mixtures consisted of5 120583L buffer B (10x) 3 120583L of PmeI and 6 120583L plasmids whiledH2O was added to a final volume of 60 120583L The linearized
plasmids were purified using the QIAquick PCR purificationkit The preparation of electrocompetent cells was doneaccording to Pichia expression kit manual (Invitrogen USA)Later about 80ndash100 120583L of the electrocompetent cells weretransferred into an overnight prechilled (kept in minus20∘C)02 cm electroporation cuvette (BioRad USA) and mixedwith 10 120583L (1ndash5 120583g) recombinant DNA The transformationmixtures (mixture of cells and plasmids) were incubated onice for 5min in the electroporation cuvette and pulsedwith anelectroporator (Gene Pulser BioRad USA) at the followingparameters charging voltage of 1500V capacitance of 25120583Fand resistance of 400Ω which generated a pulse length ofsim5ndash10ms with a field strength of sim7500Vcm Iced-cold 1Msorbitol (1mL) was added immediately after the pulse intothe cuvettes The contents were transferred into sterile bijoubottles and incubated at 30∘C for 2 hours without shakingLater 10ndash200120583L of the cells were spread on the MD agar[Minimal dextrose medium 134 (wv) yeast nitrogen base(YNB) 4 times 10minus5 (wv) biotin 2 (wv) dextrose 2 (wv)agar]The plates were incubated at 30∘C for 3ndash5 days until thetransformant colonies were formed
27 Expression and Optimization of Recombinant pPIC9Mature PI12 Protease Single colonies of the recom-binantPichia pastoris carrying the pPIC9PI12 protease were grownin 3mL of BMGY [buffered glycerol-complex medium 1(wv yeast extract) 2 (wv) peptone 134 (wv) yeastnitrogen base (YNB) 4 times 10minus5 (wv) biotin 1 (wv)glycerol 100mM potassium phosphate buffer pH 60] at
4 Enzyme Research
Table 1 List of web servers applied for full-length cDNA of PI12 protease analysis
Analysis Web serverSoftware Website(1) Open reading frame (ORF)(2) Amino acid translation(3) Intron-exon boundaries
Augustus Web server httpaugustusgobicsde
(1) Amino acid composition(2) Molecular mass(3) Isoelectric point
ProtParam httpwwwexpasyorgtool
Homology search (1) NCBI BLASTX httpwwwncbinlmnihgov(2) NCBI BLASTP
Phylogenetic tree (1) Molecular Evolutionary Genetics Analysis (MEGA 6) httpwwwmegasoftwarenet(2) CLUSTALW httpseqtoolsdsceduCGIBWcgi
Propeptide ProP httpwwwcbsdtudkservicesProP-10Open reading frame ORF Finder httpwwwncbinlmnihgovprojectsgorfSignal peptide SignalP httpwwwcbsdtudkservicesSignalPHydrophobic profile ProtScale httpauexpasyorgcgi-binMultiple sequence alignment Biology Workbench httpworkbenchsdsceduProtein motifs ScanProsite httpprositeexpasyorgscanprositeN-glycosylation NetNGlyc httpwwwcbsdtudkservicesNetNGlyc
30∘C in a shake incubator (250 rpm) overnight The nextday 1mL of the starter cultures was used as inoculumand inoculated into 10mL of BMGY in a 50mL flask andincubated in a condition as described to generate cell biomassbefore induction Cells were harvested by centrifugation at1500timesg for 10min
The cells were resuspended in 50mL BMMY medium[bufferedmethanol-complexmedium 1 (wv) yeast extract2 (wv) peptone 134 (wv) yeast nitrogen base (YNB) 4times10minus5 (wv) biotin methanol 100mM potassium phosphatebuffer pH 60] and incubated with vigorous shaking at30∘C to induce expression The expression control was doneby applying the same protein expression condition to therecombinant Pichia bearing the empty vector (pPIC9) Thegrowth of Pichia pastoris harboring recombinant plasmidGpPro2 was optimized in four different parameters in orderto enhance the protease enzyme yield The yeast was grownup to 5 days for postinduction study and the concentrationof methanol was varied from 0 to 1 Assay temperature ofactive enzyme was optimized ranging from 10∘C until 35∘Cwith 5∘C intervals The last parameter was to find out thecapability of cell density in protease production at A
600 nmfrom 1 to 10
28 Protease Assay The proteolytic activity was measured bythemodifiedmethod of Brock et al 1982 [20] using azocaseinas substrate Azocasein (05) was dissolved in 01M Tris-HCl-2mM CaCl
2 pH 7 The reaction was initiated by the
addition of 100 120583L of enzyme solution for 30min at 20∘C Anequal volume of 10 (wv) trichloroacetic acid (TCA) wasadded to terminate the reaction The absorbance was read at450 nm using UVVisible spectrophotometer Ultraspec 2100pro (Amersham Biosciences USA) One unit of azocaseinaseactivity was defined as the rate of absorbance change of 0001per min at 20∘C under the standard assay condition
29 SDS-PAGE Sodium dodecyl sulphate polyacrylamidegel electrophoresis (SDS-PAGE) was prepared according tothe Laemmli [21] method using 12 polyacrylamide gel inthe Mini PROTEAN 3 Cell apparatus (BioRad USA) Afterelectrophoresis gels were stained with Coomassie BrilliantBlue R-250 (BioRad USA)
210 Activity Staining and Cup Plate Assay Activity stainingwas performed as described elsewhere [22] The supernatantfrom recombinant Pichia clones were applied in this methodand loaded into SDS-PAGE [12 (wv)] After electrophore-sis the gel was immersed in 20 isopropanol to remove SDSand washed with distilled water (2-3 times) and the gel wastransferred into 2 skim milk yeast peptone dextrose (YPD)agar plate Protease activity onPetri dishwas also tested by theactivity ring staining or cup plate assay using 2 skim milkYPD agar plate adapted from Poza et al 2001 [23] The platewas incubated at 20∘C for 5-6 hours
3 Results and Discussion
31 Microorganism and Culture Condition After 10 daysat 4∘C the growth of G antarctica PI12 on nutrient agarsabourouddextrose agar and potato dextrose agarwas cream-coloured smooth mucous runny and glisteningThe borderwas entire circular and convex while on skim milk agar theyeast produced hydrolytic colonies (Figure 1(a)) The cellsof G antarctica PI12 occurred in oval yeast-like shape withthe size of about 2ndash4 120583m and present singly when observedvia simple and negative staining (Figures 1(b) and 1(c)) SEMphotograph showed budding yeast form of the G antarcticastrain PI12 (Figure 1(d)) All cells clumped together andpossessed smooth and oval shape appearance
Figure 2 shows TEM photographs of the thin-sectionedof the basidiomycete yeast G antarctica strain PI12 which
Enzyme Research 5
(a)
2120583m
(b)
(c) (d)
Figure 1 (a) Casein hydrolysis on skim milk agar plate (b) Negative staining with enlargement of 10000x bar = 2120583m (c) Simple stainingphotograph (d) Scanning electron microscopy (SEM) photograph with magnification 2500x Bar = 10 120583m
reveals the intracellular organelles Figures 2(a) and 2(b)show budding yeast appearance of membrane-boundednucleus and easily recognized bud scarwhile Figure 2(c) illus-trates ultrastructure comprising of a dark and thick cell walla cell membrane an irregular shape nucleus mitochondriavacuoles golgi and bud scars
32 Ribosomal RNA Identification The ITS158S rDNAITS2 region of PI12 was successfully amplified by Booet al 2013 [14] with the predicted size of 646 bp TheITS158SITS2 sequence of Glaciozyma antarctica strainPI12 had been deposited in GenBank with the accessionnumber JX896956 Amplification of ITS region showed100 identity to ITS158SITS2 sequence of Antarctic yeastCBS 8938 (AY040659) and Glaciozyma antarctica strainCBS 8943 (AY033637) National Collection of Yeast Cul-tures (NCYC) identified PI12 as an isolate of Glaciozymaantarctica based on 26S rDNA D1D2 sequencing resultwhere it displayed 99 sequence identity to Glaciozymaantarctica strain CBS 5942 [formerly known as Leucosporid-ium antarcticum strain CBS 5942 (AF444529)] Phyloge-netic tree analysis was created based on comparison of ITSsequence of this strain with the closest strains that areavailable in the database with reference to BLAST result
(Figure 3) Thirty-nine of ITS sequences from Glaciozymaclade were comparedThe phylogenetic tree constructed con-sists of three main clusters cluster I (JX896956 Glaciozymaantarctica strain PI12 AY040659 Antarctic yeast CBS 8938AY033637 Glaciozyma antarctica strain CBS 8943 EU149809Glaciozyma antarctica strain CBS 10640 AB774460 Leu-cosporidium sp KGK-2 JQ857017 Glaciozyma antarcticaisolate T21Ga EU149808 Glaciozyma antarctica strain CBS10636 AY040663 Glaciozyma antarctica strain CBS 8939EU149806Glaciozyma antarctica strainCBS 10639 EU149805Glaciozyma antarctica strain CBS 10638 AY040657 Antarcticyeast CBS 8927 AF444529 Glaciozyma antarctica strainCBS 5942 and KF934487 Glaciozyma sp KP7-5-2) isthe sequences from strains originally considered as Lantarcticum which was reclassified as Glaciozyma antarc-tica [15] Cluster II (GQ336996 Leucosporidium sp AY30EU149804 Glaciozyma watsonii strain CBS 10641 AY040661Antarctic yeast ML 4515 AY040660 Glaciozyma watsoniistrain CBS 8940 AY033638 Glaciozyma watsonii strain CBS8944 AB774462 Leucosporidium sp BSS-2 KC785578Uncul-tured Glaciozyma clone 180 AB774461 Leucosporidium spBSS-1 EU149803 Glaciozyma watsonii strain CBS 10684HQ432823 Glaciozyma watsonii DBVPG4802 HQ432818Glaciozyma watsonii DBVPG4726 HQ432820 Glaciozyma
6 Enzyme Research
2120583m
(a)
2120583m
(b)
BS
V
N
CW
CM
GM
Length 314866nm
Length 226635nm
1120583m
(c)
Figure 2 Transmission electronmicroscopy (TEM) photographs of AntarcticG antarctica strain PI12 (a) 12500xmagnification (b) 10000xmagnification (c) 27500x magnificationThe scale bars represent 2 120583m for (a) and (b) and 1 120583m for (c)
watsonii DBVPG4760 JX171176 Glaciozyma watsonii iso-late LKF08-112 DQ402535 Leucosporidium sp LC-03-120isolate 03-120 JF900360 Glaciozyma watsonii strain CBS7009 KC333171 Glaciozyma watsonii isolate AU CryS06HQ432821 Glaciozyma watsoniiDBVPG4799) is categorizedas G watsonii while cluster III (AY040664 Glaciozyma mar-tinii strain CBS 8929 AY033641 Glaciozyma martinii strain
CBS 8928 HQ432816 Glaciozyma martinii DBVPG4841KF934486 Glaciozyma sp KP7-5-1 FR682434 UnculturedBasidiomycota HF934010 Glaciozyma sp K81b JF900362Glaciozyma martinii strain CBS 9639 JF900359 Glaciozymamartinii strain CBS 6581 JF900361 Glaciozyma martiniistrain CBS 7054) is categorized as G martini Recognitionof the Glaciozyma species has been done by comparing
Enzyme Research 7
JX896956 Glaciozyma antarctica strain PI12
AY040659 Antarctic yeast CBS 8938
AY033637 Glaciozyma antarctica strain CBS 8943
EU149809 Glaciozyma antarctica strain CBS 10640
AB774460 Leucosporidium sp KGK-2
JQ857017 Glaciozyma antarctica isolate T21Ga
EU149808 Glaciozyma antarctica strain CBS 10636
AY040663 Glaciozyma antarctica strain CBS 8939
EU149806 Glaciozyma antarctica strain CBS 10639
EU149805 Glaciozyma antarctica strain CBS 10638
AY040657 Antarctic yeast CBS 8927
AF444529 Glaciozyma antarctica strain CBS 5942
KF934487 Glaciozyma sp KP7-5-2
GQ336996 Leucosporidium sp AY30
EU149804 Glaciozyma watsonii strain CBS 10641
AY040661 Antarctic yeast ML 4515
AY040660 Glaciozyma watsonii strain CBS 8940
AY033638 Glaciozyma watsonii strain CBS 8944
EU149803 Glaciozyma watsonii strain CBS 10684
JX171176 Glaciozyma watsonii isolate LKF08-112
DQ402535 Leucosporidium sp LC-03-120 isolate 03-120
AB774462 Leucosporidium sp BSS-2
KC785578 uncultured Glaciozyma clone 180
HQ432818 Glaciozyma watsonii DBVPG4726
HQ432823 Glaciozyma watsonii DBVPG4802
AB774461 Leucosporidium sp BSS-1
HQ432820 Glaciozyma watsonii DBVPG4760
KC333171 Glaciozyma watsonii isolate AU CryS06
HQ432821 Glaciozyma watsonii DBVPG4799
JF900360 Glaciozyma watsonii strain CBS 7009
AY040664 Glaciozyma martinii strain CBS 8929
AY033641 Glaciozyma martinii strain CBS 8928
HQ432816 Glaciozyma martinii DBVPG4841
KF934486 Glaciozyma sp KP7-5-1
FR682434 uncultured Basidiomycota
HF934010 Glaciozyma sp K81b
JF900362 Glaciozyma martinii strain CBS 9639
JF900359 Glaciozyma martinii strain CBS 6581
JF900361 Glaciozyma martinii strain CBS 7054
001
Clus
ter 1
Glac
iozy
ma
anta
rctic
aCl
uste
r 3
Glac
iozy
ma
mar
tinii
Clus
ter 2
Glac
iozy
ma
watso
nii
Figure 3 Neighbor-joining phylogenetic analysis of internal transcribed spacer 1 (ITS1)58S rRNA geneinternal transcribed spacer 2 (ITS2)of Glaciozyma clade Numbers indicate percentage bootstrap values calculated on 1000 repeats of the alignment The tree was constructedusingMEGA6 software [39]The scale bar represents 001 substitutions per nucleotide position GenBank accession numbers of the sequencesare indicated before strain name ITS158SITS2 sequence of Glaciozyma antarctica strain PI12 had been deposited in GenBank with theaccession number JX896956 [14]
8 Enzyme Research
their molecular phylogenetic data and physiological andmorphological characteristics [15] Phylogenetic analysis ofITS158SITS2 sequence of strain PI12 showed that thesequence was closely related to ITS158SITS2 sequenceof Glaciozyma antarctica with high homology similarity toCluster I (99-100 identity) The ITS158SITS2 sequenceof G antarctica strain PI12 was differed from G watsonii(Cluster II) by 2-3 gap in the sequence (93 identity)and G martinii by 4-5 gap in the sequence (90ndash92identity)
33 Sequence Analysis of PI12 Protease cDNA In this studyamplification of the PI12 protease gene sequence was con-ducted using genomicDNAand cDNAas a template Putativepartial PI12 protease gene with the size of 1321 bp was initiallyacquired from the G antarctica genome survey sequencelibrary (Supplementary Figure see Supplementary Mate-rial available online at httpdxdoiorg1011552014197938)However the gene was discovered as an incomplete pro-tease gene sequence DNA walking strategy successfullyamplified 1500 bp of the upstream region of partial PI12protease Both sequences of about 1500 bp (derived fromthe DNA walking reaction) and 1321 bp (partial PI12 pro-tease) were assembled and made up a total length of2687 bp protease sequence (Supplementary figure)The puta-tive PI12 protease was predicted by Augustus webserver(httpaugustusgobicsde) having intervening sequencesintronsThus reverse-transcriptase PCR (RT-PCR) and rapidamplification of cDNA ends (RACE) were accomplished toobtain the full-length cDNA As a result the ORF comprising2892 bp encoding 963 amino acid residues were successfullydeduced
Nucleotide and deduced amino acid sequences ofgenomic DNA and cDNA fragment encoding PI12 proteasewere shown in Figure 4 The size of the ORF of putative PI12protease was 3867 bp with genomic DNA as the templatewhile the ORF comprised 2892 bp encoding 963 amino acidresidues were successfully deduced through RACE strategywith cDNA as the template The ORF was predicted usingORF Finder (httpwwwncbinlmnihgovprojectsgorf)while prediction using SignalP (httpwwwcbsdtudkser-vicesSignalP) webserver allowed the identification of theputative PI12 protease prepeptide where it predicted thepresence of signal peptidase I cleavage sites [24]The putativeN-terminal signal peptide (1 to 16 the underlined amino acid)was identified as a secretion signal There is no propeptidepredicted by ProP Webserver (httpwwwcbsdtudkser-vicesProP-10) Comparison of cDNA and gene sequencesallowed determination of gene structureTherefore sequencealignment was performed between the sequence of these twotemplates and it was confirmed that the coding sequence(consisted of 19 exons) was interrupted by 18 introns withtheir length varying from 48 to 60 nucleotides However theGT and AG rule for donor-acceptor sites was not obeyedOf 18 introns 10 introns contained canonical GT and AGdinucleotides at their 51015840 and 31015840 ends while 8 introns holdnoncanonical splice site pairs with TT-GG (2 introns)AG-TG GC-GT TG-GG TC-GG TA-GG and TG-AG
dinucleotides at the splice junction Upper case letters inblue colour represent introns while lower case letters inblack colour represent exons Splice donor and acceptor werehighlighted in grey
The amino acid composition (upper case letters) withinthe ORF amino acid residues was determined by theProtParam tool of the Expasy molecular biology server(httpwwwexpasyorgtool) Translation starts at a nucle-otide position 1 and translation stop is marked with anasterisk The nucleotide sequences ATG (1 to 3) and TAG(3865 to 3867) indicate the initiation codon and termi-nal codon respectively The presumed putative sequencesof promoter CAAT (minus101 to minus98) and TATA-box (minus92to minus88) were identified in the 51015840-untranslated (UTR)region The search was achieved via Neural Network Pro-moter Prediction (Berkeley Drosophila Genome Project)(httpwwwfruitflyorgseq toolspromoterhtml) In the 31015840-untranslated (UTR) region there was a putative polyadeny-lation signal attaa (3889 to 3893) detected from the TAGstop codon to the poly (A) tail The predicted molecularmass and pI of PI12 serine protease were 10099 kDa and641 respectively Out of 963 amino acid residues in thededuced sequence of the PI12 protease 70 are negativelycharged residues (Asp + Glu) while 66 are positively chargedresidues (Arg + Lys) at neutral pH (Table 2)The total chargedresidues (Arg Asp Glu His and Lys) are 150 amino acidsin the open reading frame of PI12 protease corresponded to156 while hydrophobic residues (Ala Phe Ile Leu MetPro Val and Trp) are 441 amino acids which correspondedto 458 of the total amino acids discovered The totaluncharged residues (Asp Cys Gln Gly SerThr and Tyr) areconsisted of 372 amino acids which corresponded to 386of the total amino acids of the open reading frame of PI12protease The aliphatic index is 9290 and the grand averageof hydropathicity (GRAVY) is 0141 The instability index (II)of PI12 protease was computed to be 3414 which classify theprotein as stable
N-glycosylation sites were predicted using NetNGlyc10 Server (httpwwwcbsdtudkservicesNetNGlyc)Therewere five putative N-glycosylation sites for the PI12 proteasegene predicted using the server For N-glycosylation thesequence motif Asn-Xaa-SerThr (Xaa any amino acid butnot Pro) has been defined as a prerequisite for glycosylationThe sequencemotifs in PI12 protease occurred in the position170 NETH 365 NATV 576 NLTP 634 NDTA and 833 NSSPAmong five predicted sites only three (except positions 576and 833) showed a high potential score (averaged output ofnine neural networks) which support the prediction Theputative N-glycosylation sites were demonstrated in boxesin Figure 4 The comparison of amino acid composition ofthis cold-adapted protease fromG antarctica strain PI12 withnine other subtilisin serine proteases fromdifferent families isshown in Table 2 Analysis of this comparison depicts that thepercentage of each amino acids in PI12 protease is almost thesamewith other proteaseswhileGly (G) is themost abundantaccounting for up to 104 of the total amino acid in PI12 pro-tease PI12 protease also contained the highest percentage ofLeu (L) with 99 of the total amino acids compared to other
Enzyme Research 9
Table2Aminoacid
compo
sitionandmolecular
weighto
fproteases
acrossdifferent
families
Amino
CAQ76821
CAG25549
EDR11679
CAG38357
CAD118
98CA
L25578
EDU50747
AAW
40780
ACB3
0121
ACB3
0119
acid
Qty
Qty
Qty
Qty
Qty
Qty
Qty
Qty
Qty
Qty
Ala
(A)
97101
8191
7887
7590
8299
101
115
8189
9196
8092
105
116
Arg
(R)
4042
2730
2224
1619
1721
1821
2932
2223
2124
3134
Asn
(N)
4446
3843
4348
4149
4251
4046
5257
5154
4552
4347
Asp
(D)
4951
6269
5460
6578
5567
5158
5358
6669
5260
6370
Cys
(C)
808
506
404
405
911
809
910
909
708
1011
Gln
(Q)
2324
3438
2326
2834
3036
3742
3539
2931
1922
2629
Glu
(E)
2122
1618
1921
2024
1518
2023
2528
2627
2832
2426
Gly
(G)
100
104
89100
8493
85102
85103
7788
8493
8084
8598
8695
His
(H)
1415
1416
1314
1619
1316
1113
1618
1415
1618
1213
Ile(I)
5355
4955
5157
5364
4150
3843
5156
4446
3540
4449
Leu
(L)
9599
6876
7179
6274
6073
6675
6774
8084
7485
6875
Lys
(K)
2627
2932
4449
2935
4959
5158
4044
3032
5058
5662
Met
(M)
1212
202
202
911
1113
809
1820
808
1113
1517
Phe
(F)
4143
5966
4752
5566
2935
3641
4449
3941
3439
3438
Pro
(P)
6264
6573
6774
5870
5870
5967
5055
5154
5564
5763
ser
(S)
7780
6977
91101
6072
6680
7080
7279
111
117
6373
6167
Thr
(T)
8790
89100
7583
83100
6579
7383
7178
7276
7182
6875
Trp
(W)
505
303
607
202
1215
1214
1213
909
1214
1415
Tyr
(Y)
3334
1213
2629
708
2935
2832
3943
5053
2630
2730
val
(V)
7679
8292
8089
6679
5769
7484
5864
6872
8295
6268
Total
963
1000
893
1000
900
1000
834
1000
825
1000
878
1000
906
1000
950
1000
866
1000
906
1000
MW
(Da)
1009898
932669
944365
869455
869549
924905
979343
1009629
917209
961988
Thea
ccessio
nnu
mbersof
proteasespu
tativ
eserinep
roteaseo
fGlacio
zymaantarctica
PI12
(CAQ
76821)sub
tilisin-lik
eproteaseP
leurotuso
streatus(CA
G25549)pyrolysin
LaccariabicolorS
238N
-H82
(EDR11679)
subtilisin
-like
proteasePh
anerochaetechrysosporiu
m(C
AG38357)sub
tilisin-lik
eserinep
roteaseP
R1CMetarhiziu
maniso
pliaev
aranisopliae(CA
D11898)serinee
ndop
eptid
aseH
ypocrealixii(C
AL2
5578)peptidase
Pyrenophoratritici-repentis
(EDU50747)p
eptid
aseCr
yptococcus
neoforman
svarn
eoform
ansJEC
21(A
AW40
780)sub
tilisin-lik
eprotease
Epich
loefestucae(A
CB30121)sub
tilisin-lik
eprotease
Epich
loefestucae
(ACB
30119
)
10 Enzyme Research
atgctcttcctccccgtcctcctcctcctccttcccggcgtcactgccttcctcaaccca 60
M L F L P V L L L L L P G V T A F L N Pgttaccaaccgcgcgaccaacgccatctcctcgactcaatacctctctaacgcctatatc 120
V T N R A T N A I S S T Q Y L S N A Y IctcgaattggacctctccacccccggcctcgtcaaacgggatagcacgcccgactctGTG 180
L E L D L S T P G L V K R D S T P D SCGTCCCGCGTTCCTCCCTCGCTGTCGAAGACGAGCTAACAATGACGTGGTAGatcctaga 240
I L Eggacgtactcacgtccgtaggccgcaacggtatcaagtaccaactccgccaccgctttat 300
D V L T S V G R N G I K Y Q L R H R F Ictccccgactctgttccacggcgcttcgatcactgtcccccctggaatctcccgctccca 360
S P T L F H G A S I T V P P G I S R S QaatcgcctctctccgcggtatcaaaGTCCGTCCCTCTCTCTCCCCTACTCCCTCCGTGCT 420
I A S L R G I KAACCCTGCACGCAACAGcgcgtctggcccgttcgaaagttctcccgacccagcgcagtag 480
R V W P V R K F S R P S A Vtggacgccgatggaggaggaagcgggttctcagggtcgcctatcaaggcggcgctcatgg 540
V D A D G G G S G F S G S P I K A A L Mgggtgaaagagctcgggaagcgcgcgaacgcttatgctggagatacgtttggaccgcatg 600
G V K E L G K R A N A Y A G D T F G P HtcatgacgggggttaatgagacgcatgaggcggggttgttgggagctgggattaagGTGT 660
V M T G V N E T H E A G L L G A G I KGTTTTCGTCTTTGTTTGGGGGAGGGGGGAGGGAGGAGCTGATTAATGGGTGCAGattggg 720
I GgtgctggacactggtgAGAAGCGGTTTGGAGGGAGGAGAGGATGGAGCTGATGAGTGTGC 780
V L D T GAGGTGttgattatttgaacccgattctgggaggctgctttggacctgggtgccatatgtc 840
V D Y L N P I L G G C F G P G C H M Sgtttgggtacgacttggttggcgatgattacgatggagataacgctcctgtgccggatgt 900
F G Y D L V G D D Y D G D N A P V P D VggatccttGTGCGTCCTTCCCTCCGCGATGGGCTGGAATCTGAGCTGACGGCGTTGTTGA 960
D PTCAGttgcgagctgcgatcctcatggTTCGTCTCGCTTTCCGCTCCTCTGATGCGCTCGC 1020
F A S C D P H GTGATGCACTTGGCAATAGGaactcacgttacgggaatcattggagcgctcccgaatgcgt 1080
T H V T G I I G A L P N AttggatttactggcgtcgcacccgccgctactctgggccactaccgagtGCGCTCCCTCT 1140
F G F T G V A P A A T L G H Y R VATCCGTTCTTCCTCGCAGCTCCCACTCACTCGTGCTTCCAGGTatttggctgcactggct 1200
F G C T GtcgtcggagaagatatcGTTCGTCCTCTCTCCAGTCCCGGCTAGCGATCCTGCTTCTGAC 1260
F V G E D ITTCGCATCTCCCTCTGAGattctcgctggactcatgcgaggagtcgaggacaactgcaac 1320
I L A G L M R G V E D N C Ngtcttgaccctctctctcggaggtccaggagggtgggtcaagggcacgccggcgtccatc 1380
V L T L S L G G P G G W V K G T P A S Icttatcgaccagatcgaagcgcaaggcattctcgtcaccgtcgccactggcaactcggga 1440
L I D Q I E A Q G I L V T V A T G N S GgctgagggTGAGCTCCTCGCTACTTGCGCGTCGAGACCAGTACTAACAGGGGCGTGGACG 1500
A E GCAGGcatgttcttctccgagtctcccgcctcgacgatcaacggcctttccatcgcatcca 1560
M F F S E S P A S T I N G L S I A ScgTCCGTCTCTCCCTTCCTTCGCACGCCACTCGCTCACACTCTCTCGCAGGgacgttacc 1620
T D V Tgacctcatcgcctacaacgccaccgtctcaggccaacctgcgatcccttacctctccgcc 1680
D L I A Y N A T V S G Q P A I P Y L S A
ggggggggggtctg minus194
ctagcgcgaatcgagaaaacacaaaagaacctcccgacttggcgttgcttctgctcttct minus180
ccttctcccaaccgccgacaataaaaactataacacacacacaccccatctcgacgcctg minus120
cagacttcacgactctcctcacgctctttaccgccccacgggtcccacccacgagccacg minus60
(a)
Figure 4 Continued
Enzyme Research 11
acgcccctcaacgtcgtcgccaacagcttccgcgtccacttcacctctaccgaccccaac 1740
T P L N V V A N S F R V H F T S T D P Naaccccgtcgacgcctgctctcctcttccggctggagcgcccgacttcgccaactatgtt 1800
N P V D A C S P L P A G A P D F A N Y Vacggtcgttcagcgtgggacttgtacgttcgttaccaagtaccagaacgttctcaatgct 1860
T V V Q R G T C T F V T K Y Q N V L N AggaggGTGCGTCGGCTTCTTCTCTCCCGACTGGATTGGGCGGAGACTGACCTTCTGAGCA 1920
G GGaaagatcgtattgttgtacaactcggagggagctgggaacctcccttacctcacgccca 1980
K I V L L Y N S E G A G N L P Y L T PacggtgtcggcatcgacgccgttgcaggtcttcgtcgttccgacggactcaagGTTCGTC 2040
N G V G I D A V A G L R R S D G L KTACGCTCTTCGGTGCTGAGGCCCTTCTCTGATCTCGCTCCGACAGcttctctcgtactat 2100
L L S Y Ycagaatgccaacaagcgtctcactctgcgcttccccaagggcaagatcgtcgcaggcttg 2160
Q N A N K R L T L R F P K G K I V A G LaccgataccatcaccggcggactcatctcgTTCGTTCCTCTCCGCCTCCCTCGCTGGCCG 2220
T D T I T G G L I SATGAGCTGACGCGGTCTTTGCAGGggttactcgacgtttggtccgacgaatgacctctac 2280
G Y S T F G P T N D L Yggtcagcctaccctctctgcccctggtggcaacatcctttcgaccttccctctctccgag 2340
G Q P T L S A P G G N I L S T F P L S EggaggagtggcggtcatcagtgggacgagcatgtcgtgcccctttgtcgGTTCGTCCTTC 2400
G G V A V I S G T S M S C P F VTCCTCGTTCGTTCGCTGAGCGTCGACTGATCTTGCACGATCGCAGctggatctgcggcgg 2460
A G S A Atcctcatggccgctcgcgcttcggagaacctcacgccgcttgagatcaggagtctcctta 2520
V L M A A R A S E N L T P L E I R S L Lctaccactgcgaagcttacgccggtctcgctcttgggatcgacgcctttggtgagcgtga 2580
T T T A K L T P V S L L G S T P L V S VttcgtcaaggaggaggTACGTTCCTTTCTCGCTGCTTTGAGGATCGAGGCTGATTAGCGA 2640
I R Q G G GGCTTCGCAGGactcgttcaggttgccaaggcgctcgcggccaagacgctaatctctcctc 2700
L V Q V A K A L A A K T L I S Pacgagctcctgctcaacgacactgcgaacgcgaactacgtccagactatcaagatcaaga 2760
H E L L L N D T A N A N Y V Q T I K I Kacaccaactcgtgggcgatgaagtacaccttctcctcggccgtcgcccaaggactcggaa 2820
N T N S W A M K Y T F S S A V A Q G L Gctttcgacgcttcgggcgatatcctccctaccctcgacccagtcgccgtctctggcgcac 2880
T F D A S G D I L P T L D P V A V S G Aaggctaccgtcgcgttcaacactcggatcctcagcgtcgcgcccggcgcgacggggtccg 2940
Q A T V A F N T R I L S V A P G A T G Stcgtggcgactatcacgccgccggttcttcccgtagcggacgctgcgaggttccctatct 3000
V V A T I T P P V L P V A D A A R F P Itctctgggtggatcagggtgaatgggcaaggcgcgagggatagcagtaggaacgaggcgt 3060
F S G W I R V N G Q G A R D S S R N E AacactgGTGCGTTTGGCTCGCTTTTTGGGAGGTGCCGAGCTAATCGTTTCTCGTAGtccc 3120
Y T V PgtactttgggcttgcggcgaagatgatcgatatgcaagGTGCGTCGATGGTTCTCTCGCT 3180
Y F G L A A K M I D M QTCTCTCTCGGTTGCTCACTTCAAAGTGCAGtcctcgacaccaccgagaccatttacggtc 3240
V L D T T E T I Y Gcgggctacgcctaccccttcgtgatcgacgacgcgattggagacatccaatccaccacaa 3300
P G Y A Y P F V I D D A I G D I Q S T TcgtcgtactccaggaacctcgggcccaccgtcttcgctcgctttgccaTGCGTCGACCGT 3360
T S Y S R N L G P T V F A R F ACCTCATTTCCTGCGGCAGAGCGAGAGCTGATTCCGACCTCGAACAGctggaacccttcac 3420
T G T L Htacagcctcgatctcgtcctagccgacatcgcctttacccccacctaccccaactcctcc 3480
Y S L D L V L A D I A F T P T Y P N S S
(b)
Figure 4 Continued
12 Enzyme Research
cccgccactcgtttcgtcaagcgctccctcacgcagcacacctctgccgcttcgcacctc 3540
P A T R F V K R S L T Q H T S A A S H Lgccaagcgccgcgtctccatcgccactatcaaccccaaagccaccctcgtcgccgatcga 3600
A K R R V S I A T I N P K A T L V A D Rcagctccactcggacgtccctatcgagggcaacatcttcacccaaccctttactggaagg 3660
Q L H S D V P I E G N I F T Q P F T G Rgattacctcgtcgacgcagccccgacgggatccaccgatcgtaccgtcacttttaacggg 3720
D Y L V D A A P T G S T D R T V T F N Gcagtacgccgagaacggcctcgtgaggacggctgtgacggggacttcgtaccgcttcctc 3780
Q Y A E N G L V R T A V T G T S Y R F Lcttcgggcgttgaagatctcgggagacgcgatgtacgaggatcagtatgagagctggctc 3840
L R A L K I S G D A M Y E D Q Y E S W L3900
S L P F S F R Attttgccacctcaaaaaaaaaaaaaaaaaaaaaaaaaa 3938
lowast
tcgctaccgttctcgttccgtgcgtaggcttgttgtggacaccactgcattaagttatga
(c)
Figure 4 Nucleotide and deduced amino acid sequences of genomic DNA and cDNA encoding protease from G antarctica strain PI12 Thenucleotides are numbered from the 51015840 end of the cDNA A putative CAAT TATA box and polyadenylation signal are bold italicThe putativeN-terminal signal peptide is underlined The initiation codon is boldfaced and termination codon is represented by an asterisk (lowast) boxedpeptide sequences showed N-glycosylated asparagines residues The amino acid residues that form the catalytic triad are double underlinedand bold while introns are represented in blue font Highlighted in grey are the intron donor-acceptor splice sites
proteases Hydrophobic profile of PI12 amino acid proteasewas performed by ProtScale tools with Kyte and Doolittlemethod at httpauexpasyorgcgi-bin The hydrophobicityprofile elucidation of PI12 protease amino acid was carriedout from amino acid 1 to 963 The amino acid residuesfrom 1 to 16 (M-L-F-L-P-V-L-L-L-L-L-P-G-V-T-A) and 548 to562 (V-A-V-I-S) were hydrophobic grooves The first regionwhich consisted of 16 amino acids was very hydrophobic andwas predicted to be the signal peptide by SignalP and ProPwebserver
The bold and double-underlined amino acids showedresidues of catalytic triad aspartic acid (D) histidine (H) andserine (S) active site respectively NCBI Conserved DomainsDatabase (CDD) identified the presence of subtilase proteaseand subtilisin-like serine protease domains in the amino acidsequence The predicted PI12 protease amino acid sequencewas found sharing significant homology to the subtilisinfamily of protease The highest similarity of PI12 proteasewas observed with 42 identity to subtilisin-like proteaseof Rhodosporidium toruloides (EMS20793) 37 identity tosubtilisin-like protease of Pleurotus ostreatus (CAG25549)and 35 identity to subtilisin-like protease of Gloeophyllumtrabeum ATCC 11539 (EPQ55564) The nucleotide sequenceof G antarctica strain PI12 showed low homology to thefungus subtilisin protease family and none to other psy-chrophilic proteases The multiple sequence alignment wasillustrated in Figure 5 The consensus sequence around thecatalytic triad aspartic acid (D) histidine (H) and serine(S) of PI12 protease showed high conservation with othersubtilisin (showed in black boxes) Glycine residues G142G173 G191 G262 and G327 that were highly conserved inmost of subtilases were conserved in PI12 protease [25]Therefore PI12 protease belongs to the subtilisin subgroup ofthe subtilase serine protease superfamilyThis serine protease
of G antarctica PI12 has been deposited into GenBank withaccession number CAQ76821
34 Phylogenetic Relationship of PI12 Protease The phylo-genetic tree was constructed to demonstrate the similaritiesand relationship of protein sequence between differentspecies or organisms Figure 6 illustrates the phylogeneticdendrogram indicating the relationship of PI12 proteasewith another 25 translated amino acid sequences ofserine protease The bar scale signifies the branch lengthto approximate variance along each branch Basicallythis tree comprises two main clusters cluster I was sub-divided into subcluster a (CAG25549 EPQ55564 ETW-80907 XP 007381302 XP 001877576 CAG38357 XP007341385 CCA68627 EJU04161 XP 007369624 EIW-62675 CCO37249 XP 001830835 XP 007342189) and sub-cluster b (XP 007589863 XP 003711175 EFQ32840 XP006694395 ENH74885 EON99722) cluster II was sub-divided into subcluster c (CBQ73425 GAC97617 CCF52619)and subcluster d [CAQ76821 (PI12 protease) EMS20793]
Serine protease constructed by the phylogenetic treecomprised large size of protein with about 801 to 1032 aminoacids The majority of the translated protease sequenceswere obtained from fungus All serine proteases from cluster1 subcluster a are derived from Agaricomycetes except forEJU04161 which belongs to Dacrymycetes whereby all serineproteases from cluster 1 subcluster b are from Sordari-omycetes Cluster II subcluster c consisted of serine proteasefrom fungus belonging to Ustilaginomycetes PI12 proteasewas categorized under cluster II subcluster d It possessedhomology similarity with the subtilisin-like protease fromRhodosporidium toruloides (EMS20793) (formerly known asRhodotorula glutinis or Rhodotorula gracilis) which is a
Enzyme Research 13
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
MKG-YSWSLLQLAFTGAALATVPLSDVKRISSAS--VVPNKFVVEVESASEIPGKR--SL
MKGVLVWSILS---AGCALAVTPLSSVKRVTNLP--IVANKFIVEVDSTSDIPTKR--S-
MKLPLLSLALT---AVSVGATVPLSSVQRTTTSPNNALAGEYIVEMNNVQGLGGKR--AY
---MAHGSLASLLLLLAASAQGAAAFGGKSATGGRSVVPNSYILQVNASAPALTKRGVSV
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease ---MLFLPVLLLLLPGVTAFLNPVTNRATNAISSTQYLSNAYILELDLSTPGLVKRDSTP
lowastlowast
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
YTPHELFYRSLSARGVSYEVNKEYNS-EGLFVGAVVTVSDANDVKALAETPGVKAIRPVR
-NPHDALYESLRKRAIGFKVDREFNT-QGLFVGAALTLSDAQDVGQIQSTTGVKSIRPVR
ASTHDTLYRSLRKRGISFDVKKEYNQ-PGIMVGAALKVASPQDVQQIATMAEVKAIRPVI
FEALDHTLAAVKQNGVKYTVRQRFDAIPEAFQAVSIQVEDGASMAELAQIPGVQRVWPVS
DSILEDVLTSVGRNGIKYQLRHRFIS-PTLFHGASITVPPGISRSQIASLRGIKRVWPVR
lowastlowast
TIKSPKPVKVHVVTGKDDPEIPA-------------------------DTESTHIMTGVN
AFDRPKPVKVQVVKGPDDPALPP-------------------------DSESTHVITGVD
AVPAPERINQFTVSSPNDSRITA-------------------------DSETTHIMTGVD
LIPRPVEPTVSDFTPVSSSSSSTKSKRD-THTLEKRGTNFPPASAYLNDTFYPHVQTGID
KFSRPSAVVDADGGGSGFSGSPIKAALMGVKELGKR------ANAYAGDTFGPHVMTGVN
lowast lowast lowast lowastlowast
KLHAQGIFGAGIKIGIIDTGIDYTHPALGGALGAGHKVIGGYDFVGDAYDGTNTPVPDND
KLHAQGITGAGIKIGILDTGIDYTHPTLGGAFGPGNKVIGGFDLVGDDYDGTNTPVPDPD
KVHAAGITGEGIKIGIIDTGIDYNHPLLGAGFGPGFKVIGGYDFVGDDYTGSNNPAPDDD
VLHNKGILGQGVKIVVVDEGVDYTNPILGGCFGPGCQISFGYDFVGDNYTGTNAPVPDSD
ETHEAGLLGAGIKIGVLDTGVDYLNPILGGCFGPGCHMSFGYDLVGDDYDGDNAPVPDVD
lowast lowast lowast lowastlowastlowast lowast lowast lowastlowast lowast lowastlowast lowast lowast lowast lowast lowastlowastlowast lowast lowast lowast lowast lowastlowast lowast
PLDQCNGHGTHVAGIIGANPDNQFNISGVSYGASLSSYRVFGCSGSVSDDIIIEALLRGV
PLDQCAGHGTHVAGIIGANPDNAFNISGVAFQASLSAYRVFGCVGFVTDDVLVDALLLGF
PLDNCNGHGTHVAGIIGANPGNDFGISGVAYGSSIASYRVFGCSGFVSDDIIIDALLRGY
PFSSCSNHGTHTTGTVGALAN-EYGFSGAAPMATIGHYRVISCSGATTDEILVAALTRAL
PFASCDPHGTHVTGIIGALPN-AFGFTGVAPAATLGHYRVFGCTGFVGEDIILAGLMRGV
lowast lowastlowastlowast lowastlowast lowast lowastlowast lowast lowastlowastlowast lowast lowast lowast
QDGQDILTLSLGGTDGWTE-SSSSVVASRIAASGKIVTIAAGNEGAYGSWYSSSPGNGID
NEGQDILTLSLGGADGWTE-SVSAVVASRIAATGKVVTIAAGNDGASGAWYTSSPGNGID
SDGMDILTLSLGGVEGWTE-SSSSVVASRIADQGRVVTIAAGNDGAYGSWYASSPGTGVD
QDNVNIVSMSLGSSVGWLDDSPVQIMASYLGSKGIHVVASVGNERNEGLFAADQPAASRI
EDNCNVLTLSLGGPGGWVKGTPASILIDQIEAQGILVTVATGNSGAEGMFFSESPASTIN
lowastlowastlowast lowastlowast lowast lowast lowastlowast lowast lowast
VISVASIDNTVDNIQNATVNGVEHGPIPYFKTLPLPFTNA-LPLYVISNDTTIANDACNA
VISVASLDNTVVPLQTAQVHGVTHDPIIYQDTFPLPINDT-RPIFATSTDVTVVDDACNP
VISVASVNNIAIPVQNATVQGVEHAPIPYLEALPLNVTGS-RPIYATSTDTTVADDACNA
GTGVGGVDPTYLAAYYAYLI--KRDPVPYISPTPFNLPNN-YLLYFTSTNTSVTNDACNP
GLSIASTDVTDLIAYNATVS--GQPAIPYLSATPLNVVANSFRVHFTSTDPNNPVDACSP
lowast lowast lowast lowast lowastlowastlowast
LPDNTPDLSGFLVLVRRGSCTFVQKLTNIAAKGGNAVFIYDNGNG-----FAGIDTGKFT
LPDSTPDLSGFVVIVRRGTCTFVQKLGNVAAKGAKVTLIYDNGSG-----FGAIDTGAFV
LPNDTPDLSGYVVVIKRGTCTFVTKLQNAAAKGANTFLIYNNGGS-----FSAISVGDYT
LPSTTPNLANRVVVVQRGTCGFTVKQQNVAAAGGKIVLIYNSKGSGLIPQLDVGSTGLTA
LPAGAPDFANYVTVVQRGTCTFVTKYQNVLNAGGKIVLLYNSEGAGNLPYLTPNGVGIDA
lowastlowast lowast lowastlowast lowast lowast lowast lowast lowast lowast lowast
(a)
Figure 5 Continued
14 Enzyme Research
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
NATLIQAADGVFLAQQVAAGKSVTITFPQTGGSAQITN-PSGGLVSSFTTYGPTNDFYFK
-ASFIQAADGEFLVQQFASGQNVSLSFPQSGGLIQFPD-PAGGLISSFTSYGPSNDFFFK
-AALISETDGTFLVNQFAAKANITISFPQSGSAWNYPDDVSGGLVSDFSSYGPSYDMYFK
VGSLRYEDGLKLLSYYQSSPRGQSISFPNGPLAPYVSNTVSGGVVASYSNFGPTNELFIY
VAGLRRSDGLKLLSYYQNANKRLTLRFPKGKIVAGLTDTITGGLISGYSTFGPTNDLYGQ
lowast lowastlowast lowastlowast lowastlowast
PAFGAPGGNILSTLPVPLGSYGVESGTSMATPYVAGSAALLLSVKGKSAAVALGARTLFQ
PAVAAPGGNILSTLPVNLGLFGIESGTSMATPFVAGSAALLFQAKGTSAAVGRSARTLFE
PAVAAPGGNILSTLPIALGSYGVESGTSMATPFTAGSAALILQAKGK--SVAKSIRNLFQ
PTLAAPGTDILSTV---TGGVALMRGTSMAAPLVAGAYALLLSVGNNLKLSPEEARTILM
PTLSAPGGNILSTFPLSEGGVAVISGTSMSCPFVAGSAAVLMAARASENLTPLEIRSLLT
lowast lowastlowastlowast lowastlowastlowastlowast lowast lowastlowastlowastlowast lowast lowastlowast lowast lowast
STAQRVPSSHTDGDPLQTLSQQGAGLIDVFKALYTETIVWPTELILNDTAHYKEVHTFTV
TTAQRVPSTHTDGDPLQTLTQQGAGLINVFNAIHTTTIVSPGELILNDTAHFKGTQQFTV
TTASAVASSKTDGDPLQTVSQQGAGLIQVDRAIGTTTVVSPGQLTLNDTAHFEGFKTFNI
STSQPSPVAY-GSETLDTVVSQGAGVINMTAAIAAHTVISPAQFNLNDTQYSNNTQTLTL
TTAKLTPVSLLGSTPLVSVIRQGGGLVQVAKALAAKTLISPHELLLNDTANANYVQTIKI
VNTGKTSKQYKLTHVPAGTAQTIT---SGTIFAADGPVPLSTDYATVKLSSSSFTLRPGQ
RNAGSSAKTYTLRHIPAGTAVTVT---PGTIFPADGPVPLSTDFASVSLSTSKFTLAPGK
QNTGSKAVTYKISHVPAGTAASVQ---SGSIFAADGPVPLTADYAQVTFSLNKVTVLPGV
RNNNRFPVSYQLSWVDSTGIVTYNDGATTDIIPSTTPNYVSTSVLRVAFSQRTVTVPGGQ
KNTNSWAMKYTFSSAVAQGLGTFD--ASGDILPTLDPVAVSGAQATVAFNTRILSVAPGA
lowast lowast lowast lowast lowast lowast
SKQVTTHINAP--SGVDASTYPVFSGFIEIATAT--------ESVHVTYLGLAASLKDKQ
TQTVTARFTPP--AAADPSTFPVFSGFIQIESGT--------EQVQVSYLGLKASLKDKQ
SVPILVWFKPP--QGVDASTYPVYSGFIEIAGLG--------ETLRVAYLGVAASLKDKA
TVKVTAQFIPPNLTAQQRNQFPIYSGFVTITGQGQGAGAEQMEKYNLPFFGLGARMVDMP
TGSVVATITPPVLPVADAARFPIFSGWIRVNGQG-ARDSSRNEAYTVPYFGLAAKMIDMQ
lowast lowast lowastlowast lowast lowast lowast lowast
IVDNTNEFFGITIPAVLNNTGDAQ-LSPTNYTFSGSDFPTLLLRLAFGTPLLRVDLVDAT
VIDNTDFFFGVPTPVLTDPNGEVQ-TSPRNYSFLASDFPTLIFRLAFGSPKVVVDLVSPT
VVDNTDEYFGVKIPALLDASGDVQ-GNSTNYTLVGDDVPSLLYRFAFGSAHVRVDLVEKA
VLDTTNVALGP--NLPFIAVGQN--IQTGPTTFSESNPAVVYFRLAAGTRRLRVDLVDAS
VLDTTETIYGPGYAYPFVIDDAIGDIQSTTTSYSRNLGPTVFARFATGTLHYSLDLVLAD
lowast lowast lowast lowast lowastlowastlowastlowast lowast
IDLKPTLNSREIDDKRHVFERESFSFPHPGPKGS-------------------TFAQVKI
INFKPTLNTRALGDTHGSF----FSFPHRVKTG--------------------SFAQVPT
STVSATLSKRNWWN---------WWWPQKPSN---------------------SYDKVPI
TSYNATVPAVTNPASRLVKRSNTELTPRAAFPT--------------------LYSQVRT
IAFTPTYPNSS-PATRFVKRSLTQHTSAASHLAKRRVSIATINPKATLVADRQLHSDVPI
lowast lowast
VGTLSEQSHIPRNSDASTVDDGGYSTFAFNKP------SFANGTTIPNGTYRWLLRALKV
LGTVFELDFTSRNNDDPTDN--GFNTVAIDPP------TFANGTTIPNGQYRLLLRALRV
VGTLAEYDYVPRNSDASTIDDNGYNVFQFT-D------SFVNGTKIAAGEYKVLLRALKV
VGSLFAPSYWPPRDYLFNNGGT-YSDYEIQLDGSFTFPNGTKGMAELGRSYKVLLRALKI
EGNIFTQPFTG-RDYLVDAAPTGSTDRTVTFNGQY-AENGLVRTAVTGTSYRFLLRALKI
lowast lowast lowastlowast lowast lowast lowast lowast lowastlowastlowastlowast
lowast lowast lowastlowastlowastlowastlowast
(b)
Figure 5 Continued
Enzyme Research 15
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
TGDPTKEADFETWLSPIVGITVA
TGDPTNEADFESFLSPIIGVNAP
TGNPAEEGDYETWLSPSFSVVPA
TADPTYTSSYESWLSPSFTFTA-
SGDAMYEDQYESWLSLPFSFRA-
lowast lowastlowast
ConservedSimilar
lowast All match
(c)
Figure 5 Multiple sequence alignment between amino acid sequence of PI12 protease with other subtilisin The serine protease sequenceswere obtained from the GenBank database Their accession numbers are EMS20793 (subtilisin-like protease of Rhodosporidium toruloidesNP11) CAG25549 (subtilisin-like protease of Pleurotus ostreatus) EPQ55564 (subtilisin-like protease of Gloeophyllum trabeumATCC 11539)and ETW80907 (serine protease S8 ofHeterobasidion irregulare TC 32-1) Putative ORF of serine protease fromG antarctica PI12 was labeledas PI12 protease
CAG38357 Phanerochaete chrysosporium
EIW62675 Trametes versicolor FP-101664 SS1
XP 007369624 Dichomitus squalens LYAD-421 SS1
XP 007381302 Punctularia strigosozonata HHB-11173 SS5
CAG25549 Pleurotus ostreatus
ETW80907 Heterobasidion irregulare TC 32-1
XP 001877576 Laccaria bicolor S238N-H82
EPQ55564 Gloeophyllum trabeum ATCC 11539
XP 001830835 Coprinopsis cinerea okayama7130
XP 007341385 Auricularia delicata TFB-10046 SS5
CCA68627 Piriformospora indica DSM 11827
CCO37249 Rhizoctonia solani AG-1 IB
XP 007342189 Auricularia delicata TFB-10046 SS5
EJU04161 Dacryopinax sp DJM-731 SS1
XP 007589863 Colletotrichum fioriniae PJ7
EFQ32840 Colletotrichum graminicola M1001
ENH74885 Fusarium oxysporum f sp cubense race 1
XP 003711175 Magnaporthe oryzae 70-15
XP 006694395 Chaetomium thermophilum DSM 1495
EON99722 Togninia minima UCRPA7
CBQ73425 Sporisorium reilianum SRZ2
GAC97617 Pseudozyma hubeiensis SY62
CCF52619 Ustilago hordei
EMS20793 Rhodosporidium toruloides NP11
CAQ76821 Glaciozyma antarctica PI12
01
Clus
ter 1
Clus
ter 2
a
b
c
d
Figure 6 Phylogenetic dendrogram among 25 translated amino acid sequences of serine proteaseThe tree was constructed using a neighbor-joining algorithm with 1000 bootstrappings by MEGA 6 software [39]The scale bar represents 01 substitutions per amino acid positionTheserine protease sequences were obtained from the GenBank database GenBank accession numbers of the sequences were indicated beforethe strain name The putative serine protease of G antarctica strain PI12 has been deposited in GenBank with accession number CAQ76821
16 Enzyme Research
0
2
4
6
8
10
12
0 24 48 72 96 110
Prot
ease
activ
ity (U
mL)
Post induction time (hours)
Figure 7 Effect of postinduction time towards protease activity The highest activity was detected at 72 hours after 110 hours of incubationwith 9UmL activity followed by 48 24 and 96 hoursThe substrate (azocasein) was dissolved in Tris HCL buffer and incubated for 30min at20∘CThe absorbance was measured at 450 nm after 15min incubation A one-way ANOVAwas used to test for preference differences amongsix different postinduction times The statistical value (119875 value lt005) between hours is 0003
pink-coloured nonpathogenic oleaginous yeast (42 iden-tity) Both G antarctica PI12 and Rhodosporidium toruloidesNP11 belong to Microbotryomycetes class PI12 protease didnot show any homology similarity to other psychrophilicproteases however it showed 37 identity to hypotheticalprotein from Pseudozyma antarctica T-34 which was isolatedfrom Japan (GAC76005) (not listed in the phylogenetic tree)
35 Cloning and Expression of Cold-Adapted PI12 Protease inthe Yeast Expression System The PCR product harbouringthe mature PI12 protease with two flanking restriction sites(EcoRI and AvrII) was cloned into pPIC9 expression vectorThe recombinant plasmid harbouring PI12 protease was usedto transform both GS115 and KM71 intended to produce bothHis+ Mut+ and His+ MutS Two recombinant clones fromGS115 (His+Mut+) strain (GpPro1 andGpPro2) and one fromKM71 (His+MutS) strain (KpPro1) were successfully obtainedwhich were further analyzed for protein expression study
36 Expression and Enzyme Assay of PI12 Protease in Pichiapastoris All positive recombinants were grown in small scalecultures and assayed for the cold-adapted protease activityin the culture supernatants The clones were first grown inBMGY medium with glycerol as the sole carbon sourceFor induction of AOX1 promoter and protein productionthe growth medium was transferred to the same mediumexcept with 05 methanol in place with glycerol as thecarbon source [26] The supernatant of the recombinant Ppastoris expression cultures after 48 h of incubation time wasanalyzed to determine the recombinant protease activity bythe modified method of Brock et al (1982) [20] All testedclones secreted protease into the supernatant in differentexpression levels This might be due to the variability ofthe strength of the promoter and copy number in each cellto secrete and express the recombinant product The clonewith the highest yield was GpPro2 with 63UmL activityfollowed by KpPro1 and GpPro1 with 25UmL and 37UmLactivities respectively (data not shown) It is perceptible thatexpression systemof Pichia pastoris is the ideal system for thisprotein The preliminary assay has fruitfully produced high
protease expression at 20∘C Yet further optimization of therecombinant PI12 protease production need to be conductedin order to examine the optimum condition of this proteinReported cold-adapted proteases showed optimum activity ata range of 15∘Cndash60∘C [27ndash30] This difference was possiblyrelated to the respective genetic and physiological adaptationof the strains There are several factors that influenced thedifferent rate of expression It includes different site of inte-gration between interest gene and Pichia genome variabilityof different expression host cell to secrete and express therecombinant enzyme and different sites in the polylinker forinserting foreign genes [31]
This is the second extracellular protease reported to besynthesized by this obligate psychrophilic yeastG antarcticaThe first cold-adapted extracellular subtilase from this yeasthas been purified and characterized previously [32 33] Theextracellular serine proteinase (LAP2) from this Antarcticyeast formerly known as Leucosporidium antarcticum strain171 is specific towards synthetic substrates of chymotrypsinand subtilisin The subtilase of L antarcticum strain 171showed specific activity of 4043Umg when utilized N-SucAAPFpNA as a substrate [33]
37 Optimization of Protease Production GpPro2 was chosenfor further optimization due to its highest protease activityat the preliminary expression stage High protease activitywas detected after 72 hours of incubation (Figure 7) byusing the standard protocol method as recommended forPichia pastoris expression system (Pichia Expression KitInvitrogen USA) The expression decreased after 72 hoursand no activity was detected at 110 hours The prolongedincubation may result in the decrease of biomass productionand will subsequently reduce the expression of recombinantprotein In addition oxygen and nutrient depletion duringprolonged incubationwill trigger the stress responses in orderto survive in such harsh environment [34]
Methanol was used as a sole carbon by P pastoris Themethanol was oxidized to formaldehyde with the aid of oxy-gen as electron acceptor by an alcohol oxidase enzyme (AOXgene) [35] The optimal methanol concentration is at 05(vv) with 14UmL activity The addition of more than 05
Enzyme Research 17
02468
10121416
0 01 03 05 07 09Pr
otea
se ac
tivity
(Um
L)Methanol concentration ()
Figure 8 Effect ofmethanol concentration towards protease activityThe recombinant was cultivated for 72 hours at 30∘C in BMMYmedium05 methanol gives the highest activity with 14UmL The substrate (azocasein) was dissolved in Tris HCL buffer and incubated for 30minat 20∘CThe mixture was left for 15min and absorbance was taken at 450 nm A one-way ANOVA was used to test for preference differencesamong six different concentrations The statistical value (119875 value lt005) between concentration is 002
0
5
10
15
20
25
10 15 20 25 30 35
Prot
eas a
ctiv
ity (
Um
L)
Temperature (∘C)
Figure 9 Effect of temperature towards protease activity The reaction was carried out in temperature ranging from 10∘C to 35∘C at pH 7 for30min A one-way ANOVA was used to test for preference differences among six different temperatures The statistical value (119875 value lt005)between temperature is 0067
(vv) methanol resulted in tremendous decrease of proteaseactivity (Figure 8) Production of PI12 protease was decreasedwhen the methanol concentration increased This might bedue to the toxic effect of methanol concentration on DNAreplication and membrane synthesis Basically methanol isused to increase the permeability of the cell membrane tohelp the secretion of foreign protein [26] However if thepermeability of cell membrane is too high lowmolecular sizemetabolites would leak from the cell and such leakage willseriously damage cell activities Such condition would abusethe cellular system and might reduce the yield of proteinexpression [36]
Psychrophilic microorganism dominated in low temper-ature habitat which most other microorganism are inca-pable of withstanding due to their inability to produceenzyme that can tolerate the extreme environment con-dition Psychrophilic enzymes are always associated withhigh catalytic efficiency at low and moderate temperature(0∘C to 30∘C) compared to the mesophilic counterpart thatare generally unable to catalyze efficiently under extremely
low temperature [2] The recombinant protein assay wasdone at various temperatures ranging from 10∘C to 35∘Cusing 05 azocasein as a substrate dissolved in 50mMTris HCl in presence of 2mM CaCl
2 pH 7 In the activity
assays expressed protease showed a highest activity at 20∘C(20UmL) and decreased tremendously once it exceededmaximum temperature due to the denatured state of theenzyme (Figure 9) The active site of an enzyme is easilyaltered when the high temperature is introduced duringincubation or activity assay resulting in a low reaction rate[37]
The recombinant construct was transferred into BMMYmedia at different A
600 nm ranging from 1 to 10 The high-est activity of active enzyme was detected at A
600 nm 60(283UmL) and above (Figure 10) The slope during expo-nential phase was much higher compared to other phasesindicating an increase in the doubling time At exponentialphase the cells are adapted to the growth condition andother processes like cell growth cycle and synthesis of RNAThe yield of active enzyme during expression of P pastoris
18 Enzyme Research
05
1015202530
1 2 3 4 5 6 7 8 9 10Prot
ease
activ
ity (U
mL)
Initial inoculum A600
Figure 10 Effect of inoculums size towards protease activity The recombinant was cultivated for 72 hours at 30∘C in BMMY mediumwith 05 methanol concentration A one-way ANOVA was used to test for preference differences among six different optical densitiesThe statistical value (119875 value lt005) between optical density value is 003
(kD
a)
1160
662
450
350
250
184
144
M 1 2 3
(a) (b) (c)
Figure 11 (a) Coomassie-stained SDS-PAGE of recombinant P pastoris clone 1mL supernatants collected from the third-day culturewere concentrated 10-fold using trichloroacetic acid and 10120583L resultant products were applied to SDS-PAGE analysis Lane 1 molecularweight marker (Fermentas) Lane 2 GS115 (Control) Lane 3 GS115pPIC9 empty (Control) Lane 4 GS115pPIC9PI12prot (GpPro2)The arrow indicates the secreted recombinant PI12 protease (b) Plate activity staining of different recombinant P pastoris clones GGS115 GP GS115pPIC9 GRI GS115pPIC9PI12prot1 (GpPro1) GRII GS115pPIC9PI12prot2 (GpPro2) K KM71 KP KM71pPIC9 KRKM71pPIC9PI12prot (KpPro) (c)Gel activity staining of recombinant PI12 protease inP pastorisGpPro2 clone Lane 1 prestainedmolecularweightmarker (Fermentas) Lane 2 GS115pPIC9 empty (Control) Lane 3 GS115pPIC9PI12prot1 (GpPro2)The arrow indicates the secretedrecombinant PI12 protease
dropped after 28 hours of incubation which is at stationaryor death phase [38]
38 Detection of Recombinant Protein by SDS-PAGE andActivity Staining TheSDS-PAGE gel of the GpPro2 recombi-nant clonesrsquo supernatant showed a putative expression bandat around 993 kDa Plate staining and gel activity stainingwere executed to examine if the recombinant PI12 proteaseexpressed in P pastoris was functionally active In the platestaining the transformants showed strong proteolytic activity(Figure 11(a)) In gel staining proteolytic activity was alsodetected around 993 kDa after incubation at 20∘C This is
evidence of protease secretion into the surrounding mediawhereby it catalyzed the breakdown of casein thus formedclearing zones around the activity ring and band (Figures11(b) and 11(c)) As a conclusion the cold-adapted PI12protease was functionally expressed in P pastoris GS115(GpPro2 clone) with the size of 993 kDa
4 Conclusions
Psychrophilic yeast which was originated frommarine waterin Casey Station Antarctica was identified as Glaciozymaantarctica strain PI12 A serine protease with an open reading
Enzyme Research 19
frame of 2892 bp encoded for 963 amino acids from thisAntarctic yeast has been cloned and expressed in Pichiaexpression system This is the second extracellular pro-tease reported being synthesized by this psychrophilic yeastSequence analysis through bioinformatics studies revealedfew homology identities of this protein with other ser-ine proteases The mature PI12 protease was successfullyexpressed and secreted into the culture medium driven bythe Saccharomyces cerevisiae 120572-factor signal sequence in Ppastoris The protease production in P pastoris was bestobtained from strain GS115 (GpPro2) with 283UmL after 72hours of induction time with 05 (vv) of methanol inducerThe starting optical density at 600 nm (OD600) was 60 andthe optimum assay temperature was 20∘C The expressedprotein was detected by SDS-PAGE and activity staining witha molecular weight of 99 kDaThis cold-active enzymemightoffer an opportunity for biotechnological exploitation basedon its high catalytic activity at low temperatures and lowthermostability Structural and functional study of this cold-adapted enzyme would give insight into its adaptation to anextreme cold environment
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgment
This research was supported financially by the InitiativeBiotechnology-GMBI-MOSTI (Project number 07-05-16-MG-GMB02) andRUGS-Initiative 6 (Project number 05-02-11-1414RU)
References
[1] J W Deming ldquoPsychrophiles and polar regionsrdquo CurrentOpinion in Microbiology vol 5 no 3 pp 301ndash309 2002
[2] C Gerday M Aittaleb M Bentahir et al ldquoCold-adaptedenzymes from fundamentals to biotechnologyrdquo Trends inBiotechnology vol 18 no 3 pp 103ndash107 2000
[3] A Svingor J Kardos I Hajdu A Nemeth and P Zavodszky ldquoAbetter enzyme to cope with cold comparative flexibility studieson psychrotrophic mesophilic and thermophilic IPMDHSrdquoThe Journal of Biological Chemistry vol 276 no 30 pp 28121ndash28125 2001
[4] G Antranikian C E Vorgias and C Bertoldo ldquoExtremeenvironments as a resource for microorganisms and novel bio-catalystsrdquo Advances in Biochemical EngineeringBiotechnologyvol 96 pp 219ndash262 2005
[5] T Lonhienne C Gerday andG Feller ldquoPsychrophilic enzymesrevisiting the thermodynamic parameters of activation mayexplain local flexibilityrdquoBiochimica et Biophysica Acta vol 1543no 1 pp 1ndash10 2000
[6] R Cavicchioli and TThomas ldquoExtremophilesrdquo in Encyclopediaof Microbiology pp 317ndash337 Academic Press San Diego CalifUSA 2000
[7] G Feller E Narinx J L Arpigny et al ldquoEnzymes frompsychrophilic organismsrdquo FEMS Microbiology Reviews vol 18no 2-3 pp 189ndash202 1996
[8] J Arnorsdottir R B Smaradottir T O Magnusson S HThorbjarnardottir G Eggertsson and M M KristjanssonldquoCharacterization of a cloned subtilisin-like serine proteinasefrom a psychrotrophic Vibrio speciesrdquo European Journal ofBiochemistry vol 269 no 22 pp 5536ndash5546 2002
[9] N J Russell ldquoPsychrophilic bacteriamdashmolecular adaptations ofmembrane lipidsrdquo Comparative Biochemistry and Physiology APhysiology vol 118 no 3 pp 489ndash493 1997
[10] G Feller and C Gerday ldquoPsychrophilic enzymes hot topics incold adaptationrdquo Nature Reviews Microbiology vol 1 no 3 pp200ndash208 2003
[11] P Raspor and J Zupan ldquoYeasts in extreme environmentsrdquo inBiodiversity and Ecophysiology of Yeasts The Yeast Handbookpp 371ndash417 Springer Berlin Germany 2006
[12] M B Rao A M Tanksale M S Ghatge and V V Desh-pande ldquoMolecular and biotechnological aspects of microbialproteasesrdquoMicrobiology andMolecular Biology Reviews vol 62no 3 pp 597ndash635 1998
[13] L Polgar ldquoThe catalytic triad of serine peptidasesrdquo Cellular andMolecular Life Sciences vol 62 no 19-20 pp 2161ndash2172 2005
[14] S Y Boo C M V L Wong K F Rodrigues N NajimudinA M A Murad and N M Mahadi ldquoThermal stress responsesin Antarctic yeastGlaciozyma antarctica PI12 characterized byreal-time quantitative PCRrdquoPolar Biology vol 36 no 3 pp 381ndash389 2013
[15] B Turchetti S R T Hall L B Connell et al ldquoPsychrophilicyeasts from Antarctica and European glaciers descriptionof Glaciozyma gen nov Glaciozyma martinii sp nov andGlaciozyma watsonii sp novrdquo Extremophiles vol 15 no 5 pp573ndash586 2011
[16] N H F Hashim I Bharudin D L S Nguong et al ldquoCharac-terization of Afp1 an antifreeze protein from the psychrophilicyeast Glaciozyma antarctica PI12rdquo Extremophiles vol 17 no 1pp 63ndash73 2013
[17] A N M Ramli M A Azhar M S Shamsir et al ldquoSequenceand structural investigation of a novel psychrophilic 120572-amylasefrom Glaciozyma antarctica PI12 for cold-adaptation analysisrdquoJournal ofMolecularModeling vol 19 no 8 pp 3369ndash3383 2013
[18] A N M Ramli N M Mahadi A Rabu A M A Murad F DA Bakar and R M Illias ldquoMolecular cloning expression andbiochemical characterisation of a cold-adapted novel recombi-nant chitinase from Glaciozyma antarctica PI12rdquoMicrobial CellFactories vol 10 no 1 article 94 2011
[19] J Sambrook E Fritsch and T Maniatis Molecular Cloning ALaboratoryManual Cold SpringHarbor Laboratory Press NewYork NY USA 1989
[20] F M Brock C W Forsberg and J G Buchanan-SmithldquoProteolytic activity of rumen microorganisms and effects ofproteinase inhibitorsrdquoApplied and EnvironmentalMicrobiologyvol 44 no 3 pp 561ndash569 1982
[21] U K Laemmli ldquoCleavage of structural proteins during theassembly of the head of bacteriophage T4rdquo Nature vol 227 no5259 pp 680ndash685 1970
[22] A Henne R A Schmitz M Bomeke G Gottschalk and RDaniel ldquoScreening of environmental DNA libraries for thepresence of genes conferring lipolytic activity on Escherichiacolirdquo Applied and Environmental Microbiology vol 66 no 7 pp3113ndash3116 2000
[23] M Poza T de Miguel C Sieiro and T G Villa ldquoCharacteri-zation of a broad pH range protease of Candida caseinolyticardquoJournal of Applied Microbiology vol 91 no 5 pp 916ndash921 2001
20 Enzyme Research
[24] J D Bendtsen H Nielsen G von Heijne and S BrunakldquoImproved prediction of signal peptides SignalP 30rdquo Journalof Molecular Biology vol 340 no 4 pp 783ndash795 2004
[25] R J Siezen and J A M Leunissen ldquoSubtilases the superfamilyof subtilisin-like serine proteasesrdquo Protein Science vol 6 no 3pp 501ndash523 1997
[26] Y Katakura W Zhang G Zhuang et al ldquoEffect of methanolconcentration on the production of human 1205732- glycoprotein Idomain V by a recombinant Pichia pastoris a simple system forthe control of methanol concentration using a semiconductorgas sensorrdquo Journal of Fermentation and Bioengineering vol 86no 5 pp 482ndash487 1998
[27] Q-FWang Y-HHou Z Xu J-LMiao andG-Y Li ldquoPurifica-tion and properties of an extracellular cold-active protease fromthe psychrophilic bacterium Pseudoalteromonas sp NJ276rdquoBiochemical Engineering Journal vol 38 no 3 pp 362ndash3682008
[28] S Vazquez L Ruberto and W Mac Cormack ldquoProper-ties of extracellular proteases from three psychrotolerantStenotrophomonas maltophilia isolated from Antarctic soilrdquoPolar Biology vol 28 no 4 pp 319ndash325 2005
[29] R Zeng R Zhang J Zhao and N Lin ldquoCold-active ser-ine alkaline protease from the psychrophilic bacterium Pseu-domonas strain DY-A enzyme purification and characteriza-tionrdquo Extremophiles vol 7 no 4 pp 335ndash337 2003
[30] M Turkiewicz E Gromek H Kalinowska and M ZielinskaldquoBiosynthesis and properties of an extracellularmetalloproteasefrom the Antarctic marine bacterium Sphingomonas paucimo-bilisrdquo Journal of Biotechnology vol 70 no 1ndash3 pp 53ndash60 1999
[31] A Vassileva D A Chugh S Swaminathan and N KhannaldquoEffect of copy number on the expression levels of hepatitisB surface antigen in the methylotrophic yeast Pichia pastorisrdquoProtein Expression andPurification vol 21 no 1 pp 71ndash80 2001
[32] M Pazgier M Turkiewicz H Kalinowska and S Bielecki ldquoTheunique cold-adapted extracellular subtilase from psychrophilicyeast Leucosporidium antarcticumrdquo Journal ofMolecular Catal-ysis B Enzymatic vol 21 no 1-2 pp 39ndash42 2003
[33] M Turkiewicz M Pazgier H Kalinowska and S BieleckildquoA cold-adapted extracellular serine proteinase of the yeastLeucosporidium antarcticumrdquo Extremophiles vol 7 no 6 pp435ndash442 2003
[34] D Mattanovich B Gasser H Hohenblum and M SauerldquoStress in recombinant protein producing yeastsrdquo Journal ofBiotechnology vol 113 no 1ndash3 pp 121ndash135 2004
[35] P Secades BAlvarez and JAGuijarro ldquoPurification andprop-erties of a new psychrophilic metalloprotease (Fpp2) in the fishpathogen Flavobacterium psychrophilumrdquo FEMS MicrobiologyLetters vol 226 no 2 pp 273ndash279 2003
[36] L Y Ling I Ithoi and F M Yik ldquoOptimization for high-level expression in Pichia pastoris and purification of truncatedand full length recombinant sag2 of toxoplasma gondii fordiagnostic userdquo Southeast Asian Journal of Tropical Medicineand Public Health vol 41 no 3 pp 507ndash513 2010
[37] A Illanes ldquoStability of biocatalystsrdquo Electronic Journal ofBiotechnology vol 2 no 1 pp 1ndash9 1999
[38] B A Plantz J Sinha L Villarete K W Nickerson and VL Schlegel ldquoPichia pastoris fermentation optimization energystate and testing a growth-associated modelrdquo Applied Microbi-ology and Biotechnology vol 72 no 2 pp 297ndash305 2006
[39] K Tamura G Stecher D Peterson A Filipski and S KumarldquoMEGA6molecular evolutionary genetics analysis version 60rdquo
Molecular Biology and Evolution vol 30 no 12 pp 2725ndash27292013
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

4 Enzyme Research
Table 1 List of web servers applied for full-length cDNA of PI12 protease analysis
Analysis Web serverSoftware Website(1) Open reading frame (ORF)(2) Amino acid translation(3) Intron-exon boundaries
Augustus Web server httpaugustusgobicsde
(1) Amino acid composition(2) Molecular mass(3) Isoelectric point
ProtParam httpwwwexpasyorgtool
Homology search (1) NCBI BLASTX httpwwwncbinlmnihgov(2) NCBI BLASTP
Phylogenetic tree (1) Molecular Evolutionary Genetics Analysis (MEGA 6) httpwwwmegasoftwarenet(2) CLUSTALW httpseqtoolsdsceduCGIBWcgi
Propeptide ProP httpwwwcbsdtudkservicesProP-10Open reading frame ORF Finder httpwwwncbinlmnihgovprojectsgorfSignal peptide SignalP httpwwwcbsdtudkservicesSignalPHydrophobic profile ProtScale httpauexpasyorgcgi-binMultiple sequence alignment Biology Workbench httpworkbenchsdsceduProtein motifs ScanProsite httpprositeexpasyorgscanprositeN-glycosylation NetNGlyc httpwwwcbsdtudkservicesNetNGlyc
30∘C in a shake incubator (250 rpm) overnight The nextday 1mL of the starter cultures was used as inoculumand inoculated into 10mL of BMGY in a 50mL flask andincubated in a condition as described to generate cell biomassbefore induction Cells were harvested by centrifugation at1500timesg for 10min
The cells were resuspended in 50mL BMMY medium[bufferedmethanol-complexmedium 1 (wv) yeast extract2 (wv) peptone 134 (wv) yeast nitrogen base (YNB) 4times10minus5 (wv) biotin methanol 100mM potassium phosphatebuffer pH 60] and incubated with vigorous shaking at30∘C to induce expression The expression control was doneby applying the same protein expression condition to therecombinant Pichia bearing the empty vector (pPIC9) Thegrowth of Pichia pastoris harboring recombinant plasmidGpPro2 was optimized in four different parameters in orderto enhance the protease enzyme yield The yeast was grownup to 5 days for postinduction study and the concentrationof methanol was varied from 0 to 1 Assay temperature ofactive enzyme was optimized ranging from 10∘C until 35∘Cwith 5∘C intervals The last parameter was to find out thecapability of cell density in protease production at A
600 nmfrom 1 to 10
28 Protease Assay The proteolytic activity was measured bythemodifiedmethod of Brock et al 1982 [20] using azocaseinas substrate Azocasein (05) was dissolved in 01M Tris-HCl-2mM CaCl
2 pH 7 The reaction was initiated by the
addition of 100 120583L of enzyme solution for 30min at 20∘C Anequal volume of 10 (wv) trichloroacetic acid (TCA) wasadded to terminate the reaction The absorbance was read at450 nm using UVVisible spectrophotometer Ultraspec 2100pro (Amersham Biosciences USA) One unit of azocaseinaseactivity was defined as the rate of absorbance change of 0001per min at 20∘C under the standard assay condition
29 SDS-PAGE Sodium dodecyl sulphate polyacrylamidegel electrophoresis (SDS-PAGE) was prepared according tothe Laemmli [21] method using 12 polyacrylamide gel inthe Mini PROTEAN 3 Cell apparatus (BioRad USA) Afterelectrophoresis gels were stained with Coomassie BrilliantBlue R-250 (BioRad USA)
210 Activity Staining and Cup Plate Assay Activity stainingwas performed as described elsewhere [22] The supernatantfrom recombinant Pichia clones were applied in this methodand loaded into SDS-PAGE [12 (wv)] After electrophore-sis the gel was immersed in 20 isopropanol to remove SDSand washed with distilled water (2-3 times) and the gel wastransferred into 2 skim milk yeast peptone dextrose (YPD)agar plate Protease activity onPetri dishwas also tested by theactivity ring staining or cup plate assay using 2 skim milkYPD agar plate adapted from Poza et al 2001 [23] The platewas incubated at 20∘C for 5-6 hours
3 Results and Discussion
31 Microorganism and Culture Condition After 10 daysat 4∘C the growth of G antarctica PI12 on nutrient agarsabourouddextrose agar and potato dextrose agarwas cream-coloured smooth mucous runny and glisteningThe borderwas entire circular and convex while on skim milk agar theyeast produced hydrolytic colonies (Figure 1(a)) The cellsof G antarctica PI12 occurred in oval yeast-like shape withthe size of about 2ndash4 120583m and present singly when observedvia simple and negative staining (Figures 1(b) and 1(c)) SEMphotograph showed budding yeast form of the G antarcticastrain PI12 (Figure 1(d)) All cells clumped together andpossessed smooth and oval shape appearance
Figure 2 shows TEM photographs of the thin-sectionedof the basidiomycete yeast G antarctica strain PI12 which
Enzyme Research 5
(a)
2120583m
(b)
(c) (d)
Figure 1 (a) Casein hydrolysis on skim milk agar plate (b) Negative staining with enlargement of 10000x bar = 2120583m (c) Simple stainingphotograph (d) Scanning electron microscopy (SEM) photograph with magnification 2500x Bar = 10 120583m
reveals the intracellular organelles Figures 2(a) and 2(b)show budding yeast appearance of membrane-boundednucleus and easily recognized bud scarwhile Figure 2(c) illus-trates ultrastructure comprising of a dark and thick cell walla cell membrane an irregular shape nucleus mitochondriavacuoles golgi and bud scars
32 Ribosomal RNA Identification The ITS158S rDNAITS2 region of PI12 was successfully amplified by Booet al 2013 [14] with the predicted size of 646 bp TheITS158SITS2 sequence of Glaciozyma antarctica strainPI12 had been deposited in GenBank with the accessionnumber JX896956 Amplification of ITS region showed100 identity to ITS158SITS2 sequence of Antarctic yeastCBS 8938 (AY040659) and Glaciozyma antarctica strainCBS 8943 (AY033637) National Collection of Yeast Cul-tures (NCYC) identified PI12 as an isolate of Glaciozymaantarctica based on 26S rDNA D1D2 sequencing resultwhere it displayed 99 sequence identity to Glaciozymaantarctica strain CBS 5942 [formerly known as Leucosporid-ium antarcticum strain CBS 5942 (AF444529)] Phyloge-netic tree analysis was created based on comparison of ITSsequence of this strain with the closest strains that areavailable in the database with reference to BLAST result
(Figure 3) Thirty-nine of ITS sequences from Glaciozymaclade were comparedThe phylogenetic tree constructed con-sists of three main clusters cluster I (JX896956 Glaciozymaantarctica strain PI12 AY040659 Antarctic yeast CBS 8938AY033637 Glaciozyma antarctica strain CBS 8943 EU149809Glaciozyma antarctica strain CBS 10640 AB774460 Leu-cosporidium sp KGK-2 JQ857017 Glaciozyma antarcticaisolate T21Ga EU149808 Glaciozyma antarctica strain CBS10636 AY040663 Glaciozyma antarctica strain CBS 8939EU149806Glaciozyma antarctica strainCBS 10639 EU149805Glaciozyma antarctica strain CBS 10638 AY040657 Antarcticyeast CBS 8927 AF444529 Glaciozyma antarctica strainCBS 5942 and KF934487 Glaciozyma sp KP7-5-2) isthe sequences from strains originally considered as Lantarcticum which was reclassified as Glaciozyma antarc-tica [15] Cluster II (GQ336996 Leucosporidium sp AY30EU149804 Glaciozyma watsonii strain CBS 10641 AY040661Antarctic yeast ML 4515 AY040660 Glaciozyma watsoniistrain CBS 8940 AY033638 Glaciozyma watsonii strain CBS8944 AB774462 Leucosporidium sp BSS-2 KC785578Uncul-tured Glaciozyma clone 180 AB774461 Leucosporidium spBSS-1 EU149803 Glaciozyma watsonii strain CBS 10684HQ432823 Glaciozyma watsonii DBVPG4802 HQ432818Glaciozyma watsonii DBVPG4726 HQ432820 Glaciozyma
6 Enzyme Research
2120583m
(a)
2120583m
(b)
BS
V
N
CW
CM
GM
Length 314866nm
Length 226635nm
1120583m
(c)
Figure 2 Transmission electronmicroscopy (TEM) photographs of AntarcticG antarctica strain PI12 (a) 12500xmagnification (b) 10000xmagnification (c) 27500x magnificationThe scale bars represent 2 120583m for (a) and (b) and 1 120583m for (c)
watsonii DBVPG4760 JX171176 Glaciozyma watsonii iso-late LKF08-112 DQ402535 Leucosporidium sp LC-03-120isolate 03-120 JF900360 Glaciozyma watsonii strain CBS7009 KC333171 Glaciozyma watsonii isolate AU CryS06HQ432821 Glaciozyma watsoniiDBVPG4799) is categorizedas G watsonii while cluster III (AY040664 Glaciozyma mar-tinii strain CBS 8929 AY033641 Glaciozyma martinii strain
CBS 8928 HQ432816 Glaciozyma martinii DBVPG4841KF934486 Glaciozyma sp KP7-5-1 FR682434 UnculturedBasidiomycota HF934010 Glaciozyma sp K81b JF900362Glaciozyma martinii strain CBS 9639 JF900359 Glaciozymamartinii strain CBS 6581 JF900361 Glaciozyma martiniistrain CBS 7054) is categorized as G martini Recognitionof the Glaciozyma species has been done by comparing
Enzyme Research 7
JX896956 Glaciozyma antarctica strain PI12
AY040659 Antarctic yeast CBS 8938
AY033637 Glaciozyma antarctica strain CBS 8943
EU149809 Glaciozyma antarctica strain CBS 10640
AB774460 Leucosporidium sp KGK-2
JQ857017 Glaciozyma antarctica isolate T21Ga
EU149808 Glaciozyma antarctica strain CBS 10636
AY040663 Glaciozyma antarctica strain CBS 8939
EU149806 Glaciozyma antarctica strain CBS 10639
EU149805 Glaciozyma antarctica strain CBS 10638
AY040657 Antarctic yeast CBS 8927
AF444529 Glaciozyma antarctica strain CBS 5942
KF934487 Glaciozyma sp KP7-5-2
GQ336996 Leucosporidium sp AY30
EU149804 Glaciozyma watsonii strain CBS 10641
AY040661 Antarctic yeast ML 4515
AY040660 Glaciozyma watsonii strain CBS 8940
AY033638 Glaciozyma watsonii strain CBS 8944
EU149803 Glaciozyma watsonii strain CBS 10684
JX171176 Glaciozyma watsonii isolate LKF08-112
DQ402535 Leucosporidium sp LC-03-120 isolate 03-120
AB774462 Leucosporidium sp BSS-2
KC785578 uncultured Glaciozyma clone 180
HQ432818 Glaciozyma watsonii DBVPG4726
HQ432823 Glaciozyma watsonii DBVPG4802
AB774461 Leucosporidium sp BSS-1
HQ432820 Glaciozyma watsonii DBVPG4760
KC333171 Glaciozyma watsonii isolate AU CryS06
HQ432821 Glaciozyma watsonii DBVPG4799
JF900360 Glaciozyma watsonii strain CBS 7009
AY040664 Glaciozyma martinii strain CBS 8929
AY033641 Glaciozyma martinii strain CBS 8928
HQ432816 Glaciozyma martinii DBVPG4841
KF934486 Glaciozyma sp KP7-5-1
FR682434 uncultured Basidiomycota
HF934010 Glaciozyma sp K81b
JF900362 Glaciozyma martinii strain CBS 9639
JF900359 Glaciozyma martinii strain CBS 6581
JF900361 Glaciozyma martinii strain CBS 7054
001
Clus
ter 1
Glac
iozy
ma
anta
rctic
aCl
uste
r 3
Glac
iozy
ma
mar
tinii
Clus
ter 2
Glac
iozy
ma
watso
nii
Figure 3 Neighbor-joining phylogenetic analysis of internal transcribed spacer 1 (ITS1)58S rRNA geneinternal transcribed spacer 2 (ITS2)of Glaciozyma clade Numbers indicate percentage bootstrap values calculated on 1000 repeats of the alignment The tree was constructedusingMEGA6 software [39]The scale bar represents 001 substitutions per nucleotide position GenBank accession numbers of the sequencesare indicated before strain name ITS158SITS2 sequence of Glaciozyma antarctica strain PI12 had been deposited in GenBank with theaccession number JX896956 [14]
8 Enzyme Research
their molecular phylogenetic data and physiological andmorphological characteristics [15] Phylogenetic analysis ofITS158SITS2 sequence of strain PI12 showed that thesequence was closely related to ITS158SITS2 sequenceof Glaciozyma antarctica with high homology similarity toCluster I (99-100 identity) The ITS158SITS2 sequenceof G antarctica strain PI12 was differed from G watsonii(Cluster II) by 2-3 gap in the sequence (93 identity)and G martinii by 4-5 gap in the sequence (90ndash92identity)
33 Sequence Analysis of PI12 Protease cDNA In this studyamplification of the PI12 protease gene sequence was con-ducted using genomicDNAand cDNAas a template Putativepartial PI12 protease gene with the size of 1321 bp was initiallyacquired from the G antarctica genome survey sequencelibrary (Supplementary Figure see Supplementary Mate-rial available online at httpdxdoiorg1011552014197938)However the gene was discovered as an incomplete pro-tease gene sequence DNA walking strategy successfullyamplified 1500 bp of the upstream region of partial PI12protease Both sequences of about 1500 bp (derived fromthe DNA walking reaction) and 1321 bp (partial PI12 pro-tease) were assembled and made up a total length of2687 bp protease sequence (Supplementary figure)The puta-tive PI12 protease was predicted by Augustus webserver(httpaugustusgobicsde) having intervening sequencesintronsThus reverse-transcriptase PCR (RT-PCR) and rapidamplification of cDNA ends (RACE) were accomplished toobtain the full-length cDNA As a result the ORF comprising2892 bp encoding 963 amino acid residues were successfullydeduced
Nucleotide and deduced amino acid sequences ofgenomic DNA and cDNA fragment encoding PI12 proteasewere shown in Figure 4 The size of the ORF of putative PI12protease was 3867 bp with genomic DNA as the templatewhile the ORF comprised 2892 bp encoding 963 amino acidresidues were successfully deduced through RACE strategywith cDNA as the template The ORF was predicted usingORF Finder (httpwwwncbinlmnihgovprojectsgorf)while prediction using SignalP (httpwwwcbsdtudkser-vicesSignalP) webserver allowed the identification of theputative PI12 protease prepeptide where it predicted thepresence of signal peptidase I cleavage sites [24]The putativeN-terminal signal peptide (1 to 16 the underlined amino acid)was identified as a secretion signal There is no propeptidepredicted by ProP Webserver (httpwwwcbsdtudkser-vicesProP-10) Comparison of cDNA and gene sequencesallowed determination of gene structureTherefore sequencealignment was performed between the sequence of these twotemplates and it was confirmed that the coding sequence(consisted of 19 exons) was interrupted by 18 introns withtheir length varying from 48 to 60 nucleotides However theGT and AG rule for donor-acceptor sites was not obeyedOf 18 introns 10 introns contained canonical GT and AGdinucleotides at their 51015840 and 31015840 ends while 8 introns holdnoncanonical splice site pairs with TT-GG (2 introns)AG-TG GC-GT TG-GG TC-GG TA-GG and TG-AG
dinucleotides at the splice junction Upper case letters inblue colour represent introns while lower case letters inblack colour represent exons Splice donor and acceptor werehighlighted in grey
The amino acid composition (upper case letters) withinthe ORF amino acid residues was determined by theProtParam tool of the Expasy molecular biology server(httpwwwexpasyorgtool) Translation starts at a nucle-otide position 1 and translation stop is marked with anasterisk The nucleotide sequences ATG (1 to 3) and TAG(3865 to 3867) indicate the initiation codon and termi-nal codon respectively The presumed putative sequencesof promoter CAAT (minus101 to minus98) and TATA-box (minus92to minus88) were identified in the 51015840-untranslated (UTR)region The search was achieved via Neural Network Pro-moter Prediction (Berkeley Drosophila Genome Project)(httpwwwfruitflyorgseq toolspromoterhtml) In the 31015840-untranslated (UTR) region there was a putative polyadeny-lation signal attaa (3889 to 3893) detected from the TAGstop codon to the poly (A) tail The predicted molecularmass and pI of PI12 serine protease were 10099 kDa and641 respectively Out of 963 amino acid residues in thededuced sequence of the PI12 protease 70 are negativelycharged residues (Asp + Glu) while 66 are positively chargedresidues (Arg + Lys) at neutral pH (Table 2)The total chargedresidues (Arg Asp Glu His and Lys) are 150 amino acidsin the open reading frame of PI12 protease corresponded to156 while hydrophobic residues (Ala Phe Ile Leu MetPro Val and Trp) are 441 amino acids which correspondedto 458 of the total amino acids discovered The totaluncharged residues (Asp Cys Gln Gly SerThr and Tyr) areconsisted of 372 amino acids which corresponded to 386of the total amino acids of the open reading frame of PI12protease The aliphatic index is 9290 and the grand averageof hydropathicity (GRAVY) is 0141 The instability index (II)of PI12 protease was computed to be 3414 which classify theprotein as stable
N-glycosylation sites were predicted using NetNGlyc10 Server (httpwwwcbsdtudkservicesNetNGlyc)Therewere five putative N-glycosylation sites for the PI12 proteasegene predicted using the server For N-glycosylation thesequence motif Asn-Xaa-SerThr (Xaa any amino acid butnot Pro) has been defined as a prerequisite for glycosylationThe sequencemotifs in PI12 protease occurred in the position170 NETH 365 NATV 576 NLTP 634 NDTA and 833 NSSPAmong five predicted sites only three (except positions 576and 833) showed a high potential score (averaged output ofnine neural networks) which support the prediction Theputative N-glycosylation sites were demonstrated in boxesin Figure 4 The comparison of amino acid composition ofthis cold-adapted protease fromG antarctica strain PI12 withnine other subtilisin serine proteases fromdifferent families isshown in Table 2 Analysis of this comparison depicts that thepercentage of each amino acids in PI12 protease is almost thesamewith other proteaseswhileGly (G) is themost abundantaccounting for up to 104 of the total amino acid in PI12 pro-tease PI12 protease also contained the highest percentage ofLeu (L) with 99 of the total amino acids compared to other
Enzyme Research 9
Table2Aminoacid
compo
sitionandmolecular
weighto
fproteases
acrossdifferent
families
Amino
CAQ76821
CAG25549
EDR11679
CAG38357
CAD118
98CA
L25578
EDU50747
AAW
40780
ACB3
0121
ACB3
0119
acid
Qty
Qty
Qty
Qty
Qty
Qty
Qty
Qty
Qty
Qty
Ala
(A)
97101
8191
7887
7590
8299
101
115
8189
9196
8092
105
116
Arg
(R)
4042
2730
2224
1619
1721
1821
2932
2223
2124
3134
Asn
(N)
4446
3843
4348
4149
4251
4046
5257
5154
4552
4347
Asp
(D)
4951
6269
5460
6578
5567
5158
5358
6669
5260
6370
Cys
(C)
808
506
404
405
911
809
910
909
708
1011
Gln
(Q)
2324
3438
2326
2834
3036
3742
3539
2931
1922
2629
Glu
(E)
2122
1618
1921
2024
1518
2023
2528
2627
2832
2426
Gly
(G)
100
104
89100
8493
85102
85103
7788
8493
8084
8598
8695
His
(H)
1415
1416
1314
1619
1316
1113
1618
1415
1618
1213
Ile(I)
5355
4955
5157
5364
4150
3843
5156
4446
3540
4449
Leu
(L)
9599
6876
7179
6274
6073
6675
6774
8084
7485
6875
Lys
(K)
2627
2932
4449
2935
4959
5158
4044
3032
5058
5662
Met
(M)
1212
202
202
911
1113
809
1820
808
1113
1517
Phe
(F)
4143
5966
4752
5566
2935
3641
4449
3941
3439
3438
Pro
(P)
6264
6573
6774
5870
5870
5967
5055
5154
5564
5763
ser
(S)
7780
6977
91101
6072
6680
7080
7279
111
117
6373
6167
Thr
(T)
8790
89100
7583
83100
6579
7383
7178
7276
7182
6875
Trp
(W)
505
303
607
202
1215
1214
1213
909
1214
1415
Tyr
(Y)
3334
1213
2629
708
2935
2832
3943
5053
2630
2730
val
(V)
7679
8292
8089
6679
5769
7484
5864
6872
8295
6268
Total
963
1000
893
1000
900
1000
834
1000
825
1000
878
1000
906
1000
950
1000
866
1000
906
1000
MW
(Da)
1009898
932669
944365
869455
869549
924905
979343
1009629
917209
961988
Thea
ccessio
nnu
mbersof
proteasespu
tativ
eserinep
roteaseo
fGlacio
zymaantarctica
PI12
(CAQ
76821)sub
tilisin-lik
eproteaseP
leurotuso
streatus(CA
G25549)pyrolysin
LaccariabicolorS
238N
-H82
(EDR11679)
subtilisin
-like
proteasePh
anerochaetechrysosporiu
m(C
AG38357)sub
tilisin-lik
eserinep
roteaseP
R1CMetarhiziu
maniso
pliaev
aranisopliae(CA
D11898)serinee
ndop
eptid
aseH
ypocrealixii(C
AL2
5578)peptidase
Pyrenophoratritici-repentis
(EDU50747)p
eptid
aseCr
yptococcus
neoforman
svarn
eoform
ansJEC
21(A
AW40
780)sub
tilisin-lik
eprotease
Epich
loefestucae(A
CB30121)sub
tilisin-lik
eprotease
Epich
loefestucae
(ACB
30119
)
10 Enzyme Research
atgctcttcctccccgtcctcctcctcctccttcccggcgtcactgccttcctcaaccca 60
M L F L P V L L L L L P G V T A F L N Pgttaccaaccgcgcgaccaacgccatctcctcgactcaatacctctctaacgcctatatc 120
V T N R A T N A I S S T Q Y L S N A Y IctcgaattggacctctccacccccggcctcgtcaaacgggatagcacgcccgactctGTG 180
L E L D L S T P G L V K R D S T P D SCGTCCCGCGTTCCTCCCTCGCTGTCGAAGACGAGCTAACAATGACGTGGTAGatcctaga 240
I L Eggacgtactcacgtccgtaggccgcaacggtatcaagtaccaactccgccaccgctttat 300
D V L T S V G R N G I K Y Q L R H R F Ictccccgactctgttccacggcgcttcgatcactgtcccccctggaatctcccgctccca 360
S P T L F H G A S I T V P P G I S R S QaatcgcctctctccgcggtatcaaaGTCCGTCCCTCTCTCTCCCCTACTCCCTCCGTGCT 420
I A S L R G I KAACCCTGCACGCAACAGcgcgtctggcccgttcgaaagttctcccgacccagcgcagtag 480
R V W P V R K F S R P S A Vtggacgccgatggaggaggaagcgggttctcagggtcgcctatcaaggcggcgctcatgg 540
V D A D G G G S G F S G S P I K A A L Mgggtgaaagagctcgggaagcgcgcgaacgcttatgctggagatacgtttggaccgcatg 600
G V K E L G K R A N A Y A G D T F G P HtcatgacgggggttaatgagacgcatgaggcggggttgttgggagctgggattaagGTGT 660
V M T G V N E T H E A G L L G A G I KGTTTTCGTCTTTGTTTGGGGGAGGGGGGAGGGAGGAGCTGATTAATGGGTGCAGattggg 720
I GgtgctggacactggtgAGAAGCGGTTTGGAGGGAGGAGAGGATGGAGCTGATGAGTGTGC 780
V L D T GAGGTGttgattatttgaacccgattctgggaggctgctttggacctgggtgccatatgtc 840
V D Y L N P I L G G C F G P G C H M Sgtttgggtacgacttggttggcgatgattacgatggagataacgctcctgtgccggatgt 900
F G Y D L V G D D Y D G D N A P V P D VggatccttGTGCGTCCTTCCCTCCGCGATGGGCTGGAATCTGAGCTGACGGCGTTGTTGA 960
D PTCAGttgcgagctgcgatcctcatggTTCGTCTCGCTTTCCGCTCCTCTGATGCGCTCGC 1020
F A S C D P H GTGATGCACTTGGCAATAGGaactcacgttacgggaatcattggagcgctcccgaatgcgt 1080
T H V T G I I G A L P N AttggatttactggcgtcgcacccgccgctactctgggccactaccgagtGCGCTCCCTCT 1140
F G F T G V A P A A T L G H Y R VATCCGTTCTTCCTCGCAGCTCCCACTCACTCGTGCTTCCAGGTatttggctgcactggct 1200
F G C T GtcgtcggagaagatatcGTTCGTCCTCTCTCCAGTCCCGGCTAGCGATCCTGCTTCTGAC 1260
F V G E D ITTCGCATCTCCCTCTGAGattctcgctggactcatgcgaggagtcgaggacaactgcaac 1320
I L A G L M R G V E D N C Ngtcttgaccctctctctcggaggtccaggagggtgggtcaagggcacgccggcgtccatc 1380
V L T L S L G G P G G W V K G T P A S Icttatcgaccagatcgaagcgcaaggcattctcgtcaccgtcgccactggcaactcggga 1440
L I D Q I E A Q G I L V T V A T G N S GgctgagggTGAGCTCCTCGCTACTTGCGCGTCGAGACCAGTACTAACAGGGGCGTGGACG 1500
A E GCAGGcatgttcttctccgagtctcccgcctcgacgatcaacggcctttccatcgcatcca 1560
M F F S E S P A S T I N G L S I A ScgTCCGTCTCTCCCTTCCTTCGCACGCCACTCGCTCACACTCTCTCGCAGGgacgttacc 1620
T D V Tgacctcatcgcctacaacgccaccgtctcaggccaacctgcgatcccttacctctccgcc 1680
D L I A Y N A T V S G Q P A I P Y L S A
ggggggggggtctg minus194
ctagcgcgaatcgagaaaacacaaaagaacctcccgacttggcgttgcttctgctcttct minus180
ccttctcccaaccgccgacaataaaaactataacacacacacaccccatctcgacgcctg minus120
cagacttcacgactctcctcacgctctttaccgccccacgggtcccacccacgagccacg minus60
(a)
Figure 4 Continued
Enzyme Research 11
acgcccctcaacgtcgtcgccaacagcttccgcgtccacttcacctctaccgaccccaac 1740
T P L N V V A N S F R V H F T S T D P Naaccccgtcgacgcctgctctcctcttccggctggagcgcccgacttcgccaactatgtt 1800
N P V D A C S P L P A G A P D F A N Y Vacggtcgttcagcgtgggacttgtacgttcgttaccaagtaccagaacgttctcaatgct 1860
T V V Q R G T C T F V T K Y Q N V L N AggaggGTGCGTCGGCTTCTTCTCTCCCGACTGGATTGGGCGGAGACTGACCTTCTGAGCA 1920
G GGaaagatcgtattgttgtacaactcggagggagctgggaacctcccttacctcacgccca 1980
K I V L L Y N S E G A G N L P Y L T PacggtgtcggcatcgacgccgttgcaggtcttcgtcgttccgacggactcaagGTTCGTC 2040
N G V G I D A V A G L R R S D G L KTACGCTCTTCGGTGCTGAGGCCCTTCTCTGATCTCGCTCCGACAGcttctctcgtactat 2100
L L S Y Ycagaatgccaacaagcgtctcactctgcgcttccccaagggcaagatcgtcgcaggcttg 2160
Q N A N K R L T L R F P K G K I V A G LaccgataccatcaccggcggactcatctcgTTCGTTCCTCTCCGCCTCCCTCGCTGGCCG 2220
T D T I T G G L I SATGAGCTGACGCGGTCTTTGCAGGggttactcgacgtttggtccgacgaatgacctctac 2280
G Y S T F G P T N D L Yggtcagcctaccctctctgcccctggtggcaacatcctttcgaccttccctctctccgag 2340
G Q P T L S A P G G N I L S T F P L S EggaggagtggcggtcatcagtgggacgagcatgtcgtgcccctttgtcgGTTCGTCCTTC 2400
G G V A V I S G T S M S C P F VTCCTCGTTCGTTCGCTGAGCGTCGACTGATCTTGCACGATCGCAGctggatctgcggcgg 2460
A G S A Atcctcatggccgctcgcgcttcggagaacctcacgccgcttgagatcaggagtctcctta 2520
V L M A A R A S E N L T P L E I R S L Lctaccactgcgaagcttacgccggtctcgctcttgggatcgacgcctttggtgagcgtga 2580
T T T A K L T P V S L L G S T P L V S VttcgtcaaggaggaggTACGTTCCTTTCTCGCTGCTTTGAGGATCGAGGCTGATTAGCGA 2640
I R Q G G GGCTTCGCAGGactcgttcaggttgccaaggcgctcgcggccaagacgctaatctctcctc 2700
L V Q V A K A L A A K T L I S Pacgagctcctgctcaacgacactgcgaacgcgaactacgtccagactatcaagatcaaga 2760
H E L L L N D T A N A N Y V Q T I K I Kacaccaactcgtgggcgatgaagtacaccttctcctcggccgtcgcccaaggactcggaa 2820
N T N S W A M K Y T F S S A V A Q G L Gctttcgacgcttcgggcgatatcctccctaccctcgacccagtcgccgtctctggcgcac 2880
T F D A S G D I L P T L D P V A V S G Aaggctaccgtcgcgttcaacactcggatcctcagcgtcgcgcccggcgcgacggggtccg 2940
Q A T V A F N T R I L S V A P G A T G Stcgtggcgactatcacgccgccggttcttcccgtagcggacgctgcgaggttccctatct 3000
V V A T I T P P V L P V A D A A R F P Itctctgggtggatcagggtgaatgggcaaggcgcgagggatagcagtaggaacgaggcgt 3060
F S G W I R V N G Q G A R D S S R N E AacactgGTGCGTTTGGCTCGCTTTTTGGGAGGTGCCGAGCTAATCGTTTCTCGTAGtccc 3120
Y T V PgtactttgggcttgcggcgaagatgatcgatatgcaagGTGCGTCGATGGTTCTCTCGCT 3180
Y F G L A A K M I D M QTCTCTCTCGGTTGCTCACTTCAAAGTGCAGtcctcgacaccaccgagaccatttacggtc 3240
V L D T T E T I Y Gcgggctacgcctaccccttcgtgatcgacgacgcgattggagacatccaatccaccacaa 3300
P G Y A Y P F V I D D A I G D I Q S T TcgtcgtactccaggaacctcgggcccaccgtcttcgctcgctttgccaTGCGTCGACCGT 3360
T S Y S R N L G P T V F A R F ACCTCATTTCCTGCGGCAGAGCGAGAGCTGATTCCGACCTCGAACAGctggaacccttcac 3420
T G T L Htacagcctcgatctcgtcctagccgacatcgcctttacccccacctaccccaactcctcc 3480
Y S L D L V L A D I A F T P T Y P N S S
(b)
Figure 4 Continued
12 Enzyme Research
cccgccactcgtttcgtcaagcgctccctcacgcagcacacctctgccgcttcgcacctc 3540
P A T R F V K R S L T Q H T S A A S H Lgccaagcgccgcgtctccatcgccactatcaaccccaaagccaccctcgtcgccgatcga 3600
A K R R V S I A T I N P K A T L V A D Rcagctccactcggacgtccctatcgagggcaacatcttcacccaaccctttactggaagg 3660
Q L H S D V P I E G N I F T Q P F T G Rgattacctcgtcgacgcagccccgacgggatccaccgatcgtaccgtcacttttaacggg 3720
D Y L V D A A P T G S T D R T V T F N Gcagtacgccgagaacggcctcgtgaggacggctgtgacggggacttcgtaccgcttcctc 3780
Q Y A E N G L V R T A V T G T S Y R F Lcttcgggcgttgaagatctcgggagacgcgatgtacgaggatcagtatgagagctggctc 3840
L R A L K I S G D A M Y E D Q Y E S W L3900
S L P F S F R Attttgccacctcaaaaaaaaaaaaaaaaaaaaaaaaaa 3938
lowast
tcgctaccgttctcgttccgtgcgtaggcttgttgtggacaccactgcattaagttatga
(c)
Figure 4 Nucleotide and deduced amino acid sequences of genomic DNA and cDNA encoding protease from G antarctica strain PI12 Thenucleotides are numbered from the 51015840 end of the cDNA A putative CAAT TATA box and polyadenylation signal are bold italicThe putativeN-terminal signal peptide is underlined The initiation codon is boldfaced and termination codon is represented by an asterisk (lowast) boxedpeptide sequences showed N-glycosylated asparagines residues The amino acid residues that form the catalytic triad are double underlinedand bold while introns are represented in blue font Highlighted in grey are the intron donor-acceptor splice sites
proteases Hydrophobic profile of PI12 amino acid proteasewas performed by ProtScale tools with Kyte and Doolittlemethod at httpauexpasyorgcgi-bin The hydrophobicityprofile elucidation of PI12 protease amino acid was carriedout from amino acid 1 to 963 The amino acid residuesfrom 1 to 16 (M-L-F-L-P-V-L-L-L-L-L-P-G-V-T-A) and 548 to562 (V-A-V-I-S) were hydrophobic grooves The first regionwhich consisted of 16 amino acids was very hydrophobic andwas predicted to be the signal peptide by SignalP and ProPwebserver
The bold and double-underlined amino acids showedresidues of catalytic triad aspartic acid (D) histidine (H) andserine (S) active site respectively NCBI Conserved DomainsDatabase (CDD) identified the presence of subtilase proteaseand subtilisin-like serine protease domains in the amino acidsequence The predicted PI12 protease amino acid sequencewas found sharing significant homology to the subtilisinfamily of protease The highest similarity of PI12 proteasewas observed with 42 identity to subtilisin-like proteaseof Rhodosporidium toruloides (EMS20793) 37 identity tosubtilisin-like protease of Pleurotus ostreatus (CAG25549)and 35 identity to subtilisin-like protease of Gloeophyllumtrabeum ATCC 11539 (EPQ55564) The nucleotide sequenceof G antarctica strain PI12 showed low homology to thefungus subtilisin protease family and none to other psy-chrophilic proteases The multiple sequence alignment wasillustrated in Figure 5 The consensus sequence around thecatalytic triad aspartic acid (D) histidine (H) and serine(S) of PI12 protease showed high conservation with othersubtilisin (showed in black boxes) Glycine residues G142G173 G191 G262 and G327 that were highly conserved inmost of subtilases were conserved in PI12 protease [25]Therefore PI12 protease belongs to the subtilisin subgroup ofthe subtilase serine protease superfamilyThis serine protease
of G antarctica PI12 has been deposited into GenBank withaccession number CAQ76821
34 Phylogenetic Relationship of PI12 Protease The phylo-genetic tree was constructed to demonstrate the similaritiesand relationship of protein sequence between differentspecies or organisms Figure 6 illustrates the phylogeneticdendrogram indicating the relationship of PI12 proteasewith another 25 translated amino acid sequences ofserine protease The bar scale signifies the branch lengthto approximate variance along each branch Basicallythis tree comprises two main clusters cluster I was sub-divided into subcluster a (CAG25549 EPQ55564 ETW-80907 XP 007381302 XP 001877576 CAG38357 XP007341385 CCA68627 EJU04161 XP 007369624 EIW-62675 CCO37249 XP 001830835 XP 007342189) and sub-cluster b (XP 007589863 XP 003711175 EFQ32840 XP006694395 ENH74885 EON99722) cluster II was sub-divided into subcluster c (CBQ73425 GAC97617 CCF52619)and subcluster d [CAQ76821 (PI12 protease) EMS20793]
Serine protease constructed by the phylogenetic treecomprised large size of protein with about 801 to 1032 aminoacids The majority of the translated protease sequenceswere obtained from fungus All serine proteases from cluster1 subcluster a are derived from Agaricomycetes except forEJU04161 which belongs to Dacrymycetes whereby all serineproteases from cluster 1 subcluster b are from Sordari-omycetes Cluster II subcluster c consisted of serine proteasefrom fungus belonging to Ustilaginomycetes PI12 proteasewas categorized under cluster II subcluster d It possessedhomology similarity with the subtilisin-like protease fromRhodosporidium toruloides (EMS20793) (formerly known asRhodotorula glutinis or Rhodotorula gracilis) which is a
Enzyme Research 13
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
MKG-YSWSLLQLAFTGAALATVPLSDVKRISSAS--VVPNKFVVEVESASEIPGKR--SL
MKGVLVWSILS---AGCALAVTPLSSVKRVTNLP--IVANKFIVEVDSTSDIPTKR--S-
MKLPLLSLALT---AVSVGATVPLSSVQRTTTSPNNALAGEYIVEMNNVQGLGGKR--AY
---MAHGSLASLLLLLAASAQGAAAFGGKSATGGRSVVPNSYILQVNASAPALTKRGVSV
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease ---MLFLPVLLLLLPGVTAFLNPVTNRATNAISSTQYLSNAYILELDLSTPGLVKRDSTP
lowastlowast
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
YTPHELFYRSLSARGVSYEVNKEYNS-EGLFVGAVVTVSDANDVKALAETPGVKAIRPVR
-NPHDALYESLRKRAIGFKVDREFNT-QGLFVGAALTLSDAQDVGQIQSTTGVKSIRPVR
ASTHDTLYRSLRKRGISFDVKKEYNQ-PGIMVGAALKVASPQDVQQIATMAEVKAIRPVI
FEALDHTLAAVKQNGVKYTVRQRFDAIPEAFQAVSIQVEDGASMAELAQIPGVQRVWPVS
DSILEDVLTSVGRNGIKYQLRHRFIS-PTLFHGASITVPPGISRSQIASLRGIKRVWPVR
lowastlowast
TIKSPKPVKVHVVTGKDDPEIPA-------------------------DTESTHIMTGVN
AFDRPKPVKVQVVKGPDDPALPP-------------------------DSESTHVITGVD
AVPAPERINQFTVSSPNDSRITA-------------------------DSETTHIMTGVD
LIPRPVEPTVSDFTPVSSSSSSTKSKRD-THTLEKRGTNFPPASAYLNDTFYPHVQTGID
KFSRPSAVVDADGGGSGFSGSPIKAALMGVKELGKR------ANAYAGDTFGPHVMTGVN
lowast lowast lowast lowastlowast
KLHAQGIFGAGIKIGIIDTGIDYTHPALGGALGAGHKVIGGYDFVGDAYDGTNTPVPDND
KLHAQGITGAGIKIGILDTGIDYTHPTLGGAFGPGNKVIGGFDLVGDDYDGTNTPVPDPD
KVHAAGITGEGIKIGIIDTGIDYNHPLLGAGFGPGFKVIGGYDFVGDDYTGSNNPAPDDD
VLHNKGILGQGVKIVVVDEGVDYTNPILGGCFGPGCQISFGYDFVGDNYTGTNAPVPDSD
ETHEAGLLGAGIKIGVLDTGVDYLNPILGGCFGPGCHMSFGYDLVGDDYDGDNAPVPDVD
lowast lowast lowast lowastlowastlowast lowast lowast lowastlowast lowast lowastlowast lowast lowast lowast lowast lowastlowastlowast lowast lowast lowast lowast lowastlowast lowast
PLDQCNGHGTHVAGIIGANPDNQFNISGVSYGASLSSYRVFGCSGSVSDDIIIEALLRGV
PLDQCAGHGTHVAGIIGANPDNAFNISGVAFQASLSAYRVFGCVGFVTDDVLVDALLLGF
PLDNCNGHGTHVAGIIGANPGNDFGISGVAYGSSIASYRVFGCSGFVSDDIIIDALLRGY
PFSSCSNHGTHTTGTVGALAN-EYGFSGAAPMATIGHYRVISCSGATTDEILVAALTRAL
PFASCDPHGTHVTGIIGALPN-AFGFTGVAPAATLGHYRVFGCTGFVGEDIILAGLMRGV
lowast lowastlowastlowast lowastlowast lowast lowastlowast lowast lowastlowastlowast lowast lowast lowast
QDGQDILTLSLGGTDGWTE-SSSSVVASRIAASGKIVTIAAGNEGAYGSWYSSSPGNGID
NEGQDILTLSLGGADGWTE-SVSAVVASRIAATGKVVTIAAGNDGASGAWYTSSPGNGID
SDGMDILTLSLGGVEGWTE-SSSSVVASRIADQGRVVTIAAGNDGAYGSWYASSPGTGVD
QDNVNIVSMSLGSSVGWLDDSPVQIMASYLGSKGIHVVASVGNERNEGLFAADQPAASRI
EDNCNVLTLSLGGPGGWVKGTPASILIDQIEAQGILVTVATGNSGAEGMFFSESPASTIN
lowastlowastlowast lowastlowast lowast lowast lowastlowast lowast lowast
VISVASIDNTVDNIQNATVNGVEHGPIPYFKTLPLPFTNA-LPLYVISNDTTIANDACNA
VISVASLDNTVVPLQTAQVHGVTHDPIIYQDTFPLPINDT-RPIFATSTDVTVVDDACNP
VISVASVNNIAIPVQNATVQGVEHAPIPYLEALPLNVTGS-RPIYATSTDTTVADDACNA
GTGVGGVDPTYLAAYYAYLI--KRDPVPYISPTPFNLPNN-YLLYFTSTNTSVTNDACNP
GLSIASTDVTDLIAYNATVS--GQPAIPYLSATPLNVVANSFRVHFTSTDPNNPVDACSP
lowast lowast lowast lowast lowastlowastlowast
LPDNTPDLSGFLVLVRRGSCTFVQKLTNIAAKGGNAVFIYDNGNG-----FAGIDTGKFT
LPDSTPDLSGFVVIVRRGTCTFVQKLGNVAAKGAKVTLIYDNGSG-----FGAIDTGAFV
LPNDTPDLSGYVVVIKRGTCTFVTKLQNAAAKGANTFLIYNNGGS-----FSAISVGDYT
LPSTTPNLANRVVVVQRGTCGFTVKQQNVAAAGGKIVLIYNSKGSGLIPQLDVGSTGLTA
LPAGAPDFANYVTVVQRGTCTFVTKYQNVLNAGGKIVLLYNSEGAGNLPYLTPNGVGIDA
lowastlowast lowast lowastlowast lowast lowast lowast lowast lowast lowast lowast
(a)
Figure 5 Continued
14 Enzyme Research
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
NATLIQAADGVFLAQQVAAGKSVTITFPQTGGSAQITN-PSGGLVSSFTTYGPTNDFYFK
-ASFIQAADGEFLVQQFASGQNVSLSFPQSGGLIQFPD-PAGGLISSFTSYGPSNDFFFK
-AALISETDGTFLVNQFAAKANITISFPQSGSAWNYPDDVSGGLVSDFSSYGPSYDMYFK
VGSLRYEDGLKLLSYYQSSPRGQSISFPNGPLAPYVSNTVSGGVVASYSNFGPTNELFIY
VAGLRRSDGLKLLSYYQNANKRLTLRFPKGKIVAGLTDTITGGLISGYSTFGPTNDLYGQ
lowast lowastlowast lowastlowast lowastlowast
PAFGAPGGNILSTLPVPLGSYGVESGTSMATPYVAGSAALLLSVKGKSAAVALGARTLFQ
PAVAAPGGNILSTLPVNLGLFGIESGTSMATPFVAGSAALLFQAKGTSAAVGRSARTLFE
PAVAAPGGNILSTLPIALGSYGVESGTSMATPFTAGSAALILQAKGK--SVAKSIRNLFQ
PTLAAPGTDILSTV---TGGVALMRGTSMAAPLVAGAYALLLSVGNNLKLSPEEARTILM
PTLSAPGGNILSTFPLSEGGVAVISGTSMSCPFVAGSAAVLMAARASENLTPLEIRSLLT
lowast lowastlowastlowast lowastlowastlowastlowast lowast lowastlowastlowastlowast lowast lowastlowast lowast lowast
STAQRVPSSHTDGDPLQTLSQQGAGLIDVFKALYTETIVWPTELILNDTAHYKEVHTFTV
TTAQRVPSTHTDGDPLQTLTQQGAGLINVFNAIHTTTIVSPGELILNDTAHFKGTQQFTV
TTASAVASSKTDGDPLQTVSQQGAGLIQVDRAIGTTTVVSPGQLTLNDTAHFEGFKTFNI
STSQPSPVAY-GSETLDTVVSQGAGVINMTAAIAAHTVISPAQFNLNDTQYSNNTQTLTL
TTAKLTPVSLLGSTPLVSVIRQGGGLVQVAKALAAKTLISPHELLLNDTANANYVQTIKI
VNTGKTSKQYKLTHVPAGTAQTIT---SGTIFAADGPVPLSTDYATVKLSSSSFTLRPGQ
RNAGSSAKTYTLRHIPAGTAVTVT---PGTIFPADGPVPLSTDFASVSLSTSKFTLAPGK
QNTGSKAVTYKISHVPAGTAASVQ---SGSIFAADGPVPLTADYAQVTFSLNKVTVLPGV
RNNNRFPVSYQLSWVDSTGIVTYNDGATTDIIPSTTPNYVSTSVLRVAFSQRTVTVPGGQ
KNTNSWAMKYTFSSAVAQGLGTFD--ASGDILPTLDPVAVSGAQATVAFNTRILSVAPGA
lowast lowast lowast lowast lowast lowast
SKQVTTHINAP--SGVDASTYPVFSGFIEIATAT--------ESVHVTYLGLAASLKDKQ
TQTVTARFTPP--AAADPSTFPVFSGFIQIESGT--------EQVQVSYLGLKASLKDKQ
SVPILVWFKPP--QGVDASTYPVYSGFIEIAGLG--------ETLRVAYLGVAASLKDKA
TVKVTAQFIPPNLTAQQRNQFPIYSGFVTITGQGQGAGAEQMEKYNLPFFGLGARMVDMP
TGSVVATITPPVLPVADAARFPIFSGWIRVNGQG-ARDSSRNEAYTVPYFGLAAKMIDMQ
lowast lowast lowastlowast lowast lowast lowast lowast
IVDNTNEFFGITIPAVLNNTGDAQ-LSPTNYTFSGSDFPTLLLRLAFGTPLLRVDLVDAT
VIDNTDFFFGVPTPVLTDPNGEVQ-TSPRNYSFLASDFPTLIFRLAFGSPKVVVDLVSPT
VVDNTDEYFGVKIPALLDASGDVQ-GNSTNYTLVGDDVPSLLYRFAFGSAHVRVDLVEKA
VLDTTNVALGP--NLPFIAVGQN--IQTGPTTFSESNPAVVYFRLAAGTRRLRVDLVDAS
VLDTTETIYGPGYAYPFVIDDAIGDIQSTTTSYSRNLGPTVFARFATGTLHYSLDLVLAD
lowast lowast lowast lowast lowastlowastlowastlowast lowast
IDLKPTLNSREIDDKRHVFERESFSFPHPGPKGS-------------------TFAQVKI
INFKPTLNTRALGDTHGSF----FSFPHRVKTG--------------------SFAQVPT
STVSATLSKRNWWN---------WWWPQKPSN---------------------SYDKVPI
TSYNATVPAVTNPASRLVKRSNTELTPRAAFPT--------------------LYSQVRT
IAFTPTYPNSS-PATRFVKRSLTQHTSAASHLAKRRVSIATINPKATLVADRQLHSDVPI
lowast lowast
VGTLSEQSHIPRNSDASTVDDGGYSTFAFNKP------SFANGTTIPNGTYRWLLRALKV
LGTVFELDFTSRNNDDPTDN--GFNTVAIDPP------TFANGTTIPNGQYRLLLRALRV
VGTLAEYDYVPRNSDASTIDDNGYNVFQFT-D------SFVNGTKIAAGEYKVLLRALKV
VGSLFAPSYWPPRDYLFNNGGT-YSDYEIQLDGSFTFPNGTKGMAELGRSYKVLLRALKI
EGNIFTQPFTG-RDYLVDAAPTGSTDRTVTFNGQY-AENGLVRTAVTGTSYRFLLRALKI
lowast lowast lowastlowast lowast lowast lowast lowast lowastlowastlowastlowast
lowast lowast lowastlowastlowastlowastlowast
(b)
Figure 5 Continued
Enzyme Research 15
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
TGDPTKEADFETWLSPIVGITVA
TGDPTNEADFESFLSPIIGVNAP
TGNPAEEGDYETWLSPSFSVVPA
TADPTYTSSYESWLSPSFTFTA-
SGDAMYEDQYESWLSLPFSFRA-
lowast lowastlowast
ConservedSimilar
lowast All match
(c)
Figure 5 Multiple sequence alignment between amino acid sequence of PI12 protease with other subtilisin The serine protease sequenceswere obtained from the GenBank database Their accession numbers are EMS20793 (subtilisin-like protease of Rhodosporidium toruloidesNP11) CAG25549 (subtilisin-like protease of Pleurotus ostreatus) EPQ55564 (subtilisin-like protease of Gloeophyllum trabeumATCC 11539)and ETW80907 (serine protease S8 ofHeterobasidion irregulare TC 32-1) Putative ORF of serine protease fromG antarctica PI12 was labeledas PI12 protease
CAG38357 Phanerochaete chrysosporium
EIW62675 Trametes versicolor FP-101664 SS1
XP 007369624 Dichomitus squalens LYAD-421 SS1
XP 007381302 Punctularia strigosozonata HHB-11173 SS5
CAG25549 Pleurotus ostreatus
ETW80907 Heterobasidion irregulare TC 32-1
XP 001877576 Laccaria bicolor S238N-H82
EPQ55564 Gloeophyllum trabeum ATCC 11539
XP 001830835 Coprinopsis cinerea okayama7130
XP 007341385 Auricularia delicata TFB-10046 SS5
CCA68627 Piriformospora indica DSM 11827
CCO37249 Rhizoctonia solani AG-1 IB
XP 007342189 Auricularia delicata TFB-10046 SS5
EJU04161 Dacryopinax sp DJM-731 SS1
XP 007589863 Colletotrichum fioriniae PJ7
EFQ32840 Colletotrichum graminicola M1001
ENH74885 Fusarium oxysporum f sp cubense race 1
XP 003711175 Magnaporthe oryzae 70-15
XP 006694395 Chaetomium thermophilum DSM 1495
EON99722 Togninia minima UCRPA7
CBQ73425 Sporisorium reilianum SRZ2
GAC97617 Pseudozyma hubeiensis SY62
CCF52619 Ustilago hordei
EMS20793 Rhodosporidium toruloides NP11
CAQ76821 Glaciozyma antarctica PI12
01
Clus
ter 1
Clus
ter 2
a
b
c
d
Figure 6 Phylogenetic dendrogram among 25 translated amino acid sequences of serine proteaseThe tree was constructed using a neighbor-joining algorithm with 1000 bootstrappings by MEGA 6 software [39]The scale bar represents 01 substitutions per amino acid positionTheserine protease sequences were obtained from the GenBank database GenBank accession numbers of the sequences were indicated beforethe strain name The putative serine protease of G antarctica strain PI12 has been deposited in GenBank with accession number CAQ76821
16 Enzyme Research
0
2
4
6
8
10
12
0 24 48 72 96 110
Prot
ease
activ
ity (U
mL)
Post induction time (hours)
Figure 7 Effect of postinduction time towards protease activity The highest activity was detected at 72 hours after 110 hours of incubationwith 9UmL activity followed by 48 24 and 96 hoursThe substrate (azocasein) was dissolved in Tris HCL buffer and incubated for 30min at20∘CThe absorbance was measured at 450 nm after 15min incubation A one-way ANOVAwas used to test for preference differences amongsix different postinduction times The statistical value (119875 value lt005) between hours is 0003
pink-coloured nonpathogenic oleaginous yeast (42 iden-tity) Both G antarctica PI12 and Rhodosporidium toruloidesNP11 belong to Microbotryomycetes class PI12 protease didnot show any homology similarity to other psychrophilicproteases however it showed 37 identity to hypotheticalprotein from Pseudozyma antarctica T-34 which was isolatedfrom Japan (GAC76005) (not listed in the phylogenetic tree)
35 Cloning and Expression of Cold-Adapted PI12 Protease inthe Yeast Expression System The PCR product harbouringthe mature PI12 protease with two flanking restriction sites(EcoRI and AvrII) was cloned into pPIC9 expression vectorThe recombinant plasmid harbouring PI12 protease was usedto transform both GS115 and KM71 intended to produce bothHis+ Mut+ and His+ MutS Two recombinant clones fromGS115 (His+Mut+) strain (GpPro1 andGpPro2) and one fromKM71 (His+MutS) strain (KpPro1) were successfully obtainedwhich were further analyzed for protein expression study
36 Expression and Enzyme Assay of PI12 Protease in Pichiapastoris All positive recombinants were grown in small scalecultures and assayed for the cold-adapted protease activityin the culture supernatants The clones were first grown inBMGY medium with glycerol as the sole carbon sourceFor induction of AOX1 promoter and protein productionthe growth medium was transferred to the same mediumexcept with 05 methanol in place with glycerol as thecarbon source [26] The supernatant of the recombinant Ppastoris expression cultures after 48 h of incubation time wasanalyzed to determine the recombinant protease activity bythe modified method of Brock et al (1982) [20] All testedclones secreted protease into the supernatant in differentexpression levels This might be due to the variability ofthe strength of the promoter and copy number in each cellto secrete and express the recombinant product The clonewith the highest yield was GpPro2 with 63UmL activityfollowed by KpPro1 and GpPro1 with 25UmL and 37UmLactivities respectively (data not shown) It is perceptible thatexpression systemof Pichia pastoris is the ideal system for thisprotein The preliminary assay has fruitfully produced high
protease expression at 20∘C Yet further optimization of therecombinant PI12 protease production need to be conductedin order to examine the optimum condition of this proteinReported cold-adapted proteases showed optimum activity ata range of 15∘Cndash60∘C [27ndash30] This difference was possiblyrelated to the respective genetic and physiological adaptationof the strains There are several factors that influenced thedifferent rate of expression It includes different site of inte-gration between interest gene and Pichia genome variabilityof different expression host cell to secrete and express therecombinant enzyme and different sites in the polylinker forinserting foreign genes [31]
This is the second extracellular protease reported to besynthesized by this obligate psychrophilic yeastG antarcticaThe first cold-adapted extracellular subtilase from this yeasthas been purified and characterized previously [32 33] Theextracellular serine proteinase (LAP2) from this Antarcticyeast formerly known as Leucosporidium antarcticum strain171 is specific towards synthetic substrates of chymotrypsinand subtilisin The subtilase of L antarcticum strain 171showed specific activity of 4043Umg when utilized N-SucAAPFpNA as a substrate [33]
37 Optimization of Protease Production GpPro2 was chosenfor further optimization due to its highest protease activityat the preliminary expression stage High protease activitywas detected after 72 hours of incubation (Figure 7) byusing the standard protocol method as recommended forPichia pastoris expression system (Pichia Expression KitInvitrogen USA) The expression decreased after 72 hoursand no activity was detected at 110 hours The prolongedincubation may result in the decrease of biomass productionand will subsequently reduce the expression of recombinantprotein In addition oxygen and nutrient depletion duringprolonged incubationwill trigger the stress responses in orderto survive in such harsh environment [34]
Methanol was used as a sole carbon by P pastoris Themethanol was oxidized to formaldehyde with the aid of oxy-gen as electron acceptor by an alcohol oxidase enzyme (AOXgene) [35] The optimal methanol concentration is at 05(vv) with 14UmL activity The addition of more than 05
Enzyme Research 17
02468
10121416
0 01 03 05 07 09Pr
otea
se ac
tivity
(Um
L)Methanol concentration ()
Figure 8 Effect ofmethanol concentration towards protease activityThe recombinant was cultivated for 72 hours at 30∘C in BMMYmedium05 methanol gives the highest activity with 14UmL The substrate (azocasein) was dissolved in Tris HCL buffer and incubated for 30minat 20∘CThe mixture was left for 15min and absorbance was taken at 450 nm A one-way ANOVA was used to test for preference differencesamong six different concentrations The statistical value (119875 value lt005) between concentration is 002
0
5
10
15
20
25
10 15 20 25 30 35
Prot
eas a
ctiv
ity (
Um
L)
Temperature (∘C)
Figure 9 Effect of temperature towards protease activity The reaction was carried out in temperature ranging from 10∘C to 35∘C at pH 7 for30min A one-way ANOVA was used to test for preference differences among six different temperatures The statistical value (119875 value lt005)between temperature is 0067
(vv) methanol resulted in tremendous decrease of proteaseactivity (Figure 8) Production of PI12 protease was decreasedwhen the methanol concentration increased This might bedue to the toxic effect of methanol concentration on DNAreplication and membrane synthesis Basically methanol isused to increase the permeability of the cell membrane tohelp the secretion of foreign protein [26] However if thepermeability of cell membrane is too high lowmolecular sizemetabolites would leak from the cell and such leakage willseriously damage cell activities Such condition would abusethe cellular system and might reduce the yield of proteinexpression [36]
Psychrophilic microorganism dominated in low temper-ature habitat which most other microorganism are inca-pable of withstanding due to their inability to produceenzyme that can tolerate the extreme environment con-dition Psychrophilic enzymes are always associated withhigh catalytic efficiency at low and moderate temperature(0∘C to 30∘C) compared to the mesophilic counterpart thatare generally unable to catalyze efficiently under extremely
low temperature [2] The recombinant protein assay wasdone at various temperatures ranging from 10∘C to 35∘Cusing 05 azocasein as a substrate dissolved in 50mMTris HCl in presence of 2mM CaCl
2 pH 7 In the activity
assays expressed protease showed a highest activity at 20∘C(20UmL) and decreased tremendously once it exceededmaximum temperature due to the denatured state of theenzyme (Figure 9) The active site of an enzyme is easilyaltered when the high temperature is introduced duringincubation or activity assay resulting in a low reaction rate[37]
The recombinant construct was transferred into BMMYmedia at different A
600 nm ranging from 1 to 10 The high-est activity of active enzyme was detected at A
600 nm 60(283UmL) and above (Figure 10) The slope during expo-nential phase was much higher compared to other phasesindicating an increase in the doubling time At exponentialphase the cells are adapted to the growth condition andother processes like cell growth cycle and synthesis of RNAThe yield of active enzyme during expression of P pastoris
18 Enzyme Research
05
1015202530
1 2 3 4 5 6 7 8 9 10Prot
ease
activ
ity (U
mL)
Initial inoculum A600
Figure 10 Effect of inoculums size towards protease activity The recombinant was cultivated for 72 hours at 30∘C in BMMY mediumwith 05 methanol concentration A one-way ANOVA was used to test for preference differences among six different optical densitiesThe statistical value (119875 value lt005) between optical density value is 003
(kD
a)
1160
662
450
350
250
184
144
M 1 2 3
(a) (b) (c)
Figure 11 (a) Coomassie-stained SDS-PAGE of recombinant P pastoris clone 1mL supernatants collected from the third-day culturewere concentrated 10-fold using trichloroacetic acid and 10120583L resultant products were applied to SDS-PAGE analysis Lane 1 molecularweight marker (Fermentas) Lane 2 GS115 (Control) Lane 3 GS115pPIC9 empty (Control) Lane 4 GS115pPIC9PI12prot (GpPro2)The arrow indicates the secreted recombinant PI12 protease (b) Plate activity staining of different recombinant P pastoris clones GGS115 GP GS115pPIC9 GRI GS115pPIC9PI12prot1 (GpPro1) GRII GS115pPIC9PI12prot2 (GpPro2) K KM71 KP KM71pPIC9 KRKM71pPIC9PI12prot (KpPro) (c)Gel activity staining of recombinant PI12 protease inP pastorisGpPro2 clone Lane 1 prestainedmolecularweightmarker (Fermentas) Lane 2 GS115pPIC9 empty (Control) Lane 3 GS115pPIC9PI12prot1 (GpPro2)The arrow indicates the secretedrecombinant PI12 protease
dropped after 28 hours of incubation which is at stationaryor death phase [38]
38 Detection of Recombinant Protein by SDS-PAGE andActivity Staining TheSDS-PAGE gel of the GpPro2 recombi-nant clonesrsquo supernatant showed a putative expression bandat around 993 kDa Plate staining and gel activity stainingwere executed to examine if the recombinant PI12 proteaseexpressed in P pastoris was functionally active In the platestaining the transformants showed strong proteolytic activity(Figure 11(a)) In gel staining proteolytic activity was alsodetected around 993 kDa after incubation at 20∘C This is
evidence of protease secretion into the surrounding mediawhereby it catalyzed the breakdown of casein thus formedclearing zones around the activity ring and band (Figures11(b) and 11(c)) As a conclusion the cold-adapted PI12protease was functionally expressed in P pastoris GS115(GpPro2 clone) with the size of 993 kDa
4 Conclusions
Psychrophilic yeast which was originated frommarine waterin Casey Station Antarctica was identified as Glaciozymaantarctica strain PI12 A serine protease with an open reading
Enzyme Research 19
frame of 2892 bp encoded for 963 amino acids from thisAntarctic yeast has been cloned and expressed in Pichiaexpression system This is the second extracellular pro-tease reported being synthesized by this psychrophilic yeastSequence analysis through bioinformatics studies revealedfew homology identities of this protein with other ser-ine proteases The mature PI12 protease was successfullyexpressed and secreted into the culture medium driven bythe Saccharomyces cerevisiae 120572-factor signal sequence in Ppastoris The protease production in P pastoris was bestobtained from strain GS115 (GpPro2) with 283UmL after 72hours of induction time with 05 (vv) of methanol inducerThe starting optical density at 600 nm (OD600) was 60 andthe optimum assay temperature was 20∘C The expressedprotein was detected by SDS-PAGE and activity staining witha molecular weight of 99 kDaThis cold-active enzymemightoffer an opportunity for biotechnological exploitation basedon its high catalytic activity at low temperatures and lowthermostability Structural and functional study of this cold-adapted enzyme would give insight into its adaptation to anextreme cold environment
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgment
This research was supported financially by the InitiativeBiotechnology-GMBI-MOSTI (Project number 07-05-16-MG-GMB02) andRUGS-Initiative 6 (Project number 05-02-11-1414RU)
References
[1] J W Deming ldquoPsychrophiles and polar regionsrdquo CurrentOpinion in Microbiology vol 5 no 3 pp 301ndash309 2002
[2] C Gerday M Aittaleb M Bentahir et al ldquoCold-adaptedenzymes from fundamentals to biotechnologyrdquo Trends inBiotechnology vol 18 no 3 pp 103ndash107 2000
[3] A Svingor J Kardos I Hajdu A Nemeth and P Zavodszky ldquoAbetter enzyme to cope with cold comparative flexibility studieson psychrotrophic mesophilic and thermophilic IPMDHSrdquoThe Journal of Biological Chemistry vol 276 no 30 pp 28121ndash28125 2001
[4] G Antranikian C E Vorgias and C Bertoldo ldquoExtremeenvironments as a resource for microorganisms and novel bio-catalystsrdquo Advances in Biochemical EngineeringBiotechnologyvol 96 pp 219ndash262 2005
[5] T Lonhienne C Gerday andG Feller ldquoPsychrophilic enzymesrevisiting the thermodynamic parameters of activation mayexplain local flexibilityrdquoBiochimica et Biophysica Acta vol 1543no 1 pp 1ndash10 2000
[6] R Cavicchioli and TThomas ldquoExtremophilesrdquo in Encyclopediaof Microbiology pp 317ndash337 Academic Press San Diego CalifUSA 2000
[7] G Feller E Narinx J L Arpigny et al ldquoEnzymes frompsychrophilic organismsrdquo FEMS Microbiology Reviews vol 18no 2-3 pp 189ndash202 1996
[8] J Arnorsdottir R B Smaradottir T O Magnusson S HThorbjarnardottir G Eggertsson and M M KristjanssonldquoCharacterization of a cloned subtilisin-like serine proteinasefrom a psychrotrophic Vibrio speciesrdquo European Journal ofBiochemistry vol 269 no 22 pp 5536ndash5546 2002
[9] N J Russell ldquoPsychrophilic bacteriamdashmolecular adaptations ofmembrane lipidsrdquo Comparative Biochemistry and Physiology APhysiology vol 118 no 3 pp 489ndash493 1997
[10] G Feller and C Gerday ldquoPsychrophilic enzymes hot topics incold adaptationrdquo Nature Reviews Microbiology vol 1 no 3 pp200ndash208 2003
[11] P Raspor and J Zupan ldquoYeasts in extreme environmentsrdquo inBiodiversity and Ecophysiology of Yeasts The Yeast Handbookpp 371ndash417 Springer Berlin Germany 2006
[12] M B Rao A M Tanksale M S Ghatge and V V Desh-pande ldquoMolecular and biotechnological aspects of microbialproteasesrdquoMicrobiology andMolecular Biology Reviews vol 62no 3 pp 597ndash635 1998
[13] L Polgar ldquoThe catalytic triad of serine peptidasesrdquo Cellular andMolecular Life Sciences vol 62 no 19-20 pp 2161ndash2172 2005
[14] S Y Boo C M V L Wong K F Rodrigues N NajimudinA M A Murad and N M Mahadi ldquoThermal stress responsesin Antarctic yeastGlaciozyma antarctica PI12 characterized byreal-time quantitative PCRrdquoPolar Biology vol 36 no 3 pp 381ndash389 2013
[15] B Turchetti S R T Hall L B Connell et al ldquoPsychrophilicyeasts from Antarctica and European glaciers descriptionof Glaciozyma gen nov Glaciozyma martinii sp nov andGlaciozyma watsonii sp novrdquo Extremophiles vol 15 no 5 pp573ndash586 2011
[16] N H F Hashim I Bharudin D L S Nguong et al ldquoCharac-terization of Afp1 an antifreeze protein from the psychrophilicyeast Glaciozyma antarctica PI12rdquo Extremophiles vol 17 no 1pp 63ndash73 2013
[17] A N M Ramli M A Azhar M S Shamsir et al ldquoSequenceand structural investigation of a novel psychrophilic 120572-amylasefrom Glaciozyma antarctica PI12 for cold-adaptation analysisrdquoJournal ofMolecularModeling vol 19 no 8 pp 3369ndash3383 2013
[18] A N M Ramli N M Mahadi A Rabu A M A Murad F DA Bakar and R M Illias ldquoMolecular cloning expression andbiochemical characterisation of a cold-adapted novel recombi-nant chitinase from Glaciozyma antarctica PI12rdquoMicrobial CellFactories vol 10 no 1 article 94 2011
[19] J Sambrook E Fritsch and T Maniatis Molecular Cloning ALaboratoryManual Cold SpringHarbor Laboratory Press NewYork NY USA 1989
[20] F M Brock C W Forsberg and J G Buchanan-SmithldquoProteolytic activity of rumen microorganisms and effects ofproteinase inhibitorsrdquoApplied and EnvironmentalMicrobiologyvol 44 no 3 pp 561ndash569 1982
[21] U K Laemmli ldquoCleavage of structural proteins during theassembly of the head of bacteriophage T4rdquo Nature vol 227 no5259 pp 680ndash685 1970
[22] A Henne R A Schmitz M Bomeke G Gottschalk and RDaniel ldquoScreening of environmental DNA libraries for thepresence of genes conferring lipolytic activity on Escherichiacolirdquo Applied and Environmental Microbiology vol 66 no 7 pp3113ndash3116 2000
[23] M Poza T de Miguel C Sieiro and T G Villa ldquoCharacteri-zation of a broad pH range protease of Candida caseinolyticardquoJournal of Applied Microbiology vol 91 no 5 pp 916ndash921 2001
20 Enzyme Research
[24] J D Bendtsen H Nielsen G von Heijne and S BrunakldquoImproved prediction of signal peptides SignalP 30rdquo Journalof Molecular Biology vol 340 no 4 pp 783ndash795 2004
[25] R J Siezen and J A M Leunissen ldquoSubtilases the superfamilyof subtilisin-like serine proteasesrdquo Protein Science vol 6 no 3pp 501ndash523 1997
[26] Y Katakura W Zhang G Zhuang et al ldquoEffect of methanolconcentration on the production of human 1205732- glycoprotein Idomain V by a recombinant Pichia pastoris a simple system forthe control of methanol concentration using a semiconductorgas sensorrdquo Journal of Fermentation and Bioengineering vol 86no 5 pp 482ndash487 1998
[27] Q-FWang Y-HHou Z Xu J-LMiao andG-Y Li ldquoPurifica-tion and properties of an extracellular cold-active protease fromthe psychrophilic bacterium Pseudoalteromonas sp NJ276rdquoBiochemical Engineering Journal vol 38 no 3 pp 362ndash3682008
[28] S Vazquez L Ruberto and W Mac Cormack ldquoProper-ties of extracellular proteases from three psychrotolerantStenotrophomonas maltophilia isolated from Antarctic soilrdquoPolar Biology vol 28 no 4 pp 319ndash325 2005
[29] R Zeng R Zhang J Zhao and N Lin ldquoCold-active ser-ine alkaline protease from the psychrophilic bacterium Pseu-domonas strain DY-A enzyme purification and characteriza-tionrdquo Extremophiles vol 7 no 4 pp 335ndash337 2003
[30] M Turkiewicz E Gromek H Kalinowska and M ZielinskaldquoBiosynthesis and properties of an extracellularmetalloproteasefrom the Antarctic marine bacterium Sphingomonas paucimo-bilisrdquo Journal of Biotechnology vol 70 no 1ndash3 pp 53ndash60 1999
[31] A Vassileva D A Chugh S Swaminathan and N KhannaldquoEffect of copy number on the expression levels of hepatitisB surface antigen in the methylotrophic yeast Pichia pastorisrdquoProtein Expression andPurification vol 21 no 1 pp 71ndash80 2001
[32] M Pazgier M Turkiewicz H Kalinowska and S Bielecki ldquoTheunique cold-adapted extracellular subtilase from psychrophilicyeast Leucosporidium antarcticumrdquo Journal ofMolecular Catal-ysis B Enzymatic vol 21 no 1-2 pp 39ndash42 2003
[33] M Turkiewicz M Pazgier H Kalinowska and S BieleckildquoA cold-adapted extracellular serine proteinase of the yeastLeucosporidium antarcticumrdquo Extremophiles vol 7 no 6 pp435ndash442 2003
[34] D Mattanovich B Gasser H Hohenblum and M SauerldquoStress in recombinant protein producing yeastsrdquo Journal ofBiotechnology vol 113 no 1ndash3 pp 121ndash135 2004
[35] P Secades BAlvarez and JAGuijarro ldquoPurification andprop-erties of a new psychrophilic metalloprotease (Fpp2) in the fishpathogen Flavobacterium psychrophilumrdquo FEMS MicrobiologyLetters vol 226 no 2 pp 273ndash279 2003
[36] L Y Ling I Ithoi and F M Yik ldquoOptimization for high-level expression in Pichia pastoris and purification of truncatedand full length recombinant sag2 of toxoplasma gondii fordiagnostic userdquo Southeast Asian Journal of Tropical Medicineand Public Health vol 41 no 3 pp 507ndash513 2010
[37] A Illanes ldquoStability of biocatalystsrdquo Electronic Journal ofBiotechnology vol 2 no 1 pp 1ndash9 1999
[38] B A Plantz J Sinha L Villarete K W Nickerson and VL Schlegel ldquoPichia pastoris fermentation optimization energystate and testing a growth-associated modelrdquo Applied Microbi-ology and Biotechnology vol 72 no 2 pp 297ndash305 2006
[39] K Tamura G Stecher D Peterson A Filipski and S KumarldquoMEGA6molecular evolutionary genetics analysis version 60rdquo
Molecular Biology and Evolution vol 30 no 12 pp 2725ndash27292013
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology
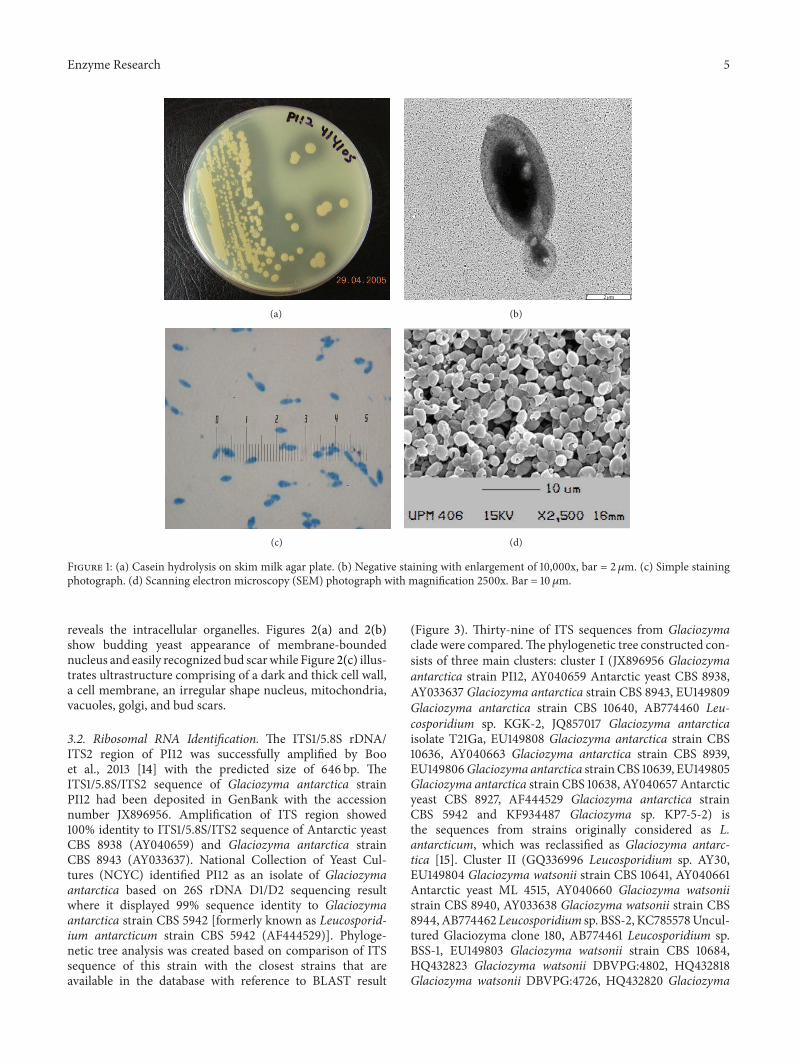
Enzyme Research 5
(a)
2120583m
(b)
(c) (d)
Figure 1 (a) Casein hydrolysis on skim milk agar plate (b) Negative staining with enlargement of 10000x bar = 2120583m (c) Simple stainingphotograph (d) Scanning electron microscopy (SEM) photograph with magnification 2500x Bar = 10 120583m
reveals the intracellular organelles Figures 2(a) and 2(b)show budding yeast appearance of membrane-boundednucleus and easily recognized bud scarwhile Figure 2(c) illus-trates ultrastructure comprising of a dark and thick cell walla cell membrane an irregular shape nucleus mitochondriavacuoles golgi and bud scars
32 Ribosomal RNA Identification The ITS158S rDNAITS2 region of PI12 was successfully amplified by Booet al 2013 [14] with the predicted size of 646 bp TheITS158SITS2 sequence of Glaciozyma antarctica strainPI12 had been deposited in GenBank with the accessionnumber JX896956 Amplification of ITS region showed100 identity to ITS158SITS2 sequence of Antarctic yeastCBS 8938 (AY040659) and Glaciozyma antarctica strainCBS 8943 (AY033637) National Collection of Yeast Cul-tures (NCYC) identified PI12 as an isolate of Glaciozymaantarctica based on 26S rDNA D1D2 sequencing resultwhere it displayed 99 sequence identity to Glaciozymaantarctica strain CBS 5942 [formerly known as Leucosporid-ium antarcticum strain CBS 5942 (AF444529)] Phyloge-netic tree analysis was created based on comparison of ITSsequence of this strain with the closest strains that areavailable in the database with reference to BLAST result
(Figure 3) Thirty-nine of ITS sequences from Glaciozymaclade were comparedThe phylogenetic tree constructed con-sists of three main clusters cluster I (JX896956 Glaciozymaantarctica strain PI12 AY040659 Antarctic yeast CBS 8938AY033637 Glaciozyma antarctica strain CBS 8943 EU149809Glaciozyma antarctica strain CBS 10640 AB774460 Leu-cosporidium sp KGK-2 JQ857017 Glaciozyma antarcticaisolate T21Ga EU149808 Glaciozyma antarctica strain CBS10636 AY040663 Glaciozyma antarctica strain CBS 8939EU149806Glaciozyma antarctica strainCBS 10639 EU149805Glaciozyma antarctica strain CBS 10638 AY040657 Antarcticyeast CBS 8927 AF444529 Glaciozyma antarctica strainCBS 5942 and KF934487 Glaciozyma sp KP7-5-2) isthe sequences from strains originally considered as Lantarcticum which was reclassified as Glaciozyma antarc-tica [15] Cluster II (GQ336996 Leucosporidium sp AY30EU149804 Glaciozyma watsonii strain CBS 10641 AY040661Antarctic yeast ML 4515 AY040660 Glaciozyma watsoniistrain CBS 8940 AY033638 Glaciozyma watsonii strain CBS8944 AB774462 Leucosporidium sp BSS-2 KC785578Uncul-tured Glaciozyma clone 180 AB774461 Leucosporidium spBSS-1 EU149803 Glaciozyma watsonii strain CBS 10684HQ432823 Glaciozyma watsonii DBVPG4802 HQ432818Glaciozyma watsonii DBVPG4726 HQ432820 Glaciozyma
6 Enzyme Research
2120583m
(a)
2120583m
(b)
BS
V
N
CW
CM
GM
Length 314866nm
Length 226635nm
1120583m
(c)
Figure 2 Transmission electronmicroscopy (TEM) photographs of AntarcticG antarctica strain PI12 (a) 12500xmagnification (b) 10000xmagnification (c) 27500x magnificationThe scale bars represent 2 120583m for (a) and (b) and 1 120583m for (c)
watsonii DBVPG4760 JX171176 Glaciozyma watsonii iso-late LKF08-112 DQ402535 Leucosporidium sp LC-03-120isolate 03-120 JF900360 Glaciozyma watsonii strain CBS7009 KC333171 Glaciozyma watsonii isolate AU CryS06HQ432821 Glaciozyma watsoniiDBVPG4799) is categorizedas G watsonii while cluster III (AY040664 Glaciozyma mar-tinii strain CBS 8929 AY033641 Glaciozyma martinii strain
CBS 8928 HQ432816 Glaciozyma martinii DBVPG4841KF934486 Glaciozyma sp KP7-5-1 FR682434 UnculturedBasidiomycota HF934010 Glaciozyma sp K81b JF900362Glaciozyma martinii strain CBS 9639 JF900359 Glaciozymamartinii strain CBS 6581 JF900361 Glaciozyma martiniistrain CBS 7054) is categorized as G martini Recognitionof the Glaciozyma species has been done by comparing
Enzyme Research 7
JX896956 Glaciozyma antarctica strain PI12
AY040659 Antarctic yeast CBS 8938
AY033637 Glaciozyma antarctica strain CBS 8943
EU149809 Glaciozyma antarctica strain CBS 10640
AB774460 Leucosporidium sp KGK-2
JQ857017 Glaciozyma antarctica isolate T21Ga
EU149808 Glaciozyma antarctica strain CBS 10636
AY040663 Glaciozyma antarctica strain CBS 8939
EU149806 Glaciozyma antarctica strain CBS 10639
EU149805 Glaciozyma antarctica strain CBS 10638
AY040657 Antarctic yeast CBS 8927
AF444529 Glaciozyma antarctica strain CBS 5942
KF934487 Glaciozyma sp KP7-5-2
GQ336996 Leucosporidium sp AY30
EU149804 Glaciozyma watsonii strain CBS 10641
AY040661 Antarctic yeast ML 4515
AY040660 Glaciozyma watsonii strain CBS 8940
AY033638 Glaciozyma watsonii strain CBS 8944
EU149803 Glaciozyma watsonii strain CBS 10684
JX171176 Glaciozyma watsonii isolate LKF08-112
DQ402535 Leucosporidium sp LC-03-120 isolate 03-120
AB774462 Leucosporidium sp BSS-2
KC785578 uncultured Glaciozyma clone 180
HQ432818 Glaciozyma watsonii DBVPG4726
HQ432823 Glaciozyma watsonii DBVPG4802
AB774461 Leucosporidium sp BSS-1
HQ432820 Glaciozyma watsonii DBVPG4760
KC333171 Glaciozyma watsonii isolate AU CryS06
HQ432821 Glaciozyma watsonii DBVPG4799
JF900360 Glaciozyma watsonii strain CBS 7009
AY040664 Glaciozyma martinii strain CBS 8929
AY033641 Glaciozyma martinii strain CBS 8928
HQ432816 Glaciozyma martinii DBVPG4841
KF934486 Glaciozyma sp KP7-5-1
FR682434 uncultured Basidiomycota
HF934010 Glaciozyma sp K81b
JF900362 Glaciozyma martinii strain CBS 9639
JF900359 Glaciozyma martinii strain CBS 6581
JF900361 Glaciozyma martinii strain CBS 7054
001
Clus
ter 1
Glac
iozy
ma
anta
rctic
aCl
uste
r 3
Glac
iozy
ma
mar
tinii
Clus
ter 2
Glac
iozy
ma
watso
nii
Figure 3 Neighbor-joining phylogenetic analysis of internal transcribed spacer 1 (ITS1)58S rRNA geneinternal transcribed spacer 2 (ITS2)of Glaciozyma clade Numbers indicate percentage bootstrap values calculated on 1000 repeats of the alignment The tree was constructedusingMEGA6 software [39]The scale bar represents 001 substitutions per nucleotide position GenBank accession numbers of the sequencesare indicated before strain name ITS158SITS2 sequence of Glaciozyma antarctica strain PI12 had been deposited in GenBank with theaccession number JX896956 [14]
8 Enzyme Research
their molecular phylogenetic data and physiological andmorphological characteristics [15] Phylogenetic analysis ofITS158SITS2 sequence of strain PI12 showed that thesequence was closely related to ITS158SITS2 sequenceof Glaciozyma antarctica with high homology similarity toCluster I (99-100 identity) The ITS158SITS2 sequenceof G antarctica strain PI12 was differed from G watsonii(Cluster II) by 2-3 gap in the sequence (93 identity)and G martinii by 4-5 gap in the sequence (90ndash92identity)
33 Sequence Analysis of PI12 Protease cDNA In this studyamplification of the PI12 protease gene sequence was con-ducted using genomicDNAand cDNAas a template Putativepartial PI12 protease gene with the size of 1321 bp was initiallyacquired from the G antarctica genome survey sequencelibrary (Supplementary Figure see Supplementary Mate-rial available online at httpdxdoiorg1011552014197938)However the gene was discovered as an incomplete pro-tease gene sequence DNA walking strategy successfullyamplified 1500 bp of the upstream region of partial PI12protease Both sequences of about 1500 bp (derived fromthe DNA walking reaction) and 1321 bp (partial PI12 pro-tease) were assembled and made up a total length of2687 bp protease sequence (Supplementary figure)The puta-tive PI12 protease was predicted by Augustus webserver(httpaugustusgobicsde) having intervening sequencesintronsThus reverse-transcriptase PCR (RT-PCR) and rapidamplification of cDNA ends (RACE) were accomplished toobtain the full-length cDNA As a result the ORF comprising2892 bp encoding 963 amino acid residues were successfullydeduced
Nucleotide and deduced amino acid sequences ofgenomic DNA and cDNA fragment encoding PI12 proteasewere shown in Figure 4 The size of the ORF of putative PI12protease was 3867 bp with genomic DNA as the templatewhile the ORF comprised 2892 bp encoding 963 amino acidresidues were successfully deduced through RACE strategywith cDNA as the template The ORF was predicted usingORF Finder (httpwwwncbinlmnihgovprojectsgorf)while prediction using SignalP (httpwwwcbsdtudkser-vicesSignalP) webserver allowed the identification of theputative PI12 protease prepeptide where it predicted thepresence of signal peptidase I cleavage sites [24]The putativeN-terminal signal peptide (1 to 16 the underlined amino acid)was identified as a secretion signal There is no propeptidepredicted by ProP Webserver (httpwwwcbsdtudkser-vicesProP-10) Comparison of cDNA and gene sequencesallowed determination of gene structureTherefore sequencealignment was performed between the sequence of these twotemplates and it was confirmed that the coding sequence(consisted of 19 exons) was interrupted by 18 introns withtheir length varying from 48 to 60 nucleotides However theGT and AG rule for donor-acceptor sites was not obeyedOf 18 introns 10 introns contained canonical GT and AGdinucleotides at their 51015840 and 31015840 ends while 8 introns holdnoncanonical splice site pairs with TT-GG (2 introns)AG-TG GC-GT TG-GG TC-GG TA-GG and TG-AG
dinucleotides at the splice junction Upper case letters inblue colour represent introns while lower case letters inblack colour represent exons Splice donor and acceptor werehighlighted in grey
The amino acid composition (upper case letters) withinthe ORF amino acid residues was determined by theProtParam tool of the Expasy molecular biology server(httpwwwexpasyorgtool) Translation starts at a nucle-otide position 1 and translation stop is marked with anasterisk The nucleotide sequences ATG (1 to 3) and TAG(3865 to 3867) indicate the initiation codon and termi-nal codon respectively The presumed putative sequencesof promoter CAAT (minus101 to minus98) and TATA-box (minus92to minus88) were identified in the 51015840-untranslated (UTR)region The search was achieved via Neural Network Pro-moter Prediction (Berkeley Drosophila Genome Project)(httpwwwfruitflyorgseq toolspromoterhtml) In the 31015840-untranslated (UTR) region there was a putative polyadeny-lation signal attaa (3889 to 3893) detected from the TAGstop codon to the poly (A) tail The predicted molecularmass and pI of PI12 serine protease were 10099 kDa and641 respectively Out of 963 amino acid residues in thededuced sequence of the PI12 protease 70 are negativelycharged residues (Asp + Glu) while 66 are positively chargedresidues (Arg + Lys) at neutral pH (Table 2)The total chargedresidues (Arg Asp Glu His and Lys) are 150 amino acidsin the open reading frame of PI12 protease corresponded to156 while hydrophobic residues (Ala Phe Ile Leu MetPro Val and Trp) are 441 amino acids which correspondedto 458 of the total amino acids discovered The totaluncharged residues (Asp Cys Gln Gly SerThr and Tyr) areconsisted of 372 amino acids which corresponded to 386of the total amino acids of the open reading frame of PI12protease The aliphatic index is 9290 and the grand averageof hydropathicity (GRAVY) is 0141 The instability index (II)of PI12 protease was computed to be 3414 which classify theprotein as stable
N-glycosylation sites were predicted using NetNGlyc10 Server (httpwwwcbsdtudkservicesNetNGlyc)Therewere five putative N-glycosylation sites for the PI12 proteasegene predicted using the server For N-glycosylation thesequence motif Asn-Xaa-SerThr (Xaa any amino acid butnot Pro) has been defined as a prerequisite for glycosylationThe sequencemotifs in PI12 protease occurred in the position170 NETH 365 NATV 576 NLTP 634 NDTA and 833 NSSPAmong five predicted sites only three (except positions 576and 833) showed a high potential score (averaged output ofnine neural networks) which support the prediction Theputative N-glycosylation sites were demonstrated in boxesin Figure 4 The comparison of amino acid composition ofthis cold-adapted protease fromG antarctica strain PI12 withnine other subtilisin serine proteases fromdifferent families isshown in Table 2 Analysis of this comparison depicts that thepercentage of each amino acids in PI12 protease is almost thesamewith other proteaseswhileGly (G) is themost abundantaccounting for up to 104 of the total amino acid in PI12 pro-tease PI12 protease also contained the highest percentage ofLeu (L) with 99 of the total amino acids compared to other
Enzyme Research 9
Table2Aminoacid
compo
sitionandmolecular
weighto
fproteases
acrossdifferent
families
Amino
CAQ76821
CAG25549
EDR11679
CAG38357
CAD118
98CA
L25578
EDU50747
AAW
40780
ACB3
0121
ACB3
0119
acid
Qty
Qty
Qty
Qty
Qty
Qty
Qty
Qty
Qty
Qty
Ala
(A)
97101
8191
7887
7590
8299
101
115
8189
9196
8092
105
116
Arg
(R)
4042
2730
2224
1619
1721
1821
2932
2223
2124
3134
Asn
(N)
4446
3843
4348
4149
4251
4046
5257
5154
4552
4347
Asp
(D)
4951
6269
5460
6578
5567
5158
5358
6669
5260
6370
Cys
(C)
808
506
404
405
911
809
910
909
708
1011
Gln
(Q)
2324
3438
2326
2834
3036
3742
3539
2931
1922
2629
Glu
(E)
2122
1618
1921
2024
1518
2023
2528
2627
2832
2426
Gly
(G)
100
104
89100
8493
85102
85103
7788
8493
8084
8598
8695
His
(H)
1415
1416
1314
1619
1316
1113
1618
1415
1618
1213
Ile(I)
5355
4955
5157
5364
4150
3843
5156
4446
3540
4449
Leu
(L)
9599
6876
7179
6274
6073
6675
6774
8084
7485
6875
Lys
(K)
2627
2932
4449
2935
4959
5158
4044
3032
5058
5662
Met
(M)
1212
202
202
911
1113
809
1820
808
1113
1517
Phe
(F)
4143
5966
4752
5566
2935
3641
4449
3941
3439
3438
Pro
(P)
6264
6573
6774
5870
5870
5967
5055
5154
5564
5763
ser
(S)
7780
6977
91101
6072
6680
7080
7279
111
117
6373
6167
Thr
(T)
8790
89100
7583
83100
6579
7383
7178
7276
7182
6875
Trp
(W)
505
303
607
202
1215
1214
1213
909
1214
1415
Tyr
(Y)
3334
1213
2629
708
2935
2832
3943
5053
2630
2730
val
(V)
7679
8292
8089
6679
5769
7484
5864
6872
8295
6268
Total
963
1000
893
1000
900
1000
834
1000
825
1000
878
1000
906
1000
950
1000
866
1000
906
1000
MW
(Da)
1009898
932669
944365
869455
869549
924905
979343
1009629
917209
961988
Thea
ccessio
nnu
mbersof
proteasespu
tativ
eserinep
roteaseo
fGlacio
zymaantarctica
PI12
(CAQ
76821)sub
tilisin-lik
eproteaseP
leurotuso
streatus(CA
G25549)pyrolysin
LaccariabicolorS
238N
-H82
(EDR11679)
subtilisin
-like
proteasePh
anerochaetechrysosporiu
m(C
AG38357)sub
tilisin-lik
eserinep
roteaseP
R1CMetarhiziu
maniso
pliaev
aranisopliae(CA
D11898)serinee
ndop
eptid
aseH
ypocrealixii(C
AL2
5578)peptidase
Pyrenophoratritici-repentis
(EDU50747)p
eptid
aseCr
yptococcus
neoforman
svarn
eoform
ansJEC
21(A
AW40
780)sub
tilisin-lik
eprotease
Epich
loefestucae(A
CB30121)sub
tilisin-lik
eprotease
Epich
loefestucae
(ACB
30119
)
10 Enzyme Research
atgctcttcctccccgtcctcctcctcctccttcccggcgtcactgccttcctcaaccca 60
M L F L P V L L L L L P G V T A F L N Pgttaccaaccgcgcgaccaacgccatctcctcgactcaatacctctctaacgcctatatc 120
V T N R A T N A I S S T Q Y L S N A Y IctcgaattggacctctccacccccggcctcgtcaaacgggatagcacgcccgactctGTG 180
L E L D L S T P G L V K R D S T P D SCGTCCCGCGTTCCTCCCTCGCTGTCGAAGACGAGCTAACAATGACGTGGTAGatcctaga 240
I L Eggacgtactcacgtccgtaggccgcaacggtatcaagtaccaactccgccaccgctttat 300
D V L T S V G R N G I K Y Q L R H R F Ictccccgactctgttccacggcgcttcgatcactgtcccccctggaatctcccgctccca 360
S P T L F H G A S I T V P P G I S R S QaatcgcctctctccgcggtatcaaaGTCCGTCCCTCTCTCTCCCCTACTCCCTCCGTGCT 420
I A S L R G I KAACCCTGCACGCAACAGcgcgtctggcccgttcgaaagttctcccgacccagcgcagtag 480
R V W P V R K F S R P S A Vtggacgccgatggaggaggaagcgggttctcagggtcgcctatcaaggcggcgctcatgg 540
V D A D G G G S G F S G S P I K A A L Mgggtgaaagagctcgggaagcgcgcgaacgcttatgctggagatacgtttggaccgcatg 600
G V K E L G K R A N A Y A G D T F G P HtcatgacgggggttaatgagacgcatgaggcggggttgttgggagctgggattaagGTGT 660
V M T G V N E T H E A G L L G A G I KGTTTTCGTCTTTGTTTGGGGGAGGGGGGAGGGAGGAGCTGATTAATGGGTGCAGattggg 720
I GgtgctggacactggtgAGAAGCGGTTTGGAGGGAGGAGAGGATGGAGCTGATGAGTGTGC 780
V L D T GAGGTGttgattatttgaacccgattctgggaggctgctttggacctgggtgccatatgtc 840
V D Y L N P I L G G C F G P G C H M Sgtttgggtacgacttggttggcgatgattacgatggagataacgctcctgtgccggatgt 900
F G Y D L V G D D Y D G D N A P V P D VggatccttGTGCGTCCTTCCCTCCGCGATGGGCTGGAATCTGAGCTGACGGCGTTGTTGA 960
D PTCAGttgcgagctgcgatcctcatggTTCGTCTCGCTTTCCGCTCCTCTGATGCGCTCGC 1020
F A S C D P H GTGATGCACTTGGCAATAGGaactcacgttacgggaatcattggagcgctcccgaatgcgt 1080
T H V T G I I G A L P N AttggatttactggcgtcgcacccgccgctactctgggccactaccgagtGCGCTCCCTCT 1140
F G F T G V A P A A T L G H Y R VATCCGTTCTTCCTCGCAGCTCCCACTCACTCGTGCTTCCAGGTatttggctgcactggct 1200
F G C T GtcgtcggagaagatatcGTTCGTCCTCTCTCCAGTCCCGGCTAGCGATCCTGCTTCTGAC 1260
F V G E D ITTCGCATCTCCCTCTGAGattctcgctggactcatgcgaggagtcgaggacaactgcaac 1320
I L A G L M R G V E D N C Ngtcttgaccctctctctcggaggtccaggagggtgggtcaagggcacgccggcgtccatc 1380
V L T L S L G G P G G W V K G T P A S Icttatcgaccagatcgaagcgcaaggcattctcgtcaccgtcgccactggcaactcggga 1440
L I D Q I E A Q G I L V T V A T G N S GgctgagggTGAGCTCCTCGCTACTTGCGCGTCGAGACCAGTACTAACAGGGGCGTGGACG 1500
A E GCAGGcatgttcttctccgagtctcccgcctcgacgatcaacggcctttccatcgcatcca 1560
M F F S E S P A S T I N G L S I A ScgTCCGTCTCTCCCTTCCTTCGCACGCCACTCGCTCACACTCTCTCGCAGGgacgttacc 1620
T D V Tgacctcatcgcctacaacgccaccgtctcaggccaacctgcgatcccttacctctccgcc 1680
D L I A Y N A T V S G Q P A I P Y L S A
ggggggggggtctg minus194
ctagcgcgaatcgagaaaacacaaaagaacctcccgacttggcgttgcttctgctcttct minus180
ccttctcccaaccgccgacaataaaaactataacacacacacaccccatctcgacgcctg minus120
cagacttcacgactctcctcacgctctttaccgccccacgggtcccacccacgagccacg minus60
(a)
Figure 4 Continued
Enzyme Research 11
acgcccctcaacgtcgtcgccaacagcttccgcgtccacttcacctctaccgaccccaac 1740
T P L N V V A N S F R V H F T S T D P Naaccccgtcgacgcctgctctcctcttccggctggagcgcccgacttcgccaactatgtt 1800
N P V D A C S P L P A G A P D F A N Y Vacggtcgttcagcgtgggacttgtacgttcgttaccaagtaccagaacgttctcaatgct 1860
T V V Q R G T C T F V T K Y Q N V L N AggaggGTGCGTCGGCTTCTTCTCTCCCGACTGGATTGGGCGGAGACTGACCTTCTGAGCA 1920
G GGaaagatcgtattgttgtacaactcggagggagctgggaacctcccttacctcacgccca 1980
K I V L L Y N S E G A G N L P Y L T PacggtgtcggcatcgacgccgttgcaggtcttcgtcgttccgacggactcaagGTTCGTC 2040
N G V G I D A V A G L R R S D G L KTACGCTCTTCGGTGCTGAGGCCCTTCTCTGATCTCGCTCCGACAGcttctctcgtactat 2100
L L S Y Ycagaatgccaacaagcgtctcactctgcgcttccccaagggcaagatcgtcgcaggcttg 2160
Q N A N K R L T L R F P K G K I V A G LaccgataccatcaccggcggactcatctcgTTCGTTCCTCTCCGCCTCCCTCGCTGGCCG 2220
T D T I T G G L I SATGAGCTGACGCGGTCTTTGCAGGggttactcgacgtttggtccgacgaatgacctctac 2280
G Y S T F G P T N D L Yggtcagcctaccctctctgcccctggtggcaacatcctttcgaccttccctctctccgag 2340
G Q P T L S A P G G N I L S T F P L S EggaggagtggcggtcatcagtgggacgagcatgtcgtgcccctttgtcgGTTCGTCCTTC 2400
G G V A V I S G T S M S C P F VTCCTCGTTCGTTCGCTGAGCGTCGACTGATCTTGCACGATCGCAGctggatctgcggcgg 2460
A G S A Atcctcatggccgctcgcgcttcggagaacctcacgccgcttgagatcaggagtctcctta 2520
V L M A A R A S E N L T P L E I R S L Lctaccactgcgaagcttacgccggtctcgctcttgggatcgacgcctttggtgagcgtga 2580
T T T A K L T P V S L L G S T P L V S VttcgtcaaggaggaggTACGTTCCTTTCTCGCTGCTTTGAGGATCGAGGCTGATTAGCGA 2640
I R Q G G GGCTTCGCAGGactcgttcaggttgccaaggcgctcgcggccaagacgctaatctctcctc 2700
L V Q V A K A L A A K T L I S Pacgagctcctgctcaacgacactgcgaacgcgaactacgtccagactatcaagatcaaga 2760
H E L L L N D T A N A N Y V Q T I K I Kacaccaactcgtgggcgatgaagtacaccttctcctcggccgtcgcccaaggactcggaa 2820
N T N S W A M K Y T F S S A V A Q G L Gctttcgacgcttcgggcgatatcctccctaccctcgacccagtcgccgtctctggcgcac 2880
T F D A S G D I L P T L D P V A V S G Aaggctaccgtcgcgttcaacactcggatcctcagcgtcgcgcccggcgcgacggggtccg 2940
Q A T V A F N T R I L S V A P G A T G Stcgtggcgactatcacgccgccggttcttcccgtagcggacgctgcgaggttccctatct 3000
V V A T I T P P V L P V A D A A R F P Itctctgggtggatcagggtgaatgggcaaggcgcgagggatagcagtaggaacgaggcgt 3060
F S G W I R V N G Q G A R D S S R N E AacactgGTGCGTTTGGCTCGCTTTTTGGGAGGTGCCGAGCTAATCGTTTCTCGTAGtccc 3120
Y T V PgtactttgggcttgcggcgaagatgatcgatatgcaagGTGCGTCGATGGTTCTCTCGCT 3180
Y F G L A A K M I D M QTCTCTCTCGGTTGCTCACTTCAAAGTGCAGtcctcgacaccaccgagaccatttacggtc 3240
V L D T T E T I Y Gcgggctacgcctaccccttcgtgatcgacgacgcgattggagacatccaatccaccacaa 3300
P G Y A Y P F V I D D A I G D I Q S T TcgtcgtactccaggaacctcgggcccaccgtcttcgctcgctttgccaTGCGTCGACCGT 3360
T S Y S R N L G P T V F A R F ACCTCATTTCCTGCGGCAGAGCGAGAGCTGATTCCGACCTCGAACAGctggaacccttcac 3420
T G T L Htacagcctcgatctcgtcctagccgacatcgcctttacccccacctaccccaactcctcc 3480
Y S L D L V L A D I A F T P T Y P N S S
(b)
Figure 4 Continued
12 Enzyme Research
cccgccactcgtttcgtcaagcgctccctcacgcagcacacctctgccgcttcgcacctc 3540
P A T R F V K R S L T Q H T S A A S H Lgccaagcgccgcgtctccatcgccactatcaaccccaaagccaccctcgtcgccgatcga 3600
A K R R V S I A T I N P K A T L V A D Rcagctccactcggacgtccctatcgagggcaacatcttcacccaaccctttactggaagg 3660
Q L H S D V P I E G N I F T Q P F T G Rgattacctcgtcgacgcagccccgacgggatccaccgatcgtaccgtcacttttaacggg 3720
D Y L V D A A P T G S T D R T V T F N Gcagtacgccgagaacggcctcgtgaggacggctgtgacggggacttcgtaccgcttcctc 3780
Q Y A E N G L V R T A V T G T S Y R F Lcttcgggcgttgaagatctcgggagacgcgatgtacgaggatcagtatgagagctggctc 3840
L R A L K I S G D A M Y E D Q Y E S W L3900
S L P F S F R Attttgccacctcaaaaaaaaaaaaaaaaaaaaaaaaaa 3938
lowast
tcgctaccgttctcgttccgtgcgtaggcttgttgtggacaccactgcattaagttatga
(c)
Figure 4 Nucleotide and deduced amino acid sequences of genomic DNA and cDNA encoding protease from G antarctica strain PI12 Thenucleotides are numbered from the 51015840 end of the cDNA A putative CAAT TATA box and polyadenylation signal are bold italicThe putativeN-terminal signal peptide is underlined The initiation codon is boldfaced and termination codon is represented by an asterisk (lowast) boxedpeptide sequences showed N-glycosylated asparagines residues The amino acid residues that form the catalytic triad are double underlinedand bold while introns are represented in blue font Highlighted in grey are the intron donor-acceptor splice sites
proteases Hydrophobic profile of PI12 amino acid proteasewas performed by ProtScale tools with Kyte and Doolittlemethod at httpauexpasyorgcgi-bin The hydrophobicityprofile elucidation of PI12 protease amino acid was carriedout from amino acid 1 to 963 The amino acid residuesfrom 1 to 16 (M-L-F-L-P-V-L-L-L-L-L-P-G-V-T-A) and 548 to562 (V-A-V-I-S) were hydrophobic grooves The first regionwhich consisted of 16 amino acids was very hydrophobic andwas predicted to be the signal peptide by SignalP and ProPwebserver
The bold and double-underlined amino acids showedresidues of catalytic triad aspartic acid (D) histidine (H) andserine (S) active site respectively NCBI Conserved DomainsDatabase (CDD) identified the presence of subtilase proteaseand subtilisin-like serine protease domains in the amino acidsequence The predicted PI12 protease amino acid sequencewas found sharing significant homology to the subtilisinfamily of protease The highest similarity of PI12 proteasewas observed with 42 identity to subtilisin-like proteaseof Rhodosporidium toruloides (EMS20793) 37 identity tosubtilisin-like protease of Pleurotus ostreatus (CAG25549)and 35 identity to subtilisin-like protease of Gloeophyllumtrabeum ATCC 11539 (EPQ55564) The nucleotide sequenceof G antarctica strain PI12 showed low homology to thefungus subtilisin protease family and none to other psy-chrophilic proteases The multiple sequence alignment wasillustrated in Figure 5 The consensus sequence around thecatalytic triad aspartic acid (D) histidine (H) and serine(S) of PI12 protease showed high conservation with othersubtilisin (showed in black boxes) Glycine residues G142G173 G191 G262 and G327 that were highly conserved inmost of subtilases were conserved in PI12 protease [25]Therefore PI12 protease belongs to the subtilisin subgroup ofthe subtilase serine protease superfamilyThis serine protease
of G antarctica PI12 has been deposited into GenBank withaccession number CAQ76821
34 Phylogenetic Relationship of PI12 Protease The phylo-genetic tree was constructed to demonstrate the similaritiesand relationship of protein sequence between differentspecies or organisms Figure 6 illustrates the phylogeneticdendrogram indicating the relationship of PI12 proteasewith another 25 translated amino acid sequences ofserine protease The bar scale signifies the branch lengthto approximate variance along each branch Basicallythis tree comprises two main clusters cluster I was sub-divided into subcluster a (CAG25549 EPQ55564 ETW-80907 XP 007381302 XP 001877576 CAG38357 XP007341385 CCA68627 EJU04161 XP 007369624 EIW-62675 CCO37249 XP 001830835 XP 007342189) and sub-cluster b (XP 007589863 XP 003711175 EFQ32840 XP006694395 ENH74885 EON99722) cluster II was sub-divided into subcluster c (CBQ73425 GAC97617 CCF52619)and subcluster d [CAQ76821 (PI12 protease) EMS20793]
Serine protease constructed by the phylogenetic treecomprised large size of protein with about 801 to 1032 aminoacids The majority of the translated protease sequenceswere obtained from fungus All serine proteases from cluster1 subcluster a are derived from Agaricomycetes except forEJU04161 which belongs to Dacrymycetes whereby all serineproteases from cluster 1 subcluster b are from Sordari-omycetes Cluster II subcluster c consisted of serine proteasefrom fungus belonging to Ustilaginomycetes PI12 proteasewas categorized under cluster II subcluster d It possessedhomology similarity with the subtilisin-like protease fromRhodosporidium toruloides (EMS20793) (formerly known asRhodotorula glutinis or Rhodotorula gracilis) which is a
Enzyme Research 13
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
MKG-YSWSLLQLAFTGAALATVPLSDVKRISSAS--VVPNKFVVEVESASEIPGKR--SL
MKGVLVWSILS---AGCALAVTPLSSVKRVTNLP--IVANKFIVEVDSTSDIPTKR--S-
MKLPLLSLALT---AVSVGATVPLSSVQRTTTSPNNALAGEYIVEMNNVQGLGGKR--AY
---MAHGSLASLLLLLAASAQGAAAFGGKSATGGRSVVPNSYILQVNASAPALTKRGVSV
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease ---MLFLPVLLLLLPGVTAFLNPVTNRATNAISSTQYLSNAYILELDLSTPGLVKRDSTP
lowastlowast
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
YTPHELFYRSLSARGVSYEVNKEYNS-EGLFVGAVVTVSDANDVKALAETPGVKAIRPVR
-NPHDALYESLRKRAIGFKVDREFNT-QGLFVGAALTLSDAQDVGQIQSTTGVKSIRPVR
ASTHDTLYRSLRKRGISFDVKKEYNQ-PGIMVGAALKVASPQDVQQIATMAEVKAIRPVI
FEALDHTLAAVKQNGVKYTVRQRFDAIPEAFQAVSIQVEDGASMAELAQIPGVQRVWPVS
DSILEDVLTSVGRNGIKYQLRHRFIS-PTLFHGASITVPPGISRSQIASLRGIKRVWPVR
lowastlowast
TIKSPKPVKVHVVTGKDDPEIPA-------------------------DTESTHIMTGVN
AFDRPKPVKVQVVKGPDDPALPP-------------------------DSESTHVITGVD
AVPAPERINQFTVSSPNDSRITA-------------------------DSETTHIMTGVD
LIPRPVEPTVSDFTPVSSSSSSTKSKRD-THTLEKRGTNFPPASAYLNDTFYPHVQTGID
KFSRPSAVVDADGGGSGFSGSPIKAALMGVKELGKR------ANAYAGDTFGPHVMTGVN
lowast lowast lowast lowastlowast
KLHAQGIFGAGIKIGIIDTGIDYTHPALGGALGAGHKVIGGYDFVGDAYDGTNTPVPDND
KLHAQGITGAGIKIGILDTGIDYTHPTLGGAFGPGNKVIGGFDLVGDDYDGTNTPVPDPD
KVHAAGITGEGIKIGIIDTGIDYNHPLLGAGFGPGFKVIGGYDFVGDDYTGSNNPAPDDD
VLHNKGILGQGVKIVVVDEGVDYTNPILGGCFGPGCQISFGYDFVGDNYTGTNAPVPDSD
ETHEAGLLGAGIKIGVLDTGVDYLNPILGGCFGPGCHMSFGYDLVGDDYDGDNAPVPDVD
lowast lowast lowast lowastlowastlowast lowast lowast lowastlowast lowast lowastlowast lowast lowast lowast lowast lowastlowastlowast lowast lowast lowast lowast lowastlowast lowast
PLDQCNGHGTHVAGIIGANPDNQFNISGVSYGASLSSYRVFGCSGSVSDDIIIEALLRGV
PLDQCAGHGTHVAGIIGANPDNAFNISGVAFQASLSAYRVFGCVGFVTDDVLVDALLLGF
PLDNCNGHGTHVAGIIGANPGNDFGISGVAYGSSIASYRVFGCSGFVSDDIIIDALLRGY
PFSSCSNHGTHTTGTVGALAN-EYGFSGAAPMATIGHYRVISCSGATTDEILVAALTRAL
PFASCDPHGTHVTGIIGALPN-AFGFTGVAPAATLGHYRVFGCTGFVGEDIILAGLMRGV
lowast lowastlowastlowast lowastlowast lowast lowastlowast lowast lowastlowastlowast lowast lowast lowast
QDGQDILTLSLGGTDGWTE-SSSSVVASRIAASGKIVTIAAGNEGAYGSWYSSSPGNGID
NEGQDILTLSLGGADGWTE-SVSAVVASRIAATGKVVTIAAGNDGASGAWYTSSPGNGID
SDGMDILTLSLGGVEGWTE-SSSSVVASRIADQGRVVTIAAGNDGAYGSWYASSPGTGVD
QDNVNIVSMSLGSSVGWLDDSPVQIMASYLGSKGIHVVASVGNERNEGLFAADQPAASRI
EDNCNVLTLSLGGPGGWVKGTPASILIDQIEAQGILVTVATGNSGAEGMFFSESPASTIN
lowastlowastlowast lowastlowast lowast lowast lowastlowast lowast lowast
VISVASIDNTVDNIQNATVNGVEHGPIPYFKTLPLPFTNA-LPLYVISNDTTIANDACNA
VISVASLDNTVVPLQTAQVHGVTHDPIIYQDTFPLPINDT-RPIFATSTDVTVVDDACNP
VISVASVNNIAIPVQNATVQGVEHAPIPYLEALPLNVTGS-RPIYATSTDTTVADDACNA
GTGVGGVDPTYLAAYYAYLI--KRDPVPYISPTPFNLPNN-YLLYFTSTNTSVTNDACNP
GLSIASTDVTDLIAYNATVS--GQPAIPYLSATPLNVVANSFRVHFTSTDPNNPVDACSP
lowast lowast lowast lowast lowastlowastlowast
LPDNTPDLSGFLVLVRRGSCTFVQKLTNIAAKGGNAVFIYDNGNG-----FAGIDTGKFT
LPDSTPDLSGFVVIVRRGTCTFVQKLGNVAAKGAKVTLIYDNGSG-----FGAIDTGAFV
LPNDTPDLSGYVVVIKRGTCTFVTKLQNAAAKGANTFLIYNNGGS-----FSAISVGDYT
LPSTTPNLANRVVVVQRGTCGFTVKQQNVAAAGGKIVLIYNSKGSGLIPQLDVGSTGLTA
LPAGAPDFANYVTVVQRGTCTFVTKYQNVLNAGGKIVLLYNSEGAGNLPYLTPNGVGIDA
lowastlowast lowast lowastlowast lowast lowast lowast lowast lowast lowast lowast
(a)
Figure 5 Continued
14 Enzyme Research
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
NATLIQAADGVFLAQQVAAGKSVTITFPQTGGSAQITN-PSGGLVSSFTTYGPTNDFYFK
-ASFIQAADGEFLVQQFASGQNVSLSFPQSGGLIQFPD-PAGGLISSFTSYGPSNDFFFK
-AALISETDGTFLVNQFAAKANITISFPQSGSAWNYPDDVSGGLVSDFSSYGPSYDMYFK
VGSLRYEDGLKLLSYYQSSPRGQSISFPNGPLAPYVSNTVSGGVVASYSNFGPTNELFIY
VAGLRRSDGLKLLSYYQNANKRLTLRFPKGKIVAGLTDTITGGLISGYSTFGPTNDLYGQ
lowast lowastlowast lowastlowast lowastlowast
PAFGAPGGNILSTLPVPLGSYGVESGTSMATPYVAGSAALLLSVKGKSAAVALGARTLFQ
PAVAAPGGNILSTLPVNLGLFGIESGTSMATPFVAGSAALLFQAKGTSAAVGRSARTLFE
PAVAAPGGNILSTLPIALGSYGVESGTSMATPFTAGSAALILQAKGK--SVAKSIRNLFQ
PTLAAPGTDILSTV---TGGVALMRGTSMAAPLVAGAYALLLSVGNNLKLSPEEARTILM
PTLSAPGGNILSTFPLSEGGVAVISGTSMSCPFVAGSAAVLMAARASENLTPLEIRSLLT
lowast lowastlowastlowast lowastlowastlowastlowast lowast lowastlowastlowastlowast lowast lowastlowast lowast lowast
STAQRVPSSHTDGDPLQTLSQQGAGLIDVFKALYTETIVWPTELILNDTAHYKEVHTFTV
TTAQRVPSTHTDGDPLQTLTQQGAGLINVFNAIHTTTIVSPGELILNDTAHFKGTQQFTV
TTASAVASSKTDGDPLQTVSQQGAGLIQVDRAIGTTTVVSPGQLTLNDTAHFEGFKTFNI
STSQPSPVAY-GSETLDTVVSQGAGVINMTAAIAAHTVISPAQFNLNDTQYSNNTQTLTL
TTAKLTPVSLLGSTPLVSVIRQGGGLVQVAKALAAKTLISPHELLLNDTANANYVQTIKI
VNTGKTSKQYKLTHVPAGTAQTIT---SGTIFAADGPVPLSTDYATVKLSSSSFTLRPGQ
RNAGSSAKTYTLRHIPAGTAVTVT---PGTIFPADGPVPLSTDFASVSLSTSKFTLAPGK
QNTGSKAVTYKISHVPAGTAASVQ---SGSIFAADGPVPLTADYAQVTFSLNKVTVLPGV
RNNNRFPVSYQLSWVDSTGIVTYNDGATTDIIPSTTPNYVSTSVLRVAFSQRTVTVPGGQ
KNTNSWAMKYTFSSAVAQGLGTFD--ASGDILPTLDPVAVSGAQATVAFNTRILSVAPGA
lowast lowast lowast lowast lowast lowast
SKQVTTHINAP--SGVDASTYPVFSGFIEIATAT--------ESVHVTYLGLAASLKDKQ
TQTVTARFTPP--AAADPSTFPVFSGFIQIESGT--------EQVQVSYLGLKASLKDKQ
SVPILVWFKPP--QGVDASTYPVYSGFIEIAGLG--------ETLRVAYLGVAASLKDKA
TVKVTAQFIPPNLTAQQRNQFPIYSGFVTITGQGQGAGAEQMEKYNLPFFGLGARMVDMP
TGSVVATITPPVLPVADAARFPIFSGWIRVNGQG-ARDSSRNEAYTVPYFGLAAKMIDMQ
lowast lowast lowastlowast lowast lowast lowast lowast
IVDNTNEFFGITIPAVLNNTGDAQ-LSPTNYTFSGSDFPTLLLRLAFGTPLLRVDLVDAT
VIDNTDFFFGVPTPVLTDPNGEVQ-TSPRNYSFLASDFPTLIFRLAFGSPKVVVDLVSPT
VVDNTDEYFGVKIPALLDASGDVQ-GNSTNYTLVGDDVPSLLYRFAFGSAHVRVDLVEKA
VLDTTNVALGP--NLPFIAVGQN--IQTGPTTFSESNPAVVYFRLAAGTRRLRVDLVDAS
VLDTTETIYGPGYAYPFVIDDAIGDIQSTTTSYSRNLGPTVFARFATGTLHYSLDLVLAD
lowast lowast lowast lowast lowastlowastlowastlowast lowast
IDLKPTLNSREIDDKRHVFERESFSFPHPGPKGS-------------------TFAQVKI
INFKPTLNTRALGDTHGSF----FSFPHRVKTG--------------------SFAQVPT
STVSATLSKRNWWN---------WWWPQKPSN---------------------SYDKVPI
TSYNATVPAVTNPASRLVKRSNTELTPRAAFPT--------------------LYSQVRT
IAFTPTYPNSS-PATRFVKRSLTQHTSAASHLAKRRVSIATINPKATLVADRQLHSDVPI
lowast lowast
VGTLSEQSHIPRNSDASTVDDGGYSTFAFNKP------SFANGTTIPNGTYRWLLRALKV
LGTVFELDFTSRNNDDPTDN--GFNTVAIDPP------TFANGTTIPNGQYRLLLRALRV
VGTLAEYDYVPRNSDASTIDDNGYNVFQFT-D------SFVNGTKIAAGEYKVLLRALKV
VGSLFAPSYWPPRDYLFNNGGT-YSDYEIQLDGSFTFPNGTKGMAELGRSYKVLLRALKI
EGNIFTQPFTG-RDYLVDAAPTGSTDRTVTFNGQY-AENGLVRTAVTGTSYRFLLRALKI
lowast lowast lowastlowast lowast lowast lowast lowast lowastlowastlowastlowast
lowast lowast lowastlowastlowastlowastlowast
(b)
Figure 5 Continued
Enzyme Research 15
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
TGDPTKEADFETWLSPIVGITVA
TGDPTNEADFESFLSPIIGVNAP
TGNPAEEGDYETWLSPSFSVVPA
TADPTYTSSYESWLSPSFTFTA-
SGDAMYEDQYESWLSLPFSFRA-
lowast lowastlowast
ConservedSimilar
lowast All match
(c)
Figure 5 Multiple sequence alignment between amino acid sequence of PI12 protease with other subtilisin The serine protease sequenceswere obtained from the GenBank database Their accession numbers are EMS20793 (subtilisin-like protease of Rhodosporidium toruloidesNP11) CAG25549 (subtilisin-like protease of Pleurotus ostreatus) EPQ55564 (subtilisin-like protease of Gloeophyllum trabeumATCC 11539)and ETW80907 (serine protease S8 ofHeterobasidion irregulare TC 32-1) Putative ORF of serine protease fromG antarctica PI12 was labeledas PI12 protease
CAG38357 Phanerochaete chrysosporium
EIW62675 Trametes versicolor FP-101664 SS1
XP 007369624 Dichomitus squalens LYAD-421 SS1
XP 007381302 Punctularia strigosozonata HHB-11173 SS5
CAG25549 Pleurotus ostreatus
ETW80907 Heterobasidion irregulare TC 32-1
XP 001877576 Laccaria bicolor S238N-H82
EPQ55564 Gloeophyllum trabeum ATCC 11539
XP 001830835 Coprinopsis cinerea okayama7130
XP 007341385 Auricularia delicata TFB-10046 SS5
CCA68627 Piriformospora indica DSM 11827
CCO37249 Rhizoctonia solani AG-1 IB
XP 007342189 Auricularia delicata TFB-10046 SS5
EJU04161 Dacryopinax sp DJM-731 SS1
XP 007589863 Colletotrichum fioriniae PJ7
EFQ32840 Colletotrichum graminicola M1001
ENH74885 Fusarium oxysporum f sp cubense race 1
XP 003711175 Magnaporthe oryzae 70-15
XP 006694395 Chaetomium thermophilum DSM 1495
EON99722 Togninia minima UCRPA7
CBQ73425 Sporisorium reilianum SRZ2
GAC97617 Pseudozyma hubeiensis SY62
CCF52619 Ustilago hordei
EMS20793 Rhodosporidium toruloides NP11
CAQ76821 Glaciozyma antarctica PI12
01
Clus
ter 1
Clus
ter 2
a
b
c
d
Figure 6 Phylogenetic dendrogram among 25 translated amino acid sequences of serine proteaseThe tree was constructed using a neighbor-joining algorithm with 1000 bootstrappings by MEGA 6 software [39]The scale bar represents 01 substitutions per amino acid positionTheserine protease sequences were obtained from the GenBank database GenBank accession numbers of the sequences were indicated beforethe strain name The putative serine protease of G antarctica strain PI12 has been deposited in GenBank with accession number CAQ76821
16 Enzyme Research
0
2
4
6
8
10
12
0 24 48 72 96 110
Prot
ease
activ
ity (U
mL)
Post induction time (hours)
Figure 7 Effect of postinduction time towards protease activity The highest activity was detected at 72 hours after 110 hours of incubationwith 9UmL activity followed by 48 24 and 96 hoursThe substrate (azocasein) was dissolved in Tris HCL buffer and incubated for 30min at20∘CThe absorbance was measured at 450 nm after 15min incubation A one-way ANOVAwas used to test for preference differences amongsix different postinduction times The statistical value (119875 value lt005) between hours is 0003
pink-coloured nonpathogenic oleaginous yeast (42 iden-tity) Both G antarctica PI12 and Rhodosporidium toruloidesNP11 belong to Microbotryomycetes class PI12 protease didnot show any homology similarity to other psychrophilicproteases however it showed 37 identity to hypotheticalprotein from Pseudozyma antarctica T-34 which was isolatedfrom Japan (GAC76005) (not listed in the phylogenetic tree)
35 Cloning and Expression of Cold-Adapted PI12 Protease inthe Yeast Expression System The PCR product harbouringthe mature PI12 protease with two flanking restriction sites(EcoRI and AvrII) was cloned into pPIC9 expression vectorThe recombinant plasmid harbouring PI12 protease was usedto transform both GS115 and KM71 intended to produce bothHis+ Mut+ and His+ MutS Two recombinant clones fromGS115 (His+Mut+) strain (GpPro1 andGpPro2) and one fromKM71 (His+MutS) strain (KpPro1) were successfully obtainedwhich were further analyzed for protein expression study
36 Expression and Enzyme Assay of PI12 Protease in Pichiapastoris All positive recombinants were grown in small scalecultures and assayed for the cold-adapted protease activityin the culture supernatants The clones were first grown inBMGY medium with glycerol as the sole carbon sourceFor induction of AOX1 promoter and protein productionthe growth medium was transferred to the same mediumexcept with 05 methanol in place with glycerol as thecarbon source [26] The supernatant of the recombinant Ppastoris expression cultures after 48 h of incubation time wasanalyzed to determine the recombinant protease activity bythe modified method of Brock et al (1982) [20] All testedclones secreted protease into the supernatant in differentexpression levels This might be due to the variability ofthe strength of the promoter and copy number in each cellto secrete and express the recombinant product The clonewith the highest yield was GpPro2 with 63UmL activityfollowed by KpPro1 and GpPro1 with 25UmL and 37UmLactivities respectively (data not shown) It is perceptible thatexpression systemof Pichia pastoris is the ideal system for thisprotein The preliminary assay has fruitfully produced high
protease expression at 20∘C Yet further optimization of therecombinant PI12 protease production need to be conductedin order to examine the optimum condition of this proteinReported cold-adapted proteases showed optimum activity ata range of 15∘Cndash60∘C [27ndash30] This difference was possiblyrelated to the respective genetic and physiological adaptationof the strains There are several factors that influenced thedifferent rate of expression It includes different site of inte-gration between interest gene and Pichia genome variabilityof different expression host cell to secrete and express therecombinant enzyme and different sites in the polylinker forinserting foreign genes [31]
This is the second extracellular protease reported to besynthesized by this obligate psychrophilic yeastG antarcticaThe first cold-adapted extracellular subtilase from this yeasthas been purified and characterized previously [32 33] Theextracellular serine proteinase (LAP2) from this Antarcticyeast formerly known as Leucosporidium antarcticum strain171 is specific towards synthetic substrates of chymotrypsinand subtilisin The subtilase of L antarcticum strain 171showed specific activity of 4043Umg when utilized N-SucAAPFpNA as a substrate [33]
37 Optimization of Protease Production GpPro2 was chosenfor further optimization due to its highest protease activityat the preliminary expression stage High protease activitywas detected after 72 hours of incubation (Figure 7) byusing the standard protocol method as recommended forPichia pastoris expression system (Pichia Expression KitInvitrogen USA) The expression decreased after 72 hoursand no activity was detected at 110 hours The prolongedincubation may result in the decrease of biomass productionand will subsequently reduce the expression of recombinantprotein In addition oxygen and nutrient depletion duringprolonged incubationwill trigger the stress responses in orderto survive in such harsh environment [34]
Methanol was used as a sole carbon by P pastoris Themethanol was oxidized to formaldehyde with the aid of oxy-gen as electron acceptor by an alcohol oxidase enzyme (AOXgene) [35] The optimal methanol concentration is at 05(vv) with 14UmL activity The addition of more than 05
Enzyme Research 17
02468
10121416
0 01 03 05 07 09Pr
otea
se ac
tivity
(Um
L)Methanol concentration ()
Figure 8 Effect ofmethanol concentration towards protease activityThe recombinant was cultivated for 72 hours at 30∘C in BMMYmedium05 methanol gives the highest activity with 14UmL The substrate (azocasein) was dissolved in Tris HCL buffer and incubated for 30minat 20∘CThe mixture was left for 15min and absorbance was taken at 450 nm A one-way ANOVA was used to test for preference differencesamong six different concentrations The statistical value (119875 value lt005) between concentration is 002
0
5
10
15
20
25
10 15 20 25 30 35
Prot
eas a
ctiv
ity (
Um
L)
Temperature (∘C)
Figure 9 Effect of temperature towards protease activity The reaction was carried out in temperature ranging from 10∘C to 35∘C at pH 7 for30min A one-way ANOVA was used to test for preference differences among six different temperatures The statistical value (119875 value lt005)between temperature is 0067
(vv) methanol resulted in tremendous decrease of proteaseactivity (Figure 8) Production of PI12 protease was decreasedwhen the methanol concentration increased This might bedue to the toxic effect of methanol concentration on DNAreplication and membrane synthesis Basically methanol isused to increase the permeability of the cell membrane tohelp the secretion of foreign protein [26] However if thepermeability of cell membrane is too high lowmolecular sizemetabolites would leak from the cell and such leakage willseriously damage cell activities Such condition would abusethe cellular system and might reduce the yield of proteinexpression [36]
Psychrophilic microorganism dominated in low temper-ature habitat which most other microorganism are inca-pable of withstanding due to their inability to produceenzyme that can tolerate the extreme environment con-dition Psychrophilic enzymes are always associated withhigh catalytic efficiency at low and moderate temperature(0∘C to 30∘C) compared to the mesophilic counterpart thatare generally unable to catalyze efficiently under extremely
low temperature [2] The recombinant protein assay wasdone at various temperatures ranging from 10∘C to 35∘Cusing 05 azocasein as a substrate dissolved in 50mMTris HCl in presence of 2mM CaCl
2 pH 7 In the activity
assays expressed protease showed a highest activity at 20∘C(20UmL) and decreased tremendously once it exceededmaximum temperature due to the denatured state of theenzyme (Figure 9) The active site of an enzyme is easilyaltered when the high temperature is introduced duringincubation or activity assay resulting in a low reaction rate[37]
The recombinant construct was transferred into BMMYmedia at different A
600 nm ranging from 1 to 10 The high-est activity of active enzyme was detected at A
600 nm 60(283UmL) and above (Figure 10) The slope during expo-nential phase was much higher compared to other phasesindicating an increase in the doubling time At exponentialphase the cells are adapted to the growth condition andother processes like cell growth cycle and synthesis of RNAThe yield of active enzyme during expression of P pastoris
18 Enzyme Research
05
1015202530
1 2 3 4 5 6 7 8 9 10Prot
ease
activ
ity (U
mL)
Initial inoculum A600
Figure 10 Effect of inoculums size towards protease activity The recombinant was cultivated for 72 hours at 30∘C in BMMY mediumwith 05 methanol concentration A one-way ANOVA was used to test for preference differences among six different optical densitiesThe statistical value (119875 value lt005) between optical density value is 003
(kD
a)
1160
662
450
350
250
184
144
M 1 2 3
(a) (b) (c)
Figure 11 (a) Coomassie-stained SDS-PAGE of recombinant P pastoris clone 1mL supernatants collected from the third-day culturewere concentrated 10-fold using trichloroacetic acid and 10120583L resultant products were applied to SDS-PAGE analysis Lane 1 molecularweight marker (Fermentas) Lane 2 GS115 (Control) Lane 3 GS115pPIC9 empty (Control) Lane 4 GS115pPIC9PI12prot (GpPro2)The arrow indicates the secreted recombinant PI12 protease (b) Plate activity staining of different recombinant P pastoris clones GGS115 GP GS115pPIC9 GRI GS115pPIC9PI12prot1 (GpPro1) GRII GS115pPIC9PI12prot2 (GpPro2) K KM71 KP KM71pPIC9 KRKM71pPIC9PI12prot (KpPro) (c)Gel activity staining of recombinant PI12 protease inP pastorisGpPro2 clone Lane 1 prestainedmolecularweightmarker (Fermentas) Lane 2 GS115pPIC9 empty (Control) Lane 3 GS115pPIC9PI12prot1 (GpPro2)The arrow indicates the secretedrecombinant PI12 protease
dropped after 28 hours of incubation which is at stationaryor death phase [38]
38 Detection of Recombinant Protein by SDS-PAGE andActivity Staining TheSDS-PAGE gel of the GpPro2 recombi-nant clonesrsquo supernatant showed a putative expression bandat around 993 kDa Plate staining and gel activity stainingwere executed to examine if the recombinant PI12 proteaseexpressed in P pastoris was functionally active In the platestaining the transformants showed strong proteolytic activity(Figure 11(a)) In gel staining proteolytic activity was alsodetected around 993 kDa after incubation at 20∘C This is
evidence of protease secretion into the surrounding mediawhereby it catalyzed the breakdown of casein thus formedclearing zones around the activity ring and band (Figures11(b) and 11(c)) As a conclusion the cold-adapted PI12protease was functionally expressed in P pastoris GS115(GpPro2 clone) with the size of 993 kDa
4 Conclusions
Psychrophilic yeast which was originated frommarine waterin Casey Station Antarctica was identified as Glaciozymaantarctica strain PI12 A serine protease with an open reading
Enzyme Research 19
frame of 2892 bp encoded for 963 amino acids from thisAntarctic yeast has been cloned and expressed in Pichiaexpression system This is the second extracellular pro-tease reported being synthesized by this psychrophilic yeastSequence analysis through bioinformatics studies revealedfew homology identities of this protein with other ser-ine proteases The mature PI12 protease was successfullyexpressed and secreted into the culture medium driven bythe Saccharomyces cerevisiae 120572-factor signal sequence in Ppastoris The protease production in P pastoris was bestobtained from strain GS115 (GpPro2) with 283UmL after 72hours of induction time with 05 (vv) of methanol inducerThe starting optical density at 600 nm (OD600) was 60 andthe optimum assay temperature was 20∘C The expressedprotein was detected by SDS-PAGE and activity staining witha molecular weight of 99 kDaThis cold-active enzymemightoffer an opportunity for biotechnological exploitation basedon its high catalytic activity at low temperatures and lowthermostability Structural and functional study of this cold-adapted enzyme would give insight into its adaptation to anextreme cold environment
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgment
This research was supported financially by the InitiativeBiotechnology-GMBI-MOSTI (Project number 07-05-16-MG-GMB02) andRUGS-Initiative 6 (Project number 05-02-11-1414RU)
References
[1] J W Deming ldquoPsychrophiles and polar regionsrdquo CurrentOpinion in Microbiology vol 5 no 3 pp 301ndash309 2002
[2] C Gerday M Aittaleb M Bentahir et al ldquoCold-adaptedenzymes from fundamentals to biotechnologyrdquo Trends inBiotechnology vol 18 no 3 pp 103ndash107 2000
[3] A Svingor J Kardos I Hajdu A Nemeth and P Zavodszky ldquoAbetter enzyme to cope with cold comparative flexibility studieson psychrotrophic mesophilic and thermophilic IPMDHSrdquoThe Journal of Biological Chemistry vol 276 no 30 pp 28121ndash28125 2001
[4] G Antranikian C E Vorgias and C Bertoldo ldquoExtremeenvironments as a resource for microorganisms and novel bio-catalystsrdquo Advances in Biochemical EngineeringBiotechnologyvol 96 pp 219ndash262 2005
[5] T Lonhienne C Gerday andG Feller ldquoPsychrophilic enzymesrevisiting the thermodynamic parameters of activation mayexplain local flexibilityrdquoBiochimica et Biophysica Acta vol 1543no 1 pp 1ndash10 2000
[6] R Cavicchioli and TThomas ldquoExtremophilesrdquo in Encyclopediaof Microbiology pp 317ndash337 Academic Press San Diego CalifUSA 2000
[7] G Feller E Narinx J L Arpigny et al ldquoEnzymes frompsychrophilic organismsrdquo FEMS Microbiology Reviews vol 18no 2-3 pp 189ndash202 1996
[8] J Arnorsdottir R B Smaradottir T O Magnusson S HThorbjarnardottir G Eggertsson and M M KristjanssonldquoCharacterization of a cloned subtilisin-like serine proteinasefrom a psychrotrophic Vibrio speciesrdquo European Journal ofBiochemistry vol 269 no 22 pp 5536ndash5546 2002
[9] N J Russell ldquoPsychrophilic bacteriamdashmolecular adaptations ofmembrane lipidsrdquo Comparative Biochemistry and Physiology APhysiology vol 118 no 3 pp 489ndash493 1997
[10] G Feller and C Gerday ldquoPsychrophilic enzymes hot topics incold adaptationrdquo Nature Reviews Microbiology vol 1 no 3 pp200ndash208 2003
[11] P Raspor and J Zupan ldquoYeasts in extreme environmentsrdquo inBiodiversity and Ecophysiology of Yeasts The Yeast Handbookpp 371ndash417 Springer Berlin Germany 2006
[12] M B Rao A M Tanksale M S Ghatge and V V Desh-pande ldquoMolecular and biotechnological aspects of microbialproteasesrdquoMicrobiology andMolecular Biology Reviews vol 62no 3 pp 597ndash635 1998
[13] L Polgar ldquoThe catalytic triad of serine peptidasesrdquo Cellular andMolecular Life Sciences vol 62 no 19-20 pp 2161ndash2172 2005
[14] S Y Boo C M V L Wong K F Rodrigues N NajimudinA M A Murad and N M Mahadi ldquoThermal stress responsesin Antarctic yeastGlaciozyma antarctica PI12 characterized byreal-time quantitative PCRrdquoPolar Biology vol 36 no 3 pp 381ndash389 2013
[15] B Turchetti S R T Hall L B Connell et al ldquoPsychrophilicyeasts from Antarctica and European glaciers descriptionof Glaciozyma gen nov Glaciozyma martinii sp nov andGlaciozyma watsonii sp novrdquo Extremophiles vol 15 no 5 pp573ndash586 2011
[16] N H F Hashim I Bharudin D L S Nguong et al ldquoCharac-terization of Afp1 an antifreeze protein from the psychrophilicyeast Glaciozyma antarctica PI12rdquo Extremophiles vol 17 no 1pp 63ndash73 2013
[17] A N M Ramli M A Azhar M S Shamsir et al ldquoSequenceand structural investigation of a novel psychrophilic 120572-amylasefrom Glaciozyma antarctica PI12 for cold-adaptation analysisrdquoJournal ofMolecularModeling vol 19 no 8 pp 3369ndash3383 2013
[18] A N M Ramli N M Mahadi A Rabu A M A Murad F DA Bakar and R M Illias ldquoMolecular cloning expression andbiochemical characterisation of a cold-adapted novel recombi-nant chitinase from Glaciozyma antarctica PI12rdquoMicrobial CellFactories vol 10 no 1 article 94 2011
[19] J Sambrook E Fritsch and T Maniatis Molecular Cloning ALaboratoryManual Cold SpringHarbor Laboratory Press NewYork NY USA 1989
[20] F M Brock C W Forsberg and J G Buchanan-SmithldquoProteolytic activity of rumen microorganisms and effects ofproteinase inhibitorsrdquoApplied and EnvironmentalMicrobiologyvol 44 no 3 pp 561ndash569 1982
[21] U K Laemmli ldquoCleavage of structural proteins during theassembly of the head of bacteriophage T4rdquo Nature vol 227 no5259 pp 680ndash685 1970
[22] A Henne R A Schmitz M Bomeke G Gottschalk and RDaniel ldquoScreening of environmental DNA libraries for thepresence of genes conferring lipolytic activity on Escherichiacolirdquo Applied and Environmental Microbiology vol 66 no 7 pp3113ndash3116 2000
[23] M Poza T de Miguel C Sieiro and T G Villa ldquoCharacteri-zation of a broad pH range protease of Candida caseinolyticardquoJournal of Applied Microbiology vol 91 no 5 pp 916ndash921 2001
20 Enzyme Research
[24] J D Bendtsen H Nielsen G von Heijne and S BrunakldquoImproved prediction of signal peptides SignalP 30rdquo Journalof Molecular Biology vol 340 no 4 pp 783ndash795 2004
[25] R J Siezen and J A M Leunissen ldquoSubtilases the superfamilyof subtilisin-like serine proteasesrdquo Protein Science vol 6 no 3pp 501ndash523 1997
[26] Y Katakura W Zhang G Zhuang et al ldquoEffect of methanolconcentration on the production of human 1205732- glycoprotein Idomain V by a recombinant Pichia pastoris a simple system forthe control of methanol concentration using a semiconductorgas sensorrdquo Journal of Fermentation and Bioengineering vol 86no 5 pp 482ndash487 1998
[27] Q-FWang Y-HHou Z Xu J-LMiao andG-Y Li ldquoPurifica-tion and properties of an extracellular cold-active protease fromthe psychrophilic bacterium Pseudoalteromonas sp NJ276rdquoBiochemical Engineering Journal vol 38 no 3 pp 362ndash3682008
[28] S Vazquez L Ruberto and W Mac Cormack ldquoProper-ties of extracellular proteases from three psychrotolerantStenotrophomonas maltophilia isolated from Antarctic soilrdquoPolar Biology vol 28 no 4 pp 319ndash325 2005
[29] R Zeng R Zhang J Zhao and N Lin ldquoCold-active ser-ine alkaline protease from the psychrophilic bacterium Pseu-domonas strain DY-A enzyme purification and characteriza-tionrdquo Extremophiles vol 7 no 4 pp 335ndash337 2003
[30] M Turkiewicz E Gromek H Kalinowska and M ZielinskaldquoBiosynthesis and properties of an extracellularmetalloproteasefrom the Antarctic marine bacterium Sphingomonas paucimo-bilisrdquo Journal of Biotechnology vol 70 no 1ndash3 pp 53ndash60 1999
[31] A Vassileva D A Chugh S Swaminathan and N KhannaldquoEffect of copy number on the expression levels of hepatitisB surface antigen in the methylotrophic yeast Pichia pastorisrdquoProtein Expression andPurification vol 21 no 1 pp 71ndash80 2001
[32] M Pazgier M Turkiewicz H Kalinowska and S Bielecki ldquoTheunique cold-adapted extracellular subtilase from psychrophilicyeast Leucosporidium antarcticumrdquo Journal ofMolecular Catal-ysis B Enzymatic vol 21 no 1-2 pp 39ndash42 2003
[33] M Turkiewicz M Pazgier H Kalinowska and S BieleckildquoA cold-adapted extracellular serine proteinase of the yeastLeucosporidium antarcticumrdquo Extremophiles vol 7 no 6 pp435ndash442 2003
[34] D Mattanovich B Gasser H Hohenblum and M SauerldquoStress in recombinant protein producing yeastsrdquo Journal ofBiotechnology vol 113 no 1ndash3 pp 121ndash135 2004
[35] P Secades BAlvarez and JAGuijarro ldquoPurification andprop-erties of a new psychrophilic metalloprotease (Fpp2) in the fishpathogen Flavobacterium psychrophilumrdquo FEMS MicrobiologyLetters vol 226 no 2 pp 273ndash279 2003
[36] L Y Ling I Ithoi and F M Yik ldquoOptimization for high-level expression in Pichia pastoris and purification of truncatedand full length recombinant sag2 of toxoplasma gondii fordiagnostic userdquo Southeast Asian Journal of Tropical Medicineand Public Health vol 41 no 3 pp 507ndash513 2010
[37] A Illanes ldquoStability of biocatalystsrdquo Electronic Journal ofBiotechnology vol 2 no 1 pp 1ndash9 1999
[38] B A Plantz J Sinha L Villarete K W Nickerson and VL Schlegel ldquoPichia pastoris fermentation optimization energystate and testing a growth-associated modelrdquo Applied Microbi-ology and Biotechnology vol 72 no 2 pp 297ndash305 2006
[39] K Tamura G Stecher D Peterson A Filipski and S KumarldquoMEGA6molecular evolutionary genetics analysis version 60rdquo
Molecular Biology and Evolution vol 30 no 12 pp 2725ndash27292013
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology
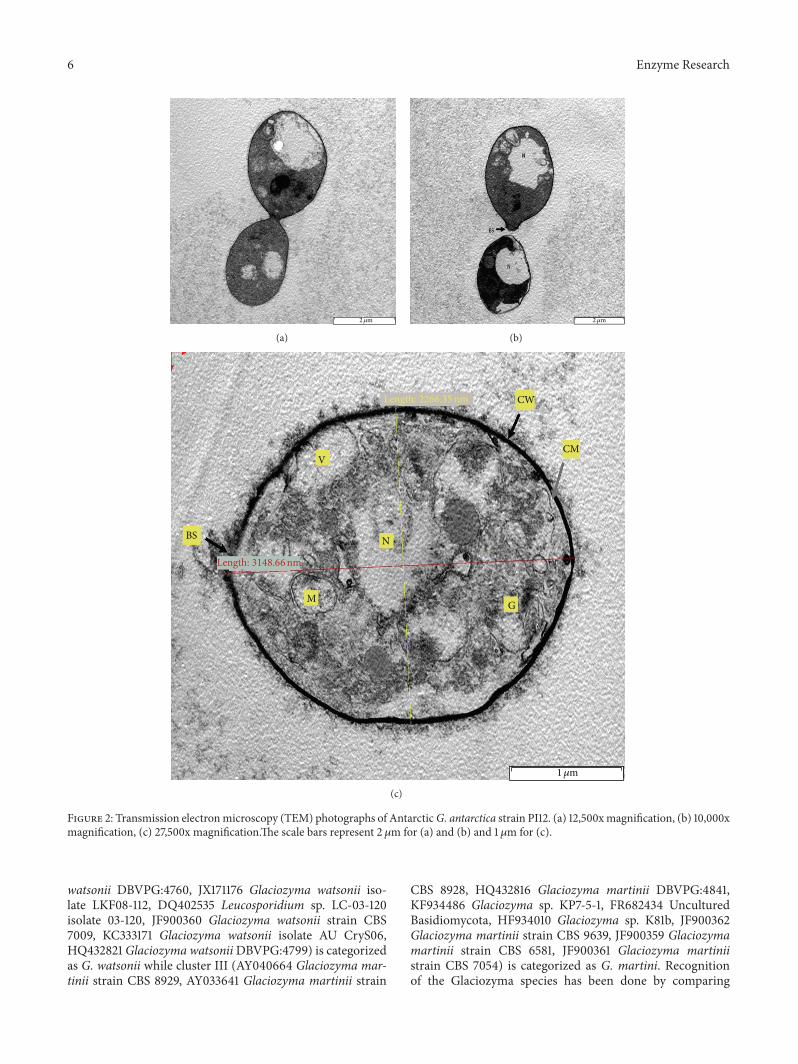
6 Enzyme Research
2120583m
(a)
2120583m
(b)
BS
V
N
CW
CM
GM
Length 314866nm
Length 226635nm
1120583m
(c)
Figure 2 Transmission electronmicroscopy (TEM) photographs of AntarcticG antarctica strain PI12 (a) 12500xmagnification (b) 10000xmagnification (c) 27500x magnificationThe scale bars represent 2 120583m for (a) and (b) and 1 120583m for (c)
watsonii DBVPG4760 JX171176 Glaciozyma watsonii iso-late LKF08-112 DQ402535 Leucosporidium sp LC-03-120isolate 03-120 JF900360 Glaciozyma watsonii strain CBS7009 KC333171 Glaciozyma watsonii isolate AU CryS06HQ432821 Glaciozyma watsoniiDBVPG4799) is categorizedas G watsonii while cluster III (AY040664 Glaciozyma mar-tinii strain CBS 8929 AY033641 Glaciozyma martinii strain
CBS 8928 HQ432816 Glaciozyma martinii DBVPG4841KF934486 Glaciozyma sp KP7-5-1 FR682434 UnculturedBasidiomycota HF934010 Glaciozyma sp K81b JF900362Glaciozyma martinii strain CBS 9639 JF900359 Glaciozymamartinii strain CBS 6581 JF900361 Glaciozyma martiniistrain CBS 7054) is categorized as G martini Recognitionof the Glaciozyma species has been done by comparing
Enzyme Research 7
JX896956 Glaciozyma antarctica strain PI12
AY040659 Antarctic yeast CBS 8938
AY033637 Glaciozyma antarctica strain CBS 8943
EU149809 Glaciozyma antarctica strain CBS 10640
AB774460 Leucosporidium sp KGK-2
JQ857017 Glaciozyma antarctica isolate T21Ga
EU149808 Glaciozyma antarctica strain CBS 10636
AY040663 Glaciozyma antarctica strain CBS 8939
EU149806 Glaciozyma antarctica strain CBS 10639
EU149805 Glaciozyma antarctica strain CBS 10638
AY040657 Antarctic yeast CBS 8927
AF444529 Glaciozyma antarctica strain CBS 5942
KF934487 Glaciozyma sp KP7-5-2
GQ336996 Leucosporidium sp AY30
EU149804 Glaciozyma watsonii strain CBS 10641
AY040661 Antarctic yeast ML 4515
AY040660 Glaciozyma watsonii strain CBS 8940
AY033638 Glaciozyma watsonii strain CBS 8944
EU149803 Glaciozyma watsonii strain CBS 10684
JX171176 Glaciozyma watsonii isolate LKF08-112
DQ402535 Leucosporidium sp LC-03-120 isolate 03-120
AB774462 Leucosporidium sp BSS-2
KC785578 uncultured Glaciozyma clone 180
HQ432818 Glaciozyma watsonii DBVPG4726
HQ432823 Glaciozyma watsonii DBVPG4802
AB774461 Leucosporidium sp BSS-1
HQ432820 Glaciozyma watsonii DBVPG4760
KC333171 Glaciozyma watsonii isolate AU CryS06
HQ432821 Glaciozyma watsonii DBVPG4799
JF900360 Glaciozyma watsonii strain CBS 7009
AY040664 Glaciozyma martinii strain CBS 8929
AY033641 Glaciozyma martinii strain CBS 8928
HQ432816 Glaciozyma martinii DBVPG4841
KF934486 Glaciozyma sp KP7-5-1
FR682434 uncultured Basidiomycota
HF934010 Glaciozyma sp K81b
JF900362 Glaciozyma martinii strain CBS 9639
JF900359 Glaciozyma martinii strain CBS 6581
JF900361 Glaciozyma martinii strain CBS 7054
001
Clus
ter 1
Glac
iozy
ma
anta
rctic
aCl
uste
r 3
Glac
iozy
ma
mar
tinii
Clus
ter 2
Glac
iozy
ma
watso
nii
Figure 3 Neighbor-joining phylogenetic analysis of internal transcribed spacer 1 (ITS1)58S rRNA geneinternal transcribed spacer 2 (ITS2)of Glaciozyma clade Numbers indicate percentage bootstrap values calculated on 1000 repeats of the alignment The tree was constructedusingMEGA6 software [39]The scale bar represents 001 substitutions per nucleotide position GenBank accession numbers of the sequencesare indicated before strain name ITS158SITS2 sequence of Glaciozyma antarctica strain PI12 had been deposited in GenBank with theaccession number JX896956 [14]
8 Enzyme Research
their molecular phylogenetic data and physiological andmorphological characteristics [15] Phylogenetic analysis ofITS158SITS2 sequence of strain PI12 showed that thesequence was closely related to ITS158SITS2 sequenceof Glaciozyma antarctica with high homology similarity toCluster I (99-100 identity) The ITS158SITS2 sequenceof G antarctica strain PI12 was differed from G watsonii(Cluster II) by 2-3 gap in the sequence (93 identity)and G martinii by 4-5 gap in the sequence (90ndash92identity)
33 Sequence Analysis of PI12 Protease cDNA In this studyamplification of the PI12 protease gene sequence was con-ducted using genomicDNAand cDNAas a template Putativepartial PI12 protease gene with the size of 1321 bp was initiallyacquired from the G antarctica genome survey sequencelibrary (Supplementary Figure see Supplementary Mate-rial available online at httpdxdoiorg1011552014197938)However the gene was discovered as an incomplete pro-tease gene sequence DNA walking strategy successfullyamplified 1500 bp of the upstream region of partial PI12protease Both sequences of about 1500 bp (derived fromthe DNA walking reaction) and 1321 bp (partial PI12 pro-tease) were assembled and made up a total length of2687 bp protease sequence (Supplementary figure)The puta-tive PI12 protease was predicted by Augustus webserver(httpaugustusgobicsde) having intervening sequencesintronsThus reverse-transcriptase PCR (RT-PCR) and rapidamplification of cDNA ends (RACE) were accomplished toobtain the full-length cDNA As a result the ORF comprising2892 bp encoding 963 amino acid residues were successfullydeduced
Nucleotide and deduced amino acid sequences ofgenomic DNA and cDNA fragment encoding PI12 proteasewere shown in Figure 4 The size of the ORF of putative PI12protease was 3867 bp with genomic DNA as the templatewhile the ORF comprised 2892 bp encoding 963 amino acidresidues were successfully deduced through RACE strategywith cDNA as the template The ORF was predicted usingORF Finder (httpwwwncbinlmnihgovprojectsgorf)while prediction using SignalP (httpwwwcbsdtudkser-vicesSignalP) webserver allowed the identification of theputative PI12 protease prepeptide where it predicted thepresence of signal peptidase I cleavage sites [24]The putativeN-terminal signal peptide (1 to 16 the underlined amino acid)was identified as a secretion signal There is no propeptidepredicted by ProP Webserver (httpwwwcbsdtudkser-vicesProP-10) Comparison of cDNA and gene sequencesallowed determination of gene structureTherefore sequencealignment was performed between the sequence of these twotemplates and it was confirmed that the coding sequence(consisted of 19 exons) was interrupted by 18 introns withtheir length varying from 48 to 60 nucleotides However theGT and AG rule for donor-acceptor sites was not obeyedOf 18 introns 10 introns contained canonical GT and AGdinucleotides at their 51015840 and 31015840 ends while 8 introns holdnoncanonical splice site pairs with TT-GG (2 introns)AG-TG GC-GT TG-GG TC-GG TA-GG and TG-AG
dinucleotides at the splice junction Upper case letters inblue colour represent introns while lower case letters inblack colour represent exons Splice donor and acceptor werehighlighted in grey
The amino acid composition (upper case letters) withinthe ORF amino acid residues was determined by theProtParam tool of the Expasy molecular biology server(httpwwwexpasyorgtool) Translation starts at a nucle-otide position 1 and translation stop is marked with anasterisk The nucleotide sequences ATG (1 to 3) and TAG(3865 to 3867) indicate the initiation codon and termi-nal codon respectively The presumed putative sequencesof promoter CAAT (minus101 to minus98) and TATA-box (minus92to minus88) were identified in the 51015840-untranslated (UTR)region The search was achieved via Neural Network Pro-moter Prediction (Berkeley Drosophila Genome Project)(httpwwwfruitflyorgseq toolspromoterhtml) In the 31015840-untranslated (UTR) region there was a putative polyadeny-lation signal attaa (3889 to 3893) detected from the TAGstop codon to the poly (A) tail The predicted molecularmass and pI of PI12 serine protease were 10099 kDa and641 respectively Out of 963 amino acid residues in thededuced sequence of the PI12 protease 70 are negativelycharged residues (Asp + Glu) while 66 are positively chargedresidues (Arg + Lys) at neutral pH (Table 2)The total chargedresidues (Arg Asp Glu His and Lys) are 150 amino acidsin the open reading frame of PI12 protease corresponded to156 while hydrophobic residues (Ala Phe Ile Leu MetPro Val and Trp) are 441 amino acids which correspondedto 458 of the total amino acids discovered The totaluncharged residues (Asp Cys Gln Gly SerThr and Tyr) areconsisted of 372 amino acids which corresponded to 386of the total amino acids of the open reading frame of PI12protease The aliphatic index is 9290 and the grand averageof hydropathicity (GRAVY) is 0141 The instability index (II)of PI12 protease was computed to be 3414 which classify theprotein as stable
N-glycosylation sites were predicted using NetNGlyc10 Server (httpwwwcbsdtudkservicesNetNGlyc)Therewere five putative N-glycosylation sites for the PI12 proteasegene predicted using the server For N-glycosylation thesequence motif Asn-Xaa-SerThr (Xaa any amino acid butnot Pro) has been defined as a prerequisite for glycosylationThe sequencemotifs in PI12 protease occurred in the position170 NETH 365 NATV 576 NLTP 634 NDTA and 833 NSSPAmong five predicted sites only three (except positions 576and 833) showed a high potential score (averaged output ofnine neural networks) which support the prediction Theputative N-glycosylation sites were demonstrated in boxesin Figure 4 The comparison of amino acid composition ofthis cold-adapted protease fromG antarctica strain PI12 withnine other subtilisin serine proteases fromdifferent families isshown in Table 2 Analysis of this comparison depicts that thepercentage of each amino acids in PI12 protease is almost thesamewith other proteaseswhileGly (G) is themost abundantaccounting for up to 104 of the total amino acid in PI12 pro-tease PI12 protease also contained the highest percentage ofLeu (L) with 99 of the total amino acids compared to other
Enzyme Research 9
Table2Aminoacid
compo
sitionandmolecular
weighto
fproteases
acrossdifferent
families
Amino
CAQ76821
CAG25549
EDR11679
CAG38357
CAD118
98CA
L25578
EDU50747
AAW
40780
ACB3
0121
ACB3
0119
acid
Qty
Qty
Qty
Qty
Qty
Qty
Qty
Qty
Qty
Qty
Ala
(A)
97101
8191
7887
7590
8299
101
115
8189
9196
8092
105
116
Arg
(R)
4042
2730
2224
1619
1721
1821
2932
2223
2124
3134
Asn
(N)
4446
3843
4348
4149
4251
4046
5257
5154
4552
4347
Asp
(D)
4951
6269
5460
6578
5567
5158
5358
6669
5260
6370
Cys
(C)
808
506
404
405
911
809
910
909
708
1011
Gln
(Q)
2324
3438
2326
2834
3036
3742
3539
2931
1922
2629
Glu
(E)
2122
1618
1921
2024
1518
2023
2528
2627
2832
2426
Gly
(G)
100
104
89100
8493
85102
85103
7788
8493
8084
8598
8695
His
(H)
1415
1416
1314
1619
1316
1113
1618
1415
1618
1213
Ile(I)
5355
4955
5157
5364
4150
3843
5156
4446
3540
4449
Leu
(L)
9599
6876
7179
6274
6073
6675
6774
8084
7485
6875
Lys
(K)
2627
2932
4449
2935
4959
5158
4044
3032
5058
5662
Met
(M)
1212
202
202
911
1113
809
1820
808
1113
1517
Phe
(F)
4143
5966
4752
5566
2935
3641
4449
3941
3439
3438
Pro
(P)
6264
6573
6774
5870
5870
5967
5055
5154
5564
5763
ser
(S)
7780
6977
91101
6072
6680
7080
7279
111
117
6373
6167
Thr
(T)
8790
89100
7583
83100
6579
7383
7178
7276
7182
6875
Trp
(W)
505
303
607
202
1215
1214
1213
909
1214
1415
Tyr
(Y)
3334
1213
2629
708
2935
2832
3943
5053
2630
2730
val
(V)
7679
8292
8089
6679
5769
7484
5864
6872
8295
6268
Total
963
1000
893
1000
900
1000
834
1000
825
1000
878
1000
906
1000
950
1000
866
1000
906
1000
MW
(Da)
1009898
932669
944365
869455
869549
924905
979343
1009629
917209
961988
Thea
ccessio
nnu
mbersof
proteasespu
tativ
eserinep
roteaseo
fGlacio
zymaantarctica
PI12
(CAQ
76821)sub
tilisin-lik
eproteaseP
leurotuso
streatus(CA
G25549)pyrolysin
LaccariabicolorS
238N
-H82
(EDR11679)
subtilisin
-like
proteasePh
anerochaetechrysosporiu
m(C
AG38357)sub
tilisin-lik
eserinep
roteaseP
R1CMetarhiziu
maniso
pliaev
aranisopliae(CA
D11898)serinee
ndop
eptid
aseH
ypocrealixii(C
AL2
5578)peptidase
Pyrenophoratritici-repentis
(EDU50747)p
eptid
aseCr
yptococcus
neoforman
svarn
eoform
ansJEC
21(A
AW40
780)sub
tilisin-lik
eprotease
Epich
loefestucae(A
CB30121)sub
tilisin-lik
eprotease
Epich
loefestucae
(ACB
30119
)
10 Enzyme Research
atgctcttcctccccgtcctcctcctcctccttcccggcgtcactgccttcctcaaccca 60
M L F L P V L L L L L P G V T A F L N Pgttaccaaccgcgcgaccaacgccatctcctcgactcaatacctctctaacgcctatatc 120
V T N R A T N A I S S T Q Y L S N A Y IctcgaattggacctctccacccccggcctcgtcaaacgggatagcacgcccgactctGTG 180
L E L D L S T P G L V K R D S T P D SCGTCCCGCGTTCCTCCCTCGCTGTCGAAGACGAGCTAACAATGACGTGGTAGatcctaga 240
I L Eggacgtactcacgtccgtaggccgcaacggtatcaagtaccaactccgccaccgctttat 300
D V L T S V G R N G I K Y Q L R H R F Ictccccgactctgttccacggcgcttcgatcactgtcccccctggaatctcccgctccca 360
S P T L F H G A S I T V P P G I S R S QaatcgcctctctccgcggtatcaaaGTCCGTCCCTCTCTCTCCCCTACTCCCTCCGTGCT 420
I A S L R G I KAACCCTGCACGCAACAGcgcgtctggcccgttcgaaagttctcccgacccagcgcagtag 480
R V W P V R K F S R P S A Vtggacgccgatggaggaggaagcgggttctcagggtcgcctatcaaggcggcgctcatgg 540
V D A D G G G S G F S G S P I K A A L Mgggtgaaagagctcgggaagcgcgcgaacgcttatgctggagatacgtttggaccgcatg 600
G V K E L G K R A N A Y A G D T F G P HtcatgacgggggttaatgagacgcatgaggcggggttgttgggagctgggattaagGTGT 660
V M T G V N E T H E A G L L G A G I KGTTTTCGTCTTTGTTTGGGGGAGGGGGGAGGGAGGAGCTGATTAATGGGTGCAGattggg 720
I GgtgctggacactggtgAGAAGCGGTTTGGAGGGAGGAGAGGATGGAGCTGATGAGTGTGC 780
V L D T GAGGTGttgattatttgaacccgattctgggaggctgctttggacctgggtgccatatgtc 840
V D Y L N P I L G G C F G P G C H M Sgtttgggtacgacttggttggcgatgattacgatggagataacgctcctgtgccggatgt 900
F G Y D L V G D D Y D G D N A P V P D VggatccttGTGCGTCCTTCCCTCCGCGATGGGCTGGAATCTGAGCTGACGGCGTTGTTGA 960
D PTCAGttgcgagctgcgatcctcatggTTCGTCTCGCTTTCCGCTCCTCTGATGCGCTCGC 1020
F A S C D P H GTGATGCACTTGGCAATAGGaactcacgttacgggaatcattggagcgctcccgaatgcgt 1080
T H V T G I I G A L P N AttggatttactggcgtcgcacccgccgctactctgggccactaccgagtGCGCTCCCTCT 1140
F G F T G V A P A A T L G H Y R VATCCGTTCTTCCTCGCAGCTCCCACTCACTCGTGCTTCCAGGTatttggctgcactggct 1200
F G C T GtcgtcggagaagatatcGTTCGTCCTCTCTCCAGTCCCGGCTAGCGATCCTGCTTCTGAC 1260
F V G E D ITTCGCATCTCCCTCTGAGattctcgctggactcatgcgaggagtcgaggacaactgcaac 1320
I L A G L M R G V E D N C Ngtcttgaccctctctctcggaggtccaggagggtgggtcaagggcacgccggcgtccatc 1380
V L T L S L G G P G G W V K G T P A S Icttatcgaccagatcgaagcgcaaggcattctcgtcaccgtcgccactggcaactcggga 1440
L I D Q I E A Q G I L V T V A T G N S GgctgagggTGAGCTCCTCGCTACTTGCGCGTCGAGACCAGTACTAACAGGGGCGTGGACG 1500
A E GCAGGcatgttcttctccgagtctcccgcctcgacgatcaacggcctttccatcgcatcca 1560
M F F S E S P A S T I N G L S I A ScgTCCGTCTCTCCCTTCCTTCGCACGCCACTCGCTCACACTCTCTCGCAGGgacgttacc 1620
T D V Tgacctcatcgcctacaacgccaccgtctcaggccaacctgcgatcccttacctctccgcc 1680
D L I A Y N A T V S G Q P A I P Y L S A
ggggggggggtctg minus194
ctagcgcgaatcgagaaaacacaaaagaacctcccgacttggcgttgcttctgctcttct minus180
ccttctcccaaccgccgacaataaaaactataacacacacacaccccatctcgacgcctg minus120
cagacttcacgactctcctcacgctctttaccgccccacgggtcccacccacgagccacg minus60
(a)
Figure 4 Continued
Enzyme Research 11
acgcccctcaacgtcgtcgccaacagcttccgcgtccacttcacctctaccgaccccaac 1740
T P L N V V A N S F R V H F T S T D P Naaccccgtcgacgcctgctctcctcttccggctggagcgcccgacttcgccaactatgtt 1800
N P V D A C S P L P A G A P D F A N Y Vacggtcgttcagcgtgggacttgtacgttcgttaccaagtaccagaacgttctcaatgct 1860
T V V Q R G T C T F V T K Y Q N V L N AggaggGTGCGTCGGCTTCTTCTCTCCCGACTGGATTGGGCGGAGACTGACCTTCTGAGCA 1920
G GGaaagatcgtattgttgtacaactcggagggagctgggaacctcccttacctcacgccca 1980
K I V L L Y N S E G A G N L P Y L T PacggtgtcggcatcgacgccgttgcaggtcttcgtcgttccgacggactcaagGTTCGTC 2040
N G V G I D A V A G L R R S D G L KTACGCTCTTCGGTGCTGAGGCCCTTCTCTGATCTCGCTCCGACAGcttctctcgtactat 2100
L L S Y Ycagaatgccaacaagcgtctcactctgcgcttccccaagggcaagatcgtcgcaggcttg 2160
Q N A N K R L T L R F P K G K I V A G LaccgataccatcaccggcggactcatctcgTTCGTTCCTCTCCGCCTCCCTCGCTGGCCG 2220
T D T I T G G L I SATGAGCTGACGCGGTCTTTGCAGGggttactcgacgtttggtccgacgaatgacctctac 2280
G Y S T F G P T N D L Yggtcagcctaccctctctgcccctggtggcaacatcctttcgaccttccctctctccgag 2340
G Q P T L S A P G G N I L S T F P L S EggaggagtggcggtcatcagtgggacgagcatgtcgtgcccctttgtcgGTTCGTCCTTC 2400
G G V A V I S G T S M S C P F VTCCTCGTTCGTTCGCTGAGCGTCGACTGATCTTGCACGATCGCAGctggatctgcggcgg 2460
A G S A Atcctcatggccgctcgcgcttcggagaacctcacgccgcttgagatcaggagtctcctta 2520
V L M A A R A S E N L T P L E I R S L Lctaccactgcgaagcttacgccggtctcgctcttgggatcgacgcctttggtgagcgtga 2580
T T T A K L T P V S L L G S T P L V S VttcgtcaaggaggaggTACGTTCCTTTCTCGCTGCTTTGAGGATCGAGGCTGATTAGCGA 2640
I R Q G G GGCTTCGCAGGactcgttcaggttgccaaggcgctcgcggccaagacgctaatctctcctc 2700
L V Q V A K A L A A K T L I S Pacgagctcctgctcaacgacactgcgaacgcgaactacgtccagactatcaagatcaaga 2760
H E L L L N D T A N A N Y V Q T I K I Kacaccaactcgtgggcgatgaagtacaccttctcctcggccgtcgcccaaggactcggaa 2820
N T N S W A M K Y T F S S A V A Q G L Gctttcgacgcttcgggcgatatcctccctaccctcgacccagtcgccgtctctggcgcac 2880
T F D A S G D I L P T L D P V A V S G Aaggctaccgtcgcgttcaacactcggatcctcagcgtcgcgcccggcgcgacggggtccg 2940
Q A T V A F N T R I L S V A P G A T G Stcgtggcgactatcacgccgccggttcttcccgtagcggacgctgcgaggttccctatct 3000
V V A T I T P P V L P V A D A A R F P Itctctgggtggatcagggtgaatgggcaaggcgcgagggatagcagtaggaacgaggcgt 3060
F S G W I R V N G Q G A R D S S R N E AacactgGTGCGTTTGGCTCGCTTTTTGGGAGGTGCCGAGCTAATCGTTTCTCGTAGtccc 3120
Y T V PgtactttgggcttgcggcgaagatgatcgatatgcaagGTGCGTCGATGGTTCTCTCGCT 3180
Y F G L A A K M I D M QTCTCTCTCGGTTGCTCACTTCAAAGTGCAGtcctcgacaccaccgagaccatttacggtc 3240
V L D T T E T I Y Gcgggctacgcctaccccttcgtgatcgacgacgcgattggagacatccaatccaccacaa 3300
P G Y A Y P F V I D D A I G D I Q S T TcgtcgtactccaggaacctcgggcccaccgtcttcgctcgctttgccaTGCGTCGACCGT 3360
T S Y S R N L G P T V F A R F ACCTCATTTCCTGCGGCAGAGCGAGAGCTGATTCCGACCTCGAACAGctggaacccttcac 3420
T G T L Htacagcctcgatctcgtcctagccgacatcgcctttacccccacctaccccaactcctcc 3480
Y S L D L V L A D I A F T P T Y P N S S
(b)
Figure 4 Continued
12 Enzyme Research
cccgccactcgtttcgtcaagcgctccctcacgcagcacacctctgccgcttcgcacctc 3540
P A T R F V K R S L T Q H T S A A S H Lgccaagcgccgcgtctccatcgccactatcaaccccaaagccaccctcgtcgccgatcga 3600
A K R R V S I A T I N P K A T L V A D Rcagctccactcggacgtccctatcgagggcaacatcttcacccaaccctttactggaagg 3660
Q L H S D V P I E G N I F T Q P F T G Rgattacctcgtcgacgcagccccgacgggatccaccgatcgtaccgtcacttttaacggg 3720
D Y L V D A A P T G S T D R T V T F N Gcagtacgccgagaacggcctcgtgaggacggctgtgacggggacttcgtaccgcttcctc 3780
Q Y A E N G L V R T A V T G T S Y R F Lcttcgggcgttgaagatctcgggagacgcgatgtacgaggatcagtatgagagctggctc 3840
L R A L K I S G D A M Y E D Q Y E S W L3900
S L P F S F R Attttgccacctcaaaaaaaaaaaaaaaaaaaaaaaaaa 3938
lowast
tcgctaccgttctcgttccgtgcgtaggcttgttgtggacaccactgcattaagttatga
(c)
Figure 4 Nucleotide and deduced amino acid sequences of genomic DNA and cDNA encoding protease from G antarctica strain PI12 Thenucleotides are numbered from the 51015840 end of the cDNA A putative CAAT TATA box and polyadenylation signal are bold italicThe putativeN-terminal signal peptide is underlined The initiation codon is boldfaced and termination codon is represented by an asterisk (lowast) boxedpeptide sequences showed N-glycosylated asparagines residues The amino acid residues that form the catalytic triad are double underlinedand bold while introns are represented in blue font Highlighted in grey are the intron donor-acceptor splice sites
proteases Hydrophobic profile of PI12 amino acid proteasewas performed by ProtScale tools with Kyte and Doolittlemethod at httpauexpasyorgcgi-bin The hydrophobicityprofile elucidation of PI12 protease amino acid was carriedout from amino acid 1 to 963 The amino acid residuesfrom 1 to 16 (M-L-F-L-P-V-L-L-L-L-L-P-G-V-T-A) and 548 to562 (V-A-V-I-S) were hydrophobic grooves The first regionwhich consisted of 16 amino acids was very hydrophobic andwas predicted to be the signal peptide by SignalP and ProPwebserver
The bold and double-underlined amino acids showedresidues of catalytic triad aspartic acid (D) histidine (H) andserine (S) active site respectively NCBI Conserved DomainsDatabase (CDD) identified the presence of subtilase proteaseand subtilisin-like serine protease domains in the amino acidsequence The predicted PI12 protease amino acid sequencewas found sharing significant homology to the subtilisinfamily of protease The highest similarity of PI12 proteasewas observed with 42 identity to subtilisin-like proteaseof Rhodosporidium toruloides (EMS20793) 37 identity tosubtilisin-like protease of Pleurotus ostreatus (CAG25549)and 35 identity to subtilisin-like protease of Gloeophyllumtrabeum ATCC 11539 (EPQ55564) The nucleotide sequenceof G antarctica strain PI12 showed low homology to thefungus subtilisin protease family and none to other psy-chrophilic proteases The multiple sequence alignment wasillustrated in Figure 5 The consensus sequence around thecatalytic triad aspartic acid (D) histidine (H) and serine(S) of PI12 protease showed high conservation with othersubtilisin (showed in black boxes) Glycine residues G142G173 G191 G262 and G327 that were highly conserved inmost of subtilases were conserved in PI12 protease [25]Therefore PI12 protease belongs to the subtilisin subgroup ofthe subtilase serine protease superfamilyThis serine protease
of G antarctica PI12 has been deposited into GenBank withaccession number CAQ76821
34 Phylogenetic Relationship of PI12 Protease The phylo-genetic tree was constructed to demonstrate the similaritiesand relationship of protein sequence between differentspecies or organisms Figure 6 illustrates the phylogeneticdendrogram indicating the relationship of PI12 proteasewith another 25 translated amino acid sequences ofserine protease The bar scale signifies the branch lengthto approximate variance along each branch Basicallythis tree comprises two main clusters cluster I was sub-divided into subcluster a (CAG25549 EPQ55564 ETW-80907 XP 007381302 XP 001877576 CAG38357 XP007341385 CCA68627 EJU04161 XP 007369624 EIW-62675 CCO37249 XP 001830835 XP 007342189) and sub-cluster b (XP 007589863 XP 003711175 EFQ32840 XP006694395 ENH74885 EON99722) cluster II was sub-divided into subcluster c (CBQ73425 GAC97617 CCF52619)and subcluster d [CAQ76821 (PI12 protease) EMS20793]
Serine protease constructed by the phylogenetic treecomprised large size of protein with about 801 to 1032 aminoacids The majority of the translated protease sequenceswere obtained from fungus All serine proteases from cluster1 subcluster a are derived from Agaricomycetes except forEJU04161 which belongs to Dacrymycetes whereby all serineproteases from cluster 1 subcluster b are from Sordari-omycetes Cluster II subcluster c consisted of serine proteasefrom fungus belonging to Ustilaginomycetes PI12 proteasewas categorized under cluster II subcluster d It possessedhomology similarity with the subtilisin-like protease fromRhodosporidium toruloides (EMS20793) (formerly known asRhodotorula glutinis or Rhodotorula gracilis) which is a
Enzyme Research 13
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
MKG-YSWSLLQLAFTGAALATVPLSDVKRISSAS--VVPNKFVVEVESASEIPGKR--SL
MKGVLVWSILS---AGCALAVTPLSSVKRVTNLP--IVANKFIVEVDSTSDIPTKR--S-
MKLPLLSLALT---AVSVGATVPLSSVQRTTTSPNNALAGEYIVEMNNVQGLGGKR--AY
---MAHGSLASLLLLLAASAQGAAAFGGKSATGGRSVVPNSYILQVNASAPALTKRGVSV
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease ---MLFLPVLLLLLPGVTAFLNPVTNRATNAISSTQYLSNAYILELDLSTPGLVKRDSTP
lowastlowast
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
YTPHELFYRSLSARGVSYEVNKEYNS-EGLFVGAVVTVSDANDVKALAETPGVKAIRPVR
-NPHDALYESLRKRAIGFKVDREFNT-QGLFVGAALTLSDAQDVGQIQSTTGVKSIRPVR
ASTHDTLYRSLRKRGISFDVKKEYNQ-PGIMVGAALKVASPQDVQQIATMAEVKAIRPVI
FEALDHTLAAVKQNGVKYTVRQRFDAIPEAFQAVSIQVEDGASMAELAQIPGVQRVWPVS
DSILEDVLTSVGRNGIKYQLRHRFIS-PTLFHGASITVPPGISRSQIASLRGIKRVWPVR
lowastlowast
TIKSPKPVKVHVVTGKDDPEIPA-------------------------DTESTHIMTGVN
AFDRPKPVKVQVVKGPDDPALPP-------------------------DSESTHVITGVD
AVPAPERINQFTVSSPNDSRITA-------------------------DSETTHIMTGVD
LIPRPVEPTVSDFTPVSSSSSSTKSKRD-THTLEKRGTNFPPASAYLNDTFYPHVQTGID
KFSRPSAVVDADGGGSGFSGSPIKAALMGVKELGKR------ANAYAGDTFGPHVMTGVN
lowast lowast lowast lowastlowast
KLHAQGIFGAGIKIGIIDTGIDYTHPALGGALGAGHKVIGGYDFVGDAYDGTNTPVPDND
KLHAQGITGAGIKIGILDTGIDYTHPTLGGAFGPGNKVIGGFDLVGDDYDGTNTPVPDPD
KVHAAGITGEGIKIGIIDTGIDYNHPLLGAGFGPGFKVIGGYDFVGDDYTGSNNPAPDDD
VLHNKGILGQGVKIVVVDEGVDYTNPILGGCFGPGCQISFGYDFVGDNYTGTNAPVPDSD
ETHEAGLLGAGIKIGVLDTGVDYLNPILGGCFGPGCHMSFGYDLVGDDYDGDNAPVPDVD
lowast lowast lowast lowastlowastlowast lowast lowast lowastlowast lowast lowastlowast lowast lowast lowast lowast lowastlowastlowast lowast lowast lowast lowast lowastlowast lowast
PLDQCNGHGTHVAGIIGANPDNQFNISGVSYGASLSSYRVFGCSGSVSDDIIIEALLRGV
PLDQCAGHGTHVAGIIGANPDNAFNISGVAFQASLSAYRVFGCVGFVTDDVLVDALLLGF
PLDNCNGHGTHVAGIIGANPGNDFGISGVAYGSSIASYRVFGCSGFVSDDIIIDALLRGY
PFSSCSNHGTHTTGTVGALAN-EYGFSGAAPMATIGHYRVISCSGATTDEILVAALTRAL
PFASCDPHGTHVTGIIGALPN-AFGFTGVAPAATLGHYRVFGCTGFVGEDIILAGLMRGV
lowast lowastlowastlowast lowastlowast lowast lowastlowast lowast lowastlowastlowast lowast lowast lowast
QDGQDILTLSLGGTDGWTE-SSSSVVASRIAASGKIVTIAAGNEGAYGSWYSSSPGNGID
NEGQDILTLSLGGADGWTE-SVSAVVASRIAATGKVVTIAAGNDGASGAWYTSSPGNGID
SDGMDILTLSLGGVEGWTE-SSSSVVASRIADQGRVVTIAAGNDGAYGSWYASSPGTGVD
QDNVNIVSMSLGSSVGWLDDSPVQIMASYLGSKGIHVVASVGNERNEGLFAADQPAASRI
EDNCNVLTLSLGGPGGWVKGTPASILIDQIEAQGILVTVATGNSGAEGMFFSESPASTIN
lowastlowastlowast lowastlowast lowast lowast lowastlowast lowast lowast
VISVASIDNTVDNIQNATVNGVEHGPIPYFKTLPLPFTNA-LPLYVISNDTTIANDACNA
VISVASLDNTVVPLQTAQVHGVTHDPIIYQDTFPLPINDT-RPIFATSTDVTVVDDACNP
VISVASVNNIAIPVQNATVQGVEHAPIPYLEALPLNVTGS-RPIYATSTDTTVADDACNA
GTGVGGVDPTYLAAYYAYLI--KRDPVPYISPTPFNLPNN-YLLYFTSTNTSVTNDACNP
GLSIASTDVTDLIAYNATVS--GQPAIPYLSATPLNVVANSFRVHFTSTDPNNPVDACSP
lowast lowast lowast lowast lowastlowastlowast
LPDNTPDLSGFLVLVRRGSCTFVQKLTNIAAKGGNAVFIYDNGNG-----FAGIDTGKFT
LPDSTPDLSGFVVIVRRGTCTFVQKLGNVAAKGAKVTLIYDNGSG-----FGAIDTGAFV
LPNDTPDLSGYVVVIKRGTCTFVTKLQNAAAKGANTFLIYNNGGS-----FSAISVGDYT
LPSTTPNLANRVVVVQRGTCGFTVKQQNVAAAGGKIVLIYNSKGSGLIPQLDVGSTGLTA
LPAGAPDFANYVTVVQRGTCTFVTKYQNVLNAGGKIVLLYNSEGAGNLPYLTPNGVGIDA
lowastlowast lowast lowastlowast lowast lowast lowast lowast lowast lowast lowast
(a)
Figure 5 Continued
14 Enzyme Research
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
NATLIQAADGVFLAQQVAAGKSVTITFPQTGGSAQITN-PSGGLVSSFTTYGPTNDFYFK
-ASFIQAADGEFLVQQFASGQNVSLSFPQSGGLIQFPD-PAGGLISSFTSYGPSNDFFFK
-AALISETDGTFLVNQFAAKANITISFPQSGSAWNYPDDVSGGLVSDFSSYGPSYDMYFK
VGSLRYEDGLKLLSYYQSSPRGQSISFPNGPLAPYVSNTVSGGVVASYSNFGPTNELFIY
VAGLRRSDGLKLLSYYQNANKRLTLRFPKGKIVAGLTDTITGGLISGYSTFGPTNDLYGQ
lowast lowastlowast lowastlowast lowastlowast
PAFGAPGGNILSTLPVPLGSYGVESGTSMATPYVAGSAALLLSVKGKSAAVALGARTLFQ
PAVAAPGGNILSTLPVNLGLFGIESGTSMATPFVAGSAALLFQAKGTSAAVGRSARTLFE
PAVAAPGGNILSTLPIALGSYGVESGTSMATPFTAGSAALILQAKGK--SVAKSIRNLFQ
PTLAAPGTDILSTV---TGGVALMRGTSMAAPLVAGAYALLLSVGNNLKLSPEEARTILM
PTLSAPGGNILSTFPLSEGGVAVISGTSMSCPFVAGSAAVLMAARASENLTPLEIRSLLT
lowast lowastlowastlowast lowastlowastlowastlowast lowast lowastlowastlowastlowast lowast lowastlowast lowast lowast
STAQRVPSSHTDGDPLQTLSQQGAGLIDVFKALYTETIVWPTELILNDTAHYKEVHTFTV
TTAQRVPSTHTDGDPLQTLTQQGAGLINVFNAIHTTTIVSPGELILNDTAHFKGTQQFTV
TTASAVASSKTDGDPLQTVSQQGAGLIQVDRAIGTTTVVSPGQLTLNDTAHFEGFKTFNI
STSQPSPVAY-GSETLDTVVSQGAGVINMTAAIAAHTVISPAQFNLNDTQYSNNTQTLTL
TTAKLTPVSLLGSTPLVSVIRQGGGLVQVAKALAAKTLISPHELLLNDTANANYVQTIKI
VNTGKTSKQYKLTHVPAGTAQTIT---SGTIFAADGPVPLSTDYATVKLSSSSFTLRPGQ
RNAGSSAKTYTLRHIPAGTAVTVT---PGTIFPADGPVPLSTDFASVSLSTSKFTLAPGK
QNTGSKAVTYKISHVPAGTAASVQ---SGSIFAADGPVPLTADYAQVTFSLNKVTVLPGV
RNNNRFPVSYQLSWVDSTGIVTYNDGATTDIIPSTTPNYVSTSVLRVAFSQRTVTVPGGQ
KNTNSWAMKYTFSSAVAQGLGTFD--ASGDILPTLDPVAVSGAQATVAFNTRILSVAPGA
lowast lowast lowast lowast lowast lowast
SKQVTTHINAP--SGVDASTYPVFSGFIEIATAT--------ESVHVTYLGLAASLKDKQ
TQTVTARFTPP--AAADPSTFPVFSGFIQIESGT--------EQVQVSYLGLKASLKDKQ
SVPILVWFKPP--QGVDASTYPVYSGFIEIAGLG--------ETLRVAYLGVAASLKDKA
TVKVTAQFIPPNLTAQQRNQFPIYSGFVTITGQGQGAGAEQMEKYNLPFFGLGARMVDMP
TGSVVATITPPVLPVADAARFPIFSGWIRVNGQG-ARDSSRNEAYTVPYFGLAAKMIDMQ
lowast lowast lowastlowast lowast lowast lowast lowast
IVDNTNEFFGITIPAVLNNTGDAQ-LSPTNYTFSGSDFPTLLLRLAFGTPLLRVDLVDAT
VIDNTDFFFGVPTPVLTDPNGEVQ-TSPRNYSFLASDFPTLIFRLAFGSPKVVVDLVSPT
VVDNTDEYFGVKIPALLDASGDVQ-GNSTNYTLVGDDVPSLLYRFAFGSAHVRVDLVEKA
VLDTTNVALGP--NLPFIAVGQN--IQTGPTTFSESNPAVVYFRLAAGTRRLRVDLVDAS
VLDTTETIYGPGYAYPFVIDDAIGDIQSTTTSYSRNLGPTVFARFATGTLHYSLDLVLAD
lowast lowast lowast lowast lowastlowastlowastlowast lowast
IDLKPTLNSREIDDKRHVFERESFSFPHPGPKGS-------------------TFAQVKI
INFKPTLNTRALGDTHGSF----FSFPHRVKTG--------------------SFAQVPT
STVSATLSKRNWWN---------WWWPQKPSN---------------------SYDKVPI
TSYNATVPAVTNPASRLVKRSNTELTPRAAFPT--------------------LYSQVRT
IAFTPTYPNSS-PATRFVKRSLTQHTSAASHLAKRRVSIATINPKATLVADRQLHSDVPI
lowast lowast
VGTLSEQSHIPRNSDASTVDDGGYSTFAFNKP------SFANGTTIPNGTYRWLLRALKV
LGTVFELDFTSRNNDDPTDN--GFNTVAIDPP------TFANGTTIPNGQYRLLLRALRV
VGTLAEYDYVPRNSDASTIDDNGYNVFQFT-D------SFVNGTKIAAGEYKVLLRALKV
VGSLFAPSYWPPRDYLFNNGGT-YSDYEIQLDGSFTFPNGTKGMAELGRSYKVLLRALKI
EGNIFTQPFTG-RDYLVDAAPTGSTDRTVTFNGQY-AENGLVRTAVTGTSYRFLLRALKI
lowast lowast lowastlowast lowast lowast lowast lowast lowastlowastlowastlowast
lowast lowast lowastlowastlowastlowastlowast
(b)
Figure 5 Continued
Enzyme Research 15
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
TGDPTKEADFETWLSPIVGITVA
TGDPTNEADFESFLSPIIGVNAP
TGNPAEEGDYETWLSPSFSVVPA
TADPTYTSSYESWLSPSFTFTA-
SGDAMYEDQYESWLSLPFSFRA-
lowast lowastlowast
ConservedSimilar
lowast All match
(c)
Figure 5 Multiple sequence alignment between amino acid sequence of PI12 protease with other subtilisin The serine protease sequenceswere obtained from the GenBank database Their accession numbers are EMS20793 (subtilisin-like protease of Rhodosporidium toruloidesNP11) CAG25549 (subtilisin-like protease of Pleurotus ostreatus) EPQ55564 (subtilisin-like protease of Gloeophyllum trabeumATCC 11539)and ETW80907 (serine protease S8 ofHeterobasidion irregulare TC 32-1) Putative ORF of serine protease fromG antarctica PI12 was labeledas PI12 protease
CAG38357 Phanerochaete chrysosporium
EIW62675 Trametes versicolor FP-101664 SS1
XP 007369624 Dichomitus squalens LYAD-421 SS1
XP 007381302 Punctularia strigosozonata HHB-11173 SS5
CAG25549 Pleurotus ostreatus
ETW80907 Heterobasidion irregulare TC 32-1
XP 001877576 Laccaria bicolor S238N-H82
EPQ55564 Gloeophyllum trabeum ATCC 11539
XP 001830835 Coprinopsis cinerea okayama7130
XP 007341385 Auricularia delicata TFB-10046 SS5
CCA68627 Piriformospora indica DSM 11827
CCO37249 Rhizoctonia solani AG-1 IB
XP 007342189 Auricularia delicata TFB-10046 SS5
EJU04161 Dacryopinax sp DJM-731 SS1
XP 007589863 Colletotrichum fioriniae PJ7
EFQ32840 Colletotrichum graminicola M1001
ENH74885 Fusarium oxysporum f sp cubense race 1
XP 003711175 Magnaporthe oryzae 70-15
XP 006694395 Chaetomium thermophilum DSM 1495
EON99722 Togninia minima UCRPA7
CBQ73425 Sporisorium reilianum SRZ2
GAC97617 Pseudozyma hubeiensis SY62
CCF52619 Ustilago hordei
EMS20793 Rhodosporidium toruloides NP11
CAQ76821 Glaciozyma antarctica PI12
01
Clus
ter 1
Clus
ter 2
a
b
c
d
Figure 6 Phylogenetic dendrogram among 25 translated amino acid sequences of serine proteaseThe tree was constructed using a neighbor-joining algorithm with 1000 bootstrappings by MEGA 6 software [39]The scale bar represents 01 substitutions per amino acid positionTheserine protease sequences were obtained from the GenBank database GenBank accession numbers of the sequences were indicated beforethe strain name The putative serine protease of G antarctica strain PI12 has been deposited in GenBank with accession number CAQ76821
16 Enzyme Research
0
2
4
6
8
10
12
0 24 48 72 96 110
Prot
ease
activ
ity (U
mL)
Post induction time (hours)
Figure 7 Effect of postinduction time towards protease activity The highest activity was detected at 72 hours after 110 hours of incubationwith 9UmL activity followed by 48 24 and 96 hoursThe substrate (azocasein) was dissolved in Tris HCL buffer and incubated for 30min at20∘CThe absorbance was measured at 450 nm after 15min incubation A one-way ANOVAwas used to test for preference differences amongsix different postinduction times The statistical value (119875 value lt005) between hours is 0003
pink-coloured nonpathogenic oleaginous yeast (42 iden-tity) Both G antarctica PI12 and Rhodosporidium toruloidesNP11 belong to Microbotryomycetes class PI12 protease didnot show any homology similarity to other psychrophilicproteases however it showed 37 identity to hypotheticalprotein from Pseudozyma antarctica T-34 which was isolatedfrom Japan (GAC76005) (not listed in the phylogenetic tree)
35 Cloning and Expression of Cold-Adapted PI12 Protease inthe Yeast Expression System The PCR product harbouringthe mature PI12 protease with two flanking restriction sites(EcoRI and AvrII) was cloned into pPIC9 expression vectorThe recombinant plasmid harbouring PI12 protease was usedto transform both GS115 and KM71 intended to produce bothHis+ Mut+ and His+ MutS Two recombinant clones fromGS115 (His+Mut+) strain (GpPro1 andGpPro2) and one fromKM71 (His+MutS) strain (KpPro1) were successfully obtainedwhich were further analyzed for protein expression study
36 Expression and Enzyme Assay of PI12 Protease in Pichiapastoris All positive recombinants were grown in small scalecultures and assayed for the cold-adapted protease activityin the culture supernatants The clones were first grown inBMGY medium with glycerol as the sole carbon sourceFor induction of AOX1 promoter and protein productionthe growth medium was transferred to the same mediumexcept with 05 methanol in place with glycerol as thecarbon source [26] The supernatant of the recombinant Ppastoris expression cultures after 48 h of incubation time wasanalyzed to determine the recombinant protease activity bythe modified method of Brock et al (1982) [20] All testedclones secreted protease into the supernatant in differentexpression levels This might be due to the variability ofthe strength of the promoter and copy number in each cellto secrete and express the recombinant product The clonewith the highest yield was GpPro2 with 63UmL activityfollowed by KpPro1 and GpPro1 with 25UmL and 37UmLactivities respectively (data not shown) It is perceptible thatexpression systemof Pichia pastoris is the ideal system for thisprotein The preliminary assay has fruitfully produced high
protease expression at 20∘C Yet further optimization of therecombinant PI12 protease production need to be conductedin order to examine the optimum condition of this proteinReported cold-adapted proteases showed optimum activity ata range of 15∘Cndash60∘C [27ndash30] This difference was possiblyrelated to the respective genetic and physiological adaptationof the strains There are several factors that influenced thedifferent rate of expression It includes different site of inte-gration between interest gene and Pichia genome variabilityof different expression host cell to secrete and express therecombinant enzyme and different sites in the polylinker forinserting foreign genes [31]
This is the second extracellular protease reported to besynthesized by this obligate psychrophilic yeastG antarcticaThe first cold-adapted extracellular subtilase from this yeasthas been purified and characterized previously [32 33] Theextracellular serine proteinase (LAP2) from this Antarcticyeast formerly known as Leucosporidium antarcticum strain171 is specific towards synthetic substrates of chymotrypsinand subtilisin The subtilase of L antarcticum strain 171showed specific activity of 4043Umg when utilized N-SucAAPFpNA as a substrate [33]
37 Optimization of Protease Production GpPro2 was chosenfor further optimization due to its highest protease activityat the preliminary expression stage High protease activitywas detected after 72 hours of incubation (Figure 7) byusing the standard protocol method as recommended forPichia pastoris expression system (Pichia Expression KitInvitrogen USA) The expression decreased after 72 hoursand no activity was detected at 110 hours The prolongedincubation may result in the decrease of biomass productionand will subsequently reduce the expression of recombinantprotein In addition oxygen and nutrient depletion duringprolonged incubationwill trigger the stress responses in orderto survive in such harsh environment [34]
Methanol was used as a sole carbon by P pastoris Themethanol was oxidized to formaldehyde with the aid of oxy-gen as electron acceptor by an alcohol oxidase enzyme (AOXgene) [35] The optimal methanol concentration is at 05(vv) with 14UmL activity The addition of more than 05
Enzyme Research 17
02468
10121416
0 01 03 05 07 09Pr
otea
se ac
tivity
(Um
L)Methanol concentration ()
Figure 8 Effect ofmethanol concentration towards protease activityThe recombinant was cultivated for 72 hours at 30∘C in BMMYmedium05 methanol gives the highest activity with 14UmL The substrate (azocasein) was dissolved in Tris HCL buffer and incubated for 30minat 20∘CThe mixture was left for 15min and absorbance was taken at 450 nm A one-way ANOVA was used to test for preference differencesamong six different concentrations The statistical value (119875 value lt005) between concentration is 002
0
5
10
15
20
25
10 15 20 25 30 35
Prot
eas a
ctiv
ity (
Um
L)
Temperature (∘C)
Figure 9 Effect of temperature towards protease activity The reaction was carried out in temperature ranging from 10∘C to 35∘C at pH 7 for30min A one-way ANOVA was used to test for preference differences among six different temperatures The statistical value (119875 value lt005)between temperature is 0067
(vv) methanol resulted in tremendous decrease of proteaseactivity (Figure 8) Production of PI12 protease was decreasedwhen the methanol concentration increased This might bedue to the toxic effect of methanol concentration on DNAreplication and membrane synthesis Basically methanol isused to increase the permeability of the cell membrane tohelp the secretion of foreign protein [26] However if thepermeability of cell membrane is too high lowmolecular sizemetabolites would leak from the cell and such leakage willseriously damage cell activities Such condition would abusethe cellular system and might reduce the yield of proteinexpression [36]
Psychrophilic microorganism dominated in low temper-ature habitat which most other microorganism are inca-pable of withstanding due to their inability to produceenzyme that can tolerate the extreme environment con-dition Psychrophilic enzymes are always associated withhigh catalytic efficiency at low and moderate temperature(0∘C to 30∘C) compared to the mesophilic counterpart thatare generally unable to catalyze efficiently under extremely
low temperature [2] The recombinant protein assay wasdone at various temperatures ranging from 10∘C to 35∘Cusing 05 azocasein as a substrate dissolved in 50mMTris HCl in presence of 2mM CaCl
2 pH 7 In the activity
assays expressed protease showed a highest activity at 20∘C(20UmL) and decreased tremendously once it exceededmaximum temperature due to the denatured state of theenzyme (Figure 9) The active site of an enzyme is easilyaltered when the high temperature is introduced duringincubation or activity assay resulting in a low reaction rate[37]
The recombinant construct was transferred into BMMYmedia at different A
600 nm ranging from 1 to 10 The high-est activity of active enzyme was detected at A
600 nm 60(283UmL) and above (Figure 10) The slope during expo-nential phase was much higher compared to other phasesindicating an increase in the doubling time At exponentialphase the cells are adapted to the growth condition andother processes like cell growth cycle and synthesis of RNAThe yield of active enzyme during expression of P pastoris
18 Enzyme Research
05
1015202530
1 2 3 4 5 6 7 8 9 10Prot
ease
activ
ity (U
mL)
Initial inoculum A600
Figure 10 Effect of inoculums size towards protease activity The recombinant was cultivated for 72 hours at 30∘C in BMMY mediumwith 05 methanol concentration A one-way ANOVA was used to test for preference differences among six different optical densitiesThe statistical value (119875 value lt005) between optical density value is 003
(kD
a)
1160
662
450
350
250
184
144
M 1 2 3
(a) (b) (c)
Figure 11 (a) Coomassie-stained SDS-PAGE of recombinant P pastoris clone 1mL supernatants collected from the third-day culturewere concentrated 10-fold using trichloroacetic acid and 10120583L resultant products were applied to SDS-PAGE analysis Lane 1 molecularweight marker (Fermentas) Lane 2 GS115 (Control) Lane 3 GS115pPIC9 empty (Control) Lane 4 GS115pPIC9PI12prot (GpPro2)The arrow indicates the secreted recombinant PI12 protease (b) Plate activity staining of different recombinant P pastoris clones GGS115 GP GS115pPIC9 GRI GS115pPIC9PI12prot1 (GpPro1) GRII GS115pPIC9PI12prot2 (GpPro2) K KM71 KP KM71pPIC9 KRKM71pPIC9PI12prot (KpPro) (c)Gel activity staining of recombinant PI12 protease inP pastorisGpPro2 clone Lane 1 prestainedmolecularweightmarker (Fermentas) Lane 2 GS115pPIC9 empty (Control) Lane 3 GS115pPIC9PI12prot1 (GpPro2)The arrow indicates the secretedrecombinant PI12 protease
dropped after 28 hours of incubation which is at stationaryor death phase [38]
38 Detection of Recombinant Protein by SDS-PAGE andActivity Staining TheSDS-PAGE gel of the GpPro2 recombi-nant clonesrsquo supernatant showed a putative expression bandat around 993 kDa Plate staining and gel activity stainingwere executed to examine if the recombinant PI12 proteaseexpressed in P pastoris was functionally active In the platestaining the transformants showed strong proteolytic activity(Figure 11(a)) In gel staining proteolytic activity was alsodetected around 993 kDa after incubation at 20∘C This is
evidence of protease secretion into the surrounding mediawhereby it catalyzed the breakdown of casein thus formedclearing zones around the activity ring and band (Figures11(b) and 11(c)) As a conclusion the cold-adapted PI12protease was functionally expressed in P pastoris GS115(GpPro2 clone) with the size of 993 kDa
4 Conclusions
Psychrophilic yeast which was originated frommarine waterin Casey Station Antarctica was identified as Glaciozymaantarctica strain PI12 A serine protease with an open reading
Enzyme Research 19
frame of 2892 bp encoded for 963 amino acids from thisAntarctic yeast has been cloned and expressed in Pichiaexpression system This is the second extracellular pro-tease reported being synthesized by this psychrophilic yeastSequence analysis through bioinformatics studies revealedfew homology identities of this protein with other ser-ine proteases The mature PI12 protease was successfullyexpressed and secreted into the culture medium driven bythe Saccharomyces cerevisiae 120572-factor signal sequence in Ppastoris The protease production in P pastoris was bestobtained from strain GS115 (GpPro2) with 283UmL after 72hours of induction time with 05 (vv) of methanol inducerThe starting optical density at 600 nm (OD600) was 60 andthe optimum assay temperature was 20∘C The expressedprotein was detected by SDS-PAGE and activity staining witha molecular weight of 99 kDaThis cold-active enzymemightoffer an opportunity for biotechnological exploitation basedon its high catalytic activity at low temperatures and lowthermostability Structural and functional study of this cold-adapted enzyme would give insight into its adaptation to anextreme cold environment
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgment
This research was supported financially by the InitiativeBiotechnology-GMBI-MOSTI (Project number 07-05-16-MG-GMB02) andRUGS-Initiative 6 (Project number 05-02-11-1414RU)
References
[1] J W Deming ldquoPsychrophiles and polar regionsrdquo CurrentOpinion in Microbiology vol 5 no 3 pp 301ndash309 2002
[2] C Gerday M Aittaleb M Bentahir et al ldquoCold-adaptedenzymes from fundamentals to biotechnologyrdquo Trends inBiotechnology vol 18 no 3 pp 103ndash107 2000
[3] A Svingor J Kardos I Hajdu A Nemeth and P Zavodszky ldquoAbetter enzyme to cope with cold comparative flexibility studieson psychrotrophic mesophilic and thermophilic IPMDHSrdquoThe Journal of Biological Chemistry vol 276 no 30 pp 28121ndash28125 2001
[4] G Antranikian C E Vorgias and C Bertoldo ldquoExtremeenvironments as a resource for microorganisms and novel bio-catalystsrdquo Advances in Biochemical EngineeringBiotechnologyvol 96 pp 219ndash262 2005
[5] T Lonhienne C Gerday andG Feller ldquoPsychrophilic enzymesrevisiting the thermodynamic parameters of activation mayexplain local flexibilityrdquoBiochimica et Biophysica Acta vol 1543no 1 pp 1ndash10 2000
[6] R Cavicchioli and TThomas ldquoExtremophilesrdquo in Encyclopediaof Microbiology pp 317ndash337 Academic Press San Diego CalifUSA 2000
[7] G Feller E Narinx J L Arpigny et al ldquoEnzymes frompsychrophilic organismsrdquo FEMS Microbiology Reviews vol 18no 2-3 pp 189ndash202 1996
[8] J Arnorsdottir R B Smaradottir T O Magnusson S HThorbjarnardottir G Eggertsson and M M KristjanssonldquoCharacterization of a cloned subtilisin-like serine proteinasefrom a psychrotrophic Vibrio speciesrdquo European Journal ofBiochemistry vol 269 no 22 pp 5536ndash5546 2002
[9] N J Russell ldquoPsychrophilic bacteriamdashmolecular adaptations ofmembrane lipidsrdquo Comparative Biochemistry and Physiology APhysiology vol 118 no 3 pp 489ndash493 1997
[10] G Feller and C Gerday ldquoPsychrophilic enzymes hot topics incold adaptationrdquo Nature Reviews Microbiology vol 1 no 3 pp200ndash208 2003
[11] P Raspor and J Zupan ldquoYeasts in extreme environmentsrdquo inBiodiversity and Ecophysiology of Yeasts The Yeast Handbookpp 371ndash417 Springer Berlin Germany 2006
[12] M B Rao A M Tanksale M S Ghatge and V V Desh-pande ldquoMolecular and biotechnological aspects of microbialproteasesrdquoMicrobiology andMolecular Biology Reviews vol 62no 3 pp 597ndash635 1998
[13] L Polgar ldquoThe catalytic triad of serine peptidasesrdquo Cellular andMolecular Life Sciences vol 62 no 19-20 pp 2161ndash2172 2005
[14] S Y Boo C M V L Wong K F Rodrigues N NajimudinA M A Murad and N M Mahadi ldquoThermal stress responsesin Antarctic yeastGlaciozyma antarctica PI12 characterized byreal-time quantitative PCRrdquoPolar Biology vol 36 no 3 pp 381ndash389 2013
[15] B Turchetti S R T Hall L B Connell et al ldquoPsychrophilicyeasts from Antarctica and European glaciers descriptionof Glaciozyma gen nov Glaciozyma martinii sp nov andGlaciozyma watsonii sp novrdquo Extremophiles vol 15 no 5 pp573ndash586 2011
[16] N H F Hashim I Bharudin D L S Nguong et al ldquoCharac-terization of Afp1 an antifreeze protein from the psychrophilicyeast Glaciozyma antarctica PI12rdquo Extremophiles vol 17 no 1pp 63ndash73 2013
[17] A N M Ramli M A Azhar M S Shamsir et al ldquoSequenceand structural investigation of a novel psychrophilic 120572-amylasefrom Glaciozyma antarctica PI12 for cold-adaptation analysisrdquoJournal ofMolecularModeling vol 19 no 8 pp 3369ndash3383 2013
[18] A N M Ramli N M Mahadi A Rabu A M A Murad F DA Bakar and R M Illias ldquoMolecular cloning expression andbiochemical characterisation of a cold-adapted novel recombi-nant chitinase from Glaciozyma antarctica PI12rdquoMicrobial CellFactories vol 10 no 1 article 94 2011
[19] J Sambrook E Fritsch and T Maniatis Molecular Cloning ALaboratoryManual Cold SpringHarbor Laboratory Press NewYork NY USA 1989
[20] F M Brock C W Forsberg and J G Buchanan-SmithldquoProteolytic activity of rumen microorganisms and effects ofproteinase inhibitorsrdquoApplied and EnvironmentalMicrobiologyvol 44 no 3 pp 561ndash569 1982
[21] U K Laemmli ldquoCleavage of structural proteins during theassembly of the head of bacteriophage T4rdquo Nature vol 227 no5259 pp 680ndash685 1970
[22] A Henne R A Schmitz M Bomeke G Gottschalk and RDaniel ldquoScreening of environmental DNA libraries for thepresence of genes conferring lipolytic activity on Escherichiacolirdquo Applied and Environmental Microbiology vol 66 no 7 pp3113ndash3116 2000
[23] M Poza T de Miguel C Sieiro and T G Villa ldquoCharacteri-zation of a broad pH range protease of Candida caseinolyticardquoJournal of Applied Microbiology vol 91 no 5 pp 916ndash921 2001
20 Enzyme Research
[24] J D Bendtsen H Nielsen G von Heijne and S BrunakldquoImproved prediction of signal peptides SignalP 30rdquo Journalof Molecular Biology vol 340 no 4 pp 783ndash795 2004
[25] R J Siezen and J A M Leunissen ldquoSubtilases the superfamilyof subtilisin-like serine proteasesrdquo Protein Science vol 6 no 3pp 501ndash523 1997
[26] Y Katakura W Zhang G Zhuang et al ldquoEffect of methanolconcentration on the production of human 1205732- glycoprotein Idomain V by a recombinant Pichia pastoris a simple system forthe control of methanol concentration using a semiconductorgas sensorrdquo Journal of Fermentation and Bioengineering vol 86no 5 pp 482ndash487 1998
[27] Q-FWang Y-HHou Z Xu J-LMiao andG-Y Li ldquoPurifica-tion and properties of an extracellular cold-active protease fromthe psychrophilic bacterium Pseudoalteromonas sp NJ276rdquoBiochemical Engineering Journal vol 38 no 3 pp 362ndash3682008
[28] S Vazquez L Ruberto and W Mac Cormack ldquoProper-ties of extracellular proteases from three psychrotolerantStenotrophomonas maltophilia isolated from Antarctic soilrdquoPolar Biology vol 28 no 4 pp 319ndash325 2005
[29] R Zeng R Zhang J Zhao and N Lin ldquoCold-active ser-ine alkaline protease from the psychrophilic bacterium Pseu-domonas strain DY-A enzyme purification and characteriza-tionrdquo Extremophiles vol 7 no 4 pp 335ndash337 2003
[30] M Turkiewicz E Gromek H Kalinowska and M ZielinskaldquoBiosynthesis and properties of an extracellularmetalloproteasefrom the Antarctic marine bacterium Sphingomonas paucimo-bilisrdquo Journal of Biotechnology vol 70 no 1ndash3 pp 53ndash60 1999
[31] A Vassileva D A Chugh S Swaminathan and N KhannaldquoEffect of copy number on the expression levels of hepatitisB surface antigen in the methylotrophic yeast Pichia pastorisrdquoProtein Expression andPurification vol 21 no 1 pp 71ndash80 2001
[32] M Pazgier M Turkiewicz H Kalinowska and S Bielecki ldquoTheunique cold-adapted extracellular subtilase from psychrophilicyeast Leucosporidium antarcticumrdquo Journal ofMolecular Catal-ysis B Enzymatic vol 21 no 1-2 pp 39ndash42 2003
[33] M Turkiewicz M Pazgier H Kalinowska and S BieleckildquoA cold-adapted extracellular serine proteinase of the yeastLeucosporidium antarcticumrdquo Extremophiles vol 7 no 6 pp435ndash442 2003
[34] D Mattanovich B Gasser H Hohenblum and M SauerldquoStress in recombinant protein producing yeastsrdquo Journal ofBiotechnology vol 113 no 1ndash3 pp 121ndash135 2004
[35] P Secades BAlvarez and JAGuijarro ldquoPurification andprop-erties of a new psychrophilic metalloprotease (Fpp2) in the fishpathogen Flavobacterium psychrophilumrdquo FEMS MicrobiologyLetters vol 226 no 2 pp 273ndash279 2003
[36] L Y Ling I Ithoi and F M Yik ldquoOptimization for high-level expression in Pichia pastoris and purification of truncatedand full length recombinant sag2 of toxoplasma gondii fordiagnostic userdquo Southeast Asian Journal of Tropical Medicineand Public Health vol 41 no 3 pp 507ndash513 2010
[37] A Illanes ldquoStability of biocatalystsrdquo Electronic Journal ofBiotechnology vol 2 no 1 pp 1ndash9 1999
[38] B A Plantz J Sinha L Villarete K W Nickerson and VL Schlegel ldquoPichia pastoris fermentation optimization energystate and testing a growth-associated modelrdquo Applied Microbi-ology and Biotechnology vol 72 no 2 pp 297ndash305 2006
[39] K Tamura G Stecher D Peterson A Filipski and S KumarldquoMEGA6molecular evolutionary genetics analysis version 60rdquo
Molecular Biology and Evolution vol 30 no 12 pp 2725ndash27292013
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

Enzyme Research 7
JX896956 Glaciozyma antarctica strain PI12
AY040659 Antarctic yeast CBS 8938
AY033637 Glaciozyma antarctica strain CBS 8943
EU149809 Glaciozyma antarctica strain CBS 10640
AB774460 Leucosporidium sp KGK-2
JQ857017 Glaciozyma antarctica isolate T21Ga
EU149808 Glaciozyma antarctica strain CBS 10636
AY040663 Glaciozyma antarctica strain CBS 8939
EU149806 Glaciozyma antarctica strain CBS 10639
EU149805 Glaciozyma antarctica strain CBS 10638
AY040657 Antarctic yeast CBS 8927
AF444529 Glaciozyma antarctica strain CBS 5942
KF934487 Glaciozyma sp KP7-5-2
GQ336996 Leucosporidium sp AY30
EU149804 Glaciozyma watsonii strain CBS 10641
AY040661 Antarctic yeast ML 4515
AY040660 Glaciozyma watsonii strain CBS 8940
AY033638 Glaciozyma watsonii strain CBS 8944
EU149803 Glaciozyma watsonii strain CBS 10684
JX171176 Glaciozyma watsonii isolate LKF08-112
DQ402535 Leucosporidium sp LC-03-120 isolate 03-120
AB774462 Leucosporidium sp BSS-2
KC785578 uncultured Glaciozyma clone 180
HQ432818 Glaciozyma watsonii DBVPG4726
HQ432823 Glaciozyma watsonii DBVPG4802
AB774461 Leucosporidium sp BSS-1
HQ432820 Glaciozyma watsonii DBVPG4760
KC333171 Glaciozyma watsonii isolate AU CryS06
HQ432821 Glaciozyma watsonii DBVPG4799
JF900360 Glaciozyma watsonii strain CBS 7009
AY040664 Glaciozyma martinii strain CBS 8929
AY033641 Glaciozyma martinii strain CBS 8928
HQ432816 Glaciozyma martinii DBVPG4841
KF934486 Glaciozyma sp KP7-5-1
FR682434 uncultured Basidiomycota
HF934010 Glaciozyma sp K81b
JF900362 Glaciozyma martinii strain CBS 9639
JF900359 Glaciozyma martinii strain CBS 6581
JF900361 Glaciozyma martinii strain CBS 7054
001
Clus
ter 1
Glac
iozy
ma
anta
rctic
aCl
uste
r 3
Glac
iozy
ma
mar
tinii
Clus
ter 2
Glac
iozy
ma
watso
nii
Figure 3 Neighbor-joining phylogenetic analysis of internal transcribed spacer 1 (ITS1)58S rRNA geneinternal transcribed spacer 2 (ITS2)of Glaciozyma clade Numbers indicate percentage bootstrap values calculated on 1000 repeats of the alignment The tree was constructedusingMEGA6 software [39]The scale bar represents 001 substitutions per nucleotide position GenBank accession numbers of the sequencesare indicated before strain name ITS158SITS2 sequence of Glaciozyma antarctica strain PI12 had been deposited in GenBank with theaccession number JX896956 [14]
8 Enzyme Research
their molecular phylogenetic data and physiological andmorphological characteristics [15] Phylogenetic analysis ofITS158SITS2 sequence of strain PI12 showed that thesequence was closely related to ITS158SITS2 sequenceof Glaciozyma antarctica with high homology similarity toCluster I (99-100 identity) The ITS158SITS2 sequenceof G antarctica strain PI12 was differed from G watsonii(Cluster II) by 2-3 gap in the sequence (93 identity)and G martinii by 4-5 gap in the sequence (90ndash92identity)
33 Sequence Analysis of PI12 Protease cDNA In this studyamplification of the PI12 protease gene sequence was con-ducted using genomicDNAand cDNAas a template Putativepartial PI12 protease gene with the size of 1321 bp was initiallyacquired from the G antarctica genome survey sequencelibrary (Supplementary Figure see Supplementary Mate-rial available online at httpdxdoiorg1011552014197938)However the gene was discovered as an incomplete pro-tease gene sequence DNA walking strategy successfullyamplified 1500 bp of the upstream region of partial PI12protease Both sequences of about 1500 bp (derived fromthe DNA walking reaction) and 1321 bp (partial PI12 pro-tease) were assembled and made up a total length of2687 bp protease sequence (Supplementary figure)The puta-tive PI12 protease was predicted by Augustus webserver(httpaugustusgobicsde) having intervening sequencesintronsThus reverse-transcriptase PCR (RT-PCR) and rapidamplification of cDNA ends (RACE) were accomplished toobtain the full-length cDNA As a result the ORF comprising2892 bp encoding 963 amino acid residues were successfullydeduced
Nucleotide and deduced amino acid sequences ofgenomic DNA and cDNA fragment encoding PI12 proteasewere shown in Figure 4 The size of the ORF of putative PI12protease was 3867 bp with genomic DNA as the templatewhile the ORF comprised 2892 bp encoding 963 amino acidresidues were successfully deduced through RACE strategywith cDNA as the template The ORF was predicted usingORF Finder (httpwwwncbinlmnihgovprojectsgorf)while prediction using SignalP (httpwwwcbsdtudkser-vicesSignalP) webserver allowed the identification of theputative PI12 protease prepeptide where it predicted thepresence of signal peptidase I cleavage sites [24]The putativeN-terminal signal peptide (1 to 16 the underlined amino acid)was identified as a secretion signal There is no propeptidepredicted by ProP Webserver (httpwwwcbsdtudkser-vicesProP-10) Comparison of cDNA and gene sequencesallowed determination of gene structureTherefore sequencealignment was performed between the sequence of these twotemplates and it was confirmed that the coding sequence(consisted of 19 exons) was interrupted by 18 introns withtheir length varying from 48 to 60 nucleotides However theGT and AG rule for donor-acceptor sites was not obeyedOf 18 introns 10 introns contained canonical GT and AGdinucleotides at their 51015840 and 31015840 ends while 8 introns holdnoncanonical splice site pairs with TT-GG (2 introns)AG-TG GC-GT TG-GG TC-GG TA-GG and TG-AG
dinucleotides at the splice junction Upper case letters inblue colour represent introns while lower case letters inblack colour represent exons Splice donor and acceptor werehighlighted in grey
The amino acid composition (upper case letters) withinthe ORF amino acid residues was determined by theProtParam tool of the Expasy molecular biology server(httpwwwexpasyorgtool) Translation starts at a nucle-otide position 1 and translation stop is marked with anasterisk The nucleotide sequences ATG (1 to 3) and TAG(3865 to 3867) indicate the initiation codon and termi-nal codon respectively The presumed putative sequencesof promoter CAAT (minus101 to minus98) and TATA-box (minus92to minus88) were identified in the 51015840-untranslated (UTR)region The search was achieved via Neural Network Pro-moter Prediction (Berkeley Drosophila Genome Project)(httpwwwfruitflyorgseq toolspromoterhtml) In the 31015840-untranslated (UTR) region there was a putative polyadeny-lation signal attaa (3889 to 3893) detected from the TAGstop codon to the poly (A) tail The predicted molecularmass and pI of PI12 serine protease were 10099 kDa and641 respectively Out of 963 amino acid residues in thededuced sequence of the PI12 protease 70 are negativelycharged residues (Asp + Glu) while 66 are positively chargedresidues (Arg + Lys) at neutral pH (Table 2)The total chargedresidues (Arg Asp Glu His and Lys) are 150 amino acidsin the open reading frame of PI12 protease corresponded to156 while hydrophobic residues (Ala Phe Ile Leu MetPro Val and Trp) are 441 amino acids which correspondedto 458 of the total amino acids discovered The totaluncharged residues (Asp Cys Gln Gly SerThr and Tyr) areconsisted of 372 amino acids which corresponded to 386of the total amino acids of the open reading frame of PI12protease The aliphatic index is 9290 and the grand averageof hydropathicity (GRAVY) is 0141 The instability index (II)of PI12 protease was computed to be 3414 which classify theprotein as stable
N-glycosylation sites were predicted using NetNGlyc10 Server (httpwwwcbsdtudkservicesNetNGlyc)Therewere five putative N-glycosylation sites for the PI12 proteasegene predicted using the server For N-glycosylation thesequence motif Asn-Xaa-SerThr (Xaa any amino acid butnot Pro) has been defined as a prerequisite for glycosylationThe sequencemotifs in PI12 protease occurred in the position170 NETH 365 NATV 576 NLTP 634 NDTA and 833 NSSPAmong five predicted sites only three (except positions 576and 833) showed a high potential score (averaged output ofnine neural networks) which support the prediction Theputative N-glycosylation sites were demonstrated in boxesin Figure 4 The comparison of amino acid composition ofthis cold-adapted protease fromG antarctica strain PI12 withnine other subtilisin serine proteases fromdifferent families isshown in Table 2 Analysis of this comparison depicts that thepercentage of each amino acids in PI12 protease is almost thesamewith other proteaseswhileGly (G) is themost abundantaccounting for up to 104 of the total amino acid in PI12 pro-tease PI12 protease also contained the highest percentage ofLeu (L) with 99 of the total amino acids compared to other
Enzyme Research 9
Table2Aminoacid
compo
sitionandmolecular
weighto
fproteases
acrossdifferent
families
Amino
CAQ76821
CAG25549
EDR11679
CAG38357
CAD118
98CA
L25578
EDU50747
AAW
40780
ACB3
0121
ACB3
0119
acid
Qty
Qty
Qty
Qty
Qty
Qty
Qty
Qty
Qty
Qty
Ala
(A)
97101
8191
7887
7590
8299
101
115
8189
9196
8092
105
116
Arg
(R)
4042
2730
2224
1619
1721
1821
2932
2223
2124
3134
Asn
(N)
4446
3843
4348
4149
4251
4046
5257
5154
4552
4347
Asp
(D)
4951
6269
5460
6578
5567
5158
5358
6669
5260
6370
Cys
(C)
808
506
404
405
911
809
910
909
708
1011
Gln
(Q)
2324
3438
2326
2834
3036
3742
3539
2931
1922
2629
Glu
(E)
2122
1618
1921
2024
1518
2023
2528
2627
2832
2426
Gly
(G)
100
104
89100
8493
85102
85103
7788
8493
8084
8598
8695
His
(H)
1415
1416
1314
1619
1316
1113
1618
1415
1618
1213
Ile(I)
5355
4955
5157
5364
4150
3843
5156
4446
3540
4449
Leu
(L)
9599
6876
7179
6274
6073
6675
6774
8084
7485
6875
Lys
(K)
2627
2932
4449
2935
4959
5158
4044
3032
5058
5662
Met
(M)
1212
202
202
911
1113
809
1820
808
1113
1517
Phe
(F)
4143
5966
4752
5566
2935
3641
4449
3941
3439
3438
Pro
(P)
6264
6573
6774
5870
5870
5967
5055
5154
5564
5763
ser
(S)
7780
6977
91101
6072
6680
7080
7279
111
117
6373
6167
Thr
(T)
8790
89100
7583
83100
6579
7383
7178
7276
7182
6875
Trp
(W)
505
303
607
202
1215
1214
1213
909
1214
1415
Tyr
(Y)
3334
1213
2629
708
2935
2832
3943
5053
2630
2730
val
(V)
7679
8292
8089
6679
5769
7484
5864
6872
8295
6268
Total
963
1000
893
1000
900
1000
834
1000
825
1000
878
1000
906
1000
950
1000
866
1000
906
1000
MW
(Da)
1009898
932669
944365
869455
869549
924905
979343
1009629
917209
961988
Thea
ccessio
nnu
mbersof
proteasespu
tativ
eserinep
roteaseo
fGlacio
zymaantarctica
PI12
(CAQ
76821)sub
tilisin-lik
eproteaseP
leurotuso
streatus(CA
G25549)pyrolysin
LaccariabicolorS
238N
-H82
(EDR11679)
subtilisin
-like
proteasePh
anerochaetechrysosporiu
m(C
AG38357)sub
tilisin-lik
eserinep
roteaseP
R1CMetarhiziu
maniso
pliaev
aranisopliae(CA
D11898)serinee
ndop
eptid
aseH
ypocrealixii(C
AL2
5578)peptidase
Pyrenophoratritici-repentis
(EDU50747)p
eptid
aseCr
yptococcus
neoforman
svarn
eoform
ansJEC
21(A
AW40
780)sub
tilisin-lik
eprotease
Epich
loefestucae(A
CB30121)sub
tilisin-lik
eprotease
Epich
loefestucae
(ACB
30119
)
10 Enzyme Research
atgctcttcctccccgtcctcctcctcctccttcccggcgtcactgccttcctcaaccca 60
M L F L P V L L L L L P G V T A F L N Pgttaccaaccgcgcgaccaacgccatctcctcgactcaatacctctctaacgcctatatc 120
V T N R A T N A I S S T Q Y L S N A Y IctcgaattggacctctccacccccggcctcgtcaaacgggatagcacgcccgactctGTG 180
L E L D L S T P G L V K R D S T P D SCGTCCCGCGTTCCTCCCTCGCTGTCGAAGACGAGCTAACAATGACGTGGTAGatcctaga 240
I L Eggacgtactcacgtccgtaggccgcaacggtatcaagtaccaactccgccaccgctttat 300
D V L T S V G R N G I K Y Q L R H R F Ictccccgactctgttccacggcgcttcgatcactgtcccccctggaatctcccgctccca 360
S P T L F H G A S I T V P P G I S R S QaatcgcctctctccgcggtatcaaaGTCCGTCCCTCTCTCTCCCCTACTCCCTCCGTGCT 420
I A S L R G I KAACCCTGCACGCAACAGcgcgtctggcccgttcgaaagttctcccgacccagcgcagtag 480
R V W P V R K F S R P S A Vtggacgccgatggaggaggaagcgggttctcagggtcgcctatcaaggcggcgctcatgg 540
V D A D G G G S G F S G S P I K A A L Mgggtgaaagagctcgggaagcgcgcgaacgcttatgctggagatacgtttggaccgcatg 600
G V K E L G K R A N A Y A G D T F G P HtcatgacgggggttaatgagacgcatgaggcggggttgttgggagctgggattaagGTGT 660
V M T G V N E T H E A G L L G A G I KGTTTTCGTCTTTGTTTGGGGGAGGGGGGAGGGAGGAGCTGATTAATGGGTGCAGattggg 720
I GgtgctggacactggtgAGAAGCGGTTTGGAGGGAGGAGAGGATGGAGCTGATGAGTGTGC 780
V L D T GAGGTGttgattatttgaacccgattctgggaggctgctttggacctgggtgccatatgtc 840
V D Y L N P I L G G C F G P G C H M Sgtttgggtacgacttggttggcgatgattacgatggagataacgctcctgtgccggatgt 900
F G Y D L V G D D Y D G D N A P V P D VggatccttGTGCGTCCTTCCCTCCGCGATGGGCTGGAATCTGAGCTGACGGCGTTGTTGA 960
D PTCAGttgcgagctgcgatcctcatggTTCGTCTCGCTTTCCGCTCCTCTGATGCGCTCGC 1020
F A S C D P H GTGATGCACTTGGCAATAGGaactcacgttacgggaatcattggagcgctcccgaatgcgt 1080
T H V T G I I G A L P N AttggatttactggcgtcgcacccgccgctactctgggccactaccgagtGCGCTCCCTCT 1140
F G F T G V A P A A T L G H Y R VATCCGTTCTTCCTCGCAGCTCCCACTCACTCGTGCTTCCAGGTatttggctgcactggct 1200
F G C T GtcgtcggagaagatatcGTTCGTCCTCTCTCCAGTCCCGGCTAGCGATCCTGCTTCTGAC 1260
F V G E D ITTCGCATCTCCCTCTGAGattctcgctggactcatgcgaggagtcgaggacaactgcaac 1320
I L A G L M R G V E D N C Ngtcttgaccctctctctcggaggtccaggagggtgggtcaagggcacgccggcgtccatc 1380
V L T L S L G G P G G W V K G T P A S Icttatcgaccagatcgaagcgcaaggcattctcgtcaccgtcgccactggcaactcggga 1440
L I D Q I E A Q G I L V T V A T G N S GgctgagggTGAGCTCCTCGCTACTTGCGCGTCGAGACCAGTACTAACAGGGGCGTGGACG 1500
A E GCAGGcatgttcttctccgagtctcccgcctcgacgatcaacggcctttccatcgcatcca 1560
M F F S E S P A S T I N G L S I A ScgTCCGTCTCTCCCTTCCTTCGCACGCCACTCGCTCACACTCTCTCGCAGGgacgttacc 1620
T D V Tgacctcatcgcctacaacgccaccgtctcaggccaacctgcgatcccttacctctccgcc 1680
D L I A Y N A T V S G Q P A I P Y L S A
ggggggggggtctg minus194
ctagcgcgaatcgagaaaacacaaaagaacctcccgacttggcgttgcttctgctcttct minus180
ccttctcccaaccgccgacaataaaaactataacacacacacaccccatctcgacgcctg minus120
cagacttcacgactctcctcacgctctttaccgccccacgggtcccacccacgagccacg minus60
(a)
Figure 4 Continued
Enzyme Research 11
acgcccctcaacgtcgtcgccaacagcttccgcgtccacttcacctctaccgaccccaac 1740
T P L N V V A N S F R V H F T S T D P Naaccccgtcgacgcctgctctcctcttccggctggagcgcccgacttcgccaactatgtt 1800
N P V D A C S P L P A G A P D F A N Y Vacggtcgttcagcgtgggacttgtacgttcgttaccaagtaccagaacgttctcaatgct 1860
T V V Q R G T C T F V T K Y Q N V L N AggaggGTGCGTCGGCTTCTTCTCTCCCGACTGGATTGGGCGGAGACTGACCTTCTGAGCA 1920
G GGaaagatcgtattgttgtacaactcggagggagctgggaacctcccttacctcacgccca 1980
K I V L L Y N S E G A G N L P Y L T PacggtgtcggcatcgacgccgttgcaggtcttcgtcgttccgacggactcaagGTTCGTC 2040
N G V G I D A V A G L R R S D G L KTACGCTCTTCGGTGCTGAGGCCCTTCTCTGATCTCGCTCCGACAGcttctctcgtactat 2100
L L S Y Ycagaatgccaacaagcgtctcactctgcgcttccccaagggcaagatcgtcgcaggcttg 2160
Q N A N K R L T L R F P K G K I V A G LaccgataccatcaccggcggactcatctcgTTCGTTCCTCTCCGCCTCCCTCGCTGGCCG 2220
T D T I T G G L I SATGAGCTGACGCGGTCTTTGCAGGggttactcgacgtttggtccgacgaatgacctctac 2280
G Y S T F G P T N D L Yggtcagcctaccctctctgcccctggtggcaacatcctttcgaccttccctctctccgag 2340
G Q P T L S A P G G N I L S T F P L S EggaggagtggcggtcatcagtgggacgagcatgtcgtgcccctttgtcgGTTCGTCCTTC 2400
G G V A V I S G T S M S C P F VTCCTCGTTCGTTCGCTGAGCGTCGACTGATCTTGCACGATCGCAGctggatctgcggcgg 2460
A G S A Atcctcatggccgctcgcgcttcggagaacctcacgccgcttgagatcaggagtctcctta 2520
V L M A A R A S E N L T P L E I R S L Lctaccactgcgaagcttacgccggtctcgctcttgggatcgacgcctttggtgagcgtga 2580
T T T A K L T P V S L L G S T P L V S VttcgtcaaggaggaggTACGTTCCTTTCTCGCTGCTTTGAGGATCGAGGCTGATTAGCGA 2640
I R Q G G GGCTTCGCAGGactcgttcaggttgccaaggcgctcgcggccaagacgctaatctctcctc 2700
L V Q V A K A L A A K T L I S Pacgagctcctgctcaacgacactgcgaacgcgaactacgtccagactatcaagatcaaga 2760
H E L L L N D T A N A N Y V Q T I K I Kacaccaactcgtgggcgatgaagtacaccttctcctcggccgtcgcccaaggactcggaa 2820
N T N S W A M K Y T F S S A V A Q G L Gctttcgacgcttcgggcgatatcctccctaccctcgacccagtcgccgtctctggcgcac 2880
T F D A S G D I L P T L D P V A V S G Aaggctaccgtcgcgttcaacactcggatcctcagcgtcgcgcccggcgcgacggggtccg 2940
Q A T V A F N T R I L S V A P G A T G Stcgtggcgactatcacgccgccggttcttcccgtagcggacgctgcgaggttccctatct 3000
V V A T I T P P V L P V A D A A R F P Itctctgggtggatcagggtgaatgggcaaggcgcgagggatagcagtaggaacgaggcgt 3060
F S G W I R V N G Q G A R D S S R N E AacactgGTGCGTTTGGCTCGCTTTTTGGGAGGTGCCGAGCTAATCGTTTCTCGTAGtccc 3120
Y T V PgtactttgggcttgcggcgaagatgatcgatatgcaagGTGCGTCGATGGTTCTCTCGCT 3180
Y F G L A A K M I D M QTCTCTCTCGGTTGCTCACTTCAAAGTGCAGtcctcgacaccaccgagaccatttacggtc 3240
V L D T T E T I Y Gcgggctacgcctaccccttcgtgatcgacgacgcgattggagacatccaatccaccacaa 3300
P G Y A Y P F V I D D A I G D I Q S T TcgtcgtactccaggaacctcgggcccaccgtcttcgctcgctttgccaTGCGTCGACCGT 3360
T S Y S R N L G P T V F A R F ACCTCATTTCCTGCGGCAGAGCGAGAGCTGATTCCGACCTCGAACAGctggaacccttcac 3420
T G T L Htacagcctcgatctcgtcctagccgacatcgcctttacccccacctaccccaactcctcc 3480
Y S L D L V L A D I A F T P T Y P N S S
(b)
Figure 4 Continued
12 Enzyme Research
cccgccactcgtttcgtcaagcgctccctcacgcagcacacctctgccgcttcgcacctc 3540
P A T R F V K R S L T Q H T S A A S H Lgccaagcgccgcgtctccatcgccactatcaaccccaaagccaccctcgtcgccgatcga 3600
A K R R V S I A T I N P K A T L V A D Rcagctccactcggacgtccctatcgagggcaacatcttcacccaaccctttactggaagg 3660
Q L H S D V P I E G N I F T Q P F T G Rgattacctcgtcgacgcagccccgacgggatccaccgatcgtaccgtcacttttaacggg 3720
D Y L V D A A P T G S T D R T V T F N Gcagtacgccgagaacggcctcgtgaggacggctgtgacggggacttcgtaccgcttcctc 3780
Q Y A E N G L V R T A V T G T S Y R F Lcttcgggcgttgaagatctcgggagacgcgatgtacgaggatcagtatgagagctggctc 3840
L R A L K I S G D A M Y E D Q Y E S W L3900
S L P F S F R Attttgccacctcaaaaaaaaaaaaaaaaaaaaaaaaaa 3938
lowast
tcgctaccgttctcgttccgtgcgtaggcttgttgtggacaccactgcattaagttatga
(c)
Figure 4 Nucleotide and deduced amino acid sequences of genomic DNA and cDNA encoding protease from G antarctica strain PI12 Thenucleotides are numbered from the 51015840 end of the cDNA A putative CAAT TATA box and polyadenylation signal are bold italicThe putativeN-terminal signal peptide is underlined The initiation codon is boldfaced and termination codon is represented by an asterisk (lowast) boxedpeptide sequences showed N-glycosylated asparagines residues The amino acid residues that form the catalytic triad are double underlinedand bold while introns are represented in blue font Highlighted in grey are the intron donor-acceptor splice sites
proteases Hydrophobic profile of PI12 amino acid proteasewas performed by ProtScale tools with Kyte and Doolittlemethod at httpauexpasyorgcgi-bin The hydrophobicityprofile elucidation of PI12 protease amino acid was carriedout from amino acid 1 to 963 The amino acid residuesfrom 1 to 16 (M-L-F-L-P-V-L-L-L-L-L-P-G-V-T-A) and 548 to562 (V-A-V-I-S) were hydrophobic grooves The first regionwhich consisted of 16 amino acids was very hydrophobic andwas predicted to be the signal peptide by SignalP and ProPwebserver
The bold and double-underlined amino acids showedresidues of catalytic triad aspartic acid (D) histidine (H) andserine (S) active site respectively NCBI Conserved DomainsDatabase (CDD) identified the presence of subtilase proteaseand subtilisin-like serine protease domains in the amino acidsequence The predicted PI12 protease amino acid sequencewas found sharing significant homology to the subtilisinfamily of protease The highest similarity of PI12 proteasewas observed with 42 identity to subtilisin-like proteaseof Rhodosporidium toruloides (EMS20793) 37 identity tosubtilisin-like protease of Pleurotus ostreatus (CAG25549)and 35 identity to subtilisin-like protease of Gloeophyllumtrabeum ATCC 11539 (EPQ55564) The nucleotide sequenceof G antarctica strain PI12 showed low homology to thefungus subtilisin protease family and none to other psy-chrophilic proteases The multiple sequence alignment wasillustrated in Figure 5 The consensus sequence around thecatalytic triad aspartic acid (D) histidine (H) and serine(S) of PI12 protease showed high conservation with othersubtilisin (showed in black boxes) Glycine residues G142G173 G191 G262 and G327 that were highly conserved inmost of subtilases were conserved in PI12 protease [25]Therefore PI12 protease belongs to the subtilisin subgroup ofthe subtilase serine protease superfamilyThis serine protease
of G antarctica PI12 has been deposited into GenBank withaccession number CAQ76821
34 Phylogenetic Relationship of PI12 Protease The phylo-genetic tree was constructed to demonstrate the similaritiesand relationship of protein sequence between differentspecies or organisms Figure 6 illustrates the phylogeneticdendrogram indicating the relationship of PI12 proteasewith another 25 translated amino acid sequences ofserine protease The bar scale signifies the branch lengthto approximate variance along each branch Basicallythis tree comprises two main clusters cluster I was sub-divided into subcluster a (CAG25549 EPQ55564 ETW-80907 XP 007381302 XP 001877576 CAG38357 XP007341385 CCA68627 EJU04161 XP 007369624 EIW-62675 CCO37249 XP 001830835 XP 007342189) and sub-cluster b (XP 007589863 XP 003711175 EFQ32840 XP006694395 ENH74885 EON99722) cluster II was sub-divided into subcluster c (CBQ73425 GAC97617 CCF52619)and subcluster d [CAQ76821 (PI12 protease) EMS20793]
Serine protease constructed by the phylogenetic treecomprised large size of protein with about 801 to 1032 aminoacids The majority of the translated protease sequenceswere obtained from fungus All serine proteases from cluster1 subcluster a are derived from Agaricomycetes except forEJU04161 which belongs to Dacrymycetes whereby all serineproteases from cluster 1 subcluster b are from Sordari-omycetes Cluster II subcluster c consisted of serine proteasefrom fungus belonging to Ustilaginomycetes PI12 proteasewas categorized under cluster II subcluster d It possessedhomology similarity with the subtilisin-like protease fromRhodosporidium toruloides (EMS20793) (formerly known asRhodotorula glutinis or Rhodotorula gracilis) which is a
Enzyme Research 13
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
MKG-YSWSLLQLAFTGAALATVPLSDVKRISSAS--VVPNKFVVEVESASEIPGKR--SL
MKGVLVWSILS---AGCALAVTPLSSVKRVTNLP--IVANKFIVEVDSTSDIPTKR--S-
MKLPLLSLALT---AVSVGATVPLSSVQRTTTSPNNALAGEYIVEMNNVQGLGGKR--AY
---MAHGSLASLLLLLAASAQGAAAFGGKSATGGRSVVPNSYILQVNASAPALTKRGVSV
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease ---MLFLPVLLLLLPGVTAFLNPVTNRATNAISSTQYLSNAYILELDLSTPGLVKRDSTP
lowastlowast
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
YTPHELFYRSLSARGVSYEVNKEYNS-EGLFVGAVVTVSDANDVKALAETPGVKAIRPVR
-NPHDALYESLRKRAIGFKVDREFNT-QGLFVGAALTLSDAQDVGQIQSTTGVKSIRPVR
ASTHDTLYRSLRKRGISFDVKKEYNQ-PGIMVGAALKVASPQDVQQIATMAEVKAIRPVI
FEALDHTLAAVKQNGVKYTVRQRFDAIPEAFQAVSIQVEDGASMAELAQIPGVQRVWPVS
DSILEDVLTSVGRNGIKYQLRHRFIS-PTLFHGASITVPPGISRSQIASLRGIKRVWPVR
lowastlowast
TIKSPKPVKVHVVTGKDDPEIPA-------------------------DTESTHIMTGVN
AFDRPKPVKVQVVKGPDDPALPP-------------------------DSESTHVITGVD
AVPAPERINQFTVSSPNDSRITA-------------------------DSETTHIMTGVD
LIPRPVEPTVSDFTPVSSSSSSTKSKRD-THTLEKRGTNFPPASAYLNDTFYPHVQTGID
KFSRPSAVVDADGGGSGFSGSPIKAALMGVKELGKR------ANAYAGDTFGPHVMTGVN
lowast lowast lowast lowastlowast
KLHAQGIFGAGIKIGIIDTGIDYTHPALGGALGAGHKVIGGYDFVGDAYDGTNTPVPDND
KLHAQGITGAGIKIGILDTGIDYTHPTLGGAFGPGNKVIGGFDLVGDDYDGTNTPVPDPD
KVHAAGITGEGIKIGIIDTGIDYNHPLLGAGFGPGFKVIGGYDFVGDDYTGSNNPAPDDD
VLHNKGILGQGVKIVVVDEGVDYTNPILGGCFGPGCQISFGYDFVGDNYTGTNAPVPDSD
ETHEAGLLGAGIKIGVLDTGVDYLNPILGGCFGPGCHMSFGYDLVGDDYDGDNAPVPDVD
lowast lowast lowast lowastlowastlowast lowast lowast lowastlowast lowast lowastlowast lowast lowast lowast lowast lowastlowastlowast lowast lowast lowast lowast lowastlowast lowast
PLDQCNGHGTHVAGIIGANPDNQFNISGVSYGASLSSYRVFGCSGSVSDDIIIEALLRGV
PLDQCAGHGTHVAGIIGANPDNAFNISGVAFQASLSAYRVFGCVGFVTDDVLVDALLLGF
PLDNCNGHGTHVAGIIGANPGNDFGISGVAYGSSIASYRVFGCSGFVSDDIIIDALLRGY
PFSSCSNHGTHTTGTVGALAN-EYGFSGAAPMATIGHYRVISCSGATTDEILVAALTRAL
PFASCDPHGTHVTGIIGALPN-AFGFTGVAPAATLGHYRVFGCTGFVGEDIILAGLMRGV
lowast lowastlowastlowast lowastlowast lowast lowastlowast lowast lowastlowastlowast lowast lowast lowast
QDGQDILTLSLGGTDGWTE-SSSSVVASRIAASGKIVTIAAGNEGAYGSWYSSSPGNGID
NEGQDILTLSLGGADGWTE-SVSAVVASRIAATGKVVTIAAGNDGASGAWYTSSPGNGID
SDGMDILTLSLGGVEGWTE-SSSSVVASRIADQGRVVTIAAGNDGAYGSWYASSPGTGVD
QDNVNIVSMSLGSSVGWLDDSPVQIMASYLGSKGIHVVASVGNERNEGLFAADQPAASRI
EDNCNVLTLSLGGPGGWVKGTPASILIDQIEAQGILVTVATGNSGAEGMFFSESPASTIN
lowastlowastlowast lowastlowast lowast lowast lowastlowast lowast lowast
VISVASIDNTVDNIQNATVNGVEHGPIPYFKTLPLPFTNA-LPLYVISNDTTIANDACNA
VISVASLDNTVVPLQTAQVHGVTHDPIIYQDTFPLPINDT-RPIFATSTDVTVVDDACNP
VISVASVNNIAIPVQNATVQGVEHAPIPYLEALPLNVTGS-RPIYATSTDTTVADDACNA
GTGVGGVDPTYLAAYYAYLI--KRDPVPYISPTPFNLPNN-YLLYFTSTNTSVTNDACNP
GLSIASTDVTDLIAYNATVS--GQPAIPYLSATPLNVVANSFRVHFTSTDPNNPVDACSP
lowast lowast lowast lowast lowastlowastlowast
LPDNTPDLSGFLVLVRRGSCTFVQKLTNIAAKGGNAVFIYDNGNG-----FAGIDTGKFT
LPDSTPDLSGFVVIVRRGTCTFVQKLGNVAAKGAKVTLIYDNGSG-----FGAIDTGAFV
LPNDTPDLSGYVVVIKRGTCTFVTKLQNAAAKGANTFLIYNNGGS-----FSAISVGDYT
LPSTTPNLANRVVVVQRGTCGFTVKQQNVAAAGGKIVLIYNSKGSGLIPQLDVGSTGLTA
LPAGAPDFANYVTVVQRGTCTFVTKYQNVLNAGGKIVLLYNSEGAGNLPYLTPNGVGIDA
lowastlowast lowast lowastlowast lowast lowast lowast lowast lowast lowast lowast
(a)
Figure 5 Continued
14 Enzyme Research
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
NATLIQAADGVFLAQQVAAGKSVTITFPQTGGSAQITN-PSGGLVSSFTTYGPTNDFYFK
-ASFIQAADGEFLVQQFASGQNVSLSFPQSGGLIQFPD-PAGGLISSFTSYGPSNDFFFK
-AALISETDGTFLVNQFAAKANITISFPQSGSAWNYPDDVSGGLVSDFSSYGPSYDMYFK
VGSLRYEDGLKLLSYYQSSPRGQSISFPNGPLAPYVSNTVSGGVVASYSNFGPTNELFIY
VAGLRRSDGLKLLSYYQNANKRLTLRFPKGKIVAGLTDTITGGLISGYSTFGPTNDLYGQ
lowast lowastlowast lowastlowast lowastlowast
PAFGAPGGNILSTLPVPLGSYGVESGTSMATPYVAGSAALLLSVKGKSAAVALGARTLFQ
PAVAAPGGNILSTLPVNLGLFGIESGTSMATPFVAGSAALLFQAKGTSAAVGRSARTLFE
PAVAAPGGNILSTLPIALGSYGVESGTSMATPFTAGSAALILQAKGK--SVAKSIRNLFQ
PTLAAPGTDILSTV---TGGVALMRGTSMAAPLVAGAYALLLSVGNNLKLSPEEARTILM
PTLSAPGGNILSTFPLSEGGVAVISGTSMSCPFVAGSAAVLMAARASENLTPLEIRSLLT
lowast lowastlowastlowast lowastlowastlowastlowast lowast lowastlowastlowastlowast lowast lowastlowast lowast lowast
STAQRVPSSHTDGDPLQTLSQQGAGLIDVFKALYTETIVWPTELILNDTAHYKEVHTFTV
TTAQRVPSTHTDGDPLQTLTQQGAGLINVFNAIHTTTIVSPGELILNDTAHFKGTQQFTV
TTASAVASSKTDGDPLQTVSQQGAGLIQVDRAIGTTTVVSPGQLTLNDTAHFEGFKTFNI
STSQPSPVAY-GSETLDTVVSQGAGVINMTAAIAAHTVISPAQFNLNDTQYSNNTQTLTL
TTAKLTPVSLLGSTPLVSVIRQGGGLVQVAKALAAKTLISPHELLLNDTANANYVQTIKI
VNTGKTSKQYKLTHVPAGTAQTIT---SGTIFAADGPVPLSTDYATVKLSSSSFTLRPGQ
RNAGSSAKTYTLRHIPAGTAVTVT---PGTIFPADGPVPLSTDFASVSLSTSKFTLAPGK
QNTGSKAVTYKISHVPAGTAASVQ---SGSIFAADGPVPLTADYAQVTFSLNKVTVLPGV
RNNNRFPVSYQLSWVDSTGIVTYNDGATTDIIPSTTPNYVSTSVLRVAFSQRTVTVPGGQ
KNTNSWAMKYTFSSAVAQGLGTFD--ASGDILPTLDPVAVSGAQATVAFNTRILSVAPGA
lowast lowast lowast lowast lowast lowast
SKQVTTHINAP--SGVDASTYPVFSGFIEIATAT--------ESVHVTYLGLAASLKDKQ
TQTVTARFTPP--AAADPSTFPVFSGFIQIESGT--------EQVQVSYLGLKASLKDKQ
SVPILVWFKPP--QGVDASTYPVYSGFIEIAGLG--------ETLRVAYLGVAASLKDKA
TVKVTAQFIPPNLTAQQRNQFPIYSGFVTITGQGQGAGAEQMEKYNLPFFGLGARMVDMP
TGSVVATITPPVLPVADAARFPIFSGWIRVNGQG-ARDSSRNEAYTVPYFGLAAKMIDMQ
lowast lowast lowastlowast lowast lowast lowast lowast
IVDNTNEFFGITIPAVLNNTGDAQ-LSPTNYTFSGSDFPTLLLRLAFGTPLLRVDLVDAT
VIDNTDFFFGVPTPVLTDPNGEVQ-TSPRNYSFLASDFPTLIFRLAFGSPKVVVDLVSPT
VVDNTDEYFGVKIPALLDASGDVQ-GNSTNYTLVGDDVPSLLYRFAFGSAHVRVDLVEKA
VLDTTNVALGP--NLPFIAVGQN--IQTGPTTFSESNPAVVYFRLAAGTRRLRVDLVDAS
VLDTTETIYGPGYAYPFVIDDAIGDIQSTTTSYSRNLGPTVFARFATGTLHYSLDLVLAD
lowast lowast lowast lowast lowastlowastlowastlowast lowast
IDLKPTLNSREIDDKRHVFERESFSFPHPGPKGS-------------------TFAQVKI
INFKPTLNTRALGDTHGSF----FSFPHRVKTG--------------------SFAQVPT
STVSATLSKRNWWN---------WWWPQKPSN---------------------SYDKVPI
TSYNATVPAVTNPASRLVKRSNTELTPRAAFPT--------------------LYSQVRT
IAFTPTYPNSS-PATRFVKRSLTQHTSAASHLAKRRVSIATINPKATLVADRQLHSDVPI
lowast lowast
VGTLSEQSHIPRNSDASTVDDGGYSTFAFNKP------SFANGTTIPNGTYRWLLRALKV
LGTVFELDFTSRNNDDPTDN--GFNTVAIDPP------TFANGTTIPNGQYRLLLRALRV
VGTLAEYDYVPRNSDASTIDDNGYNVFQFT-D------SFVNGTKIAAGEYKVLLRALKV
VGSLFAPSYWPPRDYLFNNGGT-YSDYEIQLDGSFTFPNGTKGMAELGRSYKVLLRALKI
EGNIFTQPFTG-RDYLVDAAPTGSTDRTVTFNGQY-AENGLVRTAVTGTSYRFLLRALKI
lowast lowast lowastlowast lowast lowast lowast lowast lowastlowastlowastlowast
lowast lowast lowastlowastlowastlowastlowast
(b)
Figure 5 Continued
Enzyme Research 15
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
TGDPTKEADFETWLSPIVGITVA
TGDPTNEADFESFLSPIIGVNAP
TGNPAEEGDYETWLSPSFSVVPA
TADPTYTSSYESWLSPSFTFTA-
SGDAMYEDQYESWLSLPFSFRA-
lowast lowastlowast
ConservedSimilar
lowast All match
(c)
Figure 5 Multiple sequence alignment between amino acid sequence of PI12 protease with other subtilisin The serine protease sequenceswere obtained from the GenBank database Their accession numbers are EMS20793 (subtilisin-like protease of Rhodosporidium toruloidesNP11) CAG25549 (subtilisin-like protease of Pleurotus ostreatus) EPQ55564 (subtilisin-like protease of Gloeophyllum trabeumATCC 11539)and ETW80907 (serine protease S8 ofHeterobasidion irregulare TC 32-1) Putative ORF of serine protease fromG antarctica PI12 was labeledas PI12 protease
CAG38357 Phanerochaete chrysosporium
EIW62675 Trametes versicolor FP-101664 SS1
XP 007369624 Dichomitus squalens LYAD-421 SS1
XP 007381302 Punctularia strigosozonata HHB-11173 SS5
CAG25549 Pleurotus ostreatus
ETW80907 Heterobasidion irregulare TC 32-1
XP 001877576 Laccaria bicolor S238N-H82
EPQ55564 Gloeophyllum trabeum ATCC 11539
XP 001830835 Coprinopsis cinerea okayama7130
XP 007341385 Auricularia delicata TFB-10046 SS5
CCA68627 Piriformospora indica DSM 11827
CCO37249 Rhizoctonia solani AG-1 IB
XP 007342189 Auricularia delicata TFB-10046 SS5
EJU04161 Dacryopinax sp DJM-731 SS1
XP 007589863 Colletotrichum fioriniae PJ7
EFQ32840 Colletotrichum graminicola M1001
ENH74885 Fusarium oxysporum f sp cubense race 1
XP 003711175 Magnaporthe oryzae 70-15
XP 006694395 Chaetomium thermophilum DSM 1495
EON99722 Togninia minima UCRPA7
CBQ73425 Sporisorium reilianum SRZ2
GAC97617 Pseudozyma hubeiensis SY62
CCF52619 Ustilago hordei
EMS20793 Rhodosporidium toruloides NP11
CAQ76821 Glaciozyma antarctica PI12
01
Clus
ter 1
Clus
ter 2
a
b
c
d
Figure 6 Phylogenetic dendrogram among 25 translated amino acid sequences of serine proteaseThe tree was constructed using a neighbor-joining algorithm with 1000 bootstrappings by MEGA 6 software [39]The scale bar represents 01 substitutions per amino acid positionTheserine protease sequences were obtained from the GenBank database GenBank accession numbers of the sequences were indicated beforethe strain name The putative serine protease of G antarctica strain PI12 has been deposited in GenBank with accession number CAQ76821
16 Enzyme Research
0
2
4
6
8
10
12
0 24 48 72 96 110
Prot
ease
activ
ity (U
mL)
Post induction time (hours)
Figure 7 Effect of postinduction time towards protease activity The highest activity was detected at 72 hours after 110 hours of incubationwith 9UmL activity followed by 48 24 and 96 hoursThe substrate (azocasein) was dissolved in Tris HCL buffer and incubated for 30min at20∘CThe absorbance was measured at 450 nm after 15min incubation A one-way ANOVAwas used to test for preference differences amongsix different postinduction times The statistical value (119875 value lt005) between hours is 0003
pink-coloured nonpathogenic oleaginous yeast (42 iden-tity) Both G antarctica PI12 and Rhodosporidium toruloidesNP11 belong to Microbotryomycetes class PI12 protease didnot show any homology similarity to other psychrophilicproteases however it showed 37 identity to hypotheticalprotein from Pseudozyma antarctica T-34 which was isolatedfrom Japan (GAC76005) (not listed in the phylogenetic tree)
35 Cloning and Expression of Cold-Adapted PI12 Protease inthe Yeast Expression System The PCR product harbouringthe mature PI12 protease with two flanking restriction sites(EcoRI and AvrII) was cloned into pPIC9 expression vectorThe recombinant plasmid harbouring PI12 protease was usedto transform both GS115 and KM71 intended to produce bothHis+ Mut+ and His+ MutS Two recombinant clones fromGS115 (His+Mut+) strain (GpPro1 andGpPro2) and one fromKM71 (His+MutS) strain (KpPro1) were successfully obtainedwhich were further analyzed for protein expression study
36 Expression and Enzyme Assay of PI12 Protease in Pichiapastoris All positive recombinants were grown in small scalecultures and assayed for the cold-adapted protease activityin the culture supernatants The clones were first grown inBMGY medium with glycerol as the sole carbon sourceFor induction of AOX1 promoter and protein productionthe growth medium was transferred to the same mediumexcept with 05 methanol in place with glycerol as thecarbon source [26] The supernatant of the recombinant Ppastoris expression cultures after 48 h of incubation time wasanalyzed to determine the recombinant protease activity bythe modified method of Brock et al (1982) [20] All testedclones secreted protease into the supernatant in differentexpression levels This might be due to the variability ofthe strength of the promoter and copy number in each cellto secrete and express the recombinant product The clonewith the highest yield was GpPro2 with 63UmL activityfollowed by KpPro1 and GpPro1 with 25UmL and 37UmLactivities respectively (data not shown) It is perceptible thatexpression systemof Pichia pastoris is the ideal system for thisprotein The preliminary assay has fruitfully produced high
protease expression at 20∘C Yet further optimization of therecombinant PI12 protease production need to be conductedin order to examine the optimum condition of this proteinReported cold-adapted proteases showed optimum activity ata range of 15∘Cndash60∘C [27ndash30] This difference was possiblyrelated to the respective genetic and physiological adaptationof the strains There are several factors that influenced thedifferent rate of expression It includes different site of inte-gration between interest gene and Pichia genome variabilityof different expression host cell to secrete and express therecombinant enzyme and different sites in the polylinker forinserting foreign genes [31]
This is the second extracellular protease reported to besynthesized by this obligate psychrophilic yeastG antarcticaThe first cold-adapted extracellular subtilase from this yeasthas been purified and characterized previously [32 33] Theextracellular serine proteinase (LAP2) from this Antarcticyeast formerly known as Leucosporidium antarcticum strain171 is specific towards synthetic substrates of chymotrypsinand subtilisin The subtilase of L antarcticum strain 171showed specific activity of 4043Umg when utilized N-SucAAPFpNA as a substrate [33]
37 Optimization of Protease Production GpPro2 was chosenfor further optimization due to its highest protease activityat the preliminary expression stage High protease activitywas detected after 72 hours of incubation (Figure 7) byusing the standard protocol method as recommended forPichia pastoris expression system (Pichia Expression KitInvitrogen USA) The expression decreased after 72 hoursand no activity was detected at 110 hours The prolongedincubation may result in the decrease of biomass productionand will subsequently reduce the expression of recombinantprotein In addition oxygen and nutrient depletion duringprolonged incubationwill trigger the stress responses in orderto survive in such harsh environment [34]
Methanol was used as a sole carbon by P pastoris Themethanol was oxidized to formaldehyde with the aid of oxy-gen as electron acceptor by an alcohol oxidase enzyme (AOXgene) [35] The optimal methanol concentration is at 05(vv) with 14UmL activity The addition of more than 05
Enzyme Research 17
02468
10121416
0 01 03 05 07 09Pr
otea
se ac
tivity
(Um
L)Methanol concentration ()
Figure 8 Effect ofmethanol concentration towards protease activityThe recombinant was cultivated for 72 hours at 30∘C in BMMYmedium05 methanol gives the highest activity with 14UmL The substrate (azocasein) was dissolved in Tris HCL buffer and incubated for 30minat 20∘CThe mixture was left for 15min and absorbance was taken at 450 nm A one-way ANOVA was used to test for preference differencesamong six different concentrations The statistical value (119875 value lt005) between concentration is 002
0
5
10
15
20
25
10 15 20 25 30 35
Prot
eas a
ctiv
ity (
Um
L)
Temperature (∘C)
Figure 9 Effect of temperature towards protease activity The reaction was carried out in temperature ranging from 10∘C to 35∘C at pH 7 for30min A one-way ANOVA was used to test for preference differences among six different temperatures The statistical value (119875 value lt005)between temperature is 0067
(vv) methanol resulted in tremendous decrease of proteaseactivity (Figure 8) Production of PI12 protease was decreasedwhen the methanol concentration increased This might bedue to the toxic effect of methanol concentration on DNAreplication and membrane synthesis Basically methanol isused to increase the permeability of the cell membrane tohelp the secretion of foreign protein [26] However if thepermeability of cell membrane is too high lowmolecular sizemetabolites would leak from the cell and such leakage willseriously damage cell activities Such condition would abusethe cellular system and might reduce the yield of proteinexpression [36]
Psychrophilic microorganism dominated in low temper-ature habitat which most other microorganism are inca-pable of withstanding due to their inability to produceenzyme that can tolerate the extreme environment con-dition Psychrophilic enzymes are always associated withhigh catalytic efficiency at low and moderate temperature(0∘C to 30∘C) compared to the mesophilic counterpart thatare generally unable to catalyze efficiently under extremely
low temperature [2] The recombinant protein assay wasdone at various temperatures ranging from 10∘C to 35∘Cusing 05 azocasein as a substrate dissolved in 50mMTris HCl in presence of 2mM CaCl
2 pH 7 In the activity
assays expressed protease showed a highest activity at 20∘C(20UmL) and decreased tremendously once it exceededmaximum temperature due to the denatured state of theenzyme (Figure 9) The active site of an enzyme is easilyaltered when the high temperature is introduced duringincubation or activity assay resulting in a low reaction rate[37]
The recombinant construct was transferred into BMMYmedia at different A
600 nm ranging from 1 to 10 The high-est activity of active enzyme was detected at A
600 nm 60(283UmL) and above (Figure 10) The slope during expo-nential phase was much higher compared to other phasesindicating an increase in the doubling time At exponentialphase the cells are adapted to the growth condition andother processes like cell growth cycle and synthesis of RNAThe yield of active enzyme during expression of P pastoris
18 Enzyme Research
05
1015202530
1 2 3 4 5 6 7 8 9 10Prot
ease
activ
ity (U
mL)
Initial inoculum A600
Figure 10 Effect of inoculums size towards protease activity The recombinant was cultivated for 72 hours at 30∘C in BMMY mediumwith 05 methanol concentration A one-way ANOVA was used to test for preference differences among six different optical densitiesThe statistical value (119875 value lt005) between optical density value is 003
(kD
a)
1160
662
450
350
250
184
144
M 1 2 3
(a) (b) (c)
Figure 11 (a) Coomassie-stained SDS-PAGE of recombinant P pastoris clone 1mL supernatants collected from the third-day culturewere concentrated 10-fold using trichloroacetic acid and 10120583L resultant products were applied to SDS-PAGE analysis Lane 1 molecularweight marker (Fermentas) Lane 2 GS115 (Control) Lane 3 GS115pPIC9 empty (Control) Lane 4 GS115pPIC9PI12prot (GpPro2)The arrow indicates the secreted recombinant PI12 protease (b) Plate activity staining of different recombinant P pastoris clones GGS115 GP GS115pPIC9 GRI GS115pPIC9PI12prot1 (GpPro1) GRII GS115pPIC9PI12prot2 (GpPro2) K KM71 KP KM71pPIC9 KRKM71pPIC9PI12prot (KpPro) (c)Gel activity staining of recombinant PI12 protease inP pastorisGpPro2 clone Lane 1 prestainedmolecularweightmarker (Fermentas) Lane 2 GS115pPIC9 empty (Control) Lane 3 GS115pPIC9PI12prot1 (GpPro2)The arrow indicates the secretedrecombinant PI12 protease
dropped after 28 hours of incubation which is at stationaryor death phase [38]
38 Detection of Recombinant Protein by SDS-PAGE andActivity Staining TheSDS-PAGE gel of the GpPro2 recombi-nant clonesrsquo supernatant showed a putative expression bandat around 993 kDa Plate staining and gel activity stainingwere executed to examine if the recombinant PI12 proteaseexpressed in P pastoris was functionally active In the platestaining the transformants showed strong proteolytic activity(Figure 11(a)) In gel staining proteolytic activity was alsodetected around 993 kDa after incubation at 20∘C This is
evidence of protease secretion into the surrounding mediawhereby it catalyzed the breakdown of casein thus formedclearing zones around the activity ring and band (Figures11(b) and 11(c)) As a conclusion the cold-adapted PI12protease was functionally expressed in P pastoris GS115(GpPro2 clone) with the size of 993 kDa
4 Conclusions
Psychrophilic yeast which was originated frommarine waterin Casey Station Antarctica was identified as Glaciozymaantarctica strain PI12 A serine protease with an open reading
Enzyme Research 19
frame of 2892 bp encoded for 963 amino acids from thisAntarctic yeast has been cloned and expressed in Pichiaexpression system This is the second extracellular pro-tease reported being synthesized by this psychrophilic yeastSequence analysis through bioinformatics studies revealedfew homology identities of this protein with other ser-ine proteases The mature PI12 protease was successfullyexpressed and secreted into the culture medium driven bythe Saccharomyces cerevisiae 120572-factor signal sequence in Ppastoris The protease production in P pastoris was bestobtained from strain GS115 (GpPro2) with 283UmL after 72hours of induction time with 05 (vv) of methanol inducerThe starting optical density at 600 nm (OD600) was 60 andthe optimum assay temperature was 20∘C The expressedprotein was detected by SDS-PAGE and activity staining witha molecular weight of 99 kDaThis cold-active enzymemightoffer an opportunity for biotechnological exploitation basedon its high catalytic activity at low temperatures and lowthermostability Structural and functional study of this cold-adapted enzyme would give insight into its adaptation to anextreme cold environment
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgment
This research was supported financially by the InitiativeBiotechnology-GMBI-MOSTI (Project number 07-05-16-MG-GMB02) andRUGS-Initiative 6 (Project number 05-02-11-1414RU)
References
[1] J W Deming ldquoPsychrophiles and polar regionsrdquo CurrentOpinion in Microbiology vol 5 no 3 pp 301ndash309 2002
[2] C Gerday M Aittaleb M Bentahir et al ldquoCold-adaptedenzymes from fundamentals to biotechnologyrdquo Trends inBiotechnology vol 18 no 3 pp 103ndash107 2000
[3] A Svingor J Kardos I Hajdu A Nemeth and P Zavodszky ldquoAbetter enzyme to cope with cold comparative flexibility studieson psychrotrophic mesophilic and thermophilic IPMDHSrdquoThe Journal of Biological Chemistry vol 276 no 30 pp 28121ndash28125 2001
[4] G Antranikian C E Vorgias and C Bertoldo ldquoExtremeenvironments as a resource for microorganisms and novel bio-catalystsrdquo Advances in Biochemical EngineeringBiotechnologyvol 96 pp 219ndash262 2005
[5] T Lonhienne C Gerday andG Feller ldquoPsychrophilic enzymesrevisiting the thermodynamic parameters of activation mayexplain local flexibilityrdquoBiochimica et Biophysica Acta vol 1543no 1 pp 1ndash10 2000
[6] R Cavicchioli and TThomas ldquoExtremophilesrdquo in Encyclopediaof Microbiology pp 317ndash337 Academic Press San Diego CalifUSA 2000
[7] G Feller E Narinx J L Arpigny et al ldquoEnzymes frompsychrophilic organismsrdquo FEMS Microbiology Reviews vol 18no 2-3 pp 189ndash202 1996
[8] J Arnorsdottir R B Smaradottir T O Magnusson S HThorbjarnardottir G Eggertsson and M M KristjanssonldquoCharacterization of a cloned subtilisin-like serine proteinasefrom a psychrotrophic Vibrio speciesrdquo European Journal ofBiochemistry vol 269 no 22 pp 5536ndash5546 2002
[9] N J Russell ldquoPsychrophilic bacteriamdashmolecular adaptations ofmembrane lipidsrdquo Comparative Biochemistry and Physiology APhysiology vol 118 no 3 pp 489ndash493 1997
[10] G Feller and C Gerday ldquoPsychrophilic enzymes hot topics incold adaptationrdquo Nature Reviews Microbiology vol 1 no 3 pp200ndash208 2003
[11] P Raspor and J Zupan ldquoYeasts in extreme environmentsrdquo inBiodiversity and Ecophysiology of Yeasts The Yeast Handbookpp 371ndash417 Springer Berlin Germany 2006
[12] M B Rao A M Tanksale M S Ghatge and V V Desh-pande ldquoMolecular and biotechnological aspects of microbialproteasesrdquoMicrobiology andMolecular Biology Reviews vol 62no 3 pp 597ndash635 1998
[13] L Polgar ldquoThe catalytic triad of serine peptidasesrdquo Cellular andMolecular Life Sciences vol 62 no 19-20 pp 2161ndash2172 2005
[14] S Y Boo C M V L Wong K F Rodrigues N NajimudinA M A Murad and N M Mahadi ldquoThermal stress responsesin Antarctic yeastGlaciozyma antarctica PI12 characterized byreal-time quantitative PCRrdquoPolar Biology vol 36 no 3 pp 381ndash389 2013
[15] B Turchetti S R T Hall L B Connell et al ldquoPsychrophilicyeasts from Antarctica and European glaciers descriptionof Glaciozyma gen nov Glaciozyma martinii sp nov andGlaciozyma watsonii sp novrdquo Extremophiles vol 15 no 5 pp573ndash586 2011
[16] N H F Hashim I Bharudin D L S Nguong et al ldquoCharac-terization of Afp1 an antifreeze protein from the psychrophilicyeast Glaciozyma antarctica PI12rdquo Extremophiles vol 17 no 1pp 63ndash73 2013
[17] A N M Ramli M A Azhar M S Shamsir et al ldquoSequenceand structural investigation of a novel psychrophilic 120572-amylasefrom Glaciozyma antarctica PI12 for cold-adaptation analysisrdquoJournal ofMolecularModeling vol 19 no 8 pp 3369ndash3383 2013
[18] A N M Ramli N M Mahadi A Rabu A M A Murad F DA Bakar and R M Illias ldquoMolecular cloning expression andbiochemical characterisation of a cold-adapted novel recombi-nant chitinase from Glaciozyma antarctica PI12rdquoMicrobial CellFactories vol 10 no 1 article 94 2011
[19] J Sambrook E Fritsch and T Maniatis Molecular Cloning ALaboratoryManual Cold SpringHarbor Laboratory Press NewYork NY USA 1989
[20] F M Brock C W Forsberg and J G Buchanan-SmithldquoProteolytic activity of rumen microorganisms and effects ofproteinase inhibitorsrdquoApplied and EnvironmentalMicrobiologyvol 44 no 3 pp 561ndash569 1982
[21] U K Laemmli ldquoCleavage of structural proteins during theassembly of the head of bacteriophage T4rdquo Nature vol 227 no5259 pp 680ndash685 1970
[22] A Henne R A Schmitz M Bomeke G Gottschalk and RDaniel ldquoScreening of environmental DNA libraries for thepresence of genes conferring lipolytic activity on Escherichiacolirdquo Applied and Environmental Microbiology vol 66 no 7 pp3113ndash3116 2000
[23] M Poza T de Miguel C Sieiro and T G Villa ldquoCharacteri-zation of a broad pH range protease of Candida caseinolyticardquoJournal of Applied Microbiology vol 91 no 5 pp 916ndash921 2001
20 Enzyme Research
[24] J D Bendtsen H Nielsen G von Heijne and S BrunakldquoImproved prediction of signal peptides SignalP 30rdquo Journalof Molecular Biology vol 340 no 4 pp 783ndash795 2004
[25] R J Siezen and J A M Leunissen ldquoSubtilases the superfamilyof subtilisin-like serine proteasesrdquo Protein Science vol 6 no 3pp 501ndash523 1997
[26] Y Katakura W Zhang G Zhuang et al ldquoEffect of methanolconcentration on the production of human 1205732- glycoprotein Idomain V by a recombinant Pichia pastoris a simple system forthe control of methanol concentration using a semiconductorgas sensorrdquo Journal of Fermentation and Bioengineering vol 86no 5 pp 482ndash487 1998
[27] Q-FWang Y-HHou Z Xu J-LMiao andG-Y Li ldquoPurifica-tion and properties of an extracellular cold-active protease fromthe psychrophilic bacterium Pseudoalteromonas sp NJ276rdquoBiochemical Engineering Journal vol 38 no 3 pp 362ndash3682008
[28] S Vazquez L Ruberto and W Mac Cormack ldquoProper-ties of extracellular proteases from three psychrotolerantStenotrophomonas maltophilia isolated from Antarctic soilrdquoPolar Biology vol 28 no 4 pp 319ndash325 2005
[29] R Zeng R Zhang J Zhao and N Lin ldquoCold-active ser-ine alkaline protease from the psychrophilic bacterium Pseu-domonas strain DY-A enzyme purification and characteriza-tionrdquo Extremophiles vol 7 no 4 pp 335ndash337 2003
[30] M Turkiewicz E Gromek H Kalinowska and M ZielinskaldquoBiosynthesis and properties of an extracellularmetalloproteasefrom the Antarctic marine bacterium Sphingomonas paucimo-bilisrdquo Journal of Biotechnology vol 70 no 1ndash3 pp 53ndash60 1999
[31] A Vassileva D A Chugh S Swaminathan and N KhannaldquoEffect of copy number on the expression levels of hepatitisB surface antigen in the methylotrophic yeast Pichia pastorisrdquoProtein Expression andPurification vol 21 no 1 pp 71ndash80 2001
[32] M Pazgier M Turkiewicz H Kalinowska and S Bielecki ldquoTheunique cold-adapted extracellular subtilase from psychrophilicyeast Leucosporidium antarcticumrdquo Journal ofMolecular Catal-ysis B Enzymatic vol 21 no 1-2 pp 39ndash42 2003
[33] M Turkiewicz M Pazgier H Kalinowska and S BieleckildquoA cold-adapted extracellular serine proteinase of the yeastLeucosporidium antarcticumrdquo Extremophiles vol 7 no 6 pp435ndash442 2003
[34] D Mattanovich B Gasser H Hohenblum and M SauerldquoStress in recombinant protein producing yeastsrdquo Journal ofBiotechnology vol 113 no 1ndash3 pp 121ndash135 2004
[35] P Secades BAlvarez and JAGuijarro ldquoPurification andprop-erties of a new psychrophilic metalloprotease (Fpp2) in the fishpathogen Flavobacterium psychrophilumrdquo FEMS MicrobiologyLetters vol 226 no 2 pp 273ndash279 2003
[36] L Y Ling I Ithoi and F M Yik ldquoOptimization for high-level expression in Pichia pastoris and purification of truncatedand full length recombinant sag2 of toxoplasma gondii fordiagnostic userdquo Southeast Asian Journal of Tropical Medicineand Public Health vol 41 no 3 pp 507ndash513 2010
[37] A Illanes ldquoStability of biocatalystsrdquo Electronic Journal ofBiotechnology vol 2 no 1 pp 1ndash9 1999
[38] B A Plantz J Sinha L Villarete K W Nickerson and VL Schlegel ldquoPichia pastoris fermentation optimization energystate and testing a growth-associated modelrdquo Applied Microbi-ology and Biotechnology vol 72 no 2 pp 297ndash305 2006
[39] K Tamura G Stecher D Peterson A Filipski and S KumarldquoMEGA6molecular evolutionary genetics analysis version 60rdquo
Molecular Biology and Evolution vol 30 no 12 pp 2725ndash27292013
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

8 Enzyme Research
their molecular phylogenetic data and physiological andmorphological characteristics [15] Phylogenetic analysis ofITS158SITS2 sequence of strain PI12 showed that thesequence was closely related to ITS158SITS2 sequenceof Glaciozyma antarctica with high homology similarity toCluster I (99-100 identity) The ITS158SITS2 sequenceof G antarctica strain PI12 was differed from G watsonii(Cluster II) by 2-3 gap in the sequence (93 identity)and G martinii by 4-5 gap in the sequence (90ndash92identity)
33 Sequence Analysis of PI12 Protease cDNA In this studyamplification of the PI12 protease gene sequence was con-ducted using genomicDNAand cDNAas a template Putativepartial PI12 protease gene with the size of 1321 bp was initiallyacquired from the G antarctica genome survey sequencelibrary (Supplementary Figure see Supplementary Mate-rial available online at httpdxdoiorg1011552014197938)However the gene was discovered as an incomplete pro-tease gene sequence DNA walking strategy successfullyamplified 1500 bp of the upstream region of partial PI12protease Both sequences of about 1500 bp (derived fromthe DNA walking reaction) and 1321 bp (partial PI12 pro-tease) were assembled and made up a total length of2687 bp protease sequence (Supplementary figure)The puta-tive PI12 protease was predicted by Augustus webserver(httpaugustusgobicsde) having intervening sequencesintronsThus reverse-transcriptase PCR (RT-PCR) and rapidamplification of cDNA ends (RACE) were accomplished toobtain the full-length cDNA As a result the ORF comprising2892 bp encoding 963 amino acid residues were successfullydeduced
Nucleotide and deduced amino acid sequences ofgenomic DNA and cDNA fragment encoding PI12 proteasewere shown in Figure 4 The size of the ORF of putative PI12protease was 3867 bp with genomic DNA as the templatewhile the ORF comprised 2892 bp encoding 963 amino acidresidues were successfully deduced through RACE strategywith cDNA as the template The ORF was predicted usingORF Finder (httpwwwncbinlmnihgovprojectsgorf)while prediction using SignalP (httpwwwcbsdtudkser-vicesSignalP) webserver allowed the identification of theputative PI12 protease prepeptide where it predicted thepresence of signal peptidase I cleavage sites [24]The putativeN-terminal signal peptide (1 to 16 the underlined amino acid)was identified as a secretion signal There is no propeptidepredicted by ProP Webserver (httpwwwcbsdtudkser-vicesProP-10) Comparison of cDNA and gene sequencesallowed determination of gene structureTherefore sequencealignment was performed between the sequence of these twotemplates and it was confirmed that the coding sequence(consisted of 19 exons) was interrupted by 18 introns withtheir length varying from 48 to 60 nucleotides However theGT and AG rule for donor-acceptor sites was not obeyedOf 18 introns 10 introns contained canonical GT and AGdinucleotides at their 51015840 and 31015840 ends while 8 introns holdnoncanonical splice site pairs with TT-GG (2 introns)AG-TG GC-GT TG-GG TC-GG TA-GG and TG-AG
dinucleotides at the splice junction Upper case letters inblue colour represent introns while lower case letters inblack colour represent exons Splice donor and acceptor werehighlighted in grey
The amino acid composition (upper case letters) withinthe ORF amino acid residues was determined by theProtParam tool of the Expasy molecular biology server(httpwwwexpasyorgtool) Translation starts at a nucle-otide position 1 and translation stop is marked with anasterisk The nucleotide sequences ATG (1 to 3) and TAG(3865 to 3867) indicate the initiation codon and termi-nal codon respectively The presumed putative sequencesof promoter CAAT (minus101 to minus98) and TATA-box (minus92to minus88) were identified in the 51015840-untranslated (UTR)region The search was achieved via Neural Network Pro-moter Prediction (Berkeley Drosophila Genome Project)(httpwwwfruitflyorgseq toolspromoterhtml) In the 31015840-untranslated (UTR) region there was a putative polyadeny-lation signal attaa (3889 to 3893) detected from the TAGstop codon to the poly (A) tail The predicted molecularmass and pI of PI12 serine protease were 10099 kDa and641 respectively Out of 963 amino acid residues in thededuced sequence of the PI12 protease 70 are negativelycharged residues (Asp + Glu) while 66 are positively chargedresidues (Arg + Lys) at neutral pH (Table 2)The total chargedresidues (Arg Asp Glu His and Lys) are 150 amino acidsin the open reading frame of PI12 protease corresponded to156 while hydrophobic residues (Ala Phe Ile Leu MetPro Val and Trp) are 441 amino acids which correspondedto 458 of the total amino acids discovered The totaluncharged residues (Asp Cys Gln Gly SerThr and Tyr) areconsisted of 372 amino acids which corresponded to 386of the total amino acids of the open reading frame of PI12protease The aliphatic index is 9290 and the grand averageof hydropathicity (GRAVY) is 0141 The instability index (II)of PI12 protease was computed to be 3414 which classify theprotein as stable
N-glycosylation sites were predicted using NetNGlyc10 Server (httpwwwcbsdtudkservicesNetNGlyc)Therewere five putative N-glycosylation sites for the PI12 proteasegene predicted using the server For N-glycosylation thesequence motif Asn-Xaa-SerThr (Xaa any amino acid butnot Pro) has been defined as a prerequisite for glycosylationThe sequencemotifs in PI12 protease occurred in the position170 NETH 365 NATV 576 NLTP 634 NDTA and 833 NSSPAmong five predicted sites only three (except positions 576and 833) showed a high potential score (averaged output ofnine neural networks) which support the prediction Theputative N-glycosylation sites were demonstrated in boxesin Figure 4 The comparison of amino acid composition ofthis cold-adapted protease fromG antarctica strain PI12 withnine other subtilisin serine proteases fromdifferent families isshown in Table 2 Analysis of this comparison depicts that thepercentage of each amino acids in PI12 protease is almost thesamewith other proteaseswhileGly (G) is themost abundantaccounting for up to 104 of the total amino acid in PI12 pro-tease PI12 protease also contained the highest percentage ofLeu (L) with 99 of the total amino acids compared to other
Enzyme Research 9
Table2Aminoacid
compo
sitionandmolecular
weighto
fproteases
acrossdifferent
families
Amino
CAQ76821
CAG25549
EDR11679
CAG38357
CAD118
98CA
L25578
EDU50747
AAW
40780
ACB3
0121
ACB3
0119
acid
Qty
Qty
Qty
Qty
Qty
Qty
Qty
Qty
Qty
Qty
Ala
(A)
97101
8191
7887
7590
8299
101
115
8189
9196
8092
105
116
Arg
(R)
4042
2730
2224
1619
1721
1821
2932
2223
2124
3134
Asn
(N)
4446
3843
4348
4149
4251
4046
5257
5154
4552
4347
Asp
(D)
4951
6269
5460
6578
5567
5158
5358
6669
5260
6370
Cys
(C)
808
506
404
405
911
809
910
909
708
1011
Gln
(Q)
2324
3438
2326
2834
3036
3742
3539
2931
1922
2629
Glu
(E)
2122
1618
1921
2024
1518
2023
2528
2627
2832
2426
Gly
(G)
100
104
89100
8493
85102
85103
7788
8493
8084
8598
8695
His
(H)
1415
1416
1314
1619
1316
1113
1618
1415
1618
1213
Ile(I)
5355
4955
5157
5364
4150
3843
5156
4446
3540
4449
Leu
(L)
9599
6876
7179
6274
6073
6675
6774
8084
7485
6875
Lys
(K)
2627
2932
4449
2935
4959
5158
4044
3032
5058
5662
Met
(M)
1212
202
202
911
1113
809
1820
808
1113
1517
Phe
(F)
4143
5966
4752
5566
2935
3641
4449
3941
3439
3438
Pro
(P)
6264
6573
6774
5870
5870
5967
5055
5154
5564
5763
ser
(S)
7780
6977
91101
6072
6680
7080
7279
111
117
6373
6167
Thr
(T)
8790
89100
7583
83100
6579
7383
7178
7276
7182
6875
Trp
(W)
505
303
607
202
1215
1214
1213
909
1214
1415
Tyr
(Y)
3334
1213
2629
708
2935
2832
3943
5053
2630
2730
val
(V)
7679
8292
8089
6679
5769
7484
5864
6872
8295
6268
Total
963
1000
893
1000
900
1000
834
1000
825
1000
878
1000
906
1000
950
1000
866
1000
906
1000
MW
(Da)
1009898
932669
944365
869455
869549
924905
979343
1009629
917209
961988
Thea
ccessio
nnu
mbersof
proteasespu
tativ
eserinep
roteaseo
fGlacio
zymaantarctica
PI12
(CAQ
76821)sub
tilisin-lik
eproteaseP
leurotuso
streatus(CA
G25549)pyrolysin
LaccariabicolorS
238N
-H82
(EDR11679)
subtilisin
-like
proteasePh
anerochaetechrysosporiu
m(C
AG38357)sub
tilisin-lik
eserinep
roteaseP
R1CMetarhiziu
maniso
pliaev
aranisopliae(CA
D11898)serinee
ndop
eptid
aseH
ypocrealixii(C
AL2
5578)peptidase
Pyrenophoratritici-repentis
(EDU50747)p
eptid
aseCr
yptococcus
neoforman
svarn
eoform
ansJEC
21(A
AW40
780)sub
tilisin-lik
eprotease
Epich
loefestucae(A
CB30121)sub
tilisin-lik
eprotease
Epich
loefestucae
(ACB
30119
)
10 Enzyme Research
atgctcttcctccccgtcctcctcctcctccttcccggcgtcactgccttcctcaaccca 60
M L F L P V L L L L L P G V T A F L N Pgttaccaaccgcgcgaccaacgccatctcctcgactcaatacctctctaacgcctatatc 120
V T N R A T N A I S S T Q Y L S N A Y IctcgaattggacctctccacccccggcctcgtcaaacgggatagcacgcccgactctGTG 180
L E L D L S T P G L V K R D S T P D SCGTCCCGCGTTCCTCCCTCGCTGTCGAAGACGAGCTAACAATGACGTGGTAGatcctaga 240
I L Eggacgtactcacgtccgtaggccgcaacggtatcaagtaccaactccgccaccgctttat 300
D V L T S V G R N G I K Y Q L R H R F Ictccccgactctgttccacggcgcttcgatcactgtcccccctggaatctcccgctccca 360
S P T L F H G A S I T V P P G I S R S QaatcgcctctctccgcggtatcaaaGTCCGTCCCTCTCTCTCCCCTACTCCCTCCGTGCT 420
I A S L R G I KAACCCTGCACGCAACAGcgcgtctggcccgttcgaaagttctcccgacccagcgcagtag 480
R V W P V R K F S R P S A Vtggacgccgatggaggaggaagcgggttctcagggtcgcctatcaaggcggcgctcatgg 540
V D A D G G G S G F S G S P I K A A L Mgggtgaaagagctcgggaagcgcgcgaacgcttatgctggagatacgtttggaccgcatg 600
G V K E L G K R A N A Y A G D T F G P HtcatgacgggggttaatgagacgcatgaggcggggttgttgggagctgggattaagGTGT 660
V M T G V N E T H E A G L L G A G I KGTTTTCGTCTTTGTTTGGGGGAGGGGGGAGGGAGGAGCTGATTAATGGGTGCAGattggg 720
I GgtgctggacactggtgAGAAGCGGTTTGGAGGGAGGAGAGGATGGAGCTGATGAGTGTGC 780
V L D T GAGGTGttgattatttgaacccgattctgggaggctgctttggacctgggtgccatatgtc 840
V D Y L N P I L G G C F G P G C H M Sgtttgggtacgacttggttggcgatgattacgatggagataacgctcctgtgccggatgt 900
F G Y D L V G D D Y D G D N A P V P D VggatccttGTGCGTCCTTCCCTCCGCGATGGGCTGGAATCTGAGCTGACGGCGTTGTTGA 960
D PTCAGttgcgagctgcgatcctcatggTTCGTCTCGCTTTCCGCTCCTCTGATGCGCTCGC 1020
F A S C D P H GTGATGCACTTGGCAATAGGaactcacgttacgggaatcattggagcgctcccgaatgcgt 1080
T H V T G I I G A L P N AttggatttactggcgtcgcacccgccgctactctgggccactaccgagtGCGCTCCCTCT 1140
F G F T G V A P A A T L G H Y R VATCCGTTCTTCCTCGCAGCTCCCACTCACTCGTGCTTCCAGGTatttggctgcactggct 1200
F G C T GtcgtcggagaagatatcGTTCGTCCTCTCTCCAGTCCCGGCTAGCGATCCTGCTTCTGAC 1260
F V G E D ITTCGCATCTCCCTCTGAGattctcgctggactcatgcgaggagtcgaggacaactgcaac 1320
I L A G L M R G V E D N C Ngtcttgaccctctctctcggaggtccaggagggtgggtcaagggcacgccggcgtccatc 1380
V L T L S L G G P G G W V K G T P A S Icttatcgaccagatcgaagcgcaaggcattctcgtcaccgtcgccactggcaactcggga 1440
L I D Q I E A Q G I L V T V A T G N S GgctgagggTGAGCTCCTCGCTACTTGCGCGTCGAGACCAGTACTAACAGGGGCGTGGACG 1500
A E GCAGGcatgttcttctccgagtctcccgcctcgacgatcaacggcctttccatcgcatcca 1560
M F F S E S P A S T I N G L S I A ScgTCCGTCTCTCCCTTCCTTCGCACGCCACTCGCTCACACTCTCTCGCAGGgacgttacc 1620
T D V Tgacctcatcgcctacaacgccaccgtctcaggccaacctgcgatcccttacctctccgcc 1680
D L I A Y N A T V S G Q P A I P Y L S A
ggggggggggtctg minus194
ctagcgcgaatcgagaaaacacaaaagaacctcccgacttggcgttgcttctgctcttct minus180
ccttctcccaaccgccgacaataaaaactataacacacacacaccccatctcgacgcctg minus120
cagacttcacgactctcctcacgctctttaccgccccacgggtcccacccacgagccacg minus60
(a)
Figure 4 Continued
Enzyme Research 11
acgcccctcaacgtcgtcgccaacagcttccgcgtccacttcacctctaccgaccccaac 1740
T P L N V V A N S F R V H F T S T D P Naaccccgtcgacgcctgctctcctcttccggctggagcgcccgacttcgccaactatgtt 1800
N P V D A C S P L P A G A P D F A N Y Vacggtcgttcagcgtgggacttgtacgttcgttaccaagtaccagaacgttctcaatgct 1860
T V V Q R G T C T F V T K Y Q N V L N AggaggGTGCGTCGGCTTCTTCTCTCCCGACTGGATTGGGCGGAGACTGACCTTCTGAGCA 1920
G GGaaagatcgtattgttgtacaactcggagggagctgggaacctcccttacctcacgccca 1980
K I V L L Y N S E G A G N L P Y L T PacggtgtcggcatcgacgccgttgcaggtcttcgtcgttccgacggactcaagGTTCGTC 2040
N G V G I D A V A G L R R S D G L KTACGCTCTTCGGTGCTGAGGCCCTTCTCTGATCTCGCTCCGACAGcttctctcgtactat 2100
L L S Y Ycagaatgccaacaagcgtctcactctgcgcttccccaagggcaagatcgtcgcaggcttg 2160
Q N A N K R L T L R F P K G K I V A G LaccgataccatcaccggcggactcatctcgTTCGTTCCTCTCCGCCTCCCTCGCTGGCCG 2220
T D T I T G G L I SATGAGCTGACGCGGTCTTTGCAGGggttactcgacgtttggtccgacgaatgacctctac 2280
G Y S T F G P T N D L Yggtcagcctaccctctctgcccctggtggcaacatcctttcgaccttccctctctccgag 2340
G Q P T L S A P G G N I L S T F P L S EggaggagtggcggtcatcagtgggacgagcatgtcgtgcccctttgtcgGTTCGTCCTTC 2400
G G V A V I S G T S M S C P F VTCCTCGTTCGTTCGCTGAGCGTCGACTGATCTTGCACGATCGCAGctggatctgcggcgg 2460
A G S A Atcctcatggccgctcgcgcttcggagaacctcacgccgcttgagatcaggagtctcctta 2520
V L M A A R A S E N L T P L E I R S L Lctaccactgcgaagcttacgccggtctcgctcttgggatcgacgcctttggtgagcgtga 2580
T T T A K L T P V S L L G S T P L V S VttcgtcaaggaggaggTACGTTCCTTTCTCGCTGCTTTGAGGATCGAGGCTGATTAGCGA 2640
I R Q G G GGCTTCGCAGGactcgttcaggttgccaaggcgctcgcggccaagacgctaatctctcctc 2700
L V Q V A K A L A A K T L I S Pacgagctcctgctcaacgacactgcgaacgcgaactacgtccagactatcaagatcaaga 2760
H E L L L N D T A N A N Y V Q T I K I Kacaccaactcgtgggcgatgaagtacaccttctcctcggccgtcgcccaaggactcggaa 2820
N T N S W A M K Y T F S S A V A Q G L Gctttcgacgcttcgggcgatatcctccctaccctcgacccagtcgccgtctctggcgcac 2880
T F D A S G D I L P T L D P V A V S G Aaggctaccgtcgcgttcaacactcggatcctcagcgtcgcgcccggcgcgacggggtccg 2940
Q A T V A F N T R I L S V A P G A T G Stcgtggcgactatcacgccgccggttcttcccgtagcggacgctgcgaggttccctatct 3000
V V A T I T P P V L P V A D A A R F P Itctctgggtggatcagggtgaatgggcaaggcgcgagggatagcagtaggaacgaggcgt 3060
F S G W I R V N G Q G A R D S S R N E AacactgGTGCGTTTGGCTCGCTTTTTGGGAGGTGCCGAGCTAATCGTTTCTCGTAGtccc 3120
Y T V PgtactttgggcttgcggcgaagatgatcgatatgcaagGTGCGTCGATGGTTCTCTCGCT 3180
Y F G L A A K M I D M QTCTCTCTCGGTTGCTCACTTCAAAGTGCAGtcctcgacaccaccgagaccatttacggtc 3240
V L D T T E T I Y Gcgggctacgcctaccccttcgtgatcgacgacgcgattggagacatccaatccaccacaa 3300
P G Y A Y P F V I D D A I G D I Q S T TcgtcgtactccaggaacctcgggcccaccgtcttcgctcgctttgccaTGCGTCGACCGT 3360
T S Y S R N L G P T V F A R F ACCTCATTTCCTGCGGCAGAGCGAGAGCTGATTCCGACCTCGAACAGctggaacccttcac 3420
T G T L Htacagcctcgatctcgtcctagccgacatcgcctttacccccacctaccccaactcctcc 3480
Y S L D L V L A D I A F T P T Y P N S S
(b)
Figure 4 Continued
12 Enzyme Research
cccgccactcgtttcgtcaagcgctccctcacgcagcacacctctgccgcttcgcacctc 3540
P A T R F V K R S L T Q H T S A A S H Lgccaagcgccgcgtctccatcgccactatcaaccccaaagccaccctcgtcgccgatcga 3600
A K R R V S I A T I N P K A T L V A D Rcagctccactcggacgtccctatcgagggcaacatcttcacccaaccctttactggaagg 3660
Q L H S D V P I E G N I F T Q P F T G Rgattacctcgtcgacgcagccccgacgggatccaccgatcgtaccgtcacttttaacggg 3720
D Y L V D A A P T G S T D R T V T F N Gcagtacgccgagaacggcctcgtgaggacggctgtgacggggacttcgtaccgcttcctc 3780
Q Y A E N G L V R T A V T G T S Y R F Lcttcgggcgttgaagatctcgggagacgcgatgtacgaggatcagtatgagagctggctc 3840
L R A L K I S G D A M Y E D Q Y E S W L3900
S L P F S F R Attttgccacctcaaaaaaaaaaaaaaaaaaaaaaaaaa 3938
lowast
tcgctaccgttctcgttccgtgcgtaggcttgttgtggacaccactgcattaagttatga
(c)
Figure 4 Nucleotide and deduced amino acid sequences of genomic DNA and cDNA encoding protease from G antarctica strain PI12 Thenucleotides are numbered from the 51015840 end of the cDNA A putative CAAT TATA box and polyadenylation signal are bold italicThe putativeN-terminal signal peptide is underlined The initiation codon is boldfaced and termination codon is represented by an asterisk (lowast) boxedpeptide sequences showed N-glycosylated asparagines residues The amino acid residues that form the catalytic triad are double underlinedand bold while introns are represented in blue font Highlighted in grey are the intron donor-acceptor splice sites
proteases Hydrophobic profile of PI12 amino acid proteasewas performed by ProtScale tools with Kyte and Doolittlemethod at httpauexpasyorgcgi-bin The hydrophobicityprofile elucidation of PI12 protease amino acid was carriedout from amino acid 1 to 963 The amino acid residuesfrom 1 to 16 (M-L-F-L-P-V-L-L-L-L-L-P-G-V-T-A) and 548 to562 (V-A-V-I-S) were hydrophobic grooves The first regionwhich consisted of 16 amino acids was very hydrophobic andwas predicted to be the signal peptide by SignalP and ProPwebserver
The bold and double-underlined amino acids showedresidues of catalytic triad aspartic acid (D) histidine (H) andserine (S) active site respectively NCBI Conserved DomainsDatabase (CDD) identified the presence of subtilase proteaseand subtilisin-like serine protease domains in the amino acidsequence The predicted PI12 protease amino acid sequencewas found sharing significant homology to the subtilisinfamily of protease The highest similarity of PI12 proteasewas observed with 42 identity to subtilisin-like proteaseof Rhodosporidium toruloides (EMS20793) 37 identity tosubtilisin-like protease of Pleurotus ostreatus (CAG25549)and 35 identity to subtilisin-like protease of Gloeophyllumtrabeum ATCC 11539 (EPQ55564) The nucleotide sequenceof G antarctica strain PI12 showed low homology to thefungus subtilisin protease family and none to other psy-chrophilic proteases The multiple sequence alignment wasillustrated in Figure 5 The consensus sequence around thecatalytic triad aspartic acid (D) histidine (H) and serine(S) of PI12 protease showed high conservation with othersubtilisin (showed in black boxes) Glycine residues G142G173 G191 G262 and G327 that were highly conserved inmost of subtilases were conserved in PI12 protease [25]Therefore PI12 protease belongs to the subtilisin subgroup ofthe subtilase serine protease superfamilyThis serine protease
of G antarctica PI12 has been deposited into GenBank withaccession number CAQ76821
34 Phylogenetic Relationship of PI12 Protease The phylo-genetic tree was constructed to demonstrate the similaritiesand relationship of protein sequence between differentspecies or organisms Figure 6 illustrates the phylogeneticdendrogram indicating the relationship of PI12 proteasewith another 25 translated amino acid sequences ofserine protease The bar scale signifies the branch lengthto approximate variance along each branch Basicallythis tree comprises two main clusters cluster I was sub-divided into subcluster a (CAG25549 EPQ55564 ETW-80907 XP 007381302 XP 001877576 CAG38357 XP007341385 CCA68627 EJU04161 XP 007369624 EIW-62675 CCO37249 XP 001830835 XP 007342189) and sub-cluster b (XP 007589863 XP 003711175 EFQ32840 XP006694395 ENH74885 EON99722) cluster II was sub-divided into subcluster c (CBQ73425 GAC97617 CCF52619)and subcluster d [CAQ76821 (PI12 protease) EMS20793]
Serine protease constructed by the phylogenetic treecomprised large size of protein with about 801 to 1032 aminoacids The majority of the translated protease sequenceswere obtained from fungus All serine proteases from cluster1 subcluster a are derived from Agaricomycetes except forEJU04161 which belongs to Dacrymycetes whereby all serineproteases from cluster 1 subcluster b are from Sordari-omycetes Cluster II subcluster c consisted of serine proteasefrom fungus belonging to Ustilaginomycetes PI12 proteasewas categorized under cluster II subcluster d It possessedhomology similarity with the subtilisin-like protease fromRhodosporidium toruloides (EMS20793) (formerly known asRhodotorula glutinis or Rhodotorula gracilis) which is a
Enzyme Research 13
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
MKG-YSWSLLQLAFTGAALATVPLSDVKRISSAS--VVPNKFVVEVESASEIPGKR--SL
MKGVLVWSILS---AGCALAVTPLSSVKRVTNLP--IVANKFIVEVDSTSDIPTKR--S-
MKLPLLSLALT---AVSVGATVPLSSVQRTTTSPNNALAGEYIVEMNNVQGLGGKR--AY
---MAHGSLASLLLLLAASAQGAAAFGGKSATGGRSVVPNSYILQVNASAPALTKRGVSV
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease ---MLFLPVLLLLLPGVTAFLNPVTNRATNAISSTQYLSNAYILELDLSTPGLVKRDSTP
lowastlowast
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
YTPHELFYRSLSARGVSYEVNKEYNS-EGLFVGAVVTVSDANDVKALAETPGVKAIRPVR
-NPHDALYESLRKRAIGFKVDREFNT-QGLFVGAALTLSDAQDVGQIQSTTGVKSIRPVR
ASTHDTLYRSLRKRGISFDVKKEYNQ-PGIMVGAALKVASPQDVQQIATMAEVKAIRPVI
FEALDHTLAAVKQNGVKYTVRQRFDAIPEAFQAVSIQVEDGASMAELAQIPGVQRVWPVS
DSILEDVLTSVGRNGIKYQLRHRFIS-PTLFHGASITVPPGISRSQIASLRGIKRVWPVR
lowastlowast
TIKSPKPVKVHVVTGKDDPEIPA-------------------------DTESTHIMTGVN
AFDRPKPVKVQVVKGPDDPALPP-------------------------DSESTHVITGVD
AVPAPERINQFTVSSPNDSRITA-------------------------DSETTHIMTGVD
LIPRPVEPTVSDFTPVSSSSSSTKSKRD-THTLEKRGTNFPPASAYLNDTFYPHVQTGID
KFSRPSAVVDADGGGSGFSGSPIKAALMGVKELGKR------ANAYAGDTFGPHVMTGVN
lowast lowast lowast lowastlowast
KLHAQGIFGAGIKIGIIDTGIDYTHPALGGALGAGHKVIGGYDFVGDAYDGTNTPVPDND
KLHAQGITGAGIKIGILDTGIDYTHPTLGGAFGPGNKVIGGFDLVGDDYDGTNTPVPDPD
KVHAAGITGEGIKIGIIDTGIDYNHPLLGAGFGPGFKVIGGYDFVGDDYTGSNNPAPDDD
VLHNKGILGQGVKIVVVDEGVDYTNPILGGCFGPGCQISFGYDFVGDNYTGTNAPVPDSD
ETHEAGLLGAGIKIGVLDTGVDYLNPILGGCFGPGCHMSFGYDLVGDDYDGDNAPVPDVD
lowast lowast lowast lowastlowastlowast lowast lowast lowastlowast lowast lowastlowast lowast lowast lowast lowast lowastlowastlowast lowast lowast lowast lowast lowastlowast lowast
PLDQCNGHGTHVAGIIGANPDNQFNISGVSYGASLSSYRVFGCSGSVSDDIIIEALLRGV
PLDQCAGHGTHVAGIIGANPDNAFNISGVAFQASLSAYRVFGCVGFVTDDVLVDALLLGF
PLDNCNGHGTHVAGIIGANPGNDFGISGVAYGSSIASYRVFGCSGFVSDDIIIDALLRGY
PFSSCSNHGTHTTGTVGALAN-EYGFSGAAPMATIGHYRVISCSGATTDEILVAALTRAL
PFASCDPHGTHVTGIIGALPN-AFGFTGVAPAATLGHYRVFGCTGFVGEDIILAGLMRGV
lowast lowastlowastlowast lowastlowast lowast lowastlowast lowast lowastlowastlowast lowast lowast lowast
QDGQDILTLSLGGTDGWTE-SSSSVVASRIAASGKIVTIAAGNEGAYGSWYSSSPGNGID
NEGQDILTLSLGGADGWTE-SVSAVVASRIAATGKVVTIAAGNDGASGAWYTSSPGNGID
SDGMDILTLSLGGVEGWTE-SSSSVVASRIADQGRVVTIAAGNDGAYGSWYASSPGTGVD
QDNVNIVSMSLGSSVGWLDDSPVQIMASYLGSKGIHVVASVGNERNEGLFAADQPAASRI
EDNCNVLTLSLGGPGGWVKGTPASILIDQIEAQGILVTVATGNSGAEGMFFSESPASTIN
lowastlowastlowast lowastlowast lowast lowast lowastlowast lowast lowast
VISVASIDNTVDNIQNATVNGVEHGPIPYFKTLPLPFTNA-LPLYVISNDTTIANDACNA
VISVASLDNTVVPLQTAQVHGVTHDPIIYQDTFPLPINDT-RPIFATSTDVTVVDDACNP
VISVASVNNIAIPVQNATVQGVEHAPIPYLEALPLNVTGS-RPIYATSTDTTVADDACNA
GTGVGGVDPTYLAAYYAYLI--KRDPVPYISPTPFNLPNN-YLLYFTSTNTSVTNDACNP
GLSIASTDVTDLIAYNATVS--GQPAIPYLSATPLNVVANSFRVHFTSTDPNNPVDACSP
lowast lowast lowast lowast lowastlowastlowast
LPDNTPDLSGFLVLVRRGSCTFVQKLTNIAAKGGNAVFIYDNGNG-----FAGIDTGKFT
LPDSTPDLSGFVVIVRRGTCTFVQKLGNVAAKGAKVTLIYDNGSG-----FGAIDTGAFV
LPNDTPDLSGYVVVIKRGTCTFVTKLQNAAAKGANTFLIYNNGGS-----FSAISVGDYT
LPSTTPNLANRVVVVQRGTCGFTVKQQNVAAAGGKIVLIYNSKGSGLIPQLDVGSTGLTA
LPAGAPDFANYVTVVQRGTCTFVTKYQNVLNAGGKIVLLYNSEGAGNLPYLTPNGVGIDA
lowastlowast lowast lowastlowast lowast lowast lowast lowast lowast lowast lowast
(a)
Figure 5 Continued
14 Enzyme Research
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
NATLIQAADGVFLAQQVAAGKSVTITFPQTGGSAQITN-PSGGLVSSFTTYGPTNDFYFK
-ASFIQAADGEFLVQQFASGQNVSLSFPQSGGLIQFPD-PAGGLISSFTSYGPSNDFFFK
-AALISETDGTFLVNQFAAKANITISFPQSGSAWNYPDDVSGGLVSDFSSYGPSYDMYFK
VGSLRYEDGLKLLSYYQSSPRGQSISFPNGPLAPYVSNTVSGGVVASYSNFGPTNELFIY
VAGLRRSDGLKLLSYYQNANKRLTLRFPKGKIVAGLTDTITGGLISGYSTFGPTNDLYGQ
lowast lowastlowast lowastlowast lowastlowast
PAFGAPGGNILSTLPVPLGSYGVESGTSMATPYVAGSAALLLSVKGKSAAVALGARTLFQ
PAVAAPGGNILSTLPVNLGLFGIESGTSMATPFVAGSAALLFQAKGTSAAVGRSARTLFE
PAVAAPGGNILSTLPIALGSYGVESGTSMATPFTAGSAALILQAKGK--SVAKSIRNLFQ
PTLAAPGTDILSTV---TGGVALMRGTSMAAPLVAGAYALLLSVGNNLKLSPEEARTILM
PTLSAPGGNILSTFPLSEGGVAVISGTSMSCPFVAGSAAVLMAARASENLTPLEIRSLLT
lowast lowastlowastlowast lowastlowastlowastlowast lowast lowastlowastlowastlowast lowast lowastlowast lowast lowast
STAQRVPSSHTDGDPLQTLSQQGAGLIDVFKALYTETIVWPTELILNDTAHYKEVHTFTV
TTAQRVPSTHTDGDPLQTLTQQGAGLINVFNAIHTTTIVSPGELILNDTAHFKGTQQFTV
TTASAVASSKTDGDPLQTVSQQGAGLIQVDRAIGTTTVVSPGQLTLNDTAHFEGFKTFNI
STSQPSPVAY-GSETLDTVVSQGAGVINMTAAIAAHTVISPAQFNLNDTQYSNNTQTLTL
TTAKLTPVSLLGSTPLVSVIRQGGGLVQVAKALAAKTLISPHELLLNDTANANYVQTIKI
VNTGKTSKQYKLTHVPAGTAQTIT---SGTIFAADGPVPLSTDYATVKLSSSSFTLRPGQ
RNAGSSAKTYTLRHIPAGTAVTVT---PGTIFPADGPVPLSTDFASVSLSTSKFTLAPGK
QNTGSKAVTYKISHVPAGTAASVQ---SGSIFAADGPVPLTADYAQVTFSLNKVTVLPGV
RNNNRFPVSYQLSWVDSTGIVTYNDGATTDIIPSTTPNYVSTSVLRVAFSQRTVTVPGGQ
KNTNSWAMKYTFSSAVAQGLGTFD--ASGDILPTLDPVAVSGAQATVAFNTRILSVAPGA
lowast lowast lowast lowast lowast lowast
SKQVTTHINAP--SGVDASTYPVFSGFIEIATAT--------ESVHVTYLGLAASLKDKQ
TQTVTARFTPP--AAADPSTFPVFSGFIQIESGT--------EQVQVSYLGLKASLKDKQ
SVPILVWFKPP--QGVDASTYPVYSGFIEIAGLG--------ETLRVAYLGVAASLKDKA
TVKVTAQFIPPNLTAQQRNQFPIYSGFVTITGQGQGAGAEQMEKYNLPFFGLGARMVDMP
TGSVVATITPPVLPVADAARFPIFSGWIRVNGQG-ARDSSRNEAYTVPYFGLAAKMIDMQ
lowast lowast lowastlowast lowast lowast lowast lowast
IVDNTNEFFGITIPAVLNNTGDAQ-LSPTNYTFSGSDFPTLLLRLAFGTPLLRVDLVDAT
VIDNTDFFFGVPTPVLTDPNGEVQ-TSPRNYSFLASDFPTLIFRLAFGSPKVVVDLVSPT
VVDNTDEYFGVKIPALLDASGDVQ-GNSTNYTLVGDDVPSLLYRFAFGSAHVRVDLVEKA
VLDTTNVALGP--NLPFIAVGQN--IQTGPTTFSESNPAVVYFRLAAGTRRLRVDLVDAS
VLDTTETIYGPGYAYPFVIDDAIGDIQSTTTSYSRNLGPTVFARFATGTLHYSLDLVLAD
lowast lowast lowast lowast lowastlowastlowastlowast lowast
IDLKPTLNSREIDDKRHVFERESFSFPHPGPKGS-------------------TFAQVKI
INFKPTLNTRALGDTHGSF----FSFPHRVKTG--------------------SFAQVPT
STVSATLSKRNWWN---------WWWPQKPSN---------------------SYDKVPI
TSYNATVPAVTNPASRLVKRSNTELTPRAAFPT--------------------LYSQVRT
IAFTPTYPNSS-PATRFVKRSLTQHTSAASHLAKRRVSIATINPKATLVADRQLHSDVPI
lowast lowast
VGTLSEQSHIPRNSDASTVDDGGYSTFAFNKP------SFANGTTIPNGTYRWLLRALKV
LGTVFELDFTSRNNDDPTDN--GFNTVAIDPP------TFANGTTIPNGQYRLLLRALRV
VGTLAEYDYVPRNSDASTIDDNGYNVFQFT-D------SFVNGTKIAAGEYKVLLRALKV
VGSLFAPSYWPPRDYLFNNGGT-YSDYEIQLDGSFTFPNGTKGMAELGRSYKVLLRALKI
EGNIFTQPFTG-RDYLVDAAPTGSTDRTVTFNGQY-AENGLVRTAVTGTSYRFLLRALKI
lowast lowast lowastlowast lowast lowast lowast lowast lowastlowastlowastlowast
lowast lowast lowastlowastlowastlowastlowast
(b)
Figure 5 Continued
Enzyme Research 15
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
TGDPTKEADFETWLSPIVGITVA
TGDPTNEADFESFLSPIIGVNAP
TGNPAEEGDYETWLSPSFSVVPA
TADPTYTSSYESWLSPSFTFTA-
SGDAMYEDQYESWLSLPFSFRA-
lowast lowastlowast
ConservedSimilar
lowast All match
(c)
Figure 5 Multiple sequence alignment between amino acid sequence of PI12 protease with other subtilisin The serine protease sequenceswere obtained from the GenBank database Their accession numbers are EMS20793 (subtilisin-like protease of Rhodosporidium toruloidesNP11) CAG25549 (subtilisin-like protease of Pleurotus ostreatus) EPQ55564 (subtilisin-like protease of Gloeophyllum trabeumATCC 11539)and ETW80907 (serine protease S8 ofHeterobasidion irregulare TC 32-1) Putative ORF of serine protease fromG antarctica PI12 was labeledas PI12 protease
CAG38357 Phanerochaete chrysosporium
EIW62675 Trametes versicolor FP-101664 SS1
XP 007369624 Dichomitus squalens LYAD-421 SS1
XP 007381302 Punctularia strigosozonata HHB-11173 SS5
CAG25549 Pleurotus ostreatus
ETW80907 Heterobasidion irregulare TC 32-1
XP 001877576 Laccaria bicolor S238N-H82
EPQ55564 Gloeophyllum trabeum ATCC 11539
XP 001830835 Coprinopsis cinerea okayama7130
XP 007341385 Auricularia delicata TFB-10046 SS5
CCA68627 Piriformospora indica DSM 11827
CCO37249 Rhizoctonia solani AG-1 IB
XP 007342189 Auricularia delicata TFB-10046 SS5
EJU04161 Dacryopinax sp DJM-731 SS1
XP 007589863 Colletotrichum fioriniae PJ7
EFQ32840 Colletotrichum graminicola M1001
ENH74885 Fusarium oxysporum f sp cubense race 1
XP 003711175 Magnaporthe oryzae 70-15
XP 006694395 Chaetomium thermophilum DSM 1495
EON99722 Togninia minima UCRPA7
CBQ73425 Sporisorium reilianum SRZ2
GAC97617 Pseudozyma hubeiensis SY62
CCF52619 Ustilago hordei
EMS20793 Rhodosporidium toruloides NP11
CAQ76821 Glaciozyma antarctica PI12
01
Clus
ter 1
Clus
ter 2
a
b
c
d
Figure 6 Phylogenetic dendrogram among 25 translated amino acid sequences of serine proteaseThe tree was constructed using a neighbor-joining algorithm with 1000 bootstrappings by MEGA 6 software [39]The scale bar represents 01 substitutions per amino acid positionTheserine protease sequences were obtained from the GenBank database GenBank accession numbers of the sequences were indicated beforethe strain name The putative serine protease of G antarctica strain PI12 has been deposited in GenBank with accession number CAQ76821
16 Enzyme Research
0
2
4
6
8
10
12
0 24 48 72 96 110
Prot
ease
activ
ity (U
mL)
Post induction time (hours)
Figure 7 Effect of postinduction time towards protease activity The highest activity was detected at 72 hours after 110 hours of incubationwith 9UmL activity followed by 48 24 and 96 hoursThe substrate (azocasein) was dissolved in Tris HCL buffer and incubated for 30min at20∘CThe absorbance was measured at 450 nm after 15min incubation A one-way ANOVAwas used to test for preference differences amongsix different postinduction times The statistical value (119875 value lt005) between hours is 0003
pink-coloured nonpathogenic oleaginous yeast (42 iden-tity) Both G antarctica PI12 and Rhodosporidium toruloidesNP11 belong to Microbotryomycetes class PI12 protease didnot show any homology similarity to other psychrophilicproteases however it showed 37 identity to hypotheticalprotein from Pseudozyma antarctica T-34 which was isolatedfrom Japan (GAC76005) (not listed in the phylogenetic tree)
35 Cloning and Expression of Cold-Adapted PI12 Protease inthe Yeast Expression System The PCR product harbouringthe mature PI12 protease with two flanking restriction sites(EcoRI and AvrII) was cloned into pPIC9 expression vectorThe recombinant plasmid harbouring PI12 protease was usedto transform both GS115 and KM71 intended to produce bothHis+ Mut+ and His+ MutS Two recombinant clones fromGS115 (His+Mut+) strain (GpPro1 andGpPro2) and one fromKM71 (His+MutS) strain (KpPro1) were successfully obtainedwhich were further analyzed for protein expression study
36 Expression and Enzyme Assay of PI12 Protease in Pichiapastoris All positive recombinants were grown in small scalecultures and assayed for the cold-adapted protease activityin the culture supernatants The clones were first grown inBMGY medium with glycerol as the sole carbon sourceFor induction of AOX1 promoter and protein productionthe growth medium was transferred to the same mediumexcept with 05 methanol in place with glycerol as thecarbon source [26] The supernatant of the recombinant Ppastoris expression cultures after 48 h of incubation time wasanalyzed to determine the recombinant protease activity bythe modified method of Brock et al (1982) [20] All testedclones secreted protease into the supernatant in differentexpression levels This might be due to the variability ofthe strength of the promoter and copy number in each cellto secrete and express the recombinant product The clonewith the highest yield was GpPro2 with 63UmL activityfollowed by KpPro1 and GpPro1 with 25UmL and 37UmLactivities respectively (data not shown) It is perceptible thatexpression systemof Pichia pastoris is the ideal system for thisprotein The preliminary assay has fruitfully produced high
protease expression at 20∘C Yet further optimization of therecombinant PI12 protease production need to be conductedin order to examine the optimum condition of this proteinReported cold-adapted proteases showed optimum activity ata range of 15∘Cndash60∘C [27ndash30] This difference was possiblyrelated to the respective genetic and physiological adaptationof the strains There are several factors that influenced thedifferent rate of expression It includes different site of inte-gration between interest gene and Pichia genome variabilityof different expression host cell to secrete and express therecombinant enzyme and different sites in the polylinker forinserting foreign genes [31]
This is the second extracellular protease reported to besynthesized by this obligate psychrophilic yeastG antarcticaThe first cold-adapted extracellular subtilase from this yeasthas been purified and characterized previously [32 33] Theextracellular serine proteinase (LAP2) from this Antarcticyeast formerly known as Leucosporidium antarcticum strain171 is specific towards synthetic substrates of chymotrypsinand subtilisin The subtilase of L antarcticum strain 171showed specific activity of 4043Umg when utilized N-SucAAPFpNA as a substrate [33]
37 Optimization of Protease Production GpPro2 was chosenfor further optimization due to its highest protease activityat the preliminary expression stage High protease activitywas detected after 72 hours of incubation (Figure 7) byusing the standard protocol method as recommended forPichia pastoris expression system (Pichia Expression KitInvitrogen USA) The expression decreased after 72 hoursand no activity was detected at 110 hours The prolongedincubation may result in the decrease of biomass productionand will subsequently reduce the expression of recombinantprotein In addition oxygen and nutrient depletion duringprolonged incubationwill trigger the stress responses in orderto survive in such harsh environment [34]
Methanol was used as a sole carbon by P pastoris Themethanol was oxidized to formaldehyde with the aid of oxy-gen as electron acceptor by an alcohol oxidase enzyme (AOXgene) [35] The optimal methanol concentration is at 05(vv) with 14UmL activity The addition of more than 05
Enzyme Research 17
02468
10121416
0 01 03 05 07 09Pr
otea
se ac
tivity
(Um
L)Methanol concentration ()
Figure 8 Effect ofmethanol concentration towards protease activityThe recombinant was cultivated for 72 hours at 30∘C in BMMYmedium05 methanol gives the highest activity with 14UmL The substrate (azocasein) was dissolved in Tris HCL buffer and incubated for 30minat 20∘CThe mixture was left for 15min and absorbance was taken at 450 nm A one-way ANOVA was used to test for preference differencesamong six different concentrations The statistical value (119875 value lt005) between concentration is 002
0
5
10
15
20
25
10 15 20 25 30 35
Prot
eas a
ctiv
ity (
Um
L)
Temperature (∘C)
Figure 9 Effect of temperature towards protease activity The reaction was carried out in temperature ranging from 10∘C to 35∘C at pH 7 for30min A one-way ANOVA was used to test for preference differences among six different temperatures The statistical value (119875 value lt005)between temperature is 0067
(vv) methanol resulted in tremendous decrease of proteaseactivity (Figure 8) Production of PI12 protease was decreasedwhen the methanol concentration increased This might bedue to the toxic effect of methanol concentration on DNAreplication and membrane synthesis Basically methanol isused to increase the permeability of the cell membrane tohelp the secretion of foreign protein [26] However if thepermeability of cell membrane is too high lowmolecular sizemetabolites would leak from the cell and such leakage willseriously damage cell activities Such condition would abusethe cellular system and might reduce the yield of proteinexpression [36]
Psychrophilic microorganism dominated in low temper-ature habitat which most other microorganism are inca-pable of withstanding due to their inability to produceenzyme that can tolerate the extreme environment con-dition Psychrophilic enzymes are always associated withhigh catalytic efficiency at low and moderate temperature(0∘C to 30∘C) compared to the mesophilic counterpart thatare generally unable to catalyze efficiently under extremely
low temperature [2] The recombinant protein assay wasdone at various temperatures ranging from 10∘C to 35∘Cusing 05 azocasein as a substrate dissolved in 50mMTris HCl in presence of 2mM CaCl
2 pH 7 In the activity
assays expressed protease showed a highest activity at 20∘C(20UmL) and decreased tremendously once it exceededmaximum temperature due to the denatured state of theenzyme (Figure 9) The active site of an enzyme is easilyaltered when the high temperature is introduced duringincubation or activity assay resulting in a low reaction rate[37]
The recombinant construct was transferred into BMMYmedia at different A
600 nm ranging from 1 to 10 The high-est activity of active enzyme was detected at A
600 nm 60(283UmL) and above (Figure 10) The slope during expo-nential phase was much higher compared to other phasesindicating an increase in the doubling time At exponentialphase the cells are adapted to the growth condition andother processes like cell growth cycle and synthesis of RNAThe yield of active enzyme during expression of P pastoris
18 Enzyme Research
05
1015202530
1 2 3 4 5 6 7 8 9 10Prot
ease
activ
ity (U
mL)
Initial inoculum A600
Figure 10 Effect of inoculums size towards protease activity The recombinant was cultivated for 72 hours at 30∘C in BMMY mediumwith 05 methanol concentration A one-way ANOVA was used to test for preference differences among six different optical densitiesThe statistical value (119875 value lt005) between optical density value is 003
(kD
a)
1160
662
450
350
250
184
144
M 1 2 3
(a) (b) (c)
Figure 11 (a) Coomassie-stained SDS-PAGE of recombinant P pastoris clone 1mL supernatants collected from the third-day culturewere concentrated 10-fold using trichloroacetic acid and 10120583L resultant products were applied to SDS-PAGE analysis Lane 1 molecularweight marker (Fermentas) Lane 2 GS115 (Control) Lane 3 GS115pPIC9 empty (Control) Lane 4 GS115pPIC9PI12prot (GpPro2)The arrow indicates the secreted recombinant PI12 protease (b) Plate activity staining of different recombinant P pastoris clones GGS115 GP GS115pPIC9 GRI GS115pPIC9PI12prot1 (GpPro1) GRII GS115pPIC9PI12prot2 (GpPro2) K KM71 KP KM71pPIC9 KRKM71pPIC9PI12prot (KpPro) (c)Gel activity staining of recombinant PI12 protease inP pastorisGpPro2 clone Lane 1 prestainedmolecularweightmarker (Fermentas) Lane 2 GS115pPIC9 empty (Control) Lane 3 GS115pPIC9PI12prot1 (GpPro2)The arrow indicates the secretedrecombinant PI12 protease
dropped after 28 hours of incubation which is at stationaryor death phase [38]
38 Detection of Recombinant Protein by SDS-PAGE andActivity Staining TheSDS-PAGE gel of the GpPro2 recombi-nant clonesrsquo supernatant showed a putative expression bandat around 993 kDa Plate staining and gel activity stainingwere executed to examine if the recombinant PI12 proteaseexpressed in P pastoris was functionally active In the platestaining the transformants showed strong proteolytic activity(Figure 11(a)) In gel staining proteolytic activity was alsodetected around 993 kDa after incubation at 20∘C This is
evidence of protease secretion into the surrounding mediawhereby it catalyzed the breakdown of casein thus formedclearing zones around the activity ring and band (Figures11(b) and 11(c)) As a conclusion the cold-adapted PI12protease was functionally expressed in P pastoris GS115(GpPro2 clone) with the size of 993 kDa
4 Conclusions
Psychrophilic yeast which was originated frommarine waterin Casey Station Antarctica was identified as Glaciozymaantarctica strain PI12 A serine protease with an open reading
Enzyme Research 19
frame of 2892 bp encoded for 963 amino acids from thisAntarctic yeast has been cloned and expressed in Pichiaexpression system This is the second extracellular pro-tease reported being synthesized by this psychrophilic yeastSequence analysis through bioinformatics studies revealedfew homology identities of this protein with other ser-ine proteases The mature PI12 protease was successfullyexpressed and secreted into the culture medium driven bythe Saccharomyces cerevisiae 120572-factor signal sequence in Ppastoris The protease production in P pastoris was bestobtained from strain GS115 (GpPro2) with 283UmL after 72hours of induction time with 05 (vv) of methanol inducerThe starting optical density at 600 nm (OD600) was 60 andthe optimum assay temperature was 20∘C The expressedprotein was detected by SDS-PAGE and activity staining witha molecular weight of 99 kDaThis cold-active enzymemightoffer an opportunity for biotechnological exploitation basedon its high catalytic activity at low temperatures and lowthermostability Structural and functional study of this cold-adapted enzyme would give insight into its adaptation to anextreme cold environment
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgment
This research was supported financially by the InitiativeBiotechnology-GMBI-MOSTI (Project number 07-05-16-MG-GMB02) andRUGS-Initiative 6 (Project number 05-02-11-1414RU)
References
[1] J W Deming ldquoPsychrophiles and polar regionsrdquo CurrentOpinion in Microbiology vol 5 no 3 pp 301ndash309 2002
[2] C Gerday M Aittaleb M Bentahir et al ldquoCold-adaptedenzymes from fundamentals to biotechnologyrdquo Trends inBiotechnology vol 18 no 3 pp 103ndash107 2000
[3] A Svingor J Kardos I Hajdu A Nemeth and P Zavodszky ldquoAbetter enzyme to cope with cold comparative flexibility studieson psychrotrophic mesophilic and thermophilic IPMDHSrdquoThe Journal of Biological Chemistry vol 276 no 30 pp 28121ndash28125 2001
[4] G Antranikian C E Vorgias and C Bertoldo ldquoExtremeenvironments as a resource for microorganisms and novel bio-catalystsrdquo Advances in Biochemical EngineeringBiotechnologyvol 96 pp 219ndash262 2005
[5] T Lonhienne C Gerday andG Feller ldquoPsychrophilic enzymesrevisiting the thermodynamic parameters of activation mayexplain local flexibilityrdquoBiochimica et Biophysica Acta vol 1543no 1 pp 1ndash10 2000
[6] R Cavicchioli and TThomas ldquoExtremophilesrdquo in Encyclopediaof Microbiology pp 317ndash337 Academic Press San Diego CalifUSA 2000
[7] G Feller E Narinx J L Arpigny et al ldquoEnzymes frompsychrophilic organismsrdquo FEMS Microbiology Reviews vol 18no 2-3 pp 189ndash202 1996
[8] J Arnorsdottir R B Smaradottir T O Magnusson S HThorbjarnardottir G Eggertsson and M M KristjanssonldquoCharacterization of a cloned subtilisin-like serine proteinasefrom a psychrotrophic Vibrio speciesrdquo European Journal ofBiochemistry vol 269 no 22 pp 5536ndash5546 2002
[9] N J Russell ldquoPsychrophilic bacteriamdashmolecular adaptations ofmembrane lipidsrdquo Comparative Biochemistry and Physiology APhysiology vol 118 no 3 pp 489ndash493 1997
[10] G Feller and C Gerday ldquoPsychrophilic enzymes hot topics incold adaptationrdquo Nature Reviews Microbiology vol 1 no 3 pp200ndash208 2003
[11] P Raspor and J Zupan ldquoYeasts in extreme environmentsrdquo inBiodiversity and Ecophysiology of Yeasts The Yeast Handbookpp 371ndash417 Springer Berlin Germany 2006
[12] M B Rao A M Tanksale M S Ghatge and V V Desh-pande ldquoMolecular and biotechnological aspects of microbialproteasesrdquoMicrobiology andMolecular Biology Reviews vol 62no 3 pp 597ndash635 1998
[13] L Polgar ldquoThe catalytic triad of serine peptidasesrdquo Cellular andMolecular Life Sciences vol 62 no 19-20 pp 2161ndash2172 2005
[14] S Y Boo C M V L Wong K F Rodrigues N NajimudinA M A Murad and N M Mahadi ldquoThermal stress responsesin Antarctic yeastGlaciozyma antarctica PI12 characterized byreal-time quantitative PCRrdquoPolar Biology vol 36 no 3 pp 381ndash389 2013
[15] B Turchetti S R T Hall L B Connell et al ldquoPsychrophilicyeasts from Antarctica and European glaciers descriptionof Glaciozyma gen nov Glaciozyma martinii sp nov andGlaciozyma watsonii sp novrdquo Extremophiles vol 15 no 5 pp573ndash586 2011
[16] N H F Hashim I Bharudin D L S Nguong et al ldquoCharac-terization of Afp1 an antifreeze protein from the psychrophilicyeast Glaciozyma antarctica PI12rdquo Extremophiles vol 17 no 1pp 63ndash73 2013
[17] A N M Ramli M A Azhar M S Shamsir et al ldquoSequenceand structural investigation of a novel psychrophilic 120572-amylasefrom Glaciozyma antarctica PI12 for cold-adaptation analysisrdquoJournal ofMolecularModeling vol 19 no 8 pp 3369ndash3383 2013
[18] A N M Ramli N M Mahadi A Rabu A M A Murad F DA Bakar and R M Illias ldquoMolecular cloning expression andbiochemical characterisation of a cold-adapted novel recombi-nant chitinase from Glaciozyma antarctica PI12rdquoMicrobial CellFactories vol 10 no 1 article 94 2011
[19] J Sambrook E Fritsch and T Maniatis Molecular Cloning ALaboratoryManual Cold SpringHarbor Laboratory Press NewYork NY USA 1989
[20] F M Brock C W Forsberg and J G Buchanan-SmithldquoProteolytic activity of rumen microorganisms and effects ofproteinase inhibitorsrdquoApplied and EnvironmentalMicrobiologyvol 44 no 3 pp 561ndash569 1982
[21] U K Laemmli ldquoCleavage of structural proteins during theassembly of the head of bacteriophage T4rdquo Nature vol 227 no5259 pp 680ndash685 1970
[22] A Henne R A Schmitz M Bomeke G Gottschalk and RDaniel ldquoScreening of environmental DNA libraries for thepresence of genes conferring lipolytic activity on Escherichiacolirdquo Applied and Environmental Microbiology vol 66 no 7 pp3113ndash3116 2000
[23] M Poza T de Miguel C Sieiro and T G Villa ldquoCharacteri-zation of a broad pH range protease of Candida caseinolyticardquoJournal of Applied Microbiology vol 91 no 5 pp 916ndash921 2001
20 Enzyme Research
[24] J D Bendtsen H Nielsen G von Heijne and S BrunakldquoImproved prediction of signal peptides SignalP 30rdquo Journalof Molecular Biology vol 340 no 4 pp 783ndash795 2004
[25] R J Siezen and J A M Leunissen ldquoSubtilases the superfamilyof subtilisin-like serine proteasesrdquo Protein Science vol 6 no 3pp 501ndash523 1997
[26] Y Katakura W Zhang G Zhuang et al ldquoEffect of methanolconcentration on the production of human 1205732- glycoprotein Idomain V by a recombinant Pichia pastoris a simple system forthe control of methanol concentration using a semiconductorgas sensorrdquo Journal of Fermentation and Bioengineering vol 86no 5 pp 482ndash487 1998
[27] Q-FWang Y-HHou Z Xu J-LMiao andG-Y Li ldquoPurifica-tion and properties of an extracellular cold-active protease fromthe psychrophilic bacterium Pseudoalteromonas sp NJ276rdquoBiochemical Engineering Journal vol 38 no 3 pp 362ndash3682008
[28] S Vazquez L Ruberto and W Mac Cormack ldquoProper-ties of extracellular proteases from three psychrotolerantStenotrophomonas maltophilia isolated from Antarctic soilrdquoPolar Biology vol 28 no 4 pp 319ndash325 2005
[29] R Zeng R Zhang J Zhao and N Lin ldquoCold-active ser-ine alkaline protease from the psychrophilic bacterium Pseu-domonas strain DY-A enzyme purification and characteriza-tionrdquo Extremophiles vol 7 no 4 pp 335ndash337 2003
[30] M Turkiewicz E Gromek H Kalinowska and M ZielinskaldquoBiosynthesis and properties of an extracellularmetalloproteasefrom the Antarctic marine bacterium Sphingomonas paucimo-bilisrdquo Journal of Biotechnology vol 70 no 1ndash3 pp 53ndash60 1999
[31] A Vassileva D A Chugh S Swaminathan and N KhannaldquoEffect of copy number on the expression levels of hepatitisB surface antigen in the methylotrophic yeast Pichia pastorisrdquoProtein Expression andPurification vol 21 no 1 pp 71ndash80 2001
[32] M Pazgier M Turkiewicz H Kalinowska and S Bielecki ldquoTheunique cold-adapted extracellular subtilase from psychrophilicyeast Leucosporidium antarcticumrdquo Journal ofMolecular Catal-ysis B Enzymatic vol 21 no 1-2 pp 39ndash42 2003
[33] M Turkiewicz M Pazgier H Kalinowska and S BieleckildquoA cold-adapted extracellular serine proteinase of the yeastLeucosporidium antarcticumrdquo Extremophiles vol 7 no 6 pp435ndash442 2003
[34] D Mattanovich B Gasser H Hohenblum and M SauerldquoStress in recombinant protein producing yeastsrdquo Journal ofBiotechnology vol 113 no 1ndash3 pp 121ndash135 2004
[35] P Secades BAlvarez and JAGuijarro ldquoPurification andprop-erties of a new psychrophilic metalloprotease (Fpp2) in the fishpathogen Flavobacterium psychrophilumrdquo FEMS MicrobiologyLetters vol 226 no 2 pp 273ndash279 2003
[36] L Y Ling I Ithoi and F M Yik ldquoOptimization for high-level expression in Pichia pastoris and purification of truncatedand full length recombinant sag2 of toxoplasma gondii fordiagnostic userdquo Southeast Asian Journal of Tropical Medicineand Public Health vol 41 no 3 pp 507ndash513 2010
[37] A Illanes ldquoStability of biocatalystsrdquo Electronic Journal ofBiotechnology vol 2 no 1 pp 1ndash9 1999
[38] B A Plantz J Sinha L Villarete K W Nickerson and VL Schlegel ldquoPichia pastoris fermentation optimization energystate and testing a growth-associated modelrdquo Applied Microbi-ology and Biotechnology vol 72 no 2 pp 297ndash305 2006
[39] K Tamura G Stecher D Peterson A Filipski and S KumarldquoMEGA6molecular evolutionary genetics analysis version 60rdquo
Molecular Biology and Evolution vol 30 no 12 pp 2725ndash27292013
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

Enzyme Research 9
Table2Aminoacid
compo
sitionandmolecular
weighto
fproteases
acrossdifferent
families
Amino
CAQ76821
CAG25549
EDR11679
CAG38357
CAD118
98CA
L25578
EDU50747
AAW
40780
ACB3
0121
ACB3
0119
acid
Qty
Qty
Qty
Qty
Qty
Qty
Qty
Qty
Qty
Qty
Ala
(A)
97101
8191
7887
7590
8299
101
115
8189
9196
8092
105
116
Arg
(R)
4042
2730
2224
1619
1721
1821
2932
2223
2124
3134
Asn
(N)
4446
3843
4348
4149
4251
4046
5257
5154
4552
4347
Asp
(D)
4951
6269
5460
6578
5567
5158
5358
6669
5260
6370
Cys
(C)
808
506
404
405
911
809
910
909
708
1011
Gln
(Q)
2324
3438
2326
2834
3036
3742
3539
2931
1922
2629
Glu
(E)
2122
1618
1921
2024
1518
2023
2528
2627
2832
2426
Gly
(G)
100
104
89100
8493
85102
85103
7788
8493
8084
8598
8695
His
(H)
1415
1416
1314
1619
1316
1113
1618
1415
1618
1213
Ile(I)
5355
4955
5157
5364
4150
3843
5156
4446
3540
4449
Leu
(L)
9599
6876
7179
6274
6073
6675
6774
8084
7485
6875
Lys
(K)
2627
2932
4449
2935
4959
5158
4044
3032
5058
5662
Met
(M)
1212
202
202
911
1113
809
1820
808
1113
1517
Phe
(F)
4143
5966
4752
5566
2935
3641
4449
3941
3439
3438
Pro
(P)
6264
6573
6774
5870
5870
5967
5055
5154
5564
5763
ser
(S)
7780
6977
91101
6072
6680
7080
7279
111
117
6373
6167
Thr
(T)
8790
89100
7583
83100
6579
7383
7178
7276
7182
6875
Trp
(W)
505
303
607
202
1215
1214
1213
909
1214
1415
Tyr
(Y)
3334
1213
2629
708
2935
2832
3943
5053
2630
2730
val
(V)
7679
8292
8089
6679
5769
7484
5864
6872
8295
6268
Total
963
1000
893
1000
900
1000
834
1000
825
1000
878
1000
906
1000
950
1000
866
1000
906
1000
MW
(Da)
1009898
932669
944365
869455
869549
924905
979343
1009629
917209
961988
Thea
ccessio
nnu
mbersof
proteasespu
tativ
eserinep
roteaseo
fGlacio
zymaantarctica
PI12
(CAQ
76821)sub
tilisin-lik
eproteaseP
leurotuso
streatus(CA
G25549)pyrolysin
LaccariabicolorS
238N
-H82
(EDR11679)
subtilisin
-like
proteasePh
anerochaetechrysosporiu
m(C
AG38357)sub
tilisin-lik
eserinep
roteaseP
R1CMetarhiziu
maniso
pliaev
aranisopliae(CA
D11898)serinee
ndop
eptid
aseH
ypocrealixii(C
AL2
5578)peptidase
Pyrenophoratritici-repentis
(EDU50747)p
eptid
aseCr
yptococcus
neoforman
svarn
eoform
ansJEC
21(A
AW40
780)sub
tilisin-lik
eprotease
Epich
loefestucae(A
CB30121)sub
tilisin-lik
eprotease
Epich
loefestucae
(ACB
30119
)
10 Enzyme Research
atgctcttcctccccgtcctcctcctcctccttcccggcgtcactgccttcctcaaccca 60
M L F L P V L L L L L P G V T A F L N Pgttaccaaccgcgcgaccaacgccatctcctcgactcaatacctctctaacgcctatatc 120
V T N R A T N A I S S T Q Y L S N A Y IctcgaattggacctctccacccccggcctcgtcaaacgggatagcacgcccgactctGTG 180
L E L D L S T P G L V K R D S T P D SCGTCCCGCGTTCCTCCCTCGCTGTCGAAGACGAGCTAACAATGACGTGGTAGatcctaga 240
I L Eggacgtactcacgtccgtaggccgcaacggtatcaagtaccaactccgccaccgctttat 300
D V L T S V G R N G I K Y Q L R H R F Ictccccgactctgttccacggcgcttcgatcactgtcccccctggaatctcccgctccca 360
S P T L F H G A S I T V P P G I S R S QaatcgcctctctccgcggtatcaaaGTCCGTCCCTCTCTCTCCCCTACTCCCTCCGTGCT 420
I A S L R G I KAACCCTGCACGCAACAGcgcgtctggcccgttcgaaagttctcccgacccagcgcagtag 480
R V W P V R K F S R P S A Vtggacgccgatggaggaggaagcgggttctcagggtcgcctatcaaggcggcgctcatgg 540
V D A D G G G S G F S G S P I K A A L Mgggtgaaagagctcgggaagcgcgcgaacgcttatgctggagatacgtttggaccgcatg 600
G V K E L G K R A N A Y A G D T F G P HtcatgacgggggttaatgagacgcatgaggcggggttgttgggagctgggattaagGTGT 660
V M T G V N E T H E A G L L G A G I KGTTTTCGTCTTTGTTTGGGGGAGGGGGGAGGGAGGAGCTGATTAATGGGTGCAGattggg 720
I GgtgctggacactggtgAGAAGCGGTTTGGAGGGAGGAGAGGATGGAGCTGATGAGTGTGC 780
V L D T GAGGTGttgattatttgaacccgattctgggaggctgctttggacctgggtgccatatgtc 840
V D Y L N P I L G G C F G P G C H M Sgtttgggtacgacttggttggcgatgattacgatggagataacgctcctgtgccggatgt 900
F G Y D L V G D D Y D G D N A P V P D VggatccttGTGCGTCCTTCCCTCCGCGATGGGCTGGAATCTGAGCTGACGGCGTTGTTGA 960
D PTCAGttgcgagctgcgatcctcatggTTCGTCTCGCTTTCCGCTCCTCTGATGCGCTCGC 1020
F A S C D P H GTGATGCACTTGGCAATAGGaactcacgttacgggaatcattggagcgctcccgaatgcgt 1080
T H V T G I I G A L P N AttggatttactggcgtcgcacccgccgctactctgggccactaccgagtGCGCTCCCTCT 1140
F G F T G V A P A A T L G H Y R VATCCGTTCTTCCTCGCAGCTCCCACTCACTCGTGCTTCCAGGTatttggctgcactggct 1200
F G C T GtcgtcggagaagatatcGTTCGTCCTCTCTCCAGTCCCGGCTAGCGATCCTGCTTCTGAC 1260
F V G E D ITTCGCATCTCCCTCTGAGattctcgctggactcatgcgaggagtcgaggacaactgcaac 1320
I L A G L M R G V E D N C Ngtcttgaccctctctctcggaggtccaggagggtgggtcaagggcacgccggcgtccatc 1380
V L T L S L G G P G G W V K G T P A S Icttatcgaccagatcgaagcgcaaggcattctcgtcaccgtcgccactggcaactcggga 1440
L I D Q I E A Q G I L V T V A T G N S GgctgagggTGAGCTCCTCGCTACTTGCGCGTCGAGACCAGTACTAACAGGGGCGTGGACG 1500
A E GCAGGcatgttcttctccgagtctcccgcctcgacgatcaacggcctttccatcgcatcca 1560
M F F S E S P A S T I N G L S I A ScgTCCGTCTCTCCCTTCCTTCGCACGCCACTCGCTCACACTCTCTCGCAGGgacgttacc 1620
T D V Tgacctcatcgcctacaacgccaccgtctcaggccaacctgcgatcccttacctctccgcc 1680
D L I A Y N A T V S G Q P A I P Y L S A
ggggggggggtctg minus194
ctagcgcgaatcgagaaaacacaaaagaacctcccgacttggcgttgcttctgctcttct minus180
ccttctcccaaccgccgacaataaaaactataacacacacacaccccatctcgacgcctg minus120
cagacttcacgactctcctcacgctctttaccgccccacgggtcccacccacgagccacg minus60
(a)
Figure 4 Continued
Enzyme Research 11
acgcccctcaacgtcgtcgccaacagcttccgcgtccacttcacctctaccgaccccaac 1740
T P L N V V A N S F R V H F T S T D P Naaccccgtcgacgcctgctctcctcttccggctggagcgcccgacttcgccaactatgtt 1800
N P V D A C S P L P A G A P D F A N Y Vacggtcgttcagcgtgggacttgtacgttcgttaccaagtaccagaacgttctcaatgct 1860
T V V Q R G T C T F V T K Y Q N V L N AggaggGTGCGTCGGCTTCTTCTCTCCCGACTGGATTGGGCGGAGACTGACCTTCTGAGCA 1920
G GGaaagatcgtattgttgtacaactcggagggagctgggaacctcccttacctcacgccca 1980
K I V L L Y N S E G A G N L P Y L T PacggtgtcggcatcgacgccgttgcaggtcttcgtcgttccgacggactcaagGTTCGTC 2040
N G V G I D A V A G L R R S D G L KTACGCTCTTCGGTGCTGAGGCCCTTCTCTGATCTCGCTCCGACAGcttctctcgtactat 2100
L L S Y Ycagaatgccaacaagcgtctcactctgcgcttccccaagggcaagatcgtcgcaggcttg 2160
Q N A N K R L T L R F P K G K I V A G LaccgataccatcaccggcggactcatctcgTTCGTTCCTCTCCGCCTCCCTCGCTGGCCG 2220
T D T I T G G L I SATGAGCTGACGCGGTCTTTGCAGGggttactcgacgtttggtccgacgaatgacctctac 2280
G Y S T F G P T N D L Yggtcagcctaccctctctgcccctggtggcaacatcctttcgaccttccctctctccgag 2340
G Q P T L S A P G G N I L S T F P L S EggaggagtggcggtcatcagtgggacgagcatgtcgtgcccctttgtcgGTTCGTCCTTC 2400
G G V A V I S G T S M S C P F VTCCTCGTTCGTTCGCTGAGCGTCGACTGATCTTGCACGATCGCAGctggatctgcggcgg 2460
A G S A Atcctcatggccgctcgcgcttcggagaacctcacgccgcttgagatcaggagtctcctta 2520
V L M A A R A S E N L T P L E I R S L Lctaccactgcgaagcttacgccggtctcgctcttgggatcgacgcctttggtgagcgtga 2580
T T T A K L T P V S L L G S T P L V S VttcgtcaaggaggaggTACGTTCCTTTCTCGCTGCTTTGAGGATCGAGGCTGATTAGCGA 2640
I R Q G G GGCTTCGCAGGactcgttcaggttgccaaggcgctcgcggccaagacgctaatctctcctc 2700
L V Q V A K A L A A K T L I S Pacgagctcctgctcaacgacactgcgaacgcgaactacgtccagactatcaagatcaaga 2760
H E L L L N D T A N A N Y V Q T I K I Kacaccaactcgtgggcgatgaagtacaccttctcctcggccgtcgcccaaggactcggaa 2820
N T N S W A M K Y T F S S A V A Q G L Gctttcgacgcttcgggcgatatcctccctaccctcgacccagtcgccgtctctggcgcac 2880
T F D A S G D I L P T L D P V A V S G Aaggctaccgtcgcgttcaacactcggatcctcagcgtcgcgcccggcgcgacggggtccg 2940
Q A T V A F N T R I L S V A P G A T G Stcgtggcgactatcacgccgccggttcttcccgtagcggacgctgcgaggttccctatct 3000
V V A T I T P P V L P V A D A A R F P Itctctgggtggatcagggtgaatgggcaaggcgcgagggatagcagtaggaacgaggcgt 3060
F S G W I R V N G Q G A R D S S R N E AacactgGTGCGTTTGGCTCGCTTTTTGGGAGGTGCCGAGCTAATCGTTTCTCGTAGtccc 3120
Y T V PgtactttgggcttgcggcgaagatgatcgatatgcaagGTGCGTCGATGGTTCTCTCGCT 3180
Y F G L A A K M I D M QTCTCTCTCGGTTGCTCACTTCAAAGTGCAGtcctcgacaccaccgagaccatttacggtc 3240
V L D T T E T I Y Gcgggctacgcctaccccttcgtgatcgacgacgcgattggagacatccaatccaccacaa 3300
P G Y A Y P F V I D D A I G D I Q S T TcgtcgtactccaggaacctcgggcccaccgtcttcgctcgctttgccaTGCGTCGACCGT 3360
T S Y S R N L G P T V F A R F ACCTCATTTCCTGCGGCAGAGCGAGAGCTGATTCCGACCTCGAACAGctggaacccttcac 3420
T G T L Htacagcctcgatctcgtcctagccgacatcgcctttacccccacctaccccaactcctcc 3480
Y S L D L V L A D I A F T P T Y P N S S
(b)
Figure 4 Continued
12 Enzyme Research
cccgccactcgtttcgtcaagcgctccctcacgcagcacacctctgccgcttcgcacctc 3540
P A T R F V K R S L T Q H T S A A S H Lgccaagcgccgcgtctccatcgccactatcaaccccaaagccaccctcgtcgccgatcga 3600
A K R R V S I A T I N P K A T L V A D Rcagctccactcggacgtccctatcgagggcaacatcttcacccaaccctttactggaagg 3660
Q L H S D V P I E G N I F T Q P F T G Rgattacctcgtcgacgcagccccgacgggatccaccgatcgtaccgtcacttttaacggg 3720
D Y L V D A A P T G S T D R T V T F N Gcagtacgccgagaacggcctcgtgaggacggctgtgacggggacttcgtaccgcttcctc 3780
Q Y A E N G L V R T A V T G T S Y R F Lcttcgggcgttgaagatctcgggagacgcgatgtacgaggatcagtatgagagctggctc 3840
L R A L K I S G D A M Y E D Q Y E S W L3900
S L P F S F R Attttgccacctcaaaaaaaaaaaaaaaaaaaaaaaaaa 3938
lowast
tcgctaccgttctcgttccgtgcgtaggcttgttgtggacaccactgcattaagttatga
(c)
Figure 4 Nucleotide and deduced amino acid sequences of genomic DNA and cDNA encoding protease from G antarctica strain PI12 Thenucleotides are numbered from the 51015840 end of the cDNA A putative CAAT TATA box and polyadenylation signal are bold italicThe putativeN-terminal signal peptide is underlined The initiation codon is boldfaced and termination codon is represented by an asterisk (lowast) boxedpeptide sequences showed N-glycosylated asparagines residues The amino acid residues that form the catalytic triad are double underlinedand bold while introns are represented in blue font Highlighted in grey are the intron donor-acceptor splice sites
proteases Hydrophobic profile of PI12 amino acid proteasewas performed by ProtScale tools with Kyte and Doolittlemethod at httpauexpasyorgcgi-bin The hydrophobicityprofile elucidation of PI12 protease amino acid was carriedout from amino acid 1 to 963 The amino acid residuesfrom 1 to 16 (M-L-F-L-P-V-L-L-L-L-L-P-G-V-T-A) and 548 to562 (V-A-V-I-S) were hydrophobic grooves The first regionwhich consisted of 16 amino acids was very hydrophobic andwas predicted to be the signal peptide by SignalP and ProPwebserver
The bold and double-underlined amino acids showedresidues of catalytic triad aspartic acid (D) histidine (H) andserine (S) active site respectively NCBI Conserved DomainsDatabase (CDD) identified the presence of subtilase proteaseand subtilisin-like serine protease domains in the amino acidsequence The predicted PI12 protease amino acid sequencewas found sharing significant homology to the subtilisinfamily of protease The highest similarity of PI12 proteasewas observed with 42 identity to subtilisin-like proteaseof Rhodosporidium toruloides (EMS20793) 37 identity tosubtilisin-like protease of Pleurotus ostreatus (CAG25549)and 35 identity to subtilisin-like protease of Gloeophyllumtrabeum ATCC 11539 (EPQ55564) The nucleotide sequenceof G antarctica strain PI12 showed low homology to thefungus subtilisin protease family and none to other psy-chrophilic proteases The multiple sequence alignment wasillustrated in Figure 5 The consensus sequence around thecatalytic triad aspartic acid (D) histidine (H) and serine(S) of PI12 protease showed high conservation with othersubtilisin (showed in black boxes) Glycine residues G142G173 G191 G262 and G327 that were highly conserved inmost of subtilases were conserved in PI12 protease [25]Therefore PI12 protease belongs to the subtilisin subgroup ofthe subtilase serine protease superfamilyThis serine protease
of G antarctica PI12 has been deposited into GenBank withaccession number CAQ76821
34 Phylogenetic Relationship of PI12 Protease The phylo-genetic tree was constructed to demonstrate the similaritiesand relationship of protein sequence between differentspecies or organisms Figure 6 illustrates the phylogeneticdendrogram indicating the relationship of PI12 proteasewith another 25 translated amino acid sequences ofserine protease The bar scale signifies the branch lengthto approximate variance along each branch Basicallythis tree comprises two main clusters cluster I was sub-divided into subcluster a (CAG25549 EPQ55564 ETW-80907 XP 007381302 XP 001877576 CAG38357 XP007341385 CCA68627 EJU04161 XP 007369624 EIW-62675 CCO37249 XP 001830835 XP 007342189) and sub-cluster b (XP 007589863 XP 003711175 EFQ32840 XP006694395 ENH74885 EON99722) cluster II was sub-divided into subcluster c (CBQ73425 GAC97617 CCF52619)and subcluster d [CAQ76821 (PI12 protease) EMS20793]
Serine protease constructed by the phylogenetic treecomprised large size of protein with about 801 to 1032 aminoacids The majority of the translated protease sequenceswere obtained from fungus All serine proteases from cluster1 subcluster a are derived from Agaricomycetes except forEJU04161 which belongs to Dacrymycetes whereby all serineproteases from cluster 1 subcluster b are from Sordari-omycetes Cluster II subcluster c consisted of serine proteasefrom fungus belonging to Ustilaginomycetes PI12 proteasewas categorized under cluster II subcluster d It possessedhomology similarity with the subtilisin-like protease fromRhodosporidium toruloides (EMS20793) (formerly known asRhodotorula glutinis or Rhodotorula gracilis) which is a
Enzyme Research 13
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
MKG-YSWSLLQLAFTGAALATVPLSDVKRISSAS--VVPNKFVVEVESASEIPGKR--SL
MKGVLVWSILS---AGCALAVTPLSSVKRVTNLP--IVANKFIVEVDSTSDIPTKR--S-
MKLPLLSLALT---AVSVGATVPLSSVQRTTTSPNNALAGEYIVEMNNVQGLGGKR--AY
---MAHGSLASLLLLLAASAQGAAAFGGKSATGGRSVVPNSYILQVNASAPALTKRGVSV
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease ---MLFLPVLLLLLPGVTAFLNPVTNRATNAISSTQYLSNAYILELDLSTPGLVKRDSTP
lowastlowast
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
YTPHELFYRSLSARGVSYEVNKEYNS-EGLFVGAVVTVSDANDVKALAETPGVKAIRPVR
-NPHDALYESLRKRAIGFKVDREFNT-QGLFVGAALTLSDAQDVGQIQSTTGVKSIRPVR
ASTHDTLYRSLRKRGISFDVKKEYNQ-PGIMVGAALKVASPQDVQQIATMAEVKAIRPVI
FEALDHTLAAVKQNGVKYTVRQRFDAIPEAFQAVSIQVEDGASMAELAQIPGVQRVWPVS
DSILEDVLTSVGRNGIKYQLRHRFIS-PTLFHGASITVPPGISRSQIASLRGIKRVWPVR
lowastlowast
TIKSPKPVKVHVVTGKDDPEIPA-------------------------DTESTHIMTGVN
AFDRPKPVKVQVVKGPDDPALPP-------------------------DSESTHVITGVD
AVPAPERINQFTVSSPNDSRITA-------------------------DSETTHIMTGVD
LIPRPVEPTVSDFTPVSSSSSSTKSKRD-THTLEKRGTNFPPASAYLNDTFYPHVQTGID
KFSRPSAVVDADGGGSGFSGSPIKAALMGVKELGKR------ANAYAGDTFGPHVMTGVN
lowast lowast lowast lowastlowast
KLHAQGIFGAGIKIGIIDTGIDYTHPALGGALGAGHKVIGGYDFVGDAYDGTNTPVPDND
KLHAQGITGAGIKIGILDTGIDYTHPTLGGAFGPGNKVIGGFDLVGDDYDGTNTPVPDPD
KVHAAGITGEGIKIGIIDTGIDYNHPLLGAGFGPGFKVIGGYDFVGDDYTGSNNPAPDDD
VLHNKGILGQGVKIVVVDEGVDYTNPILGGCFGPGCQISFGYDFVGDNYTGTNAPVPDSD
ETHEAGLLGAGIKIGVLDTGVDYLNPILGGCFGPGCHMSFGYDLVGDDYDGDNAPVPDVD
lowast lowast lowast lowastlowastlowast lowast lowast lowastlowast lowast lowastlowast lowast lowast lowast lowast lowastlowastlowast lowast lowast lowast lowast lowastlowast lowast
PLDQCNGHGTHVAGIIGANPDNQFNISGVSYGASLSSYRVFGCSGSVSDDIIIEALLRGV
PLDQCAGHGTHVAGIIGANPDNAFNISGVAFQASLSAYRVFGCVGFVTDDVLVDALLLGF
PLDNCNGHGTHVAGIIGANPGNDFGISGVAYGSSIASYRVFGCSGFVSDDIIIDALLRGY
PFSSCSNHGTHTTGTVGALAN-EYGFSGAAPMATIGHYRVISCSGATTDEILVAALTRAL
PFASCDPHGTHVTGIIGALPN-AFGFTGVAPAATLGHYRVFGCTGFVGEDIILAGLMRGV
lowast lowastlowastlowast lowastlowast lowast lowastlowast lowast lowastlowastlowast lowast lowast lowast
QDGQDILTLSLGGTDGWTE-SSSSVVASRIAASGKIVTIAAGNEGAYGSWYSSSPGNGID
NEGQDILTLSLGGADGWTE-SVSAVVASRIAATGKVVTIAAGNDGASGAWYTSSPGNGID
SDGMDILTLSLGGVEGWTE-SSSSVVASRIADQGRVVTIAAGNDGAYGSWYASSPGTGVD
QDNVNIVSMSLGSSVGWLDDSPVQIMASYLGSKGIHVVASVGNERNEGLFAADQPAASRI
EDNCNVLTLSLGGPGGWVKGTPASILIDQIEAQGILVTVATGNSGAEGMFFSESPASTIN
lowastlowastlowast lowastlowast lowast lowast lowastlowast lowast lowast
VISVASIDNTVDNIQNATVNGVEHGPIPYFKTLPLPFTNA-LPLYVISNDTTIANDACNA
VISVASLDNTVVPLQTAQVHGVTHDPIIYQDTFPLPINDT-RPIFATSTDVTVVDDACNP
VISVASVNNIAIPVQNATVQGVEHAPIPYLEALPLNVTGS-RPIYATSTDTTVADDACNA
GTGVGGVDPTYLAAYYAYLI--KRDPVPYISPTPFNLPNN-YLLYFTSTNTSVTNDACNP
GLSIASTDVTDLIAYNATVS--GQPAIPYLSATPLNVVANSFRVHFTSTDPNNPVDACSP
lowast lowast lowast lowast lowastlowastlowast
LPDNTPDLSGFLVLVRRGSCTFVQKLTNIAAKGGNAVFIYDNGNG-----FAGIDTGKFT
LPDSTPDLSGFVVIVRRGTCTFVQKLGNVAAKGAKVTLIYDNGSG-----FGAIDTGAFV
LPNDTPDLSGYVVVIKRGTCTFVTKLQNAAAKGANTFLIYNNGGS-----FSAISVGDYT
LPSTTPNLANRVVVVQRGTCGFTVKQQNVAAAGGKIVLIYNSKGSGLIPQLDVGSTGLTA
LPAGAPDFANYVTVVQRGTCTFVTKYQNVLNAGGKIVLLYNSEGAGNLPYLTPNGVGIDA
lowastlowast lowast lowastlowast lowast lowast lowast lowast lowast lowast lowast
(a)
Figure 5 Continued
14 Enzyme Research
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
NATLIQAADGVFLAQQVAAGKSVTITFPQTGGSAQITN-PSGGLVSSFTTYGPTNDFYFK
-ASFIQAADGEFLVQQFASGQNVSLSFPQSGGLIQFPD-PAGGLISSFTSYGPSNDFFFK
-AALISETDGTFLVNQFAAKANITISFPQSGSAWNYPDDVSGGLVSDFSSYGPSYDMYFK
VGSLRYEDGLKLLSYYQSSPRGQSISFPNGPLAPYVSNTVSGGVVASYSNFGPTNELFIY
VAGLRRSDGLKLLSYYQNANKRLTLRFPKGKIVAGLTDTITGGLISGYSTFGPTNDLYGQ
lowast lowastlowast lowastlowast lowastlowast
PAFGAPGGNILSTLPVPLGSYGVESGTSMATPYVAGSAALLLSVKGKSAAVALGARTLFQ
PAVAAPGGNILSTLPVNLGLFGIESGTSMATPFVAGSAALLFQAKGTSAAVGRSARTLFE
PAVAAPGGNILSTLPIALGSYGVESGTSMATPFTAGSAALILQAKGK--SVAKSIRNLFQ
PTLAAPGTDILSTV---TGGVALMRGTSMAAPLVAGAYALLLSVGNNLKLSPEEARTILM
PTLSAPGGNILSTFPLSEGGVAVISGTSMSCPFVAGSAAVLMAARASENLTPLEIRSLLT
lowast lowastlowastlowast lowastlowastlowastlowast lowast lowastlowastlowastlowast lowast lowastlowast lowast lowast
STAQRVPSSHTDGDPLQTLSQQGAGLIDVFKALYTETIVWPTELILNDTAHYKEVHTFTV
TTAQRVPSTHTDGDPLQTLTQQGAGLINVFNAIHTTTIVSPGELILNDTAHFKGTQQFTV
TTASAVASSKTDGDPLQTVSQQGAGLIQVDRAIGTTTVVSPGQLTLNDTAHFEGFKTFNI
STSQPSPVAY-GSETLDTVVSQGAGVINMTAAIAAHTVISPAQFNLNDTQYSNNTQTLTL
TTAKLTPVSLLGSTPLVSVIRQGGGLVQVAKALAAKTLISPHELLLNDTANANYVQTIKI
VNTGKTSKQYKLTHVPAGTAQTIT---SGTIFAADGPVPLSTDYATVKLSSSSFTLRPGQ
RNAGSSAKTYTLRHIPAGTAVTVT---PGTIFPADGPVPLSTDFASVSLSTSKFTLAPGK
QNTGSKAVTYKISHVPAGTAASVQ---SGSIFAADGPVPLTADYAQVTFSLNKVTVLPGV
RNNNRFPVSYQLSWVDSTGIVTYNDGATTDIIPSTTPNYVSTSVLRVAFSQRTVTVPGGQ
KNTNSWAMKYTFSSAVAQGLGTFD--ASGDILPTLDPVAVSGAQATVAFNTRILSVAPGA
lowast lowast lowast lowast lowast lowast
SKQVTTHINAP--SGVDASTYPVFSGFIEIATAT--------ESVHVTYLGLAASLKDKQ
TQTVTARFTPP--AAADPSTFPVFSGFIQIESGT--------EQVQVSYLGLKASLKDKQ
SVPILVWFKPP--QGVDASTYPVYSGFIEIAGLG--------ETLRVAYLGVAASLKDKA
TVKVTAQFIPPNLTAQQRNQFPIYSGFVTITGQGQGAGAEQMEKYNLPFFGLGARMVDMP
TGSVVATITPPVLPVADAARFPIFSGWIRVNGQG-ARDSSRNEAYTVPYFGLAAKMIDMQ
lowast lowast lowastlowast lowast lowast lowast lowast
IVDNTNEFFGITIPAVLNNTGDAQ-LSPTNYTFSGSDFPTLLLRLAFGTPLLRVDLVDAT
VIDNTDFFFGVPTPVLTDPNGEVQ-TSPRNYSFLASDFPTLIFRLAFGSPKVVVDLVSPT
VVDNTDEYFGVKIPALLDASGDVQ-GNSTNYTLVGDDVPSLLYRFAFGSAHVRVDLVEKA
VLDTTNVALGP--NLPFIAVGQN--IQTGPTTFSESNPAVVYFRLAAGTRRLRVDLVDAS
VLDTTETIYGPGYAYPFVIDDAIGDIQSTTTSYSRNLGPTVFARFATGTLHYSLDLVLAD
lowast lowast lowast lowast lowastlowastlowastlowast lowast
IDLKPTLNSREIDDKRHVFERESFSFPHPGPKGS-------------------TFAQVKI
INFKPTLNTRALGDTHGSF----FSFPHRVKTG--------------------SFAQVPT
STVSATLSKRNWWN---------WWWPQKPSN---------------------SYDKVPI
TSYNATVPAVTNPASRLVKRSNTELTPRAAFPT--------------------LYSQVRT
IAFTPTYPNSS-PATRFVKRSLTQHTSAASHLAKRRVSIATINPKATLVADRQLHSDVPI
lowast lowast
VGTLSEQSHIPRNSDASTVDDGGYSTFAFNKP------SFANGTTIPNGTYRWLLRALKV
LGTVFELDFTSRNNDDPTDN--GFNTVAIDPP------TFANGTTIPNGQYRLLLRALRV
VGTLAEYDYVPRNSDASTIDDNGYNVFQFT-D------SFVNGTKIAAGEYKVLLRALKV
VGSLFAPSYWPPRDYLFNNGGT-YSDYEIQLDGSFTFPNGTKGMAELGRSYKVLLRALKI
EGNIFTQPFTG-RDYLVDAAPTGSTDRTVTFNGQY-AENGLVRTAVTGTSYRFLLRALKI
lowast lowast lowastlowast lowast lowast lowast lowast lowastlowastlowastlowast
lowast lowast lowastlowastlowastlowastlowast
(b)
Figure 5 Continued
Enzyme Research 15
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
TGDPTKEADFETWLSPIVGITVA
TGDPTNEADFESFLSPIIGVNAP
TGNPAEEGDYETWLSPSFSVVPA
TADPTYTSSYESWLSPSFTFTA-
SGDAMYEDQYESWLSLPFSFRA-
lowast lowastlowast
ConservedSimilar
lowast All match
(c)
Figure 5 Multiple sequence alignment between amino acid sequence of PI12 protease with other subtilisin The serine protease sequenceswere obtained from the GenBank database Their accession numbers are EMS20793 (subtilisin-like protease of Rhodosporidium toruloidesNP11) CAG25549 (subtilisin-like protease of Pleurotus ostreatus) EPQ55564 (subtilisin-like protease of Gloeophyllum trabeumATCC 11539)and ETW80907 (serine protease S8 ofHeterobasidion irregulare TC 32-1) Putative ORF of serine protease fromG antarctica PI12 was labeledas PI12 protease
CAG38357 Phanerochaete chrysosporium
EIW62675 Trametes versicolor FP-101664 SS1
XP 007369624 Dichomitus squalens LYAD-421 SS1
XP 007381302 Punctularia strigosozonata HHB-11173 SS5
CAG25549 Pleurotus ostreatus
ETW80907 Heterobasidion irregulare TC 32-1
XP 001877576 Laccaria bicolor S238N-H82
EPQ55564 Gloeophyllum trabeum ATCC 11539
XP 001830835 Coprinopsis cinerea okayama7130
XP 007341385 Auricularia delicata TFB-10046 SS5
CCA68627 Piriformospora indica DSM 11827
CCO37249 Rhizoctonia solani AG-1 IB
XP 007342189 Auricularia delicata TFB-10046 SS5
EJU04161 Dacryopinax sp DJM-731 SS1
XP 007589863 Colletotrichum fioriniae PJ7
EFQ32840 Colletotrichum graminicola M1001
ENH74885 Fusarium oxysporum f sp cubense race 1
XP 003711175 Magnaporthe oryzae 70-15
XP 006694395 Chaetomium thermophilum DSM 1495
EON99722 Togninia minima UCRPA7
CBQ73425 Sporisorium reilianum SRZ2
GAC97617 Pseudozyma hubeiensis SY62
CCF52619 Ustilago hordei
EMS20793 Rhodosporidium toruloides NP11
CAQ76821 Glaciozyma antarctica PI12
01
Clus
ter 1
Clus
ter 2
a
b
c
d
Figure 6 Phylogenetic dendrogram among 25 translated amino acid sequences of serine proteaseThe tree was constructed using a neighbor-joining algorithm with 1000 bootstrappings by MEGA 6 software [39]The scale bar represents 01 substitutions per amino acid positionTheserine protease sequences were obtained from the GenBank database GenBank accession numbers of the sequences were indicated beforethe strain name The putative serine protease of G antarctica strain PI12 has been deposited in GenBank with accession number CAQ76821
16 Enzyme Research
0
2
4
6
8
10
12
0 24 48 72 96 110
Prot
ease
activ
ity (U
mL)
Post induction time (hours)
Figure 7 Effect of postinduction time towards protease activity The highest activity was detected at 72 hours after 110 hours of incubationwith 9UmL activity followed by 48 24 and 96 hoursThe substrate (azocasein) was dissolved in Tris HCL buffer and incubated for 30min at20∘CThe absorbance was measured at 450 nm after 15min incubation A one-way ANOVAwas used to test for preference differences amongsix different postinduction times The statistical value (119875 value lt005) between hours is 0003
pink-coloured nonpathogenic oleaginous yeast (42 iden-tity) Both G antarctica PI12 and Rhodosporidium toruloidesNP11 belong to Microbotryomycetes class PI12 protease didnot show any homology similarity to other psychrophilicproteases however it showed 37 identity to hypotheticalprotein from Pseudozyma antarctica T-34 which was isolatedfrom Japan (GAC76005) (not listed in the phylogenetic tree)
35 Cloning and Expression of Cold-Adapted PI12 Protease inthe Yeast Expression System The PCR product harbouringthe mature PI12 protease with two flanking restriction sites(EcoRI and AvrII) was cloned into pPIC9 expression vectorThe recombinant plasmid harbouring PI12 protease was usedto transform both GS115 and KM71 intended to produce bothHis+ Mut+ and His+ MutS Two recombinant clones fromGS115 (His+Mut+) strain (GpPro1 andGpPro2) and one fromKM71 (His+MutS) strain (KpPro1) were successfully obtainedwhich were further analyzed for protein expression study
36 Expression and Enzyme Assay of PI12 Protease in Pichiapastoris All positive recombinants were grown in small scalecultures and assayed for the cold-adapted protease activityin the culture supernatants The clones were first grown inBMGY medium with glycerol as the sole carbon sourceFor induction of AOX1 promoter and protein productionthe growth medium was transferred to the same mediumexcept with 05 methanol in place with glycerol as thecarbon source [26] The supernatant of the recombinant Ppastoris expression cultures after 48 h of incubation time wasanalyzed to determine the recombinant protease activity bythe modified method of Brock et al (1982) [20] All testedclones secreted protease into the supernatant in differentexpression levels This might be due to the variability ofthe strength of the promoter and copy number in each cellto secrete and express the recombinant product The clonewith the highest yield was GpPro2 with 63UmL activityfollowed by KpPro1 and GpPro1 with 25UmL and 37UmLactivities respectively (data not shown) It is perceptible thatexpression systemof Pichia pastoris is the ideal system for thisprotein The preliminary assay has fruitfully produced high
protease expression at 20∘C Yet further optimization of therecombinant PI12 protease production need to be conductedin order to examine the optimum condition of this proteinReported cold-adapted proteases showed optimum activity ata range of 15∘Cndash60∘C [27ndash30] This difference was possiblyrelated to the respective genetic and physiological adaptationof the strains There are several factors that influenced thedifferent rate of expression It includes different site of inte-gration between interest gene and Pichia genome variabilityof different expression host cell to secrete and express therecombinant enzyme and different sites in the polylinker forinserting foreign genes [31]
This is the second extracellular protease reported to besynthesized by this obligate psychrophilic yeastG antarcticaThe first cold-adapted extracellular subtilase from this yeasthas been purified and characterized previously [32 33] Theextracellular serine proteinase (LAP2) from this Antarcticyeast formerly known as Leucosporidium antarcticum strain171 is specific towards synthetic substrates of chymotrypsinand subtilisin The subtilase of L antarcticum strain 171showed specific activity of 4043Umg when utilized N-SucAAPFpNA as a substrate [33]
37 Optimization of Protease Production GpPro2 was chosenfor further optimization due to its highest protease activityat the preliminary expression stage High protease activitywas detected after 72 hours of incubation (Figure 7) byusing the standard protocol method as recommended forPichia pastoris expression system (Pichia Expression KitInvitrogen USA) The expression decreased after 72 hoursand no activity was detected at 110 hours The prolongedincubation may result in the decrease of biomass productionand will subsequently reduce the expression of recombinantprotein In addition oxygen and nutrient depletion duringprolonged incubationwill trigger the stress responses in orderto survive in such harsh environment [34]
Methanol was used as a sole carbon by P pastoris Themethanol was oxidized to formaldehyde with the aid of oxy-gen as electron acceptor by an alcohol oxidase enzyme (AOXgene) [35] The optimal methanol concentration is at 05(vv) with 14UmL activity The addition of more than 05
Enzyme Research 17
02468
10121416
0 01 03 05 07 09Pr
otea
se ac
tivity
(Um
L)Methanol concentration ()
Figure 8 Effect ofmethanol concentration towards protease activityThe recombinant was cultivated for 72 hours at 30∘C in BMMYmedium05 methanol gives the highest activity with 14UmL The substrate (azocasein) was dissolved in Tris HCL buffer and incubated for 30minat 20∘CThe mixture was left for 15min and absorbance was taken at 450 nm A one-way ANOVA was used to test for preference differencesamong six different concentrations The statistical value (119875 value lt005) between concentration is 002
0
5
10
15
20
25
10 15 20 25 30 35
Prot
eas a
ctiv
ity (
Um
L)
Temperature (∘C)
Figure 9 Effect of temperature towards protease activity The reaction was carried out in temperature ranging from 10∘C to 35∘C at pH 7 for30min A one-way ANOVA was used to test for preference differences among six different temperatures The statistical value (119875 value lt005)between temperature is 0067
(vv) methanol resulted in tremendous decrease of proteaseactivity (Figure 8) Production of PI12 protease was decreasedwhen the methanol concentration increased This might bedue to the toxic effect of methanol concentration on DNAreplication and membrane synthesis Basically methanol isused to increase the permeability of the cell membrane tohelp the secretion of foreign protein [26] However if thepermeability of cell membrane is too high lowmolecular sizemetabolites would leak from the cell and such leakage willseriously damage cell activities Such condition would abusethe cellular system and might reduce the yield of proteinexpression [36]
Psychrophilic microorganism dominated in low temper-ature habitat which most other microorganism are inca-pable of withstanding due to their inability to produceenzyme that can tolerate the extreme environment con-dition Psychrophilic enzymes are always associated withhigh catalytic efficiency at low and moderate temperature(0∘C to 30∘C) compared to the mesophilic counterpart thatare generally unable to catalyze efficiently under extremely
low temperature [2] The recombinant protein assay wasdone at various temperatures ranging from 10∘C to 35∘Cusing 05 azocasein as a substrate dissolved in 50mMTris HCl in presence of 2mM CaCl
2 pH 7 In the activity
assays expressed protease showed a highest activity at 20∘C(20UmL) and decreased tremendously once it exceededmaximum temperature due to the denatured state of theenzyme (Figure 9) The active site of an enzyme is easilyaltered when the high temperature is introduced duringincubation or activity assay resulting in a low reaction rate[37]
The recombinant construct was transferred into BMMYmedia at different A
600 nm ranging from 1 to 10 The high-est activity of active enzyme was detected at A
600 nm 60(283UmL) and above (Figure 10) The slope during expo-nential phase was much higher compared to other phasesindicating an increase in the doubling time At exponentialphase the cells are adapted to the growth condition andother processes like cell growth cycle and synthesis of RNAThe yield of active enzyme during expression of P pastoris
18 Enzyme Research
05
1015202530
1 2 3 4 5 6 7 8 9 10Prot
ease
activ
ity (U
mL)
Initial inoculum A600
Figure 10 Effect of inoculums size towards protease activity The recombinant was cultivated for 72 hours at 30∘C in BMMY mediumwith 05 methanol concentration A one-way ANOVA was used to test for preference differences among six different optical densitiesThe statistical value (119875 value lt005) between optical density value is 003
(kD
a)
1160
662
450
350
250
184
144
M 1 2 3
(a) (b) (c)
Figure 11 (a) Coomassie-stained SDS-PAGE of recombinant P pastoris clone 1mL supernatants collected from the third-day culturewere concentrated 10-fold using trichloroacetic acid and 10120583L resultant products were applied to SDS-PAGE analysis Lane 1 molecularweight marker (Fermentas) Lane 2 GS115 (Control) Lane 3 GS115pPIC9 empty (Control) Lane 4 GS115pPIC9PI12prot (GpPro2)The arrow indicates the secreted recombinant PI12 protease (b) Plate activity staining of different recombinant P pastoris clones GGS115 GP GS115pPIC9 GRI GS115pPIC9PI12prot1 (GpPro1) GRII GS115pPIC9PI12prot2 (GpPro2) K KM71 KP KM71pPIC9 KRKM71pPIC9PI12prot (KpPro) (c)Gel activity staining of recombinant PI12 protease inP pastorisGpPro2 clone Lane 1 prestainedmolecularweightmarker (Fermentas) Lane 2 GS115pPIC9 empty (Control) Lane 3 GS115pPIC9PI12prot1 (GpPro2)The arrow indicates the secretedrecombinant PI12 protease
dropped after 28 hours of incubation which is at stationaryor death phase [38]
38 Detection of Recombinant Protein by SDS-PAGE andActivity Staining TheSDS-PAGE gel of the GpPro2 recombi-nant clonesrsquo supernatant showed a putative expression bandat around 993 kDa Plate staining and gel activity stainingwere executed to examine if the recombinant PI12 proteaseexpressed in P pastoris was functionally active In the platestaining the transformants showed strong proteolytic activity(Figure 11(a)) In gel staining proteolytic activity was alsodetected around 993 kDa after incubation at 20∘C This is
evidence of protease secretion into the surrounding mediawhereby it catalyzed the breakdown of casein thus formedclearing zones around the activity ring and band (Figures11(b) and 11(c)) As a conclusion the cold-adapted PI12protease was functionally expressed in P pastoris GS115(GpPro2 clone) with the size of 993 kDa
4 Conclusions
Psychrophilic yeast which was originated frommarine waterin Casey Station Antarctica was identified as Glaciozymaantarctica strain PI12 A serine protease with an open reading
Enzyme Research 19
frame of 2892 bp encoded for 963 amino acids from thisAntarctic yeast has been cloned and expressed in Pichiaexpression system This is the second extracellular pro-tease reported being synthesized by this psychrophilic yeastSequence analysis through bioinformatics studies revealedfew homology identities of this protein with other ser-ine proteases The mature PI12 protease was successfullyexpressed and secreted into the culture medium driven bythe Saccharomyces cerevisiae 120572-factor signal sequence in Ppastoris The protease production in P pastoris was bestobtained from strain GS115 (GpPro2) with 283UmL after 72hours of induction time with 05 (vv) of methanol inducerThe starting optical density at 600 nm (OD600) was 60 andthe optimum assay temperature was 20∘C The expressedprotein was detected by SDS-PAGE and activity staining witha molecular weight of 99 kDaThis cold-active enzymemightoffer an opportunity for biotechnological exploitation basedon its high catalytic activity at low temperatures and lowthermostability Structural and functional study of this cold-adapted enzyme would give insight into its adaptation to anextreme cold environment
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgment
This research was supported financially by the InitiativeBiotechnology-GMBI-MOSTI (Project number 07-05-16-MG-GMB02) andRUGS-Initiative 6 (Project number 05-02-11-1414RU)
References
[1] J W Deming ldquoPsychrophiles and polar regionsrdquo CurrentOpinion in Microbiology vol 5 no 3 pp 301ndash309 2002
[2] C Gerday M Aittaleb M Bentahir et al ldquoCold-adaptedenzymes from fundamentals to biotechnologyrdquo Trends inBiotechnology vol 18 no 3 pp 103ndash107 2000
[3] A Svingor J Kardos I Hajdu A Nemeth and P Zavodszky ldquoAbetter enzyme to cope with cold comparative flexibility studieson psychrotrophic mesophilic and thermophilic IPMDHSrdquoThe Journal of Biological Chemistry vol 276 no 30 pp 28121ndash28125 2001
[4] G Antranikian C E Vorgias and C Bertoldo ldquoExtremeenvironments as a resource for microorganisms and novel bio-catalystsrdquo Advances in Biochemical EngineeringBiotechnologyvol 96 pp 219ndash262 2005
[5] T Lonhienne C Gerday andG Feller ldquoPsychrophilic enzymesrevisiting the thermodynamic parameters of activation mayexplain local flexibilityrdquoBiochimica et Biophysica Acta vol 1543no 1 pp 1ndash10 2000
[6] R Cavicchioli and TThomas ldquoExtremophilesrdquo in Encyclopediaof Microbiology pp 317ndash337 Academic Press San Diego CalifUSA 2000
[7] G Feller E Narinx J L Arpigny et al ldquoEnzymes frompsychrophilic organismsrdquo FEMS Microbiology Reviews vol 18no 2-3 pp 189ndash202 1996
[8] J Arnorsdottir R B Smaradottir T O Magnusson S HThorbjarnardottir G Eggertsson and M M KristjanssonldquoCharacterization of a cloned subtilisin-like serine proteinasefrom a psychrotrophic Vibrio speciesrdquo European Journal ofBiochemistry vol 269 no 22 pp 5536ndash5546 2002
[9] N J Russell ldquoPsychrophilic bacteriamdashmolecular adaptations ofmembrane lipidsrdquo Comparative Biochemistry and Physiology APhysiology vol 118 no 3 pp 489ndash493 1997
[10] G Feller and C Gerday ldquoPsychrophilic enzymes hot topics incold adaptationrdquo Nature Reviews Microbiology vol 1 no 3 pp200ndash208 2003
[11] P Raspor and J Zupan ldquoYeasts in extreme environmentsrdquo inBiodiversity and Ecophysiology of Yeasts The Yeast Handbookpp 371ndash417 Springer Berlin Germany 2006
[12] M B Rao A M Tanksale M S Ghatge and V V Desh-pande ldquoMolecular and biotechnological aspects of microbialproteasesrdquoMicrobiology andMolecular Biology Reviews vol 62no 3 pp 597ndash635 1998
[13] L Polgar ldquoThe catalytic triad of serine peptidasesrdquo Cellular andMolecular Life Sciences vol 62 no 19-20 pp 2161ndash2172 2005
[14] S Y Boo C M V L Wong K F Rodrigues N NajimudinA M A Murad and N M Mahadi ldquoThermal stress responsesin Antarctic yeastGlaciozyma antarctica PI12 characterized byreal-time quantitative PCRrdquoPolar Biology vol 36 no 3 pp 381ndash389 2013
[15] B Turchetti S R T Hall L B Connell et al ldquoPsychrophilicyeasts from Antarctica and European glaciers descriptionof Glaciozyma gen nov Glaciozyma martinii sp nov andGlaciozyma watsonii sp novrdquo Extremophiles vol 15 no 5 pp573ndash586 2011
[16] N H F Hashim I Bharudin D L S Nguong et al ldquoCharac-terization of Afp1 an antifreeze protein from the psychrophilicyeast Glaciozyma antarctica PI12rdquo Extremophiles vol 17 no 1pp 63ndash73 2013
[17] A N M Ramli M A Azhar M S Shamsir et al ldquoSequenceand structural investigation of a novel psychrophilic 120572-amylasefrom Glaciozyma antarctica PI12 for cold-adaptation analysisrdquoJournal ofMolecularModeling vol 19 no 8 pp 3369ndash3383 2013
[18] A N M Ramli N M Mahadi A Rabu A M A Murad F DA Bakar and R M Illias ldquoMolecular cloning expression andbiochemical characterisation of a cold-adapted novel recombi-nant chitinase from Glaciozyma antarctica PI12rdquoMicrobial CellFactories vol 10 no 1 article 94 2011
[19] J Sambrook E Fritsch and T Maniatis Molecular Cloning ALaboratoryManual Cold SpringHarbor Laboratory Press NewYork NY USA 1989
[20] F M Brock C W Forsberg and J G Buchanan-SmithldquoProteolytic activity of rumen microorganisms and effects ofproteinase inhibitorsrdquoApplied and EnvironmentalMicrobiologyvol 44 no 3 pp 561ndash569 1982
[21] U K Laemmli ldquoCleavage of structural proteins during theassembly of the head of bacteriophage T4rdquo Nature vol 227 no5259 pp 680ndash685 1970
[22] A Henne R A Schmitz M Bomeke G Gottschalk and RDaniel ldquoScreening of environmental DNA libraries for thepresence of genes conferring lipolytic activity on Escherichiacolirdquo Applied and Environmental Microbiology vol 66 no 7 pp3113ndash3116 2000
[23] M Poza T de Miguel C Sieiro and T G Villa ldquoCharacteri-zation of a broad pH range protease of Candida caseinolyticardquoJournal of Applied Microbiology vol 91 no 5 pp 916ndash921 2001
20 Enzyme Research
[24] J D Bendtsen H Nielsen G von Heijne and S BrunakldquoImproved prediction of signal peptides SignalP 30rdquo Journalof Molecular Biology vol 340 no 4 pp 783ndash795 2004
[25] R J Siezen and J A M Leunissen ldquoSubtilases the superfamilyof subtilisin-like serine proteasesrdquo Protein Science vol 6 no 3pp 501ndash523 1997
[26] Y Katakura W Zhang G Zhuang et al ldquoEffect of methanolconcentration on the production of human 1205732- glycoprotein Idomain V by a recombinant Pichia pastoris a simple system forthe control of methanol concentration using a semiconductorgas sensorrdquo Journal of Fermentation and Bioengineering vol 86no 5 pp 482ndash487 1998
[27] Q-FWang Y-HHou Z Xu J-LMiao andG-Y Li ldquoPurifica-tion and properties of an extracellular cold-active protease fromthe psychrophilic bacterium Pseudoalteromonas sp NJ276rdquoBiochemical Engineering Journal vol 38 no 3 pp 362ndash3682008
[28] S Vazquez L Ruberto and W Mac Cormack ldquoProper-ties of extracellular proteases from three psychrotolerantStenotrophomonas maltophilia isolated from Antarctic soilrdquoPolar Biology vol 28 no 4 pp 319ndash325 2005
[29] R Zeng R Zhang J Zhao and N Lin ldquoCold-active ser-ine alkaline protease from the psychrophilic bacterium Pseu-domonas strain DY-A enzyme purification and characteriza-tionrdquo Extremophiles vol 7 no 4 pp 335ndash337 2003
[30] M Turkiewicz E Gromek H Kalinowska and M ZielinskaldquoBiosynthesis and properties of an extracellularmetalloproteasefrom the Antarctic marine bacterium Sphingomonas paucimo-bilisrdquo Journal of Biotechnology vol 70 no 1ndash3 pp 53ndash60 1999
[31] A Vassileva D A Chugh S Swaminathan and N KhannaldquoEffect of copy number on the expression levels of hepatitisB surface antigen in the methylotrophic yeast Pichia pastorisrdquoProtein Expression andPurification vol 21 no 1 pp 71ndash80 2001
[32] M Pazgier M Turkiewicz H Kalinowska and S Bielecki ldquoTheunique cold-adapted extracellular subtilase from psychrophilicyeast Leucosporidium antarcticumrdquo Journal ofMolecular Catal-ysis B Enzymatic vol 21 no 1-2 pp 39ndash42 2003
[33] M Turkiewicz M Pazgier H Kalinowska and S BieleckildquoA cold-adapted extracellular serine proteinase of the yeastLeucosporidium antarcticumrdquo Extremophiles vol 7 no 6 pp435ndash442 2003
[34] D Mattanovich B Gasser H Hohenblum and M SauerldquoStress in recombinant protein producing yeastsrdquo Journal ofBiotechnology vol 113 no 1ndash3 pp 121ndash135 2004
[35] P Secades BAlvarez and JAGuijarro ldquoPurification andprop-erties of a new psychrophilic metalloprotease (Fpp2) in the fishpathogen Flavobacterium psychrophilumrdquo FEMS MicrobiologyLetters vol 226 no 2 pp 273ndash279 2003
[36] L Y Ling I Ithoi and F M Yik ldquoOptimization for high-level expression in Pichia pastoris and purification of truncatedand full length recombinant sag2 of toxoplasma gondii fordiagnostic userdquo Southeast Asian Journal of Tropical Medicineand Public Health vol 41 no 3 pp 507ndash513 2010
[37] A Illanes ldquoStability of biocatalystsrdquo Electronic Journal ofBiotechnology vol 2 no 1 pp 1ndash9 1999
[38] B A Plantz J Sinha L Villarete K W Nickerson and VL Schlegel ldquoPichia pastoris fermentation optimization energystate and testing a growth-associated modelrdquo Applied Microbi-ology and Biotechnology vol 72 no 2 pp 297ndash305 2006
[39] K Tamura G Stecher D Peterson A Filipski and S KumarldquoMEGA6molecular evolutionary genetics analysis version 60rdquo
Molecular Biology and Evolution vol 30 no 12 pp 2725ndash27292013
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

10 Enzyme Research
atgctcttcctccccgtcctcctcctcctccttcccggcgtcactgccttcctcaaccca 60
M L F L P V L L L L L P G V T A F L N Pgttaccaaccgcgcgaccaacgccatctcctcgactcaatacctctctaacgcctatatc 120
V T N R A T N A I S S T Q Y L S N A Y IctcgaattggacctctccacccccggcctcgtcaaacgggatagcacgcccgactctGTG 180
L E L D L S T P G L V K R D S T P D SCGTCCCGCGTTCCTCCCTCGCTGTCGAAGACGAGCTAACAATGACGTGGTAGatcctaga 240
I L Eggacgtactcacgtccgtaggccgcaacggtatcaagtaccaactccgccaccgctttat 300
D V L T S V G R N G I K Y Q L R H R F Ictccccgactctgttccacggcgcttcgatcactgtcccccctggaatctcccgctccca 360
S P T L F H G A S I T V P P G I S R S QaatcgcctctctccgcggtatcaaaGTCCGTCCCTCTCTCTCCCCTACTCCCTCCGTGCT 420
I A S L R G I KAACCCTGCACGCAACAGcgcgtctggcccgttcgaaagttctcccgacccagcgcagtag 480
R V W P V R K F S R P S A Vtggacgccgatggaggaggaagcgggttctcagggtcgcctatcaaggcggcgctcatgg 540
V D A D G G G S G F S G S P I K A A L Mgggtgaaagagctcgggaagcgcgcgaacgcttatgctggagatacgtttggaccgcatg 600
G V K E L G K R A N A Y A G D T F G P HtcatgacgggggttaatgagacgcatgaggcggggttgttgggagctgggattaagGTGT 660
V M T G V N E T H E A G L L G A G I KGTTTTCGTCTTTGTTTGGGGGAGGGGGGAGGGAGGAGCTGATTAATGGGTGCAGattggg 720
I GgtgctggacactggtgAGAAGCGGTTTGGAGGGAGGAGAGGATGGAGCTGATGAGTGTGC 780
V L D T GAGGTGttgattatttgaacccgattctgggaggctgctttggacctgggtgccatatgtc 840
V D Y L N P I L G G C F G P G C H M Sgtttgggtacgacttggttggcgatgattacgatggagataacgctcctgtgccggatgt 900
F G Y D L V G D D Y D G D N A P V P D VggatccttGTGCGTCCTTCCCTCCGCGATGGGCTGGAATCTGAGCTGACGGCGTTGTTGA 960
D PTCAGttgcgagctgcgatcctcatggTTCGTCTCGCTTTCCGCTCCTCTGATGCGCTCGC 1020
F A S C D P H GTGATGCACTTGGCAATAGGaactcacgttacgggaatcattggagcgctcccgaatgcgt 1080
T H V T G I I G A L P N AttggatttactggcgtcgcacccgccgctactctgggccactaccgagtGCGCTCCCTCT 1140
F G F T G V A P A A T L G H Y R VATCCGTTCTTCCTCGCAGCTCCCACTCACTCGTGCTTCCAGGTatttggctgcactggct 1200
F G C T GtcgtcggagaagatatcGTTCGTCCTCTCTCCAGTCCCGGCTAGCGATCCTGCTTCTGAC 1260
F V G E D ITTCGCATCTCCCTCTGAGattctcgctggactcatgcgaggagtcgaggacaactgcaac 1320
I L A G L M R G V E D N C Ngtcttgaccctctctctcggaggtccaggagggtgggtcaagggcacgccggcgtccatc 1380
V L T L S L G G P G G W V K G T P A S Icttatcgaccagatcgaagcgcaaggcattctcgtcaccgtcgccactggcaactcggga 1440
L I D Q I E A Q G I L V T V A T G N S GgctgagggTGAGCTCCTCGCTACTTGCGCGTCGAGACCAGTACTAACAGGGGCGTGGACG 1500
A E GCAGGcatgttcttctccgagtctcccgcctcgacgatcaacggcctttccatcgcatcca 1560
M F F S E S P A S T I N G L S I A ScgTCCGTCTCTCCCTTCCTTCGCACGCCACTCGCTCACACTCTCTCGCAGGgacgttacc 1620
T D V Tgacctcatcgcctacaacgccaccgtctcaggccaacctgcgatcccttacctctccgcc 1680
D L I A Y N A T V S G Q P A I P Y L S A
ggggggggggtctg minus194
ctagcgcgaatcgagaaaacacaaaagaacctcccgacttggcgttgcttctgctcttct minus180
ccttctcccaaccgccgacaataaaaactataacacacacacaccccatctcgacgcctg minus120
cagacttcacgactctcctcacgctctttaccgccccacgggtcccacccacgagccacg minus60
(a)
Figure 4 Continued
Enzyme Research 11
acgcccctcaacgtcgtcgccaacagcttccgcgtccacttcacctctaccgaccccaac 1740
T P L N V V A N S F R V H F T S T D P Naaccccgtcgacgcctgctctcctcttccggctggagcgcccgacttcgccaactatgtt 1800
N P V D A C S P L P A G A P D F A N Y Vacggtcgttcagcgtgggacttgtacgttcgttaccaagtaccagaacgttctcaatgct 1860
T V V Q R G T C T F V T K Y Q N V L N AggaggGTGCGTCGGCTTCTTCTCTCCCGACTGGATTGGGCGGAGACTGACCTTCTGAGCA 1920
G GGaaagatcgtattgttgtacaactcggagggagctgggaacctcccttacctcacgccca 1980
K I V L L Y N S E G A G N L P Y L T PacggtgtcggcatcgacgccgttgcaggtcttcgtcgttccgacggactcaagGTTCGTC 2040
N G V G I D A V A G L R R S D G L KTACGCTCTTCGGTGCTGAGGCCCTTCTCTGATCTCGCTCCGACAGcttctctcgtactat 2100
L L S Y Ycagaatgccaacaagcgtctcactctgcgcttccccaagggcaagatcgtcgcaggcttg 2160
Q N A N K R L T L R F P K G K I V A G LaccgataccatcaccggcggactcatctcgTTCGTTCCTCTCCGCCTCCCTCGCTGGCCG 2220
T D T I T G G L I SATGAGCTGACGCGGTCTTTGCAGGggttactcgacgtttggtccgacgaatgacctctac 2280
G Y S T F G P T N D L Yggtcagcctaccctctctgcccctggtggcaacatcctttcgaccttccctctctccgag 2340
G Q P T L S A P G G N I L S T F P L S EggaggagtggcggtcatcagtgggacgagcatgtcgtgcccctttgtcgGTTCGTCCTTC 2400
G G V A V I S G T S M S C P F VTCCTCGTTCGTTCGCTGAGCGTCGACTGATCTTGCACGATCGCAGctggatctgcggcgg 2460
A G S A Atcctcatggccgctcgcgcttcggagaacctcacgccgcttgagatcaggagtctcctta 2520
V L M A A R A S E N L T P L E I R S L Lctaccactgcgaagcttacgccggtctcgctcttgggatcgacgcctttggtgagcgtga 2580
T T T A K L T P V S L L G S T P L V S VttcgtcaaggaggaggTACGTTCCTTTCTCGCTGCTTTGAGGATCGAGGCTGATTAGCGA 2640
I R Q G G GGCTTCGCAGGactcgttcaggttgccaaggcgctcgcggccaagacgctaatctctcctc 2700
L V Q V A K A L A A K T L I S Pacgagctcctgctcaacgacactgcgaacgcgaactacgtccagactatcaagatcaaga 2760
H E L L L N D T A N A N Y V Q T I K I Kacaccaactcgtgggcgatgaagtacaccttctcctcggccgtcgcccaaggactcggaa 2820
N T N S W A M K Y T F S S A V A Q G L Gctttcgacgcttcgggcgatatcctccctaccctcgacccagtcgccgtctctggcgcac 2880
T F D A S G D I L P T L D P V A V S G Aaggctaccgtcgcgttcaacactcggatcctcagcgtcgcgcccggcgcgacggggtccg 2940
Q A T V A F N T R I L S V A P G A T G Stcgtggcgactatcacgccgccggttcttcccgtagcggacgctgcgaggttccctatct 3000
V V A T I T P P V L P V A D A A R F P Itctctgggtggatcagggtgaatgggcaaggcgcgagggatagcagtaggaacgaggcgt 3060
F S G W I R V N G Q G A R D S S R N E AacactgGTGCGTTTGGCTCGCTTTTTGGGAGGTGCCGAGCTAATCGTTTCTCGTAGtccc 3120
Y T V PgtactttgggcttgcggcgaagatgatcgatatgcaagGTGCGTCGATGGTTCTCTCGCT 3180
Y F G L A A K M I D M QTCTCTCTCGGTTGCTCACTTCAAAGTGCAGtcctcgacaccaccgagaccatttacggtc 3240
V L D T T E T I Y Gcgggctacgcctaccccttcgtgatcgacgacgcgattggagacatccaatccaccacaa 3300
P G Y A Y P F V I D D A I G D I Q S T TcgtcgtactccaggaacctcgggcccaccgtcttcgctcgctttgccaTGCGTCGACCGT 3360
T S Y S R N L G P T V F A R F ACCTCATTTCCTGCGGCAGAGCGAGAGCTGATTCCGACCTCGAACAGctggaacccttcac 3420
T G T L Htacagcctcgatctcgtcctagccgacatcgcctttacccccacctaccccaactcctcc 3480
Y S L D L V L A D I A F T P T Y P N S S
(b)
Figure 4 Continued
12 Enzyme Research
cccgccactcgtttcgtcaagcgctccctcacgcagcacacctctgccgcttcgcacctc 3540
P A T R F V K R S L T Q H T S A A S H Lgccaagcgccgcgtctccatcgccactatcaaccccaaagccaccctcgtcgccgatcga 3600
A K R R V S I A T I N P K A T L V A D Rcagctccactcggacgtccctatcgagggcaacatcttcacccaaccctttactggaagg 3660
Q L H S D V P I E G N I F T Q P F T G Rgattacctcgtcgacgcagccccgacgggatccaccgatcgtaccgtcacttttaacggg 3720
D Y L V D A A P T G S T D R T V T F N Gcagtacgccgagaacggcctcgtgaggacggctgtgacggggacttcgtaccgcttcctc 3780
Q Y A E N G L V R T A V T G T S Y R F Lcttcgggcgttgaagatctcgggagacgcgatgtacgaggatcagtatgagagctggctc 3840
L R A L K I S G D A M Y E D Q Y E S W L3900
S L P F S F R Attttgccacctcaaaaaaaaaaaaaaaaaaaaaaaaaa 3938
lowast
tcgctaccgttctcgttccgtgcgtaggcttgttgtggacaccactgcattaagttatga
(c)
Figure 4 Nucleotide and deduced amino acid sequences of genomic DNA and cDNA encoding protease from G antarctica strain PI12 Thenucleotides are numbered from the 51015840 end of the cDNA A putative CAAT TATA box and polyadenylation signal are bold italicThe putativeN-terminal signal peptide is underlined The initiation codon is boldfaced and termination codon is represented by an asterisk (lowast) boxedpeptide sequences showed N-glycosylated asparagines residues The amino acid residues that form the catalytic triad are double underlinedand bold while introns are represented in blue font Highlighted in grey are the intron donor-acceptor splice sites
proteases Hydrophobic profile of PI12 amino acid proteasewas performed by ProtScale tools with Kyte and Doolittlemethod at httpauexpasyorgcgi-bin The hydrophobicityprofile elucidation of PI12 protease amino acid was carriedout from amino acid 1 to 963 The amino acid residuesfrom 1 to 16 (M-L-F-L-P-V-L-L-L-L-L-P-G-V-T-A) and 548 to562 (V-A-V-I-S) were hydrophobic grooves The first regionwhich consisted of 16 amino acids was very hydrophobic andwas predicted to be the signal peptide by SignalP and ProPwebserver
The bold and double-underlined amino acids showedresidues of catalytic triad aspartic acid (D) histidine (H) andserine (S) active site respectively NCBI Conserved DomainsDatabase (CDD) identified the presence of subtilase proteaseand subtilisin-like serine protease domains in the amino acidsequence The predicted PI12 protease amino acid sequencewas found sharing significant homology to the subtilisinfamily of protease The highest similarity of PI12 proteasewas observed with 42 identity to subtilisin-like proteaseof Rhodosporidium toruloides (EMS20793) 37 identity tosubtilisin-like protease of Pleurotus ostreatus (CAG25549)and 35 identity to subtilisin-like protease of Gloeophyllumtrabeum ATCC 11539 (EPQ55564) The nucleotide sequenceof G antarctica strain PI12 showed low homology to thefungus subtilisin protease family and none to other psy-chrophilic proteases The multiple sequence alignment wasillustrated in Figure 5 The consensus sequence around thecatalytic triad aspartic acid (D) histidine (H) and serine(S) of PI12 protease showed high conservation with othersubtilisin (showed in black boxes) Glycine residues G142G173 G191 G262 and G327 that were highly conserved inmost of subtilases were conserved in PI12 protease [25]Therefore PI12 protease belongs to the subtilisin subgroup ofthe subtilase serine protease superfamilyThis serine protease
of G antarctica PI12 has been deposited into GenBank withaccession number CAQ76821
34 Phylogenetic Relationship of PI12 Protease The phylo-genetic tree was constructed to demonstrate the similaritiesand relationship of protein sequence between differentspecies or organisms Figure 6 illustrates the phylogeneticdendrogram indicating the relationship of PI12 proteasewith another 25 translated amino acid sequences ofserine protease The bar scale signifies the branch lengthto approximate variance along each branch Basicallythis tree comprises two main clusters cluster I was sub-divided into subcluster a (CAG25549 EPQ55564 ETW-80907 XP 007381302 XP 001877576 CAG38357 XP007341385 CCA68627 EJU04161 XP 007369624 EIW-62675 CCO37249 XP 001830835 XP 007342189) and sub-cluster b (XP 007589863 XP 003711175 EFQ32840 XP006694395 ENH74885 EON99722) cluster II was sub-divided into subcluster c (CBQ73425 GAC97617 CCF52619)and subcluster d [CAQ76821 (PI12 protease) EMS20793]
Serine protease constructed by the phylogenetic treecomprised large size of protein with about 801 to 1032 aminoacids The majority of the translated protease sequenceswere obtained from fungus All serine proteases from cluster1 subcluster a are derived from Agaricomycetes except forEJU04161 which belongs to Dacrymycetes whereby all serineproteases from cluster 1 subcluster b are from Sordari-omycetes Cluster II subcluster c consisted of serine proteasefrom fungus belonging to Ustilaginomycetes PI12 proteasewas categorized under cluster II subcluster d It possessedhomology similarity with the subtilisin-like protease fromRhodosporidium toruloides (EMS20793) (formerly known asRhodotorula glutinis or Rhodotorula gracilis) which is a
Enzyme Research 13
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
MKG-YSWSLLQLAFTGAALATVPLSDVKRISSAS--VVPNKFVVEVESASEIPGKR--SL
MKGVLVWSILS---AGCALAVTPLSSVKRVTNLP--IVANKFIVEVDSTSDIPTKR--S-
MKLPLLSLALT---AVSVGATVPLSSVQRTTTSPNNALAGEYIVEMNNVQGLGGKR--AY
---MAHGSLASLLLLLAASAQGAAAFGGKSATGGRSVVPNSYILQVNASAPALTKRGVSV
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease ---MLFLPVLLLLLPGVTAFLNPVTNRATNAISSTQYLSNAYILELDLSTPGLVKRDSTP
lowastlowast
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
YTPHELFYRSLSARGVSYEVNKEYNS-EGLFVGAVVTVSDANDVKALAETPGVKAIRPVR
-NPHDALYESLRKRAIGFKVDREFNT-QGLFVGAALTLSDAQDVGQIQSTTGVKSIRPVR
ASTHDTLYRSLRKRGISFDVKKEYNQ-PGIMVGAALKVASPQDVQQIATMAEVKAIRPVI
FEALDHTLAAVKQNGVKYTVRQRFDAIPEAFQAVSIQVEDGASMAELAQIPGVQRVWPVS
DSILEDVLTSVGRNGIKYQLRHRFIS-PTLFHGASITVPPGISRSQIASLRGIKRVWPVR
lowastlowast
TIKSPKPVKVHVVTGKDDPEIPA-------------------------DTESTHIMTGVN
AFDRPKPVKVQVVKGPDDPALPP-------------------------DSESTHVITGVD
AVPAPERINQFTVSSPNDSRITA-------------------------DSETTHIMTGVD
LIPRPVEPTVSDFTPVSSSSSSTKSKRD-THTLEKRGTNFPPASAYLNDTFYPHVQTGID
KFSRPSAVVDADGGGSGFSGSPIKAALMGVKELGKR------ANAYAGDTFGPHVMTGVN
lowast lowast lowast lowastlowast
KLHAQGIFGAGIKIGIIDTGIDYTHPALGGALGAGHKVIGGYDFVGDAYDGTNTPVPDND
KLHAQGITGAGIKIGILDTGIDYTHPTLGGAFGPGNKVIGGFDLVGDDYDGTNTPVPDPD
KVHAAGITGEGIKIGIIDTGIDYNHPLLGAGFGPGFKVIGGYDFVGDDYTGSNNPAPDDD
VLHNKGILGQGVKIVVVDEGVDYTNPILGGCFGPGCQISFGYDFVGDNYTGTNAPVPDSD
ETHEAGLLGAGIKIGVLDTGVDYLNPILGGCFGPGCHMSFGYDLVGDDYDGDNAPVPDVD
lowast lowast lowast lowastlowastlowast lowast lowast lowastlowast lowast lowastlowast lowast lowast lowast lowast lowastlowastlowast lowast lowast lowast lowast lowastlowast lowast
PLDQCNGHGTHVAGIIGANPDNQFNISGVSYGASLSSYRVFGCSGSVSDDIIIEALLRGV
PLDQCAGHGTHVAGIIGANPDNAFNISGVAFQASLSAYRVFGCVGFVTDDVLVDALLLGF
PLDNCNGHGTHVAGIIGANPGNDFGISGVAYGSSIASYRVFGCSGFVSDDIIIDALLRGY
PFSSCSNHGTHTTGTVGALAN-EYGFSGAAPMATIGHYRVISCSGATTDEILVAALTRAL
PFASCDPHGTHVTGIIGALPN-AFGFTGVAPAATLGHYRVFGCTGFVGEDIILAGLMRGV
lowast lowastlowastlowast lowastlowast lowast lowastlowast lowast lowastlowastlowast lowast lowast lowast
QDGQDILTLSLGGTDGWTE-SSSSVVASRIAASGKIVTIAAGNEGAYGSWYSSSPGNGID
NEGQDILTLSLGGADGWTE-SVSAVVASRIAATGKVVTIAAGNDGASGAWYTSSPGNGID
SDGMDILTLSLGGVEGWTE-SSSSVVASRIADQGRVVTIAAGNDGAYGSWYASSPGTGVD
QDNVNIVSMSLGSSVGWLDDSPVQIMASYLGSKGIHVVASVGNERNEGLFAADQPAASRI
EDNCNVLTLSLGGPGGWVKGTPASILIDQIEAQGILVTVATGNSGAEGMFFSESPASTIN
lowastlowastlowast lowastlowast lowast lowast lowastlowast lowast lowast
VISVASIDNTVDNIQNATVNGVEHGPIPYFKTLPLPFTNA-LPLYVISNDTTIANDACNA
VISVASLDNTVVPLQTAQVHGVTHDPIIYQDTFPLPINDT-RPIFATSTDVTVVDDACNP
VISVASVNNIAIPVQNATVQGVEHAPIPYLEALPLNVTGS-RPIYATSTDTTVADDACNA
GTGVGGVDPTYLAAYYAYLI--KRDPVPYISPTPFNLPNN-YLLYFTSTNTSVTNDACNP
GLSIASTDVTDLIAYNATVS--GQPAIPYLSATPLNVVANSFRVHFTSTDPNNPVDACSP
lowast lowast lowast lowast lowastlowastlowast
LPDNTPDLSGFLVLVRRGSCTFVQKLTNIAAKGGNAVFIYDNGNG-----FAGIDTGKFT
LPDSTPDLSGFVVIVRRGTCTFVQKLGNVAAKGAKVTLIYDNGSG-----FGAIDTGAFV
LPNDTPDLSGYVVVIKRGTCTFVTKLQNAAAKGANTFLIYNNGGS-----FSAISVGDYT
LPSTTPNLANRVVVVQRGTCGFTVKQQNVAAAGGKIVLIYNSKGSGLIPQLDVGSTGLTA
LPAGAPDFANYVTVVQRGTCTFVTKYQNVLNAGGKIVLLYNSEGAGNLPYLTPNGVGIDA
lowastlowast lowast lowastlowast lowast lowast lowast lowast lowast lowast lowast
(a)
Figure 5 Continued
14 Enzyme Research
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
NATLIQAADGVFLAQQVAAGKSVTITFPQTGGSAQITN-PSGGLVSSFTTYGPTNDFYFK
-ASFIQAADGEFLVQQFASGQNVSLSFPQSGGLIQFPD-PAGGLISSFTSYGPSNDFFFK
-AALISETDGTFLVNQFAAKANITISFPQSGSAWNYPDDVSGGLVSDFSSYGPSYDMYFK
VGSLRYEDGLKLLSYYQSSPRGQSISFPNGPLAPYVSNTVSGGVVASYSNFGPTNELFIY
VAGLRRSDGLKLLSYYQNANKRLTLRFPKGKIVAGLTDTITGGLISGYSTFGPTNDLYGQ
lowast lowastlowast lowastlowast lowastlowast
PAFGAPGGNILSTLPVPLGSYGVESGTSMATPYVAGSAALLLSVKGKSAAVALGARTLFQ
PAVAAPGGNILSTLPVNLGLFGIESGTSMATPFVAGSAALLFQAKGTSAAVGRSARTLFE
PAVAAPGGNILSTLPIALGSYGVESGTSMATPFTAGSAALILQAKGK--SVAKSIRNLFQ
PTLAAPGTDILSTV---TGGVALMRGTSMAAPLVAGAYALLLSVGNNLKLSPEEARTILM
PTLSAPGGNILSTFPLSEGGVAVISGTSMSCPFVAGSAAVLMAARASENLTPLEIRSLLT
lowast lowastlowastlowast lowastlowastlowastlowast lowast lowastlowastlowastlowast lowast lowastlowast lowast lowast
STAQRVPSSHTDGDPLQTLSQQGAGLIDVFKALYTETIVWPTELILNDTAHYKEVHTFTV
TTAQRVPSTHTDGDPLQTLTQQGAGLINVFNAIHTTTIVSPGELILNDTAHFKGTQQFTV
TTASAVASSKTDGDPLQTVSQQGAGLIQVDRAIGTTTVVSPGQLTLNDTAHFEGFKTFNI
STSQPSPVAY-GSETLDTVVSQGAGVINMTAAIAAHTVISPAQFNLNDTQYSNNTQTLTL
TTAKLTPVSLLGSTPLVSVIRQGGGLVQVAKALAAKTLISPHELLLNDTANANYVQTIKI
VNTGKTSKQYKLTHVPAGTAQTIT---SGTIFAADGPVPLSTDYATVKLSSSSFTLRPGQ
RNAGSSAKTYTLRHIPAGTAVTVT---PGTIFPADGPVPLSTDFASVSLSTSKFTLAPGK
QNTGSKAVTYKISHVPAGTAASVQ---SGSIFAADGPVPLTADYAQVTFSLNKVTVLPGV
RNNNRFPVSYQLSWVDSTGIVTYNDGATTDIIPSTTPNYVSTSVLRVAFSQRTVTVPGGQ
KNTNSWAMKYTFSSAVAQGLGTFD--ASGDILPTLDPVAVSGAQATVAFNTRILSVAPGA
lowast lowast lowast lowast lowast lowast
SKQVTTHINAP--SGVDASTYPVFSGFIEIATAT--------ESVHVTYLGLAASLKDKQ
TQTVTARFTPP--AAADPSTFPVFSGFIQIESGT--------EQVQVSYLGLKASLKDKQ
SVPILVWFKPP--QGVDASTYPVYSGFIEIAGLG--------ETLRVAYLGVAASLKDKA
TVKVTAQFIPPNLTAQQRNQFPIYSGFVTITGQGQGAGAEQMEKYNLPFFGLGARMVDMP
TGSVVATITPPVLPVADAARFPIFSGWIRVNGQG-ARDSSRNEAYTVPYFGLAAKMIDMQ
lowast lowast lowastlowast lowast lowast lowast lowast
IVDNTNEFFGITIPAVLNNTGDAQ-LSPTNYTFSGSDFPTLLLRLAFGTPLLRVDLVDAT
VIDNTDFFFGVPTPVLTDPNGEVQ-TSPRNYSFLASDFPTLIFRLAFGSPKVVVDLVSPT
VVDNTDEYFGVKIPALLDASGDVQ-GNSTNYTLVGDDVPSLLYRFAFGSAHVRVDLVEKA
VLDTTNVALGP--NLPFIAVGQN--IQTGPTTFSESNPAVVYFRLAAGTRRLRVDLVDAS
VLDTTETIYGPGYAYPFVIDDAIGDIQSTTTSYSRNLGPTVFARFATGTLHYSLDLVLAD
lowast lowast lowast lowast lowastlowastlowastlowast lowast
IDLKPTLNSREIDDKRHVFERESFSFPHPGPKGS-------------------TFAQVKI
INFKPTLNTRALGDTHGSF----FSFPHRVKTG--------------------SFAQVPT
STVSATLSKRNWWN---------WWWPQKPSN---------------------SYDKVPI
TSYNATVPAVTNPASRLVKRSNTELTPRAAFPT--------------------LYSQVRT
IAFTPTYPNSS-PATRFVKRSLTQHTSAASHLAKRRVSIATINPKATLVADRQLHSDVPI
lowast lowast
VGTLSEQSHIPRNSDASTVDDGGYSTFAFNKP------SFANGTTIPNGTYRWLLRALKV
LGTVFELDFTSRNNDDPTDN--GFNTVAIDPP------TFANGTTIPNGQYRLLLRALRV
VGTLAEYDYVPRNSDASTIDDNGYNVFQFT-D------SFVNGTKIAAGEYKVLLRALKV
VGSLFAPSYWPPRDYLFNNGGT-YSDYEIQLDGSFTFPNGTKGMAELGRSYKVLLRALKI
EGNIFTQPFTG-RDYLVDAAPTGSTDRTVTFNGQY-AENGLVRTAVTGTSYRFLLRALKI
lowast lowast lowastlowast lowast lowast lowast lowast lowastlowastlowastlowast
lowast lowast lowastlowastlowastlowastlowast
(b)
Figure 5 Continued
Enzyme Research 15
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
TGDPTKEADFETWLSPIVGITVA
TGDPTNEADFESFLSPIIGVNAP
TGNPAEEGDYETWLSPSFSVVPA
TADPTYTSSYESWLSPSFTFTA-
SGDAMYEDQYESWLSLPFSFRA-
lowast lowastlowast
ConservedSimilar
lowast All match
(c)
Figure 5 Multiple sequence alignment between amino acid sequence of PI12 protease with other subtilisin The serine protease sequenceswere obtained from the GenBank database Their accession numbers are EMS20793 (subtilisin-like protease of Rhodosporidium toruloidesNP11) CAG25549 (subtilisin-like protease of Pleurotus ostreatus) EPQ55564 (subtilisin-like protease of Gloeophyllum trabeumATCC 11539)and ETW80907 (serine protease S8 ofHeterobasidion irregulare TC 32-1) Putative ORF of serine protease fromG antarctica PI12 was labeledas PI12 protease
CAG38357 Phanerochaete chrysosporium
EIW62675 Trametes versicolor FP-101664 SS1
XP 007369624 Dichomitus squalens LYAD-421 SS1
XP 007381302 Punctularia strigosozonata HHB-11173 SS5
CAG25549 Pleurotus ostreatus
ETW80907 Heterobasidion irregulare TC 32-1
XP 001877576 Laccaria bicolor S238N-H82
EPQ55564 Gloeophyllum trabeum ATCC 11539
XP 001830835 Coprinopsis cinerea okayama7130
XP 007341385 Auricularia delicata TFB-10046 SS5
CCA68627 Piriformospora indica DSM 11827
CCO37249 Rhizoctonia solani AG-1 IB
XP 007342189 Auricularia delicata TFB-10046 SS5
EJU04161 Dacryopinax sp DJM-731 SS1
XP 007589863 Colletotrichum fioriniae PJ7
EFQ32840 Colletotrichum graminicola M1001
ENH74885 Fusarium oxysporum f sp cubense race 1
XP 003711175 Magnaporthe oryzae 70-15
XP 006694395 Chaetomium thermophilum DSM 1495
EON99722 Togninia minima UCRPA7
CBQ73425 Sporisorium reilianum SRZ2
GAC97617 Pseudozyma hubeiensis SY62
CCF52619 Ustilago hordei
EMS20793 Rhodosporidium toruloides NP11
CAQ76821 Glaciozyma antarctica PI12
01
Clus
ter 1
Clus
ter 2
a
b
c
d
Figure 6 Phylogenetic dendrogram among 25 translated amino acid sequences of serine proteaseThe tree was constructed using a neighbor-joining algorithm with 1000 bootstrappings by MEGA 6 software [39]The scale bar represents 01 substitutions per amino acid positionTheserine protease sequences were obtained from the GenBank database GenBank accession numbers of the sequences were indicated beforethe strain name The putative serine protease of G antarctica strain PI12 has been deposited in GenBank with accession number CAQ76821
16 Enzyme Research
0
2
4
6
8
10
12
0 24 48 72 96 110
Prot
ease
activ
ity (U
mL)
Post induction time (hours)
Figure 7 Effect of postinduction time towards protease activity The highest activity was detected at 72 hours after 110 hours of incubationwith 9UmL activity followed by 48 24 and 96 hoursThe substrate (azocasein) was dissolved in Tris HCL buffer and incubated for 30min at20∘CThe absorbance was measured at 450 nm after 15min incubation A one-way ANOVAwas used to test for preference differences amongsix different postinduction times The statistical value (119875 value lt005) between hours is 0003
pink-coloured nonpathogenic oleaginous yeast (42 iden-tity) Both G antarctica PI12 and Rhodosporidium toruloidesNP11 belong to Microbotryomycetes class PI12 protease didnot show any homology similarity to other psychrophilicproteases however it showed 37 identity to hypotheticalprotein from Pseudozyma antarctica T-34 which was isolatedfrom Japan (GAC76005) (not listed in the phylogenetic tree)
35 Cloning and Expression of Cold-Adapted PI12 Protease inthe Yeast Expression System The PCR product harbouringthe mature PI12 protease with two flanking restriction sites(EcoRI and AvrII) was cloned into pPIC9 expression vectorThe recombinant plasmid harbouring PI12 protease was usedto transform both GS115 and KM71 intended to produce bothHis+ Mut+ and His+ MutS Two recombinant clones fromGS115 (His+Mut+) strain (GpPro1 andGpPro2) and one fromKM71 (His+MutS) strain (KpPro1) were successfully obtainedwhich were further analyzed for protein expression study
36 Expression and Enzyme Assay of PI12 Protease in Pichiapastoris All positive recombinants were grown in small scalecultures and assayed for the cold-adapted protease activityin the culture supernatants The clones were first grown inBMGY medium with glycerol as the sole carbon sourceFor induction of AOX1 promoter and protein productionthe growth medium was transferred to the same mediumexcept with 05 methanol in place with glycerol as thecarbon source [26] The supernatant of the recombinant Ppastoris expression cultures after 48 h of incubation time wasanalyzed to determine the recombinant protease activity bythe modified method of Brock et al (1982) [20] All testedclones secreted protease into the supernatant in differentexpression levels This might be due to the variability ofthe strength of the promoter and copy number in each cellto secrete and express the recombinant product The clonewith the highest yield was GpPro2 with 63UmL activityfollowed by KpPro1 and GpPro1 with 25UmL and 37UmLactivities respectively (data not shown) It is perceptible thatexpression systemof Pichia pastoris is the ideal system for thisprotein The preliminary assay has fruitfully produced high
protease expression at 20∘C Yet further optimization of therecombinant PI12 protease production need to be conductedin order to examine the optimum condition of this proteinReported cold-adapted proteases showed optimum activity ata range of 15∘Cndash60∘C [27ndash30] This difference was possiblyrelated to the respective genetic and physiological adaptationof the strains There are several factors that influenced thedifferent rate of expression It includes different site of inte-gration between interest gene and Pichia genome variabilityof different expression host cell to secrete and express therecombinant enzyme and different sites in the polylinker forinserting foreign genes [31]
This is the second extracellular protease reported to besynthesized by this obligate psychrophilic yeastG antarcticaThe first cold-adapted extracellular subtilase from this yeasthas been purified and characterized previously [32 33] Theextracellular serine proteinase (LAP2) from this Antarcticyeast formerly known as Leucosporidium antarcticum strain171 is specific towards synthetic substrates of chymotrypsinand subtilisin The subtilase of L antarcticum strain 171showed specific activity of 4043Umg when utilized N-SucAAPFpNA as a substrate [33]
37 Optimization of Protease Production GpPro2 was chosenfor further optimization due to its highest protease activityat the preliminary expression stage High protease activitywas detected after 72 hours of incubation (Figure 7) byusing the standard protocol method as recommended forPichia pastoris expression system (Pichia Expression KitInvitrogen USA) The expression decreased after 72 hoursand no activity was detected at 110 hours The prolongedincubation may result in the decrease of biomass productionand will subsequently reduce the expression of recombinantprotein In addition oxygen and nutrient depletion duringprolonged incubationwill trigger the stress responses in orderto survive in such harsh environment [34]
Methanol was used as a sole carbon by P pastoris Themethanol was oxidized to formaldehyde with the aid of oxy-gen as electron acceptor by an alcohol oxidase enzyme (AOXgene) [35] The optimal methanol concentration is at 05(vv) with 14UmL activity The addition of more than 05
Enzyme Research 17
02468
10121416
0 01 03 05 07 09Pr
otea
se ac
tivity
(Um
L)Methanol concentration ()
Figure 8 Effect ofmethanol concentration towards protease activityThe recombinant was cultivated for 72 hours at 30∘C in BMMYmedium05 methanol gives the highest activity with 14UmL The substrate (azocasein) was dissolved in Tris HCL buffer and incubated for 30minat 20∘CThe mixture was left for 15min and absorbance was taken at 450 nm A one-way ANOVA was used to test for preference differencesamong six different concentrations The statistical value (119875 value lt005) between concentration is 002
0
5
10
15
20
25
10 15 20 25 30 35
Prot
eas a
ctiv
ity (
Um
L)
Temperature (∘C)
Figure 9 Effect of temperature towards protease activity The reaction was carried out in temperature ranging from 10∘C to 35∘C at pH 7 for30min A one-way ANOVA was used to test for preference differences among six different temperatures The statistical value (119875 value lt005)between temperature is 0067
(vv) methanol resulted in tremendous decrease of proteaseactivity (Figure 8) Production of PI12 protease was decreasedwhen the methanol concentration increased This might bedue to the toxic effect of methanol concentration on DNAreplication and membrane synthesis Basically methanol isused to increase the permeability of the cell membrane tohelp the secretion of foreign protein [26] However if thepermeability of cell membrane is too high lowmolecular sizemetabolites would leak from the cell and such leakage willseriously damage cell activities Such condition would abusethe cellular system and might reduce the yield of proteinexpression [36]
Psychrophilic microorganism dominated in low temper-ature habitat which most other microorganism are inca-pable of withstanding due to their inability to produceenzyme that can tolerate the extreme environment con-dition Psychrophilic enzymes are always associated withhigh catalytic efficiency at low and moderate temperature(0∘C to 30∘C) compared to the mesophilic counterpart thatare generally unable to catalyze efficiently under extremely
low temperature [2] The recombinant protein assay wasdone at various temperatures ranging from 10∘C to 35∘Cusing 05 azocasein as a substrate dissolved in 50mMTris HCl in presence of 2mM CaCl
2 pH 7 In the activity
assays expressed protease showed a highest activity at 20∘C(20UmL) and decreased tremendously once it exceededmaximum temperature due to the denatured state of theenzyme (Figure 9) The active site of an enzyme is easilyaltered when the high temperature is introduced duringincubation or activity assay resulting in a low reaction rate[37]
The recombinant construct was transferred into BMMYmedia at different A
600 nm ranging from 1 to 10 The high-est activity of active enzyme was detected at A
600 nm 60(283UmL) and above (Figure 10) The slope during expo-nential phase was much higher compared to other phasesindicating an increase in the doubling time At exponentialphase the cells are adapted to the growth condition andother processes like cell growth cycle and synthesis of RNAThe yield of active enzyme during expression of P pastoris
18 Enzyme Research
05
1015202530
1 2 3 4 5 6 7 8 9 10Prot
ease
activ
ity (U
mL)
Initial inoculum A600
Figure 10 Effect of inoculums size towards protease activity The recombinant was cultivated for 72 hours at 30∘C in BMMY mediumwith 05 methanol concentration A one-way ANOVA was used to test for preference differences among six different optical densitiesThe statistical value (119875 value lt005) between optical density value is 003
(kD
a)
1160
662
450
350
250
184
144
M 1 2 3
(a) (b) (c)
Figure 11 (a) Coomassie-stained SDS-PAGE of recombinant P pastoris clone 1mL supernatants collected from the third-day culturewere concentrated 10-fold using trichloroacetic acid and 10120583L resultant products were applied to SDS-PAGE analysis Lane 1 molecularweight marker (Fermentas) Lane 2 GS115 (Control) Lane 3 GS115pPIC9 empty (Control) Lane 4 GS115pPIC9PI12prot (GpPro2)The arrow indicates the secreted recombinant PI12 protease (b) Plate activity staining of different recombinant P pastoris clones GGS115 GP GS115pPIC9 GRI GS115pPIC9PI12prot1 (GpPro1) GRII GS115pPIC9PI12prot2 (GpPro2) K KM71 KP KM71pPIC9 KRKM71pPIC9PI12prot (KpPro) (c)Gel activity staining of recombinant PI12 protease inP pastorisGpPro2 clone Lane 1 prestainedmolecularweightmarker (Fermentas) Lane 2 GS115pPIC9 empty (Control) Lane 3 GS115pPIC9PI12prot1 (GpPro2)The arrow indicates the secretedrecombinant PI12 protease
dropped after 28 hours of incubation which is at stationaryor death phase [38]
38 Detection of Recombinant Protein by SDS-PAGE andActivity Staining TheSDS-PAGE gel of the GpPro2 recombi-nant clonesrsquo supernatant showed a putative expression bandat around 993 kDa Plate staining and gel activity stainingwere executed to examine if the recombinant PI12 proteaseexpressed in P pastoris was functionally active In the platestaining the transformants showed strong proteolytic activity(Figure 11(a)) In gel staining proteolytic activity was alsodetected around 993 kDa after incubation at 20∘C This is
evidence of protease secretion into the surrounding mediawhereby it catalyzed the breakdown of casein thus formedclearing zones around the activity ring and band (Figures11(b) and 11(c)) As a conclusion the cold-adapted PI12protease was functionally expressed in P pastoris GS115(GpPro2 clone) with the size of 993 kDa
4 Conclusions
Psychrophilic yeast which was originated frommarine waterin Casey Station Antarctica was identified as Glaciozymaantarctica strain PI12 A serine protease with an open reading
Enzyme Research 19
frame of 2892 bp encoded for 963 amino acids from thisAntarctic yeast has been cloned and expressed in Pichiaexpression system This is the second extracellular pro-tease reported being synthesized by this psychrophilic yeastSequence analysis through bioinformatics studies revealedfew homology identities of this protein with other ser-ine proteases The mature PI12 protease was successfullyexpressed and secreted into the culture medium driven bythe Saccharomyces cerevisiae 120572-factor signal sequence in Ppastoris The protease production in P pastoris was bestobtained from strain GS115 (GpPro2) with 283UmL after 72hours of induction time with 05 (vv) of methanol inducerThe starting optical density at 600 nm (OD600) was 60 andthe optimum assay temperature was 20∘C The expressedprotein was detected by SDS-PAGE and activity staining witha molecular weight of 99 kDaThis cold-active enzymemightoffer an opportunity for biotechnological exploitation basedon its high catalytic activity at low temperatures and lowthermostability Structural and functional study of this cold-adapted enzyme would give insight into its adaptation to anextreme cold environment
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgment
This research was supported financially by the InitiativeBiotechnology-GMBI-MOSTI (Project number 07-05-16-MG-GMB02) andRUGS-Initiative 6 (Project number 05-02-11-1414RU)
References
[1] J W Deming ldquoPsychrophiles and polar regionsrdquo CurrentOpinion in Microbiology vol 5 no 3 pp 301ndash309 2002
[2] C Gerday M Aittaleb M Bentahir et al ldquoCold-adaptedenzymes from fundamentals to biotechnologyrdquo Trends inBiotechnology vol 18 no 3 pp 103ndash107 2000
[3] A Svingor J Kardos I Hajdu A Nemeth and P Zavodszky ldquoAbetter enzyme to cope with cold comparative flexibility studieson psychrotrophic mesophilic and thermophilic IPMDHSrdquoThe Journal of Biological Chemistry vol 276 no 30 pp 28121ndash28125 2001
[4] G Antranikian C E Vorgias and C Bertoldo ldquoExtremeenvironments as a resource for microorganisms and novel bio-catalystsrdquo Advances in Biochemical EngineeringBiotechnologyvol 96 pp 219ndash262 2005
[5] T Lonhienne C Gerday andG Feller ldquoPsychrophilic enzymesrevisiting the thermodynamic parameters of activation mayexplain local flexibilityrdquoBiochimica et Biophysica Acta vol 1543no 1 pp 1ndash10 2000
[6] R Cavicchioli and TThomas ldquoExtremophilesrdquo in Encyclopediaof Microbiology pp 317ndash337 Academic Press San Diego CalifUSA 2000
[7] G Feller E Narinx J L Arpigny et al ldquoEnzymes frompsychrophilic organismsrdquo FEMS Microbiology Reviews vol 18no 2-3 pp 189ndash202 1996
[8] J Arnorsdottir R B Smaradottir T O Magnusson S HThorbjarnardottir G Eggertsson and M M KristjanssonldquoCharacterization of a cloned subtilisin-like serine proteinasefrom a psychrotrophic Vibrio speciesrdquo European Journal ofBiochemistry vol 269 no 22 pp 5536ndash5546 2002
[9] N J Russell ldquoPsychrophilic bacteriamdashmolecular adaptations ofmembrane lipidsrdquo Comparative Biochemistry and Physiology APhysiology vol 118 no 3 pp 489ndash493 1997
[10] G Feller and C Gerday ldquoPsychrophilic enzymes hot topics incold adaptationrdquo Nature Reviews Microbiology vol 1 no 3 pp200ndash208 2003
[11] P Raspor and J Zupan ldquoYeasts in extreme environmentsrdquo inBiodiversity and Ecophysiology of Yeasts The Yeast Handbookpp 371ndash417 Springer Berlin Germany 2006
[12] M B Rao A M Tanksale M S Ghatge and V V Desh-pande ldquoMolecular and biotechnological aspects of microbialproteasesrdquoMicrobiology andMolecular Biology Reviews vol 62no 3 pp 597ndash635 1998
[13] L Polgar ldquoThe catalytic triad of serine peptidasesrdquo Cellular andMolecular Life Sciences vol 62 no 19-20 pp 2161ndash2172 2005
[14] S Y Boo C M V L Wong K F Rodrigues N NajimudinA M A Murad and N M Mahadi ldquoThermal stress responsesin Antarctic yeastGlaciozyma antarctica PI12 characterized byreal-time quantitative PCRrdquoPolar Biology vol 36 no 3 pp 381ndash389 2013
[15] B Turchetti S R T Hall L B Connell et al ldquoPsychrophilicyeasts from Antarctica and European glaciers descriptionof Glaciozyma gen nov Glaciozyma martinii sp nov andGlaciozyma watsonii sp novrdquo Extremophiles vol 15 no 5 pp573ndash586 2011
[16] N H F Hashim I Bharudin D L S Nguong et al ldquoCharac-terization of Afp1 an antifreeze protein from the psychrophilicyeast Glaciozyma antarctica PI12rdquo Extremophiles vol 17 no 1pp 63ndash73 2013
[17] A N M Ramli M A Azhar M S Shamsir et al ldquoSequenceand structural investigation of a novel psychrophilic 120572-amylasefrom Glaciozyma antarctica PI12 for cold-adaptation analysisrdquoJournal ofMolecularModeling vol 19 no 8 pp 3369ndash3383 2013
[18] A N M Ramli N M Mahadi A Rabu A M A Murad F DA Bakar and R M Illias ldquoMolecular cloning expression andbiochemical characterisation of a cold-adapted novel recombi-nant chitinase from Glaciozyma antarctica PI12rdquoMicrobial CellFactories vol 10 no 1 article 94 2011
[19] J Sambrook E Fritsch and T Maniatis Molecular Cloning ALaboratoryManual Cold SpringHarbor Laboratory Press NewYork NY USA 1989
[20] F M Brock C W Forsberg and J G Buchanan-SmithldquoProteolytic activity of rumen microorganisms and effects ofproteinase inhibitorsrdquoApplied and EnvironmentalMicrobiologyvol 44 no 3 pp 561ndash569 1982
[21] U K Laemmli ldquoCleavage of structural proteins during theassembly of the head of bacteriophage T4rdquo Nature vol 227 no5259 pp 680ndash685 1970
[22] A Henne R A Schmitz M Bomeke G Gottschalk and RDaniel ldquoScreening of environmental DNA libraries for thepresence of genes conferring lipolytic activity on Escherichiacolirdquo Applied and Environmental Microbiology vol 66 no 7 pp3113ndash3116 2000
[23] M Poza T de Miguel C Sieiro and T G Villa ldquoCharacteri-zation of a broad pH range protease of Candida caseinolyticardquoJournal of Applied Microbiology vol 91 no 5 pp 916ndash921 2001
20 Enzyme Research
[24] J D Bendtsen H Nielsen G von Heijne and S BrunakldquoImproved prediction of signal peptides SignalP 30rdquo Journalof Molecular Biology vol 340 no 4 pp 783ndash795 2004
[25] R J Siezen and J A M Leunissen ldquoSubtilases the superfamilyof subtilisin-like serine proteasesrdquo Protein Science vol 6 no 3pp 501ndash523 1997
[26] Y Katakura W Zhang G Zhuang et al ldquoEffect of methanolconcentration on the production of human 1205732- glycoprotein Idomain V by a recombinant Pichia pastoris a simple system forthe control of methanol concentration using a semiconductorgas sensorrdquo Journal of Fermentation and Bioengineering vol 86no 5 pp 482ndash487 1998
[27] Q-FWang Y-HHou Z Xu J-LMiao andG-Y Li ldquoPurifica-tion and properties of an extracellular cold-active protease fromthe psychrophilic bacterium Pseudoalteromonas sp NJ276rdquoBiochemical Engineering Journal vol 38 no 3 pp 362ndash3682008
[28] S Vazquez L Ruberto and W Mac Cormack ldquoProper-ties of extracellular proteases from three psychrotolerantStenotrophomonas maltophilia isolated from Antarctic soilrdquoPolar Biology vol 28 no 4 pp 319ndash325 2005
[29] R Zeng R Zhang J Zhao and N Lin ldquoCold-active ser-ine alkaline protease from the psychrophilic bacterium Pseu-domonas strain DY-A enzyme purification and characteriza-tionrdquo Extremophiles vol 7 no 4 pp 335ndash337 2003
[30] M Turkiewicz E Gromek H Kalinowska and M ZielinskaldquoBiosynthesis and properties of an extracellularmetalloproteasefrom the Antarctic marine bacterium Sphingomonas paucimo-bilisrdquo Journal of Biotechnology vol 70 no 1ndash3 pp 53ndash60 1999
[31] A Vassileva D A Chugh S Swaminathan and N KhannaldquoEffect of copy number on the expression levels of hepatitisB surface antigen in the methylotrophic yeast Pichia pastorisrdquoProtein Expression andPurification vol 21 no 1 pp 71ndash80 2001
[32] M Pazgier M Turkiewicz H Kalinowska and S Bielecki ldquoTheunique cold-adapted extracellular subtilase from psychrophilicyeast Leucosporidium antarcticumrdquo Journal ofMolecular Catal-ysis B Enzymatic vol 21 no 1-2 pp 39ndash42 2003
[33] M Turkiewicz M Pazgier H Kalinowska and S BieleckildquoA cold-adapted extracellular serine proteinase of the yeastLeucosporidium antarcticumrdquo Extremophiles vol 7 no 6 pp435ndash442 2003
[34] D Mattanovich B Gasser H Hohenblum and M SauerldquoStress in recombinant protein producing yeastsrdquo Journal ofBiotechnology vol 113 no 1ndash3 pp 121ndash135 2004
[35] P Secades BAlvarez and JAGuijarro ldquoPurification andprop-erties of a new psychrophilic metalloprotease (Fpp2) in the fishpathogen Flavobacterium psychrophilumrdquo FEMS MicrobiologyLetters vol 226 no 2 pp 273ndash279 2003
[36] L Y Ling I Ithoi and F M Yik ldquoOptimization for high-level expression in Pichia pastoris and purification of truncatedand full length recombinant sag2 of toxoplasma gondii fordiagnostic userdquo Southeast Asian Journal of Tropical Medicineand Public Health vol 41 no 3 pp 507ndash513 2010
[37] A Illanes ldquoStability of biocatalystsrdquo Electronic Journal ofBiotechnology vol 2 no 1 pp 1ndash9 1999
[38] B A Plantz J Sinha L Villarete K W Nickerson and VL Schlegel ldquoPichia pastoris fermentation optimization energystate and testing a growth-associated modelrdquo Applied Microbi-ology and Biotechnology vol 72 no 2 pp 297ndash305 2006
[39] K Tamura G Stecher D Peterson A Filipski and S KumarldquoMEGA6molecular evolutionary genetics analysis version 60rdquo
Molecular Biology and Evolution vol 30 no 12 pp 2725ndash27292013
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

Enzyme Research 11
acgcccctcaacgtcgtcgccaacagcttccgcgtccacttcacctctaccgaccccaac 1740
T P L N V V A N S F R V H F T S T D P Naaccccgtcgacgcctgctctcctcttccggctggagcgcccgacttcgccaactatgtt 1800
N P V D A C S P L P A G A P D F A N Y Vacggtcgttcagcgtgggacttgtacgttcgttaccaagtaccagaacgttctcaatgct 1860
T V V Q R G T C T F V T K Y Q N V L N AggaggGTGCGTCGGCTTCTTCTCTCCCGACTGGATTGGGCGGAGACTGACCTTCTGAGCA 1920
G GGaaagatcgtattgttgtacaactcggagggagctgggaacctcccttacctcacgccca 1980
K I V L L Y N S E G A G N L P Y L T PacggtgtcggcatcgacgccgttgcaggtcttcgtcgttccgacggactcaagGTTCGTC 2040
N G V G I D A V A G L R R S D G L KTACGCTCTTCGGTGCTGAGGCCCTTCTCTGATCTCGCTCCGACAGcttctctcgtactat 2100
L L S Y Ycagaatgccaacaagcgtctcactctgcgcttccccaagggcaagatcgtcgcaggcttg 2160
Q N A N K R L T L R F P K G K I V A G LaccgataccatcaccggcggactcatctcgTTCGTTCCTCTCCGCCTCCCTCGCTGGCCG 2220
T D T I T G G L I SATGAGCTGACGCGGTCTTTGCAGGggttactcgacgtttggtccgacgaatgacctctac 2280
G Y S T F G P T N D L Yggtcagcctaccctctctgcccctggtggcaacatcctttcgaccttccctctctccgag 2340
G Q P T L S A P G G N I L S T F P L S EggaggagtggcggtcatcagtgggacgagcatgtcgtgcccctttgtcgGTTCGTCCTTC 2400
G G V A V I S G T S M S C P F VTCCTCGTTCGTTCGCTGAGCGTCGACTGATCTTGCACGATCGCAGctggatctgcggcgg 2460
A G S A Atcctcatggccgctcgcgcttcggagaacctcacgccgcttgagatcaggagtctcctta 2520
V L M A A R A S E N L T P L E I R S L Lctaccactgcgaagcttacgccggtctcgctcttgggatcgacgcctttggtgagcgtga 2580
T T T A K L T P V S L L G S T P L V S VttcgtcaaggaggaggTACGTTCCTTTCTCGCTGCTTTGAGGATCGAGGCTGATTAGCGA 2640
I R Q G G GGCTTCGCAGGactcgttcaggttgccaaggcgctcgcggccaagacgctaatctctcctc 2700
L V Q V A K A L A A K T L I S Pacgagctcctgctcaacgacactgcgaacgcgaactacgtccagactatcaagatcaaga 2760
H E L L L N D T A N A N Y V Q T I K I Kacaccaactcgtgggcgatgaagtacaccttctcctcggccgtcgcccaaggactcggaa 2820
N T N S W A M K Y T F S S A V A Q G L Gctttcgacgcttcgggcgatatcctccctaccctcgacccagtcgccgtctctggcgcac 2880
T F D A S G D I L P T L D P V A V S G Aaggctaccgtcgcgttcaacactcggatcctcagcgtcgcgcccggcgcgacggggtccg 2940
Q A T V A F N T R I L S V A P G A T G Stcgtggcgactatcacgccgccggttcttcccgtagcggacgctgcgaggttccctatct 3000
V V A T I T P P V L P V A D A A R F P Itctctgggtggatcagggtgaatgggcaaggcgcgagggatagcagtaggaacgaggcgt 3060
F S G W I R V N G Q G A R D S S R N E AacactgGTGCGTTTGGCTCGCTTTTTGGGAGGTGCCGAGCTAATCGTTTCTCGTAGtccc 3120
Y T V PgtactttgggcttgcggcgaagatgatcgatatgcaagGTGCGTCGATGGTTCTCTCGCT 3180
Y F G L A A K M I D M QTCTCTCTCGGTTGCTCACTTCAAAGTGCAGtcctcgacaccaccgagaccatttacggtc 3240
V L D T T E T I Y Gcgggctacgcctaccccttcgtgatcgacgacgcgattggagacatccaatccaccacaa 3300
P G Y A Y P F V I D D A I G D I Q S T TcgtcgtactccaggaacctcgggcccaccgtcttcgctcgctttgccaTGCGTCGACCGT 3360
T S Y S R N L G P T V F A R F ACCTCATTTCCTGCGGCAGAGCGAGAGCTGATTCCGACCTCGAACAGctggaacccttcac 3420
T G T L Htacagcctcgatctcgtcctagccgacatcgcctttacccccacctaccccaactcctcc 3480
Y S L D L V L A D I A F T P T Y P N S S
(b)
Figure 4 Continued
12 Enzyme Research
cccgccactcgtttcgtcaagcgctccctcacgcagcacacctctgccgcttcgcacctc 3540
P A T R F V K R S L T Q H T S A A S H Lgccaagcgccgcgtctccatcgccactatcaaccccaaagccaccctcgtcgccgatcga 3600
A K R R V S I A T I N P K A T L V A D Rcagctccactcggacgtccctatcgagggcaacatcttcacccaaccctttactggaagg 3660
Q L H S D V P I E G N I F T Q P F T G Rgattacctcgtcgacgcagccccgacgggatccaccgatcgtaccgtcacttttaacggg 3720
D Y L V D A A P T G S T D R T V T F N Gcagtacgccgagaacggcctcgtgaggacggctgtgacggggacttcgtaccgcttcctc 3780
Q Y A E N G L V R T A V T G T S Y R F Lcttcgggcgttgaagatctcgggagacgcgatgtacgaggatcagtatgagagctggctc 3840
L R A L K I S G D A M Y E D Q Y E S W L3900
S L P F S F R Attttgccacctcaaaaaaaaaaaaaaaaaaaaaaaaaa 3938
lowast
tcgctaccgttctcgttccgtgcgtaggcttgttgtggacaccactgcattaagttatga
(c)
Figure 4 Nucleotide and deduced amino acid sequences of genomic DNA and cDNA encoding protease from G antarctica strain PI12 Thenucleotides are numbered from the 51015840 end of the cDNA A putative CAAT TATA box and polyadenylation signal are bold italicThe putativeN-terminal signal peptide is underlined The initiation codon is boldfaced and termination codon is represented by an asterisk (lowast) boxedpeptide sequences showed N-glycosylated asparagines residues The amino acid residues that form the catalytic triad are double underlinedand bold while introns are represented in blue font Highlighted in grey are the intron donor-acceptor splice sites
proteases Hydrophobic profile of PI12 amino acid proteasewas performed by ProtScale tools with Kyte and Doolittlemethod at httpauexpasyorgcgi-bin The hydrophobicityprofile elucidation of PI12 protease amino acid was carriedout from amino acid 1 to 963 The amino acid residuesfrom 1 to 16 (M-L-F-L-P-V-L-L-L-L-L-P-G-V-T-A) and 548 to562 (V-A-V-I-S) were hydrophobic grooves The first regionwhich consisted of 16 amino acids was very hydrophobic andwas predicted to be the signal peptide by SignalP and ProPwebserver
The bold and double-underlined amino acids showedresidues of catalytic triad aspartic acid (D) histidine (H) andserine (S) active site respectively NCBI Conserved DomainsDatabase (CDD) identified the presence of subtilase proteaseand subtilisin-like serine protease domains in the amino acidsequence The predicted PI12 protease amino acid sequencewas found sharing significant homology to the subtilisinfamily of protease The highest similarity of PI12 proteasewas observed with 42 identity to subtilisin-like proteaseof Rhodosporidium toruloides (EMS20793) 37 identity tosubtilisin-like protease of Pleurotus ostreatus (CAG25549)and 35 identity to subtilisin-like protease of Gloeophyllumtrabeum ATCC 11539 (EPQ55564) The nucleotide sequenceof G antarctica strain PI12 showed low homology to thefungus subtilisin protease family and none to other psy-chrophilic proteases The multiple sequence alignment wasillustrated in Figure 5 The consensus sequence around thecatalytic triad aspartic acid (D) histidine (H) and serine(S) of PI12 protease showed high conservation with othersubtilisin (showed in black boxes) Glycine residues G142G173 G191 G262 and G327 that were highly conserved inmost of subtilases were conserved in PI12 protease [25]Therefore PI12 protease belongs to the subtilisin subgroup ofthe subtilase serine protease superfamilyThis serine protease
of G antarctica PI12 has been deposited into GenBank withaccession number CAQ76821
34 Phylogenetic Relationship of PI12 Protease The phylo-genetic tree was constructed to demonstrate the similaritiesand relationship of protein sequence between differentspecies or organisms Figure 6 illustrates the phylogeneticdendrogram indicating the relationship of PI12 proteasewith another 25 translated amino acid sequences ofserine protease The bar scale signifies the branch lengthto approximate variance along each branch Basicallythis tree comprises two main clusters cluster I was sub-divided into subcluster a (CAG25549 EPQ55564 ETW-80907 XP 007381302 XP 001877576 CAG38357 XP007341385 CCA68627 EJU04161 XP 007369624 EIW-62675 CCO37249 XP 001830835 XP 007342189) and sub-cluster b (XP 007589863 XP 003711175 EFQ32840 XP006694395 ENH74885 EON99722) cluster II was sub-divided into subcluster c (CBQ73425 GAC97617 CCF52619)and subcluster d [CAQ76821 (PI12 protease) EMS20793]
Serine protease constructed by the phylogenetic treecomprised large size of protein with about 801 to 1032 aminoacids The majority of the translated protease sequenceswere obtained from fungus All serine proteases from cluster1 subcluster a are derived from Agaricomycetes except forEJU04161 which belongs to Dacrymycetes whereby all serineproteases from cluster 1 subcluster b are from Sordari-omycetes Cluster II subcluster c consisted of serine proteasefrom fungus belonging to Ustilaginomycetes PI12 proteasewas categorized under cluster II subcluster d It possessedhomology similarity with the subtilisin-like protease fromRhodosporidium toruloides (EMS20793) (formerly known asRhodotorula glutinis or Rhodotorula gracilis) which is a
Enzyme Research 13
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
MKG-YSWSLLQLAFTGAALATVPLSDVKRISSAS--VVPNKFVVEVESASEIPGKR--SL
MKGVLVWSILS---AGCALAVTPLSSVKRVTNLP--IVANKFIVEVDSTSDIPTKR--S-
MKLPLLSLALT---AVSVGATVPLSSVQRTTTSPNNALAGEYIVEMNNVQGLGGKR--AY
---MAHGSLASLLLLLAASAQGAAAFGGKSATGGRSVVPNSYILQVNASAPALTKRGVSV
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease ---MLFLPVLLLLLPGVTAFLNPVTNRATNAISSTQYLSNAYILELDLSTPGLVKRDSTP
lowastlowast
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
YTPHELFYRSLSARGVSYEVNKEYNS-EGLFVGAVVTVSDANDVKALAETPGVKAIRPVR
-NPHDALYESLRKRAIGFKVDREFNT-QGLFVGAALTLSDAQDVGQIQSTTGVKSIRPVR
ASTHDTLYRSLRKRGISFDVKKEYNQ-PGIMVGAALKVASPQDVQQIATMAEVKAIRPVI
FEALDHTLAAVKQNGVKYTVRQRFDAIPEAFQAVSIQVEDGASMAELAQIPGVQRVWPVS
DSILEDVLTSVGRNGIKYQLRHRFIS-PTLFHGASITVPPGISRSQIASLRGIKRVWPVR
lowastlowast
TIKSPKPVKVHVVTGKDDPEIPA-------------------------DTESTHIMTGVN
AFDRPKPVKVQVVKGPDDPALPP-------------------------DSESTHVITGVD
AVPAPERINQFTVSSPNDSRITA-------------------------DSETTHIMTGVD
LIPRPVEPTVSDFTPVSSSSSSTKSKRD-THTLEKRGTNFPPASAYLNDTFYPHVQTGID
KFSRPSAVVDADGGGSGFSGSPIKAALMGVKELGKR------ANAYAGDTFGPHVMTGVN
lowast lowast lowast lowastlowast
KLHAQGIFGAGIKIGIIDTGIDYTHPALGGALGAGHKVIGGYDFVGDAYDGTNTPVPDND
KLHAQGITGAGIKIGILDTGIDYTHPTLGGAFGPGNKVIGGFDLVGDDYDGTNTPVPDPD
KVHAAGITGEGIKIGIIDTGIDYNHPLLGAGFGPGFKVIGGYDFVGDDYTGSNNPAPDDD
VLHNKGILGQGVKIVVVDEGVDYTNPILGGCFGPGCQISFGYDFVGDNYTGTNAPVPDSD
ETHEAGLLGAGIKIGVLDTGVDYLNPILGGCFGPGCHMSFGYDLVGDDYDGDNAPVPDVD
lowast lowast lowast lowastlowastlowast lowast lowast lowastlowast lowast lowastlowast lowast lowast lowast lowast lowastlowastlowast lowast lowast lowast lowast lowastlowast lowast
PLDQCNGHGTHVAGIIGANPDNQFNISGVSYGASLSSYRVFGCSGSVSDDIIIEALLRGV
PLDQCAGHGTHVAGIIGANPDNAFNISGVAFQASLSAYRVFGCVGFVTDDVLVDALLLGF
PLDNCNGHGTHVAGIIGANPGNDFGISGVAYGSSIASYRVFGCSGFVSDDIIIDALLRGY
PFSSCSNHGTHTTGTVGALAN-EYGFSGAAPMATIGHYRVISCSGATTDEILVAALTRAL
PFASCDPHGTHVTGIIGALPN-AFGFTGVAPAATLGHYRVFGCTGFVGEDIILAGLMRGV
lowast lowastlowastlowast lowastlowast lowast lowastlowast lowast lowastlowastlowast lowast lowast lowast
QDGQDILTLSLGGTDGWTE-SSSSVVASRIAASGKIVTIAAGNEGAYGSWYSSSPGNGID
NEGQDILTLSLGGADGWTE-SVSAVVASRIAATGKVVTIAAGNDGASGAWYTSSPGNGID
SDGMDILTLSLGGVEGWTE-SSSSVVASRIADQGRVVTIAAGNDGAYGSWYASSPGTGVD
QDNVNIVSMSLGSSVGWLDDSPVQIMASYLGSKGIHVVASVGNERNEGLFAADQPAASRI
EDNCNVLTLSLGGPGGWVKGTPASILIDQIEAQGILVTVATGNSGAEGMFFSESPASTIN
lowastlowastlowast lowastlowast lowast lowast lowastlowast lowast lowast
VISVASIDNTVDNIQNATVNGVEHGPIPYFKTLPLPFTNA-LPLYVISNDTTIANDACNA
VISVASLDNTVVPLQTAQVHGVTHDPIIYQDTFPLPINDT-RPIFATSTDVTVVDDACNP
VISVASVNNIAIPVQNATVQGVEHAPIPYLEALPLNVTGS-RPIYATSTDTTVADDACNA
GTGVGGVDPTYLAAYYAYLI--KRDPVPYISPTPFNLPNN-YLLYFTSTNTSVTNDACNP
GLSIASTDVTDLIAYNATVS--GQPAIPYLSATPLNVVANSFRVHFTSTDPNNPVDACSP
lowast lowast lowast lowast lowastlowastlowast
LPDNTPDLSGFLVLVRRGSCTFVQKLTNIAAKGGNAVFIYDNGNG-----FAGIDTGKFT
LPDSTPDLSGFVVIVRRGTCTFVQKLGNVAAKGAKVTLIYDNGSG-----FGAIDTGAFV
LPNDTPDLSGYVVVIKRGTCTFVTKLQNAAAKGANTFLIYNNGGS-----FSAISVGDYT
LPSTTPNLANRVVVVQRGTCGFTVKQQNVAAAGGKIVLIYNSKGSGLIPQLDVGSTGLTA
LPAGAPDFANYVTVVQRGTCTFVTKYQNVLNAGGKIVLLYNSEGAGNLPYLTPNGVGIDA
lowastlowast lowast lowastlowast lowast lowast lowast lowast lowast lowast lowast
(a)
Figure 5 Continued
14 Enzyme Research
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
NATLIQAADGVFLAQQVAAGKSVTITFPQTGGSAQITN-PSGGLVSSFTTYGPTNDFYFK
-ASFIQAADGEFLVQQFASGQNVSLSFPQSGGLIQFPD-PAGGLISSFTSYGPSNDFFFK
-AALISETDGTFLVNQFAAKANITISFPQSGSAWNYPDDVSGGLVSDFSSYGPSYDMYFK
VGSLRYEDGLKLLSYYQSSPRGQSISFPNGPLAPYVSNTVSGGVVASYSNFGPTNELFIY
VAGLRRSDGLKLLSYYQNANKRLTLRFPKGKIVAGLTDTITGGLISGYSTFGPTNDLYGQ
lowast lowastlowast lowastlowast lowastlowast
PAFGAPGGNILSTLPVPLGSYGVESGTSMATPYVAGSAALLLSVKGKSAAVALGARTLFQ
PAVAAPGGNILSTLPVNLGLFGIESGTSMATPFVAGSAALLFQAKGTSAAVGRSARTLFE
PAVAAPGGNILSTLPIALGSYGVESGTSMATPFTAGSAALILQAKGK--SVAKSIRNLFQ
PTLAAPGTDILSTV---TGGVALMRGTSMAAPLVAGAYALLLSVGNNLKLSPEEARTILM
PTLSAPGGNILSTFPLSEGGVAVISGTSMSCPFVAGSAAVLMAARASENLTPLEIRSLLT
lowast lowastlowastlowast lowastlowastlowastlowast lowast lowastlowastlowastlowast lowast lowastlowast lowast lowast
STAQRVPSSHTDGDPLQTLSQQGAGLIDVFKALYTETIVWPTELILNDTAHYKEVHTFTV
TTAQRVPSTHTDGDPLQTLTQQGAGLINVFNAIHTTTIVSPGELILNDTAHFKGTQQFTV
TTASAVASSKTDGDPLQTVSQQGAGLIQVDRAIGTTTVVSPGQLTLNDTAHFEGFKTFNI
STSQPSPVAY-GSETLDTVVSQGAGVINMTAAIAAHTVISPAQFNLNDTQYSNNTQTLTL
TTAKLTPVSLLGSTPLVSVIRQGGGLVQVAKALAAKTLISPHELLLNDTANANYVQTIKI
VNTGKTSKQYKLTHVPAGTAQTIT---SGTIFAADGPVPLSTDYATVKLSSSSFTLRPGQ
RNAGSSAKTYTLRHIPAGTAVTVT---PGTIFPADGPVPLSTDFASVSLSTSKFTLAPGK
QNTGSKAVTYKISHVPAGTAASVQ---SGSIFAADGPVPLTADYAQVTFSLNKVTVLPGV
RNNNRFPVSYQLSWVDSTGIVTYNDGATTDIIPSTTPNYVSTSVLRVAFSQRTVTVPGGQ
KNTNSWAMKYTFSSAVAQGLGTFD--ASGDILPTLDPVAVSGAQATVAFNTRILSVAPGA
lowast lowast lowast lowast lowast lowast
SKQVTTHINAP--SGVDASTYPVFSGFIEIATAT--------ESVHVTYLGLAASLKDKQ
TQTVTARFTPP--AAADPSTFPVFSGFIQIESGT--------EQVQVSYLGLKASLKDKQ
SVPILVWFKPP--QGVDASTYPVYSGFIEIAGLG--------ETLRVAYLGVAASLKDKA
TVKVTAQFIPPNLTAQQRNQFPIYSGFVTITGQGQGAGAEQMEKYNLPFFGLGARMVDMP
TGSVVATITPPVLPVADAARFPIFSGWIRVNGQG-ARDSSRNEAYTVPYFGLAAKMIDMQ
lowast lowast lowastlowast lowast lowast lowast lowast
IVDNTNEFFGITIPAVLNNTGDAQ-LSPTNYTFSGSDFPTLLLRLAFGTPLLRVDLVDAT
VIDNTDFFFGVPTPVLTDPNGEVQ-TSPRNYSFLASDFPTLIFRLAFGSPKVVVDLVSPT
VVDNTDEYFGVKIPALLDASGDVQ-GNSTNYTLVGDDVPSLLYRFAFGSAHVRVDLVEKA
VLDTTNVALGP--NLPFIAVGQN--IQTGPTTFSESNPAVVYFRLAAGTRRLRVDLVDAS
VLDTTETIYGPGYAYPFVIDDAIGDIQSTTTSYSRNLGPTVFARFATGTLHYSLDLVLAD
lowast lowast lowast lowast lowastlowastlowastlowast lowast
IDLKPTLNSREIDDKRHVFERESFSFPHPGPKGS-------------------TFAQVKI
INFKPTLNTRALGDTHGSF----FSFPHRVKTG--------------------SFAQVPT
STVSATLSKRNWWN---------WWWPQKPSN---------------------SYDKVPI
TSYNATVPAVTNPASRLVKRSNTELTPRAAFPT--------------------LYSQVRT
IAFTPTYPNSS-PATRFVKRSLTQHTSAASHLAKRRVSIATINPKATLVADRQLHSDVPI
lowast lowast
VGTLSEQSHIPRNSDASTVDDGGYSTFAFNKP------SFANGTTIPNGTYRWLLRALKV
LGTVFELDFTSRNNDDPTDN--GFNTVAIDPP------TFANGTTIPNGQYRLLLRALRV
VGTLAEYDYVPRNSDASTIDDNGYNVFQFT-D------SFVNGTKIAAGEYKVLLRALKV
VGSLFAPSYWPPRDYLFNNGGT-YSDYEIQLDGSFTFPNGTKGMAELGRSYKVLLRALKI
EGNIFTQPFTG-RDYLVDAAPTGSTDRTVTFNGQY-AENGLVRTAVTGTSYRFLLRALKI
lowast lowast lowastlowast lowast lowast lowast lowast lowastlowastlowastlowast
lowast lowast lowastlowastlowastlowastlowast
(b)
Figure 5 Continued
Enzyme Research 15
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
TGDPTKEADFETWLSPIVGITVA
TGDPTNEADFESFLSPIIGVNAP
TGNPAEEGDYETWLSPSFSVVPA
TADPTYTSSYESWLSPSFTFTA-
SGDAMYEDQYESWLSLPFSFRA-
lowast lowastlowast
ConservedSimilar
lowast All match
(c)
Figure 5 Multiple sequence alignment between amino acid sequence of PI12 protease with other subtilisin The serine protease sequenceswere obtained from the GenBank database Their accession numbers are EMS20793 (subtilisin-like protease of Rhodosporidium toruloidesNP11) CAG25549 (subtilisin-like protease of Pleurotus ostreatus) EPQ55564 (subtilisin-like protease of Gloeophyllum trabeumATCC 11539)and ETW80907 (serine protease S8 ofHeterobasidion irregulare TC 32-1) Putative ORF of serine protease fromG antarctica PI12 was labeledas PI12 protease
CAG38357 Phanerochaete chrysosporium
EIW62675 Trametes versicolor FP-101664 SS1
XP 007369624 Dichomitus squalens LYAD-421 SS1
XP 007381302 Punctularia strigosozonata HHB-11173 SS5
CAG25549 Pleurotus ostreatus
ETW80907 Heterobasidion irregulare TC 32-1
XP 001877576 Laccaria bicolor S238N-H82
EPQ55564 Gloeophyllum trabeum ATCC 11539
XP 001830835 Coprinopsis cinerea okayama7130
XP 007341385 Auricularia delicata TFB-10046 SS5
CCA68627 Piriformospora indica DSM 11827
CCO37249 Rhizoctonia solani AG-1 IB
XP 007342189 Auricularia delicata TFB-10046 SS5
EJU04161 Dacryopinax sp DJM-731 SS1
XP 007589863 Colletotrichum fioriniae PJ7
EFQ32840 Colletotrichum graminicola M1001
ENH74885 Fusarium oxysporum f sp cubense race 1
XP 003711175 Magnaporthe oryzae 70-15
XP 006694395 Chaetomium thermophilum DSM 1495
EON99722 Togninia minima UCRPA7
CBQ73425 Sporisorium reilianum SRZ2
GAC97617 Pseudozyma hubeiensis SY62
CCF52619 Ustilago hordei
EMS20793 Rhodosporidium toruloides NP11
CAQ76821 Glaciozyma antarctica PI12
01
Clus
ter 1
Clus
ter 2
a
b
c
d
Figure 6 Phylogenetic dendrogram among 25 translated amino acid sequences of serine proteaseThe tree was constructed using a neighbor-joining algorithm with 1000 bootstrappings by MEGA 6 software [39]The scale bar represents 01 substitutions per amino acid positionTheserine protease sequences were obtained from the GenBank database GenBank accession numbers of the sequences were indicated beforethe strain name The putative serine protease of G antarctica strain PI12 has been deposited in GenBank with accession number CAQ76821
16 Enzyme Research
0
2
4
6
8
10
12
0 24 48 72 96 110
Prot
ease
activ
ity (U
mL)
Post induction time (hours)
Figure 7 Effect of postinduction time towards protease activity The highest activity was detected at 72 hours after 110 hours of incubationwith 9UmL activity followed by 48 24 and 96 hoursThe substrate (azocasein) was dissolved in Tris HCL buffer and incubated for 30min at20∘CThe absorbance was measured at 450 nm after 15min incubation A one-way ANOVAwas used to test for preference differences amongsix different postinduction times The statistical value (119875 value lt005) between hours is 0003
pink-coloured nonpathogenic oleaginous yeast (42 iden-tity) Both G antarctica PI12 and Rhodosporidium toruloidesNP11 belong to Microbotryomycetes class PI12 protease didnot show any homology similarity to other psychrophilicproteases however it showed 37 identity to hypotheticalprotein from Pseudozyma antarctica T-34 which was isolatedfrom Japan (GAC76005) (not listed in the phylogenetic tree)
35 Cloning and Expression of Cold-Adapted PI12 Protease inthe Yeast Expression System The PCR product harbouringthe mature PI12 protease with two flanking restriction sites(EcoRI and AvrII) was cloned into pPIC9 expression vectorThe recombinant plasmid harbouring PI12 protease was usedto transform both GS115 and KM71 intended to produce bothHis+ Mut+ and His+ MutS Two recombinant clones fromGS115 (His+Mut+) strain (GpPro1 andGpPro2) and one fromKM71 (His+MutS) strain (KpPro1) were successfully obtainedwhich were further analyzed for protein expression study
36 Expression and Enzyme Assay of PI12 Protease in Pichiapastoris All positive recombinants were grown in small scalecultures and assayed for the cold-adapted protease activityin the culture supernatants The clones were first grown inBMGY medium with glycerol as the sole carbon sourceFor induction of AOX1 promoter and protein productionthe growth medium was transferred to the same mediumexcept with 05 methanol in place with glycerol as thecarbon source [26] The supernatant of the recombinant Ppastoris expression cultures after 48 h of incubation time wasanalyzed to determine the recombinant protease activity bythe modified method of Brock et al (1982) [20] All testedclones secreted protease into the supernatant in differentexpression levels This might be due to the variability ofthe strength of the promoter and copy number in each cellto secrete and express the recombinant product The clonewith the highest yield was GpPro2 with 63UmL activityfollowed by KpPro1 and GpPro1 with 25UmL and 37UmLactivities respectively (data not shown) It is perceptible thatexpression systemof Pichia pastoris is the ideal system for thisprotein The preliminary assay has fruitfully produced high
protease expression at 20∘C Yet further optimization of therecombinant PI12 protease production need to be conductedin order to examine the optimum condition of this proteinReported cold-adapted proteases showed optimum activity ata range of 15∘Cndash60∘C [27ndash30] This difference was possiblyrelated to the respective genetic and physiological adaptationof the strains There are several factors that influenced thedifferent rate of expression It includes different site of inte-gration between interest gene and Pichia genome variabilityof different expression host cell to secrete and express therecombinant enzyme and different sites in the polylinker forinserting foreign genes [31]
This is the second extracellular protease reported to besynthesized by this obligate psychrophilic yeastG antarcticaThe first cold-adapted extracellular subtilase from this yeasthas been purified and characterized previously [32 33] Theextracellular serine proteinase (LAP2) from this Antarcticyeast formerly known as Leucosporidium antarcticum strain171 is specific towards synthetic substrates of chymotrypsinand subtilisin The subtilase of L antarcticum strain 171showed specific activity of 4043Umg when utilized N-SucAAPFpNA as a substrate [33]
37 Optimization of Protease Production GpPro2 was chosenfor further optimization due to its highest protease activityat the preliminary expression stage High protease activitywas detected after 72 hours of incubation (Figure 7) byusing the standard protocol method as recommended forPichia pastoris expression system (Pichia Expression KitInvitrogen USA) The expression decreased after 72 hoursand no activity was detected at 110 hours The prolongedincubation may result in the decrease of biomass productionand will subsequently reduce the expression of recombinantprotein In addition oxygen and nutrient depletion duringprolonged incubationwill trigger the stress responses in orderto survive in such harsh environment [34]
Methanol was used as a sole carbon by P pastoris Themethanol was oxidized to formaldehyde with the aid of oxy-gen as electron acceptor by an alcohol oxidase enzyme (AOXgene) [35] The optimal methanol concentration is at 05(vv) with 14UmL activity The addition of more than 05
Enzyme Research 17
02468
10121416
0 01 03 05 07 09Pr
otea
se ac
tivity
(Um
L)Methanol concentration ()
Figure 8 Effect ofmethanol concentration towards protease activityThe recombinant was cultivated for 72 hours at 30∘C in BMMYmedium05 methanol gives the highest activity with 14UmL The substrate (azocasein) was dissolved in Tris HCL buffer and incubated for 30minat 20∘CThe mixture was left for 15min and absorbance was taken at 450 nm A one-way ANOVA was used to test for preference differencesamong six different concentrations The statistical value (119875 value lt005) between concentration is 002
0
5
10
15
20
25
10 15 20 25 30 35
Prot
eas a
ctiv
ity (
Um
L)
Temperature (∘C)
Figure 9 Effect of temperature towards protease activity The reaction was carried out in temperature ranging from 10∘C to 35∘C at pH 7 for30min A one-way ANOVA was used to test for preference differences among six different temperatures The statistical value (119875 value lt005)between temperature is 0067
(vv) methanol resulted in tremendous decrease of proteaseactivity (Figure 8) Production of PI12 protease was decreasedwhen the methanol concentration increased This might bedue to the toxic effect of methanol concentration on DNAreplication and membrane synthesis Basically methanol isused to increase the permeability of the cell membrane tohelp the secretion of foreign protein [26] However if thepermeability of cell membrane is too high lowmolecular sizemetabolites would leak from the cell and such leakage willseriously damage cell activities Such condition would abusethe cellular system and might reduce the yield of proteinexpression [36]
Psychrophilic microorganism dominated in low temper-ature habitat which most other microorganism are inca-pable of withstanding due to their inability to produceenzyme that can tolerate the extreme environment con-dition Psychrophilic enzymes are always associated withhigh catalytic efficiency at low and moderate temperature(0∘C to 30∘C) compared to the mesophilic counterpart thatare generally unable to catalyze efficiently under extremely
low temperature [2] The recombinant protein assay wasdone at various temperatures ranging from 10∘C to 35∘Cusing 05 azocasein as a substrate dissolved in 50mMTris HCl in presence of 2mM CaCl
2 pH 7 In the activity
assays expressed protease showed a highest activity at 20∘C(20UmL) and decreased tremendously once it exceededmaximum temperature due to the denatured state of theenzyme (Figure 9) The active site of an enzyme is easilyaltered when the high temperature is introduced duringincubation or activity assay resulting in a low reaction rate[37]
The recombinant construct was transferred into BMMYmedia at different A
600 nm ranging from 1 to 10 The high-est activity of active enzyme was detected at A
600 nm 60(283UmL) and above (Figure 10) The slope during expo-nential phase was much higher compared to other phasesindicating an increase in the doubling time At exponentialphase the cells are adapted to the growth condition andother processes like cell growth cycle and synthesis of RNAThe yield of active enzyme during expression of P pastoris
18 Enzyme Research
05
1015202530
1 2 3 4 5 6 7 8 9 10Prot
ease
activ
ity (U
mL)
Initial inoculum A600
Figure 10 Effect of inoculums size towards protease activity The recombinant was cultivated for 72 hours at 30∘C in BMMY mediumwith 05 methanol concentration A one-way ANOVA was used to test for preference differences among six different optical densitiesThe statistical value (119875 value lt005) between optical density value is 003
(kD
a)
1160
662
450
350
250
184
144
M 1 2 3
(a) (b) (c)
Figure 11 (a) Coomassie-stained SDS-PAGE of recombinant P pastoris clone 1mL supernatants collected from the third-day culturewere concentrated 10-fold using trichloroacetic acid and 10120583L resultant products were applied to SDS-PAGE analysis Lane 1 molecularweight marker (Fermentas) Lane 2 GS115 (Control) Lane 3 GS115pPIC9 empty (Control) Lane 4 GS115pPIC9PI12prot (GpPro2)The arrow indicates the secreted recombinant PI12 protease (b) Plate activity staining of different recombinant P pastoris clones GGS115 GP GS115pPIC9 GRI GS115pPIC9PI12prot1 (GpPro1) GRII GS115pPIC9PI12prot2 (GpPro2) K KM71 KP KM71pPIC9 KRKM71pPIC9PI12prot (KpPro) (c)Gel activity staining of recombinant PI12 protease inP pastorisGpPro2 clone Lane 1 prestainedmolecularweightmarker (Fermentas) Lane 2 GS115pPIC9 empty (Control) Lane 3 GS115pPIC9PI12prot1 (GpPro2)The arrow indicates the secretedrecombinant PI12 protease
dropped after 28 hours of incubation which is at stationaryor death phase [38]
38 Detection of Recombinant Protein by SDS-PAGE andActivity Staining TheSDS-PAGE gel of the GpPro2 recombi-nant clonesrsquo supernatant showed a putative expression bandat around 993 kDa Plate staining and gel activity stainingwere executed to examine if the recombinant PI12 proteaseexpressed in P pastoris was functionally active In the platestaining the transformants showed strong proteolytic activity(Figure 11(a)) In gel staining proteolytic activity was alsodetected around 993 kDa after incubation at 20∘C This is
evidence of protease secretion into the surrounding mediawhereby it catalyzed the breakdown of casein thus formedclearing zones around the activity ring and band (Figures11(b) and 11(c)) As a conclusion the cold-adapted PI12protease was functionally expressed in P pastoris GS115(GpPro2 clone) with the size of 993 kDa
4 Conclusions
Psychrophilic yeast which was originated frommarine waterin Casey Station Antarctica was identified as Glaciozymaantarctica strain PI12 A serine protease with an open reading
Enzyme Research 19
frame of 2892 bp encoded for 963 amino acids from thisAntarctic yeast has been cloned and expressed in Pichiaexpression system This is the second extracellular pro-tease reported being synthesized by this psychrophilic yeastSequence analysis through bioinformatics studies revealedfew homology identities of this protein with other ser-ine proteases The mature PI12 protease was successfullyexpressed and secreted into the culture medium driven bythe Saccharomyces cerevisiae 120572-factor signal sequence in Ppastoris The protease production in P pastoris was bestobtained from strain GS115 (GpPro2) with 283UmL after 72hours of induction time with 05 (vv) of methanol inducerThe starting optical density at 600 nm (OD600) was 60 andthe optimum assay temperature was 20∘C The expressedprotein was detected by SDS-PAGE and activity staining witha molecular weight of 99 kDaThis cold-active enzymemightoffer an opportunity for biotechnological exploitation basedon its high catalytic activity at low temperatures and lowthermostability Structural and functional study of this cold-adapted enzyme would give insight into its adaptation to anextreme cold environment
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgment
This research was supported financially by the InitiativeBiotechnology-GMBI-MOSTI (Project number 07-05-16-MG-GMB02) andRUGS-Initiative 6 (Project number 05-02-11-1414RU)
References
[1] J W Deming ldquoPsychrophiles and polar regionsrdquo CurrentOpinion in Microbiology vol 5 no 3 pp 301ndash309 2002
[2] C Gerday M Aittaleb M Bentahir et al ldquoCold-adaptedenzymes from fundamentals to biotechnologyrdquo Trends inBiotechnology vol 18 no 3 pp 103ndash107 2000
[3] A Svingor J Kardos I Hajdu A Nemeth and P Zavodszky ldquoAbetter enzyme to cope with cold comparative flexibility studieson psychrotrophic mesophilic and thermophilic IPMDHSrdquoThe Journal of Biological Chemistry vol 276 no 30 pp 28121ndash28125 2001
[4] G Antranikian C E Vorgias and C Bertoldo ldquoExtremeenvironments as a resource for microorganisms and novel bio-catalystsrdquo Advances in Biochemical EngineeringBiotechnologyvol 96 pp 219ndash262 2005
[5] T Lonhienne C Gerday andG Feller ldquoPsychrophilic enzymesrevisiting the thermodynamic parameters of activation mayexplain local flexibilityrdquoBiochimica et Biophysica Acta vol 1543no 1 pp 1ndash10 2000
[6] R Cavicchioli and TThomas ldquoExtremophilesrdquo in Encyclopediaof Microbiology pp 317ndash337 Academic Press San Diego CalifUSA 2000
[7] G Feller E Narinx J L Arpigny et al ldquoEnzymes frompsychrophilic organismsrdquo FEMS Microbiology Reviews vol 18no 2-3 pp 189ndash202 1996
[8] J Arnorsdottir R B Smaradottir T O Magnusson S HThorbjarnardottir G Eggertsson and M M KristjanssonldquoCharacterization of a cloned subtilisin-like serine proteinasefrom a psychrotrophic Vibrio speciesrdquo European Journal ofBiochemistry vol 269 no 22 pp 5536ndash5546 2002
[9] N J Russell ldquoPsychrophilic bacteriamdashmolecular adaptations ofmembrane lipidsrdquo Comparative Biochemistry and Physiology APhysiology vol 118 no 3 pp 489ndash493 1997
[10] G Feller and C Gerday ldquoPsychrophilic enzymes hot topics incold adaptationrdquo Nature Reviews Microbiology vol 1 no 3 pp200ndash208 2003
[11] P Raspor and J Zupan ldquoYeasts in extreme environmentsrdquo inBiodiversity and Ecophysiology of Yeasts The Yeast Handbookpp 371ndash417 Springer Berlin Germany 2006
[12] M B Rao A M Tanksale M S Ghatge and V V Desh-pande ldquoMolecular and biotechnological aspects of microbialproteasesrdquoMicrobiology andMolecular Biology Reviews vol 62no 3 pp 597ndash635 1998
[13] L Polgar ldquoThe catalytic triad of serine peptidasesrdquo Cellular andMolecular Life Sciences vol 62 no 19-20 pp 2161ndash2172 2005
[14] S Y Boo C M V L Wong K F Rodrigues N NajimudinA M A Murad and N M Mahadi ldquoThermal stress responsesin Antarctic yeastGlaciozyma antarctica PI12 characterized byreal-time quantitative PCRrdquoPolar Biology vol 36 no 3 pp 381ndash389 2013
[15] B Turchetti S R T Hall L B Connell et al ldquoPsychrophilicyeasts from Antarctica and European glaciers descriptionof Glaciozyma gen nov Glaciozyma martinii sp nov andGlaciozyma watsonii sp novrdquo Extremophiles vol 15 no 5 pp573ndash586 2011
[16] N H F Hashim I Bharudin D L S Nguong et al ldquoCharac-terization of Afp1 an antifreeze protein from the psychrophilicyeast Glaciozyma antarctica PI12rdquo Extremophiles vol 17 no 1pp 63ndash73 2013
[17] A N M Ramli M A Azhar M S Shamsir et al ldquoSequenceand structural investigation of a novel psychrophilic 120572-amylasefrom Glaciozyma antarctica PI12 for cold-adaptation analysisrdquoJournal ofMolecularModeling vol 19 no 8 pp 3369ndash3383 2013
[18] A N M Ramli N M Mahadi A Rabu A M A Murad F DA Bakar and R M Illias ldquoMolecular cloning expression andbiochemical characterisation of a cold-adapted novel recombi-nant chitinase from Glaciozyma antarctica PI12rdquoMicrobial CellFactories vol 10 no 1 article 94 2011
[19] J Sambrook E Fritsch and T Maniatis Molecular Cloning ALaboratoryManual Cold SpringHarbor Laboratory Press NewYork NY USA 1989
[20] F M Brock C W Forsberg and J G Buchanan-SmithldquoProteolytic activity of rumen microorganisms and effects ofproteinase inhibitorsrdquoApplied and EnvironmentalMicrobiologyvol 44 no 3 pp 561ndash569 1982
[21] U K Laemmli ldquoCleavage of structural proteins during theassembly of the head of bacteriophage T4rdquo Nature vol 227 no5259 pp 680ndash685 1970
[22] A Henne R A Schmitz M Bomeke G Gottschalk and RDaniel ldquoScreening of environmental DNA libraries for thepresence of genes conferring lipolytic activity on Escherichiacolirdquo Applied and Environmental Microbiology vol 66 no 7 pp3113ndash3116 2000
[23] M Poza T de Miguel C Sieiro and T G Villa ldquoCharacteri-zation of a broad pH range protease of Candida caseinolyticardquoJournal of Applied Microbiology vol 91 no 5 pp 916ndash921 2001
20 Enzyme Research
[24] J D Bendtsen H Nielsen G von Heijne and S BrunakldquoImproved prediction of signal peptides SignalP 30rdquo Journalof Molecular Biology vol 340 no 4 pp 783ndash795 2004
[25] R J Siezen and J A M Leunissen ldquoSubtilases the superfamilyof subtilisin-like serine proteasesrdquo Protein Science vol 6 no 3pp 501ndash523 1997
[26] Y Katakura W Zhang G Zhuang et al ldquoEffect of methanolconcentration on the production of human 1205732- glycoprotein Idomain V by a recombinant Pichia pastoris a simple system forthe control of methanol concentration using a semiconductorgas sensorrdquo Journal of Fermentation and Bioengineering vol 86no 5 pp 482ndash487 1998
[27] Q-FWang Y-HHou Z Xu J-LMiao andG-Y Li ldquoPurifica-tion and properties of an extracellular cold-active protease fromthe psychrophilic bacterium Pseudoalteromonas sp NJ276rdquoBiochemical Engineering Journal vol 38 no 3 pp 362ndash3682008
[28] S Vazquez L Ruberto and W Mac Cormack ldquoProper-ties of extracellular proteases from three psychrotolerantStenotrophomonas maltophilia isolated from Antarctic soilrdquoPolar Biology vol 28 no 4 pp 319ndash325 2005
[29] R Zeng R Zhang J Zhao and N Lin ldquoCold-active ser-ine alkaline protease from the psychrophilic bacterium Pseu-domonas strain DY-A enzyme purification and characteriza-tionrdquo Extremophiles vol 7 no 4 pp 335ndash337 2003
[30] M Turkiewicz E Gromek H Kalinowska and M ZielinskaldquoBiosynthesis and properties of an extracellularmetalloproteasefrom the Antarctic marine bacterium Sphingomonas paucimo-bilisrdquo Journal of Biotechnology vol 70 no 1ndash3 pp 53ndash60 1999
[31] A Vassileva D A Chugh S Swaminathan and N KhannaldquoEffect of copy number on the expression levels of hepatitisB surface antigen in the methylotrophic yeast Pichia pastorisrdquoProtein Expression andPurification vol 21 no 1 pp 71ndash80 2001
[32] M Pazgier M Turkiewicz H Kalinowska and S Bielecki ldquoTheunique cold-adapted extracellular subtilase from psychrophilicyeast Leucosporidium antarcticumrdquo Journal ofMolecular Catal-ysis B Enzymatic vol 21 no 1-2 pp 39ndash42 2003
[33] M Turkiewicz M Pazgier H Kalinowska and S BieleckildquoA cold-adapted extracellular serine proteinase of the yeastLeucosporidium antarcticumrdquo Extremophiles vol 7 no 6 pp435ndash442 2003
[34] D Mattanovich B Gasser H Hohenblum and M SauerldquoStress in recombinant protein producing yeastsrdquo Journal ofBiotechnology vol 113 no 1ndash3 pp 121ndash135 2004
[35] P Secades BAlvarez and JAGuijarro ldquoPurification andprop-erties of a new psychrophilic metalloprotease (Fpp2) in the fishpathogen Flavobacterium psychrophilumrdquo FEMS MicrobiologyLetters vol 226 no 2 pp 273ndash279 2003
[36] L Y Ling I Ithoi and F M Yik ldquoOptimization for high-level expression in Pichia pastoris and purification of truncatedand full length recombinant sag2 of toxoplasma gondii fordiagnostic userdquo Southeast Asian Journal of Tropical Medicineand Public Health vol 41 no 3 pp 507ndash513 2010
[37] A Illanes ldquoStability of biocatalystsrdquo Electronic Journal ofBiotechnology vol 2 no 1 pp 1ndash9 1999
[38] B A Plantz J Sinha L Villarete K W Nickerson and VL Schlegel ldquoPichia pastoris fermentation optimization energystate and testing a growth-associated modelrdquo Applied Microbi-ology and Biotechnology vol 72 no 2 pp 297ndash305 2006
[39] K Tamura G Stecher D Peterson A Filipski and S KumarldquoMEGA6molecular evolutionary genetics analysis version 60rdquo
Molecular Biology and Evolution vol 30 no 12 pp 2725ndash27292013
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

12 Enzyme Research
cccgccactcgtttcgtcaagcgctccctcacgcagcacacctctgccgcttcgcacctc 3540
P A T R F V K R S L T Q H T S A A S H Lgccaagcgccgcgtctccatcgccactatcaaccccaaagccaccctcgtcgccgatcga 3600
A K R R V S I A T I N P K A T L V A D Rcagctccactcggacgtccctatcgagggcaacatcttcacccaaccctttactggaagg 3660
Q L H S D V P I E G N I F T Q P F T G Rgattacctcgtcgacgcagccccgacgggatccaccgatcgtaccgtcacttttaacggg 3720
D Y L V D A A P T G S T D R T V T F N Gcagtacgccgagaacggcctcgtgaggacggctgtgacggggacttcgtaccgcttcctc 3780
Q Y A E N G L V R T A V T G T S Y R F Lcttcgggcgttgaagatctcgggagacgcgatgtacgaggatcagtatgagagctggctc 3840
L R A L K I S G D A M Y E D Q Y E S W L3900
S L P F S F R Attttgccacctcaaaaaaaaaaaaaaaaaaaaaaaaaa 3938
lowast
tcgctaccgttctcgttccgtgcgtaggcttgttgtggacaccactgcattaagttatga
(c)
Figure 4 Nucleotide and deduced amino acid sequences of genomic DNA and cDNA encoding protease from G antarctica strain PI12 Thenucleotides are numbered from the 51015840 end of the cDNA A putative CAAT TATA box and polyadenylation signal are bold italicThe putativeN-terminal signal peptide is underlined The initiation codon is boldfaced and termination codon is represented by an asterisk (lowast) boxedpeptide sequences showed N-glycosylated asparagines residues The amino acid residues that form the catalytic triad are double underlinedand bold while introns are represented in blue font Highlighted in grey are the intron donor-acceptor splice sites
proteases Hydrophobic profile of PI12 amino acid proteasewas performed by ProtScale tools with Kyte and Doolittlemethod at httpauexpasyorgcgi-bin The hydrophobicityprofile elucidation of PI12 protease amino acid was carriedout from amino acid 1 to 963 The amino acid residuesfrom 1 to 16 (M-L-F-L-P-V-L-L-L-L-L-P-G-V-T-A) and 548 to562 (V-A-V-I-S) were hydrophobic grooves The first regionwhich consisted of 16 amino acids was very hydrophobic andwas predicted to be the signal peptide by SignalP and ProPwebserver
The bold and double-underlined amino acids showedresidues of catalytic triad aspartic acid (D) histidine (H) andserine (S) active site respectively NCBI Conserved DomainsDatabase (CDD) identified the presence of subtilase proteaseand subtilisin-like serine protease domains in the amino acidsequence The predicted PI12 protease amino acid sequencewas found sharing significant homology to the subtilisinfamily of protease The highest similarity of PI12 proteasewas observed with 42 identity to subtilisin-like proteaseof Rhodosporidium toruloides (EMS20793) 37 identity tosubtilisin-like protease of Pleurotus ostreatus (CAG25549)and 35 identity to subtilisin-like protease of Gloeophyllumtrabeum ATCC 11539 (EPQ55564) The nucleotide sequenceof G antarctica strain PI12 showed low homology to thefungus subtilisin protease family and none to other psy-chrophilic proteases The multiple sequence alignment wasillustrated in Figure 5 The consensus sequence around thecatalytic triad aspartic acid (D) histidine (H) and serine(S) of PI12 protease showed high conservation with othersubtilisin (showed in black boxes) Glycine residues G142G173 G191 G262 and G327 that were highly conserved inmost of subtilases were conserved in PI12 protease [25]Therefore PI12 protease belongs to the subtilisin subgroup ofthe subtilase serine protease superfamilyThis serine protease
of G antarctica PI12 has been deposited into GenBank withaccession number CAQ76821
34 Phylogenetic Relationship of PI12 Protease The phylo-genetic tree was constructed to demonstrate the similaritiesand relationship of protein sequence between differentspecies or organisms Figure 6 illustrates the phylogeneticdendrogram indicating the relationship of PI12 proteasewith another 25 translated amino acid sequences ofserine protease The bar scale signifies the branch lengthto approximate variance along each branch Basicallythis tree comprises two main clusters cluster I was sub-divided into subcluster a (CAG25549 EPQ55564 ETW-80907 XP 007381302 XP 001877576 CAG38357 XP007341385 CCA68627 EJU04161 XP 007369624 EIW-62675 CCO37249 XP 001830835 XP 007342189) and sub-cluster b (XP 007589863 XP 003711175 EFQ32840 XP006694395 ENH74885 EON99722) cluster II was sub-divided into subcluster c (CBQ73425 GAC97617 CCF52619)and subcluster d [CAQ76821 (PI12 protease) EMS20793]
Serine protease constructed by the phylogenetic treecomprised large size of protein with about 801 to 1032 aminoacids The majority of the translated protease sequenceswere obtained from fungus All serine proteases from cluster1 subcluster a are derived from Agaricomycetes except forEJU04161 which belongs to Dacrymycetes whereby all serineproteases from cluster 1 subcluster b are from Sordari-omycetes Cluster II subcluster c consisted of serine proteasefrom fungus belonging to Ustilaginomycetes PI12 proteasewas categorized under cluster II subcluster d It possessedhomology similarity with the subtilisin-like protease fromRhodosporidium toruloides (EMS20793) (formerly known asRhodotorula glutinis or Rhodotorula gracilis) which is a
Enzyme Research 13
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
MKG-YSWSLLQLAFTGAALATVPLSDVKRISSAS--VVPNKFVVEVESASEIPGKR--SL
MKGVLVWSILS---AGCALAVTPLSSVKRVTNLP--IVANKFIVEVDSTSDIPTKR--S-
MKLPLLSLALT---AVSVGATVPLSSVQRTTTSPNNALAGEYIVEMNNVQGLGGKR--AY
---MAHGSLASLLLLLAASAQGAAAFGGKSATGGRSVVPNSYILQVNASAPALTKRGVSV
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease ---MLFLPVLLLLLPGVTAFLNPVTNRATNAISSTQYLSNAYILELDLSTPGLVKRDSTP
lowastlowast
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
YTPHELFYRSLSARGVSYEVNKEYNS-EGLFVGAVVTVSDANDVKALAETPGVKAIRPVR
-NPHDALYESLRKRAIGFKVDREFNT-QGLFVGAALTLSDAQDVGQIQSTTGVKSIRPVR
ASTHDTLYRSLRKRGISFDVKKEYNQ-PGIMVGAALKVASPQDVQQIATMAEVKAIRPVI
FEALDHTLAAVKQNGVKYTVRQRFDAIPEAFQAVSIQVEDGASMAELAQIPGVQRVWPVS
DSILEDVLTSVGRNGIKYQLRHRFIS-PTLFHGASITVPPGISRSQIASLRGIKRVWPVR
lowastlowast
TIKSPKPVKVHVVTGKDDPEIPA-------------------------DTESTHIMTGVN
AFDRPKPVKVQVVKGPDDPALPP-------------------------DSESTHVITGVD
AVPAPERINQFTVSSPNDSRITA-------------------------DSETTHIMTGVD
LIPRPVEPTVSDFTPVSSSSSSTKSKRD-THTLEKRGTNFPPASAYLNDTFYPHVQTGID
KFSRPSAVVDADGGGSGFSGSPIKAALMGVKELGKR------ANAYAGDTFGPHVMTGVN
lowast lowast lowast lowastlowast
KLHAQGIFGAGIKIGIIDTGIDYTHPALGGALGAGHKVIGGYDFVGDAYDGTNTPVPDND
KLHAQGITGAGIKIGILDTGIDYTHPTLGGAFGPGNKVIGGFDLVGDDYDGTNTPVPDPD
KVHAAGITGEGIKIGIIDTGIDYNHPLLGAGFGPGFKVIGGYDFVGDDYTGSNNPAPDDD
VLHNKGILGQGVKIVVVDEGVDYTNPILGGCFGPGCQISFGYDFVGDNYTGTNAPVPDSD
ETHEAGLLGAGIKIGVLDTGVDYLNPILGGCFGPGCHMSFGYDLVGDDYDGDNAPVPDVD
lowast lowast lowast lowastlowastlowast lowast lowast lowastlowast lowast lowastlowast lowast lowast lowast lowast lowastlowastlowast lowast lowast lowast lowast lowastlowast lowast
PLDQCNGHGTHVAGIIGANPDNQFNISGVSYGASLSSYRVFGCSGSVSDDIIIEALLRGV
PLDQCAGHGTHVAGIIGANPDNAFNISGVAFQASLSAYRVFGCVGFVTDDVLVDALLLGF
PLDNCNGHGTHVAGIIGANPGNDFGISGVAYGSSIASYRVFGCSGFVSDDIIIDALLRGY
PFSSCSNHGTHTTGTVGALAN-EYGFSGAAPMATIGHYRVISCSGATTDEILVAALTRAL
PFASCDPHGTHVTGIIGALPN-AFGFTGVAPAATLGHYRVFGCTGFVGEDIILAGLMRGV
lowast lowastlowastlowast lowastlowast lowast lowastlowast lowast lowastlowastlowast lowast lowast lowast
QDGQDILTLSLGGTDGWTE-SSSSVVASRIAASGKIVTIAAGNEGAYGSWYSSSPGNGID
NEGQDILTLSLGGADGWTE-SVSAVVASRIAATGKVVTIAAGNDGASGAWYTSSPGNGID
SDGMDILTLSLGGVEGWTE-SSSSVVASRIADQGRVVTIAAGNDGAYGSWYASSPGTGVD
QDNVNIVSMSLGSSVGWLDDSPVQIMASYLGSKGIHVVASVGNERNEGLFAADQPAASRI
EDNCNVLTLSLGGPGGWVKGTPASILIDQIEAQGILVTVATGNSGAEGMFFSESPASTIN
lowastlowastlowast lowastlowast lowast lowast lowastlowast lowast lowast
VISVASIDNTVDNIQNATVNGVEHGPIPYFKTLPLPFTNA-LPLYVISNDTTIANDACNA
VISVASLDNTVVPLQTAQVHGVTHDPIIYQDTFPLPINDT-RPIFATSTDVTVVDDACNP
VISVASVNNIAIPVQNATVQGVEHAPIPYLEALPLNVTGS-RPIYATSTDTTVADDACNA
GTGVGGVDPTYLAAYYAYLI--KRDPVPYISPTPFNLPNN-YLLYFTSTNTSVTNDACNP
GLSIASTDVTDLIAYNATVS--GQPAIPYLSATPLNVVANSFRVHFTSTDPNNPVDACSP
lowast lowast lowast lowast lowastlowastlowast
LPDNTPDLSGFLVLVRRGSCTFVQKLTNIAAKGGNAVFIYDNGNG-----FAGIDTGKFT
LPDSTPDLSGFVVIVRRGTCTFVQKLGNVAAKGAKVTLIYDNGSG-----FGAIDTGAFV
LPNDTPDLSGYVVVIKRGTCTFVTKLQNAAAKGANTFLIYNNGGS-----FSAISVGDYT
LPSTTPNLANRVVVVQRGTCGFTVKQQNVAAAGGKIVLIYNSKGSGLIPQLDVGSTGLTA
LPAGAPDFANYVTVVQRGTCTFVTKYQNVLNAGGKIVLLYNSEGAGNLPYLTPNGVGIDA
lowastlowast lowast lowastlowast lowast lowast lowast lowast lowast lowast lowast
(a)
Figure 5 Continued
14 Enzyme Research
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
NATLIQAADGVFLAQQVAAGKSVTITFPQTGGSAQITN-PSGGLVSSFTTYGPTNDFYFK
-ASFIQAADGEFLVQQFASGQNVSLSFPQSGGLIQFPD-PAGGLISSFTSYGPSNDFFFK
-AALISETDGTFLVNQFAAKANITISFPQSGSAWNYPDDVSGGLVSDFSSYGPSYDMYFK
VGSLRYEDGLKLLSYYQSSPRGQSISFPNGPLAPYVSNTVSGGVVASYSNFGPTNELFIY
VAGLRRSDGLKLLSYYQNANKRLTLRFPKGKIVAGLTDTITGGLISGYSTFGPTNDLYGQ
lowast lowastlowast lowastlowast lowastlowast
PAFGAPGGNILSTLPVPLGSYGVESGTSMATPYVAGSAALLLSVKGKSAAVALGARTLFQ
PAVAAPGGNILSTLPVNLGLFGIESGTSMATPFVAGSAALLFQAKGTSAAVGRSARTLFE
PAVAAPGGNILSTLPIALGSYGVESGTSMATPFTAGSAALILQAKGK--SVAKSIRNLFQ
PTLAAPGTDILSTV---TGGVALMRGTSMAAPLVAGAYALLLSVGNNLKLSPEEARTILM
PTLSAPGGNILSTFPLSEGGVAVISGTSMSCPFVAGSAAVLMAARASENLTPLEIRSLLT
lowast lowastlowastlowast lowastlowastlowastlowast lowast lowastlowastlowastlowast lowast lowastlowast lowast lowast
STAQRVPSSHTDGDPLQTLSQQGAGLIDVFKALYTETIVWPTELILNDTAHYKEVHTFTV
TTAQRVPSTHTDGDPLQTLTQQGAGLINVFNAIHTTTIVSPGELILNDTAHFKGTQQFTV
TTASAVASSKTDGDPLQTVSQQGAGLIQVDRAIGTTTVVSPGQLTLNDTAHFEGFKTFNI
STSQPSPVAY-GSETLDTVVSQGAGVINMTAAIAAHTVISPAQFNLNDTQYSNNTQTLTL
TTAKLTPVSLLGSTPLVSVIRQGGGLVQVAKALAAKTLISPHELLLNDTANANYVQTIKI
VNTGKTSKQYKLTHVPAGTAQTIT---SGTIFAADGPVPLSTDYATVKLSSSSFTLRPGQ
RNAGSSAKTYTLRHIPAGTAVTVT---PGTIFPADGPVPLSTDFASVSLSTSKFTLAPGK
QNTGSKAVTYKISHVPAGTAASVQ---SGSIFAADGPVPLTADYAQVTFSLNKVTVLPGV
RNNNRFPVSYQLSWVDSTGIVTYNDGATTDIIPSTTPNYVSTSVLRVAFSQRTVTVPGGQ
KNTNSWAMKYTFSSAVAQGLGTFD--ASGDILPTLDPVAVSGAQATVAFNTRILSVAPGA
lowast lowast lowast lowast lowast lowast
SKQVTTHINAP--SGVDASTYPVFSGFIEIATAT--------ESVHVTYLGLAASLKDKQ
TQTVTARFTPP--AAADPSTFPVFSGFIQIESGT--------EQVQVSYLGLKASLKDKQ
SVPILVWFKPP--QGVDASTYPVYSGFIEIAGLG--------ETLRVAYLGVAASLKDKA
TVKVTAQFIPPNLTAQQRNQFPIYSGFVTITGQGQGAGAEQMEKYNLPFFGLGARMVDMP
TGSVVATITPPVLPVADAARFPIFSGWIRVNGQG-ARDSSRNEAYTVPYFGLAAKMIDMQ
lowast lowast lowastlowast lowast lowast lowast lowast
IVDNTNEFFGITIPAVLNNTGDAQ-LSPTNYTFSGSDFPTLLLRLAFGTPLLRVDLVDAT
VIDNTDFFFGVPTPVLTDPNGEVQ-TSPRNYSFLASDFPTLIFRLAFGSPKVVVDLVSPT
VVDNTDEYFGVKIPALLDASGDVQ-GNSTNYTLVGDDVPSLLYRFAFGSAHVRVDLVEKA
VLDTTNVALGP--NLPFIAVGQN--IQTGPTTFSESNPAVVYFRLAAGTRRLRVDLVDAS
VLDTTETIYGPGYAYPFVIDDAIGDIQSTTTSYSRNLGPTVFARFATGTLHYSLDLVLAD
lowast lowast lowast lowast lowastlowastlowastlowast lowast
IDLKPTLNSREIDDKRHVFERESFSFPHPGPKGS-------------------TFAQVKI
INFKPTLNTRALGDTHGSF----FSFPHRVKTG--------------------SFAQVPT
STVSATLSKRNWWN---------WWWPQKPSN---------------------SYDKVPI
TSYNATVPAVTNPASRLVKRSNTELTPRAAFPT--------------------LYSQVRT
IAFTPTYPNSS-PATRFVKRSLTQHTSAASHLAKRRVSIATINPKATLVADRQLHSDVPI
lowast lowast
VGTLSEQSHIPRNSDASTVDDGGYSTFAFNKP------SFANGTTIPNGTYRWLLRALKV
LGTVFELDFTSRNNDDPTDN--GFNTVAIDPP------TFANGTTIPNGQYRLLLRALRV
VGTLAEYDYVPRNSDASTIDDNGYNVFQFT-D------SFVNGTKIAAGEYKVLLRALKV
VGSLFAPSYWPPRDYLFNNGGT-YSDYEIQLDGSFTFPNGTKGMAELGRSYKVLLRALKI
EGNIFTQPFTG-RDYLVDAAPTGSTDRTVTFNGQY-AENGLVRTAVTGTSYRFLLRALKI
lowast lowast lowastlowast lowast lowast lowast lowast lowastlowastlowastlowast
lowast lowast lowastlowastlowastlowastlowast
(b)
Figure 5 Continued
Enzyme Research 15
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
TGDPTKEADFETWLSPIVGITVA
TGDPTNEADFESFLSPIIGVNAP
TGNPAEEGDYETWLSPSFSVVPA
TADPTYTSSYESWLSPSFTFTA-
SGDAMYEDQYESWLSLPFSFRA-
lowast lowastlowast
ConservedSimilar
lowast All match
(c)
Figure 5 Multiple sequence alignment between amino acid sequence of PI12 protease with other subtilisin The serine protease sequenceswere obtained from the GenBank database Their accession numbers are EMS20793 (subtilisin-like protease of Rhodosporidium toruloidesNP11) CAG25549 (subtilisin-like protease of Pleurotus ostreatus) EPQ55564 (subtilisin-like protease of Gloeophyllum trabeumATCC 11539)and ETW80907 (serine protease S8 ofHeterobasidion irregulare TC 32-1) Putative ORF of serine protease fromG antarctica PI12 was labeledas PI12 protease
CAG38357 Phanerochaete chrysosporium
EIW62675 Trametes versicolor FP-101664 SS1
XP 007369624 Dichomitus squalens LYAD-421 SS1
XP 007381302 Punctularia strigosozonata HHB-11173 SS5
CAG25549 Pleurotus ostreatus
ETW80907 Heterobasidion irregulare TC 32-1
XP 001877576 Laccaria bicolor S238N-H82
EPQ55564 Gloeophyllum trabeum ATCC 11539
XP 001830835 Coprinopsis cinerea okayama7130
XP 007341385 Auricularia delicata TFB-10046 SS5
CCA68627 Piriformospora indica DSM 11827
CCO37249 Rhizoctonia solani AG-1 IB
XP 007342189 Auricularia delicata TFB-10046 SS5
EJU04161 Dacryopinax sp DJM-731 SS1
XP 007589863 Colletotrichum fioriniae PJ7
EFQ32840 Colletotrichum graminicola M1001
ENH74885 Fusarium oxysporum f sp cubense race 1
XP 003711175 Magnaporthe oryzae 70-15
XP 006694395 Chaetomium thermophilum DSM 1495
EON99722 Togninia minima UCRPA7
CBQ73425 Sporisorium reilianum SRZ2
GAC97617 Pseudozyma hubeiensis SY62
CCF52619 Ustilago hordei
EMS20793 Rhodosporidium toruloides NP11
CAQ76821 Glaciozyma antarctica PI12
01
Clus
ter 1
Clus
ter 2
a
b
c
d
Figure 6 Phylogenetic dendrogram among 25 translated amino acid sequences of serine proteaseThe tree was constructed using a neighbor-joining algorithm with 1000 bootstrappings by MEGA 6 software [39]The scale bar represents 01 substitutions per amino acid positionTheserine protease sequences were obtained from the GenBank database GenBank accession numbers of the sequences were indicated beforethe strain name The putative serine protease of G antarctica strain PI12 has been deposited in GenBank with accession number CAQ76821
16 Enzyme Research
0
2
4
6
8
10
12
0 24 48 72 96 110
Prot
ease
activ
ity (U
mL)
Post induction time (hours)
Figure 7 Effect of postinduction time towards protease activity The highest activity was detected at 72 hours after 110 hours of incubationwith 9UmL activity followed by 48 24 and 96 hoursThe substrate (azocasein) was dissolved in Tris HCL buffer and incubated for 30min at20∘CThe absorbance was measured at 450 nm after 15min incubation A one-way ANOVAwas used to test for preference differences amongsix different postinduction times The statistical value (119875 value lt005) between hours is 0003
pink-coloured nonpathogenic oleaginous yeast (42 iden-tity) Both G antarctica PI12 and Rhodosporidium toruloidesNP11 belong to Microbotryomycetes class PI12 protease didnot show any homology similarity to other psychrophilicproteases however it showed 37 identity to hypotheticalprotein from Pseudozyma antarctica T-34 which was isolatedfrom Japan (GAC76005) (not listed in the phylogenetic tree)
35 Cloning and Expression of Cold-Adapted PI12 Protease inthe Yeast Expression System The PCR product harbouringthe mature PI12 protease with two flanking restriction sites(EcoRI and AvrII) was cloned into pPIC9 expression vectorThe recombinant plasmid harbouring PI12 protease was usedto transform both GS115 and KM71 intended to produce bothHis+ Mut+ and His+ MutS Two recombinant clones fromGS115 (His+Mut+) strain (GpPro1 andGpPro2) and one fromKM71 (His+MutS) strain (KpPro1) were successfully obtainedwhich were further analyzed for protein expression study
36 Expression and Enzyme Assay of PI12 Protease in Pichiapastoris All positive recombinants were grown in small scalecultures and assayed for the cold-adapted protease activityin the culture supernatants The clones were first grown inBMGY medium with glycerol as the sole carbon sourceFor induction of AOX1 promoter and protein productionthe growth medium was transferred to the same mediumexcept with 05 methanol in place with glycerol as thecarbon source [26] The supernatant of the recombinant Ppastoris expression cultures after 48 h of incubation time wasanalyzed to determine the recombinant protease activity bythe modified method of Brock et al (1982) [20] All testedclones secreted protease into the supernatant in differentexpression levels This might be due to the variability ofthe strength of the promoter and copy number in each cellto secrete and express the recombinant product The clonewith the highest yield was GpPro2 with 63UmL activityfollowed by KpPro1 and GpPro1 with 25UmL and 37UmLactivities respectively (data not shown) It is perceptible thatexpression systemof Pichia pastoris is the ideal system for thisprotein The preliminary assay has fruitfully produced high
protease expression at 20∘C Yet further optimization of therecombinant PI12 protease production need to be conductedin order to examine the optimum condition of this proteinReported cold-adapted proteases showed optimum activity ata range of 15∘Cndash60∘C [27ndash30] This difference was possiblyrelated to the respective genetic and physiological adaptationof the strains There are several factors that influenced thedifferent rate of expression It includes different site of inte-gration between interest gene and Pichia genome variabilityof different expression host cell to secrete and express therecombinant enzyme and different sites in the polylinker forinserting foreign genes [31]
This is the second extracellular protease reported to besynthesized by this obligate psychrophilic yeastG antarcticaThe first cold-adapted extracellular subtilase from this yeasthas been purified and characterized previously [32 33] Theextracellular serine proteinase (LAP2) from this Antarcticyeast formerly known as Leucosporidium antarcticum strain171 is specific towards synthetic substrates of chymotrypsinand subtilisin The subtilase of L antarcticum strain 171showed specific activity of 4043Umg when utilized N-SucAAPFpNA as a substrate [33]
37 Optimization of Protease Production GpPro2 was chosenfor further optimization due to its highest protease activityat the preliminary expression stage High protease activitywas detected after 72 hours of incubation (Figure 7) byusing the standard protocol method as recommended forPichia pastoris expression system (Pichia Expression KitInvitrogen USA) The expression decreased after 72 hoursand no activity was detected at 110 hours The prolongedincubation may result in the decrease of biomass productionand will subsequently reduce the expression of recombinantprotein In addition oxygen and nutrient depletion duringprolonged incubationwill trigger the stress responses in orderto survive in such harsh environment [34]
Methanol was used as a sole carbon by P pastoris Themethanol was oxidized to formaldehyde with the aid of oxy-gen as electron acceptor by an alcohol oxidase enzyme (AOXgene) [35] The optimal methanol concentration is at 05(vv) with 14UmL activity The addition of more than 05
Enzyme Research 17
02468
10121416
0 01 03 05 07 09Pr
otea
se ac
tivity
(Um
L)Methanol concentration ()
Figure 8 Effect ofmethanol concentration towards protease activityThe recombinant was cultivated for 72 hours at 30∘C in BMMYmedium05 methanol gives the highest activity with 14UmL The substrate (azocasein) was dissolved in Tris HCL buffer and incubated for 30minat 20∘CThe mixture was left for 15min and absorbance was taken at 450 nm A one-way ANOVA was used to test for preference differencesamong six different concentrations The statistical value (119875 value lt005) between concentration is 002
0
5
10
15
20
25
10 15 20 25 30 35
Prot
eas a
ctiv
ity (
Um
L)
Temperature (∘C)
Figure 9 Effect of temperature towards protease activity The reaction was carried out in temperature ranging from 10∘C to 35∘C at pH 7 for30min A one-way ANOVA was used to test for preference differences among six different temperatures The statistical value (119875 value lt005)between temperature is 0067
(vv) methanol resulted in tremendous decrease of proteaseactivity (Figure 8) Production of PI12 protease was decreasedwhen the methanol concentration increased This might bedue to the toxic effect of methanol concentration on DNAreplication and membrane synthesis Basically methanol isused to increase the permeability of the cell membrane tohelp the secretion of foreign protein [26] However if thepermeability of cell membrane is too high lowmolecular sizemetabolites would leak from the cell and such leakage willseriously damage cell activities Such condition would abusethe cellular system and might reduce the yield of proteinexpression [36]
Psychrophilic microorganism dominated in low temper-ature habitat which most other microorganism are inca-pable of withstanding due to their inability to produceenzyme that can tolerate the extreme environment con-dition Psychrophilic enzymes are always associated withhigh catalytic efficiency at low and moderate temperature(0∘C to 30∘C) compared to the mesophilic counterpart thatare generally unable to catalyze efficiently under extremely
low temperature [2] The recombinant protein assay wasdone at various temperatures ranging from 10∘C to 35∘Cusing 05 azocasein as a substrate dissolved in 50mMTris HCl in presence of 2mM CaCl
2 pH 7 In the activity
assays expressed protease showed a highest activity at 20∘C(20UmL) and decreased tremendously once it exceededmaximum temperature due to the denatured state of theenzyme (Figure 9) The active site of an enzyme is easilyaltered when the high temperature is introduced duringincubation or activity assay resulting in a low reaction rate[37]
The recombinant construct was transferred into BMMYmedia at different A
600 nm ranging from 1 to 10 The high-est activity of active enzyme was detected at A
600 nm 60(283UmL) and above (Figure 10) The slope during expo-nential phase was much higher compared to other phasesindicating an increase in the doubling time At exponentialphase the cells are adapted to the growth condition andother processes like cell growth cycle and synthesis of RNAThe yield of active enzyme during expression of P pastoris
18 Enzyme Research
05
1015202530
1 2 3 4 5 6 7 8 9 10Prot
ease
activ
ity (U
mL)
Initial inoculum A600
Figure 10 Effect of inoculums size towards protease activity The recombinant was cultivated for 72 hours at 30∘C in BMMY mediumwith 05 methanol concentration A one-way ANOVA was used to test for preference differences among six different optical densitiesThe statistical value (119875 value lt005) between optical density value is 003
(kD
a)
1160
662
450
350
250
184
144
M 1 2 3
(a) (b) (c)
Figure 11 (a) Coomassie-stained SDS-PAGE of recombinant P pastoris clone 1mL supernatants collected from the third-day culturewere concentrated 10-fold using trichloroacetic acid and 10120583L resultant products were applied to SDS-PAGE analysis Lane 1 molecularweight marker (Fermentas) Lane 2 GS115 (Control) Lane 3 GS115pPIC9 empty (Control) Lane 4 GS115pPIC9PI12prot (GpPro2)The arrow indicates the secreted recombinant PI12 protease (b) Plate activity staining of different recombinant P pastoris clones GGS115 GP GS115pPIC9 GRI GS115pPIC9PI12prot1 (GpPro1) GRII GS115pPIC9PI12prot2 (GpPro2) K KM71 KP KM71pPIC9 KRKM71pPIC9PI12prot (KpPro) (c)Gel activity staining of recombinant PI12 protease inP pastorisGpPro2 clone Lane 1 prestainedmolecularweightmarker (Fermentas) Lane 2 GS115pPIC9 empty (Control) Lane 3 GS115pPIC9PI12prot1 (GpPro2)The arrow indicates the secretedrecombinant PI12 protease
dropped after 28 hours of incubation which is at stationaryor death phase [38]
38 Detection of Recombinant Protein by SDS-PAGE andActivity Staining TheSDS-PAGE gel of the GpPro2 recombi-nant clonesrsquo supernatant showed a putative expression bandat around 993 kDa Plate staining and gel activity stainingwere executed to examine if the recombinant PI12 proteaseexpressed in P pastoris was functionally active In the platestaining the transformants showed strong proteolytic activity(Figure 11(a)) In gel staining proteolytic activity was alsodetected around 993 kDa after incubation at 20∘C This is
evidence of protease secretion into the surrounding mediawhereby it catalyzed the breakdown of casein thus formedclearing zones around the activity ring and band (Figures11(b) and 11(c)) As a conclusion the cold-adapted PI12protease was functionally expressed in P pastoris GS115(GpPro2 clone) with the size of 993 kDa
4 Conclusions
Psychrophilic yeast which was originated frommarine waterin Casey Station Antarctica was identified as Glaciozymaantarctica strain PI12 A serine protease with an open reading
Enzyme Research 19
frame of 2892 bp encoded for 963 amino acids from thisAntarctic yeast has been cloned and expressed in Pichiaexpression system This is the second extracellular pro-tease reported being synthesized by this psychrophilic yeastSequence analysis through bioinformatics studies revealedfew homology identities of this protein with other ser-ine proteases The mature PI12 protease was successfullyexpressed and secreted into the culture medium driven bythe Saccharomyces cerevisiae 120572-factor signal sequence in Ppastoris The protease production in P pastoris was bestobtained from strain GS115 (GpPro2) with 283UmL after 72hours of induction time with 05 (vv) of methanol inducerThe starting optical density at 600 nm (OD600) was 60 andthe optimum assay temperature was 20∘C The expressedprotein was detected by SDS-PAGE and activity staining witha molecular weight of 99 kDaThis cold-active enzymemightoffer an opportunity for biotechnological exploitation basedon its high catalytic activity at low temperatures and lowthermostability Structural and functional study of this cold-adapted enzyme would give insight into its adaptation to anextreme cold environment
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgment
This research was supported financially by the InitiativeBiotechnology-GMBI-MOSTI (Project number 07-05-16-MG-GMB02) andRUGS-Initiative 6 (Project number 05-02-11-1414RU)
References
[1] J W Deming ldquoPsychrophiles and polar regionsrdquo CurrentOpinion in Microbiology vol 5 no 3 pp 301ndash309 2002
[2] C Gerday M Aittaleb M Bentahir et al ldquoCold-adaptedenzymes from fundamentals to biotechnologyrdquo Trends inBiotechnology vol 18 no 3 pp 103ndash107 2000
[3] A Svingor J Kardos I Hajdu A Nemeth and P Zavodszky ldquoAbetter enzyme to cope with cold comparative flexibility studieson psychrotrophic mesophilic and thermophilic IPMDHSrdquoThe Journal of Biological Chemistry vol 276 no 30 pp 28121ndash28125 2001
[4] G Antranikian C E Vorgias and C Bertoldo ldquoExtremeenvironments as a resource for microorganisms and novel bio-catalystsrdquo Advances in Biochemical EngineeringBiotechnologyvol 96 pp 219ndash262 2005
[5] T Lonhienne C Gerday andG Feller ldquoPsychrophilic enzymesrevisiting the thermodynamic parameters of activation mayexplain local flexibilityrdquoBiochimica et Biophysica Acta vol 1543no 1 pp 1ndash10 2000
[6] R Cavicchioli and TThomas ldquoExtremophilesrdquo in Encyclopediaof Microbiology pp 317ndash337 Academic Press San Diego CalifUSA 2000
[7] G Feller E Narinx J L Arpigny et al ldquoEnzymes frompsychrophilic organismsrdquo FEMS Microbiology Reviews vol 18no 2-3 pp 189ndash202 1996
[8] J Arnorsdottir R B Smaradottir T O Magnusson S HThorbjarnardottir G Eggertsson and M M KristjanssonldquoCharacterization of a cloned subtilisin-like serine proteinasefrom a psychrotrophic Vibrio speciesrdquo European Journal ofBiochemistry vol 269 no 22 pp 5536ndash5546 2002
[9] N J Russell ldquoPsychrophilic bacteriamdashmolecular adaptations ofmembrane lipidsrdquo Comparative Biochemistry and Physiology APhysiology vol 118 no 3 pp 489ndash493 1997
[10] G Feller and C Gerday ldquoPsychrophilic enzymes hot topics incold adaptationrdquo Nature Reviews Microbiology vol 1 no 3 pp200ndash208 2003
[11] P Raspor and J Zupan ldquoYeasts in extreme environmentsrdquo inBiodiversity and Ecophysiology of Yeasts The Yeast Handbookpp 371ndash417 Springer Berlin Germany 2006
[12] M B Rao A M Tanksale M S Ghatge and V V Desh-pande ldquoMolecular and biotechnological aspects of microbialproteasesrdquoMicrobiology andMolecular Biology Reviews vol 62no 3 pp 597ndash635 1998
[13] L Polgar ldquoThe catalytic triad of serine peptidasesrdquo Cellular andMolecular Life Sciences vol 62 no 19-20 pp 2161ndash2172 2005
[14] S Y Boo C M V L Wong K F Rodrigues N NajimudinA M A Murad and N M Mahadi ldquoThermal stress responsesin Antarctic yeastGlaciozyma antarctica PI12 characterized byreal-time quantitative PCRrdquoPolar Biology vol 36 no 3 pp 381ndash389 2013
[15] B Turchetti S R T Hall L B Connell et al ldquoPsychrophilicyeasts from Antarctica and European glaciers descriptionof Glaciozyma gen nov Glaciozyma martinii sp nov andGlaciozyma watsonii sp novrdquo Extremophiles vol 15 no 5 pp573ndash586 2011
[16] N H F Hashim I Bharudin D L S Nguong et al ldquoCharac-terization of Afp1 an antifreeze protein from the psychrophilicyeast Glaciozyma antarctica PI12rdquo Extremophiles vol 17 no 1pp 63ndash73 2013
[17] A N M Ramli M A Azhar M S Shamsir et al ldquoSequenceand structural investigation of a novel psychrophilic 120572-amylasefrom Glaciozyma antarctica PI12 for cold-adaptation analysisrdquoJournal ofMolecularModeling vol 19 no 8 pp 3369ndash3383 2013
[18] A N M Ramli N M Mahadi A Rabu A M A Murad F DA Bakar and R M Illias ldquoMolecular cloning expression andbiochemical characterisation of a cold-adapted novel recombi-nant chitinase from Glaciozyma antarctica PI12rdquoMicrobial CellFactories vol 10 no 1 article 94 2011
[19] J Sambrook E Fritsch and T Maniatis Molecular Cloning ALaboratoryManual Cold SpringHarbor Laboratory Press NewYork NY USA 1989
[20] F M Brock C W Forsberg and J G Buchanan-SmithldquoProteolytic activity of rumen microorganisms and effects ofproteinase inhibitorsrdquoApplied and EnvironmentalMicrobiologyvol 44 no 3 pp 561ndash569 1982
[21] U K Laemmli ldquoCleavage of structural proteins during theassembly of the head of bacteriophage T4rdquo Nature vol 227 no5259 pp 680ndash685 1970
[22] A Henne R A Schmitz M Bomeke G Gottschalk and RDaniel ldquoScreening of environmental DNA libraries for thepresence of genes conferring lipolytic activity on Escherichiacolirdquo Applied and Environmental Microbiology vol 66 no 7 pp3113ndash3116 2000
[23] M Poza T de Miguel C Sieiro and T G Villa ldquoCharacteri-zation of a broad pH range protease of Candida caseinolyticardquoJournal of Applied Microbiology vol 91 no 5 pp 916ndash921 2001
20 Enzyme Research
[24] J D Bendtsen H Nielsen G von Heijne and S BrunakldquoImproved prediction of signal peptides SignalP 30rdquo Journalof Molecular Biology vol 340 no 4 pp 783ndash795 2004
[25] R J Siezen and J A M Leunissen ldquoSubtilases the superfamilyof subtilisin-like serine proteasesrdquo Protein Science vol 6 no 3pp 501ndash523 1997
[26] Y Katakura W Zhang G Zhuang et al ldquoEffect of methanolconcentration on the production of human 1205732- glycoprotein Idomain V by a recombinant Pichia pastoris a simple system forthe control of methanol concentration using a semiconductorgas sensorrdquo Journal of Fermentation and Bioengineering vol 86no 5 pp 482ndash487 1998
[27] Q-FWang Y-HHou Z Xu J-LMiao andG-Y Li ldquoPurifica-tion and properties of an extracellular cold-active protease fromthe psychrophilic bacterium Pseudoalteromonas sp NJ276rdquoBiochemical Engineering Journal vol 38 no 3 pp 362ndash3682008
[28] S Vazquez L Ruberto and W Mac Cormack ldquoProper-ties of extracellular proteases from three psychrotolerantStenotrophomonas maltophilia isolated from Antarctic soilrdquoPolar Biology vol 28 no 4 pp 319ndash325 2005
[29] R Zeng R Zhang J Zhao and N Lin ldquoCold-active ser-ine alkaline protease from the psychrophilic bacterium Pseu-domonas strain DY-A enzyme purification and characteriza-tionrdquo Extremophiles vol 7 no 4 pp 335ndash337 2003
[30] M Turkiewicz E Gromek H Kalinowska and M ZielinskaldquoBiosynthesis and properties of an extracellularmetalloproteasefrom the Antarctic marine bacterium Sphingomonas paucimo-bilisrdquo Journal of Biotechnology vol 70 no 1ndash3 pp 53ndash60 1999
[31] A Vassileva D A Chugh S Swaminathan and N KhannaldquoEffect of copy number on the expression levels of hepatitisB surface antigen in the methylotrophic yeast Pichia pastorisrdquoProtein Expression andPurification vol 21 no 1 pp 71ndash80 2001
[32] M Pazgier M Turkiewicz H Kalinowska and S Bielecki ldquoTheunique cold-adapted extracellular subtilase from psychrophilicyeast Leucosporidium antarcticumrdquo Journal ofMolecular Catal-ysis B Enzymatic vol 21 no 1-2 pp 39ndash42 2003
[33] M Turkiewicz M Pazgier H Kalinowska and S BieleckildquoA cold-adapted extracellular serine proteinase of the yeastLeucosporidium antarcticumrdquo Extremophiles vol 7 no 6 pp435ndash442 2003
[34] D Mattanovich B Gasser H Hohenblum and M SauerldquoStress in recombinant protein producing yeastsrdquo Journal ofBiotechnology vol 113 no 1ndash3 pp 121ndash135 2004
[35] P Secades BAlvarez and JAGuijarro ldquoPurification andprop-erties of a new psychrophilic metalloprotease (Fpp2) in the fishpathogen Flavobacterium psychrophilumrdquo FEMS MicrobiologyLetters vol 226 no 2 pp 273ndash279 2003
[36] L Y Ling I Ithoi and F M Yik ldquoOptimization for high-level expression in Pichia pastoris and purification of truncatedand full length recombinant sag2 of toxoplasma gondii fordiagnostic userdquo Southeast Asian Journal of Tropical Medicineand Public Health vol 41 no 3 pp 507ndash513 2010
[37] A Illanes ldquoStability of biocatalystsrdquo Electronic Journal ofBiotechnology vol 2 no 1 pp 1ndash9 1999
[38] B A Plantz J Sinha L Villarete K W Nickerson and VL Schlegel ldquoPichia pastoris fermentation optimization energystate and testing a growth-associated modelrdquo Applied Microbi-ology and Biotechnology vol 72 no 2 pp 297ndash305 2006
[39] K Tamura G Stecher D Peterson A Filipski and S KumarldquoMEGA6molecular evolutionary genetics analysis version 60rdquo
Molecular Biology and Evolution vol 30 no 12 pp 2725ndash27292013
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology
Enzyme Research 13
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
MKG-YSWSLLQLAFTGAALATVPLSDVKRISSAS--VVPNKFVVEVESASEIPGKR--SL
MKGVLVWSILS---AGCALAVTPLSSVKRVTNLP--IVANKFIVEVDSTSDIPTKR--S-
MKLPLLSLALT---AVSVGATVPLSSVQRTTTSPNNALAGEYIVEMNNVQGLGGKR--AY
---MAHGSLASLLLLLAASAQGAAAFGGKSATGGRSVVPNSYILQVNASAPALTKRGVSV
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease ---MLFLPVLLLLLPGVTAFLNPVTNRATNAISSTQYLSNAYILELDLSTPGLVKRDSTP
lowastlowast
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
YTPHELFYRSLSARGVSYEVNKEYNS-EGLFVGAVVTVSDANDVKALAETPGVKAIRPVR
-NPHDALYESLRKRAIGFKVDREFNT-QGLFVGAALTLSDAQDVGQIQSTTGVKSIRPVR
ASTHDTLYRSLRKRGISFDVKKEYNQ-PGIMVGAALKVASPQDVQQIATMAEVKAIRPVI
FEALDHTLAAVKQNGVKYTVRQRFDAIPEAFQAVSIQVEDGASMAELAQIPGVQRVWPVS
DSILEDVLTSVGRNGIKYQLRHRFIS-PTLFHGASITVPPGISRSQIASLRGIKRVWPVR
lowastlowast
TIKSPKPVKVHVVTGKDDPEIPA-------------------------DTESTHIMTGVN
AFDRPKPVKVQVVKGPDDPALPP-------------------------DSESTHVITGVD
AVPAPERINQFTVSSPNDSRITA-------------------------DSETTHIMTGVD
LIPRPVEPTVSDFTPVSSSSSSTKSKRD-THTLEKRGTNFPPASAYLNDTFYPHVQTGID
KFSRPSAVVDADGGGSGFSGSPIKAALMGVKELGKR------ANAYAGDTFGPHVMTGVN
lowast lowast lowast lowastlowast
KLHAQGIFGAGIKIGIIDTGIDYTHPALGGALGAGHKVIGGYDFVGDAYDGTNTPVPDND
KLHAQGITGAGIKIGILDTGIDYTHPTLGGAFGPGNKVIGGFDLVGDDYDGTNTPVPDPD
KVHAAGITGEGIKIGIIDTGIDYNHPLLGAGFGPGFKVIGGYDFVGDDYTGSNNPAPDDD
VLHNKGILGQGVKIVVVDEGVDYTNPILGGCFGPGCQISFGYDFVGDNYTGTNAPVPDSD
ETHEAGLLGAGIKIGVLDTGVDYLNPILGGCFGPGCHMSFGYDLVGDDYDGDNAPVPDVD
lowast lowast lowast lowastlowastlowast lowast lowast lowastlowast lowast lowastlowast lowast lowast lowast lowast lowastlowastlowast lowast lowast lowast lowast lowastlowast lowast
PLDQCNGHGTHVAGIIGANPDNQFNISGVSYGASLSSYRVFGCSGSVSDDIIIEALLRGV
PLDQCAGHGTHVAGIIGANPDNAFNISGVAFQASLSAYRVFGCVGFVTDDVLVDALLLGF
PLDNCNGHGTHVAGIIGANPGNDFGISGVAYGSSIASYRVFGCSGFVSDDIIIDALLRGY
PFSSCSNHGTHTTGTVGALAN-EYGFSGAAPMATIGHYRVISCSGATTDEILVAALTRAL
PFASCDPHGTHVTGIIGALPN-AFGFTGVAPAATLGHYRVFGCTGFVGEDIILAGLMRGV
lowast lowastlowastlowast lowastlowast lowast lowastlowast lowast lowastlowastlowast lowast lowast lowast
QDGQDILTLSLGGTDGWTE-SSSSVVASRIAASGKIVTIAAGNEGAYGSWYSSSPGNGID
NEGQDILTLSLGGADGWTE-SVSAVVASRIAATGKVVTIAAGNDGASGAWYTSSPGNGID
SDGMDILTLSLGGVEGWTE-SSSSVVASRIADQGRVVTIAAGNDGAYGSWYASSPGTGVD
QDNVNIVSMSLGSSVGWLDDSPVQIMASYLGSKGIHVVASVGNERNEGLFAADQPAASRI
EDNCNVLTLSLGGPGGWVKGTPASILIDQIEAQGILVTVATGNSGAEGMFFSESPASTIN
lowastlowastlowast lowastlowast lowast lowast lowastlowast lowast lowast
VISVASIDNTVDNIQNATVNGVEHGPIPYFKTLPLPFTNA-LPLYVISNDTTIANDACNA
VISVASLDNTVVPLQTAQVHGVTHDPIIYQDTFPLPINDT-RPIFATSTDVTVVDDACNP
VISVASVNNIAIPVQNATVQGVEHAPIPYLEALPLNVTGS-RPIYATSTDTTVADDACNA
GTGVGGVDPTYLAAYYAYLI--KRDPVPYISPTPFNLPNN-YLLYFTSTNTSVTNDACNP
GLSIASTDVTDLIAYNATVS--GQPAIPYLSATPLNVVANSFRVHFTSTDPNNPVDACSP
lowast lowast lowast lowast lowastlowastlowast
LPDNTPDLSGFLVLVRRGSCTFVQKLTNIAAKGGNAVFIYDNGNG-----FAGIDTGKFT
LPDSTPDLSGFVVIVRRGTCTFVQKLGNVAAKGAKVTLIYDNGSG-----FGAIDTGAFV
LPNDTPDLSGYVVVIKRGTCTFVTKLQNAAAKGANTFLIYNNGGS-----FSAISVGDYT
LPSTTPNLANRVVVVQRGTCGFTVKQQNVAAAGGKIVLIYNSKGSGLIPQLDVGSTGLTA
LPAGAPDFANYVTVVQRGTCTFVTKYQNVLNAGGKIVLLYNSEGAGNLPYLTPNGVGIDA
lowastlowast lowast lowastlowast lowast lowast lowast lowast lowast lowast lowast
(a)
Figure 5 Continued
14 Enzyme Research
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
NATLIQAADGVFLAQQVAAGKSVTITFPQTGGSAQITN-PSGGLVSSFTTYGPTNDFYFK
-ASFIQAADGEFLVQQFASGQNVSLSFPQSGGLIQFPD-PAGGLISSFTSYGPSNDFFFK
-AALISETDGTFLVNQFAAKANITISFPQSGSAWNYPDDVSGGLVSDFSSYGPSYDMYFK
VGSLRYEDGLKLLSYYQSSPRGQSISFPNGPLAPYVSNTVSGGVVASYSNFGPTNELFIY
VAGLRRSDGLKLLSYYQNANKRLTLRFPKGKIVAGLTDTITGGLISGYSTFGPTNDLYGQ
lowast lowastlowast lowastlowast lowastlowast
PAFGAPGGNILSTLPVPLGSYGVESGTSMATPYVAGSAALLLSVKGKSAAVALGARTLFQ
PAVAAPGGNILSTLPVNLGLFGIESGTSMATPFVAGSAALLFQAKGTSAAVGRSARTLFE
PAVAAPGGNILSTLPIALGSYGVESGTSMATPFTAGSAALILQAKGK--SVAKSIRNLFQ
PTLAAPGTDILSTV---TGGVALMRGTSMAAPLVAGAYALLLSVGNNLKLSPEEARTILM
PTLSAPGGNILSTFPLSEGGVAVISGTSMSCPFVAGSAAVLMAARASENLTPLEIRSLLT
lowast lowastlowastlowast lowastlowastlowastlowast lowast lowastlowastlowastlowast lowast lowastlowast lowast lowast
STAQRVPSSHTDGDPLQTLSQQGAGLIDVFKALYTETIVWPTELILNDTAHYKEVHTFTV
TTAQRVPSTHTDGDPLQTLTQQGAGLINVFNAIHTTTIVSPGELILNDTAHFKGTQQFTV
TTASAVASSKTDGDPLQTVSQQGAGLIQVDRAIGTTTVVSPGQLTLNDTAHFEGFKTFNI
STSQPSPVAY-GSETLDTVVSQGAGVINMTAAIAAHTVISPAQFNLNDTQYSNNTQTLTL
TTAKLTPVSLLGSTPLVSVIRQGGGLVQVAKALAAKTLISPHELLLNDTANANYVQTIKI
VNTGKTSKQYKLTHVPAGTAQTIT---SGTIFAADGPVPLSTDYATVKLSSSSFTLRPGQ
RNAGSSAKTYTLRHIPAGTAVTVT---PGTIFPADGPVPLSTDFASVSLSTSKFTLAPGK
QNTGSKAVTYKISHVPAGTAASVQ---SGSIFAADGPVPLTADYAQVTFSLNKVTVLPGV
RNNNRFPVSYQLSWVDSTGIVTYNDGATTDIIPSTTPNYVSTSVLRVAFSQRTVTVPGGQ
KNTNSWAMKYTFSSAVAQGLGTFD--ASGDILPTLDPVAVSGAQATVAFNTRILSVAPGA
lowast lowast lowast lowast lowast lowast
SKQVTTHINAP--SGVDASTYPVFSGFIEIATAT--------ESVHVTYLGLAASLKDKQ
TQTVTARFTPP--AAADPSTFPVFSGFIQIESGT--------EQVQVSYLGLKASLKDKQ
SVPILVWFKPP--QGVDASTYPVYSGFIEIAGLG--------ETLRVAYLGVAASLKDKA
TVKVTAQFIPPNLTAQQRNQFPIYSGFVTITGQGQGAGAEQMEKYNLPFFGLGARMVDMP
TGSVVATITPPVLPVADAARFPIFSGWIRVNGQG-ARDSSRNEAYTVPYFGLAAKMIDMQ
lowast lowast lowastlowast lowast lowast lowast lowast
IVDNTNEFFGITIPAVLNNTGDAQ-LSPTNYTFSGSDFPTLLLRLAFGTPLLRVDLVDAT
VIDNTDFFFGVPTPVLTDPNGEVQ-TSPRNYSFLASDFPTLIFRLAFGSPKVVVDLVSPT
VVDNTDEYFGVKIPALLDASGDVQ-GNSTNYTLVGDDVPSLLYRFAFGSAHVRVDLVEKA
VLDTTNVALGP--NLPFIAVGQN--IQTGPTTFSESNPAVVYFRLAAGTRRLRVDLVDAS
VLDTTETIYGPGYAYPFVIDDAIGDIQSTTTSYSRNLGPTVFARFATGTLHYSLDLVLAD
lowast lowast lowast lowast lowastlowastlowastlowast lowast
IDLKPTLNSREIDDKRHVFERESFSFPHPGPKGS-------------------TFAQVKI
INFKPTLNTRALGDTHGSF----FSFPHRVKTG--------------------SFAQVPT
STVSATLSKRNWWN---------WWWPQKPSN---------------------SYDKVPI
TSYNATVPAVTNPASRLVKRSNTELTPRAAFPT--------------------LYSQVRT
IAFTPTYPNSS-PATRFVKRSLTQHTSAASHLAKRRVSIATINPKATLVADRQLHSDVPI
lowast lowast
VGTLSEQSHIPRNSDASTVDDGGYSTFAFNKP------SFANGTTIPNGTYRWLLRALKV
LGTVFELDFTSRNNDDPTDN--GFNTVAIDPP------TFANGTTIPNGQYRLLLRALRV
VGTLAEYDYVPRNSDASTIDDNGYNVFQFT-D------SFVNGTKIAAGEYKVLLRALKV
VGSLFAPSYWPPRDYLFNNGGT-YSDYEIQLDGSFTFPNGTKGMAELGRSYKVLLRALKI
EGNIFTQPFTG-RDYLVDAAPTGSTDRTVTFNGQY-AENGLVRTAVTGTSYRFLLRALKI
lowast lowast lowastlowast lowast lowast lowast lowast lowastlowastlowastlowast
lowast lowast lowastlowastlowastlowastlowast
(b)
Figure 5 Continued
Enzyme Research 15
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
TGDPTKEADFETWLSPIVGITVA
TGDPTNEADFESFLSPIIGVNAP
TGNPAEEGDYETWLSPSFSVVPA
TADPTYTSSYESWLSPSFTFTA-
SGDAMYEDQYESWLSLPFSFRA-
lowast lowastlowast
ConservedSimilar
lowast All match
(c)
Figure 5 Multiple sequence alignment between amino acid sequence of PI12 protease with other subtilisin The serine protease sequenceswere obtained from the GenBank database Their accession numbers are EMS20793 (subtilisin-like protease of Rhodosporidium toruloidesNP11) CAG25549 (subtilisin-like protease of Pleurotus ostreatus) EPQ55564 (subtilisin-like protease of Gloeophyllum trabeumATCC 11539)and ETW80907 (serine protease S8 ofHeterobasidion irregulare TC 32-1) Putative ORF of serine protease fromG antarctica PI12 was labeledas PI12 protease
CAG38357 Phanerochaete chrysosporium
EIW62675 Trametes versicolor FP-101664 SS1
XP 007369624 Dichomitus squalens LYAD-421 SS1
XP 007381302 Punctularia strigosozonata HHB-11173 SS5
CAG25549 Pleurotus ostreatus
ETW80907 Heterobasidion irregulare TC 32-1
XP 001877576 Laccaria bicolor S238N-H82
EPQ55564 Gloeophyllum trabeum ATCC 11539
XP 001830835 Coprinopsis cinerea okayama7130
XP 007341385 Auricularia delicata TFB-10046 SS5
CCA68627 Piriformospora indica DSM 11827
CCO37249 Rhizoctonia solani AG-1 IB
XP 007342189 Auricularia delicata TFB-10046 SS5
EJU04161 Dacryopinax sp DJM-731 SS1
XP 007589863 Colletotrichum fioriniae PJ7
EFQ32840 Colletotrichum graminicola M1001
ENH74885 Fusarium oxysporum f sp cubense race 1
XP 003711175 Magnaporthe oryzae 70-15
XP 006694395 Chaetomium thermophilum DSM 1495
EON99722 Togninia minima UCRPA7
CBQ73425 Sporisorium reilianum SRZ2
GAC97617 Pseudozyma hubeiensis SY62
CCF52619 Ustilago hordei
EMS20793 Rhodosporidium toruloides NP11
CAQ76821 Glaciozyma antarctica PI12
01
Clus
ter 1
Clus
ter 2
a
b
c
d
Figure 6 Phylogenetic dendrogram among 25 translated amino acid sequences of serine proteaseThe tree was constructed using a neighbor-joining algorithm with 1000 bootstrappings by MEGA 6 software [39]The scale bar represents 01 substitutions per amino acid positionTheserine protease sequences were obtained from the GenBank database GenBank accession numbers of the sequences were indicated beforethe strain name The putative serine protease of G antarctica strain PI12 has been deposited in GenBank with accession number CAQ76821
16 Enzyme Research
0
2
4
6
8
10
12
0 24 48 72 96 110
Prot
ease
activ
ity (U
mL)
Post induction time (hours)
Figure 7 Effect of postinduction time towards protease activity The highest activity was detected at 72 hours after 110 hours of incubationwith 9UmL activity followed by 48 24 and 96 hoursThe substrate (azocasein) was dissolved in Tris HCL buffer and incubated for 30min at20∘CThe absorbance was measured at 450 nm after 15min incubation A one-way ANOVAwas used to test for preference differences amongsix different postinduction times The statistical value (119875 value lt005) between hours is 0003
pink-coloured nonpathogenic oleaginous yeast (42 iden-tity) Both G antarctica PI12 and Rhodosporidium toruloidesNP11 belong to Microbotryomycetes class PI12 protease didnot show any homology similarity to other psychrophilicproteases however it showed 37 identity to hypotheticalprotein from Pseudozyma antarctica T-34 which was isolatedfrom Japan (GAC76005) (not listed in the phylogenetic tree)
35 Cloning and Expression of Cold-Adapted PI12 Protease inthe Yeast Expression System The PCR product harbouringthe mature PI12 protease with two flanking restriction sites(EcoRI and AvrII) was cloned into pPIC9 expression vectorThe recombinant plasmid harbouring PI12 protease was usedto transform both GS115 and KM71 intended to produce bothHis+ Mut+ and His+ MutS Two recombinant clones fromGS115 (His+Mut+) strain (GpPro1 andGpPro2) and one fromKM71 (His+MutS) strain (KpPro1) were successfully obtainedwhich were further analyzed for protein expression study
36 Expression and Enzyme Assay of PI12 Protease in Pichiapastoris All positive recombinants were grown in small scalecultures and assayed for the cold-adapted protease activityin the culture supernatants The clones were first grown inBMGY medium with glycerol as the sole carbon sourceFor induction of AOX1 promoter and protein productionthe growth medium was transferred to the same mediumexcept with 05 methanol in place with glycerol as thecarbon source [26] The supernatant of the recombinant Ppastoris expression cultures after 48 h of incubation time wasanalyzed to determine the recombinant protease activity bythe modified method of Brock et al (1982) [20] All testedclones secreted protease into the supernatant in differentexpression levels This might be due to the variability ofthe strength of the promoter and copy number in each cellto secrete and express the recombinant product The clonewith the highest yield was GpPro2 with 63UmL activityfollowed by KpPro1 and GpPro1 with 25UmL and 37UmLactivities respectively (data not shown) It is perceptible thatexpression systemof Pichia pastoris is the ideal system for thisprotein The preliminary assay has fruitfully produced high
protease expression at 20∘C Yet further optimization of therecombinant PI12 protease production need to be conductedin order to examine the optimum condition of this proteinReported cold-adapted proteases showed optimum activity ata range of 15∘Cndash60∘C [27ndash30] This difference was possiblyrelated to the respective genetic and physiological adaptationof the strains There are several factors that influenced thedifferent rate of expression It includes different site of inte-gration between interest gene and Pichia genome variabilityof different expression host cell to secrete and express therecombinant enzyme and different sites in the polylinker forinserting foreign genes [31]
This is the second extracellular protease reported to besynthesized by this obligate psychrophilic yeastG antarcticaThe first cold-adapted extracellular subtilase from this yeasthas been purified and characterized previously [32 33] Theextracellular serine proteinase (LAP2) from this Antarcticyeast formerly known as Leucosporidium antarcticum strain171 is specific towards synthetic substrates of chymotrypsinand subtilisin The subtilase of L antarcticum strain 171showed specific activity of 4043Umg when utilized N-SucAAPFpNA as a substrate [33]
37 Optimization of Protease Production GpPro2 was chosenfor further optimization due to its highest protease activityat the preliminary expression stage High protease activitywas detected after 72 hours of incubation (Figure 7) byusing the standard protocol method as recommended forPichia pastoris expression system (Pichia Expression KitInvitrogen USA) The expression decreased after 72 hoursand no activity was detected at 110 hours The prolongedincubation may result in the decrease of biomass productionand will subsequently reduce the expression of recombinantprotein In addition oxygen and nutrient depletion duringprolonged incubationwill trigger the stress responses in orderto survive in such harsh environment [34]
Methanol was used as a sole carbon by P pastoris Themethanol was oxidized to formaldehyde with the aid of oxy-gen as electron acceptor by an alcohol oxidase enzyme (AOXgene) [35] The optimal methanol concentration is at 05(vv) with 14UmL activity The addition of more than 05
Enzyme Research 17
02468
10121416
0 01 03 05 07 09Pr
otea
se ac
tivity
(Um
L)Methanol concentration ()
Figure 8 Effect ofmethanol concentration towards protease activityThe recombinant was cultivated for 72 hours at 30∘C in BMMYmedium05 methanol gives the highest activity with 14UmL The substrate (azocasein) was dissolved in Tris HCL buffer and incubated for 30minat 20∘CThe mixture was left for 15min and absorbance was taken at 450 nm A one-way ANOVA was used to test for preference differencesamong six different concentrations The statistical value (119875 value lt005) between concentration is 002
0
5
10
15
20
25
10 15 20 25 30 35
Prot
eas a
ctiv
ity (
Um
L)
Temperature (∘C)
Figure 9 Effect of temperature towards protease activity The reaction was carried out in temperature ranging from 10∘C to 35∘C at pH 7 for30min A one-way ANOVA was used to test for preference differences among six different temperatures The statistical value (119875 value lt005)between temperature is 0067
(vv) methanol resulted in tremendous decrease of proteaseactivity (Figure 8) Production of PI12 protease was decreasedwhen the methanol concentration increased This might bedue to the toxic effect of methanol concentration on DNAreplication and membrane synthesis Basically methanol isused to increase the permeability of the cell membrane tohelp the secretion of foreign protein [26] However if thepermeability of cell membrane is too high lowmolecular sizemetabolites would leak from the cell and such leakage willseriously damage cell activities Such condition would abusethe cellular system and might reduce the yield of proteinexpression [36]
Psychrophilic microorganism dominated in low temper-ature habitat which most other microorganism are inca-pable of withstanding due to their inability to produceenzyme that can tolerate the extreme environment con-dition Psychrophilic enzymes are always associated withhigh catalytic efficiency at low and moderate temperature(0∘C to 30∘C) compared to the mesophilic counterpart thatare generally unable to catalyze efficiently under extremely
low temperature [2] The recombinant protein assay wasdone at various temperatures ranging from 10∘C to 35∘Cusing 05 azocasein as a substrate dissolved in 50mMTris HCl in presence of 2mM CaCl
2 pH 7 In the activity
assays expressed protease showed a highest activity at 20∘C(20UmL) and decreased tremendously once it exceededmaximum temperature due to the denatured state of theenzyme (Figure 9) The active site of an enzyme is easilyaltered when the high temperature is introduced duringincubation or activity assay resulting in a low reaction rate[37]
The recombinant construct was transferred into BMMYmedia at different A
600 nm ranging from 1 to 10 The high-est activity of active enzyme was detected at A
600 nm 60(283UmL) and above (Figure 10) The slope during expo-nential phase was much higher compared to other phasesindicating an increase in the doubling time At exponentialphase the cells are adapted to the growth condition andother processes like cell growth cycle and synthesis of RNAThe yield of active enzyme during expression of P pastoris
18 Enzyme Research
05
1015202530
1 2 3 4 5 6 7 8 9 10Prot
ease
activ
ity (U
mL)
Initial inoculum A600
Figure 10 Effect of inoculums size towards protease activity The recombinant was cultivated for 72 hours at 30∘C in BMMY mediumwith 05 methanol concentration A one-way ANOVA was used to test for preference differences among six different optical densitiesThe statistical value (119875 value lt005) between optical density value is 003
(kD
a)
1160
662
450
350
250
184
144
M 1 2 3
(a) (b) (c)
Figure 11 (a) Coomassie-stained SDS-PAGE of recombinant P pastoris clone 1mL supernatants collected from the third-day culturewere concentrated 10-fold using trichloroacetic acid and 10120583L resultant products were applied to SDS-PAGE analysis Lane 1 molecularweight marker (Fermentas) Lane 2 GS115 (Control) Lane 3 GS115pPIC9 empty (Control) Lane 4 GS115pPIC9PI12prot (GpPro2)The arrow indicates the secreted recombinant PI12 protease (b) Plate activity staining of different recombinant P pastoris clones GGS115 GP GS115pPIC9 GRI GS115pPIC9PI12prot1 (GpPro1) GRII GS115pPIC9PI12prot2 (GpPro2) K KM71 KP KM71pPIC9 KRKM71pPIC9PI12prot (KpPro) (c)Gel activity staining of recombinant PI12 protease inP pastorisGpPro2 clone Lane 1 prestainedmolecularweightmarker (Fermentas) Lane 2 GS115pPIC9 empty (Control) Lane 3 GS115pPIC9PI12prot1 (GpPro2)The arrow indicates the secretedrecombinant PI12 protease
dropped after 28 hours of incubation which is at stationaryor death phase [38]
38 Detection of Recombinant Protein by SDS-PAGE andActivity Staining TheSDS-PAGE gel of the GpPro2 recombi-nant clonesrsquo supernatant showed a putative expression bandat around 993 kDa Plate staining and gel activity stainingwere executed to examine if the recombinant PI12 proteaseexpressed in P pastoris was functionally active In the platestaining the transformants showed strong proteolytic activity(Figure 11(a)) In gel staining proteolytic activity was alsodetected around 993 kDa after incubation at 20∘C This is
evidence of protease secretion into the surrounding mediawhereby it catalyzed the breakdown of casein thus formedclearing zones around the activity ring and band (Figures11(b) and 11(c)) As a conclusion the cold-adapted PI12protease was functionally expressed in P pastoris GS115(GpPro2 clone) with the size of 993 kDa
4 Conclusions
Psychrophilic yeast which was originated frommarine waterin Casey Station Antarctica was identified as Glaciozymaantarctica strain PI12 A serine protease with an open reading
Enzyme Research 19
frame of 2892 bp encoded for 963 amino acids from thisAntarctic yeast has been cloned and expressed in Pichiaexpression system This is the second extracellular pro-tease reported being synthesized by this psychrophilic yeastSequence analysis through bioinformatics studies revealedfew homology identities of this protein with other ser-ine proteases The mature PI12 protease was successfullyexpressed and secreted into the culture medium driven bythe Saccharomyces cerevisiae 120572-factor signal sequence in Ppastoris The protease production in P pastoris was bestobtained from strain GS115 (GpPro2) with 283UmL after 72hours of induction time with 05 (vv) of methanol inducerThe starting optical density at 600 nm (OD600) was 60 andthe optimum assay temperature was 20∘C The expressedprotein was detected by SDS-PAGE and activity staining witha molecular weight of 99 kDaThis cold-active enzymemightoffer an opportunity for biotechnological exploitation basedon its high catalytic activity at low temperatures and lowthermostability Structural and functional study of this cold-adapted enzyme would give insight into its adaptation to anextreme cold environment
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgment
This research was supported financially by the InitiativeBiotechnology-GMBI-MOSTI (Project number 07-05-16-MG-GMB02) andRUGS-Initiative 6 (Project number 05-02-11-1414RU)
References
[1] J W Deming ldquoPsychrophiles and polar regionsrdquo CurrentOpinion in Microbiology vol 5 no 3 pp 301ndash309 2002
[2] C Gerday M Aittaleb M Bentahir et al ldquoCold-adaptedenzymes from fundamentals to biotechnologyrdquo Trends inBiotechnology vol 18 no 3 pp 103ndash107 2000
[3] A Svingor J Kardos I Hajdu A Nemeth and P Zavodszky ldquoAbetter enzyme to cope with cold comparative flexibility studieson psychrotrophic mesophilic and thermophilic IPMDHSrdquoThe Journal of Biological Chemistry vol 276 no 30 pp 28121ndash28125 2001
[4] G Antranikian C E Vorgias and C Bertoldo ldquoExtremeenvironments as a resource for microorganisms and novel bio-catalystsrdquo Advances in Biochemical EngineeringBiotechnologyvol 96 pp 219ndash262 2005
[5] T Lonhienne C Gerday andG Feller ldquoPsychrophilic enzymesrevisiting the thermodynamic parameters of activation mayexplain local flexibilityrdquoBiochimica et Biophysica Acta vol 1543no 1 pp 1ndash10 2000
[6] R Cavicchioli and TThomas ldquoExtremophilesrdquo in Encyclopediaof Microbiology pp 317ndash337 Academic Press San Diego CalifUSA 2000
[7] G Feller E Narinx J L Arpigny et al ldquoEnzymes frompsychrophilic organismsrdquo FEMS Microbiology Reviews vol 18no 2-3 pp 189ndash202 1996
[8] J Arnorsdottir R B Smaradottir T O Magnusson S HThorbjarnardottir G Eggertsson and M M KristjanssonldquoCharacterization of a cloned subtilisin-like serine proteinasefrom a psychrotrophic Vibrio speciesrdquo European Journal ofBiochemistry vol 269 no 22 pp 5536ndash5546 2002
[9] N J Russell ldquoPsychrophilic bacteriamdashmolecular adaptations ofmembrane lipidsrdquo Comparative Biochemistry and Physiology APhysiology vol 118 no 3 pp 489ndash493 1997
[10] G Feller and C Gerday ldquoPsychrophilic enzymes hot topics incold adaptationrdquo Nature Reviews Microbiology vol 1 no 3 pp200ndash208 2003
[11] P Raspor and J Zupan ldquoYeasts in extreme environmentsrdquo inBiodiversity and Ecophysiology of Yeasts The Yeast Handbookpp 371ndash417 Springer Berlin Germany 2006
[12] M B Rao A M Tanksale M S Ghatge and V V Desh-pande ldquoMolecular and biotechnological aspects of microbialproteasesrdquoMicrobiology andMolecular Biology Reviews vol 62no 3 pp 597ndash635 1998
[13] L Polgar ldquoThe catalytic triad of serine peptidasesrdquo Cellular andMolecular Life Sciences vol 62 no 19-20 pp 2161ndash2172 2005
[14] S Y Boo C M V L Wong K F Rodrigues N NajimudinA M A Murad and N M Mahadi ldquoThermal stress responsesin Antarctic yeastGlaciozyma antarctica PI12 characterized byreal-time quantitative PCRrdquoPolar Biology vol 36 no 3 pp 381ndash389 2013
[15] B Turchetti S R T Hall L B Connell et al ldquoPsychrophilicyeasts from Antarctica and European glaciers descriptionof Glaciozyma gen nov Glaciozyma martinii sp nov andGlaciozyma watsonii sp novrdquo Extremophiles vol 15 no 5 pp573ndash586 2011
[16] N H F Hashim I Bharudin D L S Nguong et al ldquoCharac-terization of Afp1 an antifreeze protein from the psychrophilicyeast Glaciozyma antarctica PI12rdquo Extremophiles vol 17 no 1pp 63ndash73 2013
[17] A N M Ramli M A Azhar M S Shamsir et al ldquoSequenceand structural investigation of a novel psychrophilic 120572-amylasefrom Glaciozyma antarctica PI12 for cold-adaptation analysisrdquoJournal ofMolecularModeling vol 19 no 8 pp 3369ndash3383 2013
[18] A N M Ramli N M Mahadi A Rabu A M A Murad F DA Bakar and R M Illias ldquoMolecular cloning expression andbiochemical characterisation of a cold-adapted novel recombi-nant chitinase from Glaciozyma antarctica PI12rdquoMicrobial CellFactories vol 10 no 1 article 94 2011
[19] J Sambrook E Fritsch and T Maniatis Molecular Cloning ALaboratoryManual Cold SpringHarbor Laboratory Press NewYork NY USA 1989
[20] F M Brock C W Forsberg and J G Buchanan-SmithldquoProteolytic activity of rumen microorganisms and effects ofproteinase inhibitorsrdquoApplied and EnvironmentalMicrobiologyvol 44 no 3 pp 561ndash569 1982
[21] U K Laemmli ldquoCleavage of structural proteins during theassembly of the head of bacteriophage T4rdquo Nature vol 227 no5259 pp 680ndash685 1970
[22] A Henne R A Schmitz M Bomeke G Gottschalk and RDaniel ldquoScreening of environmental DNA libraries for thepresence of genes conferring lipolytic activity on Escherichiacolirdquo Applied and Environmental Microbiology vol 66 no 7 pp3113ndash3116 2000
[23] M Poza T de Miguel C Sieiro and T G Villa ldquoCharacteri-zation of a broad pH range protease of Candida caseinolyticardquoJournal of Applied Microbiology vol 91 no 5 pp 916ndash921 2001
20 Enzyme Research
[24] J D Bendtsen H Nielsen G von Heijne and S BrunakldquoImproved prediction of signal peptides SignalP 30rdquo Journalof Molecular Biology vol 340 no 4 pp 783ndash795 2004
[25] R J Siezen and J A M Leunissen ldquoSubtilases the superfamilyof subtilisin-like serine proteasesrdquo Protein Science vol 6 no 3pp 501ndash523 1997
[26] Y Katakura W Zhang G Zhuang et al ldquoEffect of methanolconcentration on the production of human 1205732- glycoprotein Idomain V by a recombinant Pichia pastoris a simple system forthe control of methanol concentration using a semiconductorgas sensorrdquo Journal of Fermentation and Bioengineering vol 86no 5 pp 482ndash487 1998
[27] Q-FWang Y-HHou Z Xu J-LMiao andG-Y Li ldquoPurifica-tion and properties of an extracellular cold-active protease fromthe psychrophilic bacterium Pseudoalteromonas sp NJ276rdquoBiochemical Engineering Journal vol 38 no 3 pp 362ndash3682008
[28] S Vazquez L Ruberto and W Mac Cormack ldquoProper-ties of extracellular proteases from three psychrotolerantStenotrophomonas maltophilia isolated from Antarctic soilrdquoPolar Biology vol 28 no 4 pp 319ndash325 2005
[29] R Zeng R Zhang J Zhao and N Lin ldquoCold-active ser-ine alkaline protease from the psychrophilic bacterium Pseu-domonas strain DY-A enzyme purification and characteriza-tionrdquo Extremophiles vol 7 no 4 pp 335ndash337 2003
[30] M Turkiewicz E Gromek H Kalinowska and M ZielinskaldquoBiosynthesis and properties of an extracellularmetalloproteasefrom the Antarctic marine bacterium Sphingomonas paucimo-bilisrdquo Journal of Biotechnology vol 70 no 1ndash3 pp 53ndash60 1999
[31] A Vassileva D A Chugh S Swaminathan and N KhannaldquoEffect of copy number on the expression levels of hepatitisB surface antigen in the methylotrophic yeast Pichia pastorisrdquoProtein Expression andPurification vol 21 no 1 pp 71ndash80 2001
[32] M Pazgier M Turkiewicz H Kalinowska and S Bielecki ldquoTheunique cold-adapted extracellular subtilase from psychrophilicyeast Leucosporidium antarcticumrdquo Journal ofMolecular Catal-ysis B Enzymatic vol 21 no 1-2 pp 39ndash42 2003
[33] M Turkiewicz M Pazgier H Kalinowska and S BieleckildquoA cold-adapted extracellular serine proteinase of the yeastLeucosporidium antarcticumrdquo Extremophiles vol 7 no 6 pp435ndash442 2003
[34] D Mattanovich B Gasser H Hohenblum and M SauerldquoStress in recombinant protein producing yeastsrdquo Journal ofBiotechnology vol 113 no 1ndash3 pp 121ndash135 2004
[35] P Secades BAlvarez and JAGuijarro ldquoPurification andprop-erties of a new psychrophilic metalloprotease (Fpp2) in the fishpathogen Flavobacterium psychrophilumrdquo FEMS MicrobiologyLetters vol 226 no 2 pp 273ndash279 2003
[36] L Y Ling I Ithoi and F M Yik ldquoOptimization for high-level expression in Pichia pastoris and purification of truncatedand full length recombinant sag2 of toxoplasma gondii fordiagnostic userdquo Southeast Asian Journal of Tropical Medicineand Public Health vol 41 no 3 pp 507ndash513 2010
[37] A Illanes ldquoStability of biocatalystsrdquo Electronic Journal ofBiotechnology vol 2 no 1 pp 1ndash9 1999
[38] B A Plantz J Sinha L Villarete K W Nickerson and VL Schlegel ldquoPichia pastoris fermentation optimization energystate and testing a growth-associated modelrdquo Applied Microbi-ology and Biotechnology vol 72 no 2 pp 297ndash305 2006
[39] K Tamura G Stecher D Peterson A Filipski and S KumarldquoMEGA6molecular evolutionary genetics analysis version 60rdquo
Molecular Biology and Evolution vol 30 no 12 pp 2725ndash27292013
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology
14 Enzyme Research
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
NATLIQAADGVFLAQQVAAGKSVTITFPQTGGSAQITN-PSGGLVSSFTTYGPTNDFYFK
-ASFIQAADGEFLVQQFASGQNVSLSFPQSGGLIQFPD-PAGGLISSFTSYGPSNDFFFK
-AALISETDGTFLVNQFAAKANITISFPQSGSAWNYPDDVSGGLVSDFSSYGPSYDMYFK
VGSLRYEDGLKLLSYYQSSPRGQSISFPNGPLAPYVSNTVSGGVVASYSNFGPTNELFIY
VAGLRRSDGLKLLSYYQNANKRLTLRFPKGKIVAGLTDTITGGLISGYSTFGPTNDLYGQ
lowast lowastlowast lowastlowast lowastlowast
PAFGAPGGNILSTLPVPLGSYGVESGTSMATPYVAGSAALLLSVKGKSAAVALGARTLFQ
PAVAAPGGNILSTLPVNLGLFGIESGTSMATPFVAGSAALLFQAKGTSAAVGRSARTLFE
PAVAAPGGNILSTLPIALGSYGVESGTSMATPFTAGSAALILQAKGK--SVAKSIRNLFQ
PTLAAPGTDILSTV---TGGVALMRGTSMAAPLVAGAYALLLSVGNNLKLSPEEARTILM
PTLSAPGGNILSTFPLSEGGVAVISGTSMSCPFVAGSAAVLMAARASENLTPLEIRSLLT
lowast lowastlowastlowast lowastlowastlowastlowast lowast lowastlowastlowastlowast lowast lowastlowast lowast lowast
STAQRVPSSHTDGDPLQTLSQQGAGLIDVFKALYTETIVWPTELILNDTAHYKEVHTFTV
TTAQRVPSTHTDGDPLQTLTQQGAGLINVFNAIHTTTIVSPGELILNDTAHFKGTQQFTV
TTASAVASSKTDGDPLQTVSQQGAGLIQVDRAIGTTTVVSPGQLTLNDTAHFEGFKTFNI
STSQPSPVAY-GSETLDTVVSQGAGVINMTAAIAAHTVISPAQFNLNDTQYSNNTQTLTL
TTAKLTPVSLLGSTPLVSVIRQGGGLVQVAKALAAKTLISPHELLLNDTANANYVQTIKI
VNTGKTSKQYKLTHVPAGTAQTIT---SGTIFAADGPVPLSTDYATVKLSSSSFTLRPGQ
RNAGSSAKTYTLRHIPAGTAVTVT---PGTIFPADGPVPLSTDFASVSLSTSKFTLAPGK
QNTGSKAVTYKISHVPAGTAASVQ---SGSIFAADGPVPLTADYAQVTFSLNKVTVLPGV
RNNNRFPVSYQLSWVDSTGIVTYNDGATTDIIPSTTPNYVSTSVLRVAFSQRTVTVPGGQ
KNTNSWAMKYTFSSAVAQGLGTFD--ASGDILPTLDPVAVSGAQATVAFNTRILSVAPGA
lowast lowast lowast lowast lowast lowast
SKQVTTHINAP--SGVDASTYPVFSGFIEIATAT--------ESVHVTYLGLAASLKDKQ
TQTVTARFTPP--AAADPSTFPVFSGFIQIESGT--------EQVQVSYLGLKASLKDKQ
SVPILVWFKPP--QGVDASTYPVYSGFIEIAGLG--------ETLRVAYLGVAASLKDKA
TVKVTAQFIPPNLTAQQRNQFPIYSGFVTITGQGQGAGAEQMEKYNLPFFGLGARMVDMP
TGSVVATITPPVLPVADAARFPIFSGWIRVNGQG-ARDSSRNEAYTVPYFGLAAKMIDMQ
lowast lowast lowastlowast lowast lowast lowast lowast
IVDNTNEFFGITIPAVLNNTGDAQ-LSPTNYTFSGSDFPTLLLRLAFGTPLLRVDLVDAT
VIDNTDFFFGVPTPVLTDPNGEVQ-TSPRNYSFLASDFPTLIFRLAFGSPKVVVDLVSPT
VVDNTDEYFGVKIPALLDASGDVQ-GNSTNYTLVGDDVPSLLYRFAFGSAHVRVDLVEKA
VLDTTNVALGP--NLPFIAVGQN--IQTGPTTFSESNPAVVYFRLAAGTRRLRVDLVDAS
VLDTTETIYGPGYAYPFVIDDAIGDIQSTTTSYSRNLGPTVFARFATGTLHYSLDLVLAD
lowast lowast lowast lowast lowastlowastlowastlowast lowast
IDLKPTLNSREIDDKRHVFERESFSFPHPGPKGS-------------------TFAQVKI
INFKPTLNTRALGDTHGSF----FSFPHRVKTG--------------------SFAQVPT
STVSATLSKRNWWN---------WWWPQKPSN---------------------SYDKVPI
TSYNATVPAVTNPASRLVKRSNTELTPRAAFPT--------------------LYSQVRT
IAFTPTYPNSS-PATRFVKRSLTQHTSAASHLAKRRVSIATINPKATLVADRQLHSDVPI
lowast lowast
VGTLSEQSHIPRNSDASTVDDGGYSTFAFNKP------SFANGTTIPNGTYRWLLRALKV
LGTVFELDFTSRNNDDPTDN--GFNTVAIDPP------TFANGTTIPNGQYRLLLRALRV
VGTLAEYDYVPRNSDASTIDDNGYNVFQFT-D------SFVNGTKIAAGEYKVLLRALKV
VGSLFAPSYWPPRDYLFNNGGT-YSDYEIQLDGSFTFPNGTKGMAELGRSYKVLLRALKI
EGNIFTQPFTG-RDYLVDAAPTGSTDRTVTFNGQY-AENGLVRTAVTGTSYRFLLRALKI
lowast lowast lowastlowast lowast lowast lowast lowast lowastlowastlowastlowast
lowast lowast lowastlowastlowastlowastlowast
(b)
Figure 5 Continued
Enzyme Research 15
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
TGDPTKEADFETWLSPIVGITVA
TGDPTNEADFESFLSPIIGVNAP
TGNPAEEGDYETWLSPSFSVVPA
TADPTYTSSYESWLSPSFTFTA-
SGDAMYEDQYESWLSLPFSFRA-
lowast lowastlowast
ConservedSimilar
lowast All match
(c)
Figure 5 Multiple sequence alignment between amino acid sequence of PI12 protease with other subtilisin The serine protease sequenceswere obtained from the GenBank database Their accession numbers are EMS20793 (subtilisin-like protease of Rhodosporidium toruloidesNP11) CAG25549 (subtilisin-like protease of Pleurotus ostreatus) EPQ55564 (subtilisin-like protease of Gloeophyllum trabeumATCC 11539)and ETW80907 (serine protease S8 ofHeterobasidion irregulare TC 32-1) Putative ORF of serine protease fromG antarctica PI12 was labeledas PI12 protease
CAG38357 Phanerochaete chrysosporium
EIW62675 Trametes versicolor FP-101664 SS1
XP 007369624 Dichomitus squalens LYAD-421 SS1
XP 007381302 Punctularia strigosozonata HHB-11173 SS5
CAG25549 Pleurotus ostreatus
ETW80907 Heterobasidion irregulare TC 32-1
XP 001877576 Laccaria bicolor S238N-H82
EPQ55564 Gloeophyllum trabeum ATCC 11539
XP 001830835 Coprinopsis cinerea okayama7130
XP 007341385 Auricularia delicata TFB-10046 SS5
CCA68627 Piriformospora indica DSM 11827
CCO37249 Rhizoctonia solani AG-1 IB
XP 007342189 Auricularia delicata TFB-10046 SS5
EJU04161 Dacryopinax sp DJM-731 SS1
XP 007589863 Colletotrichum fioriniae PJ7
EFQ32840 Colletotrichum graminicola M1001
ENH74885 Fusarium oxysporum f sp cubense race 1
XP 003711175 Magnaporthe oryzae 70-15
XP 006694395 Chaetomium thermophilum DSM 1495
EON99722 Togninia minima UCRPA7
CBQ73425 Sporisorium reilianum SRZ2
GAC97617 Pseudozyma hubeiensis SY62
CCF52619 Ustilago hordei
EMS20793 Rhodosporidium toruloides NP11
CAQ76821 Glaciozyma antarctica PI12
01
Clus
ter 1
Clus
ter 2
a
b
c
d
Figure 6 Phylogenetic dendrogram among 25 translated amino acid sequences of serine proteaseThe tree was constructed using a neighbor-joining algorithm with 1000 bootstrappings by MEGA 6 software [39]The scale bar represents 01 substitutions per amino acid positionTheserine protease sequences were obtained from the GenBank database GenBank accession numbers of the sequences were indicated beforethe strain name The putative serine protease of G antarctica strain PI12 has been deposited in GenBank with accession number CAQ76821
16 Enzyme Research
0
2
4
6
8
10
12
0 24 48 72 96 110
Prot
ease
activ
ity (U
mL)
Post induction time (hours)
Figure 7 Effect of postinduction time towards protease activity The highest activity was detected at 72 hours after 110 hours of incubationwith 9UmL activity followed by 48 24 and 96 hoursThe substrate (azocasein) was dissolved in Tris HCL buffer and incubated for 30min at20∘CThe absorbance was measured at 450 nm after 15min incubation A one-way ANOVAwas used to test for preference differences amongsix different postinduction times The statistical value (119875 value lt005) between hours is 0003
pink-coloured nonpathogenic oleaginous yeast (42 iden-tity) Both G antarctica PI12 and Rhodosporidium toruloidesNP11 belong to Microbotryomycetes class PI12 protease didnot show any homology similarity to other psychrophilicproteases however it showed 37 identity to hypotheticalprotein from Pseudozyma antarctica T-34 which was isolatedfrom Japan (GAC76005) (not listed in the phylogenetic tree)
35 Cloning and Expression of Cold-Adapted PI12 Protease inthe Yeast Expression System The PCR product harbouringthe mature PI12 protease with two flanking restriction sites(EcoRI and AvrII) was cloned into pPIC9 expression vectorThe recombinant plasmid harbouring PI12 protease was usedto transform both GS115 and KM71 intended to produce bothHis+ Mut+ and His+ MutS Two recombinant clones fromGS115 (His+Mut+) strain (GpPro1 andGpPro2) and one fromKM71 (His+MutS) strain (KpPro1) were successfully obtainedwhich were further analyzed for protein expression study
36 Expression and Enzyme Assay of PI12 Protease in Pichiapastoris All positive recombinants were grown in small scalecultures and assayed for the cold-adapted protease activityin the culture supernatants The clones were first grown inBMGY medium with glycerol as the sole carbon sourceFor induction of AOX1 promoter and protein productionthe growth medium was transferred to the same mediumexcept with 05 methanol in place with glycerol as thecarbon source [26] The supernatant of the recombinant Ppastoris expression cultures after 48 h of incubation time wasanalyzed to determine the recombinant protease activity bythe modified method of Brock et al (1982) [20] All testedclones secreted protease into the supernatant in differentexpression levels This might be due to the variability ofthe strength of the promoter and copy number in each cellto secrete and express the recombinant product The clonewith the highest yield was GpPro2 with 63UmL activityfollowed by KpPro1 and GpPro1 with 25UmL and 37UmLactivities respectively (data not shown) It is perceptible thatexpression systemof Pichia pastoris is the ideal system for thisprotein The preliminary assay has fruitfully produced high
protease expression at 20∘C Yet further optimization of therecombinant PI12 protease production need to be conductedin order to examine the optimum condition of this proteinReported cold-adapted proteases showed optimum activity ata range of 15∘Cndash60∘C [27ndash30] This difference was possiblyrelated to the respective genetic and physiological adaptationof the strains There are several factors that influenced thedifferent rate of expression It includes different site of inte-gration between interest gene and Pichia genome variabilityof different expression host cell to secrete and express therecombinant enzyme and different sites in the polylinker forinserting foreign genes [31]
This is the second extracellular protease reported to besynthesized by this obligate psychrophilic yeastG antarcticaThe first cold-adapted extracellular subtilase from this yeasthas been purified and characterized previously [32 33] Theextracellular serine proteinase (LAP2) from this Antarcticyeast formerly known as Leucosporidium antarcticum strain171 is specific towards synthetic substrates of chymotrypsinand subtilisin The subtilase of L antarcticum strain 171showed specific activity of 4043Umg when utilized N-SucAAPFpNA as a substrate [33]
37 Optimization of Protease Production GpPro2 was chosenfor further optimization due to its highest protease activityat the preliminary expression stage High protease activitywas detected after 72 hours of incubation (Figure 7) byusing the standard protocol method as recommended forPichia pastoris expression system (Pichia Expression KitInvitrogen USA) The expression decreased after 72 hoursand no activity was detected at 110 hours The prolongedincubation may result in the decrease of biomass productionand will subsequently reduce the expression of recombinantprotein In addition oxygen and nutrient depletion duringprolonged incubationwill trigger the stress responses in orderto survive in such harsh environment [34]
Methanol was used as a sole carbon by P pastoris Themethanol was oxidized to formaldehyde with the aid of oxy-gen as electron acceptor by an alcohol oxidase enzyme (AOXgene) [35] The optimal methanol concentration is at 05(vv) with 14UmL activity The addition of more than 05
Enzyme Research 17
02468
10121416
0 01 03 05 07 09Pr
otea
se ac
tivity
(Um
L)Methanol concentration ()
Figure 8 Effect ofmethanol concentration towards protease activityThe recombinant was cultivated for 72 hours at 30∘C in BMMYmedium05 methanol gives the highest activity with 14UmL The substrate (azocasein) was dissolved in Tris HCL buffer and incubated for 30minat 20∘CThe mixture was left for 15min and absorbance was taken at 450 nm A one-way ANOVA was used to test for preference differencesamong six different concentrations The statistical value (119875 value lt005) between concentration is 002
0
5
10
15
20
25
10 15 20 25 30 35
Prot
eas a
ctiv
ity (
Um
L)
Temperature (∘C)
Figure 9 Effect of temperature towards protease activity The reaction was carried out in temperature ranging from 10∘C to 35∘C at pH 7 for30min A one-way ANOVA was used to test for preference differences among six different temperatures The statistical value (119875 value lt005)between temperature is 0067
(vv) methanol resulted in tremendous decrease of proteaseactivity (Figure 8) Production of PI12 protease was decreasedwhen the methanol concentration increased This might bedue to the toxic effect of methanol concentration on DNAreplication and membrane synthesis Basically methanol isused to increase the permeability of the cell membrane tohelp the secretion of foreign protein [26] However if thepermeability of cell membrane is too high lowmolecular sizemetabolites would leak from the cell and such leakage willseriously damage cell activities Such condition would abusethe cellular system and might reduce the yield of proteinexpression [36]
Psychrophilic microorganism dominated in low temper-ature habitat which most other microorganism are inca-pable of withstanding due to their inability to produceenzyme that can tolerate the extreme environment con-dition Psychrophilic enzymes are always associated withhigh catalytic efficiency at low and moderate temperature(0∘C to 30∘C) compared to the mesophilic counterpart thatare generally unable to catalyze efficiently under extremely
low temperature [2] The recombinant protein assay wasdone at various temperatures ranging from 10∘C to 35∘Cusing 05 azocasein as a substrate dissolved in 50mMTris HCl in presence of 2mM CaCl
2 pH 7 In the activity
assays expressed protease showed a highest activity at 20∘C(20UmL) and decreased tremendously once it exceededmaximum temperature due to the denatured state of theenzyme (Figure 9) The active site of an enzyme is easilyaltered when the high temperature is introduced duringincubation or activity assay resulting in a low reaction rate[37]
The recombinant construct was transferred into BMMYmedia at different A
600 nm ranging from 1 to 10 The high-est activity of active enzyme was detected at A
600 nm 60(283UmL) and above (Figure 10) The slope during expo-nential phase was much higher compared to other phasesindicating an increase in the doubling time At exponentialphase the cells are adapted to the growth condition andother processes like cell growth cycle and synthesis of RNAThe yield of active enzyme during expression of P pastoris
18 Enzyme Research
05
1015202530
1 2 3 4 5 6 7 8 9 10Prot
ease
activ
ity (U
mL)
Initial inoculum A600
Figure 10 Effect of inoculums size towards protease activity The recombinant was cultivated for 72 hours at 30∘C in BMMY mediumwith 05 methanol concentration A one-way ANOVA was used to test for preference differences among six different optical densitiesThe statistical value (119875 value lt005) between optical density value is 003
(kD
a)
1160
662
450
350
250
184
144
M 1 2 3
(a) (b) (c)
Figure 11 (a) Coomassie-stained SDS-PAGE of recombinant P pastoris clone 1mL supernatants collected from the third-day culturewere concentrated 10-fold using trichloroacetic acid and 10120583L resultant products were applied to SDS-PAGE analysis Lane 1 molecularweight marker (Fermentas) Lane 2 GS115 (Control) Lane 3 GS115pPIC9 empty (Control) Lane 4 GS115pPIC9PI12prot (GpPro2)The arrow indicates the secreted recombinant PI12 protease (b) Plate activity staining of different recombinant P pastoris clones GGS115 GP GS115pPIC9 GRI GS115pPIC9PI12prot1 (GpPro1) GRII GS115pPIC9PI12prot2 (GpPro2) K KM71 KP KM71pPIC9 KRKM71pPIC9PI12prot (KpPro) (c)Gel activity staining of recombinant PI12 protease inP pastorisGpPro2 clone Lane 1 prestainedmolecularweightmarker (Fermentas) Lane 2 GS115pPIC9 empty (Control) Lane 3 GS115pPIC9PI12prot1 (GpPro2)The arrow indicates the secretedrecombinant PI12 protease
dropped after 28 hours of incubation which is at stationaryor death phase [38]
38 Detection of Recombinant Protein by SDS-PAGE andActivity Staining TheSDS-PAGE gel of the GpPro2 recombi-nant clonesrsquo supernatant showed a putative expression bandat around 993 kDa Plate staining and gel activity stainingwere executed to examine if the recombinant PI12 proteaseexpressed in P pastoris was functionally active In the platestaining the transformants showed strong proteolytic activity(Figure 11(a)) In gel staining proteolytic activity was alsodetected around 993 kDa after incubation at 20∘C This is
evidence of protease secretion into the surrounding mediawhereby it catalyzed the breakdown of casein thus formedclearing zones around the activity ring and band (Figures11(b) and 11(c)) As a conclusion the cold-adapted PI12protease was functionally expressed in P pastoris GS115(GpPro2 clone) with the size of 993 kDa
4 Conclusions
Psychrophilic yeast which was originated frommarine waterin Casey Station Antarctica was identified as Glaciozymaantarctica strain PI12 A serine protease with an open reading
Enzyme Research 19
frame of 2892 bp encoded for 963 amino acids from thisAntarctic yeast has been cloned and expressed in Pichiaexpression system This is the second extracellular pro-tease reported being synthesized by this psychrophilic yeastSequence analysis through bioinformatics studies revealedfew homology identities of this protein with other ser-ine proteases The mature PI12 protease was successfullyexpressed and secreted into the culture medium driven bythe Saccharomyces cerevisiae 120572-factor signal sequence in Ppastoris The protease production in P pastoris was bestobtained from strain GS115 (GpPro2) with 283UmL after 72hours of induction time with 05 (vv) of methanol inducerThe starting optical density at 600 nm (OD600) was 60 andthe optimum assay temperature was 20∘C The expressedprotein was detected by SDS-PAGE and activity staining witha molecular weight of 99 kDaThis cold-active enzymemightoffer an opportunity for biotechnological exploitation basedon its high catalytic activity at low temperatures and lowthermostability Structural and functional study of this cold-adapted enzyme would give insight into its adaptation to anextreme cold environment
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgment
This research was supported financially by the InitiativeBiotechnology-GMBI-MOSTI (Project number 07-05-16-MG-GMB02) andRUGS-Initiative 6 (Project number 05-02-11-1414RU)
References
[1] J W Deming ldquoPsychrophiles and polar regionsrdquo CurrentOpinion in Microbiology vol 5 no 3 pp 301ndash309 2002
[2] C Gerday M Aittaleb M Bentahir et al ldquoCold-adaptedenzymes from fundamentals to biotechnologyrdquo Trends inBiotechnology vol 18 no 3 pp 103ndash107 2000
[3] A Svingor J Kardos I Hajdu A Nemeth and P Zavodszky ldquoAbetter enzyme to cope with cold comparative flexibility studieson psychrotrophic mesophilic and thermophilic IPMDHSrdquoThe Journal of Biological Chemistry vol 276 no 30 pp 28121ndash28125 2001
[4] G Antranikian C E Vorgias and C Bertoldo ldquoExtremeenvironments as a resource for microorganisms and novel bio-catalystsrdquo Advances in Biochemical EngineeringBiotechnologyvol 96 pp 219ndash262 2005
[5] T Lonhienne C Gerday andG Feller ldquoPsychrophilic enzymesrevisiting the thermodynamic parameters of activation mayexplain local flexibilityrdquoBiochimica et Biophysica Acta vol 1543no 1 pp 1ndash10 2000
[6] R Cavicchioli and TThomas ldquoExtremophilesrdquo in Encyclopediaof Microbiology pp 317ndash337 Academic Press San Diego CalifUSA 2000
[7] G Feller E Narinx J L Arpigny et al ldquoEnzymes frompsychrophilic organismsrdquo FEMS Microbiology Reviews vol 18no 2-3 pp 189ndash202 1996
[8] J Arnorsdottir R B Smaradottir T O Magnusson S HThorbjarnardottir G Eggertsson and M M KristjanssonldquoCharacterization of a cloned subtilisin-like serine proteinasefrom a psychrotrophic Vibrio speciesrdquo European Journal ofBiochemistry vol 269 no 22 pp 5536ndash5546 2002
[9] N J Russell ldquoPsychrophilic bacteriamdashmolecular adaptations ofmembrane lipidsrdquo Comparative Biochemistry and Physiology APhysiology vol 118 no 3 pp 489ndash493 1997
[10] G Feller and C Gerday ldquoPsychrophilic enzymes hot topics incold adaptationrdquo Nature Reviews Microbiology vol 1 no 3 pp200ndash208 2003
[11] P Raspor and J Zupan ldquoYeasts in extreme environmentsrdquo inBiodiversity and Ecophysiology of Yeasts The Yeast Handbookpp 371ndash417 Springer Berlin Germany 2006
[12] M B Rao A M Tanksale M S Ghatge and V V Desh-pande ldquoMolecular and biotechnological aspects of microbialproteasesrdquoMicrobiology andMolecular Biology Reviews vol 62no 3 pp 597ndash635 1998
[13] L Polgar ldquoThe catalytic triad of serine peptidasesrdquo Cellular andMolecular Life Sciences vol 62 no 19-20 pp 2161ndash2172 2005
[14] S Y Boo C M V L Wong K F Rodrigues N NajimudinA M A Murad and N M Mahadi ldquoThermal stress responsesin Antarctic yeastGlaciozyma antarctica PI12 characterized byreal-time quantitative PCRrdquoPolar Biology vol 36 no 3 pp 381ndash389 2013
[15] B Turchetti S R T Hall L B Connell et al ldquoPsychrophilicyeasts from Antarctica and European glaciers descriptionof Glaciozyma gen nov Glaciozyma martinii sp nov andGlaciozyma watsonii sp novrdquo Extremophiles vol 15 no 5 pp573ndash586 2011
[16] N H F Hashim I Bharudin D L S Nguong et al ldquoCharac-terization of Afp1 an antifreeze protein from the psychrophilicyeast Glaciozyma antarctica PI12rdquo Extremophiles vol 17 no 1pp 63ndash73 2013
[17] A N M Ramli M A Azhar M S Shamsir et al ldquoSequenceand structural investigation of a novel psychrophilic 120572-amylasefrom Glaciozyma antarctica PI12 for cold-adaptation analysisrdquoJournal ofMolecularModeling vol 19 no 8 pp 3369ndash3383 2013
[18] A N M Ramli N M Mahadi A Rabu A M A Murad F DA Bakar and R M Illias ldquoMolecular cloning expression andbiochemical characterisation of a cold-adapted novel recombi-nant chitinase from Glaciozyma antarctica PI12rdquoMicrobial CellFactories vol 10 no 1 article 94 2011
[19] J Sambrook E Fritsch and T Maniatis Molecular Cloning ALaboratoryManual Cold SpringHarbor Laboratory Press NewYork NY USA 1989
[20] F M Brock C W Forsberg and J G Buchanan-SmithldquoProteolytic activity of rumen microorganisms and effects ofproteinase inhibitorsrdquoApplied and EnvironmentalMicrobiologyvol 44 no 3 pp 561ndash569 1982
[21] U K Laemmli ldquoCleavage of structural proteins during theassembly of the head of bacteriophage T4rdquo Nature vol 227 no5259 pp 680ndash685 1970
[22] A Henne R A Schmitz M Bomeke G Gottschalk and RDaniel ldquoScreening of environmental DNA libraries for thepresence of genes conferring lipolytic activity on Escherichiacolirdquo Applied and Environmental Microbiology vol 66 no 7 pp3113ndash3116 2000
[23] M Poza T de Miguel C Sieiro and T G Villa ldquoCharacteri-zation of a broad pH range protease of Candida caseinolyticardquoJournal of Applied Microbiology vol 91 no 5 pp 916ndash921 2001
20 Enzyme Research
[24] J D Bendtsen H Nielsen G von Heijne and S BrunakldquoImproved prediction of signal peptides SignalP 30rdquo Journalof Molecular Biology vol 340 no 4 pp 783ndash795 2004
[25] R J Siezen and J A M Leunissen ldquoSubtilases the superfamilyof subtilisin-like serine proteasesrdquo Protein Science vol 6 no 3pp 501ndash523 1997
[26] Y Katakura W Zhang G Zhuang et al ldquoEffect of methanolconcentration on the production of human 1205732- glycoprotein Idomain V by a recombinant Pichia pastoris a simple system forthe control of methanol concentration using a semiconductorgas sensorrdquo Journal of Fermentation and Bioengineering vol 86no 5 pp 482ndash487 1998
[27] Q-FWang Y-HHou Z Xu J-LMiao andG-Y Li ldquoPurifica-tion and properties of an extracellular cold-active protease fromthe psychrophilic bacterium Pseudoalteromonas sp NJ276rdquoBiochemical Engineering Journal vol 38 no 3 pp 362ndash3682008
[28] S Vazquez L Ruberto and W Mac Cormack ldquoProper-ties of extracellular proteases from three psychrotolerantStenotrophomonas maltophilia isolated from Antarctic soilrdquoPolar Biology vol 28 no 4 pp 319ndash325 2005
[29] R Zeng R Zhang J Zhao and N Lin ldquoCold-active ser-ine alkaline protease from the psychrophilic bacterium Pseu-domonas strain DY-A enzyme purification and characteriza-tionrdquo Extremophiles vol 7 no 4 pp 335ndash337 2003
[30] M Turkiewicz E Gromek H Kalinowska and M ZielinskaldquoBiosynthesis and properties of an extracellularmetalloproteasefrom the Antarctic marine bacterium Sphingomonas paucimo-bilisrdquo Journal of Biotechnology vol 70 no 1ndash3 pp 53ndash60 1999
[31] A Vassileva D A Chugh S Swaminathan and N KhannaldquoEffect of copy number on the expression levels of hepatitisB surface antigen in the methylotrophic yeast Pichia pastorisrdquoProtein Expression andPurification vol 21 no 1 pp 71ndash80 2001
[32] M Pazgier M Turkiewicz H Kalinowska and S Bielecki ldquoTheunique cold-adapted extracellular subtilase from psychrophilicyeast Leucosporidium antarcticumrdquo Journal ofMolecular Catal-ysis B Enzymatic vol 21 no 1-2 pp 39ndash42 2003
[33] M Turkiewicz M Pazgier H Kalinowska and S BieleckildquoA cold-adapted extracellular serine proteinase of the yeastLeucosporidium antarcticumrdquo Extremophiles vol 7 no 6 pp435ndash442 2003
[34] D Mattanovich B Gasser H Hohenblum and M SauerldquoStress in recombinant protein producing yeastsrdquo Journal ofBiotechnology vol 113 no 1ndash3 pp 121ndash135 2004
[35] P Secades BAlvarez and JAGuijarro ldquoPurification andprop-erties of a new psychrophilic metalloprotease (Fpp2) in the fishpathogen Flavobacterium psychrophilumrdquo FEMS MicrobiologyLetters vol 226 no 2 pp 273ndash279 2003
[36] L Y Ling I Ithoi and F M Yik ldquoOptimization for high-level expression in Pichia pastoris and purification of truncatedand full length recombinant sag2 of toxoplasma gondii fordiagnostic userdquo Southeast Asian Journal of Tropical Medicineand Public Health vol 41 no 3 pp 507ndash513 2010
[37] A Illanes ldquoStability of biocatalystsrdquo Electronic Journal ofBiotechnology vol 2 no 1 pp 1ndash9 1999
[38] B A Plantz J Sinha L Villarete K W Nickerson and VL Schlegel ldquoPichia pastoris fermentation optimization energystate and testing a growth-associated modelrdquo Applied Microbi-ology and Biotechnology vol 72 no 2 pp 297ndash305 2006
[39] K Tamura G Stecher D Peterson A Filipski and S KumarldquoMEGA6molecular evolutionary genetics analysis version 60rdquo
Molecular Biology and Evolution vol 30 no 12 pp 2725ndash27292013
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

Enzyme Research 15
ETW80907
CAG25549
EPQ55564
EMS20793PI12 protease
TGDPTKEADFETWLSPIVGITVA
TGDPTNEADFESFLSPIIGVNAP
TGNPAEEGDYETWLSPSFSVVPA
TADPTYTSSYESWLSPSFTFTA-
SGDAMYEDQYESWLSLPFSFRA-
lowast lowastlowast
ConservedSimilar
lowast All match
(c)
Figure 5 Multiple sequence alignment between amino acid sequence of PI12 protease with other subtilisin The serine protease sequenceswere obtained from the GenBank database Their accession numbers are EMS20793 (subtilisin-like protease of Rhodosporidium toruloidesNP11) CAG25549 (subtilisin-like protease of Pleurotus ostreatus) EPQ55564 (subtilisin-like protease of Gloeophyllum trabeumATCC 11539)and ETW80907 (serine protease S8 ofHeterobasidion irregulare TC 32-1) Putative ORF of serine protease fromG antarctica PI12 was labeledas PI12 protease
CAG38357 Phanerochaete chrysosporium
EIW62675 Trametes versicolor FP-101664 SS1
XP 007369624 Dichomitus squalens LYAD-421 SS1
XP 007381302 Punctularia strigosozonata HHB-11173 SS5
CAG25549 Pleurotus ostreatus
ETW80907 Heterobasidion irregulare TC 32-1
XP 001877576 Laccaria bicolor S238N-H82
EPQ55564 Gloeophyllum trabeum ATCC 11539
XP 001830835 Coprinopsis cinerea okayama7130
XP 007341385 Auricularia delicata TFB-10046 SS5
CCA68627 Piriformospora indica DSM 11827
CCO37249 Rhizoctonia solani AG-1 IB
XP 007342189 Auricularia delicata TFB-10046 SS5
EJU04161 Dacryopinax sp DJM-731 SS1
XP 007589863 Colletotrichum fioriniae PJ7
EFQ32840 Colletotrichum graminicola M1001
ENH74885 Fusarium oxysporum f sp cubense race 1
XP 003711175 Magnaporthe oryzae 70-15
XP 006694395 Chaetomium thermophilum DSM 1495
EON99722 Togninia minima UCRPA7
CBQ73425 Sporisorium reilianum SRZ2
GAC97617 Pseudozyma hubeiensis SY62
CCF52619 Ustilago hordei
EMS20793 Rhodosporidium toruloides NP11
CAQ76821 Glaciozyma antarctica PI12
01
Clus
ter 1
Clus
ter 2
a
b
c
d
Figure 6 Phylogenetic dendrogram among 25 translated amino acid sequences of serine proteaseThe tree was constructed using a neighbor-joining algorithm with 1000 bootstrappings by MEGA 6 software [39]The scale bar represents 01 substitutions per amino acid positionTheserine protease sequences were obtained from the GenBank database GenBank accession numbers of the sequences were indicated beforethe strain name The putative serine protease of G antarctica strain PI12 has been deposited in GenBank with accession number CAQ76821
16 Enzyme Research
0
2
4
6
8
10
12
0 24 48 72 96 110
Prot
ease
activ
ity (U
mL)
Post induction time (hours)
Figure 7 Effect of postinduction time towards protease activity The highest activity was detected at 72 hours after 110 hours of incubationwith 9UmL activity followed by 48 24 and 96 hoursThe substrate (azocasein) was dissolved in Tris HCL buffer and incubated for 30min at20∘CThe absorbance was measured at 450 nm after 15min incubation A one-way ANOVAwas used to test for preference differences amongsix different postinduction times The statistical value (119875 value lt005) between hours is 0003
pink-coloured nonpathogenic oleaginous yeast (42 iden-tity) Both G antarctica PI12 and Rhodosporidium toruloidesNP11 belong to Microbotryomycetes class PI12 protease didnot show any homology similarity to other psychrophilicproteases however it showed 37 identity to hypotheticalprotein from Pseudozyma antarctica T-34 which was isolatedfrom Japan (GAC76005) (not listed in the phylogenetic tree)
35 Cloning and Expression of Cold-Adapted PI12 Protease inthe Yeast Expression System The PCR product harbouringthe mature PI12 protease with two flanking restriction sites(EcoRI and AvrII) was cloned into pPIC9 expression vectorThe recombinant plasmid harbouring PI12 protease was usedto transform both GS115 and KM71 intended to produce bothHis+ Mut+ and His+ MutS Two recombinant clones fromGS115 (His+Mut+) strain (GpPro1 andGpPro2) and one fromKM71 (His+MutS) strain (KpPro1) were successfully obtainedwhich were further analyzed for protein expression study
36 Expression and Enzyme Assay of PI12 Protease in Pichiapastoris All positive recombinants were grown in small scalecultures and assayed for the cold-adapted protease activityin the culture supernatants The clones were first grown inBMGY medium with glycerol as the sole carbon sourceFor induction of AOX1 promoter and protein productionthe growth medium was transferred to the same mediumexcept with 05 methanol in place with glycerol as thecarbon source [26] The supernatant of the recombinant Ppastoris expression cultures after 48 h of incubation time wasanalyzed to determine the recombinant protease activity bythe modified method of Brock et al (1982) [20] All testedclones secreted protease into the supernatant in differentexpression levels This might be due to the variability ofthe strength of the promoter and copy number in each cellto secrete and express the recombinant product The clonewith the highest yield was GpPro2 with 63UmL activityfollowed by KpPro1 and GpPro1 with 25UmL and 37UmLactivities respectively (data not shown) It is perceptible thatexpression systemof Pichia pastoris is the ideal system for thisprotein The preliminary assay has fruitfully produced high
protease expression at 20∘C Yet further optimization of therecombinant PI12 protease production need to be conductedin order to examine the optimum condition of this proteinReported cold-adapted proteases showed optimum activity ata range of 15∘Cndash60∘C [27ndash30] This difference was possiblyrelated to the respective genetic and physiological adaptationof the strains There are several factors that influenced thedifferent rate of expression It includes different site of inte-gration between interest gene and Pichia genome variabilityof different expression host cell to secrete and express therecombinant enzyme and different sites in the polylinker forinserting foreign genes [31]
This is the second extracellular protease reported to besynthesized by this obligate psychrophilic yeastG antarcticaThe first cold-adapted extracellular subtilase from this yeasthas been purified and characterized previously [32 33] Theextracellular serine proteinase (LAP2) from this Antarcticyeast formerly known as Leucosporidium antarcticum strain171 is specific towards synthetic substrates of chymotrypsinand subtilisin The subtilase of L antarcticum strain 171showed specific activity of 4043Umg when utilized N-SucAAPFpNA as a substrate [33]
37 Optimization of Protease Production GpPro2 was chosenfor further optimization due to its highest protease activityat the preliminary expression stage High protease activitywas detected after 72 hours of incubation (Figure 7) byusing the standard protocol method as recommended forPichia pastoris expression system (Pichia Expression KitInvitrogen USA) The expression decreased after 72 hoursand no activity was detected at 110 hours The prolongedincubation may result in the decrease of biomass productionand will subsequently reduce the expression of recombinantprotein In addition oxygen and nutrient depletion duringprolonged incubationwill trigger the stress responses in orderto survive in such harsh environment [34]
Methanol was used as a sole carbon by P pastoris Themethanol was oxidized to formaldehyde with the aid of oxy-gen as electron acceptor by an alcohol oxidase enzyme (AOXgene) [35] The optimal methanol concentration is at 05(vv) with 14UmL activity The addition of more than 05
Enzyme Research 17
02468
10121416
0 01 03 05 07 09Pr
otea
se ac
tivity
(Um
L)Methanol concentration ()
Figure 8 Effect ofmethanol concentration towards protease activityThe recombinant was cultivated for 72 hours at 30∘C in BMMYmedium05 methanol gives the highest activity with 14UmL The substrate (azocasein) was dissolved in Tris HCL buffer and incubated for 30minat 20∘CThe mixture was left for 15min and absorbance was taken at 450 nm A one-way ANOVA was used to test for preference differencesamong six different concentrations The statistical value (119875 value lt005) between concentration is 002
0
5
10
15
20
25
10 15 20 25 30 35
Prot
eas a
ctiv
ity (
Um
L)
Temperature (∘C)
Figure 9 Effect of temperature towards protease activity The reaction was carried out in temperature ranging from 10∘C to 35∘C at pH 7 for30min A one-way ANOVA was used to test for preference differences among six different temperatures The statistical value (119875 value lt005)between temperature is 0067
(vv) methanol resulted in tremendous decrease of proteaseactivity (Figure 8) Production of PI12 protease was decreasedwhen the methanol concentration increased This might bedue to the toxic effect of methanol concentration on DNAreplication and membrane synthesis Basically methanol isused to increase the permeability of the cell membrane tohelp the secretion of foreign protein [26] However if thepermeability of cell membrane is too high lowmolecular sizemetabolites would leak from the cell and such leakage willseriously damage cell activities Such condition would abusethe cellular system and might reduce the yield of proteinexpression [36]
Psychrophilic microorganism dominated in low temper-ature habitat which most other microorganism are inca-pable of withstanding due to their inability to produceenzyme that can tolerate the extreme environment con-dition Psychrophilic enzymes are always associated withhigh catalytic efficiency at low and moderate temperature(0∘C to 30∘C) compared to the mesophilic counterpart thatare generally unable to catalyze efficiently under extremely
low temperature [2] The recombinant protein assay wasdone at various temperatures ranging from 10∘C to 35∘Cusing 05 azocasein as a substrate dissolved in 50mMTris HCl in presence of 2mM CaCl
2 pH 7 In the activity
assays expressed protease showed a highest activity at 20∘C(20UmL) and decreased tremendously once it exceededmaximum temperature due to the denatured state of theenzyme (Figure 9) The active site of an enzyme is easilyaltered when the high temperature is introduced duringincubation or activity assay resulting in a low reaction rate[37]
The recombinant construct was transferred into BMMYmedia at different A
600 nm ranging from 1 to 10 The high-est activity of active enzyme was detected at A
600 nm 60(283UmL) and above (Figure 10) The slope during expo-nential phase was much higher compared to other phasesindicating an increase in the doubling time At exponentialphase the cells are adapted to the growth condition andother processes like cell growth cycle and synthesis of RNAThe yield of active enzyme during expression of P pastoris
18 Enzyme Research
05
1015202530
1 2 3 4 5 6 7 8 9 10Prot
ease
activ
ity (U
mL)
Initial inoculum A600
Figure 10 Effect of inoculums size towards protease activity The recombinant was cultivated for 72 hours at 30∘C in BMMY mediumwith 05 methanol concentration A one-way ANOVA was used to test for preference differences among six different optical densitiesThe statistical value (119875 value lt005) between optical density value is 003
(kD
a)
1160
662
450
350
250
184
144
M 1 2 3
(a) (b) (c)
Figure 11 (a) Coomassie-stained SDS-PAGE of recombinant P pastoris clone 1mL supernatants collected from the third-day culturewere concentrated 10-fold using trichloroacetic acid and 10120583L resultant products were applied to SDS-PAGE analysis Lane 1 molecularweight marker (Fermentas) Lane 2 GS115 (Control) Lane 3 GS115pPIC9 empty (Control) Lane 4 GS115pPIC9PI12prot (GpPro2)The arrow indicates the secreted recombinant PI12 protease (b) Plate activity staining of different recombinant P pastoris clones GGS115 GP GS115pPIC9 GRI GS115pPIC9PI12prot1 (GpPro1) GRII GS115pPIC9PI12prot2 (GpPro2) K KM71 KP KM71pPIC9 KRKM71pPIC9PI12prot (KpPro) (c)Gel activity staining of recombinant PI12 protease inP pastorisGpPro2 clone Lane 1 prestainedmolecularweightmarker (Fermentas) Lane 2 GS115pPIC9 empty (Control) Lane 3 GS115pPIC9PI12prot1 (GpPro2)The arrow indicates the secretedrecombinant PI12 protease
dropped after 28 hours of incubation which is at stationaryor death phase [38]
38 Detection of Recombinant Protein by SDS-PAGE andActivity Staining TheSDS-PAGE gel of the GpPro2 recombi-nant clonesrsquo supernatant showed a putative expression bandat around 993 kDa Plate staining and gel activity stainingwere executed to examine if the recombinant PI12 proteaseexpressed in P pastoris was functionally active In the platestaining the transformants showed strong proteolytic activity(Figure 11(a)) In gel staining proteolytic activity was alsodetected around 993 kDa after incubation at 20∘C This is
evidence of protease secretion into the surrounding mediawhereby it catalyzed the breakdown of casein thus formedclearing zones around the activity ring and band (Figures11(b) and 11(c)) As a conclusion the cold-adapted PI12protease was functionally expressed in P pastoris GS115(GpPro2 clone) with the size of 993 kDa
4 Conclusions
Psychrophilic yeast which was originated frommarine waterin Casey Station Antarctica was identified as Glaciozymaantarctica strain PI12 A serine protease with an open reading
Enzyme Research 19
frame of 2892 bp encoded for 963 amino acids from thisAntarctic yeast has been cloned and expressed in Pichiaexpression system This is the second extracellular pro-tease reported being synthesized by this psychrophilic yeastSequence analysis through bioinformatics studies revealedfew homology identities of this protein with other ser-ine proteases The mature PI12 protease was successfullyexpressed and secreted into the culture medium driven bythe Saccharomyces cerevisiae 120572-factor signal sequence in Ppastoris The protease production in P pastoris was bestobtained from strain GS115 (GpPro2) with 283UmL after 72hours of induction time with 05 (vv) of methanol inducerThe starting optical density at 600 nm (OD600) was 60 andthe optimum assay temperature was 20∘C The expressedprotein was detected by SDS-PAGE and activity staining witha molecular weight of 99 kDaThis cold-active enzymemightoffer an opportunity for biotechnological exploitation basedon its high catalytic activity at low temperatures and lowthermostability Structural and functional study of this cold-adapted enzyme would give insight into its adaptation to anextreme cold environment
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgment
This research was supported financially by the InitiativeBiotechnology-GMBI-MOSTI (Project number 07-05-16-MG-GMB02) andRUGS-Initiative 6 (Project number 05-02-11-1414RU)
References
[1] J W Deming ldquoPsychrophiles and polar regionsrdquo CurrentOpinion in Microbiology vol 5 no 3 pp 301ndash309 2002
[2] C Gerday M Aittaleb M Bentahir et al ldquoCold-adaptedenzymes from fundamentals to biotechnologyrdquo Trends inBiotechnology vol 18 no 3 pp 103ndash107 2000
[3] A Svingor J Kardos I Hajdu A Nemeth and P Zavodszky ldquoAbetter enzyme to cope with cold comparative flexibility studieson psychrotrophic mesophilic and thermophilic IPMDHSrdquoThe Journal of Biological Chemistry vol 276 no 30 pp 28121ndash28125 2001
[4] G Antranikian C E Vorgias and C Bertoldo ldquoExtremeenvironments as a resource for microorganisms and novel bio-catalystsrdquo Advances in Biochemical EngineeringBiotechnologyvol 96 pp 219ndash262 2005
[5] T Lonhienne C Gerday andG Feller ldquoPsychrophilic enzymesrevisiting the thermodynamic parameters of activation mayexplain local flexibilityrdquoBiochimica et Biophysica Acta vol 1543no 1 pp 1ndash10 2000
[6] R Cavicchioli and TThomas ldquoExtremophilesrdquo in Encyclopediaof Microbiology pp 317ndash337 Academic Press San Diego CalifUSA 2000
[7] G Feller E Narinx J L Arpigny et al ldquoEnzymes frompsychrophilic organismsrdquo FEMS Microbiology Reviews vol 18no 2-3 pp 189ndash202 1996
[8] J Arnorsdottir R B Smaradottir T O Magnusson S HThorbjarnardottir G Eggertsson and M M KristjanssonldquoCharacterization of a cloned subtilisin-like serine proteinasefrom a psychrotrophic Vibrio speciesrdquo European Journal ofBiochemistry vol 269 no 22 pp 5536ndash5546 2002
[9] N J Russell ldquoPsychrophilic bacteriamdashmolecular adaptations ofmembrane lipidsrdquo Comparative Biochemistry and Physiology APhysiology vol 118 no 3 pp 489ndash493 1997
[10] G Feller and C Gerday ldquoPsychrophilic enzymes hot topics incold adaptationrdquo Nature Reviews Microbiology vol 1 no 3 pp200ndash208 2003
[11] P Raspor and J Zupan ldquoYeasts in extreme environmentsrdquo inBiodiversity and Ecophysiology of Yeasts The Yeast Handbookpp 371ndash417 Springer Berlin Germany 2006
[12] M B Rao A M Tanksale M S Ghatge and V V Desh-pande ldquoMolecular and biotechnological aspects of microbialproteasesrdquoMicrobiology andMolecular Biology Reviews vol 62no 3 pp 597ndash635 1998
[13] L Polgar ldquoThe catalytic triad of serine peptidasesrdquo Cellular andMolecular Life Sciences vol 62 no 19-20 pp 2161ndash2172 2005
[14] S Y Boo C M V L Wong K F Rodrigues N NajimudinA M A Murad and N M Mahadi ldquoThermal stress responsesin Antarctic yeastGlaciozyma antarctica PI12 characterized byreal-time quantitative PCRrdquoPolar Biology vol 36 no 3 pp 381ndash389 2013
[15] B Turchetti S R T Hall L B Connell et al ldquoPsychrophilicyeasts from Antarctica and European glaciers descriptionof Glaciozyma gen nov Glaciozyma martinii sp nov andGlaciozyma watsonii sp novrdquo Extremophiles vol 15 no 5 pp573ndash586 2011
[16] N H F Hashim I Bharudin D L S Nguong et al ldquoCharac-terization of Afp1 an antifreeze protein from the psychrophilicyeast Glaciozyma antarctica PI12rdquo Extremophiles vol 17 no 1pp 63ndash73 2013
[17] A N M Ramli M A Azhar M S Shamsir et al ldquoSequenceand structural investigation of a novel psychrophilic 120572-amylasefrom Glaciozyma antarctica PI12 for cold-adaptation analysisrdquoJournal ofMolecularModeling vol 19 no 8 pp 3369ndash3383 2013
[18] A N M Ramli N M Mahadi A Rabu A M A Murad F DA Bakar and R M Illias ldquoMolecular cloning expression andbiochemical characterisation of a cold-adapted novel recombi-nant chitinase from Glaciozyma antarctica PI12rdquoMicrobial CellFactories vol 10 no 1 article 94 2011
[19] J Sambrook E Fritsch and T Maniatis Molecular Cloning ALaboratoryManual Cold SpringHarbor Laboratory Press NewYork NY USA 1989
[20] F M Brock C W Forsberg and J G Buchanan-SmithldquoProteolytic activity of rumen microorganisms and effects ofproteinase inhibitorsrdquoApplied and EnvironmentalMicrobiologyvol 44 no 3 pp 561ndash569 1982
[21] U K Laemmli ldquoCleavage of structural proteins during theassembly of the head of bacteriophage T4rdquo Nature vol 227 no5259 pp 680ndash685 1970
[22] A Henne R A Schmitz M Bomeke G Gottschalk and RDaniel ldquoScreening of environmental DNA libraries for thepresence of genes conferring lipolytic activity on Escherichiacolirdquo Applied and Environmental Microbiology vol 66 no 7 pp3113ndash3116 2000
[23] M Poza T de Miguel C Sieiro and T G Villa ldquoCharacteri-zation of a broad pH range protease of Candida caseinolyticardquoJournal of Applied Microbiology vol 91 no 5 pp 916ndash921 2001
20 Enzyme Research
[24] J D Bendtsen H Nielsen G von Heijne and S BrunakldquoImproved prediction of signal peptides SignalP 30rdquo Journalof Molecular Biology vol 340 no 4 pp 783ndash795 2004
[25] R J Siezen and J A M Leunissen ldquoSubtilases the superfamilyof subtilisin-like serine proteasesrdquo Protein Science vol 6 no 3pp 501ndash523 1997
[26] Y Katakura W Zhang G Zhuang et al ldquoEffect of methanolconcentration on the production of human 1205732- glycoprotein Idomain V by a recombinant Pichia pastoris a simple system forthe control of methanol concentration using a semiconductorgas sensorrdquo Journal of Fermentation and Bioengineering vol 86no 5 pp 482ndash487 1998
[27] Q-FWang Y-HHou Z Xu J-LMiao andG-Y Li ldquoPurifica-tion and properties of an extracellular cold-active protease fromthe psychrophilic bacterium Pseudoalteromonas sp NJ276rdquoBiochemical Engineering Journal vol 38 no 3 pp 362ndash3682008
[28] S Vazquez L Ruberto and W Mac Cormack ldquoProper-ties of extracellular proteases from three psychrotolerantStenotrophomonas maltophilia isolated from Antarctic soilrdquoPolar Biology vol 28 no 4 pp 319ndash325 2005
[29] R Zeng R Zhang J Zhao and N Lin ldquoCold-active ser-ine alkaline protease from the psychrophilic bacterium Pseu-domonas strain DY-A enzyme purification and characteriza-tionrdquo Extremophiles vol 7 no 4 pp 335ndash337 2003
[30] M Turkiewicz E Gromek H Kalinowska and M ZielinskaldquoBiosynthesis and properties of an extracellularmetalloproteasefrom the Antarctic marine bacterium Sphingomonas paucimo-bilisrdquo Journal of Biotechnology vol 70 no 1ndash3 pp 53ndash60 1999
[31] A Vassileva D A Chugh S Swaminathan and N KhannaldquoEffect of copy number on the expression levels of hepatitisB surface antigen in the methylotrophic yeast Pichia pastorisrdquoProtein Expression andPurification vol 21 no 1 pp 71ndash80 2001
[32] M Pazgier M Turkiewicz H Kalinowska and S Bielecki ldquoTheunique cold-adapted extracellular subtilase from psychrophilicyeast Leucosporidium antarcticumrdquo Journal ofMolecular Catal-ysis B Enzymatic vol 21 no 1-2 pp 39ndash42 2003
[33] M Turkiewicz M Pazgier H Kalinowska and S BieleckildquoA cold-adapted extracellular serine proteinase of the yeastLeucosporidium antarcticumrdquo Extremophiles vol 7 no 6 pp435ndash442 2003
[34] D Mattanovich B Gasser H Hohenblum and M SauerldquoStress in recombinant protein producing yeastsrdquo Journal ofBiotechnology vol 113 no 1ndash3 pp 121ndash135 2004
[35] P Secades BAlvarez and JAGuijarro ldquoPurification andprop-erties of a new psychrophilic metalloprotease (Fpp2) in the fishpathogen Flavobacterium psychrophilumrdquo FEMS MicrobiologyLetters vol 226 no 2 pp 273ndash279 2003
[36] L Y Ling I Ithoi and F M Yik ldquoOptimization for high-level expression in Pichia pastoris and purification of truncatedand full length recombinant sag2 of toxoplasma gondii fordiagnostic userdquo Southeast Asian Journal of Tropical Medicineand Public Health vol 41 no 3 pp 507ndash513 2010
[37] A Illanes ldquoStability of biocatalystsrdquo Electronic Journal ofBiotechnology vol 2 no 1 pp 1ndash9 1999
[38] B A Plantz J Sinha L Villarete K W Nickerson and VL Schlegel ldquoPichia pastoris fermentation optimization energystate and testing a growth-associated modelrdquo Applied Microbi-ology and Biotechnology vol 72 no 2 pp 297ndash305 2006
[39] K Tamura G Stecher D Peterson A Filipski and S KumarldquoMEGA6molecular evolutionary genetics analysis version 60rdquo
Molecular Biology and Evolution vol 30 no 12 pp 2725ndash27292013
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology
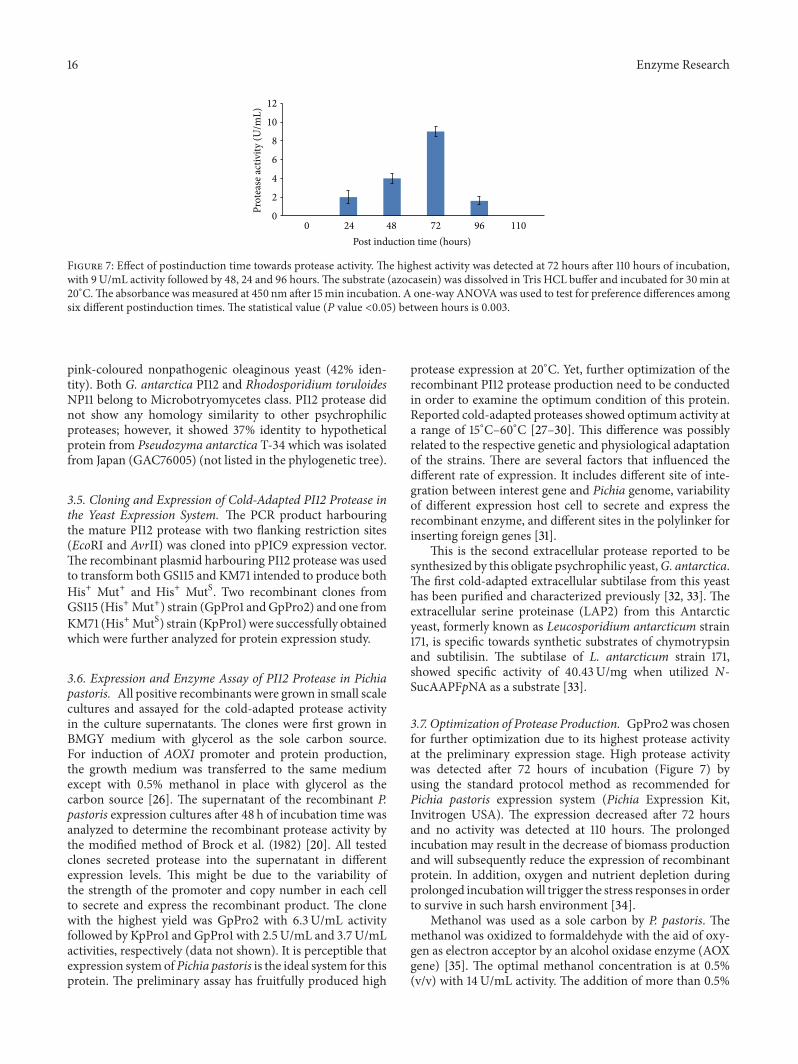
16 Enzyme Research
0
2
4
6
8
10
12
0 24 48 72 96 110
Prot
ease
activ
ity (U
mL)
Post induction time (hours)
Figure 7 Effect of postinduction time towards protease activity The highest activity was detected at 72 hours after 110 hours of incubationwith 9UmL activity followed by 48 24 and 96 hoursThe substrate (azocasein) was dissolved in Tris HCL buffer and incubated for 30min at20∘CThe absorbance was measured at 450 nm after 15min incubation A one-way ANOVAwas used to test for preference differences amongsix different postinduction times The statistical value (119875 value lt005) between hours is 0003
pink-coloured nonpathogenic oleaginous yeast (42 iden-tity) Both G antarctica PI12 and Rhodosporidium toruloidesNP11 belong to Microbotryomycetes class PI12 protease didnot show any homology similarity to other psychrophilicproteases however it showed 37 identity to hypotheticalprotein from Pseudozyma antarctica T-34 which was isolatedfrom Japan (GAC76005) (not listed in the phylogenetic tree)
35 Cloning and Expression of Cold-Adapted PI12 Protease inthe Yeast Expression System The PCR product harbouringthe mature PI12 protease with two flanking restriction sites(EcoRI and AvrII) was cloned into pPIC9 expression vectorThe recombinant plasmid harbouring PI12 protease was usedto transform both GS115 and KM71 intended to produce bothHis+ Mut+ and His+ MutS Two recombinant clones fromGS115 (His+Mut+) strain (GpPro1 andGpPro2) and one fromKM71 (His+MutS) strain (KpPro1) were successfully obtainedwhich were further analyzed for protein expression study
36 Expression and Enzyme Assay of PI12 Protease in Pichiapastoris All positive recombinants were grown in small scalecultures and assayed for the cold-adapted protease activityin the culture supernatants The clones were first grown inBMGY medium with glycerol as the sole carbon sourceFor induction of AOX1 promoter and protein productionthe growth medium was transferred to the same mediumexcept with 05 methanol in place with glycerol as thecarbon source [26] The supernatant of the recombinant Ppastoris expression cultures after 48 h of incubation time wasanalyzed to determine the recombinant protease activity bythe modified method of Brock et al (1982) [20] All testedclones secreted protease into the supernatant in differentexpression levels This might be due to the variability ofthe strength of the promoter and copy number in each cellto secrete and express the recombinant product The clonewith the highest yield was GpPro2 with 63UmL activityfollowed by KpPro1 and GpPro1 with 25UmL and 37UmLactivities respectively (data not shown) It is perceptible thatexpression systemof Pichia pastoris is the ideal system for thisprotein The preliminary assay has fruitfully produced high
protease expression at 20∘C Yet further optimization of therecombinant PI12 protease production need to be conductedin order to examine the optimum condition of this proteinReported cold-adapted proteases showed optimum activity ata range of 15∘Cndash60∘C [27ndash30] This difference was possiblyrelated to the respective genetic and physiological adaptationof the strains There are several factors that influenced thedifferent rate of expression It includes different site of inte-gration between interest gene and Pichia genome variabilityof different expression host cell to secrete and express therecombinant enzyme and different sites in the polylinker forinserting foreign genes [31]
This is the second extracellular protease reported to besynthesized by this obligate psychrophilic yeastG antarcticaThe first cold-adapted extracellular subtilase from this yeasthas been purified and characterized previously [32 33] Theextracellular serine proteinase (LAP2) from this Antarcticyeast formerly known as Leucosporidium antarcticum strain171 is specific towards synthetic substrates of chymotrypsinand subtilisin The subtilase of L antarcticum strain 171showed specific activity of 4043Umg when utilized N-SucAAPFpNA as a substrate [33]
37 Optimization of Protease Production GpPro2 was chosenfor further optimization due to its highest protease activityat the preliminary expression stage High protease activitywas detected after 72 hours of incubation (Figure 7) byusing the standard protocol method as recommended forPichia pastoris expression system (Pichia Expression KitInvitrogen USA) The expression decreased after 72 hoursand no activity was detected at 110 hours The prolongedincubation may result in the decrease of biomass productionand will subsequently reduce the expression of recombinantprotein In addition oxygen and nutrient depletion duringprolonged incubationwill trigger the stress responses in orderto survive in such harsh environment [34]
Methanol was used as a sole carbon by P pastoris Themethanol was oxidized to formaldehyde with the aid of oxy-gen as electron acceptor by an alcohol oxidase enzyme (AOXgene) [35] The optimal methanol concentration is at 05(vv) with 14UmL activity The addition of more than 05
Enzyme Research 17
02468
10121416
0 01 03 05 07 09Pr
otea
se ac
tivity
(Um
L)Methanol concentration ()
Figure 8 Effect ofmethanol concentration towards protease activityThe recombinant was cultivated for 72 hours at 30∘C in BMMYmedium05 methanol gives the highest activity with 14UmL The substrate (azocasein) was dissolved in Tris HCL buffer and incubated for 30minat 20∘CThe mixture was left for 15min and absorbance was taken at 450 nm A one-way ANOVA was used to test for preference differencesamong six different concentrations The statistical value (119875 value lt005) between concentration is 002
0
5
10
15
20
25
10 15 20 25 30 35
Prot
eas a
ctiv
ity (
Um
L)
Temperature (∘C)
Figure 9 Effect of temperature towards protease activity The reaction was carried out in temperature ranging from 10∘C to 35∘C at pH 7 for30min A one-way ANOVA was used to test for preference differences among six different temperatures The statistical value (119875 value lt005)between temperature is 0067
(vv) methanol resulted in tremendous decrease of proteaseactivity (Figure 8) Production of PI12 protease was decreasedwhen the methanol concentration increased This might bedue to the toxic effect of methanol concentration on DNAreplication and membrane synthesis Basically methanol isused to increase the permeability of the cell membrane tohelp the secretion of foreign protein [26] However if thepermeability of cell membrane is too high lowmolecular sizemetabolites would leak from the cell and such leakage willseriously damage cell activities Such condition would abusethe cellular system and might reduce the yield of proteinexpression [36]
Psychrophilic microorganism dominated in low temper-ature habitat which most other microorganism are inca-pable of withstanding due to their inability to produceenzyme that can tolerate the extreme environment con-dition Psychrophilic enzymes are always associated withhigh catalytic efficiency at low and moderate temperature(0∘C to 30∘C) compared to the mesophilic counterpart thatare generally unable to catalyze efficiently under extremely
low temperature [2] The recombinant protein assay wasdone at various temperatures ranging from 10∘C to 35∘Cusing 05 azocasein as a substrate dissolved in 50mMTris HCl in presence of 2mM CaCl
2 pH 7 In the activity
assays expressed protease showed a highest activity at 20∘C(20UmL) and decreased tremendously once it exceededmaximum temperature due to the denatured state of theenzyme (Figure 9) The active site of an enzyme is easilyaltered when the high temperature is introduced duringincubation or activity assay resulting in a low reaction rate[37]
The recombinant construct was transferred into BMMYmedia at different A
600 nm ranging from 1 to 10 The high-est activity of active enzyme was detected at A
600 nm 60(283UmL) and above (Figure 10) The slope during expo-nential phase was much higher compared to other phasesindicating an increase in the doubling time At exponentialphase the cells are adapted to the growth condition andother processes like cell growth cycle and synthesis of RNAThe yield of active enzyme during expression of P pastoris
18 Enzyme Research
05
1015202530
1 2 3 4 5 6 7 8 9 10Prot
ease
activ
ity (U
mL)
Initial inoculum A600
Figure 10 Effect of inoculums size towards protease activity The recombinant was cultivated for 72 hours at 30∘C in BMMY mediumwith 05 methanol concentration A one-way ANOVA was used to test for preference differences among six different optical densitiesThe statistical value (119875 value lt005) between optical density value is 003
(kD
a)
1160
662
450
350
250
184
144
M 1 2 3
(a) (b) (c)
Figure 11 (a) Coomassie-stained SDS-PAGE of recombinant P pastoris clone 1mL supernatants collected from the third-day culturewere concentrated 10-fold using trichloroacetic acid and 10120583L resultant products were applied to SDS-PAGE analysis Lane 1 molecularweight marker (Fermentas) Lane 2 GS115 (Control) Lane 3 GS115pPIC9 empty (Control) Lane 4 GS115pPIC9PI12prot (GpPro2)The arrow indicates the secreted recombinant PI12 protease (b) Plate activity staining of different recombinant P pastoris clones GGS115 GP GS115pPIC9 GRI GS115pPIC9PI12prot1 (GpPro1) GRII GS115pPIC9PI12prot2 (GpPro2) K KM71 KP KM71pPIC9 KRKM71pPIC9PI12prot (KpPro) (c)Gel activity staining of recombinant PI12 protease inP pastorisGpPro2 clone Lane 1 prestainedmolecularweightmarker (Fermentas) Lane 2 GS115pPIC9 empty (Control) Lane 3 GS115pPIC9PI12prot1 (GpPro2)The arrow indicates the secretedrecombinant PI12 protease
dropped after 28 hours of incubation which is at stationaryor death phase [38]
38 Detection of Recombinant Protein by SDS-PAGE andActivity Staining TheSDS-PAGE gel of the GpPro2 recombi-nant clonesrsquo supernatant showed a putative expression bandat around 993 kDa Plate staining and gel activity stainingwere executed to examine if the recombinant PI12 proteaseexpressed in P pastoris was functionally active In the platestaining the transformants showed strong proteolytic activity(Figure 11(a)) In gel staining proteolytic activity was alsodetected around 993 kDa after incubation at 20∘C This is
evidence of protease secretion into the surrounding mediawhereby it catalyzed the breakdown of casein thus formedclearing zones around the activity ring and band (Figures11(b) and 11(c)) As a conclusion the cold-adapted PI12protease was functionally expressed in P pastoris GS115(GpPro2 clone) with the size of 993 kDa
4 Conclusions
Psychrophilic yeast which was originated frommarine waterin Casey Station Antarctica was identified as Glaciozymaantarctica strain PI12 A serine protease with an open reading
Enzyme Research 19
frame of 2892 bp encoded for 963 amino acids from thisAntarctic yeast has been cloned and expressed in Pichiaexpression system This is the second extracellular pro-tease reported being synthesized by this psychrophilic yeastSequence analysis through bioinformatics studies revealedfew homology identities of this protein with other ser-ine proteases The mature PI12 protease was successfullyexpressed and secreted into the culture medium driven bythe Saccharomyces cerevisiae 120572-factor signal sequence in Ppastoris The protease production in P pastoris was bestobtained from strain GS115 (GpPro2) with 283UmL after 72hours of induction time with 05 (vv) of methanol inducerThe starting optical density at 600 nm (OD600) was 60 andthe optimum assay temperature was 20∘C The expressedprotein was detected by SDS-PAGE and activity staining witha molecular weight of 99 kDaThis cold-active enzymemightoffer an opportunity for biotechnological exploitation basedon its high catalytic activity at low temperatures and lowthermostability Structural and functional study of this cold-adapted enzyme would give insight into its adaptation to anextreme cold environment
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgment
This research was supported financially by the InitiativeBiotechnology-GMBI-MOSTI (Project number 07-05-16-MG-GMB02) andRUGS-Initiative 6 (Project number 05-02-11-1414RU)
References
[1] J W Deming ldquoPsychrophiles and polar regionsrdquo CurrentOpinion in Microbiology vol 5 no 3 pp 301ndash309 2002
[2] C Gerday M Aittaleb M Bentahir et al ldquoCold-adaptedenzymes from fundamentals to biotechnologyrdquo Trends inBiotechnology vol 18 no 3 pp 103ndash107 2000
[3] A Svingor J Kardos I Hajdu A Nemeth and P Zavodszky ldquoAbetter enzyme to cope with cold comparative flexibility studieson psychrotrophic mesophilic and thermophilic IPMDHSrdquoThe Journal of Biological Chemistry vol 276 no 30 pp 28121ndash28125 2001
[4] G Antranikian C E Vorgias and C Bertoldo ldquoExtremeenvironments as a resource for microorganisms and novel bio-catalystsrdquo Advances in Biochemical EngineeringBiotechnologyvol 96 pp 219ndash262 2005
[5] T Lonhienne C Gerday andG Feller ldquoPsychrophilic enzymesrevisiting the thermodynamic parameters of activation mayexplain local flexibilityrdquoBiochimica et Biophysica Acta vol 1543no 1 pp 1ndash10 2000
[6] R Cavicchioli and TThomas ldquoExtremophilesrdquo in Encyclopediaof Microbiology pp 317ndash337 Academic Press San Diego CalifUSA 2000
[7] G Feller E Narinx J L Arpigny et al ldquoEnzymes frompsychrophilic organismsrdquo FEMS Microbiology Reviews vol 18no 2-3 pp 189ndash202 1996
[8] J Arnorsdottir R B Smaradottir T O Magnusson S HThorbjarnardottir G Eggertsson and M M KristjanssonldquoCharacterization of a cloned subtilisin-like serine proteinasefrom a psychrotrophic Vibrio speciesrdquo European Journal ofBiochemistry vol 269 no 22 pp 5536ndash5546 2002
[9] N J Russell ldquoPsychrophilic bacteriamdashmolecular adaptations ofmembrane lipidsrdquo Comparative Biochemistry and Physiology APhysiology vol 118 no 3 pp 489ndash493 1997
[10] G Feller and C Gerday ldquoPsychrophilic enzymes hot topics incold adaptationrdquo Nature Reviews Microbiology vol 1 no 3 pp200ndash208 2003
[11] P Raspor and J Zupan ldquoYeasts in extreme environmentsrdquo inBiodiversity and Ecophysiology of Yeasts The Yeast Handbookpp 371ndash417 Springer Berlin Germany 2006
[12] M B Rao A M Tanksale M S Ghatge and V V Desh-pande ldquoMolecular and biotechnological aspects of microbialproteasesrdquoMicrobiology andMolecular Biology Reviews vol 62no 3 pp 597ndash635 1998
[13] L Polgar ldquoThe catalytic triad of serine peptidasesrdquo Cellular andMolecular Life Sciences vol 62 no 19-20 pp 2161ndash2172 2005
[14] S Y Boo C M V L Wong K F Rodrigues N NajimudinA M A Murad and N M Mahadi ldquoThermal stress responsesin Antarctic yeastGlaciozyma antarctica PI12 characterized byreal-time quantitative PCRrdquoPolar Biology vol 36 no 3 pp 381ndash389 2013
[15] B Turchetti S R T Hall L B Connell et al ldquoPsychrophilicyeasts from Antarctica and European glaciers descriptionof Glaciozyma gen nov Glaciozyma martinii sp nov andGlaciozyma watsonii sp novrdquo Extremophiles vol 15 no 5 pp573ndash586 2011
[16] N H F Hashim I Bharudin D L S Nguong et al ldquoCharac-terization of Afp1 an antifreeze protein from the psychrophilicyeast Glaciozyma antarctica PI12rdquo Extremophiles vol 17 no 1pp 63ndash73 2013
[17] A N M Ramli M A Azhar M S Shamsir et al ldquoSequenceand structural investigation of a novel psychrophilic 120572-amylasefrom Glaciozyma antarctica PI12 for cold-adaptation analysisrdquoJournal ofMolecularModeling vol 19 no 8 pp 3369ndash3383 2013
[18] A N M Ramli N M Mahadi A Rabu A M A Murad F DA Bakar and R M Illias ldquoMolecular cloning expression andbiochemical characterisation of a cold-adapted novel recombi-nant chitinase from Glaciozyma antarctica PI12rdquoMicrobial CellFactories vol 10 no 1 article 94 2011
[19] J Sambrook E Fritsch and T Maniatis Molecular Cloning ALaboratoryManual Cold SpringHarbor Laboratory Press NewYork NY USA 1989
[20] F M Brock C W Forsberg and J G Buchanan-SmithldquoProteolytic activity of rumen microorganisms and effects ofproteinase inhibitorsrdquoApplied and EnvironmentalMicrobiologyvol 44 no 3 pp 561ndash569 1982
[21] U K Laemmli ldquoCleavage of structural proteins during theassembly of the head of bacteriophage T4rdquo Nature vol 227 no5259 pp 680ndash685 1970
[22] A Henne R A Schmitz M Bomeke G Gottschalk and RDaniel ldquoScreening of environmental DNA libraries for thepresence of genes conferring lipolytic activity on Escherichiacolirdquo Applied and Environmental Microbiology vol 66 no 7 pp3113ndash3116 2000
[23] M Poza T de Miguel C Sieiro and T G Villa ldquoCharacteri-zation of a broad pH range protease of Candida caseinolyticardquoJournal of Applied Microbiology vol 91 no 5 pp 916ndash921 2001
20 Enzyme Research
[24] J D Bendtsen H Nielsen G von Heijne and S BrunakldquoImproved prediction of signal peptides SignalP 30rdquo Journalof Molecular Biology vol 340 no 4 pp 783ndash795 2004
[25] R J Siezen and J A M Leunissen ldquoSubtilases the superfamilyof subtilisin-like serine proteasesrdquo Protein Science vol 6 no 3pp 501ndash523 1997
[26] Y Katakura W Zhang G Zhuang et al ldquoEffect of methanolconcentration on the production of human 1205732- glycoprotein Idomain V by a recombinant Pichia pastoris a simple system forthe control of methanol concentration using a semiconductorgas sensorrdquo Journal of Fermentation and Bioengineering vol 86no 5 pp 482ndash487 1998
[27] Q-FWang Y-HHou Z Xu J-LMiao andG-Y Li ldquoPurifica-tion and properties of an extracellular cold-active protease fromthe psychrophilic bacterium Pseudoalteromonas sp NJ276rdquoBiochemical Engineering Journal vol 38 no 3 pp 362ndash3682008
[28] S Vazquez L Ruberto and W Mac Cormack ldquoProper-ties of extracellular proteases from three psychrotolerantStenotrophomonas maltophilia isolated from Antarctic soilrdquoPolar Biology vol 28 no 4 pp 319ndash325 2005
[29] R Zeng R Zhang J Zhao and N Lin ldquoCold-active ser-ine alkaline protease from the psychrophilic bacterium Pseu-domonas strain DY-A enzyme purification and characteriza-tionrdquo Extremophiles vol 7 no 4 pp 335ndash337 2003
[30] M Turkiewicz E Gromek H Kalinowska and M ZielinskaldquoBiosynthesis and properties of an extracellularmetalloproteasefrom the Antarctic marine bacterium Sphingomonas paucimo-bilisrdquo Journal of Biotechnology vol 70 no 1ndash3 pp 53ndash60 1999
[31] A Vassileva D A Chugh S Swaminathan and N KhannaldquoEffect of copy number on the expression levels of hepatitisB surface antigen in the methylotrophic yeast Pichia pastorisrdquoProtein Expression andPurification vol 21 no 1 pp 71ndash80 2001
[32] M Pazgier M Turkiewicz H Kalinowska and S Bielecki ldquoTheunique cold-adapted extracellular subtilase from psychrophilicyeast Leucosporidium antarcticumrdquo Journal ofMolecular Catal-ysis B Enzymatic vol 21 no 1-2 pp 39ndash42 2003
[33] M Turkiewicz M Pazgier H Kalinowska and S BieleckildquoA cold-adapted extracellular serine proteinase of the yeastLeucosporidium antarcticumrdquo Extremophiles vol 7 no 6 pp435ndash442 2003
[34] D Mattanovich B Gasser H Hohenblum and M SauerldquoStress in recombinant protein producing yeastsrdquo Journal ofBiotechnology vol 113 no 1ndash3 pp 121ndash135 2004
[35] P Secades BAlvarez and JAGuijarro ldquoPurification andprop-erties of a new psychrophilic metalloprotease (Fpp2) in the fishpathogen Flavobacterium psychrophilumrdquo FEMS MicrobiologyLetters vol 226 no 2 pp 273ndash279 2003
[36] L Y Ling I Ithoi and F M Yik ldquoOptimization for high-level expression in Pichia pastoris and purification of truncatedand full length recombinant sag2 of toxoplasma gondii fordiagnostic userdquo Southeast Asian Journal of Tropical Medicineand Public Health vol 41 no 3 pp 507ndash513 2010
[37] A Illanes ldquoStability of biocatalystsrdquo Electronic Journal ofBiotechnology vol 2 no 1 pp 1ndash9 1999
[38] B A Plantz J Sinha L Villarete K W Nickerson and VL Schlegel ldquoPichia pastoris fermentation optimization energystate and testing a growth-associated modelrdquo Applied Microbi-ology and Biotechnology vol 72 no 2 pp 297ndash305 2006
[39] K Tamura G Stecher D Peterson A Filipski and S KumarldquoMEGA6molecular evolutionary genetics analysis version 60rdquo
Molecular Biology and Evolution vol 30 no 12 pp 2725ndash27292013
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology
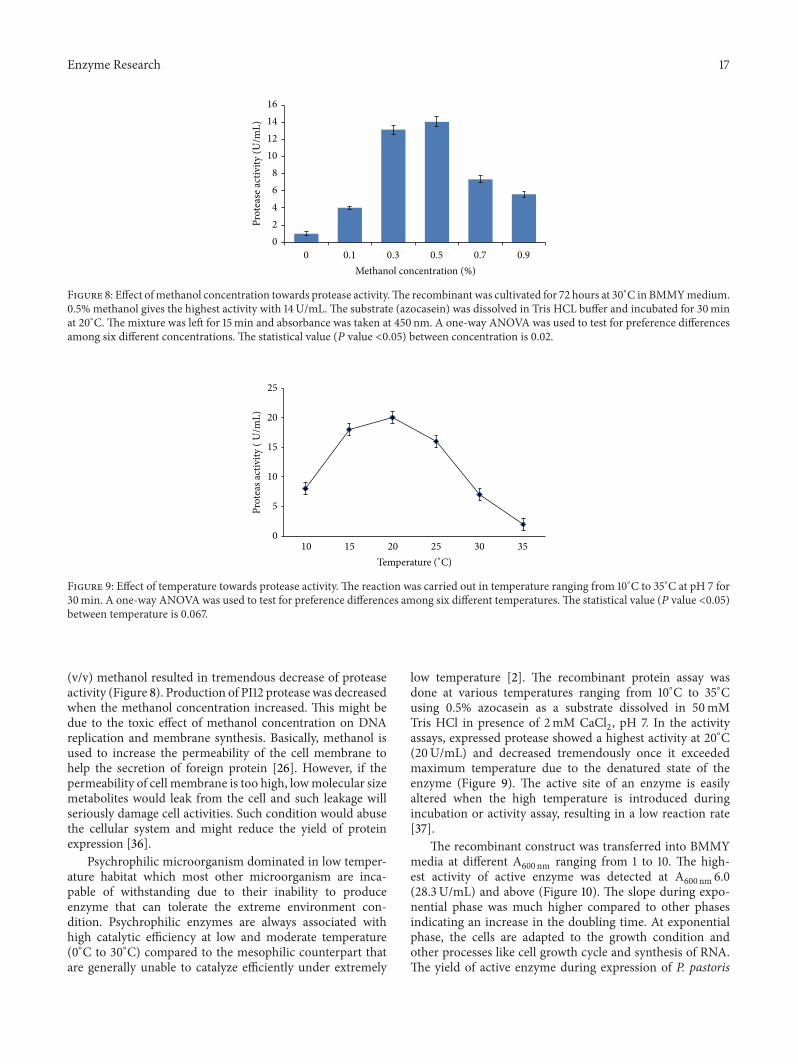
Enzyme Research 17
02468
10121416
0 01 03 05 07 09Pr
otea
se ac
tivity
(Um
L)Methanol concentration ()
Figure 8 Effect ofmethanol concentration towards protease activityThe recombinant was cultivated for 72 hours at 30∘C in BMMYmedium05 methanol gives the highest activity with 14UmL The substrate (azocasein) was dissolved in Tris HCL buffer and incubated for 30minat 20∘CThe mixture was left for 15min and absorbance was taken at 450 nm A one-way ANOVA was used to test for preference differencesamong six different concentrations The statistical value (119875 value lt005) between concentration is 002
0
5
10
15
20
25
10 15 20 25 30 35
Prot
eas a
ctiv
ity (
Um
L)
Temperature (∘C)
Figure 9 Effect of temperature towards protease activity The reaction was carried out in temperature ranging from 10∘C to 35∘C at pH 7 for30min A one-way ANOVA was used to test for preference differences among six different temperatures The statistical value (119875 value lt005)between temperature is 0067
(vv) methanol resulted in tremendous decrease of proteaseactivity (Figure 8) Production of PI12 protease was decreasedwhen the methanol concentration increased This might bedue to the toxic effect of methanol concentration on DNAreplication and membrane synthesis Basically methanol isused to increase the permeability of the cell membrane tohelp the secretion of foreign protein [26] However if thepermeability of cell membrane is too high lowmolecular sizemetabolites would leak from the cell and such leakage willseriously damage cell activities Such condition would abusethe cellular system and might reduce the yield of proteinexpression [36]
Psychrophilic microorganism dominated in low temper-ature habitat which most other microorganism are inca-pable of withstanding due to their inability to produceenzyme that can tolerate the extreme environment con-dition Psychrophilic enzymes are always associated withhigh catalytic efficiency at low and moderate temperature(0∘C to 30∘C) compared to the mesophilic counterpart thatare generally unable to catalyze efficiently under extremely
low temperature [2] The recombinant protein assay wasdone at various temperatures ranging from 10∘C to 35∘Cusing 05 azocasein as a substrate dissolved in 50mMTris HCl in presence of 2mM CaCl
2 pH 7 In the activity
assays expressed protease showed a highest activity at 20∘C(20UmL) and decreased tremendously once it exceededmaximum temperature due to the denatured state of theenzyme (Figure 9) The active site of an enzyme is easilyaltered when the high temperature is introduced duringincubation or activity assay resulting in a low reaction rate[37]
The recombinant construct was transferred into BMMYmedia at different A
600 nm ranging from 1 to 10 The high-est activity of active enzyme was detected at A
600 nm 60(283UmL) and above (Figure 10) The slope during expo-nential phase was much higher compared to other phasesindicating an increase in the doubling time At exponentialphase the cells are adapted to the growth condition andother processes like cell growth cycle and synthesis of RNAThe yield of active enzyme during expression of P pastoris
18 Enzyme Research
05
1015202530
1 2 3 4 5 6 7 8 9 10Prot
ease
activ
ity (U
mL)
Initial inoculum A600
Figure 10 Effect of inoculums size towards protease activity The recombinant was cultivated for 72 hours at 30∘C in BMMY mediumwith 05 methanol concentration A one-way ANOVA was used to test for preference differences among six different optical densitiesThe statistical value (119875 value lt005) between optical density value is 003
(kD
a)
1160
662
450
350
250
184
144
M 1 2 3
(a) (b) (c)
Figure 11 (a) Coomassie-stained SDS-PAGE of recombinant P pastoris clone 1mL supernatants collected from the third-day culturewere concentrated 10-fold using trichloroacetic acid and 10120583L resultant products were applied to SDS-PAGE analysis Lane 1 molecularweight marker (Fermentas) Lane 2 GS115 (Control) Lane 3 GS115pPIC9 empty (Control) Lane 4 GS115pPIC9PI12prot (GpPro2)The arrow indicates the secreted recombinant PI12 protease (b) Plate activity staining of different recombinant P pastoris clones GGS115 GP GS115pPIC9 GRI GS115pPIC9PI12prot1 (GpPro1) GRII GS115pPIC9PI12prot2 (GpPro2) K KM71 KP KM71pPIC9 KRKM71pPIC9PI12prot (KpPro) (c)Gel activity staining of recombinant PI12 protease inP pastorisGpPro2 clone Lane 1 prestainedmolecularweightmarker (Fermentas) Lane 2 GS115pPIC9 empty (Control) Lane 3 GS115pPIC9PI12prot1 (GpPro2)The arrow indicates the secretedrecombinant PI12 protease
dropped after 28 hours of incubation which is at stationaryor death phase [38]
38 Detection of Recombinant Protein by SDS-PAGE andActivity Staining TheSDS-PAGE gel of the GpPro2 recombi-nant clonesrsquo supernatant showed a putative expression bandat around 993 kDa Plate staining and gel activity stainingwere executed to examine if the recombinant PI12 proteaseexpressed in P pastoris was functionally active In the platestaining the transformants showed strong proteolytic activity(Figure 11(a)) In gel staining proteolytic activity was alsodetected around 993 kDa after incubation at 20∘C This is
evidence of protease secretion into the surrounding mediawhereby it catalyzed the breakdown of casein thus formedclearing zones around the activity ring and band (Figures11(b) and 11(c)) As a conclusion the cold-adapted PI12protease was functionally expressed in P pastoris GS115(GpPro2 clone) with the size of 993 kDa
4 Conclusions
Psychrophilic yeast which was originated frommarine waterin Casey Station Antarctica was identified as Glaciozymaantarctica strain PI12 A serine protease with an open reading
Enzyme Research 19
frame of 2892 bp encoded for 963 amino acids from thisAntarctic yeast has been cloned and expressed in Pichiaexpression system This is the second extracellular pro-tease reported being synthesized by this psychrophilic yeastSequence analysis through bioinformatics studies revealedfew homology identities of this protein with other ser-ine proteases The mature PI12 protease was successfullyexpressed and secreted into the culture medium driven bythe Saccharomyces cerevisiae 120572-factor signal sequence in Ppastoris The protease production in P pastoris was bestobtained from strain GS115 (GpPro2) with 283UmL after 72hours of induction time with 05 (vv) of methanol inducerThe starting optical density at 600 nm (OD600) was 60 andthe optimum assay temperature was 20∘C The expressedprotein was detected by SDS-PAGE and activity staining witha molecular weight of 99 kDaThis cold-active enzymemightoffer an opportunity for biotechnological exploitation basedon its high catalytic activity at low temperatures and lowthermostability Structural and functional study of this cold-adapted enzyme would give insight into its adaptation to anextreme cold environment
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgment
This research was supported financially by the InitiativeBiotechnology-GMBI-MOSTI (Project number 07-05-16-MG-GMB02) andRUGS-Initiative 6 (Project number 05-02-11-1414RU)
References
[1] J W Deming ldquoPsychrophiles and polar regionsrdquo CurrentOpinion in Microbiology vol 5 no 3 pp 301ndash309 2002
[2] C Gerday M Aittaleb M Bentahir et al ldquoCold-adaptedenzymes from fundamentals to biotechnologyrdquo Trends inBiotechnology vol 18 no 3 pp 103ndash107 2000
[3] A Svingor J Kardos I Hajdu A Nemeth and P Zavodszky ldquoAbetter enzyme to cope with cold comparative flexibility studieson psychrotrophic mesophilic and thermophilic IPMDHSrdquoThe Journal of Biological Chemistry vol 276 no 30 pp 28121ndash28125 2001
[4] G Antranikian C E Vorgias and C Bertoldo ldquoExtremeenvironments as a resource for microorganisms and novel bio-catalystsrdquo Advances in Biochemical EngineeringBiotechnologyvol 96 pp 219ndash262 2005
[5] T Lonhienne C Gerday andG Feller ldquoPsychrophilic enzymesrevisiting the thermodynamic parameters of activation mayexplain local flexibilityrdquoBiochimica et Biophysica Acta vol 1543no 1 pp 1ndash10 2000
[6] R Cavicchioli and TThomas ldquoExtremophilesrdquo in Encyclopediaof Microbiology pp 317ndash337 Academic Press San Diego CalifUSA 2000
[7] G Feller E Narinx J L Arpigny et al ldquoEnzymes frompsychrophilic organismsrdquo FEMS Microbiology Reviews vol 18no 2-3 pp 189ndash202 1996
[8] J Arnorsdottir R B Smaradottir T O Magnusson S HThorbjarnardottir G Eggertsson and M M KristjanssonldquoCharacterization of a cloned subtilisin-like serine proteinasefrom a psychrotrophic Vibrio speciesrdquo European Journal ofBiochemistry vol 269 no 22 pp 5536ndash5546 2002
[9] N J Russell ldquoPsychrophilic bacteriamdashmolecular adaptations ofmembrane lipidsrdquo Comparative Biochemistry and Physiology APhysiology vol 118 no 3 pp 489ndash493 1997
[10] G Feller and C Gerday ldquoPsychrophilic enzymes hot topics incold adaptationrdquo Nature Reviews Microbiology vol 1 no 3 pp200ndash208 2003
[11] P Raspor and J Zupan ldquoYeasts in extreme environmentsrdquo inBiodiversity and Ecophysiology of Yeasts The Yeast Handbookpp 371ndash417 Springer Berlin Germany 2006
[12] M B Rao A M Tanksale M S Ghatge and V V Desh-pande ldquoMolecular and biotechnological aspects of microbialproteasesrdquoMicrobiology andMolecular Biology Reviews vol 62no 3 pp 597ndash635 1998
[13] L Polgar ldquoThe catalytic triad of serine peptidasesrdquo Cellular andMolecular Life Sciences vol 62 no 19-20 pp 2161ndash2172 2005
[14] S Y Boo C M V L Wong K F Rodrigues N NajimudinA M A Murad and N M Mahadi ldquoThermal stress responsesin Antarctic yeastGlaciozyma antarctica PI12 characterized byreal-time quantitative PCRrdquoPolar Biology vol 36 no 3 pp 381ndash389 2013
[15] B Turchetti S R T Hall L B Connell et al ldquoPsychrophilicyeasts from Antarctica and European glaciers descriptionof Glaciozyma gen nov Glaciozyma martinii sp nov andGlaciozyma watsonii sp novrdquo Extremophiles vol 15 no 5 pp573ndash586 2011
[16] N H F Hashim I Bharudin D L S Nguong et al ldquoCharac-terization of Afp1 an antifreeze protein from the psychrophilicyeast Glaciozyma antarctica PI12rdquo Extremophiles vol 17 no 1pp 63ndash73 2013
[17] A N M Ramli M A Azhar M S Shamsir et al ldquoSequenceand structural investigation of a novel psychrophilic 120572-amylasefrom Glaciozyma antarctica PI12 for cold-adaptation analysisrdquoJournal ofMolecularModeling vol 19 no 8 pp 3369ndash3383 2013
[18] A N M Ramli N M Mahadi A Rabu A M A Murad F DA Bakar and R M Illias ldquoMolecular cloning expression andbiochemical characterisation of a cold-adapted novel recombi-nant chitinase from Glaciozyma antarctica PI12rdquoMicrobial CellFactories vol 10 no 1 article 94 2011
[19] J Sambrook E Fritsch and T Maniatis Molecular Cloning ALaboratoryManual Cold SpringHarbor Laboratory Press NewYork NY USA 1989
[20] F M Brock C W Forsberg and J G Buchanan-SmithldquoProteolytic activity of rumen microorganisms and effects ofproteinase inhibitorsrdquoApplied and EnvironmentalMicrobiologyvol 44 no 3 pp 561ndash569 1982
[21] U K Laemmli ldquoCleavage of structural proteins during theassembly of the head of bacteriophage T4rdquo Nature vol 227 no5259 pp 680ndash685 1970
[22] A Henne R A Schmitz M Bomeke G Gottschalk and RDaniel ldquoScreening of environmental DNA libraries for thepresence of genes conferring lipolytic activity on Escherichiacolirdquo Applied and Environmental Microbiology vol 66 no 7 pp3113ndash3116 2000
[23] M Poza T de Miguel C Sieiro and T G Villa ldquoCharacteri-zation of a broad pH range protease of Candida caseinolyticardquoJournal of Applied Microbiology vol 91 no 5 pp 916ndash921 2001
20 Enzyme Research
[24] J D Bendtsen H Nielsen G von Heijne and S BrunakldquoImproved prediction of signal peptides SignalP 30rdquo Journalof Molecular Biology vol 340 no 4 pp 783ndash795 2004
[25] R J Siezen and J A M Leunissen ldquoSubtilases the superfamilyof subtilisin-like serine proteasesrdquo Protein Science vol 6 no 3pp 501ndash523 1997
[26] Y Katakura W Zhang G Zhuang et al ldquoEffect of methanolconcentration on the production of human 1205732- glycoprotein Idomain V by a recombinant Pichia pastoris a simple system forthe control of methanol concentration using a semiconductorgas sensorrdquo Journal of Fermentation and Bioengineering vol 86no 5 pp 482ndash487 1998
[27] Q-FWang Y-HHou Z Xu J-LMiao andG-Y Li ldquoPurifica-tion and properties of an extracellular cold-active protease fromthe psychrophilic bacterium Pseudoalteromonas sp NJ276rdquoBiochemical Engineering Journal vol 38 no 3 pp 362ndash3682008
[28] S Vazquez L Ruberto and W Mac Cormack ldquoProper-ties of extracellular proteases from three psychrotolerantStenotrophomonas maltophilia isolated from Antarctic soilrdquoPolar Biology vol 28 no 4 pp 319ndash325 2005
[29] R Zeng R Zhang J Zhao and N Lin ldquoCold-active ser-ine alkaline protease from the psychrophilic bacterium Pseu-domonas strain DY-A enzyme purification and characteriza-tionrdquo Extremophiles vol 7 no 4 pp 335ndash337 2003
[30] M Turkiewicz E Gromek H Kalinowska and M ZielinskaldquoBiosynthesis and properties of an extracellularmetalloproteasefrom the Antarctic marine bacterium Sphingomonas paucimo-bilisrdquo Journal of Biotechnology vol 70 no 1ndash3 pp 53ndash60 1999
[31] A Vassileva D A Chugh S Swaminathan and N KhannaldquoEffect of copy number on the expression levels of hepatitisB surface antigen in the methylotrophic yeast Pichia pastorisrdquoProtein Expression andPurification vol 21 no 1 pp 71ndash80 2001
[32] M Pazgier M Turkiewicz H Kalinowska and S Bielecki ldquoTheunique cold-adapted extracellular subtilase from psychrophilicyeast Leucosporidium antarcticumrdquo Journal ofMolecular Catal-ysis B Enzymatic vol 21 no 1-2 pp 39ndash42 2003
[33] M Turkiewicz M Pazgier H Kalinowska and S BieleckildquoA cold-adapted extracellular serine proteinase of the yeastLeucosporidium antarcticumrdquo Extremophiles vol 7 no 6 pp435ndash442 2003
[34] D Mattanovich B Gasser H Hohenblum and M SauerldquoStress in recombinant protein producing yeastsrdquo Journal ofBiotechnology vol 113 no 1ndash3 pp 121ndash135 2004
[35] P Secades BAlvarez and JAGuijarro ldquoPurification andprop-erties of a new psychrophilic metalloprotease (Fpp2) in the fishpathogen Flavobacterium psychrophilumrdquo FEMS MicrobiologyLetters vol 226 no 2 pp 273ndash279 2003
[36] L Y Ling I Ithoi and F M Yik ldquoOptimization for high-level expression in Pichia pastoris and purification of truncatedand full length recombinant sag2 of toxoplasma gondii fordiagnostic userdquo Southeast Asian Journal of Tropical Medicineand Public Health vol 41 no 3 pp 507ndash513 2010
[37] A Illanes ldquoStability of biocatalystsrdquo Electronic Journal ofBiotechnology vol 2 no 1 pp 1ndash9 1999
[38] B A Plantz J Sinha L Villarete K W Nickerson and VL Schlegel ldquoPichia pastoris fermentation optimization energystate and testing a growth-associated modelrdquo Applied Microbi-ology and Biotechnology vol 72 no 2 pp 297ndash305 2006
[39] K Tamura G Stecher D Peterson A Filipski and S KumarldquoMEGA6molecular evolutionary genetics analysis version 60rdquo
Molecular Biology and Evolution vol 30 no 12 pp 2725ndash27292013
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology
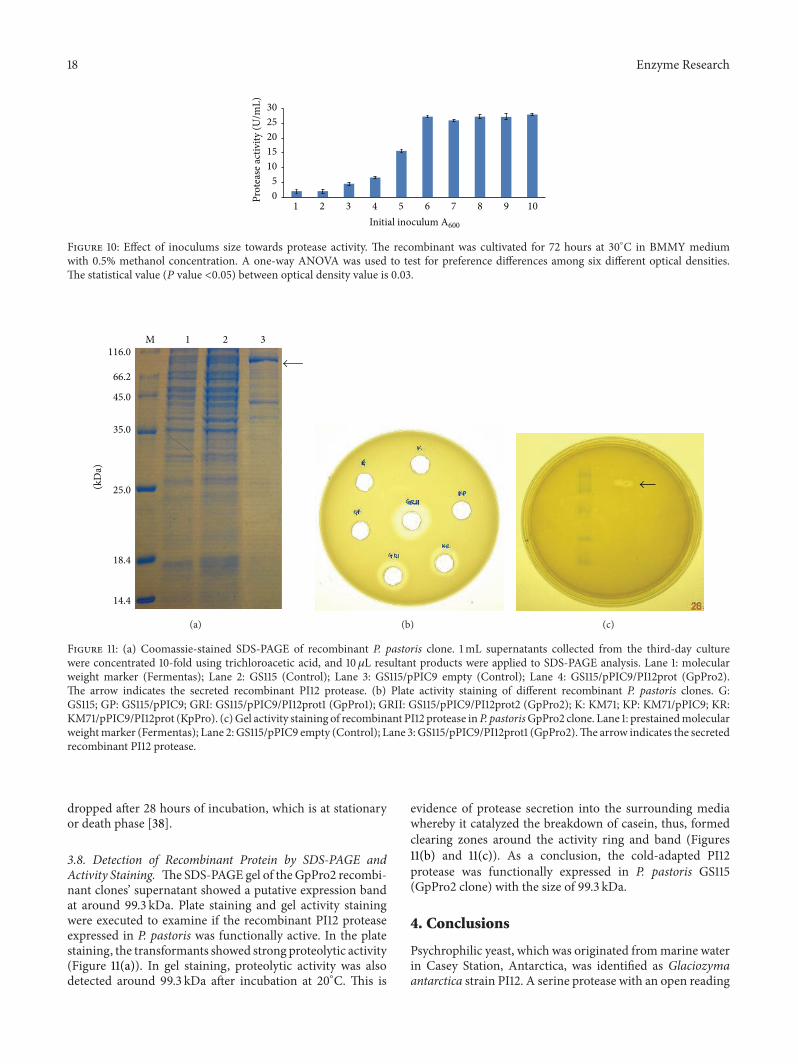
18 Enzyme Research
05
1015202530
1 2 3 4 5 6 7 8 9 10Prot
ease
activ
ity (U
mL)
Initial inoculum A600
Figure 10 Effect of inoculums size towards protease activity The recombinant was cultivated for 72 hours at 30∘C in BMMY mediumwith 05 methanol concentration A one-way ANOVA was used to test for preference differences among six different optical densitiesThe statistical value (119875 value lt005) between optical density value is 003
(kD
a)
1160
662
450
350
250
184
144
M 1 2 3
(a) (b) (c)
Figure 11 (a) Coomassie-stained SDS-PAGE of recombinant P pastoris clone 1mL supernatants collected from the third-day culturewere concentrated 10-fold using trichloroacetic acid and 10120583L resultant products were applied to SDS-PAGE analysis Lane 1 molecularweight marker (Fermentas) Lane 2 GS115 (Control) Lane 3 GS115pPIC9 empty (Control) Lane 4 GS115pPIC9PI12prot (GpPro2)The arrow indicates the secreted recombinant PI12 protease (b) Plate activity staining of different recombinant P pastoris clones GGS115 GP GS115pPIC9 GRI GS115pPIC9PI12prot1 (GpPro1) GRII GS115pPIC9PI12prot2 (GpPro2) K KM71 KP KM71pPIC9 KRKM71pPIC9PI12prot (KpPro) (c)Gel activity staining of recombinant PI12 protease inP pastorisGpPro2 clone Lane 1 prestainedmolecularweightmarker (Fermentas) Lane 2 GS115pPIC9 empty (Control) Lane 3 GS115pPIC9PI12prot1 (GpPro2)The arrow indicates the secretedrecombinant PI12 protease
dropped after 28 hours of incubation which is at stationaryor death phase [38]
38 Detection of Recombinant Protein by SDS-PAGE andActivity Staining TheSDS-PAGE gel of the GpPro2 recombi-nant clonesrsquo supernatant showed a putative expression bandat around 993 kDa Plate staining and gel activity stainingwere executed to examine if the recombinant PI12 proteaseexpressed in P pastoris was functionally active In the platestaining the transformants showed strong proteolytic activity(Figure 11(a)) In gel staining proteolytic activity was alsodetected around 993 kDa after incubation at 20∘C This is
evidence of protease secretion into the surrounding mediawhereby it catalyzed the breakdown of casein thus formedclearing zones around the activity ring and band (Figures11(b) and 11(c)) As a conclusion the cold-adapted PI12protease was functionally expressed in P pastoris GS115(GpPro2 clone) with the size of 993 kDa
4 Conclusions
Psychrophilic yeast which was originated frommarine waterin Casey Station Antarctica was identified as Glaciozymaantarctica strain PI12 A serine protease with an open reading
Enzyme Research 19
frame of 2892 bp encoded for 963 amino acids from thisAntarctic yeast has been cloned and expressed in Pichiaexpression system This is the second extracellular pro-tease reported being synthesized by this psychrophilic yeastSequence analysis through bioinformatics studies revealedfew homology identities of this protein with other ser-ine proteases The mature PI12 protease was successfullyexpressed and secreted into the culture medium driven bythe Saccharomyces cerevisiae 120572-factor signal sequence in Ppastoris The protease production in P pastoris was bestobtained from strain GS115 (GpPro2) with 283UmL after 72hours of induction time with 05 (vv) of methanol inducerThe starting optical density at 600 nm (OD600) was 60 andthe optimum assay temperature was 20∘C The expressedprotein was detected by SDS-PAGE and activity staining witha molecular weight of 99 kDaThis cold-active enzymemightoffer an opportunity for biotechnological exploitation basedon its high catalytic activity at low temperatures and lowthermostability Structural and functional study of this cold-adapted enzyme would give insight into its adaptation to anextreme cold environment
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgment
This research was supported financially by the InitiativeBiotechnology-GMBI-MOSTI (Project number 07-05-16-MG-GMB02) andRUGS-Initiative 6 (Project number 05-02-11-1414RU)
References
[1] J W Deming ldquoPsychrophiles and polar regionsrdquo CurrentOpinion in Microbiology vol 5 no 3 pp 301ndash309 2002
[2] C Gerday M Aittaleb M Bentahir et al ldquoCold-adaptedenzymes from fundamentals to biotechnologyrdquo Trends inBiotechnology vol 18 no 3 pp 103ndash107 2000
[3] A Svingor J Kardos I Hajdu A Nemeth and P Zavodszky ldquoAbetter enzyme to cope with cold comparative flexibility studieson psychrotrophic mesophilic and thermophilic IPMDHSrdquoThe Journal of Biological Chemistry vol 276 no 30 pp 28121ndash28125 2001
[4] G Antranikian C E Vorgias and C Bertoldo ldquoExtremeenvironments as a resource for microorganisms and novel bio-catalystsrdquo Advances in Biochemical EngineeringBiotechnologyvol 96 pp 219ndash262 2005
[5] T Lonhienne C Gerday andG Feller ldquoPsychrophilic enzymesrevisiting the thermodynamic parameters of activation mayexplain local flexibilityrdquoBiochimica et Biophysica Acta vol 1543no 1 pp 1ndash10 2000
[6] R Cavicchioli and TThomas ldquoExtremophilesrdquo in Encyclopediaof Microbiology pp 317ndash337 Academic Press San Diego CalifUSA 2000
[7] G Feller E Narinx J L Arpigny et al ldquoEnzymes frompsychrophilic organismsrdquo FEMS Microbiology Reviews vol 18no 2-3 pp 189ndash202 1996
[8] J Arnorsdottir R B Smaradottir T O Magnusson S HThorbjarnardottir G Eggertsson and M M KristjanssonldquoCharacterization of a cloned subtilisin-like serine proteinasefrom a psychrotrophic Vibrio speciesrdquo European Journal ofBiochemistry vol 269 no 22 pp 5536ndash5546 2002
[9] N J Russell ldquoPsychrophilic bacteriamdashmolecular adaptations ofmembrane lipidsrdquo Comparative Biochemistry and Physiology APhysiology vol 118 no 3 pp 489ndash493 1997
[10] G Feller and C Gerday ldquoPsychrophilic enzymes hot topics incold adaptationrdquo Nature Reviews Microbiology vol 1 no 3 pp200ndash208 2003
[11] P Raspor and J Zupan ldquoYeasts in extreme environmentsrdquo inBiodiversity and Ecophysiology of Yeasts The Yeast Handbookpp 371ndash417 Springer Berlin Germany 2006
[12] M B Rao A M Tanksale M S Ghatge and V V Desh-pande ldquoMolecular and biotechnological aspects of microbialproteasesrdquoMicrobiology andMolecular Biology Reviews vol 62no 3 pp 597ndash635 1998
[13] L Polgar ldquoThe catalytic triad of serine peptidasesrdquo Cellular andMolecular Life Sciences vol 62 no 19-20 pp 2161ndash2172 2005
[14] S Y Boo C M V L Wong K F Rodrigues N NajimudinA M A Murad and N M Mahadi ldquoThermal stress responsesin Antarctic yeastGlaciozyma antarctica PI12 characterized byreal-time quantitative PCRrdquoPolar Biology vol 36 no 3 pp 381ndash389 2013
[15] B Turchetti S R T Hall L B Connell et al ldquoPsychrophilicyeasts from Antarctica and European glaciers descriptionof Glaciozyma gen nov Glaciozyma martinii sp nov andGlaciozyma watsonii sp novrdquo Extremophiles vol 15 no 5 pp573ndash586 2011
[16] N H F Hashim I Bharudin D L S Nguong et al ldquoCharac-terization of Afp1 an antifreeze protein from the psychrophilicyeast Glaciozyma antarctica PI12rdquo Extremophiles vol 17 no 1pp 63ndash73 2013
[17] A N M Ramli M A Azhar M S Shamsir et al ldquoSequenceand structural investigation of a novel psychrophilic 120572-amylasefrom Glaciozyma antarctica PI12 for cold-adaptation analysisrdquoJournal ofMolecularModeling vol 19 no 8 pp 3369ndash3383 2013
[18] A N M Ramli N M Mahadi A Rabu A M A Murad F DA Bakar and R M Illias ldquoMolecular cloning expression andbiochemical characterisation of a cold-adapted novel recombi-nant chitinase from Glaciozyma antarctica PI12rdquoMicrobial CellFactories vol 10 no 1 article 94 2011
[19] J Sambrook E Fritsch and T Maniatis Molecular Cloning ALaboratoryManual Cold SpringHarbor Laboratory Press NewYork NY USA 1989
[20] F M Brock C W Forsberg and J G Buchanan-SmithldquoProteolytic activity of rumen microorganisms and effects ofproteinase inhibitorsrdquoApplied and EnvironmentalMicrobiologyvol 44 no 3 pp 561ndash569 1982
[21] U K Laemmli ldquoCleavage of structural proteins during theassembly of the head of bacteriophage T4rdquo Nature vol 227 no5259 pp 680ndash685 1970
[22] A Henne R A Schmitz M Bomeke G Gottschalk and RDaniel ldquoScreening of environmental DNA libraries for thepresence of genes conferring lipolytic activity on Escherichiacolirdquo Applied and Environmental Microbiology vol 66 no 7 pp3113ndash3116 2000
[23] M Poza T de Miguel C Sieiro and T G Villa ldquoCharacteri-zation of a broad pH range protease of Candida caseinolyticardquoJournal of Applied Microbiology vol 91 no 5 pp 916ndash921 2001
20 Enzyme Research
[24] J D Bendtsen H Nielsen G von Heijne and S BrunakldquoImproved prediction of signal peptides SignalP 30rdquo Journalof Molecular Biology vol 340 no 4 pp 783ndash795 2004
[25] R J Siezen and J A M Leunissen ldquoSubtilases the superfamilyof subtilisin-like serine proteasesrdquo Protein Science vol 6 no 3pp 501ndash523 1997
[26] Y Katakura W Zhang G Zhuang et al ldquoEffect of methanolconcentration on the production of human 1205732- glycoprotein Idomain V by a recombinant Pichia pastoris a simple system forthe control of methanol concentration using a semiconductorgas sensorrdquo Journal of Fermentation and Bioengineering vol 86no 5 pp 482ndash487 1998
[27] Q-FWang Y-HHou Z Xu J-LMiao andG-Y Li ldquoPurifica-tion and properties of an extracellular cold-active protease fromthe psychrophilic bacterium Pseudoalteromonas sp NJ276rdquoBiochemical Engineering Journal vol 38 no 3 pp 362ndash3682008
[28] S Vazquez L Ruberto and W Mac Cormack ldquoProper-ties of extracellular proteases from three psychrotolerantStenotrophomonas maltophilia isolated from Antarctic soilrdquoPolar Biology vol 28 no 4 pp 319ndash325 2005
[29] R Zeng R Zhang J Zhao and N Lin ldquoCold-active ser-ine alkaline protease from the psychrophilic bacterium Pseu-domonas strain DY-A enzyme purification and characteriza-tionrdquo Extremophiles vol 7 no 4 pp 335ndash337 2003
[30] M Turkiewicz E Gromek H Kalinowska and M ZielinskaldquoBiosynthesis and properties of an extracellularmetalloproteasefrom the Antarctic marine bacterium Sphingomonas paucimo-bilisrdquo Journal of Biotechnology vol 70 no 1ndash3 pp 53ndash60 1999
[31] A Vassileva D A Chugh S Swaminathan and N KhannaldquoEffect of copy number on the expression levels of hepatitisB surface antigen in the methylotrophic yeast Pichia pastorisrdquoProtein Expression andPurification vol 21 no 1 pp 71ndash80 2001
[32] M Pazgier M Turkiewicz H Kalinowska and S Bielecki ldquoTheunique cold-adapted extracellular subtilase from psychrophilicyeast Leucosporidium antarcticumrdquo Journal ofMolecular Catal-ysis B Enzymatic vol 21 no 1-2 pp 39ndash42 2003
[33] M Turkiewicz M Pazgier H Kalinowska and S BieleckildquoA cold-adapted extracellular serine proteinase of the yeastLeucosporidium antarcticumrdquo Extremophiles vol 7 no 6 pp435ndash442 2003
[34] D Mattanovich B Gasser H Hohenblum and M SauerldquoStress in recombinant protein producing yeastsrdquo Journal ofBiotechnology vol 113 no 1ndash3 pp 121ndash135 2004
[35] P Secades BAlvarez and JAGuijarro ldquoPurification andprop-erties of a new psychrophilic metalloprotease (Fpp2) in the fishpathogen Flavobacterium psychrophilumrdquo FEMS MicrobiologyLetters vol 226 no 2 pp 273ndash279 2003
[36] L Y Ling I Ithoi and F M Yik ldquoOptimization for high-level expression in Pichia pastoris and purification of truncatedand full length recombinant sag2 of toxoplasma gondii fordiagnostic userdquo Southeast Asian Journal of Tropical Medicineand Public Health vol 41 no 3 pp 507ndash513 2010
[37] A Illanes ldquoStability of biocatalystsrdquo Electronic Journal ofBiotechnology vol 2 no 1 pp 1ndash9 1999
[38] B A Plantz J Sinha L Villarete K W Nickerson and VL Schlegel ldquoPichia pastoris fermentation optimization energystate and testing a growth-associated modelrdquo Applied Microbi-ology and Biotechnology vol 72 no 2 pp 297ndash305 2006
[39] K Tamura G Stecher D Peterson A Filipski and S KumarldquoMEGA6molecular evolutionary genetics analysis version 60rdquo
Molecular Biology and Evolution vol 30 no 12 pp 2725ndash27292013
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

Enzyme Research 19
frame of 2892 bp encoded for 963 amino acids from thisAntarctic yeast has been cloned and expressed in Pichiaexpression system This is the second extracellular pro-tease reported being synthesized by this psychrophilic yeastSequence analysis through bioinformatics studies revealedfew homology identities of this protein with other ser-ine proteases The mature PI12 protease was successfullyexpressed and secreted into the culture medium driven bythe Saccharomyces cerevisiae 120572-factor signal sequence in Ppastoris The protease production in P pastoris was bestobtained from strain GS115 (GpPro2) with 283UmL after 72hours of induction time with 05 (vv) of methanol inducerThe starting optical density at 600 nm (OD600) was 60 andthe optimum assay temperature was 20∘C The expressedprotein was detected by SDS-PAGE and activity staining witha molecular weight of 99 kDaThis cold-active enzymemightoffer an opportunity for biotechnological exploitation basedon its high catalytic activity at low temperatures and lowthermostability Structural and functional study of this cold-adapted enzyme would give insight into its adaptation to anextreme cold environment
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgment
This research was supported financially by the InitiativeBiotechnology-GMBI-MOSTI (Project number 07-05-16-MG-GMB02) andRUGS-Initiative 6 (Project number 05-02-11-1414RU)
References
[1] J W Deming ldquoPsychrophiles and polar regionsrdquo CurrentOpinion in Microbiology vol 5 no 3 pp 301ndash309 2002
[2] C Gerday M Aittaleb M Bentahir et al ldquoCold-adaptedenzymes from fundamentals to biotechnologyrdquo Trends inBiotechnology vol 18 no 3 pp 103ndash107 2000
[3] A Svingor J Kardos I Hajdu A Nemeth and P Zavodszky ldquoAbetter enzyme to cope with cold comparative flexibility studieson psychrotrophic mesophilic and thermophilic IPMDHSrdquoThe Journal of Biological Chemistry vol 276 no 30 pp 28121ndash28125 2001
[4] G Antranikian C E Vorgias and C Bertoldo ldquoExtremeenvironments as a resource for microorganisms and novel bio-catalystsrdquo Advances in Biochemical EngineeringBiotechnologyvol 96 pp 219ndash262 2005
[5] T Lonhienne C Gerday andG Feller ldquoPsychrophilic enzymesrevisiting the thermodynamic parameters of activation mayexplain local flexibilityrdquoBiochimica et Biophysica Acta vol 1543no 1 pp 1ndash10 2000
[6] R Cavicchioli and TThomas ldquoExtremophilesrdquo in Encyclopediaof Microbiology pp 317ndash337 Academic Press San Diego CalifUSA 2000
[7] G Feller E Narinx J L Arpigny et al ldquoEnzymes frompsychrophilic organismsrdquo FEMS Microbiology Reviews vol 18no 2-3 pp 189ndash202 1996
[8] J Arnorsdottir R B Smaradottir T O Magnusson S HThorbjarnardottir G Eggertsson and M M KristjanssonldquoCharacterization of a cloned subtilisin-like serine proteinasefrom a psychrotrophic Vibrio speciesrdquo European Journal ofBiochemistry vol 269 no 22 pp 5536ndash5546 2002
[9] N J Russell ldquoPsychrophilic bacteriamdashmolecular adaptations ofmembrane lipidsrdquo Comparative Biochemistry and Physiology APhysiology vol 118 no 3 pp 489ndash493 1997
[10] G Feller and C Gerday ldquoPsychrophilic enzymes hot topics incold adaptationrdquo Nature Reviews Microbiology vol 1 no 3 pp200ndash208 2003
[11] P Raspor and J Zupan ldquoYeasts in extreme environmentsrdquo inBiodiversity and Ecophysiology of Yeasts The Yeast Handbookpp 371ndash417 Springer Berlin Germany 2006
[12] M B Rao A M Tanksale M S Ghatge and V V Desh-pande ldquoMolecular and biotechnological aspects of microbialproteasesrdquoMicrobiology andMolecular Biology Reviews vol 62no 3 pp 597ndash635 1998
[13] L Polgar ldquoThe catalytic triad of serine peptidasesrdquo Cellular andMolecular Life Sciences vol 62 no 19-20 pp 2161ndash2172 2005
[14] S Y Boo C M V L Wong K F Rodrigues N NajimudinA M A Murad and N M Mahadi ldquoThermal stress responsesin Antarctic yeastGlaciozyma antarctica PI12 characterized byreal-time quantitative PCRrdquoPolar Biology vol 36 no 3 pp 381ndash389 2013
[15] B Turchetti S R T Hall L B Connell et al ldquoPsychrophilicyeasts from Antarctica and European glaciers descriptionof Glaciozyma gen nov Glaciozyma martinii sp nov andGlaciozyma watsonii sp novrdquo Extremophiles vol 15 no 5 pp573ndash586 2011
[16] N H F Hashim I Bharudin D L S Nguong et al ldquoCharac-terization of Afp1 an antifreeze protein from the psychrophilicyeast Glaciozyma antarctica PI12rdquo Extremophiles vol 17 no 1pp 63ndash73 2013
[17] A N M Ramli M A Azhar M S Shamsir et al ldquoSequenceand structural investigation of a novel psychrophilic 120572-amylasefrom Glaciozyma antarctica PI12 for cold-adaptation analysisrdquoJournal ofMolecularModeling vol 19 no 8 pp 3369ndash3383 2013
[18] A N M Ramli N M Mahadi A Rabu A M A Murad F DA Bakar and R M Illias ldquoMolecular cloning expression andbiochemical characterisation of a cold-adapted novel recombi-nant chitinase from Glaciozyma antarctica PI12rdquoMicrobial CellFactories vol 10 no 1 article 94 2011
[19] J Sambrook E Fritsch and T Maniatis Molecular Cloning ALaboratoryManual Cold SpringHarbor Laboratory Press NewYork NY USA 1989
[20] F M Brock C W Forsberg and J G Buchanan-SmithldquoProteolytic activity of rumen microorganisms and effects ofproteinase inhibitorsrdquoApplied and EnvironmentalMicrobiologyvol 44 no 3 pp 561ndash569 1982
[21] U K Laemmli ldquoCleavage of structural proteins during theassembly of the head of bacteriophage T4rdquo Nature vol 227 no5259 pp 680ndash685 1970
[22] A Henne R A Schmitz M Bomeke G Gottschalk and RDaniel ldquoScreening of environmental DNA libraries for thepresence of genes conferring lipolytic activity on Escherichiacolirdquo Applied and Environmental Microbiology vol 66 no 7 pp3113ndash3116 2000
[23] M Poza T de Miguel C Sieiro and T G Villa ldquoCharacteri-zation of a broad pH range protease of Candida caseinolyticardquoJournal of Applied Microbiology vol 91 no 5 pp 916ndash921 2001
20 Enzyme Research
[24] J D Bendtsen H Nielsen G von Heijne and S BrunakldquoImproved prediction of signal peptides SignalP 30rdquo Journalof Molecular Biology vol 340 no 4 pp 783ndash795 2004
[25] R J Siezen and J A M Leunissen ldquoSubtilases the superfamilyof subtilisin-like serine proteasesrdquo Protein Science vol 6 no 3pp 501ndash523 1997
[26] Y Katakura W Zhang G Zhuang et al ldquoEffect of methanolconcentration on the production of human 1205732- glycoprotein Idomain V by a recombinant Pichia pastoris a simple system forthe control of methanol concentration using a semiconductorgas sensorrdquo Journal of Fermentation and Bioengineering vol 86no 5 pp 482ndash487 1998
[27] Q-FWang Y-HHou Z Xu J-LMiao andG-Y Li ldquoPurifica-tion and properties of an extracellular cold-active protease fromthe psychrophilic bacterium Pseudoalteromonas sp NJ276rdquoBiochemical Engineering Journal vol 38 no 3 pp 362ndash3682008
[28] S Vazquez L Ruberto and W Mac Cormack ldquoProper-ties of extracellular proteases from three psychrotolerantStenotrophomonas maltophilia isolated from Antarctic soilrdquoPolar Biology vol 28 no 4 pp 319ndash325 2005
[29] R Zeng R Zhang J Zhao and N Lin ldquoCold-active ser-ine alkaline protease from the psychrophilic bacterium Pseu-domonas strain DY-A enzyme purification and characteriza-tionrdquo Extremophiles vol 7 no 4 pp 335ndash337 2003
[30] M Turkiewicz E Gromek H Kalinowska and M ZielinskaldquoBiosynthesis and properties of an extracellularmetalloproteasefrom the Antarctic marine bacterium Sphingomonas paucimo-bilisrdquo Journal of Biotechnology vol 70 no 1ndash3 pp 53ndash60 1999
[31] A Vassileva D A Chugh S Swaminathan and N KhannaldquoEffect of copy number on the expression levels of hepatitisB surface antigen in the methylotrophic yeast Pichia pastorisrdquoProtein Expression andPurification vol 21 no 1 pp 71ndash80 2001
[32] M Pazgier M Turkiewicz H Kalinowska and S Bielecki ldquoTheunique cold-adapted extracellular subtilase from psychrophilicyeast Leucosporidium antarcticumrdquo Journal ofMolecular Catal-ysis B Enzymatic vol 21 no 1-2 pp 39ndash42 2003
[33] M Turkiewicz M Pazgier H Kalinowska and S BieleckildquoA cold-adapted extracellular serine proteinase of the yeastLeucosporidium antarcticumrdquo Extremophiles vol 7 no 6 pp435ndash442 2003
[34] D Mattanovich B Gasser H Hohenblum and M SauerldquoStress in recombinant protein producing yeastsrdquo Journal ofBiotechnology vol 113 no 1ndash3 pp 121ndash135 2004
[35] P Secades BAlvarez and JAGuijarro ldquoPurification andprop-erties of a new psychrophilic metalloprotease (Fpp2) in the fishpathogen Flavobacterium psychrophilumrdquo FEMS MicrobiologyLetters vol 226 no 2 pp 273ndash279 2003
[36] L Y Ling I Ithoi and F M Yik ldquoOptimization for high-level expression in Pichia pastoris and purification of truncatedand full length recombinant sag2 of toxoplasma gondii fordiagnostic userdquo Southeast Asian Journal of Tropical Medicineand Public Health vol 41 no 3 pp 507ndash513 2010
[37] A Illanes ldquoStability of biocatalystsrdquo Electronic Journal ofBiotechnology vol 2 no 1 pp 1ndash9 1999
[38] B A Plantz J Sinha L Villarete K W Nickerson and VL Schlegel ldquoPichia pastoris fermentation optimization energystate and testing a growth-associated modelrdquo Applied Microbi-ology and Biotechnology vol 72 no 2 pp 297ndash305 2006
[39] K Tamura G Stecher D Peterson A Filipski and S KumarldquoMEGA6molecular evolutionary genetics analysis version 60rdquo
Molecular Biology and Evolution vol 30 no 12 pp 2725ndash27292013
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

20 Enzyme Research
[24] J D Bendtsen H Nielsen G von Heijne and S BrunakldquoImproved prediction of signal peptides SignalP 30rdquo Journalof Molecular Biology vol 340 no 4 pp 783ndash795 2004
[25] R J Siezen and J A M Leunissen ldquoSubtilases the superfamilyof subtilisin-like serine proteasesrdquo Protein Science vol 6 no 3pp 501ndash523 1997
[26] Y Katakura W Zhang G Zhuang et al ldquoEffect of methanolconcentration on the production of human 1205732- glycoprotein Idomain V by a recombinant Pichia pastoris a simple system forthe control of methanol concentration using a semiconductorgas sensorrdquo Journal of Fermentation and Bioengineering vol 86no 5 pp 482ndash487 1998
[27] Q-FWang Y-HHou Z Xu J-LMiao andG-Y Li ldquoPurifica-tion and properties of an extracellular cold-active protease fromthe psychrophilic bacterium Pseudoalteromonas sp NJ276rdquoBiochemical Engineering Journal vol 38 no 3 pp 362ndash3682008
[28] S Vazquez L Ruberto and W Mac Cormack ldquoProper-ties of extracellular proteases from three psychrotolerantStenotrophomonas maltophilia isolated from Antarctic soilrdquoPolar Biology vol 28 no 4 pp 319ndash325 2005
[29] R Zeng R Zhang J Zhao and N Lin ldquoCold-active ser-ine alkaline protease from the psychrophilic bacterium Pseu-domonas strain DY-A enzyme purification and characteriza-tionrdquo Extremophiles vol 7 no 4 pp 335ndash337 2003
[30] M Turkiewicz E Gromek H Kalinowska and M ZielinskaldquoBiosynthesis and properties of an extracellularmetalloproteasefrom the Antarctic marine bacterium Sphingomonas paucimo-bilisrdquo Journal of Biotechnology vol 70 no 1ndash3 pp 53ndash60 1999
[31] A Vassileva D A Chugh S Swaminathan and N KhannaldquoEffect of copy number on the expression levels of hepatitisB surface antigen in the methylotrophic yeast Pichia pastorisrdquoProtein Expression andPurification vol 21 no 1 pp 71ndash80 2001
[32] M Pazgier M Turkiewicz H Kalinowska and S Bielecki ldquoTheunique cold-adapted extracellular subtilase from psychrophilicyeast Leucosporidium antarcticumrdquo Journal ofMolecular Catal-ysis B Enzymatic vol 21 no 1-2 pp 39ndash42 2003
[33] M Turkiewicz M Pazgier H Kalinowska and S BieleckildquoA cold-adapted extracellular serine proteinase of the yeastLeucosporidium antarcticumrdquo Extremophiles vol 7 no 6 pp435ndash442 2003
[34] D Mattanovich B Gasser H Hohenblum and M SauerldquoStress in recombinant protein producing yeastsrdquo Journal ofBiotechnology vol 113 no 1ndash3 pp 121ndash135 2004
[35] P Secades BAlvarez and JAGuijarro ldquoPurification andprop-erties of a new psychrophilic metalloprotease (Fpp2) in the fishpathogen Flavobacterium psychrophilumrdquo FEMS MicrobiologyLetters vol 226 no 2 pp 273ndash279 2003
[36] L Y Ling I Ithoi and F M Yik ldquoOptimization for high-level expression in Pichia pastoris and purification of truncatedand full length recombinant sag2 of toxoplasma gondii fordiagnostic userdquo Southeast Asian Journal of Tropical Medicineand Public Health vol 41 no 3 pp 507ndash513 2010
[37] A Illanes ldquoStability of biocatalystsrdquo Electronic Journal ofBiotechnology vol 2 no 1 pp 1ndash9 1999
[38] B A Plantz J Sinha L Villarete K W Nickerson and VL Schlegel ldquoPichia pastoris fermentation optimization energystate and testing a growth-associated modelrdquo Applied Microbi-ology and Biotechnology vol 72 no 2 pp 297ndash305 2006
[39] K Tamura G Stecher D Peterson A Filipski and S KumarldquoMEGA6molecular evolutionary genetics analysis version 60rdquo
Molecular Biology and Evolution vol 30 no 12 pp 2725ndash27292013
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology
Top Related