







Idiomas
Páginas
Jurídico


ASIMETRÍAS DE COMBATE Y COMPETENCIA
INTRA E INTERESPECÍFICA EN DOS ESPECIES DE
DÍPTEROS (Diptera: Richardiidae)
TESIS QUE PRESENTA FELIPE BECERRIL MORALES
PARA OBTENER EL GRADO DE DOCTOR EN CIENCIAS
ECOLOGÍA Y MANEJO DE RECURSOS NATURALES
Xalapa, Veracruz, México 2009

2
Aprobación final del documento final de tesis de grado:
“Asimetrías de combate y competencia intra e interespecífica en dos especies de dípteros
(Diptera: Richardiidae)”
Nombre Firma
Director Dr. Rogelio Macías Ordóñez ________________________
Dra. Molly Morris ________________________ Comité Tutorial
Dr. Roger Enrique Guevara Hernández ________________________
Dra. Diana Folger Pérez Staples ________________________
Dr. Mario Enrique Favila Castillo ________________________
Dr. Juan Antonio Rull Gabayet ________________________
Jurado
Dr. Francisco Díaz Fleischer ________________________

INSTITUTO DE Aprobación final del documento final de tesis de grado: ECOlOGIAj A.C.
"Asimetrías de combate y competencia ¡ntra e illterespecífica en dos especies de dípteros
(Díptera: Richardiidae)'~
Director
Comité Tutoría!
Jurado
Nombre
Dr. Rogelio Madas Ordóñez
Dra. Molly Morris
DI'. Roger Enrique Guevara Hernández
Dra. Diana Folger Pérez Staples
Dr. Mario Enrique Favila Castillo
Dr. Juan Antonio Rul\ Gabayet
Dr. Fr3J1Cisco Diaz Fleischer

3
RECONOCIMIENTOS
Terminar este tipo de trabajos suele llevar a la situación de listar e incluir en la tesis, a las personas y cosas que fueron apoyo y motivo. En correspondencia, en muchos casos, luego de revisar u hojear las tesis, solemos revisar esta sección, con ánimos de constatar y/o constatarnos, regodearnos por uno y/o por otros; quienes estuvieron allí, en el recuerdo, en el hogar, en las aulas, en el cubo itinerante, en el diamante coatepecano, en la francachela o en la fiesta, en el solar de las neblinosas tardes jalapeñas. Sea pues, que fluya la sempiterna dualidad: agradecimiento y satisfacción. Añado a la sed de gracias, la gracia de mi satisfacción, de agradecer a quien me ayudó.

4
DEDICATORIA
A mis hijas y a mi esposa, a mis padres y a mis hermanos

5
DECLARACIÓN
Excepto cuando es explícitamente indicado en el texto, el trabajo de investigación contenido en
esta tesis fue efectuado por Felipe Becerril Morales como estudiante de la carrera de Doctorado
en Ciencias (Ecología y Manejo de Recursos Naturales) entre septiembre del 2001 y febrero del
2009, bajo la supervisión del Dr. Rogelio Macías Ordóñez.
Las investigaciones reportadas en esta tesis no han sido utilizadas anteriormente para obtener
otros grados académicos, ni serán utilizadas para tales fines en el futuro.
Candidato: _____________________________
Director de tesis: _____________________________

DECLARACIÓN
Excepto cuando es explícitamente indicado en el texto, el trabajo de investigación
contenido en esta tesis fue efectuado por Felipe Becerril Morales como estudiante de la
CalTera de Doctorado en Ciencias (Ecología y Manejo de Recursos Naturales) entre
septiembre del 200 I Y febrero del 2009, bajo la supervisión del 01'. Rogelio Macías
OrdÓñez.
Las investigaciones reportadas en esta tesis no han sido uti lizadas anterionnente para
obtener otros grados académicos, ni serán utilizadas para tales fines en el futuro.
Candidato: V
Felipe Becerril Morales
Director de tesis:
Rogelio Macias Ordóñez

6
ÍNDICE
Lista de tablas .........................................................................................................................8
Lista de figuras........................................................................................................................9
Resumen ...............................................................................................................................10
I. Introducción general ............................................................................................................11
Interacciones agonísticas y territorialidad..............................................................................12
Aspectos de la historia natural de ricárdidos ..........................................................................15
Objetivos...............................................................................................................................21
Literatura citada ....................................................................................................................22
II. Territorial contests within and between two species of flies (Diptera: Richardiidae) in the
wild. ..........................................................................................................................................26
Abstract.................................................................................................................................27
Introduction...........................................................................................................................28
Material and methods ............................................................................................................30
Analysis............................................................................................................................31
Abundance and size......................................................................................................31
Fight frequency ............................................................................................................32
Resident fighting success..............................................................................................32
Concurrence Index (CX)................................................................................................33
Results ..................................................................................................................................34
Abundance and body size .................................................................................................34
Fight frequency.................................................................................................................34
Resident fighting success ..................................................................................................35
Concurrence index............................................................................................................36
Discussion.............................................................................................................................37

7
Fighting asymmetries........................................................................................................38
Acknowledgements ...............................................................................................................40
References.............................................................................................................................41
III. Competencia por territorios: exclusión de campo............................................................53
Introducción..........................................................................................................................54
Método..................................................................................................................................56
Resultados.............................................................................................................................58
Discusión ..............................................................................................................................59
Literatura citada ....................................................................................................................69
IV. Discusión general ...............................................................................................................72
Coexistencia entre O. nitens y O. ferruginea .........................................................................74
Perspectivas de estudio en la ecología de la conducta en ricárdidos .......................................76
Literatura citada ....................................................................................................................78

8
Lista de tablas
CAPÍTULO 2
Table 1 Median and range of abundance of Odontomera nitens and O. ferruginea per
census in Xalapa, Veracruz, Mexico during the two field seasons. ...................................... 46
Table 2 Fight frequencies in 2003 and 2004 among all sex-species categories in each
resident-intruder roll. .......................................................................................................... 47
Table 3 Interactions among factors affecting fight frequency among all sex-species
categories in each resident-intruder roll according to the minimum adequate model (with a
Poisson distribution) and based on contingency table arrangement of richardiids fights
frequencies in two years of survey (see Table 2). ................................................................ 48
Table 4 Factors predicting resident fighting success: Type II ANOVA (non sequential).
Binomial GLM tests with logit link. LR: likelihood ratio test. ............................................. 49
CAPÍTULO 3
Tabla 1 Número de combates en que cada categoría sexo-especie actuó como residente
según las condiciones experimentales. ................................................................................ 64
Tabla 2 Resultados de las comparaciones estadísticas según las variables medidas en cada
experimento de exclusión.................................................................................................... 65

9
Lista de figuras
CAPÍTULO 1
Figura 1. Representación de las tres especies de ricárdidos observadas en el paraje de
Coapexpan, Veracruz, durante el 2003 y 2004..................................................................... 18
Figura 2. Área de estudio. .................................................................................................. 19
CAPÍTULO 2
Figure 1 Components of contest behaviour observed in two species of richardiids flies at
the study site during 2003 and 2004. ................................................................................... 50
Figure 2 Left wing length, (median ± quartiles, and extreme values) of male and female O.
nitens and O. ferruginea...................................................................................................... 51
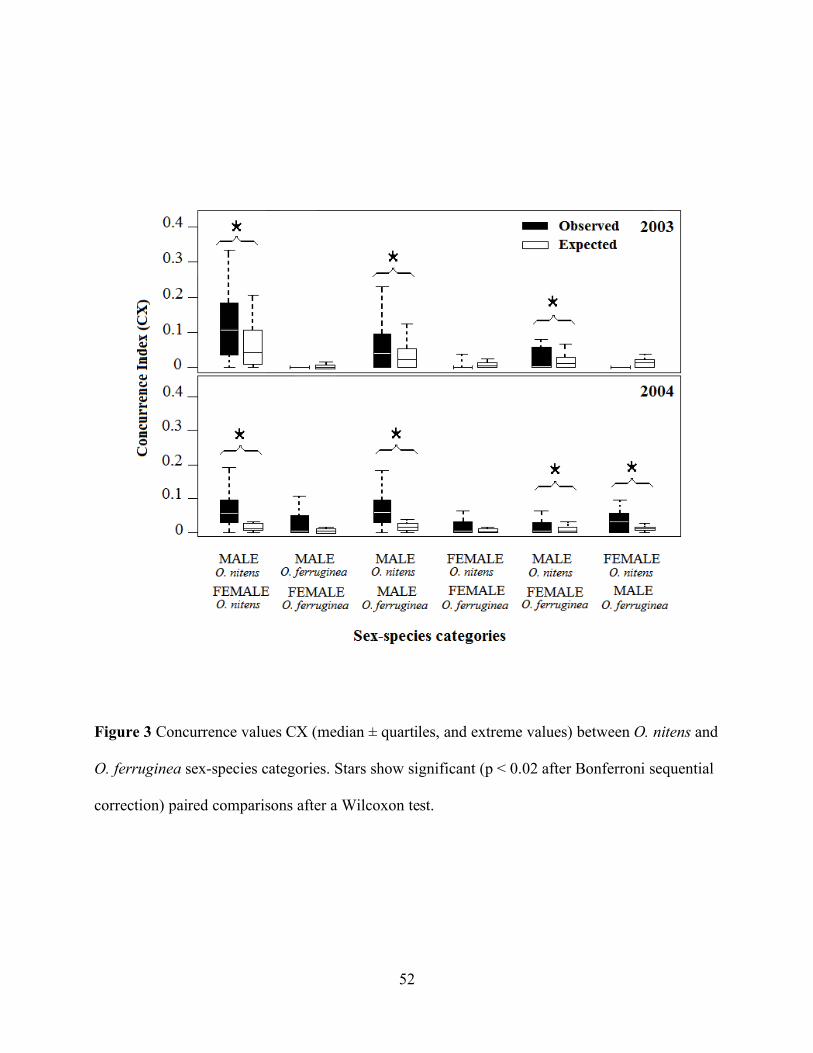
Figure 3 Concurrence values CX (median ± quartiles, and extreme values) between O.
nitens and O. ferruginea sex-species categories................................................................... 52
CAPÍTULO 3
Figura 1 Número de moscas (mediana, cuartiles 1 y 3 y valores extremos) y número de
combates (conteos de barrido) con y sin exclusión de territorios de alimentación. ............... 66
Figura 2 Abundancias (mediana, cuartiles 1 y 2 y valores máximos y mínimos) de las
categorías de ricárdidos por sexo y por especie, según los distintos tratamientos en cada
experimento. ....................................................................................................................... 67
Figura 3 Duración de los combates observados (mediana, cuantiles 1 y 3 y valores
extremos) según los tratamientos de cada experimento de exclusión. .................................. 68

10
Resumen
La coexistencia entre individuos y entre especies involucra tanto sus habilidades competitivas
como el grado en que coinciden en sus preferencias por un recurso ecológico. En este trabajo se
estudió por dos años la interacción agonística entre dos especies de mosca (Odontomera nitens y
O. ferruginea) de la familia Richardiidae, así como el efecto de sus asimetrías de combate en la
defensa de territorios de alimentación. Por otra parte, se exploró si los sitios defendidos
representan un recurso limitado, disminuyendo la disponibilidad de ellos y monitoreando la
frecuencia y duración de los combates. La disminución de los sitios a defender, aumentó la
frecuencia y duración de los combates, pero solo en el caso de las exclusiones de mayor duración.
Por primera vez se describe en detalle la conducta agresiva de este género de dípteros. Nuestros
resultados mostraron que la categoría ‘especie’, no fue una asimetría relevante en el combate. Por
otro lado, se encontró que los efectos de la residencia previa y del tamaño corporal en la
resolución del combate, dependen a su vez de la interacción entre ellos o con otros factores tales
como el sexo o la especie. Se encontró que la interacción agresiva entre individuos de ambas
especies no necesariamente surge de su concurrencia en las plantas donde se observaron, por lo
cual se resalta la necesidad de ajustar las escalas espaciales a las cuales se estudian dichas
interacciones. Lo anterior permite discutir el efecto de la magnitud y duración de la
disponibilidad de recursos en la competencia, ya sea por explotación o por interferencia y en
torno al concepto de sistemas territoriales.

11
CAPÍTULO I
I. Introducción general

12
Interacciones agonísticas y territorialidad
La conducta agresiva en los animales ha sido vista como una forma especializada de competencia
por algún recurso de acceso limitado (Archer 1988), y se ha caracterizado de acuerdo a las
circunstancias o condiciones en que los individuos intentan enfrentar diferentes presiones
ambientales, tanto en un contexto ecológico (Adams 2001) como social (Emlem & Oring 1977;
Magnhagen 2006). Los recursos por los cuales puede existir una interacción agresiva (combate)
son típicamente sitios de alimentación o sitios de agregación de parejas reproductivas. El estudio
del comportamiento agresivo se ha centrado en dos enfoques: el de territorios, el cual se basa en
interacciones de un individuo con respecto al resto de la población en un marco esencialmente
espacial (Tiebout 1996; Sirot 2000; Adams 2001); y el enfoque a nivel de díadas o pares de
individuos, mediante teoría de juegos (Parker 1974; Maynard Smith & Parker 1976; Maynard
Smith 1982; Mesterton-Gibbons & Adams 1998; Switzer et al. 2001, Magnhagen 2006).
En ocasiones los individuos delimitan y acaparan un área que contiene los recursos
necesarios para su sobrevivencia, repeliendo la intrusión de otros individuos; algunas veces
desplegando señales de advertencia y en otras mediante agresiones que implican contacto físico,
infligiendo incluso algún grado de daño. Entendemos como territorio al “área defendida” (Maher
& Lott 1995). Características tales como tamaño y permanencia de los territorios determinan
tanto el arreglo espacio-temporal de los individuos de una población (Gordon 1997) como la
demarcación de un nicho ecológico en interacción con otras especies (Gordon 2000; Adams
2001). Este último escenario, abre la posibilidad de que los individuos de dos especies que
compartan la necesidad del mismo recurso compitan por territorios (Abrams 2001).
En el conflicto por un recurso, la competencia puede establecerse de manera directa o
indirecta. La primera de ellas se identifica con la competencia por interferencia y se puede

13
manifestar al enfrentarse agresivamente dos individuos (díadas). Por otro lado, dos o más
individuos pueden competir indirectamente ejerciendo sus propias habilidades en la búsqueda y
consumo del recurso (competencia por explotación). En una interacción directa, la teoría de
combates predice que cada contendiente adopta una estrategia que lo lleve a la victoria y es
generalmente asociada a su capacidad de combate y/o a la recompensa obtenida (Hodge 1987;
Riechert 1998). Cada individuo posee atributos que le confieren capacidades de ganar un
combate, no obstante, existen combates entre contendientes que difieren muy poco en tales
atributos (combates simétricos) o bien pueden enfrentarse contendientes con francas diferencias
(combates asimétricos). Las denominadas ‘asimetrías de combate’ son un aspecto relevante en el
estudio de la conducta agresiva en díadas de organismos. A una mayor habilidad de combate,
mayor probabilidad de ganar un encuentro agresivo y es esto se le ha llamado ‘capacidad de
retención del recurso’ (RHP, por sus siglas en inglés). Se ha documentado ampliamente que el
tamaño del contendiente tiene una alta correlación con el RHP (Parker 1974), y se incluye en una
categoría denominada ‘asimetría de combate correlacionada’ (Maynard Smith & Parker 1976).
Varios ejemplos de otras asimetrías correlacionadas han sido reportados en distintos sistemas,
tales como el estado fisiológico, el uso de rasgos morfológicos o químicos especializados en la
agresión (armamento), la experiencia en combates o el sexo (ver revisión en Arnott & Elwood
2009). Por otra parte, existe una característica de los contendientes que no está correlacionada
con el RHP, no obstante puede determinar el resultado de un enfrentamiento. En muchos sistemas
territoriales se ha observado que los individuos que arriban primero al territorio (residentes)
poseen mayor probabilidad de repeler a un individuo que traspase, en segunda instancia, los
límites del territorio (intruso). Este predominio del residente se ha explicado conforme a lo que,
en teoría de combates, se ha llamado ‘estrategia del Burgués’ (Maynard Smith 1982);
aparentemente los individuos de la población obedecen una regla arbitraria de ‘respetar al

14
residente’, eludiendo potenciales enfrentamientos de alto costo. Esta explicación ha sido, por
décadas, objeto de numerosos estudios, resultando en un acerbo debate entre especialistas, en
especial respecto a su enfoque simplista y poco realista. No obstante, de tales investigaciones se
pueden reconocer al menos dos cosas: 1) el hecho de que los individuos involucrados en disputas
territoriales adquieren roles diferentes (residente e intruso) y 2) las razones del éxito del residente
podrían asociarse con asimetrías de combate correlacionadas (Kemp & Wiklund 2004).
Cuando se trata de dos especies que viven en simpatría, taxonómicamente cercanas y
morfológicamente semejantes, se espera que los individuos de cada especie coincidan en la
elección de sus recursos ecológicos, y por ello los efectos de la competencia serían más evidentes
(Juliano & Lawton 1990; Guillot 2005). Tal coexistencia tradicionalmente ha sido explicada en
concordancia al principio de Gause, de exclusión competitiva (Begon et al. 1996; Tilman 2007).
Este principio indica que dos especies no pueden ocupar el mismo nicho ecológico (Gause 1934).
Alternativamente, un reparto de recursos disminuiría la competencia y con ello permitiría la
coexistencia. Sin embargo, en recientes formulaciones de modelos de coexistencia, se han
encontrado relaciones importantes entre la competencia y la escala espacial en la que ocurren las
interacciones interespecíficas (Inouye 1999; Gordon 2000). Es decir, el criterio de coexistencia
depende de la escala (región, hábitat, etc.) a la que ocurren los encuentros entre los individuos de
dos o más especies. Sin embargo, al margen de dicho criterio, es razonable suponer que los
individuos pueden transitar en los límites de sus nichos (cualquiera que ellos sean) y coincidir en
sus requerimientos ecológicos, y con ello aumentar la probabilidad de un encuentro agresivo.
La competencia interespecífica ha sido uno de los tópicos más abordados en la ecología y
se ha documentado en una gran variedad de organismos y situaciones. Por ejemplo, pese a la
creencia generalizada de que el alimento no suele ser limitante para las especies fitófagas

15
(Lawton & Hassell 1981; Reitz & Trumble 2002), otros estudios han evidenciado la importancia
de la competencia interespecífica como un mecanismo que moldea la estructura de comunidades
de artrópodos pertenecientes a este gremio (Denno et al. 1995).
Aspectos de la historia natural de ricárdidos
Por mucho tiempo, los ricárdidos fueron ubicados como una subfamilia dentro de la
familia Otitidae (Carrejo & González, 1992). Dadas algunas diferencias en rasgos morfológicos
tanto en las alas como en el abdomen, actualmente es una familia incluida en la superfamilia
Tephritoidea, junto a otras ocho familias entre las que se cuentan Ulidiidae (= Otitidae) y
Tephritidae. Esta última familia ha sido por mucho la más estudiada, principalmente por su
importancia económica ya que varias de sus especies oviponen en frutos de especies de
importancia comercial (Díaz-Fleischer & Aluja 2000). En contraste, existe muy poca información
para la familia Richardiidae debido principalmente a que su diversidad y abundancias locales
suelen ser bajas (Pie 1998), y a la poca importancia económica que representa. De hecho
Richardiidae es señalada como la familia menos conocida de los tephritoideos (Diaz-Fleischer et
al. 2000).
En la familia Richardiidae se reconocen aproximadamente 170 especies (30 géneros
según Steyskal, 1987), restringidas al continente americano, principalmente en los trópicos
(Carrejo & González, 1992). De sus hábitos de vida se conoce muy poco, aunque se sabe que son
principalmente saprófagos. Existen reportes de sus larvas alimentándose de frutos y hojas en
descomposición (Inouye 1999; Diaz-Fleischer et al. 2000; Sivinski 2000) y también de materia
orgánica asociada a troncos caídos (Geoff Hancock, y Marcio Pie, com. pers.). También se han
observado en flores y brácteas florales de Heliconia (Richardson et al. 2000). Pie & del Claro
(2001), describieron agregaciones de no más de 10 individuos (cantidad calificada por los autores

16
como de alta densidad) en partes discretas de vegetación de pastizal, específicamente en plantas
productoras de néctar extrafloral (Qualea multiflora: Vochysiaceae), en las cuales observaron
tanto conductas de alimentación como de reproducción.
Este trabajo consistió en el estudio de la conducta territorial de dos especies del género
Odontomera (O. nitens Schiner 1868 y O. ferruginea Macquart 1844), pertenecientes a la familia
Richardiidae. La identidad de las especies se estableció a partir de especimenes colectados en
abril del 2003 con la colaboración del especialista Geoff Hancock de la Universidad de Glasgow,
quien entre los especímenes colectados en el sitio de estudio nos reportó una especie más: O.
albopilosa Hendel 1911, la cual, debido a su menor ocurrencia en el área de estudio no fue
incluida en nuestras investigaciones. Según Carrejo & González (1992), los ricárdidos son
dípteros caracterizados principalmente por la presencia de setas laterales en el segundo segmento
abdominal, fémures generalmente gruesos y espinosos, celda anal redondeada o truncada
apicalmente y alas con patrón de pocas manchas. La determinación de las dos especies estudiadas
se basó en la pigmentación presente en el escutelum, en el fémur del tercer par de patas y en la
parte dorsal de los segmentos abdominales. La especie Odontomera nitens presenta el escutelum
completamente negro, fémures y segmentos abdominales con marcas negras bien definidas. La
especie O. ferruginea presenta marcas negras en cada lado del escutelum, fémures con manchas
tenues (o ausentes) de color negro y ausencia de marcas en los segmentos abdominales. La
especie O. albopilosa es reconocida por su color negro en todo el cuerpo (Fig. 1).
En las inmediaciones de la ciudad de Xalapa, Veracruz, México, individuos adultos de
ambas especies de ricárdidos defienden territorios de alimentación en dos especies de plantas
silvestres: una planta herbácea Circium sp. (Asteracea) y principalmente una especie de planta
arbustiva Eupatorioum collinum (Asteracea). Las interacciones territoriales involucran a

17
individuos de ambos sexos y especies, e incluso a especies de otros órdenes de insectos, p. ej.
Himenoptera, Ortoptera, Coleóptera (Becerril-Morales, datos no publicados).
Otras poblaciones de las tres especies estudiadas fueron detectadas en el Jardín Botánico
Clavijero (Municipio de Xalapa), así como en los Municipios de Coatepec, Xico, Teocelo y
Apazapan, ver Fig. 2B). Sin embargo, la información obtenida se basó en el trabajo de campo
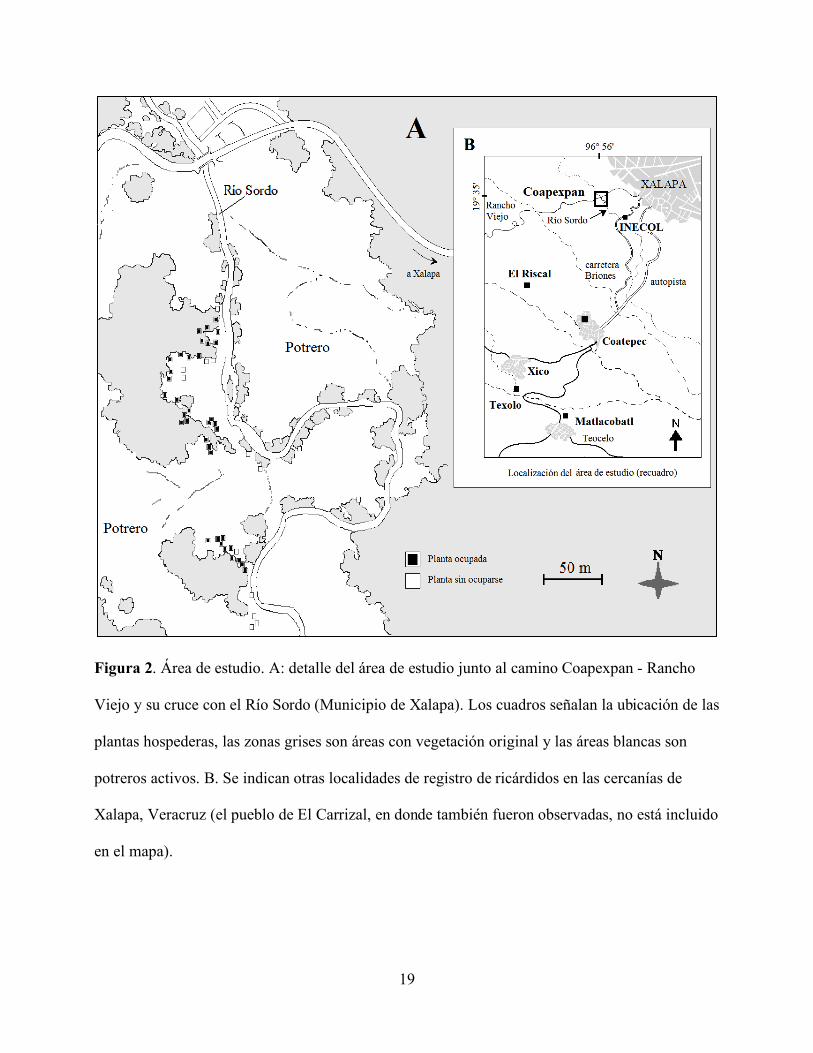
efectuado en la ribera del Río Sordo, en el paraje de Coapexpan, Municipio de Xalapa (Fig. 2A).
Dicho paraje consiste de fragmentos de bosque mesófilo de montaña y pastizales utilizados como
potreros. También otras especies de plantas fueron registradas de manera esporádica como fuente
de alimento para ambas especies de ricárdidos además de E. collinum y Circium sp., tanto en
Coapexpan como en otras localidades: Balsamina impatiens, Ricinus communis y Dioscores
alata. Ambas especies de ricárdidos se alimentan del exudado de las plantas, aparentemente
extraído mediante el contacto reiterado del labelum (del aparato bucal o probosis) sobre las
vellosidades presentes en la superficie de hojas, pecíolo y tallos, en especial de las partes jóvenes
de la planta (retoños en el caso de E. collinum).

18
Figura 1. Representación de las tres especies de ricárdidos observadas en el paraje de
Coapexpan, Veracruz, durante el 2003 y 2004.
.
19
Figura 2. Área de estudio. A: detalle del área de estudio junto al camino Coapexpan - Rancho
Viejo y su cruce con el Río Sordo (Municipio de Xalapa). Los cuadros señalan la ubicación de las
plantas hospederas, las zonas grises son áreas con vegetación original y las áreas blancas son
potreros activos. B. Se indican otras localidades de registro de ricárdidos en las cercanías de
Xalapa, Veracruz (el pueblo de El Carrizal, en donde también fueron observadas, no está incluido
en el mapa).

20
En el área de estudio fue común observar que los individuos de ambas especies de ricárdidos,
demarcaran territorios y los defendieran mediante combates que involucran contendientes de
diferente especie, sexo y tamaño. Este escenario, aunado a la facilidad para identificar los roles
de residente e intruso, representó una atractiva oportunidad de evaluar el comportamiento
territorial bajo algunos preceptos de la teoría de combates. En este caso, era factible probar en
conjunto, el efecto de asimetrías correlacionadas en la capacidad de retención del territorio por
parte del residente. Una vez que ciertos atributos, por ejemplo los propios del sexo, pueden
conferirle ventajas de combate a los individuos (Arnott & Elwood 2009), es razonable suponer
que los atributos que les confieren la identidad de especie a los individuos también puedan ser
utilizados para ganar un combate; de modo que esta condición pueda significar una asimetría de
combate. Por otra parte, en dicho escenario, es posible que los individuos se enfrenten siguiendo
una estrategia fija consistente en ahuyentar a cualquier intruso, o bien en tratar de ingresar a
cualquier territorio ya ocupado. En tal caso, la frecuencia de los combates sólo sería función de
cuántos individuos estarían presentes en la población y cuántos de ellos efectivamente
concurriesen en los sitios donde presumiblemente se concentra el recurso limitante. Más aun,
tomando en cuenta que el recurso en disputa representa un valor asociado a la decisión de iniciar,
continuar y/o terminar un combate, el restringir el acceso a dicho recurso, podría ilustrar los
mecanismos que rigen tanto la competencia como la coexistencia de ambas especies de
ricárdidos.

21
Objetivos
De acuerdo a lo anterior, en esta investigación se abordó la interacción competitiva entre las
especies O. nitens y O. ferruginea, en tres aspectos:
1) Los enfrentamientos en función de las habilidades de combate de los individuos
(asimetrías de combate) y su relación con la probabilidad de éxito del residente (capítulo
2).
2) La frecuencia de enfrentamientos y la concurrencia de potenciales contendientes desde
una perspectiva poblacional, intersexual e interespecífica dada las abundancias relativas
de cada categoría sexo-especie y el uso del recurso (capítulo 2).
3) El efecto de las fluctuaciones en la abundancia local del recurso alimenticio sobre el
comportamiento territorial (capítulo 3).

22
Literatura citada
Abrams, P.A. 2001. Describing and quantifying interspecific interactions: a commentary on
recent approaches. Oikos 94: 209-218.
Adams, E.S. 2001. Approaches to the study of territory size and shape. Annu. Rev. Ecol. Syst.
32: 277-303.
Archer, J. 1988. The behavioral biology of aggression. Cambridge University Press. Cambridge,
UK.
Arnott, G. & Elwood R.W. 2009. Assessment of fighting ability in animal contests. Anim. Behav.
77: 991-1004.
Begon, M., J.L. Harper, & C.R. Townsend. 1996. Ecology, individuals, populations and
communities. Blackwell Scientific Publications, Oxford, UK.
Carrejo, G.N.S. & O.R. González. 1992. Introducción al conocimiento de los Diptera. Centro
Editorial Universidad del Valle. Cali, Colombia.
Denno, R.F., M.S. McClure and J.R. Ott. 1995. Interspecific interactions in phytophagous
insects: competition reexamined and resurrected. Annu. Rev. Entomol. 40: 297-331.
Díaz Fleischer, F. & M. Aluja, 2000. Behavior of tephritid flies: a historical perspective. En:
Fruit flies (Tephritidae), phylogeny and evolution of behavior. Aluja M. & A.L. Norrbom
(Eds.). 39-69. CRC Press LLC. Boca Raton, USA.
Díaz Fleischer, F., D.R. Papaj, R.J. Prokopy, A.L. Norrbom & M. Aluja. 2000. Evolution of fruit
fly oviposition behaviour. In: Fruit flies (Tepritidae), phylogeny and evolution of
behavior. Aluja M. & A.L. Norrbom (Eds). 811-841. CRC Press LLC. Boca Raton, USA.

23
Emlen, S.T. & L.W. Oring. 1977. Ecology, sexual selection, and the evolution of mating systems.
Science 197: 215-223.
Gause, G.F. 1934. The strugle for existence. Williams and Wilkins. Baltymore, Maryland, USA.
Gordon, C.E. 2000. The coexistence of species. Rev. Chil. Hist. Nat. 73: 175-198.
Gordon, D.M. 1997. The population consequences of territorial behavior. TREE 2: 63-66.
Guillot, C. 2005. Entomology. Springer. Dordrecht, Netherlands.
Hodge, M. A. 1987. Agonistic interactions between female bowl and doily spiders (Araneae,
Linyphiidae): owner biased outcomes. J. Arachnol., 15:241-247.
Inouye, B.D. 1999. Integrated nested spatial scales: implications for the coexistence of
competitors on a patchy resource. J. of Anim. Ecol. 68: 150-162.
Juliano, S.A. & J.H. Lawton 1990. The relationship between competition and morphology II.
Experiments on co-occurring dysticid beetles. J. of Anim. Ecol. 59: 831-848.
Kemp, D.J. & C. Wiklund. 2004: Residency effects in animal contests. Proc. R. Soc. Lond. 271:
1707-1711.
Lawton, J.H. & M.P. Hassell. 1981. Asymmetrical competition in insects. Nature 289, 793-795.
Magnhagen, 2006. Information about an opponent can decrease aggression in male common
gobies competing for nest sites. Anim. Behav. 71: 879–884.
Maher, C.R. & D.F. Lott. 1995. Definitions of territoriality used in the study of variation in
vertebrate spacing systems. Anim. Behav. 49: 1581-1597.
Maynard Smith, J. 1982. Evolution and the theory of games. Cambridge University Press. New
York, USA.
Maynard Smith, J. & G.A. Parker. 1976. The logic of asymmetric contests. Anim. Behav. 24:
159-175.

24
Mesterton-Gibbons, M. & E.S. Adams. 1998. Animal contests as evolutionary games. Amer.
Scien. 86: 334-341.
Parker, G. A. 1974. Assessment strategy and the evolution of fighting behavior. J. Theor. Biol.
47: 223-246.
Pie, M.R. 1998. Lek behaviour as the mating strategy of Setellia sp. (Diptera: Richardiidae). J.
Insect. Behav. 11: 823–832.
Pie, M.R. & K. Del-Claro. 2001. Male-male agonistic behavior and ant-mimicry in a neotropical
richardiid (Diptera: Richardiidae). Studies on Neotrop. Fauna and Environ. 36: 1-4.
Reitz, S. R. & J. T. Trumble. 2002. Competitive displacement among insects and arachnids.
Annu. Rev. Entomol. 47: 435-465.
Richardson, B.A., C. Rogers, & M.J. Richardson. 2000. Nutrients diversity, and community
structure of two phytotelm systems in a lower montane forest, Puerto Rico. Ecol.
Entomol. 25: 348-356.
Riechert, S. E. 1998. Game theory and animal contests. – In: Game theory and animal behavior.
(Dugatkin, L.A. & Reeve, H.K. eds). Oxford University Press, New York, p. 64-93.
Sirot, E. 2000. An evolutionarily stable strategy for aggressiveness in feeding groups. Behav.
Ecol. 11: 351-356.
Sivinski, J. 2000. Breeding habits and sex in families closely related to the Tephritidae:
opportunities for comparative studies of the evolution of fruit fly behavior. En: Fruit flies
(Tephritidae), phylogeny and evolution of behavior. Aluja M. & A.L. Norrbom (Eds.). 23-
37. CRC Press LLC. Boca Raton, USA.
Steyskal, G.C. 1987. Richardiidae. In: McAlpine, J.F. (ed.) Manual of neartic diptera. Research
Branch, Agriculture, Canada.

25
Switzer, P.V., J.A. Stamps, & M. Mangel. 2001. When should a territory resident attack? Anim.
Behav. 62: 749-759.
Tiebout, H.M. III. 1996. Costs and benefits of interspecific dominance rank: are subordinates
better at finding novel locations? Anim. Behav. 51: 1375-1381.
Tilman, D. 2007. Interspecific competition and multispecies coexistence. In: May R. M. &
McLean A. R. (Eds.). Theoretical ecology principles and applications. 84 – 97 pp. Oxford
University Press Inc., New York, USA.

26
CAPÍTULO II
II. Territorial contests within and between two species of flies
(Diptera: Richardiidae) in the wild.
(Publicado en Behaviour 2009, 146: 245-262)

27
Becerril-Morales F. & Macías-Ordóñez R. Territorial contests within and between two species of
flies (Diptera: Richardiidae) in the wild.
Abstract
When two species compete for the same resources, inter and intraspecific competition also
occurs. Males and females of two syntopic phytophagous fly species (O. nitens and O.
ferruginea, Diptera: Richardiidae) compete over small ephemeral feeding territories. We assessed
fight frequency of both sexes of two species in the wild, and the simultaneous effects of potential
fight asymmetries, sex and species, on the outcome of territorial contests in relation with resident
status. Our results showed that residents won most of the fights, although the proportion of fights
won by the resident depended on the sex of the resident, the interaction between the sex of the
resident and size, and the year. Furthermore, even though some combinations of species and sex
categories could be found on the same tree more than expected by chance, this did not imply a
significantly high fight frequency for such combinations.
Key words: interspecific contests, intersexual contests fight asymmetries, resident advantage,
Odontomera.

28
Introduction
From a behavioural perspective, both intraspecific competition (including both inter and
intrasexual competition; Merilaita & Jormalainen, 1997; Grant et al., 2000) and interspecific
competition (frequently between sister species; Denno et al., 1995; Azovsky, 1996; Hart, 2003)
may be observed in the form of fights or combats; and the ability to win a disputed resource may
raise or lower the survival or reproductive chances of contenders. Models of aggressive
interactions are usually dynamic and typically focus on two contenders under a game theory
approach (Switzer et al., 2001). Differences between contenders in traits related to the probability
of winning a combat are identified as fight asymmetries (Maynard Smith & Price, 1973, Maynard
Smith & Parker, 1976; Maynard Smith, 1982; Crowley, 2000). Fight asymmetries such as
experience (Gherardi & Tiedemann, 2004) size or perceived resource value are intrinsic to the
contenders, and frequently correlated with their resource holding power (RHP, Parker, 1974).
Other fight asymmetries are intrinsic to the nature of the conflict itself (resident-intruder role)
(Mesterton-Gibbons & Adams, 1998).
It has been repeatedly reported and theoretically approached that, all other things being equal,
residents have a higher probability of wining territorial fights, frequently regardless of other
asymmetries (Kemp & Wiklund, 2004). It has also been shown however, that this resident effect
may be modified by fight asymmetries in relative or absolute size, experience, relative resource
value and information on the disputed resource. Even congeneric species and conspecific males
and females frequently differ somewhat in ecological requirements (Archer, 1988; Baird & May,
2003). Thus, when males and females of two or more species fight, inter-specific and inter-sexual
differences may also be expected to influence the outcome of fights.

29
Studies aiming to identify size asymmetries have been commonly focused on conspecific
males, and very frequently under experimental conditions. Thus, although pointed out a while
ago (Riechert & Hammerstein, 1983), they have rarely extended to a population or interespecific
perspective of the distribution and interaction of fighting asymmetries. In this work, we
documented naturally occurring territorial fights within and between males and females of two
syntopic species of phytophagous flies (Odontomera nitens, Macquart 1843 and O. ferruginea,
Schiner 1868, Diptera: Richardiidae) under field conditions. There is no published ecological
information on any species of this genus. For the whole family, the least biologically known
family of their group (Tephritoidea, Diaz-Fleischer et al., 2000), only aggression and courtship
behaviour have been described for Setellia and Sepsisoma (Pie, 1998; Pie & del Claro, 2001).
However, preliminary observations showed that individuals of both species of Odontomera
defend feeding territories on leaves of a few individuals of two common plant species in our
study site, Eupatorium collinum and Circium sp. (both Asteraceae), suggesting heterogeneity in
the quality of feeding territories.
We explored the effect of relative size, species and sex on the probability of residents to
win territorial fights. We also explored if these fights occurred randomly, according to the
relative abundance of each sex and species. In addition, we predicted that individuals of both
species would be found together (we define this as concurrence) in a larger proportion of plants
than expected by chance. As far as we know, our study could be the first to document the
relationship between this many potential fight asymmetries simultaneously, either in natural or
controlled conditions.

30
Material and methods
Fieldwork was carried out in a 2 ha area of fragmented tropical cloud forest in the central part of
the state of Veracruz, Mexico (19º 35’ N, 96º 56’ W; 1,361 m.a.s.l.). Counts, captures and
observations of territorial behaviour of both species were carried out between 09:00 and 17:00 h.
Abundance of each species and sex were recorded throughout a fixed route during two annual
seasons (30 censuses on 46 plants between May and November 2003, 23 censuses on 47 plants
between April and December of 2004). In both years, the number of occupied plants (E. collinum
and Circium sp.) increased over the season (from 11 to 20 in 2003 and from 28 to 39 in 2004).
Territories are located under the leaf base and petiole of young leaves of E. collinum, and
over the leaves of Circium sp. Intruders were defined as any individual that landed on a leaf
occupied by other fly (resident) engaged in foraging (walking, probing and bubbling on the leaf
surface, presumably feeding on plant secretions and associated bacteria, see Aluja et al. 2000)
and/or patrolling behaviour. Unlike intruders, residents were frequently forming bubbles with
fluids from the proboscis (bubbling) and probing the leaf surface (dabbing), both behaviours
associated with foraging in tephritoid flies (Aluja et al., 2000). Fights occurred above or under
leaves and were defined as unidirectional approaches of one individual to another showing
seemingly aggressive chasing display and some times scaling to intense combats. Fights were
initiated by “approaching” (see Figure 1) usually by the resident walking from the base of the leaf
towards the intruder, which usually landed near the tip of the leaf. The intruder may, however,
initiate the fight approaching the resident. If none of the flies gives up at this stage, they contact
each other frontally touching heads while moving their wings (“heading”) and/or jump forward
(“threatening”). If the contest proceeds, both flies engage in more physical interactions
(“grappling”, “boxing”, “ramming” or “wrestling”). If the contest proceeds to the wrestling stage,

31
both flies may fall down, in which case only one comes back to the leaf (the winner). Any
contender may give up at any point during the fight. Most fights lasted less than 10 seconds and
finished at the earlier stages of escalation, usually during or before “threatening”.
For each interaction we recorded species, sex, territorial status (intruder or resident),
relative size of resident (larger than, smaller than or closely matched to the intruder) and fight
outcome (resident winner – loser). The winner was defined as the fly that remained on the
territory after the fight, and the loser as the fly that abandoned the leaf. Contenders in same sex
and species interactions were distinguished by means of individual variation in colour intensity
and pigmentation of the abdomen and last abdominal segment. Those few cases in which this was
not possible were not included in the analysis. Sex was assigned based on the presence of a very
conspicuous ovipositor in females. Visual species identification was carried out based on a
taxonomic description by Geoff Hancock (University of Glasgow), who previously received
voucher material from the field site.
One hundred and eighty two flies were captured (and then released) in 2003 and 92 in
2004. Left wing length of all individuals was taken with a calliper (± 0.1 mm). Unpublished
recapture data showed low recapture rates, thus suggesting a low chance of measuring or
counting twice the same individual.
Analysis
Abundance and size
Abundance (number of flies observed per day) of each sex of each species (four sex-species
categories) were compared between 2003 and 2004 using a Wilcoxon two-sample unpaired test
(daily values as replicates). Sex-species categories were compared in terms of wing length using
Kruskal Wallis and Dunn post-hoc comparison tests (Lehner, 1996).

32
Fight frequency
Fights between all sex-species categories and roles (resident or intruder) were arranged in a
contingency table for each year. Significant interactions among factors affecting fight frequency
among all sex-species categories in each resident-intruder roll were assessed using a generalized
linear model (GLM) with a Poisson distribution by means of a simplification process of the
saturated model (Crawley, 2002). The attributes of each sex-species-role combination (each cell
in Table 2) were coded in the following grouping variables (factors in the model): (sprel) relative
species identity of contenders (inter or intraspecific); (sxrel) relative sex identity of contenders
(inter or intrasexual); (spres) species identity of the resident (O. nitens or O. ferruginea); and
(sxres) sex identity of the resident (male or female).
Resident fighting success
A logistic regression was performed on fight outcome (Hardy & Field, 1998). A quasibinomial
model with a logit link function was used since the binomial model had a slight overdispersion
(1.36) (Fox, 2002). The minimal adequate model was obtained using a stepwise elimination
process from the maximal model using a GLM analysis (Crawley, 2002). The independent
variables were species, sex, relative size and year. As a response variable, the outcome of fights
can be represented using a binary vector (0 if the resident was displaced and 1 if the resident
won) (Hardy & Field, 1998). In order to use a logit link function when all independent variables
are factors, the outcome variable must be expressed as an “odds ratio” for each factor. The odds
ratio is p / (1-p) where p = fights won by the resident and 1-p = fights won by the intruder. This is
achieved by using the “cbind” command in R, and the number of fights and resident victories in
each factor combination (Crawley, 2002). Although the overall value of p (fights won by the
resident) was predicted to be above 0.5 due to the well documented resident advantage (see

33
discussion), the main interest of this analysis was to assess the effect of each independent
variable and their interactions on the binary outcome variable. The levels of the species factor
are, therefore, the four potential combinations between O. nitens and O. ferruginea, alternating
the resident role. Resident-intruder relative size included the three categories observed in the
field: larger than, smaller than, and closely matched. The “R” statistical package version 2.1.1 and
S-plus version 4.6 were used to perform the analyses.
Concurrence Index (CX)
In this study, concurrence was defined as the fraction of host plants on a given day in which
individuals of any two sex-species categories were simultaneously observed with respect to the
total number of host plants available. A concurrence index (CX) for each sex-species category
combination (e.g., males and females of O. ferruginea) was defined daily as CX = a /(a + b + c +
d) (modified after Peng et al., 2005), in which ‘a’ is the number of host plants in that day in
which at least one individual of each of the two categories was recorded; ‘b’ is the number of
host plants in which only one category was recorded (e.g. male O. ferruginea); ‘c’ is the number
of host plants in which only the other category was recorded (e.g., females O. ferruginea) and ‘d’
is the number of host plants in which no individual was recorded on that day, but in which at least
one individual of any sex-category was observed during the whole season. This index measures
the opportunity of fighting between different sex-species categories based on how frequently they
are actually found sharing a given host plant (the fighting arena) regardless of their overall
relative frequencies. Using Wilcoxon paired test, the observed CX was compared to the expected
one (CXexp) if both sex-categories were concurring at random on host plants depending only on
their relative frequency, which is expressed as CXexp = (a+b) (a+c)/ (a+b+c+d) 2. Individuals may
remain on the same host plant for several hours, sometimes walking along the petioles and stems.

34
Even when expelled from a territory or leaving it spontaneously, flies remain flying around the
same host plant landing on several leaves, and thus a given individual may interact with several
others before abandoning each plant. Therefore, a significantly high (or low) CX reflects more (or
less) opportunities to fight than expected by their relative abundance alone.
Results
Abundance and body size
Odontomera nitens was the most abundant species, both in 2003 (around 80%, median values per
census 16.5 and 4 for each species) and 2004 (around 70%, median values per census 15 and 6
for each species, Table 1). The difference in abundance between the two species decreased in
2004, since the number of O. ferruginea males increased and approached the number of O. nitens
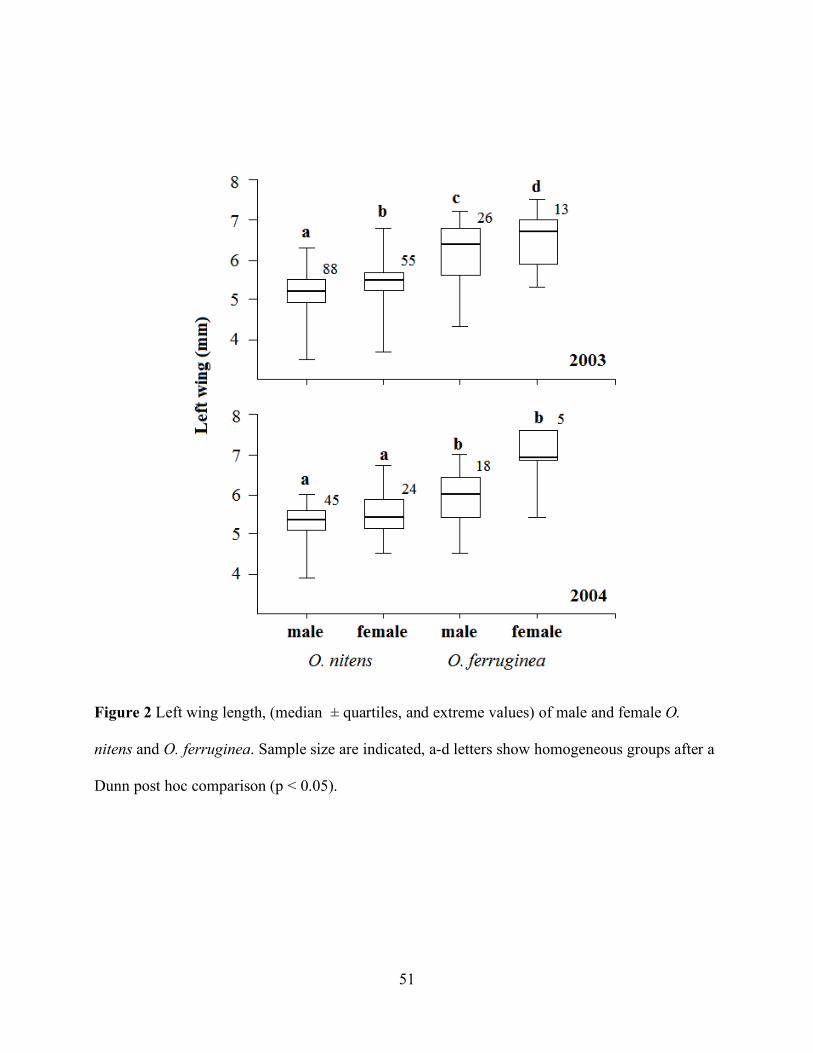
females (Table 1). The size comparison (left wing length) between sex-species categories showed
significant differences both in 2003 (Kruskal-Wallis: H (3, N = 182) = 52.7, p < 0.001) and in 2004
(H (3, N = 92) = 17.1, p < 0.001). A Dunn nonparametric multiple comparison test showed that both
O. nitens males and females were smaller than O. ferruginea males or females. Within species,
females were significantly larger than males in 2003, the year in which sample size was larger
(Figure 2).
Fight frequency
A total of 508 contests were recorded, 245 (48%) in 2003 and 263 (52%) in 2004; contenders
belonged to different species in 103 (20%) of them while in 405 (80%) they belonged to the same
species, in 370 (73%) to O. nitens and in 35 (7%) to O. ferruginea (Tables 2 and 3). Fight
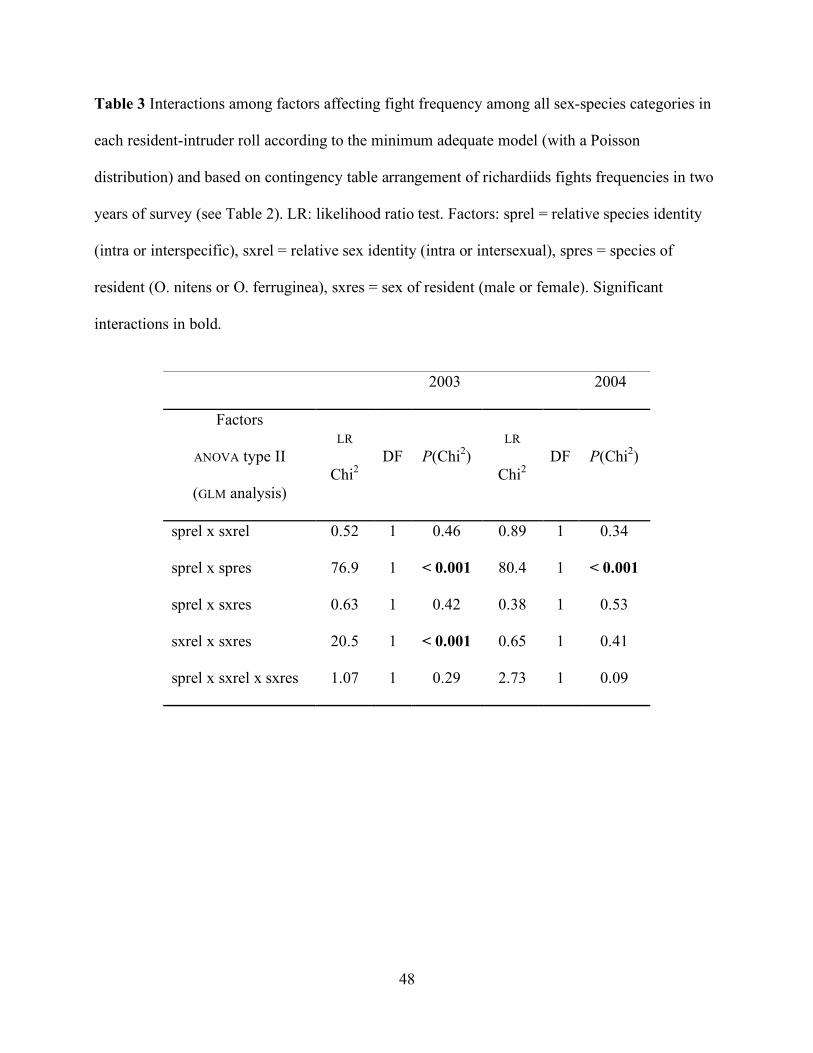
frequency was not randomly distributed among sex-species categories; the GLM analysis detected
two significant interactions. First, the interaction between relative species identity of contenders

35
(inter or intraspecific, “sprel”) and the species of the resident (“spres”) had a significant effect in
the frequency distribution of fights in both years ( p < 0.001 2003 and p < 0.001 2004) being
more frequent the fights of O. nitens as resident (table3). Second, the interaction between relative
sex identity of contenders (inter or intraspecific, “sxrel”) and the sex of the resident (“sxres”) had
a significant effect in the frequency distribution of fights, only in 2004 (p < 0.001) being more
frequent the fights of males as resident (Table 3). Thus, fights between O. nitens males were
more frequent than expected by chance. When the year was included as a factor in the model, the
frequency of intraspecific and intrasexual fights in 2004 (the year in which males of O.
ferruginea were more abundant) turned out to be significantly higher than in 2003, specially in
those cases in which males were residents (factors sprel x year, p = 0.01 and factors sxrel x sxres
x year, p = 0.002).
Resident fighting success
The proportion of fights won by residents was above 0.5 in both years (mean ± SD: 0.88 ± 0.25 in
2003; 0.67 ± 0.39 in 2004). Of the O. nitens intraspecific fights, 82% (304 / 370) were won by
residents and 80% (28 / 35) in the case of O. ferruginea. In the case of intrasexual fights, resident
males won 87% of the fights (201 / 230) and resident females won 83% (55 / 66). In the case of
intersexual fights, 81% (108 / 133) were won by resident males over intruding females, and 65%
(52 / 79) were won by resident females over intruding males. In fights in which contenders were
of different relative size, 82% (178 / 216) were won by larger residents and 73% (77 / 105) by
smaller residents. When both contenders were closely matched, 86 % (161 / 187) of the fights
were won by the resident. In the case of interspecific fights, 66% (21 / 32) were won by O. nitens
residents over O. ferruginea intruders, and 89% (63 / 71) were won by O. ferruginea residents
over O. nitens intruders.

36
Even though the proportion of the fights won by residents was slightly larger when the
intruder was smaller than the resident, it was even higher in size symmetrical fights. Relative size
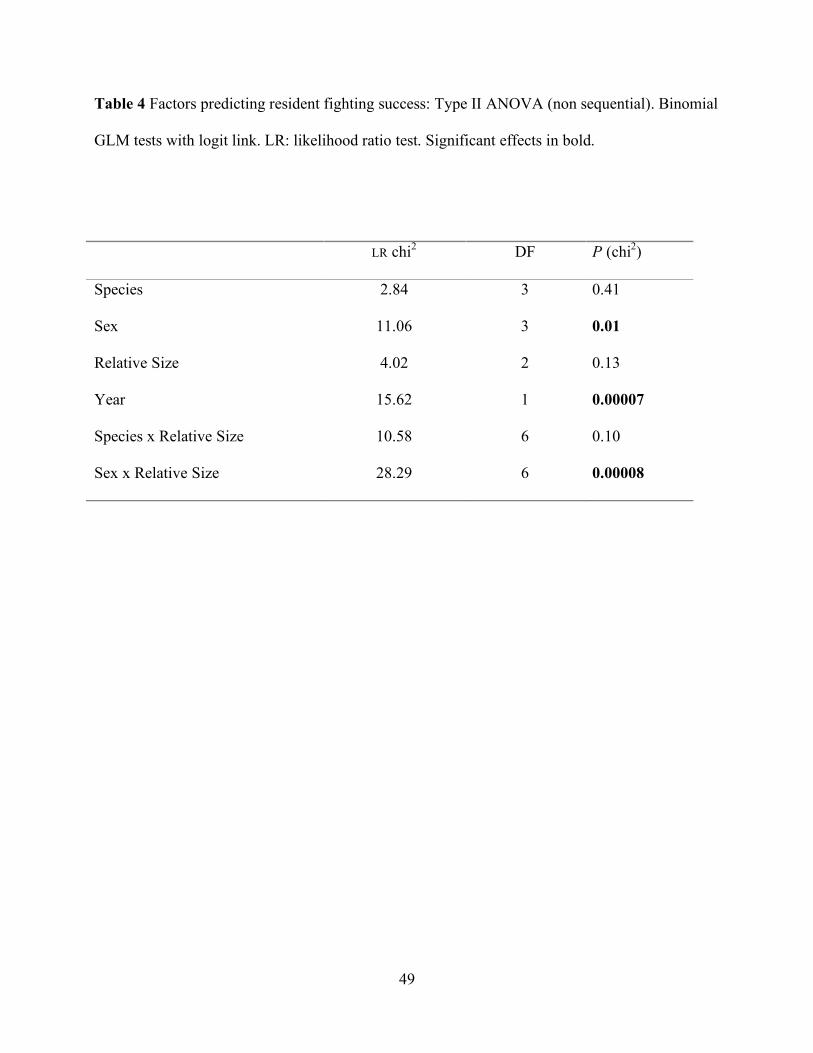
did not affect resident fighting success as a simple factor (GLM ANOVA, p = 0.13, Table 4), nor in
interactions with species in any year. As a simple factor in the model, sex significantly affected
the proportion of the fights won by residents (p = 0.01, GLM ANOVA, Table 4). Furthermore, its
interaction with relative size was also significant (p < 0.0001, GLM ANOVA, Table 4); smaller
resident males retained their territories against larger females (the only significant logistic
coefficient: β = 3.79, SE = 1.4, t = 2.6, p= 0.01). Finally, resident fighting success was different
between years (p < 0.001) and species by itself did not explain resident fighting success (p =
0.41, GLM ANOVA, Table 4).
Concurrence index
Within species, only O. nitens males and females were found in the same trees more than
expected by chance (CX observed vs expected by chance, paired Wilcoxon test: Z = 3.9, p = 0.001
in 2003 and Z = 4.0, p < 0.001 in 2004, Figure 3). Among species, O. nitens males were found in
the same trees more than expected by chance with both O. ferruginea males (CX observed vs
expected by chance, paired Wilcoxon test: Z = 2.4, p = 0.01 in 2003 and Z = 3.9, p < 0.001 in
2004) and females (CX observed vs expected by chance, paired Wilcoxon test: Z = 2.3, p = 0.01 in
2003 and Z = 2.4, p = 0.01 in 2004). Males of O. ferruginea were found in the same trees more
than expected by chance with O. nitens females (CX observed vs expected by chance, paired
Wilcoxon test: Z = 3.2, p = 0.001), but only in 2004. The remaining combinations concurred in
the same trees as expected given their relative abundance (p > 0.05 in all cases, significance
threshold was α = 0.02 after Bonferroni sequential correction).

37
Discussion
According to Díaz-Uriarte (2000), the continuous presence of intruders near feeding territories
may imply a high rate of intrusions. We expected that when difference in abundance between
potential contender categories were small (such as O. ferruginea males and O. nitens females in
2004), the concurrence values and thus the opportunities to fight would be higher than when one
of these categories was much less abundant (as in 2003) (Giraldeau & Beauchamp, 1999).
However our results were ambiguous. In 2004, interspecific encounters were less frequent, even
though interspecific concurrence values were higher. Male and female O. nitens, for instance,
showed significantly high concurrence values and also a significantly high frequency of contests,
specifically when males were residents. However, despite high concurrence values of O. nitens
males with O. ferruginea males and females, such category combinations did not show
significantly high contest frequencies.
One possibility is that territories may have different value for each species, in which case,
the resource may be worth a fight for different fractions of the flies of each species, thus the
frequency of interspecific fights would not correspond to relative abundance or to concurrence on
the same plant. The value of territories for each species could change between years, a likely
explanation for the different results in fight frequency between years, and the year effect on
fighting success. The actual value of the resource could also change drastically in a short time,
due to depletion of nutrients on the leaf surface and replenishment by plant secretions. That along
with continuous scouting by flies searching for such resource in a given host plant seem to result
in a highly dynamic system of short tenure territories disputed among those flies present at a
given moment. Results on concurrence and fight frequency suggest that some specific categories
such as O. nitens males, contest more than others for such short-tenure territories, regardless of

38
their concurrence with other categories, probably due to species-specific or sex-specific
perceived resource value (see below).
Fighting asymmetries
“Prior resident advantage” in territorial fights has been extensively reported in different
taxonomic groups (Davies, 1978; Rosenberg & Enquist, 1991; Wenseleers et al., 2002; Kemp &
Wiklund, 2004). Thus, it is not surprising that such effect was also prevalent in spite of any other
asymmetry; it is the additive or interactive effect of ownership status with other factors in natural
conditions what has been poorly explored in the past. Surprisingly however, even though size is
known to be a key factor of RHP (Parker, 1974; Petersen & Hardy, 1996), relative size was not so
as a single factor in our study. One possibility we can not rule out is that a small effect could
have gone undetected due to size categorization in a three-level relative size factor, since we did
not have the absolute size of contenders in enough cases to include it in the analysis (which
would have provide greater statistical power). Nevertheless, our results show how other
asymmetries influenced prior resident advantage. The simultaneous effect of sex and relative size
shown by their statistic interaction, for instance, did predict fight outcome. Regardless of species,
a resident advantage was observed in intraspecific fights in which resident males were not larger
but smaller than females, thus the interactive effect of relative size was contrary to that reported
in other studies in which this variable correlates positively with RHP (Maynard Smith, 1982;
Riechert, 1998).
It is important to notice that males seem to have an advantage, even though they are
smaller than females. It has been shown that males and females may differ in their fighting
strategy (Draud et al., 2004). Whether this may be responsible for an apparent female
disadvantage even against a favourable size asymmetry remains to be explored. Another

39
possibility is that the resources offered by the territories have different value for each sex.
Asymmetries in resource value have been suggested to predict the level of escalation (see Hurd
2006), and thus the probability of winning the resource in dispute. Extra floral secretions usually
contain carbohydrates more than aminoacids or lipids (Baker et al. 1978). Thus, such males may
simply be more willing to escalate as carbohydrates are a relatively more precious resource for
them (physiological energy to search for mates) than for females (for whom protein for egg
production is usually the limiting resource) (see Morrel et al. 2005). Not a single instance of
courtship, copulation or oviposition behaviour was observed on these “host” plants throughout
the study, but frequent instances of proboscis interaction with the leaf surface and bubbling (also
called post-ingestion food processing: Prokopy & Papaj 2000) performed by both males and
females of both species. The few published records on richardiid reproduction report rooting
organic material as oviposition and larval development sites (Inouye, 1999; Diaz-Fleischer et al.
2000; Sivinski 2000). All this strongly suggests only a feeding nature of these territories, but no
role in mate acquisition. It is very rare to find female competition for territories in insects,
especially against males (Sivinski, 2000). The observed female-female and female-male fights
among conspecific richardiids opens a rare opportunity to explore the role of different ecological
asymmetries in natural fights over feeding territories.
As in our study, Young (2003) found no species effect on fight outcome in co-generic fish
species competing for foraging spots. Our analyses suggest that the apparent advantage of O.
ferruginea over O. nitens in over one hundred interspecific encounters is due to specific sex and
size combinations, regardless of species identity. A question worthy of further exploration is how
the added effect of all such interactions could provide information on the competition dynamic
and coexistence between both species. Recent studies have highlighted the importance of
interspecific interactions as a mechanism shaping arthropod community structure (Reitz &

40
Trumble, 2002). It has been suggested that competition between phytophagous species is rare
(Lawton & Hassell, 1981) since their resources are rarely limited in natural conditions. However,
multiple evidence against this assumption has been put forward, especially in the case of closely
related taxa, feeding on discrete resources and living in habitats with some grade of human
perturbation (Denno et al. 1995); all three conditions are met in our study.
Most ecological models of competition for resources in the form of territorial defence are
static and focus on the defender versus the rest of the competitors (see review in Adams, 2001).
The effect of simultaneous asymmetries on fight outcome in the wild has rarely been studied. Our
study shows a complex of simultaneous factors influencing fight outcome in two congeneric
competing species in an ecological context. The next logical step is to address how individual
interactions translate into population dynamics of the interacting species.
Acknowledgements
We are grateful to Molly Morris, Roger Guevara and Glauco Machado for insightful comments
on early manuscripts. To Bill Eberhard for suggesting these flies as a study model. Geoff
Hancock kindly identified the flies. We want to express our gratitude to Roberto Munguía Steyer
for his help with the analysis and relevant comments. Our editor and two anonymous reviewers
provided challenging comments that greatly improved the manuscript. This study was partially
supported by a graduate fellowship (no. 35565) provided by Conacyt to FBM.

41
References
Adams, E.S. (2001). Approaches to the study of territory size and shape. – Annu. Rev. Ecol. Syst.
32: 277-303.
Aluja, M., Piñero, J., Jácome, I., Díaz-Fleisher, F. & Sivinski J. (2000). Behavior of flies in the
genus Anastrepha (Trypetinae: Toxotripanini). - In: Fruit flies (Tepritidae) (Aluja, M. &
Norrbom, A.L., eds). CRC Press LLC., Boca Raton, p. 375-406.
Archer, J. (1988). The behavioral biology of aggression. - Cambridge University Press,
Cambridge.
Azovsky, A.I. (1996). The effect of scale on congener coexistence: can mollusks and polychaetes
reconcile beetles to ciliates? - Oikos 77: 117-126.
Baird, J.M., & May, M.L. (2003). Fights at the dinner table: agonistic behaviour in Pachydiplaz
longipennis (Odonata: Libellulidae) at feeding sites. - J. of Insect Behav. 16: 189-216.
Baker, H.G., Opler, P.A. & Baker, I. (1978). A comparison of the amino acid complements of
floral and extrafloral nectars. - Botanical Gazette 139:322-332.
Crawley, M.J. (2002). Statistical computing, an introduction to data analysis using S-Plus. -
Wiley & Sons Ltd., Sussex.
Crowley, P.H. (2000). Hawks, doves and mixed-symmetric games. - J. Theor. Biol. 204: 543-
563.
Davies, N.B. (1978). Territorial defense in the speckled wood butterfly (Pararge aegeria), the
resident always wins. - Anim. Behav. 26: 138-147.
Denno, R.F., McClure, M.S. & Ott, J.R. (1995). Interspecific interactions in phytophagous
insects: competition reexamined and resurrected. - Annu. Rev. Entomol. 40: 297-331.

42
Díaz-Fleischer F., Papaj, D.R., Prokopy, R.J., Norrbom, A. L. & Aluja, M. (2000) Evolution of
fruit fly oviposition behavior. - In: Fruit flies (Tephritidae), Phylogeny and evolution of
behavior (Aluja M. & Norrbom A.L. eds). CRC Press LLC., Boca Raton, p. 811-841.
Díaz-Uriarte, R. (2000). Effects of aggressive interactions on antipredator behavior: empirical
and theoretical aspects. - Ph.D. Thesis, Univ. of Wisconsin, Madison.
Draud, M., Macías-Ordóñez, R., Verga, J. & Itzkowitz, M. (2004). Female and male Texas
cichlids (Herichthys cyanoguttatum) do not fight by the same rules. – Behav. Ecol. 1(15):
102–108.
Fox, J. (2002). An R and S-Plus companion to applied regression. - Sage Publications Inc.,
Thousand Oaks.
Gherardi, F. & Tiedemann, A. 2004: Agonism and interference competition in freshwater
decapods. - Behaviour 141: 1297-1324.
Giraldeau, L.A. & Beauchamp, G. (1999). Food exploitation: searching for the optimal joining
policy. - Trends Ecol. Evol. 3: 102-106.
Grant, J.W.A., Gaboury, C.L. & Levit, H.L. (2000). Competitor to resource ratio, a general
formulation of operational ser ratio, as a predictor of competitive aggression in japanese
medaka (Pisces: Oryziidae). - Behav. Ecol. 6: 670-675.
Hardy, C.W. & Field, S.A. (1998). Logistic analysis of animal contests. - Anim. Behav. 56: 787-
792.
Hart, P.J.B. (2003). Habitat use and feeding behaviour in two closely related fish species, the
three-spined and nine-spined stickleback: an experimental analysis. - J. of Anim. Ecol. 72:
777-783.

43
Hurd P.L. (2006). Resource holding potential, subjective resource value, and game theorical
model of aggressiveness signaling. - J. Theor. Biol. 241: 639-648.
Inouye, B.D. 1999. Integrated nested spatial scales: implications for the coexistence of
competitors on a patchy resource. - Journal of Animal Ecology, 68:150-162.
Kemp, D.J. & Wiklund, C. (2004). Residency effects in animal contests. - Proc. R. Soc. Lond.,
271: 1707-1711.
Lawton, J.H. & Hassell, M.P. (1981). Asymmetrical competition in insects. - Nature 289: 793-
795.
Lehner, N. P. (1996). Handbook of ethological methods. 2nd edn. - Cambridge University Press,
Cambridge.
Maynard Smith, J. (1982). Evolution and the theory of games. - Cambridge University Press,
New York.
Maynard Smith, J. & Price, G.R. (1973). The logic of animal conflict. – Nature 246: 15 – 18.
Maynard Smith J. & Parker, G.A. (1976). The logic of asymmetric contest. – Anim. Behav. 24:
159 - 175
Merilaita, S. & Jormalainen, V. (1997). Evolution of sex differences in microhabitat choice and
colour polymorphism in Idotea baltica. - Anim. Behav. 54: 769–778.
Mesterton-Gibbons, M. & Adams, E.S. (1998). Animal contests as evolutionary games. - Amer.
Sci. 86: 334-341.
Morrel, L.J., Lindström, J. & Ruxton, G.D. (2005). Why are small males aggressive? - Proc. R.
Soc. B272: 1235-1241.

44
Parker, G. A. (1974). Assessment strategy and the evolution of fighting behavior. - J. Theor. Biol.
47: 223-246.
Peng, Y.Q., Yang, D. & Wang, Q.T. (2005). Quantitative tests of interaction between pollinating
and non-pollinating fig wasps on dioecious Ficus hispida. - Ecological Entomology 30:
70-77.
Petersen G. & I.C.W. Hardy. 1996. The importance of being larger: parasitoid intruder-owner
contests and their implications for clutch size. - Anim. Behav. 51: 1363-1373.
Pie, M.R. (1998). Lek behaviour as the mating strategy of Setellia sp. (Diptera: Richardiidae). - J.
Insect. Behav. 11: 823–832.
Pie, M.R. & Del Claro, K. (2001). Male-male agonistic behavior and ant-mimicry in a neotropical
richardiid (Diptera: Richardiidae). - Studies on Neotropical Fauna and Environment 36: 1-
4.
Prokopy, R. & Papaj, D.R. (2000). Behavior of flies of the genera Ragoletis, Zonosemata and
Carpomya (Trypetinae: Carpomyina). In: Fruit flies (Tephritidae), Phylogeny and
evolution of behavior (Aluja M. & Norrbom A.L. eds). CRC Press LLC. Boca Raton,
219-252.
Reitz, S. R. & Trumble, J. T. (2002). Competitive displacement among insects and arachnids. -
Annu. Rev. Entomol. 47: 435-465.
Riechert, S. E. (1998). Game theory and animal contests. – In: Game theory and animal behavior.
(Dugatkin, L.A. & Reeve, H.K. eds). Oxford University Press, New York, p. 64-93.
Riechert, S.E. & Hammerstein, P. (1983). Game theory in the ecological context. – Annu. Rev.
Ecol. Syst. 14:377-409.

45
Rosenberg, R.H & Enquist, M. (1991). Contest behavior in Weidemeyer’s admiral butterfly
Limenitis weidemeyerii (Nymphalidae): the effect of size and residency. - Anim. Behav.
42: 805-811.
Sivinski, J. (2000). Breeding habits and sex in families closely related to the Tephritidae:
opportunities for comparative studies of the evolution of fruit fly behavior. - In: Fruit
Flies (Tepritidae), Phylogeny and Evolution of Behavior. (Aluja, M. & Norrbom, A.L.
eds). CRC Press LLC, Boca Raton, p. 23-37.
Switzer, P.V., Stamps, J.A. & Mangel, M. (2001). When should a territory resident attack? -
Anim. Behav. 62: 749-759.
Wenseleers, T., Billen J. & Hefetz, A. (2002). Territorial marking in the desert ant Cataglyphis
niger: does it pay to play bourgeois? - J. Insect. Behav. 15: 85-93.
Young, K.A. (2003). Evolution of fighting behavior under asymmetric competition: an
experimental test with juvenile salmonids. - Behav. Ecol. 14:127-134.

46
Table 1 Median and range of abundance of Odontomera nitens and O. ferruginea per census in
Xalapa, Veracruz, Mexico during the two field seasons. Significant differences in bold.
2003 2004
Species and sex Median
min - max n
Median
min - max n
p
Wilcoxon
Test
O. nitens
Males 13.0 0 – 47 30 10.0 2– 44 23 0.17
Females 3.5 0 – 27 30 4.0 0 – 24 23 0.73
Total 16.5 0 – 68 30 15.0 5 – 65 23 < 0.0001
O. ferruginea
Males 2.0 0 – 8 30 4.0 2 – 15 23 0.03
Females 1.0 0 – 7 30 1.0 0 – 7 23 0.25
Total 4.0 0 – 11 30 6 0 – 22 23 0.0008

47
Table 2 Fight frequencies in 2003 and 2004 among all sex-species categories in each resident-
intruder roll. The relative abundance of each category is shown in parenthesis. The relative
frequency of each category combination is shown at the lower right corner of each cell. The
number of observed fights for each category combination is shown at the top left corner of each
cell. Expected frequency of fights, according to the minimal model are shown at the top right
corner of each cell.
2003 Resident
MALE
O. nitens
(0.57)
FEMALE O. nitens
(0.22)
MALE O. ferruginea
(0.17)
FEMALE O. ferruginea
(0.04)
MALE O. nitens (0.57)
83 86.6
(0.32)
34 33.3
(0.12)
18 18.6
(0.09)
5 4.6
(0.02)
FEMALE O. nitens (0.31)
48 46.9
(0.17)
20 18.0
(0.06)
6 6.2
(0.05)
2 1.5
(0.01) MALE O. ferruginea (0.09)
6 5.4
(0.05)
1 1.8 (0.01)
13 9.3
(0.01)
3 3.6
(0.003)
Intruder FEMALE
O. ferruginea (0.02)
2 1.8
(0.01)
0 1.3
(0.004)
4 5.0
(0.003)
0 1.9
(0.008)
2004 Resident
MALE
O. nitens
(0.55)
FEMALE O. nitens
(0.24)
MALE O. ferruginea
(0.14)
FEMALE O. ferruginea
(0.06)
MALE O. nitens (0.47)
72 74.0
(0.3)
24 23.1
(0.13)
21 19.0
(0.07)
8 6.9
(0.03)
FEMALE O. nitens (0.38)
57 56.4
(0.02)
32 31.4
(0.09)
5 7.6
(0.05)
6 6.3
(0.02) MALE O. ferruginea (0.08)
9 10.9
(0.04)
3 4.0
(0.01)
8 6.0
(0.01)
1 1.8
(0.005)
Intruder FEMALE
O. ferruginea (0.06)
7 4.3
(0.03)
4 3.6
(0.01)
4 4.5
(0.009)
2 2.5
(0.004)
48
Table 3 Interactions among factors affecting fight frequency among all sex-species categories in
each resident-intruder roll according to the minimum adequate model (with a Poisson
distribution) and based on contingency table arrangement of richardiids fights frequencies in two
years of survey (see Table 2). LR: likelihood ratio test. Factors: sprel = relative species identity
(intra or interspecific), sxrel = relative sex identity (intra or intersexual), spres = species of
resident (O. nitens or O. ferruginea), sxres = sex of resident (male or female). Significant
interactions in bold.
2003 2004
Factors
ANOVA type II
(GLM analysis)
LR
Chi2 DF P(Chi2)
LR
Chi2 DF P(Chi2)
sprel x sxrel 0.52 1 0.46 0.89 1 0.34
sprel x spres 76.9 1 < 0.001 80.4 1 < 0.001
sprel x sxres 0.63 1 0.42 0.38 1 0.53
sxrel x sxres 20.5 1 < 0.001 0.65 1 0.41
sprel x sxrel x sxres 1.07 1 0.29 2.73 1 0.09
49
Table 4 Factors predicting resident fighting success: Type II ANOVA (non sequential). Binomial
GLM tests with logit link. LR: likelihood ratio test. Significant effects in bold.
LR chi2 DF P (chi2)
Species 2.84 3 0.41
Sex 11.06 3 0.01
Relative Size 4.02 2 0.13
Year 15.62 1 0.00007
Species x Relative Size 10.58 6 0.10
Sex x Relative Size 28.29 6 0.00008

50
Figure 1 Components of contest behaviour observed in two species of richardiids flies at the
study site during 2003 and 2004. Interactions usually occurred in the sequence indicated by the
numbers.
51
Figure 2 Left wing length, (median ± quartiles, and extreme values) of male and female O.
nitens and O. ferruginea. Sample size are indicated, a-d letters show homogeneous groups after a
Dunn post hoc comparison (p < 0.05).
52
Figure 3 Concurrence values CX (median ± quartiles, and extreme values) between O. nitens and
O. ferruginea sex-species categories. Stars show significant (p < 0.02 after Bonferroni sequential
correction) paired comparisons after a Wilcoxon test.

53
CAPÍTULO III
III. Competencia por territorios: exclusión de campo

54
Introducción
Uno de los principales retos en la investigación de los mecanismos con que se expresa la
competencia entre individuos de la misma o de distintas especies, es la identificación del recurso
en disputa (Dudley et al. 1990). En el caso de interacciones interespecíficas se ha supuesto que la
cercanía taxonómica y/o morfológica incrementa la oportunidad de comportamientos
competitivos. La razón de esto es que existiría una alta probabilidad de coincidir en la
explotación de un recurso en particular (Juliano et al. 1990; Hart 2003); la abundancia y
distribución de este modularían las formas de competencia.
La competencia por explotación implica la habilidad diferencial de uno o varios individuos de
agotar el recurso disputado, mientras que la competencia por interferencia implica la interacción
directa entre competidores por acceso al mismo (Begon et al. 1996). La defensa de territorios es
una forma recurrente en que se expresa este último tipo de competencia. Se ha propuesto que la
frecuencia e intensidad de las interacciones agresivas o combates por territorios dependen de la
densidad de los interactuantes en relación a la cantidad de territorios (revisado por Kokko et al.
2006). Estudios experimentales sugieren que la exclusión al acceso a recursos genera un aumento
en la agresión interespecífica (e.g. Petren & Case 1996) e intraespecífica (Goldber et al. 2001).
Por otro lado, desde un punto de vista de la concurrencia de dos especies que compiten por
recursos comunes, se puede esperar que entre más frecuente sea el encuentro entre individuos, la
competencia entre las especies sería más intensa (Thompson 1996).
El énfasis de la mayoría de los estudios de territorialidad en un contexto ecológico ha
consistido en relacionar atributos del territorio tales como el tamaño o la forma, con la
abundancia y distribución de los recursos (Stamps 1990; Adams 2001; Switzer et al. 2001). Sin
embargo estos estudios ignoran en gran parte las estrategias mismas de combate, el desarrollo de

55
los mismos, así como la variabilidad de atributos individuales entre intrusos y residentes (Switzer
et al. 2001).
En particular, las moscas fitófagas del género Odontomera (Familia Richardiidae), al parecer
compiten por territorios alimenticios, preferentemente en una especie arbustiva (Eupatorium
collinum, Asteracea) (Becerril-Morales & Macías-Ordóñez, 2009), no obstante se desconoce si
tal conducta está asociada a la disponibilidad de un recurso presente en tales plantas. Si bien se ha
supuesto que el alimento no suele ser un recurso limitante para las especies fitófagas (Reitz y
Trumble 2002), otros estudios han mostrado evidencia de lo contrario, en particular entre
especies filogenéticamente cercanas (e.g. Denno et al. 1995). En tales circunstancias se espera
que exista acaparamiento de recursos alimenticios (territorialidad) entre especies fitófagas, y por
tanto competencia tanto inter como intra específica por los recursos en que coinciden.
La hipótesis de que los sitios de alimentación de O. nitens Schiner 1868 y O. ferruginea
Macquart 1844, representan un recurso limitado por el cual compiten ambas especies de
ricárdidos fue puesta a prueba, manipulando la disponibilidad de territorios (hojas jóvenes de la
planta E. collinum). Es importante hacer notar que la disponibilidad de arbustos de E. collium y
hojas fue alta, y la enorme mayoría de estos no estuvieron “ocupados” ni fueron disputados. Solo
algunas hojas en ciertos árboles lo fueron, por lo cual se podría pensar que no todas las hojas
ofrecen recursos, o al menos recursos de la misma calidad, de tal manera que no se esperan
acaparamiento o combates por éstos sitios. En tal caso se predice que al disminuir
experimentalmente el acceso a las probables fuentes de alimentación, se observará un aumento
significativo en el número de disputas por los territorios restantes, así como un incremento en la
densidad de competidores por unidad de recurso. Por otro lado, se ha propuesto y documentado
que las estrategias de combate que modulan el escalamiento al valor relativo del recurso suelen

56
ser evolutivamente estables (Riechert 1998). Por lo tanto, también se espera que los combates
aumenten en duración dada la disminución en el acceso a los sitios de alimentación.
Método
El trabajo se realizó en un paraje a 1 km al suroeste de Xalapa, Veracruz (19° 35’ N, 96° 56’ O;
1,364 m.s.n.m.) y consistió en dos experimentos de exclusión de territorios de alimentación, en
una planta de E. collinum. Esta planta mostró las mayores abundancias de moscas en un estudio
paralelo (Becerril-Morales & Macías-Ordóñez 2009, Capítulo 2).
Se restringió el acceso de las moscas al recurso (territorios) mediante el uso de bolsas de tela.
El territorio defendido por una mosca fue definido como el área basal de una hoja (incluido el
pecíolo), localizada en el extremo de una rama. Sin embargo, fue frecuente observar
comportamientos de patrullaje que involucraron al resto de las hojas más extremas. En ambos
experimentos, a cada extremo de una rama se le llamó “punta” y esta fue la unidad de exclusión.
Inicialmente se identificó el número de territorios y se marcaron las puntas tanto activas (en las
que se observaron al menos un combate) como las no activas. Las marcas consistieron en
etiquetas de papel (1.0 cm2), rotuladas, enmicadas y sujetas a la rama por alambre inoxidable. Un
combate fue definido como aquel evento en que un individuo se aproximó a otro, mostrando
despliegues agresivos (particularmente proyectando las alas hacia adelante) y algunas veces
llegando al contacto físico, lo que resulta en una intensa lucha por desplazar al contendiente (ver
detalles en el capítulo 2).
Las observaciones en cada etapa de cada uno de los experimentos descritos más adelante,
consistieron en la toma de datos de forma sistemática, respecto a tres variables: 1) número de
individuos de cada sexo y especie en cada rama marcada (barrido de 30 min.); 2) la ocurrencia de

57
combates (tres observaciones de barrido de cinco min. separadas por cinco min.), registrando la
identidad de los contendientes (especie y sexo, residente o intruso); y 3) duración del combate.
El primer experimento comprendió tres etapas: 1ª) sin exclusión, 2ª) con exclusión y 3ª) sin
exclusión. La primera etapa (30 y 31 de Agosto y 1º de Septiembre del 2004, d1-d3) consistió en
la toma de datos (descritos arriba) en ramas sin exclusión. Este muestreo se repitió de manera
idéntica en tres periodos de una hora a lo largo de cada día (09:00 a 10:00 hrs.; 12:00 a 13:00
hrs.; y 15:00 a 16:00 hrs.). En la segunda etapa (2, 3 y 4 de Septiembre; d4-d6), se hicieron las
mismas mediciones que en la etapa 1, pero ahora con la mitad de los territorios excluidos
mediante embolsados de tela (tul). Las exclusiones se asignaron de manera sistemática
(alternando ramas e iniciando desde las puntas activas inferiores) tratando de distribuirlas
homogéneamente en toda la planta. Fueron excluidas también todas las plantas de E. collinum,
cercanas (en un radio de no menos de 15 m) a la planta experimental, cubriéndolas totalmente
con el mismo tipo de tela. Para la tercera etapa (7, 8 y 9 de Septiembre; d9-d11), se retiraron las
exclusiones y se procedió a tomar el mismo tipo de datos.
Para el segundo experimento se siguieron los mismos procedimientos generales de toma de
datos en la misma planta. Sin embargo, se ejecutaron dos etapas (sin exclusión y con exclusión)
de menor duración (un día) pero se realizaron el doble de sesiones de observación: seis periodos
de una hora a lo largo del día (09:00 a 10:00 hrs.; 10:30 a 11:30 hrs.; 12:00 a 13:00 hrs.; 13:30 a
14:30 hrs.; 15:00 a 16:00 hrs.; y 16:30 a 17:30 hrs.). Las observaciones sin exclusiones, se
llevaron a cabo el día 17 de Septiembre (d19) mientras que las observaciones con exclusiones se
llevaron a cabo el 18 de Septiembre (d20).
El segundo experimento pretendía determinar si una menor duración del tratamiento de
exclusión también estimularía a los individuos a exacerbar sus conductas territoriales. Para ello el
periodo de exclusión sería de un solo día. Sin embargo, no fue posible realizar los registros

58
posteriores a la exclusión debido a que hubo una disminución drástica de la abundancia de
moscas en el área de estudio. Por esta razón en este trabajo se presenta una comparación
esencialmente cualitativa entre los resultados de ambos experimentos, no obstante éstos tuvieron
un propósito común: obtener información sobre la importancia de los cambios en la
disponibilidad de territorios en la intensidad y calidad de competencia por un recurso alimenticio.
El análisis de datos consistió en el contraste de abundancias totales y abundancias por
categorías sexo-especie entre tratamientos (con exclusión o sin exclusión) de acuerdo a cada
experimento, el cual se realizó aplicando técnicas estadísticas no paramétricas que consideran
muestras pareadas: prueba de Friedman y prueba de rangos asignados de Wilcoxon (Siegel &
Castellan 1995). De modo semejante se compararon los datos del número de combates y duración
de combates para cada experimento.
Resultados
En total se observaron 92 enfrentamientos territoriales, 63 en el primer experimento y 29 en el
segundo. En ambos experimentos, la abundancia mediana del total de individuos de ambas
especies en la planta (26.5 individuos/censo, min = 7 y max = 48, ver Tabla 1) no mostró
cambios significativos en respuesta a la exclusión (Tabla 2, Fig. 1). La abundancia de ninguna de
las categorías sexo-especie fue distinta entre los tratamientos en ninguno de los dos experimentos
(Tabla 2, Fig. 2).
Se observó un aumento en el número de combates durante el periodo de exclusión en el primer
experimento pero no en el segundo (Tabla 2, Fig. 1). Los combates fueron de mayor duración
respecto a las condiciones de control en el primer experimento, pero no en el caso del segundo
(Tabla 2, Fig. 3).

59
Discusión
El primer experimento apoyó nuestras predicciones de que habría un aumento en la frecuencia y
duración de combates asociada a la exclusión de una fracción de los recursos. Se mostró también
que esto no resultó de un aumento en el número de individuos ya que las abundancias de cada
especie fueron similares. En otras palabras, no solo hay más combates por individuo cuando los
territorios disminuyen, sino que el eventual perdedor permanece por más tiempo en el combate
antes de rendirse. Sería de esperarse que no solo la duración sino el nivel de escalamiento de las
interacciones fuera mayor en los combates al disminuir la disponibilidad de territorios (Kokko et
al. 2006). Estos resultados apuntan a que en efecto los recursos ofrecidos por solo algunas hojas
de la planta hospedera son limitantes para ambas poblaciones de ricárdidos, y probablemente para
otras especies de insectos también excluidos por éstos.
Las discrepancias en los resultados de ambos experimentos ofrecen una oportunidad adicional
para especular sobre las circunstancias que podrían modular la competencia por estos territorios.
Dichas discrepancias podrían deberse a que el periodo de exclusión en el caso del segundo
experimento fue menor; por lo tanto probablemente insuficiente para promover la respuesta que
sí se observó en el primer experimento. La competencia por interferencia representa un
mecanismo más complejo que la competencia por explotación ya que la primera ocurre a través
de la interacción directa entre competidores por ejemplo, el establecimiento de territorios
(Milinsky & Parker 1991). Recientemente se ha reconocido que cambios en los beneficios que
representan el defender un recurso pueden cambiar las conductas de los individuos (Kokko et al.
2006). Es probable que los individuos compitiendo con base sólo a sus habilidades de forrajeo
(explotación) se encuentren en situación crítica una vez que el recurso limitado (en este caso,
exacerbado artificialmente) alcance niveles de escasez suficientes para optar por conductas

60
territoriales. Siendo así, los procesos involucrados en la competencia por interferencia mediante
defensa territorial probablemente requieren de un tiempo mayor al utilizado en el segundo
experimento para reflejar los efectos de un cambio en la disponibilidad del recurso. Se ha
propuesto que la duración e intensidad de la limitación de recursos puede determinar la
importancia relativa de la competencia por interferencia o explotación entre dos especies
(Milinsky & Parker 1991; Amarasekare 2002), tal como parece ser el caso en este estudio.
Un factor asociado a lo anterior y que también pudo haber contribuido a los diferentes
patrones observados en ambos experimentos es la densidad de competidores. Durante del
segundo experimento se registró una drástica caída de las abundancias de ambas especies en la
zona de estudio. Esto pudo deberse a que la noche previa del que sería el tercer día del segundo
experimento, hubo condiciones climáticas que no se habían presentado durante la temporada: una
drástica caída de temperatura al valor mínimo del verano acompañada de una tormenta y un
aumento de la presión atmosférica (datos de la estación meteorológica MMJA, El Lencero,
obtenidos en http://www47.wolframalpha.com). Se sabe que la conducta de los dípteros es
sensible a cambios en condiciones atmosféricas (Ankney 1984; Markow & O’Grady 2008), por
ejemplo en lo que se refiere a variaciones de temperatura en poblaciones de insectos estacionales
(Taylor 1981; Raghu et al. 2004).
Si bien la aparente condición atractiva de los territorios en esta planta mantuvo densidades
locales comparables a las dos primeras etapas del primer experimento, la percepción de los
competidores probablemente fue de una densidad menor de competidores a través de una menor
tasa de encuentro local. Se ha propuesto que cuando los recursos por los que se combate son
territorios, el valor relativo del territorio depende de la densidad de competidores (Kokko et al.
2006). Esto podría explicar también las diferencias entre ambos experimentos en términos del
efecto de la exclusión en la duración de los enfrentamientos. En el primer experimento las peleas

61
fueron más largas durante el periodo de exclusión, tal como se esperaría si aumentara el valor
relativo del recurso durante la exclusión ante una alta densidad de competidores. En el segundo
experimento, sin embargo, no se detectó dicho efecto probablemente debido a la percepción de
los intrusos de una menor densidad de competidores en la zona, en comparación con el primer
experimento.
Un escenario alternativo es que al tiempo del segundo experimento (previo a la baja
generalizada de abundancia de moscas en toda la zona) los territorios disponibles estaban ya
disminuyendo en cantidad y calidad del recurso que ofrecían. Por tanto, los territorios excluidos
en el segundo experimento significaron a su vez, niveles más altos de densidad de competidores
comparados incluso a los propiciados en el primer experimento, aunque su valor como recurso
podría ser menor. De acuerdo a esto, la discrepancia entre ambos experimentos podría explicarse
en términos de predicciones de otros modelos de competencia en los que sucesos de interferencia
ocurren con alta frecuencia sólo a niveles intermedios de densidad de competidores y cuando el
valor del recurso es alto (Dubois et al. 2003; Kokko & Rankin 2006).
En el primer experimento, la respuesta a las exclusiones (aumento en el número de combates)
por parte de machos de O. nitens (los cuales representaron el 70% del total de combates
observados) fue en proporción, mayor a la observada en el segundo experimento (80% (20 de 25)
vs. 28% (2 de 7) respectivamente; Z = 2.7, p = 0.006). Es probable que las variaciones en el
número de enfrentamientos observados en ambos experimentos estén asociadas a intereses
inherentes al sexo de los contendientes. En otros estudios se ha resaltado el hecho de que los
sexos difieren en importantes aspectos de los nichos ecológicos (Brecko et al. 2008). Esta
situación es consistente con uno de los resultados relevantes (importancia del sexo como
asimetría de combate) reportados en otro estudio paralelo en este sistema (Becerril-Morales &
Macías-Ordóñez 2009, Capítulo 2). Es posible además que los requerimientos energéticos

62
diferenciales de cada sexo puedan modular la decisión de contender por el recurso restringido
experimentalmente. Aún cuando nuestras observaciones de campo sugieren que los territorios no
estuvieron asociados directamente al comportamiento reproductivo, no se descarta que la defensa
de territorios de alimentación pueda ser capitalizada en un contexto reproductivo (Emlem &
Oring 1977). Se ha reconocido que la aparición de la conducta territorial no solamente obedece a
presiones selectivas asociadas a la obtención de parejas reproductivas sino que también es
aplicable a otros recursos ecológicos como el alimento (Kokko & Rankin 2006).
El escenario descrito en este ensamble de especies representa una oportunidad de explorar
una probable sinergia entre competencia por explotación y por interferencia. En un extremo, el
defender un territorio podría permitir al defensor poseer el recurso aun sin utilizarlo
inmediatamente, es decir competir por interferencia excluyendo directamente a otros aun sin
consumirlo. Sin embargo, según la definición que se adopte de territorialidad (Maher & Lott
1995), un organismo podría ocupar un territorio y consumir el recurso sin recibir intrusiones, en
cuyo caso la competencia sería solamente por explotación. Ninguno de estos escenarios extremos
parece representar los territorios estudiados en este y muchos otros trabajos ya que
simultáneamente se está consumiendo el recurso y excluyendo competidores. El considerar la
defensa territorial de recursos efímeros que se consumen y defienden simultáneamente como
casos de competencia por explotación e interferencia de manera simultánea da oportunidad a
utilizar enfoques menos restrictivos en el estudio de este tipo de mecanismos.
Los estudios sobre competencia y desplazamiento competitivo, al menos en el caso de los
insectos, se han llevado a cabo con un enfoque ecológico de gran escala, y pocas veces
observando en detalle los mecanismos de competencia, particularmente en los casos de
competencia por interferencia (revisado por Reitz & Trumble 2002). Algunos estudios en
vertebrados ofrecen modelos atractivos que combinan un enfoque ecológico-demográfico con

63
uno que analiza en detalle la cantidad y calidad de competencia por interferencia incluso a nivel
de interacciones individuales (e.g. Petren & Case 1996; Goldber et al. 2001). Las manipulaciones
en este estudio permiten aislar el efecto de la cantidad y agregación de recursos en la cantidad y
calidad de las interacciones competitivas por interferencia, intra e interespecíficas, en este caso de
dos especies de dípteros. El estudio representa una rara combinación en que las predicciones
originadas en la dinámica literatura de teoría de juegos aplicada a sistemas territoriales (Kokko et
al. 2006) se ponen a prueba en sistemas naturales de artrópodos.
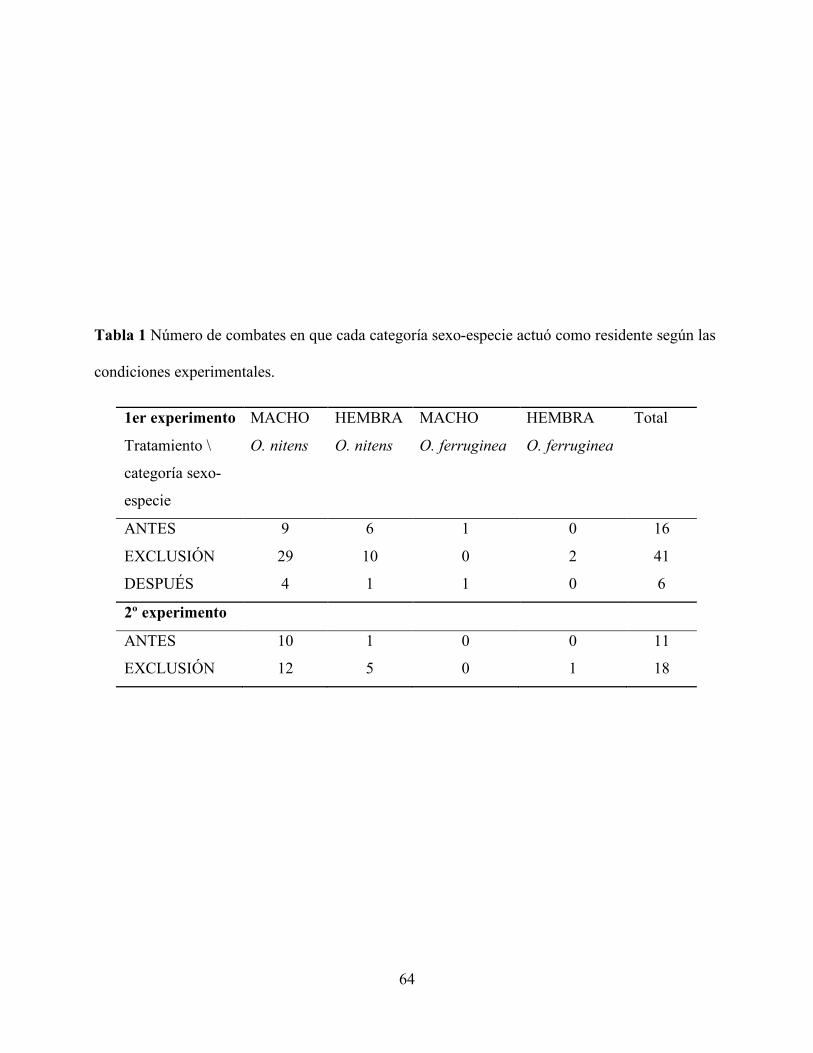
64
Tabla 1 Número de combates en que cada categoría sexo-especie actuó como residente según las
condiciones experimentales.
1er experimento
Tratamiento \
categoría sexo-
especie
MACHO
O. nitens
HEMBRA
O. nitens
MACHO
O. ferruginea
HEMBRA
O. ferruginea
Total
ANTES 9 6 1 0 16
EXCLUSIÓN 29 10 0 2 41
DESPUÉS 4 1 1 0 6
2º experimento
ANTES
EXCLUSIÓN
10
12
1
5
0
0
0
1
11
18

65
Tabla 2 Resultados de las comparaciones estadísticas según las variables medidas en cada
experimento de exclusión.
Variables Experimento Valores estadísticos Prueba
primero
Fr = 4.67, g. l. = 2, P = 0.09 Friedman
Abundancias totales
segundo
W+ = 16.5, W- = 4.5, N = 6, p ≤ 0.21
Rangos asignados de Wilcoxon
primero
Fr = 4.63, g. l. = 2, p = 0.09
Friedman
Abundancias sexo-especie
segundo
Fr = 0.33, g. l. = 1, p = 0.56 Friedman
primero
Fr = 6.94, g. l. = 2, p = 0.03
Friedman
Número de combates
segundo
W+ = 16.5, W- = 38.5, N = 10, p ≤ 0.27
Rangos asignados de Wilcoxon
primero
Fr = 16.6, g. l. = 2, p < 0.01
Friedman
Duración de combates
segundo
W+ = 6.0, W- = 15.0, N = 6, p ≤ 0.43
Rangos asignados de Wilcoxon
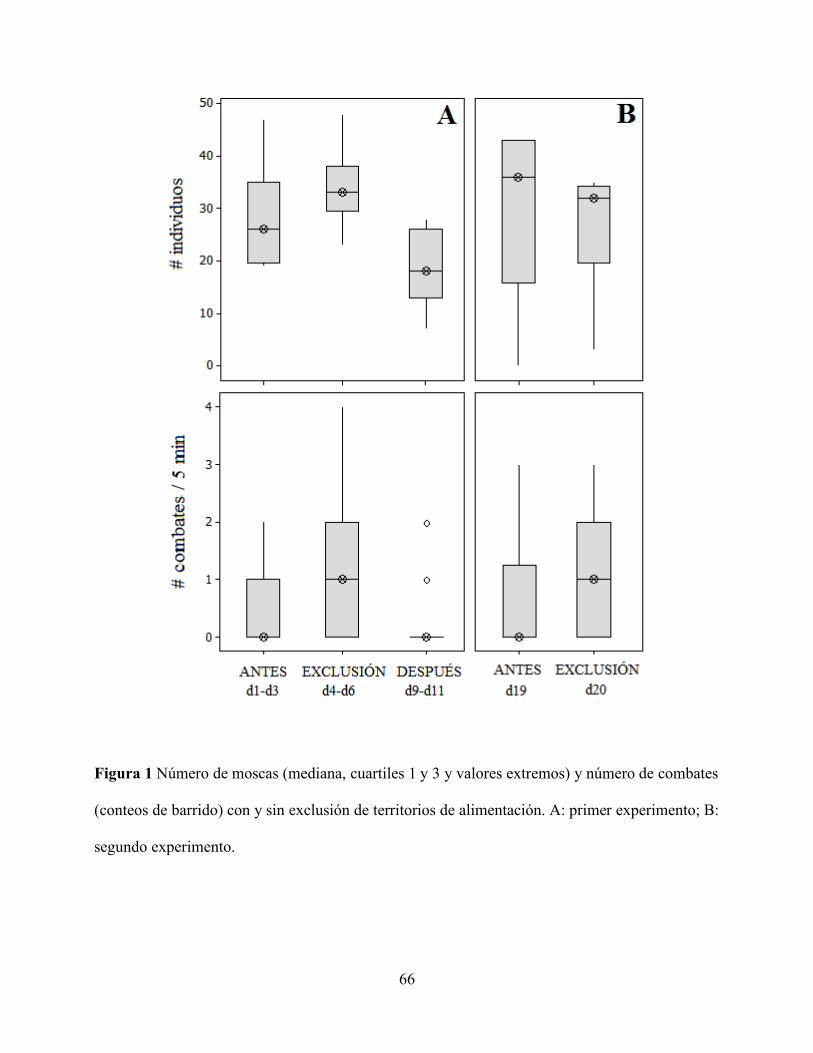
66
Figura 1 Número de moscas (mediana, cuartiles 1 y 3 y valores extremos) y número de combates
(conteos de barrido) con y sin exclusión de territorios de alimentación. A: primer experimento; B:
segundo experimento.

67
Figura 2 Abundancias (mediana, cuartiles 1 y 2 y valores máximos y mínimos) de las categorías
de ricárdidos por sexo y por especie, según los distintos tratamientos en cada experimento.

68
Figura 3 Duración de los combates observados (mediana, cuantiles 1 y 3 y valores extremos)
según los tratamientos de cada experimento de exclusión. Para el primer experimento, no se
muestra el registro de un tiempo atípicamente largo (73 s), durante el periodo previo a la
exclusión. El asterisco indica diferencias significativas (α = 0.05).

69
Literatura citada
Adams, E.S. 2001. Approaches to the study of territory size and shape. Annu. Rev. Ecol. Syst.
32: 277–303.
Amarasekare, P. 2002. Interference competition and species coexistence. Proc. Roy. Soc. Lond.
B. 269: 2541-2550.
Ankney, P.F. 1984. A note on barometric pressure and behavior in Drosophila pseudoobscura.
Behavior Genetics 14(3): 315-317.
Becerril-Morales, F. & R. Macías-Ordóñez. 2009. Territorial contests within and between two
species of flies (Diptera: Richardiidae) in the wild. Behaviour 146: 245-262.
Begon, M., Harper, J.L., & C.R. Townsend. 1996. Ecology. Individuals, populations and
communities, 3rd edn. Blackwell Scientific Publications, Oxford, UK
Brecko, J., Huyghe K., Vanhooydonck B., Herrel A., Grbac I. & R. Van Damme. 2008.
Functional and ecological relevance of intraspecific variation in body size and shape in
the lizard Podarcis melisellensis (Lacertidae). Biol. J. Linn. Soc. 94: 251-264.
Denno, R.F., McClure, M.S. & J.R.Ott. 1995. Interspecific interactions in phytophagous insects:
competition reexamined and resurrected. Annu. Rev. Entomol. 40: 297-331.
Dubois F., L.A. Giraldeau & J.W.A. Grant. 2003. Resource defense in a group-foraging context.
Behav. Ecol. 1: 2-9.
Dudley T.L., D’Antonio C.M. & S.D. Cooper. 1990. Mechanisms and consequences of
interspecific competition between two stream insects. J. Anim. Ecol. 59: 849-866.

70
Emlen S.T. & L.W. Oring. 1977. Ecology, sexual selection, and the evolution of mating systems.
Science197: 215-223.
Goldberg, J.L., Grant, J.W.A. & L. Lefebvre. 2001. Effects of the temporal predictability and
spatial clumping of food on the intensity of competitive aggression in Zenaida dove.
Behav. Ecol. 12: 490-495.
Hart, P.J.B. 2003. Habitat use and feeding behaviour in two closely related fish species, the three-
spined and nine-spined stickleback: an experimental analysis. J. Anim. Ecol. 72: 777-783.
Juliano, S.A. & J.H. Lawton. 1990. The relationship between competition and morphology II.
Experiments on co-occurring dysticid beetles. J. Anim. Ecol. 59: 831-848.
Kokko, H. & D.J. Rankin. 2006. Lonely hearts or sex in the city? Density-dependent effects in
mating systems. Phil. Trans. R. Soc. B 361: 319-334.
Kokko, H., López-Sepulcre A. & L.J. Morrell. 2006. From hawks and doves to self-consistent
games of territorial behavior. Am. Nat. 167: 901-912.
Maher, C.R. & D.F. Lott. 1995. Definitions of territoriality used in the study of variation in
vertebrate spacing systems. Anim. Behav. 49: 1581-1597.
Markow TA & P. O’Grady. 2008. Reproductive ecology of Drosophila. Functional Ecology
22(5): 747-759.
Milinsky, M. & Parker G.A. 1991. Competition for resources. In: Krebs J.R. & Davies N.B.
(Eds.). Behavioural ecology: an evolutionary approach. 3d eds. 137-168 pp. Blackwell
Scientific, Oxford. UK.
Petren K. & T.J. Case. 1996. An experimental demonstration of exploitation competition in an
ongoing invasion. Ecology 77(1): 118-132.

71
Raghu S., R.A.I. Drew & A.R. Clarke. 2004. Influence of host plant structure and microclimate
on the abundance and behavior of a tephritid fly. J. of Insect Behavior 17: 179-189.
Reitz, S. R. & J. T. Trumble. 2002. Competitive displacement among insects and arachnids.
Annu. Rev. Entomol. 47: 435-465.
Riechert, S. E. 1998. Game theory and animal contests. – In: Game theory and animal behavior.
(Dugatkin, L.A. & Reeve, H.K. eds). Oxford University Press, New York, p. 64-93.
Siegel S. & N.J. Castellan. 1995. Estadística no paramétrica, aplicada a las ciencias de la
conducta. Editorial Trillas. México DF.
Stamps J.A. 1990. The effect of contender pressure on territory size and overlap in seasonally
territorial species. Am. Nat. 135: 614-632.
Switzer P.V., Stamps J.A. & M. Mangel. 2001. When should a territory resident attack? Anim.
Behav. 62: 749-759.
Taylor F. 1981. Ecology and evolution of physiological time in insects. American Naturalist 117-
123.
Thompson, J.N. 1996. Evolutionary ecology and the conservation of biodiversity. - Trends Ecol.
Evol. 11: 300-303.

72
CAPÍTULO IV
IV. Discusión general

73
Las moscas, al igual que otros insectos como los lepidópteros, están pobremente
equipadas con estructuras especializadas en amenazar a otros individuos o infligirles algún tipo
de daño (Baker 1983). Sin embargo, una gran cantidad de especies de dípteros muestran un
amplio repertorio de conductas agonísticas y establecen territorios como estrategia competitiva
por parejas reproductivas, alimento o ambos (Dodson 1997; Headrick & Goeden 2000; Chen et
al. 2002).
Existen muchos estudios que sustentan la idea de que la competencia surge a partir tanto
de la concurrencia de al menos dos individuos (conespecíficos o no) buscando el mismo recurso,
como del hecho de que el acceso al mismo es limitado, ya sea por su escasez o por la naturaleza
de su distribución espacial y temporal. Durante los dos años de trabajo en campo sólo se
observaron conductas de defensa, alimentación, reposo y limpieza, restringidas sólo a algunas de
las plantas hospederas disponibles. Las interacciones agonísticas consistieron en delimitar un
territorio y en ahuyentar intrusos de ambos sexos, de la misma especie y de otras especies de
ricárdidos, así como de otros órdenes de insectos. En ninguna ocasión se observó alguna
interacción que sugiriera cortejo, cópula u oviposición; conductas frecuentemente descritas en
tefritoideos que defienden territorios en circunstancias similares (Sivinski 2000). Todo lo anterior
apunta a que dichas agresiones obedecen a competencia por recursos alimenticios y no por
recursos sexuales tales como sitios de cortejo, apareamiento u oviposición. A semejanza de otros
animales, los dípteros parecen resolver sus conflictos empleando combinaciones de
comportamientos tales como despliegues ritualizados y otros patrones de mayor intensidad o
“escalamiento” (Archer 1988), y los ricárdidos no son la excepción. Tal como era de esperarse
por diferentes modelos de resolución de combates (v. gr. Enquist & Leimar 1983; Hsu et al.
2006), la mayoría de los enfrentamientos observados entre ricárdidos terminaron en niveles bajos
de escalamiento.

74
Un tipo de aproximación teórica a la conducta territorial es la enfocada al residente y sus
conductas ante intrusos, sean o no de la misma especie (Adams 2001). Tal enfoque se conoce
como el “efecto del residente” (Leimar & Enquist 1984; Grafen 1987; Archer 1988; Kemp &
Wiklund 2004), que consiste en el hecho de que en la mayoría de los combates, la probabilidad
de victoria del residente es mayor a la de los intrusos. Si bien este estudio, al igual que otros
estudios previos, encontró este patrón, nuestros resultados revelan que en tal dominio intervienen
de manera interactiva otros atributos de los contendientes tales como el sexo y el tamaño. Es
decir, el modelo de residencia previa (ver “Estrategia del Burgués” en Introducción; ver Maynard
Smith 1982) parece ser sólo una arista del complejo de factores involucrados en la resolución de
conflictos territoriales. Este tipo de resultados solo es posible mediante un enfoque simultáneo de
los atributos de los combatientes y del contexto ecológico en que se desarrollan los combates,
combinación rara vez utilizada en estudios de competencia por territorios.
Coexistencia entre O. nitens y O. ferruginea
La coexistencia entre dos especies con requerimientos similares podría ser modulada por sus
habilidades competitivas (Lawlor & Maynard Smith 1976; Tiebout 1996; Gordon 2000; Young
2003). , Los individuos de O. ferruginea son de mayor tamaño que los de O. nitens, lo cual
supondría mayor capacidad competitiva en combate, sin embargo O. ferruginea fue menos
abundante. Por una parte, tal diferencia de tamaños puede implicar diferentes requerimientos
nutricionales (Persson 1985, Kindlmann et al. 1992, Adams 2001) y con ello, umbrales de
competencia-coexistencia diferentes. Individuos de la especie de talla pequeña podrían ser más
eficientes en obtener lo necesario a partir de los territorios si sus requerimientos nutricionales son
menores. Si lo anterior es cierto, O. nitens, al ser la especie de menor talla podría ser más

75
eficiente en competir en condiciones de baja disponibilidad de alimento (Wacker & von Elert
2008), aun cuando su capacidad de combate sea menor.
Muchos estudios sobre interacciones competitivas interespecíficas en insectos fitófagos
(Denno et al. 1995) y en otros taxa han sido enfocados a la de competencia por interferencia o por
explotación. La competencia por interferencia pueden ser combates en los cuales el ganador
desplaza al perdedor y monopoliza el recurso (Sirot 2000; Reitz & Trumble 2002). La
competencia por explotación implica que algunos individuos son más capaces de localizar y
consumir más eficientemente el recurso sin ningún contacto con el competidor (Reitz & Trumble
2002). Por ejemplo, en un estudio de dos especies de colibrí, la especie más pequeña compitió
sólo por explotación y fue desplazada por la especie de mayor tamaño a través de defensa del
territorio (interferencia) (Tiebout 1996). Ebersole (1985) en un estudio de competencia entre dos
especies de peces damiselas, sugirió que la competencia por explotación por alimento
aparentemente resulta en competencia por interferencia en la forma de territorialidad
interespecífica. De acuerdo a Persson (1985), en situaciones de competencia entre dos especies,
la especie de menor tamaño suele ser más resistente a la escasez del recurso en disputa y por ello
resulta ser mejor competidora por explotación. La especie de mayor tamaño puede compensar
esta desventaja basando su competencia en utilizar sus mayores habilidades para disputar
directamente el recurso (p. ej. mediante conductas territoriales), es decir, compitiendo por
interferencia. La coexistencia observada entre ambas especies en nuestro estudio podría ser
consecuencia de una relación dinámica entre las dos formas de competencia: individuos de la
especie más pequeña y abundante podrían ser favorecidos en situaciones de competencia por
explotación (Morse 1974; Persson 1985), lo cual requeriría de un estudio enfocado a explorar el
efecto de la ocupación de territorios en el consumo de los recursos que contienen.

76
Perspectivas de estudio en la ecología de la conducta en ricárdidos
El estudio del comportamiento asociado a competencia entre especies cercanas implica
considerar a la conducta agresiva como un rasgo fenotípico en aquellos individuos en conflicto.
Si bien las asimetrías de un combate parecen determinar el grado en que el residente permanece
como tal en un territorio, aun desconocemos las diferentes reglas de decisión de individuos de
cada sexo y tamaño para iniciar y/o continuar un combate. En este trabajo la mayoría de los
combates registrados fueron la suma de dos o más eventos agresivos y cada uno de estos fueron
conformados por una secuencia de elementos conductuales (ver Capítulo 2). Evaluar el efecto de
estas secuencias conductuales o estructura en “rounds” en el contexto teórico de intercambio de
información entre contendientes (Enquist & Leimar 1983; Bradbury & Vehrencamp 1998) y en
función de la categoría sexo-especie-tamaño es una línea promisoria de investigación rara vez
abordada, mucho menos en condiciones naturales, pero susceptible de llevarse a cabo en este
sistema.
Este estudio plantea también nuevas perspectivas respecto a aproximaciones de modelos
de interacción agonística en el que se involucren conflictos intersexuales (hembras expulsando a
machos de sus territorios y viceversa) e intrasexuales (hembras combatiendo contra hembras).
Estas interacciones son menos frecuentes en la naturaleza pero tampoco son raras, sin embargo,
han sido poco estudiadas en el ámbito de teoría de enfrentamientos (Draud et al. 2004). Por otro
lado, fue frecuente observar agresiones de ricárdidos dirigidas a insectos de otros órdenes. Esto
implica la posibilidad de plantear modelos novedosos de interacciones interespecíficas basados
en conducta individual en un contexto de ensambles o comunidades.
El tomar en cuenta diversas asimetrías de combate simultáneamente, incluyendo
diferencias entre especies o sexos, implica aumentar la diversidad del tipo de interacciones

77
interespecíficas tradicionalmente estudiadas en la ecología de comunidades. El dar un contexto
ecológico a estos análisis permitirá abordar de una manera mucho más dinámica el estudio de la
competencia por interferencia, la resolución de conflictos individuales, procesos coevolutivos de
aprovechamiento de recursos por especies con requerimientos similares, y finalmente los
mecanismos de composición de comunidades a diferentes escalas.

78
Literatura citada
Adams, E.S. 2001. Approaches to the study of territory size and shape. Annu. Rev. Ecol. Syst.
32: 277–303.
Archer, J., 1988. The behavioural biology of aggression. Cambridge University Press,
Cambridge, UK.
Baker, R. R. 1983. Insect Territoriality. Ann. Rev. Entomol. 28: 65-89.
Bradbury, J.W. & S.L. Vehrencamp. 1998. Principles of Animal Communication. Sinauer
Associates, Sunderland MA, USA.
Chen S., Lee A.Y., Bowens N.M., Huber R. & E.A. Kravitz. 2002. Fighting fruit flies: a model
system for the study of aggression. PNAS 99: 5664-5668.
Denno, R.F., McClure, M.S. & J.R. Ott. 1995. Interspecific interactions in phytophagous insects:
competition reexamined and resurrected. Annu. Rev. Entomol. 40: 297-331.
Dodson G.N. 1997. Resourse defence mating system in antlered flies, Phytalmia spp. (Diptera:
Tephriitidae). Ann. Entomol. Soc. Am. 90:496-504.
Draud, M., Macías-Ordóñez R., Verga J. & M. Itzkowitz. 2004. Female and male Texas cichlids
(Herichthys cyanoguttatum) do not fight by the same rules. Behav. Ecol. 15: 102-108.
Ebersole, J.P. 1985. Niche separation of two damselfish species by aggression and differential
microhabitat utilization. Ecology 66, 14-20.
Enquist, M. & O. Leimar. 1983. Evolution of fighting behavior: decision rules and assessment of
relative strength. J. Theoret. Biol., 102, 387-410.
Gordon C.E. 2000. The coexistence of species. Rev. Chil. Hist. Nat. 73: 175-198.
Grafen, A. 1987. The logic of divisively asymmetric contests: respect for ownership and
desperado effect. Anim. Behav. 35: 462-467.

79
Headrick D.H. & R.D. Goeden. 2000. Behavior of flies in the subfamily Tephritinae. In: Fruit
flies (Tephritidae), (Aluja, M. & Norrbom, A.L., eds.). pp 671-707. CRC Press LLC.
Boca Raton, USA.
Hsu, Y., Earley, R.L., & L.L. Wolf. 2006. Modulation of aggressive behaviour by fighting
experience: mechanisms and contest outcomes. Biol. Rev. 81: 33-74.
Kemp, D.J. & C. Wiklund. 2004. Residency effects in animal contests. Proc. R. Soc. Lond.
271:1707-1711.
Kindlmann, P., Dixon, A.F.G. & C.N. Brough. 1992. Intra- and interspecific relationships of
reproductive investment to body weight in insects. Oikos 64: 548-552.
Lawlor, L. R. & J. Maynard Smith. 1976. The coevolution and stability of competing species.
Am. Nat. 110: 79-98.
Leimar, O. & M. Enquist. 1984. Effects of asymmetries in owner-intruder conflicts. J. Theor.
Biol. 111: 475-491.
Maynard Smith, J. 1982. Evolution and the theory of games. Cambridge University Press. New
York, USA.
Morse, D.H. 1974. Niche breadth as a function of social dominance. Am. Nat. 108: 818-830.
Persson, L. 1985. Asymmetrical competition: are larger animals competitively superior? Am.
Nat. 2: 261-266.
Reitz, S. R. & J. T. Trumble. 2002. Competitive displacement among insects and arachnids.
Annu. Rev. Entomol. 47: 435-465.
Sirot, E. 2000. An evolutionarily stable strategy for aggressiveness in feeding groups. Behav.
Ecol. 11: 351-356.
Sivinski, J. 2000. Breeding habits and sex in families closely related to the Tephritidae:
opportunities for comparative studies of the evolution of fruit fly behavior. In: Fruit Flies

80
(Tepritidae), Phylogeny and Evolution of Behavior. (Aluja M. & Norrbom, A.L. eds). 23-
37 pp. CRC Press LLC, Boca Raton, USA.
Tiebout, H.M. III. 1996: Costs and benefits of interspecific dominance rank: are subordinates
better at finding novel locations? Anim. Behav. 51: 1375-1381.
Wacker A. & E. von Elert. 2008. Body size and food thresholds for zero growth in Dreissena
polymorpha: a mechanism underlying intraspecific competition. Freshwater Biol. 53:
2356–2363.
Young, K.A. 2003. Evolution of fighting behavior under asymmetric competition: an
experimental test with juvenile salmonids. Behav. Ecol., 1: 127-134.
Top Related