





![11 Digestion y Absorcion de Nutrientes[1] (1)](https://static.fdocuments.mx/doc/165x107/55cf9abc550346d033a32900/11-digestion-y-absorcion-de-nutrientes1-1.jpg)

Idiomas
Páginas
Jurídico


TEMA:
ABSORCION Y DIGESTION DE CARBOHIDRATOS Y LIPIDOS
CICLO DEL GLIOXILATO INTEGRANTES:
Balbín Narvaez, Deyson Paul
Ruiz Andía, Jhojan
Zuñiga Ayala, Francisco
PROFESOR:
Calderón Gómez, Javier
CURSO:
Bioquímica I
CARRERA:
FARMACIA Y BIOQUIMICA
CICLO: V

INTRODUCCIÓN
El tema siguiente de presentación de Carbohidratos, Lípidos y el Ciclo de Glioxilato, se
realizo desde el punto de vista de poder comprender los siguientes procesos que comprende en la
digestión como también el mecanismo de absorción. Tanto mediadores, enzimas y reacciones que
resultan de ello. Como también la importancia de estos Monómeros en el Organismo producto de
las reacciones como transportadores y el déficit que podría por la carencia de ellos.
Se puede observar que cada órgano como tejido o célula cumple un papel muy importante en el
proceso metabólico, por tanto debemos de tener en cuenta todo lo aprendido por lo mismo que todo
se forma una cadena de conocimiento y una cosa lleva a lo otro.

DIGESTION Y ABSORCION DE LOS CARBOHIDRATOS
LOS CARBOHIDRATOS EN LA DIETA:
Los carbohidratos forman el 60% de los alimentos ingeridos por los omnívoros y herbívoros.
El constituyente más importante es el almidón, un polisacárido que está presente en los tubérculos,
los cereales y otros vegetales. Este carbohidrato no tiene sabor pronunciado, como los
monosacáridos, a causa de que su alto peso molecular impide su penetración a las papilas
gustativas, lo cual facilita su ingestión en grandes cantidades.
El glucógeno es otro polisacárido que se ingiere en pequeña cantidad con los alimentos de
origen animal. Entre los disacáridos tiene importancia la sacarosa, que se agrega a la dieta en forma
de sustancia cristalina, y la lactosa o azúcar de leche. Los monosacáridos son principalmente la
glucosa y fructosa, que se ingieren en pequeñas cantidades con las frutas. La miel de abeja
contiene altas concentraciones de glucosa y fructosa.
DIGESTIÓN DE LOS CARBOHIDRATOS:
En los animales superiores, la digestión del almidón se inicia en la boca. La saliva,
principalmente aquella producida por la parótida, contiene una enzima, la amilasa salival, llamada
también ptialina. Esta enzima es una α- amilasa que rompe los enlaces
α- 1,4 del almidón y el glucógeno, tiene un pH óptimo de 7 y requiere ión cloruro para su actividad.
La α- amilasa, al actuar al azar sobre las cadenas lineales de almidón (amilosa), da lugar a
la formación predominante de maltosa 87% y algo de glucosa 13%. Cuando la enzima actúa sobre
un polisacárido de cadena ramificada, como el glucógeno o la amilo pectina del almidón, la ruptura al
azar de los enlaces glucosídicos α- 1,4 da lugar a maltosa, glucosa y una gran variedad de
oligosacáridos ramificados con 2 a 8 unidades de glucosa, que contiene uno o más enlaces α- 1,6.

Los enlaces α- 1,6 y α- 1,4 de las unidades de glucosa que sirven como puntos de ramificación no
son atacados por la enzima.
La enzima más importante para la digestión del almidón y glucógeno es la amilasa
pancreática, que actúa igual que la amilasa salivar. Cuando la enzima no se vierte al intestino, como
en caso de obstrucción de los conductos excretores, se encuentran granos de almidón no digeridos
en las heces.
La digestión final es llevada a cabo por enzimas específicas que están en el borde en cepillo de la
mucosa del intestino delgado: la oligo 1,6- glucosidasa, que hidroliza los enlaces α- 1,6 de la
isomaltosa y oligosacáridos ramificados, quedando de estos últimos una cadena lineal sobre la cual
actúa la α- amilasa para formar maltosa; la maltasa, que convierte la maltosa en dos glucosas; la
sucrosa , que desdobla sacarosa en glucosa y fructuosa; y la lactasa, que convierte la lactosa o
azúcar de leche en glucosa y galactosa.
Los di y oligosacáridos que escapan a la acción de la amilasa pancreática y de las enzimas
de la superficie intestinal van al tracto inferior del intestino el que, a partir de la parte inferior del hilio,
contiene bacterias que fermentan los carbohidratos formando ácidos grasos volátiles, acido láctico,
H2, metano y CO2, produciendo flatulencia y diarrea por irritación del intestino. El bien conocido
fenómeno de la flatulencia después de la ingestión de semillas de leguminosas (fríjol, soya) se debe
a la fermentación bacteriana de oligosacáridos que no pueden ser hidrolizados por las enzimas
intestinales del hombre, que son inhibidas en estos casos.
La deficiencia de disacaridasas se encuentra con relativa frecuencia en humanos. La
deficiencia puede ser de una sola enzima o de varias de ellas por una serie de razones (defecto
genético, declinación fisiológica por la edad o injuria a la mucosa intestinal)
Lo más común es la deficiencia de la lactasa, que puede ser absoluta o relativa, y que se
manifiesta por intolerancia a la leche; la lactosa no se puede absorber y es fermentada por las
bacterias intestinales con producción de ácidos y gas que originan flatulencia y diarrea.

ABSORCIÓN DE LOS CARBOHIDRATOS
Los carbohidratos se absorben en forma de monosacáridos. Algunos disacáridos, como la
sacarosa, pueden absorberse si están presentes en altas concentraciones, pero no se utilizan y son
eliminados por la orina como sustancia extraña.
Concluida la digestión, están presentes en el intestino, principalmente glucosa y cantidades
variables de fructosa y galactosa.
La absorción de la D-glucosa, D-galactosa y D-fructosa y otros monosacáridos de menor
importancia se lleva a cabo mediante transportadores que son proteínas que están en la membrana
luminal o contraluminal, que exhiben especialidad por el substrato, estereoespecificidad, cinética de
saturación y son sensibles a inhibidores. Se conoce por lo menos dos tipos de sistemas de enzimas
de transporte que catalizan la captación de monosacáridos del lumen y su transferencia a la célula:
a.- El sistema de cotransporte Na+ monosacáridos, conocido también como SGLT,
específico para la D-glucosa y D-galactosa, cataliza el transporte activo de estos azúcares
b.-Difusión facilitada, que es un sistema de transporte independiente de Na+ conocido como
GLUT – 5 , y que es específica para la D-fructosa.Adicionalmente, hay un sistema de transporte
para monosacáridos independiente del Na+, con especificidad para la D-glucosa,D-galactosa y D-
fructosa y que están presente en la membrana plasmática contraluminal. Mediante este sistema de
transporte, los monosacáridos salen de la célula.. Los sistemas de transporte luminal y contraluminal
no sólo difieren en cuanto al requerimiento de Na, sino también por la especificidad por el substrato
y la sensibilidad a inhibidores (la florida inhibe el sistema luminal y la citochalasina el sistema
contraluminal).
La glucosa y la galactosa son absorbidas a través del epitelio intestinal contra la gradiente
de concentración. La energía para este transporte activo deriva directamente de la gradiente de
concentración del Na+ o el potencial eléctrico a través de la membrana luminal. Una K+ Na+
ATPasa, presente exclusivamente en la membrana plasmática contraluminal, proporciona el
mecanismo para la transformación de la energía del ATP en energía osmótica de concentración
(química) o una combinación de concentración y gradiente electroiónica (electroquímica), a través
de la membrana plasmática . Los estudios estequiométricos demuestran que en la reacción de la
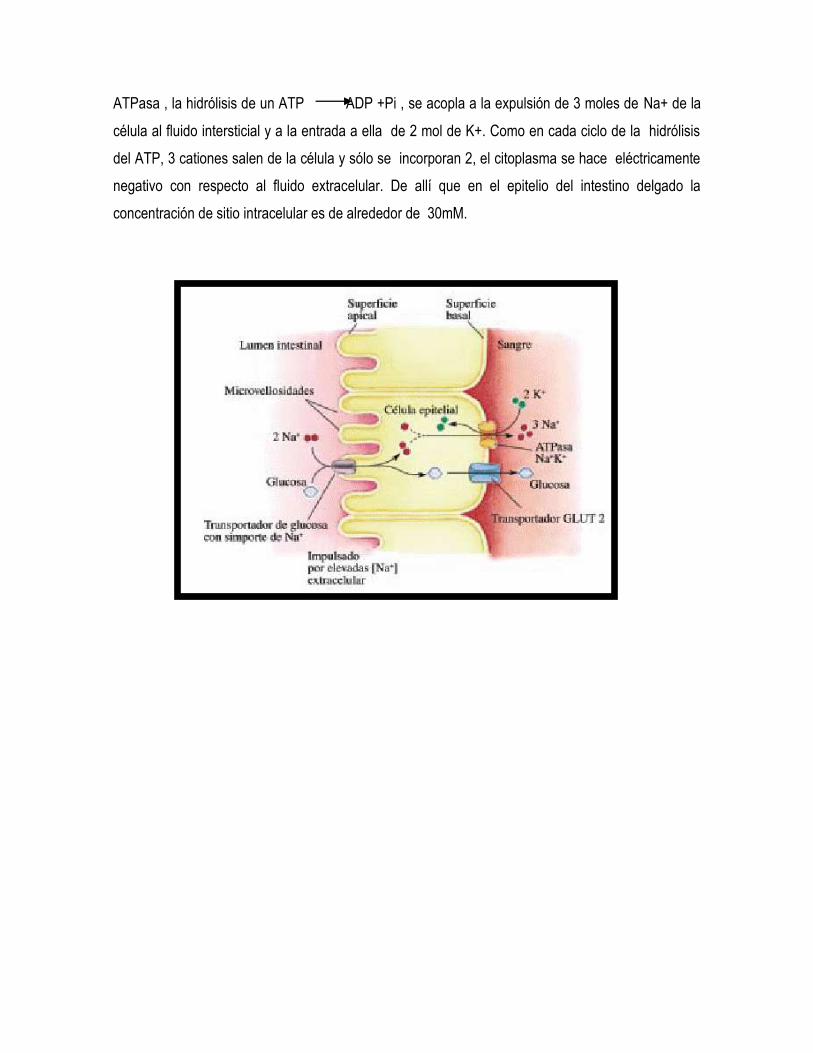
ATPasa , la hidrólisis de un ATP ADP +Pi , se acopla a la expulsión de 3 moles de Na+ de la
célula al fluido intersticial y a la entrada a ella de 2 mol de K+. Como en cada ciclo de la hidrólisis
del ATP, 3 cationes salen de la célula y sólo se incorporan 2, el citoplasma se hace eléctricamente
negativo con respecto al fluido extracelular. De allí que en el epitelio del intestino delgado la
concentración de sitio intracelular es de alrededor de 30mM.

D I G E S T I Ó N Y A B S O R C I Ó N D E L O S L Í P I D O S
DEFINICIÓN
Los lípidos constituyen alrededor del 40 al 45% de la ingestión calórica de los países
desarrollados. La mayor parte de esta energía proviene de los TRIACILGLICEROLES. Ello quiere
decir que una persona consume de 60 a 130 gramos de grasa por día y que el organismo tiene que
DIGERIR, ABSORBER, RESINTETIZAR Y TRANSPORTAR esta cuantía de lípidos.
Alrededor del 90 al 95% de los TRIGLICERIDOS de la dieta se absorben en un 50%.
DIGESTIÓN
En el estómago la digestión de los TRIACILGLICEROLES es de poca importancia, pero las
enzimas PROTEOLÍTICAS liberan los lípidos de las partículas alimenticias y se forman una emulsión
gruesa por la agitación en el estómago.
Hay una lipasa gástrica, distinta de la lipasa pancreática que se ah encontrado en varios
mamíferos y en el hombre. Esta lipasa, activa en pH 3-4, hidroliza preferentemente los ácidos grasos
de cadena corta y mediana de los triacilgliceroles, estos ácidos grasos están presentes en la leche.
La hidrólisis parcial de los triacilgliceroles a 1,2 diacilgliceroles hacen que las micelas de la
grasa de la leche sean más susceptibles a la acción subsecuente de la LIPASA PANCREÁTICA en
el intestino.
Los ácidos grasos de cadena mediana y corta pueden absorberse directamente por la
mucosa gástrica, uniéndose a la albúmina para su transporte sanguíneo hacia el hígado, donde son
OXIDADOS.
Los lípidos restantes, ingresan a la parte proximal del INTESTINO, donde hay un pH de 5.8
– 6,5. Las gruesas gotas de grasa son cubiertas por SALES BILIARES, segregadas por el hígado.
Estas sales son compuestos bifásicos que se disuelven en la interfase GRASA – AGUA, orientando
la fase HIDROFOBA hacia la grasa y la fase HIDRÓFILA hacia el medio acuoso intestinal,
favoreciendo así la DISPERSIÓN de las gotas de grasa.

Una proteína llamada COLIPASA, segregada por el páncreas, es adsorbida en la superficie
de las gotitas de grasa, y ésta proteína actúa como un gancho para fijar la LIPASA PANCREATICA
en la interfase grasa – agua.
Se piensa que las sales biliares, la colipasa y la lipasa, forman un complejo TERNARIO en
presencia de Ca+2 para hidrolizar los enlaces éster 1y 3 del TRIACILGLICEROL.
En los mamíferos NO RUMIANTES, alrededor del 85% de los triglicéridos es convertido en
2-mono-acilglicerol. Hay una hidrolasa inespecífica segregada que hace una hidrólisis muy limitada
del 2-monoacilglicerol a GLICEROL.
Los fosfolípidos son degradados por la fosfolipasa pancreática A2. La digestión de los
fosfolípidos no necesita ser completa, un 30-40% de la fosfatidilcolina de la dieta puede absorberse
como LISOFOSFATIDILCOLINA.
A medida que prosigue la digestión, los monoacilgliceroles, ácidos grasos, lisofosfolipidos,
colesterol y sales biliares, se disocian de la superficie de la gota de grasa para formar una solución
micelar.
Las micelas son agregados AMFIPATICOS que orientan sus lados hidrófobos hacia adentro
y sus grupos polares hacia el medio acuoso.

Las micro vellosidades de los entericitos contienen FOSFOLIPASAS E HIDROLASAS donde
puede proseguir la digestión de fosfolípidos y esteres de colesterol.
ABSORCIÓN DE LÍPIDOS EN EL INTESTINO
La mayor parte de los lípidos se absorben en el yeyuno, con excepción de las sales biliares
que permanecen más tiempo en el lumen para facilitar la digestión y son absorbidas en la parte
distal del ileum para ser transportadas al hígado por la vena porta (Circulación enterohepática)
La absorción ocurre cuando la solución micelar se pone en contacto con las micro
vellosidades del enterocito. Las sales biliares son esenciales no solo para la digestión sino también
para la absorción de los productos de los lípidos y de las vitaminas LIPOSOLUBLES (A, D, E y K).
Cuando hay obstrucción de los conductos biliares la absorción de los lípidos de la dieta es muy
pobre y se eliminan en forma hidrolizada por las heces (esteatorrea). El transporte de los lípidos se
realiza por una gradiente de difusión.

Dentro de las células intestinales, los ácidos grasos son ligados a una proteína de bajo peso
molecular que sirve para aumentar la efectiva solubilidad de los ácidos grasos y para proteger a la
célula de su efecto detergente.
Los productos de la digestión de los lípidos absorbidos por la mucosa intestinal son
convertidos, en este tejido, en triacilgliceroles, donde se absorbe desde el lumen intestinal el 2-
monoacilglicerol, donde se resintetiza a triacilglicerol por acción secuencial monoaclglicerol
aciltransferaza y la diacilglicerol aciltransferaza.
En cuanto a la resíntesis de la fosfatidilcolina en el enterocito se da por la esterificación de la
lisofofatidilcolina, donde esta fue formada ya por el proceso de la digestión en el intestino.
El transporte y metabolismo del colesterol es mas lento, permanece alrededor de 12 horas
en el enterocito y se incorpora a su membrana mezclándose con el colesterol preexistente.

Formación de Quilomicrones
El transporte de los lípidos del intestino delgado requiere que ellos formen un paquete
estable en medio acuoso, lo cual se consigue cubriendo los lípidos mas hidrófobos (triacilgliceroles,
esteres de colesterol) con una capa de compuestos amfipáticos constituida de fosfatidilcolina,
colesterol y varias apoproteinas, éstas últimas confieren, además marcas direccionales que van a
determinar que células han de recibir y metabolizar estos paquetes estables de lipoproteínas.
Los agregados de lipoproteínas que forma el intestino para el transporte de la grasa son los
QUILOMICRONES. En forma similar los triacilgliceroles sintetizados por el hígado son
empaquetados por lipoproteínas de muy baja densidad (VLDL). Los quilomicrones son más grandes
y tienen mayor contenido de triacilgliceroles que la VLDL.
El ensamblaje de los quilomicrones empieza en la zona apical de los entericitos con la
resíntesis de los triacilgliceroles en el retículo endoplasmático liso. Estos triacilgliceroles aparecen
como gotitas de grasa que luego son estabilizadas con una capa de fosfolípidos y las proteínas

sintetizadas por el retículo endoplasmático rugoso (RER). El retículo endoplasmico se agranda para
atrapar mas gotitas de grasa y formas vesículas las que se fusionarían con el aparato de golgi.
Enseguida, los quilomicrones son llevados a la superficie lateral del enterocito, donde ocurre la
fusión del aparato de golgi con la membrana del enterocito, segregándose los quilomicrones al
espacio intercelular, del que pasan de los vasos linfáticos para desembocar, por intermedio del
conducto toráxico, en la vena subclavia izquierda para su distribución a los diversos tejidos del
organismo.
Siguen la ruta de la vena porta, sin esterificarse, la mayoría de los ácidos grasos de cadena
corta (C2 – C10), una cantidad significativa de cadena mediana (C12) y muy poco de los que tienen
más de 12 carbonos.
Al contrario, siguen la vía linfática previa esterificación e integrando los quilomicrones, la
mayoría de los ácidos grasos de cadena larga una cantidad significativa de cadena mediana y muy
pocos de los de cadena corta.

En el lecho capilar de varios tejidos extrahepáticos (músculo esquelético, músculo cardiaco y
tejido adiposo) existe una enzima, la lipoproteína lipasa, la cual realiza la hidrólisis de los
triacilgliceroles de los quilomicrones. En los músculos, los ácidos grasos liberados son fácilmente
usados como fuente de energía, pudiendo depositarse temporalmente como triglicéridos si están en
exceso y en el tejido adiposo su suerte mayor consiste en almacenarse como sustancia de reserva.
El glicerol liberado es transportado al hígado y los riñones en donde se convierte en
DIHIDROXIACETONA FOSFATO por la acción sucesiva de la gliceroquinasa.
El remanente de los quilomicrones que han dejado los triacilgliceroles pero que aun
contienen colesterol y apolipoproteinas son llevados por el torrente sanguíneo al hígado, donde son
captados por receptores para sus apolipoproteinas por un proceso de endocitosis. Los
triacilgliceroles que ingresan al hígado por esta ruta pueden ser oxidados para proporcionar energía
o para la síntesis de cuerpos cetónicos.
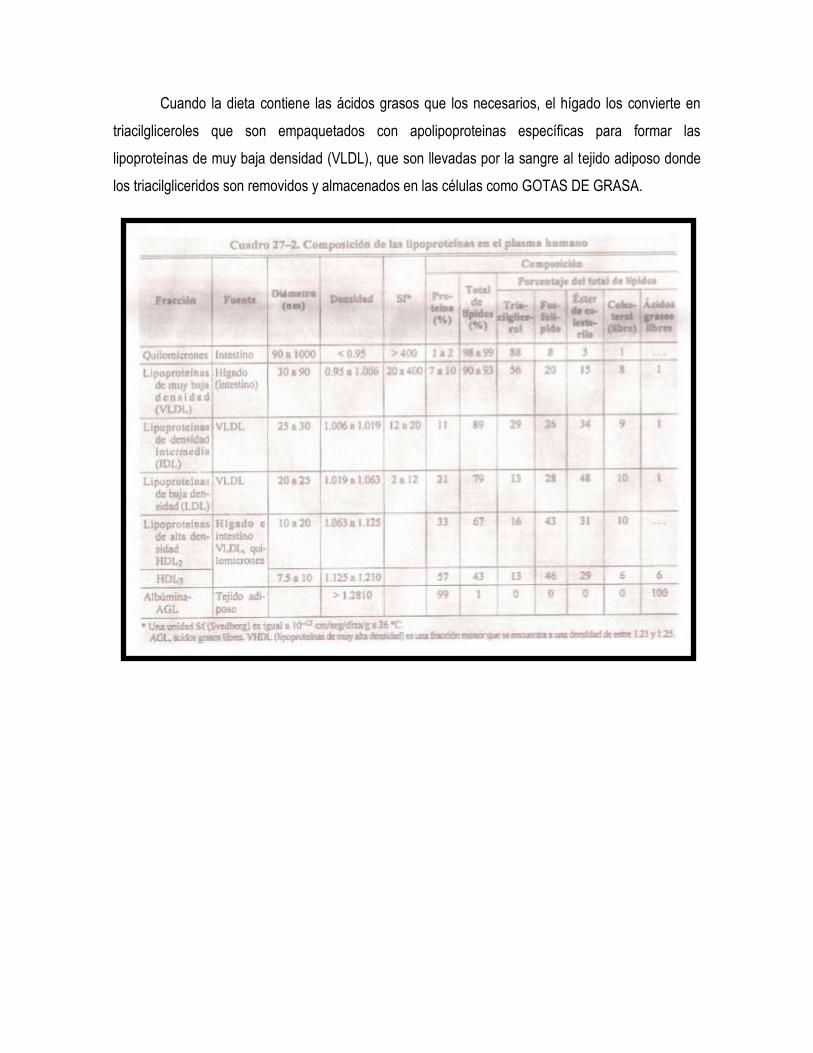
Cuando la dieta contiene las ácidos grasos que los necesarios, el hígado los convierte en
triacilgliceroles que son empaquetados con apolipoproteinas específicas para formar las
lipoproteínas de muy baja densidad (VLDL), que son llevadas por la sangre al tejido adiposo donde
los triacilgliceridos son removidos y almacenados en las células como GOTAS DE GRASA.
CICLO DE GLIOXILATO
El Ciclo de Glioxilato es una ruta anabólica asimilativa, donde: no se genera energía y se
incorpora la utilización de compuestos orgánicos. La misma es una modificación del Ciclo de Krebs
presente en: bacterias, algunas plantas y protozoas. En plantas, este ciclo ocurre en organelos
llamados: glioxisomas. Estos son necesarios para la germinación de las semillas en acetato. En el
caso de las bacterias, este ciclo se lleva a cabo cuando las fuentes de carbono son muy limitadas. A
diferencia del ciclo de Krebs, en el ciclo de glioxilato hay una serie de reacciones, que a través de la
eliminación de decarboxilaciones, forman un compuesto de cuatro carbonos; el cual ayuda a la
síntesis de glucosa. Este ciclo se caracteriza y se diferencia del Ciclo de Krebs, en que en el mismo
se produce glucosa mediante gluconeogénesis, partiendo inicialmente, de ácidos grasos y acetatos
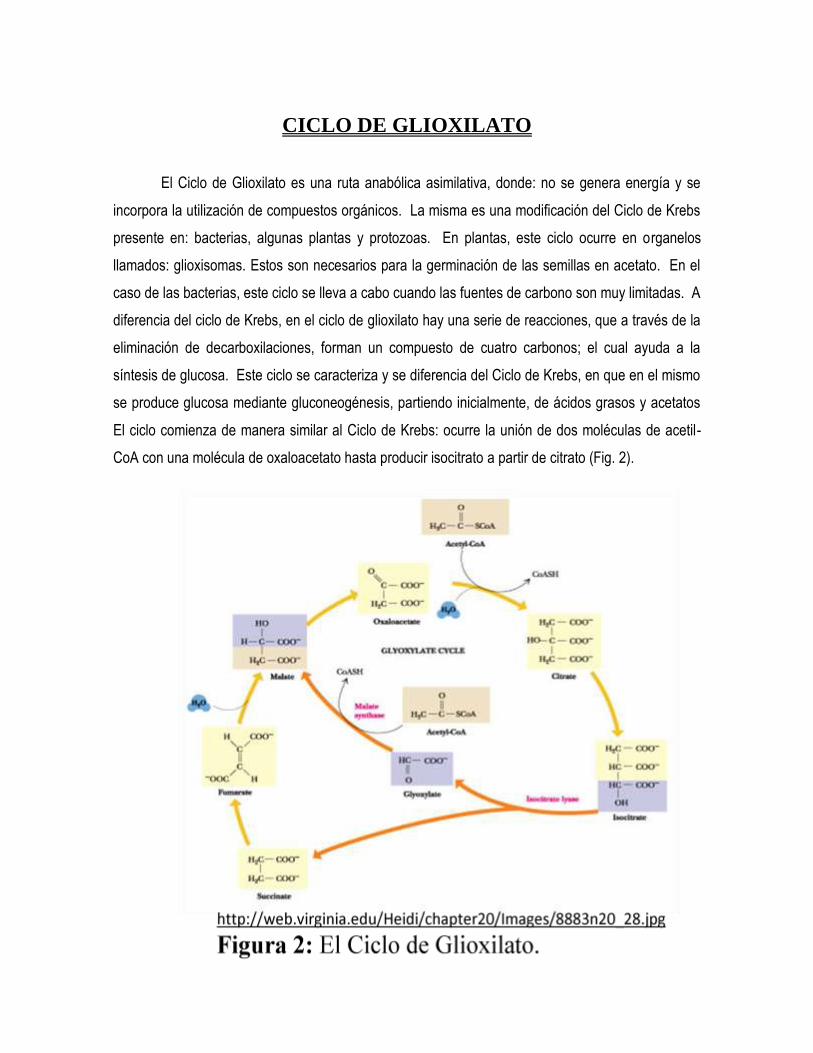
El ciclo comienza de manera similar al Ciclo de Krebs: ocurre la unión de dos moléculas de acetil-
CoA con una molécula de oxaloacetato hasta producir isocitrato a partir de citrato (Fig. 2).

Es este el punto, en donde cada uno de estos ciclos alterna sus vías, ya que el Ciclo de
Glioxilato produce succinato y glioxilato a partir de isocitrato. El glioxilato se une a acetil-CoA para
producir malato, el cual a su vez, produce oxaloacetato, siendo este último, el encargado de la
síntesis de glucosa. Este ciclo tiene dos enzimas características. Estas son: la isocitrato liasa, la
cual está encargada de degradar el isocitrato en succinato y glioxilato, y la malato sintasa, la cual
cataliza la reacción donde se une glioxilato con acetil-CoA, para producir malato.
La importancia de este ciclo recae en que le da la habilidad a ciertos organismos de crecer
en: etanol, acetato, ácidos grasos o compuestos constituído por únicamente dos carbonos. Este
ciclo es una manera alterna de sobrevivir en ambientes extremos con limitación de nutrientes, y es
regulado según las necesidades celulares de cada organismo. Algunos microoorganismos aeróbicos
como: Saccharomyces cerevisiae, Escherichia coli, Haloferax volcanii (halofilico extremo) y
Debaryomyces hansenii, llevan a cabo este ciclo.
RESUMEN DEL CICLO GLIOXILATO:
1. Incorporación de fuentes de carbonos.
2. Citrato sintasa cataliza la unión de: Oxaloacetato y Acetil-CoA, para formar citrato.
3. Si la célula lo requiere, esta molécula continúa con el Ciclo de Glioxilato.
4. Isocitrato liasa, cataliza la conversión de Isocitrato en: Glioxilato y Succinato.
5. Succinato entra de nuevo al Ciclo de Krebs y Glioxilato, y Malato sintasa cataliza su
conversión a Malato.
6. Malato deshidrogenasa, cataliza la conversión de Malato, en Oxaloacetato.

BIBLIOGRAFIA:
1) BIOQUÍMICA (Marino Villavicencio Núñez – Tomo 1 “Volumen 1,2” – 3º edición – Lima – 2007 – Fondo Editorial de la UNMSM)
2) HARPER BIOQUIMICA ILUSTRADA (Robert K. Murray – Peter A. Mayes – Daryl K.
Granner – Victor W. Rodwell)
3) BIOQUÍMICA (Marino Villavicencio Núñez – Tomo 2 “Volumen 1,2” – 3º edición – Lima – 2007 – Fondo Editorial de la UNMSM)
4) Cornah J. E., V. Germain, J. L. Ward, M. H. Beale, S. M. Smith. (2004). “Lipid
Utilization, Gluconeogenesis and Seedling Growth in Arabidopsis Mutants
Lacking the Glyoxylate Cycle Enzyme Malate Synthase”. Journal of Biological
Chemistry (279): 42916-42923.
5) Campbell M. K., S. O. Farrell. (2004). El ciclo de glioxilato: una vía
relacionada. Bioquímica. 4ta edición. (16.6): 532-533.
Top Related