Universidad Nacional de Colombia Facultad de … · SAC, e indican que la interacción social...
85
Universidad Nacional de Colombia Facultad de Ciencias Humanas Departamento de Psicología Diana Milena Cortés Patiño Efectos de la Interacción Social en el Consumo y Elección de Dosis de Etanol Bogotá, 2012
Transcript of Universidad Nacional de Colombia Facultad de … · SAC, e indican que la interacción social...

Universidad Nacional de Colombia
Facultad de Ciencias Humanas
Departamento de Psicología
Diana Milena Cortés Patiño
Efectos de la Interacción Social en el Consumo y Elección de Dosis de Etanol
Bogotá, 2012

Diana Milena Cortés Patiño
Efectos de la Interacción Social en el Consumo y Elección de Dosis de Etanol
Tesis presentada a la Maestría en Psicología
de la Universidad Nacional de Colombia
como uno de los requisitos para obtener el
grado de Magister en Psicología
Orientador: Prof. Arturo Clavijo Álvarez
Bogotá, 2012

Resumen
Efectos de la Interacción Social en el Consumo y Elección de Dosis de Etanol
Investigaciones en psicofarmacología han mostrado una asociación entre interacción social
y autoadministración de etanol (ET), sin embargo, los hallazgos experimentales son
ambiguos. Esta ambigüedad puede ser resultado de las dosis administradas y las variaciones
en los tipos de alojamiento. En este experimento expusimos tres grupos de ratas, divididas
en las condiciones de alojamiento de Interacción (GI), enriquecimiento (GE) y aislamiento
(GA) a tres dosis de ET (5%, 10% y 20%) y evaluamos su consumo de ET. En la primera
fase, evaluamos el consumo de sacarosa (SAC) con el objetivo de determinar si existían
diferencias entre el consumo de ET y de SAC en función de las condiciones de alojamiento.
En la segunda fase las ratas fueron entrenadas para auto-administrarse ET usando un
procedimiento de fading. En la tercera fase, evaluamos la elección entre diferentes pares de
soluciones de ET usando programas concurrentes VR5-VR5 (5%-10%; 10%-20%; 5-20%).
El consumo de SAC fue significativamente menor en GE, sin embargo, durante el
entrenamiento en consumo de ET y la fase de elección (fases 2 y 3) GE y GI mostraron los
mayores niveles de consumo de ET. En la tercera fase, no hubo diferencia en la tasa relativa
por ninguna dosis de ET, ni dentro, ni entre los grupos. Estos hallazgos sugieren que
condiciones grupales (GE y GI) facilitan el consumo de ET, lo que no se observa con la
SAC, e indican que la interacción social afecta diferencialmente el consumo de diversos
recursos reforzadores.
Palabras Clave: Condiciones de alojamiento, consumo de etanol, elección, ratas

Abstract
Effect of social Interaction on consumption and choice of different dosses of ethanol
Psychopharmacological research has shown an association between social interaction and
ethanol (ET) self-administration; nevertheless, experimental finding are ambiguous. This
ambiguity might result from differences in administrated doses and housing variations. We
exposed three groups of rats divided in the housing conditions of interaction (IN),
enrichment (EN) and isolation (IS) to three doses of ET (5%, 10% and 20%) and evaluated
their ET consumption. In a first phase, we evaluated sucrose (SUC) intake to determine
differences between SUC and ET consumption in function of housing conditions. In a
second phase, we trained the rats to self-administrate ET with a fading procedure. In a third
phase we assessed choice between different pairs of ET solution using concurrent schedules
VR5-VR5 (5%-10%; 10%-20%; 5-20%). SUC consumption was significantly lower in the
EN groups; however, during the ET training and choice phases (2 and 3), the EN and IN
groups shoed higher ET intake levels. In phase 3, there was no difference in relative
response rate for any dose of ET between groups. These findings suggest that group
conditions (IN and EN) facilitate ET but not SUC consumption, and indicate that social
interaction affects differentially consumption of diverse reinforcement sources.
Key-words: housing conditions, ethanol consumption, choice, rats.

LISTA DE FIGURAS
Figura 1. Relaciones económicas entre los bienes. ………………………………….….8
Figura 2. Relación de sustitución entre X y Y…………………………………………...11
Figura 3. Proceso de adicción relativa……………………………………………...……..12
Figura 4. Alojamiento del grupo de enriquecimiento……………………………………..34
Figura 5. Alojamientos individuales………………………………………………………35
Figura 6. Diferencias en la frecuencia de respuestas por SAC10% ……………….……..42
Figura 7. Box plot de consumo neto de etanol (g/kg) durante las etapas del entrenamiento
en consumo………………………………………………………………………………...44
Figura 8. Tasa de respuesta por las soluciones de ET 10%, al disminuir la cantidad de
sacarosa……………………………………………………………………………………46
Figura 9. Dispersión de los sujetos en el consumo neto por grupo de alojamiento……..48
Figura 10. Dispersión de los sujetos en el consumo neto por de tipo de elección……….49
Figura 11. Media ± SD de la tasa de respuesta por la solución con mayor concentración de
SAC (28.65%)…………………………………………………………………………..…50

LISTA DE TABLAS
Tabla 1. Organización de las fases del experimento………………………………………37
Tabla 2. Subgrupos generados para balancear el orden de elecciones a las que serían
expuestos los sujetos………………………………………………………………………39
Tabla 3. Valores p asociados al test U de Mann Whitney para la diferencia en consumo
entre pares de grupos………………………………………………………………………44

APÉNDICES
Apéndice 1. Peso corporal en los diferentes grupos durante la fase de autoadministración
de SAC
Apéndice 2. Valores asociados a las pruebas de Kruskall Wallis para consumo neto de
etanol durante la fase de entrenamiento
Apéndice 3. Número de sujetos en cada grupo clasificados por la posible fuente de control
del comportamiento de consumo durante la fase de elección.
Apéndice 4. Desempeño característico del control por la posición.
Apéndice 5. Desempeño característico del control por consumo neto de ET.
Apéndice 6. Desempeño característico del control por la dosis.

TABLA DE CONTENIDO
INTRODUCCIÓN………………………………………………………………………..1
Reforzadores alternativos y consumo de sustancias...………………………….…2
El efecto de sustitución económica y la teoría de la adicción relativa …………7
Evidencias de la adicción relativa en modelos animales …………………….13
Aislamiento y consumo de ET …………………………………………..15
Interacción y Consumo de ET……………………………………………….17
Enriquecimiento y Consumo de ET…………………………………………20
DISEÑO EXPERIMENTAL……………….…………………………………………....27
Alojamientos……………………….………………………………………..……28
Auto-administración de ET……………………………………………………....29
Elección entre dosis de ET ………………………………………………….…..30
Auto-administración de SAC……………………………………………………..30
Objetivos …………………………………………………………………………31
MÉTODO …………………………. ……………………………………...…………....32
Sujetos………………………………………….………………………………...32
Materiales e Instrumentos……………………………………………….……….33
Alojamientos………………………………………………………………....33
Cajas de Alimentación………………………………………………………36
Cajas de Condicionamiento Operante………………………………………36

Procedimiento…………………………………………………………………….36
Análisis de Datos…………………………………………………………………39
RESULTADOS…………………………………………………………………………..41
Las ratas alojadas de GE ambiental se auto-administran menos SAC…………...41
El consumo total de ET (g/kg) fue significativamente menor en GA…………….43
Las ratas mostraron indiferencia ante dosis concurrentes de ET …………… …46
DISCUSIÓN………………………………………………………………………………51
CONCLUSIONES…………………………………………………………………….….58
REFERENCIAS…………………………………………………………………………..61
APÉNDICE…………………………………………………………………………..…….69

1
Desde los inicios de las investigaciones en abuso de sustancias en los años 50,
diferentes teorías y modelos han intentado explicar cómo un organismo se hace adicto. Si
bien dichos modelos divergen en el tipo de explicación, una tesis es compartida por todos,
el uso de sustancias es una condición necesaria pero no suficiente para desarrollar una
adicción, esto es, la droga en sí misma no es responsable por la dependencia (Vuchinich y
Tucker, 1996). Surge entonces una pregunta, ¿qué tipo de variables, además de las
características farmacológicas de las sustancias, promueven y modifican el consumo?
Aproximaciones desde el análisis comportamental han intentado responder esta
cuestión vinculando variables relevantes de los fenómenos de aprendizaje a la explicación
del uso de sustancias, y han mostrado que los comportamientos de consumo son
susceptibles a variaciones en contingencias tanto operantes (Sanchis-Segura y Spanagel,
2006) como pavlovianas (Siegel, 2001). Esto demuestra que, como otros comportamientos,
la búsqueda y el uso de drogas están sujetos a las leyes de aprendizaje, y que por lo tanto,
el grado de control que una droga ejerce sobre la conducta no depende exclusivamente de
sus propiedades farmacológicas, sino del contexto específico de consumo (Higgins, Heil, y
Lussier, 2004).
Una de las áreas del análisis de la conducta que ha realizado contribuciones
importantes en la comprensión de las adicciones es la economía del comportamieto.
Aproximaciones a la dependencia desde esta área se caracterizan por estudiar el consumo
de sustancias en contextos de elección, enfatizando en los factores ambientales que
modulan la asignación del comportamiento e integrando a la explicación del consumo
conceptos provenientes de la economía, como precio, ingreso oferta o escasez (Hursh,
1991).

2
Los aportes de la economía del comportamiento a la explicación del consumo
pueden ser agrupados bajo tres grandes cuestiones (Tucker, 2005). La primera se refiere a
la variación de las preferencias por la sustancia al alterar los constreñimientos en el acceso
a esta misma, ya sea por incrementar el precio, restringir la oferta o disminuir el ingreso. La
segunda se relaciona con la variación en las preferencias al cambiar los constreñimientos en
el acceso a bienes alternativos. Finalmente, la tercera procura explicar cómo cambia la
asignación conductual cuando los consumidores deben elegir entre reforzadores que varían
en la cantidad y/o demora con que son recibidos.
Diversos estudios han sido realizados para responder a la primera cuestión (para una
revisión véase Hursh, 1991; Hursh, 1993; Hursh, 2005), demostrando que, tal como
acontece con otro tipo de reforzadores como agua o comida, el consumo de sustancias
consideradas adictivas disminuye cuando el precio aumenta; sin embargo, el incremento en
el precio de algunas sustancias psicoactivas puede llegar a ser muy alto antes de originar
una disminución en su consumo –evidencia de esto es la persistencia en el empleo de
drogas a pesar de las pérdidas económicas, laborales o familiares que puede generar el uso
de una sustancia-; por ello, la importancia de la segunda cuestión: el consumo puede ser
aumentado o disminuido variando el acceso a reforzadores alternativos.
Reforzadores alternativos y consumo de sustancias
Según la economía del comportamiento toda situación de consumo es una situación
de elección –consumir un bien implica dejar de consumir otro- por ello, el uso de una
comodidad no depende únicamente de su precio en términos monetarios o
comportamentales, depende de su costo de oportunidad (Allison, 1983). El costo de

3
oportunidad es aquello a lo que un agente económico renuncia cuando toma una decisión de
consumo (Mankiw, 2004) y depende en gran medida de la cantidad de bienes disponibles
en el mercado. Por ejemplo, en mercados con un único bien disponible, el costo de
oportunidad originado por el consumo de este bien será nulo sin importar cuál sea su
precio, pues no existen comodidades alternativas a las cuales el sujeto renuncie por su
decisión de consumo. En contraste, en mercados con múltiples posibilidades de elección, el
costo de oportunidad generado por el consumo de este bien será mayor, pues elegir este
bien significa renunciar a los demás disponibles.
Teniendo en cuenta lo anterior, una conclusión al respecto de cualquier análisis
económico de consumo es que la asignación comportamental para un bien X, cuando son
considerados los costos de oportunidad, no depende de su precio nominal (e.g. $10 por un
plato de comida, o 5 presiones de barra por un pellet en el caso de un experimentos) sino de
su precio relativo, esto es, de la razón entre el precio del bien X y el precio de los otros
bienes disponibles en el mercado (Mankiw, 2004).
La inclusión de los costos de oportunidad en el análisis del consumo puede ayudar a
entender por qué el uso de sustancias persiste a pesar de los incrementos en el precio de las
drogas. Una posible explicación es que el costo de oportunidad por el consumo de
substancias para algunos sujetos puede ser cercano de cero, es decir, no existen en el
contexto otras opciones que generen el mismo tipo de utilidad que las drogas y, en este
sentido, el precio relativo de consumir permanece bajo a pesar de los incrementos en el
precio nominal.

4
Uno de los ejemplos más claros de cómo variaciones en el precio relativo pueden
afectar el consumo de sustancias es proporcionado por el caso de Vietnam (Robins, 1975),
conocido en el área de abuso de drogas porque sólo un porcentaje de los soldados
americanos que consumieron opio durante la guerra continuaron consumiéndolo al llegar a
sus hogares y desarrollaron dependencia. En este caso –que acontece en un momento de la
historia en el que se creía que el consumo de drogas de abuso era una condición necesaria y
suficiente para desarrollar una adicción- la diferencia de consumo entre los soldados que
desarrollaron dependencia y quienes no la desarrollaron no puede ser explicada por las
propiedades farmacológicas del opio, pues todos estuvieron expuestos a la misma sustancia.
La pregunta que surgió entonces fue, ¿por qué sólo algunos soldados continuaron
consumiendo al llegar a sus hogares?
Heyman (1996) propone que la respuesta a esta pregunta estaría en la diferencia de
reforzadores disponibles para los soldados durante la guerra en Vietnam y al llegar a sus
hogares, esto es, en la diferencia en el precio relativo del opio en estos dos contextos. Así,
durante la estancia en Vietnam, los soldados tendrían fácil acceso a los opiáceos y un
acceso más restringido a otro tipo de reforzadores como, por ejemplo, el soporte social, esto
es, el precio de los opiáceos en relación al precio del soporte social sería menor en
Vietnam. Al regresar a sus hogares, con el acceso a familia y amigos, el precio del soporte
social disminuiría y, como consecuencia, aumentaría el del opio. Esta variación en los
precios relativos habría revertido la preferencia por la droga y con ello, disminuido su
consumo. Sin embargo, para algunos de los soldados el precio relativo del opio podría no
haber disminuido necesariamente, pues el contexto en sus hogares no habría garantizado la
existencia de reforzadores alternativos y estos soldados tendrían más posibilidad de

5
desarrollar una adicción. Si bien, esta es sólo una hipótesis de lo ocurrido, la evaluación de
cómo la disponibilidad de otros reforzadores en el contexto del sujeto puede afectar el
consumo, sería de gran ayuda en la comprensión del uso y abuso de psicoactivos.
Investigaciones con modelos animales han sido realizadas con el objetivo de evaluar
los efectos de la disponibilidad de otros reforzadores sobre el consumo de sustancias. En
general, estos experimentos utilizan diseños en los cuales los sujetos tienen acceso a más de
un reforzador, y se observa cómo varía la asignación del comportamiento al restringir o
facilitar el acceso a uno de los reforzadores; para ello son utilizados principalmente
programas concurrentes de razón o de intervalo, y las variaciones en el acceso a
reforzadores son hechas al alterar los valores del programa. Ejemplos de estos
experimentos serán presentados a continuación.
Carroll y Lac (1998) demostraron que la disponibilidad de comida afecta la
autoadministración de cocaína durante la fase de adquisición. Los resultados muestran que
mientras el 100% de los sujetos de los grupos con restricción alimentaria alcanzaron el
criterio de adquisición (100 infusiones durante 5 días consecutivos) sólo el 76% del grupo
sin restricción de acceso a comida lo consiguió. Adicionalmente, el número de días para
alcanzar el criterio fue significativamente mayor en el grupo con comida ad libitum. Este
efecto de la disponibilidad de comida sobre el consumo de psicoactivos también se ha
replicado en opiáceos como el etonitazeno y drogas disociativas como la feniciclidina
(Carroll y Cambpell, 2000).
La disponibilidad de sustancias dulces como la sacarosa también ha mostrado incidir
sobre la autoadministración de drogas en la fase de adquisición. En un experimento con

6
programas concurrentes, Carroll, Lac y Nygard (1989) investigaron si la
autoadministración de cocaína era afectada la por disponibilidad de una solución de
glucosa+sacarosa. Los resultados mostraron que la concurrencia de sacarosa interfirió con
la autoadministración de cocaína, de manera que cuando la sacarosa fue presentada
concurrente las ratas no alcanzaron el criterio de adquisición. Cuando la solución de
glucosa+sacarosa fue retirada las ratas aprendieron rápidamente el comportamiento de
autoadministración.
Samson, Tolliver y Roehrs (1983) reportaron resultados similares con etanol y
sacarosa en programas concurrentes de razón fija. En una fase inicial, los sujetos podían
elegir entre dos sustancias –agua ó una solución de etanol al 5% (v/v) bajo programas de
razón fija con el mismo valor. En esta fase la tasa relativa de respuesta para la solución con
etanol se encontró entre .8 y 1. En la siguiente fase, el agua fue reemplazada por una
solución de agua con sacarosa cuya concentración iba incrementando progresivamente.
Para todos los sujetos, al aumentar la concentración de sacarosa, la tasa relativa de
respuestas para la solución con etanol disminuyó. Esto es, hacer disponible la opción de
sacarosa disminuyó la preferencia por el etanol.
Un aspecto importante en relación a la disponibilidad de otros reforzadores en
situaciones de consumo es que la presencia del reforzador alternativo no debe ser
concurrente para afectar la elección de un recurso adictivo. La disponibilidad de otro
reforzador en la historia previa del animal (momentos diferentes a la situación de elección)
también afecta la tasa de elección de la sustancia psicoactiva. Carrol y Lac (1993)
demostraron que sujetos con acceso a una solución de glucosa y sacarosa durante tres
semanas precedentes a la fase de autoadministración de cocaína intravenosa demoraron

7
significativamente más para alcanzar el criterio de adquisición y sólo un 50% de los sujetos
lo consiguió, en comparación con el 75% de aquellos que únicamente recibieron agua
previamente. Este tipo de hallazgos son importantes porque brindan una caracterización
molar del contexto de elección, es decir, la asignación que un sujeto hace de su conducta no
depende únicamente de los recursos reforzadores presentes, sino de su historia de
reforzamiento.
El efecto de sustitución económica y la teoría de adicción relativa
Como fue visto anteriormente, la disponibilidad de otros reforzadores interfiere con la
auto-administración de sustancias; sin embargo, no todos los reforzadores presentes en el
contexto afectan de la misma forma los comportamientos de consumo. En el experimento
descrito de Carroll y Lac (1993), mientras el acceso a sacarosa interfirió con la
autoadministración de cocaína, la disponibilidad del agua no mostró un efecto significativo
sobre este comportamiento. Este resultado revela una característica importante de las
situaciones de consumo: la disponibilidad de otros reforzadores puede tener efectos
distintos sobre el consumo de drogas, y, el tipo de efecto observado depende de los
reforzadores presentes en el contexto y de las relaciones entre ellos.
Según la teoría económica, tres tipos de relaciones determinan cómo la concurrencia
de reforzadores afecta la asignación del comportamiento: independencia, sustituibilidad y
complementariedad. La existencia de una u otra relación depende de cómo varía el
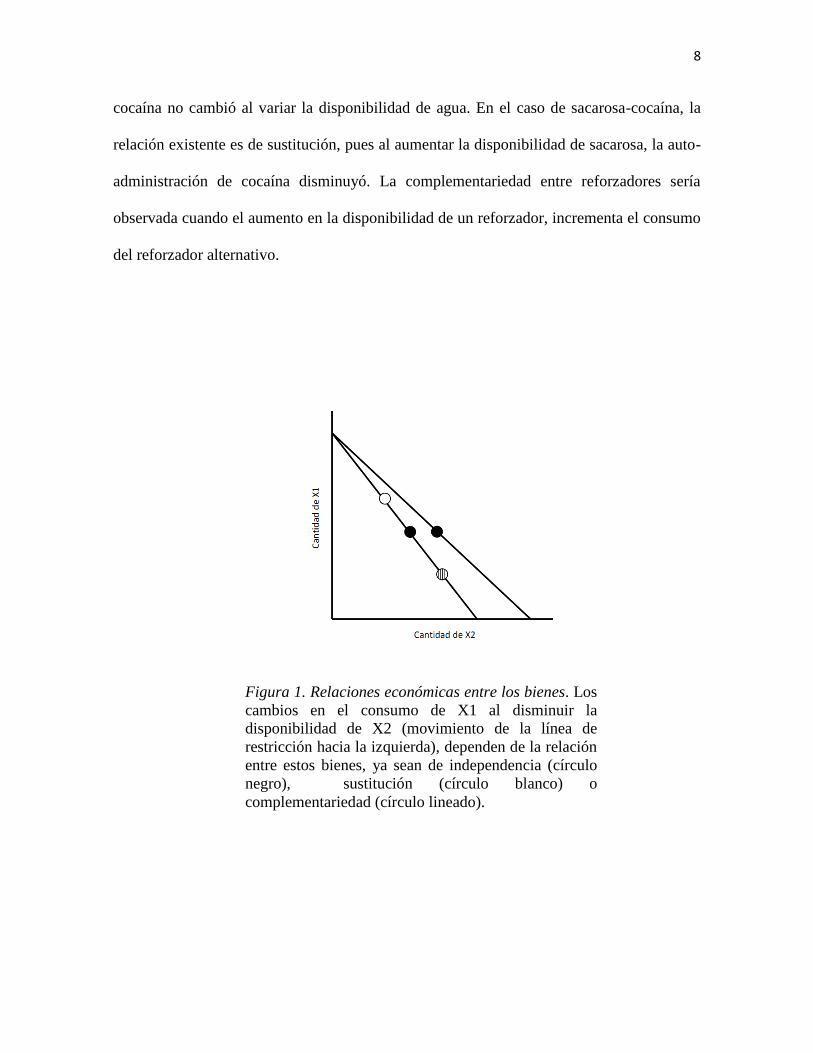
consumo de un bien al aumentar la disponibilidad de otro –o disminuir su precio (Figura 1).
En los resultados de Carroll y Lac (1993) pueden observarse dos tipos de relaciones. Para el
par de reforzadores agua-cocaína la relación es de independencia, pues el consumo de
8
cocaína no cambió al variar la disponibilidad de agua. En el caso de sacarosa-cocaína, la
relación existente es de sustitución, pues al aumentar la disponibilidad de sacarosa, la auto-
administración de cocaína disminuyó. La complementariedad entre reforzadores sería
observada cuando el aumento en la disponibilidad de un reforzador, incrementa el consumo
del reforzador alternativo.
Figura 1. Relaciones económicas entre los bienes. Los
cambios en el consumo de X1 al disminuir la
disponibilidad de X2 (movimiento de la línea de
restricción hacia la izquierda), dependen de la relación
entre estos bienes, ya sean de independencia (círculo
negro), sustitución (círculo blanco) o
complementariedad (círculo lineado).

9
Diversos estudios en economía comportamental han explorado la existencia de
relaciones de sustitución entre sustancias psicoactivas y recursos alternativos, buscando
identificar reforzadores que puedan ser utilizados en la prevención y tratamiento de la
dependencia de sustancias. En líneas generales, estos experimentos utilizan modelos
operantes en los que dos reforzadores son ofrecidos en programas concurrentes –de razón o
de intervalo-. La variable independiente es el precio de uno de los reforzadores, manipulado
al aumentar el requerimiento en un programa o la dosis, y la variable dependiente es la tasa
de respuesta para el segundo reforzador (Hursh, 2005).
Siguiendo esta línea de estudios, las investigaciones en sustitución farmacológica son
realizadas con el objetivo de identificar si relaciones de sustitución son observadas entre
sustancias con una acción farmacológica parecida o similar. En un experimento clásico con
monos Rhesus, Hursh (1993) estudió la relación de sustitución entre dos agonistas opioides
–hidromorfina y metadona-, observando cómo variaba el consumo de hidromorfina cuando
se aumentaban los valores de un programa de razón fija (RF) para autoadministración de
metadona. Los resultados mostraron que con los incrementos del RF para acceder a
metadona, disminuía su consumo y aumentaba el de la hidromorfina.
Resultados como los de Hursh (1993) son relevantes por sus implicaciones clínicas y
servir de base para las terapias de reemplazo o de sustitución farmacológica, en las que se
facilita el acceso controlado a un reforzador sustituto que comparte los mecanismos de
acción central con una sustancia objetivo, evitando con eso el consumo de la sustancia
ilegal y sus múltiples efectos adversos para el organismo (Greenwald y Hursh, 2006; Strain,
Stitzer, Leibson, y Bigelow, 1993; Vanichseni, Wongsuwan, Choopanya, y Wongpanich,
1991).

10
A pesar de la relevancia de los hallazgos en sustitución farmacológica, la relación de
sustitución no se evidencia únicamente entre recursos que comparten características
cualitativas, como la acción en el sistema nervioso central, también son encontrados entre
reforzadores que no comparten una dimensión aparente. Por ejemplo, en los experimentos
descritos de Carroll et al (1989) y de Samson et al (1983), la disminución en el precio de la
sacarosa (aumento de la dosis) generó una disminución en el consumo de la sustancia
psicoactiva (cocaína y alcohol, respectivamente), demostrando que, en situaciones de
elección, la sacarosa funcionaria como sustituto de algunas drogas a pesar de que su acción
central sea diferente. Esto muestra que la sustitución entre reforzadores es definida
funcionalmente, es decir, son los cambios comportamentales en términos de aumento o
disminución del consumo –y no las cualidades o propiedades de las sustancias usadas
como- las que determinan la existencia de esta relación.
Rachlin (2000) se basa en esta característica funcional de las relaciones de sustitución
para plantear la teoría de la adicción relativa. Según esta teoría, la adicción es el producto
de la demanda inelástica de un bien indefinido s, que está disponible únicamente a través de
las actividades de consumo X y Y. Éstas dos actividades –diferentes cualitativamente- son
mutuamente sustituibles únicamente en tanto suplen la necesidad de s, pero no son
sustitutas para ninguna otra actividad de consumo, es decir, no son sustitutos perfectos
(Figura 2). Rachlin (2000) propone que las actividades X y Y serían consumo de sustancias
e interacción social respectivamente, estos dos bienes serían sustitutos en tanto suplen una
misma necesidad s (elevación de ánimo y reducción de la ansiedad según Fisher,
Lichtenstein, y Haire-Joshu, 1993), por lo que el aumento en el precio de uno implicaría en
el aumento del consumo del otro.

11
Figura 2. Relación de sustitución entre X y Y. Aunque
no son sustitutos perfectos, ambos satisfacen la
necesidad de S.
Una de las características fundamentales de la teoría postulada por Rachlin (2000)
es que las actividades de consumo de sustancias e interacción social difieren en la forma
como varía la utilidad1 generada por su consumo repetido. Así, las conductas adictivas son
habituadas al precio, esto es, con su consumo repetido disminuye la utilidad reportada y el
sujeto debe gastar más para recibir los mismos beneficios, en otras palabras, con cada
consumo el precio aumenta. A diferencia del consumo de sustancias, la interacción social
es sensibilizada al precio, lo que implica que cuanto más se interactúa, se necesita hacer
menor para recibir determinada utilidad, por lo que podríamos decir que a mayor
interacción social, menor es el precio que debe ser pagado por la utilidad recibida.
De acuerdo con la teoría de Rachlin (2000), la dinámica entre la habituación al
precio del consumo y la sensibilización al precio de la interacción sería la responsable por
1 La utilidad es definida como el beneficio resultante del consumo de un bien

12
la generación de patrones adictivos, pues en determinadas etapas de consumo -a pesar del
aumento de precio de la droga por el efecto de habituación- sería difícil acceder a la
interacción social debido al efecto de sensibilización al precio. Por ejemplo, podríamos
plantear una situación donde inicialmente la interacción social es más barata que el
consumo de alcohol, pero una pérdida de un ser querido aumenta el precio extrínseco de la
interacción social. Aumentar el precio de la interacción se aumenta el consumo de alcohol,
producto del efecto de sustitución, razón por la cual sujeto empieza a consumir más
frecuentemente. El consumo repetido de alcohol disminuye la utilidad de ambos - el soporte
social y la ingesta de alcohol- porque al dedicar menos tiempo en la actividad social, ésta se
hace más costosa (debido al efecto de sensibilización al precio), y el alcohol también
aumenta su precio por el efecto de habituación (Figura 3).
Figura 3. Proceso de adicción relativa. Un individuo se
hace adicto cuando la tasa de sensibilización al precio es
más alta que la tasa de habituación.

13
Una de las ventajas de la teoría de la adicción relativa es que, además de proponer
una explicación de la permanencia en patrones adictivos, permitiría explicar la variabilidad
comportamental encontrada entre las personas que consumen algún tipo de droga, pues,
como señala Ahmed (2010), sólo una pequeña proporción de quienes usan psicofármacos
desarrollan dependencia. Este hecho sería explicado porque, debido a la relación de
sustitución, cualquier incremento en el precio de estos reforzadores llevaría al individuo a
distribuir su conducta para maximizar la utilidad recibida, aumentando el consumo de la
actividad cuyo precio sea menor. Este proceso de maximización puede resultar en patrones
de conducta adictivos (cuando la tasa de habituación es menor que la tasa de
sensibilización), en patrones principalmente de interacción (cuando la tasa de
sensibilización es menor que la tasa de habituación) o en un equilibrio entre estas dos
conductas, como es el caso bebedor social.
Si la hipótesis de sustitución entre estas dos conductas es cierta, se daría un paso
importante en la prevención y el tratamiento de la dependencia a drogas, pues al manipular
las variables implicadas, ya sea por precio o por disponibilidad, sería posible cambiar las
utilidades recibidas por la elección del consumo o de la interacción, facilitando una nueva
asignación conductual con un equilibrio de consumo diferente.
Evidencias de la Adicción Relativa en Modelos Animales
Estudios realizados con participantes humanos han mostrado correlaciones
negativas entre diversas medidas de relacionamiento social, como soporte social, soporte
parental, o integración social, y medidas de consumo de psicoactivos, como frecuencia de

14
uso o número de recaídas, (e.g Battistich y Hom, 1997; Brown y Riley, 2005; Frauenglass,
Routh, Pantin, y Mason, 1997). Estos encuentros ratifican la importancia de considerar
variables sociales en el análisis del abuso de drogas; sin embargo, no pueden ser tomados
como evidencia a favor de la hipótesis de la adicción relativa, pues, a partir de esos datos
correlacionales no es posible determinar la existencia de una relación de sustitución, esto
sólo es posible mediante el uso de diseños experimentales con modelos animales que
permitan especificar cómo cambian los valores de consumo cuando son manipuladas las
posibilidades de interacción
Actualmente, no se encuentran en la literatura experimental con modelos animales
estudios dirigidos a probar la hipótesis de adicción relativa; no obstante, existen múltiples
investigaciones en las que se evalúa la autoadministración de drogas en sujetos bajo
diferentes tipos de alojamiento –indivudual vs grupal-. Los resultados de estas
investigaciones permiten establecer si existen diferencias en el consumo de sustancias que
dependen de la manipulación de la interacción social, y en este sentido, serían una fuente de
evidencia para establecer la veracidad de la hipótesis postulada por Rachlin (2000).
A continuación, describiré resultados de tres líneas de estudios que evalúan cómo la
interacción social afecta el consumo de sustancias2: estudios en aislamiento, en interacción
y en enriquecimiento. A pesar de que en todas las líneas es comparado el consumo de ratas
en interacción vs ratas en aislamiento, la variable relevante al respecto de la interacción –
variable que guía la hipótesis de trabajo- es diferente en cada una, y con ella, el tipo de
interacción permitida. Así, estudios en los que el aislamiento es la variable principal no dan
2 Debido a que en el presente estudio se evaluó el consumo de alcohol, se priorizaron los estudios relativos a
esta sustancia.

15
mucha importancia a la expresión de comportamientos sociales y utilizan pocos sujetos en
el grupo control, generalmente pares. En las investigaciones de la línea de interacción, la
variable relevante es la expresión de comportamientos sociales y por ello garantizan
alojamientos grupales con un número de sujetos mayor. Los estudios en enriquecimiento
hacen énfasis en el tipo de ambiente en el que las ratas están alojadas y en la expresión de
comportamientos de la especie en condiciones naturales, por lo que son utilizadas colonias
de ratas en comparación con ratas en aislamiento.
Aislamiento y consumo de drogas
Los primeros experimentos en observar los efectos del tipo de alojamiento en el
consumo estaban interesados en determinar la acción del estrés por aislamiento en la
autoadministración de drogas. Estas investigaciones partían del supuesto según el cual
fuentes de estrés estarían asociadas a incrementos en el consumo de psicoactivos. Debido a
que pruebas fisiológicas habían mostrado que el aislamiento funciona como un evento
estresante, (Hatch, Wiberg, Balazs y Grice, 1963) se presumía que condiciones de vivienda
individual favorecerían niveles mayores de consumo. Parker y Radow (1974) evaluaron
esta hipótesis en un procedimiento en el que se observó el consumo de una solución de
etanol (ET) 25% (v/v) en ratas alojadas individualmente y en pares. La autoadministración
era realizada en sesiones de 16 horas, en las cuales los sujetos tenían acceso individual a
agua, comida y ET. Después de la sesión de autoadministración las ratas volvían a sus
respectivos alojamientos por ocho horas. Los resultados mostraron que ratas alojadas
individualmente tenían medias de consumo diario entre 10g/kg y 15g/kg, mientras que estas
medias para las ratas alojadas en pares se encontraban entre 5g/kg y 10 g/kg, evidenciando
mayores niveles de consumo en las ratas aisladas.

16
A pesar de que estos resultados parecen ir en línea con la hipótesis de la adicción
relativa, es necesario hacer ciertas consideraciones metodológicas al respecto de los
resultados. En primer lugar, resulta sorprendente que ratas se auto-administren una solución
con una concentración tan alta de ET (25% v/v) sin un entrenamiento específico de
consumo. Revisiones recientes muestran que concentraciones mayores a 12% son
aversivas para las ratas (Wolffgramm y Turyabahika-Thyen, 2006), así que no es claro bajo
qué condiciones fueron alcanzados niveles tan altos de consumo voluntario (entre 5g/kg -
15g/kg). Otra consideración está relacionada con la descripción de las viviendas, no es
descrito si las cajas de alojamiento para los dos grupos tenían las mismas medidas de área x
animal. Controlar esa variable sería fundamental, pues espacios muy restringidos podrían
generar otro tipo de estrés además del que se pretendía evaluar y podrían haber afectado los
valores de consumo.
Otros experimentos más recientes que han evaluado el efecto del aislamiento en el
consumo de alcohol muestran resultados variables y diferentes de los reportados por Parker
y Radow (1974). Por ejemplo, Thorsell, Slaweckia, Khoury, Mathe, y Ehlers (2005)
observaron que bajo las mismas condiciones de vivienda utilizadas por Parker y Radow
(1974), las ratas alojadas en pares bebieron más de una solución de ET 10% (v/v) que
aquellas alojadas individualmente cuando estaban en fases de consumo forzado; sin
embargo, no hubo diferencia en las medidas de consumo para los dos grupos cuando la
autoadministración fue voluntaria, esto es, cuando había otras fuentes de líquido
disponibles. En otro estudio cuyo objetivo era identificar diferencias en consumo de ET
(10% v/v) entre ratas de dos líneas genéticas diferentes -con preferencia por alcohol y sin
preferencia por alcohol- Ehlers, Walker, Pian, Roth, y Slawecki, (2007) observaron

17
diferencias en el consumo del ET dependientes del tipo de alojamiento (par vs individual)
en los sujetos pertenecientes al grupo con preferencia -las ratas aisladas bebieron más. Para
el grupo sin preferencia, no se observó ningún efecto del tipo de alojamiento sobre la
autoadministración de alcohol. Finalmente, Doremus, Brunell, Rajendran, y Spear (2005)
en un experimento que evaluaba los factores que influencian el consumo de ET en ratas
adolescentes y adultas, encontraron que en el grupo de adultas la autoadministración de una
solución de ET 10% (v/v) fue mayor para los sujetos alojados en pares. Esta diferencia no
se encontró en ratas adolescentes.
La variabilidad hallada tiene una implicación importante en el análisis de la
interacción social y el consumo de sustancias, el aislamiento, como privación de
interacción, no siempre favorece el consumo de psicoactivos –como era presumido en la
hipótesis del estrés por aislamiento-, pues los estudios muestran que alojamientos
individuales tienen efectos diferenciales (aumento, mantenimiento o disminución del
consumo de alcohol) que dependen de otras variables relacionadas, como la predisposición
genética, edad, o el tipo de consumo, i.e forzado vs voluntario.
Interacción Social y Consumo de drogas
A diferencia de los estudios en estrés por aislamiento, otra línea de investigaciones
hace énfasis en el comportamiento social como factor determinante en la
autoadministración de drogas. Aunque en estas investigaciones también se analiza el
consumo de sujetos en condiciones de alojamiento “individual” vs “interacción”, en los
grupos de interacción son utilizados por lo menos cuatro sujetos, esto con la finalidad de
permitir la aparición de conductas sociales como juego, exploración social y otros

18
comportamientos relacionados con la jerarquía como submisión o agresión (Wolffgramm y
Heyne, 1991)
Uno de los primeros experimentos de esta línea fue realizado por Deatherage
(1972), quien evaluó el consumo forzado de dos soluciones de ET (10% v/v y 20% v/v) en
ratas bajo condiciones de alojamiento individual y grupal (6 individuos). En su
procedimiento, el alcohol estaba disponible 24 horas en las cajas de alojamiento y no
existían otras fuentes de líquido disponibles. Los resultados mostraron que las ratas
alojadas individualmente preferían la solución más alta de ET (20%).
Resultados similares fueron obtenidos por Wolffgramm y Heyne (1991) quienes
analizaron el consumo voluntario de tres soluciones de ET (5% v/v, 10% v/v y 20%v/v) en
ratas bajo tres tipos de alojamiento: aislamiento (en el que no era posible interacción alguna
entre sujetos), contacto (en el que los sujetos interactuaban a través de una malla) y grupos
(en la que los sujetos interactuaban sin ninguna clase de restricción). Pruebas de ingesta
total de ET mostraron que las ratas aisladas bebieron significativamente más que las
agrupadas, además, y de acuerdo con los resultados de Deatherage (1972), las ratas
pertenecientes al grupo de aislamiento prefirieron la solución de alcohol 20% v/v, mientras
que las ratas en grupo prefirieron la concentración más baja (5%).
Uno de los datos más interesantes presentados por Wolffgramm y Heyne (1991) es
que el consumo de alcohol de los sujetos (g/kg de peso) variaba diferencialmente cuando
eran cambiadas las condiciones de alojamiento; así, en una fase del experimento en la cual
las condiciones de alojamiento eran revertidas, sólo se observaban incrementos en el
consumo cuando los animales pasaban de una condición de mayor interacción a una

19
situación de menor interacción (e. g. grupal para contacto, o contacto para aislamiento). En
las situaciones en las que pasaban de una situación de menor interacción a una de mayor
interacción, los niveles de consumo disminuyeron para algunos sujetos y permanecieron
constantes para otros. Daoust, Chretien, Moore, Saligaut, Lhuintre y Boismare (1985)
observaron este mismo patrón de resultados en un experimento que pretendía evaluar el
consumo de una solución de alcohol 12% al variar las condiciones de alojamiento de ratas.
Los resultados mostraron que cambios en la condición de alojamiento de individual para
grupal generaban disminuciones en el consumo de alcohol.
El efecto de condiciones grupales de alojamiento sobre el consumo de sustancias
también ha sido analizado con otros tipos de drogas como anfetaminas, opioides y cocaína
(Heyne, 1996, Heyne y Wolffgramm,1998; Ding, 2005) y se han encontrado resultados
semejantes a los descritos, señalando que alojamientos grupales –como oportunidad para
comportamientos sociales- tienen un efecto de disminución del consumo, lo que iría en
dirección de la propuesta de Rachlin (2000).
No obstante, otros estudios en los que la interacción social también es permitida en
ambientes semi-naturales no han mostrado un patrón consistente de resultados. Adams
(1996) realizó un experimento en el que ratas fueron divididas en tres grupos diferentes de
alojamiento (aislamiento, grupal y semi-natural) y se evaluó el consumo libre de una
solución de alcohol 10% (v/v). Los resultados evidenciaron que las ratas en alojamientos
semi-naturales bebieron significativamente más ET que las alojadas individualmente o en
grupo. Este hallazgo tiene una implicación en el análisis de los efectos de la interacción
social en el consumo de drogas, así como el aislamiento no siempre favorece el consumo de
drogas, la interacción social no siempre disminuye los niveles de consumo. Surge entonces

20
una pregunta ¿qué tipo de interacciones son necesarias para generar disminuciones en la
autoadministración de drogas?
Enriquecimiento Ambiental y Consumo de drogas
Los estudios sobre enriquecimiento ambiental iniciaron a mediados de los 60, cuando
Rosenzweig y su grupo de colaboradores realizaron una serie de estudios con el objetivo
de determinar si animales criados bajo condiciones diferenciales de estimulación eran
diferentes en cuanto a morfología y fisiología cerebral (Bennett, Krech y Rosenzweig,
1964; Rosenzweig y Bennett, 1969). En estos estudios, ratas alojadas en condiciones
estándares de laboratorio –criadas en aislamiento con acceso a agua y comida únicamente-
eran comparadas con ratas criadas en ambientes enriquecidos. El ambiente enriquecido era
una condición de alojamiento grupal con acceso a múltiples objetos inanimados –como
ruedas de actividad, juguetes o laberintos- que permitían que las ratas exhibieran
comportamientos asociados a la especie en condiciones naturales, como roer, corre o jugar.
Los resultados de estos experimentos demostraron que ratas mantenidas en ambientes
enriquecidos eran diferentes de ratas en ambientes pobres en medidas como el peso
cerebral, la espesura del córtex, la cantidad de conexiones sinápticas, el número de espinas
dendríticas y la actividad enzimática en diferentes áreas del cerebro.
Además de los efectos positivos en el desarrollo cerebral, reportados por los
experimentos iniciales de Rosenzweig (Bennett, Krech y Rosenzweig, 1964; Rosenzweig y
Bennett, 1969), el enriquecimiento ambiental ha demostrado tener efectos favorables en
variables comportamentales; por ejemplo, cuando son comparados con sujetos aislados, los
sujetos mantenidos en ambiente enriquecido muestran tasas de aprendizaje más rápidas y

21
respuestas de habituación más rápida a eventos estresantes (Pham, Winblad, Granholm, y
Mohammed, 2002; Mocek, Ducko, Johansson y Jezova, 2004). Otros estudios han revelado
que ratas criadas en ambientes enriquecidos exhiben menos patrones de comportamiento
anormales como estereotipia o inactividad motora en horas nocturnas (Olsson y Dahlborn,
2002).
Los efectos positivos del enriquecimiento ambiental en relación con otras condiciones
de alojamiento también se han observado cuando ratas en ambientes enriquecidos son
comparadas con animales criados en condiciones grupales, pero sin acceso a otras
actividades además de la interacción (Rosenzweig, Bennett, Hebert y Morimoto, 1978).
Este descubrimiento llevó a Rosenzweig et al (1978) a proponer que la variable relevante
de sus experimentos no era la interacción permitida entre los sujetos, sino todo el complejo
de estimulación característico de las condiciones de enriquecimiento.
Teniendo en cuenta los encuentros de Rosenzweig et al (1978) podríamos afirmar que
la interacción en condiciones pobres de alojamiento (sin otros reforzadores además de la
interacción social) es cualitativamente diferente de la interacción en situaciones de
enriquecimiento. En este sentido, es importante evaluar cómo el enriquecimiento ambiental
–diferente de otras situaciones de interacción- afecta el consumo de sustancias psicoactivas.
Rockman, Gibson, y Benarroch (1989) evaluaron los efectos del enriquecimiento
ambiental en el consumo de diferentes soluciones de ET en ratas Sprague- Dawley. En el
procedimiento, un grupo de machos fue criado en condiciones de enriquecimiento, el otro
grupo fue criado en condiciones de alojamiento individual. Después de 90 días en esas
condiciones –previo a la presentación de ET- la mitad del grupo de enriquecimiento fue

22
puesta en condiciones de aislamiento por el resto del experimento, mientras la otra mitad
permaneció en su alojamiento 8 horas al día y después fue aislada durante 16 horas para
realizar mediciones de consumo de alcohol. Los animales del grupo de aislamiento
también fueron divididos de manera que la mitad permaneció en sus condiciones originales
de alojamiento, mientras la otra mitad fue mantenida durante 8 horas en la colonia y 16
horas en aislamiento para tomar las medidas de consumo de alcohol. Esta división generó
cuatro grupos experimentales: enriquecimiento, enriquecimiento-aislamiento, aislamiento y
aislamiento-enriquecimiento para los que fueron analizadas la media de ingesta en g/kg de
ET y el porcentaje medio de consumo de ET con relación al total de ingesta de fluidos.
Los resultados mostraron que no hubo diferencias en el consumo entre los grupos
para las soluciones de ET3% y ET7%. Los sujetos de enriquecimiento-aislamiento
bebieron significativamente más de la solución de ET5% que los sujetos aislamiento-
enriquecimiento; sin embargo, esta diferencia no se encontró cuando fue presentada la
solución ET9%. Finalmente, el grupo que permaneció enriquecido durante todo el
experimento consumió significativamente más de la concentración ET 9%. Con base en
estos resultados, Rockman et. al (1989) afirman que condiciones de ambiente enriquecido
favorecen el consumo de dosis altas de ET.
Estos resultados van en contra de los hallazgos de Wolffgramm y Heyne (1991) que
muestran que ratas con acceso a interacción social consumen menos cantidades y prefieren
dosis menores de ET, por lo que el enriquecimiento ambiental sería caracterizado como una
situación de interacción diferente con efectos distintos sobre el consumo de ET. No
obstante, es necesario hacer algunas consideraciones metodológicas con respecto al
procedimiento de Rockman et. al (1989) antes de aceptar esta conclusión. En primer lugar

23
no es especificado cuantos animales habitaban en la colonia de ambiente enriquecido,
Deatherage (1972) mostró que la densidad poblacional favorece niveles altos de consumo
de ET, por eso es importante especificar el control de esta variable. La segunda –y más
importante- consideración está relacionada con el tiempo utilizado para medición de
consumo. Los investigadores realizaron las medidas de consumo en cajas individuales
durante 16 horas, esto implica que los animales de enriquecimiento eran sometidos a
cambios de condiciones de alojamiento cada vez que eran expuestos a alcohol. En
comparación con los otros grupos, aislamiento, enriquecimiento-aislamiento y aislamiento-
enriquecimiento, los sujetos que permanecieron todo el tiempo en enriquecimiento
experimentaron cambios de alojamiento durante todo el experimento y no únicamente
durante la fase previa a la presentación del alcohol, lo que pudo afectar los niveles de
consumo de ET.
Otros estudios realizados por Ellison y colaboradores en los años 80 sugieren que el
enriquecimiento ambiental puede favorecer niveles altos de consumo (Ellison, Levy y
Lorant, 1979; Ellison, Daniel y Zoraster, Ellison, 1981, Ellison, 1987). En estas
investigaciones se observó el consumo libre de ET10% y agua en ratas Long Evans bajo
condiciones de alojamiento seminatural –enriquecido- e individual. En el procedimiento los
animales tenían acceso a botellas con agua y ET10% dentro de su alojamiento y a través de
cámaras era estimado el tiempo de consumo de cada animal. Los resultados indicaron que
ratas aisladas bebían en promedio más agua y ET que ratas en colonias; sin embargo, las
ratas en colonias exhibían mayor variabilidad entre los individuos, esto es, dentro de las
colonias habían ratas que mostraban preferencia extrema por ET y otras que sólo

24
consumían agua. A diferencia de lo observado en las colonias, los animales alojados
individualmente no mostraron variaciones importantes en el consumo entre los individuos.
Los resultados de los experimentos Ellison y colaboradores tienen una implicación
importante en el análisis de la interacción social y el consumo: condiciones de interacción
pueden favorecer consumos extremos de alcohol. Ellison (1987) explica este tipo de
hallazgo refiriéndose a la organización social permitida por condiciones grupales de
alojamiento, pues el mantenimiento en colonias permite la expresión de comportamientos
sociales como los típicos de la organización jerárquica y esto genera variaciones en el
consumo de alcohol entre los individuos, de manera que ratas subordinadas beben
cantidades mayores de ET que ratas dominantes.
En un experimento más reciente, Fernández- Turuel et al (2002) crió ratas de dos
líneas genéticas seleccionadas de acuerdo con la “búsqueda por la novedad” en ambientes
enriquecidos y las comparó con ratas de esas mismas líneas criadas en duplas en cuanto a
consumo y preferencia de ET 10%. Los resultados mostraron que independientemente de la
línea genética, las ratas criadas en ambientes enriquecidos consumieron más ET que
aquellas alojadas en duplas. Este resultado también fue observado en consumo de
anfetaminas y cocaína (Bowling y Bardo, 1994; Bardo, Donohew y Harrington, 1996), por
lo que Fernánde-Turuel (2002) sugiere que el ambiente enriquecido podría favorecer
niveles altos de consumo de psicoactivos.
Hasta el momento los experimentos descritos nos llevan a las siguientes
conclusiones: (1) Los efectos del aislamiento en el consumo de sustancias son ambiguos,
algunos experimentos muestran que incrementa el consumo y otros que lo disminuye (2) La

25
interacción social –sin enriquecimiento- parece tener un efecto de disminución del
consumo (3) El enriquecimiento ambiental favorece niveles altos de consumo. A
continuación haré algunas consideraciones al respecto de estas conclusiones.
En primer lugar es necesario aclarar que los efectos del aislamiento son analizados en
relación con un grupo control (grupo de interacción). Debido a que las características del
grupo control pueden afectar diferencialmente el consumo de ET, cualquier estudio que
manipule el aislamiento debe considerar qué tipo de interacción está siendo permitida en el
grupo control para poder llegar a conclusiones válidas y replicables.
Con respecto a los estudios de interacción y enriquecimiento surgen dos
consideraciones. En general los estudios que centran su atención en la interacción social
utilizan un número pequeño de sujetos (entre tres y seis), los estudios en enriquecimiento
por su parte utilizan colonias que generalmente tienen más de 10 ratas. En este sentido las
diferencias entre estas fuentes de estudio pueden deberse al número de sujetos, y no a un
efecto especial del enriquecimiento. Como lo afirma Ellison (1987) las dinámicas de
jerarquía tienen efectos importantes en el consumo de ET, así que grupos pequeños en los
que los comportamientos de dominancia no son expresados totalmente son cualitativamente
diferentes de los grupos de enriquecimiento. Surge entonces la pregunta, qué sucede cuando
son comparados grupos de interacción y enriquecimiento con el mismo número de sujetos?
Otra consideración con respecto a los experimentos descritos es que todos utilizan
modelos de consumo libre para evaluar la autoadministración de ET. Estudios en
autoadministración de anfetaminas muestran que los efectos del enriquecimiento varían
cuando el consumo es libre y cuando el consumo es operante, de manera que, mientras en

26
situaciones de consumo libre, ratas en enriquecimiento se auto-administran más anfetamina
que ratas aisladas, cuando el consumo es operante el resultado se revierte. Teniendo estos
experimentos en cuenta es necesario evaluar como varía el consumo de ET en modelos con
autoadministración operante, pues este tipo de consumo se asemeja más al encontrado en
humanos, que deben pagar precios muy altos para acceder a psicoactivos.
En conclusión, los estudios que tuvimos en cuenta para evaluar la hipótesis de la
adicción relativa postulada por Rachlin (2000) muestran que la relación entre interacción
social y consumo de ET no es simple; por lo tanto, la evaluación de una relación de
sustitución entre estos dos comportamiento implica considerar no sólo situaciones de
interacción vs aislamiento, es necesario tener en cuenta el tipo de interacción del cual los
sujetos hacen parte para comprender los efectos que ésta pueda tener sobre el consumo. El
presente experimento pretende profundizar en las diferencias entre tipo de interacción y
consumo de alcohol.

27
DISEÑO EXPERIMENTAL
Dos características fueron observadas en las investigaciones presentadas en el
apartado anterior. En primer lugar, en la mayoría de los estudios son comparadas dos
condiciones, una condición de aislamiento y otra condición de interacción. La condición de
interacción varía entre los estudios, de manera que algunos experimentos utilizan
condiciones estándares de laboratorio con pocos animales y otros utilizan alojamientos
semi-naturales o enriquecidos en los que ratas habitan en colonias. Esta variación en los
tipos de interacción parece tener efectos diferenciales en el consumo en relación con
situaciones de aislamiento (Daoust et al, 1985; Wolfgramm y Heyne, 1991; Ellison, 1987);
sin embargo, pocos estudios comparan estas dos condiciones –interacción y
enriquecimiento- en relación con condiciones de aislamiento.
Otra característica observada en la mayoría de los estudios es que utilizan dosis
medias de ET (generalmente ET 10%), sin embargo, como muestran Wolffgramm y Heyne
(1991) las ratas muestran preferencias diferenciales por dosis altas o bajas de ET
dependiendo del tipo de alojamiento, de manera que al presentar una sola dosis para
consumo puede afectar los resultados obtenidos.
Teniendo en cuenta estas dos observaciones, el presente experimento pretendió
evaluar el consumo y la elección de dosis de ET en ratas, cuando son permitidas diferentes
condiciones de interacción. Para ello se utilizó un diseño de grupos con tres condiciones
experimentales correspondientes al tipo de interacción o alojamiento (Variable
Independiente): Grupo de Aislamiento (GA), Grupo de Enriquecimiento (GE) y Grupo de
Interacción (GI). Dos medidas fueron analizadas en cada uno de los grupos (variables

28
dependientes): el consumo total de ET (g/kg), y la preferencia por 3 diferentes dosis de ET
(5%, 10% y 20%) determinada a partir de la tasa relativa de respuesta en situaciones de
elección.
A continuación profundizaré en algunos aspectos relevantes del diseño
experimental.
Alojamientos
Debido a que el establecimiento de jerarquías lleva tiempo y a que los efectos del
enriquecimiento dependen del tiempo de exposición a este tipo de ambiente (Schenk,
Gorman, y Amit, 1990), ratas Wistar fueron colocadas en sus respectivas condiciones de
alojamiento poco tiempo después del desmame. Los alojamientos de GI y GE tenían las
mismas dimensiones y el mismo número de ratas, de manera que la única diferencia entre
estas dos condiciones eran los objetos ubicados dentro de la jaula de GE. Los alojamientos
de GA eran jaulas individuales que mantenían la misma área por rata que las otras
condiciones de manera que en esta condición las ratas tenían poca restricción de
movimiento (podían caminar y trepar en la jaula) evitando así la intervención de otras
variables que pudieran generar algún tipo de estrés (Rockman, Hall, y Glavin, 1986).
El enriquecimiento ambiental para GE fue construido de forma que permitiera que
las ratas realizaran comportamientos propios de la especie en condiciones naturales, así las
ratas en este grupo tenían material para construcción de nidos, lugares para esconderse,
materiales para roer y desgastar los dientes, una rueda para correr, y objetos varias con
varias texturas para interactuar. Con la finalidad de permitir comportamientos asociados a

29
la búsqueda de comida, los pellets eran distribuidos en diversos lugares de la jaula
(Hutchinson, Avery y Wanderwode, 2005).
Autoadministración de ET
Con base en las similitudes con el comportamiento de consumo de ET en humanos,
la autoadministración de ET en el experimento tuvo tres características: oral, operante, e
intermitente. Como Meisch (2001) señala, la auto-administración de ET por otras vías
diferentes a la oral no se asemeja al consumo humano, pues el olor y el sabor de la droga
deben hacer parte de la experiencia de consumo. En segundo lugar, se utilizó un modelo
operante en el cual las ratas debían emitir un número de respuestas para acceder a ET. Este
modelo no sólo simula las situaciones de consumo humano, además permite una mayor
precisión en las medidas de consumo individual. Finalmente, se utilizó un programa de
refuerzo intermitente (VR 5) pues como señala Hartnoll (1991) en el caso de los humanos
no todas las respuestas tienen como consecuencia alcohol, en muchas circunstancias la
exigencia comportamental para acceder a la sustancia es muy alta. En este sentido,
programas de refuerzo continuo tendrían menos validez en el estudio del consumo de
alcohol.
En general, las ratas muestran aversión a los sabores nuevos, por ello deben ser
utilizados procedimientos para facilitar la auto-administración de ET. Uno de los más
utilizados en el área de investigación en consumo es el procedimiento de fading en el que
el alcohol es presentado en una solución con sacarosa. Inicialmente el porcentaje de SAC es
mayor que el de ET y conforma avanzan las sesiones, el porcentaje de SAC en la solución
disminuye y el de ET aumenta hasta llegar al porcentaje deseado de ET con una

30
concentración mínima de SAC. Este procedimiento permite que los animales consuman
cantidades suficientes de ET para asociar el sabor, originalmente aversivo, a sus efectos. La
efectividad de este procedimiento ha sido probada incluso en situaciones sin privación de
agua y comida (Samson, Pfeffer y Tolliver, 1988).
Elección de dosis de ET
Teniendo en cuenta el resultado de Wolffggramm y Heyne (1991) que señala que en
modelos de consumo libre las ratas bajo diferentes condiciones de alojamiento muestran
preferencias diferenciales por dosis de ET, el presente experimento pretendió observar si
utilizando las mismas dosis usadas por Wolffgramm y Heyne (1991), ratas respondiendo en
programas operantes también muestran esas preferencias. La verificación de las
preferencias es importante, pues estudios muestran que la elección por sustancias
psicoactivas depende en parte del tipo de consumo (Deehan, Cain, y Kiefer, 2007), y hasta
el momento no se conocen estudios que evalúen la preferencia por dosis de ET utilizando
modelos operantes.
En este experimento se utilizaron programas concurrentes VR5-VR5 que
presentaban dos dosis diferentes de ET. En este caso, el precio en términos operantes
(número de respuestas) para ambas dosis se mantuvo constante. La preferencia por fue
determinada a partir la tasa relativa de respuesta por cada dosis en una sesión.
Autoadministración de SAC
Inicialmente, la auto-administración de SAC era parte del entrenamiento de
comportamiento operante previo a la presentación de ET, de manera que –como los datos
de moldeamiento- los resultados de esta etapa no iban a ser presentados como parte de los

31
objetivos del estudio; sin embargo, la observación temprana de los datos mostró que
existían diferencias en la tasa de respuesta por SAC dependientes del grupo de alojamiento,
por ellos se aumentó el número de sesiones y se analizó durante una semana la
autoadministración de SAC en programas VR5 para los diferentes grupos.
Objetivos
Con base en los elementos mencionados en el diseño, tres grandes objetivos fueron
planteados:
1. Determinar si existían diferencias en la tasa de respuesta por SAC entre los
grupos de alojamiento.
2. Determinar si existían diferencias en el consumo (g/kg) de ET entre los grupos de
alojamiento.
3. Determinar si existían diferencias en la elección de tres dosis de ET (5%, 10% Y
20%) dependientes de las condiciones de alojamiento.

32
MÉTODO
Sujetos
Treinta y seis ratas albinas Wistar, machos, obtenidas en el bioterio de Butantán (São
Paulo) permanecieron en el bioterio del Departamento de Psicología Experimental de la
Universidad de São Paulo en condiciones constantes de temperatura (21+2º) e iluminación.
El ciclo claro comenzaba a las 7:00 pm y finalizaba a las 7:00 am. Las sesiones
experimentales se realizaron durante la fase nocturna del ciclo de iluminación con el fin de
no interferir con los patrones de actividad de las ratas.
Después de tres días de habituación al bioterio, los animales fueron distribuidos
aleatoriamente en tres grupos: 12 sujetos fueron asignados al grupo de interacción (GI) 12
al grupo de enriquecimiento (GE) y 12 sujetos al grupo de aislamiento (GA). Durante todo
el experimento, los animales se mantuvieron en sus condiciones respectivas de alojamiento.
Los sujetos recibieron agua y comida ad lib por 45 días después de los cuales se inició la
fase de privación alimenticia. Durante la restricción, las ratas eran ubicadas
individualmente en cajas donde tenían acceso a comida y agua durante una hora al día, lo
suficiente para mantenerlas en aproximadamente el 90% de su peso. Una vez por semana,
recibían comida dentro de sus respectivos alojamientos, lo cual permitía comportamientos
de búsqueda de comida (foraging)
Antes de cada fase, el experimento era interrumpido y la privación suspendida por
una semana, esto con el objetivo de establecer nuevas líneas de base del peso y acompañar
el crecimiento de los sujetos.

33
Instrumentos
Alojamiento
Los sujetos pertenecientes a los grupos de interacción (GI, n=12) y enriquecimiento
(GE, n=12) fueron criados en jaulas de alambre con dimensiones de 100x50x50cms. Cada
alojamiento tenía cuatro bebederos localizados en el exterior de la caja. El alojamiento del
GE contenía diferentes objetos que eran reubicados y/o cambiados frecuentemente. Entre
los objetos se encontraban: juguetes de plástico y madera, escaleras de madera y metal,
cajas de cartón, escondites de plástico y cerámica, cuerdas, corchos, entre otros (Figura 4).
El piso de las cajas era cubierto con viruta y cambiado por lo menos tres veces por semana.
La única diferencia entre los alojamientos de GE y GI era la presencia de los objetos
característicos del ambiente enriquecido.
Los animales del grupo de aislamiento (GA, n=12) fueron criados en jaulas
individuales de alambre con dimensiones de 40x26x19 (Figura 5). Las cajas fueron
seleccionadas de manera tal que el área por rata se mantuviera homogénea en los tres
grupos experimentales (20.000 cm3 aproximadamente), evitando así que variables
relacionadas con el espacio, como la restricción de movimiento, intervinieran en los
resultados. Otro aspecto a señalar es que el aislamiento sólo impedía el contacto cuerpo a
cuerpo, pues, debido a las características de las cajas y su disposición en la sala, no existía
otro tipo de privación sensorial. Al igual que en los otros grupos, la viruta era cambiada por
lo menos tres veces por semana.

34
Figura 4. Alojamiento del grupo de enriquecimiento. Las
ratas tenían acceso constante a una rueda de actividad. El
alojamiento contenía objetos y juguetes que eran
cambiados frecuentemente

35
Figura 5. Alojamientos individuales.

36
Cajas de Alimentación
Las ratas se alimentaban en cajas individuales de polietileno, con dimensiones de
25x40x20cms. Las tapas de cada caja eran adaptadas para permitir la ubicación de comida
y una botella con agua. El piso de cada caja se cubría con viruta.
Cajas de condicionamiento operante
Se utilizaron 6 cajas experimentales de fabricación Med-Associates, de dimensiones
30X25X21cms, con control por microcomputador IBM-PC con la interfaz y los programas
de Med-Associates. Cada caja contaba con dos barras localizadas en los extremos derecho e
izquierdo de una de las paredes, a 8cms del piso. Bajo cada una de las barras se
encontraban los bebederos (a 2cms del piso). Sobre las barras estaba ubicada una luz blanca
que indicaba la disponibilidad del refuerzo. Cada caja tenía un ventilador que permitía la
circulación de aire y proporcionaba un ruido blanco.
Procedimiento
Todas las sesiones fueron realizadas entre las 9:00 am y la 1:00 pm, después de
iniciado el período nocturno. Las sesiones experimentales para cada sujeto ocurrían día por
medio, de forma que la mitad de cada grupo pasaba por sesión experimental un día y la otra
mitad el día siguiente. Después de las sesiones experimentales los sujetos eran ubicados en
las cajas de alimentación con acceso a agua y comida durante una hora.
La Tabla 1 muestra las diferentes fases del experimento con sus respectivas etapas.

37
Tabla 1. Organización de las fases del experimento.
Fase Etapas
1. Preparatoria y Auto-administración de SAC a. Moldeamiento
b. Auto-administración de SAC
2. Entrenamiento en consumo de ET a. SAC 10%
b. SAC 10% + ET 2,5%
c. SAC 5% + ET 5%
d. SAC 5% + ET 10%
e. SAC 2.5% + ET 10%
f. SAC 0.25% + ET 10%
3. Elección entre dosis de ET a. Elección 1
b. Elección 2*
c. Elección 3*
4. Elección entre dosis de SAC
*Los asteriscos indican las fases que no se llevaron a cabo
1. Fase preparatoria y autoadministración de sacarosa. El comportamiento de
presionar la barra fue moldeado usando un programa de razón continua. La
autoadministración de SAC se llevó a cabo durante 7 sesiones con un programa
concurrente VR5-VR5. Ambos bebederos presentaban una solución de SAC 10%
como reforzador.
2. Entrenamiento en consumo de etanol (ET)- Procedimiento de Fading. Debido
al sabor aversivo del alcohol, la autoadministración fue entrenada mediante un
procedimiento de fading en el que se aumentaba la concentración de ET conforme
se disminuía la cantidad de sacarosa en la solución. El procedimiento de fading
fue realizado manteniendo el programa concurrente VR5-VR5 de la fase anterior.

38
Una de las soluciones de SAC fue reemplazada por agua, mientras que la otra fue
sustituida gradualmente por una de ET endulzado de acuerdo con la siguiente
programación: 2 sesiones con solución de SAC 10%; 5 sesiones con solución
SAC 10%+ET 2,5%; 4 sesiones con solución de SAC 5%+ET 5%; 4 sesiones con
solución SAC 2,5%+ET10%; 8 sesiones con solución SAC 0,25%+ET 10%.
3. Fase de elección entre dosis de ET. Manteniendo el programa concurrente VR5-
VR5, cada bebedero ofrecía una de tres soluciones de ET (5%, 10% o 20%)
endulzado con SAC 0,25%; de esta manera, fueron obtenidas tres condiciones
diferentes de elección: (1) ET5%-ET10%, (2) ET5%-ET20% y (3) ET10%-
ET20%. El objetivo inicial era que todas las ratas pasaran por las tres condiciones,
por eso, cada grupo fue dividido en 3 subgrupos con el objetivo de balancear el
orden de elección al que los sujetos serían expuestos (Tabla 2).
Debido a la variabilidad intra-sujeto encontrada en la respuesta de elección
durante las primeras 10 sesiones de esta fase, se decidió aumentar a 14 el número
de sesiones con la primera condición de elección, después de las cuales se
suspendió el experimento, pues los sujetos dentro de todos los grupos no
mostraban preferencia por alguna dosis

39
Tabla 2. Subgrupos generados para balancear el orden de elecciones a las que
serían expuestos los sujetos.
Elección y número
de sesiones
Subgrupo 1
(GI=4, GE=4,
GA=4)
Subgrupo 2
(GI=4, GE=4,
GA=4)
Subgrupo 3
(GI=4, GE=4,
GA=4)
1 (10 sesiones) 5%-10% 5%-20% 10%-20%
2 (10 sesiones)* 5%-20% 10%-20% 5%-10%
3 (10 sesiones)* 10%-20% 5%-10% 5%-20%
* Los asteriscos muestran las etapas que no se llevaron a cabo
4. Fase de elección entre dosis de sacarosa. Un análisis final de preferencia
por dosis de sacarosa fue realizado con el fin de observar si la variabilidad
encontrada en la respuesta por dosis de ET se reproducía cuando se presentaban
dosis concurrentes de SAC. De esta forma, fueron presentadas en programas
concurrentes VR5-VR5 dosis de SAC 7.35% y SAC 28.65% (v/v). La
preparación de las soluciones de SAC se hizo de tal modo que una de las dosis
fuera isocalórica a una solución de ET5% y la otra a una solución de ET20%. En
esta fase todos las ratas se enfrentaron al mismo tipo de elección.
Análisis de datos
Para los datos de la fase de autoadministración de sacarosa, que cumplían con las
condiciones de normalidad y homogeneidad de varianza, se realizaron análisis factoriales
utilizando ANOVA. Las comparaciones planeadas fueron realizadas con pruebas t para
muestras independientes.

40
En las fases 2 y 3 -entrenamiento en consumo y elección de dosis de ET,
respectivamente-, el consumo de ET fue calculado en relación al peso corporal diario de los
sujetos. Debido a la varianza de los datos en estas fases, los análisis de consumo fueron
realizados con estadísticos no paramétricos. Las diferencias en consumo de ET entre los
grupos se estimaron por medio de pruebas de rangos de Kruskal Wallis. Las comparaciones
planeadas se realizaron con la prueba de Mann Whitney y las pruebas de medidas repetidas
con los estadísticos de Friedman y Wilcoxon.
Los datos de la fase de elección entre dosis de sacarosa fueron analizados por medio
de estadísticos ANOVA.

41
RESULTADOS
En relación con los objetivos determinados en el delineamiento experimental se
encontró que: (1) las ratas alojadas en condiciones de enriquecimiento se auto-
administraron menos SAC que las de los otros grupos (GA y GI). (2) El consumo total de
ET (g/kg) fue significativamente menor en GA que en los otros grupos. (3) Las ratas de
todos los grupos mostraron indiferencia ante las dosis de ET cuando fueron presentadas
concurrentes.
Las ratas alojadas en enriquecimiento ambiental se auto-administran menos SAC
La Figura 6 (a) muestra la media más error estándar del número de respuestas por
SAC para cada grupo. Un ANOVA mostró un efecto significativo de la condición
alojamiento sobre la varianza en la frecuencia de respuestas (F=7.033, p˂0.001) Análisis
post-hoc mostraron que las diferencias significativas se encontraban GE y GI (p˂0.01) y
GE y GA (p˂0.001), evidenciando niveles de consumo significativamente menores para las
ratas en la condición de enriquecimiento.
Análisis por sesión de auto-administración de SAC mostraron que en todos los días
las ratas de GE respondieron menos por SAC que las pertenecientes a GI y GA (Figura 6
b); sin embargo, comparaciones planeadas entre pares de grupo utilizando pruebas t con la
corrección de bonferroni para 7 factores mostraron que estas diferencias en el consumo
diario no fueron significativas en ningún día.
Un ANOVA reveló que no habían diferencias en el peso entre los grupos (F= 0.579,
p=0.548), de manera que las diferencias en el consumo de SAC encontradas entre las
condiciones de alojamiento no pueden ser atribuidas a esta variable. (Apéndice 1)

42
Figura 6. (a) Diferencias en la frecuencia de respuestas por SAC10% para
cada uno de los grupos. Se encontraron diferencias significativas entre GE
vs GA, y GE vs GI (F= 7.0336, p˂0.001). (b) Número de respuestas por
SAC por sesión en cada grupo. Pruebas t con corrección de bonferroni para
siete factores mostraron que las diferencias observadas no eran
significativas para ninguno de los días.
GI GE GA (a)
(b)

43
El consumo total de ET (g/kg) fue significativamente menor en GA
Análisis iniciales mostraron que los datos no cumplían con las condiciones de
normalidad y homogeneidad de varianza, por ello, se utilizaron pruebas no paramétricas de
Kruskall Wallis y U de Mann Withney, para establecer si existían diferencias entre los
grupos en esta etapa. Debido a que los todos los sujetos pasaron por las 5 etapas del
procedimiento de fading, se realizó una corrección de Bonferroni para 5 factores con el fin
de ajustar el nivel de significancia; de esta manera, fueron consideradas significativas
diferencias con p≤0.01
Los resultados mostraron diferencias en el consumo neto de ET que dependían del
tipo de alojamiento en 4 de las 5 etapas del procedimiento. En la primera etapa, con
ET2.5% v/v, no se encontraron diferencias significativas entre los grupos (H= 0.628,
p=0.73). En todas las etapas restantes, con concentraciones mayores de ET, se observaron
diferencias significativas en el consumo entre los grupos (p≤0.01, los valores del test y sus
niveles de significancia son presentados en el apéndice 2)
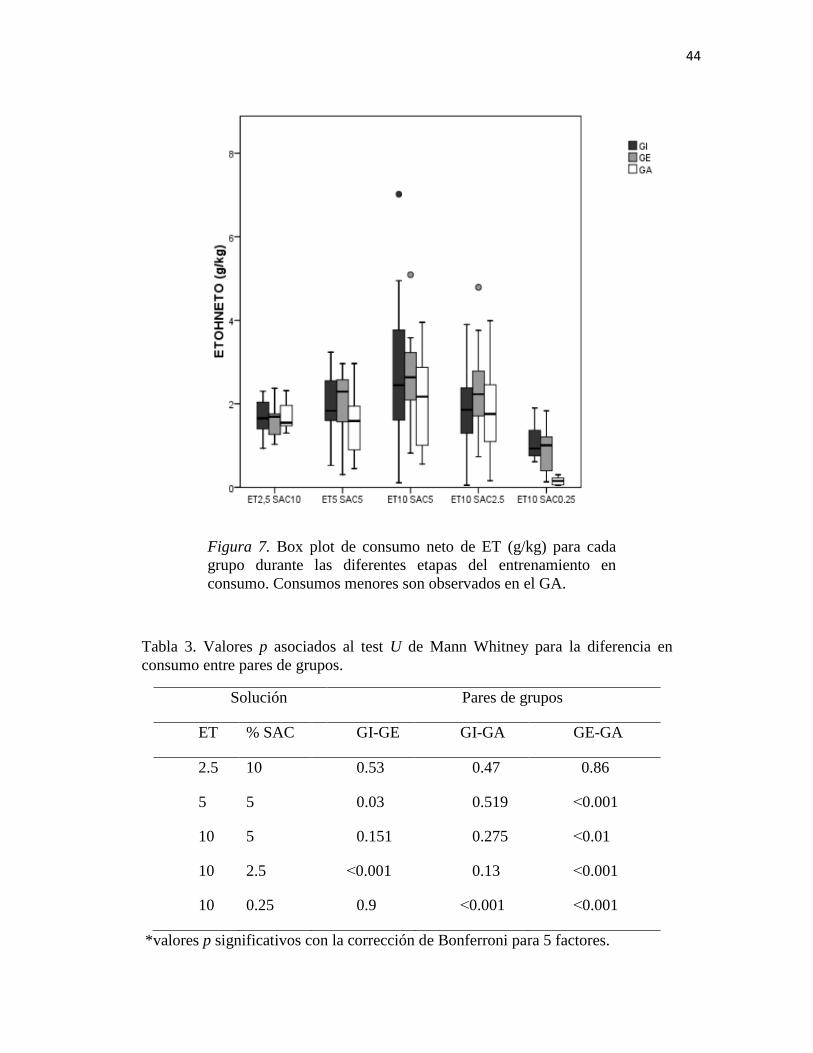
Al analizar las diferencias durante cada etapa del procedimiento de fading se encontró
que el grupo de aislamiento mostró los menores niveles de consumo durante todas las
etapas (Figura 7). Comparaciones planeadas usando la prueba U de Mann Whitney
mostraron que las mayores diferencias se encontraron entre el grupo de aislamiento (GA) y
el grupo de enriquecimiento (GE) (ver Tabla 3).
44
Figura 7. Box plot de consumo neto de ET (g/kg) para cada
grupo durante las diferentes etapas del entrenamiento en
consumo. Consumos menores son observados en el GA.
Tabla 3. Valores p asociados al test U de Mann Whitney para la diferencia en
consumo entre pares de grupos.
Solución Pares de grupos
ET % SAC GI-GE GI-GA GE-GA
2.5 10 0.53 0.47 0.86
5 5 0.03 0.519 ˂0.001
10 5 0.151 0.275 ˂0.01
10 2.5 ˂0.001 0.13 ˂0.001
10 0.25 0.9 ˂0.001 ˂0.001
*valores p significativos con la corrección de Bonferroni para 5 factores.

45
En la última etapa del procedimiento (ET 10%-SAC 0.25%), los sujetos de GA
disminuyeron significativamente la cantidad de alcohol consumida; de hecho, un análisis de
tasa relativa de respuesta, mostró que la preferencia por ET se revirtió y los sujetos
comenzaron a mostrar tasas de respuesta mayores por agua (Figura 10). Con el fin de
analizar si había diferencias en la tasa relativa de respuesta por ET dentro de los grupos
cuando era variada la concentración se sacarosa, se realizó una prueba de Friedman para
medidas repetidas. Los resultados muestran que para GI y GE no hubo diferencias (p=0.6
en ambos casos); sin embargo, para GA el valor p obtenido fue ˂0.001, lo cual indica que
dentro de este grupo hubo diferencias en la tasa de respuesta por ET dependiendo de la
concentración de sacarosa. Comparaciones pos-hoc utilizando la prueba de Wilcoxon con la
corrección de Bonferroni para 3 factores muestran que estas diferencias en el consumo de
ET ocurrieron cuando se varió la concentración de sacarosa de 2.5% a 0.25% (p˂0.001).
Una prueba adicional de Kruskal- Wallis mostró que no habían diferencias en el
consumo de ET dependiendo la localización de la barra de ET –derecha o izquierda-
(p=0.1). Tampoco se encontraron diferencias en los pesos entre los grupos (F=0.364,
p=0.69)

46
Figura 8. Tasa de respuesta por las soluciones de ET
10%, al disminuir la cantidad de sacarosa. Variaciones
mayores se encuentran en GA al cambiar la cantidad de
2.5% a 0.25%.
Las ratas mostraron indiferencia ante dosis de ET presentadas concurrentes.
Los análisis de tasa relativa de respuesta de la primera etapa de elección revelaron
que la mayoría de los sujetos era indiferente ante las dosis presentadas concurrentes. Sólo
tres de los 36 sujetos (dos de GE y uno de GI) mostraron preferencia por la dosis de ET5%.
Después de 4 sesiones adicionales con las mismas concentraciones, el experimento fue
suspendido con la finalidad de determinar porqué los sujetos no mostraban preferencias.
Un analisis caso a caso del desempeño de los sujetos durante las 14 sesiones que duró
la fase reveló que el comportamiento de autoadministración de ET estaba siendo controlado
por variables diversas, así, 16 sujetos estaban respondiendo de acuerdo con la posición de la

47
barra, 5 sujetos respondían de acuerdo con la cantidad total de ET ingerido (sin importar la
dosis) y sólo 3 respondían por la dosis. Para 11 sujetos no se consiguió establecer la fuente
de control. Los patrones de desempeño de cada tipo de control están incluídos en el
apéndice3 (apéndices 3,4,5 y 6).
Análisis de las sesiones por bloques de cinco minutos tampoco mostraron
preferencias por dosis en el inicio de la sesión. De hecho, la mayor parte de las ratas
dejaban de responder después de los primeros 5 minutos, por lo que la posibilidad de que la
indiferencia por las dosis dependiera del tiempo transcurrido de la sesión fue descartada.
Con respecto al consumo neto de ET (g/kg) en esta fase, los resultados muestran una
gran dispersión (Figura 9); no obstante, la observación realizada en la fase de
entrenamiento de que los sujetos pertenecientes al grupo de aislamiento consumían menos
ET se mantuvo en esta fase (triángulos blancos en la parte inferior de la Figura 9). Fueron
realizados análisis para confirmar esta observación. Un test de Kruskall Wallis con
corrección para 14 factores mostró que existían diferencias entre los grupos en el consumo
neto para esta fase (p˂0.001). Comparaciones planeadas usando la prueba U de Mann
Whitney evidenciaron diferencias entre GA y GE (p˂ 0.001), y entre GA y GI (p˂0.001).
La comparación entre GE y GI no reveló ninguna diferencia.
La Figura 10 muestra el consumo neto diario de todos los sujetos por tipo de elección
(10%-20%, 5%-10% y 5%-20%). A pesar de que pareciera que consumos más altos fueron
favorecidos por situaciones en las que se presentaban concurrentes las dosis 10%-20 y 5%-
3 Debido a que el experimento se detuvo en la primera etapa, sólo 4 sujetos de cada grupo pasaron por cada
tipo de elección, por ello no fue posible realizar análisis estadísticos con relación a los datos de esta fase. Los
desempeños individuales característicos de esta fase son incluídos en el apéndice.
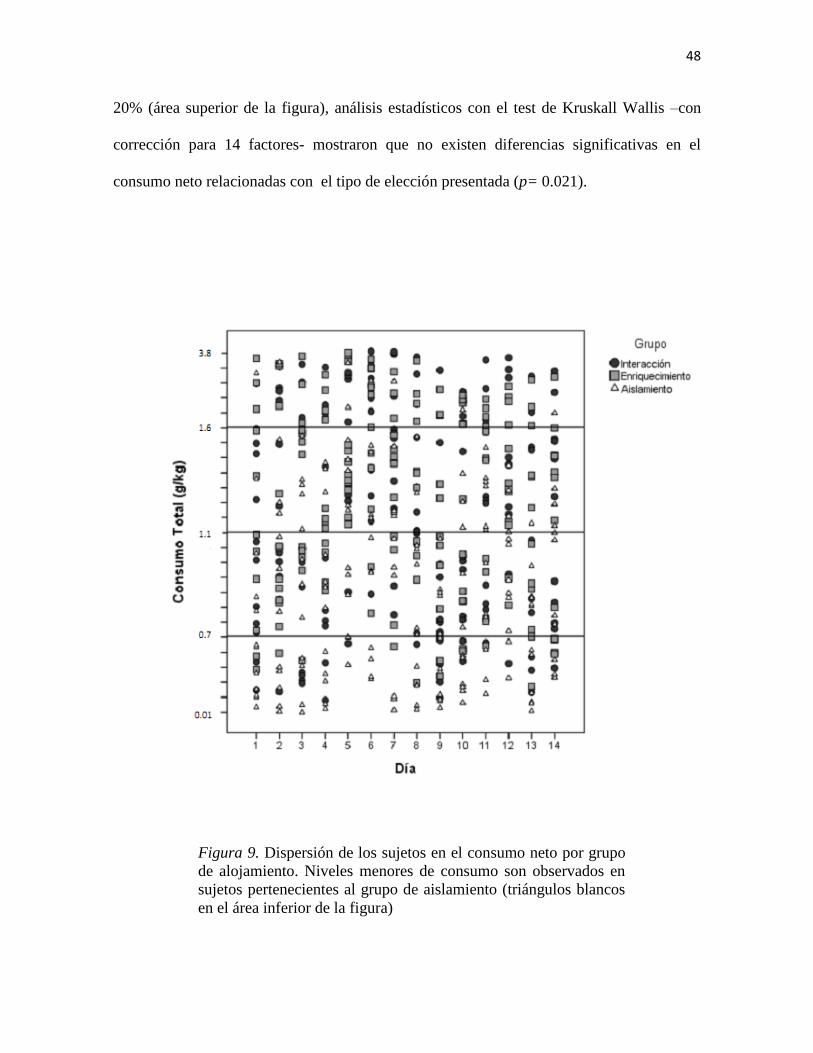
48
20% (área superior de la figura), análisis estadísticos con el test de Kruskall Wallis –con
corrección para 14 factores- mostraron que no existen diferencias significativas en el
consumo neto relacionadas con el tipo de elección presentada (p= 0.021).
Figura 9. Dispersión de los sujetos en el consumo neto por grupo
de alojamiento. Niveles menores de consumo son observados en
sujetos pertenecientes al grupo de aislamiento (triángulos blancos
en el área inferior de la figura)

49
Figura 10. Dispersión de los sujetos en el consumo neto por
de tipo de elección. No se encontraron diferencias en el
consumo dependientes de los pares de dosis presentadas
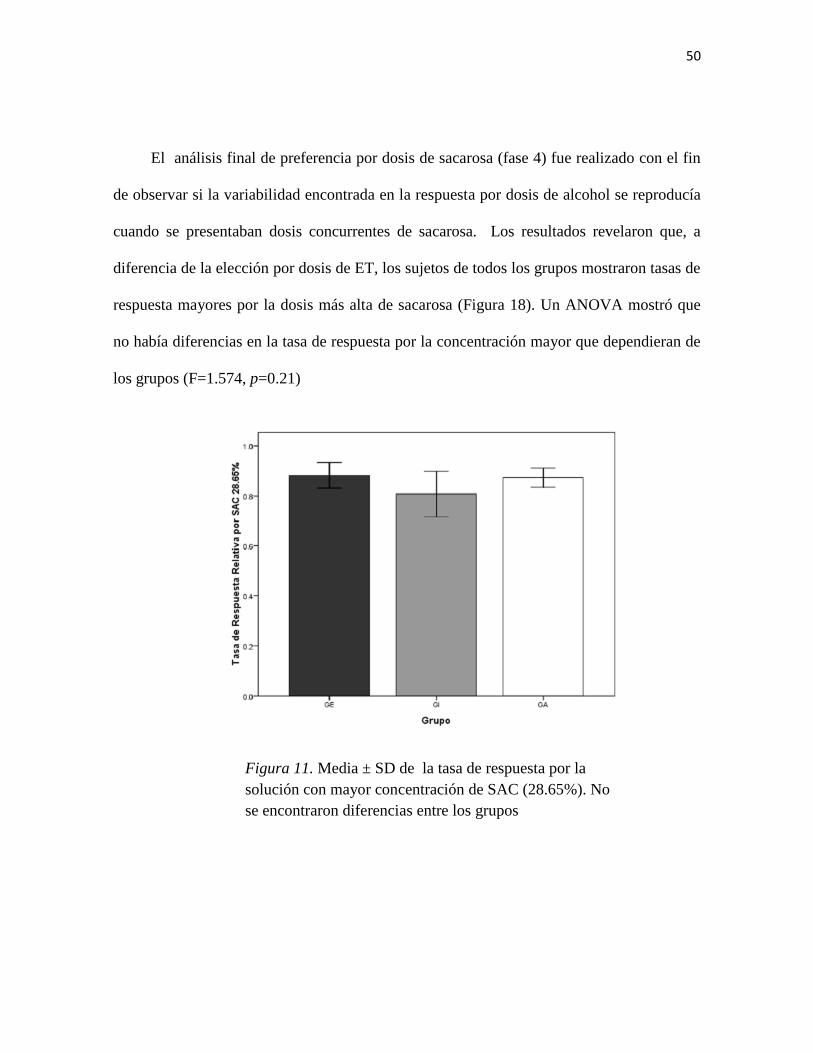
50
El análisis final de preferencia por dosis de sacarosa (fase 4) fue realizado con el fin
de observar si la variabilidad encontrada en la respuesta por dosis de alcohol se reproducía
cuando se presentaban dosis concurrentes de sacarosa. Los resultados revelaron que, a
diferencia de la elección por dosis de ET, los sujetos de todos los grupos mostraron tasas de
respuesta mayores por la dosis más alta de sacarosa (Figura 18). Un ANOVA mostró que
no había diferencias en la tasa de respuesta por la concentración mayor que dependieran de
los grupos (F=1.574, p=0.21)
Figura 11. Media ± SD de la tasa de respuesta por la
solución con mayor concentración de SAC (28.65%). No
se encontraron diferencias entre los grupos

51
Discusión
Tres grandes resultados fueron obtenidos en relación con los objetivos planteados: (1)
Las ratas de GE se auto-administran menos SAC que las ratas de GI y GA. (2) Las ratas de
GA beben significativamente menos ET que las ratas de GI y GE y (3) Las ratas mostraron
indiferencia ante dosis concurrentes de ET. Iniciaré discutiendo los resultados relacionados
con el consumo y elección de ET –que hacen parte de los objetivos originales del estudio- y
después discutiré los datos relacionados con el consumo de SAC.
Con respecto a las diferencias en el consumo de ET entre grupos, los resultados
mostraron que los sujetos aislados (GA) consumieron cantidades menores de ET que los
sujetos de GI y GE. Estos encuentros coinciden con la mayoría de los experimentos en la
línea de enriquecimiento, que sugieren que ambientes enriquecidos pueden ocasionar
niveles altos de consumo. (Rockman et al, 1989, Ellison, 1987, Fernández-Turuel, 2002);
sin embargo, divergen de los resultados de los experimentos descritos en la línea de
interacción –sin enriquecimiento-, que señalan que situaciones que favorecen
comportamientos sociales disminuyen el consumo.
Una posible explicación con respecto a esta divergencia es que la mayoría de
experimentos en los que la interacción es manipulada son realizados con grupos pequeños
de ratas (entre dos a cuatro sujetos). Alojamientos con esta cantidad de sujetos podrían no
haber permitido el establecimiento de jerarquías observado en condiciones semi-naturales
de interacción. Al no existir jerarquías de dominancia, los consumos extremos tendrían
menos probabilidad de aparecer, y la media de consumo grupal sería menor. De hecho, en

52
los experimentos de Wolffgramm (1991) con grupos de cuatro sujetos en la condición de
interacción, no se observa la variabilidad intra-grupal encontrada en otros experimentos, lo
que sería un indicio de que las jerarquías no fueron establecidas y explicaría los bajos
niveles de consumo.
En el caso del presente experimento ambos grupos (GI y GE) tenían 12 sujetos y
observaciones anecdóticas mostraron comportamientos asociados al establecimiento de
jerarquías. Si como afirma Ellison (1987), las jerarquías de dominancia favorecen
consumos extremos, deberíamos esperar que sujetos en GI y GE mostraran niveles altos
de consumo en relación a GA, que fue lo observado.
Una variable que pudo haber incrementado los niveles de consumo en GI y GE fue la
agresión entre sujetos. En diversas ocasiones fueron observados ataques agresivos en estos
grupos, de hecho, en dos ocasiones animales tuvieron que ser retirados por algunos días del
experimento por presentar heridas graves. Este tipo de observaciones indican que el tipo de
dominancia en los grupos era caracterizada por comportamientos agresivos, y las
agresiones, como fuentes de estrés social, podrían haber afectado el consumo de alcohol en
sujetos de estos grupos. Aunque esta suposición es respaldada por estudios que muestran
que situaciones de estrés social afectan la actividad dopaminérgica y el consumo de alcohol
de animales alojados grupalmente (Blanchard , McKittrick y Blanchard , 2001; Blanchard.,
Hori, y Blanchard, 1987), serían necesarios estudios adicionales con mediciones de
respuestas fisiológicas características del estrés para corroborar esta hipótesis.
Otros experimentos que observan el consumo de alcohol en condiciones de
enriquecimiento y condiciones de aislamiento han reportado niveles de consumo mayores

53
en ratas en aislamiento (Deehan, 2007; Bernardes, 2008). Para explicar la divergencia con
estos estudios fueron revisadas las medidas de las cajas de sujetos en aislamiento y de
enriquecimiento, y se encontró que las cajas de aislamiento tenían un área por rata
significativamente menor que las cajas de enriquecimiento. Por ejemplo, en el experimento
de Deehan (2007) el área por rata en las cajas de aislamiento era de 8160 cm3, mientras que
las cajas de enriquecimiento era de 32400 cm3, esto quiere decir que las ratas en ambientes
enriquecidos tenían un espacio por rata casi cuatro veces mayor que las ratas en
aislamiento. En este caso la de movimiento –utilizada en experimentos para generar estrés-
podría haber sido una variable más relevante que el aislamiento (Rockman, 1986). En el
presente experimento esta variable fue controlada de manera que el espacio por rata se
homogeneizó para todas las condiciones.
Una observación importante relacionada con el consumo (g/kg), fue que, tanto en la
fase de entrenamiento como en la fase de elección, se encontró una gran variabilidad dentro
de todos los grupos en la cantidad de ET ingerida. De esta manera, en cada condición de
alojamiento existían quienes bebían menos de 0.5 g/kg (menores a la dosis activas de ET),
y quienes bebían cantidades superiores a 4g/kg (consumo identificado como alto por
Wolffgramm, 1991). En la condición de enriquecimiento, la variabilidad en consumo de ET
fue mayor que en los otros grupos, pues se encontraban bebedores extremos que ingerían
cantidades superiores a 7g/kg al día. Este resultado coincide con las observaciones
realizadas por Ellison (1987) que señalan que condiciones de enriquecimiento –debido a la
existencia de recursos para competir y al desarrollo jerarquías de dominancia- favorecen
consumos heterogéneos de ET dentro de la población. En sus estudios, Ellison realizó
análisis etológicos que demostraban que niveles extremos de ingesta de ET eran

54
encontradas en ratas subordinadas, mientras que ratas dominantes mostraban menores
consumos de ET. En este estudio no se realizaron dichas evaluaciones etológicas, sin
embargo, es propuesto que estudios posteriores con condiciones de alojamiento grupales
incluyan análisis de jerarquía para dar cuenta de la variabilidad en la ingesta.
La variabilidad en la ingesta de ET encontrada en los grupos reproduce los datos
epidemiológicos de uso de alcohol en humanos, en los que se observan hábitos
extremamente diferentes de consumo en una misma población. En este sentido, como lo
señala Carroll (1996), la variabilidad intra-grupo es un elemento que ratifica la validez de
los modelos con animales no humanos en el estudio del consumo y la dependencia de
psicoactivos.
Con respecto a la elección de dosis de ET, los resultados mostraron que las ratas no
tenían preferencia por ninguna dosis de ET. La pregunta que surgió entonces fue, si los
sujetos son indiferentes a las dosis presentadas, ¿qué variable está controlando su
comportamiento de consumo? Como se mostró en el apartado de los resultados, posibles
fuentes de control fueron encontradas, e.g. control por posición, por consumo total de ET, o
por dosis. La variabilidad en las fuentes de control y el hecho de que estos tipos de control
se encontraran dispersos en todos los grupos fueron una prueba de que la ausencia de
preferencia no se debía a errores metodológicos (podría haber sido el caso si todos los
animales tuvieran un sesgo de posición). Las pruebas de elección entre dosis de sacarosa
también descartaron esta posibilidad, pues al cambiar el reforzador los sujetos de todos los
grupos comenzaron a responder por la dosis más alta.

55
La pregunta siguiente –que continúa vigente- fue ¿por qué ratas en una situación de
elección con dosis concurrentes de ET no desarrollan preferencias? Una primera hipótesis
para responder esta pregunta, estuvo relacionada con el tiempo transcurrido de la sesión,
así, pensamos que podrían haber preferencias por alguna dosis en los primeros minutos de
la sesión y que con el paso del tiempo y los efectos del ET, los sujetos dejarían de
responder por la dosis y estarían bajo el control de otras variables. Como se mostró en los
resultados, análisis de las sesiones por bloques de 5 minutos evidenciaron que los sujetos
paraban de responder antes de los 10 minutos de la sesión, así que las diferencias
observadas no se debían al tiempo transcurrido.
Otras posibilidades de análisis deben ser consideradas en experimentos futuros. En
primer lugar, podría ser aumentado el número de sesiones de elección con el fin de observar
si los animales desarrollarían preferencias por dosis conforme el número de sesiones
aumentaba. Los experimentos de Wolffgramm y Heyne (1991) en los que se describen
preferencias por dosis en consumo libre, los animales son expuestos a las soluciones por 70
semanas, así que es posible que las ratas demoren un tiempo específico para desarrollar
algún tipo de preferencia. El incremento del número de sesiones tendría además otra
ventaja, posibilitaría el análisis de patrones de consumo y elección en series temporales.
Estos análisis permitirían observar si el comportamiento de elección de los sujetos se veía
afectado por el consumo de los días anteriores. Esto sería útil principalmente para analizar
explicar el comportamiento de los sujetos en los que no se encontró una fuente de control.
Una posibilidad para futuras investigaciones es el aumento del valor del programa de
razón, pues existe la posibilidad de que no se presenten preferencias por dosis de alcohol
cuando son utilizados programas de refuerzo con requerimientos bajos (Meish, 2001). En

56
experimentos realizados monos reshus Meish y Lemiere (1990) encontraron que cuando los
sujetos se enfrentaban a situaciones de elección entre una solución de pentobarbital+
alcohol y una solución de pentobarbital puro en programas concurrentes de razón fija
idénticos de valores bajos, no se observaban preferencias por ninguna de las soluciones; sin
embargo, con aumentos consecutivos en los programas de razón los sujetos preferían la
solución de pentobarbital+alcohol.
Con respecto a la autoadministración de SAC, los resultados sugieren que en
condiciones de enriquecimiento disminuye el consumo de SAC pues los sujetos el grupo
GE respondieron significativamente menos por SAC que los sujetos de los grupos GA y GI.
Este resultado coincide con los de Brenes, Rodríguez, y Fornaguera (2006) que muestran
como ratas criadas en ambientes enriquecidos beben menos SAC que ratas alojadas
individualmente o en tríadas, en pruebas de preferencia entre SAC32% y agua. Ellos
explican sus resultados como un efecto antidepresivo generado por las condiciones de
enriquecimiento. Infieren el efecto antidepresivo del hecho de que ratas en aislamiento con
comportamientos asociados a estados de depresión beben más sacarosa (Van den Berg,
Pijlman, Koning, Diergaarde, Ree, y Spruijt, 1999; Brenes y Fornagueira, 2009).
La economía del comportamiento ofrece otra explicación más parsimoniosa de este
hallazgo con el concepto de precio relativo. Para ratas en enriquecimiento ambiental con
acceso a múltiples reforzadores, el precio relativo de la sacarosa es mayor que para aquellas
con menos reforzadores disponibles, por eso la tasa de respuesta por SAC es menor –están
dispuestas a pagar menos-. Para ratas con pocos reforzadores disponibles la sacarosa tiene
un precio relativo menor que para aquellas con múltiples reforzadores en su alojamiento y
por ello muestran tasas de respuesta mayor.

57
La sacarosa es considerada un reforzador poderoso porque sustenta tasas de
respuesta altas cuando es ofrecida a ratas, debido a esto, es una de las sustancias más
utilizadas como reforzadoras en estudios de motivación vs anhedonia. Sin embargo, pocos
estudios evalúan qué tipo de variables ambientales afectan el consumo de SAC. En el caso
de los efectos del enriquecimiento, son encontrados pocos estudios que evalúen como varía
la autoadministración de SAC, cuando ratas son alojadas en condiciones diferenciales, por
lo que son necesarias investigaciones que evalúen más profundamente la interacción entre
estas variables.

58
CONCLUSIONES
La Real Academia Española define a la adicción como “el hábito de quien se deja
dominar por el uso de alguna o algunas drogas tóxicas”, sin embargo, estudios desde el
análisis comportamental muestran que el control que una droga puede ejercer sobre el
organismo depende del contexto del cual el sujeto hace parte y de los reforzadores
disponibles en él. Basándose en este hallazgo, Rachlin (2000) propuso su teoría de la
adicción relativa, según la cual, la interacción social y el consumo de psicoactivos
funcionan como sustitutos, de manera que al aumentar la disponibilidad de uno, disminuye
el consumo del otro. Múltiples estudios que evalúan la relación entre interacción social y
consumo de drogas han reportado resultados ambiguos en cuanto a cómo cambia el
consumo al aumentar la interacción, de manera que la relación de sustitución entre estos
reforzadores, postulada por Rachlin (2000), no ha sido verificada.
En el presente estudio se propuso que las divergencias encontradas en la literatura
podrían provenir de las diferencias entre los estudios en cuanto al tipo de interacción
permitida (e.g. número de sujetos, condiciones ambientales de enriquecimiento). También
se consideró que si, como otros autores proponen, las ratas muestren preferencias por dosis
altas o bajas dependiendo del tipo de alojamiento, utilizar una única dosis habría afectado
los resultados de dichos estudios.
Los resultados de este experimento mostraron que ratas en condiciones de aislamiento
beben significativamente menos ET cuando son comparadas con grupos de interacción y
enriquecimiento, mientras que las ratas en estas dos últimas condiciones (GE y GI) no
difieren en los niveles de consumo de ET. Estos resultados tienen dos implicaciones cuando

59
son analizados en el contexto de estudios anteriores. En primer lugar, los efectos
diferenciales de la interacción –sin enriquecimiento- y del enriquecimiento ambiental sobre
el consumo de ET, observados en la literatura, no fueron encontrados cuando los dos tipos
de interacción fueron comparados en un mismo experimento. Esto sugiere, en línea con los
planteamientos de Ellison (1987) que en grupos grandes el establecimiento de jerarquías
favorece niveles extremos de consumo, y que este efecto no está directamente relacionado
específicamente con la situación de enriquecimiento.
En segundo lugar, el resultado de que los sujetos de GA beben menos ET contrasta
con algunos estudios que sugieren que el aislamiento como fuente de estrés aumenta el
consumo. No obstante, en estos estudios –a diferencia del presente- no se especifica el
control sobre variables diferentes al aislamiento, como la restricción de movimiento, que
pueden afectar significativamente los resultados observados.
Con respecto a la preferencia por dosis, los resultados mostraron que las ratas eran
indiferentes a concentraciones diferentes de ET presentadas concurrentes. Análisis
adicionales son necesarios para comprender el porqué de este hallazgo, sin embargo, es
probable que la indiferencia observada se deba al requerimiento de los programas
concurrentes, pues como Mesch (2001) lo señala con valores de programas bajos pueden
afectar el desarrollo de preferencias.
Los resultados encontrados ratifican el carácter relativo de los recursos que
usualmente son indicados como reforzadores. A pesar de que en muchas situaciones la
interacción social es reforzadora, esta función no es intrínseca, de modo que sus efectos
dependen del contexto del que el organismo hace parte. En ambientes sociales con muchos

60
sujetos las situaciones de interacción pueden resultar aversivas, por lo que la interacción
social no funcionaría como sustituto del consumo de sustancias, al contrario, como se
observó en el presente estudio, podría funcionar como complemento.

61
REFERENCIAS
Adams, N. y Oldham, T.D. Seminatural housing increases subsequent ethanol intake in
male Maudsley Reactive rats. J. Stud. Alcohol. 1996, 57, 349-351.
Ahmed, S., H. (2010). Validation crisis in animal models of drug addiction: Beyond non-
disordered drug use toward drug addiction. Neuroscience and Biobehavioral Reviews,
35, 172–184.
Allison, J. (1983). Behavioral Economics. New York: Praeger Publishers.
Bardo MT, Donohew RL ,y Harrington NG. (1996)Psychobiology of novelty seek-ing and
drug seeking behavior. Behav Brain Res;77:23 – 43.
Battistich, V., & Hom, A. (1997). The Relationship between students´sense of their school
as a community and their involvement in problem behaviors. American journal of
public health
Bennett EL, Diamond MC, Krech D & Rosenzweig, MR (1964) Chemical and anatomical
plasticity of brain. Science.: 146, 610–619.
Bernardes, AMT (2008) Efeitos do enriquecimento ambiental na autoadministração oral de
álcool em ratos. Tesis de maestria no publicada. Universidade de São Paulo. Brasil.
Blanchard, R.J., Hori, K., y Blanchard, D.C. (1987). Social structure and ethanol
consumption in the laboratory rat. Pharmacology Biochemistry & Behavior, 28, 437–
442.
Blanchard , R. J. McKittrick , C. R. y Blanchard , D. C. (2001). Animal models of social
stress: Effects on behavior and brain neurochemical systems. Physiology and
Behavior, 73, 261-271.
Bowling SL y Bardo MT. (1994) Locomotor and rewarding effects of amphetaminein
enriched, social and isolate reared rats. Pharmacology, Biochemistry &
Behavior;48:459 – 64.

62
Bozarth, M.A., Murray, A. y Wise, R.A., (1989). Influence of housing conditions on the
acquisition of intravenous heroin and cocaine self-administration in rats.
Pharmacology Biochernistry & Behavior, 33, 903-907.
Brenes, J.C., Rodriguez, O., y Fornaguera, J. (2006). Factor analysis of forced swimming
test, sucrose preference test and open field test on enriched, social and isolated reared
rats. Behavior and Brain Research, 169, 57–65.
Brenes, J.C. y Fornaguera, J. (2009). The effect of chronic fluoxetine on social isolation-
induced changes on sucrose consumption, immobility behavior, and serotonin and
dopamine function in hippocampus and ventral striatum. Behavior and Brain
Research, 165, 187–196.
Brown, V., & Riley, M. (2005). Social support, drug use, and employment among low-
income women. The american journal of drug and alcohol abuse , 203-223.
Carroll, M. E. y Lac, S.T. (1993). Autoshaping i.v. cocaine self-administration in rats:
effects of nondrug alternative reinforcers on acquisition. Psychopharmacology, 110,
5–12.
Carroll, M. E. y Lac, S. T.(1998). Dietary additives and the acquisition of cocaine self-
administration in rats. Psychopharmacology, 137, 81–89.
Carroll, M.E., Lac, S.T., y Nygaard, S.L. (1989). A concurrently available nondrug
reinforcer prevents the acquisition or decreases the maintenance of cocaine-reinforced
behavior. Psychopharmacology, 97, 23–29.
Daoust, M., Chretien, P., Moore, N., Saligaut, C., Lhuintre, J.P. y Boismare, F. (1985).
Isolation and striatal (3h) serotonin uptake: Role of the voluntary intake of ethanol in
rats. Pharmacology Biochemistry & Behavior, 22, 205–208.
Deatherage, G., (1972). Effect of housing density on alcohol intake in the rat. Physiology
and Behavior, 9, 55–57.

63
Deehan, G.A., Cain, M.E. y Kiefer, S.W. (2007).Differential rearing conditions alter
operant responding for ethanol in outbred rats. Alcoholism: Clinical and
Experimental Research, 31, 1692–1698.
Ding, Y., Kang, L., Li, B. y Ma, L. (2005). Enhanced cocaine self-administration in adult
rats with adolescent isolation experience. Pharmacology Biochemistry and
Behavior, 82, 673–677.
Doremus, T. L., Brunell, S. C., Rajendran, P. y Spear, L. P. (2005). Factors influencing
elevated ethanol consumption in adolescent relative to adult rats. Alcoholism:
Clinical and Experimental Research, 29, 1796–1808.
Ehlers, C.L., Walker, B.M., Pian, J.P., Roth, J.L. y Slawecki, C.J. (2007). Increased alcohol
drinking in isolate-housed alcohol-preferring rats. Behavioral Neuroscience, 121,
111–119.
Ellison, G., Levy, A. y Lorant, N. (1983). Alcohol-preferring rats in colonies show
withdrawal, inactivity, and lowered dominance. Pharmacology Biochemistry and
Behavior, 18 , 565–570.
Ellison, G., Daniel, F. y Zoraster, R. (1979). Delayed increases in Alcohol consumption
occur in rat colonies but not in isolated rats after injections of monoamine
neurotoxins. Experimental Neurology, 65, 608-615.
Ellison, G. (1981). A novel animal model of alcohol consumption based on the
development of extremes of ethanol preference in colony-housed by not isolated rats.
Behavior and Neural Biology, 31, 324-330.
Ellison, G. (1987). Stress and alcohol intake: The socio-pharmacological approach.
Physiology & Behavior, 40 , 387–392.
Fernandez-Teruel, A., Driscoll, P., Gil, L., Aguilar, R., Tobea, A., Escorihuela, R. M.
(2002). Enduring effects of environmental enrichment on novelty seeking, saccharin
and ethanol intake in two rat lines (RHA/Verh and RLA/Verh) differing in incentive-
seeking behavior. Pharmacology Biochemistry and Behavior, 73 , 225-231.

64
Fernández-Vidal, J. M. y Molina, J. C. (2004). Socially mediated alcohol preferences in
adolescent rats following interactions with an intoxicated peer. Pharmacology
Biochemistry and Behavior, 79, 229–241.
Fisher, E. B., Jr., Lichtenstein, E., y Haire-Joshu, D. (1993). Multiple determinants of
tobacco use and cessation. In Orleans, C. T. y Slade, J. D. (Eds.), Nicotine Addiction:
Principles and Management (pp. 59-88). New York: Oxford University Press.
Füllgrabe, M.W., Frauenglass, S., Routh, D., Pantin, H., & Mason, C. (1997). Family
support decreases influence of deviant peers on hispanic adolescents´substance use.
Journal of clinical child psychology , 15-23.
Greenwald, M., & Hursh, S. (2006). Behavioral economic analysis of opioid consumption
in heroin-dependent individuals: Effects of unit price and pre-session drug supply.
Drug and Alcohol Dependence , 35-48.
Hartnoll, R. (1991). The relevance of behavioural models of drug abuse and dependence
liabilities to the understanding of drug misuse in humans. In P. Willner (ed.).
Behavioural models in psychopharmacology: theoretical, industrial and clinical
perspectives. Chapter 19, 503-519. Cambridge: Cambridge University Press.
Hatch, A., G. Wiberg, T. Balazs and H. Grice (1963). Long-term isola- tion stress in
rats. Science 142: 507.
Heyman, G.M. (1996). Resolving the contradictions of addiction. Behavior and Brain
Science, 19, 561–610.
Heyne, A. (1996). The Developmentent of Opiate Addiction in the Rat. Pharmacology,
Biochemistry and Behavior , 11-25.
Heyne, A., & Wolffgramm, J. (1998). The developmente of addiction to d-anphetamine in
an animal model:same principles as for alcohol and opiate. Psychopharmacology ,
510-518
Higgins, S.T., Heil, S.H., y Lussier, J.P. (2004). Clinical implications of reinforcement as a
determinant of substance use disorders. Annual Review of Psychology, 55, 431–461.

65
Holter, S.M., Linthorst, A.C., Reul, J.M.H.M., y Spanagel, R. (2000). Withdrawal
symptoms in a long-term model of voluntary alcohol drinking in Wistar rats.
Pharmacology Biochemistry and Behavior, 66, 143-151.
Hursh, S. (1991). Behavioral Economics of Drug Self.Administration and Drug Abuse
Policy. Journal of Experimental Analysis of Behavior , 377-393.
Hursh, S. (1993). Behavioral Economics of Drug Self- Administration: An Introduction.
165-172.
Hursh, S. (2005). The Economics of Drug Abuse: A Quantitative Asessment of Drug
Demand. Molecular Interventions , 20-28.
Hutchinson, E., Avery, A & Vandewoude, S. Enviromental Enrichment for laboratory
animals.ILAR Journal 46(2) 148-161
Juárez, J. y Vázquez-Corteás, C. (2003). Alcohol intake in social housing and in isolation
before puberty and its effects on voluntary alcohol consumption in adulthood.
Developmental Psychobiology, 43, 200-207.
Mankiw, G. (2004). Principios de Economía. Madrid: McGraw-Hill/Interamérica de
España, S.A.U.
Mattick RP, Breen C, Kimber J, Davoli M. (2008). Terapia de mantenimiento con
metadona versus la no administración de tratamiento de sustitución con opiáceos para
la dependencia de opiáceos (Revisión Cochrane traducida). En: La Biblioteca
Cochrane Plus, 2008 Número 2. Oxford: Update Software Ltd. Disponible en:
http://www.update-software.com. (Traducida de The Cochrane Library, 2008 Issue 2.
Chichester, UK: John Wiley & Sons, Ltd.).
Meisch , R. A. y Lemaire , G. A. (1988). Oral self-administration of pentobarbital by rhesus
monkeys: Relative reinforcing effects under concurrent fixed-ratio schedules. Journal
of the Experimental Analysis of Behavior, 50, 75-86.

66
Meisch, R.A. y Lemaire, G.A. (1989). Oral self-administration of pentobarbital by rhesus
monkeys: maintenance of behavior by different concurrently available volumes of
drug solution. Journal of the Experimental Analysis of Behavior, 52 , 111–126.
Meisch, R. A. y Lemaire, G. A. (1990). Reinforcing effects of a pentobarbital–ethanol combination
relative to each drug alone. Pharmacology Biochemistry and Behavior, 35, 443–450.
Meisch, R. A. (2001). Oral drug self-administration: An overview of laboratory animal
studies. Alcohol, 24, 117–128.
Moncek F., Duncko R., Johansson B.B., and Jezova D. (2004). Effect of environmental
enrichment on stress related systems in rats. Journal of. Neuroendocrinoly. 16:
423-431.
Olsson, A.S. y Dahlborn, K. (2002). Improving housin conditions for laboratory mice: a
revew of „environmental enrichment‟.Laboratory Animals, 36, 243–270.
Parker, L.F. y Radow, B.L. (1974). Isolation stress and volitional ethanol consumption in
the rat. Physiology and Behavior, 12, 1–3.
Perez, C., Canal, J.R., Domingues, E. Campillo, J.E. Guillen, M. y Torres, M.D. (1997).
Individual housing influences certain biochemical parameters in the rat. Laboratory
Animals, 31, 357–361.
Pham TM, Winblad B, Granholm A, Mohammed AH. Environmental influences on brain
neurotrophins in rats. Pharmacology Biochemistry and Behavior. 2002; 73:167-175..
Rachlin, H. (2000). The Science of Sel Control. London: Harvard University Press.
Robins, L. N. (1975). Drug treatment after return in vietnam veterans. Highlights of the 20th
annual conference, veterans administration studies in mental health and behavioral
sciences. Perry point, md: central np research laboratory.
Rockman, G.E., Hall, A. y Glavin, G.B. (1986). Effects of restraint stress on voluntary
ethanol intake and ulcer proliferation in rats. Pharmacology Biochemistry and
Behavior, 25, 1083–1087.

67
Rockman , G. E. Hall , A. M. Markert , L. E. y Glavin , G. B. (1988). Influence of rearing
conditions on voluntary ethanol intake and response to stress in rats. Behavioral and
Neural Biology, 49, 184-191.
Rockman, G.E., Gibson, J.E. y Benarroch, A. (1989). Effects of environmental enrichment
on voluntary ethanol intake in rats. Pharmacology Biochemistry and Behavior, 34,
487–490.
Rosenzweig, M. R. y Bennett, E. L., Hebert, M., and Morimoto, H. (1978). Social
grouping cannot account for cerebral effect of enriched environment. Brain
Research, 153, 563-576.
Rosenzweig, M. R. y Bennett, E. L. (1969). Effects of differential environments on brain
weights and enzyme activities in gerbils, rats and mice. Develop. Psychobiology
Sanchis-Segura, C. y Spanagel, R. (2006). Behavioural assessment of drug reinforcement
and addictive features in rodents: an overview. Addiction Biology, 11, 2-38.
Samson, H. H., Pfeffer, A. O., & Tolliver, G. (1988). Oral ethanol self-administration in
rat: Models of alcohol-seeking behavior. Alcoholism: Clinical and Experimental
Research, 12, 591-598.
Samson, H. H., Tolliver, G. A., y Roehrs, T. A. (1983). Ethanol reinforced responding in
the rat: relation of ethanol introduction to later ethanol responding. Pharmacology
Biochemistry Behavior, 18, 895–900.
Schenk, S., Gorman, K., y Amit. Z (1990). Age-dependent effects of isolation housing on
the self-administration of ethanol in laboratory rats. Alcohol, 7, 321–326.
Siegel, D.J. (2001). Toward an interpersonal neurobiology of the developing mind:
attachment relationships, “mindsight” and neural integration. Infant mental health
journal, 22, 67–94.
Strain, E., Stitzer, M., Leibson, I., y Bigelow, G. (1993). Dose-response effects of
methadone in the treatment of opioid dependence. Annals of Internal Medicine, 119,
23–7.

68
Thorsell , A. Slawecki , C. J. Khoury , A. Matheé , A. A. Ehlers , C. L. (2005). Effect of
social isolation on ethanol consumption and substance P/neurokinin expression in
Wistar rats. Alcohol, 36, 91-97.
Tucker, J. (2005). Contributions of Behavioral Economics for Understanding and
Resolving Substance Abuse Disorders. 35-47.
Vanichseni, S., Wongsuwan, B., Choopanya, K., y Wongpanich, K. (1991). A controlled
trial of methadone in a population of intravenous drug users in Bangkok: implications
for prevention of HIV. International Journal of the Addictions, 26, 1313–20.
Vuchinich, R.E. y Tucker, J.A. (1996). The molar context of alcohol abuse. in: Green, L. y
Kagel, J. (Eds.), Advances in behavioral economics, vol. 3, Ablex, Norwood, NJ, 133–162.
Wolffgramm, J. y Heyne, A. (1991). Social behavior, dominance, and social deprivation of
rats determine drug choice. Pharmacology Biochemistry and Behavior, 38, 389–399.
Wolffgramm, J., & Turyabahika-Thyen, K. (2006). Loss of flexibility in alcohol taking rats:
Promoting Factors. European Addiction Research , 210-221.
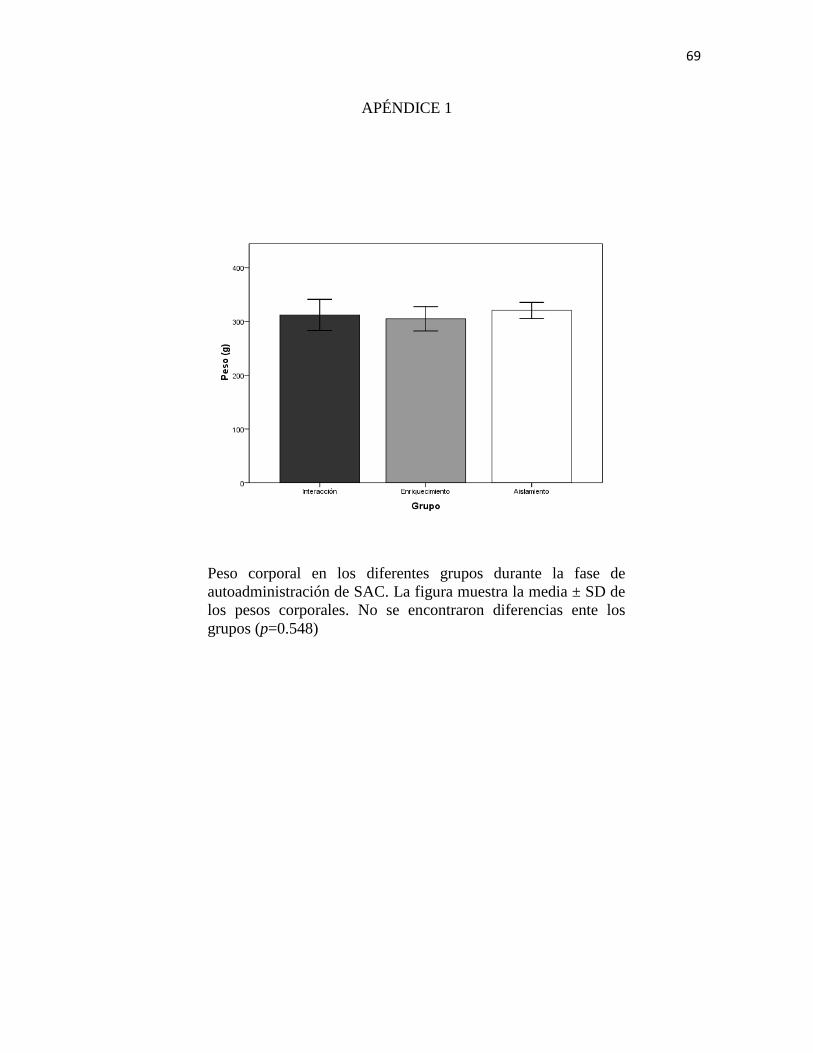
69
APÉNDICE 1
Peso corporal en los diferentes grupos durante la fase de
autoadministración de SAC. La figura muestra la media ± SD de
los pesos corporales. No se encontraron diferencias ente los
grupos (p=0.548)

70
APÉNDICE 2
Valores asociados a las pruebas de Kruskall Wallis para consumo neto de ET (g/kg)
entre los grupos de alojamiento, durante la fase de entrenamiento
% ET % SAC gl H Significancia
2,5 10 2 0.628 p=0.73
5 5 2 11.73 p˂0.01*
10 5 2 7.88 p=0.01*
10 2,5 2 27.9 p˂0.001**
10 0,25 2 63.34 p˂0.001**
*valores p significativos con la corrección de Bonferroni para 5 factores.

71
APÉNDICE 3
Número de sujetos en cada grupo clasificados por la posible fuente de control del
comportamiento de consumo durante la fase de elección.
Grupo de Alojamiento
Control Interacción Enriquecimiento Aislamiento
Posición 5 4 7
ET Neto ingerido 2 1 2
Dosis 1 2 0
Desconocido 3 5 3
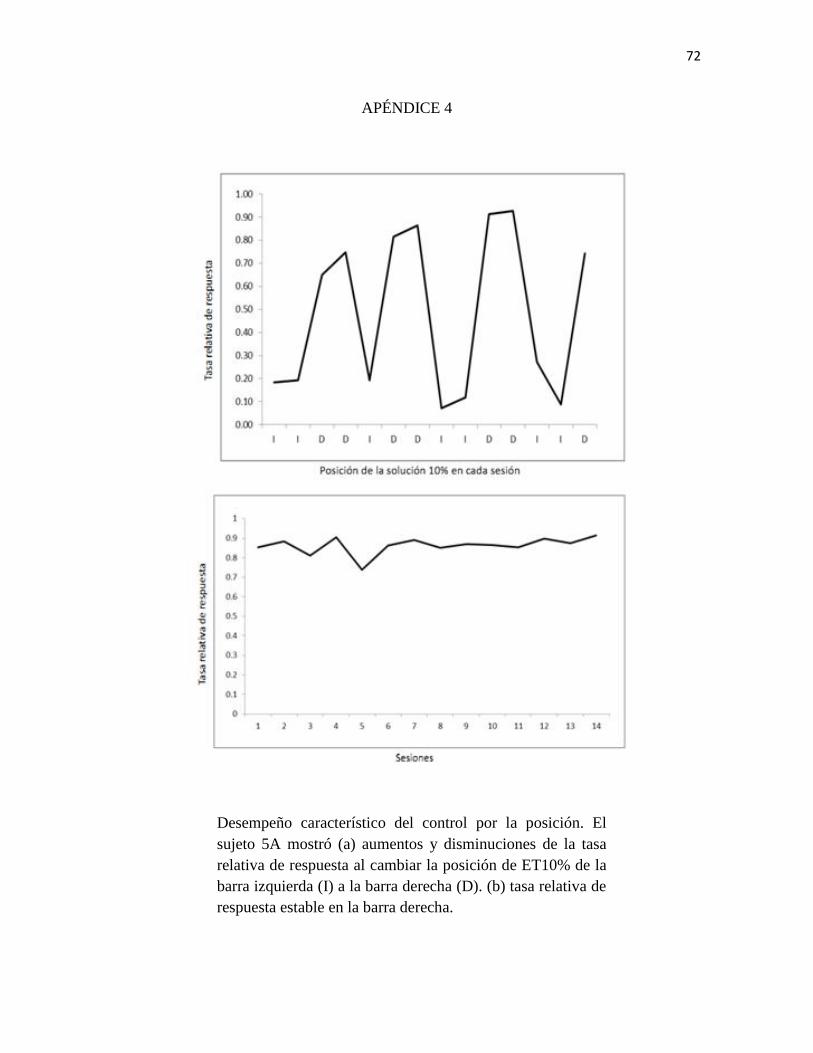
72
APÉNDICE 4
Desempeño característico del control por la posición. El
sujeto 5A mostró (a) aumentos y disminuciones de la tasa
relativa de respuesta al cambiar la posición de ET10% de la
barra izquierda (I) a la barra derecha (D). (b) tasa relativa de
respuesta estable en la barra derecha.

73
APÉNDICE 5
Desempeño típico de control por la cantidad neta de ET para
el sujeto 6E en una situación de elección ET5%-ET10%. A
pesar de las variaciones en los patrones de elección por la
dosis del 5% (a), el consumo total de ET (g/kg) se mantiene
en rangos estables (b).

74
APÉNDICE 6
Desempeño típico del control por la dosis de ET para el sujeto 8E
en una situación de elección ET5%-ET20%. (a) A partir de la sexta
sesión la tasa de respuesta relativa por ET20% está ente .8 y 1,
además el consumo neto de ET permanece estable.

75

76