
UNIVERSIDAD DE COLIMA - digeset.ucol.mxdigeset.ucol.mx/tesis_posgrado/Pdf/Raymundo Flores... ·...
54
UNIVERSIDAD DE COLIMA MAESTRÍA EN CIENCIAS, ÁREA: CIENCIAS AGRÍCOLAS Y FORESTALES “EFECTO DE LA VARIEDAD DE MAIZ SOBRE EL DESARROLLO Y SUSCEPTIBILIDAD DE LARVAS DE Spodoptera frugiperda (Lepidoptera: Noctuidae) A Bacillus thuringiensis” TESIS QUE PRESENTA RAYMUNDO FLORES HUESO COMO REQUISITO PARCIAL PARA OBTENER EL GRADO DE MAESTRO EN CIENCIAS, ÁREA: CIENCIAS AGRÍCOLAS Y FORESTALES ASESOR Dr. Roberto Lezama Gutiérrez TECOMÁN, COLIMA, MÉXICO Junio de 2000.
Transcript of UNIVERSIDAD DE COLIMA - digeset.ucol.mxdigeset.ucol.mx/tesis_posgrado/Pdf/Raymundo Flores... ·...

UNIVERSIDAD DE COLIMA
MAESTRÍA EN CIENCIAS, ÁREA: CIENCIAS AGRÍCOLAS Y FORESTALES
“EFECTO DE LA VARIEDAD DE MAIZ SOBRE EL DESARROLLO Y SUSCEPTIBILIDAD DE LARVAS DE Spodoptera frugiperda (Lepidoptera:
Noctuidae) A Bacillus thuringiensis”
TESIS
QUE PRESENTA
RAYMUNDO FLORES HUESO COMO REQUISITO PARCIAL PARA OBTENER EL GRADO DE MAESTRO
EN CIENCIAS, ÁREA: CIENCIAS AGRÍCOLAS Y FORESTALES
ASESOR Dr. Roberto Lezama Gutiérrez
TECOMÁN, COLIMA, MÉXICO Junio de 2000.

“EFECTO DE LA VARIEDAD DE MAIZ SOBRE EL DESARROLLO Y SUSCEPTIBILIDAD DE LARVAS DE
Spodoptera frugiperda (Lepidoptera: Noctuidae) A Bacillus thuringiensis”
Raymundo Flores Hueso Facultad de Ciencias Biológicas y Agropecuarias de la Universidad de Colima,
Apartado Postal No. 36, C.P. 28100, Tecomán, Colima.
RESUMEN
El gusano cogollero del maíz, Spodoptera frugiperda (J. E. Smith) Lepidoptera:
Noctuidae), es una plaga importante en el cultivo del maíz; ataca a la planta en
todas las etapas de desarrollo (crecimiento, floración y fructificación), cuando
las infestaciones son altas puede resultar la pérdida total del cultivo. El control
químico es el principal método de combate de este insecto, con sus respectivos
efectos sobre el ambiente y la salud; sin embargo, dentro de un programa de
manejo integrado de plagas, con miras al desarrollo de una agricultura
sustentable, el uso de variedades resistentes y el control microbiano,
constituyen dos principales alternativas para disminuir el uso de plaguicidas.
En este estudio se evaluó el efecto de la variedad de maíz sobre el desarrollo y
la susceptibilidad de larvas de S. frugiperda a la bacteria Bacillus thuringiensis
(Berliner), variedad Kenyae, serotipo 007. En total se evaluaron 13 variedades,
6 criollos y 7 híbridos; se encontró que cuando las larvas se alimentaron con
hojas de los criollos Uruapeño y Grano ancho, los valores de CL50 de B.
thuringiensis fueron 0.087 y 0.097 mg/ml, respectivamente y cuando las larvas
fueron alimentadas con los criollos Grano rojo, Grano pinto, Tampiqueño olote
delgado y Grano negro, los valores de CL50 oscilaron entre 0.101 y 0.180
mg/ml. Cuando las larvas se alimentaron con follaje de los híbridos V-526, P-
3288 y P-3086, los valores de CL50 de Bt fueron 0.072, 0.073 0.095 y mg/mI,
respectivamente; con los híbridos B-810, Ceres 7, H- 910 y amarillo forrajero,
los valores de CL50 encontrados fluctuaron entre 0.109 0.181 mg/ml.

Por otro lado, al evaluar la susceptibilidad de larvas, de tercero y quinto estadio
a la bacteria B. thuringiensis, alimentadas con follaje de las variedades de maíz
criollo o híbrido, en los cuales las larvas neonatas presentaron mayor y menor
susceptibilidad a Bt, se encontró que los valores de CL50 de la bacteria se
invirtieron, con respecto a la susceptibilidad encontrada en larvas neonatas, de
acuerdo a las variedades con las cuales fueron alimentadas las larvas.
También se encontró que el valor de la CL50 de la bacteria se incrementó de
acuerdo a la edad de la larva sin importar la variedad; y también, que el peso
de las larvas y pupas varía de acuerdo la variedad de la cual se alimentan.
Los resultados de esta investigación demuestran que la variedad de maíz
afecta el desarrollo y peso de larvas y pupas, así como la susceptibilidad de las
larvas a la B. thuringiensis. Así mismo, dentro de la perspectiva de utilizar a Bt
en interacción con la resistencia vegetal, dentro de un programa de manejo
integrado, se debe considerar el valor de CL50 de la bacteria, de acuerdo a la
edad de la larva y la variedad de maíz, cultivado.

CONTENIDO
Página RESUMEN……………………………………………………………………………… I
CONTENIDO……………………………………………………………………………. III
LISTA DE CUADROS………………………………………………………………….. V
I INTRODUCCIÓN……………………………………………………………………… 1
ll ANTECEDENTES…………………………………………………………………….. 7
2.1 El cultivo de maíz………………………………………………………………. 7
2. 1 . 1. Generales……………………………………………………………… 7
2. 1 . 2. Las zonas maíceras en el estado…………………………………… 8
2. 1 . 3 Plagas…………………………………………………………………… 8
2.2 El gusano cogollero S. frugiperda…………………………………………….. 9
2. 2 . 1 Posición taxonómica…………………………………………………… 9
2 . 2 . 2 Origen y distribución geográfica……………………………………... 10
2 . 2 . 3 Descripción y biología.………………………………………………... 11
2 . 2 . 4 Plantas hospederas…………………………………………………… 12
2 . 2 . 5 Daños…………………………………………………………………… 12
2 . 2 . 6 Métodos de control……………………………………………………. 13
2.3 La bacterias entornopatógenas……………………………………………….. 15
2. 3 . 1 Bacillus thuringiensis…………………………………………………... 15
2 . 3 . 2 Las toxinas del Bt……………………………………………………… 16
2 . 3 . 3 Producción comercial…………………………………………………. 17
2. 3. 4 Modo de acción…………………………………………………………. 17
2 . 3 . 5 . Resistencia varietal………………………………………………….. 18
2 . 3. 6 . El manejo integrado………………………………………………….. 20
III MATERIALES Y METODOS……………………………………………………….. 23
3.1. Lugar de la experimentación…………………………………………………. 23
3.2. Etapas del desarrollo experimental………………………………………….. 23
3.3 Material biológico……………………………………………………………….. 23
3. 3 . 1 Las variedades de maíz.………………………………………………. 23
3 . 3 . 2 La bacteria B. thuringiensis…………………………………………... 24
3 . 3 . 3 . El insecto prueba…………………………………………………….. 24
3.4 Experimentos…………………………………………………………………… 25

3. 4 . 1. Evaluación de la susceptibilidad de larvas neonatas de S.
frugiperda, alimentadas con hojas de 13 variedades de maíz, a la
bacteria B. thuringiensis variedad kenyae, serovar 007…………………….
25
3. 4 . 2 Susceptibilidad de larvas de 3º y 5º estadio de S. frugiperda, a la
bacteria B. thuringiensis, alimentadas con hojas de maíz de las
variedades seleccionadas, en base a los valores de CL50 altos y bajos de
Bt, obtenidos del bioensayo anterior…………………………………………..
25
3. 4 . 3. Evaluar el efecto de la alimentación de larvas con follaje de
maíz, de diferentes variedades, sobre el desarrollo de las
larvas……………………………………………………………………………...
26
IV RESULTADOS 28
4.1 Susceptibilidad de larvas neonatas de S. frugiperda, alimentadas con
hojas de 13 variedades de maíz, a la bacteria B. thuringiensis variedad
kenyae, serovar 007………………………………………………………………...
28
4.2 Susceptibilidad de larvas de 3º y 5º estadio de S. frugiperda, a la
bacteria B. thuringiensis, alimentadas con hojas de maíz de variedades
seleccionadas, en base a los valores de CL50 altos y bajos de Bt, obtenidos
del bioensayo anterior………………………………………………………………
29
4.3. Efecto de la alimentación de larvas con follaje de maíz, de diferentes
variedades, sobre el desarrollo de las larvas…………………………………….
31
V DISCUSION…………………………………………………………………………… 33
VI CONCLUSIONES……………………………………………………………………. 36
VII LITERATURA REVISADA…………………………………………………………. 37

LISTA DE CUADROS
Página
Cuadro 1. Características y etapa de síntesis de toxinas de B. thuringiensis…… 16
Cuadro 2. Valores de CL50 de la bacteria B. thuringiensis, en larvas de S.
frugiperda, alimentadas con cada una de las variedades
evaluadas…………………………………………………………………..
29
Cuadro 3. Valores de CL50 de B. thuringiensis en el estadio larval 3º de S.
frugiperda, alimentando a las larvas con hojas de maíz de diferentes
variedades………………………………………………………………….
30
Cuadro 4. Valores de CL50 de B. thuringiensis en el estadio larval 4º de S.
frugiperda, alimentando a las larvas con hojas de maíz de diferentes
variedades………………………………………………………………….
31
Cuadro 5. Tiempo de desarrollo y peso de larvas y pupas de S. frugiperda
alimentadas con cuatro variedades de maíz…………………………..
32

1
I INTRODUCCION
Una civilización avanzada, con sus conglomerados industriales, artes y
comodidades dependen en gran medida de la capacidad humana para
administrar el sistema ecológico, del cual forma parte, de tal manera que una
fracción pequeña del grupo pueda producir alimentación y energía para todos.
La producción de alimento suficiente, para la población mundial, exige no sólo
una aplicación más eficaz de las tecnologías actuales en la producción de
cosechas, sino la adquisición y uso efectivo, de los nuevos conocimientos en
muchos aspectos de la agricultura.
Cada vez es mayor el crecimiento de la población en el mundo y
particularmente en este país, en donde se está demandando más y mejores
alimentos. En México la dieta está constituida básicamente de maíz, cuya
producción en grano, no se es suficiente. Como es conocido, el cultivo del
maíz, es uno de los cereales de vital importancia en el mundo, tanto por el
grano como por el forraje producido, por su utilización para consumo de
humano y de diferentes especies animales. Se conoce que las reacciones de
los organismos vegetales cambian en función, de una serie de factores del
medio ambiente bióticos y abióticos, a que se hallan expuestos, lo que trae
como consecuencia una variación importante, en la calidad y rendimiento
(Huston, 1993).
Uno de los métodos de protección de cultivos es principalmente el uso de
agroquímicos (Lise et al., 1998). Además, el cambio a sistemas de cosechas
continuas o intensivas, con el uso de paquetes tecnológicos, que implica la
utilización de insumos tales como fertilizantes e insecticidas químicos, en
amplias zonas agrícolas, ha creado problemas graves de plagas; por otro lado,
la creciente demanda de productos agrícolas libres del ataque de insectos y al
mismo tiempo de residuos de plaguicidas, ha ocasionado la necesidad de
buscar alternativas de control. Actualmente, existe gran competencia en las
operaciones agrícolas y forestales y en el caso de muchos productos, la
ganancia es pequeña aunque se obtenga una máxima producción.

2
Como es conocido, durante la producción de la mayoría de los alimentos,
forrajes y/o fibras naturales, indispensables para el hombre, los insectos
destruyen una parte de dichos productos. Los insectos también son vectores de
organismos causantes de enfermedades graves en plantas, animales y el
hombre. Existen miles de especies de insectos plaga, difundidas en la mayoría
de las regiones terrestres, donde pueden vivir; su permanencia se limita a los
lugares accesibles que les proporcionan comida y condiciones climáticas
adecuadas, para su crecimiento y desarrollo, así como bióticos esenciales.
Los insectos plaga tienen una gran adaptabilidad y se han acomodado a
muchas condiciones y situaciones ecológicas del mundo; no sólo se adaptaron
para sobrevivir en épocas pasadas, sino que siguen haciéndolo a pesar de los
cambios hechos por el hombre, o de los cambios ecológicos naturales. Las
pérdidas debido a plagas y enfermedades, han sido estimadas en un 37% de la
producción agrícola mundial, con un 13% a causa de los insectos (Gatehouse
et al., 1992).
Algunas especies de insectos son plagas durante todo el año, otras sólo en
determinadas estaciones. Las plagas de insectos pueden ser activas sólo en
ciertos años y otros no. La distribución de las plagas de insectos puede
cambiar durante una cosecha. Los factores que afectan la distribución y
actividad son variables, por tanto la distribución de plagas de insectos cambia
constantemente. A pesar de ello, a menudo las mismas plagas de insectos
dañan año tras año el mismo cultivo o las mismas regiones.
En los Estados Unidos de Norteamérica, se calcula que entre 150 y 500
especies causan graves daños; sin embargo, se estima que cerca de 6,000
especies de insectos se convierten en plagas en forma ocasional, y rara vez
causan daños de importancia. También; en otros países del mundo los cultivos
sufren infestaciones por una gran diversidad de insectos. Este problema, es
uno de los más importantes que afronta la agricultura moderna en el mundo, y
esta en incremento por el empleo desmedido de los agroquímicos, que en si
mismo representan un gran peligro para la salud del hombre.

3
Uno de los objetivos del manejo y control de plagas de insectos es crear y
mantener condiciones, que impidan que los insectos causen problemas de
importancia económica. Esto se logra evitando que se establezcan o que se
diseminen, mediante la prevención o el control de las infestaciones a un nivel
que sea el mínimo posible. Lo anterior se debe condicionar al mínimo costo y
que no haya riesgos tanto para el hombre como para otros integrantes del
ambiente; o sea, que el control de las plagas se realice de manera sostenible
(Behle et al., 1997).
Los cambios culturales, agrícolas y económicos, que han ocurrido en el mundo,
han tenido una influencia profunda en el control de plagas de insectos; se
insiste en que las medidas de control no causen daño al hombre o al ambiente.
Desde el aspecto económico, un buen control se debe basar tanto en los
resultados inmediatos como en los efectos a largo plazo. Se deben evitar las
prácticas de control que impliquen dificultades futuras, aun cuando produzcan
excelentes resultados inmediatos (National Academy of Sciences (NAS), 1992).
El control biológico de plagas, mediante el uso de insectos parásitos,
depredadores y patógenos, como hongos, bacterias, virus, nematodos y
protozoarios, así como el uso de feromonas o la utilización de la resistencia
morfológica o química de las plantas, en forma integrada con otras medidas de
control, como cultural o legal, constituye una estrategia de manejo integrado de
plagas dentro de una programa de agricultura sustentable. No obstante lo
anterior, por lo general, el control biológico de plagas, se realiza con la
aplicación de un sólo agente de control y poco se ha estudiado, la asociación
de la resistencia natural del vegetal y el uso de agentes biológicos de control
(Bergman y Tingey, 1979).
Conociendo la necesidad y los inconvenientes del uso del control químico,
como única herramienta de control del gusano cogollero Spodoptera frugiperda
(J. E. Smith) en maíz, incluyendo el elevado costo de los productos, surge la
necesidad de buscar otros medios de control, como el desarrollo de variedades
de maíz resistentes al ataque de este insecto y el uso de agentes
entomopatógenos para su control, que son alternativas deseables, tanto

4
económica como ecológicamente. Se reporta, que el control biológico puede
ser mejorado con el uso concurrente de la resistencia de la planta hospedera
(Hamm y Wiseman, 1986; Wiseman y Hamm, 1993; Epsky y Capinera, 1993,
Molina et al., 1999).
El uso sólo o simultáneo de la resistencia de la planta hospedera y el control
biológico, es una efectiva táctica de manejo de plagas y pueden combinarse
con otras tácticas de control, varios autores mencionan, los efectos deletereos
de la resistencia vegetal sobre el tercer nivel trófico (el agente de control
biológico) (Painter, 1951; Bergman y Tingey, 1979; Adkisson y Dyck, 1980;
Smith, 1989; Barbosa, et al., 1991; Barbercheck, 1993; Meade y Hare, 1995).
Aun más otros reportes mencionan que el éxito de los agentes de control
biológico están relacionados con la variedad o la química de la planta (Price et
al., 1980; Boethel y Eikenbary, 1986). También, que la química de la planta
hospedera puede afectar la virulencia de patógenos de insectos, por la acción
de sus metabolitos secundarios (Hare y Andreadis, 1983; Ramoska y Todd,
1985; Reichelderfer, 1991; Schultz y Keating, 1991), o mejorar su acción
(Molina et al., 1999).
La especie S. frugiperda es susceptible a 53 especies de parasitoides, de 43
géneros y 10 familias (Ashley, 1979); también, se tienen reportes de la
susceptibilidad de este insecto a 20 especies de entomopatógenos, que
incluyen hongos, bacterias, virus, protozoarios y nematodos (Gardner y Fuxa,
1980; Gardner et al., 1984). De ellos, la bacteria Bacillus thuringiensis Berliner
es la más utilizada, en el control biológico de larvas de lepidópteros (Cannon,
1996; Behle et al., 1997) y se le considera un agente de control biológico clave
dentro de un programa de manejo integrado de plagas (Trumble, 1990;
Entwistle et al., 1993; Tabashnik et al., 1997). Bernhard, et al. (1997) afirman
que el insecticida B. thuringiensis ha sido tan eficaz, como lo son los
insecticidas químicos convencionales.
El uso combinado, de la resistencia vegetal del maíz y B. thuringiensis ha sido
estudiado, en el Ostrinia nubialis (Hübner); esta integración, de alternativas de
control, es recomendada como viable en el manejo de plagas (Bolin et al.,

5
1996). Otra de las especies de insectos que más atacan al maíz,
principalmente en América Latina, es S. frugiperda, cuando la planta se
encuentra en su fase vegetativa (Williams y Buckley 1992), etapa considerada
como crítica para la planta (Borbolla, 1981). Este insecto ocasiona una
disminución importante en la producción del cereal.
En maíz, para el control del S. frugiperda, Hamm y Wiseman (1986) y Wiseman
y Hamm (1993) demostraron que la susceptibilidad de las larvas al virus de la
polihedrosis nuclear se incrementa con el uso de estigmas de maíz
incorporados en la dieta merídica. Del mismo modo Molina et al. (1997)
reportan que el uso de estigmas de maíz y de nematodos entomopatógenos,
inhiben en desarrollo larvas, alargan el período de crecimiento, prolongan el
tiempo de exposición al ataque de nematodos y mejora la actividad de los
nematodos entomopatógenos.
La asociación de Bt y plantas resistentes para el control de plagas ha sido
estudiado en Trichoplusia ni (Hübner) y Spodoptera exigua (Hübner) en apio
(Meade y Hare, 1995) y en muchas interacciones entre Bt con otros agentes de
control y se han obtenido resultados efectivos (Creighton et al., 1975, 1981;
Bell, 1978; Kea et al., 1978; Schuster et al., 1983; Felton y Dahlman, 1984;
Ludlum et al., 1991; Meade y Hare, 1994). También, se han reportado
resultados de incompatibilidad (Krischik et al., 1988; Navon et al., 1993). Sin
embargo, la interacción de Bt con plantas resistentes y la susceptibilidad de
larvas de S. frugiperda al complejo espora-cristal de Bt, alimentadas con
diferentes genotipos de maíz, no ha sido evaluada.
Con base en lo anterior, en este trabajo se plantea la hipótesis de que, el
desarrollo de S. frugiperda y su grado de susceptibilidad a la bacteria
entomopatógena B. thuringiensis, son dependientes de las cualidades de la
planta, con la cual se les alimenta.

6
Para probar esta hipótesis, se plantearon los siguientes objetivos:
1. Evaluar la susceptibilidad de larvas de S. frugiperda, a la bacteria
entomopatógena B. thuringiensis, alimentadas con hojas de diferentes
variedades de maíz.
2. Evaluar el efecto de la alimentación de larvas, con follaje de maíz de
diferentes variedades, sobre el periodo de desarrollo.

7
II ANTECEDENTES 2.1 El cultivo de maíz
El cultivo del maíz tiene importancia especial, dado que este cereal constituye
la base de alimentación de los latinoamericanos. Su origen no se ha podido
establecer con precisión, sin embargo, se puede afirmar que el maíz ya se
cultivaba en América Latina en la época precortesiana. El maíz, actualmente,
ocupa el segundo lugar en volumen de la producción mundial, después del
trigo; se cultiva en una superficie total de 106 millones de hectáreas, con una
producción de 449 millones de toneladas con un promedio de dos toneladas
por hectárea. Se adapta ampliamente a diversas condiciones ecológicas y
edáficas, razón por la que se le cultiva en casi todo el mundo (Benz, 1993).
2.1 .1 . Generales
Tres importantes cereales, cada uno asociado estrechamente con los orígenes
de grandes civilizaciones, aparecieron en los pasados 8000 a 10,000 años. El
maíz, Z. mays L., fue domesticado en el Hemisferio Occidental; el trigo y otros
cereales de su tipo, en el Cercano Oriente, y el arroz, en el Oeste de Asia
(Figueroa et al., 1994). El maíz ha sido utilizado como alimento, moneda y
religión, por el pueblo de México. Durante siglos, la historia nacional y las
condiciones de vida de los mexicanos han estado asociadas estrechamente a
su cultivo.
La representación de plantas de maíz, o partes de ella, en códices y cerámica
prehistórica, así como los estudios de fósiles, en varias culturas, indican que
México es el centro de origen primario. Se presume que el progenitor silvestre
del maíz existió hace 8 mil años (Beadle, 1939, 1980; Figueroa et al., 1994).
Actualmente en el estado de estado de Colima, el maíz es el cultivo anual de
mayor importancia, pues en cuanto a superficie sembrada ocupa el primer lugar
dentro de los cultivos básicos (INEGI, 1991).

8
El 83% del cultivo se siembra bajo condiciones de temporal, en terrenos en
donde la precipitación pluvial se presenta mal distribuida y en muchas
ocasiones es insuficiente, durante el ciclo del cultivo, acarreando consigo
problemas al productor por la obtención de bajos rendimientos (SARH-INIFAP-
CIAPAC, 1993). En el estado, la superficie total sembrada con cultivos anuales,
en 1991, fue de 55,772 ha, de éstas, el 78.1% corresponde a maíz (Hernández,
1989; INEGI, 1991).
2.1.2. Las zonas maíceras en el estado En el estado de Colima se tienen tres zonas ecológicas, caracterizadas por su
altitud, temperatura y precipitación; una de ellas la Zona Norte, que comprende
las comunidades de Quesería, del municipio de Cuauhtémoc; Cofradía de
Suchitlán, del municipio de Comala y el municipio de Minatitlán, en donde se
tienen rendimientos promedio comprendidos entre 4.9 a 6.6 toneladas de grano
por hectárea. La Zona Centro comprende las localidades del municipio de Villa
de Álvarez, Tepames, Colima; Camotlán de Miraflores, Manzanillo y
Coquimatlán, en esta zona se obtienen rendimientos promedio comprendidos
entre 3.8 y 5 toneladas de grano por hectárea (SARH-INIFAP-CIAPAC, 1993).
La Zona Costa abarca la región del municipio de Ixtlahuacán, Rincón de López
del municipio de Armería, y Madrid del municipio de Tecomán. Los
rendimientos del cultivo alcanzan las 2.4 y 3 toneladas por hectárea (SARH-
INIFAP-CIAPAC, 1993).
2. 1. 3 Plagas
En México como en otros países, al igual que el resto de los cultivos, el maíz
guarda una relación trófica entre éste y sus herbívoros o parásitos naturales,
los cuales al encontrar un ambiente favorable, aumentan su densidad
poblacional convirtiéndose en plagas. Las plagas atacan en diferentes edades
a las plantas y si no se controlan pueden reducir considerablemente el
rendimiento, ocasionando por consecuencia, una disminución en la producción

9
potencial del cultivo. Las plagas más importantes son los insectos debido a que
además de causar daño a la planta son portadores de enfermedades.
Las plagas de mayor importancia del cultivo son la gallina ciega, Phyllophaga
spp, gusano de alambre, Elateridae, Agrotis sp, Melanotus sp, larvas de
diabrótica, Diabrotica undecimpuntata y gusano trozador, Agrotis ipsilon, y
Chorizagrotis auxiliaris, que se alimentan de las raíces de plantas jóvenes,
debilitándolas y afectando su crecimiento, además las vuelven susceptibles al
acame. También el gusano cogollero, S. frugiperda, adultos de diabróticas, D.
undecimpuntata, chicharritas, Dalbulus elimatus, trips, Frankliniella occidentalis,
F. williamsi y Hercothrips phaseoli, gusano barrenadores del tallo, Zeadiatraea
lineolata, frailecillo, Macrodactylus infuscatus, M. mexicanus y M. marinus y
gusano elotero, Helicoverpa zea Boddie, que se alimentan de hojas, tallos,
flores o frutos (SARH-INIFAP-CIAPAC, 1993).
2.2 El gusano cogollero S. frugiperda
Más de tres siglos han pasado desde que S. frugiperda fue reconocida como
una plaga, causante de serios problemas en diversos cultivos (Wiseman et al.,
1983; Ashley et al., 1989; Simmons y Wiseman 1993), limitando muchas áreas
de éstos, en el Sureste de Estados Unidos, México, Centro y Sudamérica
(Wiseman y Davis, 1979; Pashley, 1988 ). A este insecto en México se le
localiza principalmente en las regiones tropicales y subtropicales registrándose
los mayores daños en los estados de: Michoacán, Guerrero, Morelos, Oaxaca,
Veracruz, Quintana Roo, Yucatán, Coahuila, Campeche, Chiapas, Chihuahua,
Valle de México, Durango, Guanajuato, Baja California Norte, Baja California
Sur y Colima.
2. 2. 1 Posición taxonómica
Este insecto está ubicado taxonómicamente en la familia Noctuidae, dentro del
orden Lepidoptera, su posición taxonómica se presenta a continuación (Banda,
1981).

10
Phylum . . . . Anthropoda
Subphylum . . . . Mandibulata
Clase . . . . . Insecta
Subclase . . . . . Pterygota
Orden . . . . . Lepidoptera
Suborden . . . . . Frenatae
Superfamilia . . . . . Noctuoidea
Familia . . . . . Noctuidae
Tribu . . . . . Predeninii
Género . . . . . Spodoptera
Especie. . . . . S. frugiperda (J. E. Smith)
2. 2. 2 Origen y distribución geográfica
El origen de S. frugiperda no está aún bien definido, sin embargo, Walton y
Luginbill (1917), Luginbill (1928), Vickery (1929) y Borbolla (1981), coinciden al
afirmar que esta especie tiene su origen en los trópicos del Continente
Americano, especialmente en América del Sur, ya que no posee diapausa
(Carpenter et al., 1983; Carpenter y Young, 1991). En el Hemisferio Occidental
se le ha encontrado desde el Sureste de Canadá hasta Chile y Argentina;
frecuentemente es abundante en las áreas agrícolas. Es una especie de
distribución tropical aunque se le encuentra también en zonas templadas
(Sparks, 1979; Andrews, 1980; Foott y Timmins, 1982; Abbas et al., 1989).
Es una especie endémica del Continente Americano, incluyendo Las Antillas y
el Caribe, sin embargo Tood y Poole (1980) mencionan la introducción de S.
frugiperda a Israel. En México se encuentra ampliamente distribuida en todas
las zonas agrícolas, principalmente en los Estados ubicados dentro del
Territorio Costero. En los Estados con altitudes superiores a los 2000 msnm
como Puebla, Tlaxcala, Hidalgo, Guanajuato, Querétaro y el Edo. de México,
en donde éste insecto tiene menos importancia económica (Andrews, 1988).
Aparentemente, las condiciones climáticas están significativamente

11
relacionadas con las frustraciones poblacionales y la dispersión de S.
frugiperda tal como ha sido señalado por Pashley (1986).
2. 2. 3 Descripción y biología
Los adultos son palomillas que miden aproximadamente 3.75 cm de extensión
alar. En el macho son de color pardo claro, con marcas oscuras y líneas
irregulares pálidas en el centro, mientras que las de la hembra son más
oscuras y grisáceas, con diseños menos notorios (Morón y Terrón, 1988). Los
adultos presentan hábitos nocturnos y tienen una longevidad que varía de 4 a 8
días, dependiendo de las condiciones ambientales; las hembras durante su
vida son capaces de producir hasta 3,600 huevecillos (Borbolla, 1981; Silvain,
1987).
Los huevecillos son puestos en masas que varían de 40, 150 y hasta 1500
huevos por masa, colocadas en el envés de las hojas, cubiertas por escamas
de la hembra. La incubación varía de 2 a 10 días (Luginbill, 1928; Borbolla,
1981); sin embargo, en Colima el tiempo de incubación varía de 24 a 72 h y un
100% de eclosión (observación personal).
Las larvas son del tipo cruciforme, de color pardo amarillento a pardo oscuro;
en sus regiones laterales son blanquecinas y presentan líneas longitudinales
laterales pálidas y moteadas. La cabeza es parda con reticulaciones y franjas
oscuras y en el último estadio alcanzan una longitud máxima de 30-38 mm. Las
larvas neonatas viven en grupos al principio y se separan posteriormente,
debido a sus hábitos caníbales, quedando en forma general una larva por
planta de maíz (Borbolla, 1981). Inicialmente las larvas comienzan a
alimentarse en el envés de las hojas, se dispersan y se dirigen al cogollo de la
planta de maíz; aquí se alimentan de las hojas en crecimiento, las cuales
posteriormente muestran perforaciones irregulares (Sparks, 1979; Lagunes et
al., 1985).
Las larvas pasan por seis estadios en un lapso que puede durar de 2 a 3
semanas; transcurrido este tiempo se introducen en el suelo para pupar. La

12
cabeza de las larvas es de color negro en los últimos estadios, con la sutura
epicraneal bien marcada; el cuerpo es cilíndrico, de color café gris dorsalmente
y verde ventralmente, con líneas dorsales y subdorsales visibles (Borbolla,
1981).
Las pupas son de tipo obtecta y miden cerca de 2 cm de largo; son de color
pardo rojizo, con el protórax más oscuro, encontrándose normalmente
enterradas en el suelo, donde permanecen una semana aproximadamente y
luego emergen como adultos; de esta forma, se reanuda su ciclo (Lagunes et
al., 1985).
2. 2. 4 Plantas hospederas
Este insecto se alimenta de una gran diversidad de especies vegetales; en el
Continente Americano es considerada la plaga más importante, pues ataca
más de 60 especies de plantas (Hernández, 1989), en diversas familias
vegetales y se le atribuye su polifagia a una carencia en la especialización de la
alimentación larvaria (Pashley, 1986). En Colima su alimentación se limita al
maíz y sorgo exclusivamente, y en muy raras ocasiones de otras especies de
vegetales, observación personal. Así en la relación existente entre la plaga y el
cultivo es necesario considerar una serie de factores que intervienen en forma
entrelazada.
Tanto la planta como la plaga son seres vivos, en los que ocurren cambios en
todo momento. Ambos están sujetos a cambios físicos y biológicos del
ambiente que los rodea, que también es dinámico. Por lo tanto, la capacidad de
la plaga de hacer daño depende, por una parte de la densidad poblacional
existente en el cultivo. Por otro lado, las plantas de distintas especies
reaccionan de un modo diferente ante los ataques de las plagas.
2. 2. 5 Daños
En la estimación del daño que causan los insectos a las plantas se debe
considerar el tipo de cultivo, variedades, densidad de plantas, hábitos de

13
crecimiento, base de desarrollo, condición y permanencia en el campo. La
parte de la planta afectada por la plaga determina a veces la magnitud e
importancia del daño. Las larvas se alimentan del cogollo o verticilo de las
plantas de maíz y sorgo en desarrollo. En el caso del maíz diversos autores
han tratado de determinar el efecto de la defoliación, causada por S. frugiperda
sobre los rendimientos del cultivo, llegando a conclusiones muy variables y en
algunos casos contradictorias (Clavijo, 1984); sin embargo, (Hernández, 1989)
menciona que las pérdidas del área foliar causadas por el ataque de cogollero
en cultivos de maíz afecta la actividad fotosintética de las plantas,
manifestándose estos daños de manera indirecta como una disminución del
rendimiento por hectárea.
En el cultivo del maíz, el desarrollo fenológico de la planta tiene una fuerte
influencia sobre el ataque de las poblaciones S. frugiperda. Este insecto inicia
su ataque cuando las plantas tienen alrededor de 5 a 6 hojas libres y a medida
que progresa la edad de la planta, las poblaciones del insecto también
progresan hasta alcanzar el punto de máxima infestación y este se presenta
cuando la planta tiene 10 hojas libres. Sin embargo, el ataque puede llevarse a
cabo en cualquier etapa del vegetal (Wiseman et al., 1977).
Los ataques del S. frugiperda, en el Occidente de México y en particular en el
Estado de Colima, se presentan en los dos ciclos de cultivo de maíz que son
realizados anualmente; las infestaciones larvarias de esta plaga, en la Zona
Costera, aparentemente están formadas por generaciones superpuestas, que
siguen un comportamiento similar en los dos ciclos vegetativos, aunque esta
distribución puede variar en los cultivos localizados en la zona norte del Estado
de Colima (Hernández, 1989).
2. 2. 6 Métodos de control
Las medidas de control de esta plaga varían con la variedad de maíz y el
estado de desarrollo de la planta (Young, 1979). La integración de medidas
fitosanitarias aumenta la posibilidad de lograr un control efectivo de las diversas
plagas que atacan el maíz durante sus distintas etapas de crecimiento, sin

14
embargo, hasta la fecha, el control de S. frugiperda se realiza mediante varios
métodos:
a) Control mecánico. Comprende una serie de medidas, básicamente de
limpieza, con la finalidad de eliminar todo refugio que sirva de protección al
insecto, principalmente durante el invierno.
b) Control químico. Es el más comúnmente empleado y se lleva a .cabo con la
aplicación de productos químicos; sin embargo, a través del tiempo los
insectos han adquirido resistencia a varios insecticidas (Landazabal, 1973;
Brattsten et al., 1986; Roush y McKenzie, 1987; Georghiou y Lagunes-
Tejeda, 1991; Yu, 1991, 1992). Comúnmente se aplican piretroides,
organoclorados, fosforados y carbomatos; los más usados son Sevín 80%
(a dosis de 1.5 Kg por hectárea); Paratión metílico CE (1.5 litros/hectárea);
Sevín 2 % granulado (15 a 20 kg por hectárea) y Lorsban 3% granulado (15
a 20 Kg/hectárea). La aplicación de los insecticidas debe de estar dirigida al
cogollo, cuando tres de cada 10 plantas presentan daño (SARH, 1993).
c) Control biológico. En la lucha contra las plagas agrícolas, mediante el
empleo de insectos benéficos, la Dirección General de Sanidad Vegetal, de
la Secretaría de Agricultura Ganadería y Desarrollo Rural (SAGAR), ha
impulsado los programas de control biológico, para disminuir la población de
una determinada plaga.
S. frugiperda es susceptible a insectos parásitos (Ashley, 1979); a
depredadores como insectos, arañas, aves, reptiles y anfibios, entre otros; así
como, a al menos 20 de entomopatógenos, dentro de ellos, hongos, bacterias,
virus, protozoarios y nematodos (Dicke, 1972; Gardner y Fuxa, 1980; Gardner
et al., 1984; Fuxa et al., 1988; Molina et al., 1996). De ellos, la bacteria B.
thuringiensis es el agente de control biológico más utilizado en el control de
lepidópteros en cultivos de grano y hortalizas (Behle et al., 1997).

15
2.3 La bacterias entomopatógenas
Las bacterias constituyen el grupo más numeroso y quizás el más estudiado
entre los microorganismos asociados con insectos (Bucher 1960, Heimpel
1967, Heimpel y Angus 1963, Anderson y Rogoff 1966, Falcon 1971). En
general los entomopatógenos bacteriales se agrupan en: a) cristalíferas
formadoras de esporas; b) patógenos obligados; c) patógenos facultativos y d)
patógenos potenciales. Las bacterias más importantes desde el punto de vista
del control de insectos son las aeróbicas formadoras del de esporas del género
Bacillus.
El género Bacillus tiene muchos representantes con características
entomopatógenas, y pertenece a la familia Bacillaceae (González y Bruce,
1980). Dentro del Orden Lepidóptera, las especies del género Spodoptera
presentan una variada susceptibilidad a B. thuringiensis aunque en ocasiones
han mostrado cierta resistencia a cepas y variedades del bacilo. Las bacterias
patógenas de insectos son organismos unicelulares, que tienen reproducción
por fisión y tienen motilidad; algunas producen esporas con capacidad de
formación de cristales, con acción patógena contra insectos (Angus, 1954;
Falcón, 1971; Tanada y Kaya, 1993; Knowles y Dow, 1993; Bauer, 1995).
La especie B. thuringiensis es considerada entre las bacterias patógenas de
insectos, con gran potencial de uso, en el manejo de poblaciones de insectos
plaga (McClintock et al., 1995; Aronson et al., 1986). Existe otro grupo de
bacterias a la cual, se le ha considerado como patógenas potenciales. La
mayoría de ellas pueden ser desarrolladas in vitro.
2. 3. 1 Bacillus thuringiensis
La especie B. thuringiensis (Bt), se ha aislado de muchos insectos en diversos
lugares del mundo. Actualmente se reconocen 34 variedades correspondientes
a 14 serotipos diferentes (Heimpel, 1967; De Barjac y Bonnefoi, 1968; Hall et
al., 1977; De Barjac y Frachon, 1990), de las cuales las más importantes son
thuringiensis e israelensis. Las especie B. thuringiensis es tóxica ya que

16
produce protoxinas durante la fase de esporulación (Aronson et al., 1986;
Gawron-Burke and Baum, 1991) y las esporas pueden germinar lenta o
rápidamente dependiendo de la etapa en que sea utilizada (Bon et al., 1969).
2. 3. 2 Las toxinas del Bt
Durante su ciclo reproductivo Bt produce varios metabolitos tóxicos (toxinas) de
diferentes clases, como se muestra en el Cuadro 1.
Cuadro 1. Características y etapa de síntesis de toxinas de B. thuringiensis.
TOXINA ETAPA DE PRODUCCIÓN
CARACTERISTICAS REFERENCIA
Delta-Endotoxina Beta-Exotoxina Alfa-Exotoxina Gama-Exotoxina
Fase estacionaria Fase logarítmica Fase logarítmica Fase logarítmica
Glicoproteina mantenida en el cristal. Es termolábil y soluble en álcalis. Adenin nucleótido, soluble en agua y estable al calor. Proteína termolábil, soluble en agua. Enzima no identificada.
Meyer, 1986, Mohad-Salleh y, Temeyer, 1984. Lüthy, 1983. Burges, 1982, Haufler y Sindey, 1985; Ignoffo, 1974. Meyer, 1986, Mohad-Salleh y. Mohad-Salleh y.
Por lo general, para un desarrollo eficaz de los bioinsecticidas, se requieren de
pruebas de laboratorio, con el fin de seleccionar microorganismos
entomopatógenos más sobresalientes. Esta selección se realiza a través del
desarrollo de bioensayos, para conocer su actividad tóxica y su mecanismo
patogénico (Krieg y Herbert, 1984; Lidstone y Khachatourians, 1985).

17
2.3.3 Producción comercial
El desarrollo comercial de formulaciones a base de Bt se ha incrementado
ampliamente y de manera exitosa, debido a que su uso en el control de
insectos perjudiciales para la agricultura ha dado respuesta a las expectativas
de los productores (Falcon, 1971; Krieg y Herbert, 1984; Stanier et al., 1986).
Este insecticida microbiano es producido en algunos países como Francia,
Alemania, Checoslovaquia y Estados Unidos; las presentaciones del
bioinsecticida se encuentran como formulaciones en polvo humectable o
líquidos, granulados o en aerosol, los cuales contienen como ingrediente activo
esporas, cristales, células vegetativas, o mezclas de ellas.
Algunas de las formulaciones llevan como nombre de Biotrol, Thuricide, Dipel,
Lavatol, Agritol, Sporeine y Bakthane. Estas formulaciones son específicas
para insectos, no causa daño a humanos y no produce residuos tóxicos. Lo que
le permite ser utilizado en pastizales, cultivos frutícolas, hortícolas, forestales,
ornamentales, etc. (Fernández y Estefania, 1974).
2. 3. 4 Modo de acción
La principal propiedad insecticida de todas las cepas de Bt es su capacidad de
producir o sintetizar cristales proteicos de delta-endotoxinas (Krieg et al., 1968),
cuyo potencial tóxico varía, no sólo con las diferentes cepas, sino también el
medio que se utilice para su producción; además influye sobre este potencial
tóxico otros factores como temperatura, edad y vigor del insecto (Dulmage,
1971). Esta bacteria afecta al epitelio del intestino medio en el insecto tratado;
ésta es la parte inicial del proceso de infección, en donde se realiza el ataque
de la toxina (Fast, 1974; Fast et al., 1978; Huber y Luthy, 1983; Knowles et al.,
1984; Singh et al., 1986a; Gill et al., 1992), para lo que debe ser ingerida por el
insecto (Thomas y David, 1983). Las delta endotoxinas son considerados como
venenos intestinales, por lo que deben ser consumidas por el insecto
susceptible (Knowles y Dow, 1993).

18
Las proteínas son sintetizadas como protoxinas cristalinas insolubles, las
cuales se disuelven y son activadas en el intestino del insecto antes de
aparecer s us efectos. El intestino de la mayoría de los insectos tiene un pH
muy alto (Dow, 1986) y esto es esencial para solubilizar a la mayoría de las
protoxinas de Bt, las cuales son generalmente solubles solo arriba de pH 9.5.
Los intestinos medios de las larvas de estos insectos también tienen proteasas
necesarias para convertir las protoxinas a toxinas y quizás receptores sobre la
superficie de las células epiteliales del intestino medio a las cuales las toxinas o
protoxinas hieren para iniciar su acción (Fast et al., 1978; Huber y Lüthy, 1981;
Knowles et al., 1984).
Las protoxinas son activadas por las proteasas del intestino del insecto, las
cuales típicamente dividen alrededor de 500 aminoácidos provenientes del
Carbono final de protoxinas de 130kDa y 28 aminoácidos provenientes del
Nitrógeno terminal, dejando un sitio activo resistente a proteasas de 65-55 kDa
que comprime la mitad del N terminal de la protoxina (Höfte y Whiteley, 1989,
Van Rie et al., 1990). Los síntomas, más importantes que se presentan en los
insectos tratados, son una inactividad marcada, seguida por parálisis general
del cuerpo, lo cual indica que los otros órganos de la larva juegan un papel
importante en la acción entomócida de las toxinas (Singh et al., 1986b).
2. 3. 5 Resistencia varietal
Luginbill (1969) y Wiseman y Davis (1979) publicaron que el método más
efectivo e ideal de combatir insectos que atacan a las plantas es el de utilizar
variedades resistentes a los insectos. La interacción insecto-planta, entre S.
frugiperda y plantas de maíz, es uno de los temas menos estudiados en
comparación con muchos otros insectos (Wiseman y Davis, 1979).
La resistencia es un fenómeno, que refleja la interacción de la planta con el
insecto. Painter (1951) y Wiseman y Davis (1979) definen la resistencia vegetal
como la cantidad relativa de cualidades heredables, que posee una planta, con
influencia en el grado de daño que le ocasiona un insecto. Lo anterior significa,
para la agricultura práctica, la capacidad de ciertas variedades de producir una

19
mayor cosecha de mejor calidad, que las variedades comunes expuestas a la
misma especie y cantidad de insectos plaga. Con los años se han desarrollado
muchas variedades de cultivos resistentes a insectos, más notablemente en la
alfalfa, maíz, algodón, frijol, yuca, hortalizas, arroz, sorgo, soya y trigo, entre
otras especies vegetales.
Se considera que una planta es resistente cuando, bajo algunas condiciones
ambientales, por ejemplo, el medio ambiente en que se encuentra, ésta cambia
en su nivel de resistencia (Wiseman y Davis, 1979). Dado que cerca de 500
especies de insectos se les conoce como resistentes a algunos insecticidas, se
han creado una serie de medidas estratégicas para disminuir la dependencia
en el uso de sustancias químicas utilizadas para su control; dentro de estas
medidas de prevención de ataque de insectos a los cultivos, están las
variedades resistentes, que representan un método de potencial de control de
un insecto plaga, (Painter, 1968; Yang et. al., 1991, 1993).
Debido a estas características, la resistencia que posee una planta a los
insectos es, probablemente, la medida más útil de todos los factores que
incluye un manejo integrado de plagas. Se conoce que las plantas presentan
un grado diferente de sensibilidad al ataque de los insectos, es decir, los
insectos prefieren más unas variedades que otras. La razón de esta
selectividad es muy compleja, pueden intervenir en ello las características
físico-químicas de las plantas y las condiciones del medio ambiente,
especialmente el clima y el suelo (con su respectiva importancia en el estado
de nutrición de los insectos (Hernández, 1994).
Una planta puede ser resistente al ataque de insectos plaga, por numerosas
razones que incluyen, características morfológicas tales como forma, tipo de
tejido, presencia de verticilios, vaina, panoja, etc. Investigaciones recientes han
sugerido que la presencia de ciertos metabolitos secundarios de la planta
tienen un papel importante en la protección al ataque de plagas (Atkinson et al.,
1990). La nutrición del cultivo hospedero puede tener influencia sobre la
expresión de la resistencia de éste para el ataque de plagas (Wiseman y Davis,
1979).

20
Se ha demostrado que los niveles de resistencia de la planta pueden
aumentarse o disminuirse, dependiendo de la manipulación en la nutrición del
cultivo. Por otro lado es aceptable que un cultivo designado como resistente o
susceptible podría ser inducido por los nutrientes de la planta para exhibir una
mayor o menor expresión de resistencia al ataque de insectos. Painter (1951) y
Wiseman (1985) clasificaron la resistencia de plantas en tres categorías y las
define como: No preferencia, antibiosis y tolerancia. La no preferencia resulta
cuando una planta o variedad no posee las cualidades atractivas para la
oviposición, alimentación o refugio de un insecto; la antibiosis es el término
empleado para denotar los efectos adversos producidos en el insecto cuando
éste se alimenta con una planta o variedad, mientras que el tercer mecanismo
de resistencia: tolerancia, es probablemente el más desatendido se refiere
cuando las plantas pueden soportar cierta población de un insecto sin que las
pérdidas sean considerables en la producción (Wiseman, 1985; Meckenstock et
al., 1991; Diawara et al., 1991).
Las variedades de maíz presentan diferentes grados de atracción para los
adultos y las larvas de cogollero; también se reporta que el manejo que sobre
el cultivo de maíz y de otras gramíneas se tenga, también modifica el
comportamiento de selección de los adultos por un cultivo (Chang et al., 1987;
Slansky y Wheeler, 1991). Estudiándose la relación resistencia susceptibilidad
de maíces nativos en el Estado de Colima al ataque de cogollero, se observó
que tanto los adultos muestran una preferencia para ovipositar, como las larvas
para alimentarse sobre el follaje de ciertas variedades (Hernández, 1989).
2. 3. 6 El manejo integrado
El manejo integrado de plagas es una estrategia que intenta maximizar la
utilización de los factores que controlan a las plagas de manera natural. Los
enemigos naturales (parasitoides, depredadores y patógenos), la resistencia o
la tolerancia de los vegetales al ataque de las plagas y las prácticas culturales
(Isenhour y Wiseman, 1991), estas tácticas son, entre otras cosas, los

21
mecanismos que deben integrarse para lograr una mayor eficiencia en el
control de plagas.
Son varias las razones por las que se reconoce que ha aumentado en los
últimos años el interés por estas estrategias de control de plagas y
enfermedades; la mayoría asociada más bien a problemas causados por el uso
indiscriminado de plaguicidas. Se puede mencionar la aparición de resistencia
de los insectos a los agroquímicos, el impacto negativo de los mismos sobre
los organismos benéficos, los riesgos para la salud y el medio ambiente
asociados con su uso (Hernández y Benderly, 1982).
Los componentes claves, de un programa de manejo integrado de plagas de un
determinado cultivo, pueden resumirse como sigue: identificación de las
especies presentes en el agroecosistemas, reconocimiento sobre la biología y
ecología del cultivo, de las plagas y de los enemigos naturales; métodos fiables
y sencillos de muestreo de poblaciones, conocimiento de los umbrales
económicos de daños y en consecuencia de los umbrales de tolerancia o
intervención, existencia de métodos de control de plagas eficientes,
económicamente viables y compatibles (resistencia varietal, control biológico y
microbiológico, métodos culturales) (Slansky , 1992).
El control biológico es uno de los que causan menos daño ya que se utilizan
enemigos naturales para controlar la población plaga (Costa-Comelles et al.,
1992). Sin embargo, el desarrollo de variedades de maíz resistentes al ataque
de cogollero es también un método de prevención de infestaciones deseables
tanto económicamente como a favor del medio ambiente, por lo que es
conveniente buscar en primer instancia las posibles fuentes que generen
resistencia de la planta al ataque de plagas (Yang et al., 1991, 1993).
A menudo el uso de una variedad resistente en vez de insecticidas puede
alcanzar los mismos resultados, con ventajas obvias. Incluso con un nivel bajo
de resistencia, el efecto persistente y acumulativo puede facilitar el control
cuando se combina con otros factores ecológicos. La sustitución de una
variedad resistente por una susceptible puede permitir el aumento de parásitos,

22
con un menor nivel económico de daños (NAS, 1992). El binomio formado por
la resistencia varietal y el control biológico es un método barato y
ecológicamente aceptable en el manejo de poblaciones insectos.
El manejo integrado de plagas ha sido la plataforma de un nuevo concepto más
amplio, la producción integrada, que es importante no confundir con la
producción o agricultura orgánica. La producción integrada ha sido definida
como “la producción económica de frutos de alta calidad, dando prioridad a los
métodos de producción ecológicamente más seguros, minimizando los efectos
secundarios indeseables y el uso de los agroquímicos, para mejorar la
protección del medio ambiente y la sanidad humana” (Costa-Comelles et al.,
1992).
Los programas de manejo integrado de plagas de un cultivo se estructuran
generalmente alrededor de un método selectivo de control de una determinada
plaga-clave del citado cultivo. Por lo que una de las áreas importantes de
investigación es el crear variedades insecto-resistentes, dado que, la
resistencia de plantas al ataque de insectos es un fenómeno que tiene un
amplio potencial de uso en el manejo integrado de plagas (Wiseman, 1987),
puesto que la disminución de los ataques del insecto se puede transformar en
un incremento de la rentabilidad de los cultivos al tiempo que se producen
cosechas más limpias al no estar contaminadas con agroquímicos.
En maíz y sorgo, S. frugiperda se alimenta del cogollo, causando importantes
pérdidas del área foliar manifestándose estos daños como una disminución del
rendimiento por hectárea (Hernández, 1989). El desarrollo de variedades de
maíz resistentes al ataque de cogollero es un método de prevención de
infestaciones deseables tanto económicamente como a favor del medio
ambiente, por lo que es conveniente buscar en primer instancia las posibles
fuentes que generen resistencia en la planta (Yang et al., 1991, 1993).

23
III MATERIALES Y METODOS 3.1 Lugar de la experimentación
El presente trabajo se realizó en el laboratorio de Biología del Centro
Universitario de Investigación y Desarrollo Agropecuario (CUIDA) de la
Universidad de Colima, ubicado en el Crucero de Tecomán, Col. a una altitud
de 32 msnm. La temperatura media dentro del laboratorio, osciló entre 22 y
25ºC, durante el período comprendido entre los meses de febrero de 1995 a
enero de 1997.
3.2 Etapas del desarrollo experimental
El trabajo experimental se desarrolló en dos partes; en la primera se evaluó la
susceptibilidad de larvas neonatas de S. frugiperda a la bacteria B.
thuringiensis, alimentadas con follaje de 13 variedades de maíz (6 criollos del
estado de Colima y 7 híbridos comerciales). En base a los resultados obtenidos
se seleccionaron dos variedades de maíz, en las cuales se determinó una CL50
de la bacteria más baja y dos variedades, en donde las CL50 de la bacteria se
mostró alta, para la segunda etapa en la cual se evaluó la susceptibilidad de
larvas de tercero y quinto estadio S. frugiperda, a la bacteria, también en base
a sus valores de CL50.
3.3 Material biológico 3. 3. 1 Las variedades de maíz
En esta investigación se utilizaron 13 variedades de maíz (6 criollos del estado
de Colima y 7 híbridos comerciales); los criollos son Grano Ancho, Grano
Negro, Grano Pinto, Grano Rojo, Tampiqueño olote delgado, Uruapeño y los
híbridos Amarillo forrajero, Ceres 7, Pioneer 3086, Pioneer 3288, V-526, B-810
y H-910.

24
Las trece variedades de maíz, fueron sembradas en macetas de plástico de
500 ml de capacidad. Como sustrato se utilizó suelo sin esterilizar, de las áreas
verdes de las instalaciones del CUIDA. De la misma forma, se formaron camas
de 1.20 x 2.50 m, en donde se sembraron surcos con cada una de las
variedades de maíz, para contar con suficiente follaje para alimentar las larvas,
de la colonia y para los bioensayos. Cuando las plantas de maíz alcanzaron los
15 a 30 cm de altura, se les cortaron trozos de hoja, de 5 cm de longitud, para
mantenimiento de la colonia de S. frugiperda o para los experimentos. El suelo
se fertilizó y se regó de acuerdo a las necesidades de las plantas.
3. 3. 2 La bacteria B. thuringiensis (Bt)
Para este estudio se utilizó la cepa de B. thuringiensis, variedad kenyae
serovar 007, originaria del Centro de Referencia de la Organización
Internacional de Control Biológico, del Instituto Pasteur de Francia; misma que
se mantiene en formulación en polvo acetónico (T044Boo7); No. de referencia
280289, en forma de complejo espora-cristal, a -40ºC. Esta cepa se seleccionó
en base a los resultados obtenidos por Hernández (1988), con esta bacteria en
S. frugiperda.
3. 3. 3 El insecto prueba
Las larvas de S. frugiperda, fueron colectadas de campos comerciales de maíz
del estado de Colima. La técnica de cría de la colonia fue alimentando a las
larvas con hojas frescas de maíz, en vasos de plástico de 100 ml de capacidad,
mantenidos en forma individual, para evitar canibalismo. Las pupas se
mantuvieron en cámaras húmedas sobre polvo de coco estéril, hasta la
emergencia de los adultos. Los adultos se colocaron en cámaras de oviposición
de 30x30x30 cm, acondicionadas con alimento a base de miel de abeja o
sacarosa al 10% y tiras de papel plegadas en forma de acordeón, como
sustrato de oviposición (Burton y Perkins, 1989).
Cada 24 horas se colectaron las oviposturas y se cambió el alimento de los
adultos y larvas.

25
3.4 Experimentos 3. 4. 1 Evaluación de la susceptibilidad de larvas neonatas de S. frugiperda,
alimentadas con hojas de 13 variedades de maíz, a la bacteria B. thuringiensis
variedad kenyae, serovar 007.
Para este experimento, 30 trozos de hojas de maíz de 5 cm de longitud, de
cada variedad se inocularon por inmersión en una suspensión del complejo
espora y cristal de Bt con las concentraciones de 1.0, 0.1, 0.05, 0.025, 0.01 y 0
mg/ml. Las concentraciones de espora-cristal se realizaron, a partir de una
solución madre, utilizando 0.01g de la formulación de Bt, disuelto en 10 ml de
agua destilada estéril, con Tween 80 al 0.1%. Los trozos de hojas inoculadas
se dejaron 15 minutos en una campana de flujo laminar, para que el exceso de
agua se evaporara; posteriormente cada hoja se colocó en una caja Petri de
60x10 mm, sobre una capa de papel filtro húmedo, junto con cinco larvas
neonatas de S. frugiperda.
A cada caja de Petri entre ella y la tapa se les colocó una capa de papel
higiénico, para evitar el escape de las larvas. Las larvas se mantuvieron en
exposición de las hojas tratadas con Bt durante 72 horas, siguiendo la técnica
de Del Cañizo et al. (1990). Al término de éste período se contabilizaron las
larvas muertas y se calculó el porcentaje de mortalidad, para cada
concentración, en cada variedad y se procedió a los análisis estadísticos
correspondientes.
Para cada variedad de maíz se evaluaron 6 concentraciones de Bt
(tratamientos), con 5 repeticiones y 5 larvas por repetición, con un total de 150
larvas por experimento. El diseño experimental utilizado fue completamente al
azar. Los datos obtenidos se analizaron mediante análisis de varianza y la
separación de promedios mediante la prueba de rango múltiple de Duncan (P<
0.05), así como análisis Probit, para la obtención de la CL50.
3. 4. 2 Susceptibilidad de larvas de 3º y 5º estadio de S. frugiperda,
alimentadas con hojas de maíz de las variedades seleccionadas por su efecto
en los valores de CL50 de Bt.

26
Las variedades de maíz seleccionadas, que promovieron valores bajos de CL50,
fueron el Criollo uruapeño y el híbrido V-526; por otro lado, las variedades
seleccionadas que promovieron valores altos de CL50 fueron el criollo Grano
Negro (Jaripeño) y el híbrido Amarillo Forrajero. Hojas de maíz de estas cuatro
variedades fueron inoculadas por inmersión en 6 concentraciones de Bt
(tratamiento); cada una de las concentraciones fue evaluada en larvas de 3º y
5º estadio. Se utilizaron 10 larvas por cada estadio/concentración y como
repetición con un total de 60 larvas por bioensayo.
Las larvas de 3º estadio se expusieron a trozos de hojas de 10 cm2 e
inoculadas con Bt, como se indicó en los ensayos anteriores; cada larva se
conservó en un recipiente de 25 ml. En contraste, las larvas de 5º estadio
fueron expuestas a Bt con trozos de 15 cm2 y mantenidas individualmente en
recipientes de plástico de 100 ml. El tiempo de exposición fue de 72 horas.
Pasado este período se registró el número de larvas muertas en cada
concentración, para cada estadio larval y en cada variedad. La distribución de
los tratamientos y el análisis estadístico de los resultados fue similar al citado
anteriormente, mediante análisis de varianza y pruebas de medias con la
prueba de Tukey y análisis Probit para la estimación de las CL50, con P « 0.05
de probabilidad de Error Tipo I.
3. 4. 3 Evaluar el efecto de la alimentación de larvas con follaje de maíz, de
diferentes variedades, sobre el desarrollo de las larvas.
Para este experimento se seleccionaron las mismas cuatro variedades del
bioensayo anterior, en los que en el primer experimento se encontraron valores
de CL50 bajos de Bt; éstos fueron el criollo Uruapeño y el híbrido V-526 y los
seleccionados, porque en ellos la bacteria Bt presentó valores altos en sus
CL50, las cuales fueron, el criollo Grano Negro y el híbrido Amarillo Forrajero.
Con cada uno de estas cuatro variedades se alimentaron 4 grupos de 10 larvas
neonatas de S. frugiperda y se registró el período que duraron para alcanzar el
último estadio larval y el período para llegar al estado biológico de pupa y
también se registró el peso de las larvas y pupas (Molina-Ochoa et al., 1997).

27
La distribución de los tratamientos y el análisis estadístico de los resultados fue
similar al citado anteriormente, mediante análisis de varianza y pruebas de
medias por Tukey al 5%.

28
IV RESULTADOS
La hipótesis de esta investigación fue probar que existe una influencia de follaje
de maíz de diferentes variedades sobre desarrollo y la susceptibilidad de larvas
de S. frugiperda a la actividad patógena de la bacteria B. thuringiensis. Los
resultados encontrados muestraron que efectivamente, la variedad de maíz
afecta la susceptibilidad de las larvas de S. frugiperda a la bacteria, y que
además los estadios larvales tienen una preferencia marcada para alimentarse
sobre ciertas variedades.
4.1 Susceptibilidad de larvas neonatas de S. frugiperda, alimentadas con hojas
de 13 variedades de maíz, a la bacteria B. thuringiensis variedad kenyae,
serovar 007.
En el testigo no se presentó mortalidad en las larvas; mientras que en la
concentración más alta con Bt se registró un 100% de mortalidad. Los valores
de CL50 de la bacteria B. thuringiensis, en larvas de S. frugiperda, alimentadas
con cada una de las variedades se presentan en el cuadro 2. Las larvas de S.
frugiperda alimentadas con hojas de las variedades criollas Uruapeño y Grano
ancho presentaron los valores más bajos de CL50 entre las variedades criollas
(0.087 y 0.097 mg/ml, respectivamente); en el resto de las variedades criollas
los valores de CL50 oscilaron entre 0.101 a 0.180 mg/ml; en el criollo Grano
Negro se presentó el más alto valor de CL50.
En lo referente a la susceptibilidad de las larvas a Bt, alimentadas con hojas de
variedades híbridas, se encontró que sus valores de CL50, variaron entre 0.072
y 0.181 mg/ml. En los híbridos V-526 y P-3288, se presentaron los valores más
bajos y en el híbrido Amarillo forrajero se encontró el valor más alto de todos
los híbridos (0.181 mg/ml) (Cuadro 2). Se puede observar que las variedades
(una variedad sintética y un criollo), V-526 y Uruapeño presentan los valores de
CL50 más bajo y que las variedades (un híbrido y un criollo) Amarillo forrajero y
Grano Negro, por su parte presentaron valores más altos de CL50 (Cuadro 2).

29
Cuadro 2. Valores de CL50 de la bacteria B. thuringiensis, en larvas de S.
frugiperda, alimentadas con cada uno de las variedades evaluados.
Variedad CL50 (mg/ml) Intervalo de Confianza (95%)
Cr Uruapeño
Cr Grano ancho (de ocho)
Cr Grano rojo
Cr Grano pinto
CrTampiqueño olote
delgado
Cr Grano negro (Jaripeño)
HV-526
H P-3288
H P-3086
H B-810
H Ceres 7
H 910
H Amarillo forrajero
0.087 ab
0.097 ab
0.101 ab
0.119 ab
0.121 ab
0.180 ab
0.072 b
0.073 ab
0.095 ab
0.109 ab
0.118 ab
0.121 ab
0.181 a
0.053 - 0.145
0.068 - 0.145
0.075 - 0.147
0.082 - 0.172
0.065 - 0.225
0.116 - 0.281
0.044 - 0.117
0.04 - 0.131
0.059 - 0.153
0.065 - 0.184
0.084 - 0.167
0.071 - 0.205
0.127 - 0.285
* Valores entre columnas seguidos de la misma letra son iguales
estadísticamente 95% de confianza. Cr.- Criollos H.- Híbridos.
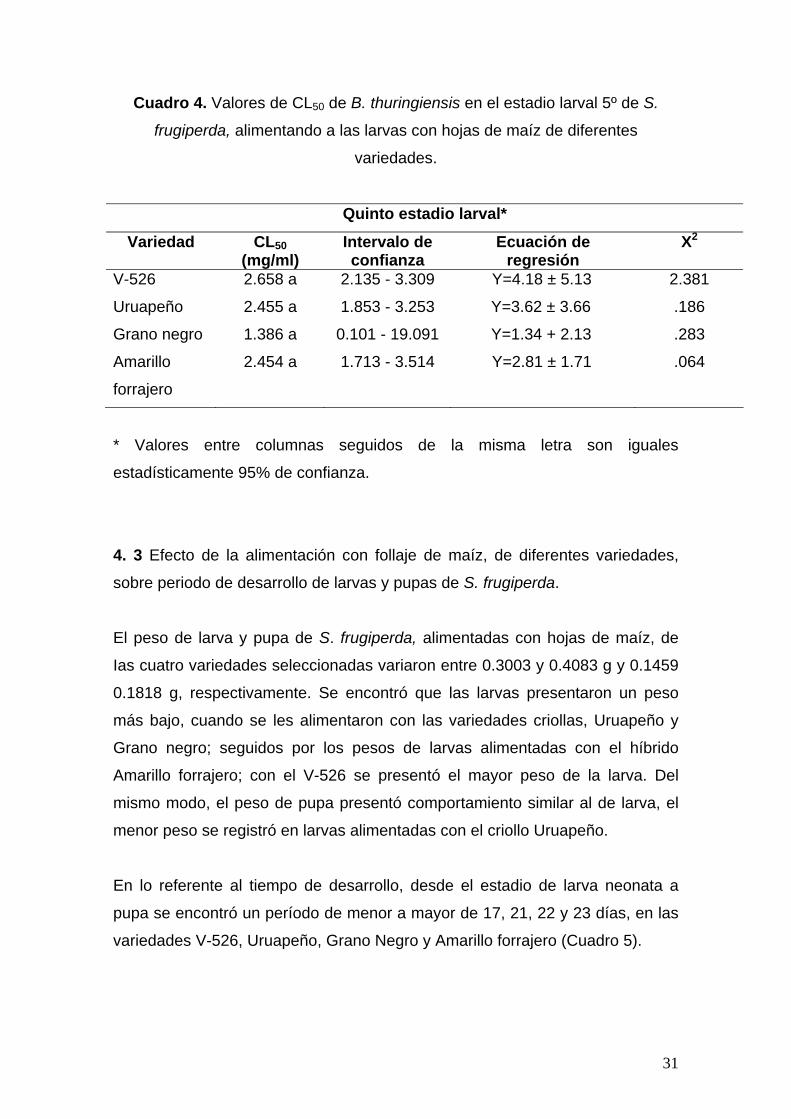
4. 2 Susceptibilidad de larvas de 3º y 5º estadio de S. frugiperda, a la bacteria
B. thuringiensis, alimentadas con hojas de maíz de variedades seleccionadas,
en base a los valores de CL50 altos y bajos de susceptibilidad a Bt.
Con los resultados obtenidos del bioensayo anterior se seleccionaron las
variedades, (una variedad sintética y un criollo), V-526 y Uruapeño, por
presentar los valores de CL50 más bajos y las variedades (un híbrido y un
criollo) Amarillo forrajero y Grano Negro, por su parte presentaron valores más

30
altos de CL50 de la Bt. Los valores de CL50 de la bacteria en estos estadios
larvales, se muestran en los Cuadros 3 y 4.
En larvas de tercer estadio se encontró que las larvas presentan de menor a
mayor susceptibilidad a Bt, cuando fueron alimentadas en las variedades V-526
< Uruapeño < Grano negro < Amarillo forrajero. En larvas de quinto estadio, la
susceptibilidad al Bt fue de V-526 < Uruapeño < Amarillo forrajero < Grano
negro. Sin embargo, en base a los entre cruzamientos de los intervalos de
confianza, de las respectivas CL50, se observa que no existen diferencias
estadísticas entre los valores obtenidos de CL50 de la bacteria, influenciado por
la variedad (Cuadro 3 y 4), pero si se encontró que los valores de CL50 de la
bacteria Bt en larvas de tercer estadio son menores, que los registrados en los
experimentos realizados con larvas de quinto estadio.
Cuadro 3. Valores de CL50 de B. thuringiensis en el estadio larval 3º de S.
frugiperda, alimentando a las larvas con hojas de maíz de diferentes
variedades.
Tercer estadio larval*
Variedad CL50 (mg/ml)
Intervalo de confianza
Ecuación de regresión
X2
V-526
Uruapeño
Grano negro
Amarillo
forrajero
1.372 a
1.059 a
0.891 a
0.677 ab
1.013 - 1.858
0.896 - 1.252
0.694 - 1.145
0.337 - 1.362
Y=3.55 ± 2.59
Y=5.78 ± 6.69
Y=4.07 ± 2.94
Y=1.23+2.75
1.544
.227
2.44
1.022
* Valores entre columnas seguidos de la misma letra son iguales
estadísticamente 95% de confianza.
31
Cuadro 4. Valores de CL50 de B. thuringiensis en el estadio larval 5º de S.
frugiperda, alimentando a las larvas con hojas de maíz de diferentes
variedades.
Quinto estadio larval*
Variedad CL50 (mg/ml)
Intervalo de confianza
Ecuación de regresión
X2
V-526
Uruapeño
Grano negro
Amarillo
forrajero
2.658 a
2.455 a
1.386 a
2.454 a
2.135 - 3.309
1.853 - 3.253
0.101 - 19.091
1.713 - 3.514
Y=4.18 ± 5.13
Y=3.62 ± 3.66
Y=1.34 + 2.13
Y=2.81 ± 1.71
2.381
.186
.283
.064
* Valores entre columnas seguidos de la misma letra son iguales
estadísticamente 95% de confianza.
4. 3 Efecto de la alimentación con follaje de maíz, de diferentes variedades,
sobre periodo de desarrollo de larvas y pupas de S. frugiperda.
El peso de larva y pupa de S. frugiperda, alimentadas con hojas de maíz, de
Ias cuatro variedades seleccionadas variaron entre 0.3003 y 0.4083 g y 0.1459
0.1818 g, respectivamente. Se encontró que las larvas presentaron un peso
más bajo, cuando se les alimentaron con las variedades criollas, Uruapeño y
Grano negro; seguidos por los pesos de larvas alimentadas con el híbrido
Amarillo forrajero; con el V-526 se presentó el mayor peso de la larva. Del
mismo modo, el peso de pupa presentó comportamiento similar al de larva, el
menor peso se registró en larvas alimentadas con el criollo Uruapeño.
En lo referente al tiempo de desarrollo, desde el estadio de larva neonata a
pupa se encontró un período de menor a mayor de 17, 21, 22 y 23 días, en las
variedades V-526, Uruapeño, Grano Negro y Amarillo forrajero (Cuadro 5).

32
Cuadro 5. Tiempo de desarrollo, peso de larva y peso de pupa de S. frugiperda
alimentadas con cuatro variedades de maíz.
Larvas Pupas
Variedad Peso (g)
Duración (días)
Peso (g)
Duración (días)
V-526
Uruapeño
Grano negro
Amarillo forrajero
0.4083
0.3003
0.3041
0.3399
15
19
20
20
0.1818
0.1459
0.1474
0.1526
17
21
22
23

33
V DISCUSION
Para la realización de la presente investigación se partió del supuesto de que al
utilizar una misma cepa o subespecie de B. thuringiensis y una sola población
de S. frugiperda, permitiría detectar el efecto de la fuente de alimento de las
larvas, sobre su susceptibilidad a Bt; lo anterior, tomando como base lo
reportado por López-Edwards et al. (1999) quienes encontraron, que las larvas
de S. frugiperda tienen diferente grado de susceptibilidad a Bt, de acuerdo al
origen geográfico de las larvas y Hernández (1988), de que las larvas de S.
frugiperda presentaba diferente susceptibilidad a Bt de acuerdo a la especie o
subespecie que se utilice.
Los resultados encontrados muestran que efectivamente, la variedad de maíz,
con el que se alimenten las larvas, afecta la susceptibilidad de las larvas de S.
frugiperda a la bacteria de B. thuringiensis y que además, las larvas se ven
afectadas en su tiempo de desarrollo larval y en el peso de las pupas;
posiblemente debido a que las larvas tienen cierto grado de preferencia entre
las variedades, ya que se observó que las larvas no querían alimentarse sobre
algunos de ellos.
Posiblemente existieron los mecanismos de resistencia vegetal de no
preferencia y/o antibiosis entre las variedades utilizadas en este estudio. Por lo
que se hacen necesarias más investigaciones, para determinar el tipo de
resistencia de estas variedades de maíz. Ya que cuando una larva ingiere
follaje que contenga alguna sustancia antibiótica, tales como el maysin y sus
análogos, ácido clorogénico, luteolina, rutina, teoapigenina, postapigenina o
premaysin (Gueldner et al., 1992) o Bt, por lo general deja de alimentarse,
debido a una parálisis intestinal, lo que por consecuencia no aumenta el peso;
en tanto que el uso de toxinas de Bt afectan el desarrollo larval y el fenómeno
de alimentación, producto de la intoxicación bacteriana, tal como lo reportan
Navon et al. (1992) y Stapel et al. (1998).

34
Los resultados obtenidos permiten explicar los siguientes fenómenos, una larva
alimentada con una variedad resistente al ataque de S. frugiperda, ya sea por
no preferencia o por antibiosis, no se desarrolla normalmente, el peso se ve
afectado y el período de desarrollo se alarga, tal como lo demostraron en sus
estudios Wiseman et al., (1976), Wiseman et al., (1992a, b) y Molina-Ochoa et
al. (1999); pero también, en este estudio se observó, que la bacteria de Bt
presentó el valor de CL50 más bajo en las larvas neonatas alimentadas con
hojas de la variedad V-526.
Con lo anterior se infiere, que las larvas mejor alimentadas están
fisiológicamente más fuertes y por ende, son menos sensibles a la actividad del
Bt. Por el contrario, en las larvas alimentadas con maíces no preferidos, se
alimentan menos, gastan más reservas (Reese y Schmidt, 1986) y son más
débiles y por lo tanto, se puede presentar una susceptibilidad y mortalidad más
altas.
Sin embargo, en los estudios del efecto de la variedad sobre el desarrollo larval
de S. frugiperda, se encontró que las larvas alimentadas con la variedad V-526
presentaron un tiempo más corto de larva y el peso fue el más alto; también los
valores de CL50 de la bacteria en larvas de 3º y 5º estadio fueron los más
elevados. Posiblemente, por que las larvas estaban más robustas y soportaron
más altas concentraciones de la bacteria; cosa que no ocurrió en larvas
alimentadas con las otras variedades (Simpson y Simpson, 1990).
Los resultados en este estudio son importantes, ya que permitieron conocer
que la bacteria utilizada en larvas neonatas, presenta valores de CL50
diferentes, de acuerdo a la variedad en que se alimentan; pero en esa edad
larval no se detecta el efecto de la planta sobre la tasa de desarrollo, ya que si
esta misma bacteria se utiliza en larvas de mayor edad (3º o 5º estadio, por
ejemplo) los valores son diferentes, de acuerdo a la interacción con la
resistencia de la planta. Tal como se detectó con el híbrido V-526, en este
estudio.

35
Lo anterior, tiene su aplicación práctica, por ejemplo, de acuerdo a la variedad
de maíz, en el que se quiere utilizar Bt, para el control de S. frugiperda, la CL50
de Bt no es la misma, por lo que de este estudio surge la recomendación de
que, no obstante que sean las mismas especies de Bt y de S. frugiperda, la
susceptibilidad de la plaga a la bacteria va a depender de la variedad del maíz
y de la edad de la larva, tal como lo reportan López-Edwards et al. (1999) y
Hernández (1988).
Resultados similares, asociados con la concentración de estigmas del maíz
resistente “Zapalote Chico” habían sido reportados por Molina-Ochoa et al.,
(1999) utilizando nematodos entomopatógenos; con los altos contenidos de
taninos y el uso del Dipel y Pydrin en contra de gusano bellotero en algodón;
además, Fuxa et al., (1988) reportaron una asociación de la edad del
hospedero y la cepa de nematodo entomopatógeno sobre S. frugiperda; y por
Schultz y Keating (1991) usando un baculovirus en contra de palomilla gitana.
Por lo que para emplear B. truringiensis, en condiciones de campo, para el
control del gusano cogollero, deberá previamente evaluarse la relación
existente entre la variedad de maíz, la población del insecto y su
susceptibilidad a B. thuringiensis.

36
VI CONCLUSIONES
El desarrollo de S. frugiperda expresado por las variables peso de larvas,
duración de los estadios larvales a pupa, peso de pupas y duración del estado
de pupa, fue afectado por las variedades de maíz.
Las variedades de maíz afectaron la susceptibilidad de los estadios larvales de
S. frugiperda a la bacteria entomopatógena B. thuringiensis.

37
VII LITERATURA REVISADA
Abbas Ali, R. G. Luttrell, H. N. Pitre, y F. M. Davis. (1989). Distribution of fall armyworm (Lepidoptera: Noctuidae) Egg Masses cotton. Environ. Entomol. 18(5): 881-885.
Adkisson, P. L., and V. A. Dyck. (1980). Resistant varietes in pest management
systems. pp. 233-251. In F. G. Maxwell and P. R. Jenninngs (Eds.). Breeding plant resistant to insect. Wiley. New York.
Anderson, R. F. y M. H. Rogoff. (1966): Crystalliferous bacteria and insect toxican. pp 67-69. In D.G. Crosby (Symposium Chairman). Natural Pest Control Agents. Advances in chem: Series N.53, Amer. Chem. Soc. Washington, D.C.
Andrews, K. L. (1980). The wholworm, Spodoptera frugiperda, in Central America and neighboring areas. Fla. Entomol. 63: 456-467.
Andrews, K. L. (1988). Latin American Research on Spodoptera frugiperda (Lepidoptera: Noctuidae). Fla. Entomol. 71(4): 630-653.
Angus, T. A. (1954). A bacterial toxin paralysing silkworm larvae. Nature 173: 54-56.
Ashley, T. R. (1979). Classification and distribution of fall armyworm parasites. Fla. Entomol. 62: 114-123.
Ashley, T. R., B. R. Wiseman, F. M. Davis, y K. L. Andrews. (1989). The fall armyworm: a bibliography. Fla. Entomol. 72: 152-202.
Aronson A. L., W. Beckman y D. Peter. (1986). Bacillus thuringiensis and related insects pathogens. Microbiol. Rev. 56: 1-24.
Atkinson J.; P. Morand y J. T. Arnason. (1990). Analogues of ciclic Hidroxamic. Acid 2,4-dihidroxy-2 H-1,4 benzoxazin-3-one: Descomposition to Benzoxazolinones and reaction with beta-Mercaotoethanol. J. Org. Chemistry. 56(5): 1788-1800.
Banda T. J. F. (1981). Importancia económica de Heliothis zea (Boddie) y determinación del umbral económico, distribución matemática y muestreo secuencial de Spodoptera frugiperda (J. E. Smith) en maíz criollo. Tesis Doctorado, ITESM Monterrey N.L: p. 74.
Barbercheck, M. E. (1993). Tritrophic level effects on entomopathogenic nematedes. Environ. Entomol. 22: 1166-1171.
Barbosa, P., V. A. Krischik, y G. C. Jones (Eds.). (1991). Microbial mediation of plant-herbivore interactions. Wiley, New York.
Bauer, L. S. (1995). Resistance: A threat to the insecticidal crystal proteins of Bacillus thuringiensis. Fla. Entomol. 78(3): 414-443.

38
Beadle, G. W. (1939). Teosinte and the origin of Zea mays. J. Heredity. 30: 245-247.
Beadle, G. W. (1980). The ancestry of corn. Scientifïc American. 242: 112-119.
Behle, R. W., M. R. McGuire, R. L. Gillespie, y B. S. Shasha. (1997). Effects of alkaline gluten on the insecticidal activity of Bacillus thuringiensis. J. Econ. Entomol. 90(2): 354-360.
Bell, J. A. (1978). Development and mortality in bollworm for resistant and susceptible soybean cultivars treated with Nomuraea rileyi or Bacillus thuringiensis. J. Ga. Entomol. Soc. 13: 50-55.
Benz B. F. (1993). Introducción en biología, ecología y conservación del género Zea. Universidad de Guadalajara: 9 -11.
Bergman, J. M., y W. M. Tingey. (1979). Aspects of interactions between plant genotypes and biological controls. Bull. Entomol. Soc. Am. 25: 275-279.
Bernhard, K., P. Jarret, M. Meadows, J. Butt, D. J. Ellis, G. M. Roberts, S. Pauli, P. Rodgers, y H. D. Burges. (1997). Natural isoletes of Bacillus thuringiensis: Worldwide distribution, characterization, and activity against Insect pests. J. Invertebr. Pothol. 70: 59-68.
Boethel, D. J., y R. D. Eikenbary (Eds.). (1986). Interactions of plant resistance and parasitoids and predators of insect. Wiley, New York.
Bolin, C. P., D. H. Hutchison, y W. D. Davis. (1996). Resistant Hybrids and Bacillus thuringiensis for Management of European Corn Borer (Lepidopteran: Pyralidae) in Sweet Corn. J. Econ. Entomol. 89(1): 82-91.
Bon, R. P. M., C. B. C. Boyce y J. Fiench. (1969). A purification and some properties of an insecticide exotoxin from Bacillus thuringiensis Berliner. Biochem. J. 144:477-488.
Borbolla, I. S. (1981). Estudio comparativo de insecticidas a diferentes dosis y número de aplicaciones para el control de gusano cogollero Spodoptera frugiperda (J.E.Smith) en maíz de temporal. Agronomía en Sinaloa-UAS 1:21-30.
Brattsten, L. B., C. W. Holyoke, Jr., J. R. Leeper, y K. F. Raffa. (1986). Insecticide resistance: challenge to pest management and basic research. Sci. 231: 1255-1260.
Bucher, G . E . (1960). Potential bacterial pathogens of insects and their characteristics. J. Insect Pathol. 2: 172-195.
Burges, H. D. (1982). Control of insects by bacteria. Parasitology. 84: 79-117.
Burton, R. L. y W. D. Perkins. (1989). Rearing the corn earworm and fall armyworm for maize resistance studies. Pp. 37-45. In Toward insect resistant maize for the third world: Proceedings of the international

39
Symposium on Methodologies for Developing host plant resistance to maize insects. Mexico, D. F.: CYMMYT.
Cannon, R. J. C. (1996). Bacillus thuringiensis use in agriculture: a molecular perspective. Biol. Rev. 71: 561-626.
Carpenter, J. E. y J. R. Young. (1991). lnteraction of inherited sterility and insecticide resistance in the fall armyworm (Lepidoptera: Noctuidae). J. Entomol. 84: 25-27.
Carpenter, J. E., J. R. Young, E. F. Knipling y A. N. Spark. (1983). Fall armyworm (Lepidoptera: Noctuidae) inheritance of gamma-induced deleterious effects and potential for pest control. J. Econ. Entomol. 76: 378-382.
Chang, N. T., R. E. Lynch, F. A. Slansky, B. R. Wiseman, y D. H. Habech. (1987). Quantitative utilization of selected grasses for fall armyworm larvae. Entomol. Exp. Apple. 45: 29-35.
Costa-Camelles, J., D. Bosch, A. Botargues, P. Cabiscol, A. Moreno, J. Portillo, N. Ris, E. Santalinas, M. J. Sarasua y j. Avila. (1992). Resultados de la aplicación en parcelas comerciales de un programa de control integrado de manzano en Lleida. Bol. San. Veg. Plagas. 18: 745-754.
Clavijo, S. (1984). Efecto de la fertilización con nitrógeno y de diferentes niveles de infestación por Spodoptera frugiperda (Lepidoptera: Noctuidae) sobre los rendimientos del maíz. Rev. Fac. Agron. (Macaray), XIII (81-4): 73-78.
Creighton, C. S., T. L. McFadden, y M. L. Robbins. (1975). Complementary influence of Bacillus thuringiensis host plant resistance and microbial-enemical control of cabbage caterpillars. HortScience 10: 487-488.
De Barjac, H. y A Bonnefoi. (1968). A classification of strains of Bacillus thuringiensis Berliner with a key to their differentiation. J. Invertebr. Pathol. 11: 335-347.
De Barjac, H. y E. Frachon. (1990). Classification of Bacillus thuringiensis strains. Entomophaga. 35: 233-240.
Del Cañizo P. J. A., V. R. Moreno, y A. C. Garijo. (1990). Guía práctica de plagas. Segunda edición. Madrid España: p. 51.
Diawara, M. M., B. R. Wiseman, y D. J. Isenhour. (1991). Bioassay for screening plant accessions for resistance to fall armyworm (Lepidoptera: Noctuidae) using artificial diets. J. Entomol. Sci. 26: 367-374.
Dicke, F. F. (1972). Philosophy on the biological control of insect pests. J. Environ. Qual. 1: 249-253.
Dow, J. A. T. (1986). Insect midgut function. Adv. Insect Physiol. 19; 187-328.

40
Dulmage, H. T. (1971). Production of delta-endotoxina by eighteen isoletes of Bacillus thuringiensis serotype 3 in fermentation media. J. Invertebr. Pathol. 18: 353-358.
Entwistle, P., M. J., Bailey, J. Cory, y S. Higgs, (Eds.) (1993). Bacillus thuringiensis: An. Environ. Biopestic. (Wiley, New York).
Epsky, N. D., y J. L. Capinera. (1993). Quantification of invasion of two strains of Steinernema carpocapsae (Weiser) into three lepidopteran larvae. J. Nematol. 25: 173-180.
Falcon, L. A. (1971). Use of bacteria for microbial control. In H.P. Burguess y N. W. Hussey (eds). Microbial Control of Insects and Mites. Academic Press, London. New York. 3: 67-95.
Fast, P. G. (1974). Bacillus thuringiensis its history and mode of action. Dev. Ind. Microbial. 15: 195-198.
Fast, P. G., D. W. Murphy, y S. S. Sohi. (1978). Bacillus thuringiensis -endotoxin: evidence that toxin acts at the surface of susceptible cells. Experientia. 34: 762-763.
Felton, G. W., and D. L. Dahlman. (1984). Allelochemical induced stress: effects of L-canavanine on the pathogenicity of Bacillus thuringiensis in Manduca sexta. J. Invertebr. Pathol. 44: 187-191.
Fernández, W. L. y V. L. Estefania (1974). Coconut water waste disposal of some desiccated coconut factories in the Philippines. Phil. Agric. 57: 359-363.
Figueroa C. J. D., B. F. Martínez, H. J. González, S. F. Sánchez., M. J. Martínez y T. M. Ruiz. (1994). Modernización tecnológica del proceso de nixtamalización. Avance y Perspectiva 13:323-329.
Foott, W. H. y P. R. Timmins. (1982). Injury to grain corn by the fall armyworm, Spodoptera frugiperda (Lepidoptera: Noctuidae), in Southuestein Ontorio, 1977-1981. Proc. Ent. Soc. Ont. 113: 17-20.
Fuxa, J. R., A. R. Ritcher, y F. Agudelo-Silva. (1988). Effect of host age and nematode strain on susceptibility of Spodoptera frugiperda to Steirnernema feltiae. J. Nematol. 20: 91-95.
Gatehouse, A. M. R., V. A. Hilder, y D. Boulter. (1992). Potential of plant-derived genes in the genetic manipulation of crops for insect resistance, in: Biotechnology in Agriculture No 7: Plant Genetic Manipulation for Crop Protection. CAB International, pp. 155-181.
Gardner, A. W. y J. R. Fuxa. (1980). Pathogens for the suppression of the fall armyworm. Fla. Entomol. 63: 439-447.

41
Gardner, A. W., N. Raymond, y D. S. Robert. (1984). The potential of microbial agents in managing populations of the fall armyworm (Lepidoptera: Noctuidae). Fla. Entomol. 67(3): 325-332.
Gawron-Burke C. and J. A. Baum. (1991). Genetic manipulation of Bacillus thuringiensis insecticidal crystal protein genes in bacteria, In: Setlow JK (Ed.). genetic engineering principles and methods, 13. Plenum Press, New York, p 237.
Georghiou, G. P. y A. Lagunes-Tejeda. (1991). The occurrence of resistance to pesticides in arthropods. Food and Agriculture Organization of the United Nations, Rome.
Gill S. S., E. A. Cowles, y P. V. Pietrantonio. (1992). The mode of action of Bacillus thuringiensis endotoxins. Ann. Rev. Entomol. 37: 615-636.
González J. M. Jr. y C. C. Bruce (1980). Patterns of plasmid DNA in crystalliferous strains of Bacillus thuringiensis. Plasmid. 3: 89-92.
Gueldner, R. C., M. E. Snook, N. W. Widstrom, y B. R. Wiseman. (1992). TLC Screen for maysin, chlorogenic acid, and other posible resistance factors to the fall armyworm and the corn earworm in Zea mays. J. Agric. Food. Chem. 40:1211-1213.
Hall, J. M., K. Aizawa, H. T. Dulmage y J. A. Correa. (1977). The pathogenicity of strains of Bacillus thuringiensis to larvae of Aedes and to Culex mosquitoes. Mosquito News. 37:246-251.
Hamm, J. J., y B. R. Wiseman. (1986). Plant resistance and nuclear polihedrosis virus for suppression of the fall armyworm (Lepidoptera: Noctuidae). Fla. Entomol. 69: 541-549.
Hare, J. D., y T. G. Andreadis. (1983). Variation in the susceptibility of Leptinotarsa decemlineata (Coleoptera: Chrysomelidae) when reared on different host plants to the fungal pathogen, Beauveria bassiana in the field and laboratory. Environ. Entomol. 12: 1892-1897.
Haufler, M. y E. K. Sindey. (1985). Laboratory evaluation of an exotoxin from Bacillus thuringiensis subsp. morrisoni to horn fly larval (Diptera: Muscidae) and mice. J. Econ. Entomol. 78: 613-616.
Heimpel, A. M. y A.T. Angus. (1963). Diseases caused by certain sporeforming bacteria. pp. 21-73 In E. A. Steinhaus (ed.). Insect oathology an Advanced Treatise Vol. 2 Academic Press, New, York.
Heimpel, A. M. (1967). A critical review of Bacillus thuringiensis Berliner and other crystalliferous bacteria. Ann. Rev. Entomol: 12:287-316.
Hernández M. J. L. (1988). Evaluation de la Toxicité de Bacillus thuringiensis sur Spodoptera frugiperda. Entomophaga. 33: 163-171.

42
Hernández M. J. L. (1989). Ecopathologie et degats de Spodopfera frugiperda (J. E. Smith) en culture de mais au Mexique (etat de Colima) posibilite de lutte a I`aide de la bacterie entomopatogene Bacillus thuringiensis. Tesis Doctorado, Escuela Nacional Superior de Agronomía, ENSAM Montpellier, Francia: 147 p.
Hernández C. J. (1994). Reproducción de lepidópteros de cereales (Diatraea spp, Heliothis zea [Boddie] y Spodoptera frugiperda [Smith]). Técnicas para la cría de insectos. Colegio de Postgrados en Ciencias Agrícolas: 49 p.
Hernández J. E. y A. Benderly. (1982). Nuevos conceptos químicos de los plaguicidas. Ciencia y desarrollo 47:33-48.
Höfte, H. y H. R. Whiteley. (1989). Insecticidal crystal proteins of Bacillus thuringiensis. Microbiol. Rev. 53: 242-255.
Huber, H. E. y P. Luthy. (1981). Bacillus thuringiensis delta endotoxin; composition and activation. pp. 209-234. In E. W. Davidson (Ed.). Pathogenesis of Invertebrate Microbial Diseases. Allenheld. Osmun Publisher. N. J.
Huston, M. (1993). Biological diversity, soils, and economics. Sci. 262: 1676-1680.
Ignoffo, C. M. (1974). Microbial control of insects: viral pathogens. In Proceedings of Summer Institute on Biological Control of Plant Insects and Diseases. Maxwell y F. A. Harris (Ed.). Univ. Press, Jackson, Miss. 541-557
INEGI. (1991). Instituto Nacional de Estadística Geografía e Informática. Atlas Agropecuario del Estado de Colima. P. 21.
Isenhour, D. J. y B. R. Wiseman. (1991). Fall armyworm resistance in progeny in maize plants regenerated via tissue culture. Fla. Entomol. 74(2): 221-228.
Kea, W. C., S. G. Turnipseed, y G. R. Carner. (1978). Influence of resistant soybeans on the susceptibility of Lepidopterous pest to insecticides. J. Econ. Entomol. 71: 58-60.
Knowles H. B., W. E. Thomas, y D. J. Ellar. (1984). Lectinlike binding of Bacillus thuringiensis var. Kurstaki lepidopteran-specific toxin is an initial step in insecticidal action. FEBS Microbiol. Lett. 168: 197-202.
Knowles H. B., y J. A. T. Dow (1993). The Crystal - endotoxins of Bacillus thuringiensis models for their mechanism of action on the insect gut. BioEssays. 15(7): 469-476.
Krieg, A., H. De Barjac y A. Bonnefoi. (1968). A new serotype of Bacillus thuringiensis var. darmstadiensis. J. Inverteb. Pathol. 10: 428-430.
Krieg. A. y G. M. Herbert. (1984). Bioinsecticides: I Bacillus thuringiensis. Adv. Biotechnol. Proc. 3: 273-290.

43
Krischik, V. A., P. Barbosa, and C. F. Reichelderfer (1988): Three trophic levels interactions: allelochemicals. Manduca sexta (L), and Bacillus thuringiensis subs. Kurstaki Berliner. Environ. Entomol. 17: 476-482.
Lagunes T. A., M. C. Rodríguez y R. R. Domínguez (1985): Plagas del maíz en la mesa central. Centro de Entomología y Acarología, Colegio de postgraduados, México: 18-21.
Landazabal, J., F. Fernández y A. Figueroa, (1973): Control biológico de Spodoptera frugiperda (J. E. Smith), con el nematodo: Neoaplectana carpocapsae en maíz Zea mayz. Acta Agron. (Colombia); 23: 41-70.
Lidstone, D. W. G. y G. G. Khachatourians. (1985). A standarized bioassay for larvicidal activity of Bacillus thuringiensis in sun flower moth Homoeosa electellum (Lepidoptera:Phycitidae). Can. Entomol. 117: 15-22.
Lise, J., M. Bonadé-Bottino, C. Girard, G. Morrot, y M. Giband. (1998). Transgenic plants for insect resistance. Plant Sci. 131: 1-11.
López-Edwards, M., J. L. Hernández-Mendoza, A. Pescador-Rubio, J. Molina-Ochoa, R. Lezama-Gutiérrez, J. J. Hamm and B. R. Wiseman. (1999). Biological diferences between five populations of fall armyworm (Lepidoptera:Noctuidae) collected from corn México. Fla. Entomol. 82(2): 254-262.
Ludlum, C. T., G. W. Felton, and S. S. Duffey. (1991). Plant defenses: chlorogenic acid and polyphenol oxidase enhance toxicity of Bacillus thuringiensis subsp. Kurstaki to Heliothis zea. J. Chem. Ecol. 17: 217-237.
Luginbill, P. (1928). The fall armyworm. USDA. Tech. Bull: 34. 92.
Luginbill, P. (1969). Developing resistant plants the ideal method of controlling insect. USDA. Pro. Res. Rep: 111. 14.
Lüthy, P. (1983). Physiology of delta-endotoxin of Bacillus thuringiensis including the ultrastructure and histopathological studies. In Basic biology of Microbial Larvacides of Vectores of Human Diseases. Michael, F. (Ed.). UNDP/WHO, Genoba. 29-36.
McClintock J. T., C. R. Schaffer y R. D. Sjoblad. (1995). A comparative Review the mammalian toxity of Bacillus thuringiensis-based pesticides. Pestic Sci. 45: 95-105.
Meade, T., y J. D. Hare. (1994). Effects of genetic and environmental host plant variation on the susceptibility of two noctuids to Bacillus thuringiensis. Entomol. Exp. Appl. 70: 165-178.
Meade, T., y J. D. Hare. (1995). Integration of host plant resistance and Bacillus thuringiensis insecticides in the management of lepidopterous pests of celery. J. Econ. Entomol. 88(6): 1787-1794.

44
Meckenstock, D. H., M. T. Castro, H. N. Pitre, y F. Gómez. (1991). Antibiosis to fall armyworm in Honduras, Landrace Sorghum. Environ. Entomol. 20: 1259-1266.
Meyer, J. A. (1986). Biological control of filth flies associated with confined livestock. Miscellaneus Publication Biological Control of Muscoid Flies. USDA-ARS 61:108-115.
Mohad-Salleh, M. B. y C. C. Beegle. (1980). Fermentation media and production of exotoxin by thee varieties of Bacillus thuringiensis. J. Invertebr. Pathol. 35: 75-83.
Molina-Ochoa J., J. J. Hamm, R. Lezama-Gutiérrez, M. Arenas-Vargas, M Arenas-Vargas y M. González-Ramírez. (1996). Virulence of six entomophogenic nematedes (Steinernematidae and Heterorabditidae) on immature stages of Spodoptera frugiperda (Lepidoptera: Noctuidae). Vedalia 3: 25-29.
Molina-Ochoa J., B. R. Wiseman, R. Lezama-Gutiérrez, J. J. Hamm, O. Rebolledo-Dominguez, M. González-Ramírez y Arenas-Vargas. (1997). Impact of resistant “Zapalote Chico” corn silks on Spodoptera frugiperda (Lepidoptera: Noctuidae) growth and development. Vedalia 4: 31-34.
Molina-Ochoa J., R. Lezama-Gutiérrez, J. J. Hamm, B. R. Wiseman y López-Edwards, M. (1999). Integrated control of fall armyworm (Lepidoptera:Noctui dae) using resistant plants and entomopathogenic nematodes (Rhabditida: Steinernematidae). Fla. Entomol. 82(2): 263-271.
Morón, A. M. y R. Terrón (1988). Entomología práctica. Instituto de Ecología. México: 12, 373, 423.
National Academy of Sciences. (1992). Control de plagas de plantas y animales. México, 1: 1-5.
Navon, A., J. D., B. A. Federici, T. S. Walsh, y U. M. Peiper. (1992). Mandibular Adduction Forte of Heliothis virescens (Lepidoptera: Noctuidae) larvae Fed the Insecticidal Crystals of Bacillus thuringiensis. J. Econ. Entomol. 85(6): 2138-2143.
Navon, A., J. D. Hare, y B. A. Federici. (1993). Interactions among Heliothis virescens larvae. Cotton condensed tannin and the CrylA(c) - endotoxin of Bacillus thuringiensis. J. Chem. Ecol. 19: 2485-2499.
Painter, R. H. (1951). Insect resistence in crop plants. Macmillan Publ. Co., New York: 520 p.
Painter, R. H. (1968). Crops that resistant insect provide a way to increase world food supply. Kansas Agric. Exp. Stn. Bull. 520: 22.
Pashley, D. P. (1986). Host-associated genetic differentiation in fall armyworm (Lepidoptera: Noctuidae) a sibling species complex?. Ann. Entomol. Soc. Am., 79: 898-904.

45
Pashley, D. P. (1988). Current status of fall armyworm host strains. Fla. Entomol. 71: 227-234.
Price, P. W., C. E. Bouton, P. Gross, B. A. McPheron, J. N. Thompson y A. E. Weiss (1980). Interactions among three trophic levels: Influence of plants on interactions between insect herbivores and natural anemies. Ann. Rev. Ecol. Syst. 11: 41-65.
Ramoska, W. A., y T. Todd. (1985). Variation in the efficacy and viability of Beauveria bassiana in the chinch bug (Hemiptera: Lygaeidae) as a result of feeding activity on selected host plants. Environ. Entomol. 14: 146-148.
Reese, J. C., y D. J. Schmidt. (1986). Physiological aspects of plant-insect interactions. Iowa State. J. Res. 60(4): 545-567.
Reichelderfer, C. F. (1991). Interactions among allelochemicals, some Lepidoptera. and Bacillus thuringiensis Berliner, pp. 507-524. In P. Barbosa, V. A. Krischilk y C. G. Jones (eds.). Microbial mediation of plant-herbivore interactions. Wiley, New York.
Roush, R. T., y J. A. McKenzie. (1987). Ecological genetics of insecticide and acaricide resistance. Ann. Rev. Entomol. 32: 361-380.
SARH. (1993). Guía para cultivar maíz de riego en el estado de Colima. Instituto Nacional de investigaciones Forestales y Agropecuarias. Centro de Investigaciones del Pacífico Centro Colima. Folleto para productores. N. 4. Colima, Colima, México.
Schultz, J. C., y S. T. Keating. (1991). Host-plant mediated interactions between the gypsy moth and a baculovirus, pp. 489-506. In P. Barbosa, V. A. Krischik and C. G. Jones (eds.) Microbial mediation of plant-herbivore interactions. Wiley, New York.
Schuster, M. F., P. D. Calvin, y W. C. Langston. (1983). Interaction of high-tannin with bollworm control by Pydrin and Dipel. pp. 72-73. In proceedings, Beltwide Cotton Production Research Conferences. National Cotton Council of America. Memphis.
Silvain, J. F. (1987). Spodoptera frugiperda. SUAD-ORSTOM, Francia, 2 p.
Simmons, A. M., y B. R. Wiseman. (1993). James Edward Smith, taxonomic author of Fall armyworm. Fla. Entomol. 76: 271-276.
Simpson, S. J. Y C. L. Simpson. (1990). The mechanisms of nutritional compensation by phytophagous insects. Pp. 111-160. In: Bernays, E. A. (Ed.). Focus on Insect-plant Interactions. 2, CRC, Boca Raton, FI.
Singh, G. J. P., P. Leo, Jr y S. G. Sarjeet. (1986a). The toxin action of Bacillus thuringiensis var. israelensis in Aedes aegypti in vivo. Pestic. Biochem. Physiol. 26: 47-55.

46
Singh, G. J. P., P. Leo, Jr y S. G. Sarjeet (1986b). Action of Bacillus subsp. israelensis delta-endotoxin on the ultraestructure of the house fly larva neuromuscular system in vitro. J. Invertebr. Pathol. 47: 155-166.
Slansky, F. Jr. (1992). Insect Nutritional Ecology as a basis for Studying Host Plant Resistance. Fla. Entomol. 73(3): 359-378.
Slansky, F. Jr. y Wheeler. (1991). Food consumption and utilization responses to dietary dilution with celluloce and water by velvetbean caterpillars, Anticarsia gemmatalis. Physiol. Entomol. 16: 99-116.
Smith, C. M. (1989). Plant resistance to insects: a fundamental approach. Wiley, New York.
Sparks, N. A. (1979). A review of the biology of the fall armyworm. Fla. Entomol. 62: 82-87.
Stanier, R. Y., E. A. Adelberg y J. L. Igraham. (1986). Microbiología. 4 (Ed.). México: 621-624.
Stapel, J. O., J. W. Deborah; J. R. Ruberson, y W. L. Joe. (1998). Development and behavior of Spodoptera exigua (Lepidoptera: Noctuidae) larvae in Choice Test with Food Substrares containg toxins of Bacillus thuringiensis. Biological control. 11(1): 29-37.
Tabashnik B. E., Y. B. Liu, N. Finson, L. Masson y D. G. Heckel. (1997). One gene in diamondback moth confers resistance to four Bacillus thuringiensis toxins. Proc. Natl. Acad. Sci. 94: 1640-1644.
Tanada y H. K. Kaya. (1993). Insect pathology. Academic Press, Inc. New York. 666 p.
Temeyer, K. B. (1984). Larvacidal activity of Bacillus thuringiensis subsp. israelensis in the diptera Haematobia irritans. Applic. Environ. Microbiol. 5: 252-255.
Thomas, W. E. y J. E. David. (1983). Bacillus thuringiensis var. Israelensis crystal delta-endotoxin: effects on insect and in vivo. J. Cell. Sci. 60: 118-197.
Tood, E. L. y R. W. Poole. (1980). Key and illustrations for the fall armyworm moths of noctuid genus Spodoptera Guenée from the Westerm Hemissphere. Ann. Entomol. Soc. Amer. 73(6): 722-738.
Trumble, J. T. (1990). Vegetable insect control with minimal use of insecticides. HortScienci. 25: 159-164.
Van Rie J., S. Jansen, H. Höfte, D. Degheele y H. Van Mellaert. (1990). Receptors on the brush border membrane of the insect midgut as determinants of the specifity of Bacillus thuringiensis d-endotoxins. Appl. Env. Microbiol. 56: 1378-1385.

47
Vickery, R. A. (1929). Studies on the fall armyworm in the Gulf Coast District of Texas. USDA Tech. Bull: 138:68
Walton W. R. y P. Luginbill. (1917). The fall armyworm, or grass worm, and its control. USDA Farmers Bull: 752: 3-12.
Williams W. P., y M. P. Buckley. (1992). Growth of Fall armyworm (Lepidoptera: Noctuidae) larvae on resistant and Susceptible Corn. J. Econ. Entomol. 85(5):2039-2042.
Wiseman B. R., W. W. McMillian y N. W. Widstrom. (1976). Feeding of corn earworm in the laboratory on excised silks of selected corn entries with notes on Orius insidiosus. Fla. Entomol. 59: 305-308.
Wiseman B. R., N. W. Widstrom, y W. W. McMillian. (1977). Ear characteristics and mechanism of resistance among selected corn earworm. Fla. Entomol. 60(2): 97-104.
Wiseman B. R. Y F. M. Davis. (1979). Plant resistence to the fall armyworm. Fla. Entomol. 62: 123-130.
Wiseman B. R., M. E. Snook, D. J. Isenhour, J. A. Mihm, y N. W. Widstrom. (1992a). Relationship between growth of corn earworm and fall armyworm larvae (Lepidoptera: Noctuidae) and maysin concentration in corn silks. J. Econ. Entomol. 85: 2473-2477.
Wiseman B. R., M. E. Snook, R. L. Wilson, y D. J. Isenhour. (1992b). Allelochemical content of selected popcorn silks-Effects on growth of corn earworm (Lepidoptera: Noctuidae). J. Econ. Entomol. 85: 2500-2504.
Wiseman B. R., F. M. Davis y W. P. Williams. (1983). Fall armyworm larval density and movement as an indication of nonpreference in resistant corn protection. Ecology 5: 135-141.
Wiseman B. R. (1985). Types and mechanisms of host plant resistance to insect attack. Insect Sci. Applic. 6: 239-242.
Wiseman B. R. (1987). Host plant resistance in crop protection in the 21st Century. pp. 505-509. In: Proceedings International Congress of plant protection, Manila, Philipines.
Wiseman B. R. y J. J. Hamm. (1993). Nuclear polyhedrosis virus and resistant corn silks enhance mortality of corn earworm (Lepidoptera: Noctuidae) larvae. Biological control 3: 337-342.
Yang G., D. J. Isenhour, y K. L. Espelie. (1991). Activity of maize leaf cuticular lipido in resistance to leaf-feeding by the fall armyworm. Fla. Entomol. 74: 229-236.
Yang G., B. R. Wiseman, D. J. Isenhour y K. L. Espelie. (1993). Chemical and ultrastructural analisys of Corn cuticular lipids and their effect on feeding by fall armiworm larvae. J. Chem. Ecol. 19(9):2055-2073.

48
Young R. J. (1979). Fall armyworm control with insecticides. Fla. Entomol. 62: 130-133.
Yu, S. J. (1991). Insecticide resistance in the fall armyworm, Spodoptera frugiperda (J. E. Smith). Pestic. Biochem. Physiol. 39: 84-91.
Yu, S. J. (1992). Detection and biochemical characterization of insecticide resistance en fall armyworm, Spodoptera frugiperda (J. E. Smith). J. Econ. Entomol. 85: 675-682.


















