
Universidad Andrés Bello EFECTO DE LA MANIPULACIÓN DEL ...
65
Universidad Andrés Bello Facultad de Ciencias Biológicas Escuela de Ingeniería en Biotecnología EFECTO DE LA MANIPULACIÓN DEL EQUILIBRIO FUENTE-SUMIDERO EN EL PERFIL DE METABOLITOS ASOCIADOS A CALIDAD EN FRUTOS DE Prunus persica (L.) BATSCH VAR `BUEN LINDO´ Tesis presentada como parte de los requisitos para optar al Grado de Magíster en Biotecnología Profesor patrocinante: Dra. Andréa Miyasaka de Almeida Facultad de Ciencias Biológicas Universidad Andrés Bello Profesor co-patrocinante: Dra. Romina Pedreschi Escuela de Agronomía Pontificia Universidad Católica de Valparaíso María Victoria Lillo Carmona Santiago, Chile Julio, 2017
Transcript of Universidad Andrés Bello EFECTO DE LA MANIPULACIÓN DEL ...

Universidad Andrés Bello
Facultad de Ciencias Biológicas
Escuela de Ingeniería en Biotecnología
EFECTO DE LA MANIPULACIÓN DEL EQUILIBRIO FUENTE-SUMIDERO EN
EL PERFIL DE METABOLITOS ASOCIADOS A CALIDAD EN FRUTOS DE
Prunus persica (L.) BATSCH VAR `BUEN LINDO´
Tesis presentada como parte de los requisitos para optar
al Grado de Magíster en Biotecnología
Profesor patrocinante: Dra. Andréa Miyasaka de Almeida
Facultad de Ciencias Biológicas
Universidad Andrés Bello
Profesor co-patrocinante: Dra. Romina Pedreschi
Escuela de Agronomía
Pontificia Universidad Católica de Valparaíso
María Victoria Lillo Carmona
Santiago, Chile
Julio, 2017

2
UNIVERSIDAD ANDRÉS BELLO
Facultad de Ciencias Biológicas
Ingeniería en Biotecnología
" EFECTO DE LA MANIPULACIÓN DEL EQUILIBRIO FUENTE-SUMIDERO
EN EL PERFIL DE METABOLITOS ASOCIADOS A CALIDAD EN FRUTOS DE
Prunus persica (L.) BATSCH VAR `BUEN LINDO´ ”
María Victoria Lillo Carmona
Este trabajo fue elaborado bajo la supervisión del Director de Tesis Dra. Andréa Miyasaka
de Almeida, en el laboratorio de Fisiología Vegetal y Genómica de frutales del Centro de
Biotecnología Vegetal, Facultad de Ciencias Biológicas, Universidad Andrés Bello,
aprobado por los miembros de la Comisión de evaluación.
SANTIAGO-CHILE
2017
Dra. Andréa Miyasaka de Almeida
Directora de Tesis
Dr. Reinaldo Campos
Comisión de Tesis
Dra. Romina Pedreschi
Co-Directora de Tesis
Dra. María Alejandra Moya
Comisión de Tesis

3
A mi estrellita en el cielo, Benny…

4
AGRADECIMIENTOS
A los primeros que quiero agradecer, además de dedicar todo este trabajo, son a mis padres,
María Elena y José. Creo que faltan palabras para agradecer todo lo que han hecho por mi
durante todos estos años, por sus palabras, motivación y enseñanzas, además de la
paciencia que tuvieron conmigo. Gracias papis.
También agradecer a la Dra. Andréa, gracias por darme la oportunidad de estar en su
laboratorio y confiar en mi trabajo, además de toda la ayuda que me brindó durante este
proceso. A la Dra. Romina, gracias por recibirme en Quillota y por sus enseñanzas. A
Claudia quien tuvo mucha paciencia en enseñarme, gracias por tu buena voluntad y
disposición, además de hacerme sentir como en casa. A Viveros El Tambo por el acceso al
material vegetal. A la Dra. María Luisa Valenzuela por su aporte al análisis de mis
resultados. A todo el grupo AM, en especial al grupo durazno y a Carlos, Lore, Esteban y
Luciano, gracias por las risas, la paciencia y la buena disposición. A la Maca, quien
escuchó todas mis tonteras y soportó el estrés mientras escribía. Agradecer a mis hermanas,
Nena, Eu y Kika por su apoyo y por escucharme siempre. A mis pequeños Fernanda y
Martín, que llegaron sólo a darme alegrías en momentos de estrés. A mi tía Margarita, por
su apoyo incondicional. A mi amigo del alma, mi BFF, Miguel. Sin ti definitivamente ésto
no habría sido igual. Gracias por enseñarme, apoyarme, escucharme, soportarme y tu
amistad incondicional. Agradezco a los amigos que conocí durante mis años en la
universidad. Camilo, gracias por tu amistad y apoyarme en cada momento. Kuky, mi
gemela perdida, gracias por escucharme y soportarme. Lily y Jossy, gracias por compartir
conmigo, aprendí tanto de ustedes, es bueno saber que aún puedo contar con su amistad.
Muri, Nico y Dany, con ustedes empezó toda esta aventura, gracias por todos los momentos
vividos, me alegra tanto saber que aún están. A Poleto y Sandri, gracias por escucharme y
contener todo mi estrés, además del cariño recibido siempre independiente del tiempo que
pase. A Mama Ru y Katy, “Show 'em what you're worth” y “Don´t f*uck it up” nunca me
hicieron mas sentido que ahora. Por último y no menos importante, mi Benny, quien me
acompañó las noches de estudio incondicionalmente, esperándome hasta la madrugada.
Gracias por tu amor, mi hermosurita.

5
FINANCIAMIENTO
Esta tesis se realizó en el Laboratorio de Fisiología Vegetal y Genómica de Frutales del
Centro de Biotecnología Vegetal de la Universidad Andrés Bello, financiado por el
proyecto FONDECYT 1130197 y FONDEQUIP EQM140074.

6
INDICE
INDICE ................................................................................................................................... 6
ABREVIATURAS ................................................................................................................. 7
RESUMEN ............................................................................................................................. 8
ABSTRACT ........................................................................................................................... 9
INTRODUCCIÓN ................................................................................................................ 10
1. Importancia económica y exportación de duraznos y nectarines ................................. 10
2. Características bioquímicas y morfológicas de Prunus persica .................................... 10
3. Desarrollo de Prunus persica ........................................................................................ 11
4. Metabolitos asociados a calidad ................................................................................... 12
5. Equilibrio fuente-sumidero ........................................................................................... 16
HIPÓTESIS .......................................................................................................................... 20
OBJETIVO GENERAL ....................................................................................................... 20
OBJETIVOS ESPECÍFICOS ............................................................................................... 20
MATERIALES Y MÉTODOS ............................................................................................. 21
1. Material vegetal ............................................................................................................ 21
2. Diseño experimental ..................................................................................................... 21
3. Fenotipificación de frutos de Prunus persica (L.) var `Buen Lindo´. ........................... 22
4. Tinción de lignina ......................................................................................................... 23
5. Elección de los puntos de muestreo para los análisis metabolómicos .......................... 23
6. Extracción de azúcares, alcoholes de azúcares, ácidos orgánicos y aminoácidos ........ 23
7. Condiciones cromatográficas ....................................................................................... 24
8. Análisis de datos metabolitos polares (GC-MS) .......................................................... 25
8.1 PLS-DA .................................................................................................................. 25
8.2 Variables importantes en la proyección (VIP) ....................................................... 25
8.3 Heat map ................................................................................................................. 26
9. Cuantificación de carotenoides totales ......................................................................... 26
RESULTADOS .................................................................................................................... 28
1. Desarrollo de los frutos de `Buen Lindo´ ..................................................................... 28
2. Fenotipificación de frutos de `Buen Lindo´ ................................................................. 30
3. Cuantificación de clorofila a, b y carotenoides totales ................................................. 32
4. Composición metabólica de los frutos.......................................................................... 34
4.1 Heat map de composición metabólica .................................................................... 34
4.2 Análisis discriminante de regresión de mínimos parciales (PLS-DA) ................... 36
4.3 Variables importantes en la proyección (VIP) ....................................................... 38
4.4 PLS- DA aplicando el VIP ..................................................................................... 41
4.5 Metabolitos correlacionados con los tratamientos de raleo .................................... 41
DISCUSIÓN ......................................................................................................................... 47
CONCLUSIONES ................................................................................................................ 53
REFERENCIAS ................................................................................................................... 54

7
ABREVIATURAS
DAB: días post-floración
HPF: hojas por fruto
PLS-DA: regresión de mínimos cuadrados parciales con análisis discriminante
S1: Etapa 1 del desarrollo del fruto
S2: Etapa 2 del desarrollo del fruto
S3: Etapa 3 del desarrollo del fruto
QC: control de calidad
GC-MS: Cromatógrafo de gases acoplado a detector de masa
VIP: variables importantes en la proyección
PF: peso fresco
SS: sólidos solubles
SSM: sólidos solubles maduros
OAA: oxaloacetato
PEP: fosfoenolpiruvato
SDH: sorbitol deshidrogenasa
SOX: sorbitol oxigenasa
CO2: dióxido de carbono
PEPC: fosfoenolpiruvato carboxilasa
TCA: ciclo de los ácidos tricarboxílicos

8
RESUMEN
Prunus persica (L.) Batsch, durazno y nectarinas, pertenece a la familia Rosaceae y es
originario de China. Es uno de los frutos más populares que se cultivan en zonas templadas
alrededor del mundo. Chile es su principal exportador del hemisferio sur en contra-estación
y Es necesario el incremento de la calidad de los frutos, atribuibles al tamaño, color,
firmeza, frescura, aroma, dulzor, acidez, jugosidad y textura para que el país se mantenga
en esa posición de liderazgo en el mercado. Metabolitos tales como azúcares, ácidos
orgánicos y aminoácidos entre otros, los cuales van cambiando dependiendo de la variedad
y el estado del desarrollo en el cual se encuentre el fruto, están directamente relacionados a
algunos de esos parámetros de calidad. Una manera de modificar el metabolismo es
cambiando el equilibrio fuente-sumidero del árbol, utilizando técnicas como el raleo. En
este trabajo se evaluó el perfil metabolómico de frutos de la variedad temprana de durazno
`Buen Lindo´ de árboles sometidos a dos tratamientos: con raleo y sin raleo. Para ello, se
realizó una extracción de metabolitos de frutos en distintos estados de desarrollo y se
analizaron mediante cromatografía de gases acoplado a un detector de masas. Los datos se
analizaron mediante análisis de regresión de mínimos cuadrados parciales con análisis
discriminante (PLS-DA), evaluando los principales metabolitos asociados a cada estado de
desarrollo del fruto, y además relacionándolos a un tratamiento de raleo. Se identificaron
los principales metabolitos involucrados en la calidad final del fruto siendo la mayor parte
azúcares y ácidos orgánicos. El efecto del raleo se observó a nivel fenotípico, incidiendo en
la producción de frutos de mayor tamaño y con mayor cantidad de sólidos solubles.
Además, el raleo modificó el perfil metabólico, encontrándose mayor cantidad relativa de
azúcares y ácidos orgánicos en el tratamiento sin raleo en madurez de consumo.

9
ABSTRACT
Prunus persica (L.) Batsch, peach and nectarines, belongs to the Rosaceae family and it is
originated from China. It is one of the most popular fruit that is grown in temperate zones
around the world. Chile is the main exporter from the Southern hemisphere in counter
season. It is necessary to increase the quality of the fruits regarding size, color, firmness,
freshness, aroma, sweetness, acidity, juiciness and texture to keep the country in this
leadership. Metabolites such as sugars, organic acids and amino acids are important, which
change depending on the variety and fruit developmental stage are directly related to some
of these quality traits. One way to modify the metabolism is changing tree source-sink
balance, using techniques such as thinning. In this researchwork the fruit metabolic profile
of the early peach variety `Buen Lindo´ from thinned and unthinned trees were evaluated.
Metabolite extraction of peach fruits at different stages of development was performed and
gas chromatography coupled to a mass spectrometry was used for the analysis. The data
were analyzed by least squares regression analysis with discriminant analysis (PLS-DA),
evaluating the main metabolites associated with each stage of development and those
related to a thinning treatment. The main metabolites involved in the final quality of the
fruit were identified and most of them were sugars and organic acids. The effect of thinning
was observed at the phenotypic level, promoting bigger size fruits and with an improved
amount of soluble solids. Indeed, thinning had a effect on the metabolic profile, with the
highest relative amount of sugars and organic acid associated with a non-thinning
treatment.

10
INTRODUCCIÓN
1. Importancia económica y exportación de duraznos y nectarines
Los duraznos y nectarines [Prunus persica (L.) Batsch] pertenecientes a la familia
de las Rosaceae son drupas de buen sabor, dulces y jugosas. La diversidad genética de
Prunus persica es muy amplia en China, existiendo alrededor de 495 cultivares
reconocidos, indicando que tiene una larga historia que se remonta desde los 1100 AC
(Huang et al., 2008). Prunus persica crece en todos los continentes, excepto la Antártica, y
la producción mundial ha incrementado continuamente en los últimos años (Jiang y Song,
2009). Es el cuarto fruto más importante en el mundo y el segundo en Europa (FAOstat,
2013). Chile aporta el 8% de las ventas mundiales de durazno, además supera el 80% de las
exportaciones del hemisferio sur, abasteciendo a los países del hemisferio norte, EE.UU.
principalmente. Sólo en el año 2015, la exportación de este fruto en Chile alcanzó las 35
mil toneladas (ODEPA, 2015).
2. Características bioquímicas y morfológicas de Prunus persica
Los frutos de Prunus persica tienen carozo, poseen un endocarpo lignificado en
donde se encierra la semilla, un mesocarpo carnoso y un exocarpo (piel) delgado (Figura 1).
Una característica que diferencia a duraznos de nectarines es la presencia de tricomas en los
duraznos. Estos frutos también se diferencian si poseen o no carozo adherido a la pulpa,
además de cómo es el proceso de ablandamiento de la pulpa: fundente o no fundente
(Colaric et al., 2005; Infante et al., 2008)

11
Figura 1: Estructura de Prunus persica (L.) Batsch, donde se observan sus distintos tejidos. Fuente:
http://www.euita.upv.es/varios/biologia/web_frutos/Estructura.htm.
3. Desarrollo de Prunus persica
El desarrollo y maduración del fruto son procesos dinámicos que involucran una
serie de cambios moleculares y bioquímicos. El desarrollo de frutos con carozo puede ser
descrito generalmente como un proceso de 4 estados observándose una curva doble
sigmoide (Chalmers y van den Ende, 1975; Tonutti et al., 1997; el-Sharkawt et al., 2007).
El primer estado (S1), corresponde a la primera fase de crecimiento exponencial, y se
caracteriza por un rápido incremento en la división y elongación celular. Durante la
segunda fase (S2), el endocarpo se endurece para formar el cuesco, proceso conocido como
endurecimiento del carozo. Además, durante esta fase no hay incremento en el tamaño del
fruto y las vías metabólicas relacionadas con la síntesis de lignina están muy activas
(Dardick et al., 2010). Posteriormente, en el tercer estado (S3), también conocido como la
segunda fase exponencial, existe un rápido incremento en el tamaño del fruto, dado por una
rápida elongación celular y entrada de agua. Finalmente, en el cuarto estado (S4) el fruto

12
llega a su tamaño final y entra a la maduración o a su estado climatérico (Figura 2). Cabe
mencionar que la fase S4 puede ser subdividida en dos fases: S4-1, en la cual el fruto llega
a su tamaño final, y S4-2, donde el fruto continua su madurez en un modo dependiente de
etileno (Trainotti et al., 2003). La fase S4-2 puede llevarse a cabo fuera del árbol y
usualmente ocurre antes del consumo humano (Borsani et al., 2009).
Figura 2: Curva de crecimiento de Prunus persica (L.) Batsch. Se observa como una curva doble sigmoidea.
Fuente: https://fisiohorticola.files.wordpress.com/2008/11/estructura-y-morfogenesis-del-fruto1.pdf.
4. Metabolitos asociados a calidad
Los atributos de calidad sensorial y valor nutritivo del fruto juegan un rol
importante en la satisfacción del consumidor e influencia futuros consumos. La calidad de
frutos de Prunus persica puede ser determinada por análisis químicos y depende
principalmente de sus componentes mayores tales como la sacarosa, ácido cítrico y málico,
carotenoides, lactonas y polifenoles (Aubert et al., 2003; Bassi y Selli, 1990; Brovelli et
al.,1999; Versari et al., 2002).
El sabor, aroma, textura y apariencia son generalmente considerados como los
atributos sensoriales más importantes dentro de los parámetros de calidad del fruto. El
sabor está relacionado con los compuestos solubles en agua, específicamente, el dulzor es
mayormente atribuible a algunos mono y disacáridos, mientras que el sabor ácido es

13
relacionado a ácidos orgánicos y pH (Colaric et al., 2005). Por otro lado, el aroma es
atribuido a compuestos que exhiben cierta volatilidad (Shepherd et al., 1993).
Prunus persica representa un modelo particular de fruta en términos de fisiología,
anatomía y metabolismo, diferente de otros modelos de frutas como el tomate, frambuesas
o uvas; y con redes metabólicas particulares durante su desarrollo, el cual está relacionado a
su morfología (una drupa con semillas cubierta por un endocarpo lignificado y un
mesocarpo jugoso) (Lombardo et al., 2011). Con esta morfología en general, hay una
extensa variabilidad en sabores, texturas y tasas de dulzor/acidez, en los cientos de
variedades que existen alrededor del mundo (Byrne, 2005; Sansavini et al., 2006).
El sabor de la pulpa de las frutas, como Prunus persica, dependen mayormente del
contenido de azúcares y ácidos, y la razón entre ellos (Esti et al., 1997; Robertson et al.,
1990; Souty y André, 1975). La sacarosa es el azúcar predominante en estos frutos
encontrándose entre un 40 y un 85% del contenido de azúcares totales, mientras que los
azúcares reductores como glucosa y fructosa están en cantidades equimolares alcanzando
hasta un 25% del contenido de azúcares totales. Sin embargo, el contenido de sorbitol es
siempre bajo (Wrolstad y Shallenberger, 1981), lo que sugiere que el sorbitol es
metabolizado a azúcares reductores (Brooks et al., 1993). Por otro lado, los ácidos
orgánicos predominantes son el ácido málico y cítrico con concentraciones en la pulpa en la
cosecha de entre 14 y 12 meq/100 g respectivamente (Souty y André, 1975). Autores
señalan que el contenido de sacarosa y ácido málico incrementan durante la maduración del
fruto cuando los azúcares reductores y el ácido cítrico disminuyen (Chapman et al., 1991;
Ishida et al., 1971).
Existe una analogía entre los azúcares reductores, sacarosa y los ácidos málico y
cítrico, en donde los azúcares reductores y el ácido cítrico son utilizados como sustratos
para el crecimiento y la respiración (Génard et al., 1994). Ambos tienen distintas
concentraciones, en donde el malato probablemente tiene una alta prioridad metabólica,
explicando la relación positiva con la tasa de crecimiento. Muchos autores han mostrado
que la concentración de sacarosa y el ácido málico incrementan durante la maduración del

14
fruto, mientras el ácido cítrico disminuye (Chapman et al., 1991; Liverani y Cangini, 1991).
Por otro lado, frutos cosechados inmaduros fisiológicamente tendrán bajas cantidades de
sacarosa y ácido málico, y alto contenido de ácido cítrico (Génard et al., 1994).
Sacarosa, glucosa, fructosa, y sorbitol son las mayores azúcares o alcoholes de
azúcares encontrados en las frutas de las mayoría de las especies de Rosaceae. La Figura 3,
simula la división de flujo de carbono desde el floema de los cuatro azúcares mencionados,
los otros compuestos de la fruta y el CO2 producido por la respiración (Génard y Souty
1996). El modelo tiene siete puntos principales: 1) la distribución de carbono que entra
como sacarosa y sorbitol es constante; 2) el carbono presente en la fruta como sorbitol es
continuamente convertido en glucosa y fructosa; 3) la transformación de carbono desde la
sacarosa en glucosa y fructosa disminuye a medida que se acumulan días post-floración; 4)
glucosa y fructosa pueden ser interconvertidas; 5) la cantidad de carbono utilizada para
sintetizar otros compuestos (no azúcares) disminuye a medida que se acumulan grados-día
y proviene de glucosa y fructosa a través de vías respiratorias; 6) el carbono utilizado para
la respiración viene desde la glucosa y la fructosa, proporcional a su cantidad en la fruta; y
7) excepto por la respiración, el flujo de carbono entre dos compuestos es proporcional a la
cantidad de carbono en la fuente (Génard y Souty, 1996).

15
Figura 3: Representación de la división del flujo de carbono desde el floema. Las flechas representan el flujo
de carbono, los círculos representan la fuente de carbono, y los rectángulos representan los carbohidratos.
Fuente: Modificado de Génard y Souty, 1996.
En frutos inmaduros de Prunus persica se guardan altos niveles de proteínas las
cuales son utilizadas en procesos que ocurren en S1 y S2. El perfil metabólico también
indica altos niveles de aminoácidos, tales como prolina, fenilalanina, tirosina y glutamina,
los cuales van disminuyendo a medida que el fruto va madurando. Una alza importante en
aminoácidos como prolina, fenilalanina, tirosina, alanina, valina ocurre en el estado S2/S3.

16
En este escenario, es probable que estos aminoácidos en estadios tempranos de desarrollo,
se conviertan en sustratos para vías de fenilpropanoides, lignina y flavonoides (Lombardo
et al., 2011).
En aspectos de sabor, el dulzor está correlacionado con el ácido cítrico, el ácido
shikímico y la razón entre azúcares/ácidos orgánicos. Aunque el ácido cítrico usualmente
está presente en bajas concentraciones en los frutos, añade más acidez a los que el ácido
málico (Génard et al., 1994). En el estudio de Collaric et al., (2005), los azúcares como la
sacarosa, fructosa y glucosa los cuales son responsables del dulzor del fruto, no se
correlacionaron con el dulzor, en cambio, la razón entre azúcares/ácidos orgánicos si lo
hicieron. Por otro lado, el azúcar más abundante, la sacarosa fue la más relacionada al
aroma. Según Collaric et al., (2005), la sacarosa y el sorbitol fueron los que tuvieron más
relación al sabor. La sacarosa como el mayor azúcar presente en frutos de Prunus persica
es importante en el dulzor, como fuente de energía y antioxidante (Huberlant y Anderson,
2003). Sorbitol, siendo alcohol de azúcar es más beneficioso que otros azúcares, aportando
en el sabor y en la textura del fruto (Rapaille et al., 2003). El ácido málico y cítrico son los
que contribuyen en mayor grado a la acidez.
Otro parámetro de calidad atractiva para el consumidor y además con múltiples
beneficios para la salud son los carotenoides. Son considerados nutrientes esenciales y no
pueden ser sintetizados por el cuerpo humano. Además son los pigmentos responsables del
color del fruto (Mayne, 1996).
5. Equilibrio fuente-sumidero
Para entender la fisiología de los frutos y la influencia de los factores limitantes de
la cosecha en la producción, es necesario el estudio de la relación fuente-sumidero y la
regulación del flujo del carbono. Las hojas son regiones de producción de asimilados
conocidas como órganos fuentes. Una hoja en desarrollo, sin embargo, puede representar
tanto como órgano fuente produciendo asimilados para exportación, así como también
puede ser sumidero, importando asimilados desde otras partes de la planta. Los sumideros

17
importan y usan asimilados durante la respiración, crecimiento y almacenaje (Guardiola y
García-Luis, 2000). Dentro del árbol, los órganos reproductivos como los frutos, compiten
por los asimilados con otros frutos, especialmente en árboles con alta carga frutal como
duraznos y manzanas (Dann et al., 1984; DeJong et al., 1987; Hansen, 1971; Maggs, 1963;
Weinberger, 1931). Es por esto, que en campos productivos es necesario reducir la carga
frutal del árbol, mediante la técnica llamada raleo para obtener frutos de mayor tamaño
(Guardiola y García-Luis, 2000; Link, 2000; Ojer y Reginato, 2002)
En árboles en plena producción el raleo debe ser llevado a cabo cada año. Las
ventajas de esta técnica han sido ampliamente descritas: la reducción del número de flores o
frutos incrementa el tamaño del fruto, mejora la calidad, y balancea la razón fruto/brote, lo
que lleva a un incremento de los asimilados hacia la fruta (Costa et al., 1978; Farley 1923;
Giulivo, 1974; Giulivo et al., 1981; Havis, 1962). La fecha de raleo es crítica para obtener
buenos resultados. Ha sido descrito que el raleo manual de frutos usualmente se debe
realizar hacia el final del estado S2 o a comienzos del estado S3. Después de la abscisión
natural, sólo el exceso de frutos en el árbol necesita ser raleado. Durante el estado S2, los
frutos incrementan en tamaño lentamente y su demanda de asimilados permanece alta
durante el endurecimiento del endocarpo (Weinberger, 1941).
El raleo de frutos afecta el tamaño del fruto y el rendimiento. La cantidad adecuada
de frutos que se van a remover depende de la genética y de la capacidad de carga. Esta
última se asocia con la edad y el tamaño del árbol, y también está influenciada por factores
externos como la poda (Guardiola y García-Luis, 2000). Independiente de la especie, un
incremento de la carga frutal, reduce el tamaño final de la fruta y el rendimiento de la
cosecha (Dennis et al., 1983). La relación fuente-sumidero y la asignación de asimilados a
los diferentes órganos juegan un rol fundamental en la determinación del rendimiento del
cultivo (Dennis et al., 1983). El tamaño del fruto depende de la razón entre el número de
hojas y el número de frutos. Reduciendo en número de frutos por árbol mediante raleo, se
incrementa la razón hoja-fruto, incrementando el tamaño del fruto (Dorsey y McMunn,
1927; Weinberger, 1931). Se ha reportado que el raleo temprano a la floración o después de
la polinización, resulta en frutos más grandes (Weinberger, 1941), indicando que el

18
crecimiento de Prunus persica es fuente-limitada durante periodos tempranos y tardíos del
desarrollo (Grossman y DeJong, 1995; Pavel y DeJong, 1993). Durante los periodos medios
del crecimiento, las limitaciones del sumidero pueden estar presentes (al menos en
cultivares que maduran tardíamente) (DeJong y Grossman, 1995). Sin embargo, el periodo
en el cual las limitaciones del crecimiento se llevan a cabo dependen de la floración y de la
fecha de cosecha del cultivar. La competencia entre frutos puede ser más evidente en frutos
de cosecha temprana y puede estar presente a través de todo el desarrollo del fruto, a pesar
de que otros factores como condiciones medioambientales (agua y disponibilidad de
carbono, nutrición de la planta, etc) pueden reprimir el crecimiento del fruto con respecto a
su potencial genético (Pavel y DeJong, 1993).
La literatura muestra que el equilibrio fuente-sumidero, la fertilización mineral, la
disponibilidad de agua y la temperatura son factores agro-medioambientales que tienen
mayor impacto en la acidez de la fruta (Etienne et al., 2013). Las prácticas de manejo en los
huertos como el raleo de frutos, la poda, o la defoliación afectan el equilibrio fuente-
sumidero de la planta. Estas prácticas también afectan la acidez de la fruta. En Prunus
persica se ha observado que un incremento en la razón fuente-sumidero incrementa el
contenido de citrato en estadios tempranos del desarrollo, y disminuye en la madurez
(Souty et al., 1999; Wu et al., 2002; Léchaudel et al., 2005). El efecto contrario ha sido
reportado para malato, con una disminución en estadios tempranos, seguido por un
incremento hacia la madurez (Wu et al., 2002; Léchaudel et al., 2005).
Se ha hipotetizado que durante los estados tempranos, grandes cantidades de
azúcares importadas desde las hojas están disponibles para la producción de malato vía
glicólisis y su conversión a citrato vía ciclo de los ácidos tricarboxílicos. Está descrito que
frutos que crecen con una alta disponibilidad de azúcares, debido a una alta razón fuente-
sumidero, son más grandes y consecuentemente poseen una tasa respiratoria más alta. En
estos estadíos, un incremento en la respiración dado por una alta disponibilidad de azúcares
puede estimular la glicólisis y la conversión de malato a citrato (Etienne et al., 2013). En
contraste, durante la maduración, las azúcares ya no están disponibles para la respiración
pues están almacenadas en la vacuola (Coombe, 1976), causando un cambio desde los

19
azúcares hasta los ácidos orgánicos (en particular el citrato) como sustrato respiratorio.
Durante esta etapa, un incremento en la respiración (dado por la gran cantidad de frutos en
respuesta a una alta razón fuente-sumidero) puede estimular la conversión de citrato a
malato para mantener el pool de los intermediarios del ciclo de los ácidos tricarboxílicos
constantes (Lobit et al., 2003, Wu et al., 2007).
Un trabajo previo de nuestro laboratorio utilizando la variedad de frutos de Prunus
persica, “Magique” (nectarin), en la etapa S3 de desarrollo mostró las primeras diferencias
en el tamaño de frutos bajo distintos tipos de raleo, y luego se incrementaron en la etapa de
postcosecha (S4), donde los frutos de árboles sin raleo tuvieron un menor diámetro respecto
de los árboles raleados. El análisis de metabolitos mediante HPAEC-PAD y HPLC
demostró que los frutos de árboles sin raleo mostraron diferencias con los frutos de árboles
raleados. Mediante análisis de componentes principales, los frutos de árboles sin raleo se
separaron de los raleados mediante el componente principal 1, con un 49,3% de
explicación. Las variables que contribuyeron a esta separación fueron glucosa, sacarosa,
trehalosa, inositol, malato, succinato y valina en la etapa final de S1; sacarosa, galactosa y
citrato durante la mitad de la S2; glucosa y fructosa al momento de la cosecha. En el
momento de la cosecha, los frutos de los árboles con raleo poseían un mayor contenido de
sólidos solubles y eran más grandes respecto a diámetro y peso (Covarrubias et al.,
manuscrito en preparación).
Con todos los datos recopilados, se podría concluir que el raleo desde estados
tempranos modifica el perfil de metabolitos asociados a calidad, por lo que es necesario
realizar un estudio más preciso durante todas las etapas del desarrollo de Prunus persica,
comparando frutos de árboles con y sin raleo.

20
HIPÓTESIS
Alteraciones en el equilibrio fuente-sumidero modifican el perfil de metabolitos de frutos
de cosecha temprana de Prunus persica (L.) Batsch.
OBJETIVO GENERAL
Evaluar el efecto de la modificación del equilibrio fuente-sumidero sobre el perfil
metabólico polar asociado a la calidad en frutos de Prunus persica (L.) Batsch var `Buen
Lindo´.
OBJETIVOS ESPECÍFICOS
1) Realizar una fenotipificación de frutos de Prunus persica (L.) Batsch var `Buen
Lindo´ sometidos a dos tratamientos: con raleo y sin raleo.
2) Identificar y cuantificar azúcares, alcoholes de azúcares, aminoácidos y ácidos
orgánicos en frutos de Prunus persica (L.) Batsch var `Buen Lindo´, sometidos a
dos tratamientos (con raleo y sin raleo) durante distintos estados de desarrollo.
3) Evaluar el perfil de los metabolitos identificados, asociándolos a un estado de
desarrollo en particular y/o calidad.

21
MATERIALES Y MÉTODOS
1. Material vegetal
El material vegetal corresponde a frutos de Prunus persica (L.) Batsch var `Buen
Lindo´ correspondiente a la temporada 2015-2016. Este fruto es un durazno, de pulpa
amarilla, de sabor dulce y cosecha temprana. El material se recolectó desde viveros El
Tambo Limitada, ubicado en la localidad de Malloa (en las coordenadas latitud -34,474581,
longitud -70,985219), en la VI Región del Libertador Bernardo O´Higgins. El portainjerto
corresponde a patrón Nemaguard y la densidad de plantación es de 4,5 x 2,2 m.
2. Diseño experimental
Se utilizaron dos árboles: uno con raleo manual con una razón de 19 hojas por fruto;
y un árbol sin raleo con una razón de 2 hojas por fruto. El raleo de frutos se realizó a los 38
días post- floración (38 DAFB). Los frutos fueron recolectados durante todo el desarrollo
desde ramillas distintas de un mismo árbol. Durante los primeros estados del desarrollo (S1,
S2 y S3), se tomaron 6 frutos desde 3 ramillas distintas en cada punto de muestreo. Estas
ramillas no volvieron a utilizarse para muestreos futuros. Para la cosecha, se recolectaron
todos los frutos del árbol. Se seleccionaron 18 frutos para el punto de madurez de cosecha y
adicionalmente 18 frutos para someterlos a 5 días a 21°C (madurez de consumo).
Para la fenotipificación, se ocuparon todos los frutos recolectados tanto para los estados
tempranos del desarrollo como para la post-cosecha. Para el análisis metabolómico se
realizaron pooles independientes. Para los estados tempranos del desarrollo, cada pool
estaba compuesto por 3 frutos de una misma ramilla, siendo que en total se analizaron 3
pooles de ramillas distintas para cada tratamiento de raleo. Para la post-cosecha, se tomaron
5 frutos individuales, en donde cada fruto era una muestra independiente. Los pooles
estaban compuestos por el mesocarpo y el exocarpo del fruto.

22
3. Fenotipificación de frutos de Prunus persica (L.) var `Buen Lindo´.
La fenotipificación de los frutos se realizó midiendo los siguientes parámetros:
- Calibre: se medió el diámetro ecuatorial en milímetros de cada fruto a través de un
medidor de calibre.
- Peso: a través de una balanza granataria se pesaron los frutos en gramos,
correspondientes a los distintos tratamientos y a los distintos estados de desarrollo.
Estos parámetros fueron medidos para todos los estados del desarrollo.
- Firmeza: fue medida a través de un penetrómetro (FT-327, EFFEGI, Italia)
expresada en libras-fuerza. Se utilizaron ambas mejillas del fruto.
- Acidez titulable: a partir de 5 mL de jugo de fruta extraído, se tituló con NaOH
0,1N hasta alcanzar un pH de 8,2. Se registró el volumen utilizado y se calculó el
porcentaje de ácido málico mediante la siguiente fórmula:
% acidez = 𝑉𝑜𝑙𝑢𝑚𝑒𝑛 𝑁𝑎𝑂𝐻 𝑢𝑡𝑖𝑙𝑖𝑧𝑎𝑑𝑜 𝑥 0,1 𝑥 0,067 𝑥 100
𝑉𝑜𝑙𝑢𝑚𝑒𝑛 𝑑𝑒 𝑗𝑢𝑔𝑜
- Sólidos solubles: se evaluó mediante la utilización de un refractómetro thermo
compensado digital (HANNA instruments inc. Woonsoket-RI-USA, Modelo HI
96811, serie 08255834). Los valores entregados corresponden a °Brix (sólidos
solubles totales)
Estos parámetros sólo fueron medidos para frutos en madurez de cosecha y
consumo.
- Porcentaje de jugo: a frutos con madurez de consumo se les midió el porcentaje de
jugo, mediante la técnica de absorción en papel descrita por Infante et al., 2008.
Este parámetro sólo fue medido para consumo.
Luego de la fenotipificación, los frutos utilizados fueron fraccionados, congelados en
nitrógeno líquido y almacenados a -80°C.

23
4. Tinción de lignina
El momento en el cual ocurre el endurecimiento se determinó mediante la tinción
específica de lignina mediante fluoroglucinol, según el protocolo descrito por Dean (1997).
Se tomaron frutos provenientes del árbol con raleo comercial y del árbol sin raleo, se les
realizó un corte transversal y al carozo se le añadió una solución de fluoroglucinol 5% en
etanol 85%. Luego se eliminó el exceso y se colocó HCl fumante hasta evidenciar un
cambio de color a rojizo, finalmente se lavó con etanol 90%.
5. Elección de los puntos de muestreo para los análisis metabolómicos
A partir de los datos entregados por la fenotipificación, se procedió a elegir los
puntos del desarrollo que mostraron diferencias significativas entre los árboles con raleo y
sin raleo. Para ello se graficó en el programa GraphPad PRISM® versión 5.0a, y luego se
realizó análisis de los datos con el programa estadístico Infostat, mediante análisis de la
varianza Tukey (p < 0.05). Los puntos de trabajo fueron los siguientes:
- 46 DAB para ambos árboles, este punto corresponde a la etapa 1 del desarrollo (S1).
- 58 DAB para raleo comercial y 64 DAB para sin raleo, correspondiente a la etapa 2
del desarrollo (S2).
- 86 DAB para raleo comercial y 93 DAB para sin raleo, correspondiente a la etapa 3
del desarrollo (S3).
- 101 DAB para raleo comercial y 110 para sin raleo, correspondiente a la cosecha de
los frutos.
- 101 DAB + 5 días a temperatura ambiente para raleo comercial y 110 DAB + 5 días
a temperatura ambiente para sin raleo, correspondiente a madurez de consumo.
6. Extracción de azúcares, alcoholes de azúcares, ácidos orgánicos y aminoácidos
El protocolo de extracción de metabolitos polares se llevó a cabo según Hatoum et
al., 2014. Los frutos almacenados previamente se molieron con mortero y nitrógeno
líquido. Se liofilizaron y de este polvo se pesaron 20 mg para la extracción. A los 20 mg se

24
les adicionó 500 µL de metanol frío y 20 µL de fenil β-D-glucopiranósido 2910 ng/µL el
cual corresponde al estándar interno. Luego se incubó a 70°C por 15 min con agitación. Se
centrifugó a máxima velocidad por 20 min. Del sobrenadante se tomaron 100 µL y se
colocaron en un tubo nuevo. Se secaron con nitrógeno gaseoso y se re-disolvieron en 120
µL de clorhidrato metoxiamina de una concentración de 20 mg/mL en piridina. Se incubó a
30°C por 90 min con agitación. Se agregaron 120 µL de N,O-
bis(trimetilsilil)trifluoroacetamida (BSTFA) y se incubaron a 37°C por 30 minutos con
agitación.
Finalmente, se inyectó 1 µL en GC-MS de cada muestra junto a un solvente
(metanol) y un control de calidad (QC). Los metabolitos detectados fueron los siguientes,
para ácidos orgánicos: oxálico, ascórbico, cítrico, quínico, succínico y málico; para
aminoácidos: arginina, alanina, leucina, valina, prolina, serina, treonina, ácido aspártico,
fenilalanina, asparagina; para azúcares y alcoholes de azúcares: glucosa, fructosa,
glucosa/manosa, sacarosa, sorbitol y mio-inositol.
7. Condiciones cromatográficas
Se utilizó un cromatógrafo de gases serie 7890B con detector selectivo de masa
serie 5977, marca Agilent Technologies, con las siguientes condiciones: el inyector estaba a
220°C, la interfase a 280°C, el flujo de helio a 1 mL/min, la fuente a 230°C, el cuadrupolo
a 150 °C, rango de scaneo 50-600 m/z y 2,66 ciclos scan/s.
Para ácidos y aminoácidos: se trabajó en modo splitless, 50°C por 1 minuto, 310°C a
10°C/min y se mantuvo por 13 minutos. Para azúcares y alcoholes de azúcares se trabajó en
modo split: 1:150, 120°C por 1 minuto, 300 °C a 10°C/min y se mantuvo por 6 minutos.

25
8. Análisis de datos metabolitos polares (GC-MS)
Al obtener los datos entregados por el cromatograma, se integraron con el algoritmo
de deconvolución. Los espectros de masa de los compuestos encontrados por la integración,
se buscaron en la librería NIST 2014 mediante el software Mass Hunter de Agilent
Technologies. La confirmación de los datos se realizó mediante comparación con
estándares externos. Se realizó la normalización de los datos utilizando el estándar interno,
el control de calidad (QC) y el peso inicial de la muestra.
8.1 PLS-DA
Una vez identificados los compuestos, se realizó un análisis discriminante de
regresión de mínimos parciales (PLS-DA) para comparar los compuestos identificados en
los distintos tratamientos y estados de desarrollo. El software que se utilizó para la
estadística fue el Unscrambler® X. Luego de esto, se graficaron los datos de cada
metabolito individualmente con el programa GraphPad PRISM® versión 5.0a, obteniendo
cantidades relativas. El análisis estadístico se realizó mediante Infostat, utilizando análisis
de la varianza Tukey (p < 0.05).
8.2 Variables importantes en la proyección (VIP)
Para ordenar los metabolitos identificados y su importancia , se calculó el VIP (variables
importantes en proyección), el cual ordena los metabolitos según su valor de correlación.
Este cálculo se realizó manualmente y a través del software Unscrambler X®.
8.2.1 Cálculo del VIP manual
Este valor se calculó mediante la siguiente fórmula:
√𝑥2 + 𝑦2
en donde “x” representa el factor 1 e “y” representa el factor 2.

26
8.2.2 VIP a través de Unscrambler X®
Este método analiza los VIP mediante observación, es decir, los metabolitos se
agrupan en clusters según su importancia, y los discrimina por factor, a diferencia del
método anterior.
8.3 Heat map
Utilizando los datos obtenidos se realizó un heat map en un ambiente de R 3.3.2 en
adición al paquete ggplot2 disponible en (www.ggplot2.org). Los datos fueron agrupados
utilizando la función hclustfun con parámetros por defecto y reorganizados considerando
las medias de cada fila.
9. Cuantificación de carotenoides totales
La cuantificación de carotenoides totales se llevó a cabo basado en el método de
Wellburn (1994). Se colocaron 0,5 mL de dimetil sulfóxido (DMSO) en un tubo Eppendorf
de 1,5 mL previamente cubierto con papel aluminio para proteger el pigmento de la luz. Se
agregaron 80 mg de polvo congelado en nitrógeno líquido de fruta, se homogenizó en
vórtex y se agregó nuevamente 0,5 mL de DMSO para dejarlo en incubación a 65°C por 2
h. Luego los tubos se dejaron durante 20 min a temperatura ambiente. Finalizado este
tiempo, el contenido del tubo se pasó por filtro miracloth, para luego depositarlo en placas
de 96 pocillos y medir en Espectrofotómetro para Microplacas Epoch a las absorbancias
descritas por Wellburn (1994) en sus fórmulas para equipos con sensibilidad entre 0,1-0,5
nm.
Las fórmulas utilizadas fueron las siguientes:
Ca= 12,47A665,1 – 3,62A649,1
Cb= 25,06A649,1 – 6,5A665,1
Cx= (100A480 – 1,29Ca – 53,78Cb)/220

27
A: Absorbancia a la longitud de onda indicada en el subíndice
Ca: Clorofila a
Cb: Clorofila b
Cx: Carotenoides totales, expresados en (μg/mL)
Estos datos se normalizaron por peso fresco de la muestra y se graficaron en
GraphPad PRISM® versión 5.0a. El análisis estadístico se realizó mediante Infostat,
utilizando análisis de la varianza Tukey (p < 0.05).

28
RESULTADOS
1. Desarrollo de los frutos de `Buen Lindo´
El desarrollo de los frutos de `Buen Lindo´ fue monitoreado a través de tinción de
lignina para definir el comienzo de la segunda etapa del desarrollo, la cual corresponde al
endurecimiento del carozo (Figura 4A y 4B). Otra forma de evidenciar el desarrollo de los
frutos fue a través de la medición de peso y diámetro (Figura 4C y 4D, respectivamente).
En la Figura 4A y 4B, se observa el desarrollo de los frutos del árbol con raleo comercial y
sin raleo respectivamente, en donde se observa que la coloración roja correspondiente a la
tinción de lignina comienza alrededor de los 58 DAB para ambos tratamientos. En la Figura
4C se observa el desarrollo de los frutos a través de todo su crecimiento, en donde los frutos
de árboles con raleo comercial muestran mayor peso con respecto a los frutos de árboles sin
raleo, llegando a obtener un mayor peso en la época de cosecha inclusive. En la Figura 4D
se observa el tamaño del fruto a través de su desarrollo, evidenciando también que los
frutos de árboles con raleo comercial son más grandes durante todas las etapas del
desarrollo que aquellos frutos de árbol sin raleo. Cabe destacar que los frutos de árbol con
raleo comercial fueron cosechados antes que los frutos de árbol sin raleo (ambos alrededor
de 12 libras-fuerza). Los frutos del árbol con raleo comercial se cosecharon a los 101 días
post-floración, y los frutos del árbol sin raleo se cosecharon a los 110 días post-floración
(DAB).

29
Figura 4: Desarrollo de frutos de árboles de `Buen Lindo´. A: Tinción de lignina durante el desarrollo del
fruto del árbol con raleo comercial, B: Tinción de lignina durante el desarrollo del fruto del árbol sin raleo, C:
Desarrollo del fruto en relación a su peso en gramos, D: Desarrollo del fruto en relación a su diámetro en
milímetros. El raleo se realizó a los 38 días post-floración (DAB), indicado en ambas imágenes con una flecha
negra. Las etapas del desarrollo fueron definidas como S1: primera etapa de desarrollo exponencial; S2:
endurecimiento del carozo; S3: segunda etapa de desarrollo exponencial; Cosecha: etapa S4 o momento
donde ocurre la cosecha del fruto; Consumo: etapa a la que corresponde la madurez de consumo del fruto (5
días posterior a su cosecha). Las líneas naranjas representan a los frutos del árbol con raleo comercial, las
líneas verdes representan a los frutos del árbol sin raleo. Se trabajó con un n= 3-5. Los asteriscos (*)
representan diferencias significativas entre los tratamientos de raleo con test Tukey (p<0.05).

30
2. Fenotipificación de frutos de `Buen Lindo´
La Figura 5A muestra la cantidad de sólidos solubles a cosecha (madurez
fisiológica) y consumo, donde se observa que ambos tratamientos no muestran diferencias a
cosecha, pero si en la madurez de consumo, donde los frutos de árbol con raleo comercial
tienen mayor cantidad de sólidos solubles que aquellos frutos de árbol sin raleo.
La Figura 5B muestra la firmeza entre frutos con los distintos tratamientos a
cosecha y madurez de consumo. Se observa la diferencia entre ambos tratamientos a
cosecha, en donde los frutos del árbol sin raleo son menos firmes que los frutos de árbol
con raleo comercial, no así a madurez de consumo, en donde no existen diferencias
significativas.
En el caso del porcentaje de acidez entre ambos tratamientos (Figura 5C), se
observa que los frutos de árbol sin raleo contienen un porcentaje de acidez titulable
expresado como ácido málico menor que los frutos de árbol con raleo comercial a cosecha.
Sin embargo, esta diferencia no es significativa en madurez de consumo.
Al comparar los porcentajes de jugo (Figura 5D), no se observan diferencias
significativas entre los tratamientos

31
Figura 5: Fenotipificación de frutos de árboles de `Buen Lindo´ . A: Cantidad de sólidos solubles a cosecha y
madurez de consumo expresados como ºBrix, B: Firmeza de los frutos a cosecha y madurez de consumo
expresados como libras-fuerza, C: Porcentaje de acidez de los frutos a cosecha y madurez de consumo
expresados como porcentaje de ácido málico, D: Porcentaje de jugo a madurez de consumo expresados como
porcentaje de jugo. Las barras naranjas representan a los frutos del árbol con raleo comercial, y las barras
verdes representan los frutos del árbol sin raleo. Las letras distintas representan diferencias significativas entre
los tratamientos de raleo con test Tukey (p<0.05).

32
3. Cuantificación de clorofila a, b y carotenoides totales
Para determinar la coloración de los frutos durante todo el desarrollo, se determinó
la concentración de clorofila a y clorofila b (Figura 6A y B respectivamente), para luego
determinar los carotenoides totales en los frutos (Figura 6C). En relación a la clorofila a
(Figura 6A), se observa que durante el desarrollo existen diferencias significativas entre los
primeros estados del desarrollo (S1 y S2) con respecto a la tercera etapa del desarrollo y la
post-cosecha, evidenciando una disminución de la concentración de clorofila a, a medida
que avanza el desarrollo, ocurriendo la misma situación para ambos tratamientos. Con
respecto a la concentración entre los tratamientos, no existen diferencias significativas en
ningún estado de desarrollo. En la clorofila b (Figura 6B), la única diferencia significativa
que existe durante el estado del desarrollo ocurre en la segunda etapa del crecimiento (S2),
donde se observa que en ambos tratamientos existe una alza en la concentración de clorofila
b. Con respecto a la concentración de clorofila b entre los tratamientos de raleo, no existen
diferencias significativas entre tratamientos para ningún estado del desarrollo. En el caso de
la cuantificación de carotenoides totales (Figura 6C), no se observaron diferencias
significativas entre los tratamientos de raleo durante ningún estado del desarrollo, sin
embargo, al comparar los estados del desarrollo durante un mismo tratamiento de raleo, se
observa que la primera fase del desarrollo (S1) para ambos tratamientos, contiene mayor
cantidad de carotenoides que desde la tercera fase del desarrollo (S3) hasta la post-cosecha.
Para la segunda etapa del desarrollo (S2) no existen diferencias significativas.

33
Figura 6: Cuantificación de Clorofila a, Clorofila b y carotenoides totales. A: Cuantificación de “clorofila a”,
a través del desarrollo del fruto de `Buen Lindo´ expresada en μg por gramo de peso fresco (PF), B:
Cuantificación de “clorofila b”, a través del desarrollo del fruto de `Buen Lindo´ expresada en μg por gramo
de peso fresco (PF), C: Cuantificación de carotenoides totales a través del desarrollo del fruto de `Buen
Lindo´ expresados como μg por gramo de peso fresco (PF). Las barras naranjas representan a los frutos del
árbol con raleo comercial, y las barras verdes representan los frutos del árbol sin raleo. Las letras mayúsculas
distintas representan diferencias significativas entre los estados del desarrollo en un mismo tratamiento, y las
letras distintas minúsculas representan diferencias significativas entre los tratamientos de raleo con test Tukey
(p<0.05).

34
4. Composición metabólica de los frutos
4.1 Heat map de composición metabólica
Para la visualización general de los metabolitos identificados y el análisis de su
comportamiento durante todos los estados de desarrollo y en los distintos tratamientos se
realizó un heat map (Figura 7). Cada corchete representa los metabolitos que se relacionan
entre sí. En el caso de la glucosa y glucosa/manosa tienen un comportamiento similar
durante todo el desarrollo en ambos tratamientos de raleo, y estos azúcares además tienen
un comportamiento similar a fructosa, principalmente en el desarrollo de frutos en el
tratamiento sin raleo. Se observa también que el ácido málico y arginina tienden a
comportarse de igual forma durante todo el estado de desarrollo, observándose mayores
cantidades relativas de estos metabolitos hacia la post-cosecha en el tratamiento sin raleo.
Ácido cítrico y sacarosa también presentan un comportamiento similar, sin embargo, se
observa que existen mayores cantidades durante la post-cosecha en el tratamiento sin raleo.

35
Figura 7: Heatmap con las cantidades relativas de los metabolitos encontrados por GC-MS. RC: Raleo
comercial, SR: Sin raleo. S1: primera etapa del desarrollo, S2: segunda etapa del desarrollo, S3: tercera etapa
del desarrollo, Cosecha: cosecha de los frutos, Consumo: frutos sometidos 5 días a 21°C.

36
4.2 Análisis discriminante de regresión de mínimos parciales (PLS-DA)
A través del análisis PLS-DA se extrajeron dos factores (Figura 8), los cuales
explican el 45% de la variabilidad de los metabolitos, y 31% de la variabilidad de las
categorías de tratamiento y estado de desarrollo. Mientras más lejos se encuentren las
variables del origen del eje, mayor variabilidad existe entre las observaciones. Por ejemplo,
el aminoácido prolina no contribuye a la discriminación del tratamiento ni del estado de
desarrollo. Por otro lado, los metabolitos fructosa, glucosa y glucosa/manosa son los que
presentan una alta variabilidad y se correlacionan positivamente con el tratamiento sin raleo
y negativamente con el raleo comercial, debido a que se encuentran en lados opuestos. Los
estados de desarrollo S1, S2 y S3 presentan contenido significativamente bajo de los
aminoácidos serina, alanina y ácido aspártico, por el contrario, estos aminoácidos se
correlacionan positivamente con la etapa de consumo. El tratamiento sin raleo en
combinación con la etapa de consumo se correlacionan positivamente con ácido aspártico,
alanina, serina y treonina.

37
Figura 8: Bi-gráfico de proyecciones latentes con análisis discriminante (PLS-DA, Partial Least Square Regression- Discriminant Analysis). Las variables
categóricas se muestran en color rojo y representan los tratamientos de raleo (raleo comercial y sin raleo), y los estados de desarrollo (S1, S2, S3, Cosecha y
Consumo). Las variables predictoras se muestran en color azul y representan a metabolitos polares como azúcares, aminoácidos y ácidos orgánicos.

38
4.3 Variables importantes en la proyección (VIP)
Para poder identificar cual de todos los metabolitos identificados tiene más
importancia en cada tratamiento, se calculó el VIP (variables importantes en proyección),
el cual ordena los metabolitos según su correlación con el tratamiento (valor de VIP
cercano a 1 está más correlacionado). En amarillo se muestran los valores de VIP más
altos, los cuales ayudan a la discriminación de las variables más importantes en la
predicción de los tratamientos (VIP más alto mayor correlación). Estos metabolitos son
ácido succínico, glucosa/manosa, ácido málico, fructosa, glucosa, ácido ascórbico, ácido
cítrico y sacarosa. Otra forma de representar el VIP se muestra en la Figura 9, en donde se
observa los metabolitos ordenados por factor. Para el factor 1 (Figura 9A), los metabolitos
más importantes se muestran al lado izquierdo, los cuales son ácido málico, ácido
succínico, ácido ascórbico, sacarosa, mioinositol y ácido cítrico. Para el factor 2 (Figura
9B) los metabolitos más importantes están en ambos costados los cuales son fenilalanina,
glucosa/manosa, fructosa, sorbitol, ácido aspártico, ácido cítrico, valina, ácido succínico y
sacarosa.

39
Tabla 1: Variables importantes en la proyección (VIP)
Metabolito Factor-1 Factor-2 VIP
Ácido succínico -0,7841022 0,3804631 0,871532231
Glucosa/manosa -0,484767 -0,7171354 0,865610898
Ácido Málico -0,8459454 0,1693879 0,862737434
Fructosa -0,5854556 -0,6129984 0,847658716
Glucosa -0,5378073 -0,6499076 0,843573696
Ácido Ascórbico -0,7908604 0,2412087 0,826826348
Ácido Cítrico -0,7843405 0,2576932 0,825588157
Sacarosa -0,7899809 0,1447987 0,803141635
Mioinositol -0,732725 0,004579464 0,73273931
Ácido L-aspártico -0,4469828 0,4843448 0,659077771
Fenilalanina -0,2721435 -0,5954436 0,654687074
Ácido oxálico -0,6095425 0,06023116 0,612511104
Asparagina -0,5740687 -0,1858971 0,603417438
L-alanina -0,3524185 0,4552709 0,575734654
Serina -0,4456698 0,3474957 0,565132579
Sorbitol -0,2403564 -0,4553068 0,514854816
L-treonina -0,4767985 0,1549843 0,501355107
L-leucina -0,4764067 -0,07881965 0,482882885
L-valina -0,03506187 0,3623419 0,364034321
Ácido Quínico 0,06633172 0,1722393 0,184570511
L-prolina -0,03200621 -0,1258789 0,12988416
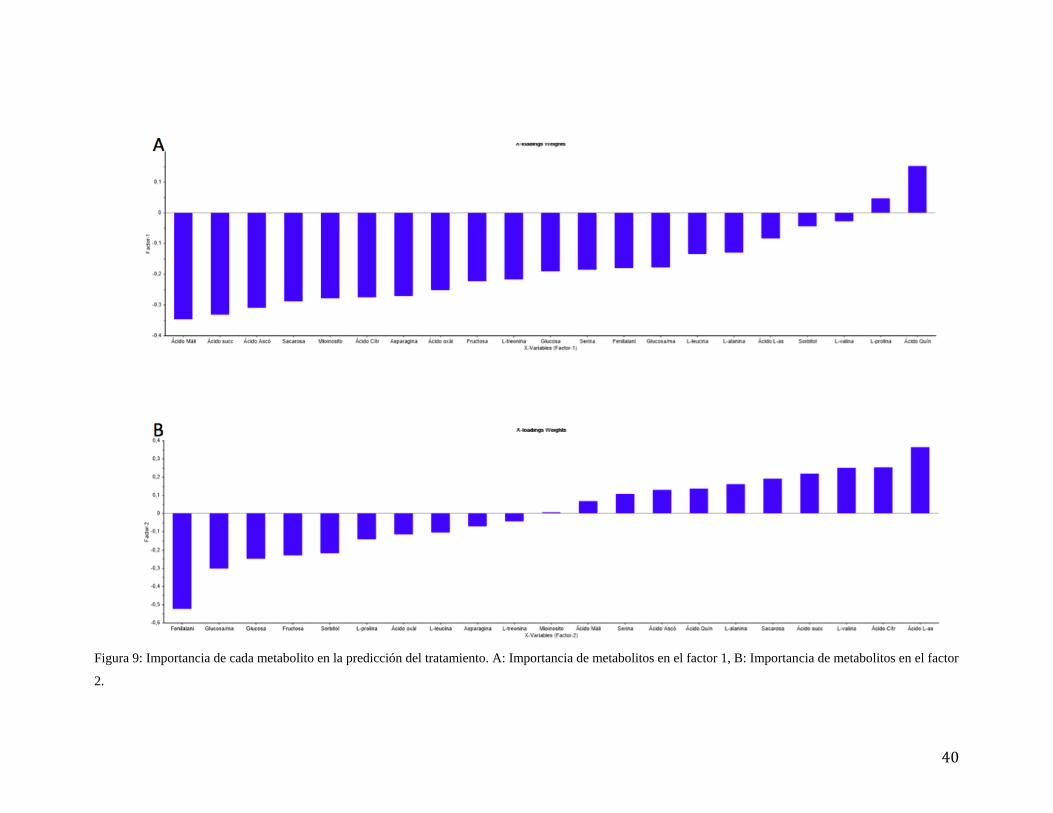
40
Figura 9: Importancia de cada metabolito en la predicción del tratamiento. A: Importancia de metabolitos en el factor 1, B: Importancia de metabolitos en el factor
2.

41
4.4 PLS- DA aplicando el VIP
Para tener una mejor modelo de predicción, se consideraron sólo los VIP más
altos (más cercanos a 1) tomando en consideración ambos criterios de discriminación y se
volvió a realizar un PLS-DA con estos datos (Figura 10). A través de este análisis se
extrajeron dos factores, los cuales explican el 82% de la variabilidad de los metabolitos, y
21% de la variabilidad de las categorías de tratamiento y estado de desarrollo. Los
metabolitos fructosa, glucosa y glucosa/manosa se correlacionan positivamente con el
tratamiento sin raleo y negativamente con el raleo comercial, debido a que se encuentran en
lados opuestos. El tratamiento sin raleo en combinación con la etapa de consumo se
correlacionan positivamente con ácido succínico, ácido málico, ácido ascórbico, ácido
cítrico, sacarosa y mioinositol.
4.5 Metabolitos correlacionados con los tratamientos de raleo
Teniendo en consideración el criterio del VIP para discriminar los metabolitos
correlacionados con los tratamiento de raleo, se graficaron estos metabolitos:

42
Figura 10: Bi-gráfico de proyecciones latentes con análisis discriminante (PLS-DA, Partial Least Square Regression- Discriminant Analysis). Las variables
categóricas se muestran en color rojo y representan los tratamientos de raleo (raleo comercial y sin raleo), y los estados de desarrollo (S1, S2, S3, Cosecha y
Consumo). Las variables predictoras se muestran en color azul y representan a metabolitos polares como azúcares, aminoácidos y ácidos orgánicos.

43
4.5.1 Azúcares
La cantidad de glucosa durante el desarrollo mostró diferencias significativas
durante la etapa S1 y la madurez de consumo, donde los frutos del árbol sin raleo tienen
mayor cantidad de glucosa que los frutos del árbol con raleo comercial (Figura 11A). No
existen diferencias significativas en los frutos del árbol sin raleo a lo largo del desarrollo,
sin embargo, para los frutos del árbol con raleo comercial, existen diferencias entre el
estado S2 con respecto a todos los otros estados del desarrollo. En relación a la cantidad de
fructosa (Figura 11B), se observaron diferencias significativas entre los frutos del árbol sin
raleo y los frutos del árbol con raleo comercial en la etapas S2 y la madurez de consumo, en
donde se observa que hay una mayor cantidad de fructosa en los frutos del árbol sin raleo.
No existen diferencias significativas entre los estados de desarrollo en un mismo
tratamiento. La cantidad relativa de glucosa/manosa (Figura 11C) fue similar durante todo
el desarrollo, sin embargo, existen diferencias significativas durante la madurez de
consumo, en donde los frutos del árbol sin raleo contienen mayor cantidad de
glucosa/manosa que los frutos del árbol sin raleo. No existen diferencias significativas entre
los estados del desarrollo en un mismo tratamiento. Se observa que la cantidad de sacarosa
(Figura 11D) aumenta a medida que avanza el desarrollo del fruto, en donde la cantidad
relativa de este metabolito es significativamente mayor durante la madurez de consumo en
los frutos del árbol sin raleo que en los frutos del árbol con raleo comercial. En los frutos en
estado de madurez de consumo del árbol con raleo comercial existen diferencias
significativas con respecto a los otros estados de desarrollo. Por otro lado, la cantidad
relativa de mioinositol (Figura 11E) también se mantiene constante durante las primeras
etapas del desarrollo, sin embargo, durante la cosecha y la madurez de consumo existen
diferencias significativas entre los tratamientos, observándose mayor cantidad de este
metabolito en los frutos del árbol sin raleo que en los frutos del árbol con raleo comercial.

44
Figura 11: Composición de azúcares en frutos de `Buen Lindo´ medida a través de cromatografía gaseosa
acoplada a espectrometría de masa (GC-MS) expresados en cantidad relativa de metabolito durante todo el
estado de desarrollo A: glucosa, B: fructosa, C: glucosa/manosa, D: sacarosa, E: mioinositol. Las barras
naranjas representan a los frutos del árbol con raleo comercial, y las barras verdes representan a los frutos del
árbol sin raleo. n= 3-5 frutos. Las letras distintas mayúsculas distintas representan diferencias significativas
entre los estados del desarrollo en un mismo tratamiento, y las letras distintas minúsculas distintas representan
diferencias significativas entre los tratamientos de raleo con test Tukey (p<0.05).

45
4.5.2 Ácidos orgánicos
La cantidad relativa de ácidos orgánicos fue medida a través de GC-MS,
normalizado por el peso inicial de la muestra. En el caso del ácido málico (Figura 12A), la
cantidad relativa se mantiene constante entre los tratamientos durante los primeros estados
del desarrollo, sin embargo, durante la cosecha y la madurez de consumo existe una
diferencia significativa entre los tratamientos de raleo, en donde los frutos del árbol sin
raleo contiene mayor cantidad relativa de ácido málico que los frutos del árbol con raleo
comercial. La diferencia también es significativa a cosecha y a madurez de consumo para
los frutos del árbol sin raleo con respecto a los primeros estados del desarrollo. La cantidad
relativa de ácido cítrico va aumentando a medida que avanza el desarrollo (Figura 12B), en
donde la segunda etapa del desarrollo (S2) y la cosecha muestran diferencias significativas
entre los tratamientos de raleo, evidenciando una mayor cantidad relativa de ácido cítrico
en los frutos del árbol sin raleo que en los frutos del árbol con raleo comercial. La
diferencia es significativa entre los estados del desarrollo para ambos tratamientos, en
donde la etapa S1 y S2 contiene menor cantidad relativa de ácido cítrico en los frutos del
árbol con raleo comercial. Para los frutos del árbol sin raleo, esta diferencia es significativa
sólo a madurez de consumo. La cantidad relativa de ácido ascórbico (Figura 12C) se
mantiene constante durante el desarrollo, sin embargo, existen diferencias significativas
entre los tratamientos durante la cosecha y la madurez de consumo, donde los frutos del
árbol sin raleo contienen mayor cantidad relativa de ácido ascórbico que los frutos del árbol
con raleo comercial. No existen diferencias significativas durante el desarrollo para un
mismo tratamiento. En el caso del ácido succínico (Figura 12D), existen diferencias
significativas entre los tratamientos de raleo durante la cosecha y la madurez de consumo,
donde los frutos del árbol sin raleo contienen mayor cantidad relativa de este metabolito
que los frutos del árbol con raleo comercial. Además existe una tendencia al aumento a
medida que avanza el estado del desarrollo en los frutos del árbol sin raleo.

46
Figura 12: Composición de ácidos orgánicos en frutos de `Buen Lindo´ medida a través de cromatografía
gaseosa acoplada a espectrometría de masa (GC-MS) expresados en cantidad relativa de metabolito durante
todo el estado de desarrollo A: ácido málico, B: ácido cítrico, C: ácido ascórbico, D: ácido succínico. Las
barras naranjas representan a los frutos del árbol con raleo comercial, y las barras verdes representan a los
frutos del árbol sin raleo. n= 3-5 frutos. Las letras distintas mayúsculas representan diferencias significativas
entre los estados del desarrollo en un mismo tratamiento, y las letras distintas minúsculas representan
diferencias significativas entre los tratamientos de raleo con test Tukey (p<0.05).

47
DISCUSIÓN
El raleo de frutos en los árboles de durazno es una práctica agrícola que se realiza
con el propósito de obtener frutos con tamaño y calidad apropiados para el mercado
(Childers, 1983). En ese trabajo se utilizó la variedad temprana de durazno `Buen
Lindo´que fue sometida a un nivel de raleo.
La cinética de crecimiento de la variedad `Buen Lindo´ presentó una tendencia a ser
una curva doble sigmoide (Figura 1) como la reportada para otras variedades de durazno
(Chalmers y van den Ende, 1975; Tonutti et al., 1997; el-Sharkawt et al., 2007). Sin
embargo, los tiempos de desarrollo para variedades tempranas como `Buen Lindo´ son más
breves que aquellos de las variedades tardías, por lo que la curva no está bien definida.
El objetivo principal del raleo es obtener frutos con mayor tamaño y calidad. Esto
concuerda con los datos obtenidos para la variedad en estudio, ya que los frutos del árbol
con raleo comercial fueron más grandes durante todo el desarrollo en comparación a los
frutos del árbol sin raleo. Inclusive la fecha de cosecha se adelantó 9 días para los frutos del
árbol con raleo comercial. Este comportamiento se debe a que la eliminación de frutos
tempranamente promueve una menor competencia por los asimilados de los frutos restantes
en el árbol (Bussi et al., 2005).
Una de las características fundamentales para que el fruto sea apetecible para el
mercado es su dulzor y su acidez, además de cómo éstos se relacionan entre sí. En el caso
de la cosecha ambos tratamientos mostraron la misma cantidad de sólidos solubles. Sin
embargo, en la madurez de consumo los frutos del árbol sin raleo fueron los que obtuvieron
menor cantidad de sólidos solubles. Se ha descrito que una vez realizado el raleo, los
fotoasimilados son destinados a los frutos que permanecen en el árbol, los cuales al tener
menor competencia entre ellos, resulta en un incremento en el contenido de sólidos solubles
en los frutos (Caruso et al., 2001; Crisosto et al., 1997). Una serie de estudios asocian los
niveles de sólidos solubles (SS) con una mayor aceptación (Bassi y Selli, 1990; Karakurt et
al., 2000). El nivel de agrado por los duraznos depende ampliamente de la concentración de
sólidos solubles maduros (SSM), que son los sólidos solubles del durazno que está listo

48
para el consumo (Crisosto et al., 2005; Crisosto et al., 2006). La Unión Europea ha
establecido un mínimo de 8°Brix para el mercado del durazno a pesar de que muchos
autores sugieren la introducción de valores más altos de SS, dependiendo de los niveles de
acidez (Kader, 1994; Testoni, 1995). Dicho esto, los frutos del árbol con raleo comercial sí
tuvieron mayor contenido de SSM, por lo tanto, serían superiores en calidad que los frutos
del árbol sin raleo.
En el caso de la acidez, la diferencia entre los tratamientos sólo ocurrió durante la
cosecha, en donde los frutos del árbol con raleo comercial tuvieron mayor porcentaje de
acidez que los frutos del árbol sin raleo.
El durazno es uno de los frutos más populares alrededor del mundo durante el
verano, debido a su alto contenido en minerales y agua, además de la presencia de
carotenoides y moléculas antioxidantes, los cuales determinan el valor nutritivo junto a los
azúcares y ácidos orgánicos (Loizzo et al., 2015). En el caso de los carotenoides, los cuales
también determinan el color del fruto, se mantuvieron sin diferencias significativas entre
los tratamientos de raleo, lo que sugiere que estos no están influenciados por la relación
fuente-sumidero.
El sabor es una combinación de atributos como el gusto, aroma y la sensación en la
boca, los cuales definen un perfil sensorial que afectan el juicio del consumidor con
respecto a la calidad del fruto. En el durazno en particular, el sabor depende principalmente
de los compuestos solubles en agua, como los azúcares y ácidos orgánicos, otorgando la
sensación de dulzor o acidez (Orazem et al., 2011).
Durante el desarrollo del fruto y su maduración existe una red de metabolitos que
actúan en conjunto y que van cambiando a medida que avanza el desarrollo. Por esta razón,
el análisis metabólico permitió identificar cuáles de ellos son los que predominan en cada
etapa de desarrollo. El azúcar glucosa/manosa, se expresó así ya que no fue posible
identificar a cuál de los dos azúcares correspondía. Estos dos azúcares son hexosas
epímeras que sólo se diferencian en un grupo hidroxilo en el carbono 2. Los azúcares

49
glucosa, glucosa/manosa y fructosa poseen un comportamiento similar durante todo el
desarrollo del fruto en ambos tratamientos. No obstante, son más abundantes hacia la post-
cosecha de frutos del árbol sin raleo.
Como ya se ha mencionado anteriormente, los azúcares representan un componente
fundamental de la calidad. Confieren el dulzor, uno de los principales atributos que
influyen en la satisfacción del consumidor. La intensidad del dulzor depende de la cantidad
global de azúcar, así como también el contenido relativo de cada azúcar individual. Dicho
esto, la fructosa, la glucosa y el sorbitol son los azúcares que predominan en el dulzor,
mucho más que la sacarosa (Kroger et al., 2006). Se ha descrito que duraznos de baja
calidad son aquellos que contienen menor fructosa y mayor sorbitol y glucosa en
comparación con los de alta calidad (Robertson y Meredith, 1988).
En el fruto investigado, altas concentraciones de hexosas han sido reportadas
durante las primeras etapas del desarrollo, cuando existe un alto requerimiento de energía
para la división celular (Moriguchi, 1990). Este es el caso de los azúcares glucosa,
glucos/manosa y fructosa; no así sacarosa.
La cantidad de azúcares es bastante baja durante los primeros estados de desarrollo,
aumentando su cantidad relativa hacia la etapa S3 para ambos tratamientos de raleo. El
aumento de sacarosa puede deberse a la translocación fotosintética desde las hojas donde
son cargadas en el floema ya sea mediante simplasto o apoplasto (Moing et al., 1997; Lo
Bianco et al., 1999; Nadwodnik y Lohaus, 2008). La baja cantidad de sacarosa durante las
etapas tempranas del desarrollo para ambos tratamientos puede deberse a que este azúcar se
esté hidrolizando a glucosa y fructosa, lo cual provee la energía necesaria para el
crecimiento del fruto y la formación del endocarpo (Lombardo et al., 2011).
La acumulación de sacarosa desde la etapa S3 del desarrollo se puede atribuir a una
síntesis de novo en el mesocarpo del fruto como resultado de la actividad de la enzima
sacarosa sintasa (Morandi et al., 2008). Sorbitol, junto a sacarosa y almidón, es el producto
final fotosintético primario y el fotoasimilado más importante que es translocado desde los

50
órganos fuente (hojas) en Prunus persica (Loescher, 1987; Layne y Flore, 1995; Zhou et
al., 2001). Este metabolito se mantiene constante durante todo el desarrollo en ambos
tratamientos. Una vez que el sorbitol es descargado a los órganos sumidero, éste es
convertido en fructosa y glucosa por la enzimas sorbitol deshidrogenasa (SDH) y sorbitol
oxigenasa (SOX) respectivamente (Morandi et al., 2008).
Los azúcares manosa y mioinositol también son de gran relevancia durante la post-
cosecha para ambos tratamientos. No obstante, la cantidad relativa es mayor para los frutos
del árbol sin raleo. Se ha descrito que manosa es un componente primario de la pared
celular asociada con el ablandamiento durante la maduración en donde sus niveles
aumentan a lo largo de este proceso y la pérdida de la firmeza (Carrari et al., 2006). Por
otro lado, mioinositol es un precursor importante en la biosíntesis de polisacáridos de pared
celular (Oms-Oliuet et al., 2011). Esto explicaría los niveles constantes de este metabolito
durante el desarrollo.
En el caso de los aminoácidos, durante la etapa S2 se ve un aumento de los
aminoácidos prolina y fenilalanina, principalmente en los frutos del árbol sin raleo. Estos
aminoácidos se utilizan como sustratos para vías de fenilpropanoides, flavonoides y lignina
(Dardick et al., 2010), lo que concuerda con la deposición de lignina durante la etapa S2.
Los aminoácidos, más que una correlación con la calidad del fruto, poseen relevancia por
ser sustratos para la síntesis de ácidos orgánicos y compuestos del metabolismo secundario
durante las etapas tempranas del desarrollo y en la post-cosecha (Lombardo et al., 2011).
En la post-cosecha, el fruto es transformado en un producto sabroso para el
consumo. Todas estas modificaciones afectan la calidad nutricional, sabor y aroma, en
donde la relación entre los azúcares y los ácidos orgánicos contribuyen al sabor final del
fruto (Borsani et al., 2009). Contrario a los estados tempranos del desarrollo, en donde
existen altas cantidades de aminoácidos - como valina, serina, alanina, fenilalanina y
prolina - éstos van disminuyendo hacia la post-cosecha del fruto, particularmente para los
frutos del árbol sin raleo. Estos aminoácidos participarían en el catabolismo de ácidos
orgánicos y como sustrato respiratorio (Lombardo et al., 2011).

51
Como se ha mencionado anteriormente, los ácidos orgánicos son importantes en el
sabor de los frutos y, como tales, contribuyen junto con los azúcares y aromas a la calidad
organoléptica de los duraznos (Sweeney et al., 1970). Los ácidos orgánicos predominantes
en la fruta madura del durazno son ácidos málico y cítrico, mientras que el ácido quínico se
acumula en cantidades más bajas (Moing et al., 1998). En el caso de los frutos del árbol sin
raleo ocurre de esta forma: existe una acumulación de ácido málico y ácido cítrico
principalmente en la madurez de consumo. El ácido málico se acumula principalmente
durante la primera fase de crecimiento rápido, la cual se produce al final de la fase de
división celular, mientras que el ácido cítrico se acumula durante la tercera fase del
desarrollo, antes del inicio de la maduración (Masia et al., 1992). En el caso de esta
investigación, la cantidad relativa de ácido cítrico aumenta durante la tercera fase del
desarrollo para ambos tratamientos. A pesar de ello, el ácido málico aumenta hacia la post-
cosecha del fruto.
La formación de acidez implica la síntesis de ácidos orgánicos, principalmente
malato, citrato y succinato que pueden almacenarse en la vacuola en grandes cantidades.
Como el citrato se produce a partir de dicarboxilatos, principalmente malato, el primer paso
en el desarrollo de la acidez es la síntesis de dicarboxilatos, como malato y oxaloacetato
(OAA). Éstos requieren la fijación de CO2 sobre un esqueleto de carbono derivado del
catabolismo de hexosas como la glucosa (Hardy, 1968; Young y Biale, 1968), que se
consigue mediante la carboxilación de fosfoenolpiruvato (PEP), catalizada por la
fosfoenolpiruvato carboxilasa (PEPC). Esta reacción tiene lugar en el citosol, ya que la PEP
es un intermedio de la vía de la glicólisis y produce OAA, que puede reducirse a malato
mediante la malato deshidrogenasa (Givan, 1999) o suministrada al ciclo de ácidos
tricarboxílicos (TCA) si la reposición es necesaria (Leegood y Walker, 2003). Estos tres
ácidos orgánicos se ven aumentados durante la post-cosecha, principalmente en los frutos
del árbol sin raleo, lo que concuerda también con la cantidad elevada de hexosas en el
mismo tratamiento.
Se ha descrito en algunos frutos que el incremento en la tasa fuente-sumidero
incrementa la cantidad de citrato en los estados tempranos del desarrollo y disminuye hacia

52
la madurez (Souty et al., 1999; Wu et al., 2002; Léchaudel et al., 2005). El efecto contrario
ha sido reportado para malato, el cual decrece en los estados tempranos del desarrollo e
incrementa hacia la madurez (Wu et al., 2002 ; Léchaudel et al., 2005). Durante los estados
tempranos, grandes cantidades de azúcares importadas desde las hojas están disponibles
para la conversión de citrato vía ciclo de los ácidos tricarboxílicos (TCA). Durante estos
estados también ocurre un aumento de la respiración en donde una alta disponibilidad de
azúcares puede estimular la glicólisis y la conversión de malato a citrato. Durante la
maduración, en contraste, los azúcares ya no están disponibles para la respiración ya que se
almacenan en la vacuola (Coombe, 1976), causando un cambio de azúcares a ácidos
orgánicos, especialmente citrato como sustrato respiratorio. Durante este estado, un
incremento en la respiración dado por la alta tasa fuente-sumidero puede estimular la
conversión de citrato a malato para mantener el pool de intermediarios del TCA constantes
(Lobit et al., 2003 ; Wu et al., 2007).
Por otro lado, el ácido ascórbico posee niveles similares durante las primeras etapas
del desarrollo, pero se ve aumentado durante la post-cosecha esencialmente en los frutos
del árbol sin raleo. Se ha descrito que el ascorbato es un antioxidante presente en
concentraciones milimolares en las hojas de las plantas y en los órganos de almacenamiento
como los frutos, lo que contribuye significativamente al estado redox celular (Noctor y
Foyer, 1998; Smirnoff, 2000; Smirnoff et al., 2001, Ishikawa et al., 2006). También se ha
informado que el ascorbato está implicado en una serie de otros procesos celulares tales
como la fotoprotección (Kiddle et al., 2003), el establecimiento de la morfología de toda la
planta, la arquitectura de las raíces y el desarrollo (Olmos et al., 2006; Horemans et al.,
2003).

53
CONCLUSIONES
El raleo promueve la obtención de frutos de mayor tamaño.
La fecha de cosecha se adelanta para los frutos provenientes del árbol con
raleo comercial.
Los metabolitos que separan los tratamientos de raleo fueron glucosa,
glucosa/manosa, sacarosa, mioinositol, fructosa, ácido málico, cítrico,
ascórbico y succínico.
Los metabolitos ácido succínico, málico, cítrico, ascórbico, sacarosa y
mioinositol además se acumulan principalmente en la etapa de madurez de
consumo.

54
REFERENCIAS
Aubert C, Gunata Z, Ambid C, Baumes R. (2003). Changes in physicochemical
characteristics and volatile constituents of yellow- and white-fleshed nectarines during
maturation and artificial ripening. Journal of Agriculture Food Chemistry 51, 3083–3091.
Bassi D, Selli R. (1990). Evaluation of fruit quality in peach and apricot. Advances in
Horticultural Science 4, 107–112.
Blanco A. (2008). Estructura y morfogénesis del fruto, recuperado de
https://fisiohorticola.files.wordpress.com/2008/11/estructura-y-morfogenesis-del-
fruto1.pdf.
Borsani J, Budde C, Porrini L, Lauxmann M, Lombardo V, Murray R, Andreo C,
Drincovich M, Lara M. (2009). Carbon metabolism of peach fruit after harvest: changes in
enzymes involved in organic acid and sugar level modifications. Jounal of Experimental
Botany 60, 1823–1837.
Brooks S, Moore J, Murphy J. (1993). Quantitative and qualitative changes in sugar content
of peach genotypes (Prunus persica L., Batsch.). Journal of the American Society for
Horticultural Science 118, 97–100.
Brovelli E, Brecht J, Sherman W, Sims C, Harrison J. (1999). Sensory and compositional
attributes of melting and non-melting-flesh peaches for the fresh market. Journal of the
Science of Food Agriculture 79, 707–712.
Bussi C, Lescourret F, Génard M, Habib R. (2005). Pruning intensity and fruit load
influence vegetative and fruit growth in an early-maturing peach tree (cv. Alexandra).
Fruits 60, 133-142.
Byrne, D. (2005). Trends in stone fruit cultivar development. HortTechnology 15, 494–500.

55
Carrari, F, Fernie, A. (2006). Metabolic regulation underlying tomato fruit development.
Journal of Experimental Botany 57, 1883–1897.
Chalmers D, van den Ende B. (1975). A reappraisal of the growth and development of
peach fruit. Australian Journal of Plant Physiology 2, 623–634.
Chapman J, Horvat, R, Forbus R. (1991). Physical and chemical changes during the
maturation of peaches (cv. Majestic). Journal of Agricultural Food Chemistry 39, 867–870.
Childers N. (1983). Peach, nectarine, apricot and almond. Modern Fruit Science 970, 203-
241
Colaric M, Veberic R, Stampar F, Hudina M. (2005). Evaluation of peach and nectarine
fruit quality and correlations between sensory and chemical attributes. Journal of Science
Food Agricultural 85, 2611–2616.
Coombe B. (1976). The development of fleshy fruits. Annual Review of Plant Physiology
27, 207–228.
Costa, G. (1978). Chemical and mechanical thinning of peach fruits. Rivista della
Ortoflorofrutticoltura Italiana 62, 63–86.
Covarrubias M, Benedetto G, Muñoz-Espinosa C, Andrade D, Maucourt M, Deborde C,
Moing A, Valenzuela M.A, Almeida AM. Primary metabolite fruit profile is altered in
response to source-sink imbalance and can be used as early quality predictors in nectarine.
Journal of Agricultural and Food Chemistry, en preparación.
Crisosto C, Johnson S, DeJong T, Day K. R. (1997). Orchard factors affecting postharvest
stone fruit quality. Hortscience 32, 820-823.

56
Crisosto C, Crisosto G. (2005). Relationship between ripe soluble solids concentration
(RSSC) and consumer acceptance of high and low acid melting flesh peach and nectarine
(Prunus persica (L.) Batsch cultivars. Postharvest Biology and Technology 38, 239–246.
Crisosto C, Crisosto G, Echeverria G, Puy J. (2006). Segregation of peach and nectarine
(Prunus persica (L.) Batsch cultivars according to their organoleptic characteristics. Post-
harvest Biology and Technology 39, 10–18.
Dann I.R., Wildes R.A., Chalmers D.J. (1984). Effects of limb girdling on growth and
development of competing fruit and vegetative tissues of peach trees. Australian Journal of
Plant Physiology 11, 49-58.
Dardick C, Callahan A, Chiozzotto R, Schaffer R, Piagnani M, Scorza R. (2010). Stone
formation in peach fruit exhibits spatial coordination of the lignin and flavonoid pathways
and similarity to Arabidopsis dehiscence. BMC Biology 8, 13
Davidovic S, Veljovic M, Pantelic M, Baosic R, Natic M, Dabic C, Pecic S, Vukosavljevic
P. (2013). Physicochemical, antioxidant and sensory properties of peach wine made from
Redhaven cultivar. Journal of Agricultural Food Chemistry 61, 1357-1363.
Dean JFD. (1997). Lignin analysis. Methods in Plant Biochemistry and Molecular Biology.
CRC Press, Boca ratón, pp 199-215.
DeJong T.M., Doyle J.F., Day K.R. (1987). Seasonal patterns of reproductive and
vegetative sink activity in early and late maturing peach (Prunus persica) cultivars.
Physiologia Platarumt 71, 83-88.
DeJong T, Grossman W. (1995). Quantifying sink and source limitations on dry matter
partitioning to fruit growth in peach trees. Physiologia Plantarum 95, 437–443.

57
Dennis F, Dilley D, Cook R. (1983) Improving fruit quality and yield. Proceeding
Michigan Society of Horticultural Science 113, 199–201.
Dorsey M, McMunn R. (1927). Relation of the time of thinning peaches to the growth of
fruit and tree. Proceeding American Society of Horticultural Science 24, 221–228.
Dziezak J. (2003). Acids . Encyclopeadia of Food Sciences and Nutrition. 2nd ed, Academic
Press, pp 7-17.
El-Sharkawy I, Kim W, El-Kereamy A, Jayasankar S, Svircev AM, Brown D. (2007)
Isolation and characterization of four ethylene signal transduction elements in plums
(Prunus salicina L.). Journal of Experimental Botany 58, 3631–3643.
Esti M, Messia M, Sinesio F, Nicotra A, Conte L, Notte E, Palleschi G. (1997). Quality
evaluation of peaches and nectarines by electrochemical and multivariate analyses:
relationships between analytical measurements and sensory attributes. Food Chemistry 60,
659–666.
Etienne A, Génard M, Lobit P, Mbeguié-A-Mbéguie D, Bugaud, C. (2013). What controls
freshy fruit acidity? A review of malate and citrate accumulation in fruit cells. Journal of
Experimental Botany 64, 4, 1451-1469.
FAOstat (2013). Food and agricultural organization statistical database, recuperado de
http://faostat3.fao.org.
Farley A. (1923). Factors that influence the effectiveness of peach thinning. Proceeding
American Society of Horticultural Science 20, 145–151.
García F. (2003). Estructura de los frutos, recuperado de
http://www.euita.upv.es/varios/biologia/web_frutos/Estructura.htm.

58
Génard M, Souty M, Holmes S, Reich M, Breuils L. (1994). Correlations among quality
parameters of peach fruit. Journal of the Science of Food and Agriculture 66, 241–245.
Génard, M, Huguet J. (1996). Modelling the response of peach fruit growth to water stress.
Tree Physiology 16, 407–415.
Génard M, Souty M. (1996). Modeling the peach sugar contents in relation to fruit growth.
Journal American Society of Horticultural Science 121, 1122–1131.
Givan CV. (1999). Evolving concepts in plant glycolysis: two centuries of progress.
Biological Reviews 74, 277–309.
Grossman Y, DeJong T. (1995). Maximum fruit growth potential and seasonal patterns of
resource dynamics during peach growth. Annals of Botany 75, 553–560.
Giulivo C. (1974) Knowledge on growth of the fruits and their contribution to the thinning
of peaches and apples technique. Incontro Frutticolo SOI, Bologna 3, 21–28.
Giulivo C, Ramina A, Costa G. (1981). The contribution of the sub-project to the
knowledge of the abscission processes and ripening fruit. Seminario su ‘I fitoregolatori nel
controllo della produzione degli alberi da frutto’ 26, 107–117.
Guardiola JL, García-Luis JL. (2000). Increasing fruit size in Citrus: Thinning and
stimulation of fruit growth. Plant Growth Regulation 31, 121–132.
Hansen P. (1970). 14C-studies on apple trees. V. Translocation of labelled compounds from
leaves to fruit and their conversion within the fruit. Physiologia Plantarum 23, 564–573.
Hansen P. (1971). The effect of cropping on the distribution of growth in apple trees.
Tidsskr. Planteavl 75, 119-127.

59
Hardy PJ. (1968). Metabolism of sugars and organic acids in immature grape berries. Plant
Physiology 43, 224–228.
Hatoum D, Annaratone C, Hertog M, Geeraerd A, Nicolai, B. (2014). Target metabolomics
study of “Braeburn” apples during long-term storage. Postharvest Biology and
Biotechnology 96, 33-41.
Havis A. (1962). Effect of time of fruit thinning of ‘Redhaven’ peach. Proceeding
American Society of Horticultural Science 80, 172–176.
Horemans N, Potters G, De Wilde L, Caubergs RJ. (2003). Dehydroascorbate uptake
activity correlates with cell growth and cell division of tobacco Bright Yellow-2 cell
cultures. Plant Physiology 133, 361–367.
Huang H, Cheng Z, Zhang Z, Wang Y. (2008). Livres. Hist. Cultiv. trends China. Peach
Bot. Prod. uses. In: Layne, D.R., Bassi, D. (Eds.), CAB Int. London, UK, 1st
ed., pp. 37.
Huberlant J, Anderson G. (2003). Sucrose. Encyclopaedia of Food Sciences and Nutrition.
2nd ed, Academic Press, pp 5536-5645.
Infante R, Meneses C, Predieri S. (2008). Sensory quality performance of two nectarine
flesh typologies exposed to distant market conditions. Journal of Food Quality 31, 526–
535.
Ishida M, Inaba A, Sobajima Y. (1971). Seasonal changes in the concentration of sugars
and organic acids in peach fruits. Science Rpt Kyoto Prefecture University Agricultural 23,
18–23.
Ishikawa T, Dowdle J, Smirnoff N. (2006). Progress in manipulating ascorbic acid
biosynthesis and accumulation in plants. Physiologia Plantarum 126, 343–355.

60
Jiang, Y, Song J. (2009). Fruits and fruit flavor: classification and biological
characterization (Chapter 1). In: Hui, Y.H. (Ed.), Handbook of Fruit and Vegetable
Flavors. A John Wiley & Sons, Inc., Publication, pp. 3-23.
Kader AA. (1994) Fruit maturity, ripening, and quality relationships. Perishable Handling
Newslett 80, 2.
Karakurt Y, Huber DJ, Sherman WB. (2000). Quality characteristics of melting and
nonmelting flesh peach genotypes. Journal of Science Food Agricultural 80, 1843–1853.
Kroger M, Meister K, Kava R. (2006). Low-calorie sweeteners and other sugar substitutes:
a review of the safety issues. Science Food Safety 5, 35–47.
Kiddle G, Pastori GM, Bernard S, Pignocchi C, Antoniw J, Verrier PJ, Foyer CH. (2003).
Effects of leaf ascorbate content on defense and photosynthesis gene expression in
Arabidopsis thaliana. Antioxidants and Redox Signaling 5, 23–32.
Layne DR, Flore JA. (1995). End-product inhibition of photosynthesis in Prunus cerasus L.
in response to wholeplant source-sink manipulation. Journal American Society
Horticultural Science 120, 583–599.
Léchaudel M, Joas J, Caro Y, Génard M, Jannoyer M. (2005). Leaf: fruit ratio and
irrigation supply affect seasonal changes in minerals, organic acids and sugars of mango
fruit. Journal of the Science of Food and Agriculture 85, 251–260.
Leegood RC, Walker RP. (2003). Regulation and roles of phosphoenolpyruvate
carboxykinase in plants. Archives of Biochemistry and Biophysics 414, 204–210.
Link H. (2000). Significance of flower and fruit thinning on fruit quality. Plant Growth
Regulation 31, 17-26.

61
Liverani A, Cangini A. (1991). Ethylene evolution and changes in carbohydrates and
organic acid during maturation of two white and two yellow fleshed peach cultivars.
Advances in Horticultural Science 5, 59–63.
Lobit P, Génard M, Wu B, Soing P, Habib R. (2003). Modelling citrate metabolism in
fruits: responses to growth and temperature. Journal of Experimental Botany 54, 2489-
2501.
Loescher, W.H. (1987). Physiology and metabolism of sugar alcohols in higher plants.
Physiologia Plantarum 70, 553–557.
Loizzo MR, Pacetti D, Lucci P. (2015). Prunus persica var. Platycarpa (Tabacchiera
peach): bioactive compounds and antioxidant activity of pulp, peel and seed ethanolic
extracts. Plant Foods for Human Nutrition 70, 331–337.
Lombardo V, Osorio S, Borsani J, Lauxmann M, Bustamante C, Budde C. (2011).
Metabolic profiling during peach fruit development and ripening reveals the metabolic
networks that underpin each developmental stage. Plant Physiology 157, 1696–1710.
Maggs D.H. (1963). The reduction in growth of apple trees brought about by fruiting.
Journal of Horticultural Science. 38, 119-128.
Masia A, Zanchin A, Rascio N, Ramina A. (1992). Some biochemical and ultrastructural
aspects of peach fruit development. Journal American Society Horticultural Science 117,
808–815.
Mayne ST. (1996). Beta-carotene, carotenoids, and disease prevention in humans. FASEB
Journal 10, 690–701.

62
Moing AF, Svanella L, Rolin D, Monet R, Gaudillere JP, Gaudillere M. (1988).
Compositional changes during fruit development of two peach cultivars differing in juice
acidity. Journal of the American Society Horticultural Science 123, 770–775.
Moriguchi T, Sanada T, Yamaki S (1990) Seasonal fluctuation of some enzymes relating to
sucrose and sorbitol metabolism in peach fruit. Journal of the American Society
Horticultural Science 115, 278–281.
Nadwodnik J, Lohaus G. (2008). Subcellular concentrations of sugar alcohols and sugars in
relation to phloem translocation in Plantago major, Plantago maritima, Prunus persica,
and Apium graveolens. Planta 227, 1079–1089.
Noctor G, Foyer CH. (1998). Ascorbate and glutathione: keeping active oxygen under
control. Annual Review of Plant Physiology and Plant Molecular Biology 49, 249–279.
ODEPA (2015). Boletín frutícola, recuperado de http://www.odepa.cl/wp-
content/files_mf/1427304052Boletinfruticola201503.pdf
Ojer M, Reginato G. (2002). Efectos de la carga frutal en durazneros. Revista de la
Facultad de Ciencias Agrarias XXXIV, 39-46.
Olmos E, Kiddle G, Pellny TK, Kumar S, Foyer CH. (2006). Modulation of plant
morphology, root architecture, and cell structure by low vitamin C in Arabidopsis thaliana.
Journal of Experimental Botany 57, 1645–1655.
Oms-Oliu G, Hertog, M, Van de Poel B, Ampofo-Asiama J, Geeraerd A, Nicolai B. (2011).
Metabolic characterization of tomato fruit during preharvestdevelopment, ripening, and
postharvest shelf-life. Postharvest Biology and Technology 62, 7–16.
Orazem P, Stampar F, Hudina M. (2011). Quality analysis of Redhaven peach fruit grafted
on 11 rootstocks of different genetic origin in a replant soil. Food Chemistry 124, 1691–
1698.

63
Pavel E, DeJong T. (1993) Source- and sink-limited growth periods of developing peach
fruits indicated by relative growth rate analysis. Journal of the American Society of
Horticultural Science 118, 820–824.
Rapaille A, Goosens J, Heume M. (2003). Sugar alcohols. Encyclopaedia of Food Sciencies
and Nutrition, 2nd ed, Academic Press, pp 7-17.
Robertson J, Meredith FI. (1988). Characteristics of fruit from high- and low-quality peach
cultivars. HortScience 23, 1032–1034.
Robertson J, Horvat R, Lyon B, Meredith J, Senter S, Okie W. (1990). Comparison of
quality characteristics of selected yellow- and white-fleshed peach cultivars. Journal Food
Science 55, 1308–1311.
Sansavini S, Gamberini A, Bassi D. (2006). Peach breeding, genetics and new cultivar
trends. Acta Horticulturae 713, 23–48.
Shepherd R, Colwill J, Daget N, Thomson D, McEwan J, Lyon D, Stone H, Sidel J,
Redlinger P, Bourne M, Mottram D, Mela D. (1993). Sensory evaluation, in Encyclopaedia
of Food Science, Food Technology and Nutrition, ed. Academic Press, pp 4023–4075.
Smirnoff N. (2000). Ascorbic acid: metabolism and functions of a multi-facetted molecule.
Current Opinion in Plant Biology 3, 229–235.
Smirnoff N, Conklin PL, Loewus FA. (2001). Biosynthesis of ascorbic acid in plants: a
renaissance. Annual Review of Plant Physiology and Plant Molecular Biology 52, 437–467.
Souty M, Génard M, Reich M, Albagnac G. (1999). Effect of assimilate supply on peach
fruit maturation and quality. Canadian Journal of Plant Science 79, 259–268.

64
Souty M, André P. (1975) Biochemical composition and quality peaches. Annales de
Technologie Agricole 24, 217–236.
Sweeney J.P., Chapman V.J., Hepner P.A. (1970). Sugar, acid and flavor in flesh fruits.
Journal American Diet Assocciation 57, 432–435.
Testoni A. (1995). Momento di raccolta, qualita´, condizionamento e confezionamento
delle pesche. Proceedings of the Symposium ‘La peschicoltura Veronese alle soglie del
2000’, 327–354.
Tonutti P, Bonghi C, Ruperti B, Tornielli G, Ramina A. (1997). Ethylene evolution and 1
aminocyclopropane-1-carboxylate oxidase gene expression during early development and
ripening of peach fruit. Journal of the American Society of Horticultural Science 122, 642–
647
Trainotti L, Zanin D, Casadoro G. (2003). A cell wall-oriented genomic approach reveals a
new and unexpected complexity of the softening in peaches. Journal of Experimental
Botany 54, 1821–1832.
Tukey H, Einset O. (1939). Effect of fruit thinning on size, color and yield of peaches and
on growth and blossoming of the trees. Proceeding American Society of Horticultural
Science 36, 314–319.
Versari A, Castellari M, Parpinello G, Riponi C, Galassi S. (2002). Characterization of
peach juices obtained from cultivars Redhaven, Suncrest and Maria Marta grown in Italy.
Food Chemistry 76, 181–185.
Wareing P.F., Patrick J. (1975). Source-sink relations and the partition of assimilate, In:
J.P. Cooper (ed.). Photosynthesis and productivity in different environments. Cambridge
University Press, Cambridge, U.K. p. 481-499.

65
Weinberger J. (1931) The relation of leaf area to size and quality of peaches. Proceedings
American Society of Horticultural Science 28, 18–22
Weinberger J. (1941) Studies on time of peach thinning from blossoming to maturity.
Proceedings American Society of Horticultural Science 38, 137–140.
Wellburn AR. (1994). The Spectral Determination of Chlorophylls a and b, as well as Total
Carotenoids, Using Various Solvents with Spectrophotometers of Different Resolution.
Plant Physiology 144, 307-313.
Wrolstad R, Shallenberger R. (1981). Free sugars and sorbitol in fruits: a compilation from
the literature.. Annals Chemistry 64, 91-103.
Wu B, Génard M, Lescourret F, Gomez L, Li S. (2002). Influence of assimilate and water
supply on seasonal variation of acids in peach (cv Suncrest). Journal of the Science of Food
and Agriculture 82, 1829–1836.
Wu B, Génard M, Lobit P, Longuenesse J, Lescourret F, Habib R, Li S. (2007). Analysis of
citrate accumulation during peach fruit development via a model approach. Journal of
Experimental Botany 58, 2583–2594.
Young RE, Biale JB. (1968). Carbon dioxide effects on fruits. Planta 81, 253–263.
Zhou R, Sicher R, Quebedeaux B. (2001). Diurnal changes in carbohydrate metabolism in
mature apple leaves. Australian Journal Plant Physiology 28, 1143–1150.


















