Unidad 3 control y regulación metabólica (1) (1) (1)

of 191
-
Upload
elva-avalos -
Category
Documents
-
view
98 -
download
1
Transcript of Unidad 3 control y regulación metabólica (1) (1) (1)
Presentacin de PowerPoint
Unidad iii.- Control y regulacin metablicaINSTITUTO TECNLOGICO DE JIQUILPAN
ALUMNAS: RUB MEDINA CORTSTANIA PAOLA ORTEGA PARRALIDIA AGUILAR OROZCOPAULINA ORTIZ MARTNEZCONTROL Y REGULACIN METABLICA3.1 CONCEPTOS DE REGULACIN DEL METABOLISMO3.2 REGULACIN A NIVEL ENZIMTICO 3.2.1 INHIBICIN 3.2.2 ACTIVACIN3.3 REGULACIN A NIVEL DNA 3.3.1 REPRESOR 3.3.2 INDUCCINInicio 9 de Abril de 2013 Final 16 de Abril de 2013.3.1 CONCEPTOS DE REGULACION DEL METABOLISMOEl metabolismo es el conjunto de reacciones bioqumicas y procesos fsico-qumicos que ocurren en una clula y en el organismo.
Estos procesos permiten:CrecerReproducirseMantener sus estructurasResponder a estmulosEtcLa metabolizacin es el proceso por el cual el organismo consigue que sustancias activas se transformen en no activas.Este proceso lo realizan en los seres humanos enzimas localizadas en el hgado.Las reacciones qumicas del metabolismo se organizan estrictamente en vas o rutas metablicas donde un sustrato es transformado en un producto y este a su vez funciona como sustrato para generar otro producto siguiendo una secuencias de reacciones bajo la intervencin de diferentes enzimas
El metabolismo se divide en dos procesos conjugados:
Catabolismo Anabolismo. CATABOLISMOFase destructiva del metabolismo en la cual se obtienen molculas sencillas, que servirn para construir las propias biomolculas, y energa para la realizacin de otras funciones celulares.
Son procesos oxidativos en los que los e-procedentes de molculas orgnicas complejas van descendiendo progresivamente de nivel energtico.
TIPOS DE CATABOLISMOCatabolismo aerbicoCatabolismo anaerbico
Si el aceptor de electrones es el oxgeno molecular la ruta o el catabolismo es aerbico y si es otra molcula es catabolismo anaerbico.
CATABOLISMO AERBICOEl catabolismo aerobio est formado por varias rutas metablicas que conducen finalmente a la obtencin de molculas de ATP. Estas molculas de ATP ms tarde sern imprescindibles para dar energa en las rutas anablicas. La energa que no se usa se disipar en forma de calor. CICLO DE KREBSCADENA RESPIRATORIACATABOLISMO ANAEROBICOCuando el catabolismo o se realiza en condiciones anaerbicas, es decir cuando el ltimo aceptor de hidrgenos o electrones no es el oxgeno, sino una molcula orgnica sencilla, las rutas de degradacin de la glucosa se llaman fermentaciones.
FERMENTACIONESLa fermentacin es un proceso catablico de oxidacin incompleta, que no requiere oxgeno, siendo el producto final un compuesto orgnico.
Estos productos finales son los que caracterizan los diversos tipos de fermentaciones.GLICLISISEs la va metablica encargada de oxidar o fermentar la glucosa y as obtener energa para la clula. sta consiste de diez reacciones enzimticas que convierten a la glucosa en dos molculas de piruvato, la cual es capaz de seguir otras vas metablicas y as continuar entregando energa al organismo. Ocurre en el citosol sin necesidad de oxigeno.
GLUCLISISCICLO DE KREBSForma parte de la respiracin aerobia, es decir, se realiza en presencia de oxgeno y se desarrolla entre los procesos de glicolisis y cadena respiratoria. Su fin es la obtencin de NADH, una molcula con poder reductor, que se utiliza para la produccin de ATP mediante la cadena respiratoria.
CICLO DE KREBS
CADENA RESPIRATORIALa energa necesaria para la sntesis del ATP por medio de la fosforilacin oxidativa se obtienen de la oxidacin del FADH+H y del NADH+H.La cadena respiratoria esta formada por transportadores y protenas que forman los complejos
Complejo I: NADH-Ubiquinona reductasaEs por donde ingresan la mayora de los electrones a la cadena. Los electrones son transferidos desde el NADH a la Co Q a travs del FMN que es parte del complejo I.
Complejo II: Succinato deshidrogenasaEs otro punto de entrada de electrones a la cadena y en su transferencia entre el FAD y la coenzima Q no libera energa suficiente para bombear protones. Por esta razon se genera un ATP menos cuando la cadena comienza por el FAD respecto a cuando comienza por en NADComplejo III: CoQCitocromo C reductasaRecibe electrones de los complejos I y II. A partir de este paso se transportan los electrones y quedan libres los H+.El complejo III involucra los citocromos y la energa aportado por este complejo para el bombeo de protones es suficiente para producir ATP
Complejo IV: Citocromo oxidasaCataliza la formacin de H2O a partir de los dos electrones: O y 2H.Este complejo contribuye con la generacin de un gradiente de protones suficiente para generar un ATP. El oxgeno se reduce a agua, mientras la energa liberada por los electrones permite la fosforilacin del ADP a ATP: el proceso se denomina fosforilacin oxidativa.
Complejo V: ATPasa o ATPsintasaAnclado a la membrana mitocondrial interna.Este complejo esta formado por los componentes F0 y F1.F0: Corresponde al canal protnico. F1: Contiene unidades catlicas de la ATPasaCadena respiratoria
VideoANABOLISMOConjunto de reacciones metablicas mediante las cuales a partir de compuestos sencillos se sintetizan molculas ms complejas.
Mediante estas reacciones se crean nuevos enlaces por lo que se requiere un aporte de energa que provendr del ATP.GLUCONEOGNESISEs una ruta metablica anablica que permite la sntesis de glucosa a partir de precursores no glucdicos. Incluye la utilizacin de varios aminocidos, lactato, piruvato, glicerol y cualquiera de los intermediarios del ciclo de los cidos tricarboxlicos.
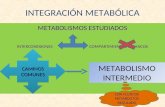
GLUCONEOGNESISEl catabolismo y el anabolismo son procesos acoplados que hacen al metabolismo en conjunto, puesto que cada uno depende del otro.La clula realiza todos los procesos con mxima eficacia y utilizando el menor consumo de energa posible. Por ello el metabolismo celular se encuentra regulado de forma precisa y constante.Se puede establecer que el ritmo del metabolismo se controla por las necesidades energticas de la clula, esto es, las clulas oxidan las molculas combustibles a la misma velocidad que requieren energa.Para entender como son controladas las vas metablicas existen dos conceptos vinculados:Regulacin ControlLa regulacin de una enzima en una ruta es cmo incrementa o disminuye su actividad en respuesta a seales o estmulos.El control llevado a cabo por esta enzima viene dado por los efectos que, dichos cambios de su actividad, tienen sobre la velocidad de la ruta.
Existen tres mecanismos por los que se regula el metabolismo en las clulas de organismos superiores.
Enzimas reguladorasControl de concentracin de una enzimaControl metablicoAPLICACIONES CLNICASUna enfermedad metablica puede derivar en la incorrecta expresin de una enzima.La determinacin de la actividad de una enzima en los lquidos biolgicos o tejidos constituye un indicador valioso de una determinada enfermedad metablica.
REGULACIN A NIVEL ENZIMTICO
Se definen como aquellas cuya actividad se puede modificar en una forma que afecte la velocidad de una reaccin catalizada por la enzima.
ENZIMAS REGULADORASFuncionamiento de la enzima Las enzimas se unen a los reactivos (sustratos)Cada enzima tiene una forma nica con un sitio o centro activo en el que se une al sustrato.
Despus de la reaccin, enzimas y productos se separan..Las molculas enzimticas no han cambiado despus de participar en la reaccin
CLASIFICACIN Enzimas alostricas
Enzimas modulados covalentemente.
39Enzima Alostricos Son aquellos que, adems del centro activo mediante el cual interactan con el sustrato, poseen otro centro de unin llamado centro alostrico mediante el cual interactan con otra molcula denominada efector o modulador
Caractersticas Pesos moleculares superiores a los de otros enzimas
Protenas oligomricas (estn formados por varias subunidades en nmero par).
Catalizan reacciones irreversible
ModuladoresEstimulan la actividad del enzima al unirse al centro alostrico, reciben el nombre de moduladores positivos o activadores.
Inhiben y se llaman moduladores negativos o inhibidores.
Presentan siempre dos formas, una activa y otra inactiva, interconvertibles por efecto del modulador
Inhibicin por producto final, retroinhibicin o control feed-back Inhibe alostricamente al enzima que cataliza la primera reaccin de dicha ruta, modulador negativo.
Una enzima sea estimulada por algn agente que se acumula en el medio. Cuando existe un exceso de sustrato, l mismo promueve su utilizacin activando a la enzima.
Modulacin Covalente Consiste en la adicin o eliminacin de un grupo qumico esencial para la catlisis (generalmente un grupo fosfato, metilo, adenilato u otros)
Modificacin de los residuos de tirosina, serina y/o treonina de las enzimas por un proceso de unin o eliminacin de grupos fosfatos.Clasificacin Reversible FosforilacinMetilacinAcetilacin ADP- ribosilacin Irreversible Activacin proteoltica
Enzimas ModuladoresSe activa cuando est unido a dicho grupoInactiva cuando ste se elimina.Caractersticas Presentan dos formas, una activa y otra inactiva, que son interconvertibles, por modificacin covalente de sus estructuras catalizada enzimas moduladores. Ejemplo glucgeno-fosforilasa, que acta en el metabolismo de los glcidos liberando unidades de glucosa-1-fosfato a partir de las cadenas polisacardicas del glucgeno. La glucgeno-fosforilasa es activada por un enzima modulador, la fosforilasa-quinasa, que une covalentemente un grupo fosfato a un resto especfico de serina en cada una de las dos subunidades del enzima modulado.
Otro enzima modulador, la fosforilasa-fosfatasa, escinde hidrolticamente dichos grupos fosfato desactivando as el enzima .
MECANISMOS DE REGULACIN Generales: Se basan en la disponibilidad de sustrato o producto.
Especficos: La regulacin de la actividad de una o varias de las enzimas de la va
54Clasificacin de los Mecanismos de RegulacinEn funcin del tiempo que tardan en cambiar la velocidad de una reaccin o de una ruta metablica:
Rpidas
Lentas
Rpidas: actan sobre la actividad de la enzima interacciones alostericas y modificaciones covalentes.
Lentas: modifican las concentraciones de enzimas por aumento (induccin) o disminucin (represin) de la sntesis o degradacin proteica.INHIBICIN A NIVEL ENZIMATICO57 Que inhiben o anulan la accin de los enzimas sin ser transformados por ellos. Inhibidores La unin de un inhibidor puede impedir la entrada del sustrato al sitio activo de la enzima y/u obstaculizar que la enzima catalice su reaccin correspondiente.Clasificacin Isostricos: ejercen su accin sobre el centro activo
Alostricos: ejercen su accin sobre otra parte de la molcula, causando un cambio conformacional con repercusin negativa en la actividad enzimtica.
Los inhibidores isostricos pueden ser de dos tipos: Inhibidor reversible: establece un equilibrio con la enzima libre, con el complejo enzima-substrato o con ambos:
Inhibidor irreversible: modifica qumicamente a la enzima
Caractersticas inhibidores ireversible Reaccionan con un grupo qumico de la enzima, modificndola covalentemente
Depende del tiempo de actuacin del inhibidor.
Utilidad se basa para identificar los grupos funcionales esenciales para la catlisis en aquellos enzimas a los que inactivan. Tipos de inhibidores irreversibles Reactivos de grupos -SH Organofosfricos Ligandos de metales Metales pesados
Aplicacin de los inhibidoresLos venenos naturales son a menudo inhibidores enzimticos que han evolucionado para defender a una planta o animal contra sus depredadores.
Aplicacin de los inhibidoresLos inhibidores artificiales son usados como medicamentos, pero tambin existen:
Insecticidas (como el malathion) Herbicidas (como el glifosato) Desinfectantes, (como el triclosn).Inhibicin Enzimtica Consiste en la disminucin o anulacin de la velocidad de la reaccin catalizada por una enzima.
Clasificacin Reversible CompetitivaNo competitivaAcompetitiva Irreversible
Inhibicin Irreversible La enzima no recobra su actividad por remocin del inhibidor libre.Esto es debido a que el inhibidor irreversible, acta por lo general modificando irreversiblemente o an destruyendo algunos de los grupos esenciales del centro activoInhibicin Reversible La enzima recobra su actividad por remocin del inhibidor libre. Clasificacin Competitivo No competitivo AcompetitivoInhibicin Competitiva El inhibidor se combina reversiblemente con la enzima en el sitio por el cual se debera unir el sustrato, impidiendo la formacin del complejo activo enzima sustrato.
Caractersticas Tanto el inhibidor como el sustrato compiten por el mismo sitio y tratan de desplazarse mutuamente de la enzima.
Muy altas concentraciones de substrato desaparece la inhibicin
Se une solo a la enzima libre
En este tipo de inhibicin los inhibidores se incluyen sustancias que estructuralmente son muy parecidas al sustrato y que por lo pueden ocupar el lugar que ocupara el mismo sobre laenzima -
La Vmx para una dada cantidad de enzima no se modifica por el agregado de inhibidor porque agregando una concentracin lo suficientemente grande de sustrato se puede desplazar completamente al inhibidorKm aumenta porque la presencia del inhibidor hace que se necesite una mayor cantidad de sustrato para saturar a la enzima que la que se necesitara si el inhibidor no estuviera presente.Inhibicin No Competitiva El inhibidor se fija a la enzima independientemente de que lohaga o no el substrato; el inhibidor, no impide la fijacin del substrato a la enzima.
Caractersticas La unin del sustrato con la enzima no es afectada por la presencia del inhibido.
No puede ser revertido por un aumento de la concentracin de sustrato
El inhibidor disminuye el valor de la Vmx sin modificar la Km ya que la unin del sustrato a la enzima no est alterada por la simultnea unin del inhibidor.
Inhibicin Acompetitiva El inhibidor se fija nicamente al complejo enzima-substratouna vez formado, impidiendo la accin cataltica
Caractersticas Se une a un lugar diferente del sitio activo de la enzima
Se une slo al complejo enzima-sustrato
Disminuye el valor de Km y tambin el de Vmx
RESUMEN DE LAINHIBICIN83
REGULACIN DE LA ACTIVIDAD ENZIMTICAUn organismo debe regular actividades catalticas de las enzimas componentes para coordinar sus numerosos procesos metablicos, responder a los cambios de medio y diferenciarse, en forma ordenada. TIPOS DE ACTIVIDAD ENZIMATICAModelo llave- cerradura:
La encima y el sustrato deben encajar.
Modelo de encaje inducido:
La enzima cambia para encajar con el sustrato
REGULACIN CELULAR DE LA ACTIVIDAD ENZIMATICA
Regulacin de la sntesis de enzimas: La clula controla cuntas molculas de cada enzima fabrica en cada momento.
Modificacin de la estructura enzimtica: Muchas enzimas tienen dos formaciones con nivel de catlisis diferente, de manera que la transicin de una a otra altera su actividad.
CONTROL DE LA ACTIVIDAD DE LA ENZIMALa actividad cataltica de la enzima puede tener regulacin directa.
Se puede controlar la actividad cataltica de una enzima por encima de la variacin de su afinidad de unin con el sustrato.
La afinidad de la unin entre una enzima y un sustrato puede variar por la unin de pequeos efectores moleculares, por lo que se modifica la actividad cataltica de la enzima.La actividad enzimtica puede ser controlada en la clula principalmente de estas cinco formas:
PRODUCCIN DE LA ENZIMALa sntesis de una enzima puede ser favorecida o desfavorecida en respuesta a determinados estmulos recibidos por la clula.
COMPARTIMENTALIZACIN DE LA ENZIMA Las enzimas pueden localizarse en diferentes compartimentos celulares, de modo que puedan tener lugar diferentes rutas metablicas de forma independiente.
ACTIVADORES ENZIMTICOS Son molculas pequeas, estimulan la actividad cataltica de una enzima. Las enzimas pueden ser activadas por ciertas molculas.
MODIFICACIN POSTRADUCCIONAL DE ENZIMASLas enzimas pueden sufrir diversas modificaciones postraduccionales como fosforilacin.La actividad enzimtica se mide por la desaparicin del sustrato o la aparicin de alguno de los productos de la reaccin, siguiendo dichos cambios en el tiempo.EFECTOS QUE AFECTAN A LA ACTIVIDAD ENZIMTICAAdems del efecto de la concentracin del sustrato en la rapidez de la reaccin enzimtica hay otros factores que afectan la actividad de las enzimas:
CONCENTRACIN DEL SUSTRATOA mayor concentracin del sustrato, a una concentracin fija de la enzima se obtiene la velocidad mxima. Despus de que se alcanza esta velocidad, un aumento en la concentracin del sustrato no tiene efecto en la velocidad de la reaccin.
CONCENTRACIN DE LA ENZIMASiempre y cuando haya sustrato disponible, un aumento en la concentracin de la enzima aumenta la velocidad enzimtica hacia cierto lmite.
TEMPERATURAHay una temperatura ptima a la que habr una mayor eficiencia enzimtica. Por lo general, entre los 50 a 60C la enzima pierde efectividad y la rapidez de la reaccin disminuye bruscamente.
El calor es un factor que desnaturaliza las protenas por lo tanto si la temperatura se eleva demasiada, la enzima pierde su actividad.
pHTodas la enzimas tienen un pH ptimo, al que habr mayor actividad o eficiencia enzimtica.El pH ptimo de la actividad enzimtica es 7, excepto las enzimas del estmago cuyo pH ptimo es cido.
PRESENCIA DE COFACTORESMuchas enzimas dependen de los cofactores, sean activadores o coenzimas para funcionar adecuadamente. Para las enzimas que tienen cofactores, la concentracin del cofactor debe ser igual o mayor que la concentracin de la enzima para obtener una actividad cataltica mxima.NOTA:Sin la presencia de iones y coenzimas no hay actividad enzimtica.CICLO DEL CIDO CTRICO110 Es una ruta metablica, es decir, una sucesin de reacciones qumicas, que forma parte de la respiracin celular en todas las clulas aerbicas. Ciclo de Krebs
PASO REACCIN Piruvato + CoA-SH+ NAD Acetil CoA + NADH + CO2 1Acetil CoA + Oxalacetato + H2O Citrato + CoA-SH + H2 Citrato Isocitrato3 Isocitrato + NAD cetoglutarato + NADH + CO2 + H4 cetoglutarato + NADH + CoA-SH Succinil- CoA + NAD + CO2 + H5 Succinil- CoA + GDP + Pi Succinato + GTP + CoA-SH6 Succinato + FAD Fumarato + FADH27 Fumarato + H2O L- malato8 L- malato + NAD Oxalacetato + NADH + H
Control de acceso o de la piruvato deshidrogenasa El complejo de piruvato deshidrogenasa (PDH) que sintetiza el acetil-CoA necesario para la primera reaccin del ciclo a partir de piruvato, procedente de la gluclisis o del catabolismo de aminocidos. Es activado por ADP, el cual abunda cuando la clula necesita energa. Inhibido por el ATP, NADH y Acetil- CoAControl del ciclo del cido ctrico Muchas de las enzimas del ciclo de Krebs son reguladas por retroalimentacin negativa, por unin alostrica del ATP.Los tres puntos de regulacin son las reacciones catalizadas por:
Citrato sintasaIsocitrato deshidrogenasaComplejo de cetoglutarato deshidrogenasaPuntos de regulacin
Inhibido por ATP, NADH, Succinil CoA citratoInhibido por ATP NADHInhibido por ATP, NADH , succinil CoACitrato sintasaLa citrato sintasa es una enzima alostrica inhibida por ATP, NADH y succinil-CoA y su propio producto, el citrato.Isocitrato deshidrogenasaEste es el segundo sitio de regulacin. En este caso el ADP y NAD son activadores alostricos de la enzima.
Puesto que el ATP y el NADH inhiben esta enzimaComplejo de cetoglutarato deshidrogenasaEste constituye el tercer sitio regulado por ATP y NADH, sin ambargo succinil-CoA tambin es un inhibidor de esta enzima. PUNTOS DE CONTROL DE LOS LPIDOSPUNTOS DE REGULACINSNTESIS:
Punto de control es el enzima que modifica el acetil-CoA, la acetil-CoA carboxilasa.
DEGRADACIN:
Enzima que activa acetil-CoA para entrar cidos grasos en la mitocondria transfirindolos a la carnitina, la carnitina aciltransferasa I.
Regulacin de la sntesis: acetil-CoA carboxilasaRegulado alostricamente:Modulador positivo: Citrato, que favorece la conformacin ms activa adems de sacar el acetil-CoA.
Modulador negativo: Palmitoil-CoA, que es el producto de la reaccin. Tambin la carga energtica, concretamente el AMP.
Regulado covalentemente Fosforilada es inactiva. Si la quinasa est favorecida bloquea la sntesis de cidos grasos y si lo es la fosfatasa activa el enzima. La insulina favorece la fosfatasa y el glucagn la quinasa.
Regulacin a largo plazo
La enzima puede variar su concentracin, se puede inducir su sntesis.
REGULACIN DE LA DEGRADACIN: CARNITINA ACILTRANSFERASA 1Es modulador el malonil-CoA que es el producto de la acetil-CoA carboxilasa. Si sube su concentracin es porque sobra energa y hay hidratos de carbono y al estar inhibido el enzima no pueden entrar los acetil-CoA en la mitocondria.
Al degradar cidos grasos se obtiene acetil-CoA que puede ir al C.A.C. si hace falta energa, pero hace falta OAA. Si faltan hidratos de carbono el acetil-CoA no puede entrar en el C.A.C. porque el OAA forma glucosa. Si no hay hidratos de carbono los niveles de OAA estn comprometidos y no podemos degradar acetil-CoA.REGULACINA nivel DNALaregulacin genticacomprende todos aquellos procesos que afectan la accin de ungena nivel de traduccin o transcripcin, regulando sus productos funcionalesGran parte de la regulacin gnica es realizada por protenas que se unen a DNA y afectan su expresin.
La regulacin implica interacciones entre el ambiente qumico de la clula y ciertas protenas codificadas por genes reguladores.
Losgenes reguladoresson aquellosgenesencargados de controlar la velocidad de sntesis de los productos de uno o de varios genes o rutas biocinticasSITIOS DE REGULACIN DE LA EXPRESIN GENTICA La regulacin de la expresin gentica ocurre en los niveles de:TranscripcinPostranscripcinTraduccinPostraduccin
La regulacin a nivel de transcripcin:Incluye todos los mecanismos que controlan la transferencia de informacin del DNA al RNA por la RNA polimerasa.Regulacin a nivel de postranscripcin:Incluye todas las modificaciones de la copia de RNA primaria antes de que se traduzca en protenas.
Regulacin a nivel de traduccin:Abarca los factores que determinan la velocidad de traduccin de las molculas de mRNA maduro.
Regulacin a nivel de Postraduccin:Incluye mecanismos que controlan el procesamiento del producto de traduccin primario en la protena madura producto. CONTROL POSITIVO Y CONTROL NEGATIVOPositivo:El control positivo es cuando el producto del gen regulador activa la expresin de los genes, acta como unactivador.Negativo:El control negativo es cuando el producto del gen regulador reprime o impide la expresin de los genes, acta como unrepresor.REGULACIN EN PROCARIOTASGenoma:Es la totalidad de lainformacin genticaque posee unorganismoo unaespecieen particular.OperonEs la unidad del cromosoma bacteriano formado por los siguientes componentes:
Operador: Controla el acceso de la ARN polimerasa al promotor.
Promotor: Donde la ARN polimerasa reconoce el sitio de inicio de la transcripcin.Gen regulador: Controla el tiempo de velocidad de transcripcin de otros genes.
Genes estructurales: Codifican las enzimas relacionadas o las protenas estructurales.
Entre los organismos procariotas como las bacterias cuyo genoma es muy pequeo los genes se organizan en forma de operones como se describi en la historia de la gentica. Es decir que varios genes involucrados en la misma ruta metablica se transcriben juntos y son controlados por el mismo promotor y la regulacin del mismo.
REGULACIN EN EUCARIOTASEn este caso el ADN posee muchas secuencias regulatorias o elementos de respuesta o secuencias internas de un promotor,algunas dentro del mismo promotor de un gen que son reconocidas por los factores de transcripcin y por protenas reguladoras aumentando la tasa de transcripcin.Regulacin a nivel de DNA
La transmisin de la informacin gentica (transcripcin), posibilita la formacin de protenas, cuyas funciones van a caracterizar la actividad y morfologa de las clulas.Laregulacin genticacomprende todos aquellos procesos que afectan la accin de ungena nivel de traduccin o transcripcin, regulando sus productos funcionales.
La regulacin de la expresin gnica se refiere al control de la cantidad y el momento de aparicin de los productos funcionales de un gen.
Gran parte de la regulacin gnica es realizada por protenas que se unen a DNA y afectan su expresin. Los dominios ms importantes de unin a DNA son:Hlice-giro hliceDedos de zincCremallera de leucina
Hlice-giro hlice
est formado por dos segmentos peptidicos en -hlice, de estructura rgida, separados por una secuencia de aminocidos, flexibles, que permite que las 2 hlices se aproximen entre s. Una de la a-hlices se encaja en el surco mayor de DNA, de forma que los residuos aminocidos de un lado de la hlice interaccionan mediante puentes de hidrogeno con las bases nitrogenadas expuestas en ese surco.
Dedos de zinc
Los dedos de Zinc coordinan iones de Zinc con una combinacin de residuos decistenaehistidina.
Cremallera de leucina
forma una hlice anfiptica en la que las leucinas de la cremallera de una de las protenas pueden sobresalir de la -hlice y trabarse o engranarse con las leucinas de la cremallera de otra protena que se encuentra situada de manera paralela para formar un dominio de bobina en espiral (coiled-coil).
REPRESIN Mediante la represin se controla la formacin de cierto numero de enzimas biosintticas de importancia capital. Es decir, las enzimas que catalizan la sntesis de un producto especfico no se sintetizan si este producto est presente en el medio. Por ejemplo: Enzimas que participan en la formacin de aminocidos arginina slo se sintetiza cuando no hay arginina en el medio de cultivo.
Una molcula pequea, frecuentemente el producto final de una ruta biosinttica, el cual se le denomina correpresor, inhibe la formacin de una enzima represible que intervine en las etapas iniciales de la biosntesis del correpresor.Regulacin de la expresin gnica en bacterias
Muchos genes bacterianos con funciones relacionadas se encuentran agrupados y estn bajo el control de un nico promotor. Esta estructura se denomina opern.
Un opern tpico incluye varios genes estructurales y una regin reguladora formada por el promotor y el operador
Los genes estructurales se transcriben a un mRNA policistrnico (codifican ms de unaprotena.) que se traduce para producir las protenas correspondientes.
Por ejemplo. Slo se forma triptfano sintetasa de E. coli cuando las clulas se desarrollan en un medio libre de triptfano.
La adicin de triptfano, que tiene carcter de correpresor, reprime completamente la formacin de sintetasa.El opern triptfano de E. coliEl opern triptfano controla la biosntesis de triptfano y es un opern reprimible.
Funcionamiento del opern triptfano:Nivel bajo de triptfano.- la protena represor no puede unirse al operador y se permite la transcripcin.
Nivel alto de triptfano - el triptfano se une al represor provocando un cambio en su estructura y permitiendo la unin al operador. Transcripcin reprimida.
Como la sntesis de la enzima cesa en forma bastante rpida, la que ya se encontraba previamente en la clula, continua catalizando la reaccin, hasta el momento en que se elimina, ya sea por degradacin o por dilucin que determina el crecimiento celular.INDUCCIN
Se refiere tambin a la activacin de la trascripcin de un gen como consecuencia de un inductor que interacta con una protena reguladora.
Esto se lleva a cabo por la adicin del sustrato de la enzima al medio en que la clula se desarrolla. El sustrato se denomina inductor, y la enzima cuya sntesis se estimula en forma notable, enzima inducible.
Comparacin Represin-InduccinRepresinInduccinPredomina en las secuencias anablicas que intervienen en la sntesis de los aminocidos y de los nucletidos.Prevalece en las secuencias enzimticas catablicas (degradacin de sustratos exgenos).Un poco de historia... En 1961, J. Monod y F. Jacob, de Francia, propusieron un mecanismo ingenioso para explicar la induccin y la represin.
En 1965, se les otorg el premio Nobel de medicina por este trabajo, y por otras contribuciones importantes en el campo de la Bioqumica.Regresando a la Represin...Para comprender el mecanismo de la represin , se examinar la regulacin de la -galactosidasa de E. coli, la cual se ha caracterizado perfectamente por medio de estudios intensivos.
H2O
E. coli utiliza indirectamente la lactosa. Primero, debe inducirse una galactsido permeasa que haga posible el paso de la lactosa al interior de la clula; despus tambin deber inducirse una - galactosidasa que hidrolice el disacrido hasta galactosa y glucosa. Asimismo se induce la formacin de una tercera enzima (tiogalactsido transacetilasa), cuya funcin an se ignora.
El sitio del DNA de E. coli en que se encuentra la clave para las enzimas que permiten la utilizacin de la lactosa se denomina lactosa opern. Este opern consta de 4 componentes principales: a) Un numero de genes estructurales que sirve como platillas para los mRNA, permiten la traduccin de la informacin para la sntesis de la enzimas que intervienen en el metabolismo de la lactosa.b) El gene operador, que se encuentra adyacente al primer gene estructural.c) La regin promotora, que aparece contigua al gene operador.d) La protena represora, que es el producto de un gene regulador ya sea topogrficamente independiente o ntimamente asociado.E la i172El opern lactosa de E. coliEl opern lactosa controla la expresin de genes responsables del metabolismo de la lactosa. Es un opern inducible.
Funcionamiento del opern lactosa: No hay lactosa en el medio - la protena represor se encuentra unida al operador y bloquea la transcripcin.
S hay lactosa en el medio - la lactosa se une al represor e induce un cambio conformacional que le impide unirse al operador. Se transcriben los genes estructurales
Se cree que el inductor modifica la conformacin de la protena represora, de manera que el enlace no ocurre en el sitio del operador. En este caso la lactosa o sus derivados, actan como inductores.Control gnico en eucariotasHay protenas de unin a DNA que modican la actividad transcripcional.No hay operones.Existencia de intrones (procesamientos alternativos), Un intrn es un fragmento de ADN que est presente en un gen pero que no codifica ningn fragmento de la protena.Separacin espacial entre transcripcin y traduccinLa estructura de la cromatina afecta a la expresin Gnica.Son ms comunes los activadores que los represores.Las modicaciones post-traduccionales son frecuentes.
Cambios en la estructura de la cromatina El nucleosoma es la unidad estructural de la cromatina y esta constituido por DNA unido a protenas denominadas histonas que rodea a un octmero de histonas:
Para que ocurra la transcripcin, la estructura de la cromatina debe modicarse para que el DNA se vuelva ms accesible a los factores de transcripcin, los activadores y la RNA polimerasa.
Modicacin de histonas
Remodelacin de la cromatina
Metilacin del DNAModicacin de histonas
Las histonas puede modificarse por fosforilacin, acetilacin, metilacin,ubiquitinacin y sumoilacin las que pueden ocurrir en mltiples residuos como Lys, Ser o Arg. Estos residuos se encuentran en el amino terminal de las histonas, el que interacciona directamente con el DNA.
Las modificaciones en las histonas son efectuadas por familias de enzimas que generalmente se asocian a complejos activadores o represores de la transcripcin.La familia de acetil tranferasas de histonas pueden acetilar residuos de Lys en las histonas H2A, H2B, H3 y H4, y disminuyen as la interaccin histona-DNA. Esto permite una movilizacin y reordenamiento del nucleosoma lo que facilita el acceso de la maquinaria de la transcripcin al DNA templado y la activacin de la expresin de un gen.Acetilacin de histonasEs llevada a cabo por una familia muy amplia de protenas denominadas histona acetil transferasas (HATs).La acetilacin en zonas reguladoras o promotoras de la transcripcin, permiten la prdida del empaquetamiento sobre el DNA y favorece la transcripcin de un gen.
Fosforilacin de histonas...Es una modificacin que involucra mltiples procesos celulares.La enzima H1 es fosforilada en Ser, H2B y H4 en Ser 1-acetilada del amino terminal de cada subunidad.Por otro lad0 se ha encontrado que la fosforilacin de la histona H3 en Ser 10 es importante en la activacin transcripcional.
Ubiquitinacin de histonas...La Ub se une al grupo -amino de residuos de Lys en la protena. Esta reaccin puede ser por una poliubiquitina, donde se ensamblan cadenas polimricas de Ub-Ub, se asocia a la destruccin va proteosoma de la protena.En las histonas la Ub es encontrada principalmente en las subunidades H2A y H2B.
Sumoilacin de histonas...En mamferos son 3 miembros de la familia de protenas SUMO: SUMO1, SUMO2 y SUMO3. Estas protenas son unidas covalentemente por un mecanismo similar al de la ubiquitinacin, donde existe una cascada de enzimas (E1-E2-E3).La protena SUMO se une se une -amino de residuos de Lys en la protena.
Metilacin de histonas...Es uno de los mecanismos de regulacin que presenta diversas funciones asociadas a la activacin o a la represin de un gen. Las enzimas que catalizan esta reaccin son las histona metil transferasas (HMTs) que metilan residuos de Lys y Arg; la metilacin aumenta la afinidad de residuos bsicos por el DNA
Remodelacin de la cromatina El trmino remodelacin de la cromatina, generalmente se refiere a cambios en la interaccin histona-DNA en el nucleosoma realizada por ciertos factores que ayudan a compactar o descompactar la cromatina, dichos factores catalizan la movilizacin y reordenamiento del nucleosoma dejando libre el templado, lo que facilita el acceso de la transcripcin.
Metilacin de DNA La metilacin del ADN es un proceso epigentico(mecanismos deregulacin genticaque no implican cambios en la secuencias deADN), que participa en la regulacin de la expresin gnica de dos maneras, directamente al impedir la unin de factores de transcripcin, e indirectamente propiciando la estructura "cerrada" de la cromatina.El ADN presenta regiones de 1000-1500 pb ricas en dinucletidos CpG ("islas CpG"), que son reconocidas por las enzimas ADN-metiltransferasas, las cuales, durante la replicacin del ADN metilan el carbono 5 de las citosinas de la cadena recin sintetizada, mantenindose as la memoria del estado metilado en la molcula hija de ADN.