
Tema 1.1- Definición de virus. Orígenes de la virología ...a_Temas... · REPLICACION VIRAL...
45
Tema 1.1- Definición de virus. Orígenes de la virología. Historia y descubrimiento de los virus. Teorías del origen de los virus. Los virus son agentes infecciosos, por lo que son parásitos intracelulares obligados. Solo se replican en las células vivas, puesto que dependen de los orgánulos y de la energía de la célula huésped. A diferencia de otros tipos celulares, un virus no viene directamente de otro virus, ya que necesita de otra célula intermediaria para poder replicarse y formar progenie viral. ESTRUCTURA BÁSICA Respecto a su genoma, puede haber virus de DNA o RNA, pero nunca de ambos a la vez. Este genoma puede ser monocatenario, bicatenario o incluso estar formado por varios fragmentos, como el virus de la gripe (Influenza). Los virus se clasifican en función de su genoma y de su transcripción a mRNA en los llamados grupos de Baltimore. Todos los virus tienen una cápside proteica que rodea y protege el material genético. Dicha cápside está formada por capsómeros, y el conjunto del genoma junto con la cápside se denomina nucleocápside. Además, algunos virus también pueden tener una envuelta lipoproteica, que en la mayoría de los casos proviene del huésped, y se adquiere a la salida de la célula por exocitosis. A los virus que no tienen envuelta se les denomina virus desnudos, mientras que a los que sí tienen envuelta se les denomina virus envueltos. Existe una gran diversidad de tamaños virales, por lo general entre 20 y 300 nm, aunque algunos pueden tener un mayor tamaño. Por lo general son más pequeños que las bacterias, pero hay excepciones, como los mimivirus, más grande que algunas bacterias (> 400nm). Viruela (360 nm), Herpesvirus (200 nm), Rabia (180 nm), VIH (120 nm), Influenza (100 nm), Parvovirus (20 nm).
Transcript of Tema 1.1- Definición de virus. Orígenes de la virología ...a_Temas... · REPLICACION VIRAL...

Tema 1.1- Definición de virus. Orígenes de la virología. Historia y descubrimiento de los virus.
Teorías del origen de los virus.
Los virus son agentes infecciosos, por lo que son parásitos intracelulares obligados. Solo se
replican en las células vivas, puesto que dependen de los orgánulos y de la energía de la célula
huésped.
A diferencia de otros tipos celulares, un virus no viene directamente de otro virus, ya que
necesita de otra célula intermediaria para poder replicarse y formar progenie viral.
ESTRUCTURA BÁSICA
Respecto a su genoma, puede haber virus de DNA o RNA, pero nunca de ambos a la vez. Este
genoma puede ser monocatenario, bicatenario o incluso estar formado por varios fragmentos,
como el virus de la gripe (Influenza). Los virus se clasifican en función de su genoma y de su
transcripción a mRNA en los llamados grupos de Baltimore.
Todos los virus tienen una cápside proteica que rodea y protege el material genético. Dicha
cápside está formada por capsómeros, y el conjunto del genoma junto con la cápside se
denomina nucleocápside.
Además, algunos virus también pueden tener una envuelta lipoproteica, que en la mayoría de
los casos proviene del huésped, y se adquiere a la salida de la célula por exocitosis.
A los virus que no tienen envuelta se les denomina virus desnudos, mientras que a los que sí
tienen envuelta se les denomina virus envueltos.
Existe una gran diversidad de tamaños virales, por lo general entre 20 y 300 nm, aunque algunos
pueden tener un mayor tamaño.
Por lo general son más pequeños que las bacterias, pero hay excepciones, como los mimivirus,
más grande que algunas bacterias (> 400nm).
Viruela (360 nm), Herpesvirus (200 nm), Rabia (180 nm), VIH (120 nm), Influenza (100 nm),
Parvovirus (20 nm).

REPLICACION VIRAL
Ciclos básicos de la replicación de un virus (pregunta de examen). SON 6¡¡¡
1) Interacción virus-célula diana: NO TODOS LOS VIRUS INFECTAN TODAS LAS CÉLULAS
(Tropismo viral), depende de las interacciones y los receptores de cada virus y de cada
tipo celular.
2) Penetración: entrada del virus en la célula que “desea” infectar (no siempre el virus
consigue penetrar en su célula diana).
3) Liberación del material genético: el material genético se libera de la cápside para
penetrar en el material genético de la célula huésped. En algunos casos, como los
bacteriófagos, la cápside se queda fuera, es decir, no llega a penetrar en la célula.
Por lo general, los virus con material genético de RNA replican en el citoplasma, mientras
que los de DNA replican en el núcleo.
4) Biosíntesis: consiste en la síntesis de múltiples copias del genoma viral dentro de la
célula huésped. Por otro lado, también se da la formación de las proteínas y la cápside
de los nuevos virus.
5) Ensamblaje: colocación y unión de los componentes sintetizados en la fase anterior,
formando nuevas partículas virales. A continuación puede darse una fase de
maduración.
6) Liberación: consiste en la salida de la célula de la progenie viral, la cual puede (o no)
conllevar la lisis de dicha célula hospedadora.
CICLO LÍTICO VS LISOGÉNICO
Los virus pueden encontrarse en dos ciclos diferentes, lítico y lisogénico. Todos los virus poseen
ciclo lítico, sin embargo, no todos poseen ciclo lisogénico. El ciclo lítico conlleva la lisis (no
siempre) de la célula, mientras que, durante el ciclo lisogénico, el material genético viral se
queda integrado en el de la célula hospedadora, como por ejemplo el VIH. En el caso del Herpes,
cuando se encuentra en ciclo lisogénico, el virus se queda latente en los núcleos de las células
neuronales de forma episomal, es decir, no se integra en el cromosoma de dichas células.

Los virus se caracterizan por poseer un estado extracelular e intracelular.
- En la fase extracelular, las partículas se denominan viriones, los cuales son
metabólicamente inertes y no realizan ninguna función biosintética.
- La fase intracelular se da a partir de la penetración.
EDAD DE LOS VIRUS
Se cree que los virus ya existían en la época de los dinosaurios. Aproximadamente, se estima
que los primeros virus eran retrovirus con más de 45 millones de años y eran virus marinos.
DONDE HAY VIDA HAY VIRUS. Se creen que los virus pueden ser igual de antiguos que las
primeras células procariotas.
En el siglo XVIII, Edward Jenner “descubrió” las vacunas al observar que los granjeros que tenían
un contacto frecuente con vacas con viruela no contraían esta enfermedad. Inoculó el líquido
de una pústula de viruela a un niño sano de 8 años y, más adelante, le introdujo el virus,
observando que el niño no se contagió.
Otros personajes importantes para el desarrollo de la microbiología en general, y de la virología
en particular, fueron Leeuwenhoek, Pasteur y Koch.
CARACTERÍSTICAS ENFERMEDADES INFECCIOSAS
- Siglo XIX → las enfermedades infecciosas están causadas por gérmenes microscópicos, que
pueden ser cultivados en medios nutritivos especiales y aislados por filtros muy finos capaces
de retener dichos gérmenes.
- Finales del siglo XIX → Ivanovski estudia el virus del mosaico del tabaco y demuestra que el
agente causal de esta enfermedad NO es una bacteria y que este no puede ser visto al
microscopio ni cultivado en medios artificiales. Postuló que podría tratarse de una toxina.
- 1898 → Beijerinck descartó que se trate de una trate toxina, ya que era capaz de multiplicarse
en los tejidos de plantas infectadas y atravesaba filtros de porcelana (Contagium vivum fluidum).
Tuvo muy poca aceptación en su tiempo.
En ese mismo año, Loeffler y Frosch describen por primera vez el origen viral de una enfermedad
animal, la fiebre aftosa o glosopeda.
Según lo visto hasta ese momento, se estipuló que los virus son agentes que no se ven al
microscopio, no son cultivables en medio de cultivo y no se retienen por filtros bacterianos.
El primer virus humano que se descubrió fue el de la fiebre amarilla (1901). Más tarde, en 1915
se descubrieron los bacteriófagos (virus que infectan a bacterias).
Entre 1950 y 1975 se sentaron las bases de la Virología moderna, tal y como la conocemos hoy
en día.
En 1966 se inició el Programa de erradicación de la viruela y en 1980 se convirtió en la primera
enfermedad viral (y hasta el momento la única) oficialmente erradicada.
En 2012 se encontró el virus de la viruela en momias de hace 300 años. En 2016 se encuentra
de nuevo en una momia de un niño, sin aparente mayor peligro en ambos casos.

Sin embargo, se intentó conectar informáticamente ambos virus secuenciándolos y se estableció
que ambos tienen un ancestro común no muy anterior al siglo XIV o XV, lo que descarta que la
viruela sea tan antigua como se creía en un principio.
¿DE DÓNDE VIENEN LOS VIRUS?
No sabemos con certeza de donde provienen los virus. No hay fósiles, puesto que son demasiado
pequeños para eso, pero si pueden quedarse insertados en otros seres vivos a los que han
infectado, y que estos sí se fosilicen. Esto hace suponer que los virus son tan antiguos como las
primeras células.
Existen dos teorías sobre el origen de los virus:
- Teoría de la coevolución (origen pre-celular): establece que los virus existían en un
mundo pre-celular como unidades auto-replicantes, por lo que podrían haber existido
antes que bacterias y eucariotas, e incluso ser sus antecesores (aunque a día de hoy esa
idea está prácticamente descartada). Se mantiene que los virus aparecieron después de
la aparición de los procariotas y que evolucionaron a partir de transposones.
- Teoría progresiva o de escape: mantiene que los virus habrían surgido a partir de
componentes celulares y que adquirieron la habilidad de replicarse autónomamente y
por tanto de evolucionar independientemente.
Lo cierto es que ninguna de las hipótesis es cierta del todo, podría ser un mix de ambas, o incluso
que existan múltiples mecanismos que no han sido descritos hasta la fecha.
TEORÍA DE LAS PANSPERMIA
Indica que un tipo de esporas y bacterias viajan por el espacio y se desarrollan si encuentran las
condiciones adecuadas en algún lugar. En el caso de los virus, estos caen sobre la tierra y la
contaminan, por lo que son los responsables del origen de la vida en la tierra. NO esta muy
aceptada por la comunidad científica, aunque cobró fuerza en los últimos años al encontrar
bacterias fosilizadas de hace millones de años en meteoritos.
MECANISMOS DE EVOLUCIÓN VIRAL (semejantes a los responsables de la evolución celular)
Estos procesos implican la generación de variantes virales nuevas mediante mutaciones,
recombinaciones y reordenamientos. Se crean variantes virales nuevas y, en ocasiones, algunas
se hacen fuertes y acaban siendo mayoritarias.
- Mutaciones:
los virus RNA son más mutables, puesto que, tanto la RNA polimerasa RNA dependiente
como la retrotranscriptasa, carecen de actividad correctora de errores en la replicación.
Las mutaciones en virus son las mismas que en el resto de organismos (deleciones,
inserciones, sustituciones…)
La tasa de mutación es la cantidad de errores que ocurren durante la replicación del
genoma.
La retrotranscriptasa del VIH comete 1 error por cada 104 bases, por lo tanto,
aproximadamente un error por cada vez que el genoma se transcribe. Esto hace que
en un enfermo de VIH se encuentren multitud de tipos del virus, en función de su
genoma (alta variabilidad genética). A estas variantes se las denomina cuasiespecies.
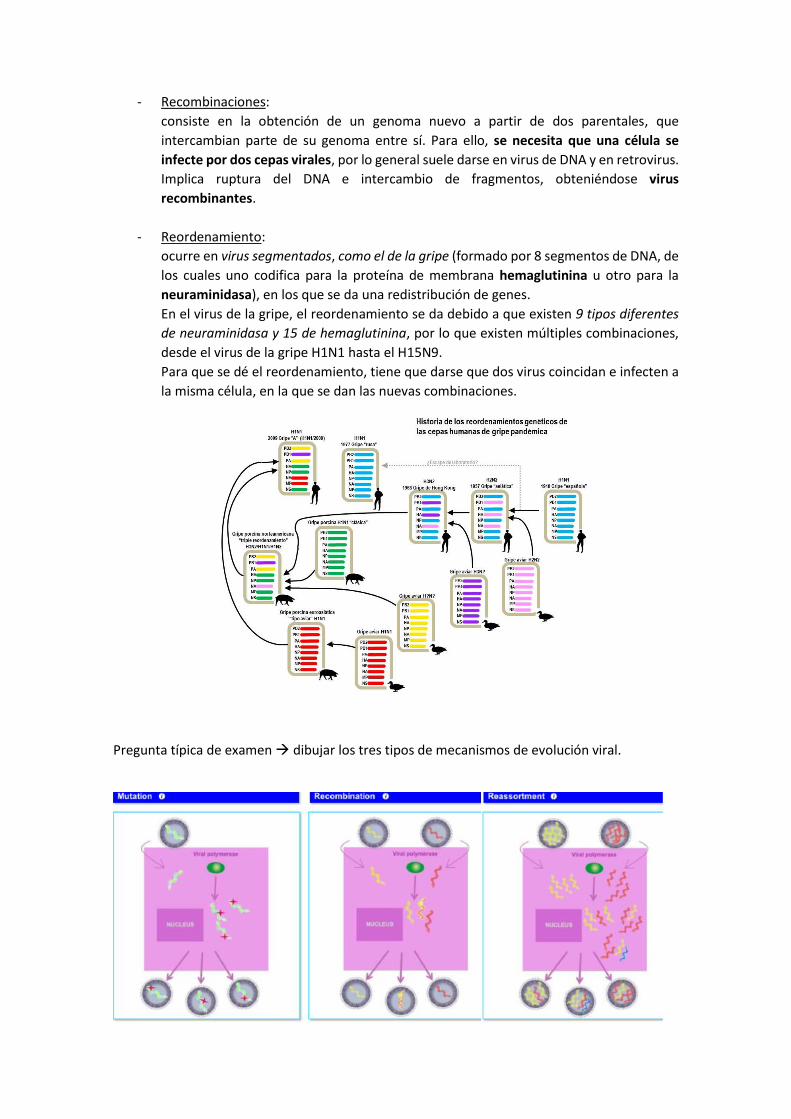
- Recombinaciones:
consiste en la obtención de un genoma nuevo a partir de dos parentales, que
intercambian parte de su genoma entre sí. Para ello, se necesita que una célula se
infecte por dos cepas virales, por lo general suele darse en virus de DNA y en retrovirus.
Implica ruptura del DNA e intercambio de fragmentos, obteniéndose virus
recombinantes.
- Reordenamiento:
ocurre en virus segmentados, como el de la gripe (formado por 8 segmentos de DNA, de
los cuales uno codifica para la proteína de membrana hemaglutinina u otro para la
neuraminidasa), en los que se da una redistribución de genes.
En el virus de la gripe, el reordenamiento se da debido a que existen 9 tipos diferentes
de neuraminidasa y 15 de hemaglutinina, por lo que existen múltiples combinaciones,
desde el virus de la gripe H1N1 hasta el H15N9.
Para que se dé el reordenamiento, tiene que darse que dos virus coincidan e infecten a
la misma célula, en la que se dan las nuevas combinaciones.
Pregunta típica de examen → dibujar los tres tipos de mecanismos de evolución viral.

Tema 1.2 – Estructura y morfología de los virus. Clasificación de Baltimore. Simetría de las
cápsides: helicoidales, icosaédricas y complejas. Virus con envuelta. Taxonomía viral.
Como ya sabemos, los virus pueden tener genoma de DNA o de RNA, pero nunca ambos al
mismo tiempo. Esto hace que existan cuatro genomas virales fundamentales: dsDNA, ssDNA,
dsRNA y ssRNA, a partir de los cuales se pueden constituir las 7 clases de genomas que engloban
todos los virus conocidos hasta la fecha (englobadas en la clasificación de Baltimore).
Clasificación de Baltimore
Todos los genomas de los virus se transcriben a mRNA, que es leído por los ribosomas para
codificar las proteínas de la cápside viral y las enzimas virales (en caso de tenerlas).
La clasificación de Baltimore depende del tipo de genoma viral y de su paso a mRNA.
(Nota: en el examen siempre hay una pregunta de la clasificación de Baltimore. Pasos que llevan
del genoma de origen al mRNA).
David Baltimore solo incluyó 6 grupos a su clasificación. El grupo VII se añadió posteriormente.
En los virus ssRNA, el genoma se puede encontrar en sentido positivo o negativo. El ssRNA (+)
tiene la misma secuencia que el mRNA, sin embargo, NO todos los RNA (+) son mRNA¡¡¡¡¡¡
(Hincapié en esto)
Incluso en grupos con el mismo tipo de genoma (ssRNA positivo en grupos IV y VI) existen
diferencias en el paso a mRNA que permiten la diferenciación de los grupos. El grupo VI genera
un intermediario de doble cadena y posee una retrotranscriptasa.
Diferentes estructuras del genoma viral
El genoma viral puede ser lineal, circular, segmentado, gapped (grupo VII), ambisense (tienen
una parte de RNA (+) y otra de RNA (-), los cuales son bastante raros), material genético con
proteínas en los extremos (generalmente en el extremo 5´) etc.
Baltimore nos permite saber la replicación y transcripción de cualquier virus conocido
La mayoría de los virus RNA replican en el citoplasma, salvo retrovirus y ortomyxovirus.
La mayoría de los virus DNA replican en el núcleo, salvo poxvirus.

Genomas virales de DNA (grupos de Baltimore I, II y VII)
Los virus de DNA necesitan una RNA polimerasa dependiente de DNA. Generalmente, los que
replican en el núcleo utilizan la polimerasa de la célula hospedadora. Si replican en el citoplasma
usan la polimerasa propia del virus (poxvirus).
• Los virus más destacados de dsDNA que replican en el núcleo son los Adenovirus
(propios de catarros y conjuntivitis), los Herpesvirus, los Papilomavirus y los
Polyomavirus.
La excepción de los virus dsDNA que replica en el citoplasma y, por lo tanto, necesita
una polimerasa propia son los Poxvirus (más grande que los virus dsDNA que replican
en el núcleo).
Todos ellos pertenecen al grupo I de la clasificación de Baltimore.
• Por otro lado, están los del grupo VII, que son gapped dsDNA. Este grupo no fue descrito
por Baltimore y fue añadido más tarde. Estos virus presentan “huecos” en su genoma
(Hepatitis B y Hepadnavirus).
Primero la DNA polimerasa celular rellena el trozo que le falta al genoma viral, y se
obtiene un genoma dsDNA “normal”.
Posteriormente se puede transcribir por dos vías, a partir de la RNA polimerasa celular
DNA dependiente. En una se genera el mRNA para generar proteínas y, en la otra, la
misma RNA polimerasa genera una hebra de ssRNA (+), sobre el cual se da la acción de
la retrotranscriptasa, que a partir de este RNA (+) genera una hebra de DNA (-), a partir
de la cual se sintetiza una dsDNA gapped con un hueco, que será introducida en la
cápside para generar la nueva progenie viral.

• El último grupo de Baltimore que presenta genoma DNA es el grupo II, cual posee un
genoma ssDNA (generalmente positivo).
Los virus de este grupo son muy pequeños, destacando los Circovirus y parvovirus
(genoma curioso), esto hace que utilicen siempre las polimerasas de la célula a la que
infectan.
La cadena inicial de ssDNA sirve de molde para generar un intermediario de dsDNA,a
partir del cual se genera el mRNA y los nuevos genomas virales (generalmente con
orientación positiva).
Genomas virales de RNA (grupos de Baltimore III, IV, V y VI)
Las células infectadas por los virus NO tienen RNA polimerasa RNA dependiente (RdRp), por lo
que SIEMPRE tienen que codificar para su propia RNA polimerasa, que se encarga de la síntesis
de nuevos genomas de RNA para la progenie viral, y de mRNA para su traducción a proteínas.
En primer lugar, tenemos los ssRNA (+), los cuales corresponden a los grupos IV y VI de la
clasificación de Baltimore.
• Los virus del grupo IV pueden ser leídos directamente por los ribosomas y utilizados
como mRNA.
En este grupo destacan los Picornavirus (Poliovirus y Rhinovirus), Coronavirus (SARS),
Calicivirus (gastroenteritis), Flavivirus (fiebre amarilla y Hepatitis C) y Togavirus
(Rubeola, el cual es peligroso en mujeres embarazadas, por su alta capacidad de
atravesar la placenta y llegar al feto, pudiendo ocasionar alteraciones graves en el feto).
Algunos de estos virus presentan en su genoma un extremo CAP y Poly(A), sin embargo,
algunos no la tienen, puesto que NO es indispensable para la lectura por lo ribosomas.
Generalmente ayudan al genoma en su salida del núcleo y en su llegada a los ribosomas,

pero como en estos virus replican en el citoplasma, no son estrictamente necesarios.
Otros, como los Picornavirus, tienen una proteína (VPg) en su extremo 5´.
• Los virus del grupo VI, también conocidos como retrovirus, tienen un genoma de ssRNA
(+), y se diferencian de los del grupo IV en que estos generan un intermediario de dsDNA
(provirus) y utilizan una retrotranscriptasa.
En este caso, el RNA(+) NO puede ser leído por los ribosomas, por lo que es utilizado
como molde para generar una cadena de DNA/RNA por la retrotranscriptasa. Este
DNA/RNA a su vez hace de molde para que se genere un intermediario de dsDNA
(provirus), capaz de penetrar y de quedarse integrado en el genoma de las células
infectadas. A partir de este dsDNA se generan las nuevas copias del genoma RNA (+) de
la progenie viral y se codifican las proteínas necesarias para su correcto funcionamiento.
Algunos de los virus de humanos más destacados en humanos son el VIH y HTLV. Otro
virus destacado en simios es el SIV, el cual se cree puede ser el precursor del VIH.
En segundo lugar tenemos los virus ssRNA(-), los cuales son característicos del grupo V de
Baltimore. Los genomas de estos virus pueden ser segmentados (Influenza) y no segmentados.
En este grupo destacan los Paramixovirus (sarampión o paperas), Rhabdovirus (rabia), Filovirus
(ébola), Orthomixovirus (Influenza)y Arenavirus (virus Lassa).
Como estos virus no pueden ser directamente traducidos a proteínas (por su orientación
negativa), generan dos intermediarios, por un lado, un mRNA que pueda ser leído por los
ribosomas y, por otro lado, nuevas cadenas de RNA(-) para rellenar las cápsides.
Algunos virus, como los Arenavirus y los Bunyavirus (virus segmentados), que presentan un RNA
ambisentido, es decir, están formados por dos fragmentos de RNA, uno largo que sirve para
generar nuevas copias de material genético viral, y uno corto, el cual contiene un tramo de
RNA(+) y otro tramo de RNA (-). A partir de este fragmento corto se generan las proteínas virales.

El tramo positivo del fragmento corto sirve de molde para la replicación, generando una
molécula negativa (glucoproteína), mientras que la negativa se transcribe directamente a mRNA
monocistrónico que codifica a las proteínas del virus (nucleoproteína).
En tercer y último lugar tenemos los virus del grupo III de Baltimore, los cuales presentan
genoma dsRNA. En este grupo destacan los Reovirus (10 segmentos) y los Rotavirus
(gastroenteritis, no incluida en el calendario de vacunación). Generalmente, presentan genomas
segmentados.
Proteínas virales
Dentro de las proteínas virales podemos destacar las proteínas estructurales y las no
estructurales.
Las proteínas estructurales son las que forman los componentes de los viriones (cápside). Sus
funciones son proteger el genoma viral, reconocer y empaquetar el genoma viral, unir al virión
con su célula diana y llevar a cabo la fusión entre ambos.
Las proteínas no estructurales son las enzimas necesarias para la replicación viral.
Por otro lado, las partículas virales tienen que ser metaestables, es decir, tienen q ser lo
suficientemente estables para no ser degradadas por el ambiente extracelular, pero no lo
bastante como para que no se dé la pérdida de la cápside una vez que está dentro de la célula
(equilibrio).
Cápside
La cápside es una capa de naturaleza proteica que rodea el genoma viral y está formada por
capsómeros.
Tiene un papel fundamental en la adsorción y penetración en la célula huésped. Determina el
tropismo viral (reconocimiento de receptores específicos de las células diana) y posee capacidad
antigénica.
Simetría: es la forma que adopta un virus en el espacio, la cual viene determinada por la
estructura de la nucleocápside. Los virus pueden tener simetría helicoidal, icosaédrica o
compleja.
• Simetría helicoidal:

Nucleocápside cilíndrica compuesta por un solo tipo de capsómeros organizados en
torno a un eje central. La longitud de la cápside depende de la longitud del ácido
nucleico. Un ejemplo es el virus del mosaico del tabaco.
• Simetría icosaédrica:
La mayoría de virus que infectan a animales y humanos
tienen este tipo de simetría.
Están formados por 20 caras idénticas y el virus icosaédrico
más sencillo posee 60 subunidades, es decir, 3 proteínas
por cada cara del icosaedro.
Además, pueden estar formadas por uno o varios tipos de
capsómeros, como los adenovirus (formados por hexones
y pentones).
• Cápsides complejas:
Los virus tienen una cápside que puede poseer estructuras adicionales como colas
proteicas o una pared exterior compleja. Algunos de los ejemplos más destacados de
virus con cápsides complejas son los bacteriófagos y los poxvirus.
Fagos → cabeza icosaédrica y cola helicoidal.
Poxvirus → tiene un sistema de protección del genoma compleja. Además, posee un
core que protege el material genético.
Envuelta viral
Está formada por una bicapa lipídica rica en colesterol y glicoproteínas, que rodea la
nucleocápside y suele proceder de la célula diana.
Hay muy pocos virus envueltos que infecten a plantas, y son muy raros en virus que infectan a
procariotas.
Algunas envueltas tienen simetría helicoidal (Influenza) y otras icosaédrica (Herpesvirus).
Ventajas y desventajas
Generalmente son más frágiles que los virus desnudos, cualquier condición que dañe la
envuelta, alterará la conformación nativa del virus y lo desestabilizará (desventaja).

Por otro lado, pueden parecer células y no ser reconocidas por el sistema inmune. Además, la
presencia de glicoproteínas aumenta cuantiosamente la afinidad del virus por la célula diana,
favoreciendo la infección (ventajas).
Asociadas a la envuelta viral puede haber otros tipos de proteínas, las cuales pueden estar N-
glicosiladas o O-glicosiladas, que consiste en la adición de azúcares a los grupos OH o a los grupos
NH de las proteínas. Estas proteínas glicosiladas pueden ser externas o transmembrana.
• Externas:
La mayor parte de la estructura es externa al virus y está anclada a él mediante dominios
transmembrana. Una de sus principales funciones es la de reconocimiento de la célula
target. Dos ejemplos de células externas son la hemaglutinina y la neuraminidasa (virus
de la gripe).
• Transmembrana:
Crean canales en la envuelta viral para el paso de iones (M2). También se altera el pH
dentro de la partícula viral para favorecer el avance del ciclo (influenza).
Algunos virus envueltos, como el virus Influenza o el VIH, tienen una serie de proteínas entre la
envoltura y la nucleocápside, llamadas proteínas de la matriz (matrix protein). Su principal
función es dirigir al virus en el momento de la salida de la célula infectada.
Ejemplos de virus con diferentes proteínas estructurales
Virus desnudos icosaédricos → Poliovirus, Adenovirus o Hepatitis A.
Virus desnudos helicoidales → Virus del mosaico del tabaco. NO se conocen virus de humanos
con esta estructura.
Virus envueltos icosaédricos → Herpesvirus, Fiebre amarilla o Rubeola.
Virus envueltos helicoidales → Rabia, Influenza, Parainfluenza, Paperas o Sarampión.
Virus complejos → Poxvirus y bacteriófagos.

Clasificación taxonómica de los virus
Los virus pueden clasificarse en función a las siguientes características:
1) Rasgos morfológicos: tamaño, envuelta…
2) Naturaleza del genoma: 7 tipos (dsDNA, ssDNA, dsRNA…)
3) Propiedades de la cápside: masa molecular, estabilidad al pH, resistencia a
detergentes…
4) Propiedades biológicas del virus: que enfermedad causa y a quien infecta, modo de
transmisión, distribución geográfica…
Los virus también pueden ordenarse por orden, familia, género y especie.
Con las técnicas de secuenciación, los virus también pueden clasificarse en base a sus
secuencias. El grado de similitud entre genomas virales se evalúa mediante programas
informáticos y se realizan árboles filogenéticos.
Los árboles filogenéticos pueden ser enraizados (cuando se estudian dos virus muy cercanos en
cuanto a secuencia, como el VIH y el SIV, o cuando existe un ancestro común entre los virus) o
no enraizados (cuando no hay indicios de un ancestro común entre los virus).
Por otro lado, los virus bacterianos se nombran utilizando códigos más sencillos (T1, T2, φX174).
Otros virus se nombran por la ciudad, río… donde el virus fue descrito por primera vez (Ébola,
Newcastle…).
Algunos virus de plantas son nombrados con el nombre del huésped y alguna característica de
la enfermedad que provoca.
Los virus de insectos a veces se nombran seguidos del nombre del insecto al que infectan,
añadido a algún efecto en el propio insecto.

Tema 1.3- Nuevos agentes infecciosos: viroides. Satélites. Priones
VIROIDES
En los años 1970 y 1971, Theodor O. Diener descubre un nuevo replicón con un genoma muy
pequeño, en torno a 10 veces más pequeño que el del más pequeño de los virus. Sin embargo,
no consiguió concentrar las partículas virales (viriones). Tras ultracentrifugar el extracto, el
“principio infeccioso” permanecía en el sobrenadante, mientras que los viriones utilizados como
control sedimentaban. Tras una centrifugación zonal, las partículas migraban hacia una zona que
correspondía a entidades de menor tamaño que los viriones. Tras incubarse con agentes
desproteinizantes, no se alteraba dicho “principio infeccioso”
Por lo tanto, con estos tres experimentos se determinó que el principio infeccioso NO era un
virión, puesto que se trataba de un ácido nucleico desnudo, además de poseer un tamaño más
pequeño que lo viriones. Se acuñó el término viroide.
Los viroides son pequeños ssRNA circulares con elevado contenido en estructura secundaria:
regiones de apareamiento intramolecular. Son los agentes infecciosos más simples de la escala
biológica. Solo infectan a plantas, no poseen cápside y son simplemente una molécula de ácido
nucleico. Algunos pueden poseer actividad ribozima (actividad autocatalítica, fundamental para
la replicación).
Existen dos grandes familias de viroides: pospviridae y avsunviroidae (actividad ribozima)
Replicación de los viroides
Los viroides NO codifican para proteínas, y su principal diferencia con los virus, además de la
ausencia cápside y las proteínas, es su capacidad para replicarse autónomamente parasitando
la maquinaria transcripcional de sus huéspedes (aunque no está claro cómo se replican).
Se cree que el mecanismo de replicación de los viroides es similar al mecanismo de círculo
rodante, provocado por una escisión autocatalítica y autoligación para producir el viroide
maduro. Los avsunviroidae son capaces de cortarse y ligarse para obtener más copias de sí
mismos (además, replican en el cloroplasto), mientras que los pospviridae utilizan enzimas
celulares y replican en el núcleo.
Patogénesis
No esta claro como causan enfermedad, pero se cree que RNA pequeños de interferencia
(siRNAs) derivados de los viroides de RNA podrían anular genes de la planta y producir la
enfermedad.
Huéspedes
Herbáceos (tomate, patata, lúpulo…) y leñosos /cítricos, palmera, vid, castaño…).
Pueden tener efectos devastadores o inapreciables. La infección solo puede darse a través de
heridas en las propias plantas (creo).
La replicación y acumulación de los viroides, además de la aparición de los síntomas que inducen
se ven favorecidos por temperaturas altas y alta intensidad lumínica- muy presentes en zonas
tropicales y subtropicales.

Alteraciones más frecuentes
Enanismo, distorsiones, clorosis, rugosidad y necrosis de las hojas, decoloraciones de los frutos,
agrietamientos del tallo y de la corteza, además de malformaciones en semillas y órganos de
reserva.
Transmisión
El principal mecanismo de transmisión es la propagación vegetativa (división de plantas
infectadas). Algunos viroides se transmiten por semilla, polen e insectos. Además, también se
cree que existe la transmisión por el uso de herramientas de poda utilizadas en una planta
infectada.
SATÉLITES
Son agentes subvirales parásitos de otros virus, es decir, necesitan un virus helper. Pueden
infectar animales, plantas o incluso bacterias (aunque la mayoría de satélites conocidos son
agentes infecciosos de plantas).
Existen dos grandes grupos de satélites en animales: Dependoparvovirus, que son satélites de
adenovirus o Herpesvirus y Deltavirus, que son satélites de HBV. Pueden ser de ssRNA o ssDNA.
Existen virus satélites capaces de codificar sus propias proteínas de cápside. También existen
virusoides o RNAs satélites, en los que la cápside es sintetizada por el helper. Ninguno de los
dos codifica para enzimas necesarias para la replicación, sino que estas son cedidas por el virus
helper, sin el que NO pueden llevar a cabo la replicación. Además, NO SON VIRUS DERIVADOS
DE LOS HELPER.
Por otro lado, pueden alterar los síntomas causados por el helper (empeorándolos o
mejorándolos).
Virus hepatitis Delta
Genoma ssRNA (-). Necesita la presencia del virus de la hepatitis B (HBV). El genoma viral utiliza
la RNA polimerasa I celular para replicar. El virus se transmite por contacto con la sangre u otros
líquidos corporales de las personas infectadas. La transmisión vertical es rara.
Patogénesis del virus hepatitis Delta
La infección con ambos virus se asocia con mayor daño hepático que la hepatitis B sola: se da
una progresión más rápida a cirrosis y muerte que en la monoinfección.
La infección por virus satélites se puede dar de dos formas: mediante coinfección (se da la
infección al mismo tiempo por el virus satélite y el helper) o superinfección (en la que el
individuo se infecta por Deltavirus cuando ya está infectado por hepatitis B). En este último caso
se da un peor pronóstico, puesto que la progresión hacia la enfermedad hepática terminal es
más rápida, mientras que en la coinfección la mayor parte de los pacientes evolucionan a la
curación con una tasa de cronificación menor del 5%.
En el mundo hay en torno a 350 millones de infectados con hepatitis B, de los cuales un 5% (unos
18 millones) presentan también el Deltavirus (HDV). Uno de los lugares donde está más
presente es en Asia, donde tiene una alta prevalencia.

PRIONES
Son responsables de las Enfermedades Espongiformes Transmisibles (EETs), que se caracterizan
por dejar el cerebro espongiforme.
Historia
Creutzfeldt y Jacob llevaron a cabo en los años 20 las primeras descripciones de la enfermedad,
donde describen los síntomas de los pacientes (síndromes neurológicos complejos que NO se
identifican con ninguna enfermedad conocida).
En 1967, Alper y col. demuestran que el responsable de esta patología es resistente a radiación
ionizante, lo que descarta que sea un virus.
En 1981, Prusiner, basándose en los trabajos de Alper y col. Y John S. Griffith, enunció la hipótesis
de solo proteína y acuña el termino prión, con el que identificaba a las partículas proteicas
infecciosas resistentes a tratamiento específicos de los ácidos nucleicos.
En 1985 tiene lugar en gran Bretaña una enfermedad en el ganado bovino, la conocida como
“enfermedad de las vacas locas”, que se denomina Encefalopatía Espongiforme Bovina (EEB).
Esta enfermedad tuvo lugar porque el ganado fue alimentado con restos de ovejas y cabras
positivas para estas encefalopatías.
Definición de prion
Es una proteína patógena cuya estructura secundaria está modificada, dando lugar a un
plegamiento alterado (también conocido como aberrante).
Ojo, importante de cara al examen.

La proteína no patológica se localiza en células del sistema inmune, estómago y
fundamentalmente en las membranas de las células neuronales. Está codificada por el gen prnp
(cromosoma 20) el cual codifica para una sialoglicoproteína, que podría intervenir en
señalización celular y en la formación sináptica.
También podrían ayudar a mantener la plasticidad de las conexiones nerviosas, al menos,
durante el desarrollo del hipocampo, una de las zonas del cerebro que primero se dañan
causando los problemas de desorientación y pérdida de memoria.
El año pasado se publicó y se demostró que la formación de espinas dendríticas en ratones WT
es mucho mayor que la de los ratones con la proteína mal plegada PrPSc. Además, se produce
la desaparición de dichas espinas y se comprueba que el daño asociado es absolutamente
dependiente del prion bueno.
Diferencias entre priones patógenos y no patógenos
- Priones no patogénicos (PrPC): tienen su estructura secundaria en forma de hélice alfa,
son solubles en detergentes y pueden ser degradados por proteasas.
- Priones patogénicos (PrPSc): tienen al menos un 40% de su estructura en forma de
lámina beta, son insolubles en detergentes y presentan cierta resistencia a la
degradación por proteasas.
Características de los priones
El sistema inmune es incapaz de atacar al prion patógeno, ya que es detectado como un mal
plegamiento de una proteína sana y no genera respuesta inmune (pueden generar una
respuesta casi inexistente).
Son agentes transmisibles no convencionales carentes de ácidos nucleicos.
Presentan una gran resistencia a la mayoría de los procedimientos de desinfección tradicionales
y no pueden ser destruidos por cocción, alcohol, ácido, radiación o métodos estándar de
autoclave.
Su sistema de seguimiento es muy complicado, porque en los tejidos accesibles (sangre, orina)
se encuentran en muy baja concentración.
Su expresión principal se da en el SNC (aunque es irregular), poseen largos periodos de
incubación y se ha visto que cerebros infectados y mantenidos en formaldehído (formol) pueden
transmitir la enfermedad durante décadas.

Hipótesis del prion
La conversión de PrPC a PrPSc constituye el evento principal de las enfermedades causadas
por priones. La forma endógena sufre un cambio conformacional que la transforma.
Su insolubilidad hace que se acumule en el citoplasma neuronal y forme agregados, causando
la muerte celular mediante neurodegeneración.
Se origina una reacción en cadena que lleva a la reducción de la forma PrPC y al acúmulo de
PrPSc en los tejidos del huésped.
Estas enfermedades son transmisibles en animales (EEF, EEB, encefalopatía transmisible del
visón…) y humanos (ECJ, kuru, GSS, IFF…).
Desde el punto de vista patológico estas enfermedades presentan una combinación variable de
pérdida neuronal, gliosis, espongiosis (vacuolización del tejido cerebral). También conlleva una
severa disfunción motora y los síntomas dependen del área cerebral dañada.
Tipos de encefalopatías espongiformes
Según etiología existen tres categorías: adquiridas, hereditarias y esporádicas.
Adquiridas (origen infeccioso)
- Vía oral:
• Creutzfeldt-Jakob variante (CJv): está asociado a la EBB por ingesta de animales
enfermos. En un principio se pensó que mediante el cocinado de la carne se
eliminaban los priones, pero no fue así. De hecho, la carne tiene poco poder
contaminante, puesto que el músculo solo posee una pequeña cantidad de
proteína priónica.
El consumo más peligroso es el del tejido nervioso (cerebro, médula espinal) y
las vísceras (bazo, intestino, ojos, amígdalas…), que son más ricos en proteína
priónica.
• Kuru (1957): se dio en Nueva Guinea, asociado al canibalismo de ciertas tribus,
que se comían los cerebros de los muertos para “adquirir su sabiduría”, lo que
hacía que el prión se transmitiera rápidamente.
- Vía iatrogénica (por procedimiento médico involuntario): puede darse a través de
sangre, cirugías o instrumental médico utilizado previamente con pacientes infectados.

Hereditarias (origen genético)
Son alrededor de un 15% y están asociadas a mutaciones en el gen prnp. Conlleva la muerte en
aproximadamente 6 meses y aparece en edades tempranas. Las más destacadas son el
Creutzfeldt-Jakob genético (CJf), el Insomnio Fatal Familiar (IFF) y el Gertsmann-Sträussler-
Scheinker (GSS).
- Insomnio familiar fatal: es una enfermedad con herencia autosómica dominante, en la
que se produce una mutación en el codón 178 del gen prnp, situado en el cromosoma
20 (20p13). Existen muy pocos casos en el mundo, de los cuales la mitad son en España
y tres cuartas partes en de los mismos en el País Vasco. Provoca una degeneración del
sistema nervioso (principalmente afecta al tálamo) y provoca una alteración severa del
ritmo circadiano (ritmo sueño-vigilia), junto con ataxia (falta de coordinación) y
trastornos endocrinológicos.
El insomnio deteriora progresivamente la salud del paciente hasta hacerle entrar en
coma y provocarle la muerte. No existe tratamiento curativo.
- Síndrome de Gertsmann-Sträussler-Scheinker (GSS): se debe a mutaciones puntuales
en la secuencia codificante para la PrPc. Aparece entre los 30 y los 60 años y posee una
incidencia muy baja (en torno a 1 y 10 casos por cada 100 millones de habitantes). La
duración de la enfermedad varía entre 3 meses a 15 años, con una media de duración
de 5 años. No existe cura.
Esporádicas
Constituye el 65% de las EETs. Tienen lugar en personas previamente sanas. Las mas destacadas
son el Creutzfeldt-Jakob esporádico(CJe) (más del 85% de los casos) y el Insomnio Fatal
esporádico (IFe).
Los casos esporádicos de ECJ ocurren en todos los países con una distribución homogénea y una
incidencia de aproximadamente 1-2 casos por cada millón de habitantes y año. No se conoce la
causa.
Llegada al sistema nervioso
Cuando se ingiere un alimento contaminado por proteínas priónicas, estas son absorbidas por
el sistema digestivo, a través del cual pueden acceder al sistema nervioso entérico, por donde
son capaces de propagarse y generar la enfermedad. Puede generarse una respuesta inmune,
pero, como hemos visto antes, esta es altamente ineficaz.
Terapias y profilaxis
A día de hoy, no existen terapias eficaces. Los estudios actuales se centran en la búsqueda de
productos que puedan impedir la conversión de la proteína normal a la forma alterada y
favorezcan la eliminación de la estructura patogénica. Para que sean eficaces, estos principios
farmacológicos han de ser capaces de atravesar la barrera hematoencefálica (BHE). Finalmente,
las terapias actuales se centran en paliar los síntomas.

Tema 1.4- Crecimiento, cuantificación y caracterización viral.
Diagnóstico de laboratorio de las infecciones virales
Podemos destacar tres grandes bloques: 1) Identificación del agente viral y aislamiento y
crecimiento del mismo. 2) Detección de componentes de la partícula viral. 3) Detección de
anticuerpos frente a la partícula viral.
Cultivo viral
Para ello, es necesario proporcionar al virus las células adecuadas en las que se puede replicar.
Por ejemplo, los bacteriófagos tienen que ser incubados con bacterias, los virus de plantas con
plantas y así sucesivamente, debido a la especificidad de los virus. Por otro lado, los virus
animales pueden cultivarse en organismos completos (como ratones o embriones de pollo).
Los embriones de pollo eran muy utilizados para la propagación viral como alternativa al uso de
organismos animales, antes del desarrollo de las técnicas de cultivo actuales. De hecho, el virus
Influenza sigue cultivándose en embriones de pollo.
Primero se lleva a cabo un primer contacto con una aguja fina, para después inocular el virus
con una más gruesa.
Cultivos celulares
En su mayor parte, los virus animales se crecen en cultivos de células animales. Estos cultivos
pueden hacerse en líneas celulares normales o en líneas tumorales.
En 1952 se obtuvo la primera línea celular continua humana: células HeLa, extraídas a partir de
un tumor de cuello de útero de una paciente afroamericana que se llamaba Henrietta Lacks.
Métodos de detección de virus en cultivo celular
El método más usual de detectar la presencia del virus es provocando la patología que causa
(efecto citopático).
El efecto citopático (ECP) es el conjunto de cambios bioquímicos, moleculares, morfológicos y
de viabilidad celular, visibles al microscopio óptico, causados durante el ciclo de replicación viral.
Solo se puede ver si las células de cultivo son target del virus en cuestión y no todos los virus lo
producen.

Dependiendo de los virus puede originar diferentes efectos como:
- Perdida de adherencia al sustrato (adenovirus, picornavirus y Herpesvirus).
- Vacuolas en el citoplasma (polyomavirus).
- Sincitios = células gigantes multinucleadas (paramyxovirus y coronavirus).
- Cuerpos de inclusión, como viriones en el núcleo (adenovirus) o en el citoplasma (Negri
bodies: virus de la rabia).
Para detectar los virus dentro de un cultivo se pueden utilizar dos métodos diferentes (directos
o indirectos).
Métodos directos
Se analiza la presencia del virus o de alguno de sus componentes. Para ello, se utilizan ensayos
de infectividad, método de dilución limitante, microscopía electrónica, hemaglutinación,
técnicas inmunológicas (ELISA) o presencia de ácidos nucleicos virales (PCR).
- Ensayo de infectividad: consiste en infectar una siembra de cultivo celular con un virus
a diferentes concentraciones (diluciones seriadas) y contar las unidades formadoras de
placa. Herpesvirus.
Es importante cubrir la monocapa celular con un medio semisólido (como el agar) para
evitar que nuevas partículas virales difundan y eliminen todo el cultivo celular.

Después de llevar a cabo las diluciones seriadas, es importante contar las unidades formadoras
de placa que se han formado por unidad de volumen (UFP/mL). Para ello, se cogen las placas
que tengas entre 10 y 100 UFP y se multiplica el número de UFP por el volumen de la dilución
por la dilución correspondiente cambiada de signo.
17*106/0,1= 1,7*107 UFP/ml
- Método de dilución limitante: este método se debe a que muchos virus no forman
placas en cultivo. Por lo tanto, se analizan otros parámetros como la TCID50, que es la
tasa infectiva 50 (dosis que hay que administrar para infectar a la mitad de la población
deseada), la LD50, que es la dosis letal 50 (cantidad que hay que añadir para matar a la
mitad de la población) y la EID50 (consigue el efecto deseado en la mitad de la población).
En otro experimento, se “tapiza” una placa de 96 pocillos con diluciones a distintas
concentraciones del cultivo, se añade el virus y se analiza la formación de sincitios.
- Microscopía electrónica: es posible observar la morfología de los virus presentes en
muestras clínicas. Presenta dos grandes limitaciones, el elevado coste de la técnica y la
gran concentración de viriones que se necesitan para poder llevarla a cabo
(aproximadamente 109 partículas virales/ml). Esto hace que sea una técnica
relativamente poco utilizada.
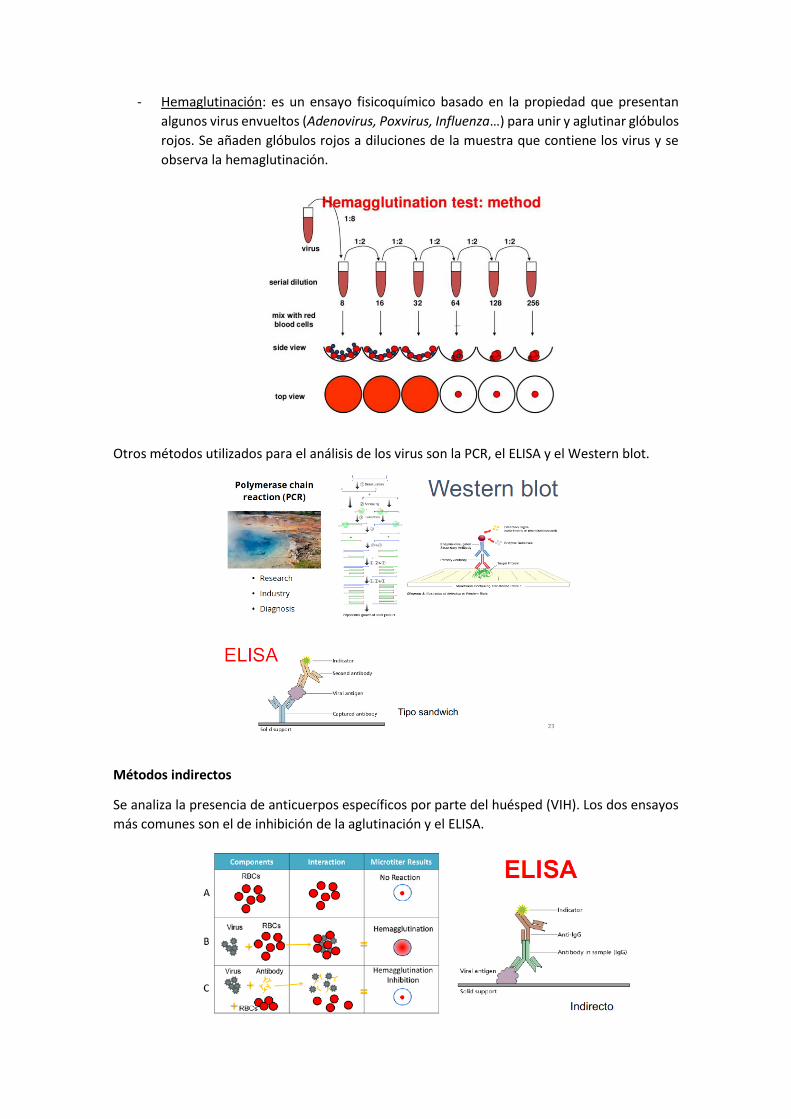
- Hemaglutinación: es un ensayo fisicoquímico basado en la propiedad que presentan
algunos virus envueltos (Adenovirus, Poxvirus, Influenza…) para unir y aglutinar glóbulos
rojos. Se añaden glóbulos rojos a diluciones de la muestra que contiene los virus y se
observa la hemaglutinación.
Otros métodos utilizados para el análisis de los virus son la PCR, el ELISA y el Western blot.
Métodos indirectos
Se analiza la presencia de anticuerpos específicos por parte del huésped (VIH). Los dos ensayos
más comunes son el de inhibición de la aglutinación y el ELISA.

Tema 2.1- Mecanismos de entrada y diseminación de la infección viral en la célula
VIRUS ANIMALES
Ciclo viral
El primer paso en el ciclo viral es el Attachment. Se trata de un proceso complejo durante el cual
las proteínas de la envuelta o la cápside (dependiendo del virus) se enlazan con factores de
attachment de las células diana, como los heparanatos, que permiten el contacto del virus con
muchos tipos celulares. Esto ocurre de forma electrostática y no requiere energía celular.
El attachment es un tipo de interacción NO específica. Las primeras colisiones se producen por
azar. El uso de componentes específicos de la superficie celular es el responsable de que los
virus sean capaces de infectar a cierto tipo de células target, lo que se conoce como tropismo
viral.
Estrategias de entrada a la célula
Existen tres grandes estrategias de entrada de los virus a las células: A) Adhesión del virión a un
único receptor de membrana celular. B) Adhesión del virión a un receptor y un correceptor. C)
Adhesión del virión a un receptor y otras proteínas de membrana de la célula target.
A- se da en los rhinovirus, que se unen a un receptor de membrana de la familia del colesterol
(LDLR).
B- se da en el VIH. El virión se une al receptor y luego puede unirse a dos correceptores distintos.
- Si se une al correceptor CCR5, el virión atacará a los monocitos.
- Si se une al correceptor CXCR4, el virión atacará a los linfocitos T.
C- se da en el virus de la hepatitis C, en el que intervienen hasta cuatro proteínas diferentes (SR-
B1, CD81, Claudina-1 y Ocludina).

Factores que intervienen en la influencia de la interacción del virus con la célula
Densidad de receptores en la superficie celular, densidad de los ligandos en la superficie del
virión, cantidad de células diana y de viriones, y otros factores como la temperatura (a 4ºC puede
darse el attachment, pero no la entrada del virión en la célula), el pH o la concentración de iones.
Receptores y correceptores celulares
Suelen ser proteínas o glicoproteínas, aunque en ciertas ocasiones también pueden ser
carbohidratos o lípidos.
Los virus se adhieren a tres tipos diferentes de moléculas receptoras de la superficie celular:
- Receptores de adhesión (baja especificidad y baja afinidad de interacción con el virus), HSV-1
y HPV usan heparanatos, Paramixoviruses y Reoviruses usan azúcar (ácido N-acetil neuramínico
-NANA).
- Receptores de entrada (se cree que son los más específicos- el reconocimiento del receptor
por el virión tiene una alta especificidad).
- Correceptores (son el segundo tipo de moléculas de superficie de las células que son
reconocidas por los viriones. En algunos casos, el enlace del virión con el receptor genera un
cambio conformacional en la proteína viral que le habilita para establecer contacto con el
correceptor y acceder a la célula).
Los virus han evolucionado de tal forma que son capaces de utilizar componentes esenciales de
las células como receptores, sin los cuales estas células no podrían existir. Esto hace que sea
muy complicado el desarrollo de antivirales que no afecten directamente a las células
infectadas.
Virus del ébola
Se trata de un caso particular, en el que el virus entra en
contacto con los respectivos receptores de membrana (DC-
SIGN/L-SIGN, TIM-1 y Axl). A continuación, se forma un
endosoma temprano, dentro del cual se encuentra la
catepsina, gracias a la cual el virus interacciona con la proteína
NPC-1, que permite la salida del genoma viral al citoplasma.
Los enfermos de Niemann Pick tipo C son inmunes al virus del
ébola, puesto que presentan un deterioro en los genes que
codifican para la NPC-1 y NPC-2, lo que provoca la acumulación
de colesterol en tejidos y órganos (puede ser peligrosa).

Sitios de attachment
- Virus desnudos: los sitios se localizan en diferentes lugares la cápside. En los Adenovirus,
se dan unas prolongaciones (knob) a partir de los pentones que permiten la interacción
con la célula diana. En los Poliovirus se dan unas invaginaciones dentro del propio virus
que permiten el acoplamiento del receptor CD155. En otros casos, la interacción puede
darse directamente con proteínas de la superficie viral (Rotavirus).
- Virus envueltos: los sitios de attachment se encuentran en las glicoproteínas presentes
en la superficie de la envuelta del virus. Un ejemplo es el virus Influenza.
Las glicoproteínas de la superficie del virus Influenza están formados por trímeros de
hemaglutinina y tetrámeros de neuraminidasa. La hemaglutinina es la responsable de
enlazar el virus con su receptor, que es ácido siálico.
La liberación de partículas del virus de la gripe desde las células requiere la actividad de
una enzima del virión: una neuraminidasa con actividad asociada a glicoproteínas de la
envuelta, es decir, la neuraminidasa participa en la salida de la nueva progenie viral.
Fuerzas de attachment
Las principales fuerzas que permiten la unión del virus con su receptor celular son los puentes
de hidrógeno, interacciones iónicas y fuerzas de Van de Waals. No se producen enlaces
covalentes, lo que hace que la unión virus-receptor sea reversible, por lo que el virus puede
separarse. Si el virus permanece unido al receptor, otras partículas virales tienen la oportunidad
de unirse a más receptores.
Entrada de los virus animales a las células
La penetración o entrada en la célula target suele ocurrir poco después del attachment.
Generalmente, este proceso es dependiente de energía, es decir, las células debes estar
metabólicamente activas para que este proceso tenga lugar.

Los virus han de atravesar la bicapa lipídica que forma la membrana plasmática, en la cual se
encuentran algunas proteínas, que son como icebergs en un mar de lípidos. Los virus adheridos
a las células pueden necesitar otros receptores antes de entrar en la célula, con los cuales puede
entrar en contacto gracias a la fluidez de la membrana. El mecanismo de entrada depende del
tipo de virus.
El mecanismo de entrada de los virus desnudos es diferente al de los virus envueltos, dentro de
los cuales también hay dos mecanismos distintos.
- Virus desnudos: la inmensa mayoría se introducen en la célula diana mediante
endocitosis, formando un endosoma. La salida del material genético de la cápside y del
endosoma es dependiente de pH, mediante la lisis o permeabilización de dicha cápside
y de la membrana del endosoma.
Algunas excepciones, como los Poliovirus, son capaces de formar un “semiendosoma”,
gracias al cual se produce la entrada del genoma viral en la célula sin que entre la cápside
(importante- ojo examen).
- Virus envueltos: existen dos modelos diferentes de entrada en las células huésped.
Ambos procesos conllevan la fusión de la envuelta del virión con la membrana celular
(las cuales tienen una naturaleza común).
Las membranas de ambos no se fusionan espontáneamente, sino que requieren de
glicoproteínas especializadas que lleven a cabo el proceso.
• Proteínas de fusión: se encuentran en la envuelta del virión en forma de dímeros o
trímeros, dependiendo de la proteína. Cada monómero tiene, al menos, dos
secuencias hidrofóbicas: una secuencia transmembrana y una secuencia de fusión,
que es responsable de la fusión mediante inserción en la membrana celular.
Los cambios conformacionales en las proteínas de fusión pueden venir dados por la
unión del virus a su receptor correspondiente o debido al bajo pH dentro del
endosoma.

Existen dos tipos distintos de fusión de membrana (en virus envueltos):
1) Fusión independiente de pH: se da la fusión de la envuelta viral con la
membrana en la superficie de la célula (Herpesvirus, VIH…).
En el caso del VIH, la zona variable de la proteína gp120 del virus entra en
contacto con el receptor CD4 de la célula. Esto hace que a continuación se una
al correceptor, ocasionando una serie de cambios que provocan la fusión de
ambas membranas.
2) Fusión desencadenada por ácido (dependiente de pH): se da entre el virus y la
membrana del endosoma (Influenza, Rhabdovirus…). Todos los virus que entran
por endosoma son dependientes de pH.
Se da una acidificación del medio progresiva debido a la acción de la bomba
H+/ATPasa, que impulsa protones dentro de la membrana mediante la hidrolisis
de ATP a ADP, acidificando el medio.
La bajada de pH provoca cambios conformacionales en las glicoproteínas, esto
hace que se exponga el péptido de fusión, que interacciona con la membrana
del endosoma y permite la liberación al citosol del virión completo.
En la siguiente imagen podemos ver detalladamente los dos mecanismos de entrada de los virus
envueltos, representados por el virus VIH y por el virus Influenza respectivamente.

Transporte intracelular
Los virus de DNA (salvo Iridovirus y Poxvirus) y algunas excepciones de virus RNA necesitan llegar
al núcleo para llevar a cabo su replicación.
En las células, las estructuras con un tamaño mayor de 20nm requieren de energía para viajar a
través del citosol. Por lo tanto, los virus encapsidados o de genoma largo utilizan los mecanismos
de motilidad celular para moverse por el citoplasma. La mayoría de estos virus utilizan los
microtúbulos.
Los microtúbulos forman parte del citoesqueleto celular y son utilizados para el transporte de
partículas. El transporte depende de proteínas motoras como quinesinas y dineínas, que son
“secuestradas” (no estoy seguro) por los virus para ser transportados al núcleo.
Por el contrario, otros virus son capaces de utilizar el citoesqueleto de actina para llevar a cabo
su movilidad. El transporte dependiente de actina es inducido por proteínas virales que
interactúan con actina o con otras proteínas motoras como miosinas.
Los virus neurotrópicos, por ejemplo, Herpesvirus, atraviesan el final de las terminaciones
nerviosas y son transportados mediante transporte axonal (transporte retrógrado asociado a
microtúbulos. La salida del núcleo se produce por transporte anterógrado).
Entrada en el núcleo
La membrana nuclear está formada por una bicapa lipídica. Esta membrana contiene entre 3000
y 5000 poros nucleares (células en crecimiento), que controlan el transporte de materiales que
entran y salen del núcleo. Cada poro nuclear está formado por un complejo de más de 50
proteínas (nucleoporinas), que controlan la entrada (importinas) y la salida (exportinas) de
componentes. En el centro de los poros se encuentra un canal mediante el cual se produce el
acceso y salida del núcleo.
Existen múltiples estrategias de entrada de los virus al núcleo, dependiendo del tipo viral.
En los virus de RNA (Orthomyxovirus y Bornavirus), el RNA entra en el núcleo a través de los
canales (y no sé que pasa- preguntar)
En los virus de DNA (Polyomavirus, Papillomavirus, Bacullovirus) y ssRNA (Lentivirus) la entrada
del virus se produce mediante unas importinas específicas.

En los virus de dsDNA (Hepadnavirus), la cápside entra parcialmente en el poro nuclear, se libera
el material genético dentro del núcleo y se degrada la cápside, que no llega a entrar en el núcleo.
En los virus ssDNA (Parvovirus, Circovirus, Geminivirus), como la cápside es pequeña atraviesa
el poro nuclear. A continuación, se libera el material genético y la cápside se degrada dentro del
núcleo.
En otros virus de dsDNA (Herpesvirus, Adenovirus), como la cápside en grande, no penetra en el
poro. De nuevo el material genético es liberado dentro y la cápside es destruida fuera del núcleo.
Pérdida de la cápside
Es la completa liberación del material genético de la cápside, con el objetivo de introducir el
genoma viral dentro del genoma. Depende del tipo de virus. Un rasgo común es que siempre se
tiene que liberar el material genético. Puede ocurrir en la membrana celular, dentro del
citoplasma, en los poros nucleares o en el propio núcleo. Es necesario un paso antes de la
replicación del genoma.
BACTERIÓFAGOS
Al igual que los virus animales, los fagos llevan a cabo interacciones con moléculas específicas
de la superficie, que funcionan como receptores y correceptores. Para muchos fagos, estas
moléculas se encuentran en la superficie celular, pero otros bacteriófagos se adhieren a otros
componentes celulares como flagelos o pili.
El attachment también es reversible en un principio y se hace irreversible después de la fijación
a otros receptores y correceptores. En el caso de los bacteriófagos la cápside no entra en la
célula huésped, sino que solo se interna el genoma viral y, en algunos casos, ciertas proteínas
virales asociadas. La cápside permanece en la superficie celular.
La entrada del genoma del fago dentro de la célula requiere la penetración de la pared celular.
Para ello, los viriones de diversos fagos poseen enzimas (como lisozima) para llevar a cabo este
proceso.

VIRUS DE PLANTAS
La presencia de pared celular en las células vegetales hace que los virus solo puedan llevar a
cabo una infección si existe un daño mecánico en la planta. Una vez dentro de las plantas, el
virus puede transmitirse de unas células a otras mediante el floema, y de unas plantas a otras
mediante plasmodesmos (no estoy seguro si es así o al reves).
Estas infecciones también puedes darse gracias a vectores, como insectos o hongos.
RESÚMEN PENETRACIÓN VIRAL EN CÉLULAS ANIMALES

Tema 2.2- Estrategias de replicación empleadas por los virus de DNA y RNA (Tema
IMPORTANTE)
Localización de la replicación viral en células eucariotas.
Generalmente, los virus de DNA replican en el núcleo, salvo los Poxvirus, que replican en el
citoplasma. Por el contrario, los virus de RNA replican en el citoplasma, salvo algunos como los
retrovirus o los Ortomyxovirus, que llevan a cabo su replicación en el núcleo.
Como ya sabemos, los virus se clasifican por su genoma mediante la clasificación de Baltimore,
como vemos en el siguiente esquema
Reglas universales de la replicación de DNA
• El DNA es sintetizado a partir de una hebra molde, mediante la incorporación de
nucleótidos al extremo 3´-OH. Además, siempre se sintetiza en sentido 5´→ 3´ de forma
semiconservativa.
• La replicación comienza en los sitios de inicio. Estos orígenes de replicación son
necesarios y sin ellos no podría llevarse a cabo este proceso.
• La replicación es llevada a cabo por DNA polimerasas dependientes de DNA (DdDp),
empleando algunas proteínas accesorias. También existen otros tipos, como las DNA
polimerasas RNA dependientes. Para que se pueda llevar a cabo se necesita siempre un
primer, que puede ser DNA, RNA o proteína.

Existen dos modelos básicos de replicación en virus: 1) Formación de una horquilla de
replicación, como los Papillomavirus, Polyomavirus, Herpesvirus y Retrovirus, todos ellos con
primers de RNA. 2) Replicación mediante desplazamiento de hebra. A diferencia de la anterior,
primero se sintetiza una de las nuevas hebras y después la siguiente. Los primers pueden ser
proteínas o el propio DNA viral, pero nunca RNA.
Orígenes de replicación viral
Dependen de cada tipo de virus. Algunos pueden llegar a tener hasta tres orígenes de
replicación, aunque como mínimo tienen uno. Cada genoma viral tiene una secuencia específica
donde comienza la replicación del material genético.
Las enzimas involucradas con la replicación del DNA son DNA polimerasas.
Los virus pequeños de DNA (dsDNA y ssDNA) no codifican para su propia maquinaria de
replicación, sino que “secuestran” y utilizan la maquinaria del hospedador (lo que les permite
tener un genoma muy pequeño) – Papillomavirus, Polyomavirus, Parvovirus…
Los virus grandes de DNA, por el contrario, codifican para su propia maquinaria de replicación –
Herpesvirus, Adenovirus, Poxvirus…
Primers
Como ya hemos dicho antes, para que se lleve a cabo la replicación necesitamos primers que
permitan la primera unión de un nucleótido a su extremo -OH, para formar las nuevas hebras
de material genético. Los primers pueden ser de RNA, DNA o proteínas.
- RNA: casi siempre son sintetizados por primasas (enzimas virales), después de que las
helicasas hayan separado las dos hebras molde del DNA.

- Proteínas: al igual que en los primers de RNA, los nucleótidos tienen que unirse al
extremo -OH. En estos casos, utilizan los grupos-OH de serinas o tirosinas.
- DNA: algunos virus de ssDNA, como los Parvovirus, utilizan su propio genoma como
primer. Al final del extremo 3´ de la secuencia del virus se encuentran secuencias
complementarias, que permiten la formación de un loop por complementariedad de
bases, permitiendo la síntesis de nuevos genomas virales.
Parvovirus (ssDNA)- Desplazamiento de hebra
Como acabamos de ver, estos virus poseen unas
secuencias específicas, complementarias entre sí, que
permiten la formación de un loop, que deja libre un
extremo -OH a partir del cual puede darse la
replicación viral (sirve como primer). A estos
fragmentos se les conoce como secuencias ITR
(Inverted Terminal Repeats).
Una vez sintetizada la nueva hebra a partir de las ITR,
aparece la enzima viral Rep 78/68, que produce un
Nick a la misma altura que el primer, pero en la
secuencia molde. Esto provoca que la secuencia que
ha servido como primer se estire, quedando libre un
extremo 3´ en la secuencia molde (la parte que NO ha
servido como primer antes), a partir del cual se
sintetiza la parte restante de la nueva hebra, que
coincidirá con la secuencia ITR del nuevo genoma
viral.
A continuación, las secuencias ITR (la molde y la recién sintetizada) se pliegan para
actuar de nuevo como primers y continuar la síntesis de la nueva hebra. Al mismo
tiempo, ambas hebras se van separando, de manera que una de ellas se introduce en
una nueva cápside y la otra repite el ciclo para generar más copias de genoma viral.
Por otro lado, la RNA polimerasa II (a partir del dsDNA lineal generado tras el
estiramiento del primer primer y después de la síntesis de su cadena complementaria-
entre los pasos 4 y 5 del esquema), sintetiza diferentes hebras de mRNA, de los cuales
algunos son traducidos a proteínas de la cápside y otros a proteínas no estructurales.

Replicación de virus dsDNA
Algunos utilizan la DNA polimerasa de la célula huésped, como los Polyomavirus (un solo origen
de replicación) o los Papillomavirus, mientras que otros son capaces de sintetizar su propia DNA
polimerasa, como los Adenovirus, los Herpesvirus (tres orígenes de replicación) y los Poxvirus
(forman loops mediante las ITR).
Adenovirus
Estos virus utilizan una proteína como primer
(precursor Terminal Protein pTP). Además,
poseen una DNA polimerasa propia, junto
con otras proteínas virales como proteínas
terminales o proteínas de enlace a ssDNA.
Presenta orígenes de replicación en ambos
extremos del genoma y la replicación del
genoma se da mediante desplazamiento de
hebra, de forma semiconservativa.
Para su replicación, se forma el complejo
pTP-Pol, en el extremo 3´-OH de la cadena
molde. A continuación, la polimerasa
comienza a sintetizar una de las nuevas
hebras de DNA, mientras que las dos hebras
de DNA inicial se van separando.
Al mismo tiempo, a la secuencia antigua del DNA que todavía no ha servido de molde (hebra
desplazada), se le unen proteínas DBP, que mantienen este DNA sin aparearse, hasta que la
polimerasa se une al primer de esta hebra y se lleva a cabo la síntesis de la nueva hebra.
Finalmente, se obtienen dos nuevas hebras que pueden repetir de nuevo el ciclo de replicación.
Herpesvirus (Herpesviridae)
Suelen encontrarse en mamíferos, animales de sangre fría y aves. Son virus envueltos de
genoma lineal dsDNA, con un tamaño de 120-200 nm y con morfología icosaédrica.
Se han aislado más de 100 Herpesvirus en diferentes huéspedes, de los cuales ocho han sido
hallados en humanos.
- Virus herpes simplex 1 (VHS-1 y VHH-1): Herpesvirus tipo 1, causantes de daños en boca
y fosas nasales. El VHS-1 es capaz de codificar al menos 74 proteínas.
- Virus herpes simplex 2 (VHS-2 y VHH-2): Herpesvirus tipo 2, causantes de daños en los
genitales. Son los más comunes después de los de tipo 1.
- Varicela Zóster (VZV, VHH-3): Herpesvirus tipo 3.
- Epstein Barr (VEB, VHH-4): Herpesvirus tipo 4.
- Citomegalovirus (CMV, VHH-5): Herpesvirus tipo 5.
- Virus herpes humano 6 (VHH-6) y virus herpes humano 7 (VHH-7).
- Virus asociado al sarcoma de Kaposi (VHH-8): puede ocasionar cáncer en pacientes
inmunodeprimidos.
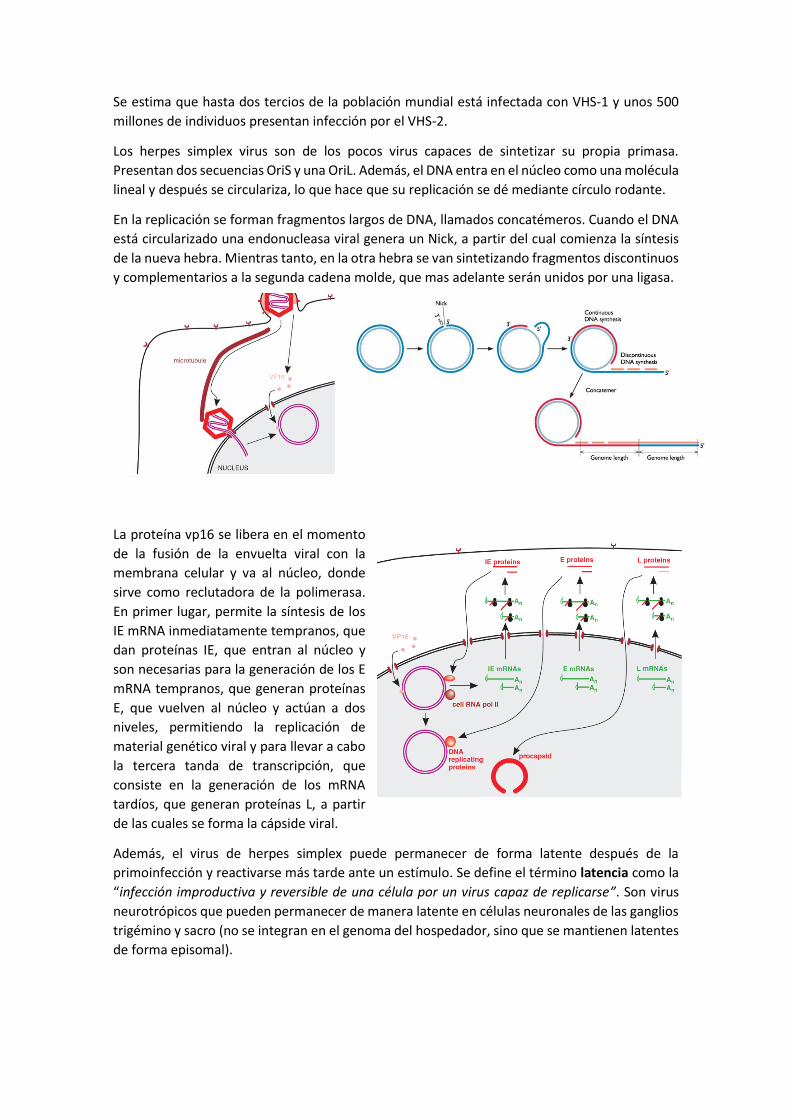
Se estima que hasta dos tercios de la población mundial está infectada con VHS-1 y unos 500
millones de individuos presentan infección por el VHS-2.
Los herpes simplex virus son de los pocos virus capaces de sintetizar su propia primasa.
Presentan dos secuencias OriS y una OriL. Además, el DNA entra en el núcleo como una molécula
lineal y después se circulariza, lo que hace que su replicación se dé mediante círculo rodante.
En la replicación se forman fragmentos largos de DNA, llamados concatémeros. Cuando el DNA
está circularizado una endonucleasa viral genera un Nick, a partir del cual comienza la síntesis
de la nueva hebra. Mientras tanto, en la otra hebra se van sintetizando fragmentos discontinuos
y complementarios a la segunda cadena molde, que mas adelante serán unidos por una ligasa.
La proteína vp16 se libera en el momento
de la fusión de la envuelta viral con la
membrana celular y va al núcleo, donde
sirve como reclutadora de la polimerasa.
En primer lugar, permite la síntesis de los
IE mRNA inmediatamente tempranos, que
dan proteínas IE, que entran al núcleo y
son necesarias para la generación de los E
mRNA tempranos, que generan proteínas
E, que vuelven al núcleo y actúan a dos
niveles, permitiendo la replicación de
material genético viral y para llevar a cabo
la tercera tanda de transcripción, que
consiste en la generación de los mRNA
tardíos, que generan proteínas L, a partir
de las cuales se forma la cápside viral.
Además, el virus de herpes simplex puede permanecer de forma latente después de la
primoinfección y reactivarse más tarde ante un estímulo. Se define el término latencia como la
“infección improductiva y reversible de una célula por un virus capaz de replicarse”. Son virus
neurotrópicos que pueden permanecer de manera latente en células neuronales de las ganglios
trigémino y sacro (no se integran en el genoma del hospedador, sino que se mantienen latentes
de forma episomal).
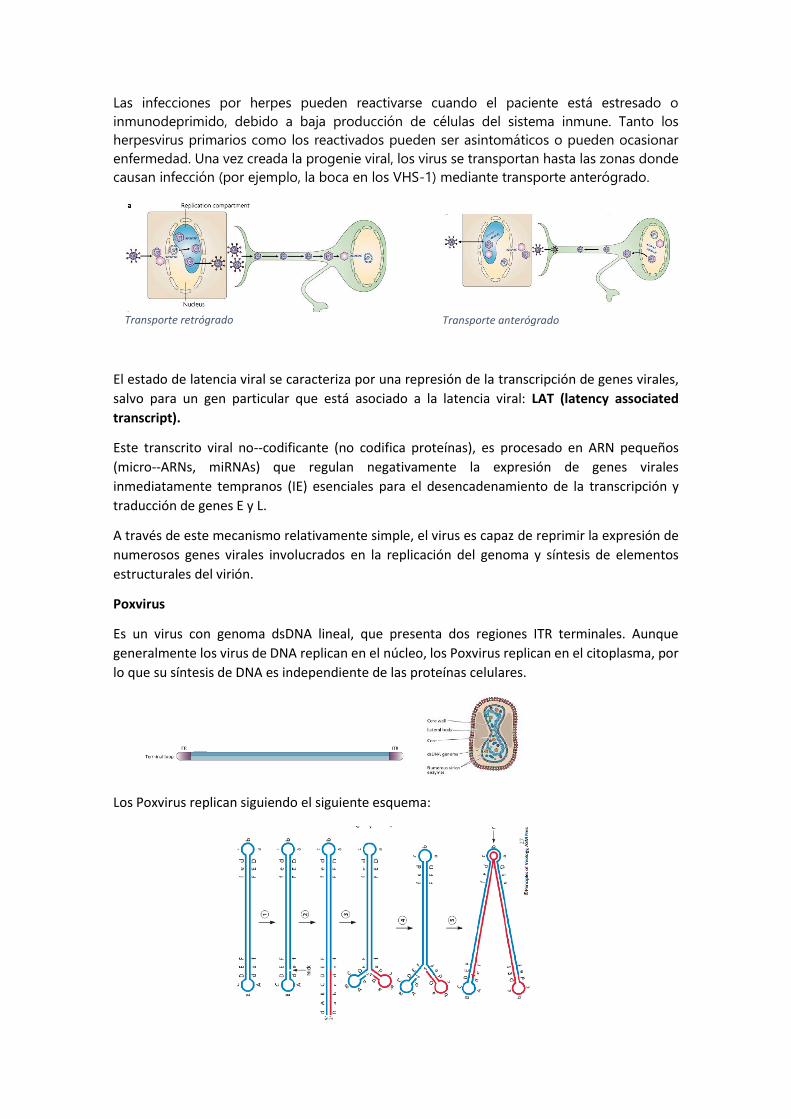
Las infecciones por herpes pueden reactivarse cuando el paciente está estresado o
inmunodeprimido, debido a baja producción de células del sistema inmune. Tanto los
herpesvirus primarios como los reactivados pueden ser asintomáticos o pueden ocasionar
enfermedad. Una vez creada la progenie viral, los virus se transportan hasta las zonas donde
causan infección (por ejemplo, la boca en los VHS-1) mediante transporte anterógrado.
El estado de latencia viral se caracteriza por una represión de la transcripción de genes virales,
salvo para un gen particular que está asociado a la latencia viral: LAT (latency associated
transcript).
Este transcrito viral no-codificante (no codifica proteínas), es procesado en ARN pequeños
(micro-ARNs, miRNAs) que regulan negativamente la expresión de genes virales
inmediatamente tempranos (IE) esenciales para el desencadenamiento de la transcripción y
traducción de genes E y L.
A través de este mecanismo relativamente simple, el virus es capaz de reprimir la expresión de
numerosos genes virales involucrados en la replicación del genoma y síntesis de elementos
estructurales del virión.
Poxvirus
Es un virus con genoma dsDNA lineal, que presenta dos regiones ITR terminales. Aunque
generalmente los virus de DNA replican en el núcleo, los Poxvirus replican en el citoplasma, por
lo que su síntesis de DNA es independiente de las proteínas celulares.
Los Poxvirus replican siguiendo el siguiente esquema:
Transporte retrógrado Transporte anterógrado

Virus de RNA
Existen tres tipos de virus de RNA 1) Aquellos cuyo genoma puede servir de molde para que la
RNA polimerasa sintetice proteínas (RNA +). 2) Aquellos que, como su genoma es de RNA (-) o
RNA (+) diferente de mRNA, tienen que sintetizar una hebra de mRNA para la síntesis proteica.
3) Aquellos que, como su genoma es de dsDNA, utilizan una de sus hebras para formar un mRNA
y sintetizar las proteínas virales.
En todos estos casos es necesaria la acción de la RNA polimerasa RNA dependiente, la cual
siempre es aportada por el virus, puesto que las células carecen de esta enzima. Las polimerasas
involucradas en la replicación suelen llamarse replicasas.
Para la síntesis del RNA hemos de seguir los siguientes criterios:
- La síntesis de RNA empieza y termina en sitios específicos.
- La RNA polimerasa RNA dependiente puede necesitar un primer para la síntesis (O NO).
- Pueden requerirse otras proteínas celulares.
- El RNA es sintetizado mediante la adición de nucleótidos, siempre en sentido 5´→3´.
Virus de ssRNA (+) → grupo IV de Baltimore
El genoma puede ser utilizado directamente como mRNA para la síntesis de proteínas virales.
Por otro lado, la replicación del genoma se da en dos pasos. En el primero, la cadena positiva es
copiada para dar una cadena negativa. En segundo lugar, esta cadena de orientación negativa
sirve para formar nuevas copias de genoma viral de orientación positiva.
Algunos ejemplos son los Picornavirus (Poliovirus, Rhinovirus), Calicivirus (gastroenteritis),
Coronavirus (SARS), Flavivirus (fiebre amarilla, virus del Nilo Occidental, Hepatitis C) y Togavirus
(Rubeola).
Picornavirus
Es un virus de pequeño tamaño y de ssRNA (+). Presenta cápside icosaédrica y es propio de
mamíferos y aves. Su peculiaridad reside en que posee una proteína asociada a su genoma (VPG)
que sirve como primer en la replicación.

Dentro de los Picornavirus se encuentras los Poliovirus.
La replicación de los Poliovirus tiene lugar en diferentes
fases. Primero, el genoma viral entra en la célula
huésped (la cápside no), para que los ribosomas lo lean
y sinteticen una poliproteína, que servirá de precursor
para la formación de otras proteínas virales.
Muchos virus replican en determinadas zonas del
citoplasma, que pueden tener una mayor facilidad,
como las vesículas de membrana inducidas por la
infección.
Virus de ssRNA (-) grupo V de Baltimore
El genoma, con orientación negativa, no puede ser utilizado directamente como mRNA. Por lo
tanto, es necesaria la síntesis de una hebra con orientación positiva, utilizando como molde el
genoma inicial. Este paso es llevado a cabo por una RNA polimerasa RNA dependiente.
A partir del genoma ssRNA (-) se sintetiza por un lado el mRNA para dar las proteínas y, por otro
lado, una hebra con orientación positiva a partir de la cual se sintetizarán las nuevas copias de
genoma viral.
Algunos ejemplos son los Paramixovirus (Paperas, sarampión), Rhabdovirus (Rabia), Bornavirus,
Filovirus (Ébola), Orthomyxovirus (Influenza), Arenavirus (virus Lassa).
Rhabdovirus
La rabia es una enfermedad neurológica aguda de consecuencias fatales. Se transmite a través
de la mordedura de animales como perros, zorros, murciélagos y mapaches. Además, si la
mordedura se da en la cabeza, la mortalidad oscila entre el 60-80%. Existe vacuna desde 1885
(Pasteur). La vacuna puede ser eficaz incluso después de la mordedura.
Se produce un daño a nivel neuronal, pero antes el virus replica en las células musculares (donde
se da la mordedura) hasta que salta a las neuronas, donde se produce el daño. Si no se trata, la
muerte se da en pocos días.
En cuanto a la replicación del virus de la rabia, a partir del genoma inicial se forman cinco mRNAs
diferentes (N, P, M, G y L), y de cada uno se codifican diferentes proteínas importantes del
virus.

Orthomyxovirus (Influenza)
Se trata de un virus segmentado, formado por ocho fragmentos de RNA que codifican para ocho
mRNAs diferentes. Este virus, como todos los fragmentado, replica en el núcleo, a pesar de ser
virus de RNA, que generalmente replican en el citoplasma. Entra en la célula mediante la
formación de un endosoma.
La maquinaria viral no puede formar sus propios mRNAs, por lo que usa RNAs celulares como
primers para iniciar la síntesis de mRNA viral. Este proceso se conoce como cap-snatching.
Virus dsRNA (replicación)
Son virus poco comunes que, al igual que los de dsDNA, tienen que ser desenrollados por una
helicasa para que se pueda llevar a cabo la replicación.
Algunos ejemplos son los Reovirus y el fago phi6, el cual replica su genoma mediante un
mecanismo semiconservativo, similar a la replicación de dsDNA, de forma que cada una de las
cadenas iniciales se utiliza para dar dos hebras hijas.
Por otro lado, los Reovirus (como otros virus dsDNA), replican mediante un mecanismo
conservativo, debido a que la molécula de doble cadena del genoma viral es conservada. Son
virus con simetría icosaédrica y con genoma segmentado (10-12 fragmentos).
En la replicación, como la hebra (+) no puede ser leída por los ribosomas (en este caso), no se
utiliza. Por lo tanto, se utiliza la hebra (-), que sirve de molde para generar RNAs (+), de los cuales
algunos serán utilizados para la síntesis proteica y otros servirán de molde para formar dobles
cadenas que serán el genoma de la progenie viral (replicación conservativa).

Transcripción inversa
Algunos virus de RNA replican sus genomas mediante un intermediario de DNA. En esta forma
de replicación se utiliza la transcripción reversa, que consiste en la síntesis de una cadena de
DNA a partir de una cadena de RNA (+), para que después se forme una doble cadena de DNA a
partir de la ssDNA (-) recién sintetizada. Ambos pasos están catalizados por la misma enzima, la
retrotranscriptasa o transcriptasa inversa. No se conocen virus procariotas que utilicen este
mecanismo. Esta enzima revolucionó el dogma central de la biología, y solo se encuentra en los
virus de las clases VI y VII de Baltimore.
Además, la transcriptasa inversa tiene tres grandes funciones: 1) DNA polimerasa RNA
dependiente. 2) Actividad RNAasa, para degradar la hebra de DNA cuando se forma el híbrido
DNA-RNA. 3) DNA polimerasa DNA dependiente, para formar el DNA de doble cadena a partir
de la hebra de DNA (-) que a su vez ha sido sintetizada a partir del RNA inicial.
La transcripción reversa tiene lugar dentro del citoplasma. Para la primera síntesis de DNA
dependiente de RNA se necesita un primer de RNA transferente. Más tarde, cuando se elimina
el RNA, un fragmento del mismo (polipurina) se mantiene intacto para ser utilizado como
cebador en la síntesis de la hebra complementaria de DNA. Como en otros casos, el primer tiene
que tener un extremo 3´-OH a partir del cual se unan los dNTS (no rNTPs), para lo cual la cadena
molde (como ya hemos visto) puede ser de DNA o RNA.
Por otro lado, la actividad RNAasa se lleva a cabo mediante la hidrólisis del grupo fosfato del
RNA. Además, solo produce escisiones en el RNA cuando se encuentra formando un dúplex, el
cual puede ser DNA:RNA o RNA:RNA. Se forman oligonucleótidos cortos con 5´-fosfato, 3´-OH.
El virus es transportado al núcleo mediante la formación de un complejo de preintegración,
antes de insertarse en el genoma del huésped en forma de provirus. Este trayecto hasta el
núcleo se realiza a través de los microtúbulos.
Para la mayoría de retrovirus este proceso ocurre durante la mitosis, por lo que solo puede darse
la infección en células en división. Sin embargo, el VIH puede infectar células que no están en
división, ya que los complejos de preintegración son capaces de entrar en el núcleo sin necesidad
de que este se encuentre facilitado por la fase de división.
Cuando el dsDNA viral se integra en el huésped, puede inducirse un estado de latencia. En
latencia el virus puede quedarse durante años, hasta que se activa la transcripción del
provirus, mediante mecanismos que aún no están muy claros. Gracias a la RNA polimerasa
celular tendremos moléculas de RNA de polaridad positiva, que servirán como genoma para
la nueva progenie, y mRNAs a partir de los cuales se sintetizarán las nuevas proteínas y virales.

Tema 2.3- Ensamblaje y salida viral
La producción de partículas virales infecciosas, que constituyen la nueva progenie viral, se
denomina maduración o morfogénesis.
Todos los viriones tienen fases comunes en el ensamblaje (formación de las subunidades
proteicas de la cápside, ensamblaje de las proteínas entre sí, empaquetamiento del genoma viral
en la cápside o liberación de los virus de la célula huésped) y, alguno virus concretos, también
presentan fases de ensamblaje específicas (como la adquisición de la envuelta o la maduración
de los viriones).
La producción de nuevas partículas virales depende de la maquinaria celular del huésped. Para
ello, los virus son capaces de utilizar proteínas celulares como chaperonas (encargadas del
plegamiento de otras proteínas), importinas y exportinas (encargadas de la entrada y salida de
moléculas del núcleo celular) y sistemas de transporte (tráfico intracelular).
Morfogénesis viral
Incluye el movimiento de los componentes virales después de su formación hacia sitios
específicos en la célula diana. La morfogénesis viral se da en dos pasos: tráfico intracelular y
ensamblaje.
- Tráfico intracelular: algunos componentes virales son sintetizados en el citoplasma y
otros en el núcleo. Estos componentes tienen que ser llevados a los sitios de ensamblaje,
para lo cual se utilizan los microtúbulos o las fibras de actina de la célula huésped.
Los sitios de ensamblaje dependen del tipo de genoma (DNA o RNA), del mecanismo de
replicación del genoma y de la presencia o ausencia de envuelta viral.
La salida del núcleo puede darse a través de exportinas (Hepadnavirus, Retrovirus,
Parvovirus, Baculovirus…), a través de la salida de la cápside (Herpesvirus, Circovirus) o
mediante ruptura de la envuelta nuclear (Papillomavirus, Polyomavirus).

Maduración y salida de los Herpesvirus
La cápside forma una especie de endosoma con la membrana nuclear, permitiendo
la salida del virus al citoplasma. A continuación, el virus es capaz de interaccionar con
el Aparato de Golgi y formar una vesícula alrededor del virus a partir del cual se forma
la envuelta viral, que le permite salir al ambiente extracelular.
- Ensamblaje viral: puede darse por dos mecanismos: 1) Ensamblaje concertado.
2) Ensamblaje secuencial.
1) Ensamblaje concertado: las proteínas de la cápside se ensamblan exclusivamente
en asociación con el material genético. Este mecanismo es propio de virus de ssRNA
(-), como Influenza, o retrovirus, como el VIH.
En los virus helicoidales el ensamblaje conlleva el revestimiento del genoma con
múltiples copias de la proteína viral correspondiente.
2) Ensamblaje secuencial: a diferencia del ensamblaje concertado, este se da por
partes. En primer lugar, las proteínas se ensamblan entre sí para formar la
procápside y, después, se da el empaquetamiento del material genético. Un ejemplo
de este tipo de ensamblaje se da en los bacteriófagos. Otro ejemplo son los
Poliovirus, que a partir del mRNA producen una poliproteína a partir de la cual se
generan las proteínas de la procápside.

Empaquetamiento del genoma
Partimos del problema de que el genoma viral debe ser distinguido del DNA o RNA celular para
el empaquetamiento. La solución reside en la presencia de señales de empaquetamiento en el
genoma viral. Aún así, en algunos casos pueden llegar a empaquetarse fragmentos del material
genético del hospedador.
Existen proteínas virales que son capaces de reconocer secuencias específicas del genoma viral
(DNA o RNA) para poder llevar a cabo correctamente el empaquetamiento. Este hecho tiene
gran importancia en el uso de vectores virales en terapia génica.
Para la producción de virus infectivos con genomas virales segmentados, se requiere el
empaquetamiento de al menos una copia de cada uno de los fragmentos de genoma viral.
Existen dos teorías para explicar el empaquetamiento de estos genomas segmentados en el virus
Influenza:
- Empaquetamiento al azar: si se diera de esta forma, la probabilidad de que coincidan
los 8 fragmentos del genoma al azar dentro de la misma cápside es de 1 por cada 400
partículas ensambladas.
- Empaquetamiento selectivo: cada uno de los 8 fragmentos de RNA presenta diferentes
tipos de señales que permiten la incorporación del genoma viral. Se cree que estas
señales están dentro de secuencias codificantes y no codificantes en los extremos 3´y 5´
del RNA viral.
Salida de la célula
Por un lado, puede darse por lisis, de forma que el virus provoca la ruptura de la membrana
celular, permitiendo la salida de las partículas virales que se encuentran en el citoplasma y
provocando la muerte de la célula huésped.
La mayoría de virus desnudos (y algunos envueltos) requieren de la lisis de la célula para poder
liberar la progenie viral. Además, todos los bacteriófagos son líticos, salvo los fagos filamentosos
y los Plasmavirus.
Para inducir la lisis se emplean virioporinas (proteínas virales con la capacidad de insertarse
en la membrana de la célula y provocar rupturas que conllevan su lisis), como los Adenovirus
y Picornavirus, y fosfolípidos líticos, que también provocan la lisis. Un ejemplo se encuentra
en los Phycodnavirus.
Por otro lado, la salida viral también puede darse mediante gemación, utilizando los complejos
ESCRT (Endosomal Sorting Complexes Required for Transport) o sin necesidad de utilizar estos
complejos, como el virus Influenza.
Existen “motivos” específicos en proteínas virales llamados dominios L (L-domains), que son responsables del reclutamiento de la maquinaria celular. Esto se da en Retrovirus, Hepadnavirus, Herpesvirus, Rhabdovirus y Poxvirus.
Como no se da la lisis, se forma una "vesícula" en el Aparato de Golgi o el Retículo Endoplásmico, que viaja por los microtúbulos y sale de la célula por exocitosis. En algunos casos, al salir de la
célula por exocitosis, la partícula viral adquiere la envuelta.

La adquisición de la envuelta se puede dar simultáneamente al ensamblaje o después del
mismo, cuando la cápside ya está formada.
Maduración viral
Es el paso del ciclo viral en el que los viriones se convierten en partículas infecciosas. A menudo,
las proteasas están involucradas en la maduración de los virus. Este es un proceso especialmente
estudiado en el virus VIH. En algunos casos, si la maduración no se completa, la partícula pierde
gran parte (o la totalidad) de su infectividad.
Una vez que ya se ha dado la maduración, la progenie viral puede salir al espacio extracelular,
pasar a otras células no infectadas o ambas a la vez. También puede inducirse la formación de
sincitios, de manera que se pueda infectar todo el conglomerado de células que se ha formado.


















