SOPORTE FISIOLOGÍA DIGESTIVA

of 38
-
Upload
willian-loor -
Category
Documents
-
view
62 -
download
0
Transcript of SOPORTE FISIOLOGÍA DIGESTIVA
DOCUMENTO DE APOYO PARA EL MDULO DE FISIOLOGA DIGESTIN. Las clulas del organismo animal requieren un aporte constante de sustancias energticas y estructurales para atender el mantenimiento de su metabolismo, crecimiento y a su multiplicacin. Estos compuestos se encuentran en los alimentos y forrajes, pero en forma directamente asimilable slo se encuentran en una muy pequea proporcin. Los alimentos se presentan, en su mayor parte, en forma de combinaciones insolubles, macromoleculares, que para ser utilizadas por el organismo deben transformarse previamente en otras formas ms sencillas para su absorcin. El conjunto de procesos que comienza con la ingestin de los alimentos, contina con su transformacin a lo largo del tracto gastrointestinal y finaliza con la eliminacin de los residuos no absorbidos, constituye la funcin denominada digestin. En el proceso digestivo, los alimentos son sometidos a una degradacin que los transforma en productos de peso molecular ms bajo, perdiendo con ello sus caractersticas especficas originales y su posible toxicidad. La digestin constituye una funcin necesaria para la vida. Esta funcin est constituida, en esencia, por una multitud de procesos fsicos y qumicos estrechamente correlacionados y finamente coordinados bajo el control del sistema nervioso autnomo y endocrino. La regulacin nerviosa de los procesos digestivos est complementada por la actividad de ciertas hormonas, cuya sntesis y secrecin depende, sobre todo, del estado de actividad funcional en que se hallen las diversas porciones del tracto gastrointestinal. De acuerdo con su origen, los alimentos pueden ser clasificados como productos vegetales, animales e inorgnicos. La mayora de los alimentos, considerados aisladamente, no cubren los requerimientos animales, tanto por lo que respecta a su composicin cualitativa como cuantitativa en principios nutritivos. Por tanto, para una alimentacin ptima y para poder proporcionar al organismo todos los factores nutritivos esenciales, es necesario recurrir a las combinaciones de alimentos. Los animales se distribuyen en herbvoros o carnvoros, segn si la parte fundamental de su alimentacin sea, respectivamente, de origen vegetal o animal. Entre los animales domsticos el perro o el gato pertenecen a los carnvoros, mientras que el caballo, la vaca, la oveja, la cabra y el conejo son herbvoros. Los omnvoros ocupan una posicin intermedia, pues se nutren de alimentos tanto de origen vegetal como animal. Sin embargo, el cerdo, que es el animal representativo de este grupo dentro de los animales domsticos, recibe habitualmente una alimentacin predominantemente vegetal. En los animales domsticos, la estructura de los diversos segmentos del tubo digestivo est claramente adaptada a la naturaleza y composicin de los alimentos que ingiere y aunque las funciones del tracto digestivo pueden ser similares en especies muy diversas, existen diferencias en la distribucin anatmica y funcional de los diferentes compartimentos digestivos.
Atendiendo a estas diferencias podemos clasificar a los animales domsticos en cuatro grandes grupos o modelos digestivos. 1. La mayora de los carnvoros y omnvoros poseen un estmago relativamente simple, denominndose no rumiantes En dichas especies el estmago consiste, en esencia, en una bolsa que contiene glndulas secretoras de cido clorhdrico y enzimas digestivas. 2. En las aves el estmago est precedido por una cmara denominada buche, y que sirve para el almacenamiento y cierto grado de fermentacin, tiene esencialmente la misma estructura del esfago, excepto en que las glndulas mucosas estn presentes solamente en la unin con el estmago. El digestivo de las aves contina con el estmago verdadero denominado pro-ventrculo, siguiendo otra "cmara" denominada molleja o estmago muscular que realiza la trituracin mecnica. Las especies herbvoras, presentan modificaciones an ms trascendentes, incluso a nivel intestinal, como veremos, estas modificaciones les permiten utilizar la celulosa y otros polisacridos como la hemicelulosa, funcin que a ellos mismos les es imposible cumplir, por no poseer enzimas capaces de hidrolizar los enlaces que unen las molculas de glucosa, existiendo para ello la simbiosis con microorganismos capaces de cumplir esta funcin. 3. El aparato digestivo de los rumiantes se ha desarrollado en un rgano pregstrico que permite una compleja fermentacin microbiana, quedando limitada esta particularidad slo a rumiantes. 4. En el equino y conejo (entre otros) se presenta a nivel intestinal una modificacin importante el "gran ciego" en el cual se produce una actividad microbiana similar a la del rumen-retculo. Adems, algunas especies como el conejo practican la cecotrofia por lo que pueden alcanzar beneficios adicionales de la actividad digestiva desarrollada en el ciego e intestino grueso, aprovechando productos que no fueron absorbidos, tales como las vitaminas del complejo B. En el Cuadro 1 se puede observar la capacidad y longitud relativa a los diversos segmentos del tracto gastrointestinal en los animales domsticos.
Cuadro 1. Capacidad y longitud relativa de los diversos segmentos del tracto gastrointestinal en los animales domsticos. Especie Parte del tubo Capacidad Capacidad Relacin
digestivo Equino Estmago Int. Delgado Ciego Coln grande Coln pequeo Total Ret, Rum, Oma, Abo Int. Delgado Ciego Coln y recto Total Estmago Int. Delgado Ciego Coln y recto Total Estmago Int. Delgado Ciego Coln y recto Total Estmago Int. Delgado Int. grueso Total
relativa absoluta % 8.5 30.2 15.9 38.4 7.0 100.0 70.8 18.5 2.8 7.9 100.0 29.2 33.5 5.6 31.7 100.0 62.3 23.3 1.3 13.1 100.0 69.5 14.6 15.9 100.0
en litros (Promedio) 17.95 63.83 33.54 81.25 14.77 211.34 252.5 66.0 9.90 28.0 356.4 8.0 9.2 1.55 8.7 27.45 4.33 1.62 0.09 0.91 6.95 0.341 0.114 0.124 0.579
long.corporal : long intestino 1:12
Bovino
1:20
Cerdo
1:14
Perro
1:6
Gato
1:4
En comparacin con la longitud corporal, el tubo digestivo de los carnvoros es relativamente corto. La alimentacin en este tipo de animales consiste principalmente en alimentos ricos en principios nutritivos, de elevada digestibilidad, que son fcilmente degradados. El tubo digestivo de los omnvoros es claramente ms largo y muestra una diferenciacin ms acusada. Finalmente, en el caso de los herbvoros, el tracto gastrointestinal presenta regiones muy dilatadas destinadas a la degradacin de hidratos de carbono complejos, de difcil degradacin y digestin a cargo de los microorganismos, ya sea en los ventrculos estomacales o cavidades pre-gstricas de los rumiantes, o en intestino grueso de los herbvoros con estmago de una sola cavidad o bolsa. GENERALIDADES SOBRE LA FORMACIN DE LOS PRINCIPIOS NUTRITIVOS
Con la excepcin de unas pocas sustancias de bajo peso molecular, como monosacridos, aminocidos, sales y agua, los elementos que integran la dieta sufren, durante su paso por el canal digestivo, una degradacin a elementos de peso molecular inferior, los cuales pueden ulteriormente ser absorbidos. Adems de las sustancias anteriormente citadas, tambin pueden ser absorbidas por la mucosa intestinal, sin previa transformacin, las vitaminas. En lneas generales podemos decir que en la digestin de los alimentos intervienen fenmenos de orden fsico y de orden qumico y en base a esto podemos dividir su estudio en tres partes o captulos. I) II) III) Transporte de los alimentos Funcin secretora Absorcin de los productos de la digestin, del agua y de los diferentes electrlitos.
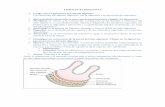
FISIOLOGIA DEL APARATO DIGESTIVO DEL RUMIANTE. Los rumiantes son animales que poseen tres compartimentos pre-gstricos encargados de la fermentacin bacteriana de los vegetales consumidos. La principal caracterstica de la digestin en estos animales es la fermentacin que ocurre en el retculo-rumen. Esta fermentacin que es provocada por microorganismos permite a los rumiantes utilizar mejor los alimentos pues en el rumen son transformados en nutrientes ms digestibles y de fcil absorcin. Antes de comenzar un estudio fisiolgico especfico para esta especie debemos mencionar algunas diferencias bsicas de estos animales con respecto a los monogstricos. El aparato digestivo de los rumiantes se diferencia anatmicamente de los monogstricos principalmente por estar formados por 4 compartimentos Rumen, Retculo, Omaso y Abomaso. Al igual que un monogstrico el aparato digestivo tiene como funcin transformar los alimentos para que puedan ser absorbidos por la mucosa y as transportados por la sangre que los moviliza hacia las clulas del organismo ya metabolizados. El animal ingiere bsicamente tres tipos de substratos, hidratos de carbono, grasas, protenas conjuntamente con agua minerales y vitaminas. El tamao de estas partculas como sabemos es inapropiado para ser absorbidas por las clulas de la pared intestinal por lo tanto deben transformarse en molculas ms simples, es decir, lograr una REDUCCIN FSICA DEL SUBSTRATO siendo la nica excepcin la absorcin de grasas no hidrolizadas siempre que sus partculas no sobrepasen las 0,5 micras. Como sabemos el mecanismo de fraccionamiento de substratos es fisicoqumico y recibe
el nombre de hidrlisis la cual se ve acelerada por las enzimas digestivas secretadas por las glndulas anexas y la pared del tubo digestivo. Las proteasas degradan protenas y en el rumiante son secretadas por el abomaso, la tripsina, quimotripsina y carboxipeptidasas por el pncreas y enzimas proteolticas intestinales que atacan pequeos pptidos. No existen proteasas en la boca ni en los primeros compartimentos de los rumiantes. El abomaso de los rumiantes es el encargado de la secrecin del HCl y proporciona un pH cido al medio para que la pepsina se active y acte, recordemos que sobre pH 5,0 es inactiva. No existe en los monogstricos, rumiantes ni aves una enzima capaz de hidrolizar la celulosa, ya que las amilasas son de tipo y no como requiere la celulosa. Su aprovechamiento como nutriente se debe a la presencia de microorganismos que poseen amilasa que se encuentra en el rumen de los rumiantes e intestino grueso de los no rumiantes. El aprovechamiento de la celulosa es de vital importancia para el rumiante. Las lipasas que degradan la grasa son las lipasas pancreticas e intestinal o entrica que funcionan luego que la grasa ha sido emulsionada por la bilis, fuera de este agente emulsionante es de mucha importancia el elemento mecnico, es decir, agitacin mecnica realizada por la contraccin de la pared intestinal, movimientos respiratorios sobre el abdomen y an por el desplazamiento del animal. Para demostrar la importancia pancretica y heptica en la absorcin de la grasa diremos que en una deficiencia pancretica se reduce la absorcin de grasas en 75% y la heptica en 50%. La importancia de la masticacin desde el punto de vista digestivo, es la de fraccionar las grandes partculas de los substratos aumentando la superficie de contacto con las enzimas. CARACTERSTICAS GENERALES DEL PROCESO DIGESTIVO DEL RUMIANTE En el rumiante el proceso digestivo tiene caractersticas que lo diferencian del monogstrico. El rumiante devuelve el alimento a la boca re-mastica y deglute el contenido rumino-reticular. Utiliza grandes cantidades de forraje. El rumen del adulto tiene una capacidad de 60-120 litros. Es capaz de sintetizar protenas a partir de Nitrgeno no proteico (urea) y su dieta no es exigente en calidad de protenas suministradas. Su proceso digestivo es constante (24 horas al da), lo que implica trabajo glandular continuo y mantenido. Como resultado del metabolismo en el rumen-retculo, hay producciones de gases, especialmente CO2 y CH4 (1 a 2 litros por minuto). Todo esto es consecuencia de la digestin realizada por la flora y fauna microbiana, que se compone aproximadamente de 5 gneros de protozoos aerbicos, en una concentracin de 103 por ml de lquido ruminal y unas 900 especies de bacterias, 98% de ellas anaerbicas, con una concentracin de 106 por ml de lquido ruminal. Estos microorganismos son capaces de digerir el 14% de celulosa, el 71% de celobiosa, el 40% de almidn y el 70% de glucosa.
MOTILIDAD DEL TUBO DIGESTIVO Su estudio tiene importancia porque es uno de los factores que determina la cuanta del aprovechamiento nutritivo de los alimentos, ya que influye en la homogenizacin y tiempo de permanencia de ellos en las diversas zonas del tubo digestivo. Mayor velocidad de trnsito aumenta la apetencia por el forraje, pero disminuye el grado de digestin y absorcin. FINALIDAD DE LOS MOVIMIENTOS Existe una necesidad de trnsito en sentido caudal para someter a los substratos que haban sido parcialmente degradados en el rumen a la accin de nuevas enzimas que se encuentran en el estmago e intestino y para llevarlos tambin a los compartimentos de absorcin representados por el intestino. La absorcin se efecta asimismo a lo largo de todo el tubo digestivo; la diferencia es cuantitativa. En los rumiantes la necesidad de colocar los substratos en el intestino para ser absorbidos en cantidades adecuadas no es tan crtica como en los monogstricos; ya que el rumen tiene una gran capacidad de absorcin. Aparte del requerimiento de trnsito, est el de homogenizar el contenido ruminoreticular y, en general, el contenido de todo el tubo digestivo. La segunda masticacin que no es indispensable para la vida normal del rumiante, contribuye a la ruptura de las membranas vegetales, facilitando mecnicamente la accin de microorganismos por el aumento de la superficie y por una mejor distribucin microorganismo - substrato. Por esta razn, la homogenizacin aumenta la velocidad de la digestin. INGESTIN DE ALIMENTOS La cantidad de alimento ingerido depende sobre todo del deseo intrnseco de alimento llamado hambre, y en el ser humano, principalmente, la variedad de alimento escogida depende del apetito. MASTICACIN Esta funcin es cumplida por los dientes, los anteriores (incisivos) permiten una accin de corte y los posteriores (molares) una accin de molienda. Todos los msculos de los maxilares trabajando juntos, pueden cerrar los dientes con una fuerza de 25 kg para los incisivos y de 90 kg para los molares (existiendo s variaciones segn la especie que se trate). En la boca tambin ocurren otros fenmenos importantes que se detallarn ms adelante. En los rumiantes la mandbula es ms estrecha que la maxila, lo cual permite llevar a cabo una molienda ms adecuada ya que la masticacin se realiza con movimientos de laterales funcionando como un molino, es tambin importante destacar que la secrecin salival es alternante (caracterstica que no ocurre en otras especies) es decir durante la rumia el flujo salival proviene del mismo lado en que el bolo est siendo masticado, mien-
tras que en el otro lado el flujo es mucho menor. DEGLUCIN Es un mecanismo complicado, principalmente porque la faringe se emplea casi todo el tiempo en otras funciones y slo se vuelve un lugar de paso de los alimentos por algunos segundos cada vez. Resulta de especial importancia que la deglucin no interfiera con la respiracin. En general se puede dividir deglucin en: 1. Etapa voluntaria que inicia el acto 2. Etapa farngea, involuntaria que constituye el paso del alimento de faringe a esfago. 3. Etapa esofgica, tambin involuntaria que corresponde al descenso del bolo de faringe a esfago. 1) ETAPA VOLUNTARIA Cuando el alimento est listo para ser deglutido se comprime o expulsa voluntariamente hacia atrs por aplicacin progresiva de la lengua contra el paladar, as la lengua empuja el bolo hacia la faringe. De aqu en adelante la deglucin se torna enteramente automtica y por lo regular no puede detenerse. 2) ETAPA FARINGEA Al ser empujado el bolo alimenticio hacia la parte posterior de la boca, estimula zonas receptoras del reflejo de deglucin que rodean el istmo de las fauces, especialmente a nivel de los pilares amigdalinos, de all salen impulsos hacia el tallo cerebral para desencadenar la siguiente serie de contracciones musculares farngeas. a) El paladar blando se eleva para ocluir las coanas, evitando as que el alimento refluya a las fosas nasales. b) Los pliegues palatofarngeos de ambos lados de la faringe se aproximan hacia la lnea media y se forma as una hendidura longitudinal que permite el paso de los alimentos masticados pero no as los trozos voluminosos, esta etapa slo dura 1 segundo. c) Las cuerdas vocales se aproximan y la epiglotis bascula hacia atrs sobre el estrecho farngeo superior, ambos movimientos impiden el paso de alimentos a la traquea. La aproximacin de las cuerdas vocales es el ms importante de estos mecanismos. d) La laringe es totalmente llevada hacia arriba y adelante por los msculos que se insertan en el hueso hioides. Este movimiento ensancha el orificio superior del esfago. Al mismo tiempo el esfnter hipofarngeo que rodea dicho orificio esofgico, y cuya funcin es impedir la entrada de aire al esfago durante la respiracin, se relaja: esto
permite paso libre y fcil del bolo de la faringe posterior a esfago superior. e) Al mismo tiempo que ocurre elevacin de la faringe y relajacin del esfnter hip farngeo, entra en accin el constrictor superior de la faringe: se produce una onda peristltica rpida que viaja hacia abajo e involucra sucesivamente al constrictor inferior y luego al esfago. Efectos de la etapa farngea sobre la respiracin. Como se mencion anteriormente esta fase no pasa de 1-2 segundos e interrumpe la respiracin durante una parte muy pequea del ciclo respiratorio normal. El centro de la deglucin inhibe especficamente el centro respiratorio bulbar durante este tiempo, y detiene la respiracin en cualquier fase para que tenga lugar el movimiento de deglucin. 3) ETAPA ESOFGICA La funcin principal del esfago es conducir los alimentos de faringe a estmago, esto lo realiza mediante dos tipos de movimientos peristlticos; PRIMARIOS Y SECUNDARIOS. El primario no es ms que la continuacin de la onda peristltica que empieza en faringe y se extiende hasta esfago en la etapa farngea. Esta onda tarda unos 5 - 10 segundos en llevar los alimentos de faringe a estmago. Pero por gravedad pueden incluso llegar antes que la onda peristltica, slo en 4 - 8 segundos. Si la onda primaria no alcanza a transportar la totalidad de los alimentos, la distensin del esfago origina una segunda onda (secundaria), que es similar a la primaria, radicando la diferencia en que su lugar de origen es la porcin distendida en lugar de ser la faringe. El contenido esofgico entra al estmago por el cardias (esfnter funcional) cuyo papel principal es evitar el reflujo de contenido gstrico. La deglucin se lleva a cabo gracias a un mecanismo activo, peristltico, lo que permite que esta se lleve a cabo incluso contra gravedad. Los lquidos pueden llegar al estmago incluso antes que la onda peristltica, algunos slidos necesitan ms que una onda peristltica para llegar a estmago. La deglucin se ve facilitada gracias a la secrecin mucosa. TIPOS DE MOVIMIENTOS ESFAGO Presenta movimientos peristlticos (25 cm/seg), que aparecen en la deglucin. Se ha comprobado la existencia de los movimientos antiperistlticos que progresan a una velocidad de 50 -100 cm/seg y aparecen durante la regurgitacin del contenido rumino reticular. El esfago es el nico segmento del tubo digestivo que muestra gran velocidad de contraccin muscular, mostrando caractersticas de musculatura estriada. En su estructura se encuentran fibras estriadas y lisas.
RUMEN. Existen movimientos con una frecuencia de 1,5 contracciones por minuto que ocurren durante las 24 horas del da, con variaciones discretas durante la noche y durante la ingestin de alimento. El ciclo motor presenta ondas peristlticas que se inician en el retculo y siguen sucesivamente al saco dorsal, ventral y de este nuevamente al saco dorsal del rumen, terminado frecuentemente en un eructo. Otro tipo de movimientos son los originados por las contracciones y relajaciones del pilar anterior del rumen, que al contraerse determina un aumento de la presin del saco ventral del rumen hacia el retculo. Los movimientos que permiten la mezcla del contenido rmino - reticular se han llamado primarios y que aquellos que van asociados al eructo. RESTO DEL TUBO DIGESTIVO Al igual que en el monogstrico se observan movimientos peristlticos, de segmentacin rtmica (que es de una alternancia de contracciones y relajaciones de pequeos segmentos de intestino), y de variaciones de tono de grandes zonas, lo que significa aumento del grado de contraccin bsica de la musculatura intestinal. CONTROL DE LA MOTILIDAD Tanto el rumen-retculo como el omaso, abomaso y el tubo intestinal de los rumiantes no difiere cualitativamente de los monogstricos. El sistema parasimptico estimula y el simptico inhibe. El control humoral es incierto; metabolitos como la glucosa inhiben "in vivo" (por su influencia sobre los centros superiores) y estimulan a trozos aislados de tejidos. Los cidos grasos inyectados "in vivo" inhiben la motilidad. PROGRESIN DE LOS ALIMENTOS EN EL TUBO DIGESTIVO DE LOS RUMIANTES Con una velocidad de pasaje mayor al apetito aumenta, en general, aceptando las variaciones individuales y aquellas derivadas del medio, incluyendo alimentacin, el 80% de los residuos integran las heces a las 100 horas y el 100% a las 160 horas. La velocidad de paso no es constante y depende de la digestibilidad del material consumido, adems de otros factores como el tamao de partcula y diferencias entre materiales consumidos y su propia digestibilidad. El 60% del tiempo permanece en el rumen, el 8% en el omaso, el 3% en el abomaso y el 29% en intestino. RUMIA La rumia empieza con una fuerte contraccin de la prensa abdominal, la abertura cardial se ensancha. La inspiracin provoca que la presin negativa intra-torxica se haga ms negativa y aumente el volumen torcico por la contraccin caudal del diafragma creando un mecanismo de succin y el contenido ruminal avanza en forma retrgrada. La rumia se inicia una hora a una hora y media luego de la ingesta de alimento. El material alimenticio poco denso o poco digerido y de mayor tamao se localiza a la altura del esfnter esofgico inferior, la rumia requiere de la coordinacin de todo el
tronco del encfalo para coordinar una serie de eventos que en conjunto logran el desarrollo de la rumia, estos incluyen la motilidad en el rumen, la apertura del esfnter esofgico inferior, onda peristltica en el rumen, masticacin, salivacin, deglucin e inhibicin de la respiracin. En las 24 horas hay 15 a 18 perodos de rumia empleando 7 a 9 horas cada da, en este perodo pasan 40 a 60 kg de alimento total (en peso hmedo) por el proceso de la rumia. El peso del bolo de rumia flucta de 80 a 120 grs y cada bolo demora 40 a 50 segundos en ser remasticado, luego de tragado y deglutido y el otro es eyectado 5 segundos despus. Si se observa por el lado izquierdo del cuello se puede ver la onda ascendente, luego hay una onda descendente que corresponde al agua deglutida que es sacada a presin contra el paladar, como dato al margen debemos decir que la rumia es el barmetro de salud de los rumiantes ya que cualquier alteracin digestiva altera la rumia. El movimiento de este material desde la porcin distal del esfago hasta la boca se realiza mediante una onda de naturaleza antiperistltica que lleva una velocidad de unos 107-112 cm/s este hecho se debe a que los msculos del esfago son estriados y por lo tanto capaces de contraccin rpida. Cualquier lesin en el tronco del encfalo puede provocar una alteracin de la rumia, no necesariamente deben presentarse alteraciones fsicas, si no tambin su inhibicin o alteracin en ritmo puede ocurrir por situaciones de estrs. El estmulo es mecnico para desencadenar la rumia es provocado por la presencia de partculas que requieren ser reducidas en tamao; las cuales se acumulan en la zona alrededor del esfago. Hay receptores nerviosos mecnicos para ese tipo de partculas (receptores de distensin) en la mucosa del retculo y rumen que rodea la zona del esfnter esofgico inferior. Aunque no se presente consumo de alimento existe en mayor o menor grado motilidad en el rumen. Durante la rumia ocurren 3 contracciones en el retculo, en lugar de 2, la contraccin extra del retculo indica que el animal est rumiando. Esa contraccin est coordinada con la apertura del esfnter esofgico inferior, antes de que comience el siguiente ciclo de rumia, el al alimento ha sido deglutido. Los ingredientes con mayor cantidad de fibra y mayor tamao de partcula son estimuladores de la rumia, BOLO DE RUMIA Se compone especialmente del contenido reticular y parte fluida del contenido del rumen (6% y 15% de peso seco respectivamente). Pesa aproximadamente 100 g y est compuesto por partculas que pueden ser regurgitadas y remasticadas dos veces o ms. Al llegar a la boca el bolo se concentra por ser exprimido contra el paladar, lo que facilita la masticacin; el agua resultante - que haba facilitado la regurgitacin a travs del esfago es deglutida inmediatamente. El bolo remasticado cae el rumen nuevamente (no sigue la gotera esofgica, que lo llevara al omaso directamente).
REGURGITACIN DEL BOLO Para que el bolo regrese a la boca desde rumen - retculo y sea remasticado, se requiere gradiente de presin intraluminal a favor del esfago. En la prctica se establecen 8,0 cm de agua, gracias a una contraccin del retculo y a una inspiracin simultnea que aumenta la presin negativa intra-torxica la cual facilita a su vez, la distensin del esfago torcico, que en rumiantes no muestra esfnter esofgico inferior. El cardias se dilata inmediatamente despus de la contraccin reticular y, al entrar el contenido del rumen - retculo al esfago, se inicia en forma refleja una rpida onda antiperistltica (50 100 cm/s) que lleva el alimento hacia la boca. La rumiacin se inicia antes de las dos semanas de edad, se mantiene durante toda la vida, y demanda 8 horas diarias. LA GOTERA ESOFGICA Funcionalmente constituye un medio de comunicacin directa entre el esfago y el omasum, evitando el rumen y retculo. En el adulto no conduce el bolo re-masticado. En el ternero cumple la funcin de llevar la leche al abomaso (que tiene las enzimas digestivas adecuadas, sin pasar por el rumen retculo), esto es importante ya que el aparato digestivo es estril en los animales recin nacidos, siendo los residuos del calostro y de la leche coagulada el sustrato que permite el establecimiento de los primeros microorganismos del pre-rumiante. Pequeas cantidades de leche que llega a estar en contacto con el rumen lleva lactobacilos, estreptococos y algunas formas de coniformes, ya esas formas microbianas son resistentes al pH cido que prevalece en los rudimentos del retculo rumen y abomaso. ERUCTO La fermentacin rumino-reticular, origina entre 1 a 2 litros de gas por minuto, especialmente CO2, derivado de la digestin de los carbohidratos y del bicarbonato de la saliva, y de otro gas CH4 (metano), resultante del metabolismo bacteriano reductivo. De los gases eructados: 66% es CO2, 26% es CH4 metano, 6% es N2 y 0,1% es H2S. Se eructan 600 litros de gas en 24 horas, pero esto ltimo est en directa relacin con el tipo de alimento ingerido. DESTINO DE LOS GASES Una parte progresa caudalmente conjuntamente con los alimentos, otra difunde hacia la sangre en forma proporcional a su presin parcial, a travs de la pared del rumen y una tercera fraccin, la ms importante, regresa por el esfago, siendo una parte eructada y entrando otra - bastante considerable - a los alvolos junto con la inspiracin. Despus de 5 segundos de eructados, los gases radioactivos marcados con C14 e inyectados en el rumen aparecen en la sangre. El CO2 a los 5 minutos aparece en la saliva (lo que indica la fuente de bicarbonato) y 3 minutos mas tarde su C se incorpora a
la lactosa constituyendo parte de su molcula. El C14 tambin es rpido para aparecer en la sangre y en la leche, pero en esta ltima se incorpora disuelto y en muy pequeas cantidades. CONTROL DE LA ERUCTACIN An no es del todo conocido. Coincide la mayora de las veces con concentraciones especficas del saco dorsal del rumen, llamadas "secundarias". Existen, aparentemente, receptores de estiramiento en el cardias. SECRECIN SALIVAL Sus funciones son: colaborar en la masticacin y deglucin, actividad enzimtica, nutrientes para microorganismos, propiedades antiespumantes y poder tampn Na+ y K+. Este mecanismo fisiolgico contribuye de manera fundamental al mantenimiento del pH ruminal, debido a la cantidad de bicarbonato que contiene la saliva, acta ajustando el pH, especialmente despus de la ingestin de alimento, que es cuando tiende a descender, a veces hasta niveles cercanos al lmite fisiolgico. Las fermentaciones que ocurren en el rumen tienden a bajar su pH. El bovino produce 50 kg de saliva en 24 horas en forma de flujo constante, y no ondulatorio como en los monogstricos que tienen fases ascendentes durante los perodos de ingestin de alimentos. Esto significa un aporte de 300 g de NaHCO3 en 24 horas, con el consiguiente efecto buffer en el lquido ruminal. En la saliva se encuentra tambin urea (6-27 mg%) en concentracin similar a la de la sangre (las bacterias pueden sintetizar protena a partir del N de la urea). Esta urea proviene del hgado y deriva del catabolismo proteico y de la dieta o de la misma saliva que ha recirculado. El exceso de urea es eliminado por los riones. IMPORTANCIA DE LA FLORA DEL RUMEN El pasto contiene aproximadamente un 20% de celulosa, que un monogstrico no puede digerir. El rumiante lo hace con un coeficiente de 50 a 90%. Las bacterias metabolizan la celulosa con produccin de cidos grasos voltiles (300 g al da). Los polisacridos intrabacterianos, al morir son, en parte, digeridos en el rumen y el resto, en el intestino. Hay dudas sobre la degradacin enzimtica inicial de la celulosa, ya que sta es muy difcil de solubilizar, por lo cual se presume que las primeras reacciones deben ser realizadas extracelularmente (fuera de los cuerpos bacterianos). Sin embargo han fracasado los intentos de encontrar, en lquidos ruminales libres de clulas, alguna muestra de celulasa extracelular. Es posible que sea muy inestable o se encuentre muy asociada a las membranas de los microorganismos. DEGRADACIN DEL ALMIDN Este es un problema ms simple, ya que hay bacterias que secretan alfa amilasa al
lquido ruminal. PROTENAS Y RUMEN Las bacterias son capaces de sintetizar protenas a partir de N no proteico, como sales de amonio o urea. Tambin pueden sintetizar protenas de mayor valor nutritivo a partir de aquellas de poco valor nutritivo. Los microorganismos desaminan aminocidos con produccin de amonio libre, cidos grasos voltiles, hidroxicidos y cetocidos. ABSORCIN EN RUMEN El grosor del epitelio o distancia que hay entre el lumen del rumen y los capilares sanguneos es de 0,05 mm con superficie queratinizada; hay espacios intracelulares de 100 nm, que estn llenos de tejido conjuntivo. No se encuentran glndulas secretoras (no se secretan enzimas digestivas). La gradiente elctrica de la pared indica que la sangre es positiva con respecto al lumen del retculo rumen 30 mV aproximadamente. Ca, Mg, PO4 se mueven muy lentamente en ambos sentidos a travs de la pared del rumen. El transporte neto es, por esto, despreciable y se dice que la pared es impermeable a ellos, an cuando se coloquen concentraciones 6 veces superiores a las normales. Su absorcin se hace en la zona intestinal. K+ Na+ Cles un ion di fusible en ambos sentidos. se transporta activamente hacia la sangre, o sea, se realiza contragrandiente. se absorbe tambin contragradiente y pasa al lquido ruminoreticular cuando en ste no existe. Cuando existe una concentracin en el lquido ruminal, de 30 milimoles/litro, lo que equivale a 2/3 de la concentracin sangunea, no hay transporte neto en ningn sentido. Se desplaza en base a gradiente elctrica y concentracin.
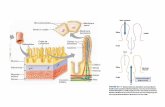
ABSORCIN DE CIDOS GRASOS Pasan a la sangre por difusin, habitualmente lo hacen desde el actico (C2) hasta el valerinico (C5). Los cidos grasos del retculo rumen se encuentran disociados o no, de acuerdo al pH del lquido ruminal; a pH neutro el 50% del acetato se absorbe como anin. El porcentaje de cidos no disociados aumenta con el medio cido, incrementndose la absorcin y transporte. TRANSPORTE DE LOS ALIMENTOS MOTILIDAD GASTROINTESTINAL En un corte tpico del tubo digestivo, de fuera hacia adentro, se aprecian las siguientes capas: 1) serosa, 2) capa muscular longitudinal, 3) capa muscular circular, 4) submucosa y 5) mucosa. Adems, se encuentra en la parte profunda de la mucosa una capa de
fibras musculares lisas, la Muscularis mucosae. Las funciones motoras del intestino las cumplen las distintas capas de msculo liso. Se observa una capa de musculatura lisa circular interna, una longitudinal externa y en determinados segmentos un engrosamiento de la circular interna formndose los esfnteres anatmicos. Las caractersticas especficas del msculo liso intestinal son las siguientes: ACTIVIDAD ELCTRICA DEL MSCULO LISO GASTROINTESTINAL El msculo liso del tubo digestivo muestra actividad elctrica en forma casi continua. Esta actividad es muy particular, pues tiende a presentar dos tipos bsicos de ondas, lentas y espigas, las ondas lentas se presentan con una frecuencia de tres a doce por minuto, y presentan oscilacin bsica continua, que tiene lugar en las membranas del msculo liso. Las ondas lentas no son potenciales de accin de tipo todo o nada; pueden presentar una intensidad de grado variable. Cuando el msculo es estimulado por distensin, mediante acetilcolina o por excitacin parasimptica, el potencial de membrana de la fibra en reposo se vuelve ms positivo, lo cual eleva todo el potencial de las ondas lentas desde su valor medio normal de -50 a -40 mV hasta valores menos negativos por un proceso de despolarizacin. Cuando la despolarizacin llega a menos de -40 mV empiezan a producirse potenciales de espiga en los mximos de las ondas lentas; la frecuencia de las espigas aumenta progresivamente a medida que el potencial va creciendo. Sin embargo, con una estimulacin muy fuerte, cuando el potencial de membrana en reposo sube hasta un valor de -15 a -20 mV, las espigas desaparecen, y la membrana queda despolarizada constantemente. La estimulacin del msculo liso por adrenalina o por excitacin simptica disminuir el potencial de membrana en reposo hasta un valor "hiperpolarizado" cercano a -70 mV, y la actividad elctrica, como la mecnica, del msculo liso prcticamente pasan a ser nulas. INICIO DE LA CONTRACCIN MUSCULAR La mayor parte de las contracciones del intestino ocurren en respuesta a los potenciales en espiga, de hecho, no suele haber contraccin en respuesta a ondas lentas cuando no tienen agujas superpuestas. En consecuencia, las espigas son comparables a los potenciales de accin en el msculo esqueltico y causan los cambios de membrana que inician la contraccin. Una contraccin tnica es una contraccin continua que dura minutos u horas; su intensidad puede aumentar o disminuir a veces, pero no se suspende la contraccin. Se cree que depende de una serie de potenciales de espiga cuya frecuencia establece el grado de contraccin tnica de los esfnteres y establece la cantidad de resistencia ofrecida en los esfnteres pilrico, ileocecal y anal interno, ayudando a regular el desplazamiento del contenido por el intestino.
En diversas partes del intestino, las contracciones rtmicas del msculo liso gastrointestinal se presentan con un ritmo tan rpido como 12 veces por minuto, o tan lento como 3 veces por minuto. Estas tambin son las frecuencias de las ondas lentas respectivas en estos segmentos; son estas ondas lentas las que establecen la frecuencia. Estas contracciones son el origen de las funciones fsicas del tubo digestivo, como la mezcla de alimentos y la propulsin peristltica de los mismos. INNERVACIN DEL INTESTINO - SISTEMA NERVIOSO INTRNSECO El sistema nervioso intrnseco est compuesto principalmente por las redes neurales: la ms externa, denominada plexo mesentrico o de Aerbach, se encuentra entre las capas musculares longitudinal y circular; la ms interna, llamada plexo submucoso o de Meissner, se encuentra en la submucosa. El plexo mesentrico controla, principalmente, los movimientos gastrointestinales en tanto que el submucoso es importante en la regulacin de la secrecin y tambin de las funciones sensoriales, recibiendo seales principalmente del epitelio intestinal y de los receptores de estiramiento de la pared intestinal. La estimulacin del plexo mesentrico estimula la actividad del intestino, causando cuatro efectos principales: 1. 2. 3. 4. Aumento de la contraccin tnica de la pared intestinal. Mayor intensidad de las contracciones rtmicas. Incremento de las contracciones rtmicas. Aumento de la velocidad de conduccin de las ondas excitadoras a lo largo de la pared intestinal.
CONTROL NEUROVEGETATIVO DEL TUBO DIGESTIVO INNERVACIN PARASIMPTICA La inervacin parasimptica se divide en aquella provista por el parasimptico craneal y por el parasimptico sacro. Con excepcin de algunas fibras que se dirigen a boca y faringe, casi todo el parasimptico craneal est incluido en los nervios vagos. El parasimptico sacro proviene de los segmentos sacro segundo, tercero y cuarto de la mdula espinal y se dirigen a la segunda mitad del intestino grueso por los nervios erectores. Las neuronas postganglionares del sistema parasimptico se encuentran en el plexo mientrico; la estimulacin de los nervios parasimpticos provoca aumento general de actividad de este plexo, que a su vez excita las paredes intestinales, y facilita la mayor parte de los reflejos intrnsecos del tubo digestivo. INNERVACIN SIMPTICA La inervacin simptica del tubo digestivo se origina en la mdula espinal entre D-8 y L-3.
En general la estimulacin del sistema nervioso simptico inhibe la actividad motora y sobre la actividad secretora tiene un efecto doble a nivel gastrointestinal, actuando de dos formas diferentes: 1. 2. por efecto directo de la noradrenalina en el msculo liso y, por accin inhibidora de la noradrenalina en las neuronas del sistema nervioso intrnseco.
La actividad motora y secretora del sistema digestivo est regulada principalmente por el sistema nervioso parasimptico que tiene un predominio 10:1 sobre la inervacin simptica. CONTROL HORMONAL DE LA MOTILIDAD GASTROINTESTINAL Se comentar ms adelante pero algunas de las hormonas ms importantes son, gastrina, colecistoquinina, secretina y pptido inhibidor gstrico.
MOVIMIENTO DEL TUBO DIGESTIVO Las dos variedades principales de movimiento son: a) Movimientos de mezcla b) Movimientos propulsores MOVIMIENTOS DE MEZCLA Dependen de contracciones rtmicas localizadas de secciones pequeas de la pared intestinal. MOVIMIENTOS PROPULSORES El principal movimiento propulsor es el peristaltismo. El peristaltismo es una propiedad inherente a todo conducto cuya pared sea un sinsitio de fibras lisas, un estmulo en cualquier sitio provoca la aparicin de un anillo de contraccin. Vemos que ocurre peristaltismo en: a) Tubo digestivo, b) conductos biliares, c) urteres d) la mayor parte de conductos formados por fibras lisas. El estmulo ms comn para la activacin del peritaltismo es la distensin, ste depende del control de nervios parasimpticos los cuales adems modifican su intensidad y velocidad. Cabe destacar que este efecto es susceptible de ser deprimido o bloqueado
totalmente con atropina la cual bloquea el plexo nervioso intramural. El porque del hecho que las ondas peristlticas se propagan hacia el ano y no hacia la boca (lo cual ocurre pero en muy pequeo grado) no est bien esclarecido an. Se cree s que esta caracterstica unidireccional se debe a una disposicin especial del plexo nervioso intramural (principalmente la porcin mesentrica) por lo cual los estmulos se transmiten con mayor facilidad en direccin caudal. MOTILIDAD GSTRICA El estmago se puede dividir en: cuerpo y antro. El fondo del estmago, en la parte alta del cuerpo, es considerado anatmicamente distinto del cuerpo; pero desde el punto de vista de su funcionamiento fisiolgico se pueden asociar. El estmago tiene tres funciones motoras distintas: a) Almacenar grandes cantidades de contenido, hasta que sea posible enviarlos a porciones inferiores; b) Mezclar los alimentos con la secrecin gstrica hasta obtener una sustancia semilquida denominada quimo; c) Vaciar progresivamente el contenido a las porciones siguientes con una velocidad tal que resulte compatible con la digestin y absorcin por el intestino delgado. 1) ALMACENAR. La pared muscular lisa del estmago es elstica, o sea que sus fibras pueden ser alargadas sin que aumente su tono; adems, mientras ms se dilata el estmago, mayor se vuelve el radio de curvatura de sus paredes. Para una cierta presin, la fuerza de distensin sobre las paredes aumenta en proporcin directa a este radio de curvatura (efecto que se conoce como ley de Laplace) por lo tanto, la presin en el estmago, aumenta poco a pesar que exista una distensin importante. 2) MEZCLAR. Las secreciones digestivas del estmago son producidas por glndulas gstricas que cubren casi toda la pared del cuerpo y del fondo. Estas secreciones, de las cuales ms adelante hablaremos en detalle, contactan con los alimentos que se encuentran cerca de la pared y a lo largo de dicha pared de producen, cada 20 segundos, ondas dbiles llamadas ondas mezcladoras u ondas tnicas, que se originan en cualquier sitio y se extienden por distancias variables hacia el ploro. Y en general son ms intensas cerca del antro lugar en que tambin se producen ondas peristlticas. Cada vez que una onda peristltica pasa por encima del antro hacia el ploro, penetra profundamente en el contenido de dicho antro. Pero la abertura del ploro es tan pequea que slo son vaciados unos pocos milmetros de contenido y as la gran masa es desplazada en direccin retrograda. Una vez que el alimento se ha mezclado con las secreciones gstricas, la mezcla que pasa al intestino se denomina quimo. Adems de las ondas tnicas y peristlticas de estmago, existe un tercer tipo, las contracciones de hambre. Normalmente son
contracciones peristlticas rtmicas, probablemente ondas de mezcla ms intensas en el cuerpo del estmago. Cuando adquieren intensidad extrema suelen fundirse para formar una contraccin tetnica continua que dura dos a tres minutos. Cuando ocurren contracciones de hambre, el sujeto percibe una sensacin dolorosa epigstrica que no suele ocurrir antes que hayan transcurrido 12 a 24 horas de la ltima ingestin. Las contracciones del estmago se dirigen siempre hacia el antro pilrico, estas son muy potentes y se visualizan en una radiografa como escotaduras llamadas lustraciones, existe un marcapaso gstrico que organiza estas contracciones. REGULACIN REFLEJA DE LAS CONTRACCIONES GSTRICAS La irritacin de la mucosa gstrica produce aumento de las contracciones gstricas del estmago. As pues, la acumulacin de secreciones gstricas que contienen grandes cantidades de cido clorhdrico aumenta la intensidad de las contracciones del estmago. En consecuencia, cuando el alimento queda saturado en exceso con secreciones gstricas, automticamente es propulsado hacia el duodeno. 3) VACIAR. El ritmo del vaciamiento gstrico est regulado por seales que provienen del estmago y el duodeno. Las primeras son de dos tipos principales: a) seales nerviosas y b) la hormona gastrina, liberada de la mucosa antral en respuesta a la presencia de ciertos tipos de alimento en el estmago. Las dos seales aumentan la fuerza de bombeo pilrico e inhiben el ploro, promoviendo as el vaciamiento gstrico. Por otra parte, las seales que provienen del duodeno deprimen la bomba pilrica y suelen aumentar el tono del ploro. En general, cuando un volumen excesivo de quimo o un exceso de ciertos tipos de quimo entran al duodeno, seales de retroalimentacin negativa enrgica, nerviosa y hormonal, deprimen la bomba pilrica y estimulan el tono del esfnter del ploro. Obviamente estas seales de retroalimentacin permiten que el quimo pase al duodeno slo con la rapidez que puede ser procesado por el intestino delgado. VOLUMEN DEL CONTENIDO GSTRICO Y EL RITMO DE VACIAMIENTO Es muy fcil ver como el aumento de volumen del alimento en el estmago puede promover un vaciamiento ms rpido del mismo. Sin embargo, esto no ocurre por los motivos que cabra esperar. No es el aumento de presin en el estmago el que provoca aumento de vaciamiento, porque en los lmites normales de volumen, este aumento no incrementa netamente la presin, segn ya estudiamos antes. Por otra parte, la distensin de la pared gstrica inicia reflejos vagales y mesentricos locales en la pared, que aumentan la actividad de la bomba pilrica y al mismo tiempo inhiben el ploro. En general el ritmo de vaciamiento de los alimentos del estmago es aproximadamente proporcional a la raz cuadrada del volumen de alimento que persiste en el estmago en todo momento.
EFECTO DE LA HORMONA GASTRINA SOBRE EL VACIAMIENTO Ms adelante veremos que la distensin, as como la presencia de algunos tipos de alimentos en el estmago, en particular las protenas, provoca la liberacin de gastrina desde la mucosa del antro. Esta ejerce un potente efecto causando secrecin del jugo gstrico cido desde las glndulas del fondo del estmago. La gastrina tambin tiene efectos estimulantes sobre las funciones motoras del estmago. Lo ms importante es que aumenta la actividad de la bomba pilrica, al mismo tiempo que relaja el ploro. Es por tanto una influencia importante en el vaciamiento de este compartimiento. Tambin tiene efecto constrictor sobre el esfnter gastro esofgico para evitar el reflujo del contenido gstrico cuando est aumentada la actividad del estmago. INHIBIDOR DEL REFLEJO ENTERO GSTRICO DEL DUODENO Y LA ACTIVIDAD PILRICA Existen varios factores que desencadenan el reflejo entero gstrico y algunos de estos factores son: 1) El grado de distensin del duodeno, 2) Irritacin duodenal, 3) Grado de acidez del quimo duodenal 4) Grado de osmolaridad del quimo, 5) Productos de degradacin proteica y de grasas. Cada vez que el pH del quimo en el duodeno cae por debajo de 3,5 a 4,0; aparece de inmediato el reflejo enterogstrico, inhibe la bomba pilrica y disminuye la liberacin de contenido gstrico cido hacia el duodeno, hasta que dicho quimo pueda ser neutralizado por las secreciones intestinales y pancreato - biliares. Los lquidos hipo o hipertnicos (sobre todo los hipertnicos), desencadenan el reflejo enterogstrico. Este efecto impide el flujo demasiado rpido de lquidos no isotnicos hacia el intestino delgado, evitando as cambios rpidos del balance de electrlitos en los lquidos corporales durante la absorcin del contenido intestinal. MOTILIDAD INTESTINAL CONTRACCIONES DE MEZCLADO A lo largo de todo el intestino delgado ocurren casi constantemente pequeas contracciones anulares. A menudo son irregulares pero las hay rtmicas, con frecuencia de unas 8 - 9 por minuto en duodeno y menor conforme se progresa distalmente. Esto tiene por consecuencia "segmentacin" del intestino delgado, en un primer tiempo, el intestino queda dividido en segmentos espaciados con regularidad, en un segundo tiempo, nuevas contracciones ocurren en distintos puntos. As estas segmentaciones "cortan" el quimo muchas veces por minuto y aseguran una buena mezcla del contenido con las secreciones del intestino delgado. Estas contracciones segmentaras slo dependen en parte del plexo nervioso intramural, pues siguen ocurriendo contracciones dbiles an al bloquear el plexo con atropina, y su intensidad aumenta por estmulo parasimptico.
Existen adems los movimientos pendulares que consisten en pequeas ondas de constriccin que se desplazan hacia adelante hacia atrs, arriba y abajo varios centmetros en el intestino. MOVIMIENTOS DE PROPULSIN (Ondas Peristlticas) Se producen en cualquier parte del intestino delgado y siempre se desplazan en direccin caudal, con una velocidad de 0,5 a 5,0 cm por segundo, pero desaparecen despus de viajar unos pocos centmetros. En consecuencia, el desplazamiento del quimo a lo largo del intestino delgado es lento, en promedio de slo 1 cm por minuto esto significa que normalmente se necesita de 3 - 10 horas para que el quimo pase desde el ploro hasta la vlvula ileocecal. REGULACIN DE LA RAPIDEZ DE DESPLAZAMIENTO DEL QUIMO POR EL INTESTINO DELGADO Depende principalmente de la velocidad de vaciamiento del estmago hacia el duodeno. Cuando el duodeno es distendido, la intensidad de las ondas peristlticas en el propio duodeno aumenta y el contenido es impulsado rpidamente a lo largo del intestino delgado para disminuir la distensin duodenal. Al desaparecer la distensin disminuye la intensidad del peristaltismo. Al alcanzar la vlvula ileocecal, el quimo queda bloqueado, a veces por varias horas hasta que el animal ingiere otra comida. Entonces el reflejo gastroentrico intensifica el peristaltismo en el leon y manda el resto del quimo a travs de la vlvula ileocecal al ciego. FUNCIN DE LA VLVULA ILEOCECAL Su funcin principal es evitar el flujo retrgrado de materias fecales de colon hacia intestino delgado, las "valvas" de dicha vlvula se proyectan en la luz del ciego, lo cual es una excelente disposicin para esta funcin. Generalmente la vlvula puede contrarrestar presiones retrgradas que alcanzan 50 a 60 cm. de agua lo que es igual a 4,4 mm de Hg. En los ltimos centmetros del leon, que preceden a la vlvula, la pared del intestino posee una capa muscular engrosada denominada esfnter ileocecal. En condiciones normales se encuentra medianamente contrado y no permite que el leon se vace en ciego, excepto inmediatamente despus de una comida, cuando el reflejo gastroileal intensifica el peristaltismo del leon. Entonces cada onda peristltica lleva al ciego unos milmetros de quimo, o sea que el ciego recibe unos 750 ml diarios de quimo. Esta resistencia al paso del quimo por la vlvula ileocecal prolonga la permanencia del quimo en intestino y facilita la absorcin. CONTROL DEL ESFNTER ILEOCECAL
El grado de control del esfnter ileocecal depende bsicamente de reflejos provenientes del ciego. Siempre que el ciego est distendido, el grado de contraccin del esfnter ileocecal se intensifica, lo cual retrasa netamente el vaciamiento de quimo adicional proveniente del leon. Cualquier irritante en el ciego provoca constriccin del esfnter ileocecal. Los reflejos del ciego al esfnter ileocecal ocurren por va del plexo mesentrico. Adems, hay que tener en cuenta que diversos reflejos vicerosimpticos iniciados por irradiacin de otras partes del tubo digestivo de los riones, o del peritoneo, tambin pueden causar contraccin intensa del esfnter, y en consecuencia, retrasar e incluso interrumpir completamente el desplazamiento del contenido intestinal a travs de la vlvula ileocecal. MOTILIDAD DE COLON Las funciones del colon son: a) Absorcin de agua y electrlitos del quimo y, b) Almacenamiento de materias fecales hasta el momento de su expulsin. La primera mitad del colon interviene principalmente en la absorcin, y la segunda mitad en el almacenamiento. Puesto que ninguna de las dos funciones requiere movimientos intensos las contracciones del colon suelen ser leves. Sin embargo, dentro de esta lentitud, los movimientos conservan las caractersticas de aquellos observados en el intestino delgado y pueden dividirse, una vez ms, en mezcladores y propulsores. En cuanto a los movimientos de propulsin, el colon no presenta ondas peristlticas parecidas a las que existen en intestino delgado, es otro tipo de movimientos que suelen ocurrir unas cuantas veces al da solamente y son ms abundantes durante unos 10 minutos en la primera hora despus del desayuno. DEFECACIN La salida continua de materia fecal a travs del ano es evitada por constriccin tnica: a) del esfnter anal interno, masa circular de msculo liso situada inmediatamente por dentro del ano (inervacin parasimptica regin sacral) y b) esfnter anal externo, compuesto de msculos estriados, voluntario y controlado por el sistema nervioso somtico. REFLEJO DE DEFECACIN Cuando las heces fecales penetran en el recto, la distensin de la pared rectal estimula los receptores de distensin e inicia las seales aferentes que pasan a travs del plexo mesentrico para iniciar ondas peristlticas reflejas en colon descendente y sigmoides, que impulsan el material fecal hacia el ano. Cuando la onda peristltica se acerca al ano el esfnter anal interno es inhibido, por el fenmeno de la "relajacin receptiva", si el esfnter anal externo se relaja, se produce la defecacin. Tiene particular importancia el hecho de poder iniciar los movimientos correspondientes tomando una inspiracin profunda para mover el diafragma hacia abajo, luego contrayendo los msculos abdominales para aumentar la presin en el vientre. Por desgracia los reflejos iniciados
en esta forma nunca son tan eficaces como los producidos naturalmente, por cuyo motivo las personas o animales que inhiben con demasiada frecuencia sus reflejos naturales acaban sufriendo estreimiento intenso. II. FUNCIN SECRETORA DEL TUBO DIGESTIVO A lo largo del tubo digestivo se encuentran glndulas que desempean dos funciones principales: en primer lugar, secretan enzimas digestivas desde la boca hasta el final del leon. Luego las glndulas mucosas, presentes desde la boca hasta el ano, producen mucus que lubrica y protege toda la superficie del tubo digestivo. La mayor parte de estas secreciones se producen slo en respuesta a la presencia de alimentos en el tracto gastrointestinal, y la cantidad secretada en cada compartimiento es casi exactamente la que se necesita para una adecuada digestin. Adems, algunas glndulas pueden modificar las caractersticas (enzimticas y otras), de su produccin en funcin del tipo de substrato presente. Por tanto, se describirn en este captulo las distintas secreciones digestivas, su papel y el control de su produccin. PRINCIPIOS GENERALES GLNDULAS DIGESTIVAS ESTMULOS LOCALES Se refiere a que la presencia de alimento en una regin del tubo digestivo suele provocar secrecin por parte de las glndulas de ese sitio o adyacentes. La mayor parte de la estimulacin local de las glndulas intestinales se deben a alguno de los tres fenmenos siguientes: 1) Estmulos tctiles o qumicos de mucosa pueden provocar reflejos que, a travs del plexo nervioso intramural, actan sobre las clulas mucosas superficiales o sobre las glndulas profundas del epitelio, 2) Los mismos reflejos pueden ser desencadenados por distensin de la pared, 3) Los estmulos tctiles o la distensin pueden aumentar la motilidad intestinal, como ya se vio en el captulo anterior, y esta hipermotilidad puede aumentar el ritmo de secrecin. ESTMULOS NERVIOSOS PARA LA SECRECIN ACTIVIDAD PARASIMPTICA La inervacin parasimptica aumenta la secrecin glandular en el tubo digestivo, en especial en las porciones altas: glndulas salivales, esofgicas, gstricas, pancreticas e incluso en las glndulas de Brnner del duodeno. En el resto del tubo digestivo, lo comn es que la secrecin responda al estmulo local, sin embargo tambin existe un ligero efecto sobre las glndulas del intestino delgado y glndulas mucosas del colon. En muchos segmentos provoca ligero aumento de la secrecin glandular. Por otra parte tambin provoca contricin de los vasos que llegan a las glndulas. Por ello, el simptico DE LA SECRECIN ESTIMULACIN DE LAS
puede tener efecto doble; si se produce aisladamente aumenta ligeramente la secrecin, pero si ya ocurre secrecin copiosa por estimulacin parasimptica, un estmulo simptico sobrepuesto disminuir la secrecin por contricin de vasos sanguneos. CONTROL HORMONAL DE LA SECRECIN DIGESTIVA Qumicamente las hormonas gastrointestinales suelen pertenecer al grupo de las protenas, polipptidos y derivados, siendo las de mayor importancia, gastrina, colecistoquinina, secretina y pptido inhibidor gstrico. PROPIEDADES LUBRICANTES Y PROTECTORAS DEL MUCUS: SU IMPORTANCIA EN EL TUBO DIGESTIVO El mucus es una secrecin espesa compuesta de agua, electrlitos y una mezcla de polisacridos, con ligeras diferencias segn su sitio de origen: pero en todas partes posee caractersticas importantes que le permiten lubricar y proteger perfectamente las paredes gastrointestinales. Es muy adherente, y forma una pelcula delgada sobre la superficie de los alimentos o partculas que estn en contacto con l, su consistencia es tal que recubre toda la pared protegindola contra alimentos mal masticados, su resistencia al desgarro permite que las partculas se deslicen fcilmente a lo largo del epitelio. El mucus causa adherencia de las partculas, unas con otras, para formar los bolos fecales, que se expulsan durante una evacuacin, resiste perfectamente la accin de enzimas digestivas, el mucopolisacrido que contiene es anftero (como cualquier otra protena), y puede amortiguar pequeas cantidades de cido o lcali. Estas diferentes propiedades explican que el mucus permita paso fcil de los alimentos y evite excoriaciones o lesiones qumicas del epitelio. El poder lubricante del mucus resulta evidente cuando se suspende la secrecin de saliva; en estas condiciones resulta muy difcil deglutir alimentos slidos, incluso con ayuda de grandes cantidades de agua. Tambin resulta evidente la importancia del mucus en la proteccin del estmago contra el cido clorhdrico. Cabe hacer notar que la proteccin de la pared gstrica contra la accin del cido clorhdrico se ve reforzada por la gran capacidad de reparacin de las clulas gstricas; las clulas maduras de la mucosa van siendo continuamente reemplazadas a un ritmo de reparacin, en el estmago humano, que alcanza a 500.000 clulas por minuto lo que en ltima instancia significa que la cubierta mucosa completa se regenera en 3 das. SECRECIN DE SALIVA Las principales glndulas salivales son las partidas, submaxilares y sublinguales. En el humano la produccin diaria de saliva oscila entre un litro y un litro y medio. La saliva consta de dos tipos de secrecin: a) Una fraccin serosa que contiene enzimas (de las cuales la de mayor importancia es la ptialina en hombre y cerdo de poco valor en
otras especies y contribuye a la digestin de almidones. En los rumiantes lactantes existe una enzima de particular importancia para la digestin de las grasas de la leche denominada lipasa bucal o estereasa pregstrica) y b) Una fraccin mucosa para la lubricacin. Las PAROTIDAS secretan fraccin serosa. Las SUBMAXILARES secretan fraccin serosa y fraccin mucosa y las SUNLINGUALES secretan fraccin mucosa. El pH de la saliva en la mayora de las especies es de 6,0 - 7,0 para accin ptima de ptialina, mientras que en los rumiantes es francamente alcalino, cercano a 8,0. CONTROL NERVIOSO DE LA SECRECIN SALIVAL Las glndulas salivales estn controladas por seales nerviosas parasimpticas de los ncleos salivales. Estos ncleos se encuentran aproximadamente en el lmite del bulbo y protuberancia, y son activados por estmulos spidos o tctiles en lengua u otras zonas de la boca, los sabores agradables tienen por resultado gran produccin de saliva. Tambin pueden aumentar o disminuir la cantidad de saliva impulsos que lleguen a los ncleos salivales desde centros superiores. As vemos que frente a un platillo apetitoso se saliva mucho. La zona del epitelio que controla estas diferencias se encuentra en el cerebro, cerca de los centros parasimpticos del hipotlamo anterior, y funciona sobre todo en respuesta a seales procedentes de las reas corticales del gusto y olfato, o de las amgdalas. Finalmente, hay salivacin por efecto de reflejos que nacen en estmago e intestino alto, en particular cuando se han deglutido alimentos muy irritantes o cuando hay nuseas, con ello al deglutir la saliva as producida se reduce la irritacin por dilucin o neutralizacin del irritante. SECRECIN ESOFGICA Esta secrecin, en la mayora de las especies, es de carcter totalmente mucoide y funciona principalmente para lubricar y favorecer la deglucin. En algunas aves se produce una secrecin denominada leche del buche la que sirve para alimentar a los polluelos. El esfago posee, en mayor proporcin, glndulas mucosas sencillas; los extremos, el inferior sobre todo, presenta glndulas compuestas. El moco formado por las glndulas compuestas de la extremidad superior impide que los alimentos lesionen la mucosa al penetrar al esfago, mientras que las glndulas compuestas cerca del cardias protegen el esfago contra la accin digestiva del jugo gstrico que a veces refluye a travs del cardias. SECRECIN GSTRICA
La mucosa del estmago posee tres clases de glndulas: 1) Glndulas Cardiales: se ubican alrededor del cardias y son de tipo tubulosa y secretan mucus y algo de bicarbonato. 2) Glndulas Pilricas: secretan jugo alcalino rico en mucus y algo de pepsingeno. 3) Glndulas principales, fndicas o ppticas: se ubican en el cuerpo del estmago y tienen tres tipos de clulas: a) Principales (zimgenas o ppticas): secretan pepsingeno b) Parietales (oxnticas): secretan cido clorhdrico c) Mucosas (del cuello): secretan mucus. MECANISMO BSICO DE LA SECRECIN DE ACIDO CLORHDRICO Las clulas parietales producen una solucin electroltica que contiene 160 milimoles de cido clorhdrico por litro, o sea, casi isotnica con los lquidos corporales. El pH de esta solucin es aproximadamente 0,8 o sea, muy cida. Para ste pH, la concentracin de iones H+ es unos cuatro millones de veces mayor que en la sangre arterial. Para lograr estas enormes concentraciones se necesitan ms de 15000 caloras de energa por litro de jugo gstrico. La secrecin de cido no es absolutamente esencial para la funcin gastrointestinal, ya que la digestin y la absorcin ocurren en el intestino delgado sin contar con el estmago: sin embargo, el cido cumple varias funciones que ayudan a la digestin. Un alto grado de acidez degrada las protenas y rompe los enlaces intermoleculares destruyendo as el tejido conjuntivo y las clulas, al liberar las molculas ionizadas que entran luego en solucin. El cido clorhdrico contina as el proceso iniciado por la masticacin, sin embargo, el cido tiene relativamente poca capacidad de degradar las protenas y polisacridos en aminocidos y glucosa, una segunda funcin es la de dar muerte a la mayora de las bacterias que entran con el alimento, obviamente nunca alcanza al 100%, pero esta funcin es bastante eficiente. Como tercera funcin del cido clorhdrico esta la de activar algunas enzimas secretadas por el estmago. Existen diferentes suposiciones sobre el mecanismo preciso de la formacin del cido clorhdrico una de ellas, la ms aceptada, es la siguiente: a) El ion cloruro es transportado activamente del citoplasma de la clula oxntica (que produce cido) a la luz de los canalculos. Ello crea un potencial negativo de -40 a -70 milivolts en los canalculos, que a su vez causa difusin pasiva de iones potasio con carga positiva del citoplasma celular hacia los canalculos. En consecuencia, a los canalculos entra cloruro de potasio. b) El agua es disociada en iones hidrgeno e hidroxilo en el citoplasma celular. Este ltimo es secretado enseguida activamente al interior de los canalculos en intercambio por iones potasio; este proceso activo est mediado por H+ - K+ ATPasa. En consecuencia, se reabsorben la mayor parte de los iones potasio que han sido secretados junto con los iones cloruro y los iones hidrgeno pasan a los canalculos. c) El agua pasa por osmosis a travs de la clula hacia los canalculos. Por tanto, la secrecin final de los canalculos es una solucin que contiene 160 milimoles por litro de cido clorhdrico y 17 milimoles/litro de cloruro de potasio. d) Finalmente, el dixido de carbono, que se ha formado
durante el metabolismo celular o a pasado de la sangre a la clula, se combina con agua por intermedio de la ANHIDRASA CARBNICA para formar cido carbnico que, a su vez, se disocia en iones bicarbonato e hidrgeno, ste ltimo se combina con el ion hidroxilo que se ha liberado en la etapa 1 para formar agua. A su vez, el ion bicarbonato sale de la clula hacia la sangre. SECRECIN DE PEPSINA Las clulas ppticas secretan sobre todo pepsina la que se forma dentro de las clulas como pepsingeno, que carece de poder digestivo, sin embargo, al penetrar en la luz de la glndula, el pepsingeno se pone en contacto con pepsina ya formada y HCl, con lo que se activa y pasa a pepsina. La pepsina es una enzima proteoltica en medio fuertemente cido (pH ptimo 2,0), pero si el pH pasa de 5,0 esta actividad disminuye rpidamente hasta desaparecer. Vemos pues, que la secrecin de cido clorhdrico es tan necesaria como la pepsina para la digestin gstrica de protenas. Tambin son eliminadas con los jugos gstricos pequeas cantidades de otras enzimas, incluyendo lipasa gstrica, amilasa gstrica y ureasa gstrica. La lipasa gstrica en realidad es una tributirasa, su principal actividad se ejerce sobre la tributirina, grasa de la mantequilla, pero casi no tiene actividad lipoltica sobre otras grasas. La amilasa gstrica desempea un papel reducido en la digestin de los almidones. La ureasa gstrica no es producida por la mucosa del estmago si no por concomitantes bacterianos del rgano, desintegra la urea y forma amoniaco y neutraliza un poco de acidez gstrica. En el caso de uremia gran cantidad de amoniaco puede eliminarse del estmago con los eructos. SECRECIN MUCOSA DEL ESTMAGO Estructuralmente las glndulas pilricas y cardiales son similares a las glndulas principales y parietales; estn formadas casi totalmente de clulas mucosas, idnticas a las clulas mucosas del cuello de las glndulas principales. Todas ellas secretan mucus claro que protege la pared gstrica de la digestin por las enzimas. Adems, la superficie de la mucosa gstrica, entre las glndulas, presenta una capa continua de clulas mucosas que producen una gran cantidad de mucus viscoso y alcalino que cubre la mucosa con una capa de gel de ms de 1 mm, suministrando as una importante proteccin para la vscera, adems de contribuir a la lubricacin para el transporte de alimentos. Una irritacin de la mucosa, incluso ligera, estimula directamente las clulas mucosas, que secretan de inmediato grandes cantidades de este mucus espeso y viscoso. REGULACIN DE LA SECRECIN GSTRICA POR MECANISMOS NERVIOSOS Y HORMONALES La regulacin nerviosa corresponde a las fibras parasimpticas del vago, y la hormonal, a
la hormona gastrina, participando tambin los reflejos del plexo mesentrico local. Vemos entonces que el mecanismo es diferente a lo que ocurre en la secrecin salival, que obedece nicamente a mecanismos nerviosos. ESTIMULACIN VAGAL DE LA SECRECIN GSTRICA Las seales nerviosas, que producen secrecin gstrica, nacen en los ncleos motores dorsales del vago, y bajan por el dcimo par al plexo mesentrico del estmago, que inerva las glndulas gstricas. Estas responden secretando grandes cantidades de pepsina y cido. Adems, las seales vgales que llegan a las glndulas pilricas, glndulas del cardias, y clulas caliciformes mucosas de las glndulas principales, aumentan considerablemente la secrecin de mucus. Otro efecto de la estimulacin vagal es que la regin antral de la mucosa gstrica produce la hormona gastrina, hormona que acta sobre las glndulas principales, aumentando la produccin de jugo gstrico muy cido. Vemos entonces que la estimulacin vagal acta sobre las glndulas del estmago directa e indirectamente, por intermedio de la gastrina. ESTIMULACIN DE LA SECRECIN GSTRICA POR MEDIO DE GASTRINA La funcin primaria de gastrina es la estimulacin de secrecin cida en el estmago, es el estimulador de la secrecin gstrica ms potente que se conoce, siendo 1500 veces ms potente que histamina. El efecto que esta hormona tenga sobre la liberacin de pepsina se ha definido como pobre pero se ha demostrado que su efecto es indirecto ya que la secrecin cida producida por estmulo de gastrina es la responsable de la secrecin de pepsina. METABOLISMO Se sabe que gastrina es una hormona trpica por ciertos tejidos de las vas digestivas superiores, bsicamente mucosa gstrica, duodenal y pncreas. SITIOS DE LOCALIZACIN El antro pilrico, o regin glandular pilrica es el nico lugar en que se encuentra una abundante cantidad de la hormona, pero tambin se han encontrado en la regin cardial del estmago, la actividad ms concentrada de gastrina est localizada en el tercio medio de la mucosa del antro. ESTIMULO PARA SU LIBERACIN La liberacin de gastrina desde el antro pilrico est normalmente mediada por reflejos neuroendocrinos, estos reflejos cortos o locales son intramurales y son activados por estmulos provocados en los receptores de la pared del estmago. Y reflejos largos que
involucran la va vagal ya sea por estimulacin vagal directa o receptores ubicados en la pared del estmago. Los reflejos cortos producen una mayor liberacin de gastrina, suficiente para producir niveles mximos de secrecin cida, los reflejos cortos o locales son estimulados por algunos elementos que componen el alimento y en menor magnitud por la distensin gstrica. INHIBICIN DE LA SECRECIN GSTRICA POR RETROALIMENTACIN Cuando la acidez del jugo gstrico baja hasta un pH de 2,0 es decir muy cida, el mecanismo de la gastrina para estimular la secrecin gstrica queda totalmente bloqueado. Este efecto probablemente resulte de dos factores diferentes. En primer lugar la acidez intensa deprime o bloquea la produccin de gastrina desde la mucosa antral. En segundo lugar, el cido podra liberar una hormona inhibidora desde la mucosa gstrica, o provocar un reflejo inhibidor que inhibe la secrecin gstrica. Este mecanismo de retroalimentacin negativa de las glndulas gstricas desempea un papel importante para la proteccin del estmago contra las secreciones cidas excesivas, adems de protector tiene importancia al mantener un pH ptimo que permite la funcin de las enzimas ppticas en el proceso digestivo. ETAPAS DE LA SECRECIN GSTRICA Existen tres etapas en la secrecin gstrica Ceflica, Gstrica e Intestinal. ETAPA CEFLICA Esta etapa tiene lugar antes que los alimentos lleguen al estmago. Se debe al estmulo que produce el aspecto, olor, recuerdo o sabor de los alimentos; mientras mayor sea el apetito, ms intensa ser la estimulacin. Las seales nerviosas que producen la etapa ceflica de la secrecin pueden originarse en la corteza cerebral o en los centros del apetito de la amgdala o el hipotlamo. Se transmite al estmago por los ncleos motores dorsales del vago. Esta etapa de la secrecin produce el 10% de la secrecin gstrica normal durante una comida. ETAPA GSTRICA Cuando los alimentos llegan al estmago, estimulan la liberacin de gastrina que a su vez inicia una secrecin menos intensa de jugo gstrico, pero que persiste varias horas, mientras el alimento permanezca en el estmago. Adems, la presencia de alimento en el estmago produce tambin: a) reflejos locales en el plexo intramural gstrico y b) reflejos vagales que ascienden hasta el tallo cerebral y vuelven al estmago; ambos tipos de reflejos producen estimulacin parasimptica de las
glndulas gstricas, y aumentan la secrecin debida al mecanismo de la gastrina. ETAPA INTESTINAL Despus que el alimento ha pasado al intestino, el estmago sigue secretando jugo; esta secrecin contina 6 a 8 hrs, mientras el quimo se encuentre en intestino delgado, y persiste an despus de seccionar todas las conexiones nerviosas del estmago. INHIBICIN DE LA SECRECIN GSTRICA POR FACTORES INTESTINALES Aunque el quimo estimula la secrecin gstrica durante la fase gstrica de la secrecin, es paradjico el hecho que frecuentemente inhiba la secrecin durante la fase intestinal, esto resulta, por lo menos, de dos influencias diferentes. a) La presencia de alimento en el intestino delgado inicia un reflejo enterogstrico que se trasmite por el plexo mientrico, los nervios simpticos y los vagos, inhibiendo la secrecin gstrica. Este reflejo puede provocarse por distensin del intestino delgado, presencia en l de cidos o productos de digestin de protenas, o irritacin de la mucosa. Es parte del mecanismo complejo que retrasa el vaciamiento, gstrico cuando el intestino est lleno. b) La presencia de un pH cido, grasas, aminocidos, lquidos hiper o hiposmticos, o cualquier factor irritante en el intestino delgado, provoca la liberacin de varias hormonas intestinales. Dos de ellas se han identificado en forma pura: SECRETINA y COLECISTOQUININA. Ambas son especialmente importantes para controlar la secrecin pancretica; la colecistoquinina es particularmente importante para estimular el vaciamiento de la vescula biliar. Sin embargo, adems de estos efectos, ambas se oponen a las acciones estimulantes de la gastrina sobre el estmago. Asimismo, otra hormona, el PPTIDO INHIBIDOR GSTRICO, tiene efecto moderado en la inhibicin de la secrecin gstrica. PPTIDO INHIBIDOR GSTRICO (GIP) No existe la seguridad que este pptido sea una hormona, fue aislado trabajando en la purificacin de colecistoquinina. De all en adelante se estudi el efecto de este pptido sobre la secrecin de cido y pepsina por el estmago y tambin su efecto sobre la motilidad de dicho rgano. El GIP se libera desde la mucosa duodenal y su liberacin presenta un comportamiento bifsico presentando dos alzas, la primera en respuesta a glucosa y la segunda se debera al contenido graso del alimento. Una vez liberado se describen 3 efectos diferentes: a) Inhibicin de la secrecin gstrica de HCl y pepsina, acompaado de una disminucin de la motilidad del cuerpo y antro estomacal (efecto enterogastrona). b) Libera insulina. c) Estimula la secrecin Intestinal. SECRECIN PANCRETICA El jugo pancretico contiene enzimas que hidrolizan carbohidratos, protenas y grasas. Las enzimas proteolticas son: tripsina, quimotripsina, carboxipeptidasas, ribonucleasas y
desoxirribonucleasas. Las tres primeras hidrolizan protenas completas y parcialmente digeridas; las carboxipeptidasas hidrolizan uniones peptdicas en pptidos menores y las nucleasas los cidos nucleicos RNA y DNA. Los carbohidratos son hidrolizados por la amilasa pancretica que acta sobre almidones, glucgeno y casi todos los dems carbohidratos (con excepcin de la celulosa) degradndolos hasta disacridos. La lipasa pancretica hidroliza las grasas y es capaz de transformar las grasas neutras en glicerina y cidos grasos. Lo que produce la clula pancretica es tripsingeno, quimotripsingeno y procarboxipeptidasas, precursoras carentes de poder enzimtico. Slo se activan despus de llegar al tubo digestivo. El tripsingeno es activado por la enzima enteroquinasa, liberada por la mucosa intestinal al contacto con el quimo. Tambin participa en esta activacin la tripsina ya formada, aunque este efecto es poco pronunciado. El quimotripsingeno es activado por la tripsina para formar quimotripsina; es probable que la activacin de la procarboxipeptidasa obedezca a un mecanismo similar. SECRECIN DE INHIBIDOR DE TRIPSINAS La importancia de que las enzimas del jugo pancretico se activen al llegar al intestino radica principalmente en que activadas en el pncreas digeriran dicho rgano. El mecanismo de inhibicin se debe a que los cinos del pncreas secretan la sustancia inhibidor de tripsina y como es la tripsina la que activa a las otras enzimas del pncreas al estar inhibida tambin se inhiben las otras. REGULACIN DE LA SECRECIN PANCRETICA Se regula por mecanismos tanto nerviosos como hormonales pero en este rgano la regulacin hormonal resulta ms importante que la nerviosa. REGULACIN NERVIOSA Durante la fase ceflica de secrecin gstrica, los vagos transmiten impulsos al pncreas; el resultado es cierta cantidad de enzimas en los cinos. Sin embargo, esta secrecin fluye muy poco hacia el intestino, porque la produccin concomitante de agua y electrlitos es escasa y el resultado es almacenamiento de enzimas en los cinos. REGULACION HORMONAL Cuando los alimentos entran al intestino delgado la secrecin pancretica aumenta
considerablemente, principalmente en respuesta a la hormona secretina. Adems otra hormona, llamada colecistoquinina eleva considerablemente la secrecin de enzimas. SECRETINA. Al margen de su importancia fisiolgica sta hormona tiene un valor histrico debido a que su descubrimiento en 1902 dio inicio a la endocrinologa (Bayliss y Starling, 1902), FUNCIONES La de mayor relevancia es la estimulacin de la secrecin pancretica (bicarbonato y fluidos), siendo la ms potente estimuladora de esta secrecin. El volumen de la secrecin pancretica es proporcional a la dosis de la secretina. La secretina tanto en perro como en gato es un potente inhibidor de gastrina. En el hombre y en el gato el efecto de secretina se traduce en la combinacin del efecto de descarga de bicarbonato desde el pncreas con la inhibicin de la secrecin cida por inhibicin de gastrina, lo que lleva a la neutralizacin del cido vaciado al intestino. Por otra parte la secretina estimula la secrecin de bicarbonato desde el hgado por ello se dice que esta hormona es el ms potente colertico entre todas las hormonas gastrointestinales (colertico que estimula a la secrecin de bilis). Sumado a este efecto tambin hay que tener en cuenta que secretina inhibe la motilidad gstrica, reduciendo con ello la cantidad de cido que entra al duodeno, teniendo adems como efecto un control en la velocidad de trnsito gastrointestinal. La secretina se produce en clulas especializadas del intestino, especialmente concentradas a nivel duodenal y en el yeyuno, conocidas como clulas S. LIBERACIN Se sabe que la llegada de cidos grasos, y aminocidos, sumados al cido clorhdrico estimulan el flujo de bicarbonato desde el pncreas y por ello se interpret que estas sustancias eran los factores liberadores, sin embargo despus pudo comprobarse que el principal estmulo para la liberacin de secretina es la llegada de HCl a duodeno. Por los estudios realizados se sabe que el principal estmulo a la liberacin de secretina es el HCl. Entre pH 4,5 y 3,0 hay un alza rpida de la secrecin pancretica, bajo pH 3,0 no aumenta la liberacin de secretina. La liberacin de secretina no es influenciada por el vago o por estimulacin colinrgica. Por accin de secretina, el pncreas produce gran cantidad de lquido que contiene mucho bicarbonato (145 mEq/l) y poco cloruro. Este flujo abundante se denomina secrecin hidroltica porque se compone de agua, electrlitos y pocas enzimas.
El mecanismo de la secretina es muy importante por dos razones. En primer lugar, la mucosa intestinal libera grandes cantidades de secretina cada vez que el pH duodenal cae bajo 4,0; inmediatamente se producen grandes cantidades de jugo pancretico rico en bicarbonato de sodio que entra en reaccin con HCl formando cido carbnico el que se disocia inmediatamente en H20 y anhdrido carbnico, ste pasa a los lquidos corporales y el duodeno termina conteniendo slo solucin neutra de NaCl. De esta forma, el quimo cido se neutraliza y la actividad digestiva del jugo gstrico desaparece (esto protege de lceras duodenales). En segundo lugar la secrecin hidroltica proporciona el pH ptimo para la accin de las enzimas pancreticas, o sea neutras o ligeramente alcalinas: la secrecin pancretica tiene pH 8,0 en promedio. COLECISTOQUININA (CCK o Pancreozimina) Cuando acta sola, en bajas dosis, CCK estimula dbilmente la secrecin de pepsina. Es sin embargo un potente competidor inhibidor de gastrina en la estimulacin de secrecin gstrica cida. CCK es una importante estimuladora de la secrecin enzimtica del pncreas, actuando sobre los cinos glandulares pancreticos. Adems, es la hormona gastrointestinal ms potente en la estimulacin de la contraccin de la vescula biliar y por lo tanto liberadora de sales biliares y bicarbonato (colagogo). A este efecto se suma la relajacin del esfnter de Oddi facilitando el flujo de bilis hacia el duodeno. La CCK se produce en clulas especializadas ubicadas en los primeros segmentos del intestino delgado, pero se debe dilucidar las dudas existentes en cuanto a su posible localizacin en otros segmentos digestivos. LIBERACION. CCK es liberada desde la mucosa del intestino delgado alto (a nivel duodenal), en respuesta a la presencia de grasas en el quimo vaciado al intestino, no se sabe como los agentes antes mencionados como estimuladores de la liberacin de CCK actan directamente en la clula endocrina u otro sitio por va de reflejos neurohumorales. A la inversa de gastrina, CCK no se libera por estmulo vagal. Las protenas no producen liberacin de CCK cuando estn en contacto con la mucosa intestinal, de modo que deben degradarse hasta aminocidos, estando en duda si pequeos pptidos pueden producir la liberacin. La liberacin de CCK est en directa relacin a la cantidad de grasas que entran al intestino, ms que a la concentracin. La cantidad de CCK liberada depende de la longitud intestinal expuesta a las grasas, en el hombre el segmento estimulable es de 80 cm. Respecto a los cidos grasos, aquellos ms cortos que C8 no producen liberacin de
CCK, en cambio los cidos grasos lurico, palmtico, esterico y oleico son fuertes estimulantes a la liberacin de CCK. Al igual que la secretina, la CCK llega al pncreas por va sangunea; esta hormona provoca la liberacin de gran cantidad de enzimas digestivas (secrecin ecblica), o sea que tiene el mismo efecto, pero de mayor potencia, que el estmulo vagal. SECRECIN DE BILIS POR EL HGADO La bilis no contiene enzimas digestivas y su importancia en la digestin se debe a su contenido en sales biliares que, aparte de activar a la lipasa pancretica, tienen una accin hidrotrpica y detergente, con ello: 1) Facilitan la emulsin de las gotas de grasa para que puedan ser digeridas por las lipasas intestinales. 2) Solubilizan los productos finales de digestin de las grasas para que puedan pasar por los linfticos a travs de la mucosa intestinal. El hgado secreta bilis constantemente, y no en forma intermitente como ocurre con la mayor parte de las secreciones digestivas; pero la bilis se almacena en la vescula biliar entre perodos digestivos, hasta que su presencia en intestino sea necesaria, entonces la vescula vaca su contenido bajo el efecto de CCK. Mientras la bilis permanece en la vescula biliar se concentra y se hace menos alcalina, adems se le agrega la mucina biliar. La intensidad de la secrecin de bilis puede modificarse por: 1) Estmulo vagal el que aumenta la secrecin fuertemente. 2) La secretina la aumenta moderadamente (80%). 3) Mayor riego sanguneo al hgado mayor secrecin. Y 4) Presencia de grandes cantidades de sales biliares en la sangre aumenta la secrecin del hgado. Luego que ejercen su funcin en el intestino, las sales biliares se reabsorben y pueden volver al hgado y cumplir su funcin, esta reabsorcin se lleva a cabo en el leon y recibe el nombre de circulacin entero-heptica de las sales biliares. La produccin diaria de bilis suele ser de 600 a 700 ml en el humano. SECRECIONES DEL INTESTINO DELGADO Secrecin de mucus por las glndulas de Brnner y clulas mucosas del epitelio. Al principio del duodeno, sobre todo entre el ploro y ampolla de Vater, se encuentran
muchas glndulas mucosas compuestas llamadas glndulas de Brnner las que producen grandes cantidades de mucus en respuesta a: a) Estmulos tctiles directos o irritantes de mucosa; b) Estmulo vagal; y c) Hormonas intestinales. La funcin principal es la proteccin de la mucosa. Cabe destacar que un estmulo simptico mantenido inhibe la secrecin de las glndulas de Brnner y se deja de producir el mucus protector. Tambin producen mucus las clulas caliciformes de las glndulas que cubren la mucosa intestinal. SECRECIN DE JUGOS DIGESTIVOS INTESTINALES, GLNDULAS DE LIEBERKHN Las secreciones son lquido extracelular puro con pH cercano al neutro y que oscila entre 6,5 a 7,5 (2000 ml diarios), estas secreciones proporcionan un lquido acuoso para la absorcin de sustancias procedentes de intestino delgado. Las enzimas presentes son ENTEROQUINASA (enzima intestinal que activa tripsingeno a tripsina) y tambin:1)
Peptidasas que llevan polipptidos aminocidos Sacarasa, maltasa, isomaltasa, lactasa/disacridos monosacridos. Lipasa intestinal: grasas neutras glicerol y cidos grasos. Amilasa intestinal: carbohidratos Disacridos.
2) 3) 4)
Cabe destacar que las clulas de las criptas de Lieberkhn tienen un ciclo de vida de 5 das lo que permite rpida reparacin de excoriaciones de la mucosa. REGULACIN DE LAS SECRECIONES El ms importante mecanismo es el reflejo local (intramural). En especial la distensin y estmulos irritantes. La regulacin nerviosa donde el parasimptico duplica o triplica la secrecin en intestino delgado y la regulacin hormonal. Algunas hormonas que promueven secreciones en otras partes del tubo digestivo tambin aumentan la secrecin en intestino delgado en especial secretina y colecistoquinina. Asimismo, algunos experimentos sugieren que otras sustancias hormonales liberadas de la mucosa del intestino delgado por el quimo pueden ayudar a controlar la secrecin. DIGESTIN Y ABSORCIN EN EL TUBO DIGESTIVO Con excepcin de algunos minerales y vitaminas se puede decir que el organismo se nutre de carbohidratos, grasas y protenas. Sin embargo, ninguno de ellos puede absorberse como tal, por lo que deben ser digeridos.
DIGESTIN DE LOS DISTINTOS ALIMENTOS LA HIDRLISIS MECANISMO PRINCIPAL DE LA DIGESTIN Casi todos los carbohidratos presentes en la dieta son grandes polisacridos o disacridos. Casi todas las grasas estn compuestas por triglicridos que son combinaciones de tres molculas de cido graso con una molcula de glicerol. Las protenas estn formadas de aminocidos unidos por enlaces peptdicos. La qumica de la digestin, es simple, el mismo principio de base, LA HIDROLISIS, se aplica en los tres tipos de alimentos, la nica diferencia estriba en las enzimas necesarias en cada caso para iniciar el proceso. Todas las enzimas digestivas son protenas aportadas por las glndulas ya mencionadas. DIGESTIN Y ASIMILACIN DE CARBOHIDRATOS, GRASAS Y PROTENAS La digestin es el desdoblamiento de los compuestos qumicos grandes de los alimentos en otros ms sencillos que puedan ser utilizados por el organismo. La palabra asimilacin incluye varias funciones que pueden enumerarse de la siguiente manera: 1. 2. 3. Absorcin de los productos finales de la digestin hacia los lquidos corporales. Transporte de los mismos a las clulas, donde sern utilizados. Transformacin qumica de algunos en otros compuestos necesarios para fines especiales. La funcin de los fenmenos digestivos y de asimilacin es proporcionar elementos nutritivos para las reacciones qumicas metablicas.
ABSORCIN INTESTINAL Una vez que los alimentos han llegado por hidrlisis a una forma molecular adecuada, comienza el proceso de absorcin por las clulas del intestino. Debe absorberse el mayor nmero de nutrientes que el organismo pueda metabolizar, a una velocidad adecuada y que guarde relacin con los requerimientos energticos y estructurales del organismo. Los nutrientes atraviesan las clulas del intestino, entrando por el lado luminal y saliendo por el lado sanguneo de las clulas por dos mecanismos generales. DIFUSIN Que consiste en que las molculas estn en constante movimiento gracias a su energa trmica, chocando continuamente entre ellas al azar. De este modo se desplazan hacia los lugares de menor concentracin (gradiente) y temperatura del medio. Con esto las molculas difunden a travs de las membranas. Este mecanismo fsico no permite concentrar contra gradiente, por esta razn, la concentracin del substrato en la sangre
debe ser inferior a la del lumen intestinal. Por este mecanismo se transportan agua, algunos iones y grasas o cuerpos resultantes de la hidrlisis de ellas. TRANSPORTE ACTIVO En el cual el substrato forma un complejo con un transportador. Este complejo se forma en la superficie luminal de las clulas y se rompe en la superficie capilar. De este modo sustancias insolubles o poco solubles son capaces de atravesar las membranas y citoplasma celular, en este proceso hay consumo de energa y se permite el transporte contra gradiente y en un solo sentido, depende de la disponibilidad de transportadores, existiendo una velocidad de transporte mximo, como ejemplo la de la glucosa es de 1 gr/hr/kg. En este tipo de mecanismo existe tambin competencia, transportndose algunos elementos preferentemente ej: la preferencia descendente Galactosa, Glucosa, Fructosa, Manosa. Por este mecanismo se transportan monosacridos y aminocidos y tambin la mayora de los minerales. DIGESTIN, ABSORCIN Y DISTRIBUCIN DE LOS CARBOHIDRATOS (CH) Los CH son elementos formados de carbono, hidrgeno y oxgeno. La unidad fundamental de un CH es un monosacrido, el ms comn es la glucosa. Otros dos monosacridos que existen frecuentemente en los alimentos son fructuosa y galactosa. La glucosa y otros monosacridos suelen polimerizarse en compuestos qumicos ms grandes, como almidones, glucgenos, pectinas y dextrinas. El carbohidrato ms abundante en la dieta es el almidn, polmero de la glucosa. Adems, de los almidones otra fuente comn son los disacridos maltosa, sacarosa y lactosa. ABSORCIN DE CARBOHIDRATOS Son absorbidos por las vellosidades intestinales que llevan la sangre a la vena porta y por ella al hgado. Los monosacridos se absorben por difusin y absorcin activa. DESTINO DE LOS MONOSACRIDOS La glucosa se transporta rpidamente por todos los lquidos corporales, sin modificarse. Sin embargo, las clulas hepticas absorben fructuosa y galactosa, la transforman en glucosa y despus van a la sangre. En consecuencia la glucosa es la base de casi todas las reacciones qumicas de los CH en el organismo, siendo su concentracin en la sangre de 90 mg por 100 ml de sangre o LEC. De aqu que en adelante los procesos que siguen son bioqumicos y a nivel celular. Participan en los procesos siguientes la insulina que se estudiar ms adelante, regulacin de la glicemia gluconeognesis etc., para llegar al final a la glucosa como fuente de Energa.
DIGESTIN, ABSORCIN Y DISTRIBUCIN DE GRASAS NEUTRAS La digestin de las grasas tambin es mediada por un fenmeno de hidrlisis, y catalizado por las lipasas. La participacin de las sales biliares ayuda mucho a su digestin pues convierten los glbulos de grasa de los alimentos en glbul