
RESPOSTA DELS ARBRES I DELS BOSCOS AL CANVI CLIMÀTIC · Els arbres vells, aïllats i lluny de la...
26
Un tast de canvi global (Cèlia Marrasé i Josep Enric Llebot, ed.) Treballs de la SCB. Vol. 54 (2003) 85-110 RESPOSTA DELS ARBRES I DELS BOSCOS AL CANVI CLIMÀTIC Emilia Gutiérrez, 1 Monserrat Ribas, 1 Octavi Planells, 1 J. Julio Camarero 2 i Jacques Tardif 3 1 Grup de Recerca de Dendroecologia i Ecologia dels Boscos. DENDRIX. Departament d’Ecologia. Facultat de Biologia. Universitat de Barcelona. 2 Unidad de Recursos Forestales. Servicio de Investigación Agroalimentaria. Gobierno de Aragón. 3 Centre for Forest Interdisciplinary Research. University of Winnipeg. Winnipeg, Manitoba. Canadà. Adreça per a la correspondència: Emilia Gutiérrez. Departament d’Ecologia. Facultat de Biologia. Universitat de Barcelona. Adreça electrònica: [email protected]. RESUM Durant el darrer segle, arbres, boscos i ecosistemes han estat responent a diversos efectes del canvi global, com són l’escalfament de la superfície terrestre, l’increment de la variabilitat climàtica o l’augment de la concentració de CO 2 . Les respostes dels arbres a aquests efectes han estat analitzades en diferents espècies de coníferes en diversos hàbitats i ambients de la península Ibèrica. Un compendi dels resultats obtinguts es mostra en el present treball. En tots els estudis presentats s’ha observat un augment de la variabilitat de les variables dendrocronològiques (gruix i composició isotòpica dels anells de creixement) en les darre- res dècades del segle xx, fet que reflecteix un increment de la variabilitat climàtica i que, al seu torn, ha restringit el ventall de respostes dels organismes a causa de l’eixamplament dels períodes de creixement i de l’augment d’esdeveniments meteorològics extrems. Des- taca, per tant, la sincronització del creixement dels arbres en detriment de les diferències explicades per causes cada cop més secundàries, com són les diferències de temperatura al llarg de gradients, altitudinals o latitudinals, o degudes a diferències d’hàbitat (orienta- ció, substrat, etc.). Els efectes es manifesten en diferents nivells temporals i espacials. Així, s’ha trobat un increment de la variabilitat intraanual en el creixement en gruix d’espècies mediterrànies com el pi blanc (Pinus halepensis) associat a un eixamplament de l’època de creixement, o augments de la variabilitat interanual i entre dècades tant a escales locals com regionals en el gruix i composició isotòpica dels anells de creixement d’espècies subalpines. Les anàlisis de les relacions entre el creixement dels arbres i el clima, d’una banda, i la composició isotòpica i el clima, de l’altra, demostren que aquest augment de la variabilitat respon a un increment de les oscil . lacions climàtiques intraanuals, interanuals i entre dèca- des. La resposta sincrònica dels arbres a un menor nombre de variables ambientals s’entén com un augment de l’estrès d’aquests organismes enfront d’aquestes variables, que esdeve-
Transcript of RESPOSTA DELS ARBRES I DELS BOSCOS AL CANVI CLIMÀTIC · Els arbres vells, aïllats i lluny de la...

Un tast de canvi global(Cèlia Marrasé i Josep Enric Llebot, ed.)
Treballs de la SCB. Vol. 54 (2003) 85-110
RESPOSTA DELS ARBRES I DELS BOSCOS AL CANVI CLIMÀTIC
Emilia Gutiérrez,1 Monserrat Ribas,1 Octavi Planells,1 J. Julio Camarero2 i JacquesTardif3
1 Grup de Recerca de Dendroecologia i Ecologia dels Boscos. DENDRIX. Departament d’Ecologia.Facultat de Biologia. Universitat de Barcelona.2 Unidad de Recursos Forestales. Servicio de Investigación Agroalimentaria. Gobierno de Aragón.3 Centre for Forest Interdisciplinary Research. University of Winnipeg. Winnipeg, Manitoba. Canadà.
Adreça per a la correspondència: Emilia Gutiérrez. Departament d’Ecologia. Facultatde Biologia. Universitat de Barcelona. Adreça electrònica: [email protected].
RESUM
Durant el darrer segle, arbres, boscos i ecosistemes han estat responent a diversos efectes delcanvi global, com són l’escalfament de la superfície terrestre, l’increment de la variabilitatclimàtica o l’augment de la concentració de CO2. Les respostes dels arbres a aquests efecteshan estat analitzades en diferents espècies de coníferes en diversos hàbitats i ambients dela península Ibèrica. Un compendi dels resultats obtinguts es mostra en el present treball.En tots els estudis presentats s’ha observat un augment de la variabilitat de les variablesdendrocronològiques (gruix i composició isotòpica dels anells de creixement) en les darre-res dècades del segle xx, fet que reflecteix un increment de la variabilitat climàtica i que,al seu torn, ha restringit el ventall de respostes dels organismes a causa de l’eixamplamentdels períodes de creixement i de l’augment d’esdeveniments meteorològics extrems. Des-taca, per tant, la sincronització del creixement dels arbres en detriment de les diferènciesexplicades per causes cada cop més secundàries, com són les diferències de temperaturaal llarg de gradients, altitudinals o latitudinals, o degudes a diferències d’hàbitat (orienta-ció, substrat, etc.). Els efectes es manifesten en diferents nivells temporals i espacials. Així,s’ha trobat un increment de la variabilitat intraanual en el creixement en gruix d’espèciesmediterrànies com el pi blanc (Pinus halepensis) associat a un eixamplament de l’època decreixement, o augments de la variabilitat interanual i entre dècades tant a escales locals comregionals en el gruix i composició isotòpica dels anells de creixement d’espècies subalpines.Les anàlisis de les relacions entre el creixement dels arbres i el clima, d’una banda, i lacomposició isotòpica i el clima, de l’altra, demostren que aquest augment de la variabilitatrespon a un increment de les oscil.lacions climàtiques intraanuals, interanuals i entre dèca-des. La resposta sincrònica dels arbres a un menor nombre de variables ambientals s’enténcom un augment de l’estrès d’aquests organismes enfront d’aquestes variables, que esdeve-

86 E. GUTIÉRREZ, M. RIBAS, O. PLANELLS, J. J. CAMARERO I J. TARDIF
nen factors limitants per al desenvolupament d’aquestes espècies. Els resultats presentatssón contrastats amb resultats similars, tant locals com globals, obtinguts en altres indretsdel planeta.
SUMMARY
Trees, forests and ecosystems have been adjusting their responses and functions to the verydiverse effects of global change: global warming, climatic variability, the increasing of theCO2 atmospheric concentrations, and changes in land use throughout the last century. Treesresponses to these effects have been analyzed for different pine species growing in differenthabitats and under different climatic regimes of the Iberian Peninsula. Here, we present aset of selected results of our research. Our results show that a variability increase in the den-drochronological series analyzed, mainly tree-ring width and isotopic concentrations sincethe second half of the 20th century as a consequence of an increase in the climatic variability.As a consequence, trees response is more synchronous over great areas and gradients due toa wider growing period and to a higher frequency of climatic extreme events. The effects ofclimatic change on tree growth and forests are significant to different spatial and temporalscales. We have found and increase in intra-annual tree-ring growth variability in Aleppopine (Pinus halepensis) in relation to a wider growing period. In the same way, there is anincrease in tree-ring width and isotopic series of the interannual and interdecadal temporalscales in locations ranging from lowland to subalpine forests. On the other hand, trees aresensitive to a lower number of climatic variables and their response is more synchronousover big areas, which could be a more stressing situation for trees. Finally, we have alsoreconstructed treeline ecotone dynamics and, as in other ecotons, treeline has remain staticfor the last fifty years while tree density has increased in the ecotone. The main reason forthis situation, again, is atributed to climatic variability. The results presented are similar toother findings in many other forests in the world.
INTRODUCCIÓ
Com més va més es donen a conèixerels resultats de les investigacions en eco-logia que posen de manifest que el canviclimàtic afecta directament tots els nivellsd’organització dels ecosistemes. L’efecte éscomplex i no és homogeni, varia segons lesregions i les espècies, ja que aquestes respo-nen de manera individual a la temperatura,a la concentració de CO2 i als factors am-bientals en general. Actualment, l’ecologiaes troba davant d’una de les situacions mésdesafiants i al mateix temps més interes-sants (Shaver et al., 2000). El Grup Intergo-
vernamental d’Experts sobre el Canvi Cli-màtic en el seu informe de 2001 proporcionade manera clara una llarga llista dels can-vis que s’estan produint en els ecosistemesterrestres com a conseqüència del canvi enl’atmosfera i el clima.
Com a fets més destacables trobem, entred’altres, que l’augment del CO2 pot produirun augment de la fotosíntesi i de la produc-ció en alguns ecosistemes (Alekseev i So-roka, 2002). No obstant això, també podendonar-se limitacions per altres factors comla manca d’aigua a causa d’una major eva-potranspiració causada per l’augment de latemperatura (Barber et al., 2000) o per una

RESPOSTA DELS ARBRES I DELS BOSCOS AL CANVI CLIMÀTIC 87
despesa metabòlica més elevada (respira-ció) dels organismes. En canvi, la velocitatde la descomposició de la materia orgànicaes veuria afavorida per una major activitatmetabòlica de fongs i bacteris. En el cas delsboscos, aquest procés es veuria afavorit peruna disminució de densitat de la fusta, talcom s’ha posat de manifest amb diverses es-pècies de coníferes al llarg d’àmplies zonesd’Euràsia i Amèrica del Nord (Briffa et al.,1998).
A la complexitat del canvi climàtic s’had’afegir un altre component: l’augment enla variabilitat del clima i de la freqüènciadels esdeveniments climàtics extrems. Enèpoques recents sembla que l’increment devariabilitat es produeix des de mitjan se-gle xx (Tullot, 1988; IPPC, 1997, 2001).
L’augment de les temperatures i el des-cens de les precipitacions determinen unamenor disponibilitat d’aigua. Aquesta cir-cumstància és més manifesta a les zones declima mediterrani (Piñol et al., 1998; Peñue-las, 2001). Concretament, al nostre país, elsecosistemes naturals i, en particular, els bos-cos, es troben restringits a les zones mun-tanyoses, de sòls poc desenvolupats, ambmarcades diferències climàtiques segons elrelleu, l’altitud i la latitud. Tot plegat, elsfa especialment sensibles al canvi climàtici a altres efectes del canvi global, com elscanvis d’usos del sòl que incideixen directa-ment en l’augment espectacular del nombred’incendis forestals, tot mantenint els eco-sistemes en fases inicials de la successió.
L’augment de temperatures que es pro-dueix des de finals del segle xix, quan acabala petita edat del gel, ha sigut molt acusaten les últimes dècades, principalment a par-tir dels anys setanta. Aquest escalfament haprovocat un allargament dels períodes decreixement i activitat dels organismes i, enconseqüència, s’han produït modificacionsen els calendaris anuals d’activitat i repòsdels organismes (fenologia), desajustos enles interaccions entre espècies i canvis sig-
nificatius en la dinàmica de les poblacions(Peñuelas et al., 2002).
Com ja s’ha assenyalat, en les zones mun-tanyenques, els intensos gradients altitu-dinals (Martín i Olcida, 2001) fan que lescondicions ecològiques variïn fortament alllarg de distàncies curtes, cosa que es reflec-teix de manera molt notable en canvis enels sòls i en la vegetació (Körner, 1999). AlsPirineus, els boscos subalpins de pi negre(Pinus uncinata), mostren zones de transició(ecotons) relativament nítides que determi-nen canvis bruscs en la diversitat d’espèciesi en el funcionament dels ecosistemes. El lí-mit superior del bosc (timberline o forest li-mit), que en gran part dels Pirineus corres-pon a una frontera entre els boscos subal-pins de pi negre (Pinus uncinata), i els pratsalpins (Caricetalia curvulae, Elyno-Seslerietea;Carreras et al., 1995) és potencialment unbon indicador dels canvis ambientals i cli-màtics (Camarero, 2000).
Dins el context de l’estudi de canvis am-bientals ràpids com el que s’està esdeveninten les darreres dècades, són necessàries sè-ries d’alta resolució (almenys anual) que en-registrin els efectes del canvi climàtic so-bre els boscos. En aquest sentit, les sèriesd’anells de creixement dels arbres són unafont de dades molt valuosa, ja que les se-ves característiques (gruix, anatomia, den-sitat, contingut isotòpic, etc.) reflecteixen lavariabilitat climàtica al llarg del temps. Perexemple, els arbres dels Pirineus que es tro-ben a altituds elevades, a més de 2.100 m,precisament a causa que es troben lluny dela influència local de les activitats humanes,enregistren de manera fiable els canvis detemperatura que es produeixen a una escalaglobal (Tardif et al., 2003).
ELS ARBRES I ELS BOSCOS
Per a tothom és ben conegut el paper queels arbres tenen com a embornals de CO2.

88 E. GUTIÉRREZ, M. RIBAS, O. PLANELLS, J. J. CAMARERO I J. TARDIF
Extracció de testimonis de fusta mitjançant una barrina
Figura 1. Els arbres vells, aïllats i lluny de la influència de les activitats locals humanes són els que millor reflecteixen elscanvis climàtics en les seves sèries d’anells de creixement. Per a obtenir les sèries d’anells no cal tallar els arbres; mitjançantuna barrina especialment dissenyada, es poden obtenir els testimonis de fusta (cores) amb els quals s’acostuma a treballar. Enles coníferes, un anell anual està format per una banda de fusta clara (fusta primerenca) i una segona banda de fusta més fosca(fusta tardana). La secció transversal d’un tronc de pi negre (Pinus uncinata) mostra una ferida causada per algun cop fort queha malmès el càmbium. L’any en què es va produir la ferida por ser datat amb precisió. Fotos ©DENDRIX.
Però a més, l’activitat del càmbium és sen-sible a les condicions de l’entorn i en elsanells queden enregistrats tots aquells es-deveniments que han afectat l’arbre al llarg
de la seva vida; també, els arbres viuenmolts anys i aquesta informació pot ser re-cuperada i, convenientment analitzada, per-met estudiar processos ecològics i ambien-

RESPOSTA DELS ARBRES I DELS BOSCOS AL CANVI CLIMÀTIC 89
Temps (any)
1750 1800 1850 1900 1950 2000
Co
nce
ntr
ació
CO
2a
tmo
sfèr
ic(p
pm
)
280
300
320
340
360
380
–11
–10
–9
–8
–7
–6
13C
PD
B c
el·lu
losa
an
ells
arb
res
(‰)
–25
–24
–23
–22
–21
–20
Concentració CO 2 atmosfèric (Friedli et al. , 1986; Keeling i Whorf, 1999)13
CPDB CO2 atmosfèric (Friedli et al., 1986; Francey i Allison, com. pers.)13
CPDB dels anells de creixement de Pinus uncinata (Castillo de Vinuesa)
i ajust corba exponencial (Feng, 1998)13
CPDB dels anells de creixement de Pinus sylvestris (Tejeros)
i ajust corba exponencial (Feng, 1998)13
CP
DB
CO
2at
mo
sfèr
ic (
‰)
Figura 2. Registres de la concentració de CO2 atmosfèric (corba vermella amb tendència exponencial ascendent) i la com-posició isotòpica en carboni 13 (corba blava, amb tendència exponencial descendent) al llarg dels darrers segles. La figuratambé mostra la δ13C dels anells de creixement del pi negre (Pinus uncinata) a Vinuesa (línia negra) i del pi roig (Pinus syl-vestris) a Tejeros (línia grisa), al llarg dels segles xix-xx i xx, respectivament. En les sèries isotòpiques dendrocronològiquess’ha ajustat una corba exponencial negativa que reprodueix la tendència decreixent de la δ13C del CO2 atmosfèric (Planells,2003).
tals amb una resolució anual (vegeu la fi-gura 1).
Els anells de creixement són sensibles ales variacions del clima, la composició at-mosfèrica, les característiques físiques i quí-miques del sòl, la competència amb els ve-ïns, les pertorbacions, etc. Tots aquests fac-tors modifiquen el nombre i la mida de lescèl.lules que componen la fusta i això esreflecteix en les taxes d’engruiximent i enles característiques físiques i químiques de
les seves cèl.lules. Altres factors produeixenferides al càmbium i, en aquest cas, els es-deveniments queden literalment marcats al’anell de l’any corresponent, tal com potsucceir, per exemple, amb ferides produïdespel foc, les allaus, els animals, les glaçades,els cops de pedres, roques i els arbres veïnsquan cauen...
En les coníferes, la formació d’un anell ésràpida al començament i s’alenteix a mesuraque avança l’estiu. Aquestes diferències de
90 E. GUTIÉRREZ, M. RIBAS, O. PLANELLS, J. J. CAMARERO I J. TARDIF
Pinus uncinata (Vinuesa)
r = 0,514; p < 0,01
Des
via
ció
est
ànd
ard
0,010
0,015
0,020
0,025
0,030
0,035
Pinus sylvestris (Tejeros)
r = 0,206; p = 0,064
Any inicial de la finestra mòbil de vint anys
1900 1910 1920 1930 1940 1950 1960 1970 1980
Des
via
ció
est
ànd
ard
0,010
0,015
0,020
0,025
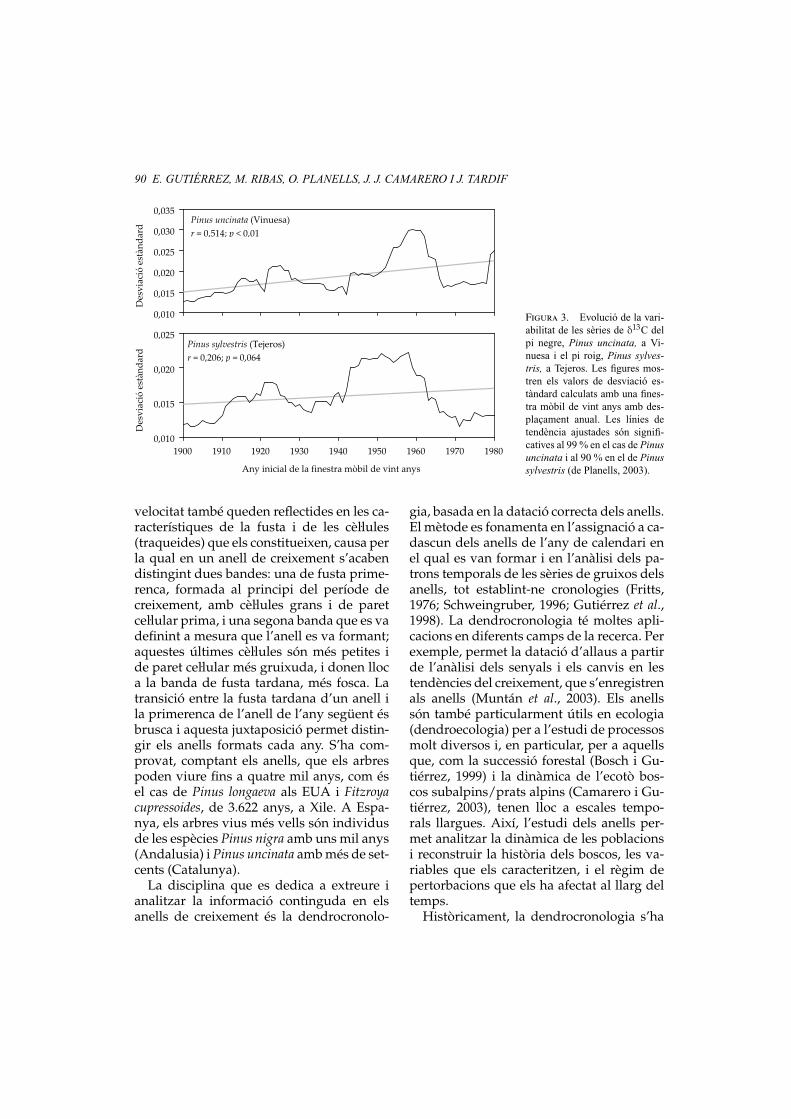
Figura 3. Evolució de la vari-abilitat de les sèries de δ13C delpi negre, Pinus uncinata, a Vi-nuesa i el pi roig, Pinus sylves-tris, a Tejeros. Les figures mos-tren els valors de desviació es-tàndard calculats amb una fines-tra mòbil de vint anys amb des-plaçament anual. Les línies detendència ajustades són signifi-catives al 99 % en el cas de Pinusuncinata i al 90 % en el de Pinussylvestris (de Planells, 2003).
velocitat també queden reflectides en les ca-racterístiques de la fusta i de les cèl.lules(traqueides) que els constitueixen, causa perla qual en un anell de creixement s’acabendistingint dues bandes: una de fusta prime-renca, formada al principi del període decreixement, amb cèl.lules grans i de paretcel.lular prima, i una segona banda que es vadefinint a mesura que l’anell es va formant;aquestes últimes cèl.lules són més petites ide paret cel.lular més gruixuda, i donen lloca la banda de fusta tardana, més fosca. Latransició entre la fusta tardana d’un anell ila primerenca de l’anell de l’any següent ésbrusca i aquesta juxtaposició permet distin-gir els anells formats cada any. S’ha com-provat, comptant els anells, que els arbrespoden viure fins a quatre mil anys, com ésel cas de Pinus longaeva als EUA i Fitzroyacupressoides, de 3.622 anys, a Xile. A Espa-nya, els arbres vius més vells són individusde les espècies Pinus nigra amb uns mil anys(Andalusia) i Pinus uncinata amb més de set-cents (Catalunya).
La disciplina que es dedica a extreure ianalitzar la informació continguda en elsanells de creixement és la dendrocronolo-
gia, basada en la datació correcta dels anells.El mètode es fonamenta en l’assignació a ca-dascun dels anells de l’any de calendari enel qual es van formar i en l’anàlisi dels pa-trons temporals de les sèries de gruixos delsanells, tot establint-ne cronologies (Fritts,1976; Schweingruber, 1996; Gutiérrez et al.,1998). La dendrocronologia té moltes apli-cacions en diferents camps de la recerca. Perexemple, permet la datació d’allaus a partirde l’anàlisi dels senyals i els canvis en lestendències del creixement, que s’enregistrenals anells (Muntán et al., 2003). Els anellssón també particularment útils en ecologia(dendroecologia) per a l’estudi de processosmolt diversos i, en particular, per a aquellsque, com la successió forestal (Bosch i Gu-tiérrez, 1999) i la dinàmica de l’ecotò bos-cos subalpins/prats alpins (Camarero i Gu-tiérrez, 2003), tenen lloc a escales tempo-rals llargues. Així, l’estudi dels anells per-met analitzar la dinàmica de les poblacionsi reconstruir la història dels boscos, les va-riables que els caracteritzen, i el règim depertorbacions que els ha afectat al llarg deltemps.
Històricament, la dendrocronologia s’ha
RESPOSTA DELS ARBRES I DELS BOSCOS AL CANVI CLIMÀTIC 91
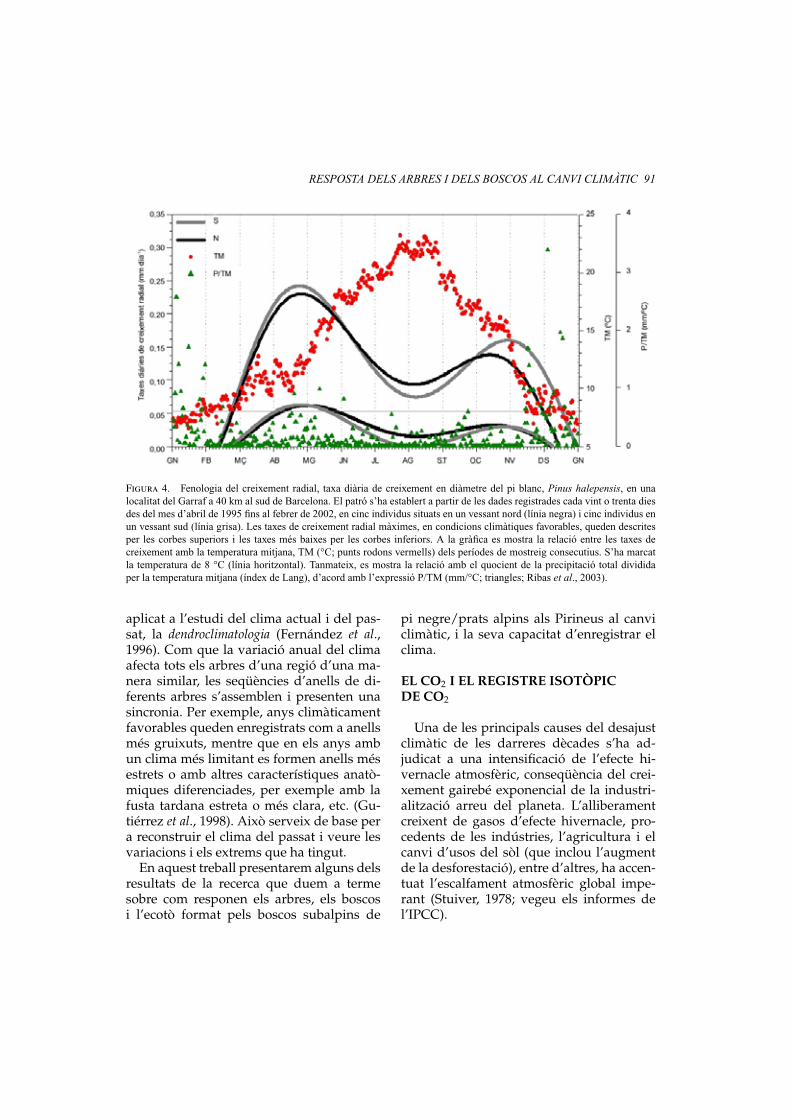
Figura 4. Fenologia del creixement radial, taxa diària de creixement en diàmetre del pi blanc, Pinus halepensis, en unalocalitat del Garraf a 40 km al sud de Barcelona. El patró s’ha establert a partir de les dades registrades cada vint o trenta diesdes del mes d’abril de 1995 fins al febrer de 2002, en cinc individus situats en un vessant nord (línia negra) i cinc individus enun vessant sud (línia grisa). Les taxes de creixement radial màximes, en condicions climàtiques favorables, queden descritesper les corbes superiors i les taxes més baixes per les corbes inferiors. A la gràfica es mostra la relació entre les taxes decreixement amb la temperatura mitjana, TM (°C; punts rodons vermells) dels períodes de mostreig consecutius. S’ha marcatla temperatura de 8 °C (línia horitzontal). Tanmateix, es mostra la relació amb el quocient de la precipitació total divididaper la temperatura mitjana (índex de Lang), d’acord amb l’expressió P/TM (mm/°C; triangles; Ribas et al., 2003).
aplicat a l’estudi del clima actual i del pas-sat, la dendroclimatologia (Fernández et al.,1996). Com que la variació anual del climaafecta tots els arbres d’una regió d’una ma-nera similar, les seqüències d’anells de di-ferents arbres s’assemblen i presenten unasincronia. Per exemple, anys climàticamentfavorables queden enregistrats com a anellsmés gruixuts, mentre que en els anys ambun clima més limitant es formen anells mésestrets o amb altres característiques anatò-miques diferenciades, per exemple amb lafusta tardana estreta o més clara, etc. (Gu-tiérrez et al., 1998). Això serveix de base pera reconstruir el clima del passat i veure lesvariacions i els extrems que ha tingut.
En aquest treball presentarem alguns delsresultats de la recerca que duem a termesobre com responen els arbres, els boscosi l’ecotò format pels boscos subalpins de
pi negre/prats alpins als Pirineus al canviclimàtic, i la seva capacitat d’enregistrar elclima.
EL CO2 I EL REGISTRE ISOTÒPICDE CO2
Una de les principals causes del desajustclimàtic de les darreres dècades s’ha ad-judicat a una intensificació de l’efecte hi-vernacle atmosfèric, conseqüència del crei-xement gairebé exponencial de la industri-alització arreu del planeta. L’alliberamentcreixent de gasos d’efecte hivernacle, pro-cedents de les indústries, l’agricultura i elcanvi d’usos del sòl (que inclou l’augmentde la desforestació), entre d’altres, ha accen-tuat l’escalfament atmosfèric global impe-rant (Stuiver, 1978; vegeu els informes del’IPCC).
92 E. GUTIÉRREZ, M. RIBAS, O. PLANELLS, J. J. CAMARERO I J. TARDIF
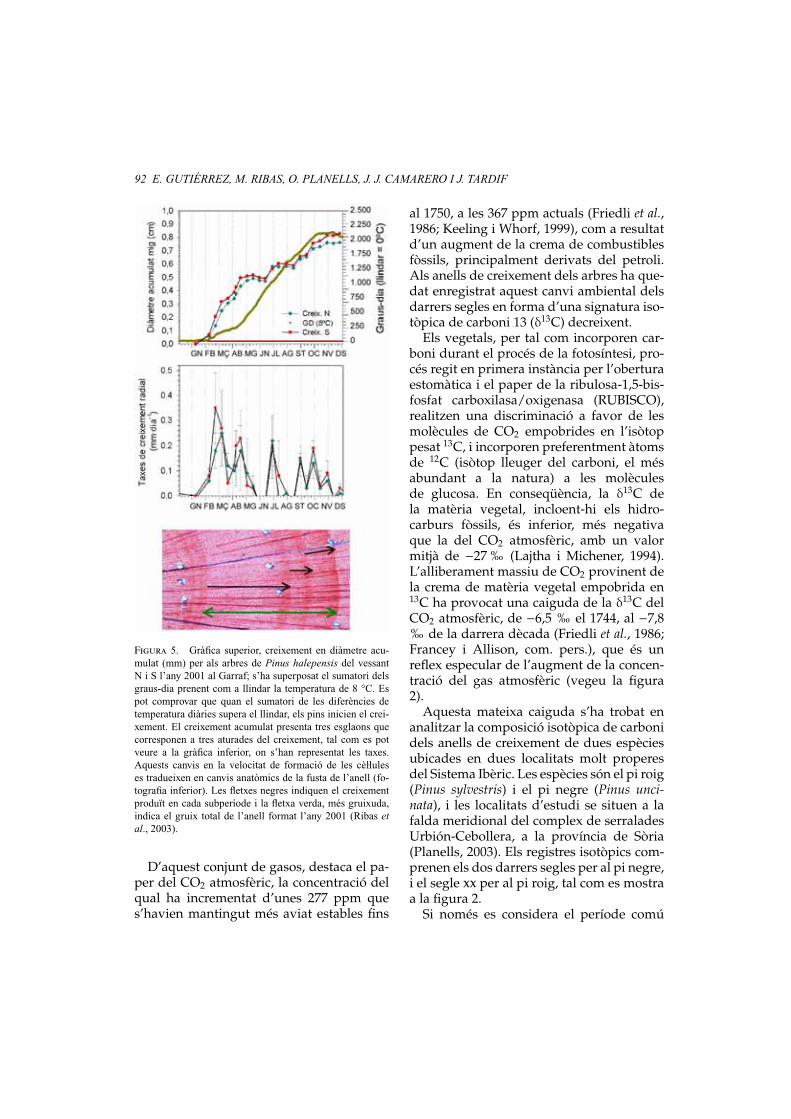
Figura 5. Gràfica superior, creixement en diàmetre acu-mulat (mm) per als arbres de Pinus halepensis del vessantN i S l’any 2001 al Garraf; s’ha superposat el sumatori delsgraus-dia prenent com a llindar la temperatura de 8 °C. Espot comprovar que quan el sumatori de les diferències detemperatura diàries supera el llindar, els pins inicien el crei-xement. El creixement acumulat presenta tres esglaons quecorresponen a tres aturades del creixement, tal com es potveure a la gràfica inferior, on s’han representat les taxes.Aquests canvis en la velocitat de formació de les cèl.luleses tradueixen en canvis anatòmics de la fusta de l’anell (fo-tografia inferior). Les fletxes negres indiquen el creixementproduït en cada subperíode i la fletxa verda, més gruixuda,indica el gruix total de l’anell format l’any 2001 (Ribas etal., 2003).
D’aquest conjunt de gasos, destaca el pa-per del CO2 atmosfèric, la concentració delqual ha incrementat d’unes 277 ppm ques’havien mantingut més aviat estables fins
al 1750, a les 367 ppm actuals (Friedli et al.,1986; Keeling i Whorf, 1999), com a resultatd’un augment de la crema de combustiblesfòssils, principalment derivats del petroli.Als anells de creixement dels arbres ha que-dat enregistrat aquest canvi ambiental delsdarrers segles en forma d’una signatura iso-tòpica de carboni 13 (δ13C) decreixent.
Els vegetals, per tal com incorporen car-boni durant el procés de la fotosíntesi, pro-cés regit en primera instància per l’oberturaestomàtica i el paper de la ribulosa-1,5-bis-fosfat carboxilasa/oxigenasa (RUBISCO),realitzen una discriminació a favor de lesmolècules de CO2 empobrides en l’isòtoppesat 13C, i incorporen preferentment àtomsde 12C (isòtop lleuger del carboni, el mésabundant a la natura) a les molèculesde glucosa. En conseqüència, la δ13C dela matèria vegetal, incloent-hi els hidro-carburs fòssils, és inferior, més negativaque la del CO2 atmosfèric, amb un valormitjà de −27 ‰ (Lajtha i Michener, 1994).L’alliberament massiu de CO2 provinent dela crema de matèria vegetal empobrida en13C ha provocat una caiguda de la δ13C delCO2 atmosfèric, de −6,5 ‰ el 1744, al −7,8‰ de la darrera dècada (Friedli et al., 1986;Francey i Allison, com. pers.), que és unreflex especular de l’augment de la concen-tració del gas atmosfèric (vegeu la figura2).
Aquesta mateixa caiguda s’ha trobat enanalitzar la composició isotòpica de carbonidels anells de creixement de dues espèciesubicades en dues localitats molt properesdel Sistema Ibèric. Les espècies són el pi roig(Pinus sylvestris) i el pi negre (Pinus unci-nata), i les localitats d’estudi se situen a lafalda meridional del complex de serraladesUrbión-Cebollera, a la província de Sòria(Planells, 2003). Els registres isotòpics com-prenen els dos darrers segles per al pi negre,i el segle xx per al pi roig, tal com es mostraa la figura 2.
Si només es considera el període comú

RESPOSTA DELS ARBRES I DELS BOSCOS AL CANVI CLIMÀTIC 93
Figura 6. Evolució de les temperatures mitjanes dels mesos de gener, febrer, març, octubre, novembre i desembre a l’estaciómeteorològica de Begues (Garraf, 40 km al sud de Barcelona) a prop de la qual s’ha dut a terme l’estudi fenològic del pi blanc,Pinus halepensis. La línia contínua horitzontal (de color verd) representa la temperatura de 8 °C, que és el llindar perquès’iniciï l’activitat del càmbium en Pinus halepensis (Garraf, 300 m snm). La línia vermella representa la tendència ascendentde la temperatura.
del segle xx, es pot observar com les cor-bes exponencials negatives ajustades (Feng,1998) a cadascuna de les sèries dendrocro-nològiques mantenen un cert paral.lelismeamb la corba de les δ13C del CO2 atmosfèric,obtinguda a partir de l’anàlisi de l’aire con-tingut en bombolles de gels polars (Friedli etal., 1986) i de mesures atmosfèriques direc-tes (Francey i Allison, com. pers.). El decre-ment de δ13C entre 1900 i 1999 per al CO2 at-mosfèric és de l’1,29 ‰. S’han trobat valorsmajors d’aquest increment per a les dues es-pècies estudiades, del 1,59 ‰ per al pi roig idel 2,47 ‰ per al pi negre. Per tant, els resul-
tats suggereixen que a l’efecte del decreixe-ment de la δ13C del CO2 atmosfèric sobre lacomposició isotòpica dels vegetals s’hi su-men altres factors, com podrien ser el discri-minat diferencial entre espècies causat perdiferències genètiques i fisiològiques i perla variació de les pressions parcials de CO2entre l’atmosfera i l’espai intercel.lular (on esprodueix la discriminació isotòpica per partde la RUBISCO). Així, hi ha un segon efectecausat directament per l’increment del CO2atmosfèric, que provoca l’empobriment iso-tòpic de la matèria vegetal (Feng i Epstein,1995).

94 E. GUTIÉRREZ, M. RIBAS, O. PLANELLS, J. J. CAMARERO I J. TARDIF
Figura 7. Representació de sèries de gruix d’anells (dades brutes), de deu arbres de pi negre, Pinus uncinata, dels qualss’ha extret una mostra al Parc Nacional d’Aigüestortes i Estany de Sant Maurici, en un rang de altituds de 2.060-2.350 m.
També, addicionalment a les tendències allarg termini descrites, degudes únicamenta l’augment de la concentració de CO2 al’atmosfera, existeix en les sèries de δ13Cdels anells de creixement una variabilitat in-teranual que respon a altres variables am-bientals, principalment a les meteorològi-ques. Com s’ha comentat més amunt, laδ13C depèn directament de la discriminacióisotòpica realitzada per la RUBISCO, la qualtambé es troba subjecta a l’obertura estomà-tica, fenomen que regula la taxa de renova-ció del CO2 a l’espai intercel.lular (Ehlerin-ger et al., 1993). Per tant, la δ13C és un reflexde les condicions d’aridesa a les quals hanestat exposats els arbres, ja que com més ele-vat és l’eixut, més temps romanen els esto-mes tancats i la RUBISCO incorpora tot elCO2 de què disposa (empobrit i enriquit), jaque a l’espai intercel.lular la taxa de renova-ció de CO2 és mínima, mentre que enfrontde condicions favorables, els estomes podenestar oberts més temps i la RUBISCO potdiscriminar en contra del 13CO2 gràcies a lacontínua disponibilitat de 12CO2 (Farquharet al., 1982).
El que s’ha observat en les sèries estan-
darditzades de δ13C mitjançant l’ajustamentexponencial (Planells, 2003) és un augmentde la variabilitat en la composició isotòpicaal llarg del segle xx (vegeu la figura 3). Lesdesviacions estàndard mòbils (amb despla-çament anual) calculades amb finestres devint anys, tant per a les sèries de pi roigcom les de pi negre, presenten una tendèn-cia creixent significativa en tots dos casos(r = 0,206 i p = 0,064 per a Pinus sylvestris;r = 0,514 i p < 0,01 per a Pinus uncinata), quepodria respondre a un increment de la va-riabilitat climàtica de les darreres dècades,és a dir, a la successió d’un major nombred’esdeveniments extrems i contrastos mete-orològics més accentuats entre anys conse-cutius.
En definitiva, en la composició isotòpicade carboni dels anells de creixement deles dues espècies estudiades, queda reflec-tit de manera bastant directa l’increment delCO2 atmosfèric. També són enregistradesles conseqüències d’aquest augment, comara els canvis en la variabilitat climàtica de-guts a les alteracions en la circulació atmos-fèrica global.

RESPOSTA DELS ARBRES I DELS BOSCOS AL CANVI CLIMÀTIC 95
Figura 8. Distribució temporal de la freqüència d’anells característics. Es representen els anells molts estrets (gruix infe-rior a 1,5 desviacions estàndard; < 1,5 SD) amb barres per sota del zero. Els anells molt gruixuts (amplada dels anells superiora 1,5 desviacions estàndard; > 1,5 SD) per sobre de la línia zero. Els anells característics han estat quantificats en les sèriesd’índexs de creixement (període 1850-1994) de pi roig, Pinus sylvestris (A); d’avet, Abies alba, (B) i pi negre, Pinus uncinata,en altituds inferiors (C) o superiors (D) de 2.125 m als Pirineus (Tardif et al., 2003).
CANVIS FENOLÒGICS SOBREEL CALENDARI D’ACTIVITATI REPÒS DELS ARBRES
La majoria d’arbres i espècies llenyosesde les zones temperades i boreals aturenel creixement durant l’època desfavorablede tardor i hivern i el reprenen quan lescondicions climàtiques tornen a ser favora-bles a la primavera i a l’estiu. Aquest pa-
tró anual d’activitat i repòs queda reflectiten l’estructura de la fusta que es va acumu-lant en forma de capes concèntriques que,en un tall transversal del tronc, es veuencom anells. En moltes espècies mediterrà-nies, com l’alzina (Quercus ilex), el garric(Quercus coccifera), el llentiscle (Pistacia len-tiscus) o el pi blanc (Pinus halepensis), mal-grat que formen anells de creixement, noacostumen a ser anuals i són molt difícils
96 E. GUTIÉRREZ, M. RIBAS, O. PLANELLS, J. J. CAMARERO I J. TARDIF
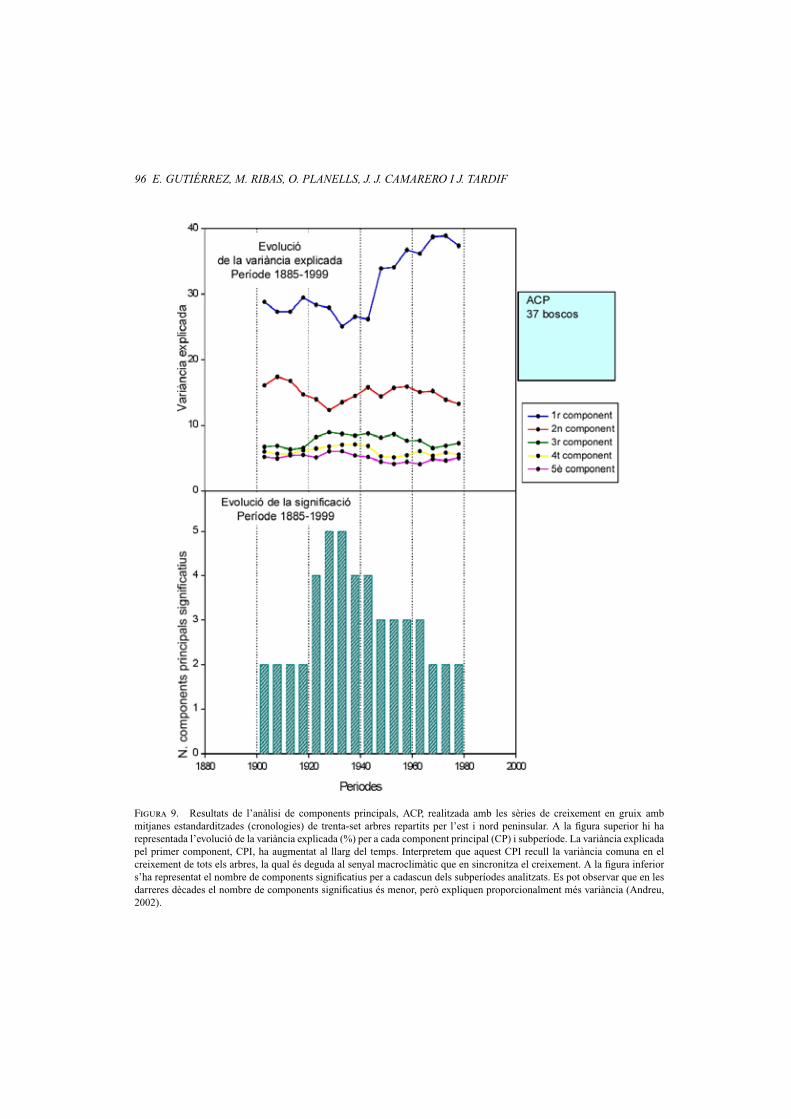
Figura 9. Resultats de l’anàlisi de components principals, ACP, realitzada amb les sèries de creixement en gruix ambmitjanes estandarditzades (cronologies) de trenta-set arbres repartits per l’est i nord peninsular. A la figura superior hi harepresentada l’evolució de la variància explicada (%) per a cada component principal (CP) i subperíode. La variància explicadapel primer component, CPI, ha augmentat al llarg del temps. Interpretem que aquest CPI recull la variància comuna en elcreixement de tots els arbres, la qual és deguda al senyal macroclimàtic que en sincronitza el creixement. A la figura inferiors’ha representat el nombre de components significatius per a cadascun dels subperíodes analitzats. Es pot observar que en lesdarreres dècades el nombre de components significatius és menor, però expliquen proporcionalment més variància (Andreu,2002).
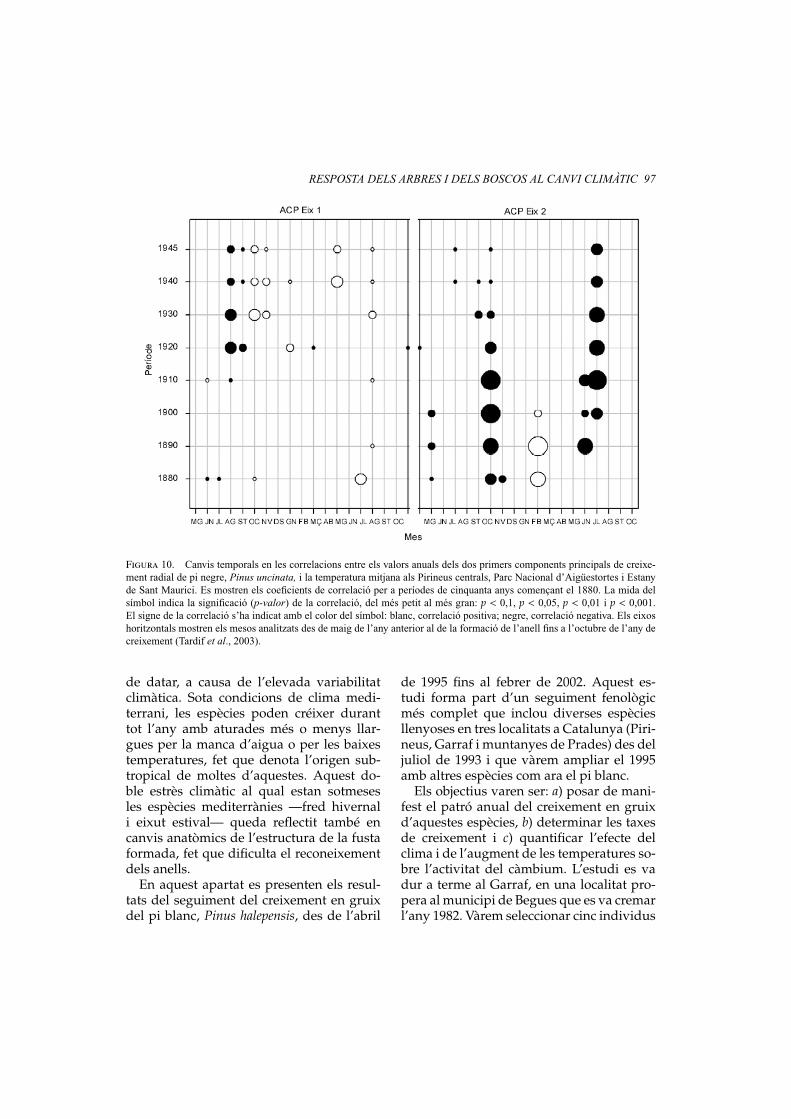
RESPOSTA DELS ARBRES I DELS BOSCOS AL CANVI CLIMÀTIC 97
Figura 10. Canvis temporals en les correlacions entre els valors anuals dels dos primers components principals de creixe-ment radial de pi negre, Pinus uncinata, i la temperatura mitjana als Pirineus centrals, Parc Nacional d’Aigüestortes i Estanyde Sant Maurici. Es mostren els coeficients de correlació per a períodes de cinquanta anys començant el 1880. La mida delsímbol indica la significació (p-valor) de la correlació, del més petit al més gran: p < 0,1, p < 0,05, p < 0,01 i p < 0,001.El signe de la correlació s’ha indicat amb el color del símbol: blanc, correlació positiva; negre, correlació negativa. Els eixoshoritzontals mostren els mesos analitzats des de maig de l’any anterior al de la formació de l’anell fins a l’octubre de l’any decreixement (Tardif et al., 2003).
de datar, a causa de l’elevada variabilitatclimàtica. Sota condicions de clima medi-terrani, les espècies poden créixer duranttot l’any amb aturades més o menys llar-gues per la manca d’aigua o per les baixestemperatures, fet que denota l’origen sub-tropical de moltes d’aquestes. Aquest do-ble estrès climàtic al qual estan sotmesesles espècies mediterrànies —fred hivernali eixut estival— queda reflectit també encanvis anatòmics de l’estructura de la fustaformada, fet que dificulta el reconeixementdels anells.
En aquest apartat es presenten els resul-tats del seguiment del creixement en gruixdel pi blanc, Pinus halepensis, des de l’abril
de 1995 fins al febrer de 2002. Aquest es-tudi forma part d’un seguiment fenològicmés complet que inclou diverses espèciesllenyoses en tres localitats a Catalunya (Piri-neus, Garraf i muntanyes de Prades) des deljuliol de 1993 i que vàrem ampliar el 1995amb altres espècies com ara el pi blanc.
Els objectius varen ser: a) posar de mani-fest el patró anual del creixement en gruixd’aquestes espècies, b) determinar les taxesde creixement i c) quantificar l’efecte delclima i de l’augment de les temperatures so-bre l’activitat del càmbium. L’estudi es vadur a terme al Garraf, en una localitat pro-pera al municipi de Begues que es va cremarl’any 1982. Vàrem seleccionar cinc individus

98 E. GUTIÉRREZ, M. RIBAS, O. PLANELLS, J. J. CAMARERO I J. TARDIF
al vessant nord i cinc més al vessant sud iels vàrem instal.lar dendròmetres de bandaa una alçada superior a 0,5 m de la base. Elsregistres de l’increment en gruix del tronc esvan fer cada vint o trenta dies, depenent del’època de l’any.
A la figura 4 es representa l’evolució anu-al de les taxes de creixement radial per atots els anys d’estudi, a les quals s’ha ajustatuna funció. Aquest ajust s’ha realitzat con-siderant el valors més alts de creixement,cosa que denota les millors condicions peral creixement, i considerant només els va-lors més baixos, és a dir, quan les condi-cions per al creixement són les pitjors delperíode. Les taxes de creixement radial mà-ximes en condicions climàtiques favorablesqueden descrites per les corbes superiors iles taxes més baixes per les corbes inferi-ors. Per tant, les corbes ajustades només re-flecteixen les condicions de mitjana per alperíode estudiat, d’abril de 1995 a febrer de2002.
Com en el cas d’altres pins de zones mésfredes però amb una clara influència medi-terrània del clima, com el pi roig i el pi ne-gre (Camarero et al., 1998), el patró de crei-xement del pi blanc és clarament bimodal:mostren dos màxims de creixement, l’una la primavera i l’altre a la tardor i cadaany s’atura a l’hivern per l’efecte de les bai-xes temperatures. Regularment, els indivi-dus del vessant sud aturen el creixement al’estiu, encara que en anys sense un estrèshídric molt sever poden continuar creixent.Al vessant nord, el creixement s’atura no-més si la sequera és molt intensa, per exem-ple, el 1995.
En aquesta mateixa figura 4 es mostrala relació de les taxes de creixement ambla temperatura mitjana (°C; cercles ver-mells) i la relació amb el quocient de laprecipitació total dividida per la tempe-ratura mitjana, P/TM (mm/°C; trianglesverds) entre dies consecutius de mostreig(Ribas et al., 2003). Aquest índex (factortermopluviomètric de Lang) resulta molt
útil per a l’estudi del clima mediterrani,especialment quan manquen dades ecofi-siològiques, com ara l’evapotranspiracióreal (Fidalgo, 1988). L’índex relaciona lesprecipitacions, i les considera com a factorfavorable per al creixement vegetal, amb latemperatura mitjana, factor que determinales pèrdues d’aigua per evapotranspiració.
Segons els nostres resultats, el pi blancinicia l’activitat del càmbium (creixementradial) quan la temperatura mitjana superaels 8 °C (vegeu la figura 4, línia horitzon-tal; vegeu la figura 5), és a dir, el llindarque determina l’inici i també el cessamentde l’activitat del càmbium és de 8 °C (vegeula figura 5; Ribas et al., 2003). Els resultatsobtinguts en aquest seguiment mostren queel pi blanc, de la mateixa manera que la ma-joria de les espècies de coníferes, té com aúnic mecanisme de control per a iniciar elseu creixement la temperatura.
En relació amb l’inici de l’activitat delcàmbium s’ha detectat a partir dels registresdel creixement radial i la seva relació amb latemperatura (dades diàries de l’estació me-teorològica de Begues, a només 8 km de dis-tància) que en els últims anys s’ha avan-çat l’inici del període de creixement. Efec-tivament, la temperatura llindar s’ha assolitabans en els darrers anys. Per tant, cal es-perar que l’augment general de les tempe-ratures afavoreixi un període de creixementmés llarg. Com es pot apreciar a la figura 6,aquesta temperatura tan sols s’atenyia en elmes de març dels anys setanta i vuitanta,i esporàdicament durant el mes de febrer.També, les temperatures dels mesos de tar-dor (octubre, novembre i desembre) han pu-jat en les últimes dècades. Aquests canvis enles temperatures afavoreixen que el períodedel creixement del pi blanc sigui més llarg.Els efectes són més evidents sobre els pro-cessos (fisiològics i de creixement) i en lesespècies que tenen una major dependènciade la temperatura que del fotoperíode.
La situació descrita per al pi blanc al Gar-raf no és excepcional. L’escalfament gene-

RESPOSTA DELS ARBRES I DELS BOSCOS AL CANVI CLIMÀTIC 99
r= 0,359 ;p < 0,01
18OPinusuncinata (Vinuesa)
Co
efic
ien
ts d
ere
gre
ssió
par
cial
0,25
0,30
0,35
0,40
r=–0,665 ;p< 0,01
Co
efic
ien
ts d
ere
gre
ssió
par
cial
–0,35
–0,30
–0,25
–0,20
–0,15
r= 0,279;p =0,055
18OPinussylvestris (Tejeros)
Co
efic
ien
ts d
ere
gre
ssió
par
cial
0,30
0,35
0,40
r= 0,397 ;p < 0,01
1900 1910 1920 1930 1940
13CPinussylvestris (Tejeros)
Co
efic
ien
ts d
ere
gre
ssió
par
cial
0,20
0,25
0,30
r=–0,046;p= 0,758
Co
efic
ien
ts d
ere
gre
ssió
par
cial
–0,30
–0,25
–0,20
–0,15
r=–0,863;p <0,01
Anyinicial delafinestramòbil decinquanta-dosanys
1900 1910 1920 1930 1940
Co
efic
ien
ts d
ere
gre
ssió
par
cial
–0,40
–0,35
–0,30
–0,25
–0,20
Figura 11. Evolució dels coeficients de regressió parcials corresponents a la temperatura mitjana (columna esquerra, líniesvermelles) i la precipitació total (columna dreta, línies blaves) del quadrimestre maig-agost. Els coeficients han estat estimatsal llarg de tot el segle xx amb regressions múltiples ortogonalitzades (vegeu el text per a més detalls) utilitzant una finestramòbil de cinquanta-dos anys, on els predictands utilitzats han estat les δ18O de Pinus uncinata i Pinus sylvestris i la δ13Cde Pinus sylvestris. Tots els valors dels coeficients de regressió parcials representats són significatius al 95 %. Les línies detendència ajustades són significatives al 99 % en tots els casos, llevat del de la δ18O per a Pinus sylvestris, que és significativaal 90 % per al cas de la temperatura, i no significativa per al de la precipitació (Planells, 2003).
ral provoca l’allargament de l’estació fa-vorable per al creixement de les plantes il’activitat dels animals i modificacions en elcalendari anual d’activitat i repòs (fenolo-gia) dels organismes (Menzel i Fabian, 1999;Inouye et al., 2000). Com a conseqüènciad’aquestes modificacions en els calendaris,en alguns ecosistemes s’han detectat desa-justos en les interaccions tròfiques (Harring-ton et al., 1999).
Al nostre país s’han detectat canvissignificatius en el creixement i la fenolo-
gia d’espècies vegetals i animals com aresposta de l’augment de temperatura il’allargament del període favorable de crei-xement (Peñuelas i Filella, 2001; Dolsa iAlbarán, 1998). En algunes espècies dioi-ques, com Pistacia lentiscus, hem detectat unaugment del nombre dels fruits no fecun-dats (de color roig) sobre els fruits fecundats(de color negre; dades no publicades). Atri-buïm aquesta desproporció a un desajust dela floració dels individus masculins i delsfemenins.

100 E. GUTIÉRREZ, M. RIBAS, O. PLANELLS, J. J. CAMARERO I J. TARDIF
r = 0,580; p < 0,01
1900 1910 1920 1930 1940
13C Pinus uncinata (Vinuesa)
Co
efic
ien
ts d
ere
gre
ssió
par
cial
0,0
0,1
0,2
r = –0,914; p < 0,01
Any inicial de la finestra mòbil de cinquanta-dos anys
1900 1910 1920 1930 1940
Co
efic
ien
ts d
ere
gre
ssió
par
cial
–0,4
–0,3
–0,2
–0,1
0,0
0,1
0,2
Figura 12. Evolució dels coeficients de regressió parcials corresponents a la temperatura mitjana (esquerra, línia vermella)i la precipitació acumulada (dreta, línia blava) del quadrimestre maig-agost. Els coeficients han estat estimats al llarg de tot elsegle xx amb regressions múltiples ortogonalitzades (vegeu el text per a més detalls) utilitzant una finestra mòbil de cinquanta-dos anys, i essent la δ13C de Pinus uncinata el predictand de les anàlisis. Els valors dels coeficients de regressió parcialscorresponents a la temperatura no són significatius en cap cas, però els coeficients de la precipitació esdevenen significatius ala segona meitat del segle xx (la línia discontínua representa el nivell de significació al 95 % per sota del qual els coeficientssón significatius). Les línies de tendència ajustades són significatives al 99 % en tots dos casos (Planells, 2003).
Figura 13. El gruix dels anells (línia discontínua) i la den-sitat de la fusta (línia més gruixuda) mostren una tendènciadecreixent en les últimes dècades. Contràriament, les tem-peratures d’abril a setembre presenten una tendència ascen-dent. Dades de l’Institut Federal Suís de Recerca Forestal,Nival i Paisatgística (WSL, Birmensdorf) corresponents a lamitjana de quatre-centes cronologies d’arbres vius de la zonaboreal de l’hemisferi nord. Des de 1960 la densitat màximade la fusta i el gruix dels anells no es corresponen amb latendència ascendent de l’augment de temperatura.
EL CREIXEMENT DELS ARBRESCOM A REGISTRES DEL CLIMA
L’efecte del canvi climàtic sobre el creixe-ment dels arbres es pot analitzar utilitzant la
informació continguda en les sèries d’anellsde creixement anual, però tal com ja s’ha co-mentat, cal extreure i destriar la informacióprèviament segons l’objectiu de l’estudi. Pertant, per a posar de manifest quin és l’efectedel clima primer hem d’obtenir un nom-bre de sèries representatives d’una locali-tat determinada, i després hem d’establirla sèrie mestra o cronologia fent compa-rables les sèries dels diferents arbres, totestandarditzant-les i, finalment, hem de po-sar de manifest quines són les variables cli-màtiques que tenen efectes significatius so-bre el creixement.
Per a l’estandardització hem fet serviruna funció spline (Cook i Peters, 1981; Cooki Kairiukstis, 1990). El grau de suavitzatde la funció spline l’hem fixat en un 50 %de la freqüència de resposta de trenta-dosanys per ressaltar la variació d’alta freqüèn-cia. Aquesta transformació de les sèries decreixement, a més d’eliminar la tendènciadeguda a l’edat, també filtra altres canvisen el creixement que no estan relacionatsamb el clima. Sobretot, és adient quan els

RESPOSTA DELS ARBRES I DELS BOSCOS AL CANVI CLIMÀTIC 101
Figura 14. Posició del límit de l’arbre (línia gruixuda decolor verd) des de 1.750 m al Tessó de Son (altitud, 2.330m, Pirineu central). Els diferents símbols representen arbresque s’han establert amb èxit a cadascun dels intervals detemps reconstruïts per a aquesta localitat. La unitat de mos-treig que hem utilitzat en aquest tipus d’estudi és una parcel-la de 140 × 30 m. El costat més llarg travessa tot l’ecotò desde l’interior del bosc (part inferior) fins als prats alpins (partsuperior) seguint l’eix altitudinal (modificat de Camarero iGutiérrez, 2003).
arbres experimenten variacions en el creixe-ment a causa de la competència amb els ve-ïns. La diferència entre els valors predits perla funció i els reals és una nova sèrie ano-menada d’índexs de creixement, amb mitjanaconstant.
Les sèries d’índexs encara conservenl’efecte que té el creixement d’un any so-bre el creixement d’anys posteriors. És adir, que no són observacions independents,condició necessària per a moltes anàlisisestadístiques (Monserud, 1986). Per aquestaraó, les sèries d’índexs han estat filtradesde nou amb models autoregressius. Aquesttractament estadístic proporciona les sè-ries residuals d’índexs de creixement que
són amitjanades per establir la cronologiade cada lloc. Hem fet servir una mitjanarobusta, per tal de ponderar l’efecte delsvalors de creixement fora de rang (mitjanarobusta biponderada; Cook, 1985).
Variabilitat de les sèries de creixement
Atès que el clima canvia al llarg deltemps, el creixement dels arbres tambémostra una variabilitat temporal, reflex dela influència del clima. Així, podem quan-tificar la variabilitat recent en les sèriesd’anells deguda al canvi climàtic actual enrelació amb els canvis o episodis climàticsdel passat. Així, un període d’estabilitat enles sèries de creixement (variància baixa)al llarg d’un període de temps pot indicaruna relaxació de l’estrès climàtic. A la fi-gura 7 es representen les sèries del gruixdels anells (dades brutes) de deu arbres. Espoden apreciar els canvis acusats en el rangde creixement.
Les anàlisis de la variabilitat de les sè-ries de creixement es poden fer de diver-ses maneres: a) quantificant els anells ca-racterístics (per exemple, anells molt prims,molt amples o anatòmicament singulars); b)calculant la variabilitat interanual o índexde sensibilitat mitjana; c) calculant la vari-ància de les sèries per períodes; d) mitjan-çant coeficients de correlació entre sèries i d)amb tècniques d’anàlisi multivariant. Ambles dues últimes tècniques es posa de mani-fest, a més, el grau de similitud (o sincro-nització) entre sèries situades en diferentspunts a l’espai (localitats) i també al llargdel temps si es fa per subperíodes.
Com a exemple de sèries d’anells carac-terístics, presentem les obtingudes per a unconjunt de cronologies establertes amb mésde dos-cents arbres corresponents a tres es-pècies: pi roig (Pinus sylvestris), avet (Abiesalba) i pi negre (Pinus uncinata) al Pirineu,al Parc Nacional d’Aigüestortes i Estany de
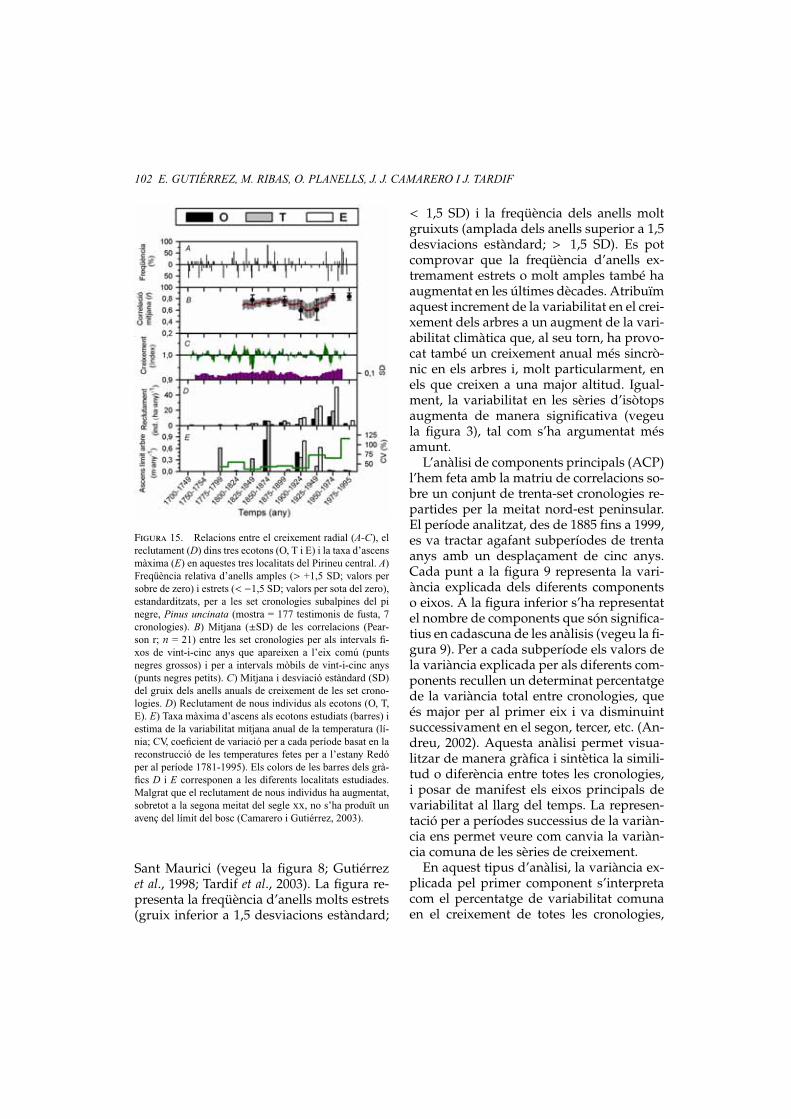
102 E. GUTIÉRREZ, M. RIBAS, O. PLANELLS, J. J. CAMARERO I J. TARDIF
Figura 15. Relacions entre el creixement radial (A-C), elreclutament (D) dins tres ecotons (O, T i E) i la taxa d’ascensmàxima (E) en aquestes tres localitats del Pirineu central. A)Freqüència relativa d’anells amples (> +1,5 SD; valors persobre de zero) i estrets (< −1,5 SD; valors per sota del zero),estandarditzats, per a les set cronologies subalpines del pinegre, Pinus uncinata (mostra = 177 testimonis de fusta, 7cronologies). B) Mitjana (±SD) de les correlacions (Pear-son r; n = 21) entre les set cronologies per als intervals fi-xos de vint-i-cinc anys que apareixen a l’eix comú (puntsnegres grossos) i per a intervals mòbils de vint-i-cinc anys(punts negres petits). C) Mitjana i desviació estàndard (SD)del gruix dels anells anuals de creixement de les set crono-logies. D) Reclutament de nous individus als ecotons (O, T,E). E) Taxa màxima d’ascens als ecotons estudiats (barres) iestima de la variabilitat mitjana anual de la temperatura (lí-nia; CV, coeficient de variació per a cada període basat en lareconstrucció de les temperatures fetes per a l’estany Redóper al període 1781-1995). Els colors de les barres dels grà-fics D i E corresponen a les diferents localitats estudiades.Malgrat que el reclutament de nous individus ha augmentat,sobretot a la segona meitat del segle xx, no s’ha produït unavenç del límit del bosc (Camarero i Gutiérrez, 2003).
Sant Maurici (vegeu la figura 8; Gutiérrezet al., 1998; Tardif et al., 2003). La figura re-presenta la freqüència d’anells molts estrets(gruix inferior a 1,5 desviacions estàndard;
< 1,5 SD) i la freqüència dels anells moltgruixuts (amplada dels anells superior a 1,5desviacions estàndard; > 1,5 SD). Es potcomprovar que la freqüència d’anells ex-tremament estrets o molt amples també haaugmentat en les últimes dècades. Atribuïmaquest increment de la variabilitat en el crei-xement dels arbres a un augment de la vari-abilitat climàtica que, al seu torn, ha provo-cat també un creixement anual més sincrò-nic en els arbres i, molt particularment, enels que creixen a una major altitud. Igual-ment, la variabilitat en les sèries d’isòtopsaugmenta de manera significativa (vegeula figura 3), tal com s’ha argumentat mésamunt.
L’anàlisi de components principals (ACP)l’hem feta amb la matriu de correlacions so-bre un conjunt de trenta-set cronologies re-partides per la meitat nord-est peninsular.El període analitzat, des de 1885 fins a 1999,es va tractar agafant subperíodes de trentaanys amb un desplaçament de cinc anys.Cada punt a la figura 9 representa la vari-ància explicada dels diferents componentso eixos. A la figura inferior s’ha representatel nombre de components que són significa-tius en cadascuna de les anàlisis (vegeu la fi-gura 9). Per a cada subperíode els valors dela variància explicada per als diferents com-ponents recullen un determinat percentatgede la variància total entre cronologies, queés major per al primer eix i va disminuintsuccessivament en el segon, tercer, etc. (An-dreu, 2002). Aquesta anàlisi permet visua-litzar de manera gràfica i sintètica la simili-tud o diferència entre totes les cronologies,i posar de manifest els eixos principals devariabilitat al llarg del temps. La represen-tació per a períodes successius de la variàn-cia ens permet veure com canvia la variàn-cia comuna de les sèries de creixement.
En aquest tipus d’anàlisi, la variància ex-plicada pel primer component s’interpretacom el percentatge de variabilitat comunaen el creixement de totes les cronologies,
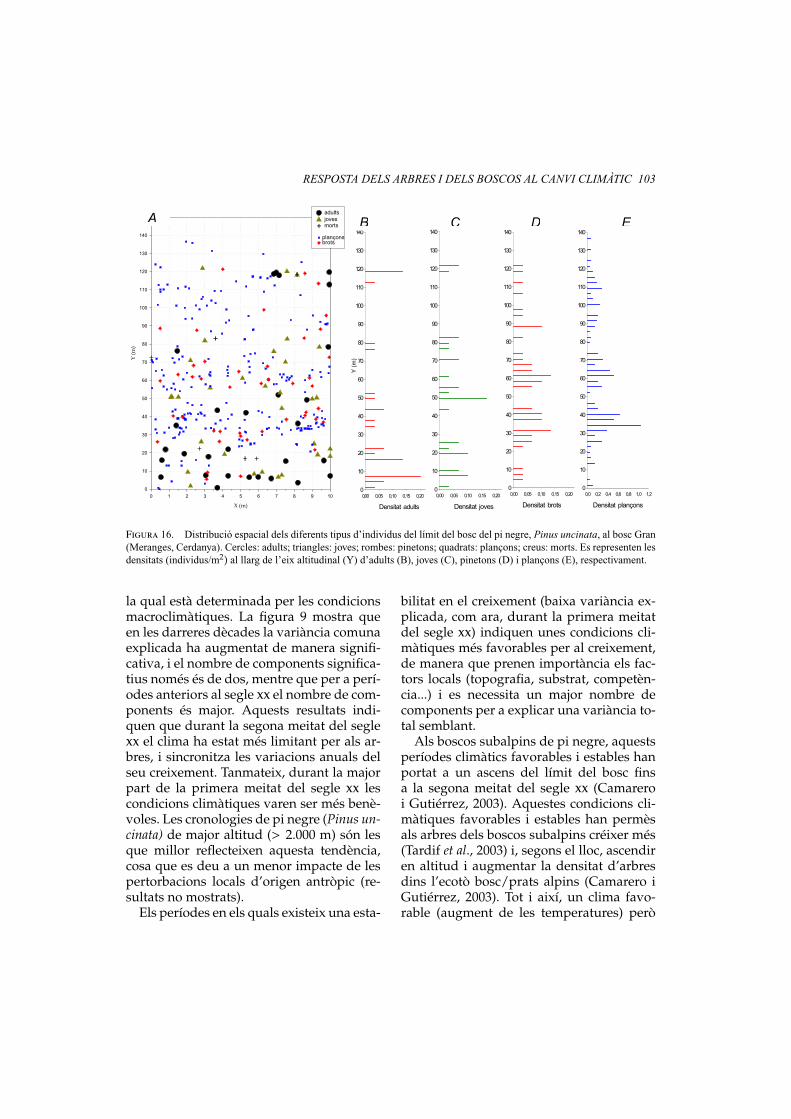
RESPOSTA DELS ARBRES I DELS BOSCOS AL CANVI CLIMÀTIC 103
X (m)
0 1 2 3 4 5 6 7 8 9 10
Y(m
)
0
10
20
30
40
50
60
70
80
90
100
110
120
130
140
150adults
jovesmorts
plançonsbrots
Densitat joves
0,00 0,05 0,10 0,15 0,200
10
20
30
40
50
60
70
80
90
100
110
120
130
140
Densitat adults
0,00 0,05 0,10 0,15 0,20
Y (
m)
0
10
20
30
40
50
60
70
80
90
100
110
120
130
140
Densitat brots
0,00 0,05 0,10 0,15 0,200
10
20
30
40
50
60
70
80
90
100
110
120
130
140
Densitat plançons
0,0 0,2 0,4 0,6 0,8 1,0 1,20
10
20
30
40
50
60
70
80
90
100
110
120
130
140
A B C D E
Figura 16. Distribució espacial dels diferents tipus d’individus del límit del bosc del pi negre, Pinus uncinata, al bosc Gran(Meranges, Cerdanya). Cercles: adults; triangles: joves; rombes: pinetons; quadrats: plançons; creus: morts. Es representen lesdensitats (individus/m2) al llarg de l’eix altitudinal (Y) d’adults (B), joves (C), pinetons (D) i plançons (E), respectivament.
la qual està determinada per les condicionsmacroclimàtiques. La figura 9 mostra queen les darreres dècades la variància comunaexplicada ha augmentat de manera signifi-cativa, i el nombre de components significa-tius només és de dos, mentre que per a perí-odes anteriors al segle xx el nombre de com-ponents és major. Aquests resultats indi-quen que durant la segona meitat del seglexx el clima ha estat més limitant per als ar-bres, i sincronitza les variacions anuals delseu creixement. Tanmateix, durant la majorpart de la primera meitat del segle xx lescondicions climàtiques varen ser més benè-voles. Les cronologies de pi negre (Pinus un-cinata) de major altitud (> 2.000 m) són lesque millor reflecteixen aquesta tendència,cosa que es deu a un menor impacte de lespertorbacions locals d’origen antròpic (re-sultats no mostrats).
Els períodes en els quals existeix una esta-
bilitat en el creixement (baixa variància ex-plicada, com ara, durant la primera meitatdel segle xx) indiquen unes condicions cli-màtiques més favorables per al creixement,de manera que prenen importància els fac-tors locals (topografia, substrat, competèn-cia...) i es necessita un major nombre decomponents per a explicar una variància to-tal semblant.
Als boscos subalpins de pi negre, aquestsperíodes climàtics favorables i estables hanportat a un ascens del límit del bosc finsa la segona meitat del segle xx (Camareroi Gutiérrez, 2003). Aquestes condicions cli-màtiques favorables i estables han permèsals arbres dels boscos subalpins créixer més(Tardif et al., 2003) i, segons el lloc, ascendiren altitud i augmentar la densitat d’arbresdins l’ecotò bosc/prats alpins (Camarero iGutiérrez, 2003). Tot i així, un clima favo-rable (augment de les temperatures) però

104 E. GUTIÉRREZ, M. RIBAS, O. PLANELLS, J. J. CAMARERO I J. TARDIF
molt inestable (augment de la variabilitatclimàtica) pot comportar un major estrès cli-màtic, tal com demostra l’aparició d’estrèsper sequera en arbres dels boscos boreals(Barber et al., 2000) o el desajust de les re-lacions clima-creixement (Briffa et al., 1998)que han constituït la base de múltiples re-construccions dendroclimàtiques.
INESTABILITAT DELS REGISTRESDENDROCRONOLÒGICS AMB ELCLIMA
Relacions creixement-clima
El canvi climàtic afecta de molt diver-ses maneres el creixement i el desenvolu-pament dels arbres. En principi, un aug-ment de temperatura pot produir un majorcreixement, atès que la temperatura exer-ceix un efecte positiu sobre les taxes decreixement dels arbres si no hi ha mancad’aigua. Així succeeix efectivament en algu-nes zones (Alekseev i Soroka, 2002), però end’altres els resultats són diferents (Barber etal., 2000).
Per posar de manifest les relacions crei-xement-clima, primer vàrem calcular vuitACP basades en les sèries de creixement dePinus uncinata al Parc Nacional d’Aigües-tortes i Estany de Sant Maurici, per a dife-rents períodes temporals de cinquanta anys,començant el 1880 i decalats deu anys, ex-cepte l’últim subperíode: 1880-1929, 1890-1939, 1900-1949, 1910-1959, 1920-1969, 1930-1979, 1940-1989 i 1945-1994 (vegeu la figura10). Per estimar les relacions creixement-clima vàrem fer servir el primer i el segoncomponents en lloc de les cronologies, jaque tots els arbres presentaven un elevatpercentage de variabilitat en comú. El pri-mer component (CPI) representa la varia-bilitat en comú deguda al senyal macrocli-màtic, i el segon separa les cronologies se-gons la seva altitud, tot seguint el gradi-
ent de temperatura associat (Gutiérrez et al.1998; Tardif et al., 2003). Després, vàrem cal-cular les correlacions dels dos primers com-ponents principals amb les temperatures,usant l’índex r de Pearson. Les temperatu-res emprades són la mitjana regional permesos obtingudes a partir de les dades deles estacions següents: Viella, Arties, Tredòs,Cabdella i pic del Migdia (vegeu la figura10).
En general, les relacions clima-creixementde Pinus uncinata als Pirineus centrals sónmolt diferents entre la primera i la segonameitats del segle xx (vegeu la figura 10).Com es pot veure, les correlacions amb elprimer component són molt baixes durantla primera meitat del segle xx però no du-rant la segona. Això vol dir que les condi-cions macroclimàtiques han pres més im-portància i s’han tornat més limitants peral creixement des de mitjan segle xx. Lescorrelacions amb el segon component delcreixement, que representa el gradient al-titudinal, es comporten justament de ma-nera contrària; a principis i mitjan segle xxel gradient altitudinal era molt fort però du-rant la segona meitat del segle xx es relaxai es perd la influència de les variables cli-màtiques sobre el creixement. Així mateix,les clàssiques relacions positives del crei-xement radial (en aquest estudi represen-tat pel primer component) i la temperaturamitjana de l’octubre anterior a l’any de crei-xement no serien importants fins al període1930-1979, és a dir, quan la majoria d’anysestudiats corresponen a la segona meitat delsegle xx. Un resultat similar s’observa ambles temperatures de maig de l’any de creixe-ment, que es tornen importants a partir de1940-1989. Les relacions amb els valors delsegon component destaquen per la presèn-cia de relacions negatives amb les tempera-tures de juliol de l’any de creixement, quepodrien indicar un cert estrès hídric, especi-alment intens a partir de 1910.
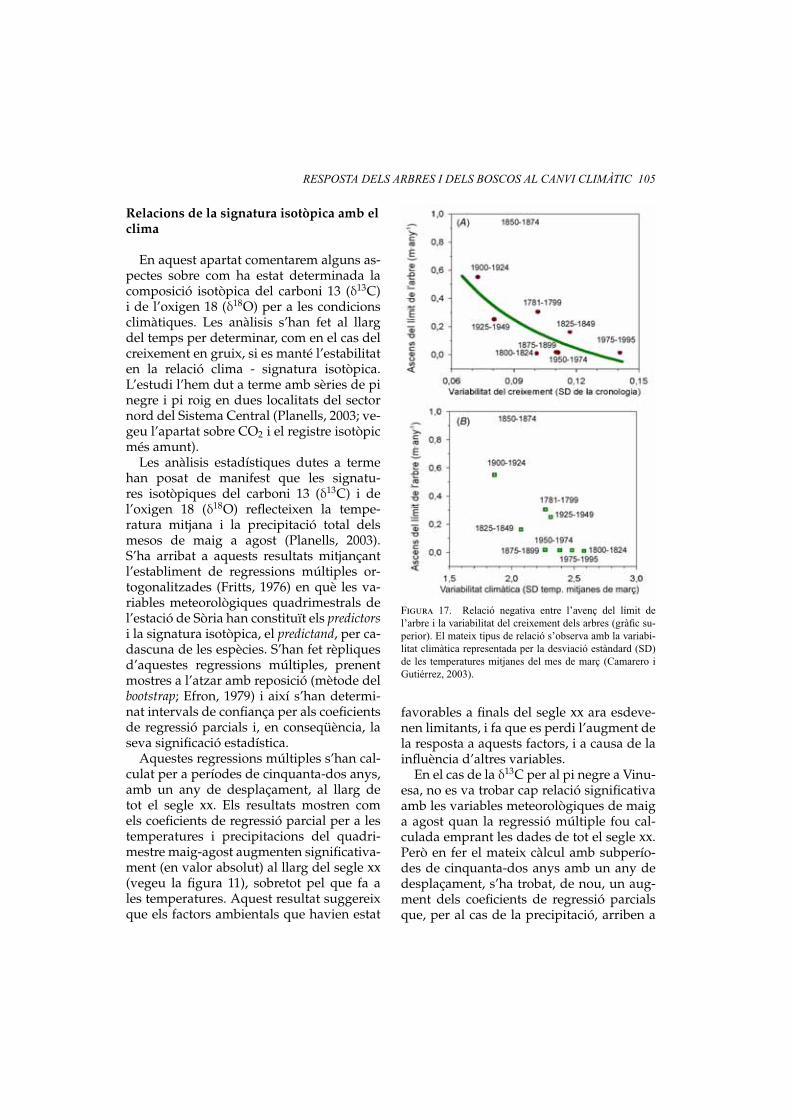
RESPOSTA DELS ARBRES I DELS BOSCOS AL CANVI CLIMÀTIC 105
Relacions de la signatura isotòpica amb elclima
En aquest apartat comentarem alguns as-pectes sobre com ha estat determinada lacomposició isotòpica del carboni 13 (δ13C)i de l’oxigen 18 (δ18O) per a les condicionsclimàtiques. Les anàlisis s’han fet al llargdel temps per determinar, com en el cas delcreixement en gruix, si es manté l’estabilitaten la relació clima - signatura isotòpica.L’estudi l’hem dut a terme amb sèries de pinegre i pi roig en dues localitats del sectornord del Sistema Central (Planells, 2003; ve-geu l’apartat sobre CO2 i el registre isotòpicmés amunt).
Les anàlisis estadístiques dutes a termehan posat de manifest que les signatu-res isotòpiques del carboni 13 (δ13C) i del’oxigen 18 (δ18O) reflecteixen la tempe-ratura mitjana i la precipitació total delsmesos de maig a agost (Planells, 2003).S’ha arribat a aquests resultats mitjançantl’establiment de regressions múltiples or-togonalitzades (Fritts, 1976) en què les va-riables meteorològiques quadrimestrals del’estació de Sòria han constituït els predictorsi la signatura isotòpica, el predictand, per ca-dascuna de les espècies. S’han fet rèpliquesd’aquestes regressions múltiples, prenentmostres a l’atzar amb reposició (mètode delbootstrap; Efron, 1979) i així s’han determi-nat intervals de confiança per als coeficientsde regressió parcials i, en conseqüència, laseva significació estadística.
Aquestes regressions múltiples s’han cal-culat per a períodes de cinquanta-dos anys,amb un any de desplaçament, al llarg detot el segle xx. Els resultats mostren comels coeficients de regressió parcial per a lestemperatures i precipitacions del quadri-mestre maig-agost augmenten significativa-ment (en valor absolut) al llarg del segle xx(vegeu la figura 11), sobretot pel que fa ales temperatures. Aquest resultat suggereixque els factors ambientals que havien estat
Figura 17. Relació negativa entre l’avenç del límit del’arbre i la variabilitat del creixement dels arbres (gràfic su-perior). El mateix tipus de relació s’observa amb la variabi-litat climàtica representada per la desviació estàndard (SD)de les temperatures mitjanes del mes de març (Camarero iGutiérrez, 2003).
favorables a finals del segle xx ara esdeve-nen limitants, i fa que es perdi l’augment dela resposta a aquests factors, i a causa de lainfluència d’altres variables.
En el cas de la δ13C per al pi negre a Vinu-esa, no es va trobar cap relació significativaamb les variables meteorològiques de maiga agost quan la regressió múltiple fou cal-culada emprant les dades de tot el segle xx.Però en fer el mateix càlcul amb subperío-des de cinquanta-dos anys amb un any dedesplaçament, s’ha trobat, de nou, un aug-ment dels coeficients de regressió parcialsque, per al cas de la precipitació, arriben a

106 E. GUTIÉRREZ, M. RIBAS, O. PLANELLS, J. J. CAMARERO I J. TARDIF
ser significatius al 95 % a finals de segle (ve-geu la figura 12). Aquest resultat corroborala hipòtesi del paràgraf anterior, i mostratambé com un canvi envers un clima mésrestrictiu provoca una major sincronitzaciódel creixement entre poblacions i espècies.Efectivament, la relació entre la δ13C de Pi-nus uncinata a Vinuesa i les variables mete-orològiques ha canviat al llarg del segle xx,atès que l’assimilació dels isòtops de δ13C tétendència a respondre a aquells factors quemés limiten el creixement (la meteorologiade maig a agost).
Així, a les dues localitats del Sistema Ibè-ric estudiades s’observa una uniformitza-ció de la resposta dels arbres al clima, quequeda reflectida en la seva composició iso-tòpica, fet que corrobora la cada vegada méselevada sincronització del creixement entreels diferents boscos de l’est i nord peninsu-lars.
EFECTES SOBRE LA DENSITATDE LA FUSTA
Segons dades de l’Institut Federal Suís deRecerca Forestal, Nival i Paisatgística, WSL(vegeu la figura 13) que concorden amb elsobtinguts per Briffa et al. (1998), la densitatde la fusta i el creixement dels arbres mos-tren tendències divergents en relació ambl’augment de les temperatures en les zo-nes temperades-fredes de l’hemisferi nord(vegeu la figura 13). A l’espai, aquesta ten-dència s’estén per àmplies zones d’Euràsia iAmèrica del Nord (Briffa et al., 1998).
Aquest fet podria explicar-se perquès’hauria sobrepassat el llindar òptim deresposta a l’escalfament; aquestes pobla-cions responen de manera negativa a unmajor estrès hídric, fins i tot a latitudselevades (Barber et al., 2000). També, ladisminució de densitat de la fusta podriatenir implicacions profundes en el funcio-nament i en l’estructura dels boscos, ja que
s’afavoriria una descomposició més ràpidaper part dels organismes. Addicionalment,l’activitat dels descomponedors també esveuria accelerada per les temperatures mésaltes.
L’ECOTÒ BOSCOS SUBALPINS/PRATSALPINS ALS PIRINEUS
Les poblacions d’arbres, als marges de laseva distribució són molt sensibles a la va-riabilitat climàtica (Tranquillini, 1979; Bru-baker, 1986). Per aquesta raó, la zona detransició o ecotò (Van der Maarel, 1990) en-tre el límit altitudinal del bosc i els prats al-pins és un bon monitor dels canvis globalsdel sistema (Kullman, 1979; Payette i Filion,1985; Lavoie i Payette, 1992: Slatyer i Noble,1992; Weisberg i Baker, 1995; Lloyd, 1996).L’ecotò, pròpiament dit, és l’àrea compresaentre el límit del bosc (zona boscosa densaa major altitud) i el límit de l’arbre (altitudmàxima a la qual es troben arbres amb untronc principal de dos metres d’alçada).
Actualment, l’interès científic està centraten la taxa d’ascens del límit del bosc degudaal canvi climàtic, als canvis d’ús del sòl, al’augment en la concentració de diòxid decarboni i a l’increment de les deposicionsde nitrogen. En els nostres estudis ens cen-trem fonamentalment en els efectes que té elcanvi climàtic (augment de les temperaturesi de la variabilitat del clima), i el canvi en elsusos del sòl a causa d’un descens de les acti-vitats ramaderes i agrícoles tradicionals alsPirineus (Bas et al., 1994; Bringue, 1995).
El canvi climàtic i els canvis en els usosdel sòl impliquen modificacions diferentsde les condicions en les quals es desen-volupen les poblacions i, per tant, es pro-dueix un canvi en la dinàmica demogrà-fica d’aquestes poblacions i, com a resul-tat, en l’estructura general del paisatge ons’ubiquen (Kullman, 1990; Lavoie i Payette,1994; Szeicz i Macdonald, 1995; Lloyd,

RESPOSTA DELS ARBRES I DELS BOSCOS AL CANVI CLIMÀTIC 107
1996). Els límits forestals altitudinals nopertorbats localment per l’home es loca-litzen on les condicions climàtiques sóntan severes que no permeten el creixementde vegetals amb port arbori. Per això, unamodificació deguda a pertorbacions locals(pastura, tales, incendis) en les condicionsen les quals es troben pot tenir efectes im-portants en la seva estructura i dinàmica.L’estudi d’aquests canvis és important per adeterminar l’efecte que aquests poden tenirsobre la biodiversitat dels ecotons alpinsi les comunitats supraforestals (Camareroi Gutiérrez, 1999). Poder predir l’evoluciódels ecotons boscos subalpins/prats alpinses converteix també en una eina de gestiómolt important per a preservar els hàbitatsi les comunitats supraforestals alpines, al-guns relictuals i restringits a pics aïllats, ique podrien ser envaïts pels boscos subal-pins.
Malgrat els augments espectaculars detemperatura que s’han produït en el Piri-neu central (Bücher i Dessens, 1991), el lí-mit de l’arbre de l’ecotò boscos/prats alpinsen diversos llocs del Pirineu no s’ha despla-çat a cotes superiors en els últims cinquantaanys (vegeu la figura 14; Camarero i Guti-érrez, 2003). Les primeres hipòtesis formu-lades pronosticaven un avenç del límit del’arbre a causa de l’escalfament global, jaque el període favorable per al creixementseria més llarg a les zones d’elevades alti-tuds i latituds. Aquesta hipòtesi s’ha com-provat en alguns llocs de latituds eleva-des on el bosc ha avançat cap a la tundra(Kullman, 2000). Els nostres resultats, des-prés d’estudiar diversos llocs no pertorbatsrecentment per activitats humanes als Piri-neus, demostren que els límits del bosc i del’arbre no s’han desplaçat a cotes superiorsdurant la segona meitat del segle xx (vegeules figures 14 i 15). El que sí que ha aug-mentat és la densitat d’arbres dins de l’ecotóbosc subalpí/prats alpins.
Les anàlisis estadístiques que hem realit-
zat amb les variables climàtiques posen demanifest que la variabilitat del clima es re-laciona de manera inversa amb l’ascens dellímit del bosc i, per tant, amb l’establimentamb èxit dels nous individus (vegeu la fi-gura 16). Així mateix, hem demostrat quehi ha una relació inversa i significativa en-tre l’ascens del límit de l’arbre i el creixe-ment (vegeu la figura 16). Els arbres ques’estableixen fora de l’àrea d’influència (mi-croclima) que generen els arbres grans o lesformes arbustives d’arbres (krummholz) mo-ririen per manca de la protecció necessà-ria. Aquesta interacció positiva (facilitació)acostuma a produir-se en ambients durscom els que es donen en cotes elevades. Siaquesta interacció positiva afecta la dinà-mica de l’ecotò en relació amb la principalpertorbació, el clima, la resposta de l’ecotòal canvi climàtic actual pot ser no lineal ipot afectar la flora alpina de manera sob-tada, atès que espècies com el pi negre sónpotencialment bones invasores.
Actualment, els nostres esforços se cen-tren a comparar l’estructura i la dinàmicad’un grup d’ecotons representatius al llargdels Pirineus, incloent-hi llocs pertorbatslocalment per l’home (ramaderia, tales, in-cendis) i llocs no pertorbats localment peròafectats pel canvi climàtic regional, perquantificar la influència del canvi climàtici d’usos del sòl sobre l’ascens del límit al-titudinal del bosc. La figura 17 mostra elsresultats preliminars del patró espacial enun ecotò no pertorbat localment situat aMeranges, a la Cerdanya. Es poden apre-ciar les ones de colonització descrites per ladensitat dels plançons de pi negre al llargde l’eix altitudinal. Aquest mateix patró java ser descrit en altres límits de boscs dis-tants i topogràficament molt diferents, peròtambé poc afectats per activitats humaneslocals, com Las Cutas (Ordesa) (Camareroi Gutiérrez, 1999). És aquest patró espacialun indicatiu de l’avenç natural del límit acausa únicament d’efectes climàtics?

108 E. GUTIÉRREZ, M. RIBAS, O. PLANELLS, J. J. CAMARERO I J. TARDIF
AGRAÏMENTS
La recerca ha estat finançada gràcies alsprojectes següents:
Respuestas fisiológicas, anatómicas y morfo-lógicas de las especies leñosas del encinar en ungradiente atlántico-mediterráneo. Núm. ref.:CICYT FOR91-0689. Ministeri d’Educació iCiència. 1992-1994.
Reconstrucciones dendroclimáticas y efectodel aumento de temperatura durante los últi-mos cien años en el límite superior del bosquede Pinus uncinata. Núm. ref.: AMB95-0160CICYT del Programa Nacional del Pla Na-cional de R+D. 1995-1998.
Projecte europeu: ForMAT Sensitivity oftree-growth to climate change and growth mo-delling from past to future. Contracte núm.:ENV4-CT97-0641. 1998-2001.
Vulnerabilidad del paisaje supraforestal pi-renaico frente al ascenso del límite superiordel bosque. Ref. REN2002-04268-02-021/ELOdel Programa Nacional del Pla Nacional deR+D+I. 2002-2005.
Projecte europeu: ISONET 400 years ofannual reconstructions of European climatevariability using a high resolution ISOtopicNETwork. Contracte núm.: EVK2-2001-00237. 2002-2005.
El doctor Jacques Tardif va treballar ambnosaltres gràcies a una beca del Programade Cooperació Catalunya-Quebec.
REFERÈNCIES
Alekseev, A. S.; Soroka, A. R. (2002). «Scots pinegrowth trends in northwestern Kola peninsula as anindicator of positive changes in the carbon cycle».Climatic Change, vol. 55, pàg. 183-196.
Andreu, L. (2002). Anàlisi de la variabilitat espai-temporaldel creixement anual de tres espècies de pi a l’est inord peninsulars. Universitat de Barcelona, Facul-tat de Biologia, Departament d’Ecologia. [Treballd’investigació realitzat per la obtenció de la Titu-lació en Estudis Avançats en Ecologia]
Barber, V. A.; Juday, G. P.; Finney, B. P. (2000). «Re-duced growth of Alaskan white spruce in the twen-
tieth century from temperature-induced droughtstress». Nature, vol. 405, pàg. 668-672.
Bas, J. (1993). Les pastures supraforestals a la Vall Ferrerai la Vall de Cardós (Pallars Sobirà). Valoració de la capa-citat ramadera de les pastures de Lladorre. Universitatde Lleida, Escola Tècnica d’Agricultura. [M. Sc. Tesi]
Bosch, O.; Gutiérrez, E. (1999). «La sucesión en losbosques de Pinus uncinata del Pirineo. De los anil-los de crecimiento a la historia del bosque». Ecología,vol. 13, pàg. 133-171.
Briffa, K. R.; Schweingruber, F. H.; Jones, P. D.; Os-born, T. J.; Shiyatov, S. G.; Vaganov, E. A. (1998).«Reduced sensitivity of recent tree-growth to tem-perature at high northen latitudes». Nature, vol. 391,pàg. 678-682.
Bringue, G. M. (1995). Comunitats i bens comunals alsPallars Sobirà, segles XV-XVIII. Barcelona: Univer-sitat Pompeu Fabra, Departament d’Història. [Tesidoctoral]
Brubaker, L. B. (1986). «Responses of tree populationsto climatic change». Vegetatio, vol. 67, pàg. 119-130.
Bücher, A.; Dessens, J. (1991). «Secular trend of sur-face temperature at an elevated observatory in thePyrenees». Journal of Climate, vol. 4, pàg. 859-868.
Camarero, J. J. (2000). Growth and regeneration patternsand processes in Pinus uncinata Ram. treeline ecotones inthe Pyrenees and in an isolated population in the Westerndistribution limit in Spain. Universitat de Barcelona.[Tesi doctoral]
Camarero, J. J.; Guerrero-Campo, J.; Gutiérrez, E.(1998). «Tree-ring growth and structure of Pinusuncinata and Pinus sylvestris in the Central Span-ish Pyrenees». Arctic, Antarctic, and Alpine Research,vol. 30, pàg. 1-10.
Camarero, J. J.; Gutiérrez, E. (1999). «Structure andrecent recruitment at alpine forest-pasture ecotonesin the Spanish Central Pyrenees». Écoscience, vol. 6,pàg. 451-464.
— (2003). «Pace and pattern of recent treeline dynam-ics: response of ecotones to climatic variability in theSpanish Pyrenees». Climatic change. [En premsa]
Carreras, J.; Carrillo, E.; Masalles, R. M.; Ninot,J. M.; Soriano, I.; Vigo, J. (1995). «Delimitation ofthe supra-forest zone in the Catalan Pyrenees». Bull.Soc. Linn. Provence, vol. 47, pàg. 27-36.
Cook, E. R. (1985). A time series analysis approach to tree-ring standardization. Tucson: University of Arizona,Lab. of Tree-Ring Research. [Tesi doctoral]
Cook, E. R.; Kairiukstis, L. A. (1990). Methods of den-drochronology. Boston: Kluwer Academic Publishers.
Cook, E. R.; Peters, K. (1981). «The smoothing spline:A new approach to standardizing forest interiortree-ring width series for dendroclimatic studies».Tree-Ring Bulletin, vol. 41, pàg. 45-53.

RESPOSTA DELS ARBRES I DELS BOSCOS AL CANVI CLIMÀTIC 109
Dolsa, A. G.; Albarán, M. T. (1998). «Aproximació ales poblacions dels lepidòpters heliòfils de les vallsde l’Escrita i Peguera». A: La investigació al Parc Na-cional d’Aigüestortes i Estany de Sant Maurici: IV Jor-nades de Recerca. Pàg. 189-207.
Efron, B. (1979). «Bootstrap methods: another look atthe jackknife». The Annals of Statistics, vol. 7, pàg. 1-26.
Ehleringer, J. R.; Farquhar, G. D.; Hall, A. E. (1993).Stable isotopes and carbon-water relations. San Diego:Academic Press.
Farquhar, G. D.; O’Leary, M. H.; Berry, J. A. (1982).«On the relationship between carbon isotope dis-crimination and the intercellular carbon dioxideconcentration of leaves». Australian Journal of PlantPhysiology, vol. 9, pàg. 121-137.
Feng, X. (1998). «Long-term ci/ca response of trees inwestern North America to atmospheric CO2 con-centration derived from carbon isotope chronolo-gies». Oecologia, vol. 117, núm. 19-25.
Feng, X.; Epstein, S. (1995). «Carbon isotopes of treesfrom arid environments and implications for recon-structing atmospheric CO2 concentration». Geochim-ica et Cosmochimica Acta, vol. 59, núm. 12, pàg. 2599-2608.
Fernández, A.; Génova, M.; Creus, J.; Gutiérrez, E.(1996). «Dendroclimatological investigation cover-ing the last 300 years in Central Spain». A: J. S.Dean; D. M. Meko; T. W. Swetnam [ed.], Tree rings,environment and humanity. Radiocarbon, special issue.Pàg. 181-190.
Fidalgo, C. (1988). Metodologia fitoclimática. Edicionesde la Universidad Autónoma de Madrid. 123 pàg.
Font Tullot, I. (1988). Historia del clima de España. Cam-bios climáticos y sus causas. Madrid: INM.
Friedli, H.; Lötscher, H.; Oeschger, H.; Siegen-thaler, U.; Stauffer, B. (1986). «13C/12C ratio ofatmospheric CO2 in the past two centuries». Nature,vol. 324, pàg. 237-238.
Fritts, H. C. (1976). Tree, rings and climate. Nova York:Academic Press.
Gutiérrez Merino, E.; Camarero Martínez, J. J.;Tardif, J.; Bosch, O.; Ribas, M. (1998). «Tendenciasrecientes del creixement i la regeneración en boscossubalpinos del Parque Nacional de Aigüestortes iEstany de Sant Maurici». Ecologia, vol. 12, pàg. 251-283.
Harrington, R.; Woiwod, I.; Sparks, T. (1999). «Cli-mate change and trophic interactions». TREE,vol. 14, núm. 4, pàg. 146-150.
Holmes, R. L. (1992). Dendrochronology program libraryversion 1992-1. Tucson: University of Arizona, Lab.of Tree-Ring Research.
Inouye, D. W.; Barr, B.; Armitage, K. B.; Inouye,B. D. (2000). «Climate change is affecting altitudinalmigrants and hibernating species». PNAS, vol. 97,núm. 4, pàg. 1630-1633.
IPCC (1997). «Special report on the regional impactsof climate change. An assessment of vulnerability.Chapter 5: Europe». <http://www.grida.no/climate/ipcc/regional/index.htm.>
— (2001). «Canvi climàtic. Informe». <http://www.ipcc.ch/pub/un/syrspanish/spm.pdf.>
Keeling, C. D.; Mook, W. G.; Tans, P. P. (1979). «Recenttrends in the 13C/12C ratio of atmospheric carbondioxide». Nature, vol. 277, pàg. 121-123.
Keeling, C. D.; Whorf, T. P. (1999). «Atmospheric CO2records from sites in the SIO air sampling network».A: Trends: A compendium of data on global change. OakRidge, Tenn: Oak Ridge National Laboratory, Car-bon Dioxide Information Analysis Center.
Körner, C. (1999). Alpine plant life: functional ecologyof high mountain ecosystems. Berlín: Springer-Verlag.338 pàg.
Kullman, L. (1979). «Change and stability in the alti-tude of the birch tree-limit in the Southern SwedishScandes 1915–1975». Acta Phytogeogr. Suec., vol. 65,pàg. 1-121.
— (1990). «Dynamics of altitudinal tree-limits in Swe-den: a review». Norsk Geografisk Tidsskrift, vol. 44,pàg. 103-116.
— (2000). «Warmer climate and tree-limit rise inthe Swedish Scandes». Fauna-och-Flora-Stockholm,vol. 95, núm. 3, pàg. 113-129.
Lajhta, K.; Michener, R. H. (1994). Stable isotopes inecology and environmental science. Oxford: BlackwellScientific Publications.
Lavoie, C.; Payette, S. (1992). «Black spruce growth-forms as a record of a changing winter environmentat tree line, Québec, Canada». Arct. Alp. Res., vol. 24,pàg. 40-49.
— (1994). «Recent fluctuations of the lichen-spruce for-est limit in subarctic Québec». J. Ecol, vol. 82, pàg.725-734.
Lloyd, A. H. (1996). Patterns and processes of tree lineresponse to late Holocene climate change in the SierraNevada, California. Tucson: University of Arizona.[Tesi doctoral]
Maarel, E. van der (1990). «Ecotones and ecoclinesare different». J. Veg. Sci., vol. 1, pàg. 135-138.
Martín, J.; Olcina, J. (2001). Climas y tiempos de Es-paña. Madrid: Alianza.
Menzel, A.; Fabian, P. (1999). «Growing season ex-tended in Europe». Nature, vol. 397, pàg. 659.
Monserud, R. A. (1986). «Time-series analyses of tree-ring chronologies». Forest Science, vol. 32, pàg. 349-372.
Muntán, E.; Andreu, L.; Oller, P.; Gutiérrez, E.;Martínez, P. (2003). «Dendrochronological study ofthe avalanche path Canal del Roc Roig. First resultsof the ALUDEX Project in the Pyrenees». Annals ofGlaciology. [En premsa]

110 E. GUTIÉRREZ, M. RIBAS, O. PLANELLS, J. J. CAMARERO I J. TARDIF
Payette, S.; Filion, L. (1985). «White spruce expansionat the tree line and recent climatic change». Can. J.For. Res., vol. 15, pàg. 241-251.
Peñuelas, J. (2001). «Cambios atmosféricos y climáti-cos y sus consecuencias sobre el funcionamiento yla estructura de los ecosistemas terrestres medite-rráneos. Ecosistemas Mediterráneos». Análisis fun-cional. CSIC-AEET, pàg. 423-455.
Peñuelas, J.; Filella, I. (2001). «Responses to a warm-ing world». Science, vol. 294, pàg. 793-795.
Peñuelas, J.; Filella, I.; Comas, P. (2002). «Changedplant and animal life cycles from 1952 to 2000 in theMediterranean region». Global Change Biol., vol. 8,pàg. 531-544.
Piñol, P.; Terradas, J.; Lloret, F. (1998). «Climatewarming, wildfire hazard, and wildfire occurrencein coastal eastern Spain». Climate Change, vol. 38,pàg. 345-357.
Planells, O. (2003). Variabilitat del gruix i de la composi-ció isotòpica dels anells de creixement de Pinus uncinatai Pinus sylvestris al Sistema Ibèric i la seva relació ambel clima. Universitat de Barcelona, Facultat de Biolo-gia, Departament d’Ecologia. [Treball d’investigaciórealitzat per a l’obtenció de la Titulació en EstudisAvançats en Ecologia]
Ribas, M.; Martín, E.; Gutiérrez, E. (2003). «Phenol-ogy of Pinus halepensis radial growth temporal pat-tern and its relationship with climatic conditions».[En revisió]
Schweingruber, F. H. (1996). Tree rings and environ-ment: dendroecology. Berna: Swiss Federal Institutefor Forest, Snow and Landscape Research, Birmens-dorf, Paul Haupt Verlag.
Shaver, G. R.; Canadell, J.; Chapin III, F. S.; Gure-vitch, J.; Harte, J.; Henry, G.; Ineson, P.; Jonas-son, S.; Melillo, J.; Pitelka, L.; Rustad, L. (2000).«Global warming and terrestrial ecosystems: a con-ceptual framework for analysis». BioScience, vol. 50,núm. 10, pàg. 871-882.
Slatyer, R. O.; Noble, I. R. (1992). «Dynamic of Mon-tane Treelines». A: A. J. Hansen; F. di Castri [ed.],Landscape boundaries: consequences for biotic diversityand ecological flows. Nova York: Springer-Verlag, pàg.346-359.
Stuiver, M. (1978). «Atmospheric carbon dioxideand carbon reservoir changes». Science, vol. 199,núm. 4326, pàg. 253-259.
Szeicz, J. M.; MacDonald, G. M. (1995). «Recent whitespruce dynamics at the Subarctic alpine tree Line ofNorth-Western Canada». J. Ecol., vol. 83, pàg. 873-885.
Tardif, J.; Camarero, J. J.; Ribas, M.; Gutiérrez, E.(2003). «Spatiotemporal variability in radial growthof trees in the Central Pyrenees: climatic and siteinfluences». Ecological Monographs, vol. 73, núm. 2,pàg. 241-257.
Tranquillini, W. (1979). Physiological ecology of theAlpine Timberline: Tree existence at high altitudes withspecial reference to the European Alps. Berlín: Springer-Verlag. 137 pàg.
Weisberg, P. J.; Baker, W. L. (1995). «Spatial variationin tree regeneration in the forest-tundra ecotone,Rocky Mountain National Park, Colorado». Can. J.For. Res., vol. 25, pàg. 1326-1339.


















