
PROMOTORES DEL CRECIMIENTO VEGETAL Y …
41
MARTÍN ESPARIZ - [email protected] BIOTECNOLOGÍA DE LOS ALIMENTOS 2019 PROMOTORES DEL CRECIMIENTO VEGETAL Y BIOINSECTICIDAS
Transcript of PROMOTORES DEL CRECIMIENTO VEGETAL Y …

M A R T Í N E S P A R I Z - E S P A R I Z @ I B R - C O N I C E T . G O V . A R
B I O T E C N O L O G Í A D E L O S A L I M E N T O S 2 0 1 9
PROMOTORES DEL CRECIMIENTO VEGETAL Y BIOINSECTICIDAS

PLANT GROWTH PROMOTINGBACTERIA
Algunas bacterias y hongos de
actúan beneficiosamente
directamente al proporcionar
un producto que mejora el
crecimiento de las plantas, y
otros actúan de manera
indirecta.
Bernard R. Glick, Capítulo 15 (p 599-620)

RIZOBACTERIAS PROMOTORAS DEL CRECIMIENTO VEGETAL (PGPR)
PGPR
La promoción directa del crecimiento de la planta generalmente implica
proporcionar a la planta un compuesto que es sintetizado por la bacteria,
como nitrógeno fijado o una hormona vegetal. Además, estas bacterias
pueden facilitar la absorción por parte de la planta de ciertos nutrientes del
medio ambiente.P R O M O C I Ó N D I R E C T A
D E L C R E C I M I E N T O
Fijación de N
Solubilización de P
Producción de fitohormonas

PLANT GROWTH PROMOTINGBACTERIA
La estimulación directa del crecimiento y desarrollo de la planta por parte de las
bacterias promotoras del crecimiento puede ocurrir de varias maneras diferentes. Las
bacterias pueden:
a) fijar el nitrógeno atmosférico al amoníaco que utiliza la planta;
b) sintetizar sideróforos que solubilizan y secuestran el hierro del suelo y lo
proporcionan a las células de las plantas.
c) sintetizar fitohormonas, como la auxina, la citoquinina o la giberelina, que mejoran
varias etapas del crecimiento de las plantas.
d) solubilizar minerales, como el fósforo, que son utilizados por la planta
e) sintetizar una enzima que pueda modular el nivel de la hormona vegetal etileno.
Bernard R. Glick, Capítulo 15 (p 599-620)

RIZOBACTERIAS PROMOTORAS DEL CRECIMIENTO VEGETAL (PGPR)
PGPR
PATÓGENOP R O M O C I Ó N I N D I R E C T A
D E L C R E C I M I E N T O
Estimulación de sistemas
de defensa de la planta
Inhibición de
patógenos
La promoción indirecta del crecimiento de la planta se produce cuando las
bacterias promotoras del crecimiento de la planta disminuyen o previenen los
efectos perjudiciales de los organismos fitopatógenos, ya sean hongos o
bacterias, es decir, actúan como agentes de control biológico.

GROWTH PROMOTION BY FREE-LIVING BACTERIA
Bernard R. Glick, Capítulo 15 (p 599-620)

GROWTH PROMOTION BY FREE-LIVING BACTERIA
Bernard R. Glick, Capítulo 15 (p 599-620)
En algunas plantas rompe la
dormancia y estimula la
germinación.
En exceso produce inhibición
elongación de la raíz,
marchitamiento, daño.

GROWTH PROMOTION BY FREE-LIVING BACTERIA
Bernard R. Glick, Capítulo 15 (p 599-620)
El IAA bacteriano puede estimular la
proliferación y el alargamiento de las células de
la planta o la actividad de la enzima ACC
sintasa, que convierte la S-adenosilmetionina
(AdoMet) en ACC.
El ACC es absorbido por la bacteria; e
hidrolizado por la enzima ACC desaminasa a
amoníaco y α-cetobutirato (α-KB).
En consecuencia, se reduce la concentración de
ACC, y por lo tanto de etileno, en la planta.
Las flechas indican pasos químicos o físicos en el
mecanismo, y el símbolo ⊥ indica la inhibición del
alargamiento de la raíz por el etileno.

GROWTH PROMOTION BY FREE-LIVING BACTERIA
Bernard R. Glick, Capítulo 15 (p 599-620)
Efecto del tratamiento de semillas de canola con bacterias promotoras
del crecimiento de plantas que contienen ACC desaminasa .
• MgSO4 (control)
• P. putida GR12-2 que
contiene ACC desaminasa
• Inhibidor químico de ´síntesis
de etileno 1-aminovinilglicina
(AVG).
Contenido de ACC de la raíz
la longitud de la raíz
Crecimiento de la planta durante 4,5 días después de la siembra de las semillas.

PGPR: DECREASING PLANT STRESS
Bernard R. Glick, Capítulo 15 (p 599-620)
Evolución temporal de la síntesis de
etileno en plantas después de estrés
ambiental o infección por patógenos
fúngicos

PGPR: DECREASING PLANT STRESS
Bernard R. Glick, Capítulo 15 (p 599-620)
Representación esquemática de una planta
en maceta inundada
El ACC que se produce en las raíces como
consecuencia del estrés es incapaz de
convertirse en etileno debido a la ausencia
de oxígeno.
El ACC se transporta a los brotes, donde el
oxígeno es abundante, y se convierte en
etileno, lo que causa epinastia y pérdida de
biomasa.

PGPR: DECREASING PLANT STRESS
Bernard R. Glick, Capítulo 15 (p 599-620)
plantas de tomate de
cincuenta y cinco días
de edad
Cultivo durante 9
días adicionales
se inundaron
durante 9 días
peso seco de las hojas y
brotes
etileno producido por
tallos de hojas de planta
de tomate de 55 días
(pecíolos)

PGPR: DECREASING PLANT STRESS
Bernard R. Glick, Capítulo 15 (p 599-620)
• Enterobacter cloacae CAL2 ACC deaminasa positiva
• P. putida UW4 ACC deaminasa positiva
• P. putida pRK415 ACC deaminasa negativa• P. Putida pRKACC es pRK415 + pRK415ACC (plásmido con ACC deaminasa)
Sin inundar Inundadas por 9 días
ACC deaminasa positivaACC deaminasa negativa

PGPR: DECREASING PLANT STRESS
Bernard R. Glick, Capítulo 15 (p 599-620)
Peso seco Etileno
Las plantas no inundadas producen aproximadamente
0.07 pmol de etileno g-1 s-1, independientemente de la
presencia o ausencia de bacterias.
Inundadas por 9 días
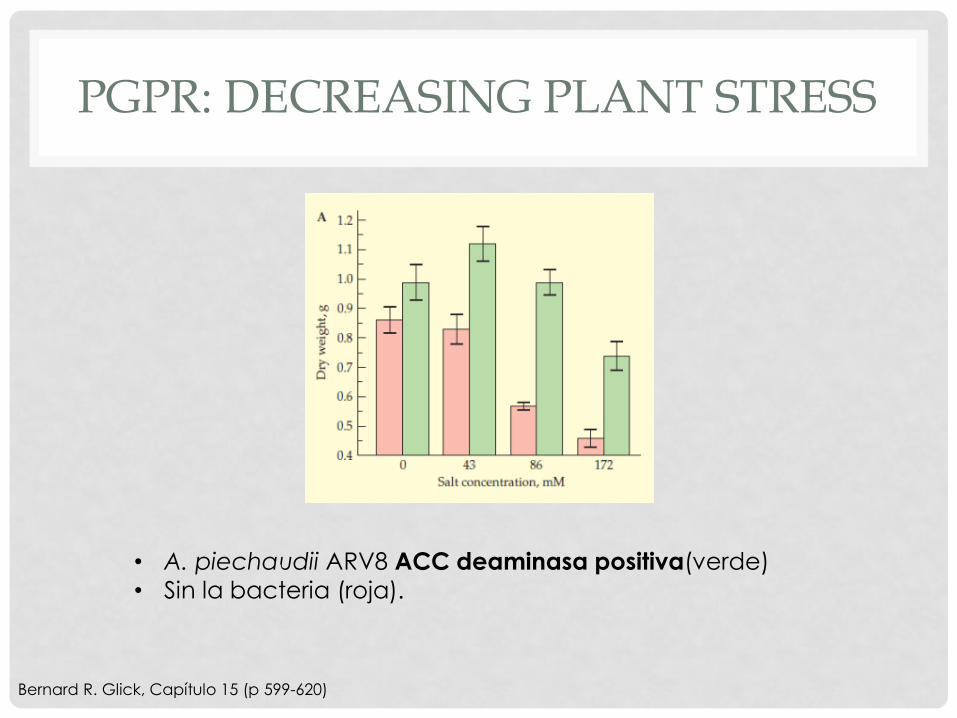
PGPR: DECREASING PLANT STRESS
Bernard R. Glick, Capítulo 15 (p 599-620)
• A. piechaudii ARV8 ACC deaminasa positiva(verde)
• Sin la bacteria (roja).

PGPR: DECREASING PLANT STRESS
Bernard R. Glick, Capítulo 15 (p 599-620)
El tratamiento de las plantas de tomate
con sal provoca un aumento en la
síntesis de estrés etileno.
La producción de etileno en presencia
de sal es parcialmente inhibida por A.
piechaudii ARV8.

PGPR: INCREASING PHOSPHORUSAVAILABILITY
Bernard R. Glick, Capítulo 15 (p 599-620)
La fitasa del hongo Aspergillus fumigatus se insertó, usando un transposón,
en el ADN cromosómico de Bacillus mucilaginosus.
La enzima fitasa puede disolver el fósforo del fosfato de calcio
Fitasa

PGPR: INCREASING PHOSPHORUSAVAILABILITY
Bernard R. Glick, Capítulo 15 (p 599-620)
La cepa bacteriana transformada puede expresar y segregar fitasa
activa.
ACC deaminasa
positivaACC deaminasa
negativa

BIOCONTROL OF PATHOGENS
Bernard R. Glick, Capítulo 15 (p 599-620)
Las bacterias promotoras del
crecimiento vegetal pueden producir
una variedad de sustancias que
limitan el daño a las plantas por los
fitopatógenos. Incluyen sideróforos,
antibióticos, otras moléculas
pequeñas y una variedad de enzimas.

BIOCONTROL OF PATHOGENS
Bernard R. Glick,
Capítulo 15 (p 599-620)

BIOCONTROL: SIDEROPHORES
Bernard R. Glick, Capítulo 15 (p 599-)
Complejo de seis coordenadas
de hierro-sideróforo.
Grupos de unión a hierro de sideróforos
microbianos.
Tres grupos funcionales bidentados en
una molécula de sideróforo se unen
con hierro férrico.

BIOCONTROL: SIDEROPHORES
Bernard R. Glick, Capítulo 15 (p 599-620)
El patógeno Pythium ultimum inhibe dramáticamente el crecimiento de
la raíz
Pseudomonas fluorescens CHA0 que posee vector que porta gen que
codifica la ARN polimerasa sigma-70 (σ70) produce mayor cantidad
de antibióticos.

BIOCONTROL: SIDEROPHORES
Bernard R. Glick, Capítulo 15 (p 599-620)
La cepa modificada fue más efectiva para proteger las plantas
de pepino contra una enfermedad de la raíz causada por el
hongo Pythium ultimum.

BIOCONTROL: ANTIBIOTICS
Bernard R. Glick, Capítulo 15 (p 599-620)
Ácido fenazina-1-carboxílico
P. fluorescens no codifica el
represor Lac cualquier gen bajo
el control del promotor tac se
expresa de forma constitutiva

BIOCONTROL: ANTIBIOTICS
Bernard R. Glick, Capítulo 15 (p 599-620)
La actividad antifúngica de la bacteria esproporcional al área de la zona deeliminación alrededor del centro de la placade Petri a la que se agrega la bacteria.
Los transconjugantes en los que la inserción cromosómica no había
inactivado ninguna función bacteriana
importante se probaron para
determinar su eficacia como cepas de
control biológico.

BIOCONTROL: ENZYMES
Bernard R. Glick, Capítulo 15 (p 599-620)
Algunas bacterias promotoras del crecimiento de las plantas
producen enzimas, como la quitinasa, la β-1,3-glucanasa, la proteasa
y la lipasa, que pueden degradar las paredes celulares de los hongos
y hacer que las células fúngicas se lisen.

BIOCONTROL: ENZYMES
Bernard R. Glick, Capítulo 15 (p 599-620)
La pared celular de los hongos se degrada por una o más enzimas
producidas por una bacteria que es agente de biocontrol. Luego, la
célula se lisa fácilmente.

NITROGEN FIXATION
Bernard R. Glick, Capítulo 15 (p 599-620)
Una amplia gama de bacterias pueden fijar nitrógeno, y varias de ellas
tienen potencial como fertilizantes para cultivos.

NITROGEN FIXATION
Bernard R. Glick, Capítulo 15 (p 599-620)
Los microorganismos más importantes
que se utilizan actualmente en la
agricultura para mejorar el contenido
de nitrógeno de las plantas incluyen
una gama de géneros y especies
rizobiales.
Estas bacterias son gram negativas,
flageladas y en forma de bastón, y
forman relaciones simbióticas con las
leguminosas.

NITROGEN FIXATION
Bernard R. Glick, Capítulo 15 (p 599-620)
En general, cada especie rizobial es
específica para un número limitado
de plantas y no interactuará con
plantas que no sean sus huéspedes
naturales.

NITROGEN FIXATION
Bernard R. Glick, Capítulo 15 (p 599-620)
Cuando no existe evidencia bioquímica o genética de la función de una proteína particular, se asigna una posible función basada en la homología de la secuencia de aminoácidos
a una proteína de secuencia conocida. Diferentes cepas rizobiales contienen diferentes subconjuntos de estas proteínas. Los genes "comunes" desempeñan la misma función en
todas las especies de rizobios.

NODULATION AND ETHYLENE
Bernard R. Glick, Capítulo 15 (p 599-620)
El etileno a menudo es producido por las plantas después de las etapas
iniciales de la infección (lo que finalmente conduce a la formación de
nódulos) por el rizobio. Este pequeño aumento en el nivel de etileno de
la planta generalmente se localiza en una porción de la raíz y puede
inhibir, y por lo tanto limitar, la subsiguiente infección y nodulación del
rizobio.

NODULATION AND ETHYLENE
Bernard R. Glick, Capítulo 15 (p 599-620)
Una forma en que algunas cepas de Rhizobium aumentan
naturalmente el número de nódulos que pueden formar en las raíces de
una leguminosa huésped es limitar el aumento de etileno que se
produce después de la infección inicial.

NODULATION AND ETHYLENE
Bernard R. Glick, Capítulo 15 (p 599-620)
• Capacidad de para nodular la
alfalfa (barras rosadas).
• Capacidad de para promover el
crecimiento de las plantas (barras
moradas).

INSECTICIDAL TOXIN OF B. THURINGIENSIS
Bernard R. Glick, Capítulo 16 (p 652-658)
Las actividades insecticidas de la bacteria Bacillus thuringiensis y los
sistemas de baculovirus de insectos se han desarrollado como insecticidas
seguros, específicos y efectivos.

INSECTICIDAL TOXIN OF B. THURINGIENSIS
Bernard R. Glick, Capítulo 16 (p 652-658)
El mercado mundial de pesticidas es enorme: actualmente supera los $ 30
mil millones por año y está creciendo rápidamente. Aunque los
bioplaguicidas, en su mayoría B. thuringiensis, representan solo alrededor
del 1% de este total, gran parte del crecimiento esperado en este campo
probablemente involucre bioplaguicidas.
invivo-group.com

INSECTICIDAL TOXIN OF B. THURINGIENSIS
Microfotografía de Bacillus thuringiensis en microscopio
electrónico de transmisión. Se muestra el cristal proteínico
romboide compuesto de toxinas Cry y una espora en
proceso.

INSECTICIDAL TOXIN OF B. THURINGIENSIS
Bernard R. Glick, Capítulo 16 (p 652-658)
Cristal parasporal de B. thuringiensis compuesto
de proteína protoxina Cry1.
Cada subunidad de proteína de 250 kDa del
cristal paraesporal contiene dos polipéptidos de
130 kDa.
La conversión de la protoxina de 130 kDa en una
toxina activa de 68 kDa requiere la combinación
de un pH ligeramente alcalino (7,5 a 8) y la
acción de una proteasas específicas, las cuales
se encuentran en el intestino del insecto.

INSECTICIDAL TOXIN OF B. THURINGIENSIS
Bernard R. Glick, Capítulo 16 (p 652-658)
Aproximadamente 15 minutos después de que se forma este canal iónico,
cesa el metabolismo celular; el insecto deja de alimentarse en unas pocas
horas, se deshidrata y finalmente muere (en aproximadamente 2 a 5 días).
En su forma activa, la proteína tóxica se
inserta en las membranas de las células
epiteliales intestinales del insecto y crea
un canal iónico, lo que conduce a una
pérdida excesiva de ATP celular.

INSECTICIDAL TOXIN OF B. THURINGIENSIS
Bernard R. Glick, Capítulo 16 (p 652-658)
Árbol filogenético de proteínas insecticidas (Cry) de B. thuringiensis. Cry1 y
Cry7 comparten menos del 45% de identidad, Cry1A y Cry1F son idénticos
entre 45% y 78%, y Cry1Ab y Cry1Ae son idénticos entre 78% y 95%.
Los diferentes colores de fondo delimitan los diferentes niveles de nomenclatura.

INSECTICIDAL TOXIN OF B. THURINGIENSIS
Bernard R. Glick, Capítulo 16 (p 652-658)


















