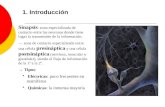
Neuroreceptores y Sinapsis

of 32
-
Upload
elvisor-visor -
Category
Documents
-
view
279 -
download
0
Transcript of Neuroreceptores y Sinapsis
-
NEUROTRANSMISORES
1. Definicin:
Un neurotransmisor (NT) es una sustancia qumica liberada selectivamente de una terminacin nerviosa por la accin de un PA, que interacciona con un receptor especfico en una estructura adyacente y que, si se recibe en cantidad suficiente, produce una determinada respuesta fisiolgica.
Para constituir un NT, una sustancia qumica debe estar presente en la terminacin nerviosa, ser liberada por un PA y, cuando se une al receptor, producir siempre el mismo efecto. Existen muchas molculas que actan como NT y se conocen al menos 18 NT mayores, varios de los cuales actan de formas ligeramente distintas.
DEFINIMOS A UN NEUROTRANSMISOR como una sustancia producida por una clula nerviosa capaz de alterar el funcionamiento de otra clula de manera breve o durable, por medio de la ocupacin de receptores especficos y por la activacin de mecanismos inicos y/o metablicos.
2. Clasificacin:
Neurotransmisores de accin rpida y molcula pequea
Son los que producen las respuestas ms inmediatas del sistema nervioso, como la transmisin de seales sensitivas hacia en encfalo y de seales motoras hacia los msculos.Este tipo de transmisores se sintetizan en el terminal presinptico, y las numerosas vesculas transmisoras presentes a este nivel los absorben por medio de transporte activo. Cada vez que llega un PA al terminal presinptico, las vesculas liberan su transmisor a la hendidura sinptica en pequeos grupos (esto tarda milisegundos o menos). Seguidamente la accin del transmisor sobre los receptores de la membrana de la neurona pstsinptica, que tambin tiene duracin aproximada de otros milisegundos. Lo ms frecuente es que el efecto consista en incrementar o disminuir la conductancia que presentan los canales inicos; p.ej., aumentar la conductancia al sodio, lo que causara una excitacin, o la del potasio o el cloruro, lo que supone una inhibicin.
Caractersticas de algunos de los ms importantes transmisores de molcula pequeaLa acetilcolina se segrega por las neuronas situadas en muchas regiones del sistema nervioso, pero especficamente en: 1) los terminales de las clulas piramidales grandes de la corteza motora; 2) diversos tipos de neuronas pertenecientes a los ganglios basales; 3) las motoneuronas que inervan los msculos esquelticos; 4) las neuronas preganglionares del sistema nervioso autnomo; 5) las neuronas posganglionares del sistema
-
nervioso parasimptico. Generalmente la acetilcolina posee un efecto excitador, pero se sabe que acta inhibiendo algunas terminaciones nerviosas parasimpticos perifricas (por ejemplo la inhibicin del corazn a cargo de los nervios vagos).La noradrenalina se segrega en los terminales de muchas neuronas cuyos somas estn situados en el tronco del encfalo y el hipotlamo. En concreto, las que estn situadas en el locus cerelus de la protuberancia envan fibras nerviosas a amplias regiones del encfalo que sirven para controlar la actividad global y el estado mental, como por ejemplo aumentar el nivel de vigilia. En la mayora de estas zonas, la noradrenalina probablemente activa receptores excitadores, pero en unas cuantas, en cambio, estimula los inhibidores. Tambin se segrega en la mayor parte de las neuronas posganglionares del sistema nervioso simptico, donde excita algunos rganos, pero inhibe otros.La dopamina se segrega en las neuronas originadas en la sustancia negra. Su terminacin se produce en la regin estriatal de los ganglios basales. El efecto que ejerce suele ser una inhibicin.La glicina se segrega en las sinapsis de la mdula espinal. Se cree que siempre acta como un transmisor inhibidor.El GABA (cido gamma-aminobutrico) es segregado en los terminales nerviosos de la mdula espinal, el cerebelo, los ganglios basales y muchas reas de la corteza. Causa inhibicin.Acetilcolina. Esta sustancia transmisora se sintetiza en el terminal presinptico a partir de la coenzima A y colina en presencia de la enzima acetiltransferasa de colina. A continuacin, se transporta a sus vesculas especficas, luego, al producirse su salida hacia la hendidura sinptica durante la transmisin de la seal nerviosa en la sinopsis, se degrada nuevamente en acetato y calina por accin de la colinesterasa (presente en el retculo formado por proteoglucano que rellena el espacio de la hendidura sinptica). Luego, una vez ms, las vesculas se reciclan en el interior del terminal presinpticos; la colina sufre un transporte activo de vuelta hacia el terminal para repetir su empleo en la sntesis de nueva acetilcolina.
Neuropptidos, transmisores de accin lenta o factores de crecimiento
A diferencia de los primeros, stos suelen provocar acciones ms prolongadas, como los cambios a largo plazo en el nmero de receptores neuronales, la apertura o cierre de ciertos canales inicos y las modificaciones persistentes en la cantidad de sinapsis o en su tamao.Por el contrario, estos neurotransmisores se sintetizan y forman en los ribosomas del soma neuronal ya como porciones ntegras de grandes molculas proteicas. Estas molculas penetran en los espacios existentes del RE del soma y luego al AG, donde suceden dos cambios. En primera instancia, la protena formadora de neuropptidos sufre una escisin enzimtica, algunos de los fragmentos (los ms pequeos) son del propio neuropptido o un precursor suyo. En segundo lugar, el AG introduce el neuropptido en minsculas vesculas transmisoras que se liberan hacia el citoplasma. A continuacin, se transportan por el axn en todas direcciones hacia el extremo de las fibras nerviosas a travs de la corriente axnica del citoplasma, viajando a una velocidad de tan slo unos centmetros al da. Finalmente estas vesculas vierten su contenido en los terminales
-
neuronales como respuesta a los PA de la misma manera que los transmisores de molcula pequea.Los neuropptidos a menudo ocasionan accionan mucho ms duraderas. Algunas de ellas consisten en cierre prolongado de los canales de calcio, los cambios persistentes en la maquinaria metablica de las clulas, en la activacin o desactivacin de gnesis especficos dentro del ncleo celular, o en alteraciones a largo plazo de la cantidad de receptores. Algunos de estos efectos duran incluso aos. 3. Mecanismo de accin:
A) ADRENALINA
La adrenalina fue descubierta en 1895 por Oliver y Schfer en extractos de glndula suprarrenal. Estos extractos tienen la propiedad de aumentar la tensin arterial y la frecuencia cardiaca. Posteriormente Stolz y Dakin identificaron su estructura qumica y finalmente fue sintetizada. La estructura de esta hormona se muestra en la figura 16. Se forma a partir del aminocido tirosina principalmente en la mdula de la glndula suprarrenal y en algunas neuronas, las llamadas neuronas simpticas. Funciona, por lo tanto, como hormona y como neurotransmisor. La noradrenalina o norepinefrina es un precursor en la biosntesis de la adrenalina. En realidad ambos compuestos se encuentran tanto en las neuronas como en las suprarrenales. Sin embargo, en general se acepta que la adrenalina es la hormona y la noradrenalina el neurotransmisor.
FIGURA 16. Estructura de adrenalina y representacin esquemtica de su mecanismo de accin.
La adrenalina ejerce importantes funciones en todo el cuerpo. Puede asegurarse con facilidad que no hay una funcin de grande o mediana importancia para el organismo en la que no participe. Por lo tanto, no es de sorprender que una enorme proporcin de las clulas de nuestro cuerpo tengan receptores adrenrgicos. Todo ello es ventajoso para quienes nos dedicamos a este campo; por lo mismo, la competencia es enorme. Es un campo apasionante, prcticamente en constante ebullicin.
Para mencionar algunas de las principales funciones de los agentes adrenrgicos dir que, dado que son de los neurotransmisores ms
-
abundantes, participan en un gran nmero de las llamadas funciones superiores, adems de que regulan la frecuencia cardiaca, la tensin arterial, la secrecin por glndulas tanto de secrecin interna como externa, el metabolismo global de la economa al regular los metabolismos especficos de rganos como el hgado, el tejido adiposo y el msculo, en fin, en casi todo tienen que ver. Sera interminable una lista de las clulas sobre las que puede actuar la adrenalina; entre ellas tenemos a la inteligente neurona, al infatigable miocito cardiaco, al humilde adipocito, a los verstiles hepatocitos, a las pequeas plaquetas, etctera.
Como resultar obvio de lo anteriormente mencionado, existe una gran cantidad de enfermedades en las que de una forma u otra estn implcitas las funciones de los agonistas adrenrgicos naturales, la adrenalina y la noradrenalina. As, se ha sugerido que en algunos trastornos mentales, como son los estados maniacos y los depresivos, existe una alteracin en el metabolismo y funcin de estas catecolaminas. En la hipertensin arterial tienen una funcin importante y de hecho algunos de los frmacos que se usan para su tratamiento son agentes con propiedades adrenrgicas. Se ha propuesto que los receptores adrenrgicos son un factor clave en el asma, y para el tratamiento de los cuadros asmticos agudos se usan agonistas adrenrgicos. Muchos de los descongestionantes nasales contienen agentes adrenrgicos.
Es fcil, entonces, imaginar por qu es importante saber qu hacen y sobre todo cmo hacen lo que hacen, los agentes adrenrgicos. Para ello hay razones cientficas bsicas: el deseo de saber ms y profundizar en los fenmenos esenciales de la vida; y razones aplicativas: lo que se descubra es importante para muchas enfermedades (desde el resfriado comn hasta las enfermedades cardiovasculares, pasando por los problemas mentales).
Existen adems razones econmicas: la industria farmacutica investiga (desafortunadamente no en nuestro pas) muy diversos agentes adrenrgicos para su posible utilizacin. Por ejemplo, la venta de un agente antiadrenrgico de uso clnico represent 50% de los ingresos de una enorme compaa trasnacional durante la dcada de los setenta. El investigador que logr el compuesto, sir James Black, contribuy adems con otro compuesto (bloqueador de la accin de la histamina) de uso clnico importantsimo y con grandes beneficios econmicos para la industria en la que trabajaba. Hace algunos aos, recibi el premio Nobel de Fisiologa y Medicina por su trabajo pionero en farmacologa molecular.
Pero dirijmonos a lo ms interesante. La adrenalina inicia sus acciones al asociarse con receptores membranales. En 1948, un investigador mexicano, Arturo Rosenblueth (quien trabajaba con el doctor Canon en el noreste de Estados Unidos), observ que la adrenalina produca contraccin en un msculo liso: la aorta; por el contrario, el msculo liso de los bronquios se relajaba. Perplejos por estos resultados contradictorios, dichos investigadores propusieron que se generaban dos tipos de mediadores, segn el tejido en el que actuaba la hormona, y que conducan a los diferentes efectos. Esta sugerencia no fue acertada, pero s constituy un inicio conceptual para dividir las acciones de la hormona. Otro investigador, Alhquist, tambin en 1948, lleg a una conclusin ms vlida y que de
-
hecho, constituye la piedra angular de lo que hoy sabemos sobre acciones adrenrgicas. Este investigador recientemente fallecido, al usar una serie de anlogos de la adrenalina, observ que la potencia relativa de estos compuestos para producir contraccin o relajacin era claramente diferente. Estas observaciones y, por supuesto, su capacidad, lo llevaron a la conclusin de que la contraccin ocurra por la activacin de un tipo de receptor al cual llam a; mientras que la relajacin se daba por la activacin de otro tipo de receptor al que denomin b. Hoy sabemos que estos tipos de receptores se subdividen an ms, y que en realidad hay tres familias de receptores para la adrenalina: los a1-, los a2- y los b-adrenrgicos. Cada una de estas familias tiene tres miembros y por lo tanto tenemos nueve receptores: a1 (A, B y D), a2 A, B y C) y b (1, 2 y 3).
Quiz parezca ms un entretenimiento que otra cosa eso de andar subdividiendo receptores para una hormona dada, pero tiene una importancia terica y prctica increble. Pondr un ejemplo: durante muchos aos se ha sabido que las catecolaminas aumentan las necesidades de oxgeno del corazn al activar a los receptores b-adrenrgicos del miocardio. Existen condiciones en las cuales es conveniente bloquear en un paciente este tipo de receptores. Para ello se han diseado diversos bloqueadores b-adrenrgicos. Por otro lado, en los cuadros asmticos se utilizan agonistas b-adrenrgicos para relajar la musculatura bronquial. Ahora bien, imaginemos a un paciente que tenga ambos tipos de problemas: cardiovasculares y asmticos. Si el galeno prescribe un bloqueador b-adrenrgico general, el paciente mejorar de su padecimiento cardiovascular, pero, qu suceder si presentara un cuadro de asma? Los receptores b de sus bronquios estaran tambin bloqueados. Afortunadamente, hoy en da la situacin no es tan complicada. Me explicar mejor: el receptor b del miocardio es b1-adrenrgico, mientras que el de los bronquios es b2 Dado que la industria farmacutica ya ha logrado sintetizar agentes selectivos para los diferentes subtipos, podemos administrar al paciente un bloqueador b1-adrenrgico selectivo y dejar en paz sus b2 para cualquier emergencia asmtica.
Deca entonces que hay tres familias o tipos bsicos de receptores adrenrgicos, todos ellos pertenecen a la familia de los siete dominios transmembranales o acoplados a protenas G. En general estos receptores se encuentran distribuidos en forma preferencial en los diferentes tejidos; por ejemplo: el corazn es rico en receptores b1-adrenrgicos, mientras que la aorta contiene mltiples receptores a1-adrenrgicos. El conocimiento de que existen tantos receptores diferentes para esta hormona y neurotransmisor es muy reciente, y an se ignora la distribucin por tipo y subtipo en cada tejido y su participacin en las acciones de este mensajero. Por otro lado, una clula puede contener ms de un tipo de receptores para la adrenalina. No hace muchos aos, en 1981, demostramos que el adipocito humano contiene receptores adrenrgicos de las tres familias principales, es decir al, a2 y b-adrenrgicos. De hecho, de la familia b, el adipocito expresa los tres subtipos: el b1, el b2 y el b3. Ahora sabemos que este hecho es bastante general y que frecuentemente las clulas expresan vanos de los subtipos para una misma hormona.
-
Otro aspecto sumamente interesante es que los receptores estn asociados a sistemas especficos de transduccin. Mi maestro, John Fain, y yo fuimos los primeros en establecer claramente este hecho. Como hemos visto, los diferentes receptores b-adrenrgicos estn acoplados activadoramente a la adenilil ciclasa a travs de Gs como hemos visto. Los receptores a2-adrenrgicos se acoplan a la ciclasa en forma inhibitoria a travs de Gi, y los receptores a1 se acoplan al recambio de fosfoinostidos a travs de Gq (ver la figura 16). Adems de la existencia de los diferentes subtipos, en los ltimos aos nos ha quedado claro que cada receptor no enciende un sealamiento lineal, sino una red, como discutimos cuando hablamos de las protenas G. As, es claro que estos receptores, adems de modular la adenilil ciclasa y el recambio de fosfoinostidos, tienen acciones sobre otros sistemas de transduccin, como son algunos canales para sodio, calcio y potasio. Ms an, el acoplamiento de un receptor no slo depende del receptor sino del repertorio de protenas G y de efectores (canales inicos, otras fosfolipasas, etc.) que expresa cada clula.
B) INSULINA
Una hormona que ha atrado el inters de miles de cientficos y del pblico en general es la insulina. Esta hormona es un polipptido con un peso molecular de 6 000 Daltones, y es producida y secretada por las clulas beta de los islotes de Langerhans del pncreas. Sin duda, gran parte del inters que se ha generado, se debe a su importancia en el mantenimiento de los niveles de glucosa en la sangre y en el tratamiento de la diabetes mellitus.
Alrededor de 1890 Mering y Minkowsky haban demostrado que la extirpacin del pncreas produce, en animales de laboratorio, un padecimiento similar a la diabetes mellitus. Fueron unos investigadores canadienses del Hospital General de Toronto, Banting y su alumno Best, un joven estudiante de medicina, quienes lograron extraer el principio activo del pncreas y demostraron su utilidad teraputica tanto en perros diabticos como en humanos; estos estudios se realizaron entre 1921 y 1922. En esos trabajos particip McLeod, en ese momento jefe del Departamento, y con quien Banting tuvo innumerables conflictos. Recibi aos despus junto con Banting el premio Nobel de Fisiologa y Medicina; Banting, por su cuenta decidi compartir su parte del premio con Best.
El primer paciente en recibir el beneficio de tales descubrimientos fue Leonard Thompson, un muchacho diabtico de 14 aos internado en el Hospital General de Toronto. La diabetes de este muchacho se encontraba totalmente descompensada y Banting y Best decidieron emplear su extracto como un tratamiento experimental. El resultado fue espectacular y abri, sin duda, una de las grandes avenidas en la historia del tratamiento de las enfermedades endocrinas en general y de la diabetes en particular. Para 1926 ya se contaba con insulina cristalina, y en 1960 Langer estableci su secuencia de aminocidos. Este investigador obtuvo el premio Nobel de Qumica.
Los estudios acerca de la insulina han continuado avanzando. Actualmente podemos cuantificar con precisin la cantidad de insulina que hay en la
-
sangre y en los diversos tejidos (metodologa que est asociada a otro premio Nobel); adems existen muy diversos preparados de insulina con diferentes velocidades de absorcin para un mejor tratamiento de los pacientes. Dos avances notables merecen tambin ser mencionados; dado que el consumo de insulina (obtenida principalment del cerdo) es muy importante, se ha considerado que su disponibilidad poda llegar a ser limitante para el tratamiento del gran nmero de pacientes que la requieren. Para evitar esto, en la dcada de los sesenta se desarrollaron los procesos de sntesis qumica completa. Estos procesos, aunque costosos, ofrecen una alternativa y se han mejorado en los ltimos aos. Otra opcin que ya ha resultado de utilidad prctica es la insercin de la informacin gentica para la sntesis de la hormona en el ADN de microorganismos. Estos microorganismos producen la hormona y es posible cultivarlos en grandes fermentadores industriales. Esta alta tecnologa de ingeniera gentica se emplea con xito en los laboratorios Lilly de Estados Unidos, que producen desde hace varios aos insulina por estos medios, para uso en pacientes.
La insulina es la principal hormona encargada de disminuir los niveles de glucosa en sangre. De los aos treinta a los setenta se descubrieron muchas de las principales acciones de la insulina. Esta hormona aumenta el transporte de glucosa al interior de las clulas y su conversin a glucgeno; adems aumenta la oxidacin del azcar. Favorece el proceso de sntesis de lpidos y disminuye tanto la movilizacin de grasa de los depsitos, como su oxidacin en el hgado; adems, aumenta el transporte de algunos aminocidos en las clulas blanco. Estas acciones, y muchas otras descritas hasta nuestros das, ocurren rpidamente, en cuestin de minutos. Adems, hay clara evidencia de que la insulina ejerce acciones ms tardas, las cuales toman de horas a das; dentro de estas ltimas est el ser un factor de crecimiento celular.
Pero, cmo hace lo que hace la insulina? Sin duda uno de los grandes avances que han ocurrido en los ltimos 15 aos es el conocimiento de la estructura del receptor de la insulina. El trabajo pionero de Pedro Cuatrecasas aclar varios aspectos. Este investigador acopl la insulina a molculas de gran tamao que no podan entrar a las clulas; la insulina, as acoplada, aun poda ejercer sus efectos. Ello estableca que el receptor se localizaba en la membrana plasmtica. Por otro lado, intent purificar el receptor, con relativo xito. Las tcnicas que se usaban en ese momento no le permitan lograr la absoluta reproducibilidad de los resultados. Su trabajo fue sujeto de mltiples criticas, justas e injustas. Hoy, ms de 20 aos despus, muchos de sus hallazgos han sido totalmente comprobados y su labor pionera es reconocida por los expertos.
Hacia 1980 trabaj en Brown University con mi maestro John Fain. En los laboratorios que estaban al final del corredor trabajaba Mike Czech, un joven y brillante ex alumno de Fain que lideraba a un grupo de entusiastas y capaces posdoctorales. Paul Pilch, qumico de corazn, haba logrado entrecruzar la insulina radiactiva con su receptor y, por medio de tcnicas refinadas (electroforesis y radioautografa), haba llegado a establecer el peso molecular del receptor. Poco despus se incorpor al grupo Joan Massagu, joven cataln quien avanz audazmente en el estudio de las
-
subunidades del receptor. As, en unos cuantos aos este grupo logr establecer la estructura del receptor de la insulina. Un sueo que durante muchos aos haban acariciado muchos investigadores. Vale la pena mencionar que nadie en este mundo est aislado, y as otros grupos, incluyendo al de Cuatrecasas, hicieron importantes contribuciones para afinar el modelo actual que se present en la figura 10. El receptor de la insulina es una protena de peso molecular aproximado de 310 000 Daltones, est formada por dos subunidades llamadas alfa con peso de 125 000 y dos beta con peso aproximado de 90 000 Daltones. Estas subunidades estn enlazadas por uniones disulfuro. Parece existir solamente un gen para el receptor de la insulina, pero por procesamiento alternativo del ARN que lo codifica da origen a dos subtipos de receptores, A y B, para la hormona. Hay evidencia de que el receptor es sintetizado como una sola protena y posteriormente es dividido y procesado. El procesamiento de este precursor del receptor no slo involucra el fraccionamiento en sus subunidades, sino que adems participan otros procesos como la incorporacin de azcares; dicho procesamiento posiblemente ocurra en vesculas especializadas del aparato de Golgi.
Las subunidades alfa contienen el sitio de fijacin de la insulina. Hay evidencia de que podra existir ms de un sitio para la hormona y de que quiz haya cierta interaccin de un sitio con el otro. Las subunidades beta han sido motivo de interesantsimos hallazgos. Kasuga y Kahn demostraron que el receptor tiene actividad de protena cinasa de tirosina. Posteriormente el gen que codifica el receptor de la insulina fue donado y ha sido expresado en muchos sistemas, as como sujeto a diferentes manipulaciones (mutaciones, formacin de quimeras, etc.) para avanzar en el mecanismo de su accin. El receptor se fosforila tanto por su propia actividad de tirosina cinasa (autofosforilacin) como por otras cinasas (fosforilacin heterloga); el mecanismo general de accin de estos receptores con actividad de protena cinasa ha sido descrito en el captulo V. Indudablemente, mucho se ha avanzado; sin embargo, faltan muchos aspectos por aclarar.
Una las principales acciones de la insulina es disminuir la concentracin de glucosa en la sangre, lo cual se logra al aumentar el transporte de azcar al interior de las clulas; este efecto puede observarse fcilmente en clulas aisladas. Durante muchos aos se ha debatido acerca de los mecanismos moleculares responsables de dicho efecto. Sabemos que el transporte de la glucosa se lleva a cabo por medio de un transportador especfico, el cual se ha aislado, e incluso reconstituido, en membranas artificiales. La pregunta fundamental ha sido, cmo aumenta la insulina el transporte del azcar? Se buscaron posibles activadores sin encontrarse ninguno. No hace mucho tiempo, dos grupos, el de Cushman y el de Kono, encontraron un hecho que ha cambiado nuestra concepcin del fenmeno. En pocas palabras, se descubri que el nmero de transportadores en la membrana plasmtica aumenta considerablemente bajo la accin de la insulina. La pregunta inmediata fue, de dnde vienen? Y surgi que, al igual que con los receptores, los transportadores se localizan no slo en la membrana plasmtica, sino tambin en vesculas intracelulares. Ms experimentos
-
confirmaron lo anterior y adems aportaron la siguiente observacin: bajo la accin de la insulina los transportadores intracelulares de glucosa se incorporan a la membrana plasmtica; por lo tanto, el nmero de transportadores disminuye en las vesculas intracelulares y aumenta en la membrana plasmtica. Adems, al terminar la accin de la insulina el proceso se revierte. Pero, cmo se relacionan la actividad de tirosina cinasa del receptor y la translocacin de transportadores de glucosa? sta es una de las grandes lagunas que quedan an en nuestro conocimiento.
C) HISTAMINA, SEROTONINA, PROSTAGLANDINAS Y ADENOSINA
Existe un amplio grupo de sustancias de intensa actividad biolgica y enorme importancia que funcionan como hormonas locales. Muchas de ellas son producidas por el organismo en respuesta a cambios o agresiones localizadas; su funcin, desde un punto de vista teleolgico, es decir finalista, es restablecer el equilibrio perdido; en este sentido son un remedio endgeno, esto es, del mismo organismo, para sus males. En 1916, sir Edward Schfer acu el trmino "autacoide" para referirse a las hormonas en general. Sin embargo, la palabra autacoide describe en forma corta y con mucha mayor propiedad a las hormonas locales; es por ello que se utiliza principalmente en este ltimo sentido. Como mencion ya en un captulo anterior, desde el punto de vista etimolgico este trmino proviene del griego autos que significa propio, y de akos que significa remedio o medicamento. Es decir, el significado etimolgico se adapta claramente a la idea de la hormona local, la cual es un automedicamento para restablecer el equilibrio del organismo. Dentro de este amplio grupo de sustancias he seleccionado a cuatro de ellas, relativamente conocidas como hormonas locales y con gran importancia fisiolgica. Ellas son: la histamina, la serotonina, la adenosina y las prostaglandinas.
i) Histamina
Este autacoide, cuya frmula presento en la figura 17, es el producto de la descarboxilacin (eliminacin de C02) de un aminocido: la histidina. Es muy interesante recordar que este producto fue aislado y sintetizado como una curiosidad qumica mucho antes de conocer su funcin en el organismo. Exista la creencia que era slo un producto de la accin de las bacterias. Para 1910 ya se saba que se produca en muchos tejidos, principalmente en el msculo liso de los vasos sanguneos; sin embargo, no fue sino hasta casi 20 aos despus, en 1927, que Best, Dale, Dualery y Thorpe la aislaron de tejidos frescos. Este hallazgo estableci, sin dudas, que la histamina es un constituyente natural de los tejidos. Es interesante adems hacer notar que la palabra histamina proviene del griego histos que quiere decir tejido y de amina: la amina de los tejidos.
La inmensa mayora de quienes no se dedican a las ciencias biomdicas, cuando oyen hablar de antihistamnicos (bloqueadores o antagonistas de la accin de la histamina), piensan exclusivamente en reacciones de tipo alrgico; lo cual no es sorprendente, ya que durante muchos aos casi toda la investigacin y por ende el conocimiento estuvieron enfocados a este aspecto. Los antihistamnicos clsicos, que todos conocemos, bloquean slo un tipo de los receptores para la histamina, los H1.
-
FIGURA 17. Estructura de la histamina y representacin esquemtica de su mecanismo de accin.
Ya en los aos cincuenta haba evidencia de que exista ms de un tipo de receptor para este autacoide, pero no fue sino hasta los setenta cuando se sintetizaron compuestos con selectividad para el otro tipo de receptores, para la histamina, los H2. En estos desarrollos fue fundamental el trabajo de sir James Black, a quien nos referimos en lneas anteriores. Poco despus de la sntesis de los antihistamnicos selectivos H2, stos fueron incorporados al arsenal de medicamentos usados en la prctica clnica. Desde hace unos dos o tres aos se han publicado trabajos que sugieren la existencia de un tercer tipo de receptor para la histamina: el H3.
Represento en la figura 17 lo que sabemos de estos receptores. Como puede apreciarse, el receptor H1 el asociado a los fenmenos alrgicos est acoplado al recambio de fosfoinostidos y al calcio, mientras que el receptor H2 est acoplado en forma activadora a la adenilil ciclasa a travs de Gs. En el caso del receptor H3 an se desconoce el sistema transductor al que se acopla. Mencionar algunas de las principales acciones de la histamina y el tipo de receptor involucrado. Se ha observado que la histamina produce dos tipos de efectos sobre los vasos sanguneos: 1) dilatacin, mediada a travs de receptores H2, y 2) contraccin, mediada por receptores H1. El enrojecimiento y el edema localizado, observados en las reacciones alrgicas se producen al activarse los receptores H1. Otro fenmeno alrgico que involucra a receptores histamnicos de este tipo es la bronco-constriccin, asociada a cuadros de tipo asmtico. El corazn tampoco escapa de tener receptores para la histamina; en este rgano, la activacin de receptores H1 disminuye la conduccin elctrica, y en algunas especies hay receptores H2 que provocan taquicardia. La histamina tambin modula la presin arterial: aparentemente la activacin de los receptores H1 la eleva y la de los H2 la disminuye. La histamina es un importante neurotransmisor en el sistema nervioso central; los tres subtipos conocidos de receptores para la histamina, parecen tener muy diversas funciones en el cerebro.
Otro efecto importante de la histamina es aumentar la secrecin gstrica. Durante muchos aos se supo que la histamina aumentaba la secrecin de cido clorhdrico en el estmago. Sin embargo, el hecho de que ninguno de
-
los antihistamnicos conocidos hasta entonces (de tipo H1) mostrara utilidad para reducir la secrecin gstrica haca dudar de la importancia fisiolgica de dicho fenmeno y se consideraba como una curiosidad con cierta utilidad diagnstica. Con la sntesis de antihistamnicos H2 result claro que la histamina s juega un papel fisiolgico en la regulacin de la secrecin gstrica. Adems, estos antagonistas H2 han resultado de enorme utilidad para el tratamiento de los pacientes con lcera gastroduodenal.
ii) Serotonina
Durante muchos aos se supo que si se dejaba coagular la sangre, en el suero se encontraba una sustancia vasoconstrictora, es decir, que aumenta el tono vascular. En 1948 Rapport y sus colaboradores aislaron y cristalizaron este compuesto, al cual le dieron el nombre de serotonina (unin de las palabras suero y tono muscular). Este compuesto, sintetizado a partir del aminocido triptofano (la serotonina es la 5-hidroxi-triptamina, 5-HT), se encuentra localizado bsicamente en tres lugares: en las clulas cromafines del intestino, en algunas neuronas del sistema nervioso central y en las plaquetas. Han sido identificados muy diversos tipos de receptores para este compuesto; por lo menos siete familias, y algunas con varios subtipos. Aparentemente los diferentes subtipos de la familia 5-HT1 estn acoplados en forma inhibitoria a la enzima adenilil ciclasa y a diversos canales inicos, a travs de Gi; mientras que los 5-HT2, estn acoplados al recambio de fosfoinostidos y al calcio. Los receptores 5-HT3 son receptores canal, los 5-HT4, 5-HT6 y 5-HT7 se acoplan activadoramente a la adenilil ciclasa, y de los 5-HT5 an se desconoce su mecanismo de acoplamiento (figura 18). Como podr observarse este es un grupo de enorme complejidad con muy diversos tipos de receptores. En este momento la tipificacin de los receptores que participan en cada una de las acciones de la serotonina es un campo de gran actividad.
Como es de esperarse, con base en su distribucin, la serotonina afecta el funcionamiento intestinal. Este autacoide aumenta notablemente la motilidad del intestino delgado. Por otro lado, produce vasoconstriccin y aumenta tanto la fuerza como la frecuencia del latido cardiaco. En la hipfisis la serotonina parece ser un importante regulador de la secrecin de algunas hormonas, como la del crecimiento, la prolactina y las gonadotrficas. Hay clara evidencia de que la serotonina es un neurotransmisor en el sistema nervioso central; se ha postulado que alteraciones en el metabolismo o en la accin de este compuesto pueden tener relevancia en algunos desequilibrios mentales como la esquizofrenia o el autismo infantil. Otro dato de importancia es que algunos compuestos alucingenos (como la psilocina y la psilocibina, productos de los hongos alucingenos y el LSD) interactan fuertemente con receptores serotonnicos. Adems este tipo de receptores parece tener una gran importancia en la modulacin del estado de nimo y por lo tanto asociado a trastornos depresivos. sta es un rea de intensa actividad.
-
FIGURA 18. Estructura de la serotonina (5-hidroxi-triptamina) y representacin esquemtica de su mecanismo de accin.
iii) Prostaglandinas
En 1930 dos gineclogos estadounidenses, Kurzrok y Lieb, descubrieron que el tero se relajaba y contraa al exponerse el semen humano. Pocos aos ms tarde, Goldblat en Inglaterra y Wuler en Suecia reportaron que el extracto de la prstata tena actividad sobre el msculo liso. El material fue identificado por Euler como un lpido, quien lo bautiz con el nombre de "prostaglandina". Fue necesario un gran avance tcnico para aislar e identificar estos compuestos, lo cual no se logr sino hasta 1960. De esa poca a la fecha el nmero de publicaciones que hablan de estos compuestos y que describen sus acciones ha aumentado en forma sorprendente. Ahora sabemos que el cido araquidnico (un cido graso polinsaturado, presente en muchos fosfolpidos de la membrana) es el precursor de un grupo de compuestos muy activos biolgicamente, entre los cuales se encuentran las prostaglandinas, los leucotrienos y los tromboxanos.
Me centrar en las prostaglandinas, no sin antes mencionar que una de las drogas ms ampliamente usadas en el mundo, la aspirina, es un inhibidor de la sntesis de prostaglandinas. Como ustedes saben, la aspirina se utiliza como analgsico (quita el dolor), antipirtico (suprime la fiebre) y antiinflamatorio. Muchas de estas acciones de la aspirina se deben a que bloquea la formacin de prostaglandinas en el organismo. En este momento hay evidencia de diversos tipos de receptores para estos compuestos con acoplamientos a la adenilil ciclasa y al recambio de fosfoinostidos. Todos estos receptores parecen corresponder a la familia de los siete dominios transmembranales.
Algunos de los efectos ms prominentes de las prostaglandinas son los siguientes: 1) inhiben la agregacin de las plaquetas al oponerse al efecto de los tromboxanos (la aspirina bloquea la sntesis de prostaglandinas pero tambin la de tromboxanos, y as ejerce su accin antitrombtica); 2) el msculo liso presenta varios tipos de respuestas: a) el msculo bronquial se
-
relaja bajo la accin de las prostaglandinas, por lo que se han usado en el tratamiento de pacientes asmticos, b) las prostaglandinas aumentan la contraccin del tero por lo mismo, se han tratado de usar como abortivos, y c) el msculo liso gastrointestinal vara en su respuesta segn la regin anatmica; 3) otra accin importante es la disminucin de la secrecin gstrica. Esta accin ha permitido tener la esperanza de disear anlogos que puedan ser tiles en el tratamiento de la lcera pptica; 4) en el sistema nervioso las prostaglandinas aumentan o disminuyen la actividad elctrica, segn la regin; 5) adems, sabemos que son importantes en la regulacin de la secrecin de algunas hormonas, y 6) desde el punto de vista metablico su accin ms importante es disminuir la movilizacin de grasa de los depsitos.
iv) Adenosina
Este compuesto es un producto del metabolismo del ATP, la moneda energtica de la clula; se libera en muchos tejidos cuando las clulas tienen una deficiencia en su aporte de oxgeno. As, se ha visto que cuando hay falta de oxigenacin en el corazn se Iibera adenosina, la cual tiende a compensar el problema aumentando el riego coronario y disminuyendo el gasto energtico.
Al igual que en el caso de las prostaglandinas, este compuesto que inhibe la agregacin plaquetaria, dilata los vasos sanguneos principalmente de las circulaciones coronaria (cardiaca) y cerebral, disminuye la liberacin de algunos neurotransmisores de tipo excitador en el sistema nervioso central, disminuye la contraccin del msculo intestinal, disminuye la liplisis en el tejido adiposo, e incluso acta como protector de la funcin heptica. Esta diversidad de acciones nuevamente nos ilustra la importancia fisiolgica que posiblemente tenga el compuesto, pero tambin nos habla de las dificultades para cualquier aplicacin teraputica. Sin embargo, se han diseado compuestos que alteran el transporte o el metabolismo de la adenosina, los cuales s se han usado ya con fines teraputicos. sta es tambin un rea activa y lo va a ser ms en los prximos aos.
Existen diversos receptores para la adenosina que junto con los del ATP constituyen la familia de los receptores purinrgicos. Todos ellos parecen tambin formar parte de la gran familia de los receptores con siete dominios transmembranales.
Otro aspecto de inters, respecto a la adenosina, es su relacin con las metilxantinas. Estas ltimas son compuestos bloqueadores de la fosfodiesterasa, la cual rompe e inactiva el AMP cclico, y adems son potentes antagonistas de las acciones de la adenosina. Las metilxantinas estn consideradas entre las drogas de mayor consumo por los seres humanos en nuestros das. La cafena, la teofilina y la teobromina son algunos de los integrantes principales de este grupo, y son constituyentes activos del caf, el t y el chocolate. Cierto es que, cada vez con mayor frecuencia, se ingiere caf descafeinado, pero para compensar se consume una gran cantidad de cafena en los refrescos de cola. Es muy posible que el efecto estimulante de estas bebidas se deba al antagonismo de la accin de la adenosina que ejercen las metilxantinas.
-
D) GLUCAGON, VASOPRESINA Y ANGIOTENSINA II
En esta seccin tratar de revisar brevemente algo de lo que sabemos acerca de tres hormonas de naturaleza peptdica, las cuales ejercen una profunda accin sobre el metabolismo: el glucagon, la vasopresina y la angiotensina II. Comenzar por el glucagon.
i) Glucagon
Esta hormona es un polipptido con un peso molecular de 3 500 Daltones aproximadamente que se produce en las clulas alfa de los islotes de Langerhans del pncreas y es una de las principales hormonas hiperglicemiantes, es decir, aquellas que aumentan los niveles de glucosa en la sangre. Recordemos que la insulina tambin es producida en los islotes pancreticos y ejerce una accin opuesta. Estas pequeas islas de clulas de secrecin interna se encuentran dentro de la estructura del pncreas rodeadas por las clulas productoras de los jugos digestivos y su funcin es regular la concentracin de glucosa en la sangre. Son un sistema de ajuste muy fino de la glucemia; cuando el nivel de glucosa en sangre se eleva, aumenta la secrecin de insulina, la cual, como ya hemos descrito, favorece la entrada de este azcar a las clulas y su posterior metabolismo, disminuyendo en consecuencia su concentracin en la sangre. Por el contrario, cuando la glucemia baja, los islotes secretan glucagon, el cual se encarga de "indicar" al hgado que aumente su produccin de glucosa, y al tejido adiposo que libere cidos grasos y glicerol como fuentes de energa y material para la sntesis heptica de ms glucosa. Visto as, nos percatamos de que estos islotes son un regulador de la glucemia; podramos llamarles un glucostato. Los islotes se pueden aislar e incubar por tiempo relativamente largo sin que se deterioren. Se ha pensado en transplantarlos a pacientes diabticos como una alternativa teraputica y miles de estudios han sido realizados con ellos.
Pero volvamos al glucagon. Esta molcula fue descubierta por Murlin y colaboradores en 1923, tan slo dos aos despus del descubrimiento de la insulina. Sin embargo, su historia ha sido un poco gris; en compensacin con la historia de la insulina, su vecina de las clulas de al lado, que es brillante, espectacular, maravillosa, como las grandes avenidas: llena de luces. El descubrimiento del glucagon recibi poco reconocimiento; para ilustrar el escaso inters que gener dir que pasaron ms de 30 aos desde su descubrimiento hasta su purificacin. Para todo existe una razn. A qu se debi el contraste de inters y avance entre insulina y glucagon? Una razn importante es la rpida aplicacin teraputica, la utilidad. La insulina poda emplearse para el tratamiento de uno de los grandes males de la humanidad, la diabetes. El glucagon no tiene la misma suerte, su utilidad en la prctica mdica es limitada: slo se utiliza para algunos casos de hipoglucemia y recientemente para algunos problemas cardiovasculares. Ni modo, una naci con estrella y la otra estrellada. Lo que ha dado popularidad al glucagon es el hecho de que su accin en algunas condiciones es adversa. Voy a explicarme. Por all en el inicio de los setenta, al desarrollarse las tcnicas de deteccin de hormonas en el suero, se observ que en algunos diabticos los niveles de insulina no estaban disminuidos como se supona; estaban normales, pero los de glucagon
-
estaban elevados. As, el grupo de Unger propuso que el glucagon quiz era responsable en parte de los problemas del diabtico. Hoy sabemos que la diabetes es una enfermedad (o grupo de enfermedades) mucho ms compleja de lo que pensbamos. Muchos factores contribuyen, y posiblemente el exceso en la secrecin de glucagon sea uno de ellos.
Para 1972 se haba logrado la sntesis qumica del glucagon y con ello se empezaron a sintetizar compuestos similares (anlogos) al glucagon con dos ideas fundamentales: conocer qu partes de su estructura son importantes para su actividad y, por otro lado, sintetizar antagonistas que pudieran bloquear las acciones de glucagon sobre el organismo y as poder, tal vez, aliviar algunos de los problemas del diabtico.
Hasta el momento slo se conoce un receptor para el glucagon, el cual est acoplado en forma activadora a la adenilil ciclasa y, en apariencia, en forma secundaria a la movilizacin de calcio, probablemente a travs del sistema fosfoinostidos-calcio. Al activar el glucagon a su receptor en el hgado, se produce un incremento rpido en los niveles de AMP cclico, el cual, a su vez, activa a la protena cinasa A y mediante la cascada de fosforilaciones, que describ captulos atrs, se activan la ruptura de glucgeno y otros procesos.
ii) Vasopresina
La vasopresina, como su nombre lo indica, es una potente hormona vasopresora, es decir, aumenta la contraccin de los vasos y la tensin arterial. Tambin recibe el nombre de hormona antidiurtica, mismo que describe otra de sus acciones principales: disminuir la diuresis, es decir, la prdida de lquidos por la orina.
La vasopresina u hormona antidiurtica es liberada por la hipfisis posterior (una glndula neurosecretora localizada en la base del cerebro) cuando la ingestin de lquidos es poca y cuando disminuye el lquido extracelular. Esta hormona es un pptido pequeo formado slo por nueve aminocidos. Su estructura fue determinada por DuVigneaud, quien adems la sintetiz, por lo que se le otorg el premio Nobel poco despus de su descubrimiento. Este investigador y su grupo estudiaron tambin la oxitocina, una hormona muy parecida a la anterior (slo cambia en dos aminocidos), responsable de la contraccin uterina durante el parto.
Esta hormona antidiurtica se utiliza para el diagnstico de la diabetes inspida (no confundir a esta enfermedad con la diabetes mellitus, la diabetes comn que todos conocemos). Se utiliza tambin como medicamento para controlar el sangrado de vrices esofgicas en los pacientes cirrticos.
La vasopresina ejerce sus efectos al activar a dos tipos de receptores: los V1 y los V2. Los efectos presores se ejercen al activar a los receptores V1, de los que existen dos subtipos (A y B) y los cuales estn acoplados a recambio de fosfoinostidos y a movilizacin de calcio. Los efectos antidiurticos se ejercen mediante receptores V2 que estn acoplados en forma activatoria a la adenilil ciclasa a travs de Gs. Actualmente contamos
-
con anlogos sintticos de la vasopresina, los cuales son selectivos hasta por 1 000 veces para algunos de los subtipos de receptores de dicha hormona. No debe pensarse que la vasopresina no tiene efectos en otros rganos; se ha observado que es un importante modulador del metabolismo heptico, adems de tener efectos sobre las plaquetas e incluso sobre algunas neuronas.
iii) Angiotensina II
Hace casi un siglo, en 1898, se descubri que el extracto del rin produce un fuerte efecto vasopresor; a este principio se le dio el nombre de renina. Posteriormente se descubri que la renina no era vasopresora por s misma, sino que era una enzima que converta a un producto inactivo del plasma, el angiotensingeno, en uno activo, la angiotensina. La angiotensina es una hormona que descubrieron el doctor Braun-Menndez y su grupo, en Argentina hacia 1939. Casi al mismo tiempo, el grupo de Page, hizo el mismo descubrimiento. Cada uno de estos grupos dio un nombre al compuesto generado en el plasma, el primero lo llam "hipertensina" y el segundo "angiotonina". Fue necesario que pasaran casi 20 aos, para que se pusieran de acuerdo los investigadores del campo en el nombre adecuado para la hormona, y en 1957 se le dio el nombre hbrido de angiotensina.
Actualmente sabemos que hay tres angiotensinas: la I, la II y la III, las cuales son productos cada vez ms pequeos; es decir, del angiotensingeno se forma la angiotensina I, de ella la angiotensina II y de sta a su vez la angiotensina III; la ms activa es la angiotensina II.
La angiotensina II es el agente vasopresor ms potente que se conoce hasta ahora; adems es un importante regulador de la secrecin de otra hormona por la corteza de la glndula suprarrenal: la aldosterona. En el corazn, la angiotensina II incrementa el flujo de calcio y la fuerza de contraccin del msculo. Otra accin importante de este pptido es favorecer la secrecin de la vasopresina. Durante los ltimos aos ha resultado evidente que este compuesto es tambin capaz de alterar el metabolismo heptico.
Pero cmo ejerce sus efectos? La angiotensina II tiene receptores especficos localizados en la membrana plasmtica de muchas clulas. Se han identificado dos tipos de receptores para la angiotensina II. El AT1 de distribucin perifrica y el que media las acciones ms conocidas del pptido. En algunas especies el gen que codifica para el receptor se ha duplicado y modificado dando origen a dos variedades de este tipo de receptor (A y B). El receptor est acoplado al recambio de fosfoinostidos a travs de Gq y a la adenilil ciclasa en forma inhibidora por medio de Gi. Adems parece estar asociado a otros sistemas de transduccin, que le permiten actuar como un poderoso factor de crecimiento en muchas clulas.
El receptor AT2 parece estar acoplado a travs de protenas G (an no bien identificadas) a una fosfatasa de protenas para residuos de tirosina. Este parece ser un nuevo sistema transductor que ahora se est estudiando con mucho inters. Por otro lado, el receptor AT2 parece expresarse en el
-
sistema nervioso y especialmente en ciertas etapas del desarrollo, lo que ha sugerido que ocupe un lugar en la diferenciacin de este importante sistema. Nuevamente otro campo en el que veremos cambios en los prximos aos.
4. Principales neurotransmisores-biosntesis:Estos extractos tienen la propiedad de aumentar la tensin arterial y la frecuencia cardiaca.La noradrenalina se segrega en los terminales de muchas neuronas cuyos somas estn situados en el tronco del encfalo y el hipotlamo. En concreto, las que estn situadas en el locus cerelus de la protuberancia envan fibras nerviosas a amplias regiones del encfalo que sirven para controlar la actividad global y el estado mental, como por ejemplo aumentar el nivel de vigilia. En la mayora de estas zonas, la noradrenalina probablemente activa receptores excitadores, pero en unas cuantas, en cambio, estimula los inhibidores. Tambin se segrega en la mayor parte de
-
las neuronas posganglionares del sistema nervioso simptico, donde excita algunos rganos, pero inhibe otros.La dopamina se segrega en las neuronas originadas en la sustancia negra. Su terminacin se produce en la regin estriatal de los ganglios basales. El efecto que ejerce suele ser una inhibicin.La glicina se segrega en las sinapsis de la mdula espinal. Se cree que siempre acta como un transmisor inhibidor.El GABA (cido gamma-aminobutrico) es segregado en los terminales nerviosos de la mdula espinal, el cerebelo, los ganglios basales y muchas reas de la corteza. Causa inhibicin.
Acetilcolina. Esta sustancia transmisora se sintetiza en el terminal presinptico a partir de la coenzima A y colina en presencia de la enzima acetiltransferasa de colina. A continuacin, se transporta a sus vesculas especficas, luego, al producirse su salida hacia la hendidura sinptica durante la transmisin de la seal nerviosa en la sinopsis, se degrada nuevamente en acetato y calina por accin de la colinesterasa (presente en el retculo formado por proteoglucano que rellena el espacio de la hendidura sinptica). Luego, una vez ms, las vesculas se reciclan en el interior del terminal presinpticos; la colina sufre un transporte activo de vuelta hacia el terminal para repetir su empleo en la sntesis de nueva acetilcolina.
SINAPSIS
Las sinapsis son uniones especializadas mediante las cuales las clulas del sistema nervioso envan seales de unas a otras y a clulas no neuronales como las musculares o glandulares. Una sinapsis entre una neurona motora y una clula muscular se denomina unin neuromuscular.
Las sinapsis permiten a las neuronas del sistema nervioso central formar una red de circuitos neuronales. Son cruciales para los procesos biolgicos que subyacen bajo la percepcin y el pensamiento. Tambin son el sistema mediante el que el sistema nervioso conecta y controla todos los sistemas del cuerpo.
El cerebro contiene un nmero inmenso de sinapsis, que en nios alcanza los 1000 billones. Este nmero disminuye con el paso de los aos, estabilizndose en la edad adulta. Se estima que un adulto puede tener entre 100 y 500 billones de sinapsis.
-
La palabra sinapsis viene de sinaptena, que Sir Charles Scott Sherrington y colaboradores formaron con las palabras griegas sin-, que significa "juntos", y hapteina, que significa "con firmeza".
Anatomia
En una sinapsis prototpica, como las que aparecen en los botones dendrticos, unas proyecciones citoplasmticas con forma de hongo desde cada clula, y en las que los extremos de ambas se aplastan uno contra otro. En esta zona, las membranas celulares de ambas clulas se juntan en una unin estrecha que permite a las molculas seal llamadas neurotransmisores pasar rpidamente de una a otra clula por difusin. Esta unin, de aproximadamente 20 nm de ancho, se conoce como hendidura sinptica.Estas sinapsis son asimtricas tanto en su estructura como en su funcionamiento. Slo la neurona presinptica segrega los neurotransmisores, que se unen a los receptores transmembrana que la clula postsinptica tiene en la hendidura. El terminal nervioso presinptico (tambin llamado botn sinptico o botn) normalmente emerge del extremo de un axn, mientras que la zona postsinptica normalmente corresponde a una dendrita, al cuerpo celular o a otras zonas celulares. La zona de la sinapsis donde se libera el neurotransmisor se denomina zona activa. En las zonas activas, las membranas de las dos clulas adyacentes estn unidas estrechamente mediante protenas de adhesin celular. Justo tras la membrana de la clula postsinptica aparece un complejo de protenas entrelazadas denominado densidad postsinptica. Las protenas de la densidad postsinptica cumplen numerosas funciones, que van desde el anclaje y movimiento de receptores de neurotransmisores de la membrana plasmtica, al anclaje de varias protenas reguladoras de la actividad de estos receptores.Tambin existe una forma menos compleja de unin llamada sinapsis elctrica, en la que las neuronas se acoplan elctricamente entre s a travs de complejos proteicos denominados uniones gap.
Sinapsis elctrica
Sinapsis elctrica es una sinapsis en la que la transmisin entre la primera neurona y la segunda no se produce por la secrecin de un neurotransmisor, como en las sinapsis qumicas, sino por el paso de iones de una clula a otra a travs de uniones gap. Las uniones gap son pequeos canales formados por el acoplamiento de complejos proteicos, basados en conexinas, en clulas estrechamente adheridas.Las sinapsis elctricas son ms rpidas que las sinapsis qumicas pero menos plsticas. En vertebrados son abundantes en la retina y en la corteza cerebral.
Sealizacin en sinapsis qumicas
La liberacin de neurotransmisores es iniciada por la llegada de un impulso nervioso (o potencial de accin), y se produce mediante un proceso muy
-
rpido de secrecin celular: en el terminal nervioso presinptico, las vesculas que contiene los neurotransmisores permanecen ancladas y preparadas junto a la membrana sinptica. Cuando llega un potencial de accin se produce una entrada de iones calcio a travs de los canales de calcio dependientes de voltaje. Los iones de calcio inician una cascada de reacciones que terminan haciendo que las membranas vesiculares se fusionen con la membrana presinptica y liberando su contenido a la hendidura sinptica. Los receptores del lado opuesto de la hendidura se unen a los neurotransmisores y fuerzan la apertura de los canales inicos cercanos de la membrana postsinptica, haciendo que los iones fluyan hacia o desde el interior, cambiando el potencial de membrana local. El resultado es excitatorio en caso de flujos de despolarizacin, o inhibitorio en caso de flujos de hiperpolarizacin. El que una sinapsis sea excitatoria o inhibitoria depende del tipo o tipos iones que se canalizan en los flujos postsinpticos, que a su vez es funcin del tipo de receptores y neurotransmisores que intervienen en la sinapsis.[editar]
Fuerza sinptica
La fuerza de una sinapsis viene dada por el cambio del potencial de membrana que ocurre cuando se activan los receptores de neurotransmisores postsinpticos. Este cambio de voltaje se denomina potencial postsinptico, y es resultado directo de los flujos inicos a travs de los canales receptores postsinpticos. Los cambios en la fuerza sinptica pueden ser a corto plazo y sin cambios permanentes en las estructuras neuronales, con una duracin de segundos o minutos, o de larga duracin (potenciacin a largo plazo o LTP), en que la activacin continuada o repetida de la sinapsis implica que los segundos mensajeros inducen la sntesis proteica en el ncleo de la neurona, alterando la estructura de la propia neurona. El aprendizaje y la memoria podran ser resultado de cambios a largo plazo en la fuerza sinptica, mediante un mecanismo de plasticidad sinptica.
Integracin de seales sinpticas
Generalmente. si una sinapsis excitatoria es fuerte, un potencial de accin en la neurona presinptica iniciar otro potencial en la clula postsinptica. En una sinapsis dbil, el potencial excitatorio postsinptico ("PEPS") no alcanzar el umbral para la iniciacin del potencial de accin. En el cerebro, cada neurona mantiene conexiones o sinapsis con muchas otras, pudiendo recibir cada una de ellas mltiples seales. Cuando se disparan potenciales de accin simultneamente en varias neuronas que se unen en sinapsis dbiles a otra neurona, pueden forzar el inicio de un impulso en esa clula a pesar de que las sinapsis son dbiles.Por otro lado, una neurona presinptica que libera neurotransmisores inhibitorios, como el GABA, puede generar un potencial inhibitorio postsinptico ("PIPS") en la neurona postsinptica, bajando su sensibilidad y la probabilidad de que se genere un potencial de accin en ella. As la respuesta de una neurona depende de las seales que recibe de otras, con las que puede tener distintos grados de influencia, dependiendo de la fuerza
-
de la sinapsis con esa neurona. John Carew Eccles realiz algunos experimentos importantes en los inicios de la investigacin sinptica, por los que recibi el Premio Nobel de Fisiologa o Medicina en 1963. Las complejas relaciones de entrada/salida conforman las bases de la computacin basada en transistores, y se cree que funcionan de forma similar en los circuitos neuronales.[editar]
Propiedades y regulacin
Tras la fusin de las vesculas sinpticas y la liberacin de las molculas transmisoras en la hendidura sinptica, el neurotransmisor es rpidamente eliminado del espacio por protenas especializadas en su reciclaje, situadas en las membranas tanto presinptica como postsinptica. Esta recaptacin evita la desensibilizacin de los receptores postsinpticos y asegura que los potenciales de accin subsiguientes generen un PEP de la misma intensidad. La necesidad de una recaptacin y el fenmeno de la desensibilizacin en los receptores y canales inicos significa que la fuerza de la sinapsis puede disminuir si un tren de potenciales de accin llega en una sucesin rpida, un fenmeno que hace que exista una dependencia de la frecuencia en las sinapsis. El sistema nervioso se aprovecha de esta propiedad para computaciones, y puede ajustar las sinapsis mediante la fosforilacin de las protenas implicadas. El tamao, nmero y tasa de reposicin de las vesculas tambin est sujeto a regulacin, as como otros muchos aspectos de la transmisin sinptica. Por ejemplo, un tipo de frmaco conocido como inhibidores selectivos de la recaptacin de serotonina o SSRI afectan a ciertas sinapsis inhibiendo la recaptacin del neurotransmisor serotonina. Por el contrario, un neurotransmisor excitatorio muy importante, la acetilcolina, no es recaptada, pero es eliminada por accin de la enzima acetilcolinesterasa.
RECEPTORES
1.- Definicion:Los receptores son estructuras proteicas que generalmente se encuentran en las membranas externas y se comportan como transductores biolgicos Los receptores son estructuras que permiten analizar todo lo que hay en el entorno y todo lo hay dentro. Cada receptor tiene una forma particular de energa. El receptor lleva a cabo la transduccin (transformacin de una forma de energa que constituye el estmulo en impulsos nerviosos) Los receptores son los componentes de una clula que tienen la capacidad de identificar una sustancia, hormona o neurotransmisor. La idea de que existen receptores, viene de principios de siglo, cuando Langley, Dale y cols., sugieren que pueden existir sustancias receptivas en la superficie de las membranas de clulas excitables: Lo primero, que al menos, dos sustancias especiales (sustancias receptivas), estn presentes
-
en la regin neural del msculo, y que los impulsos nerviosos slo pueden causar contraccin actuando en una sustancia receptora. Lo segundo que las sustancias receptoras, forman ms o menos fcilmente componentes disociables. Es una estructura que detecta las condiciones fsicas o qumicas del ambiente y transmite esta informacin al sistema nervioso, y lo hacen porque traducen los estmulos del ambiente en potenciales de accin.
PROPIEDADES
Son transductores de energa.Son sensibles slo a una forma de energa.Son sensibles a pequeas intensidades de energa amplificando la seal hacia el sistema nervioso.
2.- Mecanismo de accinEl receptor recibe el impacto del neurotransmisor y lleva a cabo la transduccin (recibe una seal y transmite otra). El ATP necesario para la sntesis de neurotransmisor es proporcionado por las mitocondrias de la Terminal presinptica. Esta sntesis debe ser muy rpida, debido a que la cantidad del mismo almacenada en las vesculas se halla limitada para unos segundos o minutos de actividad plena. Los receptores son protenas bajo control gentico. El receptor recibe el impacto del neurotransmisor y lleva a cabo la transduccin (recibe una seal y transmite otra). El ATP necesario para la sntesis de neurotransmisor es proporcionado por las mitocondrias de la terminal presinptica. Esta sntesis debe ser muy rpida, debido a que la cantidad del mismo almacenada en las vesculas se halla limitada para unos segundos o minutos de actividad plena. Los receptores son protenas bajo control gentico.
Los receptores tienen dos componentes importantes:
1. Componente de fijacin, que protruye al exterior de la membrana en el surco sinptico, y fija el neurotransmisor liberado de la terminal presinptica.
2. Componente ionforo, que penetra de la membrana al interior de la neurona y puede ser de 2 tipos: Un conducto de iones activado qumicamente -conductos activados por ligando-, cuyo paradigma es el NMDA -N-metil-D-aspartato-, verdadero conglomerado o complejo de canales inicos, que pueden ser de 3 tipos principales: Canales de sodio, Canales de potasio y Canales de cloruro. Tambin pertenecen a esta categora los receptores denominados AMPA/Kainate; y una enzima que activa un sistema metablico interno de las clulas y son receptores metabotrpicos que se unen a protenas G en el interior de las clulas. Las neuronas regulan su actividad por mecanismos de retroalimentacin que involucran a receptores de la terminacin nerviosa, conocidos como autoreceptores. Ejemplo de autoreceptor es el receptor b2-adrenrgico en las terminaciones noradrenrgicas, que media muchas de las acciones fisiolgicas de las catecolaminas endgenas Adrenalina y Noradrenalina, y es el blanco de varios agentes teraputicos. Pese a que el mecanismo de recaptura de neurotransmisores se conoce
-
desde hace unos 30 aos. La disponibilidad de frmacos selectivos que actan sobre tales receptores es bastante reciente. La clonacin de receptores y la posibilidad de expresarlos en clulas no neuronales est permitiendo un mayor conocimiento del mecanismo de accin de los diferentes transportadores, asi como la posibilidad de encontrar nuevos frmacos selectivos.
3.- Clasificacion de receptores sensitivos:
a.- De acuerdo a su localizacin
a) Perifricos
Generan una respuesta ventilatoria refleja por los cambios principalmente en la falta de oxgenoLos impulsos aferentes de estos cuerpos se unen al noveno par craneal.
b)Centrales
Se encuentran bilateralmente en la zona dorsal del bulbo raqudeoSe encuentran cerca de los centros respiratorios bulbaresModifican el patrn ventilatorio de acuerdo con los cambios en el CO2 y el H+g
b.- De acuerdo a la procedencia del estimulo
Exteroreceptores
Entregan informacin acerca del ambiente externo. Gracias a ellos los seres vivos pueden orientarse en sus desplazamientos, reconocer alimento, encontrar pareja, aprender, conocer el entorno, etc.Generalmente las sensaciones exterorreceptivas provienen de la superficie del cuerpo.
Interoreceptores
Se localizan en el interior de los rganos del cuerpo.
Son afectados por los cambios fisiolgicos de las condiciones internas como pH, temperatura, presin osmtica, etc.
Los interorreceptores incluyen a:
los mecanorreceptores sensibles a la presin sangunea,
los quimiorreceptores sensibles a las concentraciones de O2, CO2 y H+ y
-
Los sensores de temperatura del hipotlamo son tambin interorreceptores.
Habitualmente, no somos conscientes de las seales de estos receptores.
Sin embargo, en algunas ocasiones, las seales resultan en percepciones como dolor, hambre, sed, nuseas, o la sensacin, producida por receptores de tensin, de tener la vejiga o el intestino llenos.
Gracias a las sensaciones interorreceptivas el cuerpo acciona mecanismos homeostticos.
Propioreceptores
Informan acerca de la orientacin del cuerpo en el espacio y de la posicin de los miembros.
Se encuentran en el interior de los msculos, tendones y articulaciones.
Los canales semicirculares del odo son los rganos propioceptores ms importantes en muchos vertebrados.
Gracias a las sensaciones propiocpticas el cuerpo humano puede caminar y correr con la vista vendada o en la oscuridad.
c.- De acuerdo a la concesin del sistema nervioso central
Primarios (neuronas)
son terminaciones nerviosas libre que, cuando se aplica un estmulo no sensitivo producen un potencial de accin.
Secundarios: (clulas especializadas)
hacen contacto sinptico con vas nerviosas aferentes
d.- De acuerdo a la adaptacin
Fsicos
Son receptores que responden a un estmulo durante un breve periodo de tiempo, aunque el estmulo se prolongue en el tiempo. Por ejemplo, la habituacin a un olor. Son receptores que sufren un proceso de adaptacin, que es una prdida de sensibilidad, que conduce a la prdida total de la misma. Los fsicos no son buenos identificando la duracin de un estmulo
Tnicos
Son receptores que responden siempre al estmulo, aunque este se prolongue en el tiempo. En muchos receptores tnicos la intensidad de la respuesta al estmulo disminuye, pero no desaparece hasta que desaparece
-
el estmulo. La duracin del estmulo se construye segn el nivel de descarga que le llega al cerebro. Si le llega una fuerte descarga, el cerebro lo interpreta como que el estmulo acaba de empezar, y si la descarga es dbil, se interpreta como que lleva rato estimulando.
e.- De acuerdo al estimulo adecuado
Mecanoreceptores:
Reaccionan con la energa mecnica.
Responden al tacto, posicin del cuerpo, presin, gravedad, movimiento y audicin
Termorreceptores
Reaccionan con los niveles de calor del medio externo y con el de los lquidos y rganos interiores.
Detectan fro y calor.
Fotorreceptores
Reaccionan con las radiaciones luminosas.
Permiten detectar imgenes del exterior, formas y colores.
Las clulas receptoras son capaces de captar la radiacin luminosa comprendida entre la ultravioleta y la infrarroja, no siendo detectadas estas. La luz es un fenmeno ondulatorio causado por el movimiento de unas partculas llamadas fotones. Los receptores son capaces de detectar estas partculas, a veces con una sensibilidad capaz de detectar un solo fotn e interpretarlo como seal luminosa. Hay 2 tipos de receptores: conos y bastones. Para que la luz llegue a los receptores, las neuronas que estn delante se hacen transparentes, esto lo consiguen no mielinizndose. La funcin del epitelio pigmentado (de color negro para absorber la luz) es absorber toda la radiacin luminosa que no llegue a los conos ni a los bastones (para que no choque y rebote).
Quimiorreceptores:
Forman la fase de las sensaciones gustativas en la boca, olfatorias en la nariz y responden al nivel de oxigeno en sangre arterial, osmolaridad de lquidos corporales, concentracin de dixido de carbonos.
Entre los quimiorreceptores de los vertebrados terrestres, las clulas gustativas estn localizadas dentro de la boca.
-
Los receptores gustativos y las clulas de soporte que los rodean constituyen las papilas gustativas que permiten distinguir cuatro sabores primarios: dulce, cido, salado y amargo.
En los animales terrestres, el olor puede ser definido como la quimiorrecepcin de sustancias transportadas por el aire
Sin embargo, para ser detectadas, estas sustancias deben primero estar disueltas en una capa acuosa de moco que recubre a un tejido especializado, el epitelio olfativo.
En los seres humanos, este tejido est localizado en el fondo de las fosas nasales.
a) El epitelio olfativo es responsable de nuestro sentido del olfato y mucho de lo que paladeamos en la comida es el resultado de sustancias voltiles que llegan a este tejido.
b) El epitelio olfativo est compuesto por clulas de soporte, clulas basales y clulas olfativas, que son los receptores sensoriales.
Nocirreceptores: Reconocen dao tisular producido por agentes fsicos o qumicos
Son receptores especficos del dolor.
Detectan daos fsicos y qumicos en los tejidos.
Los nosiceptores son receptores que reaccionan ante algunas substancias que los tejidos daados liberan. Los tejidos al daarse liberan prostaglandinas. Estas son substancias que al liberarse activan otras clulas para que liberen histamina, la cual acta sobre los nosiceptores causando un potencial de membrana que, si alcanza el valor umbral en el primer ndulo de Ranvier, se convierte en un potencial de accin (este proceso no se lleva a cabo en la colina axnica). Estas terminaciones libres tambin estn en la dermis y en la epidermis, conduciendo estmulos dolorosos (siendo esto sensibilidad exteroceptiva). En algunos rganos internos hay terminaciones libres que actan transmitiendo sensaciones de temperatura. Son termoreceptores, los cuales estn presentes en todas las superficies del cuerpo. En estas regiones, los termoreceptores pueden ser estructuras ms especializadas llamadas corpsculos de Krause.
La actividad de los termoreceptores es relativa, es decir, el umbral de excitabilidad cambia continuamente, dependiendo siempre de la experiencia previa. Las altas temperaturas disminuyen la sensibilidad de los receptores de calor, y aumentan la de los receptores de fro, y al revs.
En la superficie del cuerpo tambin existen otros receptores, que son: corpsculos de Pacini, que son mecanoreceptores (sensibles al tacto, a la deformacin). Los corpsculos de Pacini estn formados por una terminacin nerviosa recubierta por capas de clulas fcilmente
-
deformable. Estn en la capa ms profunda de la dermis. Son receptores de adaptacin muy rpida (cuando reciben el estmulo, emiten un potencial, pero inmediatamente dejan de responder aunque el estmulo contine). Vuelven a responder, enviando una seal, cuando el estmulo cesa. Por esto son receptores especializados en captar vibraciones (por ejemplo, el tacto de una superficie rugosa).
El corpsculo de Pacini, cuando es presionado, la distancia entre capas se estrecha, y ejerce una accin de traccin sobre glcidos que se encuentran en la superficie de la terminal nerviosa. Estos glcidos (adosados a protenas canales), se estiran hacia los lados, abriendo el canal. La apertura del canal provoca la entrada de Na+ en la terminal, apareciendo el potencial receptor, el cual se transforma en potencial de accin al llegar al primer ndulo de Ranvier.
Otros tipos de receptores son los discos de Merkel. Estos son ramificaciones nerviosas con clulas en sus puntas. Son receptores de adaptacin lenta. Responden a una deformacin sostenida. Otros tipos de receptores son terminaciones libres, que son de adaptacin rpida (distintos de los discos de Merkel, no confundirlos con estos). Son de adaptacin rpida a la temperatura, y de adaptacin algo ms lenta para la recepcin de dolor.
Los corpsculos de Meissner son mecanoreceptores de adaptacin rpida. Son una fibra nerviosa simple rodeada de clulas (son distintas de los corpsculos de Pacini). Tambin estn los corpsculos de Rufini. Tienen una estructura parecida a los discos de Merkel, solo que cada terminal no est rodeada por una clula, sino que forma bulbos al enrollarse sobre s misma. Tienen adaptacin lenta, y responden a la presin sostenida. Otro tipo de receptores son las terminales nerviosas que rodean a los bulbos pilosos. Las terminales de los bulbos pilosos rodean al bulbo, informando del roce del pelo, de un cambio de posicin de este. Otro tipo son los corpsculos de Krausse.
Todos estos tipos de receptores se distribuyen de distinta forma por el cuerpo, hay zonas con ms, y zonas con menos receptores, y zonas donde no hay ninguno. La piel con pelo tiene todos los receptores. La piel sin pelo (piel glabra), tiene los de Meisser, Krausse, Pacini, Merkel, y terminales libres.
4.- Receptores de los principales neurotransmisores Receptores de la acetilcolina:
Receptor nicotnico (nAChR, llamado tambin receptor de acetilcolina ionotrpico), que logra unirse con especificidad por la nicotina, de all su nombre.
-
Receptor muscarnico (mAChR, llamado tambin receptor de acetilcolina metabotrpico), que logra unirse con especificidad por la muscarina, de all su nombre
5.- Autorreceptores
Los autorreceptores que se encuentran en cualquier neurona responden a las molculas transmisoras liberadas por la misma neurona. En el terminal nervioso, responden a las molculas transmisoras liberadas en la hendidura sinptica; sobre el cuerpo celular responderan a las molculas transmisoras liberadas por las dendritas. Funcionalmente, la mayoria de los autorreceptores parecen regular la liberacin de transmisores. Se han identificado los autorreceptores para las neuronas que contienen noradrenalina, dopamina, serotonina y GABA. Los principales tipos de autorreceptores inhibidores, descritos tanto en el sistema nervioso simptico perifrico como en el cerebro, tienen propiedades farmacolgicas similares a aquellas de los receptores ?a-adrenrgicos.
Los autorreceptores del tipo b-adrenrgico tambin han sido descritos. Esto difiere de la mayora de los otros autorreceptores conocidos, en los que la noradrenalina que acta sobre estos receptores facilita la liberacin de transmisores, y de esta forma amplifica los efectos del bombardeo neuronal. Este efecto contrasta con la accin inhibidora de los autorreceptores a-adrenrgicos y de la dopamina, los cuales ejercen un control de retroalimentacin negativa sobre la liberacin de transmisores.
6.- Receptores ionotropicos y receptores metabotropicos:
Receptores ionotrficos
Determinan la apertura o cierre de canales y producen despolarizaciones (gnesis de potenciales de respuesta excitatorios) o hiperpolarizaciones (gnesis de potenciales de respuesta inhibitorios). Es una respuesta rpida. El mecanismo de accin de estos receptores puede ser de dos formas, va sealizacin extracelular a travs de la accin de un neurotransmisor que induce, al unirse al receptor la apertura del canal, algunos canales pueden necesitar la unin de dos neurotransmisores como es el caso del receptor de Acetilcolina o el receptor NMDA que necesita glutamato y glicina. La sealizacin tambin puede ser intracelular, generalmente fosforilando en la cara citoplsmica del canal el receptor, induciendo la apertura del canal.
Receptores metabotrficos
-
Liberan mensajeros intracelulares (AMPcclico, CA y fosfolpidos). Cuando el receptor recibe el neurotransmisor, pone en funcionamiento la adenilatociclasa y el ATP se transforma en AMPcclico. Una vez ha actuado, es destruido por la fosfodiesterasa. EL AMPcclico activa una proteinquinasa, que fosforila 1 protena. Los receptores de Ca+2, cuando reciben el neurotransmisor, abren 1 canal de Ca+2, entra Ca+2 extracelular y se junta a la protena calmodulina, formando la calmodulina-Ca, que activa una proteinquinasa que fosforila una protena. El receptor de membrana, cuando recibe el neurotransmisor, activa la PLC (fosfolipasa C). La PLC acta sobre los fosfolpidos de membrana (concretamente sobre el fosfotidilinositol) y se derivan 2 productos (inositol trifosfato [IP3] y diacilglicerol). El inositol trifosfato acta sobre el retculo endoplasmtico liberando el Ca+2intracelular. El diacilglicerol, en presencia de Ca+2, activa la proteinquinasa que fosforila la protena y da lugar a la respuesta postsinptica.
1. Si el receptor es inotrpico slo abre o cierra canales. Hay receptores que ponen en marcha protenas G. El neurotransmisor acta sobre el receptor, que activa una protena G, que activa la adenil ciclasa y que transforma el ATP en AMPcclico. Este AMPcclico puede actuar sobre el canal de membrana, abrindolo. Son canales inicos operados por vas metablicas activadas por protena G. Cuando se fosforila el canal, se abre.
-
2. El neurotransmisor y el receptor provocan que la protena G active la PLC y active el fosfatidil inositol dando (IP3 y diacilglicerol), que libera el Ca+2 y la proteinquinasa fosforila la protena del canal y se abre. 3. El neurotransmisor estimula el receptor, que provoca que la protena G abra el canal. Los recientes avances sobre el conocimiento de la farmacologa de los receptores de los aminocidos excitatorios permiten la aplicacin del conocimiento fino de su papel en la etiologa de las enfermedades neurodegenerativas y su tratamiento. Los receptores ionotrpicos de los aminocidos excitatorios pueden ser divididos en dos largas familias: la familia del receptor NMDA y la familia de los receptores AMPA y KAINATO. Los estudios de clonaje de receptores han mostrado que hay un largo nmero de potenciales subtipos de receptores en ambas familias. Han sido desarrollados antagonistas para los receptores NMDA los cuales pueden interactuar como mnimo con cuatro sitios del receptor, reconocidos como drogas independientes. Para los receptores AMPA y KAINATO, dos clases de antagonistas han sido bien identificados. Razonable potencia, selectividad y penetracin cerebral son las propiedades fundamentales que presentan los antagonistas que se conocen actualmente para stos sitios y comprenden tambin la inhibicin de la liberacin del cido glutmico presinptico, como puede ser el Riluzole. La capacidad del cido glutmico para matar neuronas por su excitotoxicidad ha sido ampliamente demostrada. La acetilcolina como neurotransmisor, acta sobre los receptores nicotnicos (acta igual que la nicotina del tabaco) y sobre los receptores muscarnicos (acta por setas). Los receptores colinrgicos son Muscarnico (M1), Muscarnico (M2) y el nicotnico. Los receptores muscarnicos se pueden bloquear con atropina (se extrae de la Atropa belladona). El curare (dextrotubocuranina) bloquea la sinapsi colinrgica entre el msculo esqueltico. El mismo neurotransmisor, a veces polariza y, a veces, despolariza dependiendo del receptor y los canales que operen el receptor. En la sinapsis colinrgica, se coge cido actico y se esterifica. La colina + Co-A + cido actico dan acetilcolina, que se libera en el espacio sinptico y acta sobre el receptor nicotnico y muscarnico (M1-M2). El enzima acetilcolinesterasa hidroliza el ster de acetilcolina y libera colina y acetato. Despus es recaptado y se vuelve a formar acetilcolina. De todas las sinapsis, se tiene que conocer el neurotransmisor, la biosntesis y la degradacin del neurotransmisor. La dopamina, noradrenalina y adrenalina provienen de la fenilalanina que, mediante la fenilalaninahidroxilasa, le introduce un OH y forma la tiroxina, que mediante la tiroxinahidroxilasa le introduce otro OH y forma la dihidroxifenilalanina. Se descarboxila (se saca COO mediante la carboxilasa) y se forma la dopamina (neurotransmisor de las neuronas dopaminrgicas). Si se le introduce otro OH, se forma la noradrenalina o norepinefrina (neurotransmisor de las neuronas noradrenilaninrgicas). Por accin de una N-metil transferasa se forma la epinefrina o adrenalina (neurotransmisor de las neuronas adrenrgicas).
-
BIBLIOGRAFA
http://es.wikipedia.org/wiki/Neurotransmisor http://www.psicomag.com/neurobiologia/LOS
%20NEUROTRANSMISORES%20EN%20GENERAL.php GUYTON, Arthur... TRATADO DE FISIOLOGA MDICA. 10 Ed.
Interamericana, Mxico, 2001. GANONG, William... FISIOLOGA MDICA. 16 Ed. El Manual Moderno.
Mxico, 1998. ALBERTS, Bruce y otros BIOLOGIA MOLECULAR DE LA CELULA. 4 Ed.
Omega, Barcelona, 2004. MEYER, F... FISIOLOGA HUMANA, Ed. Salvat. Barcelona, 1986.
AnatomiaSinapsis elctricaSealizacin en sinapsis qumicasFuerza sinpticaIntegracin de seales sinpticasPropiedades y regulacin