La fotosíntesis
33
Importancia biológica de la fotosíntesis La fotosíntesis es seguramente el proceso bioquímico más importante de la Biosfera por varios motivos: 1. La síntesis de materia orgánica a partir de la inorgánica se realiza fundamentalmente mediante la fotosíntesis. Posteriormente irá pasando de unos seres vivos a otros mediante las cadenas tróficas, para ser finalmente transformada en materia propia por los diferentes seres vivos. 2. Produce la transformación de la energía luminosa en energía química, necesaria y utilizada por los seres vivos 3. En la fotosíntesis se libera oxígeno que será utilizado en la respiración aerobia como oxidante. 4. La fotosíntesis causó el cambio producido en la atmósfera primitiva, que era anaerobia y reductora. 5. De la fotosíntesis depende también la energía almacenada en combustibles fósiles como carbón, petróleo y gas natural. 6. El equilibrio necesario entre seres autótrofos y heterótrofos no sería posible sin la fotosíntesis. La fotosíntesis (del griego antiguo υώτο [foto], "luz", y σύνθεσις [síntesis], "unión") es la conversión de energía luminosa en energía química estable, siendo el adenosíntrifosfato (ATP) la primera molécula en la que queda almacenada esa energía química. Con posterioridad, el ATP se usa para sintetizar moléculas orgánicas de mayor estabilidad. Además, se debe de tener en cuenta que la vida en nuestro planeta se mantiene fundamentalmente gracias a la fotosíntesis que realizan las algas, en el medio acuático, y las plantas, en el medio terrestre, que tienen la capacidad de sintetizar materia orgánica (imprescindible para la constitución de los seres vivos ) partiendo de la luz y la materia inorgánica . De hecho, cada año los organismos fotosintetizadores fijan en forma de materia orgánica en torno a 100.000 millones de toneladas de carbono. [1][2] Los orgánulos citoplasmáticos encargados de la realización de la fotosíntesis son los cloroplastos , unas estructuras polimorfas y de color verde (esta coloración es debida a la presencia del pigmento clorofila ) propias de las células vegetales . En el interior de estos orgánulos se halla una cámara que contiene un medio interno llamado estroma, que alberga diversos componentes, entre los que cabe destacar enzimas encargadas de la transformación del dióxido de carbono en materia orgánica y unos sáculos aplastados denominados tilacoides o lamelas, cuya membrana contiene pigmentos fotosintéticos. En términos medios, una célula foliar tiene entre cincuenta y sesenta cloroplastos en su interior. [1] Los organismos que tienen la capacidad de llevar a cabo la fotosíntesis son llamados fotoautótrofos (otra nomenclatura posible es la de autótrofos , pero se debe tener en cuenta que bajo esta denominación también se engloban aquellas bacterias que realizan la quimiosíntesis ) y fijan el CO 2 atmosférico. En la actualidad se diferencian dos tipos de procesos fotosintéticos, que son la fotosíntesis oxigénica y la fotosíntesis anoxigénica. La
-
Upload
juan-carlos-1419 -
Category
Travel
-
view
5.065 -
download
1
description
se trata de la enerjia que resibe las plantas
Transcript of La fotosíntesis

Importancia biológica de la fotosíntesis
La fotosíntesis es seguramente el proceso bioquímico más importante de la Biosfera por
varios motivos:
1. La síntesis de materia orgánica a partir de la inorgánica se realiza
fundamentalmente mediante la fotosíntesis. Posteriormente irá pasando de
unos seres vivos a otros mediante las cadenas tróficas, para ser finalmente
transformada en materia propia por los diferentes seres vivos.
2. Produce la transformación de la energía luminosa en energía química,
necesaria y utilizada por los seres vivos
3. En la fotosíntesis se libera oxígeno que será utilizado en la respiración
aerobia como oxidante.
4. La fotosíntesis causó el cambio producido en la atmósfera primitiva, que era
anaerobia y reductora.
5. De la fotosíntesis depende también la energía almacenada en combustibles
fósiles como carbón, petróleo y gas natural.
6. El equilibrio necesario entre seres autótrofos y heterótrofos no sería posible
sin la fotosíntesis.
La fotosíntesis (del griego antiguoυώτο [foto], "luz", y σύνθεσις [síntesis], "unión") es la
conversión de energía luminosa en energía química estable, siendo el adenosíntrifosfato
(ATP) la primera molécula en la que queda almacenada esa energía química. Con
posterioridad, el ATP se usa para sintetizar moléculas orgánicas de mayor estabilidad.
Además, se debe de tener en cuenta que la vida en nuestro planeta se mantiene
fundamentalmente gracias a la fotosíntesis que realizan las algas, en el medio acuático, y
las plantas, en el medio terrestre, que tienen la capacidad de sintetizar materia orgánica
(imprescindible para la constitución de los seres vivos) partiendo de la luz y la materia
inorgánica. De hecho, cada año los organismos fotosintetizadores fijan en forma de materia
orgánica en torno a 100.000 millones de toneladas de carbono.[1][2]
Los orgánulos citoplasmáticos encargados de la realización de la fotosíntesis son los
cloroplastos, unas estructuras polimorfas y de color verde (esta coloración es debida a la
presencia del pigmento clorofila) propias de las células vegetales. En el interior de estos
orgánulos se halla una cámara que contiene un medio interno llamado estroma, que alberga
diversos componentes, entre los que cabe destacar enzimas encargadas de la transformación
del dióxido de carbono en materia orgánica y unos sáculos aplastados denominados
tilacoides o lamelas, cuya membrana contiene pigmentos fotosintéticos. En términos
medios, una célula foliar tiene entre cincuenta y sesenta cloroplastos en su interior.[1]
Los organismos que tienen la capacidad de llevar a cabo la fotosíntesis son llamados
fotoautótrofos (otra nomenclatura posible es la de autótrofos, pero se debe tener en cuenta
que bajo esta denominación también se engloban aquellas bacterias que realizan la
quimiosíntesis) y fijan el CO2 atmosférico. En la actualidad se diferencian dos tipos de
procesos fotosintéticos, que son la fotosíntesis oxigénica y la fotosíntesis anoxigénica. La

primera de las modalidades es la propia de las plantas superiores, las algas y las
cianobacterias, donde el dador de electrones es el agua y, como consecuencia, se desprende
oxígeno. Mientras que la segunda, también conocida con el nombre de fotosíntesis
bacteriana, la realizan las bacterias purpúreas y verdes del azufre, en las que en dador de
electrones es el sulfuro de hidrógeno, y consecuentemente, el elemento químico liberado no
será oxígeno sino azufre, que puede ser acumulado en el interior de la bacteria, o en su
defecto, expulsado al agua.[3]
A comienzos del año 2009, se publicó un artículo en la revistaNatureGeoscience en el que
científicosnorteamericanos daban a conocer el hallazgo de pequeños cristales de hematita
(en Cratón de Pilbara, en el noroeste de Australia), un mineral de hierro que data de la
época del eón Arcaico, demostrando la existencia de agua rica en oxígeno y
consecuentemente, de organismos fotosintetizadores capaces de producirlo. Gracias al
estudio realizado, se ha llegado a la conclusión de la existencia de fotosíntesis oxigénica y
de la oxigenación de la atmósfera y de los océanos hace más de 3.460 millones de años, así
como también se deduce la existencia de un número considerable de organismos capaces de
llevar a cabo la fotosíntesis para oxigenar la masa de agua mencionada, aunque sólo fuese
de manera ocasional.[4][5]
Contenido
[ocultar]
1 Historia del estudio de la fotosíntesis
o 1.1 Desde la Antigua Grecia hasta el siglo XIX
o 1.2 Siglo XX
2 El cloroplasto
o 2.1 Desarrollo
o 2.2 Estructura y abundancia
o 2.3 Función
3 Fase luminosa o fotoquímica
o 3.1 Fotofosforilaciónacíclica
o 3.2 Fase luminosa cíclica
4 Fase oscura o biosintética
5 Fotorrespiración
o 5.1 Ruta de Hatch-Slack o de las plantas C4
o 5.2 Las plantas CAM
6 Fotosistemas y pigmentos fotosintéticos
o 6.1 Los fotosistemas
6.1.1 Fotosistema I y Fotosistema II
o 6.2 Los pigmentos fotosintéticos y la absorción de la luz
7 Factores externos que influyen en el proceso
8 Fotosíntesis anoxigénica o bacteriana
9 Fotosíntesis artificial
o 9.1 Intentos de imitación de las estructura fotosintéticas
o 9.2 Célula de Grätzel

o 9.3 Disoluciones homogéneas
10 Véase también
11 Referencias
12 Bibliografía básica
13 Enlaces externos
Historia del estudio de la fotosíntesis
Desde la Antigua Grecia hasta el siglo XIX
Ya en la Antigua Grecia, el filósofoAristóteles propuso una hipótesis que sugería que la luz
solar estaba directamente relacionada con el desarrollo del color verde de las hojas de las
plantas, pero esta idea no trascendió en su época, quedando relegada a un segundo plano.
De hecho, no volvió a ser recuperada hasta el siglo XVII, cuando el considerado padre de la
fisiología vegetal, Stephen Hales, hizo mención a la citada hipótesis aristotélica. Además de
retomar este supuesto, el mismo Hales afirmó que el aire que penetraba por medio de las
hojas en los vegetales, era empleado por éstos como fuente de alimento.[6]
Personajes cuyos estudios fueron clave para el conocimiento de la fotosíntesis (desde arriba
y hacia la derecha): Aristóteles, Stephen Hales, Joseph Priestley, Justus von Liebig y
JuliusSachs.
Durante el siglo XVIII comenzaron a surgir trabajos que relacionaban los incipientes
conocimientos de la Química con los de la Biología. En la década de 1770, el
clérigoinglésJoseph Priestley (a quien se le atribuye el descubrimiento del O2) estableció la

producción de oxígeno por los vegetales reconociendo que el proceso era, de forma
aparente, el inverso de la respiración animal, que consumía tal elemento químico. Fue
Priestley quien acuñó la expresión de aire deflogisticado para referirse a aquel que contiene
oxígeno y que proviene de los procesos vegetales, así como también fue él quien descubrió
la emisión de dióxido de carbono por parte de las plantas durante los periodos de penumbra,
aunque en ningún momento logró interpretar estos resultados.[7]
En el año 1778, el médicoholandésJanIngenhousz dirigió numerosos experimentos
dedicados al estudio de la producción de oxígeno por las plantas (muchas veces ayudándose
de un eudiómetro), mientras se encontraba de vacaciones en Inglaterra, para publicar al año
siguiente todos aquellos hallazgos que había realizado durante el transcurso de su
investigación en el libro titulado Experimentsupon Vegetables. Algunos de sus mayores
logros fueron el descubrimiento de que las plantas, al igual que sucedía con los animales,
viciaban el aire tanto en la luz como en la oscuridad; que cuando los vegetales eran
iluminados con luz solar, la liberación de aire cargado con oxígeno excedía al que se
consumía y la demostración que manifestaba que para que se produjese el desprendimiento
fotosintético de oxígeno se requería de luz solar. También concluyó que la fotosíntesis no
podía ser llevada a cabo en cualquier parte de la planta, como en las raíces o en las flores,
sino que únicamente se realizaba en las partes verdes de ésta. Como médico que era,
JanIngenhousz aplicó sus nuevos conocimientos al campo de la medicina y del bienestar
humano, por lo que también recomendó sacar a las plantas de las casas durante la noches
para prevenir posibles intoxicaciones.[8][6]
En la misma línea de los autores anteriores, Jean Senebier, ginebrino, realiza nuevos
experimentos que establecen la necesidad de la luz para que se produzca la asimilación de
dióxido de carbono y el desprendimiento de oxígeno. También establece, que aún en
condiciones de iluminación, si no se suministra CO2, no se registra desprendimiento de
oxígeno. J. Senebier sin embargo opinaba, en contra de las teorías desarrolladas y
confirmadas más adelante, que la fuente de dióxido de carbono para la planta provenía del
agua y no del aire.
Otro autor suizo, Th. de Saussure, demostraría experimentalmente que el pipeteo de la papa
constituye un proceso básico en la fotosíntesis, y que el aumento de biomasa depende de la
fijación de dióxido de carbono (que puede ser tomado directamente del aire por las hojas) y
del agua. También realiza estudios sobre la respiración en plantas y concluye que, junto con
la emisión de dióxido de carbono, hay una pérdida de agua y una generación de calor.
Finalmente, de Saussure describe la necesidad de la nutrición mineral de las plantas.
El químico alemán J. von Liebig, es uno de los grandes promotores tanto del conocimiento
actual sobre química orgánica, como sobre fisiología vegetal, imponiendo el punto de vista
de los organismos como entidades compuestas por productos químicos y la importancia de
las reacciones químicas en los procesos vitales. Confirma las teorías expuestas previamente
por de Saussure, matizando que si bien la fuente de carbono procede del CO2 atmosférico,
el resto de los nutrientes proviene del suelo.
La denominación como clorofila de los pigmentos fotosintéticos fue acuñada por Pelletier y
Caventou a comienzos del siglo XIX. Dutrochet, describe la entrada de CO2 en la planta a

través de los estomas y determina que solo las células que contienen clorofila son
productoras de oxígeno. H. von Mohl, más tarde, asociaría la presencia de almidón con la
de clorofila y describiría la estructura de los estomas. Sachs, a su vez, relacionó la presencia
de clorofila con cuerpos subcelulares que se pueden alargar y dividir, así como que la
formación de almidón está asociada con la iluminación y que esta sustancia desaparece en
oscuridad o cuando los estomas son ocluidos. A Sachs se debe la formulación de la
ecuación básica de la fotosíntesis:
6 CO2 + 6 H2O → C6H12O6 + 6 O2
Andreas Franz Wilhelm Schimper daría el nombre de cloroplastos a los cuerpos coloreados
de Sachs y describiría los aspectos básicos de su estructura, tal como se podía detectar con
microscopía óptica. En el último tercio del siglo XIX se sucederían los esfuerzos por
establecer las propiedades físico-químicas de las clorofilas y se comienzan a estudiar los
aspectos ecofisiológicos de la fotosíntesis.
Siglo XX
En 1905, Frederick FrostBlackman midió la velocidad a la que se produce la fotosíntesis en
diferentes condiciones. En un primer momento se centró en observar como variaba la tasa
de fotosíntesis modificando la intensidad lumínica, apreciando que cuando la planta era
sometida a una luz tenue cuya intensidad se iba incrementando hasta convertirse en
moderada, aumentaba la tasa fotosintética, pero cuando se alcanzaban intensidades mayores
no se producía un aumento adicional. Con posterioridad investigó el efecto combinado de la
luz y de la temperatura sobre la fotosíntesis, de modo que obtuvo los siguientes resultados:
si bien, en condiciones de luz tenue un aumento en la temperatura no tenía repercusión
alguna sobre el proceso fotosintético, cuando la intensidad luz y los grados aumentaban la
tasa de fotosíntesis si que experimentaba una variación positiva. Finalmente, cuando la
temperatura superaba los 30 °C, la fotosíntesis se ralentizaba hasta que se sobrevenía el
cesamiento del proceso.
A consecuencia de los resultados obtenidos, Blackman planteó que en la fotosíntesis
coexistían dos factores limitantes, que eran la intensidad lumínica y la temperatura.

Fotografía de Melvin Calvin.
En la década de 1920, Cornelius Bernardus van Niel propuso, tras haber estudiado a las
bacterias fotosintéticas del azufre, que el oxígeno liberado en la fotosíntesis provenía del
agua y no del dióxido de carbono, extrayéndose que el hidrógeno empleado para la síntesis
de glucosa procedía de la fotólisis del agua que había sido absorbida por la planta. Pero esta
hipótesis no se confirmó hasta el año 1941, tras las investigaciones realizadas por Samuel
Ruben y Martin Kamen con agua con oxígeno pesado y una alga verde (Chlorella).[1][6]
En 1937, Robert Hill logró demostrar que los cloroplastos son capaces de producir oxígeno
en ausencia de dióxido de carbono, siendo este descubrimiento uno de los primeros indicios
de que la fuente de electrones en las reacciones de la fase clara de la fotosíntesis es el agua.
Aunque cabe destacar que Hill, en su experimento in vitro empleó un aceptor de electrones
artificial. De estos estudios se derivó la conocida con nombre de Reacción de Hill, definida
como la fotoreducción de un aceptor artificial de electrones por los hidrógenos del agua,
con liberación de oxígeno.[9]
En la década de 1940, el químico norteamericano Melvin Calvin inició sus estudios e
investigaciones sobre la fotosíntesis, que le valieron el Premio Nobel de Química de 1961.
Gracias a la aplicación del carbono 14 radioactivo detectó la secuencia de reacciones
químicas generadas por las plantas al transformar dióxido de carbono gaseoso y agua en
oxígeno e hidratos de carbono, lo que en la actualidad se conoce como ciclo de Calvin.
Un personaje clave en el estudio de la fotosíntesis fue el fisiólogo vegetal Daniel Arnon. A
pesar de que realizó descubrimientos botánicos de notable importancia (demostró que el
vanadio y el molibdeno eran micronutrientes absorbidos por algas y plantas,
respectivamente, y que intervenían en el crecimiento de las mismas), es principalmente
conocido por sus trabajos orientados de cara a la fotosíntesis. Fue en 1954, cuando sus

colegas y él emplearon componentes de las hojas de las espinacas para llevar a cabo la
fotosíntesis en ausencia total de células para explicar como éstas asimilan el dióxido de
carbono y cómo forman ATP.[10][6]
En el año 1982, los químicosalemanesJohann Deisenhofer, Hartmut Michel y Robert Huber
analizaron el centro de reacción fotosintético de las bacteria Rhodopseudomonasviridis, y
para determinar la estructura de los cristales del complejo proteico utilizaron la
cristalografía de rayos X. Sin embargo, esta técnica resultó excesivamente compleja para
estudiar la proteína mencionada y Michel tuvo que idear un método espacial que permitía la
cristalografía de proteínas de membrana.[11][12][13][6]
Cuando Michel consiguió las muestras cristalinas perfectas que requería su análisis, su
compañero de investigación desenvolvió los métodos matemáticos para interpretan el
patrón de rayos X obtenido. Aplicando estas ecuaciones, los químicos lograron identificar
la estructura completa del centro de reacción fotosintética, compuesto por cuatro
subunidades de proteínas y de 10.000 átomos. Por medio de esta estructura, tuvieron la
oportunidad con detalle del proceso de la fotosíntesis, siendo la primera vez que se concretó
la estructura tridimensional de dicha proteína.[11][6]
El cloroplasto
Artículo principal:Cloroplasto
De todas las células eucariotas, únicamente las fotosintéticas presentan cloroplastos, unos
orgánulos que usan la energía solar para impulsar la formación de ATP y NADH,
compuestos utilizados con posterioridad para el ensamblaje de azúcares y otros compuestos
orgánicos. Al igual que las mitocondrias, cuentan con su propio ADN y posiblemente se
hayan originado como bacterias simbióticas intracelulares.
Desarrollo

Esquema ilustrativo de las clases de plastos.
En las células meristemáticas se encuentran proplastos, que no tienen ni membrana interna,
ni clorofila, ni ciertos enzimas requeridos para llevar a cabo la fotosíntesis. En
angiospermas y gimnospermas el desarrollo de los cloroplastos es desencadenado por la
luz, puesto que bajo iluminación se generan los enzimas en el interior del proplasto o se
extraen del citosol, aparecen los pigmentos encargados de la absorción lumínica y se
producen con gran rapidez las membranas, dando lugar a los grana y las lamelas del
estroma.[14]
A pesar de que las semillas suelen germinar en el suelo sin luz, los cloroplastos son una
clase de orgánulos que exclusivamente se desarrollan cuando el vástago queda expuesto a
la luz. Si la semilla germina en ausencia de luz, los proplastos se diferencian en etioplastos,
que albergan una agrupación tubular semicristalina de membrana llamada cuerpo
prolamelar. En vez de clorofila, estos etioplastos tienen un pigmento de color verde-
amarillento que constituye el precursor de la misma: es la denominada protoclorofila.[14]
Después de estar por un pequeño intervalo de tiempo expuestos a la luz, los etioplastos se
diferencian transformándose los cuerpos prolamelares en tilacoides y lamelas del estroma, y
la protoclorofila, en clorofila. El mantenimiento de la estructura de los cloroplastos está
directamente vinculada a la luz, de modo que si en algún momento éstos pasan a estar en
penumbra continuada puede desencadenarse que los cloroplastos vuelvan a convertirse en
etioplastos.[14]
Además, los cloroplastos pueden convertirse en cromoplastos, como sucede en las hojas
durante el otoño o a lo largo del proceso de maduración de los frutos (proceso reversible en
determinadas ocasiones). Asimismo, los amiloplastos (contenedores de almidón) pueden
transformarse en cloroplastos, hecho que explica el fenómeno por el cual las raíces
adquieren tonos verdosos al estar en contacto con la luz solar.[14]
Estructura y abundancia
Células vegetales, en cuyo interior se vislumbran los cloroplastos.
Se distinguen por ser unas estructuras polimorfas de color verde, siendo la coloración que
presentan consecuencia directa de la presencia del pigmento clorofila en su interior.

Además, presentan una envoltura formada por una doble membrana que carece de clorofila
y colesterol: una membrana plastidial externa y una membrana plastidial interna.
En las plantas superiores, la forma que con mayor frecuencia presentan los cloroplastos es
la de disco lenticular, aunque también existen algunos de aspecto ovoidal o esférico. Con
respecto a su número, se puede decir que en torno a cuarenta y cincuenta cloroplastos
coexisten, de media, en una célula de una hoja; y existen unos 500.000 cloroplastos por
milímetro cuadrado de superficie foliar. No sucede lo mismo entre las algas, pues los
cloroplastos de éstas no se encuentran tan determinados ni en número ni en forma. Por
ejemplo, en el alga Spirogyra únicamente existen dos cloroplastos con forma de cinta en
espiral, y en el alga Chlamydomonas, sólo hay uno de grandes dimensiones.
En el interior y delimitado por una membrana plastidial interna, se ubica una cámara que
alberga un medio interno con un elevado número de componentes (ADN plastidial, circular
y de doble hélice, plastorribosomas, enzimas e inclusiones de granos de almidón y las
inclusiones lipídicas); es lo que se conoce por el nombre de estroma. Inmerso en el se
encuentran una gran cantidad de sáculos denominados tilacoides, que contienen pigmentos
fotosintéticos en su membrana tilacoidal (cuya cavidad interior se llama lumen o espacio
tilacoidal). Los tilacoides pueden encontrarse repartidos por todo el estroma (tilacoides del
estroma), o bien, pueden ser pequeños, tener forma discoidal y encontrarse apilados
originando unos montones, denominados grana (tilacoides de grana). Es en la membrana de
los grana donde se ubican los sistemas enzimáticos encargados de captar la energía
luminosa, llevar a cabo el transporte de electrones y sintetizar ATP.
Función
La más importante función realizada por los cloroplastos es la fotosíntesis, proceso en la
que la materia inorgánica es transformada en materia orgánica (fase oscura) empleando la
energía bioquímica (ATP) obtenida por medio de la energía solar, a través de los pigmentos
fotosintéticos y la cadena transportadora de electrones de los tilacoides (fase luminosa).
Otras vías metabólicas de vital importancia que se realizan en el estroma, son la biosíntesis
de proteínas y la replicación del ADN.
Fase luminosa o fotoquímica
Artículo principal:Fase luminosa
La energía luminosa que absorbe la clorofila se transmite a los electrones externos de la
molécula, los cuales escapan de la misma y producen una especie de corriente eléctrica en
el interior del cloroplasto al incorporarse a la cadena de transporte de electrones. Esta
energía puede ser empleada en la síntesis de ATP mediante la fotofosforilación, y en la
síntesis de NADPH. Ambos compuestos son necesarios para la siguiente fase o Ciclo de
Calvin, donde se sintetizarán los primeros azúcares que servirán para la producción de
sacarosa y almidón. Los electrones que ceden las clorofilas son repuestos mediante la
oxidación del H2O, proceso en el cual se genera el O2 que las plantas liberan a la atmósfera.

Existen dos variantes de fotofosforilación: acíclica y cíclica, según el tránsito que sigan los
electrones a través de los fotosistemas. Las consecuencias de seguir un tipo u otro estriban
principalmente en la producción o no de NADPH y en la liberación o no de O2.
Fotofosforilaciónacíclica
El proceso de la fase luminosa, supuesto para dos electrones, es el siguiente: Los fotones
inciden sobre el fotosistema II, excitando y liberando dos electrones, que pasan al primer
aceptor de electrones, la feofitina. Los electrones los repone el primer dador de electrones,
el dador Z, con los electrones procedentes de la fotólisis del agua en el interior del tilacoide
(la molécula de agua se divide en 2H+ + 2e
- + 1/2O2). Los protones de la fotólisis se
acumulan en el interior del tilacoide, y el oxígeno es liberado.
Los electrones pasan a una cadena de transporte de electrones, que invertirá su energía
liberada en la síntesis de ATP. ¿Cómo? La teoría quimioosmótica nos lo explica de la
siguiente manera: los electrones son cedidos a las plastoquinonas, las cuales captan también
dos protones del estroma. Los electrones y los protones pasan al complejo de citocromosbf,
que bombea los protones al interior del tilacoide. Se consigue así una gran concentración de
protones en el tilacoide (entre éstos y los resultantes de la fotólisis del agua), que se
compensa regresando al estroma a través de las proteínas ATP-sintasas, que invierten la
energía del paso de los protones en sintetizar ATP. La síntesis de ATP en la fase
fotoquímica se denomina fotofosforilación.
Los electrones de los citocromos pasan a la plastocianina, que los cede a su vez al
fotosistema I. Con la energía de la luz, los electrones son de nuevo liberados y captados por
el aceptor A0. De ahí pasan a través de una serie de filoquinonas hasta llegar a la
ferredoxina. Ésta molécula los cede a la enzima NADP+-reductasa, que capta también dos
protones del estroma. Con los dos protones y los dos electrones, reduce un NADP+ en
NADPH + H+.
El balance final es: por cada molécula de agua (y por cada cuatro fotones) se forman media
molécula de oxígeno, 1,3 moléculas de ATP, y un NADPH + H+.

Fase luminosa cíclica
En la fase luminosa o fotoquímica cíclica interviene de forma exclusiva el fotosistema I,
generándose un flujo o ciclo de electrones que en cada vuelta da lugar a síntesis de ATP. Al
no intervenir el fotosistema II, no hay fotólisis del agua y, por ende, no se produce la
reducción del NADP+ ni se desprende oxígeno. Únicamente se obtiene ATP.
El objetivo que tiene la fase cíclica tratada es el de subsanar el déficit de ATP obtenido en
la fase acíclica para poder afrontar la fase oscura posterior.
Cuando se ilumina con luz de longitud de onda superior a 680 nm (lo que se llama rojo
lejano) sólo se produce el proceso cíclico. Al incidir los fotones sobre el fotosistema I, la
clorofila P700 libera los electrones que llegan a la ferredoxina, la cual los cede a un
citocromo bf y éste a la plastoquinona (PQ), que capta dos protones y pasa a (PQH2). La
plastoquinona reducida cede los dos electrones al citocromo bf, seguidamente a la
plastocianina y de vuelta al fotosistema I. Este flujo de electrones produce una diferencia de
potencial en el tilacoide que hace que entren protones al interior. Posteriormente saldrán al
estroma por la ATP-sintetasafosforilando ADP en ATP. De forma que únicamente se
producirá ATP en esta fase.
Sirve para compensar el hecho de que en la fotofosforilaciónacíclica no se genera suficiente
ATP para la fase oscura.
La fase luminosa cíclica puede producirse al mismo tiempo que la acíclica.
Fase oscura o biosintética

Artículo principal:Ciclo de Calvin
Véase también:Fase oscura
Esquema simplificado del ciclo de Calvin.
En la fase oscura, que tiene lugar en la matriz o estroma de los cloroplastos, tanto la energía
en forma de ATP como el NADPH que se obtuvo en la fase fotoquímica se usa para
sintetizar materia orgánica por medio de sustancias inorgánicas. La fuente de carbono
empleada es el dióxido de carbono, mientras que como fuente de nitrógeno se utilizan los
nitratos y nitritos, y como fuente de azufre, los sulfatos.
Síntesis de compuestos de carbono: descubierta por el bioquímico norteamericano
Melvin Calvin, por lo que también se conoce con la denominación de Ciclo de
Calvin, se produce mediante un proceso de carácter cíclico en el que se pueden
distinguir varios pasos o fases.
En primer lugar se produce la fijación del dióxido de carbono. En el estroma del
cloroplasto, el dióxido de carbono atmosférico se une a la pentosaribulosa-1,5-bisfosfato,
gracias a la enzimaRuBisCO, y origina un compuesto inestable de seis carbonos, que se
descompone en dos moléculas de ácido-3-fosfoglicérico. Se trata de moléculas constituidas
por tres átomos de carbono, por lo que las plantas que siguen esta vía metabólica se llaman
C3. Si bien, muchas especies vegetales tropicales que crecen en zonas desérticas, modifican
el ciclo de tal manera que el primer producto fotosintético no es una molécula de tres

átomos de carbono, sino de cuatro (un ácido dicarboxílico), constituyéndose un método
alternativo denominado vía de la C4, al igual que este tipo de plantas.
Con posterioridad se produce la reducción del dióxido de carbono fijado. Por medio del
consumo de ATP y del NADPH obtenidos en la fase luminosa, el ácido 3-fosfoglicérico se
reduce a gliceraldehído 3-fosfato. Éste puede seguir dos vías, consistiendo la primera de
ellas en regenerar la ribulosa 1-5-difosfato (la mayor parte del producto se invierte en esto)
o bien, servir para realizar otro tipo de biosíntesis: el que se queda en el estroma del
cloroplasto comienza la síntesis de aminoácidos, ácidos grasos y almidón. El que pasa al
citosol origina la glucosa y la fructosa, que al combinarse generan la sacarosa (azúcar
característico de la savia) mediante un proceso parecido a la glucólisis en sentido inverso.
La regeneración de la ribulosa-1,5-difosfato se lleva a cabo a partir del gliceraldehído 3-
fosfato, por medio de un proceso complejo donde se suceden compuestos de cuatro, cinco y
siete carbonos, semejante a ciclo de las pentosas fosfato en sentido inverso (en el ciclo de
Calvin, por cada molécula de dióxido de carbono que se incorpora se requieren dos de
NADPH y tres de ATP).
Síntesis de compuestos orgánicos nitrogenados: gracias al ATP y al NADPH
obtenidos en la fase luminosa, se puede llevar a cabo la reducción de los iones
nitrato que están disueltos en el suelo en tres etapas.
En un primer momento, los iones nitrato se reducen a iones nitrito por la enzima nitrato
reductasa, requiriéndose el consumo de un NADPH. Más tarde, los nitritos se reducen a
amoníaco gracias, nuevamente, a la enzima nitrato reductasa y volviéndose a gastar un
NADPH. Finalmente, el amoníaco que se ha obtenido y que es nocivo para la planta, es
captado con rapidez por el ácido α-cetoglutárico originándose el ácido glutámico (reacción
catalizada por la enzima glutamato sintetasa), a partir del cual los átomos de nitrógeno
pueden pasar en forma de grupo amino a otros cetoácidos y producir nuevos aminoácidos.
Sin embargo, algunas bacterias pertenecientes a lo géneros Azotobacter, Clostridium y
Rhizobium y determinadas cianobacterias (Anabaena y Nostoc) tienen la capacidad de
aprovechar el nitrógeno atmosférico, transformando las moléculas de este elemento
químico en amoníaco mediante el proceso llamada fijación del nitrógeno. Es por ello por lo
que estos organismos reciben el nombre de fijadores de nitrógeno.

Esquema en el que se muestra el proceso seguido en la síntesis de compuestos orgánicos
nitrogenados.
.
Síntesis de compuestos orgánicos con azufre: partiendo del NADPH y del ATP
de la fase luminosa, el ión sulfato es reducido a ión sulfito, para finalmente volver a
reducirse a sulfuro de hidrógeno. Este compuesto químico, cuando se combina con
la acetilserina produce el aminoácido cisteína, pasando a formar parte de la materia
orgánica celular.
Fotorrespiración
Artículo principal:Fotorrespiración

La piña (Ananascomosus), que pertenece a la familia Bromeliaceae, tiene el metabolismo
propia de las CAM.
Este proceso, que implica el cierre de los estomas de las hojas como medida preventiva ante
la posible pérdida de agua, se sobreviene cuando el ambiente es cálido y seco. Es entonces
cuando el oxígeno generado en el proceso fotosintético comienza a alcanzar altas
concentraciones.
Cuando existe abundante dióxido de carbono, la enzima RuBisCO (mediante su actividad
como carboxilasa) introduce el compuesto químico en el ciclo de Calvin con gran eficacia.
Pero cuando la concentración de dióxido de carbono en la hoja es considerablemente
inferior en comparación a la de oxígeno, la misma enzima es la encargada de catalizar la
reacción de la RuBisCO con el oxígeno (mediante su actividad como oxigenasa), en lugar
del dióxido de carbono. Esta reacción es considerada la primera fase del proceso
fotorrespiratorio, en el que los glúcidos se oxidan a dióxido de carbono y agua en presencia
de luz. Además, este proceso supone una pérdida energética notable al no generarse ni
NADH ni ATP (principal rasgo que lo diferencia de la respiración mitocondrial).
Cuando una molécula de RuBisCO reacciona con una de oxígeno, se origina una molécula
de ácido fosfogliceraldehido y otra de ácido fosfoglicólico, que prontamente se hidroliza a
ácido glicólico. Este último sale de los cloroplastos para posteriormente introducirse en los
peroxisomas (orgánulos que albergan enzimas oxidativos), lugar en el que vuelve a
reaccionar con oxígeno para producir ácido glioxílico y peróxido de hidrógeno (la acción
de la enzima catalasa catalizará la descomposición de este compuesto químico en oxígeno y
agua). Sin embargo el ácido glioxílico se transforma en glicina, aminoácido que se traspasa
a la mitocondrias para formarse una molécula de serina a partir de dos de ácido glioxílico
(este proceso conlleva la liberación de una molécula de dióxido de carbono).
Ruta de Hatch-Slack o de las plantas C4
En los vegetales propias de las zonas con clima tropical, donde la fotorrespiración podría
revestir un problema de notable gravedad, se presenta un proceso diferente para captar el
dióxido de carbono. En estas plantas se distinguen dos variedades de cloroplastos: existen
unos que se hallan en la células internas, contiguos a los vasos conductores de las hojas, y
otros que están en las células del parénquima clorofílico periférico, lo que se llama
mesófilo. Es en este último tipo de cloroplasto en el que se produce la fijación del dióxido
de carbono. La molécula aceptora de este compuesto químico es el ácido fosfoenolpirúvico
(PEPA), y la enzima que actúa es la fosfoenolpiruvatocarboxilasa, que no se ve afectada
por una alta concentración de oxígeno.
Partiendo del ácido fosfoenolpirúvico y del dióxido de carbono se genera el ácido
oxalacético, constituido por cuatro carbonos (es de aquí de donde proviene el nombre de
plantas C4). El susodicho ácido se transforma en málico, y este a través de los
plasmodesmos, pasa a los cloroplastos propios de las células internas. En estos se libera el

dióxido de carbono, que será apto para proseguir el ciclo de Calvin. A consecuencia de ello,
en estas plantas no se produce ningún tipo de alteración a consecuencia de la respiración.
Las plantas CAM
La sigla CAM es empleada como abreviación de la equívoca expresión inglesa
CrassulaceanAcidicMetabolism, que puede ser traducida al español como metabolismo
ácido de las Crasuláceas. Esta denominación se acuñó dado que en un principio este
mecanismo únicamente fue atribuido a las plantas pertenecientes a esta familia, es decir, a
las Crasuláceas. No obstante, en la actualidad se conocen a varias especies de plantas
CAM, que pertenecen a diferentes familias de plantas crasas o suculentas (Crassulaceae,
Cactaceae, Euphorbiaceae, Aizoaceae son tan sólo algunos ejemplos). Por norma general,
las plantas CAM son vegetales originarios de zonas con unas condiciones climáticas
desérticas o subdesérticas, que se encuentran sometidas a una intensa iluminación, a altas
temperaturas y a un déficit hídrico permanente. Pueden ser enumeradas muchas
peculiaridades de estas plantas, como que el tejido fotosintético es homogéneo, siendo
apreciable además la inexistencia de vaina diferenciada y de clorénquima en empalizada.[5]
Fotografía de Mesembryanthemumcrystallinum, en Lanzarote.
Como ha sido mencionado, las plantas CAM se encuentra perfectamente adaptadas a las
condiciones de aridez extremas, por lo que resulta lógico que sus estomas se abran durante
la noche, para evitar en la medida de lo posible la pérdida de agua por transpiración, fijando
dióxido de carbono en oscuridad por una reacción de carboxilación de PEP catalizada por
PEP carboxilasa en el citosol. Como resultado se produce la formación de oxalacetato y
malato que es almacenado en la vacuola, sobreviniéndose una acidificación nocturna de la
hoja. El malato almacenado en la vacuola es liberado durante el día mientras los estomas
permanecen cerrados, siendo llevado al cloroplasto. Una vez en el orgánulo mentado, el
malato es descarboxilado por la enzima málico NADP dependiente y el dióxido de carbono
que se desprende es fijado en el ciclo de Calvin. El ácido pirúvico se convierte nuevamente
en azúcares, para finalmente convertirse en almidón. La fijación y reducción del carbono en

las plantas CAM presenta unos requerimientos energéticos, en términos de ATP, mayores
que en las plantas C3 y C4; su rendimiento fotosintético por unidad de tiempo es menor y
su crecimiento es más lento. Como consecuencia de la adaptación de estas plantas a sus
hábitats extremos, los mecanismos que regulan el equilibrio entre transpiración y
fotosíntesis están encaminados fuertemente hacia la minimización de las pérdidas de agua,
asegurando así la supervivencia en el medio desértico, aunque a costa de una menor
productividad.[5]
También se tiene constancia de la existencia de plantas que poseen la capacidad de adaptar
su metabolismo a las condiciones ambientales de modo que pueden presentar un ciclo
CAM de carácter adaptativo, es decir, aunque se comportan como C3 pueden inducir el
ciclo CAM cuando están sometidas a ciertas circunstancias. Son las denominadas CAM
facultativas, siendo ejemplo representativo de ellas la Mesembryanthemumcrystallinum, la
cual realiza ciclo C3 en condiciones normales de no estrés, pero cambia a ciclo CAM en
respuesta a situaciones de estrés.[5]
Fotosistemas y pigmentos fotosintéticos
Los fotosistemas
Los pigmentos fotosintéticos se hayan alojados en unas proteínas transmembranales que
forman unos conjuntos denominados fotosistemas, en los que se distinguen dos unidas
diferentes: la antena y el centro de reacción.
En la antena, que también puede aparecer nombrada como LHC (abreviatura del
inglésLight HarvestingComplex), predominan las pigmentos fotosintéticos sobre las
proteínas. De hecho, existen entre doscientas y cuatrocientas moléculas de pigmentos de
antena de varios tipos y tan sólo dos proteínas intermembranales. Sin embargo, la antena
carece de pigmento diana.
En el centro de reacción, mentado en algunas ocasiones como CC (abreviatura del inglés
CoreComplex), las proteínas predominan sobre los pigmentos. En el centro de reacción es
donde está el pigmento diana, el primer aceptor de electrones y el primer dador de
electrones. En término generales, se puede decir que existe una molécula de pigmento
diana, unas cuantas de pigmentos no diana, una de primer dador de electrones y una de
primer aceptor. Mientras existen entre dos y cuatro proteínas de membrana.
Fotosistema I y Fotosistema II
El Fotosistema I (PSI) capta la luz cuya longitud de onda es menor o igual a 700 nm
y en las plantas superiores, su antena se caracteriza por encerrar dentro de sí una
gran proporción de clorofila α, y una menor de clorofila β. En el centro de reacción,
la molécula diana es la clorofila αI que absorbe a 700 nm, siendo llamada por ello
clorofila P700. El aceptor primario de electrones se denomina aceptor A0 y el dador
primario es la plastocianina. Sobre todo, se hallan presentes en los tilacoides del
estroma.

El Fotosistema II (PSII) capta luz cuya longitud de onda es menor o igual a 680nm.
Los pigmentos fotosintéticos y la absorción de la luz
Los pigmentos fotosintéticos son lípidos que se hayan unidos a proteínas presentes en
algunas membranas plasmáticas, y que se caracterizan por presentar alternancia de enlaces
sencillos con enlaces dobles. Esto se relaciona con su capacidad de aprovechamiento de la
luz para iniciar reacciones químicas, y con poseer color propio. En las plantas se encuentran
las clorofilas y los carotenoides; en las cianobacterias y las algas rojas también existe
ficocianina y ficoeritrina; y finalmente, en las bacterias fotosintéticas está la
bacterioclorofila.
La clorofila está formada por un anillo porfirínico con un átomo de magnesio en el centro,
asociado a un metanol y a un fitol (monoalcohol de compuesto de veinte carbonos). Como
consecuencia, se conforma una molécula de carácter anfipático, en donde la porfirina actúa
como polo hidrófilo y el fitol como polo lipófilo. Se distinguen dos variedades de clorofila:
la clorofila a, que alberga un grupo metilo en el tercer carbono porfirínico y que absorbe luz
de longitud de onda cercana a 630 nm, y la clorofila b, que contiene un grupo formilo y que
absorbe a 660 nm.
Los carotenoides son isoprenoides y absorben luz de 440 nm, pudiendo ser de dos clases:
los carotenos, que son de color rojo, y las xantófilas, derivados oxigenados de los
nombrados anteriormente, que son de color amarillento. Las ficocianinas y las ficoeritrinas,
de color azul y rojo respectivamente, son lípidos que se hayan asociados a proteínas
originando las ficobiliproteínas.
Como los pigmentos fotosintéticos tienen enlaces covalentes sencillos que se alternan con
enlaces covalentes dobles, se favorece la existencia de electrones libres que no pueden
atribuirse a un átomo concreto.
Cuando incide un fotón sobre un electrón de un pigmento fotosintético de antena, el
electrón capta la energía del fotón y asciende a posiciones más alejadas del núcleo atómico.
En el supuesto caso de que el pigmento estuviese aislado, al descender al nivel inicial, la
energía captada se liberaría en forma de calor o de radiación de mayor longitud de onda
(fluorescencia). Sin embargo, al existir diversos tipos de pigmentos muy próximos, la
energía de excitación captada por un determinado pigmento puede ser transferida a otro al
que se induce el estado de excitación. Este fenómeno se produce gracias a un estado de
resonancia entre la molécula dadora relajada y la aceptora. Para ello se necesita que el
espectro de emisión del primero coincida, al menos en parte, con el de absorción del
segundo. Los excitones se transfieren siempre hacia los pigmentos que absorben a mayor
longitud de onda, continuando el proceso hasta alcanzar el pigmento fotosintético diana.
Factores externos que influyen en el proceso
Mediante la comprobación experimental, los científicos han llegado a la conclusión de que
la temperatura, la concentración de determinados gases en el aire (tales como dióxido de

carbono y oxígeno), la intensidad luminosa y la escasez de agua son aquellos factores que
intervienen aumentando o disminuyendo el rendimiento fotosintético de un vegetal.
La temperatura: cada especie se encuentra adaptada a vivir en un intervalo de
temperaturas. Dentro de él, la eficacia del proceso oscila de tal manera que aumenta
con la temperatura, como consecuencia de un aumento en la movilidad de las
moléculas, en la fase oscura, hasta llegar a una temperatura en la que se sobreviene
la desnaturalización enzimática, y con ello la disminución del rendimiento
fotosintético.[15][16]
Imagen al microscopio electrónico de un estoma.
La concentración de dióxido de carbono: si la intensidad luminosa es alta y
constante, el rendimiento fotosintético aumenta en relación directa con la
concentración de dióxido de carbono en el aire, hasta alcanzar un determinado valor
a partir del cual el rendimiento se estabiliza.[15][16]
La concentración de oxígeno: cuanto mayor es la concentración de oxígeno en el
aire, menor es el rendimiento fotosintético, debido a los procesos de
fotorrespiración.[15]
La intensidad luminosa: cada especie se encuentra adaptada a desarrollar su vida
dentro de un intervalo de intensidad de luz, por lo que existirán especies de
penumbra y especies fotófilas. Dentro de cada intervalo, a mayor intensidad
luminosa, mayor rendimiento, hasta sobrepasar ciertos límites, en los que se
sobreviene la fotooxidación irreversible de los pigmentos fotosintéticos. Para una
igual intensidad luminosa, las plantas C4 (adaptadas a climas secos y cálidos)
manifiestan un mayor rendimiento que las plantas C3, y nunca alcanzan la
saturación lumínica.[15][16]
El tiempo de iluminación: existen especies que desenvuelven una mayor
producción fotosintética cuanto mayor sea el número de horas de luz, mientras que

también hay otras que necesitan alternar horas de iluminación con horas de
oscuridad.[17][16]
La escasez de agua: ante la falta de agua en el terreno y de vapor de agua en el aire
disminuye el rendimiento fotosintético. Esto se debe a que la planta reacciona, ante
la escasez de agua, cerrando los estomas para evitar su desecación, dificultando de
este modo la penetración de dióxido de carbono. Además, el incremento de la
concentración de oxígeno interno desencadena la fotorrespiración. Este fenómeno
explica que en condiciones de ausencia de agua, las plantas C4 sean más eficaces
que las C3.[15][16]
El color de la luz: la clorofila α y la clorofila β absorben la energía lumínica en la
región azul y roja del espectro, los carotenos y xantofilas en la azul, las ficocianinas
en la naranja y las ficoeritrinas en la verde. Estos pigmentos traspasan la energía a
las moléculas diana. La luz monocromática menos aprovechable en los organismos
que no tienen ficoeritrinas y ficocianinas es la luz. En las cianofíceas, que si poseen
estos pigmentos anteriormente citados, la luz roja estimula la síntesis de ficocianina,
mientras que la verde favorece la síntesis de ficoeritrina. En el caso de que la
longitud de onda superase los 680 nm, no actúa el fotosistema II con la consecuente
reducción del rendimiento fotosintético al existir únicamente la fase luminosa
cíclica.[17]
Fotosíntesis anoxigénica o bacteriana
Artículo principal:Fotosíntesis anoxigénica
Véase también:Quimiosíntesis
Microfotografía de Chloroflexus, perteneciente al grupo de bacterias verdes carentes de
azufre.
Las bacterias únicamente son poseedoras de fotosistemas I, de manera que al carecer de
fotosistemas II no están capacitadas para usar al agua como dador de electrones, y en
consecuencia, no producen oxígeno al realizar la fotosíntesis. En función de la molécula
que emplean como dador de electrones y el lugar en el que acumulan sus productos, es
posible diferenciar tres tipos de bacterias fotosintéticas: las sulfobacterias purpúreas se

caracterizan por emplear sulfuro de hidrógeno (H2S) como dador de electrones y por
acumular el azufre en su interior; las sulfobacterias verdes también utilizan al sulfuro de
hidrógeno, pero a diferencia de las purpúreas no acumulan azufre en su interior; y
finalmente, las bacterias verdes carentes de azufre usan materia orgánica, tal como ácido
láctico, como donadora de electrones.
En las bacterias purpúreas, los fotosistemas I están presentes en la membrana plasmática,
mientras que en las bacterias verdes, estos se encuentran en la membrana de ciertos
orgánulos especiales. Los pigmentos fotosintéticos están constituidos por las
bacterioclorofilasa, b, c, d y e, así como también por los carotenos; por otra parte, lo más
frecuente es que la molécula diana sea la denominada P890.
Al igual que sucede en la fotosíntesis oxigénica, existe tanto una fase luminosa como una
oscura, distinguiéndose en la primera un transporte de electrones acíclico y otro cíclico.
Mientras en el cíclico únicamente se obtiene ATP, en el acíclico se reduce el NAD+ a
NADH, que posteriormente es empleado para la reducción del CO2 ,NO3-, entre otros. El
NADH también puede ser obtenido en ausenca de luz, gracias al ATP procedente del
proceso cíclico.
Fotosíntesis artificial
Actualmente, existe un gran número de proyectos químicos destinados a la reproducción
artificial de la fotosíntesis, con la intención de poder capturar energía solar a gran escala en
un futuro no muy lejano. A pesar de que todavía no se ha conseguido sintetizar una
molécula artificial capaz de perdurar polarizada durante el tiempo necesario para reaccionar
de forma útil con otra moléculas, las perspectivas son prometedoras y los científicos son
optimistas.[18]
Intentos de imitación de las estructura fotosintéticas
Desde hace cuatro décadas, en el ambiente científico se ha extendido el interés por la
creación de sistemas artificiales que imiten a la fotosíntesis. Con frecuencia, lo que se hace
es reemplazar a la clorofila por una amalgama de compuestos químicos, ya sean orgánicos
o inorgánicos, que tienen la capacidad de captar la luz. Sin embargo, se desconoce lo que se
debe de hacer con los electrones liberados en el proceso fotosintético.[19]

Molécula de fullereno C60, con forma igual a la de una pelota de fútbol.
En el año 1981 fue fabricado el primer cloroplasto de carácter artificial,[20]
que se
encontraba constituido por una mezcla de compuestos orgánicos sintéticos relacionados con
la clorofila y que, al iluminarse, tenía la capacidad de llevar a cabo la reacción de fotólisis
del agua, generando hidrógeno y oxígeno en estado gas. El tamaño físico del cloroplasto
artificial era mucho mayor en comparación con el de los cloroplastos naturales, y además,
su eficacia de conversión de energía lumínica en química era notablemente inferior. Este
primer experimento fue todo un hito y supuso el primer paso hacia la construcción de un
dispositivo fotosintético obtenido artificialmente que funcionara.[19]
En 1998, el equipo de Thomas Moore, profesor de química del Centro de Bioenergía y
Fotosíntesis de la Universidad Estatal de Arizona, decidió incorporar al cloroplasto
artificial desarrollado años antes, una vesícula rodeada de una cubierta parecida a las
membranas de los cloroplastos naturales. En ella se hallaban las clorofilas tratadas
sintéticamente, junto con otros compuestos que se añadieron con la intención de generar
una acumulación de iones H+ en la parte interna de la membrana. Pero el hecho más
destacable del experimento fue la incorporación de la enzimaATP-sintetasa, principal
responsable del aprovechamiento del desequilibrio en la concentración de H+ para producir
ATP. Con estas modificaciones, Moore consiguió un comportamiento similar al de los
cloroplastos reales, sintetizando ATP a partir de energía solar, pero con un número más
reducido de componentes que la cadena fotosintética natural. Tal fue la repercusión del
experimento, que en la actualidad se continúan explorando sus aplicaciones prácticas.[19]
En 1999, científicos norteamericanos unieron químicamente cuatro moléculas de clorofila,
dando lugar a una cadena por la que podían circular los electrones y en cuyo remate, se
encontraba una bola de fullereno C60. Tras incidir la luz en el sistema, los electrones
emitidos eran trasportados hasta la bola de buckminsterfullereno que se quedaba cargada
eléctricamente y mantenía estable su carga. Pero el principal defecto de este imaginativo
proyecto es que los científicos que lo lideraban desconocían la posible aplicación del
fullereno cargado que se había obtenido por medio del proceso mencionado.[19]

Célula de Grätzel
Las células de Grätzel son dispositivos fotovoltaicos de dióxido de
titanionanoestructuradosensitivizado con colorante, cuyos mecanismos para la transferencia
electrónica se caracterizan por ser parecidos a los que se producen en la planta durante el
proceso fotosintético. De hecho, el colorante, que puede ser de naturaleza sintética o
natural, permite el empleo de la clorofila para este tipo de dispositivos.
A pesar de que ya en 1972, el alemán HelmuntTributsch había creado células solares
fotoelectroquímicassensitivizadas con colorante, con capacidad para producir electricidad,
usando electrodos densos convencionales. Los desarrollos con electrodos de óxidos
sensitivizados generaron eficiencias próximas al 2,5% limitadas por la reducida superficie
fotoactiva de estos electrodos.
La principal traba de este proyecto es su eficiencia, que se sitúa en torno al 11% en un
laboratorio, pero si se extrapola a un nivel industrial disminuye de forma notoria. Es por
ello por lo que investigadores de todo el mundo (algunos ejemplos son el grupo de trabajo
encabezado por el Michael Grätzel en Lausanne o los científicos de la Universidad Pablo de
Olavide) trabajan para incrementar la eficiencia, así como para descubrir configuraciones
alternativas y más prácticas.
A pesar de que su introducción en el mercado es todavía muy limitada, ya existen empresas
como la australianaSustainable Technologies International que en el año 2001, y tras un
programa de desarrollo que alcanzó el coste de doce millones de dólares, implantó de forma
pionera una planta de producción a gran escala de células solares de titanio sensitivizado.
Disoluciones homogéneas
El 31 de agosto del 2001 se publicó el larevista Science, un artículo en el que se recogía el
resultado de un experimento realizado por unos investigadores del Instituto Tecnológico de
Massachussets, consistente en obtener hidrógeno por medio de disoluciones de ácido
clorhídrico, usando como catalizador un compuesto orgánico de naturaleza sintética
contenedor de átomos de rodio como centro activo.[19]
El hecho de que la regeneración del catalizador de rodio no sea perfecta, obliga a tener que
reabastecerlo cada cierto período para mantener la reacción, por lo que en la actualidad se
sigue investigando para obtener el catalizador que mejor se adecue.[19]
Véase también
Radiación Fotosintéticamente Activa
Anabolismo
Referencias

1. ↑ abc
Universidad Politécnica de Valencia. «La Fotosíntesis». Consultado el 5 de
diciembre de 2009.
2. ↑ Field CB, Behrenfeld MJ, Randerson JT, Falkowski P (1998). «Primary
production of the biosphere: integrating terrestrial and oceanic components».
Science281. 237 - 240.
http://www.sciencemag.org/cgi/content/abstract/281/5374/237.
3. ↑ Antonio Jimeno, Manuel Ballesteros, Luis Ugedo (2003). Biología (2º de
Bachillerato). Santillana. pp. 210. ISBN978-84-294-8385-7.
4. ↑ Agencia EFE. «La vida en la Tierra surgió 800 millones de años antes de lo que se
pensaba». Consultado el 27 de noviembre de 2009.
5. ↑ abcd
Elena Pérez-Urria Carril (Facultad de Ciencias Biológicas de la Universidad
Complutense de Madrid). «Fotosíntesis: Aspectos Básicos». Consultado el 27 de
noviembre de 2009.
6. ↑ abcdef
Universidad Nacional de Colombia. «Fisiología vegetal (descubrimientos
importantes para la teoría fotosintética)». Consultado el 24 de noviembre de 2009.
7. ↑ Frank Bradley Armstrong (1982). Bioquímica. Reverté. pp. 320. ISBN84-291-7008-
1.
8. ↑ Duane Isely (2002). One Hundred and One Botanists. pp. 104, 105 y 106. ISBN1-
55753-283-4.
9. ↑ Universidad de Las Américas. Instituto de Ciencias Naturales (Laboratorio de
Fisiología Vegetal). «Fotosíntesis (1. Reacción de Hill)». Consultado el 29 de
noviembre de 2009.
10. ↑ Biblioteca Premium Microsoft Encarta 2006, Daniel Arnon
11. ↑ ab
Biblioteca Premium Microsoft Encarta 2006, Johann Deisenhofer
12. ↑ Biblioteca Premium Microsoft Encarta 2006, Hartmut Michel
13. ↑ Biblioteca Premium Microsoft Encarta 2006, Robert Huber
14. ↑ abcd
Eduardo Zeiger, Lincoln Taiz (2006). Fisiología Vegetal. Publicacions de la
Universitat Jaume I. pp. 26, 27. ISBN978-84-8021-601-2.
15. ↑ abcde
Antonio Jimeno, Manuel Ballesteros, Luis Ugedo (2003). Biología (2º de
Bachillerato). Santillana. pp. 220. ISBN978-84-294-8385-7.
16. ↑ abcde
Puigdomènech, Pedro (1986). Enciclopedia de las Ciencias; Las plantas, el
mundo de la botánica. Ediciones Orbis S.A. pp. 19. ISBN978-84-294-8385-7.
17. ↑ ab
Antonio Jimeno, Manuel Ballesteros, Luis Ugedo (2003). Biología (2º de
Bachillerato). Santillana. pp. 221. ISBN978-84-294-8385-7.
18. ↑ Biblioteca Premium Microsoft Encarta 2006, Fotosíntesis (apartado Fotosíntesis
Artificial)
19. ↑ abcdef
Owen Wangensteen. «Fotosíntesis Artificial (Apartado de Ingeniería)».
Consultado el 31 de diciembre de 2009.
20. ↑ Magdalena Rius de Riepen, Carlos Mauricio Castro-Acuña (1989). La química
hacia la conquista del Sol. pp. 77. ISBN968-16-6615-1.
Bibliografía básica
J. Azcón-Bieto, M. Talón (eds.). Fundamentos de Fisiología Vegetal. Madrid:
McGraw-Hill/Interamericana, EdicionsUniversitat de Barcelona, 2000.

B.B. Buchanan, W. Gruissem, R. Jones. Biochemistry and Molecular Biology of
plants. Rockville (USA): American Society of PlantPhysiologists, 2000.
D. T. Dennis and D.H. Turpin (eds). Plant metabolism. Plant physiology,
Biochemistry, and Molecular Biology. Orlando, USA: AcademicPress, 1998.
H.W. Heldt. Plant Biochemistry and Molecular Biology. Oxford (U.K.): Oxford
University Press, 2004.
Frank B. Salisbury, Cleon W. Ross. Fisiología Vegetal. México: Grupo Editorial
Iberoamericana, 1994. (traducción de la 4ª edición original en inglés:
PlantPhysiology. Wadsworth, 1992; existe también una reedición de la versión
española en tres volúmenes: Madrid: Paraninfo, 2000).
L. Taiz, E. Zeiger. Plant Physiology. Sunderland, Massachussets: Sinauer
Associates Inc., 2002.

A diferencia de los animales, que necesitan digerir alimentos ya elaborados, las plantas son
capaces de producir sus propios alimentos a través de un proceso químico llamado
fotosíntesis. Para realizar la fotosíntesis las plantas disponen de un pigmento de color verde
llamado clorofila que es el encargado de absorber la luz adecuada para realizar este
proceso. Además de las plantas, la fotosíntesis también la realizan las algas verdes y ciertos
tipos de bacterias. Estos seres capaces de producir su propio alimento se conocen como
autótrofos.
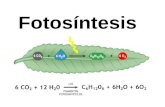
La fotosíntesis es un proceso que transforma la energía de la luz del sol en energía química.
Consiste, básicamente, en la elaboración de azúcares a partir del C02(dióxido de carbono)
minerales y agua con la ayuda de la luz solar.
¿ Qué es la fotosíntesis?

La fotosíntesis esta condicionada por cinco principales factores:
- La luz: Es necesaria para que se pueda realizar este proceso. Debe ser una luz adecuada
puesto que su eficacia depende de las diferentes longitudes de onda del espectro visible. La
más eficaz es la rojo-anaranjada. La luz azul es muy poco eficaz y prácticamente nula la
verde, aunque algunas plantas marinas son capaces de aprovecharla.
- El agua: Componente imprescindible en la reacción química de la fotosíntesis. Constituye
también el medio necesario para que se puedan disolver los elementos químicos del suelo
que la plantas deben utilizar para construir sus tejidos.
- El dióxido de carbono: Constituye el " material" que, fijado con el agua, las plantas
utilizan para sintetizar hidratos de carbono. Penetra en las hojas a través de los estomas,
aunque, en una proporción muy pequeña, puede proceder del bicarbonato disuelto en el
agua del suelo que la plantas absorben mediante sus raíces.
- Los pigmentos: Son las substancias que absorben la luz necesaria para producir la
reacción. Entre ellos, el principal es la clorofila o pigmento verde que da el color a las
plantas. La clorofila se encuentra mezclada con otros pigmentos, aunque al aparecer en una
mayor proporción, generalmente impone su color sobre el resto que queda enmascarado.
- La temperatura: Es necesaria una temperatura determinada para que puede producirse la
reacción. Se considera que la temperatura ideal para una productividad máxima se
encuentra entre los 20 y los 30 ºC, sin embargo puede producirse entre los 0 y los 50 ºC, de
acuerdo a las condiciones en que cada planta se ha ido adaptando a su medio. Es posible
incluso con una temperatura de -0,5 ºC. Por debajo del punto de congelación no puede
darse la fotosíntesis.
La fotosíntesis presenta dos fases:
Factores que condicionan la fotosíntesis
¿Fases de la fotosíntesis?

- Fase fotoquímica o reacción de Hill: Anteriormente se conocía como fase luminosa.
Para que se dé esta fase las plantas deben absorber la luz. Las plantas absorben la luz a
través de substancias llamadas pigmentos. Entre todos ellos, destaca la clorofila, que es el
pigmento de color verde que se encuentra en el interior de los cloroplastos de la célula
vegetal.
Es la gran proporción de este pigmento el que determina que las plantas presenten
principalmente su coloración verde ya que la mayor cantidad de clorofila enmascara la
menor proporción del resto de pigmentos. Las plantas las vemos verdes porque la luz verde
al no ser absorbida es captada por nuestros ojos. Sin embargo, es la luz roja -anaranjada y la
azul la que es utilizada por la mayoría de las plantas para realizar la fotosíntesis. Otras
plantas, como ciertas algas marinas rojas, , son capaces de absorber la luz verde para
realizar la fotosíntesis. Para ello utilizan pigmentos diferentes a la clorofila.
Los pigmentos deben su color a la luz que
no son capaces de absorber. Así, por
ejemplo, la clorofila absorbe prácticamente
todos los colores del espectro visible
excepto el verde. Por lo tanto, la capacidad
de absorción de la clorofila y de otros
pigmentos y la intensidad de la fotosíntesis
dependerá de los diferentes tipos de
longitud de onda lumínica. Dado que la
clorofila es el pigmento principal, la
absorción será la mayor dentro del espectro
rojo-anaranjado, inferior en el espectro azul
y prácticamente ineficaz en el espectro
verde.
Existen dos tipos de clorofila: la clorofila A
que tiene un color verde azulado y la
clorofila B que presenta un color verde
amarillento. La primera es mucho más
abundante que la segunda ya que aparece en
una proporción tres veces superior. La
clorofila A está encargada principalmente
de capturar las longitudes de onda violeta y
rojo.
Los pigmentos vegetales no se presentan
aislados sino que se combinan entre ellos.
Así, junto a la clorofila A y B , existen otros
pigmentos llamados carotenoides y
ficobilinas. Estas últimas aparecen en
La perdida del verde de la
clorofila a medida que las hojas
se van secando deja ver otros
pigmentos vegetales que antes
estaban ocultos por aquella.

organismos vegetales inferiores (algas y
cianobacterias). Los carotenoides pueden
ser carotenos, con una coloración rojiza
anaranjada y xantófilas con una coloración
amarillenta y parda. Carotenoides y
ficobilinas, junto con la clorofila B, son los
responsables de absorber aquellas
longitudes de onda que no es capaz de
absorber la clorofila A (verde y anaranjado -
rojo). De esta manera, una vez absorbida, la
transfieren a la clorofila A, para que pueda
transformarlas.
- Fase de fijación del dióxido de carbono(Ciclo de Calvin): Corresponde a lo que
anteriormente se le conocía como fase oscura. Hoy en día se prefiere omitir este término al
haberse aceptado que este proceso necesita también de la luz para poder llevarse a cabo.
Este ciclo se produce en los cloroplastos del estroma y convierte el CO2 que las plantas
absorben a través de los estomas en hidratos de carbono. Para que pueda darse este proceso
se deberán utilizar los materiales elaborados en la anterior fase.
La fotosíntesis se produce principalmente en las hojas de las plantas, aunque en menor
proporción puede producirse en los tallos, especialmente en algunas plantas que han sufrido
adaptaciones, como los cactus o las plantas crasas.
Las hojas constan fundamentalmente de las siguientes partes:
- Epidermis: La epidermis es la capa externa de la hoja que la cubre tanto por el haz como
por el envés.
- Mesófilo: El mesófilo es la capa media de la hoja.
¿Cómo se produce la fotosíntesis?

- Los haces vasculares: Son los canales que, en forma de venas, permiten el transporte de
substancias nutritivas y agua.
- Los estomas: Son una especie de agujeros o válvulas que permiten el intercambio de
gases entre el interior de la hoja y el medio exterior.
El proceso de fotosíntesis se lleva a
cabo en la capa media de la hoja o
mesófilo, en donde se hallan los
órganos especializadas en este proceso
llamados cloroplastos. Los cloroplastos
constan fundamentalmente de una
membrana externa, una membrana
interna y de una serie de sacos,
llamados tilacoides, en cuyas
membranas se forma la clorofila u
otros pigmentos. Los tilacoides. El
espacio restante interior de los
cloroplastos queda cubierto por un
fluido llamado estroma.
La reacción se produce en las
membranas de los tilacoides donde se
encuentran los pigmentos que son
capaces de absorber las diferentes
longitudes de onda de la luz. Esta
absorción de la luz produce una
reacción química cuando la energía de
los fotones descompone el agua y
libera oxígeno, protones y electrones.
Los electrones se utilizan para
sintetizar dos aparecen agrupados en
columnas verticales llamadas granas
moléculas encargadas de almacenar y
transportar energía : la ATP (
AdenosinTrifosfato o Trifosfato de
adenosina) y NADP (Nicotiamida-
Adenina Dinucleotido fosfato) .
Estas dos moléculas se utilizarán en la
siguiente fase de la fotosíntesis para
trasformar el dióxido de carbono (C02)
y el agua (H2 0) para la producción de
materia orgánica. ( hidratos de
carbono)
Foto de hoja de encina ( Quercusilex) con un
detalle ampliado en el que se pueden apreciar los
cloroplastos.

La fase de fijación del dióxido de
carbono o Ciclo de Calvin no se lleva a
cabo en los tilacoides sino en el
estroma. Durante este ciclo el dióxido
de carbono y el ATP consiguen formar
el primer compuesto orgánico en forma
de moléculas de gliceraldehido-3-
fosfato una molécula que contiene tres
átomos de carbón, a partir de las cuales
se forman los hidratos de carbono. En
la mayoría de las plantas el Ciclo de
Calvin esta ligado a la fase
fotoquímica de manera que las plantas
se regulan a través de encimas para que
ambos procesos se produzcan a la vez.
Las plantas que siguen este proceso se
denominan plantas C3
Una excepción a este tipo de plantas lo constituyen las llamadas plantas llamadas C4 y las
plantas CAM o de metabolismo ácido. Las plantas C 4 consiguen mediante una enzima
especial añadir un paso más al ciclo de Calvin y elaboran previamente al gliceraldehido-3-
fosfato una molécula que contiene 4 átomos de carbono, llamada oxaleacetato . De ahí que
se las conozca como plantas C4. Con ello consiguen superar la eficacia de la fotosíntesis en
condiciones de baja cantidad de agua disponible.
El agua es necesaria para poder metabolizar el CO2. (En el metabolismo de las plantas C3,
por cada molécula de agua y por cada cuatro fotones se forman media molécula de oxígeno,
1,3 moléculas de ATP, y un NADPH + H+.) Cuando las plantas C3 detectan la falta de agua
en el suelo, tal como ocurre en el verano, cierran los estomas y detienen el proceso de
fotosíntesis.
Las plantas C4 pueden seguir trabajando porque consiguen realizar la fotosíntesis con bajos
niveles de CO2. Pertenecen a este grupo plantas una serie de vegetales procedentes de zona
cálida y seca, tales como el maíz, el sorgo, el mijo, la caña de azúcar o la grama. Esta es la
razón por la cual la grama, por ejemplo, es tan resistente a la sequía.
Plantas C4
Plantas CAM o plantas de metabolismo
ácido

Las plantas Cam consiguen fijar el CO2 por la noche dado
que durante el día permanecen con los estomas cerrados
para evitar la pérdida de agua.
El particular proceso fotosintético que llevan a cabo las
plantas crasas, entre las que se encuentran los cactos,
explica como estas plantas han evolucionado para soportar
condiciones de sequedad ambiental extraordinarias. La
mayoría de los vegetales en el proceso de la fotosíntesis
necesitan abrir los estomas para absorber dióxido de
carbono y expulsar oxígeno,con la consecuente perdida de
agua por transpiración. Los cactus solamente abren los
estomas por la noche, par evitar la deshidratación. Así pues
el intercambio de gases se realiza en la oscuridad.
Los cactus expulsan el oxígeno a la atmósfera y absorben
dióxido de carbono, que se mantiene en forma de ácido
(generalmente ácido málico) hasta la mañana siguiente
cuando la planta , en presencia de la luz solar, realizará la
función clorofílica y extraerá el dióxido de carbono del
ácido para transformarlo en azúcar. Este proceso se
denomina C.A.M (En ingles = CrassuleanAcidMetabolism)
porque fue observado por primera vez con las crasuláceas.
Animación en flash
del metabolismo
C.A.M
Resultante de este proceso, es el oxígeno., un producto de deshecho, que proviene de
la descomposición del agua. El oxígeno, que se forma por la reacción entre el CO2 y el
agua, es expulsado de la planta a través de los estomas de las hojas.
Las plantas han tenido y tienen un papel fundamental en la historia de la vida sobre la
Tierra. Ellas son las responsables de la presencia del oxígeno, un gas necesario para la
mayoría de seres que pueblan actualmente nuestro planeta y que lo necesitan para
poder respirar. Pero esto no fue siempre así. En un principio la atmósfera de la Tierra
no tenía prácticamente oxígeno y era especialmente muy rica en dióxido de carbono
(CO2), agua en forma de vapor ( H2O) , y nitrógeno (N) . Este ambiente hubiera sido
irrespirable para la mayoría de las especies actuales que necesitan oxígeno para poder vivir.
Importancia de la fotosíntesis

Los primeros seres vivos no necesitaban oxígeno para poder respirar. Al contrario, este
gas constituía un veneno para ellos. Fueron ciertas bacterias, junto con las plantas, las
que, hace más de 2000 millones de años empezaron a iniciar el proceso de la
fotosíntesis, transformando la atmósfera y posibilitando la vida tal como se conoce en
la actualidad.