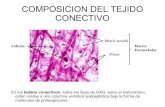
Histologia Parte 2 - Tejido Conectivo
48
Autor: Dra. Cecilia Koenig
-
Upload
janno59fifty -
Category
Documents
-
view
61 -
download
0
Transcript of Histologia Parte 2 - Tejido Conectivo

Autor: Dra. Cecilia Koenig

2
Definición y funciones Células conjuntivas Matriz extracelular Fibroblastos y fibrocitos Variedades de tejido conectivo
Definición y funciones de los tejidos conjuntivosLos tejidos conjuntivos, derivados del mesénquima, constituyen una familia de tejidos que se caracterizan porque sus células están inmersas en un abundante material intercelular, llamado la matriz extracelular.
Existen 2 variedades de células conjuntivas:
células estables, las que se originan en el mismo tejido y que sintetizan los diversos componentes de la matriz extracelular que las rodea
población de células migratorias, originadas en otros territorios del organismo, las que llegan a habitar transitoriamente el tejido conjuntivo.
La matriz extracelular es una red organizada, formada por el ensamblaje de una variedad de polisacáridos y de proteínas secretadas por las células estables, que determina las propiedades físicas de cada una de las variedades de tejido conjuntivo (Fig. 1)
Figura 1 Figura 2
Figura 3 Figura 4

3
Existen varios tipos de tejidos conjuntivos localizados en diversos sitios del organismo, adaptados a funciones específicas tales como:
mantener unidos entre sí a los otros tejidos del individuo, formando el estroma de diversos órganos (Fig. 2): TEJIDOS CONJUNTIVOS LAXOS.
contener a las células que participan en los procesos de defensa ante agenteextraños: constituyendo el sitio donde se inicia la reacción inflamatoria (Fig. 3)TEJIDOS CONJUNTIVOS LAXOS.
constituir un medio tisular adecuado para alojar células en proceso de proliferación y diferenciación para formar los elementos figurados de la sangre correspondientes a glóbulos rojos y plaquetas, y a los distintos tipos de glóbulos blancos, los que migran luego a los tejidos conjuntivos, para realizar en ellos sus funciones específicas ya sea como células cebadas, macrófagos, células plasmáticas, linfocitos y granulocitos (Fig. 4): TEJIDOS CONJUNTIVOS RETICULARES.
almacenar grasas, para su uso posterior como fuente de energía, ya sea por ellos mismos o para otros tejidos del organismo (Fig. 5) TEJIDOS ADIPOSOS.
formar láminas con una gran resistencia a la tracción, tal como ocurre en la dermis de la piel, y en los tendones y ligamentos (Fig. 6): TEJIDOS CONJUNTIVOS FIBROSOS DENSOS.
formar placas o láminas relativamente sólidas, caracterizadas por una gran resistencia a la compresión (Fig. 7): TEJIDOS CARTILAGINOSOS.
formar el principal tejido de soporte del organismo, caracterizado por su gran resistencia tanto a la tracción como a la compresión (Fig. 8): TEJIDOS ÓSEOS.
Figura 5 Figura 6
Figura 7 Figura 8

4
Células conjuntivas
Las células que están presente, normalmente, en en los tejidos conjuntivos corresponden a dos grupos distintos:
células propias de cada tipo de tejido conjuntivo, las que constituyen una población celular estable, responsable de la formación y mantención del tejido al que pertenecen, razón por la cuál se las llama células de sostén (Fig. 1) .
células conjuntivas libres, que han llegado a residir en los tejidos conjuntivos. Estas células se forman en los órganos hematopoyéticos, circulan como glóbulos blancos en la sangre, y migran luego a un tejido conjuntivo en particular, estableciéndose en él (Fig. 2)
Se distinguen entre ellas:
células de vida media larga, capaces de dividirse en el tejido conjuntivo que habitan: células cebadas y fagocitos mononucleares o macrófagos
células de vida media corta, que no se dividen, y cuya concentración en una a zona del tejido conjuntivo se relaciona con procesos de defensa del organismos: células plasmáticas, linfocitos y fagocitos porlimorfonucleares .
Células propias de los tejidos conjuntivosLas células llamadas estables o de sostén corresponden a un grupo de células diferenciadas cuyo principal rol es producir la matriz intercelular propia de cada tipo de tejido conjuntivo. Ellas se forman a partir de células mesenquimáticas localizadas en los sitios del organismo en que van a formar al tejido conjuntivo. (Fig. 1).
Estas células se caracterizan por encontrarse en proceso de activa diferenciación para sintetizar a la matriz extracelular que caracteriza al tipo de tejido conjuntivo que corresponda.
Ellas pueden diferenciarse como:
Fibroblastos Lipoblastos Osteoblastos Condroblasto
Figura 1 Figura 2

5
Se caracterizan, al microscopio de luz, por su basofilia citoplasmática . Poseen un retículo endoplásmico rugoso bien desarrollado, un aparato de Golgi definido y escasas vesículas de secreción, organelos que se relacionan con la síntesis de moléculas precursoras del colágeno, elastina, proteoglicanos y glicoproteínas de la matriz extracelular. Cuando estas células se hallan en fase de relativa quiescencia
en la matriz extracelular que han formado se las llama: fibrocitos, osteocitos y condrocitos. Los lipoblastos se diferencian posteriormente a células adiposas cuyo principal rol es almacenar grasas.
Los fibroblastos producen los tejidos conjuntivos fibrosos cuya matriz extracelular está constituida por fibras colágenas y fibras elásticas, asociadas a glicosaminoglicanos, proteoglicanos y glicoproteínas (Fig. 2, 3 y 4 )
Los condroblastos producen el tejido cartilaginoso, cuya matriz extracelular se caracteriza por la presencia de una cantidad importante de proteoglicanos asociados a ácido hialurónico y a microfibrillas de colágeno tipo II. Al quedar totalmente rodeados por la matriz cartilaginosa ellos pasan a llamarse condrocitos (Fig. 5)
Los osteoblastos producen el tejido óseo, sintetizando el componente orgánico de la matriz extracelular ósea que se caracteriza por un alto contenido en colágeno tipo I, glicosaminoglicanos y gliproteínas. Al quedar totalmente rodeados por la matriz ósea pasan a llamarse osteocitos (Fig. 6)
Figura 1 Figura 2
Figura 3 Figura 4
Figura 5 Figura 6

6
Los lipoblastos producen el tejido adiposo. Ellas se diferencian a células almacenadoras de grasa , sintetizan su matriz extracelular y se rodean de una lámina basal . Ellos pasan así a formar los adipocitos o células adiposas (Fig. 7)
Figura 7
Células conjuntivas libresEstas células se originan en la médula ósea hematopoyética y usan la circulación sanguínea como un medio de transporte hacia los tejidos conjuntivos, donde realizan sus principales funciones. (Fig. 1)
Entre ellas se encuentran las células cebadas(o mastocitos) y los macrófagos (o histiocitos) que son componentes estables del tejido conjuntivo al que llegan (Fig. 2)
Un grupo distinto lo forman las células plasmáticas, los linfocitos y los granulocitos polimorfonucleares, todos ellos células de vida media relativamente corta y que tienden a concentrarse en las zonas de tejido conjuntivo en que ocurren reacciones relacionadas con la defensa. (Fig. 3)
Figura 2 Figura 3
Figura 1

7
Células cebadas
Son células grandes de forma redondeada (20-30 um) y al microscopio de luz se caracterizan por presentar el citoplasma llena de gránulos basófilos que se tiñen metacromáticamente con azul de toluidina. Su núcleo es esférico y está situado en el centro de la célula (Fig. 1 y 2).
Ellas se ubican de preferencia vecinas a los vasos sanguíneos (Fig. 3)
Los gránulos contienen heparina, proteoglicano sulfatado de unos 750 kD, que forma la matriz de los gránulos. A esta macromolécula se asocian varias moléculas de bajo peso molecular, cargadas positivamente, como histamina; proteasas neutras, y factores quimiotácticos para eosinófilos y para neutrófilos. Su superficie muestra largas prolongaciones muy finas, su citoplasma contiene pocos organelos y sus gránulos pueden presentar un grado variable de compactación. (Fig. 4)
En la membrana de las células cebadas se ubican receptores para el fragmento Fc de las IgE, anticuerpos secretadas por las células plasmáticas. Cuando la IgE , anclada en su receptor, se une a su antígeno se produce una señal hacia el citoplasma de la célula cebada que :
estimula la síntesis de leucotrieno y su liberación hacia el extracelular induce la exocitosis de los gránulos y entre las sustancias liberadas destaca la
histamina, que aumenta la permeabilidad de las vénulas, con lo que aumenta el líquido en la matriz extracelular conjuntiva, produciendo edema.
Se origina así una reacción de hipersensibilidad inmediata a escasos minutos de haber penetrado el antígeno (alergeno) la que se manifiesta en fenómenos como edema, rinitis alérgica, asma y shock anafiláctico .
La principal función de las células cebadas es almacenar los mediadores químicos de la respuesta inflamatoria. Comparten así muchas características con los leucocitos basófilos de la sangre, pero son otra familia celular.
Figura 1 Figura 2
Figura 3 Figura 4

8
En roedores se han demostrado dos poblaciones de células cebadas : las células cebadas de tejidos conjuntivos ya descritas, y las células cebadas de mucosas de menor tamaño, cuyos gránulos contienen proteoglicanos de condroitínsulfato en lugar de heparina y con un contenido 10 veces menor de histamina. Ambos tipos de células reaccionan en forma diferente ante agentes farmacológicos que inducen la degranulación.
MacrófagosSon células mononucleadas que se caracterizan por su capacidad de fagocitar y degradar material particulado (Fig. 1)
Se originan a partir de células de la médula ósea que dan origen a los monocitos de la sangre los que luego migran desde el lumen de los capilares sanguíneos al tejido conjuntivo donde terminan su diferenciación.
Los macrófagos de los tejidos conjuntivos miden entre 10 y 30 um de diámetro y su estructura se modifica según su estado de actividad (Figs. 2 y 3)
Figura 2 Figura 3
Su superficie presenta numerosas prolongaciones digitiformes, su núcleo es indentado, y en su citoplasma presenta numerosas vacuolas endocíticas, lisosomas primarios y fagolisosomas. Tienen un retículo endoplásmico rugoso desarrollado y su aparato de Golgi es prominente. Poseen, además, microtúbulos, filamentos intermedios y microfilamentos de actina.
Los macrófagos activados tienen más prolongaciones de membrana, un mayor número de vacuolas, lisosomas, fagosomas y cuerpos residuales
Estas células se reconocen facilmente cuando cuando han
Figura 1
Figura 4 Figura 5

9
fagocitado partículas visibles al microscopio (Fig. 4 y 5)
En respuesta a ciertos procesos infecciosos los macrófagos se pueden fusionar originando células de 20 más núcleos llamadas células gigantes multinucleadas. Bajo estímulos adecuados también modifican su aspecto rodeando materiales extraños y formando las llamadas células epitelioides de cuerpo extraño.
Entre sus funciones destacan:
Su alta capacidad fagocítica les permite cumplir un rol importante en la eliminación de microorganismos, tejidos dañados y contaminantes particulados.
Su capacidad de secretar diversos factores y su participación en la respuesta inmune como células presentadoras de antígeno, se discutirán en el capítulo de Linfático y Defensa Inmune.
Células plasmáticas
Las células plasmáticas se originan en los tejidos linfáticos por diferenciación de linfocitos B activados y llegan a los tejidos conjuntivos por la circulación
sanguínea.(ver capitulo de Linfático e Inmunidad). En los tejidos conjuntivos su vida media es de 10 a 20 días.
Su función es sintetizar y secretar los anticuerpos, moléculas que pertenecen a una familia de proteínas específicas llamadas inmunoglobulinas. (Fig. 1)
Estas células tienen de 10 a 20 um de diámetro, su forma es ovalada con un citoplasma muy basófilo. El núcleo redondo y excéntrico, se caracteriza porque la heterocromatinase dispone como ruedas de carreta. Su citoplasma se caracteriza por un gran desarrollo del retículo endoplásmico rugoso. responsable de la basofilia de su citoplasma y por una aparato de Golgi yuxtanuclear muy desarrollado.(Fig. 2)
Figura 1 Figura 2

10
Los anticuerpos que está sintetizando se pueden localizar inmunocitoquímicas ubicados tanto en el espacio perinuclear, como en las cisternas del retículo endoplásmico rugoso del aparato de Golgi. Normalmente no se observan gránulos de secreción almacenados en el citoplasma de estas células. (Fig. 3)
LinfocitosSon leucocitos agranulocitos que juegan un rol fundamental en la respuesta inmune. La mayoría de los linfocitos circulantes son linfocitos pequeños (6-8 um de diámetro) (Fig. 1). Aunque
estructuralmente iguales, mediante marcadores citoquímicos es posible distinguir 3 tipos de linfocitos: linfocitos B, T y nulos.
Los linfocitos de los tejidos conectivos corresponden a linfocitos pequeños, de 6 a 8 um de diámetro. Poseen un núcleo ovoide con cromatoma muy condensada y una pequeña cantidad de citoplasma suavemente basófilo (Fig. 2).
Al MET presentan una superficie irregular debido a la presencia de numerosas proyecciones citoplasmáticas.. El citoplasma contiene escasas mitocondrias, ribosomas libres, cisternas de ergastoplasma, un Golgi pequeño y granos azurófilos (Fig. 3). Según la variedad de linfocito de que se trate su vida media celular pueden variar entre unos pocos días hasta meses y años. Su duración como células libres del tejido conjuntivo es en general de pocos días.
Cumplen un rol fundamental en la respuesta inmune. Relaciones complejas entre linfocitos B, linfocitos T y células presentadoras de antígeno generan las respuestas de defensa inmune humoral y celular. A su vez, los linfocitos nulos participan en los mecanismos de defensa dando origen a células asesinas.
Figura 3
Figura 1 Figura 2
Figura 3

11
Granulocitos polimorfonuclearesCorresponden a leucocitos que presentan un núcleo lobulado (polimorfonucleares) y contienen en su citoplasma gránulos específicos. Existen tres tipos:
polimorfonucleares neutrófilos,
eosinófilos y basófilos
Polimorfonucleares neutrófilos.
Su vida media en los tejidos conjuntivos de 1 a 4 días.
Son células de 12 a 15 um de diámetro, cuyo núcleo está formado por 2 a 5 lóbulos de cromatina condensada unidos entre sí (Figs. 1 y 2). Poseen un abundante citoplasma relativamente pobre en organelos que contiene 2 variedades de gránulos que se tiñen
poco con los colorantes usuales. Los gránulos específicos contienen fosfatasa alcalina, colagenasa, lactoferrina y proteínas básicas como lisozima y fagocitinas y los granos azurófilos a lisosomas primarios. El citoplama es en general pobre en otro tipo de organelos (Fig. 3).
Constituyen un mecanismo de defensa contra la invasión de microorganismos, especialmente bacterias. Fagocitan activamente partículas pequeñas por lo que se les ha llamado fagocitos pomorfonucleares para distinguirlos de los macrófagos.
Matriz extracelularEl análisis microscópico de la estructura del material intercelular muestra (Fig.1):
elementos fibrilares bien estructurados llamados
Figura 1 Figura 2
Figura 3
Figura 1 Figura 2

12
fibras conjuntivas, que pueden ser colágenas, reticulares o elásticas, un material poco estructurado al que se denomina sustancia fundamental
amorfa y en los sitios en que la matriz extracelular conjuntiva se asocia a células de otros
tejidos se observan las láminas basales (Fig. 2).
El análisis de la composición molecular del espacio intercelular demuestra que las tres principales clases de macromoléculas extracelulares son:
1. cadenas de polisacáridos de la clase de los glicosaminoglicanos, que pueden unirse covalentemente a proteínas, formando macromoléculas más complejas llamadas proteoglicanos. Estas moléculas forman el gel altamente hidratado que constituye la sustancia fundamental en la cuál están embebidas las células y fibras conjuntivas. La fase acuosa del gel de polisacáridos permite una rápida difusión de nutrientes, metabolitos y hormonas entre la sangre y las células tisulares
2. proteínas fibrosas que se organizan para formar estructuras bien definidas de la matriz extracelular como son las fibrillas colágenas, la lámina densa de las las láminas basales y las fibras elásticas.
3. glicoproteínas de adhesión como fibronectina que asocian entre sí a células, fibras y proteoglicanos del tejido conjuntivo y como laminina que asocia la lámina basal a las células que están rodeadas por ella.
Las variaciones en las cantidades relativas de las distintas macromoléculas presentes y en la forma en que están organizadas dan origen a variedades tan diveras de matriz como son la dura matriz extracelular del hueso y la transparente matriz extracelular de la córnea.
Fibras ConjuntivasLos principales tipos corresponden a fibras elásticas y a las estructuras formadas por fibrillas colágenas
Fibras elásticasLas fibras elásticas se estiran fácilmente y recuperan su longuitud original cuando la fuerza deformante ha desaparecido. Al microscopio electrónico aparecen formadas por un componente amorfo
rodeado y penetrado por microfibrillas de unos 10 nm de diámetro (Figs.1 y 2).
Figura 1 Figura 2

13
El principal componente de las fibras eláticas es la elastina, material proteico muy insoluble. Se caracteriza por un alto contenido en aminácidos apolares como prolina y valina y tiene un alto contenio en aminácidos no cargados como la glicina. Contiene además dos aminácidos exclusivos: desmosina e isodesmosina.
La elastina se forma por la interacciones entre moléculas solubles de tropoelastina, proteina de unos 70.000 de PM que contiene el aminacido lisina y carece del aminoácido desmosina. En la figura 3 se muestra como la interacción
entre las lisinas de 4 moléculas de tropoelastina (catalizada por la enzima lisiloxidasa, en presencia de cobre) pueden dar origen a la desmosina y unir en puntos específicos a estas hebras polipeptídicas, ricas en aminoácidos hidrofóbicos (Figs. 3 y 4).
Se ha propuesto que en estado de reposo la hebra polipeptídica rica en aminoácidos apolares se encuentra plegada sobre sí misma y que al tensionar la fibra elástica se induce el estiramiento de esta zona en cada una de las hebras polipeptidica las que son obligadas a ponerse paralelas entre sí, enfrentandose almedio acuoso del extracelular, permaneciendo unidas sólo a nivel de las desmosinas. Al desaparecer la fuerza deformante cada una de estas zonas de las hebras polipeptídicas recuperan su conformación plegada en forma espontánea.y la fibra elástica recupera su
longuitud inicial (Fig.5).
Las microfibrillas tienen como principal componente una glicoproteína llamada fibrilina, rica en aminoácidos hidrofílicos y que contiene residuos de cistina los cuales pueden formar puente disulfuro, estabilizando la estructura de las microfibrillas. En las fibras elásticas juegan un rol estructural, formando una especie de vaina alrededor de la elastina, permitiendo así la formación de estructuras fibrilares elásticas. En ausencia de microfibrillas la elastina se deposita formando láminas elásticas, como ocurre por ejemplo en la pared de las arterias.
Figura 3 Figura 4
Figura 5

14
Estructuras formadas por fibrillas colágenasLas fibrillas colágenas forman estructuras que se caracterizan por su gran resistencia a las fuerzas de tracción y el modo en que ellas se asocian entre sí varía de un tejido conjuntivo a otro. Ellas son el principal componente de:
las fibras colágenas de los tejidos conjuntivos fibrosos las fibras reticulares las laminillas colágenas del hueso.
Cada fibrilla colágena está formada por la asociación de moléculas de colágeno, proteínas fibrosas de varios tipos, que constituyen más del 25% de la masa proteica total de los mamíferos.
Las moléculas de colágeno se asocian formando polímeros que corresponden a las fibrillas colágenas visibles al microscopio electrónico (Fig.1), las cuales pueden tener cientos de m de largo y presentan una estriación transversal característica repetida cada 67 nm, la que refleja la disposición regularmente espaciada de las moléculas de colágeno (Fig.2). El diámetro de las fibrllas varía entre 10 y 300 nm, según el tipo de colágeno de que se trate.
Las moléculas del colágeno son largas y rectas (300 x1,5 nm), formadas por la asociación en una estructura helicoidal de 3 cadenas polipeptídicas, las cadenas (Fig.3).
Cada cadena contiene unos 100 aminoácidos ordenados como una hélix hacia la izquierda, con 3 aminoácidos por vuelta, correspondiendo siempre glicina al tercer residuo, de manera que cada cadena a está compuesta por una serie de secuencias de la
tripleta -Gly-X-Y- , en la cual X ó Y corresponden con gran frecuencia prolina o hydroxiprolina respectivamente. Existen más de 25 tipos distintos de cadenas a codificadas por genes diferentes , las que se asocian forman unos 15 tipos distintos de moléculas de colágeno.
Las moléculas de colageno de tipo I, II y III, son capaces de interactuar entre sí, asociándose lateralmente en forma estable por uniones covalentes entre residuos de lisina.
Figura 1 Figura 2
Figura 3

15
De hecho, en la matriz extracelular propia del cartílago las fibrillas del colágeno son muy delgadas (10 nm de diámetro) y están formadas por el ensamble de moléculas de colágeno tipo II (formadas por 3 cadenas (II) ) (ver
tejido cartilaginoso).
Las fibras colágenas, de varios mm de grosor (Figs.4), corresponden a la asociación de numerosos manojos de fibrillas de 150 a 300 nm de diámetro (Figs. 5) formadas por moléculas de colágeno tipo I (2 cadenas (I) y 1 cadena 2(I)).
Las fibras reticulares, de 0,5 a 2,0 m de grosor (visibles al ser teñidas con sales de plata) (Fig.6), están formadas por manojos de fibrillas de 50 nm diámetro formadas por moléculas de colágeno tipo III (3 cadenas
1(III)) (Fig.7).
Las laminillas colágenas óseas, están formadas por la disposición de capas sucesivas de fibrillas de colágeno tipo I . En cada capa las fibrillas están paralelas entre si, pero las capa sucesivas alternadan ordenadamente la orientación de sus fibrillas en ángulos rectos. Una disposición laminillar similar se observa en tejidos fibrosos como la córnea.
Sustancia fundamentalCorresponde a la matriz traslúcida que rodea a células y fibras conjuntivas y que no presenta una organización estructural visible al micorscopio de luz (Fig.1).
Este componente constituye un gel semisólido, bioquímicamente complejo y altamente hidratado. Las principales macromoléculas que forman la sustancia fundamental corresponden a polisacáridos de la clase de los glicosaminoglicanos, que pueden existir como tales o pueden unirse covalentemente a proteínas,
formando macromoléculas más complejas llamadas proteoglicanos.
Figura 4 Figura 5
Figura 6 Figura 7
Figura 1

16
GlicosaminoglicanosLos glicosaminoglicanos son largas cadenas de polisacáridos no ramificadas formadas por la repetición sucesiva de la unidad de disacáridos formada por: ácido urónico y hexosamina acetilada, la cual puede estar sulfatada .
El ácido hialurónico (hialuronato o hialuronano) es un glicosaminoglicano no sulfatado formado por una secuencia de 25.000 unidades sucesivas de (ácido D-glucouronico- N-acetilglucosamina) (Fig.1).
Por su riqueza en grupos ácidos está cargado negativamente y tiende a atraer cationes como el Na+ que, al ser osmóticamente activos, atraen agua disponiéndose cada molécula de hialuronato como un ovillo laxamente plegado y altamente hidratado, que ocupa un volumen importante del espacio intercelular (Figs. 1 y 2).
Los glicosaminoglicabos sulfatados no existen en general en forma libre sino asociados a proteinas.
Los principales de ellos son:
Condroitín sulfato y dermatan sulfato, que se caracterizan por estar formados por una secuencia de 70 a 200 unidades sucesivas de (N-acetil-galactosamina sulfatada y ácido D-glucourónico o L-idurónico) (Fig.3).
heparan sulfato formado por una secuencia de 20 a 100 unidades sucesivas de (N- acetil-glucosamina sulfatada y ácido glucourónico o L-idurónico)
keratan sulfato formado por la secuencia un numero variable de unidades de (D-galactosa y N-acetyl glucosamina sufatada).
Figura 1 Figura 2
Figura 3

17
Proteoglicanos
Los proteoglicanos son macromoléculas formadas por una proteina central, a lo largo de la cual se asocian, por su extremo terminal, numerosas moléculas de glicosaminoglicanos sulfatados.
Existe una enorme varidad de proteoglicanos, dependiendo del tipo y largo de la proteína central y del tipo, número y longuitud de los
glicosaminoglicanos asociados a ella. El proteoglicano decorina, que recubrela superficie de las fibrillas colágenas, contienen 1 molécula ya sea de condroitin o de derrmatansulfato (Fig. 1), mientras que el proteoglicano aggregacan que es uno de los principales componentes de la matriz extracelular del cartílago presenta alrededor de 100 moléculas de condroitinsulfato y 30 moléculas de keratansulfato unidas a una proteína central de más de 3000 aminoácidos (Fig. 1).
Existe además la posibilidad de que la sustancia fundamental se organice en agragados moleculares aún mayores ya que los proteoglicanos pueden asociarse por uno de lo extremos de la molécula de proteína central a lo largo del ácido hialurónico, por medios de proteinas globulares de unión (Fig. 2).
Se forma así la posibilidad de estabilizar la estructura de la sustancia fundamental, la cuál podrá adoptar la consistencia de un gel relativamente fluído como es el
caso de los tejidos conjuntivos laxos (Fig. 3), o de un gel practicamente sólido como en el caso del cartílago hialino (Fig. 4).
La mayoría de las funciones estructurales de los proteoglicanos se relacionan con sus características moleculares: interactúan con cationes y agua, tendiendo a inmovilizarlos, resitiendo muy bien las fuezas de compresión. El líquido que los rodea constituye el líquido tisular y a través de él fluyen los metablitos y difunden los gases respiratorios.
Figura 1 Figura 2
Figura 3 Figura 4
Figura 5

18
Las glicoproteínas de adhesiónLas glicoproteínas de adhesión de la matriz extracelular se caracterizan por ser moléculas que presentan diversos dominios que les permiten interactura simultáneamente tanto concomponentes de
la matriz extracelular comom con las células conjuntivas. En esta parte nos referiremos como ejemplo sólo a dos de ellas: la fibronectina (Fig. 1).que asocian entre sí a células, fibras y proteoglicanos del tejido conjuntivo y la laminina que asocia a células no conjuntivas con las macromoléculas de la lámina basal (Fig. 2).
Láminas basalesSon estructuras laminares que corresponden a una forma especializada de la matriz extracelular, que actúan como interfase entre las células de los tejidos epiteliales, músculares y nerviosos en los sitios en que ellas se asocian al tejido conjuntivo.
Al microscopio electrónico aparece formada por 2 capas laminares: la que está vecina a la célula es poco densa a los electrones y se denomina lamina rara (o lúcida), la segunda capa es densa a los electrones y se
denomina lamina densa , a ella se asocia una tercera capa que contiene fibrillas colágenas llamada lamina fibroreticular y que es continua con la matriz conjuntiva adyacente (Figs.1 y 2). Entre las funciones que desempeña la lámina basal se destacará en este momento sólo que ella constituye una interfase the adhesión de modo que las células poseen mecanismos de adhesión para fijarse a la membrana basal y ella a su vez está anclada en la matriz extracelular conjuntiva a las fibrillas colágenas.
Figura 1 Figura 2
Figura 1 Figura 2

19
Las membranas basales están formadas por 4 tipos de moléculas predominantes: colágeno tipo IV, perlecan (proteoglicano de heparan-sulfato), laminina y entactina (Fig.3), además de la
fibronectina y del colágeno VII que la asocia a las fibrillas de colágeno III de la lámina fibroreticular.
Las moléculas de colágeno tipo IV forman la lámina densa. Ellas difieren de las moléculas de colágeno I, II y III que forman las fibrillas colágenas. Las moléculas de colágeno IV se asocian entre primero entre sí por sus extremos carboxi terminales formando dímeros (Fig.4) los cuales a su vez se aocian entre sí lateralmente formando una lámina desda la cual se projectan perpendicularmente los extremos aminoterminal de las moléculas, los que al interactuar entre sí permiten la unión de láminas sucesivas de las redes planares formadas por los dímeros del colágeno IV.
El perlecan es un proteoglicano de heparansulfato, formado por una proteín central de unos 600.000 de PM asociada a entre 2 y 15 cadenas de heparansulfato, el cuál puede interactura directamente con colágeno IV y con laminina. Se concentra hacia la lámina rara.
La laminina es una glicoproteína de gran tamaño (850.000) (Fig.5) que presenta dominios funcionales que le permiten interactuar con: receptores para laminina de la membrana plasmática de la célula, heparansulfato, colágeno IV y entactina. Puede además interactuar con otras moléculas de laminina por los extremos de sus 4 brazo formando una estructura laminar. Se concentra hacia la lámina rara.
La entactina, que es una gliproteina pequeña se asocia a las moléculas de laminina entre sí y con colageno IV
contribuyendo a estabilizar la estructura y la asociación de las láminas rara y densa.
Figura 3 Figura 4
Figura 5

20
Fibroblastos y fibrocitos
Es la célula propia de los tejidos conjuntivos fibrosos, cuya principal función es sintetizar y mantener a la matriz extracelular propia del tejido.
De acuerdo a su actividad biosintética se pueden distinguir morfológicamente: fibroblastos que corresponden a la célula en un estado de alta actividad (Fig.1) fibrocitos que son las células poco activas o
en reposo (Fig. 2).
Los fibroblastos se distinguen estructuralmente por su aspecto fusiforme y su abundante citoplasma basófilo; los fibrocitos son más pequeños y su citoplasma es suavemente acidófilo (Figs. 3 y 4).
La organización ultraestructural de los fibroblastos diferenciado refleja el compromiso de este tipo de células en la síntesis de moléculas que forman la matriz extracelular. Poseen un retículo endoplásmico
desarrollado, un Golgi perfectamente definido y escasas vesículas de secreción, organelos que se relacionan con la síntesis de moléculas precursoras del colágeno, elastina, proteoglicanos y glicoproteínas de la MEC (Fig.5).
Presentan un desarrollado citoesqueleto de microtúbulos y de microfilamentos de actina implicados en procesos de motilidad celular. Esta propiedad es importante en la cicatrización de heridas, ya que los fibroblastos tiene la capacidad de migrar hacia la zona lesionada, proliferar y producir los componnetes de la matriz extracelular.
Figura 1 Figura 2
Figura 3 Figura 4
Figura 5 Figura 6

21
En los tejidos conjuntivos fibrosos reticulares las células propias del tejido, responsable de la sintesis y mantención del estroma reticular reciben el nombre de células reticulares. Este nombre suele inducir a confusión ya que inicialmente se usó para denominar a las células asociadas al estroma reticular, actualmente se distingue la célula reticular propiamente tal de otras células que pueden asociarse al estroma como son por ejemplo los macrófagos fijos. La estructura de las células reticulares es similar a la de los fibroblastos (Fig.6).
Fibroblastos y síntesis de microfibrillas colágenas
Los fibroblastos sintetizan y secretan el procolágeno, molécula de mayor tamaño que el colágeno, la cual presenta en ambos extremos una porción no helicoidal.
Los pasos intracelulares (Fig.1) en el proceso de sintesis de esta molécula impican las siguientes etapas:
producción del mRNA correspondiente al tipo o tipos de cadenas que se requiera, de acuerdo tipo de colágeno que se va a secretar
sintesis en ribosomas unidos a membrana e introducción hacia el lumen de cisternas del ergastoplasma de cada cadena pro-, los cuales además del peptido señal contienen una serie de aminoácidos adicionales llamados propéptidos.
En el lumen del ergastoplasma residuos precisos de prolina y lisina son hidroxilados, formando hidroprolinas e hidroxilisinas. Este paso requiere de la presencia de vitamina C. (en el escorbuto, no ocurre la hidroxilación delas prolinas y las cadenas pro- defectuosas no
pueden formas las triple helix y se degradan inmediatamente). Algunas de las hidroxilisinas son luego glicosiladas tres cadenas pro- hidroxiladas y glicosiladas se acoplan entre sí inicialmente
por los propetidos y forman las moléculas de procolágeno: estructuras helicoidal triple estabilizada por puentes de hidrógeno, cuyo extremos son más globosos por la presencia de los propéptidos. Estas moléculas se acumulan en el aparato de Golgi y son transportadas en gránulos de secreción hacia la superficie celular liberándose por exocitosis hacia el extracelular.
Para que se formen las fibrillas de colágeno a partir de las moléculas de procolágeno secretadas se requiren varios pasos (Fig. 2), que dependen de la presencia de enzimas específicas sintetizadas y secretadas por los fibroblastos en el espacio extracelular, que ocupa profundas invaginaciones en la supeficie de los fibroblastos.
Figura 1
Figura 2

22
Las etapas principales son:
remoción de los propéptidos de las moléculas de procolágeno por acción las procolágeno-peptidasas, eliminándose así los extremos globosos y quedando la molécula de colágeno, de menor peso molecular (Fig. 2)
las moléculas de colágeno son menos solubles y tienden a asociarse entre sí por interacciones entre las caras laterales de moléculas vecinas, que se disponen paralelas entre sí pero desplazadas en aproximadamente un quinto de su longitud. Esta interacción entre moléculas de colágeno se estabiliza por la formación de enlaces covalentes entre los residuos de lisina de las moléculas vecinas, en una reacción compleja que require de la enzima lisil-oxidasa (Figs. 2 y 3)La resistencia a la tracción de las fibrillas colágenas va a depender del número de enlaces covalentes que existan entre las moléculas paralelas de colágeno. Si se inhibe a la enzima lisil-oxidasa la resistencia a la tracción de la
fibrillas disminuye drásicamente y ocurren alteraciones graves en la estructura de los tejido conjuntivos.
como consecuencia de la polimerización del colágeno quedan a espacios, a largo de la fibrilla en formación, entre los extremos carboxi- y amino- terminales de las sucesivas moléculas de colágeno, Al observar las fibrillas al microscopio electrónico luego de usar tinción negativa (paso 11), se dmuestra la penetración del colorante en los sitios huecos de la fibrilla, que aparecen como un bandeo denso a los electrones (negro) mientras que los sitios de total superposición de las moleculas adyacentes no permiten la pentración del colorante y aparecen como un bandeo poco densos a los electrones (blanco). (Fig.4)
la forma en que se van a organizar las fibrillas colágenas en el extracelular depende tambien de los fibroblastos, ya que estos pueden expresar además otros tipos de colágenos como el colágeno XII (1(XII)3) el cuál no forma fibrillas sino que se asocia lateralmente en forma periódica a la superficie de las fibrillas de colágeno I y contribuye a asociarlas entre sí formando manojos paralelos de fibrillas , y las asocia también a otros elementos de la matrix extracelular.
Figura 3
Figura 4

23
Variedades de tejidos conjuntivosTejido conjuntivo laxo
Tejido conjuntivo fibroso denso
Tejido conjuntivo elástico
Tejido conjuntivo reticular
Tejido adiposo
Cartílago y hueso
Tejido conjuntivo laxoSe caracteriza por la presencia de una población relativamente alta de células residentes, ya sea propias como fibroblastos y adipocitos o migratorias como macrófagos y células cebadas, separadas
por la matriz extracelular formada por fibras colágenas y elásticas, laxamente dispuestas en una sustancia fundamental bastante fluída (Figs. 1 y 2)
Las fibras colágenas que predominan están formadas por microfibrillas de colágeno I asociadas a cantidades variables de microfibrillas de colágeno III pero en menor cantidad. Estas fibras colágenas le confieren resistencia
a la tracción (Fig.3).
Las fibras elásticas, aunque menos abundantes que las de colágeno, cumplen un rol importante ya que le otorgan al tejido la capacidad de retomar su organización original luego de ser traccionado (Fig.4).
Figura 1 Figura 2
Figura 3 Figura 4

24
La sustancia fundamental, que ocupa los espacios entre las fibras, tiene como principal componente a grandes moléculas de ácido hialurónico, el cual estabiliza una importante cantidad de agua, adoptando la
estructura de un gel poco consistente, de modo que existen innumerables espacios intermoleculares que sirven de reservorio al líquido intersticial ("fluido tisular"). Este fluído facilita la difusión del oxígeno y nutrientes desde los capilares del conjuntivo hacia células de otros tejidos (epitelios, cartíago o músculo) y de los productos de desecho del metabolismo en sentido inverso. La capacidad de la sustancia fundamental de acumular líquido es la base del proceso llamado edema. (Fig. 5).
Debido a la variedad de células que lo habitan y a la capacidad de acumular líquido proveniente del plasma sanguíneo, el conjuntivo laxo juega un rol muy importante en la iniciación del proceso de defensa orgánica a través de la generación de las respuestas inflamatoria e inmune (Fig 6).
Tejido conjuntivo fibroso densoPresenta un contenido relativamente bajo de células, las que corresponden principalmente a fibroblastos. Su matriz extracelular es muy abundante, y su principal componente son gruesas fibras colágenas. La sustancia fundamental es relativamente escasa, predominando proteoglicanos de dermatan-sulfato.
De acuerdo a la forma en que se disponen las fibras de colágeno, se distingue:
Tejido conjuntivo denso desordenado.
Las fibras colágenas forman una redtridimensional lo que le otorga resistencia en todas las
direcciones. Asociada a esta red colágena existen fibras elásticas. Entre las fibras colágenas y elásticas se ubican las células, principalmente fibroblastos y se encuentra por ejemplo en la dermis y formando la cápsula de órganos como los ganglioslinfático y el hígado (Figs.1 y 2).
Figura 5 Figura 6
Figura 1 Figura 2

25
Tejido conjuntivo denso ordenado o regular.
Las fibras de colágeno se disponen en un patrón definido que refleja una respuesta al la
direccción del requerimiento mecánico predominante. Las fibrillas de colágeno tipo I paralelas se asocian entre sí por interaccion con moléculas de colágeno tipo XII (a1(XII)3), lo que le otorga una mayor firmeza . Los fibroblastos son las únicas células presentes y se ubican entre los haces de fibras formando largas líneas paralelas. Estas células poseen escaso citoplasma y presentan un cuerpo celular alargado, con núcleos son alargados y de cromatina condensada (Fig. 3 y 4).
Las estructuras formadas por tejido conjuntivo denso ordenado están adaptadas para oponer una alta resistencia a la tracción ejercida en la dirección en que se disponen las fibras. Tal es el caso de tendones y aponeurosis.
Tejido conjuntivo elásticoEs una variedad de tejido fibroso denso en el cuál las fibras conjutivas presentes corresponden a fibras o láminas elásticas dispuestas en forma paralela. Los espacios entre las fibras elásticas están ocupados por una fina red de microfibrillas colágenas con unos pocos fibroblastos.
El tejido conjuntivo elástico forma capas en la pared de los órganos huecos sobre cuyas paredes actúan presiones desde adentro, como es el caso de los pulmones y de los vasos sanguíneos y forma algunos ligamentos como los ligamentos amarillos de la columna vertebral.
Figura 5 Figura 6
Figura 3 Figura 4

26
Tejido adiposoEs un tejido conjuntivo especializado en el que predominan las células conjuntivas llamadas adipocitos. Los lipoblastos, células precursoras de adipocitos producen cantidades importantes de
colágeno I y III, pero los adipocitos adultos secretan muy bajas cantidades de colágeno y pieden la capacidad de dividirse (Figs.1 y 2).
El tejido adiposo es uno de los tejidos más abundantes y representa alrededor del 15-20% del peso corporal del hombre y del 20-25% del peso corporal en mujeres. Los adipocitos almacenan energía en forma de triglicéridos. Debido a la baja densidad de estas moléculas y su alto valor calórico, el tejido adiposo es muy eficiente en la función de almacenaje de energía.
Los adipocitos diferenciados pierden la capacidad de dividirse; sin embargo, son células de una vida media muy larga y con capacidad de aumentar la cantidad de lípidos acumulados. Además, el tejido adiposo postnatal contiene adipocitos inmaduros y precursores de adipocitos residuales a partir de los cuales pueden diferenciarse adipocitos adicionales. Estos mecanismos se hacen operativos cuando la ingasta calórica aumenta exageradamente.
El tejido adiposo se clasifica en adiposo unilocular y el tejido adiposo multilocular, de acuerdo a las características de las células que lo constituyen.
Tejido adiposo unilocular (Fig.3).Corresponde a la variedad de tejido adiposo mas corriente en adultos. Sus células son polihédricas, miden entre 50 y 150 Um de diámetro y contienen una sola gota de lípido que llena todo el citoplasma
desplazando los organelos hacia la periferia. Al microscopio de luz cada célula aparece como un pequeño anillo de citoplasma rodeando una vacuola, resultado de la disolución de la gota lipídica, y que contiene un núcleo excéntrico y aplanado (Fig. 4).
Figura 1 Figura 2
Figura 3 Figura 4

27
El MET revela que cada célula adiposa contiene sólo una gota de lípido. En el citoplasma perinuclear se ubican un Golgi pequeño, escasas mitocondrias de forma ovalada, cisternas de RER poco desarrolladas y ribosomas libres. En el citoplasma que rodea la gota de lípido contiene vesículas de REL, algunos microtúbulos y numerosas vesículas de pinocitosis.
Tejido adiposo multilocular.
Esta variedad de tejido adiposo es de distribución restringida en el adulto. Sus células son poligonales y más pequeñas que las del tejido adiposo unilocular. Su citoplasma contiene numerosas gotas de lípido de diferente tamaño y numerosas mitocondrias conabundantes crestas. Su un núcleo está al centra y es esférico.
Este tejido adiposo se asocia con numerosos capilares sanguíneos y se conoce tambien como grasa parda. En embriones humanos y en el recién nacido, este tipo de tejido adiposo se concentra en la región interescapular y luego en individuos adultos disminuye notablemente.
Cartílago y HuesoSon tejidos conjuntivos que se caracterizan porque sus células están rodeadas de una matriz intercelular sólida y relativamente rígida. La matriz intercelular del cartílago es deformable y puede crecer por depósito de nuevo material en su interior, mientras que la matriz intercelular ósea es más bien rígida y puede crecer sólo por depósito de nuevo material sobre las superficies óseas.
CartílagoEstá formado por una abundante matriz extracelular en la cuál los condrocitos se ubican en espacios llamados lagunas (Figs. 1 y 2).
Los condrocitos sintetizan y secretan los componentes
orgánicos de la matriz extracelular que son básicamente colágeno, ácido hialurónico. proteoglicanos y glicoproteínas, y según las características de la matriz se distingue cartílago hialino y fibroso. Existe además el cartilago elástico en el cuál la elastina forma parte de la matriz extracelular.
Los vasos sanguíneos no penetran a la matriz cartilaginosa y los condrocitos se nutren con material que difunde desde de los capilares sanguíneos del tejido conjuntivo adyacente.
Figura 1 Figura 2

28
Cada placa o lámina de tejido cartilaginoso está rodeada por el pericondrio que corresponde a tejido conjuntivo denso en el cuál se distingue una capa externa fibrosa y una capa interna celular en la cual se ubican las células que pueden dar origen a los condroblastos (Fig.3), que corresponden a precursoresde los condrocitos y que difieren de ellos solo en su edad y en su mayor actividad de la sintesis de componentes de la matriz intercelular cartilaginosa.
Los condroblastos presentan un ergastoplasma y un aparato de Golgi muy desarrollados y presentan vesículas y granos de secreción, lo cuál guarda relación con su rol de sintetizar y secretar los distintos
componentes de la matriz extracelular cartilaginosa (Fig. 4) que está formada principalmente por colágeno de tipo II, proteoglicanos de condroitin y keratansulfato, ácido hialurónico y glicoproteínas.
Al disminuir su actividad de síntesis disminuye el desarrollo tanto del ergastoplasma como del aparato de Golgi, acumulan glicógeno y lípidos en su citoplasma y se les llama condrocitos (Fig. 5).
Histogénesis del cartílagoEl tejido cartilaginoso se origina en el mesenquima, a partir de células mesenquimáticas que se redondean y agrupan en conglomerados con escaso material intercelular entre ellas. Este conjunto de células precartilaginosas se llama blastema . Las células del blastema son inducidas a sintetizar matriz cartilaginosa y a partir de ese momento se les llama condroblastos, ellas se separan progresivamente a medida que aumenta la cantidad de matriz sintetizada y pasan a llamarse condrocitos. El tejido mesenquimático que rodea a la masa condrogénica pasará a constituir el pericondrio.
Crecimiento del cartílago
Las placas de tejido cartilaginoso pueden aumentar su volumen mediante dos mecanismos:
Figura 3
Figura 4 Figura 5

29
Crecimiento por aposición
Ocurre desde el pericondrio, en cuya capa celular se localizan células indiferenciadas capaces de dividirse dando origen células
que se diferenciaran a condroblastos y que producirán tejido cartilaginoso sobre la superficie del cartílago preexistente, quedando los condroblastos atrapados en la meriz que producen y pasando a ser condrocitos (Figs. 1 y 2).
Crecimiento intersticial
Ocurre porque los condrocitos son capaces de dividirse y la matriz cartlaginosa es distensible. Las células hijas ocupan inicialmente la misma laguna pero a medida que ellas secretan nueva matriz entercelular se van separando. Estas células hijas pueden volver a dividirse formándose los llamados grupos isógenos, que se
encuentran frecuentemente en cartílagos en crecimiento (Fig. 3, 4 y 5).
Figura 1 Figura 2
Figura 3 Figura 4
Figura 5

30
Tipos de CartílagoMatriz intercelular del cartílago hialino
Cartílago elástico
Cartílago fibroso o fibrocartílago
Matriz intercelular del cartílago hialinoEl colágeno corresponde a alrededor del 40% de los componentes orgánicos de la matriz cartilagiosa. Estáorganizado principalmente como fibrillas de colágeno II que se disponen como un red laxa en toda la
matriz del cartílago hialino.
A lo largo de las fibrillas de colágeno II se asocian moléculas de colágeno IX (a1( IX ) a2( IX ) a3( IX )), de modo que uno de los extremos de cada molécula se proyecta hacia la matriz extracelular circundante (Fig.1).
Los principales glicosaminoglicanos corresponden a ácido hialurónico y a proteoglicanos de condroitin y queratán-sulfato. En la matriz cartilaginosa el ácido hialurónico se asocia a entre 80 a 200 unidades de proteoglicanos, por medio de proteínas de enlace (Fig.2).
Los enormes conglomerados de hialuronato están unidos a las fibrillas colágenas por enlaces electrotáticos y puentes cruzados glicoproteicos.
Más del 60% del peso neto del cartílago hialino corresponde a agua, gran parte de la cual interactúa con los glicosaminoglicanos, encontrándose parcialmente fijada, sin embargo existen zonas en que la interacción es lo
suficientemente laxa como para permitir el flujo de solutos por la matriz cartilaginosa. La gran hidratación y la posibilidad de movimiento de agua son los dos factores que
Figura 1 Figura 2
Figura 3 Figura 4

31
permiten al cartílago su capacidad para soportar la compresión y su recuperación ante la aplicación de cargas variables, como ocurre en los cartílagos articulares.
Los proteoglicanos se concentran se en la cápsula o matriz territorial que rodea a cada laguna y de hecho los grupos isógenos están en conjunto rodeados de una zona rica en proteoglicanos, en la cuál son escasas la fibrillas de colágeno II (matriz territorial ), a diferencia de lo que ocurre en la matriz localizada entre las células cartilaginosas (matriz interterritorial ) (Figs. 3 y 4).
La adhesión entre los condrocitos y ma matriz que los rodea es estabilizada por la condronectina, glicoproteína que se asocia a receptores en la mebrana plasmpatica de las células y a los componentes de la matriz territorial.
El cartílago hialino forma el esqueleto provisional durante el desarrollo, las placa epifisiarias durante el crecimiento de los huesos (Figs. 5, 6 y 7), reviste las superficies articulares
en las articulaciones y forma parte de la pared en las grandes vías respiratorias.
Cartílago elástico
Su estructura es parecida a la del cartílago hialino, con una capa de pericondrio asociado y los condrocitos rodeados de la matriz intercelular, pero en su matriz existen
además láminas o fibras elásticas (Fig. 1) las cuales se concentranen la matriz interterritorial (Fig. 2).
Figura 5 Figura 6
Figura 7
Figura 1 Figura 2

32
Cartílago fibroso o fibrocartílagoContiene condrocitos, generalmente encapsulados en una matriz intercelular parecida a la del cartílago hialino, pero con manojos de fibrillas de colágeno I, orientados en diversas direcciones, ocupando la matriz
intercelular (Figs. 1 y 2).
HuesoEstructura y características
Organización histológica del hueso
Histogénesis del hueso
Estructura y características del tejido oseo
Figura 1 Figura 2
Figura 1 Figura 2
Figura 3 Figura 4

33
El tejido óseo es una variedad de tejido conjuntivo que se caracteriza por su rigidez y su gran resistencia tanto a la tracción como a la compresión
Está formado por la matriz ósea, que es un material
intercelular calcificado (Fig.1) y por células, que pueden corresponder a:
osteoblastos: encargados de sintetizar y secretar la parte orgánica de la matriz ósea durante su formación (Fig.2). Se ubican siempre en la superficie del tejido óseo ya que este sólo puede crecer por aposición)
osteocitos, responsables de la mantención de la mátriz ósea (Fig. 3), que se ubican en cavidades o lagunas rodeadas por el material intercelular calcificado. La nutrición de los osteocitos depende de canalículos que penetran la matriz ósea y conectan a los osteocitos vecinos entre sí y con canales vasculares que penetran al hueso (Fig. 4) o que se ubican en las membranas conjuntivas que revisten la superficies del hueso (periostio y endostio) (Fig. 5). De hecho ningun osteocito se encuentra a más de una fracción de mm de un capilar sanguíneo.
osteoclastos, células responsables de la reabsorción del tejido óseo, que participan en los procesos de remodelación de los huesos y pueden encontrarse en depresiones superficiales de la matriz ósea llamadas lagunas de Howship (Fig. 5)
La matriz intercelular oseaLa matriz intercelular ósea está formada por:
Matriz orgánica u osteoide que corresponde al 50% del peso seco del hueso (Fig.1).
Más del 90% de ella corresponde a fibrillas de colágeno I organizadas en laminillas de unos 5 um de grosor (Fig. 2).
Figura 5
Figura 1 Figura 2

34
En cada laminilla ósea, las fibrillas colágenas están paralelas entre si, pero las laminillas sucesivas alternan ordenadamente la orientación de sus fibrillas en ángulos rectos.. Esta disposición alternada de las fibrillas colágenas en laminillas sucesivas destaca particularmente al observar cortes de hueso con microscopía de luz polarizada (Fig. 3).
El resto de los componentes orgánicos son principalmente glicoproteínas como la osteonectina, proteínas ricas en ácido g-carboxiglutámico como la
osteocalcina, y proteoglicanos de condroitín y queratán-sulfato. Son moléculas ricas en grupos ácidos con gran tendencia a asociarse entre sí, capaces de unirse a calcio y que juegan un rol importante en el proceso de mineralización de la matriz ósea.
Sales minerales inorgánicas depositadas en el osteoide, que confieren al tejido su rigidez y dureza y actuan como una reserva de sales minerales, sensible a estímulos endocrinos.
Las más abundantes son fosfato de calcio amorfo y cristales de hidróxidos de calcio y de fosfato llamados hidroxiapatita (Ca10(PO4)6(OH)2). Los cristales de hidroxiapatita son aplanados (30nm por 3 nm) y se adosan a lo largo de las fibrillas colágenas, a intervalos de unos 67 nm.
La superficie del cristal está hidratada y existe una vaina de agua e iones rodeándolo, lo que facilita el intercambio de iones entre el cristal y el líquido intersticial.
Células del tejido oseoCélulas osteoprogenitoras
Osteoblastos
Osteocitos
Células de revestimiento oseo
Osteoclastos
Células osteoprogenitorasCelulas osteoprogenitoras: son células alargadas con citoplasma poco prominente, que proceden de las células mesenquimáticas primitivas y forman una población de células troncales capaces de dividirse y dar origen a células que se diferencian a osteoblastos (Figs.1 y 2).
Figura 3

35
OsteoblastosLos osteoblastos son células diferenciadas que sintetizan el colágeno y la sustancia fundamental ósea.
Cuando están en plena actividad (Fig. 1) su forma es cuboide con un
citoplasma basófilo, que refleja su ergastoplasma muy desarrollado, tienen además un prominente aparato de Golgi. Son células polarizadas que vacian sus productos de secreción por la zona del citoplasma en contacto con la matriz ósea ya formada, (matriz intercelular preósea o osteoide) (Fig. 2).
Los osteoblastos son células polihédricas con largas y delgadas prolongaciones citoplasmáticas que son rodeadas por el osteoide producido y que se asocian mediante uniones del comunicación (nexos o gap junction) con prolongaciones similares de los osteoblastos vecinos (Fig. 3).
Los osteoblastos participan también en el proceso de mineralización de la matriz órgánica ya que producen vesículas de matriz, de unos 100nm de diámetro, rodeadas de membana celular, las que acumulan Ca++
y PO4= y son ricas además en fosfatasa alcalina y pirofosfatasa, enzimas que capaces de generar iones PO4=, elevando su concentración en el medio extracelular y creando centros de nucleación para el deposito de las sales minerales.
Los osteoblastos tienen dos destinos posibles: ser rodeados por la matriz ósea que producen y pasan a ser osteocitos o permanecer en la superficie del tejido óseo recién formado, aplanándose y constituyendo las células de revestimiento óseo.
Figura 1 Figura 2
Figura 1 Figura 2
Figura 3

36
OsteocitosSon las células propias del tejido óseo formado. Su citoplasma presenta ergastoplasma, aparato de Golgi y puede presentar lisosomas.
Son capaces de sintetizar y de reabsorber, en forma limitada, a componentes de la matriz ósea ("osteolisis osteocítica"), procesos que tienen importancia en la regulación de la calcemia. Cada osteocito, bañado por una delgada capa de
líquido tisular, ocupa su laguna (Fig 1) y proyecta sus prolongaciones citoplasmáticas por los canalículos (Fig. 2), dentro de los cuales, toma contacto con prolongaciones de osteocitos vecinos mediante uniones de comunicación (Fig. 3), o con células de revestimiento óseo bañadas por el líquido tisular del tejido conjuntivo que rodea a loscapilares sanguíneos (Fig. 4).
Células de revestimiento oseo
Figura 1 Figura 2
Figura 3 Figura 4
Figura 1 Figura 2

37
Son células aplanadas que revisten las superficie óseas del interior de los huesos, y en su mayor parte corresponden a osteoblastos inactivos(Fig. 1) que están asociados entre sí y con
prolongaciones de los osteocitos (Figs. 2 y 3).
OsteoclastosSon células móviles, gigantes y multinucleadas y se localizan adosadas a la superficie de tejido óseo que debe ser removido (Fig.1).
Se originan por fusión de monocitos que han abandonado
la sangre circulante y pertenecen de hecho al sistema de fagocitos mononucleares. Contienen numerosos lisosomas y en la superficie del osteoclasto que esta en contacto con la matriz a remover se distinguen dos zonas distintas (Fig. 2): un anillo externo o zona clara que corresponde a una especialización de la superficie celular en que la membrana se asocia estrechamente al hueso que delimita el área que se va a reabsorver, y la región central o borde estriado que presenta profundos repliegues de la superficie celular bajo los cuales de concentran gran cantidad de lisosomas y mitocondrias, además de ergastoplasma y cisternas del Golgi.
La membrana de los lisosomas primarios se fusiona con la membrana celular que reviste a los repliegues del borde festoneado, liberando las enzimas lisosomales hacia el exterior y produciendo una acidificación del microambiente que baña al tejido óseo a reabsorver. El borde festoneado contiene además gran cantidad de vesículas endocíticas y lisosomas secundarios, indicando que ocurre además un proceso de fagocitosis del
Figura 3
Figura 1 Figura 2

38
material parcialmente solubilizado por el ambiente ácido y la acción lítica de la enzimas lisosomales.
Organización histológica de los huesosEn un hueso ya formado la disposición de los componentes del tejido óseo en cada zona de él está en estricta relación con las fuerzas de tracción y compresión a que esa zona está sometida habitualmente.
Sin embargo el tejido óseo que se forma primero en el feto y luego en los huesos en desarrollo no se dispone de acuerdo a estos requerimientos mecánicos, sino que constituye un tejido óseo de base llamado hueso primario o reticular ("entrelazado") que posteriormente será reemplazado por el hueso secundario o laminillar
La principal diferencia entre el hueso reticular y el laminillar radica en la disposición de las fibrillas de colágeno I, las cuales en el hueso primario se disponen en forma de manojos dispuestos en forma irregular.
De acuerdo al aspecto macroscópico que presentan se distinguen dos tipos de huesos: hueso cortical formado por tejido óseo compacto (Fig.1) y hueso esponjoso (Fig.2) en el cual el tejido óseo se dispone en trabéculas que delimitan cavidades, en las que se ubica normalmente la médula ósea.
En las superficies articulares la capa cortical de huesocompacto está cubierta por una capa de cartilago hialino: el cartílago articular (Fig. 3), mientras que el resto de la superficies del hueso están cubiertas por
membranas de tejido conjuntivo que forman el periostio (Fig. 4) y el endostio, en los que existen abundantes vasos sanguíneos que se adosan ya sea a las trabéculas del hueso esponjoso o que penetran en el hueso cortical compacto a través de los conductos de Volkman y de Havers (Fig. 5).
Figura 1 Figura 2
Figura 3 Figura 4

39
Estructura del hueso esponjoso o trabecular
Están formados por delgadas trabéculas (Fig. 1), que, en los huesos ya formados, corresponden a tejido óseo laminillar (Figs. 2 y 3) rodeadas por células de revestimiento óseo.Los osteocitos de las trabéculas óseas se mantienen desde los capilares sanguíneos vecinos ubicados en la médula ósea.
La orientación de cada trabécula y la disposición de sus laminilllas óseas
mineralizadas está determinada por las tensiones mecánicas locales con el objeto de resistir las compresiones y las tensiones a que es sometida.
Si cambia la dirección de los requerimienos mecánicos a que es sometida una trabécula (aumento de peso del individuo, crecimiento u otro) ella debe cambiar su orientación.
Figura 5
Figura 1 Figura 2
Figura 3

40
Remodelación de trabéculas en el hueso esponjoso: El cambio en la forma de una trabécula se realiza mediante dos procesos coordinados que ocurren en zonas precisas de la trabécula a remodelar:
crecimiento por aposición de la trabécula por acción de los osteoblastos los que que depositan laminillas sucesivas de tejido óseo en la zona en que existe una requerimiento mecánico, y
remoción del tejido óseo de la trabécula por acción de los osteoclastos en los sitios en que no está sometido a tracción ni compresión (Figs. 4 y 5)
Organización del hueso compacto
En la diáfisis de los hueso largos (Figs. 1, 2 y 3) el tejido óseo laminillar se organiza en tres formas distintas, formando los: sistemas de Havers, sistemas laminillares interticiales, y sistemas circunferenciales externo (subperióstico) e interno (subendóstico) (Fig. 4).
Figura 4 Figura 5
Figura 1 Figura 2
Figura 3 Figura 4

41
Cada sistema de Havers ("osteón") consiste en un largo cilindro hueco cuyas paredes están formadas por un número variable de laminillas óseas concéntricas (Fig. 5), dispuestas de tal modo que las fibrillas colágenas paralelas de cada laminilla están orientadas en forma perpendicular a la de las laminillas vecinas, (Fig. 6), entre las laminillas disponen las lagunas, que contienen los osteocitos, con los canalículos dirigidos
de preferencia en forma radial hacia la cavidad central que corresponde al conducto de Havers el cual se comunica con los conductos de Volkman y contiene vasos sanguíneos y nervios, rodeados de escaso tejido conjuntivo laxo (Figs. 7 y 8).
La forma en que se orientan los sistemas de Havers en cada zona del hueso compacto guarda estricta relación con los requerimientos mecánicos a que el hueso está sometido. Si cambian los requerimientos mecánicos se produce su reabsorción parcial o total y la formación de nuevos sistemas de Havers, proceso conocido como remodelación interna del hueso compacto o haversiano.
Los sistemas de laminillas interticiales son grupos de laminillas paralelas de forma irregular que ocupan los espacios entre sistemas de Havers y sistemas circunferenciales (Fig. 9).
Ellos corresponden a restos de sistemas deHavers que han sido parcialmente removidos durante los procesos de crecimiento y de remodelación ósea (Fig. 10).
Figura 5 Figura 6
Figura 7 Figura 8
Figura 9 Figura 10

42
Los sistemas de laminillas circunferenciales interno y externo se observan en los huesos que han terminado su proceso de crecimiento y consisten en laminillas óseas paralelas entre sí que forman bandas de grosor variable bajo el periostio y el endostio, recibiendo sus osteocitos su nutrición desde vasos sanguíneos del tejido conjuntivo (Fig. 11).
Remodelación interna del hueso compacto
En la unidad de remodelación ósea del hueso compacto existen un cono perforante que avanza y un cono de cierre que le sigue.
El cono perforante o conducto de reabsorción contiene osteoclastos activos adyacentes a los cuales avanzan removiendo el tejido óseo que no está respondiendo a tracción ni compresión y van formando un conducto de unos 200 mma de diámetro,
expensas de partes de sistemas de Havers existentes y porciones de sistemas de laminillas intersticiales el cual va siendo ocupado por un asa capilar con pericitos y rodeada de células preosteoblásticasos. El diámetro de este conducto establece el diámetro del nuevo sistema de Havers que se va a formar en él. Las células preosteoblásticas se diferencian a osteoblastos y comienzan a sintetizar la matriz orgánica del hueso (osteoide) y a depositarla sobre las paredes del conducto en laminillas sucesivas. Posteriormente la matriz ósea de cada laminilla se mineraliza. A medida que avanza el depósito de laminillas hacia el centro del conducto los osteoblastos van quedando tarapados en la matriz ósea, transformándose en osteocitos y el diámetro del conducto se va estrechando hsta alcanzar el diámetro del conducto de Havers maduro, el cuál contiene capilares, células de revestimiento óseo y escaso conjuntivo laxo (Figs. 1, 2, 3, 4).
Figura 11
Figura 1 Figura 2
Figura 3 Figura 4

43
Crecimiento de los huesosCrecimiento en grosor
Crecimiento en longitud de los huesos largos
Aumento en diámetro de la diáfisis
Crecimiento en grosor
Ocurre a partir de las células presentes en la capa interna o celular del periostio en la cual existen, adosadas a la capa externa o fibrosa, células osteoprogenitoras las cuales al dividirse originan células quese diferencian a osteoblastos. Los osteoblastos se disponen en la superficie externa del hueso y sintetizan matriz ósea orgánica, la cual poseriormente se mieraliza. Los
osteblastos atrapados pasan a formar los osteocitos del nuevo tejido óseo (Figs. 1 y 2). Mientras ocurre el crecimiento en grosor no se observa el sistema laminillar subperióstico (Fig. 3) el cual está bien desarrollado en el hueso adulto (Fig. 4)
Figura 1 Figura 2
Figura 3 Figura 4

44
Crecimiento en longitudOcurre a expensas del cartílago de la placa epifisaria (Fig.1) el cual por prolifera aumentando su longitud por crecimiento intesticial y luego se modifica y es reemplazada por hueso de acuerdo a los pasos del proceso de
osificación endocondral (Fig.2).
En la (Fig. 3) se muestran los cambios que experimenta el tejido cartilaginoso durante este proceso. El cartílago hialino normal que se observa en la zona de reposo experimenta luego (bajo en control de la hormona somatotropa) un proceso de multiplicación celular, formando columnas paralelas de células apladas en el sentido longitudinal del hueso, tal como se observa en la zona de cartílago seriado.
Posteriormente los condrocitos aumentan notablemente su tamaño, acumulando glicógeno y grasa y produciendo colágeno X y fosfatasa alcalina. La matriz cartilaginosa queda reducida a tabiques delgados, tal
como se observa en la zona de cartílago hipertrófico (Fig. 4) y posteriormente ocurre la calcificación de la matriz cartilaginosa (Fig. 5), debida a la alteración en su composición por la presencia del colágeno X y de fosfatsa alcalina.
Figura 1 Figura 2
Figura 3
Figura 4 Figura 5

45
Luego ocurre la muerte de los condrocitos hipetrofiados, aislados en la matriz calcificada. La matriz calcificada es penetrada por vasos sanguíneos que provienen desde el canal medular y que
se acompañan de osteoblastos los cuals se adosan a la supeficie de la matriz cartilaginosa calcificada y sintetizan y secretan la matrís ósea orgánica (osteoide). Los osteoclastos que también acompañan a los vasos sanguineos remueven parte de los tabiques de matriz cartilaginosa calcificada formando grandes cavidades delimitadas por espículas de matriz cartilaginosa que permanecen unidas al cartílago hepifisario y que están recubiertas por tejido óseo primario (Fig. 6) el cual se hace cada vez más grueso al continuar siendo depositado por los osteoblastos (Fig. 7).
En la Figura 8 se ilustra como este proceso permite alargar la diáfisis del hueso por desplazamiento de la epífisis. Las espículas ósea con base cartilaginosa pueden fusionarse con el hueso cortical y posteriormente experimentar remodelación ósea interna, formándose sistemas de Havers.
Figura 6 Figura 7
Figura 8

46
Aumento en diámetro de la diáfisis
Este proceso va acompañado generalmente de un aumento también en el diámetro de la cavidad medular razón por la cuál ocurre de hecho un desplazamiento lateral del hueso cortical con un aumento proporcionalmete reducido del grosor de la pared ósea misma.
Esto implica la acción coordinada de tres procesos:
crecimiento en grosor del hueso a expensan de la capa osteogénica del periostio (Fig. 1).
absorción desde el endostio, por acción de osteoclastos (Fig. 2).
remodelación interna del hueso cortical para establecer una distribución de sistemas de Havers adecuada al requerimiento mecánico del hueso en cada etapa de su vida (Figs. 3, 4 y 5).
Figura 1 Figura 2
Figura 3 Figura 4
Figura 5

47
Histogénesis del huesoEl tejido óseo se desarrolla siempre por sustitución de un tejido conjuntivo preexistente.
Osificación intramembranosa es la forma en la cual el feto forma básicamente los
huesos planos. Tiene lugar en condensaciones de tejido mesenquimatico muy vascularizado en las cuáles ciertas células mesenquimáticas se dividen formándose el blastema óseo. A partir de estas células se forman los osteoblastos que sintetizan y secretan al osteoide el cuál posteriormente se mineraliza y que va englobando a los osteoblastos los que se transforman en osteocitos (Figs. 1 y 2). Las trabéculas de hueso primario así formada son posteriormente remodeladas de acuerdo al hueso que pertenezcan.
Osificación endocondral
Es la forma en la cual el feto forma los huesos largos y cortos.
El proceso se produce en la siguiente forma (Figs. 3 y 4):
se desarrolla un modelo cartilaginoso que adopta una forma parecida a la del hueso que va a dar
origen y que está rodeado de su pericondrio en la zona media de la diáfisis se desarrollan en el pericondrio células
osteoprogenitoras y de ellas se originan osteoblastos los que producen un collar óseo subperiostico mediante un proceso de osificación directa o membranosa.
simultáneamente ocurren en el cartílago subyacente, modificaciones similares a los descritos previamente en el cartílago epifisario, formándose un centro de osificación endocondral primario (diafisario). Las epículas óseas formadas sobre restos de la matriz
Figura 1 Figura 2
Figura 3 Figura 4
Figura 5

48
cartilaginosa calcificada se unen al mango de hueso cortical que sigue engrosándose a partir de la capa osteógena del periostio.
Posteriormente se forman centros de osificación secundario a nivel del cartílago de las epífisis (Fig. 5) a partir del cual se formará el hueso esponjoso y cesará primero la osificación endocondral del cartílago hacia las epífisis.
El cartílago que permanece constituye la placa cartilaginosa epifisaria o cartilago de crecimiento de los huesos ya formados.















![Gartner, Leslie P. - Texto Atlas de Histologia, 2da Edición [6 Tejido Conectivo]](https://static.fdocuments.mx/doc/165x107/55cf863b550346484b95866e/gartner-leslie-p-texto-atlas-de-histologia-2da-edicion-6-tejido-conectivo.jpg)