
Genes en poblaciones - fcvinta.files.wordpress.com · mendeliana y extenderlos del nivel del...
12
1 CAPITULO 4 Genes en poblaciones Los principios mendelianos explican los mecanismos genéticos en individuos. Sin embargo, como criadores, nuestra labor no es cambiar a los individuos, sino las poblaciones. Entonces debemos tomar los conocimientos sobre la herencia mendeliana y extenderlos del nivel del individuo al nivel de las poblaciones. FRECUENCIAS GÉNICAS Y GENOTÍPICAS Al describir un individuo para un carácter de herencia simple, nos referimos a genes específicos que posee el individuo, o podemos describir su genotipo de uno o dos locus. Por ejemplo, podemos referirnos a la gallina Andaluza azul como si tuviera ambos alelos negro (B), o blanco (b) en un locus afectando el color de las plumas, o podemos decir que 1 pollo tiene un genotipo heterocigoto Bb en ese locus. Sin embargo ¿Cómo describimos una población? ¿Cómo describimos un lote completo de gallinas andaluzas azules? La respuesta es utilizar las frecuencias de genes y de genotipos. La frecuencia génica o frecuencia alélica es la frecuencia relativa de un alelo particular en una población. Es una medida de cuan común es el alelo relativo a otros alelos que puede haber en ese locus. Las frecuencias relativas van de cero a uno. Por ejemplo, si un alelo no existe en una población, su frecuencia es cero. Si es el único alelo en su locus en la población, su frecuencia génica es uno. Si abarca el 35 % de los genes en el locus en la población, su frecuencia génica es .35. Cuando hay solo dos alelos posibles en un locus, la frecuencia del alelo “dominante” es comúnmente representada por la letra minúscula p y la frecuencia del alelo “recesivo” por la letra minúscula q. (Los términos dominante y recesivo están puestos entre comillas aquí porque hay situaciones en que ningún alelo es dominante. En casos de ausencia de dominancia, la asignación de p o q para referirse a la frecuencia de un alelo en particular es arbitraria). Como un ejemplo considere un lote de 100 gallinas andaluzas. 36 son negras (BB), 44 son azules (Bb, en realidad gris), 20 son blancas (bb). En el locus que afecta el color de las plumas de las andaluzas hay un total de 200 genes en esta población – dos genes por cada uno de los 100 individuos. Los 36 individuos negros, cada uno tiene 2 genes, los 44 azules cada uno tiene un gen negro, y los blancos no tienen genes negros. La cantidad total de genes negros en el lote es por lo tanto 2 x 36 + 44 =116, y la frecuencia génica del alelo negro es entonces 116 de 200 o en forma decimal, 0.58. Igualmente, no hay genes blancos en las gallinas negras, 44 genes blancos en las azules y 40 genes blancos en los individuos blancos, un total de 84 genes blancos en el lote. La frecuencia génica del alelo blanco es entonces 84 de 200 o 0.42. En cada caso
Transcript of Genes en poblaciones - fcvinta.files.wordpress.com · mendeliana y extenderlos del nivel del...

1
CAPITULO 4
Genes en poblaciones
Los principios mendelianos explican los mecanismos genéticos en individuos. Sin
embargo, como criadores, nuestra labor no es cambiar a los individuos, sino las
poblaciones. Entonces debemos tomar los conocimientos sobre la herencia
mendeliana y extenderlos del nivel del individuo al nivel de las poblaciones.
FRECUENCIAS GÉNICAS Y GENOTÍPICAS
Al describir un individuo para un carácter de herencia simple, nos referimos a genes
específicos que posee el individuo, o podemos describir su genotipo de uno o dos
locus. Por ejemplo, podemos referirnos a la gallina Andaluza azul como si tuviera
ambos alelos negro (B), o blanco (b) en un locus afectando el color de las plumas, o
podemos decir que 1 pollo tiene un genotipo heterocigoto Bb en ese locus. Sin
embargo ¿Cómo describimos una población? ¿Cómo describimos un lote completo de
gallinas andaluzas azules? La respuesta es utilizar las frecuencias de genes y de
genotipos.
La frecuencia génica o frecuencia alélica es la frecuencia relativa de un alelo particular
en una población. Es una medida de cuan común es el alelo relativo a otros alelos que
puede haber en ese locus. Las frecuencias relativas van de cero a uno. Por ejemplo, si
un alelo no existe en una población, su frecuencia es cero. Si es el único alelo en su
locus en la población, su frecuencia génica es uno. Si abarca el 35 % de los genes en el
locus en la población, su frecuencia génica es .35.
Cuando hay solo dos alelos posibles en un locus, la frecuencia del alelo “dominante” es
comúnmente representada por la letra minúscula p y la frecuencia del alelo “recesivo”
por la letra minúscula q. (Los términos dominante y recesivo están puestos entre
comillas aquí porque hay situaciones en que ningún alelo es dominante. En casos de
ausencia de dominancia, la asignación de p o q para referirse a la frecuencia de un
alelo en particular es arbitraria).
Como un ejemplo considere un lote de 100 gallinas andaluzas. 36 son negras (BB), 44
son azules (Bb, en realidad gris), 20 son blancas (bb). En el locus que afecta el color de
las plumas de las andaluzas hay un total de 200 genes en esta población – dos genes
por cada uno de los 100 individuos. Los 36 individuos negros, cada uno tiene 2 genes,
los 44 azules cada uno tiene un gen negro, y los blancos no tienen genes negros. La
cantidad total de genes negros en el lote es por lo tanto 2 x 36 + 44 =116, y la
frecuencia génica del alelo negro es entonces 116 de 200 o en forma decimal, 0.58.
Igualmente, no hay genes blancos en las gallinas negras, 44 genes blancos en las azules
y 40 genes blancos en los individuos blancos, un total de 84 genes blancos en el lote.
La frecuencia génica del alelo blanco es entonces 84 de 200 o 0.42. En cada caso

2
hemos simplemente contado el número de genes de un tipo particular y dividido por el
total de genes en un locus en la población. Las ecuaciones para las frecuencias génicas
en nuestro ejemplo de las andaluzas pueden ser escritas así:
Note que p + q = 1. Esto siempre será cierto si hay solo dos alelos posibles en un locus.
Si hay múltiples alelos en un locus, entonces la suma de las frecuencias génicas debe
ser igual a 1. Por ejemplo, si los alelos posibles fueran B, b y b’ debemos llamar las
frecuencias génicas de estos alelos p,q,r, respectivamente. Entonces p + q + r = 1.
La frecuencia genotípica es la frecuencia relativa de un genotipo particular de un locus
en una población. Las letras mayúsculas son utilizadas para anotar las frecuencias
genotípicas. Con sólo dos alelos en un locus. P se refiere a la frecuencia genotípica del
genotipo homocigota “dominante”, H se refiere a la frecuencia del genotipo
heterocigoto y Q se refiere a la frecuencia del genotipo homocigota “recesivo”.
En nuestro lote de andaluzas, hay 36 genotipos BB, 44 genotipos Bb y 20 genotipos bb
de un total de 100 individuos. Por lo tanto:
Para calcular frecuencias genotípicas, simplemente cuente el número de individuos en
un genotipo particular y divide por el número total de individuos en una población.
Note que P+H+Q = 1. Esto siempre será cierto si hay solo tres genotipos posibles en un
locus. Si hay más de tres genotipos posibles, entonces la suma de las frecuencias
genotípicas de cada genotipo tiene que ser igual a 1. Por ejemplo si los alelos posibles
fueran B,b y b’ con posibles genotipos de un locus BB,bb, b’b’, Bb, Bb’ y bb, podríamos
llamar las frecuencias genotípicas P,Q,R,H (Bb) ,H( Bb’), y H (bb’),respectivamente.
Entonces P+Q+R+H (Bb)+H( Bb’)+H (bb’) = 1.
Hay varios factores que afectan las frecuencias génicas y genotípicas en una población,
entre ellos, las herramientas básicas para el mejoramiento animal: selección y sistemas
de apareamientos. El estudio de estos factores compone la rama de la genética
conocida como genética poblacional.

3
frecuencia génica o frecuencia alélica: es la frecuencia relativa de un alelo particular
en una población.
frecuencia genotípica: es la frecuencia relativa de un genotipo particular de un locus
en una población.
genética poblacional: estudio de los factores que afectan las frecuencias génicas y
genotípicas en una población.
EL EFECTO DE LA SELECCIÓN SOBRE LAS FRECUENCIAS GÉNICAS Y
GENOTÍPICAS
La selección aumenta la frecuencia génica de los alelos favorables. Cuando
seleccionamos animales de reemplazo, tratamos de seleccionar aquellos que tiene el
mejor conjunto de genes y rechazar aquellos con conjuntos de genes más pobres.
Como resultado, los miembros de la próxima generación deberían tener, en promedio,
un mejor conjunto de genes que los miembros de la población actual. Con la selección
continua, la generación siguiente debería tener incluso mejores genes, y así
sucesivamente. A través del tiempo, la selección para mejores y mejores genes causa
el incremento de la frecuencia de alelos más favorables en la población y el descenso
de alelos menos favorables.
Otra forma de decir “mejor conjunto de genes” es decir mejores valores de cría.
Cuando seleccionamos animales con mejores conjuntos de genes generación tras
generación y aumentamos la frecuencia de los alelos favorables en el proceso, lo que
estamos haciendo verdaderamente es aumentar el valor de cría promedio (y por lo
tanto, la performance promedio) de la población. Frecuencia de genes, valores de cría
medio y performance media, entonces están intrínsecamente vinculados. Si deseamos
incrementar el valor de cría promedio y performance media de una población a través
de la selección, necesariamente queremos cambiar la frecuencia génica.
El efecto inmediato de la selección es cambiar las frecuencias alélicas, pero las
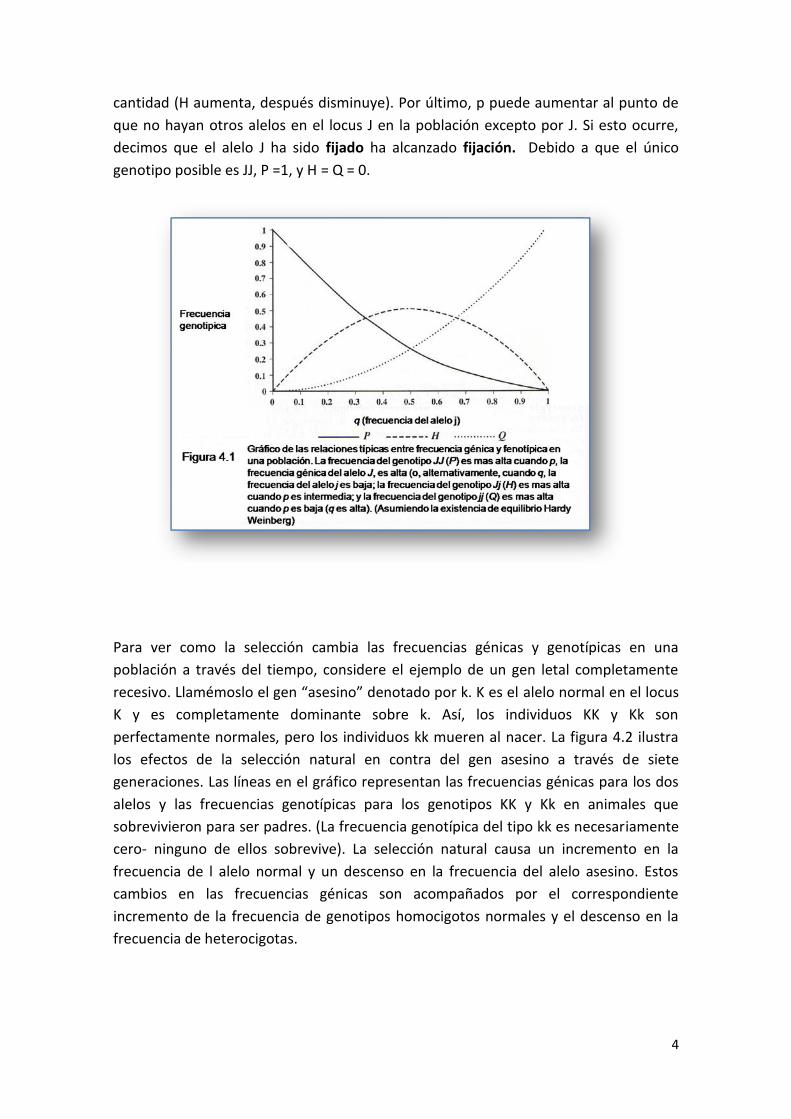
frecuencias genotípicas necesariamente lo hacen también La relación típica ente las
frecuencias génicas y genotípicas se muestran en la Figura 4.1. El eje horizontal
representa la frecuencia génica del alelo j. El eje vertical representa la frecuencia
genotípica de tres genotipos en el locus J. Hacia el lado derecho del gráfico, la
frecuencia del alelo j es alta, entonces la frecuencia del alelo J está necesariamente
baja (cercana a cero). Debido a que hay muy pocos genes J en la población, hay
también pocos genotipos JJ (P es baja), relativamente pocos genotipos Jj( H es bajo
también), y muchos genotipos jj (Q es alto). Si j es un alelo favorable, entonces con la
selección su frecuencia (q) aumentará, y a medida que nos movemos de la derecha a la
izquierda del gráfico, las frecuencias genotípicas también cambian – los genotipos JJ
serán más comunes (P aumenta), los genotipos jj serán menos comunes (Q decrece) y
los heterocigotas serán más comunes por un momento y después disminuirán en
4
cantidad (H aumenta, después disminuye). Por último, p puede aumentar al punto de
que no hayan otros alelos en el locus J en la población excepto por J. Si esto ocurre,
decimos que el alelo J ha sido fijado ha alcanzado fijación. Debido a que el único
genotipo posible es JJ, P =1, y H = Q = 0.
Para ver como la selección cambia las frecuencias génicas y genotípicas en una
población a través del tiempo, considere el ejemplo de un gen letal completamente
recesivo. Llamémoslo el gen “asesino” denotado por k. K es el alelo normal en el locus
K y es completamente dominante sobre k. Así, los individuos KK y Kk son
perfectamente normales, pero los individuos kk mueren al nacer. La figura 4.2 ilustra
los efectos de la selección natural en contra del gen asesino a través de siete
generaciones. Las líneas en el gráfico representan las frecuencias génicas para los dos
alelos y las frecuencias genotípicas para los genotipos KK y Kk en animales que
sobrevivieron para ser padres. (La frecuencia genotípica del tipo kk es necesariamente
cero- ninguno de ellos sobrevive). La selección natural causa un incremento en la
frecuencia de l alelo normal y un descenso en la frecuencia del alelo asesino. Estos
cambios en las frecuencias génicas son acompañados por el correspondiente
incremento de la frecuencia de genotipos homocigotos normales y el descenso en la
frecuencia de heterocigotas.

5
fijación: el punto en el que un alelo particular se convierte en el único alelo en su locus
en una población- la frecuencia del alelo se convierte en uno.
EL EFECTO DE LOS SISTEMAS DE APAREAMIENTO SOBRE LAS FRECUENCIAS
GÉNICAS Y GENOTÍPICAS
Los sistemas de apareamiento solos no pueden cambiar las frecuencias génicas en una
población, pero ocasionalmente un sistema de apareamiento, cuando es combinado
con la selección sí puede. Por ejemplo, los criadores que están interesados en
aumentar el tamaño corporal típicamente seleccionan los reemplazos disponibles más
grandes (machos y hembras). De esta base de animales, usualmente aparean los
machos mas grandes con la hembras más grandes para producir hijos especialmente
grandes, que luego se seleccionan para ser padres en la próxima generación. Cuando la
selección es combinada con este tipo de apareamiento las frecuencias de los alelos
que causan un tamaño corporal mayor aumentan más rápido en las generaciones
futuras, que en el caso de que la selección combinada con apareamientos
estrictamente al azar.
Sin embargo, el uso más común de los sistemas de apareamientos, es cambiar las
frecuencias genotípicas, específicamente incrementar el número de combinaciones de
genes homocigotas o el número de combinaciones heterocigotas. Las frecuencias
génicas pueden o no cambiar como resultado. Los sistemas de apareamiento

6
diseñados para afectar la homocigosis o heterocigosis caen bajo las categorías
generales de la consanguinidad y cruzamientos.
Consanguinidad
La consanguinidad, el apareamiento de parientes, aumenta la frecuencia de genotipos
homocigotos. Para ver porque, vea los pedigrí en la Figura 4.3. El de la izquierda (a) es
típico de animales con pedigrí – el pedigrí del padre compone la mitad superior, el
pedigrí de la madre la mitad inferior, y las generaciones más jóvenes aparecen más y
más a la izquierda. (En contraste, los pedigríes humanos están típicamente orientados
horizontalmente, las generaciones más viejas cercanas a la parte superior y las más
jóvenes cercanas a la parte inferior). El individuo X es consanguíneo porque el padre
(S) y la madre (D) son medio hermanos, medio hermano y media hermana. Ambos
padres tenían el mismo padre (A) y A es entonces considerado antecesor común de los
padres de X.
A (abuelo paterno) B
S (padre) B (abuela paterna) S
X X A
D (madre) A (abuelo
materno) D
C (abuela
materna) C
Figura 4.4 Pedigree y diagrama de flechas mostrando un apareamiento de medio
hermanos.
Ahora mire el pedigrí a la derecha (b). Este tipo de pedigrí es llamado diagrama de
flecha, este diagrama particular se corresponde con el pedigrí tradicional a su
izquierda. En los diagramas de flecha, los individuos pueden aparecer solo una vez, y
los ancestros que no contribuyen a la consanguinidad o relaciones genealógicas son
típicamente excluidos. (Se han incluido dos antecesores que no contribuyen, B y C, en
la figura 4.3 (b) solo para hacer la correspondencia entre dos tipos de pedigríes más
claros). Los diagramas de flecha representan esquemáticamente el flujo de genes
desde los antecesores a la descendencia.
Piense sobre el flujo de genes para un locus en particular de A hacia su descendencia.
Puede ver del diagrama de flecha que es posible para que S y D hereden copias
idénticas del mismo gen del antecesor común A, y para que X herede el gen de ambos
S y D. X entonces será homocigoto para ese gen. La chance de que esto finalmente
ocurra es uno en ocho. Considerando todos los loci, podemos esperar al menos 1/8 de
que los pares de genes del individuo consanguíneo sean homocigotos porque heredó
genes idénticos del antecesor común A de sus padres. (En realidad más de 1/8 de los
pares de genes de X deberían ser homocigotas. Muchos hubieran sido homocigotos

7
incluso sin consanguinidad). Entonces el resultado de la consanguinidad es un
incremento en la homocigosis y un descenso correspondiente de heterocigosis.
Medio hermanos: medio hermanos y hermanas.
Antecesor común: Antecesor común para más de un individuo. En el contexto del
mejoramiento, el término se refiere a un antecesor común de los padres de un
individuo consanguíneo.
Diagrama de flecha: Forma de pedigrí que representa esquemáticamente el flujo de
genes desde los antecesores hasta los descendientes.
Relaciones genealógicas: Relación entre animales debido al parentesco. Ejemplos
incluyen: hermanos completos, medio hermanos y relaciones padres hijos.
Exocría o Cruzamientos
La exocría o cruzamientos, el apareamiento de individuos no emparentados, tiene
justo el efecto opuesto de la consanguinidad. El cruzamiento aumenta la heterocigosis.
El siguiente ejemplo (bastante largo), va a demostrar el cambio en la heterocigosis
provocado por el cruzamiento, y en el proceso, va a introducir uno de los conceptos
más básicos en genética poblacional: el equilibrio Hardy- Weinberg.
Imagine dos poblaciones no emparentadas. Las frecuencias génicas en el locus B en las
poblaciones 1 y 2 son (recordar que p son los alelos dominantes y q los recesivos):
p1= 0.8 p2= 0.1
q1 = 0.2 q2 =0.9
Note que las frecuencias son muy diferentes en las dos poblaciones. Esto es evidencia
de que los dos grupos no están emparentados. En las poblaciones emparentadas se
espera que tengan frecuencias similares.
Ahora crucemos las poblaciones 1 y 2 para crear una nueva población de animales de
primera cruza – una generación F1. Los resultados de este cruzamiento se muestran
con un cuadrado de Punnett en la Figura 4.4. Las frecuencias genotípicas para los F1
aparecen en cada celda del cuadrado. Estas fueron calculadas simplemente
multiplicando las frecuencias génicas apropiadas de las poblaciones de los padres. Para
ver porqué, considere la frecuencia genotípica del genotipo homocigota BB en los
animales F1 (0.08). La frecuencia génica del alelo B en la población 1 es 0.8, y la
frecuencia génica del alelo B en la población 2 es 0.1.Por lo tanto, hay una chance del
80 % de que un hijo herede un gen B de la población 1 y un 10% de chance de que
herede un gen B similar de la población 2. La probabilidad de heredar dos genes B es el
producto de estas dos probabilidades individuales o 0.8 x0 .1 =0.08.

8
Después de combinar los valores de dos celdas heterocigotas del cuadrado de
Puennet, las frecuencias genotípicas en la población F1 son entonces
PF1 = 0.08
H F1 = 0.74 (0.72 + 0.02)
Q F1 =0.18
Las frecuencias génicas en la población F1 pueden ser derivadas de las frecuencias
genotípicas. Para hacerlo fácil, asuma que la población F1 contiene 100 individuos. Si
ocho de estos tienen dos genes B y 74 tienen un gen B, entonces de un total de 200
genes en el locus B en la población noventa (2 x 8 + 74), son genes B.
La frecuencia del aleo B es entonces 90 / 200 = 0.45. La frecuencia del alelo b es 1- 0.45
= 0.55. Así:
p F1 = 0.45
q F1 = 0.55
(El razonamiento utilizado para derivar frecuencias génicas de frecuencias genotípicas
es sintetizado en la fórmula p = P + ½ H. Se puede utilizar esta fórmula como un atajo)
Si los animales F1 son apareados entre ellos, el resultado del segundo cruzamiento o
generación F2 va a tener las frecuencias genotípicas mostradas en la Figura 4.5.
Nuevamente, sumanando los valores de las celdas heterocigotas, las frecuencias
genotípicas F2 son:
PF2 =0 .2025
H F2 = 0.495 (0.2475 + 0.2475)
Q F2 = 0.3025
Y las frecuencias génicas F2 son
p F2 = P F2 + H F2
=0 .2025 + (0.495)
= 0.45
q F2 = 1 - p F2
= 1 -0.45
= 0.55

9
Note que las frecuencias génicas no cambiaron de la generación F1 a la F2. Se
mantuvieron en 0.45 y 0.55. Debido a que las frecuencias génicas se mantuvieron
constantes, y debido a que las frecuencias genotípicas en una generación hija son una
función de la frecuencia génica en la generación paterna, esperaríamos que si los
animales F2 fueran apareados entre sí, la generación F3 resultante tendría las mismas
frecuencias genotípicas que la población F2. En otras palabras, si los apareamientos son
hechos al azar dentro de una población, las frecuencias génicas y genotípicas no
cambian.
Esta conclusión refleja lo que es el denominado equilibrio Hardy- Weinberg. La ley de
Hardy- Weinberg-llamada así por sus descubridores-dice:
En una gran población, con apareamientos al azar, en ausencia de selección, mutación o
migración las frecuencias génicas y genotípicas se mantienen constantes de generación en
generación, y las frecuencias genotípicas se relacionan con las frecuencias génicas por medio de
las fórmulas:
P = p2
H = 2 pq y
Q= q2
La ley Hardy- Weinberg esencialmente dice que si no hay fuerzas para cambiar las
frecuencias génicas y genotípicas en una población, esas frecuencias no van a cambiar.
¿Cuáles son las fuerzas que cambian las frecuencias génicas y genotípicas? La ley
menciona específicamente la selección, mutación y migración. Sabemos que la
selección modifica las frecuencias. De hecho, el propósito de la selección artificial es
cambiar las frecuencias génicas. La mutación, el proceso que altera el ADN para crear
nuevos alelos, tiene algún efecto sobre las frecuencias génicas y genotípicas, pero
debido a que la mutación es un evento raro, el efecto es pequeño. La migración, es el
movimiento de individuos hacia dentro o fuera de una población. La migración
particularmente involucra la introducción de un número grande de individuos
genéticamente diferentes en una población, puede tener grandes efectos en las
frecuencias génicas y genotípicas. La ley de Hardy- Weinberg también expone que una
población en equilibrio tiene que ser grande y apareada manera aleatoria. Las
poblaciones pequeñas se convierten en consanguíneas rápidamente, y como hemos
visto anteriormente, la consanguinidad cambia las frecuencias genotípicas
incrementando la homocigocis. Y en las pequeñas poblaciones las frecuencias génicas
pueden cambiar puramente por casualidad, fenómeno conocido como deriva
genética. El apareamiento aleatorio implica la ausencia de cualquier esquema de
apareamiento sistemático. En nuestro ejemplo del locus B, si no hubiésemos apareado
de manera aleatoria, sino que hubiéramos aplicado una regla que diga que los
genotipos BB solo pueden aparearse con otros genotipos BB, entonces podríamos

10
haber esperado frecuencias genotípicas diferentes (y no estáticas). Entonces el
apareamiento aleatorio es requerido para el equilibrio Hardy- Weinberg, y además
como sugiere nuestro ejemplo, solo se necesita una generación de apareamientos
aleatorios para alcanzar el equilibrio. El apareamiento aleatorio entre los animales F1
creó un equilibrio en la generación F2.
La derivación de las fórmulas de Hardy- Weinberg relacionando frecuencias genotípicas
con frecuencias génicas es simple. Si las frecuencias génicas en un locus en particular
en una población en equilibrio son p y q, entonces las fórmulas son las que aparecen
en el siguiente cuadro de Punnett.
p q
p p2 pq
q pq q2
Tener cuidado de no confundir p2 2pq y q2 de Hardy- Weinberg con la proporción 1:2:1
esperada cuando se aparean dos homocigotos. (Los estudiantes a menudo hacen
esto). A pesar de que la razón 1:2:1 puede ser considerada una aplicación especial de
la ley Hardy- Weinberg (cuando p=q=0.5), los dos conceptos son utilizados en
contextos muy diferentes.
¿Existen verdaderamente poblaciones en equilibrio Hardy- Weinberg? Claramente no.
Ninguna población es infinitamente grande, apareada perfectamente al azar o libre de
selección natural y mutación. Sin embargo, algunas poblaciones se encuentran muy
cercanas al equilibrio, en que que las fórmulas Hardy- Weinberg encajen bien y puede
ser de gran utilidad.
Como un ejemplo de la utilidad Hardy- Weinberg, vayamos nuevamente a las
poblaciones 1 y 2 y su descendencia F1. El objetivo de hacer este cruzamiento era
mostrar como los cruzamientos en este caso incrementan la heterocigosis. Recuerde la
Figura 4.4, que la frecuencia genotípica de los heterocigotas par el locus B en la
población F1 era 0.74. Determinamos esto multiplicando las frecuencias génicas de las
poblaciones de los progenitores. Lo que no sabemos son las frecuencias de los
heterocigotas en esas poblaciones originales. Sin embargo podemos calcularlas
asumiendo el equilibrio Hardy- Weinberg.
Las frecuencias de las poblaciones heterocigotas en las poblaciones 1 y 2, como fue
determinado por la fórmula Hardy- Weinberg son,
H1 = 2p1q1
= 2(0.8)(0.2)
= 32

11
H2 = 2p2q2
= 2(0.1)0(.9)
= 0.18
La frecuencia heterocigota promedio en las poblaciones de los padres es (H1+H2) /2 o
(0.32 +0.18)/2 =0.25.Compare esa figura con la frecuencia de los heterocigotas en los
cruzamientos F1(0.74). Cruzando las poblaciones 1 y 2, la heterocigosis casi se triplicó.
Este es un ejemplo dramático de cómo el cruzamiento aumenta la heterocigosis. Si las
poblaciones de los padres hubiesen sido más similares. (Por ejemplo que sus
frecuencias génicas en el locus B no hayan sido tan diferentes), el incremento de la
heterocigosis a partir del cruzamiento hubiese sido menos sensacional pero todavía
evidente.

12
Como usar las fórmulas Hardy- Weinberg para determinar las frecuencias génicas y genotípicas en el locus
que exhibe dominancia completa.
En una población de gallinas andaluzas (vea el ejemplo dado al comienzo del capítulo), es relativamente fácil
calcular las frecuencias génicas y genotípicas para el color de las plumas porque cada genotipo de color es
claramente identificable: los tipos BB son negros, los tipos Bb son azules, y los tipos bb son blancos. Para
determinar las frecuencias, solo necesita hacer un conteo y un poco de aritmética. ¿Pero cómo procede
cuando el locus de interés exhibe dominancia completa – es decir cuando no puede notar la diferencia entre
los genotipos heterocigotas y homocigota dominante observando sus fenotipo? Por ejemplo, ¿Cómo
determinaría las frecuencias génicas y genotípicas para el color del manto en una población de cerdos
Hampshire cuando el alelo W para la faja blanca es completamente dominante sobre el alelo w para el color
sólido? ¿Cómo sabe cuál de los cerdos con faja son Ww y cuales son WW?
El problema es pegajoso, pero sí (y solo sí) podemos asumir que la población está en (o por lo menos cerca)
equilibrio de Hardy Weinberg, una solución es posible. Con propósitos prácticos, podemos asumir el
equilibrio Hardy Weinberg si la población es razonablemente grande, si la introducción reciente de animales
del exterior es insignificante, y si, en la generación parentales más reciente, la selección y las decisiones de
apareamiento no están relacionadas al carácter de interés.
Considere una población de 1000 Hampshires de los cuales 910 tienen faja y 90 son de color sólido. Al
comienzo, la única frecuencia de la que podemos estar seguros es la frecuencia de los genotipos
homocigotas recesivos (Q), lo cual es simplemente la proporción de los cerdos de color sólido en la
población. Matemáticamente:
Tenemos una idea de la frecuencia de los genotipos heterocigotas (H) o del genotipo homocigota dominante
(P) porque ambos genotipos son parecidos – ambos tienen faja blanca. Pero si asumimos el equilibrio Hardy
Weinberg, entonces entonces,
Y
Por lo tanto


















