
ESPACIO Y TIEMPO 294 etal_Los Mamiferos de... · Discusi6n y conclusiones 312 ... con las cuales...
42
, 4. BIOGEOGRAFIA ESPACIO Y TIEMPO 294 Contexto geografico 294 Contexto temporal 299 POBLAMIENTO CONTINENTE-ISLA 301 Afinidades continentales 301 Hipotesis biogeograficas 302 Discusi6n y conclusiones 312 POBLAMIENTOISLA-ISLA 313 Dispersion 313 Vicarianza 314 IRRADIACION EVOLUCIONARlA 315 Grupos originarios de Suramerica 315 Grupos originarios de Norteamerica 318 Discusi6n y conclusiones 320 EXTINCIONES DEL CUATERNARlO 323 Cronologia y consecuencias 323 Causalidad 328 EPILOGO 334 [293]
Transcript of ESPACIO Y TIEMPO 294 etal_Los Mamiferos de... · Discusi6n y conclusiones 312 ... con las cuales...

4 BIOGEOGRAFIA
ESPACIO Y TIEMPO 294
Contexto geografico 294
Contexto temporal 299
POBLAMIENTO CONTINENTE-ISLA 301
Afinidades continentales 301
Hipotesis biogeograficas 302
Discusi6n y conclusiones 312
POBLAMIENTOISLA-ISLA 313
Dispersion 313
Vicarianza 314
IRRADIACION EVOLUCIONARlA 315
Grupos originarios de Suramerica 315
Grupos originarios de Norteamerica 318
Discusi6n y conclusiones 320
EXTINCIONES DEL CUATERNARlO 323
Cronologia y consecuencias 323
Causalidad 328
EPILOGO 334
[293]
294 BIOGEOGRAFIA
La geologia y la biologia deben com partir historias comunes Pero este afortunado suceso sera un hecho del futuro no del presente y 10 sera por aproximacion no por precision
Ernest E Williams (1989)
Dieciocho afios despues la sentencia de Williams conserva plenamente su vigencia en 10 que respecta a la biogeografia de los mamiferos terrestres (no volantes) de las Antillas subsisten incertidumbres en la compleja paleogeografia insular y el analisis filogenetico no ha alcanzado un nivel de resolucion satisfactorio para la mayoria de los taxones involucrados porotra parte el registro fosil continUa siendo deficitario no obstante haberse descubierto los primeros fosiles terciarios El presente capitulo es un intento por compendiar la informacion que existe sobre el tema (en parte incierta insatisfactoria y deficitaria) con el proposito de dar respuesta a tres preguntas esenciales (I) lCuaIes son las afinidades continentales de los mamiferos terrestres antillanos (2) lComo y cuando poblaron el territorio insular Los ancestros continentales de esa fauna (3) lQue ocurrio con los mamiferos terrestres una vez establecidos permanentemente en dicho territorio
ESPACIO Y TIEMPO
CONTEXTO GEOGRAFICO
Carecerfa de sentido abordar la biogeografia de los mamiferos cubanos at margen de las restantes Islas de las Antillas Mayores con las cuales Cuba comparte una historia comtm geologica y biotica mas inmediata que con las restantes islas caribefias Por consiguiente el ambito geognifico en el que eL tema es tratado en el presente capitulo abarca Las cuatro Islas principales de las Antillas Mayores (Cuba Jamaica La Espafiola y Puerto Rico) y sus Islas satelites asi como los grupos de Islas perifericas influidas biogeognificamente por alguna(s) de las cuatro Islas principales Islas Swan Islas Caiman Islas Bahamas Islas Virgenes y las dos isLas del norte de las Antillas Menores (cercanas a Puerto Rico) Anguila y San Martin (Fig 41) Entre alternativas de reconstruccion paleogeognifica de este segmento insular hemos seguido a lturralde-Vinent y MacPhee (1999) la version mas reciente y documentada
Para el ambito geognifico asf delimitado (que comprende 96 del territorio insular del Caribe l ) la Tabla 41 presenta la fauna conocida de mamiferos terrestres autoctonos consistente en 4 6rdenes 8 familias 28 generos (mas 2 indeterminados) y 69 especies todas endemicas del referido segmento insular 57 de las cuales (83) estan extinguidas De esta fauna 58 son roedores 20 perezosos 16 musarafias y 6 monos
lA los fines del presente capitulo las denominaciones Antill as y Caribe se emplean indistintamente como terminos equivalentes
295 ESPACIO Y TIEMPO
~) OCEANO ATLANTICO 26deg
J-~ __~~ )Bahamas laquo - ~ o km 400
---~ Florida
lt 221 Mexico ~
c~o
d _-shy ---shy P rt I Virgenes --------
u~ 0 Anguila- I
Rlco ctQ 8 Martin I c=~ -__ -------- ~I Swan ~ o ~ Qo~o
Jamaica Q
- - ------------- Y d
~ - v ~ gt o
MARCARIBE i oJ (
a
SURAMERICA
70middot
FIGURA 41 El Caribe insular_ La linea discontinua circunvala el area geografica objeto de atenci6n en el presente capitulo
TABLA 41 Marniferos terrestres aut6ctonos del segmento insular delimitado en la Fig 41 (especies en orden alfabetico las vivientes en letra negrita =especies precuaternarias) CU=Cuba JA=Jamaica LE=La Espanola PR=Puerto Rico IS=Islas Swan IC=lslas Caiman IB=Islas Bahamas IV=Islas Virgenes AS=Anguila - San Martin
lsI a S Taxonomla 1
CU JA LE PR IS IC IE IV AS
Pilosa (pcrczosos) 14 especics
MEGALONYClllDAE 2
CHOLOEPODIKAE
Acratocnus antillensls bull Acratocnus odontrigonus bull Acratocnus simorhynchus bull Acratocnus ye bull
(Continua)
296 BIOGEOGRAFiA
TABLA41 (Continuaci6n)
I s 1 a s Taxonomla 1
CD JA LE PR IS IC lB IV AS
N eocnus comes bull Neocnus dousman bull N eocnus gliriform is bull N eocnus toupiti bull Gen et sp indet bull MEGALOCNINAE
Imagocnus zazae3 bull M egalocnus rodens bull M egalocnus zile bull Parocnus browni bull P arocnus serus bull
Soricomorpha (musaraiias) 11 especles
NESOPHONTIDAE 4
Nesophontes edithae bull bull Nesophontes hypomicrus bull Nesophontes micrus bull bull N esophontes zam icrus bull N esophontes sp 1 bull Nesophontes sp 2 bull Gen et sp indet bull SOLENODONTIDAE 4
Solenodon arredondoi bull Solenodon cuhanus 5 bull Solenodon marcanoi bull Solenodon paradoxus bull
Primates (monos) 4 especies
PTIHECIIDAE 2
CALLICEBINAE
Antillothrix bernensis bull (ContinUa)
297 ESPACIO Y TIEMPO
TABLA 4 1(Continuacion)
I s a Taxonomia 1
CD JA LE PR IS IC IE IV AS
Paralouatta marianae bull Paralouatta varonai bull Xenothrix mcgregori bull
Rodentia (roedores) 40 especies
ECHllVlYIDAE 2
HETEROPSOMYINAE
Boromys offella bull Boromys torrei bull Brotomys contractus bull Brotomys voratus bull Heteropsomys antillensis bull Heteropsomys insulans bull Puertoricomys corozalus bull CAPROMYIDAE 4
PLAGIODONTINAE
Plagiodontia aedium 6 bull P lagiodontia araeum bull P lagiodontia ipnaeum bull Plagiodontia spelaeum bull Rhizoplagiodontia lemkei bull ISOLoBODONTINAE
Isolobodon portoricensis 7 bull Isolobodon montanus bull Zazamys veronicae bull HExOLOBODONTINAE
Hexolobodon phenax bull CAPROMYINAE
Capromys pilorides 67 bull Capromys sp bull
(Continua)
298 BIOGEOGRAFIA
TABLA 4 1 (Continuacion)
I s a s Taxonom1a 1
CU JA LE PR IS IC IB IV AS
Geocapromys browni 7
Geocapromys columbianus bull Geocapromys ingrahami 6
Geocapromys thoracatus
Geocapromys sp 1
Geocapromys sp 2
Macrocapromys acevedo
Macrocapromys latus
Mesocapromys angelcabrerai
Mesocapromys auritus
Mesocapromys kraglievichi
Mesocapromys nanus
Mesocapromys sanfelipensis
Mysateles melanurus
Mysateles prehensilis 6
bull bull bull bull bull bull bull bull bull
HEPTAXODONTIDAE 8
HEPTAXODONTINAE
Amblyrhiza inundata
Elasmodontomys obliquus
Quem isia grav is
Xaymacafulvopulvis 9
CLIDOMYlNAE
Clidomys osborni
Clidomys parvus
MURIDAE2
SIGMODONTlNAE
Oryzomys antillarum 10
bull
bull bull bull
bull
bull
bull bull
bull bull
bull
bull Total de especies por is1all 24 6 25 7 1 5 1 1 1
1 La taxonomia sigue las revisiones sistematicas mas recientes (Morgan 1993 1994 MacPhee y Grimaldi 1996 Horovitz y MacPhee 1999 White y MacPhee 2001 Ottenwalder 2001 Woods et aI 2001 MacPhee y Flemming 2003) con 1a adici6n de los taxones descritos posteriormente y la incorporaci6n de los cambios propuestos para las especies cubanas en el Capitulo 2 de este Compendio (Continua en pagina opuesta)
299 ESPACIO Y TIEMPO
CONTEXTO TEMPORAL
La absoluta mayoria del registro fosil de mamfferos terrestres antillanos proviene de depositos cavemarios pleistoholocenicos (escala del tiempo geol6gico en la Tabla 42) Abundan en estas cavernas restos de perezosos musarafias y roedores siendo comparativamente escasos los de monos debido quizas a la universal predileccion de los platirrinos por los ambientes densamente forestados 10 que los hace malos candidatos a la fosilizacion (MacPhee y Woods 1982)
A pesar de 10 numeroso de tales depositos en la region son escasos los fechados radiometricos del cuantioso material extraido Con excesiva frecuencia (particularmente en Cuba) estos depositos han sido asignados al Pleistoceno Superior sin fundamento suficiente para ello y es sabido que incluso en cuevas contiguas los depositos fosilfferos pueden diferir considerablemente en edad (Pregill y Olson 1981) Se superponen a esta inconveniencia la falta de estudios tafonomicos en las excavaciones y la eventual mezcla de f6siles de distintas edades como consecuencia de las multicausales alteraciones estratigraficas ocurridas en las cuevas (Nunez 1990a b Morgan y Ottenwalder 1993 MacPhee 1997 McFarlane 1999b Dfaz-Franco 2004) Sobre propuestas de tipificaci6n de los depositos paleontologicos con restos de mamiferos terrestres en Cuba vease Woloszyn y Silva (1977) Acevedo y Arredondo (1982) Gutierrez (1994) Arredondo [1999]
La Tabla 43 presenta los fechados pleistocenicos mas tempranos registrados para restos de mamfferos terrestres del Caribe insular De estos restos el roedor (Cidomys) de Jamaica con un fechado de 253 mil anos antes del presente constituye el fosH de mamifero terrestre mas viejo del Cuaternario anti11ano [Los fosiles fechados antes del surgimiento de la espectrometria de masa (aceleradora) para muestras pequefias pueden ser algo mas viejos (Jull et al 2004)]
Corresponde a MacPhee e Iturralde-Vinent (1994 1995b) e1 relevante merito de haber descubierto en la ultima decada del siglo xx los primeros fosiles terciarios de mamiferos terrestres de las Antillas extendiendo a mas de 30 millones de afios el registro f6sil de
2 Extirpada en la Subregion Antillana pero viviente en el Neotropico continental
3 Ubicacion en subfamilia s610 sugerida por White y MacPhee (200 I)
4 Endemica de la Subregion Antillana
5 Recientemente propuesta su ubicaci6n bajo el genero Atopogale (Roca et al 2004)
6 lncluye las subespecies
7 Transportada por aborigenes a otras de las islas (Rimoli 1974 Olson 1982b este trabajo)
8 Vease MacPhee y Flemming (2003) sobre la incierta situaci6n de esta familia
9 Ubicada provisionalmente en esta subfamilia por MacPhee y Flemming (2003)
10 Hallada en contexto no perturbado de edad Pleistoceno tardio (McFarlane et ai 2002) con 10 cual se descarta su supuesta introduccion por amerindios (Woods 1990)
II La correlaci6n entre el area insular y la riqueza de especies es razonablemente fuerte (R=O83) en estas islas excepto las Bahamas (Morgan 1989)
300 BIOGEOGRAFIA
TABLA 42 Escala convencional del tiempo geologico en la Era Cenozoic a y finales de la Mesozoica (Gradstein 2004) ka = kiloafios (miles de afios) Ma miJIones de afios AP = antes del presente
Periodo(Suhperiodo) Epoca Afios (aproximados) antes del presente
Era Cenozolca
Cuaternario Holoceno Pkistoceno
Presente - 11 kaAP 11 kaAP - 18 MaAP
Terciario (Ne6geno ) Plioceno Mioceno
18 - 5 MaAP 5 - 23 MaAP
oIigoceno 23 - 34 MaAP Terciario (pak6geno) Eoceno 34 - 58 MaAP
Pakoceno 58 - 65 MaAP
Era Mesozo ca
Creticico 65 - 145 -faAP
~~~------~~--- --~-
este grupo animal en la region con el hallazgo de un perezoso oligocenico en Puerto Rico (Yauco) as como un perezoso un mono y un roedor miocenicos en Cuba (Domo de Zaza) Esto unido a la presencia de huesos de una musarafia nesofontoide [referida erroneamente a Solenodon por Rivero (1997)] en ambar oligomiocenico de la Republica Dominicana (MacPhee y Grimaldi 1996) demuestra incontrovertiblemente que al menos en el Mioceno temprano (alrededor de 20 millones de afios atnis) ya estaban en las Antillas Mayores representantes de los cuatro linajes basicos de su fauna de mamiferos terrestres [La afirmacion de Abreu et al (1990 13) en cuanto a que se conocen restos fosiles de almiqui que datan de hace aproximadamente 40 millones de afios es erronea]
Pero lcuan antigua puede ser la fauna de mamiferos terrestres de las Antillas Mayores
Aunque es un hecho bien establecido por la geologia que desde finales de la Era Mesozoica y principios de la Cenozoica existieron islas en el area ocupada ahora por las Antillas Mayores la evidencia disponible indica asimismo que aquellas primeras islas sufrieron repetidas transgresiones marinas totales que habrian hecho sucumbir las biotas que pudieran haber sustentado (MacFadden 1981 Morgan y Woods 1986 MacPhee e Iturralde-Vinent 1994 Roca et al 2004) sin contar los catastroficos efectos que debio tener para el Caribe occidental el b6lido extraterrestre que impacto en Chicxulub (Yucatan) hace 65 Ma (Tada et ai 2003) Hedges (1996) ha especulado que aUn en medio de tal devastacion debe haber existido algun que otro lugar donde elementos de la fauna pudieron haber sobrevivido
301 POBLAMIENTO CONTINENTE-ISLA
TABLA 43 F echados pleistocenicos mcis tempranos (obtenidos por distintos metodos) en mamiferos terrestres del segmento insular delimitado en laFig 41 F =Fuentes (I) Jull et al 2004 (2) Goodfriend y Mitterer 1987 (3) MacPhee et al 1989 (4) Woods 1989b (5) Rega et aI 2002 (6) McFarlane 1999b (7) McFarlane 1 999a (8) McFarlane et al 1998a
Pals Localidad Taxon kaAP F
Cuba Breas de San Felipe Pilosa (Parocnus) 1l9a 1
Jamaica Cueva Coco Ree Cueva Wallingford Roadside
Primates indet Rodentia (Clidomys)
393 2530
2 3
Haiti Trouing Jeremie no5 (casimba) Rodentia (Rh izoplagiodontia) 212 4
Republica Dominicana
Cueva del Perezoso Pilosa (ficratocnus) 185 5
Puerto Rico Cueva de Ia Vaca Cueva Clara
Pilosa (Acratocnus) Soricomorpha (Nesophontes)
337 346
6 7
Anguila Cueva Cave-No-Cave Rodentia (Amblyrhiza) 2060 8
a Kulp etal (1952) reportaron un fechado radiocarbonico de gt 30000 afios para un cono de pino en el deposito fosilffero de Ciego Montero (Cuba) con restos de perezosos
(vcase tambicn Crother y Guyer 1996) pero Iturralde-Vinent y MacPhee (1999) han preguntado donde estaban esos lugares si es que existieron
Solo a partir del Eoceno Superior las Antillas Mayores en fonnacion han pennanecido emergidas hasta hoy [excepto parte de Jamaica y el sur de La Espanola que continuaron sumergidas hasta el Mioceno Medio (Iturralde-Vinent y MacPhee 1999)] Esto significa que solo durante los ultimos 40 millones de anos aproximadamente han existido pennanentemente en dicha region insular ambientes terrestres capaces de sustentar comunidades biotic as (MacPhee e Iturralde-Vinent 2000 Portell et ai 2001 MacPhee 2005) 10 cual proporciona el marco temporal de referencia para conceptuar la antigiiedad maxima posible de la fauna de mamiferos terrestres de las Antillas Mayores
POBLAMIENTO CONTINENTE-ISLA
AFINIDADES CONTINENTALES
El consenso de los autores que en las itltimas dccadas han tratado la filogenia y biogeografia de los mamiferos terrestres de las Antillas Mayores (MacFadden 1980 MacPhee y Woods 1982 Morgan 1985 1988 1993 Morgan y Woods 1986 Ford 1986 Woods 1988 1989a 1990 MacPhee e Iturralde-Vinent 1995a b Hedges 1996 Horovitz [1997] Ottenwalder 2001 Whidden y Asher 2001 MacPheeet ai 2003 Carlini y Scillatoshy
302 BIOGEOGRAFiA
Yane 2004 Davalos 2004 MacPhee y Horovitz 2004 Roca et al 2004) es practicamente unanime en cuanto a que los ancestros continentales que dieron origen a esta fauna derivaron (1) mayormente de Suramerica perezosos (Megalonychidae) monos (Pitheciidae) y roedores (Echimyidae Capromyidae y Heptaxodontidae) (2) en menor medida de Norteamerica musarafias (Nesophontidae y Solenodontidae) y (3) menos aun de Centroamerica una especie de roedor (Muridae)
HIPOTESIS BIOGEOGRAFICAS
La definicion c1asica reconoce dos tipos de islas isla continental (porcion del territorio de un continente separado de este por movimiento tectonico 0 por ascenso del nivel del mar) e isla oceanica (porcion de territorio que no ha formado parte de un continente) La definicion sigue siendo uti por 10 que implica para la formacion de las faunas insulares en uno u otro caso As en el caso de la isla continental su fauna ya estaba emplazada en la futura isla antes de producirse su separacion del continente [Vicarianza hipotesis biogeografica de fragmentacion faunal] en el caso de la isla oceanica su fauna es el resultado de la colonizacion1 por propagulos2continentales a traves de la barrera marina entre el continente y la isla [Dispersion hipotesis biogeografica de acumulacion faunal] En el primer caso el surgimiento de la barrera marina es resultado del evento vicariante en el segundo la existencia de la barrera marina es anterior al evento dispersante Ademas de operar a escala mayor (continente-isla) ambos eventos operan secundariamente a menor escala (isla-isla)
Las dos hipotesis -vicarianza y dispersion- se han disputado hasta hoy la preferencia de los biogeografos en tanto hipotesis mas viable para explicar el proceso de formacion original (continente-isla) de la fauna de mamiferos terrestres de las Antillas Mayores [Sobre la historia de las hipotesis biogeograficas relativas al Caribe insular vease RuB y Schubert 1989 Williams 1989 Pruna 2001] Yel debate continua debido ala falta de consenso sobre 10 que Williams (1989) ha llamado el problema central de la discusion a saber la ostensiblemente baja diversidad taxonomica de esta fauna a nivel de las categorias superiores (solo cuatro ordenes) en comparacion con la diversidad al mismo nivel de las faunas de islas continentales (0 de areas del continente) simi lares en tamafio y diversidad ecologica [Por ejemplo la fauna cubana de mamiferos terrestres consideradas de conjunto las especies vivientes y las extintas- es menos diversa a los niveles taxonomicos superiores (4 ordenes y 6 familias) que 10 que cabria esperar de la diversidad ecologica y extension territorial de Cuba (110 992 km2) comparada con una isla tipicamente continental como Trinidad (4828 krn2
) -veinte veces menor que Cuba y menos diversiticada ecologicamente- donde solo las especies vivientes de mamfferos terrestres representan 6 ordenes y 16 familias (Eisenberg 1989)] Este desbalance
lColonizaci6n Arribo y establecimiento de un propagulo en una isla no ocupada por la especie (MacArthur y Wilson 1967)
2Propagulo Niimero minimo de individuos de una especie capaces de reproducirse y aumentar poblacionalmente en una isla bajo condiciones favorables (MacArthur y Wilson 1967)
303 POBLAMIENTO CONTINENTE-ISLA
taxon6mico ha sido expJicado de dos maneras (1) Inicialmente hubo mas 6rdenes presentes pero la mayoria se extingui6 y s610 sobrevivieron esos cuatro [explicaci6n vicarianista] (2) S610 representantes de esos cuatro 6rdenes lograron cruzar la barrera marina [explicaci6n dispersionista]
Cinco hipot~ticos mecanismos de poblamiento continente-isla han sido invocados durante los ultimos 140 afios (y se los sigue proponiendo actualmente) para explicar la presencia de mamiferos terrestres en las Antillas Mayores A continuaci6n pasamos a resefiarlos brevemente (incluidas sus alegadas fortalezas y debilidades) en el orden aproximado en que historicamente fueron propuestos
EL PUENTE NATURAL (dispersion terrestre activa)
En el siglo XIX el puente terrestre era pnkticamente la unica hipotesis para explicar las distribuciones disyuntas de animales en buena parte del mundo y los primeros investigadores que se ocuparon de la biogeografia antillana (Fernandez [1865] 1884 Chapman [1893] Torre 1892 Lora 1895) la asumieron resueltamente en parte intrigados por el descubrimiento en Cuba en la segunda mitad del siglo de un perezoso f6sil de gran talla Y en su aplicacion a los mamiferos terrestres de las Antillas Mayores el puente sigui6 siendo a 10 largo del siglo xx la hipotesis mas favorecida por los estudiosos del tema (Vaughan 1902a Spencer 191 Oa b Torre 1910a b cAllen 1911 1917a 1918 Barbour 19141916 1945bAnthony 1918 1926 [1942] Scharff 1922 Kraglievich 1926 Beaufort 1951 Arredondo 1961 1970b Kartashov y Mayo 1974 Iturralde-Vinent 1982 1988 Arredondo y Armifiana 1996 Arredondo 1996a [1999]) quienes postularon la existencia preterita de puentes con Norte- Centro- 0 Suramerica en dependencia de las afinidades continentales de los mamiferos antillanos
Teoricamente el puente terrestre debe cumplir con una condicion esencial permitir el paso de una parte mayoritaria (y por tanto representativa de la diversidad) de la fauna continental Claro que siempre habria tipos de mamiferos menos proclives a dispersarse o para los cuales la ubicaci6n del puente resultaria inaccesible 0 las condiciones ecologicas en este fueran desfavorables pero si algunos acceden al puente no hay raz6n para pensar que el resto de esa fauna no los acompafie mayoritariamente Como dijera Simpson (1956) Los puentes no transportan faunas desbalanceadas
En consecuencia el puente terrestre como filtro es un mecanismo de poblamiento minimamente selectivo y por ende de alta diversidad inicial (sensu Iturralde-Vinent y MacPhee 1999) 10 esencial del puente es su capacidad para transportar faunas integradas (Williams 1989) Yen eno radica la objeci6n fundamental al puente terrestre surgida tempranamente en el siglo xx (Por que s610 cuatro 6rdenes de mamiferos terrestres en las Antillas si vinieron por puentes (Donde estan los fosiles de los demas ordenes que debieron venir si es que se extinguieron Los partidarios del hipotetico puente dispusieron de casi todo el resto del siglo para responder una y otra vez que otros 6rdenes de mamiferos aparecerian representados en las Antillas Mayores el dia que en estas Islas se descubrieran fosiles precuatemarios suficientemente
304 BIOGEOGRAFIA
antiguos Esto se logro por fin terminando el siglo como se explico anteriormente (vease Contexto temporal) y ique se encontro los mismos cuatro ordenes de mamiferos terrestres conocidos del Cuatemario caribefio aparentemente los Unicos que lograron llegar
Pero ademas hasta el presente no se ha demostrado inequivoca- y concluyentemente que en algun momenta durante los ultimos 40 millones de afios haya existido una conexion terrestre continua entre las Antillas Mayores (en proceso de formacion 0 plenamente formadas) y algun punto de las regiones continentales vecinas
LA BALSA NATURAL (dispersion marina pasiva)
Durante buena parte del siglo xx el puente terrestre tuvo como unico rival a la hipotesis de la balsa natural -transporte accidental de animales sobre islotes de vegetacion flotantes a la deriva despues de arrancados y expulsados mar afuera por un rio crecido- expuesta originalmente por Matthew (191319151916 1918a 1919) y desarrollada en su aplicacion a los mamfferos terreslres antillanos principal mente por Simpson (1956) asi como por olros investigadores (Darlington 1938 1957 Williams y Koopman 1952 Hershkovitz 1958 1972 Patterson 1962 Paula 1967 Patterson y Pascual 1972 Pregill 1981 Ford 1986 Woods 1989a 1990 Morgan 1993) Por su parte Hedges et al (1992) sustentaron dicha hipotesis en estimados de divergencia (basados en el reloj molecular) en la herpetofauna antillana -involucrados tangencia1mente los mamiferos [veanse objeciones a Hedges y colaboradores (Page y Lydeard 1994 Iturralde-Vinent y MacPhee 1999) respuestas a las objeciones (Hedges et al 1994 Hedges 2001) y contrarrespuestas (MacPhee e IturraldeshyVinent 2005) asi como cuestionamientos al metoda de los estimados de divergencia (Foote et al 1999 Pregill y Crother 1999 Woods et ai 2001 )]
La hip61esis de la balsa natural se origino a partir de las numerosas ocasiones en que a 10 largo de siglos tales islotes flotantes (portando 0 no vertebrados incluidos mamiferos) han sido avistados mar afuera a cientos de kilometros de la costa mas cercana en varias partes del mundo --inc1uido el Mar Caribe- segUn consta en diarios de barcos y reportes de observadores diversos (revisados por King 1962 reportes posteriores en SICSP 1970 y Censky et al 1998) Uno de estos casos acaecido en nuestra region se reporta en un informe del SICSP (Smithsonian Institution Center for Shortlived Phenomena 197050shy52)
EIB de julio de 1969 un destructor de la armada de Estados Unidos avisto un islote flotante a unos 100 km al sur de la Base Naval de Guantanamo moviendose desde el Paso de los Vientos en direccion suroeste a una velocidad de 45 kmlh islote de unos 15 m de diametro mantenfa en pie mas de 10 palmas reales de 10-15 m de altura con suelo cubierto de hierba y arbustos [la observacion con binoculares a 18 km de distancia no preciso la presencia de vida animal] La travesia fue rastreada durante 6 dfas hasta que el 19 de julio el islote se dio por perdido tras 6 horas de busqueda aerea
305 POBLAMIENTO CONTINENTE-ISLA
La colonizaci6n exitosa de una isla de las Antillas Mayores por mamiferos terrestres continentales transportados en balsas naturales a la deriva (de cada especie al menos una hembra gravida 0 dos individuos de sexo opuesto) dependeria de factores que van a operar desde el momenta mismo de formarse la balsa asi como durante su travesia marina y su arribo a la isla
a) La posibilidad de estar en la balsa Que la distribuci6n de la especie incluya areas contiguas al rio y que el animal se encuentre en ellugar y el momenta propicios para ser incorporado a la balsa antes de que el rio crecido la desprenda Por ejemplo segun Hershkovitz (1958) los equimidos -grupo ancestral de roedores antillanos- serian buenos candidatos a ser captados por la balsa porque acostumbran a anidar en huecos del tronco de los arboles a la orilla de los rios
b) El tamano y la consistencia de la balsa Muchas se desintegran en alta mar debido a su fragilidad ante la saturaci6n Algunos autores (Beaufort 1951 Arredondo 1996a [1999]) han expresado incredulidad en cuanto a que las balsas hayan podido soportar el peso de un animal como el gran perezoso cubano (Megalocnus rodens) el mayor de los mamiferos antillanos Pero esta especie evolucion6 en Cuba y el ancestro que hizo el viaje pudo ser muy pequeno [EI perezoso oligocenico de Puerto Rico es el mas pequeno de los megaloniquidos antillanos (MacPhee e Iturralde-Vinent 1995b )]
c) Las caracterfsticas fisiol6gicas de la especie Los limites fisiol6gicos de los propagulos constituyen nuestra unica objeci6n a la dispersi6n marina por balsas como mecanismo para la formaci6n de la fauna de vertebrados en las Antillas (MacPhee e IturraldeshyVinent 2005) Unas especies tienen intrinsecamente mas posibilidades que otras para sobrevivir a las vicisitudes que deben afrontar durante la travesia (deshidrataci6n inanicion insolaci6n etc) Por ejemplo los megaloniquidos han demostrado tener extraordinaria habilidad para cruzar barreras marinas fueron los primeros mamfferos suramericanos en invadir Norteamerica mucho antes de que surgiera la conexion terrestre a finales del Plioceno (Paula 1967 Olson 1978 Webb 1985b Martin y Steadman 1999) Incluso se conoce un megaloniquido acultitico del Plioceno peruano (Muizon y McDonald 1995) y otro viviente en Suramerica (Choloepus) que pnicticamente no bebe agua 10 que ha originado una leyenda aborigen que explica por que los perezosos no toman agua (Goff art 1971) Vease ademas el inciso e
d) EI regimen de las corrientes marinas Que conduzcan a la balsa en la direcci6n necesaria EI transporte de balsas de vegetacion desde los continentes hacia las islas fue durante la mayor parte del Cenozoico sumamente improbable (MacPhee e IturraldeshyVinent 2000 148) Estos autores han insistido en que hay que tener en cuenta que durante la cambiante geografia cenozoica del Caribe las corrientes marinas de superficie no favorecieron el desplazamiento de objetos fiotantes hacia las Antillas Mayores [Pero no sabemos con certeza que efecto modificador del curso de tales objetos fiotantes pudieron introducir las tormentas tropicales si previsiblemente funcionaron en el pasado como 10 hacen ahora] Dichos autores senalaron ademas (2005) que en un experimento reciente con boyas de posicionamiento satelital liberadas a la deriva desde las Antillas Menores las boyas fueron a parar mayoritariamente a Centroamerica y a otros lugares
306 B[OGEOGRAFIA
y solo unas pocas recalaron en las Antillas Mayores y las que entraron al Mar Caribe demoraron semanas 0 meses en arribar
e) La distancia a recorrer (y en consecuencia la duracion de la travesia) entre el continente y la isla (10 que repercute decisivamente en el factor c) Este es desde luego uno de los factores mas criticos En la medida que la distancia aumenta la probabilidad de alcanzar la isla disminuye drasticamente con el agravante de que la trayectoria dificilmente seria en linea recta Si tomamos como referencia la velocidad de 45 kmlh del islote ya mencionado (SICSP 1970) cubrir la distancia aerea (aproximada) de 800 km que media entre el noroeste de Suramerica y La Espafiola Ie tomaria a la balsa 74 dias Estimando el doble de este tiempo debido a desvios de la trayectoria la duracion de la travesia probablemente resultaria excesiva para la mayoria de los mamiferos Pero pudiera muy bien ocurrir que la velocidad fuera mayor 0 que la balsa portara plantas vivas que permitirian a los fitOfagos sobrevivir [Tres de los cuatro ordenes de mamiferos terrestres conocidos de las Antillas Mayores son fitOfagos] Ademas esta geologicamente bien establecido que durante etapas de variada duracion desde el Oligoceno hasta el Pleistoceno los territorios del Caribe insular y continental aumentaron considerablemente en extension con niveles del mar 100-200 m por debajo de su nivel actual oportunidades que los propagulos aprovecharian para cruzar barreras marinas significativamente menores
f) El tamafio de la isla Las islas mayores deben interceptar mas balsas (Buckley y Knedlhans 1986) 10 que repercute directamente en el factor e La balsa puede pasar entre islas pequefias y continuar viaje sin llegar a ningun lugar
g) Las condiciones (favorables 0 adversas) del desembarco en la isla y el tiempo que Ie tomaria a uno 0 mas animales depauperados encontrarun habitat apropiado Esto dependeria a su vez del tamafio y diversidad ecologica de la isla (Darlington 1938 Miller 1966)
Tal cumulo de posibles obstaculos y dificultades constituye a primera vista un desafio a la credibilidad de la hipotesis Matthew-Simpson pero en ello radica precisamente su valor explicativo bas ado en la mas absoluta improbabilidad La colonizacion por la via marina es altamente improbable pero no absolutamente imposible si se tiene en cuenta la inmensidad del tiempo geologico se vuelve probable (McKenna 1973) Basta que solo una vez entre mill ones de intentos fallidos durante ese inmenso lapso algunos de los factores anteriormente enunciados coincidan en no ser tan desfavorables para el propagulo Y eso es todo 10 que demanda la hipotesis para cada uno de los grupos monofileticos mayores de mamiferos terrestres presentes en las Antillas Mayores Las Antillas Mayores requeririan el arribo de tan solo 7 (quizas menos) propagulos fundadores a 10 largo de los ultimos 30 0 mas millones de afios (promedio de uno cada 43 mill ones) para explicar la formacion de toda su fauna de mamiferos terrestres por dispersion marina pasiva (vease Discusion y Conc1usiones)
En comparacion con la via marina el poblamiento por la via terrestre es predecible porque la mayor parte de la fauna continental de mamiferos emplazada en la region del puente va a cruzar por este una vez que el mismo se establece y los animales logran
307 POBLAMIEITO CONTINENTE-ISLA
acceder a el el poblamiento por la via marina en cambio es impredecible porque depende total mente del azar que caracteriza a la balsa Claro que en ambas vias opera el azar pero en el primer caso se trata del azar de un evento probable en el segundo el azar de un evento improbable (Simpson 1940) Asi el puente natural (mecanismo de poblamiento minimamente selectivo = alta diversidad inicial) y la balsa natural (mecanismo de poblamiento maximamente selectivo baja diversidad inicial) pueden conceptuarse como los extremos opuestos de una gama de probabilidades de dispersi6n (McKenna 1973) EI fen6meno de la dispersi6n merece la mayor atenci6n cientifica en los estudios de biogeografia insular (Cowie y Holland 2006)
Queda claro que la balsa natural es una hip6tesis consistente con el caracter de la fauna objeto de estudio puesto que el desbalance taxon6mico (sin evidencia paleontol6gica en contrario) s610 puede ser resultado de la improbabilidad del poblamiento No obstante ello no implica necesariamente que la balsa natural haya sido el imico mecanismo (ni siquiera el principal) utilizado por los mamiferos terrestres en su colonizaci6n de las Antillas Mayores La balsa natural debe satisfacer una premisa te6rica esencial dado el caracter azaroso impredecible e improbable de una colonizaci6n exitosa esta debe ocurrir a intervalos muy espaciados a traves del tiempo geol6gico 0 sea durante todo el tiempo que en el area ocupada por las Antillas Mayores ha habido ecosistemas terrestres perrnanentemente disponibles para ser colonizados (los ultimos 40 millones de afios) Sin embargo como sabemos representantes de los cuatro 6rdenes de mamiteros terrestres conocidos de las Antillas Mayores ya estaban emplazados en dicho territorio insular en el Mioceno Inferior 10 que supone un lapso de colonizaci6n limitado al Eoceno SuperiorshyOligoceno (MacPhee 2005) sugerente de un tipo de mecanismo contrario a la colonizacion alocronica exigida por la hipotesis de la balsa natural sobre todo tratandose de una fauna caracterizada por altas tasas de extincion sin reposicion aparente
La presencia en Jamaica (hasta su extinci6n en 1877) del roedor mfuido Oryzomys antillarom con estrecha afinidad centroamericana (Woods 1989a) pudiera constituir el imico caso efectivo de poblamiento (continente-isla) de las Antillas Mayores por dispersion marina dc un mamifero terrestre para el cual no es razonable invocar otro mecanismo de colonizacion
LA GUIRNALDA INSULAR (dispersion mixta)
La geologia ha establecido con suficiente certeza que durante el Cenozoico unicamente en el Pale6geno tardio y solo a traves de la Cresta de Aves (ahora sumergida excepto la isla hom6nima) pudo haber existido una cadena de islas para el transito de mamiferos terrestres entre el norte de Suramerica y las proto-Antillas Mayores [desestimados a ese efecto el Canal dc Yucatan el Elcvado de Nicaragua la Cresta de Caiman el Elevado de Beata y las Antillas Menores (Holcombe et al 1990 MacPhee e IturraldeshyVinent 1994 1995b Iturralde-Vinent y MacPhee 1999)] Holcombe y Moore (1977) y Holcombe y Edgar (1990) postularon la existencia de esta entidad y estimaron algunas de sus posibles caracteristicas Por su parte MacPhee e Iturralde-Vinent (1995b) e Iturralde-Vinent y MacPhee (1999) precisaron los extremos proximal y distal de esa
308 BIOGEOGRAFIA
estructura acotaron la duracion y la edad de la misma y Ie dieron nombre GAARlandia (Greater AntillesA vesRidgelandia)
Segun Iturralde-Vinent y MacPhee (1999) la Cresta de Aves emergio por breve tiempo (1-2 Ma) durante la transicion Eoceno-Oligoceno (35-33 MaAP) por la coincidencia de un levantamiento tectoni co y un considerable descenso eustatico del nivel del mar En ese momenta las islas septentrionales del arco de las Antillas Mayores (Cuba centroorientalshyLa Espafiola - Puerto Rico - Islas Virgenes) constituian una sola isla grande 0 una serie de islas separadas por barreras muy estrechas La Cresta de Aves emergida entre el bloque Puerto Rico - Islas Virgenes y un microcontinente noroccidental de Suramerica -separado en ese momento del resto del continente por una via marina surgida en el Eoceno tardio- dio origen a GAARlandia
Obviamente el numero de colonizaciones exitosas por mamiferos terrestres a traves de una guimalda de Islas tiende a disminuir mientras mas anchas sean las barreras marinas entre las islas que forman la guimalda y cuando esto ocurre disminuye tambien la diversidad taxonomica de los propagulos particularmente a nivel de las categorias superiores (Simpson 1956)
Vemos asi que para los mamiferos terrestres la colonizaci6n a traves de una guimalda insular puede ser tan improbable como a la deriva en una balsa natural en dependencia de la cantidad y anchura de las barreras marinas a cruzar el tamafio y la fisiografia de las islas que integran la guimalda y el regimen de las corrientes marinas y aereas entre otros factores
Resumiendo la guimalda insular puede conceptuarse como un mecanismo de poblamiento altamente selectivo (= baja diversidad inicial) que combina la improbabilidad de la balsa natural con la colonizacion restringida temporaimente del puente terrestre
En su reciente revision de la taxonomia distribuci6n y filogenia (basada en estudios moleculares) de varios grupos de mamiferos antillanos Davalos (2004) concluyo que los perezosos megalocninos los primates y los roedores histricognatos se avienen al patron de divergencia con el continente y epoca de arribo a las Antill as implicados en la hipotesis de GAARlandia pero no asi los perezosos coloepodinos que requieren una explicacion biogeognlfica distinta Esto ultimo (sospechado por Matthew 1918a) habla sido confirmado por White y MacPhee (2001 226-227)
Nuestra evidencia filogenetica no sustenta la idea de un dado unificado surgido de la invasion de un megaloniquido el analisis sugiere mas bien dos eventos de colonizaci6n independientes [y] sustenta fuertemente un origen difiletico de las subfamilias [Choloepodinae y Megalocninae] Si hubo dos migraciones a traves de GAARlandia presumiblemente poco espaciadas en el tiempo no se puede determinar en el presente
Finalmente queda claro que GAARlandia -como guirnalda insular- es la hipotesis mas robusta propuesta hasta ahora para explicar la presencia en las Antillas Mayores de
309 POBLAMIENTO CONTINENTE- ISLA
la absoluta mayona de los mamfferos terrestres de ongen suramericano porque el caracter de esa fauna supone un mecanismo de poblamiento ancestral que sea a la vez maximamente selectivo (baja diversidad ordinal) y temporalmente restringido (transicion Eoceno-Oligoceno)
Recientemente Rega et al (2002 18) expresaron A pesar de una revision exhaustiva de la paleogeografia de la cuenca del Caribe Iturralde-Vinent y MacPhee (1999) no pudieron confirmar la presencia de una conexion terrestre continua entre Suramerica y las proto-Antillas aunque hipotetizaron su existencia [Vease la Fig 42 (= figura 6 de Iturralde-Vinent y MacPhee 1999 simplificada))
En el trabajo citado por Rega y colaboradores aparece la siguiente explicacion
Actualmente no es posible ofrecer reconstrucciones paleogeograficas detalladas de la Cresta de Aves Sin embargo hay evidencia de que porciones de la cresta deben haberse elevado subaereamente en un tiempo La cuestion cntica es si la Cresta de Aves emergida pudo haber formado algun tipo de corredor entre las Antillas Mayores y el noroeste de Suramerica Perforaciones en las alturas estructurales de la cresta pudieran proveer el registro del Paleogeno tardio que se necesita para precisar la hipotesis del corredor aunque no se puede predecir si tales datos serian suficientemente detallados para determinar si en algun momento GAARlandia constituyo una sola entidad subaerea Por ejemplo pudo haber barreras marinas por corto tiempo entre las masas terrestres de la Cresta de Aves [0) ellevantamiento y la subsidencia pudieron afectar diferentes partes de GAARlandia en diferentes tiempos AItemativamente los elementos faunales pudieron haberse dispersado de manera pulsada Movimientos episOdicos de este tipo incrementarian la extincion de los elementos faunales incapaces de trasladarse a la siguiente masa terrestre en la cadena disminuyendo asi la diversidad total (Iturralde-Vinent y MacPhee 199957)
La primera parte de la explicacion deja claro que la posibilidad de que la Cresta de Aves haya sido una conexion terrestre continua no se puede descartar pero tampoco se puede afirmar La ultima parte de la explicacion describe el efecto altamente selectivo (filtrante) de la colonizacion a traves de una cadena de islas
Y continuan lturralde-Vinent y MacPhee (199957 -58) explicando
Hasta que punto el microcontinente noroccidental de Suramerica [considerado por dichos autores como el area suministradora de propagulos hacia GAARlandiaJ estuvo fisicamente separado del resto del continente no ha sido plenamente esc1arecido Si estuvo sustancialmente aislado por barreras marinas y orogenic as durante el Paleogeno tardio entonces solo la fraccion de la biota suramericana emplazada en el microcontinente estuvo en posicion de acceder a GAARlandia Esto pudiera tener que ver con el hecho de que la fauna antillana carezca de representantes de muchos grupos suramericanos [de mamfferos) que debieron haber hecho el viaje
310 BIOGEOGRAFiA
Florida o Alturas0 0
d 6 D L1anuras25deg
Yucatan
20deg
deg0
10deg
MARCARIBE
80deg 75deg
Bahamas
Cres ta de Beata
D Mares someros
D Mares profundos
OCEANO ATLANTICa
FIGURA 42 Reconstrucci6n hipotetica de GAARlandia como conexi6n terrestre continua entre las proto-Antillas Mayores y el noroeste de Suramerica en el Pale6geno tardio segun la FIgura 6 (simplificada) de Iturralde-Vinent y MacPhee (1999)
En su defensa del puente terrestre Barbour (1945b249) se refiri6 a esto - antes 10 habfa hecho Anthony [1942363]- en los terminos siguientes
El argumento de que si las Antillas hubieran estado conectadas con el continente se encontrarian los f6siles de un numero mayor de distintas criaturas me deja frio es perfectamente concebible que la conexi6n haya ocurrido en un periodo en que la parte del continente implicada no estuviera poblada por una fauna grande y variada
Y concluyen lturralde-Vinent y MacPhee (199958) su detallada explicaci6n
Desafortunadamente sabemos muy poco sobre la composici6n faunal del noroeste de Suramerica a finales del Pale6geno
Consecuentemente si se confirmara la magnitud del aislamiento del aludido microcontinente y su concomitante baja diversidad faunal GAARlandia como conexi6n terrestre continua serf a la hip6tesis mas parsimoniosa No obstante mientras no se disponga de evidencia concluyente a este respecto hipotetizar a GAARlandia como conexi6n continua (minima selectividad) la coloca en franca vulnerabilidad te6rica al tener que afrontar la objeci6n clasica al puente terrestre (baja diversidad ordinal de la
311 POBLAMIENTO CONTINENTE-TSLA
fauna) al tiempo que se la priva de ser la hip6tesis mas robusta para explicar -como guimalda de islas- la colonizaci6n de las Antillas por mamiferos procedentes de Suramerica
ELARcA DE NOE (seudovicarianza)
El primer vicarianista antillano (desde luego sin el saberlo) fue Corral (1940) con su utilizaci6n ingenua de las ideas wegenerianas de la deriva continental aparecidas la decada anterior Pero el primer (supuesto) intento de aplicar a la formaci6n de la biota caribefia el mecanismo de la vicarianza basada en la tearia de la tect6nica de placas (surgida en la sexta decada del pas ado siglo) se debe a Rosen ([1976] 1985)
Segun Rosen las Antillas Mayores se originaron en el Cremcico en la Placa Caribe aproximadamente en la posici6n que hoy dia ocupa Centroamerica y al ser desplazadas tectonicamente hacia el este interactuaron (de algun modo no precisado por Rosen) con las margenes continentales de Norte- y Suramerica de donde (en un evento unico) adquirieron toda su fauna transportada subsecuentemente por dichas islas en su transito hacia sus posiciones actuales [De ahi la denominacion metaforica dada par McKenna (1973) a este mecanismo de poblamiento] Aunque se ha presentado esta propuesta como Model0 de Vicarianza Continente-Isla de Rosen en realidad por definici6n no constituye un evento vicariante porque las proto-Antillas Mayores no resultaron del fraccionamiento y separaci6n por tectonismo de una porcion del territorio continental como exige esta hip6tesis (vease definici6n de vicarianza al comienzo de Hipotesis Biogeograficas)
El Arca de Noe de Rosen ha sido desestimada por la mayoria de los autores contemporaneos porque (1) carece de sustento geol6gico (lturralde-Vinent y MacPhee 1999) y porque (2) en el periodo en que se afrrrna ocum6 el evento muchos de los ordenes modemos de mamiferos placentarios attn no existian (Williams 1989)
EL BARCO FUNERAL VlKINGO (tanatovicarianza)
Domning et al (1997) reportaron 10 que consideraron el mamffero terrestre mas viejo de las Antillas --circunstancia que se refleja en el titulo mismo del trabajo Oldest West Indian Land Mammal- al descubrirse un perisodactilo rinocerotoide (Hyrachius sp) en una localidad de edad Eoceno Inferior a Medio en el occidente de Jamaica
Jamaica ha tenido una historia geologica y paleogeografica distinta de la historia comun a las restantes Antillas Mayores A principios del Cenozoico Jamaica occidental (a diferencia de la porcion oriental constituida por el Bloque de las Montafias Azules) formaba parte de Centro america de la cual se desgaj6 (por el desplazamiento de la Placa Caribe) para venir a integrarse despues del Eoceno Medio con 10 que serfa la futura isla de Jamaica desaparecida por transgresi6n marina en el Eoceno Superior hasta volver a emerger totalmente en el Mioceno Medio (Iturralde-Vinent y MacPhee 1999 MacPhee e Iturralde-Vinent 2000)
312 BIOGEOGRAFiA
Por consiguiente Hyrachius ---conocido de localidades de edad Eoceno Inferior a Medio en Norteamerica- debio ocupar (10 que iba a ser) Jamaica occidental mucho antes de que esta se separara de Centroamerica De hecho este fosil Uego a su destino insular contenido en las rocas de su basamento no como un ser vivo por 10 que es erroneo considerarlo un animal antillano (lturralde-Vinent y MacPhee 2004) como han supuesto algunos (Hedges 200 I Portell et al 200 I Condis [200 I D Todo 10 que hizo fue viajar tanatovicariantemente en su barco funeral vikingo -otra de las denominaciones metaforicas de McKenna (1973)
DISCUSION Y CONCLUSIONES
Hemos visto que solo disponemos de una hipotesis plausible para explicar la mayor parte del poblamiento (continente-isla) de las Antillas Mayores por mamiferos terrestres del Neotropico continental la dispersion mixta (a traves de GAARlandia) para los perezosos (excepto Choloepodinae) los monos y los roedores (excepto Oryzomys)
Por otra parte la historia biogeogrMica de las musaraiias de las Antillas Mayores (Nesophontes y Solenodon) constituye una de las situaciones mas enigmaticas entre los mamiferos de la region (Ottenwalder 2001321) Al menos cinco mecanismos de poblamiento continente-isla han sido invocados para explicar cuando como y de donde llegaron a las Antillas Mayores estas musaraiias
I EI propagulo ancestral (del Terciario de Norteamerica) coloniz6 a las Antillas Mayores por dispersion marina desde Centroamerica 0 desde la Florida (Patterson 1962)
2 El propagulo ancestral ( del Terciario) cruzo el Atlantico Sur (supuestamente mas estreshycho en aquella epoca) directamente de Africa a las Antillas Mayores (Hershkovitz 1972)
3 El propagulo ancestral (del Terciario de Norteamerica) Uego a las Antillas Mayores en el Arca de Noe de Rosen (MacFadden 1980)
4 El propagulo ancestral (del Terciario de Norteamerica) fue transportado a las Antillas desde Centroamerica en el bloque de Jamaica occidental y de alli paso por dispersion marina a las restantes Antillas Mayores (Whidden y Asher 200 I)
5 El propagulo ancestral (del Terciario de Norteamerica) lleg6 a las Antillas Mayores a traves de GAARlandia (Whidden y Asher 2001)
Aunque hay consenso entre estos (y otros) autores sobre el origen norteamericano de las musaraiias antillanas ninguno de los cinco hipoteticos mecanismos de poblamiento resiste la evidencia en contrario filogenetica yo paleogeogrMica Ademas algunas de estas hipotesis asumen que Nesophontidae y Solenodontidae comparten un antecesor comun en las Antillas pero esta relacion no ha sido confirm ada Para dilucidar el enigmatico caso de las musaraiias antillanas se requerini profundizar en la investigaci6n geologica paleogeognifica y paleontol6gica (Iturralde-Vinent y MacPhee 1999)
De acuerdo con el conocimiento filogenetico disponible sobre el conjunto de la fauna la colonizacion original de las Antillas Mayores por mamiferos terrestres implicaria el arribo
313 POBLAMIENTO ISLA-ISLA
de solo siete propagulos fundadores perezosos 2 (coloepodinos y megalocninos si como parece tienen un origen difiletico) musaraiias 2 (nesofontidos y solenodontidos si no constituyen una misma familia como consideran algunos) monos 1 (pitecidos) roedores histricognatos 1 (equimidos capromidos y heptaxodontidos) roedores muridos 1 (Oryzomys en Jamaica)
POBLAMIENTO ISLA-ISLA
La distribucion de los mamiferos terrestres a traves del area antillana demarcada al inicio del presente capitulo (vease Contexto Geogratico Fig 41) ha sido interpretada en funcion de dos hipoteticos mecanismos de poblamiento interinsular la dispersion y la vicarianza fundamentadas como se resumen seguidamente
DISPERSION
De acuerdo con la evidencia paleogeogratica mas actualizada en las principales regresiones marinas ocurridas en la cuenca caribeiia durante los ultimos 23 Ma (transiciones OligocenoshyMioceno Mioceno Superior - Plioceno y varias veces en el Pleistoceno) las masas terrestres constitutivas de la region insular en formacion aumentaron considerablemente su extension territorial disminuyendo por tanto la distancia entre elias oportunidades que los elementos faunales aprovecharian para dispersarse de unas islas a otras
Durante mucho tiempo esta ha sido la hipotesis mas favorecida por los estudiosos del tema a partir de Simpson (1956) pero en decadas recientes ha suscitado dos objeciones
1 Es dificil en tender como pudo operar la dispersion marina entre estas islas puesto que ninguna de las Antillas Mayores posee rios suficientemente caudalosos para transportar masas significativas de vegetacion flotante (MacPhee 198432)
Es oportuno recordar aqui el islote ala deriva (con mas de diez palmas reales en pie) entre las tres Antillas Mayores mas occidentales mencionado anteriormente (vease La Balsa Natural)
2 La dispersion marina es menos efectiva para explicar la asombrosa similitud de las faunas de mamiferos terrestres a traves de las Antillas Mayores septentrionales porque como proceso aleatorio no debe producir distribuciones uniformes a 10 largo de una cadena de islas 0 si las produce entonces el proceso no es aleatorio (MacPhee e lturralde-Vinent 2000 149)
Independientemente de que las aludidas distribuciones taxonomicas estan lejos de ser uniformes 0 asombrosamente similares la hipotesis de la dispersion marina contrariamente a su canicter aleatorio y altamente improbable en el caso del poblamiento continente-isla- en el caso del poblamiento isla-isla es un proceso no aleatorio (aprovecha las oportunidades brindadas puntualmente por los periodos de maxima regresion marina) y ademas altamente probable dada la incomparablemente menor distancia entre las islas caribeiias que entre estas y los continentes vecinos
314 BIOGEOGRAFiA
V ICARIANZA
La neotectonica y las transgresiones marinas del Oligoceno Medio y del Mioceno afectaron sustancialmente la paleogeografia de la Cresta de las Antillas Mayores resultando la subdivision gradual de las partes subaereas de la Cresta Asi el este de Cuba y el norte de La Espanola conectados fisicamente durante el Oligoceno Inferior se separaron al formarse el Paso de los Vientos en el Oligoceno tardio en tanto La Espanola y Puerto Rico probablemente permanecieron conectadas hasta el Mioceno tardio en que surgio el Paso de Mona (Iturralde-Vinent y MacPhee 1999)
Tales eventos geologicos habrian dividido las distribuciones de los grupos faunales emplazados previamente en el territorio continuo de las tres islas y a partir de esas disyunciones dichos autores (tambien MacPhee et al 2000 y White y MacPhee 2001) asumieron --como segunda fase del modelo de GAARlandia- la hipotesis de la vicarianza isla-isla para explicar la distribucion multiinsular de varios grupos de mamiferos terrestres a traves de las Antillas Mayores septentrionales De acuerdo con esta hipotesis cualquier diferencia en composicion faunal entre las islas (por ejemplo la ausencia de roedores heptaxodontidos en Cuba 0 de monos platirrinos en Puerto Rico) constituye evidencia de extincion local postvicariante y no de dispersion marina fallida Lo primero se puede demostrar si aparecen los fosiles pertinentes 10 segundo no se puede demostrar (MacPhee e Iturralde-Vinent 1994) Por otra parte IturraldeshyVinent y MacPhee (1999 57) han reconocido que aunque fa vicarianza isla-isla puede representar una solucion clara para las distribuciones de varios taxones cuaternarios no puede explicar todos los casos
Rega et al (2002 18) han presentado dos objeciones al modelo de la vicarianza isla-isla aplicado a los mamiferos de las Antillas Mayores
I Si la separacion de Cuba y La Espanola precedio en varios mill ones de anos la separashycion de La Espanola y Puerto Rico el modelo vicariante de lturralde-MacPhee debe predecir mayor similitud entre las faunas de La Espanola y Puerto Rico que entre las de Cuba y La Espanola De hecho es al reyes
No obstante MacPhee et al (200335) admitieron despues que la fecha mas temprana empiricamente defendible [para la apertura del Paso de Mona] debe ser el Oligoceno Medio esto invierte la secuencia de ambas disyunciones con 10 cual queda resuelta la aparente contradiccion Sin embargo en su reciente evaluacion de la congruencia entre filogenia e hipotesis geologic as en varios grupos de mamiferos terrestres de las Antillas Mayores Davalos (2004373) encontro que ninguno de los taxones cumple con la fecha y el patron de divergencia entre las masas terrestres antillanas postulados por el modelo de GAARlandia
2 La presencia de un mismo genero en las tres Islas septentrionales de las Antillas Mayores implica [por el modelo vicariante] que la diferenciacion entre estas formas durante el intervalo desde el Mioceno ha sido unicamente de nivel especifico [Dicha presencia]
315 rRRADlAcr6~ EVOLUCrONARIA
s610 puede explicarse razonablemente como el resultado de movimientos de esos animales entre las islas en el Cuaternario
De esta objecion resulta dificil disentir [El poblamiento entre islas diferenciado solo a nivel especifico no es probable que sea anterior al Pleistoceno (Simpson 1956 15)]
Debe tenerse en cuenta que la hipotesis de la vicarianza isla-isla alude a las tres islas septentrionales de las Antillas Mayores porque como se explico antes (vease El Barco Funeral Vikingo) Jamaica no existia como isla durante el lapso (Eoceno Superior shyOligoceno) consensualmente reconocido como el mas probable para la colonizacion de las proto-Antill as Mayores por la mayo ria de los mamiferos terrestres Despues de emerger como isla en el Mioceno Medio Jamaica debe haber recibido sus mamiferos terrestres por dispersion isla-isla (Davalos 2004376) y solo en el Cuaternario parece haberrecibido (por dispersion marina desde Centroamerica) al roedor murido Oryzomys
Seguramente tanto la dispersion como la vicarianza contribuyeron a que los mamiferos terrestres se irradiaran a traves de las Antillas Mayores y grupos de islas perifericas pero las maneras y tiempos en que estos mecanismos operaron a 10 largo de dicho proceso son aun en gran medida motivos de desacuerdo e incertidumbre
IRRADIACION EVOLUCIONARIA
A continuaci6n revisamos someramente las distribuciones conocidas de los diferentes grupos de mamiferos terrestres a traves de las Islas principales de las Antillas Mayores y grupos de Islas perifericas incluidas las interpretaciones y conjeturas que la literatura reciente recoge en relaci6n con la manera en que dichas distribuciones pudieron haberse conformado durante la irradiaci6n evolucionaria de los referidos grupos de mamiferos Manifestaciones de esta irradiaci6n pueden inferirse unicamente a partir de los taxones presentes en el Cuatemario pues solo 6 de las especies conocidas corresponden al Terciario
GRUPOS ORlGINARIOS DE SURAMERICA
PEREZOSOS (Megalonychidae)
Aparte de un genero miocenico de Cuba y otro del Oligoceno de Puerto Rico se conocen 4 generos (12 especies) de perezosos cuaternarios (todos extintos) restringidos a las tres islas septentrionales de las Antillas Mayores Acratocnus Neocnus Megalocnus y Parocnus Los cuatro generos estan representados en Cuba y La Espanola y Acratocnus 10 esta ademas (como unico genero de perezoso) en Puerto Rico Esta distribuci6n ha sido interpretada por MacPhee et al (2000) eomo resultado de la vicarianza oligomiocenica entre estas islas y [dado que el Banco Puerto Rico - Islas Virgenes fue la entrada de GAARlandia a las Antillas Mayores] se preguntaron si la ausencia de los otros tres generos en Puerto Rico se debe a que nunca estuvieron en ese territorio 0 a que sus
316 BIOGEOGRAFiA
fosiles atm no han sido hallados Pero como hemos visto Rega et al (2002) atribuyeron dicha distribucion a la dispersion interinsular en el Cuatemario
Por otra parte varios amilisis c1adisticos recientes ubi can a Choloepus -genero Reciente conocido solo de Centro- y Suramerica- como taxon hermano de Aeratoenus exc1uidos los demas megaloniquidos antillanos Para White y MacPhee (2001228)
tan estrecha relacion sugiere que la divergencia entre ambos [generos] ocurrio despues de que Aeratoenus divergio de Neoenus Si esta relacion implica que Aeratoenus y Neoenus se dispersaron separadamente desde Suramerica a las AntilIas mientras Choloepus permanecia en el continente (siendo necesarias al menos tres invasiones de perezosos) 0 si Choloepus invadio Suramerica desde las Antillas es algo que el material disponible no permite determinar
La diversidad adaptativa de los perezosos se manifiesta en (1) la utilizacion del habitat especies arboreas en Neoenus y Aeratoenus y terrestres en Paroenus y Megaloenus (Webb 1985a White [1993] White et al 1996 Arredondo [1999]) y en (2) la gama de tallas corporales desde los pequenos Neoenus hasta Megaloenus rodens el mayor de los mamiferos terrestres antillanos (su peso adulto estimado en 270 kg seglin Paula 19679) esto sugiere que los perezosos se irradiaron a nichos que en contextos continentales probablemente estarian ocupados por miembros de otros c1ados (MacPhee e IturraldeshyVinent 2000)
No esta claro si el abanico de tamanos entre estos perezosos enmascara alguna tendencia al gigantismo Por 10 pronto Anderson y Handley (2002) encontraron 10 contrario en perezosos (Bradypus) de las Islas de Bocas del Toro (Panama) shyseparadas secuencialmente del area continental vecina por ascensos del nivel del mar durante los ultimos 10 ka En las cinco islas los perezosos son menores que su contraparte continental y las diferencias entre las tallas medias de las cinco poblaciones expresan una reduccion lineal significativa de la talla corporal en funcion de la edad geologica de cada isla sin relacion alguna con el area de las islas 0 la distancia que las separa del continente
MONOS (Pitheciidae)
Los primates de las Antillas Mayores (cuatro especies extintas) constituyen un grupo monofiletico con Paralouatta (en Cuba) y Antillothrix (en La Espanola) como taxones hermanos seguidos en la escalera c1adistica por Xenothrix (en Jamaica) y finalmente Callieebus como el pariente continental mas cercano esto implica una sola invasion desde Suramerica antes del Mioceno Inferior (fecha mas temprana conocida para los primates fosiles antillanos) y por tanto en consonancia con GAARlandia como la via de colonizacion (Horovitz y MacPhee 1996 1999 Horovitz [1997] MacPhee y Horovitz 2004)
En desacuerdo parcial con la anterior interpretacion la filogenia de este grupo de monos obtenida por Davalos (2004) indica un antecesor comun en Cuba La Espanola y Jamaica
317 TRRADIACTON EVOLUCIONARIA
compatible con la hipotetica conexion -propuesta tentativamente por Iturralde-Vinent y MacPhee (199980-84)- entre las Montanas Azules del este de Jamaica y la peninsula suroccidental de La Espanola en la epoca de GAARlandia
A pesar de que los primates antillanos pertenecen a un grupo viviente de monos muy pequenos (los monos titles) fueron comparativamente grandes en relacion con los platirrinos modernos y sucesivamente mayores en la direccion Antillothrix Xenothrix y Paralouatta con P marianae como el mayor platirrino conocido del Mioceno neotropical (MacPhee 2005) MacPhee et al (1994) consideraron que Xenothrix debio ser un mono de andar lento y al parecer las especies de Paralouatta abandonaron la arborealidad para adoptar un modo de vida mas terrestre (MacPhee 1996a MacPhee et al 2003 MacPhee y Meldrum 2006)
ROEOORES (Echimyidae Capromyidae Heptaxodontidae)
La edad de Zazamys en Cuba demuestra que la invasion de los roedores histricognatos en las Antillas Mayores comenzo antes del Mioceno Inferior por otra parte los representantes mas antiguos de esa irradiacion evolucionaria se encuentran en La Espanola y Puerto Rico Ambas evidencias son consistentes con la hipotesis de GAARlandia (Woods 1982 1988 Woods et al 2001)
No esta claro si los capromidos evolucionaron a partir de un equimido ancestral 0 de un protocapromido 0 si los equimidos antillanos (I sub familia 3 generos y 7 especies todas extintas) forman parte de la irradiacion de los capromidos (4 subfamilias 9 generos y 27 especies 10 vivientes) Kratochvil et al (1978b) y Rodriguez et al (1979) consideraron que Capromys nan us es un eslab6n entre Echimyidae y Capromyidae pero Mesocapromys nanus esta morfologicamente mucho mas cerca de Hexolobodon y los de mas capromidos que de los equimidos (Woods 1985) Lo que si esta claro segun Woods et al (2001) es que la subfamilia Plagiodontinae endemic a de La Espanola es el linaje basal de los capromidos y que Isolobodon Hexolobodon Capromys Geocapromys Mysateles y Mesocapromys [y supuestamente Macrocapromys] forman parte de la irradiacion temprana de Capromyidae
La subfamilia Capromyinae (5 generos y 17 especies 9 vivientes) parece ser el dado de mas reciente evolucion en la familia con invasion de Cuba desde La Espanola y dispersion desde Cuba (1) por Capromys a las islas Caiman y (2) por Geocapromys a las islas Bahamas Caiman y Jamaica y desde Jamaica a las islas Swan aunque la presencia de Geocapromys en Swan pudiera deberse a introduccion aborigen (Morgan 1985 1988 1989 1993 1994 Woods 1989a 1990) Woods et al (2001) utilizaron datos moleculares para obtener estimados independientes de divergencia entre capromidos segiJn 10 cual Geocapromys divergio cerca de los 17 MaAP Mesocapromys divergio de Capromys hace 15 MaAP en tanto lvlysateles 10 hizo hace 10 MaAP Por su parte las formas de Capromys pilorides y de Mysateles prehensilis en Isla de la Juventud divergieron de sus correspondientes subespecies nominativas en la isla principal a los 24 y 08 MaAP respectivamente
318 BIOGEOGRAFiA
Woods y Howland (I979) describieron adaptaciones morfologicas de la mandibula el hioide y la faringe asociadas a mecanismos de masticacion diferentes en plagiodontinos y caprominos y Varona (1980a) llamo la atencion sobre el gran desarrollo de los molariformes (en relacion con la longitud total del cnineo) que presentan ciertos grupos de jutias fosiles en comparacion con las vivientes 10 que pudiera interpretarse como una adaptacion alimentaria a una vegetacion mas abrasiva por parte de las jutias extinguidas
Segun Woods (1990) la familia Heptaxodontidae (2 subfamilias 5 generos y 6 especies todas extintas) tambien seria parte de la irradiacion de los capromidos con origen en el este de La Espanola 0 en Puerto Rico y subsecuente dispersion al Banco de Anguila - San Martin en las Antillas Menores (pero vease MacPhee y Flemming 2003) Este banco estaria a solo 70 km del Banco Puerto Rico - Islas Virgenes en tiempos de maxima regresion marina y esta proximidad con Puerto Rico -patria de Elasmodontomys obliquus- seguramente tuvo que ver con la presencia en las islas de Anguila y San Martin del heptaxodontino gigante Amblyrhiza relacionado mas estrechamente con los roedores de las Antillas Mayores que con los del resto de las Antillas Menores (Woods 1990 1996) Aunque los roedores suelen aumentar de tamano en las islas (Case 1978) la escala del incremento de la talla en Amblyrhiza (que pudo alcanzar mas de 200 kg de peso adulto) esta fuera de toda proporcion con los patrones conocidos en otros roedores insulares (Biknevicius et 01 1993)
Clidomys (Clidomyinae) de Jamaica difiere marcadamente en su morfologia de los heptaxodontinos (vease ademas MacPhee y Flemming 2003)10 cual dificulta explicar su presencia en esa isla Como Jamaica estuvo bajo el mar hasta el Mioceno Medio el ancestro de Clidomys colonizaria esta isla por dispersion desde el extremo suroccidental de La Espanola despues que los heptaxodontinos se originaron en esta ultima isla (Woods 1989a) Pero Pascual et 01 (I 990) especularon que Clidomys pudiera ser el resultado de una invasion posterior e independiente de los capromidos
GRUPOS ORIGINARIOS DE NORTEAMERICA
El origen de las musaranas antillanas ha resultado ser hasta ahora el mayor enigma biogeografico de la fauna de mamiferos terrestres de la region aunque la mayoria de los autores coinciden en considerarlas originarias de Norteamerica (vease Discus ion y Conclusiones en Poblamiento Continente-Isla) No obstante MacPhee e Iturralde-Vinent (1994) descartaron toda posibilidad de conexion terrestre por el Estrecho de la Florida 0 por el Canal de Yucatan y si el (supuesto) evento vicari ante de Rosen (vease El Arca de Noe) -invocado por MacFadden (1980) para resolver el enigma- fuera minimamente plausible aun asi es dificil creer que las catastrofes que 10 sucedieron (vease Contexto Temporal) hayan barrido con toda esa vieja fauna continental excepto las musaranas circunstancia ya anticipada muy tempranamente por Matthew (l918a)
De otra parte la relaci6n entre Nesophontidae y Solenodontidae permanece irresuelta Si estos linajes no estan estrechamente relacionados entonces es muy posible que
319 IRRADIACION EVOLUCIONARIA
sus distribuciones sean explicadas por mecanismos diferentes (Whidden y Asher 2001)
La descomunal diferencia de talla corporal entre las musaranas pequenas (Nesophontes) y las grandes (Soienodon) seguramente impuso a ambos grupos restricciones distintas relativas a la termorregulacion y las adaptaciones energeticas En Nesophontes la seleccion debio favorecer una alta tasa de metabolismo y crecimiento camadas mas numerosas corta expectativa de vida yalta densidad poblacionaL En Soienodon en cambio la seleccion opero en la direccion de una baja tasa metabolica camadas reducidas crecimiento lento longevidad y baja densidad poblacional (Ottenwalder [1991])
MUSARANAS PEQUENAS (Nesophontidae)
Las musaranas pequenas estan representadas en las Antillas por el genero Nesophontes (y un genero miocenico indeterminado en ambar dominicano) con 6 especies cuaternarias extinguidas distribuidas en Cuba La Espanola Puerto Rico Islas Virgenes e Islas Caiman La mayor de las musaranas pequenas Nesophontes edithae evoluciono en Puerto Rico y su arribo a las Islas Virgenes supone una dispersion sumamente reciente En las Islas Caiman Nesophontes se asemeja mas a la poblacion de Cuba de donde seguramente derivo en el Plio-Pleistoceno (junto con Capromys y Geocapromys) cuando la costa sur de Cuba estuvo mucho mas cerca de las Islas Caiman (lturralde-Vinent 2003fig 4) MacPhee et ai (1994) interpretaron la distribucion de Nesophontes en Cuba y La Espanola como resultado de la vicarianza oligomiocenica ocurrida al formarse el Paso de los Vientos pero la distancia taxonomica entre los lados del Paso es solo de nivel especifico y N micros incluso vivi6 en ambas islas segtln Patterson (1962)
Allen (1918) considero que Nesophontes micrus fue una especie primitiva no modificada para la vida arb6rea 0 fos6rea y por tanto de habitos terrestres Nesophontes edithae de Puerto Rico es mucho mayor y mas robusta que las restantes especies del genero eL indice de robustez de su femur (= anchura minima de la diafisis ongitud total del hueso) muestra que este es tan robusto como el de Solenodon paradoxus un animal dos veces mayor (Morgan et ai 1980) Choate y Birney (1968) postularon un aumento cronoclinal de la taIla corporal en N edithae confirmado mas tarde por McFarlane (1999a) mediante fechados radiocarb6nicos de la secuencia estratignifica en depositos cavernarios de Puerto Rico Solenodon no vivi6 en Puerto Rico y en ausencia de una musarana de mayor tamano Nesophontes alcanzo una talla corporal mayor en esa isla que en La Espanola 0 Cuba (Woods y Eisenberg 1989)
MUSARANAS GRANDES (Solenodontidae)
Los arcaicos almiqufes (genero Soienodon) comprenden cuatro especies una viviente (S cubanus) y otra f6sil (S arredondoi) en Cuba y una viviente (S paradoxus con dos subespecies) y otra f6sil (S marcanoi) en La Espanola Como cabria esperar las dos especies cubanas se asemejan mas entre sf que a las especies de La Espanola pero
320 BIOGEOGRAFiA
enigmaticamente S marcanoi comparte caracteres con S cuban us y S paradoxus (Woods y Eisenberg 1989 Morgan y Ottenwalder 1993 Ottenwalder 200 I) Solenodon arredondoi alcanzo una talla corporal superior a la de cualquiera de las musarafias vivientes a nivel mundial Segun Morgan y Ottenwalder (1993) extrapolando las dimensiones y el peso de los almiquies vivientes S arredondoi debio tener una longitud cabeza-tronco y un peso de 450-550 mm y 1500-2000 g respectivamente en comparacion con S cubanus (360 mm y 800 g) y S paradoxus (390 mm y 1100 g)
Entre todas las especies de almiqufes Solenodon paradoxus es la que presenta los caracteres mas derivados (Otten walder 2001) Si estas relaciones son validas la distribucion de Solenodon entre Cuba y La Espafiola debio discurrir de oeste a este como propuso MacFadden (1980) quien invoco la presencia fosH de S cubanus [pero tambien S arredondoi] en Pinar del Rio para postular que el ancestro continental entr~ por Cuba opinion compartida por Perfit y Williams (1989) y por Ottenwalder ([1991] 2001) El paso de Solenodon a Puerto Rico pudo haberlo impedido la existencia de barreras ecologicas durante la conexion del este de La Espafiola con Puerto Rico 0 por la ausencia de Solenodon en esa parte de La Espafiola en aquella epoca Sin embargo las mismas condiciones ecologicas en el este de La Espafiola no parecen haber representado una barrera efectiva para Nesophontes cuyas mas altas tasa de reproduccion y densidad poblacional presumiblemente Ie permitieron expandirse flpidamente a traves de La Espafiola (Ottenwalder [1991])
Es opinion generalizada que los almiqufes antillanos constituyen el grupo de mamiferos que primero se emplazo en el territorio insular (MacPhee 2005) y en tal sentido es de interes la evidencia sobre especificidad parasito-hospedante aportada por Cruz (1980) quien encontro que la especie de acaro (genero Cubanochirus) que parasita a los almiquies es mucho mas primitiva que la especie (del mismo genero) que paras ita a las jutias y de aqui concluyo que dicho genero de acaro debio evolucionar primero en Solenodontidae y posteriormente en Capromyidae al entrar en contacto representantes de ambas familias en Cuba 0 La Espafiola
Recientemente Roca (2004) y Roca et al (2004) dieron a conocer que los resultados de su estudio genetico-molecular sobre los almiquies indican una divergencia de 25 Ma entre Solenodon cuban us y S paradoxus -compatible con la disyuncion tectonica entre Cuba y La Espafiola (Iturralde-Vinent y MacPhee 1999 McPhee et al 2003)-- y una separacion de ambas especies comparable en magnitud a la divergencia entre familias de mamlferos Consecuentemente propusieron que Solenodon cubanus fuera clasificada en el genero Atopogaie como indico Cabrera (1925) hace 82 afios
DISCUSION Y CONCLUSIONES
Las musarafias y los roedores antillanos (excepto Oryzomys) se irradiaron tanto de sus respectivos ancestros continentales que de estos 6rdenes se reconocen familias y subfamilias endemicas de la region en contraste con los perezosos (una subfamilia y tres
321 IRRADIACION EVOLUCIONARIA
tribus endemicas) y los monos (una tribu endemica) como expresion de diferentes niveles de diversificacion evolucionaria Este proceso es dificil de explicar (traUindose de c1ados que pnicticamente comparten la misma antigiiedad en las Antmas) si es correcta la interpretacion de Pascual et at (1990636)
Nosotros consideramos estos diferentes niveles de diversificacion como las diferentes etapas de evolucion alcanzadas por cada grupo [de mamiferos] Obviamente esto estuvo relacionado con las diferentes longitudes de sus historias
Pudiera ocurrir que en el futuro se compruebe que la historia biogeognifica de la region fue mucho mas complicada
La Espanola la isla mas ecol6gicamente divers a de la region debi6 ser el centro principal de evoluci6n de los mamfferos (con 87 de las familias y 73 de las subfamilias) seguida por Cuba (con 75 de las familias y 54 de las subfamilias) las dos islas de las Antillas Mayores mas relacionadas biogeograficamente (Fig 43 Tabla 44)
eL
EL
cb CAIMAN
SWAN
A Acratocnini B Cubanocnini C Megalocnini D Mesocnini E Nesophontidae F Solenodontidae G Xenothrichini
ANGUILAshyM-0 SMARTIN
H Heteropsomyinae I Plagiodontinae J Isolobodontinae
K Hexolobodontinae L Capromyinae
M Heptaxodontinae N CIidomyinae 0 Sigmodontinae
FIGURA 43 Representacion esquematica de las relaciones interinsulares (expresadas porel numero de linajes compartidos) entre las Antillas Mayores y entre estas y los grupos de islas perifericas segun las interpretaciones de los autores citados en el texto Los numeros dentro de los circulos indican la cantidad de linajes representados en cada isla relacion dudosa Contexto taxon6mico de los linajes en la Tabla 44
bull bull
bull bull
bull bull bull
bull bull
bull bull
bull
bull
bull
bull
bull bull bull bull
bull bull
bull bull
bull
bull bull bull
bull bull
bull
bull bull
bull
322 BIOGEOGRAFiA
TABLA 44 Distribuci6n de los linajcs de mamiferos terrestres a traves de las Antillas Mayores y grupos de islas perifericas LE = La Espanola CU Cuba JA Jamaica PR = Puerto Rico IC Islas Caiman IS Islas Swan IB = Islas Bahamas IV = Islas Virgenes AS = Anguila - San Martin
FamiliaSubfumiliafIribu LE CU JA PR IC IS IB IV AS
Megalonychidae
Choloepodinae
Acratocnini
Cubanocnini
Megalocninae
Megalocnini
Mesocnini
Nesophontidae
Solenodontidae
Pitheciidae
Callicebinae
Xenothrichini
Echimyidae
Heteropsomyinae
Capromyidae
Plagiodontinae
Isolobodontinae
Hexolobodontinae
Capromyinae
H e pta x 0 don tid a e
Heptaxodontinae
Clidomyinae
Muridae
Sigmodontinae
PILOSA
SORICOMORPHA
PRIMATES
RODENTIA
bull
bull
bull
bull TOTALES 12 10 4 4 2
323 EXTlICIOIES DEL CUATERNARIO
El gigantismo ha sido sin duda una tendencia evolucionaria recurrente entre los clados mayores de mamiferos antillanos Es un hecho bien conocido que algunos grupos de mamiferos han aumentado de tamafio en las islas y varios autores han intentado explicarlo a partir de enfoques diferentes (Foster 1964 Miller 1966 Sondaar 1977 Case 1978 Eisenberg y Wilson 1978 Heaney 1978 Lawlor 1982 Lomolino 1985 Angerbjom 1985 McNab 19942001)
EXTINCIONES DEL CUATERNARIO
Las islas que integran la Subregion Antillana han perdido casi 90 de sus mamiferos terrestres durante los ultimos milenios y en proporcion con su extension territorial esta perdida ha sido mayor en la Subregion que en cualquier otro lugar del planeta (MacPhee 1996b) Las extinciones del Cuatemario eliminaron mas de las tres cuartas partes de la fauna de mamiferos terrestres conocida de las Antillas Mayores sus islas satelites y grupos de islas perifericas y el examen de este proceso en sus aspectos mas generales se aborda en esta ultima parte del presente capitulo
CRONOLOGiA Y CONSECUENCIAS
Es muy improbable que un fosil represente el ultimo individuo de un taxon extinto y entonces hay que guiarse por la edad radiometrica del fosil mas reciente conocido lIamada fecha de ultima presencia- como unica aproximacion posible a la fecha de extincion del taxon
Como se explico al inicio del capitulo (vease Contexto Temporal) el registro fosH de los mamiferos terrestres de las Antillas Mayores adolece de insuficiente sustentacion radiometrica No obstante la distribucion temporal de los escasos fechados disponibles para el Cuaternario de la region abarca etapas criticas del proceso de extincion experimentado por la referida fauna a 10 largo de los ultimos 250 mil afios
De las 52 especies que es posible reconocer actualmente como miembros extintos de la fauna cuatemaria de mamiferos terrestres de las Antillas Mayores y grupos de islas perifericas los restos fosiles de 23 de elIas (442) han sido fechados radiometricamente o se han hallado asociados estratigraficamente de manera inequivoca con otro material fechado De estos 23 fechados 3 corresponden al interglacial Sangamon 4 pertenecen a la secuencia Pleistoceno terminal - Holoceno temprano 10 se ubican en ellapso entre el arribo de los aborigenes y la incursion europea y 6 son postcolombinos (a los cuales hay que sumar otras dos especies extinguidas en los siglos XIX y xx) A partir de estas 25 especies avaladas por fechas de ultima presencia distribuidas entre los cuatro intervalos temporales sefialados hemos compuesto la Tabla 45 en la que ademas aparecen las restantes 27 especies (carentes de fechado radiometrico) ubicadas en el intervalo temporal mas probable sobre la base de (1) la asociacion estratigrafica de sus restos con evidencias culturales (amerindias 0 europeas) 0 (2) la ausencia de dicha asociacion Sobre estos criterios de ubicacion temporal procede hacer dos aclaraciones
324 BIOGEOGRAFIA
TABLA 45 Intervalos temporales mas probables para las extinciones cuatemarias de los mamiferos terrestres de las Antillas Mayores y grupos de islas perifericas sobre la base de las evidencias siguientes (l) ultimo ejemplar colectado u observado en vida (2) fechado radiometrico mas tardio en material de la especie (3) asociaci6n estratigrafica mas tardia con otro material fechado (4) asociaci6n estratigrafica con restos de animales introducidos restos humanos u objetos culturales propios del intervale (5) falta de asociaci6n con evidencias culturales [Los ntimeros entre parentesis en la columna Evidencia significan adscripci6n tentativa al intervalo] Los estimados del peso corporal (en kg) estan tomados de la literatura 0 inferidos comparativamente y distribuidos entre
las clases siguientes (a) S 01 (b) 01 a lt 1 (c) 1 a lt 10 (d) 10 a lt 100 (e) 100 a lt 200 (j) ~ 200 Las
abreviaturas que designan las islas son las de la Tabla 44
F amilialEspecie Peso IsJa Evidencia Fuente
I httervalo htterg1acial Sangamon (gt80 kaAP)
HEPTAXODONTlNAE
Amblyrhiza inundata e AS 3 McFarJane et al 1998a
CLIDOMYINAE
Clidomys osborni d JA 3 Ibid
Clidomys parvus d JA 3 Ibid
II httervalo Pleistoceno terminal ~ Holoceno temprano (20-6 kaAP)
MEGALONYCHIDAE
Aeratoenus antillensis d CD 5 Arredondo 1975 a
Aeratoenus odontrigonus d PR 3 McFarJane 1999b
Aeratoenus simorhynehus d LE 2 Rega et al 2002
M egaloenus zile f LE 2 MacPhee et al 2000
SOLENODONTIDAE
Solenodon arredondoi e CD 5 Morgan y Ottenwalder 1993
PnHECIIDAE
Paralouatta varonai d CD 5 MacPhee 1996a
EcHIMYIDAE
Heteropsomys antillensis c PR 5 McFarJane 1999b
Heteropsomys insulans c PR 5 Ibid
Puertoricomys eorozalus c PR 5 Woods 1989a
CAPROMYIDAE
P lagiodontia araeum c LE 5 MacPhee et al 2000
Macrocapromys latus c CD 5 Varona y Arredondo 1979
(Continua)
325 EXTINCIONES DEL CUATERNARIO
TABLA 45 (Continuaci6n)
F ami1ia1Especie Peso Isla Evidencia Fuente
M acrocapromys latus c CU 5 Varona y Arredondo 1979
Mesocapromys kraglievichi c CU 5 Ibid
HEPTAXODONTINAE
Xaymaca fulvopulvis a JA 2 MacPhee y Flemming 2003
III Intervalo Amerindio (6-05 kaAP)
MEGALONYCHIDAE
Acratocnus ye d LE 2 MacPhee et aI 2000
Neocnus comes c LE 2 Ibid
N eocnus dousman c LE 2 Ibid
N eocnus toupiti c LE 2 Ibid
N eocnus gliriform is c CU 4 Cordova et ai 1997
M egalocnus rodens f CU 2 MacPhee et ai 2007
Parocnus browni e CU 2 Steadman et ai 2005
P arocnus serus e LE 2 MacPhee et ai 2000
N ESOPHONTIDAE
N esophontes edithae a PR 2 McFarlane 1999b
PITHECIIDAE
Antillothrix bernensis c LE 3 Rimoli 1977
EcHIMYIDAE
Brotomys contractus a LE 4 Miller 1929c
CAPROMYIDAE
P lagiodontia spelaeum c LE 4 Ibid
M acrocapromys acevedo c CU 4 NUiiez 1990a
HEPTAXODONTINAE
Elasmodontomys obliquus d PR 3 McFarlane 1999b
Iv Intervalo Postcolombino (05 kaAP - Presente)
N ESOPHONTIDAE
N esophontes hypom icrus a LE 24 MacPhee y Fle111111ing 1999
N esophontes zam icrus a LE (4) Ibid
N esophontes m icrus a CU 24 MacPhee et aI 1999
(Continua)
326 BIOGEOGRAFiA
TABLA 45 (Continuacion)
F amilialEspecie Peso Isla Evidencia Fuente
N esophontes m icrus a CD 24 MacPhee et aI 1999
N esophontes m icrus a LE 4 MacPhee et aI 2000
N esophontes sp 1 a IC 4 Morgan 1994
N esophontes sp 2 a IC 4 Ibid
Solenodon marcanoi b LE 4 Ottenwalder 2001
PITHECIIDAE
Xenothrix mcgregori c JA 4 MacPhee y Flemming 1999
EcHIMYIDAE
Boromys offeUa a CD 24 Guarch 1984
Boromys torrei a CD 24 Ibid
Brotomys voratus a LE 4 MacPhee et aI 2000
CAPROMYIDAE
P lagiodontia ipnaeum c LE 4 MacPhee y Flemming 1999
Rhizoplagiodontia lemkei c LE 4 Ibid
Isolobodon portoricensis c PR 24 Ibid
Isolobodon montanus c LE 4 Ibid
Hexolobodon phenax c LE (4) Ibid
Capromys sp c IC 24 Morgan 1994
Geocapromys columbianus c CD 4 MacPhee y Flemming 1999
Geocapromys thoracatus b IS 1 MacPhee y Marx 1997
Geocapromys sp 1 c IC 4 Morgan 1994
Geocapromys sp 2 c IC (4) Ibid
l-IEPTAXODONTINAE
Quemisia gravis c LE (1) MacPhee y Flemming 1999
MURIDAE
Oryzomys antillarum a JA 1 Ibid
1 Flemming y MacPhee (1999) alertaron en contra de la asociacion con otro material fechado Asimismo Morgan (1993) indico que la ausencia de asociacion con evidencias culturales amerindias no es necesariamente un criterio concluyente para ubicar un taxon en el intervalo temporal anterior porque hay depositos en la region que carecen de tales evidencias y son del Holoceno tardio por fechados radiocarbonicos [No obstante
327 EXTlNCIONES DEL CUATERNAR10
la ausencia de una especie en residuarios arqueol6gicos adquiere mayor certidumbre como criterio de ubicaci6n temporal de su extinci6n en la medida que aumenta el numero de residuarios estudiados que en las Antillas Mayores se cuentan por centenares] A falta de fechado absoluto de una especie consideramos que estos criterios de ubicaci6n temporal son mas utiles que la alternativa de excluir del analisis a la especie
2 En los trabajos sobre biogeografia de mamiferos antillanos los arque610gos mas citados como autoridades para establecer el inicio de la presencia aborigen en las Antillas Mayores (Kozlowski 1974 Rouse y Allaire 1978 Rouse 1989 1992 Wilson 2001) difieren al respecto entre los 6 kaAP y los 45 kaAP (pero vease Pagan et al 200523) y hemos adoptado la mas temprana de estas fechas para el comienzo del intervalo amerindio en la Tabla 45 De todos modos cualquiera que haya sido la fecha esta no es aplicable por igual al conjunto de las Antillas Mayores porque segun algunos de los autores citados el poblamiento comenz6 por Cuba y La Espanola 6 mil anos atras continu6 en Puerto Rico hace alrededor de 3 mil anos (pero vease Burney et al 1994) yen Jamaica hace menos de 2 mil e incluso a las Islas Caiman ni siquiera llegaron [Las Islas Caiman descubiertas por Col6n en 1503 carecen absolutamente de evidencia amerindia (Stokes y Keegan 1996)]
Si los datos de la Tabla 45 representan una aceptable aproximaci6n a la realidad el proceso de las extinciones cuaternarias de los mamiferos terrestres de las Antillas Mayores y grupos de islas perifericas debi6 caracterizarse entre otros por los eventos siguientes
Aunque obviamente la fauna cuaternaria debi6 ser mucho mas numerosa que 10 que refleja el registro paleontol6gico (Morgan y Woods 1986) 52 especies (81 de la fauna conocida) desaparecieron (sin reposici6n) durante los ultimos 250 mil anos
- Al parecer las extinciones se concentraron en las especies de mayor tamano Asi el peso promedio estimado para las especies de la etapa preantr6pica (intervalos I + II de la Tabla 45) es 350 kg es 328 kg para las especies del intervalo III (amerindio) y es s610 25 kg (c1ases del peso a b c) para las especies del intervalo IV (postcolombino) Ninguna especie aut6ctona de mamifero mayor de 6 kg existe hoy en la regi6n
- De acuerdo con la distribuci6n temporal de las extinciones 61 especies sobrevivieron al interglacial Sangamon (intervalo 1)48 sobrevivieron a la transici6n del Pleistoceno terminal al Holoceno temprano (intervalo II) 34 sobrevivieron al periodo amerindio (intervalo III) y s610 12 especies han sobrevivido desde la Ilegada de los europeos (intervalo IV) Las tasas de extinci6n correspondientes (exceptuado el intervalo I) son una especie cada 1167 anos en el intervalo II una cada 393 anos en el intervalo III y una cada 23 anos en el intervalo IV
- Durante todo el proceso de las extinciones cuaternarias la perdida de especies por isla fue de 91 en La Espanola 83 en Jamaica y 62 en Cuba Bahamas retuvo su unica especie (aunque extirpada en varias de las islas) Puerto Rico e Islas Caiman perdieron todas sus especies (6 y 5 respectivamente) y las Islas Swan Virgenes y Anguila - San Martin perdieron sus respectivas especies (micas
328 BIOGEOGRAFiA
De los cuatro 6rdenes de mamiferos representados en la fauna Pilosa y Primates desaparecieron totalmente Soricomorpha y Rodentia perdieron 86 y 74 de sus especies respectivamente
CAUSALIDAD
CAUSAS NATURALES
Los cicIos glaciales e interglaciales acaecidos en el Pleistoceno causaron cambios ambientales dnisticos en las Antillas con la concomitante alternancia de cIimas frios (secos) y calidos (hUmedos) asi como de descensos y ascensos del nivel del mar aumento y disminuci6n de las areas territoriales) induso (aunque con menor intensidad) en el Holoceno Estos eventos paleogeograficos han sido consistentemente documentados (Mayo y Kartashov 1972 Bonnatti y Gartner 1973 Ortega y Arcia 1982 Prance 1982 Schubert y Medina 1982 Dodge et al 1983 Ortega [1984] Haq et al 1987 Schubert 1989 Hodell et al 1991 Guilderson et al 1994 Oppo 1997 Emslie 1998 Curtis et al 2001 Iturralde-Vinent 2003) y de ellos brindamos a continuaci6n una breve resena
Finalizando el periodo interglacial anterior a la ultima glaciaci6n (Wisconsin) el mar estaba a 8-10 m por encima de su nivel actual Cuando el avance glacial del Wisconsin a1canz6 su culminaci6n (eillamado Ultimo Maximo Glacial) hace 18 ka el mar habia descendido 120 m con respecto a dicho nivel y las islas del Caribe habian incrementado considerablemente sus extensiones subaereas en comparaci6n con las que presentan hoy dia por ejemplo Puerto Rico y las Islas Virgenes se conectaron territorialmente y las Bahamas se unieron en cinco gran des islas Prevalecia un regimen frio y seco y condiciones de aridez --debidas a una reducci6n de las precipitaciones (posiblemente a menos de 440 mmlano)- y los ambientes xericos se habian extendido a traves de las islas y bancos emergidos y abundaban los pinares (los habitats mesicos estaban restringidos a parcelas relictas en 10 alto de las montanas) En el Mar Caribe y el Golfo de Mexico la temperatura superficial del mar era 5degC mas baja que la actual y en las llanuras costeras y alturas maximas eran respectivamente 2-3degC y 6degC mas bajas Los limites de las zonas ecol6gicas altitudinales descendieron varios cientos de metros y cada zona redujo su extensi6n vertical al aumentar el gradiente de temperatura Ellimite inferior de la zona periglacial
elevaci6n donde la temperatura media minima a1canza los OdegC) de la Cordillera Central de la Republica Dominicana (altura maxima del Caribe insular con evidencia de topografia glacial) se considera que estuvo entre los 1500 y 1800 mlnm (actualmente esta a 2800 mlnm) y se puede estimar que dicho limite era inferior en Cuba por estar mas al norte que la Republica Dominicana No obstante durante este periodo la temperatura experiment6 episodios cortos (unas pocas decadas) de ascensos repetidos
Un signifieativo componente bi6tieo de las referidas condiciones paleoambientales 10 constituy6 la espectacular irradiaci6n evolucionaria experimentada por un grupo numeroso de aves rapaees diurnas y nocturnas (Wetmore 1937 Brodkorb 1969 Kurotchkin y Mayo 1973 Arredondo 1976 Olson 1978 Olson y Hilgartner 1982
329 EXTINCIONES DEL ClJATERNARIO
Suarez 2000a b 2004 Suarez y Emslie 2003) Los mamiferos de las Antillas Mayores ofrecieron a estos depredadores aviares una amplia gama de tamanos de presa que en ausencia de mamiferos del orden Carnivora en la region permitio a estas aves irradiarse a este nicho alimentario con la evolucion de una variedad de especies de muy diversa tall a corporal incluidas formas gigantes Adquirir un cuerpo gigante les propicio el acceso a recursos alimentarios que no estaban siendo explotados por ningun otro depredador (Alcover y McMinn 1994) A Cuba corresponde el mejor registro fosil de rapaces cuaternarias caribenas con siete generos de especies diurnas Dentro de estas aves existieron tres gavilanes gigantes (Accipitridae) tres carairas (Falconidae) y un condor (Cathartidae) asi como cuatro generos de especies nocturnas dos lechuzas (Tytonidae) y tres Mhos (Strigidae) dos de ell os no igualados en tamano en todo el mundo Algunos de estos generos tuvieron representacion en otras islas antillanas como La Espanola Jamaica Bahamas y Gran Caiman
Al retirarse hacia el norte los hielos de la glaciacion Wisconsin el clima se tomo mas hUmedo en las Antillas durante el Holoceno temprano y el bosque mesico se expandio en la region insular El nivel del mar ascendio acompanado de mayor pluviosidad alrededor de los 7 kaAP las Islas Virgenes quedaron separadas entre si y de Puerto Rico y las Bahamas perdieron la mayor parte de su territorio Cerca de los 3 kaAP las condiciones secas retornaron a la region e hicieron desaparecer buena parte del bosque mesico Nuevamente el clima comenzo a adquirir sus condiciones actuales hacia los 25 kaAP aproximadamente y hoy transita por una fase mesica con vegetacion xerica confinada en parcelas relictas
Sin duda estos y otros eventos naturales incidieron negativamente en la sobrevivencia de mamiferos terrestres en las Antillas Mayores asi como en los grupos de pequenas islas perifericas (Pregill y Olson 1981 Olson 2006)
McFarlane et al (1998a) demostraron convincentemente como Amblyrhiza el gigante de los roedores antillanos endemico de la pequena isla de Anguila sucumbio a las repetidas y abruptas perdidas de su habitat durante el interglacial Sangamon por las nipidas y severas oscilaciones del nivel del mar (documentadas por Neumann y Hearty 1996) al no poder responder adecuadamente a tan catastroficos eventos mediante el ajuste de su masa corporal media y otro tanto debio ocurrirle en Jamaica al otro roedor gigante Clidomys (McFarlane et al 1998b) emparentado con Amblyrhiza en la misma epoca (Tabla 45 intervalo I)
El ascenso del nivel del mar seguramente promovio la extincion de mamiferos durante la transicion del Pleistoceno al Holoceno (Tabla 45 intervalo II) con la reduccion del tamano de las Islas por las transgresiones marinas (Pregill y Olson 1981 Morgan 1993) Esto ultimo haria maximamente severa la competicion asf como la presion de los depredadores [Steadman et al (2005) no yen por que el calentamiento global ocurrido durante la transicion Pleistoceno-Holoceno habria de tener un efecto negativo sobre los perezosos siendo sus descendientes actuales especies tropic ales 0 subtropicales pero no se trata del efecto
330 BIOGEOGRAFiA
directo del cambio de temperatura sobre los animales sino del efecto indirecto y progresivo que dicho cambio genera a largo plazo al reducir el territorio insular disponible]
Por esta epoca tambien desaparecieron especies de tall a mayor como Megalocnus zile en La Espanola y otras de menor tamano (en Cuba) pero que representan la tall a maxima en sus respectivos contextos taxonomicos del Cuaternario como Solenodon arredondoi Paralouatta varonai y Macrocapromys acevedo Un mamifero puede ser grande solo a costa de reducir su densidad poblacional y esto 10 hace mis vulnerable a la extinci6n sobre todo en una isla (McNab 1994 McFarlane et al 1998a) Incidentalmente Alcover et al (1998a) afirmaron que larelacion entre la talla corporal y la extincion no siempre se cumple en las especies de menor tamano y citaron como ejemplo que entre las musarafias todas las especies de Nesophontes desaparecieron mientras sobreviven especies de Solenodon pero la comparacion correcta en todo caso serfa entre los solenodones vivientes y el gigante Solenodon arredondoi y entre Nesophontes edithae -el gigante de los nesofontes extinguido en el periodo amerindio- y el resto de sus congeneres que sobrevivieron hasta los primeros siglos de la colonizacion europea (Tabla 45 intervalos III y IV)
CAUSAS ANTROPOGENICAS
Intervalo Amerindio
La mayoria de los autores que se han referido a las extinciones acaecidas durante este intervalo (entre elIos paradigmaticamente Martin 1984a b) han responsabilizado a los aborfgenes (que se supone comenzaron a poblar las Antillas Mayores hace 6 mil afios) con la desaparicion de numerosas especies de mamiferos terrestres en la region principalmente por la via de la sobrecaza y la introduccion de animales domesticos devenidos depredadores de la fauna autoctona (Morgan y Woods 1986) Este efecto negativo de la presencia indigena sobre los mamiferos de la region ha sido dramatizado por algunos autores en terminos de holocausto faunal Sin embargo pensamos que se ha exagerado Como han expresado MacPhee et al (19993) cuando uno busca evidencia dura de complicidad humana directa en [estas] extinciones no hay mucho que encontrar Las siguientes son algunas consideraciones a tener en cuenta en tal sentido
El registro radiometrico mas joven conocido hasta ahora para un perezoso antillano (MegalocnllS rodens) es de 42 kaAP (MacPhee et ai 2007) Si se acepta que los aborfgenes entraron en Cuba hace aproximadamente 6 mil anos su convivencia con los perezosos debe haber durado alrededor de 1800 afios (0 mas porque dificilmente el referfdo fechado radiometrico haya correspondido al ultimo sobreviviente de la especie) Esta cifra contradice la hipotesis del holocausto faunal segun la cualla extincion de los mamiferos insulares de mayor talla debe ocurrir con extrema rapidez -a escala de decadas 0 de centurias- a partir de su primer contacto con los humanos (Martin y Steadman 1999) A pesar de las repetidas afirmaciones en contrario (Harrington 1921 Miller 1929c Herrera [1945] Tabio 1951 Alvarez 1951a 1957 Arredondo
331 EXTINCIONES DEL CUATERNARIO
1959 1960a Veloz y Ortega 1973 Mayo 1980b Rodriguez et al 1984 Castellanos y Pino 1985 Pino y Castellanos 1985 Nunez 1990b Gutierrez 1996 Arredondo 2004) en su mayoria basadas en evidencias no exentas de posibles accidentes de deposicion (0 de reubicacion postdeposicional) en recintos cavernarios (Diaz-Franco [2006b D no se ha demostrado inequivocamente que los aborigenes antillanos se alimentaran de perezosos regularmente Sus restos en residuarios de alimentos son escasisimos en toda la region y esto contrasta con los abundantes restos de jutias y otros mamiferos de menor talla que aparecen en esos residuarios demostrativos de la preferencia de los paleoindios por estos otros mamiferos
Los mamiferos mas importantes en la economia prehistorica de las Antillas Mayores fueron las jutias Capromys y Geocapromys en Cuba Geocapromys en Jamaica e Isolobodon en La Espanola Puerto Rico e Islas Virgenes (Aguayo 1950b Tablo 1951 Rimoli 1972 Wing 2001 Diaz-Franco [2006b D Wilkins (2001) estudi61ajutia viviente de Jamaica (Geocapromys browni) en un sitio ocupado por los aborigenes durante 100 anos y no encontr6 evidencias de que durante ese tiempo la especie haya sido sobreexplotada Asimismo en Cuba las jutias conga carabali y andaraz son escasas en depositos pleistoholocenicos en cambio son muy numerosas en residuarios indigenas y de haber sido sobreexplotadas 10 mas probable es que no hubieran sobrevivido hasta hoy
La habilidad de los humanos para penetrar en profundidad el bosque tropical y mantenerse en este sustentablemente ha sido un evento comparativamente reciente (Beran y Steele 1994) En el caso antillano la caza tierra adentro ha sido explicada como un proceso tardio de desplazamiento a partir de la explotacion de abundantes recursos marinos que ofrecian condiciones 6ptimas para el sedentarismo de las comunidades aborigenes costeras (C6rdova y Arredondo 1988 Brito 2005) Peces y mariscos dominan la arqueologia costera de las Islas antillanas en tanto el consumo de recursos terrestres (mamfferos incluidos) aumenta progresivamente en la medida que los residuarios estudiados se alejan de la costa (Scudder 1991 Wilkins 2001) En las Antillas Mayores el componente terrestre de la dieta aborigen es solo 20-30 del material recobrado en los residuarios (Wing 1977 1989) Y debio ser menor alin porque el componente vegetal de la dieta no se ha preservado Por otra parte los aborigenes cubanos domesticaron la jutia conga y seguramente la reprodujeron para comer (Pose et al 1988) practica que probablemente se repitiera con otras jutias en las demas Antillas Mayores
- Fray Bartolome de las Casas hablo de 3 millones de indios en Cuba al inicio de la colonizacion pero los historiadores subsecuentes consideraron exagerada la cifra (Poey 1851 a) No obstante se dispone de dos estimados de la densidad de poblaci6n aborigen en las Antillas Mayores a la llegada de los europeos 18 indigenaslkm2 en Puerto Rico (Rouse 1956) y 1271km2 en La Espanola (Watts 1987) Extrapolando ambos valores entre las cuatro islas principales de las Antillas Mayores tenemos que en la fecha indicada la poblaci6n aborigen pudo estar entre 16 mil y 113 mil indigenas en Puerto Rico entre 21 mil y 145 mil en Jamaica entre 141 mil y 1 mill6n en La Espanola y entre 200 mil y 14 millones en Cuba Entre las cifras maximas y las minimas nos inclinamos
332 BIOGEOGRAFiA
hacia estas ultimas atendiendo a los altos indices de mortalidad infantil y morbilidad que se evidencian en las osamentas de los pobladores prehispanicos (Martinez y Rives 1990) Uno de los hechos que apuntan hacia el bajo numero de aborigenes en las Antillas Mayores es 10 rapido que desaparecieron despues de la llegada de los europeos pnkticamente sin dejar mestizaje con estos 10 cual contrasta ostensiblemente con la situacion en la America continentaL
En suma un grupo humano (1) demograiicamente irrelevante y controlado por altos indices de morbilidad (2) consumidor mayoritariamente de organismos marinos y (3) carente de la tecnologia cinegetica necesaria para poder sobreexplotar los mamiferos que Ie sirvieron de alimento (a los cuales accedio tardiamente en su desarrollo) no es concebible que haya causado e1 holocausto faunal que se Ie atribuye a partir del error de aplicar a las comunidades aborigenes antillanas un modelo cultural de la prehistoria continentaL
Tampoco hay evidencias de que los aborigenes hayan introducido carnivoros con suficiente capacidad depredadora salvo el perro Morgan y Otlenwaldder (1993) consideraron que la depredacion por perros durante el periodo amerindio es la explicacion mas plausible para la extincion del almiqui gigante de Cuba pero como se puede observar (Tabla 54) Solenodon arredondoi aparece en el intervalo II por falta de asociacion con evidencias culturales
Consecuentemente el accionar de los aborigenes y de sus animales domesticos directamente sobre la fauna no parece haber sido por sf solo un factor determinante de la extincion de mamiferos terrestres durante el periodo amerindio en las Antillas (pero vease Steadman et al 2005)
Hay que recordar en este senti do que alin en el Holoceno tardio ocurrieron cambios clirruiticos de consideracion (vease Causas Naturales) Por ejemplo Curtis et al (2001) explicaron que la extincion de algunos ani males mesofilicos de las alturas montanosas de La Espanola pudo haber antecedido a cualquier impacto humano sigrIificativo en esos lugares por coincidir dicha extincion con una tendencia a la aridez documentada isotopicamente y con evidencias de la perdida del bosque mesico ocurridas despues de 3kaAP Yen Jamaica hace solo 2shy3 mil aDos el area de la Bahia de Jackson sustentaba una gran biomasa de vertebrados en bosque hinnedo a juzgar por la cantidad y diversidad de taxones presentes en material fosil derivado de la depredacion por lechuzas hoy el area es la mas seca de la isla con matorral xerofitico y pobreza de fauna (MacPhee 1996a) Yalm en nuestros dias huracanes de gran intensidad aniquilan poblaciones de vertebrados (que no se recuperan) en is las pequenas y llanas del Caribe (Brooks y Smith 2001)
En medio de la incertidumbre sobre la verdadera causa (0 causas) de las extinciones de mamiferos en el periodo amerindio MacPhee y Marx (1997) han propuesto su hipotesis de la hipermorbilidad la cual estipula colapsos poblacionales relativamente rapidos con la introduccion de enfermedades de alta letalidad Estos autores han razonado que
Las islas son los lugares mas apropiados para experimentar perdidas catastroficas causadas por enfermedades Los mamiferos no solo serian susceptibles en virtud de su prolongado aislamiento biologico sino que en los confines de una isla la epizootia
333 EXTINCIONES DEL CUATERNARIO
tendria la potencialidad de infectar simultaneamente a la rnayoria de los individuos de vadas especies Los neonatos son inmunologicamente inmaduros y sufririan desproporcionadamente cualquier infeccion hipervirulenta La perdida excesiva de neonatos y juveniles tendria efectos severos para la sobrevivencia de la poblacion si esta se reduce por debajo de cierto nivel sobreviene la extincion porque no hay posibilidad de restitucion en el periodo postpanzootico Las especies de mayor talla corporal van a ser las mas vulnerables a la extincion dadas sus bajas tasas reproductivas largos periodos de gestacion y reducido numero de progenie Las especies de menor tamano --caracterizadas por altas tasas reproductivas periodos cortos de gestacion y mayor numero de neonatos por parto- tambien serian fuertemente diezmadas pero muchas sobrevivirian porque su estrategia reproductiva les permite perder mayor numero de individuos Las perdidas deben ocurrir relativamente rapido (a escala de decadas) mientras que otras hipotesis (digamos los cambios climaticos) permiten intervalos mucho mas largos (a escala de siglos 0 de milenios) entre la aparicion de las causas y la produccion de sus efectos Por ejemplo todo indica que los nesofontes se extinguieron muy rapidamente y seria dificil poder invocar un impacto humano directo 0 argumentar fenomenos climaticos que pudieran explicar plausiblemente perdidas tan rapidas
MacPhee y Marx (1997 191) han dicho que por el momento la hipotesis de la hipermorbilidad no pasa de ser una abstraccion en espera de adecuada sustentacion empirica Llama la atencion que linicamente en el periodo amerindio (Tabla 45 intervalo 1Il) las extinciones abarcaron todas las clases del peso corporal (de la a a laj) y que ninguna de las especies que sobrevivieron hasta el periodo postcolombino (Tabla 45 intervalo IV mas las especies vivientes) sobrepasa los 10 kg (clases del peso a b c) en clara coincidencia con los presupuestos de la hipotetica hipermorbilidad
Intervalo Postcolornbino
Por 10 menos 37 especies de mamiferos terrestres vivian en las Antillas Mayores y grupos de Islas perifericas a la lIegada de los europeos en 1492 De elIas se han extinguido 23 shymas especies que en cualquiera de los periodos precedentes (Tabla 45~ y solo subsisten 12 especies repartidas entre Cuba (8 especies) La Espanola (2) Jamaica (1) y Bahamas (1) varias de estas en peligro critico de extincion Solo dos de las especies extinguidas durante los ultimos 500 anos se conocieron en estado viviente los roedores Oryzomys antillarum de Jamaica (extinguido en 1877) y Geocapromys thoracatus de las Islas Swan (extinguido en 1955) Las causas de las extinciones ocurridas durante este periodo son sobradamente conocidas destruccion de habitats introducci6n de animales exoticos (domesticos y comensales) y demas presiones asociadas a la actividad humana entre las cuales posiblemente habria que incluir brotes de hipermorbilidad
Durante mucho tiempo se ha afirmado la posibilidad de que especies de Nesophontes hayan sobrevivido hasta las primeras decadas del siglo xx y que incluso continuen existiendo en lugares remotos de Cuba 0 La Espanola (Allen 1918 1942 Miller 1930 Aguayo 1950a Arredondo 1960b Fischer 1977 MacFadden 1980 Woods y
334 BIOGEOGRAFiA
Ottenwalder 1991) Actualmente no existe evidencia radiometrica de que alguna especie de Nesophontes haya sobrevivido al siglo xv las fechas radiocarbonicas calibradas obtenidas en muestras de Nesophontes de Cuba y La Espanola resultaron no mas tardias que los siglos XIII y XIV (MacPhee et al 1999) Por otra parte las expediciones efectuadas a fines del siglo xx con el proposito de localizar individuos vivientes del genero en La Espanola no produjeron la menor indicacion afirmativa al respecto (Woods et al 1985) Ademas a traves de las Antillas Mayores no aparecen restos de Nesophontes en los depositos formados actualmente por las lechuzas
EPiLOGO
Las Antillas Mayores son islas oceanicas no han formado parte de un continente (con la posible excepcion de una parte de Jamaica hasta el Eoceno Medio hace 45 Ma) y durante un tiempo geologico considerable (desde el Eoceno Superior hace 40 Ma) han permanecido emergidas y separadas de los continentes vecinos Consecuentemente los mamiferos terrestres (no volantes) autoctonos de estas islas constituyen una fauna oceanica que por ende cumple con todas las caracteristicas convencionales reconocidas para este tipo de entidad biogeografica
- Baja diversidad taxonomica a nivel de las categorias superiores (solo cuatro ordenes) en comparacion con islas plataformicas continentales (0 con areas continentales) de tamano y diversidad ecologica simi lares
- Alto grado de endemismo a nivel de genero (96) y familia (50) en respuesta al prolongado aislamiento
- Desbalance ecologico de la estructura trofica ausencia de mamiferos camivoros y ungulados
- Espectaculares procesos de irradiaci6n evolucionaria como resultado de la colonizacion de territorios libres de competidores y depredadores
Tendencias evolucionarias extremas gigantismo
- Control a cargo de aves rapaces con tendencia at gigantismo en ausencia de mamiferos miembros del orden Carnivora
- Mas especies de reptiles y de murcielagos que de mamiferos terrestres relaciones que en los continentes se invierten
Alta tasa de extincion de especies (84) sin reposicion
Preservacion de especies filogeneticamente arcaicas (los llamados fosiles vivientes) los almiquies

294 BIOGEOGRAFIA
La geologia y la biologia deben com partir historias comunes Pero este afortunado suceso sera un hecho del futuro no del presente y 10 sera por aproximacion no por precision
Ernest E Williams (1989)
Dieciocho afios despues la sentencia de Williams conserva plenamente su vigencia en 10 que respecta a la biogeografia de los mamiferos terrestres (no volantes) de las Antillas subsisten incertidumbres en la compleja paleogeografia insular y el analisis filogenetico no ha alcanzado un nivel de resolucion satisfactorio para la mayoria de los taxones involucrados porotra parte el registro fosil continUa siendo deficitario no obstante haberse descubierto los primeros fosiles terciarios El presente capitulo es un intento por compendiar la informacion que existe sobre el tema (en parte incierta insatisfactoria y deficitaria) con el proposito de dar respuesta a tres preguntas esenciales (I) lCuaIes son las afinidades continentales de los mamiferos terrestres antillanos (2) lComo y cuando poblaron el territorio insular Los ancestros continentales de esa fauna (3) lQue ocurrio con los mamiferos terrestres una vez establecidos permanentemente en dicho territorio
ESPACIO Y TIEMPO
CONTEXTO GEOGRAFICO
Carecerfa de sentido abordar la biogeografia de los mamiferos cubanos at margen de las restantes Islas de las Antillas Mayores con las cuales Cuba comparte una historia comtm geologica y biotica mas inmediata que con las restantes islas caribefias Por consiguiente el ambito geognifico en el que eL tema es tratado en el presente capitulo abarca Las cuatro Islas principales de las Antillas Mayores (Cuba Jamaica La Espafiola y Puerto Rico) y sus Islas satelites asi como los grupos de Islas perifericas influidas biogeognificamente por alguna(s) de las cuatro Islas principales Islas Swan Islas Caiman Islas Bahamas Islas Virgenes y las dos isLas del norte de las Antillas Menores (cercanas a Puerto Rico) Anguila y San Martin (Fig 41) Entre alternativas de reconstruccion paleogeognifica de este segmento insular hemos seguido a lturralde-Vinent y MacPhee (1999) la version mas reciente y documentada
Para el ambito geognifico asf delimitado (que comprende 96 del territorio insular del Caribe l ) la Tabla 41 presenta la fauna conocida de mamiferos terrestres autoctonos consistente en 4 6rdenes 8 familias 28 generos (mas 2 indeterminados) y 69 especies todas endemicas del referido segmento insular 57 de las cuales (83) estan extinguidas De esta fauna 58 son roedores 20 perezosos 16 musarafias y 6 monos
lA los fines del presente capitulo las denominaciones Antill as y Caribe se emplean indistintamente como terminos equivalentes
295 ESPACIO Y TIEMPO
~) OCEANO ATLANTICO 26deg
J-~ __~~ )Bahamas laquo - ~ o km 400
---~ Florida
lt 221 Mexico ~
c~o
d _-shy ---shy P rt I Virgenes --------
u~ 0 Anguila- I
Rlco ctQ 8 Martin I c=~ -__ -------- ~I Swan ~ o ~ Qo~o
Jamaica Q
- - ------------- Y d
~ - v ~ gt o
MARCARIBE i oJ (
a
SURAMERICA
70middot
FIGURA 41 El Caribe insular_ La linea discontinua circunvala el area geografica objeto de atenci6n en el presente capitulo
TABLA 41 Marniferos terrestres aut6ctonos del segmento insular delimitado en la Fig 41 (especies en orden alfabetico las vivientes en letra negrita =especies precuaternarias) CU=Cuba JA=Jamaica LE=La Espanola PR=Puerto Rico IS=Islas Swan IC=lslas Caiman IB=Islas Bahamas IV=Islas Virgenes AS=Anguila - San Martin
lsI a S Taxonomla 1
CU JA LE PR IS IC IE IV AS
Pilosa (pcrczosos) 14 especics
MEGALONYClllDAE 2
CHOLOEPODIKAE
Acratocnus antillensls bull Acratocnus odontrigonus bull Acratocnus simorhynchus bull Acratocnus ye bull
(Continua)
296 BIOGEOGRAFiA
TABLA41 (Continuaci6n)
I s 1 a s Taxonomla 1
CD JA LE PR IS IC lB IV AS
N eocnus comes bull Neocnus dousman bull N eocnus gliriform is bull N eocnus toupiti bull Gen et sp indet bull MEGALOCNINAE
Imagocnus zazae3 bull M egalocnus rodens bull M egalocnus zile bull Parocnus browni bull P arocnus serus bull
Soricomorpha (musaraiias) 11 especles
NESOPHONTIDAE 4
Nesophontes edithae bull bull Nesophontes hypomicrus bull Nesophontes micrus bull bull N esophontes zam icrus bull N esophontes sp 1 bull Nesophontes sp 2 bull Gen et sp indet bull SOLENODONTIDAE 4
Solenodon arredondoi bull Solenodon cuhanus 5 bull Solenodon marcanoi bull Solenodon paradoxus bull
Primates (monos) 4 especies
PTIHECIIDAE 2
CALLICEBINAE
Antillothrix bernensis bull (ContinUa)
297 ESPACIO Y TIEMPO
TABLA 4 1(Continuacion)
I s a Taxonomia 1
CD JA LE PR IS IC IE IV AS
Paralouatta marianae bull Paralouatta varonai bull Xenothrix mcgregori bull
Rodentia (roedores) 40 especies
ECHllVlYIDAE 2
HETEROPSOMYINAE
Boromys offella bull Boromys torrei bull Brotomys contractus bull Brotomys voratus bull Heteropsomys antillensis bull Heteropsomys insulans bull Puertoricomys corozalus bull CAPROMYIDAE 4
PLAGIODONTINAE
Plagiodontia aedium 6 bull P lagiodontia araeum bull P lagiodontia ipnaeum bull Plagiodontia spelaeum bull Rhizoplagiodontia lemkei bull ISOLoBODONTINAE
Isolobodon portoricensis 7 bull Isolobodon montanus bull Zazamys veronicae bull HExOLOBODONTINAE
Hexolobodon phenax bull CAPROMYINAE
Capromys pilorides 67 bull Capromys sp bull
(Continua)
298 BIOGEOGRAFIA
TABLA 4 1 (Continuacion)
I s a s Taxonom1a 1
CU JA LE PR IS IC IB IV AS
Geocapromys browni 7
Geocapromys columbianus bull Geocapromys ingrahami 6
Geocapromys thoracatus
Geocapromys sp 1
Geocapromys sp 2
Macrocapromys acevedo
Macrocapromys latus
Mesocapromys angelcabrerai
Mesocapromys auritus
Mesocapromys kraglievichi
Mesocapromys nanus
Mesocapromys sanfelipensis
Mysateles melanurus
Mysateles prehensilis 6
bull bull bull bull bull bull bull bull bull
HEPTAXODONTIDAE 8
HEPTAXODONTINAE
Amblyrhiza inundata
Elasmodontomys obliquus
Quem isia grav is
Xaymacafulvopulvis 9
CLIDOMYlNAE
Clidomys osborni
Clidomys parvus
MURIDAE2
SIGMODONTlNAE
Oryzomys antillarum 10
bull
bull bull bull
bull
bull
bull bull
bull bull
bull
bull Total de especies por is1all 24 6 25 7 1 5 1 1 1
1 La taxonomia sigue las revisiones sistematicas mas recientes (Morgan 1993 1994 MacPhee y Grimaldi 1996 Horovitz y MacPhee 1999 White y MacPhee 2001 Ottenwalder 2001 Woods et aI 2001 MacPhee y Flemming 2003) con 1a adici6n de los taxones descritos posteriormente y la incorporaci6n de los cambios propuestos para las especies cubanas en el Capitulo 2 de este Compendio (Continua en pagina opuesta)
299 ESPACIO Y TIEMPO
CONTEXTO TEMPORAL
La absoluta mayoria del registro fosil de mamfferos terrestres antillanos proviene de depositos cavemarios pleistoholocenicos (escala del tiempo geol6gico en la Tabla 42) Abundan en estas cavernas restos de perezosos musarafias y roedores siendo comparativamente escasos los de monos debido quizas a la universal predileccion de los platirrinos por los ambientes densamente forestados 10 que los hace malos candidatos a la fosilizacion (MacPhee y Woods 1982)
A pesar de 10 numeroso de tales depositos en la region son escasos los fechados radiometricos del cuantioso material extraido Con excesiva frecuencia (particularmente en Cuba) estos depositos han sido asignados al Pleistoceno Superior sin fundamento suficiente para ello y es sabido que incluso en cuevas contiguas los depositos fosilfferos pueden diferir considerablemente en edad (Pregill y Olson 1981) Se superponen a esta inconveniencia la falta de estudios tafonomicos en las excavaciones y la eventual mezcla de f6siles de distintas edades como consecuencia de las multicausales alteraciones estratigraficas ocurridas en las cuevas (Nunez 1990a b Morgan y Ottenwalder 1993 MacPhee 1997 McFarlane 1999b Dfaz-Franco 2004) Sobre propuestas de tipificaci6n de los depositos paleontologicos con restos de mamiferos terrestres en Cuba vease Woloszyn y Silva (1977) Acevedo y Arredondo (1982) Gutierrez (1994) Arredondo [1999]
La Tabla 43 presenta los fechados pleistocenicos mas tempranos registrados para restos de mamfferos terrestres del Caribe insular De estos restos el roedor (Cidomys) de Jamaica con un fechado de 253 mil anos antes del presente constituye el fosH de mamifero terrestre mas viejo del Cuaternario anti11ano [Los fosiles fechados antes del surgimiento de la espectrometria de masa (aceleradora) para muestras pequefias pueden ser algo mas viejos (Jull et al 2004)]
Corresponde a MacPhee e Iturralde-Vinent (1994 1995b) e1 relevante merito de haber descubierto en la ultima decada del siglo xx los primeros fosiles terciarios de mamiferos terrestres de las Antillas extendiendo a mas de 30 millones de afios el registro f6sil de
2 Extirpada en la Subregion Antillana pero viviente en el Neotropico continental
3 Ubicacion en subfamilia s610 sugerida por White y MacPhee (200 I)
4 Endemica de la Subregion Antillana
5 Recientemente propuesta su ubicaci6n bajo el genero Atopogale (Roca et al 2004)
6 lncluye las subespecies
7 Transportada por aborigenes a otras de las islas (Rimoli 1974 Olson 1982b este trabajo)
8 Vease MacPhee y Flemming (2003) sobre la incierta situaci6n de esta familia
9 Ubicada provisionalmente en esta subfamilia por MacPhee y Flemming (2003)
10 Hallada en contexto no perturbado de edad Pleistoceno tardio (McFarlane et ai 2002) con 10 cual se descarta su supuesta introduccion por amerindios (Woods 1990)
II La correlaci6n entre el area insular y la riqueza de especies es razonablemente fuerte (R=O83) en estas islas excepto las Bahamas (Morgan 1989)
300 BIOGEOGRAFIA
TABLA 42 Escala convencional del tiempo geologico en la Era Cenozoic a y finales de la Mesozoica (Gradstein 2004) ka = kiloafios (miles de afios) Ma miJIones de afios AP = antes del presente
Periodo(Suhperiodo) Epoca Afios (aproximados) antes del presente
Era Cenozolca
Cuaternario Holoceno Pkistoceno
Presente - 11 kaAP 11 kaAP - 18 MaAP
Terciario (Ne6geno ) Plioceno Mioceno
18 - 5 MaAP 5 - 23 MaAP
oIigoceno 23 - 34 MaAP Terciario (pak6geno) Eoceno 34 - 58 MaAP
Pakoceno 58 - 65 MaAP
Era Mesozo ca
Creticico 65 - 145 -faAP
~~~------~~--- --~-
este grupo animal en la region con el hallazgo de un perezoso oligocenico en Puerto Rico (Yauco) as como un perezoso un mono y un roedor miocenicos en Cuba (Domo de Zaza) Esto unido a la presencia de huesos de una musarafia nesofontoide [referida erroneamente a Solenodon por Rivero (1997)] en ambar oligomiocenico de la Republica Dominicana (MacPhee y Grimaldi 1996) demuestra incontrovertiblemente que al menos en el Mioceno temprano (alrededor de 20 millones de afios atnis) ya estaban en las Antillas Mayores representantes de los cuatro linajes basicos de su fauna de mamiferos terrestres [La afirmacion de Abreu et al (1990 13) en cuanto a que se conocen restos fosiles de almiqui que datan de hace aproximadamente 40 millones de afios es erronea]
Pero lcuan antigua puede ser la fauna de mamiferos terrestres de las Antillas Mayores
Aunque es un hecho bien establecido por la geologia que desde finales de la Era Mesozoica y principios de la Cenozoica existieron islas en el area ocupada ahora por las Antillas Mayores la evidencia disponible indica asimismo que aquellas primeras islas sufrieron repetidas transgresiones marinas totales que habrian hecho sucumbir las biotas que pudieran haber sustentado (MacFadden 1981 Morgan y Woods 1986 MacPhee e Iturralde-Vinent 1994 Roca et al 2004) sin contar los catastroficos efectos que debio tener para el Caribe occidental el b6lido extraterrestre que impacto en Chicxulub (Yucatan) hace 65 Ma (Tada et ai 2003) Hedges (1996) ha especulado que aUn en medio de tal devastacion debe haber existido algun que otro lugar donde elementos de la fauna pudieron haber sobrevivido
301 POBLAMIENTO CONTINENTE-ISLA
TABLA 43 F echados pleistocenicos mcis tempranos (obtenidos por distintos metodos) en mamiferos terrestres del segmento insular delimitado en laFig 41 F =Fuentes (I) Jull et al 2004 (2) Goodfriend y Mitterer 1987 (3) MacPhee et al 1989 (4) Woods 1989b (5) Rega et aI 2002 (6) McFarlane 1999b (7) McFarlane 1 999a (8) McFarlane et al 1998a
Pals Localidad Taxon kaAP F
Cuba Breas de San Felipe Pilosa (Parocnus) 1l9a 1
Jamaica Cueva Coco Ree Cueva Wallingford Roadside
Primates indet Rodentia (Clidomys)
393 2530
2 3
Haiti Trouing Jeremie no5 (casimba) Rodentia (Rh izoplagiodontia) 212 4
Republica Dominicana
Cueva del Perezoso Pilosa (ficratocnus) 185 5
Puerto Rico Cueva de Ia Vaca Cueva Clara
Pilosa (Acratocnus) Soricomorpha (Nesophontes)
337 346
6 7
Anguila Cueva Cave-No-Cave Rodentia (Amblyrhiza) 2060 8
a Kulp etal (1952) reportaron un fechado radiocarbonico de gt 30000 afios para un cono de pino en el deposito fosilffero de Ciego Montero (Cuba) con restos de perezosos
(vcase tambicn Crother y Guyer 1996) pero Iturralde-Vinent y MacPhee (1999) han preguntado donde estaban esos lugares si es que existieron
Solo a partir del Eoceno Superior las Antillas Mayores en fonnacion han pennanecido emergidas hasta hoy [excepto parte de Jamaica y el sur de La Espanola que continuaron sumergidas hasta el Mioceno Medio (Iturralde-Vinent y MacPhee 1999)] Esto significa que solo durante los ultimos 40 millones de anos aproximadamente han existido pennanentemente en dicha region insular ambientes terrestres capaces de sustentar comunidades biotic as (MacPhee e Iturralde-Vinent 2000 Portell et ai 2001 MacPhee 2005) 10 cual proporciona el marco temporal de referencia para conceptuar la antigiiedad maxima posible de la fauna de mamiferos terrestres de las Antillas Mayores
POBLAMIENTO CONTINENTE-ISLA
AFINIDADES CONTINENTALES
El consenso de los autores que en las itltimas dccadas han tratado la filogenia y biogeografia de los mamiferos terrestres de las Antillas Mayores (MacFadden 1980 MacPhee y Woods 1982 Morgan 1985 1988 1993 Morgan y Woods 1986 Ford 1986 Woods 1988 1989a 1990 MacPhee e Iturralde-Vinent 1995a b Hedges 1996 Horovitz [1997] Ottenwalder 2001 Whidden y Asher 2001 MacPheeet ai 2003 Carlini y Scillatoshy
302 BIOGEOGRAFiA
Yane 2004 Davalos 2004 MacPhee y Horovitz 2004 Roca et al 2004) es practicamente unanime en cuanto a que los ancestros continentales que dieron origen a esta fauna derivaron (1) mayormente de Suramerica perezosos (Megalonychidae) monos (Pitheciidae) y roedores (Echimyidae Capromyidae y Heptaxodontidae) (2) en menor medida de Norteamerica musarafias (Nesophontidae y Solenodontidae) y (3) menos aun de Centroamerica una especie de roedor (Muridae)
HIPOTESIS BIOGEOGRAFICAS
La definicion c1asica reconoce dos tipos de islas isla continental (porcion del territorio de un continente separado de este por movimiento tectonico 0 por ascenso del nivel del mar) e isla oceanica (porcion de territorio que no ha formado parte de un continente) La definicion sigue siendo uti por 10 que implica para la formacion de las faunas insulares en uno u otro caso As en el caso de la isla continental su fauna ya estaba emplazada en la futura isla antes de producirse su separacion del continente [Vicarianza hipotesis biogeografica de fragmentacion faunal] en el caso de la isla oceanica su fauna es el resultado de la colonizacion1 por propagulos2continentales a traves de la barrera marina entre el continente y la isla [Dispersion hipotesis biogeografica de acumulacion faunal] En el primer caso el surgimiento de la barrera marina es resultado del evento vicariante en el segundo la existencia de la barrera marina es anterior al evento dispersante Ademas de operar a escala mayor (continente-isla) ambos eventos operan secundariamente a menor escala (isla-isla)
Las dos hipotesis -vicarianza y dispersion- se han disputado hasta hoy la preferencia de los biogeografos en tanto hipotesis mas viable para explicar el proceso de formacion original (continente-isla) de la fauna de mamiferos terrestres de las Antillas Mayores [Sobre la historia de las hipotesis biogeograficas relativas al Caribe insular vease RuB y Schubert 1989 Williams 1989 Pruna 2001] Yel debate continua debido ala falta de consenso sobre 10 que Williams (1989) ha llamado el problema central de la discusion a saber la ostensiblemente baja diversidad taxonomica de esta fauna a nivel de las categorias superiores (solo cuatro ordenes) en comparacion con la diversidad al mismo nivel de las faunas de islas continentales (0 de areas del continente) simi lares en tamafio y diversidad ecologica [Por ejemplo la fauna cubana de mamiferos terrestres consideradas de conjunto las especies vivientes y las extintas- es menos diversa a los niveles taxonomicos superiores (4 ordenes y 6 familias) que 10 que cabria esperar de la diversidad ecologica y extension territorial de Cuba (110 992 km2) comparada con una isla tipicamente continental como Trinidad (4828 krn2
) -veinte veces menor que Cuba y menos diversiticada ecologicamente- donde solo las especies vivientes de mamfferos terrestres representan 6 ordenes y 16 familias (Eisenberg 1989)] Este desbalance
lColonizaci6n Arribo y establecimiento de un propagulo en una isla no ocupada por la especie (MacArthur y Wilson 1967)
2Propagulo Niimero minimo de individuos de una especie capaces de reproducirse y aumentar poblacionalmente en una isla bajo condiciones favorables (MacArthur y Wilson 1967)
303 POBLAMIENTO CONTINENTE-ISLA
taxon6mico ha sido expJicado de dos maneras (1) Inicialmente hubo mas 6rdenes presentes pero la mayoria se extingui6 y s610 sobrevivieron esos cuatro [explicaci6n vicarianista] (2) S610 representantes de esos cuatro 6rdenes lograron cruzar la barrera marina [explicaci6n dispersionista]
Cinco hipot~ticos mecanismos de poblamiento continente-isla han sido invocados durante los ultimos 140 afios (y se los sigue proponiendo actualmente) para explicar la presencia de mamiferos terrestres en las Antillas Mayores A continuaci6n pasamos a resefiarlos brevemente (incluidas sus alegadas fortalezas y debilidades) en el orden aproximado en que historicamente fueron propuestos
EL PUENTE NATURAL (dispersion terrestre activa)
En el siglo XIX el puente terrestre era pnkticamente la unica hipotesis para explicar las distribuciones disyuntas de animales en buena parte del mundo y los primeros investigadores que se ocuparon de la biogeografia antillana (Fernandez [1865] 1884 Chapman [1893] Torre 1892 Lora 1895) la asumieron resueltamente en parte intrigados por el descubrimiento en Cuba en la segunda mitad del siglo de un perezoso f6sil de gran talla Y en su aplicacion a los mamiferos terrestres de las Antillas Mayores el puente sigui6 siendo a 10 largo del siglo xx la hipotesis mas favorecida por los estudiosos del tema (Vaughan 1902a Spencer 191 Oa b Torre 1910a b cAllen 1911 1917a 1918 Barbour 19141916 1945bAnthony 1918 1926 [1942] Scharff 1922 Kraglievich 1926 Beaufort 1951 Arredondo 1961 1970b Kartashov y Mayo 1974 Iturralde-Vinent 1982 1988 Arredondo y Armifiana 1996 Arredondo 1996a [1999]) quienes postularon la existencia preterita de puentes con Norte- Centro- 0 Suramerica en dependencia de las afinidades continentales de los mamiferos antillanos
Teoricamente el puente terrestre debe cumplir con una condicion esencial permitir el paso de una parte mayoritaria (y por tanto representativa de la diversidad) de la fauna continental Claro que siempre habria tipos de mamiferos menos proclives a dispersarse o para los cuales la ubicaci6n del puente resultaria inaccesible 0 las condiciones ecologicas en este fueran desfavorables pero si algunos acceden al puente no hay raz6n para pensar que el resto de esa fauna no los acompafie mayoritariamente Como dijera Simpson (1956) Los puentes no transportan faunas desbalanceadas
En consecuencia el puente terrestre como filtro es un mecanismo de poblamiento minimamente selectivo y por ende de alta diversidad inicial (sensu Iturralde-Vinent y MacPhee 1999) 10 esencial del puente es su capacidad para transportar faunas integradas (Williams 1989) Yen eno radica la objeci6n fundamental al puente terrestre surgida tempranamente en el siglo xx (Por que s610 cuatro 6rdenes de mamiferos terrestres en las Antillas si vinieron por puentes (Donde estan los fosiles de los demas ordenes que debieron venir si es que se extinguieron Los partidarios del hipotetico puente dispusieron de casi todo el resto del siglo para responder una y otra vez que otros 6rdenes de mamiferos aparecerian representados en las Antillas Mayores el dia que en estas Islas se descubrieran fosiles precuatemarios suficientemente
304 BIOGEOGRAFIA
antiguos Esto se logro por fin terminando el siglo como se explico anteriormente (vease Contexto temporal) y ique se encontro los mismos cuatro ordenes de mamiferos terrestres conocidos del Cuatemario caribefio aparentemente los Unicos que lograron llegar
Pero ademas hasta el presente no se ha demostrado inequivoca- y concluyentemente que en algun momenta durante los ultimos 40 millones de afios haya existido una conexion terrestre continua entre las Antillas Mayores (en proceso de formacion 0 plenamente formadas) y algun punto de las regiones continentales vecinas
LA BALSA NATURAL (dispersion marina pasiva)
Durante buena parte del siglo xx el puente terrestre tuvo como unico rival a la hipotesis de la balsa natural -transporte accidental de animales sobre islotes de vegetacion flotantes a la deriva despues de arrancados y expulsados mar afuera por un rio crecido- expuesta originalmente por Matthew (191319151916 1918a 1919) y desarrollada en su aplicacion a los mamfferos terreslres antillanos principal mente por Simpson (1956) asi como por olros investigadores (Darlington 1938 1957 Williams y Koopman 1952 Hershkovitz 1958 1972 Patterson 1962 Paula 1967 Patterson y Pascual 1972 Pregill 1981 Ford 1986 Woods 1989a 1990 Morgan 1993) Por su parte Hedges et al (1992) sustentaron dicha hipotesis en estimados de divergencia (basados en el reloj molecular) en la herpetofauna antillana -involucrados tangencia1mente los mamiferos [veanse objeciones a Hedges y colaboradores (Page y Lydeard 1994 Iturralde-Vinent y MacPhee 1999) respuestas a las objeciones (Hedges et al 1994 Hedges 2001) y contrarrespuestas (MacPhee e IturraldeshyVinent 2005) asi como cuestionamientos al metoda de los estimados de divergencia (Foote et al 1999 Pregill y Crother 1999 Woods et ai 2001 )]
La hip61esis de la balsa natural se origino a partir de las numerosas ocasiones en que a 10 largo de siglos tales islotes flotantes (portando 0 no vertebrados incluidos mamiferos) han sido avistados mar afuera a cientos de kilometros de la costa mas cercana en varias partes del mundo --inc1uido el Mar Caribe- segUn consta en diarios de barcos y reportes de observadores diversos (revisados por King 1962 reportes posteriores en SICSP 1970 y Censky et al 1998) Uno de estos casos acaecido en nuestra region se reporta en un informe del SICSP (Smithsonian Institution Center for Shortlived Phenomena 197050shy52)
EIB de julio de 1969 un destructor de la armada de Estados Unidos avisto un islote flotante a unos 100 km al sur de la Base Naval de Guantanamo moviendose desde el Paso de los Vientos en direccion suroeste a una velocidad de 45 kmlh islote de unos 15 m de diametro mantenfa en pie mas de 10 palmas reales de 10-15 m de altura con suelo cubierto de hierba y arbustos [la observacion con binoculares a 18 km de distancia no preciso la presencia de vida animal] La travesia fue rastreada durante 6 dfas hasta que el 19 de julio el islote se dio por perdido tras 6 horas de busqueda aerea
305 POBLAMIENTO CONTINENTE-ISLA
La colonizaci6n exitosa de una isla de las Antillas Mayores por mamiferos terrestres continentales transportados en balsas naturales a la deriva (de cada especie al menos una hembra gravida 0 dos individuos de sexo opuesto) dependeria de factores que van a operar desde el momenta mismo de formarse la balsa asi como durante su travesia marina y su arribo a la isla
a) La posibilidad de estar en la balsa Que la distribuci6n de la especie incluya areas contiguas al rio y que el animal se encuentre en ellugar y el momenta propicios para ser incorporado a la balsa antes de que el rio crecido la desprenda Por ejemplo segun Hershkovitz (1958) los equimidos -grupo ancestral de roedores antillanos- serian buenos candidatos a ser captados por la balsa porque acostumbran a anidar en huecos del tronco de los arboles a la orilla de los rios
b) El tamano y la consistencia de la balsa Muchas se desintegran en alta mar debido a su fragilidad ante la saturaci6n Algunos autores (Beaufort 1951 Arredondo 1996a [1999]) han expresado incredulidad en cuanto a que las balsas hayan podido soportar el peso de un animal como el gran perezoso cubano (Megalocnus rodens) el mayor de los mamiferos antillanos Pero esta especie evolucion6 en Cuba y el ancestro que hizo el viaje pudo ser muy pequeno [EI perezoso oligocenico de Puerto Rico es el mas pequeno de los megaloniquidos antillanos (MacPhee e Iturralde-Vinent 1995b )]
c) Las caracterfsticas fisiol6gicas de la especie Los limites fisiol6gicos de los propagulos constituyen nuestra unica objeci6n a la dispersi6n marina por balsas como mecanismo para la formaci6n de la fauna de vertebrados en las Antillas (MacPhee e IturraldeshyVinent 2005) Unas especies tienen intrinsecamente mas posibilidades que otras para sobrevivir a las vicisitudes que deben afrontar durante la travesia (deshidrataci6n inanicion insolaci6n etc) Por ejemplo los megaloniquidos han demostrado tener extraordinaria habilidad para cruzar barreras marinas fueron los primeros mamfferos suramericanos en invadir Norteamerica mucho antes de que surgiera la conexion terrestre a finales del Plioceno (Paula 1967 Olson 1978 Webb 1985b Martin y Steadman 1999) Incluso se conoce un megaloniquido acultitico del Plioceno peruano (Muizon y McDonald 1995) y otro viviente en Suramerica (Choloepus) que pnicticamente no bebe agua 10 que ha originado una leyenda aborigen que explica por que los perezosos no toman agua (Goff art 1971) Vease ademas el inciso e
d) EI regimen de las corrientes marinas Que conduzcan a la balsa en la direcci6n necesaria EI transporte de balsas de vegetacion desde los continentes hacia las islas fue durante la mayor parte del Cenozoico sumamente improbable (MacPhee e IturraldeshyVinent 2000 148) Estos autores han insistido en que hay que tener en cuenta que durante la cambiante geografia cenozoica del Caribe las corrientes marinas de superficie no favorecieron el desplazamiento de objetos fiotantes hacia las Antillas Mayores [Pero no sabemos con certeza que efecto modificador del curso de tales objetos fiotantes pudieron introducir las tormentas tropicales si previsiblemente funcionaron en el pasado como 10 hacen ahora] Dichos autores senalaron ademas (2005) que en un experimento reciente con boyas de posicionamiento satelital liberadas a la deriva desde las Antillas Menores las boyas fueron a parar mayoritariamente a Centroamerica y a otros lugares
306 B[OGEOGRAFIA
y solo unas pocas recalaron en las Antillas Mayores y las que entraron al Mar Caribe demoraron semanas 0 meses en arribar
e) La distancia a recorrer (y en consecuencia la duracion de la travesia) entre el continente y la isla (10 que repercute decisivamente en el factor c) Este es desde luego uno de los factores mas criticos En la medida que la distancia aumenta la probabilidad de alcanzar la isla disminuye drasticamente con el agravante de que la trayectoria dificilmente seria en linea recta Si tomamos como referencia la velocidad de 45 kmlh del islote ya mencionado (SICSP 1970) cubrir la distancia aerea (aproximada) de 800 km que media entre el noroeste de Suramerica y La Espafiola Ie tomaria a la balsa 74 dias Estimando el doble de este tiempo debido a desvios de la trayectoria la duracion de la travesia probablemente resultaria excesiva para la mayoria de los mamiferos Pero pudiera muy bien ocurrir que la velocidad fuera mayor 0 que la balsa portara plantas vivas que permitirian a los fitOfagos sobrevivir [Tres de los cuatro ordenes de mamiferos terrestres conocidos de las Antillas Mayores son fitOfagos] Ademas esta geologicamente bien establecido que durante etapas de variada duracion desde el Oligoceno hasta el Pleistoceno los territorios del Caribe insular y continental aumentaron considerablemente en extension con niveles del mar 100-200 m por debajo de su nivel actual oportunidades que los propagulos aprovecharian para cruzar barreras marinas significativamente menores
f) El tamafio de la isla Las islas mayores deben interceptar mas balsas (Buckley y Knedlhans 1986) 10 que repercute directamente en el factor e La balsa puede pasar entre islas pequefias y continuar viaje sin llegar a ningun lugar
g) Las condiciones (favorables 0 adversas) del desembarco en la isla y el tiempo que Ie tomaria a uno 0 mas animales depauperados encontrarun habitat apropiado Esto dependeria a su vez del tamafio y diversidad ecologica de la isla (Darlington 1938 Miller 1966)
Tal cumulo de posibles obstaculos y dificultades constituye a primera vista un desafio a la credibilidad de la hipotesis Matthew-Simpson pero en ello radica precisamente su valor explicativo bas ado en la mas absoluta improbabilidad La colonizacion por la via marina es altamente improbable pero no absolutamente imposible si se tiene en cuenta la inmensidad del tiempo geologico se vuelve probable (McKenna 1973) Basta que solo una vez entre mill ones de intentos fallidos durante ese inmenso lapso algunos de los factores anteriormente enunciados coincidan en no ser tan desfavorables para el propagulo Y eso es todo 10 que demanda la hipotesis para cada uno de los grupos monofileticos mayores de mamiferos terrestres presentes en las Antillas Mayores Las Antillas Mayores requeririan el arribo de tan solo 7 (quizas menos) propagulos fundadores a 10 largo de los ultimos 30 0 mas millones de afios (promedio de uno cada 43 mill ones) para explicar la formacion de toda su fauna de mamiferos terrestres por dispersion marina pasiva (vease Discusion y Conc1usiones)
En comparacion con la via marina el poblamiento por la via terrestre es predecible porque la mayor parte de la fauna continental de mamiferos emplazada en la region del puente va a cruzar por este una vez que el mismo se establece y los animales logran
307 POBLAMIEITO CONTINENTE-ISLA
acceder a el el poblamiento por la via marina en cambio es impredecible porque depende total mente del azar que caracteriza a la balsa Claro que en ambas vias opera el azar pero en el primer caso se trata del azar de un evento probable en el segundo el azar de un evento improbable (Simpson 1940) Asi el puente natural (mecanismo de poblamiento minimamente selectivo = alta diversidad inicial) y la balsa natural (mecanismo de poblamiento maximamente selectivo baja diversidad inicial) pueden conceptuarse como los extremos opuestos de una gama de probabilidades de dispersi6n (McKenna 1973) EI fen6meno de la dispersi6n merece la mayor atenci6n cientifica en los estudios de biogeografia insular (Cowie y Holland 2006)
Queda claro que la balsa natural es una hip6tesis consistente con el caracter de la fauna objeto de estudio puesto que el desbalance taxon6mico (sin evidencia paleontol6gica en contrario) s610 puede ser resultado de la improbabilidad del poblamiento No obstante ello no implica necesariamente que la balsa natural haya sido el imico mecanismo (ni siquiera el principal) utilizado por los mamiferos terrestres en su colonizaci6n de las Antillas Mayores La balsa natural debe satisfacer una premisa te6rica esencial dado el caracter azaroso impredecible e improbable de una colonizaci6n exitosa esta debe ocurrir a intervalos muy espaciados a traves del tiempo geol6gico 0 sea durante todo el tiempo que en el area ocupada por las Antillas Mayores ha habido ecosistemas terrestres perrnanentemente disponibles para ser colonizados (los ultimos 40 millones de afios) Sin embargo como sabemos representantes de los cuatro 6rdenes de mamiteros terrestres conocidos de las Antillas Mayores ya estaban emplazados en dicho territorio insular en el Mioceno Inferior 10 que supone un lapso de colonizaci6n limitado al Eoceno SuperiorshyOligoceno (MacPhee 2005) sugerente de un tipo de mecanismo contrario a la colonizacion alocronica exigida por la hipotesis de la balsa natural sobre todo tratandose de una fauna caracterizada por altas tasas de extincion sin reposicion aparente
La presencia en Jamaica (hasta su extinci6n en 1877) del roedor mfuido Oryzomys antillarom con estrecha afinidad centroamericana (Woods 1989a) pudiera constituir el imico caso efectivo de poblamiento (continente-isla) de las Antillas Mayores por dispersion marina dc un mamifero terrestre para el cual no es razonable invocar otro mecanismo de colonizacion
LA GUIRNALDA INSULAR (dispersion mixta)
La geologia ha establecido con suficiente certeza que durante el Cenozoico unicamente en el Pale6geno tardio y solo a traves de la Cresta de Aves (ahora sumergida excepto la isla hom6nima) pudo haber existido una cadena de islas para el transito de mamiferos terrestres entre el norte de Suramerica y las proto-Antillas Mayores [desestimados a ese efecto el Canal dc Yucatan el Elcvado de Nicaragua la Cresta de Caiman el Elevado de Beata y las Antillas Menores (Holcombe et al 1990 MacPhee e IturraldeshyVinent 1994 1995b Iturralde-Vinent y MacPhee 1999)] Holcombe y Moore (1977) y Holcombe y Edgar (1990) postularon la existencia de esta entidad y estimaron algunas de sus posibles caracteristicas Por su parte MacPhee e Iturralde-Vinent (1995b) e Iturralde-Vinent y MacPhee (1999) precisaron los extremos proximal y distal de esa
308 BIOGEOGRAFIA
estructura acotaron la duracion y la edad de la misma y Ie dieron nombre GAARlandia (Greater AntillesA vesRidgelandia)
Segun Iturralde-Vinent y MacPhee (1999) la Cresta de Aves emergio por breve tiempo (1-2 Ma) durante la transicion Eoceno-Oligoceno (35-33 MaAP) por la coincidencia de un levantamiento tectoni co y un considerable descenso eustatico del nivel del mar En ese momenta las islas septentrionales del arco de las Antillas Mayores (Cuba centroorientalshyLa Espafiola - Puerto Rico - Islas Virgenes) constituian una sola isla grande 0 una serie de islas separadas por barreras muy estrechas La Cresta de Aves emergida entre el bloque Puerto Rico - Islas Virgenes y un microcontinente noroccidental de Suramerica -separado en ese momento del resto del continente por una via marina surgida en el Eoceno tardio- dio origen a GAARlandia
Obviamente el numero de colonizaciones exitosas por mamiferos terrestres a traves de una guimalda de Islas tiende a disminuir mientras mas anchas sean las barreras marinas entre las islas que forman la guimalda y cuando esto ocurre disminuye tambien la diversidad taxonomica de los propagulos particularmente a nivel de las categorias superiores (Simpson 1956)
Vemos asi que para los mamiferos terrestres la colonizaci6n a traves de una guimalda insular puede ser tan improbable como a la deriva en una balsa natural en dependencia de la cantidad y anchura de las barreras marinas a cruzar el tamafio y la fisiografia de las islas que integran la guimalda y el regimen de las corrientes marinas y aereas entre otros factores
Resumiendo la guimalda insular puede conceptuarse como un mecanismo de poblamiento altamente selectivo (= baja diversidad inicial) que combina la improbabilidad de la balsa natural con la colonizacion restringida temporaimente del puente terrestre
En su reciente revision de la taxonomia distribuci6n y filogenia (basada en estudios moleculares) de varios grupos de mamiferos antillanos Davalos (2004) concluyo que los perezosos megalocninos los primates y los roedores histricognatos se avienen al patron de divergencia con el continente y epoca de arribo a las Antill as implicados en la hipotesis de GAARlandia pero no asi los perezosos coloepodinos que requieren una explicacion biogeognlfica distinta Esto ultimo (sospechado por Matthew 1918a) habla sido confirmado por White y MacPhee (2001 226-227)
Nuestra evidencia filogenetica no sustenta la idea de un dado unificado surgido de la invasion de un megaloniquido el analisis sugiere mas bien dos eventos de colonizaci6n independientes [y] sustenta fuertemente un origen difiletico de las subfamilias [Choloepodinae y Megalocninae] Si hubo dos migraciones a traves de GAARlandia presumiblemente poco espaciadas en el tiempo no se puede determinar en el presente
Finalmente queda claro que GAARlandia -como guirnalda insular- es la hipotesis mas robusta propuesta hasta ahora para explicar la presencia en las Antillas Mayores de
309 POBLAMIENTO CONTINENTE- ISLA
la absoluta mayona de los mamfferos terrestres de ongen suramericano porque el caracter de esa fauna supone un mecanismo de poblamiento ancestral que sea a la vez maximamente selectivo (baja diversidad ordinal) y temporalmente restringido (transicion Eoceno-Oligoceno)
Recientemente Rega et al (2002 18) expresaron A pesar de una revision exhaustiva de la paleogeografia de la cuenca del Caribe Iturralde-Vinent y MacPhee (1999) no pudieron confirmar la presencia de una conexion terrestre continua entre Suramerica y las proto-Antillas aunque hipotetizaron su existencia [Vease la Fig 42 (= figura 6 de Iturralde-Vinent y MacPhee 1999 simplificada))
En el trabajo citado por Rega y colaboradores aparece la siguiente explicacion
Actualmente no es posible ofrecer reconstrucciones paleogeograficas detalladas de la Cresta de Aves Sin embargo hay evidencia de que porciones de la cresta deben haberse elevado subaereamente en un tiempo La cuestion cntica es si la Cresta de Aves emergida pudo haber formado algun tipo de corredor entre las Antillas Mayores y el noroeste de Suramerica Perforaciones en las alturas estructurales de la cresta pudieran proveer el registro del Paleogeno tardio que se necesita para precisar la hipotesis del corredor aunque no se puede predecir si tales datos serian suficientemente detallados para determinar si en algun momento GAARlandia constituyo una sola entidad subaerea Por ejemplo pudo haber barreras marinas por corto tiempo entre las masas terrestres de la Cresta de Aves [0) ellevantamiento y la subsidencia pudieron afectar diferentes partes de GAARlandia en diferentes tiempos AItemativamente los elementos faunales pudieron haberse dispersado de manera pulsada Movimientos episOdicos de este tipo incrementarian la extincion de los elementos faunales incapaces de trasladarse a la siguiente masa terrestre en la cadena disminuyendo asi la diversidad total (Iturralde-Vinent y MacPhee 199957)
La primera parte de la explicacion deja claro que la posibilidad de que la Cresta de Aves haya sido una conexion terrestre continua no se puede descartar pero tampoco se puede afirmar La ultima parte de la explicacion describe el efecto altamente selectivo (filtrante) de la colonizacion a traves de una cadena de islas
Y continuan lturralde-Vinent y MacPhee (199957 -58) explicando
Hasta que punto el microcontinente noroccidental de Suramerica [considerado por dichos autores como el area suministradora de propagulos hacia GAARlandiaJ estuvo fisicamente separado del resto del continente no ha sido plenamente esc1arecido Si estuvo sustancialmente aislado por barreras marinas y orogenic as durante el Paleogeno tardio entonces solo la fraccion de la biota suramericana emplazada en el microcontinente estuvo en posicion de acceder a GAARlandia Esto pudiera tener que ver con el hecho de que la fauna antillana carezca de representantes de muchos grupos suramericanos [de mamfferos) que debieron haber hecho el viaje
310 BIOGEOGRAFiA
Florida o Alturas0 0
d 6 D L1anuras25deg
Yucatan
20deg
deg0
10deg
MARCARIBE
80deg 75deg
Bahamas
Cres ta de Beata
D Mares someros
D Mares profundos
OCEANO ATLANTICa
FIGURA 42 Reconstrucci6n hipotetica de GAARlandia como conexi6n terrestre continua entre las proto-Antillas Mayores y el noroeste de Suramerica en el Pale6geno tardio segun la FIgura 6 (simplificada) de Iturralde-Vinent y MacPhee (1999)
En su defensa del puente terrestre Barbour (1945b249) se refiri6 a esto - antes 10 habfa hecho Anthony [1942363]- en los terminos siguientes
El argumento de que si las Antillas hubieran estado conectadas con el continente se encontrarian los f6siles de un numero mayor de distintas criaturas me deja frio es perfectamente concebible que la conexi6n haya ocurrido en un periodo en que la parte del continente implicada no estuviera poblada por una fauna grande y variada
Y concluyen lturralde-Vinent y MacPhee (199958) su detallada explicaci6n
Desafortunadamente sabemos muy poco sobre la composici6n faunal del noroeste de Suramerica a finales del Pale6geno
Consecuentemente si se confirmara la magnitud del aislamiento del aludido microcontinente y su concomitante baja diversidad faunal GAARlandia como conexi6n terrestre continua serf a la hip6tesis mas parsimoniosa No obstante mientras no se disponga de evidencia concluyente a este respecto hipotetizar a GAARlandia como conexi6n continua (minima selectividad) la coloca en franca vulnerabilidad te6rica al tener que afrontar la objeci6n clasica al puente terrestre (baja diversidad ordinal de la
311 POBLAMIENTO CONTINENTE-TSLA
fauna) al tiempo que se la priva de ser la hip6tesis mas robusta para explicar -como guimalda de islas- la colonizaci6n de las Antillas por mamiferos procedentes de Suramerica
ELARcA DE NOE (seudovicarianza)
El primer vicarianista antillano (desde luego sin el saberlo) fue Corral (1940) con su utilizaci6n ingenua de las ideas wegenerianas de la deriva continental aparecidas la decada anterior Pero el primer (supuesto) intento de aplicar a la formaci6n de la biota caribefia el mecanismo de la vicarianza basada en la tearia de la tect6nica de placas (surgida en la sexta decada del pas ado siglo) se debe a Rosen ([1976] 1985)
Segun Rosen las Antillas Mayores se originaron en el Cremcico en la Placa Caribe aproximadamente en la posici6n que hoy dia ocupa Centroamerica y al ser desplazadas tectonicamente hacia el este interactuaron (de algun modo no precisado por Rosen) con las margenes continentales de Norte- y Suramerica de donde (en un evento unico) adquirieron toda su fauna transportada subsecuentemente por dichas islas en su transito hacia sus posiciones actuales [De ahi la denominacion metaforica dada par McKenna (1973) a este mecanismo de poblamiento] Aunque se ha presentado esta propuesta como Model0 de Vicarianza Continente-Isla de Rosen en realidad por definici6n no constituye un evento vicariante porque las proto-Antillas Mayores no resultaron del fraccionamiento y separaci6n por tectonismo de una porcion del territorio continental como exige esta hip6tesis (vease definici6n de vicarianza al comienzo de Hipotesis Biogeograficas)
El Arca de Noe de Rosen ha sido desestimada por la mayoria de los autores contemporaneos porque (1) carece de sustento geol6gico (lturralde-Vinent y MacPhee 1999) y porque (2) en el periodo en que se afrrrna ocum6 el evento muchos de los ordenes modemos de mamiferos placentarios attn no existian (Williams 1989)
EL BARCO FUNERAL VlKINGO (tanatovicarianza)
Domning et al (1997) reportaron 10 que consideraron el mamffero terrestre mas viejo de las Antillas --circunstancia que se refleja en el titulo mismo del trabajo Oldest West Indian Land Mammal- al descubrirse un perisodactilo rinocerotoide (Hyrachius sp) en una localidad de edad Eoceno Inferior a Medio en el occidente de Jamaica
Jamaica ha tenido una historia geologica y paleogeografica distinta de la historia comun a las restantes Antillas Mayores A principios del Cenozoico Jamaica occidental (a diferencia de la porcion oriental constituida por el Bloque de las Montafias Azules) formaba parte de Centro america de la cual se desgaj6 (por el desplazamiento de la Placa Caribe) para venir a integrarse despues del Eoceno Medio con 10 que serfa la futura isla de Jamaica desaparecida por transgresi6n marina en el Eoceno Superior hasta volver a emerger totalmente en el Mioceno Medio (Iturralde-Vinent y MacPhee 1999 MacPhee e Iturralde-Vinent 2000)
312 BIOGEOGRAFiA
Por consiguiente Hyrachius ---conocido de localidades de edad Eoceno Inferior a Medio en Norteamerica- debio ocupar (10 que iba a ser) Jamaica occidental mucho antes de que esta se separara de Centroamerica De hecho este fosil Uego a su destino insular contenido en las rocas de su basamento no como un ser vivo por 10 que es erroneo considerarlo un animal antillano (lturralde-Vinent y MacPhee 2004) como han supuesto algunos (Hedges 200 I Portell et al 200 I Condis [200 I D Todo 10 que hizo fue viajar tanatovicariantemente en su barco funeral vikingo -otra de las denominaciones metaforicas de McKenna (1973)
DISCUSION Y CONCLUSIONES
Hemos visto que solo disponemos de una hipotesis plausible para explicar la mayor parte del poblamiento (continente-isla) de las Antillas Mayores por mamiferos terrestres del Neotropico continental la dispersion mixta (a traves de GAARlandia) para los perezosos (excepto Choloepodinae) los monos y los roedores (excepto Oryzomys)
Por otra parte la historia biogeogrMica de las musaraiias de las Antillas Mayores (Nesophontes y Solenodon) constituye una de las situaciones mas enigmaticas entre los mamiferos de la region (Ottenwalder 2001321) Al menos cinco mecanismos de poblamiento continente-isla han sido invocados para explicar cuando como y de donde llegaron a las Antillas Mayores estas musaraiias
I EI propagulo ancestral (del Terciario de Norteamerica) coloniz6 a las Antillas Mayores por dispersion marina desde Centroamerica 0 desde la Florida (Patterson 1962)
2 El propagulo ancestral ( del Terciario) cruzo el Atlantico Sur (supuestamente mas estreshycho en aquella epoca) directamente de Africa a las Antillas Mayores (Hershkovitz 1972)
3 El propagulo ancestral (del Terciario de Norteamerica) Uego a las Antillas Mayores en el Arca de Noe de Rosen (MacFadden 1980)
4 El propagulo ancestral (del Terciario de Norteamerica) fue transportado a las Antillas desde Centroamerica en el bloque de Jamaica occidental y de alli paso por dispersion marina a las restantes Antillas Mayores (Whidden y Asher 200 I)
5 El propagulo ancestral (del Terciario de Norteamerica) lleg6 a las Antillas Mayores a traves de GAARlandia (Whidden y Asher 2001)
Aunque hay consenso entre estos (y otros) autores sobre el origen norteamericano de las musaraiias antillanas ninguno de los cinco hipoteticos mecanismos de poblamiento resiste la evidencia en contrario filogenetica yo paleogeogrMica Ademas algunas de estas hipotesis asumen que Nesophontidae y Solenodontidae comparten un antecesor comun en las Antillas pero esta relacion no ha sido confirm ada Para dilucidar el enigmatico caso de las musaraiias antillanas se requerini profundizar en la investigaci6n geologica paleogeognifica y paleontol6gica (Iturralde-Vinent y MacPhee 1999)
De acuerdo con el conocimiento filogenetico disponible sobre el conjunto de la fauna la colonizacion original de las Antillas Mayores por mamiferos terrestres implicaria el arribo
313 POBLAMIENTO ISLA-ISLA
de solo siete propagulos fundadores perezosos 2 (coloepodinos y megalocninos si como parece tienen un origen difiletico) musaraiias 2 (nesofontidos y solenodontidos si no constituyen una misma familia como consideran algunos) monos 1 (pitecidos) roedores histricognatos 1 (equimidos capromidos y heptaxodontidos) roedores muridos 1 (Oryzomys en Jamaica)
POBLAMIENTO ISLA-ISLA
La distribucion de los mamiferos terrestres a traves del area antillana demarcada al inicio del presente capitulo (vease Contexto Geogratico Fig 41) ha sido interpretada en funcion de dos hipoteticos mecanismos de poblamiento interinsular la dispersion y la vicarianza fundamentadas como se resumen seguidamente
DISPERSION
De acuerdo con la evidencia paleogeogratica mas actualizada en las principales regresiones marinas ocurridas en la cuenca caribeiia durante los ultimos 23 Ma (transiciones OligocenoshyMioceno Mioceno Superior - Plioceno y varias veces en el Pleistoceno) las masas terrestres constitutivas de la region insular en formacion aumentaron considerablemente su extension territorial disminuyendo por tanto la distancia entre elias oportunidades que los elementos faunales aprovecharian para dispersarse de unas islas a otras
Durante mucho tiempo esta ha sido la hipotesis mas favorecida por los estudiosos del tema a partir de Simpson (1956) pero en decadas recientes ha suscitado dos objeciones
1 Es dificil en tender como pudo operar la dispersion marina entre estas islas puesto que ninguna de las Antillas Mayores posee rios suficientemente caudalosos para transportar masas significativas de vegetacion flotante (MacPhee 198432)
Es oportuno recordar aqui el islote ala deriva (con mas de diez palmas reales en pie) entre las tres Antillas Mayores mas occidentales mencionado anteriormente (vease La Balsa Natural)
2 La dispersion marina es menos efectiva para explicar la asombrosa similitud de las faunas de mamiferos terrestres a traves de las Antillas Mayores septentrionales porque como proceso aleatorio no debe producir distribuciones uniformes a 10 largo de una cadena de islas 0 si las produce entonces el proceso no es aleatorio (MacPhee e lturralde-Vinent 2000 149)
Independientemente de que las aludidas distribuciones taxonomicas estan lejos de ser uniformes 0 asombrosamente similares la hipotesis de la dispersion marina contrariamente a su canicter aleatorio y altamente improbable en el caso del poblamiento continente-isla- en el caso del poblamiento isla-isla es un proceso no aleatorio (aprovecha las oportunidades brindadas puntualmente por los periodos de maxima regresion marina) y ademas altamente probable dada la incomparablemente menor distancia entre las islas caribeiias que entre estas y los continentes vecinos
314 BIOGEOGRAFiA
V ICARIANZA
La neotectonica y las transgresiones marinas del Oligoceno Medio y del Mioceno afectaron sustancialmente la paleogeografia de la Cresta de las Antillas Mayores resultando la subdivision gradual de las partes subaereas de la Cresta Asi el este de Cuba y el norte de La Espanola conectados fisicamente durante el Oligoceno Inferior se separaron al formarse el Paso de los Vientos en el Oligoceno tardio en tanto La Espanola y Puerto Rico probablemente permanecieron conectadas hasta el Mioceno tardio en que surgio el Paso de Mona (Iturralde-Vinent y MacPhee 1999)
Tales eventos geologicos habrian dividido las distribuciones de los grupos faunales emplazados previamente en el territorio continuo de las tres islas y a partir de esas disyunciones dichos autores (tambien MacPhee et al 2000 y White y MacPhee 2001) asumieron --como segunda fase del modelo de GAARlandia- la hipotesis de la vicarianza isla-isla para explicar la distribucion multiinsular de varios grupos de mamiferos terrestres a traves de las Antillas Mayores septentrionales De acuerdo con esta hipotesis cualquier diferencia en composicion faunal entre las islas (por ejemplo la ausencia de roedores heptaxodontidos en Cuba 0 de monos platirrinos en Puerto Rico) constituye evidencia de extincion local postvicariante y no de dispersion marina fallida Lo primero se puede demostrar si aparecen los fosiles pertinentes 10 segundo no se puede demostrar (MacPhee e Iturralde-Vinent 1994) Por otra parte IturraldeshyVinent y MacPhee (1999 57) han reconocido que aunque fa vicarianza isla-isla puede representar una solucion clara para las distribuciones de varios taxones cuaternarios no puede explicar todos los casos
Rega et al (2002 18) han presentado dos objeciones al modelo de la vicarianza isla-isla aplicado a los mamiferos de las Antillas Mayores
I Si la separacion de Cuba y La Espanola precedio en varios mill ones de anos la separashycion de La Espanola y Puerto Rico el modelo vicariante de lturralde-MacPhee debe predecir mayor similitud entre las faunas de La Espanola y Puerto Rico que entre las de Cuba y La Espanola De hecho es al reyes
No obstante MacPhee et al (200335) admitieron despues que la fecha mas temprana empiricamente defendible [para la apertura del Paso de Mona] debe ser el Oligoceno Medio esto invierte la secuencia de ambas disyunciones con 10 cual queda resuelta la aparente contradiccion Sin embargo en su reciente evaluacion de la congruencia entre filogenia e hipotesis geologic as en varios grupos de mamiferos terrestres de las Antillas Mayores Davalos (2004373) encontro que ninguno de los taxones cumple con la fecha y el patron de divergencia entre las masas terrestres antillanas postulados por el modelo de GAARlandia
2 La presencia de un mismo genero en las tres Islas septentrionales de las Antillas Mayores implica [por el modelo vicariante] que la diferenciacion entre estas formas durante el intervalo desde el Mioceno ha sido unicamente de nivel especifico [Dicha presencia]
315 rRRADlAcr6~ EVOLUCrONARIA
s610 puede explicarse razonablemente como el resultado de movimientos de esos animales entre las islas en el Cuaternario
De esta objecion resulta dificil disentir [El poblamiento entre islas diferenciado solo a nivel especifico no es probable que sea anterior al Pleistoceno (Simpson 1956 15)]
Debe tenerse en cuenta que la hipotesis de la vicarianza isla-isla alude a las tres islas septentrionales de las Antillas Mayores porque como se explico antes (vease El Barco Funeral Vikingo) Jamaica no existia como isla durante el lapso (Eoceno Superior shyOligoceno) consensualmente reconocido como el mas probable para la colonizacion de las proto-Antill as Mayores por la mayo ria de los mamiferos terrestres Despues de emerger como isla en el Mioceno Medio Jamaica debe haber recibido sus mamiferos terrestres por dispersion isla-isla (Davalos 2004376) y solo en el Cuaternario parece haberrecibido (por dispersion marina desde Centroamerica) al roedor murido Oryzomys
Seguramente tanto la dispersion como la vicarianza contribuyeron a que los mamiferos terrestres se irradiaran a traves de las Antillas Mayores y grupos de islas perifericas pero las maneras y tiempos en que estos mecanismos operaron a 10 largo de dicho proceso son aun en gran medida motivos de desacuerdo e incertidumbre
IRRADIACION EVOLUCIONARIA
A continuaci6n revisamos someramente las distribuciones conocidas de los diferentes grupos de mamiferos terrestres a traves de las Islas principales de las Antillas Mayores y grupos de Islas perifericas incluidas las interpretaciones y conjeturas que la literatura reciente recoge en relaci6n con la manera en que dichas distribuciones pudieron haberse conformado durante la irradiaci6n evolucionaria de los referidos grupos de mamiferos Manifestaciones de esta irradiaci6n pueden inferirse unicamente a partir de los taxones presentes en el Cuatemario pues solo 6 de las especies conocidas corresponden al Terciario
GRUPOS ORlGINARIOS DE SURAMERICA
PEREZOSOS (Megalonychidae)
Aparte de un genero miocenico de Cuba y otro del Oligoceno de Puerto Rico se conocen 4 generos (12 especies) de perezosos cuaternarios (todos extintos) restringidos a las tres islas septentrionales de las Antillas Mayores Acratocnus Neocnus Megalocnus y Parocnus Los cuatro generos estan representados en Cuba y La Espanola y Acratocnus 10 esta ademas (como unico genero de perezoso) en Puerto Rico Esta distribuci6n ha sido interpretada por MacPhee et al (2000) eomo resultado de la vicarianza oligomiocenica entre estas islas y [dado que el Banco Puerto Rico - Islas Virgenes fue la entrada de GAARlandia a las Antillas Mayores] se preguntaron si la ausencia de los otros tres generos en Puerto Rico se debe a que nunca estuvieron en ese territorio 0 a que sus
316 BIOGEOGRAFiA
fosiles atm no han sido hallados Pero como hemos visto Rega et al (2002) atribuyeron dicha distribucion a la dispersion interinsular en el Cuatemario
Por otra parte varios amilisis c1adisticos recientes ubi can a Choloepus -genero Reciente conocido solo de Centro- y Suramerica- como taxon hermano de Aeratoenus exc1uidos los demas megaloniquidos antillanos Para White y MacPhee (2001228)
tan estrecha relacion sugiere que la divergencia entre ambos [generos] ocurrio despues de que Aeratoenus divergio de Neoenus Si esta relacion implica que Aeratoenus y Neoenus se dispersaron separadamente desde Suramerica a las AntilIas mientras Choloepus permanecia en el continente (siendo necesarias al menos tres invasiones de perezosos) 0 si Choloepus invadio Suramerica desde las Antillas es algo que el material disponible no permite determinar
La diversidad adaptativa de los perezosos se manifiesta en (1) la utilizacion del habitat especies arboreas en Neoenus y Aeratoenus y terrestres en Paroenus y Megaloenus (Webb 1985a White [1993] White et al 1996 Arredondo [1999]) y en (2) la gama de tallas corporales desde los pequenos Neoenus hasta Megaloenus rodens el mayor de los mamiferos terrestres antillanos (su peso adulto estimado en 270 kg seglin Paula 19679) esto sugiere que los perezosos se irradiaron a nichos que en contextos continentales probablemente estarian ocupados por miembros de otros c1ados (MacPhee e IturraldeshyVinent 2000)
No esta claro si el abanico de tamanos entre estos perezosos enmascara alguna tendencia al gigantismo Por 10 pronto Anderson y Handley (2002) encontraron 10 contrario en perezosos (Bradypus) de las Islas de Bocas del Toro (Panama) shyseparadas secuencialmente del area continental vecina por ascensos del nivel del mar durante los ultimos 10 ka En las cinco islas los perezosos son menores que su contraparte continental y las diferencias entre las tallas medias de las cinco poblaciones expresan una reduccion lineal significativa de la talla corporal en funcion de la edad geologica de cada isla sin relacion alguna con el area de las islas 0 la distancia que las separa del continente
MONOS (Pitheciidae)
Los primates de las Antillas Mayores (cuatro especies extintas) constituyen un grupo monofiletico con Paralouatta (en Cuba) y Antillothrix (en La Espanola) como taxones hermanos seguidos en la escalera c1adistica por Xenothrix (en Jamaica) y finalmente Callieebus como el pariente continental mas cercano esto implica una sola invasion desde Suramerica antes del Mioceno Inferior (fecha mas temprana conocida para los primates fosiles antillanos) y por tanto en consonancia con GAARlandia como la via de colonizacion (Horovitz y MacPhee 1996 1999 Horovitz [1997] MacPhee y Horovitz 2004)
En desacuerdo parcial con la anterior interpretacion la filogenia de este grupo de monos obtenida por Davalos (2004) indica un antecesor comun en Cuba La Espanola y Jamaica
317 TRRADIACTON EVOLUCIONARIA
compatible con la hipotetica conexion -propuesta tentativamente por Iturralde-Vinent y MacPhee (199980-84)- entre las Montanas Azules del este de Jamaica y la peninsula suroccidental de La Espanola en la epoca de GAARlandia
A pesar de que los primates antillanos pertenecen a un grupo viviente de monos muy pequenos (los monos titles) fueron comparativamente grandes en relacion con los platirrinos modernos y sucesivamente mayores en la direccion Antillothrix Xenothrix y Paralouatta con P marianae como el mayor platirrino conocido del Mioceno neotropical (MacPhee 2005) MacPhee et al (1994) consideraron que Xenothrix debio ser un mono de andar lento y al parecer las especies de Paralouatta abandonaron la arborealidad para adoptar un modo de vida mas terrestre (MacPhee 1996a MacPhee et al 2003 MacPhee y Meldrum 2006)
ROEOORES (Echimyidae Capromyidae Heptaxodontidae)
La edad de Zazamys en Cuba demuestra que la invasion de los roedores histricognatos en las Antillas Mayores comenzo antes del Mioceno Inferior por otra parte los representantes mas antiguos de esa irradiacion evolucionaria se encuentran en La Espanola y Puerto Rico Ambas evidencias son consistentes con la hipotesis de GAARlandia (Woods 1982 1988 Woods et al 2001)
No esta claro si los capromidos evolucionaron a partir de un equimido ancestral 0 de un protocapromido 0 si los equimidos antillanos (I sub familia 3 generos y 7 especies todas extintas) forman parte de la irradiacion de los capromidos (4 subfamilias 9 generos y 27 especies 10 vivientes) Kratochvil et al (1978b) y Rodriguez et al (1979) consideraron que Capromys nan us es un eslab6n entre Echimyidae y Capromyidae pero Mesocapromys nanus esta morfologicamente mucho mas cerca de Hexolobodon y los de mas capromidos que de los equimidos (Woods 1985) Lo que si esta claro segun Woods et al (2001) es que la subfamilia Plagiodontinae endemic a de La Espanola es el linaje basal de los capromidos y que Isolobodon Hexolobodon Capromys Geocapromys Mysateles y Mesocapromys [y supuestamente Macrocapromys] forman parte de la irradiacion temprana de Capromyidae
La subfamilia Capromyinae (5 generos y 17 especies 9 vivientes) parece ser el dado de mas reciente evolucion en la familia con invasion de Cuba desde La Espanola y dispersion desde Cuba (1) por Capromys a las islas Caiman y (2) por Geocapromys a las islas Bahamas Caiman y Jamaica y desde Jamaica a las islas Swan aunque la presencia de Geocapromys en Swan pudiera deberse a introduccion aborigen (Morgan 1985 1988 1989 1993 1994 Woods 1989a 1990) Woods et al (2001) utilizaron datos moleculares para obtener estimados independientes de divergencia entre capromidos segiJn 10 cual Geocapromys divergio cerca de los 17 MaAP Mesocapromys divergio de Capromys hace 15 MaAP en tanto lvlysateles 10 hizo hace 10 MaAP Por su parte las formas de Capromys pilorides y de Mysateles prehensilis en Isla de la Juventud divergieron de sus correspondientes subespecies nominativas en la isla principal a los 24 y 08 MaAP respectivamente
318 BIOGEOGRAFiA
Woods y Howland (I979) describieron adaptaciones morfologicas de la mandibula el hioide y la faringe asociadas a mecanismos de masticacion diferentes en plagiodontinos y caprominos y Varona (1980a) llamo la atencion sobre el gran desarrollo de los molariformes (en relacion con la longitud total del cnineo) que presentan ciertos grupos de jutias fosiles en comparacion con las vivientes 10 que pudiera interpretarse como una adaptacion alimentaria a una vegetacion mas abrasiva por parte de las jutias extinguidas
Segun Woods (1990) la familia Heptaxodontidae (2 subfamilias 5 generos y 6 especies todas extintas) tambien seria parte de la irradiacion de los capromidos con origen en el este de La Espanola 0 en Puerto Rico y subsecuente dispersion al Banco de Anguila - San Martin en las Antillas Menores (pero vease MacPhee y Flemming 2003) Este banco estaria a solo 70 km del Banco Puerto Rico - Islas Virgenes en tiempos de maxima regresion marina y esta proximidad con Puerto Rico -patria de Elasmodontomys obliquus- seguramente tuvo que ver con la presencia en las islas de Anguila y San Martin del heptaxodontino gigante Amblyrhiza relacionado mas estrechamente con los roedores de las Antillas Mayores que con los del resto de las Antillas Menores (Woods 1990 1996) Aunque los roedores suelen aumentar de tamano en las islas (Case 1978) la escala del incremento de la talla en Amblyrhiza (que pudo alcanzar mas de 200 kg de peso adulto) esta fuera de toda proporcion con los patrones conocidos en otros roedores insulares (Biknevicius et 01 1993)
Clidomys (Clidomyinae) de Jamaica difiere marcadamente en su morfologia de los heptaxodontinos (vease ademas MacPhee y Flemming 2003)10 cual dificulta explicar su presencia en esa isla Como Jamaica estuvo bajo el mar hasta el Mioceno Medio el ancestro de Clidomys colonizaria esta isla por dispersion desde el extremo suroccidental de La Espanola despues que los heptaxodontinos se originaron en esta ultima isla (Woods 1989a) Pero Pascual et 01 (I 990) especularon que Clidomys pudiera ser el resultado de una invasion posterior e independiente de los capromidos
GRUPOS ORIGINARIOS DE NORTEAMERICA
El origen de las musaranas antillanas ha resultado ser hasta ahora el mayor enigma biogeografico de la fauna de mamiferos terrestres de la region aunque la mayoria de los autores coinciden en considerarlas originarias de Norteamerica (vease Discus ion y Conclusiones en Poblamiento Continente-Isla) No obstante MacPhee e Iturralde-Vinent (1994) descartaron toda posibilidad de conexion terrestre por el Estrecho de la Florida 0 por el Canal de Yucatan y si el (supuesto) evento vicari ante de Rosen (vease El Arca de Noe) -invocado por MacFadden (1980) para resolver el enigma- fuera minimamente plausible aun asi es dificil creer que las catastrofes que 10 sucedieron (vease Contexto Temporal) hayan barrido con toda esa vieja fauna continental excepto las musaranas circunstancia ya anticipada muy tempranamente por Matthew (l918a)
De otra parte la relaci6n entre Nesophontidae y Solenodontidae permanece irresuelta Si estos linajes no estan estrechamente relacionados entonces es muy posible que
319 IRRADIACION EVOLUCIONARIA
sus distribuciones sean explicadas por mecanismos diferentes (Whidden y Asher 2001)
La descomunal diferencia de talla corporal entre las musaranas pequenas (Nesophontes) y las grandes (Soienodon) seguramente impuso a ambos grupos restricciones distintas relativas a la termorregulacion y las adaptaciones energeticas En Nesophontes la seleccion debio favorecer una alta tasa de metabolismo y crecimiento camadas mas numerosas corta expectativa de vida yalta densidad poblacionaL En Soienodon en cambio la seleccion opero en la direccion de una baja tasa metabolica camadas reducidas crecimiento lento longevidad y baja densidad poblacional (Ottenwalder [1991])
MUSARANAS PEQUENAS (Nesophontidae)
Las musaranas pequenas estan representadas en las Antillas por el genero Nesophontes (y un genero miocenico indeterminado en ambar dominicano) con 6 especies cuaternarias extinguidas distribuidas en Cuba La Espanola Puerto Rico Islas Virgenes e Islas Caiman La mayor de las musaranas pequenas Nesophontes edithae evoluciono en Puerto Rico y su arribo a las Islas Virgenes supone una dispersion sumamente reciente En las Islas Caiman Nesophontes se asemeja mas a la poblacion de Cuba de donde seguramente derivo en el Plio-Pleistoceno (junto con Capromys y Geocapromys) cuando la costa sur de Cuba estuvo mucho mas cerca de las Islas Caiman (lturralde-Vinent 2003fig 4) MacPhee et ai (1994) interpretaron la distribucion de Nesophontes en Cuba y La Espanola como resultado de la vicarianza oligomiocenica ocurrida al formarse el Paso de los Vientos pero la distancia taxonomica entre los lados del Paso es solo de nivel especifico y N micros incluso vivi6 en ambas islas segtln Patterson (1962)
Allen (1918) considero que Nesophontes micrus fue una especie primitiva no modificada para la vida arb6rea 0 fos6rea y por tanto de habitos terrestres Nesophontes edithae de Puerto Rico es mucho mayor y mas robusta que las restantes especies del genero eL indice de robustez de su femur (= anchura minima de la diafisis ongitud total del hueso) muestra que este es tan robusto como el de Solenodon paradoxus un animal dos veces mayor (Morgan et ai 1980) Choate y Birney (1968) postularon un aumento cronoclinal de la taIla corporal en N edithae confirmado mas tarde por McFarlane (1999a) mediante fechados radiocarb6nicos de la secuencia estratignifica en depositos cavernarios de Puerto Rico Solenodon no vivi6 en Puerto Rico y en ausencia de una musarana de mayor tamano Nesophontes alcanzo una talla corporal mayor en esa isla que en La Espanola 0 Cuba (Woods y Eisenberg 1989)
MUSARANAS GRANDES (Solenodontidae)
Los arcaicos almiqufes (genero Soienodon) comprenden cuatro especies una viviente (S cubanus) y otra f6sil (S arredondoi) en Cuba y una viviente (S paradoxus con dos subespecies) y otra f6sil (S marcanoi) en La Espanola Como cabria esperar las dos especies cubanas se asemejan mas entre sf que a las especies de La Espanola pero
320 BIOGEOGRAFiA
enigmaticamente S marcanoi comparte caracteres con S cuban us y S paradoxus (Woods y Eisenberg 1989 Morgan y Ottenwalder 1993 Ottenwalder 200 I) Solenodon arredondoi alcanzo una talla corporal superior a la de cualquiera de las musarafias vivientes a nivel mundial Segun Morgan y Ottenwalder (1993) extrapolando las dimensiones y el peso de los almiquies vivientes S arredondoi debio tener una longitud cabeza-tronco y un peso de 450-550 mm y 1500-2000 g respectivamente en comparacion con S cubanus (360 mm y 800 g) y S paradoxus (390 mm y 1100 g)
Entre todas las especies de almiqufes Solenodon paradoxus es la que presenta los caracteres mas derivados (Otten walder 2001) Si estas relaciones son validas la distribucion de Solenodon entre Cuba y La Espafiola debio discurrir de oeste a este como propuso MacFadden (1980) quien invoco la presencia fosH de S cubanus [pero tambien S arredondoi] en Pinar del Rio para postular que el ancestro continental entr~ por Cuba opinion compartida por Perfit y Williams (1989) y por Ottenwalder ([1991] 2001) El paso de Solenodon a Puerto Rico pudo haberlo impedido la existencia de barreras ecologicas durante la conexion del este de La Espafiola con Puerto Rico 0 por la ausencia de Solenodon en esa parte de La Espafiola en aquella epoca Sin embargo las mismas condiciones ecologicas en el este de La Espafiola no parecen haber representado una barrera efectiva para Nesophontes cuyas mas altas tasa de reproduccion y densidad poblacional presumiblemente Ie permitieron expandirse flpidamente a traves de La Espafiola (Ottenwalder [1991])
Es opinion generalizada que los almiqufes antillanos constituyen el grupo de mamiferos que primero se emplazo en el territorio insular (MacPhee 2005) y en tal sentido es de interes la evidencia sobre especificidad parasito-hospedante aportada por Cruz (1980) quien encontro que la especie de acaro (genero Cubanochirus) que parasita a los almiquies es mucho mas primitiva que la especie (del mismo genero) que paras ita a las jutias y de aqui concluyo que dicho genero de acaro debio evolucionar primero en Solenodontidae y posteriormente en Capromyidae al entrar en contacto representantes de ambas familias en Cuba 0 La Espafiola
Recientemente Roca (2004) y Roca et al (2004) dieron a conocer que los resultados de su estudio genetico-molecular sobre los almiquies indican una divergencia de 25 Ma entre Solenodon cuban us y S paradoxus -compatible con la disyuncion tectonica entre Cuba y La Espafiola (Iturralde-Vinent y MacPhee 1999 McPhee et al 2003)-- y una separacion de ambas especies comparable en magnitud a la divergencia entre familias de mamlferos Consecuentemente propusieron que Solenodon cubanus fuera clasificada en el genero Atopogaie como indico Cabrera (1925) hace 82 afios
DISCUSION Y CONCLUSIONES
Las musarafias y los roedores antillanos (excepto Oryzomys) se irradiaron tanto de sus respectivos ancestros continentales que de estos 6rdenes se reconocen familias y subfamilias endemicas de la region en contraste con los perezosos (una subfamilia y tres
321 IRRADIACION EVOLUCIONARIA
tribus endemicas) y los monos (una tribu endemica) como expresion de diferentes niveles de diversificacion evolucionaria Este proceso es dificil de explicar (traUindose de c1ados que pnicticamente comparten la misma antigiiedad en las Antmas) si es correcta la interpretacion de Pascual et at (1990636)
Nosotros consideramos estos diferentes niveles de diversificacion como las diferentes etapas de evolucion alcanzadas por cada grupo [de mamiferos] Obviamente esto estuvo relacionado con las diferentes longitudes de sus historias
Pudiera ocurrir que en el futuro se compruebe que la historia biogeognifica de la region fue mucho mas complicada
La Espanola la isla mas ecol6gicamente divers a de la region debi6 ser el centro principal de evoluci6n de los mamfferos (con 87 de las familias y 73 de las subfamilias) seguida por Cuba (con 75 de las familias y 54 de las subfamilias) las dos islas de las Antillas Mayores mas relacionadas biogeograficamente (Fig 43 Tabla 44)
eL
EL
cb CAIMAN
SWAN
A Acratocnini B Cubanocnini C Megalocnini D Mesocnini E Nesophontidae F Solenodontidae G Xenothrichini
ANGUILAshyM-0 SMARTIN
H Heteropsomyinae I Plagiodontinae J Isolobodontinae
K Hexolobodontinae L Capromyinae
M Heptaxodontinae N CIidomyinae 0 Sigmodontinae
FIGURA 43 Representacion esquematica de las relaciones interinsulares (expresadas porel numero de linajes compartidos) entre las Antillas Mayores y entre estas y los grupos de islas perifericas segun las interpretaciones de los autores citados en el texto Los numeros dentro de los circulos indican la cantidad de linajes representados en cada isla relacion dudosa Contexto taxon6mico de los linajes en la Tabla 44
bull bull
bull bull
bull bull bull
bull bull
bull bull
bull
bull
bull
bull
bull bull bull bull
bull bull
bull bull
bull
bull bull bull
bull bull
bull
bull bull
bull
322 BIOGEOGRAFiA
TABLA 44 Distribuci6n de los linajcs de mamiferos terrestres a traves de las Antillas Mayores y grupos de islas perifericas LE = La Espanola CU Cuba JA Jamaica PR = Puerto Rico IC Islas Caiman IS Islas Swan IB = Islas Bahamas IV = Islas Virgenes AS = Anguila - San Martin
FamiliaSubfumiliafIribu LE CU JA PR IC IS IB IV AS
Megalonychidae
Choloepodinae
Acratocnini
Cubanocnini
Megalocninae
Megalocnini
Mesocnini
Nesophontidae
Solenodontidae
Pitheciidae
Callicebinae
Xenothrichini
Echimyidae
Heteropsomyinae
Capromyidae
Plagiodontinae
Isolobodontinae
Hexolobodontinae
Capromyinae
H e pta x 0 don tid a e
Heptaxodontinae
Clidomyinae
Muridae
Sigmodontinae
PILOSA
SORICOMORPHA
PRIMATES
RODENTIA
bull
bull
bull
bull TOTALES 12 10 4 4 2
323 EXTlICIOIES DEL CUATERNARIO
El gigantismo ha sido sin duda una tendencia evolucionaria recurrente entre los clados mayores de mamiferos antillanos Es un hecho bien conocido que algunos grupos de mamiferos han aumentado de tamafio en las islas y varios autores han intentado explicarlo a partir de enfoques diferentes (Foster 1964 Miller 1966 Sondaar 1977 Case 1978 Eisenberg y Wilson 1978 Heaney 1978 Lawlor 1982 Lomolino 1985 Angerbjom 1985 McNab 19942001)
EXTINCIONES DEL CUATERNARIO
Las islas que integran la Subregion Antillana han perdido casi 90 de sus mamiferos terrestres durante los ultimos milenios y en proporcion con su extension territorial esta perdida ha sido mayor en la Subregion que en cualquier otro lugar del planeta (MacPhee 1996b) Las extinciones del Cuatemario eliminaron mas de las tres cuartas partes de la fauna de mamiferos terrestres conocida de las Antillas Mayores sus islas satelites y grupos de islas perifericas y el examen de este proceso en sus aspectos mas generales se aborda en esta ultima parte del presente capitulo
CRONOLOGiA Y CONSECUENCIAS
Es muy improbable que un fosil represente el ultimo individuo de un taxon extinto y entonces hay que guiarse por la edad radiometrica del fosil mas reciente conocido lIamada fecha de ultima presencia- como unica aproximacion posible a la fecha de extincion del taxon
Como se explico al inicio del capitulo (vease Contexto Temporal) el registro fosH de los mamiferos terrestres de las Antillas Mayores adolece de insuficiente sustentacion radiometrica No obstante la distribucion temporal de los escasos fechados disponibles para el Cuaternario de la region abarca etapas criticas del proceso de extincion experimentado por la referida fauna a 10 largo de los ultimos 250 mil afios
De las 52 especies que es posible reconocer actualmente como miembros extintos de la fauna cuatemaria de mamiferos terrestres de las Antillas Mayores y grupos de islas perifericas los restos fosiles de 23 de elIas (442) han sido fechados radiometricamente o se han hallado asociados estratigraficamente de manera inequivoca con otro material fechado De estos 23 fechados 3 corresponden al interglacial Sangamon 4 pertenecen a la secuencia Pleistoceno terminal - Holoceno temprano 10 se ubican en ellapso entre el arribo de los aborigenes y la incursion europea y 6 son postcolombinos (a los cuales hay que sumar otras dos especies extinguidas en los siglos XIX y xx) A partir de estas 25 especies avaladas por fechas de ultima presencia distribuidas entre los cuatro intervalos temporales sefialados hemos compuesto la Tabla 45 en la que ademas aparecen las restantes 27 especies (carentes de fechado radiometrico) ubicadas en el intervalo temporal mas probable sobre la base de (1) la asociacion estratigrafica de sus restos con evidencias culturales (amerindias 0 europeas) 0 (2) la ausencia de dicha asociacion Sobre estos criterios de ubicacion temporal procede hacer dos aclaraciones
324 BIOGEOGRAFIA
TABLA 45 Intervalos temporales mas probables para las extinciones cuatemarias de los mamiferos terrestres de las Antillas Mayores y grupos de islas perifericas sobre la base de las evidencias siguientes (l) ultimo ejemplar colectado u observado en vida (2) fechado radiometrico mas tardio en material de la especie (3) asociaci6n estratigrafica mas tardia con otro material fechado (4) asociaci6n estratigrafica con restos de animales introducidos restos humanos u objetos culturales propios del intervale (5) falta de asociaci6n con evidencias culturales [Los ntimeros entre parentesis en la columna Evidencia significan adscripci6n tentativa al intervalo] Los estimados del peso corporal (en kg) estan tomados de la literatura 0 inferidos comparativamente y distribuidos entre
las clases siguientes (a) S 01 (b) 01 a lt 1 (c) 1 a lt 10 (d) 10 a lt 100 (e) 100 a lt 200 (j) ~ 200 Las
abreviaturas que designan las islas son las de la Tabla 44
F amilialEspecie Peso IsJa Evidencia Fuente
I httervalo htterg1acial Sangamon (gt80 kaAP)
HEPTAXODONTlNAE
Amblyrhiza inundata e AS 3 McFarJane et al 1998a
CLIDOMYINAE
Clidomys osborni d JA 3 Ibid
Clidomys parvus d JA 3 Ibid
II httervalo Pleistoceno terminal ~ Holoceno temprano (20-6 kaAP)
MEGALONYCHIDAE
Aeratoenus antillensis d CD 5 Arredondo 1975 a
Aeratoenus odontrigonus d PR 3 McFarJane 1999b
Aeratoenus simorhynehus d LE 2 Rega et al 2002
M egaloenus zile f LE 2 MacPhee et al 2000
SOLENODONTIDAE
Solenodon arredondoi e CD 5 Morgan y Ottenwalder 1993
PnHECIIDAE
Paralouatta varonai d CD 5 MacPhee 1996a
EcHIMYIDAE
Heteropsomys antillensis c PR 5 McFarJane 1999b
Heteropsomys insulans c PR 5 Ibid
Puertoricomys eorozalus c PR 5 Woods 1989a
CAPROMYIDAE
P lagiodontia araeum c LE 5 MacPhee et al 2000
Macrocapromys latus c CD 5 Varona y Arredondo 1979
(Continua)
325 EXTINCIONES DEL CUATERNARIO
TABLA 45 (Continuaci6n)
F ami1ia1Especie Peso Isla Evidencia Fuente
M acrocapromys latus c CU 5 Varona y Arredondo 1979
Mesocapromys kraglievichi c CU 5 Ibid
HEPTAXODONTINAE
Xaymaca fulvopulvis a JA 2 MacPhee y Flemming 2003
III Intervalo Amerindio (6-05 kaAP)
MEGALONYCHIDAE
Acratocnus ye d LE 2 MacPhee et aI 2000
Neocnus comes c LE 2 Ibid
N eocnus dousman c LE 2 Ibid
N eocnus toupiti c LE 2 Ibid
N eocnus gliriform is c CU 4 Cordova et ai 1997
M egalocnus rodens f CU 2 MacPhee et ai 2007
Parocnus browni e CU 2 Steadman et ai 2005
P arocnus serus e LE 2 MacPhee et ai 2000
N ESOPHONTIDAE
N esophontes edithae a PR 2 McFarlane 1999b
PITHECIIDAE
Antillothrix bernensis c LE 3 Rimoli 1977
EcHIMYIDAE
Brotomys contractus a LE 4 Miller 1929c
CAPROMYIDAE
P lagiodontia spelaeum c LE 4 Ibid
M acrocapromys acevedo c CU 4 NUiiez 1990a
HEPTAXODONTINAE
Elasmodontomys obliquus d PR 3 McFarlane 1999b
Iv Intervalo Postcolombino (05 kaAP - Presente)
N ESOPHONTIDAE
N esophontes hypom icrus a LE 24 MacPhee y Fle111111ing 1999
N esophontes zam icrus a LE (4) Ibid
N esophontes m icrus a CU 24 MacPhee et aI 1999
(Continua)
326 BIOGEOGRAFiA
TABLA 45 (Continuacion)
F amilialEspecie Peso Isla Evidencia Fuente
N esophontes m icrus a CD 24 MacPhee et aI 1999
N esophontes m icrus a LE 4 MacPhee et aI 2000
N esophontes sp 1 a IC 4 Morgan 1994
N esophontes sp 2 a IC 4 Ibid
Solenodon marcanoi b LE 4 Ottenwalder 2001
PITHECIIDAE
Xenothrix mcgregori c JA 4 MacPhee y Flemming 1999
EcHIMYIDAE
Boromys offeUa a CD 24 Guarch 1984
Boromys torrei a CD 24 Ibid
Brotomys voratus a LE 4 MacPhee et aI 2000
CAPROMYIDAE
P lagiodontia ipnaeum c LE 4 MacPhee y Flemming 1999
Rhizoplagiodontia lemkei c LE 4 Ibid
Isolobodon portoricensis c PR 24 Ibid
Isolobodon montanus c LE 4 Ibid
Hexolobodon phenax c LE (4) Ibid
Capromys sp c IC 24 Morgan 1994
Geocapromys columbianus c CD 4 MacPhee y Flemming 1999
Geocapromys thoracatus b IS 1 MacPhee y Marx 1997
Geocapromys sp 1 c IC 4 Morgan 1994
Geocapromys sp 2 c IC (4) Ibid
l-IEPTAXODONTINAE
Quemisia gravis c LE (1) MacPhee y Flemming 1999
MURIDAE
Oryzomys antillarum a JA 1 Ibid
1 Flemming y MacPhee (1999) alertaron en contra de la asociacion con otro material fechado Asimismo Morgan (1993) indico que la ausencia de asociacion con evidencias culturales amerindias no es necesariamente un criterio concluyente para ubicar un taxon en el intervalo temporal anterior porque hay depositos en la region que carecen de tales evidencias y son del Holoceno tardio por fechados radiocarbonicos [No obstante
327 EXTlNCIONES DEL CUATERNAR10
la ausencia de una especie en residuarios arqueol6gicos adquiere mayor certidumbre como criterio de ubicaci6n temporal de su extinci6n en la medida que aumenta el numero de residuarios estudiados que en las Antillas Mayores se cuentan por centenares] A falta de fechado absoluto de una especie consideramos que estos criterios de ubicaci6n temporal son mas utiles que la alternativa de excluir del analisis a la especie
2 En los trabajos sobre biogeografia de mamiferos antillanos los arque610gos mas citados como autoridades para establecer el inicio de la presencia aborigen en las Antillas Mayores (Kozlowski 1974 Rouse y Allaire 1978 Rouse 1989 1992 Wilson 2001) difieren al respecto entre los 6 kaAP y los 45 kaAP (pero vease Pagan et al 200523) y hemos adoptado la mas temprana de estas fechas para el comienzo del intervalo amerindio en la Tabla 45 De todos modos cualquiera que haya sido la fecha esta no es aplicable por igual al conjunto de las Antillas Mayores porque segun algunos de los autores citados el poblamiento comenz6 por Cuba y La Espanola 6 mil anos atras continu6 en Puerto Rico hace alrededor de 3 mil anos (pero vease Burney et al 1994) yen Jamaica hace menos de 2 mil e incluso a las Islas Caiman ni siquiera llegaron [Las Islas Caiman descubiertas por Col6n en 1503 carecen absolutamente de evidencia amerindia (Stokes y Keegan 1996)]
Si los datos de la Tabla 45 representan una aceptable aproximaci6n a la realidad el proceso de las extinciones cuaternarias de los mamiferos terrestres de las Antillas Mayores y grupos de islas perifericas debi6 caracterizarse entre otros por los eventos siguientes
Aunque obviamente la fauna cuaternaria debi6 ser mucho mas numerosa que 10 que refleja el registro paleontol6gico (Morgan y Woods 1986) 52 especies (81 de la fauna conocida) desaparecieron (sin reposici6n) durante los ultimos 250 mil anos
- Al parecer las extinciones se concentraron en las especies de mayor tamano Asi el peso promedio estimado para las especies de la etapa preantr6pica (intervalos I + II de la Tabla 45) es 350 kg es 328 kg para las especies del intervalo III (amerindio) y es s610 25 kg (c1ases del peso a b c) para las especies del intervalo IV (postcolombino) Ninguna especie aut6ctona de mamifero mayor de 6 kg existe hoy en la regi6n
- De acuerdo con la distribuci6n temporal de las extinciones 61 especies sobrevivieron al interglacial Sangamon (intervalo 1)48 sobrevivieron a la transici6n del Pleistoceno terminal al Holoceno temprano (intervalo II) 34 sobrevivieron al periodo amerindio (intervalo III) y s610 12 especies han sobrevivido desde la Ilegada de los europeos (intervalo IV) Las tasas de extinci6n correspondientes (exceptuado el intervalo I) son una especie cada 1167 anos en el intervalo II una cada 393 anos en el intervalo III y una cada 23 anos en el intervalo IV
- Durante todo el proceso de las extinciones cuaternarias la perdida de especies por isla fue de 91 en La Espanola 83 en Jamaica y 62 en Cuba Bahamas retuvo su unica especie (aunque extirpada en varias de las islas) Puerto Rico e Islas Caiman perdieron todas sus especies (6 y 5 respectivamente) y las Islas Swan Virgenes y Anguila - San Martin perdieron sus respectivas especies (micas
328 BIOGEOGRAFiA
De los cuatro 6rdenes de mamiferos representados en la fauna Pilosa y Primates desaparecieron totalmente Soricomorpha y Rodentia perdieron 86 y 74 de sus especies respectivamente
CAUSALIDAD
CAUSAS NATURALES
Los cicIos glaciales e interglaciales acaecidos en el Pleistoceno causaron cambios ambientales dnisticos en las Antillas con la concomitante alternancia de cIimas frios (secos) y calidos (hUmedos) asi como de descensos y ascensos del nivel del mar aumento y disminuci6n de las areas territoriales) induso (aunque con menor intensidad) en el Holoceno Estos eventos paleogeograficos han sido consistentemente documentados (Mayo y Kartashov 1972 Bonnatti y Gartner 1973 Ortega y Arcia 1982 Prance 1982 Schubert y Medina 1982 Dodge et al 1983 Ortega [1984] Haq et al 1987 Schubert 1989 Hodell et al 1991 Guilderson et al 1994 Oppo 1997 Emslie 1998 Curtis et al 2001 Iturralde-Vinent 2003) y de ellos brindamos a continuaci6n una breve resena
Finalizando el periodo interglacial anterior a la ultima glaciaci6n (Wisconsin) el mar estaba a 8-10 m por encima de su nivel actual Cuando el avance glacial del Wisconsin a1canz6 su culminaci6n (eillamado Ultimo Maximo Glacial) hace 18 ka el mar habia descendido 120 m con respecto a dicho nivel y las islas del Caribe habian incrementado considerablemente sus extensiones subaereas en comparaci6n con las que presentan hoy dia por ejemplo Puerto Rico y las Islas Virgenes se conectaron territorialmente y las Bahamas se unieron en cinco gran des islas Prevalecia un regimen frio y seco y condiciones de aridez --debidas a una reducci6n de las precipitaciones (posiblemente a menos de 440 mmlano)- y los ambientes xericos se habian extendido a traves de las islas y bancos emergidos y abundaban los pinares (los habitats mesicos estaban restringidos a parcelas relictas en 10 alto de las montanas) En el Mar Caribe y el Golfo de Mexico la temperatura superficial del mar era 5degC mas baja que la actual y en las llanuras costeras y alturas maximas eran respectivamente 2-3degC y 6degC mas bajas Los limites de las zonas ecol6gicas altitudinales descendieron varios cientos de metros y cada zona redujo su extensi6n vertical al aumentar el gradiente de temperatura Ellimite inferior de la zona periglacial
elevaci6n donde la temperatura media minima a1canza los OdegC) de la Cordillera Central de la Republica Dominicana (altura maxima del Caribe insular con evidencia de topografia glacial) se considera que estuvo entre los 1500 y 1800 mlnm (actualmente esta a 2800 mlnm) y se puede estimar que dicho limite era inferior en Cuba por estar mas al norte que la Republica Dominicana No obstante durante este periodo la temperatura experiment6 episodios cortos (unas pocas decadas) de ascensos repetidos
Un signifieativo componente bi6tieo de las referidas condiciones paleoambientales 10 constituy6 la espectacular irradiaci6n evolucionaria experimentada por un grupo numeroso de aves rapaees diurnas y nocturnas (Wetmore 1937 Brodkorb 1969 Kurotchkin y Mayo 1973 Arredondo 1976 Olson 1978 Olson y Hilgartner 1982
329 EXTINCIONES DEL ClJATERNARIO
Suarez 2000a b 2004 Suarez y Emslie 2003) Los mamiferos de las Antillas Mayores ofrecieron a estos depredadores aviares una amplia gama de tamanos de presa que en ausencia de mamiferos del orden Carnivora en la region permitio a estas aves irradiarse a este nicho alimentario con la evolucion de una variedad de especies de muy diversa tall a corporal incluidas formas gigantes Adquirir un cuerpo gigante les propicio el acceso a recursos alimentarios que no estaban siendo explotados por ningun otro depredador (Alcover y McMinn 1994) A Cuba corresponde el mejor registro fosil de rapaces cuaternarias caribenas con siete generos de especies diurnas Dentro de estas aves existieron tres gavilanes gigantes (Accipitridae) tres carairas (Falconidae) y un condor (Cathartidae) asi como cuatro generos de especies nocturnas dos lechuzas (Tytonidae) y tres Mhos (Strigidae) dos de ell os no igualados en tamano en todo el mundo Algunos de estos generos tuvieron representacion en otras islas antillanas como La Espanola Jamaica Bahamas y Gran Caiman
Al retirarse hacia el norte los hielos de la glaciacion Wisconsin el clima se tomo mas hUmedo en las Antillas durante el Holoceno temprano y el bosque mesico se expandio en la region insular El nivel del mar ascendio acompanado de mayor pluviosidad alrededor de los 7 kaAP las Islas Virgenes quedaron separadas entre si y de Puerto Rico y las Bahamas perdieron la mayor parte de su territorio Cerca de los 3 kaAP las condiciones secas retornaron a la region e hicieron desaparecer buena parte del bosque mesico Nuevamente el clima comenzo a adquirir sus condiciones actuales hacia los 25 kaAP aproximadamente y hoy transita por una fase mesica con vegetacion xerica confinada en parcelas relictas
Sin duda estos y otros eventos naturales incidieron negativamente en la sobrevivencia de mamiferos terrestres en las Antillas Mayores asi como en los grupos de pequenas islas perifericas (Pregill y Olson 1981 Olson 2006)
McFarlane et al (1998a) demostraron convincentemente como Amblyrhiza el gigante de los roedores antillanos endemico de la pequena isla de Anguila sucumbio a las repetidas y abruptas perdidas de su habitat durante el interglacial Sangamon por las nipidas y severas oscilaciones del nivel del mar (documentadas por Neumann y Hearty 1996) al no poder responder adecuadamente a tan catastroficos eventos mediante el ajuste de su masa corporal media y otro tanto debio ocurrirle en Jamaica al otro roedor gigante Clidomys (McFarlane et al 1998b) emparentado con Amblyrhiza en la misma epoca (Tabla 45 intervalo I)
El ascenso del nivel del mar seguramente promovio la extincion de mamiferos durante la transicion del Pleistoceno al Holoceno (Tabla 45 intervalo II) con la reduccion del tamano de las Islas por las transgresiones marinas (Pregill y Olson 1981 Morgan 1993) Esto ultimo haria maximamente severa la competicion asf como la presion de los depredadores [Steadman et al (2005) no yen por que el calentamiento global ocurrido durante la transicion Pleistoceno-Holoceno habria de tener un efecto negativo sobre los perezosos siendo sus descendientes actuales especies tropic ales 0 subtropicales pero no se trata del efecto
330 BIOGEOGRAFiA
directo del cambio de temperatura sobre los animales sino del efecto indirecto y progresivo que dicho cambio genera a largo plazo al reducir el territorio insular disponible]
Por esta epoca tambien desaparecieron especies de tall a mayor como Megalocnus zile en La Espanola y otras de menor tamano (en Cuba) pero que representan la tall a maxima en sus respectivos contextos taxonomicos del Cuaternario como Solenodon arredondoi Paralouatta varonai y Macrocapromys acevedo Un mamifero puede ser grande solo a costa de reducir su densidad poblacional y esto 10 hace mis vulnerable a la extinci6n sobre todo en una isla (McNab 1994 McFarlane et al 1998a) Incidentalmente Alcover et al (1998a) afirmaron que larelacion entre la talla corporal y la extincion no siempre se cumple en las especies de menor tamano y citaron como ejemplo que entre las musarafias todas las especies de Nesophontes desaparecieron mientras sobreviven especies de Solenodon pero la comparacion correcta en todo caso serfa entre los solenodones vivientes y el gigante Solenodon arredondoi y entre Nesophontes edithae -el gigante de los nesofontes extinguido en el periodo amerindio- y el resto de sus congeneres que sobrevivieron hasta los primeros siglos de la colonizacion europea (Tabla 45 intervalos III y IV)
CAUSAS ANTROPOGENICAS
Intervalo Amerindio
La mayoria de los autores que se han referido a las extinciones acaecidas durante este intervalo (entre elIos paradigmaticamente Martin 1984a b) han responsabilizado a los aborfgenes (que se supone comenzaron a poblar las Antillas Mayores hace 6 mil afios) con la desaparicion de numerosas especies de mamiferos terrestres en la region principalmente por la via de la sobrecaza y la introduccion de animales domesticos devenidos depredadores de la fauna autoctona (Morgan y Woods 1986) Este efecto negativo de la presencia indigena sobre los mamiferos de la region ha sido dramatizado por algunos autores en terminos de holocausto faunal Sin embargo pensamos que se ha exagerado Como han expresado MacPhee et al (19993) cuando uno busca evidencia dura de complicidad humana directa en [estas] extinciones no hay mucho que encontrar Las siguientes son algunas consideraciones a tener en cuenta en tal sentido
El registro radiometrico mas joven conocido hasta ahora para un perezoso antillano (MegalocnllS rodens) es de 42 kaAP (MacPhee et ai 2007) Si se acepta que los aborfgenes entraron en Cuba hace aproximadamente 6 mil anos su convivencia con los perezosos debe haber durado alrededor de 1800 afios (0 mas porque dificilmente el referfdo fechado radiometrico haya correspondido al ultimo sobreviviente de la especie) Esta cifra contradice la hipotesis del holocausto faunal segun la cualla extincion de los mamiferos insulares de mayor talla debe ocurrir con extrema rapidez -a escala de decadas 0 de centurias- a partir de su primer contacto con los humanos (Martin y Steadman 1999) A pesar de las repetidas afirmaciones en contrario (Harrington 1921 Miller 1929c Herrera [1945] Tabio 1951 Alvarez 1951a 1957 Arredondo
331 EXTINCIONES DEL CUATERNARIO
1959 1960a Veloz y Ortega 1973 Mayo 1980b Rodriguez et al 1984 Castellanos y Pino 1985 Pino y Castellanos 1985 Nunez 1990b Gutierrez 1996 Arredondo 2004) en su mayoria basadas en evidencias no exentas de posibles accidentes de deposicion (0 de reubicacion postdeposicional) en recintos cavernarios (Diaz-Franco [2006b D no se ha demostrado inequivocamente que los aborigenes antillanos se alimentaran de perezosos regularmente Sus restos en residuarios de alimentos son escasisimos en toda la region y esto contrasta con los abundantes restos de jutias y otros mamiferos de menor talla que aparecen en esos residuarios demostrativos de la preferencia de los paleoindios por estos otros mamiferos
Los mamiferos mas importantes en la economia prehistorica de las Antillas Mayores fueron las jutias Capromys y Geocapromys en Cuba Geocapromys en Jamaica e Isolobodon en La Espanola Puerto Rico e Islas Virgenes (Aguayo 1950b Tablo 1951 Rimoli 1972 Wing 2001 Diaz-Franco [2006b D Wilkins (2001) estudi61ajutia viviente de Jamaica (Geocapromys browni) en un sitio ocupado por los aborigenes durante 100 anos y no encontr6 evidencias de que durante ese tiempo la especie haya sido sobreexplotada Asimismo en Cuba las jutias conga carabali y andaraz son escasas en depositos pleistoholocenicos en cambio son muy numerosas en residuarios indigenas y de haber sido sobreexplotadas 10 mas probable es que no hubieran sobrevivido hasta hoy
La habilidad de los humanos para penetrar en profundidad el bosque tropical y mantenerse en este sustentablemente ha sido un evento comparativamente reciente (Beran y Steele 1994) En el caso antillano la caza tierra adentro ha sido explicada como un proceso tardio de desplazamiento a partir de la explotacion de abundantes recursos marinos que ofrecian condiciones 6ptimas para el sedentarismo de las comunidades aborigenes costeras (C6rdova y Arredondo 1988 Brito 2005) Peces y mariscos dominan la arqueologia costera de las Islas antillanas en tanto el consumo de recursos terrestres (mamfferos incluidos) aumenta progresivamente en la medida que los residuarios estudiados se alejan de la costa (Scudder 1991 Wilkins 2001) En las Antillas Mayores el componente terrestre de la dieta aborigen es solo 20-30 del material recobrado en los residuarios (Wing 1977 1989) Y debio ser menor alin porque el componente vegetal de la dieta no se ha preservado Por otra parte los aborigenes cubanos domesticaron la jutia conga y seguramente la reprodujeron para comer (Pose et al 1988) practica que probablemente se repitiera con otras jutias en las demas Antillas Mayores
- Fray Bartolome de las Casas hablo de 3 millones de indios en Cuba al inicio de la colonizacion pero los historiadores subsecuentes consideraron exagerada la cifra (Poey 1851 a) No obstante se dispone de dos estimados de la densidad de poblaci6n aborigen en las Antillas Mayores a la llegada de los europeos 18 indigenaslkm2 en Puerto Rico (Rouse 1956) y 1271km2 en La Espanola (Watts 1987) Extrapolando ambos valores entre las cuatro islas principales de las Antillas Mayores tenemos que en la fecha indicada la poblaci6n aborigen pudo estar entre 16 mil y 113 mil indigenas en Puerto Rico entre 21 mil y 145 mil en Jamaica entre 141 mil y 1 mill6n en La Espanola y entre 200 mil y 14 millones en Cuba Entre las cifras maximas y las minimas nos inclinamos
332 BIOGEOGRAFiA
hacia estas ultimas atendiendo a los altos indices de mortalidad infantil y morbilidad que se evidencian en las osamentas de los pobladores prehispanicos (Martinez y Rives 1990) Uno de los hechos que apuntan hacia el bajo numero de aborigenes en las Antillas Mayores es 10 rapido que desaparecieron despues de la llegada de los europeos pnkticamente sin dejar mestizaje con estos 10 cual contrasta ostensiblemente con la situacion en la America continentaL
En suma un grupo humano (1) demograiicamente irrelevante y controlado por altos indices de morbilidad (2) consumidor mayoritariamente de organismos marinos y (3) carente de la tecnologia cinegetica necesaria para poder sobreexplotar los mamiferos que Ie sirvieron de alimento (a los cuales accedio tardiamente en su desarrollo) no es concebible que haya causado e1 holocausto faunal que se Ie atribuye a partir del error de aplicar a las comunidades aborigenes antillanas un modelo cultural de la prehistoria continentaL
Tampoco hay evidencias de que los aborigenes hayan introducido carnivoros con suficiente capacidad depredadora salvo el perro Morgan y Otlenwaldder (1993) consideraron que la depredacion por perros durante el periodo amerindio es la explicacion mas plausible para la extincion del almiqui gigante de Cuba pero como se puede observar (Tabla 54) Solenodon arredondoi aparece en el intervalo II por falta de asociacion con evidencias culturales
Consecuentemente el accionar de los aborigenes y de sus animales domesticos directamente sobre la fauna no parece haber sido por sf solo un factor determinante de la extincion de mamiferos terrestres durante el periodo amerindio en las Antillas (pero vease Steadman et al 2005)
Hay que recordar en este senti do que alin en el Holoceno tardio ocurrieron cambios clirruiticos de consideracion (vease Causas Naturales) Por ejemplo Curtis et al (2001) explicaron que la extincion de algunos ani males mesofilicos de las alturas montanosas de La Espanola pudo haber antecedido a cualquier impacto humano sigrIificativo en esos lugares por coincidir dicha extincion con una tendencia a la aridez documentada isotopicamente y con evidencias de la perdida del bosque mesico ocurridas despues de 3kaAP Yen Jamaica hace solo 2shy3 mil aDos el area de la Bahia de Jackson sustentaba una gran biomasa de vertebrados en bosque hinnedo a juzgar por la cantidad y diversidad de taxones presentes en material fosil derivado de la depredacion por lechuzas hoy el area es la mas seca de la isla con matorral xerofitico y pobreza de fauna (MacPhee 1996a) Yalm en nuestros dias huracanes de gran intensidad aniquilan poblaciones de vertebrados (que no se recuperan) en is las pequenas y llanas del Caribe (Brooks y Smith 2001)
En medio de la incertidumbre sobre la verdadera causa (0 causas) de las extinciones de mamiferos en el periodo amerindio MacPhee y Marx (1997) han propuesto su hipotesis de la hipermorbilidad la cual estipula colapsos poblacionales relativamente rapidos con la introduccion de enfermedades de alta letalidad Estos autores han razonado que
Las islas son los lugares mas apropiados para experimentar perdidas catastroficas causadas por enfermedades Los mamiferos no solo serian susceptibles en virtud de su prolongado aislamiento biologico sino que en los confines de una isla la epizootia
333 EXTINCIONES DEL CUATERNARIO
tendria la potencialidad de infectar simultaneamente a la rnayoria de los individuos de vadas especies Los neonatos son inmunologicamente inmaduros y sufririan desproporcionadamente cualquier infeccion hipervirulenta La perdida excesiva de neonatos y juveniles tendria efectos severos para la sobrevivencia de la poblacion si esta se reduce por debajo de cierto nivel sobreviene la extincion porque no hay posibilidad de restitucion en el periodo postpanzootico Las especies de mayor talla corporal van a ser las mas vulnerables a la extincion dadas sus bajas tasas reproductivas largos periodos de gestacion y reducido numero de progenie Las especies de menor tamano --caracterizadas por altas tasas reproductivas periodos cortos de gestacion y mayor numero de neonatos por parto- tambien serian fuertemente diezmadas pero muchas sobrevivirian porque su estrategia reproductiva les permite perder mayor numero de individuos Las perdidas deben ocurrir relativamente rapido (a escala de decadas) mientras que otras hipotesis (digamos los cambios climaticos) permiten intervalos mucho mas largos (a escala de siglos 0 de milenios) entre la aparicion de las causas y la produccion de sus efectos Por ejemplo todo indica que los nesofontes se extinguieron muy rapidamente y seria dificil poder invocar un impacto humano directo 0 argumentar fenomenos climaticos que pudieran explicar plausiblemente perdidas tan rapidas
MacPhee y Marx (1997 191) han dicho que por el momento la hipotesis de la hipermorbilidad no pasa de ser una abstraccion en espera de adecuada sustentacion empirica Llama la atencion que linicamente en el periodo amerindio (Tabla 45 intervalo 1Il) las extinciones abarcaron todas las clases del peso corporal (de la a a laj) y que ninguna de las especies que sobrevivieron hasta el periodo postcolombino (Tabla 45 intervalo IV mas las especies vivientes) sobrepasa los 10 kg (clases del peso a b c) en clara coincidencia con los presupuestos de la hipotetica hipermorbilidad
Intervalo Postcolornbino
Por 10 menos 37 especies de mamiferos terrestres vivian en las Antillas Mayores y grupos de Islas perifericas a la lIegada de los europeos en 1492 De elIas se han extinguido 23 shymas especies que en cualquiera de los periodos precedentes (Tabla 45~ y solo subsisten 12 especies repartidas entre Cuba (8 especies) La Espanola (2) Jamaica (1) y Bahamas (1) varias de estas en peligro critico de extincion Solo dos de las especies extinguidas durante los ultimos 500 anos se conocieron en estado viviente los roedores Oryzomys antillarum de Jamaica (extinguido en 1877) y Geocapromys thoracatus de las Islas Swan (extinguido en 1955) Las causas de las extinciones ocurridas durante este periodo son sobradamente conocidas destruccion de habitats introducci6n de animales exoticos (domesticos y comensales) y demas presiones asociadas a la actividad humana entre las cuales posiblemente habria que incluir brotes de hipermorbilidad
Durante mucho tiempo se ha afirmado la posibilidad de que especies de Nesophontes hayan sobrevivido hasta las primeras decadas del siglo xx y que incluso continuen existiendo en lugares remotos de Cuba 0 La Espanola (Allen 1918 1942 Miller 1930 Aguayo 1950a Arredondo 1960b Fischer 1977 MacFadden 1980 Woods y
334 BIOGEOGRAFiA
Ottenwalder 1991) Actualmente no existe evidencia radiometrica de que alguna especie de Nesophontes haya sobrevivido al siglo xv las fechas radiocarbonicas calibradas obtenidas en muestras de Nesophontes de Cuba y La Espanola resultaron no mas tardias que los siglos XIII y XIV (MacPhee et al 1999) Por otra parte las expediciones efectuadas a fines del siglo xx con el proposito de localizar individuos vivientes del genero en La Espanola no produjeron la menor indicacion afirmativa al respecto (Woods et al 1985) Ademas a traves de las Antillas Mayores no aparecen restos de Nesophontes en los depositos formados actualmente por las lechuzas
EPiLOGO
Las Antillas Mayores son islas oceanicas no han formado parte de un continente (con la posible excepcion de una parte de Jamaica hasta el Eoceno Medio hace 45 Ma) y durante un tiempo geologico considerable (desde el Eoceno Superior hace 40 Ma) han permanecido emergidas y separadas de los continentes vecinos Consecuentemente los mamiferos terrestres (no volantes) autoctonos de estas islas constituyen una fauna oceanica que por ende cumple con todas las caracteristicas convencionales reconocidas para este tipo de entidad biogeografica
- Baja diversidad taxonomica a nivel de las categorias superiores (solo cuatro ordenes) en comparacion con islas plataformicas continentales (0 con areas continentales) de tamano y diversidad ecologica simi lares
- Alto grado de endemismo a nivel de genero (96) y familia (50) en respuesta al prolongado aislamiento
- Desbalance ecologico de la estructura trofica ausencia de mamiferos camivoros y ungulados
- Espectaculares procesos de irradiaci6n evolucionaria como resultado de la colonizacion de territorios libres de competidores y depredadores
Tendencias evolucionarias extremas gigantismo
- Control a cargo de aves rapaces con tendencia at gigantismo en ausencia de mamiferos miembros del orden Carnivora
- Mas especies de reptiles y de murcielagos que de mamiferos terrestres relaciones que en los continentes se invierten
Alta tasa de extincion de especies (84) sin reposicion
Preservacion de especies filogeneticamente arcaicas (los llamados fosiles vivientes) los almiquies

295 ESPACIO Y TIEMPO
~) OCEANO ATLANTICO 26deg
J-~ __~~ )Bahamas laquo - ~ o km 400
---~ Florida
lt 221 Mexico ~
c~o
d _-shy ---shy P rt I Virgenes --------
u~ 0 Anguila- I
Rlco ctQ 8 Martin I c=~ -__ -------- ~I Swan ~ o ~ Qo~o
Jamaica Q
- - ------------- Y d
~ - v ~ gt o
MARCARIBE i oJ (
a
SURAMERICA
70middot
FIGURA 41 El Caribe insular_ La linea discontinua circunvala el area geografica objeto de atenci6n en el presente capitulo
TABLA 41 Marniferos terrestres aut6ctonos del segmento insular delimitado en la Fig 41 (especies en orden alfabetico las vivientes en letra negrita =especies precuaternarias) CU=Cuba JA=Jamaica LE=La Espanola PR=Puerto Rico IS=Islas Swan IC=lslas Caiman IB=Islas Bahamas IV=Islas Virgenes AS=Anguila - San Martin
lsI a S Taxonomla 1
CU JA LE PR IS IC IE IV AS
Pilosa (pcrczosos) 14 especics
MEGALONYClllDAE 2
CHOLOEPODIKAE
Acratocnus antillensls bull Acratocnus odontrigonus bull Acratocnus simorhynchus bull Acratocnus ye bull
(Continua)
296 BIOGEOGRAFiA
TABLA41 (Continuaci6n)
I s 1 a s Taxonomla 1
CD JA LE PR IS IC lB IV AS
N eocnus comes bull Neocnus dousman bull N eocnus gliriform is bull N eocnus toupiti bull Gen et sp indet bull MEGALOCNINAE
Imagocnus zazae3 bull M egalocnus rodens bull M egalocnus zile bull Parocnus browni bull P arocnus serus bull
Soricomorpha (musaraiias) 11 especles
NESOPHONTIDAE 4
Nesophontes edithae bull bull Nesophontes hypomicrus bull Nesophontes micrus bull bull N esophontes zam icrus bull N esophontes sp 1 bull Nesophontes sp 2 bull Gen et sp indet bull SOLENODONTIDAE 4
Solenodon arredondoi bull Solenodon cuhanus 5 bull Solenodon marcanoi bull Solenodon paradoxus bull
Primates (monos) 4 especies
PTIHECIIDAE 2
CALLICEBINAE
Antillothrix bernensis bull (ContinUa)
297 ESPACIO Y TIEMPO
TABLA 4 1(Continuacion)
I s a Taxonomia 1
CD JA LE PR IS IC IE IV AS
Paralouatta marianae bull Paralouatta varonai bull Xenothrix mcgregori bull
Rodentia (roedores) 40 especies
ECHllVlYIDAE 2
HETEROPSOMYINAE
Boromys offella bull Boromys torrei bull Brotomys contractus bull Brotomys voratus bull Heteropsomys antillensis bull Heteropsomys insulans bull Puertoricomys corozalus bull CAPROMYIDAE 4
PLAGIODONTINAE
Plagiodontia aedium 6 bull P lagiodontia araeum bull P lagiodontia ipnaeum bull Plagiodontia spelaeum bull Rhizoplagiodontia lemkei bull ISOLoBODONTINAE
Isolobodon portoricensis 7 bull Isolobodon montanus bull Zazamys veronicae bull HExOLOBODONTINAE
Hexolobodon phenax bull CAPROMYINAE
Capromys pilorides 67 bull Capromys sp bull
(Continua)
298 BIOGEOGRAFIA
TABLA 4 1 (Continuacion)
I s a s Taxonom1a 1
CU JA LE PR IS IC IB IV AS
Geocapromys browni 7
Geocapromys columbianus bull Geocapromys ingrahami 6
Geocapromys thoracatus
Geocapromys sp 1
Geocapromys sp 2
Macrocapromys acevedo
Macrocapromys latus
Mesocapromys angelcabrerai
Mesocapromys auritus
Mesocapromys kraglievichi
Mesocapromys nanus
Mesocapromys sanfelipensis
Mysateles melanurus
Mysateles prehensilis 6
bull bull bull bull bull bull bull bull bull
HEPTAXODONTIDAE 8
HEPTAXODONTINAE
Amblyrhiza inundata
Elasmodontomys obliquus
Quem isia grav is
Xaymacafulvopulvis 9
CLIDOMYlNAE
Clidomys osborni
Clidomys parvus
MURIDAE2
SIGMODONTlNAE
Oryzomys antillarum 10
bull
bull bull bull
bull
bull
bull bull
bull bull
bull
bull Total de especies por is1all 24 6 25 7 1 5 1 1 1
1 La taxonomia sigue las revisiones sistematicas mas recientes (Morgan 1993 1994 MacPhee y Grimaldi 1996 Horovitz y MacPhee 1999 White y MacPhee 2001 Ottenwalder 2001 Woods et aI 2001 MacPhee y Flemming 2003) con 1a adici6n de los taxones descritos posteriormente y la incorporaci6n de los cambios propuestos para las especies cubanas en el Capitulo 2 de este Compendio (Continua en pagina opuesta)
299 ESPACIO Y TIEMPO
CONTEXTO TEMPORAL
La absoluta mayoria del registro fosil de mamfferos terrestres antillanos proviene de depositos cavemarios pleistoholocenicos (escala del tiempo geol6gico en la Tabla 42) Abundan en estas cavernas restos de perezosos musarafias y roedores siendo comparativamente escasos los de monos debido quizas a la universal predileccion de los platirrinos por los ambientes densamente forestados 10 que los hace malos candidatos a la fosilizacion (MacPhee y Woods 1982)
A pesar de 10 numeroso de tales depositos en la region son escasos los fechados radiometricos del cuantioso material extraido Con excesiva frecuencia (particularmente en Cuba) estos depositos han sido asignados al Pleistoceno Superior sin fundamento suficiente para ello y es sabido que incluso en cuevas contiguas los depositos fosilfferos pueden diferir considerablemente en edad (Pregill y Olson 1981) Se superponen a esta inconveniencia la falta de estudios tafonomicos en las excavaciones y la eventual mezcla de f6siles de distintas edades como consecuencia de las multicausales alteraciones estratigraficas ocurridas en las cuevas (Nunez 1990a b Morgan y Ottenwalder 1993 MacPhee 1997 McFarlane 1999b Dfaz-Franco 2004) Sobre propuestas de tipificaci6n de los depositos paleontologicos con restos de mamiferos terrestres en Cuba vease Woloszyn y Silva (1977) Acevedo y Arredondo (1982) Gutierrez (1994) Arredondo [1999]
La Tabla 43 presenta los fechados pleistocenicos mas tempranos registrados para restos de mamfferos terrestres del Caribe insular De estos restos el roedor (Cidomys) de Jamaica con un fechado de 253 mil anos antes del presente constituye el fosH de mamifero terrestre mas viejo del Cuaternario anti11ano [Los fosiles fechados antes del surgimiento de la espectrometria de masa (aceleradora) para muestras pequefias pueden ser algo mas viejos (Jull et al 2004)]
Corresponde a MacPhee e Iturralde-Vinent (1994 1995b) e1 relevante merito de haber descubierto en la ultima decada del siglo xx los primeros fosiles terciarios de mamiferos terrestres de las Antillas extendiendo a mas de 30 millones de afios el registro f6sil de
2 Extirpada en la Subregion Antillana pero viviente en el Neotropico continental
3 Ubicacion en subfamilia s610 sugerida por White y MacPhee (200 I)
4 Endemica de la Subregion Antillana
5 Recientemente propuesta su ubicaci6n bajo el genero Atopogale (Roca et al 2004)
6 lncluye las subespecies
7 Transportada por aborigenes a otras de las islas (Rimoli 1974 Olson 1982b este trabajo)
8 Vease MacPhee y Flemming (2003) sobre la incierta situaci6n de esta familia
9 Ubicada provisionalmente en esta subfamilia por MacPhee y Flemming (2003)
10 Hallada en contexto no perturbado de edad Pleistoceno tardio (McFarlane et ai 2002) con 10 cual se descarta su supuesta introduccion por amerindios (Woods 1990)
II La correlaci6n entre el area insular y la riqueza de especies es razonablemente fuerte (R=O83) en estas islas excepto las Bahamas (Morgan 1989)
300 BIOGEOGRAFIA
TABLA 42 Escala convencional del tiempo geologico en la Era Cenozoic a y finales de la Mesozoica (Gradstein 2004) ka = kiloafios (miles de afios) Ma miJIones de afios AP = antes del presente
Periodo(Suhperiodo) Epoca Afios (aproximados) antes del presente
Era Cenozolca
Cuaternario Holoceno Pkistoceno
Presente - 11 kaAP 11 kaAP - 18 MaAP
Terciario (Ne6geno ) Plioceno Mioceno
18 - 5 MaAP 5 - 23 MaAP
oIigoceno 23 - 34 MaAP Terciario (pak6geno) Eoceno 34 - 58 MaAP
Pakoceno 58 - 65 MaAP
Era Mesozo ca
Creticico 65 - 145 -faAP
~~~------~~--- --~-
este grupo animal en la region con el hallazgo de un perezoso oligocenico en Puerto Rico (Yauco) as como un perezoso un mono y un roedor miocenicos en Cuba (Domo de Zaza) Esto unido a la presencia de huesos de una musarafia nesofontoide [referida erroneamente a Solenodon por Rivero (1997)] en ambar oligomiocenico de la Republica Dominicana (MacPhee y Grimaldi 1996) demuestra incontrovertiblemente que al menos en el Mioceno temprano (alrededor de 20 millones de afios atnis) ya estaban en las Antillas Mayores representantes de los cuatro linajes basicos de su fauna de mamiferos terrestres [La afirmacion de Abreu et al (1990 13) en cuanto a que se conocen restos fosiles de almiqui que datan de hace aproximadamente 40 millones de afios es erronea]
Pero lcuan antigua puede ser la fauna de mamiferos terrestres de las Antillas Mayores
Aunque es un hecho bien establecido por la geologia que desde finales de la Era Mesozoica y principios de la Cenozoica existieron islas en el area ocupada ahora por las Antillas Mayores la evidencia disponible indica asimismo que aquellas primeras islas sufrieron repetidas transgresiones marinas totales que habrian hecho sucumbir las biotas que pudieran haber sustentado (MacFadden 1981 Morgan y Woods 1986 MacPhee e Iturralde-Vinent 1994 Roca et al 2004) sin contar los catastroficos efectos que debio tener para el Caribe occidental el b6lido extraterrestre que impacto en Chicxulub (Yucatan) hace 65 Ma (Tada et ai 2003) Hedges (1996) ha especulado que aUn en medio de tal devastacion debe haber existido algun que otro lugar donde elementos de la fauna pudieron haber sobrevivido
301 POBLAMIENTO CONTINENTE-ISLA
TABLA 43 F echados pleistocenicos mcis tempranos (obtenidos por distintos metodos) en mamiferos terrestres del segmento insular delimitado en laFig 41 F =Fuentes (I) Jull et al 2004 (2) Goodfriend y Mitterer 1987 (3) MacPhee et al 1989 (4) Woods 1989b (5) Rega et aI 2002 (6) McFarlane 1999b (7) McFarlane 1 999a (8) McFarlane et al 1998a
Pals Localidad Taxon kaAP F
Cuba Breas de San Felipe Pilosa (Parocnus) 1l9a 1
Jamaica Cueva Coco Ree Cueva Wallingford Roadside
Primates indet Rodentia (Clidomys)
393 2530
2 3
Haiti Trouing Jeremie no5 (casimba) Rodentia (Rh izoplagiodontia) 212 4
Republica Dominicana
Cueva del Perezoso Pilosa (ficratocnus) 185 5
Puerto Rico Cueva de Ia Vaca Cueva Clara
Pilosa (Acratocnus) Soricomorpha (Nesophontes)
337 346
6 7
Anguila Cueva Cave-No-Cave Rodentia (Amblyrhiza) 2060 8
a Kulp etal (1952) reportaron un fechado radiocarbonico de gt 30000 afios para un cono de pino en el deposito fosilffero de Ciego Montero (Cuba) con restos de perezosos
(vcase tambicn Crother y Guyer 1996) pero Iturralde-Vinent y MacPhee (1999) han preguntado donde estaban esos lugares si es que existieron
Solo a partir del Eoceno Superior las Antillas Mayores en fonnacion han pennanecido emergidas hasta hoy [excepto parte de Jamaica y el sur de La Espanola que continuaron sumergidas hasta el Mioceno Medio (Iturralde-Vinent y MacPhee 1999)] Esto significa que solo durante los ultimos 40 millones de anos aproximadamente han existido pennanentemente en dicha region insular ambientes terrestres capaces de sustentar comunidades biotic as (MacPhee e Iturralde-Vinent 2000 Portell et ai 2001 MacPhee 2005) 10 cual proporciona el marco temporal de referencia para conceptuar la antigiiedad maxima posible de la fauna de mamiferos terrestres de las Antillas Mayores
POBLAMIENTO CONTINENTE-ISLA
AFINIDADES CONTINENTALES
El consenso de los autores que en las itltimas dccadas han tratado la filogenia y biogeografia de los mamiferos terrestres de las Antillas Mayores (MacFadden 1980 MacPhee y Woods 1982 Morgan 1985 1988 1993 Morgan y Woods 1986 Ford 1986 Woods 1988 1989a 1990 MacPhee e Iturralde-Vinent 1995a b Hedges 1996 Horovitz [1997] Ottenwalder 2001 Whidden y Asher 2001 MacPheeet ai 2003 Carlini y Scillatoshy
302 BIOGEOGRAFiA
Yane 2004 Davalos 2004 MacPhee y Horovitz 2004 Roca et al 2004) es practicamente unanime en cuanto a que los ancestros continentales que dieron origen a esta fauna derivaron (1) mayormente de Suramerica perezosos (Megalonychidae) monos (Pitheciidae) y roedores (Echimyidae Capromyidae y Heptaxodontidae) (2) en menor medida de Norteamerica musarafias (Nesophontidae y Solenodontidae) y (3) menos aun de Centroamerica una especie de roedor (Muridae)
HIPOTESIS BIOGEOGRAFICAS
La definicion c1asica reconoce dos tipos de islas isla continental (porcion del territorio de un continente separado de este por movimiento tectonico 0 por ascenso del nivel del mar) e isla oceanica (porcion de territorio que no ha formado parte de un continente) La definicion sigue siendo uti por 10 que implica para la formacion de las faunas insulares en uno u otro caso As en el caso de la isla continental su fauna ya estaba emplazada en la futura isla antes de producirse su separacion del continente [Vicarianza hipotesis biogeografica de fragmentacion faunal] en el caso de la isla oceanica su fauna es el resultado de la colonizacion1 por propagulos2continentales a traves de la barrera marina entre el continente y la isla [Dispersion hipotesis biogeografica de acumulacion faunal] En el primer caso el surgimiento de la barrera marina es resultado del evento vicariante en el segundo la existencia de la barrera marina es anterior al evento dispersante Ademas de operar a escala mayor (continente-isla) ambos eventos operan secundariamente a menor escala (isla-isla)
Las dos hipotesis -vicarianza y dispersion- se han disputado hasta hoy la preferencia de los biogeografos en tanto hipotesis mas viable para explicar el proceso de formacion original (continente-isla) de la fauna de mamiferos terrestres de las Antillas Mayores [Sobre la historia de las hipotesis biogeograficas relativas al Caribe insular vease RuB y Schubert 1989 Williams 1989 Pruna 2001] Yel debate continua debido ala falta de consenso sobre 10 que Williams (1989) ha llamado el problema central de la discusion a saber la ostensiblemente baja diversidad taxonomica de esta fauna a nivel de las categorias superiores (solo cuatro ordenes) en comparacion con la diversidad al mismo nivel de las faunas de islas continentales (0 de areas del continente) simi lares en tamafio y diversidad ecologica [Por ejemplo la fauna cubana de mamiferos terrestres consideradas de conjunto las especies vivientes y las extintas- es menos diversa a los niveles taxonomicos superiores (4 ordenes y 6 familias) que 10 que cabria esperar de la diversidad ecologica y extension territorial de Cuba (110 992 km2) comparada con una isla tipicamente continental como Trinidad (4828 krn2
) -veinte veces menor que Cuba y menos diversiticada ecologicamente- donde solo las especies vivientes de mamfferos terrestres representan 6 ordenes y 16 familias (Eisenberg 1989)] Este desbalance
lColonizaci6n Arribo y establecimiento de un propagulo en una isla no ocupada por la especie (MacArthur y Wilson 1967)
2Propagulo Niimero minimo de individuos de una especie capaces de reproducirse y aumentar poblacionalmente en una isla bajo condiciones favorables (MacArthur y Wilson 1967)
303 POBLAMIENTO CONTINENTE-ISLA
taxon6mico ha sido expJicado de dos maneras (1) Inicialmente hubo mas 6rdenes presentes pero la mayoria se extingui6 y s610 sobrevivieron esos cuatro [explicaci6n vicarianista] (2) S610 representantes de esos cuatro 6rdenes lograron cruzar la barrera marina [explicaci6n dispersionista]
Cinco hipot~ticos mecanismos de poblamiento continente-isla han sido invocados durante los ultimos 140 afios (y se los sigue proponiendo actualmente) para explicar la presencia de mamiferos terrestres en las Antillas Mayores A continuaci6n pasamos a resefiarlos brevemente (incluidas sus alegadas fortalezas y debilidades) en el orden aproximado en que historicamente fueron propuestos
EL PUENTE NATURAL (dispersion terrestre activa)
En el siglo XIX el puente terrestre era pnkticamente la unica hipotesis para explicar las distribuciones disyuntas de animales en buena parte del mundo y los primeros investigadores que se ocuparon de la biogeografia antillana (Fernandez [1865] 1884 Chapman [1893] Torre 1892 Lora 1895) la asumieron resueltamente en parte intrigados por el descubrimiento en Cuba en la segunda mitad del siglo de un perezoso f6sil de gran talla Y en su aplicacion a los mamiferos terrestres de las Antillas Mayores el puente sigui6 siendo a 10 largo del siglo xx la hipotesis mas favorecida por los estudiosos del tema (Vaughan 1902a Spencer 191 Oa b Torre 1910a b cAllen 1911 1917a 1918 Barbour 19141916 1945bAnthony 1918 1926 [1942] Scharff 1922 Kraglievich 1926 Beaufort 1951 Arredondo 1961 1970b Kartashov y Mayo 1974 Iturralde-Vinent 1982 1988 Arredondo y Armifiana 1996 Arredondo 1996a [1999]) quienes postularon la existencia preterita de puentes con Norte- Centro- 0 Suramerica en dependencia de las afinidades continentales de los mamiferos antillanos
Teoricamente el puente terrestre debe cumplir con una condicion esencial permitir el paso de una parte mayoritaria (y por tanto representativa de la diversidad) de la fauna continental Claro que siempre habria tipos de mamiferos menos proclives a dispersarse o para los cuales la ubicaci6n del puente resultaria inaccesible 0 las condiciones ecologicas en este fueran desfavorables pero si algunos acceden al puente no hay raz6n para pensar que el resto de esa fauna no los acompafie mayoritariamente Como dijera Simpson (1956) Los puentes no transportan faunas desbalanceadas
En consecuencia el puente terrestre como filtro es un mecanismo de poblamiento minimamente selectivo y por ende de alta diversidad inicial (sensu Iturralde-Vinent y MacPhee 1999) 10 esencial del puente es su capacidad para transportar faunas integradas (Williams 1989) Yen eno radica la objeci6n fundamental al puente terrestre surgida tempranamente en el siglo xx (Por que s610 cuatro 6rdenes de mamiferos terrestres en las Antillas si vinieron por puentes (Donde estan los fosiles de los demas ordenes que debieron venir si es que se extinguieron Los partidarios del hipotetico puente dispusieron de casi todo el resto del siglo para responder una y otra vez que otros 6rdenes de mamiferos aparecerian representados en las Antillas Mayores el dia que en estas Islas se descubrieran fosiles precuatemarios suficientemente
304 BIOGEOGRAFIA
antiguos Esto se logro por fin terminando el siglo como se explico anteriormente (vease Contexto temporal) y ique se encontro los mismos cuatro ordenes de mamiferos terrestres conocidos del Cuatemario caribefio aparentemente los Unicos que lograron llegar
Pero ademas hasta el presente no se ha demostrado inequivoca- y concluyentemente que en algun momenta durante los ultimos 40 millones de afios haya existido una conexion terrestre continua entre las Antillas Mayores (en proceso de formacion 0 plenamente formadas) y algun punto de las regiones continentales vecinas
LA BALSA NATURAL (dispersion marina pasiva)
Durante buena parte del siglo xx el puente terrestre tuvo como unico rival a la hipotesis de la balsa natural -transporte accidental de animales sobre islotes de vegetacion flotantes a la deriva despues de arrancados y expulsados mar afuera por un rio crecido- expuesta originalmente por Matthew (191319151916 1918a 1919) y desarrollada en su aplicacion a los mamfferos terreslres antillanos principal mente por Simpson (1956) asi como por olros investigadores (Darlington 1938 1957 Williams y Koopman 1952 Hershkovitz 1958 1972 Patterson 1962 Paula 1967 Patterson y Pascual 1972 Pregill 1981 Ford 1986 Woods 1989a 1990 Morgan 1993) Por su parte Hedges et al (1992) sustentaron dicha hipotesis en estimados de divergencia (basados en el reloj molecular) en la herpetofauna antillana -involucrados tangencia1mente los mamiferos [veanse objeciones a Hedges y colaboradores (Page y Lydeard 1994 Iturralde-Vinent y MacPhee 1999) respuestas a las objeciones (Hedges et al 1994 Hedges 2001) y contrarrespuestas (MacPhee e IturraldeshyVinent 2005) asi como cuestionamientos al metoda de los estimados de divergencia (Foote et al 1999 Pregill y Crother 1999 Woods et ai 2001 )]
La hip61esis de la balsa natural se origino a partir de las numerosas ocasiones en que a 10 largo de siglos tales islotes flotantes (portando 0 no vertebrados incluidos mamiferos) han sido avistados mar afuera a cientos de kilometros de la costa mas cercana en varias partes del mundo --inc1uido el Mar Caribe- segUn consta en diarios de barcos y reportes de observadores diversos (revisados por King 1962 reportes posteriores en SICSP 1970 y Censky et al 1998) Uno de estos casos acaecido en nuestra region se reporta en un informe del SICSP (Smithsonian Institution Center for Shortlived Phenomena 197050shy52)
EIB de julio de 1969 un destructor de la armada de Estados Unidos avisto un islote flotante a unos 100 km al sur de la Base Naval de Guantanamo moviendose desde el Paso de los Vientos en direccion suroeste a una velocidad de 45 kmlh islote de unos 15 m de diametro mantenfa en pie mas de 10 palmas reales de 10-15 m de altura con suelo cubierto de hierba y arbustos [la observacion con binoculares a 18 km de distancia no preciso la presencia de vida animal] La travesia fue rastreada durante 6 dfas hasta que el 19 de julio el islote se dio por perdido tras 6 horas de busqueda aerea
305 POBLAMIENTO CONTINENTE-ISLA
La colonizaci6n exitosa de una isla de las Antillas Mayores por mamiferos terrestres continentales transportados en balsas naturales a la deriva (de cada especie al menos una hembra gravida 0 dos individuos de sexo opuesto) dependeria de factores que van a operar desde el momenta mismo de formarse la balsa asi como durante su travesia marina y su arribo a la isla
a) La posibilidad de estar en la balsa Que la distribuci6n de la especie incluya areas contiguas al rio y que el animal se encuentre en ellugar y el momenta propicios para ser incorporado a la balsa antes de que el rio crecido la desprenda Por ejemplo segun Hershkovitz (1958) los equimidos -grupo ancestral de roedores antillanos- serian buenos candidatos a ser captados por la balsa porque acostumbran a anidar en huecos del tronco de los arboles a la orilla de los rios
b) El tamano y la consistencia de la balsa Muchas se desintegran en alta mar debido a su fragilidad ante la saturaci6n Algunos autores (Beaufort 1951 Arredondo 1996a [1999]) han expresado incredulidad en cuanto a que las balsas hayan podido soportar el peso de un animal como el gran perezoso cubano (Megalocnus rodens) el mayor de los mamiferos antillanos Pero esta especie evolucion6 en Cuba y el ancestro que hizo el viaje pudo ser muy pequeno [EI perezoso oligocenico de Puerto Rico es el mas pequeno de los megaloniquidos antillanos (MacPhee e Iturralde-Vinent 1995b )]
c) Las caracterfsticas fisiol6gicas de la especie Los limites fisiol6gicos de los propagulos constituyen nuestra unica objeci6n a la dispersi6n marina por balsas como mecanismo para la formaci6n de la fauna de vertebrados en las Antillas (MacPhee e IturraldeshyVinent 2005) Unas especies tienen intrinsecamente mas posibilidades que otras para sobrevivir a las vicisitudes que deben afrontar durante la travesia (deshidrataci6n inanicion insolaci6n etc) Por ejemplo los megaloniquidos han demostrado tener extraordinaria habilidad para cruzar barreras marinas fueron los primeros mamfferos suramericanos en invadir Norteamerica mucho antes de que surgiera la conexion terrestre a finales del Plioceno (Paula 1967 Olson 1978 Webb 1985b Martin y Steadman 1999) Incluso se conoce un megaloniquido acultitico del Plioceno peruano (Muizon y McDonald 1995) y otro viviente en Suramerica (Choloepus) que pnicticamente no bebe agua 10 que ha originado una leyenda aborigen que explica por que los perezosos no toman agua (Goff art 1971) Vease ademas el inciso e
d) EI regimen de las corrientes marinas Que conduzcan a la balsa en la direcci6n necesaria EI transporte de balsas de vegetacion desde los continentes hacia las islas fue durante la mayor parte del Cenozoico sumamente improbable (MacPhee e IturraldeshyVinent 2000 148) Estos autores han insistido en que hay que tener en cuenta que durante la cambiante geografia cenozoica del Caribe las corrientes marinas de superficie no favorecieron el desplazamiento de objetos fiotantes hacia las Antillas Mayores [Pero no sabemos con certeza que efecto modificador del curso de tales objetos fiotantes pudieron introducir las tormentas tropicales si previsiblemente funcionaron en el pasado como 10 hacen ahora] Dichos autores senalaron ademas (2005) que en un experimento reciente con boyas de posicionamiento satelital liberadas a la deriva desde las Antillas Menores las boyas fueron a parar mayoritariamente a Centroamerica y a otros lugares
306 B[OGEOGRAFIA
y solo unas pocas recalaron en las Antillas Mayores y las que entraron al Mar Caribe demoraron semanas 0 meses en arribar
e) La distancia a recorrer (y en consecuencia la duracion de la travesia) entre el continente y la isla (10 que repercute decisivamente en el factor c) Este es desde luego uno de los factores mas criticos En la medida que la distancia aumenta la probabilidad de alcanzar la isla disminuye drasticamente con el agravante de que la trayectoria dificilmente seria en linea recta Si tomamos como referencia la velocidad de 45 kmlh del islote ya mencionado (SICSP 1970) cubrir la distancia aerea (aproximada) de 800 km que media entre el noroeste de Suramerica y La Espafiola Ie tomaria a la balsa 74 dias Estimando el doble de este tiempo debido a desvios de la trayectoria la duracion de la travesia probablemente resultaria excesiva para la mayoria de los mamiferos Pero pudiera muy bien ocurrir que la velocidad fuera mayor 0 que la balsa portara plantas vivas que permitirian a los fitOfagos sobrevivir [Tres de los cuatro ordenes de mamiferos terrestres conocidos de las Antillas Mayores son fitOfagos] Ademas esta geologicamente bien establecido que durante etapas de variada duracion desde el Oligoceno hasta el Pleistoceno los territorios del Caribe insular y continental aumentaron considerablemente en extension con niveles del mar 100-200 m por debajo de su nivel actual oportunidades que los propagulos aprovecharian para cruzar barreras marinas significativamente menores
f) El tamafio de la isla Las islas mayores deben interceptar mas balsas (Buckley y Knedlhans 1986) 10 que repercute directamente en el factor e La balsa puede pasar entre islas pequefias y continuar viaje sin llegar a ningun lugar
g) Las condiciones (favorables 0 adversas) del desembarco en la isla y el tiempo que Ie tomaria a uno 0 mas animales depauperados encontrarun habitat apropiado Esto dependeria a su vez del tamafio y diversidad ecologica de la isla (Darlington 1938 Miller 1966)
Tal cumulo de posibles obstaculos y dificultades constituye a primera vista un desafio a la credibilidad de la hipotesis Matthew-Simpson pero en ello radica precisamente su valor explicativo bas ado en la mas absoluta improbabilidad La colonizacion por la via marina es altamente improbable pero no absolutamente imposible si se tiene en cuenta la inmensidad del tiempo geologico se vuelve probable (McKenna 1973) Basta que solo una vez entre mill ones de intentos fallidos durante ese inmenso lapso algunos de los factores anteriormente enunciados coincidan en no ser tan desfavorables para el propagulo Y eso es todo 10 que demanda la hipotesis para cada uno de los grupos monofileticos mayores de mamiferos terrestres presentes en las Antillas Mayores Las Antillas Mayores requeririan el arribo de tan solo 7 (quizas menos) propagulos fundadores a 10 largo de los ultimos 30 0 mas millones de afios (promedio de uno cada 43 mill ones) para explicar la formacion de toda su fauna de mamiferos terrestres por dispersion marina pasiva (vease Discusion y Conc1usiones)
En comparacion con la via marina el poblamiento por la via terrestre es predecible porque la mayor parte de la fauna continental de mamiferos emplazada en la region del puente va a cruzar por este una vez que el mismo se establece y los animales logran
307 POBLAMIEITO CONTINENTE-ISLA
acceder a el el poblamiento por la via marina en cambio es impredecible porque depende total mente del azar que caracteriza a la balsa Claro que en ambas vias opera el azar pero en el primer caso se trata del azar de un evento probable en el segundo el azar de un evento improbable (Simpson 1940) Asi el puente natural (mecanismo de poblamiento minimamente selectivo = alta diversidad inicial) y la balsa natural (mecanismo de poblamiento maximamente selectivo baja diversidad inicial) pueden conceptuarse como los extremos opuestos de una gama de probabilidades de dispersi6n (McKenna 1973) EI fen6meno de la dispersi6n merece la mayor atenci6n cientifica en los estudios de biogeografia insular (Cowie y Holland 2006)
Queda claro que la balsa natural es una hip6tesis consistente con el caracter de la fauna objeto de estudio puesto que el desbalance taxon6mico (sin evidencia paleontol6gica en contrario) s610 puede ser resultado de la improbabilidad del poblamiento No obstante ello no implica necesariamente que la balsa natural haya sido el imico mecanismo (ni siquiera el principal) utilizado por los mamiferos terrestres en su colonizaci6n de las Antillas Mayores La balsa natural debe satisfacer una premisa te6rica esencial dado el caracter azaroso impredecible e improbable de una colonizaci6n exitosa esta debe ocurrir a intervalos muy espaciados a traves del tiempo geol6gico 0 sea durante todo el tiempo que en el area ocupada por las Antillas Mayores ha habido ecosistemas terrestres perrnanentemente disponibles para ser colonizados (los ultimos 40 millones de afios) Sin embargo como sabemos representantes de los cuatro 6rdenes de mamiteros terrestres conocidos de las Antillas Mayores ya estaban emplazados en dicho territorio insular en el Mioceno Inferior 10 que supone un lapso de colonizaci6n limitado al Eoceno SuperiorshyOligoceno (MacPhee 2005) sugerente de un tipo de mecanismo contrario a la colonizacion alocronica exigida por la hipotesis de la balsa natural sobre todo tratandose de una fauna caracterizada por altas tasas de extincion sin reposicion aparente
La presencia en Jamaica (hasta su extinci6n en 1877) del roedor mfuido Oryzomys antillarom con estrecha afinidad centroamericana (Woods 1989a) pudiera constituir el imico caso efectivo de poblamiento (continente-isla) de las Antillas Mayores por dispersion marina dc un mamifero terrestre para el cual no es razonable invocar otro mecanismo de colonizacion
LA GUIRNALDA INSULAR (dispersion mixta)
La geologia ha establecido con suficiente certeza que durante el Cenozoico unicamente en el Pale6geno tardio y solo a traves de la Cresta de Aves (ahora sumergida excepto la isla hom6nima) pudo haber existido una cadena de islas para el transito de mamiferos terrestres entre el norte de Suramerica y las proto-Antillas Mayores [desestimados a ese efecto el Canal dc Yucatan el Elcvado de Nicaragua la Cresta de Caiman el Elevado de Beata y las Antillas Menores (Holcombe et al 1990 MacPhee e IturraldeshyVinent 1994 1995b Iturralde-Vinent y MacPhee 1999)] Holcombe y Moore (1977) y Holcombe y Edgar (1990) postularon la existencia de esta entidad y estimaron algunas de sus posibles caracteristicas Por su parte MacPhee e Iturralde-Vinent (1995b) e Iturralde-Vinent y MacPhee (1999) precisaron los extremos proximal y distal de esa
308 BIOGEOGRAFIA
estructura acotaron la duracion y la edad de la misma y Ie dieron nombre GAARlandia (Greater AntillesA vesRidgelandia)
Segun Iturralde-Vinent y MacPhee (1999) la Cresta de Aves emergio por breve tiempo (1-2 Ma) durante la transicion Eoceno-Oligoceno (35-33 MaAP) por la coincidencia de un levantamiento tectoni co y un considerable descenso eustatico del nivel del mar En ese momenta las islas septentrionales del arco de las Antillas Mayores (Cuba centroorientalshyLa Espafiola - Puerto Rico - Islas Virgenes) constituian una sola isla grande 0 una serie de islas separadas por barreras muy estrechas La Cresta de Aves emergida entre el bloque Puerto Rico - Islas Virgenes y un microcontinente noroccidental de Suramerica -separado en ese momento del resto del continente por una via marina surgida en el Eoceno tardio- dio origen a GAARlandia
Obviamente el numero de colonizaciones exitosas por mamiferos terrestres a traves de una guimalda de Islas tiende a disminuir mientras mas anchas sean las barreras marinas entre las islas que forman la guimalda y cuando esto ocurre disminuye tambien la diversidad taxonomica de los propagulos particularmente a nivel de las categorias superiores (Simpson 1956)
Vemos asi que para los mamiferos terrestres la colonizaci6n a traves de una guimalda insular puede ser tan improbable como a la deriva en una balsa natural en dependencia de la cantidad y anchura de las barreras marinas a cruzar el tamafio y la fisiografia de las islas que integran la guimalda y el regimen de las corrientes marinas y aereas entre otros factores
Resumiendo la guimalda insular puede conceptuarse como un mecanismo de poblamiento altamente selectivo (= baja diversidad inicial) que combina la improbabilidad de la balsa natural con la colonizacion restringida temporaimente del puente terrestre
En su reciente revision de la taxonomia distribuci6n y filogenia (basada en estudios moleculares) de varios grupos de mamiferos antillanos Davalos (2004) concluyo que los perezosos megalocninos los primates y los roedores histricognatos se avienen al patron de divergencia con el continente y epoca de arribo a las Antill as implicados en la hipotesis de GAARlandia pero no asi los perezosos coloepodinos que requieren una explicacion biogeognlfica distinta Esto ultimo (sospechado por Matthew 1918a) habla sido confirmado por White y MacPhee (2001 226-227)
Nuestra evidencia filogenetica no sustenta la idea de un dado unificado surgido de la invasion de un megaloniquido el analisis sugiere mas bien dos eventos de colonizaci6n independientes [y] sustenta fuertemente un origen difiletico de las subfamilias [Choloepodinae y Megalocninae] Si hubo dos migraciones a traves de GAARlandia presumiblemente poco espaciadas en el tiempo no se puede determinar en el presente
Finalmente queda claro que GAARlandia -como guirnalda insular- es la hipotesis mas robusta propuesta hasta ahora para explicar la presencia en las Antillas Mayores de
309 POBLAMIENTO CONTINENTE- ISLA
la absoluta mayona de los mamfferos terrestres de ongen suramericano porque el caracter de esa fauna supone un mecanismo de poblamiento ancestral que sea a la vez maximamente selectivo (baja diversidad ordinal) y temporalmente restringido (transicion Eoceno-Oligoceno)
Recientemente Rega et al (2002 18) expresaron A pesar de una revision exhaustiva de la paleogeografia de la cuenca del Caribe Iturralde-Vinent y MacPhee (1999) no pudieron confirmar la presencia de una conexion terrestre continua entre Suramerica y las proto-Antillas aunque hipotetizaron su existencia [Vease la Fig 42 (= figura 6 de Iturralde-Vinent y MacPhee 1999 simplificada))
En el trabajo citado por Rega y colaboradores aparece la siguiente explicacion
Actualmente no es posible ofrecer reconstrucciones paleogeograficas detalladas de la Cresta de Aves Sin embargo hay evidencia de que porciones de la cresta deben haberse elevado subaereamente en un tiempo La cuestion cntica es si la Cresta de Aves emergida pudo haber formado algun tipo de corredor entre las Antillas Mayores y el noroeste de Suramerica Perforaciones en las alturas estructurales de la cresta pudieran proveer el registro del Paleogeno tardio que se necesita para precisar la hipotesis del corredor aunque no se puede predecir si tales datos serian suficientemente detallados para determinar si en algun momento GAARlandia constituyo una sola entidad subaerea Por ejemplo pudo haber barreras marinas por corto tiempo entre las masas terrestres de la Cresta de Aves [0) ellevantamiento y la subsidencia pudieron afectar diferentes partes de GAARlandia en diferentes tiempos AItemativamente los elementos faunales pudieron haberse dispersado de manera pulsada Movimientos episOdicos de este tipo incrementarian la extincion de los elementos faunales incapaces de trasladarse a la siguiente masa terrestre en la cadena disminuyendo asi la diversidad total (Iturralde-Vinent y MacPhee 199957)
La primera parte de la explicacion deja claro que la posibilidad de que la Cresta de Aves haya sido una conexion terrestre continua no se puede descartar pero tampoco se puede afirmar La ultima parte de la explicacion describe el efecto altamente selectivo (filtrante) de la colonizacion a traves de una cadena de islas
Y continuan lturralde-Vinent y MacPhee (199957 -58) explicando
Hasta que punto el microcontinente noroccidental de Suramerica [considerado por dichos autores como el area suministradora de propagulos hacia GAARlandiaJ estuvo fisicamente separado del resto del continente no ha sido plenamente esc1arecido Si estuvo sustancialmente aislado por barreras marinas y orogenic as durante el Paleogeno tardio entonces solo la fraccion de la biota suramericana emplazada en el microcontinente estuvo en posicion de acceder a GAARlandia Esto pudiera tener que ver con el hecho de que la fauna antillana carezca de representantes de muchos grupos suramericanos [de mamfferos) que debieron haber hecho el viaje
310 BIOGEOGRAFiA
Florida o Alturas0 0
d 6 D L1anuras25deg
Yucatan
20deg
deg0
10deg
MARCARIBE
80deg 75deg
Bahamas
Cres ta de Beata
D Mares someros
D Mares profundos
OCEANO ATLANTICa
FIGURA 42 Reconstrucci6n hipotetica de GAARlandia como conexi6n terrestre continua entre las proto-Antillas Mayores y el noroeste de Suramerica en el Pale6geno tardio segun la FIgura 6 (simplificada) de Iturralde-Vinent y MacPhee (1999)
En su defensa del puente terrestre Barbour (1945b249) se refiri6 a esto - antes 10 habfa hecho Anthony [1942363]- en los terminos siguientes
El argumento de que si las Antillas hubieran estado conectadas con el continente se encontrarian los f6siles de un numero mayor de distintas criaturas me deja frio es perfectamente concebible que la conexi6n haya ocurrido en un periodo en que la parte del continente implicada no estuviera poblada por una fauna grande y variada
Y concluyen lturralde-Vinent y MacPhee (199958) su detallada explicaci6n
Desafortunadamente sabemos muy poco sobre la composici6n faunal del noroeste de Suramerica a finales del Pale6geno
Consecuentemente si se confirmara la magnitud del aislamiento del aludido microcontinente y su concomitante baja diversidad faunal GAARlandia como conexi6n terrestre continua serf a la hip6tesis mas parsimoniosa No obstante mientras no se disponga de evidencia concluyente a este respecto hipotetizar a GAARlandia como conexi6n continua (minima selectividad) la coloca en franca vulnerabilidad te6rica al tener que afrontar la objeci6n clasica al puente terrestre (baja diversidad ordinal de la
311 POBLAMIENTO CONTINENTE-TSLA
fauna) al tiempo que se la priva de ser la hip6tesis mas robusta para explicar -como guimalda de islas- la colonizaci6n de las Antillas por mamiferos procedentes de Suramerica
ELARcA DE NOE (seudovicarianza)
El primer vicarianista antillano (desde luego sin el saberlo) fue Corral (1940) con su utilizaci6n ingenua de las ideas wegenerianas de la deriva continental aparecidas la decada anterior Pero el primer (supuesto) intento de aplicar a la formaci6n de la biota caribefia el mecanismo de la vicarianza basada en la tearia de la tect6nica de placas (surgida en la sexta decada del pas ado siglo) se debe a Rosen ([1976] 1985)
Segun Rosen las Antillas Mayores se originaron en el Cremcico en la Placa Caribe aproximadamente en la posici6n que hoy dia ocupa Centroamerica y al ser desplazadas tectonicamente hacia el este interactuaron (de algun modo no precisado por Rosen) con las margenes continentales de Norte- y Suramerica de donde (en un evento unico) adquirieron toda su fauna transportada subsecuentemente por dichas islas en su transito hacia sus posiciones actuales [De ahi la denominacion metaforica dada par McKenna (1973) a este mecanismo de poblamiento] Aunque se ha presentado esta propuesta como Model0 de Vicarianza Continente-Isla de Rosen en realidad por definici6n no constituye un evento vicariante porque las proto-Antillas Mayores no resultaron del fraccionamiento y separaci6n por tectonismo de una porcion del territorio continental como exige esta hip6tesis (vease definici6n de vicarianza al comienzo de Hipotesis Biogeograficas)
El Arca de Noe de Rosen ha sido desestimada por la mayoria de los autores contemporaneos porque (1) carece de sustento geol6gico (lturralde-Vinent y MacPhee 1999) y porque (2) en el periodo en que se afrrrna ocum6 el evento muchos de los ordenes modemos de mamiferos placentarios attn no existian (Williams 1989)
EL BARCO FUNERAL VlKINGO (tanatovicarianza)
Domning et al (1997) reportaron 10 que consideraron el mamffero terrestre mas viejo de las Antillas --circunstancia que se refleja en el titulo mismo del trabajo Oldest West Indian Land Mammal- al descubrirse un perisodactilo rinocerotoide (Hyrachius sp) en una localidad de edad Eoceno Inferior a Medio en el occidente de Jamaica
Jamaica ha tenido una historia geologica y paleogeografica distinta de la historia comun a las restantes Antillas Mayores A principios del Cenozoico Jamaica occidental (a diferencia de la porcion oriental constituida por el Bloque de las Montafias Azules) formaba parte de Centro america de la cual se desgaj6 (por el desplazamiento de la Placa Caribe) para venir a integrarse despues del Eoceno Medio con 10 que serfa la futura isla de Jamaica desaparecida por transgresi6n marina en el Eoceno Superior hasta volver a emerger totalmente en el Mioceno Medio (Iturralde-Vinent y MacPhee 1999 MacPhee e Iturralde-Vinent 2000)
312 BIOGEOGRAFiA
Por consiguiente Hyrachius ---conocido de localidades de edad Eoceno Inferior a Medio en Norteamerica- debio ocupar (10 que iba a ser) Jamaica occidental mucho antes de que esta se separara de Centroamerica De hecho este fosil Uego a su destino insular contenido en las rocas de su basamento no como un ser vivo por 10 que es erroneo considerarlo un animal antillano (lturralde-Vinent y MacPhee 2004) como han supuesto algunos (Hedges 200 I Portell et al 200 I Condis [200 I D Todo 10 que hizo fue viajar tanatovicariantemente en su barco funeral vikingo -otra de las denominaciones metaforicas de McKenna (1973)
DISCUSION Y CONCLUSIONES
Hemos visto que solo disponemos de una hipotesis plausible para explicar la mayor parte del poblamiento (continente-isla) de las Antillas Mayores por mamiferos terrestres del Neotropico continental la dispersion mixta (a traves de GAARlandia) para los perezosos (excepto Choloepodinae) los monos y los roedores (excepto Oryzomys)
Por otra parte la historia biogeogrMica de las musaraiias de las Antillas Mayores (Nesophontes y Solenodon) constituye una de las situaciones mas enigmaticas entre los mamiferos de la region (Ottenwalder 2001321) Al menos cinco mecanismos de poblamiento continente-isla han sido invocados para explicar cuando como y de donde llegaron a las Antillas Mayores estas musaraiias
I EI propagulo ancestral (del Terciario de Norteamerica) coloniz6 a las Antillas Mayores por dispersion marina desde Centroamerica 0 desde la Florida (Patterson 1962)
2 El propagulo ancestral ( del Terciario) cruzo el Atlantico Sur (supuestamente mas estreshycho en aquella epoca) directamente de Africa a las Antillas Mayores (Hershkovitz 1972)
3 El propagulo ancestral (del Terciario de Norteamerica) Uego a las Antillas Mayores en el Arca de Noe de Rosen (MacFadden 1980)
4 El propagulo ancestral (del Terciario de Norteamerica) fue transportado a las Antillas desde Centroamerica en el bloque de Jamaica occidental y de alli paso por dispersion marina a las restantes Antillas Mayores (Whidden y Asher 200 I)
5 El propagulo ancestral (del Terciario de Norteamerica) lleg6 a las Antillas Mayores a traves de GAARlandia (Whidden y Asher 2001)
Aunque hay consenso entre estos (y otros) autores sobre el origen norteamericano de las musaraiias antillanas ninguno de los cinco hipoteticos mecanismos de poblamiento resiste la evidencia en contrario filogenetica yo paleogeogrMica Ademas algunas de estas hipotesis asumen que Nesophontidae y Solenodontidae comparten un antecesor comun en las Antillas pero esta relacion no ha sido confirm ada Para dilucidar el enigmatico caso de las musaraiias antillanas se requerini profundizar en la investigaci6n geologica paleogeognifica y paleontol6gica (Iturralde-Vinent y MacPhee 1999)
De acuerdo con el conocimiento filogenetico disponible sobre el conjunto de la fauna la colonizacion original de las Antillas Mayores por mamiferos terrestres implicaria el arribo
313 POBLAMIENTO ISLA-ISLA
de solo siete propagulos fundadores perezosos 2 (coloepodinos y megalocninos si como parece tienen un origen difiletico) musaraiias 2 (nesofontidos y solenodontidos si no constituyen una misma familia como consideran algunos) monos 1 (pitecidos) roedores histricognatos 1 (equimidos capromidos y heptaxodontidos) roedores muridos 1 (Oryzomys en Jamaica)
POBLAMIENTO ISLA-ISLA
La distribucion de los mamiferos terrestres a traves del area antillana demarcada al inicio del presente capitulo (vease Contexto Geogratico Fig 41) ha sido interpretada en funcion de dos hipoteticos mecanismos de poblamiento interinsular la dispersion y la vicarianza fundamentadas como se resumen seguidamente
DISPERSION
De acuerdo con la evidencia paleogeogratica mas actualizada en las principales regresiones marinas ocurridas en la cuenca caribeiia durante los ultimos 23 Ma (transiciones OligocenoshyMioceno Mioceno Superior - Plioceno y varias veces en el Pleistoceno) las masas terrestres constitutivas de la region insular en formacion aumentaron considerablemente su extension territorial disminuyendo por tanto la distancia entre elias oportunidades que los elementos faunales aprovecharian para dispersarse de unas islas a otras
Durante mucho tiempo esta ha sido la hipotesis mas favorecida por los estudiosos del tema a partir de Simpson (1956) pero en decadas recientes ha suscitado dos objeciones
1 Es dificil en tender como pudo operar la dispersion marina entre estas islas puesto que ninguna de las Antillas Mayores posee rios suficientemente caudalosos para transportar masas significativas de vegetacion flotante (MacPhee 198432)
Es oportuno recordar aqui el islote ala deriva (con mas de diez palmas reales en pie) entre las tres Antillas Mayores mas occidentales mencionado anteriormente (vease La Balsa Natural)
2 La dispersion marina es menos efectiva para explicar la asombrosa similitud de las faunas de mamiferos terrestres a traves de las Antillas Mayores septentrionales porque como proceso aleatorio no debe producir distribuciones uniformes a 10 largo de una cadena de islas 0 si las produce entonces el proceso no es aleatorio (MacPhee e lturralde-Vinent 2000 149)
Independientemente de que las aludidas distribuciones taxonomicas estan lejos de ser uniformes 0 asombrosamente similares la hipotesis de la dispersion marina contrariamente a su canicter aleatorio y altamente improbable en el caso del poblamiento continente-isla- en el caso del poblamiento isla-isla es un proceso no aleatorio (aprovecha las oportunidades brindadas puntualmente por los periodos de maxima regresion marina) y ademas altamente probable dada la incomparablemente menor distancia entre las islas caribeiias que entre estas y los continentes vecinos
314 BIOGEOGRAFiA
V ICARIANZA
La neotectonica y las transgresiones marinas del Oligoceno Medio y del Mioceno afectaron sustancialmente la paleogeografia de la Cresta de las Antillas Mayores resultando la subdivision gradual de las partes subaereas de la Cresta Asi el este de Cuba y el norte de La Espanola conectados fisicamente durante el Oligoceno Inferior se separaron al formarse el Paso de los Vientos en el Oligoceno tardio en tanto La Espanola y Puerto Rico probablemente permanecieron conectadas hasta el Mioceno tardio en que surgio el Paso de Mona (Iturralde-Vinent y MacPhee 1999)
Tales eventos geologicos habrian dividido las distribuciones de los grupos faunales emplazados previamente en el territorio continuo de las tres islas y a partir de esas disyunciones dichos autores (tambien MacPhee et al 2000 y White y MacPhee 2001) asumieron --como segunda fase del modelo de GAARlandia- la hipotesis de la vicarianza isla-isla para explicar la distribucion multiinsular de varios grupos de mamiferos terrestres a traves de las Antillas Mayores septentrionales De acuerdo con esta hipotesis cualquier diferencia en composicion faunal entre las islas (por ejemplo la ausencia de roedores heptaxodontidos en Cuba 0 de monos platirrinos en Puerto Rico) constituye evidencia de extincion local postvicariante y no de dispersion marina fallida Lo primero se puede demostrar si aparecen los fosiles pertinentes 10 segundo no se puede demostrar (MacPhee e Iturralde-Vinent 1994) Por otra parte IturraldeshyVinent y MacPhee (1999 57) han reconocido que aunque fa vicarianza isla-isla puede representar una solucion clara para las distribuciones de varios taxones cuaternarios no puede explicar todos los casos
Rega et al (2002 18) han presentado dos objeciones al modelo de la vicarianza isla-isla aplicado a los mamiferos de las Antillas Mayores
I Si la separacion de Cuba y La Espanola precedio en varios mill ones de anos la separashycion de La Espanola y Puerto Rico el modelo vicariante de lturralde-MacPhee debe predecir mayor similitud entre las faunas de La Espanola y Puerto Rico que entre las de Cuba y La Espanola De hecho es al reyes
No obstante MacPhee et al (200335) admitieron despues que la fecha mas temprana empiricamente defendible [para la apertura del Paso de Mona] debe ser el Oligoceno Medio esto invierte la secuencia de ambas disyunciones con 10 cual queda resuelta la aparente contradiccion Sin embargo en su reciente evaluacion de la congruencia entre filogenia e hipotesis geologic as en varios grupos de mamiferos terrestres de las Antillas Mayores Davalos (2004373) encontro que ninguno de los taxones cumple con la fecha y el patron de divergencia entre las masas terrestres antillanas postulados por el modelo de GAARlandia
2 La presencia de un mismo genero en las tres Islas septentrionales de las Antillas Mayores implica [por el modelo vicariante] que la diferenciacion entre estas formas durante el intervalo desde el Mioceno ha sido unicamente de nivel especifico [Dicha presencia]
315 rRRADlAcr6~ EVOLUCrONARIA
s610 puede explicarse razonablemente como el resultado de movimientos de esos animales entre las islas en el Cuaternario
De esta objecion resulta dificil disentir [El poblamiento entre islas diferenciado solo a nivel especifico no es probable que sea anterior al Pleistoceno (Simpson 1956 15)]
Debe tenerse en cuenta que la hipotesis de la vicarianza isla-isla alude a las tres islas septentrionales de las Antillas Mayores porque como se explico antes (vease El Barco Funeral Vikingo) Jamaica no existia como isla durante el lapso (Eoceno Superior shyOligoceno) consensualmente reconocido como el mas probable para la colonizacion de las proto-Antill as Mayores por la mayo ria de los mamiferos terrestres Despues de emerger como isla en el Mioceno Medio Jamaica debe haber recibido sus mamiferos terrestres por dispersion isla-isla (Davalos 2004376) y solo en el Cuaternario parece haberrecibido (por dispersion marina desde Centroamerica) al roedor murido Oryzomys
Seguramente tanto la dispersion como la vicarianza contribuyeron a que los mamiferos terrestres se irradiaran a traves de las Antillas Mayores y grupos de islas perifericas pero las maneras y tiempos en que estos mecanismos operaron a 10 largo de dicho proceso son aun en gran medida motivos de desacuerdo e incertidumbre
IRRADIACION EVOLUCIONARIA
A continuaci6n revisamos someramente las distribuciones conocidas de los diferentes grupos de mamiferos terrestres a traves de las Islas principales de las Antillas Mayores y grupos de Islas perifericas incluidas las interpretaciones y conjeturas que la literatura reciente recoge en relaci6n con la manera en que dichas distribuciones pudieron haberse conformado durante la irradiaci6n evolucionaria de los referidos grupos de mamiferos Manifestaciones de esta irradiaci6n pueden inferirse unicamente a partir de los taxones presentes en el Cuatemario pues solo 6 de las especies conocidas corresponden al Terciario
GRUPOS ORlGINARIOS DE SURAMERICA
PEREZOSOS (Megalonychidae)
Aparte de un genero miocenico de Cuba y otro del Oligoceno de Puerto Rico se conocen 4 generos (12 especies) de perezosos cuaternarios (todos extintos) restringidos a las tres islas septentrionales de las Antillas Mayores Acratocnus Neocnus Megalocnus y Parocnus Los cuatro generos estan representados en Cuba y La Espanola y Acratocnus 10 esta ademas (como unico genero de perezoso) en Puerto Rico Esta distribuci6n ha sido interpretada por MacPhee et al (2000) eomo resultado de la vicarianza oligomiocenica entre estas islas y [dado que el Banco Puerto Rico - Islas Virgenes fue la entrada de GAARlandia a las Antillas Mayores] se preguntaron si la ausencia de los otros tres generos en Puerto Rico se debe a que nunca estuvieron en ese territorio 0 a que sus
316 BIOGEOGRAFiA
fosiles atm no han sido hallados Pero como hemos visto Rega et al (2002) atribuyeron dicha distribucion a la dispersion interinsular en el Cuatemario
Por otra parte varios amilisis c1adisticos recientes ubi can a Choloepus -genero Reciente conocido solo de Centro- y Suramerica- como taxon hermano de Aeratoenus exc1uidos los demas megaloniquidos antillanos Para White y MacPhee (2001228)
tan estrecha relacion sugiere que la divergencia entre ambos [generos] ocurrio despues de que Aeratoenus divergio de Neoenus Si esta relacion implica que Aeratoenus y Neoenus se dispersaron separadamente desde Suramerica a las AntilIas mientras Choloepus permanecia en el continente (siendo necesarias al menos tres invasiones de perezosos) 0 si Choloepus invadio Suramerica desde las Antillas es algo que el material disponible no permite determinar
La diversidad adaptativa de los perezosos se manifiesta en (1) la utilizacion del habitat especies arboreas en Neoenus y Aeratoenus y terrestres en Paroenus y Megaloenus (Webb 1985a White [1993] White et al 1996 Arredondo [1999]) y en (2) la gama de tallas corporales desde los pequenos Neoenus hasta Megaloenus rodens el mayor de los mamiferos terrestres antillanos (su peso adulto estimado en 270 kg seglin Paula 19679) esto sugiere que los perezosos se irradiaron a nichos que en contextos continentales probablemente estarian ocupados por miembros de otros c1ados (MacPhee e IturraldeshyVinent 2000)
No esta claro si el abanico de tamanos entre estos perezosos enmascara alguna tendencia al gigantismo Por 10 pronto Anderson y Handley (2002) encontraron 10 contrario en perezosos (Bradypus) de las Islas de Bocas del Toro (Panama) shyseparadas secuencialmente del area continental vecina por ascensos del nivel del mar durante los ultimos 10 ka En las cinco islas los perezosos son menores que su contraparte continental y las diferencias entre las tallas medias de las cinco poblaciones expresan una reduccion lineal significativa de la talla corporal en funcion de la edad geologica de cada isla sin relacion alguna con el area de las islas 0 la distancia que las separa del continente
MONOS (Pitheciidae)
Los primates de las Antillas Mayores (cuatro especies extintas) constituyen un grupo monofiletico con Paralouatta (en Cuba) y Antillothrix (en La Espanola) como taxones hermanos seguidos en la escalera c1adistica por Xenothrix (en Jamaica) y finalmente Callieebus como el pariente continental mas cercano esto implica una sola invasion desde Suramerica antes del Mioceno Inferior (fecha mas temprana conocida para los primates fosiles antillanos) y por tanto en consonancia con GAARlandia como la via de colonizacion (Horovitz y MacPhee 1996 1999 Horovitz [1997] MacPhee y Horovitz 2004)
En desacuerdo parcial con la anterior interpretacion la filogenia de este grupo de monos obtenida por Davalos (2004) indica un antecesor comun en Cuba La Espanola y Jamaica
317 TRRADIACTON EVOLUCIONARIA
compatible con la hipotetica conexion -propuesta tentativamente por Iturralde-Vinent y MacPhee (199980-84)- entre las Montanas Azules del este de Jamaica y la peninsula suroccidental de La Espanola en la epoca de GAARlandia
A pesar de que los primates antillanos pertenecen a un grupo viviente de monos muy pequenos (los monos titles) fueron comparativamente grandes en relacion con los platirrinos modernos y sucesivamente mayores en la direccion Antillothrix Xenothrix y Paralouatta con P marianae como el mayor platirrino conocido del Mioceno neotropical (MacPhee 2005) MacPhee et al (1994) consideraron que Xenothrix debio ser un mono de andar lento y al parecer las especies de Paralouatta abandonaron la arborealidad para adoptar un modo de vida mas terrestre (MacPhee 1996a MacPhee et al 2003 MacPhee y Meldrum 2006)
ROEOORES (Echimyidae Capromyidae Heptaxodontidae)
La edad de Zazamys en Cuba demuestra que la invasion de los roedores histricognatos en las Antillas Mayores comenzo antes del Mioceno Inferior por otra parte los representantes mas antiguos de esa irradiacion evolucionaria se encuentran en La Espanola y Puerto Rico Ambas evidencias son consistentes con la hipotesis de GAARlandia (Woods 1982 1988 Woods et al 2001)
No esta claro si los capromidos evolucionaron a partir de un equimido ancestral 0 de un protocapromido 0 si los equimidos antillanos (I sub familia 3 generos y 7 especies todas extintas) forman parte de la irradiacion de los capromidos (4 subfamilias 9 generos y 27 especies 10 vivientes) Kratochvil et al (1978b) y Rodriguez et al (1979) consideraron que Capromys nan us es un eslab6n entre Echimyidae y Capromyidae pero Mesocapromys nanus esta morfologicamente mucho mas cerca de Hexolobodon y los de mas capromidos que de los equimidos (Woods 1985) Lo que si esta claro segun Woods et al (2001) es que la subfamilia Plagiodontinae endemic a de La Espanola es el linaje basal de los capromidos y que Isolobodon Hexolobodon Capromys Geocapromys Mysateles y Mesocapromys [y supuestamente Macrocapromys] forman parte de la irradiacion temprana de Capromyidae
La subfamilia Capromyinae (5 generos y 17 especies 9 vivientes) parece ser el dado de mas reciente evolucion en la familia con invasion de Cuba desde La Espanola y dispersion desde Cuba (1) por Capromys a las islas Caiman y (2) por Geocapromys a las islas Bahamas Caiman y Jamaica y desde Jamaica a las islas Swan aunque la presencia de Geocapromys en Swan pudiera deberse a introduccion aborigen (Morgan 1985 1988 1989 1993 1994 Woods 1989a 1990) Woods et al (2001) utilizaron datos moleculares para obtener estimados independientes de divergencia entre capromidos segiJn 10 cual Geocapromys divergio cerca de los 17 MaAP Mesocapromys divergio de Capromys hace 15 MaAP en tanto lvlysateles 10 hizo hace 10 MaAP Por su parte las formas de Capromys pilorides y de Mysateles prehensilis en Isla de la Juventud divergieron de sus correspondientes subespecies nominativas en la isla principal a los 24 y 08 MaAP respectivamente
318 BIOGEOGRAFiA
Woods y Howland (I979) describieron adaptaciones morfologicas de la mandibula el hioide y la faringe asociadas a mecanismos de masticacion diferentes en plagiodontinos y caprominos y Varona (1980a) llamo la atencion sobre el gran desarrollo de los molariformes (en relacion con la longitud total del cnineo) que presentan ciertos grupos de jutias fosiles en comparacion con las vivientes 10 que pudiera interpretarse como una adaptacion alimentaria a una vegetacion mas abrasiva por parte de las jutias extinguidas
Segun Woods (1990) la familia Heptaxodontidae (2 subfamilias 5 generos y 6 especies todas extintas) tambien seria parte de la irradiacion de los capromidos con origen en el este de La Espanola 0 en Puerto Rico y subsecuente dispersion al Banco de Anguila - San Martin en las Antillas Menores (pero vease MacPhee y Flemming 2003) Este banco estaria a solo 70 km del Banco Puerto Rico - Islas Virgenes en tiempos de maxima regresion marina y esta proximidad con Puerto Rico -patria de Elasmodontomys obliquus- seguramente tuvo que ver con la presencia en las islas de Anguila y San Martin del heptaxodontino gigante Amblyrhiza relacionado mas estrechamente con los roedores de las Antillas Mayores que con los del resto de las Antillas Menores (Woods 1990 1996) Aunque los roedores suelen aumentar de tamano en las islas (Case 1978) la escala del incremento de la talla en Amblyrhiza (que pudo alcanzar mas de 200 kg de peso adulto) esta fuera de toda proporcion con los patrones conocidos en otros roedores insulares (Biknevicius et 01 1993)
Clidomys (Clidomyinae) de Jamaica difiere marcadamente en su morfologia de los heptaxodontinos (vease ademas MacPhee y Flemming 2003)10 cual dificulta explicar su presencia en esa isla Como Jamaica estuvo bajo el mar hasta el Mioceno Medio el ancestro de Clidomys colonizaria esta isla por dispersion desde el extremo suroccidental de La Espanola despues que los heptaxodontinos se originaron en esta ultima isla (Woods 1989a) Pero Pascual et 01 (I 990) especularon que Clidomys pudiera ser el resultado de una invasion posterior e independiente de los capromidos
GRUPOS ORIGINARIOS DE NORTEAMERICA
El origen de las musaranas antillanas ha resultado ser hasta ahora el mayor enigma biogeografico de la fauna de mamiferos terrestres de la region aunque la mayoria de los autores coinciden en considerarlas originarias de Norteamerica (vease Discus ion y Conclusiones en Poblamiento Continente-Isla) No obstante MacPhee e Iturralde-Vinent (1994) descartaron toda posibilidad de conexion terrestre por el Estrecho de la Florida 0 por el Canal de Yucatan y si el (supuesto) evento vicari ante de Rosen (vease El Arca de Noe) -invocado por MacFadden (1980) para resolver el enigma- fuera minimamente plausible aun asi es dificil creer que las catastrofes que 10 sucedieron (vease Contexto Temporal) hayan barrido con toda esa vieja fauna continental excepto las musaranas circunstancia ya anticipada muy tempranamente por Matthew (l918a)
De otra parte la relaci6n entre Nesophontidae y Solenodontidae permanece irresuelta Si estos linajes no estan estrechamente relacionados entonces es muy posible que
319 IRRADIACION EVOLUCIONARIA
sus distribuciones sean explicadas por mecanismos diferentes (Whidden y Asher 2001)
La descomunal diferencia de talla corporal entre las musaranas pequenas (Nesophontes) y las grandes (Soienodon) seguramente impuso a ambos grupos restricciones distintas relativas a la termorregulacion y las adaptaciones energeticas En Nesophontes la seleccion debio favorecer una alta tasa de metabolismo y crecimiento camadas mas numerosas corta expectativa de vida yalta densidad poblacionaL En Soienodon en cambio la seleccion opero en la direccion de una baja tasa metabolica camadas reducidas crecimiento lento longevidad y baja densidad poblacional (Ottenwalder [1991])
MUSARANAS PEQUENAS (Nesophontidae)
Las musaranas pequenas estan representadas en las Antillas por el genero Nesophontes (y un genero miocenico indeterminado en ambar dominicano) con 6 especies cuaternarias extinguidas distribuidas en Cuba La Espanola Puerto Rico Islas Virgenes e Islas Caiman La mayor de las musaranas pequenas Nesophontes edithae evoluciono en Puerto Rico y su arribo a las Islas Virgenes supone una dispersion sumamente reciente En las Islas Caiman Nesophontes se asemeja mas a la poblacion de Cuba de donde seguramente derivo en el Plio-Pleistoceno (junto con Capromys y Geocapromys) cuando la costa sur de Cuba estuvo mucho mas cerca de las Islas Caiman (lturralde-Vinent 2003fig 4) MacPhee et ai (1994) interpretaron la distribucion de Nesophontes en Cuba y La Espanola como resultado de la vicarianza oligomiocenica ocurrida al formarse el Paso de los Vientos pero la distancia taxonomica entre los lados del Paso es solo de nivel especifico y N micros incluso vivi6 en ambas islas segtln Patterson (1962)
Allen (1918) considero que Nesophontes micrus fue una especie primitiva no modificada para la vida arb6rea 0 fos6rea y por tanto de habitos terrestres Nesophontes edithae de Puerto Rico es mucho mayor y mas robusta que las restantes especies del genero eL indice de robustez de su femur (= anchura minima de la diafisis ongitud total del hueso) muestra que este es tan robusto como el de Solenodon paradoxus un animal dos veces mayor (Morgan et ai 1980) Choate y Birney (1968) postularon un aumento cronoclinal de la taIla corporal en N edithae confirmado mas tarde por McFarlane (1999a) mediante fechados radiocarb6nicos de la secuencia estratignifica en depositos cavernarios de Puerto Rico Solenodon no vivi6 en Puerto Rico y en ausencia de una musarana de mayor tamano Nesophontes alcanzo una talla corporal mayor en esa isla que en La Espanola 0 Cuba (Woods y Eisenberg 1989)
MUSARANAS GRANDES (Solenodontidae)
Los arcaicos almiqufes (genero Soienodon) comprenden cuatro especies una viviente (S cubanus) y otra f6sil (S arredondoi) en Cuba y una viviente (S paradoxus con dos subespecies) y otra f6sil (S marcanoi) en La Espanola Como cabria esperar las dos especies cubanas se asemejan mas entre sf que a las especies de La Espanola pero
320 BIOGEOGRAFiA
enigmaticamente S marcanoi comparte caracteres con S cuban us y S paradoxus (Woods y Eisenberg 1989 Morgan y Ottenwalder 1993 Ottenwalder 200 I) Solenodon arredondoi alcanzo una talla corporal superior a la de cualquiera de las musarafias vivientes a nivel mundial Segun Morgan y Ottenwalder (1993) extrapolando las dimensiones y el peso de los almiquies vivientes S arredondoi debio tener una longitud cabeza-tronco y un peso de 450-550 mm y 1500-2000 g respectivamente en comparacion con S cubanus (360 mm y 800 g) y S paradoxus (390 mm y 1100 g)
Entre todas las especies de almiqufes Solenodon paradoxus es la que presenta los caracteres mas derivados (Otten walder 2001) Si estas relaciones son validas la distribucion de Solenodon entre Cuba y La Espafiola debio discurrir de oeste a este como propuso MacFadden (1980) quien invoco la presencia fosH de S cubanus [pero tambien S arredondoi] en Pinar del Rio para postular que el ancestro continental entr~ por Cuba opinion compartida por Perfit y Williams (1989) y por Ottenwalder ([1991] 2001) El paso de Solenodon a Puerto Rico pudo haberlo impedido la existencia de barreras ecologicas durante la conexion del este de La Espafiola con Puerto Rico 0 por la ausencia de Solenodon en esa parte de La Espafiola en aquella epoca Sin embargo las mismas condiciones ecologicas en el este de La Espafiola no parecen haber representado una barrera efectiva para Nesophontes cuyas mas altas tasa de reproduccion y densidad poblacional presumiblemente Ie permitieron expandirse flpidamente a traves de La Espafiola (Ottenwalder [1991])
Es opinion generalizada que los almiqufes antillanos constituyen el grupo de mamiferos que primero se emplazo en el territorio insular (MacPhee 2005) y en tal sentido es de interes la evidencia sobre especificidad parasito-hospedante aportada por Cruz (1980) quien encontro que la especie de acaro (genero Cubanochirus) que parasita a los almiquies es mucho mas primitiva que la especie (del mismo genero) que paras ita a las jutias y de aqui concluyo que dicho genero de acaro debio evolucionar primero en Solenodontidae y posteriormente en Capromyidae al entrar en contacto representantes de ambas familias en Cuba 0 La Espafiola
Recientemente Roca (2004) y Roca et al (2004) dieron a conocer que los resultados de su estudio genetico-molecular sobre los almiquies indican una divergencia de 25 Ma entre Solenodon cuban us y S paradoxus -compatible con la disyuncion tectonica entre Cuba y La Espafiola (Iturralde-Vinent y MacPhee 1999 McPhee et al 2003)-- y una separacion de ambas especies comparable en magnitud a la divergencia entre familias de mamlferos Consecuentemente propusieron que Solenodon cubanus fuera clasificada en el genero Atopogaie como indico Cabrera (1925) hace 82 afios
DISCUSION Y CONCLUSIONES
Las musarafias y los roedores antillanos (excepto Oryzomys) se irradiaron tanto de sus respectivos ancestros continentales que de estos 6rdenes se reconocen familias y subfamilias endemicas de la region en contraste con los perezosos (una subfamilia y tres
321 IRRADIACION EVOLUCIONARIA
tribus endemicas) y los monos (una tribu endemica) como expresion de diferentes niveles de diversificacion evolucionaria Este proceso es dificil de explicar (traUindose de c1ados que pnicticamente comparten la misma antigiiedad en las Antmas) si es correcta la interpretacion de Pascual et at (1990636)
Nosotros consideramos estos diferentes niveles de diversificacion como las diferentes etapas de evolucion alcanzadas por cada grupo [de mamiferos] Obviamente esto estuvo relacionado con las diferentes longitudes de sus historias
Pudiera ocurrir que en el futuro se compruebe que la historia biogeognifica de la region fue mucho mas complicada
La Espanola la isla mas ecol6gicamente divers a de la region debi6 ser el centro principal de evoluci6n de los mamfferos (con 87 de las familias y 73 de las subfamilias) seguida por Cuba (con 75 de las familias y 54 de las subfamilias) las dos islas de las Antillas Mayores mas relacionadas biogeograficamente (Fig 43 Tabla 44)
eL
EL
cb CAIMAN
SWAN
A Acratocnini B Cubanocnini C Megalocnini D Mesocnini E Nesophontidae F Solenodontidae G Xenothrichini
ANGUILAshyM-0 SMARTIN
H Heteropsomyinae I Plagiodontinae J Isolobodontinae
K Hexolobodontinae L Capromyinae
M Heptaxodontinae N CIidomyinae 0 Sigmodontinae
FIGURA 43 Representacion esquematica de las relaciones interinsulares (expresadas porel numero de linajes compartidos) entre las Antillas Mayores y entre estas y los grupos de islas perifericas segun las interpretaciones de los autores citados en el texto Los numeros dentro de los circulos indican la cantidad de linajes representados en cada isla relacion dudosa Contexto taxon6mico de los linajes en la Tabla 44
bull bull
bull bull
bull bull bull
bull bull
bull bull
bull
bull
bull
bull
bull bull bull bull
bull bull
bull bull
bull
bull bull bull
bull bull
bull
bull bull
bull
322 BIOGEOGRAFiA
TABLA 44 Distribuci6n de los linajcs de mamiferos terrestres a traves de las Antillas Mayores y grupos de islas perifericas LE = La Espanola CU Cuba JA Jamaica PR = Puerto Rico IC Islas Caiman IS Islas Swan IB = Islas Bahamas IV = Islas Virgenes AS = Anguila - San Martin
FamiliaSubfumiliafIribu LE CU JA PR IC IS IB IV AS
Megalonychidae
Choloepodinae
Acratocnini
Cubanocnini
Megalocninae
Megalocnini
Mesocnini
Nesophontidae
Solenodontidae
Pitheciidae
Callicebinae
Xenothrichini
Echimyidae
Heteropsomyinae
Capromyidae
Plagiodontinae
Isolobodontinae
Hexolobodontinae
Capromyinae
H e pta x 0 don tid a e
Heptaxodontinae
Clidomyinae
Muridae
Sigmodontinae
PILOSA
SORICOMORPHA
PRIMATES
RODENTIA
bull
bull
bull
bull TOTALES 12 10 4 4 2
323 EXTlICIOIES DEL CUATERNARIO
El gigantismo ha sido sin duda una tendencia evolucionaria recurrente entre los clados mayores de mamiferos antillanos Es un hecho bien conocido que algunos grupos de mamiferos han aumentado de tamafio en las islas y varios autores han intentado explicarlo a partir de enfoques diferentes (Foster 1964 Miller 1966 Sondaar 1977 Case 1978 Eisenberg y Wilson 1978 Heaney 1978 Lawlor 1982 Lomolino 1985 Angerbjom 1985 McNab 19942001)
EXTINCIONES DEL CUATERNARIO
Las islas que integran la Subregion Antillana han perdido casi 90 de sus mamiferos terrestres durante los ultimos milenios y en proporcion con su extension territorial esta perdida ha sido mayor en la Subregion que en cualquier otro lugar del planeta (MacPhee 1996b) Las extinciones del Cuatemario eliminaron mas de las tres cuartas partes de la fauna de mamiferos terrestres conocida de las Antillas Mayores sus islas satelites y grupos de islas perifericas y el examen de este proceso en sus aspectos mas generales se aborda en esta ultima parte del presente capitulo
CRONOLOGiA Y CONSECUENCIAS
Es muy improbable que un fosil represente el ultimo individuo de un taxon extinto y entonces hay que guiarse por la edad radiometrica del fosil mas reciente conocido lIamada fecha de ultima presencia- como unica aproximacion posible a la fecha de extincion del taxon
Como se explico al inicio del capitulo (vease Contexto Temporal) el registro fosH de los mamiferos terrestres de las Antillas Mayores adolece de insuficiente sustentacion radiometrica No obstante la distribucion temporal de los escasos fechados disponibles para el Cuaternario de la region abarca etapas criticas del proceso de extincion experimentado por la referida fauna a 10 largo de los ultimos 250 mil afios
De las 52 especies que es posible reconocer actualmente como miembros extintos de la fauna cuatemaria de mamiferos terrestres de las Antillas Mayores y grupos de islas perifericas los restos fosiles de 23 de elIas (442) han sido fechados radiometricamente o se han hallado asociados estratigraficamente de manera inequivoca con otro material fechado De estos 23 fechados 3 corresponden al interglacial Sangamon 4 pertenecen a la secuencia Pleistoceno terminal - Holoceno temprano 10 se ubican en ellapso entre el arribo de los aborigenes y la incursion europea y 6 son postcolombinos (a los cuales hay que sumar otras dos especies extinguidas en los siglos XIX y xx) A partir de estas 25 especies avaladas por fechas de ultima presencia distribuidas entre los cuatro intervalos temporales sefialados hemos compuesto la Tabla 45 en la que ademas aparecen las restantes 27 especies (carentes de fechado radiometrico) ubicadas en el intervalo temporal mas probable sobre la base de (1) la asociacion estratigrafica de sus restos con evidencias culturales (amerindias 0 europeas) 0 (2) la ausencia de dicha asociacion Sobre estos criterios de ubicacion temporal procede hacer dos aclaraciones
324 BIOGEOGRAFIA
TABLA 45 Intervalos temporales mas probables para las extinciones cuatemarias de los mamiferos terrestres de las Antillas Mayores y grupos de islas perifericas sobre la base de las evidencias siguientes (l) ultimo ejemplar colectado u observado en vida (2) fechado radiometrico mas tardio en material de la especie (3) asociaci6n estratigrafica mas tardia con otro material fechado (4) asociaci6n estratigrafica con restos de animales introducidos restos humanos u objetos culturales propios del intervale (5) falta de asociaci6n con evidencias culturales [Los ntimeros entre parentesis en la columna Evidencia significan adscripci6n tentativa al intervalo] Los estimados del peso corporal (en kg) estan tomados de la literatura 0 inferidos comparativamente y distribuidos entre
las clases siguientes (a) S 01 (b) 01 a lt 1 (c) 1 a lt 10 (d) 10 a lt 100 (e) 100 a lt 200 (j) ~ 200 Las
abreviaturas que designan las islas son las de la Tabla 44
F amilialEspecie Peso IsJa Evidencia Fuente
I httervalo htterg1acial Sangamon (gt80 kaAP)
HEPTAXODONTlNAE
Amblyrhiza inundata e AS 3 McFarJane et al 1998a
CLIDOMYINAE
Clidomys osborni d JA 3 Ibid
Clidomys parvus d JA 3 Ibid
II httervalo Pleistoceno terminal ~ Holoceno temprano (20-6 kaAP)
MEGALONYCHIDAE
Aeratoenus antillensis d CD 5 Arredondo 1975 a
Aeratoenus odontrigonus d PR 3 McFarJane 1999b
Aeratoenus simorhynehus d LE 2 Rega et al 2002
M egaloenus zile f LE 2 MacPhee et al 2000
SOLENODONTIDAE
Solenodon arredondoi e CD 5 Morgan y Ottenwalder 1993
PnHECIIDAE
Paralouatta varonai d CD 5 MacPhee 1996a
EcHIMYIDAE
Heteropsomys antillensis c PR 5 McFarJane 1999b
Heteropsomys insulans c PR 5 Ibid
Puertoricomys eorozalus c PR 5 Woods 1989a
CAPROMYIDAE
P lagiodontia araeum c LE 5 MacPhee et al 2000
Macrocapromys latus c CD 5 Varona y Arredondo 1979
(Continua)
325 EXTINCIONES DEL CUATERNARIO
TABLA 45 (Continuaci6n)
F ami1ia1Especie Peso Isla Evidencia Fuente
M acrocapromys latus c CU 5 Varona y Arredondo 1979
Mesocapromys kraglievichi c CU 5 Ibid
HEPTAXODONTINAE
Xaymaca fulvopulvis a JA 2 MacPhee y Flemming 2003
III Intervalo Amerindio (6-05 kaAP)
MEGALONYCHIDAE
Acratocnus ye d LE 2 MacPhee et aI 2000
Neocnus comes c LE 2 Ibid
N eocnus dousman c LE 2 Ibid
N eocnus toupiti c LE 2 Ibid
N eocnus gliriform is c CU 4 Cordova et ai 1997
M egalocnus rodens f CU 2 MacPhee et ai 2007
Parocnus browni e CU 2 Steadman et ai 2005
P arocnus serus e LE 2 MacPhee et ai 2000
N ESOPHONTIDAE
N esophontes edithae a PR 2 McFarlane 1999b
PITHECIIDAE
Antillothrix bernensis c LE 3 Rimoli 1977
EcHIMYIDAE
Brotomys contractus a LE 4 Miller 1929c
CAPROMYIDAE
P lagiodontia spelaeum c LE 4 Ibid
M acrocapromys acevedo c CU 4 NUiiez 1990a
HEPTAXODONTINAE
Elasmodontomys obliquus d PR 3 McFarlane 1999b
Iv Intervalo Postcolombino (05 kaAP - Presente)
N ESOPHONTIDAE
N esophontes hypom icrus a LE 24 MacPhee y Fle111111ing 1999
N esophontes zam icrus a LE (4) Ibid
N esophontes m icrus a CU 24 MacPhee et aI 1999
(Continua)
326 BIOGEOGRAFiA
TABLA 45 (Continuacion)
F amilialEspecie Peso Isla Evidencia Fuente
N esophontes m icrus a CD 24 MacPhee et aI 1999
N esophontes m icrus a LE 4 MacPhee et aI 2000
N esophontes sp 1 a IC 4 Morgan 1994
N esophontes sp 2 a IC 4 Ibid
Solenodon marcanoi b LE 4 Ottenwalder 2001
PITHECIIDAE
Xenothrix mcgregori c JA 4 MacPhee y Flemming 1999
EcHIMYIDAE
Boromys offeUa a CD 24 Guarch 1984
Boromys torrei a CD 24 Ibid
Brotomys voratus a LE 4 MacPhee et aI 2000
CAPROMYIDAE
P lagiodontia ipnaeum c LE 4 MacPhee y Flemming 1999
Rhizoplagiodontia lemkei c LE 4 Ibid
Isolobodon portoricensis c PR 24 Ibid
Isolobodon montanus c LE 4 Ibid
Hexolobodon phenax c LE (4) Ibid
Capromys sp c IC 24 Morgan 1994
Geocapromys columbianus c CD 4 MacPhee y Flemming 1999
Geocapromys thoracatus b IS 1 MacPhee y Marx 1997
Geocapromys sp 1 c IC 4 Morgan 1994
Geocapromys sp 2 c IC (4) Ibid
l-IEPTAXODONTINAE
Quemisia gravis c LE (1) MacPhee y Flemming 1999
MURIDAE
Oryzomys antillarum a JA 1 Ibid
1 Flemming y MacPhee (1999) alertaron en contra de la asociacion con otro material fechado Asimismo Morgan (1993) indico que la ausencia de asociacion con evidencias culturales amerindias no es necesariamente un criterio concluyente para ubicar un taxon en el intervalo temporal anterior porque hay depositos en la region que carecen de tales evidencias y son del Holoceno tardio por fechados radiocarbonicos [No obstante
327 EXTlNCIONES DEL CUATERNAR10
la ausencia de una especie en residuarios arqueol6gicos adquiere mayor certidumbre como criterio de ubicaci6n temporal de su extinci6n en la medida que aumenta el numero de residuarios estudiados que en las Antillas Mayores se cuentan por centenares] A falta de fechado absoluto de una especie consideramos que estos criterios de ubicaci6n temporal son mas utiles que la alternativa de excluir del analisis a la especie
2 En los trabajos sobre biogeografia de mamiferos antillanos los arque610gos mas citados como autoridades para establecer el inicio de la presencia aborigen en las Antillas Mayores (Kozlowski 1974 Rouse y Allaire 1978 Rouse 1989 1992 Wilson 2001) difieren al respecto entre los 6 kaAP y los 45 kaAP (pero vease Pagan et al 200523) y hemos adoptado la mas temprana de estas fechas para el comienzo del intervalo amerindio en la Tabla 45 De todos modos cualquiera que haya sido la fecha esta no es aplicable por igual al conjunto de las Antillas Mayores porque segun algunos de los autores citados el poblamiento comenz6 por Cuba y La Espanola 6 mil anos atras continu6 en Puerto Rico hace alrededor de 3 mil anos (pero vease Burney et al 1994) yen Jamaica hace menos de 2 mil e incluso a las Islas Caiman ni siquiera llegaron [Las Islas Caiman descubiertas por Col6n en 1503 carecen absolutamente de evidencia amerindia (Stokes y Keegan 1996)]
Si los datos de la Tabla 45 representan una aceptable aproximaci6n a la realidad el proceso de las extinciones cuaternarias de los mamiferos terrestres de las Antillas Mayores y grupos de islas perifericas debi6 caracterizarse entre otros por los eventos siguientes
Aunque obviamente la fauna cuaternaria debi6 ser mucho mas numerosa que 10 que refleja el registro paleontol6gico (Morgan y Woods 1986) 52 especies (81 de la fauna conocida) desaparecieron (sin reposici6n) durante los ultimos 250 mil anos
- Al parecer las extinciones se concentraron en las especies de mayor tamano Asi el peso promedio estimado para las especies de la etapa preantr6pica (intervalos I + II de la Tabla 45) es 350 kg es 328 kg para las especies del intervalo III (amerindio) y es s610 25 kg (c1ases del peso a b c) para las especies del intervalo IV (postcolombino) Ninguna especie aut6ctona de mamifero mayor de 6 kg existe hoy en la regi6n
- De acuerdo con la distribuci6n temporal de las extinciones 61 especies sobrevivieron al interglacial Sangamon (intervalo 1)48 sobrevivieron a la transici6n del Pleistoceno terminal al Holoceno temprano (intervalo II) 34 sobrevivieron al periodo amerindio (intervalo III) y s610 12 especies han sobrevivido desde la Ilegada de los europeos (intervalo IV) Las tasas de extinci6n correspondientes (exceptuado el intervalo I) son una especie cada 1167 anos en el intervalo II una cada 393 anos en el intervalo III y una cada 23 anos en el intervalo IV
- Durante todo el proceso de las extinciones cuaternarias la perdida de especies por isla fue de 91 en La Espanola 83 en Jamaica y 62 en Cuba Bahamas retuvo su unica especie (aunque extirpada en varias de las islas) Puerto Rico e Islas Caiman perdieron todas sus especies (6 y 5 respectivamente) y las Islas Swan Virgenes y Anguila - San Martin perdieron sus respectivas especies (micas
328 BIOGEOGRAFiA
De los cuatro 6rdenes de mamiferos representados en la fauna Pilosa y Primates desaparecieron totalmente Soricomorpha y Rodentia perdieron 86 y 74 de sus especies respectivamente
CAUSALIDAD
CAUSAS NATURALES
Los cicIos glaciales e interglaciales acaecidos en el Pleistoceno causaron cambios ambientales dnisticos en las Antillas con la concomitante alternancia de cIimas frios (secos) y calidos (hUmedos) asi como de descensos y ascensos del nivel del mar aumento y disminuci6n de las areas territoriales) induso (aunque con menor intensidad) en el Holoceno Estos eventos paleogeograficos han sido consistentemente documentados (Mayo y Kartashov 1972 Bonnatti y Gartner 1973 Ortega y Arcia 1982 Prance 1982 Schubert y Medina 1982 Dodge et al 1983 Ortega [1984] Haq et al 1987 Schubert 1989 Hodell et al 1991 Guilderson et al 1994 Oppo 1997 Emslie 1998 Curtis et al 2001 Iturralde-Vinent 2003) y de ellos brindamos a continuaci6n una breve resena
Finalizando el periodo interglacial anterior a la ultima glaciaci6n (Wisconsin) el mar estaba a 8-10 m por encima de su nivel actual Cuando el avance glacial del Wisconsin a1canz6 su culminaci6n (eillamado Ultimo Maximo Glacial) hace 18 ka el mar habia descendido 120 m con respecto a dicho nivel y las islas del Caribe habian incrementado considerablemente sus extensiones subaereas en comparaci6n con las que presentan hoy dia por ejemplo Puerto Rico y las Islas Virgenes se conectaron territorialmente y las Bahamas se unieron en cinco gran des islas Prevalecia un regimen frio y seco y condiciones de aridez --debidas a una reducci6n de las precipitaciones (posiblemente a menos de 440 mmlano)- y los ambientes xericos se habian extendido a traves de las islas y bancos emergidos y abundaban los pinares (los habitats mesicos estaban restringidos a parcelas relictas en 10 alto de las montanas) En el Mar Caribe y el Golfo de Mexico la temperatura superficial del mar era 5degC mas baja que la actual y en las llanuras costeras y alturas maximas eran respectivamente 2-3degC y 6degC mas bajas Los limites de las zonas ecol6gicas altitudinales descendieron varios cientos de metros y cada zona redujo su extensi6n vertical al aumentar el gradiente de temperatura Ellimite inferior de la zona periglacial
elevaci6n donde la temperatura media minima a1canza los OdegC) de la Cordillera Central de la Republica Dominicana (altura maxima del Caribe insular con evidencia de topografia glacial) se considera que estuvo entre los 1500 y 1800 mlnm (actualmente esta a 2800 mlnm) y se puede estimar que dicho limite era inferior en Cuba por estar mas al norte que la Republica Dominicana No obstante durante este periodo la temperatura experiment6 episodios cortos (unas pocas decadas) de ascensos repetidos
Un signifieativo componente bi6tieo de las referidas condiciones paleoambientales 10 constituy6 la espectacular irradiaci6n evolucionaria experimentada por un grupo numeroso de aves rapaees diurnas y nocturnas (Wetmore 1937 Brodkorb 1969 Kurotchkin y Mayo 1973 Arredondo 1976 Olson 1978 Olson y Hilgartner 1982
329 EXTINCIONES DEL ClJATERNARIO
Suarez 2000a b 2004 Suarez y Emslie 2003) Los mamiferos de las Antillas Mayores ofrecieron a estos depredadores aviares una amplia gama de tamanos de presa que en ausencia de mamiferos del orden Carnivora en la region permitio a estas aves irradiarse a este nicho alimentario con la evolucion de una variedad de especies de muy diversa tall a corporal incluidas formas gigantes Adquirir un cuerpo gigante les propicio el acceso a recursos alimentarios que no estaban siendo explotados por ningun otro depredador (Alcover y McMinn 1994) A Cuba corresponde el mejor registro fosil de rapaces cuaternarias caribenas con siete generos de especies diurnas Dentro de estas aves existieron tres gavilanes gigantes (Accipitridae) tres carairas (Falconidae) y un condor (Cathartidae) asi como cuatro generos de especies nocturnas dos lechuzas (Tytonidae) y tres Mhos (Strigidae) dos de ell os no igualados en tamano en todo el mundo Algunos de estos generos tuvieron representacion en otras islas antillanas como La Espanola Jamaica Bahamas y Gran Caiman
Al retirarse hacia el norte los hielos de la glaciacion Wisconsin el clima se tomo mas hUmedo en las Antillas durante el Holoceno temprano y el bosque mesico se expandio en la region insular El nivel del mar ascendio acompanado de mayor pluviosidad alrededor de los 7 kaAP las Islas Virgenes quedaron separadas entre si y de Puerto Rico y las Bahamas perdieron la mayor parte de su territorio Cerca de los 3 kaAP las condiciones secas retornaron a la region e hicieron desaparecer buena parte del bosque mesico Nuevamente el clima comenzo a adquirir sus condiciones actuales hacia los 25 kaAP aproximadamente y hoy transita por una fase mesica con vegetacion xerica confinada en parcelas relictas
Sin duda estos y otros eventos naturales incidieron negativamente en la sobrevivencia de mamiferos terrestres en las Antillas Mayores asi como en los grupos de pequenas islas perifericas (Pregill y Olson 1981 Olson 2006)
McFarlane et al (1998a) demostraron convincentemente como Amblyrhiza el gigante de los roedores antillanos endemico de la pequena isla de Anguila sucumbio a las repetidas y abruptas perdidas de su habitat durante el interglacial Sangamon por las nipidas y severas oscilaciones del nivel del mar (documentadas por Neumann y Hearty 1996) al no poder responder adecuadamente a tan catastroficos eventos mediante el ajuste de su masa corporal media y otro tanto debio ocurrirle en Jamaica al otro roedor gigante Clidomys (McFarlane et al 1998b) emparentado con Amblyrhiza en la misma epoca (Tabla 45 intervalo I)
El ascenso del nivel del mar seguramente promovio la extincion de mamiferos durante la transicion del Pleistoceno al Holoceno (Tabla 45 intervalo II) con la reduccion del tamano de las Islas por las transgresiones marinas (Pregill y Olson 1981 Morgan 1993) Esto ultimo haria maximamente severa la competicion asf como la presion de los depredadores [Steadman et al (2005) no yen por que el calentamiento global ocurrido durante la transicion Pleistoceno-Holoceno habria de tener un efecto negativo sobre los perezosos siendo sus descendientes actuales especies tropic ales 0 subtropicales pero no se trata del efecto
330 BIOGEOGRAFiA
directo del cambio de temperatura sobre los animales sino del efecto indirecto y progresivo que dicho cambio genera a largo plazo al reducir el territorio insular disponible]
Por esta epoca tambien desaparecieron especies de tall a mayor como Megalocnus zile en La Espanola y otras de menor tamano (en Cuba) pero que representan la tall a maxima en sus respectivos contextos taxonomicos del Cuaternario como Solenodon arredondoi Paralouatta varonai y Macrocapromys acevedo Un mamifero puede ser grande solo a costa de reducir su densidad poblacional y esto 10 hace mis vulnerable a la extinci6n sobre todo en una isla (McNab 1994 McFarlane et al 1998a) Incidentalmente Alcover et al (1998a) afirmaron que larelacion entre la talla corporal y la extincion no siempre se cumple en las especies de menor tamano y citaron como ejemplo que entre las musarafias todas las especies de Nesophontes desaparecieron mientras sobreviven especies de Solenodon pero la comparacion correcta en todo caso serfa entre los solenodones vivientes y el gigante Solenodon arredondoi y entre Nesophontes edithae -el gigante de los nesofontes extinguido en el periodo amerindio- y el resto de sus congeneres que sobrevivieron hasta los primeros siglos de la colonizacion europea (Tabla 45 intervalos III y IV)
CAUSAS ANTROPOGENICAS
Intervalo Amerindio
La mayoria de los autores que se han referido a las extinciones acaecidas durante este intervalo (entre elIos paradigmaticamente Martin 1984a b) han responsabilizado a los aborfgenes (que se supone comenzaron a poblar las Antillas Mayores hace 6 mil afios) con la desaparicion de numerosas especies de mamiferos terrestres en la region principalmente por la via de la sobrecaza y la introduccion de animales domesticos devenidos depredadores de la fauna autoctona (Morgan y Woods 1986) Este efecto negativo de la presencia indigena sobre los mamiferos de la region ha sido dramatizado por algunos autores en terminos de holocausto faunal Sin embargo pensamos que se ha exagerado Como han expresado MacPhee et al (19993) cuando uno busca evidencia dura de complicidad humana directa en [estas] extinciones no hay mucho que encontrar Las siguientes son algunas consideraciones a tener en cuenta en tal sentido
El registro radiometrico mas joven conocido hasta ahora para un perezoso antillano (MegalocnllS rodens) es de 42 kaAP (MacPhee et ai 2007) Si se acepta que los aborfgenes entraron en Cuba hace aproximadamente 6 mil anos su convivencia con los perezosos debe haber durado alrededor de 1800 afios (0 mas porque dificilmente el referfdo fechado radiometrico haya correspondido al ultimo sobreviviente de la especie) Esta cifra contradice la hipotesis del holocausto faunal segun la cualla extincion de los mamiferos insulares de mayor talla debe ocurrir con extrema rapidez -a escala de decadas 0 de centurias- a partir de su primer contacto con los humanos (Martin y Steadman 1999) A pesar de las repetidas afirmaciones en contrario (Harrington 1921 Miller 1929c Herrera [1945] Tabio 1951 Alvarez 1951a 1957 Arredondo
331 EXTINCIONES DEL CUATERNARIO
1959 1960a Veloz y Ortega 1973 Mayo 1980b Rodriguez et al 1984 Castellanos y Pino 1985 Pino y Castellanos 1985 Nunez 1990b Gutierrez 1996 Arredondo 2004) en su mayoria basadas en evidencias no exentas de posibles accidentes de deposicion (0 de reubicacion postdeposicional) en recintos cavernarios (Diaz-Franco [2006b D no se ha demostrado inequivocamente que los aborigenes antillanos se alimentaran de perezosos regularmente Sus restos en residuarios de alimentos son escasisimos en toda la region y esto contrasta con los abundantes restos de jutias y otros mamiferos de menor talla que aparecen en esos residuarios demostrativos de la preferencia de los paleoindios por estos otros mamiferos
Los mamiferos mas importantes en la economia prehistorica de las Antillas Mayores fueron las jutias Capromys y Geocapromys en Cuba Geocapromys en Jamaica e Isolobodon en La Espanola Puerto Rico e Islas Virgenes (Aguayo 1950b Tablo 1951 Rimoli 1972 Wing 2001 Diaz-Franco [2006b D Wilkins (2001) estudi61ajutia viviente de Jamaica (Geocapromys browni) en un sitio ocupado por los aborigenes durante 100 anos y no encontr6 evidencias de que durante ese tiempo la especie haya sido sobreexplotada Asimismo en Cuba las jutias conga carabali y andaraz son escasas en depositos pleistoholocenicos en cambio son muy numerosas en residuarios indigenas y de haber sido sobreexplotadas 10 mas probable es que no hubieran sobrevivido hasta hoy
La habilidad de los humanos para penetrar en profundidad el bosque tropical y mantenerse en este sustentablemente ha sido un evento comparativamente reciente (Beran y Steele 1994) En el caso antillano la caza tierra adentro ha sido explicada como un proceso tardio de desplazamiento a partir de la explotacion de abundantes recursos marinos que ofrecian condiciones 6ptimas para el sedentarismo de las comunidades aborigenes costeras (C6rdova y Arredondo 1988 Brito 2005) Peces y mariscos dominan la arqueologia costera de las Islas antillanas en tanto el consumo de recursos terrestres (mamfferos incluidos) aumenta progresivamente en la medida que los residuarios estudiados se alejan de la costa (Scudder 1991 Wilkins 2001) En las Antillas Mayores el componente terrestre de la dieta aborigen es solo 20-30 del material recobrado en los residuarios (Wing 1977 1989) Y debio ser menor alin porque el componente vegetal de la dieta no se ha preservado Por otra parte los aborigenes cubanos domesticaron la jutia conga y seguramente la reprodujeron para comer (Pose et al 1988) practica que probablemente se repitiera con otras jutias en las demas Antillas Mayores
- Fray Bartolome de las Casas hablo de 3 millones de indios en Cuba al inicio de la colonizacion pero los historiadores subsecuentes consideraron exagerada la cifra (Poey 1851 a) No obstante se dispone de dos estimados de la densidad de poblaci6n aborigen en las Antillas Mayores a la llegada de los europeos 18 indigenaslkm2 en Puerto Rico (Rouse 1956) y 1271km2 en La Espanola (Watts 1987) Extrapolando ambos valores entre las cuatro islas principales de las Antillas Mayores tenemos que en la fecha indicada la poblaci6n aborigen pudo estar entre 16 mil y 113 mil indigenas en Puerto Rico entre 21 mil y 145 mil en Jamaica entre 141 mil y 1 mill6n en La Espanola y entre 200 mil y 14 millones en Cuba Entre las cifras maximas y las minimas nos inclinamos
332 BIOGEOGRAFiA
hacia estas ultimas atendiendo a los altos indices de mortalidad infantil y morbilidad que se evidencian en las osamentas de los pobladores prehispanicos (Martinez y Rives 1990) Uno de los hechos que apuntan hacia el bajo numero de aborigenes en las Antillas Mayores es 10 rapido que desaparecieron despues de la llegada de los europeos pnkticamente sin dejar mestizaje con estos 10 cual contrasta ostensiblemente con la situacion en la America continentaL
En suma un grupo humano (1) demograiicamente irrelevante y controlado por altos indices de morbilidad (2) consumidor mayoritariamente de organismos marinos y (3) carente de la tecnologia cinegetica necesaria para poder sobreexplotar los mamiferos que Ie sirvieron de alimento (a los cuales accedio tardiamente en su desarrollo) no es concebible que haya causado e1 holocausto faunal que se Ie atribuye a partir del error de aplicar a las comunidades aborigenes antillanas un modelo cultural de la prehistoria continentaL
Tampoco hay evidencias de que los aborigenes hayan introducido carnivoros con suficiente capacidad depredadora salvo el perro Morgan y Otlenwaldder (1993) consideraron que la depredacion por perros durante el periodo amerindio es la explicacion mas plausible para la extincion del almiqui gigante de Cuba pero como se puede observar (Tabla 54) Solenodon arredondoi aparece en el intervalo II por falta de asociacion con evidencias culturales
Consecuentemente el accionar de los aborigenes y de sus animales domesticos directamente sobre la fauna no parece haber sido por sf solo un factor determinante de la extincion de mamiferos terrestres durante el periodo amerindio en las Antillas (pero vease Steadman et al 2005)
Hay que recordar en este senti do que alin en el Holoceno tardio ocurrieron cambios clirruiticos de consideracion (vease Causas Naturales) Por ejemplo Curtis et al (2001) explicaron que la extincion de algunos ani males mesofilicos de las alturas montanosas de La Espanola pudo haber antecedido a cualquier impacto humano sigrIificativo en esos lugares por coincidir dicha extincion con una tendencia a la aridez documentada isotopicamente y con evidencias de la perdida del bosque mesico ocurridas despues de 3kaAP Yen Jamaica hace solo 2shy3 mil aDos el area de la Bahia de Jackson sustentaba una gran biomasa de vertebrados en bosque hinnedo a juzgar por la cantidad y diversidad de taxones presentes en material fosil derivado de la depredacion por lechuzas hoy el area es la mas seca de la isla con matorral xerofitico y pobreza de fauna (MacPhee 1996a) Yalm en nuestros dias huracanes de gran intensidad aniquilan poblaciones de vertebrados (que no se recuperan) en is las pequenas y llanas del Caribe (Brooks y Smith 2001)
En medio de la incertidumbre sobre la verdadera causa (0 causas) de las extinciones de mamiferos en el periodo amerindio MacPhee y Marx (1997) han propuesto su hipotesis de la hipermorbilidad la cual estipula colapsos poblacionales relativamente rapidos con la introduccion de enfermedades de alta letalidad Estos autores han razonado que
Las islas son los lugares mas apropiados para experimentar perdidas catastroficas causadas por enfermedades Los mamiferos no solo serian susceptibles en virtud de su prolongado aislamiento biologico sino que en los confines de una isla la epizootia
333 EXTINCIONES DEL CUATERNARIO
tendria la potencialidad de infectar simultaneamente a la rnayoria de los individuos de vadas especies Los neonatos son inmunologicamente inmaduros y sufririan desproporcionadamente cualquier infeccion hipervirulenta La perdida excesiva de neonatos y juveniles tendria efectos severos para la sobrevivencia de la poblacion si esta se reduce por debajo de cierto nivel sobreviene la extincion porque no hay posibilidad de restitucion en el periodo postpanzootico Las especies de mayor talla corporal van a ser las mas vulnerables a la extincion dadas sus bajas tasas reproductivas largos periodos de gestacion y reducido numero de progenie Las especies de menor tamano --caracterizadas por altas tasas reproductivas periodos cortos de gestacion y mayor numero de neonatos por parto- tambien serian fuertemente diezmadas pero muchas sobrevivirian porque su estrategia reproductiva les permite perder mayor numero de individuos Las perdidas deben ocurrir relativamente rapido (a escala de decadas) mientras que otras hipotesis (digamos los cambios climaticos) permiten intervalos mucho mas largos (a escala de siglos 0 de milenios) entre la aparicion de las causas y la produccion de sus efectos Por ejemplo todo indica que los nesofontes se extinguieron muy rapidamente y seria dificil poder invocar un impacto humano directo 0 argumentar fenomenos climaticos que pudieran explicar plausiblemente perdidas tan rapidas
MacPhee y Marx (1997 191) han dicho que por el momento la hipotesis de la hipermorbilidad no pasa de ser una abstraccion en espera de adecuada sustentacion empirica Llama la atencion que linicamente en el periodo amerindio (Tabla 45 intervalo 1Il) las extinciones abarcaron todas las clases del peso corporal (de la a a laj) y que ninguna de las especies que sobrevivieron hasta el periodo postcolombino (Tabla 45 intervalo IV mas las especies vivientes) sobrepasa los 10 kg (clases del peso a b c) en clara coincidencia con los presupuestos de la hipotetica hipermorbilidad
Intervalo Postcolornbino
Por 10 menos 37 especies de mamiferos terrestres vivian en las Antillas Mayores y grupos de Islas perifericas a la lIegada de los europeos en 1492 De elIas se han extinguido 23 shymas especies que en cualquiera de los periodos precedentes (Tabla 45~ y solo subsisten 12 especies repartidas entre Cuba (8 especies) La Espanola (2) Jamaica (1) y Bahamas (1) varias de estas en peligro critico de extincion Solo dos de las especies extinguidas durante los ultimos 500 anos se conocieron en estado viviente los roedores Oryzomys antillarum de Jamaica (extinguido en 1877) y Geocapromys thoracatus de las Islas Swan (extinguido en 1955) Las causas de las extinciones ocurridas durante este periodo son sobradamente conocidas destruccion de habitats introducci6n de animales exoticos (domesticos y comensales) y demas presiones asociadas a la actividad humana entre las cuales posiblemente habria que incluir brotes de hipermorbilidad
Durante mucho tiempo se ha afirmado la posibilidad de que especies de Nesophontes hayan sobrevivido hasta las primeras decadas del siglo xx y que incluso continuen existiendo en lugares remotos de Cuba 0 La Espanola (Allen 1918 1942 Miller 1930 Aguayo 1950a Arredondo 1960b Fischer 1977 MacFadden 1980 Woods y
334 BIOGEOGRAFiA
Ottenwalder 1991) Actualmente no existe evidencia radiometrica de que alguna especie de Nesophontes haya sobrevivido al siglo xv las fechas radiocarbonicas calibradas obtenidas en muestras de Nesophontes de Cuba y La Espanola resultaron no mas tardias que los siglos XIII y XIV (MacPhee et al 1999) Por otra parte las expediciones efectuadas a fines del siglo xx con el proposito de localizar individuos vivientes del genero en La Espanola no produjeron la menor indicacion afirmativa al respecto (Woods et al 1985) Ademas a traves de las Antillas Mayores no aparecen restos de Nesophontes en los depositos formados actualmente por las lechuzas
EPiLOGO
Las Antillas Mayores son islas oceanicas no han formado parte de un continente (con la posible excepcion de una parte de Jamaica hasta el Eoceno Medio hace 45 Ma) y durante un tiempo geologico considerable (desde el Eoceno Superior hace 40 Ma) han permanecido emergidas y separadas de los continentes vecinos Consecuentemente los mamiferos terrestres (no volantes) autoctonos de estas islas constituyen una fauna oceanica que por ende cumple con todas las caracteristicas convencionales reconocidas para este tipo de entidad biogeografica
- Baja diversidad taxonomica a nivel de las categorias superiores (solo cuatro ordenes) en comparacion con islas plataformicas continentales (0 con areas continentales) de tamano y diversidad ecologica simi lares
- Alto grado de endemismo a nivel de genero (96) y familia (50) en respuesta al prolongado aislamiento
- Desbalance ecologico de la estructura trofica ausencia de mamiferos camivoros y ungulados
- Espectaculares procesos de irradiaci6n evolucionaria como resultado de la colonizacion de territorios libres de competidores y depredadores
Tendencias evolucionarias extremas gigantismo
- Control a cargo de aves rapaces con tendencia at gigantismo en ausencia de mamiferos miembros del orden Carnivora
- Mas especies de reptiles y de murcielagos que de mamiferos terrestres relaciones que en los continentes se invierten
Alta tasa de extincion de especies (84) sin reposicion
Preservacion de especies filogeneticamente arcaicas (los llamados fosiles vivientes) los almiquies

296 BIOGEOGRAFiA
TABLA41 (Continuaci6n)
I s 1 a s Taxonomla 1
CD JA LE PR IS IC lB IV AS
N eocnus comes bull Neocnus dousman bull N eocnus gliriform is bull N eocnus toupiti bull Gen et sp indet bull MEGALOCNINAE
Imagocnus zazae3 bull M egalocnus rodens bull M egalocnus zile bull Parocnus browni bull P arocnus serus bull
Soricomorpha (musaraiias) 11 especles
NESOPHONTIDAE 4
Nesophontes edithae bull bull Nesophontes hypomicrus bull Nesophontes micrus bull bull N esophontes zam icrus bull N esophontes sp 1 bull Nesophontes sp 2 bull Gen et sp indet bull SOLENODONTIDAE 4
Solenodon arredondoi bull Solenodon cuhanus 5 bull Solenodon marcanoi bull Solenodon paradoxus bull
Primates (monos) 4 especies
PTIHECIIDAE 2
CALLICEBINAE
Antillothrix bernensis bull (ContinUa)
297 ESPACIO Y TIEMPO
TABLA 4 1(Continuacion)
I s a Taxonomia 1
CD JA LE PR IS IC IE IV AS
Paralouatta marianae bull Paralouatta varonai bull Xenothrix mcgregori bull
Rodentia (roedores) 40 especies
ECHllVlYIDAE 2
HETEROPSOMYINAE
Boromys offella bull Boromys torrei bull Brotomys contractus bull Brotomys voratus bull Heteropsomys antillensis bull Heteropsomys insulans bull Puertoricomys corozalus bull CAPROMYIDAE 4
PLAGIODONTINAE
Plagiodontia aedium 6 bull P lagiodontia araeum bull P lagiodontia ipnaeum bull Plagiodontia spelaeum bull Rhizoplagiodontia lemkei bull ISOLoBODONTINAE
Isolobodon portoricensis 7 bull Isolobodon montanus bull Zazamys veronicae bull HExOLOBODONTINAE
Hexolobodon phenax bull CAPROMYINAE
Capromys pilorides 67 bull Capromys sp bull
(Continua)
298 BIOGEOGRAFIA
TABLA 4 1 (Continuacion)
I s a s Taxonom1a 1
CU JA LE PR IS IC IB IV AS
Geocapromys browni 7
Geocapromys columbianus bull Geocapromys ingrahami 6
Geocapromys thoracatus
Geocapromys sp 1
Geocapromys sp 2
Macrocapromys acevedo
Macrocapromys latus
Mesocapromys angelcabrerai
Mesocapromys auritus
Mesocapromys kraglievichi
Mesocapromys nanus
Mesocapromys sanfelipensis
Mysateles melanurus
Mysateles prehensilis 6
bull bull bull bull bull bull bull bull bull
HEPTAXODONTIDAE 8
HEPTAXODONTINAE
Amblyrhiza inundata
Elasmodontomys obliquus
Quem isia grav is
Xaymacafulvopulvis 9
CLIDOMYlNAE
Clidomys osborni
Clidomys parvus
MURIDAE2
SIGMODONTlNAE
Oryzomys antillarum 10
bull
bull bull bull
bull
bull
bull bull
bull bull
bull
bull Total de especies por is1all 24 6 25 7 1 5 1 1 1
1 La taxonomia sigue las revisiones sistematicas mas recientes (Morgan 1993 1994 MacPhee y Grimaldi 1996 Horovitz y MacPhee 1999 White y MacPhee 2001 Ottenwalder 2001 Woods et aI 2001 MacPhee y Flemming 2003) con 1a adici6n de los taxones descritos posteriormente y la incorporaci6n de los cambios propuestos para las especies cubanas en el Capitulo 2 de este Compendio (Continua en pagina opuesta)
299 ESPACIO Y TIEMPO
CONTEXTO TEMPORAL
La absoluta mayoria del registro fosil de mamfferos terrestres antillanos proviene de depositos cavemarios pleistoholocenicos (escala del tiempo geol6gico en la Tabla 42) Abundan en estas cavernas restos de perezosos musarafias y roedores siendo comparativamente escasos los de monos debido quizas a la universal predileccion de los platirrinos por los ambientes densamente forestados 10 que los hace malos candidatos a la fosilizacion (MacPhee y Woods 1982)
A pesar de 10 numeroso de tales depositos en la region son escasos los fechados radiometricos del cuantioso material extraido Con excesiva frecuencia (particularmente en Cuba) estos depositos han sido asignados al Pleistoceno Superior sin fundamento suficiente para ello y es sabido que incluso en cuevas contiguas los depositos fosilfferos pueden diferir considerablemente en edad (Pregill y Olson 1981) Se superponen a esta inconveniencia la falta de estudios tafonomicos en las excavaciones y la eventual mezcla de f6siles de distintas edades como consecuencia de las multicausales alteraciones estratigraficas ocurridas en las cuevas (Nunez 1990a b Morgan y Ottenwalder 1993 MacPhee 1997 McFarlane 1999b Dfaz-Franco 2004) Sobre propuestas de tipificaci6n de los depositos paleontologicos con restos de mamiferos terrestres en Cuba vease Woloszyn y Silva (1977) Acevedo y Arredondo (1982) Gutierrez (1994) Arredondo [1999]
La Tabla 43 presenta los fechados pleistocenicos mas tempranos registrados para restos de mamfferos terrestres del Caribe insular De estos restos el roedor (Cidomys) de Jamaica con un fechado de 253 mil anos antes del presente constituye el fosH de mamifero terrestre mas viejo del Cuaternario anti11ano [Los fosiles fechados antes del surgimiento de la espectrometria de masa (aceleradora) para muestras pequefias pueden ser algo mas viejos (Jull et al 2004)]
Corresponde a MacPhee e Iturralde-Vinent (1994 1995b) e1 relevante merito de haber descubierto en la ultima decada del siglo xx los primeros fosiles terciarios de mamiferos terrestres de las Antillas extendiendo a mas de 30 millones de afios el registro f6sil de
2 Extirpada en la Subregion Antillana pero viviente en el Neotropico continental
3 Ubicacion en subfamilia s610 sugerida por White y MacPhee (200 I)
4 Endemica de la Subregion Antillana
5 Recientemente propuesta su ubicaci6n bajo el genero Atopogale (Roca et al 2004)
6 lncluye las subespecies
7 Transportada por aborigenes a otras de las islas (Rimoli 1974 Olson 1982b este trabajo)
8 Vease MacPhee y Flemming (2003) sobre la incierta situaci6n de esta familia
9 Ubicada provisionalmente en esta subfamilia por MacPhee y Flemming (2003)
10 Hallada en contexto no perturbado de edad Pleistoceno tardio (McFarlane et ai 2002) con 10 cual se descarta su supuesta introduccion por amerindios (Woods 1990)
II La correlaci6n entre el area insular y la riqueza de especies es razonablemente fuerte (R=O83) en estas islas excepto las Bahamas (Morgan 1989)
300 BIOGEOGRAFIA
TABLA 42 Escala convencional del tiempo geologico en la Era Cenozoic a y finales de la Mesozoica (Gradstein 2004) ka = kiloafios (miles de afios) Ma miJIones de afios AP = antes del presente
Periodo(Suhperiodo) Epoca Afios (aproximados) antes del presente
Era Cenozolca
Cuaternario Holoceno Pkistoceno
Presente - 11 kaAP 11 kaAP - 18 MaAP
Terciario (Ne6geno ) Plioceno Mioceno
18 - 5 MaAP 5 - 23 MaAP
oIigoceno 23 - 34 MaAP Terciario (pak6geno) Eoceno 34 - 58 MaAP
Pakoceno 58 - 65 MaAP
Era Mesozo ca
Creticico 65 - 145 -faAP
~~~------~~--- --~-
este grupo animal en la region con el hallazgo de un perezoso oligocenico en Puerto Rico (Yauco) as como un perezoso un mono y un roedor miocenicos en Cuba (Domo de Zaza) Esto unido a la presencia de huesos de una musarafia nesofontoide [referida erroneamente a Solenodon por Rivero (1997)] en ambar oligomiocenico de la Republica Dominicana (MacPhee y Grimaldi 1996) demuestra incontrovertiblemente que al menos en el Mioceno temprano (alrededor de 20 millones de afios atnis) ya estaban en las Antillas Mayores representantes de los cuatro linajes basicos de su fauna de mamiferos terrestres [La afirmacion de Abreu et al (1990 13) en cuanto a que se conocen restos fosiles de almiqui que datan de hace aproximadamente 40 millones de afios es erronea]
Pero lcuan antigua puede ser la fauna de mamiferos terrestres de las Antillas Mayores
Aunque es un hecho bien establecido por la geologia que desde finales de la Era Mesozoica y principios de la Cenozoica existieron islas en el area ocupada ahora por las Antillas Mayores la evidencia disponible indica asimismo que aquellas primeras islas sufrieron repetidas transgresiones marinas totales que habrian hecho sucumbir las biotas que pudieran haber sustentado (MacFadden 1981 Morgan y Woods 1986 MacPhee e Iturralde-Vinent 1994 Roca et al 2004) sin contar los catastroficos efectos que debio tener para el Caribe occidental el b6lido extraterrestre que impacto en Chicxulub (Yucatan) hace 65 Ma (Tada et ai 2003) Hedges (1996) ha especulado que aUn en medio de tal devastacion debe haber existido algun que otro lugar donde elementos de la fauna pudieron haber sobrevivido
301 POBLAMIENTO CONTINENTE-ISLA
TABLA 43 F echados pleistocenicos mcis tempranos (obtenidos por distintos metodos) en mamiferos terrestres del segmento insular delimitado en laFig 41 F =Fuentes (I) Jull et al 2004 (2) Goodfriend y Mitterer 1987 (3) MacPhee et al 1989 (4) Woods 1989b (5) Rega et aI 2002 (6) McFarlane 1999b (7) McFarlane 1 999a (8) McFarlane et al 1998a
Pals Localidad Taxon kaAP F
Cuba Breas de San Felipe Pilosa (Parocnus) 1l9a 1
Jamaica Cueva Coco Ree Cueva Wallingford Roadside
Primates indet Rodentia (Clidomys)
393 2530
2 3
Haiti Trouing Jeremie no5 (casimba) Rodentia (Rh izoplagiodontia) 212 4
Republica Dominicana
Cueva del Perezoso Pilosa (ficratocnus) 185 5
Puerto Rico Cueva de Ia Vaca Cueva Clara
Pilosa (Acratocnus) Soricomorpha (Nesophontes)
337 346
6 7
Anguila Cueva Cave-No-Cave Rodentia (Amblyrhiza) 2060 8
a Kulp etal (1952) reportaron un fechado radiocarbonico de gt 30000 afios para un cono de pino en el deposito fosilffero de Ciego Montero (Cuba) con restos de perezosos
(vcase tambicn Crother y Guyer 1996) pero Iturralde-Vinent y MacPhee (1999) han preguntado donde estaban esos lugares si es que existieron
Solo a partir del Eoceno Superior las Antillas Mayores en fonnacion han pennanecido emergidas hasta hoy [excepto parte de Jamaica y el sur de La Espanola que continuaron sumergidas hasta el Mioceno Medio (Iturralde-Vinent y MacPhee 1999)] Esto significa que solo durante los ultimos 40 millones de anos aproximadamente han existido pennanentemente en dicha region insular ambientes terrestres capaces de sustentar comunidades biotic as (MacPhee e Iturralde-Vinent 2000 Portell et ai 2001 MacPhee 2005) 10 cual proporciona el marco temporal de referencia para conceptuar la antigiiedad maxima posible de la fauna de mamiferos terrestres de las Antillas Mayores
POBLAMIENTO CONTINENTE-ISLA
AFINIDADES CONTINENTALES
El consenso de los autores que en las itltimas dccadas han tratado la filogenia y biogeografia de los mamiferos terrestres de las Antillas Mayores (MacFadden 1980 MacPhee y Woods 1982 Morgan 1985 1988 1993 Morgan y Woods 1986 Ford 1986 Woods 1988 1989a 1990 MacPhee e Iturralde-Vinent 1995a b Hedges 1996 Horovitz [1997] Ottenwalder 2001 Whidden y Asher 2001 MacPheeet ai 2003 Carlini y Scillatoshy
302 BIOGEOGRAFiA
Yane 2004 Davalos 2004 MacPhee y Horovitz 2004 Roca et al 2004) es practicamente unanime en cuanto a que los ancestros continentales que dieron origen a esta fauna derivaron (1) mayormente de Suramerica perezosos (Megalonychidae) monos (Pitheciidae) y roedores (Echimyidae Capromyidae y Heptaxodontidae) (2) en menor medida de Norteamerica musarafias (Nesophontidae y Solenodontidae) y (3) menos aun de Centroamerica una especie de roedor (Muridae)
HIPOTESIS BIOGEOGRAFICAS
La definicion c1asica reconoce dos tipos de islas isla continental (porcion del territorio de un continente separado de este por movimiento tectonico 0 por ascenso del nivel del mar) e isla oceanica (porcion de territorio que no ha formado parte de un continente) La definicion sigue siendo uti por 10 que implica para la formacion de las faunas insulares en uno u otro caso As en el caso de la isla continental su fauna ya estaba emplazada en la futura isla antes de producirse su separacion del continente [Vicarianza hipotesis biogeografica de fragmentacion faunal] en el caso de la isla oceanica su fauna es el resultado de la colonizacion1 por propagulos2continentales a traves de la barrera marina entre el continente y la isla [Dispersion hipotesis biogeografica de acumulacion faunal] En el primer caso el surgimiento de la barrera marina es resultado del evento vicariante en el segundo la existencia de la barrera marina es anterior al evento dispersante Ademas de operar a escala mayor (continente-isla) ambos eventos operan secundariamente a menor escala (isla-isla)
Las dos hipotesis -vicarianza y dispersion- se han disputado hasta hoy la preferencia de los biogeografos en tanto hipotesis mas viable para explicar el proceso de formacion original (continente-isla) de la fauna de mamiferos terrestres de las Antillas Mayores [Sobre la historia de las hipotesis biogeograficas relativas al Caribe insular vease RuB y Schubert 1989 Williams 1989 Pruna 2001] Yel debate continua debido ala falta de consenso sobre 10 que Williams (1989) ha llamado el problema central de la discusion a saber la ostensiblemente baja diversidad taxonomica de esta fauna a nivel de las categorias superiores (solo cuatro ordenes) en comparacion con la diversidad al mismo nivel de las faunas de islas continentales (0 de areas del continente) simi lares en tamafio y diversidad ecologica [Por ejemplo la fauna cubana de mamiferos terrestres consideradas de conjunto las especies vivientes y las extintas- es menos diversa a los niveles taxonomicos superiores (4 ordenes y 6 familias) que 10 que cabria esperar de la diversidad ecologica y extension territorial de Cuba (110 992 km2) comparada con una isla tipicamente continental como Trinidad (4828 krn2
) -veinte veces menor que Cuba y menos diversiticada ecologicamente- donde solo las especies vivientes de mamfferos terrestres representan 6 ordenes y 16 familias (Eisenberg 1989)] Este desbalance
lColonizaci6n Arribo y establecimiento de un propagulo en una isla no ocupada por la especie (MacArthur y Wilson 1967)
2Propagulo Niimero minimo de individuos de una especie capaces de reproducirse y aumentar poblacionalmente en una isla bajo condiciones favorables (MacArthur y Wilson 1967)
303 POBLAMIENTO CONTINENTE-ISLA
taxon6mico ha sido expJicado de dos maneras (1) Inicialmente hubo mas 6rdenes presentes pero la mayoria se extingui6 y s610 sobrevivieron esos cuatro [explicaci6n vicarianista] (2) S610 representantes de esos cuatro 6rdenes lograron cruzar la barrera marina [explicaci6n dispersionista]
Cinco hipot~ticos mecanismos de poblamiento continente-isla han sido invocados durante los ultimos 140 afios (y se los sigue proponiendo actualmente) para explicar la presencia de mamiferos terrestres en las Antillas Mayores A continuaci6n pasamos a resefiarlos brevemente (incluidas sus alegadas fortalezas y debilidades) en el orden aproximado en que historicamente fueron propuestos
EL PUENTE NATURAL (dispersion terrestre activa)
En el siglo XIX el puente terrestre era pnkticamente la unica hipotesis para explicar las distribuciones disyuntas de animales en buena parte del mundo y los primeros investigadores que se ocuparon de la biogeografia antillana (Fernandez [1865] 1884 Chapman [1893] Torre 1892 Lora 1895) la asumieron resueltamente en parte intrigados por el descubrimiento en Cuba en la segunda mitad del siglo de un perezoso f6sil de gran talla Y en su aplicacion a los mamiferos terrestres de las Antillas Mayores el puente sigui6 siendo a 10 largo del siglo xx la hipotesis mas favorecida por los estudiosos del tema (Vaughan 1902a Spencer 191 Oa b Torre 1910a b cAllen 1911 1917a 1918 Barbour 19141916 1945bAnthony 1918 1926 [1942] Scharff 1922 Kraglievich 1926 Beaufort 1951 Arredondo 1961 1970b Kartashov y Mayo 1974 Iturralde-Vinent 1982 1988 Arredondo y Armifiana 1996 Arredondo 1996a [1999]) quienes postularon la existencia preterita de puentes con Norte- Centro- 0 Suramerica en dependencia de las afinidades continentales de los mamiferos antillanos
Teoricamente el puente terrestre debe cumplir con una condicion esencial permitir el paso de una parte mayoritaria (y por tanto representativa de la diversidad) de la fauna continental Claro que siempre habria tipos de mamiferos menos proclives a dispersarse o para los cuales la ubicaci6n del puente resultaria inaccesible 0 las condiciones ecologicas en este fueran desfavorables pero si algunos acceden al puente no hay raz6n para pensar que el resto de esa fauna no los acompafie mayoritariamente Como dijera Simpson (1956) Los puentes no transportan faunas desbalanceadas
En consecuencia el puente terrestre como filtro es un mecanismo de poblamiento minimamente selectivo y por ende de alta diversidad inicial (sensu Iturralde-Vinent y MacPhee 1999) 10 esencial del puente es su capacidad para transportar faunas integradas (Williams 1989) Yen eno radica la objeci6n fundamental al puente terrestre surgida tempranamente en el siglo xx (Por que s610 cuatro 6rdenes de mamiferos terrestres en las Antillas si vinieron por puentes (Donde estan los fosiles de los demas ordenes que debieron venir si es que se extinguieron Los partidarios del hipotetico puente dispusieron de casi todo el resto del siglo para responder una y otra vez que otros 6rdenes de mamiferos aparecerian representados en las Antillas Mayores el dia que en estas Islas se descubrieran fosiles precuatemarios suficientemente
304 BIOGEOGRAFIA
antiguos Esto se logro por fin terminando el siglo como se explico anteriormente (vease Contexto temporal) y ique se encontro los mismos cuatro ordenes de mamiferos terrestres conocidos del Cuatemario caribefio aparentemente los Unicos que lograron llegar
Pero ademas hasta el presente no se ha demostrado inequivoca- y concluyentemente que en algun momenta durante los ultimos 40 millones de afios haya existido una conexion terrestre continua entre las Antillas Mayores (en proceso de formacion 0 plenamente formadas) y algun punto de las regiones continentales vecinas
LA BALSA NATURAL (dispersion marina pasiva)
Durante buena parte del siglo xx el puente terrestre tuvo como unico rival a la hipotesis de la balsa natural -transporte accidental de animales sobre islotes de vegetacion flotantes a la deriva despues de arrancados y expulsados mar afuera por un rio crecido- expuesta originalmente por Matthew (191319151916 1918a 1919) y desarrollada en su aplicacion a los mamfferos terreslres antillanos principal mente por Simpson (1956) asi como por olros investigadores (Darlington 1938 1957 Williams y Koopman 1952 Hershkovitz 1958 1972 Patterson 1962 Paula 1967 Patterson y Pascual 1972 Pregill 1981 Ford 1986 Woods 1989a 1990 Morgan 1993) Por su parte Hedges et al (1992) sustentaron dicha hipotesis en estimados de divergencia (basados en el reloj molecular) en la herpetofauna antillana -involucrados tangencia1mente los mamiferos [veanse objeciones a Hedges y colaboradores (Page y Lydeard 1994 Iturralde-Vinent y MacPhee 1999) respuestas a las objeciones (Hedges et al 1994 Hedges 2001) y contrarrespuestas (MacPhee e IturraldeshyVinent 2005) asi como cuestionamientos al metoda de los estimados de divergencia (Foote et al 1999 Pregill y Crother 1999 Woods et ai 2001 )]
La hip61esis de la balsa natural se origino a partir de las numerosas ocasiones en que a 10 largo de siglos tales islotes flotantes (portando 0 no vertebrados incluidos mamiferos) han sido avistados mar afuera a cientos de kilometros de la costa mas cercana en varias partes del mundo --inc1uido el Mar Caribe- segUn consta en diarios de barcos y reportes de observadores diversos (revisados por King 1962 reportes posteriores en SICSP 1970 y Censky et al 1998) Uno de estos casos acaecido en nuestra region se reporta en un informe del SICSP (Smithsonian Institution Center for Shortlived Phenomena 197050shy52)
EIB de julio de 1969 un destructor de la armada de Estados Unidos avisto un islote flotante a unos 100 km al sur de la Base Naval de Guantanamo moviendose desde el Paso de los Vientos en direccion suroeste a una velocidad de 45 kmlh islote de unos 15 m de diametro mantenfa en pie mas de 10 palmas reales de 10-15 m de altura con suelo cubierto de hierba y arbustos [la observacion con binoculares a 18 km de distancia no preciso la presencia de vida animal] La travesia fue rastreada durante 6 dfas hasta que el 19 de julio el islote se dio por perdido tras 6 horas de busqueda aerea
305 POBLAMIENTO CONTINENTE-ISLA
La colonizaci6n exitosa de una isla de las Antillas Mayores por mamiferos terrestres continentales transportados en balsas naturales a la deriva (de cada especie al menos una hembra gravida 0 dos individuos de sexo opuesto) dependeria de factores que van a operar desde el momenta mismo de formarse la balsa asi como durante su travesia marina y su arribo a la isla
a) La posibilidad de estar en la balsa Que la distribuci6n de la especie incluya areas contiguas al rio y que el animal se encuentre en ellugar y el momenta propicios para ser incorporado a la balsa antes de que el rio crecido la desprenda Por ejemplo segun Hershkovitz (1958) los equimidos -grupo ancestral de roedores antillanos- serian buenos candidatos a ser captados por la balsa porque acostumbran a anidar en huecos del tronco de los arboles a la orilla de los rios
b) El tamano y la consistencia de la balsa Muchas se desintegran en alta mar debido a su fragilidad ante la saturaci6n Algunos autores (Beaufort 1951 Arredondo 1996a [1999]) han expresado incredulidad en cuanto a que las balsas hayan podido soportar el peso de un animal como el gran perezoso cubano (Megalocnus rodens) el mayor de los mamiferos antillanos Pero esta especie evolucion6 en Cuba y el ancestro que hizo el viaje pudo ser muy pequeno [EI perezoso oligocenico de Puerto Rico es el mas pequeno de los megaloniquidos antillanos (MacPhee e Iturralde-Vinent 1995b )]
c) Las caracterfsticas fisiol6gicas de la especie Los limites fisiol6gicos de los propagulos constituyen nuestra unica objeci6n a la dispersi6n marina por balsas como mecanismo para la formaci6n de la fauna de vertebrados en las Antillas (MacPhee e IturraldeshyVinent 2005) Unas especies tienen intrinsecamente mas posibilidades que otras para sobrevivir a las vicisitudes que deben afrontar durante la travesia (deshidrataci6n inanicion insolaci6n etc) Por ejemplo los megaloniquidos han demostrado tener extraordinaria habilidad para cruzar barreras marinas fueron los primeros mamfferos suramericanos en invadir Norteamerica mucho antes de que surgiera la conexion terrestre a finales del Plioceno (Paula 1967 Olson 1978 Webb 1985b Martin y Steadman 1999) Incluso se conoce un megaloniquido acultitico del Plioceno peruano (Muizon y McDonald 1995) y otro viviente en Suramerica (Choloepus) que pnicticamente no bebe agua 10 que ha originado una leyenda aborigen que explica por que los perezosos no toman agua (Goff art 1971) Vease ademas el inciso e
d) EI regimen de las corrientes marinas Que conduzcan a la balsa en la direcci6n necesaria EI transporte de balsas de vegetacion desde los continentes hacia las islas fue durante la mayor parte del Cenozoico sumamente improbable (MacPhee e IturraldeshyVinent 2000 148) Estos autores han insistido en que hay que tener en cuenta que durante la cambiante geografia cenozoica del Caribe las corrientes marinas de superficie no favorecieron el desplazamiento de objetos fiotantes hacia las Antillas Mayores [Pero no sabemos con certeza que efecto modificador del curso de tales objetos fiotantes pudieron introducir las tormentas tropicales si previsiblemente funcionaron en el pasado como 10 hacen ahora] Dichos autores senalaron ademas (2005) que en un experimento reciente con boyas de posicionamiento satelital liberadas a la deriva desde las Antillas Menores las boyas fueron a parar mayoritariamente a Centroamerica y a otros lugares
306 B[OGEOGRAFIA
y solo unas pocas recalaron en las Antillas Mayores y las que entraron al Mar Caribe demoraron semanas 0 meses en arribar
e) La distancia a recorrer (y en consecuencia la duracion de la travesia) entre el continente y la isla (10 que repercute decisivamente en el factor c) Este es desde luego uno de los factores mas criticos En la medida que la distancia aumenta la probabilidad de alcanzar la isla disminuye drasticamente con el agravante de que la trayectoria dificilmente seria en linea recta Si tomamos como referencia la velocidad de 45 kmlh del islote ya mencionado (SICSP 1970) cubrir la distancia aerea (aproximada) de 800 km que media entre el noroeste de Suramerica y La Espafiola Ie tomaria a la balsa 74 dias Estimando el doble de este tiempo debido a desvios de la trayectoria la duracion de la travesia probablemente resultaria excesiva para la mayoria de los mamiferos Pero pudiera muy bien ocurrir que la velocidad fuera mayor 0 que la balsa portara plantas vivas que permitirian a los fitOfagos sobrevivir [Tres de los cuatro ordenes de mamiferos terrestres conocidos de las Antillas Mayores son fitOfagos] Ademas esta geologicamente bien establecido que durante etapas de variada duracion desde el Oligoceno hasta el Pleistoceno los territorios del Caribe insular y continental aumentaron considerablemente en extension con niveles del mar 100-200 m por debajo de su nivel actual oportunidades que los propagulos aprovecharian para cruzar barreras marinas significativamente menores
f) El tamafio de la isla Las islas mayores deben interceptar mas balsas (Buckley y Knedlhans 1986) 10 que repercute directamente en el factor e La balsa puede pasar entre islas pequefias y continuar viaje sin llegar a ningun lugar
g) Las condiciones (favorables 0 adversas) del desembarco en la isla y el tiempo que Ie tomaria a uno 0 mas animales depauperados encontrarun habitat apropiado Esto dependeria a su vez del tamafio y diversidad ecologica de la isla (Darlington 1938 Miller 1966)
Tal cumulo de posibles obstaculos y dificultades constituye a primera vista un desafio a la credibilidad de la hipotesis Matthew-Simpson pero en ello radica precisamente su valor explicativo bas ado en la mas absoluta improbabilidad La colonizacion por la via marina es altamente improbable pero no absolutamente imposible si se tiene en cuenta la inmensidad del tiempo geologico se vuelve probable (McKenna 1973) Basta que solo una vez entre mill ones de intentos fallidos durante ese inmenso lapso algunos de los factores anteriormente enunciados coincidan en no ser tan desfavorables para el propagulo Y eso es todo 10 que demanda la hipotesis para cada uno de los grupos monofileticos mayores de mamiferos terrestres presentes en las Antillas Mayores Las Antillas Mayores requeririan el arribo de tan solo 7 (quizas menos) propagulos fundadores a 10 largo de los ultimos 30 0 mas millones de afios (promedio de uno cada 43 mill ones) para explicar la formacion de toda su fauna de mamiferos terrestres por dispersion marina pasiva (vease Discusion y Conc1usiones)
En comparacion con la via marina el poblamiento por la via terrestre es predecible porque la mayor parte de la fauna continental de mamiferos emplazada en la region del puente va a cruzar por este una vez que el mismo se establece y los animales logran
307 POBLAMIEITO CONTINENTE-ISLA
acceder a el el poblamiento por la via marina en cambio es impredecible porque depende total mente del azar que caracteriza a la balsa Claro que en ambas vias opera el azar pero en el primer caso se trata del azar de un evento probable en el segundo el azar de un evento improbable (Simpson 1940) Asi el puente natural (mecanismo de poblamiento minimamente selectivo = alta diversidad inicial) y la balsa natural (mecanismo de poblamiento maximamente selectivo baja diversidad inicial) pueden conceptuarse como los extremos opuestos de una gama de probabilidades de dispersi6n (McKenna 1973) EI fen6meno de la dispersi6n merece la mayor atenci6n cientifica en los estudios de biogeografia insular (Cowie y Holland 2006)
Queda claro que la balsa natural es una hip6tesis consistente con el caracter de la fauna objeto de estudio puesto que el desbalance taxon6mico (sin evidencia paleontol6gica en contrario) s610 puede ser resultado de la improbabilidad del poblamiento No obstante ello no implica necesariamente que la balsa natural haya sido el imico mecanismo (ni siquiera el principal) utilizado por los mamiferos terrestres en su colonizaci6n de las Antillas Mayores La balsa natural debe satisfacer una premisa te6rica esencial dado el caracter azaroso impredecible e improbable de una colonizaci6n exitosa esta debe ocurrir a intervalos muy espaciados a traves del tiempo geol6gico 0 sea durante todo el tiempo que en el area ocupada por las Antillas Mayores ha habido ecosistemas terrestres perrnanentemente disponibles para ser colonizados (los ultimos 40 millones de afios) Sin embargo como sabemos representantes de los cuatro 6rdenes de mamiteros terrestres conocidos de las Antillas Mayores ya estaban emplazados en dicho territorio insular en el Mioceno Inferior 10 que supone un lapso de colonizaci6n limitado al Eoceno SuperiorshyOligoceno (MacPhee 2005) sugerente de un tipo de mecanismo contrario a la colonizacion alocronica exigida por la hipotesis de la balsa natural sobre todo tratandose de una fauna caracterizada por altas tasas de extincion sin reposicion aparente
La presencia en Jamaica (hasta su extinci6n en 1877) del roedor mfuido Oryzomys antillarom con estrecha afinidad centroamericana (Woods 1989a) pudiera constituir el imico caso efectivo de poblamiento (continente-isla) de las Antillas Mayores por dispersion marina dc un mamifero terrestre para el cual no es razonable invocar otro mecanismo de colonizacion
LA GUIRNALDA INSULAR (dispersion mixta)
La geologia ha establecido con suficiente certeza que durante el Cenozoico unicamente en el Pale6geno tardio y solo a traves de la Cresta de Aves (ahora sumergida excepto la isla hom6nima) pudo haber existido una cadena de islas para el transito de mamiferos terrestres entre el norte de Suramerica y las proto-Antillas Mayores [desestimados a ese efecto el Canal dc Yucatan el Elcvado de Nicaragua la Cresta de Caiman el Elevado de Beata y las Antillas Menores (Holcombe et al 1990 MacPhee e IturraldeshyVinent 1994 1995b Iturralde-Vinent y MacPhee 1999)] Holcombe y Moore (1977) y Holcombe y Edgar (1990) postularon la existencia de esta entidad y estimaron algunas de sus posibles caracteristicas Por su parte MacPhee e Iturralde-Vinent (1995b) e Iturralde-Vinent y MacPhee (1999) precisaron los extremos proximal y distal de esa
308 BIOGEOGRAFIA
estructura acotaron la duracion y la edad de la misma y Ie dieron nombre GAARlandia (Greater AntillesA vesRidgelandia)
Segun Iturralde-Vinent y MacPhee (1999) la Cresta de Aves emergio por breve tiempo (1-2 Ma) durante la transicion Eoceno-Oligoceno (35-33 MaAP) por la coincidencia de un levantamiento tectoni co y un considerable descenso eustatico del nivel del mar En ese momenta las islas septentrionales del arco de las Antillas Mayores (Cuba centroorientalshyLa Espafiola - Puerto Rico - Islas Virgenes) constituian una sola isla grande 0 una serie de islas separadas por barreras muy estrechas La Cresta de Aves emergida entre el bloque Puerto Rico - Islas Virgenes y un microcontinente noroccidental de Suramerica -separado en ese momento del resto del continente por una via marina surgida en el Eoceno tardio- dio origen a GAARlandia
Obviamente el numero de colonizaciones exitosas por mamiferos terrestres a traves de una guimalda de Islas tiende a disminuir mientras mas anchas sean las barreras marinas entre las islas que forman la guimalda y cuando esto ocurre disminuye tambien la diversidad taxonomica de los propagulos particularmente a nivel de las categorias superiores (Simpson 1956)
Vemos asi que para los mamiferos terrestres la colonizaci6n a traves de una guimalda insular puede ser tan improbable como a la deriva en una balsa natural en dependencia de la cantidad y anchura de las barreras marinas a cruzar el tamafio y la fisiografia de las islas que integran la guimalda y el regimen de las corrientes marinas y aereas entre otros factores
Resumiendo la guimalda insular puede conceptuarse como un mecanismo de poblamiento altamente selectivo (= baja diversidad inicial) que combina la improbabilidad de la balsa natural con la colonizacion restringida temporaimente del puente terrestre
En su reciente revision de la taxonomia distribuci6n y filogenia (basada en estudios moleculares) de varios grupos de mamiferos antillanos Davalos (2004) concluyo que los perezosos megalocninos los primates y los roedores histricognatos se avienen al patron de divergencia con el continente y epoca de arribo a las Antill as implicados en la hipotesis de GAARlandia pero no asi los perezosos coloepodinos que requieren una explicacion biogeognlfica distinta Esto ultimo (sospechado por Matthew 1918a) habla sido confirmado por White y MacPhee (2001 226-227)
Nuestra evidencia filogenetica no sustenta la idea de un dado unificado surgido de la invasion de un megaloniquido el analisis sugiere mas bien dos eventos de colonizaci6n independientes [y] sustenta fuertemente un origen difiletico de las subfamilias [Choloepodinae y Megalocninae] Si hubo dos migraciones a traves de GAARlandia presumiblemente poco espaciadas en el tiempo no se puede determinar en el presente
Finalmente queda claro que GAARlandia -como guirnalda insular- es la hipotesis mas robusta propuesta hasta ahora para explicar la presencia en las Antillas Mayores de
309 POBLAMIENTO CONTINENTE- ISLA
la absoluta mayona de los mamfferos terrestres de ongen suramericano porque el caracter de esa fauna supone un mecanismo de poblamiento ancestral que sea a la vez maximamente selectivo (baja diversidad ordinal) y temporalmente restringido (transicion Eoceno-Oligoceno)
Recientemente Rega et al (2002 18) expresaron A pesar de una revision exhaustiva de la paleogeografia de la cuenca del Caribe Iturralde-Vinent y MacPhee (1999) no pudieron confirmar la presencia de una conexion terrestre continua entre Suramerica y las proto-Antillas aunque hipotetizaron su existencia [Vease la Fig 42 (= figura 6 de Iturralde-Vinent y MacPhee 1999 simplificada))
En el trabajo citado por Rega y colaboradores aparece la siguiente explicacion
Actualmente no es posible ofrecer reconstrucciones paleogeograficas detalladas de la Cresta de Aves Sin embargo hay evidencia de que porciones de la cresta deben haberse elevado subaereamente en un tiempo La cuestion cntica es si la Cresta de Aves emergida pudo haber formado algun tipo de corredor entre las Antillas Mayores y el noroeste de Suramerica Perforaciones en las alturas estructurales de la cresta pudieran proveer el registro del Paleogeno tardio que se necesita para precisar la hipotesis del corredor aunque no se puede predecir si tales datos serian suficientemente detallados para determinar si en algun momento GAARlandia constituyo una sola entidad subaerea Por ejemplo pudo haber barreras marinas por corto tiempo entre las masas terrestres de la Cresta de Aves [0) ellevantamiento y la subsidencia pudieron afectar diferentes partes de GAARlandia en diferentes tiempos AItemativamente los elementos faunales pudieron haberse dispersado de manera pulsada Movimientos episOdicos de este tipo incrementarian la extincion de los elementos faunales incapaces de trasladarse a la siguiente masa terrestre en la cadena disminuyendo asi la diversidad total (Iturralde-Vinent y MacPhee 199957)
La primera parte de la explicacion deja claro que la posibilidad de que la Cresta de Aves haya sido una conexion terrestre continua no se puede descartar pero tampoco se puede afirmar La ultima parte de la explicacion describe el efecto altamente selectivo (filtrante) de la colonizacion a traves de una cadena de islas
Y continuan lturralde-Vinent y MacPhee (199957 -58) explicando
Hasta que punto el microcontinente noroccidental de Suramerica [considerado por dichos autores como el area suministradora de propagulos hacia GAARlandiaJ estuvo fisicamente separado del resto del continente no ha sido plenamente esc1arecido Si estuvo sustancialmente aislado por barreras marinas y orogenic as durante el Paleogeno tardio entonces solo la fraccion de la biota suramericana emplazada en el microcontinente estuvo en posicion de acceder a GAARlandia Esto pudiera tener que ver con el hecho de que la fauna antillana carezca de representantes de muchos grupos suramericanos [de mamfferos) que debieron haber hecho el viaje
310 BIOGEOGRAFiA
Florida o Alturas0 0
d 6 D L1anuras25deg
Yucatan
20deg
deg0
10deg
MARCARIBE
80deg 75deg
Bahamas
Cres ta de Beata
D Mares someros
D Mares profundos
OCEANO ATLANTICa
FIGURA 42 Reconstrucci6n hipotetica de GAARlandia como conexi6n terrestre continua entre las proto-Antillas Mayores y el noroeste de Suramerica en el Pale6geno tardio segun la FIgura 6 (simplificada) de Iturralde-Vinent y MacPhee (1999)
En su defensa del puente terrestre Barbour (1945b249) se refiri6 a esto - antes 10 habfa hecho Anthony [1942363]- en los terminos siguientes
El argumento de que si las Antillas hubieran estado conectadas con el continente se encontrarian los f6siles de un numero mayor de distintas criaturas me deja frio es perfectamente concebible que la conexi6n haya ocurrido en un periodo en que la parte del continente implicada no estuviera poblada por una fauna grande y variada
Y concluyen lturralde-Vinent y MacPhee (199958) su detallada explicaci6n
Desafortunadamente sabemos muy poco sobre la composici6n faunal del noroeste de Suramerica a finales del Pale6geno
Consecuentemente si se confirmara la magnitud del aislamiento del aludido microcontinente y su concomitante baja diversidad faunal GAARlandia como conexi6n terrestre continua serf a la hip6tesis mas parsimoniosa No obstante mientras no se disponga de evidencia concluyente a este respecto hipotetizar a GAARlandia como conexi6n continua (minima selectividad) la coloca en franca vulnerabilidad te6rica al tener que afrontar la objeci6n clasica al puente terrestre (baja diversidad ordinal de la
311 POBLAMIENTO CONTINENTE-TSLA
fauna) al tiempo que se la priva de ser la hip6tesis mas robusta para explicar -como guimalda de islas- la colonizaci6n de las Antillas por mamiferos procedentes de Suramerica
ELARcA DE NOE (seudovicarianza)
El primer vicarianista antillano (desde luego sin el saberlo) fue Corral (1940) con su utilizaci6n ingenua de las ideas wegenerianas de la deriva continental aparecidas la decada anterior Pero el primer (supuesto) intento de aplicar a la formaci6n de la biota caribefia el mecanismo de la vicarianza basada en la tearia de la tect6nica de placas (surgida en la sexta decada del pas ado siglo) se debe a Rosen ([1976] 1985)
Segun Rosen las Antillas Mayores se originaron en el Cremcico en la Placa Caribe aproximadamente en la posici6n que hoy dia ocupa Centroamerica y al ser desplazadas tectonicamente hacia el este interactuaron (de algun modo no precisado por Rosen) con las margenes continentales de Norte- y Suramerica de donde (en un evento unico) adquirieron toda su fauna transportada subsecuentemente por dichas islas en su transito hacia sus posiciones actuales [De ahi la denominacion metaforica dada par McKenna (1973) a este mecanismo de poblamiento] Aunque se ha presentado esta propuesta como Model0 de Vicarianza Continente-Isla de Rosen en realidad por definici6n no constituye un evento vicariante porque las proto-Antillas Mayores no resultaron del fraccionamiento y separaci6n por tectonismo de una porcion del territorio continental como exige esta hip6tesis (vease definici6n de vicarianza al comienzo de Hipotesis Biogeograficas)
El Arca de Noe de Rosen ha sido desestimada por la mayoria de los autores contemporaneos porque (1) carece de sustento geol6gico (lturralde-Vinent y MacPhee 1999) y porque (2) en el periodo en que se afrrrna ocum6 el evento muchos de los ordenes modemos de mamiferos placentarios attn no existian (Williams 1989)
EL BARCO FUNERAL VlKINGO (tanatovicarianza)
Domning et al (1997) reportaron 10 que consideraron el mamffero terrestre mas viejo de las Antillas --circunstancia que se refleja en el titulo mismo del trabajo Oldest West Indian Land Mammal- al descubrirse un perisodactilo rinocerotoide (Hyrachius sp) en una localidad de edad Eoceno Inferior a Medio en el occidente de Jamaica
Jamaica ha tenido una historia geologica y paleogeografica distinta de la historia comun a las restantes Antillas Mayores A principios del Cenozoico Jamaica occidental (a diferencia de la porcion oriental constituida por el Bloque de las Montafias Azules) formaba parte de Centro america de la cual se desgaj6 (por el desplazamiento de la Placa Caribe) para venir a integrarse despues del Eoceno Medio con 10 que serfa la futura isla de Jamaica desaparecida por transgresi6n marina en el Eoceno Superior hasta volver a emerger totalmente en el Mioceno Medio (Iturralde-Vinent y MacPhee 1999 MacPhee e Iturralde-Vinent 2000)
312 BIOGEOGRAFiA
Por consiguiente Hyrachius ---conocido de localidades de edad Eoceno Inferior a Medio en Norteamerica- debio ocupar (10 que iba a ser) Jamaica occidental mucho antes de que esta se separara de Centroamerica De hecho este fosil Uego a su destino insular contenido en las rocas de su basamento no como un ser vivo por 10 que es erroneo considerarlo un animal antillano (lturralde-Vinent y MacPhee 2004) como han supuesto algunos (Hedges 200 I Portell et al 200 I Condis [200 I D Todo 10 que hizo fue viajar tanatovicariantemente en su barco funeral vikingo -otra de las denominaciones metaforicas de McKenna (1973)
DISCUSION Y CONCLUSIONES
Hemos visto que solo disponemos de una hipotesis plausible para explicar la mayor parte del poblamiento (continente-isla) de las Antillas Mayores por mamiferos terrestres del Neotropico continental la dispersion mixta (a traves de GAARlandia) para los perezosos (excepto Choloepodinae) los monos y los roedores (excepto Oryzomys)
Por otra parte la historia biogeogrMica de las musaraiias de las Antillas Mayores (Nesophontes y Solenodon) constituye una de las situaciones mas enigmaticas entre los mamiferos de la region (Ottenwalder 2001321) Al menos cinco mecanismos de poblamiento continente-isla han sido invocados para explicar cuando como y de donde llegaron a las Antillas Mayores estas musaraiias
I EI propagulo ancestral (del Terciario de Norteamerica) coloniz6 a las Antillas Mayores por dispersion marina desde Centroamerica 0 desde la Florida (Patterson 1962)
2 El propagulo ancestral ( del Terciario) cruzo el Atlantico Sur (supuestamente mas estreshycho en aquella epoca) directamente de Africa a las Antillas Mayores (Hershkovitz 1972)
3 El propagulo ancestral (del Terciario de Norteamerica) Uego a las Antillas Mayores en el Arca de Noe de Rosen (MacFadden 1980)
4 El propagulo ancestral (del Terciario de Norteamerica) fue transportado a las Antillas desde Centroamerica en el bloque de Jamaica occidental y de alli paso por dispersion marina a las restantes Antillas Mayores (Whidden y Asher 200 I)
5 El propagulo ancestral (del Terciario de Norteamerica) lleg6 a las Antillas Mayores a traves de GAARlandia (Whidden y Asher 2001)
Aunque hay consenso entre estos (y otros) autores sobre el origen norteamericano de las musaraiias antillanas ninguno de los cinco hipoteticos mecanismos de poblamiento resiste la evidencia en contrario filogenetica yo paleogeogrMica Ademas algunas de estas hipotesis asumen que Nesophontidae y Solenodontidae comparten un antecesor comun en las Antillas pero esta relacion no ha sido confirm ada Para dilucidar el enigmatico caso de las musaraiias antillanas se requerini profundizar en la investigaci6n geologica paleogeognifica y paleontol6gica (Iturralde-Vinent y MacPhee 1999)
De acuerdo con el conocimiento filogenetico disponible sobre el conjunto de la fauna la colonizacion original de las Antillas Mayores por mamiferos terrestres implicaria el arribo
313 POBLAMIENTO ISLA-ISLA
de solo siete propagulos fundadores perezosos 2 (coloepodinos y megalocninos si como parece tienen un origen difiletico) musaraiias 2 (nesofontidos y solenodontidos si no constituyen una misma familia como consideran algunos) monos 1 (pitecidos) roedores histricognatos 1 (equimidos capromidos y heptaxodontidos) roedores muridos 1 (Oryzomys en Jamaica)
POBLAMIENTO ISLA-ISLA
La distribucion de los mamiferos terrestres a traves del area antillana demarcada al inicio del presente capitulo (vease Contexto Geogratico Fig 41) ha sido interpretada en funcion de dos hipoteticos mecanismos de poblamiento interinsular la dispersion y la vicarianza fundamentadas como se resumen seguidamente
DISPERSION
De acuerdo con la evidencia paleogeogratica mas actualizada en las principales regresiones marinas ocurridas en la cuenca caribeiia durante los ultimos 23 Ma (transiciones OligocenoshyMioceno Mioceno Superior - Plioceno y varias veces en el Pleistoceno) las masas terrestres constitutivas de la region insular en formacion aumentaron considerablemente su extension territorial disminuyendo por tanto la distancia entre elias oportunidades que los elementos faunales aprovecharian para dispersarse de unas islas a otras
Durante mucho tiempo esta ha sido la hipotesis mas favorecida por los estudiosos del tema a partir de Simpson (1956) pero en decadas recientes ha suscitado dos objeciones
1 Es dificil en tender como pudo operar la dispersion marina entre estas islas puesto que ninguna de las Antillas Mayores posee rios suficientemente caudalosos para transportar masas significativas de vegetacion flotante (MacPhee 198432)
Es oportuno recordar aqui el islote ala deriva (con mas de diez palmas reales en pie) entre las tres Antillas Mayores mas occidentales mencionado anteriormente (vease La Balsa Natural)
2 La dispersion marina es menos efectiva para explicar la asombrosa similitud de las faunas de mamiferos terrestres a traves de las Antillas Mayores septentrionales porque como proceso aleatorio no debe producir distribuciones uniformes a 10 largo de una cadena de islas 0 si las produce entonces el proceso no es aleatorio (MacPhee e lturralde-Vinent 2000 149)
Independientemente de que las aludidas distribuciones taxonomicas estan lejos de ser uniformes 0 asombrosamente similares la hipotesis de la dispersion marina contrariamente a su canicter aleatorio y altamente improbable en el caso del poblamiento continente-isla- en el caso del poblamiento isla-isla es un proceso no aleatorio (aprovecha las oportunidades brindadas puntualmente por los periodos de maxima regresion marina) y ademas altamente probable dada la incomparablemente menor distancia entre las islas caribeiias que entre estas y los continentes vecinos
314 BIOGEOGRAFiA
V ICARIANZA
La neotectonica y las transgresiones marinas del Oligoceno Medio y del Mioceno afectaron sustancialmente la paleogeografia de la Cresta de las Antillas Mayores resultando la subdivision gradual de las partes subaereas de la Cresta Asi el este de Cuba y el norte de La Espanola conectados fisicamente durante el Oligoceno Inferior se separaron al formarse el Paso de los Vientos en el Oligoceno tardio en tanto La Espanola y Puerto Rico probablemente permanecieron conectadas hasta el Mioceno tardio en que surgio el Paso de Mona (Iturralde-Vinent y MacPhee 1999)
Tales eventos geologicos habrian dividido las distribuciones de los grupos faunales emplazados previamente en el territorio continuo de las tres islas y a partir de esas disyunciones dichos autores (tambien MacPhee et al 2000 y White y MacPhee 2001) asumieron --como segunda fase del modelo de GAARlandia- la hipotesis de la vicarianza isla-isla para explicar la distribucion multiinsular de varios grupos de mamiferos terrestres a traves de las Antillas Mayores septentrionales De acuerdo con esta hipotesis cualquier diferencia en composicion faunal entre las islas (por ejemplo la ausencia de roedores heptaxodontidos en Cuba 0 de monos platirrinos en Puerto Rico) constituye evidencia de extincion local postvicariante y no de dispersion marina fallida Lo primero se puede demostrar si aparecen los fosiles pertinentes 10 segundo no se puede demostrar (MacPhee e Iturralde-Vinent 1994) Por otra parte IturraldeshyVinent y MacPhee (1999 57) han reconocido que aunque fa vicarianza isla-isla puede representar una solucion clara para las distribuciones de varios taxones cuaternarios no puede explicar todos los casos
Rega et al (2002 18) han presentado dos objeciones al modelo de la vicarianza isla-isla aplicado a los mamiferos de las Antillas Mayores
I Si la separacion de Cuba y La Espanola precedio en varios mill ones de anos la separashycion de La Espanola y Puerto Rico el modelo vicariante de lturralde-MacPhee debe predecir mayor similitud entre las faunas de La Espanola y Puerto Rico que entre las de Cuba y La Espanola De hecho es al reyes
No obstante MacPhee et al (200335) admitieron despues que la fecha mas temprana empiricamente defendible [para la apertura del Paso de Mona] debe ser el Oligoceno Medio esto invierte la secuencia de ambas disyunciones con 10 cual queda resuelta la aparente contradiccion Sin embargo en su reciente evaluacion de la congruencia entre filogenia e hipotesis geologic as en varios grupos de mamiferos terrestres de las Antillas Mayores Davalos (2004373) encontro que ninguno de los taxones cumple con la fecha y el patron de divergencia entre las masas terrestres antillanas postulados por el modelo de GAARlandia
2 La presencia de un mismo genero en las tres Islas septentrionales de las Antillas Mayores implica [por el modelo vicariante] que la diferenciacion entre estas formas durante el intervalo desde el Mioceno ha sido unicamente de nivel especifico [Dicha presencia]
315 rRRADlAcr6~ EVOLUCrONARIA
s610 puede explicarse razonablemente como el resultado de movimientos de esos animales entre las islas en el Cuaternario
De esta objecion resulta dificil disentir [El poblamiento entre islas diferenciado solo a nivel especifico no es probable que sea anterior al Pleistoceno (Simpson 1956 15)]
Debe tenerse en cuenta que la hipotesis de la vicarianza isla-isla alude a las tres islas septentrionales de las Antillas Mayores porque como se explico antes (vease El Barco Funeral Vikingo) Jamaica no existia como isla durante el lapso (Eoceno Superior shyOligoceno) consensualmente reconocido como el mas probable para la colonizacion de las proto-Antill as Mayores por la mayo ria de los mamiferos terrestres Despues de emerger como isla en el Mioceno Medio Jamaica debe haber recibido sus mamiferos terrestres por dispersion isla-isla (Davalos 2004376) y solo en el Cuaternario parece haberrecibido (por dispersion marina desde Centroamerica) al roedor murido Oryzomys
Seguramente tanto la dispersion como la vicarianza contribuyeron a que los mamiferos terrestres se irradiaran a traves de las Antillas Mayores y grupos de islas perifericas pero las maneras y tiempos en que estos mecanismos operaron a 10 largo de dicho proceso son aun en gran medida motivos de desacuerdo e incertidumbre
IRRADIACION EVOLUCIONARIA
A continuaci6n revisamos someramente las distribuciones conocidas de los diferentes grupos de mamiferos terrestres a traves de las Islas principales de las Antillas Mayores y grupos de Islas perifericas incluidas las interpretaciones y conjeturas que la literatura reciente recoge en relaci6n con la manera en que dichas distribuciones pudieron haberse conformado durante la irradiaci6n evolucionaria de los referidos grupos de mamiferos Manifestaciones de esta irradiaci6n pueden inferirse unicamente a partir de los taxones presentes en el Cuatemario pues solo 6 de las especies conocidas corresponden al Terciario
GRUPOS ORlGINARIOS DE SURAMERICA
PEREZOSOS (Megalonychidae)
Aparte de un genero miocenico de Cuba y otro del Oligoceno de Puerto Rico se conocen 4 generos (12 especies) de perezosos cuaternarios (todos extintos) restringidos a las tres islas septentrionales de las Antillas Mayores Acratocnus Neocnus Megalocnus y Parocnus Los cuatro generos estan representados en Cuba y La Espanola y Acratocnus 10 esta ademas (como unico genero de perezoso) en Puerto Rico Esta distribuci6n ha sido interpretada por MacPhee et al (2000) eomo resultado de la vicarianza oligomiocenica entre estas islas y [dado que el Banco Puerto Rico - Islas Virgenes fue la entrada de GAARlandia a las Antillas Mayores] se preguntaron si la ausencia de los otros tres generos en Puerto Rico se debe a que nunca estuvieron en ese territorio 0 a que sus
316 BIOGEOGRAFiA
fosiles atm no han sido hallados Pero como hemos visto Rega et al (2002) atribuyeron dicha distribucion a la dispersion interinsular en el Cuatemario
Por otra parte varios amilisis c1adisticos recientes ubi can a Choloepus -genero Reciente conocido solo de Centro- y Suramerica- como taxon hermano de Aeratoenus exc1uidos los demas megaloniquidos antillanos Para White y MacPhee (2001228)
tan estrecha relacion sugiere que la divergencia entre ambos [generos] ocurrio despues de que Aeratoenus divergio de Neoenus Si esta relacion implica que Aeratoenus y Neoenus se dispersaron separadamente desde Suramerica a las AntilIas mientras Choloepus permanecia en el continente (siendo necesarias al menos tres invasiones de perezosos) 0 si Choloepus invadio Suramerica desde las Antillas es algo que el material disponible no permite determinar
La diversidad adaptativa de los perezosos se manifiesta en (1) la utilizacion del habitat especies arboreas en Neoenus y Aeratoenus y terrestres en Paroenus y Megaloenus (Webb 1985a White [1993] White et al 1996 Arredondo [1999]) y en (2) la gama de tallas corporales desde los pequenos Neoenus hasta Megaloenus rodens el mayor de los mamiferos terrestres antillanos (su peso adulto estimado en 270 kg seglin Paula 19679) esto sugiere que los perezosos se irradiaron a nichos que en contextos continentales probablemente estarian ocupados por miembros de otros c1ados (MacPhee e IturraldeshyVinent 2000)
No esta claro si el abanico de tamanos entre estos perezosos enmascara alguna tendencia al gigantismo Por 10 pronto Anderson y Handley (2002) encontraron 10 contrario en perezosos (Bradypus) de las Islas de Bocas del Toro (Panama) shyseparadas secuencialmente del area continental vecina por ascensos del nivel del mar durante los ultimos 10 ka En las cinco islas los perezosos son menores que su contraparte continental y las diferencias entre las tallas medias de las cinco poblaciones expresan una reduccion lineal significativa de la talla corporal en funcion de la edad geologica de cada isla sin relacion alguna con el area de las islas 0 la distancia que las separa del continente
MONOS (Pitheciidae)
Los primates de las Antillas Mayores (cuatro especies extintas) constituyen un grupo monofiletico con Paralouatta (en Cuba) y Antillothrix (en La Espanola) como taxones hermanos seguidos en la escalera c1adistica por Xenothrix (en Jamaica) y finalmente Callieebus como el pariente continental mas cercano esto implica una sola invasion desde Suramerica antes del Mioceno Inferior (fecha mas temprana conocida para los primates fosiles antillanos) y por tanto en consonancia con GAARlandia como la via de colonizacion (Horovitz y MacPhee 1996 1999 Horovitz [1997] MacPhee y Horovitz 2004)
En desacuerdo parcial con la anterior interpretacion la filogenia de este grupo de monos obtenida por Davalos (2004) indica un antecesor comun en Cuba La Espanola y Jamaica
317 TRRADIACTON EVOLUCIONARIA
compatible con la hipotetica conexion -propuesta tentativamente por Iturralde-Vinent y MacPhee (199980-84)- entre las Montanas Azules del este de Jamaica y la peninsula suroccidental de La Espanola en la epoca de GAARlandia
A pesar de que los primates antillanos pertenecen a un grupo viviente de monos muy pequenos (los monos titles) fueron comparativamente grandes en relacion con los platirrinos modernos y sucesivamente mayores en la direccion Antillothrix Xenothrix y Paralouatta con P marianae como el mayor platirrino conocido del Mioceno neotropical (MacPhee 2005) MacPhee et al (1994) consideraron que Xenothrix debio ser un mono de andar lento y al parecer las especies de Paralouatta abandonaron la arborealidad para adoptar un modo de vida mas terrestre (MacPhee 1996a MacPhee et al 2003 MacPhee y Meldrum 2006)
ROEOORES (Echimyidae Capromyidae Heptaxodontidae)
La edad de Zazamys en Cuba demuestra que la invasion de los roedores histricognatos en las Antillas Mayores comenzo antes del Mioceno Inferior por otra parte los representantes mas antiguos de esa irradiacion evolucionaria se encuentran en La Espanola y Puerto Rico Ambas evidencias son consistentes con la hipotesis de GAARlandia (Woods 1982 1988 Woods et al 2001)
No esta claro si los capromidos evolucionaron a partir de un equimido ancestral 0 de un protocapromido 0 si los equimidos antillanos (I sub familia 3 generos y 7 especies todas extintas) forman parte de la irradiacion de los capromidos (4 subfamilias 9 generos y 27 especies 10 vivientes) Kratochvil et al (1978b) y Rodriguez et al (1979) consideraron que Capromys nan us es un eslab6n entre Echimyidae y Capromyidae pero Mesocapromys nanus esta morfologicamente mucho mas cerca de Hexolobodon y los de mas capromidos que de los equimidos (Woods 1985) Lo que si esta claro segun Woods et al (2001) es que la subfamilia Plagiodontinae endemic a de La Espanola es el linaje basal de los capromidos y que Isolobodon Hexolobodon Capromys Geocapromys Mysateles y Mesocapromys [y supuestamente Macrocapromys] forman parte de la irradiacion temprana de Capromyidae
La subfamilia Capromyinae (5 generos y 17 especies 9 vivientes) parece ser el dado de mas reciente evolucion en la familia con invasion de Cuba desde La Espanola y dispersion desde Cuba (1) por Capromys a las islas Caiman y (2) por Geocapromys a las islas Bahamas Caiman y Jamaica y desde Jamaica a las islas Swan aunque la presencia de Geocapromys en Swan pudiera deberse a introduccion aborigen (Morgan 1985 1988 1989 1993 1994 Woods 1989a 1990) Woods et al (2001) utilizaron datos moleculares para obtener estimados independientes de divergencia entre capromidos segiJn 10 cual Geocapromys divergio cerca de los 17 MaAP Mesocapromys divergio de Capromys hace 15 MaAP en tanto lvlysateles 10 hizo hace 10 MaAP Por su parte las formas de Capromys pilorides y de Mysateles prehensilis en Isla de la Juventud divergieron de sus correspondientes subespecies nominativas en la isla principal a los 24 y 08 MaAP respectivamente
318 BIOGEOGRAFiA
Woods y Howland (I979) describieron adaptaciones morfologicas de la mandibula el hioide y la faringe asociadas a mecanismos de masticacion diferentes en plagiodontinos y caprominos y Varona (1980a) llamo la atencion sobre el gran desarrollo de los molariformes (en relacion con la longitud total del cnineo) que presentan ciertos grupos de jutias fosiles en comparacion con las vivientes 10 que pudiera interpretarse como una adaptacion alimentaria a una vegetacion mas abrasiva por parte de las jutias extinguidas
Segun Woods (1990) la familia Heptaxodontidae (2 subfamilias 5 generos y 6 especies todas extintas) tambien seria parte de la irradiacion de los capromidos con origen en el este de La Espanola 0 en Puerto Rico y subsecuente dispersion al Banco de Anguila - San Martin en las Antillas Menores (pero vease MacPhee y Flemming 2003) Este banco estaria a solo 70 km del Banco Puerto Rico - Islas Virgenes en tiempos de maxima regresion marina y esta proximidad con Puerto Rico -patria de Elasmodontomys obliquus- seguramente tuvo que ver con la presencia en las islas de Anguila y San Martin del heptaxodontino gigante Amblyrhiza relacionado mas estrechamente con los roedores de las Antillas Mayores que con los del resto de las Antillas Menores (Woods 1990 1996) Aunque los roedores suelen aumentar de tamano en las islas (Case 1978) la escala del incremento de la talla en Amblyrhiza (que pudo alcanzar mas de 200 kg de peso adulto) esta fuera de toda proporcion con los patrones conocidos en otros roedores insulares (Biknevicius et 01 1993)
Clidomys (Clidomyinae) de Jamaica difiere marcadamente en su morfologia de los heptaxodontinos (vease ademas MacPhee y Flemming 2003)10 cual dificulta explicar su presencia en esa isla Como Jamaica estuvo bajo el mar hasta el Mioceno Medio el ancestro de Clidomys colonizaria esta isla por dispersion desde el extremo suroccidental de La Espanola despues que los heptaxodontinos se originaron en esta ultima isla (Woods 1989a) Pero Pascual et 01 (I 990) especularon que Clidomys pudiera ser el resultado de una invasion posterior e independiente de los capromidos
GRUPOS ORIGINARIOS DE NORTEAMERICA
El origen de las musaranas antillanas ha resultado ser hasta ahora el mayor enigma biogeografico de la fauna de mamiferos terrestres de la region aunque la mayoria de los autores coinciden en considerarlas originarias de Norteamerica (vease Discus ion y Conclusiones en Poblamiento Continente-Isla) No obstante MacPhee e Iturralde-Vinent (1994) descartaron toda posibilidad de conexion terrestre por el Estrecho de la Florida 0 por el Canal de Yucatan y si el (supuesto) evento vicari ante de Rosen (vease El Arca de Noe) -invocado por MacFadden (1980) para resolver el enigma- fuera minimamente plausible aun asi es dificil creer que las catastrofes que 10 sucedieron (vease Contexto Temporal) hayan barrido con toda esa vieja fauna continental excepto las musaranas circunstancia ya anticipada muy tempranamente por Matthew (l918a)
De otra parte la relaci6n entre Nesophontidae y Solenodontidae permanece irresuelta Si estos linajes no estan estrechamente relacionados entonces es muy posible que
319 IRRADIACION EVOLUCIONARIA
sus distribuciones sean explicadas por mecanismos diferentes (Whidden y Asher 2001)
La descomunal diferencia de talla corporal entre las musaranas pequenas (Nesophontes) y las grandes (Soienodon) seguramente impuso a ambos grupos restricciones distintas relativas a la termorregulacion y las adaptaciones energeticas En Nesophontes la seleccion debio favorecer una alta tasa de metabolismo y crecimiento camadas mas numerosas corta expectativa de vida yalta densidad poblacionaL En Soienodon en cambio la seleccion opero en la direccion de una baja tasa metabolica camadas reducidas crecimiento lento longevidad y baja densidad poblacional (Ottenwalder [1991])
MUSARANAS PEQUENAS (Nesophontidae)
Las musaranas pequenas estan representadas en las Antillas por el genero Nesophontes (y un genero miocenico indeterminado en ambar dominicano) con 6 especies cuaternarias extinguidas distribuidas en Cuba La Espanola Puerto Rico Islas Virgenes e Islas Caiman La mayor de las musaranas pequenas Nesophontes edithae evoluciono en Puerto Rico y su arribo a las Islas Virgenes supone una dispersion sumamente reciente En las Islas Caiman Nesophontes se asemeja mas a la poblacion de Cuba de donde seguramente derivo en el Plio-Pleistoceno (junto con Capromys y Geocapromys) cuando la costa sur de Cuba estuvo mucho mas cerca de las Islas Caiman (lturralde-Vinent 2003fig 4) MacPhee et ai (1994) interpretaron la distribucion de Nesophontes en Cuba y La Espanola como resultado de la vicarianza oligomiocenica ocurrida al formarse el Paso de los Vientos pero la distancia taxonomica entre los lados del Paso es solo de nivel especifico y N micros incluso vivi6 en ambas islas segtln Patterson (1962)
Allen (1918) considero que Nesophontes micrus fue una especie primitiva no modificada para la vida arb6rea 0 fos6rea y por tanto de habitos terrestres Nesophontes edithae de Puerto Rico es mucho mayor y mas robusta que las restantes especies del genero eL indice de robustez de su femur (= anchura minima de la diafisis ongitud total del hueso) muestra que este es tan robusto como el de Solenodon paradoxus un animal dos veces mayor (Morgan et ai 1980) Choate y Birney (1968) postularon un aumento cronoclinal de la taIla corporal en N edithae confirmado mas tarde por McFarlane (1999a) mediante fechados radiocarb6nicos de la secuencia estratignifica en depositos cavernarios de Puerto Rico Solenodon no vivi6 en Puerto Rico y en ausencia de una musarana de mayor tamano Nesophontes alcanzo una talla corporal mayor en esa isla que en La Espanola 0 Cuba (Woods y Eisenberg 1989)
MUSARANAS GRANDES (Solenodontidae)
Los arcaicos almiqufes (genero Soienodon) comprenden cuatro especies una viviente (S cubanus) y otra f6sil (S arredondoi) en Cuba y una viviente (S paradoxus con dos subespecies) y otra f6sil (S marcanoi) en La Espanola Como cabria esperar las dos especies cubanas se asemejan mas entre sf que a las especies de La Espanola pero
320 BIOGEOGRAFiA
enigmaticamente S marcanoi comparte caracteres con S cuban us y S paradoxus (Woods y Eisenberg 1989 Morgan y Ottenwalder 1993 Ottenwalder 200 I) Solenodon arredondoi alcanzo una talla corporal superior a la de cualquiera de las musarafias vivientes a nivel mundial Segun Morgan y Ottenwalder (1993) extrapolando las dimensiones y el peso de los almiquies vivientes S arredondoi debio tener una longitud cabeza-tronco y un peso de 450-550 mm y 1500-2000 g respectivamente en comparacion con S cubanus (360 mm y 800 g) y S paradoxus (390 mm y 1100 g)
Entre todas las especies de almiqufes Solenodon paradoxus es la que presenta los caracteres mas derivados (Otten walder 2001) Si estas relaciones son validas la distribucion de Solenodon entre Cuba y La Espafiola debio discurrir de oeste a este como propuso MacFadden (1980) quien invoco la presencia fosH de S cubanus [pero tambien S arredondoi] en Pinar del Rio para postular que el ancestro continental entr~ por Cuba opinion compartida por Perfit y Williams (1989) y por Ottenwalder ([1991] 2001) El paso de Solenodon a Puerto Rico pudo haberlo impedido la existencia de barreras ecologicas durante la conexion del este de La Espafiola con Puerto Rico 0 por la ausencia de Solenodon en esa parte de La Espafiola en aquella epoca Sin embargo las mismas condiciones ecologicas en el este de La Espafiola no parecen haber representado una barrera efectiva para Nesophontes cuyas mas altas tasa de reproduccion y densidad poblacional presumiblemente Ie permitieron expandirse flpidamente a traves de La Espafiola (Ottenwalder [1991])
Es opinion generalizada que los almiqufes antillanos constituyen el grupo de mamiferos que primero se emplazo en el territorio insular (MacPhee 2005) y en tal sentido es de interes la evidencia sobre especificidad parasito-hospedante aportada por Cruz (1980) quien encontro que la especie de acaro (genero Cubanochirus) que parasita a los almiquies es mucho mas primitiva que la especie (del mismo genero) que paras ita a las jutias y de aqui concluyo que dicho genero de acaro debio evolucionar primero en Solenodontidae y posteriormente en Capromyidae al entrar en contacto representantes de ambas familias en Cuba 0 La Espafiola
Recientemente Roca (2004) y Roca et al (2004) dieron a conocer que los resultados de su estudio genetico-molecular sobre los almiquies indican una divergencia de 25 Ma entre Solenodon cuban us y S paradoxus -compatible con la disyuncion tectonica entre Cuba y La Espafiola (Iturralde-Vinent y MacPhee 1999 McPhee et al 2003)-- y una separacion de ambas especies comparable en magnitud a la divergencia entre familias de mamlferos Consecuentemente propusieron que Solenodon cubanus fuera clasificada en el genero Atopogaie como indico Cabrera (1925) hace 82 afios
DISCUSION Y CONCLUSIONES
Las musarafias y los roedores antillanos (excepto Oryzomys) se irradiaron tanto de sus respectivos ancestros continentales que de estos 6rdenes se reconocen familias y subfamilias endemicas de la region en contraste con los perezosos (una subfamilia y tres
321 IRRADIACION EVOLUCIONARIA
tribus endemicas) y los monos (una tribu endemica) como expresion de diferentes niveles de diversificacion evolucionaria Este proceso es dificil de explicar (traUindose de c1ados que pnicticamente comparten la misma antigiiedad en las Antmas) si es correcta la interpretacion de Pascual et at (1990636)
Nosotros consideramos estos diferentes niveles de diversificacion como las diferentes etapas de evolucion alcanzadas por cada grupo [de mamiferos] Obviamente esto estuvo relacionado con las diferentes longitudes de sus historias
Pudiera ocurrir que en el futuro se compruebe que la historia biogeognifica de la region fue mucho mas complicada
La Espanola la isla mas ecol6gicamente divers a de la region debi6 ser el centro principal de evoluci6n de los mamfferos (con 87 de las familias y 73 de las subfamilias) seguida por Cuba (con 75 de las familias y 54 de las subfamilias) las dos islas de las Antillas Mayores mas relacionadas biogeograficamente (Fig 43 Tabla 44)
eL
EL
cb CAIMAN
SWAN
A Acratocnini B Cubanocnini C Megalocnini D Mesocnini E Nesophontidae F Solenodontidae G Xenothrichini
ANGUILAshyM-0 SMARTIN
H Heteropsomyinae I Plagiodontinae J Isolobodontinae
K Hexolobodontinae L Capromyinae
M Heptaxodontinae N CIidomyinae 0 Sigmodontinae
FIGURA 43 Representacion esquematica de las relaciones interinsulares (expresadas porel numero de linajes compartidos) entre las Antillas Mayores y entre estas y los grupos de islas perifericas segun las interpretaciones de los autores citados en el texto Los numeros dentro de los circulos indican la cantidad de linajes representados en cada isla relacion dudosa Contexto taxon6mico de los linajes en la Tabla 44
bull bull
bull bull
bull bull bull
bull bull
bull bull
bull
bull
bull
bull
bull bull bull bull
bull bull
bull bull
bull
bull bull bull
bull bull
bull
bull bull
bull
322 BIOGEOGRAFiA
TABLA 44 Distribuci6n de los linajcs de mamiferos terrestres a traves de las Antillas Mayores y grupos de islas perifericas LE = La Espanola CU Cuba JA Jamaica PR = Puerto Rico IC Islas Caiman IS Islas Swan IB = Islas Bahamas IV = Islas Virgenes AS = Anguila - San Martin
FamiliaSubfumiliafIribu LE CU JA PR IC IS IB IV AS
Megalonychidae
Choloepodinae
Acratocnini
Cubanocnini
Megalocninae
Megalocnini
Mesocnini
Nesophontidae
Solenodontidae
Pitheciidae
Callicebinae
Xenothrichini
Echimyidae
Heteropsomyinae
Capromyidae
Plagiodontinae
Isolobodontinae
Hexolobodontinae
Capromyinae
H e pta x 0 don tid a e
Heptaxodontinae
Clidomyinae
Muridae
Sigmodontinae
PILOSA
SORICOMORPHA
PRIMATES
RODENTIA
bull
bull
bull
bull TOTALES 12 10 4 4 2
323 EXTlICIOIES DEL CUATERNARIO
El gigantismo ha sido sin duda una tendencia evolucionaria recurrente entre los clados mayores de mamiferos antillanos Es un hecho bien conocido que algunos grupos de mamiferos han aumentado de tamafio en las islas y varios autores han intentado explicarlo a partir de enfoques diferentes (Foster 1964 Miller 1966 Sondaar 1977 Case 1978 Eisenberg y Wilson 1978 Heaney 1978 Lawlor 1982 Lomolino 1985 Angerbjom 1985 McNab 19942001)
EXTINCIONES DEL CUATERNARIO
Las islas que integran la Subregion Antillana han perdido casi 90 de sus mamiferos terrestres durante los ultimos milenios y en proporcion con su extension territorial esta perdida ha sido mayor en la Subregion que en cualquier otro lugar del planeta (MacPhee 1996b) Las extinciones del Cuatemario eliminaron mas de las tres cuartas partes de la fauna de mamiferos terrestres conocida de las Antillas Mayores sus islas satelites y grupos de islas perifericas y el examen de este proceso en sus aspectos mas generales se aborda en esta ultima parte del presente capitulo
CRONOLOGiA Y CONSECUENCIAS
Es muy improbable que un fosil represente el ultimo individuo de un taxon extinto y entonces hay que guiarse por la edad radiometrica del fosil mas reciente conocido lIamada fecha de ultima presencia- como unica aproximacion posible a la fecha de extincion del taxon
Como se explico al inicio del capitulo (vease Contexto Temporal) el registro fosH de los mamiferos terrestres de las Antillas Mayores adolece de insuficiente sustentacion radiometrica No obstante la distribucion temporal de los escasos fechados disponibles para el Cuaternario de la region abarca etapas criticas del proceso de extincion experimentado por la referida fauna a 10 largo de los ultimos 250 mil afios
De las 52 especies que es posible reconocer actualmente como miembros extintos de la fauna cuatemaria de mamiferos terrestres de las Antillas Mayores y grupos de islas perifericas los restos fosiles de 23 de elIas (442) han sido fechados radiometricamente o se han hallado asociados estratigraficamente de manera inequivoca con otro material fechado De estos 23 fechados 3 corresponden al interglacial Sangamon 4 pertenecen a la secuencia Pleistoceno terminal - Holoceno temprano 10 se ubican en ellapso entre el arribo de los aborigenes y la incursion europea y 6 son postcolombinos (a los cuales hay que sumar otras dos especies extinguidas en los siglos XIX y xx) A partir de estas 25 especies avaladas por fechas de ultima presencia distribuidas entre los cuatro intervalos temporales sefialados hemos compuesto la Tabla 45 en la que ademas aparecen las restantes 27 especies (carentes de fechado radiometrico) ubicadas en el intervalo temporal mas probable sobre la base de (1) la asociacion estratigrafica de sus restos con evidencias culturales (amerindias 0 europeas) 0 (2) la ausencia de dicha asociacion Sobre estos criterios de ubicacion temporal procede hacer dos aclaraciones
324 BIOGEOGRAFIA
TABLA 45 Intervalos temporales mas probables para las extinciones cuatemarias de los mamiferos terrestres de las Antillas Mayores y grupos de islas perifericas sobre la base de las evidencias siguientes (l) ultimo ejemplar colectado u observado en vida (2) fechado radiometrico mas tardio en material de la especie (3) asociaci6n estratigrafica mas tardia con otro material fechado (4) asociaci6n estratigrafica con restos de animales introducidos restos humanos u objetos culturales propios del intervale (5) falta de asociaci6n con evidencias culturales [Los ntimeros entre parentesis en la columna Evidencia significan adscripci6n tentativa al intervalo] Los estimados del peso corporal (en kg) estan tomados de la literatura 0 inferidos comparativamente y distribuidos entre
las clases siguientes (a) S 01 (b) 01 a lt 1 (c) 1 a lt 10 (d) 10 a lt 100 (e) 100 a lt 200 (j) ~ 200 Las
abreviaturas que designan las islas son las de la Tabla 44
F amilialEspecie Peso IsJa Evidencia Fuente
I httervalo htterg1acial Sangamon (gt80 kaAP)
HEPTAXODONTlNAE
Amblyrhiza inundata e AS 3 McFarJane et al 1998a
CLIDOMYINAE
Clidomys osborni d JA 3 Ibid
Clidomys parvus d JA 3 Ibid
II httervalo Pleistoceno terminal ~ Holoceno temprano (20-6 kaAP)
MEGALONYCHIDAE
Aeratoenus antillensis d CD 5 Arredondo 1975 a
Aeratoenus odontrigonus d PR 3 McFarJane 1999b
Aeratoenus simorhynehus d LE 2 Rega et al 2002
M egaloenus zile f LE 2 MacPhee et al 2000
SOLENODONTIDAE
Solenodon arredondoi e CD 5 Morgan y Ottenwalder 1993
PnHECIIDAE
Paralouatta varonai d CD 5 MacPhee 1996a
EcHIMYIDAE
Heteropsomys antillensis c PR 5 McFarJane 1999b
Heteropsomys insulans c PR 5 Ibid
Puertoricomys eorozalus c PR 5 Woods 1989a
CAPROMYIDAE
P lagiodontia araeum c LE 5 MacPhee et al 2000
Macrocapromys latus c CD 5 Varona y Arredondo 1979
(Continua)
325 EXTINCIONES DEL CUATERNARIO
TABLA 45 (Continuaci6n)
F ami1ia1Especie Peso Isla Evidencia Fuente
M acrocapromys latus c CU 5 Varona y Arredondo 1979
Mesocapromys kraglievichi c CU 5 Ibid
HEPTAXODONTINAE
Xaymaca fulvopulvis a JA 2 MacPhee y Flemming 2003
III Intervalo Amerindio (6-05 kaAP)
MEGALONYCHIDAE
Acratocnus ye d LE 2 MacPhee et aI 2000
Neocnus comes c LE 2 Ibid
N eocnus dousman c LE 2 Ibid
N eocnus toupiti c LE 2 Ibid
N eocnus gliriform is c CU 4 Cordova et ai 1997
M egalocnus rodens f CU 2 MacPhee et ai 2007
Parocnus browni e CU 2 Steadman et ai 2005
P arocnus serus e LE 2 MacPhee et ai 2000
N ESOPHONTIDAE
N esophontes edithae a PR 2 McFarlane 1999b
PITHECIIDAE
Antillothrix bernensis c LE 3 Rimoli 1977
EcHIMYIDAE
Brotomys contractus a LE 4 Miller 1929c
CAPROMYIDAE
P lagiodontia spelaeum c LE 4 Ibid
M acrocapromys acevedo c CU 4 NUiiez 1990a
HEPTAXODONTINAE
Elasmodontomys obliquus d PR 3 McFarlane 1999b
Iv Intervalo Postcolombino (05 kaAP - Presente)
N ESOPHONTIDAE
N esophontes hypom icrus a LE 24 MacPhee y Fle111111ing 1999
N esophontes zam icrus a LE (4) Ibid
N esophontes m icrus a CU 24 MacPhee et aI 1999
(Continua)
326 BIOGEOGRAFiA
TABLA 45 (Continuacion)
F amilialEspecie Peso Isla Evidencia Fuente
N esophontes m icrus a CD 24 MacPhee et aI 1999
N esophontes m icrus a LE 4 MacPhee et aI 2000
N esophontes sp 1 a IC 4 Morgan 1994
N esophontes sp 2 a IC 4 Ibid
Solenodon marcanoi b LE 4 Ottenwalder 2001
PITHECIIDAE
Xenothrix mcgregori c JA 4 MacPhee y Flemming 1999
EcHIMYIDAE
Boromys offeUa a CD 24 Guarch 1984
Boromys torrei a CD 24 Ibid
Brotomys voratus a LE 4 MacPhee et aI 2000
CAPROMYIDAE
P lagiodontia ipnaeum c LE 4 MacPhee y Flemming 1999
Rhizoplagiodontia lemkei c LE 4 Ibid
Isolobodon portoricensis c PR 24 Ibid
Isolobodon montanus c LE 4 Ibid
Hexolobodon phenax c LE (4) Ibid
Capromys sp c IC 24 Morgan 1994
Geocapromys columbianus c CD 4 MacPhee y Flemming 1999
Geocapromys thoracatus b IS 1 MacPhee y Marx 1997
Geocapromys sp 1 c IC 4 Morgan 1994
Geocapromys sp 2 c IC (4) Ibid
l-IEPTAXODONTINAE
Quemisia gravis c LE (1) MacPhee y Flemming 1999
MURIDAE
Oryzomys antillarum a JA 1 Ibid
1 Flemming y MacPhee (1999) alertaron en contra de la asociacion con otro material fechado Asimismo Morgan (1993) indico que la ausencia de asociacion con evidencias culturales amerindias no es necesariamente un criterio concluyente para ubicar un taxon en el intervalo temporal anterior porque hay depositos en la region que carecen de tales evidencias y son del Holoceno tardio por fechados radiocarbonicos [No obstante
327 EXTlNCIONES DEL CUATERNAR10
la ausencia de una especie en residuarios arqueol6gicos adquiere mayor certidumbre como criterio de ubicaci6n temporal de su extinci6n en la medida que aumenta el numero de residuarios estudiados que en las Antillas Mayores se cuentan por centenares] A falta de fechado absoluto de una especie consideramos que estos criterios de ubicaci6n temporal son mas utiles que la alternativa de excluir del analisis a la especie
2 En los trabajos sobre biogeografia de mamiferos antillanos los arque610gos mas citados como autoridades para establecer el inicio de la presencia aborigen en las Antillas Mayores (Kozlowski 1974 Rouse y Allaire 1978 Rouse 1989 1992 Wilson 2001) difieren al respecto entre los 6 kaAP y los 45 kaAP (pero vease Pagan et al 200523) y hemos adoptado la mas temprana de estas fechas para el comienzo del intervalo amerindio en la Tabla 45 De todos modos cualquiera que haya sido la fecha esta no es aplicable por igual al conjunto de las Antillas Mayores porque segun algunos de los autores citados el poblamiento comenz6 por Cuba y La Espanola 6 mil anos atras continu6 en Puerto Rico hace alrededor de 3 mil anos (pero vease Burney et al 1994) yen Jamaica hace menos de 2 mil e incluso a las Islas Caiman ni siquiera llegaron [Las Islas Caiman descubiertas por Col6n en 1503 carecen absolutamente de evidencia amerindia (Stokes y Keegan 1996)]
Si los datos de la Tabla 45 representan una aceptable aproximaci6n a la realidad el proceso de las extinciones cuaternarias de los mamiferos terrestres de las Antillas Mayores y grupos de islas perifericas debi6 caracterizarse entre otros por los eventos siguientes
Aunque obviamente la fauna cuaternaria debi6 ser mucho mas numerosa que 10 que refleja el registro paleontol6gico (Morgan y Woods 1986) 52 especies (81 de la fauna conocida) desaparecieron (sin reposici6n) durante los ultimos 250 mil anos
- Al parecer las extinciones se concentraron en las especies de mayor tamano Asi el peso promedio estimado para las especies de la etapa preantr6pica (intervalos I + II de la Tabla 45) es 350 kg es 328 kg para las especies del intervalo III (amerindio) y es s610 25 kg (c1ases del peso a b c) para las especies del intervalo IV (postcolombino) Ninguna especie aut6ctona de mamifero mayor de 6 kg existe hoy en la regi6n
- De acuerdo con la distribuci6n temporal de las extinciones 61 especies sobrevivieron al interglacial Sangamon (intervalo 1)48 sobrevivieron a la transici6n del Pleistoceno terminal al Holoceno temprano (intervalo II) 34 sobrevivieron al periodo amerindio (intervalo III) y s610 12 especies han sobrevivido desde la Ilegada de los europeos (intervalo IV) Las tasas de extinci6n correspondientes (exceptuado el intervalo I) son una especie cada 1167 anos en el intervalo II una cada 393 anos en el intervalo III y una cada 23 anos en el intervalo IV
- Durante todo el proceso de las extinciones cuaternarias la perdida de especies por isla fue de 91 en La Espanola 83 en Jamaica y 62 en Cuba Bahamas retuvo su unica especie (aunque extirpada en varias de las islas) Puerto Rico e Islas Caiman perdieron todas sus especies (6 y 5 respectivamente) y las Islas Swan Virgenes y Anguila - San Martin perdieron sus respectivas especies (micas
328 BIOGEOGRAFiA
De los cuatro 6rdenes de mamiferos representados en la fauna Pilosa y Primates desaparecieron totalmente Soricomorpha y Rodentia perdieron 86 y 74 de sus especies respectivamente
CAUSALIDAD
CAUSAS NATURALES
Los cicIos glaciales e interglaciales acaecidos en el Pleistoceno causaron cambios ambientales dnisticos en las Antillas con la concomitante alternancia de cIimas frios (secos) y calidos (hUmedos) asi como de descensos y ascensos del nivel del mar aumento y disminuci6n de las areas territoriales) induso (aunque con menor intensidad) en el Holoceno Estos eventos paleogeograficos han sido consistentemente documentados (Mayo y Kartashov 1972 Bonnatti y Gartner 1973 Ortega y Arcia 1982 Prance 1982 Schubert y Medina 1982 Dodge et al 1983 Ortega [1984] Haq et al 1987 Schubert 1989 Hodell et al 1991 Guilderson et al 1994 Oppo 1997 Emslie 1998 Curtis et al 2001 Iturralde-Vinent 2003) y de ellos brindamos a continuaci6n una breve resena
Finalizando el periodo interglacial anterior a la ultima glaciaci6n (Wisconsin) el mar estaba a 8-10 m por encima de su nivel actual Cuando el avance glacial del Wisconsin a1canz6 su culminaci6n (eillamado Ultimo Maximo Glacial) hace 18 ka el mar habia descendido 120 m con respecto a dicho nivel y las islas del Caribe habian incrementado considerablemente sus extensiones subaereas en comparaci6n con las que presentan hoy dia por ejemplo Puerto Rico y las Islas Virgenes se conectaron territorialmente y las Bahamas se unieron en cinco gran des islas Prevalecia un regimen frio y seco y condiciones de aridez --debidas a una reducci6n de las precipitaciones (posiblemente a menos de 440 mmlano)- y los ambientes xericos se habian extendido a traves de las islas y bancos emergidos y abundaban los pinares (los habitats mesicos estaban restringidos a parcelas relictas en 10 alto de las montanas) En el Mar Caribe y el Golfo de Mexico la temperatura superficial del mar era 5degC mas baja que la actual y en las llanuras costeras y alturas maximas eran respectivamente 2-3degC y 6degC mas bajas Los limites de las zonas ecol6gicas altitudinales descendieron varios cientos de metros y cada zona redujo su extensi6n vertical al aumentar el gradiente de temperatura Ellimite inferior de la zona periglacial
elevaci6n donde la temperatura media minima a1canza los OdegC) de la Cordillera Central de la Republica Dominicana (altura maxima del Caribe insular con evidencia de topografia glacial) se considera que estuvo entre los 1500 y 1800 mlnm (actualmente esta a 2800 mlnm) y se puede estimar que dicho limite era inferior en Cuba por estar mas al norte que la Republica Dominicana No obstante durante este periodo la temperatura experiment6 episodios cortos (unas pocas decadas) de ascensos repetidos
Un signifieativo componente bi6tieo de las referidas condiciones paleoambientales 10 constituy6 la espectacular irradiaci6n evolucionaria experimentada por un grupo numeroso de aves rapaees diurnas y nocturnas (Wetmore 1937 Brodkorb 1969 Kurotchkin y Mayo 1973 Arredondo 1976 Olson 1978 Olson y Hilgartner 1982
329 EXTINCIONES DEL ClJATERNARIO
Suarez 2000a b 2004 Suarez y Emslie 2003) Los mamiferos de las Antillas Mayores ofrecieron a estos depredadores aviares una amplia gama de tamanos de presa que en ausencia de mamiferos del orden Carnivora en la region permitio a estas aves irradiarse a este nicho alimentario con la evolucion de una variedad de especies de muy diversa tall a corporal incluidas formas gigantes Adquirir un cuerpo gigante les propicio el acceso a recursos alimentarios que no estaban siendo explotados por ningun otro depredador (Alcover y McMinn 1994) A Cuba corresponde el mejor registro fosil de rapaces cuaternarias caribenas con siete generos de especies diurnas Dentro de estas aves existieron tres gavilanes gigantes (Accipitridae) tres carairas (Falconidae) y un condor (Cathartidae) asi como cuatro generos de especies nocturnas dos lechuzas (Tytonidae) y tres Mhos (Strigidae) dos de ell os no igualados en tamano en todo el mundo Algunos de estos generos tuvieron representacion en otras islas antillanas como La Espanola Jamaica Bahamas y Gran Caiman
Al retirarse hacia el norte los hielos de la glaciacion Wisconsin el clima se tomo mas hUmedo en las Antillas durante el Holoceno temprano y el bosque mesico se expandio en la region insular El nivel del mar ascendio acompanado de mayor pluviosidad alrededor de los 7 kaAP las Islas Virgenes quedaron separadas entre si y de Puerto Rico y las Bahamas perdieron la mayor parte de su territorio Cerca de los 3 kaAP las condiciones secas retornaron a la region e hicieron desaparecer buena parte del bosque mesico Nuevamente el clima comenzo a adquirir sus condiciones actuales hacia los 25 kaAP aproximadamente y hoy transita por una fase mesica con vegetacion xerica confinada en parcelas relictas
Sin duda estos y otros eventos naturales incidieron negativamente en la sobrevivencia de mamiferos terrestres en las Antillas Mayores asi como en los grupos de pequenas islas perifericas (Pregill y Olson 1981 Olson 2006)
McFarlane et al (1998a) demostraron convincentemente como Amblyrhiza el gigante de los roedores antillanos endemico de la pequena isla de Anguila sucumbio a las repetidas y abruptas perdidas de su habitat durante el interglacial Sangamon por las nipidas y severas oscilaciones del nivel del mar (documentadas por Neumann y Hearty 1996) al no poder responder adecuadamente a tan catastroficos eventos mediante el ajuste de su masa corporal media y otro tanto debio ocurrirle en Jamaica al otro roedor gigante Clidomys (McFarlane et al 1998b) emparentado con Amblyrhiza en la misma epoca (Tabla 45 intervalo I)
El ascenso del nivel del mar seguramente promovio la extincion de mamiferos durante la transicion del Pleistoceno al Holoceno (Tabla 45 intervalo II) con la reduccion del tamano de las Islas por las transgresiones marinas (Pregill y Olson 1981 Morgan 1993) Esto ultimo haria maximamente severa la competicion asf como la presion de los depredadores [Steadman et al (2005) no yen por que el calentamiento global ocurrido durante la transicion Pleistoceno-Holoceno habria de tener un efecto negativo sobre los perezosos siendo sus descendientes actuales especies tropic ales 0 subtropicales pero no se trata del efecto
330 BIOGEOGRAFiA
directo del cambio de temperatura sobre los animales sino del efecto indirecto y progresivo que dicho cambio genera a largo plazo al reducir el territorio insular disponible]
Por esta epoca tambien desaparecieron especies de tall a mayor como Megalocnus zile en La Espanola y otras de menor tamano (en Cuba) pero que representan la tall a maxima en sus respectivos contextos taxonomicos del Cuaternario como Solenodon arredondoi Paralouatta varonai y Macrocapromys acevedo Un mamifero puede ser grande solo a costa de reducir su densidad poblacional y esto 10 hace mis vulnerable a la extinci6n sobre todo en una isla (McNab 1994 McFarlane et al 1998a) Incidentalmente Alcover et al (1998a) afirmaron que larelacion entre la talla corporal y la extincion no siempre se cumple en las especies de menor tamano y citaron como ejemplo que entre las musarafias todas las especies de Nesophontes desaparecieron mientras sobreviven especies de Solenodon pero la comparacion correcta en todo caso serfa entre los solenodones vivientes y el gigante Solenodon arredondoi y entre Nesophontes edithae -el gigante de los nesofontes extinguido en el periodo amerindio- y el resto de sus congeneres que sobrevivieron hasta los primeros siglos de la colonizacion europea (Tabla 45 intervalos III y IV)
CAUSAS ANTROPOGENICAS
Intervalo Amerindio
La mayoria de los autores que se han referido a las extinciones acaecidas durante este intervalo (entre elIos paradigmaticamente Martin 1984a b) han responsabilizado a los aborfgenes (que se supone comenzaron a poblar las Antillas Mayores hace 6 mil afios) con la desaparicion de numerosas especies de mamiferos terrestres en la region principalmente por la via de la sobrecaza y la introduccion de animales domesticos devenidos depredadores de la fauna autoctona (Morgan y Woods 1986) Este efecto negativo de la presencia indigena sobre los mamiferos de la region ha sido dramatizado por algunos autores en terminos de holocausto faunal Sin embargo pensamos que se ha exagerado Como han expresado MacPhee et al (19993) cuando uno busca evidencia dura de complicidad humana directa en [estas] extinciones no hay mucho que encontrar Las siguientes son algunas consideraciones a tener en cuenta en tal sentido
El registro radiometrico mas joven conocido hasta ahora para un perezoso antillano (MegalocnllS rodens) es de 42 kaAP (MacPhee et ai 2007) Si se acepta que los aborfgenes entraron en Cuba hace aproximadamente 6 mil anos su convivencia con los perezosos debe haber durado alrededor de 1800 afios (0 mas porque dificilmente el referfdo fechado radiometrico haya correspondido al ultimo sobreviviente de la especie) Esta cifra contradice la hipotesis del holocausto faunal segun la cualla extincion de los mamiferos insulares de mayor talla debe ocurrir con extrema rapidez -a escala de decadas 0 de centurias- a partir de su primer contacto con los humanos (Martin y Steadman 1999) A pesar de las repetidas afirmaciones en contrario (Harrington 1921 Miller 1929c Herrera [1945] Tabio 1951 Alvarez 1951a 1957 Arredondo
331 EXTINCIONES DEL CUATERNARIO
1959 1960a Veloz y Ortega 1973 Mayo 1980b Rodriguez et al 1984 Castellanos y Pino 1985 Pino y Castellanos 1985 Nunez 1990b Gutierrez 1996 Arredondo 2004) en su mayoria basadas en evidencias no exentas de posibles accidentes de deposicion (0 de reubicacion postdeposicional) en recintos cavernarios (Diaz-Franco [2006b D no se ha demostrado inequivocamente que los aborigenes antillanos se alimentaran de perezosos regularmente Sus restos en residuarios de alimentos son escasisimos en toda la region y esto contrasta con los abundantes restos de jutias y otros mamiferos de menor talla que aparecen en esos residuarios demostrativos de la preferencia de los paleoindios por estos otros mamiferos
Los mamiferos mas importantes en la economia prehistorica de las Antillas Mayores fueron las jutias Capromys y Geocapromys en Cuba Geocapromys en Jamaica e Isolobodon en La Espanola Puerto Rico e Islas Virgenes (Aguayo 1950b Tablo 1951 Rimoli 1972 Wing 2001 Diaz-Franco [2006b D Wilkins (2001) estudi61ajutia viviente de Jamaica (Geocapromys browni) en un sitio ocupado por los aborigenes durante 100 anos y no encontr6 evidencias de que durante ese tiempo la especie haya sido sobreexplotada Asimismo en Cuba las jutias conga carabali y andaraz son escasas en depositos pleistoholocenicos en cambio son muy numerosas en residuarios indigenas y de haber sido sobreexplotadas 10 mas probable es que no hubieran sobrevivido hasta hoy
La habilidad de los humanos para penetrar en profundidad el bosque tropical y mantenerse en este sustentablemente ha sido un evento comparativamente reciente (Beran y Steele 1994) En el caso antillano la caza tierra adentro ha sido explicada como un proceso tardio de desplazamiento a partir de la explotacion de abundantes recursos marinos que ofrecian condiciones 6ptimas para el sedentarismo de las comunidades aborigenes costeras (C6rdova y Arredondo 1988 Brito 2005) Peces y mariscos dominan la arqueologia costera de las Islas antillanas en tanto el consumo de recursos terrestres (mamfferos incluidos) aumenta progresivamente en la medida que los residuarios estudiados se alejan de la costa (Scudder 1991 Wilkins 2001) En las Antillas Mayores el componente terrestre de la dieta aborigen es solo 20-30 del material recobrado en los residuarios (Wing 1977 1989) Y debio ser menor alin porque el componente vegetal de la dieta no se ha preservado Por otra parte los aborigenes cubanos domesticaron la jutia conga y seguramente la reprodujeron para comer (Pose et al 1988) practica que probablemente se repitiera con otras jutias en las demas Antillas Mayores
- Fray Bartolome de las Casas hablo de 3 millones de indios en Cuba al inicio de la colonizacion pero los historiadores subsecuentes consideraron exagerada la cifra (Poey 1851 a) No obstante se dispone de dos estimados de la densidad de poblaci6n aborigen en las Antillas Mayores a la llegada de los europeos 18 indigenaslkm2 en Puerto Rico (Rouse 1956) y 1271km2 en La Espanola (Watts 1987) Extrapolando ambos valores entre las cuatro islas principales de las Antillas Mayores tenemos que en la fecha indicada la poblaci6n aborigen pudo estar entre 16 mil y 113 mil indigenas en Puerto Rico entre 21 mil y 145 mil en Jamaica entre 141 mil y 1 mill6n en La Espanola y entre 200 mil y 14 millones en Cuba Entre las cifras maximas y las minimas nos inclinamos
332 BIOGEOGRAFiA
hacia estas ultimas atendiendo a los altos indices de mortalidad infantil y morbilidad que se evidencian en las osamentas de los pobladores prehispanicos (Martinez y Rives 1990) Uno de los hechos que apuntan hacia el bajo numero de aborigenes en las Antillas Mayores es 10 rapido que desaparecieron despues de la llegada de los europeos pnkticamente sin dejar mestizaje con estos 10 cual contrasta ostensiblemente con la situacion en la America continentaL
En suma un grupo humano (1) demograiicamente irrelevante y controlado por altos indices de morbilidad (2) consumidor mayoritariamente de organismos marinos y (3) carente de la tecnologia cinegetica necesaria para poder sobreexplotar los mamiferos que Ie sirvieron de alimento (a los cuales accedio tardiamente en su desarrollo) no es concebible que haya causado e1 holocausto faunal que se Ie atribuye a partir del error de aplicar a las comunidades aborigenes antillanas un modelo cultural de la prehistoria continentaL
Tampoco hay evidencias de que los aborigenes hayan introducido carnivoros con suficiente capacidad depredadora salvo el perro Morgan y Otlenwaldder (1993) consideraron que la depredacion por perros durante el periodo amerindio es la explicacion mas plausible para la extincion del almiqui gigante de Cuba pero como se puede observar (Tabla 54) Solenodon arredondoi aparece en el intervalo II por falta de asociacion con evidencias culturales
Consecuentemente el accionar de los aborigenes y de sus animales domesticos directamente sobre la fauna no parece haber sido por sf solo un factor determinante de la extincion de mamiferos terrestres durante el periodo amerindio en las Antillas (pero vease Steadman et al 2005)
Hay que recordar en este senti do que alin en el Holoceno tardio ocurrieron cambios clirruiticos de consideracion (vease Causas Naturales) Por ejemplo Curtis et al (2001) explicaron que la extincion de algunos ani males mesofilicos de las alturas montanosas de La Espanola pudo haber antecedido a cualquier impacto humano sigrIificativo en esos lugares por coincidir dicha extincion con una tendencia a la aridez documentada isotopicamente y con evidencias de la perdida del bosque mesico ocurridas despues de 3kaAP Yen Jamaica hace solo 2shy3 mil aDos el area de la Bahia de Jackson sustentaba una gran biomasa de vertebrados en bosque hinnedo a juzgar por la cantidad y diversidad de taxones presentes en material fosil derivado de la depredacion por lechuzas hoy el area es la mas seca de la isla con matorral xerofitico y pobreza de fauna (MacPhee 1996a) Yalm en nuestros dias huracanes de gran intensidad aniquilan poblaciones de vertebrados (que no se recuperan) en is las pequenas y llanas del Caribe (Brooks y Smith 2001)
En medio de la incertidumbre sobre la verdadera causa (0 causas) de las extinciones de mamiferos en el periodo amerindio MacPhee y Marx (1997) han propuesto su hipotesis de la hipermorbilidad la cual estipula colapsos poblacionales relativamente rapidos con la introduccion de enfermedades de alta letalidad Estos autores han razonado que
Las islas son los lugares mas apropiados para experimentar perdidas catastroficas causadas por enfermedades Los mamiferos no solo serian susceptibles en virtud de su prolongado aislamiento biologico sino que en los confines de una isla la epizootia
333 EXTINCIONES DEL CUATERNARIO
tendria la potencialidad de infectar simultaneamente a la rnayoria de los individuos de vadas especies Los neonatos son inmunologicamente inmaduros y sufririan desproporcionadamente cualquier infeccion hipervirulenta La perdida excesiva de neonatos y juveniles tendria efectos severos para la sobrevivencia de la poblacion si esta se reduce por debajo de cierto nivel sobreviene la extincion porque no hay posibilidad de restitucion en el periodo postpanzootico Las especies de mayor talla corporal van a ser las mas vulnerables a la extincion dadas sus bajas tasas reproductivas largos periodos de gestacion y reducido numero de progenie Las especies de menor tamano --caracterizadas por altas tasas reproductivas periodos cortos de gestacion y mayor numero de neonatos por parto- tambien serian fuertemente diezmadas pero muchas sobrevivirian porque su estrategia reproductiva les permite perder mayor numero de individuos Las perdidas deben ocurrir relativamente rapido (a escala de decadas) mientras que otras hipotesis (digamos los cambios climaticos) permiten intervalos mucho mas largos (a escala de siglos 0 de milenios) entre la aparicion de las causas y la produccion de sus efectos Por ejemplo todo indica que los nesofontes se extinguieron muy rapidamente y seria dificil poder invocar un impacto humano directo 0 argumentar fenomenos climaticos que pudieran explicar plausiblemente perdidas tan rapidas
MacPhee y Marx (1997 191) han dicho que por el momento la hipotesis de la hipermorbilidad no pasa de ser una abstraccion en espera de adecuada sustentacion empirica Llama la atencion que linicamente en el periodo amerindio (Tabla 45 intervalo 1Il) las extinciones abarcaron todas las clases del peso corporal (de la a a laj) y que ninguna de las especies que sobrevivieron hasta el periodo postcolombino (Tabla 45 intervalo IV mas las especies vivientes) sobrepasa los 10 kg (clases del peso a b c) en clara coincidencia con los presupuestos de la hipotetica hipermorbilidad
Intervalo Postcolornbino
Por 10 menos 37 especies de mamiferos terrestres vivian en las Antillas Mayores y grupos de Islas perifericas a la lIegada de los europeos en 1492 De elIas se han extinguido 23 shymas especies que en cualquiera de los periodos precedentes (Tabla 45~ y solo subsisten 12 especies repartidas entre Cuba (8 especies) La Espanola (2) Jamaica (1) y Bahamas (1) varias de estas en peligro critico de extincion Solo dos de las especies extinguidas durante los ultimos 500 anos se conocieron en estado viviente los roedores Oryzomys antillarum de Jamaica (extinguido en 1877) y Geocapromys thoracatus de las Islas Swan (extinguido en 1955) Las causas de las extinciones ocurridas durante este periodo son sobradamente conocidas destruccion de habitats introducci6n de animales exoticos (domesticos y comensales) y demas presiones asociadas a la actividad humana entre las cuales posiblemente habria que incluir brotes de hipermorbilidad
Durante mucho tiempo se ha afirmado la posibilidad de que especies de Nesophontes hayan sobrevivido hasta las primeras decadas del siglo xx y que incluso continuen existiendo en lugares remotos de Cuba 0 La Espanola (Allen 1918 1942 Miller 1930 Aguayo 1950a Arredondo 1960b Fischer 1977 MacFadden 1980 Woods y
334 BIOGEOGRAFiA
Ottenwalder 1991) Actualmente no existe evidencia radiometrica de que alguna especie de Nesophontes haya sobrevivido al siglo xv las fechas radiocarbonicas calibradas obtenidas en muestras de Nesophontes de Cuba y La Espanola resultaron no mas tardias que los siglos XIII y XIV (MacPhee et al 1999) Por otra parte las expediciones efectuadas a fines del siglo xx con el proposito de localizar individuos vivientes del genero en La Espanola no produjeron la menor indicacion afirmativa al respecto (Woods et al 1985) Ademas a traves de las Antillas Mayores no aparecen restos de Nesophontes en los depositos formados actualmente por las lechuzas
EPiLOGO
Las Antillas Mayores son islas oceanicas no han formado parte de un continente (con la posible excepcion de una parte de Jamaica hasta el Eoceno Medio hace 45 Ma) y durante un tiempo geologico considerable (desde el Eoceno Superior hace 40 Ma) han permanecido emergidas y separadas de los continentes vecinos Consecuentemente los mamiferos terrestres (no volantes) autoctonos de estas islas constituyen una fauna oceanica que por ende cumple con todas las caracteristicas convencionales reconocidas para este tipo de entidad biogeografica
- Baja diversidad taxonomica a nivel de las categorias superiores (solo cuatro ordenes) en comparacion con islas plataformicas continentales (0 con areas continentales) de tamano y diversidad ecologica simi lares
- Alto grado de endemismo a nivel de genero (96) y familia (50) en respuesta al prolongado aislamiento
- Desbalance ecologico de la estructura trofica ausencia de mamiferos camivoros y ungulados
- Espectaculares procesos de irradiaci6n evolucionaria como resultado de la colonizacion de territorios libres de competidores y depredadores
Tendencias evolucionarias extremas gigantismo
- Control a cargo de aves rapaces con tendencia at gigantismo en ausencia de mamiferos miembros del orden Carnivora
- Mas especies de reptiles y de murcielagos que de mamiferos terrestres relaciones que en los continentes se invierten
Alta tasa de extincion de especies (84) sin reposicion
Preservacion de especies filogeneticamente arcaicas (los llamados fosiles vivientes) los almiquies

297 ESPACIO Y TIEMPO
TABLA 4 1(Continuacion)
I s a Taxonomia 1
CD JA LE PR IS IC IE IV AS
Paralouatta marianae bull Paralouatta varonai bull Xenothrix mcgregori bull
Rodentia (roedores) 40 especies
ECHllVlYIDAE 2
HETEROPSOMYINAE
Boromys offella bull Boromys torrei bull Brotomys contractus bull Brotomys voratus bull Heteropsomys antillensis bull Heteropsomys insulans bull Puertoricomys corozalus bull CAPROMYIDAE 4
PLAGIODONTINAE
Plagiodontia aedium 6 bull P lagiodontia araeum bull P lagiodontia ipnaeum bull Plagiodontia spelaeum bull Rhizoplagiodontia lemkei bull ISOLoBODONTINAE
Isolobodon portoricensis 7 bull Isolobodon montanus bull Zazamys veronicae bull HExOLOBODONTINAE
Hexolobodon phenax bull CAPROMYINAE
Capromys pilorides 67 bull Capromys sp bull
(Continua)
298 BIOGEOGRAFIA
TABLA 4 1 (Continuacion)
I s a s Taxonom1a 1
CU JA LE PR IS IC IB IV AS
Geocapromys browni 7
Geocapromys columbianus bull Geocapromys ingrahami 6
Geocapromys thoracatus
Geocapromys sp 1
Geocapromys sp 2
Macrocapromys acevedo
Macrocapromys latus
Mesocapromys angelcabrerai
Mesocapromys auritus
Mesocapromys kraglievichi
Mesocapromys nanus
Mesocapromys sanfelipensis
Mysateles melanurus
Mysateles prehensilis 6
bull bull bull bull bull bull bull bull bull
HEPTAXODONTIDAE 8
HEPTAXODONTINAE
Amblyrhiza inundata
Elasmodontomys obliquus
Quem isia grav is
Xaymacafulvopulvis 9
CLIDOMYlNAE
Clidomys osborni
Clidomys parvus
MURIDAE2
SIGMODONTlNAE
Oryzomys antillarum 10
bull
bull bull bull
bull
bull
bull bull
bull bull
bull
bull Total de especies por is1all 24 6 25 7 1 5 1 1 1
1 La taxonomia sigue las revisiones sistematicas mas recientes (Morgan 1993 1994 MacPhee y Grimaldi 1996 Horovitz y MacPhee 1999 White y MacPhee 2001 Ottenwalder 2001 Woods et aI 2001 MacPhee y Flemming 2003) con 1a adici6n de los taxones descritos posteriormente y la incorporaci6n de los cambios propuestos para las especies cubanas en el Capitulo 2 de este Compendio (Continua en pagina opuesta)
299 ESPACIO Y TIEMPO
CONTEXTO TEMPORAL
La absoluta mayoria del registro fosil de mamfferos terrestres antillanos proviene de depositos cavemarios pleistoholocenicos (escala del tiempo geol6gico en la Tabla 42) Abundan en estas cavernas restos de perezosos musarafias y roedores siendo comparativamente escasos los de monos debido quizas a la universal predileccion de los platirrinos por los ambientes densamente forestados 10 que los hace malos candidatos a la fosilizacion (MacPhee y Woods 1982)
A pesar de 10 numeroso de tales depositos en la region son escasos los fechados radiometricos del cuantioso material extraido Con excesiva frecuencia (particularmente en Cuba) estos depositos han sido asignados al Pleistoceno Superior sin fundamento suficiente para ello y es sabido que incluso en cuevas contiguas los depositos fosilfferos pueden diferir considerablemente en edad (Pregill y Olson 1981) Se superponen a esta inconveniencia la falta de estudios tafonomicos en las excavaciones y la eventual mezcla de f6siles de distintas edades como consecuencia de las multicausales alteraciones estratigraficas ocurridas en las cuevas (Nunez 1990a b Morgan y Ottenwalder 1993 MacPhee 1997 McFarlane 1999b Dfaz-Franco 2004) Sobre propuestas de tipificaci6n de los depositos paleontologicos con restos de mamiferos terrestres en Cuba vease Woloszyn y Silva (1977) Acevedo y Arredondo (1982) Gutierrez (1994) Arredondo [1999]
La Tabla 43 presenta los fechados pleistocenicos mas tempranos registrados para restos de mamfferos terrestres del Caribe insular De estos restos el roedor (Cidomys) de Jamaica con un fechado de 253 mil anos antes del presente constituye el fosH de mamifero terrestre mas viejo del Cuaternario anti11ano [Los fosiles fechados antes del surgimiento de la espectrometria de masa (aceleradora) para muestras pequefias pueden ser algo mas viejos (Jull et al 2004)]
Corresponde a MacPhee e Iturralde-Vinent (1994 1995b) e1 relevante merito de haber descubierto en la ultima decada del siglo xx los primeros fosiles terciarios de mamiferos terrestres de las Antillas extendiendo a mas de 30 millones de afios el registro f6sil de
2 Extirpada en la Subregion Antillana pero viviente en el Neotropico continental
3 Ubicacion en subfamilia s610 sugerida por White y MacPhee (200 I)
4 Endemica de la Subregion Antillana
5 Recientemente propuesta su ubicaci6n bajo el genero Atopogale (Roca et al 2004)
6 lncluye las subespecies
7 Transportada por aborigenes a otras de las islas (Rimoli 1974 Olson 1982b este trabajo)
8 Vease MacPhee y Flemming (2003) sobre la incierta situaci6n de esta familia
9 Ubicada provisionalmente en esta subfamilia por MacPhee y Flemming (2003)
10 Hallada en contexto no perturbado de edad Pleistoceno tardio (McFarlane et ai 2002) con 10 cual se descarta su supuesta introduccion por amerindios (Woods 1990)
II La correlaci6n entre el area insular y la riqueza de especies es razonablemente fuerte (R=O83) en estas islas excepto las Bahamas (Morgan 1989)
300 BIOGEOGRAFIA
TABLA 42 Escala convencional del tiempo geologico en la Era Cenozoic a y finales de la Mesozoica (Gradstein 2004) ka = kiloafios (miles de afios) Ma miJIones de afios AP = antes del presente
Periodo(Suhperiodo) Epoca Afios (aproximados) antes del presente
Era Cenozolca
Cuaternario Holoceno Pkistoceno
Presente - 11 kaAP 11 kaAP - 18 MaAP
Terciario (Ne6geno ) Plioceno Mioceno
18 - 5 MaAP 5 - 23 MaAP
oIigoceno 23 - 34 MaAP Terciario (pak6geno) Eoceno 34 - 58 MaAP
Pakoceno 58 - 65 MaAP
Era Mesozo ca
Creticico 65 - 145 -faAP
~~~------~~--- --~-
este grupo animal en la region con el hallazgo de un perezoso oligocenico en Puerto Rico (Yauco) as como un perezoso un mono y un roedor miocenicos en Cuba (Domo de Zaza) Esto unido a la presencia de huesos de una musarafia nesofontoide [referida erroneamente a Solenodon por Rivero (1997)] en ambar oligomiocenico de la Republica Dominicana (MacPhee y Grimaldi 1996) demuestra incontrovertiblemente que al menos en el Mioceno temprano (alrededor de 20 millones de afios atnis) ya estaban en las Antillas Mayores representantes de los cuatro linajes basicos de su fauna de mamiferos terrestres [La afirmacion de Abreu et al (1990 13) en cuanto a que se conocen restos fosiles de almiqui que datan de hace aproximadamente 40 millones de afios es erronea]
Pero lcuan antigua puede ser la fauna de mamiferos terrestres de las Antillas Mayores
Aunque es un hecho bien establecido por la geologia que desde finales de la Era Mesozoica y principios de la Cenozoica existieron islas en el area ocupada ahora por las Antillas Mayores la evidencia disponible indica asimismo que aquellas primeras islas sufrieron repetidas transgresiones marinas totales que habrian hecho sucumbir las biotas que pudieran haber sustentado (MacFadden 1981 Morgan y Woods 1986 MacPhee e Iturralde-Vinent 1994 Roca et al 2004) sin contar los catastroficos efectos que debio tener para el Caribe occidental el b6lido extraterrestre que impacto en Chicxulub (Yucatan) hace 65 Ma (Tada et ai 2003) Hedges (1996) ha especulado que aUn en medio de tal devastacion debe haber existido algun que otro lugar donde elementos de la fauna pudieron haber sobrevivido
301 POBLAMIENTO CONTINENTE-ISLA
TABLA 43 F echados pleistocenicos mcis tempranos (obtenidos por distintos metodos) en mamiferos terrestres del segmento insular delimitado en laFig 41 F =Fuentes (I) Jull et al 2004 (2) Goodfriend y Mitterer 1987 (3) MacPhee et al 1989 (4) Woods 1989b (5) Rega et aI 2002 (6) McFarlane 1999b (7) McFarlane 1 999a (8) McFarlane et al 1998a
Pals Localidad Taxon kaAP F
Cuba Breas de San Felipe Pilosa (Parocnus) 1l9a 1
Jamaica Cueva Coco Ree Cueva Wallingford Roadside
Primates indet Rodentia (Clidomys)
393 2530
2 3
Haiti Trouing Jeremie no5 (casimba) Rodentia (Rh izoplagiodontia) 212 4
Republica Dominicana
Cueva del Perezoso Pilosa (ficratocnus) 185 5
Puerto Rico Cueva de Ia Vaca Cueva Clara
Pilosa (Acratocnus) Soricomorpha (Nesophontes)
337 346
6 7
Anguila Cueva Cave-No-Cave Rodentia (Amblyrhiza) 2060 8
a Kulp etal (1952) reportaron un fechado radiocarbonico de gt 30000 afios para un cono de pino en el deposito fosilffero de Ciego Montero (Cuba) con restos de perezosos
(vcase tambicn Crother y Guyer 1996) pero Iturralde-Vinent y MacPhee (1999) han preguntado donde estaban esos lugares si es que existieron
Solo a partir del Eoceno Superior las Antillas Mayores en fonnacion han pennanecido emergidas hasta hoy [excepto parte de Jamaica y el sur de La Espanola que continuaron sumergidas hasta el Mioceno Medio (Iturralde-Vinent y MacPhee 1999)] Esto significa que solo durante los ultimos 40 millones de anos aproximadamente han existido pennanentemente en dicha region insular ambientes terrestres capaces de sustentar comunidades biotic as (MacPhee e Iturralde-Vinent 2000 Portell et ai 2001 MacPhee 2005) 10 cual proporciona el marco temporal de referencia para conceptuar la antigiiedad maxima posible de la fauna de mamiferos terrestres de las Antillas Mayores
POBLAMIENTO CONTINENTE-ISLA
AFINIDADES CONTINENTALES
El consenso de los autores que en las itltimas dccadas han tratado la filogenia y biogeografia de los mamiferos terrestres de las Antillas Mayores (MacFadden 1980 MacPhee y Woods 1982 Morgan 1985 1988 1993 Morgan y Woods 1986 Ford 1986 Woods 1988 1989a 1990 MacPhee e Iturralde-Vinent 1995a b Hedges 1996 Horovitz [1997] Ottenwalder 2001 Whidden y Asher 2001 MacPheeet ai 2003 Carlini y Scillatoshy
302 BIOGEOGRAFiA
Yane 2004 Davalos 2004 MacPhee y Horovitz 2004 Roca et al 2004) es practicamente unanime en cuanto a que los ancestros continentales que dieron origen a esta fauna derivaron (1) mayormente de Suramerica perezosos (Megalonychidae) monos (Pitheciidae) y roedores (Echimyidae Capromyidae y Heptaxodontidae) (2) en menor medida de Norteamerica musarafias (Nesophontidae y Solenodontidae) y (3) menos aun de Centroamerica una especie de roedor (Muridae)
HIPOTESIS BIOGEOGRAFICAS
La definicion c1asica reconoce dos tipos de islas isla continental (porcion del territorio de un continente separado de este por movimiento tectonico 0 por ascenso del nivel del mar) e isla oceanica (porcion de territorio que no ha formado parte de un continente) La definicion sigue siendo uti por 10 que implica para la formacion de las faunas insulares en uno u otro caso As en el caso de la isla continental su fauna ya estaba emplazada en la futura isla antes de producirse su separacion del continente [Vicarianza hipotesis biogeografica de fragmentacion faunal] en el caso de la isla oceanica su fauna es el resultado de la colonizacion1 por propagulos2continentales a traves de la barrera marina entre el continente y la isla [Dispersion hipotesis biogeografica de acumulacion faunal] En el primer caso el surgimiento de la barrera marina es resultado del evento vicariante en el segundo la existencia de la barrera marina es anterior al evento dispersante Ademas de operar a escala mayor (continente-isla) ambos eventos operan secundariamente a menor escala (isla-isla)
Las dos hipotesis -vicarianza y dispersion- se han disputado hasta hoy la preferencia de los biogeografos en tanto hipotesis mas viable para explicar el proceso de formacion original (continente-isla) de la fauna de mamiferos terrestres de las Antillas Mayores [Sobre la historia de las hipotesis biogeograficas relativas al Caribe insular vease RuB y Schubert 1989 Williams 1989 Pruna 2001] Yel debate continua debido ala falta de consenso sobre 10 que Williams (1989) ha llamado el problema central de la discusion a saber la ostensiblemente baja diversidad taxonomica de esta fauna a nivel de las categorias superiores (solo cuatro ordenes) en comparacion con la diversidad al mismo nivel de las faunas de islas continentales (0 de areas del continente) simi lares en tamafio y diversidad ecologica [Por ejemplo la fauna cubana de mamiferos terrestres consideradas de conjunto las especies vivientes y las extintas- es menos diversa a los niveles taxonomicos superiores (4 ordenes y 6 familias) que 10 que cabria esperar de la diversidad ecologica y extension territorial de Cuba (110 992 km2) comparada con una isla tipicamente continental como Trinidad (4828 krn2
) -veinte veces menor que Cuba y menos diversiticada ecologicamente- donde solo las especies vivientes de mamfferos terrestres representan 6 ordenes y 16 familias (Eisenberg 1989)] Este desbalance
lColonizaci6n Arribo y establecimiento de un propagulo en una isla no ocupada por la especie (MacArthur y Wilson 1967)
2Propagulo Niimero minimo de individuos de una especie capaces de reproducirse y aumentar poblacionalmente en una isla bajo condiciones favorables (MacArthur y Wilson 1967)
303 POBLAMIENTO CONTINENTE-ISLA
taxon6mico ha sido expJicado de dos maneras (1) Inicialmente hubo mas 6rdenes presentes pero la mayoria se extingui6 y s610 sobrevivieron esos cuatro [explicaci6n vicarianista] (2) S610 representantes de esos cuatro 6rdenes lograron cruzar la barrera marina [explicaci6n dispersionista]
Cinco hipot~ticos mecanismos de poblamiento continente-isla han sido invocados durante los ultimos 140 afios (y se los sigue proponiendo actualmente) para explicar la presencia de mamiferos terrestres en las Antillas Mayores A continuaci6n pasamos a resefiarlos brevemente (incluidas sus alegadas fortalezas y debilidades) en el orden aproximado en que historicamente fueron propuestos
EL PUENTE NATURAL (dispersion terrestre activa)
En el siglo XIX el puente terrestre era pnkticamente la unica hipotesis para explicar las distribuciones disyuntas de animales en buena parte del mundo y los primeros investigadores que se ocuparon de la biogeografia antillana (Fernandez [1865] 1884 Chapman [1893] Torre 1892 Lora 1895) la asumieron resueltamente en parte intrigados por el descubrimiento en Cuba en la segunda mitad del siglo de un perezoso f6sil de gran talla Y en su aplicacion a los mamiferos terrestres de las Antillas Mayores el puente sigui6 siendo a 10 largo del siglo xx la hipotesis mas favorecida por los estudiosos del tema (Vaughan 1902a Spencer 191 Oa b Torre 1910a b cAllen 1911 1917a 1918 Barbour 19141916 1945bAnthony 1918 1926 [1942] Scharff 1922 Kraglievich 1926 Beaufort 1951 Arredondo 1961 1970b Kartashov y Mayo 1974 Iturralde-Vinent 1982 1988 Arredondo y Armifiana 1996 Arredondo 1996a [1999]) quienes postularon la existencia preterita de puentes con Norte- Centro- 0 Suramerica en dependencia de las afinidades continentales de los mamiferos antillanos
Teoricamente el puente terrestre debe cumplir con una condicion esencial permitir el paso de una parte mayoritaria (y por tanto representativa de la diversidad) de la fauna continental Claro que siempre habria tipos de mamiferos menos proclives a dispersarse o para los cuales la ubicaci6n del puente resultaria inaccesible 0 las condiciones ecologicas en este fueran desfavorables pero si algunos acceden al puente no hay raz6n para pensar que el resto de esa fauna no los acompafie mayoritariamente Como dijera Simpson (1956) Los puentes no transportan faunas desbalanceadas
En consecuencia el puente terrestre como filtro es un mecanismo de poblamiento minimamente selectivo y por ende de alta diversidad inicial (sensu Iturralde-Vinent y MacPhee 1999) 10 esencial del puente es su capacidad para transportar faunas integradas (Williams 1989) Yen eno radica la objeci6n fundamental al puente terrestre surgida tempranamente en el siglo xx (Por que s610 cuatro 6rdenes de mamiferos terrestres en las Antillas si vinieron por puentes (Donde estan los fosiles de los demas ordenes que debieron venir si es que se extinguieron Los partidarios del hipotetico puente dispusieron de casi todo el resto del siglo para responder una y otra vez que otros 6rdenes de mamiferos aparecerian representados en las Antillas Mayores el dia que en estas Islas se descubrieran fosiles precuatemarios suficientemente
304 BIOGEOGRAFIA
antiguos Esto se logro por fin terminando el siglo como se explico anteriormente (vease Contexto temporal) y ique se encontro los mismos cuatro ordenes de mamiferos terrestres conocidos del Cuatemario caribefio aparentemente los Unicos que lograron llegar
Pero ademas hasta el presente no se ha demostrado inequivoca- y concluyentemente que en algun momenta durante los ultimos 40 millones de afios haya existido una conexion terrestre continua entre las Antillas Mayores (en proceso de formacion 0 plenamente formadas) y algun punto de las regiones continentales vecinas
LA BALSA NATURAL (dispersion marina pasiva)
Durante buena parte del siglo xx el puente terrestre tuvo como unico rival a la hipotesis de la balsa natural -transporte accidental de animales sobre islotes de vegetacion flotantes a la deriva despues de arrancados y expulsados mar afuera por un rio crecido- expuesta originalmente por Matthew (191319151916 1918a 1919) y desarrollada en su aplicacion a los mamfferos terreslres antillanos principal mente por Simpson (1956) asi como por olros investigadores (Darlington 1938 1957 Williams y Koopman 1952 Hershkovitz 1958 1972 Patterson 1962 Paula 1967 Patterson y Pascual 1972 Pregill 1981 Ford 1986 Woods 1989a 1990 Morgan 1993) Por su parte Hedges et al (1992) sustentaron dicha hipotesis en estimados de divergencia (basados en el reloj molecular) en la herpetofauna antillana -involucrados tangencia1mente los mamiferos [veanse objeciones a Hedges y colaboradores (Page y Lydeard 1994 Iturralde-Vinent y MacPhee 1999) respuestas a las objeciones (Hedges et al 1994 Hedges 2001) y contrarrespuestas (MacPhee e IturraldeshyVinent 2005) asi como cuestionamientos al metoda de los estimados de divergencia (Foote et al 1999 Pregill y Crother 1999 Woods et ai 2001 )]
La hip61esis de la balsa natural se origino a partir de las numerosas ocasiones en que a 10 largo de siglos tales islotes flotantes (portando 0 no vertebrados incluidos mamiferos) han sido avistados mar afuera a cientos de kilometros de la costa mas cercana en varias partes del mundo --inc1uido el Mar Caribe- segUn consta en diarios de barcos y reportes de observadores diversos (revisados por King 1962 reportes posteriores en SICSP 1970 y Censky et al 1998) Uno de estos casos acaecido en nuestra region se reporta en un informe del SICSP (Smithsonian Institution Center for Shortlived Phenomena 197050shy52)
EIB de julio de 1969 un destructor de la armada de Estados Unidos avisto un islote flotante a unos 100 km al sur de la Base Naval de Guantanamo moviendose desde el Paso de los Vientos en direccion suroeste a una velocidad de 45 kmlh islote de unos 15 m de diametro mantenfa en pie mas de 10 palmas reales de 10-15 m de altura con suelo cubierto de hierba y arbustos [la observacion con binoculares a 18 km de distancia no preciso la presencia de vida animal] La travesia fue rastreada durante 6 dfas hasta que el 19 de julio el islote se dio por perdido tras 6 horas de busqueda aerea
305 POBLAMIENTO CONTINENTE-ISLA
La colonizaci6n exitosa de una isla de las Antillas Mayores por mamiferos terrestres continentales transportados en balsas naturales a la deriva (de cada especie al menos una hembra gravida 0 dos individuos de sexo opuesto) dependeria de factores que van a operar desde el momenta mismo de formarse la balsa asi como durante su travesia marina y su arribo a la isla
a) La posibilidad de estar en la balsa Que la distribuci6n de la especie incluya areas contiguas al rio y que el animal se encuentre en ellugar y el momenta propicios para ser incorporado a la balsa antes de que el rio crecido la desprenda Por ejemplo segun Hershkovitz (1958) los equimidos -grupo ancestral de roedores antillanos- serian buenos candidatos a ser captados por la balsa porque acostumbran a anidar en huecos del tronco de los arboles a la orilla de los rios
b) El tamano y la consistencia de la balsa Muchas se desintegran en alta mar debido a su fragilidad ante la saturaci6n Algunos autores (Beaufort 1951 Arredondo 1996a [1999]) han expresado incredulidad en cuanto a que las balsas hayan podido soportar el peso de un animal como el gran perezoso cubano (Megalocnus rodens) el mayor de los mamiferos antillanos Pero esta especie evolucion6 en Cuba y el ancestro que hizo el viaje pudo ser muy pequeno [EI perezoso oligocenico de Puerto Rico es el mas pequeno de los megaloniquidos antillanos (MacPhee e Iturralde-Vinent 1995b )]
c) Las caracterfsticas fisiol6gicas de la especie Los limites fisiol6gicos de los propagulos constituyen nuestra unica objeci6n a la dispersi6n marina por balsas como mecanismo para la formaci6n de la fauna de vertebrados en las Antillas (MacPhee e IturraldeshyVinent 2005) Unas especies tienen intrinsecamente mas posibilidades que otras para sobrevivir a las vicisitudes que deben afrontar durante la travesia (deshidrataci6n inanicion insolaci6n etc) Por ejemplo los megaloniquidos han demostrado tener extraordinaria habilidad para cruzar barreras marinas fueron los primeros mamfferos suramericanos en invadir Norteamerica mucho antes de que surgiera la conexion terrestre a finales del Plioceno (Paula 1967 Olson 1978 Webb 1985b Martin y Steadman 1999) Incluso se conoce un megaloniquido acultitico del Plioceno peruano (Muizon y McDonald 1995) y otro viviente en Suramerica (Choloepus) que pnicticamente no bebe agua 10 que ha originado una leyenda aborigen que explica por que los perezosos no toman agua (Goff art 1971) Vease ademas el inciso e
d) EI regimen de las corrientes marinas Que conduzcan a la balsa en la direcci6n necesaria EI transporte de balsas de vegetacion desde los continentes hacia las islas fue durante la mayor parte del Cenozoico sumamente improbable (MacPhee e IturraldeshyVinent 2000 148) Estos autores han insistido en que hay que tener en cuenta que durante la cambiante geografia cenozoica del Caribe las corrientes marinas de superficie no favorecieron el desplazamiento de objetos fiotantes hacia las Antillas Mayores [Pero no sabemos con certeza que efecto modificador del curso de tales objetos fiotantes pudieron introducir las tormentas tropicales si previsiblemente funcionaron en el pasado como 10 hacen ahora] Dichos autores senalaron ademas (2005) que en un experimento reciente con boyas de posicionamiento satelital liberadas a la deriva desde las Antillas Menores las boyas fueron a parar mayoritariamente a Centroamerica y a otros lugares
306 B[OGEOGRAFIA
y solo unas pocas recalaron en las Antillas Mayores y las que entraron al Mar Caribe demoraron semanas 0 meses en arribar
e) La distancia a recorrer (y en consecuencia la duracion de la travesia) entre el continente y la isla (10 que repercute decisivamente en el factor c) Este es desde luego uno de los factores mas criticos En la medida que la distancia aumenta la probabilidad de alcanzar la isla disminuye drasticamente con el agravante de que la trayectoria dificilmente seria en linea recta Si tomamos como referencia la velocidad de 45 kmlh del islote ya mencionado (SICSP 1970) cubrir la distancia aerea (aproximada) de 800 km que media entre el noroeste de Suramerica y La Espafiola Ie tomaria a la balsa 74 dias Estimando el doble de este tiempo debido a desvios de la trayectoria la duracion de la travesia probablemente resultaria excesiva para la mayoria de los mamiferos Pero pudiera muy bien ocurrir que la velocidad fuera mayor 0 que la balsa portara plantas vivas que permitirian a los fitOfagos sobrevivir [Tres de los cuatro ordenes de mamiferos terrestres conocidos de las Antillas Mayores son fitOfagos] Ademas esta geologicamente bien establecido que durante etapas de variada duracion desde el Oligoceno hasta el Pleistoceno los territorios del Caribe insular y continental aumentaron considerablemente en extension con niveles del mar 100-200 m por debajo de su nivel actual oportunidades que los propagulos aprovecharian para cruzar barreras marinas significativamente menores
f) El tamafio de la isla Las islas mayores deben interceptar mas balsas (Buckley y Knedlhans 1986) 10 que repercute directamente en el factor e La balsa puede pasar entre islas pequefias y continuar viaje sin llegar a ningun lugar
g) Las condiciones (favorables 0 adversas) del desembarco en la isla y el tiempo que Ie tomaria a uno 0 mas animales depauperados encontrarun habitat apropiado Esto dependeria a su vez del tamafio y diversidad ecologica de la isla (Darlington 1938 Miller 1966)
Tal cumulo de posibles obstaculos y dificultades constituye a primera vista un desafio a la credibilidad de la hipotesis Matthew-Simpson pero en ello radica precisamente su valor explicativo bas ado en la mas absoluta improbabilidad La colonizacion por la via marina es altamente improbable pero no absolutamente imposible si se tiene en cuenta la inmensidad del tiempo geologico se vuelve probable (McKenna 1973) Basta que solo una vez entre mill ones de intentos fallidos durante ese inmenso lapso algunos de los factores anteriormente enunciados coincidan en no ser tan desfavorables para el propagulo Y eso es todo 10 que demanda la hipotesis para cada uno de los grupos monofileticos mayores de mamiferos terrestres presentes en las Antillas Mayores Las Antillas Mayores requeririan el arribo de tan solo 7 (quizas menos) propagulos fundadores a 10 largo de los ultimos 30 0 mas millones de afios (promedio de uno cada 43 mill ones) para explicar la formacion de toda su fauna de mamiferos terrestres por dispersion marina pasiva (vease Discusion y Conc1usiones)
En comparacion con la via marina el poblamiento por la via terrestre es predecible porque la mayor parte de la fauna continental de mamiferos emplazada en la region del puente va a cruzar por este una vez que el mismo se establece y los animales logran
307 POBLAMIEITO CONTINENTE-ISLA
acceder a el el poblamiento por la via marina en cambio es impredecible porque depende total mente del azar que caracteriza a la balsa Claro que en ambas vias opera el azar pero en el primer caso se trata del azar de un evento probable en el segundo el azar de un evento improbable (Simpson 1940) Asi el puente natural (mecanismo de poblamiento minimamente selectivo = alta diversidad inicial) y la balsa natural (mecanismo de poblamiento maximamente selectivo baja diversidad inicial) pueden conceptuarse como los extremos opuestos de una gama de probabilidades de dispersi6n (McKenna 1973) EI fen6meno de la dispersi6n merece la mayor atenci6n cientifica en los estudios de biogeografia insular (Cowie y Holland 2006)
Queda claro que la balsa natural es una hip6tesis consistente con el caracter de la fauna objeto de estudio puesto que el desbalance taxon6mico (sin evidencia paleontol6gica en contrario) s610 puede ser resultado de la improbabilidad del poblamiento No obstante ello no implica necesariamente que la balsa natural haya sido el imico mecanismo (ni siquiera el principal) utilizado por los mamiferos terrestres en su colonizaci6n de las Antillas Mayores La balsa natural debe satisfacer una premisa te6rica esencial dado el caracter azaroso impredecible e improbable de una colonizaci6n exitosa esta debe ocurrir a intervalos muy espaciados a traves del tiempo geol6gico 0 sea durante todo el tiempo que en el area ocupada por las Antillas Mayores ha habido ecosistemas terrestres perrnanentemente disponibles para ser colonizados (los ultimos 40 millones de afios) Sin embargo como sabemos representantes de los cuatro 6rdenes de mamiteros terrestres conocidos de las Antillas Mayores ya estaban emplazados en dicho territorio insular en el Mioceno Inferior 10 que supone un lapso de colonizaci6n limitado al Eoceno SuperiorshyOligoceno (MacPhee 2005) sugerente de un tipo de mecanismo contrario a la colonizacion alocronica exigida por la hipotesis de la balsa natural sobre todo tratandose de una fauna caracterizada por altas tasas de extincion sin reposicion aparente
La presencia en Jamaica (hasta su extinci6n en 1877) del roedor mfuido Oryzomys antillarom con estrecha afinidad centroamericana (Woods 1989a) pudiera constituir el imico caso efectivo de poblamiento (continente-isla) de las Antillas Mayores por dispersion marina dc un mamifero terrestre para el cual no es razonable invocar otro mecanismo de colonizacion
LA GUIRNALDA INSULAR (dispersion mixta)
La geologia ha establecido con suficiente certeza que durante el Cenozoico unicamente en el Pale6geno tardio y solo a traves de la Cresta de Aves (ahora sumergida excepto la isla hom6nima) pudo haber existido una cadena de islas para el transito de mamiferos terrestres entre el norte de Suramerica y las proto-Antillas Mayores [desestimados a ese efecto el Canal dc Yucatan el Elcvado de Nicaragua la Cresta de Caiman el Elevado de Beata y las Antillas Menores (Holcombe et al 1990 MacPhee e IturraldeshyVinent 1994 1995b Iturralde-Vinent y MacPhee 1999)] Holcombe y Moore (1977) y Holcombe y Edgar (1990) postularon la existencia de esta entidad y estimaron algunas de sus posibles caracteristicas Por su parte MacPhee e Iturralde-Vinent (1995b) e Iturralde-Vinent y MacPhee (1999) precisaron los extremos proximal y distal de esa
308 BIOGEOGRAFIA
estructura acotaron la duracion y la edad de la misma y Ie dieron nombre GAARlandia (Greater AntillesA vesRidgelandia)
Segun Iturralde-Vinent y MacPhee (1999) la Cresta de Aves emergio por breve tiempo (1-2 Ma) durante la transicion Eoceno-Oligoceno (35-33 MaAP) por la coincidencia de un levantamiento tectoni co y un considerable descenso eustatico del nivel del mar En ese momenta las islas septentrionales del arco de las Antillas Mayores (Cuba centroorientalshyLa Espafiola - Puerto Rico - Islas Virgenes) constituian una sola isla grande 0 una serie de islas separadas por barreras muy estrechas La Cresta de Aves emergida entre el bloque Puerto Rico - Islas Virgenes y un microcontinente noroccidental de Suramerica -separado en ese momento del resto del continente por una via marina surgida en el Eoceno tardio- dio origen a GAARlandia
Obviamente el numero de colonizaciones exitosas por mamiferos terrestres a traves de una guimalda de Islas tiende a disminuir mientras mas anchas sean las barreras marinas entre las islas que forman la guimalda y cuando esto ocurre disminuye tambien la diversidad taxonomica de los propagulos particularmente a nivel de las categorias superiores (Simpson 1956)
Vemos asi que para los mamiferos terrestres la colonizaci6n a traves de una guimalda insular puede ser tan improbable como a la deriva en una balsa natural en dependencia de la cantidad y anchura de las barreras marinas a cruzar el tamafio y la fisiografia de las islas que integran la guimalda y el regimen de las corrientes marinas y aereas entre otros factores
Resumiendo la guimalda insular puede conceptuarse como un mecanismo de poblamiento altamente selectivo (= baja diversidad inicial) que combina la improbabilidad de la balsa natural con la colonizacion restringida temporaimente del puente terrestre
En su reciente revision de la taxonomia distribuci6n y filogenia (basada en estudios moleculares) de varios grupos de mamiferos antillanos Davalos (2004) concluyo que los perezosos megalocninos los primates y los roedores histricognatos se avienen al patron de divergencia con el continente y epoca de arribo a las Antill as implicados en la hipotesis de GAARlandia pero no asi los perezosos coloepodinos que requieren una explicacion biogeognlfica distinta Esto ultimo (sospechado por Matthew 1918a) habla sido confirmado por White y MacPhee (2001 226-227)
Nuestra evidencia filogenetica no sustenta la idea de un dado unificado surgido de la invasion de un megaloniquido el analisis sugiere mas bien dos eventos de colonizaci6n independientes [y] sustenta fuertemente un origen difiletico de las subfamilias [Choloepodinae y Megalocninae] Si hubo dos migraciones a traves de GAARlandia presumiblemente poco espaciadas en el tiempo no se puede determinar en el presente
Finalmente queda claro que GAARlandia -como guirnalda insular- es la hipotesis mas robusta propuesta hasta ahora para explicar la presencia en las Antillas Mayores de
309 POBLAMIENTO CONTINENTE- ISLA
la absoluta mayona de los mamfferos terrestres de ongen suramericano porque el caracter de esa fauna supone un mecanismo de poblamiento ancestral que sea a la vez maximamente selectivo (baja diversidad ordinal) y temporalmente restringido (transicion Eoceno-Oligoceno)
Recientemente Rega et al (2002 18) expresaron A pesar de una revision exhaustiva de la paleogeografia de la cuenca del Caribe Iturralde-Vinent y MacPhee (1999) no pudieron confirmar la presencia de una conexion terrestre continua entre Suramerica y las proto-Antillas aunque hipotetizaron su existencia [Vease la Fig 42 (= figura 6 de Iturralde-Vinent y MacPhee 1999 simplificada))
En el trabajo citado por Rega y colaboradores aparece la siguiente explicacion
Actualmente no es posible ofrecer reconstrucciones paleogeograficas detalladas de la Cresta de Aves Sin embargo hay evidencia de que porciones de la cresta deben haberse elevado subaereamente en un tiempo La cuestion cntica es si la Cresta de Aves emergida pudo haber formado algun tipo de corredor entre las Antillas Mayores y el noroeste de Suramerica Perforaciones en las alturas estructurales de la cresta pudieran proveer el registro del Paleogeno tardio que se necesita para precisar la hipotesis del corredor aunque no se puede predecir si tales datos serian suficientemente detallados para determinar si en algun momento GAARlandia constituyo una sola entidad subaerea Por ejemplo pudo haber barreras marinas por corto tiempo entre las masas terrestres de la Cresta de Aves [0) ellevantamiento y la subsidencia pudieron afectar diferentes partes de GAARlandia en diferentes tiempos AItemativamente los elementos faunales pudieron haberse dispersado de manera pulsada Movimientos episOdicos de este tipo incrementarian la extincion de los elementos faunales incapaces de trasladarse a la siguiente masa terrestre en la cadena disminuyendo asi la diversidad total (Iturralde-Vinent y MacPhee 199957)
La primera parte de la explicacion deja claro que la posibilidad de que la Cresta de Aves haya sido una conexion terrestre continua no se puede descartar pero tampoco se puede afirmar La ultima parte de la explicacion describe el efecto altamente selectivo (filtrante) de la colonizacion a traves de una cadena de islas
Y continuan lturralde-Vinent y MacPhee (199957 -58) explicando
Hasta que punto el microcontinente noroccidental de Suramerica [considerado por dichos autores como el area suministradora de propagulos hacia GAARlandiaJ estuvo fisicamente separado del resto del continente no ha sido plenamente esc1arecido Si estuvo sustancialmente aislado por barreras marinas y orogenic as durante el Paleogeno tardio entonces solo la fraccion de la biota suramericana emplazada en el microcontinente estuvo en posicion de acceder a GAARlandia Esto pudiera tener que ver con el hecho de que la fauna antillana carezca de representantes de muchos grupos suramericanos [de mamfferos) que debieron haber hecho el viaje
310 BIOGEOGRAFiA
Florida o Alturas0 0
d 6 D L1anuras25deg
Yucatan
20deg
deg0
10deg
MARCARIBE
80deg 75deg
Bahamas
Cres ta de Beata
D Mares someros
D Mares profundos
OCEANO ATLANTICa
FIGURA 42 Reconstrucci6n hipotetica de GAARlandia como conexi6n terrestre continua entre las proto-Antillas Mayores y el noroeste de Suramerica en el Pale6geno tardio segun la FIgura 6 (simplificada) de Iturralde-Vinent y MacPhee (1999)
En su defensa del puente terrestre Barbour (1945b249) se refiri6 a esto - antes 10 habfa hecho Anthony [1942363]- en los terminos siguientes
El argumento de que si las Antillas hubieran estado conectadas con el continente se encontrarian los f6siles de un numero mayor de distintas criaturas me deja frio es perfectamente concebible que la conexi6n haya ocurrido en un periodo en que la parte del continente implicada no estuviera poblada por una fauna grande y variada
Y concluyen lturralde-Vinent y MacPhee (199958) su detallada explicaci6n
Desafortunadamente sabemos muy poco sobre la composici6n faunal del noroeste de Suramerica a finales del Pale6geno
Consecuentemente si se confirmara la magnitud del aislamiento del aludido microcontinente y su concomitante baja diversidad faunal GAARlandia como conexi6n terrestre continua serf a la hip6tesis mas parsimoniosa No obstante mientras no se disponga de evidencia concluyente a este respecto hipotetizar a GAARlandia como conexi6n continua (minima selectividad) la coloca en franca vulnerabilidad te6rica al tener que afrontar la objeci6n clasica al puente terrestre (baja diversidad ordinal de la
311 POBLAMIENTO CONTINENTE-TSLA
fauna) al tiempo que se la priva de ser la hip6tesis mas robusta para explicar -como guimalda de islas- la colonizaci6n de las Antillas por mamiferos procedentes de Suramerica
ELARcA DE NOE (seudovicarianza)
El primer vicarianista antillano (desde luego sin el saberlo) fue Corral (1940) con su utilizaci6n ingenua de las ideas wegenerianas de la deriva continental aparecidas la decada anterior Pero el primer (supuesto) intento de aplicar a la formaci6n de la biota caribefia el mecanismo de la vicarianza basada en la tearia de la tect6nica de placas (surgida en la sexta decada del pas ado siglo) se debe a Rosen ([1976] 1985)
Segun Rosen las Antillas Mayores se originaron en el Cremcico en la Placa Caribe aproximadamente en la posici6n que hoy dia ocupa Centroamerica y al ser desplazadas tectonicamente hacia el este interactuaron (de algun modo no precisado por Rosen) con las margenes continentales de Norte- y Suramerica de donde (en un evento unico) adquirieron toda su fauna transportada subsecuentemente por dichas islas en su transito hacia sus posiciones actuales [De ahi la denominacion metaforica dada par McKenna (1973) a este mecanismo de poblamiento] Aunque se ha presentado esta propuesta como Model0 de Vicarianza Continente-Isla de Rosen en realidad por definici6n no constituye un evento vicariante porque las proto-Antillas Mayores no resultaron del fraccionamiento y separaci6n por tectonismo de una porcion del territorio continental como exige esta hip6tesis (vease definici6n de vicarianza al comienzo de Hipotesis Biogeograficas)
El Arca de Noe de Rosen ha sido desestimada por la mayoria de los autores contemporaneos porque (1) carece de sustento geol6gico (lturralde-Vinent y MacPhee 1999) y porque (2) en el periodo en que se afrrrna ocum6 el evento muchos de los ordenes modemos de mamiferos placentarios attn no existian (Williams 1989)
EL BARCO FUNERAL VlKINGO (tanatovicarianza)
Domning et al (1997) reportaron 10 que consideraron el mamffero terrestre mas viejo de las Antillas --circunstancia que se refleja en el titulo mismo del trabajo Oldest West Indian Land Mammal- al descubrirse un perisodactilo rinocerotoide (Hyrachius sp) en una localidad de edad Eoceno Inferior a Medio en el occidente de Jamaica
Jamaica ha tenido una historia geologica y paleogeografica distinta de la historia comun a las restantes Antillas Mayores A principios del Cenozoico Jamaica occidental (a diferencia de la porcion oriental constituida por el Bloque de las Montafias Azules) formaba parte de Centro america de la cual se desgaj6 (por el desplazamiento de la Placa Caribe) para venir a integrarse despues del Eoceno Medio con 10 que serfa la futura isla de Jamaica desaparecida por transgresi6n marina en el Eoceno Superior hasta volver a emerger totalmente en el Mioceno Medio (Iturralde-Vinent y MacPhee 1999 MacPhee e Iturralde-Vinent 2000)
312 BIOGEOGRAFiA
Por consiguiente Hyrachius ---conocido de localidades de edad Eoceno Inferior a Medio en Norteamerica- debio ocupar (10 que iba a ser) Jamaica occidental mucho antes de que esta se separara de Centroamerica De hecho este fosil Uego a su destino insular contenido en las rocas de su basamento no como un ser vivo por 10 que es erroneo considerarlo un animal antillano (lturralde-Vinent y MacPhee 2004) como han supuesto algunos (Hedges 200 I Portell et al 200 I Condis [200 I D Todo 10 que hizo fue viajar tanatovicariantemente en su barco funeral vikingo -otra de las denominaciones metaforicas de McKenna (1973)
DISCUSION Y CONCLUSIONES
Hemos visto que solo disponemos de una hipotesis plausible para explicar la mayor parte del poblamiento (continente-isla) de las Antillas Mayores por mamiferos terrestres del Neotropico continental la dispersion mixta (a traves de GAARlandia) para los perezosos (excepto Choloepodinae) los monos y los roedores (excepto Oryzomys)
Por otra parte la historia biogeogrMica de las musaraiias de las Antillas Mayores (Nesophontes y Solenodon) constituye una de las situaciones mas enigmaticas entre los mamiferos de la region (Ottenwalder 2001321) Al menos cinco mecanismos de poblamiento continente-isla han sido invocados para explicar cuando como y de donde llegaron a las Antillas Mayores estas musaraiias
I EI propagulo ancestral (del Terciario de Norteamerica) coloniz6 a las Antillas Mayores por dispersion marina desde Centroamerica 0 desde la Florida (Patterson 1962)
2 El propagulo ancestral ( del Terciario) cruzo el Atlantico Sur (supuestamente mas estreshycho en aquella epoca) directamente de Africa a las Antillas Mayores (Hershkovitz 1972)
3 El propagulo ancestral (del Terciario de Norteamerica) Uego a las Antillas Mayores en el Arca de Noe de Rosen (MacFadden 1980)
4 El propagulo ancestral (del Terciario de Norteamerica) fue transportado a las Antillas desde Centroamerica en el bloque de Jamaica occidental y de alli paso por dispersion marina a las restantes Antillas Mayores (Whidden y Asher 200 I)
5 El propagulo ancestral (del Terciario de Norteamerica) lleg6 a las Antillas Mayores a traves de GAARlandia (Whidden y Asher 2001)
Aunque hay consenso entre estos (y otros) autores sobre el origen norteamericano de las musaraiias antillanas ninguno de los cinco hipoteticos mecanismos de poblamiento resiste la evidencia en contrario filogenetica yo paleogeogrMica Ademas algunas de estas hipotesis asumen que Nesophontidae y Solenodontidae comparten un antecesor comun en las Antillas pero esta relacion no ha sido confirm ada Para dilucidar el enigmatico caso de las musaraiias antillanas se requerini profundizar en la investigaci6n geologica paleogeognifica y paleontol6gica (Iturralde-Vinent y MacPhee 1999)
De acuerdo con el conocimiento filogenetico disponible sobre el conjunto de la fauna la colonizacion original de las Antillas Mayores por mamiferos terrestres implicaria el arribo
313 POBLAMIENTO ISLA-ISLA
de solo siete propagulos fundadores perezosos 2 (coloepodinos y megalocninos si como parece tienen un origen difiletico) musaraiias 2 (nesofontidos y solenodontidos si no constituyen una misma familia como consideran algunos) monos 1 (pitecidos) roedores histricognatos 1 (equimidos capromidos y heptaxodontidos) roedores muridos 1 (Oryzomys en Jamaica)
POBLAMIENTO ISLA-ISLA
La distribucion de los mamiferos terrestres a traves del area antillana demarcada al inicio del presente capitulo (vease Contexto Geogratico Fig 41) ha sido interpretada en funcion de dos hipoteticos mecanismos de poblamiento interinsular la dispersion y la vicarianza fundamentadas como se resumen seguidamente
DISPERSION
De acuerdo con la evidencia paleogeogratica mas actualizada en las principales regresiones marinas ocurridas en la cuenca caribeiia durante los ultimos 23 Ma (transiciones OligocenoshyMioceno Mioceno Superior - Plioceno y varias veces en el Pleistoceno) las masas terrestres constitutivas de la region insular en formacion aumentaron considerablemente su extension territorial disminuyendo por tanto la distancia entre elias oportunidades que los elementos faunales aprovecharian para dispersarse de unas islas a otras
Durante mucho tiempo esta ha sido la hipotesis mas favorecida por los estudiosos del tema a partir de Simpson (1956) pero en decadas recientes ha suscitado dos objeciones
1 Es dificil en tender como pudo operar la dispersion marina entre estas islas puesto que ninguna de las Antillas Mayores posee rios suficientemente caudalosos para transportar masas significativas de vegetacion flotante (MacPhee 198432)
Es oportuno recordar aqui el islote ala deriva (con mas de diez palmas reales en pie) entre las tres Antillas Mayores mas occidentales mencionado anteriormente (vease La Balsa Natural)
2 La dispersion marina es menos efectiva para explicar la asombrosa similitud de las faunas de mamiferos terrestres a traves de las Antillas Mayores septentrionales porque como proceso aleatorio no debe producir distribuciones uniformes a 10 largo de una cadena de islas 0 si las produce entonces el proceso no es aleatorio (MacPhee e lturralde-Vinent 2000 149)
Independientemente de que las aludidas distribuciones taxonomicas estan lejos de ser uniformes 0 asombrosamente similares la hipotesis de la dispersion marina contrariamente a su canicter aleatorio y altamente improbable en el caso del poblamiento continente-isla- en el caso del poblamiento isla-isla es un proceso no aleatorio (aprovecha las oportunidades brindadas puntualmente por los periodos de maxima regresion marina) y ademas altamente probable dada la incomparablemente menor distancia entre las islas caribeiias que entre estas y los continentes vecinos
314 BIOGEOGRAFiA
V ICARIANZA
La neotectonica y las transgresiones marinas del Oligoceno Medio y del Mioceno afectaron sustancialmente la paleogeografia de la Cresta de las Antillas Mayores resultando la subdivision gradual de las partes subaereas de la Cresta Asi el este de Cuba y el norte de La Espanola conectados fisicamente durante el Oligoceno Inferior se separaron al formarse el Paso de los Vientos en el Oligoceno tardio en tanto La Espanola y Puerto Rico probablemente permanecieron conectadas hasta el Mioceno tardio en que surgio el Paso de Mona (Iturralde-Vinent y MacPhee 1999)
Tales eventos geologicos habrian dividido las distribuciones de los grupos faunales emplazados previamente en el territorio continuo de las tres islas y a partir de esas disyunciones dichos autores (tambien MacPhee et al 2000 y White y MacPhee 2001) asumieron --como segunda fase del modelo de GAARlandia- la hipotesis de la vicarianza isla-isla para explicar la distribucion multiinsular de varios grupos de mamiferos terrestres a traves de las Antillas Mayores septentrionales De acuerdo con esta hipotesis cualquier diferencia en composicion faunal entre las islas (por ejemplo la ausencia de roedores heptaxodontidos en Cuba 0 de monos platirrinos en Puerto Rico) constituye evidencia de extincion local postvicariante y no de dispersion marina fallida Lo primero se puede demostrar si aparecen los fosiles pertinentes 10 segundo no se puede demostrar (MacPhee e Iturralde-Vinent 1994) Por otra parte IturraldeshyVinent y MacPhee (1999 57) han reconocido que aunque fa vicarianza isla-isla puede representar una solucion clara para las distribuciones de varios taxones cuaternarios no puede explicar todos los casos
Rega et al (2002 18) han presentado dos objeciones al modelo de la vicarianza isla-isla aplicado a los mamiferos de las Antillas Mayores
I Si la separacion de Cuba y La Espanola precedio en varios mill ones de anos la separashycion de La Espanola y Puerto Rico el modelo vicariante de lturralde-MacPhee debe predecir mayor similitud entre las faunas de La Espanola y Puerto Rico que entre las de Cuba y La Espanola De hecho es al reyes
No obstante MacPhee et al (200335) admitieron despues que la fecha mas temprana empiricamente defendible [para la apertura del Paso de Mona] debe ser el Oligoceno Medio esto invierte la secuencia de ambas disyunciones con 10 cual queda resuelta la aparente contradiccion Sin embargo en su reciente evaluacion de la congruencia entre filogenia e hipotesis geologic as en varios grupos de mamiferos terrestres de las Antillas Mayores Davalos (2004373) encontro que ninguno de los taxones cumple con la fecha y el patron de divergencia entre las masas terrestres antillanas postulados por el modelo de GAARlandia
2 La presencia de un mismo genero en las tres Islas septentrionales de las Antillas Mayores implica [por el modelo vicariante] que la diferenciacion entre estas formas durante el intervalo desde el Mioceno ha sido unicamente de nivel especifico [Dicha presencia]
315 rRRADlAcr6~ EVOLUCrONARIA
s610 puede explicarse razonablemente como el resultado de movimientos de esos animales entre las islas en el Cuaternario
De esta objecion resulta dificil disentir [El poblamiento entre islas diferenciado solo a nivel especifico no es probable que sea anterior al Pleistoceno (Simpson 1956 15)]
Debe tenerse en cuenta que la hipotesis de la vicarianza isla-isla alude a las tres islas septentrionales de las Antillas Mayores porque como se explico antes (vease El Barco Funeral Vikingo) Jamaica no existia como isla durante el lapso (Eoceno Superior shyOligoceno) consensualmente reconocido como el mas probable para la colonizacion de las proto-Antill as Mayores por la mayo ria de los mamiferos terrestres Despues de emerger como isla en el Mioceno Medio Jamaica debe haber recibido sus mamiferos terrestres por dispersion isla-isla (Davalos 2004376) y solo en el Cuaternario parece haberrecibido (por dispersion marina desde Centroamerica) al roedor murido Oryzomys
Seguramente tanto la dispersion como la vicarianza contribuyeron a que los mamiferos terrestres se irradiaran a traves de las Antillas Mayores y grupos de islas perifericas pero las maneras y tiempos en que estos mecanismos operaron a 10 largo de dicho proceso son aun en gran medida motivos de desacuerdo e incertidumbre
IRRADIACION EVOLUCIONARIA
A continuaci6n revisamos someramente las distribuciones conocidas de los diferentes grupos de mamiferos terrestres a traves de las Islas principales de las Antillas Mayores y grupos de Islas perifericas incluidas las interpretaciones y conjeturas que la literatura reciente recoge en relaci6n con la manera en que dichas distribuciones pudieron haberse conformado durante la irradiaci6n evolucionaria de los referidos grupos de mamiferos Manifestaciones de esta irradiaci6n pueden inferirse unicamente a partir de los taxones presentes en el Cuatemario pues solo 6 de las especies conocidas corresponden al Terciario
GRUPOS ORlGINARIOS DE SURAMERICA
PEREZOSOS (Megalonychidae)
Aparte de un genero miocenico de Cuba y otro del Oligoceno de Puerto Rico se conocen 4 generos (12 especies) de perezosos cuaternarios (todos extintos) restringidos a las tres islas septentrionales de las Antillas Mayores Acratocnus Neocnus Megalocnus y Parocnus Los cuatro generos estan representados en Cuba y La Espanola y Acratocnus 10 esta ademas (como unico genero de perezoso) en Puerto Rico Esta distribuci6n ha sido interpretada por MacPhee et al (2000) eomo resultado de la vicarianza oligomiocenica entre estas islas y [dado que el Banco Puerto Rico - Islas Virgenes fue la entrada de GAARlandia a las Antillas Mayores] se preguntaron si la ausencia de los otros tres generos en Puerto Rico se debe a que nunca estuvieron en ese territorio 0 a que sus
316 BIOGEOGRAFiA
fosiles atm no han sido hallados Pero como hemos visto Rega et al (2002) atribuyeron dicha distribucion a la dispersion interinsular en el Cuatemario
Por otra parte varios amilisis c1adisticos recientes ubi can a Choloepus -genero Reciente conocido solo de Centro- y Suramerica- como taxon hermano de Aeratoenus exc1uidos los demas megaloniquidos antillanos Para White y MacPhee (2001228)
tan estrecha relacion sugiere que la divergencia entre ambos [generos] ocurrio despues de que Aeratoenus divergio de Neoenus Si esta relacion implica que Aeratoenus y Neoenus se dispersaron separadamente desde Suramerica a las AntilIas mientras Choloepus permanecia en el continente (siendo necesarias al menos tres invasiones de perezosos) 0 si Choloepus invadio Suramerica desde las Antillas es algo que el material disponible no permite determinar
La diversidad adaptativa de los perezosos se manifiesta en (1) la utilizacion del habitat especies arboreas en Neoenus y Aeratoenus y terrestres en Paroenus y Megaloenus (Webb 1985a White [1993] White et al 1996 Arredondo [1999]) y en (2) la gama de tallas corporales desde los pequenos Neoenus hasta Megaloenus rodens el mayor de los mamiferos terrestres antillanos (su peso adulto estimado en 270 kg seglin Paula 19679) esto sugiere que los perezosos se irradiaron a nichos que en contextos continentales probablemente estarian ocupados por miembros de otros c1ados (MacPhee e IturraldeshyVinent 2000)
No esta claro si el abanico de tamanos entre estos perezosos enmascara alguna tendencia al gigantismo Por 10 pronto Anderson y Handley (2002) encontraron 10 contrario en perezosos (Bradypus) de las Islas de Bocas del Toro (Panama) shyseparadas secuencialmente del area continental vecina por ascensos del nivel del mar durante los ultimos 10 ka En las cinco islas los perezosos son menores que su contraparte continental y las diferencias entre las tallas medias de las cinco poblaciones expresan una reduccion lineal significativa de la talla corporal en funcion de la edad geologica de cada isla sin relacion alguna con el area de las islas 0 la distancia que las separa del continente
MONOS (Pitheciidae)
Los primates de las Antillas Mayores (cuatro especies extintas) constituyen un grupo monofiletico con Paralouatta (en Cuba) y Antillothrix (en La Espanola) como taxones hermanos seguidos en la escalera c1adistica por Xenothrix (en Jamaica) y finalmente Callieebus como el pariente continental mas cercano esto implica una sola invasion desde Suramerica antes del Mioceno Inferior (fecha mas temprana conocida para los primates fosiles antillanos) y por tanto en consonancia con GAARlandia como la via de colonizacion (Horovitz y MacPhee 1996 1999 Horovitz [1997] MacPhee y Horovitz 2004)
En desacuerdo parcial con la anterior interpretacion la filogenia de este grupo de monos obtenida por Davalos (2004) indica un antecesor comun en Cuba La Espanola y Jamaica
317 TRRADIACTON EVOLUCIONARIA
compatible con la hipotetica conexion -propuesta tentativamente por Iturralde-Vinent y MacPhee (199980-84)- entre las Montanas Azules del este de Jamaica y la peninsula suroccidental de La Espanola en la epoca de GAARlandia
A pesar de que los primates antillanos pertenecen a un grupo viviente de monos muy pequenos (los monos titles) fueron comparativamente grandes en relacion con los platirrinos modernos y sucesivamente mayores en la direccion Antillothrix Xenothrix y Paralouatta con P marianae como el mayor platirrino conocido del Mioceno neotropical (MacPhee 2005) MacPhee et al (1994) consideraron que Xenothrix debio ser un mono de andar lento y al parecer las especies de Paralouatta abandonaron la arborealidad para adoptar un modo de vida mas terrestre (MacPhee 1996a MacPhee et al 2003 MacPhee y Meldrum 2006)
ROEOORES (Echimyidae Capromyidae Heptaxodontidae)
La edad de Zazamys en Cuba demuestra que la invasion de los roedores histricognatos en las Antillas Mayores comenzo antes del Mioceno Inferior por otra parte los representantes mas antiguos de esa irradiacion evolucionaria se encuentran en La Espanola y Puerto Rico Ambas evidencias son consistentes con la hipotesis de GAARlandia (Woods 1982 1988 Woods et al 2001)
No esta claro si los capromidos evolucionaron a partir de un equimido ancestral 0 de un protocapromido 0 si los equimidos antillanos (I sub familia 3 generos y 7 especies todas extintas) forman parte de la irradiacion de los capromidos (4 subfamilias 9 generos y 27 especies 10 vivientes) Kratochvil et al (1978b) y Rodriguez et al (1979) consideraron que Capromys nan us es un eslab6n entre Echimyidae y Capromyidae pero Mesocapromys nanus esta morfologicamente mucho mas cerca de Hexolobodon y los de mas capromidos que de los equimidos (Woods 1985) Lo que si esta claro segun Woods et al (2001) es que la subfamilia Plagiodontinae endemic a de La Espanola es el linaje basal de los capromidos y que Isolobodon Hexolobodon Capromys Geocapromys Mysateles y Mesocapromys [y supuestamente Macrocapromys] forman parte de la irradiacion temprana de Capromyidae
La subfamilia Capromyinae (5 generos y 17 especies 9 vivientes) parece ser el dado de mas reciente evolucion en la familia con invasion de Cuba desde La Espanola y dispersion desde Cuba (1) por Capromys a las islas Caiman y (2) por Geocapromys a las islas Bahamas Caiman y Jamaica y desde Jamaica a las islas Swan aunque la presencia de Geocapromys en Swan pudiera deberse a introduccion aborigen (Morgan 1985 1988 1989 1993 1994 Woods 1989a 1990) Woods et al (2001) utilizaron datos moleculares para obtener estimados independientes de divergencia entre capromidos segiJn 10 cual Geocapromys divergio cerca de los 17 MaAP Mesocapromys divergio de Capromys hace 15 MaAP en tanto lvlysateles 10 hizo hace 10 MaAP Por su parte las formas de Capromys pilorides y de Mysateles prehensilis en Isla de la Juventud divergieron de sus correspondientes subespecies nominativas en la isla principal a los 24 y 08 MaAP respectivamente
318 BIOGEOGRAFiA
Woods y Howland (I979) describieron adaptaciones morfologicas de la mandibula el hioide y la faringe asociadas a mecanismos de masticacion diferentes en plagiodontinos y caprominos y Varona (1980a) llamo la atencion sobre el gran desarrollo de los molariformes (en relacion con la longitud total del cnineo) que presentan ciertos grupos de jutias fosiles en comparacion con las vivientes 10 que pudiera interpretarse como una adaptacion alimentaria a una vegetacion mas abrasiva por parte de las jutias extinguidas
Segun Woods (1990) la familia Heptaxodontidae (2 subfamilias 5 generos y 6 especies todas extintas) tambien seria parte de la irradiacion de los capromidos con origen en el este de La Espanola 0 en Puerto Rico y subsecuente dispersion al Banco de Anguila - San Martin en las Antillas Menores (pero vease MacPhee y Flemming 2003) Este banco estaria a solo 70 km del Banco Puerto Rico - Islas Virgenes en tiempos de maxima regresion marina y esta proximidad con Puerto Rico -patria de Elasmodontomys obliquus- seguramente tuvo que ver con la presencia en las islas de Anguila y San Martin del heptaxodontino gigante Amblyrhiza relacionado mas estrechamente con los roedores de las Antillas Mayores que con los del resto de las Antillas Menores (Woods 1990 1996) Aunque los roedores suelen aumentar de tamano en las islas (Case 1978) la escala del incremento de la talla en Amblyrhiza (que pudo alcanzar mas de 200 kg de peso adulto) esta fuera de toda proporcion con los patrones conocidos en otros roedores insulares (Biknevicius et 01 1993)
Clidomys (Clidomyinae) de Jamaica difiere marcadamente en su morfologia de los heptaxodontinos (vease ademas MacPhee y Flemming 2003)10 cual dificulta explicar su presencia en esa isla Como Jamaica estuvo bajo el mar hasta el Mioceno Medio el ancestro de Clidomys colonizaria esta isla por dispersion desde el extremo suroccidental de La Espanola despues que los heptaxodontinos se originaron en esta ultima isla (Woods 1989a) Pero Pascual et 01 (I 990) especularon que Clidomys pudiera ser el resultado de una invasion posterior e independiente de los capromidos
GRUPOS ORIGINARIOS DE NORTEAMERICA
El origen de las musaranas antillanas ha resultado ser hasta ahora el mayor enigma biogeografico de la fauna de mamiferos terrestres de la region aunque la mayoria de los autores coinciden en considerarlas originarias de Norteamerica (vease Discus ion y Conclusiones en Poblamiento Continente-Isla) No obstante MacPhee e Iturralde-Vinent (1994) descartaron toda posibilidad de conexion terrestre por el Estrecho de la Florida 0 por el Canal de Yucatan y si el (supuesto) evento vicari ante de Rosen (vease El Arca de Noe) -invocado por MacFadden (1980) para resolver el enigma- fuera minimamente plausible aun asi es dificil creer que las catastrofes que 10 sucedieron (vease Contexto Temporal) hayan barrido con toda esa vieja fauna continental excepto las musaranas circunstancia ya anticipada muy tempranamente por Matthew (l918a)
De otra parte la relaci6n entre Nesophontidae y Solenodontidae permanece irresuelta Si estos linajes no estan estrechamente relacionados entonces es muy posible que
319 IRRADIACION EVOLUCIONARIA
sus distribuciones sean explicadas por mecanismos diferentes (Whidden y Asher 2001)
La descomunal diferencia de talla corporal entre las musaranas pequenas (Nesophontes) y las grandes (Soienodon) seguramente impuso a ambos grupos restricciones distintas relativas a la termorregulacion y las adaptaciones energeticas En Nesophontes la seleccion debio favorecer una alta tasa de metabolismo y crecimiento camadas mas numerosas corta expectativa de vida yalta densidad poblacionaL En Soienodon en cambio la seleccion opero en la direccion de una baja tasa metabolica camadas reducidas crecimiento lento longevidad y baja densidad poblacional (Ottenwalder [1991])
MUSARANAS PEQUENAS (Nesophontidae)
Las musaranas pequenas estan representadas en las Antillas por el genero Nesophontes (y un genero miocenico indeterminado en ambar dominicano) con 6 especies cuaternarias extinguidas distribuidas en Cuba La Espanola Puerto Rico Islas Virgenes e Islas Caiman La mayor de las musaranas pequenas Nesophontes edithae evoluciono en Puerto Rico y su arribo a las Islas Virgenes supone una dispersion sumamente reciente En las Islas Caiman Nesophontes se asemeja mas a la poblacion de Cuba de donde seguramente derivo en el Plio-Pleistoceno (junto con Capromys y Geocapromys) cuando la costa sur de Cuba estuvo mucho mas cerca de las Islas Caiman (lturralde-Vinent 2003fig 4) MacPhee et ai (1994) interpretaron la distribucion de Nesophontes en Cuba y La Espanola como resultado de la vicarianza oligomiocenica ocurrida al formarse el Paso de los Vientos pero la distancia taxonomica entre los lados del Paso es solo de nivel especifico y N micros incluso vivi6 en ambas islas segtln Patterson (1962)
Allen (1918) considero que Nesophontes micrus fue una especie primitiva no modificada para la vida arb6rea 0 fos6rea y por tanto de habitos terrestres Nesophontes edithae de Puerto Rico es mucho mayor y mas robusta que las restantes especies del genero eL indice de robustez de su femur (= anchura minima de la diafisis ongitud total del hueso) muestra que este es tan robusto como el de Solenodon paradoxus un animal dos veces mayor (Morgan et ai 1980) Choate y Birney (1968) postularon un aumento cronoclinal de la taIla corporal en N edithae confirmado mas tarde por McFarlane (1999a) mediante fechados radiocarb6nicos de la secuencia estratignifica en depositos cavernarios de Puerto Rico Solenodon no vivi6 en Puerto Rico y en ausencia de una musarana de mayor tamano Nesophontes alcanzo una talla corporal mayor en esa isla que en La Espanola 0 Cuba (Woods y Eisenberg 1989)
MUSARANAS GRANDES (Solenodontidae)
Los arcaicos almiqufes (genero Soienodon) comprenden cuatro especies una viviente (S cubanus) y otra f6sil (S arredondoi) en Cuba y una viviente (S paradoxus con dos subespecies) y otra f6sil (S marcanoi) en La Espanola Como cabria esperar las dos especies cubanas se asemejan mas entre sf que a las especies de La Espanola pero
320 BIOGEOGRAFiA
enigmaticamente S marcanoi comparte caracteres con S cuban us y S paradoxus (Woods y Eisenberg 1989 Morgan y Ottenwalder 1993 Ottenwalder 200 I) Solenodon arredondoi alcanzo una talla corporal superior a la de cualquiera de las musarafias vivientes a nivel mundial Segun Morgan y Ottenwalder (1993) extrapolando las dimensiones y el peso de los almiquies vivientes S arredondoi debio tener una longitud cabeza-tronco y un peso de 450-550 mm y 1500-2000 g respectivamente en comparacion con S cubanus (360 mm y 800 g) y S paradoxus (390 mm y 1100 g)
Entre todas las especies de almiqufes Solenodon paradoxus es la que presenta los caracteres mas derivados (Otten walder 2001) Si estas relaciones son validas la distribucion de Solenodon entre Cuba y La Espafiola debio discurrir de oeste a este como propuso MacFadden (1980) quien invoco la presencia fosH de S cubanus [pero tambien S arredondoi] en Pinar del Rio para postular que el ancestro continental entr~ por Cuba opinion compartida por Perfit y Williams (1989) y por Ottenwalder ([1991] 2001) El paso de Solenodon a Puerto Rico pudo haberlo impedido la existencia de barreras ecologicas durante la conexion del este de La Espafiola con Puerto Rico 0 por la ausencia de Solenodon en esa parte de La Espafiola en aquella epoca Sin embargo las mismas condiciones ecologicas en el este de La Espafiola no parecen haber representado una barrera efectiva para Nesophontes cuyas mas altas tasa de reproduccion y densidad poblacional presumiblemente Ie permitieron expandirse flpidamente a traves de La Espafiola (Ottenwalder [1991])
Es opinion generalizada que los almiqufes antillanos constituyen el grupo de mamiferos que primero se emplazo en el territorio insular (MacPhee 2005) y en tal sentido es de interes la evidencia sobre especificidad parasito-hospedante aportada por Cruz (1980) quien encontro que la especie de acaro (genero Cubanochirus) que parasita a los almiquies es mucho mas primitiva que la especie (del mismo genero) que paras ita a las jutias y de aqui concluyo que dicho genero de acaro debio evolucionar primero en Solenodontidae y posteriormente en Capromyidae al entrar en contacto representantes de ambas familias en Cuba 0 La Espafiola
Recientemente Roca (2004) y Roca et al (2004) dieron a conocer que los resultados de su estudio genetico-molecular sobre los almiquies indican una divergencia de 25 Ma entre Solenodon cuban us y S paradoxus -compatible con la disyuncion tectonica entre Cuba y La Espafiola (Iturralde-Vinent y MacPhee 1999 McPhee et al 2003)-- y una separacion de ambas especies comparable en magnitud a la divergencia entre familias de mamlferos Consecuentemente propusieron que Solenodon cubanus fuera clasificada en el genero Atopogaie como indico Cabrera (1925) hace 82 afios
DISCUSION Y CONCLUSIONES
Las musarafias y los roedores antillanos (excepto Oryzomys) se irradiaron tanto de sus respectivos ancestros continentales que de estos 6rdenes se reconocen familias y subfamilias endemicas de la region en contraste con los perezosos (una subfamilia y tres
321 IRRADIACION EVOLUCIONARIA
tribus endemicas) y los monos (una tribu endemica) como expresion de diferentes niveles de diversificacion evolucionaria Este proceso es dificil de explicar (traUindose de c1ados que pnicticamente comparten la misma antigiiedad en las Antmas) si es correcta la interpretacion de Pascual et at (1990636)
Nosotros consideramos estos diferentes niveles de diversificacion como las diferentes etapas de evolucion alcanzadas por cada grupo [de mamiferos] Obviamente esto estuvo relacionado con las diferentes longitudes de sus historias
Pudiera ocurrir que en el futuro se compruebe que la historia biogeognifica de la region fue mucho mas complicada
La Espanola la isla mas ecol6gicamente divers a de la region debi6 ser el centro principal de evoluci6n de los mamfferos (con 87 de las familias y 73 de las subfamilias) seguida por Cuba (con 75 de las familias y 54 de las subfamilias) las dos islas de las Antillas Mayores mas relacionadas biogeograficamente (Fig 43 Tabla 44)
eL
EL
cb CAIMAN
SWAN
A Acratocnini B Cubanocnini C Megalocnini D Mesocnini E Nesophontidae F Solenodontidae G Xenothrichini
ANGUILAshyM-0 SMARTIN
H Heteropsomyinae I Plagiodontinae J Isolobodontinae
K Hexolobodontinae L Capromyinae
M Heptaxodontinae N CIidomyinae 0 Sigmodontinae
FIGURA 43 Representacion esquematica de las relaciones interinsulares (expresadas porel numero de linajes compartidos) entre las Antillas Mayores y entre estas y los grupos de islas perifericas segun las interpretaciones de los autores citados en el texto Los numeros dentro de los circulos indican la cantidad de linajes representados en cada isla relacion dudosa Contexto taxon6mico de los linajes en la Tabla 44
bull bull
bull bull
bull bull bull
bull bull
bull bull
bull
bull
bull
bull
bull bull bull bull
bull bull
bull bull
bull
bull bull bull
bull bull
bull
bull bull
bull
322 BIOGEOGRAFiA
TABLA 44 Distribuci6n de los linajcs de mamiferos terrestres a traves de las Antillas Mayores y grupos de islas perifericas LE = La Espanola CU Cuba JA Jamaica PR = Puerto Rico IC Islas Caiman IS Islas Swan IB = Islas Bahamas IV = Islas Virgenes AS = Anguila - San Martin
FamiliaSubfumiliafIribu LE CU JA PR IC IS IB IV AS
Megalonychidae
Choloepodinae
Acratocnini
Cubanocnini
Megalocninae
Megalocnini
Mesocnini
Nesophontidae
Solenodontidae
Pitheciidae
Callicebinae
Xenothrichini
Echimyidae
Heteropsomyinae
Capromyidae
Plagiodontinae
Isolobodontinae
Hexolobodontinae
Capromyinae
H e pta x 0 don tid a e
Heptaxodontinae
Clidomyinae
Muridae
Sigmodontinae
PILOSA
SORICOMORPHA
PRIMATES
RODENTIA
bull
bull
bull
bull TOTALES 12 10 4 4 2
323 EXTlICIOIES DEL CUATERNARIO
El gigantismo ha sido sin duda una tendencia evolucionaria recurrente entre los clados mayores de mamiferos antillanos Es un hecho bien conocido que algunos grupos de mamiferos han aumentado de tamafio en las islas y varios autores han intentado explicarlo a partir de enfoques diferentes (Foster 1964 Miller 1966 Sondaar 1977 Case 1978 Eisenberg y Wilson 1978 Heaney 1978 Lawlor 1982 Lomolino 1985 Angerbjom 1985 McNab 19942001)
EXTINCIONES DEL CUATERNARIO
Las islas que integran la Subregion Antillana han perdido casi 90 de sus mamiferos terrestres durante los ultimos milenios y en proporcion con su extension territorial esta perdida ha sido mayor en la Subregion que en cualquier otro lugar del planeta (MacPhee 1996b) Las extinciones del Cuatemario eliminaron mas de las tres cuartas partes de la fauna de mamiferos terrestres conocida de las Antillas Mayores sus islas satelites y grupos de islas perifericas y el examen de este proceso en sus aspectos mas generales se aborda en esta ultima parte del presente capitulo
CRONOLOGiA Y CONSECUENCIAS
Es muy improbable que un fosil represente el ultimo individuo de un taxon extinto y entonces hay que guiarse por la edad radiometrica del fosil mas reciente conocido lIamada fecha de ultima presencia- como unica aproximacion posible a la fecha de extincion del taxon
Como se explico al inicio del capitulo (vease Contexto Temporal) el registro fosH de los mamiferos terrestres de las Antillas Mayores adolece de insuficiente sustentacion radiometrica No obstante la distribucion temporal de los escasos fechados disponibles para el Cuaternario de la region abarca etapas criticas del proceso de extincion experimentado por la referida fauna a 10 largo de los ultimos 250 mil afios
De las 52 especies que es posible reconocer actualmente como miembros extintos de la fauna cuatemaria de mamiferos terrestres de las Antillas Mayores y grupos de islas perifericas los restos fosiles de 23 de elIas (442) han sido fechados radiometricamente o se han hallado asociados estratigraficamente de manera inequivoca con otro material fechado De estos 23 fechados 3 corresponden al interglacial Sangamon 4 pertenecen a la secuencia Pleistoceno terminal - Holoceno temprano 10 se ubican en ellapso entre el arribo de los aborigenes y la incursion europea y 6 son postcolombinos (a los cuales hay que sumar otras dos especies extinguidas en los siglos XIX y xx) A partir de estas 25 especies avaladas por fechas de ultima presencia distribuidas entre los cuatro intervalos temporales sefialados hemos compuesto la Tabla 45 en la que ademas aparecen las restantes 27 especies (carentes de fechado radiometrico) ubicadas en el intervalo temporal mas probable sobre la base de (1) la asociacion estratigrafica de sus restos con evidencias culturales (amerindias 0 europeas) 0 (2) la ausencia de dicha asociacion Sobre estos criterios de ubicacion temporal procede hacer dos aclaraciones
324 BIOGEOGRAFIA
TABLA 45 Intervalos temporales mas probables para las extinciones cuatemarias de los mamiferos terrestres de las Antillas Mayores y grupos de islas perifericas sobre la base de las evidencias siguientes (l) ultimo ejemplar colectado u observado en vida (2) fechado radiometrico mas tardio en material de la especie (3) asociaci6n estratigrafica mas tardia con otro material fechado (4) asociaci6n estratigrafica con restos de animales introducidos restos humanos u objetos culturales propios del intervale (5) falta de asociaci6n con evidencias culturales [Los ntimeros entre parentesis en la columna Evidencia significan adscripci6n tentativa al intervalo] Los estimados del peso corporal (en kg) estan tomados de la literatura 0 inferidos comparativamente y distribuidos entre
las clases siguientes (a) S 01 (b) 01 a lt 1 (c) 1 a lt 10 (d) 10 a lt 100 (e) 100 a lt 200 (j) ~ 200 Las
abreviaturas que designan las islas son las de la Tabla 44
F amilialEspecie Peso IsJa Evidencia Fuente
I httervalo htterg1acial Sangamon (gt80 kaAP)
HEPTAXODONTlNAE
Amblyrhiza inundata e AS 3 McFarJane et al 1998a
CLIDOMYINAE
Clidomys osborni d JA 3 Ibid
Clidomys parvus d JA 3 Ibid
II httervalo Pleistoceno terminal ~ Holoceno temprano (20-6 kaAP)
MEGALONYCHIDAE
Aeratoenus antillensis d CD 5 Arredondo 1975 a
Aeratoenus odontrigonus d PR 3 McFarJane 1999b
Aeratoenus simorhynehus d LE 2 Rega et al 2002
M egaloenus zile f LE 2 MacPhee et al 2000
SOLENODONTIDAE
Solenodon arredondoi e CD 5 Morgan y Ottenwalder 1993
PnHECIIDAE
Paralouatta varonai d CD 5 MacPhee 1996a
EcHIMYIDAE
Heteropsomys antillensis c PR 5 McFarJane 1999b
Heteropsomys insulans c PR 5 Ibid
Puertoricomys eorozalus c PR 5 Woods 1989a
CAPROMYIDAE
P lagiodontia araeum c LE 5 MacPhee et al 2000
Macrocapromys latus c CD 5 Varona y Arredondo 1979
(Continua)
325 EXTINCIONES DEL CUATERNARIO
TABLA 45 (Continuaci6n)
F ami1ia1Especie Peso Isla Evidencia Fuente
M acrocapromys latus c CU 5 Varona y Arredondo 1979
Mesocapromys kraglievichi c CU 5 Ibid
HEPTAXODONTINAE
Xaymaca fulvopulvis a JA 2 MacPhee y Flemming 2003
III Intervalo Amerindio (6-05 kaAP)
MEGALONYCHIDAE
Acratocnus ye d LE 2 MacPhee et aI 2000
Neocnus comes c LE 2 Ibid
N eocnus dousman c LE 2 Ibid
N eocnus toupiti c LE 2 Ibid
N eocnus gliriform is c CU 4 Cordova et ai 1997
M egalocnus rodens f CU 2 MacPhee et ai 2007
Parocnus browni e CU 2 Steadman et ai 2005
P arocnus serus e LE 2 MacPhee et ai 2000
N ESOPHONTIDAE
N esophontes edithae a PR 2 McFarlane 1999b
PITHECIIDAE
Antillothrix bernensis c LE 3 Rimoli 1977
EcHIMYIDAE
Brotomys contractus a LE 4 Miller 1929c
CAPROMYIDAE
P lagiodontia spelaeum c LE 4 Ibid
M acrocapromys acevedo c CU 4 NUiiez 1990a
HEPTAXODONTINAE
Elasmodontomys obliquus d PR 3 McFarlane 1999b
Iv Intervalo Postcolombino (05 kaAP - Presente)
N ESOPHONTIDAE
N esophontes hypom icrus a LE 24 MacPhee y Fle111111ing 1999
N esophontes zam icrus a LE (4) Ibid
N esophontes m icrus a CU 24 MacPhee et aI 1999
(Continua)
326 BIOGEOGRAFiA
TABLA 45 (Continuacion)
F amilialEspecie Peso Isla Evidencia Fuente
N esophontes m icrus a CD 24 MacPhee et aI 1999
N esophontes m icrus a LE 4 MacPhee et aI 2000
N esophontes sp 1 a IC 4 Morgan 1994
N esophontes sp 2 a IC 4 Ibid
Solenodon marcanoi b LE 4 Ottenwalder 2001
PITHECIIDAE
Xenothrix mcgregori c JA 4 MacPhee y Flemming 1999
EcHIMYIDAE
Boromys offeUa a CD 24 Guarch 1984
Boromys torrei a CD 24 Ibid
Brotomys voratus a LE 4 MacPhee et aI 2000
CAPROMYIDAE
P lagiodontia ipnaeum c LE 4 MacPhee y Flemming 1999
Rhizoplagiodontia lemkei c LE 4 Ibid
Isolobodon portoricensis c PR 24 Ibid
Isolobodon montanus c LE 4 Ibid
Hexolobodon phenax c LE (4) Ibid
Capromys sp c IC 24 Morgan 1994
Geocapromys columbianus c CD 4 MacPhee y Flemming 1999
Geocapromys thoracatus b IS 1 MacPhee y Marx 1997
Geocapromys sp 1 c IC 4 Morgan 1994
Geocapromys sp 2 c IC (4) Ibid
l-IEPTAXODONTINAE
Quemisia gravis c LE (1) MacPhee y Flemming 1999
MURIDAE
Oryzomys antillarum a JA 1 Ibid
1 Flemming y MacPhee (1999) alertaron en contra de la asociacion con otro material fechado Asimismo Morgan (1993) indico que la ausencia de asociacion con evidencias culturales amerindias no es necesariamente un criterio concluyente para ubicar un taxon en el intervalo temporal anterior porque hay depositos en la region que carecen de tales evidencias y son del Holoceno tardio por fechados radiocarbonicos [No obstante
327 EXTlNCIONES DEL CUATERNAR10
la ausencia de una especie en residuarios arqueol6gicos adquiere mayor certidumbre como criterio de ubicaci6n temporal de su extinci6n en la medida que aumenta el numero de residuarios estudiados que en las Antillas Mayores se cuentan por centenares] A falta de fechado absoluto de una especie consideramos que estos criterios de ubicaci6n temporal son mas utiles que la alternativa de excluir del analisis a la especie
2 En los trabajos sobre biogeografia de mamiferos antillanos los arque610gos mas citados como autoridades para establecer el inicio de la presencia aborigen en las Antillas Mayores (Kozlowski 1974 Rouse y Allaire 1978 Rouse 1989 1992 Wilson 2001) difieren al respecto entre los 6 kaAP y los 45 kaAP (pero vease Pagan et al 200523) y hemos adoptado la mas temprana de estas fechas para el comienzo del intervalo amerindio en la Tabla 45 De todos modos cualquiera que haya sido la fecha esta no es aplicable por igual al conjunto de las Antillas Mayores porque segun algunos de los autores citados el poblamiento comenz6 por Cuba y La Espanola 6 mil anos atras continu6 en Puerto Rico hace alrededor de 3 mil anos (pero vease Burney et al 1994) yen Jamaica hace menos de 2 mil e incluso a las Islas Caiman ni siquiera llegaron [Las Islas Caiman descubiertas por Col6n en 1503 carecen absolutamente de evidencia amerindia (Stokes y Keegan 1996)]
Si los datos de la Tabla 45 representan una aceptable aproximaci6n a la realidad el proceso de las extinciones cuaternarias de los mamiferos terrestres de las Antillas Mayores y grupos de islas perifericas debi6 caracterizarse entre otros por los eventos siguientes
Aunque obviamente la fauna cuaternaria debi6 ser mucho mas numerosa que 10 que refleja el registro paleontol6gico (Morgan y Woods 1986) 52 especies (81 de la fauna conocida) desaparecieron (sin reposici6n) durante los ultimos 250 mil anos
- Al parecer las extinciones se concentraron en las especies de mayor tamano Asi el peso promedio estimado para las especies de la etapa preantr6pica (intervalos I + II de la Tabla 45) es 350 kg es 328 kg para las especies del intervalo III (amerindio) y es s610 25 kg (c1ases del peso a b c) para las especies del intervalo IV (postcolombino) Ninguna especie aut6ctona de mamifero mayor de 6 kg existe hoy en la regi6n
- De acuerdo con la distribuci6n temporal de las extinciones 61 especies sobrevivieron al interglacial Sangamon (intervalo 1)48 sobrevivieron a la transici6n del Pleistoceno terminal al Holoceno temprano (intervalo II) 34 sobrevivieron al periodo amerindio (intervalo III) y s610 12 especies han sobrevivido desde la Ilegada de los europeos (intervalo IV) Las tasas de extinci6n correspondientes (exceptuado el intervalo I) son una especie cada 1167 anos en el intervalo II una cada 393 anos en el intervalo III y una cada 23 anos en el intervalo IV
- Durante todo el proceso de las extinciones cuaternarias la perdida de especies por isla fue de 91 en La Espanola 83 en Jamaica y 62 en Cuba Bahamas retuvo su unica especie (aunque extirpada en varias de las islas) Puerto Rico e Islas Caiman perdieron todas sus especies (6 y 5 respectivamente) y las Islas Swan Virgenes y Anguila - San Martin perdieron sus respectivas especies (micas
328 BIOGEOGRAFiA
De los cuatro 6rdenes de mamiferos representados en la fauna Pilosa y Primates desaparecieron totalmente Soricomorpha y Rodentia perdieron 86 y 74 de sus especies respectivamente
CAUSALIDAD
CAUSAS NATURALES
Los cicIos glaciales e interglaciales acaecidos en el Pleistoceno causaron cambios ambientales dnisticos en las Antillas con la concomitante alternancia de cIimas frios (secos) y calidos (hUmedos) asi como de descensos y ascensos del nivel del mar aumento y disminuci6n de las areas territoriales) induso (aunque con menor intensidad) en el Holoceno Estos eventos paleogeograficos han sido consistentemente documentados (Mayo y Kartashov 1972 Bonnatti y Gartner 1973 Ortega y Arcia 1982 Prance 1982 Schubert y Medina 1982 Dodge et al 1983 Ortega [1984] Haq et al 1987 Schubert 1989 Hodell et al 1991 Guilderson et al 1994 Oppo 1997 Emslie 1998 Curtis et al 2001 Iturralde-Vinent 2003) y de ellos brindamos a continuaci6n una breve resena
Finalizando el periodo interglacial anterior a la ultima glaciaci6n (Wisconsin) el mar estaba a 8-10 m por encima de su nivel actual Cuando el avance glacial del Wisconsin a1canz6 su culminaci6n (eillamado Ultimo Maximo Glacial) hace 18 ka el mar habia descendido 120 m con respecto a dicho nivel y las islas del Caribe habian incrementado considerablemente sus extensiones subaereas en comparaci6n con las que presentan hoy dia por ejemplo Puerto Rico y las Islas Virgenes se conectaron territorialmente y las Bahamas se unieron en cinco gran des islas Prevalecia un regimen frio y seco y condiciones de aridez --debidas a una reducci6n de las precipitaciones (posiblemente a menos de 440 mmlano)- y los ambientes xericos se habian extendido a traves de las islas y bancos emergidos y abundaban los pinares (los habitats mesicos estaban restringidos a parcelas relictas en 10 alto de las montanas) En el Mar Caribe y el Golfo de Mexico la temperatura superficial del mar era 5degC mas baja que la actual y en las llanuras costeras y alturas maximas eran respectivamente 2-3degC y 6degC mas bajas Los limites de las zonas ecol6gicas altitudinales descendieron varios cientos de metros y cada zona redujo su extensi6n vertical al aumentar el gradiente de temperatura Ellimite inferior de la zona periglacial
elevaci6n donde la temperatura media minima a1canza los OdegC) de la Cordillera Central de la Republica Dominicana (altura maxima del Caribe insular con evidencia de topografia glacial) se considera que estuvo entre los 1500 y 1800 mlnm (actualmente esta a 2800 mlnm) y se puede estimar que dicho limite era inferior en Cuba por estar mas al norte que la Republica Dominicana No obstante durante este periodo la temperatura experiment6 episodios cortos (unas pocas decadas) de ascensos repetidos
Un signifieativo componente bi6tieo de las referidas condiciones paleoambientales 10 constituy6 la espectacular irradiaci6n evolucionaria experimentada por un grupo numeroso de aves rapaees diurnas y nocturnas (Wetmore 1937 Brodkorb 1969 Kurotchkin y Mayo 1973 Arredondo 1976 Olson 1978 Olson y Hilgartner 1982
329 EXTINCIONES DEL ClJATERNARIO
Suarez 2000a b 2004 Suarez y Emslie 2003) Los mamiferos de las Antillas Mayores ofrecieron a estos depredadores aviares una amplia gama de tamanos de presa que en ausencia de mamiferos del orden Carnivora en la region permitio a estas aves irradiarse a este nicho alimentario con la evolucion de una variedad de especies de muy diversa tall a corporal incluidas formas gigantes Adquirir un cuerpo gigante les propicio el acceso a recursos alimentarios que no estaban siendo explotados por ningun otro depredador (Alcover y McMinn 1994) A Cuba corresponde el mejor registro fosil de rapaces cuaternarias caribenas con siete generos de especies diurnas Dentro de estas aves existieron tres gavilanes gigantes (Accipitridae) tres carairas (Falconidae) y un condor (Cathartidae) asi como cuatro generos de especies nocturnas dos lechuzas (Tytonidae) y tres Mhos (Strigidae) dos de ell os no igualados en tamano en todo el mundo Algunos de estos generos tuvieron representacion en otras islas antillanas como La Espanola Jamaica Bahamas y Gran Caiman
Al retirarse hacia el norte los hielos de la glaciacion Wisconsin el clima se tomo mas hUmedo en las Antillas durante el Holoceno temprano y el bosque mesico se expandio en la region insular El nivel del mar ascendio acompanado de mayor pluviosidad alrededor de los 7 kaAP las Islas Virgenes quedaron separadas entre si y de Puerto Rico y las Bahamas perdieron la mayor parte de su territorio Cerca de los 3 kaAP las condiciones secas retornaron a la region e hicieron desaparecer buena parte del bosque mesico Nuevamente el clima comenzo a adquirir sus condiciones actuales hacia los 25 kaAP aproximadamente y hoy transita por una fase mesica con vegetacion xerica confinada en parcelas relictas
Sin duda estos y otros eventos naturales incidieron negativamente en la sobrevivencia de mamiferos terrestres en las Antillas Mayores asi como en los grupos de pequenas islas perifericas (Pregill y Olson 1981 Olson 2006)
McFarlane et al (1998a) demostraron convincentemente como Amblyrhiza el gigante de los roedores antillanos endemico de la pequena isla de Anguila sucumbio a las repetidas y abruptas perdidas de su habitat durante el interglacial Sangamon por las nipidas y severas oscilaciones del nivel del mar (documentadas por Neumann y Hearty 1996) al no poder responder adecuadamente a tan catastroficos eventos mediante el ajuste de su masa corporal media y otro tanto debio ocurrirle en Jamaica al otro roedor gigante Clidomys (McFarlane et al 1998b) emparentado con Amblyrhiza en la misma epoca (Tabla 45 intervalo I)
El ascenso del nivel del mar seguramente promovio la extincion de mamiferos durante la transicion del Pleistoceno al Holoceno (Tabla 45 intervalo II) con la reduccion del tamano de las Islas por las transgresiones marinas (Pregill y Olson 1981 Morgan 1993) Esto ultimo haria maximamente severa la competicion asf como la presion de los depredadores [Steadman et al (2005) no yen por que el calentamiento global ocurrido durante la transicion Pleistoceno-Holoceno habria de tener un efecto negativo sobre los perezosos siendo sus descendientes actuales especies tropic ales 0 subtropicales pero no se trata del efecto
330 BIOGEOGRAFiA
directo del cambio de temperatura sobre los animales sino del efecto indirecto y progresivo que dicho cambio genera a largo plazo al reducir el territorio insular disponible]
Por esta epoca tambien desaparecieron especies de tall a mayor como Megalocnus zile en La Espanola y otras de menor tamano (en Cuba) pero que representan la tall a maxima en sus respectivos contextos taxonomicos del Cuaternario como Solenodon arredondoi Paralouatta varonai y Macrocapromys acevedo Un mamifero puede ser grande solo a costa de reducir su densidad poblacional y esto 10 hace mis vulnerable a la extinci6n sobre todo en una isla (McNab 1994 McFarlane et al 1998a) Incidentalmente Alcover et al (1998a) afirmaron que larelacion entre la talla corporal y la extincion no siempre se cumple en las especies de menor tamano y citaron como ejemplo que entre las musarafias todas las especies de Nesophontes desaparecieron mientras sobreviven especies de Solenodon pero la comparacion correcta en todo caso serfa entre los solenodones vivientes y el gigante Solenodon arredondoi y entre Nesophontes edithae -el gigante de los nesofontes extinguido en el periodo amerindio- y el resto de sus congeneres que sobrevivieron hasta los primeros siglos de la colonizacion europea (Tabla 45 intervalos III y IV)
CAUSAS ANTROPOGENICAS
Intervalo Amerindio
La mayoria de los autores que se han referido a las extinciones acaecidas durante este intervalo (entre elIos paradigmaticamente Martin 1984a b) han responsabilizado a los aborfgenes (que se supone comenzaron a poblar las Antillas Mayores hace 6 mil afios) con la desaparicion de numerosas especies de mamiferos terrestres en la region principalmente por la via de la sobrecaza y la introduccion de animales domesticos devenidos depredadores de la fauna autoctona (Morgan y Woods 1986) Este efecto negativo de la presencia indigena sobre los mamiferos de la region ha sido dramatizado por algunos autores en terminos de holocausto faunal Sin embargo pensamos que se ha exagerado Como han expresado MacPhee et al (19993) cuando uno busca evidencia dura de complicidad humana directa en [estas] extinciones no hay mucho que encontrar Las siguientes son algunas consideraciones a tener en cuenta en tal sentido
El registro radiometrico mas joven conocido hasta ahora para un perezoso antillano (MegalocnllS rodens) es de 42 kaAP (MacPhee et ai 2007) Si se acepta que los aborfgenes entraron en Cuba hace aproximadamente 6 mil anos su convivencia con los perezosos debe haber durado alrededor de 1800 afios (0 mas porque dificilmente el referfdo fechado radiometrico haya correspondido al ultimo sobreviviente de la especie) Esta cifra contradice la hipotesis del holocausto faunal segun la cualla extincion de los mamiferos insulares de mayor talla debe ocurrir con extrema rapidez -a escala de decadas 0 de centurias- a partir de su primer contacto con los humanos (Martin y Steadman 1999) A pesar de las repetidas afirmaciones en contrario (Harrington 1921 Miller 1929c Herrera [1945] Tabio 1951 Alvarez 1951a 1957 Arredondo
331 EXTINCIONES DEL CUATERNARIO
1959 1960a Veloz y Ortega 1973 Mayo 1980b Rodriguez et al 1984 Castellanos y Pino 1985 Pino y Castellanos 1985 Nunez 1990b Gutierrez 1996 Arredondo 2004) en su mayoria basadas en evidencias no exentas de posibles accidentes de deposicion (0 de reubicacion postdeposicional) en recintos cavernarios (Diaz-Franco [2006b D no se ha demostrado inequivocamente que los aborigenes antillanos se alimentaran de perezosos regularmente Sus restos en residuarios de alimentos son escasisimos en toda la region y esto contrasta con los abundantes restos de jutias y otros mamiferos de menor talla que aparecen en esos residuarios demostrativos de la preferencia de los paleoindios por estos otros mamiferos
Los mamiferos mas importantes en la economia prehistorica de las Antillas Mayores fueron las jutias Capromys y Geocapromys en Cuba Geocapromys en Jamaica e Isolobodon en La Espanola Puerto Rico e Islas Virgenes (Aguayo 1950b Tablo 1951 Rimoli 1972 Wing 2001 Diaz-Franco [2006b D Wilkins (2001) estudi61ajutia viviente de Jamaica (Geocapromys browni) en un sitio ocupado por los aborigenes durante 100 anos y no encontr6 evidencias de que durante ese tiempo la especie haya sido sobreexplotada Asimismo en Cuba las jutias conga carabali y andaraz son escasas en depositos pleistoholocenicos en cambio son muy numerosas en residuarios indigenas y de haber sido sobreexplotadas 10 mas probable es que no hubieran sobrevivido hasta hoy
La habilidad de los humanos para penetrar en profundidad el bosque tropical y mantenerse en este sustentablemente ha sido un evento comparativamente reciente (Beran y Steele 1994) En el caso antillano la caza tierra adentro ha sido explicada como un proceso tardio de desplazamiento a partir de la explotacion de abundantes recursos marinos que ofrecian condiciones 6ptimas para el sedentarismo de las comunidades aborigenes costeras (C6rdova y Arredondo 1988 Brito 2005) Peces y mariscos dominan la arqueologia costera de las Islas antillanas en tanto el consumo de recursos terrestres (mamfferos incluidos) aumenta progresivamente en la medida que los residuarios estudiados se alejan de la costa (Scudder 1991 Wilkins 2001) En las Antillas Mayores el componente terrestre de la dieta aborigen es solo 20-30 del material recobrado en los residuarios (Wing 1977 1989) Y debio ser menor alin porque el componente vegetal de la dieta no se ha preservado Por otra parte los aborigenes cubanos domesticaron la jutia conga y seguramente la reprodujeron para comer (Pose et al 1988) practica que probablemente se repitiera con otras jutias en las demas Antillas Mayores
- Fray Bartolome de las Casas hablo de 3 millones de indios en Cuba al inicio de la colonizacion pero los historiadores subsecuentes consideraron exagerada la cifra (Poey 1851 a) No obstante se dispone de dos estimados de la densidad de poblaci6n aborigen en las Antillas Mayores a la llegada de los europeos 18 indigenaslkm2 en Puerto Rico (Rouse 1956) y 1271km2 en La Espanola (Watts 1987) Extrapolando ambos valores entre las cuatro islas principales de las Antillas Mayores tenemos que en la fecha indicada la poblaci6n aborigen pudo estar entre 16 mil y 113 mil indigenas en Puerto Rico entre 21 mil y 145 mil en Jamaica entre 141 mil y 1 mill6n en La Espanola y entre 200 mil y 14 millones en Cuba Entre las cifras maximas y las minimas nos inclinamos
332 BIOGEOGRAFiA
hacia estas ultimas atendiendo a los altos indices de mortalidad infantil y morbilidad que se evidencian en las osamentas de los pobladores prehispanicos (Martinez y Rives 1990) Uno de los hechos que apuntan hacia el bajo numero de aborigenes en las Antillas Mayores es 10 rapido que desaparecieron despues de la llegada de los europeos pnkticamente sin dejar mestizaje con estos 10 cual contrasta ostensiblemente con la situacion en la America continentaL
En suma un grupo humano (1) demograiicamente irrelevante y controlado por altos indices de morbilidad (2) consumidor mayoritariamente de organismos marinos y (3) carente de la tecnologia cinegetica necesaria para poder sobreexplotar los mamiferos que Ie sirvieron de alimento (a los cuales accedio tardiamente en su desarrollo) no es concebible que haya causado e1 holocausto faunal que se Ie atribuye a partir del error de aplicar a las comunidades aborigenes antillanas un modelo cultural de la prehistoria continentaL
Tampoco hay evidencias de que los aborigenes hayan introducido carnivoros con suficiente capacidad depredadora salvo el perro Morgan y Otlenwaldder (1993) consideraron que la depredacion por perros durante el periodo amerindio es la explicacion mas plausible para la extincion del almiqui gigante de Cuba pero como se puede observar (Tabla 54) Solenodon arredondoi aparece en el intervalo II por falta de asociacion con evidencias culturales
Consecuentemente el accionar de los aborigenes y de sus animales domesticos directamente sobre la fauna no parece haber sido por sf solo un factor determinante de la extincion de mamiferos terrestres durante el periodo amerindio en las Antillas (pero vease Steadman et al 2005)
Hay que recordar en este senti do que alin en el Holoceno tardio ocurrieron cambios clirruiticos de consideracion (vease Causas Naturales) Por ejemplo Curtis et al (2001) explicaron que la extincion de algunos ani males mesofilicos de las alturas montanosas de La Espanola pudo haber antecedido a cualquier impacto humano sigrIificativo en esos lugares por coincidir dicha extincion con una tendencia a la aridez documentada isotopicamente y con evidencias de la perdida del bosque mesico ocurridas despues de 3kaAP Yen Jamaica hace solo 2shy3 mil aDos el area de la Bahia de Jackson sustentaba una gran biomasa de vertebrados en bosque hinnedo a juzgar por la cantidad y diversidad de taxones presentes en material fosil derivado de la depredacion por lechuzas hoy el area es la mas seca de la isla con matorral xerofitico y pobreza de fauna (MacPhee 1996a) Yalm en nuestros dias huracanes de gran intensidad aniquilan poblaciones de vertebrados (que no se recuperan) en is las pequenas y llanas del Caribe (Brooks y Smith 2001)
En medio de la incertidumbre sobre la verdadera causa (0 causas) de las extinciones de mamiferos en el periodo amerindio MacPhee y Marx (1997) han propuesto su hipotesis de la hipermorbilidad la cual estipula colapsos poblacionales relativamente rapidos con la introduccion de enfermedades de alta letalidad Estos autores han razonado que
Las islas son los lugares mas apropiados para experimentar perdidas catastroficas causadas por enfermedades Los mamiferos no solo serian susceptibles en virtud de su prolongado aislamiento biologico sino que en los confines de una isla la epizootia
333 EXTINCIONES DEL CUATERNARIO
tendria la potencialidad de infectar simultaneamente a la rnayoria de los individuos de vadas especies Los neonatos son inmunologicamente inmaduros y sufririan desproporcionadamente cualquier infeccion hipervirulenta La perdida excesiva de neonatos y juveniles tendria efectos severos para la sobrevivencia de la poblacion si esta se reduce por debajo de cierto nivel sobreviene la extincion porque no hay posibilidad de restitucion en el periodo postpanzootico Las especies de mayor talla corporal van a ser las mas vulnerables a la extincion dadas sus bajas tasas reproductivas largos periodos de gestacion y reducido numero de progenie Las especies de menor tamano --caracterizadas por altas tasas reproductivas periodos cortos de gestacion y mayor numero de neonatos por parto- tambien serian fuertemente diezmadas pero muchas sobrevivirian porque su estrategia reproductiva les permite perder mayor numero de individuos Las perdidas deben ocurrir relativamente rapido (a escala de decadas) mientras que otras hipotesis (digamos los cambios climaticos) permiten intervalos mucho mas largos (a escala de siglos 0 de milenios) entre la aparicion de las causas y la produccion de sus efectos Por ejemplo todo indica que los nesofontes se extinguieron muy rapidamente y seria dificil poder invocar un impacto humano directo 0 argumentar fenomenos climaticos que pudieran explicar plausiblemente perdidas tan rapidas
MacPhee y Marx (1997 191) han dicho que por el momento la hipotesis de la hipermorbilidad no pasa de ser una abstraccion en espera de adecuada sustentacion empirica Llama la atencion que linicamente en el periodo amerindio (Tabla 45 intervalo 1Il) las extinciones abarcaron todas las clases del peso corporal (de la a a laj) y que ninguna de las especies que sobrevivieron hasta el periodo postcolombino (Tabla 45 intervalo IV mas las especies vivientes) sobrepasa los 10 kg (clases del peso a b c) en clara coincidencia con los presupuestos de la hipotetica hipermorbilidad
Intervalo Postcolornbino
Por 10 menos 37 especies de mamiferos terrestres vivian en las Antillas Mayores y grupos de Islas perifericas a la lIegada de los europeos en 1492 De elIas se han extinguido 23 shymas especies que en cualquiera de los periodos precedentes (Tabla 45~ y solo subsisten 12 especies repartidas entre Cuba (8 especies) La Espanola (2) Jamaica (1) y Bahamas (1) varias de estas en peligro critico de extincion Solo dos de las especies extinguidas durante los ultimos 500 anos se conocieron en estado viviente los roedores Oryzomys antillarum de Jamaica (extinguido en 1877) y Geocapromys thoracatus de las Islas Swan (extinguido en 1955) Las causas de las extinciones ocurridas durante este periodo son sobradamente conocidas destruccion de habitats introducci6n de animales exoticos (domesticos y comensales) y demas presiones asociadas a la actividad humana entre las cuales posiblemente habria que incluir brotes de hipermorbilidad
Durante mucho tiempo se ha afirmado la posibilidad de que especies de Nesophontes hayan sobrevivido hasta las primeras decadas del siglo xx y que incluso continuen existiendo en lugares remotos de Cuba 0 La Espanola (Allen 1918 1942 Miller 1930 Aguayo 1950a Arredondo 1960b Fischer 1977 MacFadden 1980 Woods y
334 BIOGEOGRAFiA
Ottenwalder 1991) Actualmente no existe evidencia radiometrica de que alguna especie de Nesophontes haya sobrevivido al siglo xv las fechas radiocarbonicas calibradas obtenidas en muestras de Nesophontes de Cuba y La Espanola resultaron no mas tardias que los siglos XIII y XIV (MacPhee et al 1999) Por otra parte las expediciones efectuadas a fines del siglo xx con el proposito de localizar individuos vivientes del genero en La Espanola no produjeron la menor indicacion afirmativa al respecto (Woods et al 1985) Ademas a traves de las Antillas Mayores no aparecen restos de Nesophontes en los depositos formados actualmente por las lechuzas
EPiLOGO
Las Antillas Mayores son islas oceanicas no han formado parte de un continente (con la posible excepcion de una parte de Jamaica hasta el Eoceno Medio hace 45 Ma) y durante un tiempo geologico considerable (desde el Eoceno Superior hace 40 Ma) han permanecido emergidas y separadas de los continentes vecinos Consecuentemente los mamiferos terrestres (no volantes) autoctonos de estas islas constituyen una fauna oceanica que por ende cumple con todas las caracteristicas convencionales reconocidas para este tipo de entidad biogeografica
- Baja diversidad taxonomica a nivel de las categorias superiores (solo cuatro ordenes) en comparacion con islas plataformicas continentales (0 con areas continentales) de tamano y diversidad ecologica simi lares
- Alto grado de endemismo a nivel de genero (96) y familia (50) en respuesta al prolongado aislamiento
- Desbalance ecologico de la estructura trofica ausencia de mamiferos camivoros y ungulados
- Espectaculares procesos de irradiaci6n evolucionaria como resultado de la colonizacion de territorios libres de competidores y depredadores
Tendencias evolucionarias extremas gigantismo
- Control a cargo de aves rapaces con tendencia at gigantismo en ausencia de mamiferos miembros del orden Carnivora
- Mas especies de reptiles y de murcielagos que de mamiferos terrestres relaciones que en los continentes se invierten
Alta tasa de extincion de especies (84) sin reposicion
Preservacion de especies filogeneticamente arcaicas (los llamados fosiles vivientes) los almiquies

298 BIOGEOGRAFIA
TABLA 4 1 (Continuacion)
I s a s Taxonom1a 1
CU JA LE PR IS IC IB IV AS
Geocapromys browni 7
Geocapromys columbianus bull Geocapromys ingrahami 6
Geocapromys thoracatus
Geocapromys sp 1
Geocapromys sp 2
Macrocapromys acevedo
Macrocapromys latus
Mesocapromys angelcabrerai
Mesocapromys auritus
Mesocapromys kraglievichi
Mesocapromys nanus
Mesocapromys sanfelipensis
Mysateles melanurus
Mysateles prehensilis 6
bull bull bull bull bull bull bull bull bull
HEPTAXODONTIDAE 8
HEPTAXODONTINAE
Amblyrhiza inundata
Elasmodontomys obliquus
Quem isia grav is
Xaymacafulvopulvis 9
CLIDOMYlNAE
Clidomys osborni
Clidomys parvus
MURIDAE2
SIGMODONTlNAE
Oryzomys antillarum 10
bull
bull bull bull
bull
bull
bull bull
bull bull
bull
bull Total de especies por is1all 24 6 25 7 1 5 1 1 1
1 La taxonomia sigue las revisiones sistematicas mas recientes (Morgan 1993 1994 MacPhee y Grimaldi 1996 Horovitz y MacPhee 1999 White y MacPhee 2001 Ottenwalder 2001 Woods et aI 2001 MacPhee y Flemming 2003) con 1a adici6n de los taxones descritos posteriormente y la incorporaci6n de los cambios propuestos para las especies cubanas en el Capitulo 2 de este Compendio (Continua en pagina opuesta)
299 ESPACIO Y TIEMPO
CONTEXTO TEMPORAL
La absoluta mayoria del registro fosil de mamfferos terrestres antillanos proviene de depositos cavemarios pleistoholocenicos (escala del tiempo geol6gico en la Tabla 42) Abundan en estas cavernas restos de perezosos musarafias y roedores siendo comparativamente escasos los de monos debido quizas a la universal predileccion de los platirrinos por los ambientes densamente forestados 10 que los hace malos candidatos a la fosilizacion (MacPhee y Woods 1982)
A pesar de 10 numeroso de tales depositos en la region son escasos los fechados radiometricos del cuantioso material extraido Con excesiva frecuencia (particularmente en Cuba) estos depositos han sido asignados al Pleistoceno Superior sin fundamento suficiente para ello y es sabido que incluso en cuevas contiguas los depositos fosilfferos pueden diferir considerablemente en edad (Pregill y Olson 1981) Se superponen a esta inconveniencia la falta de estudios tafonomicos en las excavaciones y la eventual mezcla de f6siles de distintas edades como consecuencia de las multicausales alteraciones estratigraficas ocurridas en las cuevas (Nunez 1990a b Morgan y Ottenwalder 1993 MacPhee 1997 McFarlane 1999b Dfaz-Franco 2004) Sobre propuestas de tipificaci6n de los depositos paleontologicos con restos de mamiferos terrestres en Cuba vease Woloszyn y Silva (1977) Acevedo y Arredondo (1982) Gutierrez (1994) Arredondo [1999]
La Tabla 43 presenta los fechados pleistocenicos mas tempranos registrados para restos de mamfferos terrestres del Caribe insular De estos restos el roedor (Cidomys) de Jamaica con un fechado de 253 mil anos antes del presente constituye el fosH de mamifero terrestre mas viejo del Cuaternario anti11ano [Los fosiles fechados antes del surgimiento de la espectrometria de masa (aceleradora) para muestras pequefias pueden ser algo mas viejos (Jull et al 2004)]
Corresponde a MacPhee e Iturralde-Vinent (1994 1995b) e1 relevante merito de haber descubierto en la ultima decada del siglo xx los primeros fosiles terciarios de mamiferos terrestres de las Antillas extendiendo a mas de 30 millones de afios el registro f6sil de
2 Extirpada en la Subregion Antillana pero viviente en el Neotropico continental
3 Ubicacion en subfamilia s610 sugerida por White y MacPhee (200 I)
4 Endemica de la Subregion Antillana
5 Recientemente propuesta su ubicaci6n bajo el genero Atopogale (Roca et al 2004)
6 lncluye las subespecies
7 Transportada por aborigenes a otras de las islas (Rimoli 1974 Olson 1982b este trabajo)
8 Vease MacPhee y Flemming (2003) sobre la incierta situaci6n de esta familia
9 Ubicada provisionalmente en esta subfamilia por MacPhee y Flemming (2003)
10 Hallada en contexto no perturbado de edad Pleistoceno tardio (McFarlane et ai 2002) con 10 cual se descarta su supuesta introduccion por amerindios (Woods 1990)
II La correlaci6n entre el area insular y la riqueza de especies es razonablemente fuerte (R=O83) en estas islas excepto las Bahamas (Morgan 1989)
300 BIOGEOGRAFIA
TABLA 42 Escala convencional del tiempo geologico en la Era Cenozoic a y finales de la Mesozoica (Gradstein 2004) ka = kiloafios (miles de afios) Ma miJIones de afios AP = antes del presente
Periodo(Suhperiodo) Epoca Afios (aproximados) antes del presente
Era Cenozolca
Cuaternario Holoceno Pkistoceno
Presente - 11 kaAP 11 kaAP - 18 MaAP
Terciario (Ne6geno ) Plioceno Mioceno
18 - 5 MaAP 5 - 23 MaAP
oIigoceno 23 - 34 MaAP Terciario (pak6geno) Eoceno 34 - 58 MaAP
Pakoceno 58 - 65 MaAP
Era Mesozo ca
Creticico 65 - 145 -faAP
~~~------~~--- --~-
este grupo animal en la region con el hallazgo de un perezoso oligocenico en Puerto Rico (Yauco) as como un perezoso un mono y un roedor miocenicos en Cuba (Domo de Zaza) Esto unido a la presencia de huesos de una musarafia nesofontoide [referida erroneamente a Solenodon por Rivero (1997)] en ambar oligomiocenico de la Republica Dominicana (MacPhee y Grimaldi 1996) demuestra incontrovertiblemente que al menos en el Mioceno temprano (alrededor de 20 millones de afios atnis) ya estaban en las Antillas Mayores representantes de los cuatro linajes basicos de su fauna de mamiferos terrestres [La afirmacion de Abreu et al (1990 13) en cuanto a que se conocen restos fosiles de almiqui que datan de hace aproximadamente 40 millones de afios es erronea]
Pero lcuan antigua puede ser la fauna de mamiferos terrestres de las Antillas Mayores
Aunque es un hecho bien establecido por la geologia que desde finales de la Era Mesozoica y principios de la Cenozoica existieron islas en el area ocupada ahora por las Antillas Mayores la evidencia disponible indica asimismo que aquellas primeras islas sufrieron repetidas transgresiones marinas totales que habrian hecho sucumbir las biotas que pudieran haber sustentado (MacFadden 1981 Morgan y Woods 1986 MacPhee e Iturralde-Vinent 1994 Roca et al 2004) sin contar los catastroficos efectos que debio tener para el Caribe occidental el b6lido extraterrestre que impacto en Chicxulub (Yucatan) hace 65 Ma (Tada et ai 2003) Hedges (1996) ha especulado que aUn en medio de tal devastacion debe haber existido algun que otro lugar donde elementos de la fauna pudieron haber sobrevivido
301 POBLAMIENTO CONTINENTE-ISLA
TABLA 43 F echados pleistocenicos mcis tempranos (obtenidos por distintos metodos) en mamiferos terrestres del segmento insular delimitado en laFig 41 F =Fuentes (I) Jull et al 2004 (2) Goodfriend y Mitterer 1987 (3) MacPhee et al 1989 (4) Woods 1989b (5) Rega et aI 2002 (6) McFarlane 1999b (7) McFarlane 1 999a (8) McFarlane et al 1998a
Pals Localidad Taxon kaAP F
Cuba Breas de San Felipe Pilosa (Parocnus) 1l9a 1
Jamaica Cueva Coco Ree Cueva Wallingford Roadside
Primates indet Rodentia (Clidomys)
393 2530
2 3
Haiti Trouing Jeremie no5 (casimba) Rodentia (Rh izoplagiodontia) 212 4
Republica Dominicana
Cueva del Perezoso Pilosa (ficratocnus) 185 5
Puerto Rico Cueva de Ia Vaca Cueva Clara
Pilosa (Acratocnus) Soricomorpha (Nesophontes)
337 346
6 7
Anguila Cueva Cave-No-Cave Rodentia (Amblyrhiza) 2060 8
a Kulp etal (1952) reportaron un fechado radiocarbonico de gt 30000 afios para un cono de pino en el deposito fosilffero de Ciego Montero (Cuba) con restos de perezosos
(vcase tambicn Crother y Guyer 1996) pero Iturralde-Vinent y MacPhee (1999) han preguntado donde estaban esos lugares si es que existieron
Solo a partir del Eoceno Superior las Antillas Mayores en fonnacion han pennanecido emergidas hasta hoy [excepto parte de Jamaica y el sur de La Espanola que continuaron sumergidas hasta el Mioceno Medio (Iturralde-Vinent y MacPhee 1999)] Esto significa que solo durante los ultimos 40 millones de anos aproximadamente han existido pennanentemente en dicha region insular ambientes terrestres capaces de sustentar comunidades biotic as (MacPhee e Iturralde-Vinent 2000 Portell et ai 2001 MacPhee 2005) 10 cual proporciona el marco temporal de referencia para conceptuar la antigiiedad maxima posible de la fauna de mamiferos terrestres de las Antillas Mayores
POBLAMIENTO CONTINENTE-ISLA
AFINIDADES CONTINENTALES
El consenso de los autores que en las itltimas dccadas han tratado la filogenia y biogeografia de los mamiferos terrestres de las Antillas Mayores (MacFadden 1980 MacPhee y Woods 1982 Morgan 1985 1988 1993 Morgan y Woods 1986 Ford 1986 Woods 1988 1989a 1990 MacPhee e Iturralde-Vinent 1995a b Hedges 1996 Horovitz [1997] Ottenwalder 2001 Whidden y Asher 2001 MacPheeet ai 2003 Carlini y Scillatoshy
302 BIOGEOGRAFiA
Yane 2004 Davalos 2004 MacPhee y Horovitz 2004 Roca et al 2004) es practicamente unanime en cuanto a que los ancestros continentales que dieron origen a esta fauna derivaron (1) mayormente de Suramerica perezosos (Megalonychidae) monos (Pitheciidae) y roedores (Echimyidae Capromyidae y Heptaxodontidae) (2) en menor medida de Norteamerica musarafias (Nesophontidae y Solenodontidae) y (3) menos aun de Centroamerica una especie de roedor (Muridae)
HIPOTESIS BIOGEOGRAFICAS
La definicion c1asica reconoce dos tipos de islas isla continental (porcion del territorio de un continente separado de este por movimiento tectonico 0 por ascenso del nivel del mar) e isla oceanica (porcion de territorio que no ha formado parte de un continente) La definicion sigue siendo uti por 10 que implica para la formacion de las faunas insulares en uno u otro caso As en el caso de la isla continental su fauna ya estaba emplazada en la futura isla antes de producirse su separacion del continente [Vicarianza hipotesis biogeografica de fragmentacion faunal] en el caso de la isla oceanica su fauna es el resultado de la colonizacion1 por propagulos2continentales a traves de la barrera marina entre el continente y la isla [Dispersion hipotesis biogeografica de acumulacion faunal] En el primer caso el surgimiento de la barrera marina es resultado del evento vicariante en el segundo la existencia de la barrera marina es anterior al evento dispersante Ademas de operar a escala mayor (continente-isla) ambos eventos operan secundariamente a menor escala (isla-isla)
Las dos hipotesis -vicarianza y dispersion- se han disputado hasta hoy la preferencia de los biogeografos en tanto hipotesis mas viable para explicar el proceso de formacion original (continente-isla) de la fauna de mamiferos terrestres de las Antillas Mayores [Sobre la historia de las hipotesis biogeograficas relativas al Caribe insular vease RuB y Schubert 1989 Williams 1989 Pruna 2001] Yel debate continua debido ala falta de consenso sobre 10 que Williams (1989) ha llamado el problema central de la discusion a saber la ostensiblemente baja diversidad taxonomica de esta fauna a nivel de las categorias superiores (solo cuatro ordenes) en comparacion con la diversidad al mismo nivel de las faunas de islas continentales (0 de areas del continente) simi lares en tamafio y diversidad ecologica [Por ejemplo la fauna cubana de mamiferos terrestres consideradas de conjunto las especies vivientes y las extintas- es menos diversa a los niveles taxonomicos superiores (4 ordenes y 6 familias) que 10 que cabria esperar de la diversidad ecologica y extension territorial de Cuba (110 992 km2) comparada con una isla tipicamente continental como Trinidad (4828 krn2
) -veinte veces menor que Cuba y menos diversiticada ecologicamente- donde solo las especies vivientes de mamfferos terrestres representan 6 ordenes y 16 familias (Eisenberg 1989)] Este desbalance
lColonizaci6n Arribo y establecimiento de un propagulo en una isla no ocupada por la especie (MacArthur y Wilson 1967)
2Propagulo Niimero minimo de individuos de una especie capaces de reproducirse y aumentar poblacionalmente en una isla bajo condiciones favorables (MacArthur y Wilson 1967)
303 POBLAMIENTO CONTINENTE-ISLA
taxon6mico ha sido expJicado de dos maneras (1) Inicialmente hubo mas 6rdenes presentes pero la mayoria se extingui6 y s610 sobrevivieron esos cuatro [explicaci6n vicarianista] (2) S610 representantes de esos cuatro 6rdenes lograron cruzar la barrera marina [explicaci6n dispersionista]
Cinco hipot~ticos mecanismos de poblamiento continente-isla han sido invocados durante los ultimos 140 afios (y se los sigue proponiendo actualmente) para explicar la presencia de mamiferos terrestres en las Antillas Mayores A continuaci6n pasamos a resefiarlos brevemente (incluidas sus alegadas fortalezas y debilidades) en el orden aproximado en que historicamente fueron propuestos
EL PUENTE NATURAL (dispersion terrestre activa)
En el siglo XIX el puente terrestre era pnkticamente la unica hipotesis para explicar las distribuciones disyuntas de animales en buena parte del mundo y los primeros investigadores que se ocuparon de la biogeografia antillana (Fernandez [1865] 1884 Chapman [1893] Torre 1892 Lora 1895) la asumieron resueltamente en parte intrigados por el descubrimiento en Cuba en la segunda mitad del siglo de un perezoso f6sil de gran talla Y en su aplicacion a los mamiferos terrestres de las Antillas Mayores el puente sigui6 siendo a 10 largo del siglo xx la hipotesis mas favorecida por los estudiosos del tema (Vaughan 1902a Spencer 191 Oa b Torre 1910a b cAllen 1911 1917a 1918 Barbour 19141916 1945bAnthony 1918 1926 [1942] Scharff 1922 Kraglievich 1926 Beaufort 1951 Arredondo 1961 1970b Kartashov y Mayo 1974 Iturralde-Vinent 1982 1988 Arredondo y Armifiana 1996 Arredondo 1996a [1999]) quienes postularon la existencia preterita de puentes con Norte- Centro- 0 Suramerica en dependencia de las afinidades continentales de los mamiferos antillanos
Teoricamente el puente terrestre debe cumplir con una condicion esencial permitir el paso de una parte mayoritaria (y por tanto representativa de la diversidad) de la fauna continental Claro que siempre habria tipos de mamiferos menos proclives a dispersarse o para los cuales la ubicaci6n del puente resultaria inaccesible 0 las condiciones ecologicas en este fueran desfavorables pero si algunos acceden al puente no hay raz6n para pensar que el resto de esa fauna no los acompafie mayoritariamente Como dijera Simpson (1956) Los puentes no transportan faunas desbalanceadas
En consecuencia el puente terrestre como filtro es un mecanismo de poblamiento minimamente selectivo y por ende de alta diversidad inicial (sensu Iturralde-Vinent y MacPhee 1999) 10 esencial del puente es su capacidad para transportar faunas integradas (Williams 1989) Yen eno radica la objeci6n fundamental al puente terrestre surgida tempranamente en el siglo xx (Por que s610 cuatro 6rdenes de mamiferos terrestres en las Antillas si vinieron por puentes (Donde estan los fosiles de los demas ordenes que debieron venir si es que se extinguieron Los partidarios del hipotetico puente dispusieron de casi todo el resto del siglo para responder una y otra vez que otros 6rdenes de mamiferos aparecerian representados en las Antillas Mayores el dia que en estas Islas se descubrieran fosiles precuatemarios suficientemente
304 BIOGEOGRAFIA
antiguos Esto se logro por fin terminando el siglo como se explico anteriormente (vease Contexto temporal) y ique se encontro los mismos cuatro ordenes de mamiferos terrestres conocidos del Cuatemario caribefio aparentemente los Unicos que lograron llegar
Pero ademas hasta el presente no se ha demostrado inequivoca- y concluyentemente que en algun momenta durante los ultimos 40 millones de afios haya existido una conexion terrestre continua entre las Antillas Mayores (en proceso de formacion 0 plenamente formadas) y algun punto de las regiones continentales vecinas
LA BALSA NATURAL (dispersion marina pasiva)
Durante buena parte del siglo xx el puente terrestre tuvo como unico rival a la hipotesis de la balsa natural -transporte accidental de animales sobre islotes de vegetacion flotantes a la deriva despues de arrancados y expulsados mar afuera por un rio crecido- expuesta originalmente por Matthew (191319151916 1918a 1919) y desarrollada en su aplicacion a los mamfferos terreslres antillanos principal mente por Simpson (1956) asi como por olros investigadores (Darlington 1938 1957 Williams y Koopman 1952 Hershkovitz 1958 1972 Patterson 1962 Paula 1967 Patterson y Pascual 1972 Pregill 1981 Ford 1986 Woods 1989a 1990 Morgan 1993) Por su parte Hedges et al (1992) sustentaron dicha hipotesis en estimados de divergencia (basados en el reloj molecular) en la herpetofauna antillana -involucrados tangencia1mente los mamiferos [veanse objeciones a Hedges y colaboradores (Page y Lydeard 1994 Iturralde-Vinent y MacPhee 1999) respuestas a las objeciones (Hedges et al 1994 Hedges 2001) y contrarrespuestas (MacPhee e IturraldeshyVinent 2005) asi como cuestionamientos al metoda de los estimados de divergencia (Foote et al 1999 Pregill y Crother 1999 Woods et ai 2001 )]
La hip61esis de la balsa natural se origino a partir de las numerosas ocasiones en que a 10 largo de siglos tales islotes flotantes (portando 0 no vertebrados incluidos mamiferos) han sido avistados mar afuera a cientos de kilometros de la costa mas cercana en varias partes del mundo --inc1uido el Mar Caribe- segUn consta en diarios de barcos y reportes de observadores diversos (revisados por King 1962 reportes posteriores en SICSP 1970 y Censky et al 1998) Uno de estos casos acaecido en nuestra region se reporta en un informe del SICSP (Smithsonian Institution Center for Shortlived Phenomena 197050shy52)
EIB de julio de 1969 un destructor de la armada de Estados Unidos avisto un islote flotante a unos 100 km al sur de la Base Naval de Guantanamo moviendose desde el Paso de los Vientos en direccion suroeste a una velocidad de 45 kmlh islote de unos 15 m de diametro mantenfa en pie mas de 10 palmas reales de 10-15 m de altura con suelo cubierto de hierba y arbustos [la observacion con binoculares a 18 km de distancia no preciso la presencia de vida animal] La travesia fue rastreada durante 6 dfas hasta que el 19 de julio el islote se dio por perdido tras 6 horas de busqueda aerea
305 POBLAMIENTO CONTINENTE-ISLA
La colonizaci6n exitosa de una isla de las Antillas Mayores por mamiferos terrestres continentales transportados en balsas naturales a la deriva (de cada especie al menos una hembra gravida 0 dos individuos de sexo opuesto) dependeria de factores que van a operar desde el momenta mismo de formarse la balsa asi como durante su travesia marina y su arribo a la isla
a) La posibilidad de estar en la balsa Que la distribuci6n de la especie incluya areas contiguas al rio y que el animal se encuentre en ellugar y el momenta propicios para ser incorporado a la balsa antes de que el rio crecido la desprenda Por ejemplo segun Hershkovitz (1958) los equimidos -grupo ancestral de roedores antillanos- serian buenos candidatos a ser captados por la balsa porque acostumbran a anidar en huecos del tronco de los arboles a la orilla de los rios
b) El tamano y la consistencia de la balsa Muchas se desintegran en alta mar debido a su fragilidad ante la saturaci6n Algunos autores (Beaufort 1951 Arredondo 1996a [1999]) han expresado incredulidad en cuanto a que las balsas hayan podido soportar el peso de un animal como el gran perezoso cubano (Megalocnus rodens) el mayor de los mamiferos antillanos Pero esta especie evolucion6 en Cuba y el ancestro que hizo el viaje pudo ser muy pequeno [EI perezoso oligocenico de Puerto Rico es el mas pequeno de los megaloniquidos antillanos (MacPhee e Iturralde-Vinent 1995b )]
c) Las caracterfsticas fisiol6gicas de la especie Los limites fisiol6gicos de los propagulos constituyen nuestra unica objeci6n a la dispersi6n marina por balsas como mecanismo para la formaci6n de la fauna de vertebrados en las Antillas (MacPhee e IturraldeshyVinent 2005) Unas especies tienen intrinsecamente mas posibilidades que otras para sobrevivir a las vicisitudes que deben afrontar durante la travesia (deshidrataci6n inanicion insolaci6n etc) Por ejemplo los megaloniquidos han demostrado tener extraordinaria habilidad para cruzar barreras marinas fueron los primeros mamfferos suramericanos en invadir Norteamerica mucho antes de que surgiera la conexion terrestre a finales del Plioceno (Paula 1967 Olson 1978 Webb 1985b Martin y Steadman 1999) Incluso se conoce un megaloniquido acultitico del Plioceno peruano (Muizon y McDonald 1995) y otro viviente en Suramerica (Choloepus) que pnicticamente no bebe agua 10 que ha originado una leyenda aborigen que explica por que los perezosos no toman agua (Goff art 1971) Vease ademas el inciso e
d) EI regimen de las corrientes marinas Que conduzcan a la balsa en la direcci6n necesaria EI transporte de balsas de vegetacion desde los continentes hacia las islas fue durante la mayor parte del Cenozoico sumamente improbable (MacPhee e IturraldeshyVinent 2000 148) Estos autores han insistido en que hay que tener en cuenta que durante la cambiante geografia cenozoica del Caribe las corrientes marinas de superficie no favorecieron el desplazamiento de objetos fiotantes hacia las Antillas Mayores [Pero no sabemos con certeza que efecto modificador del curso de tales objetos fiotantes pudieron introducir las tormentas tropicales si previsiblemente funcionaron en el pasado como 10 hacen ahora] Dichos autores senalaron ademas (2005) que en un experimento reciente con boyas de posicionamiento satelital liberadas a la deriva desde las Antillas Menores las boyas fueron a parar mayoritariamente a Centroamerica y a otros lugares
306 B[OGEOGRAFIA
y solo unas pocas recalaron en las Antillas Mayores y las que entraron al Mar Caribe demoraron semanas 0 meses en arribar
e) La distancia a recorrer (y en consecuencia la duracion de la travesia) entre el continente y la isla (10 que repercute decisivamente en el factor c) Este es desde luego uno de los factores mas criticos En la medida que la distancia aumenta la probabilidad de alcanzar la isla disminuye drasticamente con el agravante de que la trayectoria dificilmente seria en linea recta Si tomamos como referencia la velocidad de 45 kmlh del islote ya mencionado (SICSP 1970) cubrir la distancia aerea (aproximada) de 800 km que media entre el noroeste de Suramerica y La Espafiola Ie tomaria a la balsa 74 dias Estimando el doble de este tiempo debido a desvios de la trayectoria la duracion de la travesia probablemente resultaria excesiva para la mayoria de los mamiferos Pero pudiera muy bien ocurrir que la velocidad fuera mayor 0 que la balsa portara plantas vivas que permitirian a los fitOfagos sobrevivir [Tres de los cuatro ordenes de mamiferos terrestres conocidos de las Antillas Mayores son fitOfagos] Ademas esta geologicamente bien establecido que durante etapas de variada duracion desde el Oligoceno hasta el Pleistoceno los territorios del Caribe insular y continental aumentaron considerablemente en extension con niveles del mar 100-200 m por debajo de su nivel actual oportunidades que los propagulos aprovecharian para cruzar barreras marinas significativamente menores
f) El tamafio de la isla Las islas mayores deben interceptar mas balsas (Buckley y Knedlhans 1986) 10 que repercute directamente en el factor e La balsa puede pasar entre islas pequefias y continuar viaje sin llegar a ningun lugar
g) Las condiciones (favorables 0 adversas) del desembarco en la isla y el tiempo que Ie tomaria a uno 0 mas animales depauperados encontrarun habitat apropiado Esto dependeria a su vez del tamafio y diversidad ecologica de la isla (Darlington 1938 Miller 1966)
Tal cumulo de posibles obstaculos y dificultades constituye a primera vista un desafio a la credibilidad de la hipotesis Matthew-Simpson pero en ello radica precisamente su valor explicativo bas ado en la mas absoluta improbabilidad La colonizacion por la via marina es altamente improbable pero no absolutamente imposible si se tiene en cuenta la inmensidad del tiempo geologico se vuelve probable (McKenna 1973) Basta que solo una vez entre mill ones de intentos fallidos durante ese inmenso lapso algunos de los factores anteriormente enunciados coincidan en no ser tan desfavorables para el propagulo Y eso es todo 10 que demanda la hipotesis para cada uno de los grupos monofileticos mayores de mamiferos terrestres presentes en las Antillas Mayores Las Antillas Mayores requeririan el arribo de tan solo 7 (quizas menos) propagulos fundadores a 10 largo de los ultimos 30 0 mas millones de afios (promedio de uno cada 43 mill ones) para explicar la formacion de toda su fauna de mamiferos terrestres por dispersion marina pasiva (vease Discusion y Conc1usiones)
En comparacion con la via marina el poblamiento por la via terrestre es predecible porque la mayor parte de la fauna continental de mamiferos emplazada en la region del puente va a cruzar por este una vez que el mismo se establece y los animales logran
307 POBLAMIEITO CONTINENTE-ISLA
acceder a el el poblamiento por la via marina en cambio es impredecible porque depende total mente del azar que caracteriza a la balsa Claro que en ambas vias opera el azar pero en el primer caso se trata del azar de un evento probable en el segundo el azar de un evento improbable (Simpson 1940) Asi el puente natural (mecanismo de poblamiento minimamente selectivo = alta diversidad inicial) y la balsa natural (mecanismo de poblamiento maximamente selectivo baja diversidad inicial) pueden conceptuarse como los extremos opuestos de una gama de probabilidades de dispersi6n (McKenna 1973) EI fen6meno de la dispersi6n merece la mayor atenci6n cientifica en los estudios de biogeografia insular (Cowie y Holland 2006)
Queda claro que la balsa natural es una hip6tesis consistente con el caracter de la fauna objeto de estudio puesto que el desbalance taxon6mico (sin evidencia paleontol6gica en contrario) s610 puede ser resultado de la improbabilidad del poblamiento No obstante ello no implica necesariamente que la balsa natural haya sido el imico mecanismo (ni siquiera el principal) utilizado por los mamiferos terrestres en su colonizaci6n de las Antillas Mayores La balsa natural debe satisfacer una premisa te6rica esencial dado el caracter azaroso impredecible e improbable de una colonizaci6n exitosa esta debe ocurrir a intervalos muy espaciados a traves del tiempo geol6gico 0 sea durante todo el tiempo que en el area ocupada por las Antillas Mayores ha habido ecosistemas terrestres perrnanentemente disponibles para ser colonizados (los ultimos 40 millones de afios) Sin embargo como sabemos representantes de los cuatro 6rdenes de mamiteros terrestres conocidos de las Antillas Mayores ya estaban emplazados en dicho territorio insular en el Mioceno Inferior 10 que supone un lapso de colonizaci6n limitado al Eoceno SuperiorshyOligoceno (MacPhee 2005) sugerente de un tipo de mecanismo contrario a la colonizacion alocronica exigida por la hipotesis de la balsa natural sobre todo tratandose de una fauna caracterizada por altas tasas de extincion sin reposicion aparente
La presencia en Jamaica (hasta su extinci6n en 1877) del roedor mfuido Oryzomys antillarom con estrecha afinidad centroamericana (Woods 1989a) pudiera constituir el imico caso efectivo de poblamiento (continente-isla) de las Antillas Mayores por dispersion marina dc un mamifero terrestre para el cual no es razonable invocar otro mecanismo de colonizacion
LA GUIRNALDA INSULAR (dispersion mixta)
La geologia ha establecido con suficiente certeza que durante el Cenozoico unicamente en el Pale6geno tardio y solo a traves de la Cresta de Aves (ahora sumergida excepto la isla hom6nima) pudo haber existido una cadena de islas para el transito de mamiferos terrestres entre el norte de Suramerica y las proto-Antillas Mayores [desestimados a ese efecto el Canal dc Yucatan el Elcvado de Nicaragua la Cresta de Caiman el Elevado de Beata y las Antillas Menores (Holcombe et al 1990 MacPhee e IturraldeshyVinent 1994 1995b Iturralde-Vinent y MacPhee 1999)] Holcombe y Moore (1977) y Holcombe y Edgar (1990) postularon la existencia de esta entidad y estimaron algunas de sus posibles caracteristicas Por su parte MacPhee e Iturralde-Vinent (1995b) e Iturralde-Vinent y MacPhee (1999) precisaron los extremos proximal y distal de esa
308 BIOGEOGRAFIA
estructura acotaron la duracion y la edad de la misma y Ie dieron nombre GAARlandia (Greater AntillesA vesRidgelandia)
Segun Iturralde-Vinent y MacPhee (1999) la Cresta de Aves emergio por breve tiempo (1-2 Ma) durante la transicion Eoceno-Oligoceno (35-33 MaAP) por la coincidencia de un levantamiento tectoni co y un considerable descenso eustatico del nivel del mar En ese momenta las islas septentrionales del arco de las Antillas Mayores (Cuba centroorientalshyLa Espafiola - Puerto Rico - Islas Virgenes) constituian una sola isla grande 0 una serie de islas separadas por barreras muy estrechas La Cresta de Aves emergida entre el bloque Puerto Rico - Islas Virgenes y un microcontinente noroccidental de Suramerica -separado en ese momento del resto del continente por una via marina surgida en el Eoceno tardio- dio origen a GAARlandia
Obviamente el numero de colonizaciones exitosas por mamiferos terrestres a traves de una guimalda de Islas tiende a disminuir mientras mas anchas sean las barreras marinas entre las islas que forman la guimalda y cuando esto ocurre disminuye tambien la diversidad taxonomica de los propagulos particularmente a nivel de las categorias superiores (Simpson 1956)
Vemos asi que para los mamiferos terrestres la colonizaci6n a traves de una guimalda insular puede ser tan improbable como a la deriva en una balsa natural en dependencia de la cantidad y anchura de las barreras marinas a cruzar el tamafio y la fisiografia de las islas que integran la guimalda y el regimen de las corrientes marinas y aereas entre otros factores
Resumiendo la guimalda insular puede conceptuarse como un mecanismo de poblamiento altamente selectivo (= baja diversidad inicial) que combina la improbabilidad de la balsa natural con la colonizacion restringida temporaimente del puente terrestre
En su reciente revision de la taxonomia distribuci6n y filogenia (basada en estudios moleculares) de varios grupos de mamiferos antillanos Davalos (2004) concluyo que los perezosos megalocninos los primates y los roedores histricognatos se avienen al patron de divergencia con el continente y epoca de arribo a las Antill as implicados en la hipotesis de GAARlandia pero no asi los perezosos coloepodinos que requieren una explicacion biogeognlfica distinta Esto ultimo (sospechado por Matthew 1918a) habla sido confirmado por White y MacPhee (2001 226-227)
Nuestra evidencia filogenetica no sustenta la idea de un dado unificado surgido de la invasion de un megaloniquido el analisis sugiere mas bien dos eventos de colonizaci6n independientes [y] sustenta fuertemente un origen difiletico de las subfamilias [Choloepodinae y Megalocninae] Si hubo dos migraciones a traves de GAARlandia presumiblemente poco espaciadas en el tiempo no se puede determinar en el presente
Finalmente queda claro que GAARlandia -como guirnalda insular- es la hipotesis mas robusta propuesta hasta ahora para explicar la presencia en las Antillas Mayores de
309 POBLAMIENTO CONTINENTE- ISLA
la absoluta mayona de los mamfferos terrestres de ongen suramericano porque el caracter de esa fauna supone un mecanismo de poblamiento ancestral que sea a la vez maximamente selectivo (baja diversidad ordinal) y temporalmente restringido (transicion Eoceno-Oligoceno)
Recientemente Rega et al (2002 18) expresaron A pesar de una revision exhaustiva de la paleogeografia de la cuenca del Caribe Iturralde-Vinent y MacPhee (1999) no pudieron confirmar la presencia de una conexion terrestre continua entre Suramerica y las proto-Antillas aunque hipotetizaron su existencia [Vease la Fig 42 (= figura 6 de Iturralde-Vinent y MacPhee 1999 simplificada))
En el trabajo citado por Rega y colaboradores aparece la siguiente explicacion
Actualmente no es posible ofrecer reconstrucciones paleogeograficas detalladas de la Cresta de Aves Sin embargo hay evidencia de que porciones de la cresta deben haberse elevado subaereamente en un tiempo La cuestion cntica es si la Cresta de Aves emergida pudo haber formado algun tipo de corredor entre las Antillas Mayores y el noroeste de Suramerica Perforaciones en las alturas estructurales de la cresta pudieran proveer el registro del Paleogeno tardio que se necesita para precisar la hipotesis del corredor aunque no se puede predecir si tales datos serian suficientemente detallados para determinar si en algun momento GAARlandia constituyo una sola entidad subaerea Por ejemplo pudo haber barreras marinas por corto tiempo entre las masas terrestres de la Cresta de Aves [0) ellevantamiento y la subsidencia pudieron afectar diferentes partes de GAARlandia en diferentes tiempos AItemativamente los elementos faunales pudieron haberse dispersado de manera pulsada Movimientos episOdicos de este tipo incrementarian la extincion de los elementos faunales incapaces de trasladarse a la siguiente masa terrestre en la cadena disminuyendo asi la diversidad total (Iturralde-Vinent y MacPhee 199957)
La primera parte de la explicacion deja claro que la posibilidad de que la Cresta de Aves haya sido una conexion terrestre continua no se puede descartar pero tampoco se puede afirmar La ultima parte de la explicacion describe el efecto altamente selectivo (filtrante) de la colonizacion a traves de una cadena de islas
Y continuan lturralde-Vinent y MacPhee (199957 -58) explicando
Hasta que punto el microcontinente noroccidental de Suramerica [considerado por dichos autores como el area suministradora de propagulos hacia GAARlandiaJ estuvo fisicamente separado del resto del continente no ha sido plenamente esc1arecido Si estuvo sustancialmente aislado por barreras marinas y orogenic as durante el Paleogeno tardio entonces solo la fraccion de la biota suramericana emplazada en el microcontinente estuvo en posicion de acceder a GAARlandia Esto pudiera tener que ver con el hecho de que la fauna antillana carezca de representantes de muchos grupos suramericanos [de mamfferos) que debieron haber hecho el viaje
310 BIOGEOGRAFiA
Florida o Alturas0 0
d 6 D L1anuras25deg
Yucatan
20deg
deg0
10deg
MARCARIBE
80deg 75deg
Bahamas
Cres ta de Beata
D Mares someros
D Mares profundos
OCEANO ATLANTICa
FIGURA 42 Reconstrucci6n hipotetica de GAARlandia como conexi6n terrestre continua entre las proto-Antillas Mayores y el noroeste de Suramerica en el Pale6geno tardio segun la FIgura 6 (simplificada) de Iturralde-Vinent y MacPhee (1999)
En su defensa del puente terrestre Barbour (1945b249) se refiri6 a esto - antes 10 habfa hecho Anthony [1942363]- en los terminos siguientes
El argumento de que si las Antillas hubieran estado conectadas con el continente se encontrarian los f6siles de un numero mayor de distintas criaturas me deja frio es perfectamente concebible que la conexi6n haya ocurrido en un periodo en que la parte del continente implicada no estuviera poblada por una fauna grande y variada
Y concluyen lturralde-Vinent y MacPhee (199958) su detallada explicaci6n
Desafortunadamente sabemos muy poco sobre la composici6n faunal del noroeste de Suramerica a finales del Pale6geno
Consecuentemente si se confirmara la magnitud del aislamiento del aludido microcontinente y su concomitante baja diversidad faunal GAARlandia como conexi6n terrestre continua serf a la hip6tesis mas parsimoniosa No obstante mientras no se disponga de evidencia concluyente a este respecto hipotetizar a GAARlandia como conexi6n continua (minima selectividad) la coloca en franca vulnerabilidad te6rica al tener que afrontar la objeci6n clasica al puente terrestre (baja diversidad ordinal de la
311 POBLAMIENTO CONTINENTE-TSLA
fauna) al tiempo que se la priva de ser la hip6tesis mas robusta para explicar -como guimalda de islas- la colonizaci6n de las Antillas por mamiferos procedentes de Suramerica
ELARcA DE NOE (seudovicarianza)
El primer vicarianista antillano (desde luego sin el saberlo) fue Corral (1940) con su utilizaci6n ingenua de las ideas wegenerianas de la deriva continental aparecidas la decada anterior Pero el primer (supuesto) intento de aplicar a la formaci6n de la biota caribefia el mecanismo de la vicarianza basada en la tearia de la tect6nica de placas (surgida en la sexta decada del pas ado siglo) se debe a Rosen ([1976] 1985)
Segun Rosen las Antillas Mayores se originaron en el Cremcico en la Placa Caribe aproximadamente en la posici6n que hoy dia ocupa Centroamerica y al ser desplazadas tectonicamente hacia el este interactuaron (de algun modo no precisado por Rosen) con las margenes continentales de Norte- y Suramerica de donde (en un evento unico) adquirieron toda su fauna transportada subsecuentemente por dichas islas en su transito hacia sus posiciones actuales [De ahi la denominacion metaforica dada par McKenna (1973) a este mecanismo de poblamiento] Aunque se ha presentado esta propuesta como Model0 de Vicarianza Continente-Isla de Rosen en realidad por definici6n no constituye un evento vicariante porque las proto-Antillas Mayores no resultaron del fraccionamiento y separaci6n por tectonismo de una porcion del territorio continental como exige esta hip6tesis (vease definici6n de vicarianza al comienzo de Hipotesis Biogeograficas)
El Arca de Noe de Rosen ha sido desestimada por la mayoria de los autores contemporaneos porque (1) carece de sustento geol6gico (lturralde-Vinent y MacPhee 1999) y porque (2) en el periodo en que se afrrrna ocum6 el evento muchos de los ordenes modemos de mamiferos placentarios attn no existian (Williams 1989)
EL BARCO FUNERAL VlKINGO (tanatovicarianza)
Domning et al (1997) reportaron 10 que consideraron el mamffero terrestre mas viejo de las Antillas --circunstancia que se refleja en el titulo mismo del trabajo Oldest West Indian Land Mammal- al descubrirse un perisodactilo rinocerotoide (Hyrachius sp) en una localidad de edad Eoceno Inferior a Medio en el occidente de Jamaica
Jamaica ha tenido una historia geologica y paleogeografica distinta de la historia comun a las restantes Antillas Mayores A principios del Cenozoico Jamaica occidental (a diferencia de la porcion oriental constituida por el Bloque de las Montafias Azules) formaba parte de Centro america de la cual se desgaj6 (por el desplazamiento de la Placa Caribe) para venir a integrarse despues del Eoceno Medio con 10 que serfa la futura isla de Jamaica desaparecida por transgresi6n marina en el Eoceno Superior hasta volver a emerger totalmente en el Mioceno Medio (Iturralde-Vinent y MacPhee 1999 MacPhee e Iturralde-Vinent 2000)
312 BIOGEOGRAFiA
Por consiguiente Hyrachius ---conocido de localidades de edad Eoceno Inferior a Medio en Norteamerica- debio ocupar (10 que iba a ser) Jamaica occidental mucho antes de que esta se separara de Centroamerica De hecho este fosil Uego a su destino insular contenido en las rocas de su basamento no como un ser vivo por 10 que es erroneo considerarlo un animal antillano (lturralde-Vinent y MacPhee 2004) como han supuesto algunos (Hedges 200 I Portell et al 200 I Condis [200 I D Todo 10 que hizo fue viajar tanatovicariantemente en su barco funeral vikingo -otra de las denominaciones metaforicas de McKenna (1973)
DISCUSION Y CONCLUSIONES
Hemos visto que solo disponemos de una hipotesis plausible para explicar la mayor parte del poblamiento (continente-isla) de las Antillas Mayores por mamiferos terrestres del Neotropico continental la dispersion mixta (a traves de GAARlandia) para los perezosos (excepto Choloepodinae) los monos y los roedores (excepto Oryzomys)
Por otra parte la historia biogeogrMica de las musaraiias de las Antillas Mayores (Nesophontes y Solenodon) constituye una de las situaciones mas enigmaticas entre los mamiferos de la region (Ottenwalder 2001321) Al menos cinco mecanismos de poblamiento continente-isla han sido invocados para explicar cuando como y de donde llegaron a las Antillas Mayores estas musaraiias
I EI propagulo ancestral (del Terciario de Norteamerica) coloniz6 a las Antillas Mayores por dispersion marina desde Centroamerica 0 desde la Florida (Patterson 1962)
2 El propagulo ancestral ( del Terciario) cruzo el Atlantico Sur (supuestamente mas estreshycho en aquella epoca) directamente de Africa a las Antillas Mayores (Hershkovitz 1972)
3 El propagulo ancestral (del Terciario de Norteamerica) Uego a las Antillas Mayores en el Arca de Noe de Rosen (MacFadden 1980)
4 El propagulo ancestral (del Terciario de Norteamerica) fue transportado a las Antillas desde Centroamerica en el bloque de Jamaica occidental y de alli paso por dispersion marina a las restantes Antillas Mayores (Whidden y Asher 200 I)
5 El propagulo ancestral (del Terciario de Norteamerica) lleg6 a las Antillas Mayores a traves de GAARlandia (Whidden y Asher 2001)
Aunque hay consenso entre estos (y otros) autores sobre el origen norteamericano de las musaraiias antillanas ninguno de los cinco hipoteticos mecanismos de poblamiento resiste la evidencia en contrario filogenetica yo paleogeogrMica Ademas algunas de estas hipotesis asumen que Nesophontidae y Solenodontidae comparten un antecesor comun en las Antillas pero esta relacion no ha sido confirm ada Para dilucidar el enigmatico caso de las musaraiias antillanas se requerini profundizar en la investigaci6n geologica paleogeognifica y paleontol6gica (Iturralde-Vinent y MacPhee 1999)
De acuerdo con el conocimiento filogenetico disponible sobre el conjunto de la fauna la colonizacion original de las Antillas Mayores por mamiferos terrestres implicaria el arribo
313 POBLAMIENTO ISLA-ISLA
de solo siete propagulos fundadores perezosos 2 (coloepodinos y megalocninos si como parece tienen un origen difiletico) musaraiias 2 (nesofontidos y solenodontidos si no constituyen una misma familia como consideran algunos) monos 1 (pitecidos) roedores histricognatos 1 (equimidos capromidos y heptaxodontidos) roedores muridos 1 (Oryzomys en Jamaica)
POBLAMIENTO ISLA-ISLA
La distribucion de los mamiferos terrestres a traves del area antillana demarcada al inicio del presente capitulo (vease Contexto Geogratico Fig 41) ha sido interpretada en funcion de dos hipoteticos mecanismos de poblamiento interinsular la dispersion y la vicarianza fundamentadas como se resumen seguidamente
DISPERSION
De acuerdo con la evidencia paleogeogratica mas actualizada en las principales regresiones marinas ocurridas en la cuenca caribeiia durante los ultimos 23 Ma (transiciones OligocenoshyMioceno Mioceno Superior - Plioceno y varias veces en el Pleistoceno) las masas terrestres constitutivas de la region insular en formacion aumentaron considerablemente su extension territorial disminuyendo por tanto la distancia entre elias oportunidades que los elementos faunales aprovecharian para dispersarse de unas islas a otras
Durante mucho tiempo esta ha sido la hipotesis mas favorecida por los estudiosos del tema a partir de Simpson (1956) pero en decadas recientes ha suscitado dos objeciones
1 Es dificil en tender como pudo operar la dispersion marina entre estas islas puesto que ninguna de las Antillas Mayores posee rios suficientemente caudalosos para transportar masas significativas de vegetacion flotante (MacPhee 198432)
Es oportuno recordar aqui el islote ala deriva (con mas de diez palmas reales en pie) entre las tres Antillas Mayores mas occidentales mencionado anteriormente (vease La Balsa Natural)
2 La dispersion marina es menos efectiva para explicar la asombrosa similitud de las faunas de mamiferos terrestres a traves de las Antillas Mayores septentrionales porque como proceso aleatorio no debe producir distribuciones uniformes a 10 largo de una cadena de islas 0 si las produce entonces el proceso no es aleatorio (MacPhee e lturralde-Vinent 2000 149)
Independientemente de que las aludidas distribuciones taxonomicas estan lejos de ser uniformes 0 asombrosamente similares la hipotesis de la dispersion marina contrariamente a su canicter aleatorio y altamente improbable en el caso del poblamiento continente-isla- en el caso del poblamiento isla-isla es un proceso no aleatorio (aprovecha las oportunidades brindadas puntualmente por los periodos de maxima regresion marina) y ademas altamente probable dada la incomparablemente menor distancia entre las islas caribeiias que entre estas y los continentes vecinos
314 BIOGEOGRAFiA
V ICARIANZA
La neotectonica y las transgresiones marinas del Oligoceno Medio y del Mioceno afectaron sustancialmente la paleogeografia de la Cresta de las Antillas Mayores resultando la subdivision gradual de las partes subaereas de la Cresta Asi el este de Cuba y el norte de La Espanola conectados fisicamente durante el Oligoceno Inferior se separaron al formarse el Paso de los Vientos en el Oligoceno tardio en tanto La Espanola y Puerto Rico probablemente permanecieron conectadas hasta el Mioceno tardio en que surgio el Paso de Mona (Iturralde-Vinent y MacPhee 1999)
Tales eventos geologicos habrian dividido las distribuciones de los grupos faunales emplazados previamente en el territorio continuo de las tres islas y a partir de esas disyunciones dichos autores (tambien MacPhee et al 2000 y White y MacPhee 2001) asumieron --como segunda fase del modelo de GAARlandia- la hipotesis de la vicarianza isla-isla para explicar la distribucion multiinsular de varios grupos de mamiferos terrestres a traves de las Antillas Mayores septentrionales De acuerdo con esta hipotesis cualquier diferencia en composicion faunal entre las islas (por ejemplo la ausencia de roedores heptaxodontidos en Cuba 0 de monos platirrinos en Puerto Rico) constituye evidencia de extincion local postvicariante y no de dispersion marina fallida Lo primero se puede demostrar si aparecen los fosiles pertinentes 10 segundo no se puede demostrar (MacPhee e Iturralde-Vinent 1994) Por otra parte IturraldeshyVinent y MacPhee (1999 57) han reconocido que aunque fa vicarianza isla-isla puede representar una solucion clara para las distribuciones de varios taxones cuaternarios no puede explicar todos los casos
Rega et al (2002 18) han presentado dos objeciones al modelo de la vicarianza isla-isla aplicado a los mamiferos de las Antillas Mayores
I Si la separacion de Cuba y La Espanola precedio en varios mill ones de anos la separashycion de La Espanola y Puerto Rico el modelo vicariante de lturralde-MacPhee debe predecir mayor similitud entre las faunas de La Espanola y Puerto Rico que entre las de Cuba y La Espanola De hecho es al reyes
No obstante MacPhee et al (200335) admitieron despues que la fecha mas temprana empiricamente defendible [para la apertura del Paso de Mona] debe ser el Oligoceno Medio esto invierte la secuencia de ambas disyunciones con 10 cual queda resuelta la aparente contradiccion Sin embargo en su reciente evaluacion de la congruencia entre filogenia e hipotesis geologic as en varios grupos de mamiferos terrestres de las Antillas Mayores Davalos (2004373) encontro que ninguno de los taxones cumple con la fecha y el patron de divergencia entre las masas terrestres antillanas postulados por el modelo de GAARlandia
2 La presencia de un mismo genero en las tres Islas septentrionales de las Antillas Mayores implica [por el modelo vicariante] que la diferenciacion entre estas formas durante el intervalo desde el Mioceno ha sido unicamente de nivel especifico [Dicha presencia]
315 rRRADlAcr6~ EVOLUCrONARIA
s610 puede explicarse razonablemente como el resultado de movimientos de esos animales entre las islas en el Cuaternario
De esta objecion resulta dificil disentir [El poblamiento entre islas diferenciado solo a nivel especifico no es probable que sea anterior al Pleistoceno (Simpson 1956 15)]
Debe tenerse en cuenta que la hipotesis de la vicarianza isla-isla alude a las tres islas septentrionales de las Antillas Mayores porque como se explico antes (vease El Barco Funeral Vikingo) Jamaica no existia como isla durante el lapso (Eoceno Superior shyOligoceno) consensualmente reconocido como el mas probable para la colonizacion de las proto-Antill as Mayores por la mayo ria de los mamiferos terrestres Despues de emerger como isla en el Mioceno Medio Jamaica debe haber recibido sus mamiferos terrestres por dispersion isla-isla (Davalos 2004376) y solo en el Cuaternario parece haberrecibido (por dispersion marina desde Centroamerica) al roedor murido Oryzomys
Seguramente tanto la dispersion como la vicarianza contribuyeron a que los mamiferos terrestres se irradiaran a traves de las Antillas Mayores y grupos de islas perifericas pero las maneras y tiempos en que estos mecanismos operaron a 10 largo de dicho proceso son aun en gran medida motivos de desacuerdo e incertidumbre
IRRADIACION EVOLUCIONARIA
A continuaci6n revisamos someramente las distribuciones conocidas de los diferentes grupos de mamiferos terrestres a traves de las Islas principales de las Antillas Mayores y grupos de Islas perifericas incluidas las interpretaciones y conjeturas que la literatura reciente recoge en relaci6n con la manera en que dichas distribuciones pudieron haberse conformado durante la irradiaci6n evolucionaria de los referidos grupos de mamiferos Manifestaciones de esta irradiaci6n pueden inferirse unicamente a partir de los taxones presentes en el Cuatemario pues solo 6 de las especies conocidas corresponden al Terciario
GRUPOS ORlGINARIOS DE SURAMERICA
PEREZOSOS (Megalonychidae)
Aparte de un genero miocenico de Cuba y otro del Oligoceno de Puerto Rico se conocen 4 generos (12 especies) de perezosos cuaternarios (todos extintos) restringidos a las tres islas septentrionales de las Antillas Mayores Acratocnus Neocnus Megalocnus y Parocnus Los cuatro generos estan representados en Cuba y La Espanola y Acratocnus 10 esta ademas (como unico genero de perezoso) en Puerto Rico Esta distribuci6n ha sido interpretada por MacPhee et al (2000) eomo resultado de la vicarianza oligomiocenica entre estas islas y [dado que el Banco Puerto Rico - Islas Virgenes fue la entrada de GAARlandia a las Antillas Mayores] se preguntaron si la ausencia de los otros tres generos en Puerto Rico se debe a que nunca estuvieron en ese territorio 0 a que sus
316 BIOGEOGRAFiA
fosiles atm no han sido hallados Pero como hemos visto Rega et al (2002) atribuyeron dicha distribucion a la dispersion interinsular en el Cuatemario
Por otra parte varios amilisis c1adisticos recientes ubi can a Choloepus -genero Reciente conocido solo de Centro- y Suramerica- como taxon hermano de Aeratoenus exc1uidos los demas megaloniquidos antillanos Para White y MacPhee (2001228)
tan estrecha relacion sugiere que la divergencia entre ambos [generos] ocurrio despues de que Aeratoenus divergio de Neoenus Si esta relacion implica que Aeratoenus y Neoenus se dispersaron separadamente desde Suramerica a las AntilIas mientras Choloepus permanecia en el continente (siendo necesarias al menos tres invasiones de perezosos) 0 si Choloepus invadio Suramerica desde las Antillas es algo que el material disponible no permite determinar
La diversidad adaptativa de los perezosos se manifiesta en (1) la utilizacion del habitat especies arboreas en Neoenus y Aeratoenus y terrestres en Paroenus y Megaloenus (Webb 1985a White [1993] White et al 1996 Arredondo [1999]) y en (2) la gama de tallas corporales desde los pequenos Neoenus hasta Megaloenus rodens el mayor de los mamiferos terrestres antillanos (su peso adulto estimado en 270 kg seglin Paula 19679) esto sugiere que los perezosos se irradiaron a nichos que en contextos continentales probablemente estarian ocupados por miembros de otros c1ados (MacPhee e IturraldeshyVinent 2000)
No esta claro si el abanico de tamanos entre estos perezosos enmascara alguna tendencia al gigantismo Por 10 pronto Anderson y Handley (2002) encontraron 10 contrario en perezosos (Bradypus) de las Islas de Bocas del Toro (Panama) shyseparadas secuencialmente del area continental vecina por ascensos del nivel del mar durante los ultimos 10 ka En las cinco islas los perezosos son menores que su contraparte continental y las diferencias entre las tallas medias de las cinco poblaciones expresan una reduccion lineal significativa de la talla corporal en funcion de la edad geologica de cada isla sin relacion alguna con el area de las islas 0 la distancia que las separa del continente
MONOS (Pitheciidae)
Los primates de las Antillas Mayores (cuatro especies extintas) constituyen un grupo monofiletico con Paralouatta (en Cuba) y Antillothrix (en La Espanola) como taxones hermanos seguidos en la escalera c1adistica por Xenothrix (en Jamaica) y finalmente Callieebus como el pariente continental mas cercano esto implica una sola invasion desde Suramerica antes del Mioceno Inferior (fecha mas temprana conocida para los primates fosiles antillanos) y por tanto en consonancia con GAARlandia como la via de colonizacion (Horovitz y MacPhee 1996 1999 Horovitz [1997] MacPhee y Horovitz 2004)
En desacuerdo parcial con la anterior interpretacion la filogenia de este grupo de monos obtenida por Davalos (2004) indica un antecesor comun en Cuba La Espanola y Jamaica
317 TRRADIACTON EVOLUCIONARIA
compatible con la hipotetica conexion -propuesta tentativamente por Iturralde-Vinent y MacPhee (199980-84)- entre las Montanas Azules del este de Jamaica y la peninsula suroccidental de La Espanola en la epoca de GAARlandia
A pesar de que los primates antillanos pertenecen a un grupo viviente de monos muy pequenos (los monos titles) fueron comparativamente grandes en relacion con los platirrinos modernos y sucesivamente mayores en la direccion Antillothrix Xenothrix y Paralouatta con P marianae como el mayor platirrino conocido del Mioceno neotropical (MacPhee 2005) MacPhee et al (1994) consideraron que Xenothrix debio ser un mono de andar lento y al parecer las especies de Paralouatta abandonaron la arborealidad para adoptar un modo de vida mas terrestre (MacPhee 1996a MacPhee et al 2003 MacPhee y Meldrum 2006)
ROEOORES (Echimyidae Capromyidae Heptaxodontidae)
La edad de Zazamys en Cuba demuestra que la invasion de los roedores histricognatos en las Antillas Mayores comenzo antes del Mioceno Inferior por otra parte los representantes mas antiguos de esa irradiacion evolucionaria se encuentran en La Espanola y Puerto Rico Ambas evidencias son consistentes con la hipotesis de GAARlandia (Woods 1982 1988 Woods et al 2001)
No esta claro si los capromidos evolucionaron a partir de un equimido ancestral 0 de un protocapromido 0 si los equimidos antillanos (I sub familia 3 generos y 7 especies todas extintas) forman parte de la irradiacion de los capromidos (4 subfamilias 9 generos y 27 especies 10 vivientes) Kratochvil et al (1978b) y Rodriguez et al (1979) consideraron que Capromys nan us es un eslab6n entre Echimyidae y Capromyidae pero Mesocapromys nanus esta morfologicamente mucho mas cerca de Hexolobodon y los de mas capromidos que de los equimidos (Woods 1985) Lo que si esta claro segun Woods et al (2001) es que la subfamilia Plagiodontinae endemic a de La Espanola es el linaje basal de los capromidos y que Isolobodon Hexolobodon Capromys Geocapromys Mysateles y Mesocapromys [y supuestamente Macrocapromys] forman parte de la irradiacion temprana de Capromyidae
La subfamilia Capromyinae (5 generos y 17 especies 9 vivientes) parece ser el dado de mas reciente evolucion en la familia con invasion de Cuba desde La Espanola y dispersion desde Cuba (1) por Capromys a las islas Caiman y (2) por Geocapromys a las islas Bahamas Caiman y Jamaica y desde Jamaica a las islas Swan aunque la presencia de Geocapromys en Swan pudiera deberse a introduccion aborigen (Morgan 1985 1988 1989 1993 1994 Woods 1989a 1990) Woods et al (2001) utilizaron datos moleculares para obtener estimados independientes de divergencia entre capromidos segiJn 10 cual Geocapromys divergio cerca de los 17 MaAP Mesocapromys divergio de Capromys hace 15 MaAP en tanto lvlysateles 10 hizo hace 10 MaAP Por su parte las formas de Capromys pilorides y de Mysateles prehensilis en Isla de la Juventud divergieron de sus correspondientes subespecies nominativas en la isla principal a los 24 y 08 MaAP respectivamente
318 BIOGEOGRAFiA
Woods y Howland (I979) describieron adaptaciones morfologicas de la mandibula el hioide y la faringe asociadas a mecanismos de masticacion diferentes en plagiodontinos y caprominos y Varona (1980a) llamo la atencion sobre el gran desarrollo de los molariformes (en relacion con la longitud total del cnineo) que presentan ciertos grupos de jutias fosiles en comparacion con las vivientes 10 que pudiera interpretarse como una adaptacion alimentaria a una vegetacion mas abrasiva por parte de las jutias extinguidas
Segun Woods (1990) la familia Heptaxodontidae (2 subfamilias 5 generos y 6 especies todas extintas) tambien seria parte de la irradiacion de los capromidos con origen en el este de La Espanola 0 en Puerto Rico y subsecuente dispersion al Banco de Anguila - San Martin en las Antillas Menores (pero vease MacPhee y Flemming 2003) Este banco estaria a solo 70 km del Banco Puerto Rico - Islas Virgenes en tiempos de maxima regresion marina y esta proximidad con Puerto Rico -patria de Elasmodontomys obliquus- seguramente tuvo que ver con la presencia en las islas de Anguila y San Martin del heptaxodontino gigante Amblyrhiza relacionado mas estrechamente con los roedores de las Antillas Mayores que con los del resto de las Antillas Menores (Woods 1990 1996) Aunque los roedores suelen aumentar de tamano en las islas (Case 1978) la escala del incremento de la talla en Amblyrhiza (que pudo alcanzar mas de 200 kg de peso adulto) esta fuera de toda proporcion con los patrones conocidos en otros roedores insulares (Biknevicius et 01 1993)
Clidomys (Clidomyinae) de Jamaica difiere marcadamente en su morfologia de los heptaxodontinos (vease ademas MacPhee y Flemming 2003)10 cual dificulta explicar su presencia en esa isla Como Jamaica estuvo bajo el mar hasta el Mioceno Medio el ancestro de Clidomys colonizaria esta isla por dispersion desde el extremo suroccidental de La Espanola despues que los heptaxodontinos se originaron en esta ultima isla (Woods 1989a) Pero Pascual et 01 (I 990) especularon que Clidomys pudiera ser el resultado de una invasion posterior e independiente de los capromidos
GRUPOS ORIGINARIOS DE NORTEAMERICA
El origen de las musaranas antillanas ha resultado ser hasta ahora el mayor enigma biogeografico de la fauna de mamiferos terrestres de la region aunque la mayoria de los autores coinciden en considerarlas originarias de Norteamerica (vease Discus ion y Conclusiones en Poblamiento Continente-Isla) No obstante MacPhee e Iturralde-Vinent (1994) descartaron toda posibilidad de conexion terrestre por el Estrecho de la Florida 0 por el Canal de Yucatan y si el (supuesto) evento vicari ante de Rosen (vease El Arca de Noe) -invocado por MacFadden (1980) para resolver el enigma- fuera minimamente plausible aun asi es dificil creer que las catastrofes que 10 sucedieron (vease Contexto Temporal) hayan barrido con toda esa vieja fauna continental excepto las musaranas circunstancia ya anticipada muy tempranamente por Matthew (l918a)
De otra parte la relaci6n entre Nesophontidae y Solenodontidae permanece irresuelta Si estos linajes no estan estrechamente relacionados entonces es muy posible que
319 IRRADIACION EVOLUCIONARIA
sus distribuciones sean explicadas por mecanismos diferentes (Whidden y Asher 2001)
La descomunal diferencia de talla corporal entre las musaranas pequenas (Nesophontes) y las grandes (Soienodon) seguramente impuso a ambos grupos restricciones distintas relativas a la termorregulacion y las adaptaciones energeticas En Nesophontes la seleccion debio favorecer una alta tasa de metabolismo y crecimiento camadas mas numerosas corta expectativa de vida yalta densidad poblacionaL En Soienodon en cambio la seleccion opero en la direccion de una baja tasa metabolica camadas reducidas crecimiento lento longevidad y baja densidad poblacional (Ottenwalder [1991])
MUSARANAS PEQUENAS (Nesophontidae)
Las musaranas pequenas estan representadas en las Antillas por el genero Nesophontes (y un genero miocenico indeterminado en ambar dominicano) con 6 especies cuaternarias extinguidas distribuidas en Cuba La Espanola Puerto Rico Islas Virgenes e Islas Caiman La mayor de las musaranas pequenas Nesophontes edithae evoluciono en Puerto Rico y su arribo a las Islas Virgenes supone una dispersion sumamente reciente En las Islas Caiman Nesophontes se asemeja mas a la poblacion de Cuba de donde seguramente derivo en el Plio-Pleistoceno (junto con Capromys y Geocapromys) cuando la costa sur de Cuba estuvo mucho mas cerca de las Islas Caiman (lturralde-Vinent 2003fig 4) MacPhee et ai (1994) interpretaron la distribucion de Nesophontes en Cuba y La Espanola como resultado de la vicarianza oligomiocenica ocurrida al formarse el Paso de los Vientos pero la distancia taxonomica entre los lados del Paso es solo de nivel especifico y N micros incluso vivi6 en ambas islas segtln Patterson (1962)
Allen (1918) considero que Nesophontes micrus fue una especie primitiva no modificada para la vida arb6rea 0 fos6rea y por tanto de habitos terrestres Nesophontes edithae de Puerto Rico es mucho mayor y mas robusta que las restantes especies del genero eL indice de robustez de su femur (= anchura minima de la diafisis ongitud total del hueso) muestra que este es tan robusto como el de Solenodon paradoxus un animal dos veces mayor (Morgan et ai 1980) Choate y Birney (1968) postularon un aumento cronoclinal de la taIla corporal en N edithae confirmado mas tarde por McFarlane (1999a) mediante fechados radiocarb6nicos de la secuencia estratignifica en depositos cavernarios de Puerto Rico Solenodon no vivi6 en Puerto Rico y en ausencia de una musarana de mayor tamano Nesophontes alcanzo una talla corporal mayor en esa isla que en La Espanola 0 Cuba (Woods y Eisenberg 1989)
MUSARANAS GRANDES (Solenodontidae)
Los arcaicos almiqufes (genero Soienodon) comprenden cuatro especies una viviente (S cubanus) y otra f6sil (S arredondoi) en Cuba y una viviente (S paradoxus con dos subespecies) y otra f6sil (S marcanoi) en La Espanola Como cabria esperar las dos especies cubanas se asemejan mas entre sf que a las especies de La Espanola pero
320 BIOGEOGRAFiA
enigmaticamente S marcanoi comparte caracteres con S cuban us y S paradoxus (Woods y Eisenberg 1989 Morgan y Ottenwalder 1993 Ottenwalder 200 I) Solenodon arredondoi alcanzo una talla corporal superior a la de cualquiera de las musarafias vivientes a nivel mundial Segun Morgan y Ottenwalder (1993) extrapolando las dimensiones y el peso de los almiquies vivientes S arredondoi debio tener una longitud cabeza-tronco y un peso de 450-550 mm y 1500-2000 g respectivamente en comparacion con S cubanus (360 mm y 800 g) y S paradoxus (390 mm y 1100 g)
Entre todas las especies de almiqufes Solenodon paradoxus es la que presenta los caracteres mas derivados (Otten walder 2001) Si estas relaciones son validas la distribucion de Solenodon entre Cuba y La Espafiola debio discurrir de oeste a este como propuso MacFadden (1980) quien invoco la presencia fosH de S cubanus [pero tambien S arredondoi] en Pinar del Rio para postular que el ancestro continental entr~ por Cuba opinion compartida por Perfit y Williams (1989) y por Ottenwalder ([1991] 2001) El paso de Solenodon a Puerto Rico pudo haberlo impedido la existencia de barreras ecologicas durante la conexion del este de La Espafiola con Puerto Rico 0 por la ausencia de Solenodon en esa parte de La Espafiola en aquella epoca Sin embargo las mismas condiciones ecologicas en el este de La Espafiola no parecen haber representado una barrera efectiva para Nesophontes cuyas mas altas tasa de reproduccion y densidad poblacional presumiblemente Ie permitieron expandirse flpidamente a traves de La Espafiola (Ottenwalder [1991])
Es opinion generalizada que los almiqufes antillanos constituyen el grupo de mamiferos que primero se emplazo en el territorio insular (MacPhee 2005) y en tal sentido es de interes la evidencia sobre especificidad parasito-hospedante aportada por Cruz (1980) quien encontro que la especie de acaro (genero Cubanochirus) que parasita a los almiquies es mucho mas primitiva que la especie (del mismo genero) que paras ita a las jutias y de aqui concluyo que dicho genero de acaro debio evolucionar primero en Solenodontidae y posteriormente en Capromyidae al entrar en contacto representantes de ambas familias en Cuba 0 La Espafiola
Recientemente Roca (2004) y Roca et al (2004) dieron a conocer que los resultados de su estudio genetico-molecular sobre los almiquies indican una divergencia de 25 Ma entre Solenodon cuban us y S paradoxus -compatible con la disyuncion tectonica entre Cuba y La Espafiola (Iturralde-Vinent y MacPhee 1999 McPhee et al 2003)-- y una separacion de ambas especies comparable en magnitud a la divergencia entre familias de mamlferos Consecuentemente propusieron que Solenodon cubanus fuera clasificada en el genero Atopogaie como indico Cabrera (1925) hace 82 afios
DISCUSION Y CONCLUSIONES
Las musarafias y los roedores antillanos (excepto Oryzomys) se irradiaron tanto de sus respectivos ancestros continentales que de estos 6rdenes se reconocen familias y subfamilias endemicas de la region en contraste con los perezosos (una subfamilia y tres
321 IRRADIACION EVOLUCIONARIA
tribus endemicas) y los monos (una tribu endemica) como expresion de diferentes niveles de diversificacion evolucionaria Este proceso es dificil de explicar (traUindose de c1ados que pnicticamente comparten la misma antigiiedad en las Antmas) si es correcta la interpretacion de Pascual et at (1990636)
Nosotros consideramos estos diferentes niveles de diversificacion como las diferentes etapas de evolucion alcanzadas por cada grupo [de mamiferos] Obviamente esto estuvo relacionado con las diferentes longitudes de sus historias
Pudiera ocurrir que en el futuro se compruebe que la historia biogeognifica de la region fue mucho mas complicada
La Espanola la isla mas ecol6gicamente divers a de la region debi6 ser el centro principal de evoluci6n de los mamfferos (con 87 de las familias y 73 de las subfamilias) seguida por Cuba (con 75 de las familias y 54 de las subfamilias) las dos islas de las Antillas Mayores mas relacionadas biogeograficamente (Fig 43 Tabla 44)
eL
EL
cb CAIMAN
SWAN
A Acratocnini B Cubanocnini C Megalocnini D Mesocnini E Nesophontidae F Solenodontidae G Xenothrichini
ANGUILAshyM-0 SMARTIN
H Heteropsomyinae I Plagiodontinae J Isolobodontinae
K Hexolobodontinae L Capromyinae
M Heptaxodontinae N CIidomyinae 0 Sigmodontinae
FIGURA 43 Representacion esquematica de las relaciones interinsulares (expresadas porel numero de linajes compartidos) entre las Antillas Mayores y entre estas y los grupos de islas perifericas segun las interpretaciones de los autores citados en el texto Los numeros dentro de los circulos indican la cantidad de linajes representados en cada isla relacion dudosa Contexto taxon6mico de los linajes en la Tabla 44
bull bull
bull bull
bull bull bull
bull bull
bull bull
bull
bull
bull
bull
bull bull bull bull
bull bull
bull bull
bull
bull bull bull
bull bull
bull
bull bull
bull
322 BIOGEOGRAFiA
TABLA 44 Distribuci6n de los linajcs de mamiferos terrestres a traves de las Antillas Mayores y grupos de islas perifericas LE = La Espanola CU Cuba JA Jamaica PR = Puerto Rico IC Islas Caiman IS Islas Swan IB = Islas Bahamas IV = Islas Virgenes AS = Anguila - San Martin
FamiliaSubfumiliafIribu LE CU JA PR IC IS IB IV AS
Megalonychidae
Choloepodinae
Acratocnini
Cubanocnini
Megalocninae
Megalocnini
Mesocnini
Nesophontidae
Solenodontidae
Pitheciidae
Callicebinae
Xenothrichini
Echimyidae
Heteropsomyinae
Capromyidae
Plagiodontinae
Isolobodontinae
Hexolobodontinae
Capromyinae
H e pta x 0 don tid a e
Heptaxodontinae
Clidomyinae
Muridae
Sigmodontinae
PILOSA
SORICOMORPHA
PRIMATES
RODENTIA
bull
bull
bull
bull TOTALES 12 10 4 4 2
323 EXTlICIOIES DEL CUATERNARIO
El gigantismo ha sido sin duda una tendencia evolucionaria recurrente entre los clados mayores de mamiferos antillanos Es un hecho bien conocido que algunos grupos de mamiferos han aumentado de tamafio en las islas y varios autores han intentado explicarlo a partir de enfoques diferentes (Foster 1964 Miller 1966 Sondaar 1977 Case 1978 Eisenberg y Wilson 1978 Heaney 1978 Lawlor 1982 Lomolino 1985 Angerbjom 1985 McNab 19942001)
EXTINCIONES DEL CUATERNARIO
Las islas que integran la Subregion Antillana han perdido casi 90 de sus mamiferos terrestres durante los ultimos milenios y en proporcion con su extension territorial esta perdida ha sido mayor en la Subregion que en cualquier otro lugar del planeta (MacPhee 1996b) Las extinciones del Cuatemario eliminaron mas de las tres cuartas partes de la fauna de mamiferos terrestres conocida de las Antillas Mayores sus islas satelites y grupos de islas perifericas y el examen de este proceso en sus aspectos mas generales se aborda en esta ultima parte del presente capitulo
CRONOLOGiA Y CONSECUENCIAS
Es muy improbable que un fosil represente el ultimo individuo de un taxon extinto y entonces hay que guiarse por la edad radiometrica del fosil mas reciente conocido lIamada fecha de ultima presencia- como unica aproximacion posible a la fecha de extincion del taxon
Como se explico al inicio del capitulo (vease Contexto Temporal) el registro fosH de los mamiferos terrestres de las Antillas Mayores adolece de insuficiente sustentacion radiometrica No obstante la distribucion temporal de los escasos fechados disponibles para el Cuaternario de la region abarca etapas criticas del proceso de extincion experimentado por la referida fauna a 10 largo de los ultimos 250 mil afios
De las 52 especies que es posible reconocer actualmente como miembros extintos de la fauna cuatemaria de mamiferos terrestres de las Antillas Mayores y grupos de islas perifericas los restos fosiles de 23 de elIas (442) han sido fechados radiometricamente o se han hallado asociados estratigraficamente de manera inequivoca con otro material fechado De estos 23 fechados 3 corresponden al interglacial Sangamon 4 pertenecen a la secuencia Pleistoceno terminal - Holoceno temprano 10 se ubican en ellapso entre el arribo de los aborigenes y la incursion europea y 6 son postcolombinos (a los cuales hay que sumar otras dos especies extinguidas en los siglos XIX y xx) A partir de estas 25 especies avaladas por fechas de ultima presencia distribuidas entre los cuatro intervalos temporales sefialados hemos compuesto la Tabla 45 en la que ademas aparecen las restantes 27 especies (carentes de fechado radiometrico) ubicadas en el intervalo temporal mas probable sobre la base de (1) la asociacion estratigrafica de sus restos con evidencias culturales (amerindias 0 europeas) 0 (2) la ausencia de dicha asociacion Sobre estos criterios de ubicacion temporal procede hacer dos aclaraciones
324 BIOGEOGRAFIA
TABLA 45 Intervalos temporales mas probables para las extinciones cuatemarias de los mamiferos terrestres de las Antillas Mayores y grupos de islas perifericas sobre la base de las evidencias siguientes (l) ultimo ejemplar colectado u observado en vida (2) fechado radiometrico mas tardio en material de la especie (3) asociaci6n estratigrafica mas tardia con otro material fechado (4) asociaci6n estratigrafica con restos de animales introducidos restos humanos u objetos culturales propios del intervale (5) falta de asociaci6n con evidencias culturales [Los ntimeros entre parentesis en la columna Evidencia significan adscripci6n tentativa al intervalo] Los estimados del peso corporal (en kg) estan tomados de la literatura 0 inferidos comparativamente y distribuidos entre
las clases siguientes (a) S 01 (b) 01 a lt 1 (c) 1 a lt 10 (d) 10 a lt 100 (e) 100 a lt 200 (j) ~ 200 Las
abreviaturas que designan las islas son las de la Tabla 44
F amilialEspecie Peso IsJa Evidencia Fuente
I httervalo htterg1acial Sangamon (gt80 kaAP)
HEPTAXODONTlNAE
Amblyrhiza inundata e AS 3 McFarJane et al 1998a
CLIDOMYINAE
Clidomys osborni d JA 3 Ibid
Clidomys parvus d JA 3 Ibid
II httervalo Pleistoceno terminal ~ Holoceno temprano (20-6 kaAP)
MEGALONYCHIDAE
Aeratoenus antillensis d CD 5 Arredondo 1975 a
Aeratoenus odontrigonus d PR 3 McFarJane 1999b
Aeratoenus simorhynehus d LE 2 Rega et al 2002
M egaloenus zile f LE 2 MacPhee et al 2000
SOLENODONTIDAE
Solenodon arredondoi e CD 5 Morgan y Ottenwalder 1993
PnHECIIDAE
Paralouatta varonai d CD 5 MacPhee 1996a
EcHIMYIDAE
Heteropsomys antillensis c PR 5 McFarJane 1999b
Heteropsomys insulans c PR 5 Ibid
Puertoricomys eorozalus c PR 5 Woods 1989a
CAPROMYIDAE
P lagiodontia araeum c LE 5 MacPhee et al 2000
Macrocapromys latus c CD 5 Varona y Arredondo 1979
(Continua)
325 EXTINCIONES DEL CUATERNARIO
TABLA 45 (Continuaci6n)
F ami1ia1Especie Peso Isla Evidencia Fuente
M acrocapromys latus c CU 5 Varona y Arredondo 1979
Mesocapromys kraglievichi c CU 5 Ibid
HEPTAXODONTINAE
Xaymaca fulvopulvis a JA 2 MacPhee y Flemming 2003
III Intervalo Amerindio (6-05 kaAP)
MEGALONYCHIDAE
Acratocnus ye d LE 2 MacPhee et aI 2000
Neocnus comes c LE 2 Ibid
N eocnus dousman c LE 2 Ibid
N eocnus toupiti c LE 2 Ibid
N eocnus gliriform is c CU 4 Cordova et ai 1997
M egalocnus rodens f CU 2 MacPhee et ai 2007
Parocnus browni e CU 2 Steadman et ai 2005
P arocnus serus e LE 2 MacPhee et ai 2000
N ESOPHONTIDAE
N esophontes edithae a PR 2 McFarlane 1999b
PITHECIIDAE
Antillothrix bernensis c LE 3 Rimoli 1977
EcHIMYIDAE
Brotomys contractus a LE 4 Miller 1929c
CAPROMYIDAE
P lagiodontia spelaeum c LE 4 Ibid
M acrocapromys acevedo c CU 4 NUiiez 1990a
HEPTAXODONTINAE
Elasmodontomys obliquus d PR 3 McFarlane 1999b
Iv Intervalo Postcolombino (05 kaAP - Presente)
N ESOPHONTIDAE
N esophontes hypom icrus a LE 24 MacPhee y Fle111111ing 1999
N esophontes zam icrus a LE (4) Ibid
N esophontes m icrus a CU 24 MacPhee et aI 1999
(Continua)
326 BIOGEOGRAFiA
TABLA 45 (Continuacion)
F amilialEspecie Peso Isla Evidencia Fuente
N esophontes m icrus a CD 24 MacPhee et aI 1999
N esophontes m icrus a LE 4 MacPhee et aI 2000
N esophontes sp 1 a IC 4 Morgan 1994
N esophontes sp 2 a IC 4 Ibid
Solenodon marcanoi b LE 4 Ottenwalder 2001
PITHECIIDAE
Xenothrix mcgregori c JA 4 MacPhee y Flemming 1999
EcHIMYIDAE
Boromys offeUa a CD 24 Guarch 1984
Boromys torrei a CD 24 Ibid
Brotomys voratus a LE 4 MacPhee et aI 2000
CAPROMYIDAE
P lagiodontia ipnaeum c LE 4 MacPhee y Flemming 1999
Rhizoplagiodontia lemkei c LE 4 Ibid
Isolobodon portoricensis c PR 24 Ibid
Isolobodon montanus c LE 4 Ibid
Hexolobodon phenax c LE (4) Ibid
Capromys sp c IC 24 Morgan 1994
Geocapromys columbianus c CD 4 MacPhee y Flemming 1999
Geocapromys thoracatus b IS 1 MacPhee y Marx 1997
Geocapromys sp 1 c IC 4 Morgan 1994
Geocapromys sp 2 c IC (4) Ibid
l-IEPTAXODONTINAE
Quemisia gravis c LE (1) MacPhee y Flemming 1999
MURIDAE
Oryzomys antillarum a JA 1 Ibid
1 Flemming y MacPhee (1999) alertaron en contra de la asociacion con otro material fechado Asimismo Morgan (1993) indico que la ausencia de asociacion con evidencias culturales amerindias no es necesariamente un criterio concluyente para ubicar un taxon en el intervalo temporal anterior porque hay depositos en la region que carecen de tales evidencias y son del Holoceno tardio por fechados radiocarbonicos [No obstante
327 EXTlNCIONES DEL CUATERNAR10
la ausencia de una especie en residuarios arqueol6gicos adquiere mayor certidumbre como criterio de ubicaci6n temporal de su extinci6n en la medida que aumenta el numero de residuarios estudiados que en las Antillas Mayores se cuentan por centenares] A falta de fechado absoluto de una especie consideramos que estos criterios de ubicaci6n temporal son mas utiles que la alternativa de excluir del analisis a la especie
2 En los trabajos sobre biogeografia de mamiferos antillanos los arque610gos mas citados como autoridades para establecer el inicio de la presencia aborigen en las Antillas Mayores (Kozlowski 1974 Rouse y Allaire 1978 Rouse 1989 1992 Wilson 2001) difieren al respecto entre los 6 kaAP y los 45 kaAP (pero vease Pagan et al 200523) y hemos adoptado la mas temprana de estas fechas para el comienzo del intervalo amerindio en la Tabla 45 De todos modos cualquiera que haya sido la fecha esta no es aplicable por igual al conjunto de las Antillas Mayores porque segun algunos de los autores citados el poblamiento comenz6 por Cuba y La Espanola 6 mil anos atras continu6 en Puerto Rico hace alrededor de 3 mil anos (pero vease Burney et al 1994) yen Jamaica hace menos de 2 mil e incluso a las Islas Caiman ni siquiera llegaron [Las Islas Caiman descubiertas por Col6n en 1503 carecen absolutamente de evidencia amerindia (Stokes y Keegan 1996)]
Si los datos de la Tabla 45 representan una aceptable aproximaci6n a la realidad el proceso de las extinciones cuaternarias de los mamiferos terrestres de las Antillas Mayores y grupos de islas perifericas debi6 caracterizarse entre otros por los eventos siguientes
Aunque obviamente la fauna cuaternaria debi6 ser mucho mas numerosa que 10 que refleja el registro paleontol6gico (Morgan y Woods 1986) 52 especies (81 de la fauna conocida) desaparecieron (sin reposici6n) durante los ultimos 250 mil anos
- Al parecer las extinciones se concentraron en las especies de mayor tamano Asi el peso promedio estimado para las especies de la etapa preantr6pica (intervalos I + II de la Tabla 45) es 350 kg es 328 kg para las especies del intervalo III (amerindio) y es s610 25 kg (c1ases del peso a b c) para las especies del intervalo IV (postcolombino) Ninguna especie aut6ctona de mamifero mayor de 6 kg existe hoy en la regi6n
- De acuerdo con la distribuci6n temporal de las extinciones 61 especies sobrevivieron al interglacial Sangamon (intervalo 1)48 sobrevivieron a la transici6n del Pleistoceno terminal al Holoceno temprano (intervalo II) 34 sobrevivieron al periodo amerindio (intervalo III) y s610 12 especies han sobrevivido desde la Ilegada de los europeos (intervalo IV) Las tasas de extinci6n correspondientes (exceptuado el intervalo I) son una especie cada 1167 anos en el intervalo II una cada 393 anos en el intervalo III y una cada 23 anos en el intervalo IV
- Durante todo el proceso de las extinciones cuaternarias la perdida de especies por isla fue de 91 en La Espanola 83 en Jamaica y 62 en Cuba Bahamas retuvo su unica especie (aunque extirpada en varias de las islas) Puerto Rico e Islas Caiman perdieron todas sus especies (6 y 5 respectivamente) y las Islas Swan Virgenes y Anguila - San Martin perdieron sus respectivas especies (micas
328 BIOGEOGRAFiA
De los cuatro 6rdenes de mamiferos representados en la fauna Pilosa y Primates desaparecieron totalmente Soricomorpha y Rodentia perdieron 86 y 74 de sus especies respectivamente
CAUSALIDAD
CAUSAS NATURALES
Los cicIos glaciales e interglaciales acaecidos en el Pleistoceno causaron cambios ambientales dnisticos en las Antillas con la concomitante alternancia de cIimas frios (secos) y calidos (hUmedos) asi como de descensos y ascensos del nivel del mar aumento y disminuci6n de las areas territoriales) induso (aunque con menor intensidad) en el Holoceno Estos eventos paleogeograficos han sido consistentemente documentados (Mayo y Kartashov 1972 Bonnatti y Gartner 1973 Ortega y Arcia 1982 Prance 1982 Schubert y Medina 1982 Dodge et al 1983 Ortega [1984] Haq et al 1987 Schubert 1989 Hodell et al 1991 Guilderson et al 1994 Oppo 1997 Emslie 1998 Curtis et al 2001 Iturralde-Vinent 2003) y de ellos brindamos a continuaci6n una breve resena
Finalizando el periodo interglacial anterior a la ultima glaciaci6n (Wisconsin) el mar estaba a 8-10 m por encima de su nivel actual Cuando el avance glacial del Wisconsin a1canz6 su culminaci6n (eillamado Ultimo Maximo Glacial) hace 18 ka el mar habia descendido 120 m con respecto a dicho nivel y las islas del Caribe habian incrementado considerablemente sus extensiones subaereas en comparaci6n con las que presentan hoy dia por ejemplo Puerto Rico y las Islas Virgenes se conectaron territorialmente y las Bahamas se unieron en cinco gran des islas Prevalecia un regimen frio y seco y condiciones de aridez --debidas a una reducci6n de las precipitaciones (posiblemente a menos de 440 mmlano)- y los ambientes xericos se habian extendido a traves de las islas y bancos emergidos y abundaban los pinares (los habitats mesicos estaban restringidos a parcelas relictas en 10 alto de las montanas) En el Mar Caribe y el Golfo de Mexico la temperatura superficial del mar era 5degC mas baja que la actual y en las llanuras costeras y alturas maximas eran respectivamente 2-3degC y 6degC mas bajas Los limites de las zonas ecol6gicas altitudinales descendieron varios cientos de metros y cada zona redujo su extensi6n vertical al aumentar el gradiente de temperatura Ellimite inferior de la zona periglacial
elevaci6n donde la temperatura media minima a1canza los OdegC) de la Cordillera Central de la Republica Dominicana (altura maxima del Caribe insular con evidencia de topografia glacial) se considera que estuvo entre los 1500 y 1800 mlnm (actualmente esta a 2800 mlnm) y se puede estimar que dicho limite era inferior en Cuba por estar mas al norte que la Republica Dominicana No obstante durante este periodo la temperatura experiment6 episodios cortos (unas pocas decadas) de ascensos repetidos
Un signifieativo componente bi6tieo de las referidas condiciones paleoambientales 10 constituy6 la espectacular irradiaci6n evolucionaria experimentada por un grupo numeroso de aves rapaees diurnas y nocturnas (Wetmore 1937 Brodkorb 1969 Kurotchkin y Mayo 1973 Arredondo 1976 Olson 1978 Olson y Hilgartner 1982
329 EXTINCIONES DEL ClJATERNARIO
Suarez 2000a b 2004 Suarez y Emslie 2003) Los mamiferos de las Antillas Mayores ofrecieron a estos depredadores aviares una amplia gama de tamanos de presa que en ausencia de mamiferos del orden Carnivora en la region permitio a estas aves irradiarse a este nicho alimentario con la evolucion de una variedad de especies de muy diversa tall a corporal incluidas formas gigantes Adquirir un cuerpo gigante les propicio el acceso a recursos alimentarios que no estaban siendo explotados por ningun otro depredador (Alcover y McMinn 1994) A Cuba corresponde el mejor registro fosil de rapaces cuaternarias caribenas con siete generos de especies diurnas Dentro de estas aves existieron tres gavilanes gigantes (Accipitridae) tres carairas (Falconidae) y un condor (Cathartidae) asi como cuatro generos de especies nocturnas dos lechuzas (Tytonidae) y tres Mhos (Strigidae) dos de ell os no igualados en tamano en todo el mundo Algunos de estos generos tuvieron representacion en otras islas antillanas como La Espanola Jamaica Bahamas y Gran Caiman
Al retirarse hacia el norte los hielos de la glaciacion Wisconsin el clima se tomo mas hUmedo en las Antillas durante el Holoceno temprano y el bosque mesico se expandio en la region insular El nivel del mar ascendio acompanado de mayor pluviosidad alrededor de los 7 kaAP las Islas Virgenes quedaron separadas entre si y de Puerto Rico y las Bahamas perdieron la mayor parte de su territorio Cerca de los 3 kaAP las condiciones secas retornaron a la region e hicieron desaparecer buena parte del bosque mesico Nuevamente el clima comenzo a adquirir sus condiciones actuales hacia los 25 kaAP aproximadamente y hoy transita por una fase mesica con vegetacion xerica confinada en parcelas relictas
Sin duda estos y otros eventos naturales incidieron negativamente en la sobrevivencia de mamiferos terrestres en las Antillas Mayores asi como en los grupos de pequenas islas perifericas (Pregill y Olson 1981 Olson 2006)
McFarlane et al (1998a) demostraron convincentemente como Amblyrhiza el gigante de los roedores antillanos endemico de la pequena isla de Anguila sucumbio a las repetidas y abruptas perdidas de su habitat durante el interglacial Sangamon por las nipidas y severas oscilaciones del nivel del mar (documentadas por Neumann y Hearty 1996) al no poder responder adecuadamente a tan catastroficos eventos mediante el ajuste de su masa corporal media y otro tanto debio ocurrirle en Jamaica al otro roedor gigante Clidomys (McFarlane et al 1998b) emparentado con Amblyrhiza en la misma epoca (Tabla 45 intervalo I)
El ascenso del nivel del mar seguramente promovio la extincion de mamiferos durante la transicion del Pleistoceno al Holoceno (Tabla 45 intervalo II) con la reduccion del tamano de las Islas por las transgresiones marinas (Pregill y Olson 1981 Morgan 1993) Esto ultimo haria maximamente severa la competicion asf como la presion de los depredadores [Steadman et al (2005) no yen por que el calentamiento global ocurrido durante la transicion Pleistoceno-Holoceno habria de tener un efecto negativo sobre los perezosos siendo sus descendientes actuales especies tropic ales 0 subtropicales pero no se trata del efecto
330 BIOGEOGRAFiA
directo del cambio de temperatura sobre los animales sino del efecto indirecto y progresivo que dicho cambio genera a largo plazo al reducir el territorio insular disponible]
Por esta epoca tambien desaparecieron especies de tall a mayor como Megalocnus zile en La Espanola y otras de menor tamano (en Cuba) pero que representan la tall a maxima en sus respectivos contextos taxonomicos del Cuaternario como Solenodon arredondoi Paralouatta varonai y Macrocapromys acevedo Un mamifero puede ser grande solo a costa de reducir su densidad poblacional y esto 10 hace mis vulnerable a la extinci6n sobre todo en una isla (McNab 1994 McFarlane et al 1998a) Incidentalmente Alcover et al (1998a) afirmaron que larelacion entre la talla corporal y la extincion no siempre se cumple en las especies de menor tamano y citaron como ejemplo que entre las musarafias todas las especies de Nesophontes desaparecieron mientras sobreviven especies de Solenodon pero la comparacion correcta en todo caso serfa entre los solenodones vivientes y el gigante Solenodon arredondoi y entre Nesophontes edithae -el gigante de los nesofontes extinguido en el periodo amerindio- y el resto de sus congeneres que sobrevivieron hasta los primeros siglos de la colonizacion europea (Tabla 45 intervalos III y IV)
CAUSAS ANTROPOGENICAS
Intervalo Amerindio
La mayoria de los autores que se han referido a las extinciones acaecidas durante este intervalo (entre elIos paradigmaticamente Martin 1984a b) han responsabilizado a los aborfgenes (que se supone comenzaron a poblar las Antillas Mayores hace 6 mil afios) con la desaparicion de numerosas especies de mamiferos terrestres en la region principalmente por la via de la sobrecaza y la introduccion de animales domesticos devenidos depredadores de la fauna autoctona (Morgan y Woods 1986) Este efecto negativo de la presencia indigena sobre los mamiferos de la region ha sido dramatizado por algunos autores en terminos de holocausto faunal Sin embargo pensamos que se ha exagerado Como han expresado MacPhee et al (19993) cuando uno busca evidencia dura de complicidad humana directa en [estas] extinciones no hay mucho que encontrar Las siguientes son algunas consideraciones a tener en cuenta en tal sentido
El registro radiometrico mas joven conocido hasta ahora para un perezoso antillano (MegalocnllS rodens) es de 42 kaAP (MacPhee et ai 2007) Si se acepta que los aborfgenes entraron en Cuba hace aproximadamente 6 mil anos su convivencia con los perezosos debe haber durado alrededor de 1800 afios (0 mas porque dificilmente el referfdo fechado radiometrico haya correspondido al ultimo sobreviviente de la especie) Esta cifra contradice la hipotesis del holocausto faunal segun la cualla extincion de los mamiferos insulares de mayor talla debe ocurrir con extrema rapidez -a escala de decadas 0 de centurias- a partir de su primer contacto con los humanos (Martin y Steadman 1999) A pesar de las repetidas afirmaciones en contrario (Harrington 1921 Miller 1929c Herrera [1945] Tabio 1951 Alvarez 1951a 1957 Arredondo
331 EXTINCIONES DEL CUATERNARIO
1959 1960a Veloz y Ortega 1973 Mayo 1980b Rodriguez et al 1984 Castellanos y Pino 1985 Pino y Castellanos 1985 Nunez 1990b Gutierrez 1996 Arredondo 2004) en su mayoria basadas en evidencias no exentas de posibles accidentes de deposicion (0 de reubicacion postdeposicional) en recintos cavernarios (Diaz-Franco [2006b D no se ha demostrado inequivocamente que los aborigenes antillanos se alimentaran de perezosos regularmente Sus restos en residuarios de alimentos son escasisimos en toda la region y esto contrasta con los abundantes restos de jutias y otros mamiferos de menor talla que aparecen en esos residuarios demostrativos de la preferencia de los paleoindios por estos otros mamiferos
Los mamiferos mas importantes en la economia prehistorica de las Antillas Mayores fueron las jutias Capromys y Geocapromys en Cuba Geocapromys en Jamaica e Isolobodon en La Espanola Puerto Rico e Islas Virgenes (Aguayo 1950b Tablo 1951 Rimoli 1972 Wing 2001 Diaz-Franco [2006b D Wilkins (2001) estudi61ajutia viviente de Jamaica (Geocapromys browni) en un sitio ocupado por los aborigenes durante 100 anos y no encontr6 evidencias de que durante ese tiempo la especie haya sido sobreexplotada Asimismo en Cuba las jutias conga carabali y andaraz son escasas en depositos pleistoholocenicos en cambio son muy numerosas en residuarios indigenas y de haber sido sobreexplotadas 10 mas probable es que no hubieran sobrevivido hasta hoy
La habilidad de los humanos para penetrar en profundidad el bosque tropical y mantenerse en este sustentablemente ha sido un evento comparativamente reciente (Beran y Steele 1994) En el caso antillano la caza tierra adentro ha sido explicada como un proceso tardio de desplazamiento a partir de la explotacion de abundantes recursos marinos que ofrecian condiciones 6ptimas para el sedentarismo de las comunidades aborigenes costeras (C6rdova y Arredondo 1988 Brito 2005) Peces y mariscos dominan la arqueologia costera de las Islas antillanas en tanto el consumo de recursos terrestres (mamfferos incluidos) aumenta progresivamente en la medida que los residuarios estudiados se alejan de la costa (Scudder 1991 Wilkins 2001) En las Antillas Mayores el componente terrestre de la dieta aborigen es solo 20-30 del material recobrado en los residuarios (Wing 1977 1989) Y debio ser menor alin porque el componente vegetal de la dieta no se ha preservado Por otra parte los aborigenes cubanos domesticaron la jutia conga y seguramente la reprodujeron para comer (Pose et al 1988) practica que probablemente se repitiera con otras jutias en las demas Antillas Mayores
- Fray Bartolome de las Casas hablo de 3 millones de indios en Cuba al inicio de la colonizacion pero los historiadores subsecuentes consideraron exagerada la cifra (Poey 1851 a) No obstante se dispone de dos estimados de la densidad de poblaci6n aborigen en las Antillas Mayores a la llegada de los europeos 18 indigenaslkm2 en Puerto Rico (Rouse 1956) y 1271km2 en La Espanola (Watts 1987) Extrapolando ambos valores entre las cuatro islas principales de las Antillas Mayores tenemos que en la fecha indicada la poblaci6n aborigen pudo estar entre 16 mil y 113 mil indigenas en Puerto Rico entre 21 mil y 145 mil en Jamaica entre 141 mil y 1 mill6n en La Espanola y entre 200 mil y 14 millones en Cuba Entre las cifras maximas y las minimas nos inclinamos
332 BIOGEOGRAFiA
hacia estas ultimas atendiendo a los altos indices de mortalidad infantil y morbilidad que se evidencian en las osamentas de los pobladores prehispanicos (Martinez y Rives 1990) Uno de los hechos que apuntan hacia el bajo numero de aborigenes en las Antillas Mayores es 10 rapido que desaparecieron despues de la llegada de los europeos pnkticamente sin dejar mestizaje con estos 10 cual contrasta ostensiblemente con la situacion en la America continentaL
En suma un grupo humano (1) demograiicamente irrelevante y controlado por altos indices de morbilidad (2) consumidor mayoritariamente de organismos marinos y (3) carente de la tecnologia cinegetica necesaria para poder sobreexplotar los mamiferos que Ie sirvieron de alimento (a los cuales accedio tardiamente en su desarrollo) no es concebible que haya causado e1 holocausto faunal que se Ie atribuye a partir del error de aplicar a las comunidades aborigenes antillanas un modelo cultural de la prehistoria continentaL
Tampoco hay evidencias de que los aborigenes hayan introducido carnivoros con suficiente capacidad depredadora salvo el perro Morgan y Otlenwaldder (1993) consideraron que la depredacion por perros durante el periodo amerindio es la explicacion mas plausible para la extincion del almiqui gigante de Cuba pero como se puede observar (Tabla 54) Solenodon arredondoi aparece en el intervalo II por falta de asociacion con evidencias culturales
Consecuentemente el accionar de los aborigenes y de sus animales domesticos directamente sobre la fauna no parece haber sido por sf solo un factor determinante de la extincion de mamiferos terrestres durante el periodo amerindio en las Antillas (pero vease Steadman et al 2005)
Hay que recordar en este senti do que alin en el Holoceno tardio ocurrieron cambios clirruiticos de consideracion (vease Causas Naturales) Por ejemplo Curtis et al (2001) explicaron que la extincion de algunos ani males mesofilicos de las alturas montanosas de La Espanola pudo haber antecedido a cualquier impacto humano sigrIificativo en esos lugares por coincidir dicha extincion con una tendencia a la aridez documentada isotopicamente y con evidencias de la perdida del bosque mesico ocurridas despues de 3kaAP Yen Jamaica hace solo 2shy3 mil aDos el area de la Bahia de Jackson sustentaba una gran biomasa de vertebrados en bosque hinnedo a juzgar por la cantidad y diversidad de taxones presentes en material fosil derivado de la depredacion por lechuzas hoy el area es la mas seca de la isla con matorral xerofitico y pobreza de fauna (MacPhee 1996a) Yalm en nuestros dias huracanes de gran intensidad aniquilan poblaciones de vertebrados (que no se recuperan) en is las pequenas y llanas del Caribe (Brooks y Smith 2001)
En medio de la incertidumbre sobre la verdadera causa (0 causas) de las extinciones de mamiferos en el periodo amerindio MacPhee y Marx (1997) han propuesto su hipotesis de la hipermorbilidad la cual estipula colapsos poblacionales relativamente rapidos con la introduccion de enfermedades de alta letalidad Estos autores han razonado que
Las islas son los lugares mas apropiados para experimentar perdidas catastroficas causadas por enfermedades Los mamiferos no solo serian susceptibles en virtud de su prolongado aislamiento biologico sino que en los confines de una isla la epizootia
333 EXTINCIONES DEL CUATERNARIO
tendria la potencialidad de infectar simultaneamente a la rnayoria de los individuos de vadas especies Los neonatos son inmunologicamente inmaduros y sufririan desproporcionadamente cualquier infeccion hipervirulenta La perdida excesiva de neonatos y juveniles tendria efectos severos para la sobrevivencia de la poblacion si esta se reduce por debajo de cierto nivel sobreviene la extincion porque no hay posibilidad de restitucion en el periodo postpanzootico Las especies de mayor talla corporal van a ser las mas vulnerables a la extincion dadas sus bajas tasas reproductivas largos periodos de gestacion y reducido numero de progenie Las especies de menor tamano --caracterizadas por altas tasas reproductivas periodos cortos de gestacion y mayor numero de neonatos por parto- tambien serian fuertemente diezmadas pero muchas sobrevivirian porque su estrategia reproductiva les permite perder mayor numero de individuos Las perdidas deben ocurrir relativamente rapido (a escala de decadas) mientras que otras hipotesis (digamos los cambios climaticos) permiten intervalos mucho mas largos (a escala de siglos 0 de milenios) entre la aparicion de las causas y la produccion de sus efectos Por ejemplo todo indica que los nesofontes se extinguieron muy rapidamente y seria dificil poder invocar un impacto humano directo 0 argumentar fenomenos climaticos que pudieran explicar plausiblemente perdidas tan rapidas
MacPhee y Marx (1997 191) han dicho que por el momento la hipotesis de la hipermorbilidad no pasa de ser una abstraccion en espera de adecuada sustentacion empirica Llama la atencion que linicamente en el periodo amerindio (Tabla 45 intervalo 1Il) las extinciones abarcaron todas las clases del peso corporal (de la a a laj) y que ninguna de las especies que sobrevivieron hasta el periodo postcolombino (Tabla 45 intervalo IV mas las especies vivientes) sobrepasa los 10 kg (clases del peso a b c) en clara coincidencia con los presupuestos de la hipotetica hipermorbilidad
Intervalo Postcolornbino
Por 10 menos 37 especies de mamiferos terrestres vivian en las Antillas Mayores y grupos de Islas perifericas a la lIegada de los europeos en 1492 De elIas se han extinguido 23 shymas especies que en cualquiera de los periodos precedentes (Tabla 45~ y solo subsisten 12 especies repartidas entre Cuba (8 especies) La Espanola (2) Jamaica (1) y Bahamas (1) varias de estas en peligro critico de extincion Solo dos de las especies extinguidas durante los ultimos 500 anos se conocieron en estado viviente los roedores Oryzomys antillarum de Jamaica (extinguido en 1877) y Geocapromys thoracatus de las Islas Swan (extinguido en 1955) Las causas de las extinciones ocurridas durante este periodo son sobradamente conocidas destruccion de habitats introducci6n de animales exoticos (domesticos y comensales) y demas presiones asociadas a la actividad humana entre las cuales posiblemente habria que incluir brotes de hipermorbilidad
Durante mucho tiempo se ha afirmado la posibilidad de que especies de Nesophontes hayan sobrevivido hasta las primeras decadas del siglo xx y que incluso continuen existiendo en lugares remotos de Cuba 0 La Espanola (Allen 1918 1942 Miller 1930 Aguayo 1950a Arredondo 1960b Fischer 1977 MacFadden 1980 Woods y
334 BIOGEOGRAFiA
Ottenwalder 1991) Actualmente no existe evidencia radiometrica de que alguna especie de Nesophontes haya sobrevivido al siglo xv las fechas radiocarbonicas calibradas obtenidas en muestras de Nesophontes de Cuba y La Espanola resultaron no mas tardias que los siglos XIII y XIV (MacPhee et al 1999) Por otra parte las expediciones efectuadas a fines del siglo xx con el proposito de localizar individuos vivientes del genero en La Espanola no produjeron la menor indicacion afirmativa al respecto (Woods et al 1985) Ademas a traves de las Antillas Mayores no aparecen restos de Nesophontes en los depositos formados actualmente por las lechuzas
EPiLOGO
Las Antillas Mayores son islas oceanicas no han formado parte de un continente (con la posible excepcion de una parte de Jamaica hasta el Eoceno Medio hace 45 Ma) y durante un tiempo geologico considerable (desde el Eoceno Superior hace 40 Ma) han permanecido emergidas y separadas de los continentes vecinos Consecuentemente los mamiferos terrestres (no volantes) autoctonos de estas islas constituyen una fauna oceanica que por ende cumple con todas las caracteristicas convencionales reconocidas para este tipo de entidad biogeografica
- Baja diversidad taxonomica a nivel de las categorias superiores (solo cuatro ordenes) en comparacion con islas plataformicas continentales (0 con areas continentales) de tamano y diversidad ecologica simi lares
- Alto grado de endemismo a nivel de genero (96) y familia (50) en respuesta al prolongado aislamiento
- Desbalance ecologico de la estructura trofica ausencia de mamiferos camivoros y ungulados
- Espectaculares procesos de irradiaci6n evolucionaria como resultado de la colonizacion de territorios libres de competidores y depredadores
Tendencias evolucionarias extremas gigantismo
- Control a cargo de aves rapaces con tendencia at gigantismo en ausencia de mamiferos miembros del orden Carnivora
- Mas especies de reptiles y de murcielagos que de mamiferos terrestres relaciones que en los continentes se invierten
Alta tasa de extincion de especies (84) sin reposicion
Preservacion de especies filogeneticamente arcaicas (los llamados fosiles vivientes) los almiquies

299 ESPACIO Y TIEMPO
CONTEXTO TEMPORAL
La absoluta mayoria del registro fosil de mamfferos terrestres antillanos proviene de depositos cavemarios pleistoholocenicos (escala del tiempo geol6gico en la Tabla 42) Abundan en estas cavernas restos de perezosos musarafias y roedores siendo comparativamente escasos los de monos debido quizas a la universal predileccion de los platirrinos por los ambientes densamente forestados 10 que los hace malos candidatos a la fosilizacion (MacPhee y Woods 1982)
A pesar de 10 numeroso de tales depositos en la region son escasos los fechados radiometricos del cuantioso material extraido Con excesiva frecuencia (particularmente en Cuba) estos depositos han sido asignados al Pleistoceno Superior sin fundamento suficiente para ello y es sabido que incluso en cuevas contiguas los depositos fosilfferos pueden diferir considerablemente en edad (Pregill y Olson 1981) Se superponen a esta inconveniencia la falta de estudios tafonomicos en las excavaciones y la eventual mezcla de f6siles de distintas edades como consecuencia de las multicausales alteraciones estratigraficas ocurridas en las cuevas (Nunez 1990a b Morgan y Ottenwalder 1993 MacPhee 1997 McFarlane 1999b Dfaz-Franco 2004) Sobre propuestas de tipificaci6n de los depositos paleontologicos con restos de mamiferos terrestres en Cuba vease Woloszyn y Silva (1977) Acevedo y Arredondo (1982) Gutierrez (1994) Arredondo [1999]
La Tabla 43 presenta los fechados pleistocenicos mas tempranos registrados para restos de mamfferos terrestres del Caribe insular De estos restos el roedor (Cidomys) de Jamaica con un fechado de 253 mil anos antes del presente constituye el fosH de mamifero terrestre mas viejo del Cuaternario anti11ano [Los fosiles fechados antes del surgimiento de la espectrometria de masa (aceleradora) para muestras pequefias pueden ser algo mas viejos (Jull et al 2004)]
Corresponde a MacPhee e Iturralde-Vinent (1994 1995b) e1 relevante merito de haber descubierto en la ultima decada del siglo xx los primeros fosiles terciarios de mamiferos terrestres de las Antillas extendiendo a mas de 30 millones de afios el registro f6sil de
2 Extirpada en la Subregion Antillana pero viviente en el Neotropico continental
3 Ubicacion en subfamilia s610 sugerida por White y MacPhee (200 I)
4 Endemica de la Subregion Antillana
5 Recientemente propuesta su ubicaci6n bajo el genero Atopogale (Roca et al 2004)
6 lncluye las subespecies
7 Transportada por aborigenes a otras de las islas (Rimoli 1974 Olson 1982b este trabajo)
8 Vease MacPhee y Flemming (2003) sobre la incierta situaci6n de esta familia
9 Ubicada provisionalmente en esta subfamilia por MacPhee y Flemming (2003)
10 Hallada en contexto no perturbado de edad Pleistoceno tardio (McFarlane et ai 2002) con 10 cual se descarta su supuesta introduccion por amerindios (Woods 1990)
II La correlaci6n entre el area insular y la riqueza de especies es razonablemente fuerte (R=O83) en estas islas excepto las Bahamas (Morgan 1989)
300 BIOGEOGRAFIA
TABLA 42 Escala convencional del tiempo geologico en la Era Cenozoic a y finales de la Mesozoica (Gradstein 2004) ka = kiloafios (miles de afios) Ma miJIones de afios AP = antes del presente
Periodo(Suhperiodo) Epoca Afios (aproximados) antes del presente
Era Cenozolca
Cuaternario Holoceno Pkistoceno
Presente - 11 kaAP 11 kaAP - 18 MaAP
Terciario (Ne6geno ) Plioceno Mioceno
18 - 5 MaAP 5 - 23 MaAP
oIigoceno 23 - 34 MaAP Terciario (pak6geno) Eoceno 34 - 58 MaAP
Pakoceno 58 - 65 MaAP
Era Mesozo ca
Creticico 65 - 145 -faAP
~~~------~~--- --~-
este grupo animal en la region con el hallazgo de un perezoso oligocenico en Puerto Rico (Yauco) as como un perezoso un mono y un roedor miocenicos en Cuba (Domo de Zaza) Esto unido a la presencia de huesos de una musarafia nesofontoide [referida erroneamente a Solenodon por Rivero (1997)] en ambar oligomiocenico de la Republica Dominicana (MacPhee y Grimaldi 1996) demuestra incontrovertiblemente que al menos en el Mioceno temprano (alrededor de 20 millones de afios atnis) ya estaban en las Antillas Mayores representantes de los cuatro linajes basicos de su fauna de mamiferos terrestres [La afirmacion de Abreu et al (1990 13) en cuanto a que se conocen restos fosiles de almiqui que datan de hace aproximadamente 40 millones de afios es erronea]
Pero lcuan antigua puede ser la fauna de mamiferos terrestres de las Antillas Mayores
Aunque es un hecho bien establecido por la geologia que desde finales de la Era Mesozoica y principios de la Cenozoica existieron islas en el area ocupada ahora por las Antillas Mayores la evidencia disponible indica asimismo que aquellas primeras islas sufrieron repetidas transgresiones marinas totales que habrian hecho sucumbir las biotas que pudieran haber sustentado (MacFadden 1981 Morgan y Woods 1986 MacPhee e Iturralde-Vinent 1994 Roca et al 2004) sin contar los catastroficos efectos que debio tener para el Caribe occidental el b6lido extraterrestre que impacto en Chicxulub (Yucatan) hace 65 Ma (Tada et ai 2003) Hedges (1996) ha especulado que aUn en medio de tal devastacion debe haber existido algun que otro lugar donde elementos de la fauna pudieron haber sobrevivido
301 POBLAMIENTO CONTINENTE-ISLA
TABLA 43 F echados pleistocenicos mcis tempranos (obtenidos por distintos metodos) en mamiferos terrestres del segmento insular delimitado en laFig 41 F =Fuentes (I) Jull et al 2004 (2) Goodfriend y Mitterer 1987 (3) MacPhee et al 1989 (4) Woods 1989b (5) Rega et aI 2002 (6) McFarlane 1999b (7) McFarlane 1 999a (8) McFarlane et al 1998a
Pals Localidad Taxon kaAP F
Cuba Breas de San Felipe Pilosa (Parocnus) 1l9a 1
Jamaica Cueva Coco Ree Cueva Wallingford Roadside
Primates indet Rodentia (Clidomys)
393 2530
2 3
Haiti Trouing Jeremie no5 (casimba) Rodentia (Rh izoplagiodontia) 212 4
Republica Dominicana
Cueva del Perezoso Pilosa (ficratocnus) 185 5
Puerto Rico Cueva de Ia Vaca Cueva Clara
Pilosa (Acratocnus) Soricomorpha (Nesophontes)
337 346
6 7
Anguila Cueva Cave-No-Cave Rodentia (Amblyrhiza) 2060 8
a Kulp etal (1952) reportaron un fechado radiocarbonico de gt 30000 afios para un cono de pino en el deposito fosilffero de Ciego Montero (Cuba) con restos de perezosos
(vcase tambicn Crother y Guyer 1996) pero Iturralde-Vinent y MacPhee (1999) han preguntado donde estaban esos lugares si es que existieron
Solo a partir del Eoceno Superior las Antillas Mayores en fonnacion han pennanecido emergidas hasta hoy [excepto parte de Jamaica y el sur de La Espanola que continuaron sumergidas hasta el Mioceno Medio (Iturralde-Vinent y MacPhee 1999)] Esto significa que solo durante los ultimos 40 millones de anos aproximadamente han existido pennanentemente en dicha region insular ambientes terrestres capaces de sustentar comunidades biotic as (MacPhee e Iturralde-Vinent 2000 Portell et ai 2001 MacPhee 2005) 10 cual proporciona el marco temporal de referencia para conceptuar la antigiiedad maxima posible de la fauna de mamiferos terrestres de las Antillas Mayores
POBLAMIENTO CONTINENTE-ISLA
AFINIDADES CONTINENTALES
El consenso de los autores que en las itltimas dccadas han tratado la filogenia y biogeografia de los mamiferos terrestres de las Antillas Mayores (MacFadden 1980 MacPhee y Woods 1982 Morgan 1985 1988 1993 Morgan y Woods 1986 Ford 1986 Woods 1988 1989a 1990 MacPhee e Iturralde-Vinent 1995a b Hedges 1996 Horovitz [1997] Ottenwalder 2001 Whidden y Asher 2001 MacPheeet ai 2003 Carlini y Scillatoshy
302 BIOGEOGRAFiA
Yane 2004 Davalos 2004 MacPhee y Horovitz 2004 Roca et al 2004) es practicamente unanime en cuanto a que los ancestros continentales que dieron origen a esta fauna derivaron (1) mayormente de Suramerica perezosos (Megalonychidae) monos (Pitheciidae) y roedores (Echimyidae Capromyidae y Heptaxodontidae) (2) en menor medida de Norteamerica musarafias (Nesophontidae y Solenodontidae) y (3) menos aun de Centroamerica una especie de roedor (Muridae)
HIPOTESIS BIOGEOGRAFICAS
La definicion c1asica reconoce dos tipos de islas isla continental (porcion del territorio de un continente separado de este por movimiento tectonico 0 por ascenso del nivel del mar) e isla oceanica (porcion de territorio que no ha formado parte de un continente) La definicion sigue siendo uti por 10 que implica para la formacion de las faunas insulares en uno u otro caso As en el caso de la isla continental su fauna ya estaba emplazada en la futura isla antes de producirse su separacion del continente [Vicarianza hipotesis biogeografica de fragmentacion faunal] en el caso de la isla oceanica su fauna es el resultado de la colonizacion1 por propagulos2continentales a traves de la barrera marina entre el continente y la isla [Dispersion hipotesis biogeografica de acumulacion faunal] En el primer caso el surgimiento de la barrera marina es resultado del evento vicariante en el segundo la existencia de la barrera marina es anterior al evento dispersante Ademas de operar a escala mayor (continente-isla) ambos eventos operan secundariamente a menor escala (isla-isla)
Las dos hipotesis -vicarianza y dispersion- se han disputado hasta hoy la preferencia de los biogeografos en tanto hipotesis mas viable para explicar el proceso de formacion original (continente-isla) de la fauna de mamiferos terrestres de las Antillas Mayores [Sobre la historia de las hipotesis biogeograficas relativas al Caribe insular vease RuB y Schubert 1989 Williams 1989 Pruna 2001] Yel debate continua debido ala falta de consenso sobre 10 que Williams (1989) ha llamado el problema central de la discusion a saber la ostensiblemente baja diversidad taxonomica de esta fauna a nivel de las categorias superiores (solo cuatro ordenes) en comparacion con la diversidad al mismo nivel de las faunas de islas continentales (0 de areas del continente) simi lares en tamafio y diversidad ecologica [Por ejemplo la fauna cubana de mamiferos terrestres consideradas de conjunto las especies vivientes y las extintas- es menos diversa a los niveles taxonomicos superiores (4 ordenes y 6 familias) que 10 que cabria esperar de la diversidad ecologica y extension territorial de Cuba (110 992 km2) comparada con una isla tipicamente continental como Trinidad (4828 krn2
) -veinte veces menor que Cuba y menos diversiticada ecologicamente- donde solo las especies vivientes de mamfferos terrestres representan 6 ordenes y 16 familias (Eisenberg 1989)] Este desbalance
lColonizaci6n Arribo y establecimiento de un propagulo en una isla no ocupada por la especie (MacArthur y Wilson 1967)
2Propagulo Niimero minimo de individuos de una especie capaces de reproducirse y aumentar poblacionalmente en una isla bajo condiciones favorables (MacArthur y Wilson 1967)
303 POBLAMIENTO CONTINENTE-ISLA
taxon6mico ha sido expJicado de dos maneras (1) Inicialmente hubo mas 6rdenes presentes pero la mayoria se extingui6 y s610 sobrevivieron esos cuatro [explicaci6n vicarianista] (2) S610 representantes de esos cuatro 6rdenes lograron cruzar la barrera marina [explicaci6n dispersionista]
Cinco hipot~ticos mecanismos de poblamiento continente-isla han sido invocados durante los ultimos 140 afios (y se los sigue proponiendo actualmente) para explicar la presencia de mamiferos terrestres en las Antillas Mayores A continuaci6n pasamos a resefiarlos brevemente (incluidas sus alegadas fortalezas y debilidades) en el orden aproximado en que historicamente fueron propuestos
EL PUENTE NATURAL (dispersion terrestre activa)
En el siglo XIX el puente terrestre era pnkticamente la unica hipotesis para explicar las distribuciones disyuntas de animales en buena parte del mundo y los primeros investigadores que se ocuparon de la biogeografia antillana (Fernandez [1865] 1884 Chapman [1893] Torre 1892 Lora 1895) la asumieron resueltamente en parte intrigados por el descubrimiento en Cuba en la segunda mitad del siglo de un perezoso f6sil de gran talla Y en su aplicacion a los mamiferos terrestres de las Antillas Mayores el puente sigui6 siendo a 10 largo del siglo xx la hipotesis mas favorecida por los estudiosos del tema (Vaughan 1902a Spencer 191 Oa b Torre 1910a b cAllen 1911 1917a 1918 Barbour 19141916 1945bAnthony 1918 1926 [1942] Scharff 1922 Kraglievich 1926 Beaufort 1951 Arredondo 1961 1970b Kartashov y Mayo 1974 Iturralde-Vinent 1982 1988 Arredondo y Armifiana 1996 Arredondo 1996a [1999]) quienes postularon la existencia preterita de puentes con Norte- Centro- 0 Suramerica en dependencia de las afinidades continentales de los mamiferos antillanos
Teoricamente el puente terrestre debe cumplir con una condicion esencial permitir el paso de una parte mayoritaria (y por tanto representativa de la diversidad) de la fauna continental Claro que siempre habria tipos de mamiferos menos proclives a dispersarse o para los cuales la ubicaci6n del puente resultaria inaccesible 0 las condiciones ecologicas en este fueran desfavorables pero si algunos acceden al puente no hay raz6n para pensar que el resto de esa fauna no los acompafie mayoritariamente Como dijera Simpson (1956) Los puentes no transportan faunas desbalanceadas
En consecuencia el puente terrestre como filtro es un mecanismo de poblamiento minimamente selectivo y por ende de alta diversidad inicial (sensu Iturralde-Vinent y MacPhee 1999) 10 esencial del puente es su capacidad para transportar faunas integradas (Williams 1989) Yen eno radica la objeci6n fundamental al puente terrestre surgida tempranamente en el siglo xx (Por que s610 cuatro 6rdenes de mamiferos terrestres en las Antillas si vinieron por puentes (Donde estan los fosiles de los demas ordenes que debieron venir si es que se extinguieron Los partidarios del hipotetico puente dispusieron de casi todo el resto del siglo para responder una y otra vez que otros 6rdenes de mamiferos aparecerian representados en las Antillas Mayores el dia que en estas Islas se descubrieran fosiles precuatemarios suficientemente
304 BIOGEOGRAFIA
antiguos Esto se logro por fin terminando el siglo como se explico anteriormente (vease Contexto temporal) y ique se encontro los mismos cuatro ordenes de mamiferos terrestres conocidos del Cuatemario caribefio aparentemente los Unicos que lograron llegar
Pero ademas hasta el presente no se ha demostrado inequivoca- y concluyentemente que en algun momenta durante los ultimos 40 millones de afios haya existido una conexion terrestre continua entre las Antillas Mayores (en proceso de formacion 0 plenamente formadas) y algun punto de las regiones continentales vecinas
LA BALSA NATURAL (dispersion marina pasiva)
Durante buena parte del siglo xx el puente terrestre tuvo como unico rival a la hipotesis de la balsa natural -transporte accidental de animales sobre islotes de vegetacion flotantes a la deriva despues de arrancados y expulsados mar afuera por un rio crecido- expuesta originalmente por Matthew (191319151916 1918a 1919) y desarrollada en su aplicacion a los mamfferos terreslres antillanos principal mente por Simpson (1956) asi como por olros investigadores (Darlington 1938 1957 Williams y Koopman 1952 Hershkovitz 1958 1972 Patterson 1962 Paula 1967 Patterson y Pascual 1972 Pregill 1981 Ford 1986 Woods 1989a 1990 Morgan 1993) Por su parte Hedges et al (1992) sustentaron dicha hipotesis en estimados de divergencia (basados en el reloj molecular) en la herpetofauna antillana -involucrados tangencia1mente los mamiferos [veanse objeciones a Hedges y colaboradores (Page y Lydeard 1994 Iturralde-Vinent y MacPhee 1999) respuestas a las objeciones (Hedges et al 1994 Hedges 2001) y contrarrespuestas (MacPhee e IturraldeshyVinent 2005) asi como cuestionamientos al metoda de los estimados de divergencia (Foote et al 1999 Pregill y Crother 1999 Woods et ai 2001 )]
La hip61esis de la balsa natural se origino a partir de las numerosas ocasiones en que a 10 largo de siglos tales islotes flotantes (portando 0 no vertebrados incluidos mamiferos) han sido avistados mar afuera a cientos de kilometros de la costa mas cercana en varias partes del mundo --inc1uido el Mar Caribe- segUn consta en diarios de barcos y reportes de observadores diversos (revisados por King 1962 reportes posteriores en SICSP 1970 y Censky et al 1998) Uno de estos casos acaecido en nuestra region se reporta en un informe del SICSP (Smithsonian Institution Center for Shortlived Phenomena 197050shy52)
EIB de julio de 1969 un destructor de la armada de Estados Unidos avisto un islote flotante a unos 100 km al sur de la Base Naval de Guantanamo moviendose desde el Paso de los Vientos en direccion suroeste a una velocidad de 45 kmlh islote de unos 15 m de diametro mantenfa en pie mas de 10 palmas reales de 10-15 m de altura con suelo cubierto de hierba y arbustos [la observacion con binoculares a 18 km de distancia no preciso la presencia de vida animal] La travesia fue rastreada durante 6 dfas hasta que el 19 de julio el islote se dio por perdido tras 6 horas de busqueda aerea
305 POBLAMIENTO CONTINENTE-ISLA
La colonizaci6n exitosa de una isla de las Antillas Mayores por mamiferos terrestres continentales transportados en balsas naturales a la deriva (de cada especie al menos una hembra gravida 0 dos individuos de sexo opuesto) dependeria de factores que van a operar desde el momenta mismo de formarse la balsa asi como durante su travesia marina y su arribo a la isla
a) La posibilidad de estar en la balsa Que la distribuci6n de la especie incluya areas contiguas al rio y que el animal se encuentre en ellugar y el momenta propicios para ser incorporado a la balsa antes de que el rio crecido la desprenda Por ejemplo segun Hershkovitz (1958) los equimidos -grupo ancestral de roedores antillanos- serian buenos candidatos a ser captados por la balsa porque acostumbran a anidar en huecos del tronco de los arboles a la orilla de los rios
b) El tamano y la consistencia de la balsa Muchas se desintegran en alta mar debido a su fragilidad ante la saturaci6n Algunos autores (Beaufort 1951 Arredondo 1996a [1999]) han expresado incredulidad en cuanto a que las balsas hayan podido soportar el peso de un animal como el gran perezoso cubano (Megalocnus rodens) el mayor de los mamiferos antillanos Pero esta especie evolucion6 en Cuba y el ancestro que hizo el viaje pudo ser muy pequeno [EI perezoso oligocenico de Puerto Rico es el mas pequeno de los megaloniquidos antillanos (MacPhee e Iturralde-Vinent 1995b )]
c) Las caracterfsticas fisiol6gicas de la especie Los limites fisiol6gicos de los propagulos constituyen nuestra unica objeci6n a la dispersi6n marina por balsas como mecanismo para la formaci6n de la fauna de vertebrados en las Antillas (MacPhee e IturraldeshyVinent 2005) Unas especies tienen intrinsecamente mas posibilidades que otras para sobrevivir a las vicisitudes que deben afrontar durante la travesia (deshidrataci6n inanicion insolaci6n etc) Por ejemplo los megaloniquidos han demostrado tener extraordinaria habilidad para cruzar barreras marinas fueron los primeros mamfferos suramericanos en invadir Norteamerica mucho antes de que surgiera la conexion terrestre a finales del Plioceno (Paula 1967 Olson 1978 Webb 1985b Martin y Steadman 1999) Incluso se conoce un megaloniquido acultitico del Plioceno peruano (Muizon y McDonald 1995) y otro viviente en Suramerica (Choloepus) que pnicticamente no bebe agua 10 que ha originado una leyenda aborigen que explica por que los perezosos no toman agua (Goff art 1971) Vease ademas el inciso e
d) EI regimen de las corrientes marinas Que conduzcan a la balsa en la direcci6n necesaria EI transporte de balsas de vegetacion desde los continentes hacia las islas fue durante la mayor parte del Cenozoico sumamente improbable (MacPhee e IturraldeshyVinent 2000 148) Estos autores han insistido en que hay que tener en cuenta que durante la cambiante geografia cenozoica del Caribe las corrientes marinas de superficie no favorecieron el desplazamiento de objetos fiotantes hacia las Antillas Mayores [Pero no sabemos con certeza que efecto modificador del curso de tales objetos fiotantes pudieron introducir las tormentas tropicales si previsiblemente funcionaron en el pasado como 10 hacen ahora] Dichos autores senalaron ademas (2005) que en un experimento reciente con boyas de posicionamiento satelital liberadas a la deriva desde las Antillas Menores las boyas fueron a parar mayoritariamente a Centroamerica y a otros lugares
306 B[OGEOGRAFIA
y solo unas pocas recalaron en las Antillas Mayores y las que entraron al Mar Caribe demoraron semanas 0 meses en arribar
e) La distancia a recorrer (y en consecuencia la duracion de la travesia) entre el continente y la isla (10 que repercute decisivamente en el factor c) Este es desde luego uno de los factores mas criticos En la medida que la distancia aumenta la probabilidad de alcanzar la isla disminuye drasticamente con el agravante de que la trayectoria dificilmente seria en linea recta Si tomamos como referencia la velocidad de 45 kmlh del islote ya mencionado (SICSP 1970) cubrir la distancia aerea (aproximada) de 800 km que media entre el noroeste de Suramerica y La Espafiola Ie tomaria a la balsa 74 dias Estimando el doble de este tiempo debido a desvios de la trayectoria la duracion de la travesia probablemente resultaria excesiva para la mayoria de los mamiferos Pero pudiera muy bien ocurrir que la velocidad fuera mayor 0 que la balsa portara plantas vivas que permitirian a los fitOfagos sobrevivir [Tres de los cuatro ordenes de mamiferos terrestres conocidos de las Antillas Mayores son fitOfagos] Ademas esta geologicamente bien establecido que durante etapas de variada duracion desde el Oligoceno hasta el Pleistoceno los territorios del Caribe insular y continental aumentaron considerablemente en extension con niveles del mar 100-200 m por debajo de su nivel actual oportunidades que los propagulos aprovecharian para cruzar barreras marinas significativamente menores
f) El tamafio de la isla Las islas mayores deben interceptar mas balsas (Buckley y Knedlhans 1986) 10 que repercute directamente en el factor e La balsa puede pasar entre islas pequefias y continuar viaje sin llegar a ningun lugar
g) Las condiciones (favorables 0 adversas) del desembarco en la isla y el tiempo que Ie tomaria a uno 0 mas animales depauperados encontrarun habitat apropiado Esto dependeria a su vez del tamafio y diversidad ecologica de la isla (Darlington 1938 Miller 1966)
Tal cumulo de posibles obstaculos y dificultades constituye a primera vista un desafio a la credibilidad de la hipotesis Matthew-Simpson pero en ello radica precisamente su valor explicativo bas ado en la mas absoluta improbabilidad La colonizacion por la via marina es altamente improbable pero no absolutamente imposible si se tiene en cuenta la inmensidad del tiempo geologico se vuelve probable (McKenna 1973) Basta que solo una vez entre mill ones de intentos fallidos durante ese inmenso lapso algunos de los factores anteriormente enunciados coincidan en no ser tan desfavorables para el propagulo Y eso es todo 10 que demanda la hipotesis para cada uno de los grupos monofileticos mayores de mamiferos terrestres presentes en las Antillas Mayores Las Antillas Mayores requeririan el arribo de tan solo 7 (quizas menos) propagulos fundadores a 10 largo de los ultimos 30 0 mas millones de afios (promedio de uno cada 43 mill ones) para explicar la formacion de toda su fauna de mamiferos terrestres por dispersion marina pasiva (vease Discusion y Conc1usiones)
En comparacion con la via marina el poblamiento por la via terrestre es predecible porque la mayor parte de la fauna continental de mamiferos emplazada en la region del puente va a cruzar por este una vez que el mismo se establece y los animales logran
307 POBLAMIEITO CONTINENTE-ISLA
acceder a el el poblamiento por la via marina en cambio es impredecible porque depende total mente del azar que caracteriza a la balsa Claro que en ambas vias opera el azar pero en el primer caso se trata del azar de un evento probable en el segundo el azar de un evento improbable (Simpson 1940) Asi el puente natural (mecanismo de poblamiento minimamente selectivo = alta diversidad inicial) y la balsa natural (mecanismo de poblamiento maximamente selectivo baja diversidad inicial) pueden conceptuarse como los extremos opuestos de una gama de probabilidades de dispersi6n (McKenna 1973) EI fen6meno de la dispersi6n merece la mayor atenci6n cientifica en los estudios de biogeografia insular (Cowie y Holland 2006)
Queda claro que la balsa natural es una hip6tesis consistente con el caracter de la fauna objeto de estudio puesto que el desbalance taxon6mico (sin evidencia paleontol6gica en contrario) s610 puede ser resultado de la improbabilidad del poblamiento No obstante ello no implica necesariamente que la balsa natural haya sido el imico mecanismo (ni siquiera el principal) utilizado por los mamiferos terrestres en su colonizaci6n de las Antillas Mayores La balsa natural debe satisfacer una premisa te6rica esencial dado el caracter azaroso impredecible e improbable de una colonizaci6n exitosa esta debe ocurrir a intervalos muy espaciados a traves del tiempo geol6gico 0 sea durante todo el tiempo que en el area ocupada por las Antillas Mayores ha habido ecosistemas terrestres perrnanentemente disponibles para ser colonizados (los ultimos 40 millones de afios) Sin embargo como sabemos representantes de los cuatro 6rdenes de mamiteros terrestres conocidos de las Antillas Mayores ya estaban emplazados en dicho territorio insular en el Mioceno Inferior 10 que supone un lapso de colonizaci6n limitado al Eoceno SuperiorshyOligoceno (MacPhee 2005) sugerente de un tipo de mecanismo contrario a la colonizacion alocronica exigida por la hipotesis de la balsa natural sobre todo tratandose de una fauna caracterizada por altas tasas de extincion sin reposicion aparente
La presencia en Jamaica (hasta su extinci6n en 1877) del roedor mfuido Oryzomys antillarom con estrecha afinidad centroamericana (Woods 1989a) pudiera constituir el imico caso efectivo de poblamiento (continente-isla) de las Antillas Mayores por dispersion marina dc un mamifero terrestre para el cual no es razonable invocar otro mecanismo de colonizacion
LA GUIRNALDA INSULAR (dispersion mixta)
La geologia ha establecido con suficiente certeza que durante el Cenozoico unicamente en el Pale6geno tardio y solo a traves de la Cresta de Aves (ahora sumergida excepto la isla hom6nima) pudo haber existido una cadena de islas para el transito de mamiferos terrestres entre el norte de Suramerica y las proto-Antillas Mayores [desestimados a ese efecto el Canal dc Yucatan el Elcvado de Nicaragua la Cresta de Caiman el Elevado de Beata y las Antillas Menores (Holcombe et al 1990 MacPhee e IturraldeshyVinent 1994 1995b Iturralde-Vinent y MacPhee 1999)] Holcombe y Moore (1977) y Holcombe y Edgar (1990) postularon la existencia de esta entidad y estimaron algunas de sus posibles caracteristicas Por su parte MacPhee e Iturralde-Vinent (1995b) e Iturralde-Vinent y MacPhee (1999) precisaron los extremos proximal y distal de esa
308 BIOGEOGRAFIA
estructura acotaron la duracion y la edad de la misma y Ie dieron nombre GAARlandia (Greater AntillesA vesRidgelandia)
Segun Iturralde-Vinent y MacPhee (1999) la Cresta de Aves emergio por breve tiempo (1-2 Ma) durante la transicion Eoceno-Oligoceno (35-33 MaAP) por la coincidencia de un levantamiento tectoni co y un considerable descenso eustatico del nivel del mar En ese momenta las islas septentrionales del arco de las Antillas Mayores (Cuba centroorientalshyLa Espafiola - Puerto Rico - Islas Virgenes) constituian una sola isla grande 0 una serie de islas separadas por barreras muy estrechas La Cresta de Aves emergida entre el bloque Puerto Rico - Islas Virgenes y un microcontinente noroccidental de Suramerica -separado en ese momento del resto del continente por una via marina surgida en el Eoceno tardio- dio origen a GAARlandia
Obviamente el numero de colonizaciones exitosas por mamiferos terrestres a traves de una guimalda de Islas tiende a disminuir mientras mas anchas sean las barreras marinas entre las islas que forman la guimalda y cuando esto ocurre disminuye tambien la diversidad taxonomica de los propagulos particularmente a nivel de las categorias superiores (Simpson 1956)
Vemos asi que para los mamiferos terrestres la colonizaci6n a traves de una guimalda insular puede ser tan improbable como a la deriva en una balsa natural en dependencia de la cantidad y anchura de las barreras marinas a cruzar el tamafio y la fisiografia de las islas que integran la guimalda y el regimen de las corrientes marinas y aereas entre otros factores
Resumiendo la guimalda insular puede conceptuarse como un mecanismo de poblamiento altamente selectivo (= baja diversidad inicial) que combina la improbabilidad de la balsa natural con la colonizacion restringida temporaimente del puente terrestre
En su reciente revision de la taxonomia distribuci6n y filogenia (basada en estudios moleculares) de varios grupos de mamiferos antillanos Davalos (2004) concluyo que los perezosos megalocninos los primates y los roedores histricognatos se avienen al patron de divergencia con el continente y epoca de arribo a las Antill as implicados en la hipotesis de GAARlandia pero no asi los perezosos coloepodinos que requieren una explicacion biogeognlfica distinta Esto ultimo (sospechado por Matthew 1918a) habla sido confirmado por White y MacPhee (2001 226-227)
Nuestra evidencia filogenetica no sustenta la idea de un dado unificado surgido de la invasion de un megaloniquido el analisis sugiere mas bien dos eventos de colonizaci6n independientes [y] sustenta fuertemente un origen difiletico de las subfamilias [Choloepodinae y Megalocninae] Si hubo dos migraciones a traves de GAARlandia presumiblemente poco espaciadas en el tiempo no se puede determinar en el presente
Finalmente queda claro que GAARlandia -como guirnalda insular- es la hipotesis mas robusta propuesta hasta ahora para explicar la presencia en las Antillas Mayores de
309 POBLAMIENTO CONTINENTE- ISLA
la absoluta mayona de los mamfferos terrestres de ongen suramericano porque el caracter de esa fauna supone un mecanismo de poblamiento ancestral que sea a la vez maximamente selectivo (baja diversidad ordinal) y temporalmente restringido (transicion Eoceno-Oligoceno)
Recientemente Rega et al (2002 18) expresaron A pesar de una revision exhaustiva de la paleogeografia de la cuenca del Caribe Iturralde-Vinent y MacPhee (1999) no pudieron confirmar la presencia de una conexion terrestre continua entre Suramerica y las proto-Antillas aunque hipotetizaron su existencia [Vease la Fig 42 (= figura 6 de Iturralde-Vinent y MacPhee 1999 simplificada))
En el trabajo citado por Rega y colaboradores aparece la siguiente explicacion
Actualmente no es posible ofrecer reconstrucciones paleogeograficas detalladas de la Cresta de Aves Sin embargo hay evidencia de que porciones de la cresta deben haberse elevado subaereamente en un tiempo La cuestion cntica es si la Cresta de Aves emergida pudo haber formado algun tipo de corredor entre las Antillas Mayores y el noroeste de Suramerica Perforaciones en las alturas estructurales de la cresta pudieran proveer el registro del Paleogeno tardio que se necesita para precisar la hipotesis del corredor aunque no se puede predecir si tales datos serian suficientemente detallados para determinar si en algun momento GAARlandia constituyo una sola entidad subaerea Por ejemplo pudo haber barreras marinas por corto tiempo entre las masas terrestres de la Cresta de Aves [0) ellevantamiento y la subsidencia pudieron afectar diferentes partes de GAARlandia en diferentes tiempos AItemativamente los elementos faunales pudieron haberse dispersado de manera pulsada Movimientos episOdicos de este tipo incrementarian la extincion de los elementos faunales incapaces de trasladarse a la siguiente masa terrestre en la cadena disminuyendo asi la diversidad total (Iturralde-Vinent y MacPhee 199957)
La primera parte de la explicacion deja claro que la posibilidad de que la Cresta de Aves haya sido una conexion terrestre continua no se puede descartar pero tampoco se puede afirmar La ultima parte de la explicacion describe el efecto altamente selectivo (filtrante) de la colonizacion a traves de una cadena de islas
Y continuan lturralde-Vinent y MacPhee (199957 -58) explicando
Hasta que punto el microcontinente noroccidental de Suramerica [considerado por dichos autores como el area suministradora de propagulos hacia GAARlandiaJ estuvo fisicamente separado del resto del continente no ha sido plenamente esc1arecido Si estuvo sustancialmente aislado por barreras marinas y orogenic as durante el Paleogeno tardio entonces solo la fraccion de la biota suramericana emplazada en el microcontinente estuvo en posicion de acceder a GAARlandia Esto pudiera tener que ver con el hecho de que la fauna antillana carezca de representantes de muchos grupos suramericanos [de mamfferos) que debieron haber hecho el viaje
310 BIOGEOGRAFiA
Florida o Alturas0 0
d 6 D L1anuras25deg
Yucatan
20deg
deg0
10deg
MARCARIBE
80deg 75deg
Bahamas
Cres ta de Beata
D Mares someros
D Mares profundos
OCEANO ATLANTICa
FIGURA 42 Reconstrucci6n hipotetica de GAARlandia como conexi6n terrestre continua entre las proto-Antillas Mayores y el noroeste de Suramerica en el Pale6geno tardio segun la FIgura 6 (simplificada) de Iturralde-Vinent y MacPhee (1999)
En su defensa del puente terrestre Barbour (1945b249) se refiri6 a esto - antes 10 habfa hecho Anthony [1942363]- en los terminos siguientes
El argumento de que si las Antillas hubieran estado conectadas con el continente se encontrarian los f6siles de un numero mayor de distintas criaturas me deja frio es perfectamente concebible que la conexi6n haya ocurrido en un periodo en que la parte del continente implicada no estuviera poblada por una fauna grande y variada
Y concluyen lturralde-Vinent y MacPhee (199958) su detallada explicaci6n
Desafortunadamente sabemos muy poco sobre la composici6n faunal del noroeste de Suramerica a finales del Pale6geno
Consecuentemente si se confirmara la magnitud del aislamiento del aludido microcontinente y su concomitante baja diversidad faunal GAARlandia como conexi6n terrestre continua serf a la hip6tesis mas parsimoniosa No obstante mientras no se disponga de evidencia concluyente a este respecto hipotetizar a GAARlandia como conexi6n continua (minima selectividad) la coloca en franca vulnerabilidad te6rica al tener que afrontar la objeci6n clasica al puente terrestre (baja diversidad ordinal de la
311 POBLAMIENTO CONTINENTE-TSLA
fauna) al tiempo que se la priva de ser la hip6tesis mas robusta para explicar -como guimalda de islas- la colonizaci6n de las Antillas por mamiferos procedentes de Suramerica
ELARcA DE NOE (seudovicarianza)
El primer vicarianista antillano (desde luego sin el saberlo) fue Corral (1940) con su utilizaci6n ingenua de las ideas wegenerianas de la deriva continental aparecidas la decada anterior Pero el primer (supuesto) intento de aplicar a la formaci6n de la biota caribefia el mecanismo de la vicarianza basada en la tearia de la tect6nica de placas (surgida en la sexta decada del pas ado siglo) se debe a Rosen ([1976] 1985)
Segun Rosen las Antillas Mayores se originaron en el Cremcico en la Placa Caribe aproximadamente en la posici6n que hoy dia ocupa Centroamerica y al ser desplazadas tectonicamente hacia el este interactuaron (de algun modo no precisado por Rosen) con las margenes continentales de Norte- y Suramerica de donde (en un evento unico) adquirieron toda su fauna transportada subsecuentemente por dichas islas en su transito hacia sus posiciones actuales [De ahi la denominacion metaforica dada par McKenna (1973) a este mecanismo de poblamiento] Aunque se ha presentado esta propuesta como Model0 de Vicarianza Continente-Isla de Rosen en realidad por definici6n no constituye un evento vicariante porque las proto-Antillas Mayores no resultaron del fraccionamiento y separaci6n por tectonismo de una porcion del territorio continental como exige esta hip6tesis (vease definici6n de vicarianza al comienzo de Hipotesis Biogeograficas)
El Arca de Noe de Rosen ha sido desestimada por la mayoria de los autores contemporaneos porque (1) carece de sustento geol6gico (lturralde-Vinent y MacPhee 1999) y porque (2) en el periodo en que se afrrrna ocum6 el evento muchos de los ordenes modemos de mamiferos placentarios attn no existian (Williams 1989)
EL BARCO FUNERAL VlKINGO (tanatovicarianza)
Domning et al (1997) reportaron 10 que consideraron el mamffero terrestre mas viejo de las Antillas --circunstancia que se refleja en el titulo mismo del trabajo Oldest West Indian Land Mammal- al descubrirse un perisodactilo rinocerotoide (Hyrachius sp) en una localidad de edad Eoceno Inferior a Medio en el occidente de Jamaica
Jamaica ha tenido una historia geologica y paleogeografica distinta de la historia comun a las restantes Antillas Mayores A principios del Cenozoico Jamaica occidental (a diferencia de la porcion oriental constituida por el Bloque de las Montafias Azules) formaba parte de Centro america de la cual se desgaj6 (por el desplazamiento de la Placa Caribe) para venir a integrarse despues del Eoceno Medio con 10 que serfa la futura isla de Jamaica desaparecida por transgresi6n marina en el Eoceno Superior hasta volver a emerger totalmente en el Mioceno Medio (Iturralde-Vinent y MacPhee 1999 MacPhee e Iturralde-Vinent 2000)
312 BIOGEOGRAFiA
Por consiguiente Hyrachius ---conocido de localidades de edad Eoceno Inferior a Medio en Norteamerica- debio ocupar (10 que iba a ser) Jamaica occidental mucho antes de que esta se separara de Centroamerica De hecho este fosil Uego a su destino insular contenido en las rocas de su basamento no como un ser vivo por 10 que es erroneo considerarlo un animal antillano (lturralde-Vinent y MacPhee 2004) como han supuesto algunos (Hedges 200 I Portell et al 200 I Condis [200 I D Todo 10 que hizo fue viajar tanatovicariantemente en su barco funeral vikingo -otra de las denominaciones metaforicas de McKenna (1973)
DISCUSION Y CONCLUSIONES
Hemos visto que solo disponemos de una hipotesis plausible para explicar la mayor parte del poblamiento (continente-isla) de las Antillas Mayores por mamiferos terrestres del Neotropico continental la dispersion mixta (a traves de GAARlandia) para los perezosos (excepto Choloepodinae) los monos y los roedores (excepto Oryzomys)
Por otra parte la historia biogeogrMica de las musaraiias de las Antillas Mayores (Nesophontes y Solenodon) constituye una de las situaciones mas enigmaticas entre los mamiferos de la region (Ottenwalder 2001321) Al menos cinco mecanismos de poblamiento continente-isla han sido invocados para explicar cuando como y de donde llegaron a las Antillas Mayores estas musaraiias
I EI propagulo ancestral (del Terciario de Norteamerica) coloniz6 a las Antillas Mayores por dispersion marina desde Centroamerica 0 desde la Florida (Patterson 1962)
2 El propagulo ancestral ( del Terciario) cruzo el Atlantico Sur (supuestamente mas estreshycho en aquella epoca) directamente de Africa a las Antillas Mayores (Hershkovitz 1972)
3 El propagulo ancestral (del Terciario de Norteamerica) Uego a las Antillas Mayores en el Arca de Noe de Rosen (MacFadden 1980)
4 El propagulo ancestral (del Terciario de Norteamerica) fue transportado a las Antillas desde Centroamerica en el bloque de Jamaica occidental y de alli paso por dispersion marina a las restantes Antillas Mayores (Whidden y Asher 200 I)
5 El propagulo ancestral (del Terciario de Norteamerica) lleg6 a las Antillas Mayores a traves de GAARlandia (Whidden y Asher 2001)
Aunque hay consenso entre estos (y otros) autores sobre el origen norteamericano de las musaraiias antillanas ninguno de los cinco hipoteticos mecanismos de poblamiento resiste la evidencia en contrario filogenetica yo paleogeogrMica Ademas algunas de estas hipotesis asumen que Nesophontidae y Solenodontidae comparten un antecesor comun en las Antillas pero esta relacion no ha sido confirm ada Para dilucidar el enigmatico caso de las musaraiias antillanas se requerini profundizar en la investigaci6n geologica paleogeognifica y paleontol6gica (Iturralde-Vinent y MacPhee 1999)
De acuerdo con el conocimiento filogenetico disponible sobre el conjunto de la fauna la colonizacion original de las Antillas Mayores por mamiferos terrestres implicaria el arribo
313 POBLAMIENTO ISLA-ISLA
de solo siete propagulos fundadores perezosos 2 (coloepodinos y megalocninos si como parece tienen un origen difiletico) musaraiias 2 (nesofontidos y solenodontidos si no constituyen una misma familia como consideran algunos) monos 1 (pitecidos) roedores histricognatos 1 (equimidos capromidos y heptaxodontidos) roedores muridos 1 (Oryzomys en Jamaica)
POBLAMIENTO ISLA-ISLA
La distribucion de los mamiferos terrestres a traves del area antillana demarcada al inicio del presente capitulo (vease Contexto Geogratico Fig 41) ha sido interpretada en funcion de dos hipoteticos mecanismos de poblamiento interinsular la dispersion y la vicarianza fundamentadas como se resumen seguidamente
DISPERSION
De acuerdo con la evidencia paleogeogratica mas actualizada en las principales regresiones marinas ocurridas en la cuenca caribeiia durante los ultimos 23 Ma (transiciones OligocenoshyMioceno Mioceno Superior - Plioceno y varias veces en el Pleistoceno) las masas terrestres constitutivas de la region insular en formacion aumentaron considerablemente su extension territorial disminuyendo por tanto la distancia entre elias oportunidades que los elementos faunales aprovecharian para dispersarse de unas islas a otras
Durante mucho tiempo esta ha sido la hipotesis mas favorecida por los estudiosos del tema a partir de Simpson (1956) pero en decadas recientes ha suscitado dos objeciones
1 Es dificil en tender como pudo operar la dispersion marina entre estas islas puesto que ninguna de las Antillas Mayores posee rios suficientemente caudalosos para transportar masas significativas de vegetacion flotante (MacPhee 198432)
Es oportuno recordar aqui el islote ala deriva (con mas de diez palmas reales en pie) entre las tres Antillas Mayores mas occidentales mencionado anteriormente (vease La Balsa Natural)
2 La dispersion marina es menos efectiva para explicar la asombrosa similitud de las faunas de mamiferos terrestres a traves de las Antillas Mayores septentrionales porque como proceso aleatorio no debe producir distribuciones uniformes a 10 largo de una cadena de islas 0 si las produce entonces el proceso no es aleatorio (MacPhee e lturralde-Vinent 2000 149)
Independientemente de que las aludidas distribuciones taxonomicas estan lejos de ser uniformes 0 asombrosamente similares la hipotesis de la dispersion marina contrariamente a su canicter aleatorio y altamente improbable en el caso del poblamiento continente-isla- en el caso del poblamiento isla-isla es un proceso no aleatorio (aprovecha las oportunidades brindadas puntualmente por los periodos de maxima regresion marina) y ademas altamente probable dada la incomparablemente menor distancia entre las islas caribeiias que entre estas y los continentes vecinos
314 BIOGEOGRAFiA
V ICARIANZA
La neotectonica y las transgresiones marinas del Oligoceno Medio y del Mioceno afectaron sustancialmente la paleogeografia de la Cresta de las Antillas Mayores resultando la subdivision gradual de las partes subaereas de la Cresta Asi el este de Cuba y el norte de La Espanola conectados fisicamente durante el Oligoceno Inferior se separaron al formarse el Paso de los Vientos en el Oligoceno tardio en tanto La Espanola y Puerto Rico probablemente permanecieron conectadas hasta el Mioceno tardio en que surgio el Paso de Mona (Iturralde-Vinent y MacPhee 1999)
Tales eventos geologicos habrian dividido las distribuciones de los grupos faunales emplazados previamente en el territorio continuo de las tres islas y a partir de esas disyunciones dichos autores (tambien MacPhee et al 2000 y White y MacPhee 2001) asumieron --como segunda fase del modelo de GAARlandia- la hipotesis de la vicarianza isla-isla para explicar la distribucion multiinsular de varios grupos de mamiferos terrestres a traves de las Antillas Mayores septentrionales De acuerdo con esta hipotesis cualquier diferencia en composicion faunal entre las islas (por ejemplo la ausencia de roedores heptaxodontidos en Cuba 0 de monos platirrinos en Puerto Rico) constituye evidencia de extincion local postvicariante y no de dispersion marina fallida Lo primero se puede demostrar si aparecen los fosiles pertinentes 10 segundo no se puede demostrar (MacPhee e Iturralde-Vinent 1994) Por otra parte IturraldeshyVinent y MacPhee (1999 57) han reconocido que aunque fa vicarianza isla-isla puede representar una solucion clara para las distribuciones de varios taxones cuaternarios no puede explicar todos los casos
Rega et al (2002 18) han presentado dos objeciones al modelo de la vicarianza isla-isla aplicado a los mamiferos de las Antillas Mayores
I Si la separacion de Cuba y La Espanola precedio en varios mill ones de anos la separashycion de La Espanola y Puerto Rico el modelo vicariante de lturralde-MacPhee debe predecir mayor similitud entre las faunas de La Espanola y Puerto Rico que entre las de Cuba y La Espanola De hecho es al reyes
No obstante MacPhee et al (200335) admitieron despues que la fecha mas temprana empiricamente defendible [para la apertura del Paso de Mona] debe ser el Oligoceno Medio esto invierte la secuencia de ambas disyunciones con 10 cual queda resuelta la aparente contradiccion Sin embargo en su reciente evaluacion de la congruencia entre filogenia e hipotesis geologic as en varios grupos de mamiferos terrestres de las Antillas Mayores Davalos (2004373) encontro que ninguno de los taxones cumple con la fecha y el patron de divergencia entre las masas terrestres antillanas postulados por el modelo de GAARlandia
2 La presencia de un mismo genero en las tres Islas septentrionales de las Antillas Mayores implica [por el modelo vicariante] que la diferenciacion entre estas formas durante el intervalo desde el Mioceno ha sido unicamente de nivel especifico [Dicha presencia]
315 rRRADlAcr6~ EVOLUCrONARIA
s610 puede explicarse razonablemente como el resultado de movimientos de esos animales entre las islas en el Cuaternario
De esta objecion resulta dificil disentir [El poblamiento entre islas diferenciado solo a nivel especifico no es probable que sea anterior al Pleistoceno (Simpson 1956 15)]
Debe tenerse en cuenta que la hipotesis de la vicarianza isla-isla alude a las tres islas septentrionales de las Antillas Mayores porque como se explico antes (vease El Barco Funeral Vikingo) Jamaica no existia como isla durante el lapso (Eoceno Superior shyOligoceno) consensualmente reconocido como el mas probable para la colonizacion de las proto-Antill as Mayores por la mayo ria de los mamiferos terrestres Despues de emerger como isla en el Mioceno Medio Jamaica debe haber recibido sus mamiferos terrestres por dispersion isla-isla (Davalos 2004376) y solo en el Cuaternario parece haberrecibido (por dispersion marina desde Centroamerica) al roedor murido Oryzomys
Seguramente tanto la dispersion como la vicarianza contribuyeron a que los mamiferos terrestres se irradiaran a traves de las Antillas Mayores y grupos de islas perifericas pero las maneras y tiempos en que estos mecanismos operaron a 10 largo de dicho proceso son aun en gran medida motivos de desacuerdo e incertidumbre
IRRADIACION EVOLUCIONARIA
A continuaci6n revisamos someramente las distribuciones conocidas de los diferentes grupos de mamiferos terrestres a traves de las Islas principales de las Antillas Mayores y grupos de Islas perifericas incluidas las interpretaciones y conjeturas que la literatura reciente recoge en relaci6n con la manera en que dichas distribuciones pudieron haberse conformado durante la irradiaci6n evolucionaria de los referidos grupos de mamiferos Manifestaciones de esta irradiaci6n pueden inferirse unicamente a partir de los taxones presentes en el Cuatemario pues solo 6 de las especies conocidas corresponden al Terciario
GRUPOS ORlGINARIOS DE SURAMERICA
PEREZOSOS (Megalonychidae)
Aparte de un genero miocenico de Cuba y otro del Oligoceno de Puerto Rico se conocen 4 generos (12 especies) de perezosos cuaternarios (todos extintos) restringidos a las tres islas septentrionales de las Antillas Mayores Acratocnus Neocnus Megalocnus y Parocnus Los cuatro generos estan representados en Cuba y La Espanola y Acratocnus 10 esta ademas (como unico genero de perezoso) en Puerto Rico Esta distribuci6n ha sido interpretada por MacPhee et al (2000) eomo resultado de la vicarianza oligomiocenica entre estas islas y [dado que el Banco Puerto Rico - Islas Virgenes fue la entrada de GAARlandia a las Antillas Mayores] se preguntaron si la ausencia de los otros tres generos en Puerto Rico se debe a que nunca estuvieron en ese territorio 0 a que sus
316 BIOGEOGRAFiA
fosiles atm no han sido hallados Pero como hemos visto Rega et al (2002) atribuyeron dicha distribucion a la dispersion interinsular en el Cuatemario
Por otra parte varios amilisis c1adisticos recientes ubi can a Choloepus -genero Reciente conocido solo de Centro- y Suramerica- como taxon hermano de Aeratoenus exc1uidos los demas megaloniquidos antillanos Para White y MacPhee (2001228)
tan estrecha relacion sugiere que la divergencia entre ambos [generos] ocurrio despues de que Aeratoenus divergio de Neoenus Si esta relacion implica que Aeratoenus y Neoenus se dispersaron separadamente desde Suramerica a las AntilIas mientras Choloepus permanecia en el continente (siendo necesarias al menos tres invasiones de perezosos) 0 si Choloepus invadio Suramerica desde las Antillas es algo que el material disponible no permite determinar
La diversidad adaptativa de los perezosos se manifiesta en (1) la utilizacion del habitat especies arboreas en Neoenus y Aeratoenus y terrestres en Paroenus y Megaloenus (Webb 1985a White [1993] White et al 1996 Arredondo [1999]) y en (2) la gama de tallas corporales desde los pequenos Neoenus hasta Megaloenus rodens el mayor de los mamiferos terrestres antillanos (su peso adulto estimado en 270 kg seglin Paula 19679) esto sugiere que los perezosos se irradiaron a nichos que en contextos continentales probablemente estarian ocupados por miembros de otros c1ados (MacPhee e IturraldeshyVinent 2000)
No esta claro si el abanico de tamanos entre estos perezosos enmascara alguna tendencia al gigantismo Por 10 pronto Anderson y Handley (2002) encontraron 10 contrario en perezosos (Bradypus) de las Islas de Bocas del Toro (Panama) shyseparadas secuencialmente del area continental vecina por ascensos del nivel del mar durante los ultimos 10 ka En las cinco islas los perezosos son menores que su contraparte continental y las diferencias entre las tallas medias de las cinco poblaciones expresan una reduccion lineal significativa de la talla corporal en funcion de la edad geologica de cada isla sin relacion alguna con el area de las islas 0 la distancia que las separa del continente
MONOS (Pitheciidae)
Los primates de las Antillas Mayores (cuatro especies extintas) constituyen un grupo monofiletico con Paralouatta (en Cuba) y Antillothrix (en La Espanola) como taxones hermanos seguidos en la escalera c1adistica por Xenothrix (en Jamaica) y finalmente Callieebus como el pariente continental mas cercano esto implica una sola invasion desde Suramerica antes del Mioceno Inferior (fecha mas temprana conocida para los primates fosiles antillanos) y por tanto en consonancia con GAARlandia como la via de colonizacion (Horovitz y MacPhee 1996 1999 Horovitz [1997] MacPhee y Horovitz 2004)
En desacuerdo parcial con la anterior interpretacion la filogenia de este grupo de monos obtenida por Davalos (2004) indica un antecesor comun en Cuba La Espanola y Jamaica
317 TRRADIACTON EVOLUCIONARIA
compatible con la hipotetica conexion -propuesta tentativamente por Iturralde-Vinent y MacPhee (199980-84)- entre las Montanas Azules del este de Jamaica y la peninsula suroccidental de La Espanola en la epoca de GAARlandia
A pesar de que los primates antillanos pertenecen a un grupo viviente de monos muy pequenos (los monos titles) fueron comparativamente grandes en relacion con los platirrinos modernos y sucesivamente mayores en la direccion Antillothrix Xenothrix y Paralouatta con P marianae como el mayor platirrino conocido del Mioceno neotropical (MacPhee 2005) MacPhee et al (1994) consideraron que Xenothrix debio ser un mono de andar lento y al parecer las especies de Paralouatta abandonaron la arborealidad para adoptar un modo de vida mas terrestre (MacPhee 1996a MacPhee et al 2003 MacPhee y Meldrum 2006)
ROEOORES (Echimyidae Capromyidae Heptaxodontidae)
La edad de Zazamys en Cuba demuestra que la invasion de los roedores histricognatos en las Antillas Mayores comenzo antes del Mioceno Inferior por otra parte los representantes mas antiguos de esa irradiacion evolucionaria se encuentran en La Espanola y Puerto Rico Ambas evidencias son consistentes con la hipotesis de GAARlandia (Woods 1982 1988 Woods et al 2001)
No esta claro si los capromidos evolucionaron a partir de un equimido ancestral 0 de un protocapromido 0 si los equimidos antillanos (I sub familia 3 generos y 7 especies todas extintas) forman parte de la irradiacion de los capromidos (4 subfamilias 9 generos y 27 especies 10 vivientes) Kratochvil et al (1978b) y Rodriguez et al (1979) consideraron que Capromys nan us es un eslab6n entre Echimyidae y Capromyidae pero Mesocapromys nanus esta morfologicamente mucho mas cerca de Hexolobodon y los de mas capromidos que de los equimidos (Woods 1985) Lo que si esta claro segun Woods et al (2001) es que la subfamilia Plagiodontinae endemic a de La Espanola es el linaje basal de los capromidos y que Isolobodon Hexolobodon Capromys Geocapromys Mysateles y Mesocapromys [y supuestamente Macrocapromys] forman parte de la irradiacion temprana de Capromyidae
La subfamilia Capromyinae (5 generos y 17 especies 9 vivientes) parece ser el dado de mas reciente evolucion en la familia con invasion de Cuba desde La Espanola y dispersion desde Cuba (1) por Capromys a las islas Caiman y (2) por Geocapromys a las islas Bahamas Caiman y Jamaica y desde Jamaica a las islas Swan aunque la presencia de Geocapromys en Swan pudiera deberse a introduccion aborigen (Morgan 1985 1988 1989 1993 1994 Woods 1989a 1990) Woods et al (2001) utilizaron datos moleculares para obtener estimados independientes de divergencia entre capromidos segiJn 10 cual Geocapromys divergio cerca de los 17 MaAP Mesocapromys divergio de Capromys hace 15 MaAP en tanto lvlysateles 10 hizo hace 10 MaAP Por su parte las formas de Capromys pilorides y de Mysateles prehensilis en Isla de la Juventud divergieron de sus correspondientes subespecies nominativas en la isla principal a los 24 y 08 MaAP respectivamente
318 BIOGEOGRAFiA
Woods y Howland (I979) describieron adaptaciones morfologicas de la mandibula el hioide y la faringe asociadas a mecanismos de masticacion diferentes en plagiodontinos y caprominos y Varona (1980a) llamo la atencion sobre el gran desarrollo de los molariformes (en relacion con la longitud total del cnineo) que presentan ciertos grupos de jutias fosiles en comparacion con las vivientes 10 que pudiera interpretarse como una adaptacion alimentaria a una vegetacion mas abrasiva por parte de las jutias extinguidas
Segun Woods (1990) la familia Heptaxodontidae (2 subfamilias 5 generos y 6 especies todas extintas) tambien seria parte de la irradiacion de los capromidos con origen en el este de La Espanola 0 en Puerto Rico y subsecuente dispersion al Banco de Anguila - San Martin en las Antillas Menores (pero vease MacPhee y Flemming 2003) Este banco estaria a solo 70 km del Banco Puerto Rico - Islas Virgenes en tiempos de maxima regresion marina y esta proximidad con Puerto Rico -patria de Elasmodontomys obliquus- seguramente tuvo que ver con la presencia en las islas de Anguila y San Martin del heptaxodontino gigante Amblyrhiza relacionado mas estrechamente con los roedores de las Antillas Mayores que con los del resto de las Antillas Menores (Woods 1990 1996) Aunque los roedores suelen aumentar de tamano en las islas (Case 1978) la escala del incremento de la talla en Amblyrhiza (que pudo alcanzar mas de 200 kg de peso adulto) esta fuera de toda proporcion con los patrones conocidos en otros roedores insulares (Biknevicius et 01 1993)
Clidomys (Clidomyinae) de Jamaica difiere marcadamente en su morfologia de los heptaxodontinos (vease ademas MacPhee y Flemming 2003)10 cual dificulta explicar su presencia en esa isla Como Jamaica estuvo bajo el mar hasta el Mioceno Medio el ancestro de Clidomys colonizaria esta isla por dispersion desde el extremo suroccidental de La Espanola despues que los heptaxodontinos se originaron en esta ultima isla (Woods 1989a) Pero Pascual et 01 (I 990) especularon que Clidomys pudiera ser el resultado de una invasion posterior e independiente de los capromidos
GRUPOS ORIGINARIOS DE NORTEAMERICA
El origen de las musaranas antillanas ha resultado ser hasta ahora el mayor enigma biogeografico de la fauna de mamiferos terrestres de la region aunque la mayoria de los autores coinciden en considerarlas originarias de Norteamerica (vease Discus ion y Conclusiones en Poblamiento Continente-Isla) No obstante MacPhee e Iturralde-Vinent (1994) descartaron toda posibilidad de conexion terrestre por el Estrecho de la Florida 0 por el Canal de Yucatan y si el (supuesto) evento vicari ante de Rosen (vease El Arca de Noe) -invocado por MacFadden (1980) para resolver el enigma- fuera minimamente plausible aun asi es dificil creer que las catastrofes que 10 sucedieron (vease Contexto Temporal) hayan barrido con toda esa vieja fauna continental excepto las musaranas circunstancia ya anticipada muy tempranamente por Matthew (l918a)
De otra parte la relaci6n entre Nesophontidae y Solenodontidae permanece irresuelta Si estos linajes no estan estrechamente relacionados entonces es muy posible que
319 IRRADIACION EVOLUCIONARIA
sus distribuciones sean explicadas por mecanismos diferentes (Whidden y Asher 2001)
La descomunal diferencia de talla corporal entre las musaranas pequenas (Nesophontes) y las grandes (Soienodon) seguramente impuso a ambos grupos restricciones distintas relativas a la termorregulacion y las adaptaciones energeticas En Nesophontes la seleccion debio favorecer una alta tasa de metabolismo y crecimiento camadas mas numerosas corta expectativa de vida yalta densidad poblacionaL En Soienodon en cambio la seleccion opero en la direccion de una baja tasa metabolica camadas reducidas crecimiento lento longevidad y baja densidad poblacional (Ottenwalder [1991])
MUSARANAS PEQUENAS (Nesophontidae)
Las musaranas pequenas estan representadas en las Antillas por el genero Nesophontes (y un genero miocenico indeterminado en ambar dominicano) con 6 especies cuaternarias extinguidas distribuidas en Cuba La Espanola Puerto Rico Islas Virgenes e Islas Caiman La mayor de las musaranas pequenas Nesophontes edithae evoluciono en Puerto Rico y su arribo a las Islas Virgenes supone una dispersion sumamente reciente En las Islas Caiman Nesophontes se asemeja mas a la poblacion de Cuba de donde seguramente derivo en el Plio-Pleistoceno (junto con Capromys y Geocapromys) cuando la costa sur de Cuba estuvo mucho mas cerca de las Islas Caiman (lturralde-Vinent 2003fig 4) MacPhee et ai (1994) interpretaron la distribucion de Nesophontes en Cuba y La Espanola como resultado de la vicarianza oligomiocenica ocurrida al formarse el Paso de los Vientos pero la distancia taxonomica entre los lados del Paso es solo de nivel especifico y N micros incluso vivi6 en ambas islas segtln Patterson (1962)
Allen (1918) considero que Nesophontes micrus fue una especie primitiva no modificada para la vida arb6rea 0 fos6rea y por tanto de habitos terrestres Nesophontes edithae de Puerto Rico es mucho mayor y mas robusta que las restantes especies del genero eL indice de robustez de su femur (= anchura minima de la diafisis ongitud total del hueso) muestra que este es tan robusto como el de Solenodon paradoxus un animal dos veces mayor (Morgan et ai 1980) Choate y Birney (1968) postularon un aumento cronoclinal de la taIla corporal en N edithae confirmado mas tarde por McFarlane (1999a) mediante fechados radiocarb6nicos de la secuencia estratignifica en depositos cavernarios de Puerto Rico Solenodon no vivi6 en Puerto Rico y en ausencia de una musarana de mayor tamano Nesophontes alcanzo una talla corporal mayor en esa isla que en La Espanola 0 Cuba (Woods y Eisenberg 1989)
MUSARANAS GRANDES (Solenodontidae)
Los arcaicos almiqufes (genero Soienodon) comprenden cuatro especies una viviente (S cubanus) y otra f6sil (S arredondoi) en Cuba y una viviente (S paradoxus con dos subespecies) y otra f6sil (S marcanoi) en La Espanola Como cabria esperar las dos especies cubanas se asemejan mas entre sf que a las especies de La Espanola pero
320 BIOGEOGRAFiA
enigmaticamente S marcanoi comparte caracteres con S cuban us y S paradoxus (Woods y Eisenberg 1989 Morgan y Ottenwalder 1993 Ottenwalder 200 I) Solenodon arredondoi alcanzo una talla corporal superior a la de cualquiera de las musarafias vivientes a nivel mundial Segun Morgan y Ottenwalder (1993) extrapolando las dimensiones y el peso de los almiquies vivientes S arredondoi debio tener una longitud cabeza-tronco y un peso de 450-550 mm y 1500-2000 g respectivamente en comparacion con S cubanus (360 mm y 800 g) y S paradoxus (390 mm y 1100 g)
Entre todas las especies de almiqufes Solenodon paradoxus es la que presenta los caracteres mas derivados (Otten walder 2001) Si estas relaciones son validas la distribucion de Solenodon entre Cuba y La Espafiola debio discurrir de oeste a este como propuso MacFadden (1980) quien invoco la presencia fosH de S cubanus [pero tambien S arredondoi] en Pinar del Rio para postular que el ancestro continental entr~ por Cuba opinion compartida por Perfit y Williams (1989) y por Ottenwalder ([1991] 2001) El paso de Solenodon a Puerto Rico pudo haberlo impedido la existencia de barreras ecologicas durante la conexion del este de La Espafiola con Puerto Rico 0 por la ausencia de Solenodon en esa parte de La Espafiola en aquella epoca Sin embargo las mismas condiciones ecologicas en el este de La Espafiola no parecen haber representado una barrera efectiva para Nesophontes cuyas mas altas tasa de reproduccion y densidad poblacional presumiblemente Ie permitieron expandirse flpidamente a traves de La Espafiola (Ottenwalder [1991])
Es opinion generalizada que los almiqufes antillanos constituyen el grupo de mamiferos que primero se emplazo en el territorio insular (MacPhee 2005) y en tal sentido es de interes la evidencia sobre especificidad parasito-hospedante aportada por Cruz (1980) quien encontro que la especie de acaro (genero Cubanochirus) que parasita a los almiquies es mucho mas primitiva que la especie (del mismo genero) que paras ita a las jutias y de aqui concluyo que dicho genero de acaro debio evolucionar primero en Solenodontidae y posteriormente en Capromyidae al entrar en contacto representantes de ambas familias en Cuba 0 La Espafiola
Recientemente Roca (2004) y Roca et al (2004) dieron a conocer que los resultados de su estudio genetico-molecular sobre los almiquies indican una divergencia de 25 Ma entre Solenodon cuban us y S paradoxus -compatible con la disyuncion tectonica entre Cuba y La Espafiola (Iturralde-Vinent y MacPhee 1999 McPhee et al 2003)-- y una separacion de ambas especies comparable en magnitud a la divergencia entre familias de mamlferos Consecuentemente propusieron que Solenodon cubanus fuera clasificada en el genero Atopogaie como indico Cabrera (1925) hace 82 afios
DISCUSION Y CONCLUSIONES
Las musarafias y los roedores antillanos (excepto Oryzomys) se irradiaron tanto de sus respectivos ancestros continentales que de estos 6rdenes se reconocen familias y subfamilias endemicas de la region en contraste con los perezosos (una subfamilia y tres
321 IRRADIACION EVOLUCIONARIA
tribus endemicas) y los monos (una tribu endemica) como expresion de diferentes niveles de diversificacion evolucionaria Este proceso es dificil de explicar (traUindose de c1ados que pnicticamente comparten la misma antigiiedad en las Antmas) si es correcta la interpretacion de Pascual et at (1990636)
Nosotros consideramos estos diferentes niveles de diversificacion como las diferentes etapas de evolucion alcanzadas por cada grupo [de mamiferos] Obviamente esto estuvo relacionado con las diferentes longitudes de sus historias
Pudiera ocurrir que en el futuro se compruebe que la historia biogeognifica de la region fue mucho mas complicada
La Espanola la isla mas ecol6gicamente divers a de la region debi6 ser el centro principal de evoluci6n de los mamfferos (con 87 de las familias y 73 de las subfamilias) seguida por Cuba (con 75 de las familias y 54 de las subfamilias) las dos islas de las Antillas Mayores mas relacionadas biogeograficamente (Fig 43 Tabla 44)
eL
EL
cb CAIMAN
SWAN
A Acratocnini B Cubanocnini C Megalocnini D Mesocnini E Nesophontidae F Solenodontidae G Xenothrichini
ANGUILAshyM-0 SMARTIN
H Heteropsomyinae I Plagiodontinae J Isolobodontinae
K Hexolobodontinae L Capromyinae
M Heptaxodontinae N CIidomyinae 0 Sigmodontinae
FIGURA 43 Representacion esquematica de las relaciones interinsulares (expresadas porel numero de linajes compartidos) entre las Antillas Mayores y entre estas y los grupos de islas perifericas segun las interpretaciones de los autores citados en el texto Los numeros dentro de los circulos indican la cantidad de linajes representados en cada isla relacion dudosa Contexto taxon6mico de los linajes en la Tabla 44
bull bull
bull bull
bull bull bull
bull bull
bull bull
bull
bull
bull
bull
bull bull bull bull
bull bull
bull bull
bull
bull bull bull
bull bull
bull
bull bull
bull
322 BIOGEOGRAFiA
TABLA 44 Distribuci6n de los linajcs de mamiferos terrestres a traves de las Antillas Mayores y grupos de islas perifericas LE = La Espanola CU Cuba JA Jamaica PR = Puerto Rico IC Islas Caiman IS Islas Swan IB = Islas Bahamas IV = Islas Virgenes AS = Anguila - San Martin
FamiliaSubfumiliafIribu LE CU JA PR IC IS IB IV AS
Megalonychidae
Choloepodinae
Acratocnini
Cubanocnini
Megalocninae
Megalocnini
Mesocnini
Nesophontidae
Solenodontidae
Pitheciidae
Callicebinae
Xenothrichini
Echimyidae
Heteropsomyinae
Capromyidae
Plagiodontinae
Isolobodontinae
Hexolobodontinae
Capromyinae
H e pta x 0 don tid a e
Heptaxodontinae
Clidomyinae
Muridae
Sigmodontinae
PILOSA
SORICOMORPHA
PRIMATES
RODENTIA
bull
bull
bull
bull TOTALES 12 10 4 4 2
323 EXTlICIOIES DEL CUATERNARIO
El gigantismo ha sido sin duda una tendencia evolucionaria recurrente entre los clados mayores de mamiferos antillanos Es un hecho bien conocido que algunos grupos de mamiferos han aumentado de tamafio en las islas y varios autores han intentado explicarlo a partir de enfoques diferentes (Foster 1964 Miller 1966 Sondaar 1977 Case 1978 Eisenberg y Wilson 1978 Heaney 1978 Lawlor 1982 Lomolino 1985 Angerbjom 1985 McNab 19942001)
EXTINCIONES DEL CUATERNARIO
Las islas que integran la Subregion Antillana han perdido casi 90 de sus mamiferos terrestres durante los ultimos milenios y en proporcion con su extension territorial esta perdida ha sido mayor en la Subregion que en cualquier otro lugar del planeta (MacPhee 1996b) Las extinciones del Cuatemario eliminaron mas de las tres cuartas partes de la fauna de mamiferos terrestres conocida de las Antillas Mayores sus islas satelites y grupos de islas perifericas y el examen de este proceso en sus aspectos mas generales se aborda en esta ultima parte del presente capitulo
CRONOLOGiA Y CONSECUENCIAS
Es muy improbable que un fosil represente el ultimo individuo de un taxon extinto y entonces hay que guiarse por la edad radiometrica del fosil mas reciente conocido lIamada fecha de ultima presencia- como unica aproximacion posible a la fecha de extincion del taxon
Como se explico al inicio del capitulo (vease Contexto Temporal) el registro fosH de los mamiferos terrestres de las Antillas Mayores adolece de insuficiente sustentacion radiometrica No obstante la distribucion temporal de los escasos fechados disponibles para el Cuaternario de la region abarca etapas criticas del proceso de extincion experimentado por la referida fauna a 10 largo de los ultimos 250 mil afios
De las 52 especies que es posible reconocer actualmente como miembros extintos de la fauna cuatemaria de mamiferos terrestres de las Antillas Mayores y grupos de islas perifericas los restos fosiles de 23 de elIas (442) han sido fechados radiometricamente o se han hallado asociados estratigraficamente de manera inequivoca con otro material fechado De estos 23 fechados 3 corresponden al interglacial Sangamon 4 pertenecen a la secuencia Pleistoceno terminal - Holoceno temprano 10 se ubican en ellapso entre el arribo de los aborigenes y la incursion europea y 6 son postcolombinos (a los cuales hay que sumar otras dos especies extinguidas en los siglos XIX y xx) A partir de estas 25 especies avaladas por fechas de ultima presencia distribuidas entre los cuatro intervalos temporales sefialados hemos compuesto la Tabla 45 en la que ademas aparecen las restantes 27 especies (carentes de fechado radiometrico) ubicadas en el intervalo temporal mas probable sobre la base de (1) la asociacion estratigrafica de sus restos con evidencias culturales (amerindias 0 europeas) 0 (2) la ausencia de dicha asociacion Sobre estos criterios de ubicacion temporal procede hacer dos aclaraciones
324 BIOGEOGRAFIA
TABLA 45 Intervalos temporales mas probables para las extinciones cuatemarias de los mamiferos terrestres de las Antillas Mayores y grupos de islas perifericas sobre la base de las evidencias siguientes (l) ultimo ejemplar colectado u observado en vida (2) fechado radiometrico mas tardio en material de la especie (3) asociaci6n estratigrafica mas tardia con otro material fechado (4) asociaci6n estratigrafica con restos de animales introducidos restos humanos u objetos culturales propios del intervale (5) falta de asociaci6n con evidencias culturales [Los ntimeros entre parentesis en la columna Evidencia significan adscripci6n tentativa al intervalo] Los estimados del peso corporal (en kg) estan tomados de la literatura 0 inferidos comparativamente y distribuidos entre
las clases siguientes (a) S 01 (b) 01 a lt 1 (c) 1 a lt 10 (d) 10 a lt 100 (e) 100 a lt 200 (j) ~ 200 Las
abreviaturas que designan las islas son las de la Tabla 44
F amilialEspecie Peso IsJa Evidencia Fuente
I httervalo htterg1acial Sangamon (gt80 kaAP)
HEPTAXODONTlNAE
Amblyrhiza inundata e AS 3 McFarJane et al 1998a
CLIDOMYINAE
Clidomys osborni d JA 3 Ibid
Clidomys parvus d JA 3 Ibid
II httervalo Pleistoceno terminal ~ Holoceno temprano (20-6 kaAP)
MEGALONYCHIDAE
Aeratoenus antillensis d CD 5 Arredondo 1975 a
Aeratoenus odontrigonus d PR 3 McFarJane 1999b
Aeratoenus simorhynehus d LE 2 Rega et al 2002
M egaloenus zile f LE 2 MacPhee et al 2000
SOLENODONTIDAE
Solenodon arredondoi e CD 5 Morgan y Ottenwalder 1993
PnHECIIDAE
Paralouatta varonai d CD 5 MacPhee 1996a
EcHIMYIDAE
Heteropsomys antillensis c PR 5 McFarJane 1999b
Heteropsomys insulans c PR 5 Ibid
Puertoricomys eorozalus c PR 5 Woods 1989a
CAPROMYIDAE
P lagiodontia araeum c LE 5 MacPhee et al 2000
Macrocapromys latus c CD 5 Varona y Arredondo 1979
(Continua)
325 EXTINCIONES DEL CUATERNARIO
TABLA 45 (Continuaci6n)
F ami1ia1Especie Peso Isla Evidencia Fuente
M acrocapromys latus c CU 5 Varona y Arredondo 1979
Mesocapromys kraglievichi c CU 5 Ibid
HEPTAXODONTINAE
Xaymaca fulvopulvis a JA 2 MacPhee y Flemming 2003
III Intervalo Amerindio (6-05 kaAP)
MEGALONYCHIDAE
Acratocnus ye d LE 2 MacPhee et aI 2000
Neocnus comes c LE 2 Ibid
N eocnus dousman c LE 2 Ibid
N eocnus toupiti c LE 2 Ibid
N eocnus gliriform is c CU 4 Cordova et ai 1997
M egalocnus rodens f CU 2 MacPhee et ai 2007
Parocnus browni e CU 2 Steadman et ai 2005
P arocnus serus e LE 2 MacPhee et ai 2000
N ESOPHONTIDAE
N esophontes edithae a PR 2 McFarlane 1999b
PITHECIIDAE
Antillothrix bernensis c LE 3 Rimoli 1977
EcHIMYIDAE
Brotomys contractus a LE 4 Miller 1929c
CAPROMYIDAE
P lagiodontia spelaeum c LE 4 Ibid
M acrocapromys acevedo c CU 4 NUiiez 1990a
HEPTAXODONTINAE
Elasmodontomys obliquus d PR 3 McFarlane 1999b
Iv Intervalo Postcolombino (05 kaAP - Presente)
N ESOPHONTIDAE
N esophontes hypom icrus a LE 24 MacPhee y Fle111111ing 1999
N esophontes zam icrus a LE (4) Ibid
N esophontes m icrus a CU 24 MacPhee et aI 1999
(Continua)
326 BIOGEOGRAFiA
TABLA 45 (Continuacion)
F amilialEspecie Peso Isla Evidencia Fuente
N esophontes m icrus a CD 24 MacPhee et aI 1999
N esophontes m icrus a LE 4 MacPhee et aI 2000
N esophontes sp 1 a IC 4 Morgan 1994
N esophontes sp 2 a IC 4 Ibid
Solenodon marcanoi b LE 4 Ottenwalder 2001
PITHECIIDAE
Xenothrix mcgregori c JA 4 MacPhee y Flemming 1999
EcHIMYIDAE
Boromys offeUa a CD 24 Guarch 1984
Boromys torrei a CD 24 Ibid
Brotomys voratus a LE 4 MacPhee et aI 2000
CAPROMYIDAE
P lagiodontia ipnaeum c LE 4 MacPhee y Flemming 1999
Rhizoplagiodontia lemkei c LE 4 Ibid
Isolobodon portoricensis c PR 24 Ibid
Isolobodon montanus c LE 4 Ibid
Hexolobodon phenax c LE (4) Ibid
Capromys sp c IC 24 Morgan 1994
Geocapromys columbianus c CD 4 MacPhee y Flemming 1999
Geocapromys thoracatus b IS 1 MacPhee y Marx 1997
Geocapromys sp 1 c IC 4 Morgan 1994
Geocapromys sp 2 c IC (4) Ibid
l-IEPTAXODONTINAE
Quemisia gravis c LE (1) MacPhee y Flemming 1999
MURIDAE
Oryzomys antillarum a JA 1 Ibid
1 Flemming y MacPhee (1999) alertaron en contra de la asociacion con otro material fechado Asimismo Morgan (1993) indico que la ausencia de asociacion con evidencias culturales amerindias no es necesariamente un criterio concluyente para ubicar un taxon en el intervalo temporal anterior porque hay depositos en la region que carecen de tales evidencias y son del Holoceno tardio por fechados radiocarbonicos [No obstante
327 EXTlNCIONES DEL CUATERNAR10
la ausencia de una especie en residuarios arqueol6gicos adquiere mayor certidumbre como criterio de ubicaci6n temporal de su extinci6n en la medida que aumenta el numero de residuarios estudiados que en las Antillas Mayores se cuentan por centenares] A falta de fechado absoluto de una especie consideramos que estos criterios de ubicaci6n temporal son mas utiles que la alternativa de excluir del analisis a la especie
2 En los trabajos sobre biogeografia de mamiferos antillanos los arque610gos mas citados como autoridades para establecer el inicio de la presencia aborigen en las Antillas Mayores (Kozlowski 1974 Rouse y Allaire 1978 Rouse 1989 1992 Wilson 2001) difieren al respecto entre los 6 kaAP y los 45 kaAP (pero vease Pagan et al 200523) y hemos adoptado la mas temprana de estas fechas para el comienzo del intervalo amerindio en la Tabla 45 De todos modos cualquiera que haya sido la fecha esta no es aplicable por igual al conjunto de las Antillas Mayores porque segun algunos de los autores citados el poblamiento comenz6 por Cuba y La Espanola 6 mil anos atras continu6 en Puerto Rico hace alrededor de 3 mil anos (pero vease Burney et al 1994) yen Jamaica hace menos de 2 mil e incluso a las Islas Caiman ni siquiera llegaron [Las Islas Caiman descubiertas por Col6n en 1503 carecen absolutamente de evidencia amerindia (Stokes y Keegan 1996)]
Si los datos de la Tabla 45 representan una aceptable aproximaci6n a la realidad el proceso de las extinciones cuaternarias de los mamiferos terrestres de las Antillas Mayores y grupos de islas perifericas debi6 caracterizarse entre otros por los eventos siguientes
Aunque obviamente la fauna cuaternaria debi6 ser mucho mas numerosa que 10 que refleja el registro paleontol6gico (Morgan y Woods 1986) 52 especies (81 de la fauna conocida) desaparecieron (sin reposici6n) durante los ultimos 250 mil anos
- Al parecer las extinciones se concentraron en las especies de mayor tamano Asi el peso promedio estimado para las especies de la etapa preantr6pica (intervalos I + II de la Tabla 45) es 350 kg es 328 kg para las especies del intervalo III (amerindio) y es s610 25 kg (c1ases del peso a b c) para las especies del intervalo IV (postcolombino) Ninguna especie aut6ctona de mamifero mayor de 6 kg existe hoy en la regi6n
- De acuerdo con la distribuci6n temporal de las extinciones 61 especies sobrevivieron al interglacial Sangamon (intervalo 1)48 sobrevivieron a la transici6n del Pleistoceno terminal al Holoceno temprano (intervalo II) 34 sobrevivieron al periodo amerindio (intervalo III) y s610 12 especies han sobrevivido desde la Ilegada de los europeos (intervalo IV) Las tasas de extinci6n correspondientes (exceptuado el intervalo I) son una especie cada 1167 anos en el intervalo II una cada 393 anos en el intervalo III y una cada 23 anos en el intervalo IV
- Durante todo el proceso de las extinciones cuaternarias la perdida de especies por isla fue de 91 en La Espanola 83 en Jamaica y 62 en Cuba Bahamas retuvo su unica especie (aunque extirpada en varias de las islas) Puerto Rico e Islas Caiman perdieron todas sus especies (6 y 5 respectivamente) y las Islas Swan Virgenes y Anguila - San Martin perdieron sus respectivas especies (micas
328 BIOGEOGRAFiA
De los cuatro 6rdenes de mamiferos representados en la fauna Pilosa y Primates desaparecieron totalmente Soricomorpha y Rodentia perdieron 86 y 74 de sus especies respectivamente
CAUSALIDAD
CAUSAS NATURALES
Los cicIos glaciales e interglaciales acaecidos en el Pleistoceno causaron cambios ambientales dnisticos en las Antillas con la concomitante alternancia de cIimas frios (secos) y calidos (hUmedos) asi como de descensos y ascensos del nivel del mar aumento y disminuci6n de las areas territoriales) induso (aunque con menor intensidad) en el Holoceno Estos eventos paleogeograficos han sido consistentemente documentados (Mayo y Kartashov 1972 Bonnatti y Gartner 1973 Ortega y Arcia 1982 Prance 1982 Schubert y Medina 1982 Dodge et al 1983 Ortega [1984] Haq et al 1987 Schubert 1989 Hodell et al 1991 Guilderson et al 1994 Oppo 1997 Emslie 1998 Curtis et al 2001 Iturralde-Vinent 2003) y de ellos brindamos a continuaci6n una breve resena
Finalizando el periodo interglacial anterior a la ultima glaciaci6n (Wisconsin) el mar estaba a 8-10 m por encima de su nivel actual Cuando el avance glacial del Wisconsin a1canz6 su culminaci6n (eillamado Ultimo Maximo Glacial) hace 18 ka el mar habia descendido 120 m con respecto a dicho nivel y las islas del Caribe habian incrementado considerablemente sus extensiones subaereas en comparaci6n con las que presentan hoy dia por ejemplo Puerto Rico y las Islas Virgenes se conectaron territorialmente y las Bahamas se unieron en cinco gran des islas Prevalecia un regimen frio y seco y condiciones de aridez --debidas a una reducci6n de las precipitaciones (posiblemente a menos de 440 mmlano)- y los ambientes xericos se habian extendido a traves de las islas y bancos emergidos y abundaban los pinares (los habitats mesicos estaban restringidos a parcelas relictas en 10 alto de las montanas) En el Mar Caribe y el Golfo de Mexico la temperatura superficial del mar era 5degC mas baja que la actual y en las llanuras costeras y alturas maximas eran respectivamente 2-3degC y 6degC mas bajas Los limites de las zonas ecol6gicas altitudinales descendieron varios cientos de metros y cada zona redujo su extensi6n vertical al aumentar el gradiente de temperatura Ellimite inferior de la zona periglacial
elevaci6n donde la temperatura media minima a1canza los OdegC) de la Cordillera Central de la Republica Dominicana (altura maxima del Caribe insular con evidencia de topografia glacial) se considera que estuvo entre los 1500 y 1800 mlnm (actualmente esta a 2800 mlnm) y se puede estimar que dicho limite era inferior en Cuba por estar mas al norte que la Republica Dominicana No obstante durante este periodo la temperatura experiment6 episodios cortos (unas pocas decadas) de ascensos repetidos
Un signifieativo componente bi6tieo de las referidas condiciones paleoambientales 10 constituy6 la espectacular irradiaci6n evolucionaria experimentada por un grupo numeroso de aves rapaees diurnas y nocturnas (Wetmore 1937 Brodkorb 1969 Kurotchkin y Mayo 1973 Arredondo 1976 Olson 1978 Olson y Hilgartner 1982
329 EXTINCIONES DEL ClJATERNARIO
Suarez 2000a b 2004 Suarez y Emslie 2003) Los mamiferos de las Antillas Mayores ofrecieron a estos depredadores aviares una amplia gama de tamanos de presa que en ausencia de mamiferos del orden Carnivora en la region permitio a estas aves irradiarse a este nicho alimentario con la evolucion de una variedad de especies de muy diversa tall a corporal incluidas formas gigantes Adquirir un cuerpo gigante les propicio el acceso a recursos alimentarios que no estaban siendo explotados por ningun otro depredador (Alcover y McMinn 1994) A Cuba corresponde el mejor registro fosil de rapaces cuaternarias caribenas con siete generos de especies diurnas Dentro de estas aves existieron tres gavilanes gigantes (Accipitridae) tres carairas (Falconidae) y un condor (Cathartidae) asi como cuatro generos de especies nocturnas dos lechuzas (Tytonidae) y tres Mhos (Strigidae) dos de ell os no igualados en tamano en todo el mundo Algunos de estos generos tuvieron representacion en otras islas antillanas como La Espanola Jamaica Bahamas y Gran Caiman
Al retirarse hacia el norte los hielos de la glaciacion Wisconsin el clima se tomo mas hUmedo en las Antillas durante el Holoceno temprano y el bosque mesico se expandio en la region insular El nivel del mar ascendio acompanado de mayor pluviosidad alrededor de los 7 kaAP las Islas Virgenes quedaron separadas entre si y de Puerto Rico y las Bahamas perdieron la mayor parte de su territorio Cerca de los 3 kaAP las condiciones secas retornaron a la region e hicieron desaparecer buena parte del bosque mesico Nuevamente el clima comenzo a adquirir sus condiciones actuales hacia los 25 kaAP aproximadamente y hoy transita por una fase mesica con vegetacion xerica confinada en parcelas relictas
Sin duda estos y otros eventos naturales incidieron negativamente en la sobrevivencia de mamiferos terrestres en las Antillas Mayores asi como en los grupos de pequenas islas perifericas (Pregill y Olson 1981 Olson 2006)
McFarlane et al (1998a) demostraron convincentemente como Amblyrhiza el gigante de los roedores antillanos endemico de la pequena isla de Anguila sucumbio a las repetidas y abruptas perdidas de su habitat durante el interglacial Sangamon por las nipidas y severas oscilaciones del nivel del mar (documentadas por Neumann y Hearty 1996) al no poder responder adecuadamente a tan catastroficos eventos mediante el ajuste de su masa corporal media y otro tanto debio ocurrirle en Jamaica al otro roedor gigante Clidomys (McFarlane et al 1998b) emparentado con Amblyrhiza en la misma epoca (Tabla 45 intervalo I)
El ascenso del nivel del mar seguramente promovio la extincion de mamiferos durante la transicion del Pleistoceno al Holoceno (Tabla 45 intervalo II) con la reduccion del tamano de las Islas por las transgresiones marinas (Pregill y Olson 1981 Morgan 1993) Esto ultimo haria maximamente severa la competicion asf como la presion de los depredadores [Steadman et al (2005) no yen por que el calentamiento global ocurrido durante la transicion Pleistoceno-Holoceno habria de tener un efecto negativo sobre los perezosos siendo sus descendientes actuales especies tropic ales 0 subtropicales pero no se trata del efecto
330 BIOGEOGRAFiA
directo del cambio de temperatura sobre los animales sino del efecto indirecto y progresivo que dicho cambio genera a largo plazo al reducir el territorio insular disponible]
Por esta epoca tambien desaparecieron especies de tall a mayor como Megalocnus zile en La Espanola y otras de menor tamano (en Cuba) pero que representan la tall a maxima en sus respectivos contextos taxonomicos del Cuaternario como Solenodon arredondoi Paralouatta varonai y Macrocapromys acevedo Un mamifero puede ser grande solo a costa de reducir su densidad poblacional y esto 10 hace mis vulnerable a la extinci6n sobre todo en una isla (McNab 1994 McFarlane et al 1998a) Incidentalmente Alcover et al (1998a) afirmaron que larelacion entre la talla corporal y la extincion no siempre se cumple en las especies de menor tamano y citaron como ejemplo que entre las musarafias todas las especies de Nesophontes desaparecieron mientras sobreviven especies de Solenodon pero la comparacion correcta en todo caso serfa entre los solenodones vivientes y el gigante Solenodon arredondoi y entre Nesophontes edithae -el gigante de los nesofontes extinguido en el periodo amerindio- y el resto de sus congeneres que sobrevivieron hasta los primeros siglos de la colonizacion europea (Tabla 45 intervalos III y IV)
CAUSAS ANTROPOGENICAS
Intervalo Amerindio
La mayoria de los autores que se han referido a las extinciones acaecidas durante este intervalo (entre elIos paradigmaticamente Martin 1984a b) han responsabilizado a los aborfgenes (que se supone comenzaron a poblar las Antillas Mayores hace 6 mil afios) con la desaparicion de numerosas especies de mamiferos terrestres en la region principalmente por la via de la sobrecaza y la introduccion de animales domesticos devenidos depredadores de la fauna autoctona (Morgan y Woods 1986) Este efecto negativo de la presencia indigena sobre los mamiferos de la region ha sido dramatizado por algunos autores en terminos de holocausto faunal Sin embargo pensamos que se ha exagerado Como han expresado MacPhee et al (19993) cuando uno busca evidencia dura de complicidad humana directa en [estas] extinciones no hay mucho que encontrar Las siguientes son algunas consideraciones a tener en cuenta en tal sentido
El registro radiometrico mas joven conocido hasta ahora para un perezoso antillano (MegalocnllS rodens) es de 42 kaAP (MacPhee et ai 2007) Si se acepta que los aborfgenes entraron en Cuba hace aproximadamente 6 mil anos su convivencia con los perezosos debe haber durado alrededor de 1800 afios (0 mas porque dificilmente el referfdo fechado radiometrico haya correspondido al ultimo sobreviviente de la especie) Esta cifra contradice la hipotesis del holocausto faunal segun la cualla extincion de los mamiferos insulares de mayor talla debe ocurrir con extrema rapidez -a escala de decadas 0 de centurias- a partir de su primer contacto con los humanos (Martin y Steadman 1999) A pesar de las repetidas afirmaciones en contrario (Harrington 1921 Miller 1929c Herrera [1945] Tabio 1951 Alvarez 1951a 1957 Arredondo
331 EXTINCIONES DEL CUATERNARIO
1959 1960a Veloz y Ortega 1973 Mayo 1980b Rodriguez et al 1984 Castellanos y Pino 1985 Pino y Castellanos 1985 Nunez 1990b Gutierrez 1996 Arredondo 2004) en su mayoria basadas en evidencias no exentas de posibles accidentes de deposicion (0 de reubicacion postdeposicional) en recintos cavernarios (Diaz-Franco [2006b D no se ha demostrado inequivocamente que los aborigenes antillanos se alimentaran de perezosos regularmente Sus restos en residuarios de alimentos son escasisimos en toda la region y esto contrasta con los abundantes restos de jutias y otros mamiferos de menor talla que aparecen en esos residuarios demostrativos de la preferencia de los paleoindios por estos otros mamiferos
Los mamiferos mas importantes en la economia prehistorica de las Antillas Mayores fueron las jutias Capromys y Geocapromys en Cuba Geocapromys en Jamaica e Isolobodon en La Espanola Puerto Rico e Islas Virgenes (Aguayo 1950b Tablo 1951 Rimoli 1972 Wing 2001 Diaz-Franco [2006b D Wilkins (2001) estudi61ajutia viviente de Jamaica (Geocapromys browni) en un sitio ocupado por los aborigenes durante 100 anos y no encontr6 evidencias de que durante ese tiempo la especie haya sido sobreexplotada Asimismo en Cuba las jutias conga carabali y andaraz son escasas en depositos pleistoholocenicos en cambio son muy numerosas en residuarios indigenas y de haber sido sobreexplotadas 10 mas probable es que no hubieran sobrevivido hasta hoy
La habilidad de los humanos para penetrar en profundidad el bosque tropical y mantenerse en este sustentablemente ha sido un evento comparativamente reciente (Beran y Steele 1994) En el caso antillano la caza tierra adentro ha sido explicada como un proceso tardio de desplazamiento a partir de la explotacion de abundantes recursos marinos que ofrecian condiciones 6ptimas para el sedentarismo de las comunidades aborigenes costeras (C6rdova y Arredondo 1988 Brito 2005) Peces y mariscos dominan la arqueologia costera de las Islas antillanas en tanto el consumo de recursos terrestres (mamfferos incluidos) aumenta progresivamente en la medida que los residuarios estudiados se alejan de la costa (Scudder 1991 Wilkins 2001) En las Antillas Mayores el componente terrestre de la dieta aborigen es solo 20-30 del material recobrado en los residuarios (Wing 1977 1989) Y debio ser menor alin porque el componente vegetal de la dieta no se ha preservado Por otra parte los aborigenes cubanos domesticaron la jutia conga y seguramente la reprodujeron para comer (Pose et al 1988) practica que probablemente se repitiera con otras jutias en las demas Antillas Mayores
- Fray Bartolome de las Casas hablo de 3 millones de indios en Cuba al inicio de la colonizacion pero los historiadores subsecuentes consideraron exagerada la cifra (Poey 1851 a) No obstante se dispone de dos estimados de la densidad de poblaci6n aborigen en las Antillas Mayores a la llegada de los europeos 18 indigenaslkm2 en Puerto Rico (Rouse 1956) y 1271km2 en La Espanola (Watts 1987) Extrapolando ambos valores entre las cuatro islas principales de las Antillas Mayores tenemos que en la fecha indicada la poblaci6n aborigen pudo estar entre 16 mil y 113 mil indigenas en Puerto Rico entre 21 mil y 145 mil en Jamaica entre 141 mil y 1 mill6n en La Espanola y entre 200 mil y 14 millones en Cuba Entre las cifras maximas y las minimas nos inclinamos
332 BIOGEOGRAFiA
hacia estas ultimas atendiendo a los altos indices de mortalidad infantil y morbilidad que se evidencian en las osamentas de los pobladores prehispanicos (Martinez y Rives 1990) Uno de los hechos que apuntan hacia el bajo numero de aborigenes en las Antillas Mayores es 10 rapido que desaparecieron despues de la llegada de los europeos pnkticamente sin dejar mestizaje con estos 10 cual contrasta ostensiblemente con la situacion en la America continentaL
En suma un grupo humano (1) demograiicamente irrelevante y controlado por altos indices de morbilidad (2) consumidor mayoritariamente de organismos marinos y (3) carente de la tecnologia cinegetica necesaria para poder sobreexplotar los mamiferos que Ie sirvieron de alimento (a los cuales accedio tardiamente en su desarrollo) no es concebible que haya causado e1 holocausto faunal que se Ie atribuye a partir del error de aplicar a las comunidades aborigenes antillanas un modelo cultural de la prehistoria continentaL
Tampoco hay evidencias de que los aborigenes hayan introducido carnivoros con suficiente capacidad depredadora salvo el perro Morgan y Otlenwaldder (1993) consideraron que la depredacion por perros durante el periodo amerindio es la explicacion mas plausible para la extincion del almiqui gigante de Cuba pero como se puede observar (Tabla 54) Solenodon arredondoi aparece en el intervalo II por falta de asociacion con evidencias culturales
Consecuentemente el accionar de los aborigenes y de sus animales domesticos directamente sobre la fauna no parece haber sido por sf solo un factor determinante de la extincion de mamiferos terrestres durante el periodo amerindio en las Antillas (pero vease Steadman et al 2005)
Hay que recordar en este senti do que alin en el Holoceno tardio ocurrieron cambios clirruiticos de consideracion (vease Causas Naturales) Por ejemplo Curtis et al (2001) explicaron que la extincion de algunos ani males mesofilicos de las alturas montanosas de La Espanola pudo haber antecedido a cualquier impacto humano sigrIificativo en esos lugares por coincidir dicha extincion con una tendencia a la aridez documentada isotopicamente y con evidencias de la perdida del bosque mesico ocurridas despues de 3kaAP Yen Jamaica hace solo 2shy3 mil aDos el area de la Bahia de Jackson sustentaba una gran biomasa de vertebrados en bosque hinnedo a juzgar por la cantidad y diversidad de taxones presentes en material fosil derivado de la depredacion por lechuzas hoy el area es la mas seca de la isla con matorral xerofitico y pobreza de fauna (MacPhee 1996a) Yalm en nuestros dias huracanes de gran intensidad aniquilan poblaciones de vertebrados (que no se recuperan) en is las pequenas y llanas del Caribe (Brooks y Smith 2001)
En medio de la incertidumbre sobre la verdadera causa (0 causas) de las extinciones de mamiferos en el periodo amerindio MacPhee y Marx (1997) han propuesto su hipotesis de la hipermorbilidad la cual estipula colapsos poblacionales relativamente rapidos con la introduccion de enfermedades de alta letalidad Estos autores han razonado que
Las islas son los lugares mas apropiados para experimentar perdidas catastroficas causadas por enfermedades Los mamiferos no solo serian susceptibles en virtud de su prolongado aislamiento biologico sino que en los confines de una isla la epizootia
333 EXTINCIONES DEL CUATERNARIO
tendria la potencialidad de infectar simultaneamente a la rnayoria de los individuos de vadas especies Los neonatos son inmunologicamente inmaduros y sufririan desproporcionadamente cualquier infeccion hipervirulenta La perdida excesiva de neonatos y juveniles tendria efectos severos para la sobrevivencia de la poblacion si esta se reduce por debajo de cierto nivel sobreviene la extincion porque no hay posibilidad de restitucion en el periodo postpanzootico Las especies de mayor talla corporal van a ser las mas vulnerables a la extincion dadas sus bajas tasas reproductivas largos periodos de gestacion y reducido numero de progenie Las especies de menor tamano --caracterizadas por altas tasas reproductivas periodos cortos de gestacion y mayor numero de neonatos por parto- tambien serian fuertemente diezmadas pero muchas sobrevivirian porque su estrategia reproductiva les permite perder mayor numero de individuos Las perdidas deben ocurrir relativamente rapido (a escala de decadas) mientras que otras hipotesis (digamos los cambios climaticos) permiten intervalos mucho mas largos (a escala de siglos 0 de milenios) entre la aparicion de las causas y la produccion de sus efectos Por ejemplo todo indica que los nesofontes se extinguieron muy rapidamente y seria dificil poder invocar un impacto humano directo 0 argumentar fenomenos climaticos que pudieran explicar plausiblemente perdidas tan rapidas
MacPhee y Marx (1997 191) han dicho que por el momento la hipotesis de la hipermorbilidad no pasa de ser una abstraccion en espera de adecuada sustentacion empirica Llama la atencion que linicamente en el periodo amerindio (Tabla 45 intervalo 1Il) las extinciones abarcaron todas las clases del peso corporal (de la a a laj) y que ninguna de las especies que sobrevivieron hasta el periodo postcolombino (Tabla 45 intervalo IV mas las especies vivientes) sobrepasa los 10 kg (clases del peso a b c) en clara coincidencia con los presupuestos de la hipotetica hipermorbilidad
Intervalo Postcolornbino
Por 10 menos 37 especies de mamiferos terrestres vivian en las Antillas Mayores y grupos de Islas perifericas a la lIegada de los europeos en 1492 De elIas se han extinguido 23 shymas especies que en cualquiera de los periodos precedentes (Tabla 45~ y solo subsisten 12 especies repartidas entre Cuba (8 especies) La Espanola (2) Jamaica (1) y Bahamas (1) varias de estas en peligro critico de extincion Solo dos de las especies extinguidas durante los ultimos 500 anos se conocieron en estado viviente los roedores Oryzomys antillarum de Jamaica (extinguido en 1877) y Geocapromys thoracatus de las Islas Swan (extinguido en 1955) Las causas de las extinciones ocurridas durante este periodo son sobradamente conocidas destruccion de habitats introducci6n de animales exoticos (domesticos y comensales) y demas presiones asociadas a la actividad humana entre las cuales posiblemente habria que incluir brotes de hipermorbilidad
Durante mucho tiempo se ha afirmado la posibilidad de que especies de Nesophontes hayan sobrevivido hasta las primeras decadas del siglo xx y que incluso continuen existiendo en lugares remotos de Cuba 0 La Espanola (Allen 1918 1942 Miller 1930 Aguayo 1950a Arredondo 1960b Fischer 1977 MacFadden 1980 Woods y
334 BIOGEOGRAFiA
Ottenwalder 1991) Actualmente no existe evidencia radiometrica de que alguna especie de Nesophontes haya sobrevivido al siglo xv las fechas radiocarbonicas calibradas obtenidas en muestras de Nesophontes de Cuba y La Espanola resultaron no mas tardias que los siglos XIII y XIV (MacPhee et al 1999) Por otra parte las expediciones efectuadas a fines del siglo xx con el proposito de localizar individuos vivientes del genero en La Espanola no produjeron la menor indicacion afirmativa al respecto (Woods et al 1985) Ademas a traves de las Antillas Mayores no aparecen restos de Nesophontes en los depositos formados actualmente por las lechuzas
EPiLOGO
Las Antillas Mayores son islas oceanicas no han formado parte de un continente (con la posible excepcion de una parte de Jamaica hasta el Eoceno Medio hace 45 Ma) y durante un tiempo geologico considerable (desde el Eoceno Superior hace 40 Ma) han permanecido emergidas y separadas de los continentes vecinos Consecuentemente los mamiferos terrestres (no volantes) autoctonos de estas islas constituyen una fauna oceanica que por ende cumple con todas las caracteristicas convencionales reconocidas para este tipo de entidad biogeografica
- Baja diversidad taxonomica a nivel de las categorias superiores (solo cuatro ordenes) en comparacion con islas plataformicas continentales (0 con areas continentales) de tamano y diversidad ecologica simi lares
- Alto grado de endemismo a nivel de genero (96) y familia (50) en respuesta al prolongado aislamiento
- Desbalance ecologico de la estructura trofica ausencia de mamiferos camivoros y ungulados
- Espectaculares procesos de irradiaci6n evolucionaria como resultado de la colonizacion de territorios libres de competidores y depredadores
Tendencias evolucionarias extremas gigantismo
- Control a cargo de aves rapaces con tendencia at gigantismo en ausencia de mamiferos miembros del orden Carnivora
- Mas especies de reptiles y de murcielagos que de mamiferos terrestres relaciones que en los continentes se invierten
Alta tasa de extincion de especies (84) sin reposicion
Preservacion de especies filogeneticamente arcaicas (los llamados fosiles vivientes) los almiquies

300 BIOGEOGRAFIA
TABLA 42 Escala convencional del tiempo geologico en la Era Cenozoic a y finales de la Mesozoica (Gradstein 2004) ka = kiloafios (miles de afios) Ma miJIones de afios AP = antes del presente
Periodo(Suhperiodo) Epoca Afios (aproximados) antes del presente
Era Cenozolca
Cuaternario Holoceno Pkistoceno
Presente - 11 kaAP 11 kaAP - 18 MaAP
Terciario (Ne6geno ) Plioceno Mioceno
18 - 5 MaAP 5 - 23 MaAP
oIigoceno 23 - 34 MaAP Terciario (pak6geno) Eoceno 34 - 58 MaAP
Pakoceno 58 - 65 MaAP
Era Mesozo ca
Creticico 65 - 145 -faAP
~~~------~~--- --~-
este grupo animal en la region con el hallazgo de un perezoso oligocenico en Puerto Rico (Yauco) as como un perezoso un mono y un roedor miocenicos en Cuba (Domo de Zaza) Esto unido a la presencia de huesos de una musarafia nesofontoide [referida erroneamente a Solenodon por Rivero (1997)] en ambar oligomiocenico de la Republica Dominicana (MacPhee y Grimaldi 1996) demuestra incontrovertiblemente que al menos en el Mioceno temprano (alrededor de 20 millones de afios atnis) ya estaban en las Antillas Mayores representantes de los cuatro linajes basicos de su fauna de mamiferos terrestres [La afirmacion de Abreu et al (1990 13) en cuanto a que se conocen restos fosiles de almiqui que datan de hace aproximadamente 40 millones de afios es erronea]
Pero lcuan antigua puede ser la fauna de mamiferos terrestres de las Antillas Mayores
Aunque es un hecho bien establecido por la geologia que desde finales de la Era Mesozoica y principios de la Cenozoica existieron islas en el area ocupada ahora por las Antillas Mayores la evidencia disponible indica asimismo que aquellas primeras islas sufrieron repetidas transgresiones marinas totales que habrian hecho sucumbir las biotas que pudieran haber sustentado (MacFadden 1981 Morgan y Woods 1986 MacPhee e Iturralde-Vinent 1994 Roca et al 2004) sin contar los catastroficos efectos que debio tener para el Caribe occidental el b6lido extraterrestre que impacto en Chicxulub (Yucatan) hace 65 Ma (Tada et ai 2003) Hedges (1996) ha especulado que aUn en medio de tal devastacion debe haber existido algun que otro lugar donde elementos de la fauna pudieron haber sobrevivido
301 POBLAMIENTO CONTINENTE-ISLA
TABLA 43 F echados pleistocenicos mcis tempranos (obtenidos por distintos metodos) en mamiferos terrestres del segmento insular delimitado en laFig 41 F =Fuentes (I) Jull et al 2004 (2) Goodfriend y Mitterer 1987 (3) MacPhee et al 1989 (4) Woods 1989b (5) Rega et aI 2002 (6) McFarlane 1999b (7) McFarlane 1 999a (8) McFarlane et al 1998a
Pals Localidad Taxon kaAP F
Cuba Breas de San Felipe Pilosa (Parocnus) 1l9a 1
Jamaica Cueva Coco Ree Cueva Wallingford Roadside
Primates indet Rodentia (Clidomys)
393 2530
2 3
Haiti Trouing Jeremie no5 (casimba) Rodentia (Rh izoplagiodontia) 212 4
Republica Dominicana
Cueva del Perezoso Pilosa (ficratocnus) 185 5
Puerto Rico Cueva de Ia Vaca Cueva Clara
Pilosa (Acratocnus) Soricomorpha (Nesophontes)
337 346
6 7
Anguila Cueva Cave-No-Cave Rodentia (Amblyrhiza) 2060 8
a Kulp etal (1952) reportaron un fechado radiocarbonico de gt 30000 afios para un cono de pino en el deposito fosilffero de Ciego Montero (Cuba) con restos de perezosos
(vcase tambicn Crother y Guyer 1996) pero Iturralde-Vinent y MacPhee (1999) han preguntado donde estaban esos lugares si es que existieron
Solo a partir del Eoceno Superior las Antillas Mayores en fonnacion han pennanecido emergidas hasta hoy [excepto parte de Jamaica y el sur de La Espanola que continuaron sumergidas hasta el Mioceno Medio (Iturralde-Vinent y MacPhee 1999)] Esto significa que solo durante los ultimos 40 millones de anos aproximadamente han existido pennanentemente en dicha region insular ambientes terrestres capaces de sustentar comunidades biotic as (MacPhee e Iturralde-Vinent 2000 Portell et ai 2001 MacPhee 2005) 10 cual proporciona el marco temporal de referencia para conceptuar la antigiiedad maxima posible de la fauna de mamiferos terrestres de las Antillas Mayores
POBLAMIENTO CONTINENTE-ISLA
AFINIDADES CONTINENTALES
El consenso de los autores que en las itltimas dccadas han tratado la filogenia y biogeografia de los mamiferos terrestres de las Antillas Mayores (MacFadden 1980 MacPhee y Woods 1982 Morgan 1985 1988 1993 Morgan y Woods 1986 Ford 1986 Woods 1988 1989a 1990 MacPhee e Iturralde-Vinent 1995a b Hedges 1996 Horovitz [1997] Ottenwalder 2001 Whidden y Asher 2001 MacPheeet ai 2003 Carlini y Scillatoshy
302 BIOGEOGRAFiA
Yane 2004 Davalos 2004 MacPhee y Horovitz 2004 Roca et al 2004) es practicamente unanime en cuanto a que los ancestros continentales que dieron origen a esta fauna derivaron (1) mayormente de Suramerica perezosos (Megalonychidae) monos (Pitheciidae) y roedores (Echimyidae Capromyidae y Heptaxodontidae) (2) en menor medida de Norteamerica musarafias (Nesophontidae y Solenodontidae) y (3) menos aun de Centroamerica una especie de roedor (Muridae)
HIPOTESIS BIOGEOGRAFICAS
La definicion c1asica reconoce dos tipos de islas isla continental (porcion del territorio de un continente separado de este por movimiento tectonico 0 por ascenso del nivel del mar) e isla oceanica (porcion de territorio que no ha formado parte de un continente) La definicion sigue siendo uti por 10 que implica para la formacion de las faunas insulares en uno u otro caso As en el caso de la isla continental su fauna ya estaba emplazada en la futura isla antes de producirse su separacion del continente [Vicarianza hipotesis biogeografica de fragmentacion faunal] en el caso de la isla oceanica su fauna es el resultado de la colonizacion1 por propagulos2continentales a traves de la barrera marina entre el continente y la isla [Dispersion hipotesis biogeografica de acumulacion faunal] En el primer caso el surgimiento de la barrera marina es resultado del evento vicariante en el segundo la existencia de la barrera marina es anterior al evento dispersante Ademas de operar a escala mayor (continente-isla) ambos eventos operan secundariamente a menor escala (isla-isla)
Las dos hipotesis -vicarianza y dispersion- se han disputado hasta hoy la preferencia de los biogeografos en tanto hipotesis mas viable para explicar el proceso de formacion original (continente-isla) de la fauna de mamiferos terrestres de las Antillas Mayores [Sobre la historia de las hipotesis biogeograficas relativas al Caribe insular vease RuB y Schubert 1989 Williams 1989 Pruna 2001] Yel debate continua debido ala falta de consenso sobre 10 que Williams (1989) ha llamado el problema central de la discusion a saber la ostensiblemente baja diversidad taxonomica de esta fauna a nivel de las categorias superiores (solo cuatro ordenes) en comparacion con la diversidad al mismo nivel de las faunas de islas continentales (0 de areas del continente) simi lares en tamafio y diversidad ecologica [Por ejemplo la fauna cubana de mamiferos terrestres consideradas de conjunto las especies vivientes y las extintas- es menos diversa a los niveles taxonomicos superiores (4 ordenes y 6 familias) que 10 que cabria esperar de la diversidad ecologica y extension territorial de Cuba (110 992 km2) comparada con una isla tipicamente continental como Trinidad (4828 krn2
) -veinte veces menor que Cuba y menos diversiticada ecologicamente- donde solo las especies vivientes de mamfferos terrestres representan 6 ordenes y 16 familias (Eisenberg 1989)] Este desbalance
lColonizaci6n Arribo y establecimiento de un propagulo en una isla no ocupada por la especie (MacArthur y Wilson 1967)
2Propagulo Niimero minimo de individuos de una especie capaces de reproducirse y aumentar poblacionalmente en una isla bajo condiciones favorables (MacArthur y Wilson 1967)
303 POBLAMIENTO CONTINENTE-ISLA
taxon6mico ha sido expJicado de dos maneras (1) Inicialmente hubo mas 6rdenes presentes pero la mayoria se extingui6 y s610 sobrevivieron esos cuatro [explicaci6n vicarianista] (2) S610 representantes de esos cuatro 6rdenes lograron cruzar la barrera marina [explicaci6n dispersionista]
Cinco hipot~ticos mecanismos de poblamiento continente-isla han sido invocados durante los ultimos 140 afios (y se los sigue proponiendo actualmente) para explicar la presencia de mamiferos terrestres en las Antillas Mayores A continuaci6n pasamos a resefiarlos brevemente (incluidas sus alegadas fortalezas y debilidades) en el orden aproximado en que historicamente fueron propuestos
EL PUENTE NATURAL (dispersion terrestre activa)
En el siglo XIX el puente terrestre era pnkticamente la unica hipotesis para explicar las distribuciones disyuntas de animales en buena parte del mundo y los primeros investigadores que se ocuparon de la biogeografia antillana (Fernandez [1865] 1884 Chapman [1893] Torre 1892 Lora 1895) la asumieron resueltamente en parte intrigados por el descubrimiento en Cuba en la segunda mitad del siglo de un perezoso f6sil de gran talla Y en su aplicacion a los mamiferos terrestres de las Antillas Mayores el puente sigui6 siendo a 10 largo del siglo xx la hipotesis mas favorecida por los estudiosos del tema (Vaughan 1902a Spencer 191 Oa b Torre 1910a b cAllen 1911 1917a 1918 Barbour 19141916 1945bAnthony 1918 1926 [1942] Scharff 1922 Kraglievich 1926 Beaufort 1951 Arredondo 1961 1970b Kartashov y Mayo 1974 Iturralde-Vinent 1982 1988 Arredondo y Armifiana 1996 Arredondo 1996a [1999]) quienes postularon la existencia preterita de puentes con Norte- Centro- 0 Suramerica en dependencia de las afinidades continentales de los mamiferos antillanos
Teoricamente el puente terrestre debe cumplir con una condicion esencial permitir el paso de una parte mayoritaria (y por tanto representativa de la diversidad) de la fauna continental Claro que siempre habria tipos de mamiferos menos proclives a dispersarse o para los cuales la ubicaci6n del puente resultaria inaccesible 0 las condiciones ecologicas en este fueran desfavorables pero si algunos acceden al puente no hay raz6n para pensar que el resto de esa fauna no los acompafie mayoritariamente Como dijera Simpson (1956) Los puentes no transportan faunas desbalanceadas
En consecuencia el puente terrestre como filtro es un mecanismo de poblamiento minimamente selectivo y por ende de alta diversidad inicial (sensu Iturralde-Vinent y MacPhee 1999) 10 esencial del puente es su capacidad para transportar faunas integradas (Williams 1989) Yen eno radica la objeci6n fundamental al puente terrestre surgida tempranamente en el siglo xx (Por que s610 cuatro 6rdenes de mamiferos terrestres en las Antillas si vinieron por puentes (Donde estan los fosiles de los demas ordenes que debieron venir si es que se extinguieron Los partidarios del hipotetico puente dispusieron de casi todo el resto del siglo para responder una y otra vez que otros 6rdenes de mamiferos aparecerian representados en las Antillas Mayores el dia que en estas Islas se descubrieran fosiles precuatemarios suficientemente
304 BIOGEOGRAFIA
antiguos Esto se logro por fin terminando el siglo como se explico anteriormente (vease Contexto temporal) y ique se encontro los mismos cuatro ordenes de mamiferos terrestres conocidos del Cuatemario caribefio aparentemente los Unicos que lograron llegar
Pero ademas hasta el presente no se ha demostrado inequivoca- y concluyentemente que en algun momenta durante los ultimos 40 millones de afios haya existido una conexion terrestre continua entre las Antillas Mayores (en proceso de formacion 0 plenamente formadas) y algun punto de las regiones continentales vecinas
LA BALSA NATURAL (dispersion marina pasiva)
Durante buena parte del siglo xx el puente terrestre tuvo como unico rival a la hipotesis de la balsa natural -transporte accidental de animales sobre islotes de vegetacion flotantes a la deriva despues de arrancados y expulsados mar afuera por un rio crecido- expuesta originalmente por Matthew (191319151916 1918a 1919) y desarrollada en su aplicacion a los mamfferos terreslres antillanos principal mente por Simpson (1956) asi como por olros investigadores (Darlington 1938 1957 Williams y Koopman 1952 Hershkovitz 1958 1972 Patterson 1962 Paula 1967 Patterson y Pascual 1972 Pregill 1981 Ford 1986 Woods 1989a 1990 Morgan 1993) Por su parte Hedges et al (1992) sustentaron dicha hipotesis en estimados de divergencia (basados en el reloj molecular) en la herpetofauna antillana -involucrados tangencia1mente los mamiferos [veanse objeciones a Hedges y colaboradores (Page y Lydeard 1994 Iturralde-Vinent y MacPhee 1999) respuestas a las objeciones (Hedges et al 1994 Hedges 2001) y contrarrespuestas (MacPhee e IturraldeshyVinent 2005) asi como cuestionamientos al metoda de los estimados de divergencia (Foote et al 1999 Pregill y Crother 1999 Woods et ai 2001 )]
La hip61esis de la balsa natural se origino a partir de las numerosas ocasiones en que a 10 largo de siglos tales islotes flotantes (portando 0 no vertebrados incluidos mamiferos) han sido avistados mar afuera a cientos de kilometros de la costa mas cercana en varias partes del mundo --inc1uido el Mar Caribe- segUn consta en diarios de barcos y reportes de observadores diversos (revisados por King 1962 reportes posteriores en SICSP 1970 y Censky et al 1998) Uno de estos casos acaecido en nuestra region se reporta en un informe del SICSP (Smithsonian Institution Center for Shortlived Phenomena 197050shy52)
EIB de julio de 1969 un destructor de la armada de Estados Unidos avisto un islote flotante a unos 100 km al sur de la Base Naval de Guantanamo moviendose desde el Paso de los Vientos en direccion suroeste a una velocidad de 45 kmlh islote de unos 15 m de diametro mantenfa en pie mas de 10 palmas reales de 10-15 m de altura con suelo cubierto de hierba y arbustos [la observacion con binoculares a 18 km de distancia no preciso la presencia de vida animal] La travesia fue rastreada durante 6 dfas hasta que el 19 de julio el islote se dio por perdido tras 6 horas de busqueda aerea
305 POBLAMIENTO CONTINENTE-ISLA
La colonizaci6n exitosa de una isla de las Antillas Mayores por mamiferos terrestres continentales transportados en balsas naturales a la deriva (de cada especie al menos una hembra gravida 0 dos individuos de sexo opuesto) dependeria de factores que van a operar desde el momenta mismo de formarse la balsa asi como durante su travesia marina y su arribo a la isla
a) La posibilidad de estar en la balsa Que la distribuci6n de la especie incluya areas contiguas al rio y que el animal se encuentre en ellugar y el momenta propicios para ser incorporado a la balsa antes de que el rio crecido la desprenda Por ejemplo segun Hershkovitz (1958) los equimidos -grupo ancestral de roedores antillanos- serian buenos candidatos a ser captados por la balsa porque acostumbran a anidar en huecos del tronco de los arboles a la orilla de los rios
b) El tamano y la consistencia de la balsa Muchas se desintegran en alta mar debido a su fragilidad ante la saturaci6n Algunos autores (Beaufort 1951 Arredondo 1996a [1999]) han expresado incredulidad en cuanto a que las balsas hayan podido soportar el peso de un animal como el gran perezoso cubano (Megalocnus rodens) el mayor de los mamiferos antillanos Pero esta especie evolucion6 en Cuba y el ancestro que hizo el viaje pudo ser muy pequeno [EI perezoso oligocenico de Puerto Rico es el mas pequeno de los megaloniquidos antillanos (MacPhee e Iturralde-Vinent 1995b )]
c) Las caracterfsticas fisiol6gicas de la especie Los limites fisiol6gicos de los propagulos constituyen nuestra unica objeci6n a la dispersi6n marina por balsas como mecanismo para la formaci6n de la fauna de vertebrados en las Antillas (MacPhee e IturraldeshyVinent 2005) Unas especies tienen intrinsecamente mas posibilidades que otras para sobrevivir a las vicisitudes que deben afrontar durante la travesia (deshidrataci6n inanicion insolaci6n etc) Por ejemplo los megaloniquidos han demostrado tener extraordinaria habilidad para cruzar barreras marinas fueron los primeros mamfferos suramericanos en invadir Norteamerica mucho antes de que surgiera la conexion terrestre a finales del Plioceno (Paula 1967 Olson 1978 Webb 1985b Martin y Steadman 1999) Incluso se conoce un megaloniquido acultitico del Plioceno peruano (Muizon y McDonald 1995) y otro viviente en Suramerica (Choloepus) que pnicticamente no bebe agua 10 que ha originado una leyenda aborigen que explica por que los perezosos no toman agua (Goff art 1971) Vease ademas el inciso e
d) EI regimen de las corrientes marinas Que conduzcan a la balsa en la direcci6n necesaria EI transporte de balsas de vegetacion desde los continentes hacia las islas fue durante la mayor parte del Cenozoico sumamente improbable (MacPhee e IturraldeshyVinent 2000 148) Estos autores han insistido en que hay que tener en cuenta que durante la cambiante geografia cenozoica del Caribe las corrientes marinas de superficie no favorecieron el desplazamiento de objetos fiotantes hacia las Antillas Mayores [Pero no sabemos con certeza que efecto modificador del curso de tales objetos fiotantes pudieron introducir las tormentas tropicales si previsiblemente funcionaron en el pasado como 10 hacen ahora] Dichos autores senalaron ademas (2005) que en un experimento reciente con boyas de posicionamiento satelital liberadas a la deriva desde las Antillas Menores las boyas fueron a parar mayoritariamente a Centroamerica y a otros lugares
306 B[OGEOGRAFIA
y solo unas pocas recalaron en las Antillas Mayores y las que entraron al Mar Caribe demoraron semanas 0 meses en arribar
e) La distancia a recorrer (y en consecuencia la duracion de la travesia) entre el continente y la isla (10 que repercute decisivamente en el factor c) Este es desde luego uno de los factores mas criticos En la medida que la distancia aumenta la probabilidad de alcanzar la isla disminuye drasticamente con el agravante de que la trayectoria dificilmente seria en linea recta Si tomamos como referencia la velocidad de 45 kmlh del islote ya mencionado (SICSP 1970) cubrir la distancia aerea (aproximada) de 800 km que media entre el noroeste de Suramerica y La Espafiola Ie tomaria a la balsa 74 dias Estimando el doble de este tiempo debido a desvios de la trayectoria la duracion de la travesia probablemente resultaria excesiva para la mayoria de los mamiferos Pero pudiera muy bien ocurrir que la velocidad fuera mayor 0 que la balsa portara plantas vivas que permitirian a los fitOfagos sobrevivir [Tres de los cuatro ordenes de mamiferos terrestres conocidos de las Antillas Mayores son fitOfagos] Ademas esta geologicamente bien establecido que durante etapas de variada duracion desde el Oligoceno hasta el Pleistoceno los territorios del Caribe insular y continental aumentaron considerablemente en extension con niveles del mar 100-200 m por debajo de su nivel actual oportunidades que los propagulos aprovecharian para cruzar barreras marinas significativamente menores
f) El tamafio de la isla Las islas mayores deben interceptar mas balsas (Buckley y Knedlhans 1986) 10 que repercute directamente en el factor e La balsa puede pasar entre islas pequefias y continuar viaje sin llegar a ningun lugar
g) Las condiciones (favorables 0 adversas) del desembarco en la isla y el tiempo que Ie tomaria a uno 0 mas animales depauperados encontrarun habitat apropiado Esto dependeria a su vez del tamafio y diversidad ecologica de la isla (Darlington 1938 Miller 1966)
Tal cumulo de posibles obstaculos y dificultades constituye a primera vista un desafio a la credibilidad de la hipotesis Matthew-Simpson pero en ello radica precisamente su valor explicativo bas ado en la mas absoluta improbabilidad La colonizacion por la via marina es altamente improbable pero no absolutamente imposible si se tiene en cuenta la inmensidad del tiempo geologico se vuelve probable (McKenna 1973) Basta que solo una vez entre mill ones de intentos fallidos durante ese inmenso lapso algunos de los factores anteriormente enunciados coincidan en no ser tan desfavorables para el propagulo Y eso es todo 10 que demanda la hipotesis para cada uno de los grupos monofileticos mayores de mamiferos terrestres presentes en las Antillas Mayores Las Antillas Mayores requeririan el arribo de tan solo 7 (quizas menos) propagulos fundadores a 10 largo de los ultimos 30 0 mas millones de afios (promedio de uno cada 43 mill ones) para explicar la formacion de toda su fauna de mamiferos terrestres por dispersion marina pasiva (vease Discusion y Conc1usiones)
En comparacion con la via marina el poblamiento por la via terrestre es predecible porque la mayor parte de la fauna continental de mamiferos emplazada en la region del puente va a cruzar por este una vez que el mismo se establece y los animales logran
307 POBLAMIEITO CONTINENTE-ISLA
acceder a el el poblamiento por la via marina en cambio es impredecible porque depende total mente del azar que caracteriza a la balsa Claro que en ambas vias opera el azar pero en el primer caso se trata del azar de un evento probable en el segundo el azar de un evento improbable (Simpson 1940) Asi el puente natural (mecanismo de poblamiento minimamente selectivo = alta diversidad inicial) y la balsa natural (mecanismo de poblamiento maximamente selectivo baja diversidad inicial) pueden conceptuarse como los extremos opuestos de una gama de probabilidades de dispersi6n (McKenna 1973) EI fen6meno de la dispersi6n merece la mayor atenci6n cientifica en los estudios de biogeografia insular (Cowie y Holland 2006)
Queda claro que la balsa natural es una hip6tesis consistente con el caracter de la fauna objeto de estudio puesto que el desbalance taxon6mico (sin evidencia paleontol6gica en contrario) s610 puede ser resultado de la improbabilidad del poblamiento No obstante ello no implica necesariamente que la balsa natural haya sido el imico mecanismo (ni siquiera el principal) utilizado por los mamiferos terrestres en su colonizaci6n de las Antillas Mayores La balsa natural debe satisfacer una premisa te6rica esencial dado el caracter azaroso impredecible e improbable de una colonizaci6n exitosa esta debe ocurrir a intervalos muy espaciados a traves del tiempo geol6gico 0 sea durante todo el tiempo que en el area ocupada por las Antillas Mayores ha habido ecosistemas terrestres perrnanentemente disponibles para ser colonizados (los ultimos 40 millones de afios) Sin embargo como sabemos representantes de los cuatro 6rdenes de mamiteros terrestres conocidos de las Antillas Mayores ya estaban emplazados en dicho territorio insular en el Mioceno Inferior 10 que supone un lapso de colonizaci6n limitado al Eoceno SuperiorshyOligoceno (MacPhee 2005) sugerente de un tipo de mecanismo contrario a la colonizacion alocronica exigida por la hipotesis de la balsa natural sobre todo tratandose de una fauna caracterizada por altas tasas de extincion sin reposicion aparente
La presencia en Jamaica (hasta su extinci6n en 1877) del roedor mfuido Oryzomys antillarom con estrecha afinidad centroamericana (Woods 1989a) pudiera constituir el imico caso efectivo de poblamiento (continente-isla) de las Antillas Mayores por dispersion marina dc un mamifero terrestre para el cual no es razonable invocar otro mecanismo de colonizacion
LA GUIRNALDA INSULAR (dispersion mixta)
La geologia ha establecido con suficiente certeza que durante el Cenozoico unicamente en el Pale6geno tardio y solo a traves de la Cresta de Aves (ahora sumergida excepto la isla hom6nima) pudo haber existido una cadena de islas para el transito de mamiferos terrestres entre el norte de Suramerica y las proto-Antillas Mayores [desestimados a ese efecto el Canal dc Yucatan el Elcvado de Nicaragua la Cresta de Caiman el Elevado de Beata y las Antillas Menores (Holcombe et al 1990 MacPhee e IturraldeshyVinent 1994 1995b Iturralde-Vinent y MacPhee 1999)] Holcombe y Moore (1977) y Holcombe y Edgar (1990) postularon la existencia de esta entidad y estimaron algunas de sus posibles caracteristicas Por su parte MacPhee e Iturralde-Vinent (1995b) e Iturralde-Vinent y MacPhee (1999) precisaron los extremos proximal y distal de esa
308 BIOGEOGRAFIA
estructura acotaron la duracion y la edad de la misma y Ie dieron nombre GAARlandia (Greater AntillesA vesRidgelandia)
Segun Iturralde-Vinent y MacPhee (1999) la Cresta de Aves emergio por breve tiempo (1-2 Ma) durante la transicion Eoceno-Oligoceno (35-33 MaAP) por la coincidencia de un levantamiento tectoni co y un considerable descenso eustatico del nivel del mar En ese momenta las islas septentrionales del arco de las Antillas Mayores (Cuba centroorientalshyLa Espafiola - Puerto Rico - Islas Virgenes) constituian una sola isla grande 0 una serie de islas separadas por barreras muy estrechas La Cresta de Aves emergida entre el bloque Puerto Rico - Islas Virgenes y un microcontinente noroccidental de Suramerica -separado en ese momento del resto del continente por una via marina surgida en el Eoceno tardio- dio origen a GAARlandia
Obviamente el numero de colonizaciones exitosas por mamiferos terrestres a traves de una guimalda de Islas tiende a disminuir mientras mas anchas sean las barreras marinas entre las islas que forman la guimalda y cuando esto ocurre disminuye tambien la diversidad taxonomica de los propagulos particularmente a nivel de las categorias superiores (Simpson 1956)
Vemos asi que para los mamiferos terrestres la colonizaci6n a traves de una guimalda insular puede ser tan improbable como a la deriva en una balsa natural en dependencia de la cantidad y anchura de las barreras marinas a cruzar el tamafio y la fisiografia de las islas que integran la guimalda y el regimen de las corrientes marinas y aereas entre otros factores
Resumiendo la guimalda insular puede conceptuarse como un mecanismo de poblamiento altamente selectivo (= baja diversidad inicial) que combina la improbabilidad de la balsa natural con la colonizacion restringida temporaimente del puente terrestre
En su reciente revision de la taxonomia distribuci6n y filogenia (basada en estudios moleculares) de varios grupos de mamiferos antillanos Davalos (2004) concluyo que los perezosos megalocninos los primates y los roedores histricognatos se avienen al patron de divergencia con el continente y epoca de arribo a las Antill as implicados en la hipotesis de GAARlandia pero no asi los perezosos coloepodinos que requieren una explicacion biogeognlfica distinta Esto ultimo (sospechado por Matthew 1918a) habla sido confirmado por White y MacPhee (2001 226-227)
Nuestra evidencia filogenetica no sustenta la idea de un dado unificado surgido de la invasion de un megaloniquido el analisis sugiere mas bien dos eventos de colonizaci6n independientes [y] sustenta fuertemente un origen difiletico de las subfamilias [Choloepodinae y Megalocninae] Si hubo dos migraciones a traves de GAARlandia presumiblemente poco espaciadas en el tiempo no se puede determinar en el presente
Finalmente queda claro que GAARlandia -como guirnalda insular- es la hipotesis mas robusta propuesta hasta ahora para explicar la presencia en las Antillas Mayores de
309 POBLAMIENTO CONTINENTE- ISLA
la absoluta mayona de los mamfferos terrestres de ongen suramericano porque el caracter de esa fauna supone un mecanismo de poblamiento ancestral que sea a la vez maximamente selectivo (baja diversidad ordinal) y temporalmente restringido (transicion Eoceno-Oligoceno)
Recientemente Rega et al (2002 18) expresaron A pesar de una revision exhaustiva de la paleogeografia de la cuenca del Caribe Iturralde-Vinent y MacPhee (1999) no pudieron confirmar la presencia de una conexion terrestre continua entre Suramerica y las proto-Antillas aunque hipotetizaron su existencia [Vease la Fig 42 (= figura 6 de Iturralde-Vinent y MacPhee 1999 simplificada))
En el trabajo citado por Rega y colaboradores aparece la siguiente explicacion
Actualmente no es posible ofrecer reconstrucciones paleogeograficas detalladas de la Cresta de Aves Sin embargo hay evidencia de que porciones de la cresta deben haberse elevado subaereamente en un tiempo La cuestion cntica es si la Cresta de Aves emergida pudo haber formado algun tipo de corredor entre las Antillas Mayores y el noroeste de Suramerica Perforaciones en las alturas estructurales de la cresta pudieran proveer el registro del Paleogeno tardio que se necesita para precisar la hipotesis del corredor aunque no se puede predecir si tales datos serian suficientemente detallados para determinar si en algun momento GAARlandia constituyo una sola entidad subaerea Por ejemplo pudo haber barreras marinas por corto tiempo entre las masas terrestres de la Cresta de Aves [0) ellevantamiento y la subsidencia pudieron afectar diferentes partes de GAARlandia en diferentes tiempos AItemativamente los elementos faunales pudieron haberse dispersado de manera pulsada Movimientos episOdicos de este tipo incrementarian la extincion de los elementos faunales incapaces de trasladarse a la siguiente masa terrestre en la cadena disminuyendo asi la diversidad total (Iturralde-Vinent y MacPhee 199957)
La primera parte de la explicacion deja claro que la posibilidad de que la Cresta de Aves haya sido una conexion terrestre continua no se puede descartar pero tampoco se puede afirmar La ultima parte de la explicacion describe el efecto altamente selectivo (filtrante) de la colonizacion a traves de una cadena de islas
Y continuan lturralde-Vinent y MacPhee (199957 -58) explicando
Hasta que punto el microcontinente noroccidental de Suramerica [considerado por dichos autores como el area suministradora de propagulos hacia GAARlandiaJ estuvo fisicamente separado del resto del continente no ha sido plenamente esc1arecido Si estuvo sustancialmente aislado por barreras marinas y orogenic as durante el Paleogeno tardio entonces solo la fraccion de la biota suramericana emplazada en el microcontinente estuvo en posicion de acceder a GAARlandia Esto pudiera tener que ver con el hecho de que la fauna antillana carezca de representantes de muchos grupos suramericanos [de mamfferos) que debieron haber hecho el viaje
310 BIOGEOGRAFiA
Florida o Alturas0 0
d 6 D L1anuras25deg
Yucatan
20deg
deg0
10deg
MARCARIBE
80deg 75deg
Bahamas
Cres ta de Beata
D Mares someros
D Mares profundos
OCEANO ATLANTICa
FIGURA 42 Reconstrucci6n hipotetica de GAARlandia como conexi6n terrestre continua entre las proto-Antillas Mayores y el noroeste de Suramerica en el Pale6geno tardio segun la FIgura 6 (simplificada) de Iturralde-Vinent y MacPhee (1999)
En su defensa del puente terrestre Barbour (1945b249) se refiri6 a esto - antes 10 habfa hecho Anthony [1942363]- en los terminos siguientes
El argumento de que si las Antillas hubieran estado conectadas con el continente se encontrarian los f6siles de un numero mayor de distintas criaturas me deja frio es perfectamente concebible que la conexi6n haya ocurrido en un periodo en que la parte del continente implicada no estuviera poblada por una fauna grande y variada
Y concluyen lturralde-Vinent y MacPhee (199958) su detallada explicaci6n
Desafortunadamente sabemos muy poco sobre la composici6n faunal del noroeste de Suramerica a finales del Pale6geno
Consecuentemente si se confirmara la magnitud del aislamiento del aludido microcontinente y su concomitante baja diversidad faunal GAARlandia como conexi6n terrestre continua serf a la hip6tesis mas parsimoniosa No obstante mientras no se disponga de evidencia concluyente a este respecto hipotetizar a GAARlandia como conexi6n continua (minima selectividad) la coloca en franca vulnerabilidad te6rica al tener que afrontar la objeci6n clasica al puente terrestre (baja diversidad ordinal de la
311 POBLAMIENTO CONTINENTE-TSLA
fauna) al tiempo que se la priva de ser la hip6tesis mas robusta para explicar -como guimalda de islas- la colonizaci6n de las Antillas por mamiferos procedentes de Suramerica
ELARcA DE NOE (seudovicarianza)
El primer vicarianista antillano (desde luego sin el saberlo) fue Corral (1940) con su utilizaci6n ingenua de las ideas wegenerianas de la deriva continental aparecidas la decada anterior Pero el primer (supuesto) intento de aplicar a la formaci6n de la biota caribefia el mecanismo de la vicarianza basada en la tearia de la tect6nica de placas (surgida en la sexta decada del pas ado siglo) se debe a Rosen ([1976] 1985)
Segun Rosen las Antillas Mayores se originaron en el Cremcico en la Placa Caribe aproximadamente en la posici6n que hoy dia ocupa Centroamerica y al ser desplazadas tectonicamente hacia el este interactuaron (de algun modo no precisado por Rosen) con las margenes continentales de Norte- y Suramerica de donde (en un evento unico) adquirieron toda su fauna transportada subsecuentemente por dichas islas en su transito hacia sus posiciones actuales [De ahi la denominacion metaforica dada par McKenna (1973) a este mecanismo de poblamiento] Aunque se ha presentado esta propuesta como Model0 de Vicarianza Continente-Isla de Rosen en realidad por definici6n no constituye un evento vicariante porque las proto-Antillas Mayores no resultaron del fraccionamiento y separaci6n por tectonismo de una porcion del territorio continental como exige esta hip6tesis (vease definici6n de vicarianza al comienzo de Hipotesis Biogeograficas)
El Arca de Noe de Rosen ha sido desestimada por la mayoria de los autores contemporaneos porque (1) carece de sustento geol6gico (lturralde-Vinent y MacPhee 1999) y porque (2) en el periodo en que se afrrrna ocum6 el evento muchos de los ordenes modemos de mamiferos placentarios attn no existian (Williams 1989)
EL BARCO FUNERAL VlKINGO (tanatovicarianza)
Domning et al (1997) reportaron 10 que consideraron el mamffero terrestre mas viejo de las Antillas --circunstancia que se refleja en el titulo mismo del trabajo Oldest West Indian Land Mammal- al descubrirse un perisodactilo rinocerotoide (Hyrachius sp) en una localidad de edad Eoceno Inferior a Medio en el occidente de Jamaica
Jamaica ha tenido una historia geologica y paleogeografica distinta de la historia comun a las restantes Antillas Mayores A principios del Cenozoico Jamaica occidental (a diferencia de la porcion oriental constituida por el Bloque de las Montafias Azules) formaba parte de Centro america de la cual se desgaj6 (por el desplazamiento de la Placa Caribe) para venir a integrarse despues del Eoceno Medio con 10 que serfa la futura isla de Jamaica desaparecida por transgresi6n marina en el Eoceno Superior hasta volver a emerger totalmente en el Mioceno Medio (Iturralde-Vinent y MacPhee 1999 MacPhee e Iturralde-Vinent 2000)
312 BIOGEOGRAFiA
Por consiguiente Hyrachius ---conocido de localidades de edad Eoceno Inferior a Medio en Norteamerica- debio ocupar (10 que iba a ser) Jamaica occidental mucho antes de que esta se separara de Centroamerica De hecho este fosil Uego a su destino insular contenido en las rocas de su basamento no como un ser vivo por 10 que es erroneo considerarlo un animal antillano (lturralde-Vinent y MacPhee 2004) como han supuesto algunos (Hedges 200 I Portell et al 200 I Condis [200 I D Todo 10 que hizo fue viajar tanatovicariantemente en su barco funeral vikingo -otra de las denominaciones metaforicas de McKenna (1973)
DISCUSION Y CONCLUSIONES
Hemos visto que solo disponemos de una hipotesis plausible para explicar la mayor parte del poblamiento (continente-isla) de las Antillas Mayores por mamiferos terrestres del Neotropico continental la dispersion mixta (a traves de GAARlandia) para los perezosos (excepto Choloepodinae) los monos y los roedores (excepto Oryzomys)
Por otra parte la historia biogeogrMica de las musaraiias de las Antillas Mayores (Nesophontes y Solenodon) constituye una de las situaciones mas enigmaticas entre los mamiferos de la region (Ottenwalder 2001321) Al menos cinco mecanismos de poblamiento continente-isla han sido invocados para explicar cuando como y de donde llegaron a las Antillas Mayores estas musaraiias
I EI propagulo ancestral (del Terciario de Norteamerica) coloniz6 a las Antillas Mayores por dispersion marina desde Centroamerica 0 desde la Florida (Patterson 1962)
2 El propagulo ancestral ( del Terciario) cruzo el Atlantico Sur (supuestamente mas estreshycho en aquella epoca) directamente de Africa a las Antillas Mayores (Hershkovitz 1972)
3 El propagulo ancestral (del Terciario de Norteamerica) Uego a las Antillas Mayores en el Arca de Noe de Rosen (MacFadden 1980)
4 El propagulo ancestral (del Terciario de Norteamerica) fue transportado a las Antillas desde Centroamerica en el bloque de Jamaica occidental y de alli paso por dispersion marina a las restantes Antillas Mayores (Whidden y Asher 200 I)
5 El propagulo ancestral (del Terciario de Norteamerica) lleg6 a las Antillas Mayores a traves de GAARlandia (Whidden y Asher 2001)
Aunque hay consenso entre estos (y otros) autores sobre el origen norteamericano de las musaraiias antillanas ninguno de los cinco hipoteticos mecanismos de poblamiento resiste la evidencia en contrario filogenetica yo paleogeogrMica Ademas algunas de estas hipotesis asumen que Nesophontidae y Solenodontidae comparten un antecesor comun en las Antillas pero esta relacion no ha sido confirm ada Para dilucidar el enigmatico caso de las musaraiias antillanas se requerini profundizar en la investigaci6n geologica paleogeognifica y paleontol6gica (Iturralde-Vinent y MacPhee 1999)
De acuerdo con el conocimiento filogenetico disponible sobre el conjunto de la fauna la colonizacion original de las Antillas Mayores por mamiferos terrestres implicaria el arribo
313 POBLAMIENTO ISLA-ISLA
de solo siete propagulos fundadores perezosos 2 (coloepodinos y megalocninos si como parece tienen un origen difiletico) musaraiias 2 (nesofontidos y solenodontidos si no constituyen una misma familia como consideran algunos) monos 1 (pitecidos) roedores histricognatos 1 (equimidos capromidos y heptaxodontidos) roedores muridos 1 (Oryzomys en Jamaica)
POBLAMIENTO ISLA-ISLA
La distribucion de los mamiferos terrestres a traves del area antillana demarcada al inicio del presente capitulo (vease Contexto Geogratico Fig 41) ha sido interpretada en funcion de dos hipoteticos mecanismos de poblamiento interinsular la dispersion y la vicarianza fundamentadas como se resumen seguidamente
DISPERSION
De acuerdo con la evidencia paleogeogratica mas actualizada en las principales regresiones marinas ocurridas en la cuenca caribeiia durante los ultimos 23 Ma (transiciones OligocenoshyMioceno Mioceno Superior - Plioceno y varias veces en el Pleistoceno) las masas terrestres constitutivas de la region insular en formacion aumentaron considerablemente su extension territorial disminuyendo por tanto la distancia entre elias oportunidades que los elementos faunales aprovecharian para dispersarse de unas islas a otras
Durante mucho tiempo esta ha sido la hipotesis mas favorecida por los estudiosos del tema a partir de Simpson (1956) pero en decadas recientes ha suscitado dos objeciones
1 Es dificil en tender como pudo operar la dispersion marina entre estas islas puesto que ninguna de las Antillas Mayores posee rios suficientemente caudalosos para transportar masas significativas de vegetacion flotante (MacPhee 198432)
Es oportuno recordar aqui el islote ala deriva (con mas de diez palmas reales en pie) entre las tres Antillas Mayores mas occidentales mencionado anteriormente (vease La Balsa Natural)
2 La dispersion marina es menos efectiva para explicar la asombrosa similitud de las faunas de mamiferos terrestres a traves de las Antillas Mayores septentrionales porque como proceso aleatorio no debe producir distribuciones uniformes a 10 largo de una cadena de islas 0 si las produce entonces el proceso no es aleatorio (MacPhee e lturralde-Vinent 2000 149)
Independientemente de que las aludidas distribuciones taxonomicas estan lejos de ser uniformes 0 asombrosamente similares la hipotesis de la dispersion marina contrariamente a su canicter aleatorio y altamente improbable en el caso del poblamiento continente-isla- en el caso del poblamiento isla-isla es un proceso no aleatorio (aprovecha las oportunidades brindadas puntualmente por los periodos de maxima regresion marina) y ademas altamente probable dada la incomparablemente menor distancia entre las islas caribeiias que entre estas y los continentes vecinos
314 BIOGEOGRAFiA
V ICARIANZA
La neotectonica y las transgresiones marinas del Oligoceno Medio y del Mioceno afectaron sustancialmente la paleogeografia de la Cresta de las Antillas Mayores resultando la subdivision gradual de las partes subaereas de la Cresta Asi el este de Cuba y el norte de La Espanola conectados fisicamente durante el Oligoceno Inferior se separaron al formarse el Paso de los Vientos en el Oligoceno tardio en tanto La Espanola y Puerto Rico probablemente permanecieron conectadas hasta el Mioceno tardio en que surgio el Paso de Mona (Iturralde-Vinent y MacPhee 1999)
Tales eventos geologicos habrian dividido las distribuciones de los grupos faunales emplazados previamente en el territorio continuo de las tres islas y a partir de esas disyunciones dichos autores (tambien MacPhee et al 2000 y White y MacPhee 2001) asumieron --como segunda fase del modelo de GAARlandia- la hipotesis de la vicarianza isla-isla para explicar la distribucion multiinsular de varios grupos de mamiferos terrestres a traves de las Antillas Mayores septentrionales De acuerdo con esta hipotesis cualquier diferencia en composicion faunal entre las islas (por ejemplo la ausencia de roedores heptaxodontidos en Cuba 0 de monos platirrinos en Puerto Rico) constituye evidencia de extincion local postvicariante y no de dispersion marina fallida Lo primero se puede demostrar si aparecen los fosiles pertinentes 10 segundo no se puede demostrar (MacPhee e Iturralde-Vinent 1994) Por otra parte IturraldeshyVinent y MacPhee (1999 57) han reconocido que aunque fa vicarianza isla-isla puede representar una solucion clara para las distribuciones de varios taxones cuaternarios no puede explicar todos los casos
Rega et al (2002 18) han presentado dos objeciones al modelo de la vicarianza isla-isla aplicado a los mamiferos de las Antillas Mayores
I Si la separacion de Cuba y La Espanola precedio en varios mill ones de anos la separashycion de La Espanola y Puerto Rico el modelo vicariante de lturralde-MacPhee debe predecir mayor similitud entre las faunas de La Espanola y Puerto Rico que entre las de Cuba y La Espanola De hecho es al reyes
No obstante MacPhee et al (200335) admitieron despues que la fecha mas temprana empiricamente defendible [para la apertura del Paso de Mona] debe ser el Oligoceno Medio esto invierte la secuencia de ambas disyunciones con 10 cual queda resuelta la aparente contradiccion Sin embargo en su reciente evaluacion de la congruencia entre filogenia e hipotesis geologic as en varios grupos de mamiferos terrestres de las Antillas Mayores Davalos (2004373) encontro que ninguno de los taxones cumple con la fecha y el patron de divergencia entre las masas terrestres antillanas postulados por el modelo de GAARlandia
2 La presencia de un mismo genero en las tres Islas septentrionales de las Antillas Mayores implica [por el modelo vicariante] que la diferenciacion entre estas formas durante el intervalo desde el Mioceno ha sido unicamente de nivel especifico [Dicha presencia]
315 rRRADlAcr6~ EVOLUCrONARIA
s610 puede explicarse razonablemente como el resultado de movimientos de esos animales entre las islas en el Cuaternario
De esta objecion resulta dificil disentir [El poblamiento entre islas diferenciado solo a nivel especifico no es probable que sea anterior al Pleistoceno (Simpson 1956 15)]
Debe tenerse en cuenta que la hipotesis de la vicarianza isla-isla alude a las tres islas septentrionales de las Antillas Mayores porque como se explico antes (vease El Barco Funeral Vikingo) Jamaica no existia como isla durante el lapso (Eoceno Superior shyOligoceno) consensualmente reconocido como el mas probable para la colonizacion de las proto-Antill as Mayores por la mayo ria de los mamiferos terrestres Despues de emerger como isla en el Mioceno Medio Jamaica debe haber recibido sus mamiferos terrestres por dispersion isla-isla (Davalos 2004376) y solo en el Cuaternario parece haberrecibido (por dispersion marina desde Centroamerica) al roedor murido Oryzomys
Seguramente tanto la dispersion como la vicarianza contribuyeron a que los mamiferos terrestres se irradiaran a traves de las Antillas Mayores y grupos de islas perifericas pero las maneras y tiempos en que estos mecanismos operaron a 10 largo de dicho proceso son aun en gran medida motivos de desacuerdo e incertidumbre
IRRADIACION EVOLUCIONARIA
A continuaci6n revisamos someramente las distribuciones conocidas de los diferentes grupos de mamiferos terrestres a traves de las Islas principales de las Antillas Mayores y grupos de Islas perifericas incluidas las interpretaciones y conjeturas que la literatura reciente recoge en relaci6n con la manera en que dichas distribuciones pudieron haberse conformado durante la irradiaci6n evolucionaria de los referidos grupos de mamiferos Manifestaciones de esta irradiaci6n pueden inferirse unicamente a partir de los taxones presentes en el Cuatemario pues solo 6 de las especies conocidas corresponden al Terciario
GRUPOS ORlGINARIOS DE SURAMERICA
PEREZOSOS (Megalonychidae)
Aparte de un genero miocenico de Cuba y otro del Oligoceno de Puerto Rico se conocen 4 generos (12 especies) de perezosos cuaternarios (todos extintos) restringidos a las tres islas septentrionales de las Antillas Mayores Acratocnus Neocnus Megalocnus y Parocnus Los cuatro generos estan representados en Cuba y La Espanola y Acratocnus 10 esta ademas (como unico genero de perezoso) en Puerto Rico Esta distribuci6n ha sido interpretada por MacPhee et al (2000) eomo resultado de la vicarianza oligomiocenica entre estas islas y [dado que el Banco Puerto Rico - Islas Virgenes fue la entrada de GAARlandia a las Antillas Mayores] se preguntaron si la ausencia de los otros tres generos en Puerto Rico se debe a que nunca estuvieron en ese territorio 0 a que sus
316 BIOGEOGRAFiA
fosiles atm no han sido hallados Pero como hemos visto Rega et al (2002) atribuyeron dicha distribucion a la dispersion interinsular en el Cuatemario
Por otra parte varios amilisis c1adisticos recientes ubi can a Choloepus -genero Reciente conocido solo de Centro- y Suramerica- como taxon hermano de Aeratoenus exc1uidos los demas megaloniquidos antillanos Para White y MacPhee (2001228)
tan estrecha relacion sugiere que la divergencia entre ambos [generos] ocurrio despues de que Aeratoenus divergio de Neoenus Si esta relacion implica que Aeratoenus y Neoenus se dispersaron separadamente desde Suramerica a las AntilIas mientras Choloepus permanecia en el continente (siendo necesarias al menos tres invasiones de perezosos) 0 si Choloepus invadio Suramerica desde las Antillas es algo que el material disponible no permite determinar
La diversidad adaptativa de los perezosos se manifiesta en (1) la utilizacion del habitat especies arboreas en Neoenus y Aeratoenus y terrestres en Paroenus y Megaloenus (Webb 1985a White [1993] White et al 1996 Arredondo [1999]) y en (2) la gama de tallas corporales desde los pequenos Neoenus hasta Megaloenus rodens el mayor de los mamiferos terrestres antillanos (su peso adulto estimado en 270 kg seglin Paula 19679) esto sugiere que los perezosos se irradiaron a nichos que en contextos continentales probablemente estarian ocupados por miembros de otros c1ados (MacPhee e IturraldeshyVinent 2000)
No esta claro si el abanico de tamanos entre estos perezosos enmascara alguna tendencia al gigantismo Por 10 pronto Anderson y Handley (2002) encontraron 10 contrario en perezosos (Bradypus) de las Islas de Bocas del Toro (Panama) shyseparadas secuencialmente del area continental vecina por ascensos del nivel del mar durante los ultimos 10 ka En las cinco islas los perezosos son menores que su contraparte continental y las diferencias entre las tallas medias de las cinco poblaciones expresan una reduccion lineal significativa de la talla corporal en funcion de la edad geologica de cada isla sin relacion alguna con el area de las islas 0 la distancia que las separa del continente
MONOS (Pitheciidae)
Los primates de las Antillas Mayores (cuatro especies extintas) constituyen un grupo monofiletico con Paralouatta (en Cuba) y Antillothrix (en La Espanola) como taxones hermanos seguidos en la escalera c1adistica por Xenothrix (en Jamaica) y finalmente Callieebus como el pariente continental mas cercano esto implica una sola invasion desde Suramerica antes del Mioceno Inferior (fecha mas temprana conocida para los primates fosiles antillanos) y por tanto en consonancia con GAARlandia como la via de colonizacion (Horovitz y MacPhee 1996 1999 Horovitz [1997] MacPhee y Horovitz 2004)
En desacuerdo parcial con la anterior interpretacion la filogenia de este grupo de monos obtenida por Davalos (2004) indica un antecesor comun en Cuba La Espanola y Jamaica
317 TRRADIACTON EVOLUCIONARIA
compatible con la hipotetica conexion -propuesta tentativamente por Iturralde-Vinent y MacPhee (199980-84)- entre las Montanas Azules del este de Jamaica y la peninsula suroccidental de La Espanola en la epoca de GAARlandia
A pesar de que los primates antillanos pertenecen a un grupo viviente de monos muy pequenos (los monos titles) fueron comparativamente grandes en relacion con los platirrinos modernos y sucesivamente mayores en la direccion Antillothrix Xenothrix y Paralouatta con P marianae como el mayor platirrino conocido del Mioceno neotropical (MacPhee 2005) MacPhee et al (1994) consideraron que Xenothrix debio ser un mono de andar lento y al parecer las especies de Paralouatta abandonaron la arborealidad para adoptar un modo de vida mas terrestre (MacPhee 1996a MacPhee et al 2003 MacPhee y Meldrum 2006)
ROEOORES (Echimyidae Capromyidae Heptaxodontidae)
La edad de Zazamys en Cuba demuestra que la invasion de los roedores histricognatos en las Antillas Mayores comenzo antes del Mioceno Inferior por otra parte los representantes mas antiguos de esa irradiacion evolucionaria se encuentran en La Espanola y Puerto Rico Ambas evidencias son consistentes con la hipotesis de GAARlandia (Woods 1982 1988 Woods et al 2001)
No esta claro si los capromidos evolucionaron a partir de un equimido ancestral 0 de un protocapromido 0 si los equimidos antillanos (I sub familia 3 generos y 7 especies todas extintas) forman parte de la irradiacion de los capromidos (4 subfamilias 9 generos y 27 especies 10 vivientes) Kratochvil et al (1978b) y Rodriguez et al (1979) consideraron que Capromys nan us es un eslab6n entre Echimyidae y Capromyidae pero Mesocapromys nanus esta morfologicamente mucho mas cerca de Hexolobodon y los de mas capromidos que de los equimidos (Woods 1985) Lo que si esta claro segun Woods et al (2001) es que la subfamilia Plagiodontinae endemic a de La Espanola es el linaje basal de los capromidos y que Isolobodon Hexolobodon Capromys Geocapromys Mysateles y Mesocapromys [y supuestamente Macrocapromys] forman parte de la irradiacion temprana de Capromyidae
La subfamilia Capromyinae (5 generos y 17 especies 9 vivientes) parece ser el dado de mas reciente evolucion en la familia con invasion de Cuba desde La Espanola y dispersion desde Cuba (1) por Capromys a las islas Caiman y (2) por Geocapromys a las islas Bahamas Caiman y Jamaica y desde Jamaica a las islas Swan aunque la presencia de Geocapromys en Swan pudiera deberse a introduccion aborigen (Morgan 1985 1988 1989 1993 1994 Woods 1989a 1990) Woods et al (2001) utilizaron datos moleculares para obtener estimados independientes de divergencia entre capromidos segiJn 10 cual Geocapromys divergio cerca de los 17 MaAP Mesocapromys divergio de Capromys hace 15 MaAP en tanto lvlysateles 10 hizo hace 10 MaAP Por su parte las formas de Capromys pilorides y de Mysateles prehensilis en Isla de la Juventud divergieron de sus correspondientes subespecies nominativas en la isla principal a los 24 y 08 MaAP respectivamente
318 BIOGEOGRAFiA
Woods y Howland (I979) describieron adaptaciones morfologicas de la mandibula el hioide y la faringe asociadas a mecanismos de masticacion diferentes en plagiodontinos y caprominos y Varona (1980a) llamo la atencion sobre el gran desarrollo de los molariformes (en relacion con la longitud total del cnineo) que presentan ciertos grupos de jutias fosiles en comparacion con las vivientes 10 que pudiera interpretarse como una adaptacion alimentaria a una vegetacion mas abrasiva por parte de las jutias extinguidas
Segun Woods (1990) la familia Heptaxodontidae (2 subfamilias 5 generos y 6 especies todas extintas) tambien seria parte de la irradiacion de los capromidos con origen en el este de La Espanola 0 en Puerto Rico y subsecuente dispersion al Banco de Anguila - San Martin en las Antillas Menores (pero vease MacPhee y Flemming 2003) Este banco estaria a solo 70 km del Banco Puerto Rico - Islas Virgenes en tiempos de maxima regresion marina y esta proximidad con Puerto Rico -patria de Elasmodontomys obliquus- seguramente tuvo que ver con la presencia en las islas de Anguila y San Martin del heptaxodontino gigante Amblyrhiza relacionado mas estrechamente con los roedores de las Antillas Mayores que con los del resto de las Antillas Menores (Woods 1990 1996) Aunque los roedores suelen aumentar de tamano en las islas (Case 1978) la escala del incremento de la talla en Amblyrhiza (que pudo alcanzar mas de 200 kg de peso adulto) esta fuera de toda proporcion con los patrones conocidos en otros roedores insulares (Biknevicius et 01 1993)
Clidomys (Clidomyinae) de Jamaica difiere marcadamente en su morfologia de los heptaxodontinos (vease ademas MacPhee y Flemming 2003)10 cual dificulta explicar su presencia en esa isla Como Jamaica estuvo bajo el mar hasta el Mioceno Medio el ancestro de Clidomys colonizaria esta isla por dispersion desde el extremo suroccidental de La Espanola despues que los heptaxodontinos se originaron en esta ultima isla (Woods 1989a) Pero Pascual et 01 (I 990) especularon que Clidomys pudiera ser el resultado de una invasion posterior e independiente de los capromidos
GRUPOS ORIGINARIOS DE NORTEAMERICA
El origen de las musaranas antillanas ha resultado ser hasta ahora el mayor enigma biogeografico de la fauna de mamiferos terrestres de la region aunque la mayoria de los autores coinciden en considerarlas originarias de Norteamerica (vease Discus ion y Conclusiones en Poblamiento Continente-Isla) No obstante MacPhee e Iturralde-Vinent (1994) descartaron toda posibilidad de conexion terrestre por el Estrecho de la Florida 0 por el Canal de Yucatan y si el (supuesto) evento vicari ante de Rosen (vease El Arca de Noe) -invocado por MacFadden (1980) para resolver el enigma- fuera minimamente plausible aun asi es dificil creer que las catastrofes que 10 sucedieron (vease Contexto Temporal) hayan barrido con toda esa vieja fauna continental excepto las musaranas circunstancia ya anticipada muy tempranamente por Matthew (l918a)
De otra parte la relaci6n entre Nesophontidae y Solenodontidae permanece irresuelta Si estos linajes no estan estrechamente relacionados entonces es muy posible que
319 IRRADIACION EVOLUCIONARIA
sus distribuciones sean explicadas por mecanismos diferentes (Whidden y Asher 2001)
La descomunal diferencia de talla corporal entre las musaranas pequenas (Nesophontes) y las grandes (Soienodon) seguramente impuso a ambos grupos restricciones distintas relativas a la termorregulacion y las adaptaciones energeticas En Nesophontes la seleccion debio favorecer una alta tasa de metabolismo y crecimiento camadas mas numerosas corta expectativa de vida yalta densidad poblacionaL En Soienodon en cambio la seleccion opero en la direccion de una baja tasa metabolica camadas reducidas crecimiento lento longevidad y baja densidad poblacional (Ottenwalder [1991])
MUSARANAS PEQUENAS (Nesophontidae)
Las musaranas pequenas estan representadas en las Antillas por el genero Nesophontes (y un genero miocenico indeterminado en ambar dominicano) con 6 especies cuaternarias extinguidas distribuidas en Cuba La Espanola Puerto Rico Islas Virgenes e Islas Caiman La mayor de las musaranas pequenas Nesophontes edithae evoluciono en Puerto Rico y su arribo a las Islas Virgenes supone una dispersion sumamente reciente En las Islas Caiman Nesophontes se asemeja mas a la poblacion de Cuba de donde seguramente derivo en el Plio-Pleistoceno (junto con Capromys y Geocapromys) cuando la costa sur de Cuba estuvo mucho mas cerca de las Islas Caiman (lturralde-Vinent 2003fig 4) MacPhee et ai (1994) interpretaron la distribucion de Nesophontes en Cuba y La Espanola como resultado de la vicarianza oligomiocenica ocurrida al formarse el Paso de los Vientos pero la distancia taxonomica entre los lados del Paso es solo de nivel especifico y N micros incluso vivi6 en ambas islas segtln Patterson (1962)
Allen (1918) considero que Nesophontes micrus fue una especie primitiva no modificada para la vida arb6rea 0 fos6rea y por tanto de habitos terrestres Nesophontes edithae de Puerto Rico es mucho mayor y mas robusta que las restantes especies del genero eL indice de robustez de su femur (= anchura minima de la diafisis ongitud total del hueso) muestra que este es tan robusto como el de Solenodon paradoxus un animal dos veces mayor (Morgan et ai 1980) Choate y Birney (1968) postularon un aumento cronoclinal de la taIla corporal en N edithae confirmado mas tarde por McFarlane (1999a) mediante fechados radiocarb6nicos de la secuencia estratignifica en depositos cavernarios de Puerto Rico Solenodon no vivi6 en Puerto Rico y en ausencia de una musarana de mayor tamano Nesophontes alcanzo una talla corporal mayor en esa isla que en La Espanola 0 Cuba (Woods y Eisenberg 1989)
MUSARANAS GRANDES (Solenodontidae)
Los arcaicos almiqufes (genero Soienodon) comprenden cuatro especies una viviente (S cubanus) y otra f6sil (S arredondoi) en Cuba y una viviente (S paradoxus con dos subespecies) y otra f6sil (S marcanoi) en La Espanola Como cabria esperar las dos especies cubanas se asemejan mas entre sf que a las especies de La Espanola pero
320 BIOGEOGRAFiA
enigmaticamente S marcanoi comparte caracteres con S cuban us y S paradoxus (Woods y Eisenberg 1989 Morgan y Ottenwalder 1993 Ottenwalder 200 I) Solenodon arredondoi alcanzo una talla corporal superior a la de cualquiera de las musarafias vivientes a nivel mundial Segun Morgan y Ottenwalder (1993) extrapolando las dimensiones y el peso de los almiquies vivientes S arredondoi debio tener una longitud cabeza-tronco y un peso de 450-550 mm y 1500-2000 g respectivamente en comparacion con S cubanus (360 mm y 800 g) y S paradoxus (390 mm y 1100 g)
Entre todas las especies de almiqufes Solenodon paradoxus es la que presenta los caracteres mas derivados (Otten walder 2001) Si estas relaciones son validas la distribucion de Solenodon entre Cuba y La Espafiola debio discurrir de oeste a este como propuso MacFadden (1980) quien invoco la presencia fosH de S cubanus [pero tambien S arredondoi] en Pinar del Rio para postular que el ancestro continental entr~ por Cuba opinion compartida por Perfit y Williams (1989) y por Ottenwalder ([1991] 2001) El paso de Solenodon a Puerto Rico pudo haberlo impedido la existencia de barreras ecologicas durante la conexion del este de La Espafiola con Puerto Rico 0 por la ausencia de Solenodon en esa parte de La Espafiola en aquella epoca Sin embargo las mismas condiciones ecologicas en el este de La Espafiola no parecen haber representado una barrera efectiva para Nesophontes cuyas mas altas tasa de reproduccion y densidad poblacional presumiblemente Ie permitieron expandirse flpidamente a traves de La Espafiola (Ottenwalder [1991])
Es opinion generalizada que los almiqufes antillanos constituyen el grupo de mamiferos que primero se emplazo en el territorio insular (MacPhee 2005) y en tal sentido es de interes la evidencia sobre especificidad parasito-hospedante aportada por Cruz (1980) quien encontro que la especie de acaro (genero Cubanochirus) que parasita a los almiquies es mucho mas primitiva que la especie (del mismo genero) que paras ita a las jutias y de aqui concluyo que dicho genero de acaro debio evolucionar primero en Solenodontidae y posteriormente en Capromyidae al entrar en contacto representantes de ambas familias en Cuba 0 La Espafiola
Recientemente Roca (2004) y Roca et al (2004) dieron a conocer que los resultados de su estudio genetico-molecular sobre los almiquies indican una divergencia de 25 Ma entre Solenodon cuban us y S paradoxus -compatible con la disyuncion tectonica entre Cuba y La Espafiola (Iturralde-Vinent y MacPhee 1999 McPhee et al 2003)-- y una separacion de ambas especies comparable en magnitud a la divergencia entre familias de mamlferos Consecuentemente propusieron que Solenodon cubanus fuera clasificada en el genero Atopogaie como indico Cabrera (1925) hace 82 afios
DISCUSION Y CONCLUSIONES
Las musarafias y los roedores antillanos (excepto Oryzomys) se irradiaron tanto de sus respectivos ancestros continentales que de estos 6rdenes se reconocen familias y subfamilias endemicas de la region en contraste con los perezosos (una subfamilia y tres
321 IRRADIACION EVOLUCIONARIA
tribus endemicas) y los monos (una tribu endemica) como expresion de diferentes niveles de diversificacion evolucionaria Este proceso es dificil de explicar (traUindose de c1ados que pnicticamente comparten la misma antigiiedad en las Antmas) si es correcta la interpretacion de Pascual et at (1990636)
Nosotros consideramos estos diferentes niveles de diversificacion como las diferentes etapas de evolucion alcanzadas por cada grupo [de mamiferos] Obviamente esto estuvo relacionado con las diferentes longitudes de sus historias
Pudiera ocurrir que en el futuro se compruebe que la historia biogeognifica de la region fue mucho mas complicada
La Espanola la isla mas ecol6gicamente divers a de la region debi6 ser el centro principal de evoluci6n de los mamfferos (con 87 de las familias y 73 de las subfamilias) seguida por Cuba (con 75 de las familias y 54 de las subfamilias) las dos islas de las Antillas Mayores mas relacionadas biogeograficamente (Fig 43 Tabla 44)
eL
EL
cb CAIMAN
SWAN
A Acratocnini B Cubanocnini C Megalocnini D Mesocnini E Nesophontidae F Solenodontidae G Xenothrichini
ANGUILAshyM-0 SMARTIN
H Heteropsomyinae I Plagiodontinae J Isolobodontinae
K Hexolobodontinae L Capromyinae
M Heptaxodontinae N CIidomyinae 0 Sigmodontinae
FIGURA 43 Representacion esquematica de las relaciones interinsulares (expresadas porel numero de linajes compartidos) entre las Antillas Mayores y entre estas y los grupos de islas perifericas segun las interpretaciones de los autores citados en el texto Los numeros dentro de los circulos indican la cantidad de linajes representados en cada isla relacion dudosa Contexto taxon6mico de los linajes en la Tabla 44
bull bull
bull bull
bull bull bull
bull bull
bull bull
bull
bull
bull
bull
bull bull bull bull
bull bull
bull bull
bull
bull bull bull
bull bull
bull
bull bull
bull
322 BIOGEOGRAFiA
TABLA 44 Distribuci6n de los linajcs de mamiferos terrestres a traves de las Antillas Mayores y grupos de islas perifericas LE = La Espanola CU Cuba JA Jamaica PR = Puerto Rico IC Islas Caiman IS Islas Swan IB = Islas Bahamas IV = Islas Virgenes AS = Anguila - San Martin
FamiliaSubfumiliafIribu LE CU JA PR IC IS IB IV AS
Megalonychidae
Choloepodinae
Acratocnini
Cubanocnini
Megalocninae
Megalocnini
Mesocnini
Nesophontidae
Solenodontidae
Pitheciidae
Callicebinae
Xenothrichini
Echimyidae
Heteropsomyinae
Capromyidae
Plagiodontinae
Isolobodontinae
Hexolobodontinae
Capromyinae
H e pta x 0 don tid a e
Heptaxodontinae
Clidomyinae
Muridae
Sigmodontinae
PILOSA
SORICOMORPHA
PRIMATES
RODENTIA
bull
bull
bull
bull TOTALES 12 10 4 4 2
323 EXTlICIOIES DEL CUATERNARIO
El gigantismo ha sido sin duda una tendencia evolucionaria recurrente entre los clados mayores de mamiferos antillanos Es un hecho bien conocido que algunos grupos de mamiferos han aumentado de tamafio en las islas y varios autores han intentado explicarlo a partir de enfoques diferentes (Foster 1964 Miller 1966 Sondaar 1977 Case 1978 Eisenberg y Wilson 1978 Heaney 1978 Lawlor 1982 Lomolino 1985 Angerbjom 1985 McNab 19942001)
EXTINCIONES DEL CUATERNARIO
Las islas que integran la Subregion Antillana han perdido casi 90 de sus mamiferos terrestres durante los ultimos milenios y en proporcion con su extension territorial esta perdida ha sido mayor en la Subregion que en cualquier otro lugar del planeta (MacPhee 1996b) Las extinciones del Cuatemario eliminaron mas de las tres cuartas partes de la fauna de mamiferos terrestres conocida de las Antillas Mayores sus islas satelites y grupos de islas perifericas y el examen de este proceso en sus aspectos mas generales se aborda en esta ultima parte del presente capitulo
CRONOLOGiA Y CONSECUENCIAS
Es muy improbable que un fosil represente el ultimo individuo de un taxon extinto y entonces hay que guiarse por la edad radiometrica del fosil mas reciente conocido lIamada fecha de ultima presencia- como unica aproximacion posible a la fecha de extincion del taxon
Como se explico al inicio del capitulo (vease Contexto Temporal) el registro fosH de los mamiferos terrestres de las Antillas Mayores adolece de insuficiente sustentacion radiometrica No obstante la distribucion temporal de los escasos fechados disponibles para el Cuaternario de la region abarca etapas criticas del proceso de extincion experimentado por la referida fauna a 10 largo de los ultimos 250 mil afios
De las 52 especies que es posible reconocer actualmente como miembros extintos de la fauna cuatemaria de mamiferos terrestres de las Antillas Mayores y grupos de islas perifericas los restos fosiles de 23 de elIas (442) han sido fechados radiometricamente o se han hallado asociados estratigraficamente de manera inequivoca con otro material fechado De estos 23 fechados 3 corresponden al interglacial Sangamon 4 pertenecen a la secuencia Pleistoceno terminal - Holoceno temprano 10 se ubican en ellapso entre el arribo de los aborigenes y la incursion europea y 6 son postcolombinos (a los cuales hay que sumar otras dos especies extinguidas en los siglos XIX y xx) A partir de estas 25 especies avaladas por fechas de ultima presencia distribuidas entre los cuatro intervalos temporales sefialados hemos compuesto la Tabla 45 en la que ademas aparecen las restantes 27 especies (carentes de fechado radiometrico) ubicadas en el intervalo temporal mas probable sobre la base de (1) la asociacion estratigrafica de sus restos con evidencias culturales (amerindias 0 europeas) 0 (2) la ausencia de dicha asociacion Sobre estos criterios de ubicacion temporal procede hacer dos aclaraciones
324 BIOGEOGRAFIA
TABLA 45 Intervalos temporales mas probables para las extinciones cuatemarias de los mamiferos terrestres de las Antillas Mayores y grupos de islas perifericas sobre la base de las evidencias siguientes (l) ultimo ejemplar colectado u observado en vida (2) fechado radiometrico mas tardio en material de la especie (3) asociaci6n estratigrafica mas tardia con otro material fechado (4) asociaci6n estratigrafica con restos de animales introducidos restos humanos u objetos culturales propios del intervale (5) falta de asociaci6n con evidencias culturales [Los ntimeros entre parentesis en la columna Evidencia significan adscripci6n tentativa al intervalo] Los estimados del peso corporal (en kg) estan tomados de la literatura 0 inferidos comparativamente y distribuidos entre
las clases siguientes (a) S 01 (b) 01 a lt 1 (c) 1 a lt 10 (d) 10 a lt 100 (e) 100 a lt 200 (j) ~ 200 Las
abreviaturas que designan las islas son las de la Tabla 44
F amilialEspecie Peso IsJa Evidencia Fuente
I httervalo htterg1acial Sangamon (gt80 kaAP)
HEPTAXODONTlNAE
Amblyrhiza inundata e AS 3 McFarJane et al 1998a
CLIDOMYINAE
Clidomys osborni d JA 3 Ibid
Clidomys parvus d JA 3 Ibid
II httervalo Pleistoceno terminal ~ Holoceno temprano (20-6 kaAP)
MEGALONYCHIDAE
Aeratoenus antillensis d CD 5 Arredondo 1975 a
Aeratoenus odontrigonus d PR 3 McFarJane 1999b
Aeratoenus simorhynehus d LE 2 Rega et al 2002
M egaloenus zile f LE 2 MacPhee et al 2000
SOLENODONTIDAE
Solenodon arredondoi e CD 5 Morgan y Ottenwalder 1993
PnHECIIDAE
Paralouatta varonai d CD 5 MacPhee 1996a
EcHIMYIDAE
Heteropsomys antillensis c PR 5 McFarJane 1999b
Heteropsomys insulans c PR 5 Ibid
Puertoricomys eorozalus c PR 5 Woods 1989a
CAPROMYIDAE
P lagiodontia araeum c LE 5 MacPhee et al 2000
Macrocapromys latus c CD 5 Varona y Arredondo 1979
(Continua)
325 EXTINCIONES DEL CUATERNARIO
TABLA 45 (Continuaci6n)
F ami1ia1Especie Peso Isla Evidencia Fuente
M acrocapromys latus c CU 5 Varona y Arredondo 1979
Mesocapromys kraglievichi c CU 5 Ibid
HEPTAXODONTINAE
Xaymaca fulvopulvis a JA 2 MacPhee y Flemming 2003
III Intervalo Amerindio (6-05 kaAP)
MEGALONYCHIDAE
Acratocnus ye d LE 2 MacPhee et aI 2000
Neocnus comes c LE 2 Ibid
N eocnus dousman c LE 2 Ibid
N eocnus toupiti c LE 2 Ibid
N eocnus gliriform is c CU 4 Cordova et ai 1997
M egalocnus rodens f CU 2 MacPhee et ai 2007
Parocnus browni e CU 2 Steadman et ai 2005
P arocnus serus e LE 2 MacPhee et ai 2000
N ESOPHONTIDAE
N esophontes edithae a PR 2 McFarlane 1999b
PITHECIIDAE
Antillothrix bernensis c LE 3 Rimoli 1977
EcHIMYIDAE
Brotomys contractus a LE 4 Miller 1929c
CAPROMYIDAE
P lagiodontia spelaeum c LE 4 Ibid
M acrocapromys acevedo c CU 4 NUiiez 1990a
HEPTAXODONTINAE
Elasmodontomys obliquus d PR 3 McFarlane 1999b
Iv Intervalo Postcolombino (05 kaAP - Presente)
N ESOPHONTIDAE
N esophontes hypom icrus a LE 24 MacPhee y Fle111111ing 1999
N esophontes zam icrus a LE (4) Ibid
N esophontes m icrus a CU 24 MacPhee et aI 1999
(Continua)
326 BIOGEOGRAFiA
TABLA 45 (Continuacion)
F amilialEspecie Peso Isla Evidencia Fuente
N esophontes m icrus a CD 24 MacPhee et aI 1999
N esophontes m icrus a LE 4 MacPhee et aI 2000
N esophontes sp 1 a IC 4 Morgan 1994
N esophontes sp 2 a IC 4 Ibid
Solenodon marcanoi b LE 4 Ottenwalder 2001
PITHECIIDAE
Xenothrix mcgregori c JA 4 MacPhee y Flemming 1999
EcHIMYIDAE
Boromys offeUa a CD 24 Guarch 1984
Boromys torrei a CD 24 Ibid
Brotomys voratus a LE 4 MacPhee et aI 2000
CAPROMYIDAE
P lagiodontia ipnaeum c LE 4 MacPhee y Flemming 1999
Rhizoplagiodontia lemkei c LE 4 Ibid
Isolobodon portoricensis c PR 24 Ibid
Isolobodon montanus c LE 4 Ibid
Hexolobodon phenax c LE (4) Ibid
Capromys sp c IC 24 Morgan 1994
Geocapromys columbianus c CD 4 MacPhee y Flemming 1999
Geocapromys thoracatus b IS 1 MacPhee y Marx 1997
Geocapromys sp 1 c IC 4 Morgan 1994
Geocapromys sp 2 c IC (4) Ibid
l-IEPTAXODONTINAE
Quemisia gravis c LE (1) MacPhee y Flemming 1999
MURIDAE
Oryzomys antillarum a JA 1 Ibid
1 Flemming y MacPhee (1999) alertaron en contra de la asociacion con otro material fechado Asimismo Morgan (1993) indico que la ausencia de asociacion con evidencias culturales amerindias no es necesariamente un criterio concluyente para ubicar un taxon en el intervalo temporal anterior porque hay depositos en la region que carecen de tales evidencias y son del Holoceno tardio por fechados radiocarbonicos [No obstante
327 EXTlNCIONES DEL CUATERNAR10
la ausencia de una especie en residuarios arqueol6gicos adquiere mayor certidumbre como criterio de ubicaci6n temporal de su extinci6n en la medida que aumenta el numero de residuarios estudiados que en las Antillas Mayores se cuentan por centenares] A falta de fechado absoluto de una especie consideramos que estos criterios de ubicaci6n temporal son mas utiles que la alternativa de excluir del analisis a la especie
2 En los trabajos sobre biogeografia de mamiferos antillanos los arque610gos mas citados como autoridades para establecer el inicio de la presencia aborigen en las Antillas Mayores (Kozlowski 1974 Rouse y Allaire 1978 Rouse 1989 1992 Wilson 2001) difieren al respecto entre los 6 kaAP y los 45 kaAP (pero vease Pagan et al 200523) y hemos adoptado la mas temprana de estas fechas para el comienzo del intervalo amerindio en la Tabla 45 De todos modos cualquiera que haya sido la fecha esta no es aplicable por igual al conjunto de las Antillas Mayores porque segun algunos de los autores citados el poblamiento comenz6 por Cuba y La Espanola 6 mil anos atras continu6 en Puerto Rico hace alrededor de 3 mil anos (pero vease Burney et al 1994) yen Jamaica hace menos de 2 mil e incluso a las Islas Caiman ni siquiera llegaron [Las Islas Caiman descubiertas por Col6n en 1503 carecen absolutamente de evidencia amerindia (Stokes y Keegan 1996)]
Si los datos de la Tabla 45 representan una aceptable aproximaci6n a la realidad el proceso de las extinciones cuaternarias de los mamiferos terrestres de las Antillas Mayores y grupos de islas perifericas debi6 caracterizarse entre otros por los eventos siguientes
Aunque obviamente la fauna cuaternaria debi6 ser mucho mas numerosa que 10 que refleja el registro paleontol6gico (Morgan y Woods 1986) 52 especies (81 de la fauna conocida) desaparecieron (sin reposici6n) durante los ultimos 250 mil anos
- Al parecer las extinciones se concentraron en las especies de mayor tamano Asi el peso promedio estimado para las especies de la etapa preantr6pica (intervalos I + II de la Tabla 45) es 350 kg es 328 kg para las especies del intervalo III (amerindio) y es s610 25 kg (c1ases del peso a b c) para las especies del intervalo IV (postcolombino) Ninguna especie aut6ctona de mamifero mayor de 6 kg existe hoy en la regi6n
- De acuerdo con la distribuci6n temporal de las extinciones 61 especies sobrevivieron al interglacial Sangamon (intervalo 1)48 sobrevivieron a la transici6n del Pleistoceno terminal al Holoceno temprano (intervalo II) 34 sobrevivieron al periodo amerindio (intervalo III) y s610 12 especies han sobrevivido desde la Ilegada de los europeos (intervalo IV) Las tasas de extinci6n correspondientes (exceptuado el intervalo I) son una especie cada 1167 anos en el intervalo II una cada 393 anos en el intervalo III y una cada 23 anos en el intervalo IV
- Durante todo el proceso de las extinciones cuaternarias la perdida de especies por isla fue de 91 en La Espanola 83 en Jamaica y 62 en Cuba Bahamas retuvo su unica especie (aunque extirpada en varias de las islas) Puerto Rico e Islas Caiman perdieron todas sus especies (6 y 5 respectivamente) y las Islas Swan Virgenes y Anguila - San Martin perdieron sus respectivas especies (micas
328 BIOGEOGRAFiA
De los cuatro 6rdenes de mamiferos representados en la fauna Pilosa y Primates desaparecieron totalmente Soricomorpha y Rodentia perdieron 86 y 74 de sus especies respectivamente
CAUSALIDAD
CAUSAS NATURALES
Los cicIos glaciales e interglaciales acaecidos en el Pleistoceno causaron cambios ambientales dnisticos en las Antillas con la concomitante alternancia de cIimas frios (secos) y calidos (hUmedos) asi como de descensos y ascensos del nivel del mar aumento y disminuci6n de las areas territoriales) induso (aunque con menor intensidad) en el Holoceno Estos eventos paleogeograficos han sido consistentemente documentados (Mayo y Kartashov 1972 Bonnatti y Gartner 1973 Ortega y Arcia 1982 Prance 1982 Schubert y Medina 1982 Dodge et al 1983 Ortega [1984] Haq et al 1987 Schubert 1989 Hodell et al 1991 Guilderson et al 1994 Oppo 1997 Emslie 1998 Curtis et al 2001 Iturralde-Vinent 2003) y de ellos brindamos a continuaci6n una breve resena
Finalizando el periodo interglacial anterior a la ultima glaciaci6n (Wisconsin) el mar estaba a 8-10 m por encima de su nivel actual Cuando el avance glacial del Wisconsin a1canz6 su culminaci6n (eillamado Ultimo Maximo Glacial) hace 18 ka el mar habia descendido 120 m con respecto a dicho nivel y las islas del Caribe habian incrementado considerablemente sus extensiones subaereas en comparaci6n con las que presentan hoy dia por ejemplo Puerto Rico y las Islas Virgenes se conectaron territorialmente y las Bahamas se unieron en cinco gran des islas Prevalecia un regimen frio y seco y condiciones de aridez --debidas a una reducci6n de las precipitaciones (posiblemente a menos de 440 mmlano)- y los ambientes xericos se habian extendido a traves de las islas y bancos emergidos y abundaban los pinares (los habitats mesicos estaban restringidos a parcelas relictas en 10 alto de las montanas) En el Mar Caribe y el Golfo de Mexico la temperatura superficial del mar era 5degC mas baja que la actual y en las llanuras costeras y alturas maximas eran respectivamente 2-3degC y 6degC mas bajas Los limites de las zonas ecol6gicas altitudinales descendieron varios cientos de metros y cada zona redujo su extensi6n vertical al aumentar el gradiente de temperatura Ellimite inferior de la zona periglacial
elevaci6n donde la temperatura media minima a1canza los OdegC) de la Cordillera Central de la Republica Dominicana (altura maxima del Caribe insular con evidencia de topografia glacial) se considera que estuvo entre los 1500 y 1800 mlnm (actualmente esta a 2800 mlnm) y se puede estimar que dicho limite era inferior en Cuba por estar mas al norte que la Republica Dominicana No obstante durante este periodo la temperatura experiment6 episodios cortos (unas pocas decadas) de ascensos repetidos
Un signifieativo componente bi6tieo de las referidas condiciones paleoambientales 10 constituy6 la espectacular irradiaci6n evolucionaria experimentada por un grupo numeroso de aves rapaees diurnas y nocturnas (Wetmore 1937 Brodkorb 1969 Kurotchkin y Mayo 1973 Arredondo 1976 Olson 1978 Olson y Hilgartner 1982
329 EXTINCIONES DEL ClJATERNARIO
Suarez 2000a b 2004 Suarez y Emslie 2003) Los mamiferos de las Antillas Mayores ofrecieron a estos depredadores aviares una amplia gama de tamanos de presa que en ausencia de mamiferos del orden Carnivora en la region permitio a estas aves irradiarse a este nicho alimentario con la evolucion de una variedad de especies de muy diversa tall a corporal incluidas formas gigantes Adquirir un cuerpo gigante les propicio el acceso a recursos alimentarios que no estaban siendo explotados por ningun otro depredador (Alcover y McMinn 1994) A Cuba corresponde el mejor registro fosil de rapaces cuaternarias caribenas con siete generos de especies diurnas Dentro de estas aves existieron tres gavilanes gigantes (Accipitridae) tres carairas (Falconidae) y un condor (Cathartidae) asi como cuatro generos de especies nocturnas dos lechuzas (Tytonidae) y tres Mhos (Strigidae) dos de ell os no igualados en tamano en todo el mundo Algunos de estos generos tuvieron representacion en otras islas antillanas como La Espanola Jamaica Bahamas y Gran Caiman
Al retirarse hacia el norte los hielos de la glaciacion Wisconsin el clima se tomo mas hUmedo en las Antillas durante el Holoceno temprano y el bosque mesico se expandio en la region insular El nivel del mar ascendio acompanado de mayor pluviosidad alrededor de los 7 kaAP las Islas Virgenes quedaron separadas entre si y de Puerto Rico y las Bahamas perdieron la mayor parte de su territorio Cerca de los 3 kaAP las condiciones secas retornaron a la region e hicieron desaparecer buena parte del bosque mesico Nuevamente el clima comenzo a adquirir sus condiciones actuales hacia los 25 kaAP aproximadamente y hoy transita por una fase mesica con vegetacion xerica confinada en parcelas relictas
Sin duda estos y otros eventos naturales incidieron negativamente en la sobrevivencia de mamiferos terrestres en las Antillas Mayores asi como en los grupos de pequenas islas perifericas (Pregill y Olson 1981 Olson 2006)
McFarlane et al (1998a) demostraron convincentemente como Amblyrhiza el gigante de los roedores antillanos endemico de la pequena isla de Anguila sucumbio a las repetidas y abruptas perdidas de su habitat durante el interglacial Sangamon por las nipidas y severas oscilaciones del nivel del mar (documentadas por Neumann y Hearty 1996) al no poder responder adecuadamente a tan catastroficos eventos mediante el ajuste de su masa corporal media y otro tanto debio ocurrirle en Jamaica al otro roedor gigante Clidomys (McFarlane et al 1998b) emparentado con Amblyrhiza en la misma epoca (Tabla 45 intervalo I)
El ascenso del nivel del mar seguramente promovio la extincion de mamiferos durante la transicion del Pleistoceno al Holoceno (Tabla 45 intervalo II) con la reduccion del tamano de las Islas por las transgresiones marinas (Pregill y Olson 1981 Morgan 1993) Esto ultimo haria maximamente severa la competicion asf como la presion de los depredadores [Steadman et al (2005) no yen por que el calentamiento global ocurrido durante la transicion Pleistoceno-Holoceno habria de tener un efecto negativo sobre los perezosos siendo sus descendientes actuales especies tropic ales 0 subtropicales pero no se trata del efecto
330 BIOGEOGRAFiA
directo del cambio de temperatura sobre los animales sino del efecto indirecto y progresivo que dicho cambio genera a largo plazo al reducir el territorio insular disponible]
Por esta epoca tambien desaparecieron especies de tall a mayor como Megalocnus zile en La Espanola y otras de menor tamano (en Cuba) pero que representan la tall a maxima en sus respectivos contextos taxonomicos del Cuaternario como Solenodon arredondoi Paralouatta varonai y Macrocapromys acevedo Un mamifero puede ser grande solo a costa de reducir su densidad poblacional y esto 10 hace mis vulnerable a la extinci6n sobre todo en una isla (McNab 1994 McFarlane et al 1998a) Incidentalmente Alcover et al (1998a) afirmaron que larelacion entre la talla corporal y la extincion no siempre se cumple en las especies de menor tamano y citaron como ejemplo que entre las musarafias todas las especies de Nesophontes desaparecieron mientras sobreviven especies de Solenodon pero la comparacion correcta en todo caso serfa entre los solenodones vivientes y el gigante Solenodon arredondoi y entre Nesophontes edithae -el gigante de los nesofontes extinguido en el periodo amerindio- y el resto de sus congeneres que sobrevivieron hasta los primeros siglos de la colonizacion europea (Tabla 45 intervalos III y IV)
CAUSAS ANTROPOGENICAS
Intervalo Amerindio
La mayoria de los autores que se han referido a las extinciones acaecidas durante este intervalo (entre elIos paradigmaticamente Martin 1984a b) han responsabilizado a los aborfgenes (que se supone comenzaron a poblar las Antillas Mayores hace 6 mil afios) con la desaparicion de numerosas especies de mamiferos terrestres en la region principalmente por la via de la sobrecaza y la introduccion de animales domesticos devenidos depredadores de la fauna autoctona (Morgan y Woods 1986) Este efecto negativo de la presencia indigena sobre los mamiferos de la region ha sido dramatizado por algunos autores en terminos de holocausto faunal Sin embargo pensamos que se ha exagerado Como han expresado MacPhee et al (19993) cuando uno busca evidencia dura de complicidad humana directa en [estas] extinciones no hay mucho que encontrar Las siguientes son algunas consideraciones a tener en cuenta en tal sentido
El registro radiometrico mas joven conocido hasta ahora para un perezoso antillano (MegalocnllS rodens) es de 42 kaAP (MacPhee et ai 2007) Si se acepta que los aborfgenes entraron en Cuba hace aproximadamente 6 mil anos su convivencia con los perezosos debe haber durado alrededor de 1800 afios (0 mas porque dificilmente el referfdo fechado radiometrico haya correspondido al ultimo sobreviviente de la especie) Esta cifra contradice la hipotesis del holocausto faunal segun la cualla extincion de los mamiferos insulares de mayor talla debe ocurrir con extrema rapidez -a escala de decadas 0 de centurias- a partir de su primer contacto con los humanos (Martin y Steadman 1999) A pesar de las repetidas afirmaciones en contrario (Harrington 1921 Miller 1929c Herrera [1945] Tabio 1951 Alvarez 1951a 1957 Arredondo
331 EXTINCIONES DEL CUATERNARIO
1959 1960a Veloz y Ortega 1973 Mayo 1980b Rodriguez et al 1984 Castellanos y Pino 1985 Pino y Castellanos 1985 Nunez 1990b Gutierrez 1996 Arredondo 2004) en su mayoria basadas en evidencias no exentas de posibles accidentes de deposicion (0 de reubicacion postdeposicional) en recintos cavernarios (Diaz-Franco [2006b D no se ha demostrado inequivocamente que los aborigenes antillanos se alimentaran de perezosos regularmente Sus restos en residuarios de alimentos son escasisimos en toda la region y esto contrasta con los abundantes restos de jutias y otros mamiferos de menor talla que aparecen en esos residuarios demostrativos de la preferencia de los paleoindios por estos otros mamiferos
Los mamiferos mas importantes en la economia prehistorica de las Antillas Mayores fueron las jutias Capromys y Geocapromys en Cuba Geocapromys en Jamaica e Isolobodon en La Espanola Puerto Rico e Islas Virgenes (Aguayo 1950b Tablo 1951 Rimoli 1972 Wing 2001 Diaz-Franco [2006b D Wilkins (2001) estudi61ajutia viviente de Jamaica (Geocapromys browni) en un sitio ocupado por los aborigenes durante 100 anos y no encontr6 evidencias de que durante ese tiempo la especie haya sido sobreexplotada Asimismo en Cuba las jutias conga carabali y andaraz son escasas en depositos pleistoholocenicos en cambio son muy numerosas en residuarios indigenas y de haber sido sobreexplotadas 10 mas probable es que no hubieran sobrevivido hasta hoy
La habilidad de los humanos para penetrar en profundidad el bosque tropical y mantenerse en este sustentablemente ha sido un evento comparativamente reciente (Beran y Steele 1994) En el caso antillano la caza tierra adentro ha sido explicada como un proceso tardio de desplazamiento a partir de la explotacion de abundantes recursos marinos que ofrecian condiciones 6ptimas para el sedentarismo de las comunidades aborigenes costeras (C6rdova y Arredondo 1988 Brito 2005) Peces y mariscos dominan la arqueologia costera de las Islas antillanas en tanto el consumo de recursos terrestres (mamfferos incluidos) aumenta progresivamente en la medida que los residuarios estudiados se alejan de la costa (Scudder 1991 Wilkins 2001) En las Antillas Mayores el componente terrestre de la dieta aborigen es solo 20-30 del material recobrado en los residuarios (Wing 1977 1989) Y debio ser menor alin porque el componente vegetal de la dieta no se ha preservado Por otra parte los aborigenes cubanos domesticaron la jutia conga y seguramente la reprodujeron para comer (Pose et al 1988) practica que probablemente se repitiera con otras jutias en las demas Antillas Mayores
- Fray Bartolome de las Casas hablo de 3 millones de indios en Cuba al inicio de la colonizacion pero los historiadores subsecuentes consideraron exagerada la cifra (Poey 1851 a) No obstante se dispone de dos estimados de la densidad de poblaci6n aborigen en las Antillas Mayores a la llegada de los europeos 18 indigenaslkm2 en Puerto Rico (Rouse 1956) y 1271km2 en La Espanola (Watts 1987) Extrapolando ambos valores entre las cuatro islas principales de las Antillas Mayores tenemos que en la fecha indicada la poblaci6n aborigen pudo estar entre 16 mil y 113 mil indigenas en Puerto Rico entre 21 mil y 145 mil en Jamaica entre 141 mil y 1 mill6n en La Espanola y entre 200 mil y 14 millones en Cuba Entre las cifras maximas y las minimas nos inclinamos
332 BIOGEOGRAFiA
hacia estas ultimas atendiendo a los altos indices de mortalidad infantil y morbilidad que se evidencian en las osamentas de los pobladores prehispanicos (Martinez y Rives 1990) Uno de los hechos que apuntan hacia el bajo numero de aborigenes en las Antillas Mayores es 10 rapido que desaparecieron despues de la llegada de los europeos pnkticamente sin dejar mestizaje con estos 10 cual contrasta ostensiblemente con la situacion en la America continentaL
En suma un grupo humano (1) demograiicamente irrelevante y controlado por altos indices de morbilidad (2) consumidor mayoritariamente de organismos marinos y (3) carente de la tecnologia cinegetica necesaria para poder sobreexplotar los mamiferos que Ie sirvieron de alimento (a los cuales accedio tardiamente en su desarrollo) no es concebible que haya causado e1 holocausto faunal que se Ie atribuye a partir del error de aplicar a las comunidades aborigenes antillanas un modelo cultural de la prehistoria continentaL
Tampoco hay evidencias de que los aborigenes hayan introducido carnivoros con suficiente capacidad depredadora salvo el perro Morgan y Otlenwaldder (1993) consideraron que la depredacion por perros durante el periodo amerindio es la explicacion mas plausible para la extincion del almiqui gigante de Cuba pero como se puede observar (Tabla 54) Solenodon arredondoi aparece en el intervalo II por falta de asociacion con evidencias culturales
Consecuentemente el accionar de los aborigenes y de sus animales domesticos directamente sobre la fauna no parece haber sido por sf solo un factor determinante de la extincion de mamiferos terrestres durante el periodo amerindio en las Antillas (pero vease Steadman et al 2005)
Hay que recordar en este senti do que alin en el Holoceno tardio ocurrieron cambios clirruiticos de consideracion (vease Causas Naturales) Por ejemplo Curtis et al (2001) explicaron que la extincion de algunos ani males mesofilicos de las alturas montanosas de La Espanola pudo haber antecedido a cualquier impacto humano sigrIificativo en esos lugares por coincidir dicha extincion con una tendencia a la aridez documentada isotopicamente y con evidencias de la perdida del bosque mesico ocurridas despues de 3kaAP Yen Jamaica hace solo 2shy3 mil aDos el area de la Bahia de Jackson sustentaba una gran biomasa de vertebrados en bosque hinnedo a juzgar por la cantidad y diversidad de taxones presentes en material fosil derivado de la depredacion por lechuzas hoy el area es la mas seca de la isla con matorral xerofitico y pobreza de fauna (MacPhee 1996a) Yalm en nuestros dias huracanes de gran intensidad aniquilan poblaciones de vertebrados (que no se recuperan) en is las pequenas y llanas del Caribe (Brooks y Smith 2001)
En medio de la incertidumbre sobre la verdadera causa (0 causas) de las extinciones de mamiferos en el periodo amerindio MacPhee y Marx (1997) han propuesto su hipotesis de la hipermorbilidad la cual estipula colapsos poblacionales relativamente rapidos con la introduccion de enfermedades de alta letalidad Estos autores han razonado que
Las islas son los lugares mas apropiados para experimentar perdidas catastroficas causadas por enfermedades Los mamiferos no solo serian susceptibles en virtud de su prolongado aislamiento biologico sino que en los confines de una isla la epizootia
333 EXTINCIONES DEL CUATERNARIO
tendria la potencialidad de infectar simultaneamente a la rnayoria de los individuos de vadas especies Los neonatos son inmunologicamente inmaduros y sufririan desproporcionadamente cualquier infeccion hipervirulenta La perdida excesiva de neonatos y juveniles tendria efectos severos para la sobrevivencia de la poblacion si esta se reduce por debajo de cierto nivel sobreviene la extincion porque no hay posibilidad de restitucion en el periodo postpanzootico Las especies de mayor talla corporal van a ser las mas vulnerables a la extincion dadas sus bajas tasas reproductivas largos periodos de gestacion y reducido numero de progenie Las especies de menor tamano --caracterizadas por altas tasas reproductivas periodos cortos de gestacion y mayor numero de neonatos por parto- tambien serian fuertemente diezmadas pero muchas sobrevivirian porque su estrategia reproductiva les permite perder mayor numero de individuos Las perdidas deben ocurrir relativamente rapido (a escala de decadas) mientras que otras hipotesis (digamos los cambios climaticos) permiten intervalos mucho mas largos (a escala de siglos 0 de milenios) entre la aparicion de las causas y la produccion de sus efectos Por ejemplo todo indica que los nesofontes se extinguieron muy rapidamente y seria dificil poder invocar un impacto humano directo 0 argumentar fenomenos climaticos que pudieran explicar plausiblemente perdidas tan rapidas
MacPhee y Marx (1997 191) han dicho que por el momento la hipotesis de la hipermorbilidad no pasa de ser una abstraccion en espera de adecuada sustentacion empirica Llama la atencion que linicamente en el periodo amerindio (Tabla 45 intervalo 1Il) las extinciones abarcaron todas las clases del peso corporal (de la a a laj) y que ninguna de las especies que sobrevivieron hasta el periodo postcolombino (Tabla 45 intervalo IV mas las especies vivientes) sobrepasa los 10 kg (clases del peso a b c) en clara coincidencia con los presupuestos de la hipotetica hipermorbilidad
Intervalo Postcolornbino
Por 10 menos 37 especies de mamiferos terrestres vivian en las Antillas Mayores y grupos de Islas perifericas a la lIegada de los europeos en 1492 De elIas se han extinguido 23 shymas especies que en cualquiera de los periodos precedentes (Tabla 45~ y solo subsisten 12 especies repartidas entre Cuba (8 especies) La Espanola (2) Jamaica (1) y Bahamas (1) varias de estas en peligro critico de extincion Solo dos de las especies extinguidas durante los ultimos 500 anos se conocieron en estado viviente los roedores Oryzomys antillarum de Jamaica (extinguido en 1877) y Geocapromys thoracatus de las Islas Swan (extinguido en 1955) Las causas de las extinciones ocurridas durante este periodo son sobradamente conocidas destruccion de habitats introducci6n de animales exoticos (domesticos y comensales) y demas presiones asociadas a la actividad humana entre las cuales posiblemente habria que incluir brotes de hipermorbilidad
Durante mucho tiempo se ha afirmado la posibilidad de que especies de Nesophontes hayan sobrevivido hasta las primeras decadas del siglo xx y que incluso continuen existiendo en lugares remotos de Cuba 0 La Espanola (Allen 1918 1942 Miller 1930 Aguayo 1950a Arredondo 1960b Fischer 1977 MacFadden 1980 Woods y
334 BIOGEOGRAFiA
Ottenwalder 1991) Actualmente no existe evidencia radiometrica de que alguna especie de Nesophontes haya sobrevivido al siglo xv las fechas radiocarbonicas calibradas obtenidas en muestras de Nesophontes de Cuba y La Espanola resultaron no mas tardias que los siglos XIII y XIV (MacPhee et al 1999) Por otra parte las expediciones efectuadas a fines del siglo xx con el proposito de localizar individuos vivientes del genero en La Espanola no produjeron la menor indicacion afirmativa al respecto (Woods et al 1985) Ademas a traves de las Antillas Mayores no aparecen restos de Nesophontes en los depositos formados actualmente por las lechuzas
EPiLOGO
Las Antillas Mayores son islas oceanicas no han formado parte de un continente (con la posible excepcion de una parte de Jamaica hasta el Eoceno Medio hace 45 Ma) y durante un tiempo geologico considerable (desde el Eoceno Superior hace 40 Ma) han permanecido emergidas y separadas de los continentes vecinos Consecuentemente los mamiferos terrestres (no volantes) autoctonos de estas islas constituyen una fauna oceanica que por ende cumple con todas las caracteristicas convencionales reconocidas para este tipo de entidad biogeografica
- Baja diversidad taxonomica a nivel de las categorias superiores (solo cuatro ordenes) en comparacion con islas plataformicas continentales (0 con areas continentales) de tamano y diversidad ecologica simi lares
- Alto grado de endemismo a nivel de genero (96) y familia (50) en respuesta al prolongado aislamiento
- Desbalance ecologico de la estructura trofica ausencia de mamiferos camivoros y ungulados
- Espectaculares procesos de irradiaci6n evolucionaria como resultado de la colonizacion de territorios libres de competidores y depredadores
Tendencias evolucionarias extremas gigantismo
- Control a cargo de aves rapaces con tendencia at gigantismo en ausencia de mamiferos miembros del orden Carnivora
- Mas especies de reptiles y de murcielagos que de mamiferos terrestres relaciones que en los continentes se invierten
Alta tasa de extincion de especies (84) sin reposicion
Preservacion de especies filogeneticamente arcaicas (los llamados fosiles vivientes) los almiquies

301 POBLAMIENTO CONTINENTE-ISLA
TABLA 43 F echados pleistocenicos mcis tempranos (obtenidos por distintos metodos) en mamiferos terrestres del segmento insular delimitado en laFig 41 F =Fuentes (I) Jull et al 2004 (2) Goodfriend y Mitterer 1987 (3) MacPhee et al 1989 (4) Woods 1989b (5) Rega et aI 2002 (6) McFarlane 1999b (7) McFarlane 1 999a (8) McFarlane et al 1998a
Pals Localidad Taxon kaAP F
Cuba Breas de San Felipe Pilosa (Parocnus) 1l9a 1
Jamaica Cueva Coco Ree Cueva Wallingford Roadside
Primates indet Rodentia (Clidomys)
393 2530
2 3
Haiti Trouing Jeremie no5 (casimba) Rodentia (Rh izoplagiodontia) 212 4
Republica Dominicana
Cueva del Perezoso Pilosa (ficratocnus) 185 5
Puerto Rico Cueva de Ia Vaca Cueva Clara
Pilosa (Acratocnus) Soricomorpha (Nesophontes)
337 346
6 7
Anguila Cueva Cave-No-Cave Rodentia (Amblyrhiza) 2060 8
a Kulp etal (1952) reportaron un fechado radiocarbonico de gt 30000 afios para un cono de pino en el deposito fosilffero de Ciego Montero (Cuba) con restos de perezosos
(vcase tambicn Crother y Guyer 1996) pero Iturralde-Vinent y MacPhee (1999) han preguntado donde estaban esos lugares si es que existieron
Solo a partir del Eoceno Superior las Antillas Mayores en fonnacion han pennanecido emergidas hasta hoy [excepto parte de Jamaica y el sur de La Espanola que continuaron sumergidas hasta el Mioceno Medio (Iturralde-Vinent y MacPhee 1999)] Esto significa que solo durante los ultimos 40 millones de anos aproximadamente han existido pennanentemente en dicha region insular ambientes terrestres capaces de sustentar comunidades biotic as (MacPhee e Iturralde-Vinent 2000 Portell et ai 2001 MacPhee 2005) 10 cual proporciona el marco temporal de referencia para conceptuar la antigiiedad maxima posible de la fauna de mamiferos terrestres de las Antillas Mayores
POBLAMIENTO CONTINENTE-ISLA
AFINIDADES CONTINENTALES
El consenso de los autores que en las itltimas dccadas han tratado la filogenia y biogeografia de los mamiferos terrestres de las Antillas Mayores (MacFadden 1980 MacPhee y Woods 1982 Morgan 1985 1988 1993 Morgan y Woods 1986 Ford 1986 Woods 1988 1989a 1990 MacPhee e Iturralde-Vinent 1995a b Hedges 1996 Horovitz [1997] Ottenwalder 2001 Whidden y Asher 2001 MacPheeet ai 2003 Carlini y Scillatoshy
302 BIOGEOGRAFiA
Yane 2004 Davalos 2004 MacPhee y Horovitz 2004 Roca et al 2004) es practicamente unanime en cuanto a que los ancestros continentales que dieron origen a esta fauna derivaron (1) mayormente de Suramerica perezosos (Megalonychidae) monos (Pitheciidae) y roedores (Echimyidae Capromyidae y Heptaxodontidae) (2) en menor medida de Norteamerica musarafias (Nesophontidae y Solenodontidae) y (3) menos aun de Centroamerica una especie de roedor (Muridae)
HIPOTESIS BIOGEOGRAFICAS
La definicion c1asica reconoce dos tipos de islas isla continental (porcion del territorio de un continente separado de este por movimiento tectonico 0 por ascenso del nivel del mar) e isla oceanica (porcion de territorio que no ha formado parte de un continente) La definicion sigue siendo uti por 10 que implica para la formacion de las faunas insulares en uno u otro caso As en el caso de la isla continental su fauna ya estaba emplazada en la futura isla antes de producirse su separacion del continente [Vicarianza hipotesis biogeografica de fragmentacion faunal] en el caso de la isla oceanica su fauna es el resultado de la colonizacion1 por propagulos2continentales a traves de la barrera marina entre el continente y la isla [Dispersion hipotesis biogeografica de acumulacion faunal] En el primer caso el surgimiento de la barrera marina es resultado del evento vicariante en el segundo la existencia de la barrera marina es anterior al evento dispersante Ademas de operar a escala mayor (continente-isla) ambos eventos operan secundariamente a menor escala (isla-isla)
Las dos hipotesis -vicarianza y dispersion- se han disputado hasta hoy la preferencia de los biogeografos en tanto hipotesis mas viable para explicar el proceso de formacion original (continente-isla) de la fauna de mamiferos terrestres de las Antillas Mayores [Sobre la historia de las hipotesis biogeograficas relativas al Caribe insular vease RuB y Schubert 1989 Williams 1989 Pruna 2001] Yel debate continua debido ala falta de consenso sobre 10 que Williams (1989) ha llamado el problema central de la discusion a saber la ostensiblemente baja diversidad taxonomica de esta fauna a nivel de las categorias superiores (solo cuatro ordenes) en comparacion con la diversidad al mismo nivel de las faunas de islas continentales (0 de areas del continente) simi lares en tamafio y diversidad ecologica [Por ejemplo la fauna cubana de mamiferos terrestres consideradas de conjunto las especies vivientes y las extintas- es menos diversa a los niveles taxonomicos superiores (4 ordenes y 6 familias) que 10 que cabria esperar de la diversidad ecologica y extension territorial de Cuba (110 992 km2) comparada con una isla tipicamente continental como Trinidad (4828 krn2
) -veinte veces menor que Cuba y menos diversiticada ecologicamente- donde solo las especies vivientes de mamfferos terrestres representan 6 ordenes y 16 familias (Eisenberg 1989)] Este desbalance
lColonizaci6n Arribo y establecimiento de un propagulo en una isla no ocupada por la especie (MacArthur y Wilson 1967)
2Propagulo Niimero minimo de individuos de una especie capaces de reproducirse y aumentar poblacionalmente en una isla bajo condiciones favorables (MacArthur y Wilson 1967)
303 POBLAMIENTO CONTINENTE-ISLA
taxon6mico ha sido expJicado de dos maneras (1) Inicialmente hubo mas 6rdenes presentes pero la mayoria se extingui6 y s610 sobrevivieron esos cuatro [explicaci6n vicarianista] (2) S610 representantes de esos cuatro 6rdenes lograron cruzar la barrera marina [explicaci6n dispersionista]
Cinco hipot~ticos mecanismos de poblamiento continente-isla han sido invocados durante los ultimos 140 afios (y se los sigue proponiendo actualmente) para explicar la presencia de mamiferos terrestres en las Antillas Mayores A continuaci6n pasamos a resefiarlos brevemente (incluidas sus alegadas fortalezas y debilidades) en el orden aproximado en que historicamente fueron propuestos
EL PUENTE NATURAL (dispersion terrestre activa)
En el siglo XIX el puente terrestre era pnkticamente la unica hipotesis para explicar las distribuciones disyuntas de animales en buena parte del mundo y los primeros investigadores que se ocuparon de la biogeografia antillana (Fernandez [1865] 1884 Chapman [1893] Torre 1892 Lora 1895) la asumieron resueltamente en parte intrigados por el descubrimiento en Cuba en la segunda mitad del siglo de un perezoso f6sil de gran talla Y en su aplicacion a los mamiferos terrestres de las Antillas Mayores el puente sigui6 siendo a 10 largo del siglo xx la hipotesis mas favorecida por los estudiosos del tema (Vaughan 1902a Spencer 191 Oa b Torre 1910a b cAllen 1911 1917a 1918 Barbour 19141916 1945bAnthony 1918 1926 [1942] Scharff 1922 Kraglievich 1926 Beaufort 1951 Arredondo 1961 1970b Kartashov y Mayo 1974 Iturralde-Vinent 1982 1988 Arredondo y Armifiana 1996 Arredondo 1996a [1999]) quienes postularon la existencia preterita de puentes con Norte- Centro- 0 Suramerica en dependencia de las afinidades continentales de los mamiferos antillanos
Teoricamente el puente terrestre debe cumplir con una condicion esencial permitir el paso de una parte mayoritaria (y por tanto representativa de la diversidad) de la fauna continental Claro que siempre habria tipos de mamiferos menos proclives a dispersarse o para los cuales la ubicaci6n del puente resultaria inaccesible 0 las condiciones ecologicas en este fueran desfavorables pero si algunos acceden al puente no hay raz6n para pensar que el resto de esa fauna no los acompafie mayoritariamente Como dijera Simpson (1956) Los puentes no transportan faunas desbalanceadas
En consecuencia el puente terrestre como filtro es un mecanismo de poblamiento minimamente selectivo y por ende de alta diversidad inicial (sensu Iturralde-Vinent y MacPhee 1999) 10 esencial del puente es su capacidad para transportar faunas integradas (Williams 1989) Yen eno radica la objeci6n fundamental al puente terrestre surgida tempranamente en el siglo xx (Por que s610 cuatro 6rdenes de mamiferos terrestres en las Antillas si vinieron por puentes (Donde estan los fosiles de los demas ordenes que debieron venir si es que se extinguieron Los partidarios del hipotetico puente dispusieron de casi todo el resto del siglo para responder una y otra vez que otros 6rdenes de mamiferos aparecerian representados en las Antillas Mayores el dia que en estas Islas se descubrieran fosiles precuatemarios suficientemente
304 BIOGEOGRAFIA
antiguos Esto se logro por fin terminando el siglo como se explico anteriormente (vease Contexto temporal) y ique se encontro los mismos cuatro ordenes de mamiferos terrestres conocidos del Cuatemario caribefio aparentemente los Unicos que lograron llegar
Pero ademas hasta el presente no se ha demostrado inequivoca- y concluyentemente que en algun momenta durante los ultimos 40 millones de afios haya existido una conexion terrestre continua entre las Antillas Mayores (en proceso de formacion 0 plenamente formadas) y algun punto de las regiones continentales vecinas
LA BALSA NATURAL (dispersion marina pasiva)
Durante buena parte del siglo xx el puente terrestre tuvo como unico rival a la hipotesis de la balsa natural -transporte accidental de animales sobre islotes de vegetacion flotantes a la deriva despues de arrancados y expulsados mar afuera por un rio crecido- expuesta originalmente por Matthew (191319151916 1918a 1919) y desarrollada en su aplicacion a los mamfferos terreslres antillanos principal mente por Simpson (1956) asi como por olros investigadores (Darlington 1938 1957 Williams y Koopman 1952 Hershkovitz 1958 1972 Patterson 1962 Paula 1967 Patterson y Pascual 1972 Pregill 1981 Ford 1986 Woods 1989a 1990 Morgan 1993) Por su parte Hedges et al (1992) sustentaron dicha hipotesis en estimados de divergencia (basados en el reloj molecular) en la herpetofauna antillana -involucrados tangencia1mente los mamiferos [veanse objeciones a Hedges y colaboradores (Page y Lydeard 1994 Iturralde-Vinent y MacPhee 1999) respuestas a las objeciones (Hedges et al 1994 Hedges 2001) y contrarrespuestas (MacPhee e IturraldeshyVinent 2005) asi como cuestionamientos al metoda de los estimados de divergencia (Foote et al 1999 Pregill y Crother 1999 Woods et ai 2001 )]
La hip61esis de la balsa natural se origino a partir de las numerosas ocasiones en que a 10 largo de siglos tales islotes flotantes (portando 0 no vertebrados incluidos mamiferos) han sido avistados mar afuera a cientos de kilometros de la costa mas cercana en varias partes del mundo --inc1uido el Mar Caribe- segUn consta en diarios de barcos y reportes de observadores diversos (revisados por King 1962 reportes posteriores en SICSP 1970 y Censky et al 1998) Uno de estos casos acaecido en nuestra region se reporta en un informe del SICSP (Smithsonian Institution Center for Shortlived Phenomena 197050shy52)
EIB de julio de 1969 un destructor de la armada de Estados Unidos avisto un islote flotante a unos 100 km al sur de la Base Naval de Guantanamo moviendose desde el Paso de los Vientos en direccion suroeste a una velocidad de 45 kmlh islote de unos 15 m de diametro mantenfa en pie mas de 10 palmas reales de 10-15 m de altura con suelo cubierto de hierba y arbustos [la observacion con binoculares a 18 km de distancia no preciso la presencia de vida animal] La travesia fue rastreada durante 6 dfas hasta que el 19 de julio el islote se dio por perdido tras 6 horas de busqueda aerea
305 POBLAMIENTO CONTINENTE-ISLA
La colonizaci6n exitosa de una isla de las Antillas Mayores por mamiferos terrestres continentales transportados en balsas naturales a la deriva (de cada especie al menos una hembra gravida 0 dos individuos de sexo opuesto) dependeria de factores que van a operar desde el momenta mismo de formarse la balsa asi como durante su travesia marina y su arribo a la isla
a) La posibilidad de estar en la balsa Que la distribuci6n de la especie incluya areas contiguas al rio y que el animal se encuentre en ellugar y el momenta propicios para ser incorporado a la balsa antes de que el rio crecido la desprenda Por ejemplo segun Hershkovitz (1958) los equimidos -grupo ancestral de roedores antillanos- serian buenos candidatos a ser captados por la balsa porque acostumbran a anidar en huecos del tronco de los arboles a la orilla de los rios
b) El tamano y la consistencia de la balsa Muchas se desintegran en alta mar debido a su fragilidad ante la saturaci6n Algunos autores (Beaufort 1951 Arredondo 1996a [1999]) han expresado incredulidad en cuanto a que las balsas hayan podido soportar el peso de un animal como el gran perezoso cubano (Megalocnus rodens) el mayor de los mamiferos antillanos Pero esta especie evolucion6 en Cuba y el ancestro que hizo el viaje pudo ser muy pequeno [EI perezoso oligocenico de Puerto Rico es el mas pequeno de los megaloniquidos antillanos (MacPhee e Iturralde-Vinent 1995b )]
c) Las caracterfsticas fisiol6gicas de la especie Los limites fisiol6gicos de los propagulos constituyen nuestra unica objeci6n a la dispersi6n marina por balsas como mecanismo para la formaci6n de la fauna de vertebrados en las Antillas (MacPhee e IturraldeshyVinent 2005) Unas especies tienen intrinsecamente mas posibilidades que otras para sobrevivir a las vicisitudes que deben afrontar durante la travesia (deshidrataci6n inanicion insolaci6n etc) Por ejemplo los megaloniquidos han demostrado tener extraordinaria habilidad para cruzar barreras marinas fueron los primeros mamfferos suramericanos en invadir Norteamerica mucho antes de que surgiera la conexion terrestre a finales del Plioceno (Paula 1967 Olson 1978 Webb 1985b Martin y Steadman 1999) Incluso se conoce un megaloniquido acultitico del Plioceno peruano (Muizon y McDonald 1995) y otro viviente en Suramerica (Choloepus) que pnicticamente no bebe agua 10 que ha originado una leyenda aborigen que explica por que los perezosos no toman agua (Goff art 1971) Vease ademas el inciso e
d) EI regimen de las corrientes marinas Que conduzcan a la balsa en la direcci6n necesaria EI transporte de balsas de vegetacion desde los continentes hacia las islas fue durante la mayor parte del Cenozoico sumamente improbable (MacPhee e IturraldeshyVinent 2000 148) Estos autores han insistido en que hay que tener en cuenta que durante la cambiante geografia cenozoica del Caribe las corrientes marinas de superficie no favorecieron el desplazamiento de objetos fiotantes hacia las Antillas Mayores [Pero no sabemos con certeza que efecto modificador del curso de tales objetos fiotantes pudieron introducir las tormentas tropicales si previsiblemente funcionaron en el pasado como 10 hacen ahora] Dichos autores senalaron ademas (2005) que en un experimento reciente con boyas de posicionamiento satelital liberadas a la deriva desde las Antillas Menores las boyas fueron a parar mayoritariamente a Centroamerica y a otros lugares
306 B[OGEOGRAFIA
y solo unas pocas recalaron en las Antillas Mayores y las que entraron al Mar Caribe demoraron semanas 0 meses en arribar
e) La distancia a recorrer (y en consecuencia la duracion de la travesia) entre el continente y la isla (10 que repercute decisivamente en el factor c) Este es desde luego uno de los factores mas criticos En la medida que la distancia aumenta la probabilidad de alcanzar la isla disminuye drasticamente con el agravante de que la trayectoria dificilmente seria en linea recta Si tomamos como referencia la velocidad de 45 kmlh del islote ya mencionado (SICSP 1970) cubrir la distancia aerea (aproximada) de 800 km que media entre el noroeste de Suramerica y La Espafiola Ie tomaria a la balsa 74 dias Estimando el doble de este tiempo debido a desvios de la trayectoria la duracion de la travesia probablemente resultaria excesiva para la mayoria de los mamiferos Pero pudiera muy bien ocurrir que la velocidad fuera mayor 0 que la balsa portara plantas vivas que permitirian a los fitOfagos sobrevivir [Tres de los cuatro ordenes de mamiferos terrestres conocidos de las Antillas Mayores son fitOfagos] Ademas esta geologicamente bien establecido que durante etapas de variada duracion desde el Oligoceno hasta el Pleistoceno los territorios del Caribe insular y continental aumentaron considerablemente en extension con niveles del mar 100-200 m por debajo de su nivel actual oportunidades que los propagulos aprovecharian para cruzar barreras marinas significativamente menores
f) El tamafio de la isla Las islas mayores deben interceptar mas balsas (Buckley y Knedlhans 1986) 10 que repercute directamente en el factor e La balsa puede pasar entre islas pequefias y continuar viaje sin llegar a ningun lugar
g) Las condiciones (favorables 0 adversas) del desembarco en la isla y el tiempo que Ie tomaria a uno 0 mas animales depauperados encontrarun habitat apropiado Esto dependeria a su vez del tamafio y diversidad ecologica de la isla (Darlington 1938 Miller 1966)
Tal cumulo de posibles obstaculos y dificultades constituye a primera vista un desafio a la credibilidad de la hipotesis Matthew-Simpson pero en ello radica precisamente su valor explicativo bas ado en la mas absoluta improbabilidad La colonizacion por la via marina es altamente improbable pero no absolutamente imposible si se tiene en cuenta la inmensidad del tiempo geologico se vuelve probable (McKenna 1973) Basta que solo una vez entre mill ones de intentos fallidos durante ese inmenso lapso algunos de los factores anteriormente enunciados coincidan en no ser tan desfavorables para el propagulo Y eso es todo 10 que demanda la hipotesis para cada uno de los grupos monofileticos mayores de mamiferos terrestres presentes en las Antillas Mayores Las Antillas Mayores requeririan el arribo de tan solo 7 (quizas menos) propagulos fundadores a 10 largo de los ultimos 30 0 mas millones de afios (promedio de uno cada 43 mill ones) para explicar la formacion de toda su fauna de mamiferos terrestres por dispersion marina pasiva (vease Discusion y Conc1usiones)
En comparacion con la via marina el poblamiento por la via terrestre es predecible porque la mayor parte de la fauna continental de mamiferos emplazada en la region del puente va a cruzar por este una vez que el mismo se establece y los animales logran
307 POBLAMIEITO CONTINENTE-ISLA
acceder a el el poblamiento por la via marina en cambio es impredecible porque depende total mente del azar que caracteriza a la balsa Claro que en ambas vias opera el azar pero en el primer caso se trata del azar de un evento probable en el segundo el azar de un evento improbable (Simpson 1940) Asi el puente natural (mecanismo de poblamiento minimamente selectivo = alta diversidad inicial) y la balsa natural (mecanismo de poblamiento maximamente selectivo baja diversidad inicial) pueden conceptuarse como los extremos opuestos de una gama de probabilidades de dispersi6n (McKenna 1973) EI fen6meno de la dispersi6n merece la mayor atenci6n cientifica en los estudios de biogeografia insular (Cowie y Holland 2006)
Queda claro que la balsa natural es una hip6tesis consistente con el caracter de la fauna objeto de estudio puesto que el desbalance taxon6mico (sin evidencia paleontol6gica en contrario) s610 puede ser resultado de la improbabilidad del poblamiento No obstante ello no implica necesariamente que la balsa natural haya sido el imico mecanismo (ni siquiera el principal) utilizado por los mamiferos terrestres en su colonizaci6n de las Antillas Mayores La balsa natural debe satisfacer una premisa te6rica esencial dado el caracter azaroso impredecible e improbable de una colonizaci6n exitosa esta debe ocurrir a intervalos muy espaciados a traves del tiempo geol6gico 0 sea durante todo el tiempo que en el area ocupada por las Antillas Mayores ha habido ecosistemas terrestres perrnanentemente disponibles para ser colonizados (los ultimos 40 millones de afios) Sin embargo como sabemos representantes de los cuatro 6rdenes de mamiteros terrestres conocidos de las Antillas Mayores ya estaban emplazados en dicho territorio insular en el Mioceno Inferior 10 que supone un lapso de colonizaci6n limitado al Eoceno SuperiorshyOligoceno (MacPhee 2005) sugerente de un tipo de mecanismo contrario a la colonizacion alocronica exigida por la hipotesis de la balsa natural sobre todo tratandose de una fauna caracterizada por altas tasas de extincion sin reposicion aparente
La presencia en Jamaica (hasta su extinci6n en 1877) del roedor mfuido Oryzomys antillarom con estrecha afinidad centroamericana (Woods 1989a) pudiera constituir el imico caso efectivo de poblamiento (continente-isla) de las Antillas Mayores por dispersion marina dc un mamifero terrestre para el cual no es razonable invocar otro mecanismo de colonizacion
LA GUIRNALDA INSULAR (dispersion mixta)
La geologia ha establecido con suficiente certeza que durante el Cenozoico unicamente en el Pale6geno tardio y solo a traves de la Cresta de Aves (ahora sumergida excepto la isla hom6nima) pudo haber existido una cadena de islas para el transito de mamiferos terrestres entre el norte de Suramerica y las proto-Antillas Mayores [desestimados a ese efecto el Canal dc Yucatan el Elcvado de Nicaragua la Cresta de Caiman el Elevado de Beata y las Antillas Menores (Holcombe et al 1990 MacPhee e IturraldeshyVinent 1994 1995b Iturralde-Vinent y MacPhee 1999)] Holcombe y Moore (1977) y Holcombe y Edgar (1990) postularon la existencia de esta entidad y estimaron algunas de sus posibles caracteristicas Por su parte MacPhee e Iturralde-Vinent (1995b) e Iturralde-Vinent y MacPhee (1999) precisaron los extremos proximal y distal de esa
308 BIOGEOGRAFIA
estructura acotaron la duracion y la edad de la misma y Ie dieron nombre GAARlandia (Greater AntillesA vesRidgelandia)
Segun Iturralde-Vinent y MacPhee (1999) la Cresta de Aves emergio por breve tiempo (1-2 Ma) durante la transicion Eoceno-Oligoceno (35-33 MaAP) por la coincidencia de un levantamiento tectoni co y un considerable descenso eustatico del nivel del mar En ese momenta las islas septentrionales del arco de las Antillas Mayores (Cuba centroorientalshyLa Espafiola - Puerto Rico - Islas Virgenes) constituian una sola isla grande 0 una serie de islas separadas por barreras muy estrechas La Cresta de Aves emergida entre el bloque Puerto Rico - Islas Virgenes y un microcontinente noroccidental de Suramerica -separado en ese momento del resto del continente por una via marina surgida en el Eoceno tardio- dio origen a GAARlandia
Obviamente el numero de colonizaciones exitosas por mamiferos terrestres a traves de una guimalda de Islas tiende a disminuir mientras mas anchas sean las barreras marinas entre las islas que forman la guimalda y cuando esto ocurre disminuye tambien la diversidad taxonomica de los propagulos particularmente a nivel de las categorias superiores (Simpson 1956)
Vemos asi que para los mamiferos terrestres la colonizaci6n a traves de una guimalda insular puede ser tan improbable como a la deriva en una balsa natural en dependencia de la cantidad y anchura de las barreras marinas a cruzar el tamafio y la fisiografia de las islas que integran la guimalda y el regimen de las corrientes marinas y aereas entre otros factores
Resumiendo la guimalda insular puede conceptuarse como un mecanismo de poblamiento altamente selectivo (= baja diversidad inicial) que combina la improbabilidad de la balsa natural con la colonizacion restringida temporaimente del puente terrestre
En su reciente revision de la taxonomia distribuci6n y filogenia (basada en estudios moleculares) de varios grupos de mamiferos antillanos Davalos (2004) concluyo que los perezosos megalocninos los primates y los roedores histricognatos se avienen al patron de divergencia con el continente y epoca de arribo a las Antill as implicados en la hipotesis de GAARlandia pero no asi los perezosos coloepodinos que requieren una explicacion biogeognlfica distinta Esto ultimo (sospechado por Matthew 1918a) habla sido confirmado por White y MacPhee (2001 226-227)
Nuestra evidencia filogenetica no sustenta la idea de un dado unificado surgido de la invasion de un megaloniquido el analisis sugiere mas bien dos eventos de colonizaci6n independientes [y] sustenta fuertemente un origen difiletico de las subfamilias [Choloepodinae y Megalocninae] Si hubo dos migraciones a traves de GAARlandia presumiblemente poco espaciadas en el tiempo no se puede determinar en el presente
Finalmente queda claro que GAARlandia -como guirnalda insular- es la hipotesis mas robusta propuesta hasta ahora para explicar la presencia en las Antillas Mayores de
309 POBLAMIENTO CONTINENTE- ISLA
la absoluta mayona de los mamfferos terrestres de ongen suramericano porque el caracter de esa fauna supone un mecanismo de poblamiento ancestral que sea a la vez maximamente selectivo (baja diversidad ordinal) y temporalmente restringido (transicion Eoceno-Oligoceno)
Recientemente Rega et al (2002 18) expresaron A pesar de una revision exhaustiva de la paleogeografia de la cuenca del Caribe Iturralde-Vinent y MacPhee (1999) no pudieron confirmar la presencia de una conexion terrestre continua entre Suramerica y las proto-Antillas aunque hipotetizaron su existencia [Vease la Fig 42 (= figura 6 de Iturralde-Vinent y MacPhee 1999 simplificada))
En el trabajo citado por Rega y colaboradores aparece la siguiente explicacion
Actualmente no es posible ofrecer reconstrucciones paleogeograficas detalladas de la Cresta de Aves Sin embargo hay evidencia de que porciones de la cresta deben haberse elevado subaereamente en un tiempo La cuestion cntica es si la Cresta de Aves emergida pudo haber formado algun tipo de corredor entre las Antillas Mayores y el noroeste de Suramerica Perforaciones en las alturas estructurales de la cresta pudieran proveer el registro del Paleogeno tardio que se necesita para precisar la hipotesis del corredor aunque no se puede predecir si tales datos serian suficientemente detallados para determinar si en algun momento GAARlandia constituyo una sola entidad subaerea Por ejemplo pudo haber barreras marinas por corto tiempo entre las masas terrestres de la Cresta de Aves [0) ellevantamiento y la subsidencia pudieron afectar diferentes partes de GAARlandia en diferentes tiempos AItemativamente los elementos faunales pudieron haberse dispersado de manera pulsada Movimientos episOdicos de este tipo incrementarian la extincion de los elementos faunales incapaces de trasladarse a la siguiente masa terrestre en la cadena disminuyendo asi la diversidad total (Iturralde-Vinent y MacPhee 199957)
La primera parte de la explicacion deja claro que la posibilidad de que la Cresta de Aves haya sido una conexion terrestre continua no se puede descartar pero tampoco se puede afirmar La ultima parte de la explicacion describe el efecto altamente selectivo (filtrante) de la colonizacion a traves de una cadena de islas
Y continuan lturralde-Vinent y MacPhee (199957 -58) explicando
Hasta que punto el microcontinente noroccidental de Suramerica [considerado por dichos autores como el area suministradora de propagulos hacia GAARlandiaJ estuvo fisicamente separado del resto del continente no ha sido plenamente esc1arecido Si estuvo sustancialmente aislado por barreras marinas y orogenic as durante el Paleogeno tardio entonces solo la fraccion de la biota suramericana emplazada en el microcontinente estuvo en posicion de acceder a GAARlandia Esto pudiera tener que ver con el hecho de que la fauna antillana carezca de representantes de muchos grupos suramericanos [de mamfferos) que debieron haber hecho el viaje
310 BIOGEOGRAFiA
Florida o Alturas0 0
d 6 D L1anuras25deg
Yucatan
20deg
deg0
10deg
MARCARIBE
80deg 75deg
Bahamas
Cres ta de Beata
D Mares someros
D Mares profundos
OCEANO ATLANTICa
FIGURA 42 Reconstrucci6n hipotetica de GAARlandia como conexi6n terrestre continua entre las proto-Antillas Mayores y el noroeste de Suramerica en el Pale6geno tardio segun la FIgura 6 (simplificada) de Iturralde-Vinent y MacPhee (1999)
En su defensa del puente terrestre Barbour (1945b249) se refiri6 a esto - antes 10 habfa hecho Anthony [1942363]- en los terminos siguientes
El argumento de que si las Antillas hubieran estado conectadas con el continente se encontrarian los f6siles de un numero mayor de distintas criaturas me deja frio es perfectamente concebible que la conexi6n haya ocurrido en un periodo en que la parte del continente implicada no estuviera poblada por una fauna grande y variada
Y concluyen lturralde-Vinent y MacPhee (199958) su detallada explicaci6n
Desafortunadamente sabemos muy poco sobre la composici6n faunal del noroeste de Suramerica a finales del Pale6geno
Consecuentemente si se confirmara la magnitud del aislamiento del aludido microcontinente y su concomitante baja diversidad faunal GAARlandia como conexi6n terrestre continua serf a la hip6tesis mas parsimoniosa No obstante mientras no se disponga de evidencia concluyente a este respecto hipotetizar a GAARlandia como conexi6n continua (minima selectividad) la coloca en franca vulnerabilidad te6rica al tener que afrontar la objeci6n clasica al puente terrestre (baja diversidad ordinal de la
311 POBLAMIENTO CONTINENTE-TSLA
fauna) al tiempo que se la priva de ser la hip6tesis mas robusta para explicar -como guimalda de islas- la colonizaci6n de las Antillas por mamiferos procedentes de Suramerica
ELARcA DE NOE (seudovicarianza)
El primer vicarianista antillano (desde luego sin el saberlo) fue Corral (1940) con su utilizaci6n ingenua de las ideas wegenerianas de la deriva continental aparecidas la decada anterior Pero el primer (supuesto) intento de aplicar a la formaci6n de la biota caribefia el mecanismo de la vicarianza basada en la tearia de la tect6nica de placas (surgida en la sexta decada del pas ado siglo) se debe a Rosen ([1976] 1985)
Segun Rosen las Antillas Mayores se originaron en el Cremcico en la Placa Caribe aproximadamente en la posici6n que hoy dia ocupa Centroamerica y al ser desplazadas tectonicamente hacia el este interactuaron (de algun modo no precisado por Rosen) con las margenes continentales de Norte- y Suramerica de donde (en un evento unico) adquirieron toda su fauna transportada subsecuentemente por dichas islas en su transito hacia sus posiciones actuales [De ahi la denominacion metaforica dada par McKenna (1973) a este mecanismo de poblamiento] Aunque se ha presentado esta propuesta como Model0 de Vicarianza Continente-Isla de Rosen en realidad por definici6n no constituye un evento vicariante porque las proto-Antillas Mayores no resultaron del fraccionamiento y separaci6n por tectonismo de una porcion del territorio continental como exige esta hip6tesis (vease definici6n de vicarianza al comienzo de Hipotesis Biogeograficas)
El Arca de Noe de Rosen ha sido desestimada por la mayoria de los autores contemporaneos porque (1) carece de sustento geol6gico (lturralde-Vinent y MacPhee 1999) y porque (2) en el periodo en que se afrrrna ocum6 el evento muchos de los ordenes modemos de mamiferos placentarios attn no existian (Williams 1989)
EL BARCO FUNERAL VlKINGO (tanatovicarianza)
Domning et al (1997) reportaron 10 que consideraron el mamffero terrestre mas viejo de las Antillas --circunstancia que se refleja en el titulo mismo del trabajo Oldest West Indian Land Mammal- al descubrirse un perisodactilo rinocerotoide (Hyrachius sp) en una localidad de edad Eoceno Inferior a Medio en el occidente de Jamaica
Jamaica ha tenido una historia geologica y paleogeografica distinta de la historia comun a las restantes Antillas Mayores A principios del Cenozoico Jamaica occidental (a diferencia de la porcion oriental constituida por el Bloque de las Montafias Azules) formaba parte de Centro america de la cual se desgaj6 (por el desplazamiento de la Placa Caribe) para venir a integrarse despues del Eoceno Medio con 10 que serfa la futura isla de Jamaica desaparecida por transgresi6n marina en el Eoceno Superior hasta volver a emerger totalmente en el Mioceno Medio (Iturralde-Vinent y MacPhee 1999 MacPhee e Iturralde-Vinent 2000)
312 BIOGEOGRAFiA
Por consiguiente Hyrachius ---conocido de localidades de edad Eoceno Inferior a Medio en Norteamerica- debio ocupar (10 que iba a ser) Jamaica occidental mucho antes de que esta se separara de Centroamerica De hecho este fosil Uego a su destino insular contenido en las rocas de su basamento no como un ser vivo por 10 que es erroneo considerarlo un animal antillano (lturralde-Vinent y MacPhee 2004) como han supuesto algunos (Hedges 200 I Portell et al 200 I Condis [200 I D Todo 10 que hizo fue viajar tanatovicariantemente en su barco funeral vikingo -otra de las denominaciones metaforicas de McKenna (1973)
DISCUSION Y CONCLUSIONES
Hemos visto que solo disponemos de una hipotesis plausible para explicar la mayor parte del poblamiento (continente-isla) de las Antillas Mayores por mamiferos terrestres del Neotropico continental la dispersion mixta (a traves de GAARlandia) para los perezosos (excepto Choloepodinae) los monos y los roedores (excepto Oryzomys)
Por otra parte la historia biogeogrMica de las musaraiias de las Antillas Mayores (Nesophontes y Solenodon) constituye una de las situaciones mas enigmaticas entre los mamiferos de la region (Ottenwalder 2001321) Al menos cinco mecanismos de poblamiento continente-isla han sido invocados para explicar cuando como y de donde llegaron a las Antillas Mayores estas musaraiias
I EI propagulo ancestral (del Terciario de Norteamerica) coloniz6 a las Antillas Mayores por dispersion marina desde Centroamerica 0 desde la Florida (Patterson 1962)
2 El propagulo ancestral ( del Terciario) cruzo el Atlantico Sur (supuestamente mas estreshycho en aquella epoca) directamente de Africa a las Antillas Mayores (Hershkovitz 1972)
3 El propagulo ancestral (del Terciario de Norteamerica) Uego a las Antillas Mayores en el Arca de Noe de Rosen (MacFadden 1980)
4 El propagulo ancestral (del Terciario de Norteamerica) fue transportado a las Antillas desde Centroamerica en el bloque de Jamaica occidental y de alli paso por dispersion marina a las restantes Antillas Mayores (Whidden y Asher 200 I)
5 El propagulo ancestral (del Terciario de Norteamerica) lleg6 a las Antillas Mayores a traves de GAARlandia (Whidden y Asher 2001)
Aunque hay consenso entre estos (y otros) autores sobre el origen norteamericano de las musaraiias antillanas ninguno de los cinco hipoteticos mecanismos de poblamiento resiste la evidencia en contrario filogenetica yo paleogeogrMica Ademas algunas de estas hipotesis asumen que Nesophontidae y Solenodontidae comparten un antecesor comun en las Antillas pero esta relacion no ha sido confirm ada Para dilucidar el enigmatico caso de las musaraiias antillanas se requerini profundizar en la investigaci6n geologica paleogeognifica y paleontol6gica (Iturralde-Vinent y MacPhee 1999)
De acuerdo con el conocimiento filogenetico disponible sobre el conjunto de la fauna la colonizacion original de las Antillas Mayores por mamiferos terrestres implicaria el arribo
313 POBLAMIENTO ISLA-ISLA
de solo siete propagulos fundadores perezosos 2 (coloepodinos y megalocninos si como parece tienen un origen difiletico) musaraiias 2 (nesofontidos y solenodontidos si no constituyen una misma familia como consideran algunos) monos 1 (pitecidos) roedores histricognatos 1 (equimidos capromidos y heptaxodontidos) roedores muridos 1 (Oryzomys en Jamaica)
POBLAMIENTO ISLA-ISLA
La distribucion de los mamiferos terrestres a traves del area antillana demarcada al inicio del presente capitulo (vease Contexto Geogratico Fig 41) ha sido interpretada en funcion de dos hipoteticos mecanismos de poblamiento interinsular la dispersion y la vicarianza fundamentadas como se resumen seguidamente
DISPERSION
De acuerdo con la evidencia paleogeogratica mas actualizada en las principales regresiones marinas ocurridas en la cuenca caribeiia durante los ultimos 23 Ma (transiciones OligocenoshyMioceno Mioceno Superior - Plioceno y varias veces en el Pleistoceno) las masas terrestres constitutivas de la region insular en formacion aumentaron considerablemente su extension territorial disminuyendo por tanto la distancia entre elias oportunidades que los elementos faunales aprovecharian para dispersarse de unas islas a otras
Durante mucho tiempo esta ha sido la hipotesis mas favorecida por los estudiosos del tema a partir de Simpson (1956) pero en decadas recientes ha suscitado dos objeciones
1 Es dificil en tender como pudo operar la dispersion marina entre estas islas puesto que ninguna de las Antillas Mayores posee rios suficientemente caudalosos para transportar masas significativas de vegetacion flotante (MacPhee 198432)
Es oportuno recordar aqui el islote ala deriva (con mas de diez palmas reales en pie) entre las tres Antillas Mayores mas occidentales mencionado anteriormente (vease La Balsa Natural)
2 La dispersion marina es menos efectiva para explicar la asombrosa similitud de las faunas de mamiferos terrestres a traves de las Antillas Mayores septentrionales porque como proceso aleatorio no debe producir distribuciones uniformes a 10 largo de una cadena de islas 0 si las produce entonces el proceso no es aleatorio (MacPhee e lturralde-Vinent 2000 149)
Independientemente de que las aludidas distribuciones taxonomicas estan lejos de ser uniformes 0 asombrosamente similares la hipotesis de la dispersion marina contrariamente a su canicter aleatorio y altamente improbable en el caso del poblamiento continente-isla- en el caso del poblamiento isla-isla es un proceso no aleatorio (aprovecha las oportunidades brindadas puntualmente por los periodos de maxima regresion marina) y ademas altamente probable dada la incomparablemente menor distancia entre las islas caribeiias que entre estas y los continentes vecinos
314 BIOGEOGRAFiA
V ICARIANZA
La neotectonica y las transgresiones marinas del Oligoceno Medio y del Mioceno afectaron sustancialmente la paleogeografia de la Cresta de las Antillas Mayores resultando la subdivision gradual de las partes subaereas de la Cresta Asi el este de Cuba y el norte de La Espanola conectados fisicamente durante el Oligoceno Inferior se separaron al formarse el Paso de los Vientos en el Oligoceno tardio en tanto La Espanola y Puerto Rico probablemente permanecieron conectadas hasta el Mioceno tardio en que surgio el Paso de Mona (Iturralde-Vinent y MacPhee 1999)
Tales eventos geologicos habrian dividido las distribuciones de los grupos faunales emplazados previamente en el territorio continuo de las tres islas y a partir de esas disyunciones dichos autores (tambien MacPhee et al 2000 y White y MacPhee 2001) asumieron --como segunda fase del modelo de GAARlandia- la hipotesis de la vicarianza isla-isla para explicar la distribucion multiinsular de varios grupos de mamiferos terrestres a traves de las Antillas Mayores septentrionales De acuerdo con esta hipotesis cualquier diferencia en composicion faunal entre las islas (por ejemplo la ausencia de roedores heptaxodontidos en Cuba 0 de monos platirrinos en Puerto Rico) constituye evidencia de extincion local postvicariante y no de dispersion marina fallida Lo primero se puede demostrar si aparecen los fosiles pertinentes 10 segundo no se puede demostrar (MacPhee e Iturralde-Vinent 1994) Por otra parte IturraldeshyVinent y MacPhee (1999 57) han reconocido que aunque fa vicarianza isla-isla puede representar una solucion clara para las distribuciones de varios taxones cuaternarios no puede explicar todos los casos
Rega et al (2002 18) han presentado dos objeciones al modelo de la vicarianza isla-isla aplicado a los mamiferos de las Antillas Mayores
I Si la separacion de Cuba y La Espanola precedio en varios mill ones de anos la separashycion de La Espanola y Puerto Rico el modelo vicariante de lturralde-MacPhee debe predecir mayor similitud entre las faunas de La Espanola y Puerto Rico que entre las de Cuba y La Espanola De hecho es al reyes
No obstante MacPhee et al (200335) admitieron despues que la fecha mas temprana empiricamente defendible [para la apertura del Paso de Mona] debe ser el Oligoceno Medio esto invierte la secuencia de ambas disyunciones con 10 cual queda resuelta la aparente contradiccion Sin embargo en su reciente evaluacion de la congruencia entre filogenia e hipotesis geologic as en varios grupos de mamiferos terrestres de las Antillas Mayores Davalos (2004373) encontro que ninguno de los taxones cumple con la fecha y el patron de divergencia entre las masas terrestres antillanas postulados por el modelo de GAARlandia
2 La presencia de un mismo genero en las tres Islas septentrionales de las Antillas Mayores implica [por el modelo vicariante] que la diferenciacion entre estas formas durante el intervalo desde el Mioceno ha sido unicamente de nivel especifico [Dicha presencia]
315 rRRADlAcr6~ EVOLUCrONARIA
s610 puede explicarse razonablemente como el resultado de movimientos de esos animales entre las islas en el Cuaternario
De esta objecion resulta dificil disentir [El poblamiento entre islas diferenciado solo a nivel especifico no es probable que sea anterior al Pleistoceno (Simpson 1956 15)]
Debe tenerse en cuenta que la hipotesis de la vicarianza isla-isla alude a las tres islas septentrionales de las Antillas Mayores porque como se explico antes (vease El Barco Funeral Vikingo) Jamaica no existia como isla durante el lapso (Eoceno Superior shyOligoceno) consensualmente reconocido como el mas probable para la colonizacion de las proto-Antill as Mayores por la mayo ria de los mamiferos terrestres Despues de emerger como isla en el Mioceno Medio Jamaica debe haber recibido sus mamiferos terrestres por dispersion isla-isla (Davalos 2004376) y solo en el Cuaternario parece haberrecibido (por dispersion marina desde Centroamerica) al roedor murido Oryzomys
Seguramente tanto la dispersion como la vicarianza contribuyeron a que los mamiferos terrestres se irradiaran a traves de las Antillas Mayores y grupos de islas perifericas pero las maneras y tiempos en que estos mecanismos operaron a 10 largo de dicho proceso son aun en gran medida motivos de desacuerdo e incertidumbre
IRRADIACION EVOLUCIONARIA
A continuaci6n revisamos someramente las distribuciones conocidas de los diferentes grupos de mamiferos terrestres a traves de las Islas principales de las Antillas Mayores y grupos de Islas perifericas incluidas las interpretaciones y conjeturas que la literatura reciente recoge en relaci6n con la manera en que dichas distribuciones pudieron haberse conformado durante la irradiaci6n evolucionaria de los referidos grupos de mamiferos Manifestaciones de esta irradiaci6n pueden inferirse unicamente a partir de los taxones presentes en el Cuatemario pues solo 6 de las especies conocidas corresponden al Terciario
GRUPOS ORlGINARIOS DE SURAMERICA
PEREZOSOS (Megalonychidae)
Aparte de un genero miocenico de Cuba y otro del Oligoceno de Puerto Rico se conocen 4 generos (12 especies) de perezosos cuaternarios (todos extintos) restringidos a las tres islas septentrionales de las Antillas Mayores Acratocnus Neocnus Megalocnus y Parocnus Los cuatro generos estan representados en Cuba y La Espanola y Acratocnus 10 esta ademas (como unico genero de perezoso) en Puerto Rico Esta distribuci6n ha sido interpretada por MacPhee et al (2000) eomo resultado de la vicarianza oligomiocenica entre estas islas y [dado que el Banco Puerto Rico - Islas Virgenes fue la entrada de GAARlandia a las Antillas Mayores] se preguntaron si la ausencia de los otros tres generos en Puerto Rico se debe a que nunca estuvieron en ese territorio 0 a que sus
316 BIOGEOGRAFiA
fosiles atm no han sido hallados Pero como hemos visto Rega et al (2002) atribuyeron dicha distribucion a la dispersion interinsular en el Cuatemario
Por otra parte varios amilisis c1adisticos recientes ubi can a Choloepus -genero Reciente conocido solo de Centro- y Suramerica- como taxon hermano de Aeratoenus exc1uidos los demas megaloniquidos antillanos Para White y MacPhee (2001228)
tan estrecha relacion sugiere que la divergencia entre ambos [generos] ocurrio despues de que Aeratoenus divergio de Neoenus Si esta relacion implica que Aeratoenus y Neoenus se dispersaron separadamente desde Suramerica a las AntilIas mientras Choloepus permanecia en el continente (siendo necesarias al menos tres invasiones de perezosos) 0 si Choloepus invadio Suramerica desde las Antillas es algo que el material disponible no permite determinar
La diversidad adaptativa de los perezosos se manifiesta en (1) la utilizacion del habitat especies arboreas en Neoenus y Aeratoenus y terrestres en Paroenus y Megaloenus (Webb 1985a White [1993] White et al 1996 Arredondo [1999]) y en (2) la gama de tallas corporales desde los pequenos Neoenus hasta Megaloenus rodens el mayor de los mamiferos terrestres antillanos (su peso adulto estimado en 270 kg seglin Paula 19679) esto sugiere que los perezosos se irradiaron a nichos que en contextos continentales probablemente estarian ocupados por miembros de otros c1ados (MacPhee e IturraldeshyVinent 2000)
No esta claro si el abanico de tamanos entre estos perezosos enmascara alguna tendencia al gigantismo Por 10 pronto Anderson y Handley (2002) encontraron 10 contrario en perezosos (Bradypus) de las Islas de Bocas del Toro (Panama) shyseparadas secuencialmente del area continental vecina por ascensos del nivel del mar durante los ultimos 10 ka En las cinco islas los perezosos son menores que su contraparte continental y las diferencias entre las tallas medias de las cinco poblaciones expresan una reduccion lineal significativa de la talla corporal en funcion de la edad geologica de cada isla sin relacion alguna con el area de las islas 0 la distancia que las separa del continente
MONOS (Pitheciidae)
Los primates de las Antillas Mayores (cuatro especies extintas) constituyen un grupo monofiletico con Paralouatta (en Cuba) y Antillothrix (en La Espanola) como taxones hermanos seguidos en la escalera c1adistica por Xenothrix (en Jamaica) y finalmente Callieebus como el pariente continental mas cercano esto implica una sola invasion desde Suramerica antes del Mioceno Inferior (fecha mas temprana conocida para los primates fosiles antillanos) y por tanto en consonancia con GAARlandia como la via de colonizacion (Horovitz y MacPhee 1996 1999 Horovitz [1997] MacPhee y Horovitz 2004)
En desacuerdo parcial con la anterior interpretacion la filogenia de este grupo de monos obtenida por Davalos (2004) indica un antecesor comun en Cuba La Espanola y Jamaica
317 TRRADIACTON EVOLUCIONARIA
compatible con la hipotetica conexion -propuesta tentativamente por Iturralde-Vinent y MacPhee (199980-84)- entre las Montanas Azules del este de Jamaica y la peninsula suroccidental de La Espanola en la epoca de GAARlandia
A pesar de que los primates antillanos pertenecen a un grupo viviente de monos muy pequenos (los monos titles) fueron comparativamente grandes en relacion con los platirrinos modernos y sucesivamente mayores en la direccion Antillothrix Xenothrix y Paralouatta con P marianae como el mayor platirrino conocido del Mioceno neotropical (MacPhee 2005) MacPhee et al (1994) consideraron que Xenothrix debio ser un mono de andar lento y al parecer las especies de Paralouatta abandonaron la arborealidad para adoptar un modo de vida mas terrestre (MacPhee 1996a MacPhee et al 2003 MacPhee y Meldrum 2006)
ROEOORES (Echimyidae Capromyidae Heptaxodontidae)
La edad de Zazamys en Cuba demuestra que la invasion de los roedores histricognatos en las Antillas Mayores comenzo antes del Mioceno Inferior por otra parte los representantes mas antiguos de esa irradiacion evolucionaria se encuentran en La Espanola y Puerto Rico Ambas evidencias son consistentes con la hipotesis de GAARlandia (Woods 1982 1988 Woods et al 2001)
No esta claro si los capromidos evolucionaron a partir de un equimido ancestral 0 de un protocapromido 0 si los equimidos antillanos (I sub familia 3 generos y 7 especies todas extintas) forman parte de la irradiacion de los capromidos (4 subfamilias 9 generos y 27 especies 10 vivientes) Kratochvil et al (1978b) y Rodriguez et al (1979) consideraron que Capromys nan us es un eslab6n entre Echimyidae y Capromyidae pero Mesocapromys nanus esta morfologicamente mucho mas cerca de Hexolobodon y los de mas capromidos que de los equimidos (Woods 1985) Lo que si esta claro segun Woods et al (2001) es que la subfamilia Plagiodontinae endemic a de La Espanola es el linaje basal de los capromidos y que Isolobodon Hexolobodon Capromys Geocapromys Mysateles y Mesocapromys [y supuestamente Macrocapromys] forman parte de la irradiacion temprana de Capromyidae
La subfamilia Capromyinae (5 generos y 17 especies 9 vivientes) parece ser el dado de mas reciente evolucion en la familia con invasion de Cuba desde La Espanola y dispersion desde Cuba (1) por Capromys a las islas Caiman y (2) por Geocapromys a las islas Bahamas Caiman y Jamaica y desde Jamaica a las islas Swan aunque la presencia de Geocapromys en Swan pudiera deberse a introduccion aborigen (Morgan 1985 1988 1989 1993 1994 Woods 1989a 1990) Woods et al (2001) utilizaron datos moleculares para obtener estimados independientes de divergencia entre capromidos segiJn 10 cual Geocapromys divergio cerca de los 17 MaAP Mesocapromys divergio de Capromys hace 15 MaAP en tanto lvlysateles 10 hizo hace 10 MaAP Por su parte las formas de Capromys pilorides y de Mysateles prehensilis en Isla de la Juventud divergieron de sus correspondientes subespecies nominativas en la isla principal a los 24 y 08 MaAP respectivamente
318 BIOGEOGRAFiA
Woods y Howland (I979) describieron adaptaciones morfologicas de la mandibula el hioide y la faringe asociadas a mecanismos de masticacion diferentes en plagiodontinos y caprominos y Varona (1980a) llamo la atencion sobre el gran desarrollo de los molariformes (en relacion con la longitud total del cnineo) que presentan ciertos grupos de jutias fosiles en comparacion con las vivientes 10 que pudiera interpretarse como una adaptacion alimentaria a una vegetacion mas abrasiva por parte de las jutias extinguidas
Segun Woods (1990) la familia Heptaxodontidae (2 subfamilias 5 generos y 6 especies todas extintas) tambien seria parte de la irradiacion de los capromidos con origen en el este de La Espanola 0 en Puerto Rico y subsecuente dispersion al Banco de Anguila - San Martin en las Antillas Menores (pero vease MacPhee y Flemming 2003) Este banco estaria a solo 70 km del Banco Puerto Rico - Islas Virgenes en tiempos de maxima regresion marina y esta proximidad con Puerto Rico -patria de Elasmodontomys obliquus- seguramente tuvo que ver con la presencia en las islas de Anguila y San Martin del heptaxodontino gigante Amblyrhiza relacionado mas estrechamente con los roedores de las Antillas Mayores que con los del resto de las Antillas Menores (Woods 1990 1996) Aunque los roedores suelen aumentar de tamano en las islas (Case 1978) la escala del incremento de la talla en Amblyrhiza (que pudo alcanzar mas de 200 kg de peso adulto) esta fuera de toda proporcion con los patrones conocidos en otros roedores insulares (Biknevicius et 01 1993)
Clidomys (Clidomyinae) de Jamaica difiere marcadamente en su morfologia de los heptaxodontinos (vease ademas MacPhee y Flemming 2003)10 cual dificulta explicar su presencia en esa isla Como Jamaica estuvo bajo el mar hasta el Mioceno Medio el ancestro de Clidomys colonizaria esta isla por dispersion desde el extremo suroccidental de La Espanola despues que los heptaxodontinos se originaron en esta ultima isla (Woods 1989a) Pero Pascual et 01 (I 990) especularon que Clidomys pudiera ser el resultado de una invasion posterior e independiente de los capromidos
GRUPOS ORIGINARIOS DE NORTEAMERICA
El origen de las musaranas antillanas ha resultado ser hasta ahora el mayor enigma biogeografico de la fauna de mamiferos terrestres de la region aunque la mayoria de los autores coinciden en considerarlas originarias de Norteamerica (vease Discus ion y Conclusiones en Poblamiento Continente-Isla) No obstante MacPhee e Iturralde-Vinent (1994) descartaron toda posibilidad de conexion terrestre por el Estrecho de la Florida 0 por el Canal de Yucatan y si el (supuesto) evento vicari ante de Rosen (vease El Arca de Noe) -invocado por MacFadden (1980) para resolver el enigma- fuera minimamente plausible aun asi es dificil creer que las catastrofes que 10 sucedieron (vease Contexto Temporal) hayan barrido con toda esa vieja fauna continental excepto las musaranas circunstancia ya anticipada muy tempranamente por Matthew (l918a)
De otra parte la relaci6n entre Nesophontidae y Solenodontidae permanece irresuelta Si estos linajes no estan estrechamente relacionados entonces es muy posible que
319 IRRADIACION EVOLUCIONARIA
sus distribuciones sean explicadas por mecanismos diferentes (Whidden y Asher 2001)
La descomunal diferencia de talla corporal entre las musaranas pequenas (Nesophontes) y las grandes (Soienodon) seguramente impuso a ambos grupos restricciones distintas relativas a la termorregulacion y las adaptaciones energeticas En Nesophontes la seleccion debio favorecer una alta tasa de metabolismo y crecimiento camadas mas numerosas corta expectativa de vida yalta densidad poblacionaL En Soienodon en cambio la seleccion opero en la direccion de una baja tasa metabolica camadas reducidas crecimiento lento longevidad y baja densidad poblacional (Ottenwalder [1991])
MUSARANAS PEQUENAS (Nesophontidae)
Las musaranas pequenas estan representadas en las Antillas por el genero Nesophontes (y un genero miocenico indeterminado en ambar dominicano) con 6 especies cuaternarias extinguidas distribuidas en Cuba La Espanola Puerto Rico Islas Virgenes e Islas Caiman La mayor de las musaranas pequenas Nesophontes edithae evoluciono en Puerto Rico y su arribo a las Islas Virgenes supone una dispersion sumamente reciente En las Islas Caiman Nesophontes se asemeja mas a la poblacion de Cuba de donde seguramente derivo en el Plio-Pleistoceno (junto con Capromys y Geocapromys) cuando la costa sur de Cuba estuvo mucho mas cerca de las Islas Caiman (lturralde-Vinent 2003fig 4) MacPhee et ai (1994) interpretaron la distribucion de Nesophontes en Cuba y La Espanola como resultado de la vicarianza oligomiocenica ocurrida al formarse el Paso de los Vientos pero la distancia taxonomica entre los lados del Paso es solo de nivel especifico y N micros incluso vivi6 en ambas islas segtln Patterson (1962)
Allen (1918) considero que Nesophontes micrus fue una especie primitiva no modificada para la vida arb6rea 0 fos6rea y por tanto de habitos terrestres Nesophontes edithae de Puerto Rico es mucho mayor y mas robusta que las restantes especies del genero eL indice de robustez de su femur (= anchura minima de la diafisis ongitud total del hueso) muestra que este es tan robusto como el de Solenodon paradoxus un animal dos veces mayor (Morgan et ai 1980) Choate y Birney (1968) postularon un aumento cronoclinal de la taIla corporal en N edithae confirmado mas tarde por McFarlane (1999a) mediante fechados radiocarb6nicos de la secuencia estratignifica en depositos cavernarios de Puerto Rico Solenodon no vivi6 en Puerto Rico y en ausencia de una musarana de mayor tamano Nesophontes alcanzo una talla corporal mayor en esa isla que en La Espanola 0 Cuba (Woods y Eisenberg 1989)
MUSARANAS GRANDES (Solenodontidae)
Los arcaicos almiqufes (genero Soienodon) comprenden cuatro especies una viviente (S cubanus) y otra f6sil (S arredondoi) en Cuba y una viviente (S paradoxus con dos subespecies) y otra f6sil (S marcanoi) en La Espanola Como cabria esperar las dos especies cubanas se asemejan mas entre sf que a las especies de La Espanola pero
320 BIOGEOGRAFiA
enigmaticamente S marcanoi comparte caracteres con S cuban us y S paradoxus (Woods y Eisenberg 1989 Morgan y Ottenwalder 1993 Ottenwalder 200 I) Solenodon arredondoi alcanzo una talla corporal superior a la de cualquiera de las musarafias vivientes a nivel mundial Segun Morgan y Ottenwalder (1993) extrapolando las dimensiones y el peso de los almiquies vivientes S arredondoi debio tener una longitud cabeza-tronco y un peso de 450-550 mm y 1500-2000 g respectivamente en comparacion con S cubanus (360 mm y 800 g) y S paradoxus (390 mm y 1100 g)
Entre todas las especies de almiqufes Solenodon paradoxus es la que presenta los caracteres mas derivados (Otten walder 2001) Si estas relaciones son validas la distribucion de Solenodon entre Cuba y La Espafiola debio discurrir de oeste a este como propuso MacFadden (1980) quien invoco la presencia fosH de S cubanus [pero tambien S arredondoi] en Pinar del Rio para postular que el ancestro continental entr~ por Cuba opinion compartida por Perfit y Williams (1989) y por Ottenwalder ([1991] 2001) El paso de Solenodon a Puerto Rico pudo haberlo impedido la existencia de barreras ecologicas durante la conexion del este de La Espafiola con Puerto Rico 0 por la ausencia de Solenodon en esa parte de La Espafiola en aquella epoca Sin embargo las mismas condiciones ecologicas en el este de La Espafiola no parecen haber representado una barrera efectiva para Nesophontes cuyas mas altas tasa de reproduccion y densidad poblacional presumiblemente Ie permitieron expandirse flpidamente a traves de La Espafiola (Ottenwalder [1991])
Es opinion generalizada que los almiqufes antillanos constituyen el grupo de mamiferos que primero se emplazo en el territorio insular (MacPhee 2005) y en tal sentido es de interes la evidencia sobre especificidad parasito-hospedante aportada por Cruz (1980) quien encontro que la especie de acaro (genero Cubanochirus) que parasita a los almiquies es mucho mas primitiva que la especie (del mismo genero) que paras ita a las jutias y de aqui concluyo que dicho genero de acaro debio evolucionar primero en Solenodontidae y posteriormente en Capromyidae al entrar en contacto representantes de ambas familias en Cuba 0 La Espafiola
Recientemente Roca (2004) y Roca et al (2004) dieron a conocer que los resultados de su estudio genetico-molecular sobre los almiquies indican una divergencia de 25 Ma entre Solenodon cuban us y S paradoxus -compatible con la disyuncion tectonica entre Cuba y La Espafiola (Iturralde-Vinent y MacPhee 1999 McPhee et al 2003)-- y una separacion de ambas especies comparable en magnitud a la divergencia entre familias de mamlferos Consecuentemente propusieron que Solenodon cubanus fuera clasificada en el genero Atopogaie como indico Cabrera (1925) hace 82 afios
DISCUSION Y CONCLUSIONES
Las musarafias y los roedores antillanos (excepto Oryzomys) se irradiaron tanto de sus respectivos ancestros continentales que de estos 6rdenes se reconocen familias y subfamilias endemicas de la region en contraste con los perezosos (una subfamilia y tres
321 IRRADIACION EVOLUCIONARIA
tribus endemicas) y los monos (una tribu endemica) como expresion de diferentes niveles de diversificacion evolucionaria Este proceso es dificil de explicar (traUindose de c1ados que pnicticamente comparten la misma antigiiedad en las Antmas) si es correcta la interpretacion de Pascual et at (1990636)
Nosotros consideramos estos diferentes niveles de diversificacion como las diferentes etapas de evolucion alcanzadas por cada grupo [de mamiferos] Obviamente esto estuvo relacionado con las diferentes longitudes de sus historias
Pudiera ocurrir que en el futuro se compruebe que la historia biogeognifica de la region fue mucho mas complicada
La Espanola la isla mas ecol6gicamente divers a de la region debi6 ser el centro principal de evoluci6n de los mamfferos (con 87 de las familias y 73 de las subfamilias) seguida por Cuba (con 75 de las familias y 54 de las subfamilias) las dos islas de las Antillas Mayores mas relacionadas biogeograficamente (Fig 43 Tabla 44)
eL
EL
cb CAIMAN
SWAN
A Acratocnini B Cubanocnini C Megalocnini D Mesocnini E Nesophontidae F Solenodontidae G Xenothrichini
ANGUILAshyM-0 SMARTIN
H Heteropsomyinae I Plagiodontinae J Isolobodontinae
K Hexolobodontinae L Capromyinae
M Heptaxodontinae N CIidomyinae 0 Sigmodontinae
FIGURA 43 Representacion esquematica de las relaciones interinsulares (expresadas porel numero de linajes compartidos) entre las Antillas Mayores y entre estas y los grupos de islas perifericas segun las interpretaciones de los autores citados en el texto Los numeros dentro de los circulos indican la cantidad de linajes representados en cada isla relacion dudosa Contexto taxon6mico de los linajes en la Tabla 44
bull bull
bull bull
bull bull bull
bull bull
bull bull
bull
bull
bull
bull
bull bull bull bull
bull bull
bull bull
bull
bull bull bull
bull bull
bull
bull bull
bull
322 BIOGEOGRAFiA
TABLA 44 Distribuci6n de los linajcs de mamiferos terrestres a traves de las Antillas Mayores y grupos de islas perifericas LE = La Espanola CU Cuba JA Jamaica PR = Puerto Rico IC Islas Caiman IS Islas Swan IB = Islas Bahamas IV = Islas Virgenes AS = Anguila - San Martin
FamiliaSubfumiliafIribu LE CU JA PR IC IS IB IV AS
Megalonychidae
Choloepodinae
Acratocnini
Cubanocnini
Megalocninae
Megalocnini
Mesocnini
Nesophontidae
Solenodontidae
Pitheciidae
Callicebinae
Xenothrichini
Echimyidae
Heteropsomyinae
Capromyidae
Plagiodontinae
Isolobodontinae
Hexolobodontinae
Capromyinae
H e pta x 0 don tid a e
Heptaxodontinae
Clidomyinae
Muridae
Sigmodontinae
PILOSA
SORICOMORPHA
PRIMATES
RODENTIA
bull
bull
bull
bull TOTALES 12 10 4 4 2
323 EXTlICIOIES DEL CUATERNARIO
El gigantismo ha sido sin duda una tendencia evolucionaria recurrente entre los clados mayores de mamiferos antillanos Es un hecho bien conocido que algunos grupos de mamiferos han aumentado de tamafio en las islas y varios autores han intentado explicarlo a partir de enfoques diferentes (Foster 1964 Miller 1966 Sondaar 1977 Case 1978 Eisenberg y Wilson 1978 Heaney 1978 Lawlor 1982 Lomolino 1985 Angerbjom 1985 McNab 19942001)
EXTINCIONES DEL CUATERNARIO
Las islas que integran la Subregion Antillana han perdido casi 90 de sus mamiferos terrestres durante los ultimos milenios y en proporcion con su extension territorial esta perdida ha sido mayor en la Subregion que en cualquier otro lugar del planeta (MacPhee 1996b) Las extinciones del Cuatemario eliminaron mas de las tres cuartas partes de la fauna de mamiferos terrestres conocida de las Antillas Mayores sus islas satelites y grupos de islas perifericas y el examen de este proceso en sus aspectos mas generales se aborda en esta ultima parte del presente capitulo
CRONOLOGiA Y CONSECUENCIAS
Es muy improbable que un fosil represente el ultimo individuo de un taxon extinto y entonces hay que guiarse por la edad radiometrica del fosil mas reciente conocido lIamada fecha de ultima presencia- como unica aproximacion posible a la fecha de extincion del taxon
Como se explico al inicio del capitulo (vease Contexto Temporal) el registro fosH de los mamiferos terrestres de las Antillas Mayores adolece de insuficiente sustentacion radiometrica No obstante la distribucion temporal de los escasos fechados disponibles para el Cuaternario de la region abarca etapas criticas del proceso de extincion experimentado por la referida fauna a 10 largo de los ultimos 250 mil afios
De las 52 especies que es posible reconocer actualmente como miembros extintos de la fauna cuatemaria de mamiferos terrestres de las Antillas Mayores y grupos de islas perifericas los restos fosiles de 23 de elIas (442) han sido fechados radiometricamente o se han hallado asociados estratigraficamente de manera inequivoca con otro material fechado De estos 23 fechados 3 corresponden al interglacial Sangamon 4 pertenecen a la secuencia Pleistoceno terminal - Holoceno temprano 10 se ubican en ellapso entre el arribo de los aborigenes y la incursion europea y 6 son postcolombinos (a los cuales hay que sumar otras dos especies extinguidas en los siglos XIX y xx) A partir de estas 25 especies avaladas por fechas de ultima presencia distribuidas entre los cuatro intervalos temporales sefialados hemos compuesto la Tabla 45 en la que ademas aparecen las restantes 27 especies (carentes de fechado radiometrico) ubicadas en el intervalo temporal mas probable sobre la base de (1) la asociacion estratigrafica de sus restos con evidencias culturales (amerindias 0 europeas) 0 (2) la ausencia de dicha asociacion Sobre estos criterios de ubicacion temporal procede hacer dos aclaraciones
324 BIOGEOGRAFIA
TABLA 45 Intervalos temporales mas probables para las extinciones cuatemarias de los mamiferos terrestres de las Antillas Mayores y grupos de islas perifericas sobre la base de las evidencias siguientes (l) ultimo ejemplar colectado u observado en vida (2) fechado radiometrico mas tardio en material de la especie (3) asociaci6n estratigrafica mas tardia con otro material fechado (4) asociaci6n estratigrafica con restos de animales introducidos restos humanos u objetos culturales propios del intervale (5) falta de asociaci6n con evidencias culturales [Los ntimeros entre parentesis en la columna Evidencia significan adscripci6n tentativa al intervalo] Los estimados del peso corporal (en kg) estan tomados de la literatura 0 inferidos comparativamente y distribuidos entre
las clases siguientes (a) S 01 (b) 01 a lt 1 (c) 1 a lt 10 (d) 10 a lt 100 (e) 100 a lt 200 (j) ~ 200 Las
abreviaturas que designan las islas son las de la Tabla 44
F amilialEspecie Peso IsJa Evidencia Fuente
I httervalo htterg1acial Sangamon (gt80 kaAP)
HEPTAXODONTlNAE
Amblyrhiza inundata e AS 3 McFarJane et al 1998a
CLIDOMYINAE
Clidomys osborni d JA 3 Ibid
Clidomys parvus d JA 3 Ibid
II httervalo Pleistoceno terminal ~ Holoceno temprano (20-6 kaAP)
MEGALONYCHIDAE
Aeratoenus antillensis d CD 5 Arredondo 1975 a
Aeratoenus odontrigonus d PR 3 McFarJane 1999b
Aeratoenus simorhynehus d LE 2 Rega et al 2002
M egaloenus zile f LE 2 MacPhee et al 2000
SOLENODONTIDAE
Solenodon arredondoi e CD 5 Morgan y Ottenwalder 1993
PnHECIIDAE
Paralouatta varonai d CD 5 MacPhee 1996a
EcHIMYIDAE
Heteropsomys antillensis c PR 5 McFarJane 1999b
Heteropsomys insulans c PR 5 Ibid
Puertoricomys eorozalus c PR 5 Woods 1989a
CAPROMYIDAE
P lagiodontia araeum c LE 5 MacPhee et al 2000
Macrocapromys latus c CD 5 Varona y Arredondo 1979
(Continua)
325 EXTINCIONES DEL CUATERNARIO
TABLA 45 (Continuaci6n)
F ami1ia1Especie Peso Isla Evidencia Fuente
M acrocapromys latus c CU 5 Varona y Arredondo 1979
Mesocapromys kraglievichi c CU 5 Ibid
HEPTAXODONTINAE
Xaymaca fulvopulvis a JA 2 MacPhee y Flemming 2003
III Intervalo Amerindio (6-05 kaAP)
MEGALONYCHIDAE
Acratocnus ye d LE 2 MacPhee et aI 2000
Neocnus comes c LE 2 Ibid
N eocnus dousman c LE 2 Ibid
N eocnus toupiti c LE 2 Ibid
N eocnus gliriform is c CU 4 Cordova et ai 1997
M egalocnus rodens f CU 2 MacPhee et ai 2007
Parocnus browni e CU 2 Steadman et ai 2005
P arocnus serus e LE 2 MacPhee et ai 2000
N ESOPHONTIDAE
N esophontes edithae a PR 2 McFarlane 1999b
PITHECIIDAE
Antillothrix bernensis c LE 3 Rimoli 1977
EcHIMYIDAE
Brotomys contractus a LE 4 Miller 1929c
CAPROMYIDAE
P lagiodontia spelaeum c LE 4 Ibid
M acrocapromys acevedo c CU 4 NUiiez 1990a
HEPTAXODONTINAE
Elasmodontomys obliquus d PR 3 McFarlane 1999b
Iv Intervalo Postcolombino (05 kaAP - Presente)
N ESOPHONTIDAE
N esophontes hypom icrus a LE 24 MacPhee y Fle111111ing 1999
N esophontes zam icrus a LE (4) Ibid
N esophontes m icrus a CU 24 MacPhee et aI 1999
(Continua)
326 BIOGEOGRAFiA
TABLA 45 (Continuacion)
F amilialEspecie Peso Isla Evidencia Fuente
N esophontes m icrus a CD 24 MacPhee et aI 1999
N esophontes m icrus a LE 4 MacPhee et aI 2000
N esophontes sp 1 a IC 4 Morgan 1994
N esophontes sp 2 a IC 4 Ibid
Solenodon marcanoi b LE 4 Ottenwalder 2001
PITHECIIDAE
Xenothrix mcgregori c JA 4 MacPhee y Flemming 1999
EcHIMYIDAE
Boromys offeUa a CD 24 Guarch 1984
Boromys torrei a CD 24 Ibid
Brotomys voratus a LE 4 MacPhee et aI 2000
CAPROMYIDAE
P lagiodontia ipnaeum c LE 4 MacPhee y Flemming 1999
Rhizoplagiodontia lemkei c LE 4 Ibid
Isolobodon portoricensis c PR 24 Ibid
Isolobodon montanus c LE 4 Ibid
Hexolobodon phenax c LE (4) Ibid
Capromys sp c IC 24 Morgan 1994
Geocapromys columbianus c CD 4 MacPhee y Flemming 1999
Geocapromys thoracatus b IS 1 MacPhee y Marx 1997
Geocapromys sp 1 c IC 4 Morgan 1994
Geocapromys sp 2 c IC (4) Ibid
l-IEPTAXODONTINAE
Quemisia gravis c LE (1) MacPhee y Flemming 1999
MURIDAE
Oryzomys antillarum a JA 1 Ibid
1 Flemming y MacPhee (1999) alertaron en contra de la asociacion con otro material fechado Asimismo Morgan (1993) indico que la ausencia de asociacion con evidencias culturales amerindias no es necesariamente un criterio concluyente para ubicar un taxon en el intervalo temporal anterior porque hay depositos en la region que carecen de tales evidencias y son del Holoceno tardio por fechados radiocarbonicos [No obstante
327 EXTlNCIONES DEL CUATERNAR10
la ausencia de una especie en residuarios arqueol6gicos adquiere mayor certidumbre como criterio de ubicaci6n temporal de su extinci6n en la medida que aumenta el numero de residuarios estudiados que en las Antillas Mayores se cuentan por centenares] A falta de fechado absoluto de una especie consideramos que estos criterios de ubicaci6n temporal son mas utiles que la alternativa de excluir del analisis a la especie
2 En los trabajos sobre biogeografia de mamiferos antillanos los arque610gos mas citados como autoridades para establecer el inicio de la presencia aborigen en las Antillas Mayores (Kozlowski 1974 Rouse y Allaire 1978 Rouse 1989 1992 Wilson 2001) difieren al respecto entre los 6 kaAP y los 45 kaAP (pero vease Pagan et al 200523) y hemos adoptado la mas temprana de estas fechas para el comienzo del intervalo amerindio en la Tabla 45 De todos modos cualquiera que haya sido la fecha esta no es aplicable por igual al conjunto de las Antillas Mayores porque segun algunos de los autores citados el poblamiento comenz6 por Cuba y La Espanola 6 mil anos atras continu6 en Puerto Rico hace alrededor de 3 mil anos (pero vease Burney et al 1994) yen Jamaica hace menos de 2 mil e incluso a las Islas Caiman ni siquiera llegaron [Las Islas Caiman descubiertas por Col6n en 1503 carecen absolutamente de evidencia amerindia (Stokes y Keegan 1996)]
Si los datos de la Tabla 45 representan una aceptable aproximaci6n a la realidad el proceso de las extinciones cuaternarias de los mamiferos terrestres de las Antillas Mayores y grupos de islas perifericas debi6 caracterizarse entre otros por los eventos siguientes
Aunque obviamente la fauna cuaternaria debi6 ser mucho mas numerosa que 10 que refleja el registro paleontol6gico (Morgan y Woods 1986) 52 especies (81 de la fauna conocida) desaparecieron (sin reposici6n) durante los ultimos 250 mil anos
- Al parecer las extinciones se concentraron en las especies de mayor tamano Asi el peso promedio estimado para las especies de la etapa preantr6pica (intervalos I + II de la Tabla 45) es 350 kg es 328 kg para las especies del intervalo III (amerindio) y es s610 25 kg (c1ases del peso a b c) para las especies del intervalo IV (postcolombino) Ninguna especie aut6ctona de mamifero mayor de 6 kg existe hoy en la regi6n
- De acuerdo con la distribuci6n temporal de las extinciones 61 especies sobrevivieron al interglacial Sangamon (intervalo 1)48 sobrevivieron a la transici6n del Pleistoceno terminal al Holoceno temprano (intervalo II) 34 sobrevivieron al periodo amerindio (intervalo III) y s610 12 especies han sobrevivido desde la Ilegada de los europeos (intervalo IV) Las tasas de extinci6n correspondientes (exceptuado el intervalo I) son una especie cada 1167 anos en el intervalo II una cada 393 anos en el intervalo III y una cada 23 anos en el intervalo IV
- Durante todo el proceso de las extinciones cuaternarias la perdida de especies por isla fue de 91 en La Espanola 83 en Jamaica y 62 en Cuba Bahamas retuvo su unica especie (aunque extirpada en varias de las islas) Puerto Rico e Islas Caiman perdieron todas sus especies (6 y 5 respectivamente) y las Islas Swan Virgenes y Anguila - San Martin perdieron sus respectivas especies (micas
328 BIOGEOGRAFiA
De los cuatro 6rdenes de mamiferos representados en la fauna Pilosa y Primates desaparecieron totalmente Soricomorpha y Rodentia perdieron 86 y 74 de sus especies respectivamente
CAUSALIDAD
CAUSAS NATURALES
Los cicIos glaciales e interglaciales acaecidos en el Pleistoceno causaron cambios ambientales dnisticos en las Antillas con la concomitante alternancia de cIimas frios (secos) y calidos (hUmedos) asi como de descensos y ascensos del nivel del mar aumento y disminuci6n de las areas territoriales) induso (aunque con menor intensidad) en el Holoceno Estos eventos paleogeograficos han sido consistentemente documentados (Mayo y Kartashov 1972 Bonnatti y Gartner 1973 Ortega y Arcia 1982 Prance 1982 Schubert y Medina 1982 Dodge et al 1983 Ortega [1984] Haq et al 1987 Schubert 1989 Hodell et al 1991 Guilderson et al 1994 Oppo 1997 Emslie 1998 Curtis et al 2001 Iturralde-Vinent 2003) y de ellos brindamos a continuaci6n una breve resena
Finalizando el periodo interglacial anterior a la ultima glaciaci6n (Wisconsin) el mar estaba a 8-10 m por encima de su nivel actual Cuando el avance glacial del Wisconsin a1canz6 su culminaci6n (eillamado Ultimo Maximo Glacial) hace 18 ka el mar habia descendido 120 m con respecto a dicho nivel y las islas del Caribe habian incrementado considerablemente sus extensiones subaereas en comparaci6n con las que presentan hoy dia por ejemplo Puerto Rico y las Islas Virgenes se conectaron territorialmente y las Bahamas se unieron en cinco gran des islas Prevalecia un regimen frio y seco y condiciones de aridez --debidas a una reducci6n de las precipitaciones (posiblemente a menos de 440 mmlano)- y los ambientes xericos se habian extendido a traves de las islas y bancos emergidos y abundaban los pinares (los habitats mesicos estaban restringidos a parcelas relictas en 10 alto de las montanas) En el Mar Caribe y el Golfo de Mexico la temperatura superficial del mar era 5degC mas baja que la actual y en las llanuras costeras y alturas maximas eran respectivamente 2-3degC y 6degC mas bajas Los limites de las zonas ecol6gicas altitudinales descendieron varios cientos de metros y cada zona redujo su extensi6n vertical al aumentar el gradiente de temperatura Ellimite inferior de la zona periglacial
elevaci6n donde la temperatura media minima a1canza los OdegC) de la Cordillera Central de la Republica Dominicana (altura maxima del Caribe insular con evidencia de topografia glacial) se considera que estuvo entre los 1500 y 1800 mlnm (actualmente esta a 2800 mlnm) y se puede estimar que dicho limite era inferior en Cuba por estar mas al norte que la Republica Dominicana No obstante durante este periodo la temperatura experiment6 episodios cortos (unas pocas decadas) de ascensos repetidos
Un signifieativo componente bi6tieo de las referidas condiciones paleoambientales 10 constituy6 la espectacular irradiaci6n evolucionaria experimentada por un grupo numeroso de aves rapaees diurnas y nocturnas (Wetmore 1937 Brodkorb 1969 Kurotchkin y Mayo 1973 Arredondo 1976 Olson 1978 Olson y Hilgartner 1982
329 EXTINCIONES DEL ClJATERNARIO
Suarez 2000a b 2004 Suarez y Emslie 2003) Los mamiferos de las Antillas Mayores ofrecieron a estos depredadores aviares una amplia gama de tamanos de presa que en ausencia de mamiferos del orden Carnivora en la region permitio a estas aves irradiarse a este nicho alimentario con la evolucion de una variedad de especies de muy diversa tall a corporal incluidas formas gigantes Adquirir un cuerpo gigante les propicio el acceso a recursos alimentarios que no estaban siendo explotados por ningun otro depredador (Alcover y McMinn 1994) A Cuba corresponde el mejor registro fosil de rapaces cuaternarias caribenas con siete generos de especies diurnas Dentro de estas aves existieron tres gavilanes gigantes (Accipitridae) tres carairas (Falconidae) y un condor (Cathartidae) asi como cuatro generos de especies nocturnas dos lechuzas (Tytonidae) y tres Mhos (Strigidae) dos de ell os no igualados en tamano en todo el mundo Algunos de estos generos tuvieron representacion en otras islas antillanas como La Espanola Jamaica Bahamas y Gran Caiman
Al retirarse hacia el norte los hielos de la glaciacion Wisconsin el clima se tomo mas hUmedo en las Antillas durante el Holoceno temprano y el bosque mesico se expandio en la region insular El nivel del mar ascendio acompanado de mayor pluviosidad alrededor de los 7 kaAP las Islas Virgenes quedaron separadas entre si y de Puerto Rico y las Bahamas perdieron la mayor parte de su territorio Cerca de los 3 kaAP las condiciones secas retornaron a la region e hicieron desaparecer buena parte del bosque mesico Nuevamente el clima comenzo a adquirir sus condiciones actuales hacia los 25 kaAP aproximadamente y hoy transita por una fase mesica con vegetacion xerica confinada en parcelas relictas
Sin duda estos y otros eventos naturales incidieron negativamente en la sobrevivencia de mamiferos terrestres en las Antillas Mayores asi como en los grupos de pequenas islas perifericas (Pregill y Olson 1981 Olson 2006)
McFarlane et al (1998a) demostraron convincentemente como Amblyrhiza el gigante de los roedores antillanos endemico de la pequena isla de Anguila sucumbio a las repetidas y abruptas perdidas de su habitat durante el interglacial Sangamon por las nipidas y severas oscilaciones del nivel del mar (documentadas por Neumann y Hearty 1996) al no poder responder adecuadamente a tan catastroficos eventos mediante el ajuste de su masa corporal media y otro tanto debio ocurrirle en Jamaica al otro roedor gigante Clidomys (McFarlane et al 1998b) emparentado con Amblyrhiza en la misma epoca (Tabla 45 intervalo I)
El ascenso del nivel del mar seguramente promovio la extincion de mamiferos durante la transicion del Pleistoceno al Holoceno (Tabla 45 intervalo II) con la reduccion del tamano de las Islas por las transgresiones marinas (Pregill y Olson 1981 Morgan 1993) Esto ultimo haria maximamente severa la competicion asf como la presion de los depredadores [Steadman et al (2005) no yen por que el calentamiento global ocurrido durante la transicion Pleistoceno-Holoceno habria de tener un efecto negativo sobre los perezosos siendo sus descendientes actuales especies tropic ales 0 subtropicales pero no se trata del efecto
330 BIOGEOGRAFiA
directo del cambio de temperatura sobre los animales sino del efecto indirecto y progresivo que dicho cambio genera a largo plazo al reducir el territorio insular disponible]
Por esta epoca tambien desaparecieron especies de tall a mayor como Megalocnus zile en La Espanola y otras de menor tamano (en Cuba) pero que representan la tall a maxima en sus respectivos contextos taxonomicos del Cuaternario como Solenodon arredondoi Paralouatta varonai y Macrocapromys acevedo Un mamifero puede ser grande solo a costa de reducir su densidad poblacional y esto 10 hace mis vulnerable a la extinci6n sobre todo en una isla (McNab 1994 McFarlane et al 1998a) Incidentalmente Alcover et al (1998a) afirmaron que larelacion entre la talla corporal y la extincion no siempre se cumple en las especies de menor tamano y citaron como ejemplo que entre las musarafias todas las especies de Nesophontes desaparecieron mientras sobreviven especies de Solenodon pero la comparacion correcta en todo caso serfa entre los solenodones vivientes y el gigante Solenodon arredondoi y entre Nesophontes edithae -el gigante de los nesofontes extinguido en el periodo amerindio- y el resto de sus congeneres que sobrevivieron hasta los primeros siglos de la colonizacion europea (Tabla 45 intervalos III y IV)
CAUSAS ANTROPOGENICAS
Intervalo Amerindio
La mayoria de los autores que se han referido a las extinciones acaecidas durante este intervalo (entre elIos paradigmaticamente Martin 1984a b) han responsabilizado a los aborfgenes (que se supone comenzaron a poblar las Antillas Mayores hace 6 mil afios) con la desaparicion de numerosas especies de mamiferos terrestres en la region principalmente por la via de la sobrecaza y la introduccion de animales domesticos devenidos depredadores de la fauna autoctona (Morgan y Woods 1986) Este efecto negativo de la presencia indigena sobre los mamiferos de la region ha sido dramatizado por algunos autores en terminos de holocausto faunal Sin embargo pensamos que se ha exagerado Como han expresado MacPhee et al (19993) cuando uno busca evidencia dura de complicidad humana directa en [estas] extinciones no hay mucho que encontrar Las siguientes son algunas consideraciones a tener en cuenta en tal sentido
El registro radiometrico mas joven conocido hasta ahora para un perezoso antillano (MegalocnllS rodens) es de 42 kaAP (MacPhee et ai 2007) Si se acepta que los aborfgenes entraron en Cuba hace aproximadamente 6 mil anos su convivencia con los perezosos debe haber durado alrededor de 1800 afios (0 mas porque dificilmente el referfdo fechado radiometrico haya correspondido al ultimo sobreviviente de la especie) Esta cifra contradice la hipotesis del holocausto faunal segun la cualla extincion de los mamiferos insulares de mayor talla debe ocurrir con extrema rapidez -a escala de decadas 0 de centurias- a partir de su primer contacto con los humanos (Martin y Steadman 1999) A pesar de las repetidas afirmaciones en contrario (Harrington 1921 Miller 1929c Herrera [1945] Tabio 1951 Alvarez 1951a 1957 Arredondo
331 EXTINCIONES DEL CUATERNARIO
1959 1960a Veloz y Ortega 1973 Mayo 1980b Rodriguez et al 1984 Castellanos y Pino 1985 Pino y Castellanos 1985 Nunez 1990b Gutierrez 1996 Arredondo 2004) en su mayoria basadas en evidencias no exentas de posibles accidentes de deposicion (0 de reubicacion postdeposicional) en recintos cavernarios (Diaz-Franco [2006b D no se ha demostrado inequivocamente que los aborigenes antillanos se alimentaran de perezosos regularmente Sus restos en residuarios de alimentos son escasisimos en toda la region y esto contrasta con los abundantes restos de jutias y otros mamiferos de menor talla que aparecen en esos residuarios demostrativos de la preferencia de los paleoindios por estos otros mamiferos
Los mamiferos mas importantes en la economia prehistorica de las Antillas Mayores fueron las jutias Capromys y Geocapromys en Cuba Geocapromys en Jamaica e Isolobodon en La Espanola Puerto Rico e Islas Virgenes (Aguayo 1950b Tablo 1951 Rimoli 1972 Wing 2001 Diaz-Franco [2006b D Wilkins (2001) estudi61ajutia viviente de Jamaica (Geocapromys browni) en un sitio ocupado por los aborigenes durante 100 anos y no encontr6 evidencias de que durante ese tiempo la especie haya sido sobreexplotada Asimismo en Cuba las jutias conga carabali y andaraz son escasas en depositos pleistoholocenicos en cambio son muy numerosas en residuarios indigenas y de haber sido sobreexplotadas 10 mas probable es que no hubieran sobrevivido hasta hoy
La habilidad de los humanos para penetrar en profundidad el bosque tropical y mantenerse en este sustentablemente ha sido un evento comparativamente reciente (Beran y Steele 1994) En el caso antillano la caza tierra adentro ha sido explicada como un proceso tardio de desplazamiento a partir de la explotacion de abundantes recursos marinos que ofrecian condiciones 6ptimas para el sedentarismo de las comunidades aborigenes costeras (C6rdova y Arredondo 1988 Brito 2005) Peces y mariscos dominan la arqueologia costera de las Islas antillanas en tanto el consumo de recursos terrestres (mamfferos incluidos) aumenta progresivamente en la medida que los residuarios estudiados se alejan de la costa (Scudder 1991 Wilkins 2001) En las Antillas Mayores el componente terrestre de la dieta aborigen es solo 20-30 del material recobrado en los residuarios (Wing 1977 1989) Y debio ser menor alin porque el componente vegetal de la dieta no se ha preservado Por otra parte los aborigenes cubanos domesticaron la jutia conga y seguramente la reprodujeron para comer (Pose et al 1988) practica que probablemente se repitiera con otras jutias en las demas Antillas Mayores
- Fray Bartolome de las Casas hablo de 3 millones de indios en Cuba al inicio de la colonizacion pero los historiadores subsecuentes consideraron exagerada la cifra (Poey 1851 a) No obstante se dispone de dos estimados de la densidad de poblaci6n aborigen en las Antillas Mayores a la llegada de los europeos 18 indigenaslkm2 en Puerto Rico (Rouse 1956) y 1271km2 en La Espanola (Watts 1987) Extrapolando ambos valores entre las cuatro islas principales de las Antillas Mayores tenemos que en la fecha indicada la poblaci6n aborigen pudo estar entre 16 mil y 113 mil indigenas en Puerto Rico entre 21 mil y 145 mil en Jamaica entre 141 mil y 1 mill6n en La Espanola y entre 200 mil y 14 millones en Cuba Entre las cifras maximas y las minimas nos inclinamos
332 BIOGEOGRAFiA
hacia estas ultimas atendiendo a los altos indices de mortalidad infantil y morbilidad que se evidencian en las osamentas de los pobladores prehispanicos (Martinez y Rives 1990) Uno de los hechos que apuntan hacia el bajo numero de aborigenes en las Antillas Mayores es 10 rapido que desaparecieron despues de la llegada de los europeos pnkticamente sin dejar mestizaje con estos 10 cual contrasta ostensiblemente con la situacion en la America continentaL
En suma un grupo humano (1) demograiicamente irrelevante y controlado por altos indices de morbilidad (2) consumidor mayoritariamente de organismos marinos y (3) carente de la tecnologia cinegetica necesaria para poder sobreexplotar los mamiferos que Ie sirvieron de alimento (a los cuales accedio tardiamente en su desarrollo) no es concebible que haya causado e1 holocausto faunal que se Ie atribuye a partir del error de aplicar a las comunidades aborigenes antillanas un modelo cultural de la prehistoria continentaL
Tampoco hay evidencias de que los aborigenes hayan introducido carnivoros con suficiente capacidad depredadora salvo el perro Morgan y Otlenwaldder (1993) consideraron que la depredacion por perros durante el periodo amerindio es la explicacion mas plausible para la extincion del almiqui gigante de Cuba pero como se puede observar (Tabla 54) Solenodon arredondoi aparece en el intervalo II por falta de asociacion con evidencias culturales
Consecuentemente el accionar de los aborigenes y de sus animales domesticos directamente sobre la fauna no parece haber sido por sf solo un factor determinante de la extincion de mamiferos terrestres durante el periodo amerindio en las Antillas (pero vease Steadman et al 2005)
Hay que recordar en este senti do que alin en el Holoceno tardio ocurrieron cambios clirruiticos de consideracion (vease Causas Naturales) Por ejemplo Curtis et al (2001) explicaron que la extincion de algunos ani males mesofilicos de las alturas montanosas de La Espanola pudo haber antecedido a cualquier impacto humano sigrIificativo en esos lugares por coincidir dicha extincion con una tendencia a la aridez documentada isotopicamente y con evidencias de la perdida del bosque mesico ocurridas despues de 3kaAP Yen Jamaica hace solo 2shy3 mil aDos el area de la Bahia de Jackson sustentaba una gran biomasa de vertebrados en bosque hinnedo a juzgar por la cantidad y diversidad de taxones presentes en material fosil derivado de la depredacion por lechuzas hoy el area es la mas seca de la isla con matorral xerofitico y pobreza de fauna (MacPhee 1996a) Yalm en nuestros dias huracanes de gran intensidad aniquilan poblaciones de vertebrados (que no se recuperan) en is las pequenas y llanas del Caribe (Brooks y Smith 2001)
En medio de la incertidumbre sobre la verdadera causa (0 causas) de las extinciones de mamiferos en el periodo amerindio MacPhee y Marx (1997) han propuesto su hipotesis de la hipermorbilidad la cual estipula colapsos poblacionales relativamente rapidos con la introduccion de enfermedades de alta letalidad Estos autores han razonado que
Las islas son los lugares mas apropiados para experimentar perdidas catastroficas causadas por enfermedades Los mamiferos no solo serian susceptibles en virtud de su prolongado aislamiento biologico sino que en los confines de una isla la epizootia
333 EXTINCIONES DEL CUATERNARIO
tendria la potencialidad de infectar simultaneamente a la rnayoria de los individuos de vadas especies Los neonatos son inmunologicamente inmaduros y sufririan desproporcionadamente cualquier infeccion hipervirulenta La perdida excesiva de neonatos y juveniles tendria efectos severos para la sobrevivencia de la poblacion si esta se reduce por debajo de cierto nivel sobreviene la extincion porque no hay posibilidad de restitucion en el periodo postpanzootico Las especies de mayor talla corporal van a ser las mas vulnerables a la extincion dadas sus bajas tasas reproductivas largos periodos de gestacion y reducido numero de progenie Las especies de menor tamano --caracterizadas por altas tasas reproductivas periodos cortos de gestacion y mayor numero de neonatos por parto- tambien serian fuertemente diezmadas pero muchas sobrevivirian porque su estrategia reproductiva les permite perder mayor numero de individuos Las perdidas deben ocurrir relativamente rapido (a escala de decadas) mientras que otras hipotesis (digamos los cambios climaticos) permiten intervalos mucho mas largos (a escala de siglos 0 de milenios) entre la aparicion de las causas y la produccion de sus efectos Por ejemplo todo indica que los nesofontes se extinguieron muy rapidamente y seria dificil poder invocar un impacto humano directo 0 argumentar fenomenos climaticos que pudieran explicar plausiblemente perdidas tan rapidas
MacPhee y Marx (1997 191) han dicho que por el momento la hipotesis de la hipermorbilidad no pasa de ser una abstraccion en espera de adecuada sustentacion empirica Llama la atencion que linicamente en el periodo amerindio (Tabla 45 intervalo 1Il) las extinciones abarcaron todas las clases del peso corporal (de la a a laj) y que ninguna de las especies que sobrevivieron hasta el periodo postcolombino (Tabla 45 intervalo IV mas las especies vivientes) sobrepasa los 10 kg (clases del peso a b c) en clara coincidencia con los presupuestos de la hipotetica hipermorbilidad
Intervalo Postcolornbino
Por 10 menos 37 especies de mamiferos terrestres vivian en las Antillas Mayores y grupos de Islas perifericas a la lIegada de los europeos en 1492 De elIas se han extinguido 23 shymas especies que en cualquiera de los periodos precedentes (Tabla 45~ y solo subsisten 12 especies repartidas entre Cuba (8 especies) La Espanola (2) Jamaica (1) y Bahamas (1) varias de estas en peligro critico de extincion Solo dos de las especies extinguidas durante los ultimos 500 anos se conocieron en estado viviente los roedores Oryzomys antillarum de Jamaica (extinguido en 1877) y Geocapromys thoracatus de las Islas Swan (extinguido en 1955) Las causas de las extinciones ocurridas durante este periodo son sobradamente conocidas destruccion de habitats introducci6n de animales exoticos (domesticos y comensales) y demas presiones asociadas a la actividad humana entre las cuales posiblemente habria que incluir brotes de hipermorbilidad
Durante mucho tiempo se ha afirmado la posibilidad de que especies de Nesophontes hayan sobrevivido hasta las primeras decadas del siglo xx y que incluso continuen existiendo en lugares remotos de Cuba 0 La Espanola (Allen 1918 1942 Miller 1930 Aguayo 1950a Arredondo 1960b Fischer 1977 MacFadden 1980 Woods y
334 BIOGEOGRAFiA
Ottenwalder 1991) Actualmente no existe evidencia radiometrica de que alguna especie de Nesophontes haya sobrevivido al siglo xv las fechas radiocarbonicas calibradas obtenidas en muestras de Nesophontes de Cuba y La Espanola resultaron no mas tardias que los siglos XIII y XIV (MacPhee et al 1999) Por otra parte las expediciones efectuadas a fines del siglo xx con el proposito de localizar individuos vivientes del genero en La Espanola no produjeron la menor indicacion afirmativa al respecto (Woods et al 1985) Ademas a traves de las Antillas Mayores no aparecen restos de Nesophontes en los depositos formados actualmente por las lechuzas
EPiLOGO
Las Antillas Mayores son islas oceanicas no han formado parte de un continente (con la posible excepcion de una parte de Jamaica hasta el Eoceno Medio hace 45 Ma) y durante un tiempo geologico considerable (desde el Eoceno Superior hace 40 Ma) han permanecido emergidas y separadas de los continentes vecinos Consecuentemente los mamiferos terrestres (no volantes) autoctonos de estas islas constituyen una fauna oceanica que por ende cumple con todas las caracteristicas convencionales reconocidas para este tipo de entidad biogeografica
- Baja diversidad taxonomica a nivel de las categorias superiores (solo cuatro ordenes) en comparacion con islas plataformicas continentales (0 con areas continentales) de tamano y diversidad ecologica simi lares
- Alto grado de endemismo a nivel de genero (96) y familia (50) en respuesta al prolongado aislamiento
- Desbalance ecologico de la estructura trofica ausencia de mamiferos camivoros y ungulados
- Espectaculares procesos de irradiaci6n evolucionaria como resultado de la colonizacion de territorios libres de competidores y depredadores
Tendencias evolucionarias extremas gigantismo
- Control a cargo de aves rapaces con tendencia at gigantismo en ausencia de mamiferos miembros del orden Carnivora
- Mas especies de reptiles y de murcielagos que de mamiferos terrestres relaciones que en los continentes se invierten
Alta tasa de extincion de especies (84) sin reposicion
Preservacion de especies filogeneticamente arcaicas (los llamados fosiles vivientes) los almiquies

302 BIOGEOGRAFiA
Yane 2004 Davalos 2004 MacPhee y Horovitz 2004 Roca et al 2004) es practicamente unanime en cuanto a que los ancestros continentales que dieron origen a esta fauna derivaron (1) mayormente de Suramerica perezosos (Megalonychidae) monos (Pitheciidae) y roedores (Echimyidae Capromyidae y Heptaxodontidae) (2) en menor medida de Norteamerica musarafias (Nesophontidae y Solenodontidae) y (3) menos aun de Centroamerica una especie de roedor (Muridae)
HIPOTESIS BIOGEOGRAFICAS
La definicion c1asica reconoce dos tipos de islas isla continental (porcion del territorio de un continente separado de este por movimiento tectonico 0 por ascenso del nivel del mar) e isla oceanica (porcion de territorio que no ha formado parte de un continente) La definicion sigue siendo uti por 10 que implica para la formacion de las faunas insulares en uno u otro caso As en el caso de la isla continental su fauna ya estaba emplazada en la futura isla antes de producirse su separacion del continente [Vicarianza hipotesis biogeografica de fragmentacion faunal] en el caso de la isla oceanica su fauna es el resultado de la colonizacion1 por propagulos2continentales a traves de la barrera marina entre el continente y la isla [Dispersion hipotesis biogeografica de acumulacion faunal] En el primer caso el surgimiento de la barrera marina es resultado del evento vicariante en el segundo la existencia de la barrera marina es anterior al evento dispersante Ademas de operar a escala mayor (continente-isla) ambos eventos operan secundariamente a menor escala (isla-isla)
Las dos hipotesis -vicarianza y dispersion- se han disputado hasta hoy la preferencia de los biogeografos en tanto hipotesis mas viable para explicar el proceso de formacion original (continente-isla) de la fauna de mamiferos terrestres de las Antillas Mayores [Sobre la historia de las hipotesis biogeograficas relativas al Caribe insular vease RuB y Schubert 1989 Williams 1989 Pruna 2001] Yel debate continua debido ala falta de consenso sobre 10 que Williams (1989) ha llamado el problema central de la discusion a saber la ostensiblemente baja diversidad taxonomica de esta fauna a nivel de las categorias superiores (solo cuatro ordenes) en comparacion con la diversidad al mismo nivel de las faunas de islas continentales (0 de areas del continente) simi lares en tamafio y diversidad ecologica [Por ejemplo la fauna cubana de mamiferos terrestres consideradas de conjunto las especies vivientes y las extintas- es menos diversa a los niveles taxonomicos superiores (4 ordenes y 6 familias) que 10 que cabria esperar de la diversidad ecologica y extension territorial de Cuba (110 992 km2) comparada con una isla tipicamente continental como Trinidad (4828 krn2
) -veinte veces menor que Cuba y menos diversiticada ecologicamente- donde solo las especies vivientes de mamfferos terrestres representan 6 ordenes y 16 familias (Eisenberg 1989)] Este desbalance
lColonizaci6n Arribo y establecimiento de un propagulo en una isla no ocupada por la especie (MacArthur y Wilson 1967)
2Propagulo Niimero minimo de individuos de una especie capaces de reproducirse y aumentar poblacionalmente en una isla bajo condiciones favorables (MacArthur y Wilson 1967)
303 POBLAMIENTO CONTINENTE-ISLA
taxon6mico ha sido expJicado de dos maneras (1) Inicialmente hubo mas 6rdenes presentes pero la mayoria se extingui6 y s610 sobrevivieron esos cuatro [explicaci6n vicarianista] (2) S610 representantes de esos cuatro 6rdenes lograron cruzar la barrera marina [explicaci6n dispersionista]
Cinco hipot~ticos mecanismos de poblamiento continente-isla han sido invocados durante los ultimos 140 afios (y se los sigue proponiendo actualmente) para explicar la presencia de mamiferos terrestres en las Antillas Mayores A continuaci6n pasamos a resefiarlos brevemente (incluidas sus alegadas fortalezas y debilidades) en el orden aproximado en que historicamente fueron propuestos
EL PUENTE NATURAL (dispersion terrestre activa)
En el siglo XIX el puente terrestre era pnkticamente la unica hipotesis para explicar las distribuciones disyuntas de animales en buena parte del mundo y los primeros investigadores que se ocuparon de la biogeografia antillana (Fernandez [1865] 1884 Chapman [1893] Torre 1892 Lora 1895) la asumieron resueltamente en parte intrigados por el descubrimiento en Cuba en la segunda mitad del siglo de un perezoso f6sil de gran talla Y en su aplicacion a los mamiferos terrestres de las Antillas Mayores el puente sigui6 siendo a 10 largo del siglo xx la hipotesis mas favorecida por los estudiosos del tema (Vaughan 1902a Spencer 191 Oa b Torre 1910a b cAllen 1911 1917a 1918 Barbour 19141916 1945bAnthony 1918 1926 [1942] Scharff 1922 Kraglievich 1926 Beaufort 1951 Arredondo 1961 1970b Kartashov y Mayo 1974 Iturralde-Vinent 1982 1988 Arredondo y Armifiana 1996 Arredondo 1996a [1999]) quienes postularon la existencia preterita de puentes con Norte- Centro- 0 Suramerica en dependencia de las afinidades continentales de los mamiferos antillanos
Teoricamente el puente terrestre debe cumplir con una condicion esencial permitir el paso de una parte mayoritaria (y por tanto representativa de la diversidad) de la fauna continental Claro que siempre habria tipos de mamiferos menos proclives a dispersarse o para los cuales la ubicaci6n del puente resultaria inaccesible 0 las condiciones ecologicas en este fueran desfavorables pero si algunos acceden al puente no hay raz6n para pensar que el resto de esa fauna no los acompafie mayoritariamente Como dijera Simpson (1956) Los puentes no transportan faunas desbalanceadas
En consecuencia el puente terrestre como filtro es un mecanismo de poblamiento minimamente selectivo y por ende de alta diversidad inicial (sensu Iturralde-Vinent y MacPhee 1999) 10 esencial del puente es su capacidad para transportar faunas integradas (Williams 1989) Yen eno radica la objeci6n fundamental al puente terrestre surgida tempranamente en el siglo xx (Por que s610 cuatro 6rdenes de mamiferos terrestres en las Antillas si vinieron por puentes (Donde estan los fosiles de los demas ordenes que debieron venir si es que se extinguieron Los partidarios del hipotetico puente dispusieron de casi todo el resto del siglo para responder una y otra vez que otros 6rdenes de mamiferos aparecerian representados en las Antillas Mayores el dia que en estas Islas se descubrieran fosiles precuatemarios suficientemente
304 BIOGEOGRAFIA
antiguos Esto se logro por fin terminando el siglo como se explico anteriormente (vease Contexto temporal) y ique se encontro los mismos cuatro ordenes de mamiferos terrestres conocidos del Cuatemario caribefio aparentemente los Unicos que lograron llegar
Pero ademas hasta el presente no se ha demostrado inequivoca- y concluyentemente que en algun momenta durante los ultimos 40 millones de afios haya existido una conexion terrestre continua entre las Antillas Mayores (en proceso de formacion 0 plenamente formadas) y algun punto de las regiones continentales vecinas
LA BALSA NATURAL (dispersion marina pasiva)
Durante buena parte del siglo xx el puente terrestre tuvo como unico rival a la hipotesis de la balsa natural -transporte accidental de animales sobre islotes de vegetacion flotantes a la deriva despues de arrancados y expulsados mar afuera por un rio crecido- expuesta originalmente por Matthew (191319151916 1918a 1919) y desarrollada en su aplicacion a los mamfferos terreslres antillanos principal mente por Simpson (1956) asi como por olros investigadores (Darlington 1938 1957 Williams y Koopman 1952 Hershkovitz 1958 1972 Patterson 1962 Paula 1967 Patterson y Pascual 1972 Pregill 1981 Ford 1986 Woods 1989a 1990 Morgan 1993) Por su parte Hedges et al (1992) sustentaron dicha hipotesis en estimados de divergencia (basados en el reloj molecular) en la herpetofauna antillana -involucrados tangencia1mente los mamiferos [veanse objeciones a Hedges y colaboradores (Page y Lydeard 1994 Iturralde-Vinent y MacPhee 1999) respuestas a las objeciones (Hedges et al 1994 Hedges 2001) y contrarrespuestas (MacPhee e IturraldeshyVinent 2005) asi como cuestionamientos al metoda de los estimados de divergencia (Foote et al 1999 Pregill y Crother 1999 Woods et ai 2001 )]
La hip61esis de la balsa natural se origino a partir de las numerosas ocasiones en que a 10 largo de siglos tales islotes flotantes (portando 0 no vertebrados incluidos mamiferos) han sido avistados mar afuera a cientos de kilometros de la costa mas cercana en varias partes del mundo --inc1uido el Mar Caribe- segUn consta en diarios de barcos y reportes de observadores diversos (revisados por King 1962 reportes posteriores en SICSP 1970 y Censky et al 1998) Uno de estos casos acaecido en nuestra region se reporta en un informe del SICSP (Smithsonian Institution Center for Shortlived Phenomena 197050shy52)
EIB de julio de 1969 un destructor de la armada de Estados Unidos avisto un islote flotante a unos 100 km al sur de la Base Naval de Guantanamo moviendose desde el Paso de los Vientos en direccion suroeste a una velocidad de 45 kmlh islote de unos 15 m de diametro mantenfa en pie mas de 10 palmas reales de 10-15 m de altura con suelo cubierto de hierba y arbustos [la observacion con binoculares a 18 km de distancia no preciso la presencia de vida animal] La travesia fue rastreada durante 6 dfas hasta que el 19 de julio el islote se dio por perdido tras 6 horas de busqueda aerea
305 POBLAMIENTO CONTINENTE-ISLA
La colonizaci6n exitosa de una isla de las Antillas Mayores por mamiferos terrestres continentales transportados en balsas naturales a la deriva (de cada especie al menos una hembra gravida 0 dos individuos de sexo opuesto) dependeria de factores que van a operar desde el momenta mismo de formarse la balsa asi como durante su travesia marina y su arribo a la isla
a) La posibilidad de estar en la balsa Que la distribuci6n de la especie incluya areas contiguas al rio y que el animal se encuentre en ellugar y el momenta propicios para ser incorporado a la balsa antes de que el rio crecido la desprenda Por ejemplo segun Hershkovitz (1958) los equimidos -grupo ancestral de roedores antillanos- serian buenos candidatos a ser captados por la balsa porque acostumbran a anidar en huecos del tronco de los arboles a la orilla de los rios
b) El tamano y la consistencia de la balsa Muchas se desintegran en alta mar debido a su fragilidad ante la saturaci6n Algunos autores (Beaufort 1951 Arredondo 1996a [1999]) han expresado incredulidad en cuanto a que las balsas hayan podido soportar el peso de un animal como el gran perezoso cubano (Megalocnus rodens) el mayor de los mamiferos antillanos Pero esta especie evolucion6 en Cuba y el ancestro que hizo el viaje pudo ser muy pequeno [EI perezoso oligocenico de Puerto Rico es el mas pequeno de los megaloniquidos antillanos (MacPhee e Iturralde-Vinent 1995b )]
c) Las caracterfsticas fisiol6gicas de la especie Los limites fisiol6gicos de los propagulos constituyen nuestra unica objeci6n a la dispersi6n marina por balsas como mecanismo para la formaci6n de la fauna de vertebrados en las Antillas (MacPhee e IturraldeshyVinent 2005) Unas especies tienen intrinsecamente mas posibilidades que otras para sobrevivir a las vicisitudes que deben afrontar durante la travesia (deshidrataci6n inanicion insolaci6n etc) Por ejemplo los megaloniquidos han demostrado tener extraordinaria habilidad para cruzar barreras marinas fueron los primeros mamfferos suramericanos en invadir Norteamerica mucho antes de que surgiera la conexion terrestre a finales del Plioceno (Paula 1967 Olson 1978 Webb 1985b Martin y Steadman 1999) Incluso se conoce un megaloniquido acultitico del Plioceno peruano (Muizon y McDonald 1995) y otro viviente en Suramerica (Choloepus) que pnicticamente no bebe agua 10 que ha originado una leyenda aborigen que explica por que los perezosos no toman agua (Goff art 1971) Vease ademas el inciso e
d) EI regimen de las corrientes marinas Que conduzcan a la balsa en la direcci6n necesaria EI transporte de balsas de vegetacion desde los continentes hacia las islas fue durante la mayor parte del Cenozoico sumamente improbable (MacPhee e IturraldeshyVinent 2000 148) Estos autores han insistido en que hay que tener en cuenta que durante la cambiante geografia cenozoica del Caribe las corrientes marinas de superficie no favorecieron el desplazamiento de objetos fiotantes hacia las Antillas Mayores [Pero no sabemos con certeza que efecto modificador del curso de tales objetos fiotantes pudieron introducir las tormentas tropicales si previsiblemente funcionaron en el pasado como 10 hacen ahora] Dichos autores senalaron ademas (2005) que en un experimento reciente con boyas de posicionamiento satelital liberadas a la deriva desde las Antillas Menores las boyas fueron a parar mayoritariamente a Centroamerica y a otros lugares
306 B[OGEOGRAFIA
y solo unas pocas recalaron en las Antillas Mayores y las que entraron al Mar Caribe demoraron semanas 0 meses en arribar
e) La distancia a recorrer (y en consecuencia la duracion de la travesia) entre el continente y la isla (10 que repercute decisivamente en el factor c) Este es desde luego uno de los factores mas criticos En la medida que la distancia aumenta la probabilidad de alcanzar la isla disminuye drasticamente con el agravante de que la trayectoria dificilmente seria en linea recta Si tomamos como referencia la velocidad de 45 kmlh del islote ya mencionado (SICSP 1970) cubrir la distancia aerea (aproximada) de 800 km que media entre el noroeste de Suramerica y La Espafiola Ie tomaria a la balsa 74 dias Estimando el doble de este tiempo debido a desvios de la trayectoria la duracion de la travesia probablemente resultaria excesiva para la mayoria de los mamiferos Pero pudiera muy bien ocurrir que la velocidad fuera mayor 0 que la balsa portara plantas vivas que permitirian a los fitOfagos sobrevivir [Tres de los cuatro ordenes de mamiferos terrestres conocidos de las Antillas Mayores son fitOfagos] Ademas esta geologicamente bien establecido que durante etapas de variada duracion desde el Oligoceno hasta el Pleistoceno los territorios del Caribe insular y continental aumentaron considerablemente en extension con niveles del mar 100-200 m por debajo de su nivel actual oportunidades que los propagulos aprovecharian para cruzar barreras marinas significativamente menores
f) El tamafio de la isla Las islas mayores deben interceptar mas balsas (Buckley y Knedlhans 1986) 10 que repercute directamente en el factor e La balsa puede pasar entre islas pequefias y continuar viaje sin llegar a ningun lugar
g) Las condiciones (favorables 0 adversas) del desembarco en la isla y el tiempo que Ie tomaria a uno 0 mas animales depauperados encontrarun habitat apropiado Esto dependeria a su vez del tamafio y diversidad ecologica de la isla (Darlington 1938 Miller 1966)
Tal cumulo de posibles obstaculos y dificultades constituye a primera vista un desafio a la credibilidad de la hipotesis Matthew-Simpson pero en ello radica precisamente su valor explicativo bas ado en la mas absoluta improbabilidad La colonizacion por la via marina es altamente improbable pero no absolutamente imposible si se tiene en cuenta la inmensidad del tiempo geologico se vuelve probable (McKenna 1973) Basta que solo una vez entre mill ones de intentos fallidos durante ese inmenso lapso algunos de los factores anteriormente enunciados coincidan en no ser tan desfavorables para el propagulo Y eso es todo 10 que demanda la hipotesis para cada uno de los grupos monofileticos mayores de mamiferos terrestres presentes en las Antillas Mayores Las Antillas Mayores requeririan el arribo de tan solo 7 (quizas menos) propagulos fundadores a 10 largo de los ultimos 30 0 mas millones de afios (promedio de uno cada 43 mill ones) para explicar la formacion de toda su fauna de mamiferos terrestres por dispersion marina pasiva (vease Discusion y Conc1usiones)
En comparacion con la via marina el poblamiento por la via terrestre es predecible porque la mayor parte de la fauna continental de mamiferos emplazada en la region del puente va a cruzar por este una vez que el mismo se establece y los animales logran
307 POBLAMIEITO CONTINENTE-ISLA
acceder a el el poblamiento por la via marina en cambio es impredecible porque depende total mente del azar que caracteriza a la balsa Claro que en ambas vias opera el azar pero en el primer caso se trata del azar de un evento probable en el segundo el azar de un evento improbable (Simpson 1940) Asi el puente natural (mecanismo de poblamiento minimamente selectivo = alta diversidad inicial) y la balsa natural (mecanismo de poblamiento maximamente selectivo baja diversidad inicial) pueden conceptuarse como los extremos opuestos de una gama de probabilidades de dispersi6n (McKenna 1973) EI fen6meno de la dispersi6n merece la mayor atenci6n cientifica en los estudios de biogeografia insular (Cowie y Holland 2006)
Queda claro que la balsa natural es una hip6tesis consistente con el caracter de la fauna objeto de estudio puesto que el desbalance taxon6mico (sin evidencia paleontol6gica en contrario) s610 puede ser resultado de la improbabilidad del poblamiento No obstante ello no implica necesariamente que la balsa natural haya sido el imico mecanismo (ni siquiera el principal) utilizado por los mamiferos terrestres en su colonizaci6n de las Antillas Mayores La balsa natural debe satisfacer una premisa te6rica esencial dado el caracter azaroso impredecible e improbable de una colonizaci6n exitosa esta debe ocurrir a intervalos muy espaciados a traves del tiempo geol6gico 0 sea durante todo el tiempo que en el area ocupada por las Antillas Mayores ha habido ecosistemas terrestres perrnanentemente disponibles para ser colonizados (los ultimos 40 millones de afios) Sin embargo como sabemos representantes de los cuatro 6rdenes de mamiteros terrestres conocidos de las Antillas Mayores ya estaban emplazados en dicho territorio insular en el Mioceno Inferior 10 que supone un lapso de colonizaci6n limitado al Eoceno SuperiorshyOligoceno (MacPhee 2005) sugerente de un tipo de mecanismo contrario a la colonizacion alocronica exigida por la hipotesis de la balsa natural sobre todo tratandose de una fauna caracterizada por altas tasas de extincion sin reposicion aparente
La presencia en Jamaica (hasta su extinci6n en 1877) del roedor mfuido Oryzomys antillarom con estrecha afinidad centroamericana (Woods 1989a) pudiera constituir el imico caso efectivo de poblamiento (continente-isla) de las Antillas Mayores por dispersion marina dc un mamifero terrestre para el cual no es razonable invocar otro mecanismo de colonizacion
LA GUIRNALDA INSULAR (dispersion mixta)
La geologia ha establecido con suficiente certeza que durante el Cenozoico unicamente en el Pale6geno tardio y solo a traves de la Cresta de Aves (ahora sumergida excepto la isla hom6nima) pudo haber existido una cadena de islas para el transito de mamiferos terrestres entre el norte de Suramerica y las proto-Antillas Mayores [desestimados a ese efecto el Canal dc Yucatan el Elcvado de Nicaragua la Cresta de Caiman el Elevado de Beata y las Antillas Menores (Holcombe et al 1990 MacPhee e IturraldeshyVinent 1994 1995b Iturralde-Vinent y MacPhee 1999)] Holcombe y Moore (1977) y Holcombe y Edgar (1990) postularon la existencia de esta entidad y estimaron algunas de sus posibles caracteristicas Por su parte MacPhee e Iturralde-Vinent (1995b) e Iturralde-Vinent y MacPhee (1999) precisaron los extremos proximal y distal de esa
308 BIOGEOGRAFIA
estructura acotaron la duracion y la edad de la misma y Ie dieron nombre GAARlandia (Greater AntillesA vesRidgelandia)
Segun Iturralde-Vinent y MacPhee (1999) la Cresta de Aves emergio por breve tiempo (1-2 Ma) durante la transicion Eoceno-Oligoceno (35-33 MaAP) por la coincidencia de un levantamiento tectoni co y un considerable descenso eustatico del nivel del mar En ese momenta las islas septentrionales del arco de las Antillas Mayores (Cuba centroorientalshyLa Espafiola - Puerto Rico - Islas Virgenes) constituian una sola isla grande 0 una serie de islas separadas por barreras muy estrechas La Cresta de Aves emergida entre el bloque Puerto Rico - Islas Virgenes y un microcontinente noroccidental de Suramerica -separado en ese momento del resto del continente por una via marina surgida en el Eoceno tardio- dio origen a GAARlandia
Obviamente el numero de colonizaciones exitosas por mamiferos terrestres a traves de una guimalda de Islas tiende a disminuir mientras mas anchas sean las barreras marinas entre las islas que forman la guimalda y cuando esto ocurre disminuye tambien la diversidad taxonomica de los propagulos particularmente a nivel de las categorias superiores (Simpson 1956)
Vemos asi que para los mamiferos terrestres la colonizaci6n a traves de una guimalda insular puede ser tan improbable como a la deriva en una balsa natural en dependencia de la cantidad y anchura de las barreras marinas a cruzar el tamafio y la fisiografia de las islas que integran la guimalda y el regimen de las corrientes marinas y aereas entre otros factores
Resumiendo la guimalda insular puede conceptuarse como un mecanismo de poblamiento altamente selectivo (= baja diversidad inicial) que combina la improbabilidad de la balsa natural con la colonizacion restringida temporaimente del puente terrestre
En su reciente revision de la taxonomia distribuci6n y filogenia (basada en estudios moleculares) de varios grupos de mamiferos antillanos Davalos (2004) concluyo que los perezosos megalocninos los primates y los roedores histricognatos se avienen al patron de divergencia con el continente y epoca de arribo a las Antill as implicados en la hipotesis de GAARlandia pero no asi los perezosos coloepodinos que requieren una explicacion biogeognlfica distinta Esto ultimo (sospechado por Matthew 1918a) habla sido confirmado por White y MacPhee (2001 226-227)
Nuestra evidencia filogenetica no sustenta la idea de un dado unificado surgido de la invasion de un megaloniquido el analisis sugiere mas bien dos eventos de colonizaci6n independientes [y] sustenta fuertemente un origen difiletico de las subfamilias [Choloepodinae y Megalocninae] Si hubo dos migraciones a traves de GAARlandia presumiblemente poco espaciadas en el tiempo no se puede determinar en el presente
Finalmente queda claro que GAARlandia -como guirnalda insular- es la hipotesis mas robusta propuesta hasta ahora para explicar la presencia en las Antillas Mayores de
309 POBLAMIENTO CONTINENTE- ISLA
la absoluta mayona de los mamfferos terrestres de ongen suramericano porque el caracter de esa fauna supone un mecanismo de poblamiento ancestral que sea a la vez maximamente selectivo (baja diversidad ordinal) y temporalmente restringido (transicion Eoceno-Oligoceno)
Recientemente Rega et al (2002 18) expresaron A pesar de una revision exhaustiva de la paleogeografia de la cuenca del Caribe Iturralde-Vinent y MacPhee (1999) no pudieron confirmar la presencia de una conexion terrestre continua entre Suramerica y las proto-Antillas aunque hipotetizaron su existencia [Vease la Fig 42 (= figura 6 de Iturralde-Vinent y MacPhee 1999 simplificada))
En el trabajo citado por Rega y colaboradores aparece la siguiente explicacion
Actualmente no es posible ofrecer reconstrucciones paleogeograficas detalladas de la Cresta de Aves Sin embargo hay evidencia de que porciones de la cresta deben haberse elevado subaereamente en un tiempo La cuestion cntica es si la Cresta de Aves emergida pudo haber formado algun tipo de corredor entre las Antillas Mayores y el noroeste de Suramerica Perforaciones en las alturas estructurales de la cresta pudieran proveer el registro del Paleogeno tardio que se necesita para precisar la hipotesis del corredor aunque no se puede predecir si tales datos serian suficientemente detallados para determinar si en algun momento GAARlandia constituyo una sola entidad subaerea Por ejemplo pudo haber barreras marinas por corto tiempo entre las masas terrestres de la Cresta de Aves [0) ellevantamiento y la subsidencia pudieron afectar diferentes partes de GAARlandia en diferentes tiempos AItemativamente los elementos faunales pudieron haberse dispersado de manera pulsada Movimientos episOdicos de este tipo incrementarian la extincion de los elementos faunales incapaces de trasladarse a la siguiente masa terrestre en la cadena disminuyendo asi la diversidad total (Iturralde-Vinent y MacPhee 199957)
La primera parte de la explicacion deja claro que la posibilidad de que la Cresta de Aves haya sido una conexion terrestre continua no se puede descartar pero tampoco se puede afirmar La ultima parte de la explicacion describe el efecto altamente selectivo (filtrante) de la colonizacion a traves de una cadena de islas
Y continuan lturralde-Vinent y MacPhee (199957 -58) explicando
Hasta que punto el microcontinente noroccidental de Suramerica [considerado por dichos autores como el area suministradora de propagulos hacia GAARlandiaJ estuvo fisicamente separado del resto del continente no ha sido plenamente esc1arecido Si estuvo sustancialmente aislado por barreras marinas y orogenic as durante el Paleogeno tardio entonces solo la fraccion de la biota suramericana emplazada en el microcontinente estuvo en posicion de acceder a GAARlandia Esto pudiera tener que ver con el hecho de que la fauna antillana carezca de representantes de muchos grupos suramericanos [de mamfferos) que debieron haber hecho el viaje
310 BIOGEOGRAFiA
Florida o Alturas0 0
d 6 D L1anuras25deg
Yucatan
20deg
deg0
10deg
MARCARIBE
80deg 75deg
Bahamas
Cres ta de Beata
D Mares someros
D Mares profundos
OCEANO ATLANTICa
FIGURA 42 Reconstrucci6n hipotetica de GAARlandia como conexi6n terrestre continua entre las proto-Antillas Mayores y el noroeste de Suramerica en el Pale6geno tardio segun la FIgura 6 (simplificada) de Iturralde-Vinent y MacPhee (1999)
En su defensa del puente terrestre Barbour (1945b249) se refiri6 a esto - antes 10 habfa hecho Anthony [1942363]- en los terminos siguientes
El argumento de que si las Antillas hubieran estado conectadas con el continente se encontrarian los f6siles de un numero mayor de distintas criaturas me deja frio es perfectamente concebible que la conexi6n haya ocurrido en un periodo en que la parte del continente implicada no estuviera poblada por una fauna grande y variada
Y concluyen lturralde-Vinent y MacPhee (199958) su detallada explicaci6n
Desafortunadamente sabemos muy poco sobre la composici6n faunal del noroeste de Suramerica a finales del Pale6geno
Consecuentemente si se confirmara la magnitud del aislamiento del aludido microcontinente y su concomitante baja diversidad faunal GAARlandia como conexi6n terrestre continua serf a la hip6tesis mas parsimoniosa No obstante mientras no se disponga de evidencia concluyente a este respecto hipotetizar a GAARlandia como conexi6n continua (minima selectividad) la coloca en franca vulnerabilidad te6rica al tener que afrontar la objeci6n clasica al puente terrestre (baja diversidad ordinal de la
311 POBLAMIENTO CONTINENTE-TSLA
fauna) al tiempo que se la priva de ser la hip6tesis mas robusta para explicar -como guimalda de islas- la colonizaci6n de las Antillas por mamiferos procedentes de Suramerica
ELARcA DE NOE (seudovicarianza)
El primer vicarianista antillano (desde luego sin el saberlo) fue Corral (1940) con su utilizaci6n ingenua de las ideas wegenerianas de la deriva continental aparecidas la decada anterior Pero el primer (supuesto) intento de aplicar a la formaci6n de la biota caribefia el mecanismo de la vicarianza basada en la tearia de la tect6nica de placas (surgida en la sexta decada del pas ado siglo) se debe a Rosen ([1976] 1985)
Segun Rosen las Antillas Mayores se originaron en el Cremcico en la Placa Caribe aproximadamente en la posici6n que hoy dia ocupa Centroamerica y al ser desplazadas tectonicamente hacia el este interactuaron (de algun modo no precisado por Rosen) con las margenes continentales de Norte- y Suramerica de donde (en un evento unico) adquirieron toda su fauna transportada subsecuentemente por dichas islas en su transito hacia sus posiciones actuales [De ahi la denominacion metaforica dada par McKenna (1973) a este mecanismo de poblamiento] Aunque se ha presentado esta propuesta como Model0 de Vicarianza Continente-Isla de Rosen en realidad por definici6n no constituye un evento vicariante porque las proto-Antillas Mayores no resultaron del fraccionamiento y separaci6n por tectonismo de una porcion del territorio continental como exige esta hip6tesis (vease definici6n de vicarianza al comienzo de Hipotesis Biogeograficas)
El Arca de Noe de Rosen ha sido desestimada por la mayoria de los autores contemporaneos porque (1) carece de sustento geol6gico (lturralde-Vinent y MacPhee 1999) y porque (2) en el periodo en que se afrrrna ocum6 el evento muchos de los ordenes modemos de mamiferos placentarios attn no existian (Williams 1989)
EL BARCO FUNERAL VlKINGO (tanatovicarianza)
Domning et al (1997) reportaron 10 que consideraron el mamffero terrestre mas viejo de las Antillas --circunstancia que se refleja en el titulo mismo del trabajo Oldest West Indian Land Mammal- al descubrirse un perisodactilo rinocerotoide (Hyrachius sp) en una localidad de edad Eoceno Inferior a Medio en el occidente de Jamaica
Jamaica ha tenido una historia geologica y paleogeografica distinta de la historia comun a las restantes Antillas Mayores A principios del Cenozoico Jamaica occidental (a diferencia de la porcion oriental constituida por el Bloque de las Montafias Azules) formaba parte de Centro america de la cual se desgaj6 (por el desplazamiento de la Placa Caribe) para venir a integrarse despues del Eoceno Medio con 10 que serfa la futura isla de Jamaica desaparecida por transgresi6n marina en el Eoceno Superior hasta volver a emerger totalmente en el Mioceno Medio (Iturralde-Vinent y MacPhee 1999 MacPhee e Iturralde-Vinent 2000)
312 BIOGEOGRAFiA
Por consiguiente Hyrachius ---conocido de localidades de edad Eoceno Inferior a Medio en Norteamerica- debio ocupar (10 que iba a ser) Jamaica occidental mucho antes de que esta se separara de Centroamerica De hecho este fosil Uego a su destino insular contenido en las rocas de su basamento no como un ser vivo por 10 que es erroneo considerarlo un animal antillano (lturralde-Vinent y MacPhee 2004) como han supuesto algunos (Hedges 200 I Portell et al 200 I Condis [200 I D Todo 10 que hizo fue viajar tanatovicariantemente en su barco funeral vikingo -otra de las denominaciones metaforicas de McKenna (1973)
DISCUSION Y CONCLUSIONES
Hemos visto que solo disponemos de una hipotesis plausible para explicar la mayor parte del poblamiento (continente-isla) de las Antillas Mayores por mamiferos terrestres del Neotropico continental la dispersion mixta (a traves de GAARlandia) para los perezosos (excepto Choloepodinae) los monos y los roedores (excepto Oryzomys)
Por otra parte la historia biogeogrMica de las musaraiias de las Antillas Mayores (Nesophontes y Solenodon) constituye una de las situaciones mas enigmaticas entre los mamiferos de la region (Ottenwalder 2001321) Al menos cinco mecanismos de poblamiento continente-isla han sido invocados para explicar cuando como y de donde llegaron a las Antillas Mayores estas musaraiias
I EI propagulo ancestral (del Terciario de Norteamerica) coloniz6 a las Antillas Mayores por dispersion marina desde Centroamerica 0 desde la Florida (Patterson 1962)
2 El propagulo ancestral ( del Terciario) cruzo el Atlantico Sur (supuestamente mas estreshycho en aquella epoca) directamente de Africa a las Antillas Mayores (Hershkovitz 1972)
3 El propagulo ancestral (del Terciario de Norteamerica) Uego a las Antillas Mayores en el Arca de Noe de Rosen (MacFadden 1980)
4 El propagulo ancestral (del Terciario de Norteamerica) fue transportado a las Antillas desde Centroamerica en el bloque de Jamaica occidental y de alli paso por dispersion marina a las restantes Antillas Mayores (Whidden y Asher 200 I)
5 El propagulo ancestral (del Terciario de Norteamerica) lleg6 a las Antillas Mayores a traves de GAARlandia (Whidden y Asher 2001)
Aunque hay consenso entre estos (y otros) autores sobre el origen norteamericano de las musaraiias antillanas ninguno de los cinco hipoteticos mecanismos de poblamiento resiste la evidencia en contrario filogenetica yo paleogeogrMica Ademas algunas de estas hipotesis asumen que Nesophontidae y Solenodontidae comparten un antecesor comun en las Antillas pero esta relacion no ha sido confirm ada Para dilucidar el enigmatico caso de las musaraiias antillanas se requerini profundizar en la investigaci6n geologica paleogeognifica y paleontol6gica (Iturralde-Vinent y MacPhee 1999)
De acuerdo con el conocimiento filogenetico disponible sobre el conjunto de la fauna la colonizacion original de las Antillas Mayores por mamiferos terrestres implicaria el arribo
313 POBLAMIENTO ISLA-ISLA
de solo siete propagulos fundadores perezosos 2 (coloepodinos y megalocninos si como parece tienen un origen difiletico) musaraiias 2 (nesofontidos y solenodontidos si no constituyen una misma familia como consideran algunos) monos 1 (pitecidos) roedores histricognatos 1 (equimidos capromidos y heptaxodontidos) roedores muridos 1 (Oryzomys en Jamaica)
POBLAMIENTO ISLA-ISLA
La distribucion de los mamiferos terrestres a traves del area antillana demarcada al inicio del presente capitulo (vease Contexto Geogratico Fig 41) ha sido interpretada en funcion de dos hipoteticos mecanismos de poblamiento interinsular la dispersion y la vicarianza fundamentadas como se resumen seguidamente
DISPERSION
De acuerdo con la evidencia paleogeogratica mas actualizada en las principales regresiones marinas ocurridas en la cuenca caribeiia durante los ultimos 23 Ma (transiciones OligocenoshyMioceno Mioceno Superior - Plioceno y varias veces en el Pleistoceno) las masas terrestres constitutivas de la region insular en formacion aumentaron considerablemente su extension territorial disminuyendo por tanto la distancia entre elias oportunidades que los elementos faunales aprovecharian para dispersarse de unas islas a otras
Durante mucho tiempo esta ha sido la hipotesis mas favorecida por los estudiosos del tema a partir de Simpson (1956) pero en decadas recientes ha suscitado dos objeciones
1 Es dificil en tender como pudo operar la dispersion marina entre estas islas puesto que ninguna de las Antillas Mayores posee rios suficientemente caudalosos para transportar masas significativas de vegetacion flotante (MacPhee 198432)
Es oportuno recordar aqui el islote ala deriva (con mas de diez palmas reales en pie) entre las tres Antillas Mayores mas occidentales mencionado anteriormente (vease La Balsa Natural)
2 La dispersion marina es menos efectiva para explicar la asombrosa similitud de las faunas de mamiferos terrestres a traves de las Antillas Mayores septentrionales porque como proceso aleatorio no debe producir distribuciones uniformes a 10 largo de una cadena de islas 0 si las produce entonces el proceso no es aleatorio (MacPhee e lturralde-Vinent 2000 149)
Independientemente de que las aludidas distribuciones taxonomicas estan lejos de ser uniformes 0 asombrosamente similares la hipotesis de la dispersion marina contrariamente a su canicter aleatorio y altamente improbable en el caso del poblamiento continente-isla- en el caso del poblamiento isla-isla es un proceso no aleatorio (aprovecha las oportunidades brindadas puntualmente por los periodos de maxima regresion marina) y ademas altamente probable dada la incomparablemente menor distancia entre las islas caribeiias que entre estas y los continentes vecinos
314 BIOGEOGRAFiA
V ICARIANZA
La neotectonica y las transgresiones marinas del Oligoceno Medio y del Mioceno afectaron sustancialmente la paleogeografia de la Cresta de las Antillas Mayores resultando la subdivision gradual de las partes subaereas de la Cresta Asi el este de Cuba y el norte de La Espanola conectados fisicamente durante el Oligoceno Inferior se separaron al formarse el Paso de los Vientos en el Oligoceno tardio en tanto La Espanola y Puerto Rico probablemente permanecieron conectadas hasta el Mioceno tardio en que surgio el Paso de Mona (Iturralde-Vinent y MacPhee 1999)
Tales eventos geologicos habrian dividido las distribuciones de los grupos faunales emplazados previamente en el territorio continuo de las tres islas y a partir de esas disyunciones dichos autores (tambien MacPhee et al 2000 y White y MacPhee 2001) asumieron --como segunda fase del modelo de GAARlandia- la hipotesis de la vicarianza isla-isla para explicar la distribucion multiinsular de varios grupos de mamiferos terrestres a traves de las Antillas Mayores septentrionales De acuerdo con esta hipotesis cualquier diferencia en composicion faunal entre las islas (por ejemplo la ausencia de roedores heptaxodontidos en Cuba 0 de monos platirrinos en Puerto Rico) constituye evidencia de extincion local postvicariante y no de dispersion marina fallida Lo primero se puede demostrar si aparecen los fosiles pertinentes 10 segundo no se puede demostrar (MacPhee e Iturralde-Vinent 1994) Por otra parte IturraldeshyVinent y MacPhee (1999 57) han reconocido que aunque fa vicarianza isla-isla puede representar una solucion clara para las distribuciones de varios taxones cuaternarios no puede explicar todos los casos
Rega et al (2002 18) han presentado dos objeciones al modelo de la vicarianza isla-isla aplicado a los mamiferos de las Antillas Mayores
I Si la separacion de Cuba y La Espanola precedio en varios mill ones de anos la separashycion de La Espanola y Puerto Rico el modelo vicariante de lturralde-MacPhee debe predecir mayor similitud entre las faunas de La Espanola y Puerto Rico que entre las de Cuba y La Espanola De hecho es al reyes
No obstante MacPhee et al (200335) admitieron despues que la fecha mas temprana empiricamente defendible [para la apertura del Paso de Mona] debe ser el Oligoceno Medio esto invierte la secuencia de ambas disyunciones con 10 cual queda resuelta la aparente contradiccion Sin embargo en su reciente evaluacion de la congruencia entre filogenia e hipotesis geologic as en varios grupos de mamiferos terrestres de las Antillas Mayores Davalos (2004373) encontro que ninguno de los taxones cumple con la fecha y el patron de divergencia entre las masas terrestres antillanas postulados por el modelo de GAARlandia
2 La presencia de un mismo genero en las tres Islas septentrionales de las Antillas Mayores implica [por el modelo vicariante] que la diferenciacion entre estas formas durante el intervalo desde el Mioceno ha sido unicamente de nivel especifico [Dicha presencia]
315 rRRADlAcr6~ EVOLUCrONARIA
s610 puede explicarse razonablemente como el resultado de movimientos de esos animales entre las islas en el Cuaternario
De esta objecion resulta dificil disentir [El poblamiento entre islas diferenciado solo a nivel especifico no es probable que sea anterior al Pleistoceno (Simpson 1956 15)]
Debe tenerse en cuenta que la hipotesis de la vicarianza isla-isla alude a las tres islas septentrionales de las Antillas Mayores porque como se explico antes (vease El Barco Funeral Vikingo) Jamaica no existia como isla durante el lapso (Eoceno Superior shyOligoceno) consensualmente reconocido como el mas probable para la colonizacion de las proto-Antill as Mayores por la mayo ria de los mamiferos terrestres Despues de emerger como isla en el Mioceno Medio Jamaica debe haber recibido sus mamiferos terrestres por dispersion isla-isla (Davalos 2004376) y solo en el Cuaternario parece haberrecibido (por dispersion marina desde Centroamerica) al roedor murido Oryzomys
Seguramente tanto la dispersion como la vicarianza contribuyeron a que los mamiferos terrestres se irradiaran a traves de las Antillas Mayores y grupos de islas perifericas pero las maneras y tiempos en que estos mecanismos operaron a 10 largo de dicho proceso son aun en gran medida motivos de desacuerdo e incertidumbre
IRRADIACION EVOLUCIONARIA
A continuaci6n revisamos someramente las distribuciones conocidas de los diferentes grupos de mamiferos terrestres a traves de las Islas principales de las Antillas Mayores y grupos de Islas perifericas incluidas las interpretaciones y conjeturas que la literatura reciente recoge en relaci6n con la manera en que dichas distribuciones pudieron haberse conformado durante la irradiaci6n evolucionaria de los referidos grupos de mamiferos Manifestaciones de esta irradiaci6n pueden inferirse unicamente a partir de los taxones presentes en el Cuatemario pues solo 6 de las especies conocidas corresponden al Terciario
GRUPOS ORlGINARIOS DE SURAMERICA
PEREZOSOS (Megalonychidae)
Aparte de un genero miocenico de Cuba y otro del Oligoceno de Puerto Rico se conocen 4 generos (12 especies) de perezosos cuaternarios (todos extintos) restringidos a las tres islas septentrionales de las Antillas Mayores Acratocnus Neocnus Megalocnus y Parocnus Los cuatro generos estan representados en Cuba y La Espanola y Acratocnus 10 esta ademas (como unico genero de perezoso) en Puerto Rico Esta distribuci6n ha sido interpretada por MacPhee et al (2000) eomo resultado de la vicarianza oligomiocenica entre estas islas y [dado que el Banco Puerto Rico - Islas Virgenes fue la entrada de GAARlandia a las Antillas Mayores] se preguntaron si la ausencia de los otros tres generos en Puerto Rico se debe a que nunca estuvieron en ese territorio 0 a que sus
316 BIOGEOGRAFiA
fosiles atm no han sido hallados Pero como hemos visto Rega et al (2002) atribuyeron dicha distribucion a la dispersion interinsular en el Cuatemario
Por otra parte varios amilisis c1adisticos recientes ubi can a Choloepus -genero Reciente conocido solo de Centro- y Suramerica- como taxon hermano de Aeratoenus exc1uidos los demas megaloniquidos antillanos Para White y MacPhee (2001228)
tan estrecha relacion sugiere que la divergencia entre ambos [generos] ocurrio despues de que Aeratoenus divergio de Neoenus Si esta relacion implica que Aeratoenus y Neoenus se dispersaron separadamente desde Suramerica a las AntilIas mientras Choloepus permanecia en el continente (siendo necesarias al menos tres invasiones de perezosos) 0 si Choloepus invadio Suramerica desde las Antillas es algo que el material disponible no permite determinar
La diversidad adaptativa de los perezosos se manifiesta en (1) la utilizacion del habitat especies arboreas en Neoenus y Aeratoenus y terrestres en Paroenus y Megaloenus (Webb 1985a White [1993] White et al 1996 Arredondo [1999]) y en (2) la gama de tallas corporales desde los pequenos Neoenus hasta Megaloenus rodens el mayor de los mamiferos terrestres antillanos (su peso adulto estimado en 270 kg seglin Paula 19679) esto sugiere que los perezosos se irradiaron a nichos que en contextos continentales probablemente estarian ocupados por miembros de otros c1ados (MacPhee e IturraldeshyVinent 2000)
No esta claro si el abanico de tamanos entre estos perezosos enmascara alguna tendencia al gigantismo Por 10 pronto Anderson y Handley (2002) encontraron 10 contrario en perezosos (Bradypus) de las Islas de Bocas del Toro (Panama) shyseparadas secuencialmente del area continental vecina por ascensos del nivel del mar durante los ultimos 10 ka En las cinco islas los perezosos son menores que su contraparte continental y las diferencias entre las tallas medias de las cinco poblaciones expresan una reduccion lineal significativa de la talla corporal en funcion de la edad geologica de cada isla sin relacion alguna con el area de las islas 0 la distancia que las separa del continente
MONOS (Pitheciidae)
Los primates de las Antillas Mayores (cuatro especies extintas) constituyen un grupo monofiletico con Paralouatta (en Cuba) y Antillothrix (en La Espanola) como taxones hermanos seguidos en la escalera c1adistica por Xenothrix (en Jamaica) y finalmente Callieebus como el pariente continental mas cercano esto implica una sola invasion desde Suramerica antes del Mioceno Inferior (fecha mas temprana conocida para los primates fosiles antillanos) y por tanto en consonancia con GAARlandia como la via de colonizacion (Horovitz y MacPhee 1996 1999 Horovitz [1997] MacPhee y Horovitz 2004)
En desacuerdo parcial con la anterior interpretacion la filogenia de este grupo de monos obtenida por Davalos (2004) indica un antecesor comun en Cuba La Espanola y Jamaica
317 TRRADIACTON EVOLUCIONARIA
compatible con la hipotetica conexion -propuesta tentativamente por Iturralde-Vinent y MacPhee (199980-84)- entre las Montanas Azules del este de Jamaica y la peninsula suroccidental de La Espanola en la epoca de GAARlandia
A pesar de que los primates antillanos pertenecen a un grupo viviente de monos muy pequenos (los monos titles) fueron comparativamente grandes en relacion con los platirrinos modernos y sucesivamente mayores en la direccion Antillothrix Xenothrix y Paralouatta con P marianae como el mayor platirrino conocido del Mioceno neotropical (MacPhee 2005) MacPhee et al (1994) consideraron que Xenothrix debio ser un mono de andar lento y al parecer las especies de Paralouatta abandonaron la arborealidad para adoptar un modo de vida mas terrestre (MacPhee 1996a MacPhee et al 2003 MacPhee y Meldrum 2006)
ROEOORES (Echimyidae Capromyidae Heptaxodontidae)
La edad de Zazamys en Cuba demuestra que la invasion de los roedores histricognatos en las Antillas Mayores comenzo antes del Mioceno Inferior por otra parte los representantes mas antiguos de esa irradiacion evolucionaria se encuentran en La Espanola y Puerto Rico Ambas evidencias son consistentes con la hipotesis de GAARlandia (Woods 1982 1988 Woods et al 2001)
No esta claro si los capromidos evolucionaron a partir de un equimido ancestral 0 de un protocapromido 0 si los equimidos antillanos (I sub familia 3 generos y 7 especies todas extintas) forman parte de la irradiacion de los capromidos (4 subfamilias 9 generos y 27 especies 10 vivientes) Kratochvil et al (1978b) y Rodriguez et al (1979) consideraron que Capromys nan us es un eslab6n entre Echimyidae y Capromyidae pero Mesocapromys nanus esta morfologicamente mucho mas cerca de Hexolobodon y los de mas capromidos que de los equimidos (Woods 1985) Lo que si esta claro segun Woods et al (2001) es que la subfamilia Plagiodontinae endemic a de La Espanola es el linaje basal de los capromidos y que Isolobodon Hexolobodon Capromys Geocapromys Mysateles y Mesocapromys [y supuestamente Macrocapromys] forman parte de la irradiacion temprana de Capromyidae
La subfamilia Capromyinae (5 generos y 17 especies 9 vivientes) parece ser el dado de mas reciente evolucion en la familia con invasion de Cuba desde La Espanola y dispersion desde Cuba (1) por Capromys a las islas Caiman y (2) por Geocapromys a las islas Bahamas Caiman y Jamaica y desde Jamaica a las islas Swan aunque la presencia de Geocapromys en Swan pudiera deberse a introduccion aborigen (Morgan 1985 1988 1989 1993 1994 Woods 1989a 1990) Woods et al (2001) utilizaron datos moleculares para obtener estimados independientes de divergencia entre capromidos segiJn 10 cual Geocapromys divergio cerca de los 17 MaAP Mesocapromys divergio de Capromys hace 15 MaAP en tanto lvlysateles 10 hizo hace 10 MaAP Por su parte las formas de Capromys pilorides y de Mysateles prehensilis en Isla de la Juventud divergieron de sus correspondientes subespecies nominativas en la isla principal a los 24 y 08 MaAP respectivamente
318 BIOGEOGRAFiA
Woods y Howland (I979) describieron adaptaciones morfologicas de la mandibula el hioide y la faringe asociadas a mecanismos de masticacion diferentes en plagiodontinos y caprominos y Varona (1980a) llamo la atencion sobre el gran desarrollo de los molariformes (en relacion con la longitud total del cnineo) que presentan ciertos grupos de jutias fosiles en comparacion con las vivientes 10 que pudiera interpretarse como una adaptacion alimentaria a una vegetacion mas abrasiva por parte de las jutias extinguidas
Segun Woods (1990) la familia Heptaxodontidae (2 subfamilias 5 generos y 6 especies todas extintas) tambien seria parte de la irradiacion de los capromidos con origen en el este de La Espanola 0 en Puerto Rico y subsecuente dispersion al Banco de Anguila - San Martin en las Antillas Menores (pero vease MacPhee y Flemming 2003) Este banco estaria a solo 70 km del Banco Puerto Rico - Islas Virgenes en tiempos de maxima regresion marina y esta proximidad con Puerto Rico -patria de Elasmodontomys obliquus- seguramente tuvo que ver con la presencia en las islas de Anguila y San Martin del heptaxodontino gigante Amblyrhiza relacionado mas estrechamente con los roedores de las Antillas Mayores que con los del resto de las Antillas Menores (Woods 1990 1996) Aunque los roedores suelen aumentar de tamano en las islas (Case 1978) la escala del incremento de la talla en Amblyrhiza (que pudo alcanzar mas de 200 kg de peso adulto) esta fuera de toda proporcion con los patrones conocidos en otros roedores insulares (Biknevicius et 01 1993)
Clidomys (Clidomyinae) de Jamaica difiere marcadamente en su morfologia de los heptaxodontinos (vease ademas MacPhee y Flemming 2003)10 cual dificulta explicar su presencia en esa isla Como Jamaica estuvo bajo el mar hasta el Mioceno Medio el ancestro de Clidomys colonizaria esta isla por dispersion desde el extremo suroccidental de La Espanola despues que los heptaxodontinos se originaron en esta ultima isla (Woods 1989a) Pero Pascual et 01 (I 990) especularon que Clidomys pudiera ser el resultado de una invasion posterior e independiente de los capromidos
GRUPOS ORIGINARIOS DE NORTEAMERICA
El origen de las musaranas antillanas ha resultado ser hasta ahora el mayor enigma biogeografico de la fauna de mamiferos terrestres de la region aunque la mayoria de los autores coinciden en considerarlas originarias de Norteamerica (vease Discus ion y Conclusiones en Poblamiento Continente-Isla) No obstante MacPhee e Iturralde-Vinent (1994) descartaron toda posibilidad de conexion terrestre por el Estrecho de la Florida 0 por el Canal de Yucatan y si el (supuesto) evento vicari ante de Rosen (vease El Arca de Noe) -invocado por MacFadden (1980) para resolver el enigma- fuera minimamente plausible aun asi es dificil creer que las catastrofes que 10 sucedieron (vease Contexto Temporal) hayan barrido con toda esa vieja fauna continental excepto las musaranas circunstancia ya anticipada muy tempranamente por Matthew (l918a)
De otra parte la relaci6n entre Nesophontidae y Solenodontidae permanece irresuelta Si estos linajes no estan estrechamente relacionados entonces es muy posible que
319 IRRADIACION EVOLUCIONARIA
sus distribuciones sean explicadas por mecanismos diferentes (Whidden y Asher 2001)
La descomunal diferencia de talla corporal entre las musaranas pequenas (Nesophontes) y las grandes (Soienodon) seguramente impuso a ambos grupos restricciones distintas relativas a la termorregulacion y las adaptaciones energeticas En Nesophontes la seleccion debio favorecer una alta tasa de metabolismo y crecimiento camadas mas numerosas corta expectativa de vida yalta densidad poblacionaL En Soienodon en cambio la seleccion opero en la direccion de una baja tasa metabolica camadas reducidas crecimiento lento longevidad y baja densidad poblacional (Ottenwalder [1991])
MUSARANAS PEQUENAS (Nesophontidae)
Las musaranas pequenas estan representadas en las Antillas por el genero Nesophontes (y un genero miocenico indeterminado en ambar dominicano) con 6 especies cuaternarias extinguidas distribuidas en Cuba La Espanola Puerto Rico Islas Virgenes e Islas Caiman La mayor de las musaranas pequenas Nesophontes edithae evoluciono en Puerto Rico y su arribo a las Islas Virgenes supone una dispersion sumamente reciente En las Islas Caiman Nesophontes se asemeja mas a la poblacion de Cuba de donde seguramente derivo en el Plio-Pleistoceno (junto con Capromys y Geocapromys) cuando la costa sur de Cuba estuvo mucho mas cerca de las Islas Caiman (lturralde-Vinent 2003fig 4) MacPhee et ai (1994) interpretaron la distribucion de Nesophontes en Cuba y La Espanola como resultado de la vicarianza oligomiocenica ocurrida al formarse el Paso de los Vientos pero la distancia taxonomica entre los lados del Paso es solo de nivel especifico y N micros incluso vivi6 en ambas islas segtln Patterson (1962)
Allen (1918) considero que Nesophontes micrus fue una especie primitiva no modificada para la vida arb6rea 0 fos6rea y por tanto de habitos terrestres Nesophontes edithae de Puerto Rico es mucho mayor y mas robusta que las restantes especies del genero eL indice de robustez de su femur (= anchura minima de la diafisis ongitud total del hueso) muestra que este es tan robusto como el de Solenodon paradoxus un animal dos veces mayor (Morgan et ai 1980) Choate y Birney (1968) postularon un aumento cronoclinal de la taIla corporal en N edithae confirmado mas tarde por McFarlane (1999a) mediante fechados radiocarb6nicos de la secuencia estratignifica en depositos cavernarios de Puerto Rico Solenodon no vivi6 en Puerto Rico y en ausencia de una musarana de mayor tamano Nesophontes alcanzo una talla corporal mayor en esa isla que en La Espanola 0 Cuba (Woods y Eisenberg 1989)
MUSARANAS GRANDES (Solenodontidae)
Los arcaicos almiqufes (genero Soienodon) comprenden cuatro especies una viviente (S cubanus) y otra f6sil (S arredondoi) en Cuba y una viviente (S paradoxus con dos subespecies) y otra f6sil (S marcanoi) en La Espanola Como cabria esperar las dos especies cubanas se asemejan mas entre sf que a las especies de La Espanola pero
320 BIOGEOGRAFiA
enigmaticamente S marcanoi comparte caracteres con S cuban us y S paradoxus (Woods y Eisenberg 1989 Morgan y Ottenwalder 1993 Ottenwalder 200 I) Solenodon arredondoi alcanzo una talla corporal superior a la de cualquiera de las musarafias vivientes a nivel mundial Segun Morgan y Ottenwalder (1993) extrapolando las dimensiones y el peso de los almiquies vivientes S arredondoi debio tener una longitud cabeza-tronco y un peso de 450-550 mm y 1500-2000 g respectivamente en comparacion con S cubanus (360 mm y 800 g) y S paradoxus (390 mm y 1100 g)
Entre todas las especies de almiqufes Solenodon paradoxus es la que presenta los caracteres mas derivados (Otten walder 2001) Si estas relaciones son validas la distribucion de Solenodon entre Cuba y La Espafiola debio discurrir de oeste a este como propuso MacFadden (1980) quien invoco la presencia fosH de S cubanus [pero tambien S arredondoi] en Pinar del Rio para postular que el ancestro continental entr~ por Cuba opinion compartida por Perfit y Williams (1989) y por Ottenwalder ([1991] 2001) El paso de Solenodon a Puerto Rico pudo haberlo impedido la existencia de barreras ecologicas durante la conexion del este de La Espafiola con Puerto Rico 0 por la ausencia de Solenodon en esa parte de La Espafiola en aquella epoca Sin embargo las mismas condiciones ecologicas en el este de La Espafiola no parecen haber representado una barrera efectiva para Nesophontes cuyas mas altas tasa de reproduccion y densidad poblacional presumiblemente Ie permitieron expandirse flpidamente a traves de La Espafiola (Ottenwalder [1991])
Es opinion generalizada que los almiqufes antillanos constituyen el grupo de mamiferos que primero se emplazo en el territorio insular (MacPhee 2005) y en tal sentido es de interes la evidencia sobre especificidad parasito-hospedante aportada por Cruz (1980) quien encontro que la especie de acaro (genero Cubanochirus) que parasita a los almiquies es mucho mas primitiva que la especie (del mismo genero) que paras ita a las jutias y de aqui concluyo que dicho genero de acaro debio evolucionar primero en Solenodontidae y posteriormente en Capromyidae al entrar en contacto representantes de ambas familias en Cuba 0 La Espafiola
Recientemente Roca (2004) y Roca et al (2004) dieron a conocer que los resultados de su estudio genetico-molecular sobre los almiquies indican una divergencia de 25 Ma entre Solenodon cuban us y S paradoxus -compatible con la disyuncion tectonica entre Cuba y La Espafiola (Iturralde-Vinent y MacPhee 1999 McPhee et al 2003)-- y una separacion de ambas especies comparable en magnitud a la divergencia entre familias de mamlferos Consecuentemente propusieron que Solenodon cubanus fuera clasificada en el genero Atopogaie como indico Cabrera (1925) hace 82 afios
DISCUSION Y CONCLUSIONES
Las musarafias y los roedores antillanos (excepto Oryzomys) se irradiaron tanto de sus respectivos ancestros continentales que de estos 6rdenes se reconocen familias y subfamilias endemicas de la region en contraste con los perezosos (una subfamilia y tres
321 IRRADIACION EVOLUCIONARIA
tribus endemicas) y los monos (una tribu endemica) como expresion de diferentes niveles de diversificacion evolucionaria Este proceso es dificil de explicar (traUindose de c1ados que pnicticamente comparten la misma antigiiedad en las Antmas) si es correcta la interpretacion de Pascual et at (1990636)
Nosotros consideramos estos diferentes niveles de diversificacion como las diferentes etapas de evolucion alcanzadas por cada grupo [de mamiferos] Obviamente esto estuvo relacionado con las diferentes longitudes de sus historias
Pudiera ocurrir que en el futuro se compruebe que la historia biogeognifica de la region fue mucho mas complicada
La Espanola la isla mas ecol6gicamente divers a de la region debi6 ser el centro principal de evoluci6n de los mamfferos (con 87 de las familias y 73 de las subfamilias) seguida por Cuba (con 75 de las familias y 54 de las subfamilias) las dos islas de las Antillas Mayores mas relacionadas biogeograficamente (Fig 43 Tabla 44)
eL
EL
cb CAIMAN
SWAN
A Acratocnini B Cubanocnini C Megalocnini D Mesocnini E Nesophontidae F Solenodontidae G Xenothrichini
ANGUILAshyM-0 SMARTIN
H Heteropsomyinae I Plagiodontinae J Isolobodontinae
K Hexolobodontinae L Capromyinae
M Heptaxodontinae N CIidomyinae 0 Sigmodontinae
FIGURA 43 Representacion esquematica de las relaciones interinsulares (expresadas porel numero de linajes compartidos) entre las Antillas Mayores y entre estas y los grupos de islas perifericas segun las interpretaciones de los autores citados en el texto Los numeros dentro de los circulos indican la cantidad de linajes representados en cada isla relacion dudosa Contexto taxon6mico de los linajes en la Tabla 44
bull bull
bull bull
bull bull bull
bull bull
bull bull
bull
bull
bull
bull
bull bull bull bull
bull bull
bull bull
bull
bull bull bull
bull bull
bull
bull bull
bull
322 BIOGEOGRAFiA
TABLA 44 Distribuci6n de los linajcs de mamiferos terrestres a traves de las Antillas Mayores y grupos de islas perifericas LE = La Espanola CU Cuba JA Jamaica PR = Puerto Rico IC Islas Caiman IS Islas Swan IB = Islas Bahamas IV = Islas Virgenes AS = Anguila - San Martin
FamiliaSubfumiliafIribu LE CU JA PR IC IS IB IV AS
Megalonychidae
Choloepodinae
Acratocnini
Cubanocnini
Megalocninae
Megalocnini
Mesocnini
Nesophontidae
Solenodontidae
Pitheciidae
Callicebinae
Xenothrichini
Echimyidae
Heteropsomyinae
Capromyidae
Plagiodontinae
Isolobodontinae
Hexolobodontinae
Capromyinae
H e pta x 0 don tid a e
Heptaxodontinae
Clidomyinae
Muridae
Sigmodontinae
PILOSA
SORICOMORPHA
PRIMATES
RODENTIA
bull
bull
bull
bull TOTALES 12 10 4 4 2
323 EXTlICIOIES DEL CUATERNARIO
El gigantismo ha sido sin duda una tendencia evolucionaria recurrente entre los clados mayores de mamiferos antillanos Es un hecho bien conocido que algunos grupos de mamiferos han aumentado de tamafio en las islas y varios autores han intentado explicarlo a partir de enfoques diferentes (Foster 1964 Miller 1966 Sondaar 1977 Case 1978 Eisenberg y Wilson 1978 Heaney 1978 Lawlor 1982 Lomolino 1985 Angerbjom 1985 McNab 19942001)
EXTINCIONES DEL CUATERNARIO
Las islas que integran la Subregion Antillana han perdido casi 90 de sus mamiferos terrestres durante los ultimos milenios y en proporcion con su extension territorial esta perdida ha sido mayor en la Subregion que en cualquier otro lugar del planeta (MacPhee 1996b) Las extinciones del Cuatemario eliminaron mas de las tres cuartas partes de la fauna de mamiferos terrestres conocida de las Antillas Mayores sus islas satelites y grupos de islas perifericas y el examen de este proceso en sus aspectos mas generales se aborda en esta ultima parte del presente capitulo
CRONOLOGiA Y CONSECUENCIAS
Es muy improbable que un fosil represente el ultimo individuo de un taxon extinto y entonces hay que guiarse por la edad radiometrica del fosil mas reciente conocido lIamada fecha de ultima presencia- como unica aproximacion posible a la fecha de extincion del taxon
Como se explico al inicio del capitulo (vease Contexto Temporal) el registro fosH de los mamiferos terrestres de las Antillas Mayores adolece de insuficiente sustentacion radiometrica No obstante la distribucion temporal de los escasos fechados disponibles para el Cuaternario de la region abarca etapas criticas del proceso de extincion experimentado por la referida fauna a 10 largo de los ultimos 250 mil afios
De las 52 especies que es posible reconocer actualmente como miembros extintos de la fauna cuatemaria de mamiferos terrestres de las Antillas Mayores y grupos de islas perifericas los restos fosiles de 23 de elIas (442) han sido fechados radiometricamente o se han hallado asociados estratigraficamente de manera inequivoca con otro material fechado De estos 23 fechados 3 corresponden al interglacial Sangamon 4 pertenecen a la secuencia Pleistoceno terminal - Holoceno temprano 10 se ubican en ellapso entre el arribo de los aborigenes y la incursion europea y 6 son postcolombinos (a los cuales hay que sumar otras dos especies extinguidas en los siglos XIX y xx) A partir de estas 25 especies avaladas por fechas de ultima presencia distribuidas entre los cuatro intervalos temporales sefialados hemos compuesto la Tabla 45 en la que ademas aparecen las restantes 27 especies (carentes de fechado radiometrico) ubicadas en el intervalo temporal mas probable sobre la base de (1) la asociacion estratigrafica de sus restos con evidencias culturales (amerindias 0 europeas) 0 (2) la ausencia de dicha asociacion Sobre estos criterios de ubicacion temporal procede hacer dos aclaraciones
324 BIOGEOGRAFIA
TABLA 45 Intervalos temporales mas probables para las extinciones cuatemarias de los mamiferos terrestres de las Antillas Mayores y grupos de islas perifericas sobre la base de las evidencias siguientes (l) ultimo ejemplar colectado u observado en vida (2) fechado radiometrico mas tardio en material de la especie (3) asociaci6n estratigrafica mas tardia con otro material fechado (4) asociaci6n estratigrafica con restos de animales introducidos restos humanos u objetos culturales propios del intervale (5) falta de asociaci6n con evidencias culturales [Los ntimeros entre parentesis en la columna Evidencia significan adscripci6n tentativa al intervalo] Los estimados del peso corporal (en kg) estan tomados de la literatura 0 inferidos comparativamente y distribuidos entre
las clases siguientes (a) S 01 (b) 01 a lt 1 (c) 1 a lt 10 (d) 10 a lt 100 (e) 100 a lt 200 (j) ~ 200 Las
abreviaturas que designan las islas son las de la Tabla 44
F amilialEspecie Peso IsJa Evidencia Fuente
I httervalo htterg1acial Sangamon (gt80 kaAP)
HEPTAXODONTlNAE
Amblyrhiza inundata e AS 3 McFarJane et al 1998a
CLIDOMYINAE
Clidomys osborni d JA 3 Ibid
Clidomys parvus d JA 3 Ibid
II httervalo Pleistoceno terminal ~ Holoceno temprano (20-6 kaAP)
MEGALONYCHIDAE
Aeratoenus antillensis d CD 5 Arredondo 1975 a
Aeratoenus odontrigonus d PR 3 McFarJane 1999b
Aeratoenus simorhynehus d LE 2 Rega et al 2002
M egaloenus zile f LE 2 MacPhee et al 2000
SOLENODONTIDAE
Solenodon arredondoi e CD 5 Morgan y Ottenwalder 1993
PnHECIIDAE
Paralouatta varonai d CD 5 MacPhee 1996a
EcHIMYIDAE
Heteropsomys antillensis c PR 5 McFarJane 1999b
Heteropsomys insulans c PR 5 Ibid
Puertoricomys eorozalus c PR 5 Woods 1989a
CAPROMYIDAE
P lagiodontia araeum c LE 5 MacPhee et al 2000
Macrocapromys latus c CD 5 Varona y Arredondo 1979
(Continua)
325 EXTINCIONES DEL CUATERNARIO
TABLA 45 (Continuaci6n)
F ami1ia1Especie Peso Isla Evidencia Fuente
M acrocapromys latus c CU 5 Varona y Arredondo 1979
Mesocapromys kraglievichi c CU 5 Ibid
HEPTAXODONTINAE
Xaymaca fulvopulvis a JA 2 MacPhee y Flemming 2003
III Intervalo Amerindio (6-05 kaAP)
MEGALONYCHIDAE
Acratocnus ye d LE 2 MacPhee et aI 2000
Neocnus comes c LE 2 Ibid
N eocnus dousman c LE 2 Ibid
N eocnus toupiti c LE 2 Ibid
N eocnus gliriform is c CU 4 Cordova et ai 1997
M egalocnus rodens f CU 2 MacPhee et ai 2007
Parocnus browni e CU 2 Steadman et ai 2005
P arocnus serus e LE 2 MacPhee et ai 2000
N ESOPHONTIDAE
N esophontes edithae a PR 2 McFarlane 1999b
PITHECIIDAE
Antillothrix bernensis c LE 3 Rimoli 1977
EcHIMYIDAE
Brotomys contractus a LE 4 Miller 1929c
CAPROMYIDAE
P lagiodontia spelaeum c LE 4 Ibid
M acrocapromys acevedo c CU 4 NUiiez 1990a
HEPTAXODONTINAE
Elasmodontomys obliquus d PR 3 McFarlane 1999b
Iv Intervalo Postcolombino (05 kaAP - Presente)
N ESOPHONTIDAE
N esophontes hypom icrus a LE 24 MacPhee y Fle111111ing 1999
N esophontes zam icrus a LE (4) Ibid
N esophontes m icrus a CU 24 MacPhee et aI 1999
(Continua)
326 BIOGEOGRAFiA
TABLA 45 (Continuacion)
F amilialEspecie Peso Isla Evidencia Fuente
N esophontes m icrus a CD 24 MacPhee et aI 1999
N esophontes m icrus a LE 4 MacPhee et aI 2000
N esophontes sp 1 a IC 4 Morgan 1994
N esophontes sp 2 a IC 4 Ibid
Solenodon marcanoi b LE 4 Ottenwalder 2001
PITHECIIDAE
Xenothrix mcgregori c JA 4 MacPhee y Flemming 1999
EcHIMYIDAE
Boromys offeUa a CD 24 Guarch 1984
Boromys torrei a CD 24 Ibid
Brotomys voratus a LE 4 MacPhee et aI 2000
CAPROMYIDAE
P lagiodontia ipnaeum c LE 4 MacPhee y Flemming 1999
Rhizoplagiodontia lemkei c LE 4 Ibid
Isolobodon portoricensis c PR 24 Ibid
Isolobodon montanus c LE 4 Ibid
Hexolobodon phenax c LE (4) Ibid
Capromys sp c IC 24 Morgan 1994
Geocapromys columbianus c CD 4 MacPhee y Flemming 1999
Geocapromys thoracatus b IS 1 MacPhee y Marx 1997
Geocapromys sp 1 c IC 4 Morgan 1994
Geocapromys sp 2 c IC (4) Ibid
l-IEPTAXODONTINAE
Quemisia gravis c LE (1) MacPhee y Flemming 1999
MURIDAE
Oryzomys antillarum a JA 1 Ibid
1 Flemming y MacPhee (1999) alertaron en contra de la asociacion con otro material fechado Asimismo Morgan (1993) indico que la ausencia de asociacion con evidencias culturales amerindias no es necesariamente un criterio concluyente para ubicar un taxon en el intervalo temporal anterior porque hay depositos en la region que carecen de tales evidencias y son del Holoceno tardio por fechados radiocarbonicos [No obstante
327 EXTlNCIONES DEL CUATERNAR10
la ausencia de una especie en residuarios arqueol6gicos adquiere mayor certidumbre como criterio de ubicaci6n temporal de su extinci6n en la medida que aumenta el numero de residuarios estudiados que en las Antillas Mayores se cuentan por centenares] A falta de fechado absoluto de una especie consideramos que estos criterios de ubicaci6n temporal son mas utiles que la alternativa de excluir del analisis a la especie
2 En los trabajos sobre biogeografia de mamiferos antillanos los arque610gos mas citados como autoridades para establecer el inicio de la presencia aborigen en las Antillas Mayores (Kozlowski 1974 Rouse y Allaire 1978 Rouse 1989 1992 Wilson 2001) difieren al respecto entre los 6 kaAP y los 45 kaAP (pero vease Pagan et al 200523) y hemos adoptado la mas temprana de estas fechas para el comienzo del intervalo amerindio en la Tabla 45 De todos modos cualquiera que haya sido la fecha esta no es aplicable por igual al conjunto de las Antillas Mayores porque segun algunos de los autores citados el poblamiento comenz6 por Cuba y La Espanola 6 mil anos atras continu6 en Puerto Rico hace alrededor de 3 mil anos (pero vease Burney et al 1994) yen Jamaica hace menos de 2 mil e incluso a las Islas Caiman ni siquiera llegaron [Las Islas Caiman descubiertas por Col6n en 1503 carecen absolutamente de evidencia amerindia (Stokes y Keegan 1996)]
Si los datos de la Tabla 45 representan una aceptable aproximaci6n a la realidad el proceso de las extinciones cuaternarias de los mamiferos terrestres de las Antillas Mayores y grupos de islas perifericas debi6 caracterizarse entre otros por los eventos siguientes
Aunque obviamente la fauna cuaternaria debi6 ser mucho mas numerosa que 10 que refleja el registro paleontol6gico (Morgan y Woods 1986) 52 especies (81 de la fauna conocida) desaparecieron (sin reposici6n) durante los ultimos 250 mil anos
- Al parecer las extinciones se concentraron en las especies de mayor tamano Asi el peso promedio estimado para las especies de la etapa preantr6pica (intervalos I + II de la Tabla 45) es 350 kg es 328 kg para las especies del intervalo III (amerindio) y es s610 25 kg (c1ases del peso a b c) para las especies del intervalo IV (postcolombino) Ninguna especie aut6ctona de mamifero mayor de 6 kg existe hoy en la regi6n
- De acuerdo con la distribuci6n temporal de las extinciones 61 especies sobrevivieron al interglacial Sangamon (intervalo 1)48 sobrevivieron a la transici6n del Pleistoceno terminal al Holoceno temprano (intervalo II) 34 sobrevivieron al periodo amerindio (intervalo III) y s610 12 especies han sobrevivido desde la Ilegada de los europeos (intervalo IV) Las tasas de extinci6n correspondientes (exceptuado el intervalo I) son una especie cada 1167 anos en el intervalo II una cada 393 anos en el intervalo III y una cada 23 anos en el intervalo IV
- Durante todo el proceso de las extinciones cuaternarias la perdida de especies por isla fue de 91 en La Espanola 83 en Jamaica y 62 en Cuba Bahamas retuvo su unica especie (aunque extirpada en varias de las islas) Puerto Rico e Islas Caiman perdieron todas sus especies (6 y 5 respectivamente) y las Islas Swan Virgenes y Anguila - San Martin perdieron sus respectivas especies (micas
328 BIOGEOGRAFiA
De los cuatro 6rdenes de mamiferos representados en la fauna Pilosa y Primates desaparecieron totalmente Soricomorpha y Rodentia perdieron 86 y 74 de sus especies respectivamente
CAUSALIDAD
CAUSAS NATURALES
Los cicIos glaciales e interglaciales acaecidos en el Pleistoceno causaron cambios ambientales dnisticos en las Antillas con la concomitante alternancia de cIimas frios (secos) y calidos (hUmedos) asi como de descensos y ascensos del nivel del mar aumento y disminuci6n de las areas territoriales) induso (aunque con menor intensidad) en el Holoceno Estos eventos paleogeograficos han sido consistentemente documentados (Mayo y Kartashov 1972 Bonnatti y Gartner 1973 Ortega y Arcia 1982 Prance 1982 Schubert y Medina 1982 Dodge et al 1983 Ortega [1984] Haq et al 1987 Schubert 1989 Hodell et al 1991 Guilderson et al 1994 Oppo 1997 Emslie 1998 Curtis et al 2001 Iturralde-Vinent 2003) y de ellos brindamos a continuaci6n una breve resena
Finalizando el periodo interglacial anterior a la ultima glaciaci6n (Wisconsin) el mar estaba a 8-10 m por encima de su nivel actual Cuando el avance glacial del Wisconsin a1canz6 su culminaci6n (eillamado Ultimo Maximo Glacial) hace 18 ka el mar habia descendido 120 m con respecto a dicho nivel y las islas del Caribe habian incrementado considerablemente sus extensiones subaereas en comparaci6n con las que presentan hoy dia por ejemplo Puerto Rico y las Islas Virgenes se conectaron territorialmente y las Bahamas se unieron en cinco gran des islas Prevalecia un regimen frio y seco y condiciones de aridez --debidas a una reducci6n de las precipitaciones (posiblemente a menos de 440 mmlano)- y los ambientes xericos se habian extendido a traves de las islas y bancos emergidos y abundaban los pinares (los habitats mesicos estaban restringidos a parcelas relictas en 10 alto de las montanas) En el Mar Caribe y el Golfo de Mexico la temperatura superficial del mar era 5degC mas baja que la actual y en las llanuras costeras y alturas maximas eran respectivamente 2-3degC y 6degC mas bajas Los limites de las zonas ecol6gicas altitudinales descendieron varios cientos de metros y cada zona redujo su extensi6n vertical al aumentar el gradiente de temperatura Ellimite inferior de la zona periglacial
elevaci6n donde la temperatura media minima a1canza los OdegC) de la Cordillera Central de la Republica Dominicana (altura maxima del Caribe insular con evidencia de topografia glacial) se considera que estuvo entre los 1500 y 1800 mlnm (actualmente esta a 2800 mlnm) y se puede estimar que dicho limite era inferior en Cuba por estar mas al norte que la Republica Dominicana No obstante durante este periodo la temperatura experiment6 episodios cortos (unas pocas decadas) de ascensos repetidos
Un signifieativo componente bi6tieo de las referidas condiciones paleoambientales 10 constituy6 la espectacular irradiaci6n evolucionaria experimentada por un grupo numeroso de aves rapaees diurnas y nocturnas (Wetmore 1937 Brodkorb 1969 Kurotchkin y Mayo 1973 Arredondo 1976 Olson 1978 Olson y Hilgartner 1982
329 EXTINCIONES DEL ClJATERNARIO
Suarez 2000a b 2004 Suarez y Emslie 2003) Los mamiferos de las Antillas Mayores ofrecieron a estos depredadores aviares una amplia gama de tamanos de presa que en ausencia de mamiferos del orden Carnivora en la region permitio a estas aves irradiarse a este nicho alimentario con la evolucion de una variedad de especies de muy diversa tall a corporal incluidas formas gigantes Adquirir un cuerpo gigante les propicio el acceso a recursos alimentarios que no estaban siendo explotados por ningun otro depredador (Alcover y McMinn 1994) A Cuba corresponde el mejor registro fosil de rapaces cuaternarias caribenas con siete generos de especies diurnas Dentro de estas aves existieron tres gavilanes gigantes (Accipitridae) tres carairas (Falconidae) y un condor (Cathartidae) asi como cuatro generos de especies nocturnas dos lechuzas (Tytonidae) y tres Mhos (Strigidae) dos de ell os no igualados en tamano en todo el mundo Algunos de estos generos tuvieron representacion en otras islas antillanas como La Espanola Jamaica Bahamas y Gran Caiman
Al retirarse hacia el norte los hielos de la glaciacion Wisconsin el clima se tomo mas hUmedo en las Antillas durante el Holoceno temprano y el bosque mesico se expandio en la region insular El nivel del mar ascendio acompanado de mayor pluviosidad alrededor de los 7 kaAP las Islas Virgenes quedaron separadas entre si y de Puerto Rico y las Bahamas perdieron la mayor parte de su territorio Cerca de los 3 kaAP las condiciones secas retornaron a la region e hicieron desaparecer buena parte del bosque mesico Nuevamente el clima comenzo a adquirir sus condiciones actuales hacia los 25 kaAP aproximadamente y hoy transita por una fase mesica con vegetacion xerica confinada en parcelas relictas
Sin duda estos y otros eventos naturales incidieron negativamente en la sobrevivencia de mamiferos terrestres en las Antillas Mayores asi como en los grupos de pequenas islas perifericas (Pregill y Olson 1981 Olson 2006)
McFarlane et al (1998a) demostraron convincentemente como Amblyrhiza el gigante de los roedores antillanos endemico de la pequena isla de Anguila sucumbio a las repetidas y abruptas perdidas de su habitat durante el interglacial Sangamon por las nipidas y severas oscilaciones del nivel del mar (documentadas por Neumann y Hearty 1996) al no poder responder adecuadamente a tan catastroficos eventos mediante el ajuste de su masa corporal media y otro tanto debio ocurrirle en Jamaica al otro roedor gigante Clidomys (McFarlane et al 1998b) emparentado con Amblyrhiza en la misma epoca (Tabla 45 intervalo I)
El ascenso del nivel del mar seguramente promovio la extincion de mamiferos durante la transicion del Pleistoceno al Holoceno (Tabla 45 intervalo II) con la reduccion del tamano de las Islas por las transgresiones marinas (Pregill y Olson 1981 Morgan 1993) Esto ultimo haria maximamente severa la competicion asf como la presion de los depredadores [Steadman et al (2005) no yen por que el calentamiento global ocurrido durante la transicion Pleistoceno-Holoceno habria de tener un efecto negativo sobre los perezosos siendo sus descendientes actuales especies tropic ales 0 subtropicales pero no se trata del efecto
330 BIOGEOGRAFiA
directo del cambio de temperatura sobre los animales sino del efecto indirecto y progresivo que dicho cambio genera a largo plazo al reducir el territorio insular disponible]
Por esta epoca tambien desaparecieron especies de tall a mayor como Megalocnus zile en La Espanola y otras de menor tamano (en Cuba) pero que representan la tall a maxima en sus respectivos contextos taxonomicos del Cuaternario como Solenodon arredondoi Paralouatta varonai y Macrocapromys acevedo Un mamifero puede ser grande solo a costa de reducir su densidad poblacional y esto 10 hace mis vulnerable a la extinci6n sobre todo en una isla (McNab 1994 McFarlane et al 1998a) Incidentalmente Alcover et al (1998a) afirmaron que larelacion entre la talla corporal y la extincion no siempre se cumple en las especies de menor tamano y citaron como ejemplo que entre las musarafias todas las especies de Nesophontes desaparecieron mientras sobreviven especies de Solenodon pero la comparacion correcta en todo caso serfa entre los solenodones vivientes y el gigante Solenodon arredondoi y entre Nesophontes edithae -el gigante de los nesofontes extinguido en el periodo amerindio- y el resto de sus congeneres que sobrevivieron hasta los primeros siglos de la colonizacion europea (Tabla 45 intervalos III y IV)
CAUSAS ANTROPOGENICAS
Intervalo Amerindio
La mayoria de los autores que se han referido a las extinciones acaecidas durante este intervalo (entre elIos paradigmaticamente Martin 1984a b) han responsabilizado a los aborfgenes (que se supone comenzaron a poblar las Antillas Mayores hace 6 mil afios) con la desaparicion de numerosas especies de mamiferos terrestres en la region principalmente por la via de la sobrecaza y la introduccion de animales domesticos devenidos depredadores de la fauna autoctona (Morgan y Woods 1986) Este efecto negativo de la presencia indigena sobre los mamiferos de la region ha sido dramatizado por algunos autores en terminos de holocausto faunal Sin embargo pensamos que se ha exagerado Como han expresado MacPhee et al (19993) cuando uno busca evidencia dura de complicidad humana directa en [estas] extinciones no hay mucho que encontrar Las siguientes son algunas consideraciones a tener en cuenta en tal sentido
El registro radiometrico mas joven conocido hasta ahora para un perezoso antillano (MegalocnllS rodens) es de 42 kaAP (MacPhee et ai 2007) Si se acepta que los aborfgenes entraron en Cuba hace aproximadamente 6 mil anos su convivencia con los perezosos debe haber durado alrededor de 1800 afios (0 mas porque dificilmente el referfdo fechado radiometrico haya correspondido al ultimo sobreviviente de la especie) Esta cifra contradice la hipotesis del holocausto faunal segun la cualla extincion de los mamiferos insulares de mayor talla debe ocurrir con extrema rapidez -a escala de decadas 0 de centurias- a partir de su primer contacto con los humanos (Martin y Steadman 1999) A pesar de las repetidas afirmaciones en contrario (Harrington 1921 Miller 1929c Herrera [1945] Tabio 1951 Alvarez 1951a 1957 Arredondo
331 EXTINCIONES DEL CUATERNARIO
1959 1960a Veloz y Ortega 1973 Mayo 1980b Rodriguez et al 1984 Castellanos y Pino 1985 Pino y Castellanos 1985 Nunez 1990b Gutierrez 1996 Arredondo 2004) en su mayoria basadas en evidencias no exentas de posibles accidentes de deposicion (0 de reubicacion postdeposicional) en recintos cavernarios (Diaz-Franco [2006b D no se ha demostrado inequivocamente que los aborigenes antillanos se alimentaran de perezosos regularmente Sus restos en residuarios de alimentos son escasisimos en toda la region y esto contrasta con los abundantes restos de jutias y otros mamiferos de menor talla que aparecen en esos residuarios demostrativos de la preferencia de los paleoindios por estos otros mamiferos
Los mamiferos mas importantes en la economia prehistorica de las Antillas Mayores fueron las jutias Capromys y Geocapromys en Cuba Geocapromys en Jamaica e Isolobodon en La Espanola Puerto Rico e Islas Virgenes (Aguayo 1950b Tablo 1951 Rimoli 1972 Wing 2001 Diaz-Franco [2006b D Wilkins (2001) estudi61ajutia viviente de Jamaica (Geocapromys browni) en un sitio ocupado por los aborigenes durante 100 anos y no encontr6 evidencias de que durante ese tiempo la especie haya sido sobreexplotada Asimismo en Cuba las jutias conga carabali y andaraz son escasas en depositos pleistoholocenicos en cambio son muy numerosas en residuarios indigenas y de haber sido sobreexplotadas 10 mas probable es que no hubieran sobrevivido hasta hoy
La habilidad de los humanos para penetrar en profundidad el bosque tropical y mantenerse en este sustentablemente ha sido un evento comparativamente reciente (Beran y Steele 1994) En el caso antillano la caza tierra adentro ha sido explicada como un proceso tardio de desplazamiento a partir de la explotacion de abundantes recursos marinos que ofrecian condiciones 6ptimas para el sedentarismo de las comunidades aborigenes costeras (C6rdova y Arredondo 1988 Brito 2005) Peces y mariscos dominan la arqueologia costera de las Islas antillanas en tanto el consumo de recursos terrestres (mamfferos incluidos) aumenta progresivamente en la medida que los residuarios estudiados se alejan de la costa (Scudder 1991 Wilkins 2001) En las Antillas Mayores el componente terrestre de la dieta aborigen es solo 20-30 del material recobrado en los residuarios (Wing 1977 1989) Y debio ser menor alin porque el componente vegetal de la dieta no se ha preservado Por otra parte los aborigenes cubanos domesticaron la jutia conga y seguramente la reprodujeron para comer (Pose et al 1988) practica que probablemente se repitiera con otras jutias en las demas Antillas Mayores
- Fray Bartolome de las Casas hablo de 3 millones de indios en Cuba al inicio de la colonizacion pero los historiadores subsecuentes consideraron exagerada la cifra (Poey 1851 a) No obstante se dispone de dos estimados de la densidad de poblaci6n aborigen en las Antillas Mayores a la llegada de los europeos 18 indigenaslkm2 en Puerto Rico (Rouse 1956) y 1271km2 en La Espanola (Watts 1987) Extrapolando ambos valores entre las cuatro islas principales de las Antillas Mayores tenemos que en la fecha indicada la poblaci6n aborigen pudo estar entre 16 mil y 113 mil indigenas en Puerto Rico entre 21 mil y 145 mil en Jamaica entre 141 mil y 1 mill6n en La Espanola y entre 200 mil y 14 millones en Cuba Entre las cifras maximas y las minimas nos inclinamos
332 BIOGEOGRAFiA
hacia estas ultimas atendiendo a los altos indices de mortalidad infantil y morbilidad que se evidencian en las osamentas de los pobladores prehispanicos (Martinez y Rives 1990) Uno de los hechos que apuntan hacia el bajo numero de aborigenes en las Antillas Mayores es 10 rapido que desaparecieron despues de la llegada de los europeos pnkticamente sin dejar mestizaje con estos 10 cual contrasta ostensiblemente con la situacion en la America continentaL
En suma un grupo humano (1) demograiicamente irrelevante y controlado por altos indices de morbilidad (2) consumidor mayoritariamente de organismos marinos y (3) carente de la tecnologia cinegetica necesaria para poder sobreexplotar los mamiferos que Ie sirvieron de alimento (a los cuales accedio tardiamente en su desarrollo) no es concebible que haya causado e1 holocausto faunal que se Ie atribuye a partir del error de aplicar a las comunidades aborigenes antillanas un modelo cultural de la prehistoria continentaL
Tampoco hay evidencias de que los aborigenes hayan introducido carnivoros con suficiente capacidad depredadora salvo el perro Morgan y Otlenwaldder (1993) consideraron que la depredacion por perros durante el periodo amerindio es la explicacion mas plausible para la extincion del almiqui gigante de Cuba pero como se puede observar (Tabla 54) Solenodon arredondoi aparece en el intervalo II por falta de asociacion con evidencias culturales
Consecuentemente el accionar de los aborigenes y de sus animales domesticos directamente sobre la fauna no parece haber sido por sf solo un factor determinante de la extincion de mamiferos terrestres durante el periodo amerindio en las Antillas (pero vease Steadman et al 2005)
Hay que recordar en este senti do que alin en el Holoceno tardio ocurrieron cambios clirruiticos de consideracion (vease Causas Naturales) Por ejemplo Curtis et al (2001) explicaron que la extincion de algunos ani males mesofilicos de las alturas montanosas de La Espanola pudo haber antecedido a cualquier impacto humano sigrIificativo en esos lugares por coincidir dicha extincion con una tendencia a la aridez documentada isotopicamente y con evidencias de la perdida del bosque mesico ocurridas despues de 3kaAP Yen Jamaica hace solo 2shy3 mil aDos el area de la Bahia de Jackson sustentaba una gran biomasa de vertebrados en bosque hinnedo a juzgar por la cantidad y diversidad de taxones presentes en material fosil derivado de la depredacion por lechuzas hoy el area es la mas seca de la isla con matorral xerofitico y pobreza de fauna (MacPhee 1996a) Yalm en nuestros dias huracanes de gran intensidad aniquilan poblaciones de vertebrados (que no se recuperan) en is las pequenas y llanas del Caribe (Brooks y Smith 2001)
En medio de la incertidumbre sobre la verdadera causa (0 causas) de las extinciones de mamiferos en el periodo amerindio MacPhee y Marx (1997) han propuesto su hipotesis de la hipermorbilidad la cual estipula colapsos poblacionales relativamente rapidos con la introduccion de enfermedades de alta letalidad Estos autores han razonado que
Las islas son los lugares mas apropiados para experimentar perdidas catastroficas causadas por enfermedades Los mamiferos no solo serian susceptibles en virtud de su prolongado aislamiento biologico sino que en los confines de una isla la epizootia
333 EXTINCIONES DEL CUATERNARIO
tendria la potencialidad de infectar simultaneamente a la rnayoria de los individuos de vadas especies Los neonatos son inmunologicamente inmaduros y sufririan desproporcionadamente cualquier infeccion hipervirulenta La perdida excesiva de neonatos y juveniles tendria efectos severos para la sobrevivencia de la poblacion si esta se reduce por debajo de cierto nivel sobreviene la extincion porque no hay posibilidad de restitucion en el periodo postpanzootico Las especies de mayor talla corporal van a ser las mas vulnerables a la extincion dadas sus bajas tasas reproductivas largos periodos de gestacion y reducido numero de progenie Las especies de menor tamano --caracterizadas por altas tasas reproductivas periodos cortos de gestacion y mayor numero de neonatos por parto- tambien serian fuertemente diezmadas pero muchas sobrevivirian porque su estrategia reproductiva les permite perder mayor numero de individuos Las perdidas deben ocurrir relativamente rapido (a escala de decadas) mientras que otras hipotesis (digamos los cambios climaticos) permiten intervalos mucho mas largos (a escala de siglos 0 de milenios) entre la aparicion de las causas y la produccion de sus efectos Por ejemplo todo indica que los nesofontes se extinguieron muy rapidamente y seria dificil poder invocar un impacto humano directo 0 argumentar fenomenos climaticos que pudieran explicar plausiblemente perdidas tan rapidas
MacPhee y Marx (1997 191) han dicho que por el momento la hipotesis de la hipermorbilidad no pasa de ser una abstraccion en espera de adecuada sustentacion empirica Llama la atencion que linicamente en el periodo amerindio (Tabla 45 intervalo 1Il) las extinciones abarcaron todas las clases del peso corporal (de la a a laj) y que ninguna de las especies que sobrevivieron hasta el periodo postcolombino (Tabla 45 intervalo IV mas las especies vivientes) sobrepasa los 10 kg (clases del peso a b c) en clara coincidencia con los presupuestos de la hipotetica hipermorbilidad
Intervalo Postcolornbino
Por 10 menos 37 especies de mamiferos terrestres vivian en las Antillas Mayores y grupos de Islas perifericas a la lIegada de los europeos en 1492 De elIas se han extinguido 23 shymas especies que en cualquiera de los periodos precedentes (Tabla 45~ y solo subsisten 12 especies repartidas entre Cuba (8 especies) La Espanola (2) Jamaica (1) y Bahamas (1) varias de estas en peligro critico de extincion Solo dos de las especies extinguidas durante los ultimos 500 anos se conocieron en estado viviente los roedores Oryzomys antillarum de Jamaica (extinguido en 1877) y Geocapromys thoracatus de las Islas Swan (extinguido en 1955) Las causas de las extinciones ocurridas durante este periodo son sobradamente conocidas destruccion de habitats introducci6n de animales exoticos (domesticos y comensales) y demas presiones asociadas a la actividad humana entre las cuales posiblemente habria que incluir brotes de hipermorbilidad
Durante mucho tiempo se ha afirmado la posibilidad de que especies de Nesophontes hayan sobrevivido hasta las primeras decadas del siglo xx y que incluso continuen existiendo en lugares remotos de Cuba 0 La Espanola (Allen 1918 1942 Miller 1930 Aguayo 1950a Arredondo 1960b Fischer 1977 MacFadden 1980 Woods y
334 BIOGEOGRAFiA
Ottenwalder 1991) Actualmente no existe evidencia radiometrica de que alguna especie de Nesophontes haya sobrevivido al siglo xv las fechas radiocarbonicas calibradas obtenidas en muestras de Nesophontes de Cuba y La Espanola resultaron no mas tardias que los siglos XIII y XIV (MacPhee et al 1999) Por otra parte las expediciones efectuadas a fines del siglo xx con el proposito de localizar individuos vivientes del genero en La Espanola no produjeron la menor indicacion afirmativa al respecto (Woods et al 1985) Ademas a traves de las Antillas Mayores no aparecen restos de Nesophontes en los depositos formados actualmente por las lechuzas
EPiLOGO
Las Antillas Mayores son islas oceanicas no han formado parte de un continente (con la posible excepcion de una parte de Jamaica hasta el Eoceno Medio hace 45 Ma) y durante un tiempo geologico considerable (desde el Eoceno Superior hace 40 Ma) han permanecido emergidas y separadas de los continentes vecinos Consecuentemente los mamiferos terrestres (no volantes) autoctonos de estas islas constituyen una fauna oceanica que por ende cumple con todas las caracteristicas convencionales reconocidas para este tipo de entidad biogeografica
- Baja diversidad taxonomica a nivel de las categorias superiores (solo cuatro ordenes) en comparacion con islas plataformicas continentales (0 con areas continentales) de tamano y diversidad ecologica simi lares
- Alto grado de endemismo a nivel de genero (96) y familia (50) en respuesta al prolongado aislamiento
- Desbalance ecologico de la estructura trofica ausencia de mamiferos camivoros y ungulados
- Espectaculares procesos de irradiaci6n evolucionaria como resultado de la colonizacion de territorios libres de competidores y depredadores
Tendencias evolucionarias extremas gigantismo
- Control a cargo de aves rapaces con tendencia at gigantismo en ausencia de mamiferos miembros del orden Carnivora
- Mas especies de reptiles y de murcielagos que de mamiferos terrestres relaciones que en los continentes se invierten
Alta tasa de extincion de especies (84) sin reposicion
Preservacion de especies filogeneticamente arcaicas (los llamados fosiles vivientes) los almiquies

303 POBLAMIENTO CONTINENTE-ISLA
taxon6mico ha sido expJicado de dos maneras (1) Inicialmente hubo mas 6rdenes presentes pero la mayoria se extingui6 y s610 sobrevivieron esos cuatro [explicaci6n vicarianista] (2) S610 representantes de esos cuatro 6rdenes lograron cruzar la barrera marina [explicaci6n dispersionista]
Cinco hipot~ticos mecanismos de poblamiento continente-isla han sido invocados durante los ultimos 140 afios (y se los sigue proponiendo actualmente) para explicar la presencia de mamiferos terrestres en las Antillas Mayores A continuaci6n pasamos a resefiarlos brevemente (incluidas sus alegadas fortalezas y debilidades) en el orden aproximado en que historicamente fueron propuestos
EL PUENTE NATURAL (dispersion terrestre activa)
En el siglo XIX el puente terrestre era pnkticamente la unica hipotesis para explicar las distribuciones disyuntas de animales en buena parte del mundo y los primeros investigadores que se ocuparon de la biogeografia antillana (Fernandez [1865] 1884 Chapman [1893] Torre 1892 Lora 1895) la asumieron resueltamente en parte intrigados por el descubrimiento en Cuba en la segunda mitad del siglo de un perezoso f6sil de gran talla Y en su aplicacion a los mamiferos terrestres de las Antillas Mayores el puente sigui6 siendo a 10 largo del siglo xx la hipotesis mas favorecida por los estudiosos del tema (Vaughan 1902a Spencer 191 Oa b Torre 1910a b cAllen 1911 1917a 1918 Barbour 19141916 1945bAnthony 1918 1926 [1942] Scharff 1922 Kraglievich 1926 Beaufort 1951 Arredondo 1961 1970b Kartashov y Mayo 1974 Iturralde-Vinent 1982 1988 Arredondo y Armifiana 1996 Arredondo 1996a [1999]) quienes postularon la existencia preterita de puentes con Norte- Centro- 0 Suramerica en dependencia de las afinidades continentales de los mamiferos antillanos
Teoricamente el puente terrestre debe cumplir con una condicion esencial permitir el paso de una parte mayoritaria (y por tanto representativa de la diversidad) de la fauna continental Claro que siempre habria tipos de mamiferos menos proclives a dispersarse o para los cuales la ubicaci6n del puente resultaria inaccesible 0 las condiciones ecologicas en este fueran desfavorables pero si algunos acceden al puente no hay raz6n para pensar que el resto de esa fauna no los acompafie mayoritariamente Como dijera Simpson (1956) Los puentes no transportan faunas desbalanceadas
En consecuencia el puente terrestre como filtro es un mecanismo de poblamiento minimamente selectivo y por ende de alta diversidad inicial (sensu Iturralde-Vinent y MacPhee 1999) 10 esencial del puente es su capacidad para transportar faunas integradas (Williams 1989) Yen eno radica la objeci6n fundamental al puente terrestre surgida tempranamente en el siglo xx (Por que s610 cuatro 6rdenes de mamiferos terrestres en las Antillas si vinieron por puentes (Donde estan los fosiles de los demas ordenes que debieron venir si es que se extinguieron Los partidarios del hipotetico puente dispusieron de casi todo el resto del siglo para responder una y otra vez que otros 6rdenes de mamiferos aparecerian representados en las Antillas Mayores el dia que en estas Islas se descubrieran fosiles precuatemarios suficientemente
304 BIOGEOGRAFIA
antiguos Esto se logro por fin terminando el siglo como se explico anteriormente (vease Contexto temporal) y ique se encontro los mismos cuatro ordenes de mamiferos terrestres conocidos del Cuatemario caribefio aparentemente los Unicos que lograron llegar
Pero ademas hasta el presente no se ha demostrado inequivoca- y concluyentemente que en algun momenta durante los ultimos 40 millones de afios haya existido una conexion terrestre continua entre las Antillas Mayores (en proceso de formacion 0 plenamente formadas) y algun punto de las regiones continentales vecinas
LA BALSA NATURAL (dispersion marina pasiva)
Durante buena parte del siglo xx el puente terrestre tuvo como unico rival a la hipotesis de la balsa natural -transporte accidental de animales sobre islotes de vegetacion flotantes a la deriva despues de arrancados y expulsados mar afuera por un rio crecido- expuesta originalmente por Matthew (191319151916 1918a 1919) y desarrollada en su aplicacion a los mamfferos terreslres antillanos principal mente por Simpson (1956) asi como por olros investigadores (Darlington 1938 1957 Williams y Koopman 1952 Hershkovitz 1958 1972 Patterson 1962 Paula 1967 Patterson y Pascual 1972 Pregill 1981 Ford 1986 Woods 1989a 1990 Morgan 1993) Por su parte Hedges et al (1992) sustentaron dicha hipotesis en estimados de divergencia (basados en el reloj molecular) en la herpetofauna antillana -involucrados tangencia1mente los mamiferos [veanse objeciones a Hedges y colaboradores (Page y Lydeard 1994 Iturralde-Vinent y MacPhee 1999) respuestas a las objeciones (Hedges et al 1994 Hedges 2001) y contrarrespuestas (MacPhee e IturraldeshyVinent 2005) asi como cuestionamientos al metoda de los estimados de divergencia (Foote et al 1999 Pregill y Crother 1999 Woods et ai 2001 )]
La hip61esis de la balsa natural se origino a partir de las numerosas ocasiones en que a 10 largo de siglos tales islotes flotantes (portando 0 no vertebrados incluidos mamiferos) han sido avistados mar afuera a cientos de kilometros de la costa mas cercana en varias partes del mundo --inc1uido el Mar Caribe- segUn consta en diarios de barcos y reportes de observadores diversos (revisados por King 1962 reportes posteriores en SICSP 1970 y Censky et al 1998) Uno de estos casos acaecido en nuestra region se reporta en un informe del SICSP (Smithsonian Institution Center for Shortlived Phenomena 197050shy52)
EIB de julio de 1969 un destructor de la armada de Estados Unidos avisto un islote flotante a unos 100 km al sur de la Base Naval de Guantanamo moviendose desde el Paso de los Vientos en direccion suroeste a una velocidad de 45 kmlh islote de unos 15 m de diametro mantenfa en pie mas de 10 palmas reales de 10-15 m de altura con suelo cubierto de hierba y arbustos [la observacion con binoculares a 18 km de distancia no preciso la presencia de vida animal] La travesia fue rastreada durante 6 dfas hasta que el 19 de julio el islote se dio por perdido tras 6 horas de busqueda aerea
305 POBLAMIENTO CONTINENTE-ISLA
La colonizaci6n exitosa de una isla de las Antillas Mayores por mamiferos terrestres continentales transportados en balsas naturales a la deriva (de cada especie al menos una hembra gravida 0 dos individuos de sexo opuesto) dependeria de factores que van a operar desde el momenta mismo de formarse la balsa asi como durante su travesia marina y su arribo a la isla
a) La posibilidad de estar en la balsa Que la distribuci6n de la especie incluya areas contiguas al rio y que el animal se encuentre en ellugar y el momenta propicios para ser incorporado a la balsa antes de que el rio crecido la desprenda Por ejemplo segun Hershkovitz (1958) los equimidos -grupo ancestral de roedores antillanos- serian buenos candidatos a ser captados por la balsa porque acostumbran a anidar en huecos del tronco de los arboles a la orilla de los rios
b) El tamano y la consistencia de la balsa Muchas se desintegran en alta mar debido a su fragilidad ante la saturaci6n Algunos autores (Beaufort 1951 Arredondo 1996a [1999]) han expresado incredulidad en cuanto a que las balsas hayan podido soportar el peso de un animal como el gran perezoso cubano (Megalocnus rodens) el mayor de los mamiferos antillanos Pero esta especie evolucion6 en Cuba y el ancestro que hizo el viaje pudo ser muy pequeno [EI perezoso oligocenico de Puerto Rico es el mas pequeno de los megaloniquidos antillanos (MacPhee e Iturralde-Vinent 1995b )]
c) Las caracterfsticas fisiol6gicas de la especie Los limites fisiol6gicos de los propagulos constituyen nuestra unica objeci6n a la dispersi6n marina por balsas como mecanismo para la formaci6n de la fauna de vertebrados en las Antillas (MacPhee e IturraldeshyVinent 2005) Unas especies tienen intrinsecamente mas posibilidades que otras para sobrevivir a las vicisitudes que deben afrontar durante la travesia (deshidrataci6n inanicion insolaci6n etc) Por ejemplo los megaloniquidos han demostrado tener extraordinaria habilidad para cruzar barreras marinas fueron los primeros mamfferos suramericanos en invadir Norteamerica mucho antes de que surgiera la conexion terrestre a finales del Plioceno (Paula 1967 Olson 1978 Webb 1985b Martin y Steadman 1999) Incluso se conoce un megaloniquido acultitico del Plioceno peruano (Muizon y McDonald 1995) y otro viviente en Suramerica (Choloepus) que pnicticamente no bebe agua 10 que ha originado una leyenda aborigen que explica por que los perezosos no toman agua (Goff art 1971) Vease ademas el inciso e
d) EI regimen de las corrientes marinas Que conduzcan a la balsa en la direcci6n necesaria EI transporte de balsas de vegetacion desde los continentes hacia las islas fue durante la mayor parte del Cenozoico sumamente improbable (MacPhee e IturraldeshyVinent 2000 148) Estos autores han insistido en que hay que tener en cuenta que durante la cambiante geografia cenozoica del Caribe las corrientes marinas de superficie no favorecieron el desplazamiento de objetos fiotantes hacia las Antillas Mayores [Pero no sabemos con certeza que efecto modificador del curso de tales objetos fiotantes pudieron introducir las tormentas tropicales si previsiblemente funcionaron en el pasado como 10 hacen ahora] Dichos autores senalaron ademas (2005) que en un experimento reciente con boyas de posicionamiento satelital liberadas a la deriva desde las Antillas Menores las boyas fueron a parar mayoritariamente a Centroamerica y a otros lugares
306 B[OGEOGRAFIA
y solo unas pocas recalaron en las Antillas Mayores y las que entraron al Mar Caribe demoraron semanas 0 meses en arribar
e) La distancia a recorrer (y en consecuencia la duracion de la travesia) entre el continente y la isla (10 que repercute decisivamente en el factor c) Este es desde luego uno de los factores mas criticos En la medida que la distancia aumenta la probabilidad de alcanzar la isla disminuye drasticamente con el agravante de que la trayectoria dificilmente seria en linea recta Si tomamos como referencia la velocidad de 45 kmlh del islote ya mencionado (SICSP 1970) cubrir la distancia aerea (aproximada) de 800 km que media entre el noroeste de Suramerica y La Espafiola Ie tomaria a la balsa 74 dias Estimando el doble de este tiempo debido a desvios de la trayectoria la duracion de la travesia probablemente resultaria excesiva para la mayoria de los mamiferos Pero pudiera muy bien ocurrir que la velocidad fuera mayor 0 que la balsa portara plantas vivas que permitirian a los fitOfagos sobrevivir [Tres de los cuatro ordenes de mamiferos terrestres conocidos de las Antillas Mayores son fitOfagos] Ademas esta geologicamente bien establecido que durante etapas de variada duracion desde el Oligoceno hasta el Pleistoceno los territorios del Caribe insular y continental aumentaron considerablemente en extension con niveles del mar 100-200 m por debajo de su nivel actual oportunidades que los propagulos aprovecharian para cruzar barreras marinas significativamente menores
f) El tamafio de la isla Las islas mayores deben interceptar mas balsas (Buckley y Knedlhans 1986) 10 que repercute directamente en el factor e La balsa puede pasar entre islas pequefias y continuar viaje sin llegar a ningun lugar
g) Las condiciones (favorables 0 adversas) del desembarco en la isla y el tiempo que Ie tomaria a uno 0 mas animales depauperados encontrarun habitat apropiado Esto dependeria a su vez del tamafio y diversidad ecologica de la isla (Darlington 1938 Miller 1966)
Tal cumulo de posibles obstaculos y dificultades constituye a primera vista un desafio a la credibilidad de la hipotesis Matthew-Simpson pero en ello radica precisamente su valor explicativo bas ado en la mas absoluta improbabilidad La colonizacion por la via marina es altamente improbable pero no absolutamente imposible si se tiene en cuenta la inmensidad del tiempo geologico se vuelve probable (McKenna 1973) Basta que solo una vez entre mill ones de intentos fallidos durante ese inmenso lapso algunos de los factores anteriormente enunciados coincidan en no ser tan desfavorables para el propagulo Y eso es todo 10 que demanda la hipotesis para cada uno de los grupos monofileticos mayores de mamiferos terrestres presentes en las Antillas Mayores Las Antillas Mayores requeririan el arribo de tan solo 7 (quizas menos) propagulos fundadores a 10 largo de los ultimos 30 0 mas millones de afios (promedio de uno cada 43 mill ones) para explicar la formacion de toda su fauna de mamiferos terrestres por dispersion marina pasiva (vease Discusion y Conc1usiones)
En comparacion con la via marina el poblamiento por la via terrestre es predecible porque la mayor parte de la fauna continental de mamiferos emplazada en la region del puente va a cruzar por este una vez que el mismo se establece y los animales logran
307 POBLAMIEITO CONTINENTE-ISLA
acceder a el el poblamiento por la via marina en cambio es impredecible porque depende total mente del azar que caracteriza a la balsa Claro que en ambas vias opera el azar pero en el primer caso se trata del azar de un evento probable en el segundo el azar de un evento improbable (Simpson 1940) Asi el puente natural (mecanismo de poblamiento minimamente selectivo = alta diversidad inicial) y la balsa natural (mecanismo de poblamiento maximamente selectivo baja diversidad inicial) pueden conceptuarse como los extremos opuestos de una gama de probabilidades de dispersi6n (McKenna 1973) EI fen6meno de la dispersi6n merece la mayor atenci6n cientifica en los estudios de biogeografia insular (Cowie y Holland 2006)
Queda claro que la balsa natural es una hip6tesis consistente con el caracter de la fauna objeto de estudio puesto que el desbalance taxon6mico (sin evidencia paleontol6gica en contrario) s610 puede ser resultado de la improbabilidad del poblamiento No obstante ello no implica necesariamente que la balsa natural haya sido el imico mecanismo (ni siquiera el principal) utilizado por los mamiferos terrestres en su colonizaci6n de las Antillas Mayores La balsa natural debe satisfacer una premisa te6rica esencial dado el caracter azaroso impredecible e improbable de una colonizaci6n exitosa esta debe ocurrir a intervalos muy espaciados a traves del tiempo geol6gico 0 sea durante todo el tiempo que en el area ocupada por las Antillas Mayores ha habido ecosistemas terrestres perrnanentemente disponibles para ser colonizados (los ultimos 40 millones de afios) Sin embargo como sabemos representantes de los cuatro 6rdenes de mamiteros terrestres conocidos de las Antillas Mayores ya estaban emplazados en dicho territorio insular en el Mioceno Inferior 10 que supone un lapso de colonizaci6n limitado al Eoceno SuperiorshyOligoceno (MacPhee 2005) sugerente de un tipo de mecanismo contrario a la colonizacion alocronica exigida por la hipotesis de la balsa natural sobre todo tratandose de una fauna caracterizada por altas tasas de extincion sin reposicion aparente
La presencia en Jamaica (hasta su extinci6n en 1877) del roedor mfuido Oryzomys antillarom con estrecha afinidad centroamericana (Woods 1989a) pudiera constituir el imico caso efectivo de poblamiento (continente-isla) de las Antillas Mayores por dispersion marina dc un mamifero terrestre para el cual no es razonable invocar otro mecanismo de colonizacion
LA GUIRNALDA INSULAR (dispersion mixta)
La geologia ha establecido con suficiente certeza que durante el Cenozoico unicamente en el Pale6geno tardio y solo a traves de la Cresta de Aves (ahora sumergida excepto la isla hom6nima) pudo haber existido una cadena de islas para el transito de mamiferos terrestres entre el norte de Suramerica y las proto-Antillas Mayores [desestimados a ese efecto el Canal dc Yucatan el Elcvado de Nicaragua la Cresta de Caiman el Elevado de Beata y las Antillas Menores (Holcombe et al 1990 MacPhee e IturraldeshyVinent 1994 1995b Iturralde-Vinent y MacPhee 1999)] Holcombe y Moore (1977) y Holcombe y Edgar (1990) postularon la existencia de esta entidad y estimaron algunas de sus posibles caracteristicas Por su parte MacPhee e Iturralde-Vinent (1995b) e Iturralde-Vinent y MacPhee (1999) precisaron los extremos proximal y distal de esa
308 BIOGEOGRAFIA
estructura acotaron la duracion y la edad de la misma y Ie dieron nombre GAARlandia (Greater AntillesA vesRidgelandia)
Segun Iturralde-Vinent y MacPhee (1999) la Cresta de Aves emergio por breve tiempo (1-2 Ma) durante la transicion Eoceno-Oligoceno (35-33 MaAP) por la coincidencia de un levantamiento tectoni co y un considerable descenso eustatico del nivel del mar En ese momenta las islas septentrionales del arco de las Antillas Mayores (Cuba centroorientalshyLa Espafiola - Puerto Rico - Islas Virgenes) constituian una sola isla grande 0 una serie de islas separadas por barreras muy estrechas La Cresta de Aves emergida entre el bloque Puerto Rico - Islas Virgenes y un microcontinente noroccidental de Suramerica -separado en ese momento del resto del continente por una via marina surgida en el Eoceno tardio- dio origen a GAARlandia
Obviamente el numero de colonizaciones exitosas por mamiferos terrestres a traves de una guimalda de Islas tiende a disminuir mientras mas anchas sean las barreras marinas entre las islas que forman la guimalda y cuando esto ocurre disminuye tambien la diversidad taxonomica de los propagulos particularmente a nivel de las categorias superiores (Simpson 1956)
Vemos asi que para los mamiferos terrestres la colonizaci6n a traves de una guimalda insular puede ser tan improbable como a la deriva en una balsa natural en dependencia de la cantidad y anchura de las barreras marinas a cruzar el tamafio y la fisiografia de las islas que integran la guimalda y el regimen de las corrientes marinas y aereas entre otros factores
Resumiendo la guimalda insular puede conceptuarse como un mecanismo de poblamiento altamente selectivo (= baja diversidad inicial) que combina la improbabilidad de la balsa natural con la colonizacion restringida temporaimente del puente terrestre
En su reciente revision de la taxonomia distribuci6n y filogenia (basada en estudios moleculares) de varios grupos de mamiferos antillanos Davalos (2004) concluyo que los perezosos megalocninos los primates y los roedores histricognatos se avienen al patron de divergencia con el continente y epoca de arribo a las Antill as implicados en la hipotesis de GAARlandia pero no asi los perezosos coloepodinos que requieren una explicacion biogeognlfica distinta Esto ultimo (sospechado por Matthew 1918a) habla sido confirmado por White y MacPhee (2001 226-227)
Nuestra evidencia filogenetica no sustenta la idea de un dado unificado surgido de la invasion de un megaloniquido el analisis sugiere mas bien dos eventos de colonizaci6n independientes [y] sustenta fuertemente un origen difiletico de las subfamilias [Choloepodinae y Megalocninae] Si hubo dos migraciones a traves de GAARlandia presumiblemente poco espaciadas en el tiempo no se puede determinar en el presente
Finalmente queda claro que GAARlandia -como guirnalda insular- es la hipotesis mas robusta propuesta hasta ahora para explicar la presencia en las Antillas Mayores de
309 POBLAMIENTO CONTINENTE- ISLA
la absoluta mayona de los mamfferos terrestres de ongen suramericano porque el caracter de esa fauna supone un mecanismo de poblamiento ancestral que sea a la vez maximamente selectivo (baja diversidad ordinal) y temporalmente restringido (transicion Eoceno-Oligoceno)
Recientemente Rega et al (2002 18) expresaron A pesar de una revision exhaustiva de la paleogeografia de la cuenca del Caribe Iturralde-Vinent y MacPhee (1999) no pudieron confirmar la presencia de una conexion terrestre continua entre Suramerica y las proto-Antillas aunque hipotetizaron su existencia [Vease la Fig 42 (= figura 6 de Iturralde-Vinent y MacPhee 1999 simplificada))
En el trabajo citado por Rega y colaboradores aparece la siguiente explicacion
Actualmente no es posible ofrecer reconstrucciones paleogeograficas detalladas de la Cresta de Aves Sin embargo hay evidencia de que porciones de la cresta deben haberse elevado subaereamente en un tiempo La cuestion cntica es si la Cresta de Aves emergida pudo haber formado algun tipo de corredor entre las Antillas Mayores y el noroeste de Suramerica Perforaciones en las alturas estructurales de la cresta pudieran proveer el registro del Paleogeno tardio que se necesita para precisar la hipotesis del corredor aunque no se puede predecir si tales datos serian suficientemente detallados para determinar si en algun momento GAARlandia constituyo una sola entidad subaerea Por ejemplo pudo haber barreras marinas por corto tiempo entre las masas terrestres de la Cresta de Aves [0) ellevantamiento y la subsidencia pudieron afectar diferentes partes de GAARlandia en diferentes tiempos AItemativamente los elementos faunales pudieron haberse dispersado de manera pulsada Movimientos episOdicos de este tipo incrementarian la extincion de los elementos faunales incapaces de trasladarse a la siguiente masa terrestre en la cadena disminuyendo asi la diversidad total (Iturralde-Vinent y MacPhee 199957)
La primera parte de la explicacion deja claro que la posibilidad de que la Cresta de Aves haya sido una conexion terrestre continua no se puede descartar pero tampoco se puede afirmar La ultima parte de la explicacion describe el efecto altamente selectivo (filtrante) de la colonizacion a traves de una cadena de islas
Y continuan lturralde-Vinent y MacPhee (199957 -58) explicando
Hasta que punto el microcontinente noroccidental de Suramerica [considerado por dichos autores como el area suministradora de propagulos hacia GAARlandiaJ estuvo fisicamente separado del resto del continente no ha sido plenamente esc1arecido Si estuvo sustancialmente aislado por barreras marinas y orogenic as durante el Paleogeno tardio entonces solo la fraccion de la biota suramericana emplazada en el microcontinente estuvo en posicion de acceder a GAARlandia Esto pudiera tener que ver con el hecho de que la fauna antillana carezca de representantes de muchos grupos suramericanos [de mamfferos) que debieron haber hecho el viaje
310 BIOGEOGRAFiA
Florida o Alturas0 0
d 6 D L1anuras25deg
Yucatan
20deg
deg0
10deg
MARCARIBE
80deg 75deg
Bahamas
Cres ta de Beata
D Mares someros
D Mares profundos
OCEANO ATLANTICa
FIGURA 42 Reconstrucci6n hipotetica de GAARlandia como conexi6n terrestre continua entre las proto-Antillas Mayores y el noroeste de Suramerica en el Pale6geno tardio segun la FIgura 6 (simplificada) de Iturralde-Vinent y MacPhee (1999)
En su defensa del puente terrestre Barbour (1945b249) se refiri6 a esto - antes 10 habfa hecho Anthony [1942363]- en los terminos siguientes
El argumento de que si las Antillas hubieran estado conectadas con el continente se encontrarian los f6siles de un numero mayor de distintas criaturas me deja frio es perfectamente concebible que la conexi6n haya ocurrido en un periodo en que la parte del continente implicada no estuviera poblada por una fauna grande y variada
Y concluyen lturralde-Vinent y MacPhee (199958) su detallada explicaci6n
Desafortunadamente sabemos muy poco sobre la composici6n faunal del noroeste de Suramerica a finales del Pale6geno
Consecuentemente si se confirmara la magnitud del aislamiento del aludido microcontinente y su concomitante baja diversidad faunal GAARlandia como conexi6n terrestre continua serf a la hip6tesis mas parsimoniosa No obstante mientras no se disponga de evidencia concluyente a este respecto hipotetizar a GAARlandia como conexi6n continua (minima selectividad) la coloca en franca vulnerabilidad te6rica al tener que afrontar la objeci6n clasica al puente terrestre (baja diversidad ordinal de la
311 POBLAMIENTO CONTINENTE-TSLA
fauna) al tiempo que se la priva de ser la hip6tesis mas robusta para explicar -como guimalda de islas- la colonizaci6n de las Antillas por mamiferos procedentes de Suramerica
ELARcA DE NOE (seudovicarianza)
El primer vicarianista antillano (desde luego sin el saberlo) fue Corral (1940) con su utilizaci6n ingenua de las ideas wegenerianas de la deriva continental aparecidas la decada anterior Pero el primer (supuesto) intento de aplicar a la formaci6n de la biota caribefia el mecanismo de la vicarianza basada en la tearia de la tect6nica de placas (surgida en la sexta decada del pas ado siglo) se debe a Rosen ([1976] 1985)
Segun Rosen las Antillas Mayores se originaron en el Cremcico en la Placa Caribe aproximadamente en la posici6n que hoy dia ocupa Centroamerica y al ser desplazadas tectonicamente hacia el este interactuaron (de algun modo no precisado por Rosen) con las margenes continentales de Norte- y Suramerica de donde (en un evento unico) adquirieron toda su fauna transportada subsecuentemente por dichas islas en su transito hacia sus posiciones actuales [De ahi la denominacion metaforica dada par McKenna (1973) a este mecanismo de poblamiento] Aunque se ha presentado esta propuesta como Model0 de Vicarianza Continente-Isla de Rosen en realidad por definici6n no constituye un evento vicariante porque las proto-Antillas Mayores no resultaron del fraccionamiento y separaci6n por tectonismo de una porcion del territorio continental como exige esta hip6tesis (vease definici6n de vicarianza al comienzo de Hipotesis Biogeograficas)
El Arca de Noe de Rosen ha sido desestimada por la mayoria de los autores contemporaneos porque (1) carece de sustento geol6gico (lturralde-Vinent y MacPhee 1999) y porque (2) en el periodo en que se afrrrna ocum6 el evento muchos de los ordenes modemos de mamiferos placentarios attn no existian (Williams 1989)
EL BARCO FUNERAL VlKINGO (tanatovicarianza)
Domning et al (1997) reportaron 10 que consideraron el mamffero terrestre mas viejo de las Antillas --circunstancia que se refleja en el titulo mismo del trabajo Oldest West Indian Land Mammal- al descubrirse un perisodactilo rinocerotoide (Hyrachius sp) en una localidad de edad Eoceno Inferior a Medio en el occidente de Jamaica
Jamaica ha tenido una historia geologica y paleogeografica distinta de la historia comun a las restantes Antillas Mayores A principios del Cenozoico Jamaica occidental (a diferencia de la porcion oriental constituida por el Bloque de las Montafias Azules) formaba parte de Centro america de la cual se desgaj6 (por el desplazamiento de la Placa Caribe) para venir a integrarse despues del Eoceno Medio con 10 que serfa la futura isla de Jamaica desaparecida por transgresi6n marina en el Eoceno Superior hasta volver a emerger totalmente en el Mioceno Medio (Iturralde-Vinent y MacPhee 1999 MacPhee e Iturralde-Vinent 2000)
312 BIOGEOGRAFiA
Por consiguiente Hyrachius ---conocido de localidades de edad Eoceno Inferior a Medio en Norteamerica- debio ocupar (10 que iba a ser) Jamaica occidental mucho antes de que esta se separara de Centroamerica De hecho este fosil Uego a su destino insular contenido en las rocas de su basamento no como un ser vivo por 10 que es erroneo considerarlo un animal antillano (lturralde-Vinent y MacPhee 2004) como han supuesto algunos (Hedges 200 I Portell et al 200 I Condis [200 I D Todo 10 que hizo fue viajar tanatovicariantemente en su barco funeral vikingo -otra de las denominaciones metaforicas de McKenna (1973)
DISCUSION Y CONCLUSIONES
Hemos visto que solo disponemos de una hipotesis plausible para explicar la mayor parte del poblamiento (continente-isla) de las Antillas Mayores por mamiferos terrestres del Neotropico continental la dispersion mixta (a traves de GAARlandia) para los perezosos (excepto Choloepodinae) los monos y los roedores (excepto Oryzomys)
Por otra parte la historia biogeogrMica de las musaraiias de las Antillas Mayores (Nesophontes y Solenodon) constituye una de las situaciones mas enigmaticas entre los mamiferos de la region (Ottenwalder 2001321) Al menos cinco mecanismos de poblamiento continente-isla han sido invocados para explicar cuando como y de donde llegaron a las Antillas Mayores estas musaraiias
I EI propagulo ancestral (del Terciario de Norteamerica) coloniz6 a las Antillas Mayores por dispersion marina desde Centroamerica 0 desde la Florida (Patterson 1962)
2 El propagulo ancestral ( del Terciario) cruzo el Atlantico Sur (supuestamente mas estreshycho en aquella epoca) directamente de Africa a las Antillas Mayores (Hershkovitz 1972)
3 El propagulo ancestral (del Terciario de Norteamerica) Uego a las Antillas Mayores en el Arca de Noe de Rosen (MacFadden 1980)
4 El propagulo ancestral (del Terciario de Norteamerica) fue transportado a las Antillas desde Centroamerica en el bloque de Jamaica occidental y de alli paso por dispersion marina a las restantes Antillas Mayores (Whidden y Asher 200 I)
5 El propagulo ancestral (del Terciario de Norteamerica) lleg6 a las Antillas Mayores a traves de GAARlandia (Whidden y Asher 2001)
Aunque hay consenso entre estos (y otros) autores sobre el origen norteamericano de las musaraiias antillanas ninguno de los cinco hipoteticos mecanismos de poblamiento resiste la evidencia en contrario filogenetica yo paleogeogrMica Ademas algunas de estas hipotesis asumen que Nesophontidae y Solenodontidae comparten un antecesor comun en las Antillas pero esta relacion no ha sido confirm ada Para dilucidar el enigmatico caso de las musaraiias antillanas se requerini profundizar en la investigaci6n geologica paleogeognifica y paleontol6gica (Iturralde-Vinent y MacPhee 1999)
De acuerdo con el conocimiento filogenetico disponible sobre el conjunto de la fauna la colonizacion original de las Antillas Mayores por mamiferos terrestres implicaria el arribo
313 POBLAMIENTO ISLA-ISLA
de solo siete propagulos fundadores perezosos 2 (coloepodinos y megalocninos si como parece tienen un origen difiletico) musaraiias 2 (nesofontidos y solenodontidos si no constituyen una misma familia como consideran algunos) monos 1 (pitecidos) roedores histricognatos 1 (equimidos capromidos y heptaxodontidos) roedores muridos 1 (Oryzomys en Jamaica)
POBLAMIENTO ISLA-ISLA
La distribucion de los mamiferos terrestres a traves del area antillana demarcada al inicio del presente capitulo (vease Contexto Geogratico Fig 41) ha sido interpretada en funcion de dos hipoteticos mecanismos de poblamiento interinsular la dispersion y la vicarianza fundamentadas como se resumen seguidamente
DISPERSION
De acuerdo con la evidencia paleogeogratica mas actualizada en las principales regresiones marinas ocurridas en la cuenca caribeiia durante los ultimos 23 Ma (transiciones OligocenoshyMioceno Mioceno Superior - Plioceno y varias veces en el Pleistoceno) las masas terrestres constitutivas de la region insular en formacion aumentaron considerablemente su extension territorial disminuyendo por tanto la distancia entre elias oportunidades que los elementos faunales aprovecharian para dispersarse de unas islas a otras
Durante mucho tiempo esta ha sido la hipotesis mas favorecida por los estudiosos del tema a partir de Simpson (1956) pero en decadas recientes ha suscitado dos objeciones
1 Es dificil en tender como pudo operar la dispersion marina entre estas islas puesto que ninguna de las Antillas Mayores posee rios suficientemente caudalosos para transportar masas significativas de vegetacion flotante (MacPhee 198432)
Es oportuno recordar aqui el islote ala deriva (con mas de diez palmas reales en pie) entre las tres Antillas Mayores mas occidentales mencionado anteriormente (vease La Balsa Natural)
2 La dispersion marina es menos efectiva para explicar la asombrosa similitud de las faunas de mamiferos terrestres a traves de las Antillas Mayores septentrionales porque como proceso aleatorio no debe producir distribuciones uniformes a 10 largo de una cadena de islas 0 si las produce entonces el proceso no es aleatorio (MacPhee e lturralde-Vinent 2000 149)
Independientemente de que las aludidas distribuciones taxonomicas estan lejos de ser uniformes 0 asombrosamente similares la hipotesis de la dispersion marina contrariamente a su canicter aleatorio y altamente improbable en el caso del poblamiento continente-isla- en el caso del poblamiento isla-isla es un proceso no aleatorio (aprovecha las oportunidades brindadas puntualmente por los periodos de maxima regresion marina) y ademas altamente probable dada la incomparablemente menor distancia entre las islas caribeiias que entre estas y los continentes vecinos
314 BIOGEOGRAFiA
V ICARIANZA
La neotectonica y las transgresiones marinas del Oligoceno Medio y del Mioceno afectaron sustancialmente la paleogeografia de la Cresta de las Antillas Mayores resultando la subdivision gradual de las partes subaereas de la Cresta Asi el este de Cuba y el norte de La Espanola conectados fisicamente durante el Oligoceno Inferior se separaron al formarse el Paso de los Vientos en el Oligoceno tardio en tanto La Espanola y Puerto Rico probablemente permanecieron conectadas hasta el Mioceno tardio en que surgio el Paso de Mona (Iturralde-Vinent y MacPhee 1999)
Tales eventos geologicos habrian dividido las distribuciones de los grupos faunales emplazados previamente en el territorio continuo de las tres islas y a partir de esas disyunciones dichos autores (tambien MacPhee et al 2000 y White y MacPhee 2001) asumieron --como segunda fase del modelo de GAARlandia- la hipotesis de la vicarianza isla-isla para explicar la distribucion multiinsular de varios grupos de mamiferos terrestres a traves de las Antillas Mayores septentrionales De acuerdo con esta hipotesis cualquier diferencia en composicion faunal entre las islas (por ejemplo la ausencia de roedores heptaxodontidos en Cuba 0 de monos platirrinos en Puerto Rico) constituye evidencia de extincion local postvicariante y no de dispersion marina fallida Lo primero se puede demostrar si aparecen los fosiles pertinentes 10 segundo no se puede demostrar (MacPhee e Iturralde-Vinent 1994) Por otra parte IturraldeshyVinent y MacPhee (1999 57) han reconocido que aunque fa vicarianza isla-isla puede representar una solucion clara para las distribuciones de varios taxones cuaternarios no puede explicar todos los casos
Rega et al (2002 18) han presentado dos objeciones al modelo de la vicarianza isla-isla aplicado a los mamiferos de las Antillas Mayores
I Si la separacion de Cuba y La Espanola precedio en varios mill ones de anos la separashycion de La Espanola y Puerto Rico el modelo vicariante de lturralde-MacPhee debe predecir mayor similitud entre las faunas de La Espanola y Puerto Rico que entre las de Cuba y La Espanola De hecho es al reyes
No obstante MacPhee et al (200335) admitieron despues que la fecha mas temprana empiricamente defendible [para la apertura del Paso de Mona] debe ser el Oligoceno Medio esto invierte la secuencia de ambas disyunciones con 10 cual queda resuelta la aparente contradiccion Sin embargo en su reciente evaluacion de la congruencia entre filogenia e hipotesis geologic as en varios grupos de mamiferos terrestres de las Antillas Mayores Davalos (2004373) encontro que ninguno de los taxones cumple con la fecha y el patron de divergencia entre las masas terrestres antillanas postulados por el modelo de GAARlandia
2 La presencia de un mismo genero en las tres Islas septentrionales de las Antillas Mayores implica [por el modelo vicariante] que la diferenciacion entre estas formas durante el intervalo desde el Mioceno ha sido unicamente de nivel especifico [Dicha presencia]
315 rRRADlAcr6~ EVOLUCrONARIA
s610 puede explicarse razonablemente como el resultado de movimientos de esos animales entre las islas en el Cuaternario
De esta objecion resulta dificil disentir [El poblamiento entre islas diferenciado solo a nivel especifico no es probable que sea anterior al Pleistoceno (Simpson 1956 15)]
Debe tenerse en cuenta que la hipotesis de la vicarianza isla-isla alude a las tres islas septentrionales de las Antillas Mayores porque como se explico antes (vease El Barco Funeral Vikingo) Jamaica no existia como isla durante el lapso (Eoceno Superior shyOligoceno) consensualmente reconocido como el mas probable para la colonizacion de las proto-Antill as Mayores por la mayo ria de los mamiferos terrestres Despues de emerger como isla en el Mioceno Medio Jamaica debe haber recibido sus mamiferos terrestres por dispersion isla-isla (Davalos 2004376) y solo en el Cuaternario parece haberrecibido (por dispersion marina desde Centroamerica) al roedor murido Oryzomys
Seguramente tanto la dispersion como la vicarianza contribuyeron a que los mamiferos terrestres se irradiaran a traves de las Antillas Mayores y grupos de islas perifericas pero las maneras y tiempos en que estos mecanismos operaron a 10 largo de dicho proceso son aun en gran medida motivos de desacuerdo e incertidumbre
IRRADIACION EVOLUCIONARIA
A continuaci6n revisamos someramente las distribuciones conocidas de los diferentes grupos de mamiferos terrestres a traves de las Islas principales de las Antillas Mayores y grupos de Islas perifericas incluidas las interpretaciones y conjeturas que la literatura reciente recoge en relaci6n con la manera en que dichas distribuciones pudieron haberse conformado durante la irradiaci6n evolucionaria de los referidos grupos de mamiferos Manifestaciones de esta irradiaci6n pueden inferirse unicamente a partir de los taxones presentes en el Cuatemario pues solo 6 de las especies conocidas corresponden al Terciario
GRUPOS ORlGINARIOS DE SURAMERICA
PEREZOSOS (Megalonychidae)
Aparte de un genero miocenico de Cuba y otro del Oligoceno de Puerto Rico se conocen 4 generos (12 especies) de perezosos cuaternarios (todos extintos) restringidos a las tres islas septentrionales de las Antillas Mayores Acratocnus Neocnus Megalocnus y Parocnus Los cuatro generos estan representados en Cuba y La Espanola y Acratocnus 10 esta ademas (como unico genero de perezoso) en Puerto Rico Esta distribuci6n ha sido interpretada por MacPhee et al (2000) eomo resultado de la vicarianza oligomiocenica entre estas islas y [dado que el Banco Puerto Rico - Islas Virgenes fue la entrada de GAARlandia a las Antillas Mayores] se preguntaron si la ausencia de los otros tres generos en Puerto Rico se debe a que nunca estuvieron en ese territorio 0 a que sus
316 BIOGEOGRAFiA
fosiles atm no han sido hallados Pero como hemos visto Rega et al (2002) atribuyeron dicha distribucion a la dispersion interinsular en el Cuatemario
Por otra parte varios amilisis c1adisticos recientes ubi can a Choloepus -genero Reciente conocido solo de Centro- y Suramerica- como taxon hermano de Aeratoenus exc1uidos los demas megaloniquidos antillanos Para White y MacPhee (2001228)
tan estrecha relacion sugiere que la divergencia entre ambos [generos] ocurrio despues de que Aeratoenus divergio de Neoenus Si esta relacion implica que Aeratoenus y Neoenus se dispersaron separadamente desde Suramerica a las AntilIas mientras Choloepus permanecia en el continente (siendo necesarias al menos tres invasiones de perezosos) 0 si Choloepus invadio Suramerica desde las Antillas es algo que el material disponible no permite determinar
La diversidad adaptativa de los perezosos se manifiesta en (1) la utilizacion del habitat especies arboreas en Neoenus y Aeratoenus y terrestres en Paroenus y Megaloenus (Webb 1985a White [1993] White et al 1996 Arredondo [1999]) y en (2) la gama de tallas corporales desde los pequenos Neoenus hasta Megaloenus rodens el mayor de los mamiferos terrestres antillanos (su peso adulto estimado en 270 kg seglin Paula 19679) esto sugiere que los perezosos se irradiaron a nichos que en contextos continentales probablemente estarian ocupados por miembros de otros c1ados (MacPhee e IturraldeshyVinent 2000)
No esta claro si el abanico de tamanos entre estos perezosos enmascara alguna tendencia al gigantismo Por 10 pronto Anderson y Handley (2002) encontraron 10 contrario en perezosos (Bradypus) de las Islas de Bocas del Toro (Panama) shyseparadas secuencialmente del area continental vecina por ascensos del nivel del mar durante los ultimos 10 ka En las cinco islas los perezosos son menores que su contraparte continental y las diferencias entre las tallas medias de las cinco poblaciones expresan una reduccion lineal significativa de la talla corporal en funcion de la edad geologica de cada isla sin relacion alguna con el area de las islas 0 la distancia que las separa del continente
MONOS (Pitheciidae)
Los primates de las Antillas Mayores (cuatro especies extintas) constituyen un grupo monofiletico con Paralouatta (en Cuba) y Antillothrix (en La Espanola) como taxones hermanos seguidos en la escalera c1adistica por Xenothrix (en Jamaica) y finalmente Callieebus como el pariente continental mas cercano esto implica una sola invasion desde Suramerica antes del Mioceno Inferior (fecha mas temprana conocida para los primates fosiles antillanos) y por tanto en consonancia con GAARlandia como la via de colonizacion (Horovitz y MacPhee 1996 1999 Horovitz [1997] MacPhee y Horovitz 2004)
En desacuerdo parcial con la anterior interpretacion la filogenia de este grupo de monos obtenida por Davalos (2004) indica un antecesor comun en Cuba La Espanola y Jamaica
317 TRRADIACTON EVOLUCIONARIA
compatible con la hipotetica conexion -propuesta tentativamente por Iturralde-Vinent y MacPhee (199980-84)- entre las Montanas Azules del este de Jamaica y la peninsula suroccidental de La Espanola en la epoca de GAARlandia
A pesar de que los primates antillanos pertenecen a un grupo viviente de monos muy pequenos (los monos titles) fueron comparativamente grandes en relacion con los platirrinos modernos y sucesivamente mayores en la direccion Antillothrix Xenothrix y Paralouatta con P marianae como el mayor platirrino conocido del Mioceno neotropical (MacPhee 2005) MacPhee et al (1994) consideraron que Xenothrix debio ser un mono de andar lento y al parecer las especies de Paralouatta abandonaron la arborealidad para adoptar un modo de vida mas terrestre (MacPhee 1996a MacPhee et al 2003 MacPhee y Meldrum 2006)
ROEOORES (Echimyidae Capromyidae Heptaxodontidae)
La edad de Zazamys en Cuba demuestra que la invasion de los roedores histricognatos en las Antillas Mayores comenzo antes del Mioceno Inferior por otra parte los representantes mas antiguos de esa irradiacion evolucionaria se encuentran en La Espanola y Puerto Rico Ambas evidencias son consistentes con la hipotesis de GAARlandia (Woods 1982 1988 Woods et al 2001)
No esta claro si los capromidos evolucionaron a partir de un equimido ancestral 0 de un protocapromido 0 si los equimidos antillanos (I sub familia 3 generos y 7 especies todas extintas) forman parte de la irradiacion de los capromidos (4 subfamilias 9 generos y 27 especies 10 vivientes) Kratochvil et al (1978b) y Rodriguez et al (1979) consideraron que Capromys nan us es un eslab6n entre Echimyidae y Capromyidae pero Mesocapromys nanus esta morfologicamente mucho mas cerca de Hexolobodon y los de mas capromidos que de los equimidos (Woods 1985) Lo que si esta claro segun Woods et al (2001) es que la subfamilia Plagiodontinae endemic a de La Espanola es el linaje basal de los capromidos y que Isolobodon Hexolobodon Capromys Geocapromys Mysateles y Mesocapromys [y supuestamente Macrocapromys] forman parte de la irradiacion temprana de Capromyidae
La subfamilia Capromyinae (5 generos y 17 especies 9 vivientes) parece ser el dado de mas reciente evolucion en la familia con invasion de Cuba desde La Espanola y dispersion desde Cuba (1) por Capromys a las islas Caiman y (2) por Geocapromys a las islas Bahamas Caiman y Jamaica y desde Jamaica a las islas Swan aunque la presencia de Geocapromys en Swan pudiera deberse a introduccion aborigen (Morgan 1985 1988 1989 1993 1994 Woods 1989a 1990) Woods et al (2001) utilizaron datos moleculares para obtener estimados independientes de divergencia entre capromidos segiJn 10 cual Geocapromys divergio cerca de los 17 MaAP Mesocapromys divergio de Capromys hace 15 MaAP en tanto lvlysateles 10 hizo hace 10 MaAP Por su parte las formas de Capromys pilorides y de Mysateles prehensilis en Isla de la Juventud divergieron de sus correspondientes subespecies nominativas en la isla principal a los 24 y 08 MaAP respectivamente
318 BIOGEOGRAFiA
Woods y Howland (I979) describieron adaptaciones morfologicas de la mandibula el hioide y la faringe asociadas a mecanismos de masticacion diferentes en plagiodontinos y caprominos y Varona (1980a) llamo la atencion sobre el gran desarrollo de los molariformes (en relacion con la longitud total del cnineo) que presentan ciertos grupos de jutias fosiles en comparacion con las vivientes 10 que pudiera interpretarse como una adaptacion alimentaria a una vegetacion mas abrasiva por parte de las jutias extinguidas
Segun Woods (1990) la familia Heptaxodontidae (2 subfamilias 5 generos y 6 especies todas extintas) tambien seria parte de la irradiacion de los capromidos con origen en el este de La Espanola 0 en Puerto Rico y subsecuente dispersion al Banco de Anguila - San Martin en las Antillas Menores (pero vease MacPhee y Flemming 2003) Este banco estaria a solo 70 km del Banco Puerto Rico - Islas Virgenes en tiempos de maxima regresion marina y esta proximidad con Puerto Rico -patria de Elasmodontomys obliquus- seguramente tuvo que ver con la presencia en las islas de Anguila y San Martin del heptaxodontino gigante Amblyrhiza relacionado mas estrechamente con los roedores de las Antillas Mayores que con los del resto de las Antillas Menores (Woods 1990 1996) Aunque los roedores suelen aumentar de tamano en las islas (Case 1978) la escala del incremento de la talla en Amblyrhiza (que pudo alcanzar mas de 200 kg de peso adulto) esta fuera de toda proporcion con los patrones conocidos en otros roedores insulares (Biknevicius et 01 1993)
Clidomys (Clidomyinae) de Jamaica difiere marcadamente en su morfologia de los heptaxodontinos (vease ademas MacPhee y Flemming 2003)10 cual dificulta explicar su presencia en esa isla Como Jamaica estuvo bajo el mar hasta el Mioceno Medio el ancestro de Clidomys colonizaria esta isla por dispersion desde el extremo suroccidental de La Espanola despues que los heptaxodontinos se originaron en esta ultima isla (Woods 1989a) Pero Pascual et 01 (I 990) especularon que Clidomys pudiera ser el resultado de una invasion posterior e independiente de los capromidos
GRUPOS ORIGINARIOS DE NORTEAMERICA
El origen de las musaranas antillanas ha resultado ser hasta ahora el mayor enigma biogeografico de la fauna de mamiferos terrestres de la region aunque la mayoria de los autores coinciden en considerarlas originarias de Norteamerica (vease Discus ion y Conclusiones en Poblamiento Continente-Isla) No obstante MacPhee e Iturralde-Vinent (1994) descartaron toda posibilidad de conexion terrestre por el Estrecho de la Florida 0 por el Canal de Yucatan y si el (supuesto) evento vicari ante de Rosen (vease El Arca de Noe) -invocado por MacFadden (1980) para resolver el enigma- fuera minimamente plausible aun asi es dificil creer que las catastrofes que 10 sucedieron (vease Contexto Temporal) hayan barrido con toda esa vieja fauna continental excepto las musaranas circunstancia ya anticipada muy tempranamente por Matthew (l918a)
De otra parte la relaci6n entre Nesophontidae y Solenodontidae permanece irresuelta Si estos linajes no estan estrechamente relacionados entonces es muy posible que
319 IRRADIACION EVOLUCIONARIA
sus distribuciones sean explicadas por mecanismos diferentes (Whidden y Asher 2001)
La descomunal diferencia de talla corporal entre las musaranas pequenas (Nesophontes) y las grandes (Soienodon) seguramente impuso a ambos grupos restricciones distintas relativas a la termorregulacion y las adaptaciones energeticas En Nesophontes la seleccion debio favorecer una alta tasa de metabolismo y crecimiento camadas mas numerosas corta expectativa de vida yalta densidad poblacionaL En Soienodon en cambio la seleccion opero en la direccion de una baja tasa metabolica camadas reducidas crecimiento lento longevidad y baja densidad poblacional (Ottenwalder [1991])
MUSARANAS PEQUENAS (Nesophontidae)
Las musaranas pequenas estan representadas en las Antillas por el genero Nesophontes (y un genero miocenico indeterminado en ambar dominicano) con 6 especies cuaternarias extinguidas distribuidas en Cuba La Espanola Puerto Rico Islas Virgenes e Islas Caiman La mayor de las musaranas pequenas Nesophontes edithae evoluciono en Puerto Rico y su arribo a las Islas Virgenes supone una dispersion sumamente reciente En las Islas Caiman Nesophontes se asemeja mas a la poblacion de Cuba de donde seguramente derivo en el Plio-Pleistoceno (junto con Capromys y Geocapromys) cuando la costa sur de Cuba estuvo mucho mas cerca de las Islas Caiman (lturralde-Vinent 2003fig 4) MacPhee et ai (1994) interpretaron la distribucion de Nesophontes en Cuba y La Espanola como resultado de la vicarianza oligomiocenica ocurrida al formarse el Paso de los Vientos pero la distancia taxonomica entre los lados del Paso es solo de nivel especifico y N micros incluso vivi6 en ambas islas segtln Patterson (1962)
Allen (1918) considero que Nesophontes micrus fue una especie primitiva no modificada para la vida arb6rea 0 fos6rea y por tanto de habitos terrestres Nesophontes edithae de Puerto Rico es mucho mayor y mas robusta que las restantes especies del genero eL indice de robustez de su femur (= anchura minima de la diafisis ongitud total del hueso) muestra que este es tan robusto como el de Solenodon paradoxus un animal dos veces mayor (Morgan et ai 1980) Choate y Birney (1968) postularon un aumento cronoclinal de la taIla corporal en N edithae confirmado mas tarde por McFarlane (1999a) mediante fechados radiocarb6nicos de la secuencia estratignifica en depositos cavernarios de Puerto Rico Solenodon no vivi6 en Puerto Rico y en ausencia de una musarana de mayor tamano Nesophontes alcanzo una talla corporal mayor en esa isla que en La Espanola 0 Cuba (Woods y Eisenberg 1989)
MUSARANAS GRANDES (Solenodontidae)
Los arcaicos almiqufes (genero Soienodon) comprenden cuatro especies una viviente (S cubanus) y otra f6sil (S arredondoi) en Cuba y una viviente (S paradoxus con dos subespecies) y otra f6sil (S marcanoi) en La Espanola Como cabria esperar las dos especies cubanas se asemejan mas entre sf que a las especies de La Espanola pero
320 BIOGEOGRAFiA
enigmaticamente S marcanoi comparte caracteres con S cuban us y S paradoxus (Woods y Eisenberg 1989 Morgan y Ottenwalder 1993 Ottenwalder 200 I) Solenodon arredondoi alcanzo una talla corporal superior a la de cualquiera de las musarafias vivientes a nivel mundial Segun Morgan y Ottenwalder (1993) extrapolando las dimensiones y el peso de los almiquies vivientes S arredondoi debio tener una longitud cabeza-tronco y un peso de 450-550 mm y 1500-2000 g respectivamente en comparacion con S cubanus (360 mm y 800 g) y S paradoxus (390 mm y 1100 g)
Entre todas las especies de almiqufes Solenodon paradoxus es la que presenta los caracteres mas derivados (Otten walder 2001) Si estas relaciones son validas la distribucion de Solenodon entre Cuba y La Espafiola debio discurrir de oeste a este como propuso MacFadden (1980) quien invoco la presencia fosH de S cubanus [pero tambien S arredondoi] en Pinar del Rio para postular que el ancestro continental entr~ por Cuba opinion compartida por Perfit y Williams (1989) y por Ottenwalder ([1991] 2001) El paso de Solenodon a Puerto Rico pudo haberlo impedido la existencia de barreras ecologicas durante la conexion del este de La Espafiola con Puerto Rico 0 por la ausencia de Solenodon en esa parte de La Espafiola en aquella epoca Sin embargo las mismas condiciones ecologicas en el este de La Espafiola no parecen haber representado una barrera efectiva para Nesophontes cuyas mas altas tasa de reproduccion y densidad poblacional presumiblemente Ie permitieron expandirse flpidamente a traves de La Espafiola (Ottenwalder [1991])
Es opinion generalizada que los almiqufes antillanos constituyen el grupo de mamiferos que primero se emplazo en el territorio insular (MacPhee 2005) y en tal sentido es de interes la evidencia sobre especificidad parasito-hospedante aportada por Cruz (1980) quien encontro que la especie de acaro (genero Cubanochirus) que parasita a los almiquies es mucho mas primitiva que la especie (del mismo genero) que paras ita a las jutias y de aqui concluyo que dicho genero de acaro debio evolucionar primero en Solenodontidae y posteriormente en Capromyidae al entrar en contacto representantes de ambas familias en Cuba 0 La Espafiola
Recientemente Roca (2004) y Roca et al (2004) dieron a conocer que los resultados de su estudio genetico-molecular sobre los almiquies indican una divergencia de 25 Ma entre Solenodon cuban us y S paradoxus -compatible con la disyuncion tectonica entre Cuba y La Espafiola (Iturralde-Vinent y MacPhee 1999 McPhee et al 2003)-- y una separacion de ambas especies comparable en magnitud a la divergencia entre familias de mamlferos Consecuentemente propusieron que Solenodon cubanus fuera clasificada en el genero Atopogaie como indico Cabrera (1925) hace 82 afios
DISCUSION Y CONCLUSIONES
Las musarafias y los roedores antillanos (excepto Oryzomys) se irradiaron tanto de sus respectivos ancestros continentales que de estos 6rdenes se reconocen familias y subfamilias endemicas de la region en contraste con los perezosos (una subfamilia y tres
321 IRRADIACION EVOLUCIONARIA
tribus endemicas) y los monos (una tribu endemica) como expresion de diferentes niveles de diversificacion evolucionaria Este proceso es dificil de explicar (traUindose de c1ados que pnicticamente comparten la misma antigiiedad en las Antmas) si es correcta la interpretacion de Pascual et at (1990636)
Nosotros consideramos estos diferentes niveles de diversificacion como las diferentes etapas de evolucion alcanzadas por cada grupo [de mamiferos] Obviamente esto estuvo relacionado con las diferentes longitudes de sus historias
Pudiera ocurrir que en el futuro se compruebe que la historia biogeognifica de la region fue mucho mas complicada
La Espanola la isla mas ecol6gicamente divers a de la region debi6 ser el centro principal de evoluci6n de los mamfferos (con 87 de las familias y 73 de las subfamilias) seguida por Cuba (con 75 de las familias y 54 de las subfamilias) las dos islas de las Antillas Mayores mas relacionadas biogeograficamente (Fig 43 Tabla 44)
eL
EL
cb CAIMAN
SWAN
A Acratocnini B Cubanocnini C Megalocnini D Mesocnini E Nesophontidae F Solenodontidae G Xenothrichini
ANGUILAshyM-0 SMARTIN
H Heteropsomyinae I Plagiodontinae J Isolobodontinae
K Hexolobodontinae L Capromyinae
M Heptaxodontinae N CIidomyinae 0 Sigmodontinae
FIGURA 43 Representacion esquematica de las relaciones interinsulares (expresadas porel numero de linajes compartidos) entre las Antillas Mayores y entre estas y los grupos de islas perifericas segun las interpretaciones de los autores citados en el texto Los numeros dentro de los circulos indican la cantidad de linajes representados en cada isla relacion dudosa Contexto taxon6mico de los linajes en la Tabla 44
bull bull
bull bull
bull bull bull
bull bull
bull bull
bull
bull
bull
bull
bull bull bull bull
bull bull
bull bull
bull
bull bull bull
bull bull
bull
bull bull
bull
322 BIOGEOGRAFiA
TABLA 44 Distribuci6n de los linajcs de mamiferos terrestres a traves de las Antillas Mayores y grupos de islas perifericas LE = La Espanola CU Cuba JA Jamaica PR = Puerto Rico IC Islas Caiman IS Islas Swan IB = Islas Bahamas IV = Islas Virgenes AS = Anguila - San Martin
FamiliaSubfumiliafIribu LE CU JA PR IC IS IB IV AS
Megalonychidae
Choloepodinae
Acratocnini
Cubanocnini
Megalocninae
Megalocnini
Mesocnini
Nesophontidae
Solenodontidae
Pitheciidae
Callicebinae
Xenothrichini
Echimyidae
Heteropsomyinae
Capromyidae
Plagiodontinae
Isolobodontinae
Hexolobodontinae
Capromyinae
H e pta x 0 don tid a e
Heptaxodontinae
Clidomyinae
Muridae
Sigmodontinae
PILOSA
SORICOMORPHA
PRIMATES
RODENTIA
bull
bull
bull
bull TOTALES 12 10 4 4 2
323 EXTlICIOIES DEL CUATERNARIO
El gigantismo ha sido sin duda una tendencia evolucionaria recurrente entre los clados mayores de mamiferos antillanos Es un hecho bien conocido que algunos grupos de mamiferos han aumentado de tamafio en las islas y varios autores han intentado explicarlo a partir de enfoques diferentes (Foster 1964 Miller 1966 Sondaar 1977 Case 1978 Eisenberg y Wilson 1978 Heaney 1978 Lawlor 1982 Lomolino 1985 Angerbjom 1985 McNab 19942001)
EXTINCIONES DEL CUATERNARIO
Las islas que integran la Subregion Antillana han perdido casi 90 de sus mamiferos terrestres durante los ultimos milenios y en proporcion con su extension territorial esta perdida ha sido mayor en la Subregion que en cualquier otro lugar del planeta (MacPhee 1996b) Las extinciones del Cuatemario eliminaron mas de las tres cuartas partes de la fauna de mamiferos terrestres conocida de las Antillas Mayores sus islas satelites y grupos de islas perifericas y el examen de este proceso en sus aspectos mas generales se aborda en esta ultima parte del presente capitulo
CRONOLOGiA Y CONSECUENCIAS
Es muy improbable que un fosil represente el ultimo individuo de un taxon extinto y entonces hay que guiarse por la edad radiometrica del fosil mas reciente conocido lIamada fecha de ultima presencia- como unica aproximacion posible a la fecha de extincion del taxon
Como se explico al inicio del capitulo (vease Contexto Temporal) el registro fosH de los mamiferos terrestres de las Antillas Mayores adolece de insuficiente sustentacion radiometrica No obstante la distribucion temporal de los escasos fechados disponibles para el Cuaternario de la region abarca etapas criticas del proceso de extincion experimentado por la referida fauna a 10 largo de los ultimos 250 mil afios
De las 52 especies que es posible reconocer actualmente como miembros extintos de la fauna cuatemaria de mamiferos terrestres de las Antillas Mayores y grupos de islas perifericas los restos fosiles de 23 de elIas (442) han sido fechados radiometricamente o se han hallado asociados estratigraficamente de manera inequivoca con otro material fechado De estos 23 fechados 3 corresponden al interglacial Sangamon 4 pertenecen a la secuencia Pleistoceno terminal - Holoceno temprano 10 se ubican en ellapso entre el arribo de los aborigenes y la incursion europea y 6 son postcolombinos (a los cuales hay que sumar otras dos especies extinguidas en los siglos XIX y xx) A partir de estas 25 especies avaladas por fechas de ultima presencia distribuidas entre los cuatro intervalos temporales sefialados hemos compuesto la Tabla 45 en la que ademas aparecen las restantes 27 especies (carentes de fechado radiometrico) ubicadas en el intervalo temporal mas probable sobre la base de (1) la asociacion estratigrafica de sus restos con evidencias culturales (amerindias 0 europeas) 0 (2) la ausencia de dicha asociacion Sobre estos criterios de ubicacion temporal procede hacer dos aclaraciones
324 BIOGEOGRAFIA
TABLA 45 Intervalos temporales mas probables para las extinciones cuatemarias de los mamiferos terrestres de las Antillas Mayores y grupos de islas perifericas sobre la base de las evidencias siguientes (l) ultimo ejemplar colectado u observado en vida (2) fechado radiometrico mas tardio en material de la especie (3) asociaci6n estratigrafica mas tardia con otro material fechado (4) asociaci6n estratigrafica con restos de animales introducidos restos humanos u objetos culturales propios del intervale (5) falta de asociaci6n con evidencias culturales [Los ntimeros entre parentesis en la columna Evidencia significan adscripci6n tentativa al intervalo] Los estimados del peso corporal (en kg) estan tomados de la literatura 0 inferidos comparativamente y distribuidos entre
las clases siguientes (a) S 01 (b) 01 a lt 1 (c) 1 a lt 10 (d) 10 a lt 100 (e) 100 a lt 200 (j) ~ 200 Las
abreviaturas que designan las islas son las de la Tabla 44
F amilialEspecie Peso IsJa Evidencia Fuente
I httervalo htterg1acial Sangamon (gt80 kaAP)
HEPTAXODONTlNAE
Amblyrhiza inundata e AS 3 McFarJane et al 1998a
CLIDOMYINAE
Clidomys osborni d JA 3 Ibid
Clidomys parvus d JA 3 Ibid
II httervalo Pleistoceno terminal ~ Holoceno temprano (20-6 kaAP)
MEGALONYCHIDAE
Aeratoenus antillensis d CD 5 Arredondo 1975 a
Aeratoenus odontrigonus d PR 3 McFarJane 1999b
Aeratoenus simorhynehus d LE 2 Rega et al 2002
M egaloenus zile f LE 2 MacPhee et al 2000
SOLENODONTIDAE
Solenodon arredondoi e CD 5 Morgan y Ottenwalder 1993
PnHECIIDAE
Paralouatta varonai d CD 5 MacPhee 1996a
EcHIMYIDAE
Heteropsomys antillensis c PR 5 McFarJane 1999b
Heteropsomys insulans c PR 5 Ibid
Puertoricomys eorozalus c PR 5 Woods 1989a
CAPROMYIDAE
P lagiodontia araeum c LE 5 MacPhee et al 2000
Macrocapromys latus c CD 5 Varona y Arredondo 1979
(Continua)
325 EXTINCIONES DEL CUATERNARIO
TABLA 45 (Continuaci6n)
F ami1ia1Especie Peso Isla Evidencia Fuente
M acrocapromys latus c CU 5 Varona y Arredondo 1979
Mesocapromys kraglievichi c CU 5 Ibid
HEPTAXODONTINAE
Xaymaca fulvopulvis a JA 2 MacPhee y Flemming 2003
III Intervalo Amerindio (6-05 kaAP)
MEGALONYCHIDAE
Acratocnus ye d LE 2 MacPhee et aI 2000
Neocnus comes c LE 2 Ibid
N eocnus dousman c LE 2 Ibid
N eocnus toupiti c LE 2 Ibid
N eocnus gliriform is c CU 4 Cordova et ai 1997
M egalocnus rodens f CU 2 MacPhee et ai 2007
Parocnus browni e CU 2 Steadman et ai 2005
P arocnus serus e LE 2 MacPhee et ai 2000
N ESOPHONTIDAE
N esophontes edithae a PR 2 McFarlane 1999b
PITHECIIDAE
Antillothrix bernensis c LE 3 Rimoli 1977
EcHIMYIDAE
Brotomys contractus a LE 4 Miller 1929c
CAPROMYIDAE
P lagiodontia spelaeum c LE 4 Ibid
M acrocapromys acevedo c CU 4 NUiiez 1990a
HEPTAXODONTINAE
Elasmodontomys obliquus d PR 3 McFarlane 1999b
Iv Intervalo Postcolombino (05 kaAP - Presente)
N ESOPHONTIDAE
N esophontes hypom icrus a LE 24 MacPhee y Fle111111ing 1999
N esophontes zam icrus a LE (4) Ibid
N esophontes m icrus a CU 24 MacPhee et aI 1999
(Continua)
326 BIOGEOGRAFiA
TABLA 45 (Continuacion)
F amilialEspecie Peso Isla Evidencia Fuente
N esophontes m icrus a CD 24 MacPhee et aI 1999
N esophontes m icrus a LE 4 MacPhee et aI 2000
N esophontes sp 1 a IC 4 Morgan 1994
N esophontes sp 2 a IC 4 Ibid
Solenodon marcanoi b LE 4 Ottenwalder 2001
PITHECIIDAE
Xenothrix mcgregori c JA 4 MacPhee y Flemming 1999
EcHIMYIDAE
Boromys offeUa a CD 24 Guarch 1984
Boromys torrei a CD 24 Ibid
Brotomys voratus a LE 4 MacPhee et aI 2000
CAPROMYIDAE
P lagiodontia ipnaeum c LE 4 MacPhee y Flemming 1999
Rhizoplagiodontia lemkei c LE 4 Ibid
Isolobodon portoricensis c PR 24 Ibid
Isolobodon montanus c LE 4 Ibid
Hexolobodon phenax c LE (4) Ibid
Capromys sp c IC 24 Morgan 1994
Geocapromys columbianus c CD 4 MacPhee y Flemming 1999
Geocapromys thoracatus b IS 1 MacPhee y Marx 1997
Geocapromys sp 1 c IC 4 Morgan 1994
Geocapromys sp 2 c IC (4) Ibid
l-IEPTAXODONTINAE
Quemisia gravis c LE (1) MacPhee y Flemming 1999
MURIDAE
Oryzomys antillarum a JA 1 Ibid
1 Flemming y MacPhee (1999) alertaron en contra de la asociacion con otro material fechado Asimismo Morgan (1993) indico que la ausencia de asociacion con evidencias culturales amerindias no es necesariamente un criterio concluyente para ubicar un taxon en el intervalo temporal anterior porque hay depositos en la region que carecen de tales evidencias y son del Holoceno tardio por fechados radiocarbonicos [No obstante
327 EXTlNCIONES DEL CUATERNAR10
la ausencia de una especie en residuarios arqueol6gicos adquiere mayor certidumbre como criterio de ubicaci6n temporal de su extinci6n en la medida que aumenta el numero de residuarios estudiados que en las Antillas Mayores se cuentan por centenares] A falta de fechado absoluto de una especie consideramos que estos criterios de ubicaci6n temporal son mas utiles que la alternativa de excluir del analisis a la especie
2 En los trabajos sobre biogeografia de mamiferos antillanos los arque610gos mas citados como autoridades para establecer el inicio de la presencia aborigen en las Antillas Mayores (Kozlowski 1974 Rouse y Allaire 1978 Rouse 1989 1992 Wilson 2001) difieren al respecto entre los 6 kaAP y los 45 kaAP (pero vease Pagan et al 200523) y hemos adoptado la mas temprana de estas fechas para el comienzo del intervalo amerindio en la Tabla 45 De todos modos cualquiera que haya sido la fecha esta no es aplicable por igual al conjunto de las Antillas Mayores porque segun algunos de los autores citados el poblamiento comenz6 por Cuba y La Espanola 6 mil anos atras continu6 en Puerto Rico hace alrededor de 3 mil anos (pero vease Burney et al 1994) yen Jamaica hace menos de 2 mil e incluso a las Islas Caiman ni siquiera llegaron [Las Islas Caiman descubiertas por Col6n en 1503 carecen absolutamente de evidencia amerindia (Stokes y Keegan 1996)]
Si los datos de la Tabla 45 representan una aceptable aproximaci6n a la realidad el proceso de las extinciones cuaternarias de los mamiferos terrestres de las Antillas Mayores y grupos de islas perifericas debi6 caracterizarse entre otros por los eventos siguientes
Aunque obviamente la fauna cuaternaria debi6 ser mucho mas numerosa que 10 que refleja el registro paleontol6gico (Morgan y Woods 1986) 52 especies (81 de la fauna conocida) desaparecieron (sin reposici6n) durante los ultimos 250 mil anos
- Al parecer las extinciones se concentraron en las especies de mayor tamano Asi el peso promedio estimado para las especies de la etapa preantr6pica (intervalos I + II de la Tabla 45) es 350 kg es 328 kg para las especies del intervalo III (amerindio) y es s610 25 kg (c1ases del peso a b c) para las especies del intervalo IV (postcolombino) Ninguna especie aut6ctona de mamifero mayor de 6 kg existe hoy en la regi6n
- De acuerdo con la distribuci6n temporal de las extinciones 61 especies sobrevivieron al interglacial Sangamon (intervalo 1)48 sobrevivieron a la transici6n del Pleistoceno terminal al Holoceno temprano (intervalo II) 34 sobrevivieron al periodo amerindio (intervalo III) y s610 12 especies han sobrevivido desde la Ilegada de los europeos (intervalo IV) Las tasas de extinci6n correspondientes (exceptuado el intervalo I) son una especie cada 1167 anos en el intervalo II una cada 393 anos en el intervalo III y una cada 23 anos en el intervalo IV
- Durante todo el proceso de las extinciones cuaternarias la perdida de especies por isla fue de 91 en La Espanola 83 en Jamaica y 62 en Cuba Bahamas retuvo su unica especie (aunque extirpada en varias de las islas) Puerto Rico e Islas Caiman perdieron todas sus especies (6 y 5 respectivamente) y las Islas Swan Virgenes y Anguila - San Martin perdieron sus respectivas especies (micas
328 BIOGEOGRAFiA
De los cuatro 6rdenes de mamiferos representados en la fauna Pilosa y Primates desaparecieron totalmente Soricomorpha y Rodentia perdieron 86 y 74 de sus especies respectivamente
CAUSALIDAD
CAUSAS NATURALES
Los cicIos glaciales e interglaciales acaecidos en el Pleistoceno causaron cambios ambientales dnisticos en las Antillas con la concomitante alternancia de cIimas frios (secos) y calidos (hUmedos) asi como de descensos y ascensos del nivel del mar aumento y disminuci6n de las areas territoriales) induso (aunque con menor intensidad) en el Holoceno Estos eventos paleogeograficos han sido consistentemente documentados (Mayo y Kartashov 1972 Bonnatti y Gartner 1973 Ortega y Arcia 1982 Prance 1982 Schubert y Medina 1982 Dodge et al 1983 Ortega [1984] Haq et al 1987 Schubert 1989 Hodell et al 1991 Guilderson et al 1994 Oppo 1997 Emslie 1998 Curtis et al 2001 Iturralde-Vinent 2003) y de ellos brindamos a continuaci6n una breve resena
Finalizando el periodo interglacial anterior a la ultima glaciaci6n (Wisconsin) el mar estaba a 8-10 m por encima de su nivel actual Cuando el avance glacial del Wisconsin a1canz6 su culminaci6n (eillamado Ultimo Maximo Glacial) hace 18 ka el mar habia descendido 120 m con respecto a dicho nivel y las islas del Caribe habian incrementado considerablemente sus extensiones subaereas en comparaci6n con las que presentan hoy dia por ejemplo Puerto Rico y las Islas Virgenes se conectaron territorialmente y las Bahamas se unieron en cinco gran des islas Prevalecia un regimen frio y seco y condiciones de aridez --debidas a una reducci6n de las precipitaciones (posiblemente a menos de 440 mmlano)- y los ambientes xericos se habian extendido a traves de las islas y bancos emergidos y abundaban los pinares (los habitats mesicos estaban restringidos a parcelas relictas en 10 alto de las montanas) En el Mar Caribe y el Golfo de Mexico la temperatura superficial del mar era 5degC mas baja que la actual y en las llanuras costeras y alturas maximas eran respectivamente 2-3degC y 6degC mas bajas Los limites de las zonas ecol6gicas altitudinales descendieron varios cientos de metros y cada zona redujo su extensi6n vertical al aumentar el gradiente de temperatura Ellimite inferior de la zona periglacial
elevaci6n donde la temperatura media minima a1canza los OdegC) de la Cordillera Central de la Republica Dominicana (altura maxima del Caribe insular con evidencia de topografia glacial) se considera que estuvo entre los 1500 y 1800 mlnm (actualmente esta a 2800 mlnm) y se puede estimar que dicho limite era inferior en Cuba por estar mas al norte que la Republica Dominicana No obstante durante este periodo la temperatura experiment6 episodios cortos (unas pocas decadas) de ascensos repetidos
Un signifieativo componente bi6tieo de las referidas condiciones paleoambientales 10 constituy6 la espectacular irradiaci6n evolucionaria experimentada por un grupo numeroso de aves rapaees diurnas y nocturnas (Wetmore 1937 Brodkorb 1969 Kurotchkin y Mayo 1973 Arredondo 1976 Olson 1978 Olson y Hilgartner 1982
329 EXTINCIONES DEL ClJATERNARIO
Suarez 2000a b 2004 Suarez y Emslie 2003) Los mamiferos de las Antillas Mayores ofrecieron a estos depredadores aviares una amplia gama de tamanos de presa que en ausencia de mamiferos del orden Carnivora en la region permitio a estas aves irradiarse a este nicho alimentario con la evolucion de una variedad de especies de muy diversa tall a corporal incluidas formas gigantes Adquirir un cuerpo gigante les propicio el acceso a recursos alimentarios que no estaban siendo explotados por ningun otro depredador (Alcover y McMinn 1994) A Cuba corresponde el mejor registro fosil de rapaces cuaternarias caribenas con siete generos de especies diurnas Dentro de estas aves existieron tres gavilanes gigantes (Accipitridae) tres carairas (Falconidae) y un condor (Cathartidae) asi como cuatro generos de especies nocturnas dos lechuzas (Tytonidae) y tres Mhos (Strigidae) dos de ell os no igualados en tamano en todo el mundo Algunos de estos generos tuvieron representacion en otras islas antillanas como La Espanola Jamaica Bahamas y Gran Caiman
Al retirarse hacia el norte los hielos de la glaciacion Wisconsin el clima se tomo mas hUmedo en las Antillas durante el Holoceno temprano y el bosque mesico se expandio en la region insular El nivel del mar ascendio acompanado de mayor pluviosidad alrededor de los 7 kaAP las Islas Virgenes quedaron separadas entre si y de Puerto Rico y las Bahamas perdieron la mayor parte de su territorio Cerca de los 3 kaAP las condiciones secas retornaron a la region e hicieron desaparecer buena parte del bosque mesico Nuevamente el clima comenzo a adquirir sus condiciones actuales hacia los 25 kaAP aproximadamente y hoy transita por una fase mesica con vegetacion xerica confinada en parcelas relictas
Sin duda estos y otros eventos naturales incidieron negativamente en la sobrevivencia de mamiferos terrestres en las Antillas Mayores asi como en los grupos de pequenas islas perifericas (Pregill y Olson 1981 Olson 2006)
McFarlane et al (1998a) demostraron convincentemente como Amblyrhiza el gigante de los roedores antillanos endemico de la pequena isla de Anguila sucumbio a las repetidas y abruptas perdidas de su habitat durante el interglacial Sangamon por las nipidas y severas oscilaciones del nivel del mar (documentadas por Neumann y Hearty 1996) al no poder responder adecuadamente a tan catastroficos eventos mediante el ajuste de su masa corporal media y otro tanto debio ocurrirle en Jamaica al otro roedor gigante Clidomys (McFarlane et al 1998b) emparentado con Amblyrhiza en la misma epoca (Tabla 45 intervalo I)
El ascenso del nivel del mar seguramente promovio la extincion de mamiferos durante la transicion del Pleistoceno al Holoceno (Tabla 45 intervalo II) con la reduccion del tamano de las Islas por las transgresiones marinas (Pregill y Olson 1981 Morgan 1993) Esto ultimo haria maximamente severa la competicion asf como la presion de los depredadores [Steadman et al (2005) no yen por que el calentamiento global ocurrido durante la transicion Pleistoceno-Holoceno habria de tener un efecto negativo sobre los perezosos siendo sus descendientes actuales especies tropic ales 0 subtropicales pero no se trata del efecto
330 BIOGEOGRAFiA
directo del cambio de temperatura sobre los animales sino del efecto indirecto y progresivo que dicho cambio genera a largo plazo al reducir el territorio insular disponible]
Por esta epoca tambien desaparecieron especies de tall a mayor como Megalocnus zile en La Espanola y otras de menor tamano (en Cuba) pero que representan la tall a maxima en sus respectivos contextos taxonomicos del Cuaternario como Solenodon arredondoi Paralouatta varonai y Macrocapromys acevedo Un mamifero puede ser grande solo a costa de reducir su densidad poblacional y esto 10 hace mis vulnerable a la extinci6n sobre todo en una isla (McNab 1994 McFarlane et al 1998a) Incidentalmente Alcover et al (1998a) afirmaron que larelacion entre la talla corporal y la extincion no siempre se cumple en las especies de menor tamano y citaron como ejemplo que entre las musarafias todas las especies de Nesophontes desaparecieron mientras sobreviven especies de Solenodon pero la comparacion correcta en todo caso serfa entre los solenodones vivientes y el gigante Solenodon arredondoi y entre Nesophontes edithae -el gigante de los nesofontes extinguido en el periodo amerindio- y el resto de sus congeneres que sobrevivieron hasta los primeros siglos de la colonizacion europea (Tabla 45 intervalos III y IV)
CAUSAS ANTROPOGENICAS
Intervalo Amerindio
La mayoria de los autores que se han referido a las extinciones acaecidas durante este intervalo (entre elIos paradigmaticamente Martin 1984a b) han responsabilizado a los aborfgenes (que se supone comenzaron a poblar las Antillas Mayores hace 6 mil afios) con la desaparicion de numerosas especies de mamiferos terrestres en la region principalmente por la via de la sobrecaza y la introduccion de animales domesticos devenidos depredadores de la fauna autoctona (Morgan y Woods 1986) Este efecto negativo de la presencia indigena sobre los mamiferos de la region ha sido dramatizado por algunos autores en terminos de holocausto faunal Sin embargo pensamos que se ha exagerado Como han expresado MacPhee et al (19993) cuando uno busca evidencia dura de complicidad humana directa en [estas] extinciones no hay mucho que encontrar Las siguientes son algunas consideraciones a tener en cuenta en tal sentido
El registro radiometrico mas joven conocido hasta ahora para un perezoso antillano (MegalocnllS rodens) es de 42 kaAP (MacPhee et ai 2007) Si se acepta que los aborfgenes entraron en Cuba hace aproximadamente 6 mil anos su convivencia con los perezosos debe haber durado alrededor de 1800 afios (0 mas porque dificilmente el referfdo fechado radiometrico haya correspondido al ultimo sobreviviente de la especie) Esta cifra contradice la hipotesis del holocausto faunal segun la cualla extincion de los mamiferos insulares de mayor talla debe ocurrir con extrema rapidez -a escala de decadas 0 de centurias- a partir de su primer contacto con los humanos (Martin y Steadman 1999) A pesar de las repetidas afirmaciones en contrario (Harrington 1921 Miller 1929c Herrera [1945] Tabio 1951 Alvarez 1951a 1957 Arredondo
331 EXTINCIONES DEL CUATERNARIO
1959 1960a Veloz y Ortega 1973 Mayo 1980b Rodriguez et al 1984 Castellanos y Pino 1985 Pino y Castellanos 1985 Nunez 1990b Gutierrez 1996 Arredondo 2004) en su mayoria basadas en evidencias no exentas de posibles accidentes de deposicion (0 de reubicacion postdeposicional) en recintos cavernarios (Diaz-Franco [2006b D no se ha demostrado inequivocamente que los aborigenes antillanos se alimentaran de perezosos regularmente Sus restos en residuarios de alimentos son escasisimos en toda la region y esto contrasta con los abundantes restos de jutias y otros mamiferos de menor talla que aparecen en esos residuarios demostrativos de la preferencia de los paleoindios por estos otros mamiferos
Los mamiferos mas importantes en la economia prehistorica de las Antillas Mayores fueron las jutias Capromys y Geocapromys en Cuba Geocapromys en Jamaica e Isolobodon en La Espanola Puerto Rico e Islas Virgenes (Aguayo 1950b Tablo 1951 Rimoli 1972 Wing 2001 Diaz-Franco [2006b D Wilkins (2001) estudi61ajutia viviente de Jamaica (Geocapromys browni) en un sitio ocupado por los aborigenes durante 100 anos y no encontr6 evidencias de que durante ese tiempo la especie haya sido sobreexplotada Asimismo en Cuba las jutias conga carabali y andaraz son escasas en depositos pleistoholocenicos en cambio son muy numerosas en residuarios indigenas y de haber sido sobreexplotadas 10 mas probable es que no hubieran sobrevivido hasta hoy
La habilidad de los humanos para penetrar en profundidad el bosque tropical y mantenerse en este sustentablemente ha sido un evento comparativamente reciente (Beran y Steele 1994) En el caso antillano la caza tierra adentro ha sido explicada como un proceso tardio de desplazamiento a partir de la explotacion de abundantes recursos marinos que ofrecian condiciones 6ptimas para el sedentarismo de las comunidades aborigenes costeras (C6rdova y Arredondo 1988 Brito 2005) Peces y mariscos dominan la arqueologia costera de las Islas antillanas en tanto el consumo de recursos terrestres (mamfferos incluidos) aumenta progresivamente en la medida que los residuarios estudiados se alejan de la costa (Scudder 1991 Wilkins 2001) En las Antillas Mayores el componente terrestre de la dieta aborigen es solo 20-30 del material recobrado en los residuarios (Wing 1977 1989) Y debio ser menor alin porque el componente vegetal de la dieta no se ha preservado Por otra parte los aborigenes cubanos domesticaron la jutia conga y seguramente la reprodujeron para comer (Pose et al 1988) practica que probablemente se repitiera con otras jutias en las demas Antillas Mayores
- Fray Bartolome de las Casas hablo de 3 millones de indios en Cuba al inicio de la colonizacion pero los historiadores subsecuentes consideraron exagerada la cifra (Poey 1851 a) No obstante se dispone de dos estimados de la densidad de poblaci6n aborigen en las Antillas Mayores a la llegada de los europeos 18 indigenaslkm2 en Puerto Rico (Rouse 1956) y 1271km2 en La Espanola (Watts 1987) Extrapolando ambos valores entre las cuatro islas principales de las Antillas Mayores tenemos que en la fecha indicada la poblaci6n aborigen pudo estar entre 16 mil y 113 mil indigenas en Puerto Rico entre 21 mil y 145 mil en Jamaica entre 141 mil y 1 mill6n en La Espanola y entre 200 mil y 14 millones en Cuba Entre las cifras maximas y las minimas nos inclinamos
332 BIOGEOGRAFiA
hacia estas ultimas atendiendo a los altos indices de mortalidad infantil y morbilidad que se evidencian en las osamentas de los pobladores prehispanicos (Martinez y Rives 1990) Uno de los hechos que apuntan hacia el bajo numero de aborigenes en las Antillas Mayores es 10 rapido que desaparecieron despues de la llegada de los europeos pnkticamente sin dejar mestizaje con estos 10 cual contrasta ostensiblemente con la situacion en la America continentaL
En suma un grupo humano (1) demograiicamente irrelevante y controlado por altos indices de morbilidad (2) consumidor mayoritariamente de organismos marinos y (3) carente de la tecnologia cinegetica necesaria para poder sobreexplotar los mamiferos que Ie sirvieron de alimento (a los cuales accedio tardiamente en su desarrollo) no es concebible que haya causado e1 holocausto faunal que se Ie atribuye a partir del error de aplicar a las comunidades aborigenes antillanas un modelo cultural de la prehistoria continentaL
Tampoco hay evidencias de que los aborigenes hayan introducido carnivoros con suficiente capacidad depredadora salvo el perro Morgan y Otlenwaldder (1993) consideraron que la depredacion por perros durante el periodo amerindio es la explicacion mas plausible para la extincion del almiqui gigante de Cuba pero como se puede observar (Tabla 54) Solenodon arredondoi aparece en el intervalo II por falta de asociacion con evidencias culturales
Consecuentemente el accionar de los aborigenes y de sus animales domesticos directamente sobre la fauna no parece haber sido por sf solo un factor determinante de la extincion de mamiferos terrestres durante el periodo amerindio en las Antillas (pero vease Steadman et al 2005)
Hay que recordar en este senti do que alin en el Holoceno tardio ocurrieron cambios clirruiticos de consideracion (vease Causas Naturales) Por ejemplo Curtis et al (2001) explicaron que la extincion de algunos ani males mesofilicos de las alturas montanosas de La Espanola pudo haber antecedido a cualquier impacto humano sigrIificativo en esos lugares por coincidir dicha extincion con una tendencia a la aridez documentada isotopicamente y con evidencias de la perdida del bosque mesico ocurridas despues de 3kaAP Yen Jamaica hace solo 2shy3 mil aDos el area de la Bahia de Jackson sustentaba una gran biomasa de vertebrados en bosque hinnedo a juzgar por la cantidad y diversidad de taxones presentes en material fosil derivado de la depredacion por lechuzas hoy el area es la mas seca de la isla con matorral xerofitico y pobreza de fauna (MacPhee 1996a) Yalm en nuestros dias huracanes de gran intensidad aniquilan poblaciones de vertebrados (que no se recuperan) en is las pequenas y llanas del Caribe (Brooks y Smith 2001)
En medio de la incertidumbre sobre la verdadera causa (0 causas) de las extinciones de mamiferos en el periodo amerindio MacPhee y Marx (1997) han propuesto su hipotesis de la hipermorbilidad la cual estipula colapsos poblacionales relativamente rapidos con la introduccion de enfermedades de alta letalidad Estos autores han razonado que
Las islas son los lugares mas apropiados para experimentar perdidas catastroficas causadas por enfermedades Los mamiferos no solo serian susceptibles en virtud de su prolongado aislamiento biologico sino que en los confines de una isla la epizootia
333 EXTINCIONES DEL CUATERNARIO
tendria la potencialidad de infectar simultaneamente a la rnayoria de los individuos de vadas especies Los neonatos son inmunologicamente inmaduros y sufririan desproporcionadamente cualquier infeccion hipervirulenta La perdida excesiva de neonatos y juveniles tendria efectos severos para la sobrevivencia de la poblacion si esta se reduce por debajo de cierto nivel sobreviene la extincion porque no hay posibilidad de restitucion en el periodo postpanzootico Las especies de mayor talla corporal van a ser las mas vulnerables a la extincion dadas sus bajas tasas reproductivas largos periodos de gestacion y reducido numero de progenie Las especies de menor tamano --caracterizadas por altas tasas reproductivas periodos cortos de gestacion y mayor numero de neonatos por parto- tambien serian fuertemente diezmadas pero muchas sobrevivirian porque su estrategia reproductiva les permite perder mayor numero de individuos Las perdidas deben ocurrir relativamente rapido (a escala de decadas) mientras que otras hipotesis (digamos los cambios climaticos) permiten intervalos mucho mas largos (a escala de siglos 0 de milenios) entre la aparicion de las causas y la produccion de sus efectos Por ejemplo todo indica que los nesofontes se extinguieron muy rapidamente y seria dificil poder invocar un impacto humano directo 0 argumentar fenomenos climaticos que pudieran explicar plausiblemente perdidas tan rapidas
MacPhee y Marx (1997 191) han dicho que por el momento la hipotesis de la hipermorbilidad no pasa de ser una abstraccion en espera de adecuada sustentacion empirica Llama la atencion que linicamente en el periodo amerindio (Tabla 45 intervalo 1Il) las extinciones abarcaron todas las clases del peso corporal (de la a a laj) y que ninguna de las especies que sobrevivieron hasta el periodo postcolombino (Tabla 45 intervalo IV mas las especies vivientes) sobrepasa los 10 kg (clases del peso a b c) en clara coincidencia con los presupuestos de la hipotetica hipermorbilidad
Intervalo Postcolornbino
Por 10 menos 37 especies de mamiferos terrestres vivian en las Antillas Mayores y grupos de Islas perifericas a la lIegada de los europeos en 1492 De elIas se han extinguido 23 shymas especies que en cualquiera de los periodos precedentes (Tabla 45~ y solo subsisten 12 especies repartidas entre Cuba (8 especies) La Espanola (2) Jamaica (1) y Bahamas (1) varias de estas en peligro critico de extincion Solo dos de las especies extinguidas durante los ultimos 500 anos se conocieron en estado viviente los roedores Oryzomys antillarum de Jamaica (extinguido en 1877) y Geocapromys thoracatus de las Islas Swan (extinguido en 1955) Las causas de las extinciones ocurridas durante este periodo son sobradamente conocidas destruccion de habitats introducci6n de animales exoticos (domesticos y comensales) y demas presiones asociadas a la actividad humana entre las cuales posiblemente habria que incluir brotes de hipermorbilidad
Durante mucho tiempo se ha afirmado la posibilidad de que especies de Nesophontes hayan sobrevivido hasta las primeras decadas del siglo xx y que incluso continuen existiendo en lugares remotos de Cuba 0 La Espanola (Allen 1918 1942 Miller 1930 Aguayo 1950a Arredondo 1960b Fischer 1977 MacFadden 1980 Woods y
334 BIOGEOGRAFiA
Ottenwalder 1991) Actualmente no existe evidencia radiometrica de que alguna especie de Nesophontes haya sobrevivido al siglo xv las fechas radiocarbonicas calibradas obtenidas en muestras de Nesophontes de Cuba y La Espanola resultaron no mas tardias que los siglos XIII y XIV (MacPhee et al 1999) Por otra parte las expediciones efectuadas a fines del siglo xx con el proposito de localizar individuos vivientes del genero en La Espanola no produjeron la menor indicacion afirmativa al respecto (Woods et al 1985) Ademas a traves de las Antillas Mayores no aparecen restos de Nesophontes en los depositos formados actualmente por las lechuzas
EPiLOGO
Las Antillas Mayores son islas oceanicas no han formado parte de un continente (con la posible excepcion de una parte de Jamaica hasta el Eoceno Medio hace 45 Ma) y durante un tiempo geologico considerable (desde el Eoceno Superior hace 40 Ma) han permanecido emergidas y separadas de los continentes vecinos Consecuentemente los mamiferos terrestres (no volantes) autoctonos de estas islas constituyen una fauna oceanica que por ende cumple con todas las caracteristicas convencionales reconocidas para este tipo de entidad biogeografica
- Baja diversidad taxonomica a nivel de las categorias superiores (solo cuatro ordenes) en comparacion con islas plataformicas continentales (0 con areas continentales) de tamano y diversidad ecologica simi lares
- Alto grado de endemismo a nivel de genero (96) y familia (50) en respuesta al prolongado aislamiento
- Desbalance ecologico de la estructura trofica ausencia de mamiferos camivoros y ungulados
- Espectaculares procesos de irradiaci6n evolucionaria como resultado de la colonizacion de territorios libres de competidores y depredadores
Tendencias evolucionarias extremas gigantismo
- Control a cargo de aves rapaces con tendencia at gigantismo en ausencia de mamiferos miembros del orden Carnivora
- Mas especies de reptiles y de murcielagos que de mamiferos terrestres relaciones que en los continentes se invierten
Alta tasa de extincion de especies (84) sin reposicion
Preservacion de especies filogeneticamente arcaicas (los llamados fosiles vivientes) los almiquies

304 BIOGEOGRAFIA
antiguos Esto se logro por fin terminando el siglo como se explico anteriormente (vease Contexto temporal) y ique se encontro los mismos cuatro ordenes de mamiferos terrestres conocidos del Cuatemario caribefio aparentemente los Unicos que lograron llegar
Pero ademas hasta el presente no se ha demostrado inequivoca- y concluyentemente que en algun momenta durante los ultimos 40 millones de afios haya existido una conexion terrestre continua entre las Antillas Mayores (en proceso de formacion 0 plenamente formadas) y algun punto de las regiones continentales vecinas
LA BALSA NATURAL (dispersion marina pasiva)
Durante buena parte del siglo xx el puente terrestre tuvo como unico rival a la hipotesis de la balsa natural -transporte accidental de animales sobre islotes de vegetacion flotantes a la deriva despues de arrancados y expulsados mar afuera por un rio crecido- expuesta originalmente por Matthew (191319151916 1918a 1919) y desarrollada en su aplicacion a los mamfferos terreslres antillanos principal mente por Simpson (1956) asi como por olros investigadores (Darlington 1938 1957 Williams y Koopman 1952 Hershkovitz 1958 1972 Patterson 1962 Paula 1967 Patterson y Pascual 1972 Pregill 1981 Ford 1986 Woods 1989a 1990 Morgan 1993) Por su parte Hedges et al (1992) sustentaron dicha hipotesis en estimados de divergencia (basados en el reloj molecular) en la herpetofauna antillana -involucrados tangencia1mente los mamiferos [veanse objeciones a Hedges y colaboradores (Page y Lydeard 1994 Iturralde-Vinent y MacPhee 1999) respuestas a las objeciones (Hedges et al 1994 Hedges 2001) y contrarrespuestas (MacPhee e IturraldeshyVinent 2005) asi como cuestionamientos al metoda de los estimados de divergencia (Foote et al 1999 Pregill y Crother 1999 Woods et ai 2001 )]
La hip61esis de la balsa natural se origino a partir de las numerosas ocasiones en que a 10 largo de siglos tales islotes flotantes (portando 0 no vertebrados incluidos mamiferos) han sido avistados mar afuera a cientos de kilometros de la costa mas cercana en varias partes del mundo --inc1uido el Mar Caribe- segUn consta en diarios de barcos y reportes de observadores diversos (revisados por King 1962 reportes posteriores en SICSP 1970 y Censky et al 1998) Uno de estos casos acaecido en nuestra region se reporta en un informe del SICSP (Smithsonian Institution Center for Shortlived Phenomena 197050shy52)
EIB de julio de 1969 un destructor de la armada de Estados Unidos avisto un islote flotante a unos 100 km al sur de la Base Naval de Guantanamo moviendose desde el Paso de los Vientos en direccion suroeste a una velocidad de 45 kmlh islote de unos 15 m de diametro mantenfa en pie mas de 10 palmas reales de 10-15 m de altura con suelo cubierto de hierba y arbustos [la observacion con binoculares a 18 km de distancia no preciso la presencia de vida animal] La travesia fue rastreada durante 6 dfas hasta que el 19 de julio el islote se dio por perdido tras 6 horas de busqueda aerea
305 POBLAMIENTO CONTINENTE-ISLA
La colonizaci6n exitosa de una isla de las Antillas Mayores por mamiferos terrestres continentales transportados en balsas naturales a la deriva (de cada especie al menos una hembra gravida 0 dos individuos de sexo opuesto) dependeria de factores que van a operar desde el momenta mismo de formarse la balsa asi como durante su travesia marina y su arribo a la isla
a) La posibilidad de estar en la balsa Que la distribuci6n de la especie incluya areas contiguas al rio y que el animal se encuentre en ellugar y el momenta propicios para ser incorporado a la balsa antes de que el rio crecido la desprenda Por ejemplo segun Hershkovitz (1958) los equimidos -grupo ancestral de roedores antillanos- serian buenos candidatos a ser captados por la balsa porque acostumbran a anidar en huecos del tronco de los arboles a la orilla de los rios
b) El tamano y la consistencia de la balsa Muchas se desintegran en alta mar debido a su fragilidad ante la saturaci6n Algunos autores (Beaufort 1951 Arredondo 1996a [1999]) han expresado incredulidad en cuanto a que las balsas hayan podido soportar el peso de un animal como el gran perezoso cubano (Megalocnus rodens) el mayor de los mamiferos antillanos Pero esta especie evolucion6 en Cuba y el ancestro que hizo el viaje pudo ser muy pequeno [EI perezoso oligocenico de Puerto Rico es el mas pequeno de los megaloniquidos antillanos (MacPhee e Iturralde-Vinent 1995b )]
c) Las caracterfsticas fisiol6gicas de la especie Los limites fisiol6gicos de los propagulos constituyen nuestra unica objeci6n a la dispersi6n marina por balsas como mecanismo para la formaci6n de la fauna de vertebrados en las Antillas (MacPhee e IturraldeshyVinent 2005) Unas especies tienen intrinsecamente mas posibilidades que otras para sobrevivir a las vicisitudes que deben afrontar durante la travesia (deshidrataci6n inanicion insolaci6n etc) Por ejemplo los megaloniquidos han demostrado tener extraordinaria habilidad para cruzar barreras marinas fueron los primeros mamfferos suramericanos en invadir Norteamerica mucho antes de que surgiera la conexion terrestre a finales del Plioceno (Paula 1967 Olson 1978 Webb 1985b Martin y Steadman 1999) Incluso se conoce un megaloniquido acultitico del Plioceno peruano (Muizon y McDonald 1995) y otro viviente en Suramerica (Choloepus) que pnicticamente no bebe agua 10 que ha originado una leyenda aborigen que explica por que los perezosos no toman agua (Goff art 1971) Vease ademas el inciso e
d) EI regimen de las corrientes marinas Que conduzcan a la balsa en la direcci6n necesaria EI transporte de balsas de vegetacion desde los continentes hacia las islas fue durante la mayor parte del Cenozoico sumamente improbable (MacPhee e IturraldeshyVinent 2000 148) Estos autores han insistido en que hay que tener en cuenta que durante la cambiante geografia cenozoica del Caribe las corrientes marinas de superficie no favorecieron el desplazamiento de objetos fiotantes hacia las Antillas Mayores [Pero no sabemos con certeza que efecto modificador del curso de tales objetos fiotantes pudieron introducir las tormentas tropicales si previsiblemente funcionaron en el pasado como 10 hacen ahora] Dichos autores senalaron ademas (2005) que en un experimento reciente con boyas de posicionamiento satelital liberadas a la deriva desde las Antillas Menores las boyas fueron a parar mayoritariamente a Centroamerica y a otros lugares
306 B[OGEOGRAFIA
y solo unas pocas recalaron en las Antillas Mayores y las que entraron al Mar Caribe demoraron semanas 0 meses en arribar
e) La distancia a recorrer (y en consecuencia la duracion de la travesia) entre el continente y la isla (10 que repercute decisivamente en el factor c) Este es desde luego uno de los factores mas criticos En la medida que la distancia aumenta la probabilidad de alcanzar la isla disminuye drasticamente con el agravante de que la trayectoria dificilmente seria en linea recta Si tomamos como referencia la velocidad de 45 kmlh del islote ya mencionado (SICSP 1970) cubrir la distancia aerea (aproximada) de 800 km que media entre el noroeste de Suramerica y La Espafiola Ie tomaria a la balsa 74 dias Estimando el doble de este tiempo debido a desvios de la trayectoria la duracion de la travesia probablemente resultaria excesiva para la mayoria de los mamiferos Pero pudiera muy bien ocurrir que la velocidad fuera mayor 0 que la balsa portara plantas vivas que permitirian a los fitOfagos sobrevivir [Tres de los cuatro ordenes de mamiferos terrestres conocidos de las Antillas Mayores son fitOfagos] Ademas esta geologicamente bien establecido que durante etapas de variada duracion desde el Oligoceno hasta el Pleistoceno los territorios del Caribe insular y continental aumentaron considerablemente en extension con niveles del mar 100-200 m por debajo de su nivel actual oportunidades que los propagulos aprovecharian para cruzar barreras marinas significativamente menores
f) El tamafio de la isla Las islas mayores deben interceptar mas balsas (Buckley y Knedlhans 1986) 10 que repercute directamente en el factor e La balsa puede pasar entre islas pequefias y continuar viaje sin llegar a ningun lugar
g) Las condiciones (favorables 0 adversas) del desembarco en la isla y el tiempo que Ie tomaria a uno 0 mas animales depauperados encontrarun habitat apropiado Esto dependeria a su vez del tamafio y diversidad ecologica de la isla (Darlington 1938 Miller 1966)
Tal cumulo de posibles obstaculos y dificultades constituye a primera vista un desafio a la credibilidad de la hipotesis Matthew-Simpson pero en ello radica precisamente su valor explicativo bas ado en la mas absoluta improbabilidad La colonizacion por la via marina es altamente improbable pero no absolutamente imposible si se tiene en cuenta la inmensidad del tiempo geologico se vuelve probable (McKenna 1973) Basta que solo una vez entre mill ones de intentos fallidos durante ese inmenso lapso algunos de los factores anteriormente enunciados coincidan en no ser tan desfavorables para el propagulo Y eso es todo 10 que demanda la hipotesis para cada uno de los grupos monofileticos mayores de mamiferos terrestres presentes en las Antillas Mayores Las Antillas Mayores requeririan el arribo de tan solo 7 (quizas menos) propagulos fundadores a 10 largo de los ultimos 30 0 mas millones de afios (promedio de uno cada 43 mill ones) para explicar la formacion de toda su fauna de mamiferos terrestres por dispersion marina pasiva (vease Discusion y Conc1usiones)
En comparacion con la via marina el poblamiento por la via terrestre es predecible porque la mayor parte de la fauna continental de mamiferos emplazada en la region del puente va a cruzar por este una vez que el mismo se establece y los animales logran
307 POBLAMIEITO CONTINENTE-ISLA
acceder a el el poblamiento por la via marina en cambio es impredecible porque depende total mente del azar que caracteriza a la balsa Claro que en ambas vias opera el azar pero en el primer caso se trata del azar de un evento probable en el segundo el azar de un evento improbable (Simpson 1940) Asi el puente natural (mecanismo de poblamiento minimamente selectivo = alta diversidad inicial) y la balsa natural (mecanismo de poblamiento maximamente selectivo baja diversidad inicial) pueden conceptuarse como los extremos opuestos de una gama de probabilidades de dispersi6n (McKenna 1973) EI fen6meno de la dispersi6n merece la mayor atenci6n cientifica en los estudios de biogeografia insular (Cowie y Holland 2006)
Queda claro que la balsa natural es una hip6tesis consistente con el caracter de la fauna objeto de estudio puesto que el desbalance taxon6mico (sin evidencia paleontol6gica en contrario) s610 puede ser resultado de la improbabilidad del poblamiento No obstante ello no implica necesariamente que la balsa natural haya sido el imico mecanismo (ni siquiera el principal) utilizado por los mamiferos terrestres en su colonizaci6n de las Antillas Mayores La balsa natural debe satisfacer una premisa te6rica esencial dado el caracter azaroso impredecible e improbable de una colonizaci6n exitosa esta debe ocurrir a intervalos muy espaciados a traves del tiempo geol6gico 0 sea durante todo el tiempo que en el area ocupada por las Antillas Mayores ha habido ecosistemas terrestres perrnanentemente disponibles para ser colonizados (los ultimos 40 millones de afios) Sin embargo como sabemos representantes de los cuatro 6rdenes de mamiteros terrestres conocidos de las Antillas Mayores ya estaban emplazados en dicho territorio insular en el Mioceno Inferior 10 que supone un lapso de colonizaci6n limitado al Eoceno SuperiorshyOligoceno (MacPhee 2005) sugerente de un tipo de mecanismo contrario a la colonizacion alocronica exigida por la hipotesis de la balsa natural sobre todo tratandose de una fauna caracterizada por altas tasas de extincion sin reposicion aparente
La presencia en Jamaica (hasta su extinci6n en 1877) del roedor mfuido Oryzomys antillarom con estrecha afinidad centroamericana (Woods 1989a) pudiera constituir el imico caso efectivo de poblamiento (continente-isla) de las Antillas Mayores por dispersion marina dc un mamifero terrestre para el cual no es razonable invocar otro mecanismo de colonizacion
LA GUIRNALDA INSULAR (dispersion mixta)
La geologia ha establecido con suficiente certeza que durante el Cenozoico unicamente en el Pale6geno tardio y solo a traves de la Cresta de Aves (ahora sumergida excepto la isla hom6nima) pudo haber existido una cadena de islas para el transito de mamiferos terrestres entre el norte de Suramerica y las proto-Antillas Mayores [desestimados a ese efecto el Canal dc Yucatan el Elcvado de Nicaragua la Cresta de Caiman el Elevado de Beata y las Antillas Menores (Holcombe et al 1990 MacPhee e IturraldeshyVinent 1994 1995b Iturralde-Vinent y MacPhee 1999)] Holcombe y Moore (1977) y Holcombe y Edgar (1990) postularon la existencia de esta entidad y estimaron algunas de sus posibles caracteristicas Por su parte MacPhee e Iturralde-Vinent (1995b) e Iturralde-Vinent y MacPhee (1999) precisaron los extremos proximal y distal de esa
308 BIOGEOGRAFIA
estructura acotaron la duracion y la edad de la misma y Ie dieron nombre GAARlandia (Greater AntillesA vesRidgelandia)
Segun Iturralde-Vinent y MacPhee (1999) la Cresta de Aves emergio por breve tiempo (1-2 Ma) durante la transicion Eoceno-Oligoceno (35-33 MaAP) por la coincidencia de un levantamiento tectoni co y un considerable descenso eustatico del nivel del mar En ese momenta las islas septentrionales del arco de las Antillas Mayores (Cuba centroorientalshyLa Espafiola - Puerto Rico - Islas Virgenes) constituian una sola isla grande 0 una serie de islas separadas por barreras muy estrechas La Cresta de Aves emergida entre el bloque Puerto Rico - Islas Virgenes y un microcontinente noroccidental de Suramerica -separado en ese momento del resto del continente por una via marina surgida en el Eoceno tardio- dio origen a GAARlandia
Obviamente el numero de colonizaciones exitosas por mamiferos terrestres a traves de una guimalda de Islas tiende a disminuir mientras mas anchas sean las barreras marinas entre las islas que forman la guimalda y cuando esto ocurre disminuye tambien la diversidad taxonomica de los propagulos particularmente a nivel de las categorias superiores (Simpson 1956)
Vemos asi que para los mamiferos terrestres la colonizaci6n a traves de una guimalda insular puede ser tan improbable como a la deriva en una balsa natural en dependencia de la cantidad y anchura de las barreras marinas a cruzar el tamafio y la fisiografia de las islas que integran la guimalda y el regimen de las corrientes marinas y aereas entre otros factores
Resumiendo la guimalda insular puede conceptuarse como un mecanismo de poblamiento altamente selectivo (= baja diversidad inicial) que combina la improbabilidad de la balsa natural con la colonizacion restringida temporaimente del puente terrestre
En su reciente revision de la taxonomia distribuci6n y filogenia (basada en estudios moleculares) de varios grupos de mamiferos antillanos Davalos (2004) concluyo que los perezosos megalocninos los primates y los roedores histricognatos se avienen al patron de divergencia con el continente y epoca de arribo a las Antill as implicados en la hipotesis de GAARlandia pero no asi los perezosos coloepodinos que requieren una explicacion biogeognlfica distinta Esto ultimo (sospechado por Matthew 1918a) habla sido confirmado por White y MacPhee (2001 226-227)
Nuestra evidencia filogenetica no sustenta la idea de un dado unificado surgido de la invasion de un megaloniquido el analisis sugiere mas bien dos eventos de colonizaci6n independientes [y] sustenta fuertemente un origen difiletico de las subfamilias [Choloepodinae y Megalocninae] Si hubo dos migraciones a traves de GAARlandia presumiblemente poco espaciadas en el tiempo no se puede determinar en el presente
Finalmente queda claro que GAARlandia -como guirnalda insular- es la hipotesis mas robusta propuesta hasta ahora para explicar la presencia en las Antillas Mayores de
309 POBLAMIENTO CONTINENTE- ISLA
la absoluta mayona de los mamfferos terrestres de ongen suramericano porque el caracter de esa fauna supone un mecanismo de poblamiento ancestral que sea a la vez maximamente selectivo (baja diversidad ordinal) y temporalmente restringido (transicion Eoceno-Oligoceno)
Recientemente Rega et al (2002 18) expresaron A pesar de una revision exhaustiva de la paleogeografia de la cuenca del Caribe Iturralde-Vinent y MacPhee (1999) no pudieron confirmar la presencia de una conexion terrestre continua entre Suramerica y las proto-Antillas aunque hipotetizaron su existencia [Vease la Fig 42 (= figura 6 de Iturralde-Vinent y MacPhee 1999 simplificada))
En el trabajo citado por Rega y colaboradores aparece la siguiente explicacion
Actualmente no es posible ofrecer reconstrucciones paleogeograficas detalladas de la Cresta de Aves Sin embargo hay evidencia de que porciones de la cresta deben haberse elevado subaereamente en un tiempo La cuestion cntica es si la Cresta de Aves emergida pudo haber formado algun tipo de corredor entre las Antillas Mayores y el noroeste de Suramerica Perforaciones en las alturas estructurales de la cresta pudieran proveer el registro del Paleogeno tardio que se necesita para precisar la hipotesis del corredor aunque no se puede predecir si tales datos serian suficientemente detallados para determinar si en algun momento GAARlandia constituyo una sola entidad subaerea Por ejemplo pudo haber barreras marinas por corto tiempo entre las masas terrestres de la Cresta de Aves [0) ellevantamiento y la subsidencia pudieron afectar diferentes partes de GAARlandia en diferentes tiempos AItemativamente los elementos faunales pudieron haberse dispersado de manera pulsada Movimientos episOdicos de este tipo incrementarian la extincion de los elementos faunales incapaces de trasladarse a la siguiente masa terrestre en la cadena disminuyendo asi la diversidad total (Iturralde-Vinent y MacPhee 199957)
La primera parte de la explicacion deja claro que la posibilidad de que la Cresta de Aves haya sido una conexion terrestre continua no se puede descartar pero tampoco se puede afirmar La ultima parte de la explicacion describe el efecto altamente selectivo (filtrante) de la colonizacion a traves de una cadena de islas
Y continuan lturralde-Vinent y MacPhee (199957 -58) explicando
Hasta que punto el microcontinente noroccidental de Suramerica [considerado por dichos autores como el area suministradora de propagulos hacia GAARlandiaJ estuvo fisicamente separado del resto del continente no ha sido plenamente esc1arecido Si estuvo sustancialmente aislado por barreras marinas y orogenic as durante el Paleogeno tardio entonces solo la fraccion de la biota suramericana emplazada en el microcontinente estuvo en posicion de acceder a GAARlandia Esto pudiera tener que ver con el hecho de que la fauna antillana carezca de representantes de muchos grupos suramericanos [de mamfferos) que debieron haber hecho el viaje
310 BIOGEOGRAFiA
Florida o Alturas0 0
d 6 D L1anuras25deg
Yucatan
20deg
deg0
10deg
MARCARIBE
80deg 75deg
Bahamas
Cres ta de Beata
D Mares someros
D Mares profundos
OCEANO ATLANTICa
FIGURA 42 Reconstrucci6n hipotetica de GAARlandia como conexi6n terrestre continua entre las proto-Antillas Mayores y el noroeste de Suramerica en el Pale6geno tardio segun la FIgura 6 (simplificada) de Iturralde-Vinent y MacPhee (1999)
En su defensa del puente terrestre Barbour (1945b249) se refiri6 a esto - antes 10 habfa hecho Anthony [1942363]- en los terminos siguientes
El argumento de que si las Antillas hubieran estado conectadas con el continente se encontrarian los f6siles de un numero mayor de distintas criaturas me deja frio es perfectamente concebible que la conexi6n haya ocurrido en un periodo en que la parte del continente implicada no estuviera poblada por una fauna grande y variada
Y concluyen lturralde-Vinent y MacPhee (199958) su detallada explicaci6n
Desafortunadamente sabemos muy poco sobre la composici6n faunal del noroeste de Suramerica a finales del Pale6geno
Consecuentemente si se confirmara la magnitud del aislamiento del aludido microcontinente y su concomitante baja diversidad faunal GAARlandia como conexi6n terrestre continua serf a la hip6tesis mas parsimoniosa No obstante mientras no se disponga de evidencia concluyente a este respecto hipotetizar a GAARlandia como conexi6n continua (minima selectividad) la coloca en franca vulnerabilidad te6rica al tener que afrontar la objeci6n clasica al puente terrestre (baja diversidad ordinal de la
311 POBLAMIENTO CONTINENTE-TSLA
fauna) al tiempo que se la priva de ser la hip6tesis mas robusta para explicar -como guimalda de islas- la colonizaci6n de las Antillas por mamiferos procedentes de Suramerica
ELARcA DE NOE (seudovicarianza)
El primer vicarianista antillano (desde luego sin el saberlo) fue Corral (1940) con su utilizaci6n ingenua de las ideas wegenerianas de la deriva continental aparecidas la decada anterior Pero el primer (supuesto) intento de aplicar a la formaci6n de la biota caribefia el mecanismo de la vicarianza basada en la tearia de la tect6nica de placas (surgida en la sexta decada del pas ado siglo) se debe a Rosen ([1976] 1985)
Segun Rosen las Antillas Mayores se originaron en el Cremcico en la Placa Caribe aproximadamente en la posici6n que hoy dia ocupa Centroamerica y al ser desplazadas tectonicamente hacia el este interactuaron (de algun modo no precisado por Rosen) con las margenes continentales de Norte- y Suramerica de donde (en un evento unico) adquirieron toda su fauna transportada subsecuentemente por dichas islas en su transito hacia sus posiciones actuales [De ahi la denominacion metaforica dada par McKenna (1973) a este mecanismo de poblamiento] Aunque se ha presentado esta propuesta como Model0 de Vicarianza Continente-Isla de Rosen en realidad por definici6n no constituye un evento vicariante porque las proto-Antillas Mayores no resultaron del fraccionamiento y separaci6n por tectonismo de una porcion del territorio continental como exige esta hip6tesis (vease definici6n de vicarianza al comienzo de Hipotesis Biogeograficas)
El Arca de Noe de Rosen ha sido desestimada por la mayoria de los autores contemporaneos porque (1) carece de sustento geol6gico (lturralde-Vinent y MacPhee 1999) y porque (2) en el periodo en que se afrrrna ocum6 el evento muchos de los ordenes modemos de mamiferos placentarios attn no existian (Williams 1989)
EL BARCO FUNERAL VlKINGO (tanatovicarianza)
Domning et al (1997) reportaron 10 que consideraron el mamffero terrestre mas viejo de las Antillas --circunstancia que se refleja en el titulo mismo del trabajo Oldest West Indian Land Mammal- al descubrirse un perisodactilo rinocerotoide (Hyrachius sp) en una localidad de edad Eoceno Inferior a Medio en el occidente de Jamaica
Jamaica ha tenido una historia geologica y paleogeografica distinta de la historia comun a las restantes Antillas Mayores A principios del Cenozoico Jamaica occidental (a diferencia de la porcion oriental constituida por el Bloque de las Montafias Azules) formaba parte de Centro america de la cual se desgaj6 (por el desplazamiento de la Placa Caribe) para venir a integrarse despues del Eoceno Medio con 10 que serfa la futura isla de Jamaica desaparecida por transgresi6n marina en el Eoceno Superior hasta volver a emerger totalmente en el Mioceno Medio (Iturralde-Vinent y MacPhee 1999 MacPhee e Iturralde-Vinent 2000)
312 BIOGEOGRAFiA
Por consiguiente Hyrachius ---conocido de localidades de edad Eoceno Inferior a Medio en Norteamerica- debio ocupar (10 que iba a ser) Jamaica occidental mucho antes de que esta se separara de Centroamerica De hecho este fosil Uego a su destino insular contenido en las rocas de su basamento no como un ser vivo por 10 que es erroneo considerarlo un animal antillano (lturralde-Vinent y MacPhee 2004) como han supuesto algunos (Hedges 200 I Portell et al 200 I Condis [200 I D Todo 10 que hizo fue viajar tanatovicariantemente en su barco funeral vikingo -otra de las denominaciones metaforicas de McKenna (1973)
DISCUSION Y CONCLUSIONES
Hemos visto que solo disponemos de una hipotesis plausible para explicar la mayor parte del poblamiento (continente-isla) de las Antillas Mayores por mamiferos terrestres del Neotropico continental la dispersion mixta (a traves de GAARlandia) para los perezosos (excepto Choloepodinae) los monos y los roedores (excepto Oryzomys)
Por otra parte la historia biogeogrMica de las musaraiias de las Antillas Mayores (Nesophontes y Solenodon) constituye una de las situaciones mas enigmaticas entre los mamiferos de la region (Ottenwalder 2001321) Al menos cinco mecanismos de poblamiento continente-isla han sido invocados para explicar cuando como y de donde llegaron a las Antillas Mayores estas musaraiias
I EI propagulo ancestral (del Terciario de Norteamerica) coloniz6 a las Antillas Mayores por dispersion marina desde Centroamerica 0 desde la Florida (Patterson 1962)
2 El propagulo ancestral ( del Terciario) cruzo el Atlantico Sur (supuestamente mas estreshycho en aquella epoca) directamente de Africa a las Antillas Mayores (Hershkovitz 1972)
3 El propagulo ancestral (del Terciario de Norteamerica) Uego a las Antillas Mayores en el Arca de Noe de Rosen (MacFadden 1980)
4 El propagulo ancestral (del Terciario de Norteamerica) fue transportado a las Antillas desde Centroamerica en el bloque de Jamaica occidental y de alli paso por dispersion marina a las restantes Antillas Mayores (Whidden y Asher 200 I)
5 El propagulo ancestral (del Terciario de Norteamerica) lleg6 a las Antillas Mayores a traves de GAARlandia (Whidden y Asher 2001)
Aunque hay consenso entre estos (y otros) autores sobre el origen norteamericano de las musaraiias antillanas ninguno de los cinco hipoteticos mecanismos de poblamiento resiste la evidencia en contrario filogenetica yo paleogeogrMica Ademas algunas de estas hipotesis asumen que Nesophontidae y Solenodontidae comparten un antecesor comun en las Antillas pero esta relacion no ha sido confirm ada Para dilucidar el enigmatico caso de las musaraiias antillanas se requerini profundizar en la investigaci6n geologica paleogeognifica y paleontol6gica (Iturralde-Vinent y MacPhee 1999)
De acuerdo con el conocimiento filogenetico disponible sobre el conjunto de la fauna la colonizacion original de las Antillas Mayores por mamiferos terrestres implicaria el arribo
313 POBLAMIENTO ISLA-ISLA
de solo siete propagulos fundadores perezosos 2 (coloepodinos y megalocninos si como parece tienen un origen difiletico) musaraiias 2 (nesofontidos y solenodontidos si no constituyen una misma familia como consideran algunos) monos 1 (pitecidos) roedores histricognatos 1 (equimidos capromidos y heptaxodontidos) roedores muridos 1 (Oryzomys en Jamaica)
POBLAMIENTO ISLA-ISLA
La distribucion de los mamiferos terrestres a traves del area antillana demarcada al inicio del presente capitulo (vease Contexto Geogratico Fig 41) ha sido interpretada en funcion de dos hipoteticos mecanismos de poblamiento interinsular la dispersion y la vicarianza fundamentadas como se resumen seguidamente
DISPERSION
De acuerdo con la evidencia paleogeogratica mas actualizada en las principales regresiones marinas ocurridas en la cuenca caribeiia durante los ultimos 23 Ma (transiciones OligocenoshyMioceno Mioceno Superior - Plioceno y varias veces en el Pleistoceno) las masas terrestres constitutivas de la region insular en formacion aumentaron considerablemente su extension territorial disminuyendo por tanto la distancia entre elias oportunidades que los elementos faunales aprovecharian para dispersarse de unas islas a otras
Durante mucho tiempo esta ha sido la hipotesis mas favorecida por los estudiosos del tema a partir de Simpson (1956) pero en decadas recientes ha suscitado dos objeciones
1 Es dificil en tender como pudo operar la dispersion marina entre estas islas puesto que ninguna de las Antillas Mayores posee rios suficientemente caudalosos para transportar masas significativas de vegetacion flotante (MacPhee 198432)
Es oportuno recordar aqui el islote ala deriva (con mas de diez palmas reales en pie) entre las tres Antillas Mayores mas occidentales mencionado anteriormente (vease La Balsa Natural)
2 La dispersion marina es menos efectiva para explicar la asombrosa similitud de las faunas de mamiferos terrestres a traves de las Antillas Mayores septentrionales porque como proceso aleatorio no debe producir distribuciones uniformes a 10 largo de una cadena de islas 0 si las produce entonces el proceso no es aleatorio (MacPhee e lturralde-Vinent 2000 149)
Independientemente de que las aludidas distribuciones taxonomicas estan lejos de ser uniformes 0 asombrosamente similares la hipotesis de la dispersion marina contrariamente a su canicter aleatorio y altamente improbable en el caso del poblamiento continente-isla- en el caso del poblamiento isla-isla es un proceso no aleatorio (aprovecha las oportunidades brindadas puntualmente por los periodos de maxima regresion marina) y ademas altamente probable dada la incomparablemente menor distancia entre las islas caribeiias que entre estas y los continentes vecinos
314 BIOGEOGRAFiA
V ICARIANZA
La neotectonica y las transgresiones marinas del Oligoceno Medio y del Mioceno afectaron sustancialmente la paleogeografia de la Cresta de las Antillas Mayores resultando la subdivision gradual de las partes subaereas de la Cresta Asi el este de Cuba y el norte de La Espanola conectados fisicamente durante el Oligoceno Inferior se separaron al formarse el Paso de los Vientos en el Oligoceno tardio en tanto La Espanola y Puerto Rico probablemente permanecieron conectadas hasta el Mioceno tardio en que surgio el Paso de Mona (Iturralde-Vinent y MacPhee 1999)
Tales eventos geologicos habrian dividido las distribuciones de los grupos faunales emplazados previamente en el territorio continuo de las tres islas y a partir de esas disyunciones dichos autores (tambien MacPhee et al 2000 y White y MacPhee 2001) asumieron --como segunda fase del modelo de GAARlandia- la hipotesis de la vicarianza isla-isla para explicar la distribucion multiinsular de varios grupos de mamiferos terrestres a traves de las Antillas Mayores septentrionales De acuerdo con esta hipotesis cualquier diferencia en composicion faunal entre las islas (por ejemplo la ausencia de roedores heptaxodontidos en Cuba 0 de monos platirrinos en Puerto Rico) constituye evidencia de extincion local postvicariante y no de dispersion marina fallida Lo primero se puede demostrar si aparecen los fosiles pertinentes 10 segundo no se puede demostrar (MacPhee e Iturralde-Vinent 1994) Por otra parte IturraldeshyVinent y MacPhee (1999 57) han reconocido que aunque fa vicarianza isla-isla puede representar una solucion clara para las distribuciones de varios taxones cuaternarios no puede explicar todos los casos
Rega et al (2002 18) han presentado dos objeciones al modelo de la vicarianza isla-isla aplicado a los mamiferos de las Antillas Mayores
I Si la separacion de Cuba y La Espanola precedio en varios mill ones de anos la separashycion de La Espanola y Puerto Rico el modelo vicariante de lturralde-MacPhee debe predecir mayor similitud entre las faunas de La Espanola y Puerto Rico que entre las de Cuba y La Espanola De hecho es al reyes
No obstante MacPhee et al (200335) admitieron despues que la fecha mas temprana empiricamente defendible [para la apertura del Paso de Mona] debe ser el Oligoceno Medio esto invierte la secuencia de ambas disyunciones con 10 cual queda resuelta la aparente contradiccion Sin embargo en su reciente evaluacion de la congruencia entre filogenia e hipotesis geologic as en varios grupos de mamiferos terrestres de las Antillas Mayores Davalos (2004373) encontro que ninguno de los taxones cumple con la fecha y el patron de divergencia entre las masas terrestres antillanas postulados por el modelo de GAARlandia
2 La presencia de un mismo genero en las tres Islas septentrionales de las Antillas Mayores implica [por el modelo vicariante] que la diferenciacion entre estas formas durante el intervalo desde el Mioceno ha sido unicamente de nivel especifico [Dicha presencia]
315 rRRADlAcr6~ EVOLUCrONARIA
s610 puede explicarse razonablemente como el resultado de movimientos de esos animales entre las islas en el Cuaternario
De esta objecion resulta dificil disentir [El poblamiento entre islas diferenciado solo a nivel especifico no es probable que sea anterior al Pleistoceno (Simpson 1956 15)]
Debe tenerse en cuenta que la hipotesis de la vicarianza isla-isla alude a las tres islas septentrionales de las Antillas Mayores porque como se explico antes (vease El Barco Funeral Vikingo) Jamaica no existia como isla durante el lapso (Eoceno Superior shyOligoceno) consensualmente reconocido como el mas probable para la colonizacion de las proto-Antill as Mayores por la mayo ria de los mamiferos terrestres Despues de emerger como isla en el Mioceno Medio Jamaica debe haber recibido sus mamiferos terrestres por dispersion isla-isla (Davalos 2004376) y solo en el Cuaternario parece haberrecibido (por dispersion marina desde Centroamerica) al roedor murido Oryzomys
Seguramente tanto la dispersion como la vicarianza contribuyeron a que los mamiferos terrestres se irradiaran a traves de las Antillas Mayores y grupos de islas perifericas pero las maneras y tiempos en que estos mecanismos operaron a 10 largo de dicho proceso son aun en gran medida motivos de desacuerdo e incertidumbre
IRRADIACION EVOLUCIONARIA
A continuaci6n revisamos someramente las distribuciones conocidas de los diferentes grupos de mamiferos terrestres a traves de las Islas principales de las Antillas Mayores y grupos de Islas perifericas incluidas las interpretaciones y conjeturas que la literatura reciente recoge en relaci6n con la manera en que dichas distribuciones pudieron haberse conformado durante la irradiaci6n evolucionaria de los referidos grupos de mamiferos Manifestaciones de esta irradiaci6n pueden inferirse unicamente a partir de los taxones presentes en el Cuatemario pues solo 6 de las especies conocidas corresponden al Terciario
GRUPOS ORlGINARIOS DE SURAMERICA
PEREZOSOS (Megalonychidae)
Aparte de un genero miocenico de Cuba y otro del Oligoceno de Puerto Rico se conocen 4 generos (12 especies) de perezosos cuaternarios (todos extintos) restringidos a las tres islas septentrionales de las Antillas Mayores Acratocnus Neocnus Megalocnus y Parocnus Los cuatro generos estan representados en Cuba y La Espanola y Acratocnus 10 esta ademas (como unico genero de perezoso) en Puerto Rico Esta distribuci6n ha sido interpretada por MacPhee et al (2000) eomo resultado de la vicarianza oligomiocenica entre estas islas y [dado que el Banco Puerto Rico - Islas Virgenes fue la entrada de GAARlandia a las Antillas Mayores] se preguntaron si la ausencia de los otros tres generos en Puerto Rico se debe a que nunca estuvieron en ese territorio 0 a que sus
316 BIOGEOGRAFiA
fosiles atm no han sido hallados Pero como hemos visto Rega et al (2002) atribuyeron dicha distribucion a la dispersion interinsular en el Cuatemario
Por otra parte varios amilisis c1adisticos recientes ubi can a Choloepus -genero Reciente conocido solo de Centro- y Suramerica- como taxon hermano de Aeratoenus exc1uidos los demas megaloniquidos antillanos Para White y MacPhee (2001228)
tan estrecha relacion sugiere que la divergencia entre ambos [generos] ocurrio despues de que Aeratoenus divergio de Neoenus Si esta relacion implica que Aeratoenus y Neoenus se dispersaron separadamente desde Suramerica a las AntilIas mientras Choloepus permanecia en el continente (siendo necesarias al menos tres invasiones de perezosos) 0 si Choloepus invadio Suramerica desde las Antillas es algo que el material disponible no permite determinar
La diversidad adaptativa de los perezosos se manifiesta en (1) la utilizacion del habitat especies arboreas en Neoenus y Aeratoenus y terrestres en Paroenus y Megaloenus (Webb 1985a White [1993] White et al 1996 Arredondo [1999]) y en (2) la gama de tallas corporales desde los pequenos Neoenus hasta Megaloenus rodens el mayor de los mamiferos terrestres antillanos (su peso adulto estimado en 270 kg seglin Paula 19679) esto sugiere que los perezosos se irradiaron a nichos que en contextos continentales probablemente estarian ocupados por miembros de otros c1ados (MacPhee e IturraldeshyVinent 2000)
No esta claro si el abanico de tamanos entre estos perezosos enmascara alguna tendencia al gigantismo Por 10 pronto Anderson y Handley (2002) encontraron 10 contrario en perezosos (Bradypus) de las Islas de Bocas del Toro (Panama) shyseparadas secuencialmente del area continental vecina por ascensos del nivel del mar durante los ultimos 10 ka En las cinco islas los perezosos son menores que su contraparte continental y las diferencias entre las tallas medias de las cinco poblaciones expresan una reduccion lineal significativa de la talla corporal en funcion de la edad geologica de cada isla sin relacion alguna con el area de las islas 0 la distancia que las separa del continente
MONOS (Pitheciidae)
Los primates de las Antillas Mayores (cuatro especies extintas) constituyen un grupo monofiletico con Paralouatta (en Cuba) y Antillothrix (en La Espanola) como taxones hermanos seguidos en la escalera c1adistica por Xenothrix (en Jamaica) y finalmente Callieebus como el pariente continental mas cercano esto implica una sola invasion desde Suramerica antes del Mioceno Inferior (fecha mas temprana conocida para los primates fosiles antillanos) y por tanto en consonancia con GAARlandia como la via de colonizacion (Horovitz y MacPhee 1996 1999 Horovitz [1997] MacPhee y Horovitz 2004)
En desacuerdo parcial con la anterior interpretacion la filogenia de este grupo de monos obtenida por Davalos (2004) indica un antecesor comun en Cuba La Espanola y Jamaica
317 TRRADIACTON EVOLUCIONARIA
compatible con la hipotetica conexion -propuesta tentativamente por Iturralde-Vinent y MacPhee (199980-84)- entre las Montanas Azules del este de Jamaica y la peninsula suroccidental de La Espanola en la epoca de GAARlandia
A pesar de que los primates antillanos pertenecen a un grupo viviente de monos muy pequenos (los monos titles) fueron comparativamente grandes en relacion con los platirrinos modernos y sucesivamente mayores en la direccion Antillothrix Xenothrix y Paralouatta con P marianae como el mayor platirrino conocido del Mioceno neotropical (MacPhee 2005) MacPhee et al (1994) consideraron que Xenothrix debio ser un mono de andar lento y al parecer las especies de Paralouatta abandonaron la arborealidad para adoptar un modo de vida mas terrestre (MacPhee 1996a MacPhee et al 2003 MacPhee y Meldrum 2006)
ROEOORES (Echimyidae Capromyidae Heptaxodontidae)
La edad de Zazamys en Cuba demuestra que la invasion de los roedores histricognatos en las Antillas Mayores comenzo antes del Mioceno Inferior por otra parte los representantes mas antiguos de esa irradiacion evolucionaria se encuentran en La Espanola y Puerto Rico Ambas evidencias son consistentes con la hipotesis de GAARlandia (Woods 1982 1988 Woods et al 2001)
No esta claro si los capromidos evolucionaron a partir de un equimido ancestral 0 de un protocapromido 0 si los equimidos antillanos (I sub familia 3 generos y 7 especies todas extintas) forman parte de la irradiacion de los capromidos (4 subfamilias 9 generos y 27 especies 10 vivientes) Kratochvil et al (1978b) y Rodriguez et al (1979) consideraron que Capromys nan us es un eslab6n entre Echimyidae y Capromyidae pero Mesocapromys nanus esta morfologicamente mucho mas cerca de Hexolobodon y los de mas capromidos que de los equimidos (Woods 1985) Lo que si esta claro segun Woods et al (2001) es que la subfamilia Plagiodontinae endemic a de La Espanola es el linaje basal de los capromidos y que Isolobodon Hexolobodon Capromys Geocapromys Mysateles y Mesocapromys [y supuestamente Macrocapromys] forman parte de la irradiacion temprana de Capromyidae
La subfamilia Capromyinae (5 generos y 17 especies 9 vivientes) parece ser el dado de mas reciente evolucion en la familia con invasion de Cuba desde La Espanola y dispersion desde Cuba (1) por Capromys a las islas Caiman y (2) por Geocapromys a las islas Bahamas Caiman y Jamaica y desde Jamaica a las islas Swan aunque la presencia de Geocapromys en Swan pudiera deberse a introduccion aborigen (Morgan 1985 1988 1989 1993 1994 Woods 1989a 1990) Woods et al (2001) utilizaron datos moleculares para obtener estimados independientes de divergencia entre capromidos segiJn 10 cual Geocapromys divergio cerca de los 17 MaAP Mesocapromys divergio de Capromys hace 15 MaAP en tanto lvlysateles 10 hizo hace 10 MaAP Por su parte las formas de Capromys pilorides y de Mysateles prehensilis en Isla de la Juventud divergieron de sus correspondientes subespecies nominativas en la isla principal a los 24 y 08 MaAP respectivamente
318 BIOGEOGRAFiA
Woods y Howland (I979) describieron adaptaciones morfologicas de la mandibula el hioide y la faringe asociadas a mecanismos de masticacion diferentes en plagiodontinos y caprominos y Varona (1980a) llamo la atencion sobre el gran desarrollo de los molariformes (en relacion con la longitud total del cnineo) que presentan ciertos grupos de jutias fosiles en comparacion con las vivientes 10 que pudiera interpretarse como una adaptacion alimentaria a una vegetacion mas abrasiva por parte de las jutias extinguidas
Segun Woods (1990) la familia Heptaxodontidae (2 subfamilias 5 generos y 6 especies todas extintas) tambien seria parte de la irradiacion de los capromidos con origen en el este de La Espanola 0 en Puerto Rico y subsecuente dispersion al Banco de Anguila - San Martin en las Antillas Menores (pero vease MacPhee y Flemming 2003) Este banco estaria a solo 70 km del Banco Puerto Rico - Islas Virgenes en tiempos de maxima regresion marina y esta proximidad con Puerto Rico -patria de Elasmodontomys obliquus- seguramente tuvo que ver con la presencia en las islas de Anguila y San Martin del heptaxodontino gigante Amblyrhiza relacionado mas estrechamente con los roedores de las Antillas Mayores que con los del resto de las Antillas Menores (Woods 1990 1996) Aunque los roedores suelen aumentar de tamano en las islas (Case 1978) la escala del incremento de la talla en Amblyrhiza (que pudo alcanzar mas de 200 kg de peso adulto) esta fuera de toda proporcion con los patrones conocidos en otros roedores insulares (Biknevicius et 01 1993)
Clidomys (Clidomyinae) de Jamaica difiere marcadamente en su morfologia de los heptaxodontinos (vease ademas MacPhee y Flemming 2003)10 cual dificulta explicar su presencia en esa isla Como Jamaica estuvo bajo el mar hasta el Mioceno Medio el ancestro de Clidomys colonizaria esta isla por dispersion desde el extremo suroccidental de La Espanola despues que los heptaxodontinos se originaron en esta ultima isla (Woods 1989a) Pero Pascual et 01 (I 990) especularon que Clidomys pudiera ser el resultado de una invasion posterior e independiente de los capromidos
GRUPOS ORIGINARIOS DE NORTEAMERICA
El origen de las musaranas antillanas ha resultado ser hasta ahora el mayor enigma biogeografico de la fauna de mamiferos terrestres de la region aunque la mayoria de los autores coinciden en considerarlas originarias de Norteamerica (vease Discus ion y Conclusiones en Poblamiento Continente-Isla) No obstante MacPhee e Iturralde-Vinent (1994) descartaron toda posibilidad de conexion terrestre por el Estrecho de la Florida 0 por el Canal de Yucatan y si el (supuesto) evento vicari ante de Rosen (vease El Arca de Noe) -invocado por MacFadden (1980) para resolver el enigma- fuera minimamente plausible aun asi es dificil creer que las catastrofes que 10 sucedieron (vease Contexto Temporal) hayan barrido con toda esa vieja fauna continental excepto las musaranas circunstancia ya anticipada muy tempranamente por Matthew (l918a)
De otra parte la relaci6n entre Nesophontidae y Solenodontidae permanece irresuelta Si estos linajes no estan estrechamente relacionados entonces es muy posible que
319 IRRADIACION EVOLUCIONARIA
sus distribuciones sean explicadas por mecanismos diferentes (Whidden y Asher 2001)
La descomunal diferencia de talla corporal entre las musaranas pequenas (Nesophontes) y las grandes (Soienodon) seguramente impuso a ambos grupos restricciones distintas relativas a la termorregulacion y las adaptaciones energeticas En Nesophontes la seleccion debio favorecer una alta tasa de metabolismo y crecimiento camadas mas numerosas corta expectativa de vida yalta densidad poblacionaL En Soienodon en cambio la seleccion opero en la direccion de una baja tasa metabolica camadas reducidas crecimiento lento longevidad y baja densidad poblacional (Ottenwalder [1991])
MUSARANAS PEQUENAS (Nesophontidae)
Las musaranas pequenas estan representadas en las Antillas por el genero Nesophontes (y un genero miocenico indeterminado en ambar dominicano) con 6 especies cuaternarias extinguidas distribuidas en Cuba La Espanola Puerto Rico Islas Virgenes e Islas Caiman La mayor de las musaranas pequenas Nesophontes edithae evoluciono en Puerto Rico y su arribo a las Islas Virgenes supone una dispersion sumamente reciente En las Islas Caiman Nesophontes se asemeja mas a la poblacion de Cuba de donde seguramente derivo en el Plio-Pleistoceno (junto con Capromys y Geocapromys) cuando la costa sur de Cuba estuvo mucho mas cerca de las Islas Caiman (lturralde-Vinent 2003fig 4) MacPhee et ai (1994) interpretaron la distribucion de Nesophontes en Cuba y La Espanola como resultado de la vicarianza oligomiocenica ocurrida al formarse el Paso de los Vientos pero la distancia taxonomica entre los lados del Paso es solo de nivel especifico y N micros incluso vivi6 en ambas islas segtln Patterson (1962)
Allen (1918) considero que Nesophontes micrus fue una especie primitiva no modificada para la vida arb6rea 0 fos6rea y por tanto de habitos terrestres Nesophontes edithae de Puerto Rico es mucho mayor y mas robusta que las restantes especies del genero eL indice de robustez de su femur (= anchura minima de la diafisis ongitud total del hueso) muestra que este es tan robusto como el de Solenodon paradoxus un animal dos veces mayor (Morgan et ai 1980) Choate y Birney (1968) postularon un aumento cronoclinal de la taIla corporal en N edithae confirmado mas tarde por McFarlane (1999a) mediante fechados radiocarb6nicos de la secuencia estratignifica en depositos cavernarios de Puerto Rico Solenodon no vivi6 en Puerto Rico y en ausencia de una musarana de mayor tamano Nesophontes alcanzo una talla corporal mayor en esa isla que en La Espanola 0 Cuba (Woods y Eisenberg 1989)
MUSARANAS GRANDES (Solenodontidae)
Los arcaicos almiqufes (genero Soienodon) comprenden cuatro especies una viviente (S cubanus) y otra f6sil (S arredondoi) en Cuba y una viviente (S paradoxus con dos subespecies) y otra f6sil (S marcanoi) en La Espanola Como cabria esperar las dos especies cubanas se asemejan mas entre sf que a las especies de La Espanola pero
320 BIOGEOGRAFiA
enigmaticamente S marcanoi comparte caracteres con S cuban us y S paradoxus (Woods y Eisenberg 1989 Morgan y Ottenwalder 1993 Ottenwalder 200 I) Solenodon arredondoi alcanzo una talla corporal superior a la de cualquiera de las musarafias vivientes a nivel mundial Segun Morgan y Ottenwalder (1993) extrapolando las dimensiones y el peso de los almiquies vivientes S arredondoi debio tener una longitud cabeza-tronco y un peso de 450-550 mm y 1500-2000 g respectivamente en comparacion con S cubanus (360 mm y 800 g) y S paradoxus (390 mm y 1100 g)
Entre todas las especies de almiqufes Solenodon paradoxus es la que presenta los caracteres mas derivados (Otten walder 2001) Si estas relaciones son validas la distribucion de Solenodon entre Cuba y La Espafiola debio discurrir de oeste a este como propuso MacFadden (1980) quien invoco la presencia fosH de S cubanus [pero tambien S arredondoi] en Pinar del Rio para postular que el ancestro continental entr~ por Cuba opinion compartida por Perfit y Williams (1989) y por Ottenwalder ([1991] 2001) El paso de Solenodon a Puerto Rico pudo haberlo impedido la existencia de barreras ecologicas durante la conexion del este de La Espafiola con Puerto Rico 0 por la ausencia de Solenodon en esa parte de La Espafiola en aquella epoca Sin embargo las mismas condiciones ecologicas en el este de La Espafiola no parecen haber representado una barrera efectiva para Nesophontes cuyas mas altas tasa de reproduccion y densidad poblacional presumiblemente Ie permitieron expandirse flpidamente a traves de La Espafiola (Ottenwalder [1991])
Es opinion generalizada que los almiqufes antillanos constituyen el grupo de mamiferos que primero se emplazo en el territorio insular (MacPhee 2005) y en tal sentido es de interes la evidencia sobre especificidad parasito-hospedante aportada por Cruz (1980) quien encontro que la especie de acaro (genero Cubanochirus) que parasita a los almiquies es mucho mas primitiva que la especie (del mismo genero) que paras ita a las jutias y de aqui concluyo que dicho genero de acaro debio evolucionar primero en Solenodontidae y posteriormente en Capromyidae al entrar en contacto representantes de ambas familias en Cuba 0 La Espafiola
Recientemente Roca (2004) y Roca et al (2004) dieron a conocer que los resultados de su estudio genetico-molecular sobre los almiquies indican una divergencia de 25 Ma entre Solenodon cuban us y S paradoxus -compatible con la disyuncion tectonica entre Cuba y La Espafiola (Iturralde-Vinent y MacPhee 1999 McPhee et al 2003)-- y una separacion de ambas especies comparable en magnitud a la divergencia entre familias de mamlferos Consecuentemente propusieron que Solenodon cubanus fuera clasificada en el genero Atopogaie como indico Cabrera (1925) hace 82 afios
DISCUSION Y CONCLUSIONES
Las musarafias y los roedores antillanos (excepto Oryzomys) se irradiaron tanto de sus respectivos ancestros continentales que de estos 6rdenes se reconocen familias y subfamilias endemicas de la region en contraste con los perezosos (una subfamilia y tres
321 IRRADIACION EVOLUCIONARIA
tribus endemicas) y los monos (una tribu endemica) como expresion de diferentes niveles de diversificacion evolucionaria Este proceso es dificil de explicar (traUindose de c1ados que pnicticamente comparten la misma antigiiedad en las Antmas) si es correcta la interpretacion de Pascual et at (1990636)
Nosotros consideramos estos diferentes niveles de diversificacion como las diferentes etapas de evolucion alcanzadas por cada grupo [de mamiferos] Obviamente esto estuvo relacionado con las diferentes longitudes de sus historias
Pudiera ocurrir que en el futuro se compruebe que la historia biogeognifica de la region fue mucho mas complicada
La Espanola la isla mas ecol6gicamente divers a de la region debi6 ser el centro principal de evoluci6n de los mamfferos (con 87 de las familias y 73 de las subfamilias) seguida por Cuba (con 75 de las familias y 54 de las subfamilias) las dos islas de las Antillas Mayores mas relacionadas biogeograficamente (Fig 43 Tabla 44)
eL
EL
cb CAIMAN
SWAN
A Acratocnini B Cubanocnini C Megalocnini D Mesocnini E Nesophontidae F Solenodontidae G Xenothrichini
ANGUILAshyM-0 SMARTIN
H Heteropsomyinae I Plagiodontinae J Isolobodontinae
K Hexolobodontinae L Capromyinae
M Heptaxodontinae N CIidomyinae 0 Sigmodontinae
FIGURA 43 Representacion esquematica de las relaciones interinsulares (expresadas porel numero de linajes compartidos) entre las Antillas Mayores y entre estas y los grupos de islas perifericas segun las interpretaciones de los autores citados en el texto Los numeros dentro de los circulos indican la cantidad de linajes representados en cada isla relacion dudosa Contexto taxon6mico de los linajes en la Tabla 44
bull bull
bull bull
bull bull bull
bull bull
bull bull
bull
bull
bull
bull
bull bull bull bull
bull bull
bull bull
bull
bull bull bull
bull bull
bull
bull bull
bull
322 BIOGEOGRAFiA
TABLA 44 Distribuci6n de los linajcs de mamiferos terrestres a traves de las Antillas Mayores y grupos de islas perifericas LE = La Espanola CU Cuba JA Jamaica PR = Puerto Rico IC Islas Caiman IS Islas Swan IB = Islas Bahamas IV = Islas Virgenes AS = Anguila - San Martin
FamiliaSubfumiliafIribu LE CU JA PR IC IS IB IV AS
Megalonychidae
Choloepodinae
Acratocnini
Cubanocnini
Megalocninae
Megalocnini
Mesocnini
Nesophontidae
Solenodontidae
Pitheciidae
Callicebinae
Xenothrichini
Echimyidae
Heteropsomyinae
Capromyidae
Plagiodontinae
Isolobodontinae
Hexolobodontinae
Capromyinae
H e pta x 0 don tid a e
Heptaxodontinae
Clidomyinae
Muridae
Sigmodontinae
PILOSA
SORICOMORPHA
PRIMATES
RODENTIA
bull
bull
bull
bull TOTALES 12 10 4 4 2
323 EXTlICIOIES DEL CUATERNARIO
El gigantismo ha sido sin duda una tendencia evolucionaria recurrente entre los clados mayores de mamiferos antillanos Es un hecho bien conocido que algunos grupos de mamiferos han aumentado de tamafio en las islas y varios autores han intentado explicarlo a partir de enfoques diferentes (Foster 1964 Miller 1966 Sondaar 1977 Case 1978 Eisenberg y Wilson 1978 Heaney 1978 Lawlor 1982 Lomolino 1985 Angerbjom 1985 McNab 19942001)
EXTINCIONES DEL CUATERNARIO
Las islas que integran la Subregion Antillana han perdido casi 90 de sus mamiferos terrestres durante los ultimos milenios y en proporcion con su extension territorial esta perdida ha sido mayor en la Subregion que en cualquier otro lugar del planeta (MacPhee 1996b) Las extinciones del Cuatemario eliminaron mas de las tres cuartas partes de la fauna de mamiferos terrestres conocida de las Antillas Mayores sus islas satelites y grupos de islas perifericas y el examen de este proceso en sus aspectos mas generales se aborda en esta ultima parte del presente capitulo
CRONOLOGiA Y CONSECUENCIAS
Es muy improbable que un fosil represente el ultimo individuo de un taxon extinto y entonces hay que guiarse por la edad radiometrica del fosil mas reciente conocido lIamada fecha de ultima presencia- como unica aproximacion posible a la fecha de extincion del taxon
Como se explico al inicio del capitulo (vease Contexto Temporal) el registro fosH de los mamiferos terrestres de las Antillas Mayores adolece de insuficiente sustentacion radiometrica No obstante la distribucion temporal de los escasos fechados disponibles para el Cuaternario de la region abarca etapas criticas del proceso de extincion experimentado por la referida fauna a 10 largo de los ultimos 250 mil afios
De las 52 especies que es posible reconocer actualmente como miembros extintos de la fauna cuatemaria de mamiferos terrestres de las Antillas Mayores y grupos de islas perifericas los restos fosiles de 23 de elIas (442) han sido fechados radiometricamente o se han hallado asociados estratigraficamente de manera inequivoca con otro material fechado De estos 23 fechados 3 corresponden al interglacial Sangamon 4 pertenecen a la secuencia Pleistoceno terminal - Holoceno temprano 10 se ubican en ellapso entre el arribo de los aborigenes y la incursion europea y 6 son postcolombinos (a los cuales hay que sumar otras dos especies extinguidas en los siglos XIX y xx) A partir de estas 25 especies avaladas por fechas de ultima presencia distribuidas entre los cuatro intervalos temporales sefialados hemos compuesto la Tabla 45 en la que ademas aparecen las restantes 27 especies (carentes de fechado radiometrico) ubicadas en el intervalo temporal mas probable sobre la base de (1) la asociacion estratigrafica de sus restos con evidencias culturales (amerindias 0 europeas) 0 (2) la ausencia de dicha asociacion Sobre estos criterios de ubicacion temporal procede hacer dos aclaraciones
324 BIOGEOGRAFIA
TABLA 45 Intervalos temporales mas probables para las extinciones cuatemarias de los mamiferos terrestres de las Antillas Mayores y grupos de islas perifericas sobre la base de las evidencias siguientes (l) ultimo ejemplar colectado u observado en vida (2) fechado radiometrico mas tardio en material de la especie (3) asociaci6n estratigrafica mas tardia con otro material fechado (4) asociaci6n estratigrafica con restos de animales introducidos restos humanos u objetos culturales propios del intervale (5) falta de asociaci6n con evidencias culturales [Los ntimeros entre parentesis en la columna Evidencia significan adscripci6n tentativa al intervalo] Los estimados del peso corporal (en kg) estan tomados de la literatura 0 inferidos comparativamente y distribuidos entre
las clases siguientes (a) S 01 (b) 01 a lt 1 (c) 1 a lt 10 (d) 10 a lt 100 (e) 100 a lt 200 (j) ~ 200 Las
abreviaturas que designan las islas son las de la Tabla 44
F amilialEspecie Peso IsJa Evidencia Fuente
I httervalo htterg1acial Sangamon (gt80 kaAP)
HEPTAXODONTlNAE
Amblyrhiza inundata e AS 3 McFarJane et al 1998a
CLIDOMYINAE
Clidomys osborni d JA 3 Ibid
Clidomys parvus d JA 3 Ibid
II httervalo Pleistoceno terminal ~ Holoceno temprano (20-6 kaAP)
MEGALONYCHIDAE
Aeratoenus antillensis d CD 5 Arredondo 1975 a
Aeratoenus odontrigonus d PR 3 McFarJane 1999b
Aeratoenus simorhynehus d LE 2 Rega et al 2002
M egaloenus zile f LE 2 MacPhee et al 2000
SOLENODONTIDAE
Solenodon arredondoi e CD 5 Morgan y Ottenwalder 1993
PnHECIIDAE
Paralouatta varonai d CD 5 MacPhee 1996a
EcHIMYIDAE
Heteropsomys antillensis c PR 5 McFarJane 1999b
Heteropsomys insulans c PR 5 Ibid
Puertoricomys eorozalus c PR 5 Woods 1989a
CAPROMYIDAE
P lagiodontia araeum c LE 5 MacPhee et al 2000
Macrocapromys latus c CD 5 Varona y Arredondo 1979
(Continua)
325 EXTINCIONES DEL CUATERNARIO
TABLA 45 (Continuaci6n)
F ami1ia1Especie Peso Isla Evidencia Fuente
M acrocapromys latus c CU 5 Varona y Arredondo 1979
Mesocapromys kraglievichi c CU 5 Ibid
HEPTAXODONTINAE
Xaymaca fulvopulvis a JA 2 MacPhee y Flemming 2003
III Intervalo Amerindio (6-05 kaAP)
MEGALONYCHIDAE
Acratocnus ye d LE 2 MacPhee et aI 2000
Neocnus comes c LE 2 Ibid
N eocnus dousman c LE 2 Ibid
N eocnus toupiti c LE 2 Ibid
N eocnus gliriform is c CU 4 Cordova et ai 1997
M egalocnus rodens f CU 2 MacPhee et ai 2007
Parocnus browni e CU 2 Steadman et ai 2005
P arocnus serus e LE 2 MacPhee et ai 2000
N ESOPHONTIDAE
N esophontes edithae a PR 2 McFarlane 1999b
PITHECIIDAE
Antillothrix bernensis c LE 3 Rimoli 1977
EcHIMYIDAE
Brotomys contractus a LE 4 Miller 1929c
CAPROMYIDAE
P lagiodontia spelaeum c LE 4 Ibid
M acrocapromys acevedo c CU 4 NUiiez 1990a
HEPTAXODONTINAE
Elasmodontomys obliquus d PR 3 McFarlane 1999b
Iv Intervalo Postcolombino (05 kaAP - Presente)
N ESOPHONTIDAE
N esophontes hypom icrus a LE 24 MacPhee y Fle111111ing 1999
N esophontes zam icrus a LE (4) Ibid
N esophontes m icrus a CU 24 MacPhee et aI 1999
(Continua)
326 BIOGEOGRAFiA
TABLA 45 (Continuacion)
F amilialEspecie Peso Isla Evidencia Fuente
N esophontes m icrus a CD 24 MacPhee et aI 1999
N esophontes m icrus a LE 4 MacPhee et aI 2000
N esophontes sp 1 a IC 4 Morgan 1994
N esophontes sp 2 a IC 4 Ibid
Solenodon marcanoi b LE 4 Ottenwalder 2001
PITHECIIDAE
Xenothrix mcgregori c JA 4 MacPhee y Flemming 1999
EcHIMYIDAE
Boromys offeUa a CD 24 Guarch 1984
Boromys torrei a CD 24 Ibid
Brotomys voratus a LE 4 MacPhee et aI 2000
CAPROMYIDAE
P lagiodontia ipnaeum c LE 4 MacPhee y Flemming 1999
Rhizoplagiodontia lemkei c LE 4 Ibid
Isolobodon portoricensis c PR 24 Ibid
Isolobodon montanus c LE 4 Ibid
Hexolobodon phenax c LE (4) Ibid
Capromys sp c IC 24 Morgan 1994
Geocapromys columbianus c CD 4 MacPhee y Flemming 1999
Geocapromys thoracatus b IS 1 MacPhee y Marx 1997
Geocapromys sp 1 c IC 4 Morgan 1994
Geocapromys sp 2 c IC (4) Ibid
l-IEPTAXODONTINAE
Quemisia gravis c LE (1) MacPhee y Flemming 1999
MURIDAE
Oryzomys antillarum a JA 1 Ibid
1 Flemming y MacPhee (1999) alertaron en contra de la asociacion con otro material fechado Asimismo Morgan (1993) indico que la ausencia de asociacion con evidencias culturales amerindias no es necesariamente un criterio concluyente para ubicar un taxon en el intervalo temporal anterior porque hay depositos en la region que carecen de tales evidencias y son del Holoceno tardio por fechados radiocarbonicos [No obstante
327 EXTlNCIONES DEL CUATERNAR10
la ausencia de una especie en residuarios arqueol6gicos adquiere mayor certidumbre como criterio de ubicaci6n temporal de su extinci6n en la medida que aumenta el numero de residuarios estudiados que en las Antillas Mayores se cuentan por centenares] A falta de fechado absoluto de una especie consideramos que estos criterios de ubicaci6n temporal son mas utiles que la alternativa de excluir del analisis a la especie
2 En los trabajos sobre biogeografia de mamiferos antillanos los arque610gos mas citados como autoridades para establecer el inicio de la presencia aborigen en las Antillas Mayores (Kozlowski 1974 Rouse y Allaire 1978 Rouse 1989 1992 Wilson 2001) difieren al respecto entre los 6 kaAP y los 45 kaAP (pero vease Pagan et al 200523) y hemos adoptado la mas temprana de estas fechas para el comienzo del intervalo amerindio en la Tabla 45 De todos modos cualquiera que haya sido la fecha esta no es aplicable por igual al conjunto de las Antillas Mayores porque segun algunos de los autores citados el poblamiento comenz6 por Cuba y La Espanola 6 mil anos atras continu6 en Puerto Rico hace alrededor de 3 mil anos (pero vease Burney et al 1994) yen Jamaica hace menos de 2 mil e incluso a las Islas Caiman ni siquiera llegaron [Las Islas Caiman descubiertas por Col6n en 1503 carecen absolutamente de evidencia amerindia (Stokes y Keegan 1996)]
Si los datos de la Tabla 45 representan una aceptable aproximaci6n a la realidad el proceso de las extinciones cuaternarias de los mamiferos terrestres de las Antillas Mayores y grupos de islas perifericas debi6 caracterizarse entre otros por los eventos siguientes
Aunque obviamente la fauna cuaternaria debi6 ser mucho mas numerosa que 10 que refleja el registro paleontol6gico (Morgan y Woods 1986) 52 especies (81 de la fauna conocida) desaparecieron (sin reposici6n) durante los ultimos 250 mil anos
- Al parecer las extinciones se concentraron en las especies de mayor tamano Asi el peso promedio estimado para las especies de la etapa preantr6pica (intervalos I + II de la Tabla 45) es 350 kg es 328 kg para las especies del intervalo III (amerindio) y es s610 25 kg (c1ases del peso a b c) para las especies del intervalo IV (postcolombino) Ninguna especie aut6ctona de mamifero mayor de 6 kg existe hoy en la regi6n
- De acuerdo con la distribuci6n temporal de las extinciones 61 especies sobrevivieron al interglacial Sangamon (intervalo 1)48 sobrevivieron a la transici6n del Pleistoceno terminal al Holoceno temprano (intervalo II) 34 sobrevivieron al periodo amerindio (intervalo III) y s610 12 especies han sobrevivido desde la Ilegada de los europeos (intervalo IV) Las tasas de extinci6n correspondientes (exceptuado el intervalo I) son una especie cada 1167 anos en el intervalo II una cada 393 anos en el intervalo III y una cada 23 anos en el intervalo IV
- Durante todo el proceso de las extinciones cuaternarias la perdida de especies por isla fue de 91 en La Espanola 83 en Jamaica y 62 en Cuba Bahamas retuvo su unica especie (aunque extirpada en varias de las islas) Puerto Rico e Islas Caiman perdieron todas sus especies (6 y 5 respectivamente) y las Islas Swan Virgenes y Anguila - San Martin perdieron sus respectivas especies (micas
328 BIOGEOGRAFiA
De los cuatro 6rdenes de mamiferos representados en la fauna Pilosa y Primates desaparecieron totalmente Soricomorpha y Rodentia perdieron 86 y 74 de sus especies respectivamente
CAUSALIDAD
CAUSAS NATURALES
Los cicIos glaciales e interglaciales acaecidos en el Pleistoceno causaron cambios ambientales dnisticos en las Antillas con la concomitante alternancia de cIimas frios (secos) y calidos (hUmedos) asi como de descensos y ascensos del nivel del mar aumento y disminuci6n de las areas territoriales) induso (aunque con menor intensidad) en el Holoceno Estos eventos paleogeograficos han sido consistentemente documentados (Mayo y Kartashov 1972 Bonnatti y Gartner 1973 Ortega y Arcia 1982 Prance 1982 Schubert y Medina 1982 Dodge et al 1983 Ortega [1984] Haq et al 1987 Schubert 1989 Hodell et al 1991 Guilderson et al 1994 Oppo 1997 Emslie 1998 Curtis et al 2001 Iturralde-Vinent 2003) y de ellos brindamos a continuaci6n una breve resena
Finalizando el periodo interglacial anterior a la ultima glaciaci6n (Wisconsin) el mar estaba a 8-10 m por encima de su nivel actual Cuando el avance glacial del Wisconsin a1canz6 su culminaci6n (eillamado Ultimo Maximo Glacial) hace 18 ka el mar habia descendido 120 m con respecto a dicho nivel y las islas del Caribe habian incrementado considerablemente sus extensiones subaereas en comparaci6n con las que presentan hoy dia por ejemplo Puerto Rico y las Islas Virgenes se conectaron territorialmente y las Bahamas se unieron en cinco gran des islas Prevalecia un regimen frio y seco y condiciones de aridez --debidas a una reducci6n de las precipitaciones (posiblemente a menos de 440 mmlano)- y los ambientes xericos se habian extendido a traves de las islas y bancos emergidos y abundaban los pinares (los habitats mesicos estaban restringidos a parcelas relictas en 10 alto de las montanas) En el Mar Caribe y el Golfo de Mexico la temperatura superficial del mar era 5degC mas baja que la actual y en las llanuras costeras y alturas maximas eran respectivamente 2-3degC y 6degC mas bajas Los limites de las zonas ecol6gicas altitudinales descendieron varios cientos de metros y cada zona redujo su extensi6n vertical al aumentar el gradiente de temperatura Ellimite inferior de la zona periglacial
elevaci6n donde la temperatura media minima a1canza los OdegC) de la Cordillera Central de la Republica Dominicana (altura maxima del Caribe insular con evidencia de topografia glacial) se considera que estuvo entre los 1500 y 1800 mlnm (actualmente esta a 2800 mlnm) y se puede estimar que dicho limite era inferior en Cuba por estar mas al norte que la Republica Dominicana No obstante durante este periodo la temperatura experiment6 episodios cortos (unas pocas decadas) de ascensos repetidos
Un signifieativo componente bi6tieo de las referidas condiciones paleoambientales 10 constituy6 la espectacular irradiaci6n evolucionaria experimentada por un grupo numeroso de aves rapaees diurnas y nocturnas (Wetmore 1937 Brodkorb 1969 Kurotchkin y Mayo 1973 Arredondo 1976 Olson 1978 Olson y Hilgartner 1982
329 EXTINCIONES DEL ClJATERNARIO
Suarez 2000a b 2004 Suarez y Emslie 2003) Los mamiferos de las Antillas Mayores ofrecieron a estos depredadores aviares una amplia gama de tamanos de presa que en ausencia de mamiferos del orden Carnivora en la region permitio a estas aves irradiarse a este nicho alimentario con la evolucion de una variedad de especies de muy diversa tall a corporal incluidas formas gigantes Adquirir un cuerpo gigante les propicio el acceso a recursos alimentarios que no estaban siendo explotados por ningun otro depredador (Alcover y McMinn 1994) A Cuba corresponde el mejor registro fosil de rapaces cuaternarias caribenas con siete generos de especies diurnas Dentro de estas aves existieron tres gavilanes gigantes (Accipitridae) tres carairas (Falconidae) y un condor (Cathartidae) asi como cuatro generos de especies nocturnas dos lechuzas (Tytonidae) y tres Mhos (Strigidae) dos de ell os no igualados en tamano en todo el mundo Algunos de estos generos tuvieron representacion en otras islas antillanas como La Espanola Jamaica Bahamas y Gran Caiman
Al retirarse hacia el norte los hielos de la glaciacion Wisconsin el clima se tomo mas hUmedo en las Antillas durante el Holoceno temprano y el bosque mesico se expandio en la region insular El nivel del mar ascendio acompanado de mayor pluviosidad alrededor de los 7 kaAP las Islas Virgenes quedaron separadas entre si y de Puerto Rico y las Bahamas perdieron la mayor parte de su territorio Cerca de los 3 kaAP las condiciones secas retornaron a la region e hicieron desaparecer buena parte del bosque mesico Nuevamente el clima comenzo a adquirir sus condiciones actuales hacia los 25 kaAP aproximadamente y hoy transita por una fase mesica con vegetacion xerica confinada en parcelas relictas
Sin duda estos y otros eventos naturales incidieron negativamente en la sobrevivencia de mamiferos terrestres en las Antillas Mayores asi como en los grupos de pequenas islas perifericas (Pregill y Olson 1981 Olson 2006)
McFarlane et al (1998a) demostraron convincentemente como Amblyrhiza el gigante de los roedores antillanos endemico de la pequena isla de Anguila sucumbio a las repetidas y abruptas perdidas de su habitat durante el interglacial Sangamon por las nipidas y severas oscilaciones del nivel del mar (documentadas por Neumann y Hearty 1996) al no poder responder adecuadamente a tan catastroficos eventos mediante el ajuste de su masa corporal media y otro tanto debio ocurrirle en Jamaica al otro roedor gigante Clidomys (McFarlane et al 1998b) emparentado con Amblyrhiza en la misma epoca (Tabla 45 intervalo I)
El ascenso del nivel del mar seguramente promovio la extincion de mamiferos durante la transicion del Pleistoceno al Holoceno (Tabla 45 intervalo II) con la reduccion del tamano de las Islas por las transgresiones marinas (Pregill y Olson 1981 Morgan 1993) Esto ultimo haria maximamente severa la competicion asf como la presion de los depredadores [Steadman et al (2005) no yen por que el calentamiento global ocurrido durante la transicion Pleistoceno-Holoceno habria de tener un efecto negativo sobre los perezosos siendo sus descendientes actuales especies tropic ales 0 subtropicales pero no se trata del efecto
330 BIOGEOGRAFiA
directo del cambio de temperatura sobre los animales sino del efecto indirecto y progresivo que dicho cambio genera a largo plazo al reducir el territorio insular disponible]
Por esta epoca tambien desaparecieron especies de tall a mayor como Megalocnus zile en La Espanola y otras de menor tamano (en Cuba) pero que representan la tall a maxima en sus respectivos contextos taxonomicos del Cuaternario como Solenodon arredondoi Paralouatta varonai y Macrocapromys acevedo Un mamifero puede ser grande solo a costa de reducir su densidad poblacional y esto 10 hace mis vulnerable a la extinci6n sobre todo en una isla (McNab 1994 McFarlane et al 1998a) Incidentalmente Alcover et al (1998a) afirmaron que larelacion entre la talla corporal y la extincion no siempre se cumple en las especies de menor tamano y citaron como ejemplo que entre las musarafias todas las especies de Nesophontes desaparecieron mientras sobreviven especies de Solenodon pero la comparacion correcta en todo caso serfa entre los solenodones vivientes y el gigante Solenodon arredondoi y entre Nesophontes edithae -el gigante de los nesofontes extinguido en el periodo amerindio- y el resto de sus congeneres que sobrevivieron hasta los primeros siglos de la colonizacion europea (Tabla 45 intervalos III y IV)
CAUSAS ANTROPOGENICAS
Intervalo Amerindio
La mayoria de los autores que se han referido a las extinciones acaecidas durante este intervalo (entre elIos paradigmaticamente Martin 1984a b) han responsabilizado a los aborfgenes (que se supone comenzaron a poblar las Antillas Mayores hace 6 mil afios) con la desaparicion de numerosas especies de mamiferos terrestres en la region principalmente por la via de la sobrecaza y la introduccion de animales domesticos devenidos depredadores de la fauna autoctona (Morgan y Woods 1986) Este efecto negativo de la presencia indigena sobre los mamiferos de la region ha sido dramatizado por algunos autores en terminos de holocausto faunal Sin embargo pensamos que se ha exagerado Como han expresado MacPhee et al (19993) cuando uno busca evidencia dura de complicidad humana directa en [estas] extinciones no hay mucho que encontrar Las siguientes son algunas consideraciones a tener en cuenta en tal sentido
El registro radiometrico mas joven conocido hasta ahora para un perezoso antillano (MegalocnllS rodens) es de 42 kaAP (MacPhee et ai 2007) Si se acepta que los aborfgenes entraron en Cuba hace aproximadamente 6 mil anos su convivencia con los perezosos debe haber durado alrededor de 1800 afios (0 mas porque dificilmente el referfdo fechado radiometrico haya correspondido al ultimo sobreviviente de la especie) Esta cifra contradice la hipotesis del holocausto faunal segun la cualla extincion de los mamiferos insulares de mayor talla debe ocurrir con extrema rapidez -a escala de decadas 0 de centurias- a partir de su primer contacto con los humanos (Martin y Steadman 1999) A pesar de las repetidas afirmaciones en contrario (Harrington 1921 Miller 1929c Herrera [1945] Tabio 1951 Alvarez 1951a 1957 Arredondo
331 EXTINCIONES DEL CUATERNARIO
1959 1960a Veloz y Ortega 1973 Mayo 1980b Rodriguez et al 1984 Castellanos y Pino 1985 Pino y Castellanos 1985 Nunez 1990b Gutierrez 1996 Arredondo 2004) en su mayoria basadas en evidencias no exentas de posibles accidentes de deposicion (0 de reubicacion postdeposicional) en recintos cavernarios (Diaz-Franco [2006b D no se ha demostrado inequivocamente que los aborigenes antillanos se alimentaran de perezosos regularmente Sus restos en residuarios de alimentos son escasisimos en toda la region y esto contrasta con los abundantes restos de jutias y otros mamiferos de menor talla que aparecen en esos residuarios demostrativos de la preferencia de los paleoindios por estos otros mamiferos
Los mamiferos mas importantes en la economia prehistorica de las Antillas Mayores fueron las jutias Capromys y Geocapromys en Cuba Geocapromys en Jamaica e Isolobodon en La Espanola Puerto Rico e Islas Virgenes (Aguayo 1950b Tablo 1951 Rimoli 1972 Wing 2001 Diaz-Franco [2006b D Wilkins (2001) estudi61ajutia viviente de Jamaica (Geocapromys browni) en un sitio ocupado por los aborigenes durante 100 anos y no encontr6 evidencias de que durante ese tiempo la especie haya sido sobreexplotada Asimismo en Cuba las jutias conga carabali y andaraz son escasas en depositos pleistoholocenicos en cambio son muy numerosas en residuarios indigenas y de haber sido sobreexplotadas 10 mas probable es que no hubieran sobrevivido hasta hoy
La habilidad de los humanos para penetrar en profundidad el bosque tropical y mantenerse en este sustentablemente ha sido un evento comparativamente reciente (Beran y Steele 1994) En el caso antillano la caza tierra adentro ha sido explicada como un proceso tardio de desplazamiento a partir de la explotacion de abundantes recursos marinos que ofrecian condiciones 6ptimas para el sedentarismo de las comunidades aborigenes costeras (C6rdova y Arredondo 1988 Brito 2005) Peces y mariscos dominan la arqueologia costera de las Islas antillanas en tanto el consumo de recursos terrestres (mamfferos incluidos) aumenta progresivamente en la medida que los residuarios estudiados se alejan de la costa (Scudder 1991 Wilkins 2001) En las Antillas Mayores el componente terrestre de la dieta aborigen es solo 20-30 del material recobrado en los residuarios (Wing 1977 1989) Y debio ser menor alin porque el componente vegetal de la dieta no se ha preservado Por otra parte los aborigenes cubanos domesticaron la jutia conga y seguramente la reprodujeron para comer (Pose et al 1988) practica que probablemente se repitiera con otras jutias en las demas Antillas Mayores
- Fray Bartolome de las Casas hablo de 3 millones de indios en Cuba al inicio de la colonizacion pero los historiadores subsecuentes consideraron exagerada la cifra (Poey 1851 a) No obstante se dispone de dos estimados de la densidad de poblaci6n aborigen en las Antillas Mayores a la llegada de los europeos 18 indigenaslkm2 en Puerto Rico (Rouse 1956) y 1271km2 en La Espanola (Watts 1987) Extrapolando ambos valores entre las cuatro islas principales de las Antillas Mayores tenemos que en la fecha indicada la poblaci6n aborigen pudo estar entre 16 mil y 113 mil indigenas en Puerto Rico entre 21 mil y 145 mil en Jamaica entre 141 mil y 1 mill6n en La Espanola y entre 200 mil y 14 millones en Cuba Entre las cifras maximas y las minimas nos inclinamos
332 BIOGEOGRAFiA
hacia estas ultimas atendiendo a los altos indices de mortalidad infantil y morbilidad que se evidencian en las osamentas de los pobladores prehispanicos (Martinez y Rives 1990) Uno de los hechos que apuntan hacia el bajo numero de aborigenes en las Antillas Mayores es 10 rapido que desaparecieron despues de la llegada de los europeos pnkticamente sin dejar mestizaje con estos 10 cual contrasta ostensiblemente con la situacion en la America continentaL
En suma un grupo humano (1) demograiicamente irrelevante y controlado por altos indices de morbilidad (2) consumidor mayoritariamente de organismos marinos y (3) carente de la tecnologia cinegetica necesaria para poder sobreexplotar los mamiferos que Ie sirvieron de alimento (a los cuales accedio tardiamente en su desarrollo) no es concebible que haya causado e1 holocausto faunal que se Ie atribuye a partir del error de aplicar a las comunidades aborigenes antillanas un modelo cultural de la prehistoria continentaL
Tampoco hay evidencias de que los aborigenes hayan introducido carnivoros con suficiente capacidad depredadora salvo el perro Morgan y Otlenwaldder (1993) consideraron que la depredacion por perros durante el periodo amerindio es la explicacion mas plausible para la extincion del almiqui gigante de Cuba pero como se puede observar (Tabla 54) Solenodon arredondoi aparece en el intervalo II por falta de asociacion con evidencias culturales
Consecuentemente el accionar de los aborigenes y de sus animales domesticos directamente sobre la fauna no parece haber sido por sf solo un factor determinante de la extincion de mamiferos terrestres durante el periodo amerindio en las Antillas (pero vease Steadman et al 2005)
Hay que recordar en este senti do que alin en el Holoceno tardio ocurrieron cambios clirruiticos de consideracion (vease Causas Naturales) Por ejemplo Curtis et al (2001) explicaron que la extincion de algunos ani males mesofilicos de las alturas montanosas de La Espanola pudo haber antecedido a cualquier impacto humano sigrIificativo en esos lugares por coincidir dicha extincion con una tendencia a la aridez documentada isotopicamente y con evidencias de la perdida del bosque mesico ocurridas despues de 3kaAP Yen Jamaica hace solo 2shy3 mil aDos el area de la Bahia de Jackson sustentaba una gran biomasa de vertebrados en bosque hinnedo a juzgar por la cantidad y diversidad de taxones presentes en material fosil derivado de la depredacion por lechuzas hoy el area es la mas seca de la isla con matorral xerofitico y pobreza de fauna (MacPhee 1996a) Yalm en nuestros dias huracanes de gran intensidad aniquilan poblaciones de vertebrados (que no se recuperan) en is las pequenas y llanas del Caribe (Brooks y Smith 2001)
En medio de la incertidumbre sobre la verdadera causa (0 causas) de las extinciones de mamiferos en el periodo amerindio MacPhee y Marx (1997) han propuesto su hipotesis de la hipermorbilidad la cual estipula colapsos poblacionales relativamente rapidos con la introduccion de enfermedades de alta letalidad Estos autores han razonado que
Las islas son los lugares mas apropiados para experimentar perdidas catastroficas causadas por enfermedades Los mamiferos no solo serian susceptibles en virtud de su prolongado aislamiento biologico sino que en los confines de una isla la epizootia
333 EXTINCIONES DEL CUATERNARIO
tendria la potencialidad de infectar simultaneamente a la rnayoria de los individuos de vadas especies Los neonatos son inmunologicamente inmaduros y sufririan desproporcionadamente cualquier infeccion hipervirulenta La perdida excesiva de neonatos y juveniles tendria efectos severos para la sobrevivencia de la poblacion si esta se reduce por debajo de cierto nivel sobreviene la extincion porque no hay posibilidad de restitucion en el periodo postpanzootico Las especies de mayor talla corporal van a ser las mas vulnerables a la extincion dadas sus bajas tasas reproductivas largos periodos de gestacion y reducido numero de progenie Las especies de menor tamano --caracterizadas por altas tasas reproductivas periodos cortos de gestacion y mayor numero de neonatos por parto- tambien serian fuertemente diezmadas pero muchas sobrevivirian porque su estrategia reproductiva les permite perder mayor numero de individuos Las perdidas deben ocurrir relativamente rapido (a escala de decadas) mientras que otras hipotesis (digamos los cambios climaticos) permiten intervalos mucho mas largos (a escala de siglos 0 de milenios) entre la aparicion de las causas y la produccion de sus efectos Por ejemplo todo indica que los nesofontes se extinguieron muy rapidamente y seria dificil poder invocar un impacto humano directo 0 argumentar fenomenos climaticos que pudieran explicar plausiblemente perdidas tan rapidas
MacPhee y Marx (1997 191) han dicho que por el momento la hipotesis de la hipermorbilidad no pasa de ser una abstraccion en espera de adecuada sustentacion empirica Llama la atencion que linicamente en el periodo amerindio (Tabla 45 intervalo 1Il) las extinciones abarcaron todas las clases del peso corporal (de la a a laj) y que ninguna de las especies que sobrevivieron hasta el periodo postcolombino (Tabla 45 intervalo IV mas las especies vivientes) sobrepasa los 10 kg (clases del peso a b c) en clara coincidencia con los presupuestos de la hipotetica hipermorbilidad
Intervalo Postcolornbino
Por 10 menos 37 especies de mamiferos terrestres vivian en las Antillas Mayores y grupos de Islas perifericas a la lIegada de los europeos en 1492 De elIas se han extinguido 23 shymas especies que en cualquiera de los periodos precedentes (Tabla 45~ y solo subsisten 12 especies repartidas entre Cuba (8 especies) La Espanola (2) Jamaica (1) y Bahamas (1) varias de estas en peligro critico de extincion Solo dos de las especies extinguidas durante los ultimos 500 anos se conocieron en estado viviente los roedores Oryzomys antillarum de Jamaica (extinguido en 1877) y Geocapromys thoracatus de las Islas Swan (extinguido en 1955) Las causas de las extinciones ocurridas durante este periodo son sobradamente conocidas destruccion de habitats introducci6n de animales exoticos (domesticos y comensales) y demas presiones asociadas a la actividad humana entre las cuales posiblemente habria que incluir brotes de hipermorbilidad
Durante mucho tiempo se ha afirmado la posibilidad de que especies de Nesophontes hayan sobrevivido hasta las primeras decadas del siglo xx y que incluso continuen existiendo en lugares remotos de Cuba 0 La Espanola (Allen 1918 1942 Miller 1930 Aguayo 1950a Arredondo 1960b Fischer 1977 MacFadden 1980 Woods y
334 BIOGEOGRAFiA
Ottenwalder 1991) Actualmente no existe evidencia radiometrica de que alguna especie de Nesophontes haya sobrevivido al siglo xv las fechas radiocarbonicas calibradas obtenidas en muestras de Nesophontes de Cuba y La Espanola resultaron no mas tardias que los siglos XIII y XIV (MacPhee et al 1999) Por otra parte las expediciones efectuadas a fines del siglo xx con el proposito de localizar individuos vivientes del genero en La Espanola no produjeron la menor indicacion afirmativa al respecto (Woods et al 1985) Ademas a traves de las Antillas Mayores no aparecen restos de Nesophontes en los depositos formados actualmente por las lechuzas
EPiLOGO
Las Antillas Mayores son islas oceanicas no han formado parte de un continente (con la posible excepcion de una parte de Jamaica hasta el Eoceno Medio hace 45 Ma) y durante un tiempo geologico considerable (desde el Eoceno Superior hace 40 Ma) han permanecido emergidas y separadas de los continentes vecinos Consecuentemente los mamiferos terrestres (no volantes) autoctonos de estas islas constituyen una fauna oceanica que por ende cumple con todas las caracteristicas convencionales reconocidas para este tipo de entidad biogeografica
- Baja diversidad taxonomica a nivel de las categorias superiores (solo cuatro ordenes) en comparacion con islas plataformicas continentales (0 con areas continentales) de tamano y diversidad ecologica simi lares
- Alto grado de endemismo a nivel de genero (96) y familia (50) en respuesta al prolongado aislamiento
- Desbalance ecologico de la estructura trofica ausencia de mamiferos camivoros y ungulados
- Espectaculares procesos de irradiaci6n evolucionaria como resultado de la colonizacion de territorios libres de competidores y depredadores
Tendencias evolucionarias extremas gigantismo
- Control a cargo de aves rapaces con tendencia at gigantismo en ausencia de mamiferos miembros del orden Carnivora
- Mas especies de reptiles y de murcielagos que de mamiferos terrestres relaciones que en los continentes se invierten
Alta tasa de extincion de especies (84) sin reposicion
Preservacion de especies filogeneticamente arcaicas (los llamados fosiles vivientes) los almiquies

305 POBLAMIENTO CONTINENTE-ISLA
La colonizaci6n exitosa de una isla de las Antillas Mayores por mamiferos terrestres continentales transportados en balsas naturales a la deriva (de cada especie al menos una hembra gravida 0 dos individuos de sexo opuesto) dependeria de factores que van a operar desde el momenta mismo de formarse la balsa asi como durante su travesia marina y su arribo a la isla
a) La posibilidad de estar en la balsa Que la distribuci6n de la especie incluya areas contiguas al rio y que el animal se encuentre en ellugar y el momenta propicios para ser incorporado a la balsa antes de que el rio crecido la desprenda Por ejemplo segun Hershkovitz (1958) los equimidos -grupo ancestral de roedores antillanos- serian buenos candidatos a ser captados por la balsa porque acostumbran a anidar en huecos del tronco de los arboles a la orilla de los rios
b) El tamano y la consistencia de la balsa Muchas se desintegran en alta mar debido a su fragilidad ante la saturaci6n Algunos autores (Beaufort 1951 Arredondo 1996a [1999]) han expresado incredulidad en cuanto a que las balsas hayan podido soportar el peso de un animal como el gran perezoso cubano (Megalocnus rodens) el mayor de los mamiferos antillanos Pero esta especie evolucion6 en Cuba y el ancestro que hizo el viaje pudo ser muy pequeno [EI perezoso oligocenico de Puerto Rico es el mas pequeno de los megaloniquidos antillanos (MacPhee e Iturralde-Vinent 1995b )]
c) Las caracterfsticas fisiol6gicas de la especie Los limites fisiol6gicos de los propagulos constituyen nuestra unica objeci6n a la dispersi6n marina por balsas como mecanismo para la formaci6n de la fauna de vertebrados en las Antillas (MacPhee e IturraldeshyVinent 2005) Unas especies tienen intrinsecamente mas posibilidades que otras para sobrevivir a las vicisitudes que deben afrontar durante la travesia (deshidrataci6n inanicion insolaci6n etc) Por ejemplo los megaloniquidos han demostrado tener extraordinaria habilidad para cruzar barreras marinas fueron los primeros mamfferos suramericanos en invadir Norteamerica mucho antes de que surgiera la conexion terrestre a finales del Plioceno (Paula 1967 Olson 1978 Webb 1985b Martin y Steadman 1999) Incluso se conoce un megaloniquido acultitico del Plioceno peruano (Muizon y McDonald 1995) y otro viviente en Suramerica (Choloepus) que pnicticamente no bebe agua 10 que ha originado una leyenda aborigen que explica por que los perezosos no toman agua (Goff art 1971) Vease ademas el inciso e
d) EI regimen de las corrientes marinas Que conduzcan a la balsa en la direcci6n necesaria EI transporte de balsas de vegetacion desde los continentes hacia las islas fue durante la mayor parte del Cenozoico sumamente improbable (MacPhee e IturraldeshyVinent 2000 148) Estos autores han insistido en que hay que tener en cuenta que durante la cambiante geografia cenozoica del Caribe las corrientes marinas de superficie no favorecieron el desplazamiento de objetos fiotantes hacia las Antillas Mayores [Pero no sabemos con certeza que efecto modificador del curso de tales objetos fiotantes pudieron introducir las tormentas tropicales si previsiblemente funcionaron en el pasado como 10 hacen ahora] Dichos autores senalaron ademas (2005) que en un experimento reciente con boyas de posicionamiento satelital liberadas a la deriva desde las Antillas Menores las boyas fueron a parar mayoritariamente a Centroamerica y a otros lugares
306 B[OGEOGRAFIA
y solo unas pocas recalaron en las Antillas Mayores y las que entraron al Mar Caribe demoraron semanas 0 meses en arribar
e) La distancia a recorrer (y en consecuencia la duracion de la travesia) entre el continente y la isla (10 que repercute decisivamente en el factor c) Este es desde luego uno de los factores mas criticos En la medida que la distancia aumenta la probabilidad de alcanzar la isla disminuye drasticamente con el agravante de que la trayectoria dificilmente seria en linea recta Si tomamos como referencia la velocidad de 45 kmlh del islote ya mencionado (SICSP 1970) cubrir la distancia aerea (aproximada) de 800 km que media entre el noroeste de Suramerica y La Espafiola Ie tomaria a la balsa 74 dias Estimando el doble de este tiempo debido a desvios de la trayectoria la duracion de la travesia probablemente resultaria excesiva para la mayoria de los mamiferos Pero pudiera muy bien ocurrir que la velocidad fuera mayor 0 que la balsa portara plantas vivas que permitirian a los fitOfagos sobrevivir [Tres de los cuatro ordenes de mamiferos terrestres conocidos de las Antillas Mayores son fitOfagos] Ademas esta geologicamente bien establecido que durante etapas de variada duracion desde el Oligoceno hasta el Pleistoceno los territorios del Caribe insular y continental aumentaron considerablemente en extension con niveles del mar 100-200 m por debajo de su nivel actual oportunidades que los propagulos aprovecharian para cruzar barreras marinas significativamente menores
f) El tamafio de la isla Las islas mayores deben interceptar mas balsas (Buckley y Knedlhans 1986) 10 que repercute directamente en el factor e La balsa puede pasar entre islas pequefias y continuar viaje sin llegar a ningun lugar
g) Las condiciones (favorables 0 adversas) del desembarco en la isla y el tiempo que Ie tomaria a uno 0 mas animales depauperados encontrarun habitat apropiado Esto dependeria a su vez del tamafio y diversidad ecologica de la isla (Darlington 1938 Miller 1966)
Tal cumulo de posibles obstaculos y dificultades constituye a primera vista un desafio a la credibilidad de la hipotesis Matthew-Simpson pero en ello radica precisamente su valor explicativo bas ado en la mas absoluta improbabilidad La colonizacion por la via marina es altamente improbable pero no absolutamente imposible si se tiene en cuenta la inmensidad del tiempo geologico se vuelve probable (McKenna 1973) Basta que solo una vez entre mill ones de intentos fallidos durante ese inmenso lapso algunos de los factores anteriormente enunciados coincidan en no ser tan desfavorables para el propagulo Y eso es todo 10 que demanda la hipotesis para cada uno de los grupos monofileticos mayores de mamiferos terrestres presentes en las Antillas Mayores Las Antillas Mayores requeririan el arribo de tan solo 7 (quizas menos) propagulos fundadores a 10 largo de los ultimos 30 0 mas millones de afios (promedio de uno cada 43 mill ones) para explicar la formacion de toda su fauna de mamiferos terrestres por dispersion marina pasiva (vease Discusion y Conc1usiones)
En comparacion con la via marina el poblamiento por la via terrestre es predecible porque la mayor parte de la fauna continental de mamiferos emplazada en la region del puente va a cruzar por este una vez que el mismo se establece y los animales logran
307 POBLAMIEITO CONTINENTE-ISLA
acceder a el el poblamiento por la via marina en cambio es impredecible porque depende total mente del azar que caracteriza a la balsa Claro que en ambas vias opera el azar pero en el primer caso se trata del azar de un evento probable en el segundo el azar de un evento improbable (Simpson 1940) Asi el puente natural (mecanismo de poblamiento minimamente selectivo = alta diversidad inicial) y la balsa natural (mecanismo de poblamiento maximamente selectivo baja diversidad inicial) pueden conceptuarse como los extremos opuestos de una gama de probabilidades de dispersi6n (McKenna 1973) EI fen6meno de la dispersi6n merece la mayor atenci6n cientifica en los estudios de biogeografia insular (Cowie y Holland 2006)
Queda claro que la balsa natural es una hip6tesis consistente con el caracter de la fauna objeto de estudio puesto que el desbalance taxon6mico (sin evidencia paleontol6gica en contrario) s610 puede ser resultado de la improbabilidad del poblamiento No obstante ello no implica necesariamente que la balsa natural haya sido el imico mecanismo (ni siquiera el principal) utilizado por los mamiferos terrestres en su colonizaci6n de las Antillas Mayores La balsa natural debe satisfacer una premisa te6rica esencial dado el caracter azaroso impredecible e improbable de una colonizaci6n exitosa esta debe ocurrir a intervalos muy espaciados a traves del tiempo geol6gico 0 sea durante todo el tiempo que en el area ocupada por las Antillas Mayores ha habido ecosistemas terrestres perrnanentemente disponibles para ser colonizados (los ultimos 40 millones de afios) Sin embargo como sabemos representantes de los cuatro 6rdenes de mamiteros terrestres conocidos de las Antillas Mayores ya estaban emplazados en dicho territorio insular en el Mioceno Inferior 10 que supone un lapso de colonizaci6n limitado al Eoceno SuperiorshyOligoceno (MacPhee 2005) sugerente de un tipo de mecanismo contrario a la colonizacion alocronica exigida por la hipotesis de la balsa natural sobre todo tratandose de una fauna caracterizada por altas tasas de extincion sin reposicion aparente
La presencia en Jamaica (hasta su extinci6n en 1877) del roedor mfuido Oryzomys antillarom con estrecha afinidad centroamericana (Woods 1989a) pudiera constituir el imico caso efectivo de poblamiento (continente-isla) de las Antillas Mayores por dispersion marina dc un mamifero terrestre para el cual no es razonable invocar otro mecanismo de colonizacion
LA GUIRNALDA INSULAR (dispersion mixta)
La geologia ha establecido con suficiente certeza que durante el Cenozoico unicamente en el Pale6geno tardio y solo a traves de la Cresta de Aves (ahora sumergida excepto la isla hom6nima) pudo haber existido una cadena de islas para el transito de mamiferos terrestres entre el norte de Suramerica y las proto-Antillas Mayores [desestimados a ese efecto el Canal dc Yucatan el Elcvado de Nicaragua la Cresta de Caiman el Elevado de Beata y las Antillas Menores (Holcombe et al 1990 MacPhee e IturraldeshyVinent 1994 1995b Iturralde-Vinent y MacPhee 1999)] Holcombe y Moore (1977) y Holcombe y Edgar (1990) postularon la existencia de esta entidad y estimaron algunas de sus posibles caracteristicas Por su parte MacPhee e Iturralde-Vinent (1995b) e Iturralde-Vinent y MacPhee (1999) precisaron los extremos proximal y distal de esa
308 BIOGEOGRAFIA
estructura acotaron la duracion y la edad de la misma y Ie dieron nombre GAARlandia (Greater AntillesA vesRidgelandia)
Segun Iturralde-Vinent y MacPhee (1999) la Cresta de Aves emergio por breve tiempo (1-2 Ma) durante la transicion Eoceno-Oligoceno (35-33 MaAP) por la coincidencia de un levantamiento tectoni co y un considerable descenso eustatico del nivel del mar En ese momenta las islas septentrionales del arco de las Antillas Mayores (Cuba centroorientalshyLa Espafiola - Puerto Rico - Islas Virgenes) constituian una sola isla grande 0 una serie de islas separadas por barreras muy estrechas La Cresta de Aves emergida entre el bloque Puerto Rico - Islas Virgenes y un microcontinente noroccidental de Suramerica -separado en ese momento del resto del continente por una via marina surgida en el Eoceno tardio- dio origen a GAARlandia
Obviamente el numero de colonizaciones exitosas por mamiferos terrestres a traves de una guimalda de Islas tiende a disminuir mientras mas anchas sean las barreras marinas entre las islas que forman la guimalda y cuando esto ocurre disminuye tambien la diversidad taxonomica de los propagulos particularmente a nivel de las categorias superiores (Simpson 1956)
Vemos asi que para los mamiferos terrestres la colonizaci6n a traves de una guimalda insular puede ser tan improbable como a la deriva en una balsa natural en dependencia de la cantidad y anchura de las barreras marinas a cruzar el tamafio y la fisiografia de las islas que integran la guimalda y el regimen de las corrientes marinas y aereas entre otros factores
Resumiendo la guimalda insular puede conceptuarse como un mecanismo de poblamiento altamente selectivo (= baja diversidad inicial) que combina la improbabilidad de la balsa natural con la colonizacion restringida temporaimente del puente terrestre
En su reciente revision de la taxonomia distribuci6n y filogenia (basada en estudios moleculares) de varios grupos de mamiferos antillanos Davalos (2004) concluyo que los perezosos megalocninos los primates y los roedores histricognatos se avienen al patron de divergencia con el continente y epoca de arribo a las Antill as implicados en la hipotesis de GAARlandia pero no asi los perezosos coloepodinos que requieren una explicacion biogeognlfica distinta Esto ultimo (sospechado por Matthew 1918a) habla sido confirmado por White y MacPhee (2001 226-227)
Nuestra evidencia filogenetica no sustenta la idea de un dado unificado surgido de la invasion de un megaloniquido el analisis sugiere mas bien dos eventos de colonizaci6n independientes [y] sustenta fuertemente un origen difiletico de las subfamilias [Choloepodinae y Megalocninae] Si hubo dos migraciones a traves de GAARlandia presumiblemente poco espaciadas en el tiempo no se puede determinar en el presente
Finalmente queda claro que GAARlandia -como guirnalda insular- es la hipotesis mas robusta propuesta hasta ahora para explicar la presencia en las Antillas Mayores de
309 POBLAMIENTO CONTINENTE- ISLA
la absoluta mayona de los mamfferos terrestres de ongen suramericano porque el caracter de esa fauna supone un mecanismo de poblamiento ancestral que sea a la vez maximamente selectivo (baja diversidad ordinal) y temporalmente restringido (transicion Eoceno-Oligoceno)
Recientemente Rega et al (2002 18) expresaron A pesar de una revision exhaustiva de la paleogeografia de la cuenca del Caribe Iturralde-Vinent y MacPhee (1999) no pudieron confirmar la presencia de una conexion terrestre continua entre Suramerica y las proto-Antillas aunque hipotetizaron su existencia [Vease la Fig 42 (= figura 6 de Iturralde-Vinent y MacPhee 1999 simplificada))
En el trabajo citado por Rega y colaboradores aparece la siguiente explicacion
Actualmente no es posible ofrecer reconstrucciones paleogeograficas detalladas de la Cresta de Aves Sin embargo hay evidencia de que porciones de la cresta deben haberse elevado subaereamente en un tiempo La cuestion cntica es si la Cresta de Aves emergida pudo haber formado algun tipo de corredor entre las Antillas Mayores y el noroeste de Suramerica Perforaciones en las alturas estructurales de la cresta pudieran proveer el registro del Paleogeno tardio que se necesita para precisar la hipotesis del corredor aunque no se puede predecir si tales datos serian suficientemente detallados para determinar si en algun momento GAARlandia constituyo una sola entidad subaerea Por ejemplo pudo haber barreras marinas por corto tiempo entre las masas terrestres de la Cresta de Aves [0) ellevantamiento y la subsidencia pudieron afectar diferentes partes de GAARlandia en diferentes tiempos AItemativamente los elementos faunales pudieron haberse dispersado de manera pulsada Movimientos episOdicos de este tipo incrementarian la extincion de los elementos faunales incapaces de trasladarse a la siguiente masa terrestre en la cadena disminuyendo asi la diversidad total (Iturralde-Vinent y MacPhee 199957)
La primera parte de la explicacion deja claro que la posibilidad de que la Cresta de Aves haya sido una conexion terrestre continua no se puede descartar pero tampoco se puede afirmar La ultima parte de la explicacion describe el efecto altamente selectivo (filtrante) de la colonizacion a traves de una cadena de islas
Y continuan lturralde-Vinent y MacPhee (199957 -58) explicando
Hasta que punto el microcontinente noroccidental de Suramerica [considerado por dichos autores como el area suministradora de propagulos hacia GAARlandiaJ estuvo fisicamente separado del resto del continente no ha sido plenamente esc1arecido Si estuvo sustancialmente aislado por barreras marinas y orogenic as durante el Paleogeno tardio entonces solo la fraccion de la biota suramericana emplazada en el microcontinente estuvo en posicion de acceder a GAARlandia Esto pudiera tener que ver con el hecho de que la fauna antillana carezca de representantes de muchos grupos suramericanos [de mamfferos) que debieron haber hecho el viaje
310 BIOGEOGRAFiA
Florida o Alturas0 0
d 6 D L1anuras25deg
Yucatan
20deg
deg0
10deg
MARCARIBE
80deg 75deg
Bahamas
Cres ta de Beata
D Mares someros
D Mares profundos
OCEANO ATLANTICa
FIGURA 42 Reconstrucci6n hipotetica de GAARlandia como conexi6n terrestre continua entre las proto-Antillas Mayores y el noroeste de Suramerica en el Pale6geno tardio segun la FIgura 6 (simplificada) de Iturralde-Vinent y MacPhee (1999)
En su defensa del puente terrestre Barbour (1945b249) se refiri6 a esto - antes 10 habfa hecho Anthony [1942363]- en los terminos siguientes
El argumento de que si las Antillas hubieran estado conectadas con el continente se encontrarian los f6siles de un numero mayor de distintas criaturas me deja frio es perfectamente concebible que la conexi6n haya ocurrido en un periodo en que la parte del continente implicada no estuviera poblada por una fauna grande y variada
Y concluyen lturralde-Vinent y MacPhee (199958) su detallada explicaci6n
Desafortunadamente sabemos muy poco sobre la composici6n faunal del noroeste de Suramerica a finales del Pale6geno
Consecuentemente si se confirmara la magnitud del aislamiento del aludido microcontinente y su concomitante baja diversidad faunal GAARlandia como conexi6n terrestre continua serf a la hip6tesis mas parsimoniosa No obstante mientras no se disponga de evidencia concluyente a este respecto hipotetizar a GAARlandia como conexi6n continua (minima selectividad) la coloca en franca vulnerabilidad te6rica al tener que afrontar la objeci6n clasica al puente terrestre (baja diversidad ordinal de la
311 POBLAMIENTO CONTINENTE-TSLA
fauna) al tiempo que se la priva de ser la hip6tesis mas robusta para explicar -como guimalda de islas- la colonizaci6n de las Antillas por mamiferos procedentes de Suramerica
ELARcA DE NOE (seudovicarianza)
El primer vicarianista antillano (desde luego sin el saberlo) fue Corral (1940) con su utilizaci6n ingenua de las ideas wegenerianas de la deriva continental aparecidas la decada anterior Pero el primer (supuesto) intento de aplicar a la formaci6n de la biota caribefia el mecanismo de la vicarianza basada en la tearia de la tect6nica de placas (surgida en la sexta decada del pas ado siglo) se debe a Rosen ([1976] 1985)
Segun Rosen las Antillas Mayores se originaron en el Cremcico en la Placa Caribe aproximadamente en la posici6n que hoy dia ocupa Centroamerica y al ser desplazadas tectonicamente hacia el este interactuaron (de algun modo no precisado por Rosen) con las margenes continentales de Norte- y Suramerica de donde (en un evento unico) adquirieron toda su fauna transportada subsecuentemente por dichas islas en su transito hacia sus posiciones actuales [De ahi la denominacion metaforica dada par McKenna (1973) a este mecanismo de poblamiento] Aunque se ha presentado esta propuesta como Model0 de Vicarianza Continente-Isla de Rosen en realidad por definici6n no constituye un evento vicariante porque las proto-Antillas Mayores no resultaron del fraccionamiento y separaci6n por tectonismo de una porcion del territorio continental como exige esta hip6tesis (vease definici6n de vicarianza al comienzo de Hipotesis Biogeograficas)
El Arca de Noe de Rosen ha sido desestimada por la mayoria de los autores contemporaneos porque (1) carece de sustento geol6gico (lturralde-Vinent y MacPhee 1999) y porque (2) en el periodo en que se afrrrna ocum6 el evento muchos de los ordenes modemos de mamiferos placentarios attn no existian (Williams 1989)
EL BARCO FUNERAL VlKINGO (tanatovicarianza)
Domning et al (1997) reportaron 10 que consideraron el mamffero terrestre mas viejo de las Antillas --circunstancia que se refleja en el titulo mismo del trabajo Oldest West Indian Land Mammal- al descubrirse un perisodactilo rinocerotoide (Hyrachius sp) en una localidad de edad Eoceno Inferior a Medio en el occidente de Jamaica
Jamaica ha tenido una historia geologica y paleogeografica distinta de la historia comun a las restantes Antillas Mayores A principios del Cenozoico Jamaica occidental (a diferencia de la porcion oriental constituida por el Bloque de las Montafias Azules) formaba parte de Centro america de la cual se desgaj6 (por el desplazamiento de la Placa Caribe) para venir a integrarse despues del Eoceno Medio con 10 que serfa la futura isla de Jamaica desaparecida por transgresi6n marina en el Eoceno Superior hasta volver a emerger totalmente en el Mioceno Medio (Iturralde-Vinent y MacPhee 1999 MacPhee e Iturralde-Vinent 2000)
312 BIOGEOGRAFiA
Por consiguiente Hyrachius ---conocido de localidades de edad Eoceno Inferior a Medio en Norteamerica- debio ocupar (10 que iba a ser) Jamaica occidental mucho antes de que esta se separara de Centroamerica De hecho este fosil Uego a su destino insular contenido en las rocas de su basamento no como un ser vivo por 10 que es erroneo considerarlo un animal antillano (lturralde-Vinent y MacPhee 2004) como han supuesto algunos (Hedges 200 I Portell et al 200 I Condis [200 I D Todo 10 que hizo fue viajar tanatovicariantemente en su barco funeral vikingo -otra de las denominaciones metaforicas de McKenna (1973)
DISCUSION Y CONCLUSIONES
Hemos visto que solo disponemos de una hipotesis plausible para explicar la mayor parte del poblamiento (continente-isla) de las Antillas Mayores por mamiferos terrestres del Neotropico continental la dispersion mixta (a traves de GAARlandia) para los perezosos (excepto Choloepodinae) los monos y los roedores (excepto Oryzomys)
Por otra parte la historia biogeogrMica de las musaraiias de las Antillas Mayores (Nesophontes y Solenodon) constituye una de las situaciones mas enigmaticas entre los mamiferos de la region (Ottenwalder 2001321) Al menos cinco mecanismos de poblamiento continente-isla han sido invocados para explicar cuando como y de donde llegaron a las Antillas Mayores estas musaraiias
I EI propagulo ancestral (del Terciario de Norteamerica) coloniz6 a las Antillas Mayores por dispersion marina desde Centroamerica 0 desde la Florida (Patterson 1962)
2 El propagulo ancestral ( del Terciario) cruzo el Atlantico Sur (supuestamente mas estreshycho en aquella epoca) directamente de Africa a las Antillas Mayores (Hershkovitz 1972)
3 El propagulo ancestral (del Terciario de Norteamerica) Uego a las Antillas Mayores en el Arca de Noe de Rosen (MacFadden 1980)
4 El propagulo ancestral (del Terciario de Norteamerica) fue transportado a las Antillas desde Centroamerica en el bloque de Jamaica occidental y de alli paso por dispersion marina a las restantes Antillas Mayores (Whidden y Asher 200 I)
5 El propagulo ancestral (del Terciario de Norteamerica) lleg6 a las Antillas Mayores a traves de GAARlandia (Whidden y Asher 2001)
Aunque hay consenso entre estos (y otros) autores sobre el origen norteamericano de las musaraiias antillanas ninguno de los cinco hipoteticos mecanismos de poblamiento resiste la evidencia en contrario filogenetica yo paleogeogrMica Ademas algunas de estas hipotesis asumen que Nesophontidae y Solenodontidae comparten un antecesor comun en las Antillas pero esta relacion no ha sido confirm ada Para dilucidar el enigmatico caso de las musaraiias antillanas se requerini profundizar en la investigaci6n geologica paleogeognifica y paleontol6gica (Iturralde-Vinent y MacPhee 1999)
De acuerdo con el conocimiento filogenetico disponible sobre el conjunto de la fauna la colonizacion original de las Antillas Mayores por mamiferos terrestres implicaria el arribo
313 POBLAMIENTO ISLA-ISLA
de solo siete propagulos fundadores perezosos 2 (coloepodinos y megalocninos si como parece tienen un origen difiletico) musaraiias 2 (nesofontidos y solenodontidos si no constituyen una misma familia como consideran algunos) monos 1 (pitecidos) roedores histricognatos 1 (equimidos capromidos y heptaxodontidos) roedores muridos 1 (Oryzomys en Jamaica)
POBLAMIENTO ISLA-ISLA
La distribucion de los mamiferos terrestres a traves del area antillana demarcada al inicio del presente capitulo (vease Contexto Geogratico Fig 41) ha sido interpretada en funcion de dos hipoteticos mecanismos de poblamiento interinsular la dispersion y la vicarianza fundamentadas como se resumen seguidamente
DISPERSION
De acuerdo con la evidencia paleogeogratica mas actualizada en las principales regresiones marinas ocurridas en la cuenca caribeiia durante los ultimos 23 Ma (transiciones OligocenoshyMioceno Mioceno Superior - Plioceno y varias veces en el Pleistoceno) las masas terrestres constitutivas de la region insular en formacion aumentaron considerablemente su extension territorial disminuyendo por tanto la distancia entre elias oportunidades que los elementos faunales aprovecharian para dispersarse de unas islas a otras
Durante mucho tiempo esta ha sido la hipotesis mas favorecida por los estudiosos del tema a partir de Simpson (1956) pero en decadas recientes ha suscitado dos objeciones
1 Es dificil en tender como pudo operar la dispersion marina entre estas islas puesto que ninguna de las Antillas Mayores posee rios suficientemente caudalosos para transportar masas significativas de vegetacion flotante (MacPhee 198432)
Es oportuno recordar aqui el islote ala deriva (con mas de diez palmas reales en pie) entre las tres Antillas Mayores mas occidentales mencionado anteriormente (vease La Balsa Natural)
2 La dispersion marina es menos efectiva para explicar la asombrosa similitud de las faunas de mamiferos terrestres a traves de las Antillas Mayores septentrionales porque como proceso aleatorio no debe producir distribuciones uniformes a 10 largo de una cadena de islas 0 si las produce entonces el proceso no es aleatorio (MacPhee e lturralde-Vinent 2000 149)
Independientemente de que las aludidas distribuciones taxonomicas estan lejos de ser uniformes 0 asombrosamente similares la hipotesis de la dispersion marina contrariamente a su canicter aleatorio y altamente improbable en el caso del poblamiento continente-isla- en el caso del poblamiento isla-isla es un proceso no aleatorio (aprovecha las oportunidades brindadas puntualmente por los periodos de maxima regresion marina) y ademas altamente probable dada la incomparablemente menor distancia entre las islas caribeiias que entre estas y los continentes vecinos
314 BIOGEOGRAFiA
V ICARIANZA
La neotectonica y las transgresiones marinas del Oligoceno Medio y del Mioceno afectaron sustancialmente la paleogeografia de la Cresta de las Antillas Mayores resultando la subdivision gradual de las partes subaereas de la Cresta Asi el este de Cuba y el norte de La Espanola conectados fisicamente durante el Oligoceno Inferior se separaron al formarse el Paso de los Vientos en el Oligoceno tardio en tanto La Espanola y Puerto Rico probablemente permanecieron conectadas hasta el Mioceno tardio en que surgio el Paso de Mona (Iturralde-Vinent y MacPhee 1999)
Tales eventos geologicos habrian dividido las distribuciones de los grupos faunales emplazados previamente en el territorio continuo de las tres islas y a partir de esas disyunciones dichos autores (tambien MacPhee et al 2000 y White y MacPhee 2001) asumieron --como segunda fase del modelo de GAARlandia- la hipotesis de la vicarianza isla-isla para explicar la distribucion multiinsular de varios grupos de mamiferos terrestres a traves de las Antillas Mayores septentrionales De acuerdo con esta hipotesis cualquier diferencia en composicion faunal entre las islas (por ejemplo la ausencia de roedores heptaxodontidos en Cuba 0 de monos platirrinos en Puerto Rico) constituye evidencia de extincion local postvicariante y no de dispersion marina fallida Lo primero se puede demostrar si aparecen los fosiles pertinentes 10 segundo no se puede demostrar (MacPhee e Iturralde-Vinent 1994) Por otra parte IturraldeshyVinent y MacPhee (1999 57) han reconocido que aunque fa vicarianza isla-isla puede representar una solucion clara para las distribuciones de varios taxones cuaternarios no puede explicar todos los casos
Rega et al (2002 18) han presentado dos objeciones al modelo de la vicarianza isla-isla aplicado a los mamiferos de las Antillas Mayores
I Si la separacion de Cuba y La Espanola precedio en varios mill ones de anos la separashycion de La Espanola y Puerto Rico el modelo vicariante de lturralde-MacPhee debe predecir mayor similitud entre las faunas de La Espanola y Puerto Rico que entre las de Cuba y La Espanola De hecho es al reyes
No obstante MacPhee et al (200335) admitieron despues que la fecha mas temprana empiricamente defendible [para la apertura del Paso de Mona] debe ser el Oligoceno Medio esto invierte la secuencia de ambas disyunciones con 10 cual queda resuelta la aparente contradiccion Sin embargo en su reciente evaluacion de la congruencia entre filogenia e hipotesis geologic as en varios grupos de mamiferos terrestres de las Antillas Mayores Davalos (2004373) encontro que ninguno de los taxones cumple con la fecha y el patron de divergencia entre las masas terrestres antillanas postulados por el modelo de GAARlandia
2 La presencia de un mismo genero en las tres Islas septentrionales de las Antillas Mayores implica [por el modelo vicariante] que la diferenciacion entre estas formas durante el intervalo desde el Mioceno ha sido unicamente de nivel especifico [Dicha presencia]
315 rRRADlAcr6~ EVOLUCrONARIA
s610 puede explicarse razonablemente como el resultado de movimientos de esos animales entre las islas en el Cuaternario
De esta objecion resulta dificil disentir [El poblamiento entre islas diferenciado solo a nivel especifico no es probable que sea anterior al Pleistoceno (Simpson 1956 15)]
Debe tenerse en cuenta que la hipotesis de la vicarianza isla-isla alude a las tres islas septentrionales de las Antillas Mayores porque como se explico antes (vease El Barco Funeral Vikingo) Jamaica no existia como isla durante el lapso (Eoceno Superior shyOligoceno) consensualmente reconocido como el mas probable para la colonizacion de las proto-Antill as Mayores por la mayo ria de los mamiferos terrestres Despues de emerger como isla en el Mioceno Medio Jamaica debe haber recibido sus mamiferos terrestres por dispersion isla-isla (Davalos 2004376) y solo en el Cuaternario parece haberrecibido (por dispersion marina desde Centroamerica) al roedor murido Oryzomys
Seguramente tanto la dispersion como la vicarianza contribuyeron a que los mamiferos terrestres se irradiaran a traves de las Antillas Mayores y grupos de islas perifericas pero las maneras y tiempos en que estos mecanismos operaron a 10 largo de dicho proceso son aun en gran medida motivos de desacuerdo e incertidumbre
IRRADIACION EVOLUCIONARIA
A continuaci6n revisamos someramente las distribuciones conocidas de los diferentes grupos de mamiferos terrestres a traves de las Islas principales de las Antillas Mayores y grupos de Islas perifericas incluidas las interpretaciones y conjeturas que la literatura reciente recoge en relaci6n con la manera en que dichas distribuciones pudieron haberse conformado durante la irradiaci6n evolucionaria de los referidos grupos de mamiferos Manifestaciones de esta irradiaci6n pueden inferirse unicamente a partir de los taxones presentes en el Cuatemario pues solo 6 de las especies conocidas corresponden al Terciario
GRUPOS ORlGINARIOS DE SURAMERICA
PEREZOSOS (Megalonychidae)
Aparte de un genero miocenico de Cuba y otro del Oligoceno de Puerto Rico se conocen 4 generos (12 especies) de perezosos cuaternarios (todos extintos) restringidos a las tres islas septentrionales de las Antillas Mayores Acratocnus Neocnus Megalocnus y Parocnus Los cuatro generos estan representados en Cuba y La Espanola y Acratocnus 10 esta ademas (como unico genero de perezoso) en Puerto Rico Esta distribuci6n ha sido interpretada por MacPhee et al (2000) eomo resultado de la vicarianza oligomiocenica entre estas islas y [dado que el Banco Puerto Rico - Islas Virgenes fue la entrada de GAARlandia a las Antillas Mayores] se preguntaron si la ausencia de los otros tres generos en Puerto Rico se debe a que nunca estuvieron en ese territorio 0 a que sus
316 BIOGEOGRAFiA
fosiles atm no han sido hallados Pero como hemos visto Rega et al (2002) atribuyeron dicha distribucion a la dispersion interinsular en el Cuatemario
Por otra parte varios amilisis c1adisticos recientes ubi can a Choloepus -genero Reciente conocido solo de Centro- y Suramerica- como taxon hermano de Aeratoenus exc1uidos los demas megaloniquidos antillanos Para White y MacPhee (2001228)
tan estrecha relacion sugiere que la divergencia entre ambos [generos] ocurrio despues de que Aeratoenus divergio de Neoenus Si esta relacion implica que Aeratoenus y Neoenus se dispersaron separadamente desde Suramerica a las AntilIas mientras Choloepus permanecia en el continente (siendo necesarias al menos tres invasiones de perezosos) 0 si Choloepus invadio Suramerica desde las Antillas es algo que el material disponible no permite determinar
La diversidad adaptativa de los perezosos se manifiesta en (1) la utilizacion del habitat especies arboreas en Neoenus y Aeratoenus y terrestres en Paroenus y Megaloenus (Webb 1985a White [1993] White et al 1996 Arredondo [1999]) y en (2) la gama de tallas corporales desde los pequenos Neoenus hasta Megaloenus rodens el mayor de los mamiferos terrestres antillanos (su peso adulto estimado en 270 kg seglin Paula 19679) esto sugiere que los perezosos se irradiaron a nichos que en contextos continentales probablemente estarian ocupados por miembros de otros c1ados (MacPhee e IturraldeshyVinent 2000)
No esta claro si el abanico de tamanos entre estos perezosos enmascara alguna tendencia al gigantismo Por 10 pronto Anderson y Handley (2002) encontraron 10 contrario en perezosos (Bradypus) de las Islas de Bocas del Toro (Panama) shyseparadas secuencialmente del area continental vecina por ascensos del nivel del mar durante los ultimos 10 ka En las cinco islas los perezosos son menores que su contraparte continental y las diferencias entre las tallas medias de las cinco poblaciones expresan una reduccion lineal significativa de la talla corporal en funcion de la edad geologica de cada isla sin relacion alguna con el area de las islas 0 la distancia que las separa del continente
MONOS (Pitheciidae)
Los primates de las Antillas Mayores (cuatro especies extintas) constituyen un grupo monofiletico con Paralouatta (en Cuba) y Antillothrix (en La Espanola) como taxones hermanos seguidos en la escalera c1adistica por Xenothrix (en Jamaica) y finalmente Callieebus como el pariente continental mas cercano esto implica una sola invasion desde Suramerica antes del Mioceno Inferior (fecha mas temprana conocida para los primates fosiles antillanos) y por tanto en consonancia con GAARlandia como la via de colonizacion (Horovitz y MacPhee 1996 1999 Horovitz [1997] MacPhee y Horovitz 2004)
En desacuerdo parcial con la anterior interpretacion la filogenia de este grupo de monos obtenida por Davalos (2004) indica un antecesor comun en Cuba La Espanola y Jamaica
317 TRRADIACTON EVOLUCIONARIA
compatible con la hipotetica conexion -propuesta tentativamente por Iturralde-Vinent y MacPhee (199980-84)- entre las Montanas Azules del este de Jamaica y la peninsula suroccidental de La Espanola en la epoca de GAARlandia
A pesar de que los primates antillanos pertenecen a un grupo viviente de monos muy pequenos (los monos titles) fueron comparativamente grandes en relacion con los platirrinos modernos y sucesivamente mayores en la direccion Antillothrix Xenothrix y Paralouatta con P marianae como el mayor platirrino conocido del Mioceno neotropical (MacPhee 2005) MacPhee et al (1994) consideraron que Xenothrix debio ser un mono de andar lento y al parecer las especies de Paralouatta abandonaron la arborealidad para adoptar un modo de vida mas terrestre (MacPhee 1996a MacPhee et al 2003 MacPhee y Meldrum 2006)
ROEOORES (Echimyidae Capromyidae Heptaxodontidae)
La edad de Zazamys en Cuba demuestra que la invasion de los roedores histricognatos en las Antillas Mayores comenzo antes del Mioceno Inferior por otra parte los representantes mas antiguos de esa irradiacion evolucionaria se encuentran en La Espanola y Puerto Rico Ambas evidencias son consistentes con la hipotesis de GAARlandia (Woods 1982 1988 Woods et al 2001)
No esta claro si los capromidos evolucionaron a partir de un equimido ancestral 0 de un protocapromido 0 si los equimidos antillanos (I sub familia 3 generos y 7 especies todas extintas) forman parte de la irradiacion de los capromidos (4 subfamilias 9 generos y 27 especies 10 vivientes) Kratochvil et al (1978b) y Rodriguez et al (1979) consideraron que Capromys nan us es un eslab6n entre Echimyidae y Capromyidae pero Mesocapromys nanus esta morfologicamente mucho mas cerca de Hexolobodon y los de mas capromidos que de los equimidos (Woods 1985) Lo que si esta claro segun Woods et al (2001) es que la subfamilia Plagiodontinae endemic a de La Espanola es el linaje basal de los capromidos y que Isolobodon Hexolobodon Capromys Geocapromys Mysateles y Mesocapromys [y supuestamente Macrocapromys] forman parte de la irradiacion temprana de Capromyidae
La subfamilia Capromyinae (5 generos y 17 especies 9 vivientes) parece ser el dado de mas reciente evolucion en la familia con invasion de Cuba desde La Espanola y dispersion desde Cuba (1) por Capromys a las islas Caiman y (2) por Geocapromys a las islas Bahamas Caiman y Jamaica y desde Jamaica a las islas Swan aunque la presencia de Geocapromys en Swan pudiera deberse a introduccion aborigen (Morgan 1985 1988 1989 1993 1994 Woods 1989a 1990) Woods et al (2001) utilizaron datos moleculares para obtener estimados independientes de divergencia entre capromidos segiJn 10 cual Geocapromys divergio cerca de los 17 MaAP Mesocapromys divergio de Capromys hace 15 MaAP en tanto lvlysateles 10 hizo hace 10 MaAP Por su parte las formas de Capromys pilorides y de Mysateles prehensilis en Isla de la Juventud divergieron de sus correspondientes subespecies nominativas en la isla principal a los 24 y 08 MaAP respectivamente
318 BIOGEOGRAFiA
Woods y Howland (I979) describieron adaptaciones morfologicas de la mandibula el hioide y la faringe asociadas a mecanismos de masticacion diferentes en plagiodontinos y caprominos y Varona (1980a) llamo la atencion sobre el gran desarrollo de los molariformes (en relacion con la longitud total del cnineo) que presentan ciertos grupos de jutias fosiles en comparacion con las vivientes 10 que pudiera interpretarse como una adaptacion alimentaria a una vegetacion mas abrasiva por parte de las jutias extinguidas
Segun Woods (1990) la familia Heptaxodontidae (2 subfamilias 5 generos y 6 especies todas extintas) tambien seria parte de la irradiacion de los capromidos con origen en el este de La Espanola 0 en Puerto Rico y subsecuente dispersion al Banco de Anguila - San Martin en las Antillas Menores (pero vease MacPhee y Flemming 2003) Este banco estaria a solo 70 km del Banco Puerto Rico - Islas Virgenes en tiempos de maxima regresion marina y esta proximidad con Puerto Rico -patria de Elasmodontomys obliquus- seguramente tuvo que ver con la presencia en las islas de Anguila y San Martin del heptaxodontino gigante Amblyrhiza relacionado mas estrechamente con los roedores de las Antillas Mayores que con los del resto de las Antillas Menores (Woods 1990 1996) Aunque los roedores suelen aumentar de tamano en las islas (Case 1978) la escala del incremento de la talla en Amblyrhiza (que pudo alcanzar mas de 200 kg de peso adulto) esta fuera de toda proporcion con los patrones conocidos en otros roedores insulares (Biknevicius et 01 1993)
Clidomys (Clidomyinae) de Jamaica difiere marcadamente en su morfologia de los heptaxodontinos (vease ademas MacPhee y Flemming 2003)10 cual dificulta explicar su presencia en esa isla Como Jamaica estuvo bajo el mar hasta el Mioceno Medio el ancestro de Clidomys colonizaria esta isla por dispersion desde el extremo suroccidental de La Espanola despues que los heptaxodontinos se originaron en esta ultima isla (Woods 1989a) Pero Pascual et 01 (I 990) especularon que Clidomys pudiera ser el resultado de una invasion posterior e independiente de los capromidos
GRUPOS ORIGINARIOS DE NORTEAMERICA
El origen de las musaranas antillanas ha resultado ser hasta ahora el mayor enigma biogeografico de la fauna de mamiferos terrestres de la region aunque la mayoria de los autores coinciden en considerarlas originarias de Norteamerica (vease Discus ion y Conclusiones en Poblamiento Continente-Isla) No obstante MacPhee e Iturralde-Vinent (1994) descartaron toda posibilidad de conexion terrestre por el Estrecho de la Florida 0 por el Canal de Yucatan y si el (supuesto) evento vicari ante de Rosen (vease El Arca de Noe) -invocado por MacFadden (1980) para resolver el enigma- fuera minimamente plausible aun asi es dificil creer que las catastrofes que 10 sucedieron (vease Contexto Temporal) hayan barrido con toda esa vieja fauna continental excepto las musaranas circunstancia ya anticipada muy tempranamente por Matthew (l918a)
De otra parte la relaci6n entre Nesophontidae y Solenodontidae permanece irresuelta Si estos linajes no estan estrechamente relacionados entonces es muy posible que
319 IRRADIACION EVOLUCIONARIA
sus distribuciones sean explicadas por mecanismos diferentes (Whidden y Asher 2001)
La descomunal diferencia de talla corporal entre las musaranas pequenas (Nesophontes) y las grandes (Soienodon) seguramente impuso a ambos grupos restricciones distintas relativas a la termorregulacion y las adaptaciones energeticas En Nesophontes la seleccion debio favorecer una alta tasa de metabolismo y crecimiento camadas mas numerosas corta expectativa de vida yalta densidad poblacionaL En Soienodon en cambio la seleccion opero en la direccion de una baja tasa metabolica camadas reducidas crecimiento lento longevidad y baja densidad poblacional (Ottenwalder [1991])
MUSARANAS PEQUENAS (Nesophontidae)
Las musaranas pequenas estan representadas en las Antillas por el genero Nesophontes (y un genero miocenico indeterminado en ambar dominicano) con 6 especies cuaternarias extinguidas distribuidas en Cuba La Espanola Puerto Rico Islas Virgenes e Islas Caiman La mayor de las musaranas pequenas Nesophontes edithae evoluciono en Puerto Rico y su arribo a las Islas Virgenes supone una dispersion sumamente reciente En las Islas Caiman Nesophontes se asemeja mas a la poblacion de Cuba de donde seguramente derivo en el Plio-Pleistoceno (junto con Capromys y Geocapromys) cuando la costa sur de Cuba estuvo mucho mas cerca de las Islas Caiman (lturralde-Vinent 2003fig 4) MacPhee et ai (1994) interpretaron la distribucion de Nesophontes en Cuba y La Espanola como resultado de la vicarianza oligomiocenica ocurrida al formarse el Paso de los Vientos pero la distancia taxonomica entre los lados del Paso es solo de nivel especifico y N micros incluso vivi6 en ambas islas segtln Patterson (1962)
Allen (1918) considero que Nesophontes micrus fue una especie primitiva no modificada para la vida arb6rea 0 fos6rea y por tanto de habitos terrestres Nesophontes edithae de Puerto Rico es mucho mayor y mas robusta que las restantes especies del genero eL indice de robustez de su femur (= anchura minima de la diafisis ongitud total del hueso) muestra que este es tan robusto como el de Solenodon paradoxus un animal dos veces mayor (Morgan et ai 1980) Choate y Birney (1968) postularon un aumento cronoclinal de la taIla corporal en N edithae confirmado mas tarde por McFarlane (1999a) mediante fechados radiocarb6nicos de la secuencia estratignifica en depositos cavernarios de Puerto Rico Solenodon no vivi6 en Puerto Rico y en ausencia de una musarana de mayor tamano Nesophontes alcanzo una talla corporal mayor en esa isla que en La Espanola 0 Cuba (Woods y Eisenberg 1989)
MUSARANAS GRANDES (Solenodontidae)
Los arcaicos almiqufes (genero Soienodon) comprenden cuatro especies una viviente (S cubanus) y otra f6sil (S arredondoi) en Cuba y una viviente (S paradoxus con dos subespecies) y otra f6sil (S marcanoi) en La Espanola Como cabria esperar las dos especies cubanas se asemejan mas entre sf que a las especies de La Espanola pero
320 BIOGEOGRAFiA
enigmaticamente S marcanoi comparte caracteres con S cuban us y S paradoxus (Woods y Eisenberg 1989 Morgan y Ottenwalder 1993 Ottenwalder 200 I) Solenodon arredondoi alcanzo una talla corporal superior a la de cualquiera de las musarafias vivientes a nivel mundial Segun Morgan y Ottenwalder (1993) extrapolando las dimensiones y el peso de los almiquies vivientes S arredondoi debio tener una longitud cabeza-tronco y un peso de 450-550 mm y 1500-2000 g respectivamente en comparacion con S cubanus (360 mm y 800 g) y S paradoxus (390 mm y 1100 g)
Entre todas las especies de almiqufes Solenodon paradoxus es la que presenta los caracteres mas derivados (Otten walder 2001) Si estas relaciones son validas la distribucion de Solenodon entre Cuba y La Espafiola debio discurrir de oeste a este como propuso MacFadden (1980) quien invoco la presencia fosH de S cubanus [pero tambien S arredondoi] en Pinar del Rio para postular que el ancestro continental entr~ por Cuba opinion compartida por Perfit y Williams (1989) y por Ottenwalder ([1991] 2001) El paso de Solenodon a Puerto Rico pudo haberlo impedido la existencia de barreras ecologicas durante la conexion del este de La Espafiola con Puerto Rico 0 por la ausencia de Solenodon en esa parte de La Espafiola en aquella epoca Sin embargo las mismas condiciones ecologicas en el este de La Espafiola no parecen haber representado una barrera efectiva para Nesophontes cuyas mas altas tasa de reproduccion y densidad poblacional presumiblemente Ie permitieron expandirse flpidamente a traves de La Espafiola (Ottenwalder [1991])
Es opinion generalizada que los almiqufes antillanos constituyen el grupo de mamiferos que primero se emplazo en el territorio insular (MacPhee 2005) y en tal sentido es de interes la evidencia sobre especificidad parasito-hospedante aportada por Cruz (1980) quien encontro que la especie de acaro (genero Cubanochirus) que parasita a los almiquies es mucho mas primitiva que la especie (del mismo genero) que paras ita a las jutias y de aqui concluyo que dicho genero de acaro debio evolucionar primero en Solenodontidae y posteriormente en Capromyidae al entrar en contacto representantes de ambas familias en Cuba 0 La Espafiola
Recientemente Roca (2004) y Roca et al (2004) dieron a conocer que los resultados de su estudio genetico-molecular sobre los almiquies indican una divergencia de 25 Ma entre Solenodon cuban us y S paradoxus -compatible con la disyuncion tectonica entre Cuba y La Espafiola (Iturralde-Vinent y MacPhee 1999 McPhee et al 2003)-- y una separacion de ambas especies comparable en magnitud a la divergencia entre familias de mamlferos Consecuentemente propusieron que Solenodon cubanus fuera clasificada en el genero Atopogaie como indico Cabrera (1925) hace 82 afios
DISCUSION Y CONCLUSIONES
Las musarafias y los roedores antillanos (excepto Oryzomys) se irradiaron tanto de sus respectivos ancestros continentales que de estos 6rdenes se reconocen familias y subfamilias endemicas de la region en contraste con los perezosos (una subfamilia y tres
321 IRRADIACION EVOLUCIONARIA
tribus endemicas) y los monos (una tribu endemica) como expresion de diferentes niveles de diversificacion evolucionaria Este proceso es dificil de explicar (traUindose de c1ados que pnicticamente comparten la misma antigiiedad en las Antmas) si es correcta la interpretacion de Pascual et at (1990636)
Nosotros consideramos estos diferentes niveles de diversificacion como las diferentes etapas de evolucion alcanzadas por cada grupo [de mamiferos] Obviamente esto estuvo relacionado con las diferentes longitudes de sus historias
Pudiera ocurrir que en el futuro se compruebe que la historia biogeognifica de la region fue mucho mas complicada
La Espanola la isla mas ecol6gicamente divers a de la region debi6 ser el centro principal de evoluci6n de los mamfferos (con 87 de las familias y 73 de las subfamilias) seguida por Cuba (con 75 de las familias y 54 de las subfamilias) las dos islas de las Antillas Mayores mas relacionadas biogeograficamente (Fig 43 Tabla 44)
eL
EL
cb CAIMAN
SWAN
A Acratocnini B Cubanocnini C Megalocnini D Mesocnini E Nesophontidae F Solenodontidae G Xenothrichini
ANGUILAshyM-0 SMARTIN
H Heteropsomyinae I Plagiodontinae J Isolobodontinae
K Hexolobodontinae L Capromyinae
M Heptaxodontinae N CIidomyinae 0 Sigmodontinae
FIGURA 43 Representacion esquematica de las relaciones interinsulares (expresadas porel numero de linajes compartidos) entre las Antillas Mayores y entre estas y los grupos de islas perifericas segun las interpretaciones de los autores citados en el texto Los numeros dentro de los circulos indican la cantidad de linajes representados en cada isla relacion dudosa Contexto taxon6mico de los linajes en la Tabla 44
bull bull
bull bull
bull bull bull
bull bull
bull bull
bull
bull
bull
bull
bull bull bull bull
bull bull
bull bull
bull
bull bull bull
bull bull
bull
bull bull
bull
322 BIOGEOGRAFiA
TABLA 44 Distribuci6n de los linajcs de mamiferos terrestres a traves de las Antillas Mayores y grupos de islas perifericas LE = La Espanola CU Cuba JA Jamaica PR = Puerto Rico IC Islas Caiman IS Islas Swan IB = Islas Bahamas IV = Islas Virgenes AS = Anguila - San Martin
FamiliaSubfumiliafIribu LE CU JA PR IC IS IB IV AS
Megalonychidae
Choloepodinae
Acratocnini
Cubanocnini
Megalocninae
Megalocnini
Mesocnini
Nesophontidae
Solenodontidae
Pitheciidae
Callicebinae
Xenothrichini
Echimyidae
Heteropsomyinae
Capromyidae
Plagiodontinae
Isolobodontinae
Hexolobodontinae
Capromyinae
H e pta x 0 don tid a e
Heptaxodontinae
Clidomyinae
Muridae
Sigmodontinae
PILOSA
SORICOMORPHA
PRIMATES
RODENTIA
bull
bull
bull
bull TOTALES 12 10 4 4 2
323 EXTlICIOIES DEL CUATERNARIO
El gigantismo ha sido sin duda una tendencia evolucionaria recurrente entre los clados mayores de mamiferos antillanos Es un hecho bien conocido que algunos grupos de mamiferos han aumentado de tamafio en las islas y varios autores han intentado explicarlo a partir de enfoques diferentes (Foster 1964 Miller 1966 Sondaar 1977 Case 1978 Eisenberg y Wilson 1978 Heaney 1978 Lawlor 1982 Lomolino 1985 Angerbjom 1985 McNab 19942001)
EXTINCIONES DEL CUATERNARIO
Las islas que integran la Subregion Antillana han perdido casi 90 de sus mamiferos terrestres durante los ultimos milenios y en proporcion con su extension territorial esta perdida ha sido mayor en la Subregion que en cualquier otro lugar del planeta (MacPhee 1996b) Las extinciones del Cuatemario eliminaron mas de las tres cuartas partes de la fauna de mamiferos terrestres conocida de las Antillas Mayores sus islas satelites y grupos de islas perifericas y el examen de este proceso en sus aspectos mas generales se aborda en esta ultima parte del presente capitulo
CRONOLOGiA Y CONSECUENCIAS
Es muy improbable que un fosil represente el ultimo individuo de un taxon extinto y entonces hay que guiarse por la edad radiometrica del fosil mas reciente conocido lIamada fecha de ultima presencia- como unica aproximacion posible a la fecha de extincion del taxon
Como se explico al inicio del capitulo (vease Contexto Temporal) el registro fosH de los mamiferos terrestres de las Antillas Mayores adolece de insuficiente sustentacion radiometrica No obstante la distribucion temporal de los escasos fechados disponibles para el Cuaternario de la region abarca etapas criticas del proceso de extincion experimentado por la referida fauna a 10 largo de los ultimos 250 mil afios
De las 52 especies que es posible reconocer actualmente como miembros extintos de la fauna cuatemaria de mamiferos terrestres de las Antillas Mayores y grupos de islas perifericas los restos fosiles de 23 de elIas (442) han sido fechados radiometricamente o se han hallado asociados estratigraficamente de manera inequivoca con otro material fechado De estos 23 fechados 3 corresponden al interglacial Sangamon 4 pertenecen a la secuencia Pleistoceno terminal - Holoceno temprano 10 se ubican en ellapso entre el arribo de los aborigenes y la incursion europea y 6 son postcolombinos (a los cuales hay que sumar otras dos especies extinguidas en los siglos XIX y xx) A partir de estas 25 especies avaladas por fechas de ultima presencia distribuidas entre los cuatro intervalos temporales sefialados hemos compuesto la Tabla 45 en la que ademas aparecen las restantes 27 especies (carentes de fechado radiometrico) ubicadas en el intervalo temporal mas probable sobre la base de (1) la asociacion estratigrafica de sus restos con evidencias culturales (amerindias 0 europeas) 0 (2) la ausencia de dicha asociacion Sobre estos criterios de ubicacion temporal procede hacer dos aclaraciones
324 BIOGEOGRAFIA
TABLA 45 Intervalos temporales mas probables para las extinciones cuatemarias de los mamiferos terrestres de las Antillas Mayores y grupos de islas perifericas sobre la base de las evidencias siguientes (l) ultimo ejemplar colectado u observado en vida (2) fechado radiometrico mas tardio en material de la especie (3) asociaci6n estratigrafica mas tardia con otro material fechado (4) asociaci6n estratigrafica con restos de animales introducidos restos humanos u objetos culturales propios del intervale (5) falta de asociaci6n con evidencias culturales [Los ntimeros entre parentesis en la columna Evidencia significan adscripci6n tentativa al intervalo] Los estimados del peso corporal (en kg) estan tomados de la literatura 0 inferidos comparativamente y distribuidos entre
las clases siguientes (a) S 01 (b) 01 a lt 1 (c) 1 a lt 10 (d) 10 a lt 100 (e) 100 a lt 200 (j) ~ 200 Las
abreviaturas que designan las islas son las de la Tabla 44
F amilialEspecie Peso IsJa Evidencia Fuente
I httervalo htterg1acial Sangamon (gt80 kaAP)
HEPTAXODONTlNAE
Amblyrhiza inundata e AS 3 McFarJane et al 1998a
CLIDOMYINAE
Clidomys osborni d JA 3 Ibid
Clidomys parvus d JA 3 Ibid
II httervalo Pleistoceno terminal ~ Holoceno temprano (20-6 kaAP)
MEGALONYCHIDAE
Aeratoenus antillensis d CD 5 Arredondo 1975 a
Aeratoenus odontrigonus d PR 3 McFarJane 1999b
Aeratoenus simorhynehus d LE 2 Rega et al 2002
M egaloenus zile f LE 2 MacPhee et al 2000
SOLENODONTIDAE
Solenodon arredondoi e CD 5 Morgan y Ottenwalder 1993
PnHECIIDAE
Paralouatta varonai d CD 5 MacPhee 1996a
EcHIMYIDAE
Heteropsomys antillensis c PR 5 McFarJane 1999b
Heteropsomys insulans c PR 5 Ibid
Puertoricomys eorozalus c PR 5 Woods 1989a
CAPROMYIDAE
P lagiodontia araeum c LE 5 MacPhee et al 2000
Macrocapromys latus c CD 5 Varona y Arredondo 1979
(Continua)
325 EXTINCIONES DEL CUATERNARIO
TABLA 45 (Continuaci6n)
F ami1ia1Especie Peso Isla Evidencia Fuente
M acrocapromys latus c CU 5 Varona y Arredondo 1979
Mesocapromys kraglievichi c CU 5 Ibid
HEPTAXODONTINAE
Xaymaca fulvopulvis a JA 2 MacPhee y Flemming 2003
III Intervalo Amerindio (6-05 kaAP)
MEGALONYCHIDAE
Acratocnus ye d LE 2 MacPhee et aI 2000
Neocnus comes c LE 2 Ibid
N eocnus dousman c LE 2 Ibid
N eocnus toupiti c LE 2 Ibid
N eocnus gliriform is c CU 4 Cordova et ai 1997
M egalocnus rodens f CU 2 MacPhee et ai 2007
Parocnus browni e CU 2 Steadman et ai 2005
P arocnus serus e LE 2 MacPhee et ai 2000
N ESOPHONTIDAE
N esophontes edithae a PR 2 McFarlane 1999b
PITHECIIDAE
Antillothrix bernensis c LE 3 Rimoli 1977
EcHIMYIDAE
Brotomys contractus a LE 4 Miller 1929c
CAPROMYIDAE
P lagiodontia spelaeum c LE 4 Ibid
M acrocapromys acevedo c CU 4 NUiiez 1990a
HEPTAXODONTINAE
Elasmodontomys obliquus d PR 3 McFarlane 1999b
Iv Intervalo Postcolombino (05 kaAP - Presente)
N ESOPHONTIDAE
N esophontes hypom icrus a LE 24 MacPhee y Fle111111ing 1999
N esophontes zam icrus a LE (4) Ibid
N esophontes m icrus a CU 24 MacPhee et aI 1999
(Continua)
326 BIOGEOGRAFiA
TABLA 45 (Continuacion)
F amilialEspecie Peso Isla Evidencia Fuente
N esophontes m icrus a CD 24 MacPhee et aI 1999
N esophontes m icrus a LE 4 MacPhee et aI 2000
N esophontes sp 1 a IC 4 Morgan 1994
N esophontes sp 2 a IC 4 Ibid
Solenodon marcanoi b LE 4 Ottenwalder 2001
PITHECIIDAE
Xenothrix mcgregori c JA 4 MacPhee y Flemming 1999
EcHIMYIDAE
Boromys offeUa a CD 24 Guarch 1984
Boromys torrei a CD 24 Ibid
Brotomys voratus a LE 4 MacPhee et aI 2000
CAPROMYIDAE
P lagiodontia ipnaeum c LE 4 MacPhee y Flemming 1999
Rhizoplagiodontia lemkei c LE 4 Ibid
Isolobodon portoricensis c PR 24 Ibid
Isolobodon montanus c LE 4 Ibid
Hexolobodon phenax c LE (4) Ibid
Capromys sp c IC 24 Morgan 1994
Geocapromys columbianus c CD 4 MacPhee y Flemming 1999
Geocapromys thoracatus b IS 1 MacPhee y Marx 1997
Geocapromys sp 1 c IC 4 Morgan 1994
Geocapromys sp 2 c IC (4) Ibid
l-IEPTAXODONTINAE
Quemisia gravis c LE (1) MacPhee y Flemming 1999
MURIDAE
Oryzomys antillarum a JA 1 Ibid
1 Flemming y MacPhee (1999) alertaron en contra de la asociacion con otro material fechado Asimismo Morgan (1993) indico que la ausencia de asociacion con evidencias culturales amerindias no es necesariamente un criterio concluyente para ubicar un taxon en el intervalo temporal anterior porque hay depositos en la region que carecen de tales evidencias y son del Holoceno tardio por fechados radiocarbonicos [No obstante
327 EXTlNCIONES DEL CUATERNAR10
la ausencia de una especie en residuarios arqueol6gicos adquiere mayor certidumbre como criterio de ubicaci6n temporal de su extinci6n en la medida que aumenta el numero de residuarios estudiados que en las Antillas Mayores se cuentan por centenares] A falta de fechado absoluto de una especie consideramos que estos criterios de ubicaci6n temporal son mas utiles que la alternativa de excluir del analisis a la especie
2 En los trabajos sobre biogeografia de mamiferos antillanos los arque610gos mas citados como autoridades para establecer el inicio de la presencia aborigen en las Antillas Mayores (Kozlowski 1974 Rouse y Allaire 1978 Rouse 1989 1992 Wilson 2001) difieren al respecto entre los 6 kaAP y los 45 kaAP (pero vease Pagan et al 200523) y hemos adoptado la mas temprana de estas fechas para el comienzo del intervalo amerindio en la Tabla 45 De todos modos cualquiera que haya sido la fecha esta no es aplicable por igual al conjunto de las Antillas Mayores porque segun algunos de los autores citados el poblamiento comenz6 por Cuba y La Espanola 6 mil anos atras continu6 en Puerto Rico hace alrededor de 3 mil anos (pero vease Burney et al 1994) yen Jamaica hace menos de 2 mil e incluso a las Islas Caiman ni siquiera llegaron [Las Islas Caiman descubiertas por Col6n en 1503 carecen absolutamente de evidencia amerindia (Stokes y Keegan 1996)]
Si los datos de la Tabla 45 representan una aceptable aproximaci6n a la realidad el proceso de las extinciones cuaternarias de los mamiferos terrestres de las Antillas Mayores y grupos de islas perifericas debi6 caracterizarse entre otros por los eventos siguientes
Aunque obviamente la fauna cuaternaria debi6 ser mucho mas numerosa que 10 que refleja el registro paleontol6gico (Morgan y Woods 1986) 52 especies (81 de la fauna conocida) desaparecieron (sin reposici6n) durante los ultimos 250 mil anos
- Al parecer las extinciones se concentraron en las especies de mayor tamano Asi el peso promedio estimado para las especies de la etapa preantr6pica (intervalos I + II de la Tabla 45) es 350 kg es 328 kg para las especies del intervalo III (amerindio) y es s610 25 kg (c1ases del peso a b c) para las especies del intervalo IV (postcolombino) Ninguna especie aut6ctona de mamifero mayor de 6 kg existe hoy en la regi6n
- De acuerdo con la distribuci6n temporal de las extinciones 61 especies sobrevivieron al interglacial Sangamon (intervalo 1)48 sobrevivieron a la transici6n del Pleistoceno terminal al Holoceno temprano (intervalo II) 34 sobrevivieron al periodo amerindio (intervalo III) y s610 12 especies han sobrevivido desde la Ilegada de los europeos (intervalo IV) Las tasas de extinci6n correspondientes (exceptuado el intervalo I) son una especie cada 1167 anos en el intervalo II una cada 393 anos en el intervalo III y una cada 23 anos en el intervalo IV
- Durante todo el proceso de las extinciones cuaternarias la perdida de especies por isla fue de 91 en La Espanola 83 en Jamaica y 62 en Cuba Bahamas retuvo su unica especie (aunque extirpada en varias de las islas) Puerto Rico e Islas Caiman perdieron todas sus especies (6 y 5 respectivamente) y las Islas Swan Virgenes y Anguila - San Martin perdieron sus respectivas especies (micas
328 BIOGEOGRAFiA
De los cuatro 6rdenes de mamiferos representados en la fauna Pilosa y Primates desaparecieron totalmente Soricomorpha y Rodentia perdieron 86 y 74 de sus especies respectivamente
CAUSALIDAD
CAUSAS NATURALES
Los cicIos glaciales e interglaciales acaecidos en el Pleistoceno causaron cambios ambientales dnisticos en las Antillas con la concomitante alternancia de cIimas frios (secos) y calidos (hUmedos) asi como de descensos y ascensos del nivel del mar aumento y disminuci6n de las areas territoriales) induso (aunque con menor intensidad) en el Holoceno Estos eventos paleogeograficos han sido consistentemente documentados (Mayo y Kartashov 1972 Bonnatti y Gartner 1973 Ortega y Arcia 1982 Prance 1982 Schubert y Medina 1982 Dodge et al 1983 Ortega [1984] Haq et al 1987 Schubert 1989 Hodell et al 1991 Guilderson et al 1994 Oppo 1997 Emslie 1998 Curtis et al 2001 Iturralde-Vinent 2003) y de ellos brindamos a continuaci6n una breve resena
Finalizando el periodo interglacial anterior a la ultima glaciaci6n (Wisconsin) el mar estaba a 8-10 m por encima de su nivel actual Cuando el avance glacial del Wisconsin a1canz6 su culminaci6n (eillamado Ultimo Maximo Glacial) hace 18 ka el mar habia descendido 120 m con respecto a dicho nivel y las islas del Caribe habian incrementado considerablemente sus extensiones subaereas en comparaci6n con las que presentan hoy dia por ejemplo Puerto Rico y las Islas Virgenes se conectaron territorialmente y las Bahamas se unieron en cinco gran des islas Prevalecia un regimen frio y seco y condiciones de aridez --debidas a una reducci6n de las precipitaciones (posiblemente a menos de 440 mmlano)- y los ambientes xericos se habian extendido a traves de las islas y bancos emergidos y abundaban los pinares (los habitats mesicos estaban restringidos a parcelas relictas en 10 alto de las montanas) En el Mar Caribe y el Golfo de Mexico la temperatura superficial del mar era 5degC mas baja que la actual y en las llanuras costeras y alturas maximas eran respectivamente 2-3degC y 6degC mas bajas Los limites de las zonas ecol6gicas altitudinales descendieron varios cientos de metros y cada zona redujo su extensi6n vertical al aumentar el gradiente de temperatura Ellimite inferior de la zona periglacial
elevaci6n donde la temperatura media minima a1canza los OdegC) de la Cordillera Central de la Republica Dominicana (altura maxima del Caribe insular con evidencia de topografia glacial) se considera que estuvo entre los 1500 y 1800 mlnm (actualmente esta a 2800 mlnm) y se puede estimar que dicho limite era inferior en Cuba por estar mas al norte que la Republica Dominicana No obstante durante este periodo la temperatura experiment6 episodios cortos (unas pocas decadas) de ascensos repetidos
Un signifieativo componente bi6tieo de las referidas condiciones paleoambientales 10 constituy6 la espectacular irradiaci6n evolucionaria experimentada por un grupo numeroso de aves rapaees diurnas y nocturnas (Wetmore 1937 Brodkorb 1969 Kurotchkin y Mayo 1973 Arredondo 1976 Olson 1978 Olson y Hilgartner 1982
329 EXTINCIONES DEL ClJATERNARIO
Suarez 2000a b 2004 Suarez y Emslie 2003) Los mamiferos de las Antillas Mayores ofrecieron a estos depredadores aviares una amplia gama de tamanos de presa que en ausencia de mamiferos del orden Carnivora en la region permitio a estas aves irradiarse a este nicho alimentario con la evolucion de una variedad de especies de muy diversa tall a corporal incluidas formas gigantes Adquirir un cuerpo gigante les propicio el acceso a recursos alimentarios que no estaban siendo explotados por ningun otro depredador (Alcover y McMinn 1994) A Cuba corresponde el mejor registro fosil de rapaces cuaternarias caribenas con siete generos de especies diurnas Dentro de estas aves existieron tres gavilanes gigantes (Accipitridae) tres carairas (Falconidae) y un condor (Cathartidae) asi como cuatro generos de especies nocturnas dos lechuzas (Tytonidae) y tres Mhos (Strigidae) dos de ell os no igualados en tamano en todo el mundo Algunos de estos generos tuvieron representacion en otras islas antillanas como La Espanola Jamaica Bahamas y Gran Caiman
Al retirarse hacia el norte los hielos de la glaciacion Wisconsin el clima se tomo mas hUmedo en las Antillas durante el Holoceno temprano y el bosque mesico se expandio en la region insular El nivel del mar ascendio acompanado de mayor pluviosidad alrededor de los 7 kaAP las Islas Virgenes quedaron separadas entre si y de Puerto Rico y las Bahamas perdieron la mayor parte de su territorio Cerca de los 3 kaAP las condiciones secas retornaron a la region e hicieron desaparecer buena parte del bosque mesico Nuevamente el clima comenzo a adquirir sus condiciones actuales hacia los 25 kaAP aproximadamente y hoy transita por una fase mesica con vegetacion xerica confinada en parcelas relictas
Sin duda estos y otros eventos naturales incidieron negativamente en la sobrevivencia de mamiferos terrestres en las Antillas Mayores asi como en los grupos de pequenas islas perifericas (Pregill y Olson 1981 Olson 2006)
McFarlane et al (1998a) demostraron convincentemente como Amblyrhiza el gigante de los roedores antillanos endemico de la pequena isla de Anguila sucumbio a las repetidas y abruptas perdidas de su habitat durante el interglacial Sangamon por las nipidas y severas oscilaciones del nivel del mar (documentadas por Neumann y Hearty 1996) al no poder responder adecuadamente a tan catastroficos eventos mediante el ajuste de su masa corporal media y otro tanto debio ocurrirle en Jamaica al otro roedor gigante Clidomys (McFarlane et al 1998b) emparentado con Amblyrhiza en la misma epoca (Tabla 45 intervalo I)
El ascenso del nivel del mar seguramente promovio la extincion de mamiferos durante la transicion del Pleistoceno al Holoceno (Tabla 45 intervalo II) con la reduccion del tamano de las Islas por las transgresiones marinas (Pregill y Olson 1981 Morgan 1993) Esto ultimo haria maximamente severa la competicion asf como la presion de los depredadores [Steadman et al (2005) no yen por que el calentamiento global ocurrido durante la transicion Pleistoceno-Holoceno habria de tener un efecto negativo sobre los perezosos siendo sus descendientes actuales especies tropic ales 0 subtropicales pero no se trata del efecto
330 BIOGEOGRAFiA
directo del cambio de temperatura sobre los animales sino del efecto indirecto y progresivo que dicho cambio genera a largo plazo al reducir el territorio insular disponible]
Por esta epoca tambien desaparecieron especies de tall a mayor como Megalocnus zile en La Espanola y otras de menor tamano (en Cuba) pero que representan la tall a maxima en sus respectivos contextos taxonomicos del Cuaternario como Solenodon arredondoi Paralouatta varonai y Macrocapromys acevedo Un mamifero puede ser grande solo a costa de reducir su densidad poblacional y esto 10 hace mis vulnerable a la extinci6n sobre todo en una isla (McNab 1994 McFarlane et al 1998a) Incidentalmente Alcover et al (1998a) afirmaron que larelacion entre la talla corporal y la extincion no siempre se cumple en las especies de menor tamano y citaron como ejemplo que entre las musarafias todas las especies de Nesophontes desaparecieron mientras sobreviven especies de Solenodon pero la comparacion correcta en todo caso serfa entre los solenodones vivientes y el gigante Solenodon arredondoi y entre Nesophontes edithae -el gigante de los nesofontes extinguido en el periodo amerindio- y el resto de sus congeneres que sobrevivieron hasta los primeros siglos de la colonizacion europea (Tabla 45 intervalos III y IV)
CAUSAS ANTROPOGENICAS
Intervalo Amerindio
La mayoria de los autores que se han referido a las extinciones acaecidas durante este intervalo (entre elIos paradigmaticamente Martin 1984a b) han responsabilizado a los aborfgenes (que se supone comenzaron a poblar las Antillas Mayores hace 6 mil afios) con la desaparicion de numerosas especies de mamiferos terrestres en la region principalmente por la via de la sobrecaza y la introduccion de animales domesticos devenidos depredadores de la fauna autoctona (Morgan y Woods 1986) Este efecto negativo de la presencia indigena sobre los mamiferos de la region ha sido dramatizado por algunos autores en terminos de holocausto faunal Sin embargo pensamos que se ha exagerado Como han expresado MacPhee et al (19993) cuando uno busca evidencia dura de complicidad humana directa en [estas] extinciones no hay mucho que encontrar Las siguientes son algunas consideraciones a tener en cuenta en tal sentido
El registro radiometrico mas joven conocido hasta ahora para un perezoso antillano (MegalocnllS rodens) es de 42 kaAP (MacPhee et ai 2007) Si se acepta que los aborfgenes entraron en Cuba hace aproximadamente 6 mil anos su convivencia con los perezosos debe haber durado alrededor de 1800 afios (0 mas porque dificilmente el referfdo fechado radiometrico haya correspondido al ultimo sobreviviente de la especie) Esta cifra contradice la hipotesis del holocausto faunal segun la cualla extincion de los mamiferos insulares de mayor talla debe ocurrir con extrema rapidez -a escala de decadas 0 de centurias- a partir de su primer contacto con los humanos (Martin y Steadman 1999) A pesar de las repetidas afirmaciones en contrario (Harrington 1921 Miller 1929c Herrera [1945] Tabio 1951 Alvarez 1951a 1957 Arredondo
331 EXTINCIONES DEL CUATERNARIO
1959 1960a Veloz y Ortega 1973 Mayo 1980b Rodriguez et al 1984 Castellanos y Pino 1985 Pino y Castellanos 1985 Nunez 1990b Gutierrez 1996 Arredondo 2004) en su mayoria basadas en evidencias no exentas de posibles accidentes de deposicion (0 de reubicacion postdeposicional) en recintos cavernarios (Diaz-Franco [2006b D no se ha demostrado inequivocamente que los aborigenes antillanos se alimentaran de perezosos regularmente Sus restos en residuarios de alimentos son escasisimos en toda la region y esto contrasta con los abundantes restos de jutias y otros mamiferos de menor talla que aparecen en esos residuarios demostrativos de la preferencia de los paleoindios por estos otros mamiferos
Los mamiferos mas importantes en la economia prehistorica de las Antillas Mayores fueron las jutias Capromys y Geocapromys en Cuba Geocapromys en Jamaica e Isolobodon en La Espanola Puerto Rico e Islas Virgenes (Aguayo 1950b Tablo 1951 Rimoli 1972 Wing 2001 Diaz-Franco [2006b D Wilkins (2001) estudi61ajutia viviente de Jamaica (Geocapromys browni) en un sitio ocupado por los aborigenes durante 100 anos y no encontr6 evidencias de que durante ese tiempo la especie haya sido sobreexplotada Asimismo en Cuba las jutias conga carabali y andaraz son escasas en depositos pleistoholocenicos en cambio son muy numerosas en residuarios indigenas y de haber sido sobreexplotadas 10 mas probable es que no hubieran sobrevivido hasta hoy
La habilidad de los humanos para penetrar en profundidad el bosque tropical y mantenerse en este sustentablemente ha sido un evento comparativamente reciente (Beran y Steele 1994) En el caso antillano la caza tierra adentro ha sido explicada como un proceso tardio de desplazamiento a partir de la explotacion de abundantes recursos marinos que ofrecian condiciones 6ptimas para el sedentarismo de las comunidades aborigenes costeras (C6rdova y Arredondo 1988 Brito 2005) Peces y mariscos dominan la arqueologia costera de las Islas antillanas en tanto el consumo de recursos terrestres (mamfferos incluidos) aumenta progresivamente en la medida que los residuarios estudiados se alejan de la costa (Scudder 1991 Wilkins 2001) En las Antillas Mayores el componente terrestre de la dieta aborigen es solo 20-30 del material recobrado en los residuarios (Wing 1977 1989) Y debio ser menor alin porque el componente vegetal de la dieta no se ha preservado Por otra parte los aborigenes cubanos domesticaron la jutia conga y seguramente la reprodujeron para comer (Pose et al 1988) practica que probablemente se repitiera con otras jutias en las demas Antillas Mayores
- Fray Bartolome de las Casas hablo de 3 millones de indios en Cuba al inicio de la colonizacion pero los historiadores subsecuentes consideraron exagerada la cifra (Poey 1851 a) No obstante se dispone de dos estimados de la densidad de poblaci6n aborigen en las Antillas Mayores a la llegada de los europeos 18 indigenaslkm2 en Puerto Rico (Rouse 1956) y 1271km2 en La Espanola (Watts 1987) Extrapolando ambos valores entre las cuatro islas principales de las Antillas Mayores tenemos que en la fecha indicada la poblaci6n aborigen pudo estar entre 16 mil y 113 mil indigenas en Puerto Rico entre 21 mil y 145 mil en Jamaica entre 141 mil y 1 mill6n en La Espanola y entre 200 mil y 14 millones en Cuba Entre las cifras maximas y las minimas nos inclinamos
332 BIOGEOGRAFiA
hacia estas ultimas atendiendo a los altos indices de mortalidad infantil y morbilidad que se evidencian en las osamentas de los pobladores prehispanicos (Martinez y Rives 1990) Uno de los hechos que apuntan hacia el bajo numero de aborigenes en las Antillas Mayores es 10 rapido que desaparecieron despues de la llegada de los europeos pnkticamente sin dejar mestizaje con estos 10 cual contrasta ostensiblemente con la situacion en la America continentaL
En suma un grupo humano (1) demograiicamente irrelevante y controlado por altos indices de morbilidad (2) consumidor mayoritariamente de organismos marinos y (3) carente de la tecnologia cinegetica necesaria para poder sobreexplotar los mamiferos que Ie sirvieron de alimento (a los cuales accedio tardiamente en su desarrollo) no es concebible que haya causado e1 holocausto faunal que se Ie atribuye a partir del error de aplicar a las comunidades aborigenes antillanas un modelo cultural de la prehistoria continentaL
Tampoco hay evidencias de que los aborigenes hayan introducido carnivoros con suficiente capacidad depredadora salvo el perro Morgan y Otlenwaldder (1993) consideraron que la depredacion por perros durante el periodo amerindio es la explicacion mas plausible para la extincion del almiqui gigante de Cuba pero como se puede observar (Tabla 54) Solenodon arredondoi aparece en el intervalo II por falta de asociacion con evidencias culturales
Consecuentemente el accionar de los aborigenes y de sus animales domesticos directamente sobre la fauna no parece haber sido por sf solo un factor determinante de la extincion de mamiferos terrestres durante el periodo amerindio en las Antillas (pero vease Steadman et al 2005)
Hay que recordar en este senti do que alin en el Holoceno tardio ocurrieron cambios clirruiticos de consideracion (vease Causas Naturales) Por ejemplo Curtis et al (2001) explicaron que la extincion de algunos ani males mesofilicos de las alturas montanosas de La Espanola pudo haber antecedido a cualquier impacto humano sigrIificativo en esos lugares por coincidir dicha extincion con una tendencia a la aridez documentada isotopicamente y con evidencias de la perdida del bosque mesico ocurridas despues de 3kaAP Yen Jamaica hace solo 2shy3 mil aDos el area de la Bahia de Jackson sustentaba una gran biomasa de vertebrados en bosque hinnedo a juzgar por la cantidad y diversidad de taxones presentes en material fosil derivado de la depredacion por lechuzas hoy el area es la mas seca de la isla con matorral xerofitico y pobreza de fauna (MacPhee 1996a) Yalm en nuestros dias huracanes de gran intensidad aniquilan poblaciones de vertebrados (que no se recuperan) en is las pequenas y llanas del Caribe (Brooks y Smith 2001)
En medio de la incertidumbre sobre la verdadera causa (0 causas) de las extinciones de mamiferos en el periodo amerindio MacPhee y Marx (1997) han propuesto su hipotesis de la hipermorbilidad la cual estipula colapsos poblacionales relativamente rapidos con la introduccion de enfermedades de alta letalidad Estos autores han razonado que
Las islas son los lugares mas apropiados para experimentar perdidas catastroficas causadas por enfermedades Los mamiferos no solo serian susceptibles en virtud de su prolongado aislamiento biologico sino que en los confines de una isla la epizootia
333 EXTINCIONES DEL CUATERNARIO
tendria la potencialidad de infectar simultaneamente a la rnayoria de los individuos de vadas especies Los neonatos son inmunologicamente inmaduros y sufririan desproporcionadamente cualquier infeccion hipervirulenta La perdida excesiva de neonatos y juveniles tendria efectos severos para la sobrevivencia de la poblacion si esta se reduce por debajo de cierto nivel sobreviene la extincion porque no hay posibilidad de restitucion en el periodo postpanzootico Las especies de mayor talla corporal van a ser las mas vulnerables a la extincion dadas sus bajas tasas reproductivas largos periodos de gestacion y reducido numero de progenie Las especies de menor tamano --caracterizadas por altas tasas reproductivas periodos cortos de gestacion y mayor numero de neonatos por parto- tambien serian fuertemente diezmadas pero muchas sobrevivirian porque su estrategia reproductiva les permite perder mayor numero de individuos Las perdidas deben ocurrir relativamente rapido (a escala de decadas) mientras que otras hipotesis (digamos los cambios climaticos) permiten intervalos mucho mas largos (a escala de siglos 0 de milenios) entre la aparicion de las causas y la produccion de sus efectos Por ejemplo todo indica que los nesofontes se extinguieron muy rapidamente y seria dificil poder invocar un impacto humano directo 0 argumentar fenomenos climaticos que pudieran explicar plausiblemente perdidas tan rapidas
MacPhee y Marx (1997 191) han dicho que por el momento la hipotesis de la hipermorbilidad no pasa de ser una abstraccion en espera de adecuada sustentacion empirica Llama la atencion que linicamente en el periodo amerindio (Tabla 45 intervalo 1Il) las extinciones abarcaron todas las clases del peso corporal (de la a a laj) y que ninguna de las especies que sobrevivieron hasta el periodo postcolombino (Tabla 45 intervalo IV mas las especies vivientes) sobrepasa los 10 kg (clases del peso a b c) en clara coincidencia con los presupuestos de la hipotetica hipermorbilidad
Intervalo Postcolornbino
Por 10 menos 37 especies de mamiferos terrestres vivian en las Antillas Mayores y grupos de Islas perifericas a la lIegada de los europeos en 1492 De elIas se han extinguido 23 shymas especies que en cualquiera de los periodos precedentes (Tabla 45~ y solo subsisten 12 especies repartidas entre Cuba (8 especies) La Espanola (2) Jamaica (1) y Bahamas (1) varias de estas en peligro critico de extincion Solo dos de las especies extinguidas durante los ultimos 500 anos se conocieron en estado viviente los roedores Oryzomys antillarum de Jamaica (extinguido en 1877) y Geocapromys thoracatus de las Islas Swan (extinguido en 1955) Las causas de las extinciones ocurridas durante este periodo son sobradamente conocidas destruccion de habitats introducci6n de animales exoticos (domesticos y comensales) y demas presiones asociadas a la actividad humana entre las cuales posiblemente habria que incluir brotes de hipermorbilidad
Durante mucho tiempo se ha afirmado la posibilidad de que especies de Nesophontes hayan sobrevivido hasta las primeras decadas del siglo xx y que incluso continuen existiendo en lugares remotos de Cuba 0 La Espanola (Allen 1918 1942 Miller 1930 Aguayo 1950a Arredondo 1960b Fischer 1977 MacFadden 1980 Woods y
334 BIOGEOGRAFiA
Ottenwalder 1991) Actualmente no existe evidencia radiometrica de que alguna especie de Nesophontes haya sobrevivido al siglo xv las fechas radiocarbonicas calibradas obtenidas en muestras de Nesophontes de Cuba y La Espanola resultaron no mas tardias que los siglos XIII y XIV (MacPhee et al 1999) Por otra parte las expediciones efectuadas a fines del siglo xx con el proposito de localizar individuos vivientes del genero en La Espanola no produjeron la menor indicacion afirmativa al respecto (Woods et al 1985) Ademas a traves de las Antillas Mayores no aparecen restos de Nesophontes en los depositos formados actualmente por las lechuzas
EPiLOGO
Las Antillas Mayores son islas oceanicas no han formado parte de un continente (con la posible excepcion de una parte de Jamaica hasta el Eoceno Medio hace 45 Ma) y durante un tiempo geologico considerable (desde el Eoceno Superior hace 40 Ma) han permanecido emergidas y separadas de los continentes vecinos Consecuentemente los mamiferos terrestres (no volantes) autoctonos de estas islas constituyen una fauna oceanica que por ende cumple con todas las caracteristicas convencionales reconocidas para este tipo de entidad biogeografica
- Baja diversidad taxonomica a nivel de las categorias superiores (solo cuatro ordenes) en comparacion con islas plataformicas continentales (0 con areas continentales) de tamano y diversidad ecologica simi lares
- Alto grado de endemismo a nivel de genero (96) y familia (50) en respuesta al prolongado aislamiento
- Desbalance ecologico de la estructura trofica ausencia de mamiferos camivoros y ungulados
- Espectaculares procesos de irradiaci6n evolucionaria como resultado de la colonizacion de territorios libres de competidores y depredadores
Tendencias evolucionarias extremas gigantismo
- Control a cargo de aves rapaces con tendencia at gigantismo en ausencia de mamiferos miembros del orden Carnivora
- Mas especies de reptiles y de murcielagos que de mamiferos terrestres relaciones que en los continentes se invierten
Alta tasa de extincion de especies (84) sin reposicion
Preservacion de especies filogeneticamente arcaicas (los llamados fosiles vivientes) los almiquies

306 B[OGEOGRAFIA
y solo unas pocas recalaron en las Antillas Mayores y las que entraron al Mar Caribe demoraron semanas 0 meses en arribar
e) La distancia a recorrer (y en consecuencia la duracion de la travesia) entre el continente y la isla (10 que repercute decisivamente en el factor c) Este es desde luego uno de los factores mas criticos En la medida que la distancia aumenta la probabilidad de alcanzar la isla disminuye drasticamente con el agravante de que la trayectoria dificilmente seria en linea recta Si tomamos como referencia la velocidad de 45 kmlh del islote ya mencionado (SICSP 1970) cubrir la distancia aerea (aproximada) de 800 km que media entre el noroeste de Suramerica y La Espafiola Ie tomaria a la balsa 74 dias Estimando el doble de este tiempo debido a desvios de la trayectoria la duracion de la travesia probablemente resultaria excesiva para la mayoria de los mamiferos Pero pudiera muy bien ocurrir que la velocidad fuera mayor 0 que la balsa portara plantas vivas que permitirian a los fitOfagos sobrevivir [Tres de los cuatro ordenes de mamiferos terrestres conocidos de las Antillas Mayores son fitOfagos] Ademas esta geologicamente bien establecido que durante etapas de variada duracion desde el Oligoceno hasta el Pleistoceno los territorios del Caribe insular y continental aumentaron considerablemente en extension con niveles del mar 100-200 m por debajo de su nivel actual oportunidades que los propagulos aprovecharian para cruzar barreras marinas significativamente menores
f) El tamafio de la isla Las islas mayores deben interceptar mas balsas (Buckley y Knedlhans 1986) 10 que repercute directamente en el factor e La balsa puede pasar entre islas pequefias y continuar viaje sin llegar a ningun lugar
g) Las condiciones (favorables 0 adversas) del desembarco en la isla y el tiempo que Ie tomaria a uno 0 mas animales depauperados encontrarun habitat apropiado Esto dependeria a su vez del tamafio y diversidad ecologica de la isla (Darlington 1938 Miller 1966)
Tal cumulo de posibles obstaculos y dificultades constituye a primera vista un desafio a la credibilidad de la hipotesis Matthew-Simpson pero en ello radica precisamente su valor explicativo bas ado en la mas absoluta improbabilidad La colonizacion por la via marina es altamente improbable pero no absolutamente imposible si se tiene en cuenta la inmensidad del tiempo geologico se vuelve probable (McKenna 1973) Basta que solo una vez entre mill ones de intentos fallidos durante ese inmenso lapso algunos de los factores anteriormente enunciados coincidan en no ser tan desfavorables para el propagulo Y eso es todo 10 que demanda la hipotesis para cada uno de los grupos monofileticos mayores de mamiferos terrestres presentes en las Antillas Mayores Las Antillas Mayores requeririan el arribo de tan solo 7 (quizas menos) propagulos fundadores a 10 largo de los ultimos 30 0 mas millones de afios (promedio de uno cada 43 mill ones) para explicar la formacion de toda su fauna de mamiferos terrestres por dispersion marina pasiva (vease Discusion y Conc1usiones)
En comparacion con la via marina el poblamiento por la via terrestre es predecible porque la mayor parte de la fauna continental de mamiferos emplazada en la region del puente va a cruzar por este una vez que el mismo se establece y los animales logran
307 POBLAMIEITO CONTINENTE-ISLA
acceder a el el poblamiento por la via marina en cambio es impredecible porque depende total mente del azar que caracteriza a la balsa Claro que en ambas vias opera el azar pero en el primer caso se trata del azar de un evento probable en el segundo el azar de un evento improbable (Simpson 1940) Asi el puente natural (mecanismo de poblamiento minimamente selectivo = alta diversidad inicial) y la balsa natural (mecanismo de poblamiento maximamente selectivo baja diversidad inicial) pueden conceptuarse como los extremos opuestos de una gama de probabilidades de dispersi6n (McKenna 1973) EI fen6meno de la dispersi6n merece la mayor atenci6n cientifica en los estudios de biogeografia insular (Cowie y Holland 2006)
Queda claro que la balsa natural es una hip6tesis consistente con el caracter de la fauna objeto de estudio puesto que el desbalance taxon6mico (sin evidencia paleontol6gica en contrario) s610 puede ser resultado de la improbabilidad del poblamiento No obstante ello no implica necesariamente que la balsa natural haya sido el imico mecanismo (ni siquiera el principal) utilizado por los mamiferos terrestres en su colonizaci6n de las Antillas Mayores La balsa natural debe satisfacer una premisa te6rica esencial dado el caracter azaroso impredecible e improbable de una colonizaci6n exitosa esta debe ocurrir a intervalos muy espaciados a traves del tiempo geol6gico 0 sea durante todo el tiempo que en el area ocupada por las Antillas Mayores ha habido ecosistemas terrestres perrnanentemente disponibles para ser colonizados (los ultimos 40 millones de afios) Sin embargo como sabemos representantes de los cuatro 6rdenes de mamiteros terrestres conocidos de las Antillas Mayores ya estaban emplazados en dicho territorio insular en el Mioceno Inferior 10 que supone un lapso de colonizaci6n limitado al Eoceno SuperiorshyOligoceno (MacPhee 2005) sugerente de un tipo de mecanismo contrario a la colonizacion alocronica exigida por la hipotesis de la balsa natural sobre todo tratandose de una fauna caracterizada por altas tasas de extincion sin reposicion aparente
La presencia en Jamaica (hasta su extinci6n en 1877) del roedor mfuido Oryzomys antillarom con estrecha afinidad centroamericana (Woods 1989a) pudiera constituir el imico caso efectivo de poblamiento (continente-isla) de las Antillas Mayores por dispersion marina dc un mamifero terrestre para el cual no es razonable invocar otro mecanismo de colonizacion
LA GUIRNALDA INSULAR (dispersion mixta)
La geologia ha establecido con suficiente certeza que durante el Cenozoico unicamente en el Pale6geno tardio y solo a traves de la Cresta de Aves (ahora sumergida excepto la isla hom6nima) pudo haber existido una cadena de islas para el transito de mamiferos terrestres entre el norte de Suramerica y las proto-Antillas Mayores [desestimados a ese efecto el Canal dc Yucatan el Elcvado de Nicaragua la Cresta de Caiman el Elevado de Beata y las Antillas Menores (Holcombe et al 1990 MacPhee e IturraldeshyVinent 1994 1995b Iturralde-Vinent y MacPhee 1999)] Holcombe y Moore (1977) y Holcombe y Edgar (1990) postularon la existencia de esta entidad y estimaron algunas de sus posibles caracteristicas Por su parte MacPhee e Iturralde-Vinent (1995b) e Iturralde-Vinent y MacPhee (1999) precisaron los extremos proximal y distal de esa
308 BIOGEOGRAFIA
estructura acotaron la duracion y la edad de la misma y Ie dieron nombre GAARlandia (Greater AntillesA vesRidgelandia)
Segun Iturralde-Vinent y MacPhee (1999) la Cresta de Aves emergio por breve tiempo (1-2 Ma) durante la transicion Eoceno-Oligoceno (35-33 MaAP) por la coincidencia de un levantamiento tectoni co y un considerable descenso eustatico del nivel del mar En ese momenta las islas septentrionales del arco de las Antillas Mayores (Cuba centroorientalshyLa Espafiola - Puerto Rico - Islas Virgenes) constituian una sola isla grande 0 una serie de islas separadas por barreras muy estrechas La Cresta de Aves emergida entre el bloque Puerto Rico - Islas Virgenes y un microcontinente noroccidental de Suramerica -separado en ese momento del resto del continente por una via marina surgida en el Eoceno tardio- dio origen a GAARlandia
Obviamente el numero de colonizaciones exitosas por mamiferos terrestres a traves de una guimalda de Islas tiende a disminuir mientras mas anchas sean las barreras marinas entre las islas que forman la guimalda y cuando esto ocurre disminuye tambien la diversidad taxonomica de los propagulos particularmente a nivel de las categorias superiores (Simpson 1956)
Vemos asi que para los mamiferos terrestres la colonizaci6n a traves de una guimalda insular puede ser tan improbable como a la deriva en una balsa natural en dependencia de la cantidad y anchura de las barreras marinas a cruzar el tamafio y la fisiografia de las islas que integran la guimalda y el regimen de las corrientes marinas y aereas entre otros factores
Resumiendo la guimalda insular puede conceptuarse como un mecanismo de poblamiento altamente selectivo (= baja diversidad inicial) que combina la improbabilidad de la balsa natural con la colonizacion restringida temporaimente del puente terrestre
En su reciente revision de la taxonomia distribuci6n y filogenia (basada en estudios moleculares) de varios grupos de mamiferos antillanos Davalos (2004) concluyo que los perezosos megalocninos los primates y los roedores histricognatos se avienen al patron de divergencia con el continente y epoca de arribo a las Antill as implicados en la hipotesis de GAARlandia pero no asi los perezosos coloepodinos que requieren una explicacion biogeognlfica distinta Esto ultimo (sospechado por Matthew 1918a) habla sido confirmado por White y MacPhee (2001 226-227)
Nuestra evidencia filogenetica no sustenta la idea de un dado unificado surgido de la invasion de un megaloniquido el analisis sugiere mas bien dos eventos de colonizaci6n independientes [y] sustenta fuertemente un origen difiletico de las subfamilias [Choloepodinae y Megalocninae] Si hubo dos migraciones a traves de GAARlandia presumiblemente poco espaciadas en el tiempo no se puede determinar en el presente
Finalmente queda claro que GAARlandia -como guirnalda insular- es la hipotesis mas robusta propuesta hasta ahora para explicar la presencia en las Antillas Mayores de
309 POBLAMIENTO CONTINENTE- ISLA
la absoluta mayona de los mamfferos terrestres de ongen suramericano porque el caracter de esa fauna supone un mecanismo de poblamiento ancestral que sea a la vez maximamente selectivo (baja diversidad ordinal) y temporalmente restringido (transicion Eoceno-Oligoceno)
Recientemente Rega et al (2002 18) expresaron A pesar de una revision exhaustiva de la paleogeografia de la cuenca del Caribe Iturralde-Vinent y MacPhee (1999) no pudieron confirmar la presencia de una conexion terrestre continua entre Suramerica y las proto-Antillas aunque hipotetizaron su existencia [Vease la Fig 42 (= figura 6 de Iturralde-Vinent y MacPhee 1999 simplificada))
En el trabajo citado por Rega y colaboradores aparece la siguiente explicacion
Actualmente no es posible ofrecer reconstrucciones paleogeograficas detalladas de la Cresta de Aves Sin embargo hay evidencia de que porciones de la cresta deben haberse elevado subaereamente en un tiempo La cuestion cntica es si la Cresta de Aves emergida pudo haber formado algun tipo de corredor entre las Antillas Mayores y el noroeste de Suramerica Perforaciones en las alturas estructurales de la cresta pudieran proveer el registro del Paleogeno tardio que se necesita para precisar la hipotesis del corredor aunque no se puede predecir si tales datos serian suficientemente detallados para determinar si en algun momento GAARlandia constituyo una sola entidad subaerea Por ejemplo pudo haber barreras marinas por corto tiempo entre las masas terrestres de la Cresta de Aves [0) ellevantamiento y la subsidencia pudieron afectar diferentes partes de GAARlandia en diferentes tiempos AItemativamente los elementos faunales pudieron haberse dispersado de manera pulsada Movimientos episOdicos de este tipo incrementarian la extincion de los elementos faunales incapaces de trasladarse a la siguiente masa terrestre en la cadena disminuyendo asi la diversidad total (Iturralde-Vinent y MacPhee 199957)
La primera parte de la explicacion deja claro que la posibilidad de que la Cresta de Aves haya sido una conexion terrestre continua no se puede descartar pero tampoco se puede afirmar La ultima parte de la explicacion describe el efecto altamente selectivo (filtrante) de la colonizacion a traves de una cadena de islas
Y continuan lturralde-Vinent y MacPhee (199957 -58) explicando
Hasta que punto el microcontinente noroccidental de Suramerica [considerado por dichos autores como el area suministradora de propagulos hacia GAARlandiaJ estuvo fisicamente separado del resto del continente no ha sido plenamente esc1arecido Si estuvo sustancialmente aislado por barreras marinas y orogenic as durante el Paleogeno tardio entonces solo la fraccion de la biota suramericana emplazada en el microcontinente estuvo en posicion de acceder a GAARlandia Esto pudiera tener que ver con el hecho de que la fauna antillana carezca de representantes de muchos grupos suramericanos [de mamfferos) que debieron haber hecho el viaje
310 BIOGEOGRAFiA
Florida o Alturas0 0
d 6 D L1anuras25deg
Yucatan
20deg
deg0
10deg
MARCARIBE
80deg 75deg
Bahamas
Cres ta de Beata
D Mares someros
D Mares profundos
OCEANO ATLANTICa
FIGURA 42 Reconstrucci6n hipotetica de GAARlandia como conexi6n terrestre continua entre las proto-Antillas Mayores y el noroeste de Suramerica en el Pale6geno tardio segun la FIgura 6 (simplificada) de Iturralde-Vinent y MacPhee (1999)
En su defensa del puente terrestre Barbour (1945b249) se refiri6 a esto - antes 10 habfa hecho Anthony [1942363]- en los terminos siguientes
El argumento de que si las Antillas hubieran estado conectadas con el continente se encontrarian los f6siles de un numero mayor de distintas criaturas me deja frio es perfectamente concebible que la conexi6n haya ocurrido en un periodo en que la parte del continente implicada no estuviera poblada por una fauna grande y variada
Y concluyen lturralde-Vinent y MacPhee (199958) su detallada explicaci6n
Desafortunadamente sabemos muy poco sobre la composici6n faunal del noroeste de Suramerica a finales del Pale6geno
Consecuentemente si se confirmara la magnitud del aislamiento del aludido microcontinente y su concomitante baja diversidad faunal GAARlandia como conexi6n terrestre continua serf a la hip6tesis mas parsimoniosa No obstante mientras no se disponga de evidencia concluyente a este respecto hipotetizar a GAARlandia como conexi6n continua (minima selectividad) la coloca en franca vulnerabilidad te6rica al tener que afrontar la objeci6n clasica al puente terrestre (baja diversidad ordinal de la
311 POBLAMIENTO CONTINENTE-TSLA
fauna) al tiempo que se la priva de ser la hip6tesis mas robusta para explicar -como guimalda de islas- la colonizaci6n de las Antillas por mamiferos procedentes de Suramerica
ELARcA DE NOE (seudovicarianza)
El primer vicarianista antillano (desde luego sin el saberlo) fue Corral (1940) con su utilizaci6n ingenua de las ideas wegenerianas de la deriva continental aparecidas la decada anterior Pero el primer (supuesto) intento de aplicar a la formaci6n de la biota caribefia el mecanismo de la vicarianza basada en la tearia de la tect6nica de placas (surgida en la sexta decada del pas ado siglo) se debe a Rosen ([1976] 1985)
Segun Rosen las Antillas Mayores se originaron en el Cremcico en la Placa Caribe aproximadamente en la posici6n que hoy dia ocupa Centroamerica y al ser desplazadas tectonicamente hacia el este interactuaron (de algun modo no precisado por Rosen) con las margenes continentales de Norte- y Suramerica de donde (en un evento unico) adquirieron toda su fauna transportada subsecuentemente por dichas islas en su transito hacia sus posiciones actuales [De ahi la denominacion metaforica dada par McKenna (1973) a este mecanismo de poblamiento] Aunque se ha presentado esta propuesta como Model0 de Vicarianza Continente-Isla de Rosen en realidad por definici6n no constituye un evento vicariante porque las proto-Antillas Mayores no resultaron del fraccionamiento y separaci6n por tectonismo de una porcion del territorio continental como exige esta hip6tesis (vease definici6n de vicarianza al comienzo de Hipotesis Biogeograficas)
El Arca de Noe de Rosen ha sido desestimada por la mayoria de los autores contemporaneos porque (1) carece de sustento geol6gico (lturralde-Vinent y MacPhee 1999) y porque (2) en el periodo en que se afrrrna ocum6 el evento muchos de los ordenes modemos de mamiferos placentarios attn no existian (Williams 1989)
EL BARCO FUNERAL VlKINGO (tanatovicarianza)
Domning et al (1997) reportaron 10 que consideraron el mamffero terrestre mas viejo de las Antillas --circunstancia que se refleja en el titulo mismo del trabajo Oldest West Indian Land Mammal- al descubrirse un perisodactilo rinocerotoide (Hyrachius sp) en una localidad de edad Eoceno Inferior a Medio en el occidente de Jamaica
Jamaica ha tenido una historia geologica y paleogeografica distinta de la historia comun a las restantes Antillas Mayores A principios del Cenozoico Jamaica occidental (a diferencia de la porcion oriental constituida por el Bloque de las Montafias Azules) formaba parte de Centro america de la cual se desgaj6 (por el desplazamiento de la Placa Caribe) para venir a integrarse despues del Eoceno Medio con 10 que serfa la futura isla de Jamaica desaparecida por transgresi6n marina en el Eoceno Superior hasta volver a emerger totalmente en el Mioceno Medio (Iturralde-Vinent y MacPhee 1999 MacPhee e Iturralde-Vinent 2000)
312 BIOGEOGRAFiA
Por consiguiente Hyrachius ---conocido de localidades de edad Eoceno Inferior a Medio en Norteamerica- debio ocupar (10 que iba a ser) Jamaica occidental mucho antes de que esta se separara de Centroamerica De hecho este fosil Uego a su destino insular contenido en las rocas de su basamento no como un ser vivo por 10 que es erroneo considerarlo un animal antillano (lturralde-Vinent y MacPhee 2004) como han supuesto algunos (Hedges 200 I Portell et al 200 I Condis [200 I D Todo 10 que hizo fue viajar tanatovicariantemente en su barco funeral vikingo -otra de las denominaciones metaforicas de McKenna (1973)
DISCUSION Y CONCLUSIONES
Hemos visto que solo disponemos de una hipotesis plausible para explicar la mayor parte del poblamiento (continente-isla) de las Antillas Mayores por mamiferos terrestres del Neotropico continental la dispersion mixta (a traves de GAARlandia) para los perezosos (excepto Choloepodinae) los monos y los roedores (excepto Oryzomys)
Por otra parte la historia biogeogrMica de las musaraiias de las Antillas Mayores (Nesophontes y Solenodon) constituye una de las situaciones mas enigmaticas entre los mamiferos de la region (Ottenwalder 2001321) Al menos cinco mecanismos de poblamiento continente-isla han sido invocados para explicar cuando como y de donde llegaron a las Antillas Mayores estas musaraiias
I EI propagulo ancestral (del Terciario de Norteamerica) coloniz6 a las Antillas Mayores por dispersion marina desde Centroamerica 0 desde la Florida (Patterson 1962)
2 El propagulo ancestral ( del Terciario) cruzo el Atlantico Sur (supuestamente mas estreshycho en aquella epoca) directamente de Africa a las Antillas Mayores (Hershkovitz 1972)
3 El propagulo ancestral (del Terciario de Norteamerica) Uego a las Antillas Mayores en el Arca de Noe de Rosen (MacFadden 1980)
4 El propagulo ancestral (del Terciario de Norteamerica) fue transportado a las Antillas desde Centroamerica en el bloque de Jamaica occidental y de alli paso por dispersion marina a las restantes Antillas Mayores (Whidden y Asher 200 I)
5 El propagulo ancestral (del Terciario de Norteamerica) lleg6 a las Antillas Mayores a traves de GAARlandia (Whidden y Asher 2001)
Aunque hay consenso entre estos (y otros) autores sobre el origen norteamericano de las musaraiias antillanas ninguno de los cinco hipoteticos mecanismos de poblamiento resiste la evidencia en contrario filogenetica yo paleogeogrMica Ademas algunas de estas hipotesis asumen que Nesophontidae y Solenodontidae comparten un antecesor comun en las Antillas pero esta relacion no ha sido confirm ada Para dilucidar el enigmatico caso de las musaraiias antillanas se requerini profundizar en la investigaci6n geologica paleogeognifica y paleontol6gica (Iturralde-Vinent y MacPhee 1999)
De acuerdo con el conocimiento filogenetico disponible sobre el conjunto de la fauna la colonizacion original de las Antillas Mayores por mamiferos terrestres implicaria el arribo
313 POBLAMIENTO ISLA-ISLA
de solo siete propagulos fundadores perezosos 2 (coloepodinos y megalocninos si como parece tienen un origen difiletico) musaraiias 2 (nesofontidos y solenodontidos si no constituyen una misma familia como consideran algunos) monos 1 (pitecidos) roedores histricognatos 1 (equimidos capromidos y heptaxodontidos) roedores muridos 1 (Oryzomys en Jamaica)
POBLAMIENTO ISLA-ISLA
La distribucion de los mamiferos terrestres a traves del area antillana demarcada al inicio del presente capitulo (vease Contexto Geogratico Fig 41) ha sido interpretada en funcion de dos hipoteticos mecanismos de poblamiento interinsular la dispersion y la vicarianza fundamentadas como se resumen seguidamente
DISPERSION
De acuerdo con la evidencia paleogeogratica mas actualizada en las principales regresiones marinas ocurridas en la cuenca caribeiia durante los ultimos 23 Ma (transiciones OligocenoshyMioceno Mioceno Superior - Plioceno y varias veces en el Pleistoceno) las masas terrestres constitutivas de la region insular en formacion aumentaron considerablemente su extension territorial disminuyendo por tanto la distancia entre elias oportunidades que los elementos faunales aprovecharian para dispersarse de unas islas a otras
Durante mucho tiempo esta ha sido la hipotesis mas favorecida por los estudiosos del tema a partir de Simpson (1956) pero en decadas recientes ha suscitado dos objeciones
1 Es dificil en tender como pudo operar la dispersion marina entre estas islas puesto que ninguna de las Antillas Mayores posee rios suficientemente caudalosos para transportar masas significativas de vegetacion flotante (MacPhee 198432)
Es oportuno recordar aqui el islote ala deriva (con mas de diez palmas reales en pie) entre las tres Antillas Mayores mas occidentales mencionado anteriormente (vease La Balsa Natural)
2 La dispersion marina es menos efectiva para explicar la asombrosa similitud de las faunas de mamiferos terrestres a traves de las Antillas Mayores septentrionales porque como proceso aleatorio no debe producir distribuciones uniformes a 10 largo de una cadena de islas 0 si las produce entonces el proceso no es aleatorio (MacPhee e lturralde-Vinent 2000 149)
Independientemente de que las aludidas distribuciones taxonomicas estan lejos de ser uniformes 0 asombrosamente similares la hipotesis de la dispersion marina contrariamente a su canicter aleatorio y altamente improbable en el caso del poblamiento continente-isla- en el caso del poblamiento isla-isla es un proceso no aleatorio (aprovecha las oportunidades brindadas puntualmente por los periodos de maxima regresion marina) y ademas altamente probable dada la incomparablemente menor distancia entre las islas caribeiias que entre estas y los continentes vecinos
314 BIOGEOGRAFiA
V ICARIANZA
La neotectonica y las transgresiones marinas del Oligoceno Medio y del Mioceno afectaron sustancialmente la paleogeografia de la Cresta de las Antillas Mayores resultando la subdivision gradual de las partes subaereas de la Cresta Asi el este de Cuba y el norte de La Espanola conectados fisicamente durante el Oligoceno Inferior se separaron al formarse el Paso de los Vientos en el Oligoceno tardio en tanto La Espanola y Puerto Rico probablemente permanecieron conectadas hasta el Mioceno tardio en que surgio el Paso de Mona (Iturralde-Vinent y MacPhee 1999)
Tales eventos geologicos habrian dividido las distribuciones de los grupos faunales emplazados previamente en el territorio continuo de las tres islas y a partir de esas disyunciones dichos autores (tambien MacPhee et al 2000 y White y MacPhee 2001) asumieron --como segunda fase del modelo de GAARlandia- la hipotesis de la vicarianza isla-isla para explicar la distribucion multiinsular de varios grupos de mamiferos terrestres a traves de las Antillas Mayores septentrionales De acuerdo con esta hipotesis cualquier diferencia en composicion faunal entre las islas (por ejemplo la ausencia de roedores heptaxodontidos en Cuba 0 de monos platirrinos en Puerto Rico) constituye evidencia de extincion local postvicariante y no de dispersion marina fallida Lo primero se puede demostrar si aparecen los fosiles pertinentes 10 segundo no se puede demostrar (MacPhee e Iturralde-Vinent 1994) Por otra parte IturraldeshyVinent y MacPhee (1999 57) han reconocido que aunque fa vicarianza isla-isla puede representar una solucion clara para las distribuciones de varios taxones cuaternarios no puede explicar todos los casos
Rega et al (2002 18) han presentado dos objeciones al modelo de la vicarianza isla-isla aplicado a los mamiferos de las Antillas Mayores
I Si la separacion de Cuba y La Espanola precedio en varios mill ones de anos la separashycion de La Espanola y Puerto Rico el modelo vicariante de lturralde-MacPhee debe predecir mayor similitud entre las faunas de La Espanola y Puerto Rico que entre las de Cuba y La Espanola De hecho es al reyes
No obstante MacPhee et al (200335) admitieron despues que la fecha mas temprana empiricamente defendible [para la apertura del Paso de Mona] debe ser el Oligoceno Medio esto invierte la secuencia de ambas disyunciones con 10 cual queda resuelta la aparente contradiccion Sin embargo en su reciente evaluacion de la congruencia entre filogenia e hipotesis geologic as en varios grupos de mamiferos terrestres de las Antillas Mayores Davalos (2004373) encontro que ninguno de los taxones cumple con la fecha y el patron de divergencia entre las masas terrestres antillanas postulados por el modelo de GAARlandia
2 La presencia de un mismo genero en las tres Islas septentrionales de las Antillas Mayores implica [por el modelo vicariante] que la diferenciacion entre estas formas durante el intervalo desde el Mioceno ha sido unicamente de nivel especifico [Dicha presencia]
315 rRRADlAcr6~ EVOLUCrONARIA
s610 puede explicarse razonablemente como el resultado de movimientos de esos animales entre las islas en el Cuaternario
De esta objecion resulta dificil disentir [El poblamiento entre islas diferenciado solo a nivel especifico no es probable que sea anterior al Pleistoceno (Simpson 1956 15)]
Debe tenerse en cuenta que la hipotesis de la vicarianza isla-isla alude a las tres islas septentrionales de las Antillas Mayores porque como se explico antes (vease El Barco Funeral Vikingo) Jamaica no existia como isla durante el lapso (Eoceno Superior shyOligoceno) consensualmente reconocido como el mas probable para la colonizacion de las proto-Antill as Mayores por la mayo ria de los mamiferos terrestres Despues de emerger como isla en el Mioceno Medio Jamaica debe haber recibido sus mamiferos terrestres por dispersion isla-isla (Davalos 2004376) y solo en el Cuaternario parece haberrecibido (por dispersion marina desde Centroamerica) al roedor murido Oryzomys
Seguramente tanto la dispersion como la vicarianza contribuyeron a que los mamiferos terrestres se irradiaran a traves de las Antillas Mayores y grupos de islas perifericas pero las maneras y tiempos en que estos mecanismos operaron a 10 largo de dicho proceso son aun en gran medida motivos de desacuerdo e incertidumbre
IRRADIACION EVOLUCIONARIA
A continuaci6n revisamos someramente las distribuciones conocidas de los diferentes grupos de mamiferos terrestres a traves de las Islas principales de las Antillas Mayores y grupos de Islas perifericas incluidas las interpretaciones y conjeturas que la literatura reciente recoge en relaci6n con la manera en que dichas distribuciones pudieron haberse conformado durante la irradiaci6n evolucionaria de los referidos grupos de mamiferos Manifestaciones de esta irradiaci6n pueden inferirse unicamente a partir de los taxones presentes en el Cuatemario pues solo 6 de las especies conocidas corresponden al Terciario
GRUPOS ORlGINARIOS DE SURAMERICA
PEREZOSOS (Megalonychidae)
Aparte de un genero miocenico de Cuba y otro del Oligoceno de Puerto Rico se conocen 4 generos (12 especies) de perezosos cuaternarios (todos extintos) restringidos a las tres islas septentrionales de las Antillas Mayores Acratocnus Neocnus Megalocnus y Parocnus Los cuatro generos estan representados en Cuba y La Espanola y Acratocnus 10 esta ademas (como unico genero de perezoso) en Puerto Rico Esta distribuci6n ha sido interpretada por MacPhee et al (2000) eomo resultado de la vicarianza oligomiocenica entre estas islas y [dado que el Banco Puerto Rico - Islas Virgenes fue la entrada de GAARlandia a las Antillas Mayores] se preguntaron si la ausencia de los otros tres generos en Puerto Rico se debe a que nunca estuvieron en ese territorio 0 a que sus
316 BIOGEOGRAFiA
fosiles atm no han sido hallados Pero como hemos visto Rega et al (2002) atribuyeron dicha distribucion a la dispersion interinsular en el Cuatemario
Por otra parte varios amilisis c1adisticos recientes ubi can a Choloepus -genero Reciente conocido solo de Centro- y Suramerica- como taxon hermano de Aeratoenus exc1uidos los demas megaloniquidos antillanos Para White y MacPhee (2001228)
tan estrecha relacion sugiere que la divergencia entre ambos [generos] ocurrio despues de que Aeratoenus divergio de Neoenus Si esta relacion implica que Aeratoenus y Neoenus se dispersaron separadamente desde Suramerica a las AntilIas mientras Choloepus permanecia en el continente (siendo necesarias al menos tres invasiones de perezosos) 0 si Choloepus invadio Suramerica desde las Antillas es algo que el material disponible no permite determinar
La diversidad adaptativa de los perezosos se manifiesta en (1) la utilizacion del habitat especies arboreas en Neoenus y Aeratoenus y terrestres en Paroenus y Megaloenus (Webb 1985a White [1993] White et al 1996 Arredondo [1999]) y en (2) la gama de tallas corporales desde los pequenos Neoenus hasta Megaloenus rodens el mayor de los mamiferos terrestres antillanos (su peso adulto estimado en 270 kg seglin Paula 19679) esto sugiere que los perezosos se irradiaron a nichos que en contextos continentales probablemente estarian ocupados por miembros de otros c1ados (MacPhee e IturraldeshyVinent 2000)
No esta claro si el abanico de tamanos entre estos perezosos enmascara alguna tendencia al gigantismo Por 10 pronto Anderson y Handley (2002) encontraron 10 contrario en perezosos (Bradypus) de las Islas de Bocas del Toro (Panama) shyseparadas secuencialmente del area continental vecina por ascensos del nivel del mar durante los ultimos 10 ka En las cinco islas los perezosos son menores que su contraparte continental y las diferencias entre las tallas medias de las cinco poblaciones expresan una reduccion lineal significativa de la talla corporal en funcion de la edad geologica de cada isla sin relacion alguna con el area de las islas 0 la distancia que las separa del continente
MONOS (Pitheciidae)
Los primates de las Antillas Mayores (cuatro especies extintas) constituyen un grupo monofiletico con Paralouatta (en Cuba) y Antillothrix (en La Espanola) como taxones hermanos seguidos en la escalera c1adistica por Xenothrix (en Jamaica) y finalmente Callieebus como el pariente continental mas cercano esto implica una sola invasion desde Suramerica antes del Mioceno Inferior (fecha mas temprana conocida para los primates fosiles antillanos) y por tanto en consonancia con GAARlandia como la via de colonizacion (Horovitz y MacPhee 1996 1999 Horovitz [1997] MacPhee y Horovitz 2004)
En desacuerdo parcial con la anterior interpretacion la filogenia de este grupo de monos obtenida por Davalos (2004) indica un antecesor comun en Cuba La Espanola y Jamaica
317 TRRADIACTON EVOLUCIONARIA
compatible con la hipotetica conexion -propuesta tentativamente por Iturralde-Vinent y MacPhee (199980-84)- entre las Montanas Azules del este de Jamaica y la peninsula suroccidental de La Espanola en la epoca de GAARlandia
A pesar de que los primates antillanos pertenecen a un grupo viviente de monos muy pequenos (los monos titles) fueron comparativamente grandes en relacion con los platirrinos modernos y sucesivamente mayores en la direccion Antillothrix Xenothrix y Paralouatta con P marianae como el mayor platirrino conocido del Mioceno neotropical (MacPhee 2005) MacPhee et al (1994) consideraron que Xenothrix debio ser un mono de andar lento y al parecer las especies de Paralouatta abandonaron la arborealidad para adoptar un modo de vida mas terrestre (MacPhee 1996a MacPhee et al 2003 MacPhee y Meldrum 2006)
ROEOORES (Echimyidae Capromyidae Heptaxodontidae)
La edad de Zazamys en Cuba demuestra que la invasion de los roedores histricognatos en las Antillas Mayores comenzo antes del Mioceno Inferior por otra parte los representantes mas antiguos de esa irradiacion evolucionaria se encuentran en La Espanola y Puerto Rico Ambas evidencias son consistentes con la hipotesis de GAARlandia (Woods 1982 1988 Woods et al 2001)
No esta claro si los capromidos evolucionaron a partir de un equimido ancestral 0 de un protocapromido 0 si los equimidos antillanos (I sub familia 3 generos y 7 especies todas extintas) forman parte de la irradiacion de los capromidos (4 subfamilias 9 generos y 27 especies 10 vivientes) Kratochvil et al (1978b) y Rodriguez et al (1979) consideraron que Capromys nan us es un eslab6n entre Echimyidae y Capromyidae pero Mesocapromys nanus esta morfologicamente mucho mas cerca de Hexolobodon y los de mas capromidos que de los equimidos (Woods 1985) Lo que si esta claro segun Woods et al (2001) es que la subfamilia Plagiodontinae endemic a de La Espanola es el linaje basal de los capromidos y que Isolobodon Hexolobodon Capromys Geocapromys Mysateles y Mesocapromys [y supuestamente Macrocapromys] forman parte de la irradiacion temprana de Capromyidae
La subfamilia Capromyinae (5 generos y 17 especies 9 vivientes) parece ser el dado de mas reciente evolucion en la familia con invasion de Cuba desde La Espanola y dispersion desde Cuba (1) por Capromys a las islas Caiman y (2) por Geocapromys a las islas Bahamas Caiman y Jamaica y desde Jamaica a las islas Swan aunque la presencia de Geocapromys en Swan pudiera deberse a introduccion aborigen (Morgan 1985 1988 1989 1993 1994 Woods 1989a 1990) Woods et al (2001) utilizaron datos moleculares para obtener estimados independientes de divergencia entre capromidos segiJn 10 cual Geocapromys divergio cerca de los 17 MaAP Mesocapromys divergio de Capromys hace 15 MaAP en tanto lvlysateles 10 hizo hace 10 MaAP Por su parte las formas de Capromys pilorides y de Mysateles prehensilis en Isla de la Juventud divergieron de sus correspondientes subespecies nominativas en la isla principal a los 24 y 08 MaAP respectivamente
318 BIOGEOGRAFiA
Woods y Howland (I979) describieron adaptaciones morfologicas de la mandibula el hioide y la faringe asociadas a mecanismos de masticacion diferentes en plagiodontinos y caprominos y Varona (1980a) llamo la atencion sobre el gran desarrollo de los molariformes (en relacion con la longitud total del cnineo) que presentan ciertos grupos de jutias fosiles en comparacion con las vivientes 10 que pudiera interpretarse como una adaptacion alimentaria a una vegetacion mas abrasiva por parte de las jutias extinguidas
Segun Woods (1990) la familia Heptaxodontidae (2 subfamilias 5 generos y 6 especies todas extintas) tambien seria parte de la irradiacion de los capromidos con origen en el este de La Espanola 0 en Puerto Rico y subsecuente dispersion al Banco de Anguila - San Martin en las Antillas Menores (pero vease MacPhee y Flemming 2003) Este banco estaria a solo 70 km del Banco Puerto Rico - Islas Virgenes en tiempos de maxima regresion marina y esta proximidad con Puerto Rico -patria de Elasmodontomys obliquus- seguramente tuvo que ver con la presencia en las islas de Anguila y San Martin del heptaxodontino gigante Amblyrhiza relacionado mas estrechamente con los roedores de las Antillas Mayores que con los del resto de las Antillas Menores (Woods 1990 1996) Aunque los roedores suelen aumentar de tamano en las islas (Case 1978) la escala del incremento de la talla en Amblyrhiza (que pudo alcanzar mas de 200 kg de peso adulto) esta fuera de toda proporcion con los patrones conocidos en otros roedores insulares (Biknevicius et 01 1993)
Clidomys (Clidomyinae) de Jamaica difiere marcadamente en su morfologia de los heptaxodontinos (vease ademas MacPhee y Flemming 2003)10 cual dificulta explicar su presencia en esa isla Como Jamaica estuvo bajo el mar hasta el Mioceno Medio el ancestro de Clidomys colonizaria esta isla por dispersion desde el extremo suroccidental de La Espanola despues que los heptaxodontinos se originaron en esta ultima isla (Woods 1989a) Pero Pascual et 01 (I 990) especularon que Clidomys pudiera ser el resultado de una invasion posterior e independiente de los capromidos
GRUPOS ORIGINARIOS DE NORTEAMERICA
El origen de las musaranas antillanas ha resultado ser hasta ahora el mayor enigma biogeografico de la fauna de mamiferos terrestres de la region aunque la mayoria de los autores coinciden en considerarlas originarias de Norteamerica (vease Discus ion y Conclusiones en Poblamiento Continente-Isla) No obstante MacPhee e Iturralde-Vinent (1994) descartaron toda posibilidad de conexion terrestre por el Estrecho de la Florida 0 por el Canal de Yucatan y si el (supuesto) evento vicari ante de Rosen (vease El Arca de Noe) -invocado por MacFadden (1980) para resolver el enigma- fuera minimamente plausible aun asi es dificil creer que las catastrofes que 10 sucedieron (vease Contexto Temporal) hayan barrido con toda esa vieja fauna continental excepto las musaranas circunstancia ya anticipada muy tempranamente por Matthew (l918a)
De otra parte la relaci6n entre Nesophontidae y Solenodontidae permanece irresuelta Si estos linajes no estan estrechamente relacionados entonces es muy posible que
319 IRRADIACION EVOLUCIONARIA
sus distribuciones sean explicadas por mecanismos diferentes (Whidden y Asher 2001)
La descomunal diferencia de talla corporal entre las musaranas pequenas (Nesophontes) y las grandes (Soienodon) seguramente impuso a ambos grupos restricciones distintas relativas a la termorregulacion y las adaptaciones energeticas En Nesophontes la seleccion debio favorecer una alta tasa de metabolismo y crecimiento camadas mas numerosas corta expectativa de vida yalta densidad poblacionaL En Soienodon en cambio la seleccion opero en la direccion de una baja tasa metabolica camadas reducidas crecimiento lento longevidad y baja densidad poblacional (Ottenwalder [1991])
MUSARANAS PEQUENAS (Nesophontidae)
Las musaranas pequenas estan representadas en las Antillas por el genero Nesophontes (y un genero miocenico indeterminado en ambar dominicano) con 6 especies cuaternarias extinguidas distribuidas en Cuba La Espanola Puerto Rico Islas Virgenes e Islas Caiman La mayor de las musaranas pequenas Nesophontes edithae evoluciono en Puerto Rico y su arribo a las Islas Virgenes supone una dispersion sumamente reciente En las Islas Caiman Nesophontes se asemeja mas a la poblacion de Cuba de donde seguramente derivo en el Plio-Pleistoceno (junto con Capromys y Geocapromys) cuando la costa sur de Cuba estuvo mucho mas cerca de las Islas Caiman (lturralde-Vinent 2003fig 4) MacPhee et ai (1994) interpretaron la distribucion de Nesophontes en Cuba y La Espanola como resultado de la vicarianza oligomiocenica ocurrida al formarse el Paso de los Vientos pero la distancia taxonomica entre los lados del Paso es solo de nivel especifico y N micros incluso vivi6 en ambas islas segtln Patterson (1962)
Allen (1918) considero que Nesophontes micrus fue una especie primitiva no modificada para la vida arb6rea 0 fos6rea y por tanto de habitos terrestres Nesophontes edithae de Puerto Rico es mucho mayor y mas robusta que las restantes especies del genero eL indice de robustez de su femur (= anchura minima de la diafisis ongitud total del hueso) muestra que este es tan robusto como el de Solenodon paradoxus un animal dos veces mayor (Morgan et ai 1980) Choate y Birney (1968) postularon un aumento cronoclinal de la taIla corporal en N edithae confirmado mas tarde por McFarlane (1999a) mediante fechados radiocarb6nicos de la secuencia estratignifica en depositos cavernarios de Puerto Rico Solenodon no vivi6 en Puerto Rico y en ausencia de una musarana de mayor tamano Nesophontes alcanzo una talla corporal mayor en esa isla que en La Espanola 0 Cuba (Woods y Eisenberg 1989)
MUSARANAS GRANDES (Solenodontidae)
Los arcaicos almiqufes (genero Soienodon) comprenden cuatro especies una viviente (S cubanus) y otra f6sil (S arredondoi) en Cuba y una viviente (S paradoxus con dos subespecies) y otra f6sil (S marcanoi) en La Espanola Como cabria esperar las dos especies cubanas se asemejan mas entre sf que a las especies de La Espanola pero
320 BIOGEOGRAFiA
enigmaticamente S marcanoi comparte caracteres con S cuban us y S paradoxus (Woods y Eisenberg 1989 Morgan y Ottenwalder 1993 Ottenwalder 200 I) Solenodon arredondoi alcanzo una talla corporal superior a la de cualquiera de las musarafias vivientes a nivel mundial Segun Morgan y Ottenwalder (1993) extrapolando las dimensiones y el peso de los almiquies vivientes S arredondoi debio tener una longitud cabeza-tronco y un peso de 450-550 mm y 1500-2000 g respectivamente en comparacion con S cubanus (360 mm y 800 g) y S paradoxus (390 mm y 1100 g)
Entre todas las especies de almiqufes Solenodon paradoxus es la que presenta los caracteres mas derivados (Otten walder 2001) Si estas relaciones son validas la distribucion de Solenodon entre Cuba y La Espafiola debio discurrir de oeste a este como propuso MacFadden (1980) quien invoco la presencia fosH de S cubanus [pero tambien S arredondoi] en Pinar del Rio para postular que el ancestro continental entr~ por Cuba opinion compartida por Perfit y Williams (1989) y por Ottenwalder ([1991] 2001) El paso de Solenodon a Puerto Rico pudo haberlo impedido la existencia de barreras ecologicas durante la conexion del este de La Espafiola con Puerto Rico 0 por la ausencia de Solenodon en esa parte de La Espafiola en aquella epoca Sin embargo las mismas condiciones ecologicas en el este de La Espafiola no parecen haber representado una barrera efectiva para Nesophontes cuyas mas altas tasa de reproduccion y densidad poblacional presumiblemente Ie permitieron expandirse flpidamente a traves de La Espafiola (Ottenwalder [1991])
Es opinion generalizada que los almiqufes antillanos constituyen el grupo de mamiferos que primero se emplazo en el territorio insular (MacPhee 2005) y en tal sentido es de interes la evidencia sobre especificidad parasito-hospedante aportada por Cruz (1980) quien encontro que la especie de acaro (genero Cubanochirus) que parasita a los almiquies es mucho mas primitiva que la especie (del mismo genero) que paras ita a las jutias y de aqui concluyo que dicho genero de acaro debio evolucionar primero en Solenodontidae y posteriormente en Capromyidae al entrar en contacto representantes de ambas familias en Cuba 0 La Espafiola
Recientemente Roca (2004) y Roca et al (2004) dieron a conocer que los resultados de su estudio genetico-molecular sobre los almiquies indican una divergencia de 25 Ma entre Solenodon cuban us y S paradoxus -compatible con la disyuncion tectonica entre Cuba y La Espafiola (Iturralde-Vinent y MacPhee 1999 McPhee et al 2003)-- y una separacion de ambas especies comparable en magnitud a la divergencia entre familias de mamlferos Consecuentemente propusieron que Solenodon cubanus fuera clasificada en el genero Atopogaie como indico Cabrera (1925) hace 82 afios
DISCUSION Y CONCLUSIONES
Las musarafias y los roedores antillanos (excepto Oryzomys) se irradiaron tanto de sus respectivos ancestros continentales que de estos 6rdenes se reconocen familias y subfamilias endemicas de la region en contraste con los perezosos (una subfamilia y tres
321 IRRADIACION EVOLUCIONARIA
tribus endemicas) y los monos (una tribu endemica) como expresion de diferentes niveles de diversificacion evolucionaria Este proceso es dificil de explicar (traUindose de c1ados que pnicticamente comparten la misma antigiiedad en las Antmas) si es correcta la interpretacion de Pascual et at (1990636)
Nosotros consideramos estos diferentes niveles de diversificacion como las diferentes etapas de evolucion alcanzadas por cada grupo [de mamiferos] Obviamente esto estuvo relacionado con las diferentes longitudes de sus historias
Pudiera ocurrir que en el futuro se compruebe que la historia biogeognifica de la region fue mucho mas complicada
La Espanola la isla mas ecol6gicamente divers a de la region debi6 ser el centro principal de evoluci6n de los mamfferos (con 87 de las familias y 73 de las subfamilias) seguida por Cuba (con 75 de las familias y 54 de las subfamilias) las dos islas de las Antillas Mayores mas relacionadas biogeograficamente (Fig 43 Tabla 44)
eL
EL
cb CAIMAN
SWAN
A Acratocnini B Cubanocnini C Megalocnini D Mesocnini E Nesophontidae F Solenodontidae G Xenothrichini
ANGUILAshyM-0 SMARTIN
H Heteropsomyinae I Plagiodontinae J Isolobodontinae
K Hexolobodontinae L Capromyinae
M Heptaxodontinae N CIidomyinae 0 Sigmodontinae
FIGURA 43 Representacion esquematica de las relaciones interinsulares (expresadas porel numero de linajes compartidos) entre las Antillas Mayores y entre estas y los grupos de islas perifericas segun las interpretaciones de los autores citados en el texto Los numeros dentro de los circulos indican la cantidad de linajes representados en cada isla relacion dudosa Contexto taxon6mico de los linajes en la Tabla 44
bull bull
bull bull
bull bull bull
bull bull
bull bull
bull
bull
bull
bull
bull bull bull bull
bull bull
bull bull
bull
bull bull bull
bull bull
bull
bull bull
bull
322 BIOGEOGRAFiA
TABLA 44 Distribuci6n de los linajcs de mamiferos terrestres a traves de las Antillas Mayores y grupos de islas perifericas LE = La Espanola CU Cuba JA Jamaica PR = Puerto Rico IC Islas Caiman IS Islas Swan IB = Islas Bahamas IV = Islas Virgenes AS = Anguila - San Martin
FamiliaSubfumiliafIribu LE CU JA PR IC IS IB IV AS
Megalonychidae
Choloepodinae
Acratocnini
Cubanocnini
Megalocninae
Megalocnini
Mesocnini
Nesophontidae
Solenodontidae
Pitheciidae
Callicebinae
Xenothrichini
Echimyidae
Heteropsomyinae
Capromyidae
Plagiodontinae
Isolobodontinae
Hexolobodontinae
Capromyinae
H e pta x 0 don tid a e
Heptaxodontinae
Clidomyinae
Muridae
Sigmodontinae
PILOSA
SORICOMORPHA
PRIMATES
RODENTIA
bull
bull
bull
bull TOTALES 12 10 4 4 2
323 EXTlICIOIES DEL CUATERNARIO
El gigantismo ha sido sin duda una tendencia evolucionaria recurrente entre los clados mayores de mamiferos antillanos Es un hecho bien conocido que algunos grupos de mamiferos han aumentado de tamafio en las islas y varios autores han intentado explicarlo a partir de enfoques diferentes (Foster 1964 Miller 1966 Sondaar 1977 Case 1978 Eisenberg y Wilson 1978 Heaney 1978 Lawlor 1982 Lomolino 1985 Angerbjom 1985 McNab 19942001)
EXTINCIONES DEL CUATERNARIO
Las islas que integran la Subregion Antillana han perdido casi 90 de sus mamiferos terrestres durante los ultimos milenios y en proporcion con su extension territorial esta perdida ha sido mayor en la Subregion que en cualquier otro lugar del planeta (MacPhee 1996b) Las extinciones del Cuatemario eliminaron mas de las tres cuartas partes de la fauna de mamiferos terrestres conocida de las Antillas Mayores sus islas satelites y grupos de islas perifericas y el examen de este proceso en sus aspectos mas generales se aborda en esta ultima parte del presente capitulo
CRONOLOGiA Y CONSECUENCIAS
Es muy improbable que un fosil represente el ultimo individuo de un taxon extinto y entonces hay que guiarse por la edad radiometrica del fosil mas reciente conocido lIamada fecha de ultima presencia- como unica aproximacion posible a la fecha de extincion del taxon
Como se explico al inicio del capitulo (vease Contexto Temporal) el registro fosH de los mamiferos terrestres de las Antillas Mayores adolece de insuficiente sustentacion radiometrica No obstante la distribucion temporal de los escasos fechados disponibles para el Cuaternario de la region abarca etapas criticas del proceso de extincion experimentado por la referida fauna a 10 largo de los ultimos 250 mil afios
De las 52 especies que es posible reconocer actualmente como miembros extintos de la fauna cuatemaria de mamiferos terrestres de las Antillas Mayores y grupos de islas perifericas los restos fosiles de 23 de elIas (442) han sido fechados radiometricamente o se han hallado asociados estratigraficamente de manera inequivoca con otro material fechado De estos 23 fechados 3 corresponden al interglacial Sangamon 4 pertenecen a la secuencia Pleistoceno terminal - Holoceno temprano 10 se ubican en ellapso entre el arribo de los aborigenes y la incursion europea y 6 son postcolombinos (a los cuales hay que sumar otras dos especies extinguidas en los siglos XIX y xx) A partir de estas 25 especies avaladas por fechas de ultima presencia distribuidas entre los cuatro intervalos temporales sefialados hemos compuesto la Tabla 45 en la que ademas aparecen las restantes 27 especies (carentes de fechado radiometrico) ubicadas en el intervalo temporal mas probable sobre la base de (1) la asociacion estratigrafica de sus restos con evidencias culturales (amerindias 0 europeas) 0 (2) la ausencia de dicha asociacion Sobre estos criterios de ubicacion temporal procede hacer dos aclaraciones
324 BIOGEOGRAFIA
TABLA 45 Intervalos temporales mas probables para las extinciones cuatemarias de los mamiferos terrestres de las Antillas Mayores y grupos de islas perifericas sobre la base de las evidencias siguientes (l) ultimo ejemplar colectado u observado en vida (2) fechado radiometrico mas tardio en material de la especie (3) asociaci6n estratigrafica mas tardia con otro material fechado (4) asociaci6n estratigrafica con restos de animales introducidos restos humanos u objetos culturales propios del intervale (5) falta de asociaci6n con evidencias culturales [Los ntimeros entre parentesis en la columna Evidencia significan adscripci6n tentativa al intervalo] Los estimados del peso corporal (en kg) estan tomados de la literatura 0 inferidos comparativamente y distribuidos entre
las clases siguientes (a) S 01 (b) 01 a lt 1 (c) 1 a lt 10 (d) 10 a lt 100 (e) 100 a lt 200 (j) ~ 200 Las
abreviaturas que designan las islas son las de la Tabla 44
F amilialEspecie Peso IsJa Evidencia Fuente
I httervalo htterg1acial Sangamon (gt80 kaAP)
HEPTAXODONTlNAE
Amblyrhiza inundata e AS 3 McFarJane et al 1998a
CLIDOMYINAE
Clidomys osborni d JA 3 Ibid
Clidomys parvus d JA 3 Ibid
II httervalo Pleistoceno terminal ~ Holoceno temprano (20-6 kaAP)
MEGALONYCHIDAE
Aeratoenus antillensis d CD 5 Arredondo 1975 a
Aeratoenus odontrigonus d PR 3 McFarJane 1999b
Aeratoenus simorhynehus d LE 2 Rega et al 2002
M egaloenus zile f LE 2 MacPhee et al 2000
SOLENODONTIDAE
Solenodon arredondoi e CD 5 Morgan y Ottenwalder 1993
PnHECIIDAE
Paralouatta varonai d CD 5 MacPhee 1996a
EcHIMYIDAE
Heteropsomys antillensis c PR 5 McFarJane 1999b
Heteropsomys insulans c PR 5 Ibid
Puertoricomys eorozalus c PR 5 Woods 1989a
CAPROMYIDAE
P lagiodontia araeum c LE 5 MacPhee et al 2000
Macrocapromys latus c CD 5 Varona y Arredondo 1979
(Continua)
325 EXTINCIONES DEL CUATERNARIO
TABLA 45 (Continuaci6n)
F ami1ia1Especie Peso Isla Evidencia Fuente
M acrocapromys latus c CU 5 Varona y Arredondo 1979
Mesocapromys kraglievichi c CU 5 Ibid
HEPTAXODONTINAE
Xaymaca fulvopulvis a JA 2 MacPhee y Flemming 2003
III Intervalo Amerindio (6-05 kaAP)
MEGALONYCHIDAE
Acratocnus ye d LE 2 MacPhee et aI 2000
Neocnus comes c LE 2 Ibid
N eocnus dousman c LE 2 Ibid
N eocnus toupiti c LE 2 Ibid
N eocnus gliriform is c CU 4 Cordova et ai 1997
M egalocnus rodens f CU 2 MacPhee et ai 2007
Parocnus browni e CU 2 Steadman et ai 2005
P arocnus serus e LE 2 MacPhee et ai 2000
N ESOPHONTIDAE
N esophontes edithae a PR 2 McFarlane 1999b
PITHECIIDAE
Antillothrix bernensis c LE 3 Rimoli 1977
EcHIMYIDAE
Brotomys contractus a LE 4 Miller 1929c
CAPROMYIDAE
P lagiodontia spelaeum c LE 4 Ibid
M acrocapromys acevedo c CU 4 NUiiez 1990a
HEPTAXODONTINAE
Elasmodontomys obliquus d PR 3 McFarlane 1999b
Iv Intervalo Postcolombino (05 kaAP - Presente)
N ESOPHONTIDAE
N esophontes hypom icrus a LE 24 MacPhee y Fle111111ing 1999
N esophontes zam icrus a LE (4) Ibid
N esophontes m icrus a CU 24 MacPhee et aI 1999
(Continua)
326 BIOGEOGRAFiA
TABLA 45 (Continuacion)
F amilialEspecie Peso Isla Evidencia Fuente
N esophontes m icrus a CD 24 MacPhee et aI 1999
N esophontes m icrus a LE 4 MacPhee et aI 2000
N esophontes sp 1 a IC 4 Morgan 1994
N esophontes sp 2 a IC 4 Ibid
Solenodon marcanoi b LE 4 Ottenwalder 2001
PITHECIIDAE
Xenothrix mcgregori c JA 4 MacPhee y Flemming 1999
EcHIMYIDAE
Boromys offeUa a CD 24 Guarch 1984
Boromys torrei a CD 24 Ibid
Brotomys voratus a LE 4 MacPhee et aI 2000
CAPROMYIDAE
P lagiodontia ipnaeum c LE 4 MacPhee y Flemming 1999
Rhizoplagiodontia lemkei c LE 4 Ibid
Isolobodon portoricensis c PR 24 Ibid
Isolobodon montanus c LE 4 Ibid
Hexolobodon phenax c LE (4) Ibid
Capromys sp c IC 24 Morgan 1994
Geocapromys columbianus c CD 4 MacPhee y Flemming 1999
Geocapromys thoracatus b IS 1 MacPhee y Marx 1997
Geocapromys sp 1 c IC 4 Morgan 1994
Geocapromys sp 2 c IC (4) Ibid
l-IEPTAXODONTINAE
Quemisia gravis c LE (1) MacPhee y Flemming 1999
MURIDAE
Oryzomys antillarum a JA 1 Ibid
1 Flemming y MacPhee (1999) alertaron en contra de la asociacion con otro material fechado Asimismo Morgan (1993) indico que la ausencia de asociacion con evidencias culturales amerindias no es necesariamente un criterio concluyente para ubicar un taxon en el intervalo temporal anterior porque hay depositos en la region que carecen de tales evidencias y son del Holoceno tardio por fechados radiocarbonicos [No obstante
327 EXTlNCIONES DEL CUATERNAR10
la ausencia de una especie en residuarios arqueol6gicos adquiere mayor certidumbre como criterio de ubicaci6n temporal de su extinci6n en la medida que aumenta el numero de residuarios estudiados que en las Antillas Mayores se cuentan por centenares] A falta de fechado absoluto de una especie consideramos que estos criterios de ubicaci6n temporal son mas utiles que la alternativa de excluir del analisis a la especie
2 En los trabajos sobre biogeografia de mamiferos antillanos los arque610gos mas citados como autoridades para establecer el inicio de la presencia aborigen en las Antillas Mayores (Kozlowski 1974 Rouse y Allaire 1978 Rouse 1989 1992 Wilson 2001) difieren al respecto entre los 6 kaAP y los 45 kaAP (pero vease Pagan et al 200523) y hemos adoptado la mas temprana de estas fechas para el comienzo del intervalo amerindio en la Tabla 45 De todos modos cualquiera que haya sido la fecha esta no es aplicable por igual al conjunto de las Antillas Mayores porque segun algunos de los autores citados el poblamiento comenz6 por Cuba y La Espanola 6 mil anos atras continu6 en Puerto Rico hace alrededor de 3 mil anos (pero vease Burney et al 1994) yen Jamaica hace menos de 2 mil e incluso a las Islas Caiman ni siquiera llegaron [Las Islas Caiman descubiertas por Col6n en 1503 carecen absolutamente de evidencia amerindia (Stokes y Keegan 1996)]
Si los datos de la Tabla 45 representan una aceptable aproximaci6n a la realidad el proceso de las extinciones cuaternarias de los mamiferos terrestres de las Antillas Mayores y grupos de islas perifericas debi6 caracterizarse entre otros por los eventos siguientes
Aunque obviamente la fauna cuaternaria debi6 ser mucho mas numerosa que 10 que refleja el registro paleontol6gico (Morgan y Woods 1986) 52 especies (81 de la fauna conocida) desaparecieron (sin reposici6n) durante los ultimos 250 mil anos
- Al parecer las extinciones se concentraron en las especies de mayor tamano Asi el peso promedio estimado para las especies de la etapa preantr6pica (intervalos I + II de la Tabla 45) es 350 kg es 328 kg para las especies del intervalo III (amerindio) y es s610 25 kg (c1ases del peso a b c) para las especies del intervalo IV (postcolombino) Ninguna especie aut6ctona de mamifero mayor de 6 kg existe hoy en la regi6n
- De acuerdo con la distribuci6n temporal de las extinciones 61 especies sobrevivieron al interglacial Sangamon (intervalo 1)48 sobrevivieron a la transici6n del Pleistoceno terminal al Holoceno temprano (intervalo II) 34 sobrevivieron al periodo amerindio (intervalo III) y s610 12 especies han sobrevivido desde la Ilegada de los europeos (intervalo IV) Las tasas de extinci6n correspondientes (exceptuado el intervalo I) son una especie cada 1167 anos en el intervalo II una cada 393 anos en el intervalo III y una cada 23 anos en el intervalo IV
- Durante todo el proceso de las extinciones cuaternarias la perdida de especies por isla fue de 91 en La Espanola 83 en Jamaica y 62 en Cuba Bahamas retuvo su unica especie (aunque extirpada en varias de las islas) Puerto Rico e Islas Caiman perdieron todas sus especies (6 y 5 respectivamente) y las Islas Swan Virgenes y Anguila - San Martin perdieron sus respectivas especies (micas
328 BIOGEOGRAFiA
De los cuatro 6rdenes de mamiferos representados en la fauna Pilosa y Primates desaparecieron totalmente Soricomorpha y Rodentia perdieron 86 y 74 de sus especies respectivamente
CAUSALIDAD
CAUSAS NATURALES
Los cicIos glaciales e interglaciales acaecidos en el Pleistoceno causaron cambios ambientales dnisticos en las Antillas con la concomitante alternancia de cIimas frios (secos) y calidos (hUmedos) asi como de descensos y ascensos del nivel del mar aumento y disminuci6n de las areas territoriales) induso (aunque con menor intensidad) en el Holoceno Estos eventos paleogeograficos han sido consistentemente documentados (Mayo y Kartashov 1972 Bonnatti y Gartner 1973 Ortega y Arcia 1982 Prance 1982 Schubert y Medina 1982 Dodge et al 1983 Ortega [1984] Haq et al 1987 Schubert 1989 Hodell et al 1991 Guilderson et al 1994 Oppo 1997 Emslie 1998 Curtis et al 2001 Iturralde-Vinent 2003) y de ellos brindamos a continuaci6n una breve resena
Finalizando el periodo interglacial anterior a la ultima glaciaci6n (Wisconsin) el mar estaba a 8-10 m por encima de su nivel actual Cuando el avance glacial del Wisconsin a1canz6 su culminaci6n (eillamado Ultimo Maximo Glacial) hace 18 ka el mar habia descendido 120 m con respecto a dicho nivel y las islas del Caribe habian incrementado considerablemente sus extensiones subaereas en comparaci6n con las que presentan hoy dia por ejemplo Puerto Rico y las Islas Virgenes se conectaron territorialmente y las Bahamas se unieron en cinco gran des islas Prevalecia un regimen frio y seco y condiciones de aridez --debidas a una reducci6n de las precipitaciones (posiblemente a menos de 440 mmlano)- y los ambientes xericos se habian extendido a traves de las islas y bancos emergidos y abundaban los pinares (los habitats mesicos estaban restringidos a parcelas relictas en 10 alto de las montanas) En el Mar Caribe y el Golfo de Mexico la temperatura superficial del mar era 5degC mas baja que la actual y en las llanuras costeras y alturas maximas eran respectivamente 2-3degC y 6degC mas bajas Los limites de las zonas ecol6gicas altitudinales descendieron varios cientos de metros y cada zona redujo su extensi6n vertical al aumentar el gradiente de temperatura Ellimite inferior de la zona periglacial
elevaci6n donde la temperatura media minima a1canza los OdegC) de la Cordillera Central de la Republica Dominicana (altura maxima del Caribe insular con evidencia de topografia glacial) se considera que estuvo entre los 1500 y 1800 mlnm (actualmente esta a 2800 mlnm) y se puede estimar que dicho limite era inferior en Cuba por estar mas al norte que la Republica Dominicana No obstante durante este periodo la temperatura experiment6 episodios cortos (unas pocas decadas) de ascensos repetidos
Un signifieativo componente bi6tieo de las referidas condiciones paleoambientales 10 constituy6 la espectacular irradiaci6n evolucionaria experimentada por un grupo numeroso de aves rapaees diurnas y nocturnas (Wetmore 1937 Brodkorb 1969 Kurotchkin y Mayo 1973 Arredondo 1976 Olson 1978 Olson y Hilgartner 1982
329 EXTINCIONES DEL ClJATERNARIO
Suarez 2000a b 2004 Suarez y Emslie 2003) Los mamiferos de las Antillas Mayores ofrecieron a estos depredadores aviares una amplia gama de tamanos de presa que en ausencia de mamiferos del orden Carnivora en la region permitio a estas aves irradiarse a este nicho alimentario con la evolucion de una variedad de especies de muy diversa tall a corporal incluidas formas gigantes Adquirir un cuerpo gigante les propicio el acceso a recursos alimentarios que no estaban siendo explotados por ningun otro depredador (Alcover y McMinn 1994) A Cuba corresponde el mejor registro fosil de rapaces cuaternarias caribenas con siete generos de especies diurnas Dentro de estas aves existieron tres gavilanes gigantes (Accipitridae) tres carairas (Falconidae) y un condor (Cathartidae) asi como cuatro generos de especies nocturnas dos lechuzas (Tytonidae) y tres Mhos (Strigidae) dos de ell os no igualados en tamano en todo el mundo Algunos de estos generos tuvieron representacion en otras islas antillanas como La Espanola Jamaica Bahamas y Gran Caiman
Al retirarse hacia el norte los hielos de la glaciacion Wisconsin el clima se tomo mas hUmedo en las Antillas durante el Holoceno temprano y el bosque mesico se expandio en la region insular El nivel del mar ascendio acompanado de mayor pluviosidad alrededor de los 7 kaAP las Islas Virgenes quedaron separadas entre si y de Puerto Rico y las Bahamas perdieron la mayor parte de su territorio Cerca de los 3 kaAP las condiciones secas retornaron a la region e hicieron desaparecer buena parte del bosque mesico Nuevamente el clima comenzo a adquirir sus condiciones actuales hacia los 25 kaAP aproximadamente y hoy transita por una fase mesica con vegetacion xerica confinada en parcelas relictas
Sin duda estos y otros eventos naturales incidieron negativamente en la sobrevivencia de mamiferos terrestres en las Antillas Mayores asi como en los grupos de pequenas islas perifericas (Pregill y Olson 1981 Olson 2006)
McFarlane et al (1998a) demostraron convincentemente como Amblyrhiza el gigante de los roedores antillanos endemico de la pequena isla de Anguila sucumbio a las repetidas y abruptas perdidas de su habitat durante el interglacial Sangamon por las nipidas y severas oscilaciones del nivel del mar (documentadas por Neumann y Hearty 1996) al no poder responder adecuadamente a tan catastroficos eventos mediante el ajuste de su masa corporal media y otro tanto debio ocurrirle en Jamaica al otro roedor gigante Clidomys (McFarlane et al 1998b) emparentado con Amblyrhiza en la misma epoca (Tabla 45 intervalo I)
El ascenso del nivel del mar seguramente promovio la extincion de mamiferos durante la transicion del Pleistoceno al Holoceno (Tabla 45 intervalo II) con la reduccion del tamano de las Islas por las transgresiones marinas (Pregill y Olson 1981 Morgan 1993) Esto ultimo haria maximamente severa la competicion asf como la presion de los depredadores [Steadman et al (2005) no yen por que el calentamiento global ocurrido durante la transicion Pleistoceno-Holoceno habria de tener un efecto negativo sobre los perezosos siendo sus descendientes actuales especies tropic ales 0 subtropicales pero no se trata del efecto
330 BIOGEOGRAFiA
directo del cambio de temperatura sobre los animales sino del efecto indirecto y progresivo que dicho cambio genera a largo plazo al reducir el territorio insular disponible]
Por esta epoca tambien desaparecieron especies de tall a mayor como Megalocnus zile en La Espanola y otras de menor tamano (en Cuba) pero que representan la tall a maxima en sus respectivos contextos taxonomicos del Cuaternario como Solenodon arredondoi Paralouatta varonai y Macrocapromys acevedo Un mamifero puede ser grande solo a costa de reducir su densidad poblacional y esto 10 hace mis vulnerable a la extinci6n sobre todo en una isla (McNab 1994 McFarlane et al 1998a) Incidentalmente Alcover et al (1998a) afirmaron que larelacion entre la talla corporal y la extincion no siempre se cumple en las especies de menor tamano y citaron como ejemplo que entre las musarafias todas las especies de Nesophontes desaparecieron mientras sobreviven especies de Solenodon pero la comparacion correcta en todo caso serfa entre los solenodones vivientes y el gigante Solenodon arredondoi y entre Nesophontes edithae -el gigante de los nesofontes extinguido en el periodo amerindio- y el resto de sus congeneres que sobrevivieron hasta los primeros siglos de la colonizacion europea (Tabla 45 intervalos III y IV)
CAUSAS ANTROPOGENICAS
Intervalo Amerindio
La mayoria de los autores que se han referido a las extinciones acaecidas durante este intervalo (entre elIos paradigmaticamente Martin 1984a b) han responsabilizado a los aborfgenes (que se supone comenzaron a poblar las Antillas Mayores hace 6 mil afios) con la desaparicion de numerosas especies de mamiferos terrestres en la region principalmente por la via de la sobrecaza y la introduccion de animales domesticos devenidos depredadores de la fauna autoctona (Morgan y Woods 1986) Este efecto negativo de la presencia indigena sobre los mamiferos de la region ha sido dramatizado por algunos autores en terminos de holocausto faunal Sin embargo pensamos que se ha exagerado Como han expresado MacPhee et al (19993) cuando uno busca evidencia dura de complicidad humana directa en [estas] extinciones no hay mucho que encontrar Las siguientes son algunas consideraciones a tener en cuenta en tal sentido
El registro radiometrico mas joven conocido hasta ahora para un perezoso antillano (MegalocnllS rodens) es de 42 kaAP (MacPhee et ai 2007) Si se acepta que los aborfgenes entraron en Cuba hace aproximadamente 6 mil anos su convivencia con los perezosos debe haber durado alrededor de 1800 afios (0 mas porque dificilmente el referfdo fechado radiometrico haya correspondido al ultimo sobreviviente de la especie) Esta cifra contradice la hipotesis del holocausto faunal segun la cualla extincion de los mamiferos insulares de mayor talla debe ocurrir con extrema rapidez -a escala de decadas 0 de centurias- a partir de su primer contacto con los humanos (Martin y Steadman 1999) A pesar de las repetidas afirmaciones en contrario (Harrington 1921 Miller 1929c Herrera [1945] Tabio 1951 Alvarez 1951a 1957 Arredondo
331 EXTINCIONES DEL CUATERNARIO
1959 1960a Veloz y Ortega 1973 Mayo 1980b Rodriguez et al 1984 Castellanos y Pino 1985 Pino y Castellanos 1985 Nunez 1990b Gutierrez 1996 Arredondo 2004) en su mayoria basadas en evidencias no exentas de posibles accidentes de deposicion (0 de reubicacion postdeposicional) en recintos cavernarios (Diaz-Franco [2006b D no se ha demostrado inequivocamente que los aborigenes antillanos se alimentaran de perezosos regularmente Sus restos en residuarios de alimentos son escasisimos en toda la region y esto contrasta con los abundantes restos de jutias y otros mamiferos de menor talla que aparecen en esos residuarios demostrativos de la preferencia de los paleoindios por estos otros mamiferos
Los mamiferos mas importantes en la economia prehistorica de las Antillas Mayores fueron las jutias Capromys y Geocapromys en Cuba Geocapromys en Jamaica e Isolobodon en La Espanola Puerto Rico e Islas Virgenes (Aguayo 1950b Tablo 1951 Rimoli 1972 Wing 2001 Diaz-Franco [2006b D Wilkins (2001) estudi61ajutia viviente de Jamaica (Geocapromys browni) en un sitio ocupado por los aborigenes durante 100 anos y no encontr6 evidencias de que durante ese tiempo la especie haya sido sobreexplotada Asimismo en Cuba las jutias conga carabali y andaraz son escasas en depositos pleistoholocenicos en cambio son muy numerosas en residuarios indigenas y de haber sido sobreexplotadas 10 mas probable es que no hubieran sobrevivido hasta hoy
La habilidad de los humanos para penetrar en profundidad el bosque tropical y mantenerse en este sustentablemente ha sido un evento comparativamente reciente (Beran y Steele 1994) En el caso antillano la caza tierra adentro ha sido explicada como un proceso tardio de desplazamiento a partir de la explotacion de abundantes recursos marinos que ofrecian condiciones 6ptimas para el sedentarismo de las comunidades aborigenes costeras (C6rdova y Arredondo 1988 Brito 2005) Peces y mariscos dominan la arqueologia costera de las Islas antillanas en tanto el consumo de recursos terrestres (mamfferos incluidos) aumenta progresivamente en la medida que los residuarios estudiados se alejan de la costa (Scudder 1991 Wilkins 2001) En las Antillas Mayores el componente terrestre de la dieta aborigen es solo 20-30 del material recobrado en los residuarios (Wing 1977 1989) Y debio ser menor alin porque el componente vegetal de la dieta no se ha preservado Por otra parte los aborigenes cubanos domesticaron la jutia conga y seguramente la reprodujeron para comer (Pose et al 1988) practica que probablemente se repitiera con otras jutias en las demas Antillas Mayores
- Fray Bartolome de las Casas hablo de 3 millones de indios en Cuba al inicio de la colonizacion pero los historiadores subsecuentes consideraron exagerada la cifra (Poey 1851 a) No obstante se dispone de dos estimados de la densidad de poblaci6n aborigen en las Antillas Mayores a la llegada de los europeos 18 indigenaslkm2 en Puerto Rico (Rouse 1956) y 1271km2 en La Espanola (Watts 1987) Extrapolando ambos valores entre las cuatro islas principales de las Antillas Mayores tenemos que en la fecha indicada la poblaci6n aborigen pudo estar entre 16 mil y 113 mil indigenas en Puerto Rico entre 21 mil y 145 mil en Jamaica entre 141 mil y 1 mill6n en La Espanola y entre 200 mil y 14 millones en Cuba Entre las cifras maximas y las minimas nos inclinamos
332 BIOGEOGRAFiA
hacia estas ultimas atendiendo a los altos indices de mortalidad infantil y morbilidad que se evidencian en las osamentas de los pobladores prehispanicos (Martinez y Rives 1990) Uno de los hechos que apuntan hacia el bajo numero de aborigenes en las Antillas Mayores es 10 rapido que desaparecieron despues de la llegada de los europeos pnkticamente sin dejar mestizaje con estos 10 cual contrasta ostensiblemente con la situacion en la America continentaL
En suma un grupo humano (1) demograiicamente irrelevante y controlado por altos indices de morbilidad (2) consumidor mayoritariamente de organismos marinos y (3) carente de la tecnologia cinegetica necesaria para poder sobreexplotar los mamiferos que Ie sirvieron de alimento (a los cuales accedio tardiamente en su desarrollo) no es concebible que haya causado e1 holocausto faunal que se Ie atribuye a partir del error de aplicar a las comunidades aborigenes antillanas un modelo cultural de la prehistoria continentaL
Tampoco hay evidencias de que los aborigenes hayan introducido carnivoros con suficiente capacidad depredadora salvo el perro Morgan y Otlenwaldder (1993) consideraron que la depredacion por perros durante el periodo amerindio es la explicacion mas plausible para la extincion del almiqui gigante de Cuba pero como se puede observar (Tabla 54) Solenodon arredondoi aparece en el intervalo II por falta de asociacion con evidencias culturales
Consecuentemente el accionar de los aborigenes y de sus animales domesticos directamente sobre la fauna no parece haber sido por sf solo un factor determinante de la extincion de mamiferos terrestres durante el periodo amerindio en las Antillas (pero vease Steadman et al 2005)
Hay que recordar en este senti do que alin en el Holoceno tardio ocurrieron cambios clirruiticos de consideracion (vease Causas Naturales) Por ejemplo Curtis et al (2001) explicaron que la extincion de algunos ani males mesofilicos de las alturas montanosas de La Espanola pudo haber antecedido a cualquier impacto humano sigrIificativo en esos lugares por coincidir dicha extincion con una tendencia a la aridez documentada isotopicamente y con evidencias de la perdida del bosque mesico ocurridas despues de 3kaAP Yen Jamaica hace solo 2shy3 mil aDos el area de la Bahia de Jackson sustentaba una gran biomasa de vertebrados en bosque hinnedo a juzgar por la cantidad y diversidad de taxones presentes en material fosil derivado de la depredacion por lechuzas hoy el area es la mas seca de la isla con matorral xerofitico y pobreza de fauna (MacPhee 1996a) Yalm en nuestros dias huracanes de gran intensidad aniquilan poblaciones de vertebrados (que no se recuperan) en is las pequenas y llanas del Caribe (Brooks y Smith 2001)
En medio de la incertidumbre sobre la verdadera causa (0 causas) de las extinciones de mamiferos en el periodo amerindio MacPhee y Marx (1997) han propuesto su hipotesis de la hipermorbilidad la cual estipula colapsos poblacionales relativamente rapidos con la introduccion de enfermedades de alta letalidad Estos autores han razonado que
Las islas son los lugares mas apropiados para experimentar perdidas catastroficas causadas por enfermedades Los mamiferos no solo serian susceptibles en virtud de su prolongado aislamiento biologico sino que en los confines de una isla la epizootia
333 EXTINCIONES DEL CUATERNARIO
tendria la potencialidad de infectar simultaneamente a la rnayoria de los individuos de vadas especies Los neonatos son inmunologicamente inmaduros y sufririan desproporcionadamente cualquier infeccion hipervirulenta La perdida excesiva de neonatos y juveniles tendria efectos severos para la sobrevivencia de la poblacion si esta se reduce por debajo de cierto nivel sobreviene la extincion porque no hay posibilidad de restitucion en el periodo postpanzootico Las especies de mayor talla corporal van a ser las mas vulnerables a la extincion dadas sus bajas tasas reproductivas largos periodos de gestacion y reducido numero de progenie Las especies de menor tamano --caracterizadas por altas tasas reproductivas periodos cortos de gestacion y mayor numero de neonatos por parto- tambien serian fuertemente diezmadas pero muchas sobrevivirian porque su estrategia reproductiva les permite perder mayor numero de individuos Las perdidas deben ocurrir relativamente rapido (a escala de decadas) mientras que otras hipotesis (digamos los cambios climaticos) permiten intervalos mucho mas largos (a escala de siglos 0 de milenios) entre la aparicion de las causas y la produccion de sus efectos Por ejemplo todo indica que los nesofontes se extinguieron muy rapidamente y seria dificil poder invocar un impacto humano directo 0 argumentar fenomenos climaticos que pudieran explicar plausiblemente perdidas tan rapidas
MacPhee y Marx (1997 191) han dicho que por el momento la hipotesis de la hipermorbilidad no pasa de ser una abstraccion en espera de adecuada sustentacion empirica Llama la atencion que linicamente en el periodo amerindio (Tabla 45 intervalo 1Il) las extinciones abarcaron todas las clases del peso corporal (de la a a laj) y que ninguna de las especies que sobrevivieron hasta el periodo postcolombino (Tabla 45 intervalo IV mas las especies vivientes) sobrepasa los 10 kg (clases del peso a b c) en clara coincidencia con los presupuestos de la hipotetica hipermorbilidad
Intervalo Postcolornbino
Por 10 menos 37 especies de mamiferos terrestres vivian en las Antillas Mayores y grupos de Islas perifericas a la lIegada de los europeos en 1492 De elIas se han extinguido 23 shymas especies que en cualquiera de los periodos precedentes (Tabla 45~ y solo subsisten 12 especies repartidas entre Cuba (8 especies) La Espanola (2) Jamaica (1) y Bahamas (1) varias de estas en peligro critico de extincion Solo dos de las especies extinguidas durante los ultimos 500 anos se conocieron en estado viviente los roedores Oryzomys antillarum de Jamaica (extinguido en 1877) y Geocapromys thoracatus de las Islas Swan (extinguido en 1955) Las causas de las extinciones ocurridas durante este periodo son sobradamente conocidas destruccion de habitats introducci6n de animales exoticos (domesticos y comensales) y demas presiones asociadas a la actividad humana entre las cuales posiblemente habria que incluir brotes de hipermorbilidad
Durante mucho tiempo se ha afirmado la posibilidad de que especies de Nesophontes hayan sobrevivido hasta las primeras decadas del siglo xx y que incluso continuen existiendo en lugares remotos de Cuba 0 La Espanola (Allen 1918 1942 Miller 1930 Aguayo 1950a Arredondo 1960b Fischer 1977 MacFadden 1980 Woods y
334 BIOGEOGRAFiA
Ottenwalder 1991) Actualmente no existe evidencia radiometrica de que alguna especie de Nesophontes haya sobrevivido al siglo xv las fechas radiocarbonicas calibradas obtenidas en muestras de Nesophontes de Cuba y La Espanola resultaron no mas tardias que los siglos XIII y XIV (MacPhee et al 1999) Por otra parte las expediciones efectuadas a fines del siglo xx con el proposito de localizar individuos vivientes del genero en La Espanola no produjeron la menor indicacion afirmativa al respecto (Woods et al 1985) Ademas a traves de las Antillas Mayores no aparecen restos de Nesophontes en los depositos formados actualmente por las lechuzas
EPiLOGO
Las Antillas Mayores son islas oceanicas no han formado parte de un continente (con la posible excepcion de una parte de Jamaica hasta el Eoceno Medio hace 45 Ma) y durante un tiempo geologico considerable (desde el Eoceno Superior hace 40 Ma) han permanecido emergidas y separadas de los continentes vecinos Consecuentemente los mamiferos terrestres (no volantes) autoctonos de estas islas constituyen una fauna oceanica que por ende cumple con todas las caracteristicas convencionales reconocidas para este tipo de entidad biogeografica
- Baja diversidad taxonomica a nivel de las categorias superiores (solo cuatro ordenes) en comparacion con islas plataformicas continentales (0 con areas continentales) de tamano y diversidad ecologica simi lares
- Alto grado de endemismo a nivel de genero (96) y familia (50) en respuesta al prolongado aislamiento
- Desbalance ecologico de la estructura trofica ausencia de mamiferos camivoros y ungulados
- Espectaculares procesos de irradiaci6n evolucionaria como resultado de la colonizacion de territorios libres de competidores y depredadores
Tendencias evolucionarias extremas gigantismo
- Control a cargo de aves rapaces con tendencia at gigantismo en ausencia de mamiferos miembros del orden Carnivora
- Mas especies de reptiles y de murcielagos que de mamiferos terrestres relaciones que en los continentes se invierten
Alta tasa de extincion de especies (84) sin reposicion
Preservacion de especies filogeneticamente arcaicas (los llamados fosiles vivientes) los almiquies

307 POBLAMIEITO CONTINENTE-ISLA
acceder a el el poblamiento por la via marina en cambio es impredecible porque depende total mente del azar que caracteriza a la balsa Claro que en ambas vias opera el azar pero en el primer caso se trata del azar de un evento probable en el segundo el azar de un evento improbable (Simpson 1940) Asi el puente natural (mecanismo de poblamiento minimamente selectivo = alta diversidad inicial) y la balsa natural (mecanismo de poblamiento maximamente selectivo baja diversidad inicial) pueden conceptuarse como los extremos opuestos de una gama de probabilidades de dispersi6n (McKenna 1973) EI fen6meno de la dispersi6n merece la mayor atenci6n cientifica en los estudios de biogeografia insular (Cowie y Holland 2006)
Queda claro que la balsa natural es una hip6tesis consistente con el caracter de la fauna objeto de estudio puesto que el desbalance taxon6mico (sin evidencia paleontol6gica en contrario) s610 puede ser resultado de la improbabilidad del poblamiento No obstante ello no implica necesariamente que la balsa natural haya sido el imico mecanismo (ni siquiera el principal) utilizado por los mamiferos terrestres en su colonizaci6n de las Antillas Mayores La balsa natural debe satisfacer una premisa te6rica esencial dado el caracter azaroso impredecible e improbable de una colonizaci6n exitosa esta debe ocurrir a intervalos muy espaciados a traves del tiempo geol6gico 0 sea durante todo el tiempo que en el area ocupada por las Antillas Mayores ha habido ecosistemas terrestres perrnanentemente disponibles para ser colonizados (los ultimos 40 millones de afios) Sin embargo como sabemos representantes de los cuatro 6rdenes de mamiteros terrestres conocidos de las Antillas Mayores ya estaban emplazados en dicho territorio insular en el Mioceno Inferior 10 que supone un lapso de colonizaci6n limitado al Eoceno SuperiorshyOligoceno (MacPhee 2005) sugerente de un tipo de mecanismo contrario a la colonizacion alocronica exigida por la hipotesis de la balsa natural sobre todo tratandose de una fauna caracterizada por altas tasas de extincion sin reposicion aparente
La presencia en Jamaica (hasta su extinci6n en 1877) del roedor mfuido Oryzomys antillarom con estrecha afinidad centroamericana (Woods 1989a) pudiera constituir el imico caso efectivo de poblamiento (continente-isla) de las Antillas Mayores por dispersion marina dc un mamifero terrestre para el cual no es razonable invocar otro mecanismo de colonizacion
LA GUIRNALDA INSULAR (dispersion mixta)
La geologia ha establecido con suficiente certeza que durante el Cenozoico unicamente en el Pale6geno tardio y solo a traves de la Cresta de Aves (ahora sumergida excepto la isla hom6nima) pudo haber existido una cadena de islas para el transito de mamiferos terrestres entre el norte de Suramerica y las proto-Antillas Mayores [desestimados a ese efecto el Canal dc Yucatan el Elcvado de Nicaragua la Cresta de Caiman el Elevado de Beata y las Antillas Menores (Holcombe et al 1990 MacPhee e IturraldeshyVinent 1994 1995b Iturralde-Vinent y MacPhee 1999)] Holcombe y Moore (1977) y Holcombe y Edgar (1990) postularon la existencia de esta entidad y estimaron algunas de sus posibles caracteristicas Por su parte MacPhee e Iturralde-Vinent (1995b) e Iturralde-Vinent y MacPhee (1999) precisaron los extremos proximal y distal de esa
308 BIOGEOGRAFIA
estructura acotaron la duracion y la edad de la misma y Ie dieron nombre GAARlandia (Greater AntillesA vesRidgelandia)
Segun Iturralde-Vinent y MacPhee (1999) la Cresta de Aves emergio por breve tiempo (1-2 Ma) durante la transicion Eoceno-Oligoceno (35-33 MaAP) por la coincidencia de un levantamiento tectoni co y un considerable descenso eustatico del nivel del mar En ese momenta las islas septentrionales del arco de las Antillas Mayores (Cuba centroorientalshyLa Espafiola - Puerto Rico - Islas Virgenes) constituian una sola isla grande 0 una serie de islas separadas por barreras muy estrechas La Cresta de Aves emergida entre el bloque Puerto Rico - Islas Virgenes y un microcontinente noroccidental de Suramerica -separado en ese momento del resto del continente por una via marina surgida en el Eoceno tardio- dio origen a GAARlandia
Obviamente el numero de colonizaciones exitosas por mamiferos terrestres a traves de una guimalda de Islas tiende a disminuir mientras mas anchas sean las barreras marinas entre las islas que forman la guimalda y cuando esto ocurre disminuye tambien la diversidad taxonomica de los propagulos particularmente a nivel de las categorias superiores (Simpson 1956)
Vemos asi que para los mamiferos terrestres la colonizaci6n a traves de una guimalda insular puede ser tan improbable como a la deriva en una balsa natural en dependencia de la cantidad y anchura de las barreras marinas a cruzar el tamafio y la fisiografia de las islas que integran la guimalda y el regimen de las corrientes marinas y aereas entre otros factores
Resumiendo la guimalda insular puede conceptuarse como un mecanismo de poblamiento altamente selectivo (= baja diversidad inicial) que combina la improbabilidad de la balsa natural con la colonizacion restringida temporaimente del puente terrestre
En su reciente revision de la taxonomia distribuci6n y filogenia (basada en estudios moleculares) de varios grupos de mamiferos antillanos Davalos (2004) concluyo que los perezosos megalocninos los primates y los roedores histricognatos se avienen al patron de divergencia con el continente y epoca de arribo a las Antill as implicados en la hipotesis de GAARlandia pero no asi los perezosos coloepodinos que requieren una explicacion biogeognlfica distinta Esto ultimo (sospechado por Matthew 1918a) habla sido confirmado por White y MacPhee (2001 226-227)
Nuestra evidencia filogenetica no sustenta la idea de un dado unificado surgido de la invasion de un megaloniquido el analisis sugiere mas bien dos eventos de colonizaci6n independientes [y] sustenta fuertemente un origen difiletico de las subfamilias [Choloepodinae y Megalocninae] Si hubo dos migraciones a traves de GAARlandia presumiblemente poco espaciadas en el tiempo no se puede determinar en el presente
Finalmente queda claro que GAARlandia -como guirnalda insular- es la hipotesis mas robusta propuesta hasta ahora para explicar la presencia en las Antillas Mayores de
309 POBLAMIENTO CONTINENTE- ISLA
la absoluta mayona de los mamfferos terrestres de ongen suramericano porque el caracter de esa fauna supone un mecanismo de poblamiento ancestral que sea a la vez maximamente selectivo (baja diversidad ordinal) y temporalmente restringido (transicion Eoceno-Oligoceno)
Recientemente Rega et al (2002 18) expresaron A pesar de una revision exhaustiva de la paleogeografia de la cuenca del Caribe Iturralde-Vinent y MacPhee (1999) no pudieron confirmar la presencia de una conexion terrestre continua entre Suramerica y las proto-Antillas aunque hipotetizaron su existencia [Vease la Fig 42 (= figura 6 de Iturralde-Vinent y MacPhee 1999 simplificada))
En el trabajo citado por Rega y colaboradores aparece la siguiente explicacion
Actualmente no es posible ofrecer reconstrucciones paleogeograficas detalladas de la Cresta de Aves Sin embargo hay evidencia de que porciones de la cresta deben haberse elevado subaereamente en un tiempo La cuestion cntica es si la Cresta de Aves emergida pudo haber formado algun tipo de corredor entre las Antillas Mayores y el noroeste de Suramerica Perforaciones en las alturas estructurales de la cresta pudieran proveer el registro del Paleogeno tardio que se necesita para precisar la hipotesis del corredor aunque no se puede predecir si tales datos serian suficientemente detallados para determinar si en algun momento GAARlandia constituyo una sola entidad subaerea Por ejemplo pudo haber barreras marinas por corto tiempo entre las masas terrestres de la Cresta de Aves [0) ellevantamiento y la subsidencia pudieron afectar diferentes partes de GAARlandia en diferentes tiempos AItemativamente los elementos faunales pudieron haberse dispersado de manera pulsada Movimientos episOdicos de este tipo incrementarian la extincion de los elementos faunales incapaces de trasladarse a la siguiente masa terrestre en la cadena disminuyendo asi la diversidad total (Iturralde-Vinent y MacPhee 199957)
La primera parte de la explicacion deja claro que la posibilidad de que la Cresta de Aves haya sido una conexion terrestre continua no se puede descartar pero tampoco se puede afirmar La ultima parte de la explicacion describe el efecto altamente selectivo (filtrante) de la colonizacion a traves de una cadena de islas
Y continuan lturralde-Vinent y MacPhee (199957 -58) explicando
Hasta que punto el microcontinente noroccidental de Suramerica [considerado por dichos autores como el area suministradora de propagulos hacia GAARlandiaJ estuvo fisicamente separado del resto del continente no ha sido plenamente esc1arecido Si estuvo sustancialmente aislado por barreras marinas y orogenic as durante el Paleogeno tardio entonces solo la fraccion de la biota suramericana emplazada en el microcontinente estuvo en posicion de acceder a GAARlandia Esto pudiera tener que ver con el hecho de que la fauna antillana carezca de representantes de muchos grupos suramericanos [de mamfferos) que debieron haber hecho el viaje
310 BIOGEOGRAFiA
Florida o Alturas0 0
d 6 D L1anuras25deg
Yucatan
20deg
deg0
10deg
MARCARIBE
80deg 75deg
Bahamas
Cres ta de Beata
D Mares someros
D Mares profundos
OCEANO ATLANTICa
FIGURA 42 Reconstrucci6n hipotetica de GAARlandia como conexi6n terrestre continua entre las proto-Antillas Mayores y el noroeste de Suramerica en el Pale6geno tardio segun la FIgura 6 (simplificada) de Iturralde-Vinent y MacPhee (1999)
En su defensa del puente terrestre Barbour (1945b249) se refiri6 a esto - antes 10 habfa hecho Anthony [1942363]- en los terminos siguientes
El argumento de que si las Antillas hubieran estado conectadas con el continente se encontrarian los f6siles de un numero mayor de distintas criaturas me deja frio es perfectamente concebible que la conexi6n haya ocurrido en un periodo en que la parte del continente implicada no estuviera poblada por una fauna grande y variada
Y concluyen lturralde-Vinent y MacPhee (199958) su detallada explicaci6n
Desafortunadamente sabemos muy poco sobre la composici6n faunal del noroeste de Suramerica a finales del Pale6geno
Consecuentemente si se confirmara la magnitud del aislamiento del aludido microcontinente y su concomitante baja diversidad faunal GAARlandia como conexi6n terrestre continua serf a la hip6tesis mas parsimoniosa No obstante mientras no se disponga de evidencia concluyente a este respecto hipotetizar a GAARlandia como conexi6n continua (minima selectividad) la coloca en franca vulnerabilidad te6rica al tener que afrontar la objeci6n clasica al puente terrestre (baja diversidad ordinal de la
311 POBLAMIENTO CONTINENTE-TSLA
fauna) al tiempo que se la priva de ser la hip6tesis mas robusta para explicar -como guimalda de islas- la colonizaci6n de las Antillas por mamiferos procedentes de Suramerica
ELARcA DE NOE (seudovicarianza)
El primer vicarianista antillano (desde luego sin el saberlo) fue Corral (1940) con su utilizaci6n ingenua de las ideas wegenerianas de la deriva continental aparecidas la decada anterior Pero el primer (supuesto) intento de aplicar a la formaci6n de la biota caribefia el mecanismo de la vicarianza basada en la tearia de la tect6nica de placas (surgida en la sexta decada del pas ado siglo) se debe a Rosen ([1976] 1985)
Segun Rosen las Antillas Mayores se originaron en el Cremcico en la Placa Caribe aproximadamente en la posici6n que hoy dia ocupa Centroamerica y al ser desplazadas tectonicamente hacia el este interactuaron (de algun modo no precisado por Rosen) con las margenes continentales de Norte- y Suramerica de donde (en un evento unico) adquirieron toda su fauna transportada subsecuentemente por dichas islas en su transito hacia sus posiciones actuales [De ahi la denominacion metaforica dada par McKenna (1973) a este mecanismo de poblamiento] Aunque se ha presentado esta propuesta como Model0 de Vicarianza Continente-Isla de Rosen en realidad por definici6n no constituye un evento vicariante porque las proto-Antillas Mayores no resultaron del fraccionamiento y separaci6n por tectonismo de una porcion del territorio continental como exige esta hip6tesis (vease definici6n de vicarianza al comienzo de Hipotesis Biogeograficas)
El Arca de Noe de Rosen ha sido desestimada por la mayoria de los autores contemporaneos porque (1) carece de sustento geol6gico (lturralde-Vinent y MacPhee 1999) y porque (2) en el periodo en que se afrrrna ocum6 el evento muchos de los ordenes modemos de mamiferos placentarios attn no existian (Williams 1989)
EL BARCO FUNERAL VlKINGO (tanatovicarianza)
Domning et al (1997) reportaron 10 que consideraron el mamffero terrestre mas viejo de las Antillas --circunstancia que se refleja en el titulo mismo del trabajo Oldest West Indian Land Mammal- al descubrirse un perisodactilo rinocerotoide (Hyrachius sp) en una localidad de edad Eoceno Inferior a Medio en el occidente de Jamaica
Jamaica ha tenido una historia geologica y paleogeografica distinta de la historia comun a las restantes Antillas Mayores A principios del Cenozoico Jamaica occidental (a diferencia de la porcion oriental constituida por el Bloque de las Montafias Azules) formaba parte de Centro america de la cual se desgaj6 (por el desplazamiento de la Placa Caribe) para venir a integrarse despues del Eoceno Medio con 10 que serfa la futura isla de Jamaica desaparecida por transgresi6n marina en el Eoceno Superior hasta volver a emerger totalmente en el Mioceno Medio (Iturralde-Vinent y MacPhee 1999 MacPhee e Iturralde-Vinent 2000)
312 BIOGEOGRAFiA
Por consiguiente Hyrachius ---conocido de localidades de edad Eoceno Inferior a Medio en Norteamerica- debio ocupar (10 que iba a ser) Jamaica occidental mucho antes de que esta se separara de Centroamerica De hecho este fosil Uego a su destino insular contenido en las rocas de su basamento no como un ser vivo por 10 que es erroneo considerarlo un animal antillano (lturralde-Vinent y MacPhee 2004) como han supuesto algunos (Hedges 200 I Portell et al 200 I Condis [200 I D Todo 10 que hizo fue viajar tanatovicariantemente en su barco funeral vikingo -otra de las denominaciones metaforicas de McKenna (1973)
DISCUSION Y CONCLUSIONES
Hemos visto que solo disponemos de una hipotesis plausible para explicar la mayor parte del poblamiento (continente-isla) de las Antillas Mayores por mamiferos terrestres del Neotropico continental la dispersion mixta (a traves de GAARlandia) para los perezosos (excepto Choloepodinae) los monos y los roedores (excepto Oryzomys)
Por otra parte la historia biogeogrMica de las musaraiias de las Antillas Mayores (Nesophontes y Solenodon) constituye una de las situaciones mas enigmaticas entre los mamiferos de la region (Ottenwalder 2001321) Al menos cinco mecanismos de poblamiento continente-isla han sido invocados para explicar cuando como y de donde llegaron a las Antillas Mayores estas musaraiias
I EI propagulo ancestral (del Terciario de Norteamerica) coloniz6 a las Antillas Mayores por dispersion marina desde Centroamerica 0 desde la Florida (Patterson 1962)
2 El propagulo ancestral ( del Terciario) cruzo el Atlantico Sur (supuestamente mas estreshycho en aquella epoca) directamente de Africa a las Antillas Mayores (Hershkovitz 1972)
3 El propagulo ancestral (del Terciario de Norteamerica) Uego a las Antillas Mayores en el Arca de Noe de Rosen (MacFadden 1980)
4 El propagulo ancestral (del Terciario de Norteamerica) fue transportado a las Antillas desde Centroamerica en el bloque de Jamaica occidental y de alli paso por dispersion marina a las restantes Antillas Mayores (Whidden y Asher 200 I)
5 El propagulo ancestral (del Terciario de Norteamerica) lleg6 a las Antillas Mayores a traves de GAARlandia (Whidden y Asher 2001)
Aunque hay consenso entre estos (y otros) autores sobre el origen norteamericano de las musaraiias antillanas ninguno de los cinco hipoteticos mecanismos de poblamiento resiste la evidencia en contrario filogenetica yo paleogeogrMica Ademas algunas de estas hipotesis asumen que Nesophontidae y Solenodontidae comparten un antecesor comun en las Antillas pero esta relacion no ha sido confirm ada Para dilucidar el enigmatico caso de las musaraiias antillanas se requerini profundizar en la investigaci6n geologica paleogeognifica y paleontol6gica (Iturralde-Vinent y MacPhee 1999)
De acuerdo con el conocimiento filogenetico disponible sobre el conjunto de la fauna la colonizacion original de las Antillas Mayores por mamiferos terrestres implicaria el arribo
313 POBLAMIENTO ISLA-ISLA
de solo siete propagulos fundadores perezosos 2 (coloepodinos y megalocninos si como parece tienen un origen difiletico) musaraiias 2 (nesofontidos y solenodontidos si no constituyen una misma familia como consideran algunos) monos 1 (pitecidos) roedores histricognatos 1 (equimidos capromidos y heptaxodontidos) roedores muridos 1 (Oryzomys en Jamaica)
POBLAMIENTO ISLA-ISLA
La distribucion de los mamiferos terrestres a traves del area antillana demarcada al inicio del presente capitulo (vease Contexto Geogratico Fig 41) ha sido interpretada en funcion de dos hipoteticos mecanismos de poblamiento interinsular la dispersion y la vicarianza fundamentadas como se resumen seguidamente
DISPERSION
De acuerdo con la evidencia paleogeogratica mas actualizada en las principales regresiones marinas ocurridas en la cuenca caribeiia durante los ultimos 23 Ma (transiciones OligocenoshyMioceno Mioceno Superior - Plioceno y varias veces en el Pleistoceno) las masas terrestres constitutivas de la region insular en formacion aumentaron considerablemente su extension territorial disminuyendo por tanto la distancia entre elias oportunidades que los elementos faunales aprovecharian para dispersarse de unas islas a otras
Durante mucho tiempo esta ha sido la hipotesis mas favorecida por los estudiosos del tema a partir de Simpson (1956) pero en decadas recientes ha suscitado dos objeciones
1 Es dificil en tender como pudo operar la dispersion marina entre estas islas puesto que ninguna de las Antillas Mayores posee rios suficientemente caudalosos para transportar masas significativas de vegetacion flotante (MacPhee 198432)
Es oportuno recordar aqui el islote ala deriva (con mas de diez palmas reales en pie) entre las tres Antillas Mayores mas occidentales mencionado anteriormente (vease La Balsa Natural)
2 La dispersion marina es menos efectiva para explicar la asombrosa similitud de las faunas de mamiferos terrestres a traves de las Antillas Mayores septentrionales porque como proceso aleatorio no debe producir distribuciones uniformes a 10 largo de una cadena de islas 0 si las produce entonces el proceso no es aleatorio (MacPhee e lturralde-Vinent 2000 149)
Independientemente de que las aludidas distribuciones taxonomicas estan lejos de ser uniformes 0 asombrosamente similares la hipotesis de la dispersion marina contrariamente a su canicter aleatorio y altamente improbable en el caso del poblamiento continente-isla- en el caso del poblamiento isla-isla es un proceso no aleatorio (aprovecha las oportunidades brindadas puntualmente por los periodos de maxima regresion marina) y ademas altamente probable dada la incomparablemente menor distancia entre las islas caribeiias que entre estas y los continentes vecinos
314 BIOGEOGRAFiA
V ICARIANZA
La neotectonica y las transgresiones marinas del Oligoceno Medio y del Mioceno afectaron sustancialmente la paleogeografia de la Cresta de las Antillas Mayores resultando la subdivision gradual de las partes subaereas de la Cresta Asi el este de Cuba y el norte de La Espanola conectados fisicamente durante el Oligoceno Inferior se separaron al formarse el Paso de los Vientos en el Oligoceno tardio en tanto La Espanola y Puerto Rico probablemente permanecieron conectadas hasta el Mioceno tardio en que surgio el Paso de Mona (Iturralde-Vinent y MacPhee 1999)
Tales eventos geologicos habrian dividido las distribuciones de los grupos faunales emplazados previamente en el territorio continuo de las tres islas y a partir de esas disyunciones dichos autores (tambien MacPhee et al 2000 y White y MacPhee 2001) asumieron --como segunda fase del modelo de GAARlandia- la hipotesis de la vicarianza isla-isla para explicar la distribucion multiinsular de varios grupos de mamiferos terrestres a traves de las Antillas Mayores septentrionales De acuerdo con esta hipotesis cualquier diferencia en composicion faunal entre las islas (por ejemplo la ausencia de roedores heptaxodontidos en Cuba 0 de monos platirrinos en Puerto Rico) constituye evidencia de extincion local postvicariante y no de dispersion marina fallida Lo primero se puede demostrar si aparecen los fosiles pertinentes 10 segundo no se puede demostrar (MacPhee e Iturralde-Vinent 1994) Por otra parte IturraldeshyVinent y MacPhee (1999 57) han reconocido que aunque fa vicarianza isla-isla puede representar una solucion clara para las distribuciones de varios taxones cuaternarios no puede explicar todos los casos
Rega et al (2002 18) han presentado dos objeciones al modelo de la vicarianza isla-isla aplicado a los mamiferos de las Antillas Mayores
I Si la separacion de Cuba y La Espanola precedio en varios mill ones de anos la separashycion de La Espanola y Puerto Rico el modelo vicariante de lturralde-MacPhee debe predecir mayor similitud entre las faunas de La Espanola y Puerto Rico que entre las de Cuba y La Espanola De hecho es al reyes
No obstante MacPhee et al (200335) admitieron despues que la fecha mas temprana empiricamente defendible [para la apertura del Paso de Mona] debe ser el Oligoceno Medio esto invierte la secuencia de ambas disyunciones con 10 cual queda resuelta la aparente contradiccion Sin embargo en su reciente evaluacion de la congruencia entre filogenia e hipotesis geologic as en varios grupos de mamiferos terrestres de las Antillas Mayores Davalos (2004373) encontro que ninguno de los taxones cumple con la fecha y el patron de divergencia entre las masas terrestres antillanas postulados por el modelo de GAARlandia
2 La presencia de un mismo genero en las tres Islas septentrionales de las Antillas Mayores implica [por el modelo vicariante] que la diferenciacion entre estas formas durante el intervalo desde el Mioceno ha sido unicamente de nivel especifico [Dicha presencia]
315 rRRADlAcr6~ EVOLUCrONARIA
s610 puede explicarse razonablemente como el resultado de movimientos de esos animales entre las islas en el Cuaternario
De esta objecion resulta dificil disentir [El poblamiento entre islas diferenciado solo a nivel especifico no es probable que sea anterior al Pleistoceno (Simpson 1956 15)]
Debe tenerse en cuenta que la hipotesis de la vicarianza isla-isla alude a las tres islas septentrionales de las Antillas Mayores porque como se explico antes (vease El Barco Funeral Vikingo) Jamaica no existia como isla durante el lapso (Eoceno Superior shyOligoceno) consensualmente reconocido como el mas probable para la colonizacion de las proto-Antill as Mayores por la mayo ria de los mamiferos terrestres Despues de emerger como isla en el Mioceno Medio Jamaica debe haber recibido sus mamiferos terrestres por dispersion isla-isla (Davalos 2004376) y solo en el Cuaternario parece haberrecibido (por dispersion marina desde Centroamerica) al roedor murido Oryzomys
Seguramente tanto la dispersion como la vicarianza contribuyeron a que los mamiferos terrestres se irradiaran a traves de las Antillas Mayores y grupos de islas perifericas pero las maneras y tiempos en que estos mecanismos operaron a 10 largo de dicho proceso son aun en gran medida motivos de desacuerdo e incertidumbre
IRRADIACION EVOLUCIONARIA
A continuaci6n revisamos someramente las distribuciones conocidas de los diferentes grupos de mamiferos terrestres a traves de las Islas principales de las Antillas Mayores y grupos de Islas perifericas incluidas las interpretaciones y conjeturas que la literatura reciente recoge en relaci6n con la manera en que dichas distribuciones pudieron haberse conformado durante la irradiaci6n evolucionaria de los referidos grupos de mamiferos Manifestaciones de esta irradiaci6n pueden inferirse unicamente a partir de los taxones presentes en el Cuatemario pues solo 6 de las especies conocidas corresponden al Terciario
GRUPOS ORlGINARIOS DE SURAMERICA
PEREZOSOS (Megalonychidae)
Aparte de un genero miocenico de Cuba y otro del Oligoceno de Puerto Rico se conocen 4 generos (12 especies) de perezosos cuaternarios (todos extintos) restringidos a las tres islas septentrionales de las Antillas Mayores Acratocnus Neocnus Megalocnus y Parocnus Los cuatro generos estan representados en Cuba y La Espanola y Acratocnus 10 esta ademas (como unico genero de perezoso) en Puerto Rico Esta distribuci6n ha sido interpretada por MacPhee et al (2000) eomo resultado de la vicarianza oligomiocenica entre estas islas y [dado que el Banco Puerto Rico - Islas Virgenes fue la entrada de GAARlandia a las Antillas Mayores] se preguntaron si la ausencia de los otros tres generos en Puerto Rico se debe a que nunca estuvieron en ese territorio 0 a que sus
316 BIOGEOGRAFiA
fosiles atm no han sido hallados Pero como hemos visto Rega et al (2002) atribuyeron dicha distribucion a la dispersion interinsular en el Cuatemario
Por otra parte varios amilisis c1adisticos recientes ubi can a Choloepus -genero Reciente conocido solo de Centro- y Suramerica- como taxon hermano de Aeratoenus exc1uidos los demas megaloniquidos antillanos Para White y MacPhee (2001228)
tan estrecha relacion sugiere que la divergencia entre ambos [generos] ocurrio despues de que Aeratoenus divergio de Neoenus Si esta relacion implica que Aeratoenus y Neoenus se dispersaron separadamente desde Suramerica a las AntilIas mientras Choloepus permanecia en el continente (siendo necesarias al menos tres invasiones de perezosos) 0 si Choloepus invadio Suramerica desde las Antillas es algo que el material disponible no permite determinar
La diversidad adaptativa de los perezosos se manifiesta en (1) la utilizacion del habitat especies arboreas en Neoenus y Aeratoenus y terrestres en Paroenus y Megaloenus (Webb 1985a White [1993] White et al 1996 Arredondo [1999]) y en (2) la gama de tallas corporales desde los pequenos Neoenus hasta Megaloenus rodens el mayor de los mamiferos terrestres antillanos (su peso adulto estimado en 270 kg seglin Paula 19679) esto sugiere que los perezosos se irradiaron a nichos que en contextos continentales probablemente estarian ocupados por miembros de otros c1ados (MacPhee e IturraldeshyVinent 2000)
No esta claro si el abanico de tamanos entre estos perezosos enmascara alguna tendencia al gigantismo Por 10 pronto Anderson y Handley (2002) encontraron 10 contrario en perezosos (Bradypus) de las Islas de Bocas del Toro (Panama) shyseparadas secuencialmente del area continental vecina por ascensos del nivel del mar durante los ultimos 10 ka En las cinco islas los perezosos son menores que su contraparte continental y las diferencias entre las tallas medias de las cinco poblaciones expresan una reduccion lineal significativa de la talla corporal en funcion de la edad geologica de cada isla sin relacion alguna con el area de las islas 0 la distancia que las separa del continente
MONOS (Pitheciidae)
Los primates de las Antillas Mayores (cuatro especies extintas) constituyen un grupo monofiletico con Paralouatta (en Cuba) y Antillothrix (en La Espanola) como taxones hermanos seguidos en la escalera c1adistica por Xenothrix (en Jamaica) y finalmente Callieebus como el pariente continental mas cercano esto implica una sola invasion desde Suramerica antes del Mioceno Inferior (fecha mas temprana conocida para los primates fosiles antillanos) y por tanto en consonancia con GAARlandia como la via de colonizacion (Horovitz y MacPhee 1996 1999 Horovitz [1997] MacPhee y Horovitz 2004)
En desacuerdo parcial con la anterior interpretacion la filogenia de este grupo de monos obtenida por Davalos (2004) indica un antecesor comun en Cuba La Espanola y Jamaica
317 TRRADIACTON EVOLUCIONARIA
compatible con la hipotetica conexion -propuesta tentativamente por Iturralde-Vinent y MacPhee (199980-84)- entre las Montanas Azules del este de Jamaica y la peninsula suroccidental de La Espanola en la epoca de GAARlandia
A pesar de que los primates antillanos pertenecen a un grupo viviente de monos muy pequenos (los monos titles) fueron comparativamente grandes en relacion con los platirrinos modernos y sucesivamente mayores en la direccion Antillothrix Xenothrix y Paralouatta con P marianae como el mayor platirrino conocido del Mioceno neotropical (MacPhee 2005) MacPhee et al (1994) consideraron que Xenothrix debio ser un mono de andar lento y al parecer las especies de Paralouatta abandonaron la arborealidad para adoptar un modo de vida mas terrestre (MacPhee 1996a MacPhee et al 2003 MacPhee y Meldrum 2006)
ROEOORES (Echimyidae Capromyidae Heptaxodontidae)
La edad de Zazamys en Cuba demuestra que la invasion de los roedores histricognatos en las Antillas Mayores comenzo antes del Mioceno Inferior por otra parte los representantes mas antiguos de esa irradiacion evolucionaria se encuentran en La Espanola y Puerto Rico Ambas evidencias son consistentes con la hipotesis de GAARlandia (Woods 1982 1988 Woods et al 2001)
No esta claro si los capromidos evolucionaron a partir de un equimido ancestral 0 de un protocapromido 0 si los equimidos antillanos (I sub familia 3 generos y 7 especies todas extintas) forman parte de la irradiacion de los capromidos (4 subfamilias 9 generos y 27 especies 10 vivientes) Kratochvil et al (1978b) y Rodriguez et al (1979) consideraron que Capromys nan us es un eslab6n entre Echimyidae y Capromyidae pero Mesocapromys nanus esta morfologicamente mucho mas cerca de Hexolobodon y los de mas capromidos que de los equimidos (Woods 1985) Lo que si esta claro segun Woods et al (2001) es que la subfamilia Plagiodontinae endemic a de La Espanola es el linaje basal de los capromidos y que Isolobodon Hexolobodon Capromys Geocapromys Mysateles y Mesocapromys [y supuestamente Macrocapromys] forman parte de la irradiacion temprana de Capromyidae
La subfamilia Capromyinae (5 generos y 17 especies 9 vivientes) parece ser el dado de mas reciente evolucion en la familia con invasion de Cuba desde La Espanola y dispersion desde Cuba (1) por Capromys a las islas Caiman y (2) por Geocapromys a las islas Bahamas Caiman y Jamaica y desde Jamaica a las islas Swan aunque la presencia de Geocapromys en Swan pudiera deberse a introduccion aborigen (Morgan 1985 1988 1989 1993 1994 Woods 1989a 1990) Woods et al (2001) utilizaron datos moleculares para obtener estimados independientes de divergencia entre capromidos segiJn 10 cual Geocapromys divergio cerca de los 17 MaAP Mesocapromys divergio de Capromys hace 15 MaAP en tanto lvlysateles 10 hizo hace 10 MaAP Por su parte las formas de Capromys pilorides y de Mysateles prehensilis en Isla de la Juventud divergieron de sus correspondientes subespecies nominativas en la isla principal a los 24 y 08 MaAP respectivamente
318 BIOGEOGRAFiA
Woods y Howland (I979) describieron adaptaciones morfologicas de la mandibula el hioide y la faringe asociadas a mecanismos de masticacion diferentes en plagiodontinos y caprominos y Varona (1980a) llamo la atencion sobre el gran desarrollo de los molariformes (en relacion con la longitud total del cnineo) que presentan ciertos grupos de jutias fosiles en comparacion con las vivientes 10 que pudiera interpretarse como una adaptacion alimentaria a una vegetacion mas abrasiva por parte de las jutias extinguidas
Segun Woods (1990) la familia Heptaxodontidae (2 subfamilias 5 generos y 6 especies todas extintas) tambien seria parte de la irradiacion de los capromidos con origen en el este de La Espanola 0 en Puerto Rico y subsecuente dispersion al Banco de Anguila - San Martin en las Antillas Menores (pero vease MacPhee y Flemming 2003) Este banco estaria a solo 70 km del Banco Puerto Rico - Islas Virgenes en tiempos de maxima regresion marina y esta proximidad con Puerto Rico -patria de Elasmodontomys obliquus- seguramente tuvo que ver con la presencia en las islas de Anguila y San Martin del heptaxodontino gigante Amblyrhiza relacionado mas estrechamente con los roedores de las Antillas Mayores que con los del resto de las Antillas Menores (Woods 1990 1996) Aunque los roedores suelen aumentar de tamano en las islas (Case 1978) la escala del incremento de la talla en Amblyrhiza (que pudo alcanzar mas de 200 kg de peso adulto) esta fuera de toda proporcion con los patrones conocidos en otros roedores insulares (Biknevicius et 01 1993)
Clidomys (Clidomyinae) de Jamaica difiere marcadamente en su morfologia de los heptaxodontinos (vease ademas MacPhee y Flemming 2003)10 cual dificulta explicar su presencia en esa isla Como Jamaica estuvo bajo el mar hasta el Mioceno Medio el ancestro de Clidomys colonizaria esta isla por dispersion desde el extremo suroccidental de La Espanola despues que los heptaxodontinos se originaron en esta ultima isla (Woods 1989a) Pero Pascual et 01 (I 990) especularon que Clidomys pudiera ser el resultado de una invasion posterior e independiente de los capromidos
GRUPOS ORIGINARIOS DE NORTEAMERICA
El origen de las musaranas antillanas ha resultado ser hasta ahora el mayor enigma biogeografico de la fauna de mamiferos terrestres de la region aunque la mayoria de los autores coinciden en considerarlas originarias de Norteamerica (vease Discus ion y Conclusiones en Poblamiento Continente-Isla) No obstante MacPhee e Iturralde-Vinent (1994) descartaron toda posibilidad de conexion terrestre por el Estrecho de la Florida 0 por el Canal de Yucatan y si el (supuesto) evento vicari ante de Rosen (vease El Arca de Noe) -invocado por MacFadden (1980) para resolver el enigma- fuera minimamente plausible aun asi es dificil creer que las catastrofes que 10 sucedieron (vease Contexto Temporal) hayan barrido con toda esa vieja fauna continental excepto las musaranas circunstancia ya anticipada muy tempranamente por Matthew (l918a)
De otra parte la relaci6n entre Nesophontidae y Solenodontidae permanece irresuelta Si estos linajes no estan estrechamente relacionados entonces es muy posible que
319 IRRADIACION EVOLUCIONARIA
sus distribuciones sean explicadas por mecanismos diferentes (Whidden y Asher 2001)
La descomunal diferencia de talla corporal entre las musaranas pequenas (Nesophontes) y las grandes (Soienodon) seguramente impuso a ambos grupos restricciones distintas relativas a la termorregulacion y las adaptaciones energeticas En Nesophontes la seleccion debio favorecer una alta tasa de metabolismo y crecimiento camadas mas numerosas corta expectativa de vida yalta densidad poblacionaL En Soienodon en cambio la seleccion opero en la direccion de una baja tasa metabolica camadas reducidas crecimiento lento longevidad y baja densidad poblacional (Ottenwalder [1991])
MUSARANAS PEQUENAS (Nesophontidae)
Las musaranas pequenas estan representadas en las Antillas por el genero Nesophontes (y un genero miocenico indeterminado en ambar dominicano) con 6 especies cuaternarias extinguidas distribuidas en Cuba La Espanola Puerto Rico Islas Virgenes e Islas Caiman La mayor de las musaranas pequenas Nesophontes edithae evoluciono en Puerto Rico y su arribo a las Islas Virgenes supone una dispersion sumamente reciente En las Islas Caiman Nesophontes se asemeja mas a la poblacion de Cuba de donde seguramente derivo en el Plio-Pleistoceno (junto con Capromys y Geocapromys) cuando la costa sur de Cuba estuvo mucho mas cerca de las Islas Caiman (lturralde-Vinent 2003fig 4) MacPhee et ai (1994) interpretaron la distribucion de Nesophontes en Cuba y La Espanola como resultado de la vicarianza oligomiocenica ocurrida al formarse el Paso de los Vientos pero la distancia taxonomica entre los lados del Paso es solo de nivel especifico y N micros incluso vivi6 en ambas islas segtln Patterson (1962)
Allen (1918) considero que Nesophontes micrus fue una especie primitiva no modificada para la vida arb6rea 0 fos6rea y por tanto de habitos terrestres Nesophontes edithae de Puerto Rico es mucho mayor y mas robusta que las restantes especies del genero eL indice de robustez de su femur (= anchura minima de la diafisis ongitud total del hueso) muestra que este es tan robusto como el de Solenodon paradoxus un animal dos veces mayor (Morgan et ai 1980) Choate y Birney (1968) postularon un aumento cronoclinal de la taIla corporal en N edithae confirmado mas tarde por McFarlane (1999a) mediante fechados radiocarb6nicos de la secuencia estratignifica en depositos cavernarios de Puerto Rico Solenodon no vivi6 en Puerto Rico y en ausencia de una musarana de mayor tamano Nesophontes alcanzo una talla corporal mayor en esa isla que en La Espanola 0 Cuba (Woods y Eisenberg 1989)
MUSARANAS GRANDES (Solenodontidae)
Los arcaicos almiqufes (genero Soienodon) comprenden cuatro especies una viviente (S cubanus) y otra f6sil (S arredondoi) en Cuba y una viviente (S paradoxus con dos subespecies) y otra f6sil (S marcanoi) en La Espanola Como cabria esperar las dos especies cubanas se asemejan mas entre sf que a las especies de La Espanola pero
320 BIOGEOGRAFiA
enigmaticamente S marcanoi comparte caracteres con S cuban us y S paradoxus (Woods y Eisenberg 1989 Morgan y Ottenwalder 1993 Ottenwalder 200 I) Solenodon arredondoi alcanzo una talla corporal superior a la de cualquiera de las musarafias vivientes a nivel mundial Segun Morgan y Ottenwalder (1993) extrapolando las dimensiones y el peso de los almiquies vivientes S arredondoi debio tener una longitud cabeza-tronco y un peso de 450-550 mm y 1500-2000 g respectivamente en comparacion con S cubanus (360 mm y 800 g) y S paradoxus (390 mm y 1100 g)
Entre todas las especies de almiqufes Solenodon paradoxus es la que presenta los caracteres mas derivados (Otten walder 2001) Si estas relaciones son validas la distribucion de Solenodon entre Cuba y La Espafiola debio discurrir de oeste a este como propuso MacFadden (1980) quien invoco la presencia fosH de S cubanus [pero tambien S arredondoi] en Pinar del Rio para postular que el ancestro continental entr~ por Cuba opinion compartida por Perfit y Williams (1989) y por Ottenwalder ([1991] 2001) El paso de Solenodon a Puerto Rico pudo haberlo impedido la existencia de barreras ecologicas durante la conexion del este de La Espafiola con Puerto Rico 0 por la ausencia de Solenodon en esa parte de La Espafiola en aquella epoca Sin embargo las mismas condiciones ecologicas en el este de La Espafiola no parecen haber representado una barrera efectiva para Nesophontes cuyas mas altas tasa de reproduccion y densidad poblacional presumiblemente Ie permitieron expandirse flpidamente a traves de La Espafiola (Ottenwalder [1991])
Es opinion generalizada que los almiqufes antillanos constituyen el grupo de mamiferos que primero se emplazo en el territorio insular (MacPhee 2005) y en tal sentido es de interes la evidencia sobre especificidad parasito-hospedante aportada por Cruz (1980) quien encontro que la especie de acaro (genero Cubanochirus) que parasita a los almiquies es mucho mas primitiva que la especie (del mismo genero) que paras ita a las jutias y de aqui concluyo que dicho genero de acaro debio evolucionar primero en Solenodontidae y posteriormente en Capromyidae al entrar en contacto representantes de ambas familias en Cuba 0 La Espafiola
Recientemente Roca (2004) y Roca et al (2004) dieron a conocer que los resultados de su estudio genetico-molecular sobre los almiquies indican una divergencia de 25 Ma entre Solenodon cuban us y S paradoxus -compatible con la disyuncion tectonica entre Cuba y La Espafiola (Iturralde-Vinent y MacPhee 1999 McPhee et al 2003)-- y una separacion de ambas especies comparable en magnitud a la divergencia entre familias de mamlferos Consecuentemente propusieron que Solenodon cubanus fuera clasificada en el genero Atopogaie como indico Cabrera (1925) hace 82 afios
DISCUSION Y CONCLUSIONES
Las musarafias y los roedores antillanos (excepto Oryzomys) se irradiaron tanto de sus respectivos ancestros continentales que de estos 6rdenes se reconocen familias y subfamilias endemicas de la region en contraste con los perezosos (una subfamilia y tres
321 IRRADIACION EVOLUCIONARIA
tribus endemicas) y los monos (una tribu endemica) como expresion de diferentes niveles de diversificacion evolucionaria Este proceso es dificil de explicar (traUindose de c1ados que pnicticamente comparten la misma antigiiedad en las Antmas) si es correcta la interpretacion de Pascual et at (1990636)
Nosotros consideramos estos diferentes niveles de diversificacion como las diferentes etapas de evolucion alcanzadas por cada grupo [de mamiferos] Obviamente esto estuvo relacionado con las diferentes longitudes de sus historias
Pudiera ocurrir que en el futuro se compruebe que la historia biogeognifica de la region fue mucho mas complicada
La Espanola la isla mas ecol6gicamente divers a de la region debi6 ser el centro principal de evoluci6n de los mamfferos (con 87 de las familias y 73 de las subfamilias) seguida por Cuba (con 75 de las familias y 54 de las subfamilias) las dos islas de las Antillas Mayores mas relacionadas biogeograficamente (Fig 43 Tabla 44)
eL
EL
cb CAIMAN
SWAN
A Acratocnini B Cubanocnini C Megalocnini D Mesocnini E Nesophontidae F Solenodontidae G Xenothrichini
ANGUILAshyM-0 SMARTIN
H Heteropsomyinae I Plagiodontinae J Isolobodontinae
K Hexolobodontinae L Capromyinae
M Heptaxodontinae N CIidomyinae 0 Sigmodontinae
FIGURA 43 Representacion esquematica de las relaciones interinsulares (expresadas porel numero de linajes compartidos) entre las Antillas Mayores y entre estas y los grupos de islas perifericas segun las interpretaciones de los autores citados en el texto Los numeros dentro de los circulos indican la cantidad de linajes representados en cada isla relacion dudosa Contexto taxon6mico de los linajes en la Tabla 44
bull bull
bull bull
bull bull bull
bull bull
bull bull
bull
bull
bull
bull
bull bull bull bull
bull bull
bull bull
bull
bull bull bull
bull bull
bull
bull bull
bull
322 BIOGEOGRAFiA
TABLA 44 Distribuci6n de los linajcs de mamiferos terrestres a traves de las Antillas Mayores y grupos de islas perifericas LE = La Espanola CU Cuba JA Jamaica PR = Puerto Rico IC Islas Caiman IS Islas Swan IB = Islas Bahamas IV = Islas Virgenes AS = Anguila - San Martin
FamiliaSubfumiliafIribu LE CU JA PR IC IS IB IV AS
Megalonychidae
Choloepodinae
Acratocnini
Cubanocnini
Megalocninae
Megalocnini
Mesocnini
Nesophontidae
Solenodontidae
Pitheciidae
Callicebinae
Xenothrichini
Echimyidae
Heteropsomyinae
Capromyidae
Plagiodontinae
Isolobodontinae
Hexolobodontinae
Capromyinae
H e pta x 0 don tid a e
Heptaxodontinae
Clidomyinae
Muridae
Sigmodontinae
PILOSA
SORICOMORPHA
PRIMATES
RODENTIA
bull
bull
bull
bull TOTALES 12 10 4 4 2
323 EXTlICIOIES DEL CUATERNARIO
El gigantismo ha sido sin duda una tendencia evolucionaria recurrente entre los clados mayores de mamiferos antillanos Es un hecho bien conocido que algunos grupos de mamiferos han aumentado de tamafio en las islas y varios autores han intentado explicarlo a partir de enfoques diferentes (Foster 1964 Miller 1966 Sondaar 1977 Case 1978 Eisenberg y Wilson 1978 Heaney 1978 Lawlor 1982 Lomolino 1985 Angerbjom 1985 McNab 19942001)
EXTINCIONES DEL CUATERNARIO
Las islas que integran la Subregion Antillana han perdido casi 90 de sus mamiferos terrestres durante los ultimos milenios y en proporcion con su extension territorial esta perdida ha sido mayor en la Subregion que en cualquier otro lugar del planeta (MacPhee 1996b) Las extinciones del Cuatemario eliminaron mas de las tres cuartas partes de la fauna de mamiferos terrestres conocida de las Antillas Mayores sus islas satelites y grupos de islas perifericas y el examen de este proceso en sus aspectos mas generales se aborda en esta ultima parte del presente capitulo
CRONOLOGiA Y CONSECUENCIAS
Es muy improbable que un fosil represente el ultimo individuo de un taxon extinto y entonces hay que guiarse por la edad radiometrica del fosil mas reciente conocido lIamada fecha de ultima presencia- como unica aproximacion posible a la fecha de extincion del taxon
Como se explico al inicio del capitulo (vease Contexto Temporal) el registro fosH de los mamiferos terrestres de las Antillas Mayores adolece de insuficiente sustentacion radiometrica No obstante la distribucion temporal de los escasos fechados disponibles para el Cuaternario de la region abarca etapas criticas del proceso de extincion experimentado por la referida fauna a 10 largo de los ultimos 250 mil afios
De las 52 especies que es posible reconocer actualmente como miembros extintos de la fauna cuatemaria de mamiferos terrestres de las Antillas Mayores y grupos de islas perifericas los restos fosiles de 23 de elIas (442) han sido fechados radiometricamente o se han hallado asociados estratigraficamente de manera inequivoca con otro material fechado De estos 23 fechados 3 corresponden al interglacial Sangamon 4 pertenecen a la secuencia Pleistoceno terminal - Holoceno temprano 10 se ubican en ellapso entre el arribo de los aborigenes y la incursion europea y 6 son postcolombinos (a los cuales hay que sumar otras dos especies extinguidas en los siglos XIX y xx) A partir de estas 25 especies avaladas por fechas de ultima presencia distribuidas entre los cuatro intervalos temporales sefialados hemos compuesto la Tabla 45 en la que ademas aparecen las restantes 27 especies (carentes de fechado radiometrico) ubicadas en el intervalo temporal mas probable sobre la base de (1) la asociacion estratigrafica de sus restos con evidencias culturales (amerindias 0 europeas) 0 (2) la ausencia de dicha asociacion Sobre estos criterios de ubicacion temporal procede hacer dos aclaraciones
324 BIOGEOGRAFIA
TABLA 45 Intervalos temporales mas probables para las extinciones cuatemarias de los mamiferos terrestres de las Antillas Mayores y grupos de islas perifericas sobre la base de las evidencias siguientes (l) ultimo ejemplar colectado u observado en vida (2) fechado radiometrico mas tardio en material de la especie (3) asociaci6n estratigrafica mas tardia con otro material fechado (4) asociaci6n estratigrafica con restos de animales introducidos restos humanos u objetos culturales propios del intervale (5) falta de asociaci6n con evidencias culturales [Los ntimeros entre parentesis en la columna Evidencia significan adscripci6n tentativa al intervalo] Los estimados del peso corporal (en kg) estan tomados de la literatura 0 inferidos comparativamente y distribuidos entre
las clases siguientes (a) S 01 (b) 01 a lt 1 (c) 1 a lt 10 (d) 10 a lt 100 (e) 100 a lt 200 (j) ~ 200 Las
abreviaturas que designan las islas son las de la Tabla 44
F amilialEspecie Peso IsJa Evidencia Fuente
I httervalo htterg1acial Sangamon (gt80 kaAP)
HEPTAXODONTlNAE
Amblyrhiza inundata e AS 3 McFarJane et al 1998a
CLIDOMYINAE
Clidomys osborni d JA 3 Ibid
Clidomys parvus d JA 3 Ibid
II httervalo Pleistoceno terminal ~ Holoceno temprano (20-6 kaAP)
MEGALONYCHIDAE
Aeratoenus antillensis d CD 5 Arredondo 1975 a
Aeratoenus odontrigonus d PR 3 McFarJane 1999b
Aeratoenus simorhynehus d LE 2 Rega et al 2002
M egaloenus zile f LE 2 MacPhee et al 2000
SOLENODONTIDAE
Solenodon arredondoi e CD 5 Morgan y Ottenwalder 1993
PnHECIIDAE
Paralouatta varonai d CD 5 MacPhee 1996a
EcHIMYIDAE
Heteropsomys antillensis c PR 5 McFarJane 1999b
Heteropsomys insulans c PR 5 Ibid
Puertoricomys eorozalus c PR 5 Woods 1989a
CAPROMYIDAE
P lagiodontia araeum c LE 5 MacPhee et al 2000
Macrocapromys latus c CD 5 Varona y Arredondo 1979
(Continua)
325 EXTINCIONES DEL CUATERNARIO
TABLA 45 (Continuaci6n)
F ami1ia1Especie Peso Isla Evidencia Fuente
M acrocapromys latus c CU 5 Varona y Arredondo 1979
Mesocapromys kraglievichi c CU 5 Ibid
HEPTAXODONTINAE
Xaymaca fulvopulvis a JA 2 MacPhee y Flemming 2003
III Intervalo Amerindio (6-05 kaAP)
MEGALONYCHIDAE
Acratocnus ye d LE 2 MacPhee et aI 2000
Neocnus comes c LE 2 Ibid
N eocnus dousman c LE 2 Ibid
N eocnus toupiti c LE 2 Ibid
N eocnus gliriform is c CU 4 Cordova et ai 1997
M egalocnus rodens f CU 2 MacPhee et ai 2007
Parocnus browni e CU 2 Steadman et ai 2005
P arocnus serus e LE 2 MacPhee et ai 2000
N ESOPHONTIDAE
N esophontes edithae a PR 2 McFarlane 1999b
PITHECIIDAE
Antillothrix bernensis c LE 3 Rimoli 1977
EcHIMYIDAE
Brotomys contractus a LE 4 Miller 1929c
CAPROMYIDAE
P lagiodontia spelaeum c LE 4 Ibid
M acrocapromys acevedo c CU 4 NUiiez 1990a
HEPTAXODONTINAE
Elasmodontomys obliquus d PR 3 McFarlane 1999b
Iv Intervalo Postcolombino (05 kaAP - Presente)
N ESOPHONTIDAE
N esophontes hypom icrus a LE 24 MacPhee y Fle111111ing 1999
N esophontes zam icrus a LE (4) Ibid
N esophontes m icrus a CU 24 MacPhee et aI 1999
(Continua)
326 BIOGEOGRAFiA
TABLA 45 (Continuacion)
F amilialEspecie Peso Isla Evidencia Fuente
N esophontes m icrus a CD 24 MacPhee et aI 1999
N esophontes m icrus a LE 4 MacPhee et aI 2000
N esophontes sp 1 a IC 4 Morgan 1994
N esophontes sp 2 a IC 4 Ibid
Solenodon marcanoi b LE 4 Ottenwalder 2001
PITHECIIDAE
Xenothrix mcgregori c JA 4 MacPhee y Flemming 1999
EcHIMYIDAE
Boromys offeUa a CD 24 Guarch 1984
Boromys torrei a CD 24 Ibid
Brotomys voratus a LE 4 MacPhee et aI 2000
CAPROMYIDAE
P lagiodontia ipnaeum c LE 4 MacPhee y Flemming 1999
Rhizoplagiodontia lemkei c LE 4 Ibid
Isolobodon portoricensis c PR 24 Ibid
Isolobodon montanus c LE 4 Ibid
Hexolobodon phenax c LE (4) Ibid
Capromys sp c IC 24 Morgan 1994
Geocapromys columbianus c CD 4 MacPhee y Flemming 1999
Geocapromys thoracatus b IS 1 MacPhee y Marx 1997
Geocapromys sp 1 c IC 4 Morgan 1994
Geocapromys sp 2 c IC (4) Ibid
l-IEPTAXODONTINAE
Quemisia gravis c LE (1) MacPhee y Flemming 1999
MURIDAE
Oryzomys antillarum a JA 1 Ibid
1 Flemming y MacPhee (1999) alertaron en contra de la asociacion con otro material fechado Asimismo Morgan (1993) indico que la ausencia de asociacion con evidencias culturales amerindias no es necesariamente un criterio concluyente para ubicar un taxon en el intervalo temporal anterior porque hay depositos en la region que carecen de tales evidencias y son del Holoceno tardio por fechados radiocarbonicos [No obstante
327 EXTlNCIONES DEL CUATERNAR10
la ausencia de una especie en residuarios arqueol6gicos adquiere mayor certidumbre como criterio de ubicaci6n temporal de su extinci6n en la medida que aumenta el numero de residuarios estudiados que en las Antillas Mayores se cuentan por centenares] A falta de fechado absoluto de una especie consideramos que estos criterios de ubicaci6n temporal son mas utiles que la alternativa de excluir del analisis a la especie
2 En los trabajos sobre biogeografia de mamiferos antillanos los arque610gos mas citados como autoridades para establecer el inicio de la presencia aborigen en las Antillas Mayores (Kozlowski 1974 Rouse y Allaire 1978 Rouse 1989 1992 Wilson 2001) difieren al respecto entre los 6 kaAP y los 45 kaAP (pero vease Pagan et al 200523) y hemos adoptado la mas temprana de estas fechas para el comienzo del intervalo amerindio en la Tabla 45 De todos modos cualquiera que haya sido la fecha esta no es aplicable por igual al conjunto de las Antillas Mayores porque segun algunos de los autores citados el poblamiento comenz6 por Cuba y La Espanola 6 mil anos atras continu6 en Puerto Rico hace alrededor de 3 mil anos (pero vease Burney et al 1994) yen Jamaica hace menos de 2 mil e incluso a las Islas Caiman ni siquiera llegaron [Las Islas Caiman descubiertas por Col6n en 1503 carecen absolutamente de evidencia amerindia (Stokes y Keegan 1996)]
Si los datos de la Tabla 45 representan una aceptable aproximaci6n a la realidad el proceso de las extinciones cuaternarias de los mamiferos terrestres de las Antillas Mayores y grupos de islas perifericas debi6 caracterizarse entre otros por los eventos siguientes
Aunque obviamente la fauna cuaternaria debi6 ser mucho mas numerosa que 10 que refleja el registro paleontol6gico (Morgan y Woods 1986) 52 especies (81 de la fauna conocida) desaparecieron (sin reposici6n) durante los ultimos 250 mil anos
- Al parecer las extinciones se concentraron en las especies de mayor tamano Asi el peso promedio estimado para las especies de la etapa preantr6pica (intervalos I + II de la Tabla 45) es 350 kg es 328 kg para las especies del intervalo III (amerindio) y es s610 25 kg (c1ases del peso a b c) para las especies del intervalo IV (postcolombino) Ninguna especie aut6ctona de mamifero mayor de 6 kg existe hoy en la regi6n
- De acuerdo con la distribuci6n temporal de las extinciones 61 especies sobrevivieron al interglacial Sangamon (intervalo 1)48 sobrevivieron a la transici6n del Pleistoceno terminal al Holoceno temprano (intervalo II) 34 sobrevivieron al periodo amerindio (intervalo III) y s610 12 especies han sobrevivido desde la Ilegada de los europeos (intervalo IV) Las tasas de extinci6n correspondientes (exceptuado el intervalo I) son una especie cada 1167 anos en el intervalo II una cada 393 anos en el intervalo III y una cada 23 anos en el intervalo IV
- Durante todo el proceso de las extinciones cuaternarias la perdida de especies por isla fue de 91 en La Espanola 83 en Jamaica y 62 en Cuba Bahamas retuvo su unica especie (aunque extirpada en varias de las islas) Puerto Rico e Islas Caiman perdieron todas sus especies (6 y 5 respectivamente) y las Islas Swan Virgenes y Anguila - San Martin perdieron sus respectivas especies (micas
328 BIOGEOGRAFiA
De los cuatro 6rdenes de mamiferos representados en la fauna Pilosa y Primates desaparecieron totalmente Soricomorpha y Rodentia perdieron 86 y 74 de sus especies respectivamente
CAUSALIDAD
CAUSAS NATURALES
Los cicIos glaciales e interglaciales acaecidos en el Pleistoceno causaron cambios ambientales dnisticos en las Antillas con la concomitante alternancia de cIimas frios (secos) y calidos (hUmedos) asi como de descensos y ascensos del nivel del mar aumento y disminuci6n de las areas territoriales) induso (aunque con menor intensidad) en el Holoceno Estos eventos paleogeograficos han sido consistentemente documentados (Mayo y Kartashov 1972 Bonnatti y Gartner 1973 Ortega y Arcia 1982 Prance 1982 Schubert y Medina 1982 Dodge et al 1983 Ortega [1984] Haq et al 1987 Schubert 1989 Hodell et al 1991 Guilderson et al 1994 Oppo 1997 Emslie 1998 Curtis et al 2001 Iturralde-Vinent 2003) y de ellos brindamos a continuaci6n una breve resena
Finalizando el periodo interglacial anterior a la ultima glaciaci6n (Wisconsin) el mar estaba a 8-10 m por encima de su nivel actual Cuando el avance glacial del Wisconsin a1canz6 su culminaci6n (eillamado Ultimo Maximo Glacial) hace 18 ka el mar habia descendido 120 m con respecto a dicho nivel y las islas del Caribe habian incrementado considerablemente sus extensiones subaereas en comparaci6n con las que presentan hoy dia por ejemplo Puerto Rico y las Islas Virgenes se conectaron territorialmente y las Bahamas se unieron en cinco gran des islas Prevalecia un regimen frio y seco y condiciones de aridez --debidas a una reducci6n de las precipitaciones (posiblemente a menos de 440 mmlano)- y los ambientes xericos se habian extendido a traves de las islas y bancos emergidos y abundaban los pinares (los habitats mesicos estaban restringidos a parcelas relictas en 10 alto de las montanas) En el Mar Caribe y el Golfo de Mexico la temperatura superficial del mar era 5degC mas baja que la actual y en las llanuras costeras y alturas maximas eran respectivamente 2-3degC y 6degC mas bajas Los limites de las zonas ecol6gicas altitudinales descendieron varios cientos de metros y cada zona redujo su extensi6n vertical al aumentar el gradiente de temperatura Ellimite inferior de la zona periglacial
elevaci6n donde la temperatura media minima a1canza los OdegC) de la Cordillera Central de la Republica Dominicana (altura maxima del Caribe insular con evidencia de topografia glacial) se considera que estuvo entre los 1500 y 1800 mlnm (actualmente esta a 2800 mlnm) y se puede estimar que dicho limite era inferior en Cuba por estar mas al norte que la Republica Dominicana No obstante durante este periodo la temperatura experiment6 episodios cortos (unas pocas decadas) de ascensos repetidos
Un signifieativo componente bi6tieo de las referidas condiciones paleoambientales 10 constituy6 la espectacular irradiaci6n evolucionaria experimentada por un grupo numeroso de aves rapaees diurnas y nocturnas (Wetmore 1937 Brodkorb 1969 Kurotchkin y Mayo 1973 Arredondo 1976 Olson 1978 Olson y Hilgartner 1982
329 EXTINCIONES DEL ClJATERNARIO
Suarez 2000a b 2004 Suarez y Emslie 2003) Los mamiferos de las Antillas Mayores ofrecieron a estos depredadores aviares una amplia gama de tamanos de presa que en ausencia de mamiferos del orden Carnivora en la region permitio a estas aves irradiarse a este nicho alimentario con la evolucion de una variedad de especies de muy diversa tall a corporal incluidas formas gigantes Adquirir un cuerpo gigante les propicio el acceso a recursos alimentarios que no estaban siendo explotados por ningun otro depredador (Alcover y McMinn 1994) A Cuba corresponde el mejor registro fosil de rapaces cuaternarias caribenas con siete generos de especies diurnas Dentro de estas aves existieron tres gavilanes gigantes (Accipitridae) tres carairas (Falconidae) y un condor (Cathartidae) asi como cuatro generos de especies nocturnas dos lechuzas (Tytonidae) y tres Mhos (Strigidae) dos de ell os no igualados en tamano en todo el mundo Algunos de estos generos tuvieron representacion en otras islas antillanas como La Espanola Jamaica Bahamas y Gran Caiman
Al retirarse hacia el norte los hielos de la glaciacion Wisconsin el clima se tomo mas hUmedo en las Antillas durante el Holoceno temprano y el bosque mesico se expandio en la region insular El nivel del mar ascendio acompanado de mayor pluviosidad alrededor de los 7 kaAP las Islas Virgenes quedaron separadas entre si y de Puerto Rico y las Bahamas perdieron la mayor parte de su territorio Cerca de los 3 kaAP las condiciones secas retornaron a la region e hicieron desaparecer buena parte del bosque mesico Nuevamente el clima comenzo a adquirir sus condiciones actuales hacia los 25 kaAP aproximadamente y hoy transita por una fase mesica con vegetacion xerica confinada en parcelas relictas
Sin duda estos y otros eventos naturales incidieron negativamente en la sobrevivencia de mamiferos terrestres en las Antillas Mayores asi como en los grupos de pequenas islas perifericas (Pregill y Olson 1981 Olson 2006)
McFarlane et al (1998a) demostraron convincentemente como Amblyrhiza el gigante de los roedores antillanos endemico de la pequena isla de Anguila sucumbio a las repetidas y abruptas perdidas de su habitat durante el interglacial Sangamon por las nipidas y severas oscilaciones del nivel del mar (documentadas por Neumann y Hearty 1996) al no poder responder adecuadamente a tan catastroficos eventos mediante el ajuste de su masa corporal media y otro tanto debio ocurrirle en Jamaica al otro roedor gigante Clidomys (McFarlane et al 1998b) emparentado con Amblyrhiza en la misma epoca (Tabla 45 intervalo I)
El ascenso del nivel del mar seguramente promovio la extincion de mamiferos durante la transicion del Pleistoceno al Holoceno (Tabla 45 intervalo II) con la reduccion del tamano de las Islas por las transgresiones marinas (Pregill y Olson 1981 Morgan 1993) Esto ultimo haria maximamente severa la competicion asf como la presion de los depredadores [Steadman et al (2005) no yen por que el calentamiento global ocurrido durante la transicion Pleistoceno-Holoceno habria de tener un efecto negativo sobre los perezosos siendo sus descendientes actuales especies tropic ales 0 subtropicales pero no se trata del efecto
330 BIOGEOGRAFiA
directo del cambio de temperatura sobre los animales sino del efecto indirecto y progresivo que dicho cambio genera a largo plazo al reducir el territorio insular disponible]
Por esta epoca tambien desaparecieron especies de tall a mayor como Megalocnus zile en La Espanola y otras de menor tamano (en Cuba) pero que representan la tall a maxima en sus respectivos contextos taxonomicos del Cuaternario como Solenodon arredondoi Paralouatta varonai y Macrocapromys acevedo Un mamifero puede ser grande solo a costa de reducir su densidad poblacional y esto 10 hace mis vulnerable a la extinci6n sobre todo en una isla (McNab 1994 McFarlane et al 1998a) Incidentalmente Alcover et al (1998a) afirmaron que larelacion entre la talla corporal y la extincion no siempre se cumple en las especies de menor tamano y citaron como ejemplo que entre las musarafias todas las especies de Nesophontes desaparecieron mientras sobreviven especies de Solenodon pero la comparacion correcta en todo caso serfa entre los solenodones vivientes y el gigante Solenodon arredondoi y entre Nesophontes edithae -el gigante de los nesofontes extinguido en el periodo amerindio- y el resto de sus congeneres que sobrevivieron hasta los primeros siglos de la colonizacion europea (Tabla 45 intervalos III y IV)
CAUSAS ANTROPOGENICAS
Intervalo Amerindio
La mayoria de los autores que se han referido a las extinciones acaecidas durante este intervalo (entre elIos paradigmaticamente Martin 1984a b) han responsabilizado a los aborfgenes (que se supone comenzaron a poblar las Antillas Mayores hace 6 mil afios) con la desaparicion de numerosas especies de mamiferos terrestres en la region principalmente por la via de la sobrecaza y la introduccion de animales domesticos devenidos depredadores de la fauna autoctona (Morgan y Woods 1986) Este efecto negativo de la presencia indigena sobre los mamiferos de la region ha sido dramatizado por algunos autores en terminos de holocausto faunal Sin embargo pensamos que se ha exagerado Como han expresado MacPhee et al (19993) cuando uno busca evidencia dura de complicidad humana directa en [estas] extinciones no hay mucho que encontrar Las siguientes son algunas consideraciones a tener en cuenta en tal sentido
El registro radiometrico mas joven conocido hasta ahora para un perezoso antillano (MegalocnllS rodens) es de 42 kaAP (MacPhee et ai 2007) Si se acepta que los aborfgenes entraron en Cuba hace aproximadamente 6 mil anos su convivencia con los perezosos debe haber durado alrededor de 1800 afios (0 mas porque dificilmente el referfdo fechado radiometrico haya correspondido al ultimo sobreviviente de la especie) Esta cifra contradice la hipotesis del holocausto faunal segun la cualla extincion de los mamiferos insulares de mayor talla debe ocurrir con extrema rapidez -a escala de decadas 0 de centurias- a partir de su primer contacto con los humanos (Martin y Steadman 1999) A pesar de las repetidas afirmaciones en contrario (Harrington 1921 Miller 1929c Herrera [1945] Tabio 1951 Alvarez 1951a 1957 Arredondo
331 EXTINCIONES DEL CUATERNARIO
1959 1960a Veloz y Ortega 1973 Mayo 1980b Rodriguez et al 1984 Castellanos y Pino 1985 Pino y Castellanos 1985 Nunez 1990b Gutierrez 1996 Arredondo 2004) en su mayoria basadas en evidencias no exentas de posibles accidentes de deposicion (0 de reubicacion postdeposicional) en recintos cavernarios (Diaz-Franco [2006b D no se ha demostrado inequivocamente que los aborigenes antillanos se alimentaran de perezosos regularmente Sus restos en residuarios de alimentos son escasisimos en toda la region y esto contrasta con los abundantes restos de jutias y otros mamiferos de menor talla que aparecen en esos residuarios demostrativos de la preferencia de los paleoindios por estos otros mamiferos
Los mamiferos mas importantes en la economia prehistorica de las Antillas Mayores fueron las jutias Capromys y Geocapromys en Cuba Geocapromys en Jamaica e Isolobodon en La Espanola Puerto Rico e Islas Virgenes (Aguayo 1950b Tablo 1951 Rimoli 1972 Wing 2001 Diaz-Franco [2006b D Wilkins (2001) estudi61ajutia viviente de Jamaica (Geocapromys browni) en un sitio ocupado por los aborigenes durante 100 anos y no encontr6 evidencias de que durante ese tiempo la especie haya sido sobreexplotada Asimismo en Cuba las jutias conga carabali y andaraz son escasas en depositos pleistoholocenicos en cambio son muy numerosas en residuarios indigenas y de haber sido sobreexplotadas 10 mas probable es que no hubieran sobrevivido hasta hoy
La habilidad de los humanos para penetrar en profundidad el bosque tropical y mantenerse en este sustentablemente ha sido un evento comparativamente reciente (Beran y Steele 1994) En el caso antillano la caza tierra adentro ha sido explicada como un proceso tardio de desplazamiento a partir de la explotacion de abundantes recursos marinos que ofrecian condiciones 6ptimas para el sedentarismo de las comunidades aborigenes costeras (C6rdova y Arredondo 1988 Brito 2005) Peces y mariscos dominan la arqueologia costera de las Islas antillanas en tanto el consumo de recursos terrestres (mamfferos incluidos) aumenta progresivamente en la medida que los residuarios estudiados se alejan de la costa (Scudder 1991 Wilkins 2001) En las Antillas Mayores el componente terrestre de la dieta aborigen es solo 20-30 del material recobrado en los residuarios (Wing 1977 1989) Y debio ser menor alin porque el componente vegetal de la dieta no se ha preservado Por otra parte los aborigenes cubanos domesticaron la jutia conga y seguramente la reprodujeron para comer (Pose et al 1988) practica que probablemente se repitiera con otras jutias en las demas Antillas Mayores
- Fray Bartolome de las Casas hablo de 3 millones de indios en Cuba al inicio de la colonizacion pero los historiadores subsecuentes consideraron exagerada la cifra (Poey 1851 a) No obstante se dispone de dos estimados de la densidad de poblaci6n aborigen en las Antillas Mayores a la llegada de los europeos 18 indigenaslkm2 en Puerto Rico (Rouse 1956) y 1271km2 en La Espanola (Watts 1987) Extrapolando ambos valores entre las cuatro islas principales de las Antillas Mayores tenemos que en la fecha indicada la poblaci6n aborigen pudo estar entre 16 mil y 113 mil indigenas en Puerto Rico entre 21 mil y 145 mil en Jamaica entre 141 mil y 1 mill6n en La Espanola y entre 200 mil y 14 millones en Cuba Entre las cifras maximas y las minimas nos inclinamos
332 BIOGEOGRAFiA
hacia estas ultimas atendiendo a los altos indices de mortalidad infantil y morbilidad que se evidencian en las osamentas de los pobladores prehispanicos (Martinez y Rives 1990) Uno de los hechos que apuntan hacia el bajo numero de aborigenes en las Antillas Mayores es 10 rapido que desaparecieron despues de la llegada de los europeos pnkticamente sin dejar mestizaje con estos 10 cual contrasta ostensiblemente con la situacion en la America continentaL
En suma un grupo humano (1) demograiicamente irrelevante y controlado por altos indices de morbilidad (2) consumidor mayoritariamente de organismos marinos y (3) carente de la tecnologia cinegetica necesaria para poder sobreexplotar los mamiferos que Ie sirvieron de alimento (a los cuales accedio tardiamente en su desarrollo) no es concebible que haya causado e1 holocausto faunal que se Ie atribuye a partir del error de aplicar a las comunidades aborigenes antillanas un modelo cultural de la prehistoria continentaL
Tampoco hay evidencias de que los aborigenes hayan introducido carnivoros con suficiente capacidad depredadora salvo el perro Morgan y Otlenwaldder (1993) consideraron que la depredacion por perros durante el periodo amerindio es la explicacion mas plausible para la extincion del almiqui gigante de Cuba pero como se puede observar (Tabla 54) Solenodon arredondoi aparece en el intervalo II por falta de asociacion con evidencias culturales
Consecuentemente el accionar de los aborigenes y de sus animales domesticos directamente sobre la fauna no parece haber sido por sf solo un factor determinante de la extincion de mamiferos terrestres durante el periodo amerindio en las Antillas (pero vease Steadman et al 2005)
Hay que recordar en este senti do que alin en el Holoceno tardio ocurrieron cambios clirruiticos de consideracion (vease Causas Naturales) Por ejemplo Curtis et al (2001) explicaron que la extincion de algunos ani males mesofilicos de las alturas montanosas de La Espanola pudo haber antecedido a cualquier impacto humano sigrIificativo en esos lugares por coincidir dicha extincion con una tendencia a la aridez documentada isotopicamente y con evidencias de la perdida del bosque mesico ocurridas despues de 3kaAP Yen Jamaica hace solo 2shy3 mil aDos el area de la Bahia de Jackson sustentaba una gran biomasa de vertebrados en bosque hinnedo a juzgar por la cantidad y diversidad de taxones presentes en material fosil derivado de la depredacion por lechuzas hoy el area es la mas seca de la isla con matorral xerofitico y pobreza de fauna (MacPhee 1996a) Yalm en nuestros dias huracanes de gran intensidad aniquilan poblaciones de vertebrados (que no se recuperan) en is las pequenas y llanas del Caribe (Brooks y Smith 2001)
En medio de la incertidumbre sobre la verdadera causa (0 causas) de las extinciones de mamiferos en el periodo amerindio MacPhee y Marx (1997) han propuesto su hipotesis de la hipermorbilidad la cual estipula colapsos poblacionales relativamente rapidos con la introduccion de enfermedades de alta letalidad Estos autores han razonado que
Las islas son los lugares mas apropiados para experimentar perdidas catastroficas causadas por enfermedades Los mamiferos no solo serian susceptibles en virtud de su prolongado aislamiento biologico sino que en los confines de una isla la epizootia
333 EXTINCIONES DEL CUATERNARIO
tendria la potencialidad de infectar simultaneamente a la rnayoria de los individuos de vadas especies Los neonatos son inmunologicamente inmaduros y sufririan desproporcionadamente cualquier infeccion hipervirulenta La perdida excesiva de neonatos y juveniles tendria efectos severos para la sobrevivencia de la poblacion si esta se reduce por debajo de cierto nivel sobreviene la extincion porque no hay posibilidad de restitucion en el periodo postpanzootico Las especies de mayor talla corporal van a ser las mas vulnerables a la extincion dadas sus bajas tasas reproductivas largos periodos de gestacion y reducido numero de progenie Las especies de menor tamano --caracterizadas por altas tasas reproductivas periodos cortos de gestacion y mayor numero de neonatos por parto- tambien serian fuertemente diezmadas pero muchas sobrevivirian porque su estrategia reproductiva les permite perder mayor numero de individuos Las perdidas deben ocurrir relativamente rapido (a escala de decadas) mientras que otras hipotesis (digamos los cambios climaticos) permiten intervalos mucho mas largos (a escala de siglos 0 de milenios) entre la aparicion de las causas y la produccion de sus efectos Por ejemplo todo indica que los nesofontes se extinguieron muy rapidamente y seria dificil poder invocar un impacto humano directo 0 argumentar fenomenos climaticos que pudieran explicar plausiblemente perdidas tan rapidas
MacPhee y Marx (1997 191) han dicho que por el momento la hipotesis de la hipermorbilidad no pasa de ser una abstraccion en espera de adecuada sustentacion empirica Llama la atencion que linicamente en el periodo amerindio (Tabla 45 intervalo 1Il) las extinciones abarcaron todas las clases del peso corporal (de la a a laj) y que ninguna de las especies que sobrevivieron hasta el periodo postcolombino (Tabla 45 intervalo IV mas las especies vivientes) sobrepasa los 10 kg (clases del peso a b c) en clara coincidencia con los presupuestos de la hipotetica hipermorbilidad
Intervalo Postcolornbino
Por 10 menos 37 especies de mamiferos terrestres vivian en las Antillas Mayores y grupos de Islas perifericas a la lIegada de los europeos en 1492 De elIas se han extinguido 23 shymas especies que en cualquiera de los periodos precedentes (Tabla 45~ y solo subsisten 12 especies repartidas entre Cuba (8 especies) La Espanola (2) Jamaica (1) y Bahamas (1) varias de estas en peligro critico de extincion Solo dos de las especies extinguidas durante los ultimos 500 anos se conocieron en estado viviente los roedores Oryzomys antillarum de Jamaica (extinguido en 1877) y Geocapromys thoracatus de las Islas Swan (extinguido en 1955) Las causas de las extinciones ocurridas durante este periodo son sobradamente conocidas destruccion de habitats introducci6n de animales exoticos (domesticos y comensales) y demas presiones asociadas a la actividad humana entre las cuales posiblemente habria que incluir brotes de hipermorbilidad
Durante mucho tiempo se ha afirmado la posibilidad de que especies de Nesophontes hayan sobrevivido hasta las primeras decadas del siglo xx y que incluso continuen existiendo en lugares remotos de Cuba 0 La Espanola (Allen 1918 1942 Miller 1930 Aguayo 1950a Arredondo 1960b Fischer 1977 MacFadden 1980 Woods y
334 BIOGEOGRAFiA
Ottenwalder 1991) Actualmente no existe evidencia radiometrica de que alguna especie de Nesophontes haya sobrevivido al siglo xv las fechas radiocarbonicas calibradas obtenidas en muestras de Nesophontes de Cuba y La Espanola resultaron no mas tardias que los siglos XIII y XIV (MacPhee et al 1999) Por otra parte las expediciones efectuadas a fines del siglo xx con el proposito de localizar individuos vivientes del genero en La Espanola no produjeron la menor indicacion afirmativa al respecto (Woods et al 1985) Ademas a traves de las Antillas Mayores no aparecen restos de Nesophontes en los depositos formados actualmente por las lechuzas
EPiLOGO
Las Antillas Mayores son islas oceanicas no han formado parte de un continente (con la posible excepcion de una parte de Jamaica hasta el Eoceno Medio hace 45 Ma) y durante un tiempo geologico considerable (desde el Eoceno Superior hace 40 Ma) han permanecido emergidas y separadas de los continentes vecinos Consecuentemente los mamiferos terrestres (no volantes) autoctonos de estas islas constituyen una fauna oceanica que por ende cumple con todas las caracteristicas convencionales reconocidas para este tipo de entidad biogeografica
- Baja diversidad taxonomica a nivel de las categorias superiores (solo cuatro ordenes) en comparacion con islas plataformicas continentales (0 con areas continentales) de tamano y diversidad ecologica simi lares
- Alto grado de endemismo a nivel de genero (96) y familia (50) en respuesta al prolongado aislamiento
- Desbalance ecologico de la estructura trofica ausencia de mamiferos camivoros y ungulados
- Espectaculares procesos de irradiaci6n evolucionaria como resultado de la colonizacion de territorios libres de competidores y depredadores
Tendencias evolucionarias extremas gigantismo
- Control a cargo de aves rapaces con tendencia at gigantismo en ausencia de mamiferos miembros del orden Carnivora
- Mas especies de reptiles y de murcielagos que de mamiferos terrestres relaciones que en los continentes se invierten
Alta tasa de extincion de especies (84) sin reposicion
Preservacion de especies filogeneticamente arcaicas (los llamados fosiles vivientes) los almiquies

308 BIOGEOGRAFIA
estructura acotaron la duracion y la edad de la misma y Ie dieron nombre GAARlandia (Greater AntillesA vesRidgelandia)
Segun Iturralde-Vinent y MacPhee (1999) la Cresta de Aves emergio por breve tiempo (1-2 Ma) durante la transicion Eoceno-Oligoceno (35-33 MaAP) por la coincidencia de un levantamiento tectoni co y un considerable descenso eustatico del nivel del mar En ese momenta las islas septentrionales del arco de las Antillas Mayores (Cuba centroorientalshyLa Espafiola - Puerto Rico - Islas Virgenes) constituian una sola isla grande 0 una serie de islas separadas por barreras muy estrechas La Cresta de Aves emergida entre el bloque Puerto Rico - Islas Virgenes y un microcontinente noroccidental de Suramerica -separado en ese momento del resto del continente por una via marina surgida en el Eoceno tardio- dio origen a GAARlandia
Obviamente el numero de colonizaciones exitosas por mamiferos terrestres a traves de una guimalda de Islas tiende a disminuir mientras mas anchas sean las barreras marinas entre las islas que forman la guimalda y cuando esto ocurre disminuye tambien la diversidad taxonomica de los propagulos particularmente a nivel de las categorias superiores (Simpson 1956)
Vemos asi que para los mamiferos terrestres la colonizaci6n a traves de una guimalda insular puede ser tan improbable como a la deriva en una balsa natural en dependencia de la cantidad y anchura de las barreras marinas a cruzar el tamafio y la fisiografia de las islas que integran la guimalda y el regimen de las corrientes marinas y aereas entre otros factores
Resumiendo la guimalda insular puede conceptuarse como un mecanismo de poblamiento altamente selectivo (= baja diversidad inicial) que combina la improbabilidad de la balsa natural con la colonizacion restringida temporaimente del puente terrestre
En su reciente revision de la taxonomia distribuci6n y filogenia (basada en estudios moleculares) de varios grupos de mamiferos antillanos Davalos (2004) concluyo que los perezosos megalocninos los primates y los roedores histricognatos se avienen al patron de divergencia con el continente y epoca de arribo a las Antill as implicados en la hipotesis de GAARlandia pero no asi los perezosos coloepodinos que requieren una explicacion biogeognlfica distinta Esto ultimo (sospechado por Matthew 1918a) habla sido confirmado por White y MacPhee (2001 226-227)
Nuestra evidencia filogenetica no sustenta la idea de un dado unificado surgido de la invasion de un megaloniquido el analisis sugiere mas bien dos eventos de colonizaci6n independientes [y] sustenta fuertemente un origen difiletico de las subfamilias [Choloepodinae y Megalocninae] Si hubo dos migraciones a traves de GAARlandia presumiblemente poco espaciadas en el tiempo no se puede determinar en el presente
Finalmente queda claro que GAARlandia -como guirnalda insular- es la hipotesis mas robusta propuesta hasta ahora para explicar la presencia en las Antillas Mayores de
309 POBLAMIENTO CONTINENTE- ISLA
la absoluta mayona de los mamfferos terrestres de ongen suramericano porque el caracter de esa fauna supone un mecanismo de poblamiento ancestral que sea a la vez maximamente selectivo (baja diversidad ordinal) y temporalmente restringido (transicion Eoceno-Oligoceno)
Recientemente Rega et al (2002 18) expresaron A pesar de una revision exhaustiva de la paleogeografia de la cuenca del Caribe Iturralde-Vinent y MacPhee (1999) no pudieron confirmar la presencia de una conexion terrestre continua entre Suramerica y las proto-Antillas aunque hipotetizaron su existencia [Vease la Fig 42 (= figura 6 de Iturralde-Vinent y MacPhee 1999 simplificada))
En el trabajo citado por Rega y colaboradores aparece la siguiente explicacion
Actualmente no es posible ofrecer reconstrucciones paleogeograficas detalladas de la Cresta de Aves Sin embargo hay evidencia de que porciones de la cresta deben haberse elevado subaereamente en un tiempo La cuestion cntica es si la Cresta de Aves emergida pudo haber formado algun tipo de corredor entre las Antillas Mayores y el noroeste de Suramerica Perforaciones en las alturas estructurales de la cresta pudieran proveer el registro del Paleogeno tardio que se necesita para precisar la hipotesis del corredor aunque no se puede predecir si tales datos serian suficientemente detallados para determinar si en algun momento GAARlandia constituyo una sola entidad subaerea Por ejemplo pudo haber barreras marinas por corto tiempo entre las masas terrestres de la Cresta de Aves [0) ellevantamiento y la subsidencia pudieron afectar diferentes partes de GAARlandia en diferentes tiempos AItemativamente los elementos faunales pudieron haberse dispersado de manera pulsada Movimientos episOdicos de este tipo incrementarian la extincion de los elementos faunales incapaces de trasladarse a la siguiente masa terrestre en la cadena disminuyendo asi la diversidad total (Iturralde-Vinent y MacPhee 199957)
La primera parte de la explicacion deja claro que la posibilidad de que la Cresta de Aves haya sido una conexion terrestre continua no se puede descartar pero tampoco se puede afirmar La ultima parte de la explicacion describe el efecto altamente selectivo (filtrante) de la colonizacion a traves de una cadena de islas
Y continuan lturralde-Vinent y MacPhee (199957 -58) explicando
Hasta que punto el microcontinente noroccidental de Suramerica [considerado por dichos autores como el area suministradora de propagulos hacia GAARlandiaJ estuvo fisicamente separado del resto del continente no ha sido plenamente esc1arecido Si estuvo sustancialmente aislado por barreras marinas y orogenic as durante el Paleogeno tardio entonces solo la fraccion de la biota suramericana emplazada en el microcontinente estuvo en posicion de acceder a GAARlandia Esto pudiera tener que ver con el hecho de que la fauna antillana carezca de representantes de muchos grupos suramericanos [de mamfferos) que debieron haber hecho el viaje
310 BIOGEOGRAFiA
Florida o Alturas0 0
d 6 D L1anuras25deg
Yucatan
20deg
deg0
10deg
MARCARIBE
80deg 75deg
Bahamas
Cres ta de Beata
D Mares someros
D Mares profundos
OCEANO ATLANTICa
FIGURA 42 Reconstrucci6n hipotetica de GAARlandia como conexi6n terrestre continua entre las proto-Antillas Mayores y el noroeste de Suramerica en el Pale6geno tardio segun la FIgura 6 (simplificada) de Iturralde-Vinent y MacPhee (1999)
En su defensa del puente terrestre Barbour (1945b249) se refiri6 a esto - antes 10 habfa hecho Anthony [1942363]- en los terminos siguientes
El argumento de que si las Antillas hubieran estado conectadas con el continente se encontrarian los f6siles de un numero mayor de distintas criaturas me deja frio es perfectamente concebible que la conexi6n haya ocurrido en un periodo en que la parte del continente implicada no estuviera poblada por una fauna grande y variada
Y concluyen lturralde-Vinent y MacPhee (199958) su detallada explicaci6n
Desafortunadamente sabemos muy poco sobre la composici6n faunal del noroeste de Suramerica a finales del Pale6geno
Consecuentemente si se confirmara la magnitud del aislamiento del aludido microcontinente y su concomitante baja diversidad faunal GAARlandia como conexi6n terrestre continua serf a la hip6tesis mas parsimoniosa No obstante mientras no se disponga de evidencia concluyente a este respecto hipotetizar a GAARlandia como conexi6n continua (minima selectividad) la coloca en franca vulnerabilidad te6rica al tener que afrontar la objeci6n clasica al puente terrestre (baja diversidad ordinal de la
311 POBLAMIENTO CONTINENTE-TSLA
fauna) al tiempo que se la priva de ser la hip6tesis mas robusta para explicar -como guimalda de islas- la colonizaci6n de las Antillas por mamiferos procedentes de Suramerica
ELARcA DE NOE (seudovicarianza)
El primer vicarianista antillano (desde luego sin el saberlo) fue Corral (1940) con su utilizaci6n ingenua de las ideas wegenerianas de la deriva continental aparecidas la decada anterior Pero el primer (supuesto) intento de aplicar a la formaci6n de la biota caribefia el mecanismo de la vicarianza basada en la tearia de la tect6nica de placas (surgida en la sexta decada del pas ado siglo) se debe a Rosen ([1976] 1985)
Segun Rosen las Antillas Mayores se originaron en el Cremcico en la Placa Caribe aproximadamente en la posici6n que hoy dia ocupa Centroamerica y al ser desplazadas tectonicamente hacia el este interactuaron (de algun modo no precisado por Rosen) con las margenes continentales de Norte- y Suramerica de donde (en un evento unico) adquirieron toda su fauna transportada subsecuentemente por dichas islas en su transito hacia sus posiciones actuales [De ahi la denominacion metaforica dada par McKenna (1973) a este mecanismo de poblamiento] Aunque se ha presentado esta propuesta como Model0 de Vicarianza Continente-Isla de Rosen en realidad por definici6n no constituye un evento vicariante porque las proto-Antillas Mayores no resultaron del fraccionamiento y separaci6n por tectonismo de una porcion del territorio continental como exige esta hip6tesis (vease definici6n de vicarianza al comienzo de Hipotesis Biogeograficas)
El Arca de Noe de Rosen ha sido desestimada por la mayoria de los autores contemporaneos porque (1) carece de sustento geol6gico (lturralde-Vinent y MacPhee 1999) y porque (2) en el periodo en que se afrrrna ocum6 el evento muchos de los ordenes modemos de mamiferos placentarios attn no existian (Williams 1989)
EL BARCO FUNERAL VlKINGO (tanatovicarianza)
Domning et al (1997) reportaron 10 que consideraron el mamffero terrestre mas viejo de las Antillas --circunstancia que se refleja en el titulo mismo del trabajo Oldest West Indian Land Mammal- al descubrirse un perisodactilo rinocerotoide (Hyrachius sp) en una localidad de edad Eoceno Inferior a Medio en el occidente de Jamaica
Jamaica ha tenido una historia geologica y paleogeografica distinta de la historia comun a las restantes Antillas Mayores A principios del Cenozoico Jamaica occidental (a diferencia de la porcion oriental constituida por el Bloque de las Montafias Azules) formaba parte de Centro america de la cual se desgaj6 (por el desplazamiento de la Placa Caribe) para venir a integrarse despues del Eoceno Medio con 10 que serfa la futura isla de Jamaica desaparecida por transgresi6n marina en el Eoceno Superior hasta volver a emerger totalmente en el Mioceno Medio (Iturralde-Vinent y MacPhee 1999 MacPhee e Iturralde-Vinent 2000)
312 BIOGEOGRAFiA
Por consiguiente Hyrachius ---conocido de localidades de edad Eoceno Inferior a Medio en Norteamerica- debio ocupar (10 que iba a ser) Jamaica occidental mucho antes de que esta se separara de Centroamerica De hecho este fosil Uego a su destino insular contenido en las rocas de su basamento no como un ser vivo por 10 que es erroneo considerarlo un animal antillano (lturralde-Vinent y MacPhee 2004) como han supuesto algunos (Hedges 200 I Portell et al 200 I Condis [200 I D Todo 10 que hizo fue viajar tanatovicariantemente en su barco funeral vikingo -otra de las denominaciones metaforicas de McKenna (1973)
DISCUSION Y CONCLUSIONES
Hemos visto que solo disponemos de una hipotesis plausible para explicar la mayor parte del poblamiento (continente-isla) de las Antillas Mayores por mamiferos terrestres del Neotropico continental la dispersion mixta (a traves de GAARlandia) para los perezosos (excepto Choloepodinae) los monos y los roedores (excepto Oryzomys)
Por otra parte la historia biogeogrMica de las musaraiias de las Antillas Mayores (Nesophontes y Solenodon) constituye una de las situaciones mas enigmaticas entre los mamiferos de la region (Ottenwalder 2001321) Al menos cinco mecanismos de poblamiento continente-isla han sido invocados para explicar cuando como y de donde llegaron a las Antillas Mayores estas musaraiias
I EI propagulo ancestral (del Terciario de Norteamerica) coloniz6 a las Antillas Mayores por dispersion marina desde Centroamerica 0 desde la Florida (Patterson 1962)
2 El propagulo ancestral ( del Terciario) cruzo el Atlantico Sur (supuestamente mas estreshycho en aquella epoca) directamente de Africa a las Antillas Mayores (Hershkovitz 1972)
3 El propagulo ancestral (del Terciario de Norteamerica) Uego a las Antillas Mayores en el Arca de Noe de Rosen (MacFadden 1980)
4 El propagulo ancestral (del Terciario de Norteamerica) fue transportado a las Antillas desde Centroamerica en el bloque de Jamaica occidental y de alli paso por dispersion marina a las restantes Antillas Mayores (Whidden y Asher 200 I)
5 El propagulo ancestral (del Terciario de Norteamerica) lleg6 a las Antillas Mayores a traves de GAARlandia (Whidden y Asher 2001)
Aunque hay consenso entre estos (y otros) autores sobre el origen norteamericano de las musaraiias antillanas ninguno de los cinco hipoteticos mecanismos de poblamiento resiste la evidencia en contrario filogenetica yo paleogeogrMica Ademas algunas de estas hipotesis asumen que Nesophontidae y Solenodontidae comparten un antecesor comun en las Antillas pero esta relacion no ha sido confirm ada Para dilucidar el enigmatico caso de las musaraiias antillanas se requerini profundizar en la investigaci6n geologica paleogeognifica y paleontol6gica (Iturralde-Vinent y MacPhee 1999)
De acuerdo con el conocimiento filogenetico disponible sobre el conjunto de la fauna la colonizacion original de las Antillas Mayores por mamiferos terrestres implicaria el arribo
313 POBLAMIENTO ISLA-ISLA
de solo siete propagulos fundadores perezosos 2 (coloepodinos y megalocninos si como parece tienen un origen difiletico) musaraiias 2 (nesofontidos y solenodontidos si no constituyen una misma familia como consideran algunos) monos 1 (pitecidos) roedores histricognatos 1 (equimidos capromidos y heptaxodontidos) roedores muridos 1 (Oryzomys en Jamaica)
POBLAMIENTO ISLA-ISLA
La distribucion de los mamiferos terrestres a traves del area antillana demarcada al inicio del presente capitulo (vease Contexto Geogratico Fig 41) ha sido interpretada en funcion de dos hipoteticos mecanismos de poblamiento interinsular la dispersion y la vicarianza fundamentadas como se resumen seguidamente
DISPERSION
De acuerdo con la evidencia paleogeogratica mas actualizada en las principales regresiones marinas ocurridas en la cuenca caribeiia durante los ultimos 23 Ma (transiciones OligocenoshyMioceno Mioceno Superior - Plioceno y varias veces en el Pleistoceno) las masas terrestres constitutivas de la region insular en formacion aumentaron considerablemente su extension territorial disminuyendo por tanto la distancia entre elias oportunidades que los elementos faunales aprovecharian para dispersarse de unas islas a otras
Durante mucho tiempo esta ha sido la hipotesis mas favorecida por los estudiosos del tema a partir de Simpson (1956) pero en decadas recientes ha suscitado dos objeciones
1 Es dificil en tender como pudo operar la dispersion marina entre estas islas puesto que ninguna de las Antillas Mayores posee rios suficientemente caudalosos para transportar masas significativas de vegetacion flotante (MacPhee 198432)
Es oportuno recordar aqui el islote ala deriva (con mas de diez palmas reales en pie) entre las tres Antillas Mayores mas occidentales mencionado anteriormente (vease La Balsa Natural)
2 La dispersion marina es menos efectiva para explicar la asombrosa similitud de las faunas de mamiferos terrestres a traves de las Antillas Mayores septentrionales porque como proceso aleatorio no debe producir distribuciones uniformes a 10 largo de una cadena de islas 0 si las produce entonces el proceso no es aleatorio (MacPhee e lturralde-Vinent 2000 149)
Independientemente de que las aludidas distribuciones taxonomicas estan lejos de ser uniformes 0 asombrosamente similares la hipotesis de la dispersion marina contrariamente a su canicter aleatorio y altamente improbable en el caso del poblamiento continente-isla- en el caso del poblamiento isla-isla es un proceso no aleatorio (aprovecha las oportunidades brindadas puntualmente por los periodos de maxima regresion marina) y ademas altamente probable dada la incomparablemente menor distancia entre las islas caribeiias que entre estas y los continentes vecinos
314 BIOGEOGRAFiA
V ICARIANZA
La neotectonica y las transgresiones marinas del Oligoceno Medio y del Mioceno afectaron sustancialmente la paleogeografia de la Cresta de las Antillas Mayores resultando la subdivision gradual de las partes subaereas de la Cresta Asi el este de Cuba y el norte de La Espanola conectados fisicamente durante el Oligoceno Inferior se separaron al formarse el Paso de los Vientos en el Oligoceno tardio en tanto La Espanola y Puerto Rico probablemente permanecieron conectadas hasta el Mioceno tardio en que surgio el Paso de Mona (Iturralde-Vinent y MacPhee 1999)
Tales eventos geologicos habrian dividido las distribuciones de los grupos faunales emplazados previamente en el territorio continuo de las tres islas y a partir de esas disyunciones dichos autores (tambien MacPhee et al 2000 y White y MacPhee 2001) asumieron --como segunda fase del modelo de GAARlandia- la hipotesis de la vicarianza isla-isla para explicar la distribucion multiinsular de varios grupos de mamiferos terrestres a traves de las Antillas Mayores septentrionales De acuerdo con esta hipotesis cualquier diferencia en composicion faunal entre las islas (por ejemplo la ausencia de roedores heptaxodontidos en Cuba 0 de monos platirrinos en Puerto Rico) constituye evidencia de extincion local postvicariante y no de dispersion marina fallida Lo primero se puede demostrar si aparecen los fosiles pertinentes 10 segundo no se puede demostrar (MacPhee e Iturralde-Vinent 1994) Por otra parte IturraldeshyVinent y MacPhee (1999 57) han reconocido que aunque fa vicarianza isla-isla puede representar una solucion clara para las distribuciones de varios taxones cuaternarios no puede explicar todos los casos
Rega et al (2002 18) han presentado dos objeciones al modelo de la vicarianza isla-isla aplicado a los mamiferos de las Antillas Mayores
I Si la separacion de Cuba y La Espanola precedio en varios mill ones de anos la separashycion de La Espanola y Puerto Rico el modelo vicariante de lturralde-MacPhee debe predecir mayor similitud entre las faunas de La Espanola y Puerto Rico que entre las de Cuba y La Espanola De hecho es al reyes
No obstante MacPhee et al (200335) admitieron despues que la fecha mas temprana empiricamente defendible [para la apertura del Paso de Mona] debe ser el Oligoceno Medio esto invierte la secuencia de ambas disyunciones con 10 cual queda resuelta la aparente contradiccion Sin embargo en su reciente evaluacion de la congruencia entre filogenia e hipotesis geologic as en varios grupos de mamiferos terrestres de las Antillas Mayores Davalos (2004373) encontro que ninguno de los taxones cumple con la fecha y el patron de divergencia entre las masas terrestres antillanas postulados por el modelo de GAARlandia
2 La presencia de un mismo genero en las tres Islas septentrionales de las Antillas Mayores implica [por el modelo vicariante] que la diferenciacion entre estas formas durante el intervalo desde el Mioceno ha sido unicamente de nivel especifico [Dicha presencia]
315 rRRADlAcr6~ EVOLUCrONARIA
s610 puede explicarse razonablemente como el resultado de movimientos de esos animales entre las islas en el Cuaternario
De esta objecion resulta dificil disentir [El poblamiento entre islas diferenciado solo a nivel especifico no es probable que sea anterior al Pleistoceno (Simpson 1956 15)]
Debe tenerse en cuenta que la hipotesis de la vicarianza isla-isla alude a las tres islas septentrionales de las Antillas Mayores porque como se explico antes (vease El Barco Funeral Vikingo) Jamaica no existia como isla durante el lapso (Eoceno Superior shyOligoceno) consensualmente reconocido como el mas probable para la colonizacion de las proto-Antill as Mayores por la mayo ria de los mamiferos terrestres Despues de emerger como isla en el Mioceno Medio Jamaica debe haber recibido sus mamiferos terrestres por dispersion isla-isla (Davalos 2004376) y solo en el Cuaternario parece haberrecibido (por dispersion marina desde Centroamerica) al roedor murido Oryzomys
Seguramente tanto la dispersion como la vicarianza contribuyeron a que los mamiferos terrestres se irradiaran a traves de las Antillas Mayores y grupos de islas perifericas pero las maneras y tiempos en que estos mecanismos operaron a 10 largo de dicho proceso son aun en gran medida motivos de desacuerdo e incertidumbre
IRRADIACION EVOLUCIONARIA
A continuaci6n revisamos someramente las distribuciones conocidas de los diferentes grupos de mamiferos terrestres a traves de las Islas principales de las Antillas Mayores y grupos de Islas perifericas incluidas las interpretaciones y conjeturas que la literatura reciente recoge en relaci6n con la manera en que dichas distribuciones pudieron haberse conformado durante la irradiaci6n evolucionaria de los referidos grupos de mamiferos Manifestaciones de esta irradiaci6n pueden inferirse unicamente a partir de los taxones presentes en el Cuatemario pues solo 6 de las especies conocidas corresponden al Terciario
GRUPOS ORlGINARIOS DE SURAMERICA
PEREZOSOS (Megalonychidae)
Aparte de un genero miocenico de Cuba y otro del Oligoceno de Puerto Rico se conocen 4 generos (12 especies) de perezosos cuaternarios (todos extintos) restringidos a las tres islas septentrionales de las Antillas Mayores Acratocnus Neocnus Megalocnus y Parocnus Los cuatro generos estan representados en Cuba y La Espanola y Acratocnus 10 esta ademas (como unico genero de perezoso) en Puerto Rico Esta distribuci6n ha sido interpretada por MacPhee et al (2000) eomo resultado de la vicarianza oligomiocenica entre estas islas y [dado que el Banco Puerto Rico - Islas Virgenes fue la entrada de GAARlandia a las Antillas Mayores] se preguntaron si la ausencia de los otros tres generos en Puerto Rico se debe a que nunca estuvieron en ese territorio 0 a que sus
316 BIOGEOGRAFiA
fosiles atm no han sido hallados Pero como hemos visto Rega et al (2002) atribuyeron dicha distribucion a la dispersion interinsular en el Cuatemario
Por otra parte varios amilisis c1adisticos recientes ubi can a Choloepus -genero Reciente conocido solo de Centro- y Suramerica- como taxon hermano de Aeratoenus exc1uidos los demas megaloniquidos antillanos Para White y MacPhee (2001228)
tan estrecha relacion sugiere que la divergencia entre ambos [generos] ocurrio despues de que Aeratoenus divergio de Neoenus Si esta relacion implica que Aeratoenus y Neoenus se dispersaron separadamente desde Suramerica a las AntilIas mientras Choloepus permanecia en el continente (siendo necesarias al menos tres invasiones de perezosos) 0 si Choloepus invadio Suramerica desde las Antillas es algo que el material disponible no permite determinar
La diversidad adaptativa de los perezosos se manifiesta en (1) la utilizacion del habitat especies arboreas en Neoenus y Aeratoenus y terrestres en Paroenus y Megaloenus (Webb 1985a White [1993] White et al 1996 Arredondo [1999]) y en (2) la gama de tallas corporales desde los pequenos Neoenus hasta Megaloenus rodens el mayor de los mamiferos terrestres antillanos (su peso adulto estimado en 270 kg seglin Paula 19679) esto sugiere que los perezosos se irradiaron a nichos que en contextos continentales probablemente estarian ocupados por miembros de otros c1ados (MacPhee e IturraldeshyVinent 2000)
No esta claro si el abanico de tamanos entre estos perezosos enmascara alguna tendencia al gigantismo Por 10 pronto Anderson y Handley (2002) encontraron 10 contrario en perezosos (Bradypus) de las Islas de Bocas del Toro (Panama) shyseparadas secuencialmente del area continental vecina por ascensos del nivel del mar durante los ultimos 10 ka En las cinco islas los perezosos son menores que su contraparte continental y las diferencias entre las tallas medias de las cinco poblaciones expresan una reduccion lineal significativa de la talla corporal en funcion de la edad geologica de cada isla sin relacion alguna con el area de las islas 0 la distancia que las separa del continente
MONOS (Pitheciidae)
Los primates de las Antillas Mayores (cuatro especies extintas) constituyen un grupo monofiletico con Paralouatta (en Cuba) y Antillothrix (en La Espanola) como taxones hermanos seguidos en la escalera c1adistica por Xenothrix (en Jamaica) y finalmente Callieebus como el pariente continental mas cercano esto implica una sola invasion desde Suramerica antes del Mioceno Inferior (fecha mas temprana conocida para los primates fosiles antillanos) y por tanto en consonancia con GAARlandia como la via de colonizacion (Horovitz y MacPhee 1996 1999 Horovitz [1997] MacPhee y Horovitz 2004)
En desacuerdo parcial con la anterior interpretacion la filogenia de este grupo de monos obtenida por Davalos (2004) indica un antecesor comun en Cuba La Espanola y Jamaica
317 TRRADIACTON EVOLUCIONARIA
compatible con la hipotetica conexion -propuesta tentativamente por Iturralde-Vinent y MacPhee (199980-84)- entre las Montanas Azules del este de Jamaica y la peninsula suroccidental de La Espanola en la epoca de GAARlandia
A pesar de que los primates antillanos pertenecen a un grupo viviente de monos muy pequenos (los monos titles) fueron comparativamente grandes en relacion con los platirrinos modernos y sucesivamente mayores en la direccion Antillothrix Xenothrix y Paralouatta con P marianae como el mayor platirrino conocido del Mioceno neotropical (MacPhee 2005) MacPhee et al (1994) consideraron que Xenothrix debio ser un mono de andar lento y al parecer las especies de Paralouatta abandonaron la arborealidad para adoptar un modo de vida mas terrestre (MacPhee 1996a MacPhee et al 2003 MacPhee y Meldrum 2006)
ROEOORES (Echimyidae Capromyidae Heptaxodontidae)
La edad de Zazamys en Cuba demuestra que la invasion de los roedores histricognatos en las Antillas Mayores comenzo antes del Mioceno Inferior por otra parte los representantes mas antiguos de esa irradiacion evolucionaria se encuentran en La Espanola y Puerto Rico Ambas evidencias son consistentes con la hipotesis de GAARlandia (Woods 1982 1988 Woods et al 2001)
No esta claro si los capromidos evolucionaron a partir de un equimido ancestral 0 de un protocapromido 0 si los equimidos antillanos (I sub familia 3 generos y 7 especies todas extintas) forman parte de la irradiacion de los capromidos (4 subfamilias 9 generos y 27 especies 10 vivientes) Kratochvil et al (1978b) y Rodriguez et al (1979) consideraron que Capromys nan us es un eslab6n entre Echimyidae y Capromyidae pero Mesocapromys nanus esta morfologicamente mucho mas cerca de Hexolobodon y los de mas capromidos que de los equimidos (Woods 1985) Lo que si esta claro segun Woods et al (2001) es que la subfamilia Plagiodontinae endemic a de La Espanola es el linaje basal de los capromidos y que Isolobodon Hexolobodon Capromys Geocapromys Mysateles y Mesocapromys [y supuestamente Macrocapromys] forman parte de la irradiacion temprana de Capromyidae
La subfamilia Capromyinae (5 generos y 17 especies 9 vivientes) parece ser el dado de mas reciente evolucion en la familia con invasion de Cuba desde La Espanola y dispersion desde Cuba (1) por Capromys a las islas Caiman y (2) por Geocapromys a las islas Bahamas Caiman y Jamaica y desde Jamaica a las islas Swan aunque la presencia de Geocapromys en Swan pudiera deberse a introduccion aborigen (Morgan 1985 1988 1989 1993 1994 Woods 1989a 1990) Woods et al (2001) utilizaron datos moleculares para obtener estimados independientes de divergencia entre capromidos segiJn 10 cual Geocapromys divergio cerca de los 17 MaAP Mesocapromys divergio de Capromys hace 15 MaAP en tanto lvlysateles 10 hizo hace 10 MaAP Por su parte las formas de Capromys pilorides y de Mysateles prehensilis en Isla de la Juventud divergieron de sus correspondientes subespecies nominativas en la isla principal a los 24 y 08 MaAP respectivamente
318 BIOGEOGRAFiA
Woods y Howland (I979) describieron adaptaciones morfologicas de la mandibula el hioide y la faringe asociadas a mecanismos de masticacion diferentes en plagiodontinos y caprominos y Varona (1980a) llamo la atencion sobre el gran desarrollo de los molariformes (en relacion con la longitud total del cnineo) que presentan ciertos grupos de jutias fosiles en comparacion con las vivientes 10 que pudiera interpretarse como una adaptacion alimentaria a una vegetacion mas abrasiva por parte de las jutias extinguidas
Segun Woods (1990) la familia Heptaxodontidae (2 subfamilias 5 generos y 6 especies todas extintas) tambien seria parte de la irradiacion de los capromidos con origen en el este de La Espanola 0 en Puerto Rico y subsecuente dispersion al Banco de Anguila - San Martin en las Antillas Menores (pero vease MacPhee y Flemming 2003) Este banco estaria a solo 70 km del Banco Puerto Rico - Islas Virgenes en tiempos de maxima regresion marina y esta proximidad con Puerto Rico -patria de Elasmodontomys obliquus- seguramente tuvo que ver con la presencia en las islas de Anguila y San Martin del heptaxodontino gigante Amblyrhiza relacionado mas estrechamente con los roedores de las Antillas Mayores que con los del resto de las Antillas Menores (Woods 1990 1996) Aunque los roedores suelen aumentar de tamano en las islas (Case 1978) la escala del incremento de la talla en Amblyrhiza (que pudo alcanzar mas de 200 kg de peso adulto) esta fuera de toda proporcion con los patrones conocidos en otros roedores insulares (Biknevicius et 01 1993)
Clidomys (Clidomyinae) de Jamaica difiere marcadamente en su morfologia de los heptaxodontinos (vease ademas MacPhee y Flemming 2003)10 cual dificulta explicar su presencia en esa isla Como Jamaica estuvo bajo el mar hasta el Mioceno Medio el ancestro de Clidomys colonizaria esta isla por dispersion desde el extremo suroccidental de La Espanola despues que los heptaxodontinos se originaron en esta ultima isla (Woods 1989a) Pero Pascual et 01 (I 990) especularon que Clidomys pudiera ser el resultado de una invasion posterior e independiente de los capromidos
GRUPOS ORIGINARIOS DE NORTEAMERICA
El origen de las musaranas antillanas ha resultado ser hasta ahora el mayor enigma biogeografico de la fauna de mamiferos terrestres de la region aunque la mayoria de los autores coinciden en considerarlas originarias de Norteamerica (vease Discus ion y Conclusiones en Poblamiento Continente-Isla) No obstante MacPhee e Iturralde-Vinent (1994) descartaron toda posibilidad de conexion terrestre por el Estrecho de la Florida 0 por el Canal de Yucatan y si el (supuesto) evento vicari ante de Rosen (vease El Arca de Noe) -invocado por MacFadden (1980) para resolver el enigma- fuera minimamente plausible aun asi es dificil creer que las catastrofes que 10 sucedieron (vease Contexto Temporal) hayan barrido con toda esa vieja fauna continental excepto las musaranas circunstancia ya anticipada muy tempranamente por Matthew (l918a)
De otra parte la relaci6n entre Nesophontidae y Solenodontidae permanece irresuelta Si estos linajes no estan estrechamente relacionados entonces es muy posible que
319 IRRADIACION EVOLUCIONARIA
sus distribuciones sean explicadas por mecanismos diferentes (Whidden y Asher 2001)
La descomunal diferencia de talla corporal entre las musaranas pequenas (Nesophontes) y las grandes (Soienodon) seguramente impuso a ambos grupos restricciones distintas relativas a la termorregulacion y las adaptaciones energeticas En Nesophontes la seleccion debio favorecer una alta tasa de metabolismo y crecimiento camadas mas numerosas corta expectativa de vida yalta densidad poblacionaL En Soienodon en cambio la seleccion opero en la direccion de una baja tasa metabolica camadas reducidas crecimiento lento longevidad y baja densidad poblacional (Ottenwalder [1991])
MUSARANAS PEQUENAS (Nesophontidae)
Las musaranas pequenas estan representadas en las Antillas por el genero Nesophontes (y un genero miocenico indeterminado en ambar dominicano) con 6 especies cuaternarias extinguidas distribuidas en Cuba La Espanola Puerto Rico Islas Virgenes e Islas Caiman La mayor de las musaranas pequenas Nesophontes edithae evoluciono en Puerto Rico y su arribo a las Islas Virgenes supone una dispersion sumamente reciente En las Islas Caiman Nesophontes se asemeja mas a la poblacion de Cuba de donde seguramente derivo en el Plio-Pleistoceno (junto con Capromys y Geocapromys) cuando la costa sur de Cuba estuvo mucho mas cerca de las Islas Caiman (lturralde-Vinent 2003fig 4) MacPhee et ai (1994) interpretaron la distribucion de Nesophontes en Cuba y La Espanola como resultado de la vicarianza oligomiocenica ocurrida al formarse el Paso de los Vientos pero la distancia taxonomica entre los lados del Paso es solo de nivel especifico y N micros incluso vivi6 en ambas islas segtln Patterson (1962)
Allen (1918) considero que Nesophontes micrus fue una especie primitiva no modificada para la vida arb6rea 0 fos6rea y por tanto de habitos terrestres Nesophontes edithae de Puerto Rico es mucho mayor y mas robusta que las restantes especies del genero eL indice de robustez de su femur (= anchura minima de la diafisis ongitud total del hueso) muestra que este es tan robusto como el de Solenodon paradoxus un animal dos veces mayor (Morgan et ai 1980) Choate y Birney (1968) postularon un aumento cronoclinal de la taIla corporal en N edithae confirmado mas tarde por McFarlane (1999a) mediante fechados radiocarb6nicos de la secuencia estratignifica en depositos cavernarios de Puerto Rico Solenodon no vivi6 en Puerto Rico y en ausencia de una musarana de mayor tamano Nesophontes alcanzo una talla corporal mayor en esa isla que en La Espanola 0 Cuba (Woods y Eisenberg 1989)
MUSARANAS GRANDES (Solenodontidae)
Los arcaicos almiqufes (genero Soienodon) comprenden cuatro especies una viviente (S cubanus) y otra f6sil (S arredondoi) en Cuba y una viviente (S paradoxus con dos subespecies) y otra f6sil (S marcanoi) en La Espanola Como cabria esperar las dos especies cubanas se asemejan mas entre sf que a las especies de La Espanola pero
320 BIOGEOGRAFiA
enigmaticamente S marcanoi comparte caracteres con S cuban us y S paradoxus (Woods y Eisenberg 1989 Morgan y Ottenwalder 1993 Ottenwalder 200 I) Solenodon arredondoi alcanzo una talla corporal superior a la de cualquiera de las musarafias vivientes a nivel mundial Segun Morgan y Ottenwalder (1993) extrapolando las dimensiones y el peso de los almiquies vivientes S arredondoi debio tener una longitud cabeza-tronco y un peso de 450-550 mm y 1500-2000 g respectivamente en comparacion con S cubanus (360 mm y 800 g) y S paradoxus (390 mm y 1100 g)
Entre todas las especies de almiqufes Solenodon paradoxus es la que presenta los caracteres mas derivados (Otten walder 2001) Si estas relaciones son validas la distribucion de Solenodon entre Cuba y La Espafiola debio discurrir de oeste a este como propuso MacFadden (1980) quien invoco la presencia fosH de S cubanus [pero tambien S arredondoi] en Pinar del Rio para postular que el ancestro continental entr~ por Cuba opinion compartida por Perfit y Williams (1989) y por Ottenwalder ([1991] 2001) El paso de Solenodon a Puerto Rico pudo haberlo impedido la existencia de barreras ecologicas durante la conexion del este de La Espafiola con Puerto Rico 0 por la ausencia de Solenodon en esa parte de La Espafiola en aquella epoca Sin embargo las mismas condiciones ecologicas en el este de La Espafiola no parecen haber representado una barrera efectiva para Nesophontes cuyas mas altas tasa de reproduccion y densidad poblacional presumiblemente Ie permitieron expandirse flpidamente a traves de La Espafiola (Ottenwalder [1991])
Es opinion generalizada que los almiqufes antillanos constituyen el grupo de mamiferos que primero se emplazo en el territorio insular (MacPhee 2005) y en tal sentido es de interes la evidencia sobre especificidad parasito-hospedante aportada por Cruz (1980) quien encontro que la especie de acaro (genero Cubanochirus) que parasita a los almiquies es mucho mas primitiva que la especie (del mismo genero) que paras ita a las jutias y de aqui concluyo que dicho genero de acaro debio evolucionar primero en Solenodontidae y posteriormente en Capromyidae al entrar en contacto representantes de ambas familias en Cuba 0 La Espafiola
Recientemente Roca (2004) y Roca et al (2004) dieron a conocer que los resultados de su estudio genetico-molecular sobre los almiquies indican una divergencia de 25 Ma entre Solenodon cuban us y S paradoxus -compatible con la disyuncion tectonica entre Cuba y La Espafiola (Iturralde-Vinent y MacPhee 1999 McPhee et al 2003)-- y una separacion de ambas especies comparable en magnitud a la divergencia entre familias de mamlferos Consecuentemente propusieron que Solenodon cubanus fuera clasificada en el genero Atopogaie como indico Cabrera (1925) hace 82 afios
DISCUSION Y CONCLUSIONES
Las musarafias y los roedores antillanos (excepto Oryzomys) se irradiaron tanto de sus respectivos ancestros continentales que de estos 6rdenes se reconocen familias y subfamilias endemicas de la region en contraste con los perezosos (una subfamilia y tres
321 IRRADIACION EVOLUCIONARIA
tribus endemicas) y los monos (una tribu endemica) como expresion de diferentes niveles de diversificacion evolucionaria Este proceso es dificil de explicar (traUindose de c1ados que pnicticamente comparten la misma antigiiedad en las Antmas) si es correcta la interpretacion de Pascual et at (1990636)
Nosotros consideramos estos diferentes niveles de diversificacion como las diferentes etapas de evolucion alcanzadas por cada grupo [de mamiferos] Obviamente esto estuvo relacionado con las diferentes longitudes de sus historias
Pudiera ocurrir que en el futuro se compruebe que la historia biogeognifica de la region fue mucho mas complicada
La Espanola la isla mas ecol6gicamente divers a de la region debi6 ser el centro principal de evoluci6n de los mamfferos (con 87 de las familias y 73 de las subfamilias) seguida por Cuba (con 75 de las familias y 54 de las subfamilias) las dos islas de las Antillas Mayores mas relacionadas biogeograficamente (Fig 43 Tabla 44)
eL
EL
cb CAIMAN
SWAN
A Acratocnini B Cubanocnini C Megalocnini D Mesocnini E Nesophontidae F Solenodontidae G Xenothrichini
ANGUILAshyM-0 SMARTIN
H Heteropsomyinae I Plagiodontinae J Isolobodontinae
K Hexolobodontinae L Capromyinae
M Heptaxodontinae N CIidomyinae 0 Sigmodontinae
FIGURA 43 Representacion esquematica de las relaciones interinsulares (expresadas porel numero de linajes compartidos) entre las Antillas Mayores y entre estas y los grupos de islas perifericas segun las interpretaciones de los autores citados en el texto Los numeros dentro de los circulos indican la cantidad de linajes representados en cada isla relacion dudosa Contexto taxon6mico de los linajes en la Tabla 44
bull bull
bull bull
bull bull bull
bull bull
bull bull
bull
bull
bull
bull
bull bull bull bull
bull bull
bull bull
bull
bull bull bull
bull bull
bull
bull bull
bull
322 BIOGEOGRAFiA
TABLA 44 Distribuci6n de los linajcs de mamiferos terrestres a traves de las Antillas Mayores y grupos de islas perifericas LE = La Espanola CU Cuba JA Jamaica PR = Puerto Rico IC Islas Caiman IS Islas Swan IB = Islas Bahamas IV = Islas Virgenes AS = Anguila - San Martin
FamiliaSubfumiliafIribu LE CU JA PR IC IS IB IV AS
Megalonychidae
Choloepodinae
Acratocnini
Cubanocnini
Megalocninae
Megalocnini
Mesocnini
Nesophontidae
Solenodontidae
Pitheciidae
Callicebinae
Xenothrichini
Echimyidae
Heteropsomyinae
Capromyidae
Plagiodontinae
Isolobodontinae
Hexolobodontinae
Capromyinae
H e pta x 0 don tid a e
Heptaxodontinae
Clidomyinae
Muridae
Sigmodontinae
PILOSA
SORICOMORPHA
PRIMATES
RODENTIA
bull
bull
bull
bull TOTALES 12 10 4 4 2
323 EXTlICIOIES DEL CUATERNARIO
El gigantismo ha sido sin duda una tendencia evolucionaria recurrente entre los clados mayores de mamiferos antillanos Es un hecho bien conocido que algunos grupos de mamiferos han aumentado de tamafio en las islas y varios autores han intentado explicarlo a partir de enfoques diferentes (Foster 1964 Miller 1966 Sondaar 1977 Case 1978 Eisenberg y Wilson 1978 Heaney 1978 Lawlor 1982 Lomolino 1985 Angerbjom 1985 McNab 19942001)
EXTINCIONES DEL CUATERNARIO
Las islas que integran la Subregion Antillana han perdido casi 90 de sus mamiferos terrestres durante los ultimos milenios y en proporcion con su extension territorial esta perdida ha sido mayor en la Subregion que en cualquier otro lugar del planeta (MacPhee 1996b) Las extinciones del Cuatemario eliminaron mas de las tres cuartas partes de la fauna de mamiferos terrestres conocida de las Antillas Mayores sus islas satelites y grupos de islas perifericas y el examen de este proceso en sus aspectos mas generales se aborda en esta ultima parte del presente capitulo
CRONOLOGiA Y CONSECUENCIAS
Es muy improbable que un fosil represente el ultimo individuo de un taxon extinto y entonces hay que guiarse por la edad radiometrica del fosil mas reciente conocido lIamada fecha de ultima presencia- como unica aproximacion posible a la fecha de extincion del taxon
Como se explico al inicio del capitulo (vease Contexto Temporal) el registro fosH de los mamiferos terrestres de las Antillas Mayores adolece de insuficiente sustentacion radiometrica No obstante la distribucion temporal de los escasos fechados disponibles para el Cuaternario de la region abarca etapas criticas del proceso de extincion experimentado por la referida fauna a 10 largo de los ultimos 250 mil afios
De las 52 especies que es posible reconocer actualmente como miembros extintos de la fauna cuatemaria de mamiferos terrestres de las Antillas Mayores y grupos de islas perifericas los restos fosiles de 23 de elIas (442) han sido fechados radiometricamente o se han hallado asociados estratigraficamente de manera inequivoca con otro material fechado De estos 23 fechados 3 corresponden al interglacial Sangamon 4 pertenecen a la secuencia Pleistoceno terminal - Holoceno temprano 10 se ubican en ellapso entre el arribo de los aborigenes y la incursion europea y 6 son postcolombinos (a los cuales hay que sumar otras dos especies extinguidas en los siglos XIX y xx) A partir de estas 25 especies avaladas por fechas de ultima presencia distribuidas entre los cuatro intervalos temporales sefialados hemos compuesto la Tabla 45 en la que ademas aparecen las restantes 27 especies (carentes de fechado radiometrico) ubicadas en el intervalo temporal mas probable sobre la base de (1) la asociacion estratigrafica de sus restos con evidencias culturales (amerindias 0 europeas) 0 (2) la ausencia de dicha asociacion Sobre estos criterios de ubicacion temporal procede hacer dos aclaraciones
324 BIOGEOGRAFIA
TABLA 45 Intervalos temporales mas probables para las extinciones cuatemarias de los mamiferos terrestres de las Antillas Mayores y grupos de islas perifericas sobre la base de las evidencias siguientes (l) ultimo ejemplar colectado u observado en vida (2) fechado radiometrico mas tardio en material de la especie (3) asociaci6n estratigrafica mas tardia con otro material fechado (4) asociaci6n estratigrafica con restos de animales introducidos restos humanos u objetos culturales propios del intervale (5) falta de asociaci6n con evidencias culturales [Los ntimeros entre parentesis en la columna Evidencia significan adscripci6n tentativa al intervalo] Los estimados del peso corporal (en kg) estan tomados de la literatura 0 inferidos comparativamente y distribuidos entre
las clases siguientes (a) S 01 (b) 01 a lt 1 (c) 1 a lt 10 (d) 10 a lt 100 (e) 100 a lt 200 (j) ~ 200 Las
abreviaturas que designan las islas son las de la Tabla 44
F amilialEspecie Peso IsJa Evidencia Fuente
I httervalo htterg1acial Sangamon (gt80 kaAP)
HEPTAXODONTlNAE
Amblyrhiza inundata e AS 3 McFarJane et al 1998a
CLIDOMYINAE
Clidomys osborni d JA 3 Ibid
Clidomys parvus d JA 3 Ibid
II httervalo Pleistoceno terminal ~ Holoceno temprano (20-6 kaAP)
MEGALONYCHIDAE
Aeratoenus antillensis d CD 5 Arredondo 1975 a
Aeratoenus odontrigonus d PR 3 McFarJane 1999b
Aeratoenus simorhynehus d LE 2 Rega et al 2002
M egaloenus zile f LE 2 MacPhee et al 2000
SOLENODONTIDAE
Solenodon arredondoi e CD 5 Morgan y Ottenwalder 1993
PnHECIIDAE
Paralouatta varonai d CD 5 MacPhee 1996a
EcHIMYIDAE
Heteropsomys antillensis c PR 5 McFarJane 1999b
Heteropsomys insulans c PR 5 Ibid
Puertoricomys eorozalus c PR 5 Woods 1989a
CAPROMYIDAE
P lagiodontia araeum c LE 5 MacPhee et al 2000
Macrocapromys latus c CD 5 Varona y Arredondo 1979
(Continua)
325 EXTINCIONES DEL CUATERNARIO
TABLA 45 (Continuaci6n)
F ami1ia1Especie Peso Isla Evidencia Fuente
M acrocapromys latus c CU 5 Varona y Arredondo 1979
Mesocapromys kraglievichi c CU 5 Ibid
HEPTAXODONTINAE
Xaymaca fulvopulvis a JA 2 MacPhee y Flemming 2003
III Intervalo Amerindio (6-05 kaAP)
MEGALONYCHIDAE
Acratocnus ye d LE 2 MacPhee et aI 2000
Neocnus comes c LE 2 Ibid
N eocnus dousman c LE 2 Ibid
N eocnus toupiti c LE 2 Ibid
N eocnus gliriform is c CU 4 Cordova et ai 1997
M egalocnus rodens f CU 2 MacPhee et ai 2007
Parocnus browni e CU 2 Steadman et ai 2005
P arocnus serus e LE 2 MacPhee et ai 2000
N ESOPHONTIDAE
N esophontes edithae a PR 2 McFarlane 1999b
PITHECIIDAE
Antillothrix bernensis c LE 3 Rimoli 1977
EcHIMYIDAE
Brotomys contractus a LE 4 Miller 1929c
CAPROMYIDAE
P lagiodontia spelaeum c LE 4 Ibid
M acrocapromys acevedo c CU 4 NUiiez 1990a
HEPTAXODONTINAE
Elasmodontomys obliquus d PR 3 McFarlane 1999b
Iv Intervalo Postcolombino (05 kaAP - Presente)
N ESOPHONTIDAE
N esophontes hypom icrus a LE 24 MacPhee y Fle111111ing 1999
N esophontes zam icrus a LE (4) Ibid
N esophontes m icrus a CU 24 MacPhee et aI 1999
(Continua)
326 BIOGEOGRAFiA
TABLA 45 (Continuacion)
F amilialEspecie Peso Isla Evidencia Fuente
N esophontes m icrus a CD 24 MacPhee et aI 1999
N esophontes m icrus a LE 4 MacPhee et aI 2000
N esophontes sp 1 a IC 4 Morgan 1994
N esophontes sp 2 a IC 4 Ibid
Solenodon marcanoi b LE 4 Ottenwalder 2001
PITHECIIDAE
Xenothrix mcgregori c JA 4 MacPhee y Flemming 1999
EcHIMYIDAE
Boromys offeUa a CD 24 Guarch 1984
Boromys torrei a CD 24 Ibid
Brotomys voratus a LE 4 MacPhee et aI 2000
CAPROMYIDAE
P lagiodontia ipnaeum c LE 4 MacPhee y Flemming 1999
Rhizoplagiodontia lemkei c LE 4 Ibid
Isolobodon portoricensis c PR 24 Ibid
Isolobodon montanus c LE 4 Ibid
Hexolobodon phenax c LE (4) Ibid
Capromys sp c IC 24 Morgan 1994
Geocapromys columbianus c CD 4 MacPhee y Flemming 1999
Geocapromys thoracatus b IS 1 MacPhee y Marx 1997
Geocapromys sp 1 c IC 4 Morgan 1994
Geocapromys sp 2 c IC (4) Ibid
l-IEPTAXODONTINAE
Quemisia gravis c LE (1) MacPhee y Flemming 1999
MURIDAE
Oryzomys antillarum a JA 1 Ibid
1 Flemming y MacPhee (1999) alertaron en contra de la asociacion con otro material fechado Asimismo Morgan (1993) indico que la ausencia de asociacion con evidencias culturales amerindias no es necesariamente un criterio concluyente para ubicar un taxon en el intervalo temporal anterior porque hay depositos en la region que carecen de tales evidencias y son del Holoceno tardio por fechados radiocarbonicos [No obstante
327 EXTlNCIONES DEL CUATERNAR10
la ausencia de una especie en residuarios arqueol6gicos adquiere mayor certidumbre como criterio de ubicaci6n temporal de su extinci6n en la medida que aumenta el numero de residuarios estudiados que en las Antillas Mayores se cuentan por centenares] A falta de fechado absoluto de una especie consideramos que estos criterios de ubicaci6n temporal son mas utiles que la alternativa de excluir del analisis a la especie
2 En los trabajos sobre biogeografia de mamiferos antillanos los arque610gos mas citados como autoridades para establecer el inicio de la presencia aborigen en las Antillas Mayores (Kozlowski 1974 Rouse y Allaire 1978 Rouse 1989 1992 Wilson 2001) difieren al respecto entre los 6 kaAP y los 45 kaAP (pero vease Pagan et al 200523) y hemos adoptado la mas temprana de estas fechas para el comienzo del intervalo amerindio en la Tabla 45 De todos modos cualquiera que haya sido la fecha esta no es aplicable por igual al conjunto de las Antillas Mayores porque segun algunos de los autores citados el poblamiento comenz6 por Cuba y La Espanola 6 mil anos atras continu6 en Puerto Rico hace alrededor de 3 mil anos (pero vease Burney et al 1994) yen Jamaica hace menos de 2 mil e incluso a las Islas Caiman ni siquiera llegaron [Las Islas Caiman descubiertas por Col6n en 1503 carecen absolutamente de evidencia amerindia (Stokes y Keegan 1996)]
Si los datos de la Tabla 45 representan una aceptable aproximaci6n a la realidad el proceso de las extinciones cuaternarias de los mamiferos terrestres de las Antillas Mayores y grupos de islas perifericas debi6 caracterizarse entre otros por los eventos siguientes
Aunque obviamente la fauna cuaternaria debi6 ser mucho mas numerosa que 10 que refleja el registro paleontol6gico (Morgan y Woods 1986) 52 especies (81 de la fauna conocida) desaparecieron (sin reposici6n) durante los ultimos 250 mil anos
- Al parecer las extinciones se concentraron en las especies de mayor tamano Asi el peso promedio estimado para las especies de la etapa preantr6pica (intervalos I + II de la Tabla 45) es 350 kg es 328 kg para las especies del intervalo III (amerindio) y es s610 25 kg (c1ases del peso a b c) para las especies del intervalo IV (postcolombino) Ninguna especie aut6ctona de mamifero mayor de 6 kg existe hoy en la regi6n
- De acuerdo con la distribuci6n temporal de las extinciones 61 especies sobrevivieron al interglacial Sangamon (intervalo 1)48 sobrevivieron a la transici6n del Pleistoceno terminal al Holoceno temprano (intervalo II) 34 sobrevivieron al periodo amerindio (intervalo III) y s610 12 especies han sobrevivido desde la Ilegada de los europeos (intervalo IV) Las tasas de extinci6n correspondientes (exceptuado el intervalo I) son una especie cada 1167 anos en el intervalo II una cada 393 anos en el intervalo III y una cada 23 anos en el intervalo IV
- Durante todo el proceso de las extinciones cuaternarias la perdida de especies por isla fue de 91 en La Espanola 83 en Jamaica y 62 en Cuba Bahamas retuvo su unica especie (aunque extirpada en varias de las islas) Puerto Rico e Islas Caiman perdieron todas sus especies (6 y 5 respectivamente) y las Islas Swan Virgenes y Anguila - San Martin perdieron sus respectivas especies (micas
328 BIOGEOGRAFiA
De los cuatro 6rdenes de mamiferos representados en la fauna Pilosa y Primates desaparecieron totalmente Soricomorpha y Rodentia perdieron 86 y 74 de sus especies respectivamente
CAUSALIDAD
CAUSAS NATURALES
Los cicIos glaciales e interglaciales acaecidos en el Pleistoceno causaron cambios ambientales dnisticos en las Antillas con la concomitante alternancia de cIimas frios (secos) y calidos (hUmedos) asi como de descensos y ascensos del nivel del mar aumento y disminuci6n de las areas territoriales) induso (aunque con menor intensidad) en el Holoceno Estos eventos paleogeograficos han sido consistentemente documentados (Mayo y Kartashov 1972 Bonnatti y Gartner 1973 Ortega y Arcia 1982 Prance 1982 Schubert y Medina 1982 Dodge et al 1983 Ortega [1984] Haq et al 1987 Schubert 1989 Hodell et al 1991 Guilderson et al 1994 Oppo 1997 Emslie 1998 Curtis et al 2001 Iturralde-Vinent 2003) y de ellos brindamos a continuaci6n una breve resena
Finalizando el periodo interglacial anterior a la ultima glaciaci6n (Wisconsin) el mar estaba a 8-10 m por encima de su nivel actual Cuando el avance glacial del Wisconsin a1canz6 su culminaci6n (eillamado Ultimo Maximo Glacial) hace 18 ka el mar habia descendido 120 m con respecto a dicho nivel y las islas del Caribe habian incrementado considerablemente sus extensiones subaereas en comparaci6n con las que presentan hoy dia por ejemplo Puerto Rico y las Islas Virgenes se conectaron territorialmente y las Bahamas se unieron en cinco gran des islas Prevalecia un regimen frio y seco y condiciones de aridez --debidas a una reducci6n de las precipitaciones (posiblemente a menos de 440 mmlano)- y los ambientes xericos se habian extendido a traves de las islas y bancos emergidos y abundaban los pinares (los habitats mesicos estaban restringidos a parcelas relictas en 10 alto de las montanas) En el Mar Caribe y el Golfo de Mexico la temperatura superficial del mar era 5degC mas baja que la actual y en las llanuras costeras y alturas maximas eran respectivamente 2-3degC y 6degC mas bajas Los limites de las zonas ecol6gicas altitudinales descendieron varios cientos de metros y cada zona redujo su extensi6n vertical al aumentar el gradiente de temperatura Ellimite inferior de la zona periglacial
elevaci6n donde la temperatura media minima a1canza los OdegC) de la Cordillera Central de la Republica Dominicana (altura maxima del Caribe insular con evidencia de topografia glacial) se considera que estuvo entre los 1500 y 1800 mlnm (actualmente esta a 2800 mlnm) y se puede estimar que dicho limite era inferior en Cuba por estar mas al norte que la Republica Dominicana No obstante durante este periodo la temperatura experiment6 episodios cortos (unas pocas decadas) de ascensos repetidos
Un signifieativo componente bi6tieo de las referidas condiciones paleoambientales 10 constituy6 la espectacular irradiaci6n evolucionaria experimentada por un grupo numeroso de aves rapaees diurnas y nocturnas (Wetmore 1937 Brodkorb 1969 Kurotchkin y Mayo 1973 Arredondo 1976 Olson 1978 Olson y Hilgartner 1982
329 EXTINCIONES DEL ClJATERNARIO
Suarez 2000a b 2004 Suarez y Emslie 2003) Los mamiferos de las Antillas Mayores ofrecieron a estos depredadores aviares una amplia gama de tamanos de presa que en ausencia de mamiferos del orden Carnivora en la region permitio a estas aves irradiarse a este nicho alimentario con la evolucion de una variedad de especies de muy diversa tall a corporal incluidas formas gigantes Adquirir un cuerpo gigante les propicio el acceso a recursos alimentarios que no estaban siendo explotados por ningun otro depredador (Alcover y McMinn 1994) A Cuba corresponde el mejor registro fosil de rapaces cuaternarias caribenas con siete generos de especies diurnas Dentro de estas aves existieron tres gavilanes gigantes (Accipitridae) tres carairas (Falconidae) y un condor (Cathartidae) asi como cuatro generos de especies nocturnas dos lechuzas (Tytonidae) y tres Mhos (Strigidae) dos de ell os no igualados en tamano en todo el mundo Algunos de estos generos tuvieron representacion en otras islas antillanas como La Espanola Jamaica Bahamas y Gran Caiman
Al retirarse hacia el norte los hielos de la glaciacion Wisconsin el clima se tomo mas hUmedo en las Antillas durante el Holoceno temprano y el bosque mesico se expandio en la region insular El nivel del mar ascendio acompanado de mayor pluviosidad alrededor de los 7 kaAP las Islas Virgenes quedaron separadas entre si y de Puerto Rico y las Bahamas perdieron la mayor parte de su territorio Cerca de los 3 kaAP las condiciones secas retornaron a la region e hicieron desaparecer buena parte del bosque mesico Nuevamente el clima comenzo a adquirir sus condiciones actuales hacia los 25 kaAP aproximadamente y hoy transita por una fase mesica con vegetacion xerica confinada en parcelas relictas
Sin duda estos y otros eventos naturales incidieron negativamente en la sobrevivencia de mamiferos terrestres en las Antillas Mayores asi como en los grupos de pequenas islas perifericas (Pregill y Olson 1981 Olson 2006)
McFarlane et al (1998a) demostraron convincentemente como Amblyrhiza el gigante de los roedores antillanos endemico de la pequena isla de Anguila sucumbio a las repetidas y abruptas perdidas de su habitat durante el interglacial Sangamon por las nipidas y severas oscilaciones del nivel del mar (documentadas por Neumann y Hearty 1996) al no poder responder adecuadamente a tan catastroficos eventos mediante el ajuste de su masa corporal media y otro tanto debio ocurrirle en Jamaica al otro roedor gigante Clidomys (McFarlane et al 1998b) emparentado con Amblyrhiza en la misma epoca (Tabla 45 intervalo I)
El ascenso del nivel del mar seguramente promovio la extincion de mamiferos durante la transicion del Pleistoceno al Holoceno (Tabla 45 intervalo II) con la reduccion del tamano de las Islas por las transgresiones marinas (Pregill y Olson 1981 Morgan 1993) Esto ultimo haria maximamente severa la competicion asf como la presion de los depredadores [Steadman et al (2005) no yen por que el calentamiento global ocurrido durante la transicion Pleistoceno-Holoceno habria de tener un efecto negativo sobre los perezosos siendo sus descendientes actuales especies tropic ales 0 subtropicales pero no se trata del efecto
330 BIOGEOGRAFiA
directo del cambio de temperatura sobre los animales sino del efecto indirecto y progresivo que dicho cambio genera a largo plazo al reducir el territorio insular disponible]
Por esta epoca tambien desaparecieron especies de tall a mayor como Megalocnus zile en La Espanola y otras de menor tamano (en Cuba) pero que representan la tall a maxima en sus respectivos contextos taxonomicos del Cuaternario como Solenodon arredondoi Paralouatta varonai y Macrocapromys acevedo Un mamifero puede ser grande solo a costa de reducir su densidad poblacional y esto 10 hace mis vulnerable a la extinci6n sobre todo en una isla (McNab 1994 McFarlane et al 1998a) Incidentalmente Alcover et al (1998a) afirmaron que larelacion entre la talla corporal y la extincion no siempre se cumple en las especies de menor tamano y citaron como ejemplo que entre las musarafias todas las especies de Nesophontes desaparecieron mientras sobreviven especies de Solenodon pero la comparacion correcta en todo caso serfa entre los solenodones vivientes y el gigante Solenodon arredondoi y entre Nesophontes edithae -el gigante de los nesofontes extinguido en el periodo amerindio- y el resto de sus congeneres que sobrevivieron hasta los primeros siglos de la colonizacion europea (Tabla 45 intervalos III y IV)
CAUSAS ANTROPOGENICAS
Intervalo Amerindio
La mayoria de los autores que se han referido a las extinciones acaecidas durante este intervalo (entre elIos paradigmaticamente Martin 1984a b) han responsabilizado a los aborfgenes (que se supone comenzaron a poblar las Antillas Mayores hace 6 mil afios) con la desaparicion de numerosas especies de mamiferos terrestres en la region principalmente por la via de la sobrecaza y la introduccion de animales domesticos devenidos depredadores de la fauna autoctona (Morgan y Woods 1986) Este efecto negativo de la presencia indigena sobre los mamiferos de la region ha sido dramatizado por algunos autores en terminos de holocausto faunal Sin embargo pensamos que se ha exagerado Como han expresado MacPhee et al (19993) cuando uno busca evidencia dura de complicidad humana directa en [estas] extinciones no hay mucho que encontrar Las siguientes son algunas consideraciones a tener en cuenta en tal sentido
El registro radiometrico mas joven conocido hasta ahora para un perezoso antillano (MegalocnllS rodens) es de 42 kaAP (MacPhee et ai 2007) Si se acepta que los aborfgenes entraron en Cuba hace aproximadamente 6 mil anos su convivencia con los perezosos debe haber durado alrededor de 1800 afios (0 mas porque dificilmente el referfdo fechado radiometrico haya correspondido al ultimo sobreviviente de la especie) Esta cifra contradice la hipotesis del holocausto faunal segun la cualla extincion de los mamiferos insulares de mayor talla debe ocurrir con extrema rapidez -a escala de decadas 0 de centurias- a partir de su primer contacto con los humanos (Martin y Steadman 1999) A pesar de las repetidas afirmaciones en contrario (Harrington 1921 Miller 1929c Herrera [1945] Tabio 1951 Alvarez 1951a 1957 Arredondo
331 EXTINCIONES DEL CUATERNARIO
1959 1960a Veloz y Ortega 1973 Mayo 1980b Rodriguez et al 1984 Castellanos y Pino 1985 Pino y Castellanos 1985 Nunez 1990b Gutierrez 1996 Arredondo 2004) en su mayoria basadas en evidencias no exentas de posibles accidentes de deposicion (0 de reubicacion postdeposicional) en recintos cavernarios (Diaz-Franco [2006b D no se ha demostrado inequivocamente que los aborigenes antillanos se alimentaran de perezosos regularmente Sus restos en residuarios de alimentos son escasisimos en toda la region y esto contrasta con los abundantes restos de jutias y otros mamiferos de menor talla que aparecen en esos residuarios demostrativos de la preferencia de los paleoindios por estos otros mamiferos
Los mamiferos mas importantes en la economia prehistorica de las Antillas Mayores fueron las jutias Capromys y Geocapromys en Cuba Geocapromys en Jamaica e Isolobodon en La Espanola Puerto Rico e Islas Virgenes (Aguayo 1950b Tablo 1951 Rimoli 1972 Wing 2001 Diaz-Franco [2006b D Wilkins (2001) estudi61ajutia viviente de Jamaica (Geocapromys browni) en un sitio ocupado por los aborigenes durante 100 anos y no encontr6 evidencias de que durante ese tiempo la especie haya sido sobreexplotada Asimismo en Cuba las jutias conga carabali y andaraz son escasas en depositos pleistoholocenicos en cambio son muy numerosas en residuarios indigenas y de haber sido sobreexplotadas 10 mas probable es que no hubieran sobrevivido hasta hoy
La habilidad de los humanos para penetrar en profundidad el bosque tropical y mantenerse en este sustentablemente ha sido un evento comparativamente reciente (Beran y Steele 1994) En el caso antillano la caza tierra adentro ha sido explicada como un proceso tardio de desplazamiento a partir de la explotacion de abundantes recursos marinos que ofrecian condiciones 6ptimas para el sedentarismo de las comunidades aborigenes costeras (C6rdova y Arredondo 1988 Brito 2005) Peces y mariscos dominan la arqueologia costera de las Islas antillanas en tanto el consumo de recursos terrestres (mamfferos incluidos) aumenta progresivamente en la medida que los residuarios estudiados se alejan de la costa (Scudder 1991 Wilkins 2001) En las Antillas Mayores el componente terrestre de la dieta aborigen es solo 20-30 del material recobrado en los residuarios (Wing 1977 1989) Y debio ser menor alin porque el componente vegetal de la dieta no se ha preservado Por otra parte los aborigenes cubanos domesticaron la jutia conga y seguramente la reprodujeron para comer (Pose et al 1988) practica que probablemente se repitiera con otras jutias en las demas Antillas Mayores
- Fray Bartolome de las Casas hablo de 3 millones de indios en Cuba al inicio de la colonizacion pero los historiadores subsecuentes consideraron exagerada la cifra (Poey 1851 a) No obstante se dispone de dos estimados de la densidad de poblaci6n aborigen en las Antillas Mayores a la llegada de los europeos 18 indigenaslkm2 en Puerto Rico (Rouse 1956) y 1271km2 en La Espanola (Watts 1987) Extrapolando ambos valores entre las cuatro islas principales de las Antillas Mayores tenemos que en la fecha indicada la poblaci6n aborigen pudo estar entre 16 mil y 113 mil indigenas en Puerto Rico entre 21 mil y 145 mil en Jamaica entre 141 mil y 1 mill6n en La Espanola y entre 200 mil y 14 millones en Cuba Entre las cifras maximas y las minimas nos inclinamos
332 BIOGEOGRAFiA
hacia estas ultimas atendiendo a los altos indices de mortalidad infantil y morbilidad que se evidencian en las osamentas de los pobladores prehispanicos (Martinez y Rives 1990) Uno de los hechos que apuntan hacia el bajo numero de aborigenes en las Antillas Mayores es 10 rapido que desaparecieron despues de la llegada de los europeos pnkticamente sin dejar mestizaje con estos 10 cual contrasta ostensiblemente con la situacion en la America continentaL
En suma un grupo humano (1) demograiicamente irrelevante y controlado por altos indices de morbilidad (2) consumidor mayoritariamente de organismos marinos y (3) carente de la tecnologia cinegetica necesaria para poder sobreexplotar los mamiferos que Ie sirvieron de alimento (a los cuales accedio tardiamente en su desarrollo) no es concebible que haya causado e1 holocausto faunal que se Ie atribuye a partir del error de aplicar a las comunidades aborigenes antillanas un modelo cultural de la prehistoria continentaL
Tampoco hay evidencias de que los aborigenes hayan introducido carnivoros con suficiente capacidad depredadora salvo el perro Morgan y Otlenwaldder (1993) consideraron que la depredacion por perros durante el periodo amerindio es la explicacion mas plausible para la extincion del almiqui gigante de Cuba pero como se puede observar (Tabla 54) Solenodon arredondoi aparece en el intervalo II por falta de asociacion con evidencias culturales
Consecuentemente el accionar de los aborigenes y de sus animales domesticos directamente sobre la fauna no parece haber sido por sf solo un factor determinante de la extincion de mamiferos terrestres durante el periodo amerindio en las Antillas (pero vease Steadman et al 2005)
Hay que recordar en este senti do que alin en el Holoceno tardio ocurrieron cambios clirruiticos de consideracion (vease Causas Naturales) Por ejemplo Curtis et al (2001) explicaron que la extincion de algunos ani males mesofilicos de las alturas montanosas de La Espanola pudo haber antecedido a cualquier impacto humano sigrIificativo en esos lugares por coincidir dicha extincion con una tendencia a la aridez documentada isotopicamente y con evidencias de la perdida del bosque mesico ocurridas despues de 3kaAP Yen Jamaica hace solo 2shy3 mil aDos el area de la Bahia de Jackson sustentaba una gran biomasa de vertebrados en bosque hinnedo a juzgar por la cantidad y diversidad de taxones presentes en material fosil derivado de la depredacion por lechuzas hoy el area es la mas seca de la isla con matorral xerofitico y pobreza de fauna (MacPhee 1996a) Yalm en nuestros dias huracanes de gran intensidad aniquilan poblaciones de vertebrados (que no se recuperan) en is las pequenas y llanas del Caribe (Brooks y Smith 2001)
En medio de la incertidumbre sobre la verdadera causa (0 causas) de las extinciones de mamiferos en el periodo amerindio MacPhee y Marx (1997) han propuesto su hipotesis de la hipermorbilidad la cual estipula colapsos poblacionales relativamente rapidos con la introduccion de enfermedades de alta letalidad Estos autores han razonado que
Las islas son los lugares mas apropiados para experimentar perdidas catastroficas causadas por enfermedades Los mamiferos no solo serian susceptibles en virtud de su prolongado aislamiento biologico sino que en los confines de una isla la epizootia
333 EXTINCIONES DEL CUATERNARIO
tendria la potencialidad de infectar simultaneamente a la rnayoria de los individuos de vadas especies Los neonatos son inmunologicamente inmaduros y sufririan desproporcionadamente cualquier infeccion hipervirulenta La perdida excesiva de neonatos y juveniles tendria efectos severos para la sobrevivencia de la poblacion si esta se reduce por debajo de cierto nivel sobreviene la extincion porque no hay posibilidad de restitucion en el periodo postpanzootico Las especies de mayor talla corporal van a ser las mas vulnerables a la extincion dadas sus bajas tasas reproductivas largos periodos de gestacion y reducido numero de progenie Las especies de menor tamano --caracterizadas por altas tasas reproductivas periodos cortos de gestacion y mayor numero de neonatos por parto- tambien serian fuertemente diezmadas pero muchas sobrevivirian porque su estrategia reproductiva les permite perder mayor numero de individuos Las perdidas deben ocurrir relativamente rapido (a escala de decadas) mientras que otras hipotesis (digamos los cambios climaticos) permiten intervalos mucho mas largos (a escala de siglos 0 de milenios) entre la aparicion de las causas y la produccion de sus efectos Por ejemplo todo indica que los nesofontes se extinguieron muy rapidamente y seria dificil poder invocar un impacto humano directo 0 argumentar fenomenos climaticos que pudieran explicar plausiblemente perdidas tan rapidas
MacPhee y Marx (1997 191) han dicho que por el momento la hipotesis de la hipermorbilidad no pasa de ser una abstraccion en espera de adecuada sustentacion empirica Llama la atencion que linicamente en el periodo amerindio (Tabla 45 intervalo 1Il) las extinciones abarcaron todas las clases del peso corporal (de la a a laj) y que ninguna de las especies que sobrevivieron hasta el periodo postcolombino (Tabla 45 intervalo IV mas las especies vivientes) sobrepasa los 10 kg (clases del peso a b c) en clara coincidencia con los presupuestos de la hipotetica hipermorbilidad
Intervalo Postcolornbino
Por 10 menos 37 especies de mamiferos terrestres vivian en las Antillas Mayores y grupos de Islas perifericas a la lIegada de los europeos en 1492 De elIas se han extinguido 23 shymas especies que en cualquiera de los periodos precedentes (Tabla 45~ y solo subsisten 12 especies repartidas entre Cuba (8 especies) La Espanola (2) Jamaica (1) y Bahamas (1) varias de estas en peligro critico de extincion Solo dos de las especies extinguidas durante los ultimos 500 anos se conocieron en estado viviente los roedores Oryzomys antillarum de Jamaica (extinguido en 1877) y Geocapromys thoracatus de las Islas Swan (extinguido en 1955) Las causas de las extinciones ocurridas durante este periodo son sobradamente conocidas destruccion de habitats introducci6n de animales exoticos (domesticos y comensales) y demas presiones asociadas a la actividad humana entre las cuales posiblemente habria que incluir brotes de hipermorbilidad
Durante mucho tiempo se ha afirmado la posibilidad de que especies de Nesophontes hayan sobrevivido hasta las primeras decadas del siglo xx y que incluso continuen existiendo en lugares remotos de Cuba 0 La Espanola (Allen 1918 1942 Miller 1930 Aguayo 1950a Arredondo 1960b Fischer 1977 MacFadden 1980 Woods y
334 BIOGEOGRAFiA
Ottenwalder 1991) Actualmente no existe evidencia radiometrica de que alguna especie de Nesophontes haya sobrevivido al siglo xv las fechas radiocarbonicas calibradas obtenidas en muestras de Nesophontes de Cuba y La Espanola resultaron no mas tardias que los siglos XIII y XIV (MacPhee et al 1999) Por otra parte las expediciones efectuadas a fines del siglo xx con el proposito de localizar individuos vivientes del genero en La Espanola no produjeron la menor indicacion afirmativa al respecto (Woods et al 1985) Ademas a traves de las Antillas Mayores no aparecen restos de Nesophontes en los depositos formados actualmente por las lechuzas
EPiLOGO
Las Antillas Mayores son islas oceanicas no han formado parte de un continente (con la posible excepcion de una parte de Jamaica hasta el Eoceno Medio hace 45 Ma) y durante un tiempo geologico considerable (desde el Eoceno Superior hace 40 Ma) han permanecido emergidas y separadas de los continentes vecinos Consecuentemente los mamiferos terrestres (no volantes) autoctonos de estas islas constituyen una fauna oceanica que por ende cumple con todas las caracteristicas convencionales reconocidas para este tipo de entidad biogeografica
- Baja diversidad taxonomica a nivel de las categorias superiores (solo cuatro ordenes) en comparacion con islas plataformicas continentales (0 con areas continentales) de tamano y diversidad ecologica simi lares
- Alto grado de endemismo a nivel de genero (96) y familia (50) en respuesta al prolongado aislamiento
- Desbalance ecologico de la estructura trofica ausencia de mamiferos camivoros y ungulados
- Espectaculares procesos de irradiaci6n evolucionaria como resultado de la colonizacion de territorios libres de competidores y depredadores
Tendencias evolucionarias extremas gigantismo
- Control a cargo de aves rapaces con tendencia at gigantismo en ausencia de mamiferos miembros del orden Carnivora
- Mas especies de reptiles y de murcielagos que de mamiferos terrestres relaciones que en los continentes se invierten
Alta tasa de extincion de especies (84) sin reposicion
Preservacion de especies filogeneticamente arcaicas (los llamados fosiles vivientes) los almiquies

309 POBLAMIENTO CONTINENTE- ISLA
la absoluta mayona de los mamfferos terrestres de ongen suramericano porque el caracter de esa fauna supone un mecanismo de poblamiento ancestral que sea a la vez maximamente selectivo (baja diversidad ordinal) y temporalmente restringido (transicion Eoceno-Oligoceno)
Recientemente Rega et al (2002 18) expresaron A pesar de una revision exhaustiva de la paleogeografia de la cuenca del Caribe Iturralde-Vinent y MacPhee (1999) no pudieron confirmar la presencia de una conexion terrestre continua entre Suramerica y las proto-Antillas aunque hipotetizaron su existencia [Vease la Fig 42 (= figura 6 de Iturralde-Vinent y MacPhee 1999 simplificada))
En el trabajo citado por Rega y colaboradores aparece la siguiente explicacion
Actualmente no es posible ofrecer reconstrucciones paleogeograficas detalladas de la Cresta de Aves Sin embargo hay evidencia de que porciones de la cresta deben haberse elevado subaereamente en un tiempo La cuestion cntica es si la Cresta de Aves emergida pudo haber formado algun tipo de corredor entre las Antillas Mayores y el noroeste de Suramerica Perforaciones en las alturas estructurales de la cresta pudieran proveer el registro del Paleogeno tardio que se necesita para precisar la hipotesis del corredor aunque no se puede predecir si tales datos serian suficientemente detallados para determinar si en algun momento GAARlandia constituyo una sola entidad subaerea Por ejemplo pudo haber barreras marinas por corto tiempo entre las masas terrestres de la Cresta de Aves [0) ellevantamiento y la subsidencia pudieron afectar diferentes partes de GAARlandia en diferentes tiempos AItemativamente los elementos faunales pudieron haberse dispersado de manera pulsada Movimientos episOdicos de este tipo incrementarian la extincion de los elementos faunales incapaces de trasladarse a la siguiente masa terrestre en la cadena disminuyendo asi la diversidad total (Iturralde-Vinent y MacPhee 199957)
La primera parte de la explicacion deja claro que la posibilidad de que la Cresta de Aves haya sido una conexion terrestre continua no se puede descartar pero tampoco se puede afirmar La ultima parte de la explicacion describe el efecto altamente selectivo (filtrante) de la colonizacion a traves de una cadena de islas
Y continuan lturralde-Vinent y MacPhee (199957 -58) explicando
Hasta que punto el microcontinente noroccidental de Suramerica [considerado por dichos autores como el area suministradora de propagulos hacia GAARlandiaJ estuvo fisicamente separado del resto del continente no ha sido plenamente esc1arecido Si estuvo sustancialmente aislado por barreras marinas y orogenic as durante el Paleogeno tardio entonces solo la fraccion de la biota suramericana emplazada en el microcontinente estuvo en posicion de acceder a GAARlandia Esto pudiera tener que ver con el hecho de que la fauna antillana carezca de representantes de muchos grupos suramericanos [de mamfferos) que debieron haber hecho el viaje
310 BIOGEOGRAFiA
Florida o Alturas0 0
d 6 D L1anuras25deg
Yucatan
20deg
deg0
10deg
MARCARIBE
80deg 75deg
Bahamas
Cres ta de Beata
D Mares someros
D Mares profundos
OCEANO ATLANTICa
FIGURA 42 Reconstrucci6n hipotetica de GAARlandia como conexi6n terrestre continua entre las proto-Antillas Mayores y el noroeste de Suramerica en el Pale6geno tardio segun la FIgura 6 (simplificada) de Iturralde-Vinent y MacPhee (1999)
En su defensa del puente terrestre Barbour (1945b249) se refiri6 a esto - antes 10 habfa hecho Anthony [1942363]- en los terminos siguientes
El argumento de que si las Antillas hubieran estado conectadas con el continente se encontrarian los f6siles de un numero mayor de distintas criaturas me deja frio es perfectamente concebible que la conexi6n haya ocurrido en un periodo en que la parte del continente implicada no estuviera poblada por una fauna grande y variada
Y concluyen lturralde-Vinent y MacPhee (199958) su detallada explicaci6n
Desafortunadamente sabemos muy poco sobre la composici6n faunal del noroeste de Suramerica a finales del Pale6geno
Consecuentemente si se confirmara la magnitud del aislamiento del aludido microcontinente y su concomitante baja diversidad faunal GAARlandia como conexi6n terrestre continua serf a la hip6tesis mas parsimoniosa No obstante mientras no se disponga de evidencia concluyente a este respecto hipotetizar a GAARlandia como conexi6n continua (minima selectividad) la coloca en franca vulnerabilidad te6rica al tener que afrontar la objeci6n clasica al puente terrestre (baja diversidad ordinal de la
311 POBLAMIENTO CONTINENTE-TSLA
fauna) al tiempo que se la priva de ser la hip6tesis mas robusta para explicar -como guimalda de islas- la colonizaci6n de las Antillas por mamiferos procedentes de Suramerica
ELARcA DE NOE (seudovicarianza)
El primer vicarianista antillano (desde luego sin el saberlo) fue Corral (1940) con su utilizaci6n ingenua de las ideas wegenerianas de la deriva continental aparecidas la decada anterior Pero el primer (supuesto) intento de aplicar a la formaci6n de la biota caribefia el mecanismo de la vicarianza basada en la tearia de la tect6nica de placas (surgida en la sexta decada del pas ado siglo) se debe a Rosen ([1976] 1985)
Segun Rosen las Antillas Mayores se originaron en el Cremcico en la Placa Caribe aproximadamente en la posici6n que hoy dia ocupa Centroamerica y al ser desplazadas tectonicamente hacia el este interactuaron (de algun modo no precisado por Rosen) con las margenes continentales de Norte- y Suramerica de donde (en un evento unico) adquirieron toda su fauna transportada subsecuentemente por dichas islas en su transito hacia sus posiciones actuales [De ahi la denominacion metaforica dada par McKenna (1973) a este mecanismo de poblamiento] Aunque se ha presentado esta propuesta como Model0 de Vicarianza Continente-Isla de Rosen en realidad por definici6n no constituye un evento vicariante porque las proto-Antillas Mayores no resultaron del fraccionamiento y separaci6n por tectonismo de una porcion del territorio continental como exige esta hip6tesis (vease definici6n de vicarianza al comienzo de Hipotesis Biogeograficas)
El Arca de Noe de Rosen ha sido desestimada por la mayoria de los autores contemporaneos porque (1) carece de sustento geol6gico (lturralde-Vinent y MacPhee 1999) y porque (2) en el periodo en que se afrrrna ocum6 el evento muchos de los ordenes modemos de mamiferos placentarios attn no existian (Williams 1989)
EL BARCO FUNERAL VlKINGO (tanatovicarianza)
Domning et al (1997) reportaron 10 que consideraron el mamffero terrestre mas viejo de las Antillas --circunstancia que se refleja en el titulo mismo del trabajo Oldest West Indian Land Mammal- al descubrirse un perisodactilo rinocerotoide (Hyrachius sp) en una localidad de edad Eoceno Inferior a Medio en el occidente de Jamaica
Jamaica ha tenido una historia geologica y paleogeografica distinta de la historia comun a las restantes Antillas Mayores A principios del Cenozoico Jamaica occidental (a diferencia de la porcion oriental constituida por el Bloque de las Montafias Azules) formaba parte de Centro america de la cual se desgaj6 (por el desplazamiento de la Placa Caribe) para venir a integrarse despues del Eoceno Medio con 10 que serfa la futura isla de Jamaica desaparecida por transgresi6n marina en el Eoceno Superior hasta volver a emerger totalmente en el Mioceno Medio (Iturralde-Vinent y MacPhee 1999 MacPhee e Iturralde-Vinent 2000)
312 BIOGEOGRAFiA
Por consiguiente Hyrachius ---conocido de localidades de edad Eoceno Inferior a Medio en Norteamerica- debio ocupar (10 que iba a ser) Jamaica occidental mucho antes de que esta se separara de Centroamerica De hecho este fosil Uego a su destino insular contenido en las rocas de su basamento no como un ser vivo por 10 que es erroneo considerarlo un animal antillano (lturralde-Vinent y MacPhee 2004) como han supuesto algunos (Hedges 200 I Portell et al 200 I Condis [200 I D Todo 10 que hizo fue viajar tanatovicariantemente en su barco funeral vikingo -otra de las denominaciones metaforicas de McKenna (1973)
DISCUSION Y CONCLUSIONES
Hemos visto que solo disponemos de una hipotesis plausible para explicar la mayor parte del poblamiento (continente-isla) de las Antillas Mayores por mamiferos terrestres del Neotropico continental la dispersion mixta (a traves de GAARlandia) para los perezosos (excepto Choloepodinae) los monos y los roedores (excepto Oryzomys)
Por otra parte la historia biogeogrMica de las musaraiias de las Antillas Mayores (Nesophontes y Solenodon) constituye una de las situaciones mas enigmaticas entre los mamiferos de la region (Ottenwalder 2001321) Al menos cinco mecanismos de poblamiento continente-isla han sido invocados para explicar cuando como y de donde llegaron a las Antillas Mayores estas musaraiias
I EI propagulo ancestral (del Terciario de Norteamerica) coloniz6 a las Antillas Mayores por dispersion marina desde Centroamerica 0 desde la Florida (Patterson 1962)
2 El propagulo ancestral ( del Terciario) cruzo el Atlantico Sur (supuestamente mas estreshycho en aquella epoca) directamente de Africa a las Antillas Mayores (Hershkovitz 1972)
3 El propagulo ancestral (del Terciario de Norteamerica) Uego a las Antillas Mayores en el Arca de Noe de Rosen (MacFadden 1980)
4 El propagulo ancestral (del Terciario de Norteamerica) fue transportado a las Antillas desde Centroamerica en el bloque de Jamaica occidental y de alli paso por dispersion marina a las restantes Antillas Mayores (Whidden y Asher 200 I)
5 El propagulo ancestral (del Terciario de Norteamerica) lleg6 a las Antillas Mayores a traves de GAARlandia (Whidden y Asher 2001)
Aunque hay consenso entre estos (y otros) autores sobre el origen norteamericano de las musaraiias antillanas ninguno de los cinco hipoteticos mecanismos de poblamiento resiste la evidencia en contrario filogenetica yo paleogeogrMica Ademas algunas de estas hipotesis asumen que Nesophontidae y Solenodontidae comparten un antecesor comun en las Antillas pero esta relacion no ha sido confirm ada Para dilucidar el enigmatico caso de las musaraiias antillanas se requerini profundizar en la investigaci6n geologica paleogeognifica y paleontol6gica (Iturralde-Vinent y MacPhee 1999)
De acuerdo con el conocimiento filogenetico disponible sobre el conjunto de la fauna la colonizacion original de las Antillas Mayores por mamiferos terrestres implicaria el arribo
313 POBLAMIENTO ISLA-ISLA
de solo siete propagulos fundadores perezosos 2 (coloepodinos y megalocninos si como parece tienen un origen difiletico) musaraiias 2 (nesofontidos y solenodontidos si no constituyen una misma familia como consideran algunos) monos 1 (pitecidos) roedores histricognatos 1 (equimidos capromidos y heptaxodontidos) roedores muridos 1 (Oryzomys en Jamaica)
POBLAMIENTO ISLA-ISLA
La distribucion de los mamiferos terrestres a traves del area antillana demarcada al inicio del presente capitulo (vease Contexto Geogratico Fig 41) ha sido interpretada en funcion de dos hipoteticos mecanismos de poblamiento interinsular la dispersion y la vicarianza fundamentadas como se resumen seguidamente
DISPERSION
De acuerdo con la evidencia paleogeogratica mas actualizada en las principales regresiones marinas ocurridas en la cuenca caribeiia durante los ultimos 23 Ma (transiciones OligocenoshyMioceno Mioceno Superior - Plioceno y varias veces en el Pleistoceno) las masas terrestres constitutivas de la region insular en formacion aumentaron considerablemente su extension territorial disminuyendo por tanto la distancia entre elias oportunidades que los elementos faunales aprovecharian para dispersarse de unas islas a otras
Durante mucho tiempo esta ha sido la hipotesis mas favorecida por los estudiosos del tema a partir de Simpson (1956) pero en decadas recientes ha suscitado dos objeciones
1 Es dificil en tender como pudo operar la dispersion marina entre estas islas puesto que ninguna de las Antillas Mayores posee rios suficientemente caudalosos para transportar masas significativas de vegetacion flotante (MacPhee 198432)
Es oportuno recordar aqui el islote ala deriva (con mas de diez palmas reales en pie) entre las tres Antillas Mayores mas occidentales mencionado anteriormente (vease La Balsa Natural)
2 La dispersion marina es menos efectiva para explicar la asombrosa similitud de las faunas de mamiferos terrestres a traves de las Antillas Mayores septentrionales porque como proceso aleatorio no debe producir distribuciones uniformes a 10 largo de una cadena de islas 0 si las produce entonces el proceso no es aleatorio (MacPhee e lturralde-Vinent 2000 149)
Independientemente de que las aludidas distribuciones taxonomicas estan lejos de ser uniformes 0 asombrosamente similares la hipotesis de la dispersion marina contrariamente a su canicter aleatorio y altamente improbable en el caso del poblamiento continente-isla- en el caso del poblamiento isla-isla es un proceso no aleatorio (aprovecha las oportunidades brindadas puntualmente por los periodos de maxima regresion marina) y ademas altamente probable dada la incomparablemente menor distancia entre las islas caribeiias que entre estas y los continentes vecinos
314 BIOGEOGRAFiA
V ICARIANZA
La neotectonica y las transgresiones marinas del Oligoceno Medio y del Mioceno afectaron sustancialmente la paleogeografia de la Cresta de las Antillas Mayores resultando la subdivision gradual de las partes subaereas de la Cresta Asi el este de Cuba y el norte de La Espanola conectados fisicamente durante el Oligoceno Inferior se separaron al formarse el Paso de los Vientos en el Oligoceno tardio en tanto La Espanola y Puerto Rico probablemente permanecieron conectadas hasta el Mioceno tardio en que surgio el Paso de Mona (Iturralde-Vinent y MacPhee 1999)
Tales eventos geologicos habrian dividido las distribuciones de los grupos faunales emplazados previamente en el territorio continuo de las tres islas y a partir de esas disyunciones dichos autores (tambien MacPhee et al 2000 y White y MacPhee 2001) asumieron --como segunda fase del modelo de GAARlandia- la hipotesis de la vicarianza isla-isla para explicar la distribucion multiinsular de varios grupos de mamiferos terrestres a traves de las Antillas Mayores septentrionales De acuerdo con esta hipotesis cualquier diferencia en composicion faunal entre las islas (por ejemplo la ausencia de roedores heptaxodontidos en Cuba 0 de monos platirrinos en Puerto Rico) constituye evidencia de extincion local postvicariante y no de dispersion marina fallida Lo primero se puede demostrar si aparecen los fosiles pertinentes 10 segundo no se puede demostrar (MacPhee e Iturralde-Vinent 1994) Por otra parte IturraldeshyVinent y MacPhee (1999 57) han reconocido que aunque fa vicarianza isla-isla puede representar una solucion clara para las distribuciones de varios taxones cuaternarios no puede explicar todos los casos
Rega et al (2002 18) han presentado dos objeciones al modelo de la vicarianza isla-isla aplicado a los mamiferos de las Antillas Mayores
I Si la separacion de Cuba y La Espanola precedio en varios mill ones de anos la separashycion de La Espanola y Puerto Rico el modelo vicariante de lturralde-MacPhee debe predecir mayor similitud entre las faunas de La Espanola y Puerto Rico que entre las de Cuba y La Espanola De hecho es al reyes
No obstante MacPhee et al (200335) admitieron despues que la fecha mas temprana empiricamente defendible [para la apertura del Paso de Mona] debe ser el Oligoceno Medio esto invierte la secuencia de ambas disyunciones con 10 cual queda resuelta la aparente contradiccion Sin embargo en su reciente evaluacion de la congruencia entre filogenia e hipotesis geologic as en varios grupos de mamiferos terrestres de las Antillas Mayores Davalos (2004373) encontro que ninguno de los taxones cumple con la fecha y el patron de divergencia entre las masas terrestres antillanas postulados por el modelo de GAARlandia
2 La presencia de un mismo genero en las tres Islas septentrionales de las Antillas Mayores implica [por el modelo vicariante] que la diferenciacion entre estas formas durante el intervalo desde el Mioceno ha sido unicamente de nivel especifico [Dicha presencia]
315 rRRADlAcr6~ EVOLUCrONARIA
s610 puede explicarse razonablemente como el resultado de movimientos de esos animales entre las islas en el Cuaternario
De esta objecion resulta dificil disentir [El poblamiento entre islas diferenciado solo a nivel especifico no es probable que sea anterior al Pleistoceno (Simpson 1956 15)]
Debe tenerse en cuenta que la hipotesis de la vicarianza isla-isla alude a las tres islas septentrionales de las Antillas Mayores porque como se explico antes (vease El Barco Funeral Vikingo) Jamaica no existia como isla durante el lapso (Eoceno Superior shyOligoceno) consensualmente reconocido como el mas probable para la colonizacion de las proto-Antill as Mayores por la mayo ria de los mamiferos terrestres Despues de emerger como isla en el Mioceno Medio Jamaica debe haber recibido sus mamiferos terrestres por dispersion isla-isla (Davalos 2004376) y solo en el Cuaternario parece haberrecibido (por dispersion marina desde Centroamerica) al roedor murido Oryzomys
Seguramente tanto la dispersion como la vicarianza contribuyeron a que los mamiferos terrestres se irradiaran a traves de las Antillas Mayores y grupos de islas perifericas pero las maneras y tiempos en que estos mecanismos operaron a 10 largo de dicho proceso son aun en gran medida motivos de desacuerdo e incertidumbre
IRRADIACION EVOLUCIONARIA
A continuaci6n revisamos someramente las distribuciones conocidas de los diferentes grupos de mamiferos terrestres a traves de las Islas principales de las Antillas Mayores y grupos de Islas perifericas incluidas las interpretaciones y conjeturas que la literatura reciente recoge en relaci6n con la manera en que dichas distribuciones pudieron haberse conformado durante la irradiaci6n evolucionaria de los referidos grupos de mamiferos Manifestaciones de esta irradiaci6n pueden inferirse unicamente a partir de los taxones presentes en el Cuatemario pues solo 6 de las especies conocidas corresponden al Terciario
GRUPOS ORlGINARIOS DE SURAMERICA
PEREZOSOS (Megalonychidae)
Aparte de un genero miocenico de Cuba y otro del Oligoceno de Puerto Rico se conocen 4 generos (12 especies) de perezosos cuaternarios (todos extintos) restringidos a las tres islas septentrionales de las Antillas Mayores Acratocnus Neocnus Megalocnus y Parocnus Los cuatro generos estan representados en Cuba y La Espanola y Acratocnus 10 esta ademas (como unico genero de perezoso) en Puerto Rico Esta distribuci6n ha sido interpretada por MacPhee et al (2000) eomo resultado de la vicarianza oligomiocenica entre estas islas y [dado que el Banco Puerto Rico - Islas Virgenes fue la entrada de GAARlandia a las Antillas Mayores] se preguntaron si la ausencia de los otros tres generos en Puerto Rico se debe a que nunca estuvieron en ese territorio 0 a que sus
316 BIOGEOGRAFiA
fosiles atm no han sido hallados Pero como hemos visto Rega et al (2002) atribuyeron dicha distribucion a la dispersion interinsular en el Cuatemario
Por otra parte varios amilisis c1adisticos recientes ubi can a Choloepus -genero Reciente conocido solo de Centro- y Suramerica- como taxon hermano de Aeratoenus exc1uidos los demas megaloniquidos antillanos Para White y MacPhee (2001228)
tan estrecha relacion sugiere que la divergencia entre ambos [generos] ocurrio despues de que Aeratoenus divergio de Neoenus Si esta relacion implica que Aeratoenus y Neoenus se dispersaron separadamente desde Suramerica a las AntilIas mientras Choloepus permanecia en el continente (siendo necesarias al menos tres invasiones de perezosos) 0 si Choloepus invadio Suramerica desde las Antillas es algo que el material disponible no permite determinar
La diversidad adaptativa de los perezosos se manifiesta en (1) la utilizacion del habitat especies arboreas en Neoenus y Aeratoenus y terrestres en Paroenus y Megaloenus (Webb 1985a White [1993] White et al 1996 Arredondo [1999]) y en (2) la gama de tallas corporales desde los pequenos Neoenus hasta Megaloenus rodens el mayor de los mamiferos terrestres antillanos (su peso adulto estimado en 270 kg seglin Paula 19679) esto sugiere que los perezosos se irradiaron a nichos que en contextos continentales probablemente estarian ocupados por miembros de otros c1ados (MacPhee e IturraldeshyVinent 2000)
No esta claro si el abanico de tamanos entre estos perezosos enmascara alguna tendencia al gigantismo Por 10 pronto Anderson y Handley (2002) encontraron 10 contrario en perezosos (Bradypus) de las Islas de Bocas del Toro (Panama) shyseparadas secuencialmente del area continental vecina por ascensos del nivel del mar durante los ultimos 10 ka En las cinco islas los perezosos son menores que su contraparte continental y las diferencias entre las tallas medias de las cinco poblaciones expresan una reduccion lineal significativa de la talla corporal en funcion de la edad geologica de cada isla sin relacion alguna con el area de las islas 0 la distancia que las separa del continente
MONOS (Pitheciidae)
Los primates de las Antillas Mayores (cuatro especies extintas) constituyen un grupo monofiletico con Paralouatta (en Cuba) y Antillothrix (en La Espanola) como taxones hermanos seguidos en la escalera c1adistica por Xenothrix (en Jamaica) y finalmente Callieebus como el pariente continental mas cercano esto implica una sola invasion desde Suramerica antes del Mioceno Inferior (fecha mas temprana conocida para los primates fosiles antillanos) y por tanto en consonancia con GAARlandia como la via de colonizacion (Horovitz y MacPhee 1996 1999 Horovitz [1997] MacPhee y Horovitz 2004)
En desacuerdo parcial con la anterior interpretacion la filogenia de este grupo de monos obtenida por Davalos (2004) indica un antecesor comun en Cuba La Espanola y Jamaica
317 TRRADIACTON EVOLUCIONARIA
compatible con la hipotetica conexion -propuesta tentativamente por Iturralde-Vinent y MacPhee (199980-84)- entre las Montanas Azules del este de Jamaica y la peninsula suroccidental de La Espanola en la epoca de GAARlandia
A pesar de que los primates antillanos pertenecen a un grupo viviente de monos muy pequenos (los monos titles) fueron comparativamente grandes en relacion con los platirrinos modernos y sucesivamente mayores en la direccion Antillothrix Xenothrix y Paralouatta con P marianae como el mayor platirrino conocido del Mioceno neotropical (MacPhee 2005) MacPhee et al (1994) consideraron que Xenothrix debio ser un mono de andar lento y al parecer las especies de Paralouatta abandonaron la arborealidad para adoptar un modo de vida mas terrestre (MacPhee 1996a MacPhee et al 2003 MacPhee y Meldrum 2006)
ROEOORES (Echimyidae Capromyidae Heptaxodontidae)
La edad de Zazamys en Cuba demuestra que la invasion de los roedores histricognatos en las Antillas Mayores comenzo antes del Mioceno Inferior por otra parte los representantes mas antiguos de esa irradiacion evolucionaria se encuentran en La Espanola y Puerto Rico Ambas evidencias son consistentes con la hipotesis de GAARlandia (Woods 1982 1988 Woods et al 2001)
No esta claro si los capromidos evolucionaron a partir de un equimido ancestral 0 de un protocapromido 0 si los equimidos antillanos (I sub familia 3 generos y 7 especies todas extintas) forman parte de la irradiacion de los capromidos (4 subfamilias 9 generos y 27 especies 10 vivientes) Kratochvil et al (1978b) y Rodriguez et al (1979) consideraron que Capromys nan us es un eslab6n entre Echimyidae y Capromyidae pero Mesocapromys nanus esta morfologicamente mucho mas cerca de Hexolobodon y los de mas capromidos que de los equimidos (Woods 1985) Lo que si esta claro segun Woods et al (2001) es que la subfamilia Plagiodontinae endemic a de La Espanola es el linaje basal de los capromidos y que Isolobodon Hexolobodon Capromys Geocapromys Mysateles y Mesocapromys [y supuestamente Macrocapromys] forman parte de la irradiacion temprana de Capromyidae
La subfamilia Capromyinae (5 generos y 17 especies 9 vivientes) parece ser el dado de mas reciente evolucion en la familia con invasion de Cuba desde La Espanola y dispersion desde Cuba (1) por Capromys a las islas Caiman y (2) por Geocapromys a las islas Bahamas Caiman y Jamaica y desde Jamaica a las islas Swan aunque la presencia de Geocapromys en Swan pudiera deberse a introduccion aborigen (Morgan 1985 1988 1989 1993 1994 Woods 1989a 1990) Woods et al (2001) utilizaron datos moleculares para obtener estimados independientes de divergencia entre capromidos segiJn 10 cual Geocapromys divergio cerca de los 17 MaAP Mesocapromys divergio de Capromys hace 15 MaAP en tanto lvlysateles 10 hizo hace 10 MaAP Por su parte las formas de Capromys pilorides y de Mysateles prehensilis en Isla de la Juventud divergieron de sus correspondientes subespecies nominativas en la isla principal a los 24 y 08 MaAP respectivamente
318 BIOGEOGRAFiA
Woods y Howland (I979) describieron adaptaciones morfologicas de la mandibula el hioide y la faringe asociadas a mecanismos de masticacion diferentes en plagiodontinos y caprominos y Varona (1980a) llamo la atencion sobre el gran desarrollo de los molariformes (en relacion con la longitud total del cnineo) que presentan ciertos grupos de jutias fosiles en comparacion con las vivientes 10 que pudiera interpretarse como una adaptacion alimentaria a una vegetacion mas abrasiva por parte de las jutias extinguidas
Segun Woods (1990) la familia Heptaxodontidae (2 subfamilias 5 generos y 6 especies todas extintas) tambien seria parte de la irradiacion de los capromidos con origen en el este de La Espanola 0 en Puerto Rico y subsecuente dispersion al Banco de Anguila - San Martin en las Antillas Menores (pero vease MacPhee y Flemming 2003) Este banco estaria a solo 70 km del Banco Puerto Rico - Islas Virgenes en tiempos de maxima regresion marina y esta proximidad con Puerto Rico -patria de Elasmodontomys obliquus- seguramente tuvo que ver con la presencia en las islas de Anguila y San Martin del heptaxodontino gigante Amblyrhiza relacionado mas estrechamente con los roedores de las Antillas Mayores que con los del resto de las Antillas Menores (Woods 1990 1996) Aunque los roedores suelen aumentar de tamano en las islas (Case 1978) la escala del incremento de la talla en Amblyrhiza (que pudo alcanzar mas de 200 kg de peso adulto) esta fuera de toda proporcion con los patrones conocidos en otros roedores insulares (Biknevicius et 01 1993)
Clidomys (Clidomyinae) de Jamaica difiere marcadamente en su morfologia de los heptaxodontinos (vease ademas MacPhee y Flemming 2003)10 cual dificulta explicar su presencia en esa isla Como Jamaica estuvo bajo el mar hasta el Mioceno Medio el ancestro de Clidomys colonizaria esta isla por dispersion desde el extremo suroccidental de La Espanola despues que los heptaxodontinos se originaron en esta ultima isla (Woods 1989a) Pero Pascual et 01 (I 990) especularon que Clidomys pudiera ser el resultado de una invasion posterior e independiente de los capromidos
GRUPOS ORIGINARIOS DE NORTEAMERICA
El origen de las musaranas antillanas ha resultado ser hasta ahora el mayor enigma biogeografico de la fauna de mamiferos terrestres de la region aunque la mayoria de los autores coinciden en considerarlas originarias de Norteamerica (vease Discus ion y Conclusiones en Poblamiento Continente-Isla) No obstante MacPhee e Iturralde-Vinent (1994) descartaron toda posibilidad de conexion terrestre por el Estrecho de la Florida 0 por el Canal de Yucatan y si el (supuesto) evento vicari ante de Rosen (vease El Arca de Noe) -invocado por MacFadden (1980) para resolver el enigma- fuera minimamente plausible aun asi es dificil creer que las catastrofes que 10 sucedieron (vease Contexto Temporal) hayan barrido con toda esa vieja fauna continental excepto las musaranas circunstancia ya anticipada muy tempranamente por Matthew (l918a)
De otra parte la relaci6n entre Nesophontidae y Solenodontidae permanece irresuelta Si estos linajes no estan estrechamente relacionados entonces es muy posible que
319 IRRADIACION EVOLUCIONARIA
sus distribuciones sean explicadas por mecanismos diferentes (Whidden y Asher 2001)
La descomunal diferencia de talla corporal entre las musaranas pequenas (Nesophontes) y las grandes (Soienodon) seguramente impuso a ambos grupos restricciones distintas relativas a la termorregulacion y las adaptaciones energeticas En Nesophontes la seleccion debio favorecer una alta tasa de metabolismo y crecimiento camadas mas numerosas corta expectativa de vida yalta densidad poblacionaL En Soienodon en cambio la seleccion opero en la direccion de una baja tasa metabolica camadas reducidas crecimiento lento longevidad y baja densidad poblacional (Ottenwalder [1991])
MUSARANAS PEQUENAS (Nesophontidae)
Las musaranas pequenas estan representadas en las Antillas por el genero Nesophontes (y un genero miocenico indeterminado en ambar dominicano) con 6 especies cuaternarias extinguidas distribuidas en Cuba La Espanola Puerto Rico Islas Virgenes e Islas Caiman La mayor de las musaranas pequenas Nesophontes edithae evoluciono en Puerto Rico y su arribo a las Islas Virgenes supone una dispersion sumamente reciente En las Islas Caiman Nesophontes se asemeja mas a la poblacion de Cuba de donde seguramente derivo en el Plio-Pleistoceno (junto con Capromys y Geocapromys) cuando la costa sur de Cuba estuvo mucho mas cerca de las Islas Caiman (lturralde-Vinent 2003fig 4) MacPhee et ai (1994) interpretaron la distribucion de Nesophontes en Cuba y La Espanola como resultado de la vicarianza oligomiocenica ocurrida al formarse el Paso de los Vientos pero la distancia taxonomica entre los lados del Paso es solo de nivel especifico y N micros incluso vivi6 en ambas islas segtln Patterson (1962)
Allen (1918) considero que Nesophontes micrus fue una especie primitiva no modificada para la vida arb6rea 0 fos6rea y por tanto de habitos terrestres Nesophontes edithae de Puerto Rico es mucho mayor y mas robusta que las restantes especies del genero eL indice de robustez de su femur (= anchura minima de la diafisis ongitud total del hueso) muestra que este es tan robusto como el de Solenodon paradoxus un animal dos veces mayor (Morgan et ai 1980) Choate y Birney (1968) postularon un aumento cronoclinal de la taIla corporal en N edithae confirmado mas tarde por McFarlane (1999a) mediante fechados radiocarb6nicos de la secuencia estratignifica en depositos cavernarios de Puerto Rico Solenodon no vivi6 en Puerto Rico y en ausencia de una musarana de mayor tamano Nesophontes alcanzo una talla corporal mayor en esa isla que en La Espanola 0 Cuba (Woods y Eisenberg 1989)
MUSARANAS GRANDES (Solenodontidae)
Los arcaicos almiqufes (genero Soienodon) comprenden cuatro especies una viviente (S cubanus) y otra f6sil (S arredondoi) en Cuba y una viviente (S paradoxus con dos subespecies) y otra f6sil (S marcanoi) en La Espanola Como cabria esperar las dos especies cubanas se asemejan mas entre sf que a las especies de La Espanola pero
320 BIOGEOGRAFiA
enigmaticamente S marcanoi comparte caracteres con S cuban us y S paradoxus (Woods y Eisenberg 1989 Morgan y Ottenwalder 1993 Ottenwalder 200 I) Solenodon arredondoi alcanzo una talla corporal superior a la de cualquiera de las musarafias vivientes a nivel mundial Segun Morgan y Ottenwalder (1993) extrapolando las dimensiones y el peso de los almiquies vivientes S arredondoi debio tener una longitud cabeza-tronco y un peso de 450-550 mm y 1500-2000 g respectivamente en comparacion con S cubanus (360 mm y 800 g) y S paradoxus (390 mm y 1100 g)
Entre todas las especies de almiqufes Solenodon paradoxus es la que presenta los caracteres mas derivados (Otten walder 2001) Si estas relaciones son validas la distribucion de Solenodon entre Cuba y La Espafiola debio discurrir de oeste a este como propuso MacFadden (1980) quien invoco la presencia fosH de S cubanus [pero tambien S arredondoi] en Pinar del Rio para postular que el ancestro continental entr~ por Cuba opinion compartida por Perfit y Williams (1989) y por Ottenwalder ([1991] 2001) El paso de Solenodon a Puerto Rico pudo haberlo impedido la existencia de barreras ecologicas durante la conexion del este de La Espafiola con Puerto Rico 0 por la ausencia de Solenodon en esa parte de La Espafiola en aquella epoca Sin embargo las mismas condiciones ecologicas en el este de La Espafiola no parecen haber representado una barrera efectiva para Nesophontes cuyas mas altas tasa de reproduccion y densidad poblacional presumiblemente Ie permitieron expandirse flpidamente a traves de La Espafiola (Ottenwalder [1991])
Es opinion generalizada que los almiqufes antillanos constituyen el grupo de mamiferos que primero se emplazo en el territorio insular (MacPhee 2005) y en tal sentido es de interes la evidencia sobre especificidad parasito-hospedante aportada por Cruz (1980) quien encontro que la especie de acaro (genero Cubanochirus) que parasita a los almiquies es mucho mas primitiva que la especie (del mismo genero) que paras ita a las jutias y de aqui concluyo que dicho genero de acaro debio evolucionar primero en Solenodontidae y posteriormente en Capromyidae al entrar en contacto representantes de ambas familias en Cuba 0 La Espafiola
Recientemente Roca (2004) y Roca et al (2004) dieron a conocer que los resultados de su estudio genetico-molecular sobre los almiquies indican una divergencia de 25 Ma entre Solenodon cuban us y S paradoxus -compatible con la disyuncion tectonica entre Cuba y La Espafiola (Iturralde-Vinent y MacPhee 1999 McPhee et al 2003)-- y una separacion de ambas especies comparable en magnitud a la divergencia entre familias de mamlferos Consecuentemente propusieron que Solenodon cubanus fuera clasificada en el genero Atopogaie como indico Cabrera (1925) hace 82 afios
DISCUSION Y CONCLUSIONES
Las musarafias y los roedores antillanos (excepto Oryzomys) se irradiaron tanto de sus respectivos ancestros continentales que de estos 6rdenes se reconocen familias y subfamilias endemicas de la region en contraste con los perezosos (una subfamilia y tres
321 IRRADIACION EVOLUCIONARIA
tribus endemicas) y los monos (una tribu endemica) como expresion de diferentes niveles de diversificacion evolucionaria Este proceso es dificil de explicar (traUindose de c1ados que pnicticamente comparten la misma antigiiedad en las Antmas) si es correcta la interpretacion de Pascual et at (1990636)
Nosotros consideramos estos diferentes niveles de diversificacion como las diferentes etapas de evolucion alcanzadas por cada grupo [de mamiferos] Obviamente esto estuvo relacionado con las diferentes longitudes de sus historias
Pudiera ocurrir que en el futuro se compruebe que la historia biogeognifica de la region fue mucho mas complicada
La Espanola la isla mas ecol6gicamente divers a de la region debi6 ser el centro principal de evoluci6n de los mamfferos (con 87 de las familias y 73 de las subfamilias) seguida por Cuba (con 75 de las familias y 54 de las subfamilias) las dos islas de las Antillas Mayores mas relacionadas biogeograficamente (Fig 43 Tabla 44)
eL
EL
cb CAIMAN
SWAN
A Acratocnini B Cubanocnini C Megalocnini D Mesocnini E Nesophontidae F Solenodontidae G Xenothrichini
ANGUILAshyM-0 SMARTIN
H Heteropsomyinae I Plagiodontinae J Isolobodontinae
K Hexolobodontinae L Capromyinae
M Heptaxodontinae N CIidomyinae 0 Sigmodontinae
FIGURA 43 Representacion esquematica de las relaciones interinsulares (expresadas porel numero de linajes compartidos) entre las Antillas Mayores y entre estas y los grupos de islas perifericas segun las interpretaciones de los autores citados en el texto Los numeros dentro de los circulos indican la cantidad de linajes representados en cada isla relacion dudosa Contexto taxon6mico de los linajes en la Tabla 44
bull bull
bull bull
bull bull bull
bull bull
bull bull
bull
bull
bull
bull
bull bull bull bull
bull bull
bull bull
bull
bull bull bull
bull bull
bull
bull bull
bull
322 BIOGEOGRAFiA
TABLA 44 Distribuci6n de los linajcs de mamiferos terrestres a traves de las Antillas Mayores y grupos de islas perifericas LE = La Espanola CU Cuba JA Jamaica PR = Puerto Rico IC Islas Caiman IS Islas Swan IB = Islas Bahamas IV = Islas Virgenes AS = Anguila - San Martin
FamiliaSubfumiliafIribu LE CU JA PR IC IS IB IV AS
Megalonychidae
Choloepodinae
Acratocnini
Cubanocnini
Megalocninae
Megalocnini
Mesocnini
Nesophontidae
Solenodontidae
Pitheciidae
Callicebinae
Xenothrichini
Echimyidae
Heteropsomyinae
Capromyidae
Plagiodontinae
Isolobodontinae
Hexolobodontinae
Capromyinae
H e pta x 0 don tid a e
Heptaxodontinae
Clidomyinae
Muridae
Sigmodontinae
PILOSA
SORICOMORPHA
PRIMATES
RODENTIA
bull
bull
bull
bull TOTALES 12 10 4 4 2
323 EXTlICIOIES DEL CUATERNARIO
El gigantismo ha sido sin duda una tendencia evolucionaria recurrente entre los clados mayores de mamiferos antillanos Es un hecho bien conocido que algunos grupos de mamiferos han aumentado de tamafio en las islas y varios autores han intentado explicarlo a partir de enfoques diferentes (Foster 1964 Miller 1966 Sondaar 1977 Case 1978 Eisenberg y Wilson 1978 Heaney 1978 Lawlor 1982 Lomolino 1985 Angerbjom 1985 McNab 19942001)
EXTINCIONES DEL CUATERNARIO
Las islas que integran la Subregion Antillana han perdido casi 90 de sus mamiferos terrestres durante los ultimos milenios y en proporcion con su extension territorial esta perdida ha sido mayor en la Subregion que en cualquier otro lugar del planeta (MacPhee 1996b) Las extinciones del Cuatemario eliminaron mas de las tres cuartas partes de la fauna de mamiferos terrestres conocida de las Antillas Mayores sus islas satelites y grupos de islas perifericas y el examen de este proceso en sus aspectos mas generales se aborda en esta ultima parte del presente capitulo
CRONOLOGiA Y CONSECUENCIAS
Es muy improbable que un fosil represente el ultimo individuo de un taxon extinto y entonces hay que guiarse por la edad radiometrica del fosil mas reciente conocido lIamada fecha de ultima presencia- como unica aproximacion posible a la fecha de extincion del taxon
Como se explico al inicio del capitulo (vease Contexto Temporal) el registro fosH de los mamiferos terrestres de las Antillas Mayores adolece de insuficiente sustentacion radiometrica No obstante la distribucion temporal de los escasos fechados disponibles para el Cuaternario de la region abarca etapas criticas del proceso de extincion experimentado por la referida fauna a 10 largo de los ultimos 250 mil afios
De las 52 especies que es posible reconocer actualmente como miembros extintos de la fauna cuatemaria de mamiferos terrestres de las Antillas Mayores y grupos de islas perifericas los restos fosiles de 23 de elIas (442) han sido fechados radiometricamente o se han hallado asociados estratigraficamente de manera inequivoca con otro material fechado De estos 23 fechados 3 corresponden al interglacial Sangamon 4 pertenecen a la secuencia Pleistoceno terminal - Holoceno temprano 10 se ubican en ellapso entre el arribo de los aborigenes y la incursion europea y 6 son postcolombinos (a los cuales hay que sumar otras dos especies extinguidas en los siglos XIX y xx) A partir de estas 25 especies avaladas por fechas de ultima presencia distribuidas entre los cuatro intervalos temporales sefialados hemos compuesto la Tabla 45 en la que ademas aparecen las restantes 27 especies (carentes de fechado radiometrico) ubicadas en el intervalo temporal mas probable sobre la base de (1) la asociacion estratigrafica de sus restos con evidencias culturales (amerindias 0 europeas) 0 (2) la ausencia de dicha asociacion Sobre estos criterios de ubicacion temporal procede hacer dos aclaraciones
324 BIOGEOGRAFIA
TABLA 45 Intervalos temporales mas probables para las extinciones cuatemarias de los mamiferos terrestres de las Antillas Mayores y grupos de islas perifericas sobre la base de las evidencias siguientes (l) ultimo ejemplar colectado u observado en vida (2) fechado radiometrico mas tardio en material de la especie (3) asociaci6n estratigrafica mas tardia con otro material fechado (4) asociaci6n estratigrafica con restos de animales introducidos restos humanos u objetos culturales propios del intervale (5) falta de asociaci6n con evidencias culturales [Los ntimeros entre parentesis en la columna Evidencia significan adscripci6n tentativa al intervalo] Los estimados del peso corporal (en kg) estan tomados de la literatura 0 inferidos comparativamente y distribuidos entre
las clases siguientes (a) S 01 (b) 01 a lt 1 (c) 1 a lt 10 (d) 10 a lt 100 (e) 100 a lt 200 (j) ~ 200 Las
abreviaturas que designan las islas son las de la Tabla 44
F amilialEspecie Peso IsJa Evidencia Fuente
I httervalo htterg1acial Sangamon (gt80 kaAP)
HEPTAXODONTlNAE
Amblyrhiza inundata e AS 3 McFarJane et al 1998a
CLIDOMYINAE
Clidomys osborni d JA 3 Ibid
Clidomys parvus d JA 3 Ibid
II httervalo Pleistoceno terminal ~ Holoceno temprano (20-6 kaAP)
MEGALONYCHIDAE
Aeratoenus antillensis d CD 5 Arredondo 1975 a
Aeratoenus odontrigonus d PR 3 McFarJane 1999b
Aeratoenus simorhynehus d LE 2 Rega et al 2002
M egaloenus zile f LE 2 MacPhee et al 2000
SOLENODONTIDAE
Solenodon arredondoi e CD 5 Morgan y Ottenwalder 1993
PnHECIIDAE
Paralouatta varonai d CD 5 MacPhee 1996a
EcHIMYIDAE
Heteropsomys antillensis c PR 5 McFarJane 1999b
Heteropsomys insulans c PR 5 Ibid
Puertoricomys eorozalus c PR 5 Woods 1989a
CAPROMYIDAE
P lagiodontia araeum c LE 5 MacPhee et al 2000
Macrocapromys latus c CD 5 Varona y Arredondo 1979
(Continua)
325 EXTINCIONES DEL CUATERNARIO
TABLA 45 (Continuaci6n)
F ami1ia1Especie Peso Isla Evidencia Fuente
M acrocapromys latus c CU 5 Varona y Arredondo 1979
Mesocapromys kraglievichi c CU 5 Ibid
HEPTAXODONTINAE
Xaymaca fulvopulvis a JA 2 MacPhee y Flemming 2003
III Intervalo Amerindio (6-05 kaAP)
MEGALONYCHIDAE
Acratocnus ye d LE 2 MacPhee et aI 2000
Neocnus comes c LE 2 Ibid
N eocnus dousman c LE 2 Ibid
N eocnus toupiti c LE 2 Ibid
N eocnus gliriform is c CU 4 Cordova et ai 1997
M egalocnus rodens f CU 2 MacPhee et ai 2007
Parocnus browni e CU 2 Steadman et ai 2005
P arocnus serus e LE 2 MacPhee et ai 2000
N ESOPHONTIDAE
N esophontes edithae a PR 2 McFarlane 1999b
PITHECIIDAE
Antillothrix bernensis c LE 3 Rimoli 1977
EcHIMYIDAE
Brotomys contractus a LE 4 Miller 1929c
CAPROMYIDAE
P lagiodontia spelaeum c LE 4 Ibid
M acrocapromys acevedo c CU 4 NUiiez 1990a
HEPTAXODONTINAE
Elasmodontomys obliquus d PR 3 McFarlane 1999b
Iv Intervalo Postcolombino (05 kaAP - Presente)
N ESOPHONTIDAE
N esophontes hypom icrus a LE 24 MacPhee y Fle111111ing 1999
N esophontes zam icrus a LE (4) Ibid
N esophontes m icrus a CU 24 MacPhee et aI 1999
(Continua)
326 BIOGEOGRAFiA
TABLA 45 (Continuacion)
F amilialEspecie Peso Isla Evidencia Fuente
N esophontes m icrus a CD 24 MacPhee et aI 1999
N esophontes m icrus a LE 4 MacPhee et aI 2000
N esophontes sp 1 a IC 4 Morgan 1994
N esophontes sp 2 a IC 4 Ibid
Solenodon marcanoi b LE 4 Ottenwalder 2001
PITHECIIDAE
Xenothrix mcgregori c JA 4 MacPhee y Flemming 1999
EcHIMYIDAE
Boromys offeUa a CD 24 Guarch 1984
Boromys torrei a CD 24 Ibid
Brotomys voratus a LE 4 MacPhee et aI 2000
CAPROMYIDAE
P lagiodontia ipnaeum c LE 4 MacPhee y Flemming 1999
Rhizoplagiodontia lemkei c LE 4 Ibid
Isolobodon portoricensis c PR 24 Ibid
Isolobodon montanus c LE 4 Ibid
Hexolobodon phenax c LE (4) Ibid
Capromys sp c IC 24 Morgan 1994
Geocapromys columbianus c CD 4 MacPhee y Flemming 1999
Geocapromys thoracatus b IS 1 MacPhee y Marx 1997
Geocapromys sp 1 c IC 4 Morgan 1994
Geocapromys sp 2 c IC (4) Ibid
l-IEPTAXODONTINAE
Quemisia gravis c LE (1) MacPhee y Flemming 1999
MURIDAE
Oryzomys antillarum a JA 1 Ibid
1 Flemming y MacPhee (1999) alertaron en contra de la asociacion con otro material fechado Asimismo Morgan (1993) indico que la ausencia de asociacion con evidencias culturales amerindias no es necesariamente un criterio concluyente para ubicar un taxon en el intervalo temporal anterior porque hay depositos en la region que carecen de tales evidencias y son del Holoceno tardio por fechados radiocarbonicos [No obstante
327 EXTlNCIONES DEL CUATERNAR10
la ausencia de una especie en residuarios arqueol6gicos adquiere mayor certidumbre como criterio de ubicaci6n temporal de su extinci6n en la medida que aumenta el numero de residuarios estudiados que en las Antillas Mayores se cuentan por centenares] A falta de fechado absoluto de una especie consideramos que estos criterios de ubicaci6n temporal son mas utiles que la alternativa de excluir del analisis a la especie
2 En los trabajos sobre biogeografia de mamiferos antillanos los arque610gos mas citados como autoridades para establecer el inicio de la presencia aborigen en las Antillas Mayores (Kozlowski 1974 Rouse y Allaire 1978 Rouse 1989 1992 Wilson 2001) difieren al respecto entre los 6 kaAP y los 45 kaAP (pero vease Pagan et al 200523) y hemos adoptado la mas temprana de estas fechas para el comienzo del intervalo amerindio en la Tabla 45 De todos modos cualquiera que haya sido la fecha esta no es aplicable por igual al conjunto de las Antillas Mayores porque segun algunos de los autores citados el poblamiento comenz6 por Cuba y La Espanola 6 mil anos atras continu6 en Puerto Rico hace alrededor de 3 mil anos (pero vease Burney et al 1994) yen Jamaica hace menos de 2 mil e incluso a las Islas Caiman ni siquiera llegaron [Las Islas Caiman descubiertas por Col6n en 1503 carecen absolutamente de evidencia amerindia (Stokes y Keegan 1996)]
Si los datos de la Tabla 45 representan una aceptable aproximaci6n a la realidad el proceso de las extinciones cuaternarias de los mamiferos terrestres de las Antillas Mayores y grupos de islas perifericas debi6 caracterizarse entre otros por los eventos siguientes
Aunque obviamente la fauna cuaternaria debi6 ser mucho mas numerosa que 10 que refleja el registro paleontol6gico (Morgan y Woods 1986) 52 especies (81 de la fauna conocida) desaparecieron (sin reposici6n) durante los ultimos 250 mil anos
- Al parecer las extinciones se concentraron en las especies de mayor tamano Asi el peso promedio estimado para las especies de la etapa preantr6pica (intervalos I + II de la Tabla 45) es 350 kg es 328 kg para las especies del intervalo III (amerindio) y es s610 25 kg (c1ases del peso a b c) para las especies del intervalo IV (postcolombino) Ninguna especie aut6ctona de mamifero mayor de 6 kg existe hoy en la regi6n
- De acuerdo con la distribuci6n temporal de las extinciones 61 especies sobrevivieron al interglacial Sangamon (intervalo 1)48 sobrevivieron a la transici6n del Pleistoceno terminal al Holoceno temprano (intervalo II) 34 sobrevivieron al periodo amerindio (intervalo III) y s610 12 especies han sobrevivido desde la Ilegada de los europeos (intervalo IV) Las tasas de extinci6n correspondientes (exceptuado el intervalo I) son una especie cada 1167 anos en el intervalo II una cada 393 anos en el intervalo III y una cada 23 anos en el intervalo IV
- Durante todo el proceso de las extinciones cuaternarias la perdida de especies por isla fue de 91 en La Espanola 83 en Jamaica y 62 en Cuba Bahamas retuvo su unica especie (aunque extirpada en varias de las islas) Puerto Rico e Islas Caiman perdieron todas sus especies (6 y 5 respectivamente) y las Islas Swan Virgenes y Anguila - San Martin perdieron sus respectivas especies (micas
328 BIOGEOGRAFiA
De los cuatro 6rdenes de mamiferos representados en la fauna Pilosa y Primates desaparecieron totalmente Soricomorpha y Rodentia perdieron 86 y 74 de sus especies respectivamente
CAUSALIDAD
CAUSAS NATURALES
Los cicIos glaciales e interglaciales acaecidos en el Pleistoceno causaron cambios ambientales dnisticos en las Antillas con la concomitante alternancia de cIimas frios (secos) y calidos (hUmedos) asi como de descensos y ascensos del nivel del mar aumento y disminuci6n de las areas territoriales) induso (aunque con menor intensidad) en el Holoceno Estos eventos paleogeograficos han sido consistentemente documentados (Mayo y Kartashov 1972 Bonnatti y Gartner 1973 Ortega y Arcia 1982 Prance 1982 Schubert y Medina 1982 Dodge et al 1983 Ortega [1984] Haq et al 1987 Schubert 1989 Hodell et al 1991 Guilderson et al 1994 Oppo 1997 Emslie 1998 Curtis et al 2001 Iturralde-Vinent 2003) y de ellos brindamos a continuaci6n una breve resena
Finalizando el periodo interglacial anterior a la ultima glaciaci6n (Wisconsin) el mar estaba a 8-10 m por encima de su nivel actual Cuando el avance glacial del Wisconsin a1canz6 su culminaci6n (eillamado Ultimo Maximo Glacial) hace 18 ka el mar habia descendido 120 m con respecto a dicho nivel y las islas del Caribe habian incrementado considerablemente sus extensiones subaereas en comparaci6n con las que presentan hoy dia por ejemplo Puerto Rico y las Islas Virgenes se conectaron territorialmente y las Bahamas se unieron en cinco gran des islas Prevalecia un regimen frio y seco y condiciones de aridez --debidas a una reducci6n de las precipitaciones (posiblemente a menos de 440 mmlano)- y los ambientes xericos se habian extendido a traves de las islas y bancos emergidos y abundaban los pinares (los habitats mesicos estaban restringidos a parcelas relictas en 10 alto de las montanas) En el Mar Caribe y el Golfo de Mexico la temperatura superficial del mar era 5degC mas baja que la actual y en las llanuras costeras y alturas maximas eran respectivamente 2-3degC y 6degC mas bajas Los limites de las zonas ecol6gicas altitudinales descendieron varios cientos de metros y cada zona redujo su extensi6n vertical al aumentar el gradiente de temperatura Ellimite inferior de la zona periglacial
elevaci6n donde la temperatura media minima a1canza los OdegC) de la Cordillera Central de la Republica Dominicana (altura maxima del Caribe insular con evidencia de topografia glacial) se considera que estuvo entre los 1500 y 1800 mlnm (actualmente esta a 2800 mlnm) y se puede estimar que dicho limite era inferior en Cuba por estar mas al norte que la Republica Dominicana No obstante durante este periodo la temperatura experiment6 episodios cortos (unas pocas decadas) de ascensos repetidos
Un signifieativo componente bi6tieo de las referidas condiciones paleoambientales 10 constituy6 la espectacular irradiaci6n evolucionaria experimentada por un grupo numeroso de aves rapaees diurnas y nocturnas (Wetmore 1937 Brodkorb 1969 Kurotchkin y Mayo 1973 Arredondo 1976 Olson 1978 Olson y Hilgartner 1982
329 EXTINCIONES DEL ClJATERNARIO
Suarez 2000a b 2004 Suarez y Emslie 2003) Los mamiferos de las Antillas Mayores ofrecieron a estos depredadores aviares una amplia gama de tamanos de presa que en ausencia de mamiferos del orden Carnivora en la region permitio a estas aves irradiarse a este nicho alimentario con la evolucion de una variedad de especies de muy diversa tall a corporal incluidas formas gigantes Adquirir un cuerpo gigante les propicio el acceso a recursos alimentarios que no estaban siendo explotados por ningun otro depredador (Alcover y McMinn 1994) A Cuba corresponde el mejor registro fosil de rapaces cuaternarias caribenas con siete generos de especies diurnas Dentro de estas aves existieron tres gavilanes gigantes (Accipitridae) tres carairas (Falconidae) y un condor (Cathartidae) asi como cuatro generos de especies nocturnas dos lechuzas (Tytonidae) y tres Mhos (Strigidae) dos de ell os no igualados en tamano en todo el mundo Algunos de estos generos tuvieron representacion en otras islas antillanas como La Espanola Jamaica Bahamas y Gran Caiman
Al retirarse hacia el norte los hielos de la glaciacion Wisconsin el clima se tomo mas hUmedo en las Antillas durante el Holoceno temprano y el bosque mesico se expandio en la region insular El nivel del mar ascendio acompanado de mayor pluviosidad alrededor de los 7 kaAP las Islas Virgenes quedaron separadas entre si y de Puerto Rico y las Bahamas perdieron la mayor parte de su territorio Cerca de los 3 kaAP las condiciones secas retornaron a la region e hicieron desaparecer buena parte del bosque mesico Nuevamente el clima comenzo a adquirir sus condiciones actuales hacia los 25 kaAP aproximadamente y hoy transita por una fase mesica con vegetacion xerica confinada en parcelas relictas
Sin duda estos y otros eventos naturales incidieron negativamente en la sobrevivencia de mamiferos terrestres en las Antillas Mayores asi como en los grupos de pequenas islas perifericas (Pregill y Olson 1981 Olson 2006)
McFarlane et al (1998a) demostraron convincentemente como Amblyrhiza el gigante de los roedores antillanos endemico de la pequena isla de Anguila sucumbio a las repetidas y abruptas perdidas de su habitat durante el interglacial Sangamon por las nipidas y severas oscilaciones del nivel del mar (documentadas por Neumann y Hearty 1996) al no poder responder adecuadamente a tan catastroficos eventos mediante el ajuste de su masa corporal media y otro tanto debio ocurrirle en Jamaica al otro roedor gigante Clidomys (McFarlane et al 1998b) emparentado con Amblyrhiza en la misma epoca (Tabla 45 intervalo I)
El ascenso del nivel del mar seguramente promovio la extincion de mamiferos durante la transicion del Pleistoceno al Holoceno (Tabla 45 intervalo II) con la reduccion del tamano de las Islas por las transgresiones marinas (Pregill y Olson 1981 Morgan 1993) Esto ultimo haria maximamente severa la competicion asf como la presion de los depredadores [Steadman et al (2005) no yen por que el calentamiento global ocurrido durante la transicion Pleistoceno-Holoceno habria de tener un efecto negativo sobre los perezosos siendo sus descendientes actuales especies tropic ales 0 subtropicales pero no se trata del efecto
330 BIOGEOGRAFiA
directo del cambio de temperatura sobre los animales sino del efecto indirecto y progresivo que dicho cambio genera a largo plazo al reducir el territorio insular disponible]
Por esta epoca tambien desaparecieron especies de tall a mayor como Megalocnus zile en La Espanola y otras de menor tamano (en Cuba) pero que representan la tall a maxima en sus respectivos contextos taxonomicos del Cuaternario como Solenodon arredondoi Paralouatta varonai y Macrocapromys acevedo Un mamifero puede ser grande solo a costa de reducir su densidad poblacional y esto 10 hace mis vulnerable a la extinci6n sobre todo en una isla (McNab 1994 McFarlane et al 1998a) Incidentalmente Alcover et al (1998a) afirmaron que larelacion entre la talla corporal y la extincion no siempre se cumple en las especies de menor tamano y citaron como ejemplo que entre las musarafias todas las especies de Nesophontes desaparecieron mientras sobreviven especies de Solenodon pero la comparacion correcta en todo caso serfa entre los solenodones vivientes y el gigante Solenodon arredondoi y entre Nesophontes edithae -el gigante de los nesofontes extinguido en el periodo amerindio- y el resto de sus congeneres que sobrevivieron hasta los primeros siglos de la colonizacion europea (Tabla 45 intervalos III y IV)
CAUSAS ANTROPOGENICAS
Intervalo Amerindio
La mayoria de los autores que se han referido a las extinciones acaecidas durante este intervalo (entre elIos paradigmaticamente Martin 1984a b) han responsabilizado a los aborfgenes (que se supone comenzaron a poblar las Antillas Mayores hace 6 mil afios) con la desaparicion de numerosas especies de mamiferos terrestres en la region principalmente por la via de la sobrecaza y la introduccion de animales domesticos devenidos depredadores de la fauna autoctona (Morgan y Woods 1986) Este efecto negativo de la presencia indigena sobre los mamiferos de la region ha sido dramatizado por algunos autores en terminos de holocausto faunal Sin embargo pensamos que se ha exagerado Como han expresado MacPhee et al (19993) cuando uno busca evidencia dura de complicidad humana directa en [estas] extinciones no hay mucho que encontrar Las siguientes son algunas consideraciones a tener en cuenta en tal sentido
El registro radiometrico mas joven conocido hasta ahora para un perezoso antillano (MegalocnllS rodens) es de 42 kaAP (MacPhee et ai 2007) Si se acepta que los aborfgenes entraron en Cuba hace aproximadamente 6 mil anos su convivencia con los perezosos debe haber durado alrededor de 1800 afios (0 mas porque dificilmente el referfdo fechado radiometrico haya correspondido al ultimo sobreviviente de la especie) Esta cifra contradice la hipotesis del holocausto faunal segun la cualla extincion de los mamiferos insulares de mayor talla debe ocurrir con extrema rapidez -a escala de decadas 0 de centurias- a partir de su primer contacto con los humanos (Martin y Steadman 1999) A pesar de las repetidas afirmaciones en contrario (Harrington 1921 Miller 1929c Herrera [1945] Tabio 1951 Alvarez 1951a 1957 Arredondo
331 EXTINCIONES DEL CUATERNARIO
1959 1960a Veloz y Ortega 1973 Mayo 1980b Rodriguez et al 1984 Castellanos y Pino 1985 Pino y Castellanos 1985 Nunez 1990b Gutierrez 1996 Arredondo 2004) en su mayoria basadas en evidencias no exentas de posibles accidentes de deposicion (0 de reubicacion postdeposicional) en recintos cavernarios (Diaz-Franco [2006b D no se ha demostrado inequivocamente que los aborigenes antillanos se alimentaran de perezosos regularmente Sus restos en residuarios de alimentos son escasisimos en toda la region y esto contrasta con los abundantes restos de jutias y otros mamiferos de menor talla que aparecen en esos residuarios demostrativos de la preferencia de los paleoindios por estos otros mamiferos
Los mamiferos mas importantes en la economia prehistorica de las Antillas Mayores fueron las jutias Capromys y Geocapromys en Cuba Geocapromys en Jamaica e Isolobodon en La Espanola Puerto Rico e Islas Virgenes (Aguayo 1950b Tablo 1951 Rimoli 1972 Wing 2001 Diaz-Franco [2006b D Wilkins (2001) estudi61ajutia viviente de Jamaica (Geocapromys browni) en un sitio ocupado por los aborigenes durante 100 anos y no encontr6 evidencias de que durante ese tiempo la especie haya sido sobreexplotada Asimismo en Cuba las jutias conga carabali y andaraz son escasas en depositos pleistoholocenicos en cambio son muy numerosas en residuarios indigenas y de haber sido sobreexplotadas 10 mas probable es que no hubieran sobrevivido hasta hoy
La habilidad de los humanos para penetrar en profundidad el bosque tropical y mantenerse en este sustentablemente ha sido un evento comparativamente reciente (Beran y Steele 1994) En el caso antillano la caza tierra adentro ha sido explicada como un proceso tardio de desplazamiento a partir de la explotacion de abundantes recursos marinos que ofrecian condiciones 6ptimas para el sedentarismo de las comunidades aborigenes costeras (C6rdova y Arredondo 1988 Brito 2005) Peces y mariscos dominan la arqueologia costera de las Islas antillanas en tanto el consumo de recursos terrestres (mamfferos incluidos) aumenta progresivamente en la medida que los residuarios estudiados se alejan de la costa (Scudder 1991 Wilkins 2001) En las Antillas Mayores el componente terrestre de la dieta aborigen es solo 20-30 del material recobrado en los residuarios (Wing 1977 1989) Y debio ser menor alin porque el componente vegetal de la dieta no se ha preservado Por otra parte los aborigenes cubanos domesticaron la jutia conga y seguramente la reprodujeron para comer (Pose et al 1988) practica que probablemente se repitiera con otras jutias en las demas Antillas Mayores
- Fray Bartolome de las Casas hablo de 3 millones de indios en Cuba al inicio de la colonizacion pero los historiadores subsecuentes consideraron exagerada la cifra (Poey 1851 a) No obstante se dispone de dos estimados de la densidad de poblaci6n aborigen en las Antillas Mayores a la llegada de los europeos 18 indigenaslkm2 en Puerto Rico (Rouse 1956) y 1271km2 en La Espanola (Watts 1987) Extrapolando ambos valores entre las cuatro islas principales de las Antillas Mayores tenemos que en la fecha indicada la poblaci6n aborigen pudo estar entre 16 mil y 113 mil indigenas en Puerto Rico entre 21 mil y 145 mil en Jamaica entre 141 mil y 1 mill6n en La Espanola y entre 200 mil y 14 millones en Cuba Entre las cifras maximas y las minimas nos inclinamos
332 BIOGEOGRAFiA
hacia estas ultimas atendiendo a los altos indices de mortalidad infantil y morbilidad que se evidencian en las osamentas de los pobladores prehispanicos (Martinez y Rives 1990) Uno de los hechos que apuntan hacia el bajo numero de aborigenes en las Antillas Mayores es 10 rapido que desaparecieron despues de la llegada de los europeos pnkticamente sin dejar mestizaje con estos 10 cual contrasta ostensiblemente con la situacion en la America continentaL
En suma un grupo humano (1) demograiicamente irrelevante y controlado por altos indices de morbilidad (2) consumidor mayoritariamente de organismos marinos y (3) carente de la tecnologia cinegetica necesaria para poder sobreexplotar los mamiferos que Ie sirvieron de alimento (a los cuales accedio tardiamente en su desarrollo) no es concebible que haya causado e1 holocausto faunal que se Ie atribuye a partir del error de aplicar a las comunidades aborigenes antillanas un modelo cultural de la prehistoria continentaL
Tampoco hay evidencias de que los aborigenes hayan introducido carnivoros con suficiente capacidad depredadora salvo el perro Morgan y Otlenwaldder (1993) consideraron que la depredacion por perros durante el periodo amerindio es la explicacion mas plausible para la extincion del almiqui gigante de Cuba pero como se puede observar (Tabla 54) Solenodon arredondoi aparece en el intervalo II por falta de asociacion con evidencias culturales
Consecuentemente el accionar de los aborigenes y de sus animales domesticos directamente sobre la fauna no parece haber sido por sf solo un factor determinante de la extincion de mamiferos terrestres durante el periodo amerindio en las Antillas (pero vease Steadman et al 2005)
Hay que recordar en este senti do que alin en el Holoceno tardio ocurrieron cambios clirruiticos de consideracion (vease Causas Naturales) Por ejemplo Curtis et al (2001) explicaron que la extincion de algunos ani males mesofilicos de las alturas montanosas de La Espanola pudo haber antecedido a cualquier impacto humano sigrIificativo en esos lugares por coincidir dicha extincion con una tendencia a la aridez documentada isotopicamente y con evidencias de la perdida del bosque mesico ocurridas despues de 3kaAP Yen Jamaica hace solo 2shy3 mil aDos el area de la Bahia de Jackson sustentaba una gran biomasa de vertebrados en bosque hinnedo a juzgar por la cantidad y diversidad de taxones presentes en material fosil derivado de la depredacion por lechuzas hoy el area es la mas seca de la isla con matorral xerofitico y pobreza de fauna (MacPhee 1996a) Yalm en nuestros dias huracanes de gran intensidad aniquilan poblaciones de vertebrados (que no se recuperan) en is las pequenas y llanas del Caribe (Brooks y Smith 2001)
En medio de la incertidumbre sobre la verdadera causa (0 causas) de las extinciones de mamiferos en el periodo amerindio MacPhee y Marx (1997) han propuesto su hipotesis de la hipermorbilidad la cual estipula colapsos poblacionales relativamente rapidos con la introduccion de enfermedades de alta letalidad Estos autores han razonado que
Las islas son los lugares mas apropiados para experimentar perdidas catastroficas causadas por enfermedades Los mamiferos no solo serian susceptibles en virtud de su prolongado aislamiento biologico sino que en los confines de una isla la epizootia
333 EXTINCIONES DEL CUATERNARIO
tendria la potencialidad de infectar simultaneamente a la rnayoria de los individuos de vadas especies Los neonatos son inmunologicamente inmaduros y sufririan desproporcionadamente cualquier infeccion hipervirulenta La perdida excesiva de neonatos y juveniles tendria efectos severos para la sobrevivencia de la poblacion si esta se reduce por debajo de cierto nivel sobreviene la extincion porque no hay posibilidad de restitucion en el periodo postpanzootico Las especies de mayor talla corporal van a ser las mas vulnerables a la extincion dadas sus bajas tasas reproductivas largos periodos de gestacion y reducido numero de progenie Las especies de menor tamano --caracterizadas por altas tasas reproductivas periodos cortos de gestacion y mayor numero de neonatos por parto- tambien serian fuertemente diezmadas pero muchas sobrevivirian porque su estrategia reproductiva les permite perder mayor numero de individuos Las perdidas deben ocurrir relativamente rapido (a escala de decadas) mientras que otras hipotesis (digamos los cambios climaticos) permiten intervalos mucho mas largos (a escala de siglos 0 de milenios) entre la aparicion de las causas y la produccion de sus efectos Por ejemplo todo indica que los nesofontes se extinguieron muy rapidamente y seria dificil poder invocar un impacto humano directo 0 argumentar fenomenos climaticos que pudieran explicar plausiblemente perdidas tan rapidas
MacPhee y Marx (1997 191) han dicho que por el momento la hipotesis de la hipermorbilidad no pasa de ser una abstraccion en espera de adecuada sustentacion empirica Llama la atencion que linicamente en el periodo amerindio (Tabla 45 intervalo 1Il) las extinciones abarcaron todas las clases del peso corporal (de la a a laj) y que ninguna de las especies que sobrevivieron hasta el periodo postcolombino (Tabla 45 intervalo IV mas las especies vivientes) sobrepasa los 10 kg (clases del peso a b c) en clara coincidencia con los presupuestos de la hipotetica hipermorbilidad
Intervalo Postcolornbino
Por 10 menos 37 especies de mamiferos terrestres vivian en las Antillas Mayores y grupos de Islas perifericas a la lIegada de los europeos en 1492 De elIas se han extinguido 23 shymas especies que en cualquiera de los periodos precedentes (Tabla 45~ y solo subsisten 12 especies repartidas entre Cuba (8 especies) La Espanola (2) Jamaica (1) y Bahamas (1) varias de estas en peligro critico de extincion Solo dos de las especies extinguidas durante los ultimos 500 anos se conocieron en estado viviente los roedores Oryzomys antillarum de Jamaica (extinguido en 1877) y Geocapromys thoracatus de las Islas Swan (extinguido en 1955) Las causas de las extinciones ocurridas durante este periodo son sobradamente conocidas destruccion de habitats introducci6n de animales exoticos (domesticos y comensales) y demas presiones asociadas a la actividad humana entre las cuales posiblemente habria que incluir brotes de hipermorbilidad
Durante mucho tiempo se ha afirmado la posibilidad de que especies de Nesophontes hayan sobrevivido hasta las primeras decadas del siglo xx y que incluso continuen existiendo en lugares remotos de Cuba 0 La Espanola (Allen 1918 1942 Miller 1930 Aguayo 1950a Arredondo 1960b Fischer 1977 MacFadden 1980 Woods y
334 BIOGEOGRAFiA
Ottenwalder 1991) Actualmente no existe evidencia radiometrica de que alguna especie de Nesophontes haya sobrevivido al siglo xv las fechas radiocarbonicas calibradas obtenidas en muestras de Nesophontes de Cuba y La Espanola resultaron no mas tardias que los siglos XIII y XIV (MacPhee et al 1999) Por otra parte las expediciones efectuadas a fines del siglo xx con el proposito de localizar individuos vivientes del genero en La Espanola no produjeron la menor indicacion afirmativa al respecto (Woods et al 1985) Ademas a traves de las Antillas Mayores no aparecen restos de Nesophontes en los depositos formados actualmente por las lechuzas
EPiLOGO
Las Antillas Mayores son islas oceanicas no han formado parte de un continente (con la posible excepcion de una parte de Jamaica hasta el Eoceno Medio hace 45 Ma) y durante un tiempo geologico considerable (desde el Eoceno Superior hace 40 Ma) han permanecido emergidas y separadas de los continentes vecinos Consecuentemente los mamiferos terrestres (no volantes) autoctonos de estas islas constituyen una fauna oceanica que por ende cumple con todas las caracteristicas convencionales reconocidas para este tipo de entidad biogeografica
- Baja diversidad taxonomica a nivel de las categorias superiores (solo cuatro ordenes) en comparacion con islas plataformicas continentales (0 con areas continentales) de tamano y diversidad ecologica simi lares
- Alto grado de endemismo a nivel de genero (96) y familia (50) en respuesta al prolongado aislamiento
- Desbalance ecologico de la estructura trofica ausencia de mamiferos camivoros y ungulados
- Espectaculares procesos de irradiaci6n evolucionaria como resultado de la colonizacion de territorios libres de competidores y depredadores
Tendencias evolucionarias extremas gigantismo
- Control a cargo de aves rapaces con tendencia at gigantismo en ausencia de mamiferos miembros del orden Carnivora
- Mas especies de reptiles y de murcielagos que de mamiferos terrestres relaciones que en los continentes se invierten
Alta tasa de extincion de especies (84) sin reposicion
Preservacion de especies filogeneticamente arcaicas (los llamados fosiles vivientes) los almiquies
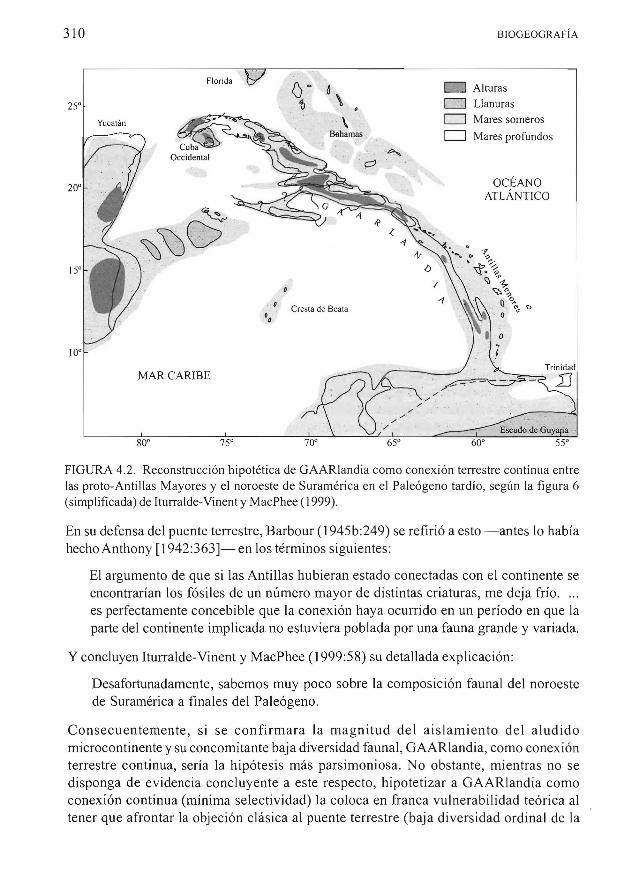
310 BIOGEOGRAFiA
Florida o Alturas0 0
d 6 D L1anuras25deg
Yucatan
20deg
deg0
10deg
MARCARIBE
80deg 75deg
Bahamas
Cres ta de Beata
D Mares someros
D Mares profundos
OCEANO ATLANTICa
FIGURA 42 Reconstrucci6n hipotetica de GAARlandia como conexi6n terrestre continua entre las proto-Antillas Mayores y el noroeste de Suramerica en el Pale6geno tardio segun la FIgura 6 (simplificada) de Iturralde-Vinent y MacPhee (1999)
En su defensa del puente terrestre Barbour (1945b249) se refiri6 a esto - antes 10 habfa hecho Anthony [1942363]- en los terminos siguientes
El argumento de que si las Antillas hubieran estado conectadas con el continente se encontrarian los f6siles de un numero mayor de distintas criaturas me deja frio es perfectamente concebible que la conexi6n haya ocurrido en un periodo en que la parte del continente implicada no estuviera poblada por una fauna grande y variada
Y concluyen lturralde-Vinent y MacPhee (199958) su detallada explicaci6n
Desafortunadamente sabemos muy poco sobre la composici6n faunal del noroeste de Suramerica a finales del Pale6geno
Consecuentemente si se confirmara la magnitud del aislamiento del aludido microcontinente y su concomitante baja diversidad faunal GAARlandia como conexi6n terrestre continua serf a la hip6tesis mas parsimoniosa No obstante mientras no se disponga de evidencia concluyente a este respecto hipotetizar a GAARlandia como conexi6n continua (minima selectividad) la coloca en franca vulnerabilidad te6rica al tener que afrontar la objeci6n clasica al puente terrestre (baja diversidad ordinal de la
311 POBLAMIENTO CONTINENTE-TSLA
fauna) al tiempo que se la priva de ser la hip6tesis mas robusta para explicar -como guimalda de islas- la colonizaci6n de las Antillas por mamiferos procedentes de Suramerica
ELARcA DE NOE (seudovicarianza)
El primer vicarianista antillano (desde luego sin el saberlo) fue Corral (1940) con su utilizaci6n ingenua de las ideas wegenerianas de la deriva continental aparecidas la decada anterior Pero el primer (supuesto) intento de aplicar a la formaci6n de la biota caribefia el mecanismo de la vicarianza basada en la tearia de la tect6nica de placas (surgida en la sexta decada del pas ado siglo) se debe a Rosen ([1976] 1985)
Segun Rosen las Antillas Mayores se originaron en el Cremcico en la Placa Caribe aproximadamente en la posici6n que hoy dia ocupa Centroamerica y al ser desplazadas tectonicamente hacia el este interactuaron (de algun modo no precisado por Rosen) con las margenes continentales de Norte- y Suramerica de donde (en un evento unico) adquirieron toda su fauna transportada subsecuentemente por dichas islas en su transito hacia sus posiciones actuales [De ahi la denominacion metaforica dada par McKenna (1973) a este mecanismo de poblamiento] Aunque se ha presentado esta propuesta como Model0 de Vicarianza Continente-Isla de Rosen en realidad por definici6n no constituye un evento vicariante porque las proto-Antillas Mayores no resultaron del fraccionamiento y separaci6n por tectonismo de una porcion del territorio continental como exige esta hip6tesis (vease definici6n de vicarianza al comienzo de Hipotesis Biogeograficas)
El Arca de Noe de Rosen ha sido desestimada por la mayoria de los autores contemporaneos porque (1) carece de sustento geol6gico (lturralde-Vinent y MacPhee 1999) y porque (2) en el periodo en que se afrrrna ocum6 el evento muchos de los ordenes modemos de mamiferos placentarios attn no existian (Williams 1989)
EL BARCO FUNERAL VlKINGO (tanatovicarianza)
Domning et al (1997) reportaron 10 que consideraron el mamffero terrestre mas viejo de las Antillas --circunstancia que se refleja en el titulo mismo del trabajo Oldest West Indian Land Mammal- al descubrirse un perisodactilo rinocerotoide (Hyrachius sp) en una localidad de edad Eoceno Inferior a Medio en el occidente de Jamaica
Jamaica ha tenido una historia geologica y paleogeografica distinta de la historia comun a las restantes Antillas Mayores A principios del Cenozoico Jamaica occidental (a diferencia de la porcion oriental constituida por el Bloque de las Montafias Azules) formaba parte de Centro america de la cual se desgaj6 (por el desplazamiento de la Placa Caribe) para venir a integrarse despues del Eoceno Medio con 10 que serfa la futura isla de Jamaica desaparecida por transgresi6n marina en el Eoceno Superior hasta volver a emerger totalmente en el Mioceno Medio (Iturralde-Vinent y MacPhee 1999 MacPhee e Iturralde-Vinent 2000)
312 BIOGEOGRAFiA
Por consiguiente Hyrachius ---conocido de localidades de edad Eoceno Inferior a Medio en Norteamerica- debio ocupar (10 que iba a ser) Jamaica occidental mucho antes de que esta se separara de Centroamerica De hecho este fosil Uego a su destino insular contenido en las rocas de su basamento no como un ser vivo por 10 que es erroneo considerarlo un animal antillano (lturralde-Vinent y MacPhee 2004) como han supuesto algunos (Hedges 200 I Portell et al 200 I Condis [200 I D Todo 10 que hizo fue viajar tanatovicariantemente en su barco funeral vikingo -otra de las denominaciones metaforicas de McKenna (1973)
DISCUSION Y CONCLUSIONES
Hemos visto que solo disponemos de una hipotesis plausible para explicar la mayor parte del poblamiento (continente-isla) de las Antillas Mayores por mamiferos terrestres del Neotropico continental la dispersion mixta (a traves de GAARlandia) para los perezosos (excepto Choloepodinae) los monos y los roedores (excepto Oryzomys)
Por otra parte la historia biogeogrMica de las musaraiias de las Antillas Mayores (Nesophontes y Solenodon) constituye una de las situaciones mas enigmaticas entre los mamiferos de la region (Ottenwalder 2001321) Al menos cinco mecanismos de poblamiento continente-isla han sido invocados para explicar cuando como y de donde llegaron a las Antillas Mayores estas musaraiias
I EI propagulo ancestral (del Terciario de Norteamerica) coloniz6 a las Antillas Mayores por dispersion marina desde Centroamerica 0 desde la Florida (Patterson 1962)
2 El propagulo ancestral ( del Terciario) cruzo el Atlantico Sur (supuestamente mas estreshycho en aquella epoca) directamente de Africa a las Antillas Mayores (Hershkovitz 1972)
3 El propagulo ancestral (del Terciario de Norteamerica) Uego a las Antillas Mayores en el Arca de Noe de Rosen (MacFadden 1980)
4 El propagulo ancestral (del Terciario de Norteamerica) fue transportado a las Antillas desde Centroamerica en el bloque de Jamaica occidental y de alli paso por dispersion marina a las restantes Antillas Mayores (Whidden y Asher 200 I)
5 El propagulo ancestral (del Terciario de Norteamerica) lleg6 a las Antillas Mayores a traves de GAARlandia (Whidden y Asher 2001)
Aunque hay consenso entre estos (y otros) autores sobre el origen norteamericano de las musaraiias antillanas ninguno de los cinco hipoteticos mecanismos de poblamiento resiste la evidencia en contrario filogenetica yo paleogeogrMica Ademas algunas de estas hipotesis asumen que Nesophontidae y Solenodontidae comparten un antecesor comun en las Antillas pero esta relacion no ha sido confirm ada Para dilucidar el enigmatico caso de las musaraiias antillanas se requerini profundizar en la investigaci6n geologica paleogeognifica y paleontol6gica (Iturralde-Vinent y MacPhee 1999)
De acuerdo con el conocimiento filogenetico disponible sobre el conjunto de la fauna la colonizacion original de las Antillas Mayores por mamiferos terrestres implicaria el arribo
313 POBLAMIENTO ISLA-ISLA
de solo siete propagulos fundadores perezosos 2 (coloepodinos y megalocninos si como parece tienen un origen difiletico) musaraiias 2 (nesofontidos y solenodontidos si no constituyen una misma familia como consideran algunos) monos 1 (pitecidos) roedores histricognatos 1 (equimidos capromidos y heptaxodontidos) roedores muridos 1 (Oryzomys en Jamaica)
POBLAMIENTO ISLA-ISLA
La distribucion de los mamiferos terrestres a traves del area antillana demarcada al inicio del presente capitulo (vease Contexto Geogratico Fig 41) ha sido interpretada en funcion de dos hipoteticos mecanismos de poblamiento interinsular la dispersion y la vicarianza fundamentadas como se resumen seguidamente
DISPERSION
De acuerdo con la evidencia paleogeogratica mas actualizada en las principales regresiones marinas ocurridas en la cuenca caribeiia durante los ultimos 23 Ma (transiciones OligocenoshyMioceno Mioceno Superior - Plioceno y varias veces en el Pleistoceno) las masas terrestres constitutivas de la region insular en formacion aumentaron considerablemente su extension territorial disminuyendo por tanto la distancia entre elias oportunidades que los elementos faunales aprovecharian para dispersarse de unas islas a otras
Durante mucho tiempo esta ha sido la hipotesis mas favorecida por los estudiosos del tema a partir de Simpson (1956) pero en decadas recientes ha suscitado dos objeciones
1 Es dificil en tender como pudo operar la dispersion marina entre estas islas puesto que ninguna de las Antillas Mayores posee rios suficientemente caudalosos para transportar masas significativas de vegetacion flotante (MacPhee 198432)
Es oportuno recordar aqui el islote ala deriva (con mas de diez palmas reales en pie) entre las tres Antillas Mayores mas occidentales mencionado anteriormente (vease La Balsa Natural)
2 La dispersion marina es menos efectiva para explicar la asombrosa similitud de las faunas de mamiferos terrestres a traves de las Antillas Mayores septentrionales porque como proceso aleatorio no debe producir distribuciones uniformes a 10 largo de una cadena de islas 0 si las produce entonces el proceso no es aleatorio (MacPhee e lturralde-Vinent 2000 149)
Independientemente de que las aludidas distribuciones taxonomicas estan lejos de ser uniformes 0 asombrosamente similares la hipotesis de la dispersion marina contrariamente a su canicter aleatorio y altamente improbable en el caso del poblamiento continente-isla- en el caso del poblamiento isla-isla es un proceso no aleatorio (aprovecha las oportunidades brindadas puntualmente por los periodos de maxima regresion marina) y ademas altamente probable dada la incomparablemente menor distancia entre las islas caribeiias que entre estas y los continentes vecinos
314 BIOGEOGRAFiA
V ICARIANZA
La neotectonica y las transgresiones marinas del Oligoceno Medio y del Mioceno afectaron sustancialmente la paleogeografia de la Cresta de las Antillas Mayores resultando la subdivision gradual de las partes subaereas de la Cresta Asi el este de Cuba y el norte de La Espanola conectados fisicamente durante el Oligoceno Inferior se separaron al formarse el Paso de los Vientos en el Oligoceno tardio en tanto La Espanola y Puerto Rico probablemente permanecieron conectadas hasta el Mioceno tardio en que surgio el Paso de Mona (Iturralde-Vinent y MacPhee 1999)
Tales eventos geologicos habrian dividido las distribuciones de los grupos faunales emplazados previamente en el territorio continuo de las tres islas y a partir de esas disyunciones dichos autores (tambien MacPhee et al 2000 y White y MacPhee 2001) asumieron --como segunda fase del modelo de GAARlandia- la hipotesis de la vicarianza isla-isla para explicar la distribucion multiinsular de varios grupos de mamiferos terrestres a traves de las Antillas Mayores septentrionales De acuerdo con esta hipotesis cualquier diferencia en composicion faunal entre las islas (por ejemplo la ausencia de roedores heptaxodontidos en Cuba 0 de monos platirrinos en Puerto Rico) constituye evidencia de extincion local postvicariante y no de dispersion marina fallida Lo primero se puede demostrar si aparecen los fosiles pertinentes 10 segundo no se puede demostrar (MacPhee e Iturralde-Vinent 1994) Por otra parte IturraldeshyVinent y MacPhee (1999 57) han reconocido que aunque fa vicarianza isla-isla puede representar una solucion clara para las distribuciones de varios taxones cuaternarios no puede explicar todos los casos
Rega et al (2002 18) han presentado dos objeciones al modelo de la vicarianza isla-isla aplicado a los mamiferos de las Antillas Mayores
I Si la separacion de Cuba y La Espanola precedio en varios mill ones de anos la separashycion de La Espanola y Puerto Rico el modelo vicariante de lturralde-MacPhee debe predecir mayor similitud entre las faunas de La Espanola y Puerto Rico que entre las de Cuba y La Espanola De hecho es al reyes
No obstante MacPhee et al (200335) admitieron despues que la fecha mas temprana empiricamente defendible [para la apertura del Paso de Mona] debe ser el Oligoceno Medio esto invierte la secuencia de ambas disyunciones con 10 cual queda resuelta la aparente contradiccion Sin embargo en su reciente evaluacion de la congruencia entre filogenia e hipotesis geologic as en varios grupos de mamiferos terrestres de las Antillas Mayores Davalos (2004373) encontro que ninguno de los taxones cumple con la fecha y el patron de divergencia entre las masas terrestres antillanas postulados por el modelo de GAARlandia
2 La presencia de un mismo genero en las tres Islas septentrionales de las Antillas Mayores implica [por el modelo vicariante] que la diferenciacion entre estas formas durante el intervalo desde el Mioceno ha sido unicamente de nivel especifico [Dicha presencia]
315 rRRADlAcr6~ EVOLUCrONARIA
s610 puede explicarse razonablemente como el resultado de movimientos de esos animales entre las islas en el Cuaternario
De esta objecion resulta dificil disentir [El poblamiento entre islas diferenciado solo a nivel especifico no es probable que sea anterior al Pleistoceno (Simpson 1956 15)]
Debe tenerse en cuenta que la hipotesis de la vicarianza isla-isla alude a las tres islas septentrionales de las Antillas Mayores porque como se explico antes (vease El Barco Funeral Vikingo) Jamaica no existia como isla durante el lapso (Eoceno Superior shyOligoceno) consensualmente reconocido como el mas probable para la colonizacion de las proto-Antill as Mayores por la mayo ria de los mamiferos terrestres Despues de emerger como isla en el Mioceno Medio Jamaica debe haber recibido sus mamiferos terrestres por dispersion isla-isla (Davalos 2004376) y solo en el Cuaternario parece haberrecibido (por dispersion marina desde Centroamerica) al roedor murido Oryzomys
Seguramente tanto la dispersion como la vicarianza contribuyeron a que los mamiferos terrestres se irradiaran a traves de las Antillas Mayores y grupos de islas perifericas pero las maneras y tiempos en que estos mecanismos operaron a 10 largo de dicho proceso son aun en gran medida motivos de desacuerdo e incertidumbre
IRRADIACION EVOLUCIONARIA
A continuaci6n revisamos someramente las distribuciones conocidas de los diferentes grupos de mamiferos terrestres a traves de las Islas principales de las Antillas Mayores y grupos de Islas perifericas incluidas las interpretaciones y conjeturas que la literatura reciente recoge en relaci6n con la manera en que dichas distribuciones pudieron haberse conformado durante la irradiaci6n evolucionaria de los referidos grupos de mamiferos Manifestaciones de esta irradiaci6n pueden inferirse unicamente a partir de los taxones presentes en el Cuatemario pues solo 6 de las especies conocidas corresponden al Terciario
GRUPOS ORlGINARIOS DE SURAMERICA
PEREZOSOS (Megalonychidae)
Aparte de un genero miocenico de Cuba y otro del Oligoceno de Puerto Rico se conocen 4 generos (12 especies) de perezosos cuaternarios (todos extintos) restringidos a las tres islas septentrionales de las Antillas Mayores Acratocnus Neocnus Megalocnus y Parocnus Los cuatro generos estan representados en Cuba y La Espanola y Acratocnus 10 esta ademas (como unico genero de perezoso) en Puerto Rico Esta distribuci6n ha sido interpretada por MacPhee et al (2000) eomo resultado de la vicarianza oligomiocenica entre estas islas y [dado que el Banco Puerto Rico - Islas Virgenes fue la entrada de GAARlandia a las Antillas Mayores] se preguntaron si la ausencia de los otros tres generos en Puerto Rico se debe a que nunca estuvieron en ese territorio 0 a que sus
316 BIOGEOGRAFiA
fosiles atm no han sido hallados Pero como hemos visto Rega et al (2002) atribuyeron dicha distribucion a la dispersion interinsular en el Cuatemario
Por otra parte varios amilisis c1adisticos recientes ubi can a Choloepus -genero Reciente conocido solo de Centro- y Suramerica- como taxon hermano de Aeratoenus exc1uidos los demas megaloniquidos antillanos Para White y MacPhee (2001228)
tan estrecha relacion sugiere que la divergencia entre ambos [generos] ocurrio despues de que Aeratoenus divergio de Neoenus Si esta relacion implica que Aeratoenus y Neoenus se dispersaron separadamente desde Suramerica a las AntilIas mientras Choloepus permanecia en el continente (siendo necesarias al menos tres invasiones de perezosos) 0 si Choloepus invadio Suramerica desde las Antillas es algo que el material disponible no permite determinar
La diversidad adaptativa de los perezosos se manifiesta en (1) la utilizacion del habitat especies arboreas en Neoenus y Aeratoenus y terrestres en Paroenus y Megaloenus (Webb 1985a White [1993] White et al 1996 Arredondo [1999]) y en (2) la gama de tallas corporales desde los pequenos Neoenus hasta Megaloenus rodens el mayor de los mamiferos terrestres antillanos (su peso adulto estimado en 270 kg seglin Paula 19679) esto sugiere que los perezosos se irradiaron a nichos que en contextos continentales probablemente estarian ocupados por miembros de otros c1ados (MacPhee e IturraldeshyVinent 2000)
No esta claro si el abanico de tamanos entre estos perezosos enmascara alguna tendencia al gigantismo Por 10 pronto Anderson y Handley (2002) encontraron 10 contrario en perezosos (Bradypus) de las Islas de Bocas del Toro (Panama) shyseparadas secuencialmente del area continental vecina por ascensos del nivel del mar durante los ultimos 10 ka En las cinco islas los perezosos son menores que su contraparte continental y las diferencias entre las tallas medias de las cinco poblaciones expresan una reduccion lineal significativa de la talla corporal en funcion de la edad geologica de cada isla sin relacion alguna con el area de las islas 0 la distancia que las separa del continente
MONOS (Pitheciidae)
Los primates de las Antillas Mayores (cuatro especies extintas) constituyen un grupo monofiletico con Paralouatta (en Cuba) y Antillothrix (en La Espanola) como taxones hermanos seguidos en la escalera c1adistica por Xenothrix (en Jamaica) y finalmente Callieebus como el pariente continental mas cercano esto implica una sola invasion desde Suramerica antes del Mioceno Inferior (fecha mas temprana conocida para los primates fosiles antillanos) y por tanto en consonancia con GAARlandia como la via de colonizacion (Horovitz y MacPhee 1996 1999 Horovitz [1997] MacPhee y Horovitz 2004)
En desacuerdo parcial con la anterior interpretacion la filogenia de este grupo de monos obtenida por Davalos (2004) indica un antecesor comun en Cuba La Espanola y Jamaica
317 TRRADIACTON EVOLUCIONARIA
compatible con la hipotetica conexion -propuesta tentativamente por Iturralde-Vinent y MacPhee (199980-84)- entre las Montanas Azules del este de Jamaica y la peninsula suroccidental de La Espanola en la epoca de GAARlandia
A pesar de que los primates antillanos pertenecen a un grupo viviente de monos muy pequenos (los monos titles) fueron comparativamente grandes en relacion con los platirrinos modernos y sucesivamente mayores en la direccion Antillothrix Xenothrix y Paralouatta con P marianae como el mayor platirrino conocido del Mioceno neotropical (MacPhee 2005) MacPhee et al (1994) consideraron que Xenothrix debio ser un mono de andar lento y al parecer las especies de Paralouatta abandonaron la arborealidad para adoptar un modo de vida mas terrestre (MacPhee 1996a MacPhee et al 2003 MacPhee y Meldrum 2006)
ROEOORES (Echimyidae Capromyidae Heptaxodontidae)
La edad de Zazamys en Cuba demuestra que la invasion de los roedores histricognatos en las Antillas Mayores comenzo antes del Mioceno Inferior por otra parte los representantes mas antiguos de esa irradiacion evolucionaria se encuentran en La Espanola y Puerto Rico Ambas evidencias son consistentes con la hipotesis de GAARlandia (Woods 1982 1988 Woods et al 2001)
No esta claro si los capromidos evolucionaron a partir de un equimido ancestral 0 de un protocapromido 0 si los equimidos antillanos (I sub familia 3 generos y 7 especies todas extintas) forman parte de la irradiacion de los capromidos (4 subfamilias 9 generos y 27 especies 10 vivientes) Kratochvil et al (1978b) y Rodriguez et al (1979) consideraron que Capromys nan us es un eslab6n entre Echimyidae y Capromyidae pero Mesocapromys nanus esta morfologicamente mucho mas cerca de Hexolobodon y los de mas capromidos que de los equimidos (Woods 1985) Lo que si esta claro segun Woods et al (2001) es que la subfamilia Plagiodontinae endemic a de La Espanola es el linaje basal de los capromidos y que Isolobodon Hexolobodon Capromys Geocapromys Mysateles y Mesocapromys [y supuestamente Macrocapromys] forman parte de la irradiacion temprana de Capromyidae
La subfamilia Capromyinae (5 generos y 17 especies 9 vivientes) parece ser el dado de mas reciente evolucion en la familia con invasion de Cuba desde La Espanola y dispersion desde Cuba (1) por Capromys a las islas Caiman y (2) por Geocapromys a las islas Bahamas Caiman y Jamaica y desde Jamaica a las islas Swan aunque la presencia de Geocapromys en Swan pudiera deberse a introduccion aborigen (Morgan 1985 1988 1989 1993 1994 Woods 1989a 1990) Woods et al (2001) utilizaron datos moleculares para obtener estimados independientes de divergencia entre capromidos segiJn 10 cual Geocapromys divergio cerca de los 17 MaAP Mesocapromys divergio de Capromys hace 15 MaAP en tanto lvlysateles 10 hizo hace 10 MaAP Por su parte las formas de Capromys pilorides y de Mysateles prehensilis en Isla de la Juventud divergieron de sus correspondientes subespecies nominativas en la isla principal a los 24 y 08 MaAP respectivamente
318 BIOGEOGRAFiA
Woods y Howland (I979) describieron adaptaciones morfologicas de la mandibula el hioide y la faringe asociadas a mecanismos de masticacion diferentes en plagiodontinos y caprominos y Varona (1980a) llamo la atencion sobre el gran desarrollo de los molariformes (en relacion con la longitud total del cnineo) que presentan ciertos grupos de jutias fosiles en comparacion con las vivientes 10 que pudiera interpretarse como una adaptacion alimentaria a una vegetacion mas abrasiva por parte de las jutias extinguidas
Segun Woods (1990) la familia Heptaxodontidae (2 subfamilias 5 generos y 6 especies todas extintas) tambien seria parte de la irradiacion de los capromidos con origen en el este de La Espanola 0 en Puerto Rico y subsecuente dispersion al Banco de Anguila - San Martin en las Antillas Menores (pero vease MacPhee y Flemming 2003) Este banco estaria a solo 70 km del Banco Puerto Rico - Islas Virgenes en tiempos de maxima regresion marina y esta proximidad con Puerto Rico -patria de Elasmodontomys obliquus- seguramente tuvo que ver con la presencia en las islas de Anguila y San Martin del heptaxodontino gigante Amblyrhiza relacionado mas estrechamente con los roedores de las Antillas Mayores que con los del resto de las Antillas Menores (Woods 1990 1996) Aunque los roedores suelen aumentar de tamano en las islas (Case 1978) la escala del incremento de la talla en Amblyrhiza (que pudo alcanzar mas de 200 kg de peso adulto) esta fuera de toda proporcion con los patrones conocidos en otros roedores insulares (Biknevicius et 01 1993)
Clidomys (Clidomyinae) de Jamaica difiere marcadamente en su morfologia de los heptaxodontinos (vease ademas MacPhee y Flemming 2003)10 cual dificulta explicar su presencia en esa isla Como Jamaica estuvo bajo el mar hasta el Mioceno Medio el ancestro de Clidomys colonizaria esta isla por dispersion desde el extremo suroccidental de La Espanola despues que los heptaxodontinos se originaron en esta ultima isla (Woods 1989a) Pero Pascual et 01 (I 990) especularon que Clidomys pudiera ser el resultado de una invasion posterior e independiente de los capromidos
GRUPOS ORIGINARIOS DE NORTEAMERICA
El origen de las musaranas antillanas ha resultado ser hasta ahora el mayor enigma biogeografico de la fauna de mamiferos terrestres de la region aunque la mayoria de los autores coinciden en considerarlas originarias de Norteamerica (vease Discus ion y Conclusiones en Poblamiento Continente-Isla) No obstante MacPhee e Iturralde-Vinent (1994) descartaron toda posibilidad de conexion terrestre por el Estrecho de la Florida 0 por el Canal de Yucatan y si el (supuesto) evento vicari ante de Rosen (vease El Arca de Noe) -invocado por MacFadden (1980) para resolver el enigma- fuera minimamente plausible aun asi es dificil creer que las catastrofes que 10 sucedieron (vease Contexto Temporal) hayan barrido con toda esa vieja fauna continental excepto las musaranas circunstancia ya anticipada muy tempranamente por Matthew (l918a)
De otra parte la relaci6n entre Nesophontidae y Solenodontidae permanece irresuelta Si estos linajes no estan estrechamente relacionados entonces es muy posible que
319 IRRADIACION EVOLUCIONARIA
sus distribuciones sean explicadas por mecanismos diferentes (Whidden y Asher 2001)
La descomunal diferencia de talla corporal entre las musaranas pequenas (Nesophontes) y las grandes (Soienodon) seguramente impuso a ambos grupos restricciones distintas relativas a la termorregulacion y las adaptaciones energeticas En Nesophontes la seleccion debio favorecer una alta tasa de metabolismo y crecimiento camadas mas numerosas corta expectativa de vida yalta densidad poblacionaL En Soienodon en cambio la seleccion opero en la direccion de una baja tasa metabolica camadas reducidas crecimiento lento longevidad y baja densidad poblacional (Ottenwalder [1991])
MUSARANAS PEQUENAS (Nesophontidae)
Las musaranas pequenas estan representadas en las Antillas por el genero Nesophontes (y un genero miocenico indeterminado en ambar dominicano) con 6 especies cuaternarias extinguidas distribuidas en Cuba La Espanola Puerto Rico Islas Virgenes e Islas Caiman La mayor de las musaranas pequenas Nesophontes edithae evoluciono en Puerto Rico y su arribo a las Islas Virgenes supone una dispersion sumamente reciente En las Islas Caiman Nesophontes se asemeja mas a la poblacion de Cuba de donde seguramente derivo en el Plio-Pleistoceno (junto con Capromys y Geocapromys) cuando la costa sur de Cuba estuvo mucho mas cerca de las Islas Caiman (lturralde-Vinent 2003fig 4) MacPhee et ai (1994) interpretaron la distribucion de Nesophontes en Cuba y La Espanola como resultado de la vicarianza oligomiocenica ocurrida al formarse el Paso de los Vientos pero la distancia taxonomica entre los lados del Paso es solo de nivel especifico y N micros incluso vivi6 en ambas islas segtln Patterson (1962)
Allen (1918) considero que Nesophontes micrus fue una especie primitiva no modificada para la vida arb6rea 0 fos6rea y por tanto de habitos terrestres Nesophontes edithae de Puerto Rico es mucho mayor y mas robusta que las restantes especies del genero eL indice de robustez de su femur (= anchura minima de la diafisis ongitud total del hueso) muestra que este es tan robusto como el de Solenodon paradoxus un animal dos veces mayor (Morgan et ai 1980) Choate y Birney (1968) postularon un aumento cronoclinal de la taIla corporal en N edithae confirmado mas tarde por McFarlane (1999a) mediante fechados radiocarb6nicos de la secuencia estratignifica en depositos cavernarios de Puerto Rico Solenodon no vivi6 en Puerto Rico y en ausencia de una musarana de mayor tamano Nesophontes alcanzo una talla corporal mayor en esa isla que en La Espanola 0 Cuba (Woods y Eisenberg 1989)
MUSARANAS GRANDES (Solenodontidae)
Los arcaicos almiqufes (genero Soienodon) comprenden cuatro especies una viviente (S cubanus) y otra f6sil (S arredondoi) en Cuba y una viviente (S paradoxus con dos subespecies) y otra f6sil (S marcanoi) en La Espanola Como cabria esperar las dos especies cubanas se asemejan mas entre sf que a las especies de La Espanola pero
320 BIOGEOGRAFiA
enigmaticamente S marcanoi comparte caracteres con S cuban us y S paradoxus (Woods y Eisenberg 1989 Morgan y Ottenwalder 1993 Ottenwalder 200 I) Solenodon arredondoi alcanzo una talla corporal superior a la de cualquiera de las musarafias vivientes a nivel mundial Segun Morgan y Ottenwalder (1993) extrapolando las dimensiones y el peso de los almiquies vivientes S arredondoi debio tener una longitud cabeza-tronco y un peso de 450-550 mm y 1500-2000 g respectivamente en comparacion con S cubanus (360 mm y 800 g) y S paradoxus (390 mm y 1100 g)
Entre todas las especies de almiqufes Solenodon paradoxus es la que presenta los caracteres mas derivados (Otten walder 2001) Si estas relaciones son validas la distribucion de Solenodon entre Cuba y La Espafiola debio discurrir de oeste a este como propuso MacFadden (1980) quien invoco la presencia fosH de S cubanus [pero tambien S arredondoi] en Pinar del Rio para postular que el ancestro continental entr~ por Cuba opinion compartida por Perfit y Williams (1989) y por Ottenwalder ([1991] 2001) El paso de Solenodon a Puerto Rico pudo haberlo impedido la existencia de barreras ecologicas durante la conexion del este de La Espafiola con Puerto Rico 0 por la ausencia de Solenodon en esa parte de La Espafiola en aquella epoca Sin embargo las mismas condiciones ecologicas en el este de La Espafiola no parecen haber representado una barrera efectiva para Nesophontes cuyas mas altas tasa de reproduccion y densidad poblacional presumiblemente Ie permitieron expandirse flpidamente a traves de La Espafiola (Ottenwalder [1991])
Es opinion generalizada que los almiqufes antillanos constituyen el grupo de mamiferos que primero se emplazo en el territorio insular (MacPhee 2005) y en tal sentido es de interes la evidencia sobre especificidad parasito-hospedante aportada por Cruz (1980) quien encontro que la especie de acaro (genero Cubanochirus) que parasita a los almiquies es mucho mas primitiva que la especie (del mismo genero) que paras ita a las jutias y de aqui concluyo que dicho genero de acaro debio evolucionar primero en Solenodontidae y posteriormente en Capromyidae al entrar en contacto representantes de ambas familias en Cuba 0 La Espafiola
Recientemente Roca (2004) y Roca et al (2004) dieron a conocer que los resultados de su estudio genetico-molecular sobre los almiquies indican una divergencia de 25 Ma entre Solenodon cuban us y S paradoxus -compatible con la disyuncion tectonica entre Cuba y La Espafiola (Iturralde-Vinent y MacPhee 1999 McPhee et al 2003)-- y una separacion de ambas especies comparable en magnitud a la divergencia entre familias de mamlferos Consecuentemente propusieron que Solenodon cubanus fuera clasificada en el genero Atopogaie como indico Cabrera (1925) hace 82 afios
DISCUSION Y CONCLUSIONES
Las musarafias y los roedores antillanos (excepto Oryzomys) se irradiaron tanto de sus respectivos ancestros continentales que de estos 6rdenes se reconocen familias y subfamilias endemicas de la region en contraste con los perezosos (una subfamilia y tres
321 IRRADIACION EVOLUCIONARIA
tribus endemicas) y los monos (una tribu endemica) como expresion de diferentes niveles de diversificacion evolucionaria Este proceso es dificil de explicar (traUindose de c1ados que pnicticamente comparten la misma antigiiedad en las Antmas) si es correcta la interpretacion de Pascual et at (1990636)
Nosotros consideramos estos diferentes niveles de diversificacion como las diferentes etapas de evolucion alcanzadas por cada grupo [de mamiferos] Obviamente esto estuvo relacionado con las diferentes longitudes de sus historias
Pudiera ocurrir que en el futuro se compruebe que la historia biogeognifica de la region fue mucho mas complicada
La Espanola la isla mas ecol6gicamente divers a de la region debi6 ser el centro principal de evoluci6n de los mamfferos (con 87 de las familias y 73 de las subfamilias) seguida por Cuba (con 75 de las familias y 54 de las subfamilias) las dos islas de las Antillas Mayores mas relacionadas biogeograficamente (Fig 43 Tabla 44)
eL
EL
cb CAIMAN
SWAN
A Acratocnini B Cubanocnini C Megalocnini D Mesocnini E Nesophontidae F Solenodontidae G Xenothrichini
ANGUILAshyM-0 SMARTIN
H Heteropsomyinae I Plagiodontinae J Isolobodontinae
K Hexolobodontinae L Capromyinae
M Heptaxodontinae N CIidomyinae 0 Sigmodontinae
FIGURA 43 Representacion esquematica de las relaciones interinsulares (expresadas porel numero de linajes compartidos) entre las Antillas Mayores y entre estas y los grupos de islas perifericas segun las interpretaciones de los autores citados en el texto Los numeros dentro de los circulos indican la cantidad de linajes representados en cada isla relacion dudosa Contexto taxon6mico de los linajes en la Tabla 44
bull bull
bull bull
bull bull bull
bull bull
bull bull
bull
bull
bull
bull
bull bull bull bull
bull bull
bull bull
bull
bull bull bull
bull bull
bull
bull bull
bull
322 BIOGEOGRAFiA
TABLA 44 Distribuci6n de los linajcs de mamiferos terrestres a traves de las Antillas Mayores y grupos de islas perifericas LE = La Espanola CU Cuba JA Jamaica PR = Puerto Rico IC Islas Caiman IS Islas Swan IB = Islas Bahamas IV = Islas Virgenes AS = Anguila - San Martin
FamiliaSubfumiliafIribu LE CU JA PR IC IS IB IV AS
Megalonychidae
Choloepodinae
Acratocnini
Cubanocnini
Megalocninae
Megalocnini
Mesocnini
Nesophontidae
Solenodontidae
Pitheciidae
Callicebinae
Xenothrichini
Echimyidae
Heteropsomyinae
Capromyidae
Plagiodontinae
Isolobodontinae
Hexolobodontinae
Capromyinae
H e pta x 0 don tid a e
Heptaxodontinae
Clidomyinae
Muridae
Sigmodontinae
PILOSA
SORICOMORPHA
PRIMATES
RODENTIA
bull
bull
bull
bull TOTALES 12 10 4 4 2
323 EXTlICIOIES DEL CUATERNARIO
El gigantismo ha sido sin duda una tendencia evolucionaria recurrente entre los clados mayores de mamiferos antillanos Es un hecho bien conocido que algunos grupos de mamiferos han aumentado de tamafio en las islas y varios autores han intentado explicarlo a partir de enfoques diferentes (Foster 1964 Miller 1966 Sondaar 1977 Case 1978 Eisenberg y Wilson 1978 Heaney 1978 Lawlor 1982 Lomolino 1985 Angerbjom 1985 McNab 19942001)
EXTINCIONES DEL CUATERNARIO
Las islas que integran la Subregion Antillana han perdido casi 90 de sus mamiferos terrestres durante los ultimos milenios y en proporcion con su extension territorial esta perdida ha sido mayor en la Subregion que en cualquier otro lugar del planeta (MacPhee 1996b) Las extinciones del Cuatemario eliminaron mas de las tres cuartas partes de la fauna de mamiferos terrestres conocida de las Antillas Mayores sus islas satelites y grupos de islas perifericas y el examen de este proceso en sus aspectos mas generales se aborda en esta ultima parte del presente capitulo
CRONOLOGiA Y CONSECUENCIAS
Es muy improbable que un fosil represente el ultimo individuo de un taxon extinto y entonces hay que guiarse por la edad radiometrica del fosil mas reciente conocido lIamada fecha de ultima presencia- como unica aproximacion posible a la fecha de extincion del taxon
Como se explico al inicio del capitulo (vease Contexto Temporal) el registro fosH de los mamiferos terrestres de las Antillas Mayores adolece de insuficiente sustentacion radiometrica No obstante la distribucion temporal de los escasos fechados disponibles para el Cuaternario de la region abarca etapas criticas del proceso de extincion experimentado por la referida fauna a 10 largo de los ultimos 250 mil afios
De las 52 especies que es posible reconocer actualmente como miembros extintos de la fauna cuatemaria de mamiferos terrestres de las Antillas Mayores y grupos de islas perifericas los restos fosiles de 23 de elIas (442) han sido fechados radiometricamente o se han hallado asociados estratigraficamente de manera inequivoca con otro material fechado De estos 23 fechados 3 corresponden al interglacial Sangamon 4 pertenecen a la secuencia Pleistoceno terminal - Holoceno temprano 10 se ubican en ellapso entre el arribo de los aborigenes y la incursion europea y 6 son postcolombinos (a los cuales hay que sumar otras dos especies extinguidas en los siglos XIX y xx) A partir de estas 25 especies avaladas por fechas de ultima presencia distribuidas entre los cuatro intervalos temporales sefialados hemos compuesto la Tabla 45 en la que ademas aparecen las restantes 27 especies (carentes de fechado radiometrico) ubicadas en el intervalo temporal mas probable sobre la base de (1) la asociacion estratigrafica de sus restos con evidencias culturales (amerindias 0 europeas) 0 (2) la ausencia de dicha asociacion Sobre estos criterios de ubicacion temporal procede hacer dos aclaraciones
324 BIOGEOGRAFIA
TABLA 45 Intervalos temporales mas probables para las extinciones cuatemarias de los mamiferos terrestres de las Antillas Mayores y grupos de islas perifericas sobre la base de las evidencias siguientes (l) ultimo ejemplar colectado u observado en vida (2) fechado radiometrico mas tardio en material de la especie (3) asociaci6n estratigrafica mas tardia con otro material fechado (4) asociaci6n estratigrafica con restos de animales introducidos restos humanos u objetos culturales propios del intervale (5) falta de asociaci6n con evidencias culturales [Los ntimeros entre parentesis en la columna Evidencia significan adscripci6n tentativa al intervalo] Los estimados del peso corporal (en kg) estan tomados de la literatura 0 inferidos comparativamente y distribuidos entre
las clases siguientes (a) S 01 (b) 01 a lt 1 (c) 1 a lt 10 (d) 10 a lt 100 (e) 100 a lt 200 (j) ~ 200 Las
abreviaturas que designan las islas son las de la Tabla 44
F amilialEspecie Peso IsJa Evidencia Fuente
I httervalo htterg1acial Sangamon (gt80 kaAP)
HEPTAXODONTlNAE
Amblyrhiza inundata e AS 3 McFarJane et al 1998a
CLIDOMYINAE
Clidomys osborni d JA 3 Ibid
Clidomys parvus d JA 3 Ibid
II httervalo Pleistoceno terminal ~ Holoceno temprano (20-6 kaAP)
MEGALONYCHIDAE
Aeratoenus antillensis d CD 5 Arredondo 1975 a
Aeratoenus odontrigonus d PR 3 McFarJane 1999b
Aeratoenus simorhynehus d LE 2 Rega et al 2002
M egaloenus zile f LE 2 MacPhee et al 2000
SOLENODONTIDAE
Solenodon arredondoi e CD 5 Morgan y Ottenwalder 1993
PnHECIIDAE
Paralouatta varonai d CD 5 MacPhee 1996a
EcHIMYIDAE
Heteropsomys antillensis c PR 5 McFarJane 1999b
Heteropsomys insulans c PR 5 Ibid
Puertoricomys eorozalus c PR 5 Woods 1989a
CAPROMYIDAE
P lagiodontia araeum c LE 5 MacPhee et al 2000
Macrocapromys latus c CD 5 Varona y Arredondo 1979
(Continua)
325 EXTINCIONES DEL CUATERNARIO
TABLA 45 (Continuaci6n)
F ami1ia1Especie Peso Isla Evidencia Fuente
M acrocapromys latus c CU 5 Varona y Arredondo 1979
Mesocapromys kraglievichi c CU 5 Ibid
HEPTAXODONTINAE
Xaymaca fulvopulvis a JA 2 MacPhee y Flemming 2003
III Intervalo Amerindio (6-05 kaAP)
MEGALONYCHIDAE
Acratocnus ye d LE 2 MacPhee et aI 2000
Neocnus comes c LE 2 Ibid
N eocnus dousman c LE 2 Ibid
N eocnus toupiti c LE 2 Ibid
N eocnus gliriform is c CU 4 Cordova et ai 1997
M egalocnus rodens f CU 2 MacPhee et ai 2007
Parocnus browni e CU 2 Steadman et ai 2005
P arocnus serus e LE 2 MacPhee et ai 2000
N ESOPHONTIDAE
N esophontes edithae a PR 2 McFarlane 1999b
PITHECIIDAE
Antillothrix bernensis c LE 3 Rimoli 1977
EcHIMYIDAE
Brotomys contractus a LE 4 Miller 1929c
CAPROMYIDAE
P lagiodontia spelaeum c LE 4 Ibid
M acrocapromys acevedo c CU 4 NUiiez 1990a
HEPTAXODONTINAE
Elasmodontomys obliquus d PR 3 McFarlane 1999b
Iv Intervalo Postcolombino (05 kaAP - Presente)
N ESOPHONTIDAE
N esophontes hypom icrus a LE 24 MacPhee y Fle111111ing 1999
N esophontes zam icrus a LE (4) Ibid
N esophontes m icrus a CU 24 MacPhee et aI 1999
(Continua)
326 BIOGEOGRAFiA
TABLA 45 (Continuacion)
F amilialEspecie Peso Isla Evidencia Fuente
N esophontes m icrus a CD 24 MacPhee et aI 1999
N esophontes m icrus a LE 4 MacPhee et aI 2000
N esophontes sp 1 a IC 4 Morgan 1994
N esophontes sp 2 a IC 4 Ibid
Solenodon marcanoi b LE 4 Ottenwalder 2001
PITHECIIDAE
Xenothrix mcgregori c JA 4 MacPhee y Flemming 1999
EcHIMYIDAE
Boromys offeUa a CD 24 Guarch 1984
Boromys torrei a CD 24 Ibid
Brotomys voratus a LE 4 MacPhee et aI 2000
CAPROMYIDAE
P lagiodontia ipnaeum c LE 4 MacPhee y Flemming 1999
Rhizoplagiodontia lemkei c LE 4 Ibid
Isolobodon portoricensis c PR 24 Ibid
Isolobodon montanus c LE 4 Ibid
Hexolobodon phenax c LE (4) Ibid
Capromys sp c IC 24 Morgan 1994
Geocapromys columbianus c CD 4 MacPhee y Flemming 1999
Geocapromys thoracatus b IS 1 MacPhee y Marx 1997
Geocapromys sp 1 c IC 4 Morgan 1994
Geocapromys sp 2 c IC (4) Ibid
l-IEPTAXODONTINAE
Quemisia gravis c LE (1) MacPhee y Flemming 1999
MURIDAE
Oryzomys antillarum a JA 1 Ibid
1 Flemming y MacPhee (1999) alertaron en contra de la asociacion con otro material fechado Asimismo Morgan (1993) indico que la ausencia de asociacion con evidencias culturales amerindias no es necesariamente un criterio concluyente para ubicar un taxon en el intervalo temporal anterior porque hay depositos en la region que carecen de tales evidencias y son del Holoceno tardio por fechados radiocarbonicos [No obstante
327 EXTlNCIONES DEL CUATERNAR10
la ausencia de una especie en residuarios arqueol6gicos adquiere mayor certidumbre como criterio de ubicaci6n temporal de su extinci6n en la medida que aumenta el numero de residuarios estudiados que en las Antillas Mayores se cuentan por centenares] A falta de fechado absoluto de una especie consideramos que estos criterios de ubicaci6n temporal son mas utiles que la alternativa de excluir del analisis a la especie
2 En los trabajos sobre biogeografia de mamiferos antillanos los arque610gos mas citados como autoridades para establecer el inicio de la presencia aborigen en las Antillas Mayores (Kozlowski 1974 Rouse y Allaire 1978 Rouse 1989 1992 Wilson 2001) difieren al respecto entre los 6 kaAP y los 45 kaAP (pero vease Pagan et al 200523) y hemos adoptado la mas temprana de estas fechas para el comienzo del intervalo amerindio en la Tabla 45 De todos modos cualquiera que haya sido la fecha esta no es aplicable por igual al conjunto de las Antillas Mayores porque segun algunos de los autores citados el poblamiento comenz6 por Cuba y La Espanola 6 mil anos atras continu6 en Puerto Rico hace alrededor de 3 mil anos (pero vease Burney et al 1994) yen Jamaica hace menos de 2 mil e incluso a las Islas Caiman ni siquiera llegaron [Las Islas Caiman descubiertas por Col6n en 1503 carecen absolutamente de evidencia amerindia (Stokes y Keegan 1996)]
Si los datos de la Tabla 45 representan una aceptable aproximaci6n a la realidad el proceso de las extinciones cuaternarias de los mamiferos terrestres de las Antillas Mayores y grupos de islas perifericas debi6 caracterizarse entre otros por los eventos siguientes
Aunque obviamente la fauna cuaternaria debi6 ser mucho mas numerosa que 10 que refleja el registro paleontol6gico (Morgan y Woods 1986) 52 especies (81 de la fauna conocida) desaparecieron (sin reposici6n) durante los ultimos 250 mil anos
- Al parecer las extinciones se concentraron en las especies de mayor tamano Asi el peso promedio estimado para las especies de la etapa preantr6pica (intervalos I + II de la Tabla 45) es 350 kg es 328 kg para las especies del intervalo III (amerindio) y es s610 25 kg (c1ases del peso a b c) para las especies del intervalo IV (postcolombino) Ninguna especie aut6ctona de mamifero mayor de 6 kg existe hoy en la regi6n
- De acuerdo con la distribuci6n temporal de las extinciones 61 especies sobrevivieron al interglacial Sangamon (intervalo 1)48 sobrevivieron a la transici6n del Pleistoceno terminal al Holoceno temprano (intervalo II) 34 sobrevivieron al periodo amerindio (intervalo III) y s610 12 especies han sobrevivido desde la Ilegada de los europeos (intervalo IV) Las tasas de extinci6n correspondientes (exceptuado el intervalo I) son una especie cada 1167 anos en el intervalo II una cada 393 anos en el intervalo III y una cada 23 anos en el intervalo IV
- Durante todo el proceso de las extinciones cuaternarias la perdida de especies por isla fue de 91 en La Espanola 83 en Jamaica y 62 en Cuba Bahamas retuvo su unica especie (aunque extirpada en varias de las islas) Puerto Rico e Islas Caiman perdieron todas sus especies (6 y 5 respectivamente) y las Islas Swan Virgenes y Anguila - San Martin perdieron sus respectivas especies (micas
328 BIOGEOGRAFiA
De los cuatro 6rdenes de mamiferos representados en la fauna Pilosa y Primates desaparecieron totalmente Soricomorpha y Rodentia perdieron 86 y 74 de sus especies respectivamente
CAUSALIDAD
CAUSAS NATURALES
Los cicIos glaciales e interglaciales acaecidos en el Pleistoceno causaron cambios ambientales dnisticos en las Antillas con la concomitante alternancia de cIimas frios (secos) y calidos (hUmedos) asi como de descensos y ascensos del nivel del mar aumento y disminuci6n de las areas territoriales) induso (aunque con menor intensidad) en el Holoceno Estos eventos paleogeograficos han sido consistentemente documentados (Mayo y Kartashov 1972 Bonnatti y Gartner 1973 Ortega y Arcia 1982 Prance 1982 Schubert y Medina 1982 Dodge et al 1983 Ortega [1984] Haq et al 1987 Schubert 1989 Hodell et al 1991 Guilderson et al 1994 Oppo 1997 Emslie 1998 Curtis et al 2001 Iturralde-Vinent 2003) y de ellos brindamos a continuaci6n una breve resena
Finalizando el periodo interglacial anterior a la ultima glaciaci6n (Wisconsin) el mar estaba a 8-10 m por encima de su nivel actual Cuando el avance glacial del Wisconsin a1canz6 su culminaci6n (eillamado Ultimo Maximo Glacial) hace 18 ka el mar habia descendido 120 m con respecto a dicho nivel y las islas del Caribe habian incrementado considerablemente sus extensiones subaereas en comparaci6n con las que presentan hoy dia por ejemplo Puerto Rico y las Islas Virgenes se conectaron territorialmente y las Bahamas se unieron en cinco gran des islas Prevalecia un regimen frio y seco y condiciones de aridez --debidas a una reducci6n de las precipitaciones (posiblemente a menos de 440 mmlano)- y los ambientes xericos se habian extendido a traves de las islas y bancos emergidos y abundaban los pinares (los habitats mesicos estaban restringidos a parcelas relictas en 10 alto de las montanas) En el Mar Caribe y el Golfo de Mexico la temperatura superficial del mar era 5degC mas baja que la actual y en las llanuras costeras y alturas maximas eran respectivamente 2-3degC y 6degC mas bajas Los limites de las zonas ecol6gicas altitudinales descendieron varios cientos de metros y cada zona redujo su extensi6n vertical al aumentar el gradiente de temperatura Ellimite inferior de la zona periglacial
elevaci6n donde la temperatura media minima a1canza los OdegC) de la Cordillera Central de la Republica Dominicana (altura maxima del Caribe insular con evidencia de topografia glacial) se considera que estuvo entre los 1500 y 1800 mlnm (actualmente esta a 2800 mlnm) y se puede estimar que dicho limite era inferior en Cuba por estar mas al norte que la Republica Dominicana No obstante durante este periodo la temperatura experiment6 episodios cortos (unas pocas decadas) de ascensos repetidos
Un signifieativo componente bi6tieo de las referidas condiciones paleoambientales 10 constituy6 la espectacular irradiaci6n evolucionaria experimentada por un grupo numeroso de aves rapaees diurnas y nocturnas (Wetmore 1937 Brodkorb 1969 Kurotchkin y Mayo 1973 Arredondo 1976 Olson 1978 Olson y Hilgartner 1982
329 EXTINCIONES DEL ClJATERNARIO
Suarez 2000a b 2004 Suarez y Emslie 2003) Los mamiferos de las Antillas Mayores ofrecieron a estos depredadores aviares una amplia gama de tamanos de presa que en ausencia de mamiferos del orden Carnivora en la region permitio a estas aves irradiarse a este nicho alimentario con la evolucion de una variedad de especies de muy diversa tall a corporal incluidas formas gigantes Adquirir un cuerpo gigante les propicio el acceso a recursos alimentarios que no estaban siendo explotados por ningun otro depredador (Alcover y McMinn 1994) A Cuba corresponde el mejor registro fosil de rapaces cuaternarias caribenas con siete generos de especies diurnas Dentro de estas aves existieron tres gavilanes gigantes (Accipitridae) tres carairas (Falconidae) y un condor (Cathartidae) asi como cuatro generos de especies nocturnas dos lechuzas (Tytonidae) y tres Mhos (Strigidae) dos de ell os no igualados en tamano en todo el mundo Algunos de estos generos tuvieron representacion en otras islas antillanas como La Espanola Jamaica Bahamas y Gran Caiman
Al retirarse hacia el norte los hielos de la glaciacion Wisconsin el clima se tomo mas hUmedo en las Antillas durante el Holoceno temprano y el bosque mesico se expandio en la region insular El nivel del mar ascendio acompanado de mayor pluviosidad alrededor de los 7 kaAP las Islas Virgenes quedaron separadas entre si y de Puerto Rico y las Bahamas perdieron la mayor parte de su territorio Cerca de los 3 kaAP las condiciones secas retornaron a la region e hicieron desaparecer buena parte del bosque mesico Nuevamente el clima comenzo a adquirir sus condiciones actuales hacia los 25 kaAP aproximadamente y hoy transita por una fase mesica con vegetacion xerica confinada en parcelas relictas
Sin duda estos y otros eventos naturales incidieron negativamente en la sobrevivencia de mamiferos terrestres en las Antillas Mayores asi como en los grupos de pequenas islas perifericas (Pregill y Olson 1981 Olson 2006)
McFarlane et al (1998a) demostraron convincentemente como Amblyrhiza el gigante de los roedores antillanos endemico de la pequena isla de Anguila sucumbio a las repetidas y abruptas perdidas de su habitat durante el interglacial Sangamon por las nipidas y severas oscilaciones del nivel del mar (documentadas por Neumann y Hearty 1996) al no poder responder adecuadamente a tan catastroficos eventos mediante el ajuste de su masa corporal media y otro tanto debio ocurrirle en Jamaica al otro roedor gigante Clidomys (McFarlane et al 1998b) emparentado con Amblyrhiza en la misma epoca (Tabla 45 intervalo I)
El ascenso del nivel del mar seguramente promovio la extincion de mamiferos durante la transicion del Pleistoceno al Holoceno (Tabla 45 intervalo II) con la reduccion del tamano de las Islas por las transgresiones marinas (Pregill y Olson 1981 Morgan 1993) Esto ultimo haria maximamente severa la competicion asf como la presion de los depredadores [Steadman et al (2005) no yen por que el calentamiento global ocurrido durante la transicion Pleistoceno-Holoceno habria de tener un efecto negativo sobre los perezosos siendo sus descendientes actuales especies tropic ales 0 subtropicales pero no se trata del efecto
330 BIOGEOGRAFiA
directo del cambio de temperatura sobre los animales sino del efecto indirecto y progresivo que dicho cambio genera a largo plazo al reducir el territorio insular disponible]
Por esta epoca tambien desaparecieron especies de tall a mayor como Megalocnus zile en La Espanola y otras de menor tamano (en Cuba) pero que representan la tall a maxima en sus respectivos contextos taxonomicos del Cuaternario como Solenodon arredondoi Paralouatta varonai y Macrocapromys acevedo Un mamifero puede ser grande solo a costa de reducir su densidad poblacional y esto 10 hace mis vulnerable a la extinci6n sobre todo en una isla (McNab 1994 McFarlane et al 1998a) Incidentalmente Alcover et al (1998a) afirmaron que larelacion entre la talla corporal y la extincion no siempre se cumple en las especies de menor tamano y citaron como ejemplo que entre las musarafias todas las especies de Nesophontes desaparecieron mientras sobreviven especies de Solenodon pero la comparacion correcta en todo caso serfa entre los solenodones vivientes y el gigante Solenodon arredondoi y entre Nesophontes edithae -el gigante de los nesofontes extinguido en el periodo amerindio- y el resto de sus congeneres que sobrevivieron hasta los primeros siglos de la colonizacion europea (Tabla 45 intervalos III y IV)
CAUSAS ANTROPOGENICAS
Intervalo Amerindio
La mayoria de los autores que se han referido a las extinciones acaecidas durante este intervalo (entre elIos paradigmaticamente Martin 1984a b) han responsabilizado a los aborfgenes (que se supone comenzaron a poblar las Antillas Mayores hace 6 mil afios) con la desaparicion de numerosas especies de mamiferos terrestres en la region principalmente por la via de la sobrecaza y la introduccion de animales domesticos devenidos depredadores de la fauna autoctona (Morgan y Woods 1986) Este efecto negativo de la presencia indigena sobre los mamiferos de la region ha sido dramatizado por algunos autores en terminos de holocausto faunal Sin embargo pensamos que se ha exagerado Como han expresado MacPhee et al (19993) cuando uno busca evidencia dura de complicidad humana directa en [estas] extinciones no hay mucho que encontrar Las siguientes son algunas consideraciones a tener en cuenta en tal sentido
El registro radiometrico mas joven conocido hasta ahora para un perezoso antillano (MegalocnllS rodens) es de 42 kaAP (MacPhee et ai 2007) Si se acepta que los aborfgenes entraron en Cuba hace aproximadamente 6 mil anos su convivencia con los perezosos debe haber durado alrededor de 1800 afios (0 mas porque dificilmente el referfdo fechado radiometrico haya correspondido al ultimo sobreviviente de la especie) Esta cifra contradice la hipotesis del holocausto faunal segun la cualla extincion de los mamiferos insulares de mayor talla debe ocurrir con extrema rapidez -a escala de decadas 0 de centurias- a partir de su primer contacto con los humanos (Martin y Steadman 1999) A pesar de las repetidas afirmaciones en contrario (Harrington 1921 Miller 1929c Herrera [1945] Tabio 1951 Alvarez 1951a 1957 Arredondo
331 EXTINCIONES DEL CUATERNARIO
1959 1960a Veloz y Ortega 1973 Mayo 1980b Rodriguez et al 1984 Castellanos y Pino 1985 Pino y Castellanos 1985 Nunez 1990b Gutierrez 1996 Arredondo 2004) en su mayoria basadas en evidencias no exentas de posibles accidentes de deposicion (0 de reubicacion postdeposicional) en recintos cavernarios (Diaz-Franco [2006b D no se ha demostrado inequivocamente que los aborigenes antillanos se alimentaran de perezosos regularmente Sus restos en residuarios de alimentos son escasisimos en toda la region y esto contrasta con los abundantes restos de jutias y otros mamiferos de menor talla que aparecen en esos residuarios demostrativos de la preferencia de los paleoindios por estos otros mamiferos
Los mamiferos mas importantes en la economia prehistorica de las Antillas Mayores fueron las jutias Capromys y Geocapromys en Cuba Geocapromys en Jamaica e Isolobodon en La Espanola Puerto Rico e Islas Virgenes (Aguayo 1950b Tablo 1951 Rimoli 1972 Wing 2001 Diaz-Franco [2006b D Wilkins (2001) estudi61ajutia viviente de Jamaica (Geocapromys browni) en un sitio ocupado por los aborigenes durante 100 anos y no encontr6 evidencias de que durante ese tiempo la especie haya sido sobreexplotada Asimismo en Cuba las jutias conga carabali y andaraz son escasas en depositos pleistoholocenicos en cambio son muy numerosas en residuarios indigenas y de haber sido sobreexplotadas 10 mas probable es que no hubieran sobrevivido hasta hoy
La habilidad de los humanos para penetrar en profundidad el bosque tropical y mantenerse en este sustentablemente ha sido un evento comparativamente reciente (Beran y Steele 1994) En el caso antillano la caza tierra adentro ha sido explicada como un proceso tardio de desplazamiento a partir de la explotacion de abundantes recursos marinos que ofrecian condiciones 6ptimas para el sedentarismo de las comunidades aborigenes costeras (C6rdova y Arredondo 1988 Brito 2005) Peces y mariscos dominan la arqueologia costera de las Islas antillanas en tanto el consumo de recursos terrestres (mamfferos incluidos) aumenta progresivamente en la medida que los residuarios estudiados se alejan de la costa (Scudder 1991 Wilkins 2001) En las Antillas Mayores el componente terrestre de la dieta aborigen es solo 20-30 del material recobrado en los residuarios (Wing 1977 1989) Y debio ser menor alin porque el componente vegetal de la dieta no se ha preservado Por otra parte los aborigenes cubanos domesticaron la jutia conga y seguramente la reprodujeron para comer (Pose et al 1988) practica que probablemente se repitiera con otras jutias en las demas Antillas Mayores
- Fray Bartolome de las Casas hablo de 3 millones de indios en Cuba al inicio de la colonizacion pero los historiadores subsecuentes consideraron exagerada la cifra (Poey 1851 a) No obstante se dispone de dos estimados de la densidad de poblaci6n aborigen en las Antillas Mayores a la llegada de los europeos 18 indigenaslkm2 en Puerto Rico (Rouse 1956) y 1271km2 en La Espanola (Watts 1987) Extrapolando ambos valores entre las cuatro islas principales de las Antillas Mayores tenemos que en la fecha indicada la poblaci6n aborigen pudo estar entre 16 mil y 113 mil indigenas en Puerto Rico entre 21 mil y 145 mil en Jamaica entre 141 mil y 1 mill6n en La Espanola y entre 200 mil y 14 millones en Cuba Entre las cifras maximas y las minimas nos inclinamos
332 BIOGEOGRAFiA
hacia estas ultimas atendiendo a los altos indices de mortalidad infantil y morbilidad que se evidencian en las osamentas de los pobladores prehispanicos (Martinez y Rives 1990) Uno de los hechos que apuntan hacia el bajo numero de aborigenes en las Antillas Mayores es 10 rapido que desaparecieron despues de la llegada de los europeos pnkticamente sin dejar mestizaje con estos 10 cual contrasta ostensiblemente con la situacion en la America continentaL
En suma un grupo humano (1) demograiicamente irrelevante y controlado por altos indices de morbilidad (2) consumidor mayoritariamente de organismos marinos y (3) carente de la tecnologia cinegetica necesaria para poder sobreexplotar los mamiferos que Ie sirvieron de alimento (a los cuales accedio tardiamente en su desarrollo) no es concebible que haya causado e1 holocausto faunal que se Ie atribuye a partir del error de aplicar a las comunidades aborigenes antillanas un modelo cultural de la prehistoria continentaL
Tampoco hay evidencias de que los aborigenes hayan introducido carnivoros con suficiente capacidad depredadora salvo el perro Morgan y Otlenwaldder (1993) consideraron que la depredacion por perros durante el periodo amerindio es la explicacion mas plausible para la extincion del almiqui gigante de Cuba pero como se puede observar (Tabla 54) Solenodon arredondoi aparece en el intervalo II por falta de asociacion con evidencias culturales
Consecuentemente el accionar de los aborigenes y de sus animales domesticos directamente sobre la fauna no parece haber sido por sf solo un factor determinante de la extincion de mamiferos terrestres durante el periodo amerindio en las Antillas (pero vease Steadman et al 2005)
Hay que recordar en este senti do que alin en el Holoceno tardio ocurrieron cambios clirruiticos de consideracion (vease Causas Naturales) Por ejemplo Curtis et al (2001) explicaron que la extincion de algunos ani males mesofilicos de las alturas montanosas de La Espanola pudo haber antecedido a cualquier impacto humano sigrIificativo en esos lugares por coincidir dicha extincion con una tendencia a la aridez documentada isotopicamente y con evidencias de la perdida del bosque mesico ocurridas despues de 3kaAP Yen Jamaica hace solo 2shy3 mil aDos el area de la Bahia de Jackson sustentaba una gran biomasa de vertebrados en bosque hinnedo a juzgar por la cantidad y diversidad de taxones presentes en material fosil derivado de la depredacion por lechuzas hoy el area es la mas seca de la isla con matorral xerofitico y pobreza de fauna (MacPhee 1996a) Yalm en nuestros dias huracanes de gran intensidad aniquilan poblaciones de vertebrados (que no se recuperan) en is las pequenas y llanas del Caribe (Brooks y Smith 2001)
En medio de la incertidumbre sobre la verdadera causa (0 causas) de las extinciones de mamiferos en el periodo amerindio MacPhee y Marx (1997) han propuesto su hipotesis de la hipermorbilidad la cual estipula colapsos poblacionales relativamente rapidos con la introduccion de enfermedades de alta letalidad Estos autores han razonado que
Las islas son los lugares mas apropiados para experimentar perdidas catastroficas causadas por enfermedades Los mamiferos no solo serian susceptibles en virtud de su prolongado aislamiento biologico sino que en los confines de una isla la epizootia
333 EXTINCIONES DEL CUATERNARIO
tendria la potencialidad de infectar simultaneamente a la rnayoria de los individuos de vadas especies Los neonatos son inmunologicamente inmaduros y sufririan desproporcionadamente cualquier infeccion hipervirulenta La perdida excesiva de neonatos y juveniles tendria efectos severos para la sobrevivencia de la poblacion si esta se reduce por debajo de cierto nivel sobreviene la extincion porque no hay posibilidad de restitucion en el periodo postpanzootico Las especies de mayor talla corporal van a ser las mas vulnerables a la extincion dadas sus bajas tasas reproductivas largos periodos de gestacion y reducido numero de progenie Las especies de menor tamano --caracterizadas por altas tasas reproductivas periodos cortos de gestacion y mayor numero de neonatos por parto- tambien serian fuertemente diezmadas pero muchas sobrevivirian porque su estrategia reproductiva les permite perder mayor numero de individuos Las perdidas deben ocurrir relativamente rapido (a escala de decadas) mientras que otras hipotesis (digamos los cambios climaticos) permiten intervalos mucho mas largos (a escala de siglos 0 de milenios) entre la aparicion de las causas y la produccion de sus efectos Por ejemplo todo indica que los nesofontes se extinguieron muy rapidamente y seria dificil poder invocar un impacto humano directo 0 argumentar fenomenos climaticos que pudieran explicar plausiblemente perdidas tan rapidas
MacPhee y Marx (1997 191) han dicho que por el momento la hipotesis de la hipermorbilidad no pasa de ser una abstraccion en espera de adecuada sustentacion empirica Llama la atencion que linicamente en el periodo amerindio (Tabla 45 intervalo 1Il) las extinciones abarcaron todas las clases del peso corporal (de la a a laj) y que ninguna de las especies que sobrevivieron hasta el periodo postcolombino (Tabla 45 intervalo IV mas las especies vivientes) sobrepasa los 10 kg (clases del peso a b c) en clara coincidencia con los presupuestos de la hipotetica hipermorbilidad
Intervalo Postcolornbino
Por 10 menos 37 especies de mamiferos terrestres vivian en las Antillas Mayores y grupos de Islas perifericas a la lIegada de los europeos en 1492 De elIas se han extinguido 23 shymas especies que en cualquiera de los periodos precedentes (Tabla 45~ y solo subsisten 12 especies repartidas entre Cuba (8 especies) La Espanola (2) Jamaica (1) y Bahamas (1) varias de estas en peligro critico de extincion Solo dos de las especies extinguidas durante los ultimos 500 anos se conocieron en estado viviente los roedores Oryzomys antillarum de Jamaica (extinguido en 1877) y Geocapromys thoracatus de las Islas Swan (extinguido en 1955) Las causas de las extinciones ocurridas durante este periodo son sobradamente conocidas destruccion de habitats introducci6n de animales exoticos (domesticos y comensales) y demas presiones asociadas a la actividad humana entre las cuales posiblemente habria que incluir brotes de hipermorbilidad
Durante mucho tiempo se ha afirmado la posibilidad de que especies de Nesophontes hayan sobrevivido hasta las primeras decadas del siglo xx y que incluso continuen existiendo en lugares remotos de Cuba 0 La Espanola (Allen 1918 1942 Miller 1930 Aguayo 1950a Arredondo 1960b Fischer 1977 MacFadden 1980 Woods y
334 BIOGEOGRAFiA
Ottenwalder 1991) Actualmente no existe evidencia radiometrica de que alguna especie de Nesophontes haya sobrevivido al siglo xv las fechas radiocarbonicas calibradas obtenidas en muestras de Nesophontes de Cuba y La Espanola resultaron no mas tardias que los siglos XIII y XIV (MacPhee et al 1999) Por otra parte las expediciones efectuadas a fines del siglo xx con el proposito de localizar individuos vivientes del genero en La Espanola no produjeron la menor indicacion afirmativa al respecto (Woods et al 1985) Ademas a traves de las Antillas Mayores no aparecen restos de Nesophontes en los depositos formados actualmente por las lechuzas
EPiLOGO
Las Antillas Mayores son islas oceanicas no han formado parte de un continente (con la posible excepcion de una parte de Jamaica hasta el Eoceno Medio hace 45 Ma) y durante un tiempo geologico considerable (desde el Eoceno Superior hace 40 Ma) han permanecido emergidas y separadas de los continentes vecinos Consecuentemente los mamiferos terrestres (no volantes) autoctonos de estas islas constituyen una fauna oceanica que por ende cumple con todas las caracteristicas convencionales reconocidas para este tipo de entidad biogeografica
- Baja diversidad taxonomica a nivel de las categorias superiores (solo cuatro ordenes) en comparacion con islas plataformicas continentales (0 con areas continentales) de tamano y diversidad ecologica simi lares
- Alto grado de endemismo a nivel de genero (96) y familia (50) en respuesta al prolongado aislamiento
- Desbalance ecologico de la estructura trofica ausencia de mamiferos camivoros y ungulados
- Espectaculares procesos de irradiaci6n evolucionaria como resultado de la colonizacion de territorios libres de competidores y depredadores
Tendencias evolucionarias extremas gigantismo
- Control a cargo de aves rapaces con tendencia at gigantismo en ausencia de mamiferos miembros del orden Carnivora
- Mas especies de reptiles y de murcielagos que de mamiferos terrestres relaciones que en los continentes se invierten
Alta tasa de extincion de especies (84) sin reposicion
Preservacion de especies filogeneticamente arcaicas (los llamados fosiles vivientes) los almiquies

311 POBLAMIENTO CONTINENTE-TSLA
fauna) al tiempo que se la priva de ser la hip6tesis mas robusta para explicar -como guimalda de islas- la colonizaci6n de las Antillas por mamiferos procedentes de Suramerica
ELARcA DE NOE (seudovicarianza)
El primer vicarianista antillano (desde luego sin el saberlo) fue Corral (1940) con su utilizaci6n ingenua de las ideas wegenerianas de la deriva continental aparecidas la decada anterior Pero el primer (supuesto) intento de aplicar a la formaci6n de la biota caribefia el mecanismo de la vicarianza basada en la tearia de la tect6nica de placas (surgida en la sexta decada del pas ado siglo) se debe a Rosen ([1976] 1985)
Segun Rosen las Antillas Mayores se originaron en el Cremcico en la Placa Caribe aproximadamente en la posici6n que hoy dia ocupa Centroamerica y al ser desplazadas tectonicamente hacia el este interactuaron (de algun modo no precisado por Rosen) con las margenes continentales de Norte- y Suramerica de donde (en un evento unico) adquirieron toda su fauna transportada subsecuentemente por dichas islas en su transito hacia sus posiciones actuales [De ahi la denominacion metaforica dada par McKenna (1973) a este mecanismo de poblamiento] Aunque se ha presentado esta propuesta como Model0 de Vicarianza Continente-Isla de Rosen en realidad por definici6n no constituye un evento vicariante porque las proto-Antillas Mayores no resultaron del fraccionamiento y separaci6n por tectonismo de una porcion del territorio continental como exige esta hip6tesis (vease definici6n de vicarianza al comienzo de Hipotesis Biogeograficas)
El Arca de Noe de Rosen ha sido desestimada por la mayoria de los autores contemporaneos porque (1) carece de sustento geol6gico (lturralde-Vinent y MacPhee 1999) y porque (2) en el periodo en que se afrrrna ocum6 el evento muchos de los ordenes modemos de mamiferos placentarios attn no existian (Williams 1989)
EL BARCO FUNERAL VlKINGO (tanatovicarianza)
Domning et al (1997) reportaron 10 que consideraron el mamffero terrestre mas viejo de las Antillas --circunstancia que se refleja en el titulo mismo del trabajo Oldest West Indian Land Mammal- al descubrirse un perisodactilo rinocerotoide (Hyrachius sp) en una localidad de edad Eoceno Inferior a Medio en el occidente de Jamaica
Jamaica ha tenido una historia geologica y paleogeografica distinta de la historia comun a las restantes Antillas Mayores A principios del Cenozoico Jamaica occidental (a diferencia de la porcion oriental constituida por el Bloque de las Montafias Azules) formaba parte de Centro america de la cual se desgaj6 (por el desplazamiento de la Placa Caribe) para venir a integrarse despues del Eoceno Medio con 10 que serfa la futura isla de Jamaica desaparecida por transgresi6n marina en el Eoceno Superior hasta volver a emerger totalmente en el Mioceno Medio (Iturralde-Vinent y MacPhee 1999 MacPhee e Iturralde-Vinent 2000)
312 BIOGEOGRAFiA
Por consiguiente Hyrachius ---conocido de localidades de edad Eoceno Inferior a Medio en Norteamerica- debio ocupar (10 que iba a ser) Jamaica occidental mucho antes de que esta se separara de Centroamerica De hecho este fosil Uego a su destino insular contenido en las rocas de su basamento no como un ser vivo por 10 que es erroneo considerarlo un animal antillano (lturralde-Vinent y MacPhee 2004) como han supuesto algunos (Hedges 200 I Portell et al 200 I Condis [200 I D Todo 10 que hizo fue viajar tanatovicariantemente en su barco funeral vikingo -otra de las denominaciones metaforicas de McKenna (1973)
DISCUSION Y CONCLUSIONES
Hemos visto que solo disponemos de una hipotesis plausible para explicar la mayor parte del poblamiento (continente-isla) de las Antillas Mayores por mamiferos terrestres del Neotropico continental la dispersion mixta (a traves de GAARlandia) para los perezosos (excepto Choloepodinae) los monos y los roedores (excepto Oryzomys)
Por otra parte la historia biogeogrMica de las musaraiias de las Antillas Mayores (Nesophontes y Solenodon) constituye una de las situaciones mas enigmaticas entre los mamiferos de la region (Ottenwalder 2001321) Al menos cinco mecanismos de poblamiento continente-isla han sido invocados para explicar cuando como y de donde llegaron a las Antillas Mayores estas musaraiias
I EI propagulo ancestral (del Terciario de Norteamerica) coloniz6 a las Antillas Mayores por dispersion marina desde Centroamerica 0 desde la Florida (Patterson 1962)
2 El propagulo ancestral ( del Terciario) cruzo el Atlantico Sur (supuestamente mas estreshycho en aquella epoca) directamente de Africa a las Antillas Mayores (Hershkovitz 1972)
3 El propagulo ancestral (del Terciario de Norteamerica) Uego a las Antillas Mayores en el Arca de Noe de Rosen (MacFadden 1980)
4 El propagulo ancestral (del Terciario de Norteamerica) fue transportado a las Antillas desde Centroamerica en el bloque de Jamaica occidental y de alli paso por dispersion marina a las restantes Antillas Mayores (Whidden y Asher 200 I)
5 El propagulo ancestral (del Terciario de Norteamerica) lleg6 a las Antillas Mayores a traves de GAARlandia (Whidden y Asher 2001)
Aunque hay consenso entre estos (y otros) autores sobre el origen norteamericano de las musaraiias antillanas ninguno de los cinco hipoteticos mecanismos de poblamiento resiste la evidencia en contrario filogenetica yo paleogeogrMica Ademas algunas de estas hipotesis asumen que Nesophontidae y Solenodontidae comparten un antecesor comun en las Antillas pero esta relacion no ha sido confirm ada Para dilucidar el enigmatico caso de las musaraiias antillanas se requerini profundizar en la investigaci6n geologica paleogeognifica y paleontol6gica (Iturralde-Vinent y MacPhee 1999)
De acuerdo con el conocimiento filogenetico disponible sobre el conjunto de la fauna la colonizacion original de las Antillas Mayores por mamiferos terrestres implicaria el arribo
313 POBLAMIENTO ISLA-ISLA
de solo siete propagulos fundadores perezosos 2 (coloepodinos y megalocninos si como parece tienen un origen difiletico) musaraiias 2 (nesofontidos y solenodontidos si no constituyen una misma familia como consideran algunos) monos 1 (pitecidos) roedores histricognatos 1 (equimidos capromidos y heptaxodontidos) roedores muridos 1 (Oryzomys en Jamaica)
POBLAMIENTO ISLA-ISLA
La distribucion de los mamiferos terrestres a traves del area antillana demarcada al inicio del presente capitulo (vease Contexto Geogratico Fig 41) ha sido interpretada en funcion de dos hipoteticos mecanismos de poblamiento interinsular la dispersion y la vicarianza fundamentadas como se resumen seguidamente
DISPERSION
De acuerdo con la evidencia paleogeogratica mas actualizada en las principales regresiones marinas ocurridas en la cuenca caribeiia durante los ultimos 23 Ma (transiciones OligocenoshyMioceno Mioceno Superior - Plioceno y varias veces en el Pleistoceno) las masas terrestres constitutivas de la region insular en formacion aumentaron considerablemente su extension territorial disminuyendo por tanto la distancia entre elias oportunidades que los elementos faunales aprovecharian para dispersarse de unas islas a otras
Durante mucho tiempo esta ha sido la hipotesis mas favorecida por los estudiosos del tema a partir de Simpson (1956) pero en decadas recientes ha suscitado dos objeciones
1 Es dificil en tender como pudo operar la dispersion marina entre estas islas puesto que ninguna de las Antillas Mayores posee rios suficientemente caudalosos para transportar masas significativas de vegetacion flotante (MacPhee 198432)
Es oportuno recordar aqui el islote ala deriva (con mas de diez palmas reales en pie) entre las tres Antillas Mayores mas occidentales mencionado anteriormente (vease La Balsa Natural)
2 La dispersion marina es menos efectiva para explicar la asombrosa similitud de las faunas de mamiferos terrestres a traves de las Antillas Mayores septentrionales porque como proceso aleatorio no debe producir distribuciones uniformes a 10 largo de una cadena de islas 0 si las produce entonces el proceso no es aleatorio (MacPhee e lturralde-Vinent 2000 149)
Independientemente de que las aludidas distribuciones taxonomicas estan lejos de ser uniformes 0 asombrosamente similares la hipotesis de la dispersion marina contrariamente a su canicter aleatorio y altamente improbable en el caso del poblamiento continente-isla- en el caso del poblamiento isla-isla es un proceso no aleatorio (aprovecha las oportunidades brindadas puntualmente por los periodos de maxima regresion marina) y ademas altamente probable dada la incomparablemente menor distancia entre las islas caribeiias que entre estas y los continentes vecinos
314 BIOGEOGRAFiA
V ICARIANZA
La neotectonica y las transgresiones marinas del Oligoceno Medio y del Mioceno afectaron sustancialmente la paleogeografia de la Cresta de las Antillas Mayores resultando la subdivision gradual de las partes subaereas de la Cresta Asi el este de Cuba y el norte de La Espanola conectados fisicamente durante el Oligoceno Inferior se separaron al formarse el Paso de los Vientos en el Oligoceno tardio en tanto La Espanola y Puerto Rico probablemente permanecieron conectadas hasta el Mioceno tardio en que surgio el Paso de Mona (Iturralde-Vinent y MacPhee 1999)
Tales eventos geologicos habrian dividido las distribuciones de los grupos faunales emplazados previamente en el territorio continuo de las tres islas y a partir de esas disyunciones dichos autores (tambien MacPhee et al 2000 y White y MacPhee 2001) asumieron --como segunda fase del modelo de GAARlandia- la hipotesis de la vicarianza isla-isla para explicar la distribucion multiinsular de varios grupos de mamiferos terrestres a traves de las Antillas Mayores septentrionales De acuerdo con esta hipotesis cualquier diferencia en composicion faunal entre las islas (por ejemplo la ausencia de roedores heptaxodontidos en Cuba 0 de monos platirrinos en Puerto Rico) constituye evidencia de extincion local postvicariante y no de dispersion marina fallida Lo primero se puede demostrar si aparecen los fosiles pertinentes 10 segundo no se puede demostrar (MacPhee e Iturralde-Vinent 1994) Por otra parte IturraldeshyVinent y MacPhee (1999 57) han reconocido que aunque fa vicarianza isla-isla puede representar una solucion clara para las distribuciones de varios taxones cuaternarios no puede explicar todos los casos
Rega et al (2002 18) han presentado dos objeciones al modelo de la vicarianza isla-isla aplicado a los mamiferos de las Antillas Mayores
I Si la separacion de Cuba y La Espanola precedio en varios mill ones de anos la separashycion de La Espanola y Puerto Rico el modelo vicariante de lturralde-MacPhee debe predecir mayor similitud entre las faunas de La Espanola y Puerto Rico que entre las de Cuba y La Espanola De hecho es al reyes
No obstante MacPhee et al (200335) admitieron despues que la fecha mas temprana empiricamente defendible [para la apertura del Paso de Mona] debe ser el Oligoceno Medio esto invierte la secuencia de ambas disyunciones con 10 cual queda resuelta la aparente contradiccion Sin embargo en su reciente evaluacion de la congruencia entre filogenia e hipotesis geologic as en varios grupos de mamiferos terrestres de las Antillas Mayores Davalos (2004373) encontro que ninguno de los taxones cumple con la fecha y el patron de divergencia entre las masas terrestres antillanas postulados por el modelo de GAARlandia
2 La presencia de un mismo genero en las tres Islas septentrionales de las Antillas Mayores implica [por el modelo vicariante] que la diferenciacion entre estas formas durante el intervalo desde el Mioceno ha sido unicamente de nivel especifico [Dicha presencia]
315 rRRADlAcr6~ EVOLUCrONARIA
s610 puede explicarse razonablemente como el resultado de movimientos de esos animales entre las islas en el Cuaternario
De esta objecion resulta dificil disentir [El poblamiento entre islas diferenciado solo a nivel especifico no es probable que sea anterior al Pleistoceno (Simpson 1956 15)]
Debe tenerse en cuenta que la hipotesis de la vicarianza isla-isla alude a las tres islas septentrionales de las Antillas Mayores porque como se explico antes (vease El Barco Funeral Vikingo) Jamaica no existia como isla durante el lapso (Eoceno Superior shyOligoceno) consensualmente reconocido como el mas probable para la colonizacion de las proto-Antill as Mayores por la mayo ria de los mamiferos terrestres Despues de emerger como isla en el Mioceno Medio Jamaica debe haber recibido sus mamiferos terrestres por dispersion isla-isla (Davalos 2004376) y solo en el Cuaternario parece haberrecibido (por dispersion marina desde Centroamerica) al roedor murido Oryzomys
Seguramente tanto la dispersion como la vicarianza contribuyeron a que los mamiferos terrestres se irradiaran a traves de las Antillas Mayores y grupos de islas perifericas pero las maneras y tiempos en que estos mecanismos operaron a 10 largo de dicho proceso son aun en gran medida motivos de desacuerdo e incertidumbre
IRRADIACION EVOLUCIONARIA
A continuaci6n revisamos someramente las distribuciones conocidas de los diferentes grupos de mamiferos terrestres a traves de las Islas principales de las Antillas Mayores y grupos de Islas perifericas incluidas las interpretaciones y conjeturas que la literatura reciente recoge en relaci6n con la manera en que dichas distribuciones pudieron haberse conformado durante la irradiaci6n evolucionaria de los referidos grupos de mamiferos Manifestaciones de esta irradiaci6n pueden inferirse unicamente a partir de los taxones presentes en el Cuatemario pues solo 6 de las especies conocidas corresponden al Terciario
GRUPOS ORlGINARIOS DE SURAMERICA
PEREZOSOS (Megalonychidae)
Aparte de un genero miocenico de Cuba y otro del Oligoceno de Puerto Rico se conocen 4 generos (12 especies) de perezosos cuaternarios (todos extintos) restringidos a las tres islas septentrionales de las Antillas Mayores Acratocnus Neocnus Megalocnus y Parocnus Los cuatro generos estan representados en Cuba y La Espanola y Acratocnus 10 esta ademas (como unico genero de perezoso) en Puerto Rico Esta distribuci6n ha sido interpretada por MacPhee et al (2000) eomo resultado de la vicarianza oligomiocenica entre estas islas y [dado que el Banco Puerto Rico - Islas Virgenes fue la entrada de GAARlandia a las Antillas Mayores] se preguntaron si la ausencia de los otros tres generos en Puerto Rico se debe a que nunca estuvieron en ese territorio 0 a que sus
316 BIOGEOGRAFiA
fosiles atm no han sido hallados Pero como hemos visto Rega et al (2002) atribuyeron dicha distribucion a la dispersion interinsular en el Cuatemario
Por otra parte varios amilisis c1adisticos recientes ubi can a Choloepus -genero Reciente conocido solo de Centro- y Suramerica- como taxon hermano de Aeratoenus exc1uidos los demas megaloniquidos antillanos Para White y MacPhee (2001228)
tan estrecha relacion sugiere que la divergencia entre ambos [generos] ocurrio despues de que Aeratoenus divergio de Neoenus Si esta relacion implica que Aeratoenus y Neoenus se dispersaron separadamente desde Suramerica a las AntilIas mientras Choloepus permanecia en el continente (siendo necesarias al menos tres invasiones de perezosos) 0 si Choloepus invadio Suramerica desde las Antillas es algo que el material disponible no permite determinar
La diversidad adaptativa de los perezosos se manifiesta en (1) la utilizacion del habitat especies arboreas en Neoenus y Aeratoenus y terrestres en Paroenus y Megaloenus (Webb 1985a White [1993] White et al 1996 Arredondo [1999]) y en (2) la gama de tallas corporales desde los pequenos Neoenus hasta Megaloenus rodens el mayor de los mamiferos terrestres antillanos (su peso adulto estimado en 270 kg seglin Paula 19679) esto sugiere que los perezosos se irradiaron a nichos que en contextos continentales probablemente estarian ocupados por miembros de otros c1ados (MacPhee e IturraldeshyVinent 2000)
No esta claro si el abanico de tamanos entre estos perezosos enmascara alguna tendencia al gigantismo Por 10 pronto Anderson y Handley (2002) encontraron 10 contrario en perezosos (Bradypus) de las Islas de Bocas del Toro (Panama) shyseparadas secuencialmente del area continental vecina por ascensos del nivel del mar durante los ultimos 10 ka En las cinco islas los perezosos son menores que su contraparte continental y las diferencias entre las tallas medias de las cinco poblaciones expresan una reduccion lineal significativa de la talla corporal en funcion de la edad geologica de cada isla sin relacion alguna con el area de las islas 0 la distancia que las separa del continente
MONOS (Pitheciidae)
Los primates de las Antillas Mayores (cuatro especies extintas) constituyen un grupo monofiletico con Paralouatta (en Cuba) y Antillothrix (en La Espanola) como taxones hermanos seguidos en la escalera c1adistica por Xenothrix (en Jamaica) y finalmente Callieebus como el pariente continental mas cercano esto implica una sola invasion desde Suramerica antes del Mioceno Inferior (fecha mas temprana conocida para los primates fosiles antillanos) y por tanto en consonancia con GAARlandia como la via de colonizacion (Horovitz y MacPhee 1996 1999 Horovitz [1997] MacPhee y Horovitz 2004)
En desacuerdo parcial con la anterior interpretacion la filogenia de este grupo de monos obtenida por Davalos (2004) indica un antecesor comun en Cuba La Espanola y Jamaica
317 TRRADIACTON EVOLUCIONARIA
compatible con la hipotetica conexion -propuesta tentativamente por Iturralde-Vinent y MacPhee (199980-84)- entre las Montanas Azules del este de Jamaica y la peninsula suroccidental de La Espanola en la epoca de GAARlandia
A pesar de que los primates antillanos pertenecen a un grupo viviente de monos muy pequenos (los monos titles) fueron comparativamente grandes en relacion con los platirrinos modernos y sucesivamente mayores en la direccion Antillothrix Xenothrix y Paralouatta con P marianae como el mayor platirrino conocido del Mioceno neotropical (MacPhee 2005) MacPhee et al (1994) consideraron que Xenothrix debio ser un mono de andar lento y al parecer las especies de Paralouatta abandonaron la arborealidad para adoptar un modo de vida mas terrestre (MacPhee 1996a MacPhee et al 2003 MacPhee y Meldrum 2006)
ROEOORES (Echimyidae Capromyidae Heptaxodontidae)
La edad de Zazamys en Cuba demuestra que la invasion de los roedores histricognatos en las Antillas Mayores comenzo antes del Mioceno Inferior por otra parte los representantes mas antiguos de esa irradiacion evolucionaria se encuentran en La Espanola y Puerto Rico Ambas evidencias son consistentes con la hipotesis de GAARlandia (Woods 1982 1988 Woods et al 2001)
No esta claro si los capromidos evolucionaron a partir de un equimido ancestral 0 de un protocapromido 0 si los equimidos antillanos (I sub familia 3 generos y 7 especies todas extintas) forman parte de la irradiacion de los capromidos (4 subfamilias 9 generos y 27 especies 10 vivientes) Kratochvil et al (1978b) y Rodriguez et al (1979) consideraron que Capromys nan us es un eslab6n entre Echimyidae y Capromyidae pero Mesocapromys nanus esta morfologicamente mucho mas cerca de Hexolobodon y los de mas capromidos que de los equimidos (Woods 1985) Lo que si esta claro segun Woods et al (2001) es que la subfamilia Plagiodontinae endemic a de La Espanola es el linaje basal de los capromidos y que Isolobodon Hexolobodon Capromys Geocapromys Mysateles y Mesocapromys [y supuestamente Macrocapromys] forman parte de la irradiacion temprana de Capromyidae
La subfamilia Capromyinae (5 generos y 17 especies 9 vivientes) parece ser el dado de mas reciente evolucion en la familia con invasion de Cuba desde La Espanola y dispersion desde Cuba (1) por Capromys a las islas Caiman y (2) por Geocapromys a las islas Bahamas Caiman y Jamaica y desde Jamaica a las islas Swan aunque la presencia de Geocapromys en Swan pudiera deberse a introduccion aborigen (Morgan 1985 1988 1989 1993 1994 Woods 1989a 1990) Woods et al (2001) utilizaron datos moleculares para obtener estimados independientes de divergencia entre capromidos segiJn 10 cual Geocapromys divergio cerca de los 17 MaAP Mesocapromys divergio de Capromys hace 15 MaAP en tanto lvlysateles 10 hizo hace 10 MaAP Por su parte las formas de Capromys pilorides y de Mysateles prehensilis en Isla de la Juventud divergieron de sus correspondientes subespecies nominativas en la isla principal a los 24 y 08 MaAP respectivamente
318 BIOGEOGRAFiA
Woods y Howland (I979) describieron adaptaciones morfologicas de la mandibula el hioide y la faringe asociadas a mecanismos de masticacion diferentes en plagiodontinos y caprominos y Varona (1980a) llamo la atencion sobre el gran desarrollo de los molariformes (en relacion con la longitud total del cnineo) que presentan ciertos grupos de jutias fosiles en comparacion con las vivientes 10 que pudiera interpretarse como una adaptacion alimentaria a una vegetacion mas abrasiva por parte de las jutias extinguidas
Segun Woods (1990) la familia Heptaxodontidae (2 subfamilias 5 generos y 6 especies todas extintas) tambien seria parte de la irradiacion de los capromidos con origen en el este de La Espanola 0 en Puerto Rico y subsecuente dispersion al Banco de Anguila - San Martin en las Antillas Menores (pero vease MacPhee y Flemming 2003) Este banco estaria a solo 70 km del Banco Puerto Rico - Islas Virgenes en tiempos de maxima regresion marina y esta proximidad con Puerto Rico -patria de Elasmodontomys obliquus- seguramente tuvo que ver con la presencia en las islas de Anguila y San Martin del heptaxodontino gigante Amblyrhiza relacionado mas estrechamente con los roedores de las Antillas Mayores que con los del resto de las Antillas Menores (Woods 1990 1996) Aunque los roedores suelen aumentar de tamano en las islas (Case 1978) la escala del incremento de la talla en Amblyrhiza (que pudo alcanzar mas de 200 kg de peso adulto) esta fuera de toda proporcion con los patrones conocidos en otros roedores insulares (Biknevicius et 01 1993)
Clidomys (Clidomyinae) de Jamaica difiere marcadamente en su morfologia de los heptaxodontinos (vease ademas MacPhee y Flemming 2003)10 cual dificulta explicar su presencia en esa isla Como Jamaica estuvo bajo el mar hasta el Mioceno Medio el ancestro de Clidomys colonizaria esta isla por dispersion desde el extremo suroccidental de La Espanola despues que los heptaxodontinos se originaron en esta ultima isla (Woods 1989a) Pero Pascual et 01 (I 990) especularon que Clidomys pudiera ser el resultado de una invasion posterior e independiente de los capromidos
GRUPOS ORIGINARIOS DE NORTEAMERICA
El origen de las musaranas antillanas ha resultado ser hasta ahora el mayor enigma biogeografico de la fauna de mamiferos terrestres de la region aunque la mayoria de los autores coinciden en considerarlas originarias de Norteamerica (vease Discus ion y Conclusiones en Poblamiento Continente-Isla) No obstante MacPhee e Iturralde-Vinent (1994) descartaron toda posibilidad de conexion terrestre por el Estrecho de la Florida 0 por el Canal de Yucatan y si el (supuesto) evento vicari ante de Rosen (vease El Arca de Noe) -invocado por MacFadden (1980) para resolver el enigma- fuera minimamente plausible aun asi es dificil creer que las catastrofes que 10 sucedieron (vease Contexto Temporal) hayan barrido con toda esa vieja fauna continental excepto las musaranas circunstancia ya anticipada muy tempranamente por Matthew (l918a)
De otra parte la relaci6n entre Nesophontidae y Solenodontidae permanece irresuelta Si estos linajes no estan estrechamente relacionados entonces es muy posible que
319 IRRADIACION EVOLUCIONARIA
sus distribuciones sean explicadas por mecanismos diferentes (Whidden y Asher 2001)
La descomunal diferencia de talla corporal entre las musaranas pequenas (Nesophontes) y las grandes (Soienodon) seguramente impuso a ambos grupos restricciones distintas relativas a la termorregulacion y las adaptaciones energeticas En Nesophontes la seleccion debio favorecer una alta tasa de metabolismo y crecimiento camadas mas numerosas corta expectativa de vida yalta densidad poblacionaL En Soienodon en cambio la seleccion opero en la direccion de una baja tasa metabolica camadas reducidas crecimiento lento longevidad y baja densidad poblacional (Ottenwalder [1991])
MUSARANAS PEQUENAS (Nesophontidae)
Las musaranas pequenas estan representadas en las Antillas por el genero Nesophontes (y un genero miocenico indeterminado en ambar dominicano) con 6 especies cuaternarias extinguidas distribuidas en Cuba La Espanola Puerto Rico Islas Virgenes e Islas Caiman La mayor de las musaranas pequenas Nesophontes edithae evoluciono en Puerto Rico y su arribo a las Islas Virgenes supone una dispersion sumamente reciente En las Islas Caiman Nesophontes se asemeja mas a la poblacion de Cuba de donde seguramente derivo en el Plio-Pleistoceno (junto con Capromys y Geocapromys) cuando la costa sur de Cuba estuvo mucho mas cerca de las Islas Caiman (lturralde-Vinent 2003fig 4) MacPhee et ai (1994) interpretaron la distribucion de Nesophontes en Cuba y La Espanola como resultado de la vicarianza oligomiocenica ocurrida al formarse el Paso de los Vientos pero la distancia taxonomica entre los lados del Paso es solo de nivel especifico y N micros incluso vivi6 en ambas islas segtln Patterson (1962)
Allen (1918) considero que Nesophontes micrus fue una especie primitiva no modificada para la vida arb6rea 0 fos6rea y por tanto de habitos terrestres Nesophontes edithae de Puerto Rico es mucho mayor y mas robusta que las restantes especies del genero eL indice de robustez de su femur (= anchura minima de la diafisis ongitud total del hueso) muestra que este es tan robusto como el de Solenodon paradoxus un animal dos veces mayor (Morgan et ai 1980) Choate y Birney (1968) postularon un aumento cronoclinal de la taIla corporal en N edithae confirmado mas tarde por McFarlane (1999a) mediante fechados radiocarb6nicos de la secuencia estratignifica en depositos cavernarios de Puerto Rico Solenodon no vivi6 en Puerto Rico y en ausencia de una musarana de mayor tamano Nesophontes alcanzo una talla corporal mayor en esa isla que en La Espanola 0 Cuba (Woods y Eisenberg 1989)
MUSARANAS GRANDES (Solenodontidae)
Los arcaicos almiqufes (genero Soienodon) comprenden cuatro especies una viviente (S cubanus) y otra f6sil (S arredondoi) en Cuba y una viviente (S paradoxus con dos subespecies) y otra f6sil (S marcanoi) en La Espanola Como cabria esperar las dos especies cubanas se asemejan mas entre sf que a las especies de La Espanola pero
320 BIOGEOGRAFiA
enigmaticamente S marcanoi comparte caracteres con S cuban us y S paradoxus (Woods y Eisenberg 1989 Morgan y Ottenwalder 1993 Ottenwalder 200 I) Solenodon arredondoi alcanzo una talla corporal superior a la de cualquiera de las musarafias vivientes a nivel mundial Segun Morgan y Ottenwalder (1993) extrapolando las dimensiones y el peso de los almiquies vivientes S arredondoi debio tener una longitud cabeza-tronco y un peso de 450-550 mm y 1500-2000 g respectivamente en comparacion con S cubanus (360 mm y 800 g) y S paradoxus (390 mm y 1100 g)
Entre todas las especies de almiqufes Solenodon paradoxus es la que presenta los caracteres mas derivados (Otten walder 2001) Si estas relaciones son validas la distribucion de Solenodon entre Cuba y La Espafiola debio discurrir de oeste a este como propuso MacFadden (1980) quien invoco la presencia fosH de S cubanus [pero tambien S arredondoi] en Pinar del Rio para postular que el ancestro continental entr~ por Cuba opinion compartida por Perfit y Williams (1989) y por Ottenwalder ([1991] 2001) El paso de Solenodon a Puerto Rico pudo haberlo impedido la existencia de barreras ecologicas durante la conexion del este de La Espafiola con Puerto Rico 0 por la ausencia de Solenodon en esa parte de La Espafiola en aquella epoca Sin embargo las mismas condiciones ecologicas en el este de La Espafiola no parecen haber representado una barrera efectiva para Nesophontes cuyas mas altas tasa de reproduccion y densidad poblacional presumiblemente Ie permitieron expandirse flpidamente a traves de La Espafiola (Ottenwalder [1991])
Es opinion generalizada que los almiqufes antillanos constituyen el grupo de mamiferos que primero se emplazo en el territorio insular (MacPhee 2005) y en tal sentido es de interes la evidencia sobre especificidad parasito-hospedante aportada por Cruz (1980) quien encontro que la especie de acaro (genero Cubanochirus) que parasita a los almiquies es mucho mas primitiva que la especie (del mismo genero) que paras ita a las jutias y de aqui concluyo que dicho genero de acaro debio evolucionar primero en Solenodontidae y posteriormente en Capromyidae al entrar en contacto representantes de ambas familias en Cuba 0 La Espafiola
Recientemente Roca (2004) y Roca et al (2004) dieron a conocer que los resultados de su estudio genetico-molecular sobre los almiquies indican una divergencia de 25 Ma entre Solenodon cuban us y S paradoxus -compatible con la disyuncion tectonica entre Cuba y La Espafiola (Iturralde-Vinent y MacPhee 1999 McPhee et al 2003)-- y una separacion de ambas especies comparable en magnitud a la divergencia entre familias de mamlferos Consecuentemente propusieron que Solenodon cubanus fuera clasificada en el genero Atopogaie como indico Cabrera (1925) hace 82 afios
DISCUSION Y CONCLUSIONES
Las musarafias y los roedores antillanos (excepto Oryzomys) se irradiaron tanto de sus respectivos ancestros continentales que de estos 6rdenes se reconocen familias y subfamilias endemicas de la region en contraste con los perezosos (una subfamilia y tres
321 IRRADIACION EVOLUCIONARIA
tribus endemicas) y los monos (una tribu endemica) como expresion de diferentes niveles de diversificacion evolucionaria Este proceso es dificil de explicar (traUindose de c1ados que pnicticamente comparten la misma antigiiedad en las Antmas) si es correcta la interpretacion de Pascual et at (1990636)
Nosotros consideramos estos diferentes niveles de diversificacion como las diferentes etapas de evolucion alcanzadas por cada grupo [de mamiferos] Obviamente esto estuvo relacionado con las diferentes longitudes de sus historias
Pudiera ocurrir que en el futuro se compruebe que la historia biogeognifica de la region fue mucho mas complicada
La Espanola la isla mas ecol6gicamente divers a de la region debi6 ser el centro principal de evoluci6n de los mamfferos (con 87 de las familias y 73 de las subfamilias) seguida por Cuba (con 75 de las familias y 54 de las subfamilias) las dos islas de las Antillas Mayores mas relacionadas biogeograficamente (Fig 43 Tabla 44)
eL
EL
cb CAIMAN
SWAN
A Acratocnini B Cubanocnini C Megalocnini D Mesocnini E Nesophontidae F Solenodontidae G Xenothrichini
ANGUILAshyM-0 SMARTIN
H Heteropsomyinae I Plagiodontinae J Isolobodontinae
K Hexolobodontinae L Capromyinae
M Heptaxodontinae N CIidomyinae 0 Sigmodontinae
FIGURA 43 Representacion esquematica de las relaciones interinsulares (expresadas porel numero de linajes compartidos) entre las Antillas Mayores y entre estas y los grupos de islas perifericas segun las interpretaciones de los autores citados en el texto Los numeros dentro de los circulos indican la cantidad de linajes representados en cada isla relacion dudosa Contexto taxon6mico de los linajes en la Tabla 44
bull bull
bull bull
bull bull bull
bull bull
bull bull
bull
bull
bull
bull
bull bull bull bull
bull bull
bull bull
bull
bull bull bull
bull bull
bull
bull bull
bull
322 BIOGEOGRAFiA
TABLA 44 Distribuci6n de los linajcs de mamiferos terrestres a traves de las Antillas Mayores y grupos de islas perifericas LE = La Espanola CU Cuba JA Jamaica PR = Puerto Rico IC Islas Caiman IS Islas Swan IB = Islas Bahamas IV = Islas Virgenes AS = Anguila - San Martin
FamiliaSubfumiliafIribu LE CU JA PR IC IS IB IV AS
Megalonychidae
Choloepodinae
Acratocnini
Cubanocnini
Megalocninae
Megalocnini
Mesocnini
Nesophontidae
Solenodontidae
Pitheciidae
Callicebinae
Xenothrichini
Echimyidae
Heteropsomyinae
Capromyidae
Plagiodontinae
Isolobodontinae
Hexolobodontinae
Capromyinae
H e pta x 0 don tid a e
Heptaxodontinae
Clidomyinae
Muridae
Sigmodontinae
PILOSA
SORICOMORPHA
PRIMATES
RODENTIA
bull
bull
bull
bull TOTALES 12 10 4 4 2
323 EXTlICIOIES DEL CUATERNARIO
El gigantismo ha sido sin duda una tendencia evolucionaria recurrente entre los clados mayores de mamiferos antillanos Es un hecho bien conocido que algunos grupos de mamiferos han aumentado de tamafio en las islas y varios autores han intentado explicarlo a partir de enfoques diferentes (Foster 1964 Miller 1966 Sondaar 1977 Case 1978 Eisenberg y Wilson 1978 Heaney 1978 Lawlor 1982 Lomolino 1985 Angerbjom 1985 McNab 19942001)
EXTINCIONES DEL CUATERNARIO
Las islas que integran la Subregion Antillana han perdido casi 90 de sus mamiferos terrestres durante los ultimos milenios y en proporcion con su extension territorial esta perdida ha sido mayor en la Subregion que en cualquier otro lugar del planeta (MacPhee 1996b) Las extinciones del Cuatemario eliminaron mas de las tres cuartas partes de la fauna de mamiferos terrestres conocida de las Antillas Mayores sus islas satelites y grupos de islas perifericas y el examen de este proceso en sus aspectos mas generales se aborda en esta ultima parte del presente capitulo
CRONOLOGiA Y CONSECUENCIAS
Es muy improbable que un fosil represente el ultimo individuo de un taxon extinto y entonces hay que guiarse por la edad radiometrica del fosil mas reciente conocido lIamada fecha de ultima presencia- como unica aproximacion posible a la fecha de extincion del taxon
Como se explico al inicio del capitulo (vease Contexto Temporal) el registro fosH de los mamiferos terrestres de las Antillas Mayores adolece de insuficiente sustentacion radiometrica No obstante la distribucion temporal de los escasos fechados disponibles para el Cuaternario de la region abarca etapas criticas del proceso de extincion experimentado por la referida fauna a 10 largo de los ultimos 250 mil afios
De las 52 especies que es posible reconocer actualmente como miembros extintos de la fauna cuatemaria de mamiferos terrestres de las Antillas Mayores y grupos de islas perifericas los restos fosiles de 23 de elIas (442) han sido fechados radiometricamente o se han hallado asociados estratigraficamente de manera inequivoca con otro material fechado De estos 23 fechados 3 corresponden al interglacial Sangamon 4 pertenecen a la secuencia Pleistoceno terminal - Holoceno temprano 10 se ubican en ellapso entre el arribo de los aborigenes y la incursion europea y 6 son postcolombinos (a los cuales hay que sumar otras dos especies extinguidas en los siglos XIX y xx) A partir de estas 25 especies avaladas por fechas de ultima presencia distribuidas entre los cuatro intervalos temporales sefialados hemos compuesto la Tabla 45 en la que ademas aparecen las restantes 27 especies (carentes de fechado radiometrico) ubicadas en el intervalo temporal mas probable sobre la base de (1) la asociacion estratigrafica de sus restos con evidencias culturales (amerindias 0 europeas) 0 (2) la ausencia de dicha asociacion Sobre estos criterios de ubicacion temporal procede hacer dos aclaraciones
324 BIOGEOGRAFIA
TABLA 45 Intervalos temporales mas probables para las extinciones cuatemarias de los mamiferos terrestres de las Antillas Mayores y grupos de islas perifericas sobre la base de las evidencias siguientes (l) ultimo ejemplar colectado u observado en vida (2) fechado radiometrico mas tardio en material de la especie (3) asociaci6n estratigrafica mas tardia con otro material fechado (4) asociaci6n estratigrafica con restos de animales introducidos restos humanos u objetos culturales propios del intervale (5) falta de asociaci6n con evidencias culturales [Los ntimeros entre parentesis en la columna Evidencia significan adscripci6n tentativa al intervalo] Los estimados del peso corporal (en kg) estan tomados de la literatura 0 inferidos comparativamente y distribuidos entre
las clases siguientes (a) S 01 (b) 01 a lt 1 (c) 1 a lt 10 (d) 10 a lt 100 (e) 100 a lt 200 (j) ~ 200 Las
abreviaturas que designan las islas son las de la Tabla 44
F amilialEspecie Peso IsJa Evidencia Fuente
I httervalo htterg1acial Sangamon (gt80 kaAP)
HEPTAXODONTlNAE
Amblyrhiza inundata e AS 3 McFarJane et al 1998a
CLIDOMYINAE
Clidomys osborni d JA 3 Ibid
Clidomys parvus d JA 3 Ibid
II httervalo Pleistoceno terminal ~ Holoceno temprano (20-6 kaAP)
MEGALONYCHIDAE
Aeratoenus antillensis d CD 5 Arredondo 1975 a
Aeratoenus odontrigonus d PR 3 McFarJane 1999b
Aeratoenus simorhynehus d LE 2 Rega et al 2002
M egaloenus zile f LE 2 MacPhee et al 2000
SOLENODONTIDAE
Solenodon arredondoi e CD 5 Morgan y Ottenwalder 1993
PnHECIIDAE
Paralouatta varonai d CD 5 MacPhee 1996a
EcHIMYIDAE
Heteropsomys antillensis c PR 5 McFarJane 1999b
Heteropsomys insulans c PR 5 Ibid
Puertoricomys eorozalus c PR 5 Woods 1989a
CAPROMYIDAE
P lagiodontia araeum c LE 5 MacPhee et al 2000
Macrocapromys latus c CD 5 Varona y Arredondo 1979
(Continua)
325 EXTINCIONES DEL CUATERNARIO
TABLA 45 (Continuaci6n)
F ami1ia1Especie Peso Isla Evidencia Fuente
M acrocapromys latus c CU 5 Varona y Arredondo 1979
Mesocapromys kraglievichi c CU 5 Ibid
HEPTAXODONTINAE
Xaymaca fulvopulvis a JA 2 MacPhee y Flemming 2003
III Intervalo Amerindio (6-05 kaAP)
MEGALONYCHIDAE
Acratocnus ye d LE 2 MacPhee et aI 2000
Neocnus comes c LE 2 Ibid
N eocnus dousman c LE 2 Ibid
N eocnus toupiti c LE 2 Ibid
N eocnus gliriform is c CU 4 Cordova et ai 1997
M egalocnus rodens f CU 2 MacPhee et ai 2007
Parocnus browni e CU 2 Steadman et ai 2005
P arocnus serus e LE 2 MacPhee et ai 2000
N ESOPHONTIDAE
N esophontes edithae a PR 2 McFarlane 1999b
PITHECIIDAE
Antillothrix bernensis c LE 3 Rimoli 1977
EcHIMYIDAE
Brotomys contractus a LE 4 Miller 1929c
CAPROMYIDAE
P lagiodontia spelaeum c LE 4 Ibid
M acrocapromys acevedo c CU 4 NUiiez 1990a
HEPTAXODONTINAE
Elasmodontomys obliquus d PR 3 McFarlane 1999b
Iv Intervalo Postcolombino (05 kaAP - Presente)
N ESOPHONTIDAE
N esophontes hypom icrus a LE 24 MacPhee y Fle111111ing 1999
N esophontes zam icrus a LE (4) Ibid
N esophontes m icrus a CU 24 MacPhee et aI 1999
(Continua)
326 BIOGEOGRAFiA
TABLA 45 (Continuacion)
F amilialEspecie Peso Isla Evidencia Fuente
N esophontes m icrus a CD 24 MacPhee et aI 1999
N esophontes m icrus a LE 4 MacPhee et aI 2000
N esophontes sp 1 a IC 4 Morgan 1994
N esophontes sp 2 a IC 4 Ibid
Solenodon marcanoi b LE 4 Ottenwalder 2001
PITHECIIDAE
Xenothrix mcgregori c JA 4 MacPhee y Flemming 1999
EcHIMYIDAE
Boromys offeUa a CD 24 Guarch 1984
Boromys torrei a CD 24 Ibid
Brotomys voratus a LE 4 MacPhee et aI 2000
CAPROMYIDAE
P lagiodontia ipnaeum c LE 4 MacPhee y Flemming 1999
Rhizoplagiodontia lemkei c LE 4 Ibid
Isolobodon portoricensis c PR 24 Ibid
Isolobodon montanus c LE 4 Ibid
Hexolobodon phenax c LE (4) Ibid
Capromys sp c IC 24 Morgan 1994
Geocapromys columbianus c CD 4 MacPhee y Flemming 1999
Geocapromys thoracatus b IS 1 MacPhee y Marx 1997
Geocapromys sp 1 c IC 4 Morgan 1994
Geocapromys sp 2 c IC (4) Ibid
l-IEPTAXODONTINAE
Quemisia gravis c LE (1) MacPhee y Flemming 1999
MURIDAE
Oryzomys antillarum a JA 1 Ibid
1 Flemming y MacPhee (1999) alertaron en contra de la asociacion con otro material fechado Asimismo Morgan (1993) indico que la ausencia de asociacion con evidencias culturales amerindias no es necesariamente un criterio concluyente para ubicar un taxon en el intervalo temporal anterior porque hay depositos en la region que carecen de tales evidencias y son del Holoceno tardio por fechados radiocarbonicos [No obstante
327 EXTlNCIONES DEL CUATERNAR10
la ausencia de una especie en residuarios arqueol6gicos adquiere mayor certidumbre como criterio de ubicaci6n temporal de su extinci6n en la medida que aumenta el numero de residuarios estudiados que en las Antillas Mayores se cuentan por centenares] A falta de fechado absoluto de una especie consideramos que estos criterios de ubicaci6n temporal son mas utiles que la alternativa de excluir del analisis a la especie
2 En los trabajos sobre biogeografia de mamiferos antillanos los arque610gos mas citados como autoridades para establecer el inicio de la presencia aborigen en las Antillas Mayores (Kozlowski 1974 Rouse y Allaire 1978 Rouse 1989 1992 Wilson 2001) difieren al respecto entre los 6 kaAP y los 45 kaAP (pero vease Pagan et al 200523) y hemos adoptado la mas temprana de estas fechas para el comienzo del intervalo amerindio en la Tabla 45 De todos modos cualquiera que haya sido la fecha esta no es aplicable por igual al conjunto de las Antillas Mayores porque segun algunos de los autores citados el poblamiento comenz6 por Cuba y La Espanola 6 mil anos atras continu6 en Puerto Rico hace alrededor de 3 mil anos (pero vease Burney et al 1994) yen Jamaica hace menos de 2 mil e incluso a las Islas Caiman ni siquiera llegaron [Las Islas Caiman descubiertas por Col6n en 1503 carecen absolutamente de evidencia amerindia (Stokes y Keegan 1996)]
Si los datos de la Tabla 45 representan una aceptable aproximaci6n a la realidad el proceso de las extinciones cuaternarias de los mamiferos terrestres de las Antillas Mayores y grupos de islas perifericas debi6 caracterizarse entre otros por los eventos siguientes
Aunque obviamente la fauna cuaternaria debi6 ser mucho mas numerosa que 10 que refleja el registro paleontol6gico (Morgan y Woods 1986) 52 especies (81 de la fauna conocida) desaparecieron (sin reposici6n) durante los ultimos 250 mil anos
- Al parecer las extinciones se concentraron en las especies de mayor tamano Asi el peso promedio estimado para las especies de la etapa preantr6pica (intervalos I + II de la Tabla 45) es 350 kg es 328 kg para las especies del intervalo III (amerindio) y es s610 25 kg (c1ases del peso a b c) para las especies del intervalo IV (postcolombino) Ninguna especie aut6ctona de mamifero mayor de 6 kg existe hoy en la regi6n
- De acuerdo con la distribuci6n temporal de las extinciones 61 especies sobrevivieron al interglacial Sangamon (intervalo 1)48 sobrevivieron a la transici6n del Pleistoceno terminal al Holoceno temprano (intervalo II) 34 sobrevivieron al periodo amerindio (intervalo III) y s610 12 especies han sobrevivido desde la Ilegada de los europeos (intervalo IV) Las tasas de extinci6n correspondientes (exceptuado el intervalo I) son una especie cada 1167 anos en el intervalo II una cada 393 anos en el intervalo III y una cada 23 anos en el intervalo IV
- Durante todo el proceso de las extinciones cuaternarias la perdida de especies por isla fue de 91 en La Espanola 83 en Jamaica y 62 en Cuba Bahamas retuvo su unica especie (aunque extirpada en varias de las islas) Puerto Rico e Islas Caiman perdieron todas sus especies (6 y 5 respectivamente) y las Islas Swan Virgenes y Anguila - San Martin perdieron sus respectivas especies (micas
328 BIOGEOGRAFiA
De los cuatro 6rdenes de mamiferos representados en la fauna Pilosa y Primates desaparecieron totalmente Soricomorpha y Rodentia perdieron 86 y 74 de sus especies respectivamente
CAUSALIDAD
CAUSAS NATURALES
Los cicIos glaciales e interglaciales acaecidos en el Pleistoceno causaron cambios ambientales dnisticos en las Antillas con la concomitante alternancia de cIimas frios (secos) y calidos (hUmedos) asi como de descensos y ascensos del nivel del mar aumento y disminuci6n de las areas territoriales) induso (aunque con menor intensidad) en el Holoceno Estos eventos paleogeograficos han sido consistentemente documentados (Mayo y Kartashov 1972 Bonnatti y Gartner 1973 Ortega y Arcia 1982 Prance 1982 Schubert y Medina 1982 Dodge et al 1983 Ortega [1984] Haq et al 1987 Schubert 1989 Hodell et al 1991 Guilderson et al 1994 Oppo 1997 Emslie 1998 Curtis et al 2001 Iturralde-Vinent 2003) y de ellos brindamos a continuaci6n una breve resena
Finalizando el periodo interglacial anterior a la ultima glaciaci6n (Wisconsin) el mar estaba a 8-10 m por encima de su nivel actual Cuando el avance glacial del Wisconsin a1canz6 su culminaci6n (eillamado Ultimo Maximo Glacial) hace 18 ka el mar habia descendido 120 m con respecto a dicho nivel y las islas del Caribe habian incrementado considerablemente sus extensiones subaereas en comparaci6n con las que presentan hoy dia por ejemplo Puerto Rico y las Islas Virgenes se conectaron territorialmente y las Bahamas se unieron en cinco gran des islas Prevalecia un regimen frio y seco y condiciones de aridez --debidas a una reducci6n de las precipitaciones (posiblemente a menos de 440 mmlano)- y los ambientes xericos se habian extendido a traves de las islas y bancos emergidos y abundaban los pinares (los habitats mesicos estaban restringidos a parcelas relictas en 10 alto de las montanas) En el Mar Caribe y el Golfo de Mexico la temperatura superficial del mar era 5degC mas baja que la actual y en las llanuras costeras y alturas maximas eran respectivamente 2-3degC y 6degC mas bajas Los limites de las zonas ecol6gicas altitudinales descendieron varios cientos de metros y cada zona redujo su extensi6n vertical al aumentar el gradiente de temperatura Ellimite inferior de la zona periglacial
elevaci6n donde la temperatura media minima a1canza los OdegC) de la Cordillera Central de la Republica Dominicana (altura maxima del Caribe insular con evidencia de topografia glacial) se considera que estuvo entre los 1500 y 1800 mlnm (actualmente esta a 2800 mlnm) y se puede estimar que dicho limite era inferior en Cuba por estar mas al norte que la Republica Dominicana No obstante durante este periodo la temperatura experiment6 episodios cortos (unas pocas decadas) de ascensos repetidos
Un signifieativo componente bi6tieo de las referidas condiciones paleoambientales 10 constituy6 la espectacular irradiaci6n evolucionaria experimentada por un grupo numeroso de aves rapaees diurnas y nocturnas (Wetmore 1937 Brodkorb 1969 Kurotchkin y Mayo 1973 Arredondo 1976 Olson 1978 Olson y Hilgartner 1982
329 EXTINCIONES DEL ClJATERNARIO
Suarez 2000a b 2004 Suarez y Emslie 2003) Los mamiferos de las Antillas Mayores ofrecieron a estos depredadores aviares una amplia gama de tamanos de presa que en ausencia de mamiferos del orden Carnivora en la region permitio a estas aves irradiarse a este nicho alimentario con la evolucion de una variedad de especies de muy diversa tall a corporal incluidas formas gigantes Adquirir un cuerpo gigante les propicio el acceso a recursos alimentarios que no estaban siendo explotados por ningun otro depredador (Alcover y McMinn 1994) A Cuba corresponde el mejor registro fosil de rapaces cuaternarias caribenas con siete generos de especies diurnas Dentro de estas aves existieron tres gavilanes gigantes (Accipitridae) tres carairas (Falconidae) y un condor (Cathartidae) asi como cuatro generos de especies nocturnas dos lechuzas (Tytonidae) y tres Mhos (Strigidae) dos de ell os no igualados en tamano en todo el mundo Algunos de estos generos tuvieron representacion en otras islas antillanas como La Espanola Jamaica Bahamas y Gran Caiman
Al retirarse hacia el norte los hielos de la glaciacion Wisconsin el clima se tomo mas hUmedo en las Antillas durante el Holoceno temprano y el bosque mesico se expandio en la region insular El nivel del mar ascendio acompanado de mayor pluviosidad alrededor de los 7 kaAP las Islas Virgenes quedaron separadas entre si y de Puerto Rico y las Bahamas perdieron la mayor parte de su territorio Cerca de los 3 kaAP las condiciones secas retornaron a la region e hicieron desaparecer buena parte del bosque mesico Nuevamente el clima comenzo a adquirir sus condiciones actuales hacia los 25 kaAP aproximadamente y hoy transita por una fase mesica con vegetacion xerica confinada en parcelas relictas
Sin duda estos y otros eventos naturales incidieron negativamente en la sobrevivencia de mamiferos terrestres en las Antillas Mayores asi como en los grupos de pequenas islas perifericas (Pregill y Olson 1981 Olson 2006)
McFarlane et al (1998a) demostraron convincentemente como Amblyrhiza el gigante de los roedores antillanos endemico de la pequena isla de Anguila sucumbio a las repetidas y abruptas perdidas de su habitat durante el interglacial Sangamon por las nipidas y severas oscilaciones del nivel del mar (documentadas por Neumann y Hearty 1996) al no poder responder adecuadamente a tan catastroficos eventos mediante el ajuste de su masa corporal media y otro tanto debio ocurrirle en Jamaica al otro roedor gigante Clidomys (McFarlane et al 1998b) emparentado con Amblyrhiza en la misma epoca (Tabla 45 intervalo I)
El ascenso del nivel del mar seguramente promovio la extincion de mamiferos durante la transicion del Pleistoceno al Holoceno (Tabla 45 intervalo II) con la reduccion del tamano de las Islas por las transgresiones marinas (Pregill y Olson 1981 Morgan 1993) Esto ultimo haria maximamente severa la competicion asf como la presion de los depredadores [Steadman et al (2005) no yen por que el calentamiento global ocurrido durante la transicion Pleistoceno-Holoceno habria de tener un efecto negativo sobre los perezosos siendo sus descendientes actuales especies tropic ales 0 subtropicales pero no se trata del efecto
330 BIOGEOGRAFiA
directo del cambio de temperatura sobre los animales sino del efecto indirecto y progresivo que dicho cambio genera a largo plazo al reducir el territorio insular disponible]
Por esta epoca tambien desaparecieron especies de tall a mayor como Megalocnus zile en La Espanola y otras de menor tamano (en Cuba) pero que representan la tall a maxima en sus respectivos contextos taxonomicos del Cuaternario como Solenodon arredondoi Paralouatta varonai y Macrocapromys acevedo Un mamifero puede ser grande solo a costa de reducir su densidad poblacional y esto 10 hace mis vulnerable a la extinci6n sobre todo en una isla (McNab 1994 McFarlane et al 1998a) Incidentalmente Alcover et al (1998a) afirmaron que larelacion entre la talla corporal y la extincion no siempre se cumple en las especies de menor tamano y citaron como ejemplo que entre las musarafias todas las especies de Nesophontes desaparecieron mientras sobreviven especies de Solenodon pero la comparacion correcta en todo caso serfa entre los solenodones vivientes y el gigante Solenodon arredondoi y entre Nesophontes edithae -el gigante de los nesofontes extinguido en el periodo amerindio- y el resto de sus congeneres que sobrevivieron hasta los primeros siglos de la colonizacion europea (Tabla 45 intervalos III y IV)
CAUSAS ANTROPOGENICAS
Intervalo Amerindio
La mayoria de los autores que se han referido a las extinciones acaecidas durante este intervalo (entre elIos paradigmaticamente Martin 1984a b) han responsabilizado a los aborfgenes (que se supone comenzaron a poblar las Antillas Mayores hace 6 mil afios) con la desaparicion de numerosas especies de mamiferos terrestres en la region principalmente por la via de la sobrecaza y la introduccion de animales domesticos devenidos depredadores de la fauna autoctona (Morgan y Woods 1986) Este efecto negativo de la presencia indigena sobre los mamiferos de la region ha sido dramatizado por algunos autores en terminos de holocausto faunal Sin embargo pensamos que se ha exagerado Como han expresado MacPhee et al (19993) cuando uno busca evidencia dura de complicidad humana directa en [estas] extinciones no hay mucho que encontrar Las siguientes son algunas consideraciones a tener en cuenta en tal sentido
El registro radiometrico mas joven conocido hasta ahora para un perezoso antillano (MegalocnllS rodens) es de 42 kaAP (MacPhee et ai 2007) Si se acepta que los aborfgenes entraron en Cuba hace aproximadamente 6 mil anos su convivencia con los perezosos debe haber durado alrededor de 1800 afios (0 mas porque dificilmente el referfdo fechado radiometrico haya correspondido al ultimo sobreviviente de la especie) Esta cifra contradice la hipotesis del holocausto faunal segun la cualla extincion de los mamiferos insulares de mayor talla debe ocurrir con extrema rapidez -a escala de decadas 0 de centurias- a partir de su primer contacto con los humanos (Martin y Steadman 1999) A pesar de las repetidas afirmaciones en contrario (Harrington 1921 Miller 1929c Herrera [1945] Tabio 1951 Alvarez 1951a 1957 Arredondo
331 EXTINCIONES DEL CUATERNARIO
1959 1960a Veloz y Ortega 1973 Mayo 1980b Rodriguez et al 1984 Castellanos y Pino 1985 Pino y Castellanos 1985 Nunez 1990b Gutierrez 1996 Arredondo 2004) en su mayoria basadas en evidencias no exentas de posibles accidentes de deposicion (0 de reubicacion postdeposicional) en recintos cavernarios (Diaz-Franco [2006b D no se ha demostrado inequivocamente que los aborigenes antillanos se alimentaran de perezosos regularmente Sus restos en residuarios de alimentos son escasisimos en toda la region y esto contrasta con los abundantes restos de jutias y otros mamiferos de menor talla que aparecen en esos residuarios demostrativos de la preferencia de los paleoindios por estos otros mamiferos
Los mamiferos mas importantes en la economia prehistorica de las Antillas Mayores fueron las jutias Capromys y Geocapromys en Cuba Geocapromys en Jamaica e Isolobodon en La Espanola Puerto Rico e Islas Virgenes (Aguayo 1950b Tablo 1951 Rimoli 1972 Wing 2001 Diaz-Franco [2006b D Wilkins (2001) estudi61ajutia viviente de Jamaica (Geocapromys browni) en un sitio ocupado por los aborigenes durante 100 anos y no encontr6 evidencias de que durante ese tiempo la especie haya sido sobreexplotada Asimismo en Cuba las jutias conga carabali y andaraz son escasas en depositos pleistoholocenicos en cambio son muy numerosas en residuarios indigenas y de haber sido sobreexplotadas 10 mas probable es que no hubieran sobrevivido hasta hoy
La habilidad de los humanos para penetrar en profundidad el bosque tropical y mantenerse en este sustentablemente ha sido un evento comparativamente reciente (Beran y Steele 1994) En el caso antillano la caza tierra adentro ha sido explicada como un proceso tardio de desplazamiento a partir de la explotacion de abundantes recursos marinos que ofrecian condiciones 6ptimas para el sedentarismo de las comunidades aborigenes costeras (C6rdova y Arredondo 1988 Brito 2005) Peces y mariscos dominan la arqueologia costera de las Islas antillanas en tanto el consumo de recursos terrestres (mamfferos incluidos) aumenta progresivamente en la medida que los residuarios estudiados se alejan de la costa (Scudder 1991 Wilkins 2001) En las Antillas Mayores el componente terrestre de la dieta aborigen es solo 20-30 del material recobrado en los residuarios (Wing 1977 1989) Y debio ser menor alin porque el componente vegetal de la dieta no se ha preservado Por otra parte los aborigenes cubanos domesticaron la jutia conga y seguramente la reprodujeron para comer (Pose et al 1988) practica que probablemente se repitiera con otras jutias en las demas Antillas Mayores
- Fray Bartolome de las Casas hablo de 3 millones de indios en Cuba al inicio de la colonizacion pero los historiadores subsecuentes consideraron exagerada la cifra (Poey 1851 a) No obstante se dispone de dos estimados de la densidad de poblaci6n aborigen en las Antillas Mayores a la llegada de los europeos 18 indigenaslkm2 en Puerto Rico (Rouse 1956) y 1271km2 en La Espanola (Watts 1987) Extrapolando ambos valores entre las cuatro islas principales de las Antillas Mayores tenemos que en la fecha indicada la poblaci6n aborigen pudo estar entre 16 mil y 113 mil indigenas en Puerto Rico entre 21 mil y 145 mil en Jamaica entre 141 mil y 1 mill6n en La Espanola y entre 200 mil y 14 millones en Cuba Entre las cifras maximas y las minimas nos inclinamos
332 BIOGEOGRAFiA
hacia estas ultimas atendiendo a los altos indices de mortalidad infantil y morbilidad que se evidencian en las osamentas de los pobladores prehispanicos (Martinez y Rives 1990) Uno de los hechos que apuntan hacia el bajo numero de aborigenes en las Antillas Mayores es 10 rapido que desaparecieron despues de la llegada de los europeos pnkticamente sin dejar mestizaje con estos 10 cual contrasta ostensiblemente con la situacion en la America continentaL
En suma un grupo humano (1) demograiicamente irrelevante y controlado por altos indices de morbilidad (2) consumidor mayoritariamente de organismos marinos y (3) carente de la tecnologia cinegetica necesaria para poder sobreexplotar los mamiferos que Ie sirvieron de alimento (a los cuales accedio tardiamente en su desarrollo) no es concebible que haya causado e1 holocausto faunal que se Ie atribuye a partir del error de aplicar a las comunidades aborigenes antillanas un modelo cultural de la prehistoria continentaL
Tampoco hay evidencias de que los aborigenes hayan introducido carnivoros con suficiente capacidad depredadora salvo el perro Morgan y Otlenwaldder (1993) consideraron que la depredacion por perros durante el periodo amerindio es la explicacion mas plausible para la extincion del almiqui gigante de Cuba pero como se puede observar (Tabla 54) Solenodon arredondoi aparece en el intervalo II por falta de asociacion con evidencias culturales
Consecuentemente el accionar de los aborigenes y de sus animales domesticos directamente sobre la fauna no parece haber sido por sf solo un factor determinante de la extincion de mamiferos terrestres durante el periodo amerindio en las Antillas (pero vease Steadman et al 2005)
Hay que recordar en este senti do que alin en el Holoceno tardio ocurrieron cambios clirruiticos de consideracion (vease Causas Naturales) Por ejemplo Curtis et al (2001) explicaron que la extincion de algunos ani males mesofilicos de las alturas montanosas de La Espanola pudo haber antecedido a cualquier impacto humano sigrIificativo en esos lugares por coincidir dicha extincion con una tendencia a la aridez documentada isotopicamente y con evidencias de la perdida del bosque mesico ocurridas despues de 3kaAP Yen Jamaica hace solo 2shy3 mil aDos el area de la Bahia de Jackson sustentaba una gran biomasa de vertebrados en bosque hinnedo a juzgar por la cantidad y diversidad de taxones presentes en material fosil derivado de la depredacion por lechuzas hoy el area es la mas seca de la isla con matorral xerofitico y pobreza de fauna (MacPhee 1996a) Yalm en nuestros dias huracanes de gran intensidad aniquilan poblaciones de vertebrados (que no se recuperan) en is las pequenas y llanas del Caribe (Brooks y Smith 2001)
En medio de la incertidumbre sobre la verdadera causa (0 causas) de las extinciones de mamiferos en el periodo amerindio MacPhee y Marx (1997) han propuesto su hipotesis de la hipermorbilidad la cual estipula colapsos poblacionales relativamente rapidos con la introduccion de enfermedades de alta letalidad Estos autores han razonado que
Las islas son los lugares mas apropiados para experimentar perdidas catastroficas causadas por enfermedades Los mamiferos no solo serian susceptibles en virtud de su prolongado aislamiento biologico sino que en los confines de una isla la epizootia
333 EXTINCIONES DEL CUATERNARIO
tendria la potencialidad de infectar simultaneamente a la rnayoria de los individuos de vadas especies Los neonatos son inmunologicamente inmaduros y sufririan desproporcionadamente cualquier infeccion hipervirulenta La perdida excesiva de neonatos y juveniles tendria efectos severos para la sobrevivencia de la poblacion si esta se reduce por debajo de cierto nivel sobreviene la extincion porque no hay posibilidad de restitucion en el periodo postpanzootico Las especies de mayor talla corporal van a ser las mas vulnerables a la extincion dadas sus bajas tasas reproductivas largos periodos de gestacion y reducido numero de progenie Las especies de menor tamano --caracterizadas por altas tasas reproductivas periodos cortos de gestacion y mayor numero de neonatos por parto- tambien serian fuertemente diezmadas pero muchas sobrevivirian porque su estrategia reproductiva les permite perder mayor numero de individuos Las perdidas deben ocurrir relativamente rapido (a escala de decadas) mientras que otras hipotesis (digamos los cambios climaticos) permiten intervalos mucho mas largos (a escala de siglos 0 de milenios) entre la aparicion de las causas y la produccion de sus efectos Por ejemplo todo indica que los nesofontes se extinguieron muy rapidamente y seria dificil poder invocar un impacto humano directo 0 argumentar fenomenos climaticos que pudieran explicar plausiblemente perdidas tan rapidas
MacPhee y Marx (1997 191) han dicho que por el momento la hipotesis de la hipermorbilidad no pasa de ser una abstraccion en espera de adecuada sustentacion empirica Llama la atencion que linicamente en el periodo amerindio (Tabla 45 intervalo 1Il) las extinciones abarcaron todas las clases del peso corporal (de la a a laj) y que ninguna de las especies que sobrevivieron hasta el periodo postcolombino (Tabla 45 intervalo IV mas las especies vivientes) sobrepasa los 10 kg (clases del peso a b c) en clara coincidencia con los presupuestos de la hipotetica hipermorbilidad
Intervalo Postcolornbino
Por 10 menos 37 especies de mamiferos terrestres vivian en las Antillas Mayores y grupos de Islas perifericas a la lIegada de los europeos en 1492 De elIas se han extinguido 23 shymas especies que en cualquiera de los periodos precedentes (Tabla 45~ y solo subsisten 12 especies repartidas entre Cuba (8 especies) La Espanola (2) Jamaica (1) y Bahamas (1) varias de estas en peligro critico de extincion Solo dos de las especies extinguidas durante los ultimos 500 anos se conocieron en estado viviente los roedores Oryzomys antillarum de Jamaica (extinguido en 1877) y Geocapromys thoracatus de las Islas Swan (extinguido en 1955) Las causas de las extinciones ocurridas durante este periodo son sobradamente conocidas destruccion de habitats introducci6n de animales exoticos (domesticos y comensales) y demas presiones asociadas a la actividad humana entre las cuales posiblemente habria que incluir brotes de hipermorbilidad
Durante mucho tiempo se ha afirmado la posibilidad de que especies de Nesophontes hayan sobrevivido hasta las primeras decadas del siglo xx y que incluso continuen existiendo en lugares remotos de Cuba 0 La Espanola (Allen 1918 1942 Miller 1930 Aguayo 1950a Arredondo 1960b Fischer 1977 MacFadden 1980 Woods y
334 BIOGEOGRAFiA
Ottenwalder 1991) Actualmente no existe evidencia radiometrica de que alguna especie de Nesophontes haya sobrevivido al siglo xv las fechas radiocarbonicas calibradas obtenidas en muestras de Nesophontes de Cuba y La Espanola resultaron no mas tardias que los siglos XIII y XIV (MacPhee et al 1999) Por otra parte las expediciones efectuadas a fines del siglo xx con el proposito de localizar individuos vivientes del genero en La Espanola no produjeron la menor indicacion afirmativa al respecto (Woods et al 1985) Ademas a traves de las Antillas Mayores no aparecen restos de Nesophontes en los depositos formados actualmente por las lechuzas
EPiLOGO
Las Antillas Mayores son islas oceanicas no han formado parte de un continente (con la posible excepcion de una parte de Jamaica hasta el Eoceno Medio hace 45 Ma) y durante un tiempo geologico considerable (desde el Eoceno Superior hace 40 Ma) han permanecido emergidas y separadas de los continentes vecinos Consecuentemente los mamiferos terrestres (no volantes) autoctonos de estas islas constituyen una fauna oceanica que por ende cumple con todas las caracteristicas convencionales reconocidas para este tipo de entidad biogeografica
- Baja diversidad taxonomica a nivel de las categorias superiores (solo cuatro ordenes) en comparacion con islas plataformicas continentales (0 con areas continentales) de tamano y diversidad ecologica simi lares
- Alto grado de endemismo a nivel de genero (96) y familia (50) en respuesta al prolongado aislamiento
- Desbalance ecologico de la estructura trofica ausencia de mamiferos camivoros y ungulados
- Espectaculares procesos de irradiaci6n evolucionaria como resultado de la colonizacion de territorios libres de competidores y depredadores
Tendencias evolucionarias extremas gigantismo
- Control a cargo de aves rapaces con tendencia at gigantismo en ausencia de mamiferos miembros del orden Carnivora
- Mas especies de reptiles y de murcielagos que de mamiferos terrestres relaciones que en los continentes se invierten
Alta tasa de extincion de especies (84) sin reposicion
Preservacion de especies filogeneticamente arcaicas (los llamados fosiles vivientes) los almiquies

312 BIOGEOGRAFiA
Por consiguiente Hyrachius ---conocido de localidades de edad Eoceno Inferior a Medio en Norteamerica- debio ocupar (10 que iba a ser) Jamaica occidental mucho antes de que esta se separara de Centroamerica De hecho este fosil Uego a su destino insular contenido en las rocas de su basamento no como un ser vivo por 10 que es erroneo considerarlo un animal antillano (lturralde-Vinent y MacPhee 2004) como han supuesto algunos (Hedges 200 I Portell et al 200 I Condis [200 I D Todo 10 que hizo fue viajar tanatovicariantemente en su barco funeral vikingo -otra de las denominaciones metaforicas de McKenna (1973)
DISCUSION Y CONCLUSIONES
Hemos visto que solo disponemos de una hipotesis plausible para explicar la mayor parte del poblamiento (continente-isla) de las Antillas Mayores por mamiferos terrestres del Neotropico continental la dispersion mixta (a traves de GAARlandia) para los perezosos (excepto Choloepodinae) los monos y los roedores (excepto Oryzomys)
Por otra parte la historia biogeogrMica de las musaraiias de las Antillas Mayores (Nesophontes y Solenodon) constituye una de las situaciones mas enigmaticas entre los mamiferos de la region (Ottenwalder 2001321) Al menos cinco mecanismos de poblamiento continente-isla han sido invocados para explicar cuando como y de donde llegaron a las Antillas Mayores estas musaraiias
I EI propagulo ancestral (del Terciario de Norteamerica) coloniz6 a las Antillas Mayores por dispersion marina desde Centroamerica 0 desde la Florida (Patterson 1962)
2 El propagulo ancestral ( del Terciario) cruzo el Atlantico Sur (supuestamente mas estreshycho en aquella epoca) directamente de Africa a las Antillas Mayores (Hershkovitz 1972)
3 El propagulo ancestral (del Terciario de Norteamerica) Uego a las Antillas Mayores en el Arca de Noe de Rosen (MacFadden 1980)
4 El propagulo ancestral (del Terciario de Norteamerica) fue transportado a las Antillas desde Centroamerica en el bloque de Jamaica occidental y de alli paso por dispersion marina a las restantes Antillas Mayores (Whidden y Asher 200 I)
5 El propagulo ancestral (del Terciario de Norteamerica) lleg6 a las Antillas Mayores a traves de GAARlandia (Whidden y Asher 2001)
Aunque hay consenso entre estos (y otros) autores sobre el origen norteamericano de las musaraiias antillanas ninguno de los cinco hipoteticos mecanismos de poblamiento resiste la evidencia en contrario filogenetica yo paleogeogrMica Ademas algunas de estas hipotesis asumen que Nesophontidae y Solenodontidae comparten un antecesor comun en las Antillas pero esta relacion no ha sido confirm ada Para dilucidar el enigmatico caso de las musaraiias antillanas se requerini profundizar en la investigaci6n geologica paleogeognifica y paleontol6gica (Iturralde-Vinent y MacPhee 1999)
De acuerdo con el conocimiento filogenetico disponible sobre el conjunto de la fauna la colonizacion original de las Antillas Mayores por mamiferos terrestres implicaria el arribo
313 POBLAMIENTO ISLA-ISLA
de solo siete propagulos fundadores perezosos 2 (coloepodinos y megalocninos si como parece tienen un origen difiletico) musaraiias 2 (nesofontidos y solenodontidos si no constituyen una misma familia como consideran algunos) monos 1 (pitecidos) roedores histricognatos 1 (equimidos capromidos y heptaxodontidos) roedores muridos 1 (Oryzomys en Jamaica)
POBLAMIENTO ISLA-ISLA
La distribucion de los mamiferos terrestres a traves del area antillana demarcada al inicio del presente capitulo (vease Contexto Geogratico Fig 41) ha sido interpretada en funcion de dos hipoteticos mecanismos de poblamiento interinsular la dispersion y la vicarianza fundamentadas como se resumen seguidamente
DISPERSION
De acuerdo con la evidencia paleogeogratica mas actualizada en las principales regresiones marinas ocurridas en la cuenca caribeiia durante los ultimos 23 Ma (transiciones OligocenoshyMioceno Mioceno Superior - Plioceno y varias veces en el Pleistoceno) las masas terrestres constitutivas de la region insular en formacion aumentaron considerablemente su extension territorial disminuyendo por tanto la distancia entre elias oportunidades que los elementos faunales aprovecharian para dispersarse de unas islas a otras
Durante mucho tiempo esta ha sido la hipotesis mas favorecida por los estudiosos del tema a partir de Simpson (1956) pero en decadas recientes ha suscitado dos objeciones
1 Es dificil en tender como pudo operar la dispersion marina entre estas islas puesto que ninguna de las Antillas Mayores posee rios suficientemente caudalosos para transportar masas significativas de vegetacion flotante (MacPhee 198432)
Es oportuno recordar aqui el islote ala deriva (con mas de diez palmas reales en pie) entre las tres Antillas Mayores mas occidentales mencionado anteriormente (vease La Balsa Natural)
2 La dispersion marina es menos efectiva para explicar la asombrosa similitud de las faunas de mamiferos terrestres a traves de las Antillas Mayores septentrionales porque como proceso aleatorio no debe producir distribuciones uniformes a 10 largo de una cadena de islas 0 si las produce entonces el proceso no es aleatorio (MacPhee e lturralde-Vinent 2000 149)
Independientemente de que las aludidas distribuciones taxonomicas estan lejos de ser uniformes 0 asombrosamente similares la hipotesis de la dispersion marina contrariamente a su canicter aleatorio y altamente improbable en el caso del poblamiento continente-isla- en el caso del poblamiento isla-isla es un proceso no aleatorio (aprovecha las oportunidades brindadas puntualmente por los periodos de maxima regresion marina) y ademas altamente probable dada la incomparablemente menor distancia entre las islas caribeiias que entre estas y los continentes vecinos
314 BIOGEOGRAFiA
V ICARIANZA
La neotectonica y las transgresiones marinas del Oligoceno Medio y del Mioceno afectaron sustancialmente la paleogeografia de la Cresta de las Antillas Mayores resultando la subdivision gradual de las partes subaereas de la Cresta Asi el este de Cuba y el norte de La Espanola conectados fisicamente durante el Oligoceno Inferior se separaron al formarse el Paso de los Vientos en el Oligoceno tardio en tanto La Espanola y Puerto Rico probablemente permanecieron conectadas hasta el Mioceno tardio en que surgio el Paso de Mona (Iturralde-Vinent y MacPhee 1999)
Tales eventos geologicos habrian dividido las distribuciones de los grupos faunales emplazados previamente en el territorio continuo de las tres islas y a partir de esas disyunciones dichos autores (tambien MacPhee et al 2000 y White y MacPhee 2001) asumieron --como segunda fase del modelo de GAARlandia- la hipotesis de la vicarianza isla-isla para explicar la distribucion multiinsular de varios grupos de mamiferos terrestres a traves de las Antillas Mayores septentrionales De acuerdo con esta hipotesis cualquier diferencia en composicion faunal entre las islas (por ejemplo la ausencia de roedores heptaxodontidos en Cuba 0 de monos platirrinos en Puerto Rico) constituye evidencia de extincion local postvicariante y no de dispersion marina fallida Lo primero se puede demostrar si aparecen los fosiles pertinentes 10 segundo no se puede demostrar (MacPhee e Iturralde-Vinent 1994) Por otra parte IturraldeshyVinent y MacPhee (1999 57) han reconocido que aunque fa vicarianza isla-isla puede representar una solucion clara para las distribuciones de varios taxones cuaternarios no puede explicar todos los casos
Rega et al (2002 18) han presentado dos objeciones al modelo de la vicarianza isla-isla aplicado a los mamiferos de las Antillas Mayores
I Si la separacion de Cuba y La Espanola precedio en varios mill ones de anos la separashycion de La Espanola y Puerto Rico el modelo vicariante de lturralde-MacPhee debe predecir mayor similitud entre las faunas de La Espanola y Puerto Rico que entre las de Cuba y La Espanola De hecho es al reyes
No obstante MacPhee et al (200335) admitieron despues que la fecha mas temprana empiricamente defendible [para la apertura del Paso de Mona] debe ser el Oligoceno Medio esto invierte la secuencia de ambas disyunciones con 10 cual queda resuelta la aparente contradiccion Sin embargo en su reciente evaluacion de la congruencia entre filogenia e hipotesis geologic as en varios grupos de mamiferos terrestres de las Antillas Mayores Davalos (2004373) encontro que ninguno de los taxones cumple con la fecha y el patron de divergencia entre las masas terrestres antillanas postulados por el modelo de GAARlandia
2 La presencia de un mismo genero en las tres Islas septentrionales de las Antillas Mayores implica [por el modelo vicariante] que la diferenciacion entre estas formas durante el intervalo desde el Mioceno ha sido unicamente de nivel especifico [Dicha presencia]
315 rRRADlAcr6~ EVOLUCrONARIA
s610 puede explicarse razonablemente como el resultado de movimientos de esos animales entre las islas en el Cuaternario
De esta objecion resulta dificil disentir [El poblamiento entre islas diferenciado solo a nivel especifico no es probable que sea anterior al Pleistoceno (Simpson 1956 15)]
Debe tenerse en cuenta que la hipotesis de la vicarianza isla-isla alude a las tres islas septentrionales de las Antillas Mayores porque como se explico antes (vease El Barco Funeral Vikingo) Jamaica no existia como isla durante el lapso (Eoceno Superior shyOligoceno) consensualmente reconocido como el mas probable para la colonizacion de las proto-Antill as Mayores por la mayo ria de los mamiferos terrestres Despues de emerger como isla en el Mioceno Medio Jamaica debe haber recibido sus mamiferos terrestres por dispersion isla-isla (Davalos 2004376) y solo en el Cuaternario parece haberrecibido (por dispersion marina desde Centroamerica) al roedor murido Oryzomys
Seguramente tanto la dispersion como la vicarianza contribuyeron a que los mamiferos terrestres se irradiaran a traves de las Antillas Mayores y grupos de islas perifericas pero las maneras y tiempos en que estos mecanismos operaron a 10 largo de dicho proceso son aun en gran medida motivos de desacuerdo e incertidumbre
IRRADIACION EVOLUCIONARIA
A continuaci6n revisamos someramente las distribuciones conocidas de los diferentes grupos de mamiferos terrestres a traves de las Islas principales de las Antillas Mayores y grupos de Islas perifericas incluidas las interpretaciones y conjeturas que la literatura reciente recoge en relaci6n con la manera en que dichas distribuciones pudieron haberse conformado durante la irradiaci6n evolucionaria de los referidos grupos de mamiferos Manifestaciones de esta irradiaci6n pueden inferirse unicamente a partir de los taxones presentes en el Cuatemario pues solo 6 de las especies conocidas corresponden al Terciario
GRUPOS ORlGINARIOS DE SURAMERICA
PEREZOSOS (Megalonychidae)
Aparte de un genero miocenico de Cuba y otro del Oligoceno de Puerto Rico se conocen 4 generos (12 especies) de perezosos cuaternarios (todos extintos) restringidos a las tres islas septentrionales de las Antillas Mayores Acratocnus Neocnus Megalocnus y Parocnus Los cuatro generos estan representados en Cuba y La Espanola y Acratocnus 10 esta ademas (como unico genero de perezoso) en Puerto Rico Esta distribuci6n ha sido interpretada por MacPhee et al (2000) eomo resultado de la vicarianza oligomiocenica entre estas islas y [dado que el Banco Puerto Rico - Islas Virgenes fue la entrada de GAARlandia a las Antillas Mayores] se preguntaron si la ausencia de los otros tres generos en Puerto Rico se debe a que nunca estuvieron en ese territorio 0 a que sus
316 BIOGEOGRAFiA
fosiles atm no han sido hallados Pero como hemos visto Rega et al (2002) atribuyeron dicha distribucion a la dispersion interinsular en el Cuatemario
Por otra parte varios amilisis c1adisticos recientes ubi can a Choloepus -genero Reciente conocido solo de Centro- y Suramerica- como taxon hermano de Aeratoenus exc1uidos los demas megaloniquidos antillanos Para White y MacPhee (2001228)
tan estrecha relacion sugiere que la divergencia entre ambos [generos] ocurrio despues de que Aeratoenus divergio de Neoenus Si esta relacion implica que Aeratoenus y Neoenus se dispersaron separadamente desde Suramerica a las AntilIas mientras Choloepus permanecia en el continente (siendo necesarias al menos tres invasiones de perezosos) 0 si Choloepus invadio Suramerica desde las Antillas es algo que el material disponible no permite determinar
La diversidad adaptativa de los perezosos se manifiesta en (1) la utilizacion del habitat especies arboreas en Neoenus y Aeratoenus y terrestres en Paroenus y Megaloenus (Webb 1985a White [1993] White et al 1996 Arredondo [1999]) y en (2) la gama de tallas corporales desde los pequenos Neoenus hasta Megaloenus rodens el mayor de los mamiferos terrestres antillanos (su peso adulto estimado en 270 kg seglin Paula 19679) esto sugiere que los perezosos se irradiaron a nichos que en contextos continentales probablemente estarian ocupados por miembros de otros c1ados (MacPhee e IturraldeshyVinent 2000)
No esta claro si el abanico de tamanos entre estos perezosos enmascara alguna tendencia al gigantismo Por 10 pronto Anderson y Handley (2002) encontraron 10 contrario en perezosos (Bradypus) de las Islas de Bocas del Toro (Panama) shyseparadas secuencialmente del area continental vecina por ascensos del nivel del mar durante los ultimos 10 ka En las cinco islas los perezosos son menores que su contraparte continental y las diferencias entre las tallas medias de las cinco poblaciones expresan una reduccion lineal significativa de la talla corporal en funcion de la edad geologica de cada isla sin relacion alguna con el area de las islas 0 la distancia que las separa del continente
MONOS (Pitheciidae)
Los primates de las Antillas Mayores (cuatro especies extintas) constituyen un grupo monofiletico con Paralouatta (en Cuba) y Antillothrix (en La Espanola) como taxones hermanos seguidos en la escalera c1adistica por Xenothrix (en Jamaica) y finalmente Callieebus como el pariente continental mas cercano esto implica una sola invasion desde Suramerica antes del Mioceno Inferior (fecha mas temprana conocida para los primates fosiles antillanos) y por tanto en consonancia con GAARlandia como la via de colonizacion (Horovitz y MacPhee 1996 1999 Horovitz [1997] MacPhee y Horovitz 2004)
En desacuerdo parcial con la anterior interpretacion la filogenia de este grupo de monos obtenida por Davalos (2004) indica un antecesor comun en Cuba La Espanola y Jamaica
317 TRRADIACTON EVOLUCIONARIA
compatible con la hipotetica conexion -propuesta tentativamente por Iturralde-Vinent y MacPhee (199980-84)- entre las Montanas Azules del este de Jamaica y la peninsula suroccidental de La Espanola en la epoca de GAARlandia
A pesar de que los primates antillanos pertenecen a un grupo viviente de monos muy pequenos (los monos titles) fueron comparativamente grandes en relacion con los platirrinos modernos y sucesivamente mayores en la direccion Antillothrix Xenothrix y Paralouatta con P marianae como el mayor platirrino conocido del Mioceno neotropical (MacPhee 2005) MacPhee et al (1994) consideraron que Xenothrix debio ser un mono de andar lento y al parecer las especies de Paralouatta abandonaron la arborealidad para adoptar un modo de vida mas terrestre (MacPhee 1996a MacPhee et al 2003 MacPhee y Meldrum 2006)
ROEOORES (Echimyidae Capromyidae Heptaxodontidae)
La edad de Zazamys en Cuba demuestra que la invasion de los roedores histricognatos en las Antillas Mayores comenzo antes del Mioceno Inferior por otra parte los representantes mas antiguos de esa irradiacion evolucionaria se encuentran en La Espanola y Puerto Rico Ambas evidencias son consistentes con la hipotesis de GAARlandia (Woods 1982 1988 Woods et al 2001)
No esta claro si los capromidos evolucionaron a partir de un equimido ancestral 0 de un protocapromido 0 si los equimidos antillanos (I sub familia 3 generos y 7 especies todas extintas) forman parte de la irradiacion de los capromidos (4 subfamilias 9 generos y 27 especies 10 vivientes) Kratochvil et al (1978b) y Rodriguez et al (1979) consideraron que Capromys nan us es un eslab6n entre Echimyidae y Capromyidae pero Mesocapromys nanus esta morfologicamente mucho mas cerca de Hexolobodon y los de mas capromidos que de los equimidos (Woods 1985) Lo que si esta claro segun Woods et al (2001) es que la subfamilia Plagiodontinae endemic a de La Espanola es el linaje basal de los capromidos y que Isolobodon Hexolobodon Capromys Geocapromys Mysateles y Mesocapromys [y supuestamente Macrocapromys] forman parte de la irradiacion temprana de Capromyidae
La subfamilia Capromyinae (5 generos y 17 especies 9 vivientes) parece ser el dado de mas reciente evolucion en la familia con invasion de Cuba desde La Espanola y dispersion desde Cuba (1) por Capromys a las islas Caiman y (2) por Geocapromys a las islas Bahamas Caiman y Jamaica y desde Jamaica a las islas Swan aunque la presencia de Geocapromys en Swan pudiera deberse a introduccion aborigen (Morgan 1985 1988 1989 1993 1994 Woods 1989a 1990) Woods et al (2001) utilizaron datos moleculares para obtener estimados independientes de divergencia entre capromidos segiJn 10 cual Geocapromys divergio cerca de los 17 MaAP Mesocapromys divergio de Capromys hace 15 MaAP en tanto lvlysateles 10 hizo hace 10 MaAP Por su parte las formas de Capromys pilorides y de Mysateles prehensilis en Isla de la Juventud divergieron de sus correspondientes subespecies nominativas en la isla principal a los 24 y 08 MaAP respectivamente
318 BIOGEOGRAFiA
Woods y Howland (I979) describieron adaptaciones morfologicas de la mandibula el hioide y la faringe asociadas a mecanismos de masticacion diferentes en plagiodontinos y caprominos y Varona (1980a) llamo la atencion sobre el gran desarrollo de los molariformes (en relacion con la longitud total del cnineo) que presentan ciertos grupos de jutias fosiles en comparacion con las vivientes 10 que pudiera interpretarse como una adaptacion alimentaria a una vegetacion mas abrasiva por parte de las jutias extinguidas
Segun Woods (1990) la familia Heptaxodontidae (2 subfamilias 5 generos y 6 especies todas extintas) tambien seria parte de la irradiacion de los capromidos con origen en el este de La Espanola 0 en Puerto Rico y subsecuente dispersion al Banco de Anguila - San Martin en las Antillas Menores (pero vease MacPhee y Flemming 2003) Este banco estaria a solo 70 km del Banco Puerto Rico - Islas Virgenes en tiempos de maxima regresion marina y esta proximidad con Puerto Rico -patria de Elasmodontomys obliquus- seguramente tuvo que ver con la presencia en las islas de Anguila y San Martin del heptaxodontino gigante Amblyrhiza relacionado mas estrechamente con los roedores de las Antillas Mayores que con los del resto de las Antillas Menores (Woods 1990 1996) Aunque los roedores suelen aumentar de tamano en las islas (Case 1978) la escala del incremento de la talla en Amblyrhiza (que pudo alcanzar mas de 200 kg de peso adulto) esta fuera de toda proporcion con los patrones conocidos en otros roedores insulares (Biknevicius et 01 1993)
Clidomys (Clidomyinae) de Jamaica difiere marcadamente en su morfologia de los heptaxodontinos (vease ademas MacPhee y Flemming 2003)10 cual dificulta explicar su presencia en esa isla Como Jamaica estuvo bajo el mar hasta el Mioceno Medio el ancestro de Clidomys colonizaria esta isla por dispersion desde el extremo suroccidental de La Espanola despues que los heptaxodontinos se originaron en esta ultima isla (Woods 1989a) Pero Pascual et 01 (I 990) especularon que Clidomys pudiera ser el resultado de una invasion posterior e independiente de los capromidos
GRUPOS ORIGINARIOS DE NORTEAMERICA
El origen de las musaranas antillanas ha resultado ser hasta ahora el mayor enigma biogeografico de la fauna de mamiferos terrestres de la region aunque la mayoria de los autores coinciden en considerarlas originarias de Norteamerica (vease Discus ion y Conclusiones en Poblamiento Continente-Isla) No obstante MacPhee e Iturralde-Vinent (1994) descartaron toda posibilidad de conexion terrestre por el Estrecho de la Florida 0 por el Canal de Yucatan y si el (supuesto) evento vicari ante de Rosen (vease El Arca de Noe) -invocado por MacFadden (1980) para resolver el enigma- fuera minimamente plausible aun asi es dificil creer que las catastrofes que 10 sucedieron (vease Contexto Temporal) hayan barrido con toda esa vieja fauna continental excepto las musaranas circunstancia ya anticipada muy tempranamente por Matthew (l918a)
De otra parte la relaci6n entre Nesophontidae y Solenodontidae permanece irresuelta Si estos linajes no estan estrechamente relacionados entonces es muy posible que
319 IRRADIACION EVOLUCIONARIA
sus distribuciones sean explicadas por mecanismos diferentes (Whidden y Asher 2001)
La descomunal diferencia de talla corporal entre las musaranas pequenas (Nesophontes) y las grandes (Soienodon) seguramente impuso a ambos grupos restricciones distintas relativas a la termorregulacion y las adaptaciones energeticas En Nesophontes la seleccion debio favorecer una alta tasa de metabolismo y crecimiento camadas mas numerosas corta expectativa de vida yalta densidad poblacionaL En Soienodon en cambio la seleccion opero en la direccion de una baja tasa metabolica camadas reducidas crecimiento lento longevidad y baja densidad poblacional (Ottenwalder [1991])
MUSARANAS PEQUENAS (Nesophontidae)
Las musaranas pequenas estan representadas en las Antillas por el genero Nesophontes (y un genero miocenico indeterminado en ambar dominicano) con 6 especies cuaternarias extinguidas distribuidas en Cuba La Espanola Puerto Rico Islas Virgenes e Islas Caiman La mayor de las musaranas pequenas Nesophontes edithae evoluciono en Puerto Rico y su arribo a las Islas Virgenes supone una dispersion sumamente reciente En las Islas Caiman Nesophontes se asemeja mas a la poblacion de Cuba de donde seguramente derivo en el Plio-Pleistoceno (junto con Capromys y Geocapromys) cuando la costa sur de Cuba estuvo mucho mas cerca de las Islas Caiman (lturralde-Vinent 2003fig 4) MacPhee et ai (1994) interpretaron la distribucion de Nesophontes en Cuba y La Espanola como resultado de la vicarianza oligomiocenica ocurrida al formarse el Paso de los Vientos pero la distancia taxonomica entre los lados del Paso es solo de nivel especifico y N micros incluso vivi6 en ambas islas segtln Patterson (1962)
Allen (1918) considero que Nesophontes micrus fue una especie primitiva no modificada para la vida arb6rea 0 fos6rea y por tanto de habitos terrestres Nesophontes edithae de Puerto Rico es mucho mayor y mas robusta que las restantes especies del genero eL indice de robustez de su femur (= anchura minima de la diafisis ongitud total del hueso) muestra que este es tan robusto como el de Solenodon paradoxus un animal dos veces mayor (Morgan et ai 1980) Choate y Birney (1968) postularon un aumento cronoclinal de la taIla corporal en N edithae confirmado mas tarde por McFarlane (1999a) mediante fechados radiocarb6nicos de la secuencia estratignifica en depositos cavernarios de Puerto Rico Solenodon no vivi6 en Puerto Rico y en ausencia de una musarana de mayor tamano Nesophontes alcanzo una talla corporal mayor en esa isla que en La Espanola 0 Cuba (Woods y Eisenberg 1989)
MUSARANAS GRANDES (Solenodontidae)
Los arcaicos almiqufes (genero Soienodon) comprenden cuatro especies una viviente (S cubanus) y otra f6sil (S arredondoi) en Cuba y una viviente (S paradoxus con dos subespecies) y otra f6sil (S marcanoi) en La Espanola Como cabria esperar las dos especies cubanas se asemejan mas entre sf que a las especies de La Espanola pero
320 BIOGEOGRAFiA
enigmaticamente S marcanoi comparte caracteres con S cuban us y S paradoxus (Woods y Eisenberg 1989 Morgan y Ottenwalder 1993 Ottenwalder 200 I) Solenodon arredondoi alcanzo una talla corporal superior a la de cualquiera de las musarafias vivientes a nivel mundial Segun Morgan y Ottenwalder (1993) extrapolando las dimensiones y el peso de los almiquies vivientes S arredondoi debio tener una longitud cabeza-tronco y un peso de 450-550 mm y 1500-2000 g respectivamente en comparacion con S cubanus (360 mm y 800 g) y S paradoxus (390 mm y 1100 g)
Entre todas las especies de almiqufes Solenodon paradoxus es la que presenta los caracteres mas derivados (Otten walder 2001) Si estas relaciones son validas la distribucion de Solenodon entre Cuba y La Espafiola debio discurrir de oeste a este como propuso MacFadden (1980) quien invoco la presencia fosH de S cubanus [pero tambien S arredondoi] en Pinar del Rio para postular que el ancestro continental entr~ por Cuba opinion compartida por Perfit y Williams (1989) y por Ottenwalder ([1991] 2001) El paso de Solenodon a Puerto Rico pudo haberlo impedido la existencia de barreras ecologicas durante la conexion del este de La Espafiola con Puerto Rico 0 por la ausencia de Solenodon en esa parte de La Espafiola en aquella epoca Sin embargo las mismas condiciones ecologicas en el este de La Espafiola no parecen haber representado una barrera efectiva para Nesophontes cuyas mas altas tasa de reproduccion y densidad poblacional presumiblemente Ie permitieron expandirse flpidamente a traves de La Espafiola (Ottenwalder [1991])
Es opinion generalizada que los almiqufes antillanos constituyen el grupo de mamiferos que primero se emplazo en el territorio insular (MacPhee 2005) y en tal sentido es de interes la evidencia sobre especificidad parasito-hospedante aportada por Cruz (1980) quien encontro que la especie de acaro (genero Cubanochirus) que parasita a los almiquies es mucho mas primitiva que la especie (del mismo genero) que paras ita a las jutias y de aqui concluyo que dicho genero de acaro debio evolucionar primero en Solenodontidae y posteriormente en Capromyidae al entrar en contacto representantes de ambas familias en Cuba 0 La Espafiola
Recientemente Roca (2004) y Roca et al (2004) dieron a conocer que los resultados de su estudio genetico-molecular sobre los almiquies indican una divergencia de 25 Ma entre Solenodon cuban us y S paradoxus -compatible con la disyuncion tectonica entre Cuba y La Espafiola (Iturralde-Vinent y MacPhee 1999 McPhee et al 2003)-- y una separacion de ambas especies comparable en magnitud a la divergencia entre familias de mamlferos Consecuentemente propusieron que Solenodon cubanus fuera clasificada en el genero Atopogaie como indico Cabrera (1925) hace 82 afios
DISCUSION Y CONCLUSIONES
Las musarafias y los roedores antillanos (excepto Oryzomys) se irradiaron tanto de sus respectivos ancestros continentales que de estos 6rdenes se reconocen familias y subfamilias endemicas de la region en contraste con los perezosos (una subfamilia y tres
321 IRRADIACION EVOLUCIONARIA
tribus endemicas) y los monos (una tribu endemica) como expresion de diferentes niveles de diversificacion evolucionaria Este proceso es dificil de explicar (traUindose de c1ados que pnicticamente comparten la misma antigiiedad en las Antmas) si es correcta la interpretacion de Pascual et at (1990636)
Nosotros consideramos estos diferentes niveles de diversificacion como las diferentes etapas de evolucion alcanzadas por cada grupo [de mamiferos] Obviamente esto estuvo relacionado con las diferentes longitudes de sus historias
Pudiera ocurrir que en el futuro se compruebe que la historia biogeognifica de la region fue mucho mas complicada
La Espanola la isla mas ecol6gicamente divers a de la region debi6 ser el centro principal de evoluci6n de los mamfferos (con 87 de las familias y 73 de las subfamilias) seguida por Cuba (con 75 de las familias y 54 de las subfamilias) las dos islas de las Antillas Mayores mas relacionadas biogeograficamente (Fig 43 Tabla 44)
eL
EL
cb CAIMAN
SWAN
A Acratocnini B Cubanocnini C Megalocnini D Mesocnini E Nesophontidae F Solenodontidae G Xenothrichini
ANGUILAshyM-0 SMARTIN
H Heteropsomyinae I Plagiodontinae J Isolobodontinae
K Hexolobodontinae L Capromyinae
M Heptaxodontinae N CIidomyinae 0 Sigmodontinae
FIGURA 43 Representacion esquematica de las relaciones interinsulares (expresadas porel numero de linajes compartidos) entre las Antillas Mayores y entre estas y los grupos de islas perifericas segun las interpretaciones de los autores citados en el texto Los numeros dentro de los circulos indican la cantidad de linajes representados en cada isla relacion dudosa Contexto taxon6mico de los linajes en la Tabla 44
bull bull
bull bull
bull bull bull
bull bull
bull bull
bull
bull
bull
bull
bull bull bull bull
bull bull
bull bull
bull
bull bull bull
bull bull
bull
bull bull
bull
322 BIOGEOGRAFiA
TABLA 44 Distribuci6n de los linajcs de mamiferos terrestres a traves de las Antillas Mayores y grupos de islas perifericas LE = La Espanola CU Cuba JA Jamaica PR = Puerto Rico IC Islas Caiman IS Islas Swan IB = Islas Bahamas IV = Islas Virgenes AS = Anguila - San Martin
FamiliaSubfumiliafIribu LE CU JA PR IC IS IB IV AS
Megalonychidae
Choloepodinae
Acratocnini
Cubanocnini
Megalocninae
Megalocnini
Mesocnini
Nesophontidae
Solenodontidae
Pitheciidae
Callicebinae
Xenothrichini
Echimyidae
Heteropsomyinae
Capromyidae
Plagiodontinae
Isolobodontinae
Hexolobodontinae
Capromyinae
H e pta x 0 don tid a e
Heptaxodontinae
Clidomyinae
Muridae
Sigmodontinae
PILOSA
SORICOMORPHA
PRIMATES
RODENTIA
bull
bull
bull
bull TOTALES 12 10 4 4 2
323 EXTlICIOIES DEL CUATERNARIO
El gigantismo ha sido sin duda una tendencia evolucionaria recurrente entre los clados mayores de mamiferos antillanos Es un hecho bien conocido que algunos grupos de mamiferos han aumentado de tamafio en las islas y varios autores han intentado explicarlo a partir de enfoques diferentes (Foster 1964 Miller 1966 Sondaar 1977 Case 1978 Eisenberg y Wilson 1978 Heaney 1978 Lawlor 1982 Lomolino 1985 Angerbjom 1985 McNab 19942001)
EXTINCIONES DEL CUATERNARIO
Las islas que integran la Subregion Antillana han perdido casi 90 de sus mamiferos terrestres durante los ultimos milenios y en proporcion con su extension territorial esta perdida ha sido mayor en la Subregion que en cualquier otro lugar del planeta (MacPhee 1996b) Las extinciones del Cuatemario eliminaron mas de las tres cuartas partes de la fauna de mamiferos terrestres conocida de las Antillas Mayores sus islas satelites y grupos de islas perifericas y el examen de este proceso en sus aspectos mas generales se aborda en esta ultima parte del presente capitulo
CRONOLOGiA Y CONSECUENCIAS
Es muy improbable que un fosil represente el ultimo individuo de un taxon extinto y entonces hay que guiarse por la edad radiometrica del fosil mas reciente conocido lIamada fecha de ultima presencia- como unica aproximacion posible a la fecha de extincion del taxon
Como se explico al inicio del capitulo (vease Contexto Temporal) el registro fosH de los mamiferos terrestres de las Antillas Mayores adolece de insuficiente sustentacion radiometrica No obstante la distribucion temporal de los escasos fechados disponibles para el Cuaternario de la region abarca etapas criticas del proceso de extincion experimentado por la referida fauna a 10 largo de los ultimos 250 mil afios
De las 52 especies que es posible reconocer actualmente como miembros extintos de la fauna cuatemaria de mamiferos terrestres de las Antillas Mayores y grupos de islas perifericas los restos fosiles de 23 de elIas (442) han sido fechados radiometricamente o se han hallado asociados estratigraficamente de manera inequivoca con otro material fechado De estos 23 fechados 3 corresponden al interglacial Sangamon 4 pertenecen a la secuencia Pleistoceno terminal - Holoceno temprano 10 se ubican en ellapso entre el arribo de los aborigenes y la incursion europea y 6 son postcolombinos (a los cuales hay que sumar otras dos especies extinguidas en los siglos XIX y xx) A partir de estas 25 especies avaladas por fechas de ultima presencia distribuidas entre los cuatro intervalos temporales sefialados hemos compuesto la Tabla 45 en la que ademas aparecen las restantes 27 especies (carentes de fechado radiometrico) ubicadas en el intervalo temporal mas probable sobre la base de (1) la asociacion estratigrafica de sus restos con evidencias culturales (amerindias 0 europeas) 0 (2) la ausencia de dicha asociacion Sobre estos criterios de ubicacion temporal procede hacer dos aclaraciones
324 BIOGEOGRAFIA
TABLA 45 Intervalos temporales mas probables para las extinciones cuatemarias de los mamiferos terrestres de las Antillas Mayores y grupos de islas perifericas sobre la base de las evidencias siguientes (l) ultimo ejemplar colectado u observado en vida (2) fechado radiometrico mas tardio en material de la especie (3) asociaci6n estratigrafica mas tardia con otro material fechado (4) asociaci6n estratigrafica con restos de animales introducidos restos humanos u objetos culturales propios del intervale (5) falta de asociaci6n con evidencias culturales [Los ntimeros entre parentesis en la columna Evidencia significan adscripci6n tentativa al intervalo] Los estimados del peso corporal (en kg) estan tomados de la literatura 0 inferidos comparativamente y distribuidos entre
las clases siguientes (a) S 01 (b) 01 a lt 1 (c) 1 a lt 10 (d) 10 a lt 100 (e) 100 a lt 200 (j) ~ 200 Las
abreviaturas que designan las islas son las de la Tabla 44
F amilialEspecie Peso IsJa Evidencia Fuente
I httervalo htterg1acial Sangamon (gt80 kaAP)
HEPTAXODONTlNAE
Amblyrhiza inundata e AS 3 McFarJane et al 1998a
CLIDOMYINAE
Clidomys osborni d JA 3 Ibid
Clidomys parvus d JA 3 Ibid
II httervalo Pleistoceno terminal ~ Holoceno temprano (20-6 kaAP)
MEGALONYCHIDAE
Aeratoenus antillensis d CD 5 Arredondo 1975 a
Aeratoenus odontrigonus d PR 3 McFarJane 1999b
Aeratoenus simorhynehus d LE 2 Rega et al 2002
M egaloenus zile f LE 2 MacPhee et al 2000
SOLENODONTIDAE
Solenodon arredondoi e CD 5 Morgan y Ottenwalder 1993
PnHECIIDAE
Paralouatta varonai d CD 5 MacPhee 1996a
EcHIMYIDAE
Heteropsomys antillensis c PR 5 McFarJane 1999b
Heteropsomys insulans c PR 5 Ibid
Puertoricomys eorozalus c PR 5 Woods 1989a
CAPROMYIDAE
P lagiodontia araeum c LE 5 MacPhee et al 2000
Macrocapromys latus c CD 5 Varona y Arredondo 1979
(Continua)
325 EXTINCIONES DEL CUATERNARIO
TABLA 45 (Continuaci6n)
F ami1ia1Especie Peso Isla Evidencia Fuente
M acrocapromys latus c CU 5 Varona y Arredondo 1979
Mesocapromys kraglievichi c CU 5 Ibid
HEPTAXODONTINAE
Xaymaca fulvopulvis a JA 2 MacPhee y Flemming 2003
III Intervalo Amerindio (6-05 kaAP)
MEGALONYCHIDAE
Acratocnus ye d LE 2 MacPhee et aI 2000
Neocnus comes c LE 2 Ibid
N eocnus dousman c LE 2 Ibid
N eocnus toupiti c LE 2 Ibid
N eocnus gliriform is c CU 4 Cordova et ai 1997
M egalocnus rodens f CU 2 MacPhee et ai 2007
Parocnus browni e CU 2 Steadman et ai 2005
P arocnus serus e LE 2 MacPhee et ai 2000
N ESOPHONTIDAE
N esophontes edithae a PR 2 McFarlane 1999b
PITHECIIDAE
Antillothrix bernensis c LE 3 Rimoli 1977
EcHIMYIDAE
Brotomys contractus a LE 4 Miller 1929c
CAPROMYIDAE
P lagiodontia spelaeum c LE 4 Ibid
M acrocapromys acevedo c CU 4 NUiiez 1990a
HEPTAXODONTINAE
Elasmodontomys obliquus d PR 3 McFarlane 1999b
Iv Intervalo Postcolombino (05 kaAP - Presente)
N ESOPHONTIDAE
N esophontes hypom icrus a LE 24 MacPhee y Fle111111ing 1999
N esophontes zam icrus a LE (4) Ibid
N esophontes m icrus a CU 24 MacPhee et aI 1999
(Continua)
326 BIOGEOGRAFiA
TABLA 45 (Continuacion)
F amilialEspecie Peso Isla Evidencia Fuente
N esophontes m icrus a CD 24 MacPhee et aI 1999
N esophontes m icrus a LE 4 MacPhee et aI 2000
N esophontes sp 1 a IC 4 Morgan 1994
N esophontes sp 2 a IC 4 Ibid
Solenodon marcanoi b LE 4 Ottenwalder 2001
PITHECIIDAE
Xenothrix mcgregori c JA 4 MacPhee y Flemming 1999
EcHIMYIDAE
Boromys offeUa a CD 24 Guarch 1984
Boromys torrei a CD 24 Ibid
Brotomys voratus a LE 4 MacPhee et aI 2000
CAPROMYIDAE
P lagiodontia ipnaeum c LE 4 MacPhee y Flemming 1999
Rhizoplagiodontia lemkei c LE 4 Ibid
Isolobodon portoricensis c PR 24 Ibid
Isolobodon montanus c LE 4 Ibid
Hexolobodon phenax c LE (4) Ibid
Capromys sp c IC 24 Morgan 1994
Geocapromys columbianus c CD 4 MacPhee y Flemming 1999
Geocapromys thoracatus b IS 1 MacPhee y Marx 1997
Geocapromys sp 1 c IC 4 Morgan 1994
Geocapromys sp 2 c IC (4) Ibid
l-IEPTAXODONTINAE
Quemisia gravis c LE (1) MacPhee y Flemming 1999
MURIDAE
Oryzomys antillarum a JA 1 Ibid
1 Flemming y MacPhee (1999) alertaron en contra de la asociacion con otro material fechado Asimismo Morgan (1993) indico que la ausencia de asociacion con evidencias culturales amerindias no es necesariamente un criterio concluyente para ubicar un taxon en el intervalo temporal anterior porque hay depositos en la region que carecen de tales evidencias y son del Holoceno tardio por fechados radiocarbonicos [No obstante
327 EXTlNCIONES DEL CUATERNAR10
la ausencia de una especie en residuarios arqueol6gicos adquiere mayor certidumbre como criterio de ubicaci6n temporal de su extinci6n en la medida que aumenta el numero de residuarios estudiados que en las Antillas Mayores se cuentan por centenares] A falta de fechado absoluto de una especie consideramos que estos criterios de ubicaci6n temporal son mas utiles que la alternativa de excluir del analisis a la especie
2 En los trabajos sobre biogeografia de mamiferos antillanos los arque610gos mas citados como autoridades para establecer el inicio de la presencia aborigen en las Antillas Mayores (Kozlowski 1974 Rouse y Allaire 1978 Rouse 1989 1992 Wilson 2001) difieren al respecto entre los 6 kaAP y los 45 kaAP (pero vease Pagan et al 200523) y hemos adoptado la mas temprana de estas fechas para el comienzo del intervalo amerindio en la Tabla 45 De todos modos cualquiera que haya sido la fecha esta no es aplicable por igual al conjunto de las Antillas Mayores porque segun algunos de los autores citados el poblamiento comenz6 por Cuba y La Espanola 6 mil anos atras continu6 en Puerto Rico hace alrededor de 3 mil anos (pero vease Burney et al 1994) yen Jamaica hace menos de 2 mil e incluso a las Islas Caiman ni siquiera llegaron [Las Islas Caiman descubiertas por Col6n en 1503 carecen absolutamente de evidencia amerindia (Stokes y Keegan 1996)]
Si los datos de la Tabla 45 representan una aceptable aproximaci6n a la realidad el proceso de las extinciones cuaternarias de los mamiferos terrestres de las Antillas Mayores y grupos de islas perifericas debi6 caracterizarse entre otros por los eventos siguientes
Aunque obviamente la fauna cuaternaria debi6 ser mucho mas numerosa que 10 que refleja el registro paleontol6gico (Morgan y Woods 1986) 52 especies (81 de la fauna conocida) desaparecieron (sin reposici6n) durante los ultimos 250 mil anos
- Al parecer las extinciones se concentraron en las especies de mayor tamano Asi el peso promedio estimado para las especies de la etapa preantr6pica (intervalos I + II de la Tabla 45) es 350 kg es 328 kg para las especies del intervalo III (amerindio) y es s610 25 kg (c1ases del peso a b c) para las especies del intervalo IV (postcolombino) Ninguna especie aut6ctona de mamifero mayor de 6 kg existe hoy en la regi6n
- De acuerdo con la distribuci6n temporal de las extinciones 61 especies sobrevivieron al interglacial Sangamon (intervalo 1)48 sobrevivieron a la transici6n del Pleistoceno terminal al Holoceno temprano (intervalo II) 34 sobrevivieron al periodo amerindio (intervalo III) y s610 12 especies han sobrevivido desde la Ilegada de los europeos (intervalo IV) Las tasas de extinci6n correspondientes (exceptuado el intervalo I) son una especie cada 1167 anos en el intervalo II una cada 393 anos en el intervalo III y una cada 23 anos en el intervalo IV
- Durante todo el proceso de las extinciones cuaternarias la perdida de especies por isla fue de 91 en La Espanola 83 en Jamaica y 62 en Cuba Bahamas retuvo su unica especie (aunque extirpada en varias de las islas) Puerto Rico e Islas Caiman perdieron todas sus especies (6 y 5 respectivamente) y las Islas Swan Virgenes y Anguila - San Martin perdieron sus respectivas especies (micas
328 BIOGEOGRAFiA
De los cuatro 6rdenes de mamiferos representados en la fauna Pilosa y Primates desaparecieron totalmente Soricomorpha y Rodentia perdieron 86 y 74 de sus especies respectivamente
CAUSALIDAD
CAUSAS NATURALES
Los cicIos glaciales e interglaciales acaecidos en el Pleistoceno causaron cambios ambientales dnisticos en las Antillas con la concomitante alternancia de cIimas frios (secos) y calidos (hUmedos) asi como de descensos y ascensos del nivel del mar aumento y disminuci6n de las areas territoriales) induso (aunque con menor intensidad) en el Holoceno Estos eventos paleogeograficos han sido consistentemente documentados (Mayo y Kartashov 1972 Bonnatti y Gartner 1973 Ortega y Arcia 1982 Prance 1982 Schubert y Medina 1982 Dodge et al 1983 Ortega [1984] Haq et al 1987 Schubert 1989 Hodell et al 1991 Guilderson et al 1994 Oppo 1997 Emslie 1998 Curtis et al 2001 Iturralde-Vinent 2003) y de ellos brindamos a continuaci6n una breve resena
Finalizando el periodo interglacial anterior a la ultima glaciaci6n (Wisconsin) el mar estaba a 8-10 m por encima de su nivel actual Cuando el avance glacial del Wisconsin a1canz6 su culminaci6n (eillamado Ultimo Maximo Glacial) hace 18 ka el mar habia descendido 120 m con respecto a dicho nivel y las islas del Caribe habian incrementado considerablemente sus extensiones subaereas en comparaci6n con las que presentan hoy dia por ejemplo Puerto Rico y las Islas Virgenes se conectaron territorialmente y las Bahamas se unieron en cinco gran des islas Prevalecia un regimen frio y seco y condiciones de aridez --debidas a una reducci6n de las precipitaciones (posiblemente a menos de 440 mmlano)- y los ambientes xericos se habian extendido a traves de las islas y bancos emergidos y abundaban los pinares (los habitats mesicos estaban restringidos a parcelas relictas en 10 alto de las montanas) En el Mar Caribe y el Golfo de Mexico la temperatura superficial del mar era 5degC mas baja que la actual y en las llanuras costeras y alturas maximas eran respectivamente 2-3degC y 6degC mas bajas Los limites de las zonas ecol6gicas altitudinales descendieron varios cientos de metros y cada zona redujo su extensi6n vertical al aumentar el gradiente de temperatura Ellimite inferior de la zona periglacial
elevaci6n donde la temperatura media minima a1canza los OdegC) de la Cordillera Central de la Republica Dominicana (altura maxima del Caribe insular con evidencia de topografia glacial) se considera que estuvo entre los 1500 y 1800 mlnm (actualmente esta a 2800 mlnm) y se puede estimar que dicho limite era inferior en Cuba por estar mas al norte que la Republica Dominicana No obstante durante este periodo la temperatura experiment6 episodios cortos (unas pocas decadas) de ascensos repetidos
Un signifieativo componente bi6tieo de las referidas condiciones paleoambientales 10 constituy6 la espectacular irradiaci6n evolucionaria experimentada por un grupo numeroso de aves rapaees diurnas y nocturnas (Wetmore 1937 Brodkorb 1969 Kurotchkin y Mayo 1973 Arredondo 1976 Olson 1978 Olson y Hilgartner 1982
329 EXTINCIONES DEL ClJATERNARIO
Suarez 2000a b 2004 Suarez y Emslie 2003) Los mamiferos de las Antillas Mayores ofrecieron a estos depredadores aviares una amplia gama de tamanos de presa que en ausencia de mamiferos del orden Carnivora en la region permitio a estas aves irradiarse a este nicho alimentario con la evolucion de una variedad de especies de muy diversa tall a corporal incluidas formas gigantes Adquirir un cuerpo gigante les propicio el acceso a recursos alimentarios que no estaban siendo explotados por ningun otro depredador (Alcover y McMinn 1994) A Cuba corresponde el mejor registro fosil de rapaces cuaternarias caribenas con siete generos de especies diurnas Dentro de estas aves existieron tres gavilanes gigantes (Accipitridae) tres carairas (Falconidae) y un condor (Cathartidae) asi como cuatro generos de especies nocturnas dos lechuzas (Tytonidae) y tres Mhos (Strigidae) dos de ell os no igualados en tamano en todo el mundo Algunos de estos generos tuvieron representacion en otras islas antillanas como La Espanola Jamaica Bahamas y Gran Caiman
Al retirarse hacia el norte los hielos de la glaciacion Wisconsin el clima se tomo mas hUmedo en las Antillas durante el Holoceno temprano y el bosque mesico se expandio en la region insular El nivel del mar ascendio acompanado de mayor pluviosidad alrededor de los 7 kaAP las Islas Virgenes quedaron separadas entre si y de Puerto Rico y las Bahamas perdieron la mayor parte de su territorio Cerca de los 3 kaAP las condiciones secas retornaron a la region e hicieron desaparecer buena parte del bosque mesico Nuevamente el clima comenzo a adquirir sus condiciones actuales hacia los 25 kaAP aproximadamente y hoy transita por una fase mesica con vegetacion xerica confinada en parcelas relictas
Sin duda estos y otros eventos naturales incidieron negativamente en la sobrevivencia de mamiferos terrestres en las Antillas Mayores asi como en los grupos de pequenas islas perifericas (Pregill y Olson 1981 Olson 2006)
McFarlane et al (1998a) demostraron convincentemente como Amblyrhiza el gigante de los roedores antillanos endemico de la pequena isla de Anguila sucumbio a las repetidas y abruptas perdidas de su habitat durante el interglacial Sangamon por las nipidas y severas oscilaciones del nivel del mar (documentadas por Neumann y Hearty 1996) al no poder responder adecuadamente a tan catastroficos eventos mediante el ajuste de su masa corporal media y otro tanto debio ocurrirle en Jamaica al otro roedor gigante Clidomys (McFarlane et al 1998b) emparentado con Amblyrhiza en la misma epoca (Tabla 45 intervalo I)
El ascenso del nivel del mar seguramente promovio la extincion de mamiferos durante la transicion del Pleistoceno al Holoceno (Tabla 45 intervalo II) con la reduccion del tamano de las Islas por las transgresiones marinas (Pregill y Olson 1981 Morgan 1993) Esto ultimo haria maximamente severa la competicion asf como la presion de los depredadores [Steadman et al (2005) no yen por que el calentamiento global ocurrido durante la transicion Pleistoceno-Holoceno habria de tener un efecto negativo sobre los perezosos siendo sus descendientes actuales especies tropic ales 0 subtropicales pero no se trata del efecto
330 BIOGEOGRAFiA
directo del cambio de temperatura sobre los animales sino del efecto indirecto y progresivo que dicho cambio genera a largo plazo al reducir el territorio insular disponible]
Por esta epoca tambien desaparecieron especies de tall a mayor como Megalocnus zile en La Espanola y otras de menor tamano (en Cuba) pero que representan la tall a maxima en sus respectivos contextos taxonomicos del Cuaternario como Solenodon arredondoi Paralouatta varonai y Macrocapromys acevedo Un mamifero puede ser grande solo a costa de reducir su densidad poblacional y esto 10 hace mis vulnerable a la extinci6n sobre todo en una isla (McNab 1994 McFarlane et al 1998a) Incidentalmente Alcover et al (1998a) afirmaron que larelacion entre la talla corporal y la extincion no siempre se cumple en las especies de menor tamano y citaron como ejemplo que entre las musarafias todas las especies de Nesophontes desaparecieron mientras sobreviven especies de Solenodon pero la comparacion correcta en todo caso serfa entre los solenodones vivientes y el gigante Solenodon arredondoi y entre Nesophontes edithae -el gigante de los nesofontes extinguido en el periodo amerindio- y el resto de sus congeneres que sobrevivieron hasta los primeros siglos de la colonizacion europea (Tabla 45 intervalos III y IV)
CAUSAS ANTROPOGENICAS
Intervalo Amerindio
La mayoria de los autores que se han referido a las extinciones acaecidas durante este intervalo (entre elIos paradigmaticamente Martin 1984a b) han responsabilizado a los aborfgenes (que se supone comenzaron a poblar las Antillas Mayores hace 6 mil afios) con la desaparicion de numerosas especies de mamiferos terrestres en la region principalmente por la via de la sobrecaza y la introduccion de animales domesticos devenidos depredadores de la fauna autoctona (Morgan y Woods 1986) Este efecto negativo de la presencia indigena sobre los mamiferos de la region ha sido dramatizado por algunos autores en terminos de holocausto faunal Sin embargo pensamos que se ha exagerado Como han expresado MacPhee et al (19993) cuando uno busca evidencia dura de complicidad humana directa en [estas] extinciones no hay mucho que encontrar Las siguientes son algunas consideraciones a tener en cuenta en tal sentido
El registro radiometrico mas joven conocido hasta ahora para un perezoso antillano (MegalocnllS rodens) es de 42 kaAP (MacPhee et ai 2007) Si se acepta que los aborfgenes entraron en Cuba hace aproximadamente 6 mil anos su convivencia con los perezosos debe haber durado alrededor de 1800 afios (0 mas porque dificilmente el referfdo fechado radiometrico haya correspondido al ultimo sobreviviente de la especie) Esta cifra contradice la hipotesis del holocausto faunal segun la cualla extincion de los mamiferos insulares de mayor talla debe ocurrir con extrema rapidez -a escala de decadas 0 de centurias- a partir de su primer contacto con los humanos (Martin y Steadman 1999) A pesar de las repetidas afirmaciones en contrario (Harrington 1921 Miller 1929c Herrera [1945] Tabio 1951 Alvarez 1951a 1957 Arredondo
331 EXTINCIONES DEL CUATERNARIO
1959 1960a Veloz y Ortega 1973 Mayo 1980b Rodriguez et al 1984 Castellanos y Pino 1985 Pino y Castellanos 1985 Nunez 1990b Gutierrez 1996 Arredondo 2004) en su mayoria basadas en evidencias no exentas de posibles accidentes de deposicion (0 de reubicacion postdeposicional) en recintos cavernarios (Diaz-Franco [2006b D no se ha demostrado inequivocamente que los aborigenes antillanos se alimentaran de perezosos regularmente Sus restos en residuarios de alimentos son escasisimos en toda la region y esto contrasta con los abundantes restos de jutias y otros mamiferos de menor talla que aparecen en esos residuarios demostrativos de la preferencia de los paleoindios por estos otros mamiferos
Los mamiferos mas importantes en la economia prehistorica de las Antillas Mayores fueron las jutias Capromys y Geocapromys en Cuba Geocapromys en Jamaica e Isolobodon en La Espanola Puerto Rico e Islas Virgenes (Aguayo 1950b Tablo 1951 Rimoli 1972 Wing 2001 Diaz-Franco [2006b D Wilkins (2001) estudi61ajutia viviente de Jamaica (Geocapromys browni) en un sitio ocupado por los aborigenes durante 100 anos y no encontr6 evidencias de que durante ese tiempo la especie haya sido sobreexplotada Asimismo en Cuba las jutias conga carabali y andaraz son escasas en depositos pleistoholocenicos en cambio son muy numerosas en residuarios indigenas y de haber sido sobreexplotadas 10 mas probable es que no hubieran sobrevivido hasta hoy
La habilidad de los humanos para penetrar en profundidad el bosque tropical y mantenerse en este sustentablemente ha sido un evento comparativamente reciente (Beran y Steele 1994) En el caso antillano la caza tierra adentro ha sido explicada como un proceso tardio de desplazamiento a partir de la explotacion de abundantes recursos marinos que ofrecian condiciones 6ptimas para el sedentarismo de las comunidades aborigenes costeras (C6rdova y Arredondo 1988 Brito 2005) Peces y mariscos dominan la arqueologia costera de las Islas antillanas en tanto el consumo de recursos terrestres (mamfferos incluidos) aumenta progresivamente en la medida que los residuarios estudiados se alejan de la costa (Scudder 1991 Wilkins 2001) En las Antillas Mayores el componente terrestre de la dieta aborigen es solo 20-30 del material recobrado en los residuarios (Wing 1977 1989) Y debio ser menor alin porque el componente vegetal de la dieta no se ha preservado Por otra parte los aborigenes cubanos domesticaron la jutia conga y seguramente la reprodujeron para comer (Pose et al 1988) practica que probablemente se repitiera con otras jutias en las demas Antillas Mayores
- Fray Bartolome de las Casas hablo de 3 millones de indios en Cuba al inicio de la colonizacion pero los historiadores subsecuentes consideraron exagerada la cifra (Poey 1851 a) No obstante se dispone de dos estimados de la densidad de poblaci6n aborigen en las Antillas Mayores a la llegada de los europeos 18 indigenaslkm2 en Puerto Rico (Rouse 1956) y 1271km2 en La Espanola (Watts 1987) Extrapolando ambos valores entre las cuatro islas principales de las Antillas Mayores tenemos que en la fecha indicada la poblaci6n aborigen pudo estar entre 16 mil y 113 mil indigenas en Puerto Rico entre 21 mil y 145 mil en Jamaica entre 141 mil y 1 mill6n en La Espanola y entre 200 mil y 14 millones en Cuba Entre las cifras maximas y las minimas nos inclinamos
332 BIOGEOGRAFiA
hacia estas ultimas atendiendo a los altos indices de mortalidad infantil y morbilidad que se evidencian en las osamentas de los pobladores prehispanicos (Martinez y Rives 1990) Uno de los hechos que apuntan hacia el bajo numero de aborigenes en las Antillas Mayores es 10 rapido que desaparecieron despues de la llegada de los europeos pnkticamente sin dejar mestizaje con estos 10 cual contrasta ostensiblemente con la situacion en la America continentaL
En suma un grupo humano (1) demograiicamente irrelevante y controlado por altos indices de morbilidad (2) consumidor mayoritariamente de organismos marinos y (3) carente de la tecnologia cinegetica necesaria para poder sobreexplotar los mamiferos que Ie sirvieron de alimento (a los cuales accedio tardiamente en su desarrollo) no es concebible que haya causado e1 holocausto faunal que se Ie atribuye a partir del error de aplicar a las comunidades aborigenes antillanas un modelo cultural de la prehistoria continentaL
Tampoco hay evidencias de que los aborigenes hayan introducido carnivoros con suficiente capacidad depredadora salvo el perro Morgan y Otlenwaldder (1993) consideraron que la depredacion por perros durante el periodo amerindio es la explicacion mas plausible para la extincion del almiqui gigante de Cuba pero como se puede observar (Tabla 54) Solenodon arredondoi aparece en el intervalo II por falta de asociacion con evidencias culturales
Consecuentemente el accionar de los aborigenes y de sus animales domesticos directamente sobre la fauna no parece haber sido por sf solo un factor determinante de la extincion de mamiferos terrestres durante el periodo amerindio en las Antillas (pero vease Steadman et al 2005)
Hay que recordar en este senti do que alin en el Holoceno tardio ocurrieron cambios clirruiticos de consideracion (vease Causas Naturales) Por ejemplo Curtis et al (2001) explicaron que la extincion de algunos ani males mesofilicos de las alturas montanosas de La Espanola pudo haber antecedido a cualquier impacto humano sigrIificativo en esos lugares por coincidir dicha extincion con una tendencia a la aridez documentada isotopicamente y con evidencias de la perdida del bosque mesico ocurridas despues de 3kaAP Yen Jamaica hace solo 2shy3 mil aDos el area de la Bahia de Jackson sustentaba una gran biomasa de vertebrados en bosque hinnedo a juzgar por la cantidad y diversidad de taxones presentes en material fosil derivado de la depredacion por lechuzas hoy el area es la mas seca de la isla con matorral xerofitico y pobreza de fauna (MacPhee 1996a) Yalm en nuestros dias huracanes de gran intensidad aniquilan poblaciones de vertebrados (que no se recuperan) en is las pequenas y llanas del Caribe (Brooks y Smith 2001)
En medio de la incertidumbre sobre la verdadera causa (0 causas) de las extinciones de mamiferos en el periodo amerindio MacPhee y Marx (1997) han propuesto su hipotesis de la hipermorbilidad la cual estipula colapsos poblacionales relativamente rapidos con la introduccion de enfermedades de alta letalidad Estos autores han razonado que
Las islas son los lugares mas apropiados para experimentar perdidas catastroficas causadas por enfermedades Los mamiferos no solo serian susceptibles en virtud de su prolongado aislamiento biologico sino que en los confines de una isla la epizootia
333 EXTINCIONES DEL CUATERNARIO
tendria la potencialidad de infectar simultaneamente a la rnayoria de los individuos de vadas especies Los neonatos son inmunologicamente inmaduros y sufririan desproporcionadamente cualquier infeccion hipervirulenta La perdida excesiva de neonatos y juveniles tendria efectos severos para la sobrevivencia de la poblacion si esta se reduce por debajo de cierto nivel sobreviene la extincion porque no hay posibilidad de restitucion en el periodo postpanzootico Las especies de mayor talla corporal van a ser las mas vulnerables a la extincion dadas sus bajas tasas reproductivas largos periodos de gestacion y reducido numero de progenie Las especies de menor tamano --caracterizadas por altas tasas reproductivas periodos cortos de gestacion y mayor numero de neonatos por parto- tambien serian fuertemente diezmadas pero muchas sobrevivirian porque su estrategia reproductiva les permite perder mayor numero de individuos Las perdidas deben ocurrir relativamente rapido (a escala de decadas) mientras que otras hipotesis (digamos los cambios climaticos) permiten intervalos mucho mas largos (a escala de siglos 0 de milenios) entre la aparicion de las causas y la produccion de sus efectos Por ejemplo todo indica que los nesofontes se extinguieron muy rapidamente y seria dificil poder invocar un impacto humano directo 0 argumentar fenomenos climaticos que pudieran explicar plausiblemente perdidas tan rapidas
MacPhee y Marx (1997 191) han dicho que por el momento la hipotesis de la hipermorbilidad no pasa de ser una abstraccion en espera de adecuada sustentacion empirica Llama la atencion que linicamente en el periodo amerindio (Tabla 45 intervalo 1Il) las extinciones abarcaron todas las clases del peso corporal (de la a a laj) y que ninguna de las especies que sobrevivieron hasta el periodo postcolombino (Tabla 45 intervalo IV mas las especies vivientes) sobrepasa los 10 kg (clases del peso a b c) en clara coincidencia con los presupuestos de la hipotetica hipermorbilidad
Intervalo Postcolornbino
Por 10 menos 37 especies de mamiferos terrestres vivian en las Antillas Mayores y grupos de Islas perifericas a la lIegada de los europeos en 1492 De elIas se han extinguido 23 shymas especies que en cualquiera de los periodos precedentes (Tabla 45~ y solo subsisten 12 especies repartidas entre Cuba (8 especies) La Espanola (2) Jamaica (1) y Bahamas (1) varias de estas en peligro critico de extincion Solo dos de las especies extinguidas durante los ultimos 500 anos se conocieron en estado viviente los roedores Oryzomys antillarum de Jamaica (extinguido en 1877) y Geocapromys thoracatus de las Islas Swan (extinguido en 1955) Las causas de las extinciones ocurridas durante este periodo son sobradamente conocidas destruccion de habitats introducci6n de animales exoticos (domesticos y comensales) y demas presiones asociadas a la actividad humana entre las cuales posiblemente habria que incluir brotes de hipermorbilidad
Durante mucho tiempo se ha afirmado la posibilidad de que especies de Nesophontes hayan sobrevivido hasta las primeras decadas del siglo xx y que incluso continuen existiendo en lugares remotos de Cuba 0 La Espanola (Allen 1918 1942 Miller 1930 Aguayo 1950a Arredondo 1960b Fischer 1977 MacFadden 1980 Woods y
334 BIOGEOGRAFiA
Ottenwalder 1991) Actualmente no existe evidencia radiometrica de que alguna especie de Nesophontes haya sobrevivido al siglo xv las fechas radiocarbonicas calibradas obtenidas en muestras de Nesophontes de Cuba y La Espanola resultaron no mas tardias que los siglos XIII y XIV (MacPhee et al 1999) Por otra parte las expediciones efectuadas a fines del siglo xx con el proposito de localizar individuos vivientes del genero en La Espanola no produjeron la menor indicacion afirmativa al respecto (Woods et al 1985) Ademas a traves de las Antillas Mayores no aparecen restos de Nesophontes en los depositos formados actualmente por las lechuzas
EPiLOGO
Las Antillas Mayores son islas oceanicas no han formado parte de un continente (con la posible excepcion de una parte de Jamaica hasta el Eoceno Medio hace 45 Ma) y durante un tiempo geologico considerable (desde el Eoceno Superior hace 40 Ma) han permanecido emergidas y separadas de los continentes vecinos Consecuentemente los mamiferos terrestres (no volantes) autoctonos de estas islas constituyen una fauna oceanica que por ende cumple con todas las caracteristicas convencionales reconocidas para este tipo de entidad biogeografica
- Baja diversidad taxonomica a nivel de las categorias superiores (solo cuatro ordenes) en comparacion con islas plataformicas continentales (0 con areas continentales) de tamano y diversidad ecologica simi lares
- Alto grado de endemismo a nivel de genero (96) y familia (50) en respuesta al prolongado aislamiento
- Desbalance ecologico de la estructura trofica ausencia de mamiferos camivoros y ungulados
- Espectaculares procesos de irradiaci6n evolucionaria como resultado de la colonizacion de territorios libres de competidores y depredadores
Tendencias evolucionarias extremas gigantismo
- Control a cargo de aves rapaces con tendencia at gigantismo en ausencia de mamiferos miembros del orden Carnivora
- Mas especies de reptiles y de murcielagos que de mamiferos terrestres relaciones que en los continentes se invierten
Alta tasa de extincion de especies (84) sin reposicion
Preservacion de especies filogeneticamente arcaicas (los llamados fosiles vivientes) los almiquies

313 POBLAMIENTO ISLA-ISLA
de solo siete propagulos fundadores perezosos 2 (coloepodinos y megalocninos si como parece tienen un origen difiletico) musaraiias 2 (nesofontidos y solenodontidos si no constituyen una misma familia como consideran algunos) monos 1 (pitecidos) roedores histricognatos 1 (equimidos capromidos y heptaxodontidos) roedores muridos 1 (Oryzomys en Jamaica)
POBLAMIENTO ISLA-ISLA
La distribucion de los mamiferos terrestres a traves del area antillana demarcada al inicio del presente capitulo (vease Contexto Geogratico Fig 41) ha sido interpretada en funcion de dos hipoteticos mecanismos de poblamiento interinsular la dispersion y la vicarianza fundamentadas como se resumen seguidamente
DISPERSION
De acuerdo con la evidencia paleogeogratica mas actualizada en las principales regresiones marinas ocurridas en la cuenca caribeiia durante los ultimos 23 Ma (transiciones OligocenoshyMioceno Mioceno Superior - Plioceno y varias veces en el Pleistoceno) las masas terrestres constitutivas de la region insular en formacion aumentaron considerablemente su extension territorial disminuyendo por tanto la distancia entre elias oportunidades que los elementos faunales aprovecharian para dispersarse de unas islas a otras
Durante mucho tiempo esta ha sido la hipotesis mas favorecida por los estudiosos del tema a partir de Simpson (1956) pero en decadas recientes ha suscitado dos objeciones
1 Es dificil en tender como pudo operar la dispersion marina entre estas islas puesto que ninguna de las Antillas Mayores posee rios suficientemente caudalosos para transportar masas significativas de vegetacion flotante (MacPhee 198432)
Es oportuno recordar aqui el islote ala deriva (con mas de diez palmas reales en pie) entre las tres Antillas Mayores mas occidentales mencionado anteriormente (vease La Balsa Natural)
2 La dispersion marina es menos efectiva para explicar la asombrosa similitud de las faunas de mamiferos terrestres a traves de las Antillas Mayores septentrionales porque como proceso aleatorio no debe producir distribuciones uniformes a 10 largo de una cadena de islas 0 si las produce entonces el proceso no es aleatorio (MacPhee e lturralde-Vinent 2000 149)
Independientemente de que las aludidas distribuciones taxonomicas estan lejos de ser uniformes 0 asombrosamente similares la hipotesis de la dispersion marina contrariamente a su canicter aleatorio y altamente improbable en el caso del poblamiento continente-isla- en el caso del poblamiento isla-isla es un proceso no aleatorio (aprovecha las oportunidades brindadas puntualmente por los periodos de maxima regresion marina) y ademas altamente probable dada la incomparablemente menor distancia entre las islas caribeiias que entre estas y los continentes vecinos
314 BIOGEOGRAFiA
V ICARIANZA
La neotectonica y las transgresiones marinas del Oligoceno Medio y del Mioceno afectaron sustancialmente la paleogeografia de la Cresta de las Antillas Mayores resultando la subdivision gradual de las partes subaereas de la Cresta Asi el este de Cuba y el norte de La Espanola conectados fisicamente durante el Oligoceno Inferior se separaron al formarse el Paso de los Vientos en el Oligoceno tardio en tanto La Espanola y Puerto Rico probablemente permanecieron conectadas hasta el Mioceno tardio en que surgio el Paso de Mona (Iturralde-Vinent y MacPhee 1999)
Tales eventos geologicos habrian dividido las distribuciones de los grupos faunales emplazados previamente en el territorio continuo de las tres islas y a partir de esas disyunciones dichos autores (tambien MacPhee et al 2000 y White y MacPhee 2001) asumieron --como segunda fase del modelo de GAARlandia- la hipotesis de la vicarianza isla-isla para explicar la distribucion multiinsular de varios grupos de mamiferos terrestres a traves de las Antillas Mayores septentrionales De acuerdo con esta hipotesis cualquier diferencia en composicion faunal entre las islas (por ejemplo la ausencia de roedores heptaxodontidos en Cuba 0 de monos platirrinos en Puerto Rico) constituye evidencia de extincion local postvicariante y no de dispersion marina fallida Lo primero se puede demostrar si aparecen los fosiles pertinentes 10 segundo no se puede demostrar (MacPhee e Iturralde-Vinent 1994) Por otra parte IturraldeshyVinent y MacPhee (1999 57) han reconocido que aunque fa vicarianza isla-isla puede representar una solucion clara para las distribuciones de varios taxones cuaternarios no puede explicar todos los casos
Rega et al (2002 18) han presentado dos objeciones al modelo de la vicarianza isla-isla aplicado a los mamiferos de las Antillas Mayores
I Si la separacion de Cuba y La Espanola precedio en varios mill ones de anos la separashycion de La Espanola y Puerto Rico el modelo vicariante de lturralde-MacPhee debe predecir mayor similitud entre las faunas de La Espanola y Puerto Rico que entre las de Cuba y La Espanola De hecho es al reyes
No obstante MacPhee et al (200335) admitieron despues que la fecha mas temprana empiricamente defendible [para la apertura del Paso de Mona] debe ser el Oligoceno Medio esto invierte la secuencia de ambas disyunciones con 10 cual queda resuelta la aparente contradiccion Sin embargo en su reciente evaluacion de la congruencia entre filogenia e hipotesis geologic as en varios grupos de mamiferos terrestres de las Antillas Mayores Davalos (2004373) encontro que ninguno de los taxones cumple con la fecha y el patron de divergencia entre las masas terrestres antillanas postulados por el modelo de GAARlandia
2 La presencia de un mismo genero en las tres Islas septentrionales de las Antillas Mayores implica [por el modelo vicariante] que la diferenciacion entre estas formas durante el intervalo desde el Mioceno ha sido unicamente de nivel especifico [Dicha presencia]
315 rRRADlAcr6~ EVOLUCrONARIA
s610 puede explicarse razonablemente como el resultado de movimientos de esos animales entre las islas en el Cuaternario
De esta objecion resulta dificil disentir [El poblamiento entre islas diferenciado solo a nivel especifico no es probable que sea anterior al Pleistoceno (Simpson 1956 15)]
Debe tenerse en cuenta que la hipotesis de la vicarianza isla-isla alude a las tres islas septentrionales de las Antillas Mayores porque como se explico antes (vease El Barco Funeral Vikingo) Jamaica no existia como isla durante el lapso (Eoceno Superior shyOligoceno) consensualmente reconocido como el mas probable para la colonizacion de las proto-Antill as Mayores por la mayo ria de los mamiferos terrestres Despues de emerger como isla en el Mioceno Medio Jamaica debe haber recibido sus mamiferos terrestres por dispersion isla-isla (Davalos 2004376) y solo en el Cuaternario parece haberrecibido (por dispersion marina desde Centroamerica) al roedor murido Oryzomys
Seguramente tanto la dispersion como la vicarianza contribuyeron a que los mamiferos terrestres se irradiaran a traves de las Antillas Mayores y grupos de islas perifericas pero las maneras y tiempos en que estos mecanismos operaron a 10 largo de dicho proceso son aun en gran medida motivos de desacuerdo e incertidumbre
IRRADIACION EVOLUCIONARIA
A continuaci6n revisamos someramente las distribuciones conocidas de los diferentes grupos de mamiferos terrestres a traves de las Islas principales de las Antillas Mayores y grupos de Islas perifericas incluidas las interpretaciones y conjeturas que la literatura reciente recoge en relaci6n con la manera en que dichas distribuciones pudieron haberse conformado durante la irradiaci6n evolucionaria de los referidos grupos de mamiferos Manifestaciones de esta irradiaci6n pueden inferirse unicamente a partir de los taxones presentes en el Cuatemario pues solo 6 de las especies conocidas corresponden al Terciario
GRUPOS ORlGINARIOS DE SURAMERICA
PEREZOSOS (Megalonychidae)
Aparte de un genero miocenico de Cuba y otro del Oligoceno de Puerto Rico se conocen 4 generos (12 especies) de perezosos cuaternarios (todos extintos) restringidos a las tres islas septentrionales de las Antillas Mayores Acratocnus Neocnus Megalocnus y Parocnus Los cuatro generos estan representados en Cuba y La Espanola y Acratocnus 10 esta ademas (como unico genero de perezoso) en Puerto Rico Esta distribuci6n ha sido interpretada por MacPhee et al (2000) eomo resultado de la vicarianza oligomiocenica entre estas islas y [dado que el Banco Puerto Rico - Islas Virgenes fue la entrada de GAARlandia a las Antillas Mayores] se preguntaron si la ausencia de los otros tres generos en Puerto Rico se debe a que nunca estuvieron en ese territorio 0 a que sus
316 BIOGEOGRAFiA
fosiles atm no han sido hallados Pero como hemos visto Rega et al (2002) atribuyeron dicha distribucion a la dispersion interinsular en el Cuatemario
Por otra parte varios amilisis c1adisticos recientes ubi can a Choloepus -genero Reciente conocido solo de Centro- y Suramerica- como taxon hermano de Aeratoenus exc1uidos los demas megaloniquidos antillanos Para White y MacPhee (2001228)
tan estrecha relacion sugiere que la divergencia entre ambos [generos] ocurrio despues de que Aeratoenus divergio de Neoenus Si esta relacion implica que Aeratoenus y Neoenus se dispersaron separadamente desde Suramerica a las AntilIas mientras Choloepus permanecia en el continente (siendo necesarias al menos tres invasiones de perezosos) 0 si Choloepus invadio Suramerica desde las Antillas es algo que el material disponible no permite determinar
La diversidad adaptativa de los perezosos se manifiesta en (1) la utilizacion del habitat especies arboreas en Neoenus y Aeratoenus y terrestres en Paroenus y Megaloenus (Webb 1985a White [1993] White et al 1996 Arredondo [1999]) y en (2) la gama de tallas corporales desde los pequenos Neoenus hasta Megaloenus rodens el mayor de los mamiferos terrestres antillanos (su peso adulto estimado en 270 kg seglin Paula 19679) esto sugiere que los perezosos se irradiaron a nichos que en contextos continentales probablemente estarian ocupados por miembros de otros c1ados (MacPhee e IturraldeshyVinent 2000)
No esta claro si el abanico de tamanos entre estos perezosos enmascara alguna tendencia al gigantismo Por 10 pronto Anderson y Handley (2002) encontraron 10 contrario en perezosos (Bradypus) de las Islas de Bocas del Toro (Panama) shyseparadas secuencialmente del area continental vecina por ascensos del nivel del mar durante los ultimos 10 ka En las cinco islas los perezosos son menores que su contraparte continental y las diferencias entre las tallas medias de las cinco poblaciones expresan una reduccion lineal significativa de la talla corporal en funcion de la edad geologica de cada isla sin relacion alguna con el area de las islas 0 la distancia que las separa del continente
MONOS (Pitheciidae)
Los primates de las Antillas Mayores (cuatro especies extintas) constituyen un grupo monofiletico con Paralouatta (en Cuba) y Antillothrix (en La Espanola) como taxones hermanos seguidos en la escalera c1adistica por Xenothrix (en Jamaica) y finalmente Callieebus como el pariente continental mas cercano esto implica una sola invasion desde Suramerica antes del Mioceno Inferior (fecha mas temprana conocida para los primates fosiles antillanos) y por tanto en consonancia con GAARlandia como la via de colonizacion (Horovitz y MacPhee 1996 1999 Horovitz [1997] MacPhee y Horovitz 2004)
En desacuerdo parcial con la anterior interpretacion la filogenia de este grupo de monos obtenida por Davalos (2004) indica un antecesor comun en Cuba La Espanola y Jamaica
317 TRRADIACTON EVOLUCIONARIA
compatible con la hipotetica conexion -propuesta tentativamente por Iturralde-Vinent y MacPhee (199980-84)- entre las Montanas Azules del este de Jamaica y la peninsula suroccidental de La Espanola en la epoca de GAARlandia
A pesar de que los primates antillanos pertenecen a un grupo viviente de monos muy pequenos (los monos titles) fueron comparativamente grandes en relacion con los platirrinos modernos y sucesivamente mayores en la direccion Antillothrix Xenothrix y Paralouatta con P marianae como el mayor platirrino conocido del Mioceno neotropical (MacPhee 2005) MacPhee et al (1994) consideraron que Xenothrix debio ser un mono de andar lento y al parecer las especies de Paralouatta abandonaron la arborealidad para adoptar un modo de vida mas terrestre (MacPhee 1996a MacPhee et al 2003 MacPhee y Meldrum 2006)
ROEOORES (Echimyidae Capromyidae Heptaxodontidae)
La edad de Zazamys en Cuba demuestra que la invasion de los roedores histricognatos en las Antillas Mayores comenzo antes del Mioceno Inferior por otra parte los representantes mas antiguos de esa irradiacion evolucionaria se encuentran en La Espanola y Puerto Rico Ambas evidencias son consistentes con la hipotesis de GAARlandia (Woods 1982 1988 Woods et al 2001)
No esta claro si los capromidos evolucionaron a partir de un equimido ancestral 0 de un protocapromido 0 si los equimidos antillanos (I sub familia 3 generos y 7 especies todas extintas) forman parte de la irradiacion de los capromidos (4 subfamilias 9 generos y 27 especies 10 vivientes) Kratochvil et al (1978b) y Rodriguez et al (1979) consideraron que Capromys nan us es un eslab6n entre Echimyidae y Capromyidae pero Mesocapromys nanus esta morfologicamente mucho mas cerca de Hexolobodon y los de mas capromidos que de los equimidos (Woods 1985) Lo que si esta claro segun Woods et al (2001) es que la subfamilia Plagiodontinae endemic a de La Espanola es el linaje basal de los capromidos y que Isolobodon Hexolobodon Capromys Geocapromys Mysateles y Mesocapromys [y supuestamente Macrocapromys] forman parte de la irradiacion temprana de Capromyidae
La subfamilia Capromyinae (5 generos y 17 especies 9 vivientes) parece ser el dado de mas reciente evolucion en la familia con invasion de Cuba desde La Espanola y dispersion desde Cuba (1) por Capromys a las islas Caiman y (2) por Geocapromys a las islas Bahamas Caiman y Jamaica y desde Jamaica a las islas Swan aunque la presencia de Geocapromys en Swan pudiera deberse a introduccion aborigen (Morgan 1985 1988 1989 1993 1994 Woods 1989a 1990) Woods et al (2001) utilizaron datos moleculares para obtener estimados independientes de divergencia entre capromidos segiJn 10 cual Geocapromys divergio cerca de los 17 MaAP Mesocapromys divergio de Capromys hace 15 MaAP en tanto lvlysateles 10 hizo hace 10 MaAP Por su parte las formas de Capromys pilorides y de Mysateles prehensilis en Isla de la Juventud divergieron de sus correspondientes subespecies nominativas en la isla principal a los 24 y 08 MaAP respectivamente
318 BIOGEOGRAFiA
Woods y Howland (I979) describieron adaptaciones morfologicas de la mandibula el hioide y la faringe asociadas a mecanismos de masticacion diferentes en plagiodontinos y caprominos y Varona (1980a) llamo la atencion sobre el gran desarrollo de los molariformes (en relacion con la longitud total del cnineo) que presentan ciertos grupos de jutias fosiles en comparacion con las vivientes 10 que pudiera interpretarse como una adaptacion alimentaria a una vegetacion mas abrasiva por parte de las jutias extinguidas
Segun Woods (1990) la familia Heptaxodontidae (2 subfamilias 5 generos y 6 especies todas extintas) tambien seria parte de la irradiacion de los capromidos con origen en el este de La Espanola 0 en Puerto Rico y subsecuente dispersion al Banco de Anguila - San Martin en las Antillas Menores (pero vease MacPhee y Flemming 2003) Este banco estaria a solo 70 km del Banco Puerto Rico - Islas Virgenes en tiempos de maxima regresion marina y esta proximidad con Puerto Rico -patria de Elasmodontomys obliquus- seguramente tuvo que ver con la presencia en las islas de Anguila y San Martin del heptaxodontino gigante Amblyrhiza relacionado mas estrechamente con los roedores de las Antillas Mayores que con los del resto de las Antillas Menores (Woods 1990 1996) Aunque los roedores suelen aumentar de tamano en las islas (Case 1978) la escala del incremento de la talla en Amblyrhiza (que pudo alcanzar mas de 200 kg de peso adulto) esta fuera de toda proporcion con los patrones conocidos en otros roedores insulares (Biknevicius et 01 1993)
Clidomys (Clidomyinae) de Jamaica difiere marcadamente en su morfologia de los heptaxodontinos (vease ademas MacPhee y Flemming 2003)10 cual dificulta explicar su presencia en esa isla Como Jamaica estuvo bajo el mar hasta el Mioceno Medio el ancestro de Clidomys colonizaria esta isla por dispersion desde el extremo suroccidental de La Espanola despues que los heptaxodontinos se originaron en esta ultima isla (Woods 1989a) Pero Pascual et 01 (I 990) especularon que Clidomys pudiera ser el resultado de una invasion posterior e independiente de los capromidos
GRUPOS ORIGINARIOS DE NORTEAMERICA
El origen de las musaranas antillanas ha resultado ser hasta ahora el mayor enigma biogeografico de la fauna de mamiferos terrestres de la region aunque la mayoria de los autores coinciden en considerarlas originarias de Norteamerica (vease Discus ion y Conclusiones en Poblamiento Continente-Isla) No obstante MacPhee e Iturralde-Vinent (1994) descartaron toda posibilidad de conexion terrestre por el Estrecho de la Florida 0 por el Canal de Yucatan y si el (supuesto) evento vicari ante de Rosen (vease El Arca de Noe) -invocado por MacFadden (1980) para resolver el enigma- fuera minimamente plausible aun asi es dificil creer que las catastrofes que 10 sucedieron (vease Contexto Temporal) hayan barrido con toda esa vieja fauna continental excepto las musaranas circunstancia ya anticipada muy tempranamente por Matthew (l918a)
De otra parte la relaci6n entre Nesophontidae y Solenodontidae permanece irresuelta Si estos linajes no estan estrechamente relacionados entonces es muy posible que
319 IRRADIACION EVOLUCIONARIA
sus distribuciones sean explicadas por mecanismos diferentes (Whidden y Asher 2001)
La descomunal diferencia de talla corporal entre las musaranas pequenas (Nesophontes) y las grandes (Soienodon) seguramente impuso a ambos grupos restricciones distintas relativas a la termorregulacion y las adaptaciones energeticas En Nesophontes la seleccion debio favorecer una alta tasa de metabolismo y crecimiento camadas mas numerosas corta expectativa de vida yalta densidad poblacionaL En Soienodon en cambio la seleccion opero en la direccion de una baja tasa metabolica camadas reducidas crecimiento lento longevidad y baja densidad poblacional (Ottenwalder [1991])
MUSARANAS PEQUENAS (Nesophontidae)
Las musaranas pequenas estan representadas en las Antillas por el genero Nesophontes (y un genero miocenico indeterminado en ambar dominicano) con 6 especies cuaternarias extinguidas distribuidas en Cuba La Espanola Puerto Rico Islas Virgenes e Islas Caiman La mayor de las musaranas pequenas Nesophontes edithae evoluciono en Puerto Rico y su arribo a las Islas Virgenes supone una dispersion sumamente reciente En las Islas Caiman Nesophontes se asemeja mas a la poblacion de Cuba de donde seguramente derivo en el Plio-Pleistoceno (junto con Capromys y Geocapromys) cuando la costa sur de Cuba estuvo mucho mas cerca de las Islas Caiman (lturralde-Vinent 2003fig 4) MacPhee et ai (1994) interpretaron la distribucion de Nesophontes en Cuba y La Espanola como resultado de la vicarianza oligomiocenica ocurrida al formarse el Paso de los Vientos pero la distancia taxonomica entre los lados del Paso es solo de nivel especifico y N micros incluso vivi6 en ambas islas segtln Patterson (1962)
Allen (1918) considero que Nesophontes micrus fue una especie primitiva no modificada para la vida arb6rea 0 fos6rea y por tanto de habitos terrestres Nesophontes edithae de Puerto Rico es mucho mayor y mas robusta que las restantes especies del genero eL indice de robustez de su femur (= anchura minima de la diafisis ongitud total del hueso) muestra que este es tan robusto como el de Solenodon paradoxus un animal dos veces mayor (Morgan et ai 1980) Choate y Birney (1968) postularon un aumento cronoclinal de la taIla corporal en N edithae confirmado mas tarde por McFarlane (1999a) mediante fechados radiocarb6nicos de la secuencia estratignifica en depositos cavernarios de Puerto Rico Solenodon no vivi6 en Puerto Rico y en ausencia de una musarana de mayor tamano Nesophontes alcanzo una talla corporal mayor en esa isla que en La Espanola 0 Cuba (Woods y Eisenberg 1989)
MUSARANAS GRANDES (Solenodontidae)
Los arcaicos almiqufes (genero Soienodon) comprenden cuatro especies una viviente (S cubanus) y otra f6sil (S arredondoi) en Cuba y una viviente (S paradoxus con dos subespecies) y otra f6sil (S marcanoi) en La Espanola Como cabria esperar las dos especies cubanas se asemejan mas entre sf que a las especies de La Espanola pero
320 BIOGEOGRAFiA
enigmaticamente S marcanoi comparte caracteres con S cuban us y S paradoxus (Woods y Eisenberg 1989 Morgan y Ottenwalder 1993 Ottenwalder 200 I) Solenodon arredondoi alcanzo una talla corporal superior a la de cualquiera de las musarafias vivientes a nivel mundial Segun Morgan y Ottenwalder (1993) extrapolando las dimensiones y el peso de los almiquies vivientes S arredondoi debio tener una longitud cabeza-tronco y un peso de 450-550 mm y 1500-2000 g respectivamente en comparacion con S cubanus (360 mm y 800 g) y S paradoxus (390 mm y 1100 g)
Entre todas las especies de almiqufes Solenodon paradoxus es la que presenta los caracteres mas derivados (Otten walder 2001) Si estas relaciones son validas la distribucion de Solenodon entre Cuba y La Espafiola debio discurrir de oeste a este como propuso MacFadden (1980) quien invoco la presencia fosH de S cubanus [pero tambien S arredondoi] en Pinar del Rio para postular que el ancestro continental entr~ por Cuba opinion compartida por Perfit y Williams (1989) y por Ottenwalder ([1991] 2001) El paso de Solenodon a Puerto Rico pudo haberlo impedido la existencia de barreras ecologicas durante la conexion del este de La Espafiola con Puerto Rico 0 por la ausencia de Solenodon en esa parte de La Espafiola en aquella epoca Sin embargo las mismas condiciones ecologicas en el este de La Espafiola no parecen haber representado una barrera efectiva para Nesophontes cuyas mas altas tasa de reproduccion y densidad poblacional presumiblemente Ie permitieron expandirse flpidamente a traves de La Espafiola (Ottenwalder [1991])
Es opinion generalizada que los almiqufes antillanos constituyen el grupo de mamiferos que primero se emplazo en el territorio insular (MacPhee 2005) y en tal sentido es de interes la evidencia sobre especificidad parasito-hospedante aportada por Cruz (1980) quien encontro que la especie de acaro (genero Cubanochirus) que parasita a los almiquies es mucho mas primitiva que la especie (del mismo genero) que paras ita a las jutias y de aqui concluyo que dicho genero de acaro debio evolucionar primero en Solenodontidae y posteriormente en Capromyidae al entrar en contacto representantes de ambas familias en Cuba 0 La Espafiola
Recientemente Roca (2004) y Roca et al (2004) dieron a conocer que los resultados de su estudio genetico-molecular sobre los almiquies indican una divergencia de 25 Ma entre Solenodon cuban us y S paradoxus -compatible con la disyuncion tectonica entre Cuba y La Espafiola (Iturralde-Vinent y MacPhee 1999 McPhee et al 2003)-- y una separacion de ambas especies comparable en magnitud a la divergencia entre familias de mamlferos Consecuentemente propusieron que Solenodon cubanus fuera clasificada en el genero Atopogaie como indico Cabrera (1925) hace 82 afios
DISCUSION Y CONCLUSIONES
Las musarafias y los roedores antillanos (excepto Oryzomys) se irradiaron tanto de sus respectivos ancestros continentales que de estos 6rdenes se reconocen familias y subfamilias endemicas de la region en contraste con los perezosos (una subfamilia y tres
321 IRRADIACION EVOLUCIONARIA
tribus endemicas) y los monos (una tribu endemica) como expresion de diferentes niveles de diversificacion evolucionaria Este proceso es dificil de explicar (traUindose de c1ados que pnicticamente comparten la misma antigiiedad en las Antmas) si es correcta la interpretacion de Pascual et at (1990636)
Nosotros consideramos estos diferentes niveles de diversificacion como las diferentes etapas de evolucion alcanzadas por cada grupo [de mamiferos] Obviamente esto estuvo relacionado con las diferentes longitudes de sus historias
Pudiera ocurrir que en el futuro se compruebe que la historia biogeognifica de la region fue mucho mas complicada
La Espanola la isla mas ecol6gicamente divers a de la region debi6 ser el centro principal de evoluci6n de los mamfferos (con 87 de las familias y 73 de las subfamilias) seguida por Cuba (con 75 de las familias y 54 de las subfamilias) las dos islas de las Antillas Mayores mas relacionadas biogeograficamente (Fig 43 Tabla 44)
eL
EL
cb CAIMAN
SWAN
A Acratocnini B Cubanocnini C Megalocnini D Mesocnini E Nesophontidae F Solenodontidae G Xenothrichini
ANGUILAshyM-0 SMARTIN
H Heteropsomyinae I Plagiodontinae J Isolobodontinae
K Hexolobodontinae L Capromyinae
M Heptaxodontinae N CIidomyinae 0 Sigmodontinae
FIGURA 43 Representacion esquematica de las relaciones interinsulares (expresadas porel numero de linajes compartidos) entre las Antillas Mayores y entre estas y los grupos de islas perifericas segun las interpretaciones de los autores citados en el texto Los numeros dentro de los circulos indican la cantidad de linajes representados en cada isla relacion dudosa Contexto taxon6mico de los linajes en la Tabla 44
bull bull
bull bull
bull bull bull
bull bull
bull bull
bull
bull
bull
bull
bull bull bull bull
bull bull
bull bull
bull
bull bull bull
bull bull
bull
bull bull
bull
322 BIOGEOGRAFiA
TABLA 44 Distribuci6n de los linajcs de mamiferos terrestres a traves de las Antillas Mayores y grupos de islas perifericas LE = La Espanola CU Cuba JA Jamaica PR = Puerto Rico IC Islas Caiman IS Islas Swan IB = Islas Bahamas IV = Islas Virgenes AS = Anguila - San Martin
FamiliaSubfumiliafIribu LE CU JA PR IC IS IB IV AS
Megalonychidae
Choloepodinae
Acratocnini
Cubanocnini
Megalocninae
Megalocnini
Mesocnini
Nesophontidae
Solenodontidae
Pitheciidae
Callicebinae
Xenothrichini
Echimyidae
Heteropsomyinae
Capromyidae
Plagiodontinae
Isolobodontinae
Hexolobodontinae
Capromyinae
H e pta x 0 don tid a e
Heptaxodontinae
Clidomyinae
Muridae
Sigmodontinae
PILOSA
SORICOMORPHA
PRIMATES
RODENTIA
bull
bull
bull
bull TOTALES 12 10 4 4 2
323 EXTlICIOIES DEL CUATERNARIO
El gigantismo ha sido sin duda una tendencia evolucionaria recurrente entre los clados mayores de mamiferos antillanos Es un hecho bien conocido que algunos grupos de mamiferos han aumentado de tamafio en las islas y varios autores han intentado explicarlo a partir de enfoques diferentes (Foster 1964 Miller 1966 Sondaar 1977 Case 1978 Eisenberg y Wilson 1978 Heaney 1978 Lawlor 1982 Lomolino 1985 Angerbjom 1985 McNab 19942001)
EXTINCIONES DEL CUATERNARIO
Las islas que integran la Subregion Antillana han perdido casi 90 de sus mamiferos terrestres durante los ultimos milenios y en proporcion con su extension territorial esta perdida ha sido mayor en la Subregion que en cualquier otro lugar del planeta (MacPhee 1996b) Las extinciones del Cuatemario eliminaron mas de las tres cuartas partes de la fauna de mamiferos terrestres conocida de las Antillas Mayores sus islas satelites y grupos de islas perifericas y el examen de este proceso en sus aspectos mas generales se aborda en esta ultima parte del presente capitulo
CRONOLOGiA Y CONSECUENCIAS
Es muy improbable que un fosil represente el ultimo individuo de un taxon extinto y entonces hay que guiarse por la edad radiometrica del fosil mas reciente conocido lIamada fecha de ultima presencia- como unica aproximacion posible a la fecha de extincion del taxon
Como se explico al inicio del capitulo (vease Contexto Temporal) el registro fosH de los mamiferos terrestres de las Antillas Mayores adolece de insuficiente sustentacion radiometrica No obstante la distribucion temporal de los escasos fechados disponibles para el Cuaternario de la region abarca etapas criticas del proceso de extincion experimentado por la referida fauna a 10 largo de los ultimos 250 mil afios
De las 52 especies que es posible reconocer actualmente como miembros extintos de la fauna cuatemaria de mamiferos terrestres de las Antillas Mayores y grupos de islas perifericas los restos fosiles de 23 de elIas (442) han sido fechados radiometricamente o se han hallado asociados estratigraficamente de manera inequivoca con otro material fechado De estos 23 fechados 3 corresponden al interglacial Sangamon 4 pertenecen a la secuencia Pleistoceno terminal - Holoceno temprano 10 se ubican en ellapso entre el arribo de los aborigenes y la incursion europea y 6 son postcolombinos (a los cuales hay que sumar otras dos especies extinguidas en los siglos XIX y xx) A partir de estas 25 especies avaladas por fechas de ultima presencia distribuidas entre los cuatro intervalos temporales sefialados hemos compuesto la Tabla 45 en la que ademas aparecen las restantes 27 especies (carentes de fechado radiometrico) ubicadas en el intervalo temporal mas probable sobre la base de (1) la asociacion estratigrafica de sus restos con evidencias culturales (amerindias 0 europeas) 0 (2) la ausencia de dicha asociacion Sobre estos criterios de ubicacion temporal procede hacer dos aclaraciones
324 BIOGEOGRAFIA
TABLA 45 Intervalos temporales mas probables para las extinciones cuatemarias de los mamiferos terrestres de las Antillas Mayores y grupos de islas perifericas sobre la base de las evidencias siguientes (l) ultimo ejemplar colectado u observado en vida (2) fechado radiometrico mas tardio en material de la especie (3) asociaci6n estratigrafica mas tardia con otro material fechado (4) asociaci6n estratigrafica con restos de animales introducidos restos humanos u objetos culturales propios del intervale (5) falta de asociaci6n con evidencias culturales [Los ntimeros entre parentesis en la columna Evidencia significan adscripci6n tentativa al intervalo] Los estimados del peso corporal (en kg) estan tomados de la literatura 0 inferidos comparativamente y distribuidos entre
las clases siguientes (a) S 01 (b) 01 a lt 1 (c) 1 a lt 10 (d) 10 a lt 100 (e) 100 a lt 200 (j) ~ 200 Las
abreviaturas que designan las islas son las de la Tabla 44
F amilialEspecie Peso IsJa Evidencia Fuente
I httervalo htterg1acial Sangamon (gt80 kaAP)
HEPTAXODONTlNAE
Amblyrhiza inundata e AS 3 McFarJane et al 1998a
CLIDOMYINAE
Clidomys osborni d JA 3 Ibid
Clidomys parvus d JA 3 Ibid
II httervalo Pleistoceno terminal ~ Holoceno temprano (20-6 kaAP)
MEGALONYCHIDAE
Aeratoenus antillensis d CD 5 Arredondo 1975 a
Aeratoenus odontrigonus d PR 3 McFarJane 1999b
Aeratoenus simorhynehus d LE 2 Rega et al 2002
M egaloenus zile f LE 2 MacPhee et al 2000
SOLENODONTIDAE
Solenodon arredondoi e CD 5 Morgan y Ottenwalder 1993
PnHECIIDAE
Paralouatta varonai d CD 5 MacPhee 1996a
EcHIMYIDAE
Heteropsomys antillensis c PR 5 McFarJane 1999b
Heteropsomys insulans c PR 5 Ibid
Puertoricomys eorozalus c PR 5 Woods 1989a
CAPROMYIDAE
P lagiodontia araeum c LE 5 MacPhee et al 2000
Macrocapromys latus c CD 5 Varona y Arredondo 1979
(Continua)
325 EXTINCIONES DEL CUATERNARIO
TABLA 45 (Continuaci6n)
F ami1ia1Especie Peso Isla Evidencia Fuente
M acrocapromys latus c CU 5 Varona y Arredondo 1979
Mesocapromys kraglievichi c CU 5 Ibid
HEPTAXODONTINAE
Xaymaca fulvopulvis a JA 2 MacPhee y Flemming 2003
III Intervalo Amerindio (6-05 kaAP)
MEGALONYCHIDAE
Acratocnus ye d LE 2 MacPhee et aI 2000
Neocnus comes c LE 2 Ibid
N eocnus dousman c LE 2 Ibid
N eocnus toupiti c LE 2 Ibid
N eocnus gliriform is c CU 4 Cordova et ai 1997
M egalocnus rodens f CU 2 MacPhee et ai 2007
Parocnus browni e CU 2 Steadman et ai 2005
P arocnus serus e LE 2 MacPhee et ai 2000
N ESOPHONTIDAE
N esophontes edithae a PR 2 McFarlane 1999b
PITHECIIDAE
Antillothrix bernensis c LE 3 Rimoli 1977
EcHIMYIDAE
Brotomys contractus a LE 4 Miller 1929c
CAPROMYIDAE
P lagiodontia spelaeum c LE 4 Ibid
M acrocapromys acevedo c CU 4 NUiiez 1990a
HEPTAXODONTINAE
Elasmodontomys obliquus d PR 3 McFarlane 1999b
Iv Intervalo Postcolombino (05 kaAP - Presente)
N ESOPHONTIDAE
N esophontes hypom icrus a LE 24 MacPhee y Fle111111ing 1999
N esophontes zam icrus a LE (4) Ibid
N esophontes m icrus a CU 24 MacPhee et aI 1999
(Continua)
326 BIOGEOGRAFiA
TABLA 45 (Continuacion)
F amilialEspecie Peso Isla Evidencia Fuente
N esophontes m icrus a CD 24 MacPhee et aI 1999
N esophontes m icrus a LE 4 MacPhee et aI 2000
N esophontes sp 1 a IC 4 Morgan 1994
N esophontes sp 2 a IC 4 Ibid
Solenodon marcanoi b LE 4 Ottenwalder 2001
PITHECIIDAE
Xenothrix mcgregori c JA 4 MacPhee y Flemming 1999
EcHIMYIDAE
Boromys offeUa a CD 24 Guarch 1984
Boromys torrei a CD 24 Ibid
Brotomys voratus a LE 4 MacPhee et aI 2000
CAPROMYIDAE
P lagiodontia ipnaeum c LE 4 MacPhee y Flemming 1999
Rhizoplagiodontia lemkei c LE 4 Ibid
Isolobodon portoricensis c PR 24 Ibid
Isolobodon montanus c LE 4 Ibid
Hexolobodon phenax c LE (4) Ibid
Capromys sp c IC 24 Morgan 1994
Geocapromys columbianus c CD 4 MacPhee y Flemming 1999
Geocapromys thoracatus b IS 1 MacPhee y Marx 1997
Geocapromys sp 1 c IC 4 Morgan 1994
Geocapromys sp 2 c IC (4) Ibid
l-IEPTAXODONTINAE
Quemisia gravis c LE (1) MacPhee y Flemming 1999
MURIDAE
Oryzomys antillarum a JA 1 Ibid
1 Flemming y MacPhee (1999) alertaron en contra de la asociacion con otro material fechado Asimismo Morgan (1993) indico que la ausencia de asociacion con evidencias culturales amerindias no es necesariamente un criterio concluyente para ubicar un taxon en el intervalo temporal anterior porque hay depositos en la region que carecen de tales evidencias y son del Holoceno tardio por fechados radiocarbonicos [No obstante
327 EXTlNCIONES DEL CUATERNAR10
la ausencia de una especie en residuarios arqueol6gicos adquiere mayor certidumbre como criterio de ubicaci6n temporal de su extinci6n en la medida que aumenta el numero de residuarios estudiados que en las Antillas Mayores se cuentan por centenares] A falta de fechado absoluto de una especie consideramos que estos criterios de ubicaci6n temporal son mas utiles que la alternativa de excluir del analisis a la especie
2 En los trabajos sobre biogeografia de mamiferos antillanos los arque610gos mas citados como autoridades para establecer el inicio de la presencia aborigen en las Antillas Mayores (Kozlowski 1974 Rouse y Allaire 1978 Rouse 1989 1992 Wilson 2001) difieren al respecto entre los 6 kaAP y los 45 kaAP (pero vease Pagan et al 200523) y hemos adoptado la mas temprana de estas fechas para el comienzo del intervalo amerindio en la Tabla 45 De todos modos cualquiera que haya sido la fecha esta no es aplicable por igual al conjunto de las Antillas Mayores porque segun algunos de los autores citados el poblamiento comenz6 por Cuba y La Espanola 6 mil anos atras continu6 en Puerto Rico hace alrededor de 3 mil anos (pero vease Burney et al 1994) yen Jamaica hace menos de 2 mil e incluso a las Islas Caiman ni siquiera llegaron [Las Islas Caiman descubiertas por Col6n en 1503 carecen absolutamente de evidencia amerindia (Stokes y Keegan 1996)]
Si los datos de la Tabla 45 representan una aceptable aproximaci6n a la realidad el proceso de las extinciones cuaternarias de los mamiferos terrestres de las Antillas Mayores y grupos de islas perifericas debi6 caracterizarse entre otros por los eventos siguientes
Aunque obviamente la fauna cuaternaria debi6 ser mucho mas numerosa que 10 que refleja el registro paleontol6gico (Morgan y Woods 1986) 52 especies (81 de la fauna conocida) desaparecieron (sin reposici6n) durante los ultimos 250 mil anos
- Al parecer las extinciones se concentraron en las especies de mayor tamano Asi el peso promedio estimado para las especies de la etapa preantr6pica (intervalos I + II de la Tabla 45) es 350 kg es 328 kg para las especies del intervalo III (amerindio) y es s610 25 kg (c1ases del peso a b c) para las especies del intervalo IV (postcolombino) Ninguna especie aut6ctona de mamifero mayor de 6 kg existe hoy en la regi6n
- De acuerdo con la distribuci6n temporal de las extinciones 61 especies sobrevivieron al interglacial Sangamon (intervalo 1)48 sobrevivieron a la transici6n del Pleistoceno terminal al Holoceno temprano (intervalo II) 34 sobrevivieron al periodo amerindio (intervalo III) y s610 12 especies han sobrevivido desde la Ilegada de los europeos (intervalo IV) Las tasas de extinci6n correspondientes (exceptuado el intervalo I) son una especie cada 1167 anos en el intervalo II una cada 393 anos en el intervalo III y una cada 23 anos en el intervalo IV
- Durante todo el proceso de las extinciones cuaternarias la perdida de especies por isla fue de 91 en La Espanola 83 en Jamaica y 62 en Cuba Bahamas retuvo su unica especie (aunque extirpada en varias de las islas) Puerto Rico e Islas Caiman perdieron todas sus especies (6 y 5 respectivamente) y las Islas Swan Virgenes y Anguila - San Martin perdieron sus respectivas especies (micas
328 BIOGEOGRAFiA
De los cuatro 6rdenes de mamiferos representados en la fauna Pilosa y Primates desaparecieron totalmente Soricomorpha y Rodentia perdieron 86 y 74 de sus especies respectivamente
CAUSALIDAD
CAUSAS NATURALES
Los cicIos glaciales e interglaciales acaecidos en el Pleistoceno causaron cambios ambientales dnisticos en las Antillas con la concomitante alternancia de cIimas frios (secos) y calidos (hUmedos) asi como de descensos y ascensos del nivel del mar aumento y disminuci6n de las areas territoriales) induso (aunque con menor intensidad) en el Holoceno Estos eventos paleogeograficos han sido consistentemente documentados (Mayo y Kartashov 1972 Bonnatti y Gartner 1973 Ortega y Arcia 1982 Prance 1982 Schubert y Medina 1982 Dodge et al 1983 Ortega [1984] Haq et al 1987 Schubert 1989 Hodell et al 1991 Guilderson et al 1994 Oppo 1997 Emslie 1998 Curtis et al 2001 Iturralde-Vinent 2003) y de ellos brindamos a continuaci6n una breve resena
Finalizando el periodo interglacial anterior a la ultima glaciaci6n (Wisconsin) el mar estaba a 8-10 m por encima de su nivel actual Cuando el avance glacial del Wisconsin a1canz6 su culminaci6n (eillamado Ultimo Maximo Glacial) hace 18 ka el mar habia descendido 120 m con respecto a dicho nivel y las islas del Caribe habian incrementado considerablemente sus extensiones subaereas en comparaci6n con las que presentan hoy dia por ejemplo Puerto Rico y las Islas Virgenes se conectaron territorialmente y las Bahamas se unieron en cinco gran des islas Prevalecia un regimen frio y seco y condiciones de aridez --debidas a una reducci6n de las precipitaciones (posiblemente a menos de 440 mmlano)- y los ambientes xericos se habian extendido a traves de las islas y bancos emergidos y abundaban los pinares (los habitats mesicos estaban restringidos a parcelas relictas en 10 alto de las montanas) En el Mar Caribe y el Golfo de Mexico la temperatura superficial del mar era 5degC mas baja que la actual y en las llanuras costeras y alturas maximas eran respectivamente 2-3degC y 6degC mas bajas Los limites de las zonas ecol6gicas altitudinales descendieron varios cientos de metros y cada zona redujo su extensi6n vertical al aumentar el gradiente de temperatura Ellimite inferior de la zona periglacial
elevaci6n donde la temperatura media minima a1canza los OdegC) de la Cordillera Central de la Republica Dominicana (altura maxima del Caribe insular con evidencia de topografia glacial) se considera que estuvo entre los 1500 y 1800 mlnm (actualmente esta a 2800 mlnm) y se puede estimar que dicho limite era inferior en Cuba por estar mas al norte que la Republica Dominicana No obstante durante este periodo la temperatura experiment6 episodios cortos (unas pocas decadas) de ascensos repetidos
Un signifieativo componente bi6tieo de las referidas condiciones paleoambientales 10 constituy6 la espectacular irradiaci6n evolucionaria experimentada por un grupo numeroso de aves rapaees diurnas y nocturnas (Wetmore 1937 Brodkorb 1969 Kurotchkin y Mayo 1973 Arredondo 1976 Olson 1978 Olson y Hilgartner 1982
329 EXTINCIONES DEL ClJATERNARIO
Suarez 2000a b 2004 Suarez y Emslie 2003) Los mamiferos de las Antillas Mayores ofrecieron a estos depredadores aviares una amplia gama de tamanos de presa que en ausencia de mamiferos del orden Carnivora en la region permitio a estas aves irradiarse a este nicho alimentario con la evolucion de una variedad de especies de muy diversa tall a corporal incluidas formas gigantes Adquirir un cuerpo gigante les propicio el acceso a recursos alimentarios que no estaban siendo explotados por ningun otro depredador (Alcover y McMinn 1994) A Cuba corresponde el mejor registro fosil de rapaces cuaternarias caribenas con siete generos de especies diurnas Dentro de estas aves existieron tres gavilanes gigantes (Accipitridae) tres carairas (Falconidae) y un condor (Cathartidae) asi como cuatro generos de especies nocturnas dos lechuzas (Tytonidae) y tres Mhos (Strigidae) dos de ell os no igualados en tamano en todo el mundo Algunos de estos generos tuvieron representacion en otras islas antillanas como La Espanola Jamaica Bahamas y Gran Caiman
Al retirarse hacia el norte los hielos de la glaciacion Wisconsin el clima se tomo mas hUmedo en las Antillas durante el Holoceno temprano y el bosque mesico se expandio en la region insular El nivel del mar ascendio acompanado de mayor pluviosidad alrededor de los 7 kaAP las Islas Virgenes quedaron separadas entre si y de Puerto Rico y las Bahamas perdieron la mayor parte de su territorio Cerca de los 3 kaAP las condiciones secas retornaron a la region e hicieron desaparecer buena parte del bosque mesico Nuevamente el clima comenzo a adquirir sus condiciones actuales hacia los 25 kaAP aproximadamente y hoy transita por una fase mesica con vegetacion xerica confinada en parcelas relictas
Sin duda estos y otros eventos naturales incidieron negativamente en la sobrevivencia de mamiferos terrestres en las Antillas Mayores asi como en los grupos de pequenas islas perifericas (Pregill y Olson 1981 Olson 2006)
McFarlane et al (1998a) demostraron convincentemente como Amblyrhiza el gigante de los roedores antillanos endemico de la pequena isla de Anguila sucumbio a las repetidas y abruptas perdidas de su habitat durante el interglacial Sangamon por las nipidas y severas oscilaciones del nivel del mar (documentadas por Neumann y Hearty 1996) al no poder responder adecuadamente a tan catastroficos eventos mediante el ajuste de su masa corporal media y otro tanto debio ocurrirle en Jamaica al otro roedor gigante Clidomys (McFarlane et al 1998b) emparentado con Amblyrhiza en la misma epoca (Tabla 45 intervalo I)
El ascenso del nivel del mar seguramente promovio la extincion de mamiferos durante la transicion del Pleistoceno al Holoceno (Tabla 45 intervalo II) con la reduccion del tamano de las Islas por las transgresiones marinas (Pregill y Olson 1981 Morgan 1993) Esto ultimo haria maximamente severa la competicion asf como la presion de los depredadores [Steadman et al (2005) no yen por que el calentamiento global ocurrido durante la transicion Pleistoceno-Holoceno habria de tener un efecto negativo sobre los perezosos siendo sus descendientes actuales especies tropic ales 0 subtropicales pero no se trata del efecto
330 BIOGEOGRAFiA
directo del cambio de temperatura sobre los animales sino del efecto indirecto y progresivo que dicho cambio genera a largo plazo al reducir el territorio insular disponible]
Por esta epoca tambien desaparecieron especies de tall a mayor como Megalocnus zile en La Espanola y otras de menor tamano (en Cuba) pero que representan la tall a maxima en sus respectivos contextos taxonomicos del Cuaternario como Solenodon arredondoi Paralouatta varonai y Macrocapromys acevedo Un mamifero puede ser grande solo a costa de reducir su densidad poblacional y esto 10 hace mis vulnerable a la extinci6n sobre todo en una isla (McNab 1994 McFarlane et al 1998a) Incidentalmente Alcover et al (1998a) afirmaron que larelacion entre la talla corporal y la extincion no siempre se cumple en las especies de menor tamano y citaron como ejemplo que entre las musarafias todas las especies de Nesophontes desaparecieron mientras sobreviven especies de Solenodon pero la comparacion correcta en todo caso serfa entre los solenodones vivientes y el gigante Solenodon arredondoi y entre Nesophontes edithae -el gigante de los nesofontes extinguido en el periodo amerindio- y el resto de sus congeneres que sobrevivieron hasta los primeros siglos de la colonizacion europea (Tabla 45 intervalos III y IV)
CAUSAS ANTROPOGENICAS
Intervalo Amerindio
La mayoria de los autores que se han referido a las extinciones acaecidas durante este intervalo (entre elIos paradigmaticamente Martin 1984a b) han responsabilizado a los aborfgenes (que se supone comenzaron a poblar las Antillas Mayores hace 6 mil afios) con la desaparicion de numerosas especies de mamiferos terrestres en la region principalmente por la via de la sobrecaza y la introduccion de animales domesticos devenidos depredadores de la fauna autoctona (Morgan y Woods 1986) Este efecto negativo de la presencia indigena sobre los mamiferos de la region ha sido dramatizado por algunos autores en terminos de holocausto faunal Sin embargo pensamos que se ha exagerado Como han expresado MacPhee et al (19993) cuando uno busca evidencia dura de complicidad humana directa en [estas] extinciones no hay mucho que encontrar Las siguientes son algunas consideraciones a tener en cuenta en tal sentido
El registro radiometrico mas joven conocido hasta ahora para un perezoso antillano (MegalocnllS rodens) es de 42 kaAP (MacPhee et ai 2007) Si se acepta que los aborfgenes entraron en Cuba hace aproximadamente 6 mil anos su convivencia con los perezosos debe haber durado alrededor de 1800 afios (0 mas porque dificilmente el referfdo fechado radiometrico haya correspondido al ultimo sobreviviente de la especie) Esta cifra contradice la hipotesis del holocausto faunal segun la cualla extincion de los mamiferos insulares de mayor talla debe ocurrir con extrema rapidez -a escala de decadas 0 de centurias- a partir de su primer contacto con los humanos (Martin y Steadman 1999) A pesar de las repetidas afirmaciones en contrario (Harrington 1921 Miller 1929c Herrera [1945] Tabio 1951 Alvarez 1951a 1957 Arredondo
331 EXTINCIONES DEL CUATERNARIO
1959 1960a Veloz y Ortega 1973 Mayo 1980b Rodriguez et al 1984 Castellanos y Pino 1985 Pino y Castellanos 1985 Nunez 1990b Gutierrez 1996 Arredondo 2004) en su mayoria basadas en evidencias no exentas de posibles accidentes de deposicion (0 de reubicacion postdeposicional) en recintos cavernarios (Diaz-Franco [2006b D no se ha demostrado inequivocamente que los aborigenes antillanos se alimentaran de perezosos regularmente Sus restos en residuarios de alimentos son escasisimos en toda la region y esto contrasta con los abundantes restos de jutias y otros mamiferos de menor talla que aparecen en esos residuarios demostrativos de la preferencia de los paleoindios por estos otros mamiferos
Los mamiferos mas importantes en la economia prehistorica de las Antillas Mayores fueron las jutias Capromys y Geocapromys en Cuba Geocapromys en Jamaica e Isolobodon en La Espanola Puerto Rico e Islas Virgenes (Aguayo 1950b Tablo 1951 Rimoli 1972 Wing 2001 Diaz-Franco [2006b D Wilkins (2001) estudi61ajutia viviente de Jamaica (Geocapromys browni) en un sitio ocupado por los aborigenes durante 100 anos y no encontr6 evidencias de que durante ese tiempo la especie haya sido sobreexplotada Asimismo en Cuba las jutias conga carabali y andaraz son escasas en depositos pleistoholocenicos en cambio son muy numerosas en residuarios indigenas y de haber sido sobreexplotadas 10 mas probable es que no hubieran sobrevivido hasta hoy
La habilidad de los humanos para penetrar en profundidad el bosque tropical y mantenerse en este sustentablemente ha sido un evento comparativamente reciente (Beran y Steele 1994) En el caso antillano la caza tierra adentro ha sido explicada como un proceso tardio de desplazamiento a partir de la explotacion de abundantes recursos marinos que ofrecian condiciones 6ptimas para el sedentarismo de las comunidades aborigenes costeras (C6rdova y Arredondo 1988 Brito 2005) Peces y mariscos dominan la arqueologia costera de las Islas antillanas en tanto el consumo de recursos terrestres (mamfferos incluidos) aumenta progresivamente en la medida que los residuarios estudiados se alejan de la costa (Scudder 1991 Wilkins 2001) En las Antillas Mayores el componente terrestre de la dieta aborigen es solo 20-30 del material recobrado en los residuarios (Wing 1977 1989) Y debio ser menor alin porque el componente vegetal de la dieta no se ha preservado Por otra parte los aborigenes cubanos domesticaron la jutia conga y seguramente la reprodujeron para comer (Pose et al 1988) practica que probablemente se repitiera con otras jutias en las demas Antillas Mayores
- Fray Bartolome de las Casas hablo de 3 millones de indios en Cuba al inicio de la colonizacion pero los historiadores subsecuentes consideraron exagerada la cifra (Poey 1851 a) No obstante se dispone de dos estimados de la densidad de poblaci6n aborigen en las Antillas Mayores a la llegada de los europeos 18 indigenaslkm2 en Puerto Rico (Rouse 1956) y 1271km2 en La Espanola (Watts 1987) Extrapolando ambos valores entre las cuatro islas principales de las Antillas Mayores tenemos que en la fecha indicada la poblaci6n aborigen pudo estar entre 16 mil y 113 mil indigenas en Puerto Rico entre 21 mil y 145 mil en Jamaica entre 141 mil y 1 mill6n en La Espanola y entre 200 mil y 14 millones en Cuba Entre las cifras maximas y las minimas nos inclinamos
332 BIOGEOGRAFiA
hacia estas ultimas atendiendo a los altos indices de mortalidad infantil y morbilidad que se evidencian en las osamentas de los pobladores prehispanicos (Martinez y Rives 1990) Uno de los hechos que apuntan hacia el bajo numero de aborigenes en las Antillas Mayores es 10 rapido que desaparecieron despues de la llegada de los europeos pnkticamente sin dejar mestizaje con estos 10 cual contrasta ostensiblemente con la situacion en la America continentaL
En suma un grupo humano (1) demograiicamente irrelevante y controlado por altos indices de morbilidad (2) consumidor mayoritariamente de organismos marinos y (3) carente de la tecnologia cinegetica necesaria para poder sobreexplotar los mamiferos que Ie sirvieron de alimento (a los cuales accedio tardiamente en su desarrollo) no es concebible que haya causado e1 holocausto faunal que se Ie atribuye a partir del error de aplicar a las comunidades aborigenes antillanas un modelo cultural de la prehistoria continentaL
Tampoco hay evidencias de que los aborigenes hayan introducido carnivoros con suficiente capacidad depredadora salvo el perro Morgan y Otlenwaldder (1993) consideraron que la depredacion por perros durante el periodo amerindio es la explicacion mas plausible para la extincion del almiqui gigante de Cuba pero como se puede observar (Tabla 54) Solenodon arredondoi aparece en el intervalo II por falta de asociacion con evidencias culturales
Consecuentemente el accionar de los aborigenes y de sus animales domesticos directamente sobre la fauna no parece haber sido por sf solo un factor determinante de la extincion de mamiferos terrestres durante el periodo amerindio en las Antillas (pero vease Steadman et al 2005)
Hay que recordar en este senti do que alin en el Holoceno tardio ocurrieron cambios clirruiticos de consideracion (vease Causas Naturales) Por ejemplo Curtis et al (2001) explicaron que la extincion de algunos ani males mesofilicos de las alturas montanosas de La Espanola pudo haber antecedido a cualquier impacto humano sigrIificativo en esos lugares por coincidir dicha extincion con una tendencia a la aridez documentada isotopicamente y con evidencias de la perdida del bosque mesico ocurridas despues de 3kaAP Yen Jamaica hace solo 2shy3 mil aDos el area de la Bahia de Jackson sustentaba una gran biomasa de vertebrados en bosque hinnedo a juzgar por la cantidad y diversidad de taxones presentes en material fosil derivado de la depredacion por lechuzas hoy el area es la mas seca de la isla con matorral xerofitico y pobreza de fauna (MacPhee 1996a) Yalm en nuestros dias huracanes de gran intensidad aniquilan poblaciones de vertebrados (que no se recuperan) en is las pequenas y llanas del Caribe (Brooks y Smith 2001)
En medio de la incertidumbre sobre la verdadera causa (0 causas) de las extinciones de mamiferos en el periodo amerindio MacPhee y Marx (1997) han propuesto su hipotesis de la hipermorbilidad la cual estipula colapsos poblacionales relativamente rapidos con la introduccion de enfermedades de alta letalidad Estos autores han razonado que
Las islas son los lugares mas apropiados para experimentar perdidas catastroficas causadas por enfermedades Los mamiferos no solo serian susceptibles en virtud de su prolongado aislamiento biologico sino que en los confines de una isla la epizootia
333 EXTINCIONES DEL CUATERNARIO
tendria la potencialidad de infectar simultaneamente a la rnayoria de los individuos de vadas especies Los neonatos son inmunologicamente inmaduros y sufririan desproporcionadamente cualquier infeccion hipervirulenta La perdida excesiva de neonatos y juveniles tendria efectos severos para la sobrevivencia de la poblacion si esta se reduce por debajo de cierto nivel sobreviene la extincion porque no hay posibilidad de restitucion en el periodo postpanzootico Las especies de mayor talla corporal van a ser las mas vulnerables a la extincion dadas sus bajas tasas reproductivas largos periodos de gestacion y reducido numero de progenie Las especies de menor tamano --caracterizadas por altas tasas reproductivas periodos cortos de gestacion y mayor numero de neonatos por parto- tambien serian fuertemente diezmadas pero muchas sobrevivirian porque su estrategia reproductiva les permite perder mayor numero de individuos Las perdidas deben ocurrir relativamente rapido (a escala de decadas) mientras que otras hipotesis (digamos los cambios climaticos) permiten intervalos mucho mas largos (a escala de siglos 0 de milenios) entre la aparicion de las causas y la produccion de sus efectos Por ejemplo todo indica que los nesofontes se extinguieron muy rapidamente y seria dificil poder invocar un impacto humano directo 0 argumentar fenomenos climaticos que pudieran explicar plausiblemente perdidas tan rapidas
MacPhee y Marx (1997 191) han dicho que por el momento la hipotesis de la hipermorbilidad no pasa de ser una abstraccion en espera de adecuada sustentacion empirica Llama la atencion que linicamente en el periodo amerindio (Tabla 45 intervalo 1Il) las extinciones abarcaron todas las clases del peso corporal (de la a a laj) y que ninguna de las especies que sobrevivieron hasta el periodo postcolombino (Tabla 45 intervalo IV mas las especies vivientes) sobrepasa los 10 kg (clases del peso a b c) en clara coincidencia con los presupuestos de la hipotetica hipermorbilidad
Intervalo Postcolornbino
Por 10 menos 37 especies de mamiferos terrestres vivian en las Antillas Mayores y grupos de Islas perifericas a la lIegada de los europeos en 1492 De elIas se han extinguido 23 shymas especies que en cualquiera de los periodos precedentes (Tabla 45~ y solo subsisten 12 especies repartidas entre Cuba (8 especies) La Espanola (2) Jamaica (1) y Bahamas (1) varias de estas en peligro critico de extincion Solo dos de las especies extinguidas durante los ultimos 500 anos se conocieron en estado viviente los roedores Oryzomys antillarum de Jamaica (extinguido en 1877) y Geocapromys thoracatus de las Islas Swan (extinguido en 1955) Las causas de las extinciones ocurridas durante este periodo son sobradamente conocidas destruccion de habitats introducci6n de animales exoticos (domesticos y comensales) y demas presiones asociadas a la actividad humana entre las cuales posiblemente habria que incluir brotes de hipermorbilidad
Durante mucho tiempo se ha afirmado la posibilidad de que especies de Nesophontes hayan sobrevivido hasta las primeras decadas del siglo xx y que incluso continuen existiendo en lugares remotos de Cuba 0 La Espanola (Allen 1918 1942 Miller 1930 Aguayo 1950a Arredondo 1960b Fischer 1977 MacFadden 1980 Woods y
334 BIOGEOGRAFiA
Ottenwalder 1991) Actualmente no existe evidencia radiometrica de que alguna especie de Nesophontes haya sobrevivido al siglo xv las fechas radiocarbonicas calibradas obtenidas en muestras de Nesophontes de Cuba y La Espanola resultaron no mas tardias que los siglos XIII y XIV (MacPhee et al 1999) Por otra parte las expediciones efectuadas a fines del siglo xx con el proposito de localizar individuos vivientes del genero en La Espanola no produjeron la menor indicacion afirmativa al respecto (Woods et al 1985) Ademas a traves de las Antillas Mayores no aparecen restos de Nesophontes en los depositos formados actualmente por las lechuzas
EPiLOGO
Las Antillas Mayores son islas oceanicas no han formado parte de un continente (con la posible excepcion de una parte de Jamaica hasta el Eoceno Medio hace 45 Ma) y durante un tiempo geologico considerable (desde el Eoceno Superior hace 40 Ma) han permanecido emergidas y separadas de los continentes vecinos Consecuentemente los mamiferos terrestres (no volantes) autoctonos de estas islas constituyen una fauna oceanica que por ende cumple con todas las caracteristicas convencionales reconocidas para este tipo de entidad biogeografica
- Baja diversidad taxonomica a nivel de las categorias superiores (solo cuatro ordenes) en comparacion con islas plataformicas continentales (0 con areas continentales) de tamano y diversidad ecologica simi lares
- Alto grado de endemismo a nivel de genero (96) y familia (50) en respuesta al prolongado aislamiento
- Desbalance ecologico de la estructura trofica ausencia de mamiferos camivoros y ungulados
- Espectaculares procesos de irradiaci6n evolucionaria como resultado de la colonizacion de territorios libres de competidores y depredadores
Tendencias evolucionarias extremas gigantismo
- Control a cargo de aves rapaces con tendencia at gigantismo en ausencia de mamiferos miembros del orden Carnivora
- Mas especies de reptiles y de murcielagos que de mamiferos terrestres relaciones que en los continentes se invierten
Alta tasa de extincion de especies (84) sin reposicion
Preservacion de especies filogeneticamente arcaicas (los llamados fosiles vivientes) los almiquies

314 BIOGEOGRAFiA
V ICARIANZA
La neotectonica y las transgresiones marinas del Oligoceno Medio y del Mioceno afectaron sustancialmente la paleogeografia de la Cresta de las Antillas Mayores resultando la subdivision gradual de las partes subaereas de la Cresta Asi el este de Cuba y el norte de La Espanola conectados fisicamente durante el Oligoceno Inferior se separaron al formarse el Paso de los Vientos en el Oligoceno tardio en tanto La Espanola y Puerto Rico probablemente permanecieron conectadas hasta el Mioceno tardio en que surgio el Paso de Mona (Iturralde-Vinent y MacPhee 1999)
Tales eventos geologicos habrian dividido las distribuciones de los grupos faunales emplazados previamente en el territorio continuo de las tres islas y a partir de esas disyunciones dichos autores (tambien MacPhee et al 2000 y White y MacPhee 2001) asumieron --como segunda fase del modelo de GAARlandia- la hipotesis de la vicarianza isla-isla para explicar la distribucion multiinsular de varios grupos de mamiferos terrestres a traves de las Antillas Mayores septentrionales De acuerdo con esta hipotesis cualquier diferencia en composicion faunal entre las islas (por ejemplo la ausencia de roedores heptaxodontidos en Cuba 0 de monos platirrinos en Puerto Rico) constituye evidencia de extincion local postvicariante y no de dispersion marina fallida Lo primero se puede demostrar si aparecen los fosiles pertinentes 10 segundo no se puede demostrar (MacPhee e Iturralde-Vinent 1994) Por otra parte IturraldeshyVinent y MacPhee (1999 57) han reconocido que aunque fa vicarianza isla-isla puede representar una solucion clara para las distribuciones de varios taxones cuaternarios no puede explicar todos los casos
Rega et al (2002 18) han presentado dos objeciones al modelo de la vicarianza isla-isla aplicado a los mamiferos de las Antillas Mayores
I Si la separacion de Cuba y La Espanola precedio en varios mill ones de anos la separashycion de La Espanola y Puerto Rico el modelo vicariante de lturralde-MacPhee debe predecir mayor similitud entre las faunas de La Espanola y Puerto Rico que entre las de Cuba y La Espanola De hecho es al reyes
No obstante MacPhee et al (200335) admitieron despues que la fecha mas temprana empiricamente defendible [para la apertura del Paso de Mona] debe ser el Oligoceno Medio esto invierte la secuencia de ambas disyunciones con 10 cual queda resuelta la aparente contradiccion Sin embargo en su reciente evaluacion de la congruencia entre filogenia e hipotesis geologic as en varios grupos de mamiferos terrestres de las Antillas Mayores Davalos (2004373) encontro que ninguno de los taxones cumple con la fecha y el patron de divergencia entre las masas terrestres antillanas postulados por el modelo de GAARlandia
2 La presencia de un mismo genero en las tres Islas septentrionales de las Antillas Mayores implica [por el modelo vicariante] que la diferenciacion entre estas formas durante el intervalo desde el Mioceno ha sido unicamente de nivel especifico [Dicha presencia]
315 rRRADlAcr6~ EVOLUCrONARIA
s610 puede explicarse razonablemente como el resultado de movimientos de esos animales entre las islas en el Cuaternario
De esta objecion resulta dificil disentir [El poblamiento entre islas diferenciado solo a nivel especifico no es probable que sea anterior al Pleistoceno (Simpson 1956 15)]
Debe tenerse en cuenta que la hipotesis de la vicarianza isla-isla alude a las tres islas septentrionales de las Antillas Mayores porque como se explico antes (vease El Barco Funeral Vikingo) Jamaica no existia como isla durante el lapso (Eoceno Superior shyOligoceno) consensualmente reconocido como el mas probable para la colonizacion de las proto-Antill as Mayores por la mayo ria de los mamiferos terrestres Despues de emerger como isla en el Mioceno Medio Jamaica debe haber recibido sus mamiferos terrestres por dispersion isla-isla (Davalos 2004376) y solo en el Cuaternario parece haberrecibido (por dispersion marina desde Centroamerica) al roedor murido Oryzomys
Seguramente tanto la dispersion como la vicarianza contribuyeron a que los mamiferos terrestres se irradiaran a traves de las Antillas Mayores y grupos de islas perifericas pero las maneras y tiempos en que estos mecanismos operaron a 10 largo de dicho proceso son aun en gran medida motivos de desacuerdo e incertidumbre
IRRADIACION EVOLUCIONARIA
A continuaci6n revisamos someramente las distribuciones conocidas de los diferentes grupos de mamiferos terrestres a traves de las Islas principales de las Antillas Mayores y grupos de Islas perifericas incluidas las interpretaciones y conjeturas que la literatura reciente recoge en relaci6n con la manera en que dichas distribuciones pudieron haberse conformado durante la irradiaci6n evolucionaria de los referidos grupos de mamiferos Manifestaciones de esta irradiaci6n pueden inferirse unicamente a partir de los taxones presentes en el Cuatemario pues solo 6 de las especies conocidas corresponden al Terciario
GRUPOS ORlGINARIOS DE SURAMERICA
PEREZOSOS (Megalonychidae)
Aparte de un genero miocenico de Cuba y otro del Oligoceno de Puerto Rico se conocen 4 generos (12 especies) de perezosos cuaternarios (todos extintos) restringidos a las tres islas septentrionales de las Antillas Mayores Acratocnus Neocnus Megalocnus y Parocnus Los cuatro generos estan representados en Cuba y La Espanola y Acratocnus 10 esta ademas (como unico genero de perezoso) en Puerto Rico Esta distribuci6n ha sido interpretada por MacPhee et al (2000) eomo resultado de la vicarianza oligomiocenica entre estas islas y [dado que el Banco Puerto Rico - Islas Virgenes fue la entrada de GAARlandia a las Antillas Mayores] se preguntaron si la ausencia de los otros tres generos en Puerto Rico se debe a que nunca estuvieron en ese territorio 0 a que sus
316 BIOGEOGRAFiA
fosiles atm no han sido hallados Pero como hemos visto Rega et al (2002) atribuyeron dicha distribucion a la dispersion interinsular en el Cuatemario
Por otra parte varios amilisis c1adisticos recientes ubi can a Choloepus -genero Reciente conocido solo de Centro- y Suramerica- como taxon hermano de Aeratoenus exc1uidos los demas megaloniquidos antillanos Para White y MacPhee (2001228)
tan estrecha relacion sugiere que la divergencia entre ambos [generos] ocurrio despues de que Aeratoenus divergio de Neoenus Si esta relacion implica que Aeratoenus y Neoenus se dispersaron separadamente desde Suramerica a las AntilIas mientras Choloepus permanecia en el continente (siendo necesarias al menos tres invasiones de perezosos) 0 si Choloepus invadio Suramerica desde las Antillas es algo que el material disponible no permite determinar
La diversidad adaptativa de los perezosos se manifiesta en (1) la utilizacion del habitat especies arboreas en Neoenus y Aeratoenus y terrestres en Paroenus y Megaloenus (Webb 1985a White [1993] White et al 1996 Arredondo [1999]) y en (2) la gama de tallas corporales desde los pequenos Neoenus hasta Megaloenus rodens el mayor de los mamiferos terrestres antillanos (su peso adulto estimado en 270 kg seglin Paula 19679) esto sugiere que los perezosos se irradiaron a nichos que en contextos continentales probablemente estarian ocupados por miembros de otros c1ados (MacPhee e IturraldeshyVinent 2000)
No esta claro si el abanico de tamanos entre estos perezosos enmascara alguna tendencia al gigantismo Por 10 pronto Anderson y Handley (2002) encontraron 10 contrario en perezosos (Bradypus) de las Islas de Bocas del Toro (Panama) shyseparadas secuencialmente del area continental vecina por ascensos del nivel del mar durante los ultimos 10 ka En las cinco islas los perezosos son menores que su contraparte continental y las diferencias entre las tallas medias de las cinco poblaciones expresan una reduccion lineal significativa de la talla corporal en funcion de la edad geologica de cada isla sin relacion alguna con el area de las islas 0 la distancia que las separa del continente
MONOS (Pitheciidae)
Los primates de las Antillas Mayores (cuatro especies extintas) constituyen un grupo monofiletico con Paralouatta (en Cuba) y Antillothrix (en La Espanola) como taxones hermanos seguidos en la escalera c1adistica por Xenothrix (en Jamaica) y finalmente Callieebus como el pariente continental mas cercano esto implica una sola invasion desde Suramerica antes del Mioceno Inferior (fecha mas temprana conocida para los primates fosiles antillanos) y por tanto en consonancia con GAARlandia como la via de colonizacion (Horovitz y MacPhee 1996 1999 Horovitz [1997] MacPhee y Horovitz 2004)
En desacuerdo parcial con la anterior interpretacion la filogenia de este grupo de monos obtenida por Davalos (2004) indica un antecesor comun en Cuba La Espanola y Jamaica
317 TRRADIACTON EVOLUCIONARIA
compatible con la hipotetica conexion -propuesta tentativamente por Iturralde-Vinent y MacPhee (199980-84)- entre las Montanas Azules del este de Jamaica y la peninsula suroccidental de La Espanola en la epoca de GAARlandia
A pesar de que los primates antillanos pertenecen a un grupo viviente de monos muy pequenos (los monos titles) fueron comparativamente grandes en relacion con los platirrinos modernos y sucesivamente mayores en la direccion Antillothrix Xenothrix y Paralouatta con P marianae como el mayor platirrino conocido del Mioceno neotropical (MacPhee 2005) MacPhee et al (1994) consideraron que Xenothrix debio ser un mono de andar lento y al parecer las especies de Paralouatta abandonaron la arborealidad para adoptar un modo de vida mas terrestre (MacPhee 1996a MacPhee et al 2003 MacPhee y Meldrum 2006)
ROEOORES (Echimyidae Capromyidae Heptaxodontidae)
La edad de Zazamys en Cuba demuestra que la invasion de los roedores histricognatos en las Antillas Mayores comenzo antes del Mioceno Inferior por otra parte los representantes mas antiguos de esa irradiacion evolucionaria se encuentran en La Espanola y Puerto Rico Ambas evidencias son consistentes con la hipotesis de GAARlandia (Woods 1982 1988 Woods et al 2001)
No esta claro si los capromidos evolucionaron a partir de un equimido ancestral 0 de un protocapromido 0 si los equimidos antillanos (I sub familia 3 generos y 7 especies todas extintas) forman parte de la irradiacion de los capromidos (4 subfamilias 9 generos y 27 especies 10 vivientes) Kratochvil et al (1978b) y Rodriguez et al (1979) consideraron que Capromys nan us es un eslab6n entre Echimyidae y Capromyidae pero Mesocapromys nanus esta morfologicamente mucho mas cerca de Hexolobodon y los de mas capromidos que de los equimidos (Woods 1985) Lo que si esta claro segun Woods et al (2001) es que la subfamilia Plagiodontinae endemic a de La Espanola es el linaje basal de los capromidos y que Isolobodon Hexolobodon Capromys Geocapromys Mysateles y Mesocapromys [y supuestamente Macrocapromys] forman parte de la irradiacion temprana de Capromyidae
La subfamilia Capromyinae (5 generos y 17 especies 9 vivientes) parece ser el dado de mas reciente evolucion en la familia con invasion de Cuba desde La Espanola y dispersion desde Cuba (1) por Capromys a las islas Caiman y (2) por Geocapromys a las islas Bahamas Caiman y Jamaica y desde Jamaica a las islas Swan aunque la presencia de Geocapromys en Swan pudiera deberse a introduccion aborigen (Morgan 1985 1988 1989 1993 1994 Woods 1989a 1990) Woods et al (2001) utilizaron datos moleculares para obtener estimados independientes de divergencia entre capromidos segiJn 10 cual Geocapromys divergio cerca de los 17 MaAP Mesocapromys divergio de Capromys hace 15 MaAP en tanto lvlysateles 10 hizo hace 10 MaAP Por su parte las formas de Capromys pilorides y de Mysateles prehensilis en Isla de la Juventud divergieron de sus correspondientes subespecies nominativas en la isla principal a los 24 y 08 MaAP respectivamente
318 BIOGEOGRAFiA
Woods y Howland (I979) describieron adaptaciones morfologicas de la mandibula el hioide y la faringe asociadas a mecanismos de masticacion diferentes en plagiodontinos y caprominos y Varona (1980a) llamo la atencion sobre el gran desarrollo de los molariformes (en relacion con la longitud total del cnineo) que presentan ciertos grupos de jutias fosiles en comparacion con las vivientes 10 que pudiera interpretarse como una adaptacion alimentaria a una vegetacion mas abrasiva por parte de las jutias extinguidas
Segun Woods (1990) la familia Heptaxodontidae (2 subfamilias 5 generos y 6 especies todas extintas) tambien seria parte de la irradiacion de los capromidos con origen en el este de La Espanola 0 en Puerto Rico y subsecuente dispersion al Banco de Anguila - San Martin en las Antillas Menores (pero vease MacPhee y Flemming 2003) Este banco estaria a solo 70 km del Banco Puerto Rico - Islas Virgenes en tiempos de maxima regresion marina y esta proximidad con Puerto Rico -patria de Elasmodontomys obliquus- seguramente tuvo que ver con la presencia en las islas de Anguila y San Martin del heptaxodontino gigante Amblyrhiza relacionado mas estrechamente con los roedores de las Antillas Mayores que con los del resto de las Antillas Menores (Woods 1990 1996) Aunque los roedores suelen aumentar de tamano en las islas (Case 1978) la escala del incremento de la talla en Amblyrhiza (que pudo alcanzar mas de 200 kg de peso adulto) esta fuera de toda proporcion con los patrones conocidos en otros roedores insulares (Biknevicius et 01 1993)
Clidomys (Clidomyinae) de Jamaica difiere marcadamente en su morfologia de los heptaxodontinos (vease ademas MacPhee y Flemming 2003)10 cual dificulta explicar su presencia en esa isla Como Jamaica estuvo bajo el mar hasta el Mioceno Medio el ancestro de Clidomys colonizaria esta isla por dispersion desde el extremo suroccidental de La Espanola despues que los heptaxodontinos se originaron en esta ultima isla (Woods 1989a) Pero Pascual et 01 (I 990) especularon que Clidomys pudiera ser el resultado de una invasion posterior e independiente de los capromidos
GRUPOS ORIGINARIOS DE NORTEAMERICA
El origen de las musaranas antillanas ha resultado ser hasta ahora el mayor enigma biogeografico de la fauna de mamiferos terrestres de la region aunque la mayoria de los autores coinciden en considerarlas originarias de Norteamerica (vease Discus ion y Conclusiones en Poblamiento Continente-Isla) No obstante MacPhee e Iturralde-Vinent (1994) descartaron toda posibilidad de conexion terrestre por el Estrecho de la Florida 0 por el Canal de Yucatan y si el (supuesto) evento vicari ante de Rosen (vease El Arca de Noe) -invocado por MacFadden (1980) para resolver el enigma- fuera minimamente plausible aun asi es dificil creer que las catastrofes que 10 sucedieron (vease Contexto Temporal) hayan barrido con toda esa vieja fauna continental excepto las musaranas circunstancia ya anticipada muy tempranamente por Matthew (l918a)
De otra parte la relaci6n entre Nesophontidae y Solenodontidae permanece irresuelta Si estos linajes no estan estrechamente relacionados entonces es muy posible que
319 IRRADIACION EVOLUCIONARIA
sus distribuciones sean explicadas por mecanismos diferentes (Whidden y Asher 2001)
La descomunal diferencia de talla corporal entre las musaranas pequenas (Nesophontes) y las grandes (Soienodon) seguramente impuso a ambos grupos restricciones distintas relativas a la termorregulacion y las adaptaciones energeticas En Nesophontes la seleccion debio favorecer una alta tasa de metabolismo y crecimiento camadas mas numerosas corta expectativa de vida yalta densidad poblacionaL En Soienodon en cambio la seleccion opero en la direccion de una baja tasa metabolica camadas reducidas crecimiento lento longevidad y baja densidad poblacional (Ottenwalder [1991])
MUSARANAS PEQUENAS (Nesophontidae)
Las musaranas pequenas estan representadas en las Antillas por el genero Nesophontes (y un genero miocenico indeterminado en ambar dominicano) con 6 especies cuaternarias extinguidas distribuidas en Cuba La Espanola Puerto Rico Islas Virgenes e Islas Caiman La mayor de las musaranas pequenas Nesophontes edithae evoluciono en Puerto Rico y su arribo a las Islas Virgenes supone una dispersion sumamente reciente En las Islas Caiman Nesophontes se asemeja mas a la poblacion de Cuba de donde seguramente derivo en el Plio-Pleistoceno (junto con Capromys y Geocapromys) cuando la costa sur de Cuba estuvo mucho mas cerca de las Islas Caiman (lturralde-Vinent 2003fig 4) MacPhee et ai (1994) interpretaron la distribucion de Nesophontes en Cuba y La Espanola como resultado de la vicarianza oligomiocenica ocurrida al formarse el Paso de los Vientos pero la distancia taxonomica entre los lados del Paso es solo de nivel especifico y N micros incluso vivi6 en ambas islas segtln Patterson (1962)
Allen (1918) considero que Nesophontes micrus fue una especie primitiva no modificada para la vida arb6rea 0 fos6rea y por tanto de habitos terrestres Nesophontes edithae de Puerto Rico es mucho mayor y mas robusta que las restantes especies del genero eL indice de robustez de su femur (= anchura minima de la diafisis ongitud total del hueso) muestra que este es tan robusto como el de Solenodon paradoxus un animal dos veces mayor (Morgan et ai 1980) Choate y Birney (1968) postularon un aumento cronoclinal de la taIla corporal en N edithae confirmado mas tarde por McFarlane (1999a) mediante fechados radiocarb6nicos de la secuencia estratignifica en depositos cavernarios de Puerto Rico Solenodon no vivi6 en Puerto Rico y en ausencia de una musarana de mayor tamano Nesophontes alcanzo una talla corporal mayor en esa isla que en La Espanola 0 Cuba (Woods y Eisenberg 1989)
MUSARANAS GRANDES (Solenodontidae)
Los arcaicos almiqufes (genero Soienodon) comprenden cuatro especies una viviente (S cubanus) y otra f6sil (S arredondoi) en Cuba y una viviente (S paradoxus con dos subespecies) y otra f6sil (S marcanoi) en La Espanola Como cabria esperar las dos especies cubanas se asemejan mas entre sf que a las especies de La Espanola pero
320 BIOGEOGRAFiA
enigmaticamente S marcanoi comparte caracteres con S cuban us y S paradoxus (Woods y Eisenberg 1989 Morgan y Ottenwalder 1993 Ottenwalder 200 I) Solenodon arredondoi alcanzo una talla corporal superior a la de cualquiera de las musarafias vivientes a nivel mundial Segun Morgan y Ottenwalder (1993) extrapolando las dimensiones y el peso de los almiquies vivientes S arredondoi debio tener una longitud cabeza-tronco y un peso de 450-550 mm y 1500-2000 g respectivamente en comparacion con S cubanus (360 mm y 800 g) y S paradoxus (390 mm y 1100 g)
Entre todas las especies de almiqufes Solenodon paradoxus es la que presenta los caracteres mas derivados (Otten walder 2001) Si estas relaciones son validas la distribucion de Solenodon entre Cuba y La Espafiola debio discurrir de oeste a este como propuso MacFadden (1980) quien invoco la presencia fosH de S cubanus [pero tambien S arredondoi] en Pinar del Rio para postular que el ancestro continental entr~ por Cuba opinion compartida por Perfit y Williams (1989) y por Ottenwalder ([1991] 2001) El paso de Solenodon a Puerto Rico pudo haberlo impedido la existencia de barreras ecologicas durante la conexion del este de La Espafiola con Puerto Rico 0 por la ausencia de Solenodon en esa parte de La Espafiola en aquella epoca Sin embargo las mismas condiciones ecologicas en el este de La Espafiola no parecen haber representado una barrera efectiva para Nesophontes cuyas mas altas tasa de reproduccion y densidad poblacional presumiblemente Ie permitieron expandirse flpidamente a traves de La Espafiola (Ottenwalder [1991])
Es opinion generalizada que los almiqufes antillanos constituyen el grupo de mamiferos que primero se emplazo en el territorio insular (MacPhee 2005) y en tal sentido es de interes la evidencia sobre especificidad parasito-hospedante aportada por Cruz (1980) quien encontro que la especie de acaro (genero Cubanochirus) que parasita a los almiquies es mucho mas primitiva que la especie (del mismo genero) que paras ita a las jutias y de aqui concluyo que dicho genero de acaro debio evolucionar primero en Solenodontidae y posteriormente en Capromyidae al entrar en contacto representantes de ambas familias en Cuba 0 La Espafiola
Recientemente Roca (2004) y Roca et al (2004) dieron a conocer que los resultados de su estudio genetico-molecular sobre los almiquies indican una divergencia de 25 Ma entre Solenodon cuban us y S paradoxus -compatible con la disyuncion tectonica entre Cuba y La Espafiola (Iturralde-Vinent y MacPhee 1999 McPhee et al 2003)-- y una separacion de ambas especies comparable en magnitud a la divergencia entre familias de mamlferos Consecuentemente propusieron que Solenodon cubanus fuera clasificada en el genero Atopogaie como indico Cabrera (1925) hace 82 afios
DISCUSION Y CONCLUSIONES
Las musarafias y los roedores antillanos (excepto Oryzomys) se irradiaron tanto de sus respectivos ancestros continentales que de estos 6rdenes se reconocen familias y subfamilias endemicas de la region en contraste con los perezosos (una subfamilia y tres
321 IRRADIACION EVOLUCIONARIA
tribus endemicas) y los monos (una tribu endemica) como expresion de diferentes niveles de diversificacion evolucionaria Este proceso es dificil de explicar (traUindose de c1ados que pnicticamente comparten la misma antigiiedad en las Antmas) si es correcta la interpretacion de Pascual et at (1990636)
Nosotros consideramos estos diferentes niveles de diversificacion como las diferentes etapas de evolucion alcanzadas por cada grupo [de mamiferos] Obviamente esto estuvo relacionado con las diferentes longitudes de sus historias
Pudiera ocurrir que en el futuro se compruebe que la historia biogeognifica de la region fue mucho mas complicada
La Espanola la isla mas ecol6gicamente divers a de la region debi6 ser el centro principal de evoluci6n de los mamfferos (con 87 de las familias y 73 de las subfamilias) seguida por Cuba (con 75 de las familias y 54 de las subfamilias) las dos islas de las Antillas Mayores mas relacionadas biogeograficamente (Fig 43 Tabla 44)
eL
EL
cb CAIMAN
SWAN
A Acratocnini B Cubanocnini C Megalocnini D Mesocnini E Nesophontidae F Solenodontidae G Xenothrichini
ANGUILAshyM-0 SMARTIN
H Heteropsomyinae I Plagiodontinae J Isolobodontinae
K Hexolobodontinae L Capromyinae
M Heptaxodontinae N CIidomyinae 0 Sigmodontinae
FIGURA 43 Representacion esquematica de las relaciones interinsulares (expresadas porel numero de linajes compartidos) entre las Antillas Mayores y entre estas y los grupos de islas perifericas segun las interpretaciones de los autores citados en el texto Los numeros dentro de los circulos indican la cantidad de linajes representados en cada isla relacion dudosa Contexto taxon6mico de los linajes en la Tabla 44
bull bull
bull bull
bull bull bull
bull bull
bull bull
bull
bull
bull
bull
bull bull bull bull
bull bull
bull bull
bull
bull bull bull
bull bull
bull
bull bull
bull
322 BIOGEOGRAFiA
TABLA 44 Distribuci6n de los linajcs de mamiferos terrestres a traves de las Antillas Mayores y grupos de islas perifericas LE = La Espanola CU Cuba JA Jamaica PR = Puerto Rico IC Islas Caiman IS Islas Swan IB = Islas Bahamas IV = Islas Virgenes AS = Anguila - San Martin
FamiliaSubfumiliafIribu LE CU JA PR IC IS IB IV AS
Megalonychidae
Choloepodinae
Acratocnini
Cubanocnini
Megalocninae
Megalocnini
Mesocnini
Nesophontidae
Solenodontidae
Pitheciidae
Callicebinae
Xenothrichini
Echimyidae
Heteropsomyinae
Capromyidae
Plagiodontinae
Isolobodontinae
Hexolobodontinae
Capromyinae
H e pta x 0 don tid a e
Heptaxodontinae
Clidomyinae
Muridae
Sigmodontinae
PILOSA
SORICOMORPHA
PRIMATES
RODENTIA
bull
bull
bull
bull TOTALES 12 10 4 4 2
323 EXTlICIOIES DEL CUATERNARIO
El gigantismo ha sido sin duda una tendencia evolucionaria recurrente entre los clados mayores de mamiferos antillanos Es un hecho bien conocido que algunos grupos de mamiferos han aumentado de tamafio en las islas y varios autores han intentado explicarlo a partir de enfoques diferentes (Foster 1964 Miller 1966 Sondaar 1977 Case 1978 Eisenberg y Wilson 1978 Heaney 1978 Lawlor 1982 Lomolino 1985 Angerbjom 1985 McNab 19942001)
EXTINCIONES DEL CUATERNARIO
Las islas que integran la Subregion Antillana han perdido casi 90 de sus mamiferos terrestres durante los ultimos milenios y en proporcion con su extension territorial esta perdida ha sido mayor en la Subregion que en cualquier otro lugar del planeta (MacPhee 1996b) Las extinciones del Cuatemario eliminaron mas de las tres cuartas partes de la fauna de mamiferos terrestres conocida de las Antillas Mayores sus islas satelites y grupos de islas perifericas y el examen de este proceso en sus aspectos mas generales se aborda en esta ultima parte del presente capitulo
CRONOLOGiA Y CONSECUENCIAS
Es muy improbable que un fosil represente el ultimo individuo de un taxon extinto y entonces hay que guiarse por la edad radiometrica del fosil mas reciente conocido lIamada fecha de ultima presencia- como unica aproximacion posible a la fecha de extincion del taxon
Como se explico al inicio del capitulo (vease Contexto Temporal) el registro fosH de los mamiferos terrestres de las Antillas Mayores adolece de insuficiente sustentacion radiometrica No obstante la distribucion temporal de los escasos fechados disponibles para el Cuaternario de la region abarca etapas criticas del proceso de extincion experimentado por la referida fauna a 10 largo de los ultimos 250 mil afios
De las 52 especies que es posible reconocer actualmente como miembros extintos de la fauna cuatemaria de mamiferos terrestres de las Antillas Mayores y grupos de islas perifericas los restos fosiles de 23 de elIas (442) han sido fechados radiometricamente o se han hallado asociados estratigraficamente de manera inequivoca con otro material fechado De estos 23 fechados 3 corresponden al interglacial Sangamon 4 pertenecen a la secuencia Pleistoceno terminal - Holoceno temprano 10 se ubican en ellapso entre el arribo de los aborigenes y la incursion europea y 6 son postcolombinos (a los cuales hay que sumar otras dos especies extinguidas en los siglos XIX y xx) A partir de estas 25 especies avaladas por fechas de ultima presencia distribuidas entre los cuatro intervalos temporales sefialados hemos compuesto la Tabla 45 en la que ademas aparecen las restantes 27 especies (carentes de fechado radiometrico) ubicadas en el intervalo temporal mas probable sobre la base de (1) la asociacion estratigrafica de sus restos con evidencias culturales (amerindias 0 europeas) 0 (2) la ausencia de dicha asociacion Sobre estos criterios de ubicacion temporal procede hacer dos aclaraciones
324 BIOGEOGRAFIA
TABLA 45 Intervalos temporales mas probables para las extinciones cuatemarias de los mamiferos terrestres de las Antillas Mayores y grupos de islas perifericas sobre la base de las evidencias siguientes (l) ultimo ejemplar colectado u observado en vida (2) fechado radiometrico mas tardio en material de la especie (3) asociaci6n estratigrafica mas tardia con otro material fechado (4) asociaci6n estratigrafica con restos de animales introducidos restos humanos u objetos culturales propios del intervale (5) falta de asociaci6n con evidencias culturales [Los ntimeros entre parentesis en la columna Evidencia significan adscripci6n tentativa al intervalo] Los estimados del peso corporal (en kg) estan tomados de la literatura 0 inferidos comparativamente y distribuidos entre
las clases siguientes (a) S 01 (b) 01 a lt 1 (c) 1 a lt 10 (d) 10 a lt 100 (e) 100 a lt 200 (j) ~ 200 Las
abreviaturas que designan las islas son las de la Tabla 44
F amilialEspecie Peso IsJa Evidencia Fuente
I httervalo htterg1acial Sangamon (gt80 kaAP)
HEPTAXODONTlNAE
Amblyrhiza inundata e AS 3 McFarJane et al 1998a
CLIDOMYINAE
Clidomys osborni d JA 3 Ibid
Clidomys parvus d JA 3 Ibid
II httervalo Pleistoceno terminal ~ Holoceno temprano (20-6 kaAP)
MEGALONYCHIDAE
Aeratoenus antillensis d CD 5 Arredondo 1975 a
Aeratoenus odontrigonus d PR 3 McFarJane 1999b
Aeratoenus simorhynehus d LE 2 Rega et al 2002
M egaloenus zile f LE 2 MacPhee et al 2000
SOLENODONTIDAE
Solenodon arredondoi e CD 5 Morgan y Ottenwalder 1993
PnHECIIDAE
Paralouatta varonai d CD 5 MacPhee 1996a
EcHIMYIDAE
Heteropsomys antillensis c PR 5 McFarJane 1999b
Heteropsomys insulans c PR 5 Ibid
Puertoricomys eorozalus c PR 5 Woods 1989a
CAPROMYIDAE
P lagiodontia araeum c LE 5 MacPhee et al 2000
Macrocapromys latus c CD 5 Varona y Arredondo 1979
(Continua)
325 EXTINCIONES DEL CUATERNARIO
TABLA 45 (Continuaci6n)
F ami1ia1Especie Peso Isla Evidencia Fuente
M acrocapromys latus c CU 5 Varona y Arredondo 1979
Mesocapromys kraglievichi c CU 5 Ibid
HEPTAXODONTINAE
Xaymaca fulvopulvis a JA 2 MacPhee y Flemming 2003
III Intervalo Amerindio (6-05 kaAP)
MEGALONYCHIDAE
Acratocnus ye d LE 2 MacPhee et aI 2000
Neocnus comes c LE 2 Ibid
N eocnus dousman c LE 2 Ibid
N eocnus toupiti c LE 2 Ibid
N eocnus gliriform is c CU 4 Cordova et ai 1997
M egalocnus rodens f CU 2 MacPhee et ai 2007
Parocnus browni e CU 2 Steadman et ai 2005
P arocnus serus e LE 2 MacPhee et ai 2000
N ESOPHONTIDAE
N esophontes edithae a PR 2 McFarlane 1999b
PITHECIIDAE
Antillothrix bernensis c LE 3 Rimoli 1977
EcHIMYIDAE
Brotomys contractus a LE 4 Miller 1929c
CAPROMYIDAE
P lagiodontia spelaeum c LE 4 Ibid
M acrocapromys acevedo c CU 4 NUiiez 1990a
HEPTAXODONTINAE
Elasmodontomys obliquus d PR 3 McFarlane 1999b
Iv Intervalo Postcolombino (05 kaAP - Presente)
N ESOPHONTIDAE
N esophontes hypom icrus a LE 24 MacPhee y Fle111111ing 1999
N esophontes zam icrus a LE (4) Ibid
N esophontes m icrus a CU 24 MacPhee et aI 1999
(Continua)
326 BIOGEOGRAFiA
TABLA 45 (Continuacion)
F amilialEspecie Peso Isla Evidencia Fuente
N esophontes m icrus a CD 24 MacPhee et aI 1999
N esophontes m icrus a LE 4 MacPhee et aI 2000
N esophontes sp 1 a IC 4 Morgan 1994
N esophontes sp 2 a IC 4 Ibid
Solenodon marcanoi b LE 4 Ottenwalder 2001
PITHECIIDAE
Xenothrix mcgregori c JA 4 MacPhee y Flemming 1999
EcHIMYIDAE
Boromys offeUa a CD 24 Guarch 1984
Boromys torrei a CD 24 Ibid
Brotomys voratus a LE 4 MacPhee et aI 2000
CAPROMYIDAE
P lagiodontia ipnaeum c LE 4 MacPhee y Flemming 1999
Rhizoplagiodontia lemkei c LE 4 Ibid
Isolobodon portoricensis c PR 24 Ibid
Isolobodon montanus c LE 4 Ibid
Hexolobodon phenax c LE (4) Ibid
Capromys sp c IC 24 Morgan 1994
Geocapromys columbianus c CD 4 MacPhee y Flemming 1999
Geocapromys thoracatus b IS 1 MacPhee y Marx 1997
Geocapromys sp 1 c IC 4 Morgan 1994
Geocapromys sp 2 c IC (4) Ibid
l-IEPTAXODONTINAE
Quemisia gravis c LE (1) MacPhee y Flemming 1999
MURIDAE
Oryzomys antillarum a JA 1 Ibid
1 Flemming y MacPhee (1999) alertaron en contra de la asociacion con otro material fechado Asimismo Morgan (1993) indico que la ausencia de asociacion con evidencias culturales amerindias no es necesariamente un criterio concluyente para ubicar un taxon en el intervalo temporal anterior porque hay depositos en la region que carecen de tales evidencias y son del Holoceno tardio por fechados radiocarbonicos [No obstante
327 EXTlNCIONES DEL CUATERNAR10
la ausencia de una especie en residuarios arqueol6gicos adquiere mayor certidumbre como criterio de ubicaci6n temporal de su extinci6n en la medida que aumenta el numero de residuarios estudiados que en las Antillas Mayores se cuentan por centenares] A falta de fechado absoluto de una especie consideramos que estos criterios de ubicaci6n temporal son mas utiles que la alternativa de excluir del analisis a la especie
2 En los trabajos sobre biogeografia de mamiferos antillanos los arque610gos mas citados como autoridades para establecer el inicio de la presencia aborigen en las Antillas Mayores (Kozlowski 1974 Rouse y Allaire 1978 Rouse 1989 1992 Wilson 2001) difieren al respecto entre los 6 kaAP y los 45 kaAP (pero vease Pagan et al 200523) y hemos adoptado la mas temprana de estas fechas para el comienzo del intervalo amerindio en la Tabla 45 De todos modos cualquiera que haya sido la fecha esta no es aplicable por igual al conjunto de las Antillas Mayores porque segun algunos de los autores citados el poblamiento comenz6 por Cuba y La Espanola 6 mil anos atras continu6 en Puerto Rico hace alrededor de 3 mil anos (pero vease Burney et al 1994) yen Jamaica hace menos de 2 mil e incluso a las Islas Caiman ni siquiera llegaron [Las Islas Caiman descubiertas por Col6n en 1503 carecen absolutamente de evidencia amerindia (Stokes y Keegan 1996)]
Si los datos de la Tabla 45 representan una aceptable aproximaci6n a la realidad el proceso de las extinciones cuaternarias de los mamiferos terrestres de las Antillas Mayores y grupos de islas perifericas debi6 caracterizarse entre otros por los eventos siguientes
Aunque obviamente la fauna cuaternaria debi6 ser mucho mas numerosa que 10 que refleja el registro paleontol6gico (Morgan y Woods 1986) 52 especies (81 de la fauna conocida) desaparecieron (sin reposici6n) durante los ultimos 250 mil anos
- Al parecer las extinciones se concentraron en las especies de mayor tamano Asi el peso promedio estimado para las especies de la etapa preantr6pica (intervalos I + II de la Tabla 45) es 350 kg es 328 kg para las especies del intervalo III (amerindio) y es s610 25 kg (c1ases del peso a b c) para las especies del intervalo IV (postcolombino) Ninguna especie aut6ctona de mamifero mayor de 6 kg existe hoy en la regi6n
- De acuerdo con la distribuci6n temporal de las extinciones 61 especies sobrevivieron al interglacial Sangamon (intervalo 1)48 sobrevivieron a la transici6n del Pleistoceno terminal al Holoceno temprano (intervalo II) 34 sobrevivieron al periodo amerindio (intervalo III) y s610 12 especies han sobrevivido desde la Ilegada de los europeos (intervalo IV) Las tasas de extinci6n correspondientes (exceptuado el intervalo I) son una especie cada 1167 anos en el intervalo II una cada 393 anos en el intervalo III y una cada 23 anos en el intervalo IV
- Durante todo el proceso de las extinciones cuaternarias la perdida de especies por isla fue de 91 en La Espanola 83 en Jamaica y 62 en Cuba Bahamas retuvo su unica especie (aunque extirpada en varias de las islas) Puerto Rico e Islas Caiman perdieron todas sus especies (6 y 5 respectivamente) y las Islas Swan Virgenes y Anguila - San Martin perdieron sus respectivas especies (micas
328 BIOGEOGRAFiA
De los cuatro 6rdenes de mamiferos representados en la fauna Pilosa y Primates desaparecieron totalmente Soricomorpha y Rodentia perdieron 86 y 74 de sus especies respectivamente
CAUSALIDAD
CAUSAS NATURALES
Los cicIos glaciales e interglaciales acaecidos en el Pleistoceno causaron cambios ambientales dnisticos en las Antillas con la concomitante alternancia de cIimas frios (secos) y calidos (hUmedos) asi como de descensos y ascensos del nivel del mar aumento y disminuci6n de las areas territoriales) induso (aunque con menor intensidad) en el Holoceno Estos eventos paleogeograficos han sido consistentemente documentados (Mayo y Kartashov 1972 Bonnatti y Gartner 1973 Ortega y Arcia 1982 Prance 1982 Schubert y Medina 1982 Dodge et al 1983 Ortega [1984] Haq et al 1987 Schubert 1989 Hodell et al 1991 Guilderson et al 1994 Oppo 1997 Emslie 1998 Curtis et al 2001 Iturralde-Vinent 2003) y de ellos brindamos a continuaci6n una breve resena
Finalizando el periodo interglacial anterior a la ultima glaciaci6n (Wisconsin) el mar estaba a 8-10 m por encima de su nivel actual Cuando el avance glacial del Wisconsin a1canz6 su culminaci6n (eillamado Ultimo Maximo Glacial) hace 18 ka el mar habia descendido 120 m con respecto a dicho nivel y las islas del Caribe habian incrementado considerablemente sus extensiones subaereas en comparaci6n con las que presentan hoy dia por ejemplo Puerto Rico y las Islas Virgenes se conectaron territorialmente y las Bahamas se unieron en cinco gran des islas Prevalecia un regimen frio y seco y condiciones de aridez --debidas a una reducci6n de las precipitaciones (posiblemente a menos de 440 mmlano)- y los ambientes xericos se habian extendido a traves de las islas y bancos emergidos y abundaban los pinares (los habitats mesicos estaban restringidos a parcelas relictas en 10 alto de las montanas) En el Mar Caribe y el Golfo de Mexico la temperatura superficial del mar era 5degC mas baja que la actual y en las llanuras costeras y alturas maximas eran respectivamente 2-3degC y 6degC mas bajas Los limites de las zonas ecol6gicas altitudinales descendieron varios cientos de metros y cada zona redujo su extensi6n vertical al aumentar el gradiente de temperatura Ellimite inferior de la zona periglacial
elevaci6n donde la temperatura media minima a1canza los OdegC) de la Cordillera Central de la Republica Dominicana (altura maxima del Caribe insular con evidencia de topografia glacial) se considera que estuvo entre los 1500 y 1800 mlnm (actualmente esta a 2800 mlnm) y se puede estimar que dicho limite era inferior en Cuba por estar mas al norte que la Republica Dominicana No obstante durante este periodo la temperatura experiment6 episodios cortos (unas pocas decadas) de ascensos repetidos
Un signifieativo componente bi6tieo de las referidas condiciones paleoambientales 10 constituy6 la espectacular irradiaci6n evolucionaria experimentada por un grupo numeroso de aves rapaees diurnas y nocturnas (Wetmore 1937 Brodkorb 1969 Kurotchkin y Mayo 1973 Arredondo 1976 Olson 1978 Olson y Hilgartner 1982
329 EXTINCIONES DEL ClJATERNARIO
Suarez 2000a b 2004 Suarez y Emslie 2003) Los mamiferos de las Antillas Mayores ofrecieron a estos depredadores aviares una amplia gama de tamanos de presa que en ausencia de mamiferos del orden Carnivora en la region permitio a estas aves irradiarse a este nicho alimentario con la evolucion de una variedad de especies de muy diversa tall a corporal incluidas formas gigantes Adquirir un cuerpo gigante les propicio el acceso a recursos alimentarios que no estaban siendo explotados por ningun otro depredador (Alcover y McMinn 1994) A Cuba corresponde el mejor registro fosil de rapaces cuaternarias caribenas con siete generos de especies diurnas Dentro de estas aves existieron tres gavilanes gigantes (Accipitridae) tres carairas (Falconidae) y un condor (Cathartidae) asi como cuatro generos de especies nocturnas dos lechuzas (Tytonidae) y tres Mhos (Strigidae) dos de ell os no igualados en tamano en todo el mundo Algunos de estos generos tuvieron representacion en otras islas antillanas como La Espanola Jamaica Bahamas y Gran Caiman
Al retirarse hacia el norte los hielos de la glaciacion Wisconsin el clima se tomo mas hUmedo en las Antillas durante el Holoceno temprano y el bosque mesico se expandio en la region insular El nivel del mar ascendio acompanado de mayor pluviosidad alrededor de los 7 kaAP las Islas Virgenes quedaron separadas entre si y de Puerto Rico y las Bahamas perdieron la mayor parte de su territorio Cerca de los 3 kaAP las condiciones secas retornaron a la region e hicieron desaparecer buena parte del bosque mesico Nuevamente el clima comenzo a adquirir sus condiciones actuales hacia los 25 kaAP aproximadamente y hoy transita por una fase mesica con vegetacion xerica confinada en parcelas relictas
Sin duda estos y otros eventos naturales incidieron negativamente en la sobrevivencia de mamiferos terrestres en las Antillas Mayores asi como en los grupos de pequenas islas perifericas (Pregill y Olson 1981 Olson 2006)
McFarlane et al (1998a) demostraron convincentemente como Amblyrhiza el gigante de los roedores antillanos endemico de la pequena isla de Anguila sucumbio a las repetidas y abruptas perdidas de su habitat durante el interglacial Sangamon por las nipidas y severas oscilaciones del nivel del mar (documentadas por Neumann y Hearty 1996) al no poder responder adecuadamente a tan catastroficos eventos mediante el ajuste de su masa corporal media y otro tanto debio ocurrirle en Jamaica al otro roedor gigante Clidomys (McFarlane et al 1998b) emparentado con Amblyrhiza en la misma epoca (Tabla 45 intervalo I)
El ascenso del nivel del mar seguramente promovio la extincion de mamiferos durante la transicion del Pleistoceno al Holoceno (Tabla 45 intervalo II) con la reduccion del tamano de las Islas por las transgresiones marinas (Pregill y Olson 1981 Morgan 1993) Esto ultimo haria maximamente severa la competicion asf como la presion de los depredadores [Steadman et al (2005) no yen por que el calentamiento global ocurrido durante la transicion Pleistoceno-Holoceno habria de tener un efecto negativo sobre los perezosos siendo sus descendientes actuales especies tropic ales 0 subtropicales pero no se trata del efecto
330 BIOGEOGRAFiA
directo del cambio de temperatura sobre los animales sino del efecto indirecto y progresivo que dicho cambio genera a largo plazo al reducir el territorio insular disponible]
Por esta epoca tambien desaparecieron especies de tall a mayor como Megalocnus zile en La Espanola y otras de menor tamano (en Cuba) pero que representan la tall a maxima en sus respectivos contextos taxonomicos del Cuaternario como Solenodon arredondoi Paralouatta varonai y Macrocapromys acevedo Un mamifero puede ser grande solo a costa de reducir su densidad poblacional y esto 10 hace mis vulnerable a la extinci6n sobre todo en una isla (McNab 1994 McFarlane et al 1998a) Incidentalmente Alcover et al (1998a) afirmaron que larelacion entre la talla corporal y la extincion no siempre se cumple en las especies de menor tamano y citaron como ejemplo que entre las musarafias todas las especies de Nesophontes desaparecieron mientras sobreviven especies de Solenodon pero la comparacion correcta en todo caso serfa entre los solenodones vivientes y el gigante Solenodon arredondoi y entre Nesophontes edithae -el gigante de los nesofontes extinguido en el periodo amerindio- y el resto de sus congeneres que sobrevivieron hasta los primeros siglos de la colonizacion europea (Tabla 45 intervalos III y IV)
CAUSAS ANTROPOGENICAS
Intervalo Amerindio
La mayoria de los autores que se han referido a las extinciones acaecidas durante este intervalo (entre elIos paradigmaticamente Martin 1984a b) han responsabilizado a los aborfgenes (que se supone comenzaron a poblar las Antillas Mayores hace 6 mil afios) con la desaparicion de numerosas especies de mamiferos terrestres en la region principalmente por la via de la sobrecaza y la introduccion de animales domesticos devenidos depredadores de la fauna autoctona (Morgan y Woods 1986) Este efecto negativo de la presencia indigena sobre los mamiferos de la region ha sido dramatizado por algunos autores en terminos de holocausto faunal Sin embargo pensamos que se ha exagerado Como han expresado MacPhee et al (19993) cuando uno busca evidencia dura de complicidad humana directa en [estas] extinciones no hay mucho que encontrar Las siguientes son algunas consideraciones a tener en cuenta en tal sentido
El registro radiometrico mas joven conocido hasta ahora para un perezoso antillano (MegalocnllS rodens) es de 42 kaAP (MacPhee et ai 2007) Si se acepta que los aborfgenes entraron en Cuba hace aproximadamente 6 mil anos su convivencia con los perezosos debe haber durado alrededor de 1800 afios (0 mas porque dificilmente el referfdo fechado radiometrico haya correspondido al ultimo sobreviviente de la especie) Esta cifra contradice la hipotesis del holocausto faunal segun la cualla extincion de los mamiferos insulares de mayor talla debe ocurrir con extrema rapidez -a escala de decadas 0 de centurias- a partir de su primer contacto con los humanos (Martin y Steadman 1999) A pesar de las repetidas afirmaciones en contrario (Harrington 1921 Miller 1929c Herrera [1945] Tabio 1951 Alvarez 1951a 1957 Arredondo
331 EXTINCIONES DEL CUATERNARIO
1959 1960a Veloz y Ortega 1973 Mayo 1980b Rodriguez et al 1984 Castellanos y Pino 1985 Pino y Castellanos 1985 Nunez 1990b Gutierrez 1996 Arredondo 2004) en su mayoria basadas en evidencias no exentas de posibles accidentes de deposicion (0 de reubicacion postdeposicional) en recintos cavernarios (Diaz-Franco [2006b D no se ha demostrado inequivocamente que los aborigenes antillanos se alimentaran de perezosos regularmente Sus restos en residuarios de alimentos son escasisimos en toda la region y esto contrasta con los abundantes restos de jutias y otros mamiferos de menor talla que aparecen en esos residuarios demostrativos de la preferencia de los paleoindios por estos otros mamiferos
Los mamiferos mas importantes en la economia prehistorica de las Antillas Mayores fueron las jutias Capromys y Geocapromys en Cuba Geocapromys en Jamaica e Isolobodon en La Espanola Puerto Rico e Islas Virgenes (Aguayo 1950b Tablo 1951 Rimoli 1972 Wing 2001 Diaz-Franco [2006b D Wilkins (2001) estudi61ajutia viviente de Jamaica (Geocapromys browni) en un sitio ocupado por los aborigenes durante 100 anos y no encontr6 evidencias de que durante ese tiempo la especie haya sido sobreexplotada Asimismo en Cuba las jutias conga carabali y andaraz son escasas en depositos pleistoholocenicos en cambio son muy numerosas en residuarios indigenas y de haber sido sobreexplotadas 10 mas probable es que no hubieran sobrevivido hasta hoy
La habilidad de los humanos para penetrar en profundidad el bosque tropical y mantenerse en este sustentablemente ha sido un evento comparativamente reciente (Beran y Steele 1994) En el caso antillano la caza tierra adentro ha sido explicada como un proceso tardio de desplazamiento a partir de la explotacion de abundantes recursos marinos que ofrecian condiciones 6ptimas para el sedentarismo de las comunidades aborigenes costeras (C6rdova y Arredondo 1988 Brito 2005) Peces y mariscos dominan la arqueologia costera de las Islas antillanas en tanto el consumo de recursos terrestres (mamfferos incluidos) aumenta progresivamente en la medida que los residuarios estudiados se alejan de la costa (Scudder 1991 Wilkins 2001) En las Antillas Mayores el componente terrestre de la dieta aborigen es solo 20-30 del material recobrado en los residuarios (Wing 1977 1989) Y debio ser menor alin porque el componente vegetal de la dieta no se ha preservado Por otra parte los aborigenes cubanos domesticaron la jutia conga y seguramente la reprodujeron para comer (Pose et al 1988) practica que probablemente se repitiera con otras jutias en las demas Antillas Mayores
- Fray Bartolome de las Casas hablo de 3 millones de indios en Cuba al inicio de la colonizacion pero los historiadores subsecuentes consideraron exagerada la cifra (Poey 1851 a) No obstante se dispone de dos estimados de la densidad de poblaci6n aborigen en las Antillas Mayores a la llegada de los europeos 18 indigenaslkm2 en Puerto Rico (Rouse 1956) y 1271km2 en La Espanola (Watts 1987) Extrapolando ambos valores entre las cuatro islas principales de las Antillas Mayores tenemos que en la fecha indicada la poblaci6n aborigen pudo estar entre 16 mil y 113 mil indigenas en Puerto Rico entre 21 mil y 145 mil en Jamaica entre 141 mil y 1 mill6n en La Espanola y entre 200 mil y 14 millones en Cuba Entre las cifras maximas y las minimas nos inclinamos
332 BIOGEOGRAFiA
hacia estas ultimas atendiendo a los altos indices de mortalidad infantil y morbilidad que se evidencian en las osamentas de los pobladores prehispanicos (Martinez y Rives 1990) Uno de los hechos que apuntan hacia el bajo numero de aborigenes en las Antillas Mayores es 10 rapido que desaparecieron despues de la llegada de los europeos pnkticamente sin dejar mestizaje con estos 10 cual contrasta ostensiblemente con la situacion en la America continentaL
En suma un grupo humano (1) demograiicamente irrelevante y controlado por altos indices de morbilidad (2) consumidor mayoritariamente de organismos marinos y (3) carente de la tecnologia cinegetica necesaria para poder sobreexplotar los mamiferos que Ie sirvieron de alimento (a los cuales accedio tardiamente en su desarrollo) no es concebible que haya causado e1 holocausto faunal que se Ie atribuye a partir del error de aplicar a las comunidades aborigenes antillanas un modelo cultural de la prehistoria continentaL
Tampoco hay evidencias de que los aborigenes hayan introducido carnivoros con suficiente capacidad depredadora salvo el perro Morgan y Otlenwaldder (1993) consideraron que la depredacion por perros durante el periodo amerindio es la explicacion mas plausible para la extincion del almiqui gigante de Cuba pero como se puede observar (Tabla 54) Solenodon arredondoi aparece en el intervalo II por falta de asociacion con evidencias culturales
Consecuentemente el accionar de los aborigenes y de sus animales domesticos directamente sobre la fauna no parece haber sido por sf solo un factor determinante de la extincion de mamiferos terrestres durante el periodo amerindio en las Antillas (pero vease Steadman et al 2005)
Hay que recordar en este senti do que alin en el Holoceno tardio ocurrieron cambios clirruiticos de consideracion (vease Causas Naturales) Por ejemplo Curtis et al (2001) explicaron que la extincion de algunos ani males mesofilicos de las alturas montanosas de La Espanola pudo haber antecedido a cualquier impacto humano sigrIificativo en esos lugares por coincidir dicha extincion con una tendencia a la aridez documentada isotopicamente y con evidencias de la perdida del bosque mesico ocurridas despues de 3kaAP Yen Jamaica hace solo 2shy3 mil aDos el area de la Bahia de Jackson sustentaba una gran biomasa de vertebrados en bosque hinnedo a juzgar por la cantidad y diversidad de taxones presentes en material fosil derivado de la depredacion por lechuzas hoy el area es la mas seca de la isla con matorral xerofitico y pobreza de fauna (MacPhee 1996a) Yalm en nuestros dias huracanes de gran intensidad aniquilan poblaciones de vertebrados (que no se recuperan) en is las pequenas y llanas del Caribe (Brooks y Smith 2001)
En medio de la incertidumbre sobre la verdadera causa (0 causas) de las extinciones de mamiferos en el periodo amerindio MacPhee y Marx (1997) han propuesto su hipotesis de la hipermorbilidad la cual estipula colapsos poblacionales relativamente rapidos con la introduccion de enfermedades de alta letalidad Estos autores han razonado que
Las islas son los lugares mas apropiados para experimentar perdidas catastroficas causadas por enfermedades Los mamiferos no solo serian susceptibles en virtud de su prolongado aislamiento biologico sino que en los confines de una isla la epizootia
333 EXTINCIONES DEL CUATERNARIO
tendria la potencialidad de infectar simultaneamente a la rnayoria de los individuos de vadas especies Los neonatos son inmunologicamente inmaduros y sufririan desproporcionadamente cualquier infeccion hipervirulenta La perdida excesiva de neonatos y juveniles tendria efectos severos para la sobrevivencia de la poblacion si esta se reduce por debajo de cierto nivel sobreviene la extincion porque no hay posibilidad de restitucion en el periodo postpanzootico Las especies de mayor talla corporal van a ser las mas vulnerables a la extincion dadas sus bajas tasas reproductivas largos periodos de gestacion y reducido numero de progenie Las especies de menor tamano --caracterizadas por altas tasas reproductivas periodos cortos de gestacion y mayor numero de neonatos por parto- tambien serian fuertemente diezmadas pero muchas sobrevivirian porque su estrategia reproductiva les permite perder mayor numero de individuos Las perdidas deben ocurrir relativamente rapido (a escala de decadas) mientras que otras hipotesis (digamos los cambios climaticos) permiten intervalos mucho mas largos (a escala de siglos 0 de milenios) entre la aparicion de las causas y la produccion de sus efectos Por ejemplo todo indica que los nesofontes se extinguieron muy rapidamente y seria dificil poder invocar un impacto humano directo 0 argumentar fenomenos climaticos que pudieran explicar plausiblemente perdidas tan rapidas
MacPhee y Marx (1997 191) han dicho que por el momento la hipotesis de la hipermorbilidad no pasa de ser una abstraccion en espera de adecuada sustentacion empirica Llama la atencion que linicamente en el periodo amerindio (Tabla 45 intervalo 1Il) las extinciones abarcaron todas las clases del peso corporal (de la a a laj) y que ninguna de las especies que sobrevivieron hasta el periodo postcolombino (Tabla 45 intervalo IV mas las especies vivientes) sobrepasa los 10 kg (clases del peso a b c) en clara coincidencia con los presupuestos de la hipotetica hipermorbilidad
Intervalo Postcolornbino
Por 10 menos 37 especies de mamiferos terrestres vivian en las Antillas Mayores y grupos de Islas perifericas a la lIegada de los europeos en 1492 De elIas se han extinguido 23 shymas especies que en cualquiera de los periodos precedentes (Tabla 45~ y solo subsisten 12 especies repartidas entre Cuba (8 especies) La Espanola (2) Jamaica (1) y Bahamas (1) varias de estas en peligro critico de extincion Solo dos de las especies extinguidas durante los ultimos 500 anos se conocieron en estado viviente los roedores Oryzomys antillarum de Jamaica (extinguido en 1877) y Geocapromys thoracatus de las Islas Swan (extinguido en 1955) Las causas de las extinciones ocurridas durante este periodo son sobradamente conocidas destruccion de habitats introducci6n de animales exoticos (domesticos y comensales) y demas presiones asociadas a la actividad humana entre las cuales posiblemente habria que incluir brotes de hipermorbilidad
Durante mucho tiempo se ha afirmado la posibilidad de que especies de Nesophontes hayan sobrevivido hasta las primeras decadas del siglo xx y que incluso continuen existiendo en lugares remotos de Cuba 0 La Espanola (Allen 1918 1942 Miller 1930 Aguayo 1950a Arredondo 1960b Fischer 1977 MacFadden 1980 Woods y
334 BIOGEOGRAFiA
Ottenwalder 1991) Actualmente no existe evidencia radiometrica de que alguna especie de Nesophontes haya sobrevivido al siglo xv las fechas radiocarbonicas calibradas obtenidas en muestras de Nesophontes de Cuba y La Espanola resultaron no mas tardias que los siglos XIII y XIV (MacPhee et al 1999) Por otra parte las expediciones efectuadas a fines del siglo xx con el proposito de localizar individuos vivientes del genero en La Espanola no produjeron la menor indicacion afirmativa al respecto (Woods et al 1985) Ademas a traves de las Antillas Mayores no aparecen restos de Nesophontes en los depositos formados actualmente por las lechuzas
EPiLOGO
Las Antillas Mayores son islas oceanicas no han formado parte de un continente (con la posible excepcion de una parte de Jamaica hasta el Eoceno Medio hace 45 Ma) y durante un tiempo geologico considerable (desde el Eoceno Superior hace 40 Ma) han permanecido emergidas y separadas de los continentes vecinos Consecuentemente los mamiferos terrestres (no volantes) autoctonos de estas islas constituyen una fauna oceanica que por ende cumple con todas las caracteristicas convencionales reconocidas para este tipo de entidad biogeografica
- Baja diversidad taxonomica a nivel de las categorias superiores (solo cuatro ordenes) en comparacion con islas plataformicas continentales (0 con areas continentales) de tamano y diversidad ecologica simi lares
- Alto grado de endemismo a nivel de genero (96) y familia (50) en respuesta al prolongado aislamiento
- Desbalance ecologico de la estructura trofica ausencia de mamiferos camivoros y ungulados
- Espectaculares procesos de irradiaci6n evolucionaria como resultado de la colonizacion de territorios libres de competidores y depredadores
Tendencias evolucionarias extremas gigantismo
- Control a cargo de aves rapaces con tendencia at gigantismo en ausencia de mamiferos miembros del orden Carnivora
- Mas especies de reptiles y de murcielagos que de mamiferos terrestres relaciones que en los continentes se invierten
Alta tasa de extincion de especies (84) sin reposicion
Preservacion de especies filogeneticamente arcaicas (los llamados fosiles vivientes) los almiquies

315 rRRADlAcr6~ EVOLUCrONARIA
s610 puede explicarse razonablemente como el resultado de movimientos de esos animales entre las islas en el Cuaternario
De esta objecion resulta dificil disentir [El poblamiento entre islas diferenciado solo a nivel especifico no es probable que sea anterior al Pleistoceno (Simpson 1956 15)]
Debe tenerse en cuenta que la hipotesis de la vicarianza isla-isla alude a las tres islas septentrionales de las Antillas Mayores porque como se explico antes (vease El Barco Funeral Vikingo) Jamaica no existia como isla durante el lapso (Eoceno Superior shyOligoceno) consensualmente reconocido como el mas probable para la colonizacion de las proto-Antill as Mayores por la mayo ria de los mamiferos terrestres Despues de emerger como isla en el Mioceno Medio Jamaica debe haber recibido sus mamiferos terrestres por dispersion isla-isla (Davalos 2004376) y solo en el Cuaternario parece haberrecibido (por dispersion marina desde Centroamerica) al roedor murido Oryzomys
Seguramente tanto la dispersion como la vicarianza contribuyeron a que los mamiferos terrestres se irradiaran a traves de las Antillas Mayores y grupos de islas perifericas pero las maneras y tiempos en que estos mecanismos operaron a 10 largo de dicho proceso son aun en gran medida motivos de desacuerdo e incertidumbre
IRRADIACION EVOLUCIONARIA
A continuaci6n revisamos someramente las distribuciones conocidas de los diferentes grupos de mamiferos terrestres a traves de las Islas principales de las Antillas Mayores y grupos de Islas perifericas incluidas las interpretaciones y conjeturas que la literatura reciente recoge en relaci6n con la manera en que dichas distribuciones pudieron haberse conformado durante la irradiaci6n evolucionaria de los referidos grupos de mamiferos Manifestaciones de esta irradiaci6n pueden inferirse unicamente a partir de los taxones presentes en el Cuatemario pues solo 6 de las especies conocidas corresponden al Terciario
GRUPOS ORlGINARIOS DE SURAMERICA
PEREZOSOS (Megalonychidae)
Aparte de un genero miocenico de Cuba y otro del Oligoceno de Puerto Rico se conocen 4 generos (12 especies) de perezosos cuaternarios (todos extintos) restringidos a las tres islas septentrionales de las Antillas Mayores Acratocnus Neocnus Megalocnus y Parocnus Los cuatro generos estan representados en Cuba y La Espanola y Acratocnus 10 esta ademas (como unico genero de perezoso) en Puerto Rico Esta distribuci6n ha sido interpretada por MacPhee et al (2000) eomo resultado de la vicarianza oligomiocenica entre estas islas y [dado que el Banco Puerto Rico - Islas Virgenes fue la entrada de GAARlandia a las Antillas Mayores] se preguntaron si la ausencia de los otros tres generos en Puerto Rico se debe a que nunca estuvieron en ese territorio 0 a que sus
316 BIOGEOGRAFiA
fosiles atm no han sido hallados Pero como hemos visto Rega et al (2002) atribuyeron dicha distribucion a la dispersion interinsular en el Cuatemario
Por otra parte varios amilisis c1adisticos recientes ubi can a Choloepus -genero Reciente conocido solo de Centro- y Suramerica- como taxon hermano de Aeratoenus exc1uidos los demas megaloniquidos antillanos Para White y MacPhee (2001228)
tan estrecha relacion sugiere que la divergencia entre ambos [generos] ocurrio despues de que Aeratoenus divergio de Neoenus Si esta relacion implica que Aeratoenus y Neoenus se dispersaron separadamente desde Suramerica a las AntilIas mientras Choloepus permanecia en el continente (siendo necesarias al menos tres invasiones de perezosos) 0 si Choloepus invadio Suramerica desde las Antillas es algo que el material disponible no permite determinar
La diversidad adaptativa de los perezosos se manifiesta en (1) la utilizacion del habitat especies arboreas en Neoenus y Aeratoenus y terrestres en Paroenus y Megaloenus (Webb 1985a White [1993] White et al 1996 Arredondo [1999]) y en (2) la gama de tallas corporales desde los pequenos Neoenus hasta Megaloenus rodens el mayor de los mamiferos terrestres antillanos (su peso adulto estimado en 270 kg seglin Paula 19679) esto sugiere que los perezosos se irradiaron a nichos que en contextos continentales probablemente estarian ocupados por miembros de otros c1ados (MacPhee e IturraldeshyVinent 2000)
No esta claro si el abanico de tamanos entre estos perezosos enmascara alguna tendencia al gigantismo Por 10 pronto Anderson y Handley (2002) encontraron 10 contrario en perezosos (Bradypus) de las Islas de Bocas del Toro (Panama) shyseparadas secuencialmente del area continental vecina por ascensos del nivel del mar durante los ultimos 10 ka En las cinco islas los perezosos son menores que su contraparte continental y las diferencias entre las tallas medias de las cinco poblaciones expresan una reduccion lineal significativa de la talla corporal en funcion de la edad geologica de cada isla sin relacion alguna con el area de las islas 0 la distancia que las separa del continente
MONOS (Pitheciidae)
Los primates de las Antillas Mayores (cuatro especies extintas) constituyen un grupo monofiletico con Paralouatta (en Cuba) y Antillothrix (en La Espanola) como taxones hermanos seguidos en la escalera c1adistica por Xenothrix (en Jamaica) y finalmente Callieebus como el pariente continental mas cercano esto implica una sola invasion desde Suramerica antes del Mioceno Inferior (fecha mas temprana conocida para los primates fosiles antillanos) y por tanto en consonancia con GAARlandia como la via de colonizacion (Horovitz y MacPhee 1996 1999 Horovitz [1997] MacPhee y Horovitz 2004)
En desacuerdo parcial con la anterior interpretacion la filogenia de este grupo de monos obtenida por Davalos (2004) indica un antecesor comun en Cuba La Espanola y Jamaica
317 TRRADIACTON EVOLUCIONARIA
compatible con la hipotetica conexion -propuesta tentativamente por Iturralde-Vinent y MacPhee (199980-84)- entre las Montanas Azules del este de Jamaica y la peninsula suroccidental de La Espanola en la epoca de GAARlandia
A pesar de que los primates antillanos pertenecen a un grupo viviente de monos muy pequenos (los monos titles) fueron comparativamente grandes en relacion con los platirrinos modernos y sucesivamente mayores en la direccion Antillothrix Xenothrix y Paralouatta con P marianae como el mayor platirrino conocido del Mioceno neotropical (MacPhee 2005) MacPhee et al (1994) consideraron que Xenothrix debio ser un mono de andar lento y al parecer las especies de Paralouatta abandonaron la arborealidad para adoptar un modo de vida mas terrestre (MacPhee 1996a MacPhee et al 2003 MacPhee y Meldrum 2006)
ROEOORES (Echimyidae Capromyidae Heptaxodontidae)
La edad de Zazamys en Cuba demuestra que la invasion de los roedores histricognatos en las Antillas Mayores comenzo antes del Mioceno Inferior por otra parte los representantes mas antiguos de esa irradiacion evolucionaria se encuentran en La Espanola y Puerto Rico Ambas evidencias son consistentes con la hipotesis de GAARlandia (Woods 1982 1988 Woods et al 2001)
No esta claro si los capromidos evolucionaron a partir de un equimido ancestral 0 de un protocapromido 0 si los equimidos antillanos (I sub familia 3 generos y 7 especies todas extintas) forman parte de la irradiacion de los capromidos (4 subfamilias 9 generos y 27 especies 10 vivientes) Kratochvil et al (1978b) y Rodriguez et al (1979) consideraron que Capromys nan us es un eslab6n entre Echimyidae y Capromyidae pero Mesocapromys nanus esta morfologicamente mucho mas cerca de Hexolobodon y los de mas capromidos que de los equimidos (Woods 1985) Lo que si esta claro segun Woods et al (2001) es que la subfamilia Plagiodontinae endemic a de La Espanola es el linaje basal de los capromidos y que Isolobodon Hexolobodon Capromys Geocapromys Mysateles y Mesocapromys [y supuestamente Macrocapromys] forman parte de la irradiacion temprana de Capromyidae
La subfamilia Capromyinae (5 generos y 17 especies 9 vivientes) parece ser el dado de mas reciente evolucion en la familia con invasion de Cuba desde La Espanola y dispersion desde Cuba (1) por Capromys a las islas Caiman y (2) por Geocapromys a las islas Bahamas Caiman y Jamaica y desde Jamaica a las islas Swan aunque la presencia de Geocapromys en Swan pudiera deberse a introduccion aborigen (Morgan 1985 1988 1989 1993 1994 Woods 1989a 1990) Woods et al (2001) utilizaron datos moleculares para obtener estimados independientes de divergencia entre capromidos segiJn 10 cual Geocapromys divergio cerca de los 17 MaAP Mesocapromys divergio de Capromys hace 15 MaAP en tanto lvlysateles 10 hizo hace 10 MaAP Por su parte las formas de Capromys pilorides y de Mysateles prehensilis en Isla de la Juventud divergieron de sus correspondientes subespecies nominativas en la isla principal a los 24 y 08 MaAP respectivamente
318 BIOGEOGRAFiA
Woods y Howland (I979) describieron adaptaciones morfologicas de la mandibula el hioide y la faringe asociadas a mecanismos de masticacion diferentes en plagiodontinos y caprominos y Varona (1980a) llamo la atencion sobre el gran desarrollo de los molariformes (en relacion con la longitud total del cnineo) que presentan ciertos grupos de jutias fosiles en comparacion con las vivientes 10 que pudiera interpretarse como una adaptacion alimentaria a una vegetacion mas abrasiva por parte de las jutias extinguidas
Segun Woods (1990) la familia Heptaxodontidae (2 subfamilias 5 generos y 6 especies todas extintas) tambien seria parte de la irradiacion de los capromidos con origen en el este de La Espanola 0 en Puerto Rico y subsecuente dispersion al Banco de Anguila - San Martin en las Antillas Menores (pero vease MacPhee y Flemming 2003) Este banco estaria a solo 70 km del Banco Puerto Rico - Islas Virgenes en tiempos de maxima regresion marina y esta proximidad con Puerto Rico -patria de Elasmodontomys obliquus- seguramente tuvo que ver con la presencia en las islas de Anguila y San Martin del heptaxodontino gigante Amblyrhiza relacionado mas estrechamente con los roedores de las Antillas Mayores que con los del resto de las Antillas Menores (Woods 1990 1996) Aunque los roedores suelen aumentar de tamano en las islas (Case 1978) la escala del incremento de la talla en Amblyrhiza (que pudo alcanzar mas de 200 kg de peso adulto) esta fuera de toda proporcion con los patrones conocidos en otros roedores insulares (Biknevicius et 01 1993)
Clidomys (Clidomyinae) de Jamaica difiere marcadamente en su morfologia de los heptaxodontinos (vease ademas MacPhee y Flemming 2003)10 cual dificulta explicar su presencia en esa isla Como Jamaica estuvo bajo el mar hasta el Mioceno Medio el ancestro de Clidomys colonizaria esta isla por dispersion desde el extremo suroccidental de La Espanola despues que los heptaxodontinos se originaron en esta ultima isla (Woods 1989a) Pero Pascual et 01 (I 990) especularon que Clidomys pudiera ser el resultado de una invasion posterior e independiente de los capromidos
GRUPOS ORIGINARIOS DE NORTEAMERICA
El origen de las musaranas antillanas ha resultado ser hasta ahora el mayor enigma biogeografico de la fauna de mamiferos terrestres de la region aunque la mayoria de los autores coinciden en considerarlas originarias de Norteamerica (vease Discus ion y Conclusiones en Poblamiento Continente-Isla) No obstante MacPhee e Iturralde-Vinent (1994) descartaron toda posibilidad de conexion terrestre por el Estrecho de la Florida 0 por el Canal de Yucatan y si el (supuesto) evento vicari ante de Rosen (vease El Arca de Noe) -invocado por MacFadden (1980) para resolver el enigma- fuera minimamente plausible aun asi es dificil creer que las catastrofes que 10 sucedieron (vease Contexto Temporal) hayan barrido con toda esa vieja fauna continental excepto las musaranas circunstancia ya anticipada muy tempranamente por Matthew (l918a)
De otra parte la relaci6n entre Nesophontidae y Solenodontidae permanece irresuelta Si estos linajes no estan estrechamente relacionados entonces es muy posible que
319 IRRADIACION EVOLUCIONARIA
sus distribuciones sean explicadas por mecanismos diferentes (Whidden y Asher 2001)
La descomunal diferencia de talla corporal entre las musaranas pequenas (Nesophontes) y las grandes (Soienodon) seguramente impuso a ambos grupos restricciones distintas relativas a la termorregulacion y las adaptaciones energeticas En Nesophontes la seleccion debio favorecer una alta tasa de metabolismo y crecimiento camadas mas numerosas corta expectativa de vida yalta densidad poblacionaL En Soienodon en cambio la seleccion opero en la direccion de una baja tasa metabolica camadas reducidas crecimiento lento longevidad y baja densidad poblacional (Ottenwalder [1991])
MUSARANAS PEQUENAS (Nesophontidae)
Las musaranas pequenas estan representadas en las Antillas por el genero Nesophontes (y un genero miocenico indeterminado en ambar dominicano) con 6 especies cuaternarias extinguidas distribuidas en Cuba La Espanola Puerto Rico Islas Virgenes e Islas Caiman La mayor de las musaranas pequenas Nesophontes edithae evoluciono en Puerto Rico y su arribo a las Islas Virgenes supone una dispersion sumamente reciente En las Islas Caiman Nesophontes se asemeja mas a la poblacion de Cuba de donde seguramente derivo en el Plio-Pleistoceno (junto con Capromys y Geocapromys) cuando la costa sur de Cuba estuvo mucho mas cerca de las Islas Caiman (lturralde-Vinent 2003fig 4) MacPhee et ai (1994) interpretaron la distribucion de Nesophontes en Cuba y La Espanola como resultado de la vicarianza oligomiocenica ocurrida al formarse el Paso de los Vientos pero la distancia taxonomica entre los lados del Paso es solo de nivel especifico y N micros incluso vivi6 en ambas islas segtln Patterson (1962)
Allen (1918) considero que Nesophontes micrus fue una especie primitiva no modificada para la vida arb6rea 0 fos6rea y por tanto de habitos terrestres Nesophontes edithae de Puerto Rico es mucho mayor y mas robusta que las restantes especies del genero eL indice de robustez de su femur (= anchura minima de la diafisis ongitud total del hueso) muestra que este es tan robusto como el de Solenodon paradoxus un animal dos veces mayor (Morgan et ai 1980) Choate y Birney (1968) postularon un aumento cronoclinal de la taIla corporal en N edithae confirmado mas tarde por McFarlane (1999a) mediante fechados radiocarb6nicos de la secuencia estratignifica en depositos cavernarios de Puerto Rico Solenodon no vivi6 en Puerto Rico y en ausencia de una musarana de mayor tamano Nesophontes alcanzo una talla corporal mayor en esa isla que en La Espanola 0 Cuba (Woods y Eisenberg 1989)
MUSARANAS GRANDES (Solenodontidae)
Los arcaicos almiqufes (genero Soienodon) comprenden cuatro especies una viviente (S cubanus) y otra f6sil (S arredondoi) en Cuba y una viviente (S paradoxus con dos subespecies) y otra f6sil (S marcanoi) en La Espanola Como cabria esperar las dos especies cubanas se asemejan mas entre sf que a las especies de La Espanola pero
320 BIOGEOGRAFiA
enigmaticamente S marcanoi comparte caracteres con S cuban us y S paradoxus (Woods y Eisenberg 1989 Morgan y Ottenwalder 1993 Ottenwalder 200 I) Solenodon arredondoi alcanzo una talla corporal superior a la de cualquiera de las musarafias vivientes a nivel mundial Segun Morgan y Ottenwalder (1993) extrapolando las dimensiones y el peso de los almiquies vivientes S arredondoi debio tener una longitud cabeza-tronco y un peso de 450-550 mm y 1500-2000 g respectivamente en comparacion con S cubanus (360 mm y 800 g) y S paradoxus (390 mm y 1100 g)
Entre todas las especies de almiqufes Solenodon paradoxus es la que presenta los caracteres mas derivados (Otten walder 2001) Si estas relaciones son validas la distribucion de Solenodon entre Cuba y La Espafiola debio discurrir de oeste a este como propuso MacFadden (1980) quien invoco la presencia fosH de S cubanus [pero tambien S arredondoi] en Pinar del Rio para postular que el ancestro continental entr~ por Cuba opinion compartida por Perfit y Williams (1989) y por Ottenwalder ([1991] 2001) El paso de Solenodon a Puerto Rico pudo haberlo impedido la existencia de barreras ecologicas durante la conexion del este de La Espafiola con Puerto Rico 0 por la ausencia de Solenodon en esa parte de La Espafiola en aquella epoca Sin embargo las mismas condiciones ecologicas en el este de La Espafiola no parecen haber representado una barrera efectiva para Nesophontes cuyas mas altas tasa de reproduccion y densidad poblacional presumiblemente Ie permitieron expandirse flpidamente a traves de La Espafiola (Ottenwalder [1991])
Es opinion generalizada que los almiqufes antillanos constituyen el grupo de mamiferos que primero se emplazo en el territorio insular (MacPhee 2005) y en tal sentido es de interes la evidencia sobre especificidad parasito-hospedante aportada por Cruz (1980) quien encontro que la especie de acaro (genero Cubanochirus) que parasita a los almiquies es mucho mas primitiva que la especie (del mismo genero) que paras ita a las jutias y de aqui concluyo que dicho genero de acaro debio evolucionar primero en Solenodontidae y posteriormente en Capromyidae al entrar en contacto representantes de ambas familias en Cuba 0 La Espafiola
Recientemente Roca (2004) y Roca et al (2004) dieron a conocer que los resultados de su estudio genetico-molecular sobre los almiquies indican una divergencia de 25 Ma entre Solenodon cuban us y S paradoxus -compatible con la disyuncion tectonica entre Cuba y La Espafiola (Iturralde-Vinent y MacPhee 1999 McPhee et al 2003)-- y una separacion de ambas especies comparable en magnitud a la divergencia entre familias de mamlferos Consecuentemente propusieron que Solenodon cubanus fuera clasificada en el genero Atopogaie como indico Cabrera (1925) hace 82 afios
DISCUSION Y CONCLUSIONES
Las musarafias y los roedores antillanos (excepto Oryzomys) se irradiaron tanto de sus respectivos ancestros continentales que de estos 6rdenes se reconocen familias y subfamilias endemicas de la region en contraste con los perezosos (una subfamilia y tres
321 IRRADIACION EVOLUCIONARIA
tribus endemicas) y los monos (una tribu endemica) como expresion de diferentes niveles de diversificacion evolucionaria Este proceso es dificil de explicar (traUindose de c1ados que pnicticamente comparten la misma antigiiedad en las Antmas) si es correcta la interpretacion de Pascual et at (1990636)
Nosotros consideramos estos diferentes niveles de diversificacion como las diferentes etapas de evolucion alcanzadas por cada grupo [de mamiferos] Obviamente esto estuvo relacionado con las diferentes longitudes de sus historias
Pudiera ocurrir que en el futuro se compruebe que la historia biogeognifica de la region fue mucho mas complicada
La Espanola la isla mas ecol6gicamente divers a de la region debi6 ser el centro principal de evoluci6n de los mamfferos (con 87 de las familias y 73 de las subfamilias) seguida por Cuba (con 75 de las familias y 54 de las subfamilias) las dos islas de las Antillas Mayores mas relacionadas biogeograficamente (Fig 43 Tabla 44)
eL
EL
cb CAIMAN
SWAN
A Acratocnini B Cubanocnini C Megalocnini D Mesocnini E Nesophontidae F Solenodontidae G Xenothrichini
ANGUILAshyM-0 SMARTIN
H Heteropsomyinae I Plagiodontinae J Isolobodontinae
K Hexolobodontinae L Capromyinae
M Heptaxodontinae N CIidomyinae 0 Sigmodontinae
FIGURA 43 Representacion esquematica de las relaciones interinsulares (expresadas porel numero de linajes compartidos) entre las Antillas Mayores y entre estas y los grupos de islas perifericas segun las interpretaciones de los autores citados en el texto Los numeros dentro de los circulos indican la cantidad de linajes representados en cada isla relacion dudosa Contexto taxon6mico de los linajes en la Tabla 44
bull bull
bull bull
bull bull bull
bull bull
bull bull
bull
bull
bull
bull
bull bull bull bull
bull bull
bull bull
bull
bull bull bull
bull bull
bull
bull bull
bull
322 BIOGEOGRAFiA
TABLA 44 Distribuci6n de los linajcs de mamiferos terrestres a traves de las Antillas Mayores y grupos de islas perifericas LE = La Espanola CU Cuba JA Jamaica PR = Puerto Rico IC Islas Caiman IS Islas Swan IB = Islas Bahamas IV = Islas Virgenes AS = Anguila - San Martin
FamiliaSubfumiliafIribu LE CU JA PR IC IS IB IV AS
Megalonychidae
Choloepodinae
Acratocnini
Cubanocnini
Megalocninae
Megalocnini
Mesocnini
Nesophontidae
Solenodontidae
Pitheciidae
Callicebinae
Xenothrichini
Echimyidae
Heteropsomyinae
Capromyidae
Plagiodontinae
Isolobodontinae
Hexolobodontinae
Capromyinae
H e pta x 0 don tid a e
Heptaxodontinae
Clidomyinae
Muridae
Sigmodontinae
PILOSA
SORICOMORPHA
PRIMATES
RODENTIA
bull
bull
bull
bull TOTALES 12 10 4 4 2
323 EXTlICIOIES DEL CUATERNARIO
El gigantismo ha sido sin duda una tendencia evolucionaria recurrente entre los clados mayores de mamiferos antillanos Es un hecho bien conocido que algunos grupos de mamiferos han aumentado de tamafio en las islas y varios autores han intentado explicarlo a partir de enfoques diferentes (Foster 1964 Miller 1966 Sondaar 1977 Case 1978 Eisenberg y Wilson 1978 Heaney 1978 Lawlor 1982 Lomolino 1985 Angerbjom 1985 McNab 19942001)
EXTINCIONES DEL CUATERNARIO
Las islas que integran la Subregion Antillana han perdido casi 90 de sus mamiferos terrestres durante los ultimos milenios y en proporcion con su extension territorial esta perdida ha sido mayor en la Subregion que en cualquier otro lugar del planeta (MacPhee 1996b) Las extinciones del Cuatemario eliminaron mas de las tres cuartas partes de la fauna de mamiferos terrestres conocida de las Antillas Mayores sus islas satelites y grupos de islas perifericas y el examen de este proceso en sus aspectos mas generales se aborda en esta ultima parte del presente capitulo
CRONOLOGiA Y CONSECUENCIAS
Es muy improbable que un fosil represente el ultimo individuo de un taxon extinto y entonces hay que guiarse por la edad radiometrica del fosil mas reciente conocido lIamada fecha de ultima presencia- como unica aproximacion posible a la fecha de extincion del taxon
Como se explico al inicio del capitulo (vease Contexto Temporal) el registro fosH de los mamiferos terrestres de las Antillas Mayores adolece de insuficiente sustentacion radiometrica No obstante la distribucion temporal de los escasos fechados disponibles para el Cuaternario de la region abarca etapas criticas del proceso de extincion experimentado por la referida fauna a 10 largo de los ultimos 250 mil afios
De las 52 especies que es posible reconocer actualmente como miembros extintos de la fauna cuatemaria de mamiferos terrestres de las Antillas Mayores y grupos de islas perifericas los restos fosiles de 23 de elIas (442) han sido fechados radiometricamente o se han hallado asociados estratigraficamente de manera inequivoca con otro material fechado De estos 23 fechados 3 corresponden al interglacial Sangamon 4 pertenecen a la secuencia Pleistoceno terminal - Holoceno temprano 10 se ubican en ellapso entre el arribo de los aborigenes y la incursion europea y 6 son postcolombinos (a los cuales hay que sumar otras dos especies extinguidas en los siglos XIX y xx) A partir de estas 25 especies avaladas por fechas de ultima presencia distribuidas entre los cuatro intervalos temporales sefialados hemos compuesto la Tabla 45 en la que ademas aparecen las restantes 27 especies (carentes de fechado radiometrico) ubicadas en el intervalo temporal mas probable sobre la base de (1) la asociacion estratigrafica de sus restos con evidencias culturales (amerindias 0 europeas) 0 (2) la ausencia de dicha asociacion Sobre estos criterios de ubicacion temporal procede hacer dos aclaraciones
324 BIOGEOGRAFIA
TABLA 45 Intervalos temporales mas probables para las extinciones cuatemarias de los mamiferos terrestres de las Antillas Mayores y grupos de islas perifericas sobre la base de las evidencias siguientes (l) ultimo ejemplar colectado u observado en vida (2) fechado radiometrico mas tardio en material de la especie (3) asociaci6n estratigrafica mas tardia con otro material fechado (4) asociaci6n estratigrafica con restos de animales introducidos restos humanos u objetos culturales propios del intervale (5) falta de asociaci6n con evidencias culturales [Los ntimeros entre parentesis en la columna Evidencia significan adscripci6n tentativa al intervalo] Los estimados del peso corporal (en kg) estan tomados de la literatura 0 inferidos comparativamente y distribuidos entre
las clases siguientes (a) S 01 (b) 01 a lt 1 (c) 1 a lt 10 (d) 10 a lt 100 (e) 100 a lt 200 (j) ~ 200 Las
abreviaturas que designan las islas son las de la Tabla 44
F amilialEspecie Peso IsJa Evidencia Fuente
I httervalo htterg1acial Sangamon (gt80 kaAP)
HEPTAXODONTlNAE
Amblyrhiza inundata e AS 3 McFarJane et al 1998a
CLIDOMYINAE
Clidomys osborni d JA 3 Ibid
Clidomys parvus d JA 3 Ibid
II httervalo Pleistoceno terminal ~ Holoceno temprano (20-6 kaAP)
MEGALONYCHIDAE
Aeratoenus antillensis d CD 5 Arredondo 1975 a
Aeratoenus odontrigonus d PR 3 McFarJane 1999b
Aeratoenus simorhynehus d LE 2 Rega et al 2002
M egaloenus zile f LE 2 MacPhee et al 2000
SOLENODONTIDAE
Solenodon arredondoi e CD 5 Morgan y Ottenwalder 1993
PnHECIIDAE
Paralouatta varonai d CD 5 MacPhee 1996a
EcHIMYIDAE
Heteropsomys antillensis c PR 5 McFarJane 1999b
Heteropsomys insulans c PR 5 Ibid
Puertoricomys eorozalus c PR 5 Woods 1989a
CAPROMYIDAE
P lagiodontia araeum c LE 5 MacPhee et al 2000
Macrocapromys latus c CD 5 Varona y Arredondo 1979
(Continua)
325 EXTINCIONES DEL CUATERNARIO
TABLA 45 (Continuaci6n)
F ami1ia1Especie Peso Isla Evidencia Fuente
M acrocapromys latus c CU 5 Varona y Arredondo 1979
Mesocapromys kraglievichi c CU 5 Ibid
HEPTAXODONTINAE
Xaymaca fulvopulvis a JA 2 MacPhee y Flemming 2003
III Intervalo Amerindio (6-05 kaAP)
MEGALONYCHIDAE
Acratocnus ye d LE 2 MacPhee et aI 2000
Neocnus comes c LE 2 Ibid
N eocnus dousman c LE 2 Ibid
N eocnus toupiti c LE 2 Ibid
N eocnus gliriform is c CU 4 Cordova et ai 1997
M egalocnus rodens f CU 2 MacPhee et ai 2007
Parocnus browni e CU 2 Steadman et ai 2005
P arocnus serus e LE 2 MacPhee et ai 2000
N ESOPHONTIDAE
N esophontes edithae a PR 2 McFarlane 1999b
PITHECIIDAE
Antillothrix bernensis c LE 3 Rimoli 1977
EcHIMYIDAE
Brotomys contractus a LE 4 Miller 1929c
CAPROMYIDAE
P lagiodontia spelaeum c LE 4 Ibid
M acrocapromys acevedo c CU 4 NUiiez 1990a
HEPTAXODONTINAE
Elasmodontomys obliquus d PR 3 McFarlane 1999b
Iv Intervalo Postcolombino (05 kaAP - Presente)
N ESOPHONTIDAE
N esophontes hypom icrus a LE 24 MacPhee y Fle111111ing 1999
N esophontes zam icrus a LE (4) Ibid
N esophontes m icrus a CU 24 MacPhee et aI 1999
(Continua)
326 BIOGEOGRAFiA
TABLA 45 (Continuacion)
F amilialEspecie Peso Isla Evidencia Fuente
N esophontes m icrus a CD 24 MacPhee et aI 1999
N esophontes m icrus a LE 4 MacPhee et aI 2000
N esophontes sp 1 a IC 4 Morgan 1994
N esophontes sp 2 a IC 4 Ibid
Solenodon marcanoi b LE 4 Ottenwalder 2001
PITHECIIDAE
Xenothrix mcgregori c JA 4 MacPhee y Flemming 1999
EcHIMYIDAE
Boromys offeUa a CD 24 Guarch 1984
Boromys torrei a CD 24 Ibid
Brotomys voratus a LE 4 MacPhee et aI 2000
CAPROMYIDAE
P lagiodontia ipnaeum c LE 4 MacPhee y Flemming 1999
Rhizoplagiodontia lemkei c LE 4 Ibid
Isolobodon portoricensis c PR 24 Ibid
Isolobodon montanus c LE 4 Ibid
Hexolobodon phenax c LE (4) Ibid
Capromys sp c IC 24 Morgan 1994
Geocapromys columbianus c CD 4 MacPhee y Flemming 1999
Geocapromys thoracatus b IS 1 MacPhee y Marx 1997
Geocapromys sp 1 c IC 4 Morgan 1994
Geocapromys sp 2 c IC (4) Ibid
l-IEPTAXODONTINAE
Quemisia gravis c LE (1) MacPhee y Flemming 1999
MURIDAE
Oryzomys antillarum a JA 1 Ibid
1 Flemming y MacPhee (1999) alertaron en contra de la asociacion con otro material fechado Asimismo Morgan (1993) indico que la ausencia de asociacion con evidencias culturales amerindias no es necesariamente un criterio concluyente para ubicar un taxon en el intervalo temporal anterior porque hay depositos en la region que carecen de tales evidencias y son del Holoceno tardio por fechados radiocarbonicos [No obstante
327 EXTlNCIONES DEL CUATERNAR10
la ausencia de una especie en residuarios arqueol6gicos adquiere mayor certidumbre como criterio de ubicaci6n temporal de su extinci6n en la medida que aumenta el numero de residuarios estudiados que en las Antillas Mayores se cuentan por centenares] A falta de fechado absoluto de una especie consideramos que estos criterios de ubicaci6n temporal son mas utiles que la alternativa de excluir del analisis a la especie
2 En los trabajos sobre biogeografia de mamiferos antillanos los arque610gos mas citados como autoridades para establecer el inicio de la presencia aborigen en las Antillas Mayores (Kozlowski 1974 Rouse y Allaire 1978 Rouse 1989 1992 Wilson 2001) difieren al respecto entre los 6 kaAP y los 45 kaAP (pero vease Pagan et al 200523) y hemos adoptado la mas temprana de estas fechas para el comienzo del intervalo amerindio en la Tabla 45 De todos modos cualquiera que haya sido la fecha esta no es aplicable por igual al conjunto de las Antillas Mayores porque segun algunos de los autores citados el poblamiento comenz6 por Cuba y La Espanola 6 mil anos atras continu6 en Puerto Rico hace alrededor de 3 mil anos (pero vease Burney et al 1994) yen Jamaica hace menos de 2 mil e incluso a las Islas Caiman ni siquiera llegaron [Las Islas Caiman descubiertas por Col6n en 1503 carecen absolutamente de evidencia amerindia (Stokes y Keegan 1996)]
Si los datos de la Tabla 45 representan una aceptable aproximaci6n a la realidad el proceso de las extinciones cuaternarias de los mamiferos terrestres de las Antillas Mayores y grupos de islas perifericas debi6 caracterizarse entre otros por los eventos siguientes
Aunque obviamente la fauna cuaternaria debi6 ser mucho mas numerosa que 10 que refleja el registro paleontol6gico (Morgan y Woods 1986) 52 especies (81 de la fauna conocida) desaparecieron (sin reposici6n) durante los ultimos 250 mil anos
- Al parecer las extinciones se concentraron en las especies de mayor tamano Asi el peso promedio estimado para las especies de la etapa preantr6pica (intervalos I + II de la Tabla 45) es 350 kg es 328 kg para las especies del intervalo III (amerindio) y es s610 25 kg (c1ases del peso a b c) para las especies del intervalo IV (postcolombino) Ninguna especie aut6ctona de mamifero mayor de 6 kg existe hoy en la regi6n
- De acuerdo con la distribuci6n temporal de las extinciones 61 especies sobrevivieron al interglacial Sangamon (intervalo 1)48 sobrevivieron a la transici6n del Pleistoceno terminal al Holoceno temprano (intervalo II) 34 sobrevivieron al periodo amerindio (intervalo III) y s610 12 especies han sobrevivido desde la Ilegada de los europeos (intervalo IV) Las tasas de extinci6n correspondientes (exceptuado el intervalo I) son una especie cada 1167 anos en el intervalo II una cada 393 anos en el intervalo III y una cada 23 anos en el intervalo IV
- Durante todo el proceso de las extinciones cuaternarias la perdida de especies por isla fue de 91 en La Espanola 83 en Jamaica y 62 en Cuba Bahamas retuvo su unica especie (aunque extirpada en varias de las islas) Puerto Rico e Islas Caiman perdieron todas sus especies (6 y 5 respectivamente) y las Islas Swan Virgenes y Anguila - San Martin perdieron sus respectivas especies (micas
328 BIOGEOGRAFiA
De los cuatro 6rdenes de mamiferos representados en la fauna Pilosa y Primates desaparecieron totalmente Soricomorpha y Rodentia perdieron 86 y 74 de sus especies respectivamente
CAUSALIDAD
CAUSAS NATURALES
Los cicIos glaciales e interglaciales acaecidos en el Pleistoceno causaron cambios ambientales dnisticos en las Antillas con la concomitante alternancia de cIimas frios (secos) y calidos (hUmedos) asi como de descensos y ascensos del nivel del mar aumento y disminuci6n de las areas territoriales) induso (aunque con menor intensidad) en el Holoceno Estos eventos paleogeograficos han sido consistentemente documentados (Mayo y Kartashov 1972 Bonnatti y Gartner 1973 Ortega y Arcia 1982 Prance 1982 Schubert y Medina 1982 Dodge et al 1983 Ortega [1984] Haq et al 1987 Schubert 1989 Hodell et al 1991 Guilderson et al 1994 Oppo 1997 Emslie 1998 Curtis et al 2001 Iturralde-Vinent 2003) y de ellos brindamos a continuaci6n una breve resena
Finalizando el periodo interglacial anterior a la ultima glaciaci6n (Wisconsin) el mar estaba a 8-10 m por encima de su nivel actual Cuando el avance glacial del Wisconsin a1canz6 su culminaci6n (eillamado Ultimo Maximo Glacial) hace 18 ka el mar habia descendido 120 m con respecto a dicho nivel y las islas del Caribe habian incrementado considerablemente sus extensiones subaereas en comparaci6n con las que presentan hoy dia por ejemplo Puerto Rico y las Islas Virgenes se conectaron territorialmente y las Bahamas se unieron en cinco gran des islas Prevalecia un regimen frio y seco y condiciones de aridez --debidas a una reducci6n de las precipitaciones (posiblemente a menos de 440 mmlano)- y los ambientes xericos se habian extendido a traves de las islas y bancos emergidos y abundaban los pinares (los habitats mesicos estaban restringidos a parcelas relictas en 10 alto de las montanas) En el Mar Caribe y el Golfo de Mexico la temperatura superficial del mar era 5degC mas baja que la actual y en las llanuras costeras y alturas maximas eran respectivamente 2-3degC y 6degC mas bajas Los limites de las zonas ecol6gicas altitudinales descendieron varios cientos de metros y cada zona redujo su extensi6n vertical al aumentar el gradiente de temperatura Ellimite inferior de la zona periglacial
elevaci6n donde la temperatura media minima a1canza los OdegC) de la Cordillera Central de la Republica Dominicana (altura maxima del Caribe insular con evidencia de topografia glacial) se considera que estuvo entre los 1500 y 1800 mlnm (actualmente esta a 2800 mlnm) y se puede estimar que dicho limite era inferior en Cuba por estar mas al norte que la Republica Dominicana No obstante durante este periodo la temperatura experiment6 episodios cortos (unas pocas decadas) de ascensos repetidos
Un signifieativo componente bi6tieo de las referidas condiciones paleoambientales 10 constituy6 la espectacular irradiaci6n evolucionaria experimentada por un grupo numeroso de aves rapaees diurnas y nocturnas (Wetmore 1937 Brodkorb 1969 Kurotchkin y Mayo 1973 Arredondo 1976 Olson 1978 Olson y Hilgartner 1982
329 EXTINCIONES DEL ClJATERNARIO
Suarez 2000a b 2004 Suarez y Emslie 2003) Los mamiferos de las Antillas Mayores ofrecieron a estos depredadores aviares una amplia gama de tamanos de presa que en ausencia de mamiferos del orden Carnivora en la region permitio a estas aves irradiarse a este nicho alimentario con la evolucion de una variedad de especies de muy diversa tall a corporal incluidas formas gigantes Adquirir un cuerpo gigante les propicio el acceso a recursos alimentarios que no estaban siendo explotados por ningun otro depredador (Alcover y McMinn 1994) A Cuba corresponde el mejor registro fosil de rapaces cuaternarias caribenas con siete generos de especies diurnas Dentro de estas aves existieron tres gavilanes gigantes (Accipitridae) tres carairas (Falconidae) y un condor (Cathartidae) asi como cuatro generos de especies nocturnas dos lechuzas (Tytonidae) y tres Mhos (Strigidae) dos de ell os no igualados en tamano en todo el mundo Algunos de estos generos tuvieron representacion en otras islas antillanas como La Espanola Jamaica Bahamas y Gran Caiman
Al retirarse hacia el norte los hielos de la glaciacion Wisconsin el clima se tomo mas hUmedo en las Antillas durante el Holoceno temprano y el bosque mesico se expandio en la region insular El nivel del mar ascendio acompanado de mayor pluviosidad alrededor de los 7 kaAP las Islas Virgenes quedaron separadas entre si y de Puerto Rico y las Bahamas perdieron la mayor parte de su territorio Cerca de los 3 kaAP las condiciones secas retornaron a la region e hicieron desaparecer buena parte del bosque mesico Nuevamente el clima comenzo a adquirir sus condiciones actuales hacia los 25 kaAP aproximadamente y hoy transita por una fase mesica con vegetacion xerica confinada en parcelas relictas
Sin duda estos y otros eventos naturales incidieron negativamente en la sobrevivencia de mamiferos terrestres en las Antillas Mayores asi como en los grupos de pequenas islas perifericas (Pregill y Olson 1981 Olson 2006)
McFarlane et al (1998a) demostraron convincentemente como Amblyrhiza el gigante de los roedores antillanos endemico de la pequena isla de Anguila sucumbio a las repetidas y abruptas perdidas de su habitat durante el interglacial Sangamon por las nipidas y severas oscilaciones del nivel del mar (documentadas por Neumann y Hearty 1996) al no poder responder adecuadamente a tan catastroficos eventos mediante el ajuste de su masa corporal media y otro tanto debio ocurrirle en Jamaica al otro roedor gigante Clidomys (McFarlane et al 1998b) emparentado con Amblyrhiza en la misma epoca (Tabla 45 intervalo I)
El ascenso del nivel del mar seguramente promovio la extincion de mamiferos durante la transicion del Pleistoceno al Holoceno (Tabla 45 intervalo II) con la reduccion del tamano de las Islas por las transgresiones marinas (Pregill y Olson 1981 Morgan 1993) Esto ultimo haria maximamente severa la competicion asf como la presion de los depredadores [Steadman et al (2005) no yen por que el calentamiento global ocurrido durante la transicion Pleistoceno-Holoceno habria de tener un efecto negativo sobre los perezosos siendo sus descendientes actuales especies tropic ales 0 subtropicales pero no se trata del efecto
330 BIOGEOGRAFiA
directo del cambio de temperatura sobre los animales sino del efecto indirecto y progresivo que dicho cambio genera a largo plazo al reducir el territorio insular disponible]
Por esta epoca tambien desaparecieron especies de tall a mayor como Megalocnus zile en La Espanola y otras de menor tamano (en Cuba) pero que representan la tall a maxima en sus respectivos contextos taxonomicos del Cuaternario como Solenodon arredondoi Paralouatta varonai y Macrocapromys acevedo Un mamifero puede ser grande solo a costa de reducir su densidad poblacional y esto 10 hace mis vulnerable a la extinci6n sobre todo en una isla (McNab 1994 McFarlane et al 1998a) Incidentalmente Alcover et al (1998a) afirmaron que larelacion entre la talla corporal y la extincion no siempre se cumple en las especies de menor tamano y citaron como ejemplo que entre las musarafias todas las especies de Nesophontes desaparecieron mientras sobreviven especies de Solenodon pero la comparacion correcta en todo caso serfa entre los solenodones vivientes y el gigante Solenodon arredondoi y entre Nesophontes edithae -el gigante de los nesofontes extinguido en el periodo amerindio- y el resto de sus congeneres que sobrevivieron hasta los primeros siglos de la colonizacion europea (Tabla 45 intervalos III y IV)
CAUSAS ANTROPOGENICAS
Intervalo Amerindio
La mayoria de los autores que se han referido a las extinciones acaecidas durante este intervalo (entre elIos paradigmaticamente Martin 1984a b) han responsabilizado a los aborfgenes (que se supone comenzaron a poblar las Antillas Mayores hace 6 mil afios) con la desaparicion de numerosas especies de mamiferos terrestres en la region principalmente por la via de la sobrecaza y la introduccion de animales domesticos devenidos depredadores de la fauna autoctona (Morgan y Woods 1986) Este efecto negativo de la presencia indigena sobre los mamiferos de la region ha sido dramatizado por algunos autores en terminos de holocausto faunal Sin embargo pensamos que se ha exagerado Como han expresado MacPhee et al (19993) cuando uno busca evidencia dura de complicidad humana directa en [estas] extinciones no hay mucho que encontrar Las siguientes son algunas consideraciones a tener en cuenta en tal sentido
El registro radiometrico mas joven conocido hasta ahora para un perezoso antillano (MegalocnllS rodens) es de 42 kaAP (MacPhee et ai 2007) Si se acepta que los aborfgenes entraron en Cuba hace aproximadamente 6 mil anos su convivencia con los perezosos debe haber durado alrededor de 1800 afios (0 mas porque dificilmente el referfdo fechado radiometrico haya correspondido al ultimo sobreviviente de la especie) Esta cifra contradice la hipotesis del holocausto faunal segun la cualla extincion de los mamiferos insulares de mayor talla debe ocurrir con extrema rapidez -a escala de decadas 0 de centurias- a partir de su primer contacto con los humanos (Martin y Steadman 1999) A pesar de las repetidas afirmaciones en contrario (Harrington 1921 Miller 1929c Herrera [1945] Tabio 1951 Alvarez 1951a 1957 Arredondo
331 EXTINCIONES DEL CUATERNARIO
1959 1960a Veloz y Ortega 1973 Mayo 1980b Rodriguez et al 1984 Castellanos y Pino 1985 Pino y Castellanos 1985 Nunez 1990b Gutierrez 1996 Arredondo 2004) en su mayoria basadas en evidencias no exentas de posibles accidentes de deposicion (0 de reubicacion postdeposicional) en recintos cavernarios (Diaz-Franco [2006b D no se ha demostrado inequivocamente que los aborigenes antillanos se alimentaran de perezosos regularmente Sus restos en residuarios de alimentos son escasisimos en toda la region y esto contrasta con los abundantes restos de jutias y otros mamiferos de menor talla que aparecen en esos residuarios demostrativos de la preferencia de los paleoindios por estos otros mamiferos
Los mamiferos mas importantes en la economia prehistorica de las Antillas Mayores fueron las jutias Capromys y Geocapromys en Cuba Geocapromys en Jamaica e Isolobodon en La Espanola Puerto Rico e Islas Virgenes (Aguayo 1950b Tablo 1951 Rimoli 1972 Wing 2001 Diaz-Franco [2006b D Wilkins (2001) estudi61ajutia viviente de Jamaica (Geocapromys browni) en un sitio ocupado por los aborigenes durante 100 anos y no encontr6 evidencias de que durante ese tiempo la especie haya sido sobreexplotada Asimismo en Cuba las jutias conga carabali y andaraz son escasas en depositos pleistoholocenicos en cambio son muy numerosas en residuarios indigenas y de haber sido sobreexplotadas 10 mas probable es que no hubieran sobrevivido hasta hoy
La habilidad de los humanos para penetrar en profundidad el bosque tropical y mantenerse en este sustentablemente ha sido un evento comparativamente reciente (Beran y Steele 1994) En el caso antillano la caza tierra adentro ha sido explicada como un proceso tardio de desplazamiento a partir de la explotacion de abundantes recursos marinos que ofrecian condiciones 6ptimas para el sedentarismo de las comunidades aborigenes costeras (C6rdova y Arredondo 1988 Brito 2005) Peces y mariscos dominan la arqueologia costera de las Islas antillanas en tanto el consumo de recursos terrestres (mamfferos incluidos) aumenta progresivamente en la medida que los residuarios estudiados se alejan de la costa (Scudder 1991 Wilkins 2001) En las Antillas Mayores el componente terrestre de la dieta aborigen es solo 20-30 del material recobrado en los residuarios (Wing 1977 1989) Y debio ser menor alin porque el componente vegetal de la dieta no se ha preservado Por otra parte los aborigenes cubanos domesticaron la jutia conga y seguramente la reprodujeron para comer (Pose et al 1988) practica que probablemente se repitiera con otras jutias en las demas Antillas Mayores
- Fray Bartolome de las Casas hablo de 3 millones de indios en Cuba al inicio de la colonizacion pero los historiadores subsecuentes consideraron exagerada la cifra (Poey 1851 a) No obstante se dispone de dos estimados de la densidad de poblaci6n aborigen en las Antillas Mayores a la llegada de los europeos 18 indigenaslkm2 en Puerto Rico (Rouse 1956) y 1271km2 en La Espanola (Watts 1987) Extrapolando ambos valores entre las cuatro islas principales de las Antillas Mayores tenemos que en la fecha indicada la poblaci6n aborigen pudo estar entre 16 mil y 113 mil indigenas en Puerto Rico entre 21 mil y 145 mil en Jamaica entre 141 mil y 1 mill6n en La Espanola y entre 200 mil y 14 millones en Cuba Entre las cifras maximas y las minimas nos inclinamos
332 BIOGEOGRAFiA
hacia estas ultimas atendiendo a los altos indices de mortalidad infantil y morbilidad que se evidencian en las osamentas de los pobladores prehispanicos (Martinez y Rives 1990) Uno de los hechos que apuntan hacia el bajo numero de aborigenes en las Antillas Mayores es 10 rapido que desaparecieron despues de la llegada de los europeos pnkticamente sin dejar mestizaje con estos 10 cual contrasta ostensiblemente con la situacion en la America continentaL
En suma un grupo humano (1) demograiicamente irrelevante y controlado por altos indices de morbilidad (2) consumidor mayoritariamente de organismos marinos y (3) carente de la tecnologia cinegetica necesaria para poder sobreexplotar los mamiferos que Ie sirvieron de alimento (a los cuales accedio tardiamente en su desarrollo) no es concebible que haya causado e1 holocausto faunal que se Ie atribuye a partir del error de aplicar a las comunidades aborigenes antillanas un modelo cultural de la prehistoria continentaL
Tampoco hay evidencias de que los aborigenes hayan introducido carnivoros con suficiente capacidad depredadora salvo el perro Morgan y Otlenwaldder (1993) consideraron que la depredacion por perros durante el periodo amerindio es la explicacion mas plausible para la extincion del almiqui gigante de Cuba pero como se puede observar (Tabla 54) Solenodon arredondoi aparece en el intervalo II por falta de asociacion con evidencias culturales
Consecuentemente el accionar de los aborigenes y de sus animales domesticos directamente sobre la fauna no parece haber sido por sf solo un factor determinante de la extincion de mamiferos terrestres durante el periodo amerindio en las Antillas (pero vease Steadman et al 2005)
Hay que recordar en este senti do que alin en el Holoceno tardio ocurrieron cambios clirruiticos de consideracion (vease Causas Naturales) Por ejemplo Curtis et al (2001) explicaron que la extincion de algunos ani males mesofilicos de las alturas montanosas de La Espanola pudo haber antecedido a cualquier impacto humano sigrIificativo en esos lugares por coincidir dicha extincion con una tendencia a la aridez documentada isotopicamente y con evidencias de la perdida del bosque mesico ocurridas despues de 3kaAP Yen Jamaica hace solo 2shy3 mil aDos el area de la Bahia de Jackson sustentaba una gran biomasa de vertebrados en bosque hinnedo a juzgar por la cantidad y diversidad de taxones presentes en material fosil derivado de la depredacion por lechuzas hoy el area es la mas seca de la isla con matorral xerofitico y pobreza de fauna (MacPhee 1996a) Yalm en nuestros dias huracanes de gran intensidad aniquilan poblaciones de vertebrados (que no se recuperan) en is las pequenas y llanas del Caribe (Brooks y Smith 2001)
En medio de la incertidumbre sobre la verdadera causa (0 causas) de las extinciones de mamiferos en el periodo amerindio MacPhee y Marx (1997) han propuesto su hipotesis de la hipermorbilidad la cual estipula colapsos poblacionales relativamente rapidos con la introduccion de enfermedades de alta letalidad Estos autores han razonado que
Las islas son los lugares mas apropiados para experimentar perdidas catastroficas causadas por enfermedades Los mamiferos no solo serian susceptibles en virtud de su prolongado aislamiento biologico sino que en los confines de una isla la epizootia
333 EXTINCIONES DEL CUATERNARIO
tendria la potencialidad de infectar simultaneamente a la rnayoria de los individuos de vadas especies Los neonatos son inmunologicamente inmaduros y sufririan desproporcionadamente cualquier infeccion hipervirulenta La perdida excesiva de neonatos y juveniles tendria efectos severos para la sobrevivencia de la poblacion si esta se reduce por debajo de cierto nivel sobreviene la extincion porque no hay posibilidad de restitucion en el periodo postpanzootico Las especies de mayor talla corporal van a ser las mas vulnerables a la extincion dadas sus bajas tasas reproductivas largos periodos de gestacion y reducido numero de progenie Las especies de menor tamano --caracterizadas por altas tasas reproductivas periodos cortos de gestacion y mayor numero de neonatos por parto- tambien serian fuertemente diezmadas pero muchas sobrevivirian porque su estrategia reproductiva les permite perder mayor numero de individuos Las perdidas deben ocurrir relativamente rapido (a escala de decadas) mientras que otras hipotesis (digamos los cambios climaticos) permiten intervalos mucho mas largos (a escala de siglos 0 de milenios) entre la aparicion de las causas y la produccion de sus efectos Por ejemplo todo indica que los nesofontes se extinguieron muy rapidamente y seria dificil poder invocar un impacto humano directo 0 argumentar fenomenos climaticos que pudieran explicar plausiblemente perdidas tan rapidas
MacPhee y Marx (1997 191) han dicho que por el momento la hipotesis de la hipermorbilidad no pasa de ser una abstraccion en espera de adecuada sustentacion empirica Llama la atencion que linicamente en el periodo amerindio (Tabla 45 intervalo 1Il) las extinciones abarcaron todas las clases del peso corporal (de la a a laj) y que ninguna de las especies que sobrevivieron hasta el periodo postcolombino (Tabla 45 intervalo IV mas las especies vivientes) sobrepasa los 10 kg (clases del peso a b c) en clara coincidencia con los presupuestos de la hipotetica hipermorbilidad
Intervalo Postcolornbino
Por 10 menos 37 especies de mamiferos terrestres vivian en las Antillas Mayores y grupos de Islas perifericas a la lIegada de los europeos en 1492 De elIas se han extinguido 23 shymas especies que en cualquiera de los periodos precedentes (Tabla 45~ y solo subsisten 12 especies repartidas entre Cuba (8 especies) La Espanola (2) Jamaica (1) y Bahamas (1) varias de estas en peligro critico de extincion Solo dos de las especies extinguidas durante los ultimos 500 anos se conocieron en estado viviente los roedores Oryzomys antillarum de Jamaica (extinguido en 1877) y Geocapromys thoracatus de las Islas Swan (extinguido en 1955) Las causas de las extinciones ocurridas durante este periodo son sobradamente conocidas destruccion de habitats introducci6n de animales exoticos (domesticos y comensales) y demas presiones asociadas a la actividad humana entre las cuales posiblemente habria que incluir brotes de hipermorbilidad
Durante mucho tiempo se ha afirmado la posibilidad de que especies de Nesophontes hayan sobrevivido hasta las primeras decadas del siglo xx y que incluso continuen existiendo en lugares remotos de Cuba 0 La Espanola (Allen 1918 1942 Miller 1930 Aguayo 1950a Arredondo 1960b Fischer 1977 MacFadden 1980 Woods y
334 BIOGEOGRAFiA
Ottenwalder 1991) Actualmente no existe evidencia radiometrica de que alguna especie de Nesophontes haya sobrevivido al siglo xv las fechas radiocarbonicas calibradas obtenidas en muestras de Nesophontes de Cuba y La Espanola resultaron no mas tardias que los siglos XIII y XIV (MacPhee et al 1999) Por otra parte las expediciones efectuadas a fines del siglo xx con el proposito de localizar individuos vivientes del genero en La Espanola no produjeron la menor indicacion afirmativa al respecto (Woods et al 1985) Ademas a traves de las Antillas Mayores no aparecen restos de Nesophontes en los depositos formados actualmente por las lechuzas
EPiLOGO
Las Antillas Mayores son islas oceanicas no han formado parte de un continente (con la posible excepcion de una parte de Jamaica hasta el Eoceno Medio hace 45 Ma) y durante un tiempo geologico considerable (desde el Eoceno Superior hace 40 Ma) han permanecido emergidas y separadas de los continentes vecinos Consecuentemente los mamiferos terrestres (no volantes) autoctonos de estas islas constituyen una fauna oceanica que por ende cumple con todas las caracteristicas convencionales reconocidas para este tipo de entidad biogeografica
- Baja diversidad taxonomica a nivel de las categorias superiores (solo cuatro ordenes) en comparacion con islas plataformicas continentales (0 con areas continentales) de tamano y diversidad ecologica simi lares
- Alto grado de endemismo a nivel de genero (96) y familia (50) en respuesta al prolongado aislamiento
- Desbalance ecologico de la estructura trofica ausencia de mamiferos camivoros y ungulados
- Espectaculares procesos de irradiaci6n evolucionaria como resultado de la colonizacion de territorios libres de competidores y depredadores
Tendencias evolucionarias extremas gigantismo
- Control a cargo de aves rapaces con tendencia at gigantismo en ausencia de mamiferos miembros del orden Carnivora
- Mas especies de reptiles y de murcielagos que de mamiferos terrestres relaciones que en los continentes se invierten
Alta tasa de extincion de especies (84) sin reposicion
Preservacion de especies filogeneticamente arcaicas (los llamados fosiles vivientes) los almiquies

316 BIOGEOGRAFiA
fosiles atm no han sido hallados Pero como hemos visto Rega et al (2002) atribuyeron dicha distribucion a la dispersion interinsular en el Cuatemario
Por otra parte varios amilisis c1adisticos recientes ubi can a Choloepus -genero Reciente conocido solo de Centro- y Suramerica- como taxon hermano de Aeratoenus exc1uidos los demas megaloniquidos antillanos Para White y MacPhee (2001228)
tan estrecha relacion sugiere que la divergencia entre ambos [generos] ocurrio despues de que Aeratoenus divergio de Neoenus Si esta relacion implica que Aeratoenus y Neoenus se dispersaron separadamente desde Suramerica a las AntilIas mientras Choloepus permanecia en el continente (siendo necesarias al menos tres invasiones de perezosos) 0 si Choloepus invadio Suramerica desde las Antillas es algo que el material disponible no permite determinar
La diversidad adaptativa de los perezosos se manifiesta en (1) la utilizacion del habitat especies arboreas en Neoenus y Aeratoenus y terrestres en Paroenus y Megaloenus (Webb 1985a White [1993] White et al 1996 Arredondo [1999]) y en (2) la gama de tallas corporales desde los pequenos Neoenus hasta Megaloenus rodens el mayor de los mamiferos terrestres antillanos (su peso adulto estimado en 270 kg seglin Paula 19679) esto sugiere que los perezosos se irradiaron a nichos que en contextos continentales probablemente estarian ocupados por miembros de otros c1ados (MacPhee e IturraldeshyVinent 2000)
No esta claro si el abanico de tamanos entre estos perezosos enmascara alguna tendencia al gigantismo Por 10 pronto Anderson y Handley (2002) encontraron 10 contrario en perezosos (Bradypus) de las Islas de Bocas del Toro (Panama) shyseparadas secuencialmente del area continental vecina por ascensos del nivel del mar durante los ultimos 10 ka En las cinco islas los perezosos son menores que su contraparte continental y las diferencias entre las tallas medias de las cinco poblaciones expresan una reduccion lineal significativa de la talla corporal en funcion de la edad geologica de cada isla sin relacion alguna con el area de las islas 0 la distancia que las separa del continente
MONOS (Pitheciidae)
Los primates de las Antillas Mayores (cuatro especies extintas) constituyen un grupo monofiletico con Paralouatta (en Cuba) y Antillothrix (en La Espanola) como taxones hermanos seguidos en la escalera c1adistica por Xenothrix (en Jamaica) y finalmente Callieebus como el pariente continental mas cercano esto implica una sola invasion desde Suramerica antes del Mioceno Inferior (fecha mas temprana conocida para los primates fosiles antillanos) y por tanto en consonancia con GAARlandia como la via de colonizacion (Horovitz y MacPhee 1996 1999 Horovitz [1997] MacPhee y Horovitz 2004)
En desacuerdo parcial con la anterior interpretacion la filogenia de este grupo de monos obtenida por Davalos (2004) indica un antecesor comun en Cuba La Espanola y Jamaica
317 TRRADIACTON EVOLUCIONARIA
compatible con la hipotetica conexion -propuesta tentativamente por Iturralde-Vinent y MacPhee (199980-84)- entre las Montanas Azules del este de Jamaica y la peninsula suroccidental de La Espanola en la epoca de GAARlandia
A pesar de que los primates antillanos pertenecen a un grupo viviente de monos muy pequenos (los monos titles) fueron comparativamente grandes en relacion con los platirrinos modernos y sucesivamente mayores en la direccion Antillothrix Xenothrix y Paralouatta con P marianae como el mayor platirrino conocido del Mioceno neotropical (MacPhee 2005) MacPhee et al (1994) consideraron que Xenothrix debio ser un mono de andar lento y al parecer las especies de Paralouatta abandonaron la arborealidad para adoptar un modo de vida mas terrestre (MacPhee 1996a MacPhee et al 2003 MacPhee y Meldrum 2006)
ROEOORES (Echimyidae Capromyidae Heptaxodontidae)
La edad de Zazamys en Cuba demuestra que la invasion de los roedores histricognatos en las Antillas Mayores comenzo antes del Mioceno Inferior por otra parte los representantes mas antiguos de esa irradiacion evolucionaria se encuentran en La Espanola y Puerto Rico Ambas evidencias son consistentes con la hipotesis de GAARlandia (Woods 1982 1988 Woods et al 2001)
No esta claro si los capromidos evolucionaron a partir de un equimido ancestral 0 de un protocapromido 0 si los equimidos antillanos (I sub familia 3 generos y 7 especies todas extintas) forman parte de la irradiacion de los capromidos (4 subfamilias 9 generos y 27 especies 10 vivientes) Kratochvil et al (1978b) y Rodriguez et al (1979) consideraron que Capromys nan us es un eslab6n entre Echimyidae y Capromyidae pero Mesocapromys nanus esta morfologicamente mucho mas cerca de Hexolobodon y los de mas capromidos que de los equimidos (Woods 1985) Lo que si esta claro segun Woods et al (2001) es que la subfamilia Plagiodontinae endemic a de La Espanola es el linaje basal de los capromidos y que Isolobodon Hexolobodon Capromys Geocapromys Mysateles y Mesocapromys [y supuestamente Macrocapromys] forman parte de la irradiacion temprana de Capromyidae
La subfamilia Capromyinae (5 generos y 17 especies 9 vivientes) parece ser el dado de mas reciente evolucion en la familia con invasion de Cuba desde La Espanola y dispersion desde Cuba (1) por Capromys a las islas Caiman y (2) por Geocapromys a las islas Bahamas Caiman y Jamaica y desde Jamaica a las islas Swan aunque la presencia de Geocapromys en Swan pudiera deberse a introduccion aborigen (Morgan 1985 1988 1989 1993 1994 Woods 1989a 1990) Woods et al (2001) utilizaron datos moleculares para obtener estimados independientes de divergencia entre capromidos segiJn 10 cual Geocapromys divergio cerca de los 17 MaAP Mesocapromys divergio de Capromys hace 15 MaAP en tanto lvlysateles 10 hizo hace 10 MaAP Por su parte las formas de Capromys pilorides y de Mysateles prehensilis en Isla de la Juventud divergieron de sus correspondientes subespecies nominativas en la isla principal a los 24 y 08 MaAP respectivamente
318 BIOGEOGRAFiA
Woods y Howland (I979) describieron adaptaciones morfologicas de la mandibula el hioide y la faringe asociadas a mecanismos de masticacion diferentes en plagiodontinos y caprominos y Varona (1980a) llamo la atencion sobre el gran desarrollo de los molariformes (en relacion con la longitud total del cnineo) que presentan ciertos grupos de jutias fosiles en comparacion con las vivientes 10 que pudiera interpretarse como una adaptacion alimentaria a una vegetacion mas abrasiva por parte de las jutias extinguidas
Segun Woods (1990) la familia Heptaxodontidae (2 subfamilias 5 generos y 6 especies todas extintas) tambien seria parte de la irradiacion de los capromidos con origen en el este de La Espanola 0 en Puerto Rico y subsecuente dispersion al Banco de Anguila - San Martin en las Antillas Menores (pero vease MacPhee y Flemming 2003) Este banco estaria a solo 70 km del Banco Puerto Rico - Islas Virgenes en tiempos de maxima regresion marina y esta proximidad con Puerto Rico -patria de Elasmodontomys obliquus- seguramente tuvo que ver con la presencia en las islas de Anguila y San Martin del heptaxodontino gigante Amblyrhiza relacionado mas estrechamente con los roedores de las Antillas Mayores que con los del resto de las Antillas Menores (Woods 1990 1996) Aunque los roedores suelen aumentar de tamano en las islas (Case 1978) la escala del incremento de la talla en Amblyrhiza (que pudo alcanzar mas de 200 kg de peso adulto) esta fuera de toda proporcion con los patrones conocidos en otros roedores insulares (Biknevicius et 01 1993)
Clidomys (Clidomyinae) de Jamaica difiere marcadamente en su morfologia de los heptaxodontinos (vease ademas MacPhee y Flemming 2003)10 cual dificulta explicar su presencia en esa isla Como Jamaica estuvo bajo el mar hasta el Mioceno Medio el ancestro de Clidomys colonizaria esta isla por dispersion desde el extremo suroccidental de La Espanola despues que los heptaxodontinos se originaron en esta ultima isla (Woods 1989a) Pero Pascual et 01 (I 990) especularon que Clidomys pudiera ser el resultado de una invasion posterior e independiente de los capromidos
GRUPOS ORIGINARIOS DE NORTEAMERICA
El origen de las musaranas antillanas ha resultado ser hasta ahora el mayor enigma biogeografico de la fauna de mamiferos terrestres de la region aunque la mayoria de los autores coinciden en considerarlas originarias de Norteamerica (vease Discus ion y Conclusiones en Poblamiento Continente-Isla) No obstante MacPhee e Iturralde-Vinent (1994) descartaron toda posibilidad de conexion terrestre por el Estrecho de la Florida 0 por el Canal de Yucatan y si el (supuesto) evento vicari ante de Rosen (vease El Arca de Noe) -invocado por MacFadden (1980) para resolver el enigma- fuera minimamente plausible aun asi es dificil creer que las catastrofes que 10 sucedieron (vease Contexto Temporal) hayan barrido con toda esa vieja fauna continental excepto las musaranas circunstancia ya anticipada muy tempranamente por Matthew (l918a)
De otra parte la relaci6n entre Nesophontidae y Solenodontidae permanece irresuelta Si estos linajes no estan estrechamente relacionados entonces es muy posible que
319 IRRADIACION EVOLUCIONARIA
sus distribuciones sean explicadas por mecanismos diferentes (Whidden y Asher 2001)
La descomunal diferencia de talla corporal entre las musaranas pequenas (Nesophontes) y las grandes (Soienodon) seguramente impuso a ambos grupos restricciones distintas relativas a la termorregulacion y las adaptaciones energeticas En Nesophontes la seleccion debio favorecer una alta tasa de metabolismo y crecimiento camadas mas numerosas corta expectativa de vida yalta densidad poblacionaL En Soienodon en cambio la seleccion opero en la direccion de una baja tasa metabolica camadas reducidas crecimiento lento longevidad y baja densidad poblacional (Ottenwalder [1991])
MUSARANAS PEQUENAS (Nesophontidae)
Las musaranas pequenas estan representadas en las Antillas por el genero Nesophontes (y un genero miocenico indeterminado en ambar dominicano) con 6 especies cuaternarias extinguidas distribuidas en Cuba La Espanola Puerto Rico Islas Virgenes e Islas Caiman La mayor de las musaranas pequenas Nesophontes edithae evoluciono en Puerto Rico y su arribo a las Islas Virgenes supone una dispersion sumamente reciente En las Islas Caiman Nesophontes se asemeja mas a la poblacion de Cuba de donde seguramente derivo en el Plio-Pleistoceno (junto con Capromys y Geocapromys) cuando la costa sur de Cuba estuvo mucho mas cerca de las Islas Caiman (lturralde-Vinent 2003fig 4) MacPhee et ai (1994) interpretaron la distribucion de Nesophontes en Cuba y La Espanola como resultado de la vicarianza oligomiocenica ocurrida al formarse el Paso de los Vientos pero la distancia taxonomica entre los lados del Paso es solo de nivel especifico y N micros incluso vivi6 en ambas islas segtln Patterson (1962)
Allen (1918) considero que Nesophontes micrus fue una especie primitiva no modificada para la vida arb6rea 0 fos6rea y por tanto de habitos terrestres Nesophontes edithae de Puerto Rico es mucho mayor y mas robusta que las restantes especies del genero eL indice de robustez de su femur (= anchura minima de la diafisis ongitud total del hueso) muestra que este es tan robusto como el de Solenodon paradoxus un animal dos veces mayor (Morgan et ai 1980) Choate y Birney (1968) postularon un aumento cronoclinal de la taIla corporal en N edithae confirmado mas tarde por McFarlane (1999a) mediante fechados radiocarb6nicos de la secuencia estratignifica en depositos cavernarios de Puerto Rico Solenodon no vivi6 en Puerto Rico y en ausencia de una musarana de mayor tamano Nesophontes alcanzo una talla corporal mayor en esa isla que en La Espanola 0 Cuba (Woods y Eisenberg 1989)
MUSARANAS GRANDES (Solenodontidae)
Los arcaicos almiqufes (genero Soienodon) comprenden cuatro especies una viviente (S cubanus) y otra f6sil (S arredondoi) en Cuba y una viviente (S paradoxus con dos subespecies) y otra f6sil (S marcanoi) en La Espanola Como cabria esperar las dos especies cubanas se asemejan mas entre sf que a las especies de La Espanola pero
320 BIOGEOGRAFiA
enigmaticamente S marcanoi comparte caracteres con S cuban us y S paradoxus (Woods y Eisenberg 1989 Morgan y Ottenwalder 1993 Ottenwalder 200 I) Solenodon arredondoi alcanzo una talla corporal superior a la de cualquiera de las musarafias vivientes a nivel mundial Segun Morgan y Ottenwalder (1993) extrapolando las dimensiones y el peso de los almiquies vivientes S arredondoi debio tener una longitud cabeza-tronco y un peso de 450-550 mm y 1500-2000 g respectivamente en comparacion con S cubanus (360 mm y 800 g) y S paradoxus (390 mm y 1100 g)
Entre todas las especies de almiqufes Solenodon paradoxus es la que presenta los caracteres mas derivados (Otten walder 2001) Si estas relaciones son validas la distribucion de Solenodon entre Cuba y La Espafiola debio discurrir de oeste a este como propuso MacFadden (1980) quien invoco la presencia fosH de S cubanus [pero tambien S arredondoi] en Pinar del Rio para postular que el ancestro continental entr~ por Cuba opinion compartida por Perfit y Williams (1989) y por Ottenwalder ([1991] 2001) El paso de Solenodon a Puerto Rico pudo haberlo impedido la existencia de barreras ecologicas durante la conexion del este de La Espafiola con Puerto Rico 0 por la ausencia de Solenodon en esa parte de La Espafiola en aquella epoca Sin embargo las mismas condiciones ecologicas en el este de La Espafiola no parecen haber representado una barrera efectiva para Nesophontes cuyas mas altas tasa de reproduccion y densidad poblacional presumiblemente Ie permitieron expandirse flpidamente a traves de La Espafiola (Ottenwalder [1991])
Es opinion generalizada que los almiqufes antillanos constituyen el grupo de mamiferos que primero se emplazo en el territorio insular (MacPhee 2005) y en tal sentido es de interes la evidencia sobre especificidad parasito-hospedante aportada por Cruz (1980) quien encontro que la especie de acaro (genero Cubanochirus) que parasita a los almiquies es mucho mas primitiva que la especie (del mismo genero) que paras ita a las jutias y de aqui concluyo que dicho genero de acaro debio evolucionar primero en Solenodontidae y posteriormente en Capromyidae al entrar en contacto representantes de ambas familias en Cuba 0 La Espafiola
Recientemente Roca (2004) y Roca et al (2004) dieron a conocer que los resultados de su estudio genetico-molecular sobre los almiquies indican una divergencia de 25 Ma entre Solenodon cuban us y S paradoxus -compatible con la disyuncion tectonica entre Cuba y La Espafiola (Iturralde-Vinent y MacPhee 1999 McPhee et al 2003)-- y una separacion de ambas especies comparable en magnitud a la divergencia entre familias de mamlferos Consecuentemente propusieron que Solenodon cubanus fuera clasificada en el genero Atopogaie como indico Cabrera (1925) hace 82 afios
DISCUSION Y CONCLUSIONES
Las musarafias y los roedores antillanos (excepto Oryzomys) se irradiaron tanto de sus respectivos ancestros continentales que de estos 6rdenes se reconocen familias y subfamilias endemicas de la region en contraste con los perezosos (una subfamilia y tres
321 IRRADIACION EVOLUCIONARIA
tribus endemicas) y los monos (una tribu endemica) como expresion de diferentes niveles de diversificacion evolucionaria Este proceso es dificil de explicar (traUindose de c1ados que pnicticamente comparten la misma antigiiedad en las Antmas) si es correcta la interpretacion de Pascual et at (1990636)
Nosotros consideramos estos diferentes niveles de diversificacion como las diferentes etapas de evolucion alcanzadas por cada grupo [de mamiferos] Obviamente esto estuvo relacionado con las diferentes longitudes de sus historias
Pudiera ocurrir que en el futuro se compruebe que la historia biogeognifica de la region fue mucho mas complicada
La Espanola la isla mas ecol6gicamente divers a de la region debi6 ser el centro principal de evoluci6n de los mamfferos (con 87 de las familias y 73 de las subfamilias) seguida por Cuba (con 75 de las familias y 54 de las subfamilias) las dos islas de las Antillas Mayores mas relacionadas biogeograficamente (Fig 43 Tabla 44)
eL
EL
cb CAIMAN
SWAN
A Acratocnini B Cubanocnini C Megalocnini D Mesocnini E Nesophontidae F Solenodontidae G Xenothrichini
ANGUILAshyM-0 SMARTIN
H Heteropsomyinae I Plagiodontinae J Isolobodontinae
K Hexolobodontinae L Capromyinae
M Heptaxodontinae N CIidomyinae 0 Sigmodontinae
FIGURA 43 Representacion esquematica de las relaciones interinsulares (expresadas porel numero de linajes compartidos) entre las Antillas Mayores y entre estas y los grupos de islas perifericas segun las interpretaciones de los autores citados en el texto Los numeros dentro de los circulos indican la cantidad de linajes representados en cada isla relacion dudosa Contexto taxon6mico de los linajes en la Tabla 44
bull bull
bull bull
bull bull bull
bull bull
bull bull
bull
bull
bull
bull
bull bull bull bull
bull bull
bull bull
bull
bull bull bull
bull bull
bull
bull bull
bull
322 BIOGEOGRAFiA
TABLA 44 Distribuci6n de los linajcs de mamiferos terrestres a traves de las Antillas Mayores y grupos de islas perifericas LE = La Espanola CU Cuba JA Jamaica PR = Puerto Rico IC Islas Caiman IS Islas Swan IB = Islas Bahamas IV = Islas Virgenes AS = Anguila - San Martin
FamiliaSubfumiliafIribu LE CU JA PR IC IS IB IV AS
Megalonychidae
Choloepodinae
Acratocnini
Cubanocnini
Megalocninae
Megalocnini
Mesocnini
Nesophontidae
Solenodontidae
Pitheciidae
Callicebinae
Xenothrichini
Echimyidae
Heteropsomyinae
Capromyidae
Plagiodontinae
Isolobodontinae
Hexolobodontinae
Capromyinae
H e pta x 0 don tid a e
Heptaxodontinae
Clidomyinae
Muridae
Sigmodontinae
PILOSA
SORICOMORPHA
PRIMATES
RODENTIA
bull
bull
bull
bull TOTALES 12 10 4 4 2
323 EXTlICIOIES DEL CUATERNARIO
El gigantismo ha sido sin duda una tendencia evolucionaria recurrente entre los clados mayores de mamiferos antillanos Es un hecho bien conocido que algunos grupos de mamiferos han aumentado de tamafio en las islas y varios autores han intentado explicarlo a partir de enfoques diferentes (Foster 1964 Miller 1966 Sondaar 1977 Case 1978 Eisenberg y Wilson 1978 Heaney 1978 Lawlor 1982 Lomolino 1985 Angerbjom 1985 McNab 19942001)
EXTINCIONES DEL CUATERNARIO
Las islas que integran la Subregion Antillana han perdido casi 90 de sus mamiferos terrestres durante los ultimos milenios y en proporcion con su extension territorial esta perdida ha sido mayor en la Subregion que en cualquier otro lugar del planeta (MacPhee 1996b) Las extinciones del Cuatemario eliminaron mas de las tres cuartas partes de la fauna de mamiferos terrestres conocida de las Antillas Mayores sus islas satelites y grupos de islas perifericas y el examen de este proceso en sus aspectos mas generales se aborda en esta ultima parte del presente capitulo
CRONOLOGiA Y CONSECUENCIAS
Es muy improbable que un fosil represente el ultimo individuo de un taxon extinto y entonces hay que guiarse por la edad radiometrica del fosil mas reciente conocido lIamada fecha de ultima presencia- como unica aproximacion posible a la fecha de extincion del taxon
Como se explico al inicio del capitulo (vease Contexto Temporal) el registro fosH de los mamiferos terrestres de las Antillas Mayores adolece de insuficiente sustentacion radiometrica No obstante la distribucion temporal de los escasos fechados disponibles para el Cuaternario de la region abarca etapas criticas del proceso de extincion experimentado por la referida fauna a 10 largo de los ultimos 250 mil afios
De las 52 especies que es posible reconocer actualmente como miembros extintos de la fauna cuatemaria de mamiferos terrestres de las Antillas Mayores y grupos de islas perifericas los restos fosiles de 23 de elIas (442) han sido fechados radiometricamente o se han hallado asociados estratigraficamente de manera inequivoca con otro material fechado De estos 23 fechados 3 corresponden al interglacial Sangamon 4 pertenecen a la secuencia Pleistoceno terminal - Holoceno temprano 10 se ubican en ellapso entre el arribo de los aborigenes y la incursion europea y 6 son postcolombinos (a los cuales hay que sumar otras dos especies extinguidas en los siglos XIX y xx) A partir de estas 25 especies avaladas por fechas de ultima presencia distribuidas entre los cuatro intervalos temporales sefialados hemos compuesto la Tabla 45 en la que ademas aparecen las restantes 27 especies (carentes de fechado radiometrico) ubicadas en el intervalo temporal mas probable sobre la base de (1) la asociacion estratigrafica de sus restos con evidencias culturales (amerindias 0 europeas) 0 (2) la ausencia de dicha asociacion Sobre estos criterios de ubicacion temporal procede hacer dos aclaraciones
324 BIOGEOGRAFIA
TABLA 45 Intervalos temporales mas probables para las extinciones cuatemarias de los mamiferos terrestres de las Antillas Mayores y grupos de islas perifericas sobre la base de las evidencias siguientes (l) ultimo ejemplar colectado u observado en vida (2) fechado radiometrico mas tardio en material de la especie (3) asociaci6n estratigrafica mas tardia con otro material fechado (4) asociaci6n estratigrafica con restos de animales introducidos restos humanos u objetos culturales propios del intervale (5) falta de asociaci6n con evidencias culturales [Los ntimeros entre parentesis en la columna Evidencia significan adscripci6n tentativa al intervalo] Los estimados del peso corporal (en kg) estan tomados de la literatura 0 inferidos comparativamente y distribuidos entre
las clases siguientes (a) S 01 (b) 01 a lt 1 (c) 1 a lt 10 (d) 10 a lt 100 (e) 100 a lt 200 (j) ~ 200 Las
abreviaturas que designan las islas son las de la Tabla 44
F amilialEspecie Peso IsJa Evidencia Fuente
I httervalo htterg1acial Sangamon (gt80 kaAP)
HEPTAXODONTlNAE
Amblyrhiza inundata e AS 3 McFarJane et al 1998a
CLIDOMYINAE
Clidomys osborni d JA 3 Ibid
Clidomys parvus d JA 3 Ibid
II httervalo Pleistoceno terminal ~ Holoceno temprano (20-6 kaAP)
MEGALONYCHIDAE
Aeratoenus antillensis d CD 5 Arredondo 1975 a
Aeratoenus odontrigonus d PR 3 McFarJane 1999b
Aeratoenus simorhynehus d LE 2 Rega et al 2002
M egaloenus zile f LE 2 MacPhee et al 2000
SOLENODONTIDAE
Solenodon arredondoi e CD 5 Morgan y Ottenwalder 1993
PnHECIIDAE
Paralouatta varonai d CD 5 MacPhee 1996a
EcHIMYIDAE
Heteropsomys antillensis c PR 5 McFarJane 1999b
Heteropsomys insulans c PR 5 Ibid
Puertoricomys eorozalus c PR 5 Woods 1989a
CAPROMYIDAE
P lagiodontia araeum c LE 5 MacPhee et al 2000
Macrocapromys latus c CD 5 Varona y Arredondo 1979
(Continua)
325 EXTINCIONES DEL CUATERNARIO
TABLA 45 (Continuaci6n)
F ami1ia1Especie Peso Isla Evidencia Fuente
M acrocapromys latus c CU 5 Varona y Arredondo 1979
Mesocapromys kraglievichi c CU 5 Ibid
HEPTAXODONTINAE
Xaymaca fulvopulvis a JA 2 MacPhee y Flemming 2003
III Intervalo Amerindio (6-05 kaAP)
MEGALONYCHIDAE
Acratocnus ye d LE 2 MacPhee et aI 2000
Neocnus comes c LE 2 Ibid
N eocnus dousman c LE 2 Ibid
N eocnus toupiti c LE 2 Ibid
N eocnus gliriform is c CU 4 Cordova et ai 1997
M egalocnus rodens f CU 2 MacPhee et ai 2007
Parocnus browni e CU 2 Steadman et ai 2005
P arocnus serus e LE 2 MacPhee et ai 2000
N ESOPHONTIDAE
N esophontes edithae a PR 2 McFarlane 1999b
PITHECIIDAE
Antillothrix bernensis c LE 3 Rimoli 1977
EcHIMYIDAE
Brotomys contractus a LE 4 Miller 1929c
CAPROMYIDAE
P lagiodontia spelaeum c LE 4 Ibid
M acrocapromys acevedo c CU 4 NUiiez 1990a
HEPTAXODONTINAE
Elasmodontomys obliquus d PR 3 McFarlane 1999b
Iv Intervalo Postcolombino (05 kaAP - Presente)
N ESOPHONTIDAE
N esophontes hypom icrus a LE 24 MacPhee y Fle111111ing 1999
N esophontes zam icrus a LE (4) Ibid
N esophontes m icrus a CU 24 MacPhee et aI 1999
(Continua)
326 BIOGEOGRAFiA
TABLA 45 (Continuacion)
F amilialEspecie Peso Isla Evidencia Fuente
N esophontes m icrus a CD 24 MacPhee et aI 1999
N esophontes m icrus a LE 4 MacPhee et aI 2000
N esophontes sp 1 a IC 4 Morgan 1994
N esophontes sp 2 a IC 4 Ibid
Solenodon marcanoi b LE 4 Ottenwalder 2001
PITHECIIDAE
Xenothrix mcgregori c JA 4 MacPhee y Flemming 1999
EcHIMYIDAE
Boromys offeUa a CD 24 Guarch 1984
Boromys torrei a CD 24 Ibid
Brotomys voratus a LE 4 MacPhee et aI 2000
CAPROMYIDAE
P lagiodontia ipnaeum c LE 4 MacPhee y Flemming 1999
Rhizoplagiodontia lemkei c LE 4 Ibid
Isolobodon portoricensis c PR 24 Ibid
Isolobodon montanus c LE 4 Ibid
Hexolobodon phenax c LE (4) Ibid
Capromys sp c IC 24 Morgan 1994
Geocapromys columbianus c CD 4 MacPhee y Flemming 1999
Geocapromys thoracatus b IS 1 MacPhee y Marx 1997
Geocapromys sp 1 c IC 4 Morgan 1994
Geocapromys sp 2 c IC (4) Ibid
l-IEPTAXODONTINAE
Quemisia gravis c LE (1) MacPhee y Flemming 1999
MURIDAE
Oryzomys antillarum a JA 1 Ibid
1 Flemming y MacPhee (1999) alertaron en contra de la asociacion con otro material fechado Asimismo Morgan (1993) indico que la ausencia de asociacion con evidencias culturales amerindias no es necesariamente un criterio concluyente para ubicar un taxon en el intervalo temporal anterior porque hay depositos en la region que carecen de tales evidencias y son del Holoceno tardio por fechados radiocarbonicos [No obstante
327 EXTlNCIONES DEL CUATERNAR10
la ausencia de una especie en residuarios arqueol6gicos adquiere mayor certidumbre como criterio de ubicaci6n temporal de su extinci6n en la medida que aumenta el numero de residuarios estudiados que en las Antillas Mayores se cuentan por centenares] A falta de fechado absoluto de una especie consideramos que estos criterios de ubicaci6n temporal son mas utiles que la alternativa de excluir del analisis a la especie
2 En los trabajos sobre biogeografia de mamiferos antillanos los arque610gos mas citados como autoridades para establecer el inicio de la presencia aborigen en las Antillas Mayores (Kozlowski 1974 Rouse y Allaire 1978 Rouse 1989 1992 Wilson 2001) difieren al respecto entre los 6 kaAP y los 45 kaAP (pero vease Pagan et al 200523) y hemos adoptado la mas temprana de estas fechas para el comienzo del intervalo amerindio en la Tabla 45 De todos modos cualquiera que haya sido la fecha esta no es aplicable por igual al conjunto de las Antillas Mayores porque segun algunos de los autores citados el poblamiento comenz6 por Cuba y La Espanola 6 mil anos atras continu6 en Puerto Rico hace alrededor de 3 mil anos (pero vease Burney et al 1994) yen Jamaica hace menos de 2 mil e incluso a las Islas Caiman ni siquiera llegaron [Las Islas Caiman descubiertas por Col6n en 1503 carecen absolutamente de evidencia amerindia (Stokes y Keegan 1996)]
Si los datos de la Tabla 45 representan una aceptable aproximaci6n a la realidad el proceso de las extinciones cuaternarias de los mamiferos terrestres de las Antillas Mayores y grupos de islas perifericas debi6 caracterizarse entre otros por los eventos siguientes
Aunque obviamente la fauna cuaternaria debi6 ser mucho mas numerosa que 10 que refleja el registro paleontol6gico (Morgan y Woods 1986) 52 especies (81 de la fauna conocida) desaparecieron (sin reposici6n) durante los ultimos 250 mil anos
- Al parecer las extinciones se concentraron en las especies de mayor tamano Asi el peso promedio estimado para las especies de la etapa preantr6pica (intervalos I + II de la Tabla 45) es 350 kg es 328 kg para las especies del intervalo III (amerindio) y es s610 25 kg (c1ases del peso a b c) para las especies del intervalo IV (postcolombino) Ninguna especie aut6ctona de mamifero mayor de 6 kg existe hoy en la regi6n
- De acuerdo con la distribuci6n temporal de las extinciones 61 especies sobrevivieron al interglacial Sangamon (intervalo 1)48 sobrevivieron a la transici6n del Pleistoceno terminal al Holoceno temprano (intervalo II) 34 sobrevivieron al periodo amerindio (intervalo III) y s610 12 especies han sobrevivido desde la Ilegada de los europeos (intervalo IV) Las tasas de extinci6n correspondientes (exceptuado el intervalo I) son una especie cada 1167 anos en el intervalo II una cada 393 anos en el intervalo III y una cada 23 anos en el intervalo IV
- Durante todo el proceso de las extinciones cuaternarias la perdida de especies por isla fue de 91 en La Espanola 83 en Jamaica y 62 en Cuba Bahamas retuvo su unica especie (aunque extirpada en varias de las islas) Puerto Rico e Islas Caiman perdieron todas sus especies (6 y 5 respectivamente) y las Islas Swan Virgenes y Anguila - San Martin perdieron sus respectivas especies (micas
328 BIOGEOGRAFiA
De los cuatro 6rdenes de mamiferos representados en la fauna Pilosa y Primates desaparecieron totalmente Soricomorpha y Rodentia perdieron 86 y 74 de sus especies respectivamente
CAUSALIDAD
CAUSAS NATURALES
Los cicIos glaciales e interglaciales acaecidos en el Pleistoceno causaron cambios ambientales dnisticos en las Antillas con la concomitante alternancia de cIimas frios (secos) y calidos (hUmedos) asi como de descensos y ascensos del nivel del mar aumento y disminuci6n de las areas territoriales) induso (aunque con menor intensidad) en el Holoceno Estos eventos paleogeograficos han sido consistentemente documentados (Mayo y Kartashov 1972 Bonnatti y Gartner 1973 Ortega y Arcia 1982 Prance 1982 Schubert y Medina 1982 Dodge et al 1983 Ortega [1984] Haq et al 1987 Schubert 1989 Hodell et al 1991 Guilderson et al 1994 Oppo 1997 Emslie 1998 Curtis et al 2001 Iturralde-Vinent 2003) y de ellos brindamos a continuaci6n una breve resena
Finalizando el periodo interglacial anterior a la ultima glaciaci6n (Wisconsin) el mar estaba a 8-10 m por encima de su nivel actual Cuando el avance glacial del Wisconsin a1canz6 su culminaci6n (eillamado Ultimo Maximo Glacial) hace 18 ka el mar habia descendido 120 m con respecto a dicho nivel y las islas del Caribe habian incrementado considerablemente sus extensiones subaereas en comparaci6n con las que presentan hoy dia por ejemplo Puerto Rico y las Islas Virgenes se conectaron territorialmente y las Bahamas se unieron en cinco gran des islas Prevalecia un regimen frio y seco y condiciones de aridez --debidas a una reducci6n de las precipitaciones (posiblemente a menos de 440 mmlano)- y los ambientes xericos se habian extendido a traves de las islas y bancos emergidos y abundaban los pinares (los habitats mesicos estaban restringidos a parcelas relictas en 10 alto de las montanas) En el Mar Caribe y el Golfo de Mexico la temperatura superficial del mar era 5degC mas baja que la actual y en las llanuras costeras y alturas maximas eran respectivamente 2-3degC y 6degC mas bajas Los limites de las zonas ecol6gicas altitudinales descendieron varios cientos de metros y cada zona redujo su extensi6n vertical al aumentar el gradiente de temperatura Ellimite inferior de la zona periglacial
elevaci6n donde la temperatura media minima a1canza los OdegC) de la Cordillera Central de la Republica Dominicana (altura maxima del Caribe insular con evidencia de topografia glacial) se considera que estuvo entre los 1500 y 1800 mlnm (actualmente esta a 2800 mlnm) y se puede estimar que dicho limite era inferior en Cuba por estar mas al norte que la Republica Dominicana No obstante durante este periodo la temperatura experiment6 episodios cortos (unas pocas decadas) de ascensos repetidos
Un signifieativo componente bi6tieo de las referidas condiciones paleoambientales 10 constituy6 la espectacular irradiaci6n evolucionaria experimentada por un grupo numeroso de aves rapaees diurnas y nocturnas (Wetmore 1937 Brodkorb 1969 Kurotchkin y Mayo 1973 Arredondo 1976 Olson 1978 Olson y Hilgartner 1982
329 EXTINCIONES DEL ClJATERNARIO
Suarez 2000a b 2004 Suarez y Emslie 2003) Los mamiferos de las Antillas Mayores ofrecieron a estos depredadores aviares una amplia gama de tamanos de presa que en ausencia de mamiferos del orden Carnivora en la region permitio a estas aves irradiarse a este nicho alimentario con la evolucion de una variedad de especies de muy diversa tall a corporal incluidas formas gigantes Adquirir un cuerpo gigante les propicio el acceso a recursos alimentarios que no estaban siendo explotados por ningun otro depredador (Alcover y McMinn 1994) A Cuba corresponde el mejor registro fosil de rapaces cuaternarias caribenas con siete generos de especies diurnas Dentro de estas aves existieron tres gavilanes gigantes (Accipitridae) tres carairas (Falconidae) y un condor (Cathartidae) asi como cuatro generos de especies nocturnas dos lechuzas (Tytonidae) y tres Mhos (Strigidae) dos de ell os no igualados en tamano en todo el mundo Algunos de estos generos tuvieron representacion en otras islas antillanas como La Espanola Jamaica Bahamas y Gran Caiman
Al retirarse hacia el norte los hielos de la glaciacion Wisconsin el clima se tomo mas hUmedo en las Antillas durante el Holoceno temprano y el bosque mesico se expandio en la region insular El nivel del mar ascendio acompanado de mayor pluviosidad alrededor de los 7 kaAP las Islas Virgenes quedaron separadas entre si y de Puerto Rico y las Bahamas perdieron la mayor parte de su territorio Cerca de los 3 kaAP las condiciones secas retornaron a la region e hicieron desaparecer buena parte del bosque mesico Nuevamente el clima comenzo a adquirir sus condiciones actuales hacia los 25 kaAP aproximadamente y hoy transita por una fase mesica con vegetacion xerica confinada en parcelas relictas
Sin duda estos y otros eventos naturales incidieron negativamente en la sobrevivencia de mamiferos terrestres en las Antillas Mayores asi como en los grupos de pequenas islas perifericas (Pregill y Olson 1981 Olson 2006)
McFarlane et al (1998a) demostraron convincentemente como Amblyrhiza el gigante de los roedores antillanos endemico de la pequena isla de Anguila sucumbio a las repetidas y abruptas perdidas de su habitat durante el interglacial Sangamon por las nipidas y severas oscilaciones del nivel del mar (documentadas por Neumann y Hearty 1996) al no poder responder adecuadamente a tan catastroficos eventos mediante el ajuste de su masa corporal media y otro tanto debio ocurrirle en Jamaica al otro roedor gigante Clidomys (McFarlane et al 1998b) emparentado con Amblyrhiza en la misma epoca (Tabla 45 intervalo I)
El ascenso del nivel del mar seguramente promovio la extincion de mamiferos durante la transicion del Pleistoceno al Holoceno (Tabla 45 intervalo II) con la reduccion del tamano de las Islas por las transgresiones marinas (Pregill y Olson 1981 Morgan 1993) Esto ultimo haria maximamente severa la competicion asf como la presion de los depredadores [Steadman et al (2005) no yen por que el calentamiento global ocurrido durante la transicion Pleistoceno-Holoceno habria de tener un efecto negativo sobre los perezosos siendo sus descendientes actuales especies tropic ales 0 subtropicales pero no se trata del efecto
330 BIOGEOGRAFiA
directo del cambio de temperatura sobre los animales sino del efecto indirecto y progresivo que dicho cambio genera a largo plazo al reducir el territorio insular disponible]
Por esta epoca tambien desaparecieron especies de tall a mayor como Megalocnus zile en La Espanola y otras de menor tamano (en Cuba) pero que representan la tall a maxima en sus respectivos contextos taxonomicos del Cuaternario como Solenodon arredondoi Paralouatta varonai y Macrocapromys acevedo Un mamifero puede ser grande solo a costa de reducir su densidad poblacional y esto 10 hace mis vulnerable a la extinci6n sobre todo en una isla (McNab 1994 McFarlane et al 1998a) Incidentalmente Alcover et al (1998a) afirmaron que larelacion entre la talla corporal y la extincion no siempre se cumple en las especies de menor tamano y citaron como ejemplo que entre las musarafias todas las especies de Nesophontes desaparecieron mientras sobreviven especies de Solenodon pero la comparacion correcta en todo caso serfa entre los solenodones vivientes y el gigante Solenodon arredondoi y entre Nesophontes edithae -el gigante de los nesofontes extinguido en el periodo amerindio- y el resto de sus congeneres que sobrevivieron hasta los primeros siglos de la colonizacion europea (Tabla 45 intervalos III y IV)
CAUSAS ANTROPOGENICAS
Intervalo Amerindio
La mayoria de los autores que se han referido a las extinciones acaecidas durante este intervalo (entre elIos paradigmaticamente Martin 1984a b) han responsabilizado a los aborfgenes (que se supone comenzaron a poblar las Antillas Mayores hace 6 mil afios) con la desaparicion de numerosas especies de mamiferos terrestres en la region principalmente por la via de la sobrecaza y la introduccion de animales domesticos devenidos depredadores de la fauna autoctona (Morgan y Woods 1986) Este efecto negativo de la presencia indigena sobre los mamiferos de la region ha sido dramatizado por algunos autores en terminos de holocausto faunal Sin embargo pensamos que se ha exagerado Como han expresado MacPhee et al (19993) cuando uno busca evidencia dura de complicidad humana directa en [estas] extinciones no hay mucho que encontrar Las siguientes son algunas consideraciones a tener en cuenta en tal sentido
El registro radiometrico mas joven conocido hasta ahora para un perezoso antillano (MegalocnllS rodens) es de 42 kaAP (MacPhee et ai 2007) Si se acepta que los aborfgenes entraron en Cuba hace aproximadamente 6 mil anos su convivencia con los perezosos debe haber durado alrededor de 1800 afios (0 mas porque dificilmente el referfdo fechado radiometrico haya correspondido al ultimo sobreviviente de la especie) Esta cifra contradice la hipotesis del holocausto faunal segun la cualla extincion de los mamiferos insulares de mayor talla debe ocurrir con extrema rapidez -a escala de decadas 0 de centurias- a partir de su primer contacto con los humanos (Martin y Steadman 1999) A pesar de las repetidas afirmaciones en contrario (Harrington 1921 Miller 1929c Herrera [1945] Tabio 1951 Alvarez 1951a 1957 Arredondo
331 EXTINCIONES DEL CUATERNARIO
1959 1960a Veloz y Ortega 1973 Mayo 1980b Rodriguez et al 1984 Castellanos y Pino 1985 Pino y Castellanos 1985 Nunez 1990b Gutierrez 1996 Arredondo 2004) en su mayoria basadas en evidencias no exentas de posibles accidentes de deposicion (0 de reubicacion postdeposicional) en recintos cavernarios (Diaz-Franco [2006b D no se ha demostrado inequivocamente que los aborigenes antillanos se alimentaran de perezosos regularmente Sus restos en residuarios de alimentos son escasisimos en toda la region y esto contrasta con los abundantes restos de jutias y otros mamiferos de menor talla que aparecen en esos residuarios demostrativos de la preferencia de los paleoindios por estos otros mamiferos
Los mamiferos mas importantes en la economia prehistorica de las Antillas Mayores fueron las jutias Capromys y Geocapromys en Cuba Geocapromys en Jamaica e Isolobodon en La Espanola Puerto Rico e Islas Virgenes (Aguayo 1950b Tablo 1951 Rimoli 1972 Wing 2001 Diaz-Franco [2006b D Wilkins (2001) estudi61ajutia viviente de Jamaica (Geocapromys browni) en un sitio ocupado por los aborigenes durante 100 anos y no encontr6 evidencias de que durante ese tiempo la especie haya sido sobreexplotada Asimismo en Cuba las jutias conga carabali y andaraz son escasas en depositos pleistoholocenicos en cambio son muy numerosas en residuarios indigenas y de haber sido sobreexplotadas 10 mas probable es que no hubieran sobrevivido hasta hoy
La habilidad de los humanos para penetrar en profundidad el bosque tropical y mantenerse en este sustentablemente ha sido un evento comparativamente reciente (Beran y Steele 1994) En el caso antillano la caza tierra adentro ha sido explicada como un proceso tardio de desplazamiento a partir de la explotacion de abundantes recursos marinos que ofrecian condiciones 6ptimas para el sedentarismo de las comunidades aborigenes costeras (C6rdova y Arredondo 1988 Brito 2005) Peces y mariscos dominan la arqueologia costera de las Islas antillanas en tanto el consumo de recursos terrestres (mamfferos incluidos) aumenta progresivamente en la medida que los residuarios estudiados se alejan de la costa (Scudder 1991 Wilkins 2001) En las Antillas Mayores el componente terrestre de la dieta aborigen es solo 20-30 del material recobrado en los residuarios (Wing 1977 1989) Y debio ser menor alin porque el componente vegetal de la dieta no se ha preservado Por otra parte los aborigenes cubanos domesticaron la jutia conga y seguramente la reprodujeron para comer (Pose et al 1988) practica que probablemente se repitiera con otras jutias en las demas Antillas Mayores
- Fray Bartolome de las Casas hablo de 3 millones de indios en Cuba al inicio de la colonizacion pero los historiadores subsecuentes consideraron exagerada la cifra (Poey 1851 a) No obstante se dispone de dos estimados de la densidad de poblaci6n aborigen en las Antillas Mayores a la llegada de los europeos 18 indigenaslkm2 en Puerto Rico (Rouse 1956) y 1271km2 en La Espanola (Watts 1987) Extrapolando ambos valores entre las cuatro islas principales de las Antillas Mayores tenemos que en la fecha indicada la poblaci6n aborigen pudo estar entre 16 mil y 113 mil indigenas en Puerto Rico entre 21 mil y 145 mil en Jamaica entre 141 mil y 1 mill6n en La Espanola y entre 200 mil y 14 millones en Cuba Entre las cifras maximas y las minimas nos inclinamos
332 BIOGEOGRAFiA
hacia estas ultimas atendiendo a los altos indices de mortalidad infantil y morbilidad que se evidencian en las osamentas de los pobladores prehispanicos (Martinez y Rives 1990) Uno de los hechos que apuntan hacia el bajo numero de aborigenes en las Antillas Mayores es 10 rapido que desaparecieron despues de la llegada de los europeos pnkticamente sin dejar mestizaje con estos 10 cual contrasta ostensiblemente con la situacion en la America continentaL
En suma un grupo humano (1) demograiicamente irrelevante y controlado por altos indices de morbilidad (2) consumidor mayoritariamente de organismos marinos y (3) carente de la tecnologia cinegetica necesaria para poder sobreexplotar los mamiferos que Ie sirvieron de alimento (a los cuales accedio tardiamente en su desarrollo) no es concebible que haya causado e1 holocausto faunal que se Ie atribuye a partir del error de aplicar a las comunidades aborigenes antillanas un modelo cultural de la prehistoria continentaL
Tampoco hay evidencias de que los aborigenes hayan introducido carnivoros con suficiente capacidad depredadora salvo el perro Morgan y Otlenwaldder (1993) consideraron que la depredacion por perros durante el periodo amerindio es la explicacion mas plausible para la extincion del almiqui gigante de Cuba pero como se puede observar (Tabla 54) Solenodon arredondoi aparece en el intervalo II por falta de asociacion con evidencias culturales
Consecuentemente el accionar de los aborigenes y de sus animales domesticos directamente sobre la fauna no parece haber sido por sf solo un factor determinante de la extincion de mamiferos terrestres durante el periodo amerindio en las Antillas (pero vease Steadman et al 2005)
Hay que recordar en este senti do que alin en el Holoceno tardio ocurrieron cambios clirruiticos de consideracion (vease Causas Naturales) Por ejemplo Curtis et al (2001) explicaron que la extincion de algunos ani males mesofilicos de las alturas montanosas de La Espanola pudo haber antecedido a cualquier impacto humano sigrIificativo en esos lugares por coincidir dicha extincion con una tendencia a la aridez documentada isotopicamente y con evidencias de la perdida del bosque mesico ocurridas despues de 3kaAP Yen Jamaica hace solo 2shy3 mil aDos el area de la Bahia de Jackson sustentaba una gran biomasa de vertebrados en bosque hinnedo a juzgar por la cantidad y diversidad de taxones presentes en material fosil derivado de la depredacion por lechuzas hoy el area es la mas seca de la isla con matorral xerofitico y pobreza de fauna (MacPhee 1996a) Yalm en nuestros dias huracanes de gran intensidad aniquilan poblaciones de vertebrados (que no se recuperan) en is las pequenas y llanas del Caribe (Brooks y Smith 2001)
En medio de la incertidumbre sobre la verdadera causa (0 causas) de las extinciones de mamiferos en el periodo amerindio MacPhee y Marx (1997) han propuesto su hipotesis de la hipermorbilidad la cual estipula colapsos poblacionales relativamente rapidos con la introduccion de enfermedades de alta letalidad Estos autores han razonado que
Las islas son los lugares mas apropiados para experimentar perdidas catastroficas causadas por enfermedades Los mamiferos no solo serian susceptibles en virtud de su prolongado aislamiento biologico sino que en los confines de una isla la epizootia
333 EXTINCIONES DEL CUATERNARIO
tendria la potencialidad de infectar simultaneamente a la rnayoria de los individuos de vadas especies Los neonatos son inmunologicamente inmaduros y sufririan desproporcionadamente cualquier infeccion hipervirulenta La perdida excesiva de neonatos y juveniles tendria efectos severos para la sobrevivencia de la poblacion si esta se reduce por debajo de cierto nivel sobreviene la extincion porque no hay posibilidad de restitucion en el periodo postpanzootico Las especies de mayor talla corporal van a ser las mas vulnerables a la extincion dadas sus bajas tasas reproductivas largos periodos de gestacion y reducido numero de progenie Las especies de menor tamano --caracterizadas por altas tasas reproductivas periodos cortos de gestacion y mayor numero de neonatos por parto- tambien serian fuertemente diezmadas pero muchas sobrevivirian porque su estrategia reproductiva les permite perder mayor numero de individuos Las perdidas deben ocurrir relativamente rapido (a escala de decadas) mientras que otras hipotesis (digamos los cambios climaticos) permiten intervalos mucho mas largos (a escala de siglos 0 de milenios) entre la aparicion de las causas y la produccion de sus efectos Por ejemplo todo indica que los nesofontes se extinguieron muy rapidamente y seria dificil poder invocar un impacto humano directo 0 argumentar fenomenos climaticos que pudieran explicar plausiblemente perdidas tan rapidas
MacPhee y Marx (1997 191) han dicho que por el momento la hipotesis de la hipermorbilidad no pasa de ser una abstraccion en espera de adecuada sustentacion empirica Llama la atencion que linicamente en el periodo amerindio (Tabla 45 intervalo 1Il) las extinciones abarcaron todas las clases del peso corporal (de la a a laj) y que ninguna de las especies que sobrevivieron hasta el periodo postcolombino (Tabla 45 intervalo IV mas las especies vivientes) sobrepasa los 10 kg (clases del peso a b c) en clara coincidencia con los presupuestos de la hipotetica hipermorbilidad
Intervalo Postcolornbino
Por 10 menos 37 especies de mamiferos terrestres vivian en las Antillas Mayores y grupos de Islas perifericas a la lIegada de los europeos en 1492 De elIas se han extinguido 23 shymas especies que en cualquiera de los periodos precedentes (Tabla 45~ y solo subsisten 12 especies repartidas entre Cuba (8 especies) La Espanola (2) Jamaica (1) y Bahamas (1) varias de estas en peligro critico de extincion Solo dos de las especies extinguidas durante los ultimos 500 anos se conocieron en estado viviente los roedores Oryzomys antillarum de Jamaica (extinguido en 1877) y Geocapromys thoracatus de las Islas Swan (extinguido en 1955) Las causas de las extinciones ocurridas durante este periodo son sobradamente conocidas destruccion de habitats introducci6n de animales exoticos (domesticos y comensales) y demas presiones asociadas a la actividad humana entre las cuales posiblemente habria que incluir brotes de hipermorbilidad
Durante mucho tiempo se ha afirmado la posibilidad de que especies de Nesophontes hayan sobrevivido hasta las primeras decadas del siglo xx y que incluso continuen existiendo en lugares remotos de Cuba 0 La Espanola (Allen 1918 1942 Miller 1930 Aguayo 1950a Arredondo 1960b Fischer 1977 MacFadden 1980 Woods y
334 BIOGEOGRAFiA
Ottenwalder 1991) Actualmente no existe evidencia radiometrica de que alguna especie de Nesophontes haya sobrevivido al siglo xv las fechas radiocarbonicas calibradas obtenidas en muestras de Nesophontes de Cuba y La Espanola resultaron no mas tardias que los siglos XIII y XIV (MacPhee et al 1999) Por otra parte las expediciones efectuadas a fines del siglo xx con el proposito de localizar individuos vivientes del genero en La Espanola no produjeron la menor indicacion afirmativa al respecto (Woods et al 1985) Ademas a traves de las Antillas Mayores no aparecen restos de Nesophontes en los depositos formados actualmente por las lechuzas
EPiLOGO
Las Antillas Mayores son islas oceanicas no han formado parte de un continente (con la posible excepcion de una parte de Jamaica hasta el Eoceno Medio hace 45 Ma) y durante un tiempo geologico considerable (desde el Eoceno Superior hace 40 Ma) han permanecido emergidas y separadas de los continentes vecinos Consecuentemente los mamiferos terrestres (no volantes) autoctonos de estas islas constituyen una fauna oceanica que por ende cumple con todas las caracteristicas convencionales reconocidas para este tipo de entidad biogeografica
- Baja diversidad taxonomica a nivel de las categorias superiores (solo cuatro ordenes) en comparacion con islas plataformicas continentales (0 con areas continentales) de tamano y diversidad ecologica simi lares
- Alto grado de endemismo a nivel de genero (96) y familia (50) en respuesta al prolongado aislamiento
- Desbalance ecologico de la estructura trofica ausencia de mamiferos camivoros y ungulados
- Espectaculares procesos de irradiaci6n evolucionaria como resultado de la colonizacion de territorios libres de competidores y depredadores
Tendencias evolucionarias extremas gigantismo
- Control a cargo de aves rapaces con tendencia at gigantismo en ausencia de mamiferos miembros del orden Carnivora
- Mas especies de reptiles y de murcielagos que de mamiferos terrestres relaciones que en los continentes se invierten
Alta tasa de extincion de especies (84) sin reposicion
Preservacion de especies filogeneticamente arcaicas (los llamados fosiles vivientes) los almiquies

317 TRRADIACTON EVOLUCIONARIA
compatible con la hipotetica conexion -propuesta tentativamente por Iturralde-Vinent y MacPhee (199980-84)- entre las Montanas Azules del este de Jamaica y la peninsula suroccidental de La Espanola en la epoca de GAARlandia
A pesar de que los primates antillanos pertenecen a un grupo viviente de monos muy pequenos (los monos titles) fueron comparativamente grandes en relacion con los platirrinos modernos y sucesivamente mayores en la direccion Antillothrix Xenothrix y Paralouatta con P marianae como el mayor platirrino conocido del Mioceno neotropical (MacPhee 2005) MacPhee et al (1994) consideraron que Xenothrix debio ser un mono de andar lento y al parecer las especies de Paralouatta abandonaron la arborealidad para adoptar un modo de vida mas terrestre (MacPhee 1996a MacPhee et al 2003 MacPhee y Meldrum 2006)
ROEOORES (Echimyidae Capromyidae Heptaxodontidae)
La edad de Zazamys en Cuba demuestra que la invasion de los roedores histricognatos en las Antillas Mayores comenzo antes del Mioceno Inferior por otra parte los representantes mas antiguos de esa irradiacion evolucionaria se encuentran en La Espanola y Puerto Rico Ambas evidencias son consistentes con la hipotesis de GAARlandia (Woods 1982 1988 Woods et al 2001)
No esta claro si los capromidos evolucionaron a partir de un equimido ancestral 0 de un protocapromido 0 si los equimidos antillanos (I sub familia 3 generos y 7 especies todas extintas) forman parte de la irradiacion de los capromidos (4 subfamilias 9 generos y 27 especies 10 vivientes) Kratochvil et al (1978b) y Rodriguez et al (1979) consideraron que Capromys nan us es un eslab6n entre Echimyidae y Capromyidae pero Mesocapromys nanus esta morfologicamente mucho mas cerca de Hexolobodon y los de mas capromidos que de los equimidos (Woods 1985) Lo que si esta claro segun Woods et al (2001) es que la subfamilia Plagiodontinae endemic a de La Espanola es el linaje basal de los capromidos y que Isolobodon Hexolobodon Capromys Geocapromys Mysateles y Mesocapromys [y supuestamente Macrocapromys] forman parte de la irradiacion temprana de Capromyidae
La subfamilia Capromyinae (5 generos y 17 especies 9 vivientes) parece ser el dado de mas reciente evolucion en la familia con invasion de Cuba desde La Espanola y dispersion desde Cuba (1) por Capromys a las islas Caiman y (2) por Geocapromys a las islas Bahamas Caiman y Jamaica y desde Jamaica a las islas Swan aunque la presencia de Geocapromys en Swan pudiera deberse a introduccion aborigen (Morgan 1985 1988 1989 1993 1994 Woods 1989a 1990) Woods et al (2001) utilizaron datos moleculares para obtener estimados independientes de divergencia entre capromidos segiJn 10 cual Geocapromys divergio cerca de los 17 MaAP Mesocapromys divergio de Capromys hace 15 MaAP en tanto lvlysateles 10 hizo hace 10 MaAP Por su parte las formas de Capromys pilorides y de Mysateles prehensilis en Isla de la Juventud divergieron de sus correspondientes subespecies nominativas en la isla principal a los 24 y 08 MaAP respectivamente
318 BIOGEOGRAFiA
Woods y Howland (I979) describieron adaptaciones morfologicas de la mandibula el hioide y la faringe asociadas a mecanismos de masticacion diferentes en plagiodontinos y caprominos y Varona (1980a) llamo la atencion sobre el gran desarrollo de los molariformes (en relacion con la longitud total del cnineo) que presentan ciertos grupos de jutias fosiles en comparacion con las vivientes 10 que pudiera interpretarse como una adaptacion alimentaria a una vegetacion mas abrasiva por parte de las jutias extinguidas
Segun Woods (1990) la familia Heptaxodontidae (2 subfamilias 5 generos y 6 especies todas extintas) tambien seria parte de la irradiacion de los capromidos con origen en el este de La Espanola 0 en Puerto Rico y subsecuente dispersion al Banco de Anguila - San Martin en las Antillas Menores (pero vease MacPhee y Flemming 2003) Este banco estaria a solo 70 km del Banco Puerto Rico - Islas Virgenes en tiempos de maxima regresion marina y esta proximidad con Puerto Rico -patria de Elasmodontomys obliquus- seguramente tuvo que ver con la presencia en las islas de Anguila y San Martin del heptaxodontino gigante Amblyrhiza relacionado mas estrechamente con los roedores de las Antillas Mayores que con los del resto de las Antillas Menores (Woods 1990 1996) Aunque los roedores suelen aumentar de tamano en las islas (Case 1978) la escala del incremento de la talla en Amblyrhiza (que pudo alcanzar mas de 200 kg de peso adulto) esta fuera de toda proporcion con los patrones conocidos en otros roedores insulares (Biknevicius et 01 1993)
Clidomys (Clidomyinae) de Jamaica difiere marcadamente en su morfologia de los heptaxodontinos (vease ademas MacPhee y Flemming 2003)10 cual dificulta explicar su presencia en esa isla Como Jamaica estuvo bajo el mar hasta el Mioceno Medio el ancestro de Clidomys colonizaria esta isla por dispersion desde el extremo suroccidental de La Espanola despues que los heptaxodontinos se originaron en esta ultima isla (Woods 1989a) Pero Pascual et 01 (I 990) especularon que Clidomys pudiera ser el resultado de una invasion posterior e independiente de los capromidos
GRUPOS ORIGINARIOS DE NORTEAMERICA
El origen de las musaranas antillanas ha resultado ser hasta ahora el mayor enigma biogeografico de la fauna de mamiferos terrestres de la region aunque la mayoria de los autores coinciden en considerarlas originarias de Norteamerica (vease Discus ion y Conclusiones en Poblamiento Continente-Isla) No obstante MacPhee e Iturralde-Vinent (1994) descartaron toda posibilidad de conexion terrestre por el Estrecho de la Florida 0 por el Canal de Yucatan y si el (supuesto) evento vicari ante de Rosen (vease El Arca de Noe) -invocado por MacFadden (1980) para resolver el enigma- fuera minimamente plausible aun asi es dificil creer que las catastrofes que 10 sucedieron (vease Contexto Temporal) hayan barrido con toda esa vieja fauna continental excepto las musaranas circunstancia ya anticipada muy tempranamente por Matthew (l918a)
De otra parte la relaci6n entre Nesophontidae y Solenodontidae permanece irresuelta Si estos linajes no estan estrechamente relacionados entonces es muy posible que
319 IRRADIACION EVOLUCIONARIA
sus distribuciones sean explicadas por mecanismos diferentes (Whidden y Asher 2001)
La descomunal diferencia de talla corporal entre las musaranas pequenas (Nesophontes) y las grandes (Soienodon) seguramente impuso a ambos grupos restricciones distintas relativas a la termorregulacion y las adaptaciones energeticas En Nesophontes la seleccion debio favorecer una alta tasa de metabolismo y crecimiento camadas mas numerosas corta expectativa de vida yalta densidad poblacionaL En Soienodon en cambio la seleccion opero en la direccion de una baja tasa metabolica camadas reducidas crecimiento lento longevidad y baja densidad poblacional (Ottenwalder [1991])
MUSARANAS PEQUENAS (Nesophontidae)
Las musaranas pequenas estan representadas en las Antillas por el genero Nesophontes (y un genero miocenico indeterminado en ambar dominicano) con 6 especies cuaternarias extinguidas distribuidas en Cuba La Espanola Puerto Rico Islas Virgenes e Islas Caiman La mayor de las musaranas pequenas Nesophontes edithae evoluciono en Puerto Rico y su arribo a las Islas Virgenes supone una dispersion sumamente reciente En las Islas Caiman Nesophontes se asemeja mas a la poblacion de Cuba de donde seguramente derivo en el Plio-Pleistoceno (junto con Capromys y Geocapromys) cuando la costa sur de Cuba estuvo mucho mas cerca de las Islas Caiman (lturralde-Vinent 2003fig 4) MacPhee et ai (1994) interpretaron la distribucion de Nesophontes en Cuba y La Espanola como resultado de la vicarianza oligomiocenica ocurrida al formarse el Paso de los Vientos pero la distancia taxonomica entre los lados del Paso es solo de nivel especifico y N micros incluso vivi6 en ambas islas segtln Patterson (1962)
Allen (1918) considero que Nesophontes micrus fue una especie primitiva no modificada para la vida arb6rea 0 fos6rea y por tanto de habitos terrestres Nesophontes edithae de Puerto Rico es mucho mayor y mas robusta que las restantes especies del genero eL indice de robustez de su femur (= anchura minima de la diafisis ongitud total del hueso) muestra que este es tan robusto como el de Solenodon paradoxus un animal dos veces mayor (Morgan et ai 1980) Choate y Birney (1968) postularon un aumento cronoclinal de la taIla corporal en N edithae confirmado mas tarde por McFarlane (1999a) mediante fechados radiocarb6nicos de la secuencia estratignifica en depositos cavernarios de Puerto Rico Solenodon no vivi6 en Puerto Rico y en ausencia de una musarana de mayor tamano Nesophontes alcanzo una talla corporal mayor en esa isla que en La Espanola 0 Cuba (Woods y Eisenberg 1989)
MUSARANAS GRANDES (Solenodontidae)
Los arcaicos almiqufes (genero Soienodon) comprenden cuatro especies una viviente (S cubanus) y otra f6sil (S arredondoi) en Cuba y una viviente (S paradoxus con dos subespecies) y otra f6sil (S marcanoi) en La Espanola Como cabria esperar las dos especies cubanas se asemejan mas entre sf que a las especies de La Espanola pero
320 BIOGEOGRAFiA
enigmaticamente S marcanoi comparte caracteres con S cuban us y S paradoxus (Woods y Eisenberg 1989 Morgan y Ottenwalder 1993 Ottenwalder 200 I) Solenodon arredondoi alcanzo una talla corporal superior a la de cualquiera de las musarafias vivientes a nivel mundial Segun Morgan y Ottenwalder (1993) extrapolando las dimensiones y el peso de los almiquies vivientes S arredondoi debio tener una longitud cabeza-tronco y un peso de 450-550 mm y 1500-2000 g respectivamente en comparacion con S cubanus (360 mm y 800 g) y S paradoxus (390 mm y 1100 g)
Entre todas las especies de almiqufes Solenodon paradoxus es la que presenta los caracteres mas derivados (Otten walder 2001) Si estas relaciones son validas la distribucion de Solenodon entre Cuba y La Espafiola debio discurrir de oeste a este como propuso MacFadden (1980) quien invoco la presencia fosH de S cubanus [pero tambien S arredondoi] en Pinar del Rio para postular que el ancestro continental entr~ por Cuba opinion compartida por Perfit y Williams (1989) y por Ottenwalder ([1991] 2001) El paso de Solenodon a Puerto Rico pudo haberlo impedido la existencia de barreras ecologicas durante la conexion del este de La Espafiola con Puerto Rico 0 por la ausencia de Solenodon en esa parte de La Espafiola en aquella epoca Sin embargo las mismas condiciones ecologicas en el este de La Espafiola no parecen haber representado una barrera efectiva para Nesophontes cuyas mas altas tasa de reproduccion y densidad poblacional presumiblemente Ie permitieron expandirse flpidamente a traves de La Espafiola (Ottenwalder [1991])
Es opinion generalizada que los almiqufes antillanos constituyen el grupo de mamiferos que primero se emplazo en el territorio insular (MacPhee 2005) y en tal sentido es de interes la evidencia sobre especificidad parasito-hospedante aportada por Cruz (1980) quien encontro que la especie de acaro (genero Cubanochirus) que parasita a los almiquies es mucho mas primitiva que la especie (del mismo genero) que paras ita a las jutias y de aqui concluyo que dicho genero de acaro debio evolucionar primero en Solenodontidae y posteriormente en Capromyidae al entrar en contacto representantes de ambas familias en Cuba 0 La Espafiola
Recientemente Roca (2004) y Roca et al (2004) dieron a conocer que los resultados de su estudio genetico-molecular sobre los almiquies indican una divergencia de 25 Ma entre Solenodon cuban us y S paradoxus -compatible con la disyuncion tectonica entre Cuba y La Espafiola (Iturralde-Vinent y MacPhee 1999 McPhee et al 2003)-- y una separacion de ambas especies comparable en magnitud a la divergencia entre familias de mamlferos Consecuentemente propusieron que Solenodon cubanus fuera clasificada en el genero Atopogaie como indico Cabrera (1925) hace 82 afios
DISCUSION Y CONCLUSIONES
Las musarafias y los roedores antillanos (excepto Oryzomys) se irradiaron tanto de sus respectivos ancestros continentales que de estos 6rdenes se reconocen familias y subfamilias endemicas de la region en contraste con los perezosos (una subfamilia y tres
321 IRRADIACION EVOLUCIONARIA
tribus endemicas) y los monos (una tribu endemica) como expresion de diferentes niveles de diversificacion evolucionaria Este proceso es dificil de explicar (traUindose de c1ados que pnicticamente comparten la misma antigiiedad en las Antmas) si es correcta la interpretacion de Pascual et at (1990636)
Nosotros consideramos estos diferentes niveles de diversificacion como las diferentes etapas de evolucion alcanzadas por cada grupo [de mamiferos] Obviamente esto estuvo relacionado con las diferentes longitudes de sus historias
Pudiera ocurrir que en el futuro se compruebe que la historia biogeognifica de la region fue mucho mas complicada
La Espanola la isla mas ecol6gicamente divers a de la region debi6 ser el centro principal de evoluci6n de los mamfferos (con 87 de las familias y 73 de las subfamilias) seguida por Cuba (con 75 de las familias y 54 de las subfamilias) las dos islas de las Antillas Mayores mas relacionadas biogeograficamente (Fig 43 Tabla 44)
eL
EL
cb CAIMAN
SWAN
A Acratocnini B Cubanocnini C Megalocnini D Mesocnini E Nesophontidae F Solenodontidae G Xenothrichini
ANGUILAshyM-0 SMARTIN
H Heteropsomyinae I Plagiodontinae J Isolobodontinae
K Hexolobodontinae L Capromyinae
M Heptaxodontinae N CIidomyinae 0 Sigmodontinae
FIGURA 43 Representacion esquematica de las relaciones interinsulares (expresadas porel numero de linajes compartidos) entre las Antillas Mayores y entre estas y los grupos de islas perifericas segun las interpretaciones de los autores citados en el texto Los numeros dentro de los circulos indican la cantidad de linajes representados en cada isla relacion dudosa Contexto taxon6mico de los linajes en la Tabla 44
bull bull
bull bull
bull bull bull
bull bull
bull bull
bull
bull
bull
bull
bull bull bull bull
bull bull
bull bull
bull
bull bull bull
bull bull
bull
bull bull
bull
322 BIOGEOGRAFiA
TABLA 44 Distribuci6n de los linajcs de mamiferos terrestres a traves de las Antillas Mayores y grupos de islas perifericas LE = La Espanola CU Cuba JA Jamaica PR = Puerto Rico IC Islas Caiman IS Islas Swan IB = Islas Bahamas IV = Islas Virgenes AS = Anguila - San Martin
FamiliaSubfumiliafIribu LE CU JA PR IC IS IB IV AS
Megalonychidae
Choloepodinae
Acratocnini
Cubanocnini
Megalocninae
Megalocnini
Mesocnini
Nesophontidae
Solenodontidae
Pitheciidae
Callicebinae
Xenothrichini
Echimyidae
Heteropsomyinae
Capromyidae
Plagiodontinae
Isolobodontinae
Hexolobodontinae
Capromyinae
H e pta x 0 don tid a e
Heptaxodontinae
Clidomyinae
Muridae
Sigmodontinae
PILOSA
SORICOMORPHA
PRIMATES
RODENTIA
bull
bull
bull
bull TOTALES 12 10 4 4 2
323 EXTlICIOIES DEL CUATERNARIO
El gigantismo ha sido sin duda una tendencia evolucionaria recurrente entre los clados mayores de mamiferos antillanos Es un hecho bien conocido que algunos grupos de mamiferos han aumentado de tamafio en las islas y varios autores han intentado explicarlo a partir de enfoques diferentes (Foster 1964 Miller 1966 Sondaar 1977 Case 1978 Eisenberg y Wilson 1978 Heaney 1978 Lawlor 1982 Lomolino 1985 Angerbjom 1985 McNab 19942001)
EXTINCIONES DEL CUATERNARIO
Las islas que integran la Subregion Antillana han perdido casi 90 de sus mamiferos terrestres durante los ultimos milenios y en proporcion con su extension territorial esta perdida ha sido mayor en la Subregion que en cualquier otro lugar del planeta (MacPhee 1996b) Las extinciones del Cuatemario eliminaron mas de las tres cuartas partes de la fauna de mamiferos terrestres conocida de las Antillas Mayores sus islas satelites y grupos de islas perifericas y el examen de este proceso en sus aspectos mas generales se aborda en esta ultima parte del presente capitulo
CRONOLOGiA Y CONSECUENCIAS
Es muy improbable que un fosil represente el ultimo individuo de un taxon extinto y entonces hay que guiarse por la edad radiometrica del fosil mas reciente conocido lIamada fecha de ultima presencia- como unica aproximacion posible a la fecha de extincion del taxon
Como se explico al inicio del capitulo (vease Contexto Temporal) el registro fosH de los mamiferos terrestres de las Antillas Mayores adolece de insuficiente sustentacion radiometrica No obstante la distribucion temporal de los escasos fechados disponibles para el Cuaternario de la region abarca etapas criticas del proceso de extincion experimentado por la referida fauna a 10 largo de los ultimos 250 mil afios
De las 52 especies que es posible reconocer actualmente como miembros extintos de la fauna cuatemaria de mamiferos terrestres de las Antillas Mayores y grupos de islas perifericas los restos fosiles de 23 de elIas (442) han sido fechados radiometricamente o se han hallado asociados estratigraficamente de manera inequivoca con otro material fechado De estos 23 fechados 3 corresponden al interglacial Sangamon 4 pertenecen a la secuencia Pleistoceno terminal - Holoceno temprano 10 se ubican en ellapso entre el arribo de los aborigenes y la incursion europea y 6 son postcolombinos (a los cuales hay que sumar otras dos especies extinguidas en los siglos XIX y xx) A partir de estas 25 especies avaladas por fechas de ultima presencia distribuidas entre los cuatro intervalos temporales sefialados hemos compuesto la Tabla 45 en la que ademas aparecen las restantes 27 especies (carentes de fechado radiometrico) ubicadas en el intervalo temporal mas probable sobre la base de (1) la asociacion estratigrafica de sus restos con evidencias culturales (amerindias 0 europeas) 0 (2) la ausencia de dicha asociacion Sobre estos criterios de ubicacion temporal procede hacer dos aclaraciones
324 BIOGEOGRAFIA
TABLA 45 Intervalos temporales mas probables para las extinciones cuatemarias de los mamiferos terrestres de las Antillas Mayores y grupos de islas perifericas sobre la base de las evidencias siguientes (l) ultimo ejemplar colectado u observado en vida (2) fechado radiometrico mas tardio en material de la especie (3) asociaci6n estratigrafica mas tardia con otro material fechado (4) asociaci6n estratigrafica con restos de animales introducidos restos humanos u objetos culturales propios del intervale (5) falta de asociaci6n con evidencias culturales [Los ntimeros entre parentesis en la columna Evidencia significan adscripci6n tentativa al intervalo] Los estimados del peso corporal (en kg) estan tomados de la literatura 0 inferidos comparativamente y distribuidos entre
las clases siguientes (a) S 01 (b) 01 a lt 1 (c) 1 a lt 10 (d) 10 a lt 100 (e) 100 a lt 200 (j) ~ 200 Las
abreviaturas que designan las islas son las de la Tabla 44
F amilialEspecie Peso IsJa Evidencia Fuente
I httervalo htterg1acial Sangamon (gt80 kaAP)
HEPTAXODONTlNAE
Amblyrhiza inundata e AS 3 McFarJane et al 1998a
CLIDOMYINAE
Clidomys osborni d JA 3 Ibid
Clidomys parvus d JA 3 Ibid
II httervalo Pleistoceno terminal ~ Holoceno temprano (20-6 kaAP)
MEGALONYCHIDAE
Aeratoenus antillensis d CD 5 Arredondo 1975 a
Aeratoenus odontrigonus d PR 3 McFarJane 1999b
Aeratoenus simorhynehus d LE 2 Rega et al 2002
M egaloenus zile f LE 2 MacPhee et al 2000
SOLENODONTIDAE
Solenodon arredondoi e CD 5 Morgan y Ottenwalder 1993
PnHECIIDAE
Paralouatta varonai d CD 5 MacPhee 1996a
EcHIMYIDAE
Heteropsomys antillensis c PR 5 McFarJane 1999b
Heteropsomys insulans c PR 5 Ibid
Puertoricomys eorozalus c PR 5 Woods 1989a
CAPROMYIDAE
P lagiodontia araeum c LE 5 MacPhee et al 2000
Macrocapromys latus c CD 5 Varona y Arredondo 1979
(Continua)
325 EXTINCIONES DEL CUATERNARIO
TABLA 45 (Continuaci6n)
F ami1ia1Especie Peso Isla Evidencia Fuente
M acrocapromys latus c CU 5 Varona y Arredondo 1979
Mesocapromys kraglievichi c CU 5 Ibid
HEPTAXODONTINAE
Xaymaca fulvopulvis a JA 2 MacPhee y Flemming 2003
III Intervalo Amerindio (6-05 kaAP)
MEGALONYCHIDAE
Acratocnus ye d LE 2 MacPhee et aI 2000
Neocnus comes c LE 2 Ibid
N eocnus dousman c LE 2 Ibid
N eocnus toupiti c LE 2 Ibid
N eocnus gliriform is c CU 4 Cordova et ai 1997
M egalocnus rodens f CU 2 MacPhee et ai 2007
Parocnus browni e CU 2 Steadman et ai 2005
P arocnus serus e LE 2 MacPhee et ai 2000
N ESOPHONTIDAE
N esophontes edithae a PR 2 McFarlane 1999b
PITHECIIDAE
Antillothrix bernensis c LE 3 Rimoli 1977
EcHIMYIDAE
Brotomys contractus a LE 4 Miller 1929c
CAPROMYIDAE
P lagiodontia spelaeum c LE 4 Ibid
M acrocapromys acevedo c CU 4 NUiiez 1990a
HEPTAXODONTINAE
Elasmodontomys obliquus d PR 3 McFarlane 1999b
Iv Intervalo Postcolombino (05 kaAP - Presente)
N ESOPHONTIDAE
N esophontes hypom icrus a LE 24 MacPhee y Fle111111ing 1999
N esophontes zam icrus a LE (4) Ibid
N esophontes m icrus a CU 24 MacPhee et aI 1999
(Continua)
326 BIOGEOGRAFiA
TABLA 45 (Continuacion)
F amilialEspecie Peso Isla Evidencia Fuente
N esophontes m icrus a CD 24 MacPhee et aI 1999
N esophontes m icrus a LE 4 MacPhee et aI 2000
N esophontes sp 1 a IC 4 Morgan 1994
N esophontes sp 2 a IC 4 Ibid
Solenodon marcanoi b LE 4 Ottenwalder 2001
PITHECIIDAE
Xenothrix mcgregori c JA 4 MacPhee y Flemming 1999
EcHIMYIDAE
Boromys offeUa a CD 24 Guarch 1984
Boromys torrei a CD 24 Ibid
Brotomys voratus a LE 4 MacPhee et aI 2000
CAPROMYIDAE
P lagiodontia ipnaeum c LE 4 MacPhee y Flemming 1999
Rhizoplagiodontia lemkei c LE 4 Ibid
Isolobodon portoricensis c PR 24 Ibid
Isolobodon montanus c LE 4 Ibid
Hexolobodon phenax c LE (4) Ibid
Capromys sp c IC 24 Morgan 1994
Geocapromys columbianus c CD 4 MacPhee y Flemming 1999
Geocapromys thoracatus b IS 1 MacPhee y Marx 1997
Geocapromys sp 1 c IC 4 Morgan 1994
Geocapromys sp 2 c IC (4) Ibid
l-IEPTAXODONTINAE
Quemisia gravis c LE (1) MacPhee y Flemming 1999
MURIDAE
Oryzomys antillarum a JA 1 Ibid
1 Flemming y MacPhee (1999) alertaron en contra de la asociacion con otro material fechado Asimismo Morgan (1993) indico que la ausencia de asociacion con evidencias culturales amerindias no es necesariamente un criterio concluyente para ubicar un taxon en el intervalo temporal anterior porque hay depositos en la region que carecen de tales evidencias y son del Holoceno tardio por fechados radiocarbonicos [No obstante
327 EXTlNCIONES DEL CUATERNAR10
la ausencia de una especie en residuarios arqueol6gicos adquiere mayor certidumbre como criterio de ubicaci6n temporal de su extinci6n en la medida que aumenta el numero de residuarios estudiados que en las Antillas Mayores se cuentan por centenares] A falta de fechado absoluto de una especie consideramos que estos criterios de ubicaci6n temporal son mas utiles que la alternativa de excluir del analisis a la especie
2 En los trabajos sobre biogeografia de mamiferos antillanos los arque610gos mas citados como autoridades para establecer el inicio de la presencia aborigen en las Antillas Mayores (Kozlowski 1974 Rouse y Allaire 1978 Rouse 1989 1992 Wilson 2001) difieren al respecto entre los 6 kaAP y los 45 kaAP (pero vease Pagan et al 200523) y hemos adoptado la mas temprana de estas fechas para el comienzo del intervalo amerindio en la Tabla 45 De todos modos cualquiera que haya sido la fecha esta no es aplicable por igual al conjunto de las Antillas Mayores porque segun algunos de los autores citados el poblamiento comenz6 por Cuba y La Espanola 6 mil anos atras continu6 en Puerto Rico hace alrededor de 3 mil anos (pero vease Burney et al 1994) yen Jamaica hace menos de 2 mil e incluso a las Islas Caiman ni siquiera llegaron [Las Islas Caiman descubiertas por Col6n en 1503 carecen absolutamente de evidencia amerindia (Stokes y Keegan 1996)]
Si los datos de la Tabla 45 representan una aceptable aproximaci6n a la realidad el proceso de las extinciones cuaternarias de los mamiferos terrestres de las Antillas Mayores y grupos de islas perifericas debi6 caracterizarse entre otros por los eventos siguientes
Aunque obviamente la fauna cuaternaria debi6 ser mucho mas numerosa que 10 que refleja el registro paleontol6gico (Morgan y Woods 1986) 52 especies (81 de la fauna conocida) desaparecieron (sin reposici6n) durante los ultimos 250 mil anos
- Al parecer las extinciones se concentraron en las especies de mayor tamano Asi el peso promedio estimado para las especies de la etapa preantr6pica (intervalos I + II de la Tabla 45) es 350 kg es 328 kg para las especies del intervalo III (amerindio) y es s610 25 kg (c1ases del peso a b c) para las especies del intervalo IV (postcolombino) Ninguna especie aut6ctona de mamifero mayor de 6 kg existe hoy en la regi6n
- De acuerdo con la distribuci6n temporal de las extinciones 61 especies sobrevivieron al interglacial Sangamon (intervalo 1)48 sobrevivieron a la transici6n del Pleistoceno terminal al Holoceno temprano (intervalo II) 34 sobrevivieron al periodo amerindio (intervalo III) y s610 12 especies han sobrevivido desde la Ilegada de los europeos (intervalo IV) Las tasas de extinci6n correspondientes (exceptuado el intervalo I) son una especie cada 1167 anos en el intervalo II una cada 393 anos en el intervalo III y una cada 23 anos en el intervalo IV
- Durante todo el proceso de las extinciones cuaternarias la perdida de especies por isla fue de 91 en La Espanola 83 en Jamaica y 62 en Cuba Bahamas retuvo su unica especie (aunque extirpada en varias de las islas) Puerto Rico e Islas Caiman perdieron todas sus especies (6 y 5 respectivamente) y las Islas Swan Virgenes y Anguila - San Martin perdieron sus respectivas especies (micas
328 BIOGEOGRAFiA
De los cuatro 6rdenes de mamiferos representados en la fauna Pilosa y Primates desaparecieron totalmente Soricomorpha y Rodentia perdieron 86 y 74 de sus especies respectivamente
CAUSALIDAD
CAUSAS NATURALES
Los cicIos glaciales e interglaciales acaecidos en el Pleistoceno causaron cambios ambientales dnisticos en las Antillas con la concomitante alternancia de cIimas frios (secos) y calidos (hUmedos) asi como de descensos y ascensos del nivel del mar aumento y disminuci6n de las areas territoriales) induso (aunque con menor intensidad) en el Holoceno Estos eventos paleogeograficos han sido consistentemente documentados (Mayo y Kartashov 1972 Bonnatti y Gartner 1973 Ortega y Arcia 1982 Prance 1982 Schubert y Medina 1982 Dodge et al 1983 Ortega [1984] Haq et al 1987 Schubert 1989 Hodell et al 1991 Guilderson et al 1994 Oppo 1997 Emslie 1998 Curtis et al 2001 Iturralde-Vinent 2003) y de ellos brindamos a continuaci6n una breve resena
Finalizando el periodo interglacial anterior a la ultima glaciaci6n (Wisconsin) el mar estaba a 8-10 m por encima de su nivel actual Cuando el avance glacial del Wisconsin a1canz6 su culminaci6n (eillamado Ultimo Maximo Glacial) hace 18 ka el mar habia descendido 120 m con respecto a dicho nivel y las islas del Caribe habian incrementado considerablemente sus extensiones subaereas en comparaci6n con las que presentan hoy dia por ejemplo Puerto Rico y las Islas Virgenes se conectaron territorialmente y las Bahamas se unieron en cinco gran des islas Prevalecia un regimen frio y seco y condiciones de aridez --debidas a una reducci6n de las precipitaciones (posiblemente a menos de 440 mmlano)- y los ambientes xericos se habian extendido a traves de las islas y bancos emergidos y abundaban los pinares (los habitats mesicos estaban restringidos a parcelas relictas en 10 alto de las montanas) En el Mar Caribe y el Golfo de Mexico la temperatura superficial del mar era 5degC mas baja que la actual y en las llanuras costeras y alturas maximas eran respectivamente 2-3degC y 6degC mas bajas Los limites de las zonas ecol6gicas altitudinales descendieron varios cientos de metros y cada zona redujo su extensi6n vertical al aumentar el gradiente de temperatura Ellimite inferior de la zona periglacial
elevaci6n donde la temperatura media minima a1canza los OdegC) de la Cordillera Central de la Republica Dominicana (altura maxima del Caribe insular con evidencia de topografia glacial) se considera que estuvo entre los 1500 y 1800 mlnm (actualmente esta a 2800 mlnm) y se puede estimar que dicho limite era inferior en Cuba por estar mas al norte que la Republica Dominicana No obstante durante este periodo la temperatura experiment6 episodios cortos (unas pocas decadas) de ascensos repetidos
Un signifieativo componente bi6tieo de las referidas condiciones paleoambientales 10 constituy6 la espectacular irradiaci6n evolucionaria experimentada por un grupo numeroso de aves rapaees diurnas y nocturnas (Wetmore 1937 Brodkorb 1969 Kurotchkin y Mayo 1973 Arredondo 1976 Olson 1978 Olson y Hilgartner 1982
329 EXTINCIONES DEL ClJATERNARIO
Suarez 2000a b 2004 Suarez y Emslie 2003) Los mamiferos de las Antillas Mayores ofrecieron a estos depredadores aviares una amplia gama de tamanos de presa que en ausencia de mamiferos del orden Carnivora en la region permitio a estas aves irradiarse a este nicho alimentario con la evolucion de una variedad de especies de muy diversa tall a corporal incluidas formas gigantes Adquirir un cuerpo gigante les propicio el acceso a recursos alimentarios que no estaban siendo explotados por ningun otro depredador (Alcover y McMinn 1994) A Cuba corresponde el mejor registro fosil de rapaces cuaternarias caribenas con siete generos de especies diurnas Dentro de estas aves existieron tres gavilanes gigantes (Accipitridae) tres carairas (Falconidae) y un condor (Cathartidae) asi como cuatro generos de especies nocturnas dos lechuzas (Tytonidae) y tres Mhos (Strigidae) dos de ell os no igualados en tamano en todo el mundo Algunos de estos generos tuvieron representacion en otras islas antillanas como La Espanola Jamaica Bahamas y Gran Caiman
Al retirarse hacia el norte los hielos de la glaciacion Wisconsin el clima se tomo mas hUmedo en las Antillas durante el Holoceno temprano y el bosque mesico se expandio en la region insular El nivel del mar ascendio acompanado de mayor pluviosidad alrededor de los 7 kaAP las Islas Virgenes quedaron separadas entre si y de Puerto Rico y las Bahamas perdieron la mayor parte de su territorio Cerca de los 3 kaAP las condiciones secas retornaron a la region e hicieron desaparecer buena parte del bosque mesico Nuevamente el clima comenzo a adquirir sus condiciones actuales hacia los 25 kaAP aproximadamente y hoy transita por una fase mesica con vegetacion xerica confinada en parcelas relictas
Sin duda estos y otros eventos naturales incidieron negativamente en la sobrevivencia de mamiferos terrestres en las Antillas Mayores asi como en los grupos de pequenas islas perifericas (Pregill y Olson 1981 Olson 2006)
McFarlane et al (1998a) demostraron convincentemente como Amblyrhiza el gigante de los roedores antillanos endemico de la pequena isla de Anguila sucumbio a las repetidas y abruptas perdidas de su habitat durante el interglacial Sangamon por las nipidas y severas oscilaciones del nivel del mar (documentadas por Neumann y Hearty 1996) al no poder responder adecuadamente a tan catastroficos eventos mediante el ajuste de su masa corporal media y otro tanto debio ocurrirle en Jamaica al otro roedor gigante Clidomys (McFarlane et al 1998b) emparentado con Amblyrhiza en la misma epoca (Tabla 45 intervalo I)
El ascenso del nivel del mar seguramente promovio la extincion de mamiferos durante la transicion del Pleistoceno al Holoceno (Tabla 45 intervalo II) con la reduccion del tamano de las Islas por las transgresiones marinas (Pregill y Olson 1981 Morgan 1993) Esto ultimo haria maximamente severa la competicion asf como la presion de los depredadores [Steadman et al (2005) no yen por que el calentamiento global ocurrido durante la transicion Pleistoceno-Holoceno habria de tener un efecto negativo sobre los perezosos siendo sus descendientes actuales especies tropic ales 0 subtropicales pero no se trata del efecto
330 BIOGEOGRAFiA
directo del cambio de temperatura sobre los animales sino del efecto indirecto y progresivo que dicho cambio genera a largo plazo al reducir el territorio insular disponible]
Por esta epoca tambien desaparecieron especies de tall a mayor como Megalocnus zile en La Espanola y otras de menor tamano (en Cuba) pero que representan la tall a maxima en sus respectivos contextos taxonomicos del Cuaternario como Solenodon arredondoi Paralouatta varonai y Macrocapromys acevedo Un mamifero puede ser grande solo a costa de reducir su densidad poblacional y esto 10 hace mis vulnerable a la extinci6n sobre todo en una isla (McNab 1994 McFarlane et al 1998a) Incidentalmente Alcover et al (1998a) afirmaron que larelacion entre la talla corporal y la extincion no siempre se cumple en las especies de menor tamano y citaron como ejemplo que entre las musarafias todas las especies de Nesophontes desaparecieron mientras sobreviven especies de Solenodon pero la comparacion correcta en todo caso serfa entre los solenodones vivientes y el gigante Solenodon arredondoi y entre Nesophontes edithae -el gigante de los nesofontes extinguido en el periodo amerindio- y el resto de sus congeneres que sobrevivieron hasta los primeros siglos de la colonizacion europea (Tabla 45 intervalos III y IV)
CAUSAS ANTROPOGENICAS
Intervalo Amerindio
La mayoria de los autores que se han referido a las extinciones acaecidas durante este intervalo (entre elIos paradigmaticamente Martin 1984a b) han responsabilizado a los aborfgenes (que se supone comenzaron a poblar las Antillas Mayores hace 6 mil afios) con la desaparicion de numerosas especies de mamiferos terrestres en la region principalmente por la via de la sobrecaza y la introduccion de animales domesticos devenidos depredadores de la fauna autoctona (Morgan y Woods 1986) Este efecto negativo de la presencia indigena sobre los mamiferos de la region ha sido dramatizado por algunos autores en terminos de holocausto faunal Sin embargo pensamos que se ha exagerado Como han expresado MacPhee et al (19993) cuando uno busca evidencia dura de complicidad humana directa en [estas] extinciones no hay mucho que encontrar Las siguientes son algunas consideraciones a tener en cuenta en tal sentido
El registro radiometrico mas joven conocido hasta ahora para un perezoso antillano (MegalocnllS rodens) es de 42 kaAP (MacPhee et ai 2007) Si se acepta que los aborfgenes entraron en Cuba hace aproximadamente 6 mil anos su convivencia con los perezosos debe haber durado alrededor de 1800 afios (0 mas porque dificilmente el referfdo fechado radiometrico haya correspondido al ultimo sobreviviente de la especie) Esta cifra contradice la hipotesis del holocausto faunal segun la cualla extincion de los mamiferos insulares de mayor talla debe ocurrir con extrema rapidez -a escala de decadas 0 de centurias- a partir de su primer contacto con los humanos (Martin y Steadman 1999) A pesar de las repetidas afirmaciones en contrario (Harrington 1921 Miller 1929c Herrera [1945] Tabio 1951 Alvarez 1951a 1957 Arredondo
331 EXTINCIONES DEL CUATERNARIO
1959 1960a Veloz y Ortega 1973 Mayo 1980b Rodriguez et al 1984 Castellanos y Pino 1985 Pino y Castellanos 1985 Nunez 1990b Gutierrez 1996 Arredondo 2004) en su mayoria basadas en evidencias no exentas de posibles accidentes de deposicion (0 de reubicacion postdeposicional) en recintos cavernarios (Diaz-Franco [2006b D no se ha demostrado inequivocamente que los aborigenes antillanos se alimentaran de perezosos regularmente Sus restos en residuarios de alimentos son escasisimos en toda la region y esto contrasta con los abundantes restos de jutias y otros mamiferos de menor talla que aparecen en esos residuarios demostrativos de la preferencia de los paleoindios por estos otros mamiferos
Los mamiferos mas importantes en la economia prehistorica de las Antillas Mayores fueron las jutias Capromys y Geocapromys en Cuba Geocapromys en Jamaica e Isolobodon en La Espanola Puerto Rico e Islas Virgenes (Aguayo 1950b Tablo 1951 Rimoli 1972 Wing 2001 Diaz-Franco [2006b D Wilkins (2001) estudi61ajutia viviente de Jamaica (Geocapromys browni) en un sitio ocupado por los aborigenes durante 100 anos y no encontr6 evidencias de que durante ese tiempo la especie haya sido sobreexplotada Asimismo en Cuba las jutias conga carabali y andaraz son escasas en depositos pleistoholocenicos en cambio son muy numerosas en residuarios indigenas y de haber sido sobreexplotadas 10 mas probable es que no hubieran sobrevivido hasta hoy
La habilidad de los humanos para penetrar en profundidad el bosque tropical y mantenerse en este sustentablemente ha sido un evento comparativamente reciente (Beran y Steele 1994) En el caso antillano la caza tierra adentro ha sido explicada como un proceso tardio de desplazamiento a partir de la explotacion de abundantes recursos marinos que ofrecian condiciones 6ptimas para el sedentarismo de las comunidades aborigenes costeras (C6rdova y Arredondo 1988 Brito 2005) Peces y mariscos dominan la arqueologia costera de las Islas antillanas en tanto el consumo de recursos terrestres (mamfferos incluidos) aumenta progresivamente en la medida que los residuarios estudiados se alejan de la costa (Scudder 1991 Wilkins 2001) En las Antillas Mayores el componente terrestre de la dieta aborigen es solo 20-30 del material recobrado en los residuarios (Wing 1977 1989) Y debio ser menor alin porque el componente vegetal de la dieta no se ha preservado Por otra parte los aborigenes cubanos domesticaron la jutia conga y seguramente la reprodujeron para comer (Pose et al 1988) practica que probablemente se repitiera con otras jutias en las demas Antillas Mayores
- Fray Bartolome de las Casas hablo de 3 millones de indios en Cuba al inicio de la colonizacion pero los historiadores subsecuentes consideraron exagerada la cifra (Poey 1851 a) No obstante se dispone de dos estimados de la densidad de poblaci6n aborigen en las Antillas Mayores a la llegada de los europeos 18 indigenaslkm2 en Puerto Rico (Rouse 1956) y 1271km2 en La Espanola (Watts 1987) Extrapolando ambos valores entre las cuatro islas principales de las Antillas Mayores tenemos que en la fecha indicada la poblaci6n aborigen pudo estar entre 16 mil y 113 mil indigenas en Puerto Rico entre 21 mil y 145 mil en Jamaica entre 141 mil y 1 mill6n en La Espanola y entre 200 mil y 14 millones en Cuba Entre las cifras maximas y las minimas nos inclinamos
332 BIOGEOGRAFiA
hacia estas ultimas atendiendo a los altos indices de mortalidad infantil y morbilidad que se evidencian en las osamentas de los pobladores prehispanicos (Martinez y Rives 1990) Uno de los hechos que apuntan hacia el bajo numero de aborigenes en las Antillas Mayores es 10 rapido que desaparecieron despues de la llegada de los europeos pnkticamente sin dejar mestizaje con estos 10 cual contrasta ostensiblemente con la situacion en la America continentaL
En suma un grupo humano (1) demograiicamente irrelevante y controlado por altos indices de morbilidad (2) consumidor mayoritariamente de organismos marinos y (3) carente de la tecnologia cinegetica necesaria para poder sobreexplotar los mamiferos que Ie sirvieron de alimento (a los cuales accedio tardiamente en su desarrollo) no es concebible que haya causado e1 holocausto faunal que se Ie atribuye a partir del error de aplicar a las comunidades aborigenes antillanas un modelo cultural de la prehistoria continentaL
Tampoco hay evidencias de que los aborigenes hayan introducido carnivoros con suficiente capacidad depredadora salvo el perro Morgan y Otlenwaldder (1993) consideraron que la depredacion por perros durante el periodo amerindio es la explicacion mas plausible para la extincion del almiqui gigante de Cuba pero como se puede observar (Tabla 54) Solenodon arredondoi aparece en el intervalo II por falta de asociacion con evidencias culturales
Consecuentemente el accionar de los aborigenes y de sus animales domesticos directamente sobre la fauna no parece haber sido por sf solo un factor determinante de la extincion de mamiferos terrestres durante el periodo amerindio en las Antillas (pero vease Steadman et al 2005)
Hay que recordar en este senti do que alin en el Holoceno tardio ocurrieron cambios clirruiticos de consideracion (vease Causas Naturales) Por ejemplo Curtis et al (2001) explicaron que la extincion de algunos ani males mesofilicos de las alturas montanosas de La Espanola pudo haber antecedido a cualquier impacto humano sigrIificativo en esos lugares por coincidir dicha extincion con una tendencia a la aridez documentada isotopicamente y con evidencias de la perdida del bosque mesico ocurridas despues de 3kaAP Yen Jamaica hace solo 2shy3 mil aDos el area de la Bahia de Jackson sustentaba una gran biomasa de vertebrados en bosque hinnedo a juzgar por la cantidad y diversidad de taxones presentes en material fosil derivado de la depredacion por lechuzas hoy el area es la mas seca de la isla con matorral xerofitico y pobreza de fauna (MacPhee 1996a) Yalm en nuestros dias huracanes de gran intensidad aniquilan poblaciones de vertebrados (que no se recuperan) en is las pequenas y llanas del Caribe (Brooks y Smith 2001)
En medio de la incertidumbre sobre la verdadera causa (0 causas) de las extinciones de mamiferos en el periodo amerindio MacPhee y Marx (1997) han propuesto su hipotesis de la hipermorbilidad la cual estipula colapsos poblacionales relativamente rapidos con la introduccion de enfermedades de alta letalidad Estos autores han razonado que
Las islas son los lugares mas apropiados para experimentar perdidas catastroficas causadas por enfermedades Los mamiferos no solo serian susceptibles en virtud de su prolongado aislamiento biologico sino que en los confines de una isla la epizootia
333 EXTINCIONES DEL CUATERNARIO
tendria la potencialidad de infectar simultaneamente a la rnayoria de los individuos de vadas especies Los neonatos son inmunologicamente inmaduros y sufririan desproporcionadamente cualquier infeccion hipervirulenta La perdida excesiva de neonatos y juveniles tendria efectos severos para la sobrevivencia de la poblacion si esta se reduce por debajo de cierto nivel sobreviene la extincion porque no hay posibilidad de restitucion en el periodo postpanzootico Las especies de mayor talla corporal van a ser las mas vulnerables a la extincion dadas sus bajas tasas reproductivas largos periodos de gestacion y reducido numero de progenie Las especies de menor tamano --caracterizadas por altas tasas reproductivas periodos cortos de gestacion y mayor numero de neonatos por parto- tambien serian fuertemente diezmadas pero muchas sobrevivirian porque su estrategia reproductiva les permite perder mayor numero de individuos Las perdidas deben ocurrir relativamente rapido (a escala de decadas) mientras que otras hipotesis (digamos los cambios climaticos) permiten intervalos mucho mas largos (a escala de siglos 0 de milenios) entre la aparicion de las causas y la produccion de sus efectos Por ejemplo todo indica que los nesofontes se extinguieron muy rapidamente y seria dificil poder invocar un impacto humano directo 0 argumentar fenomenos climaticos que pudieran explicar plausiblemente perdidas tan rapidas
MacPhee y Marx (1997 191) han dicho que por el momento la hipotesis de la hipermorbilidad no pasa de ser una abstraccion en espera de adecuada sustentacion empirica Llama la atencion que linicamente en el periodo amerindio (Tabla 45 intervalo 1Il) las extinciones abarcaron todas las clases del peso corporal (de la a a laj) y que ninguna de las especies que sobrevivieron hasta el periodo postcolombino (Tabla 45 intervalo IV mas las especies vivientes) sobrepasa los 10 kg (clases del peso a b c) en clara coincidencia con los presupuestos de la hipotetica hipermorbilidad
Intervalo Postcolornbino
Por 10 menos 37 especies de mamiferos terrestres vivian en las Antillas Mayores y grupos de Islas perifericas a la lIegada de los europeos en 1492 De elIas se han extinguido 23 shymas especies que en cualquiera de los periodos precedentes (Tabla 45~ y solo subsisten 12 especies repartidas entre Cuba (8 especies) La Espanola (2) Jamaica (1) y Bahamas (1) varias de estas en peligro critico de extincion Solo dos de las especies extinguidas durante los ultimos 500 anos se conocieron en estado viviente los roedores Oryzomys antillarum de Jamaica (extinguido en 1877) y Geocapromys thoracatus de las Islas Swan (extinguido en 1955) Las causas de las extinciones ocurridas durante este periodo son sobradamente conocidas destruccion de habitats introducci6n de animales exoticos (domesticos y comensales) y demas presiones asociadas a la actividad humana entre las cuales posiblemente habria que incluir brotes de hipermorbilidad
Durante mucho tiempo se ha afirmado la posibilidad de que especies de Nesophontes hayan sobrevivido hasta las primeras decadas del siglo xx y que incluso continuen existiendo en lugares remotos de Cuba 0 La Espanola (Allen 1918 1942 Miller 1930 Aguayo 1950a Arredondo 1960b Fischer 1977 MacFadden 1980 Woods y
334 BIOGEOGRAFiA
Ottenwalder 1991) Actualmente no existe evidencia radiometrica de que alguna especie de Nesophontes haya sobrevivido al siglo xv las fechas radiocarbonicas calibradas obtenidas en muestras de Nesophontes de Cuba y La Espanola resultaron no mas tardias que los siglos XIII y XIV (MacPhee et al 1999) Por otra parte las expediciones efectuadas a fines del siglo xx con el proposito de localizar individuos vivientes del genero en La Espanola no produjeron la menor indicacion afirmativa al respecto (Woods et al 1985) Ademas a traves de las Antillas Mayores no aparecen restos de Nesophontes en los depositos formados actualmente por las lechuzas
EPiLOGO
Las Antillas Mayores son islas oceanicas no han formado parte de un continente (con la posible excepcion de una parte de Jamaica hasta el Eoceno Medio hace 45 Ma) y durante un tiempo geologico considerable (desde el Eoceno Superior hace 40 Ma) han permanecido emergidas y separadas de los continentes vecinos Consecuentemente los mamiferos terrestres (no volantes) autoctonos de estas islas constituyen una fauna oceanica que por ende cumple con todas las caracteristicas convencionales reconocidas para este tipo de entidad biogeografica
- Baja diversidad taxonomica a nivel de las categorias superiores (solo cuatro ordenes) en comparacion con islas plataformicas continentales (0 con areas continentales) de tamano y diversidad ecologica simi lares
- Alto grado de endemismo a nivel de genero (96) y familia (50) en respuesta al prolongado aislamiento
- Desbalance ecologico de la estructura trofica ausencia de mamiferos camivoros y ungulados
- Espectaculares procesos de irradiaci6n evolucionaria como resultado de la colonizacion de territorios libres de competidores y depredadores
Tendencias evolucionarias extremas gigantismo
- Control a cargo de aves rapaces con tendencia at gigantismo en ausencia de mamiferos miembros del orden Carnivora
- Mas especies de reptiles y de murcielagos que de mamiferos terrestres relaciones que en los continentes se invierten
Alta tasa de extincion de especies (84) sin reposicion
Preservacion de especies filogeneticamente arcaicas (los llamados fosiles vivientes) los almiquies

318 BIOGEOGRAFiA
Woods y Howland (I979) describieron adaptaciones morfologicas de la mandibula el hioide y la faringe asociadas a mecanismos de masticacion diferentes en plagiodontinos y caprominos y Varona (1980a) llamo la atencion sobre el gran desarrollo de los molariformes (en relacion con la longitud total del cnineo) que presentan ciertos grupos de jutias fosiles en comparacion con las vivientes 10 que pudiera interpretarse como una adaptacion alimentaria a una vegetacion mas abrasiva por parte de las jutias extinguidas
Segun Woods (1990) la familia Heptaxodontidae (2 subfamilias 5 generos y 6 especies todas extintas) tambien seria parte de la irradiacion de los capromidos con origen en el este de La Espanola 0 en Puerto Rico y subsecuente dispersion al Banco de Anguila - San Martin en las Antillas Menores (pero vease MacPhee y Flemming 2003) Este banco estaria a solo 70 km del Banco Puerto Rico - Islas Virgenes en tiempos de maxima regresion marina y esta proximidad con Puerto Rico -patria de Elasmodontomys obliquus- seguramente tuvo que ver con la presencia en las islas de Anguila y San Martin del heptaxodontino gigante Amblyrhiza relacionado mas estrechamente con los roedores de las Antillas Mayores que con los del resto de las Antillas Menores (Woods 1990 1996) Aunque los roedores suelen aumentar de tamano en las islas (Case 1978) la escala del incremento de la talla en Amblyrhiza (que pudo alcanzar mas de 200 kg de peso adulto) esta fuera de toda proporcion con los patrones conocidos en otros roedores insulares (Biknevicius et 01 1993)
Clidomys (Clidomyinae) de Jamaica difiere marcadamente en su morfologia de los heptaxodontinos (vease ademas MacPhee y Flemming 2003)10 cual dificulta explicar su presencia en esa isla Como Jamaica estuvo bajo el mar hasta el Mioceno Medio el ancestro de Clidomys colonizaria esta isla por dispersion desde el extremo suroccidental de La Espanola despues que los heptaxodontinos se originaron en esta ultima isla (Woods 1989a) Pero Pascual et 01 (I 990) especularon que Clidomys pudiera ser el resultado de una invasion posterior e independiente de los capromidos
GRUPOS ORIGINARIOS DE NORTEAMERICA
El origen de las musaranas antillanas ha resultado ser hasta ahora el mayor enigma biogeografico de la fauna de mamiferos terrestres de la region aunque la mayoria de los autores coinciden en considerarlas originarias de Norteamerica (vease Discus ion y Conclusiones en Poblamiento Continente-Isla) No obstante MacPhee e Iturralde-Vinent (1994) descartaron toda posibilidad de conexion terrestre por el Estrecho de la Florida 0 por el Canal de Yucatan y si el (supuesto) evento vicari ante de Rosen (vease El Arca de Noe) -invocado por MacFadden (1980) para resolver el enigma- fuera minimamente plausible aun asi es dificil creer que las catastrofes que 10 sucedieron (vease Contexto Temporal) hayan barrido con toda esa vieja fauna continental excepto las musaranas circunstancia ya anticipada muy tempranamente por Matthew (l918a)
De otra parte la relaci6n entre Nesophontidae y Solenodontidae permanece irresuelta Si estos linajes no estan estrechamente relacionados entonces es muy posible que
319 IRRADIACION EVOLUCIONARIA
sus distribuciones sean explicadas por mecanismos diferentes (Whidden y Asher 2001)
La descomunal diferencia de talla corporal entre las musaranas pequenas (Nesophontes) y las grandes (Soienodon) seguramente impuso a ambos grupos restricciones distintas relativas a la termorregulacion y las adaptaciones energeticas En Nesophontes la seleccion debio favorecer una alta tasa de metabolismo y crecimiento camadas mas numerosas corta expectativa de vida yalta densidad poblacionaL En Soienodon en cambio la seleccion opero en la direccion de una baja tasa metabolica camadas reducidas crecimiento lento longevidad y baja densidad poblacional (Ottenwalder [1991])
MUSARANAS PEQUENAS (Nesophontidae)
Las musaranas pequenas estan representadas en las Antillas por el genero Nesophontes (y un genero miocenico indeterminado en ambar dominicano) con 6 especies cuaternarias extinguidas distribuidas en Cuba La Espanola Puerto Rico Islas Virgenes e Islas Caiman La mayor de las musaranas pequenas Nesophontes edithae evoluciono en Puerto Rico y su arribo a las Islas Virgenes supone una dispersion sumamente reciente En las Islas Caiman Nesophontes se asemeja mas a la poblacion de Cuba de donde seguramente derivo en el Plio-Pleistoceno (junto con Capromys y Geocapromys) cuando la costa sur de Cuba estuvo mucho mas cerca de las Islas Caiman (lturralde-Vinent 2003fig 4) MacPhee et ai (1994) interpretaron la distribucion de Nesophontes en Cuba y La Espanola como resultado de la vicarianza oligomiocenica ocurrida al formarse el Paso de los Vientos pero la distancia taxonomica entre los lados del Paso es solo de nivel especifico y N micros incluso vivi6 en ambas islas segtln Patterson (1962)
Allen (1918) considero que Nesophontes micrus fue una especie primitiva no modificada para la vida arb6rea 0 fos6rea y por tanto de habitos terrestres Nesophontes edithae de Puerto Rico es mucho mayor y mas robusta que las restantes especies del genero eL indice de robustez de su femur (= anchura minima de la diafisis ongitud total del hueso) muestra que este es tan robusto como el de Solenodon paradoxus un animal dos veces mayor (Morgan et ai 1980) Choate y Birney (1968) postularon un aumento cronoclinal de la taIla corporal en N edithae confirmado mas tarde por McFarlane (1999a) mediante fechados radiocarb6nicos de la secuencia estratignifica en depositos cavernarios de Puerto Rico Solenodon no vivi6 en Puerto Rico y en ausencia de una musarana de mayor tamano Nesophontes alcanzo una talla corporal mayor en esa isla que en La Espanola 0 Cuba (Woods y Eisenberg 1989)
MUSARANAS GRANDES (Solenodontidae)
Los arcaicos almiqufes (genero Soienodon) comprenden cuatro especies una viviente (S cubanus) y otra f6sil (S arredondoi) en Cuba y una viviente (S paradoxus con dos subespecies) y otra f6sil (S marcanoi) en La Espanola Como cabria esperar las dos especies cubanas se asemejan mas entre sf que a las especies de La Espanola pero
320 BIOGEOGRAFiA
enigmaticamente S marcanoi comparte caracteres con S cuban us y S paradoxus (Woods y Eisenberg 1989 Morgan y Ottenwalder 1993 Ottenwalder 200 I) Solenodon arredondoi alcanzo una talla corporal superior a la de cualquiera de las musarafias vivientes a nivel mundial Segun Morgan y Ottenwalder (1993) extrapolando las dimensiones y el peso de los almiquies vivientes S arredondoi debio tener una longitud cabeza-tronco y un peso de 450-550 mm y 1500-2000 g respectivamente en comparacion con S cubanus (360 mm y 800 g) y S paradoxus (390 mm y 1100 g)
Entre todas las especies de almiqufes Solenodon paradoxus es la que presenta los caracteres mas derivados (Otten walder 2001) Si estas relaciones son validas la distribucion de Solenodon entre Cuba y La Espafiola debio discurrir de oeste a este como propuso MacFadden (1980) quien invoco la presencia fosH de S cubanus [pero tambien S arredondoi] en Pinar del Rio para postular que el ancestro continental entr~ por Cuba opinion compartida por Perfit y Williams (1989) y por Ottenwalder ([1991] 2001) El paso de Solenodon a Puerto Rico pudo haberlo impedido la existencia de barreras ecologicas durante la conexion del este de La Espafiola con Puerto Rico 0 por la ausencia de Solenodon en esa parte de La Espafiola en aquella epoca Sin embargo las mismas condiciones ecologicas en el este de La Espafiola no parecen haber representado una barrera efectiva para Nesophontes cuyas mas altas tasa de reproduccion y densidad poblacional presumiblemente Ie permitieron expandirse flpidamente a traves de La Espafiola (Ottenwalder [1991])
Es opinion generalizada que los almiqufes antillanos constituyen el grupo de mamiferos que primero se emplazo en el territorio insular (MacPhee 2005) y en tal sentido es de interes la evidencia sobre especificidad parasito-hospedante aportada por Cruz (1980) quien encontro que la especie de acaro (genero Cubanochirus) que parasita a los almiquies es mucho mas primitiva que la especie (del mismo genero) que paras ita a las jutias y de aqui concluyo que dicho genero de acaro debio evolucionar primero en Solenodontidae y posteriormente en Capromyidae al entrar en contacto representantes de ambas familias en Cuba 0 La Espafiola
Recientemente Roca (2004) y Roca et al (2004) dieron a conocer que los resultados de su estudio genetico-molecular sobre los almiquies indican una divergencia de 25 Ma entre Solenodon cuban us y S paradoxus -compatible con la disyuncion tectonica entre Cuba y La Espafiola (Iturralde-Vinent y MacPhee 1999 McPhee et al 2003)-- y una separacion de ambas especies comparable en magnitud a la divergencia entre familias de mamlferos Consecuentemente propusieron que Solenodon cubanus fuera clasificada en el genero Atopogaie como indico Cabrera (1925) hace 82 afios
DISCUSION Y CONCLUSIONES
Las musarafias y los roedores antillanos (excepto Oryzomys) se irradiaron tanto de sus respectivos ancestros continentales que de estos 6rdenes se reconocen familias y subfamilias endemicas de la region en contraste con los perezosos (una subfamilia y tres
321 IRRADIACION EVOLUCIONARIA
tribus endemicas) y los monos (una tribu endemica) como expresion de diferentes niveles de diversificacion evolucionaria Este proceso es dificil de explicar (traUindose de c1ados que pnicticamente comparten la misma antigiiedad en las Antmas) si es correcta la interpretacion de Pascual et at (1990636)
Nosotros consideramos estos diferentes niveles de diversificacion como las diferentes etapas de evolucion alcanzadas por cada grupo [de mamiferos] Obviamente esto estuvo relacionado con las diferentes longitudes de sus historias
Pudiera ocurrir que en el futuro se compruebe que la historia biogeognifica de la region fue mucho mas complicada
La Espanola la isla mas ecol6gicamente divers a de la region debi6 ser el centro principal de evoluci6n de los mamfferos (con 87 de las familias y 73 de las subfamilias) seguida por Cuba (con 75 de las familias y 54 de las subfamilias) las dos islas de las Antillas Mayores mas relacionadas biogeograficamente (Fig 43 Tabla 44)
eL
EL
cb CAIMAN
SWAN
A Acratocnini B Cubanocnini C Megalocnini D Mesocnini E Nesophontidae F Solenodontidae G Xenothrichini
ANGUILAshyM-0 SMARTIN
H Heteropsomyinae I Plagiodontinae J Isolobodontinae
K Hexolobodontinae L Capromyinae
M Heptaxodontinae N CIidomyinae 0 Sigmodontinae
FIGURA 43 Representacion esquematica de las relaciones interinsulares (expresadas porel numero de linajes compartidos) entre las Antillas Mayores y entre estas y los grupos de islas perifericas segun las interpretaciones de los autores citados en el texto Los numeros dentro de los circulos indican la cantidad de linajes representados en cada isla relacion dudosa Contexto taxon6mico de los linajes en la Tabla 44
bull bull
bull bull
bull bull bull
bull bull
bull bull
bull
bull
bull
bull
bull bull bull bull
bull bull
bull bull
bull
bull bull bull
bull bull
bull
bull bull
bull
322 BIOGEOGRAFiA
TABLA 44 Distribuci6n de los linajcs de mamiferos terrestres a traves de las Antillas Mayores y grupos de islas perifericas LE = La Espanola CU Cuba JA Jamaica PR = Puerto Rico IC Islas Caiman IS Islas Swan IB = Islas Bahamas IV = Islas Virgenes AS = Anguila - San Martin
FamiliaSubfumiliafIribu LE CU JA PR IC IS IB IV AS
Megalonychidae
Choloepodinae
Acratocnini
Cubanocnini
Megalocninae
Megalocnini
Mesocnini
Nesophontidae
Solenodontidae
Pitheciidae
Callicebinae
Xenothrichini
Echimyidae
Heteropsomyinae
Capromyidae
Plagiodontinae
Isolobodontinae
Hexolobodontinae
Capromyinae
H e pta x 0 don tid a e
Heptaxodontinae
Clidomyinae
Muridae
Sigmodontinae
PILOSA
SORICOMORPHA
PRIMATES
RODENTIA
bull
bull
bull
bull TOTALES 12 10 4 4 2
323 EXTlICIOIES DEL CUATERNARIO
El gigantismo ha sido sin duda una tendencia evolucionaria recurrente entre los clados mayores de mamiferos antillanos Es un hecho bien conocido que algunos grupos de mamiferos han aumentado de tamafio en las islas y varios autores han intentado explicarlo a partir de enfoques diferentes (Foster 1964 Miller 1966 Sondaar 1977 Case 1978 Eisenberg y Wilson 1978 Heaney 1978 Lawlor 1982 Lomolino 1985 Angerbjom 1985 McNab 19942001)
EXTINCIONES DEL CUATERNARIO
Las islas que integran la Subregion Antillana han perdido casi 90 de sus mamiferos terrestres durante los ultimos milenios y en proporcion con su extension territorial esta perdida ha sido mayor en la Subregion que en cualquier otro lugar del planeta (MacPhee 1996b) Las extinciones del Cuatemario eliminaron mas de las tres cuartas partes de la fauna de mamiferos terrestres conocida de las Antillas Mayores sus islas satelites y grupos de islas perifericas y el examen de este proceso en sus aspectos mas generales se aborda en esta ultima parte del presente capitulo
CRONOLOGiA Y CONSECUENCIAS
Es muy improbable que un fosil represente el ultimo individuo de un taxon extinto y entonces hay que guiarse por la edad radiometrica del fosil mas reciente conocido lIamada fecha de ultima presencia- como unica aproximacion posible a la fecha de extincion del taxon
Como se explico al inicio del capitulo (vease Contexto Temporal) el registro fosH de los mamiferos terrestres de las Antillas Mayores adolece de insuficiente sustentacion radiometrica No obstante la distribucion temporal de los escasos fechados disponibles para el Cuaternario de la region abarca etapas criticas del proceso de extincion experimentado por la referida fauna a 10 largo de los ultimos 250 mil afios
De las 52 especies que es posible reconocer actualmente como miembros extintos de la fauna cuatemaria de mamiferos terrestres de las Antillas Mayores y grupos de islas perifericas los restos fosiles de 23 de elIas (442) han sido fechados radiometricamente o se han hallado asociados estratigraficamente de manera inequivoca con otro material fechado De estos 23 fechados 3 corresponden al interglacial Sangamon 4 pertenecen a la secuencia Pleistoceno terminal - Holoceno temprano 10 se ubican en ellapso entre el arribo de los aborigenes y la incursion europea y 6 son postcolombinos (a los cuales hay que sumar otras dos especies extinguidas en los siglos XIX y xx) A partir de estas 25 especies avaladas por fechas de ultima presencia distribuidas entre los cuatro intervalos temporales sefialados hemos compuesto la Tabla 45 en la que ademas aparecen las restantes 27 especies (carentes de fechado radiometrico) ubicadas en el intervalo temporal mas probable sobre la base de (1) la asociacion estratigrafica de sus restos con evidencias culturales (amerindias 0 europeas) 0 (2) la ausencia de dicha asociacion Sobre estos criterios de ubicacion temporal procede hacer dos aclaraciones
324 BIOGEOGRAFIA
TABLA 45 Intervalos temporales mas probables para las extinciones cuatemarias de los mamiferos terrestres de las Antillas Mayores y grupos de islas perifericas sobre la base de las evidencias siguientes (l) ultimo ejemplar colectado u observado en vida (2) fechado radiometrico mas tardio en material de la especie (3) asociaci6n estratigrafica mas tardia con otro material fechado (4) asociaci6n estratigrafica con restos de animales introducidos restos humanos u objetos culturales propios del intervale (5) falta de asociaci6n con evidencias culturales [Los ntimeros entre parentesis en la columna Evidencia significan adscripci6n tentativa al intervalo] Los estimados del peso corporal (en kg) estan tomados de la literatura 0 inferidos comparativamente y distribuidos entre
las clases siguientes (a) S 01 (b) 01 a lt 1 (c) 1 a lt 10 (d) 10 a lt 100 (e) 100 a lt 200 (j) ~ 200 Las
abreviaturas que designan las islas son las de la Tabla 44
F amilialEspecie Peso IsJa Evidencia Fuente
I httervalo htterg1acial Sangamon (gt80 kaAP)
HEPTAXODONTlNAE
Amblyrhiza inundata e AS 3 McFarJane et al 1998a
CLIDOMYINAE
Clidomys osborni d JA 3 Ibid
Clidomys parvus d JA 3 Ibid
II httervalo Pleistoceno terminal ~ Holoceno temprano (20-6 kaAP)
MEGALONYCHIDAE
Aeratoenus antillensis d CD 5 Arredondo 1975 a
Aeratoenus odontrigonus d PR 3 McFarJane 1999b
Aeratoenus simorhynehus d LE 2 Rega et al 2002
M egaloenus zile f LE 2 MacPhee et al 2000
SOLENODONTIDAE
Solenodon arredondoi e CD 5 Morgan y Ottenwalder 1993
PnHECIIDAE
Paralouatta varonai d CD 5 MacPhee 1996a
EcHIMYIDAE
Heteropsomys antillensis c PR 5 McFarJane 1999b
Heteropsomys insulans c PR 5 Ibid
Puertoricomys eorozalus c PR 5 Woods 1989a
CAPROMYIDAE
P lagiodontia araeum c LE 5 MacPhee et al 2000
Macrocapromys latus c CD 5 Varona y Arredondo 1979
(Continua)
325 EXTINCIONES DEL CUATERNARIO
TABLA 45 (Continuaci6n)
F ami1ia1Especie Peso Isla Evidencia Fuente
M acrocapromys latus c CU 5 Varona y Arredondo 1979
Mesocapromys kraglievichi c CU 5 Ibid
HEPTAXODONTINAE
Xaymaca fulvopulvis a JA 2 MacPhee y Flemming 2003
III Intervalo Amerindio (6-05 kaAP)
MEGALONYCHIDAE
Acratocnus ye d LE 2 MacPhee et aI 2000
Neocnus comes c LE 2 Ibid
N eocnus dousman c LE 2 Ibid
N eocnus toupiti c LE 2 Ibid
N eocnus gliriform is c CU 4 Cordova et ai 1997
M egalocnus rodens f CU 2 MacPhee et ai 2007
Parocnus browni e CU 2 Steadman et ai 2005
P arocnus serus e LE 2 MacPhee et ai 2000
N ESOPHONTIDAE
N esophontes edithae a PR 2 McFarlane 1999b
PITHECIIDAE
Antillothrix bernensis c LE 3 Rimoli 1977
EcHIMYIDAE
Brotomys contractus a LE 4 Miller 1929c
CAPROMYIDAE
P lagiodontia spelaeum c LE 4 Ibid
M acrocapromys acevedo c CU 4 NUiiez 1990a
HEPTAXODONTINAE
Elasmodontomys obliquus d PR 3 McFarlane 1999b
Iv Intervalo Postcolombino (05 kaAP - Presente)
N ESOPHONTIDAE
N esophontes hypom icrus a LE 24 MacPhee y Fle111111ing 1999
N esophontes zam icrus a LE (4) Ibid
N esophontes m icrus a CU 24 MacPhee et aI 1999
(Continua)
326 BIOGEOGRAFiA
TABLA 45 (Continuacion)
F amilialEspecie Peso Isla Evidencia Fuente
N esophontes m icrus a CD 24 MacPhee et aI 1999
N esophontes m icrus a LE 4 MacPhee et aI 2000
N esophontes sp 1 a IC 4 Morgan 1994
N esophontes sp 2 a IC 4 Ibid
Solenodon marcanoi b LE 4 Ottenwalder 2001
PITHECIIDAE
Xenothrix mcgregori c JA 4 MacPhee y Flemming 1999
EcHIMYIDAE
Boromys offeUa a CD 24 Guarch 1984
Boromys torrei a CD 24 Ibid
Brotomys voratus a LE 4 MacPhee et aI 2000
CAPROMYIDAE
P lagiodontia ipnaeum c LE 4 MacPhee y Flemming 1999
Rhizoplagiodontia lemkei c LE 4 Ibid
Isolobodon portoricensis c PR 24 Ibid
Isolobodon montanus c LE 4 Ibid
Hexolobodon phenax c LE (4) Ibid
Capromys sp c IC 24 Morgan 1994
Geocapromys columbianus c CD 4 MacPhee y Flemming 1999
Geocapromys thoracatus b IS 1 MacPhee y Marx 1997
Geocapromys sp 1 c IC 4 Morgan 1994
Geocapromys sp 2 c IC (4) Ibid
l-IEPTAXODONTINAE
Quemisia gravis c LE (1) MacPhee y Flemming 1999
MURIDAE
Oryzomys antillarum a JA 1 Ibid
1 Flemming y MacPhee (1999) alertaron en contra de la asociacion con otro material fechado Asimismo Morgan (1993) indico que la ausencia de asociacion con evidencias culturales amerindias no es necesariamente un criterio concluyente para ubicar un taxon en el intervalo temporal anterior porque hay depositos en la region que carecen de tales evidencias y son del Holoceno tardio por fechados radiocarbonicos [No obstante
327 EXTlNCIONES DEL CUATERNAR10
la ausencia de una especie en residuarios arqueol6gicos adquiere mayor certidumbre como criterio de ubicaci6n temporal de su extinci6n en la medida que aumenta el numero de residuarios estudiados que en las Antillas Mayores se cuentan por centenares] A falta de fechado absoluto de una especie consideramos que estos criterios de ubicaci6n temporal son mas utiles que la alternativa de excluir del analisis a la especie
2 En los trabajos sobre biogeografia de mamiferos antillanos los arque610gos mas citados como autoridades para establecer el inicio de la presencia aborigen en las Antillas Mayores (Kozlowski 1974 Rouse y Allaire 1978 Rouse 1989 1992 Wilson 2001) difieren al respecto entre los 6 kaAP y los 45 kaAP (pero vease Pagan et al 200523) y hemos adoptado la mas temprana de estas fechas para el comienzo del intervalo amerindio en la Tabla 45 De todos modos cualquiera que haya sido la fecha esta no es aplicable por igual al conjunto de las Antillas Mayores porque segun algunos de los autores citados el poblamiento comenz6 por Cuba y La Espanola 6 mil anos atras continu6 en Puerto Rico hace alrededor de 3 mil anos (pero vease Burney et al 1994) yen Jamaica hace menos de 2 mil e incluso a las Islas Caiman ni siquiera llegaron [Las Islas Caiman descubiertas por Col6n en 1503 carecen absolutamente de evidencia amerindia (Stokes y Keegan 1996)]
Si los datos de la Tabla 45 representan una aceptable aproximaci6n a la realidad el proceso de las extinciones cuaternarias de los mamiferos terrestres de las Antillas Mayores y grupos de islas perifericas debi6 caracterizarse entre otros por los eventos siguientes
Aunque obviamente la fauna cuaternaria debi6 ser mucho mas numerosa que 10 que refleja el registro paleontol6gico (Morgan y Woods 1986) 52 especies (81 de la fauna conocida) desaparecieron (sin reposici6n) durante los ultimos 250 mil anos
- Al parecer las extinciones se concentraron en las especies de mayor tamano Asi el peso promedio estimado para las especies de la etapa preantr6pica (intervalos I + II de la Tabla 45) es 350 kg es 328 kg para las especies del intervalo III (amerindio) y es s610 25 kg (c1ases del peso a b c) para las especies del intervalo IV (postcolombino) Ninguna especie aut6ctona de mamifero mayor de 6 kg existe hoy en la regi6n
- De acuerdo con la distribuci6n temporal de las extinciones 61 especies sobrevivieron al interglacial Sangamon (intervalo 1)48 sobrevivieron a la transici6n del Pleistoceno terminal al Holoceno temprano (intervalo II) 34 sobrevivieron al periodo amerindio (intervalo III) y s610 12 especies han sobrevivido desde la Ilegada de los europeos (intervalo IV) Las tasas de extinci6n correspondientes (exceptuado el intervalo I) son una especie cada 1167 anos en el intervalo II una cada 393 anos en el intervalo III y una cada 23 anos en el intervalo IV
- Durante todo el proceso de las extinciones cuaternarias la perdida de especies por isla fue de 91 en La Espanola 83 en Jamaica y 62 en Cuba Bahamas retuvo su unica especie (aunque extirpada en varias de las islas) Puerto Rico e Islas Caiman perdieron todas sus especies (6 y 5 respectivamente) y las Islas Swan Virgenes y Anguila - San Martin perdieron sus respectivas especies (micas
328 BIOGEOGRAFiA
De los cuatro 6rdenes de mamiferos representados en la fauna Pilosa y Primates desaparecieron totalmente Soricomorpha y Rodentia perdieron 86 y 74 de sus especies respectivamente
CAUSALIDAD
CAUSAS NATURALES
Los cicIos glaciales e interglaciales acaecidos en el Pleistoceno causaron cambios ambientales dnisticos en las Antillas con la concomitante alternancia de cIimas frios (secos) y calidos (hUmedos) asi como de descensos y ascensos del nivel del mar aumento y disminuci6n de las areas territoriales) induso (aunque con menor intensidad) en el Holoceno Estos eventos paleogeograficos han sido consistentemente documentados (Mayo y Kartashov 1972 Bonnatti y Gartner 1973 Ortega y Arcia 1982 Prance 1982 Schubert y Medina 1982 Dodge et al 1983 Ortega [1984] Haq et al 1987 Schubert 1989 Hodell et al 1991 Guilderson et al 1994 Oppo 1997 Emslie 1998 Curtis et al 2001 Iturralde-Vinent 2003) y de ellos brindamos a continuaci6n una breve resena
Finalizando el periodo interglacial anterior a la ultima glaciaci6n (Wisconsin) el mar estaba a 8-10 m por encima de su nivel actual Cuando el avance glacial del Wisconsin a1canz6 su culminaci6n (eillamado Ultimo Maximo Glacial) hace 18 ka el mar habia descendido 120 m con respecto a dicho nivel y las islas del Caribe habian incrementado considerablemente sus extensiones subaereas en comparaci6n con las que presentan hoy dia por ejemplo Puerto Rico y las Islas Virgenes se conectaron territorialmente y las Bahamas se unieron en cinco gran des islas Prevalecia un regimen frio y seco y condiciones de aridez --debidas a una reducci6n de las precipitaciones (posiblemente a menos de 440 mmlano)- y los ambientes xericos se habian extendido a traves de las islas y bancos emergidos y abundaban los pinares (los habitats mesicos estaban restringidos a parcelas relictas en 10 alto de las montanas) En el Mar Caribe y el Golfo de Mexico la temperatura superficial del mar era 5degC mas baja que la actual y en las llanuras costeras y alturas maximas eran respectivamente 2-3degC y 6degC mas bajas Los limites de las zonas ecol6gicas altitudinales descendieron varios cientos de metros y cada zona redujo su extensi6n vertical al aumentar el gradiente de temperatura Ellimite inferior de la zona periglacial
elevaci6n donde la temperatura media minima a1canza los OdegC) de la Cordillera Central de la Republica Dominicana (altura maxima del Caribe insular con evidencia de topografia glacial) se considera que estuvo entre los 1500 y 1800 mlnm (actualmente esta a 2800 mlnm) y se puede estimar que dicho limite era inferior en Cuba por estar mas al norte que la Republica Dominicana No obstante durante este periodo la temperatura experiment6 episodios cortos (unas pocas decadas) de ascensos repetidos
Un signifieativo componente bi6tieo de las referidas condiciones paleoambientales 10 constituy6 la espectacular irradiaci6n evolucionaria experimentada por un grupo numeroso de aves rapaees diurnas y nocturnas (Wetmore 1937 Brodkorb 1969 Kurotchkin y Mayo 1973 Arredondo 1976 Olson 1978 Olson y Hilgartner 1982
329 EXTINCIONES DEL ClJATERNARIO
Suarez 2000a b 2004 Suarez y Emslie 2003) Los mamiferos de las Antillas Mayores ofrecieron a estos depredadores aviares una amplia gama de tamanos de presa que en ausencia de mamiferos del orden Carnivora en la region permitio a estas aves irradiarse a este nicho alimentario con la evolucion de una variedad de especies de muy diversa tall a corporal incluidas formas gigantes Adquirir un cuerpo gigante les propicio el acceso a recursos alimentarios que no estaban siendo explotados por ningun otro depredador (Alcover y McMinn 1994) A Cuba corresponde el mejor registro fosil de rapaces cuaternarias caribenas con siete generos de especies diurnas Dentro de estas aves existieron tres gavilanes gigantes (Accipitridae) tres carairas (Falconidae) y un condor (Cathartidae) asi como cuatro generos de especies nocturnas dos lechuzas (Tytonidae) y tres Mhos (Strigidae) dos de ell os no igualados en tamano en todo el mundo Algunos de estos generos tuvieron representacion en otras islas antillanas como La Espanola Jamaica Bahamas y Gran Caiman
Al retirarse hacia el norte los hielos de la glaciacion Wisconsin el clima se tomo mas hUmedo en las Antillas durante el Holoceno temprano y el bosque mesico se expandio en la region insular El nivel del mar ascendio acompanado de mayor pluviosidad alrededor de los 7 kaAP las Islas Virgenes quedaron separadas entre si y de Puerto Rico y las Bahamas perdieron la mayor parte de su territorio Cerca de los 3 kaAP las condiciones secas retornaron a la region e hicieron desaparecer buena parte del bosque mesico Nuevamente el clima comenzo a adquirir sus condiciones actuales hacia los 25 kaAP aproximadamente y hoy transita por una fase mesica con vegetacion xerica confinada en parcelas relictas
Sin duda estos y otros eventos naturales incidieron negativamente en la sobrevivencia de mamiferos terrestres en las Antillas Mayores asi como en los grupos de pequenas islas perifericas (Pregill y Olson 1981 Olson 2006)
McFarlane et al (1998a) demostraron convincentemente como Amblyrhiza el gigante de los roedores antillanos endemico de la pequena isla de Anguila sucumbio a las repetidas y abruptas perdidas de su habitat durante el interglacial Sangamon por las nipidas y severas oscilaciones del nivel del mar (documentadas por Neumann y Hearty 1996) al no poder responder adecuadamente a tan catastroficos eventos mediante el ajuste de su masa corporal media y otro tanto debio ocurrirle en Jamaica al otro roedor gigante Clidomys (McFarlane et al 1998b) emparentado con Amblyrhiza en la misma epoca (Tabla 45 intervalo I)
El ascenso del nivel del mar seguramente promovio la extincion de mamiferos durante la transicion del Pleistoceno al Holoceno (Tabla 45 intervalo II) con la reduccion del tamano de las Islas por las transgresiones marinas (Pregill y Olson 1981 Morgan 1993) Esto ultimo haria maximamente severa la competicion asf como la presion de los depredadores [Steadman et al (2005) no yen por que el calentamiento global ocurrido durante la transicion Pleistoceno-Holoceno habria de tener un efecto negativo sobre los perezosos siendo sus descendientes actuales especies tropic ales 0 subtropicales pero no se trata del efecto
330 BIOGEOGRAFiA
directo del cambio de temperatura sobre los animales sino del efecto indirecto y progresivo que dicho cambio genera a largo plazo al reducir el territorio insular disponible]
Por esta epoca tambien desaparecieron especies de tall a mayor como Megalocnus zile en La Espanola y otras de menor tamano (en Cuba) pero que representan la tall a maxima en sus respectivos contextos taxonomicos del Cuaternario como Solenodon arredondoi Paralouatta varonai y Macrocapromys acevedo Un mamifero puede ser grande solo a costa de reducir su densidad poblacional y esto 10 hace mis vulnerable a la extinci6n sobre todo en una isla (McNab 1994 McFarlane et al 1998a) Incidentalmente Alcover et al (1998a) afirmaron que larelacion entre la talla corporal y la extincion no siempre se cumple en las especies de menor tamano y citaron como ejemplo que entre las musarafias todas las especies de Nesophontes desaparecieron mientras sobreviven especies de Solenodon pero la comparacion correcta en todo caso serfa entre los solenodones vivientes y el gigante Solenodon arredondoi y entre Nesophontes edithae -el gigante de los nesofontes extinguido en el periodo amerindio- y el resto de sus congeneres que sobrevivieron hasta los primeros siglos de la colonizacion europea (Tabla 45 intervalos III y IV)
CAUSAS ANTROPOGENICAS
Intervalo Amerindio
La mayoria de los autores que se han referido a las extinciones acaecidas durante este intervalo (entre elIos paradigmaticamente Martin 1984a b) han responsabilizado a los aborfgenes (que se supone comenzaron a poblar las Antillas Mayores hace 6 mil afios) con la desaparicion de numerosas especies de mamiferos terrestres en la region principalmente por la via de la sobrecaza y la introduccion de animales domesticos devenidos depredadores de la fauna autoctona (Morgan y Woods 1986) Este efecto negativo de la presencia indigena sobre los mamiferos de la region ha sido dramatizado por algunos autores en terminos de holocausto faunal Sin embargo pensamos que se ha exagerado Como han expresado MacPhee et al (19993) cuando uno busca evidencia dura de complicidad humana directa en [estas] extinciones no hay mucho que encontrar Las siguientes son algunas consideraciones a tener en cuenta en tal sentido
El registro radiometrico mas joven conocido hasta ahora para un perezoso antillano (MegalocnllS rodens) es de 42 kaAP (MacPhee et ai 2007) Si se acepta que los aborfgenes entraron en Cuba hace aproximadamente 6 mil anos su convivencia con los perezosos debe haber durado alrededor de 1800 afios (0 mas porque dificilmente el referfdo fechado radiometrico haya correspondido al ultimo sobreviviente de la especie) Esta cifra contradice la hipotesis del holocausto faunal segun la cualla extincion de los mamiferos insulares de mayor talla debe ocurrir con extrema rapidez -a escala de decadas 0 de centurias- a partir de su primer contacto con los humanos (Martin y Steadman 1999) A pesar de las repetidas afirmaciones en contrario (Harrington 1921 Miller 1929c Herrera [1945] Tabio 1951 Alvarez 1951a 1957 Arredondo
331 EXTINCIONES DEL CUATERNARIO
1959 1960a Veloz y Ortega 1973 Mayo 1980b Rodriguez et al 1984 Castellanos y Pino 1985 Pino y Castellanos 1985 Nunez 1990b Gutierrez 1996 Arredondo 2004) en su mayoria basadas en evidencias no exentas de posibles accidentes de deposicion (0 de reubicacion postdeposicional) en recintos cavernarios (Diaz-Franco [2006b D no se ha demostrado inequivocamente que los aborigenes antillanos se alimentaran de perezosos regularmente Sus restos en residuarios de alimentos son escasisimos en toda la region y esto contrasta con los abundantes restos de jutias y otros mamiferos de menor talla que aparecen en esos residuarios demostrativos de la preferencia de los paleoindios por estos otros mamiferos
Los mamiferos mas importantes en la economia prehistorica de las Antillas Mayores fueron las jutias Capromys y Geocapromys en Cuba Geocapromys en Jamaica e Isolobodon en La Espanola Puerto Rico e Islas Virgenes (Aguayo 1950b Tablo 1951 Rimoli 1972 Wing 2001 Diaz-Franco [2006b D Wilkins (2001) estudi61ajutia viviente de Jamaica (Geocapromys browni) en un sitio ocupado por los aborigenes durante 100 anos y no encontr6 evidencias de que durante ese tiempo la especie haya sido sobreexplotada Asimismo en Cuba las jutias conga carabali y andaraz son escasas en depositos pleistoholocenicos en cambio son muy numerosas en residuarios indigenas y de haber sido sobreexplotadas 10 mas probable es que no hubieran sobrevivido hasta hoy
La habilidad de los humanos para penetrar en profundidad el bosque tropical y mantenerse en este sustentablemente ha sido un evento comparativamente reciente (Beran y Steele 1994) En el caso antillano la caza tierra adentro ha sido explicada como un proceso tardio de desplazamiento a partir de la explotacion de abundantes recursos marinos que ofrecian condiciones 6ptimas para el sedentarismo de las comunidades aborigenes costeras (C6rdova y Arredondo 1988 Brito 2005) Peces y mariscos dominan la arqueologia costera de las Islas antillanas en tanto el consumo de recursos terrestres (mamfferos incluidos) aumenta progresivamente en la medida que los residuarios estudiados se alejan de la costa (Scudder 1991 Wilkins 2001) En las Antillas Mayores el componente terrestre de la dieta aborigen es solo 20-30 del material recobrado en los residuarios (Wing 1977 1989) Y debio ser menor alin porque el componente vegetal de la dieta no se ha preservado Por otra parte los aborigenes cubanos domesticaron la jutia conga y seguramente la reprodujeron para comer (Pose et al 1988) practica que probablemente se repitiera con otras jutias en las demas Antillas Mayores
- Fray Bartolome de las Casas hablo de 3 millones de indios en Cuba al inicio de la colonizacion pero los historiadores subsecuentes consideraron exagerada la cifra (Poey 1851 a) No obstante se dispone de dos estimados de la densidad de poblaci6n aborigen en las Antillas Mayores a la llegada de los europeos 18 indigenaslkm2 en Puerto Rico (Rouse 1956) y 1271km2 en La Espanola (Watts 1987) Extrapolando ambos valores entre las cuatro islas principales de las Antillas Mayores tenemos que en la fecha indicada la poblaci6n aborigen pudo estar entre 16 mil y 113 mil indigenas en Puerto Rico entre 21 mil y 145 mil en Jamaica entre 141 mil y 1 mill6n en La Espanola y entre 200 mil y 14 millones en Cuba Entre las cifras maximas y las minimas nos inclinamos
332 BIOGEOGRAFiA
hacia estas ultimas atendiendo a los altos indices de mortalidad infantil y morbilidad que se evidencian en las osamentas de los pobladores prehispanicos (Martinez y Rives 1990) Uno de los hechos que apuntan hacia el bajo numero de aborigenes en las Antillas Mayores es 10 rapido que desaparecieron despues de la llegada de los europeos pnkticamente sin dejar mestizaje con estos 10 cual contrasta ostensiblemente con la situacion en la America continentaL
En suma un grupo humano (1) demograiicamente irrelevante y controlado por altos indices de morbilidad (2) consumidor mayoritariamente de organismos marinos y (3) carente de la tecnologia cinegetica necesaria para poder sobreexplotar los mamiferos que Ie sirvieron de alimento (a los cuales accedio tardiamente en su desarrollo) no es concebible que haya causado e1 holocausto faunal que se Ie atribuye a partir del error de aplicar a las comunidades aborigenes antillanas un modelo cultural de la prehistoria continentaL
Tampoco hay evidencias de que los aborigenes hayan introducido carnivoros con suficiente capacidad depredadora salvo el perro Morgan y Otlenwaldder (1993) consideraron que la depredacion por perros durante el periodo amerindio es la explicacion mas plausible para la extincion del almiqui gigante de Cuba pero como se puede observar (Tabla 54) Solenodon arredondoi aparece en el intervalo II por falta de asociacion con evidencias culturales
Consecuentemente el accionar de los aborigenes y de sus animales domesticos directamente sobre la fauna no parece haber sido por sf solo un factor determinante de la extincion de mamiferos terrestres durante el periodo amerindio en las Antillas (pero vease Steadman et al 2005)
Hay que recordar en este senti do que alin en el Holoceno tardio ocurrieron cambios clirruiticos de consideracion (vease Causas Naturales) Por ejemplo Curtis et al (2001) explicaron que la extincion de algunos ani males mesofilicos de las alturas montanosas de La Espanola pudo haber antecedido a cualquier impacto humano sigrIificativo en esos lugares por coincidir dicha extincion con una tendencia a la aridez documentada isotopicamente y con evidencias de la perdida del bosque mesico ocurridas despues de 3kaAP Yen Jamaica hace solo 2shy3 mil aDos el area de la Bahia de Jackson sustentaba una gran biomasa de vertebrados en bosque hinnedo a juzgar por la cantidad y diversidad de taxones presentes en material fosil derivado de la depredacion por lechuzas hoy el area es la mas seca de la isla con matorral xerofitico y pobreza de fauna (MacPhee 1996a) Yalm en nuestros dias huracanes de gran intensidad aniquilan poblaciones de vertebrados (que no se recuperan) en is las pequenas y llanas del Caribe (Brooks y Smith 2001)
En medio de la incertidumbre sobre la verdadera causa (0 causas) de las extinciones de mamiferos en el periodo amerindio MacPhee y Marx (1997) han propuesto su hipotesis de la hipermorbilidad la cual estipula colapsos poblacionales relativamente rapidos con la introduccion de enfermedades de alta letalidad Estos autores han razonado que
Las islas son los lugares mas apropiados para experimentar perdidas catastroficas causadas por enfermedades Los mamiferos no solo serian susceptibles en virtud de su prolongado aislamiento biologico sino que en los confines de una isla la epizootia
333 EXTINCIONES DEL CUATERNARIO
tendria la potencialidad de infectar simultaneamente a la rnayoria de los individuos de vadas especies Los neonatos son inmunologicamente inmaduros y sufririan desproporcionadamente cualquier infeccion hipervirulenta La perdida excesiva de neonatos y juveniles tendria efectos severos para la sobrevivencia de la poblacion si esta se reduce por debajo de cierto nivel sobreviene la extincion porque no hay posibilidad de restitucion en el periodo postpanzootico Las especies de mayor talla corporal van a ser las mas vulnerables a la extincion dadas sus bajas tasas reproductivas largos periodos de gestacion y reducido numero de progenie Las especies de menor tamano --caracterizadas por altas tasas reproductivas periodos cortos de gestacion y mayor numero de neonatos por parto- tambien serian fuertemente diezmadas pero muchas sobrevivirian porque su estrategia reproductiva les permite perder mayor numero de individuos Las perdidas deben ocurrir relativamente rapido (a escala de decadas) mientras que otras hipotesis (digamos los cambios climaticos) permiten intervalos mucho mas largos (a escala de siglos 0 de milenios) entre la aparicion de las causas y la produccion de sus efectos Por ejemplo todo indica que los nesofontes se extinguieron muy rapidamente y seria dificil poder invocar un impacto humano directo 0 argumentar fenomenos climaticos que pudieran explicar plausiblemente perdidas tan rapidas
MacPhee y Marx (1997 191) han dicho que por el momento la hipotesis de la hipermorbilidad no pasa de ser una abstraccion en espera de adecuada sustentacion empirica Llama la atencion que linicamente en el periodo amerindio (Tabla 45 intervalo 1Il) las extinciones abarcaron todas las clases del peso corporal (de la a a laj) y que ninguna de las especies que sobrevivieron hasta el periodo postcolombino (Tabla 45 intervalo IV mas las especies vivientes) sobrepasa los 10 kg (clases del peso a b c) en clara coincidencia con los presupuestos de la hipotetica hipermorbilidad
Intervalo Postcolornbino
Por 10 menos 37 especies de mamiferos terrestres vivian en las Antillas Mayores y grupos de Islas perifericas a la lIegada de los europeos en 1492 De elIas se han extinguido 23 shymas especies que en cualquiera de los periodos precedentes (Tabla 45~ y solo subsisten 12 especies repartidas entre Cuba (8 especies) La Espanola (2) Jamaica (1) y Bahamas (1) varias de estas en peligro critico de extincion Solo dos de las especies extinguidas durante los ultimos 500 anos se conocieron en estado viviente los roedores Oryzomys antillarum de Jamaica (extinguido en 1877) y Geocapromys thoracatus de las Islas Swan (extinguido en 1955) Las causas de las extinciones ocurridas durante este periodo son sobradamente conocidas destruccion de habitats introducci6n de animales exoticos (domesticos y comensales) y demas presiones asociadas a la actividad humana entre las cuales posiblemente habria que incluir brotes de hipermorbilidad
Durante mucho tiempo se ha afirmado la posibilidad de que especies de Nesophontes hayan sobrevivido hasta las primeras decadas del siglo xx y que incluso continuen existiendo en lugares remotos de Cuba 0 La Espanola (Allen 1918 1942 Miller 1930 Aguayo 1950a Arredondo 1960b Fischer 1977 MacFadden 1980 Woods y
334 BIOGEOGRAFiA
Ottenwalder 1991) Actualmente no existe evidencia radiometrica de que alguna especie de Nesophontes haya sobrevivido al siglo xv las fechas radiocarbonicas calibradas obtenidas en muestras de Nesophontes de Cuba y La Espanola resultaron no mas tardias que los siglos XIII y XIV (MacPhee et al 1999) Por otra parte las expediciones efectuadas a fines del siglo xx con el proposito de localizar individuos vivientes del genero en La Espanola no produjeron la menor indicacion afirmativa al respecto (Woods et al 1985) Ademas a traves de las Antillas Mayores no aparecen restos de Nesophontes en los depositos formados actualmente por las lechuzas
EPiLOGO
Las Antillas Mayores son islas oceanicas no han formado parte de un continente (con la posible excepcion de una parte de Jamaica hasta el Eoceno Medio hace 45 Ma) y durante un tiempo geologico considerable (desde el Eoceno Superior hace 40 Ma) han permanecido emergidas y separadas de los continentes vecinos Consecuentemente los mamiferos terrestres (no volantes) autoctonos de estas islas constituyen una fauna oceanica que por ende cumple con todas las caracteristicas convencionales reconocidas para este tipo de entidad biogeografica
- Baja diversidad taxonomica a nivel de las categorias superiores (solo cuatro ordenes) en comparacion con islas plataformicas continentales (0 con areas continentales) de tamano y diversidad ecologica simi lares
- Alto grado de endemismo a nivel de genero (96) y familia (50) en respuesta al prolongado aislamiento
- Desbalance ecologico de la estructura trofica ausencia de mamiferos camivoros y ungulados
- Espectaculares procesos de irradiaci6n evolucionaria como resultado de la colonizacion de territorios libres de competidores y depredadores
Tendencias evolucionarias extremas gigantismo
- Control a cargo de aves rapaces con tendencia at gigantismo en ausencia de mamiferos miembros del orden Carnivora
- Mas especies de reptiles y de murcielagos que de mamiferos terrestres relaciones que en los continentes se invierten
Alta tasa de extincion de especies (84) sin reposicion
Preservacion de especies filogeneticamente arcaicas (los llamados fosiles vivientes) los almiquies

319 IRRADIACION EVOLUCIONARIA
sus distribuciones sean explicadas por mecanismos diferentes (Whidden y Asher 2001)
La descomunal diferencia de talla corporal entre las musaranas pequenas (Nesophontes) y las grandes (Soienodon) seguramente impuso a ambos grupos restricciones distintas relativas a la termorregulacion y las adaptaciones energeticas En Nesophontes la seleccion debio favorecer una alta tasa de metabolismo y crecimiento camadas mas numerosas corta expectativa de vida yalta densidad poblacionaL En Soienodon en cambio la seleccion opero en la direccion de una baja tasa metabolica camadas reducidas crecimiento lento longevidad y baja densidad poblacional (Ottenwalder [1991])
MUSARANAS PEQUENAS (Nesophontidae)
Las musaranas pequenas estan representadas en las Antillas por el genero Nesophontes (y un genero miocenico indeterminado en ambar dominicano) con 6 especies cuaternarias extinguidas distribuidas en Cuba La Espanola Puerto Rico Islas Virgenes e Islas Caiman La mayor de las musaranas pequenas Nesophontes edithae evoluciono en Puerto Rico y su arribo a las Islas Virgenes supone una dispersion sumamente reciente En las Islas Caiman Nesophontes se asemeja mas a la poblacion de Cuba de donde seguramente derivo en el Plio-Pleistoceno (junto con Capromys y Geocapromys) cuando la costa sur de Cuba estuvo mucho mas cerca de las Islas Caiman (lturralde-Vinent 2003fig 4) MacPhee et ai (1994) interpretaron la distribucion de Nesophontes en Cuba y La Espanola como resultado de la vicarianza oligomiocenica ocurrida al formarse el Paso de los Vientos pero la distancia taxonomica entre los lados del Paso es solo de nivel especifico y N micros incluso vivi6 en ambas islas segtln Patterson (1962)
Allen (1918) considero que Nesophontes micrus fue una especie primitiva no modificada para la vida arb6rea 0 fos6rea y por tanto de habitos terrestres Nesophontes edithae de Puerto Rico es mucho mayor y mas robusta que las restantes especies del genero eL indice de robustez de su femur (= anchura minima de la diafisis ongitud total del hueso) muestra que este es tan robusto como el de Solenodon paradoxus un animal dos veces mayor (Morgan et ai 1980) Choate y Birney (1968) postularon un aumento cronoclinal de la taIla corporal en N edithae confirmado mas tarde por McFarlane (1999a) mediante fechados radiocarb6nicos de la secuencia estratignifica en depositos cavernarios de Puerto Rico Solenodon no vivi6 en Puerto Rico y en ausencia de una musarana de mayor tamano Nesophontes alcanzo una talla corporal mayor en esa isla que en La Espanola 0 Cuba (Woods y Eisenberg 1989)
MUSARANAS GRANDES (Solenodontidae)
Los arcaicos almiqufes (genero Soienodon) comprenden cuatro especies una viviente (S cubanus) y otra f6sil (S arredondoi) en Cuba y una viviente (S paradoxus con dos subespecies) y otra f6sil (S marcanoi) en La Espanola Como cabria esperar las dos especies cubanas se asemejan mas entre sf que a las especies de La Espanola pero
320 BIOGEOGRAFiA
enigmaticamente S marcanoi comparte caracteres con S cuban us y S paradoxus (Woods y Eisenberg 1989 Morgan y Ottenwalder 1993 Ottenwalder 200 I) Solenodon arredondoi alcanzo una talla corporal superior a la de cualquiera de las musarafias vivientes a nivel mundial Segun Morgan y Ottenwalder (1993) extrapolando las dimensiones y el peso de los almiquies vivientes S arredondoi debio tener una longitud cabeza-tronco y un peso de 450-550 mm y 1500-2000 g respectivamente en comparacion con S cubanus (360 mm y 800 g) y S paradoxus (390 mm y 1100 g)
Entre todas las especies de almiqufes Solenodon paradoxus es la que presenta los caracteres mas derivados (Otten walder 2001) Si estas relaciones son validas la distribucion de Solenodon entre Cuba y La Espafiola debio discurrir de oeste a este como propuso MacFadden (1980) quien invoco la presencia fosH de S cubanus [pero tambien S arredondoi] en Pinar del Rio para postular que el ancestro continental entr~ por Cuba opinion compartida por Perfit y Williams (1989) y por Ottenwalder ([1991] 2001) El paso de Solenodon a Puerto Rico pudo haberlo impedido la existencia de barreras ecologicas durante la conexion del este de La Espafiola con Puerto Rico 0 por la ausencia de Solenodon en esa parte de La Espafiola en aquella epoca Sin embargo las mismas condiciones ecologicas en el este de La Espafiola no parecen haber representado una barrera efectiva para Nesophontes cuyas mas altas tasa de reproduccion y densidad poblacional presumiblemente Ie permitieron expandirse flpidamente a traves de La Espafiola (Ottenwalder [1991])
Es opinion generalizada que los almiqufes antillanos constituyen el grupo de mamiferos que primero se emplazo en el territorio insular (MacPhee 2005) y en tal sentido es de interes la evidencia sobre especificidad parasito-hospedante aportada por Cruz (1980) quien encontro que la especie de acaro (genero Cubanochirus) que parasita a los almiquies es mucho mas primitiva que la especie (del mismo genero) que paras ita a las jutias y de aqui concluyo que dicho genero de acaro debio evolucionar primero en Solenodontidae y posteriormente en Capromyidae al entrar en contacto representantes de ambas familias en Cuba 0 La Espafiola
Recientemente Roca (2004) y Roca et al (2004) dieron a conocer que los resultados de su estudio genetico-molecular sobre los almiquies indican una divergencia de 25 Ma entre Solenodon cuban us y S paradoxus -compatible con la disyuncion tectonica entre Cuba y La Espafiola (Iturralde-Vinent y MacPhee 1999 McPhee et al 2003)-- y una separacion de ambas especies comparable en magnitud a la divergencia entre familias de mamlferos Consecuentemente propusieron que Solenodon cubanus fuera clasificada en el genero Atopogaie como indico Cabrera (1925) hace 82 afios
DISCUSION Y CONCLUSIONES
Las musarafias y los roedores antillanos (excepto Oryzomys) se irradiaron tanto de sus respectivos ancestros continentales que de estos 6rdenes se reconocen familias y subfamilias endemicas de la region en contraste con los perezosos (una subfamilia y tres
321 IRRADIACION EVOLUCIONARIA
tribus endemicas) y los monos (una tribu endemica) como expresion de diferentes niveles de diversificacion evolucionaria Este proceso es dificil de explicar (traUindose de c1ados que pnicticamente comparten la misma antigiiedad en las Antmas) si es correcta la interpretacion de Pascual et at (1990636)
Nosotros consideramos estos diferentes niveles de diversificacion como las diferentes etapas de evolucion alcanzadas por cada grupo [de mamiferos] Obviamente esto estuvo relacionado con las diferentes longitudes de sus historias
Pudiera ocurrir que en el futuro se compruebe que la historia biogeognifica de la region fue mucho mas complicada
La Espanola la isla mas ecol6gicamente divers a de la region debi6 ser el centro principal de evoluci6n de los mamfferos (con 87 de las familias y 73 de las subfamilias) seguida por Cuba (con 75 de las familias y 54 de las subfamilias) las dos islas de las Antillas Mayores mas relacionadas biogeograficamente (Fig 43 Tabla 44)
eL
EL
cb CAIMAN
SWAN
A Acratocnini B Cubanocnini C Megalocnini D Mesocnini E Nesophontidae F Solenodontidae G Xenothrichini
ANGUILAshyM-0 SMARTIN
H Heteropsomyinae I Plagiodontinae J Isolobodontinae
K Hexolobodontinae L Capromyinae
M Heptaxodontinae N CIidomyinae 0 Sigmodontinae
FIGURA 43 Representacion esquematica de las relaciones interinsulares (expresadas porel numero de linajes compartidos) entre las Antillas Mayores y entre estas y los grupos de islas perifericas segun las interpretaciones de los autores citados en el texto Los numeros dentro de los circulos indican la cantidad de linajes representados en cada isla relacion dudosa Contexto taxon6mico de los linajes en la Tabla 44
bull bull
bull bull
bull bull bull
bull bull
bull bull
bull
bull
bull
bull
bull bull bull bull
bull bull
bull bull
bull
bull bull bull
bull bull
bull
bull bull
bull
322 BIOGEOGRAFiA
TABLA 44 Distribuci6n de los linajcs de mamiferos terrestres a traves de las Antillas Mayores y grupos de islas perifericas LE = La Espanola CU Cuba JA Jamaica PR = Puerto Rico IC Islas Caiman IS Islas Swan IB = Islas Bahamas IV = Islas Virgenes AS = Anguila - San Martin
FamiliaSubfumiliafIribu LE CU JA PR IC IS IB IV AS
Megalonychidae
Choloepodinae
Acratocnini
Cubanocnini
Megalocninae
Megalocnini
Mesocnini
Nesophontidae
Solenodontidae
Pitheciidae
Callicebinae
Xenothrichini
Echimyidae
Heteropsomyinae
Capromyidae
Plagiodontinae
Isolobodontinae
Hexolobodontinae
Capromyinae
H e pta x 0 don tid a e
Heptaxodontinae
Clidomyinae
Muridae
Sigmodontinae
PILOSA
SORICOMORPHA
PRIMATES
RODENTIA
bull
bull
bull
bull TOTALES 12 10 4 4 2
323 EXTlICIOIES DEL CUATERNARIO
El gigantismo ha sido sin duda una tendencia evolucionaria recurrente entre los clados mayores de mamiferos antillanos Es un hecho bien conocido que algunos grupos de mamiferos han aumentado de tamafio en las islas y varios autores han intentado explicarlo a partir de enfoques diferentes (Foster 1964 Miller 1966 Sondaar 1977 Case 1978 Eisenberg y Wilson 1978 Heaney 1978 Lawlor 1982 Lomolino 1985 Angerbjom 1985 McNab 19942001)
EXTINCIONES DEL CUATERNARIO
Las islas que integran la Subregion Antillana han perdido casi 90 de sus mamiferos terrestres durante los ultimos milenios y en proporcion con su extension territorial esta perdida ha sido mayor en la Subregion que en cualquier otro lugar del planeta (MacPhee 1996b) Las extinciones del Cuatemario eliminaron mas de las tres cuartas partes de la fauna de mamiferos terrestres conocida de las Antillas Mayores sus islas satelites y grupos de islas perifericas y el examen de este proceso en sus aspectos mas generales se aborda en esta ultima parte del presente capitulo
CRONOLOGiA Y CONSECUENCIAS
Es muy improbable que un fosil represente el ultimo individuo de un taxon extinto y entonces hay que guiarse por la edad radiometrica del fosil mas reciente conocido lIamada fecha de ultima presencia- como unica aproximacion posible a la fecha de extincion del taxon
Como se explico al inicio del capitulo (vease Contexto Temporal) el registro fosH de los mamiferos terrestres de las Antillas Mayores adolece de insuficiente sustentacion radiometrica No obstante la distribucion temporal de los escasos fechados disponibles para el Cuaternario de la region abarca etapas criticas del proceso de extincion experimentado por la referida fauna a 10 largo de los ultimos 250 mil afios
De las 52 especies que es posible reconocer actualmente como miembros extintos de la fauna cuatemaria de mamiferos terrestres de las Antillas Mayores y grupos de islas perifericas los restos fosiles de 23 de elIas (442) han sido fechados radiometricamente o se han hallado asociados estratigraficamente de manera inequivoca con otro material fechado De estos 23 fechados 3 corresponden al interglacial Sangamon 4 pertenecen a la secuencia Pleistoceno terminal - Holoceno temprano 10 se ubican en ellapso entre el arribo de los aborigenes y la incursion europea y 6 son postcolombinos (a los cuales hay que sumar otras dos especies extinguidas en los siglos XIX y xx) A partir de estas 25 especies avaladas por fechas de ultima presencia distribuidas entre los cuatro intervalos temporales sefialados hemos compuesto la Tabla 45 en la que ademas aparecen las restantes 27 especies (carentes de fechado radiometrico) ubicadas en el intervalo temporal mas probable sobre la base de (1) la asociacion estratigrafica de sus restos con evidencias culturales (amerindias 0 europeas) 0 (2) la ausencia de dicha asociacion Sobre estos criterios de ubicacion temporal procede hacer dos aclaraciones
324 BIOGEOGRAFIA
TABLA 45 Intervalos temporales mas probables para las extinciones cuatemarias de los mamiferos terrestres de las Antillas Mayores y grupos de islas perifericas sobre la base de las evidencias siguientes (l) ultimo ejemplar colectado u observado en vida (2) fechado radiometrico mas tardio en material de la especie (3) asociaci6n estratigrafica mas tardia con otro material fechado (4) asociaci6n estratigrafica con restos de animales introducidos restos humanos u objetos culturales propios del intervale (5) falta de asociaci6n con evidencias culturales [Los ntimeros entre parentesis en la columna Evidencia significan adscripci6n tentativa al intervalo] Los estimados del peso corporal (en kg) estan tomados de la literatura 0 inferidos comparativamente y distribuidos entre
las clases siguientes (a) S 01 (b) 01 a lt 1 (c) 1 a lt 10 (d) 10 a lt 100 (e) 100 a lt 200 (j) ~ 200 Las
abreviaturas que designan las islas son las de la Tabla 44
F amilialEspecie Peso IsJa Evidencia Fuente
I httervalo htterg1acial Sangamon (gt80 kaAP)
HEPTAXODONTlNAE
Amblyrhiza inundata e AS 3 McFarJane et al 1998a
CLIDOMYINAE
Clidomys osborni d JA 3 Ibid
Clidomys parvus d JA 3 Ibid
II httervalo Pleistoceno terminal ~ Holoceno temprano (20-6 kaAP)
MEGALONYCHIDAE
Aeratoenus antillensis d CD 5 Arredondo 1975 a
Aeratoenus odontrigonus d PR 3 McFarJane 1999b
Aeratoenus simorhynehus d LE 2 Rega et al 2002
M egaloenus zile f LE 2 MacPhee et al 2000
SOLENODONTIDAE
Solenodon arredondoi e CD 5 Morgan y Ottenwalder 1993
PnHECIIDAE
Paralouatta varonai d CD 5 MacPhee 1996a
EcHIMYIDAE
Heteropsomys antillensis c PR 5 McFarJane 1999b
Heteropsomys insulans c PR 5 Ibid
Puertoricomys eorozalus c PR 5 Woods 1989a
CAPROMYIDAE
P lagiodontia araeum c LE 5 MacPhee et al 2000
Macrocapromys latus c CD 5 Varona y Arredondo 1979
(Continua)
325 EXTINCIONES DEL CUATERNARIO
TABLA 45 (Continuaci6n)
F ami1ia1Especie Peso Isla Evidencia Fuente
M acrocapromys latus c CU 5 Varona y Arredondo 1979
Mesocapromys kraglievichi c CU 5 Ibid
HEPTAXODONTINAE
Xaymaca fulvopulvis a JA 2 MacPhee y Flemming 2003
III Intervalo Amerindio (6-05 kaAP)
MEGALONYCHIDAE
Acratocnus ye d LE 2 MacPhee et aI 2000
Neocnus comes c LE 2 Ibid
N eocnus dousman c LE 2 Ibid
N eocnus toupiti c LE 2 Ibid
N eocnus gliriform is c CU 4 Cordova et ai 1997
M egalocnus rodens f CU 2 MacPhee et ai 2007
Parocnus browni e CU 2 Steadman et ai 2005
P arocnus serus e LE 2 MacPhee et ai 2000
N ESOPHONTIDAE
N esophontes edithae a PR 2 McFarlane 1999b
PITHECIIDAE
Antillothrix bernensis c LE 3 Rimoli 1977
EcHIMYIDAE
Brotomys contractus a LE 4 Miller 1929c
CAPROMYIDAE
P lagiodontia spelaeum c LE 4 Ibid
M acrocapromys acevedo c CU 4 NUiiez 1990a
HEPTAXODONTINAE
Elasmodontomys obliquus d PR 3 McFarlane 1999b
Iv Intervalo Postcolombino (05 kaAP - Presente)
N ESOPHONTIDAE
N esophontes hypom icrus a LE 24 MacPhee y Fle111111ing 1999
N esophontes zam icrus a LE (4) Ibid
N esophontes m icrus a CU 24 MacPhee et aI 1999
(Continua)
326 BIOGEOGRAFiA
TABLA 45 (Continuacion)
F amilialEspecie Peso Isla Evidencia Fuente
N esophontes m icrus a CD 24 MacPhee et aI 1999
N esophontes m icrus a LE 4 MacPhee et aI 2000
N esophontes sp 1 a IC 4 Morgan 1994
N esophontes sp 2 a IC 4 Ibid
Solenodon marcanoi b LE 4 Ottenwalder 2001
PITHECIIDAE
Xenothrix mcgregori c JA 4 MacPhee y Flemming 1999
EcHIMYIDAE
Boromys offeUa a CD 24 Guarch 1984
Boromys torrei a CD 24 Ibid
Brotomys voratus a LE 4 MacPhee et aI 2000
CAPROMYIDAE
P lagiodontia ipnaeum c LE 4 MacPhee y Flemming 1999
Rhizoplagiodontia lemkei c LE 4 Ibid
Isolobodon portoricensis c PR 24 Ibid
Isolobodon montanus c LE 4 Ibid
Hexolobodon phenax c LE (4) Ibid
Capromys sp c IC 24 Morgan 1994
Geocapromys columbianus c CD 4 MacPhee y Flemming 1999
Geocapromys thoracatus b IS 1 MacPhee y Marx 1997
Geocapromys sp 1 c IC 4 Morgan 1994
Geocapromys sp 2 c IC (4) Ibid
l-IEPTAXODONTINAE
Quemisia gravis c LE (1) MacPhee y Flemming 1999
MURIDAE
Oryzomys antillarum a JA 1 Ibid
1 Flemming y MacPhee (1999) alertaron en contra de la asociacion con otro material fechado Asimismo Morgan (1993) indico que la ausencia de asociacion con evidencias culturales amerindias no es necesariamente un criterio concluyente para ubicar un taxon en el intervalo temporal anterior porque hay depositos en la region que carecen de tales evidencias y son del Holoceno tardio por fechados radiocarbonicos [No obstante
327 EXTlNCIONES DEL CUATERNAR10
la ausencia de una especie en residuarios arqueol6gicos adquiere mayor certidumbre como criterio de ubicaci6n temporal de su extinci6n en la medida que aumenta el numero de residuarios estudiados que en las Antillas Mayores se cuentan por centenares] A falta de fechado absoluto de una especie consideramos que estos criterios de ubicaci6n temporal son mas utiles que la alternativa de excluir del analisis a la especie
2 En los trabajos sobre biogeografia de mamiferos antillanos los arque610gos mas citados como autoridades para establecer el inicio de la presencia aborigen en las Antillas Mayores (Kozlowski 1974 Rouse y Allaire 1978 Rouse 1989 1992 Wilson 2001) difieren al respecto entre los 6 kaAP y los 45 kaAP (pero vease Pagan et al 200523) y hemos adoptado la mas temprana de estas fechas para el comienzo del intervalo amerindio en la Tabla 45 De todos modos cualquiera que haya sido la fecha esta no es aplicable por igual al conjunto de las Antillas Mayores porque segun algunos de los autores citados el poblamiento comenz6 por Cuba y La Espanola 6 mil anos atras continu6 en Puerto Rico hace alrededor de 3 mil anos (pero vease Burney et al 1994) yen Jamaica hace menos de 2 mil e incluso a las Islas Caiman ni siquiera llegaron [Las Islas Caiman descubiertas por Col6n en 1503 carecen absolutamente de evidencia amerindia (Stokes y Keegan 1996)]
Si los datos de la Tabla 45 representan una aceptable aproximaci6n a la realidad el proceso de las extinciones cuaternarias de los mamiferos terrestres de las Antillas Mayores y grupos de islas perifericas debi6 caracterizarse entre otros por los eventos siguientes
Aunque obviamente la fauna cuaternaria debi6 ser mucho mas numerosa que 10 que refleja el registro paleontol6gico (Morgan y Woods 1986) 52 especies (81 de la fauna conocida) desaparecieron (sin reposici6n) durante los ultimos 250 mil anos
- Al parecer las extinciones se concentraron en las especies de mayor tamano Asi el peso promedio estimado para las especies de la etapa preantr6pica (intervalos I + II de la Tabla 45) es 350 kg es 328 kg para las especies del intervalo III (amerindio) y es s610 25 kg (c1ases del peso a b c) para las especies del intervalo IV (postcolombino) Ninguna especie aut6ctona de mamifero mayor de 6 kg existe hoy en la regi6n
- De acuerdo con la distribuci6n temporal de las extinciones 61 especies sobrevivieron al interglacial Sangamon (intervalo 1)48 sobrevivieron a la transici6n del Pleistoceno terminal al Holoceno temprano (intervalo II) 34 sobrevivieron al periodo amerindio (intervalo III) y s610 12 especies han sobrevivido desde la Ilegada de los europeos (intervalo IV) Las tasas de extinci6n correspondientes (exceptuado el intervalo I) son una especie cada 1167 anos en el intervalo II una cada 393 anos en el intervalo III y una cada 23 anos en el intervalo IV
- Durante todo el proceso de las extinciones cuaternarias la perdida de especies por isla fue de 91 en La Espanola 83 en Jamaica y 62 en Cuba Bahamas retuvo su unica especie (aunque extirpada en varias de las islas) Puerto Rico e Islas Caiman perdieron todas sus especies (6 y 5 respectivamente) y las Islas Swan Virgenes y Anguila - San Martin perdieron sus respectivas especies (micas
328 BIOGEOGRAFiA
De los cuatro 6rdenes de mamiferos representados en la fauna Pilosa y Primates desaparecieron totalmente Soricomorpha y Rodentia perdieron 86 y 74 de sus especies respectivamente
CAUSALIDAD
CAUSAS NATURALES
Los cicIos glaciales e interglaciales acaecidos en el Pleistoceno causaron cambios ambientales dnisticos en las Antillas con la concomitante alternancia de cIimas frios (secos) y calidos (hUmedos) asi como de descensos y ascensos del nivel del mar aumento y disminuci6n de las areas territoriales) induso (aunque con menor intensidad) en el Holoceno Estos eventos paleogeograficos han sido consistentemente documentados (Mayo y Kartashov 1972 Bonnatti y Gartner 1973 Ortega y Arcia 1982 Prance 1982 Schubert y Medina 1982 Dodge et al 1983 Ortega [1984] Haq et al 1987 Schubert 1989 Hodell et al 1991 Guilderson et al 1994 Oppo 1997 Emslie 1998 Curtis et al 2001 Iturralde-Vinent 2003) y de ellos brindamos a continuaci6n una breve resena
Finalizando el periodo interglacial anterior a la ultima glaciaci6n (Wisconsin) el mar estaba a 8-10 m por encima de su nivel actual Cuando el avance glacial del Wisconsin a1canz6 su culminaci6n (eillamado Ultimo Maximo Glacial) hace 18 ka el mar habia descendido 120 m con respecto a dicho nivel y las islas del Caribe habian incrementado considerablemente sus extensiones subaereas en comparaci6n con las que presentan hoy dia por ejemplo Puerto Rico y las Islas Virgenes se conectaron territorialmente y las Bahamas se unieron en cinco gran des islas Prevalecia un regimen frio y seco y condiciones de aridez --debidas a una reducci6n de las precipitaciones (posiblemente a menos de 440 mmlano)- y los ambientes xericos se habian extendido a traves de las islas y bancos emergidos y abundaban los pinares (los habitats mesicos estaban restringidos a parcelas relictas en 10 alto de las montanas) En el Mar Caribe y el Golfo de Mexico la temperatura superficial del mar era 5degC mas baja que la actual y en las llanuras costeras y alturas maximas eran respectivamente 2-3degC y 6degC mas bajas Los limites de las zonas ecol6gicas altitudinales descendieron varios cientos de metros y cada zona redujo su extensi6n vertical al aumentar el gradiente de temperatura Ellimite inferior de la zona periglacial
elevaci6n donde la temperatura media minima a1canza los OdegC) de la Cordillera Central de la Republica Dominicana (altura maxima del Caribe insular con evidencia de topografia glacial) se considera que estuvo entre los 1500 y 1800 mlnm (actualmente esta a 2800 mlnm) y se puede estimar que dicho limite era inferior en Cuba por estar mas al norte que la Republica Dominicana No obstante durante este periodo la temperatura experiment6 episodios cortos (unas pocas decadas) de ascensos repetidos
Un signifieativo componente bi6tieo de las referidas condiciones paleoambientales 10 constituy6 la espectacular irradiaci6n evolucionaria experimentada por un grupo numeroso de aves rapaees diurnas y nocturnas (Wetmore 1937 Brodkorb 1969 Kurotchkin y Mayo 1973 Arredondo 1976 Olson 1978 Olson y Hilgartner 1982
329 EXTINCIONES DEL ClJATERNARIO
Suarez 2000a b 2004 Suarez y Emslie 2003) Los mamiferos de las Antillas Mayores ofrecieron a estos depredadores aviares una amplia gama de tamanos de presa que en ausencia de mamiferos del orden Carnivora en la region permitio a estas aves irradiarse a este nicho alimentario con la evolucion de una variedad de especies de muy diversa tall a corporal incluidas formas gigantes Adquirir un cuerpo gigante les propicio el acceso a recursos alimentarios que no estaban siendo explotados por ningun otro depredador (Alcover y McMinn 1994) A Cuba corresponde el mejor registro fosil de rapaces cuaternarias caribenas con siete generos de especies diurnas Dentro de estas aves existieron tres gavilanes gigantes (Accipitridae) tres carairas (Falconidae) y un condor (Cathartidae) asi como cuatro generos de especies nocturnas dos lechuzas (Tytonidae) y tres Mhos (Strigidae) dos de ell os no igualados en tamano en todo el mundo Algunos de estos generos tuvieron representacion en otras islas antillanas como La Espanola Jamaica Bahamas y Gran Caiman
Al retirarse hacia el norte los hielos de la glaciacion Wisconsin el clima se tomo mas hUmedo en las Antillas durante el Holoceno temprano y el bosque mesico se expandio en la region insular El nivel del mar ascendio acompanado de mayor pluviosidad alrededor de los 7 kaAP las Islas Virgenes quedaron separadas entre si y de Puerto Rico y las Bahamas perdieron la mayor parte de su territorio Cerca de los 3 kaAP las condiciones secas retornaron a la region e hicieron desaparecer buena parte del bosque mesico Nuevamente el clima comenzo a adquirir sus condiciones actuales hacia los 25 kaAP aproximadamente y hoy transita por una fase mesica con vegetacion xerica confinada en parcelas relictas
Sin duda estos y otros eventos naturales incidieron negativamente en la sobrevivencia de mamiferos terrestres en las Antillas Mayores asi como en los grupos de pequenas islas perifericas (Pregill y Olson 1981 Olson 2006)
McFarlane et al (1998a) demostraron convincentemente como Amblyrhiza el gigante de los roedores antillanos endemico de la pequena isla de Anguila sucumbio a las repetidas y abruptas perdidas de su habitat durante el interglacial Sangamon por las nipidas y severas oscilaciones del nivel del mar (documentadas por Neumann y Hearty 1996) al no poder responder adecuadamente a tan catastroficos eventos mediante el ajuste de su masa corporal media y otro tanto debio ocurrirle en Jamaica al otro roedor gigante Clidomys (McFarlane et al 1998b) emparentado con Amblyrhiza en la misma epoca (Tabla 45 intervalo I)
El ascenso del nivel del mar seguramente promovio la extincion de mamiferos durante la transicion del Pleistoceno al Holoceno (Tabla 45 intervalo II) con la reduccion del tamano de las Islas por las transgresiones marinas (Pregill y Olson 1981 Morgan 1993) Esto ultimo haria maximamente severa la competicion asf como la presion de los depredadores [Steadman et al (2005) no yen por que el calentamiento global ocurrido durante la transicion Pleistoceno-Holoceno habria de tener un efecto negativo sobre los perezosos siendo sus descendientes actuales especies tropic ales 0 subtropicales pero no se trata del efecto
330 BIOGEOGRAFiA
directo del cambio de temperatura sobre los animales sino del efecto indirecto y progresivo que dicho cambio genera a largo plazo al reducir el territorio insular disponible]
Por esta epoca tambien desaparecieron especies de tall a mayor como Megalocnus zile en La Espanola y otras de menor tamano (en Cuba) pero que representan la tall a maxima en sus respectivos contextos taxonomicos del Cuaternario como Solenodon arredondoi Paralouatta varonai y Macrocapromys acevedo Un mamifero puede ser grande solo a costa de reducir su densidad poblacional y esto 10 hace mis vulnerable a la extinci6n sobre todo en una isla (McNab 1994 McFarlane et al 1998a) Incidentalmente Alcover et al (1998a) afirmaron que larelacion entre la talla corporal y la extincion no siempre se cumple en las especies de menor tamano y citaron como ejemplo que entre las musarafias todas las especies de Nesophontes desaparecieron mientras sobreviven especies de Solenodon pero la comparacion correcta en todo caso serfa entre los solenodones vivientes y el gigante Solenodon arredondoi y entre Nesophontes edithae -el gigante de los nesofontes extinguido en el periodo amerindio- y el resto de sus congeneres que sobrevivieron hasta los primeros siglos de la colonizacion europea (Tabla 45 intervalos III y IV)
CAUSAS ANTROPOGENICAS
Intervalo Amerindio
La mayoria de los autores que se han referido a las extinciones acaecidas durante este intervalo (entre elIos paradigmaticamente Martin 1984a b) han responsabilizado a los aborfgenes (que se supone comenzaron a poblar las Antillas Mayores hace 6 mil afios) con la desaparicion de numerosas especies de mamiferos terrestres en la region principalmente por la via de la sobrecaza y la introduccion de animales domesticos devenidos depredadores de la fauna autoctona (Morgan y Woods 1986) Este efecto negativo de la presencia indigena sobre los mamiferos de la region ha sido dramatizado por algunos autores en terminos de holocausto faunal Sin embargo pensamos que se ha exagerado Como han expresado MacPhee et al (19993) cuando uno busca evidencia dura de complicidad humana directa en [estas] extinciones no hay mucho que encontrar Las siguientes son algunas consideraciones a tener en cuenta en tal sentido
El registro radiometrico mas joven conocido hasta ahora para un perezoso antillano (MegalocnllS rodens) es de 42 kaAP (MacPhee et ai 2007) Si se acepta que los aborfgenes entraron en Cuba hace aproximadamente 6 mil anos su convivencia con los perezosos debe haber durado alrededor de 1800 afios (0 mas porque dificilmente el referfdo fechado radiometrico haya correspondido al ultimo sobreviviente de la especie) Esta cifra contradice la hipotesis del holocausto faunal segun la cualla extincion de los mamiferos insulares de mayor talla debe ocurrir con extrema rapidez -a escala de decadas 0 de centurias- a partir de su primer contacto con los humanos (Martin y Steadman 1999) A pesar de las repetidas afirmaciones en contrario (Harrington 1921 Miller 1929c Herrera [1945] Tabio 1951 Alvarez 1951a 1957 Arredondo
331 EXTINCIONES DEL CUATERNARIO
1959 1960a Veloz y Ortega 1973 Mayo 1980b Rodriguez et al 1984 Castellanos y Pino 1985 Pino y Castellanos 1985 Nunez 1990b Gutierrez 1996 Arredondo 2004) en su mayoria basadas en evidencias no exentas de posibles accidentes de deposicion (0 de reubicacion postdeposicional) en recintos cavernarios (Diaz-Franco [2006b D no se ha demostrado inequivocamente que los aborigenes antillanos se alimentaran de perezosos regularmente Sus restos en residuarios de alimentos son escasisimos en toda la region y esto contrasta con los abundantes restos de jutias y otros mamiferos de menor talla que aparecen en esos residuarios demostrativos de la preferencia de los paleoindios por estos otros mamiferos
Los mamiferos mas importantes en la economia prehistorica de las Antillas Mayores fueron las jutias Capromys y Geocapromys en Cuba Geocapromys en Jamaica e Isolobodon en La Espanola Puerto Rico e Islas Virgenes (Aguayo 1950b Tablo 1951 Rimoli 1972 Wing 2001 Diaz-Franco [2006b D Wilkins (2001) estudi61ajutia viviente de Jamaica (Geocapromys browni) en un sitio ocupado por los aborigenes durante 100 anos y no encontr6 evidencias de que durante ese tiempo la especie haya sido sobreexplotada Asimismo en Cuba las jutias conga carabali y andaraz son escasas en depositos pleistoholocenicos en cambio son muy numerosas en residuarios indigenas y de haber sido sobreexplotadas 10 mas probable es que no hubieran sobrevivido hasta hoy
La habilidad de los humanos para penetrar en profundidad el bosque tropical y mantenerse en este sustentablemente ha sido un evento comparativamente reciente (Beran y Steele 1994) En el caso antillano la caza tierra adentro ha sido explicada como un proceso tardio de desplazamiento a partir de la explotacion de abundantes recursos marinos que ofrecian condiciones 6ptimas para el sedentarismo de las comunidades aborigenes costeras (C6rdova y Arredondo 1988 Brito 2005) Peces y mariscos dominan la arqueologia costera de las Islas antillanas en tanto el consumo de recursos terrestres (mamfferos incluidos) aumenta progresivamente en la medida que los residuarios estudiados se alejan de la costa (Scudder 1991 Wilkins 2001) En las Antillas Mayores el componente terrestre de la dieta aborigen es solo 20-30 del material recobrado en los residuarios (Wing 1977 1989) Y debio ser menor alin porque el componente vegetal de la dieta no se ha preservado Por otra parte los aborigenes cubanos domesticaron la jutia conga y seguramente la reprodujeron para comer (Pose et al 1988) practica que probablemente se repitiera con otras jutias en las demas Antillas Mayores
- Fray Bartolome de las Casas hablo de 3 millones de indios en Cuba al inicio de la colonizacion pero los historiadores subsecuentes consideraron exagerada la cifra (Poey 1851 a) No obstante se dispone de dos estimados de la densidad de poblaci6n aborigen en las Antillas Mayores a la llegada de los europeos 18 indigenaslkm2 en Puerto Rico (Rouse 1956) y 1271km2 en La Espanola (Watts 1987) Extrapolando ambos valores entre las cuatro islas principales de las Antillas Mayores tenemos que en la fecha indicada la poblaci6n aborigen pudo estar entre 16 mil y 113 mil indigenas en Puerto Rico entre 21 mil y 145 mil en Jamaica entre 141 mil y 1 mill6n en La Espanola y entre 200 mil y 14 millones en Cuba Entre las cifras maximas y las minimas nos inclinamos
332 BIOGEOGRAFiA
hacia estas ultimas atendiendo a los altos indices de mortalidad infantil y morbilidad que se evidencian en las osamentas de los pobladores prehispanicos (Martinez y Rives 1990) Uno de los hechos que apuntan hacia el bajo numero de aborigenes en las Antillas Mayores es 10 rapido que desaparecieron despues de la llegada de los europeos pnkticamente sin dejar mestizaje con estos 10 cual contrasta ostensiblemente con la situacion en la America continentaL
En suma un grupo humano (1) demograiicamente irrelevante y controlado por altos indices de morbilidad (2) consumidor mayoritariamente de organismos marinos y (3) carente de la tecnologia cinegetica necesaria para poder sobreexplotar los mamiferos que Ie sirvieron de alimento (a los cuales accedio tardiamente en su desarrollo) no es concebible que haya causado e1 holocausto faunal que se Ie atribuye a partir del error de aplicar a las comunidades aborigenes antillanas un modelo cultural de la prehistoria continentaL
Tampoco hay evidencias de que los aborigenes hayan introducido carnivoros con suficiente capacidad depredadora salvo el perro Morgan y Otlenwaldder (1993) consideraron que la depredacion por perros durante el periodo amerindio es la explicacion mas plausible para la extincion del almiqui gigante de Cuba pero como se puede observar (Tabla 54) Solenodon arredondoi aparece en el intervalo II por falta de asociacion con evidencias culturales
Consecuentemente el accionar de los aborigenes y de sus animales domesticos directamente sobre la fauna no parece haber sido por sf solo un factor determinante de la extincion de mamiferos terrestres durante el periodo amerindio en las Antillas (pero vease Steadman et al 2005)
Hay que recordar en este senti do que alin en el Holoceno tardio ocurrieron cambios clirruiticos de consideracion (vease Causas Naturales) Por ejemplo Curtis et al (2001) explicaron que la extincion de algunos ani males mesofilicos de las alturas montanosas de La Espanola pudo haber antecedido a cualquier impacto humano sigrIificativo en esos lugares por coincidir dicha extincion con una tendencia a la aridez documentada isotopicamente y con evidencias de la perdida del bosque mesico ocurridas despues de 3kaAP Yen Jamaica hace solo 2shy3 mil aDos el area de la Bahia de Jackson sustentaba una gran biomasa de vertebrados en bosque hinnedo a juzgar por la cantidad y diversidad de taxones presentes en material fosil derivado de la depredacion por lechuzas hoy el area es la mas seca de la isla con matorral xerofitico y pobreza de fauna (MacPhee 1996a) Yalm en nuestros dias huracanes de gran intensidad aniquilan poblaciones de vertebrados (que no se recuperan) en is las pequenas y llanas del Caribe (Brooks y Smith 2001)
En medio de la incertidumbre sobre la verdadera causa (0 causas) de las extinciones de mamiferos en el periodo amerindio MacPhee y Marx (1997) han propuesto su hipotesis de la hipermorbilidad la cual estipula colapsos poblacionales relativamente rapidos con la introduccion de enfermedades de alta letalidad Estos autores han razonado que
Las islas son los lugares mas apropiados para experimentar perdidas catastroficas causadas por enfermedades Los mamiferos no solo serian susceptibles en virtud de su prolongado aislamiento biologico sino que en los confines de una isla la epizootia
333 EXTINCIONES DEL CUATERNARIO
tendria la potencialidad de infectar simultaneamente a la rnayoria de los individuos de vadas especies Los neonatos son inmunologicamente inmaduros y sufririan desproporcionadamente cualquier infeccion hipervirulenta La perdida excesiva de neonatos y juveniles tendria efectos severos para la sobrevivencia de la poblacion si esta se reduce por debajo de cierto nivel sobreviene la extincion porque no hay posibilidad de restitucion en el periodo postpanzootico Las especies de mayor talla corporal van a ser las mas vulnerables a la extincion dadas sus bajas tasas reproductivas largos periodos de gestacion y reducido numero de progenie Las especies de menor tamano --caracterizadas por altas tasas reproductivas periodos cortos de gestacion y mayor numero de neonatos por parto- tambien serian fuertemente diezmadas pero muchas sobrevivirian porque su estrategia reproductiva les permite perder mayor numero de individuos Las perdidas deben ocurrir relativamente rapido (a escala de decadas) mientras que otras hipotesis (digamos los cambios climaticos) permiten intervalos mucho mas largos (a escala de siglos 0 de milenios) entre la aparicion de las causas y la produccion de sus efectos Por ejemplo todo indica que los nesofontes se extinguieron muy rapidamente y seria dificil poder invocar un impacto humano directo 0 argumentar fenomenos climaticos que pudieran explicar plausiblemente perdidas tan rapidas
MacPhee y Marx (1997 191) han dicho que por el momento la hipotesis de la hipermorbilidad no pasa de ser una abstraccion en espera de adecuada sustentacion empirica Llama la atencion que linicamente en el periodo amerindio (Tabla 45 intervalo 1Il) las extinciones abarcaron todas las clases del peso corporal (de la a a laj) y que ninguna de las especies que sobrevivieron hasta el periodo postcolombino (Tabla 45 intervalo IV mas las especies vivientes) sobrepasa los 10 kg (clases del peso a b c) en clara coincidencia con los presupuestos de la hipotetica hipermorbilidad
Intervalo Postcolornbino
Por 10 menos 37 especies de mamiferos terrestres vivian en las Antillas Mayores y grupos de Islas perifericas a la lIegada de los europeos en 1492 De elIas se han extinguido 23 shymas especies que en cualquiera de los periodos precedentes (Tabla 45~ y solo subsisten 12 especies repartidas entre Cuba (8 especies) La Espanola (2) Jamaica (1) y Bahamas (1) varias de estas en peligro critico de extincion Solo dos de las especies extinguidas durante los ultimos 500 anos se conocieron en estado viviente los roedores Oryzomys antillarum de Jamaica (extinguido en 1877) y Geocapromys thoracatus de las Islas Swan (extinguido en 1955) Las causas de las extinciones ocurridas durante este periodo son sobradamente conocidas destruccion de habitats introducci6n de animales exoticos (domesticos y comensales) y demas presiones asociadas a la actividad humana entre las cuales posiblemente habria que incluir brotes de hipermorbilidad
Durante mucho tiempo se ha afirmado la posibilidad de que especies de Nesophontes hayan sobrevivido hasta las primeras decadas del siglo xx y que incluso continuen existiendo en lugares remotos de Cuba 0 La Espanola (Allen 1918 1942 Miller 1930 Aguayo 1950a Arredondo 1960b Fischer 1977 MacFadden 1980 Woods y
334 BIOGEOGRAFiA
Ottenwalder 1991) Actualmente no existe evidencia radiometrica de que alguna especie de Nesophontes haya sobrevivido al siglo xv las fechas radiocarbonicas calibradas obtenidas en muestras de Nesophontes de Cuba y La Espanola resultaron no mas tardias que los siglos XIII y XIV (MacPhee et al 1999) Por otra parte las expediciones efectuadas a fines del siglo xx con el proposito de localizar individuos vivientes del genero en La Espanola no produjeron la menor indicacion afirmativa al respecto (Woods et al 1985) Ademas a traves de las Antillas Mayores no aparecen restos de Nesophontes en los depositos formados actualmente por las lechuzas
EPiLOGO
Las Antillas Mayores son islas oceanicas no han formado parte de un continente (con la posible excepcion de una parte de Jamaica hasta el Eoceno Medio hace 45 Ma) y durante un tiempo geologico considerable (desde el Eoceno Superior hace 40 Ma) han permanecido emergidas y separadas de los continentes vecinos Consecuentemente los mamiferos terrestres (no volantes) autoctonos de estas islas constituyen una fauna oceanica que por ende cumple con todas las caracteristicas convencionales reconocidas para este tipo de entidad biogeografica
- Baja diversidad taxonomica a nivel de las categorias superiores (solo cuatro ordenes) en comparacion con islas plataformicas continentales (0 con areas continentales) de tamano y diversidad ecologica simi lares
- Alto grado de endemismo a nivel de genero (96) y familia (50) en respuesta al prolongado aislamiento
- Desbalance ecologico de la estructura trofica ausencia de mamiferos camivoros y ungulados
- Espectaculares procesos de irradiaci6n evolucionaria como resultado de la colonizacion de territorios libres de competidores y depredadores
Tendencias evolucionarias extremas gigantismo
- Control a cargo de aves rapaces con tendencia at gigantismo en ausencia de mamiferos miembros del orden Carnivora
- Mas especies de reptiles y de murcielagos que de mamiferos terrestres relaciones que en los continentes se invierten
Alta tasa de extincion de especies (84) sin reposicion
Preservacion de especies filogeneticamente arcaicas (los llamados fosiles vivientes) los almiquies

320 BIOGEOGRAFiA
enigmaticamente S marcanoi comparte caracteres con S cuban us y S paradoxus (Woods y Eisenberg 1989 Morgan y Ottenwalder 1993 Ottenwalder 200 I) Solenodon arredondoi alcanzo una talla corporal superior a la de cualquiera de las musarafias vivientes a nivel mundial Segun Morgan y Ottenwalder (1993) extrapolando las dimensiones y el peso de los almiquies vivientes S arredondoi debio tener una longitud cabeza-tronco y un peso de 450-550 mm y 1500-2000 g respectivamente en comparacion con S cubanus (360 mm y 800 g) y S paradoxus (390 mm y 1100 g)
Entre todas las especies de almiqufes Solenodon paradoxus es la que presenta los caracteres mas derivados (Otten walder 2001) Si estas relaciones son validas la distribucion de Solenodon entre Cuba y La Espafiola debio discurrir de oeste a este como propuso MacFadden (1980) quien invoco la presencia fosH de S cubanus [pero tambien S arredondoi] en Pinar del Rio para postular que el ancestro continental entr~ por Cuba opinion compartida por Perfit y Williams (1989) y por Ottenwalder ([1991] 2001) El paso de Solenodon a Puerto Rico pudo haberlo impedido la existencia de barreras ecologicas durante la conexion del este de La Espafiola con Puerto Rico 0 por la ausencia de Solenodon en esa parte de La Espafiola en aquella epoca Sin embargo las mismas condiciones ecologicas en el este de La Espafiola no parecen haber representado una barrera efectiva para Nesophontes cuyas mas altas tasa de reproduccion y densidad poblacional presumiblemente Ie permitieron expandirse flpidamente a traves de La Espafiola (Ottenwalder [1991])
Es opinion generalizada que los almiqufes antillanos constituyen el grupo de mamiferos que primero se emplazo en el territorio insular (MacPhee 2005) y en tal sentido es de interes la evidencia sobre especificidad parasito-hospedante aportada por Cruz (1980) quien encontro que la especie de acaro (genero Cubanochirus) que parasita a los almiquies es mucho mas primitiva que la especie (del mismo genero) que paras ita a las jutias y de aqui concluyo que dicho genero de acaro debio evolucionar primero en Solenodontidae y posteriormente en Capromyidae al entrar en contacto representantes de ambas familias en Cuba 0 La Espafiola
Recientemente Roca (2004) y Roca et al (2004) dieron a conocer que los resultados de su estudio genetico-molecular sobre los almiquies indican una divergencia de 25 Ma entre Solenodon cuban us y S paradoxus -compatible con la disyuncion tectonica entre Cuba y La Espafiola (Iturralde-Vinent y MacPhee 1999 McPhee et al 2003)-- y una separacion de ambas especies comparable en magnitud a la divergencia entre familias de mamlferos Consecuentemente propusieron que Solenodon cubanus fuera clasificada en el genero Atopogaie como indico Cabrera (1925) hace 82 afios
DISCUSION Y CONCLUSIONES
Las musarafias y los roedores antillanos (excepto Oryzomys) se irradiaron tanto de sus respectivos ancestros continentales que de estos 6rdenes se reconocen familias y subfamilias endemicas de la region en contraste con los perezosos (una subfamilia y tres
321 IRRADIACION EVOLUCIONARIA
tribus endemicas) y los monos (una tribu endemica) como expresion de diferentes niveles de diversificacion evolucionaria Este proceso es dificil de explicar (traUindose de c1ados que pnicticamente comparten la misma antigiiedad en las Antmas) si es correcta la interpretacion de Pascual et at (1990636)
Nosotros consideramos estos diferentes niveles de diversificacion como las diferentes etapas de evolucion alcanzadas por cada grupo [de mamiferos] Obviamente esto estuvo relacionado con las diferentes longitudes de sus historias
Pudiera ocurrir que en el futuro se compruebe que la historia biogeognifica de la region fue mucho mas complicada
La Espanola la isla mas ecol6gicamente divers a de la region debi6 ser el centro principal de evoluci6n de los mamfferos (con 87 de las familias y 73 de las subfamilias) seguida por Cuba (con 75 de las familias y 54 de las subfamilias) las dos islas de las Antillas Mayores mas relacionadas biogeograficamente (Fig 43 Tabla 44)
eL
EL
cb CAIMAN
SWAN
A Acratocnini B Cubanocnini C Megalocnini D Mesocnini E Nesophontidae F Solenodontidae G Xenothrichini
ANGUILAshyM-0 SMARTIN
H Heteropsomyinae I Plagiodontinae J Isolobodontinae
K Hexolobodontinae L Capromyinae
M Heptaxodontinae N CIidomyinae 0 Sigmodontinae
FIGURA 43 Representacion esquematica de las relaciones interinsulares (expresadas porel numero de linajes compartidos) entre las Antillas Mayores y entre estas y los grupos de islas perifericas segun las interpretaciones de los autores citados en el texto Los numeros dentro de los circulos indican la cantidad de linajes representados en cada isla relacion dudosa Contexto taxon6mico de los linajes en la Tabla 44
bull bull
bull bull
bull bull bull
bull bull
bull bull
bull
bull
bull
bull
bull bull bull bull
bull bull
bull bull
bull
bull bull bull
bull bull
bull
bull bull
bull
322 BIOGEOGRAFiA
TABLA 44 Distribuci6n de los linajcs de mamiferos terrestres a traves de las Antillas Mayores y grupos de islas perifericas LE = La Espanola CU Cuba JA Jamaica PR = Puerto Rico IC Islas Caiman IS Islas Swan IB = Islas Bahamas IV = Islas Virgenes AS = Anguila - San Martin
FamiliaSubfumiliafIribu LE CU JA PR IC IS IB IV AS
Megalonychidae
Choloepodinae
Acratocnini
Cubanocnini
Megalocninae
Megalocnini
Mesocnini
Nesophontidae
Solenodontidae
Pitheciidae
Callicebinae
Xenothrichini
Echimyidae
Heteropsomyinae
Capromyidae
Plagiodontinae
Isolobodontinae
Hexolobodontinae
Capromyinae
H e pta x 0 don tid a e
Heptaxodontinae
Clidomyinae
Muridae
Sigmodontinae
PILOSA
SORICOMORPHA
PRIMATES
RODENTIA
bull
bull
bull
bull TOTALES 12 10 4 4 2
323 EXTlICIOIES DEL CUATERNARIO
El gigantismo ha sido sin duda una tendencia evolucionaria recurrente entre los clados mayores de mamiferos antillanos Es un hecho bien conocido que algunos grupos de mamiferos han aumentado de tamafio en las islas y varios autores han intentado explicarlo a partir de enfoques diferentes (Foster 1964 Miller 1966 Sondaar 1977 Case 1978 Eisenberg y Wilson 1978 Heaney 1978 Lawlor 1982 Lomolino 1985 Angerbjom 1985 McNab 19942001)
EXTINCIONES DEL CUATERNARIO
Las islas que integran la Subregion Antillana han perdido casi 90 de sus mamiferos terrestres durante los ultimos milenios y en proporcion con su extension territorial esta perdida ha sido mayor en la Subregion que en cualquier otro lugar del planeta (MacPhee 1996b) Las extinciones del Cuatemario eliminaron mas de las tres cuartas partes de la fauna de mamiferos terrestres conocida de las Antillas Mayores sus islas satelites y grupos de islas perifericas y el examen de este proceso en sus aspectos mas generales se aborda en esta ultima parte del presente capitulo
CRONOLOGiA Y CONSECUENCIAS
Es muy improbable que un fosil represente el ultimo individuo de un taxon extinto y entonces hay que guiarse por la edad radiometrica del fosil mas reciente conocido lIamada fecha de ultima presencia- como unica aproximacion posible a la fecha de extincion del taxon
Como se explico al inicio del capitulo (vease Contexto Temporal) el registro fosH de los mamiferos terrestres de las Antillas Mayores adolece de insuficiente sustentacion radiometrica No obstante la distribucion temporal de los escasos fechados disponibles para el Cuaternario de la region abarca etapas criticas del proceso de extincion experimentado por la referida fauna a 10 largo de los ultimos 250 mil afios
De las 52 especies que es posible reconocer actualmente como miembros extintos de la fauna cuatemaria de mamiferos terrestres de las Antillas Mayores y grupos de islas perifericas los restos fosiles de 23 de elIas (442) han sido fechados radiometricamente o se han hallado asociados estratigraficamente de manera inequivoca con otro material fechado De estos 23 fechados 3 corresponden al interglacial Sangamon 4 pertenecen a la secuencia Pleistoceno terminal - Holoceno temprano 10 se ubican en ellapso entre el arribo de los aborigenes y la incursion europea y 6 son postcolombinos (a los cuales hay que sumar otras dos especies extinguidas en los siglos XIX y xx) A partir de estas 25 especies avaladas por fechas de ultima presencia distribuidas entre los cuatro intervalos temporales sefialados hemos compuesto la Tabla 45 en la que ademas aparecen las restantes 27 especies (carentes de fechado radiometrico) ubicadas en el intervalo temporal mas probable sobre la base de (1) la asociacion estratigrafica de sus restos con evidencias culturales (amerindias 0 europeas) 0 (2) la ausencia de dicha asociacion Sobre estos criterios de ubicacion temporal procede hacer dos aclaraciones
324 BIOGEOGRAFIA
TABLA 45 Intervalos temporales mas probables para las extinciones cuatemarias de los mamiferos terrestres de las Antillas Mayores y grupos de islas perifericas sobre la base de las evidencias siguientes (l) ultimo ejemplar colectado u observado en vida (2) fechado radiometrico mas tardio en material de la especie (3) asociaci6n estratigrafica mas tardia con otro material fechado (4) asociaci6n estratigrafica con restos de animales introducidos restos humanos u objetos culturales propios del intervale (5) falta de asociaci6n con evidencias culturales [Los ntimeros entre parentesis en la columna Evidencia significan adscripci6n tentativa al intervalo] Los estimados del peso corporal (en kg) estan tomados de la literatura 0 inferidos comparativamente y distribuidos entre
las clases siguientes (a) S 01 (b) 01 a lt 1 (c) 1 a lt 10 (d) 10 a lt 100 (e) 100 a lt 200 (j) ~ 200 Las
abreviaturas que designan las islas son las de la Tabla 44
F amilialEspecie Peso IsJa Evidencia Fuente
I httervalo htterg1acial Sangamon (gt80 kaAP)
HEPTAXODONTlNAE
Amblyrhiza inundata e AS 3 McFarJane et al 1998a
CLIDOMYINAE
Clidomys osborni d JA 3 Ibid
Clidomys parvus d JA 3 Ibid
II httervalo Pleistoceno terminal ~ Holoceno temprano (20-6 kaAP)
MEGALONYCHIDAE
Aeratoenus antillensis d CD 5 Arredondo 1975 a
Aeratoenus odontrigonus d PR 3 McFarJane 1999b
Aeratoenus simorhynehus d LE 2 Rega et al 2002
M egaloenus zile f LE 2 MacPhee et al 2000
SOLENODONTIDAE
Solenodon arredondoi e CD 5 Morgan y Ottenwalder 1993
PnHECIIDAE
Paralouatta varonai d CD 5 MacPhee 1996a
EcHIMYIDAE
Heteropsomys antillensis c PR 5 McFarJane 1999b
Heteropsomys insulans c PR 5 Ibid
Puertoricomys eorozalus c PR 5 Woods 1989a
CAPROMYIDAE
P lagiodontia araeum c LE 5 MacPhee et al 2000
Macrocapromys latus c CD 5 Varona y Arredondo 1979
(Continua)
325 EXTINCIONES DEL CUATERNARIO
TABLA 45 (Continuaci6n)
F ami1ia1Especie Peso Isla Evidencia Fuente
M acrocapromys latus c CU 5 Varona y Arredondo 1979
Mesocapromys kraglievichi c CU 5 Ibid
HEPTAXODONTINAE
Xaymaca fulvopulvis a JA 2 MacPhee y Flemming 2003
III Intervalo Amerindio (6-05 kaAP)
MEGALONYCHIDAE
Acratocnus ye d LE 2 MacPhee et aI 2000
Neocnus comes c LE 2 Ibid
N eocnus dousman c LE 2 Ibid
N eocnus toupiti c LE 2 Ibid
N eocnus gliriform is c CU 4 Cordova et ai 1997
M egalocnus rodens f CU 2 MacPhee et ai 2007
Parocnus browni e CU 2 Steadman et ai 2005
P arocnus serus e LE 2 MacPhee et ai 2000
N ESOPHONTIDAE
N esophontes edithae a PR 2 McFarlane 1999b
PITHECIIDAE
Antillothrix bernensis c LE 3 Rimoli 1977
EcHIMYIDAE
Brotomys contractus a LE 4 Miller 1929c
CAPROMYIDAE
P lagiodontia spelaeum c LE 4 Ibid
M acrocapromys acevedo c CU 4 NUiiez 1990a
HEPTAXODONTINAE
Elasmodontomys obliquus d PR 3 McFarlane 1999b
Iv Intervalo Postcolombino (05 kaAP - Presente)
N ESOPHONTIDAE
N esophontes hypom icrus a LE 24 MacPhee y Fle111111ing 1999
N esophontes zam icrus a LE (4) Ibid
N esophontes m icrus a CU 24 MacPhee et aI 1999
(Continua)
326 BIOGEOGRAFiA
TABLA 45 (Continuacion)
F amilialEspecie Peso Isla Evidencia Fuente
N esophontes m icrus a CD 24 MacPhee et aI 1999
N esophontes m icrus a LE 4 MacPhee et aI 2000
N esophontes sp 1 a IC 4 Morgan 1994
N esophontes sp 2 a IC 4 Ibid
Solenodon marcanoi b LE 4 Ottenwalder 2001
PITHECIIDAE
Xenothrix mcgregori c JA 4 MacPhee y Flemming 1999
EcHIMYIDAE
Boromys offeUa a CD 24 Guarch 1984
Boromys torrei a CD 24 Ibid
Brotomys voratus a LE 4 MacPhee et aI 2000
CAPROMYIDAE
P lagiodontia ipnaeum c LE 4 MacPhee y Flemming 1999
Rhizoplagiodontia lemkei c LE 4 Ibid
Isolobodon portoricensis c PR 24 Ibid
Isolobodon montanus c LE 4 Ibid
Hexolobodon phenax c LE (4) Ibid
Capromys sp c IC 24 Morgan 1994
Geocapromys columbianus c CD 4 MacPhee y Flemming 1999
Geocapromys thoracatus b IS 1 MacPhee y Marx 1997
Geocapromys sp 1 c IC 4 Morgan 1994
Geocapromys sp 2 c IC (4) Ibid
l-IEPTAXODONTINAE
Quemisia gravis c LE (1) MacPhee y Flemming 1999
MURIDAE
Oryzomys antillarum a JA 1 Ibid
1 Flemming y MacPhee (1999) alertaron en contra de la asociacion con otro material fechado Asimismo Morgan (1993) indico que la ausencia de asociacion con evidencias culturales amerindias no es necesariamente un criterio concluyente para ubicar un taxon en el intervalo temporal anterior porque hay depositos en la region que carecen de tales evidencias y son del Holoceno tardio por fechados radiocarbonicos [No obstante
327 EXTlNCIONES DEL CUATERNAR10
la ausencia de una especie en residuarios arqueol6gicos adquiere mayor certidumbre como criterio de ubicaci6n temporal de su extinci6n en la medida que aumenta el numero de residuarios estudiados que en las Antillas Mayores se cuentan por centenares] A falta de fechado absoluto de una especie consideramos que estos criterios de ubicaci6n temporal son mas utiles que la alternativa de excluir del analisis a la especie
2 En los trabajos sobre biogeografia de mamiferos antillanos los arque610gos mas citados como autoridades para establecer el inicio de la presencia aborigen en las Antillas Mayores (Kozlowski 1974 Rouse y Allaire 1978 Rouse 1989 1992 Wilson 2001) difieren al respecto entre los 6 kaAP y los 45 kaAP (pero vease Pagan et al 200523) y hemos adoptado la mas temprana de estas fechas para el comienzo del intervalo amerindio en la Tabla 45 De todos modos cualquiera que haya sido la fecha esta no es aplicable por igual al conjunto de las Antillas Mayores porque segun algunos de los autores citados el poblamiento comenz6 por Cuba y La Espanola 6 mil anos atras continu6 en Puerto Rico hace alrededor de 3 mil anos (pero vease Burney et al 1994) yen Jamaica hace menos de 2 mil e incluso a las Islas Caiman ni siquiera llegaron [Las Islas Caiman descubiertas por Col6n en 1503 carecen absolutamente de evidencia amerindia (Stokes y Keegan 1996)]
Si los datos de la Tabla 45 representan una aceptable aproximaci6n a la realidad el proceso de las extinciones cuaternarias de los mamiferos terrestres de las Antillas Mayores y grupos de islas perifericas debi6 caracterizarse entre otros por los eventos siguientes
Aunque obviamente la fauna cuaternaria debi6 ser mucho mas numerosa que 10 que refleja el registro paleontol6gico (Morgan y Woods 1986) 52 especies (81 de la fauna conocida) desaparecieron (sin reposici6n) durante los ultimos 250 mil anos
- Al parecer las extinciones se concentraron en las especies de mayor tamano Asi el peso promedio estimado para las especies de la etapa preantr6pica (intervalos I + II de la Tabla 45) es 350 kg es 328 kg para las especies del intervalo III (amerindio) y es s610 25 kg (c1ases del peso a b c) para las especies del intervalo IV (postcolombino) Ninguna especie aut6ctona de mamifero mayor de 6 kg existe hoy en la regi6n
- De acuerdo con la distribuci6n temporal de las extinciones 61 especies sobrevivieron al interglacial Sangamon (intervalo 1)48 sobrevivieron a la transici6n del Pleistoceno terminal al Holoceno temprano (intervalo II) 34 sobrevivieron al periodo amerindio (intervalo III) y s610 12 especies han sobrevivido desde la Ilegada de los europeos (intervalo IV) Las tasas de extinci6n correspondientes (exceptuado el intervalo I) son una especie cada 1167 anos en el intervalo II una cada 393 anos en el intervalo III y una cada 23 anos en el intervalo IV
- Durante todo el proceso de las extinciones cuaternarias la perdida de especies por isla fue de 91 en La Espanola 83 en Jamaica y 62 en Cuba Bahamas retuvo su unica especie (aunque extirpada en varias de las islas) Puerto Rico e Islas Caiman perdieron todas sus especies (6 y 5 respectivamente) y las Islas Swan Virgenes y Anguila - San Martin perdieron sus respectivas especies (micas
328 BIOGEOGRAFiA
De los cuatro 6rdenes de mamiferos representados en la fauna Pilosa y Primates desaparecieron totalmente Soricomorpha y Rodentia perdieron 86 y 74 de sus especies respectivamente
CAUSALIDAD
CAUSAS NATURALES
Los cicIos glaciales e interglaciales acaecidos en el Pleistoceno causaron cambios ambientales dnisticos en las Antillas con la concomitante alternancia de cIimas frios (secos) y calidos (hUmedos) asi como de descensos y ascensos del nivel del mar aumento y disminuci6n de las areas territoriales) induso (aunque con menor intensidad) en el Holoceno Estos eventos paleogeograficos han sido consistentemente documentados (Mayo y Kartashov 1972 Bonnatti y Gartner 1973 Ortega y Arcia 1982 Prance 1982 Schubert y Medina 1982 Dodge et al 1983 Ortega [1984] Haq et al 1987 Schubert 1989 Hodell et al 1991 Guilderson et al 1994 Oppo 1997 Emslie 1998 Curtis et al 2001 Iturralde-Vinent 2003) y de ellos brindamos a continuaci6n una breve resena
Finalizando el periodo interglacial anterior a la ultima glaciaci6n (Wisconsin) el mar estaba a 8-10 m por encima de su nivel actual Cuando el avance glacial del Wisconsin a1canz6 su culminaci6n (eillamado Ultimo Maximo Glacial) hace 18 ka el mar habia descendido 120 m con respecto a dicho nivel y las islas del Caribe habian incrementado considerablemente sus extensiones subaereas en comparaci6n con las que presentan hoy dia por ejemplo Puerto Rico y las Islas Virgenes se conectaron territorialmente y las Bahamas se unieron en cinco gran des islas Prevalecia un regimen frio y seco y condiciones de aridez --debidas a una reducci6n de las precipitaciones (posiblemente a menos de 440 mmlano)- y los ambientes xericos se habian extendido a traves de las islas y bancos emergidos y abundaban los pinares (los habitats mesicos estaban restringidos a parcelas relictas en 10 alto de las montanas) En el Mar Caribe y el Golfo de Mexico la temperatura superficial del mar era 5degC mas baja que la actual y en las llanuras costeras y alturas maximas eran respectivamente 2-3degC y 6degC mas bajas Los limites de las zonas ecol6gicas altitudinales descendieron varios cientos de metros y cada zona redujo su extensi6n vertical al aumentar el gradiente de temperatura Ellimite inferior de la zona periglacial
elevaci6n donde la temperatura media minima a1canza los OdegC) de la Cordillera Central de la Republica Dominicana (altura maxima del Caribe insular con evidencia de topografia glacial) se considera que estuvo entre los 1500 y 1800 mlnm (actualmente esta a 2800 mlnm) y se puede estimar que dicho limite era inferior en Cuba por estar mas al norte que la Republica Dominicana No obstante durante este periodo la temperatura experiment6 episodios cortos (unas pocas decadas) de ascensos repetidos
Un signifieativo componente bi6tieo de las referidas condiciones paleoambientales 10 constituy6 la espectacular irradiaci6n evolucionaria experimentada por un grupo numeroso de aves rapaees diurnas y nocturnas (Wetmore 1937 Brodkorb 1969 Kurotchkin y Mayo 1973 Arredondo 1976 Olson 1978 Olson y Hilgartner 1982
329 EXTINCIONES DEL ClJATERNARIO
Suarez 2000a b 2004 Suarez y Emslie 2003) Los mamiferos de las Antillas Mayores ofrecieron a estos depredadores aviares una amplia gama de tamanos de presa que en ausencia de mamiferos del orden Carnivora en la region permitio a estas aves irradiarse a este nicho alimentario con la evolucion de una variedad de especies de muy diversa tall a corporal incluidas formas gigantes Adquirir un cuerpo gigante les propicio el acceso a recursos alimentarios que no estaban siendo explotados por ningun otro depredador (Alcover y McMinn 1994) A Cuba corresponde el mejor registro fosil de rapaces cuaternarias caribenas con siete generos de especies diurnas Dentro de estas aves existieron tres gavilanes gigantes (Accipitridae) tres carairas (Falconidae) y un condor (Cathartidae) asi como cuatro generos de especies nocturnas dos lechuzas (Tytonidae) y tres Mhos (Strigidae) dos de ell os no igualados en tamano en todo el mundo Algunos de estos generos tuvieron representacion en otras islas antillanas como La Espanola Jamaica Bahamas y Gran Caiman
Al retirarse hacia el norte los hielos de la glaciacion Wisconsin el clima se tomo mas hUmedo en las Antillas durante el Holoceno temprano y el bosque mesico se expandio en la region insular El nivel del mar ascendio acompanado de mayor pluviosidad alrededor de los 7 kaAP las Islas Virgenes quedaron separadas entre si y de Puerto Rico y las Bahamas perdieron la mayor parte de su territorio Cerca de los 3 kaAP las condiciones secas retornaron a la region e hicieron desaparecer buena parte del bosque mesico Nuevamente el clima comenzo a adquirir sus condiciones actuales hacia los 25 kaAP aproximadamente y hoy transita por una fase mesica con vegetacion xerica confinada en parcelas relictas
Sin duda estos y otros eventos naturales incidieron negativamente en la sobrevivencia de mamiferos terrestres en las Antillas Mayores asi como en los grupos de pequenas islas perifericas (Pregill y Olson 1981 Olson 2006)
McFarlane et al (1998a) demostraron convincentemente como Amblyrhiza el gigante de los roedores antillanos endemico de la pequena isla de Anguila sucumbio a las repetidas y abruptas perdidas de su habitat durante el interglacial Sangamon por las nipidas y severas oscilaciones del nivel del mar (documentadas por Neumann y Hearty 1996) al no poder responder adecuadamente a tan catastroficos eventos mediante el ajuste de su masa corporal media y otro tanto debio ocurrirle en Jamaica al otro roedor gigante Clidomys (McFarlane et al 1998b) emparentado con Amblyrhiza en la misma epoca (Tabla 45 intervalo I)
El ascenso del nivel del mar seguramente promovio la extincion de mamiferos durante la transicion del Pleistoceno al Holoceno (Tabla 45 intervalo II) con la reduccion del tamano de las Islas por las transgresiones marinas (Pregill y Olson 1981 Morgan 1993) Esto ultimo haria maximamente severa la competicion asf como la presion de los depredadores [Steadman et al (2005) no yen por que el calentamiento global ocurrido durante la transicion Pleistoceno-Holoceno habria de tener un efecto negativo sobre los perezosos siendo sus descendientes actuales especies tropic ales 0 subtropicales pero no se trata del efecto
330 BIOGEOGRAFiA
directo del cambio de temperatura sobre los animales sino del efecto indirecto y progresivo que dicho cambio genera a largo plazo al reducir el territorio insular disponible]
Por esta epoca tambien desaparecieron especies de tall a mayor como Megalocnus zile en La Espanola y otras de menor tamano (en Cuba) pero que representan la tall a maxima en sus respectivos contextos taxonomicos del Cuaternario como Solenodon arredondoi Paralouatta varonai y Macrocapromys acevedo Un mamifero puede ser grande solo a costa de reducir su densidad poblacional y esto 10 hace mis vulnerable a la extinci6n sobre todo en una isla (McNab 1994 McFarlane et al 1998a) Incidentalmente Alcover et al (1998a) afirmaron que larelacion entre la talla corporal y la extincion no siempre se cumple en las especies de menor tamano y citaron como ejemplo que entre las musarafias todas las especies de Nesophontes desaparecieron mientras sobreviven especies de Solenodon pero la comparacion correcta en todo caso serfa entre los solenodones vivientes y el gigante Solenodon arredondoi y entre Nesophontes edithae -el gigante de los nesofontes extinguido en el periodo amerindio- y el resto de sus congeneres que sobrevivieron hasta los primeros siglos de la colonizacion europea (Tabla 45 intervalos III y IV)
CAUSAS ANTROPOGENICAS
Intervalo Amerindio
La mayoria de los autores que se han referido a las extinciones acaecidas durante este intervalo (entre elIos paradigmaticamente Martin 1984a b) han responsabilizado a los aborfgenes (que se supone comenzaron a poblar las Antillas Mayores hace 6 mil afios) con la desaparicion de numerosas especies de mamiferos terrestres en la region principalmente por la via de la sobrecaza y la introduccion de animales domesticos devenidos depredadores de la fauna autoctona (Morgan y Woods 1986) Este efecto negativo de la presencia indigena sobre los mamiferos de la region ha sido dramatizado por algunos autores en terminos de holocausto faunal Sin embargo pensamos que se ha exagerado Como han expresado MacPhee et al (19993) cuando uno busca evidencia dura de complicidad humana directa en [estas] extinciones no hay mucho que encontrar Las siguientes son algunas consideraciones a tener en cuenta en tal sentido
El registro radiometrico mas joven conocido hasta ahora para un perezoso antillano (MegalocnllS rodens) es de 42 kaAP (MacPhee et ai 2007) Si se acepta que los aborfgenes entraron en Cuba hace aproximadamente 6 mil anos su convivencia con los perezosos debe haber durado alrededor de 1800 afios (0 mas porque dificilmente el referfdo fechado radiometrico haya correspondido al ultimo sobreviviente de la especie) Esta cifra contradice la hipotesis del holocausto faunal segun la cualla extincion de los mamiferos insulares de mayor talla debe ocurrir con extrema rapidez -a escala de decadas 0 de centurias- a partir de su primer contacto con los humanos (Martin y Steadman 1999) A pesar de las repetidas afirmaciones en contrario (Harrington 1921 Miller 1929c Herrera [1945] Tabio 1951 Alvarez 1951a 1957 Arredondo
331 EXTINCIONES DEL CUATERNARIO
1959 1960a Veloz y Ortega 1973 Mayo 1980b Rodriguez et al 1984 Castellanos y Pino 1985 Pino y Castellanos 1985 Nunez 1990b Gutierrez 1996 Arredondo 2004) en su mayoria basadas en evidencias no exentas de posibles accidentes de deposicion (0 de reubicacion postdeposicional) en recintos cavernarios (Diaz-Franco [2006b D no se ha demostrado inequivocamente que los aborigenes antillanos se alimentaran de perezosos regularmente Sus restos en residuarios de alimentos son escasisimos en toda la region y esto contrasta con los abundantes restos de jutias y otros mamiferos de menor talla que aparecen en esos residuarios demostrativos de la preferencia de los paleoindios por estos otros mamiferos
Los mamiferos mas importantes en la economia prehistorica de las Antillas Mayores fueron las jutias Capromys y Geocapromys en Cuba Geocapromys en Jamaica e Isolobodon en La Espanola Puerto Rico e Islas Virgenes (Aguayo 1950b Tablo 1951 Rimoli 1972 Wing 2001 Diaz-Franco [2006b D Wilkins (2001) estudi61ajutia viviente de Jamaica (Geocapromys browni) en un sitio ocupado por los aborigenes durante 100 anos y no encontr6 evidencias de que durante ese tiempo la especie haya sido sobreexplotada Asimismo en Cuba las jutias conga carabali y andaraz son escasas en depositos pleistoholocenicos en cambio son muy numerosas en residuarios indigenas y de haber sido sobreexplotadas 10 mas probable es que no hubieran sobrevivido hasta hoy
La habilidad de los humanos para penetrar en profundidad el bosque tropical y mantenerse en este sustentablemente ha sido un evento comparativamente reciente (Beran y Steele 1994) En el caso antillano la caza tierra adentro ha sido explicada como un proceso tardio de desplazamiento a partir de la explotacion de abundantes recursos marinos que ofrecian condiciones 6ptimas para el sedentarismo de las comunidades aborigenes costeras (C6rdova y Arredondo 1988 Brito 2005) Peces y mariscos dominan la arqueologia costera de las Islas antillanas en tanto el consumo de recursos terrestres (mamfferos incluidos) aumenta progresivamente en la medida que los residuarios estudiados se alejan de la costa (Scudder 1991 Wilkins 2001) En las Antillas Mayores el componente terrestre de la dieta aborigen es solo 20-30 del material recobrado en los residuarios (Wing 1977 1989) Y debio ser menor alin porque el componente vegetal de la dieta no se ha preservado Por otra parte los aborigenes cubanos domesticaron la jutia conga y seguramente la reprodujeron para comer (Pose et al 1988) practica que probablemente se repitiera con otras jutias en las demas Antillas Mayores
- Fray Bartolome de las Casas hablo de 3 millones de indios en Cuba al inicio de la colonizacion pero los historiadores subsecuentes consideraron exagerada la cifra (Poey 1851 a) No obstante se dispone de dos estimados de la densidad de poblaci6n aborigen en las Antillas Mayores a la llegada de los europeos 18 indigenaslkm2 en Puerto Rico (Rouse 1956) y 1271km2 en La Espanola (Watts 1987) Extrapolando ambos valores entre las cuatro islas principales de las Antillas Mayores tenemos que en la fecha indicada la poblaci6n aborigen pudo estar entre 16 mil y 113 mil indigenas en Puerto Rico entre 21 mil y 145 mil en Jamaica entre 141 mil y 1 mill6n en La Espanola y entre 200 mil y 14 millones en Cuba Entre las cifras maximas y las minimas nos inclinamos
332 BIOGEOGRAFiA
hacia estas ultimas atendiendo a los altos indices de mortalidad infantil y morbilidad que se evidencian en las osamentas de los pobladores prehispanicos (Martinez y Rives 1990) Uno de los hechos que apuntan hacia el bajo numero de aborigenes en las Antillas Mayores es 10 rapido que desaparecieron despues de la llegada de los europeos pnkticamente sin dejar mestizaje con estos 10 cual contrasta ostensiblemente con la situacion en la America continentaL
En suma un grupo humano (1) demograiicamente irrelevante y controlado por altos indices de morbilidad (2) consumidor mayoritariamente de organismos marinos y (3) carente de la tecnologia cinegetica necesaria para poder sobreexplotar los mamiferos que Ie sirvieron de alimento (a los cuales accedio tardiamente en su desarrollo) no es concebible que haya causado e1 holocausto faunal que se Ie atribuye a partir del error de aplicar a las comunidades aborigenes antillanas un modelo cultural de la prehistoria continentaL
Tampoco hay evidencias de que los aborigenes hayan introducido carnivoros con suficiente capacidad depredadora salvo el perro Morgan y Otlenwaldder (1993) consideraron que la depredacion por perros durante el periodo amerindio es la explicacion mas plausible para la extincion del almiqui gigante de Cuba pero como se puede observar (Tabla 54) Solenodon arredondoi aparece en el intervalo II por falta de asociacion con evidencias culturales
Consecuentemente el accionar de los aborigenes y de sus animales domesticos directamente sobre la fauna no parece haber sido por sf solo un factor determinante de la extincion de mamiferos terrestres durante el periodo amerindio en las Antillas (pero vease Steadman et al 2005)
Hay que recordar en este senti do que alin en el Holoceno tardio ocurrieron cambios clirruiticos de consideracion (vease Causas Naturales) Por ejemplo Curtis et al (2001) explicaron que la extincion de algunos ani males mesofilicos de las alturas montanosas de La Espanola pudo haber antecedido a cualquier impacto humano sigrIificativo en esos lugares por coincidir dicha extincion con una tendencia a la aridez documentada isotopicamente y con evidencias de la perdida del bosque mesico ocurridas despues de 3kaAP Yen Jamaica hace solo 2shy3 mil aDos el area de la Bahia de Jackson sustentaba una gran biomasa de vertebrados en bosque hinnedo a juzgar por la cantidad y diversidad de taxones presentes en material fosil derivado de la depredacion por lechuzas hoy el area es la mas seca de la isla con matorral xerofitico y pobreza de fauna (MacPhee 1996a) Yalm en nuestros dias huracanes de gran intensidad aniquilan poblaciones de vertebrados (que no se recuperan) en is las pequenas y llanas del Caribe (Brooks y Smith 2001)
En medio de la incertidumbre sobre la verdadera causa (0 causas) de las extinciones de mamiferos en el periodo amerindio MacPhee y Marx (1997) han propuesto su hipotesis de la hipermorbilidad la cual estipula colapsos poblacionales relativamente rapidos con la introduccion de enfermedades de alta letalidad Estos autores han razonado que
Las islas son los lugares mas apropiados para experimentar perdidas catastroficas causadas por enfermedades Los mamiferos no solo serian susceptibles en virtud de su prolongado aislamiento biologico sino que en los confines de una isla la epizootia
333 EXTINCIONES DEL CUATERNARIO
tendria la potencialidad de infectar simultaneamente a la rnayoria de los individuos de vadas especies Los neonatos son inmunologicamente inmaduros y sufririan desproporcionadamente cualquier infeccion hipervirulenta La perdida excesiva de neonatos y juveniles tendria efectos severos para la sobrevivencia de la poblacion si esta se reduce por debajo de cierto nivel sobreviene la extincion porque no hay posibilidad de restitucion en el periodo postpanzootico Las especies de mayor talla corporal van a ser las mas vulnerables a la extincion dadas sus bajas tasas reproductivas largos periodos de gestacion y reducido numero de progenie Las especies de menor tamano --caracterizadas por altas tasas reproductivas periodos cortos de gestacion y mayor numero de neonatos por parto- tambien serian fuertemente diezmadas pero muchas sobrevivirian porque su estrategia reproductiva les permite perder mayor numero de individuos Las perdidas deben ocurrir relativamente rapido (a escala de decadas) mientras que otras hipotesis (digamos los cambios climaticos) permiten intervalos mucho mas largos (a escala de siglos 0 de milenios) entre la aparicion de las causas y la produccion de sus efectos Por ejemplo todo indica que los nesofontes se extinguieron muy rapidamente y seria dificil poder invocar un impacto humano directo 0 argumentar fenomenos climaticos que pudieran explicar plausiblemente perdidas tan rapidas
MacPhee y Marx (1997 191) han dicho que por el momento la hipotesis de la hipermorbilidad no pasa de ser una abstraccion en espera de adecuada sustentacion empirica Llama la atencion que linicamente en el periodo amerindio (Tabla 45 intervalo 1Il) las extinciones abarcaron todas las clases del peso corporal (de la a a laj) y que ninguna de las especies que sobrevivieron hasta el periodo postcolombino (Tabla 45 intervalo IV mas las especies vivientes) sobrepasa los 10 kg (clases del peso a b c) en clara coincidencia con los presupuestos de la hipotetica hipermorbilidad
Intervalo Postcolornbino
Por 10 menos 37 especies de mamiferos terrestres vivian en las Antillas Mayores y grupos de Islas perifericas a la lIegada de los europeos en 1492 De elIas se han extinguido 23 shymas especies que en cualquiera de los periodos precedentes (Tabla 45~ y solo subsisten 12 especies repartidas entre Cuba (8 especies) La Espanola (2) Jamaica (1) y Bahamas (1) varias de estas en peligro critico de extincion Solo dos de las especies extinguidas durante los ultimos 500 anos se conocieron en estado viviente los roedores Oryzomys antillarum de Jamaica (extinguido en 1877) y Geocapromys thoracatus de las Islas Swan (extinguido en 1955) Las causas de las extinciones ocurridas durante este periodo son sobradamente conocidas destruccion de habitats introducci6n de animales exoticos (domesticos y comensales) y demas presiones asociadas a la actividad humana entre las cuales posiblemente habria que incluir brotes de hipermorbilidad
Durante mucho tiempo se ha afirmado la posibilidad de que especies de Nesophontes hayan sobrevivido hasta las primeras decadas del siglo xx y que incluso continuen existiendo en lugares remotos de Cuba 0 La Espanola (Allen 1918 1942 Miller 1930 Aguayo 1950a Arredondo 1960b Fischer 1977 MacFadden 1980 Woods y
334 BIOGEOGRAFiA
Ottenwalder 1991) Actualmente no existe evidencia radiometrica de que alguna especie de Nesophontes haya sobrevivido al siglo xv las fechas radiocarbonicas calibradas obtenidas en muestras de Nesophontes de Cuba y La Espanola resultaron no mas tardias que los siglos XIII y XIV (MacPhee et al 1999) Por otra parte las expediciones efectuadas a fines del siglo xx con el proposito de localizar individuos vivientes del genero en La Espanola no produjeron la menor indicacion afirmativa al respecto (Woods et al 1985) Ademas a traves de las Antillas Mayores no aparecen restos de Nesophontes en los depositos formados actualmente por las lechuzas
EPiLOGO
Las Antillas Mayores son islas oceanicas no han formado parte de un continente (con la posible excepcion de una parte de Jamaica hasta el Eoceno Medio hace 45 Ma) y durante un tiempo geologico considerable (desde el Eoceno Superior hace 40 Ma) han permanecido emergidas y separadas de los continentes vecinos Consecuentemente los mamiferos terrestres (no volantes) autoctonos de estas islas constituyen una fauna oceanica que por ende cumple con todas las caracteristicas convencionales reconocidas para este tipo de entidad biogeografica
- Baja diversidad taxonomica a nivel de las categorias superiores (solo cuatro ordenes) en comparacion con islas plataformicas continentales (0 con areas continentales) de tamano y diversidad ecologica simi lares
- Alto grado de endemismo a nivel de genero (96) y familia (50) en respuesta al prolongado aislamiento
- Desbalance ecologico de la estructura trofica ausencia de mamiferos camivoros y ungulados
- Espectaculares procesos de irradiaci6n evolucionaria como resultado de la colonizacion de territorios libres de competidores y depredadores
Tendencias evolucionarias extremas gigantismo
- Control a cargo de aves rapaces con tendencia at gigantismo en ausencia de mamiferos miembros del orden Carnivora
- Mas especies de reptiles y de murcielagos que de mamiferos terrestres relaciones que en los continentes se invierten
Alta tasa de extincion de especies (84) sin reposicion
Preservacion de especies filogeneticamente arcaicas (los llamados fosiles vivientes) los almiquies
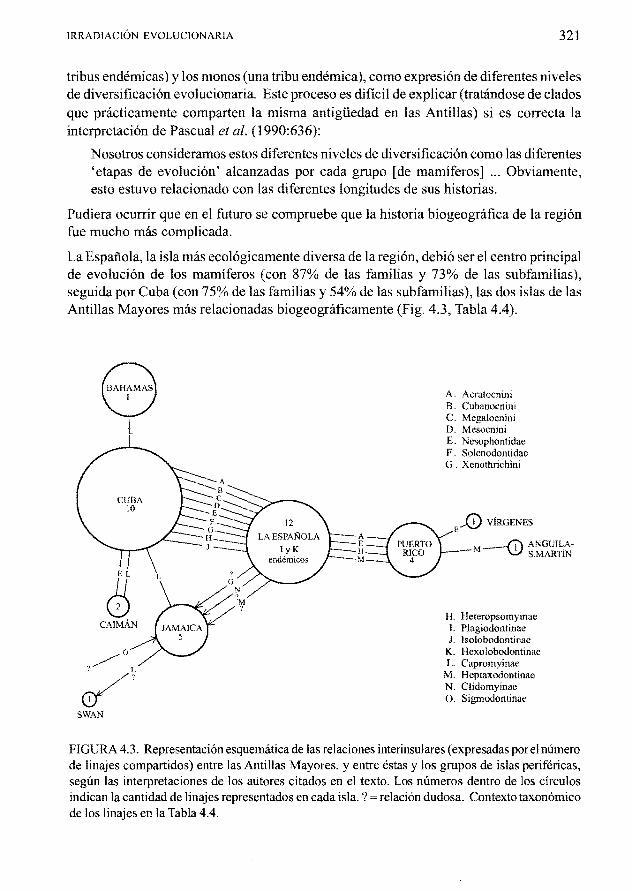
321 IRRADIACION EVOLUCIONARIA
tribus endemicas) y los monos (una tribu endemica) como expresion de diferentes niveles de diversificacion evolucionaria Este proceso es dificil de explicar (traUindose de c1ados que pnicticamente comparten la misma antigiiedad en las Antmas) si es correcta la interpretacion de Pascual et at (1990636)
Nosotros consideramos estos diferentes niveles de diversificacion como las diferentes etapas de evolucion alcanzadas por cada grupo [de mamiferos] Obviamente esto estuvo relacionado con las diferentes longitudes de sus historias
Pudiera ocurrir que en el futuro se compruebe que la historia biogeognifica de la region fue mucho mas complicada
La Espanola la isla mas ecol6gicamente divers a de la region debi6 ser el centro principal de evoluci6n de los mamfferos (con 87 de las familias y 73 de las subfamilias) seguida por Cuba (con 75 de las familias y 54 de las subfamilias) las dos islas de las Antillas Mayores mas relacionadas biogeograficamente (Fig 43 Tabla 44)
eL
EL
cb CAIMAN
SWAN
A Acratocnini B Cubanocnini C Megalocnini D Mesocnini E Nesophontidae F Solenodontidae G Xenothrichini
ANGUILAshyM-0 SMARTIN
H Heteropsomyinae I Plagiodontinae J Isolobodontinae
K Hexolobodontinae L Capromyinae
M Heptaxodontinae N CIidomyinae 0 Sigmodontinae
FIGURA 43 Representacion esquematica de las relaciones interinsulares (expresadas porel numero de linajes compartidos) entre las Antillas Mayores y entre estas y los grupos de islas perifericas segun las interpretaciones de los autores citados en el texto Los numeros dentro de los circulos indican la cantidad de linajes representados en cada isla relacion dudosa Contexto taxon6mico de los linajes en la Tabla 44
bull bull
bull bull
bull bull bull
bull bull
bull bull
bull
bull
bull
bull
bull bull bull bull
bull bull
bull bull
bull
bull bull bull
bull bull
bull
bull bull
bull
322 BIOGEOGRAFiA
TABLA 44 Distribuci6n de los linajcs de mamiferos terrestres a traves de las Antillas Mayores y grupos de islas perifericas LE = La Espanola CU Cuba JA Jamaica PR = Puerto Rico IC Islas Caiman IS Islas Swan IB = Islas Bahamas IV = Islas Virgenes AS = Anguila - San Martin
FamiliaSubfumiliafIribu LE CU JA PR IC IS IB IV AS
Megalonychidae
Choloepodinae
Acratocnini
Cubanocnini
Megalocninae
Megalocnini
Mesocnini
Nesophontidae
Solenodontidae
Pitheciidae
Callicebinae
Xenothrichini
Echimyidae
Heteropsomyinae
Capromyidae
Plagiodontinae
Isolobodontinae
Hexolobodontinae
Capromyinae
H e pta x 0 don tid a e
Heptaxodontinae
Clidomyinae
Muridae
Sigmodontinae
PILOSA
SORICOMORPHA
PRIMATES
RODENTIA
bull
bull
bull
bull TOTALES 12 10 4 4 2
323 EXTlICIOIES DEL CUATERNARIO
El gigantismo ha sido sin duda una tendencia evolucionaria recurrente entre los clados mayores de mamiferos antillanos Es un hecho bien conocido que algunos grupos de mamiferos han aumentado de tamafio en las islas y varios autores han intentado explicarlo a partir de enfoques diferentes (Foster 1964 Miller 1966 Sondaar 1977 Case 1978 Eisenberg y Wilson 1978 Heaney 1978 Lawlor 1982 Lomolino 1985 Angerbjom 1985 McNab 19942001)
EXTINCIONES DEL CUATERNARIO
Las islas que integran la Subregion Antillana han perdido casi 90 de sus mamiferos terrestres durante los ultimos milenios y en proporcion con su extension territorial esta perdida ha sido mayor en la Subregion que en cualquier otro lugar del planeta (MacPhee 1996b) Las extinciones del Cuatemario eliminaron mas de las tres cuartas partes de la fauna de mamiferos terrestres conocida de las Antillas Mayores sus islas satelites y grupos de islas perifericas y el examen de este proceso en sus aspectos mas generales se aborda en esta ultima parte del presente capitulo
CRONOLOGiA Y CONSECUENCIAS
Es muy improbable que un fosil represente el ultimo individuo de un taxon extinto y entonces hay que guiarse por la edad radiometrica del fosil mas reciente conocido lIamada fecha de ultima presencia- como unica aproximacion posible a la fecha de extincion del taxon
Como se explico al inicio del capitulo (vease Contexto Temporal) el registro fosH de los mamiferos terrestres de las Antillas Mayores adolece de insuficiente sustentacion radiometrica No obstante la distribucion temporal de los escasos fechados disponibles para el Cuaternario de la region abarca etapas criticas del proceso de extincion experimentado por la referida fauna a 10 largo de los ultimos 250 mil afios
De las 52 especies que es posible reconocer actualmente como miembros extintos de la fauna cuatemaria de mamiferos terrestres de las Antillas Mayores y grupos de islas perifericas los restos fosiles de 23 de elIas (442) han sido fechados radiometricamente o se han hallado asociados estratigraficamente de manera inequivoca con otro material fechado De estos 23 fechados 3 corresponden al interglacial Sangamon 4 pertenecen a la secuencia Pleistoceno terminal - Holoceno temprano 10 se ubican en ellapso entre el arribo de los aborigenes y la incursion europea y 6 son postcolombinos (a los cuales hay que sumar otras dos especies extinguidas en los siglos XIX y xx) A partir de estas 25 especies avaladas por fechas de ultima presencia distribuidas entre los cuatro intervalos temporales sefialados hemos compuesto la Tabla 45 en la que ademas aparecen las restantes 27 especies (carentes de fechado radiometrico) ubicadas en el intervalo temporal mas probable sobre la base de (1) la asociacion estratigrafica de sus restos con evidencias culturales (amerindias 0 europeas) 0 (2) la ausencia de dicha asociacion Sobre estos criterios de ubicacion temporal procede hacer dos aclaraciones
324 BIOGEOGRAFIA
TABLA 45 Intervalos temporales mas probables para las extinciones cuatemarias de los mamiferos terrestres de las Antillas Mayores y grupos de islas perifericas sobre la base de las evidencias siguientes (l) ultimo ejemplar colectado u observado en vida (2) fechado radiometrico mas tardio en material de la especie (3) asociaci6n estratigrafica mas tardia con otro material fechado (4) asociaci6n estratigrafica con restos de animales introducidos restos humanos u objetos culturales propios del intervale (5) falta de asociaci6n con evidencias culturales [Los ntimeros entre parentesis en la columna Evidencia significan adscripci6n tentativa al intervalo] Los estimados del peso corporal (en kg) estan tomados de la literatura 0 inferidos comparativamente y distribuidos entre
las clases siguientes (a) S 01 (b) 01 a lt 1 (c) 1 a lt 10 (d) 10 a lt 100 (e) 100 a lt 200 (j) ~ 200 Las
abreviaturas que designan las islas son las de la Tabla 44
F amilialEspecie Peso IsJa Evidencia Fuente
I httervalo htterg1acial Sangamon (gt80 kaAP)
HEPTAXODONTlNAE
Amblyrhiza inundata e AS 3 McFarJane et al 1998a
CLIDOMYINAE
Clidomys osborni d JA 3 Ibid
Clidomys parvus d JA 3 Ibid
II httervalo Pleistoceno terminal ~ Holoceno temprano (20-6 kaAP)
MEGALONYCHIDAE
Aeratoenus antillensis d CD 5 Arredondo 1975 a
Aeratoenus odontrigonus d PR 3 McFarJane 1999b
Aeratoenus simorhynehus d LE 2 Rega et al 2002
M egaloenus zile f LE 2 MacPhee et al 2000
SOLENODONTIDAE
Solenodon arredondoi e CD 5 Morgan y Ottenwalder 1993
PnHECIIDAE
Paralouatta varonai d CD 5 MacPhee 1996a
EcHIMYIDAE
Heteropsomys antillensis c PR 5 McFarJane 1999b
Heteropsomys insulans c PR 5 Ibid
Puertoricomys eorozalus c PR 5 Woods 1989a
CAPROMYIDAE
P lagiodontia araeum c LE 5 MacPhee et al 2000
Macrocapromys latus c CD 5 Varona y Arredondo 1979
(Continua)
325 EXTINCIONES DEL CUATERNARIO
TABLA 45 (Continuaci6n)
F ami1ia1Especie Peso Isla Evidencia Fuente
M acrocapromys latus c CU 5 Varona y Arredondo 1979
Mesocapromys kraglievichi c CU 5 Ibid
HEPTAXODONTINAE
Xaymaca fulvopulvis a JA 2 MacPhee y Flemming 2003
III Intervalo Amerindio (6-05 kaAP)
MEGALONYCHIDAE
Acratocnus ye d LE 2 MacPhee et aI 2000
Neocnus comes c LE 2 Ibid
N eocnus dousman c LE 2 Ibid
N eocnus toupiti c LE 2 Ibid
N eocnus gliriform is c CU 4 Cordova et ai 1997
M egalocnus rodens f CU 2 MacPhee et ai 2007
Parocnus browni e CU 2 Steadman et ai 2005
P arocnus serus e LE 2 MacPhee et ai 2000
N ESOPHONTIDAE
N esophontes edithae a PR 2 McFarlane 1999b
PITHECIIDAE
Antillothrix bernensis c LE 3 Rimoli 1977
EcHIMYIDAE
Brotomys contractus a LE 4 Miller 1929c
CAPROMYIDAE
P lagiodontia spelaeum c LE 4 Ibid
M acrocapromys acevedo c CU 4 NUiiez 1990a
HEPTAXODONTINAE
Elasmodontomys obliquus d PR 3 McFarlane 1999b
Iv Intervalo Postcolombino (05 kaAP - Presente)
N ESOPHONTIDAE
N esophontes hypom icrus a LE 24 MacPhee y Fle111111ing 1999
N esophontes zam icrus a LE (4) Ibid
N esophontes m icrus a CU 24 MacPhee et aI 1999
(Continua)
326 BIOGEOGRAFiA
TABLA 45 (Continuacion)
F amilialEspecie Peso Isla Evidencia Fuente
N esophontes m icrus a CD 24 MacPhee et aI 1999
N esophontes m icrus a LE 4 MacPhee et aI 2000
N esophontes sp 1 a IC 4 Morgan 1994
N esophontes sp 2 a IC 4 Ibid
Solenodon marcanoi b LE 4 Ottenwalder 2001
PITHECIIDAE
Xenothrix mcgregori c JA 4 MacPhee y Flemming 1999
EcHIMYIDAE
Boromys offeUa a CD 24 Guarch 1984
Boromys torrei a CD 24 Ibid
Brotomys voratus a LE 4 MacPhee et aI 2000
CAPROMYIDAE
P lagiodontia ipnaeum c LE 4 MacPhee y Flemming 1999
Rhizoplagiodontia lemkei c LE 4 Ibid
Isolobodon portoricensis c PR 24 Ibid
Isolobodon montanus c LE 4 Ibid
Hexolobodon phenax c LE (4) Ibid
Capromys sp c IC 24 Morgan 1994
Geocapromys columbianus c CD 4 MacPhee y Flemming 1999
Geocapromys thoracatus b IS 1 MacPhee y Marx 1997
Geocapromys sp 1 c IC 4 Morgan 1994
Geocapromys sp 2 c IC (4) Ibid
l-IEPTAXODONTINAE
Quemisia gravis c LE (1) MacPhee y Flemming 1999
MURIDAE
Oryzomys antillarum a JA 1 Ibid
1 Flemming y MacPhee (1999) alertaron en contra de la asociacion con otro material fechado Asimismo Morgan (1993) indico que la ausencia de asociacion con evidencias culturales amerindias no es necesariamente un criterio concluyente para ubicar un taxon en el intervalo temporal anterior porque hay depositos en la region que carecen de tales evidencias y son del Holoceno tardio por fechados radiocarbonicos [No obstante
327 EXTlNCIONES DEL CUATERNAR10
la ausencia de una especie en residuarios arqueol6gicos adquiere mayor certidumbre como criterio de ubicaci6n temporal de su extinci6n en la medida que aumenta el numero de residuarios estudiados que en las Antillas Mayores se cuentan por centenares] A falta de fechado absoluto de una especie consideramos que estos criterios de ubicaci6n temporal son mas utiles que la alternativa de excluir del analisis a la especie
2 En los trabajos sobre biogeografia de mamiferos antillanos los arque610gos mas citados como autoridades para establecer el inicio de la presencia aborigen en las Antillas Mayores (Kozlowski 1974 Rouse y Allaire 1978 Rouse 1989 1992 Wilson 2001) difieren al respecto entre los 6 kaAP y los 45 kaAP (pero vease Pagan et al 200523) y hemos adoptado la mas temprana de estas fechas para el comienzo del intervalo amerindio en la Tabla 45 De todos modos cualquiera que haya sido la fecha esta no es aplicable por igual al conjunto de las Antillas Mayores porque segun algunos de los autores citados el poblamiento comenz6 por Cuba y La Espanola 6 mil anos atras continu6 en Puerto Rico hace alrededor de 3 mil anos (pero vease Burney et al 1994) yen Jamaica hace menos de 2 mil e incluso a las Islas Caiman ni siquiera llegaron [Las Islas Caiman descubiertas por Col6n en 1503 carecen absolutamente de evidencia amerindia (Stokes y Keegan 1996)]
Si los datos de la Tabla 45 representan una aceptable aproximaci6n a la realidad el proceso de las extinciones cuaternarias de los mamiferos terrestres de las Antillas Mayores y grupos de islas perifericas debi6 caracterizarse entre otros por los eventos siguientes
Aunque obviamente la fauna cuaternaria debi6 ser mucho mas numerosa que 10 que refleja el registro paleontol6gico (Morgan y Woods 1986) 52 especies (81 de la fauna conocida) desaparecieron (sin reposici6n) durante los ultimos 250 mil anos
- Al parecer las extinciones se concentraron en las especies de mayor tamano Asi el peso promedio estimado para las especies de la etapa preantr6pica (intervalos I + II de la Tabla 45) es 350 kg es 328 kg para las especies del intervalo III (amerindio) y es s610 25 kg (c1ases del peso a b c) para las especies del intervalo IV (postcolombino) Ninguna especie aut6ctona de mamifero mayor de 6 kg existe hoy en la regi6n
- De acuerdo con la distribuci6n temporal de las extinciones 61 especies sobrevivieron al interglacial Sangamon (intervalo 1)48 sobrevivieron a la transici6n del Pleistoceno terminal al Holoceno temprano (intervalo II) 34 sobrevivieron al periodo amerindio (intervalo III) y s610 12 especies han sobrevivido desde la Ilegada de los europeos (intervalo IV) Las tasas de extinci6n correspondientes (exceptuado el intervalo I) son una especie cada 1167 anos en el intervalo II una cada 393 anos en el intervalo III y una cada 23 anos en el intervalo IV
- Durante todo el proceso de las extinciones cuaternarias la perdida de especies por isla fue de 91 en La Espanola 83 en Jamaica y 62 en Cuba Bahamas retuvo su unica especie (aunque extirpada en varias de las islas) Puerto Rico e Islas Caiman perdieron todas sus especies (6 y 5 respectivamente) y las Islas Swan Virgenes y Anguila - San Martin perdieron sus respectivas especies (micas
328 BIOGEOGRAFiA
De los cuatro 6rdenes de mamiferos representados en la fauna Pilosa y Primates desaparecieron totalmente Soricomorpha y Rodentia perdieron 86 y 74 de sus especies respectivamente
CAUSALIDAD
CAUSAS NATURALES
Los cicIos glaciales e interglaciales acaecidos en el Pleistoceno causaron cambios ambientales dnisticos en las Antillas con la concomitante alternancia de cIimas frios (secos) y calidos (hUmedos) asi como de descensos y ascensos del nivel del mar aumento y disminuci6n de las areas territoriales) induso (aunque con menor intensidad) en el Holoceno Estos eventos paleogeograficos han sido consistentemente documentados (Mayo y Kartashov 1972 Bonnatti y Gartner 1973 Ortega y Arcia 1982 Prance 1982 Schubert y Medina 1982 Dodge et al 1983 Ortega [1984] Haq et al 1987 Schubert 1989 Hodell et al 1991 Guilderson et al 1994 Oppo 1997 Emslie 1998 Curtis et al 2001 Iturralde-Vinent 2003) y de ellos brindamos a continuaci6n una breve resena
Finalizando el periodo interglacial anterior a la ultima glaciaci6n (Wisconsin) el mar estaba a 8-10 m por encima de su nivel actual Cuando el avance glacial del Wisconsin a1canz6 su culminaci6n (eillamado Ultimo Maximo Glacial) hace 18 ka el mar habia descendido 120 m con respecto a dicho nivel y las islas del Caribe habian incrementado considerablemente sus extensiones subaereas en comparaci6n con las que presentan hoy dia por ejemplo Puerto Rico y las Islas Virgenes se conectaron territorialmente y las Bahamas se unieron en cinco gran des islas Prevalecia un regimen frio y seco y condiciones de aridez --debidas a una reducci6n de las precipitaciones (posiblemente a menos de 440 mmlano)- y los ambientes xericos se habian extendido a traves de las islas y bancos emergidos y abundaban los pinares (los habitats mesicos estaban restringidos a parcelas relictas en 10 alto de las montanas) En el Mar Caribe y el Golfo de Mexico la temperatura superficial del mar era 5degC mas baja que la actual y en las llanuras costeras y alturas maximas eran respectivamente 2-3degC y 6degC mas bajas Los limites de las zonas ecol6gicas altitudinales descendieron varios cientos de metros y cada zona redujo su extensi6n vertical al aumentar el gradiente de temperatura Ellimite inferior de la zona periglacial
elevaci6n donde la temperatura media minima a1canza los OdegC) de la Cordillera Central de la Republica Dominicana (altura maxima del Caribe insular con evidencia de topografia glacial) se considera que estuvo entre los 1500 y 1800 mlnm (actualmente esta a 2800 mlnm) y se puede estimar que dicho limite era inferior en Cuba por estar mas al norte que la Republica Dominicana No obstante durante este periodo la temperatura experiment6 episodios cortos (unas pocas decadas) de ascensos repetidos
Un signifieativo componente bi6tieo de las referidas condiciones paleoambientales 10 constituy6 la espectacular irradiaci6n evolucionaria experimentada por un grupo numeroso de aves rapaees diurnas y nocturnas (Wetmore 1937 Brodkorb 1969 Kurotchkin y Mayo 1973 Arredondo 1976 Olson 1978 Olson y Hilgartner 1982
329 EXTINCIONES DEL ClJATERNARIO
Suarez 2000a b 2004 Suarez y Emslie 2003) Los mamiferos de las Antillas Mayores ofrecieron a estos depredadores aviares una amplia gama de tamanos de presa que en ausencia de mamiferos del orden Carnivora en la region permitio a estas aves irradiarse a este nicho alimentario con la evolucion de una variedad de especies de muy diversa tall a corporal incluidas formas gigantes Adquirir un cuerpo gigante les propicio el acceso a recursos alimentarios que no estaban siendo explotados por ningun otro depredador (Alcover y McMinn 1994) A Cuba corresponde el mejor registro fosil de rapaces cuaternarias caribenas con siete generos de especies diurnas Dentro de estas aves existieron tres gavilanes gigantes (Accipitridae) tres carairas (Falconidae) y un condor (Cathartidae) asi como cuatro generos de especies nocturnas dos lechuzas (Tytonidae) y tres Mhos (Strigidae) dos de ell os no igualados en tamano en todo el mundo Algunos de estos generos tuvieron representacion en otras islas antillanas como La Espanola Jamaica Bahamas y Gran Caiman
Al retirarse hacia el norte los hielos de la glaciacion Wisconsin el clima se tomo mas hUmedo en las Antillas durante el Holoceno temprano y el bosque mesico se expandio en la region insular El nivel del mar ascendio acompanado de mayor pluviosidad alrededor de los 7 kaAP las Islas Virgenes quedaron separadas entre si y de Puerto Rico y las Bahamas perdieron la mayor parte de su territorio Cerca de los 3 kaAP las condiciones secas retornaron a la region e hicieron desaparecer buena parte del bosque mesico Nuevamente el clima comenzo a adquirir sus condiciones actuales hacia los 25 kaAP aproximadamente y hoy transita por una fase mesica con vegetacion xerica confinada en parcelas relictas
Sin duda estos y otros eventos naturales incidieron negativamente en la sobrevivencia de mamiferos terrestres en las Antillas Mayores asi como en los grupos de pequenas islas perifericas (Pregill y Olson 1981 Olson 2006)
McFarlane et al (1998a) demostraron convincentemente como Amblyrhiza el gigante de los roedores antillanos endemico de la pequena isla de Anguila sucumbio a las repetidas y abruptas perdidas de su habitat durante el interglacial Sangamon por las nipidas y severas oscilaciones del nivel del mar (documentadas por Neumann y Hearty 1996) al no poder responder adecuadamente a tan catastroficos eventos mediante el ajuste de su masa corporal media y otro tanto debio ocurrirle en Jamaica al otro roedor gigante Clidomys (McFarlane et al 1998b) emparentado con Amblyrhiza en la misma epoca (Tabla 45 intervalo I)
El ascenso del nivel del mar seguramente promovio la extincion de mamiferos durante la transicion del Pleistoceno al Holoceno (Tabla 45 intervalo II) con la reduccion del tamano de las Islas por las transgresiones marinas (Pregill y Olson 1981 Morgan 1993) Esto ultimo haria maximamente severa la competicion asf como la presion de los depredadores [Steadman et al (2005) no yen por que el calentamiento global ocurrido durante la transicion Pleistoceno-Holoceno habria de tener un efecto negativo sobre los perezosos siendo sus descendientes actuales especies tropic ales 0 subtropicales pero no se trata del efecto
330 BIOGEOGRAFiA
directo del cambio de temperatura sobre los animales sino del efecto indirecto y progresivo que dicho cambio genera a largo plazo al reducir el territorio insular disponible]
Por esta epoca tambien desaparecieron especies de tall a mayor como Megalocnus zile en La Espanola y otras de menor tamano (en Cuba) pero que representan la tall a maxima en sus respectivos contextos taxonomicos del Cuaternario como Solenodon arredondoi Paralouatta varonai y Macrocapromys acevedo Un mamifero puede ser grande solo a costa de reducir su densidad poblacional y esto 10 hace mis vulnerable a la extinci6n sobre todo en una isla (McNab 1994 McFarlane et al 1998a) Incidentalmente Alcover et al (1998a) afirmaron que larelacion entre la talla corporal y la extincion no siempre se cumple en las especies de menor tamano y citaron como ejemplo que entre las musarafias todas las especies de Nesophontes desaparecieron mientras sobreviven especies de Solenodon pero la comparacion correcta en todo caso serfa entre los solenodones vivientes y el gigante Solenodon arredondoi y entre Nesophontes edithae -el gigante de los nesofontes extinguido en el periodo amerindio- y el resto de sus congeneres que sobrevivieron hasta los primeros siglos de la colonizacion europea (Tabla 45 intervalos III y IV)
CAUSAS ANTROPOGENICAS
Intervalo Amerindio
La mayoria de los autores que se han referido a las extinciones acaecidas durante este intervalo (entre elIos paradigmaticamente Martin 1984a b) han responsabilizado a los aborfgenes (que se supone comenzaron a poblar las Antillas Mayores hace 6 mil afios) con la desaparicion de numerosas especies de mamiferos terrestres en la region principalmente por la via de la sobrecaza y la introduccion de animales domesticos devenidos depredadores de la fauna autoctona (Morgan y Woods 1986) Este efecto negativo de la presencia indigena sobre los mamiferos de la region ha sido dramatizado por algunos autores en terminos de holocausto faunal Sin embargo pensamos que se ha exagerado Como han expresado MacPhee et al (19993) cuando uno busca evidencia dura de complicidad humana directa en [estas] extinciones no hay mucho que encontrar Las siguientes son algunas consideraciones a tener en cuenta en tal sentido
El registro radiometrico mas joven conocido hasta ahora para un perezoso antillano (MegalocnllS rodens) es de 42 kaAP (MacPhee et ai 2007) Si se acepta que los aborfgenes entraron en Cuba hace aproximadamente 6 mil anos su convivencia con los perezosos debe haber durado alrededor de 1800 afios (0 mas porque dificilmente el referfdo fechado radiometrico haya correspondido al ultimo sobreviviente de la especie) Esta cifra contradice la hipotesis del holocausto faunal segun la cualla extincion de los mamiferos insulares de mayor talla debe ocurrir con extrema rapidez -a escala de decadas 0 de centurias- a partir de su primer contacto con los humanos (Martin y Steadman 1999) A pesar de las repetidas afirmaciones en contrario (Harrington 1921 Miller 1929c Herrera [1945] Tabio 1951 Alvarez 1951a 1957 Arredondo
331 EXTINCIONES DEL CUATERNARIO
1959 1960a Veloz y Ortega 1973 Mayo 1980b Rodriguez et al 1984 Castellanos y Pino 1985 Pino y Castellanos 1985 Nunez 1990b Gutierrez 1996 Arredondo 2004) en su mayoria basadas en evidencias no exentas de posibles accidentes de deposicion (0 de reubicacion postdeposicional) en recintos cavernarios (Diaz-Franco [2006b D no se ha demostrado inequivocamente que los aborigenes antillanos se alimentaran de perezosos regularmente Sus restos en residuarios de alimentos son escasisimos en toda la region y esto contrasta con los abundantes restos de jutias y otros mamiferos de menor talla que aparecen en esos residuarios demostrativos de la preferencia de los paleoindios por estos otros mamiferos
Los mamiferos mas importantes en la economia prehistorica de las Antillas Mayores fueron las jutias Capromys y Geocapromys en Cuba Geocapromys en Jamaica e Isolobodon en La Espanola Puerto Rico e Islas Virgenes (Aguayo 1950b Tablo 1951 Rimoli 1972 Wing 2001 Diaz-Franco [2006b D Wilkins (2001) estudi61ajutia viviente de Jamaica (Geocapromys browni) en un sitio ocupado por los aborigenes durante 100 anos y no encontr6 evidencias de que durante ese tiempo la especie haya sido sobreexplotada Asimismo en Cuba las jutias conga carabali y andaraz son escasas en depositos pleistoholocenicos en cambio son muy numerosas en residuarios indigenas y de haber sido sobreexplotadas 10 mas probable es que no hubieran sobrevivido hasta hoy
La habilidad de los humanos para penetrar en profundidad el bosque tropical y mantenerse en este sustentablemente ha sido un evento comparativamente reciente (Beran y Steele 1994) En el caso antillano la caza tierra adentro ha sido explicada como un proceso tardio de desplazamiento a partir de la explotacion de abundantes recursos marinos que ofrecian condiciones 6ptimas para el sedentarismo de las comunidades aborigenes costeras (C6rdova y Arredondo 1988 Brito 2005) Peces y mariscos dominan la arqueologia costera de las Islas antillanas en tanto el consumo de recursos terrestres (mamfferos incluidos) aumenta progresivamente en la medida que los residuarios estudiados se alejan de la costa (Scudder 1991 Wilkins 2001) En las Antillas Mayores el componente terrestre de la dieta aborigen es solo 20-30 del material recobrado en los residuarios (Wing 1977 1989) Y debio ser menor alin porque el componente vegetal de la dieta no se ha preservado Por otra parte los aborigenes cubanos domesticaron la jutia conga y seguramente la reprodujeron para comer (Pose et al 1988) practica que probablemente se repitiera con otras jutias en las demas Antillas Mayores
- Fray Bartolome de las Casas hablo de 3 millones de indios en Cuba al inicio de la colonizacion pero los historiadores subsecuentes consideraron exagerada la cifra (Poey 1851 a) No obstante se dispone de dos estimados de la densidad de poblaci6n aborigen en las Antillas Mayores a la llegada de los europeos 18 indigenaslkm2 en Puerto Rico (Rouse 1956) y 1271km2 en La Espanola (Watts 1987) Extrapolando ambos valores entre las cuatro islas principales de las Antillas Mayores tenemos que en la fecha indicada la poblaci6n aborigen pudo estar entre 16 mil y 113 mil indigenas en Puerto Rico entre 21 mil y 145 mil en Jamaica entre 141 mil y 1 mill6n en La Espanola y entre 200 mil y 14 millones en Cuba Entre las cifras maximas y las minimas nos inclinamos
332 BIOGEOGRAFiA
hacia estas ultimas atendiendo a los altos indices de mortalidad infantil y morbilidad que se evidencian en las osamentas de los pobladores prehispanicos (Martinez y Rives 1990) Uno de los hechos que apuntan hacia el bajo numero de aborigenes en las Antillas Mayores es 10 rapido que desaparecieron despues de la llegada de los europeos pnkticamente sin dejar mestizaje con estos 10 cual contrasta ostensiblemente con la situacion en la America continentaL
En suma un grupo humano (1) demograiicamente irrelevante y controlado por altos indices de morbilidad (2) consumidor mayoritariamente de organismos marinos y (3) carente de la tecnologia cinegetica necesaria para poder sobreexplotar los mamiferos que Ie sirvieron de alimento (a los cuales accedio tardiamente en su desarrollo) no es concebible que haya causado e1 holocausto faunal que se Ie atribuye a partir del error de aplicar a las comunidades aborigenes antillanas un modelo cultural de la prehistoria continentaL
Tampoco hay evidencias de que los aborigenes hayan introducido carnivoros con suficiente capacidad depredadora salvo el perro Morgan y Otlenwaldder (1993) consideraron que la depredacion por perros durante el periodo amerindio es la explicacion mas plausible para la extincion del almiqui gigante de Cuba pero como se puede observar (Tabla 54) Solenodon arredondoi aparece en el intervalo II por falta de asociacion con evidencias culturales
Consecuentemente el accionar de los aborigenes y de sus animales domesticos directamente sobre la fauna no parece haber sido por sf solo un factor determinante de la extincion de mamiferos terrestres durante el periodo amerindio en las Antillas (pero vease Steadman et al 2005)
Hay que recordar en este senti do que alin en el Holoceno tardio ocurrieron cambios clirruiticos de consideracion (vease Causas Naturales) Por ejemplo Curtis et al (2001) explicaron que la extincion de algunos ani males mesofilicos de las alturas montanosas de La Espanola pudo haber antecedido a cualquier impacto humano sigrIificativo en esos lugares por coincidir dicha extincion con una tendencia a la aridez documentada isotopicamente y con evidencias de la perdida del bosque mesico ocurridas despues de 3kaAP Yen Jamaica hace solo 2shy3 mil aDos el area de la Bahia de Jackson sustentaba una gran biomasa de vertebrados en bosque hinnedo a juzgar por la cantidad y diversidad de taxones presentes en material fosil derivado de la depredacion por lechuzas hoy el area es la mas seca de la isla con matorral xerofitico y pobreza de fauna (MacPhee 1996a) Yalm en nuestros dias huracanes de gran intensidad aniquilan poblaciones de vertebrados (que no se recuperan) en is las pequenas y llanas del Caribe (Brooks y Smith 2001)
En medio de la incertidumbre sobre la verdadera causa (0 causas) de las extinciones de mamiferos en el periodo amerindio MacPhee y Marx (1997) han propuesto su hipotesis de la hipermorbilidad la cual estipula colapsos poblacionales relativamente rapidos con la introduccion de enfermedades de alta letalidad Estos autores han razonado que
Las islas son los lugares mas apropiados para experimentar perdidas catastroficas causadas por enfermedades Los mamiferos no solo serian susceptibles en virtud de su prolongado aislamiento biologico sino que en los confines de una isla la epizootia
333 EXTINCIONES DEL CUATERNARIO
tendria la potencialidad de infectar simultaneamente a la rnayoria de los individuos de vadas especies Los neonatos son inmunologicamente inmaduros y sufririan desproporcionadamente cualquier infeccion hipervirulenta La perdida excesiva de neonatos y juveniles tendria efectos severos para la sobrevivencia de la poblacion si esta se reduce por debajo de cierto nivel sobreviene la extincion porque no hay posibilidad de restitucion en el periodo postpanzootico Las especies de mayor talla corporal van a ser las mas vulnerables a la extincion dadas sus bajas tasas reproductivas largos periodos de gestacion y reducido numero de progenie Las especies de menor tamano --caracterizadas por altas tasas reproductivas periodos cortos de gestacion y mayor numero de neonatos por parto- tambien serian fuertemente diezmadas pero muchas sobrevivirian porque su estrategia reproductiva les permite perder mayor numero de individuos Las perdidas deben ocurrir relativamente rapido (a escala de decadas) mientras que otras hipotesis (digamos los cambios climaticos) permiten intervalos mucho mas largos (a escala de siglos 0 de milenios) entre la aparicion de las causas y la produccion de sus efectos Por ejemplo todo indica que los nesofontes se extinguieron muy rapidamente y seria dificil poder invocar un impacto humano directo 0 argumentar fenomenos climaticos que pudieran explicar plausiblemente perdidas tan rapidas
MacPhee y Marx (1997 191) han dicho que por el momento la hipotesis de la hipermorbilidad no pasa de ser una abstraccion en espera de adecuada sustentacion empirica Llama la atencion que linicamente en el periodo amerindio (Tabla 45 intervalo 1Il) las extinciones abarcaron todas las clases del peso corporal (de la a a laj) y que ninguna de las especies que sobrevivieron hasta el periodo postcolombino (Tabla 45 intervalo IV mas las especies vivientes) sobrepasa los 10 kg (clases del peso a b c) en clara coincidencia con los presupuestos de la hipotetica hipermorbilidad
Intervalo Postcolornbino
Por 10 menos 37 especies de mamiferos terrestres vivian en las Antillas Mayores y grupos de Islas perifericas a la lIegada de los europeos en 1492 De elIas se han extinguido 23 shymas especies que en cualquiera de los periodos precedentes (Tabla 45~ y solo subsisten 12 especies repartidas entre Cuba (8 especies) La Espanola (2) Jamaica (1) y Bahamas (1) varias de estas en peligro critico de extincion Solo dos de las especies extinguidas durante los ultimos 500 anos se conocieron en estado viviente los roedores Oryzomys antillarum de Jamaica (extinguido en 1877) y Geocapromys thoracatus de las Islas Swan (extinguido en 1955) Las causas de las extinciones ocurridas durante este periodo son sobradamente conocidas destruccion de habitats introducci6n de animales exoticos (domesticos y comensales) y demas presiones asociadas a la actividad humana entre las cuales posiblemente habria que incluir brotes de hipermorbilidad
Durante mucho tiempo se ha afirmado la posibilidad de que especies de Nesophontes hayan sobrevivido hasta las primeras decadas del siglo xx y que incluso continuen existiendo en lugares remotos de Cuba 0 La Espanola (Allen 1918 1942 Miller 1930 Aguayo 1950a Arredondo 1960b Fischer 1977 MacFadden 1980 Woods y
334 BIOGEOGRAFiA
Ottenwalder 1991) Actualmente no existe evidencia radiometrica de que alguna especie de Nesophontes haya sobrevivido al siglo xv las fechas radiocarbonicas calibradas obtenidas en muestras de Nesophontes de Cuba y La Espanola resultaron no mas tardias que los siglos XIII y XIV (MacPhee et al 1999) Por otra parte las expediciones efectuadas a fines del siglo xx con el proposito de localizar individuos vivientes del genero en La Espanola no produjeron la menor indicacion afirmativa al respecto (Woods et al 1985) Ademas a traves de las Antillas Mayores no aparecen restos de Nesophontes en los depositos formados actualmente por las lechuzas
EPiLOGO
Las Antillas Mayores son islas oceanicas no han formado parte de un continente (con la posible excepcion de una parte de Jamaica hasta el Eoceno Medio hace 45 Ma) y durante un tiempo geologico considerable (desde el Eoceno Superior hace 40 Ma) han permanecido emergidas y separadas de los continentes vecinos Consecuentemente los mamiferos terrestres (no volantes) autoctonos de estas islas constituyen una fauna oceanica que por ende cumple con todas las caracteristicas convencionales reconocidas para este tipo de entidad biogeografica
- Baja diversidad taxonomica a nivel de las categorias superiores (solo cuatro ordenes) en comparacion con islas plataformicas continentales (0 con areas continentales) de tamano y diversidad ecologica simi lares
- Alto grado de endemismo a nivel de genero (96) y familia (50) en respuesta al prolongado aislamiento
- Desbalance ecologico de la estructura trofica ausencia de mamiferos camivoros y ungulados
- Espectaculares procesos de irradiaci6n evolucionaria como resultado de la colonizacion de territorios libres de competidores y depredadores
Tendencias evolucionarias extremas gigantismo
- Control a cargo de aves rapaces con tendencia at gigantismo en ausencia de mamiferos miembros del orden Carnivora
- Mas especies de reptiles y de murcielagos que de mamiferos terrestres relaciones que en los continentes se invierten
Alta tasa de extincion de especies (84) sin reposicion
Preservacion de especies filogeneticamente arcaicas (los llamados fosiles vivientes) los almiquies

bull bull
bull bull
bull bull bull
bull bull
bull bull
bull
bull
bull
bull
bull bull bull bull
bull bull
bull bull
bull
bull bull bull
bull bull
bull
bull bull
bull
322 BIOGEOGRAFiA
TABLA 44 Distribuci6n de los linajcs de mamiferos terrestres a traves de las Antillas Mayores y grupos de islas perifericas LE = La Espanola CU Cuba JA Jamaica PR = Puerto Rico IC Islas Caiman IS Islas Swan IB = Islas Bahamas IV = Islas Virgenes AS = Anguila - San Martin
FamiliaSubfumiliafIribu LE CU JA PR IC IS IB IV AS
Megalonychidae
Choloepodinae
Acratocnini
Cubanocnini
Megalocninae
Megalocnini
Mesocnini
Nesophontidae
Solenodontidae
Pitheciidae
Callicebinae
Xenothrichini
Echimyidae
Heteropsomyinae
Capromyidae
Plagiodontinae
Isolobodontinae
Hexolobodontinae
Capromyinae
H e pta x 0 don tid a e
Heptaxodontinae
Clidomyinae
Muridae
Sigmodontinae
PILOSA
SORICOMORPHA
PRIMATES
RODENTIA
bull
bull
bull
bull TOTALES 12 10 4 4 2
323 EXTlICIOIES DEL CUATERNARIO
El gigantismo ha sido sin duda una tendencia evolucionaria recurrente entre los clados mayores de mamiferos antillanos Es un hecho bien conocido que algunos grupos de mamiferos han aumentado de tamafio en las islas y varios autores han intentado explicarlo a partir de enfoques diferentes (Foster 1964 Miller 1966 Sondaar 1977 Case 1978 Eisenberg y Wilson 1978 Heaney 1978 Lawlor 1982 Lomolino 1985 Angerbjom 1985 McNab 19942001)
EXTINCIONES DEL CUATERNARIO
Las islas que integran la Subregion Antillana han perdido casi 90 de sus mamiferos terrestres durante los ultimos milenios y en proporcion con su extension territorial esta perdida ha sido mayor en la Subregion que en cualquier otro lugar del planeta (MacPhee 1996b) Las extinciones del Cuatemario eliminaron mas de las tres cuartas partes de la fauna de mamiferos terrestres conocida de las Antillas Mayores sus islas satelites y grupos de islas perifericas y el examen de este proceso en sus aspectos mas generales se aborda en esta ultima parte del presente capitulo
CRONOLOGiA Y CONSECUENCIAS
Es muy improbable que un fosil represente el ultimo individuo de un taxon extinto y entonces hay que guiarse por la edad radiometrica del fosil mas reciente conocido lIamada fecha de ultima presencia- como unica aproximacion posible a la fecha de extincion del taxon
Como se explico al inicio del capitulo (vease Contexto Temporal) el registro fosH de los mamiferos terrestres de las Antillas Mayores adolece de insuficiente sustentacion radiometrica No obstante la distribucion temporal de los escasos fechados disponibles para el Cuaternario de la region abarca etapas criticas del proceso de extincion experimentado por la referida fauna a 10 largo de los ultimos 250 mil afios
De las 52 especies que es posible reconocer actualmente como miembros extintos de la fauna cuatemaria de mamiferos terrestres de las Antillas Mayores y grupos de islas perifericas los restos fosiles de 23 de elIas (442) han sido fechados radiometricamente o se han hallado asociados estratigraficamente de manera inequivoca con otro material fechado De estos 23 fechados 3 corresponden al interglacial Sangamon 4 pertenecen a la secuencia Pleistoceno terminal - Holoceno temprano 10 se ubican en ellapso entre el arribo de los aborigenes y la incursion europea y 6 son postcolombinos (a los cuales hay que sumar otras dos especies extinguidas en los siglos XIX y xx) A partir de estas 25 especies avaladas por fechas de ultima presencia distribuidas entre los cuatro intervalos temporales sefialados hemos compuesto la Tabla 45 en la que ademas aparecen las restantes 27 especies (carentes de fechado radiometrico) ubicadas en el intervalo temporal mas probable sobre la base de (1) la asociacion estratigrafica de sus restos con evidencias culturales (amerindias 0 europeas) 0 (2) la ausencia de dicha asociacion Sobre estos criterios de ubicacion temporal procede hacer dos aclaraciones
324 BIOGEOGRAFIA
TABLA 45 Intervalos temporales mas probables para las extinciones cuatemarias de los mamiferos terrestres de las Antillas Mayores y grupos de islas perifericas sobre la base de las evidencias siguientes (l) ultimo ejemplar colectado u observado en vida (2) fechado radiometrico mas tardio en material de la especie (3) asociaci6n estratigrafica mas tardia con otro material fechado (4) asociaci6n estratigrafica con restos de animales introducidos restos humanos u objetos culturales propios del intervale (5) falta de asociaci6n con evidencias culturales [Los ntimeros entre parentesis en la columna Evidencia significan adscripci6n tentativa al intervalo] Los estimados del peso corporal (en kg) estan tomados de la literatura 0 inferidos comparativamente y distribuidos entre
las clases siguientes (a) S 01 (b) 01 a lt 1 (c) 1 a lt 10 (d) 10 a lt 100 (e) 100 a lt 200 (j) ~ 200 Las
abreviaturas que designan las islas son las de la Tabla 44
F amilialEspecie Peso IsJa Evidencia Fuente
I httervalo htterg1acial Sangamon (gt80 kaAP)
HEPTAXODONTlNAE
Amblyrhiza inundata e AS 3 McFarJane et al 1998a
CLIDOMYINAE
Clidomys osborni d JA 3 Ibid
Clidomys parvus d JA 3 Ibid
II httervalo Pleistoceno terminal ~ Holoceno temprano (20-6 kaAP)
MEGALONYCHIDAE
Aeratoenus antillensis d CD 5 Arredondo 1975 a
Aeratoenus odontrigonus d PR 3 McFarJane 1999b
Aeratoenus simorhynehus d LE 2 Rega et al 2002
M egaloenus zile f LE 2 MacPhee et al 2000
SOLENODONTIDAE
Solenodon arredondoi e CD 5 Morgan y Ottenwalder 1993
PnHECIIDAE
Paralouatta varonai d CD 5 MacPhee 1996a
EcHIMYIDAE
Heteropsomys antillensis c PR 5 McFarJane 1999b
Heteropsomys insulans c PR 5 Ibid
Puertoricomys eorozalus c PR 5 Woods 1989a
CAPROMYIDAE
P lagiodontia araeum c LE 5 MacPhee et al 2000
Macrocapromys latus c CD 5 Varona y Arredondo 1979
(Continua)
325 EXTINCIONES DEL CUATERNARIO
TABLA 45 (Continuaci6n)
F ami1ia1Especie Peso Isla Evidencia Fuente
M acrocapromys latus c CU 5 Varona y Arredondo 1979
Mesocapromys kraglievichi c CU 5 Ibid
HEPTAXODONTINAE
Xaymaca fulvopulvis a JA 2 MacPhee y Flemming 2003
III Intervalo Amerindio (6-05 kaAP)
MEGALONYCHIDAE
Acratocnus ye d LE 2 MacPhee et aI 2000
Neocnus comes c LE 2 Ibid
N eocnus dousman c LE 2 Ibid
N eocnus toupiti c LE 2 Ibid
N eocnus gliriform is c CU 4 Cordova et ai 1997
M egalocnus rodens f CU 2 MacPhee et ai 2007
Parocnus browni e CU 2 Steadman et ai 2005
P arocnus serus e LE 2 MacPhee et ai 2000
N ESOPHONTIDAE
N esophontes edithae a PR 2 McFarlane 1999b
PITHECIIDAE
Antillothrix bernensis c LE 3 Rimoli 1977
EcHIMYIDAE
Brotomys contractus a LE 4 Miller 1929c
CAPROMYIDAE
P lagiodontia spelaeum c LE 4 Ibid
M acrocapromys acevedo c CU 4 NUiiez 1990a
HEPTAXODONTINAE
Elasmodontomys obliquus d PR 3 McFarlane 1999b
Iv Intervalo Postcolombino (05 kaAP - Presente)
N ESOPHONTIDAE
N esophontes hypom icrus a LE 24 MacPhee y Fle111111ing 1999
N esophontes zam icrus a LE (4) Ibid
N esophontes m icrus a CU 24 MacPhee et aI 1999
(Continua)
326 BIOGEOGRAFiA
TABLA 45 (Continuacion)
F amilialEspecie Peso Isla Evidencia Fuente
N esophontes m icrus a CD 24 MacPhee et aI 1999
N esophontes m icrus a LE 4 MacPhee et aI 2000
N esophontes sp 1 a IC 4 Morgan 1994
N esophontes sp 2 a IC 4 Ibid
Solenodon marcanoi b LE 4 Ottenwalder 2001
PITHECIIDAE
Xenothrix mcgregori c JA 4 MacPhee y Flemming 1999
EcHIMYIDAE
Boromys offeUa a CD 24 Guarch 1984
Boromys torrei a CD 24 Ibid
Brotomys voratus a LE 4 MacPhee et aI 2000
CAPROMYIDAE
P lagiodontia ipnaeum c LE 4 MacPhee y Flemming 1999
Rhizoplagiodontia lemkei c LE 4 Ibid
Isolobodon portoricensis c PR 24 Ibid
Isolobodon montanus c LE 4 Ibid
Hexolobodon phenax c LE (4) Ibid
Capromys sp c IC 24 Morgan 1994
Geocapromys columbianus c CD 4 MacPhee y Flemming 1999
Geocapromys thoracatus b IS 1 MacPhee y Marx 1997
Geocapromys sp 1 c IC 4 Morgan 1994
Geocapromys sp 2 c IC (4) Ibid
l-IEPTAXODONTINAE
Quemisia gravis c LE (1) MacPhee y Flemming 1999
MURIDAE
Oryzomys antillarum a JA 1 Ibid
1 Flemming y MacPhee (1999) alertaron en contra de la asociacion con otro material fechado Asimismo Morgan (1993) indico que la ausencia de asociacion con evidencias culturales amerindias no es necesariamente un criterio concluyente para ubicar un taxon en el intervalo temporal anterior porque hay depositos en la region que carecen de tales evidencias y son del Holoceno tardio por fechados radiocarbonicos [No obstante
327 EXTlNCIONES DEL CUATERNAR10
la ausencia de una especie en residuarios arqueol6gicos adquiere mayor certidumbre como criterio de ubicaci6n temporal de su extinci6n en la medida que aumenta el numero de residuarios estudiados que en las Antillas Mayores se cuentan por centenares] A falta de fechado absoluto de una especie consideramos que estos criterios de ubicaci6n temporal son mas utiles que la alternativa de excluir del analisis a la especie
2 En los trabajos sobre biogeografia de mamiferos antillanos los arque610gos mas citados como autoridades para establecer el inicio de la presencia aborigen en las Antillas Mayores (Kozlowski 1974 Rouse y Allaire 1978 Rouse 1989 1992 Wilson 2001) difieren al respecto entre los 6 kaAP y los 45 kaAP (pero vease Pagan et al 200523) y hemos adoptado la mas temprana de estas fechas para el comienzo del intervalo amerindio en la Tabla 45 De todos modos cualquiera que haya sido la fecha esta no es aplicable por igual al conjunto de las Antillas Mayores porque segun algunos de los autores citados el poblamiento comenz6 por Cuba y La Espanola 6 mil anos atras continu6 en Puerto Rico hace alrededor de 3 mil anos (pero vease Burney et al 1994) yen Jamaica hace menos de 2 mil e incluso a las Islas Caiman ni siquiera llegaron [Las Islas Caiman descubiertas por Col6n en 1503 carecen absolutamente de evidencia amerindia (Stokes y Keegan 1996)]
Si los datos de la Tabla 45 representan una aceptable aproximaci6n a la realidad el proceso de las extinciones cuaternarias de los mamiferos terrestres de las Antillas Mayores y grupos de islas perifericas debi6 caracterizarse entre otros por los eventos siguientes
Aunque obviamente la fauna cuaternaria debi6 ser mucho mas numerosa que 10 que refleja el registro paleontol6gico (Morgan y Woods 1986) 52 especies (81 de la fauna conocida) desaparecieron (sin reposici6n) durante los ultimos 250 mil anos
- Al parecer las extinciones se concentraron en las especies de mayor tamano Asi el peso promedio estimado para las especies de la etapa preantr6pica (intervalos I + II de la Tabla 45) es 350 kg es 328 kg para las especies del intervalo III (amerindio) y es s610 25 kg (c1ases del peso a b c) para las especies del intervalo IV (postcolombino) Ninguna especie aut6ctona de mamifero mayor de 6 kg existe hoy en la regi6n
- De acuerdo con la distribuci6n temporal de las extinciones 61 especies sobrevivieron al interglacial Sangamon (intervalo 1)48 sobrevivieron a la transici6n del Pleistoceno terminal al Holoceno temprano (intervalo II) 34 sobrevivieron al periodo amerindio (intervalo III) y s610 12 especies han sobrevivido desde la Ilegada de los europeos (intervalo IV) Las tasas de extinci6n correspondientes (exceptuado el intervalo I) son una especie cada 1167 anos en el intervalo II una cada 393 anos en el intervalo III y una cada 23 anos en el intervalo IV
- Durante todo el proceso de las extinciones cuaternarias la perdida de especies por isla fue de 91 en La Espanola 83 en Jamaica y 62 en Cuba Bahamas retuvo su unica especie (aunque extirpada en varias de las islas) Puerto Rico e Islas Caiman perdieron todas sus especies (6 y 5 respectivamente) y las Islas Swan Virgenes y Anguila - San Martin perdieron sus respectivas especies (micas
328 BIOGEOGRAFiA
De los cuatro 6rdenes de mamiferos representados en la fauna Pilosa y Primates desaparecieron totalmente Soricomorpha y Rodentia perdieron 86 y 74 de sus especies respectivamente
CAUSALIDAD
CAUSAS NATURALES
Los cicIos glaciales e interglaciales acaecidos en el Pleistoceno causaron cambios ambientales dnisticos en las Antillas con la concomitante alternancia de cIimas frios (secos) y calidos (hUmedos) asi como de descensos y ascensos del nivel del mar aumento y disminuci6n de las areas territoriales) induso (aunque con menor intensidad) en el Holoceno Estos eventos paleogeograficos han sido consistentemente documentados (Mayo y Kartashov 1972 Bonnatti y Gartner 1973 Ortega y Arcia 1982 Prance 1982 Schubert y Medina 1982 Dodge et al 1983 Ortega [1984] Haq et al 1987 Schubert 1989 Hodell et al 1991 Guilderson et al 1994 Oppo 1997 Emslie 1998 Curtis et al 2001 Iturralde-Vinent 2003) y de ellos brindamos a continuaci6n una breve resena
Finalizando el periodo interglacial anterior a la ultima glaciaci6n (Wisconsin) el mar estaba a 8-10 m por encima de su nivel actual Cuando el avance glacial del Wisconsin a1canz6 su culminaci6n (eillamado Ultimo Maximo Glacial) hace 18 ka el mar habia descendido 120 m con respecto a dicho nivel y las islas del Caribe habian incrementado considerablemente sus extensiones subaereas en comparaci6n con las que presentan hoy dia por ejemplo Puerto Rico y las Islas Virgenes se conectaron territorialmente y las Bahamas se unieron en cinco gran des islas Prevalecia un regimen frio y seco y condiciones de aridez --debidas a una reducci6n de las precipitaciones (posiblemente a menos de 440 mmlano)- y los ambientes xericos se habian extendido a traves de las islas y bancos emergidos y abundaban los pinares (los habitats mesicos estaban restringidos a parcelas relictas en 10 alto de las montanas) En el Mar Caribe y el Golfo de Mexico la temperatura superficial del mar era 5degC mas baja que la actual y en las llanuras costeras y alturas maximas eran respectivamente 2-3degC y 6degC mas bajas Los limites de las zonas ecol6gicas altitudinales descendieron varios cientos de metros y cada zona redujo su extensi6n vertical al aumentar el gradiente de temperatura Ellimite inferior de la zona periglacial
elevaci6n donde la temperatura media minima a1canza los OdegC) de la Cordillera Central de la Republica Dominicana (altura maxima del Caribe insular con evidencia de topografia glacial) se considera que estuvo entre los 1500 y 1800 mlnm (actualmente esta a 2800 mlnm) y se puede estimar que dicho limite era inferior en Cuba por estar mas al norte que la Republica Dominicana No obstante durante este periodo la temperatura experiment6 episodios cortos (unas pocas decadas) de ascensos repetidos
Un signifieativo componente bi6tieo de las referidas condiciones paleoambientales 10 constituy6 la espectacular irradiaci6n evolucionaria experimentada por un grupo numeroso de aves rapaees diurnas y nocturnas (Wetmore 1937 Brodkorb 1969 Kurotchkin y Mayo 1973 Arredondo 1976 Olson 1978 Olson y Hilgartner 1982
329 EXTINCIONES DEL ClJATERNARIO
Suarez 2000a b 2004 Suarez y Emslie 2003) Los mamiferos de las Antillas Mayores ofrecieron a estos depredadores aviares una amplia gama de tamanos de presa que en ausencia de mamiferos del orden Carnivora en la region permitio a estas aves irradiarse a este nicho alimentario con la evolucion de una variedad de especies de muy diversa tall a corporal incluidas formas gigantes Adquirir un cuerpo gigante les propicio el acceso a recursos alimentarios que no estaban siendo explotados por ningun otro depredador (Alcover y McMinn 1994) A Cuba corresponde el mejor registro fosil de rapaces cuaternarias caribenas con siete generos de especies diurnas Dentro de estas aves existieron tres gavilanes gigantes (Accipitridae) tres carairas (Falconidae) y un condor (Cathartidae) asi como cuatro generos de especies nocturnas dos lechuzas (Tytonidae) y tres Mhos (Strigidae) dos de ell os no igualados en tamano en todo el mundo Algunos de estos generos tuvieron representacion en otras islas antillanas como La Espanola Jamaica Bahamas y Gran Caiman
Al retirarse hacia el norte los hielos de la glaciacion Wisconsin el clima se tomo mas hUmedo en las Antillas durante el Holoceno temprano y el bosque mesico se expandio en la region insular El nivel del mar ascendio acompanado de mayor pluviosidad alrededor de los 7 kaAP las Islas Virgenes quedaron separadas entre si y de Puerto Rico y las Bahamas perdieron la mayor parte de su territorio Cerca de los 3 kaAP las condiciones secas retornaron a la region e hicieron desaparecer buena parte del bosque mesico Nuevamente el clima comenzo a adquirir sus condiciones actuales hacia los 25 kaAP aproximadamente y hoy transita por una fase mesica con vegetacion xerica confinada en parcelas relictas
Sin duda estos y otros eventos naturales incidieron negativamente en la sobrevivencia de mamiferos terrestres en las Antillas Mayores asi como en los grupos de pequenas islas perifericas (Pregill y Olson 1981 Olson 2006)
McFarlane et al (1998a) demostraron convincentemente como Amblyrhiza el gigante de los roedores antillanos endemico de la pequena isla de Anguila sucumbio a las repetidas y abruptas perdidas de su habitat durante el interglacial Sangamon por las nipidas y severas oscilaciones del nivel del mar (documentadas por Neumann y Hearty 1996) al no poder responder adecuadamente a tan catastroficos eventos mediante el ajuste de su masa corporal media y otro tanto debio ocurrirle en Jamaica al otro roedor gigante Clidomys (McFarlane et al 1998b) emparentado con Amblyrhiza en la misma epoca (Tabla 45 intervalo I)
El ascenso del nivel del mar seguramente promovio la extincion de mamiferos durante la transicion del Pleistoceno al Holoceno (Tabla 45 intervalo II) con la reduccion del tamano de las Islas por las transgresiones marinas (Pregill y Olson 1981 Morgan 1993) Esto ultimo haria maximamente severa la competicion asf como la presion de los depredadores [Steadman et al (2005) no yen por que el calentamiento global ocurrido durante la transicion Pleistoceno-Holoceno habria de tener un efecto negativo sobre los perezosos siendo sus descendientes actuales especies tropic ales 0 subtropicales pero no se trata del efecto
330 BIOGEOGRAFiA
directo del cambio de temperatura sobre los animales sino del efecto indirecto y progresivo que dicho cambio genera a largo plazo al reducir el territorio insular disponible]
Por esta epoca tambien desaparecieron especies de tall a mayor como Megalocnus zile en La Espanola y otras de menor tamano (en Cuba) pero que representan la tall a maxima en sus respectivos contextos taxonomicos del Cuaternario como Solenodon arredondoi Paralouatta varonai y Macrocapromys acevedo Un mamifero puede ser grande solo a costa de reducir su densidad poblacional y esto 10 hace mis vulnerable a la extinci6n sobre todo en una isla (McNab 1994 McFarlane et al 1998a) Incidentalmente Alcover et al (1998a) afirmaron que larelacion entre la talla corporal y la extincion no siempre se cumple en las especies de menor tamano y citaron como ejemplo que entre las musarafias todas las especies de Nesophontes desaparecieron mientras sobreviven especies de Solenodon pero la comparacion correcta en todo caso serfa entre los solenodones vivientes y el gigante Solenodon arredondoi y entre Nesophontes edithae -el gigante de los nesofontes extinguido en el periodo amerindio- y el resto de sus congeneres que sobrevivieron hasta los primeros siglos de la colonizacion europea (Tabla 45 intervalos III y IV)
CAUSAS ANTROPOGENICAS
Intervalo Amerindio
La mayoria de los autores que se han referido a las extinciones acaecidas durante este intervalo (entre elIos paradigmaticamente Martin 1984a b) han responsabilizado a los aborfgenes (que se supone comenzaron a poblar las Antillas Mayores hace 6 mil afios) con la desaparicion de numerosas especies de mamiferos terrestres en la region principalmente por la via de la sobrecaza y la introduccion de animales domesticos devenidos depredadores de la fauna autoctona (Morgan y Woods 1986) Este efecto negativo de la presencia indigena sobre los mamiferos de la region ha sido dramatizado por algunos autores en terminos de holocausto faunal Sin embargo pensamos que se ha exagerado Como han expresado MacPhee et al (19993) cuando uno busca evidencia dura de complicidad humana directa en [estas] extinciones no hay mucho que encontrar Las siguientes son algunas consideraciones a tener en cuenta en tal sentido
El registro radiometrico mas joven conocido hasta ahora para un perezoso antillano (MegalocnllS rodens) es de 42 kaAP (MacPhee et ai 2007) Si se acepta que los aborfgenes entraron en Cuba hace aproximadamente 6 mil anos su convivencia con los perezosos debe haber durado alrededor de 1800 afios (0 mas porque dificilmente el referfdo fechado radiometrico haya correspondido al ultimo sobreviviente de la especie) Esta cifra contradice la hipotesis del holocausto faunal segun la cualla extincion de los mamiferos insulares de mayor talla debe ocurrir con extrema rapidez -a escala de decadas 0 de centurias- a partir de su primer contacto con los humanos (Martin y Steadman 1999) A pesar de las repetidas afirmaciones en contrario (Harrington 1921 Miller 1929c Herrera [1945] Tabio 1951 Alvarez 1951a 1957 Arredondo
331 EXTINCIONES DEL CUATERNARIO
1959 1960a Veloz y Ortega 1973 Mayo 1980b Rodriguez et al 1984 Castellanos y Pino 1985 Pino y Castellanos 1985 Nunez 1990b Gutierrez 1996 Arredondo 2004) en su mayoria basadas en evidencias no exentas de posibles accidentes de deposicion (0 de reubicacion postdeposicional) en recintos cavernarios (Diaz-Franco [2006b D no se ha demostrado inequivocamente que los aborigenes antillanos se alimentaran de perezosos regularmente Sus restos en residuarios de alimentos son escasisimos en toda la region y esto contrasta con los abundantes restos de jutias y otros mamiferos de menor talla que aparecen en esos residuarios demostrativos de la preferencia de los paleoindios por estos otros mamiferos
Los mamiferos mas importantes en la economia prehistorica de las Antillas Mayores fueron las jutias Capromys y Geocapromys en Cuba Geocapromys en Jamaica e Isolobodon en La Espanola Puerto Rico e Islas Virgenes (Aguayo 1950b Tablo 1951 Rimoli 1972 Wing 2001 Diaz-Franco [2006b D Wilkins (2001) estudi61ajutia viviente de Jamaica (Geocapromys browni) en un sitio ocupado por los aborigenes durante 100 anos y no encontr6 evidencias de que durante ese tiempo la especie haya sido sobreexplotada Asimismo en Cuba las jutias conga carabali y andaraz son escasas en depositos pleistoholocenicos en cambio son muy numerosas en residuarios indigenas y de haber sido sobreexplotadas 10 mas probable es que no hubieran sobrevivido hasta hoy
La habilidad de los humanos para penetrar en profundidad el bosque tropical y mantenerse en este sustentablemente ha sido un evento comparativamente reciente (Beran y Steele 1994) En el caso antillano la caza tierra adentro ha sido explicada como un proceso tardio de desplazamiento a partir de la explotacion de abundantes recursos marinos que ofrecian condiciones 6ptimas para el sedentarismo de las comunidades aborigenes costeras (C6rdova y Arredondo 1988 Brito 2005) Peces y mariscos dominan la arqueologia costera de las Islas antillanas en tanto el consumo de recursos terrestres (mamfferos incluidos) aumenta progresivamente en la medida que los residuarios estudiados se alejan de la costa (Scudder 1991 Wilkins 2001) En las Antillas Mayores el componente terrestre de la dieta aborigen es solo 20-30 del material recobrado en los residuarios (Wing 1977 1989) Y debio ser menor alin porque el componente vegetal de la dieta no se ha preservado Por otra parte los aborigenes cubanos domesticaron la jutia conga y seguramente la reprodujeron para comer (Pose et al 1988) practica que probablemente se repitiera con otras jutias en las demas Antillas Mayores
- Fray Bartolome de las Casas hablo de 3 millones de indios en Cuba al inicio de la colonizacion pero los historiadores subsecuentes consideraron exagerada la cifra (Poey 1851 a) No obstante se dispone de dos estimados de la densidad de poblaci6n aborigen en las Antillas Mayores a la llegada de los europeos 18 indigenaslkm2 en Puerto Rico (Rouse 1956) y 1271km2 en La Espanola (Watts 1987) Extrapolando ambos valores entre las cuatro islas principales de las Antillas Mayores tenemos que en la fecha indicada la poblaci6n aborigen pudo estar entre 16 mil y 113 mil indigenas en Puerto Rico entre 21 mil y 145 mil en Jamaica entre 141 mil y 1 mill6n en La Espanola y entre 200 mil y 14 millones en Cuba Entre las cifras maximas y las minimas nos inclinamos
332 BIOGEOGRAFiA
hacia estas ultimas atendiendo a los altos indices de mortalidad infantil y morbilidad que se evidencian en las osamentas de los pobladores prehispanicos (Martinez y Rives 1990) Uno de los hechos que apuntan hacia el bajo numero de aborigenes en las Antillas Mayores es 10 rapido que desaparecieron despues de la llegada de los europeos pnkticamente sin dejar mestizaje con estos 10 cual contrasta ostensiblemente con la situacion en la America continentaL
En suma un grupo humano (1) demograiicamente irrelevante y controlado por altos indices de morbilidad (2) consumidor mayoritariamente de organismos marinos y (3) carente de la tecnologia cinegetica necesaria para poder sobreexplotar los mamiferos que Ie sirvieron de alimento (a los cuales accedio tardiamente en su desarrollo) no es concebible que haya causado e1 holocausto faunal que se Ie atribuye a partir del error de aplicar a las comunidades aborigenes antillanas un modelo cultural de la prehistoria continentaL
Tampoco hay evidencias de que los aborigenes hayan introducido carnivoros con suficiente capacidad depredadora salvo el perro Morgan y Otlenwaldder (1993) consideraron que la depredacion por perros durante el periodo amerindio es la explicacion mas plausible para la extincion del almiqui gigante de Cuba pero como se puede observar (Tabla 54) Solenodon arredondoi aparece en el intervalo II por falta de asociacion con evidencias culturales
Consecuentemente el accionar de los aborigenes y de sus animales domesticos directamente sobre la fauna no parece haber sido por sf solo un factor determinante de la extincion de mamiferos terrestres durante el periodo amerindio en las Antillas (pero vease Steadman et al 2005)
Hay que recordar en este senti do que alin en el Holoceno tardio ocurrieron cambios clirruiticos de consideracion (vease Causas Naturales) Por ejemplo Curtis et al (2001) explicaron que la extincion de algunos ani males mesofilicos de las alturas montanosas de La Espanola pudo haber antecedido a cualquier impacto humano sigrIificativo en esos lugares por coincidir dicha extincion con una tendencia a la aridez documentada isotopicamente y con evidencias de la perdida del bosque mesico ocurridas despues de 3kaAP Yen Jamaica hace solo 2shy3 mil aDos el area de la Bahia de Jackson sustentaba una gran biomasa de vertebrados en bosque hinnedo a juzgar por la cantidad y diversidad de taxones presentes en material fosil derivado de la depredacion por lechuzas hoy el area es la mas seca de la isla con matorral xerofitico y pobreza de fauna (MacPhee 1996a) Yalm en nuestros dias huracanes de gran intensidad aniquilan poblaciones de vertebrados (que no se recuperan) en is las pequenas y llanas del Caribe (Brooks y Smith 2001)
En medio de la incertidumbre sobre la verdadera causa (0 causas) de las extinciones de mamiferos en el periodo amerindio MacPhee y Marx (1997) han propuesto su hipotesis de la hipermorbilidad la cual estipula colapsos poblacionales relativamente rapidos con la introduccion de enfermedades de alta letalidad Estos autores han razonado que
Las islas son los lugares mas apropiados para experimentar perdidas catastroficas causadas por enfermedades Los mamiferos no solo serian susceptibles en virtud de su prolongado aislamiento biologico sino que en los confines de una isla la epizootia
333 EXTINCIONES DEL CUATERNARIO
tendria la potencialidad de infectar simultaneamente a la rnayoria de los individuos de vadas especies Los neonatos son inmunologicamente inmaduros y sufririan desproporcionadamente cualquier infeccion hipervirulenta La perdida excesiva de neonatos y juveniles tendria efectos severos para la sobrevivencia de la poblacion si esta se reduce por debajo de cierto nivel sobreviene la extincion porque no hay posibilidad de restitucion en el periodo postpanzootico Las especies de mayor talla corporal van a ser las mas vulnerables a la extincion dadas sus bajas tasas reproductivas largos periodos de gestacion y reducido numero de progenie Las especies de menor tamano --caracterizadas por altas tasas reproductivas periodos cortos de gestacion y mayor numero de neonatos por parto- tambien serian fuertemente diezmadas pero muchas sobrevivirian porque su estrategia reproductiva les permite perder mayor numero de individuos Las perdidas deben ocurrir relativamente rapido (a escala de decadas) mientras que otras hipotesis (digamos los cambios climaticos) permiten intervalos mucho mas largos (a escala de siglos 0 de milenios) entre la aparicion de las causas y la produccion de sus efectos Por ejemplo todo indica que los nesofontes se extinguieron muy rapidamente y seria dificil poder invocar un impacto humano directo 0 argumentar fenomenos climaticos que pudieran explicar plausiblemente perdidas tan rapidas
MacPhee y Marx (1997 191) han dicho que por el momento la hipotesis de la hipermorbilidad no pasa de ser una abstraccion en espera de adecuada sustentacion empirica Llama la atencion que linicamente en el periodo amerindio (Tabla 45 intervalo 1Il) las extinciones abarcaron todas las clases del peso corporal (de la a a laj) y que ninguna de las especies que sobrevivieron hasta el periodo postcolombino (Tabla 45 intervalo IV mas las especies vivientes) sobrepasa los 10 kg (clases del peso a b c) en clara coincidencia con los presupuestos de la hipotetica hipermorbilidad
Intervalo Postcolornbino
Por 10 menos 37 especies de mamiferos terrestres vivian en las Antillas Mayores y grupos de Islas perifericas a la lIegada de los europeos en 1492 De elIas se han extinguido 23 shymas especies que en cualquiera de los periodos precedentes (Tabla 45~ y solo subsisten 12 especies repartidas entre Cuba (8 especies) La Espanola (2) Jamaica (1) y Bahamas (1) varias de estas en peligro critico de extincion Solo dos de las especies extinguidas durante los ultimos 500 anos se conocieron en estado viviente los roedores Oryzomys antillarum de Jamaica (extinguido en 1877) y Geocapromys thoracatus de las Islas Swan (extinguido en 1955) Las causas de las extinciones ocurridas durante este periodo son sobradamente conocidas destruccion de habitats introducci6n de animales exoticos (domesticos y comensales) y demas presiones asociadas a la actividad humana entre las cuales posiblemente habria que incluir brotes de hipermorbilidad
Durante mucho tiempo se ha afirmado la posibilidad de que especies de Nesophontes hayan sobrevivido hasta las primeras decadas del siglo xx y que incluso continuen existiendo en lugares remotos de Cuba 0 La Espanola (Allen 1918 1942 Miller 1930 Aguayo 1950a Arredondo 1960b Fischer 1977 MacFadden 1980 Woods y
334 BIOGEOGRAFiA
Ottenwalder 1991) Actualmente no existe evidencia radiometrica de que alguna especie de Nesophontes haya sobrevivido al siglo xv las fechas radiocarbonicas calibradas obtenidas en muestras de Nesophontes de Cuba y La Espanola resultaron no mas tardias que los siglos XIII y XIV (MacPhee et al 1999) Por otra parte las expediciones efectuadas a fines del siglo xx con el proposito de localizar individuos vivientes del genero en La Espanola no produjeron la menor indicacion afirmativa al respecto (Woods et al 1985) Ademas a traves de las Antillas Mayores no aparecen restos de Nesophontes en los depositos formados actualmente por las lechuzas
EPiLOGO
Las Antillas Mayores son islas oceanicas no han formado parte de un continente (con la posible excepcion de una parte de Jamaica hasta el Eoceno Medio hace 45 Ma) y durante un tiempo geologico considerable (desde el Eoceno Superior hace 40 Ma) han permanecido emergidas y separadas de los continentes vecinos Consecuentemente los mamiferos terrestres (no volantes) autoctonos de estas islas constituyen una fauna oceanica que por ende cumple con todas las caracteristicas convencionales reconocidas para este tipo de entidad biogeografica
- Baja diversidad taxonomica a nivel de las categorias superiores (solo cuatro ordenes) en comparacion con islas plataformicas continentales (0 con areas continentales) de tamano y diversidad ecologica simi lares
- Alto grado de endemismo a nivel de genero (96) y familia (50) en respuesta al prolongado aislamiento
- Desbalance ecologico de la estructura trofica ausencia de mamiferos camivoros y ungulados
- Espectaculares procesos de irradiaci6n evolucionaria como resultado de la colonizacion de territorios libres de competidores y depredadores
Tendencias evolucionarias extremas gigantismo
- Control a cargo de aves rapaces con tendencia at gigantismo en ausencia de mamiferos miembros del orden Carnivora
- Mas especies de reptiles y de murcielagos que de mamiferos terrestres relaciones que en los continentes se invierten
Alta tasa de extincion de especies (84) sin reposicion
Preservacion de especies filogeneticamente arcaicas (los llamados fosiles vivientes) los almiquies

323 EXTlICIOIES DEL CUATERNARIO
El gigantismo ha sido sin duda una tendencia evolucionaria recurrente entre los clados mayores de mamiferos antillanos Es un hecho bien conocido que algunos grupos de mamiferos han aumentado de tamafio en las islas y varios autores han intentado explicarlo a partir de enfoques diferentes (Foster 1964 Miller 1966 Sondaar 1977 Case 1978 Eisenberg y Wilson 1978 Heaney 1978 Lawlor 1982 Lomolino 1985 Angerbjom 1985 McNab 19942001)
EXTINCIONES DEL CUATERNARIO
Las islas que integran la Subregion Antillana han perdido casi 90 de sus mamiferos terrestres durante los ultimos milenios y en proporcion con su extension territorial esta perdida ha sido mayor en la Subregion que en cualquier otro lugar del planeta (MacPhee 1996b) Las extinciones del Cuatemario eliminaron mas de las tres cuartas partes de la fauna de mamiferos terrestres conocida de las Antillas Mayores sus islas satelites y grupos de islas perifericas y el examen de este proceso en sus aspectos mas generales se aborda en esta ultima parte del presente capitulo
CRONOLOGiA Y CONSECUENCIAS
Es muy improbable que un fosil represente el ultimo individuo de un taxon extinto y entonces hay que guiarse por la edad radiometrica del fosil mas reciente conocido lIamada fecha de ultima presencia- como unica aproximacion posible a la fecha de extincion del taxon
Como se explico al inicio del capitulo (vease Contexto Temporal) el registro fosH de los mamiferos terrestres de las Antillas Mayores adolece de insuficiente sustentacion radiometrica No obstante la distribucion temporal de los escasos fechados disponibles para el Cuaternario de la region abarca etapas criticas del proceso de extincion experimentado por la referida fauna a 10 largo de los ultimos 250 mil afios
De las 52 especies que es posible reconocer actualmente como miembros extintos de la fauna cuatemaria de mamiferos terrestres de las Antillas Mayores y grupos de islas perifericas los restos fosiles de 23 de elIas (442) han sido fechados radiometricamente o se han hallado asociados estratigraficamente de manera inequivoca con otro material fechado De estos 23 fechados 3 corresponden al interglacial Sangamon 4 pertenecen a la secuencia Pleistoceno terminal - Holoceno temprano 10 se ubican en ellapso entre el arribo de los aborigenes y la incursion europea y 6 son postcolombinos (a los cuales hay que sumar otras dos especies extinguidas en los siglos XIX y xx) A partir de estas 25 especies avaladas por fechas de ultima presencia distribuidas entre los cuatro intervalos temporales sefialados hemos compuesto la Tabla 45 en la que ademas aparecen las restantes 27 especies (carentes de fechado radiometrico) ubicadas en el intervalo temporal mas probable sobre la base de (1) la asociacion estratigrafica de sus restos con evidencias culturales (amerindias 0 europeas) 0 (2) la ausencia de dicha asociacion Sobre estos criterios de ubicacion temporal procede hacer dos aclaraciones
324 BIOGEOGRAFIA
TABLA 45 Intervalos temporales mas probables para las extinciones cuatemarias de los mamiferos terrestres de las Antillas Mayores y grupos de islas perifericas sobre la base de las evidencias siguientes (l) ultimo ejemplar colectado u observado en vida (2) fechado radiometrico mas tardio en material de la especie (3) asociaci6n estratigrafica mas tardia con otro material fechado (4) asociaci6n estratigrafica con restos de animales introducidos restos humanos u objetos culturales propios del intervale (5) falta de asociaci6n con evidencias culturales [Los ntimeros entre parentesis en la columna Evidencia significan adscripci6n tentativa al intervalo] Los estimados del peso corporal (en kg) estan tomados de la literatura 0 inferidos comparativamente y distribuidos entre
las clases siguientes (a) S 01 (b) 01 a lt 1 (c) 1 a lt 10 (d) 10 a lt 100 (e) 100 a lt 200 (j) ~ 200 Las
abreviaturas que designan las islas son las de la Tabla 44
F amilialEspecie Peso IsJa Evidencia Fuente
I httervalo htterg1acial Sangamon (gt80 kaAP)
HEPTAXODONTlNAE
Amblyrhiza inundata e AS 3 McFarJane et al 1998a
CLIDOMYINAE
Clidomys osborni d JA 3 Ibid
Clidomys parvus d JA 3 Ibid
II httervalo Pleistoceno terminal ~ Holoceno temprano (20-6 kaAP)
MEGALONYCHIDAE
Aeratoenus antillensis d CD 5 Arredondo 1975 a
Aeratoenus odontrigonus d PR 3 McFarJane 1999b
Aeratoenus simorhynehus d LE 2 Rega et al 2002
M egaloenus zile f LE 2 MacPhee et al 2000
SOLENODONTIDAE
Solenodon arredondoi e CD 5 Morgan y Ottenwalder 1993
PnHECIIDAE
Paralouatta varonai d CD 5 MacPhee 1996a
EcHIMYIDAE
Heteropsomys antillensis c PR 5 McFarJane 1999b
Heteropsomys insulans c PR 5 Ibid
Puertoricomys eorozalus c PR 5 Woods 1989a
CAPROMYIDAE
P lagiodontia araeum c LE 5 MacPhee et al 2000
Macrocapromys latus c CD 5 Varona y Arredondo 1979
(Continua)
325 EXTINCIONES DEL CUATERNARIO
TABLA 45 (Continuaci6n)
F ami1ia1Especie Peso Isla Evidencia Fuente
M acrocapromys latus c CU 5 Varona y Arredondo 1979
Mesocapromys kraglievichi c CU 5 Ibid
HEPTAXODONTINAE
Xaymaca fulvopulvis a JA 2 MacPhee y Flemming 2003
III Intervalo Amerindio (6-05 kaAP)
MEGALONYCHIDAE
Acratocnus ye d LE 2 MacPhee et aI 2000
Neocnus comes c LE 2 Ibid
N eocnus dousman c LE 2 Ibid
N eocnus toupiti c LE 2 Ibid
N eocnus gliriform is c CU 4 Cordova et ai 1997
M egalocnus rodens f CU 2 MacPhee et ai 2007
Parocnus browni e CU 2 Steadman et ai 2005
P arocnus serus e LE 2 MacPhee et ai 2000
N ESOPHONTIDAE
N esophontes edithae a PR 2 McFarlane 1999b
PITHECIIDAE
Antillothrix bernensis c LE 3 Rimoli 1977
EcHIMYIDAE
Brotomys contractus a LE 4 Miller 1929c
CAPROMYIDAE
P lagiodontia spelaeum c LE 4 Ibid
M acrocapromys acevedo c CU 4 NUiiez 1990a
HEPTAXODONTINAE
Elasmodontomys obliquus d PR 3 McFarlane 1999b
Iv Intervalo Postcolombino (05 kaAP - Presente)
N ESOPHONTIDAE
N esophontes hypom icrus a LE 24 MacPhee y Fle111111ing 1999
N esophontes zam icrus a LE (4) Ibid
N esophontes m icrus a CU 24 MacPhee et aI 1999
(Continua)
326 BIOGEOGRAFiA
TABLA 45 (Continuacion)
F amilialEspecie Peso Isla Evidencia Fuente
N esophontes m icrus a CD 24 MacPhee et aI 1999
N esophontes m icrus a LE 4 MacPhee et aI 2000
N esophontes sp 1 a IC 4 Morgan 1994
N esophontes sp 2 a IC 4 Ibid
Solenodon marcanoi b LE 4 Ottenwalder 2001
PITHECIIDAE
Xenothrix mcgregori c JA 4 MacPhee y Flemming 1999
EcHIMYIDAE
Boromys offeUa a CD 24 Guarch 1984
Boromys torrei a CD 24 Ibid
Brotomys voratus a LE 4 MacPhee et aI 2000
CAPROMYIDAE
P lagiodontia ipnaeum c LE 4 MacPhee y Flemming 1999
Rhizoplagiodontia lemkei c LE 4 Ibid
Isolobodon portoricensis c PR 24 Ibid
Isolobodon montanus c LE 4 Ibid
Hexolobodon phenax c LE (4) Ibid
Capromys sp c IC 24 Morgan 1994
Geocapromys columbianus c CD 4 MacPhee y Flemming 1999
Geocapromys thoracatus b IS 1 MacPhee y Marx 1997
Geocapromys sp 1 c IC 4 Morgan 1994
Geocapromys sp 2 c IC (4) Ibid
l-IEPTAXODONTINAE
Quemisia gravis c LE (1) MacPhee y Flemming 1999
MURIDAE
Oryzomys antillarum a JA 1 Ibid
1 Flemming y MacPhee (1999) alertaron en contra de la asociacion con otro material fechado Asimismo Morgan (1993) indico que la ausencia de asociacion con evidencias culturales amerindias no es necesariamente un criterio concluyente para ubicar un taxon en el intervalo temporal anterior porque hay depositos en la region que carecen de tales evidencias y son del Holoceno tardio por fechados radiocarbonicos [No obstante
327 EXTlNCIONES DEL CUATERNAR10
la ausencia de una especie en residuarios arqueol6gicos adquiere mayor certidumbre como criterio de ubicaci6n temporal de su extinci6n en la medida que aumenta el numero de residuarios estudiados que en las Antillas Mayores se cuentan por centenares] A falta de fechado absoluto de una especie consideramos que estos criterios de ubicaci6n temporal son mas utiles que la alternativa de excluir del analisis a la especie
2 En los trabajos sobre biogeografia de mamiferos antillanos los arque610gos mas citados como autoridades para establecer el inicio de la presencia aborigen en las Antillas Mayores (Kozlowski 1974 Rouse y Allaire 1978 Rouse 1989 1992 Wilson 2001) difieren al respecto entre los 6 kaAP y los 45 kaAP (pero vease Pagan et al 200523) y hemos adoptado la mas temprana de estas fechas para el comienzo del intervalo amerindio en la Tabla 45 De todos modos cualquiera que haya sido la fecha esta no es aplicable por igual al conjunto de las Antillas Mayores porque segun algunos de los autores citados el poblamiento comenz6 por Cuba y La Espanola 6 mil anos atras continu6 en Puerto Rico hace alrededor de 3 mil anos (pero vease Burney et al 1994) yen Jamaica hace menos de 2 mil e incluso a las Islas Caiman ni siquiera llegaron [Las Islas Caiman descubiertas por Col6n en 1503 carecen absolutamente de evidencia amerindia (Stokes y Keegan 1996)]
Si los datos de la Tabla 45 representan una aceptable aproximaci6n a la realidad el proceso de las extinciones cuaternarias de los mamiferos terrestres de las Antillas Mayores y grupos de islas perifericas debi6 caracterizarse entre otros por los eventos siguientes
Aunque obviamente la fauna cuaternaria debi6 ser mucho mas numerosa que 10 que refleja el registro paleontol6gico (Morgan y Woods 1986) 52 especies (81 de la fauna conocida) desaparecieron (sin reposici6n) durante los ultimos 250 mil anos
- Al parecer las extinciones se concentraron en las especies de mayor tamano Asi el peso promedio estimado para las especies de la etapa preantr6pica (intervalos I + II de la Tabla 45) es 350 kg es 328 kg para las especies del intervalo III (amerindio) y es s610 25 kg (c1ases del peso a b c) para las especies del intervalo IV (postcolombino) Ninguna especie aut6ctona de mamifero mayor de 6 kg existe hoy en la regi6n
- De acuerdo con la distribuci6n temporal de las extinciones 61 especies sobrevivieron al interglacial Sangamon (intervalo 1)48 sobrevivieron a la transici6n del Pleistoceno terminal al Holoceno temprano (intervalo II) 34 sobrevivieron al periodo amerindio (intervalo III) y s610 12 especies han sobrevivido desde la Ilegada de los europeos (intervalo IV) Las tasas de extinci6n correspondientes (exceptuado el intervalo I) son una especie cada 1167 anos en el intervalo II una cada 393 anos en el intervalo III y una cada 23 anos en el intervalo IV
- Durante todo el proceso de las extinciones cuaternarias la perdida de especies por isla fue de 91 en La Espanola 83 en Jamaica y 62 en Cuba Bahamas retuvo su unica especie (aunque extirpada en varias de las islas) Puerto Rico e Islas Caiman perdieron todas sus especies (6 y 5 respectivamente) y las Islas Swan Virgenes y Anguila - San Martin perdieron sus respectivas especies (micas
328 BIOGEOGRAFiA
De los cuatro 6rdenes de mamiferos representados en la fauna Pilosa y Primates desaparecieron totalmente Soricomorpha y Rodentia perdieron 86 y 74 de sus especies respectivamente
CAUSALIDAD
CAUSAS NATURALES
Los cicIos glaciales e interglaciales acaecidos en el Pleistoceno causaron cambios ambientales dnisticos en las Antillas con la concomitante alternancia de cIimas frios (secos) y calidos (hUmedos) asi como de descensos y ascensos del nivel del mar aumento y disminuci6n de las areas territoriales) induso (aunque con menor intensidad) en el Holoceno Estos eventos paleogeograficos han sido consistentemente documentados (Mayo y Kartashov 1972 Bonnatti y Gartner 1973 Ortega y Arcia 1982 Prance 1982 Schubert y Medina 1982 Dodge et al 1983 Ortega [1984] Haq et al 1987 Schubert 1989 Hodell et al 1991 Guilderson et al 1994 Oppo 1997 Emslie 1998 Curtis et al 2001 Iturralde-Vinent 2003) y de ellos brindamos a continuaci6n una breve resena
Finalizando el periodo interglacial anterior a la ultima glaciaci6n (Wisconsin) el mar estaba a 8-10 m por encima de su nivel actual Cuando el avance glacial del Wisconsin a1canz6 su culminaci6n (eillamado Ultimo Maximo Glacial) hace 18 ka el mar habia descendido 120 m con respecto a dicho nivel y las islas del Caribe habian incrementado considerablemente sus extensiones subaereas en comparaci6n con las que presentan hoy dia por ejemplo Puerto Rico y las Islas Virgenes se conectaron territorialmente y las Bahamas se unieron en cinco gran des islas Prevalecia un regimen frio y seco y condiciones de aridez --debidas a una reducci6n de las precipitaciones (posiblemente a menos de 440 mmlano)- y los ambientes xericos se habian extendido a traves de las islas y bancos emergidos y abundaban los pinares (los habitats mesicos estaban restringidos a parcelas relictas en 10 alto de las montanas) En el Mar Caribe y el Golfo de Mexico la temperatura superficial del mar era 5degC mas baja que la actual y en las llanuras costeras y alturas maximas eran respectivamente 2-3degC y 6degC mas bajas Los limites de las zonas ecol6gicas altitudinales descendieron varios cientos de metros y cada zona redujo su extensi6n vertical al aumentar el gradiente de temperatura Ellimite inferior de la zona periglacial
elevaci6n donde la temperatura media minima a1canza los OdegC) de la Cordillera Central de la Republica Dominicana (altura maxima del Caribe insular con evidencia de topografia glacial) se considera que estuvo entre los 1500 y 1800 mlnm (actualmente esta a 2800 mlnm) y se puede estimar que dicho limite era inferior en Cuba por estar mas al norte que la Republica Dominicana No obstante durante este periodo la temperatura experiment6 episodios cortos (unas pocas decadas) de ascensos repetidos
Un signifieativo componente bi6tieo de las referidas condiciones paleoambientales 10 constituy6 la espectacular irradiaci6n evolucionaria experimentada por un grupo numeroso de aves rapaees diurnas y nocturnas (Wetmore 1937 Brodkorb 1969 Kurotchkin y Mayo 1973 Arredondo 1976 Olson 1978 Olson y Hilgartner 1982
329 EXTINCIONES DEL ClJATERNARIO
Suarez 2000a b 2004 Suarez y Emslie 2003) Los mamiferos de las Antillas Mayores ofrecieron a estos depredadores aviares una amplia gama de tamanos de presa que en ausencia de mamiferos del orden Carnivora en la region permitio a estas aves irradiarse a este nicho alimentario con la evolucion de una variedad de especies de muy diversa tall a corporal incluidas formas gigantes Adquirir un cuerpo gigante les propicio el acceso a recursos alimentarios que no estaban siendo explotados por ningun otro depredador (Alcover y McMinn 1994) A Cuba corresponde el mejor registro fosil de rapaces cuaternarias caribenas con siete generos de especies diurnas Dentro de estas aves existieron tres gavilanes gigantes (Accipitridae) tres carairas (Falconidae) y un condor (Cathartidae) asi como cuatro generos de especies nocturnas dos lechuzas (Tytonidae) y tres Mhos (Strigidae) dos de ell os no igualados en tamano en todo el mundo Algunos de estos generos tuvieron representacion en otras islas antillanas como La Espanola Jamaica Bahamas y Gran Caiman
Al retirarse hacia el norte los hielos de la glaciacion Wisconsin el clima se tomo mas hUmedo en las Antillas durante el Holoceno temprano y el bosque mesico se expandio en la region insular El nivel del mar ascendio acompanado de mayor pluviosidad alrededor de los 7 kaAP las Islas Virgenes quedaron separadas entre si y de Puerto Rico y las Bahamas perdieron la mayor parte de su territorio Cerca de los 3 kaAP las condiciones secas retornaron a la region e hicieron desaparecer buena parte del bosque mesico Nuevamente el clima comenzo a adquirir sus condiciones actuales hacia los 25 kaAP aproximadamente y hoy transita por una fase mesica con vegetacion xerica confinada en parcelas relictas
Sin duda estos y otros eventos naturales incidieron negativamente en la sobrevivencia de mamiferos terrestres en las Antillas Mayores asi como en los grupos de pequenas islas perifericas (Pregill y Olson 1981 Olson 2006)
McFarlane et al (1998a) demostraron convincentemente como Amblyrhiza el gigante de los roedores antillanos endemico de la pequena isla de Anguila sucumbio a las repetidas y abruptas perdidas de su habitat durante el interglacial Sangamon por las nipidas y severas oscilaciones del nivel del mar (documentadas por Neumann y Hearty 1996) al no poder responder adecuadamente a tan catastroficos eventos mediante el ajuste de su masa corporal media y otro tanto debio ocurrirle en Jamaica al otro roedor gigante Clidomys (McFarlane et al 1998b) emparentado con Amblyrhiza en la misma epoca (Tabla 45 intervalo I)
El ascenso del nivel del mar seguramente promovio la extincion de mamiferos durante la transicion del Pleistoceno al Holoceno (Tabla 45 intervalo II) con la reduccion del tamano de las Islas por las transgresiones marinas (Pregill y Olson 1981 Morgan 1993) Esto ultimo haria maximamente severa la competicion asf como la presion de los depredadores [Steadman et al (2005) no yen por que el calentamiento global ocurrido durante la transicion Pleistoceno-Holoceno habria de tener un efecto negativo sobre los perezosos siendo sus descendientes actuales especies tropic ales 0 subtropicales pero no se trata del efecto
330 BIOGEOGRAFiA
directo del cambio de temperatura sobre los animales sino del efecto indirecto y progresivo que dicho cambio genera a largo plazo al reducir el territorio insular disponible]
Por esta epoca tambien desaparecieron especies de tall a mayor como Megalocnus zile en La Espanola y otras de menor tamano (en Cuba) pero que representan la tall a maxima en sus respectivos contextos taxonomicos del Cuaternario como Solenodon arredondoi Paralouatta varonai y Macrocapromys acevedo Un mamifero puede ser grande solo a costa de reducir su densidad poblacional y esto 10 hace mis vulnerable a la extinci6n sobre todo en una isla (McNab 1994 McFarlane et al 1998a) Incidentalmente Alcover et al (1998a) afirmaron que larelacion entre la talla corporal y la extincion no siempre se cumple en las especies de menor tamano y citaron como ejemplo que entre las musarafias todas las especies de Nesophontes desaparecieron mientras sobreviven especies de Solenodon pero la comparacion correcta en todo caso serfa entre los solenodones vivientes y el gigante Solenodon arredondoi y entre Nesophontes edithae -el gigante de los nesofontes extinguido en el periodo amerindio- y el resto de sus congeneres que sobrevivieron hasta los primeros siglos de la colonizacion europea (Tabla 45 intervalos III y IV)
CAUSAS ANTROPOGENICAS
Intervalo Amerindio
La mayoria de los autores que se han referido a las extinciones acaecidas durante este intervalo (entre elIos paradigmaticamente Martin 1984a b) han responsabilizado a los aborfgenes (que se supone comenzaron a poblar las Antillas Mayores hace 6 mil afios) con la desaparicion de numerosas especies de mamiferos terrestres en la region principalmente por la via de la sobrecaza y la introduccion de animales domesticos devenidos depredadores de la fauna autoctona (Morgan y Woods 1986) Este efecto negativo de la presencia indigena sobre los mamiferos de la region ha sido dramatizado por algunos autores en terminos de holocausto faunal Sin embargo pensamos que se ha exagerado Como han expresado MacPhee et al (19993) cuando uno busca evidencia dura de complicidad humana directa en [estas] extinciones no hay mucho que encontrar Las siguientes son algunas consideraciones a tener en cuenta en tal sentido
El registro radiometrico mas joven conocido hasta ahora para un perezoso antillano (MegalocnllS rodens) es de 42 kaAP (MacPhee et ai 2007) Si se acepta que los aborfgenes entraron en Cuba hace aproximadamente 6 mil anos su convivencia con los perezosos debe haber durado alrededor de 1800 afios (0 mas porque dificilmente el referfdo fechado radiometrico haya correspondido al ultimo sobreviviente de la especie) Esta cifra contradice la hipotesis del holocausto faunal segun la cualla extincion de los mamiferos insulares de mayor talla debe ocurrir con extrema rapidez -a escala de decadas 0 de centurias- a partir de su primer contacto con los humanos (Martin y Steadman 1999) A pesar de las repetidas afirmaciones en contrario (Harrington 1921 Miller 1929c Herrera [1945] Tabio 1951 Alvarez 1951a 1957 Arredondo
331 EXTINCIONES DEL CUATERNARIO
1959 1960a Veloz y Ortega 1973 Mayo 1980b Rodriguez et al 1984 Castellanos y Pino 1985 Pino y Castellanos 1985 Nunez 1990b Gutierrez 1996 Arredondo 2004) en su mayoria basadas en evidencias no exentas de posibles accidentes de deposicion (0 de reubicacion postdeposicional) en recintos cavernarios (Diaz-Franco [2006b D no se ha demostrado inequivocamente que los aborigenes antillanos se alimentaran de perezosos regularmente Sus restos en residuarios de alimentos son escasisimos en toda la region y esto contrasta con los abundantes restos de jutias y otros mamiferos de menor talla que aparecen en esos residuarios demostrativos de la preferencia de los paleoindios por estos otros mamiferos
Los mamiferos mas importantes en la economia prehistorica de las Antillas Mayores fueron las jutias Capromys y Geocapromys en Cuba Geocapromys en Jamaica e Isolobodon en La Espanola Puerto Rico e Islas Virgenes (Aguayo 1950b Tablo 1951 Rimoli 1972 Wing 2001 Diaz-Franco [2006b D Wilkins (2001) estudi61ajutia viviente de Jamaica (Geocapromys browni) en un sitio ocupado por los aborigenes durante 100 anos y no encontr6 evidencias de que durante ese tiempo la especie haya sido sobreexplotada Asimismo en Cuba las jutias conga carabali y andaraz son escasas en depositos pleistoholocenicos en cambio son muy numerosas en residuarios indigenas y de haber sido sobreexplotadas 10 mas probable es que no hubieran sobrevivido hasta hoy
La habilidad de los humanos para penetrar en profundidad el bosque tropical y mantenerse en este sustentablemente ha sido un evento comparativamente reciente (Beran y Steele 1994) En el caso antillano la caza tierra adentro ha sido explicada como un proceso tardio de desplazamiento a partir de la explotacion de abundantes recursos marinos que ofrecian condiciones 6ptimas para el sedentarismo de las comunidades aborigenes costeras (C6rdova y Arredondo 1988 Brito 2005) Peces y mariscos dominan la arqueologia costera de las Islas antillanas en tanto el consumo de recursos terrestres (mamfferos incluidos) aumenta progresivamente en la medida que los residuarios estudiados se alejan de la costa (Scudder 1991 Wilkins 2001) En las Antillas Mayores el componente terrestre de la dieta aborigen es solo 20-30 del material recobrado en los residuarios (Wing 1977 1989) Y debio ser menor alin porque el componente vegetal de la dieta no se ha preservado Por otra parte los aborigenes cubanos domesticaron la jutia conga y seguramente la reprodujeron para comer (Pose et al 1988) practica que probablemente se repitiera con otras jutias en las demas Antillas Mayores
- Fray Bartolome de las Casas hablo de 3 millones de indios en Cuba al inicio de la colonizacion pero los historiadores subsecuentes consideraron exagerada la cifra (Poey 1851 a) No obstante se dispone de dos estimados de la densidad de poblaci6n aborigen en las Antillas Mayores a la llegada de los europeos 18 indigenaslkm2 en Puerto Rico (Rouse 1956) y 1271km2 en La Espanola (Watts 1987) Extrapolando ambos valores entre las cuatro islas principales de las Antillas Mayores tenemos que en la fecha indicada la poblaci6n aborigen pudo estar entre 16 mil y 113 mil indigenas en Puerto Rico entre 21 mil y 145 mil en Jamaica entre 141 mil y 1 mill6n en La Espanola y entre 200 mil y 14 millones en Cuba Entre las cifras maximas y las minimas nos inclinamos
332 BIOGEOGRAFiA
hacia estas ultimas atendiendo a los altos indices de mortalidad infantil y morbilidad que se evidencian en las osamentas de los pobladores prehispanicos (Martinez y Rives 1990) Uno de los hechos que apuntan hacia el bajo numero de aborigenes en las Antillas Mayores es 10 rapido que desaparecieron despues de la llegada de los europeos pnkticamente sin dejar mestizaje con estos 10 cual contrasta ostensiblemente con la situacion en la America continentaL
En suma un grupo humano (1) demograiicamente irrelevante y controlado por altos indices de morbilidad (2) consumidor mayoritariamente de organismos marinos y (3) carente de la tecnologia cinegetica necesaria para poder sobreexplotar los mamiferos que Ie sirvieron de alimento (a los cuales accedio tardiamente en su desarrollo) no es concebible que haya causado e1 holocausto faunal que se Ie atribuye a partir del error de aplicar a las comunidades aborigenes antillanas un modelo cultural de la prehistoria continentaL
Tampoco hay evidencias de que los aborigenes hayan introducido carnivoros con suficiente capacidad depredadora salvo el perro Morgan y Otlenwaldder (1993) consideraron que la depredacion por perros durante el periodo amerindio es la explicacion mas plausible para la extincion del almiqui gigante de Cuba pero como se puede observar (Tabla 54) Solenodon arredondoi aparece en el intervalo II por falta de asociacion con evidencias culturales
Consecuentemente el accionar de los aborigenes y de sus animales domesticos directamente sobre la fauna no parece haber sido por sf solo un factor determinante de la extincion de mamiferos terrestres durante el periodo amerindio en las Antillas (pero vease Steadman et al 2005)
Hay que recordar en este senti do que alin en el Holoceno tardio ocurrieron cambios clirruiticos de consideracion (vease Causas Naturales) Por ejemplo Curtis et al (2001) explicaron que la extincion de algunos ani males mesofilicos de las alturas montanosas de La Espanola pudo haber antecedido a cualquier impacto humano sigrIificativo en esos lugares por coincidir dicha extincion con una tendencia a la aridez documentada isotopicamente y con evidencias de la perdida del bosque mesico ocurridas despues de 3kaAP Yen Jamaica hace solo 2shy3 mil aDos el area de la Bahia de Jackson sustentaba una gran biomasa de vertebrados en bosque hinnedo a juzgar por la cantidad y diversidad de taxones presentes en material fosil derivado de la depredacion por lechuzas hoy el area es la mas seca de la isla con matorral xerofitico y pobreza de fauna (MacPhee 1996a) Yalm en nuestros dias huracanes de gran intensidad aniquilan poblaciones de vertebrados (que no se recuperan) en is las pequenas y llanas del Caribe (Brooks y Smith 2001)
En medio de la incertidumbre sobre la verdadera causa (0 causas) de las extinciones de mamiferos en el periodo amerindio MacPhee y Marx (1997) han propuesto su hipotesis de la hipermorbilidad la cual estipula colapsos poblacionales relativamente rapidos con la introduccion de enfermedades de alta letalidad Estos autores han razonado que
Las islas son los lugares mas apropiados para experimentar perdidas catastroficas causadas por enfermedades Los mamiferos no solo serian susceptibles en virtud de su prolongado aislamiento biologico sino que en los confines de una isla la epizootia
333 EXTINCIONES DEL CUATERNARIO
tendria la potencialidad de infectar simultaneamente a la rnayoria de los individuos de vadas especies Los neonatos son inmunologicamente inmaduros y sufririan desproporcionadamente cualquier infeccion hipervirulenta La perdida excesiva de neonatos y juveniles tendria efectos severos para la sobrevivencia de la poblacion si esta se reduce por debajo de cierto nivel sobreviene la extincion porque no hay posibilidad de restitucion en el periodo postpanzootico Las especies de mayor talla corporal van a ser las mas vulnerables a la extincion dadas sus bajas tasas reproductivas largos periodos de gestacion y reducido numero de progenie Las especies de menor tamano --caracterizadas por altas tasas reproductivas periodos cortos de gestacion y mayor numero de neonatos por parto- tambien serian fuertemente diezmadas pero muchas sobrevivirian porque su estrategia reproductiva les permite perder mayor numero de individuos Las perdidas deben ocurrir relativamente rapido (a escala de decadas) mientras que otras hipotesis (digamos los cambios climaticos) permiten intervalos mucho mas largos (a escala de siglos 0 de milenios) entre la aparicion de las causas y la produccion de sus efectos Por ejemplo todo indica que los nesofontes se extinguieron muy rapidamente y seria dificil poder invocar un impacto humano directo 0 argumentar fenomenos climaticos que pudieran explicar plausiblemente perdidas tan rapidas
MacPhee y Marx (1997 191) han dicho que por el momento la hipotesis de la hipermorbilidad no pasa de ser una abstraccion en espera de adecuada sustentacion empirica Llama la atencion que linicamente en el periodo amerindio (Tabla 45 intervalo 1Il) las extinciones abarcaron todas las clases del peso corporal (de la a a laj) y que ninguna de las especies que sobrevivieron hasta el periodo postcolombino (Tabla 45 intervalo IV mas las especies vivientes) sobrepasa los 10 kg (clases del peso a b c) en clara coincidencia con los presupuestos de la hipotetica hipermorbilidad
Intervalo Postcolornbino
Por 10 menos 37 especies de mamiferos terrestres vivian en las Antillas Mayores y grupos de Islas perifericas a la lIegada de los europeos en 1492 De elIas se han extinguido 23 shymas especies que en cualquiera de los periodos precedentes (Tabla 45~ y solo subsisten 12 especies repartidas entre Cuba (8 especies) La Espanola (2) Jamaica (1) y Bahamas (1) varias de estas en peligro critico de extincion Solo dos de las especies extinguidas durante los ultimos 500 anos se conocieron en estado viviente los roedores Oryzomys antillarum de Jamaica (extinguido en 1877) y Geocapromys thoracatus de las Islas Swan (extinguido en 1955) Las causas de las extinciones ocurridas durante este periodo son sobradamente conocidas destruccion de habitats introducci6n de animales exoticos (domesticos y comensales) y demas presiones asociadas a la actividad humana entre las cuales posiblemente habria que incluir brotes de hipermorbilidad
Durante mucho tiempo se ha afirmado la posibilidad de que especies de Nesophontes hayan sobrevivido hasta las primeras decadas del siglo xx y que incluso continuen existiendo en lugares remotos de Cuba 0 La Espanola (Allen 1918 1942 Miller 1930 Aguayo 1950a Arredondo 1960b Fischer 1977 MacFadden 1980 Woods y
334 BIOGEOGRAFiA
Ottenwalder 1991) Actualmente no existe evidencia radiometrica de que alguna especie de Nesophontes haya sobrevivido al siglo xv las fechas radiocarbonicas calibradas obtenidas en muestras de Nesophontes de Cuba y La Espanola resultaron no mas tardias que los siglos XIII y XIV (MacPhee et al 1999) Por otra parte las expediciones efectuadas a fines del siglo xx con el proposito de localizar individuos vivientes del genero en La Espanola no produjeron la menor indicacion afirmativa al respecto (Woods et al 1985) Ademas a traves de las Antillas Mayores no aparecen restos de Nesophontes en los depositos formados actualmente por las lechuzas
EPiLOGO
Las Antillas Mayores son islas oceanicas no han formado parte de un continente (con la posible excepcion de una parte de Jamaica hasta el Eoceno Medio hace 45 Ma) y durante un tiempo geologico considerable (desde el Eoceno Superior hace 40 Ma) han permanecido emergidas y separadas de los continentes vecinos Consecuentemente los mamiferos terrestres (no volantes) autoctonos de estas islas constituyen una fauna oceanica que por ende cumple con todas las caracteristicas convencionales reconocidas para este tipo de entidad biogeografica
- Baja diversidad taxonomica a nivel de las categorias superiores (solo cuatro ordenes) en comparacion con islas plataformicas continentales (0 con areas continentales) de tamano y diversidad ecologica simi lares
- Alto grado de endemismo a nivel de genero (96) y familia (50) en respuesta al prolongado aislamiento
- Desbalance ecologico de la estructura trofica ausencia de mamiferos camivoros y ungulados
- Espectaculares procesos de irradiaci6n evolucionaria como resultado de la colonizacion de territorios libres de competidores y depredadores
Tendencias evolucionarias extremas gigantismo
- Control a cargo de aves rapaces con tendencia at gigantismo en ausencia de mamiferos miembros del orden Carnivora
- Mas especies de reptiles y de murcielagos que de mamiferos terrestres relaciones que en los continentes se invierten
Alta tasa de extincion de especies (84) sin reposicion
Preservacion de especies filogeneticamente arcaicas (los llamados fosiles vivientes) los almiquies
324 BIOGEOGRAFIA
TABLA 45 Intervalos temporales mas probables para las extinciones cuatemarias de los mamiferos terrestres de las Antillas Mayores y grupos de islas perifericas sobre la base de las evidencias siguientes (l) ultimo ejemplar colectado u observado en vida (2) fechado radiometrico mas tardio en material de la especie (3) asociaci6n estratigrafica mas tardia con otro material fechado (4) asociaci6n estratigrafica con restos de animales introducidos restos humanos u objetos culturales propios del intervale (5) falta de asociaci6n con evidencias culturales [Los ntimeros entre parentesis en la columna Evidencia significan adscripci6n tentativa al intervalo] Los estimados del peso corporal (en kg) estan tomados de la literatura 0 inferidos comparativamente y distribuidos entre
las clases siguientes (a) S 01 (b) 01 a lt 1 (c) 1 a lt 10 (d) 10 a lt 100 (e) 100 a lt 200 (j) ~ 200 Las
abreviaturas que designan las islas son las de la Tabla 44
F amilialEspecie Peso IsJa Evidencia Fuente
I httervalo htterg1acial Sangamon (gt80 kaAP)
HEPTAXODONTlNAE
Amblyrhiza inundata e AS 3 McFarJane et al 1998a
CLIDOMYINAE
Clidomys osborni d JA 3 Ibid
Clidomys parvus d JA 3 Ibid
II httervalo Pleistoceno terminal ~ Holoceno temprano (20-6 kaAP)
MEGALONYCHIDAE
Aeratoenus antillensis d CD 5 Arredondo 1975 a
Aeratoenus odontrigonus d PR 3 McFarJane 1999b
Aeratoenus simorhynehus d LE 2 Rega et al 2002
M egaloenus zile f LE 2 MacPhee et al 2000
SOLENODONTIDAE
Solenodon arredondoi e CD 5 Morgan y Ottenwalder 1993
PnHECIIDAE
Paralouatta varonai d CD 5 MacPhee 1996a
EcHIMYIDAE
Heteropsomys antillensis c PR 5 McFarJane 1999b
Heteropsomys insulans c PR 5 Ibid
Puertoricomys eorozalus c PR 5 Woods 1989a
CAPROMYIDAE
P lagiodontia araeum c LE 5 MacPhee et al 2000
Macrocapromys latus c CD 5 Varona y Arredondo 1979
(Continua)
325 EXTINCIONES DEL CUATERNARIO
TABLA 45 (Continuaci6n)
F ami1ia1Especie Peso Isla Evidencia Fuente
M acrocapromys latus c CU 5 Varona y Arredondo 1979
Mesocapromys kraglievichi c CU 5 Ibid
HEPTAXODONTINAE
Xaymaca fulvopulvis a JA 2 MacPhee y Flemming 2003
III Intervalo Amerindio (6-05 kaAP)
MEGALONYCHIDAE
Acratocnus ye d LE 2 MacPhee et aI 2000
Neocnus comes c LE 2 Ibid
N eocnus dousman c LE 2 Ibid
N eocnus toupiti c LE 2 Ibid
N eocnus gliriform is c CU 4 Cordova et ai 1997
M egalocnus rodens f CU 2 MacPhee et ai 2007
Parocnus browni e CU 2 Steadman et ai 2005
P arocnus serus e LE 2 MacPhee et ai 2000
N ESOPHONTIDAE
N esophontes edithae a PR 2 McFarlane 1999b
PITHECIIDAE
Antillothrix bernensis c LE 3 Rimoli 1977
EcHIMYIDAE
Brotomys contractus a LE 4 Miller 1929c
CAPROMYIDAE
P lagiodontia spelaeum c LE 4 Ibid
M acrocapromys acevedo c CU 4 NUiiez 1990a
HEPTAXODONTINAE
Elasmodontomys obliquus d PR 3 McFarlane 1999b
Iv Intervalo Postcolombino (05 kaAP - Presente)
N ESOPHONTIDAE
N esophontes hypom icrus a LE 24 MacPhee y Fle111111ing 1999
N esophontes zam icrus a LE (4) Ibid
N esophontes m icrus a CU 24 MacPhee et aI 1999
(Continua)
326 BIOGEOGRAFiA
TABLA 45 (Continuacion)
F amilialEspecie Peso Isla Evidencia Fuente
N esophontes m icrus a CD 24 MacPhee et aI 1999
N esophontes m icrus a LE 4 MacPhee et aI 2000
N esophontes sp 1 a IC 4 Morgan 1994
N esophontes sp 2 a IC 4 Ibid
Solenodon marcanoi b LE 4 Ottenwalder 2001
PITHECIIDAE
Xenothrix mcgregori c JA 4 MacPhee y Flemming 1999
EcHIMYIDAE
Boromys offeUa a CD 24 Guarch 1984
Boromys torrei a CD 24 Ibid
Brotomys voratus a LE 4 MacPhee et aI 2000
CAPROMYIDAE
P lagiodontia ipnaeum c LE 4 MacPhee y Flemming 1999
Rhizoplagiodontia lemkei c LE 4 Ibid
Isolobodon portoricensis c PR 24 Ibid
Isolobodon montanus c LE 4 Ibid
Hexolobodon phenax c LE (4) Ibid
Capromys sp c IC 24 Morgan 1994
Geocapromys columbianus c CD 4 MacPhee y Flemming 1999
Geocapromys thoracatus b IS 1 MacPhee y Marx 1997
Geocapromys sp 1 c IC 4 Morgan 1994
Geocapromys sp 2 c IC (4) Ibid
l-IEPTAXODONTINAE
Quemisia gravis c LE (1) MacPhee y Flemming 1999
MURIDAE
Oryzomys antillarum a JA 1 Ibid
1 Flemming y MacPhee (1999) alertaron en contra de la asociacion con otro material fechado Asimismo Morgan (1993) indico que la ausencia de asociacion con evidencias culturales amerindias no es necesariamente un criterio concluyente para ubicar un taxon en el intervalo temporal anterior porque hay depositos en la region que carecen de tales evidencias y son del Holoceno tardio por fechados radiocarbonicos [No obstante
327 EXTlNCIONES DEL CUATERNAR10
la ausencia de una especie en residuarios arqueol6gicos adquiere mayor certidumbre como criterio de ubicaci6n temporal de su extinci6n en la medida que aumenta el numero de residuarios estudiados que en las Antillas Mayores se cuentan por centenares] A falta de fechado absoluto de una especie consideramos que estos criterios de ubicaci6n temporal son mas utiles que la alternativa de excluir del analisis a la especie
2 En los trabajos sobre biogeografia de mamiferos antillanos los arque610gos mas citados como autoridades para establecer el inicio de la presencia aborigen en las Antillas Mayores (Kozlowski 1974 Rouse y Allaire 1978 Rouse 1989 1992 Wilson 2001) difieren al respecto entre los 6 kaAP y los 45 kaAP (pero vease Pagan et al 200523) y hemos adoptado la mas temprana de estas fechas para el comienzo del intervalo amerindio en la Tabla 45 De todos modos cualquiera que haya sido la fecha esta no es aplicable por igual al conjunto de las Antillas Mayores porque segun algunos de los autores citados el poblamiento comenz6 por Cuba y La Espanola 6 mil anos atras continu6 en Puerto Rico hace alrededor de 3 mil anos (pero vease Burney et al 1994) yen Jamaica hace menos de 2 mil e incluso a las Islas Caiman ni siquiera llegaron [Las Islas Caiman descubiertas por Col6n en 1503 carecen absolutamente de evidencia amerindia (Stokes y Keegan 1996)]
Si los datos de la Tabla 45 representan una aceptable aproximaci6n a la realidad el proceso de las extinciones cuaternarias de los mamiferos terrestres de las Antillas Mayores y grupos de islas perifericas debi6 caracterizarse entre otros por los eventos siguientes
Aunque obviamente la fauna cuaternaria debi6 ser mucho mas numerosa que 10 que refleja el registro paleontol6gico (Morgan y Woods 1986) 52 especies (81 de la fauna conocida) desaparecieron (sin reposici6n) durante los ultimos 250 mil anos
- Al parecer las extinciones se concentraron en las especies de mayor tamano Asi el peso promedio estimado para las especies de la etapa preantr6pica (intervalos I + II de la Tabla 45) es 350 kg es 328 kg para las especies del intervalo III (amerindio) y es s610 25 kg (c1ases del peso a b c) para las especies del intervalo IV (postcolombino) Ninguna especie aut6ctona de mamifero mayor de 6 kg existe hoy en la regi6n
- De acuerdo con la distribuci6n temporal de las extinciones 61 especies sobrevivieron al interglacial Sangamon (intervalo 1)48 sobrevivieron a la transici6n del Pleistoceno terminal al Holoceno temprano (intervalo II) 34 sobrevivieron al periodo amerindio (intervalo III) y s610 12 especies han sobrevivido desde la Ilegada de los europeos (intervalo IV) Las tasas de extinci6n correspondientes (exceptuado el intervalo I) son una especie cada 1167 anos en el intervalo II una cada 393 anos en el intervalo III y una cada 23 anos en el intervalo IV
- Durante todo el proceso de las extinciones cuaternarias la perdida de especies por isla fue de 91 en La Espanola 83 en Jamaica y 62 en Cuba Bahamas retuvo su unica especie (aunque extirpada en varias de las islas) Puerto Rico e Islas Caiman perdieron todas sus especies (6 y 5 respectivamente) y las Islas Swan Virgenes y Anguila - San Martin perdieron sus respectivas especies (micas
328 BIOGEOGRAFiA
De los cuatro 6rdenes de mamiferos representados en la fauna Pilosa y Primates desaparecieron totalmente Soricomorpha y Rodentia perdieron 86 y 74 de sus especies respectivamente
CAUSALIDAD
CAUSAS NATURALES
Los cicIos glaciales e interglaciales acaecidos en el Pleistoceno causaron cambios ambientales dnisticos en las Antillas con la concomitante alternancia de cIimas frios (secos) y calidos (hUmedos) asi como de descensos y ascensos del nivel del mar aumento y disminuci6n de las areas territoriales) induso (aunque con menor intensidad) en el Holoceno Estos eventos paleogeograficos han sido consistentemente documentados (Mayo y Kartashov 1972 Bonnatti y Gartner 1973 Ortega y Arcia 1982 Prance 1982 Schubert y Medina 1982 Dodge et al 1983 Ortega [1984] Haq et al 1987 Schubert 1989 Hodell et al 1991 Guilderson et al 1994 Oppo 1997 Emslie 1998 Curtis et al 2001 Iturralde-Vinent 2003) y de ellos brindamos a continuaci6n una breve resena
Finalizando el periodo interglacial anterior a la ultima glaciaci6n (Wisconsin) el mar estaba a 8-10 m por encima de su nivel actual Cuando el avance glacial del Wisconsin a1canz6 su culminaci6n (eillamado Ultimo Maximo Glacial) hace 18 ka el mar habia descendido 120 m con respecto a dicho nivel y las islas del Caribe habian incrementado considerablemente sus extensiones subaereas en comparaci6n con las que presentan hoy dia por ejemplo Puerto Rico y las Islas Virgenes se conectaron territorialmente y las Bahamas se unieron en cinco gran des islas Prevalecia un regimen frio y seco y condiciones de aridez --debidas a una reducci6n de las precipitaciones (posiblemente a menos de 440 mmlano)- y los ambientes xericos se habian extendido a traves de las islas y bancos emergidos y abundaban los pinares (los habitats mesicos estaban restringidos a parcelas relictas en 10 alto de las montanas) En el Mar Caribe y el Golfo de Mexico la temperatura superficial del mar era 5degC mas baja que la actual y en las llanuras costeras y alturas maximas eran respectivamente 2-3degC y 6degC mas bajas Los limites de las zonas ecol6gicas altitudinales descendieron varios cientos de metros y cada zona redujo su extensi6n vertical al aumentar el gradiente de temperatura Ellimite inferior de la zona periglacial
elevaci6n donde la temperatura media minima a1canza los OdegC) de la Cordillera Central de la Republica Dominicana (altura maxima del Caribe insular con evidencia de topografia glacial) se considera que estuvo entre los 1500 y 1800 mlnm (actualmente esta a 2800 mlnm) y se puede estimar que dicho limite era inferior en Cuba por estar mas al norte que la Republica Dominicana No obstante durante este periodo la temperatura experiment6 episodios cortos (unas pocas decadas) de ascensos repetidos
Un signifieativo componente bi6tieo de las referidas condiciones paleoambientales 10 constituy6 la espectacular irradiaci6n evolucionaria experimentada por un grupo numeroso de aves rapaees diurnas y nocturnas (Wetmore 1937 Brodkorb 1969 Kurotchkin y Mayo 1973 Arredondo 1976 Olson 1978 Olson y Hilgartner 1982
329 EXTINCIONES DEL ClJATERNARIO
Suarez 2000a b 2004 Suarez y Emslie 2003) Los mamiferos de las Antillas Mayores ofrecieron a estos depredadores aviares una amplia gama de tamanos de presa que en ausencia de mamiferos del orden Carnivora en la region permitio a estas aves irradiarse a este nicho alimentario con la evolucion de una variedad de especies de muy diversa tall a corporal incluidas formas gigantes Adquirir un cuerpo gigante les propicio el acceso a recursos alimentarios que no estaban siendo explotados por ningun otro depredador (Alcover y McMinn 1994) A Cuba corresponde el mejor registro fosil de rapaces cuaternarias caribenas con siete generos de especies diurnas Dentro de estas aves existieron tres gavilanes gigantes (Accipitridae) tres carairas (Falconidae) y un condor (Cathartidae) asi como cuatro generos de especies nocturnas dos lechuzas (Tytonidae) y tres Mhos (Strigidae) dos de ell os no igualados en tamano en todo el mundo Algunos de estos generos tuvieron representacion en otras islas antillanas como La Espanola Jamaica Bahamas y Gran Caiman
Al retirarse hacia el norte los hielos de la glaciacion Wisconsin el clima se tomo mas hUmedo en las Antillas durante el Holoceno temprano y el bosque mesico se expandio en la region insular El nivel del mar ascendio acompanado de mayor pluviosidad alrededor de los 7 kaAP las Islas Virgenes quedaron separadas entre si y de Puerto Rico y las Bahamas perdieron la mayor parte de su territorio Cerca de los 3 kaAP las condiciones secas retornaron a la region e hicieron desaparecer buena parte del bosque mesico Nuevamente el clima comenzo a adquirir sus condiciones actuales hacia los 25 kaAP aproximadamente y hoy transita por una fase mesica con vegetacion xerica confinada en parcelas relictas
Sin duda estos y otros eventos naturales incidieron negativamente en la sobrevivencia de mamiferos terrestres en las Antillas Mayores asi como en los grupos de pequenas islas perifericas (Pregill y Olson 1981 Olson 2006)
McFarlane et al (1998a) demostraron convincentemente como Amblyrhiza el gigante de los roedores antillanos endemico de la pequena isla de Anguila sucumbio a las repetidas y abruptas perdidas de su habitat durante el interglacial Sangamon por las nipidas y severas oscilaciones del nivel del mar (documentadas por Neumann y Hearty 1996) al no poder responder adecuadamente a tan catastroficos eventos mediante el ajuste de su masa corporal media y otro tanto debio ocurrirle en Jamaica al otro roedor gigante Clidomys (McFarlane et al 1998b) emparentado con Amblyrhiza en la misma epoca (Tabla 45 intervalo I)
El ascenso del nivel del mar seguramente promovio la extincion de mamiferos durante la transicion del Pleistoceno al Holoceno (Tabla 45 intervalo II) con la reduccion del tamano de las Islas por las transgresiones marinas (Pregill y Olson 1981 Morgan 1993) Esto ultimo haria maximamente severa la competicion asf como la presion de los depredadores [Steadman et al (2005) no yen por que el calentamiento global ocurrido durante la transicion Pleistoceno-Holoceno habria de tener un efecto negativo sobre los perezosos siendo sus descendientes actuales especies tropic ales 0 subtropicales pero no se trata del efecto
330 BIOGEOGRAFiA
directo del cambio de temperatura sobre los animales sino del efecto indirecto y progresivo que dicho cambio genera a largo plazo al reducir el territorio insular disponible]
Por esta epoca tambien desaparecieron especies de tall a mayor como Megalocnus zile en La Espanola y otras de menor tamano (en Cuba) pero que representan la tall a maxima en sus respectivos contextos taxonomicos del Cuaternario como Solenodon arredondoi Paralouatta varonai y Macrocapromys acevedo Un mamifero puede ser grande solo a costa de reducir su densidad poblacional y esto 10 hace mis vulnerable a la extinci6n sobre todo en una isla (McNab 1994 McFarlane et al 1998a) Incidentalmente Alcover et al (1998a) afirmaron que larelacion entre la talla corporal y la extincion no siempre se cumple en las especies de menor tamano y citaron como ejemplo que entre las musarafias todas las especies de Nesophontes desaparecieron mientras sobreviven especies de Solenodon pero la comparacion correcta en todo caso serfa entre los solenodones vivientes y el gigante Solenodon arredondoi y entre Nesophontes edithae -el gigante de los nesofontes extinguido en el periodo amerindio- y el resto de sus congeneres que sobrevivieron hasta los primeros siglos de la colonizacion europea (Tabla 45 intervalos III y IV)
CAUSAS ANTROPOGENICAS
Intervalo Amerindio
La mayoria de los autores que se han referido a las extinciones acaecidas durante este intervalo (entre elIos paradigmaticamente Martin 1984a b) han responsabilizado a los aborfgenes (que se supone comenzaron a poblar las Antillas Mayores hace 6 mil afios) con la desaparicion de numerosas especies de mamiferos terrestres en la region principalmente por la via de la sobrecaza y la introduccion de animales domesticos devenidos depredadores de la fauna autoctona (Morgan y Woods 1986) Este efecto negativo de la presencia indigena sobre los mamiferos de la region ha sido dramatizado por algunos autores en terminos de holocausto faunal Sin embargo pensamos que se ha exagerado Como han expresado MacPhee et al (19993) cuando uno busca evidencia dura de complicidad humana directa en [estas] extinciones no hay mucho que encontrar Las siguientes son algunas consideraciones a tener en cuenta en tal sentido
El registro radiometrico mas joven conocido hasta ahora para un perezoso antillano (MegalocnllS rodens) es de 42 kaAP (MacPhee et ai 2007) Si se acepta que los aborfgenes entraron en Cuba hace aproximadamente 6 mil anos su convivencia con los perezosos debe haber durado alrededor de 1800 afios (0 mas porque dificilmente el referfdo fechado radiometrico haya correspondido al ultimo sobreviviente de la especie) Esta cifra contradice la hipotesis del holocausto faunal segun la cualla extincion de los mamiferos insulares de mayor talla debe ocurrir con extrema rapidez -a escala de decadas 0 de centurias- a partir de su primer contacto con los humanos (Martin y Steadman 1999) A pesar de las repetidas afirmaciones en contrario (Harrington 1921 Miller 1929c Herrera [1945] Tabio 1951 Alvarez 1951a 1957 Arredondo
331 EXTINCIONES DEL CUATERNARIO
1959 1960a Veloz y Ortega 1973 Mayo 1980b Rodriguez et al 1984 Castellanos y Pino 1985 Pino y Castellanos 1985 Nunez 1990b Gutierrez 1996 Arredondo 2004) en su mayoria basadas en evidencias no exentas de posibles accidentes de deposicion (0 de reubicacion postdeposicional) en recintos cavernarios (Diaz-Franco [2006b D no se ha demostrado inequivocamente que los aborigenes antillanos se alimentaran de perezosos regularmente Sus restos en residuarios de alimentos son escasisimos en toda la region y esto contrasta con los abundantes restos de jutias y otros mamiferos de menor talla que aparecen en esos residuarios demostrativos de la preferencia de los paleoindios por estos otros mamiferos
Los mamiferos mas importantes en la economia prehistorica de las Antillas Mayores fueron las jutias Capromys y Geocapromys en Cuba Geocapromys en Jamaica e Isolobodon en La Espanola Puerto Rico e Islas Virgenes (Aguayo 1950b Tablo 1951 Rimoli 1972 Wing 2001 Diaz-Franco [2006b D Wilkins (2001) estudi61ajutia viviente de Jamaica (Geocapromys browni) en un sitio ocupado por los aborigenes durante 100 anos y no encontr6 evidencias de que durante ese tiempo la especie haya sido sobreexplotada Asimismo en Cuba las jutias conga carabali y andaraz son escasas en depositos pleistoholocenicos en cambio son muy numerosas en residuarios indigenas y de haber sido sobreexplotadas 10 mas probable es que no hubieran sobrevivido hasta hoy
La habilidad de los humanos para penetrar en profundidad el bosque tropical y mantenerse en este sustentablemente ha sido un evento comparativamente reciente (Beran y Steele 1994) En el caso antillano la caza tierra adentro ha sido explicada como un proceso tardio de desplazamiento a partir de la explotacion de abundantes recursos marinos que ofrecian condiciones 6ptimas para el sedentarismo de las comunidades aborigenes costeras (C6rdova y Arredondo 1988 Brito 2005) Peces y mariscos dominan la arqueologia costera de las Islas antillanas en tanto el consumo de recursos terrestres (mamfferos incluidos) aumenta progresivamente en la medida que los residuarios estudiados se alejan de la costa (Scudder 1991 Wilkins 2001) En las Antillas Mayores el componente terrestre de la dieta aborigen es solo 20-30 del material recobrado en los residuarios (Wing 1977 1989) Y debio ser menor alin porque el componente vegetal de la dieta no se ha preservado Por otra parte los aborigenes cubanos domesticaron la jutia conga y seguramente la reprodujeron para comer (Pose et al 1988) practica que probablemente se repitiera con otras jutias en las demas Antillas Mayores
- Fray Bartolome de las Casas hablo de 3 millones de indios en Cuba al inicio de la colonizacion pero los historiadores subsecuentes consideraron exagerada la cifra (Poey 1851 a) No obstante se dispone de dos estimados de la densidad de poblaci6n aborigen en las Antillas Mayores a la llegada de los europeos 18 indigenaslkm2 en Puerto Rico (Rouse 1956) y 1271km2 en La Espanola (Watts 1987) Extrapolando ambos valores entre las cuatro islas principales de las Antillas Mayores tenemos que en la fecha indicada la poblaci6n aborigen pudo estar entre 16 mil y 113 mil indigenas en Puerto Rico entre 21 mil y 145 mil en Jamaica entre 141 mil y 1 mill6n en La Espanola y entre 200 mil y 14 millones en Cuba Entre las cifras maximas y las minimas nos inclinamos
332 BIOGEOGRAFiA
hacia estas ultimas atendiendo a los altos indices de mortalidad infantil y morbilidad que se evidencian en las osamentas de los pobladores prehispanicos (Martinez y Rives 1990) Uno de los hechos que apuntan hacia el bajo numero de aborigenes en las Antillas Mayores es 10 rapido que desaparecieron despues de la llegada de los europeos pnkticamente sin dejar mestizaje con estos 10 cual contrasta ostensiblemente con la situacion en la America continentaL
En suma un grupo humano (1) demograiicamente irrelevante y controlado por altos indices de morbilidad (2) consumidor mayoritariamente de organismos marinos y (3) carente de la tecnologia cinegetica necesaria para poder sobreexplotar los mamiferos que Ie sirvieron de alimento (a los cuales accedio tardiamente en su desarrollo) no es concebible que haya causado e1 holocausto faunal que se Ie atribuye a partir del error de aplicar a las comunidades aborigenes antillanas un modelo cultural de la prehistoria continentaL
Tampoco hay evidencias de que los aborigenes hayan introducido carnivoros con suficiente capacidad depredadora salvo el perro Morgan y Otlenwaldder (1993) consideraron que la depredacion por perros durante el periodo amerindio es la explicacion mas plausible para la extincion del almiqui gigante de Cuba pero como se puede observar (Tabla 54) Solenodon arredondoi aparece en el intervalo II por falta de asociacion con evidencias culturales
Consecuentemente el accionar de los aborigenes y de sus animales domesticos directamente sobre la fauna no parece haber sido por sf solo un factor determinante de la extincion de mamiferos terrestres durante el periodo amerindio en las Antillas (pero vease Steadman et al 2005)
Hay que recordar en este senti do que alin en el Holoceno tardio ocurrieron cambios clirruiticos de consideracion (vease Causas Naturales) Por ejemplo Curtis et al (2001) explicaron que la extincion de algunos ani males mesofilicos de las alturas montanosas de La Espanola pudo haber antecedido a cualquier impacto humano sigrIificativo en esos lugares por coincidir dicha extincion con una tendencia a la aridez documentada isotopicamente y con evidencias de la perdida del bosque mesico ocurridas despues de 3kaAP Yen Jamaica hace solo 2shy3 mil aDos el area de la Bahia de Jackson sustentaba una gran biomasa de vertebrados en bosque hinnedo a juzgar por la cantidad y diversidad de taxones presentes en material fosil derivado de la depredacion por lechuzas hoy el area es la mas seca de la isla con matorral xerofitico y pobreza de fauna (MacPhee 1996a) Yalm en nuestros dias huracanes de gran intensidad aniquilan poblaciones de vertebrados (que no se recuperan) en is las pequenas y llanas del Caribe (Brooks y Smith 2001)
En medio de la incertidumbre sobre la verdadera causa (0 causas) de las extinciones de mamiferos en el periodo amerindio MacPhee y Marx (1997) han propuesto su hipotesis de la hipermorbilidad la cual estipula colapsos poblacionales relativamente rapidos con la introduccion de enfermedades de alta letalidad Estos autores han razonado que
Las islas son los lugares mas apropiados para experimentar perdidas catastroficas causadas por enfermedades Los mamiferos no solo serian susceptibles en virtud de su prolongado aislamiento biologico sino que en los confines de una isla la epizootia
333 EXTINCIONES DEL CUATERNARIO
tendria la potencialidad de infectar simultaneamente a la rnayoria de los individuos de vadas especies Los neonatos son inmunologicamente inmaduros y sufririan desproporcionadamente cualquier infeccion hipervirulenta La perdida excesiva de neonatos y juveniles tendria efectos severos para la sobrevivencia de la poblacion si esta se reduce por debajo de cierto nivel sobreviene la extincion porque no hay posibilidad de restitucion en el periodo postpanzootico Las especies de mayor talla corporal van a ser las mas vulnerables a la extincion dadas sus bajas tasas reproductivas largos periodos de gestacion y reducido numero de progenie Las especies de menor tamano --caracterizadas por altas tasas reproductivas periodos cortos de gestacion y mayor numero de neonatos por parto- tambien serian fuertemente diezmadas pero muchas sobrevivirian porque su estrategia reproductiva les permite perder mayor numero de individuos Las perdidas deben ocurrir relativamente rapido (a escala de decadas) mientras que otras hipotesis (digamos los cambios climaticos) permiten intervalos mucho mas largos (a escala de siglos 0 de milenios) entre la aparicion de las causas y la produccion de sus efectos Por ejemplo todo indica que los nesofontes se extinguieron muy rapidamente y seria dificil poder invocar un impacto humano directo 0 argumentar fenomenos climaticos que pudieran explicar plausiblemente perdidas tan rapidas
MacPhee y Marx (1997 191) han dicho que por el momento la hipotesis de la hipermorbilidad no pasa de ser una abstraccion en espera de adecuada sustentacion empirica Llama la atencion que linicamente en el periodo amerindio (Tabla 45 intervalo 1Il) las extinciones abarcaron todas las clases del peso corporal (de la a a laj) y que ninguna de las especies que sobrevivieron hasta el periodo postcolombino (Tabla 45 intervalo IV mas las especies vivientes) sobrepasa los 10 kg (clases del peso a b c) en clara coincidencia con los presupuestos de la hipotetica hipermorbilidad
Intervalo Postcolornbino
Por 10 menos 37 especies de mamiferos terrestres vivian en las Antillas Mayores y grupos de Islas perifericas a la lIegada de los europeos en 1492 De elIas se han extinguido 23 shymas especies que en cualquiera de los periodos precedentes (Tabla 45~ y solo subsisten 12 especies repartidas entre Cuba (8 especies) La Espanola (2) Jamaica (1) y Bahamas (1) varias de estas en peligro critico de extincion Solo dos de las especies extinguidas durante los ultimos 500 anos se conocieron en estado viviente los roedores Oryzomys antillarum de Jamaica (extinguido en 1877) y Geocapromys thoracatus de las Islas Swan (extinguido en 1955) Las causas de las extinciones ocurridas durante este periodo son sobradamente conocidas destruccion de habitats introducci6n de animales exoticos (domesticos y comensales) y demas presiones asociadas a la actividad humana entre las cuales posiblemente habria que incluir brotes de hipermorbilidad
Durante mucho tiempo se ha afirmado la posibilidad de que especies de Nesophontes hayan sobrevivido hasta las primeras decadas del siglo xx y que incluso continuen existiendo en lugares remotos de Cuba 0 La Espanola (Allen 1918 1942 Miller 1930 Aguayo 1950a Arredondo 1960b Fischer 1977 MacFadden 1980 Woods y
334 BIOGEOGRAFiA
Ottenwalder 1991) Actualmente no existe evidencia radiometrica de que alguna especie de Nesophontes haya sobrevivido al siglo xv las fechas radiocarbonicas calibradas obtenidas en muestras de Nesophontes de Cuba y La Espanola resultaron no mas tardias que los siglos XIII y XIV (MacPhee et al 1999) Por otra parte las expediciones efectuadas a fines del siglo xx con el proposito de localizar individuos vivientes del genero en La Espanola no produjeron la menor indicacion afirmativa al respecto (Woods et al 1985) Ademas a traves de las Antillas Mayores no aparecen restos de Nesophontes en los depositos formados actualmente por las lechuzas
EPiLOGO
Las Antillas Mayores son islas oceanicas no han formado parte de un continente (con la posible excepcion de una parte de Jamaica hasta el Eoceno Medio hace 45 Ma) y durante un tiempo geologico considerable (desde el Eoceno Superior hace 40 Ma) han permanecido emergidas y separadas de los continentes vecinos Consecuentemente los mamiferos terrestres (no volantes) autoctonos de estas islas constituyen una fauna oceanica que por ende cumple con todas las caracteristicas convencionales reconocidas para este tipo de entidad biogeografica
- Baja diversidad taxonomica a nivel de las categorias superiores (solo cuatro ordenes) en comparacion con islas plataformicas continentales (0 con areas continentales) de tamano y diversidad ecologica simi lares
- Alto grado de endemismo a nivel de genero (96) y familia (50) en respuesta al prolongado aislamiento
- Desbalance ecologico de la estructura trofica ausencia de mamiferos camivoros y ungulados
- Espectaculares procesos de irradiaci6n evolucionaria como resultado de la colonizacion de territorios libres de competidores y depredadores
Tendencias evolucionarias extremas gigantismo
- Control a cargo de aves rapaces con tendencia at gigantismo en ausencia de mamiferos miembros del orden Carnivora
- Mas especies de reptiles y de murcielagos que de mamiferos terrestres relaciones que en los continentes se invierten
Alta tasa de extincion de especies (84) sin reposicion
Preservacion de especies filogeneticamente arcaicas (los llamados fosiles vivientes) los almiquies

325 EXTINCIONES DEL CUATERNARIO
TABLA 45 (Continuaci6n)
F ami1ia1Especie Peso Isla Evidencia Fuente
M acrocapromys latus c CU 5 Varona y Arredondo 1979
Mesocapromys kraglievichi c CU 5 Ibid
HEPTAXODONTINAE
Xaymaca fulvopulvis a JA 2 MacPhee y Flemming 2003
III Intervalo Amerindio (6-05 kaAP)
MEGALONYCHIDAE
Acratocnus ye d LE 2 MacPhee et aI 2000
Neocnus comes c LE 2 Ibid
N eocnus dousman c LE 2 Ibid
N eocnus toupiti c LE 2 Ibid
N eocnus gliriform is c CU 4 Cordova et ai 1997
M egalocnus rodens f CU 2 MacPhee et ai 2007
Parocnus browni e CU 2 Steadman et ai 2005
P arocnus serus e LE 2 MacPhee et ai 2000
N ESOPHONTIDAE
N esophontes edithae a PR 2 McFarlane 1999b
PITHECIIDAE
Antillothrix bernensis c LE 3 Rimoli 1977
EcHIMYIDAE
Brotomys contractus a LE 4 Miller 1929c
CAPROMYIDAE
P lagiodontia spelaeum c LE 4 Ibid
M acrocapromys acevedo c CU 4 NUiiez 1990a
HEPTAXODONTINAE
Elasmodontomys obliquus d PR 3 McFarlane 1999b
Iv Intervalo Postcolombino (05 kaAP - Presente)
N ESOPHONTIDAE
N esophontes hypom icrus a LE 24 MacPhee y Fle111111ing 1999
N esophontes zam icrus a LE (4) Ibid
N esophontes m icrus a CU 24 MacPhee et aI 1999
(Continua)
326 BIOGEOGRAFiA
TABLA 45 (Continuacion)
F amilialEspecie Peso Isla Evidencia Fuente
N esophontes m icrus a CD 24 MacPhee et aI 1999
N esophontes m icrus a LE 4 MacPhee et aI 2000
N esophontes sp 1 a IC 4 Morgan 1994
N esophontes sp 2 a IC 4 Ibid
Solenodon marcanoi b LE 4 Ottenwalder 2001
PITHECIIDAE
Xenothrix mcgregori c JA 4 MacPhee y Flemming 1999
EcHIMYIDAE
Boromys offeUa a CD 24 Guarch 1984
Boromys torrei a CD 24 Ibid
Brotomys voratus a LE 4 MacPhee et aI 2000
CAPROMYIDAE
P lagiodontia ipnaeum c LE 4 MacPhee y Flemming 1999
Rhizoplagiodontia lemkei c LE 4 Ibid
Isolobodon portoricensis c PR 24 Ibid
Isolobodon montanus c LE 4 Ibid
Hexolobodon phenax c LE (4) Ibid
Capromys sp c IC 24 Morgan 1994
Geocapromys columbianus c CD 4 MacPhee y Flemming 1999
Geocapromys thoracatus b IS 1 MacPhee y Marx 1997
Geocapromys sp 1 c IC 4 Morgan 1994
Geocapromys sp 2 c IC (4) Ibid
l-IEPTAXODONTINAE
Quemisia gravis c LE (1) MacPhee y Flemming 1999
MURIDAE
Oryzomys antillarum a JA 1 Ibid
1 Flemming y MacPhee (1999) alertaron en contra de la asociacion con otro material fechado Asimismo Morgan (1993) indico que la ausencia de asociacion con evidencias culturales amerindias no es necesariamente un criterio concluyente para ubicar un taxon en el intervalo temporal anterior porque hay depositos en la region que carecen de tales evidencias y son del Holoceno tardio por fechados radiocarbonicos [No obstante
327 EXTlNCIONES DEL CUATERNAR10
la ausencia de una especie en residuarios arqueol6gicos adquiere mayor certidumbre como criterio de ubicaci6n temporal de su extinci6n en la medida que aumenta el numero de residuarios estudiados que en las Antillas Mayores se cuentan por centenares] A falta de fechado absoluto de una especie consideramos que estos criterios de ubicaci6n temporal son mas utiles que la alternativa de excluir del analisis a la especie
2 En los trabajos sobre biogeografia de mamiferos antillanos los arque610gos mas citados como autoridades para establecer el inicio de la presencia aborigen en las Antillas Mayores (Kozlowski 1974 Rouse y Allaire 1978 Rouse 1989 1992 Wilson 2001) difieren al respecto entre los 6 kaAP y los 45 kaAP (pero vease Pagan et al 200523) y hemos adoptado la mas temprana de estas fechas para el comienzo del intervalo amerindio en la Tabla 45 De todos modos cualquiera que haya sido la fecha esta no es aplicable por igual al conjunto de las Antillas Mayores porque segun algunos de los autores citados el poblamiento comenz6 por Cuba y La Espanola 6 mil anos atras continu6 en Puerto Rico hace alrededor de 3 mil anos (pero vease Burney et al 1994) yen Jamaica hace menos de 2 mil e incluso a las Islas Caiman ni siquiera llegaron [Las Islas Caiman descubiertas por Col6n en 1503 carecen absolutamente de evidencia amerindia (Stokes y Keegan 1996)]
Si los datos de la Tabla 45 representan una aceptable aproximaci6n a la realidad el proceso de las extinciones cuaternarias de los mamiferos terrestres de las Antillas Mayores y grupos de islas perifericas debi6 caracterizarse entre otros por los eventos siguientes
Aunque obviamente la fauna cuaternaria debi6 ser mucho mas numerosa que 10 que refleja el registro paleontol6gico (Morgan y Woods 1986) 52 especies (81 de la fauna conocida) desaparecieron (sin reposici6n) durante los ultimos 250 mil anos
- Al parecer las extinciones se concentraron en las especies de mayor tamano Asi el peso promedio estimado para las especies de la etapa preantr6pica (intervalos I + II de la Tabla 45) es 350 kg es 328 kg para las especies del intervalo III (amerindio) y es s610 25 kg (c1ases del peso a b c) para las especies del intervalo IV (postcolombino) Ninguna especie aut6ctona de mamifero mayor de 6 kg existe hoy en la regi6n
- De acuerdo con la distribuci6n temporal de las extinciones 61 especies sobrevivieron al interglacial Sangamon (intervalo 1)48 sobrevivieron a la transici6n del Pleistoceno terminal al Holoceno temprano (intervalo II) 34 sobrevivieron al periodo amerindio (intervalo III) y s610 12 especies han sobrevivido desde la Ilegada de los europeos (intervalo IV) Las tasas de extinci6n correspondientes (exceptuado el intervalo I) son una especie cada 1167 anos en el intervalo II una cada 393 anos en el intervalo III y una cada 23 anos en el intervalo IV
- Durante todo el proceso de las extinciones cuaternarias la perdida de especies por isla fue de 91 en La Espanola 83 en Jamaica y 62 en Cuba Bahamas retuvo su unica especie (aunque extirpada en varias de las islas) Puerto Rico e Islas Caiman perdieron todas sus especies (6 y 5 respectivamente) y las Islas Swan Virgenes y Anguila - San Martin perdieron sus respectivas especies (micas
328 BIOGEOGRAFiA
De los cuatro 6rdenes de mamiferos representados en la fauna Pilosa y Primates desaparecieron totalmente Soricomorpha y Rodentia perdieron 86 y 74 de sus especies respectivamente
CAUSALIDAD
CAUSAS NATURALES
Los cicIos glaciales e interglaciales acaecidos en el Pleistoceno causaron cambios ambientales dnisticos en las Antillas con la concomitante alternancia de cIimas frios (secos) y calidos (hUmedos) asi como de descensos y ascensos del nivel del mar aumento y disminuci6n de las areas territoriales) induso (aunque con menor intensidad) en el Holoceno Estos eventos paleogeograficos han sido consistentemente documentados (Mayo y Kartashov 1972 Bonnatti y Gartner 1973 Ortega y Arcia 1982 Prance 1982 Schubert y Medina 1982 Dodge et al 1983 Ortega [1984] Haq et al 1987 Schubert 1989 Hodell et al 1991 Guilderson et al 1994 Oppo 1997 Emslie 1998 Curtis et al 2001 Iturralde-Vinent 2003) y de ellos brindamos a continuaci6n una breve resena
Finalizando el periodo interglacial anterior a la ultima glaciaci6n (Wisconsin) el mar estaba a 8-10 m por encima de su nivel actual Cuando el avance glacial del Wisconsin a1canz6 su culminaci6n (eillamado Ultimo Maximo Glacial) hace 18 ka el mar habia descendido 120 m con respecto a dicho nivel y las islas del Caribe habian incrementado considerablemente sus extensiones subaereas en comparaci6n con las que presentan hoy dia por ejemplo Puerto Rico y las Islas Virgenes se conectaron territorialmente y las Bahamas se unieron en cinco gran des islas Prevalecia un regimen frio y seco y condiciones de aridez --debidas a una reducci6n de las precipitaciones (posiblemente a menos de 440 mmlano)- y los ambientes xericos se habian extendido a traves de las islas y bancos emergidos y abundaban los pinares (los habitats mesicos estaban restringidos a parcelas relictas en 10 alto de las montanas) En el Mar Caribe y el Golfo de Mexico la temperatura superficial del mar era 5degC mas baja que la actual y en las llanuras costeras y alturas maximas eran respectivamente 2-3degC y 6degC mas bajas Los limites de las zonas ecol6gicas altitudinales descendieron varios cientos de metros y cada zona redujo su extensi6n vertical al aumentar el gradiente de temperatura Ellimite inferior de la zona periglacial
elevaci6n donde la temperatura media minima a1canza los OdegC) de la Cordillera Central de la Republica Dominicana (altura maxima del Caribe insular con evidencia de topografia glacial) se considera que estuvo entre los 1500 y 1800 mlnm (actualmente esta a 2800 mlnm) y se puede estimar que dicho limite era inferior en Cuba por estar mas al norte que la Republica Dominicana No obstante durante este periodo la temperatura experiment6 episodios cortos (unas pocas decadas) de ascensos repetidos
Un signifieativo componente bi6tieo de las referidas condiciones paleoambientales 10 constituy6 la espectacular irradiaci6n evolucionaria experimentada por un grupo numeroso de aves rapaees diurnas y nocturnas (Wetmore 1937 Brodkorb 1969 Kurotchkin y Mayo 1973 Arredondo 1976 Olson 1978 Olson y Hilgartner 1982
329 EXTINCIONES DEL ClJATERNARIO
Suarez 2000a b 2004 Suarez y Emslie 2003) Los mamiferos de las Antillas Mayores ofrecieron a estos depredadores aviares una amplia gama de tamanos de presa que en ausencia de mamiferos del orden Carnivora en la region permitio a estas aves irradiarse a este nicho alimentario con la evolucion de una variedad de especies de muy diversa tall a corporal incluidas formas gigantes Adquirir un cuerpo gigante les propicio el acceso a recursos alimentarios que no estaban siendo explotados por ningun otro depredador (Alcover y McMinn 1994) A Cuba corresponde el mejor registro fosil de rapaces cuaternarias caribenas con siete generos de especies diurnas Dentro de estas aves existieron tres gavilanes gigantes (Accipitridae) tres carairas (Falconidae) y un condor (Cathartidae) asi como cuatro generos de especies nocturnas dos lechuzas (Tytonidae) y tres Mhos (Strigidae) dos de ell os no igualados en tamano en todo el mundo Algunos de estos generos tuvieron representacion en otras islas antillanas como La Espanola Jamaica Bahamas y Gran Caiman
Al retirarse hacia el norte los hielos de la glaciacion Wisconsin el clima se tomo mas hUmedo en las Antillas durante el Holoceno temprano y el bosque mesico se expandio en la region insular El nivel del mar ascendio acompanado de mayor pluviosidad alrededor de los 7 kaAP las Islas Virgenes quedaron separadas entre si y de Puerto Rico y las Bahamas perdieron la mayor parte de su territorio Cerca de los 3 kaAP las condiciones secas retornaron a la region e hicieron desaparecer buena parte del bosque mesico Nuevamente el clima comenzo a adquirir sus condiciones actuales hacia los 25 kaAP aproximadamente y hoy transita por una fase mesica con vegetacion xerica confinada en parcelas relictas
Sin duda estos y otros eventos naturales incidieron negativamente en la sobrevivencia de mamiferos terrestres en las Antillas Mayores asi como en los grupos de pequenas islas perifericas (Pregill y Olson 1981 Olson 2006)
McFarlane et al (1998a) demostraron convincentemente como Amblyrhiza el gigante de los roedores antillanos endemico de la pequena isla de Anguila sucumbio a las repetidas y abruptas perdidas de su habitat durante el interglacial Sangamon por las nipidas y severas oscilaciones del nivel del mar (documentadas por Neumann y Hearty 1996) al no poder responder adecuadamente a tan catastroficos eventos mediante el ajuste de su masa corporal media y otro tanto debio ocurrirle en Jamaica al otro roedor gigante Clidomys (McFarlane et al 1998b) emparentado con Amblyrhiza en la misma epoca (Tabla 45 intervalo I)
El ascenso del nivel del mar seguramente promovio la extincion de mamiferos durante la transicion del Pleistoceno al Holoceno (Tabla 45 intervalo II) con la reduccion del tamano de las Islas por las transgresiones marinas (Pregill y Olson 1981 Morgan 1993) Esto ultimo haria maximamente severa la competicion asf como la presion de los depredadores [Steadman et al (2005) no yen por que el calentamiento global ocurrido durante la transicion Pleistoceno-Holoceno habria de tener un efecto negativo sobre los perezosos siendo sus descendientes actuales especies tropic ales 0 subtropicales pero no se trata del efecto
330 BIOGEOGRAFiA
directo del cambio de temperatura sobre los animales sino del efecto indirecto y progresivo que dicho cambio genera a largo plazo al reducir el territorio insular disponible]
Por esta epoca tambien desaparecieron especies de tall a mayor como Megalocnus zile en La Espanola y otras de menor tamano (en Cuba) pero que representan la tall a maxima en sus respectivos contextos taxonomicos del Cuaternario como Solenodon arredondoi Paralouatta varonai y Macrocapromys acevedo Un mamifero puede ser grande solo a costa de reducir su densidad poblacional y esto 10 hace mis vulnerable a la extinci6n sobre todo en una isla (McNab 1994 McFarlane et al 1998a) Incidentalmente Alcover et al (1998a) afirmaron que larelacion entre la talla corporal y la extincion no siempre se cumple en las especies de menor tamano y citaron como ejemplo que entre las musarafias todas las especies de Nesophontes desaparecieron mientras sobreviven especies de Solenodon pero la comparacion correcta en todo caso serfa entre los solenodones vivientes y el gigante Solenodon arredondoi y entre Nesophontes edithae -el gigante de los nesofontes extinguido en el periodo amerindio- y el resto de sus congeneres que sobrevivieron hasta los primeros siglos de la colonizacion europea (Tabla 45 intervalos III y IV)
CAUSAS ANTROPOGENICAS
Intervalo Amerindio
La mayoria de los autores que se han referido a las extinciones acaecidas durante este intervalo (entre elIos paradigmaticamente Martin 1984a b) han responsabilizado a los aborfgenes (que se supone comenzaron a poblar las Antillas Mayores hace 6 mil afios) con la desaparicion de numerosas especies de mamiferos terrestres en la region principalmente por la via de la sobrecaza y la introduccion de animales domesticos devenidos depredadores de la fauna autoctona (Morgan y Woods 1986) Este efecto negativo de la presencia indigena sobre los mamiferos de la region ha sido dramatizado por algunos autores en terminos de holocausto faunal Sin embargo pensamos que se ha exagerado Como han expresado MacPhee et al (19993) cuando uno busca evidencia dura de complicidad humana directa en [estas] extinciones no hay mucho que encontrar Las siguientes son algunas consideraciones a tener en cuenta en tal sentido
El registro radiometrico mas joven conocido hasta ahora para un perezoso antillano (MegalocnllS rodens) es de 42 kaAP (MacPhee et ai 2007) Si se acepta que los aborfgenes entraron en Cuba hace aproximadamente 6 mil anos su convivencia con los perezosos debe haber durado alrededor de 1800 afios (0 mas porque dificilmente el referfdo fechado radiometrico haya correspondido al ultimo sobreviviente de la especie) Esta cifra contradice la hipotesis del holocausto faunal segun la cualla extincion de los mamiferos insulares de mayor talla debe ocurrir con extrema rapidez -a escala de decadas 0 de centurias- a partir de su primer contacto con los humanos (Martin y Steadman 1999) A pesar de las repetidas afirmaciones en contrario (Harrington 1921 Miller 1929c Herrera [1945] Tabio 1951 Alvarez 1951a 1957 Arredondo
331 EXTINCIONES DEL CUATERNARIO
1959 1960a Veloz y Ortega 1973 Mayo 1980b Rodriguez et al 1984 Castellanos y Pino 1985 Pino y Castellanos 1985 Nunez 1990b Gutierrez 1996 Arredondo 2004) en su mayoria basadas en evidencias no exentas de posibles accidentes de deposicion (0 de reubicacion postdeposicional) en recintos cavernarios (Diaz-Franco [2006b D no se ha demostrado inequivocamente que los aborigenes antillanos se alimentaran de perezosos regularmente Sus restos en residuarios de alimentos son escasisimos en toda la region y esto contrasta con los abundantes restos de jutias y otros mamiferos de menor talla que aparecen en esos residuarios demostrativos de la preferencia de los paleoindios por estos otros mamiferos
Los mamiferos mas importantes en la economia prehistorica de las Antillas Mayores fueron las jutias Capromys y Geocapromys en Cuba Geocapromys en Jamaica e Isolobodon en La Espanola Puerto Rico e Islas Virgenes (Aguayo 1950b Tablo 1951 Rimoli 1972 Wing 2001 Diaz-Franco [2006b D Wilkins (2001) estudi61ajutia viviente de Jamaica (Geocapromys browni) en un sitio ocupado por los aborigenes durante 100 anos y no encontr6 evidencias de que durante ese tiempo la especie haya sido sobreexplotada Asimismo en Cuba las jutias conga carabali y andaraz son escasas en depositos pleistoholocenicos en cambio son muy numerosas en residuarios indigenas y de haber sido sobreexplotadas 10 mas probable es que no hubieran sobrevivido hasta hoy
La habilidad de los humanos para penetrar en profundidad el bosque tropical y mantenerse en este sustentablemente ha sido un evento comparativamente reciente (Beran y Steele 1994) En el caso antillano la caza tierra adentro ha sido explicada como un proceso tardio de desplazamiento a partir de la explotacion de abundantes recursos marinos que ofrecian condiciones 6ptimas para el sedentarismo de las comunidades aborigenes costeras (C6rdova y Arredondo 1988 Brito 2005) Peces y mariscos dominan la arqueologia costera de las Islas antillanas en tanto el consumo de recursos terrestres (mamfferos incluidos) aumenta progresivamente en la medida que los residuarios estudiados se alejan de la costa (Scudder 1991 Wilkins 2001) En las Antillas Mayores el componente terrestre de la dieta aborigen es solo 20-30 del material recobrado en los residuarios (Wing 1977 1989) Y debio ser menor alin porque el componente vegetal de la dieta no se ha preservado Por otra parte los aborigenes cubanos domesticaron la jutia conga y seguramente la reprodujeron para comer (Pose et al 1988) practica que probablemente se repitiera con otras jutias en las demas Antillas Mayores
- Fray Bartolome de las Casas hablo de 3 millones de indios en Cuba al inicio de la colonizacion pero los historiadores subsecuentes consideraron exagerada la cifra (Poey 1851 a) No obstante se dispone de dos estimados de la densidad de poblaci6n aborigen en las Antillas Mayores a la llegada de los europeos 18 indigenaslkm2 en Puerto Rico (Rouse 1956) y 1271km2 en La Espanola (Watts 1987) Extrapolando ambos valores entre las cuatro islas principales de las Antillas Mayores tenemos que en la fecha indicada la poblaci6n aborigen pudo estar entre 16 mil y 113 mil indigenas en Puerto Rico entre 21 mil y 145 mil en Jamaica entre 141 mil y 1 mill6n en La Espanola y entre 200 mil y 14 millones en Cuba Entre las cifras maximas y las minimas nos inclinamos
332 BIOGEOGRAFiA
hacia estas ultimas atendiendo a los altos indices de mortalidad infantil y morbilidad que se evidencian en las osamentas de los pobladores prehispanicos (Martinez y Rives 1990) Uno de los hechos que apuntan hacia el bajo numero de aborigenes en las Antillas Mayores es 10 rapido que desaparecieron despues de la llegada de los europeos pnkticamente sin dejar mestizaje con estos 10 cual contrasta ostensiblemente con la situacion en la America continentaL
En suma un grupo humano (1) demograiicamente irrelevante y controlado por altos indices de morbilidad (2) consumidor mayoritariamente de organismos marinos y (3) carente de la tecnologia cinegetica necesaria para poder sobreexplotar los mamiferos que Ie sirvieron de alimento (a los cuales accedio tardiamente en su desarrollo) no es concebible que haya causado e1 holocausto faunal que se Ie atribuye a partir del error de aplicar a las comunidades aborigenes antillanas un modelo cultural de la prehistoria continentaL
Tampoco hay evidencias de que los aborigenes hayan introducido carnivoros con suficiente capacidad depredadora salvo el perro Morgan y Otlenwaldder (1993) consideraron que la depredacion por perros durante el periodo amerindio es la explicacion mas plausible para la extincion del almiqui gigante de Cuba pero como se puede observar (Tabla 54) Solenodon arredondoi aparece en el intervalo II por falta de asociacion con evidencias culturales
Consecuentemente el accionar de los aborigenes y de sus animales domesticos directamente sobre la fauna no parece haber sido por sf solo un factor determinante de la extincion de mamiferos terrestres durante el periodo amerindio en las Antillas (pero vease Steadman et al 2005)
Hay que recordar en este senti do que alin en el Holoceno tardio ocurrieron cambios clirruiticos de consideracion (vease Causas Naturales) Por ejemplo Curtis et al (2001) explicaron que la extincion de algunos ani males mesofilicos de las alturas montanosas de La Espanola pudo haber antecedido a cualquier impacto humano sigrIificativo en esos lugares por coincidir dicha extincion con una tendencia a la aridez documentada isotopicamente y con evidencias de la perdida del bosque mesico ocurridas despues de 3kaAP Yen Jamaica hace solo 2shy3 mil aDos el area de la Bahia de Jackson sustentaba una gran biomasa de vertebrados en bosque hinnedo a juzgar por la cantidad y diversidad de taxones presentes en material fosil derivado de la depredacion por lechuzas hoy el area es la mas seca de la isla con matorral xerofitico y pobreza de fauna (MacPhee 1996a) Yalm en nuestros dias huracanes de gran intensidad aniquilan poblaciones de vertebrados (que no se recuperan) en is las pequenas y llanas del Caribe (Brooks y Smith 2001)
En medio de la incertidumbre sobre la verdadera causa (0 causas) de las extinciones de mamiferos en el periodo amerindio MacPhee y Marx (1997) han propuesto su hipotesis de la hipermorbilidad la cual estipula colapsos poblacionales relativamente rapidos con la introduccion de enfermedades de alta letalidad Estos autores han razonado que
Las islas son los lugares mas apropiados para experimentar perdidas catastroficas causadas por enfermedades Los mamiferos no solo serian susceptibles en virtud de su prolongado aislamiento biologico sino que en los confines de una isla la epizootia
333 EXTINCIONES DEL CUATERNARIO
tendria la potencialidad de infectar simultaneamente a la rnayoria de los individuos de vadas especies Los neonatos son inmunologicamente inmaduros y sufririan desproporcionadamente cualquier infeccion hipervirulenta La perdida excesiva de neonatos y juveniles tendria efectos severos para la sobrevivencia de la poblacion si esta se reduce por debajo de cierto nivel sobreviene la extincion porque no hay posibilidad de restitucion en el periodo postpanzootico Las especies de mayor talla corporal van a ser las mas vulnerables a la extincion dadas sus bajas tasas reproductivas largos periodos de gestacion y reducido numero de progenie Las especies de menor tamano --caracterizadas por altas tasas reproductivas periodos cortos de gestacion y mayor numero de neonatos por parto- tambien serian fuertemente diezmadas pero muchas sobrevivirian porque su estrategia reproductiva les permite perder mayor numero de individuos Las perdidas deben ocurrir relativamente rapido (a escala de decadas) mientras que otras hipotesis (digamos los cambios climaticos) permiten intervalos mucho mas largos (a escala de siglos 0 de milenios) entre la aparicion de las causas y la produccion de sus efectos Por ejemplo todo indica que los nesofontes se extinguieron muy rapidamente y seria dificil poder invocar un impacto humano directo 0 argumentar fenomenos climaticos que pudieran explicar plausiblemente perdidas tan rapidas
MacPhee y Marx (1997 191) han dicho que por el momento la hipotesis de la hipermorbilidad no pasa de ser una abstraccion en espera de adecuada sustentacion empirica Llama la atencion que linicamente en el periodo amerindio (Tabla 45 intervalo 1Il) las extinciones abarcaron todas las clases del peso corporal (de la a a laj) y que ninguna de las especies que sobrevivieron hasta el periodo postcolombino (Tabla 45 intervalo IV mas las especies vivientes) sobrepasa los 10 kg (clases del peso a b c) en clara coincidencia con los presupuestos de la hipotetica hipermorbilidad
Intervalo Postcolornbino
Por 10 menos 37 especies de mamiferos terrestres vivian en las Antillas Mayores y grupos de Islas perifericas a la lIegada de los europeos en 1492 De elIas se han extinguido 23 shymas especies que en cualquiera de los periodos precedentes (Tabla 45~ y solo subsisten 12 especies repartidas entre Cuba (8 especies) La Espanola (2) Jamaica (1) y Bahamas (1) varias de estas en peligro critico de extincion Solo dos de las especies extinguidas durante los ultimos 500 anos se conocieron en estado viviente los roedores Oryzomys antillarum de Jamaica (extinguido en 1877) y Geocapromys thoracatus de las Islas Swan (extinguido en 1955) Las causas de las extinciones ocurridas durante este periodo son sobradamente conocidas destruccion de habitats introducci6n de animales exoticos (domesticos y comensales) y demas presiones asociadas a la actividad humana entre las cuales posiblemente habria que incluir brotes de hipermorbilidad
Durante mucho tiempo se ha afirmado la posibilidad de que especies de Nesophontes hayan sobrevivido hasta las primeras decadas del siglo xx y que incluso continuen existiendo en lugares remotos de Cuba 0 La Espanola (Allen 1918 1942 Miller 1930 Aguayo 1950a Arredondo 1960b Fischer 1977 MacFadden 1980 Woods y
334 BIOGEOGRAFiA
Ottenwalder 1991) Actualmente no existe evidencia radiometrica de que alguna especie de Nesophontes haya sobrevivido al siglo xv las fechas radiocarbonicas calibradas obtenidas en muestras de Nesophontes de Cuba y La Espanola resultaron no mas tardias que los siglos XIII y XIV (MacPhee et al 1999) Por otra parte las expediciones efectuadas a fines del siglo xx con el proposito de localizar individuos vivientes del genero en La Espanola no produjeron la menor indicacion afirmativa al respecto (Woods et al 1985) Ademas a traves de las Antillas Mayores no aparecen restos de Nesophontes en los depositos formados actualmente por las lechuzas
EPiLOGO
Las Antillas Mayores son islas oceanicas no han formado parte de un continente (con la posible excepcion de una parte de Jamaica hasta el Eoceno Medio hace 45 Ma) y durante un tiempo geologico considerable (desde el Eoceno Superior hace 40 Ma) han permanecido emergidas y separadas de los continentes vecinos Consecuentemente los mamiferos terrestres (no volantes) autoctonos de estas islas constituyen una fauna oceanica que por ende cumple con todas las caracteristicas convencionales reconocidas para este tipo de entidad biogeografica
- Baja diversidad taxonomica a nivel de las categorias superiores (solo cuatro ordenes) en comparacion con islas plataformicas continentales (0 con areas continentales) de tamano y diversidad ecologica simi lares
- Alto grado de endemismo a nivel de genero (96) y familia (50) en respuesta al prolongado aislamiento
- Desbalance ecologico de la estructura trofica ausencia de mamiferos camivoros y ungulados
- Espectaculares procesos de irradiaci6n evolucionaria como resultado de la colonizacion de territorios libres de competidores y depredadores
Tendencias evolucionarias extremas gigantismo
- Control a cargo de aves rapaces con tendencia at gigantismo en ausencia de mamiferos miembros del orden Carnivora
- Mas especies de reptiles y de murcielagos que de mamiferos terrestres relaciones que en los continentes se invierten
Alta tasa de extincion de especies (84) sin reposicion
Preservacion de especies filogeneticamente arcaicas (los llamados fosiles vivientes) los almiquies
326 BIOGEOGRAFiA
TABLA 45 (Continuacion)
F amilialEspecie Peso Isla Evidencia Fuente
N esophontes m icrus a CD 24 MacPhee et aI 1999
N esophontes m icrus a LE 4 MacPhee et aI 2000
N esophontes sp 1 a IC 4 Morgan 1994
N esophontes sp 2 a IC 4 Ibid
Solenodon marcanoi b LE 4 Ottenwalder 2001
PITHECIIDAE
Xenothrix mcgregori c JA 4 MacPhee y Flemming 1999
EcHIMYIDAE
Boromys offeUa a CD 24 Guarch 1984
Boromys torrei a CD 24 Ibid
Brotomys voratus a LE 4 MacPhee et aI 2000
CAPROMYIDAE
P lagiodontia ipnaeum c LE 4 MacPhee y Flemming 1999
Rhizoplagiodontia lemkei c LE 4 Ibid
Isolobodon portoricensis c PR 24 Ibid
Isolobodon montanus c LE 4 Ibid
Hexolobodon phenax c LE (4) Ibid
Capromys sp c IC 24 Morgan 1994
Geocapromys columbianus c CD 4 MacPhee y Flemming 1999
Geocapromys thoracatus b IS 1 MacPhee y Marx 1997
Geocapromys sp 1 c IC 4 Morgan 1994
Geocapromys sp 2 c IC (4) Ibid
l-IEPTAXODONTINAE
Quemisia gravis c LE (1) MacPhee y Flemming 1999
MURIDAE
Oryzomys antillarum a JA 1 Ibid
1 Flemming y MacPhee (1999) alertaron en contra de la asociacion con otro material fechado Asimismo Morgan (1993) indico que la ausencia de asociacion con evidencias culturales amerindias no es necesariamente un criterio concluyente para ubicar un taxon en el intervalo temporal anterior porque hay depositos en la region que carecen de tales evidencias y son del Holoceno tardio por fechados radiocarbonicos [No obstante
327 EXTlNCIONES DEL CUATERNAR10
la ausencia de una especie en residuarios arqueol6gicos adquiere mayor certidumbre como criterio de ubicaci6n temporal de su extinci6n en la medida que aumenta el numero de residuarios estudiados que en las Antillas Mayores se cuentan por centenares] A falta de fechado absoluto de una especie consideramos que estos criterios de ubicaci6n temporal son mas utiles que la alternativa de excluir del analisis a la especie
2 En los trabajos sobre biogeografia de mamiferos antillanos los arque610gos mas citados como autoridades para establecer el inicio de la presencia aborigen en las Antillas Mayores (Kozlowski 1974 Rouse y Allaire 1978 Rouse 1989 1992 Wilson 2001) difieren al respecto entre los 6 kaAP y los 45 kaAP (pero vease Pagan et al 200523) y hemos adoptado la mas temprana de estas fechas para el comienzo del intervalo amerindio en la Tabla 45 De todos modos cualquiera que haya sido la fecha esta no es aplicable por igual al conjunto de las Antillas Mayores porque segun algunos de los autores citados el poblamiento comenz6 por Cuba y La Espanola 6 mil anos atras continu6 en Puerto Rico hace alrededor de 3 mil anos (pero vease Burney et al 1994) yen Jamaica hace menos de 2 mil e incluso a las Islas Caiman ni siquiera llegaron [Las Islas Caiman descubiertas por Col6n en 1503 carecen absolutamente de evidencia amerindia (Stokes y Keegan 1996)]
Si los datos de la Tabla 45 representan una aceptable aproximaci6n a la realidad el proceso de las extinciones cuaternarias de los mamiferos terrestres de las Antillas Mayores y grupos de islas perifericas debi6 caracterizarse entre otros por los eventos siguientes
Aunque obviamente la fauna cuaternaria debi6 ser mucho mas numerosa que 10 que refleja el registro paleontol6gico (Morgan y Woods 1986) 52 especies (81 de la fauna conocida) desaparecieron (sin reposici6n) durante los ultimos 250 mil anos
- Al parecer las extinciones se concentraron en las especies de mayor tamano Asi el peso promedio estimado para las especies de la etapa preantr6pica (intervalos I + II de la Tabla 45) es 350 kg es 328 kg para las especies del intervalo III (amerindio) y es s610 25 kg (c1ases del peso a b c) para las especies del intervalo IV (postcolombino) Ninguna especie aut6ctona de mamifero mayor de 6 kg existe hoy en la regi6n
- De acuerdo con la distribuci6n temporal de las extinciones 61 especies sobrevivieron al interglacial Sangamon (intervalo 1)48 sobrevivieron a la transici6n del Pleistoceno terminal al Holoceno temprano (intervalo II) 34 sobrevivieron al periodo amerindio (intervalo III) y s610 12 especies han sobrevivido desde la Ilegada de los europeos (intervalo IV) Las tasas de extinci6n correspondientes (exceptuado el intervalo I) son una especie cada 1167 anos en el intervalo II una cada 393 anos en el intervalo III y una cada 23 anos en el intervalo IV
- Durante todo el proceso de las extinciones cuaternarias la perdida de especies por isla fue de 91 en La Espanola 83 en Jamaica y 62 en Cuba Bahamas retuvo su unica especie (aunque extirpada en varias de las islas) Puerto Rico e Islas Caiman perdieron todas sus especies (6 y 5 respectivamente) y las Islas Swan Virgenes y Anguila - San Martin perdieron sus respectivas especies (micas
328 BIOGEOGRAFiA
De los cuatro 6rdenes de mamiferos representados en la fauna Pilosa y Primates desaparecieron totalmente Soricomorpha y Rodentia perdieron 86 y 74 de sus especies respectivamente
CAUSALIDAD
CAUSAS NATURALES
Los cicIos glaciales e interglaciales acaecidos en el Pleistoceno causaron cambios ambientales dnisticos en las Antillas con la concomitante alternancia de cIimas frios (secos) y calidos (hUmedos) asi como de descensos y ascensos del nivel del mar aumento y disminuci6n de las areas territoriales) induso (aunque con menor intensidad) en el Holoceno Estos eventos paleogeograficos han sido consistentemente documentados (Mayo y Kartashov 1972 Bonnatti y Gartner 1973 Ortega y Arcia 1982 Prance 1982 Schubert y Medina 1982 Dodge et al 1983 Ortega [1984] Haq et al 1987 Schubert 1989 Hodell et al 1991 Guilderson et al 1994 Oppo 1997 Emslie 1998 Curtis et al 2001 Iturralde-Vinent 2003) y de ellos brindamos a continuaci6n una breve resena
Finalizando el periodo interglacial anterior a la ultima glaciaci6n (Wisconsin) el mar estaba a 8-10 m por encima de su nivel actual Cuando el avance glacial del Wisconsin a1canz6 su culminaci6n (eillamado Ultimo Maximo Glacial) hace 18 ka el mar habia descendido 120 m con respecto a dicho nivel y las islas del Caribe habian incrementado considerablemente sus extensiones subaereas en comparaci6n con las que presentan hoy dia por ejemplo Puerto Rico y las Islas Virgenes se conectaron territorialmente y las Bahamas se unieron en cinco gran des islas Prevalecia un regimen frio y seco y condiciones de aridez --debidas a una reducci6n de las precipitaciones (posiblemente a menos de 440 mmlano)- y los ambientes xericos se habian extendido a traves de las islas y bancos emergidos y abundaban los pinares (los habitats mesicos estaban restringidos a parcelas relictas en 10 alto de las montanas) En el Mar Caribe y el Golfo de Mexico la temperatura superficial del mar era 5degC mas baja que la actual y en las llanuras costeras y alturas maximas eran respectivamente 2-3degC y 6degC mas bajas Los limites de las zonas ecol6gicas altitudinales descendieron varios cientos de metros y cada zona redujo su extensi6n vertical al aumentar el gradiente de temperatura Ellimite inferior de la zona periglacial
elevaci6n donde la temperatura media minima a1canza los OdegC) de la Cordillera Central de la Republica Dominicana (altura maxima del Caribe insular con evidencia de topografia glacial) se considera que estuvo entre los 1500 y 1800 mlnm (actualmente esta a 2800 mlnm) y se puede estimar que dicho limite era inferior en Cuba por estar mas al norte que la Republica Dominicana No obstante durante este periodo la temperatura experiment6 episodios cortos (unas pocas decadas) de ascensos repetidos
Un signifieativo componente bi6tieo de las referidas condiciones paleoambientales 10 constituy6 la espectacular irradiaci6n evolucionaria experimentada por un grupo numeroso de aves rapaees diurnas y nocturnas (Wetmore 1937 Brodkorb 1969 Kurotchkin y Mayo 1973 Arredondo 1976 Olson 1978 Olson y Hilgartner 1982
329 EXTINCIONES DEL ClJATERNARIO
Suarez 2000a b 2004 Suarez y Emslie 2003) Los mamiferos de las Antillas Mayores ofrecieron a estos depredadores aviares una amplia gama de tamanos de presa que en ausencia de mamiferos del orden Carnivora en la region permitio a estas aves irradiarse a este nicho alimentario con la evolucion de una variedad de especies de muy diversa tall a corporal incluidas formas gigantes Adquirir un cuerpo gigante les propicio el acceso a recursos alimentarios que no estaban siendo explotados por ningun otro depredador (Alcover y McMinn 1994) A Cuba corresponde el mejor registro fosil de rapaces cuaternarias caribenas con siete generos de especies diurnas Dentro de estas aves existieron tres gavilanes gigantes (Accipitridae) tres carairas (Falconidae) y un condor (Cathartidae) asi como cuatro generos de especies nocturnas dos lechuzas (Tytonidae) y tres Mhos (Strigidae) dos de ell os no igualados en tamano en todo el mundo Algunos de estos generos tuvieron representacion en otras islas antillanas como La Espanola Jamaica Bahamas y Gran Caiman
Al retirarse hacia el norte los hielos de la glaciacion Wisconsin el clima se tomo mas hUmedo en las Antillas durante el Holoceno temprano y el bosque mesico se expandio en la region insular El nivel del mar ascendio acompanado de mayor pluviosidad alrededor de los 7 kaAP las Islas Virgenes quedaron separadas entre si y de Puerto Rico y las Bahamas perdieron la mayor parte de su territorio Cerca de los 3 kaAP las condiciones secas retornaron a la region e hicieron desaparecer buena parte del bosque mesico Nuevamente el clima comenzo a adquirir sus condiciones actuales hacia los 25 kaAP aproximadamente y hoy transita por una fase mesica con vegetacion xerica confinada en parcelas relictas
Sin duda estos y otros eventos naturales incidieron negativamente en la sobrevivencia de mamiferos terrestres en las Antillas Mayores asi como en los grupos de pequenas islas perifericas (Pregill y Olson 1981 Olson 2006)
McFarlane et al (1998a) demostraron convincentemente como Amblyrhiza el gigante de los roedores antillanos endemico de la pequena isla de Anguila sucumbio a las repetidas y abruptas perdidas de su habitat durante el interglacial Sangamon por las nipidas y severas oscilaciones del nivel del mar (documentadas por Neumann y Hearty 1996) al no poder responder adecuadamente a tan catastroficos eventos mediante el ajuste de su masa corporal media y otro tanto debio ocurrirle en Jamaica al otro roedor gigante Clidomys (McFarlane et al 1998b) emparentado con Amblyrhiza en la misma epoca (Tabla 45 intervalo I)
El ascenso del nivel del mar seguramente promovio la extincion de mamiferos durante la transicion del Pleistoceno al Holoceno (Tabla 45 intervalo II) con la reduccion del tamano de las Islas por las transgresiones marinas (Pregill y Olson 1981 Morgan 1993) Esto ultimo haria maximamente severa la competicion asf como la presion de los depredadores [Steadman et al (2005) no yen por que el calentamiento global ocurrido durante la transicion Pleistoceno-Holoceno habria de tener un efecto negativo sobre los perezosos siendo sus descendientes actuales especies tropic ales 0 subtropicales pero no se trata del efecto
330 BIOGEOGRAFiA
directo del cambio de temperatura sobre los animales sino del efecto indirecto y progresivo que dicho cambio genera a largo plazo al reducir el territorio insular disponible]
Por esta epoca tambien desaparecieron especies de tall a mayor como Megalocnus zile en La Espanola y otras de menor tamano (en Cuba) pero que representan la tall a maxima en sus respectivos contextos taxonomicos del Cuaternario como Solenodon arredondoi Paralouatta varonai y Macrocapromys acevedo Un mamifero puede ser grande solo a costa de reducir su densidad poblacional y esto 10 hace mis vulnerable a la extinci6n sobre todo en una isla (McNab 1994 McFarlane et al 1998a) Incidentalmente Alcover et al (1998a) afirmaron que larelacion entre la talla corporal y la extincion no siempre se cumple en las especies de menor tamano y citaron como ejemplo que entre las musarafias todas las especies de Nesophontes desaparecieron mientras sobreviven especies de Solenodon pero la comparacion correcta en todo caso serfa entre los solenodones vivientes y el gigante Solenodon arredondoi y entre Nesophontes edithae -el gigante de los nesofontes extinguido en el periodo amerindio- y el resto de sus congeneres que sobrevivieron hasta los primeros siglos de la colonizacion europea (Tabla 45 intervalos III y IV)
CAUSAS ANTROPOGENICAS
Intervalo Amerindio
La mayoria de los autores que se han referido a las extinciones acaecidas durante este intervalo (entre elIos paradigmaticamente Martin 1984a b) han responsabilizado a los aborfgenes (que se supone comenzaron a poblar las Antillas Mayores hace 6 mil afios) con la desaparicion de numerosas especies de mamiferos terrestres en la region principalmente por la via de la sobrecaza y la introduccion de animales domesticos devenidos depredadores de la fauna autoctona (Morgan y Woods 1986) Este efecto negativo de la presencia indigena sobre los mamiferos de la region ha sido dramatizado por algunos autores en terminos de holocausto faunal Sin embargo pensamos que se ha exagerado Como han expresado MacPhee et al (19993) cuando uno busca evidencia dura de complicidad humana directa en [estas] extinciones no hay mucho que encontrar Las siguientes son algunas consideraciones a tener en cuenta en tal sentido
El registro radiometrico mas joven conocido hasta ahora para un perezoso antillano (MegalocnllS rodens) es de 42 kaAP (MacPhee et ai 2007) Si se acepta que los aborfgenes entraron en Cuba hace aproximadamente 6 mil anos su convivencia con los perezosos debe haber durado alrededor de 1800 afios (0 mas porque dificilmente el referfdo fechado radiometrico haya correspondido al ultimo sobreviviente de la especie) Esta cifra contradice la hipotesis del holocausto faunal segun la cualla extincion de los mamiferos insulares de mayor talla debe ocurrir con extrema rapidez -a escala de decadas 0 de centurias- a partir de su primer contacto con los humanos (Martin y Steadman 1999) A pesar de las repetidas afirmaciones en contrario (Harrington 1921 Miller 1929c Herrera [1945] Tabio 1951 Alvarez 1951a 1957 Arredondo
331 EXTINCIONES DEL CUATERNARIO
1959 1960a Veloz y Ortega 1973 Mayo 1980b Rodriguez et al 1984 Castellanos y Pino 1985 Pino y Castellanos 1985 Nunez 1990b Gutierrez 1996 Arredondo 2004) en su mayoria basadas en evidencias no exentas de posibles accidentes de deposicion (0 de reubicacion postdeposicional) en recintos cavernarios (Diaz-Franco [2006b D no se ha demostrado inequivocamente que los aborigenes antillanos se alimentaran de perezosos regularmente Sus restos en residuarios de alimentos son escasisimos en toda la region y esto contrasta con los abundantes restos de jutias y otros mamiferos de menor talla que aparecen en esos residuarios demostrativos de la preferencia de los paleoindios por estos otros mamiferos
Los mamiferos mas importantes en la economia prehistorica de las Antillas Mayores fueron las jutias Capromys y Geocapromys en Cuba Geocapromys en Jamaica e Isolobodon en La Espanola Puerto Rico e Islas Virgenes (Aguayo 1950b Tablo 1951 Rimoli 1972 Wing 2001 Diaz-Franco [2006b D Wilkins (2001) estudi61ajutia viviente de Jamaica (Geocapromys browni) en un sitio ocupado por los aborigenes durante 100 anos y no encontr6 evidencias de que durante ese tiempo la especie haya sido sobreexplotada Asimismo en Cuba las jutias conga carabali y andaraz son escasas en depositos pleistoholocenicos en cambio son muy numerosas en residuarios indigenas y de haber sido sobreexplotadas 10 mas probable es que no hubieran sobrevivido hasta hoy
La habilidad de los humanos para penetrar en profundidad el bosque tropical y mantenerse en este sustentablemente ha sido un evento comparativamente reciente (Beran y Steele 1994) En el caso antillano la caza tierra adentro ha sido explicada como un proceso tardio de desplazamiento a partir de la explotacion de abundantes recursos marinos que ofrecian condiciones 6ptimas para el sedentarismo de las comunidades aborigenes costeras (C6rdova y Arredondo 1988 Brito 2005) Peces y mariscos dominan la arqueologia costera de las Islas antillanas en tanto el consumo de recursos terrestres (mamfferos incluidos) aumenta progresivamente en la medida que los residuarios estudiados se alejan de la costa (Scudder 1991 Wilkins 2001) En las Antillas Mayores el componente terrestre de la dieta aborigen es solo 20-30 del material recobrado en los residuarios (Wing 1977 1989) Y debio ser menor alin porque el componente vegetal de la dieta no se ha preservado Por otra parte los aborigenes cubanos domesticaron la jutia conga y seguramente la reprodujeron para comer (Pose et al 1988) practica que probablemente se repitiera con otras jutias en las demas Antillas Mayores
- Fray Bartolome de las Casas hablo de 3 millones de indios en Cuba al inicio de la colonizacion pero los historiadores subsecuentes consideraron exagerada la cifra (Poey 1851 a) No obstante se dispone de dos estimados de la densidad de poblaci6n aborigen en las Antillas Mayores a la llegada de los europeos 18 indigenaslkm2 en Puerto Rico (Rouse 1956) y 1271km2 en La Espanola (Watts 1987) Extrapolando ambos valores entre las cuatro islas principales de las Antillas Mayores tenemos que en la fecha indicada la poblaci6n aborigen pudo estar entre 16 mil y 113 mil indigenas en Puerto Rico entre 21 mil y 145 mil en Jamaica entre 141 mil y 1 mill6n en La Espanola y entre 200 mil y 14 millones en Cuba Entre las cifras maximas y las minimas nos inclinamos
332 BIOGEOGRAFiA
hacia estas ultimas atendiendo a los altos indices de mortalidad infantil y morbilidad que se evidencian en las osamentas de los pobladores prehispanicos (Martinez y Rives 1990) Uno de los hechos que apuntan hacia el bajo numero de aborigenes en las Antillas Mayores es 10 rapido que desaparecieron despues de la llegada de los europeos pnkticamente sin dejar mestizaje con estos 10 cual contrasta ostensiblemente con la situacion en la America continentaL
En suma un grupo humano (1) demograiicamente irrelevante y controlado por altos indices de morbilidad (2) consumidor mayoritariamente de organismos marinos y (3) carente de la tecnologia cinegetica necesaria para poder sobreexplotar los mamiferos que Ie sirvieron de alimento (a los cuales accedio tardiamente en su desarrollo) no es concebible que haya causado e1 holocausto faunal que se Ie atribuye a partir del error de aplicar a las comunidades aborigenes antillanas un modelo cultural de la prehistoria continentaL
Tampoco hay evidencias de que los aborigenes hayan introducido carnivoros con suficiente capacidad depredadora salvo el perro Morgan y Otlenwaldder (1993) consideraron que la depredacion por perros durante el periodo amerindio es la explicacion mas plausible para la extincion del almiqui gigante de Cuba pero como se puede observar (Tabla 54) Solenodon arredondoi aparece en el intervalo II por falta de asociacion con evidencias culturales
Consecuentemente el accionar de los aborigenes y de sus animales domesticos directamente sobre la fauna no parece haber sido por sf solo un factor determinante de la extincion de mamiferos terrestres durante el periodo amerindio en las Antillas (pero vease Steadman et al 2005)
Hay que recordar en este senti do que alin en el Holoceno tardio ocurrieron cambios clirruiticos de consideracion (vease Causas Naturales) Por ejemplo Curtis et al (2001) explicaron que la extincion de algunos ani males mesofilicos de las alturas montanosas de La Espanola pudo haber antecedido a cualquier impacto humano sigrIificativo en esos lugares por coincidir dicha extincion con una tendencia a la aridez documentada isotopicamente y con evidencias de la perdida del bosque mesico ocurridas despues de 3kaAP Yen Jamaica hace solo 2shy3 mil aDos el area de la Bahia de Jackson sustentaba una gran biomasa de vertebrados en bosque hinnedo a juzgar por la cantidad y diversidad de taxones presentes en material fosil derivado de la depredacion por lechuzas hoy el area es la mas seca de la isla con matorral xerofitico y pobreza de fauna (MacPhee 1996a) Yalm en nuestros dias huracanes de gran intensidad aniquilan poblaciones de vertebrados (que no se recuperan) en is las pequenas y llanas del Caribe (Brooks y Smith 2001)
En medio de la incertidumbre sobre la verdadera causa (0 causas) de las extinciones de mamiferos en el periodo amerindio MacPhee y Marx (1997) han propuesto su hipotesis de la hipermorbilidad la cual estipula colapsos poblacionales relativamente rapidos con la introduccion de enfermedades de alta letalidad Estos autores han razonado que
Las islas son los lugares mas apropiados para experimentar perdidas catastroficas causadas por enfermedades Los mamiferos no solo serian susceptibles en virtud de su prolongado aislamiento biologico sino que en los confines de una isla la epizootia
333 EXTINCIONES DEL CUATERNARIO
tendria la potencialidad de infectar simultaneamente a la rnayoria de los individuos de vadas especies Los neonatos son inmunologicamente inmaduros y sufririan desproporcionadamente cualquier infeccion hipervirulenta La perdida excesiva de neonatos y juveniles tendria efectos severos para la sobrevivencia de la poblacion si esta se reduce por debajo de cierto nivel sobreviene la extincion porque no hay posibilidad de restitucion en el periodo postpanzootico Las especies de mayor talla corporal van a ser las mas vulnerables a la extincion dadas sus bajas tasas reproductivas largos periodos de gestacion y reducido numero de progenie Las especies de menor tamano --caracterizadas por altas tasas reproductivas periodos cortos de gestacion y mayor numero de neonatos por parto- tambien serian fuertemente diezmadas pero muchas sobrevivirian porque su estrategia reproductiva les permite perder mayor numero de individuos Las perdidas deben ocurrir relativamente rapido (a escala de decadas) mientras que otras hipotesis (digamos los cambios climaticos) permiten intervalos mucho mas largos (a escala de siglos 0 de milenios) entre la aparicion de las causas y la produccion de sus efectos Por ejemplo todo indica que los nesofontes se extinguieron muy rapidamente y seria dificil poder invocar un impacto humano directo 0 argumentar fenomenos climaticos que pudieran explicar plausiblemente perdidas tan rapidas
MacPhee y Marx (1997 191) han dicho que por el momento la hipotesis de la hipermorbilidad no pasa de ser una abstraccion en espera de adecuada sustentacion empirica Llama la atencion que linicamente en el periodo amerindio (Tabla 45 intervalo 1Il) las extinciones abarcaron todas las clases del peso corporal (de la a a laj) y que ninguna de las especies que sobrevivieron hasta el periodo postcolombino (Tabla 45 intervalo IV mas las especies vivientes) sobrepasa los 10 kg (clases del peso a b c) en clara coincidencia con los presupuestos de la hipotetica hipermorbilidad
Intervalo Postcolornbino
Por 10 menos 37 especies de mamiferos terrestres vivian en las Antillas Mayores y grupos de Islas perifericas a la lIegada de los europeos en 1492 De elIas se han extinguido 23 shymas especies que en cualquiera de los periodos precedentes (Tabla 45~ y solo subsisten 12 especies repartidas entre Cuba (8 especies) La Espanola (2) Jamaica (1) y Bahamas (1) varias de estas en peligro critico de extincion Solo dos de las especies extinguidas durante los ultimos 500 anos se conocieron en estado viviente los roedores Oryzomys antillarum de Jamaica (extinguido en 1877) y Geocapromys thoracatus de las Islas Swan (extinguido en 1955) Las causas de las extinciones ocurridas durante este periodo son sobradamente conocidas destruccion de habitats introducci6n de animales exoticos (domesticos y comensales) y demas presiones asociadas a la actividad humana entre las cuales posiblemente habria que incluir brotes de hipermorbilidad
Durante mucho tiempo se ha afirmado la posibilidad de que especies de Nesophontes hayan sobrevivido hasta las primeras decadas del siglo xx y que incluso continuen existiendo en lugares remotos de Cuba 0 La Espanola (Allen 1918 1942 Miller 1930 Aguayo 1950a Arredondo 1960b Fischer 1977 MacFadden 1980 Woods y
334 BIOGEOGRAFiA
Ottenwalder 1991) Actualmente no existe evidencia radiometrica de que alguna especie de Nesophontes haya sobrevivido al siglo xv las fechas radiocarbonicas calibradas obtenidas en muestras de Nesophontes de Cuba y La Espanola resultaron no mas tardias que los siglos XIII y XIV (MacPhee et al 1999) Por otra parte las expediciones efectuadas a fines del siglo xx con el proposito de localizar individuos vivientes del genero en La Espanola no produjeron la menor indicacion afirmativa al respecto (Woods et al 1985) Ademas a traves de las Antillas Mayores no aparecen restos de Nesophontes en los depositos formados actualmente por las lechuzas
EPiLOGO
Las Antillas Mayores son islas oceanicas no han formado parte de un continente (con la posible excepcion de una parte de Jamaica hasta el Eoceno Medio hace 45 Ma) y durante un tiempo geologico considerable (desde el Eoceno Superior hace 40 Ma) han permanecido emergidas y separadas de los continentes vecinos Consecuentemente los mamiferos terrestres (no volantes) autoctonos de estas islas constituyen una fauna oceanica que por ende cumple con todas las caracteristicas convencionales reconocidas para este tipo de entidad biogeografica
- Baja diversidad taxonomica a nivel de las categorias superiores (solo cuatro ordenes) en comparacion con islas plataformicas continentales (0 con areas continentales) de tamano y diversidad ecologica simi lares
- Alto grado de endemismo a nivel de genero (96) y familia (50) en respuesta al prolongado aislamiento
- Desbalance ecologico de la estructura trofica ausencia de mamiferos camivoros y ungulados
- Espectaculares procesos de irradiaci6n evolucionaria como resultado de la colonizacion de territorios libres de competidores y depredadores
Tendencias evolucionarias extremas gigantismo
- Control a cargo de aves rapaces con tendencia at gigantismo en ausencia de mamiferos miembros del orden Carnivora
- Mas especies de reptiles y de murcielagos que de mamiferos terrestres relaciones que en los continentes se invierten
Alta tasa de extincion de especies (84) sin reposicion
Preservacion de especies filogeneticamente arcaicas (los llamados fosiles vivientes) los almiquies

327 EXTlNCIONES DEL CUATERNAR10
la ausencia de una especie en residuarios arqueol6gicos adquiere mayor certidumbre como criterio de ubicaci6n temporal de su extinci6n en la medida que aumenta el numero de residuarios estudiados que en las Antillas Mayores se cuentan por centenares] A falta de fechado absoluto de una especie consideramos que estos criterios de ubicaci6n temporal son mas utiles que la alternativa de excluir del analisis a la especie
2 En los trabajos sobre biogeografia de mamiferos antillanos los arque610gos mas citados como autoridades para establecer el inicio de la presencia aborigen en las Antillas Mayores (Kozlowski 1974 Rouse y Allaire 1978 Rouse 1989 1992 Wilson 2001) difieren al respecto entre los 6 kaAP y los 45 kaAP (pero vease Pagan et al 200523) y hemos adoptado la mas temprana de estas fechas para el comienzo del intervalo amerindio en la Tabla 45 De todos modos cualquiera que haya sido la fecha esta no es aplicable por igual al conjunto de las Antillas Mayores porque segun algunos de los autores citados el poblamiento comenz6 por Cuba y La Espanola 6 mil anos atras continu6 en Puerto Rico hace alrededor de 3 mil anos (pero vease Burney et al 1994) yen Jamaica hace menos de 2 mil e incluso a las Islas Caiman ni siquiera llegaron [Las Islas Caiman descubiertas por Col6n en 1503 carecen absolutamente de evidencia amerindia (Stokes y Keegan 1996)]
Si los datos de la Tabla 45 representan una aceptable aproximaci6n a la realidad el proceso de las extinciones cuaternarias de los mamiferos terrestres de las Antillas Mayores y grupos de islas perifericas debi6 caracterizarse entre otros por los eventos siguientes
Aunque obviamente la fauna cuaternaria debi6 ser mucho mas numerosa que 10 que refleja el registro paleontol6gico (Morgan y Woods 1986) 52 especies (81 de la fauna conocida) desaparecieron (sin reposici6n) durante los ultimos 250 mil anos
- Al parecer las extinciones se concentraron en las especies de mayor tamano Asi el peso promedio estimado para las especies de la etapa preantr6pica (intervalos I + II de la Tabla 45) es 350 kg es 328 kg para las especies del intervalo III (amerindio) y es s610 25 kg (c1ases del peso a b c) para las especies del intervalo IV (postcolombino) Ninguna especie aut6ctona de mamifero mayor de 6 kg existe hoy en la regi6n
- De acuerdo con la distribuci6n temporal de las extinciones 61 especies sobrevivieron al interglacial Sangamon (intervalo 1)48 sobrevivieron a la transici6n del Pleistoceno terminal al Holoceno temprano (intervalo II) 34 sobrevivieron al periodo amerindio (intervalo III) y s610 12 especies han sobrevivido desde la Ilegada de los europeos (intervalo IV) Las tasas de extinci6n correspondientes (exceptuado el intervalo I) son una especie cada 1167 anos en el intervalo II una cada 393 anos en el intervalo III y una cada 23 anos en el intervalo IV
- Durante todo el proceso de las extinciones cuaternarias la perdida de especies por isla fue de 91 en La Espanola 83 en Jamaica y 62 en Cuba Bahamas retuvo su unica especie (aunque extirpada en varias de las islas) Puerto Rico e Islas Caiman perdieron todas sus especies (6 y 5 respectivamente) y las Islas Swan Virgenes y Anguila - San Martin perdieron sus respectivas especies (micas
328 BIOGEOGRAFiA
De los cuatro 6rdenes de mamiferos representados en la fauna Pilosa y Primates desaparecieron totalmente Soricomorpha y Rodentia perdieron 86 y 74 de sus especies respectivamente
CAUSALIDAD
CAUSAS NATURALES
Los cicIos glaciales e interglaciales acaecidos en el Pleistoceno causaron cambios ambientales dnisticos en las Antillas con la concomitante alternancia de cIimas frios (secos) y calidos (hUmedos) asi como de descensos y ascensos del nivel del mar aumento y disminuci6n de las areas territoriales) induso (aunque con menor intensidad) en el Holoceno Estos eventos paleogeograficos han sido consistentemente documentados (Mayo y Kartashov 1972 Bonnatti y Gartner 1973 Ortega y Arcia 1982 Prance 1982 Schubert y Medina 1982 Dodge et al 1983 Ortega [1984] Haq et al 1987 Schubert 1989 Hodell et al 1991 Guilderson et al 1994 Oppo 1997 Emslie 1998 Curtis et al 2001 Iturralde-Vinent 2003) y de ellos brindamos a continuaci6n una breve resena
Finalizando el periodo interglacial anterior a la ultima glaciaci6n (Wisconsin) el mar estaba a 8-10 m por encima de su nivel actual Cuando el avance glacial del Wisconsin a1canz6 su culminaci6n (eillamado Ultimo Maximo Glacial) hace 18 ka el mar habia descendido 120 m con respecto a dicho nivel y las islas del Caribe habian incrementado considerablemente sus extensiones subaereas en comparaci6n con las que presentan hoy dia por ejemplo Puerto Rico y las Islas Virgenes se conectaron territorialmente y las Bahamas se unieron en cinco gran des islas Prevalecia un regimen frio y seco y condiciones de aridez --debidas a una reducci6n de las precipitaciones (posiblemente a menos de 440 mmlano)- y los ambientes xericos se habian extendido a traves de las islas y bancos emergidos y abundaban los pinares (los habitats mesicos estaban restringidos a parcelas relictas en 10 alto de las montanas) En el Mar Caribe y el Golfo de Mexico la temperatura superficial del mar era 5degC mas baja que la actual y en las llanuras costeras y alturas maximas eran respectivamente 2-3degC y 6degC mas bajas Los limites de las zonas ecol6gicas altitudinales descendieron varios cientos de metros y cada zona redujo su extensi6n vertical al aumentar el gradiente de temperatura Ellimite inferior de la zona periglacial
elevaci6n donde la temperatura media minima a1canza los OdegC) de la Cordillera Central de la Republica Dominicana (altura maxima del Caribe insular con evidencia de topografia glacial) se considera que estuvo entre los 1500 y 1800 mlnm (actualmente esta a 2800 mlnm) y se puede estimar que dicho limite era inferior en Cuba por estar mas al norte que la Republica Dominicana No obstante durante este periodo la temperatura experiment6 episodios cortos (unas pocas decadas) de ascensos repetidos
Un signifieativo componente bi6tieo de las referidas condiciones paleoambientales 10 constituy6 la espectacular irradiaci6n evolucionaria experimentada por un grupo numeroso de aves rapaees diurnas y nocturnas (Wetmore 1937 Brodkorb 1969 Kurotchkin y Mayo 1973 Arredondo 1976 Olson 1978 Olson y Hilgartner 1982
329 EXTINCIONES DEL ClJATERNARIO
Suarez 2000a b 2004 Suarez y Emslie 2003) Los mamiferos de las Antillas Mayores ofrecieron a estos depredadores aviares una amplia gama de tamanos de presa que en ausencia de mamiferos del orden Carnivora en la region permitio a estas aves irradiarse a este nicho alimentario con la evolucion de una variedad de especies de muy diversa tall a corporal incluidas formas gigantes Adquirir un cuerpo gigante les propicio el acceso a recursos alimentarios que no estaban siendo explotados por ningun otro depredador (Alcover y McMinn 1994) A Cuba corresponde el mejor registro fosil de rapaces cuaternarias caribenas con siete generos de especies diurnas Dentro de estas aves existieron tres gavilanes gigantes (Accipitridae) tres carairas (Falconidae) y un condor (Cathartidae) asi como cuatro generos de especies nocturnas dos lechuzas (Tytonidae) y tres Mhos (Strigidae) dos de ell os no igualados en tamano en todo el mundo Algunos de estos generos tuvieron representacion en otras islas antillanas como La Espanola Jamaica Bahamas y Gran Caiman
Al retirarse hacia el norte los hielos de la glaciacion Wisconsin el clima se tomo mas hUmedo en las Antillas durante el Holoceno temprano y el bosque mesico se expandio en la region insular El nivel del mar ascendio acompanado de mayor pluviosidad alrededor de los 7 kaAP las Islas Virgenes quedaron separadas entre si y de Puerto Rico y las Bahamas perdieron la mayor parte de su territorio Cerca de los 3 kaAP las condiciones secas retornaron a la region e hicieron desaparecer buena parte del bosque mesico Nuevamente el clima comenzo a adquirir sus condiciones actuales hacia los 25 kaAP aproximadamente y hoy transita por una fase mesica con vegetacion xerica confinada en parcelas relictas
Sin duda estos y otros eventos naturales incidieron negativamente en la sobrevivencia de mamiferos terrestres en las Antillas Mayores asi como en los grupos de pequenas islas perifericas (Pregill y Olson 1981 Olson 2006)
McFarlane et al (1998a) demostraron convincentemente como Amblyrhiza el gigante de los roedores antillanos endemico de la pequena isla de Anguila sucumbio a las repetidas y abruptas perdidas de su habitat durante el interglacial Sangamon por las nipidas y severas oscilaciones del nivel del mar (documentadas por Neumann y Hearty 1996) al no poder responder adecuadamente a tan catastroficos eventos mediante el ajuste de su masa corporal media y otro tanto debio ocurrirle en Jamaica al otro roedor gigante Clidomys (McFarlane et al 1998b) emparentado con Amblyrhiza en la misma epoca (Tabla 45 intervalo I)
El ascenso del nivel del mar seguramente promovio la extincion de mamiferos durante la transicion del Pleistoceno al Holoceno (Tabla 45 intervalo II) con la reduccion del tamano de las Islas por las transgresiones marinas (Pregill y Olson 1981 Morgan 1993) Esto ultimo haria maximamente severa la competicion asf como la presion de los depredadores [Steadman et al (2005) no yen por que el calentamiento global ocurrido durante la transicion Pleistoceno-Holoceno habria de tener un efecto negativo sobre los perezosos siendo sus descendientes actuales especies tropic ales 0 subtropicales pero no se trata del efecto
330 BIOGEOGRAFiA
directo del cambio de temperatura sobre los animales sino del efecto indirecto y progresivo que dicho cambio genera a largo plazo al reducir el territorio insular disponible]
Por esta epoca tambien desaparecieron especies de tall a mayor como Megalocnus zile en La Espanola y otras de menor tamano (en Cuba) pero que representan la tall a maxima en sus respectivos contextos taxonomicos del Cuaternario como Solenodon arredondoi Paralouatta varonai y Macrocapromys acevedo Un mamifero puede ser grande solo a costa de reducir su densidad poblacional y esto 10 hace mis vulnerable a la extinci6n sobre todo en una isla (McNab 1994 McFarlane et al 1998a) Incidentalmente Alcover et al (1998a) afirmaron que larelacion entre la talla corporal y la extincion no siempre se cumple en las especies de menor tamano y citaron como ejemplo que entre las musarafias todas las especies de Nesophontes desaparecieron mientras sobreviven especies de Solenodon pero la comparacion correcta en todo caso serfa entre los solenodones vivientes y el gigante Solenodon arredondoi y entre Nesophontes edithae -el gigante de los nesofontes extinguido en el periodo amerindio- y el resto de sus congeneres que sobrevivieron hasta los primeros siglos de la colonizacion europea (Tabla 45 intervalos III y IV)
CAUSAS ANTROPOGENICAS
Intervalo Amerindio
La mayoria de los autores que se han referido a las extinciones acaecidas durante este intervalo (entre elIos paradigmaticamente Martin 1984a b) han responsabilizado a los aborfgenes (que se supone comenzaron a poblar las Antillas Mayores hace 6 mil afios) con la desaparicion de numerosas especies de mamiferos terrestres en la region principalmente por la via de la sobrecaza y la introduccion de animales domesticos devenidos depredadores de la fauna autoctona (Morgan y Woods 1986) Este efecto negativo de la presencia indigena sobre los mamiferos de la region ha sido dramatizado por algunos autores en terminos de holocausto faunal Sin embargo pensamos que se ha exagerado Como han expresado MacPhee et al (19993) cuando uno busca evidencia dura de complicidad humana directa en [estas] extinciones no hay mucho que encontrar Las siguientes son algunas consideraciones a tener en cuenta en tal sentido
El registro radiometrico mas joven conocido hasta ahora para un perezoso antillano (MegalocnllS rodens) es de 42 kaAP (MacPhee et ai 2007) Si se acepta que los aborfgenes entraron en Cuba hace aproximadamente 6 mil anos su convivencia con los perezosos debe haber durado alrededor de 1800 afios (0 mas porque dificilmente el referfdo fechado radiometrico haya correspondido al ultimo sobreviviente de la especie) Esta cifra contradice la hipotesis del holocausto faunal segun la cualla extincion de los mamiferos insulares de mayor talla debe ocurrir con extrema rapidez -a escala de decadas 0 de centurias- a partir de su primer contacto con los humanos (Martin y Steadman 1999) A pesar de las repetidas afirmaciones en contrario (Harrington 1921 Miller 1929c Herrera [1945] Tabio 1951 Alvarez 1951a 1957 Arredondo
331 EXTINCIONES DEL CUATERNARIO
1959 1960a Veloz y Ortega 1973 Mayo 1980b Rodriguez et al 1984 Castellanos y Pino 1985 Pino y Castellanos 1985 Nunez 1990b Gutierrez 1996 Arredondo 2004) en su mayoria basadas en evidencias no exentas de posibles accidentes de deposicion (0 de reubicacion postdeposicional) en recintos cavernarios (Diaz-Franco [2006b D no se ha demostrado inequivocamente que los aborigenes antillanos se alimentaran de perezosos regularmente Sus restos en residuarios de alimentos son escasisimos en toda la region y esto contrasta con los abundantes restos de jutias y otros mamiferos de menor talla que aparecen en esos residuarios demostrativos de la preferencia de los paleoindios por estos otros mamiferos
Los mamiferos mas importantes en la economia prehistorica de las Antillas Mayores fueron las jutias Capromys y Geocapromys en Cuba Geocapromys en Jamaica e Isolobodon en La Espanola Puerto Rico e Islas Virgenes (Aguayo 1950b Tablo 1951 Rimoli 1972 Wing 2001 Diaz-Franco [2006b D Wilkins (2001) estudi61ajutia viviente de Jamaica (Geocapromys browni) en un sitio ocupado por los aborigenes durante 100 anos y no encontr6 evidencias de que durante ese tiempo la especie haya sido sobreexplotada Asimismo en Cuba las jutias conga carabali y andaraz son escasas en depositos pleistoholocenicos en cambio son muy numerosas en residuarios indigenas y de haber sido sobreexplotadas 10 mas probable es que no hubieran sobrevivido hasta hoy
La habilidad de los humanos para penetrar en profundidad el bosque tropical y mantenerse en este sustentablemente ha sido un evento comparativamente reciente (Beran y Steele 1994) En el caso antillano la caza tierra adentro ha sido explicada como un proceso tardio de desplazamiento a partir de la explotacion de abundantes recursos marinos que ofrecian condiciones 6ptimas para el sedentarismo de las comunidades aborigenes costeras (C6rdova y Arredondo 1988 Brito 2005) Peces y mariscos dominan la arqueologia costera de las Islas antillanas en tanto el consumo de recursos terrestres (mamfferos incluidos) aumenta progresivamente en la medida que los residuarios estudiados se alejan de la costa (Scudder 1991 Wilkins 2001) En las Antillas Mayores el componente terrestre de la dieta aborigen es solo 20-30 del material recobrado en los residuarios (Wing 1977 1989) Y debio ser menor alin porque el componente vegetal de la dieta no se ha preservado Por otra parte los aborigenes cubanos domesticaron la jutia conga y seguramente la reprodujeron para comer (Pose et al 1988) practica que probablemente se repitiera con otras jutias en las demas Antillas Mayores
- Fray Bartolome de las Casas hablo de 3 millones de indios en Cuba al inicio de la colonizacion pero los historiadores subsecuentes consideraron exagerada la cifra (Poey 1851 a) No obstante se dispone de dos estimados de la densidad de poblaci6n aborigen en las Antillas Mayores a la llegada de los europeos 18 indigenaslkm2 en Puerto Rico (Rouse 1956) y 1271km2 en La Espanola (Watts 1987) Extrapolando ambos valores entre las cuatro islas principales de las Antillas Mayores tenemos que en la fecha indicada la poblaci6n aborigen pudo estar entre 16 mil y 113 mil indigenas en Puerto Rico entre 21 mil y 145 mil en Jamaica entre 141 mil y 1 mill6n en La Espanola y entre 200 mil y 14 millones en Cuba Entre las cifras maximas y las minimas nos inclinamos
332 BIOGEOGRAFiA
hacia estas ultimas atendiendo a los altos indices de mortalidad infantil y morbilidad que se evidencian en las osamentas de los pobladores prehispanicos (Martinez y Rives 1990) Uno de los hechos que apuntan hacia el bajo numero de aborigenes en las Antillas Mayores es 10 rapido que desaparecieron despues de la llegada de los europeos pnkticamente sin dejar mestizaje con estos 10 cual contrasta ostensiblemente con la situacion en la America continentaL
En suma un grupo humano (1) demograiicamente irrelevante y controlado por altos indices de morbilidad (2) consumidor mayoritariamente de organismos marinos y (3) carente de la tecnologia cinegetica necesaria para poder sobreexplotar los mamiferos que Ie sirvieron de alimento (a los cuales accedio tardiamente en su desarrollo) no es concebible que haya causado e1 holocausto faunal que se Ie atribuye a partir del error de aplicar a las comunidades aborigenes antillanas un modelo cultural de la prehistoria continentaL
Tampoco hay evidencias de que los aborigenes hayan introducido carnivoros con suficiente capacidad depredadora salvo el perro Morgan y Otlenwaldder (1993) consideraron que la depredacion por perros durante el periodo amerindio es la explicacion mas plausible para la extincion del almiqui gigante de Cuba pero como se puede observar (Tabla 54) Solenodon arredondoi aparece en el intervalo II por falta de asociacion con evidencias culturales
Consecuentemente el accionar de los aborigenes y de sus animales domesticos directamente sobre la fauna no parece haber sido por sf solo un factor determinante de la extincion de mamiferos terrestres durante el periodo amerindio en las Antillas (pero vease Steadman et al 2005)
Hay que recordar en este senti do que alin en el Holoceno tardio ocurrieron cambios clirruiticos de consideracion (vease Causas Naturales) Por ejemplo Curtis et al (2001) explicaron que la extincion de algunos ani males mesofilicos de las alturas montanosas de La Espanola pudo haber antecedido a cualquier impacto humano sigrIificativo en esos lugares por coincidir dicha extincion con una tendencia a la aridez documentada isotopicamente y con evidencias de la perdida del bosque mesico ocurridas despues de 3kaAP Yen Jamaica hace solo 2shy3 mil aDos el area de la Bahia de Jackson sustentaba una gran biomasa de vertebrados en bosque hinnedo a juzgar por la cantidad y diversidad de taxones presentes en material fosil derivado de la depredacion por lechuzas hoy el area es la mas seca de la isla con matorral xerofitico y pobreza de fauna (MacPhee 1996a) Yalm en nuestros dias huracanes de gran intensidad aniquilan poblaciones de vertebrados (que no se recuperan) en is las pequenas y llanas del Caribe (Brooks y Smith 2001)
En medio de la incertidumbre sobre la verdadera causa (0 causas) de las extinciones de mamiferos en el periodo amerindio MacPhee y Marx (1997) han propuesto su hipotesis de la hipermorbilidad la cual estipula colapsos poblacionales relativamente rapidos con la introduccion de enfermedades de alta letalidad Estos autores han razonado que
Las islas son los lugares mas apropiados para experimentar perdidas catastroficas causadas por enfermedades Los mamiferos no solo serian susceptibles en virtud de su prolongado aislamiento biologico sino que en los confines de una isla la epizootia
333 EXTINCIONES DEL CUATERNARIO
tendria la potencialidad de infectar simultaneamente a la rnayoria de los individuos de vadas especies Los neonatos son inmunologicamente inmaduros y sufririan desproporcionadamente cualquier infeccion hipervirulenta La perdida excesiva de neonatos y juveniles tendria efectos severos para la sobrevivencia de la poblacion si esta se reduce por debajo de cierto nivel sobreviene la extincion porque no hay posibilidad de restitucion en el periodo postpanzootico Las especies de mayor talla corporal van a ser las mas vulnerables a la extincion dadas sus bajas tasas reproductivas largos periodos de gestacion y reducido numero de progenie Las especies de menor tamano --caracterizadas por altas tasas reproductivas periodos cortos de gestacion y mayor numero de neonatos por parto- tambien serian fuertemente diezmadas pero muchas sobrevivirian porque su estrategia reproductiva les permite perder mayor numero de individuos Las perdidas deben ocurrir relativamente rapido (a escala de decadas) mientras que otras hipotesis (digamos los cambios climaticos) permiten intervalos mucho mas largos (a escala de siglos 0 de milenios) entre la aparicion de las causas y la produccion de sus efectos Por ejemplo todo indica que los nesofontes se extinguieron muy rapidamente y seria dificil poder invocar un impacto humano directo 0 argumentar fenomenos climaticos que pudieran explicar plausiblemente perdidas tan rapidas
MacPhee y Marx (1997 191) han dicho que por el momento la hipotesis de la hipermorbilidad no pasa de ser una abstraccion en espera de adecuada sustentacion empirica Llama la atencion que linicamente en el periodo amerindio (Tabla 45 intervalo 1Il) las extinciones abarcaron todas las clases del peso corporal (de la a a laj) y que ninguna de las especies que sobrevivieron hasta el periodo postcolombino (Tabla 45 intervalo IV mas las especies vivientes) sobrepasa los 10 kg (clases del peso a b c) en clara coincidencia con los presupuestos de la hipotetica hipermorbilidad
Intervalo Postcolornbino
Por 10 menos 37 especies de mamiferos terrestres vivian en las Antillas Mayores y grupos de Islas perifericas a la lIegada de los europeos en 1492 De elIas se han extinguido 23 shymas especies que en cualquiera de los periodos precedentes (Tabla 45~ y solo subsisten 12 especies repartidas entre Cuba (8 especies) La Espanola (2) Jamaica (1) y Bahamas (1) varias de estas en peligro critico de extincion Solo dos de las especies extinguidas durante los ultimos 500 anos se conocieron en estado viviente los roedores Oryzomys antillarum de Jamaica (extinguido en 1877) y Geocapromys thoracatus de las Islas Swan (extinguido en 1955) Las causas de las extinciones ocurridas durante este periodo son sobradamente conocidas destruccion de habitats introducci6n de animales exoticos (domesticos y comensales) y demas presiones asociadas a la actividad humana entre las cuales posiblemente habria que incluir brotes de hipermorbilidad
Durante mucho tiempo se ha afirmado la posibilidad de que especies de Nesophontes hayan sobrevivido hasta las primeras decadas del siglo xx y que incluso continuen existiendo en lugares remotos de Cuba 0 La Espanola (Allen 1918 1942 Miller 1930 Aguayo 1950a Arredondo 1960b Fischer 1977 MacFadden 1980 Woods y
334 BIOGEOGRAFiA
Ottenwalder 1991) Actualmente no existe evidencia radiometrica de que alguna especie de Nesophontes haya sobrevivido al siglo xv las fechas radiocarbonicas calibradas obtenidas en muestras de Nesophontes de Cuba y La Espanola resultaron no mas tardias que los siglos XIII y XIV (MacPhee et al 1999) Por otra parte las expediciones efectuadas a fines del siglo xx con el proposito de localizar individuos vivientes del genero en La Espanola no produjeron la menor indicacion afirmativa al respecto (Woods et al 1985) Ademas a traves de las Antillas Mayores no aparecen restos de Nesophontes en los depositos formados actualmente por las lechuzas
EPiLOGO
Las Antillas Mayores son islas oceanicas no han formado parte de un continente (con la posible excepcion de una parte de Jamaica hasta el Eoceno Medio hace 45 Ma) y durante un tiempo geologico considerable (desde el Eoceno Superior hace 40 Ma) han permanecido emergidas y separadas de los continentes vecinos Consecuentemente los mamiferos terrestres (no volantes) autoctonos de estas islas constituyen una fauna oceanica que por ende cumple con todas las caracteristicas convencionales reconocidas para este tipo de entidad biogeografica
- Baja diversidad taxonomica a nivel de las categorias superiores (solo cuatro ordenes) en comparacion con islas plataformicas continentales (0 con areas continentales) de tamano y diversidad ecologica simi lares
- Alto grado de endemismo a nivel de genero (96) y familia (50) en respuesta al prolongado aislamiento
- Desbalance ecologico de la estructura trofica ausencia de mamiferos camivoros y ungulados
- Espectaculares procesos de irradiaci6n evolucionaria como resultado de la colonizacion de territorios libres de competidores y depredadores
Tendencias evolucionarias extremas gigantismo
- Control a cargo de aves rapaces con tendencia at gigantismo en ausencia de mamiferos miembros del orden Carnivora
- Mas especies de reptiles y de murcielagos que de mamiferos terrestres relaciones que en los continentes se invierten
Alta tasa de extincion de especies (84) sin reposicion
Preservacion de especies filogeneticamente arcaicas (los llamados fosiles vivientes) los almiquies

328 BIOGEOGRAFiA
De los cuatro 6rdenes de mamiferos representados en la fauna Pilosa y Primates desaparecieron totalmente Soricomorpha y Rodentia perdieron 86 y 74 de sus especies respectivamente
CAUSALIDAD
CAUSAS NATURALES
Los cicIos glaciales e interglaciales acaecidos en el Pleistoceno causaron cambios ambientales dnisticos en las Antillas con la concomitante alternancia de cIimas frios (secos) y calidos (hUmedos) asi como de descensos y ascensos del nivel del mar aumento y disminuci6n de las areas territoriales) induso (aunque con menor intensidad) en el Holoceno Estos eventos paleogeograficos han sido consistentemente documentados (Mayo y Kartashov 1972 Bonnatti y Gartner 1973 Ortega y Arcia 1982 Prance 1982 Schubert y Medina 1982 Dodge et al 1983 Ortega [1984] Haq et al 1987 Schubert 1989 Hodell et al 1991 Guilderson et al 1994 Oppo 1997 Emslie 1998 Curtis et al 2001 Iturralde-Vinent 2003) y de ellos brindamos a continuaci6n una breve resena
Finalizando el periodo interglacial anterior a la ultima glaciaci6n (Wisconsin) el mar estaba a 8-10 m por encima de su nivel actual Cuando el avance glacial del Wisconsin a1canz6 su culminaci6n (eillamado Ultimo Maximo Glacial) hace 18 ka el mar habia descendido 120 m con respecto a dicho nivel y las islas del Caribe habian incrementado considerablemente sus extensiones subaereas en comparaci6n con las que presentan hoy dia por ejemplo Puerto Rico y las Islas Virgenes se conectaron territorialmente y las Bahamas se unieron en cinco gran des islas Prevalecia un regimen frio y seco y condiciones de aridez --debidas a una reducci6n de las precipitaciones (posiblemente a menos de 440 mmlano)- y los ambientes xericos se habian extendido a traves de las islas y bancos emergidos y abundaban los pinares (los habitats mesicos estaban restringidos a parcelas relictas en 10 alto de las montanas) En el Mar Caribe y el Golfo de Mexico la temperatura superficial del mar era 5degC mas baja que la actual y en las llanuras costeras y alturas maximas eran respectivamente 2-3degC y 6degC mas bajas Los limites de las zonas ecol6gicas altitudinales descendieron varios cientos de metros y cada zona redujo su extensi6n vertical al aumentar el gradiente de temperatura Ellimite inferior de la zona periglacial
elevaci6n donde la temperatura media minima a1canza los OdegC) de la Cordillera Central de la Republica Dominicana (altura maxima del Caribe insular con evidencia de topografia glacial) se considera que estuvo entre los 1500 y 1800 mlnm (actualmente esta a 2800 mlnm) y se puede estimar que dicho limite era inferior en Cuba por estar mas al norte que la Republica Dominicana No obstante durante este periodo la temperatura experiment6 episodios cortos (unas pocas decadas) de ascensos repetidos
Un signifieativo componente bi6tieo de las referidas condiciones paleoambientales 10 constituy6 la espectacular irradiaci6n evolucionaria experimentada por un grupo numeroso de aves rapaees diurnas y nocturnas (Wetmore 1937 Brodkorb 1969 Kurotchkin y Mayo 1973 Arredondo 1976 Olson 1978 Olson y Hilgartner 1982
329 EXTINCIONES DEL ClJATERNARIO
Suarez 2000a b 2004 Suarez y Emslie 2003) Los mamiferos de las Antillas Mayores ofrecieron a estos depredadores aviares una amplia gama de tamanos de presa que en ausencia de mamiferos del orden Carnivora en la region permitio a estas aves irradiarse a este nicho alimentario con la evolucion de una variedad de especies de muy diversa tall a corporal incluidas formas gigantes Adquirir un cuerpo gigante les propicio el acceso a recursos alimentarios que no estaban siendo explotados por ningun otro depredador (Alcover y McMinn 1994) A Cuba corresponde el mejor registro fosil de rapaces cuaternarias caribenas con siete generos de especies diurnas Dentro de estas aves existieron tres gavilanes gigantes (Accipitridae) tres carairas (Falconidae) y un condor (Cathartidae) asi como cuatro generos de especies nocturnas dos lechuzas (Tytonidae) y tres Mhos (Strigidae) dos de ell os no igualados en tamano en todo el mundo Algunos de estos generos tuvieron representacion en otras islas antillanas como La Espanola Jamaica Bahamas y Gran Caiman
Al retirarse hacia el norte los hielos de la glaciacion Wisconsin el clima se tomo mas hUmedo en las Antillas durante el Holoceno temprano y el bosque mesico se expandio en la region insular El nivel del mar ascendio acompanado de mayor pluviosidad alrededor de los 7 kaAP las Islas Virgenes quedaron separadas entre si y de Puerto Rico y las Bahamas perdieron la mayor parte de su territorio Cerca de los 3 kaAP las condiciones secas retornaron a la region e hicieron desaparecer buena parte del bosque mesico Nuevamente el clima comenzo a adquirir sus condiciones actuales hacia los 25 kaAP aproximadamente y hoy transita por una fase mesica con vegetacion xerica confinada en parcelas relictas
Sin duda estos y otros eventos naturales incidieron negativamente en la sobrevivencia de mamiferos terrestres en las Antillas Mayores asi como en los grupos de pequenas islas perifericas (Pregill y Olson 1981 Olson 2006)
McFarlane et al (1998a) demostraron convincentemente como Amblyrhiza el gigante de los roedores antillanos endemico de la pequena isla de Anguila sucumbio a las repetidas y abruptas perdidas de su habitat durante el interglacial Sangamon por las nipidas y severas oscilaciones del nivel del mar (documentadas por Neumann y Hearty 1996) al no poder responder adecuadamente a tan catastroficos eventos mediante el ajuste de su masa corporal media y otro tanto debio ocurrirle en Jamaica al otro roedor gigante Clidomys (McFarlane et al 1998b) emparentado con Amblyrhiza en la misma epoca (Tabla 45 intervalo I)
El ascenso del nivel del mar seguramente promovio la extincion de mamiferos durante la transicion del Pleistoceno al Holoceno (Tabla 45 intervalo II) con la reduccion del tamano de las Islas por las transgresiones marinas (Pregill y Olson 1981 Morgan 1993) Esto ultimo haria maximamente severa la competicion asf como la presion de los depredadores [Steadman et al (2005) no yen por que el calentamiento global ocurrido durante la transicion Pleistoceno-Holoceno habria de tener un efecto negativo sobre los perezosos siendo sus descendientes actuales especies tropic ales 0 subtropicales pero no se trata del efecto
330 BIOGEOGRAFiA
directo del cambio de temperatura sobre los animales sino del efecto indirecto y progresivo que dicho cambio genera a largo plazo al reducir el territorio insular disponible]
Por esta epoca tambien desaparecieron especies de tall a mayor como Megalocnus zile en La Espanola y otras de menor tamano (en Cuba) pero que representan la tall a maxima en sus respectivos contextos taxonomicos del Cuaternario como Solenodon arredondoi Paralouatta varonai y Macrocapromys acevedo Un mamifero puede ser grande solo a costa de reducir su densidad poblacional y esto 10 hace mis vulnerable a la extinci6n sobre todo en una isla (McNab 1994 McFarlane et al 1998a) Incidentalmente Alcover et al (1998a) afirmaron que larelacion entre la talla corporal y la extincion no siempre se cumple en las especies de menor tamano y citaron como ejemplo que entre las musarafias todas las especies de Nesophontes desaparecieron mientras sobreviven especies de Solenodon pero la comparacion correcta en todo caso serfa entre los solenodones vivientes y el gigante Solenodon arredondoi y entre Nesophontes edithae -el gigante de los nesofontes extinguido en el periodo amerindio- y el resto de sus congeneres que sobrevivieron hasta los primeros siglos de la colonizacion europea (Tabla 45 intervalos III y IV)
CAUSAS ANTROPOGENICAS
Intervalo Amerindio
La mayoria de los autores que se han referido a las extinciones acaecidas durante este intervalo (entre elIos paradigmaticamente Martin 1984a b) han responsabilizado a los aborfgenes (que se supone comenzaron a poblar las Antillas Mayores hace 6 mil afios) con la desaparicion de numerosas especies de mamiferos terrestres en la region principalmente por la via de la sobrecaza y la introduccion de animales domesticos devenidos depredadores de la fauna autoctona (Morgan y Woods 1986) Este efecto negativo de la presencia indigena sobre los mamiferos de la region ha sido dramatizado por algunos autores en terminos de holocausto faunal Sin embargo pensamos que se ha exagerado Como han expresado MacPhee et al (19993) cuando uno busca evidencia dura de complicidad humana directa en [estas] extinciones no hay mucho que encontrar Las siguientes son algunas consideraciones a tener en cuenta en tal sentido
El registro radiometrico mas joven conocido hasta ahora para un perezoso antillano (MegalocnllS rodens) es de 42 kaAP (MacPhee et ai 2007) Si se acepta que los aborfgenes entraron en Cuba hace aproximadamente 6 mil anos su convivencia con los perezosos debe haber durado alrededor de 1800 afios (0 mas porque dificilmente el referfdo fechado radiometrico haya correspondido al ultimo sobreviviente de la especie) Esta cifra contradice la hipotesis del holocausto faunal segun la cualla extincion de los mamiferos insulares de mayor talla debe ocurrir con extrema rapidez -a escala de decadas 0 de centurias- a partir de su primer contacto con los humanos (Martin y Steadman 1999) A pesar de las repetidas afirmaciones en contrario (Harrington 1921 Miller 1929c Herrera [1945] Tabio 1951 Alvarez 1951a 1957 Arredondo
331 EXTINCIONES DEL CUATERNARIO
1959 1960a Veloz y Ortega 1973 Mayo 1980b Rodriguez et al 1984 Castellanos y Pino 1985 Pino y Castellanos 1985 Nunez 1990b Gutierrez 1996 Arredondo 2004) en su mayoria basadas en evidencias no exentas de posibles accidentes de deposicion (0 de reubicacion postdeposicional) en recintos cavernarios (Diaz-Franco [2006b D no se ha demostrado inequivocamente que los aborigenes antillanos se alimentaran de perezosos regularmente Sus restos en residuarios de alimentos son escasisimos en toda la region y esto contrasta con los abundantes restos de jutias y otros mamiferos de menor talla que aparecen en esos residuarios demostrativos de la preferencia de los paleoindios por estos otros mamiferos
Los mamiferos mas importantes en la economia prehistorica de las Antillas Mayores fueron las jutias Capromys y Geocapromys en Cuba Geocapromys en Jamaica e Isolobodon en La Espanola Puerto Rico e Islas Virgenes (Aguayo 1950b Tablo 1951 Rimoli 1972 Wing 2001 Diaz-Franco [2006b D Wilkins (2001) estudi61ajutia viviente de Jamaica (Geocapromys browni) en un sitio ocupado por los aborigenes durante 100 anos y no encontr6 evidencias de que durante ese tiempo la especie haya sido sobreexplotada Asimismo en Cuba las jutias conga carabali y andaraz son escasas en depositos pleistoholocenicos en cambio son muy numerosas en residuarios indigenas y de haber sido sobreexplotadas 10 mas probable es que no hubieran sobrevivido hasta hoy
La habilidad de los humanos para penetrar en profundidad el bosque tropical y mantenerse en este sustentablemente ha sido un evento comparativamente reciente (Beran y Steele 1994) En el caso antillano la caza tierra adentro ha sido explicada como un proceso tardio de desplazamiento a partir de la explotacion de abundantes recursos marinos que ofrecian condiciones 6ptimas para el sedentarismo de las comunidades aborigenes costeras (C6rdova y Arredondo 1988 Brito 2005) Peces y mariscos dominan la arqueologia costera de las Islas antillanas en tanto el consumo de recursos terrestres (mamfferos incluidos) aumenta progresivamente en la medida que los residuarios estudiados se alejan de la costa (Scudder 1991 Wilkins 2001) En las Antillas Mayores el componente terrestre de la dieta aborigen es solo 20-30 del material recobrado en los residuarios (Wing 1977 1989) Y debio ser menor alin porque el componente vegetal de la dieta no se ha preservado Por otra parte los aborigenes cubanos domesticaron la jutia conga y seguramente la reprodujeron para comer (Pose et al 1988) practica que probablemente se repitiera con otras jutias en las demas Antillas Mayores
- Fray Bartolome de las Casas hablo de 3 millones de indios en Cuba al inicio de la colonizacion pero los historiadores subsecuentes consideraron exagerada la cifra (Poey 1851 a) No obstante se dispone de dos estimados de la densidad de poblaci6n aborigen en las Antillas Mayores a la llegada de los europeos 18 indigenaslkm2 en Puerto Rico (Rouse 1956) y 1271km2 en La Espanola (Watts 1987) Extrapolando ambos valores entre las cuatro islas principales de las Antillas Mayores tenemos que en la fecha indicada la poblaci6n aborigen pudo estar entre 16 mil y 113 mil indigenas en Puerto Rico entre 21 mil y 145 mil en Jamaica entre 141 mil y 1 mill6n en La Espanola y entre 200 mil y 14 millones en Cuba Entre las cifras maximas y las minimas nos inclinamos
332 BIOGEOGRAFiA
hacia estas ultimas atendiendo a los altos indices de mortalidad infantil y morbilidad que se evidencian en las osamentas de los pobladores prehispanicos (Martinez y Rives 1990) Uno de los hechos que apuntan hacia el bajo numero de aborigenes en las Antillas Mayores es 10 rapido que desaparecieron despues de la llegada de los europeos pnkticamente sin dejar mestizaje con estos 10 cual contrasta ostensiblemente con la situacion en la America continentaL
En suma un grupo humano (1) demograiicamente irrelevante y controlado por altos indices de morbilidad (2) consumidor mayoritariamente de organismos marinos y (3) carente de la tecnologia cinegetica necesaria para poder sobreexplotar los mamiferos que Ie sirvieron de alimento (a los cuales accedio tardiamente en su desarrollo) no es concebible que haya causado e1 holocausto faunal que se Ie atribuye a partir del error de aplicar a las comunidades aborigenes antillanas un modelo cultural de la prehistoria continentaL
Tampoco hay evidencias de que los aborigenes hayan introducido carnivoros con suficiente capacidad depredadora salvo el perro Morgan y Otlenwaldder (1993) consideraron que la depredacion por perros durante el periodo amerindio es la explicacion mas plausible para la extincion del almiqui gigante de Cuba pero como se puede observar (Tabla 54) Solenodon arredondoi aparece en el intervalo II por falta de asociacion con evidencias culturales
Consecuentemente el accionar de los aborigenes y de sus animales domesticos directamente sobre la fauna no parece haber sido por sf solo un factor determinante de la extincion de mamiferos terrestres durante el periodo amerindio en las Antillas (pero vease Steadman et al 2005)
Hay que recordar en este senti do que alin en el Holoceno tardio ocurrieron cambios clirruiticos de consideracion (vease Causas Naturales) Por ejemplo Curtis et al (2001) explicaron que la extincion de algunos ani males mesofilicos de las alturas montanosas de La Espanola pudo haber antecedido a cualquier impacto humano sigrIificativo en esos lugares por coincidir dicha extincion con una tendencia a la aridez documentada isotopicamente y con evidencias de la perdida del bosque mesico ocurridas despues de 3kaAP Yen Jamaica hace solo 2shy3 mil aDos el area de la Bahia de Jackson sustentaba una gran biomasa de vertebrados en bosque hinnedo a juzgar por la cantidad y diversidad de taxones presentes en material fosil derivado de la depredacion por lechuzas hoy el area es la mas seca de la isla con matorral xerofitico y pobreza de fauna (MacPhee 1996a) Yalm en nuestros dias huracanes de gran intensidad aniquilan poblaciones de vertebrados (que no se recuperan) en is las pequenas y llanas del Caribe (Brooks y Smith 2001)
En medio de la incertidumbre sobre la verdadera causa (0 causas) de las extinciones de mamiferos en el periodo amerindio MacPhee y Marx (1997) han propuesto su hipotesis de la hipermorbilidad la cual estipula colapsos poblacionales relativamente rapidos con la introduccion de enfermedades de alta letalidad Estos autores han razonado que
Las islas son los lugares mas apropiados para experimentar perdidas catastroficas causadas por enfermedades Los mamiferos no solo serian susceptibles en virtud de su prolongado aislamiento biologico sino que en los confines de una isla la epizootia
333 EXTINCIONES DEL CUATERNARIO
tendria la potencialidad de infectar simultaneamente a la rnayoria de los individuos de vadas especies Los neonatos son inmunologicamente inmaduros y sufririan desproporcionadamente cualquier infeccion hipervirulenta La perdida excesiva de neonatos y juveniles tendria efectos severos para la sobrevivencia de la poblacion si esta se reduce por debajo de cierto nivel sobreviene la extincion porque no hay posibilidad de restitucion en el periodo postpanzootico Las especies de mayor talla corporal van a ser las mas vulnerables a la extincion dadas sus bajas tasas reproductivas largos periodos de gestacion y reducido numero de progenie Las especies de menor tamano --caracterizadas por altas tasas reproductivas periodos cortos de gestacion y mayor numero de neonatos por parto- tambien serian fuertemente diezmadas pero muchas sobrevivirian porque su estrategia reproductiva les permite perder mayor numero de individuos Las perdidas deben ocurrir relativamente rapido (a escala de decadas) mientras que otras hipotesis (digamos los cambios climaticos) permiten intervalos mucho mas largos (a escala de siglos 0 de milenios) entre la aparicion de las causas y la produccion de sus efectos Por ejemplo todo indica que los nesofontes se extinguieron muy rapidamente y seria dificil poder invocar un impacto humano directo 0 argumentar fenomenos climaticos que pudieran explicar plausiblemente perdidas tan rapidas
MacPhee y Marx (1997 191) han dicho que por el momento la hipotesis de la hipermorbilidad no pasa de ser una abstraccion en espera de adecuada sustentacion empirica Llama la atencion que linicamente en el periodo amerindio (Tabla 45 intervalo 1Il) las extinciones abarcaron todas las clases del peso corporal (de la a a laj) y que ninguna de las especies que sobrevivieron hasta el periodo postcolombino (Tabla 45 intervalo IV mas las especies vivientes) sobrepasa los 10 kg (clases del peso a b c) en clara coincidencia con los presupuestos de la hipotetica hipermorbilidad
Intervalo Postcolornbino
Por 10 menos 37 especies de mamiferos terrestres vivian en las Antillas Mayores y grupos de Islas perifericas a la lIegada de los europeos en 1492 De elIas se han extinguido 23 shymas especies que en cualquiera de los periodos precedentes (Tabla 45~ y solo subsisten 12 especies repartidas entre Cuba (8 especies) La Espanola (2) Jamaica (1) y Bahamas (1) varias de estas en peligro critico de extincion Solo dos de las especies extinguidas durante los ultimos 500 anos se conocieron en estado viviente los roedores Oryzomys antillarum de Jamaica (extinguido en 1877) y Geocapromys thoracatus de las Islas Swan (extinguido en 1955) Las causas de las extinciones ocurridas durante este periodo son sobradamente conocidas destruccion de habitats introducci6n de animales exoticos (domesticos y comensales) y demas presiones asociadas a la actividad humana entre las cuales posiblemente habria que incluir brotes de hipermorbilidad
Durante mucho tiempo se ha afirmado la posibilidad de que especies de Nesophontes hayan sobrevivido hasta las primeras decadas del siglo xx y que incluso continuen existiendo en lugares remotos de Cuba 0 La Espanola (Allen 1918 1942 Miller 1930 Aguayo 1950a Arredondo 1960b Fischer 1977 MacFadden 1980 Woods y
334 BIOGEOGRAFiA
Ottenwalder 1991) Actualmente no existe evidencia radiometrica de que alguna especie de Nesophontes haya sobrevivido al siglo xv las fechas radiocarbonicas calibradas obtenidas en muestras de Nesophontes de Cuba y La Espanola resultaron no mas tardias que los siglos XIII y XIV (MacPhee et al 1999) Por otra parte las expediciones efectuadas a fines del siglo xx con el proposito de localizar individuos vivientes del genero en La Espanola no produjeron la menor indicacion afirmativa al respecto (Woods et al 1985) Ademas a traves de las Antillas Mayores no aparecen restos de Nesophontes en los depositos formados actualmente por las lechuzas
EPiLOGO
Las Antillas Mayores son islas oceanicas no han formado parte de un continente (con la posible excepcion de una parte de Jamaica hasta el Eoceno Medio hace 45 Ma) y durante un tiempo geologico considerable (desde el Eoceno Superior hace 40 Ma) han permanecido emergidas y separadas de los continentes vecinos Consecuentemente los mamiferos terrestres (no volantes) autoctonos de estas islas constituyen una fauna oceanica que por ende cumple con todas las caracteristicas convencionales reconocidas para este tipo de entidad biogeografica
- Baja diversidad taxonomica a nivel de las categorias superiores (solo cuatro ordenes) en comparacion con islas plataformicas continentales (0 con areas continentales) de tamano y diversidad ecologica simi lares
- Alto grado de endemismo a nivel de genero (96) y familia (50) en respuesta al prolongado aislamiento
- Desbalance ecologico de la estructura trofica ausencia de mamiferos camivoros y ungulados
- Espectaculares procesos de irradiaci6n evolucionaria como resultado de la colonizacion de territorios libres de competidores y depredadores
Tendencias evolucionarias extremas gigantismo
- Control a cargo de aves rapaces con tendencia at gigantismo en ausencia de mamiferos miembros del orden Carnivora
- Mas especies de reptiles y de murcielagos que de mamiferos terrestres relaciones que en los continentes se invierten
Alta tasa de extincion de especies (84) sin reposicion
Preservacion de especies filogeneticamente arcaicas (los llamados fosiles vivientes) los almiquies

329 EXTINCIONES DEL ClJATERNARIO
Suarez 2000a b 2004 Suarez y Emslie 2003) Los mamiferos de las Antillas Mayores ofrecieron a estos depredadores aviares una amplia gama de tamanos de presa que en ausencia de mamiferos del orden Carnivora en la region permitio a estas aves irradiarse a este nicho alimentario con la evolucion de una variedad de especies de muy diversa tall a corporal incluidas formas gigantes Adquirir un cuerpo gigante les propicio el acceso a recursos alimentarios que no estaban siendo explotados por ningun otro depredador (Alcover y McMinn 1994) A Cuba corresponde el mejor registro fosil de rapaces cuaternarias caribenas con siete generos de especies diurnas Dentro de estas aves existieron tres gavilanes gigantes (Accipitridae) tres carairas (Falconidae) y un condor (Cathartidae) asi como cuatro generos de especies nocturnas dos lechuzas (Tytonidae) y tres Mhos (Strigidae) dos de ell os no igualados en tamano en todo el mundo Algunos de estos generos tuvieron representacion en otras islas antillanas como La Espanola Jamaica Bahamas y Gran Caiman
Al retirarse hacia el norte los hielos de la glaciacion Wisconsin el clima se tomo mas hUmedo en las Antillas durante el Holoceno temprano y el bosque mesico se expandio en la region insular El nivel del mar ascendio acompanado de mayor pluviosidad alrededor de los 7 kaAP las Islas Virgenes quedaron separadas entre si y de Puerto Rico y las Bahamas perdieron la mayor parte de su territorio Cerca de los 3 kaAP las condiciones secas retornaron a la region e hicieron desaparecer buena parte del bosque mesico Nuevamente el clima comenzo a adquirir sus condiciones actuales hacia los 25 kaAP aproximadamente y hoy transita por una fase mesica con vegetacion xerica confinada en parcelas relictas
Sin duda estos y otros eventos naturales incidieron negativamente en la sobrevivencia de mamiferos terrestres en las Antillas Mayores asi como en los grupos de pequenas islas perifericas (Pregill y Olson 1981 Olson 2006)
McFarlane et al (1998a) demostraron convincentemente como Amblyrhiza el gigante de los roedores antillanos endemico de la pequena isla de Anguila sucumbio a las repetidas y abruptas perdidas de su habitat durante el interglacial Sangamon por las nipidas y severas oscilaciones del nivel del mar (documentadas por Neumann y Hearty 1996) al no poder responder adecuadamente a tan catastroficos eventos mediante el ajuste de su masa corporal media y otro tanto debio ocurrirle en Jamaica al otro roedor gigante Clidomys (McFarlane et al 1998b) emparentado con Amblyrhiza en la misma epoca (Tabla 45 intervalo I)
El ascenso del nivel del mar seguramente promovio la extincion de mamiferos durante la transicion del Pleistoceno al Holoceno (Tabla 45 intervalo II) con la reduccion del tamano de las Islas por las transgresiones marinas (Pregill y Olson 1981 Morgan 1993) Esto ultimo haria maximamente severa la competicion asf como la presion de los depredadores [Steadman et al (2005) no yen por que el calentamiento global ocurrido durante la transicion Pleistoceno-Holoceno habria de tener un efecto negativo sobre los perezosos siendo sus descendientes actuales especies tropic ales 0 subtropicales pero no se trata del efecto
330 BIOGEOGRAFiA
directo del cambio de temperatura sobre los animales sino del efecto indirecto y progresivo que dicho cambio genera a largo plazo al reducir el territorio insular disponible]
Por esta epoca tambien desaparecieron especies de tall a mayor como Megalocnus zile en La Espanola y otras de menor tamano (en Cuba) pero que representan la tall a maxima en sus respectivos contextos taxonomicos del Cuaternario como Solenodon arredondoi Paralouatta varonai y Macrocapromys acevedo Un mamifero puede ser grande solo a costa de reducir su densidad poblacional y esto 10 hace mis vulnerable a la extinci6n sobre todo en una isla (McNab 1994 McFarlane et al 1998a) Incidentalmente Alcover et al (1998a) afirmaron que larelacion entre la talla corporal y la extincion no siempre se cumple en las especies de menor tamano y citaron como ejemplo que entre las musarafias todas las especies de Nesophontes desaparecieron mientras sobreviven especies de Solenodon pero la comparacion correcta en todo caso serfa entre los solenodones vivientes y el gigante Solenodon arredondoi y entre Nesophontes edithae -el gigante de los nesofontes extinguido en el periodo amerindio- y el resto de sus congeneres que sobrevivieron hasta los primeros siglos de la colonizacion europea (Tabla 45 intervalos III y IV)
CAUSAS ANTROPOGENICAS
Intervalo Amerindio
La mayoria de los autores que se han referido a las extinciones acaecidas durante este intervalo (entre elIos paradigmaticamente Martin 1984a b) han responsabilizado a los aborfgenes (que se supone comenzaron a poblar las Antillas Mayores hace 6 mil afios) con la desaparicion de numerosas especies de mamiferos terrestres en la region principalmente por la via de la sobrecaza y la introduccion de animales domesticos devenidos depredadores de la fauna autoctona (Morgan y Woods 1986) Este efecto negativo de la presencia indigena sobre los mamiferos de la region ha sido dramatizado por algunos autores en terminos de holocausto faunal Sin embargo pensamos que se ha exagerado Como han expresado MacPhee et al (19993) cuando uno busca evidencia dura de complicidad humana directa en [estas] extinciones no hay mucho que encontrar Las siguientes son algunas consideraciones a tener en cuenta en tal sentido
El registro radiometrico mas joven conocido hasta ahora para un perezoso antillano (MegalocnllS rodens) es de 42 kaAP (MacPhee et ai 2007) Si se acepta que los aborfgenes entraron en Cuba hace aproximadamente 6 mil anos su convivencia con los perezosos debe haber durado alrededor de 1800 afios (0 mas porque dificilmente el referfdo fechado radiometrico haya correspondido al ultimo sobreviviente de la especie) Esta cifra contradice la hipotesis del holocausto faunal segun la cualla extincion de los mamiferos insulares de mayor talla debe ocurrir con extrema rapidez -a escala de decadas 0 de centurias- a partir de su primer contacto con los humanos (Martin y Steadman 1999) A pesar de las repetidas afirmaciones en contrario (Harrington 1921 Miller 1929c Herrera [1945] Tabio 1951 Alvarez 1951a 1957 Arredondo
331 EXTINCIONES DEL CUATERNARIO
1959 1960a Veloz y Ortega 1973 Mayo 1980b Rodriguez et al 1984 Castellanos y Pino 1985 Pino y Castellanos 1985 Nunez 1990b Gutierrez 1996 Arredondo 2004) en su mayoria basadas en evidencias no exentas de posibles accidentes de deposicion (0 de reubicacion postdeposicional) en recintos cavernarios (Diaz-Franco [2006b D no se ha demostrado inequivocamente que los aborigenes antillanos se alimentaran de perezosos regularmente Sus restos en residuarios de alimentos son escasisimos en toda la region y esto contrasta con los abundantes restos de jutias y otros mamiferos de menor talla que aparecen en esos residuarios demostrativos de la preferencia de los paleoindios por estos otros mamiferos
Los mamiferos mas importantes en la economia prehistorica de las Antillas Mayores fueron las jutias Capromys y Geocapromys en Cuba Geocapromys en Jamaica e Isolobodon en La Espanola Puerto Rico e Islas Virgenes (Aguayo 1950b Tablo 1951 Rimoli 1972 Wing 2001 Diaz-Franco [2006b D Wilkins (2001) estudi61ajutia viviente de Jamaica (Geocapromys browni) en un sitio ocupado por los aborigenes durante 100 anos y no encontr6 evidencias de que durante ese tiempo la especie haya sido sobreexplotada Asimismo en Cuba las jutias conga carabali y andaraz son escasas en depositos pleistoholocenicos en cambio son muy numerosas en residuarios indigenas y de haber sido sobreexplotadas 10 mas probable es que no hubieran sobrevivido hasta hoy
La habilidad de los humanos para penetrar en profundidad el bosque tropical y mantenerse en este sustentablemente ha sido un evento comparativamente reciente (Beran y Steele 1994) En el caso antillano la caza tierra adentro ha sido explicada como un proceso tardio de desplazamiento a partir de la explotacion de abundantes recursos marinos que ofrecian condiciones 6ptimas para el sedentarismo de las comunidades aborigenes costeras (C6rdova y Arredondo 1988 Brito 2005) Peces y mariscos dominan la arqueologia costera de las Islas antillanas en tanto el consumo de recursos terrestres (mamfferos incluidos) aumenta progresivamente en la medida que los residuarios estudiados se alejan de la costa (Scudder 1991 Wilkins 2001) En las Antillas Mayores el componente terrestre de la dieta aborigen es solo 20-30 del material recobrado en los residuarios (Wing 1977 1989) Y debio ser menor alin porque el componente vegetal de la dieta no se ha preservado Por otra parte los aborigenes cubanos domesticaron la jutia conga y seguramente la reprodujeron para comer (Pose et al 1988) practica que probablemente se repitiera con otras jutias en las demas Antillas Mayores
- Fray Bartolome de las Casas hablo de 3 millones de indios en Cuba al inicio de la colonizacion pero los historiadores subsecuentes consideraron exagerada la cifra (Poey 1851 a) No obstante se dispone de dos estimados de la densidad de poblaci6n aborigen en las Antillas Mayores a la llegada de los europeos 18 indigenaslkm2 en Puerto Rico (Rouse 1956) y 1271km2 en La Espanola (Watts 1987) Extrapolando ambos valores entre las cuatro islas principales de las Antillas Mayores tenemos que en la fecha indicada la poblaci6n aborigen pudo estar entre 16 mil y 113 mil indigenas en Puerto Rico entre 21 mil y 145 mil en Jamaica entre 141 mil y 1 mill6n en La Espanola y entre 200 mil y 14 millones en Cuba Entre las cifras maximas y las minimas nos inclinamos
332 BIOGEOGRAFiA
hacia estas ultimas atendiendo a los altos indices de mortalidad infantil y morbilidad que se evidencian en las osamentas de los pobladores prehispanicos (Martinez y Rives 1990) Uno de los hechos que apuntan hacia el bajo numero de aborigenes en las Antillas Mayores es 10 rapido que desaparecieron despues de la llegada de los europeos pnkticamente sin dejar mestizaje con estos 10 cual contrasta ostensiblemente con la situacion en la America continentaL
En suma un grupo humano (1) demograiicamente irrelevante y controlado por altos indices de morbilidad (2) consumidor mayoritariamente de organismos marinos y (3) carente de la tecnologia cinegetica necesaria para poder sobreexplotar los mamiferos que Ie sirvieron de alimento (a los cuales accedio tardiamente en su desarrollo) no es concebible que haya causado e1 holocausto faunal que se Ie atribuye a partir del error de aplicar a las comunidades aborigenes antillanas un modelo cultural de la prehistoria continentaL
Tampoco hay evidencias de que los aborigenes hayan introducido carnivoros con suficiente capacidad depredadora salvo el perro Morgan y Otlenwaldder (1993) consideraron que la depredacion por perros durante el periodo amerindio es la explicacion mas plausible para la extincion del almiqui gigante de Cuba pero como se puede observar (Tabla 54) Solenodon arredondoi aparece en el intervalo II por falta de asociacion con evidencias culturales
Consecuentemente el accionar de los aborigenes y de sus animales domesticos directamente sobre la fauna no parece haber sido por sf solo un factor determinante de la extincion de mamiferos terrestres durante el periodo amerindio en las Antillas (pero vease Steadman et al 2005)
Hay que recordar en este senti do que alin en el Holoceno tardio ocurrieron cambios clirruiticos de consideracion (vease Causas Naturales) Por ejemplo Curtis et al (2001) explicaron que la extincion de algunos ani males mesofilicos de las alturas montanosas de La Espanola pudo haber antecedido a cualquier impacto humano sigrIificativo en esos lugares por coincidir dicha extincion con una tendencia a la aridez documentada isotopicamente y con evidencias de la perdida del bosque mesico ocurridas despues de 3kaAP Yen Jamaica hace solo 2shy3 mil aDos el area de la Bahia de Jackson sustentaba una gran biomasa de vertebrados en bosque hinnedo a juzgar por la cantidad y diversidad de taxones presentes en material fosil derivado de la depredacion por lechuzas hoy el area es la mas seca de la isla con matorral xerofitico y pobreza de fauna (MacPhee 1996a) Yalm en nuestros dias huracanes de gran intensidad aniquilan poblaciones de vertebrados (que no se recuperan) en is las pequenas y llanas del Caribe (Brooks y Smith 2001)
En medio de la incertidumbre sobre la verdadera causa (0 causas) de las extinciones de mamiferos en el periodo amerindio MacPhee y Marx (1997) han propuesto su hipotesis de la hipermorbilidad la cual estipula colapsos poblacionales relativamente rapidos con la introduccion de enfermedades de alta letalidad Estos autores han razonado que
Las islas son los lugares mas apropiados para experimentar perdidas catastroficas causadas por enfermedades Los mamiferos no solo serian susceptibles en virtud de su prolongado aislamiento biologico sino que en los confines de una isla la epizootia
333 EXTINCIONES DEL CUATERNARIO
tendria la potencialidad de infectar simultaneamente a la rnayoria de los individuos de vadas especies Los neonatos son inmunologicamente inmaduros y sufririan desproporcionadamente cualquier infeccion hipervirulenta La perdida excesiva de neonatos y juveniles tendria efectos severos para la sobrevivencia de la poblacion si esta se reduce por debajo de cierto nivel sobreviene la extincion porque no hay posibilidad de restitucion en el periodo postpanzootico Las especies de mayor talla corporal van a ser las mas vulnerables a la extincion dadas sus bajas tasas reproductivas largos periodos de gestacion y reducido numero de progenie Las especies de menor tamano --caracterizadas por altas tasas reproductivas periodos cortos de gestacion y mayor numero de neonatos por parto- tambien serian fuertemente diezmadas pero muchas sobrevivirian porque su estrategia reproductiva les permite perder mayor numero de individuos Las perdidas deben ocurrir relativamente rapido (a escala de decadas) mientras que otras hipotesis (digamos los cambios climaticos) permiten intervalos mucho mas largos (a escala de siglos 0 de milenios) entre la aparicion de las causas y la produccion de sus efectos Por ejemplo todo indica que los nesofontes se extinguieron muy rapidamente y seria dificil poder invocar un impacto humano directo 0 argumentar fenomenos climaticos que pudieran explicar plausiblemente perdidas tan rapidas
MacPhee y Marx (1997 191) han dicho que por el momento la hipotesis de la hipermorbilidad no pasa de ser una abstraccion en espera de adecuada sustentacion empirica Llama la atencion que linicamente en el periodo amerindio (Tabla 45 intervalo 1Il) las extinciones abarcaron todas las clases del peso corporal (de la a a laj) y que ninguna de las especies que sobrevivieron hasta el periodo postcolombino (Tabla 45 intervalo IV mas las especies vivientes) sobrepasa los 10 kg (clases del peso a b c) en clara coincidencia con los presupuestos de la hipotetica hipermorbilidad
Intervalo Postcolornbino
Por 10 menos 37 especies de mamiferos terrestres vivian en las Antillas Mayores y grupos de Islas perifericas a la lIegada de los europeos en 1492 De elIas se han extinguido 23 shymas especies que en cualquiera de los periodos precedentes (Tabla 45~ y solo subsisten 12 especies repartidas entre Cuba (8 especies) La Espanola (2) Jamaica (1) y Bahamas (1) varias de estas en peligro critico de extincion Solo dos de las especies extinguidas durante los ultimos 500 anos se conocieron en estado viviente los roedores Oryzomys antillarum de Jamaica (extinguido en 1877) y Geocapromys thoracatus de las Islas Swan (extinguido en 1955) Las causas de las extinciones ocurridas durante este periodo son sobradamente conocidas destruccion de habitats introducci6n de animales exoticos (domesticos y comensales) y demas presiones asociadas a la actividad humana entre las cuales posiblemente habria que incluir brotes de hipermorbilidad
Durante mucho tiempo se ha afirmado la posibilidad de que especies de Nesophontes hayan sobrevivido hasta las primeras decadas del siglo xx y que incluso continuen existiendo en lugares remotos de Cuba 0 La Espanola (Allen 1918 1942 Miller 1930 Aguayo 1950a Arredondo 1960b Fischer 1977 MacFadden 1980 Woods y
334 BIOGEOGRAFiA
Ottenwalder 1991) Actualmente no existe evidencia radiometrica de que alguna especie de Nesophontes haya sobrevivido al siglo xv las fechas radiocarbonicas calibradas obtenidas en muestras de Nesophontes de Cuba y La Espanola resultaron no mas tardias que los siglos XIII y XIV (MacPhee et al 1999) Por otra parte las expediciones efectuadas a fines del siglo xx con el proposito de localizar individuos vivientes del genero en La Espanola no produjeron la menor indicacion afirmativa al respecto (Woods et al 1985) Ademas a traves de las Antillas Mayores no aparecen restos de Nesophontes en los depositos formados actualmente por las lechuzas
EPiLOGO
Las Antillas Mayores son islas oceanicas no han formado parte de un continente (con la posible excepcion de una parte de Jamaica hasta el Eoceno Medio hace 45 Ma) y durante un tiempo geologico considerable (desde el Eoceno Superior hace 40 Ma) han permanecido emergidas y separadas de los continentes vecinos Consecuentemente los mamiferos terrestres (no volantes) autoctonos de estas islas constituyen una fauna oceanica que por ende cumple con todas las caracteristicas convencionales reconocidas para este tipo de entidad biogeografica
- Baja diversidad taxonomica a nivel de las categorias superiores (solo cuatro ordenes) en comparacion con islas plataformicas continentales (0 con areas continentales) de tamano y diversidad ecologica simi lares
- Alto grado de endemismo a nivel de genero (96) y familia (50) en respuesta al prolongado aislamiento
- Desbalance ecologico de la estructura trofica ausencia de mamiferos camivoros y ungulados
- Espectaculares procesos de irradiaci6n evolucionaria como resultado de la colonizacion de territorios libres de competidores y depredadores
Tendencias evolucionarias extremas gigantismo
- Control a cargo de aves rapaces con tendencia at gigantismo en ausencia de mamiferos miembros del orden Carnivora
- Mas especies de reptiles y de murcielagos que de mamiferos terrestres relaciones que en los continentes se invierten
Alta tasa de extincion de especies (84) sin reposicion
Preservacion de especies filogeneticamente arcaicas (los llamados fosiles vivientes) los almiquies

330 BIOGEOGRAFiA
directo del cambio de temperatura sobre los animales sino del efecto indirecto y progresivo que dicho cambio genera a largo plazo al reducir el territorio insular disponible]
Por esta epoca tambien desaparecieron especies de tall a mayor como Megalocnus zile en La Espanola y otras de menor tamano (en Cuba) pero que representan la tall a maxima en sus respectivos contextos taxonomicos del Cuaternario como Solenodon arredondoi Paralouatta varonai y Macrocapromys acevedo Un mamifero puede ser grande solo a costa de reducir su densidad poblacional y esto 10 hace mis vulnerable a la extinci6n sobre todo en una isla (McNab 1994 McFarlane et al 1998a) Incidentalmente Alcover et al (1998a) afirmaron que larelacion entre la talla corporal y la extincion no siempre se cumple en las especies de menor tamano y citaron como ejemplo que entre las musarafias todas las especies de Nesophontes desaparecieron mientras sobreviven especies de Solenodon pero la comparacion correcta en todo caso serfa entre los solenodones vivientes y el gigante Solenodon arredondoi y entre Nesophontes edithae -el gigante de los nesofontes extinguido en el periodo amerindio- y el resto de sus congeneres que sobrevivieron hasta los primeros siglos de la colonizacion europea (Tabla 45 intervalos III y IV)
CAUSAS ANTROPOGENICAS
Intervalo Amerindio
La mayoria de los autores que se han referido a las extinciones acaecidas durante este intervalo (entre elIos paradigmaticamente Martin 1984a b) han responsabilizado a los aborfgenes (que se supone comenzaron a poblar las Antillas Mayores hace 6 mil afios) con la desaparicion de numerosas especies de mamiferos terrestres en la region principalmente por la via de la sobrecaza y la introduccion de animales domesticos devenidos depredadores de la fauna autoctona (Morgan y Woods 1986) Este efecto negativo de la presencia indigena sobre los mamiferos de la region ha sido dramatizado por algunos autores en terminos de holocausto faunal Sin embargo pensamos que se ha exagerado Como han expresado MacPhee et al (19993) cuando uno busca evidencia dura de complicidad humana directa en [estas] extinciones no hay mucho que encontrar Las siguientes son algunas consideraciones a tener en cuenta en tal sentido
El registro radiometrico mas joven conocido hasta ahora para un perezoso antillano (MegalocnllS rodens) es de 42 kaAP (MacPhee et ai 2007) Si se acepta que los aborfgenes entraron en Cuba hace aproximadamente 6 mil anos su convivencia con los perezosos debe haber durado alrededor de 1800 afios (0 mas porque dificilmente el referfdo fechado radiometrico haya correspondido al ultimo sobreviviente de la especie) Esta cifra contradice la hipotesis del holocausto faunal segun la cualla extincion de los mamiferos insulares de mayor talla debe ocurrir con extrema rapidez -a escala de decadas 0 de centurias- a partir de su primer contacto con los humanos (Martin y Steadman 1999) A pesar de las repetidas afirmaciones en contrario (Harrington 1921 Miller 1929c Herrera [1945] Tabio 1951 Alvarez 1951a 1957 Arredondo
331 EXTINCIONES DEL CUATERNARIO
1959 1960a Veloz y Ortega 1973 Mayo 1980b Rodriguez et al 1984 Castellanos y Pino 1985 Pino y Castellanos 1985 Nunez 1990b Gutierrez 1996 Arredondo 2004) en su mayoria basadas en evidencias no exentas de posibles accidentes de deposicion (0 de reubicacion postdeposicional) en recintos cavernarios (Diaz-Franco [2006b D no se ha demostrado inequivocamente que los aborigenes antillanos se alimentaran de perezosos regularmente Sus restos en residuarios de alimentos son escasisimos en toda la region y esto contrasta con los abundantes restos de jutias y otros mamiferos de menor talla que aparecen en esos residuarios demostrativos de la preferencia de los paleoindios por estos otros mamiferos
Los mamiferos mas importantes en la economia prehistorica de las Antillas Mayores fueron las jutias Capromys y Geocapromys en Cuba Geocapromys en Jamaica e Isolobodon en La Espanola Puerto Rico e Islas Virgenes (Aguayo 1950b Tablo 1951 Rimoli 1972 Wing 2001 Diaz-Franco [2006b D Wilkins (2001) estudi61ajutia viviente de Jamaica (Geocapromys browni) en un sitio ocupado por los aborigenes durante 100 anos y no encontr6 evidencias de que durante ese tiempo la especie haya sido sobreexplotada Asimismo en Cuba las jutias conga carabali y andaraz son escasas en depositos pleistoholocenicos en cambio son muy numerosas en residuarios indigenas y de haber sido sobreexplotadas 10 mas probable es que no hubieran sobrevivido hasta hoy
La habilidad de los humanos para penetrar en profundidad el bosque tropical y mantenerse en este sustentablemente ha sido un evento comparativamente reciente (Beran y Steele 1994) En el caso antillano la caza tierra adentro ha sido explicada como un proceso tardio de desplazamiento a partir de la explotacion de abundantes recursos marinos que ofrecian condiciones 6ptimas para el sedentarismo de las comunidades aborigenes costeras (C6rdova y Arredondo 1988 Brito 2005) Peces y mariscos dominan la arqueologia costera de las Islas antillanas en tanto el consumo de recursos terrestres (mamfferos incluidos) aumenta progresivamente en la medida que los residuarios estudiados se alejan de la costa (Scudder 1991 Wilkins 2001) En las Antillas Mayores el componente terrestre de la dieta aborigen es solo 20-30 del material recobrado en los residuarios (Wing 1977 1989) Y debio ser menor alin porque el componente vegetal de la dieta no se ha preservado Por otra parte los aborigenes cubanos domesticaron la jutia conga y seguramente la reprodujeron para comer (Pose et al 1988) practica que probablemente se repitiera con otras jutias en las demas Antillas Mayores
- Fray Bartolome de las Casas hablo de 3 millones de indios en Cuba al inicio de la colonizacion pero los historiadores subsecuentes consideraron exagerada la cifra (Poey 1851 a) No obstante se dispone de dos estimados de la densidad de poblaci6n aborigen en las Antillas Mayores a la llegada de los europeos 18 indigenaslkm2 en Puerto Rico (Rouse 1956) y 1271km2 en La Espanola (Watts 1987) Extrapolando ambos valores entre las cuatro islas principales de las Antillas Mayores tenemos que en la fecha indicada la poblaci6n aborigen pudo estar entre 16 mil y 113 mil indigenas en Puerto Rico entre 21 mil y 145 mil en Jamaica entre 141 mil y 1 mill6n en La Espanola y entre 200 mil y 14 millones en Cuba Entre las cifras maximas y las minimas nos inclinamos
332 BIOGEOGRAFiA
hacia estas ultimas atendiendo a los altos indices de mortalidad infantil y morbilidad que se evidencian en las osamentas de los pobladores prehispanicos (Martinez y Rives 1990) Uno de los hechos que apuntan hacia el bajo numero de aborigenes en las Antillas Mayores es 10 rapido que desaparecieron despues de la llegada de los europeos pnkticamente sin dejar mestizaje con estos 10 cual contrasta ostensiblemente con la situacion en la America continentaL
En suma un grupo humano (1) demograiicamente irrelevante y controlado por altos indices de morbilidad (2) consumidor mayoritariamente de organismos marinos y (3) carente de la tecnologia cinegetica necesaria para poder sobreexplotar los mamiferos que Ie sirvieron de alimento (a los cuales accedio tardiamente en su desarrollo) no es concebible que haya causado e1 holocausto faunal que se Ie atribuye a partir del error de aplicar a las comunidades aborigenes antillanas un modelo cultural de la prehistoria continentaL
Tampoco hay evidencias de que los aborigenes hayan introducido carnivoros con suficiente capacidad depredadora salvo el perro Morgan y Otlenwaldder (1993) consideraron que la depredacion por perros durante el periodo amerindio es la explicacion mas plausible para la extincion del almiqui gigante de Cuba pero como se puede observar (Tabla 54) Solenodon arredondoi aparece en el intervalo II por falta de asociacion con evidencias culturales
Consecuentemente el accionar de los aborigenes y de sus animales domesticos directamente sobre la fauna no parece haber sido por sf solo un factor determinante de la extincion de mamiferos terrestres durante el periodo amerindio en las Antillas (pero vease Steadman et al 2005)
Hay que recordar en este senti do que alin en el Holoceno tardio ocurrieron cambios clirruiticos de consideracion (vease Causas Naturales) Por ejemplo Curtis et al (2001) explicaron que la extincion de algunos ani males mesofilicos de las alturas montanosas de La Espanola pudo haber antecedido a cualquier impacto humano sigrIificativo en esos lugares por coincidir dicha extincion con una tendencia a la aridez documentada isotopicamente y con evidencias de la perdida del bosque mesico ocurridas despues de 3kaAP Yen Jamaica hace solo 2shy3 mil aDos el area de la Bahia de Jackson sustentaba una gran biomasa de vertebrados en bosque hinnedo a juzgar por la cantidad y diversidad de taxones presentes en material fosil derivado de la depredacion por lechuzas hoy el area es la mas seca de la isla con matorral xerofitico y pobreza de fauna (MacPhee 1996a) Yalm en nuestros dias huracanes de gran intensidad aniquilan poblaciones de vertebrados (que no se recuperan) en is las pequenas y llanas del Caribe (Brooks y Smith 2001)
En medio de la incertidumbre sobre la verdadera causa (0 causas) de las extinciones de mamiferos en el periodo amerindio MacPhee y Marx (1997) han propuesto su hipotesis de la hipermorbilidad la cual estipula colapsos poblacionales relativamente rapidos con la introduccion de enfermedades de alta letalidad Estos autores han razonado que
Las islas son los lugares mas apropiados para experimentar perdidas catastroficas causadas por enfermedades Los mamiferos no solo serian susceptibles en virtud de su prolongado aislamiento biologico sino que en los confines de una isla la epizootia
333 EXTINCIONES DEL CUATERNARIO
tendria la potencialidad de infectar simultaneamente a la rnayoria de los individuos de vadas especies Los neonatos son inmunologicamente inmaduros y sufririan desproporcionadamente cualquier infeccion hipervirulenta La perdida excesiva de neonatos y juveniles tendria efectos severos para la sobrevivencia de la poblacion si esta se reduce por debajo de cierto nivel sobreviene la extincion porque no hay posibilidad de restitucion en el periodo postpanzootico Las especies de mayor talla corporal van a ser las mas vulnerables a la extincion dadas sus bajas tasas reproductivas largos periodos de gestacion y reducido numero de progenie Las especies de menor tamano --caracterizadas por altas tasas reproductivas periodos cortos de gestacion y mayor numero de neonatos por parto- tambien serian fuertemente diezmadas pero muchas sobrevivirian porque su estrategia reproductiva les permite perder mayor numero de individuos Las perdidas deben ocurrir relativamente rapido (a escala de decadas) mientras que otras hipotesis (digamos los cambios climaticos) permiten intervalos mucho mas largos (a escala de siglos 0 de milenios) entre la aparicion de las causas y la produccion de sus efectos Por ejemplo todo indica que los nesofontes se extinguieron muy rapidamente y seria dificil poder invocar un impacto humano directo 0 argumentar fenomenos climaticos que pudieran explicar plausiblemente perdidas tan rapidas
MacPhee y Marx (1997 191) han dicho que por el momento la hipotesis de la hipermorbilidad no pasa de ser una abstraccion en espera de adecuada sustentacion empirica Llama la atencion que linicamente en el periodo amerindio (Tabla 45 intervalo 1Il) las extinciones abarcaron todas las clases del peso corporal (de la a a laj) y que ninguna de las especies que sobrevivieron hasta el periodo postcolombino (Tabla 45 intervalo IV mas las especies vivientes) sobrepasa los 10 kg (clases del peso a b c) en clara coincidencia con los presupuestos de la hipotetica hipermorbilidad
Intervalo Postcolornbino
Por 10 menos 37 especies de mamiferos terrestres vivian en las Antillas Mayores y grupos de Islas perifericas a la lIegada de los europeos en 1492 De elIas se han extinguido 23 shymas especies que en cualquiera de los periodos precedentes (Tabla 45~ y solo subsisten 12 especies repartidas entre Cuba (8 especies) La Espanola (2) Jamaica (1) y Bahamas (1) varias de estas en peligro critico de extincion Solo dos de las especies extinguidas durante los ultimos 500 anos se conocieron en estado viviente los roedores Oryzomys antillarum de Jamaica (extinguido en 1877) y Geocapromys thoracatus de las Islas Swan (extinguido en 1955) Las causas de las extinciones ocurridas durante este periodo son sobradamente conocidas destruccion de habitats introducci6n de animales exoticos (domesticos y comensales) y demas presiones asociadas a la actividad humana entre las cuales posiblemente habria que incluir brotes de hipermorbilidad
Durante mucho tiempo se ha afirmado la posibilidad de que especies de Nesophontes hayan sobrevivido hasta las primeras decadas del siglo xx y que incluso continuen existiendo en lugares remotos de Cuba 0 La Espanola (Allen 1918 1942 Miller 1930 Aguayo 1950a Arredondo 1960b Fischer 1977 MacFadden 1980 Woods y
334 BIOGEOGRAFiA
Ottenwalder 1991) Actualmente no existe evidencia radiometrica de que alguna especie de Nesophontes haya sobrevivido al siglo xv las fechas radiocarbonicas calibradas obtenidas en muestras de Nesophontes de Cuba y La Espanola resultaron no mas tardias que los siglos XIII y XIV (MacPhee et al 1999) Por otra parte las expediciones efectuadas a fines del siglo xx con el proposito de localizar individuos vivientes del genero en La Espanola no produjeron la menor indicacion afirmativa al respecto (Woods et al 1985) Ademas a traves de las Antillas Mayores no aparecen restos de Nesophontes en los depositos formados actualmente por las lechuzas
EPiLOGO
Las Antillas Mayores son islas oceanicas no han formado parte de un continente (con la posible excepcion de una parte de Jamaica hasta el Eoceno Medio hace 45 Ma) y durante un tiempo geologico considerable (desde el Eoceno Superior hace 40 Ma) han permanecido emergidas y separadas de los continentes vecinos Consecuentemente los mamiferos terrestres (no volantes) autoctonos de estas islas constituyen una fauna oceanica que por ende cumple con todas las caracteristicas convencionales reconocidas para este tipo de entidad biogeografica
- Baja diversidad taxonomica a nivel de las categorias superiores (solo cuatro ordenes) en comparacion con islas plataformicas continentales (0 con areas continentales) de tamano y diversidad ecologica simi lares
- Alto grado de endemismo a nivel de genero (96) y familia (50) en respuesta al prolongado aislamiento
- Desbalance ecologico de la estructura trofica ausencia de mamiferos camivoros y ungulados
- Espectaculares procesos de irradiaci6n evolucionaria como resultado de la colonizacion de territorios libres de competidores y depredadores
Tendencias evolucionarias extremas gigantismo
- Control a cargo de aves rapaces con tendencia at gigantismo en ausencia de mamiferos miembros del orden Carnivora
- Mas especies de reptiles y de murcielagos que de mamiferos terrestres relaciones que en los continentes se invierten
Alta tasa de extincion de especies (84) sin reposicion
Preservacion de especies filogeneticamente arcaicas (los llamados fosiles vivientes) los almiquies

331 EXTINCIONES DEL CUATERNARIO
1959 1960a Veloz y Ortega 1973 Mayo 1980b Rodriguez et al 1984 Castellanos y Pino 1985 Pino y Castellanos 1985 Nunez 1990b Gutierrez 1996 Arredondo 2004) en su mayoria basadas en evidencias no exentas de posibles accidentes de deposicion (0 de reubicacion postdeposicional) en recintos cavernarios (Diaz-Franco [2006b D no se ha demostrado inequivocamente que los aborigenes antillanos se alimentaran de perezosos regularmente Sus restos en residuarios de alimentos son escasisimos en toda la region y esto contrasta con los abundantes restos de jutias y otros mamiferos de menor talla que aparecen en esos residuarios demostrativos de la preferencia de los paleoindios por estos otros mamiferos
Los mamiferos mas importantes en la economia prehistorica de las Antillas Mayores fueron las jutias Capromys y Geocapromys en Cuba Geocapromys en Jamaica e Isolobodon en La Espanola Puerto Rico e Islas Virgenes (Aguayo 1950b Tablo 1951 Rimoli 1972 Wing 2001 Diaz-Franco [2006b D Wilkins (2001) estudi61ajutia viviente de Jamaica (Geocapromys browni) en un sitio ocupado por los aborigenes durante 100 anos y no encontr6 evidencias de que durante ese tiempo la especie haya sido sobreexplotada Asimismo en Cuba las jutias conga carabali y andaraz son escasas en depositos pleistoholocenicos en cambio son muy numerosas en residuarios indigenas y de haber sido sobreexplotadas 10 mas probable es que no hubieran sobrevivido hasta hoy
La habilidad de los humanos para penetrar en profundidad el bosque tropical y mantenerse en este sustentablemente ha sido un evento comparativamente reciente (Beran y Steele 1994) En el caso antillano la caza tierra adentro ha sido explicada como un proceso tardio de desplazamiento a partir de la explotacion de abundantes recursos marinos que ofrecian condiciones 6ptimas para el sedentarismo de las comunidades aborigenes costeras (C6rdova y Arredondo 1988 Brito 2005) Peces y mariscos dominan la arqueologia costera de las Islas antillanas en tanto el consumo de recursos terrestres (mamfferos incluidos) aumenta progresivamente en la medida que los residuarios estudiados se alejan de la costa (Scudder 1991 Wilkins 2001) En las Antillas Mayores el componente terrestre de la dieta aborigen es solo 20-30 del material recobrado en los residuarios (Wing 1977 1989) Y debio ser menor alin porque el componente vegetal de la dieta no se ha preservado Por otra parte los aborigenes cubanos domesticaron la jutia conga y seguramente la reprodujeron para comer (Pose et al 1988) practica que probablemente se repitiera con otras jutias en las demas Antillas Mayores
- Fray Bartolome de las Casas hablo de 3 millones de indios en Cuba al inicio de la colonizacion pero los historiadores subsecuentes consideraron exagerada la cifra (Poey 1851 a) No obstante se dispone de dos estimados de la densidad de poblaci6n aborigen en las Antillas Mayores a la llegada de los europeos 18 indigenaslkm2 en Puerto Rico (Rouse 1956) y 1271km2 en La Espanola (Watts 1987) Extrapolando ambos valores entre las cuatro islas principales de las Antillas Mayores tenemos que en la fecha indicada la poblaci6n aborigen pudo estar entre 16 mil y 113 mil indigenas en Puerto Rico entre 21 mil y 145 mil en Jamaica entre 141 mil y 1 mill6n en La Espanola y entre 200 mil y 14 millones en Cuba Entre las cifras maximas y las minimas nos inclinamos
332 BIOGEOGRAFiA
hacia estas ultimas atendiendo a los altos indices de mortalidad infantil y morbilidad que se evidencian en las osamentas de los pobladores prehispanicos (Martinez y Rives 1990) Uno de los hechos que apuntan hacia el bajo numero de aborigenes en las Antillas Mayores es 10 rapido que desaparecieron despues de la llegada de los europeos pnkticamente sin dejar mestizaje con estos 10 cual contrasta ostensiblemente con la situacion en la America continentaL
En suma un grupo humano (1) demograiicamente irrelevante y controlado por altos indices de morbilidad (2) consumidor mayoritariamente de organismos marinos y (3) carente de la tecnologia cinegetica necesaria para poder sobreexplotar los mamiferos que Ie sirvieron de alimento (a los cuales accedio tardiamente en su desarrollo) no es concebible que haya causado e1 holocausto faunal que se Ie atribuye a partir del error de aplicar a las comunidades aborigenes antillanas un modelo cultural de la prehistoria continentaL
Tampoco hay evidencias de que los aborigenes hayan introducido carnivoros con suficiente capacidad depredadora salvo el perro Morgan y Otlenwaldder (1993) consideraron que la depredacion por perros durante el periodo amerindio es la explicacion mas plausible para la extincion del almiqui gigante de Cuba pero como se puede observar (Tabla 54) Solenodon arredondoi aparece en el intervalo II por falta de asociacion con evidencias culturales
Consecuentemente el accionar de los aborigenes y de sus animales domesticos directamente sobre la fauna no parece haber sido por sf solo un factor determinante de la extincion de mamiferos terrestres durante el periodo amerindio en las Antillas (pero vease Steadman et al 2005)
Hay que recordar en este senti do que alin en el Holoceno tardio ocurrieron cambios clirruiticos de consideracion (vease Causas Naturales) Por ejemplo Curtis et al (2001) explicaron que la extincion de algunos ani males mesofilicos de las alturas montanosas de La Espanola pudo haber antecedido a cualquier impacto humano sigrIificativo en esos lugares por coincidir dicha extincion con una tendencia a la aridez documentada isotopicamente y con evidencias de la perdida del bosque mesico ocurridas despues de 3kaAP Yen Jamaica hace solo 2shy3 mil aDos el area de la Bahia de Jackson sustentaba una gran biomasa de vertebrados en bosque hinnedo a juzgar por la cantidad y diversidad de taxones presentes en material fosil derivado de la depredacion por lechuzas hoy el area es la mas seca de la isla con matorral xerofitico y pobreza de fauna (MacPhee 1996a) Yalm en nuestros dias huracanes de gran intensidad aniquilan poblaciones de vertebrados (que no se recuperan) en is las pequenas y llanas del Caribe (Brooks y Smith 2001)
En medio de la incertidumbre sobre la verdadera causa (0 causas) de las extinciones de mamiferos en el periodo amerindio MacPhee y Marx (1997) han propuesto su hipotesis de la hipermorbilidad la cual estipula colapsos poblacionales relativamente rapidos con la introduccion de enfermedades de alta letalidad Estos autores han razonado que
Las islas son los lugares mas apropiados para experimentar perdidas catastroficas causadas por enfermedades Los mamiferos no solo serian susceptibles en virtud de su prolongado aislamiento biologico sino que en los confines de una isla la epizootia
333 EXTINCIONES DEL CUATERNARIO
tendria la potencialidad de infectar simultaneamente a la rnayoria de los individuos de vadas especies Los neonatos son inmunologicamente inmaduros y sufririan desproporcionadamente cualquier infeccion hipervirulenta La perdida excesiva de neonatos y juveniles tendria efectos severos para la sobrevivencia de la poblacion si esta se reduce por debajo de cierto nivel sobreviene la extincion porque no hay posibilidad de restitucion en el periodo postpanzootico Las especies de mayor talla corporal van a ser las mas vulnerables a la extincion dadas sus bajas tasas reproductivas largos periodos de gestacion y reducido numero de progenie Las especies de menor tamano --caracterizadas por altas tasas reproductivas periodos cortos de gestacion y mayor numero de neonatos por parto- tambien serian fuertemente diezmadas pero muchas sobrevivirian porque su estrategia reproductiva les permite perder mayor numero de individuos Las perdidas deben ocurrir relativamente rapido (a escala de decadas) mientras que otras hipotesis (digamos los cambios climaticos) permiten intervalos mucho mas largos (a escala de siglos 0 de milenios) entre la aparicion de las causas y la produccion de sus efectos Por ejemplo todo indica que los nesofontes se extinguieron muy rapidamente y seria dificil poder invocar un impacto humano directo 0 argumentar fenomenos climaticos que pudieran explicar plausiblemente perdidas tan rapidas
MacPhee y Marx (1997 191) han dicho que por el momento la hipotesis de la hipermorbilidad no pasa de ser una abstraccion en espera de adecuada sustentacion empirica Llama la atencion que linicamente en el periodo amerindio (Tabla 45 intervalo 1Il) las extinciones abarcaron todas las clases del peso corporal (de la a a laj) y que ninguna de las especies que sobrevivieron hasta el periodo postcolombino (Tabla 45 intervalo IV mas las especies vivientes) sobrepasa los 10 kg (clases del peso a b c) en clara coincidencia con los presupuestos de la hipotetica hipermorbilidad
Intervalo Postcolornbino
Por 10 menos 37 especies de mamiferos terrestres vivian en las Antillas Mayores y grupos de Islas perifericas a la lIegada de los europeos en 1492 De elIas se han extinguido 23 shymas especies que en cualquiera de los periodos precedentes (Tabla 45~ y solo subsisten 12 especies repartidas entre Cuba (8 especies) La Espanola (2) Jamaica (1) y Bahamas (1) varias de estas en peligro critico de extincion Solo dos de las especies extinguidas durante los ultimos 500 anos se conocieron en estado viviente los roedores Oryzomys antillarum de Jamaica (extinguido en 1877) y Geocapromys thoracatus de las Islas Swan (extinguido en 1955) Las causas de las extinciones ocurridas durante este periodo son sobradamente conocidas destruccion de habitats introducci6n de animales exoticos (domesticos y comensales) y demas presiones asociadas a la actividad humana entre las cuales posiblemente habria que incluir brotes de hipermorbilidad
Durante mucho tiempo se ha afirmado la posibilidad de que especies de Nesophontes hayan sobrevivido hasta las primeras decadas del siglo xx y que incluso continuen existiendo en lugares remotos de Cuba 0 La Espanola (Allen 1918 1942 Miller 1930 Aguayo 1950a Arredondo 1960b Fischer 1977 MacFadden 1980 Woods y
334 BIOGEOGRAFiA
Ottenwalder 1991) Actualmente no existe evidencia radiometrica de que alguna especie de Nesophontes haya sobrevivido al siglo xv las fechas radiocarbonicas calibradas obtenidas en muestras de Nesophontes de Cuba y La Espanola resultaron no mas tardias que los siglos XIII y XIV (MacPhee et al 1999) Por otra parte las expediciones efectuadas a fines del siglo xx con el proposito de localizar individuos vivientes del genero en La Espanola no produjeron la menor indicacion afirmativa al respecto (Woods et al 1985) Ademas a traves de las Antillas Mayores no aparecen restos de Nesophontes en los depositos formados actualmente por las lechuzas
EPiLOGO
Las Antillas Mayores son islas oceanicas no han formado parte de un continente (con la posible excepcion de una parte de Jamaica hasta el Eoceno Medio hace 45 Ma) y durante un tiempo geologico considerable (desde el Eoceno Superior hace 40 Ma) han permanecido emergidas y separadas de los continentes vecinos Consecuentemente los mamiferos terrestres (no volantes) autoctonos de estas islas constituyen una fauna oceanica que por ende cumple con todas las caracteristicas convencionales reconocidas para este tipo de entidad biogeografica
- Baja diversidad taxonomica a nivel de las categorias superiores (solo cuatro ordenes) en comparacion con islas plataformicas continentales (0 con areas continentales) de tamano y diversidad ecologica simi lares
- Alto grado de endemismo a nivel de genero (96) y familia (50) en respuesta al prolongado aislamiento
- Desbalance ecologico de la estructura trofica ausencia de mamiferos camivoros y ungulados
- Espectaculares procesos de irradiaci6n evolucionaria como resultado de la colonizacion de territorios libres de competidores y depredadores
Tendencias evolucionarias extremas gigantismo
- Control a cargo de aves rapaces con tendencia at gigantismo en ausencia de mamiferos miembros del orden Carnivora
- Mas especies de reptiles y de murcielagos que de mamiferos terrestres relaciones que en los continentes se invierten
Alta tasa de extincion de especies (84) sin reposicion
Preservacion de especies filogeneticamente arcaicas (los llamados fosiles vivientes) los almiquies

332 BIOGEOGRAFiA
hacia estas ultimas atendiendo a los altos indices de mortalidad infantil y morbilidad que se evidencian en las osamentas de los pobladores prehispanicos (Martinez y Rives 1990) Uno de los hechos que apuntan hacia el bajo numero de aborigenes en las Antillas Mayores es 10 rapido que desaparecieron despues de la llegada de los europeos pnkticamente sin dejar mestizaje con estos 10 cual contrasta ostensiblemente con la situacion en la America continentaL
En suma un grupo humano (1) demograiicamente irrelevante y controlado por altos indices de morbilidad (2) consumidor mayoritariamente de organismos marinos y (3) carente de la tecnologia cinegetica necesaria para poder sobreexplotar los mamiferos que Ie sirvieron de alimento (a los cuales accedio tardiamente en su desarrollo) no es concebible que haya causado e1 holocausto faunal que se Ie atribuye a partir del error de aplicar a las comunidades aborigenes antillanas un modelo cultural de la prehistoria continentaL
Tampoco hay evidencias de que los aborigenes hayan introducido carnivoros con suficiente capacidad depredadora salvo el perro Morgan y Otlenwaldder (1993) consideraron que la depredacion por perros durante el periodo amerindio es la explicacion mas plausible para la extincion del almiqui gigante de Cuba pero como se puede observar (Tabla 54) Solenodon arredondoi aparece en el intervalo II por falta de asociacion con evidencias culturales
Consecuentemente el accionar de los aborigenes y de sus animales domesticos directamente sobre la fauna no parece haber sido por sf solo un factor determinante de la extincion de mamiferos terrestres durante el periodo amerindio en las Antillas (pero vease Steadman et al 2005)
Hay que recordar en este senti do que alin en el Holoceno tardio ocurrieron cambios clirruiticos de consideracion (vease Causas Naturales) Por ejemplo Curtis et al (2001) explicaron que la extincion de algunos ani males mesofilicos de las alturas montanosas de La Espanola pudo haber antecedido a cualquier impacto humano sigrIificativo en esos lugares por coincidir dicha extincion con una tendencia a la aridez documentada isotopicamente y con evidencias de la perdida del bosque mesico ocurridas despues de 3kaAP Yen Jamaica hace solo 2shy3 mil aDos el area de la Bahia de Jackson sustentaba una gran biomasa de vertebrados en bosque hinnedo a juzgar por la cantidad y diversidad de taxones presentes en material fosil derivado de la depredacion por lechuzas hoy el area es la mas seca de la isla con matorral xerofitico y pobreza de fauna (MacPhee 1996a) Yalm en nuestros dias huracanes de gran intensidad aniquilan poblaciones de vertebrados (que no se recuperan) en is las pequenas y llanas del Caribe (Brooks y Smith 2001)
En medio de la incertidumbre sobre la verdadera causa (0 causas) de las extinciones de mamiferos en el periodo amerindio MacPhee y Marx (1997) han propuesto su hipotesis de la hipermorbilidad la cual estipula colapsos poblacionales relativamente rapidos con la introduccion de enfermedades de alta letalidad Estos autores han razonado que
Las islas son los lugares mas apropiados para experimentar perdidas catastroficas causadas por enfermedades Los mamiferos no solo serian susceptibles en virtud de su prolongado aislamiento biologico sino que en los confines de una isla la epizootia
333 EXTINCIONES DEL CUATERNARIO
tendria la potencialidad de infectar simultaneamente a la rnayoria de los individuos de vadas especies Los neonatos son inmunologicamente inmaduros y sufririan desproporcionadamente cualquier infeccion hipervirulenta La perdida excesiva de neonatos y juveniles tendria efectos severos para la sobrevivencia de la poblacion si esta se reduce por debajo de cierto nivel sobreviene la extincion porque no hay posibilidad de restitucion en el periodo postpanzootico Las especies de mayor talla corporal van a ser las mas vulnerables a la extincion dadas sus bajas tasas reproductivas largos periodos de gestacion y reducido numero de progenie Las especies de menor tamano --caracterizadas por altas tasas reproductivas periodos cortos de gestacion y mayor numero de neonatos por parto- tambien serian fuertemente diezmadas pero muchas sobrevivirian porque su estrategia reproductiva les permite perder mayor numero de individuos Las perdidas deben ocurrir relativamente rapido (a escala de decadas) mientras que otras hipotesis (digamos los cambios climaticos) permiten intervalos mucho mas largos (a escala de siglos 0 de milenios) entre la aparicion de las causas y la produccion de sus efectos Por ejemplo todo indica que los nesofontes se extinguieron muy rapidamente y seria dificil poder invocar un impacto humano directo 0 argumentar fenomenos climaticos que pudieran explicar plausiblemente perdidas tan rapidas
MacPhee y Marx (1997 191) han dicho que por el momento la hipotesis de la hipermorbilidad no pasa de ser una abstraccion en espera de adecuada sustentacion empirica Llama la atencion que linicamente en el periodo amerindio (Tabla 45 intervalo 1Il) las extinciones abarcaron todas las clases del peso corporal (de la a a laj) y que ninguna de las especies que sobrevivieron hasta el periodo postcolombino (Tabla 45 intervalo IV mas las especies vivientes) sobrepasa los 10 kg (clases del peso a b c) en clara coincidencia con los presupuestos de la hipotetica hipermorbilidad
Intervalo Postcolornbino
Por 10 menos 37 especies de mamiferos terrestres vivian en las Antillas Mayores y grupos de Islas perifericas a la lIegada de los europeos en 1492 De elIas se han extinguido 23 shymas especies que en cualquiera de los periodos precedentes (Tabla 45~ y solo subsisten 12 especies repartidas entre Cuba (8 especies) La Espanola (2) Jamaica (1) y Bahamas (1) varias de estas en peligro critico de extincion Solo dos de las especies extinguidas durante los ultimos 500 anos se conocieron en estado viviente los roedores Oryzomys antillarum de Jamaica (extinguido en 1877) y Geocapromys thoracatus de las Islas Swan (extinguido en 1955) Las causas de las extinciones ocurridas durante este periodo son sobradamente conocidas destruccion de habitats introducci6n de animales exoticos (domesticos y comensales) y demas presiones asociadas a la actividad humana entre las cuales posiblemente habria que incluir brotes de hipermorbilidad
Durante mucho tiempo se ha afirmado la posibilidad de que especies de Nesophontes hayan sobrevivido hasta las primeras decadas del siglo xx y que incluso continuen existiendo en lugares remotos de Cuba 0 La Espanola (Allen 1918 1942 Miller 1930 Aguayo 1950a Arredondo 1960b Fischer 1977 MacFadden 1980 Woods y
334 BIOGEOGRAFiA
Ottenwalder 1991) Actualmente no existe evidencia radiometrica de que alguna especie de Nesophontes haya sobrevivido al siglo xv las fechas radiocarbonicas calibradas obtenidas en muestras de Nesophontes de Cuba y La Espanola resultaron no mas tardias que los siglos XIII y XIV (MacPhee et al 1999) Por otra parte las expediciones efectuadas a fines del siglo xx con el proposito de localizar individuos vivientes del genero en La Espanola no produjeron la menor indicacion afirmativa al respecto (Woods et al 1985) Ademas a traves de las Antillas Mayores no aparecen restos de Nesophontes en los depositos formados actualmente por las lechuzas
EPiLOGO
Las Antillas Mayores son islas oceanicas no han formado parte de un continente (con la posible excepcion de una parte de Jamaica hasta el Eoceno Medio hace 45 Ma) y durante un tiempo geologico considerable (desde el Eoceno Superior hace 40 Ma) han permanecido emergidas y separadas de los continentes vecinos Consecuentemente los mamiferos terrestres (no volantes) autoctonos de estas islas constituyen una fauna oceanica que por ende cumple con todas las caracteristicas convencionales reconocidas para este tipo de entidad biogeografica
- Baja diversidad taxonomica a nivel de las categorias superiores (solo cuatro ordenes) en comparacion con islas plataformicas continentales (0 con areas continentales) de tamano y diversidad ecologica simi lares
- Alto grado de endemismo a nivel de genero (96) y familia (50) en respuesta al prolongado aislamiento
- Desbalance ecologico de la estructura trofica ausencia de mamiferos camivoros y ungulados
- Espectaculares procesos de irradiaci6n evolucionaria como resultado de la colonizacion de territorios libres de competidores y depredadores
Tendencias evolucionarias extremas gigantismo
- Control a cargo de aves rapaces con tendencia at gigantismo en ausencia de mamiferos miembros del orden Carnivora
- Mas especies de reptiles y de murcielagos que de mamiferos terrestres relaciones que en los continentes se invierten
Alta tasa de extincion de especies (84) sin reposicion
Preservacion de especies filogeneticamente arcaicas (los llamados fosiles vivientes) los almiquies

333 EXTINCIONES DEL CUATERNARIO
tendria la potencialidad de infectar simultaneamente a la rnayoria de los individuos de vadas especies Los neonatos son inmunologicamente inmaduros y sufririan desproporcionadamente cualquier infeccion hipervirulenta La perdida excesiva de neonatos y juveniles tendria efectos severos para la sobrevivencia de la poblacion si esta se reduce por debajo de cierto nivel sobreviene la extincion porque no hay posibilidad de restitucion en el periodo postpanzootico Las especies de mayor talla corporal van a ser las mas vulnerables a la extincion dadas sus bajas tasas reproductivas largos periodos de gestacion y reducido numero de progenie Las especies de menor tamano --caracterizadas por altas tasas reproductivas periodos cortos de gestacion y mayor numero de neonatos por parto- tambien serian fuertemente diezmadas pero muchas sobrevivirian porque su estrategia reproductiva les permite perder mayor numero de individuos Las perdidas deben ocurrir relativamente rapido (a escala de decadas) mientras que otras hipotesis (digamos los cambios climaticos) permiten intervalos mucho mas largos (a escala de siglos 0 de milenios) entre la aparicion de las causas y la produccion de sus efectos Por ejemplo todo indica que los nesofontes se extinguieron muy rapidamente y seria dificil poder invocar un impacto humano directo 0 argumentar fenomenos climaticos que pudieran explicar plausiblemente perdidas tan rapidas
MacPhee y Marx (1997 191) han dicho que por el momento la hipotesis de la hipermorbilidad no pasa de ser una abstraccion en espera de adecuada sustentacion empirica Llama la atencion que linicamente en el periodo amerindio (Tabla 45 intervalo 1Il) las extinciones abarcaron todas las clases del peso corporal (de la a a laj) y que ninguna de las especies que sobrevivieron hasta el periodo postcolombino (Tabla 45 intervalo IV mas las especies vivientes) sobrepasa los 10 kg (clases del peso a b c) en clara coincidencia con los presupuestos de la hipotetica hipermorbilidad
Intervalo Postcolornbino
Por 10 menos 37 especies de mamiferos terrestres vivian en las Antillas Mayores y grupos de Islas perifericas a la lIegada de los europeos en 1492 De elIas se han extinguido 23 shymas especies que en cualquiera de los periodos precedentes (Tabla 45~ y solo subsisten 12 especies repartidas entre Cuba (8 especies) La Espanola (2) Jamaica (1) y Bahamas (1) varias de estas en peligro critico de extincion Solo dos de las especies extinguidas durante los ultimos 500 anos se conocieron en estado viviente los roedores Oryzomys antillarum de Jamaica (extinguido en 1877) y Geocapromys thoracatus de las Islas Swan (extinguido en 1955) Las causas de las extinciones ocurridas durante este periodo son sobradamente conocidas destruccion de habitats introducci6n de animales exoticos (domesticos y comensales) y demas presiones asociadas a la actividad humana entre las cuales posiblemente habria que incluir brotes de hipermorbilidad
Durante mucho tiempo se ha afirmado la posibilidad de que especies de Nesophontes hayan sobrevivido hasta las primeras decadas del siglo xx y que incluso continuen existiendo en lugares remotos de Cuba 0 La Espanola (Allen 1918 1942 Miller 1930 Aguayo 1950a Arredondo 1960b Fischer 1977 MacFadden 1980 Woods y
334 BIOGEOGRAFiA
Ottenwalder 1991) Actualmente no existe evidencia radiometrica de que alguna especie de Nesophontes haya sobrevivido al siglo xv las fechas radiocarbonicas calibradas obtenidas en muestras de Nesophontes de Cuba y La Espanola resultaron no mas tardias que los siglos XIII y XIV (MacPhee et al 1999) Por otra parte las expediciones efectuadas a fines del siglo xx con el proposito de localizar individuos vivientes del genero en La Espanola no produjeron la menor indicacion afirmativa al respecto (Woods et al 1985) Ademas a traves de las Antillas Mayores no aparecen restos de Nesophontes en los depositos formados actualmente por las lechuzas
EPiLOGO
Las Antillas Mayores son islas oceanicas no han formado parte de un continente (con la posible excepcion de una parte de Jamaica hasta el Eoceno Medio hace 45 Ma) y durante un tiempo geologico considerable (desde el Eoceno Superior hace 40 Ma) han permanecido emergidas y separadas de los continentes vecinos Consecuentemente los mamiferos terrestres (no volantes) autoctonos de estas islas constituyen una fauna oceanica que por ende cumple con todas las caracteristicas convencionales reconocidas para este tipo de entidad biogeografica
- Baja diversidad taxonomica a nivel de las categorias superiores (solo cuatro ordenes) en comparacion con islas plataformicas continentales (0 con areas continentales) de tamano y diversidad ecologica simi lares
- Alto grado de endemismo a nivel de genero (96) y familia (50) en respuesta al prolongado aislamiento
- Desbalance ecologico de la estructura trofica ausencia de mamiferos camivoros y ungulados
- Espectaculares procesos de irradiaci6n evolucionaria como resultado de la colonizacion de territorios libres de competidores y depredadores
Tendencias evolucionarias extremas gigantismo
- Control a cargo de aves rapaces con tendencia at gigantismo en ausencia de mamiferos miembros del orden Carnivora
- Mas especies de reptiles y de murcielagos que de mamiferos terrestres relaciones que en los continentes se invierten
Alta tasa de extincion de especies (84) sin reposicion
Preservacion de especies filogeneticamente arcaicas (los llamados fosiles vivientes) los almiquies

334 BIOGEOGRAFiA
Ottenwalder 1991) Actualmente no existe evidencia radiometrica de que alguna especie de Nesophontes haya sobrevivido al siglo xv las fechas radiocarbonicas calibradas obtenidas en muestras de Nesophontes de Cuba y La Espanola resultaron no mas tardias que los siglos XIII y XIV (MacPhee et al 1999) Por otra parte las expediciones efectuadas a fines del siglo xx con el proposito de localizar individuos vivientes del genero en La Espanola no produjeron la menor indicacion afirmativa al respecto (Woods et al 1985) Ademas a traves de las Antillas Mayores no aparecen restos de Nesophontes en los depositos formados actualmente por las lechuzas
EPiLOGO
Las Antillas Mayores son islas oceanicas no han formado parte de un continente (con la posible excepcion de una parte de Jamaica hasta el Eoceno Medio hace 45 Ma) y durante un tiempo geologico considerable (desde el Eoceno Superior hace 40 Ma) han permanecido emergidas y separadas de los continentes vecinos Consecuentemente los mamiferos terrestres (no volantes) autoctonos de estas islas constituyen una fauna oceanica que por ende cumple con todas las caracteristicas convencionales reconocidas para este tipo de entidad biogeografica
- Baja diversidad taxonomica a nivel de las categorias superiores (solo cuatro ordenes) en comparacion con islas plataformicas continentales (0 con areas continentales) de tamano y diversidad ecologica simi lares
- Alto grado de endemismo a nivel de genero (96) y familia (50) en respuesta al prolongado aislamiento
- Desbalance ecologico de la estructura trofica ausencia de mamiferos camivoros y ungulados
- Espectaculares procesos de irradiaci6n evolucionaria como resultado de la colonizacion de territorios libres de competidores y depredadores
Tendencias evolucionarias extremas gigantismo
- Control a cargo de aves rapaces con tendencia at gigantismo en ausencia de mamiferos miembros del orden Carnivora
- Mas especies de reptiles y de murcielagos que de mamiferos terrestres relaciones que en los continentes se invierten
Alta tasa de extincion de especies (84) sin reposicion
Preservacion de especies filogeneticamente arcaicas (los llamados fosiles vivientes) los almiquies


















