EFECTO DEL CONSUMO DE NITRÓGENO DE LA MICROALGA ... · de Ciencias de la UNAM, por la...
90
Programa de Estudios de Posgrado TESIS Que para obtener el grado de Maestro en Ciencias Uso, Manejo y Preservación de los Recursos Naturales P r e s e n t a (Orientación en Biotecnología) EFECTO DEL CONSUMO DE NITRÓGENO DE LA MICROALGA Desmodesmus communis SOBRE LA COMPOSICIÓN BIOQUÍMICA, PRODUCTIVIDAD DE LA BIOMASA, COMUNIDAD BACTERIANA Y LONGITUD DE LOS TELÓMEROS La Paz, Baja California Sur, mayo 2018. Jessica Guadalupe Elias Castelo
Transcript of EFECTO DEL CONSUMO DE NITRÓGENO DE LA MICROALGA ... · de Ciencias de la UNAM, por la...

Programa de Estudios de Posgrado
TESIS
Que para obtener el grado de
Maestro en Ciencias
Uso, Manejo y Preservación de los Recursos Naturales
P r e s e n t a
(Orientación en Biotecnología)
EFECTO DEL CONSUMO DE NITRÓGENO DE LA
MICROALGA Desmodesmus communis SOBRE LA
COMPOSICIÓN BIOQUÍMICA, PRODUCTIVIDAD
DE LA BIOMASA, COMUNIDAD BACTERIANA Y
LONGITUD DE LOS TELÓMEROS
EFECTO DEL CONSUMO DE NITROGENO EN LA MICROALGA Desmodesmus communis
SOBRE LA COMPOSICIÓN BIOQUÍMICA, PRODUCTIVIDAD DE LA BIOMASA,
COMUNIDAD BACTERIANA Y LONGITUD DE LOS TELÓMEROS
La Paz, Baja California Sur, mayo 2018.
Jessica Guadalupe Elias Castelo


CONFORMACIÓN DE COMITÉS
Comité tutorial
Co-Director de tesis Dra. Bertha Olivia Arredondo Vega - CIBNOR
Co-Director de tesis Dr. Juan Pedro Luna Arias - CINVESTAV
Tutor de tesis Dra. Thelma Rosa Castellanos Cervantes - CIBNOR
Comité revisor
Dra. Bertha Olivia Arredondo Vega
Dr. Juan Pedro Luna Arias
Dra. Thelma Rosa Castellanos Cervantes
Jurado de examen
Dra. Bertha Olivia Arredondo Vega
Dr. Juan Pedro Luna Arias
Dra. Thelma Rosa Castellanos Cervantes
Suplente
Dr. Dariel Tovar Ramírez

i
Resumen
Las microalgas son organismos fotosintéticos que contienen clorofila y acumulan compuestos de interés biotecnológico. En condiciones de estrés como la disminución de la concentración de nitrógeno en el medio, incrementan la concentración de especies reactivas de oxígeno (ERO). Las ERO causan daño a biomoléculas como los lípidos, el ADN y sus telómeros, siendo estos responsables de la senescencia celular. La ausencia de daño estructural se debe a moléculas con capacidad antioxidante como los carotenoides. En el presente trabajo se evaluó el efecto del consumo de nitrógeno en el medio de cultivo y nitrógeno elemental interno así como sus razones isotópicas durante el crecimiento Desmodesmus communis sobre la tasa de crecimiento, productividad de la biomasa, la composición bioquímica, comunidad bacteriana y longitud de los telómeros. Las condiciones de cultivo batch fueron: medio F/2, fotoperiodo 12:12h luz: oscuridad, aireación continua, intensidad luminosa 120 µmol fotón m-2s-1, pH 7.8. La biomasa se cosechó por centrifugación a los días 0, 5, 10, 12 y 17. Los resultados obtenidos se analizaron estadísticamente con un ANOVA de una vía y un test Duncan con un α de 0.05. Los resultados fueron: el nitrógeno externo disminuyó con respecto al tiempo, obteniendo el agotamiento total en el día 10 con diferencias significativas entre el día 0 y los días 5,10,12 y 17. La tasa de crecimiento, concentración celular y productividad de la biomasa máxima se obtuvieron a los días 2, 3 y 5, respectivamente. Las proteínas (10.7%), carbohidratos (39.2%) y lípidos totales (33%) a los días 0, 12, 5 respectivamente. Las proteínas disminuyeron significativamente del día 0 al día 10 y se mantuvieron en concentración similar los días 12 y 17 mientras que los carbohidratos aumentaron significativamente del día 0 al 5, 10, 12 y17. En cuanto a los ácidos grasos los más abundantes fueron 18:1 ω9 y 16:0, siendo este último el más abundante en el día 17 presentando diferencias significativas con el día 0. La clorofila a disminuyó significativamente del día 0 al 10. Se identificaron distintas bacterias en donde la mayoría fueron α- proteobacterias, un bacteroide y una β- proteobacteria. En cuanto los telómeros se encontró que la longitud de éstos permaneció constante. Los mecanismos que desarrolla la microalga para vivir bajo los efectos del estrés como la modificación de ácidos grasos y la interacción de las bacterias presentes constituyen una herramienta biotecnológica en el ámbito de los biocombustibles y la biorremediación respectivamente.
Palabras clave: Telómeros, bacterias, nitrógeno, lípidos, carbohidratos, proteínas, y pigmentos.

Summary
The microalgae are photosynthetic organisms that contain chlorophyll and accumulate compounds of biotechnological interest. Under conditions of stress such as the decrease in the concentration of nitrogen in the medium, they increase the concentration of reactive oxygen species (ROS). ROS cause damage to biomolecules such as lipids, DNA and their telomeres, which are responsible for cellular senescence. The absence of structural damage is due to molecules with antioxidant capacity such as carotenoids. In the present work, the effect of nitrogen consumption on the culture medium and internal elemental nitrogen as well as its isotopic ratios during Desmodesmus communis growth on the growth rate, biomass productivity, biochemical composition, bacterial community and length was evaluated. Of the telomeres. The batch culture conditions were: medium F / 2, photoperiod 12: 12h light: dark, continuous aeration, luminous intensity 120 μmol photon m-2s-1, pH 7.8. The biomass was harvested by centrifugation at days 0, 5, 10, 12 and 17. The results obtained were analyzed statistically with a one-way ANOVA and a Duncan test with an α of 0.05. The results were: external nitrogen decreased with respect to time, obtaining total depletion on day 10 with significant differences between day 0 and days 5,10,12 and 17. The growth rate, cellular concentration and productivity of the Maximum biomass were obtained at days 2, 3 and 5, respectively. Proteins (10.7%), carbohydrates (39.2%) and total lipids (33%) at days 0, 12, 5 respectively. The proteins decreased significantly from day 0 to day 10 and were maintained at similar concentration on days 12 and 17 while carbohydrates increased significantly from day 0 to 5, 10, 12 and 17. Regarding fatty acids, the most abundant were 18: 1 ω9 and 16: 0, the latter being the most abundant on day 17, presenting significant differences with day 0. Chlorophyll a decreased significantly from day 0 to 10. They were identified different bacteria where the majority were α-proteobacteria, a bacteroid and a β-proteobacteria. Once the telomeres were found, the length of these remained constant. The mechanisms developed by the microalga to live under the effects of stress such as the modification of fatty acids and the interaction of the bacteria present constitute a biotechnological tool in the field of biofuels and bioremediation, respectively. Keywords : Telomers, bacteria, nitrogen, lipids, carbohydrates, proteins, and pigments.

Dedicatoria
A mi hermosa hija Sofía por su espera, por alentarme con su gran
amor a ser el mejor ejemplo para ella.
A mis padres Jorge Elias y Guadalupe Castelo por apoyarme en
todos los aspectos de mi vida, para poder hacer posible alcanzar
mis metas y por ser mí más grande ejemplo de perseverancia.
A mis amigos de maestría por el gusto de haberlos conocido.

Agradecimientos
Agradezco al Centro de Investigaciones Biológicas de Noroeste, S.C. por
brindarme la oportunidad de realizar mis estudios de posgrado y
proporcionar la infraestructura necesaria para llevar a cabo este trabajo de
maestría.
A CONACYT por la beca otorgada número 589407.
Al CINVESTAV por permitirme realizar una estancia de investigación y
proporcionar la infraestructura para llevarla a cabo.
A mi comité tutorial, Dra. Bertha Olivia Arredondo Vega, Dr. Juan Pedro
Luna Arias y Dra. Thelma Rosa Castellanos Cervantes, por su apoyo,
paciencia y valiosos consejos.
A la técnico responsable del Laboratorio de Biotecnología de Microalgas,
la Dra. Laura Carreón Palau por enseñarme y guiarme en la realización
de las técnicas en el laboratorio y por ser una guía.
Al técnico responsable del Laboratorio de Ecología Microbiana Molecular
el M. en C. Ángel Edgardo Carrillo García por guiarme en la ejecución de
las técnicas realizadas en el laboratorio.
Al Dr. Iban Murillo Murillo y la IBQ. Celina Beltrán Camacho personal del
Laboratorio de Análisis Químico de Aguas por guiarme en la realización de
las técnicas realizadas en el laboratorio.
Al Dr. Eberto Novelo Maldonado del Laboratorio de Algas Continentales.
Ecología y Taxonomía, Departamento de Biología Comparada, Facultad
de Ciencias de la UNAM, por la identificación de la cepa de Desmodesmus
communis mediante herramientas de microscopía electrónica, y a la Ing.
en Biotecnología Daniela Bárcenas Pérez del Laboratorio de Biotecnología
de Microalgas, por la comprobación taxonómica de la cepa mediante
marcadores moleculares 18S y Rbcl.

A la ayudante de investigador la Bióloga María Luisa Labra Barrios por su
asistencia técnica en la realización de los experimentos llevados a cabo
en el Laboratorio de Biología Celular del CINVESTAV.
A Refugio Contreras Contreras por ser un apoyo emocional durante mi
estancia en CINVESTAV, gracias por acompañarme.
Al Lic. Horacio Sandoval Gómez responsable del Laboratorio de Cómputo
del CIBNOR por su asistencia técnica en el uso de equipo informático.
A mis maestros de las asignaturas de maestría.
A mis compañeros de maestría a los cuales siempre llevare en mi corazón,
en especial a Caroli, Carmen y Jeb que me acompañaron cercanamente
en esta hermosa etapa.
A todos muchas gracias ♥.

Contenido
Resumen ............................................................................................................ ii
Summary ............................................................................................................ ii
Dedicatoria ........................................................................................................ iii
Agradecimientos .............................................................................................. iv
Contenido .......................................................................................................... vi
Lista de figuras ............................................................................................... viii
Lista de tablas .................................................................................................. ix
1. INTRODUCCIÓN ......................................................................................... 1
1.1.1 Usos de las microalgas ....................................................................... 3
1.2 Comunidad de bacterias con las microalgas ............................................. 3
1.2.1 Relación bacteria - microalga ............................................................. 4
1.3 Nutrición de las microalgas y mecanismos de obtención de energía ........ 5
1.4 Nitrógeno .................................................................................................. 6
1.4.1 El metabolismo del nitrógeno y el del carbono están interconectados y
se regulan recíprocamente .......................................................................... 8
1.5 Efecto del oxígeno en la oxidación de las biomoléculas ........................... 9
1.5.1 Efecto de los radicales libres en el ADN y en la longitud de los
telómeros .................................................................................................. 10
1.6 Antioxidantes........................................................................................... 11
1.7 Microalga utilizada en este trabajo .......................................................... 13
1.7.1 Generalidades .................................................................................. 13
1.7.2 Descripción ....................................................................................... 14
1.7.3 Clasificación ..................................................................................... 14
1.7.4 Ciclo celular ...................................................................................... 15
2. ANTECEDENTES ..................................................................................... 16
2.1 Respuesta bioquímica a condiciones de consumo de nitrógeno ......... 16
2.2 Lípidos y ácidos grasos en microalgas ................................................ 17
2.2.1 Biosíntesis de ácidos grasos ..................................................... 19
2.3 Telómeros y microalgas ...................................................................... 20
3. JUSTIFICACIÓN ....................................................................................... 22
4. HIPÓTESIS ................................................................................................ 23
5. OBJETIVOS .............................................................................................. 24
6. MATERIAL Y MÉTODOS .......................................................................... 25

6.1 Material biológico y condiciones de cultivo .......................................... 25
6.2 Análisis experimental .......................................................................... 26
6.2.1 Determinación de peso seco, orgánico e inorgánico ................. 26
6.2.2 Determinación de nitratos .......................................................... 27
6.2.3 Determinación de la relación carbono: nitrógeno elemental y la
relación entre 13C:12C y 15N:14N ................................................................ 28
6.2.4 Determinación de proteínas ...................................................... 29
6.2.5 Determinación de carbohidratos ............................................... 30
6.2.6 Determinación de pigmentos, lípidos totales y ácidos grasos ... 31
6.2.7 Pigmentos ................................................................................. 31
6.2.8 Lípidos totales ........................................................................... 32
6.2.9 Ácidos grasos ............................................................................ 33
6.2.10 Determinación de bacterias presentes ...................................... 34
6.2.10.1 Extracción de ADN .................................................................... 34
6.2.10.2 Electroforesis ............................................................................. 35
6.2.10.3 SSCP ......................................................................................... 35
6.2.11 Determinación de la longitud de los telómeros .......................... 39
6.3 Análisis estadístico .............................................................................. 42
7. RESULTADOS .......................................................................................... 43
8. DISCUSIÓN ............................................................................................... 59
9. CONCLUSIONES ...................................................................................... 64
10. LITERATURA CITADA ............................................................................. 65

Lista de figuras
Figura 1. Blancos de oxidación de especies reactivas y radicales libres.. ....... 10
Figura 2. Representación del ciclo celular Cn (fisión múltiple) ....................... 16
Figura 3. Desmodesmus communis identificación de la cepa por Dr. Eberto
Novelo M. ......................................................................................................... 25
Figura 4. Curva de crecimiento D. communis. ................................................ 43
Figura 5. Rendimiento de la biomasa del cultivo de D. communis mg L-1,. ..... 44
Figura 6. Productividad de la biomasa del cultivo de D. communis mg L-1 d-1..44
Figura 7. Porcentaje de peso orgánico y peso inorgánico de la biomasa del
cultivo de D. communis .. ................................................................................. 45
Figura 8. Porcentaje de proteínas con respecto al peso seco de D. communis
......................................................................................................................... 48
Figura 9. Porcentaje de carbohidratos de D. communis con respecto al peso
seco .. ............................................................................................................... 49
Figura 10. Perfil cuantitativo de pigmentos de D. communis µg mg-1. ............. 50
Figura 11. Porcentaje de lípidos de D. communis con respecto al peso seco. 51
Figura 12. Porcentaje de ácidos grasos con respecto al total de ácidos grasos .
......................................................................................................................... 54
Figura 13. Electroforesis en gel de acrilamida técnica SSCP.. ........................ 56
Figura 14. Técnica TRF. .................................................................................. 58

Lista de tablas
Tabla I. Aplicaciones de los pigmentos carotenoides. ..................................... 13
Tabla II. Comportamiento de biomoléculas en respuesta al consumo de
nitrógeno. ......................................................................................................... 17
Tabla III. Contenido de lípidos totales en la familia Scenedesmaceae. ........... 18
Tabla IV. Presencia de ácidos grasos en microalgas. ..................................... 20
Tabla V. Antecedentes del estudio de telómeros en microalgas...................... 21
Tabla VI. Programa de PCR (para técnica SSCP). .......................................... 36
Tabla VII. Mezcla de reacción gel de acrilamida. ............................................. 37
Tabla VIII. Mezcla de reacción para digestión. ................................................ 38
Tabla IX. Mezcla de reacción para marcado de la sonda. ............................... 41
Tabla X. Tasa de crecimiento y tiempo de generación. ................................... 43
Tabla XI. Concentración de nitratos (µM) en medio de cultivo de D. communis.
......................................................................................................................... 46
Tabla XII. Relación carbono: nitrógeno intracelular de la biomasa del cultivo de
D. communis. ................................................................................................... 47
Tabla XIII. Relación 15N: 14N de la biomasa del cultivo de D. communis.. ....... 47
Tabla XIV. Relación 13C: 12C de la biomasa del cultivo de D. communis.. ....... 48
Tabla XV. Porcentaje de ácidos grasos saturados, monoinsaturados y
poliinsaturados con respecto a los ácidos grasos totales ................................. 52
Tabla XVI. Bacterias cultivables presentes en el cultivo de D. communis. ...... 55
Tabla XVII. Bacterias identificadas mediante la técnica SSCP. ...................... 57

1
1. INTRODUCCIÓN
Las microalgas del género Desmodesmus (antes Scenedesmus) se encuentran
en cuerpos de agua dulce, en las aguas residuales de las plantas de tratamiento
y en aguas contaminadas con metales pesados (Perales-Vela et al., 2006). La
capacidad de sobrevivir en dichos ambientes se ha explicado por su plasticidad
fenotípica ya que puede crecer como células independientes o como cenobios
(agrupación de células). Su capacidad de responder a ambientes estresantes
hacen a las especies de este género buenas candidatas para su uso
biotecnológico. Por ejemplo, S. almeriensis puede ser inducida por la irradiancia
para producir luteína, un pigmento que evita la degeneración retinal (Sánchez et
al., 2008) y S. incrassatulus se utiliza en el tratamiento de aguas residuales para
la remoción de metales pesados (Peña-Castro et al., 2004). Esta capacidad de
adaptarse a ambientes estresantes como la deficiencia de nitrógeno en donde
se incrementa la concentración de especies reactivas de oxígeno (Fujita et al.,
2006), para contrarrestarlas, las células presentan mecanismos de protección
(Bumbak et al., 2011; Vilar-Rojas et al., 1996) para contrarrestarlas, es decir, los
antioxidantes (Sahnoun et al., 1997). Cuando las especies reactivas de oxígeno
no son contrarrestadas de manera eficaz pueden causar daños, al ADN,
telómeros, lípidos membranales, así como proteínas y carbohidratos a nivel de
los organelos. La senescencia celular es un proceso irreversible de declinación
de la proliferación celular en relación con la edad. Es un proceso activo,
genéticamente programado, que responde a una inducción dada por el
acortamiento telomérico, generando una señal semejante a la producida por el
daño en el ADN (Jeyapalan y Sedivy, 2008).
Debido a que las microalgas pueden almacenar el nitrógeno que consumen del
medio (Lavín y Lourenço, 2005), es importante para poder entender las
respuestas, medir el nitrógeno intracelular y de esta manera, determinar si las
células se encuentran en condición de deficiencia interna. En este trabajo se
determinó el nitrógeno elemental (Lourenço et al., 2004) así como relación
carbono:nitrógeno.

Considerando que las microalgas presentan plasticidad metabólica, el presente
trabajo se enfocó en evaluar el efecto del estrés generado por la deficiencia de
nitrógeno en diferentes biomoléculas: proteínas, carbohidratos, lípidos, ácidos
grasos y pigmentos, y a nivel del ADN se midió la longitud de los telómeros de la
microalga Desmodesmus communis. Así mismo, se identificaron las bacterias
presentes en el cultivo de la microalga, haciendo énfasis en aquellas que tienen
capacidad para fijar nitrógeno.
1.1 Generalidades
Las microalgas son organismos fotosintéticos, unicelulares que pueden formar
cadenas, colonias o cenobios, y se les puede encontrar en agua marina, dulce,
salobre, en el suelo o en ambientes hipersalinos y nevados (Metting, 1996;
Vaulot et al., 2008; Holzinger et al., 2016). Se les consideran responsables de la
síntesis de material orgánico en los ecosistemas acuáticos, así como del 40% de
la fotosíntesis que se lleva a cabo en el planeta (Margalef, 1981).
Las microalgas son de gran importancia ambiental ya que fijan más del 40% del
carbono de la tierra, además de ofrecer a la biósfera una considerable proporción
de oxígeno. Su importancia ecológica radica en su abundancia, su extrema
biodiversidad y la habilidad de sobrevivir en una variedad de ambientes, desde
los muy extremos como los suelos desérticos, hasta ambientes moderados como
lagos de agua dulce y océanos (Norton et al., 1996). Son un grupo diverso de
microorganismos fotosintéticos con una estructura simple (unicelulares) que
permite el rápido crecimiento celular y por lo tanto una mayor producción de
biomasa (Li et al., 2009). Las microalgas son organismos fotoautótrofos, siendo
la luz su principal fuente de energía y el dióxido de carbono (CO2) su principal
fuente de carbono (Martin, 2010); pero también pueden utilizar una fuente de
carbono orgánico (glucosa, acetato de sodio) por lo que se convierten en
organismos mixotróficos (Arias Peñarada et al., 2013).

1.1.1 Usos de las microalgas
En los últimos años se han logrado avances importantes en el uso de las
microalgas para diversos fines como salud humana, cosmetología, purificación
de aguas residuales, prevención de contaminación acuática, industria
farmacéutica, acuicultura, producción de pigmentos y antibióticos, entre otros
(Jasso et al., 2012), por esto se han vuelto de gran interés para la producción de
compuestos de alto valor agregado como la astaxantina, el β-caroteno, la luteína,
la cantaxantina y clorofila (Qin et al., 2008). También son una fuente de biodiesel
y otros productos de gran interés comercial (Bitog et al., 2011). Teniendo en
cuenta la enorme biodiversidad de las microalgas y los recientes desarrollos en
ingeniería genética y metabólica, este grupo de organismos representa una
fuente prometedora para generar nuevos productos y aplicaciones. Con el
desarrollo de sistemas de cultivo y cosecha, la biotecnología de microalgas
puede satisfacer las altas demandas de las industrias alimentaria, energética y
farmacéutica (Harun et al., 2010).
1.2 Comunidad de bacterias con las microalgas
Las microalgas y las bacterias habitan juntas en casi todos los ambientes
acuáticos y desempeñan un papel crucial en el ciclo de nutrientes y el flujo de
energía (Ramanan et al., 2016). Las bacterias tienen una gran influencia en el
crecimiento de microalgas en condiciones autotróficas, ya sea positiva o negativa
(Ethier et al., 2011; Ortiz-Marquez et al., 2012). En general, las bacterias
promueven el crecimiento de microalgas reduciendo la concentración de oxígeno
disuelto en el medio de cultivo, consumiendo los materiales orgánicos
excretados por algas (Mouget et al., 1995) y secretando biotina, cobalamina y
tiamina (Croft et al., 2005). A su vez, las microalgas le retribuyen a las bacterias
oxígeno y compuestos extracelulares. Tal reciprocidad implica que el crecimiento
de microalgas puede ser potenciado por bacterias específicas(de-Bashan et al.,
2002; Ethier et al., 2011). Recientemente, se ha encontrado que las moléculas
de señalización de "quorum” sensing producidas por bacterias pueden afectar
no solo a las cianobacterias, sino también a las microalgas eucariotas, y que a

su vez, las algas eucariotas también pueden interferir con la señalización química
de las bacterias (Borowitzka et al., 2016).
1.2.1 Relación bacteria - microalga
Las microalgas crecen asociadas a bacterias de forma natural. El concepto de
“anillo microbiano” introducido por Azam et al. (1998) cambió notablemente la
percepción del papel que tienen los microorganismos en los ecosistemas
acuáticos. Este concepto plantea la importancia de la interacción bacteria-
microalga como componentes básicos del ecosistema, proponiendo que las
bacterias no sólo son recicladoras de la materia orgánica, sino que además
tienen la capacidad de absorber nutrientes desde el océano y hacer circular
fuentes de carbono mediante diversas interacciones ecológicas con los otros
componentes del plancton (Azam, 1998).
Dentro de los componentes del fitoplancton, uno de los organismos con los
cuales las bacterias interactúan directamente son las microalgas. Diversas
investigaciones a nivel de laboratorio y en campo han demostrado fenómenos
de estímulo e inhibición de crecimiento en microalgas y/o bacterias (Borowitzka
et al., 2016; Sieburth, 1959, 1971). Las bacterias pueden incrementar el
crecimiento microalgal (Dae-Hyun et al., 2014) mediante la producción de
algunas vitaminas como la cobalamina, cofactor de la enzima metionina sintasa
(Croft et al., 2005). También, se ha demostrado que las sustancias orgánicas
derivadas del fitoplancton en ecosistemas naturales, son utilizadas por las
bacterias como sustrato de crecimiento (Ohara et al., 1993; Mouget et al., 1995).
Las interacciones microbianas son el núcleo del éxito o fracaso de las especies.
Estas interacciones pueden ser de tipo mutualista, simbiótica, comensalista o
parasitaria, y a menudo son consideradas especies específicas (Ashen y Goff,
2000; Sapp et al., 2007).
La producción de vitaminas por bacterias, como resultado de la interacción entre
bacterias y fitoplancton, ha sido frecuentemente investigada. Diversos estudios
sugieren una fuerte correlación entre el agotamiento de la vitamina en el agua y
la terminación de la floculación de diatomeas (Ohwada, 1973), siendo la vitamina
B12 la más estudiada (Carlucci et al., 1969; Ryther y Guillard, 1962). Esta es

necesaria en organismos que carecen de la enzima metionina sintasa
independiente de vitamina B12.
El estudio más importante de las interacciones entre bacterias y microalgas
debido a la producción de vitamina B12 por las bacterias se llevó a cabo en 1974,
cuando varias especies de diatomeas se cultivaron axénicamente en presencia
o ausencia de vitamina B12, y en presencia de bacterias marinas heterotróficas
productoras de vitamina B12. En casi todos los casos, las bacterias parecían
mejorar el rendimiento de las células de diatomeas en relación con los cultivos
limitados con vitamina B12, pero no con el nivel de los cultivos suplementados
con B12 (Haines y Guillard, 1974).
Croft et al. (2005) realizaron una revisión bibliográfica sobre los requerimientos
de vitamina B12 en 326 especies de microalgas, y la confirmaron como requisito.
También demostraron que una bacteria del género Halomonas aumentó la
producción de vitamina B12 cuando se les proporcionó un extracto comercial de
microalga; éste fue interpretado como evidencia de que las bacterias tienen una
correlación siendo suplementos vitamínicos (y particularmente vitaminas B12) a
escala global a la mayoría del fitoplancton B12 auxotrófico, a cambio de carbono
orgánico (Droop, 2007).
1.3 Nutrición de las microalgas y mecanismos de obtención de energía
En las microalgas su nutrición es por absorción iónica de C, H, O, N, P, S, K, Ca,
Fe y Mg, así como de los elementos traza: Mn, B, Co, Cu, Zn y Mo. De los grupos
que las conforman, las cianofitas, clorofitas, crisófitas, euglenofitas y diatomeas
pennales son más comunes en agua dulce (Margalef, 1981).
La diversidad de especies en un ambiente acuático se encuentra determinada
por la proporción que existe entre los nutrientes disponibles en el medio y los
requerimientos nutricionales de cada especie. De esta forma, diferentes
especies creciendo en una misma comunidad pueden estar limitadas por
distintos recursos (Rouzic y Bertru, 1997).

La composición química de las microalgas se encuentra directamente
determinada por la toma de material inorgánico (cationes y aniones) del medio
acuático circundante. La asimilación de nutrientes inorgánicos por las microalgas
es importante ya que estos organismos representan la principal fuente de
nutrientes en ecosistemas acuáticos continentales y marinos. La entrada de
materia inorgánica en las células microalgales constituye la principal ruta de
entrada en la cadena alimenticia con la subsecuente transformación en biomasa
de herbívoros y carnívoros (Krivtsov y Sigee, 2005).
1.4 Nitrógeno
El nitrógeno es uno de los componentes esenciales de muchas biomoléculas
clave, como aminoácidos y nucleótidos. La síntesis de proteínas normalmente
depende de un adecuado suministro de nitrógeno (Sze, 1998). Un incremento en
la disponibilidad de nitrógeno inorgánico generalmente deriva en un incremento
de la abundancia de productores primarios. Sin embargo, altos niveles de
nitrógeno inorgánico que no pueden ser asimilados por los sistemas ecológicos
pueden causar efectos adversos en los organismos menos tolerantes (Camargo
y Alonso, 2006).
El nitrógeno se presenta de diferentes formas en el agua, incluyendo nitrógeno
molecular (N2), amonio (NH4+), nitrito (NO2
-), nitrato (NO3-), otros óxidos de
nitrógeno y nitrógeno en compuestos orgánicos. El amonio normalmente se
encuentra en baja concentración en aguas oxigenadas donde las bacterias lo
oxidan a nitrato (Sze, 1998).
En los cultivos, el nitrógeno se aporta en la forma de sales de amonio o nitratos,
ya que son éstas las principales formas de nitrógeno que usan las microalgas,
aunque la mayoría puede utilizar también nitritos, urea y otras formas de
nitrógeno orgánico. El nitrógeno es aceptado en las vías metabólicas de las
microalgas en forma de amonio, mientras que la asimilación de nitrato requiere
de la transformación de nitrato a nitrito y luego a amonio mediante cuatro pasos
de reducción que requieren energía, resultando más favorable para las
microalgas usar el amonio cuando está disponible (Sze, 1998).

La asimilación del nitrato consta de tres etapas: 1) absorción, 2) reducción del
nitrato a amonio, y 3) incorporación del amonio a esqueletos carbonados para la
síntesis de aminoácidos, proceso que recibe el nombre específico de asimilación
del amonio (Maldonado, 2008).
En una primera reacción, el nitrato es reducido a nitrito por la enzima nitrato
reductasa. Dicha reacción requiere dos electrones suministrados por una
molécula de piridín nucleótido reducido. A continuación, el nitrito es reducido a
amonio por la enzima nitrito reductasa, en una reacción que requiere seis
electrones donados por la ferredoxina reducida. La reducción del nitrato a
amonio consume, un total de ocho electrones. El poder reductor requerido se
genera en la fase luminosa de la fotosíntesis (Foyer y Noctor, 2003). Las
vacuolas constituyen el principal sitio de almacenamiento del nitrato (Maldonado,
2008).
La deficiencia de nitrógeno influye directamente en la formación de aminoácidos,
lo cual a su vez limita la traducción del mRNA y por lo tanto se reduce la síntesis
de proteínas. Por otra parte, la eficiencia del fotosistema II disminuye inicialmente
como una consecuencia de la disipación térmica de la energía de excitación
absorbida en el lecho pigmentario, derivando en una reducción de la tasa de
fotosíntesis, lo cual a su vez lleva a una reducción en la tasa de respiración
(Wehr, 2007).
La tasa de crecimiento microalgal es una función de las reservas intracelulares
del nutriente limitante: cuando está disponible, la abundancia del nutriente
determina su tasa de asimilación y su almacenamiento intracelular (Droop,
1973). Si hay producción continua, la cantidad de reservas intracelulares
contenidas en cada organismo disminuye. La reducción del nutriente limitante
lleva a un decremento en la tasa de crecimiento hasta que alcanza su valor
mínimo, momento en el cual no puede haber más crecimiento (fase estacionaria)
(Lampert y Sommer, 2007).
El nitrógeno presenta dos isótopos estables: 14N y 15N. El estado de valencia del
nitrógeno varia de +5 (NO3-) a -3 (NH4
+), dando como resultado una amplia
variación natural de composiciones isotópicas. El contenido en 15N de una
sustancia se expresa como su desviación isotópica, δ15N, respecto al estándar

del aire atmosférico. El reservorio de nitrógeno atmosférico, corresponde al
estándar de referencia, por tanto su valor será de 0%. Al fabricarse los
fertilizantes agrícolas a partir del nitrógeno atmosférico, y presentar este proceso
un fraccionamiento muy bajo, el valor isotópico del nitrógeno de estos se
encuentra comprendido alrededor del valor del N del aire (entre +5% a – %). De
manera similar, el nitrógeno procedente de efluentes ganaderos o de fosas
sépticas o redes de alcantarillado, al proceder de amonio el cual ha sufrido
procesos de volatilización importantes, tiene valores de δ15N isotópicamente más
pesados, con valores comprendidos entre 10 y 20% (Vitòria et al., 2004). En el
presente trabajo el δ15N se utilizó como indicador de mixotrofía.
La asimilación del nitrógeno requiere poder reductor, ATP y esqueletos
carbonados. Los esqueletos carbonados son proporcionados por la oxidación de
carbohidratos. La fijación de nitrógeno y el metabolismo de carbohidratos está
íntimamente ligado (Coruzzi y Zhou, 2001; O’Brien et al., 2016) . Se ha
observado que las mitocondrias son sumamente sensibles a la presión oxidativa
en todos los organismos aerobios, lo que se refleja en cantidades elevadas de
oxidación en lípidos y proteínas, y en mutaciones del ADN mitocondrial (Lenaz,
1998; Ríos, 2003). Debido al daño oxidativo acumulado se puede originar la
pérdida de la capacidad funcional de la célula, lo que conduce al envejecimiento
y a la muerte (Hayflick, 1985; Medvedev, 1990; Von Zglinicki, 2002).
1.4.1 El metabolismo del nitrógeno y el del carbono están interconectados
y se regulan recíprocamente
La asimilación del nitrógeno es un proceso íntimamente conectado al
metabolismo del carbono. La reducción del nitrato a amonio y su posterior
asimilación en glutamato requieren diez electrones, de los cuales dos son usados
en el citosol como NADH por la enzima nitrato reductasa, y los ocho restantes
son utilizados en el cloroplasto en forma de ferredoxina reducida (seis por la
enzima nitrito reductasa y dos por la ferredoxina-glutamato sintasa). El NADH
necesario para la reducción del nitrato a nitrito proviene, en su mayor parte, del
NADPH generado fotosintéticamente en los cloroplastos, y que es exportado al

citosol mediante las lanzaderas triosa fosfato-fosfoglicerato y malato-oxalacetato
(Coruzzi y Zhou, 2001).
La asimilación del nitrógeno origina un flujo de carbono desde carbohidratos
hasta aminoácidos. En ausencia de fotosíntesis, el almidón o la sacarosa pueden
suministrar dicho carbono. En la luz, los azúcares que se sintetizan en el ciclo
reductivo de las pentosas fosfato constituyen la fuente última de esqueletos
carbonados para sustentar la síntesis de aminoácidos. Así pues, en los
organismos fotosintéticos, el carbono asimilado se reparte entre la formación de
carbohidratos y la síntesis de aminoácidos. El flujo de carbono orgánico hacia
una u otra vía se modula a través de la regulación de enzimas claves de ambas
rutas, como respuesta, principalmente, a la luz y a la disponibilidad de carbono
y nitrógeno (Coruzzi y Zhou, 2001).
1.5 Efecto del oxígeno en la oxidación de las biomoléculas
El oxígeno es un compuesto esencial en el metabolismo de todos los organismos
aerobios, ya que participa en diversas reacciones de oxidación, incluyendo la
respiración. Durante estos procesos el oxígeno molecular se reduce, dando
origen a las llamadas especies reactivas de oxígeno, las que en su mayoría son
radicales libres. Los radicales libres se pueden formar en el interior de las células
como producto de sus actividades fisiológicas normales (Bunker, 1992). La figura
1 muestra el efecto de los radicales libres en las biomoléculas; proteínas, ácidos
nucleicos, carbohidratos y lípidos.

1.5.1 Efecto de los radicales libres en el ADN y en la longitud de los
telómeros
El daño provocado a nivel del ADN por los radicales libres puede generar
mutaciones somáticas que llevarían a la síntesis de proteínas defectuosas y
posiblemente a la generación de transformaciones malignas, como es el caso de
la desoxirribosa, la que al oxidarse puede inducir el rompimiento del enlace entre
este azúcar y el grupo fosfato del siguiente nucleótido, mecanismo mediante el
cual se forman rompimientos de cadena sencilla (Breen y Murphy, 1995;
Dizdaroglu y Jaruga, 2012), convirtiendo a los telómeros en un blanco
susceptible a daño. Los radicales hidroxilos tienen la capacidad de reaccionar
también con las bases nitrogenadas del ADN. El tipo predominante de alteración
que puede observarse a este nivel son las sustituciones, aunque también se
pueden observar deleciones y con menor frecuencia inserciones. Se ha visto que
las sustituciones frecuentemente involucran al par guanina-citocina, con el que
los radicales hidroxilos y el oxígeno simple reaccionan directamente. Las
mutaciones se concentran en regiones específicas del ADN, que se denominan
Figura 1. Blancos de oxidación de especies reactivas y radicales libres. Modificado de Dizdaroglu et al. (2002); Valko et al. (2004); Benov y Beema (2003), Halliwell y Chirico, (1993); y Lobo et al. (2010).Tomado de Carocho y Ferreira (2013).

“puntos calientes”, lo que indica que están relacionadas con las secuencias de
bases púricas y pirimídicas (Imlay y Linn, 1988).
Los telómeros son regiones de ADN no codificante ubicadas en los extremos de
los cromosomas eucariónticos. Están constituidos por secuencias de ADN
altamente conservadas, repetidas en tándem (TTAGGG)n, y proteínas
asociadas. Presentan una estructura especial que impide su unión a los
extremos de otros cromosomas, previniendo la fusión telomérica (Lu et al., 2013;
Zakian, 1995). Cumplen un papel esencial en la preservación de la integridad
cromosómica, protegiendo al ADN codificante de deleción por fenómeno de fin
de replicación y la degradación, contribuyendo al mantenimiento de la estabilidad
cromosómica. Moderan importantes interacciones entre los cromosomas y la
matriz nuclear, pudiendo además ejercer efectos sobre la transcripción de genes
situados en regiones subteloméricas e interactúan con los mecanismos
regulatorios del ciclo celular (Blackburn, 1991).
Distintas evidencias indican que el acortamiento telomérico durante el
envejecimiento de células somáticas normales in vitro juega un papel causal en
la senescencia celular. Una longitud telomérica crítica estaría asociada con un
bloqueo en la replicación característica de las células senescentes (Mondello y
Scovassi, 2004).
La senescencia celular es un proceso irreversible de declinación de la
proliferación celular en relación con la edad. Es un proceso activo,
genéticamente programado que responde a una inducción dada por el
acortamiento telomérico, generando una señal semejante a la producida por el
daño en el ADN (Jeyapalan y Sedivy, 2008).
1.6 Antioxidantes
Las células presentan mecanismos de protección, de manera que los radicales
libres pueden ser transformados a productos menos tóxicos o no tóxicos. La
protección de las células contra los radicales libres derivados del oxígeno
comprende no solo la captura de estos intermediarios agresivos, sino también la

prevención de su formación, la inhibición de su propagación y la reparación de
las lesiones (Gavia García y González Martínez, 2015).
La célula cuenta con dos niveles de defensa para actuar en contra de los
radicales libres, la primera es la prevención; esto implica la acción de
procedimientos que bloquean su formación, como sería la presencia de
proteínas que se unen a metales (en particular hierro y cobre) lo que controla
eficientemente la lipoperoxidación y la fragmentación del ADN, ya que de esta
manera se evita la participación de estos metales en las reacciones donde se
producen las diferentes especies reactivas de oxígeno (Vilar-Rojas et al., 1996).
En un segundo nivel de protección está la acción de los antioxidantes, que
eliminan a los radicales para suprimir su actividad nociva en la célula. Estos
agentes pueden dividirse en dos categorías: en enzimáticos y no enzimáticos
(Pietta, 2000; Ratnam et al., 2006.).
Los antioxidantes no enzimáticos se unen a los radicales libres, y los transfieren
de sitios donde pueden provocar graves daños (membranas) a compartimentos
celulares donde sus efectos sean menos drásticos (citoplasma), o bien, los
transforman en radicales menos agresivos (Halliwell, 1991).
Dentro de los antioxidantes no enzimáticos están a los pigmentos carotenoides
cuya principal función es captar energía luminosa, para luego ser transferida a la
clorofila y transformarse durante la fotosíntesis. Los carotenoides siempre
acompañan a la clorofila en una relación de tres a cuatro partes de clorofila por
una parte de carotenoide (Meléndez-Martínez et al., 2007). Gran parte de la
importancia nutricional de estos pigmentos radica en el hecho de que algunos de
ellos poseen actividad pro vitamina A. Durante la última década se ha
demostrado que juegan un papel importante en la prevención de diversas
enfermedades degenerativas humanas (Meléndez-Martínez et al., 2007).
La actividad antioxidante de estos pigmentos depende de una serie de factores,
como su estructura química (tamaño, número de sustituyentes, configuración cis
o trans), su concentración, la presión parcial de oxígeno o su interacción con
otros antioxidantes, sobre todo las vitaminas C y E. En un principio estos estudios
se llevaron a cabo basándose principalmente en el -caroteno. El mecanismo de

la actividad antioxidante de este compuesto está relacionado con su carácter
hidrofóbico y con su capacidad para "retirar" el oxígeno singulete y desactivar
radicales libres (Jialal et al., 1991). En la tabla I se muestran las aplicaciones de
distintos pigmentos carotenoides.
Tabla I. Aplicaciones de los pigmentos carotenoides Bhosale, (2004).
Carotenoide Utilidad
Licopeno Medicamentos contra enfermedades cardiovasculares, cáncer de próstata.
Preparaciones cosméticas.
β-caroteno Agente anticancerígeno, fuente de vitamina A. Colorante en alimentos.
Fotoprotector.
Preparaciones cosméticas.
Astaxantina Complemento alimenticio principal en acuacultura.
Colorante en alimentos.
Luteína Prevención de la degeneración macular senil (AMD por sus siglas en inglés).
Preparaciones cosméticas.
Zeaxantina Prevención de la degeneración muscular senil (AMD por sus siglas en inglés).
1.7 Microalga utilizada en este trabajo
Desmodesmus communis
1.7.1 Generalidades
Desmodesmus es un microalga de la clase Chlorophyceae, orden
Sphaeropleales, familia Scenedesmaceae, subfamilia Desmodesmoideae.
Originalmente este género fue nombrado como Scenedesmus por Mayen 1823,

pero después Hegewald et al. (1997) reubicaron todas las especies del
subgénero Desmodesmus de Scenedesmus al género Desmodesmus utilizando
caracteres genéticos. Este es probablemente el género de Chlorococcales más
comúnmente reportado y frecuentemente abundante en aguas ricas en
nutrientes, especialmente con un alto contenido de nitrógeno (Wehr, 2007).
1.7.2 Descripción
Las células en la colonia se organizan en múltiplos de dos, aunque las especies
más comunes presentan cuatro u ocho células (Lee, 2008). Estas se unen entre
sí en toda su longitud. Las células externas del cenobio con espinas polares con
simetría bilateral, no tangencial y márgenes rectos o ligeramente convexos. Las
espinas son rectas o ligeramente sinuosas. En general son células cilíndricas,
las células interiores con los polos redondeados y las exteriores con polos
ligeramente alargados y curvos en la dirección de la espina. Las espinas se
desarrollan apicalmente y sin engrosamientos basales. Entre las células puede
observarse una membrana de unión que no sobrepasa la curva de los ápices de
las células. En la superficie de las células se observan rosetas apicales o
cercanas a los polos. Las células contienen un cloroplasto parietal laminar y
lateral con un pirenoide excéntrico. En las células adultas se presentan gotas de
aceite amarillento. Sus dimensiones son: células de 4.7 a 6.3 µm de diámetro,
12-18 µm de largo, espinas hasta 12.5 µm de largo (Guiry y Guiry, 2018).
1.7.3 Clasificación (Guiry y Guiry, 2018)
Clasificación:
Imperio Eukaryota
Reino Plantae
Subreino Viridiplantae
Infrareino Chlorophyta
Phylum Chlorophyta
Subphylum Chlorophytina

Clase Chlorophyceae
Orden Sphaeropleales
Familia Scenedesmaceae
Subfamilia Desmodesmoideae
Género Desmodesmus
1.7.4 Ciclo celular
Una característica que promueve el interés biotecnológico en esta microalga es
su ciclo celular (Ševčíková et al., 2013) , el cual se caracteriza porque se dividen
en más de dos células hijas en un ciclo celular (denotado como ciclo de fisión
múltiple). Generalmente, cualquier división ocurrirá en 2n células hijas (ciclo tipo
Cn), donde “n” es un número entero de 1 a 15. Los tipos de ciclo celular C1 y Cn
son intercambiables en algunas especies. Las células cultivadas bajo
condiciones de crecimiento desfavorables, con una tasa de crecimiento baja, se
dividirán en sólo 2 (n = 1, C1); células hijas, mientras que las mismas células,
cuando crecen en condiciones óptimas, se pueden dividir en 8 (n = 3, Cn) o 16
(n = 4, Cn) células hijas.
Estas microalgas son populares en los estudios del ciclo celular (Prison y
Lorenzen, 1966; Tamiya, 1966) porque pueden sincronizarse fácilmente
alternando los períodos de luz y oscuridad, un procedimiento que se considera
natural y donde la sincronía inducida es muy alta. Debido a la presencia de
múltiples replicaciones de ADN, división nuclear y celular, el ciclo es mucho más
complejo que el esquema clásico y tiene una serie de modificaciones. Es
importante destacar que hay una amplia superposición de la duplicación del
genoma por la replicación del ADN, la separación del genoma por la división
nuclear, y la división celular, y es aún más complejo ya que los procesos del ciclo
celular se coordinan con procesos equivalentes tanto en las mitocondrias como
en los cloroplastos (Ševčíková et al., 2013).
Aunque el ciclo celular de tipo Cn también se producen en otros organismos, en
muchos grupos taxonómicos de microalgas es único. Los ciclos Cn son
característicos para la mayoría de las células en los órdenes de microalgas

Chlorococcales y Volvocales como Chlorella, Desmodesmus, Scenedesmus y
Chlamydomonas. En la figura 2 se muestra una representación del ciclo Cn
(fisión múltiple) y sus etapas G1, CP, pS, S, G2, G3 y C.
Figura 2 . Representación del ciclo celular Cn (fisión múltiple) (Borowitzka et al., 2016) donde se representa G1: fase del ciclo celular, en la que existe crecimiento celular con síntesis de proteínas y de ARN, CP: etapa en el ciclo celular en el que la célula se compromete a desencadenar una terminación de la secuencia de procesos que conduce a la duplicación de estructuras reproductivas, la cual consiste de pS: fase de pre-replicación entre el punto de restricción y el comienzo de la replicación del ADN. S: fase durante la cual la replicación de ADN toma lugar, G2: fase entre la terminación de la replicación del ADN y el inicio de la mitosis, M: fase durante la cual sucede la división nuclear, G3: fase entre la división nuclear y la división celular. Los procesos que conducen a la división celular se supone que tienen lugar durante esta fase, y C: fase durante la cual se produce la división celular y la formación de células hijas.
2. ANTECEDENTES
2.1 Respuesta bioquímica a condiciones de consumo de nitrógeno
Se han realizado diversos trabajos en microalgas con relación a la disminución
(consumo) de nitrógeno del medio de cultivo y el efecto en la concentración de
diferentes biomoléculas (Tabla II).

Tabla II. Comportamiento de biomoléculas en respuesta al consumo de nitrógeno.
Biomolécula Respuesta Referencia
ARNm Disminución Doucette y Harrison, 1991
Proteínas Disminución en su
concentración
Doucette y Harrison, 1991; Li et al., 2012; Simionato et al.,
2013
Lípidos Aumento en su concentración
Pirastru et al., 2012;
Thompson, 1996
Carbohidratos Aumento en su concentración
Coruzzi y Zhou, 2001; Pirastru et al., 2012
Carotenoides Aumentan Ben-Amotz et al., 1982; Qin et
al., 2008
Clorofila Baja su concentración ,
disminuyendo la fotosíntesis Lazar, 2003
2.2 Lípidos y ácidos grasos en microalgas
Las microalgas han atraído la atención de la comunidad científica, en particular
los bioquímicos lipídicos y los biotecnólogos de microalgas, ya que se les
considera una fuente de valiosos ingredientes nutricionales, tales como ácidos
grasos altamente insaturados de cadena larga (LC-PUFA) de la familia ω3 y
carotenoides, así como los ácidos grasos saturados que son utilizados para la
producción de biodiesel (Imahara et al., 2007).
Las microalgas bajo condiciones normales de cultivo presentan un contenido de
lípidos que por lo general varía entre el 20 y el 50 % de su peso seco, sin
embargo, cuando son sometidas a situaciones de estrés, frecuentemente
incrementan el contenido de lípidos totales, por lo que se han reportado valores
en intervalos más amplios como se señala en la tabla III.

Tabla III. Contenido de lípidos totales en la familia Scenedesmaceae.
Microalga % Lípidos (peso seco) Referencia
Scenedesmus dimorphus 31 Shen et al., 2009
Scenedesmus incrassatulus
8 -12 Arias-Peñaranda et al.,
2013
Scenedesmus obliquus 10 – 43 Ho et al., 2010
Scenedesmus rubescens 27 - 43 Lin y Lin, 2011; Tan y
Lin, 2011
Scenedesmus sp. 7 – 53 Xin et al., 2010
Los ácidos grasos son compuestos de carbono alifático que varían en longitud,
grado de insaturación y estructura. Dada la enorme diversidad de microalgas y
sus hábitats en la naturaleza (Guschina y Harwood, 2006), estos organismos
presentan una mayor variedad de ácidos grasos, que los que se encuentran en
plantas superiores. En plantas superiores hay lípidos, donde son más comunes
los ácidos grasos saturados e insaturados de 16 carbonos (C16, C18) de cadena
lineal, con hasta tres insaturaciones.
En ciertas clases de microalga son abundantes los ácidos grasos poliinsaturados
de cadena larga (LC-PUFA), con cadenas de C20 y C22 y un mayor número de
insaturaciones (hasta seis). La composición de ácidos grasos de microalgas
varía sustancialmente entre los representantes de los diferentes grupos
filogenéticos (Borowitzka y Borowitzka, 1988; Hu et al., 2008; Lang et al., 2011).
Los ácidos grasos suelen contener número par de carbonos, pero también se ha
descrito que los ácidos grasos con cadenas impares existen en microalgas (Lang
et al., 2011).

2.2.1 Biosíntesis de ácidos grasos
En microalgas verdes, la ruta metabólica para la síntesis de ácidos grasos
empieza por la unión del complejo luz-biomasa, el cual está mediado por la
clorofila y carotenoides, los cuales capturan la energía lumínica en forma de
fotones. La energía es empleada por el fotosistema II en la oxidación catalítica
del agua, formando protones, electrones y O2 molecular. Los electrones se
transportan mediante la cadena de transporte fotosintética para promover la
producción de NADPH mediante la reducción de ferredoxina. Dentro del lumen
tilacoidal se genera un gradiente electroquímico debido a la liberación de
protones después de la oxidación de agua, el cual se emplea para la formación
de ATP mediante la vía ATP sintasa. Los productos de la fotosíntesis NADPH y
ATP son sustratos del ciclo de Calvin (Beer et al., 2009), en donde la enzima
ribulosa biofosfato carboxilasa (Rubisco) cataliza la conversión de CO2 y
ribulosa-5-fosfato en dos triosas, las cuales son convertidas subsecuentemente
en piruvato y acetil-CoA (Fischer et al., 2008). Las moléculas de acetil-CoA son
carboxiladas dentro del cloroplasto por la acetil-CoA carboxilasa (ACC) para la
síntesis de malonil-CoA (Hu et al., 2008), molécula que es transferida a la
proteína acarreadora de grupos acil (ACP) del complejo multienzimático sintasa
de ácidos grasos (SAG). La subunidad cetoacil-ACP sintasa (CAS) cataliza la
condensación de malonil-ACP mediante tres reacciones cíclicas: reducción,
deshidratación y reducción; condensando el producto con otra molécula de
malonil-CoA (Guschina y Harwood, 2006). El ciclo se repite hasta formar
cadenas saturadas de ácido palmítico (C16:0) o esteárico (C18:0). La enzima
ACP-sintasa abre la cadena acil y libera el ácido graso (Scott et al., 2010).
En la tabla IV se muestran algunos ácidos grasos presentes en microalgas.

Tabla IV. Presencia de ácidos grasos en microalgas.
Ácido graso Presencia Referencia
C16 y C18 (saturados y monoinsaturados)
Variación en abundancias en
distintos grupos de microalgas
Borowitzka y Borowitzka, 1988; Hu et
al., 2008
C16:0 y C18:1 ω9 Abundantes en la
división Chlorophyta Lang et al., 2011
ácido α-linolénico
ácido γ-linolénico
Presentes en la clase Chlorophyceae
Lang et al., 2011
ARA, EPA y DHA Frecuencia del 5%
aprox. En 900 cepas de la clase Chlorophyceae
Lang et al., 2011
2.3 Telómeros y microalgas
En cuanto a telómeros se ha demostrado que las especies reactivas de oxígeno
ocasionan una actividad telomerasa defectuosa (Caradonna y Mauro, 2016),
siendo de interés ya que se ha visto que la actividad de la telomerasa está
presente en la mayoría de los tumores (D’Souza et al., 2013). Una disminución
en su actividad causa acortamiento de los telómeros iniciando la senescencia
prematura, siendo la interacción de los antioxidantes de interés ya que al reducir
las ERO se reducen la pérdida de la actividad telomerasa (Zeiher et al., 2004).
Los telómeros en microalgas han sido poco estudiados; sin embargo se reporta
que extractos crudos de microalgas han presentado actividad antiproliferativa
sobre líneas celulares de cáncer (HeLa), lo que sugiere la existencia de
metabolitos secundarios específicos que podrían interferir con la mitosis
celular, probablemente como resultado de la inhibición o regulación sobre la
actividad de la telomerasa (Moo-Puc et al., 2009). En la tabla V se muestran los
estudios realizado en microalgas en cuanto a la longitud de los telómeros.

Tabla V. Antecedentes del estudio de telómeros en microalgas.
Referencia Organismo Hallazgo
Ševčíková et al., 2013
Desmodesmus quadricauda
Alta actividad de telomerasa, con aumentos finalizando la fase oscura.
Fulnečková et al., 2012
Orden Chlamydomonadales
Secuencia telomérica de tipo Arabidopsis es ancestral para las
microalgas verdes.

3. JUSTIFICACIÓN
El estudio de la microalga Desmodesmus.communis es importante para generar
conocimiento básico, ya que su plasticidad metabólica le permite en condiciones
de estrés como el consumo de nitrógeno evaluar el potencial para la producción
de compuestos bioactivos y su posterior escalamiento biotecnológico.

4. HIPÓTESIS
Si el consumo de nitrógeno durante el crecimiento de la microalga
Desmodesmus communis provoca una disminución del contenido de proteínas,
clorofila, ácidos grasos y el nitrógeno intracelular, entonces la relación C:N
aumentará y se espera que la longitud de los telómeros se acorten.
En el cultivo de Desmodesmus communis las bacterias asociadas a la microalga
contribuirán aportando nitrógeno orgánico.

5. OBJETIVOS
a. Objetivo general
Evaluar el efecto del consumo de nitrógeno en la respuesta de la microalga
Desmodesmus communis en cuanto a nitrógeno intracelular, contenido de
proteínas, carbohidratos, lípidos, ácidos grasos, pigmentos, bacterias presentes
en el cultivo de la microalga y la longitud de los telómeros.
b. Objetivos específicos
1. Determinar si la microalga Desmodesmus communis se encuentra en
condición de deficiencia de nitrógeno mediante la medición de la relación
C: N y el contenido de nitrógeno elemental.
2. Evaluar el efecto de la deficiencia de nitrógeno sobre la tasa de
crecimiento, productividad y composición bioquímica de la biomasa de la
microalga Desmodesmus communis en diferentes fases de crecimiento.
3. Evaluar el efecto de la deficiencia de nitrógeno sobre el perfil de ácidos
grasos y pigmentos de la biomasa microalgal en diferentes fases de
crecimiento de la microalga Desmodesmus communis.
4. Determinar la presencia de bacterias asociadas al crecimiento de la
microalga Desmodesmus communis en diferentes fases de crecimiento.
5. Evaluar el efecto de la deficiencia de nitrógeno sobre la longitud de los
telómeros de la microalga Desmodesmus communis en diferentes fases
de crecimiento.

6. MATERIAL Y MÉTODOS
6.1 Material biológico y condiciones de cultivo
Desmodesmus communis fue colectada y aislada de la presa Miguel Hidalgo de
Ciudad Victoria, Tamaulipas en el año 2009 por la M. en C. Mónica Rodríguez
Palacios del Laboratorio de Fitoplancton de la UAM-Iztapalapa. Fue donada al
cepario del CIBNOR en donde se mantiene con la clave CIB53 desde el año
2009. La cepa fue identificada mediante microscopía electrónica de transmisión
por el Dr. Eberto Novelo Maldonado del Laboratorio de Algas Continentales.
Ecología y Taxonomía. Departamento de Biología Comparada de la Facultad de
Ciencias de la UNAM. En la figura 3 se muestra Desmodesmus communis
imagen obtenida con un microscopio Nikon Eclipse E600, con contraste
diferencial de interferencias (DIC) y cámara digital Nikon DXM1200E.
Figura 3. Desmodesmus communis identificada por el Dr. Eberto Novelo M.
La cepa de D. communis se cultivó en medio f/2 (Guillard y Ryther, 1962)
adicionada de 10% de agua de mar, en matraces Erlenmeyer a de vidrio en un
volumen final de 1.7 L. El cultivo se inició con un inóculo de 1 x 106 cél. mL-1. Las
condiciones de cultivo fueron: temperatura 24±1°C, irradiancia incidente de 120
µmol fotón m-2 s-1, fotoperiodo: 12 h luz: 12 h oscuridad, aireación continua, pH
inicial de 7.8.
En las condiciones anteriores, se realizó la curva de crecimiento, tomando
diariamente bajo condiciones de esterilidad, una alícuota de 5 mL de cultivo, la
cual se contó en la cámara de neubauer en un microscopio (Primo Star ZESS),

como lo indica Arredondo-Vega y Voltolina (2017). Una vez conocida la curva de
crecimiento, se seleccionaron los tiempos de muestreo (día 0, 5, 10, 12 y 17), en
los cuales se tomaron 50 mL de cultivo, que se centrifugaron a 1428 x g, 10 °C
durante 5 min en la centrifuga refrigerada (Allegra® X-15R). El sobrenadante de
cada muestra, así como una parte de la biomasa fresca se mantuvieron en
congelación a -20 ºC y -80 °C respectivamente. Otra parte de la biomasa se
liofilizó (liofilizadora LABCONCO Free Zone 1).
6.2 Análisis experimental
6.2.1 Determinación de peso seco, orgánico e inorgánico (Sorokin,
1973)
El peso seco total y cenizas se realizó de acuerdo a Sorokin (1973), la cual
consiste en la separación de la biomasa del medio de cultivo mediante filtración
al vacío en membranas de fibra de vidrio Whatman GF/F de tamaño de poro 0.7
µm previamente a peso constante. La biomasa filtrada se lavó con 10 mL de
agua destilada. Primeramente, se ajustaron a peso constante los filtros a utilizar,
esto se logró de la siguiente manera: se lavaron las membranas de fibra de vidrio
con agua destilada y se dejaron secar a temperatura ambiente en un área en la
que se encontraran protegidos de contaminantes. Una vez secos, se transfirieron
a sobres de papel aluminio y se pre-quemaron a 490 °C por 6 h. Los filtros pre-
quemados se colocaron en un desecador durante 1.5 h y se pesaron con una
balanza analítica de cuatro dígitos (OHAUS PIONEER™), posteriormente se
colocaron en un horno de secado (TERLAB TE-H35D) a 65 °C por 1 h y
posteriormente en un desecador 1.5 h, esto último se repitió hasta verificar que
el peso permaneciera constante.
Luego se tomó un volumen conocido del cultivo de la microalga y se filtró al vacío
en las membranas de fibra de vidrio a peso constante. Las muestras de
Desmodesmus communis se lavaron con 10 mL de agua destilada. Se guardaron
los filtros en los sobres. Los sobres (entreabiertos) con el filtro se colocaron en
el horno de secado (TERLAB TE-H35D) a 65 °C durante al menos 12 h. Después
del secado, se colocaron los filtros en el desecador durante 1.5 h y se pesaron

rápidamente. Se introdujeron de nuevo los filtros al horno de secado a 65 ºC por
1 h y enseguida 1.5 h en el desecador y se pesaron, esto se repitió hasta
asegurarse que el peso permaneciera constante.
Por diferencia entre el peso del filtro vacío y el peso del filtro con la muestra y
dividido entre el volumen de cultivo filtrado, se obtuvo el dato de peso seco total
(µg mL-1 o mg mL-1). El filtro con la biomasa seca y pesada se incineró en una
mufla (TERLAB MR) a 490 °C 6 h. Se introdujeron de nuevo los filtros al horno
de secado 1 h a 65 ºC, enseguida 1.5 h en el desecador y se pesaron, esto se
repitió hasta asegurarse que el peso permaneciera constante. Por diferencia de
peso entre el filtro vacío y el filtro con las cenizas, se obtuvo el peso inorgánico
o peso de cenizas (µg mL-1 o mg mL-1). Por diferencia entre el peso del filtro con
la muestra seca y el filtro con las cenizas, se obtuvo el peso orgánico.
6.2.2 Determinación de nitratos (Strickland y Parsons, 1972)
Del sobrenadante resultante de la cosecha del cultivo de cada tiempo
muestreado, se destinaron 50 mL para determinar la concentración de nitrógeno
presente en forma de nitratos. Para lo cual se realizó la metodología que se
describe a continuación:
a) Primeramente, se realizaron diluciones de las muestras con agua
desionizada obteniendo un volumen total de 10 mL para para muestra,
posteriormente se agregaron 200 µL de cloruro de amonio concentrado
(125 g cloruro de amonio en 500 mL de agua desionizada).
b) Después se procedió a la preparación de la columna de cadmio la cual
fue empacada con 25 g de cadmio, el cual se lavó previamente tres veces
con HCl al 10% hasta eliminar impurezas, y se enjuagó con agua corriente
hasta que el pH de la solución fue superior a 6.
c) Posteriormente se lavó con una solución diluida de cloruro de amonio (25
mL de la solución de amonio concentrado en 1 L de agua desionizada)
por 5 veces y se escurrió.
d) Enseguida se agregaron 50 mL de solución de sulfato de cobre al 2% y
se agitó hasta que el color azul desapareció.

e) Se enjuagó 3 veces con cloruro de amonio diluido.
f) Posterior a la activación del cadmio se empacó la columna, se colocó en
un equipo Quikchem FIA + y se procedió a colocar las muestras en el auto
muestreador para su análisis. La curva de calibración de nitritos fue de 0.1
µM a 5 µM NO3-1.
6.2.3 Determinación de la relación carbono: nitrógeno elemental y la
relación entre 13C:12C y 15N:14N (Peterson y Fry,1987)
La biomasa previamente liofilizada se pulverizó y homogenizó con un agitador
de vórtice. Se pesaron en cada tiempo de cosecha y por duplicado 2 mg con una
balanza RADWAG MYA 2.3Y, y se empaquetaron en cápsulas de estaño, para
ser analizado posteriormente por un Analizador Elemental de Combustión,
marca Costech, modelo ECS 4010, y con un Espectrómetro de Masas de
Razones Isotópicas marca Thermo Scientific, modelo Delta V Plus. El análisis se
llevó a cabo de la siguiente manera:
a) Las cápsulas de estaño se introdujeron en el reactor de combustión.
b) El gas portador de helio se conmuta a un volumen de oxígeno que es
elegido por el operador dependiendo del tamaño y composición de la
muestra.
a) La muestra y la cápsula de estaño reaccionan con el oxígeno y se queman
a 1020 ºC, así es como la muestra se descompone en sus componentes
elementales (N2: nitrógeno molecular y CO2: dióxido de carbono).
b) Posteriormente estos elementos acarreados por el gas fluyen a través de
la columna de separación por cromatografía de gases.
c) Los gases se separan y son detectados secuencialmente por el detector
de conductividad térmica (TCD). El TCD genera una señal que es
proporcional a la cantidad de elemento en la muestra.
d) El análisis de los datos se realizó con el software ISOdata 3.0 utilizando
como estándar BBOT el cual tiene un porcentaje de nitrógeno elemental
de 6.51% y de carbono elemental 72.53%. El software compara el pico
elemental y calcula la relación isotópica con respecto al estándar
conocido.

6.2.4 Determinación de proteínas (Lowry et al., 1951)
Se pesaron 5 mg de muestra liofilizada y pulverizada, se colocó en un tubo de
vidrio limpio previamente lavado con Micro 90, enjuagado con agua destilada y
secado y se siguieron los pasos que se describen a continuación:
a) Se agregó 1 mL de NaOH 0.1 N y se colocaron los tubos en un baño
ultrasónico (VWR ultrasonic cleaner) durante 5 min.
b) Se agregaron 4 mL de NaOH 0.1 N y se colocó papel aluminio en la boca
del tubo, se tapó con una tapa sin septo, se sometió a hidrólisis alcalina
en un baño a 100°C durante 1 h, transcurrido el tiempo de hidrólisis se
enfriaron los tubos a temperatura ambiente.
c) El extracto alcalino se filtró pasando éste a través de una pipeta Pasteur
limpia con un tapón de fibra de vidrio y recibiendo el extracto libre de
partículas en otro tubo limpio y seco.
Posteriormente se procedió a realizar los cálculos de la alícuota para la
cuantificación de proteínas (Lowry et al., 1951). La alícuota calculada se
completó a 1 mL con NaOH 0.1 N, y se agitó en un vortex para su total
homogenización y se procedió a realizar los pasos que se describen a
continuación:
a) Se agregó 5 mL de la solución C, la cual está compuesta de la solución
A (Na2CO3 al 2 % (p/v) en NaOH 0.1 N) y la solución B (mezcla 1:1 de
CuSO4:5H2O al 5 % y tartrato de K al 1 %) en una proporción (50:1). Se
mezcló en el vortex y dejó reposar 15 min.
b) Transcurrido el tiempo de reposo se agregaron 500 μL de la solución 1:1
de Folin: H2O destilada (solución D), se agitó en el vortex y dejó reposar
en la oscuridad por 40 min.
c) Después se leyó en un espectrofotómetro (HACH DR 5000) a 750 nm
calibrando el equipo con un blanco preparado usando el mismo
tratamiento que el de las muestras, pero utilizando 1 mL de NaOH 0.1 N
en lugar de muestra. Previamente se realizó una curva de calibración
con una solución concentrada de 300 µg mL-1 de albúmina sérica bovina

(BSA por sus siglas en inglés), a partir de la cual se preparó un gradiente
de concentración de 0 a 150 µg mL-1.
6.2.5 Determinación de carbohidratos (DuBois et al., 1956)
La determinación de carbohidratos se realizó utilizando el método colorimétrico
del fenol-ácido sulfúrico (DuBois et al., 1956). Primeramente, se pesó 5 mg de
muestra liofilizada, se colocó en un tubo de vidrio limpio previamente lavado con
Micro 90, enjuagado con agua destilada y secado. Y se procedió a realizar los
pasos que se describen a continuación:
a) Se agregó 1 mL de H2SO4 1 M, se colocaron los tubos en un baño
ultrasónico (VWR ultrasonic cleaner) durante 5 min y se agregaron 4 mL
de H2SO4 1 M.
b) Se colocó papel aluminio en la boca del tubo y se tapó con una tapa sin
septo, luego se introdujeron los tubos a un termo baño (TERLAB) a 100
°C durante 1 h, transcurrido el tiempo de hidrólisis se enfriaron los tubos
a temperatura ambiente.
c) El extracto ácido se filtró pasando éste a través de una pipeta Pasteur
limpia con un tapón de fibra de vidrio y recibiendo el extracto libre de
partículas en otro tubo limpio y seco.
d) Se agregó 1 mL de fenol al 5 % y se mezcló, se dejó reposar 40 min,
enseguida se agregó lentamente 5 mL de H2SO4 concentrado todo esto
en campana de extracción, se enfrió a temperatura ambiente y se leyó a
485 nm en un espectrofotómetro (Spectronic® 20 GENESYS™),
calibrando el espectrofotómetro con un blanco que se prepara de la
misma manera, sustituyendo el extracto ácido de la muestra con 1 mL de
H2SO4 1M. Previamente se realizó una curva de calibración usando un
gradiente de concentraciones de glucosa anhidra preparadas a partir de
una solución de 120 µg mL-1, el gradiente de concentración fue de 0 a
120 µg mL-1 .

6.2.6 Determinación de pigmentos, lípidos totales y ácidos grasos
Para la determinación de pigmentos se utilizó la técnica de Lamers et al. (2010).
A partir de un volumen conocido del cultivo, se centrifugó a 1850 x g por 10 min
a 5 ºC y la pastilla celular obtenida se sometió a los siguientes pasos:
a) Se agregó 4 mL de la mezcla cloroformo: metanol (2:2.5 v/v) y 5 µL
antioxidante butil-hydroxi tolueno (BHT). Se homogenizó con vortex por 1
min y se introdujeron los tubos en baño un baño ultrasónico (VWR
ultrasonic cleaner) con hielo por 20 min.
b) Posteriormente se agregaron 2.5 mL de la mezcla TRIS ácido: TRIS
base: NaCl 1M (3:0.4:5 v/v) con pH de 7.5, para facilitar la separación de
la fase acuosa y orgánica (Mulders et al., 2014). Se homogenizó con
vortex 1 min, se colocaron los tubos en un baño ultrasónico (VWR
ultrasonic cleaner) con hielo 20 min y se centrifugó a 1850 x g por 15 min
a 10°C.
c) Se recuperó la fase inferior con pipeta Pasteur nueva mediante burbujeo
y se colocó en un tubo de vidrio con tapón de rosca limpio.
d) Para recuperar los lípidos y pigmentos remanentes, se agregó 1 mL de
cloroformo al tubo anterior, y se repitieron los pasos de homogenizar,
sonicar y centrifugar.
e) Finalmente se recupera el segundo extracto (lípidos y pigmentos) con una
pipeta Pasteur nueva mediante burbujeo y se colocaron en el tubo en
donde está el primer extracto.
6.2.7 Pigmentos (Lamers et al., 2010)
Para recuperar solamente los pigmentos, del extracto anterior se tomó una
alícuota calculada primeramente con los valores anteriormente reportados para
esta microalga en el Laboratorio de Biotecnología de Microalgas en CIBNOR por
la estudiante de licenciatura Diana Kareli Valenzuela y se realizó la metodología
que se describe a continuación:

a) Se utilizó una alícuota de 0.5 mL, se colocó en un tubo limpio y se secó
con nitrógeno gaseoso.
b) Después del secado se agregaron 3 mL de acetona al 100 % grado HPLC
y se procedió a realizar el cálculo del volumen de inyección, el cual se
realizó utilizando los valores determinados mediante espectrofotometría
de clorofila a clorofila b y carotenoides totales obtenidos midiendo la
absorbancia a 470 nm, 645 nm y 662 nm, esto con la finalidad de que
cada pigmento se encuentre dentro del intervalo lineal de la curva de
calibración (2 a 10 ng) que se realizó con siete estándares de pigmentos
(fucoxantina, violaxantina, diadinoxantina, astaxantina trans, zeaxantina,
luteína, cantaxantina, clorofila b, xantafila, β-caroteno cis, criptoxantina,
clorofila a y β-caroteno trans).
c) Las alícuotas (entre 1 y 8 µL) se inyectaron en un HPLC 1200 Infinity
series Agilent Technologies.
6.2.8 Lípidos totales (Marsh y Weinstein, 1966)
Se calculó la alícuota correspondiente del extracto clorofórmico a utilizar para
que se encuentre en el intervalo lineal de la curva de calibración que va de 30 a
180 μg de lípidos. Se utilizaron los datos de concentración de lípidos
previamente obtenidos en el Laboratorio de Biotecnología de Microalgas en
CIBNOR por la estudiante Diana Kareli Valenzuela y se procedió a realizar la
metodología que se describe a continuación:
a) El volumen calculado se encontró entre 50 y 70 µL de extracto.
b) Se colocó en un tubo limpio con tapón de rosca, se secó con nitrógeno,
se agregó 2 mL de H2SO4 concentrado y se tapó la boca del tubo con
papel aluminio y se cerró el tubo hasta el primer tope, sin apretar
demasiado.
c) Posteriormente se colocó el tubo en un horno TERLAB TE-H35D a 200°C
por 15 min, después de trascurrido el tiempo se dejó enfriar a temperatura
ambiente.

d) Se agregaron lentamente con el tubo inclinado 3 mL de agua destilada,
se homogenizó con vortex hasta que se mezcló el agua y el ácido y dejar
reposar en baño de hielo por 5 min.
e) Enseguida se leyó la absorbancia a 375 nm en un espectrofotómetro
Spectronic® 20 GENESYS™, calibrando previamente el equipo con un
blanco de H2SO4 que se trató de igual manera que las muestras.
6.2.9 Ácidos grasos (Sato y Murata, 1988)
Se colocó el resto del extracto total de cloroformo en un tubo de tapón de rosca
limpio, se secó con nitrógeno gaseoso y se procedió a realizar la metodología
que se describe a continuación:
a) Se agregaron 2.5 mL de la mezcla de HCl:CH3OH (5:95 v/v). El tubo se
tapó y se cerró apretando fuerte para evitar la evaporación de la muestra
se selló con cinta teflón por dentro y fuera de la tapa, la cual debe tener
una cubierta de teflón para evitar que los solventes disuelvan los plásticos
de la parte interior de la tapa. Una vez selladas las tapas, lo tubos se
colocaron en un termo baño (TERLAB) a 85 °C durante 2.5 h.
b) Los metilésteres obtenidos de la reacción de derivatización se extraen
con 1.5 mL de hexano grado HPLC. Se repite la extracción dos veces. El
extracto total de ácidos grasos en hexano grado HPLC se recupera en un
nuevo tubo limpio. Para inyectar se calculó el volumen de extracto en
hexano para tener una concentración de ácidos grasos totales de 1200
μg mL-1 y se inyectaron entre 1 y 3 μL en el cromatógrafo de gases-
espectrómetro de masas HP-GCD1800B con detector de ionización
electrónica. La curva de calibración se realizó una mezcla de estándares
de 31 ácidos grasos metil-esterificados marca Sigma.

6.2.10 Determinación de bacterias presentes
Para cultivar las bacterias presentes, cultivables por medios convencionales se
tomaron 300 mL del sobrenadante del medio de cultivo resultante de la cosecha
de biomasa a los diferentes tiempos (0, 5, 10, 12 y 17 días). Para detectar
bacterias que no se desarrollen en medios de cultivo, se aisló ADN, se amplifico
con oligonucleótidos dominio bacteria y se enviaron a secuenciación parcial.
De los 300 mL separados del sobrenadante, se centrifugaron 250 mL a 13 000 x
g durante 5 min en una centrifuga eppendorf 5804 R, y la pastilla celular
resultante fue suspendida en 1 mL de sobrenadante, de este se tomaron 100 µL
y se inocularon por siembra masiva en placas Petri con el mismo medio en el
que se cultivaron las algas y en Agar nutritivo para cada tiempo de cosecha. Se
incubó a 35 °C hasta observar crecimiento. Posteriormente, de las colonias
obtenidas se sembraron por estría cruzada para determinar pureza y se
determinó su morfología colonial.
Con los 50 mL restantes de medio se centrifugaron a 13 000 x g durante 5 min
(eppendorf 5804 R) para formar una pastilla celular suspendida en 1 mL de
sobrenadante, el cual fue utilizado para realizar la extracción de ADN de la
comunidad bacteriana.
6.2.10.1 Extracción de ADN
La extracción de ADN, tanto de las colonias obtenidas como de los pellets
obtenidos a los diferentes tiempos de cosecha, se llevó a cabo utilizando
FastDNA®SPIN kit for soil, como se describe a continuación:
a) Se agregaron al pellet 978 µL de Buffer de fosfato de sodio y 122 µL de
buffer MT, se realizó la ruptura celular con el uso de FastPrep® a 6.0 m/s
durante 40 s.
b) Se centrifugó a 13 000 x g durante 10 min en una microcentrifuga (VWR
2416), se transfirió el sobrenadante a tubos de 2.0 mL y se agregaron 250
µL de PPS y se mezcló 10 veces.
c) Posteriormente, se centrifugó a 13 000 x g durante 5 min y se transfirió el
sobrenadante a tubos de 15 mL, se agregó 1 mL de Binding matrix, se

invirtió por 2 min y se dejó precipitar durante 10 min a temperatura
ambiente.
d) Se descartó el sobrenadante hasta quedar aprox. 1.2 mL, se resuspendió
la Binding matrix y se trasfirieron 600 µL a un SPIN filtre y se centrifugo
13 000 x g por 1.5 min, posteriormente se repitió la trasferencia de 600 µL
al SPIN filtre y se centrifugo nuevamente.
e) Enseguida se procedió a hacer un lavado para el que se agregaron 500
µL de SEWS- M y se centrifugo a 13 000 g por 1.5 min, para eliminar todo
el alcohol presente se centrifugó nuevamente a 13 000 x g durante 2.5
min y se pasó el SPIN filtre a un tubo nuevo, se dejó secar 5 min a
temperatura ambiente y luego se procedió a eluir el ADN de la matriz con
50 µL de DES, se resuspendió, se incubó por 5 min a 55°C y se centrifugó
a 13 000 x g por 1 min el ADN extraído se resguardo a -20°C.
6.2.10.2 Electroforesis
Después de la extracción de ADN se realizó una electroforesis en agarosa al 1%,
con bromuro de etidio, a 100V durante 30 min. El patrón electroforético se
documentó en un equipo Molecular Imagen® Gel DOC TM XR+.
Una vez obtenido el ADN de las bacterias y corroborado una buena
concentración en gel se procedió a amplificar con primers de dominio bacteria y
enviar a secuenciar.
En cuanto al ADN de la fracción del sobrenadante después de comprobar una
buena concentración por medio de electroforesis y nanodrop, se inició el
procedimiento para obtener el perfil de la comunidad bacteriana por medio de
SSCP de sus siglas en inglés (Single Stranded Conformational Polymorphism).
6.2.10.3 SSCP (Schwieger y Tebbe, 1998)
La técnica comprende los siguientes pasos
1. Extracción de ADN
2. Amplificación por PCR
3. Digestión de una cadena de ADN

4. Electroforesis en gel de poliacrilamida
5. Tinción del gel
Reacción en cadena de la polimerasa (PCR)
Para el ADN correspondiente a los tiempos de cosecha se procedió a realizar
reacción de PCR en un termociclador TECHNE TC - 312. La reacción de PCR
se preparó de la siguiente manera:
Reactivos 1Rx
Buffer 5x Gotaq (7.5 mM HgCl2) 5 µL
dNTP’s (c/u 10 mM. 40mM Total) 0.5 µL
F (10 mM) 0.25 µL
R (10 mM) 0.25 µL
Go Taq (5 u/ µL) 0.125 µL
Templado 0.5 µL
H2O 18.375 µL
Tabla VI. Programa de PCR (para técnica SSCP).
Etapa Temperatura Tiempo Número de
ciclos
Inicio 94°C 3 min
Desnaturalización 94°C 1 min
30 ciclos Alineamiento 50°C 1 min
Extensión 72°C 1 min
Elongación final 72°C 5 min
Conservación 26°C

Se comprobaron los productos de PCR mediante electroforesis como se
describe en el apartado 6.2.10.2.
Después, procedimos a purificar los productos de PCR con el kit QIAquickspin
column como de describe a continuación:
a) En un tubo colector de 2.0 mL, se centrifugó por 1min a 9 000 x g, se
descartó lo obtenido en el tubo colector y se colocó la columna de nuevo
en el tubo
b) Para el lavado se adiciono 0.75 mL de buffer PE y se centrifugó a 9 000
x g durante 1 min, se descartó lo obtenido en el tubo colector y se
centrifugó adicionalmente 9 000 x g durante 1 min para eliminar
totalmente el etanol residual proveniente del buffer PE.
c) Posteriormente se colocó la columna en un tubo de microcentrifuga
limpio con capacidad de 1.5 mL. Para eluir el ADN se utilizaron 30µL de
buffer EB (10mM Tris – Cl, pH 8.5), se dejó reposar por 1 min y luego se
centrifugo a 13 000 x g por 1 min.
Electroforesis acrilamida
Se prepararon las placas, se limpiaron con etanol y se adiciono a la placa grande
Repel Silano y a la chica Bind Silano.
El gel se preparó a una concentración de 0.625% en las proporciones a
continuación:
Tabla VII. Mezcla de reacción gel de acrilamida.
Componente Volumen
Volumen final: 20 mL
Solución MDE 2x 6.25 mL
TBE 10x 2 mL
Agua bidestilada 11.752 mL
TEMED 8 µL
APS 40% 20 µL

Digestión de productos de PCR
La digestión de los productos de PCR se realizó como se muestra la mezcla de
reacción:
Tabla VIII. Mezcla de reacción para digestión.
Componente Volumen
ʎ- exonucleasa 1 µL
ʎ- exonucleasa Buffer 3 µL
H2O desionizada 6 µL
Productos de PCR 20 µL
Una vez preparada la reacción se mezcló e incubó a 37 °C por 45 min. Una vez
concluido el tiempo de incubación se procedió a purificar inmediatamente con el
kit MiniElute, Qiagen.
Posteriormente se adicionó a la muestra 9 µL de Buffer de carga para SSCP, se
incubo a 95 °C durante 2 min, enseguida se incubó en hielo 3 min y se cargó el
gel con el estándar y las muestras. Las condiciones de la electroforesis fueron:
350 V, 85 mA, 20 °C durante 18 h.
Tinción del gel con nitrato plata
Una vez terminada la electroforesis se colocó el gel en una charola de acero
inoxidable y se siguieron los pasos que se describen a continuación:
a) Se adiciono la solución fijadora y se incubó durante 30 min a temperatura
ambiente.
b) Posteriormente se enjuagó con agua desionizada, se eliminó el agua y se
adicionó la solución de nitrito de plata y se incubó durante 30 min a
temperatura ambiente.
c) Se eliminó la solución de nitrito de plata, se enjuagó con agua desionizada
y se transfirió el gel a otra charola en donde se adicionó la solución

reveladora agitando suavemente hasta que las manchas tuvieron la
intensidad deseada.
d) Se desechó la solución reveladora y se paró el revelado con solución
fijadora.
e) Para finalizar enjuagando el gel con agua desionizada.
6.2.11 Determinación de la longitud de los telómeros
Primeramente, se realizó la digestión enzimática de las muestras de DNA
genómico (1-5 µg) con las endonucleasas Rsa I y Taq I. Posteriormente se
realizó una electroforesis en un gel de agarosa al 1% en buffer TAE 1X.
Southern blot
Primeramente, se realizó lavado del gel con agua destilada y se colocó sobre un
recipiente que contenía 300 mL (10 veces el volumen del gel) de HCl 0.25M. Se
agitó por 30 min a temperatura ambiente muy suavemente para evitar romperlo.
Esto se hace para la despurinación parcial de los fragmentos de DNA. Se
descartó el HCl y el gel se enjuagó rápidamente con 300 mL de agua destilada.
Se adicionaron 300 mL solución de desnaturalización (NaCl 1.5 M/NaOH 0.5 M)
y se incubó durante 20 min a temperatura ambiente. Se reemplazó esta solución
por otros 300 mL de la misma solución fresca a temperatura ambiente y se
incubó por otros 20 min. En este paso el DNA queda de cadena sencilla para el
subsecuente análisis de hibridación. Se descartó la solución de
desnaturalización y el gel se enjuagó con agua destilada. Se adicionaron 300 mL
de solución neutralizante NaCl 1.5 M/Tris-HCl 0.5 M pH 7.0 fresca y se agitó a
temperatura ambiente por otros 20 min. Se reemplazó por otros 300 mL de
solución de neutralización fresca y se agitó por otros 20 min. Es necesario
mantener un pH menor a 9.0 para la hibridación.
Para realizar la trasferencia, en un recipiente de plástico rectangular de
aproximadamente 500 mL se adicionaron 300 mL de solución SSC 20X (NaCl 3
M, citrato de sodio dihidratado 0.3 M pH 7.0) y dentro de este recipiente se colocó
una superficie de acrílico sobre la cual depositó una tira de papel filtro 3 veces
más largo que el tamaño del gel y se humedeció con solución SSC 20X. En
seguida, se colocaron 3 piezas de papel filtro del mismo tamaño del gel y se

humedecieron con solución SSC 20X. Se colocó el gel sobre el papel filtro
descartando todas las burbujas (usando una pipeta como rodillo) y
posteriormente una pieza de membrana de nylon con carga positiva,
previamente hidratada en agua destilada durante 5 min. Se evitó la formación de
burbujas. Sobre la membrana se colocaron 3 piezas del mismo tamaño de la
membrana de papel filtro humedecidas en la misma solución. Sobre el papel filtro
se colocó una base de 10 centímetros de papel sanitas del mismo tamaño de la
membrana. Finalmente, se colocó una placa de acrílico o cristal para dar
estabilidad a la torre, se colocó un peso de 0.5 kg y se dejó a temperatura
ambiente durante 2 días.
Desensamble de la transferencia.
Se retiraron el papel sanita y filtro que recubrían la membrana y se marcó con un
lápiz la orientación. Se enjuagó la membrana con SSC 2X y se colocó sobre
papel Whatmann 3MM y se dejó secar a temperatura ambiente. Se realizó la
tinción del gel transferido con bromuro de etidio para valorar la eficiencia de la
transferencia.
Inmovilización o fijado.
La inmovilización se realizó con luz UV en un UV Cross-linker 1800 de
Stratagene con la membrana de nylon completamente seca antes de la
irradiación. Se aplicó una potencia de 1200 Joules/cm2.

Marcado de la sonda.
Para esto se preparó la siguiente mezcla de reacción:
Tabla IX. Mezcla de reacción para marcado de la sonda.
Reactivos Concentración de stock Volumen por c/u
Agua Desionizada 14 µL
Buffer de reacción para T4 polinucleótido cinasa
(NEB) 10X 2 µL
Sonda TELPR-3A 100 ng 1 µL
T4 polinucleótido cinasa 10 Unidades/µL 1 µL
[-32P] ATP 9.25MBq (250 µCi/25 µL)
10 µCi 2 µL
Se dio una centrifugación rápida para bajar toda la mezcla de reacción, se incubó
a 37 ºC por 60 min y se agregaron 180 µL de buffer TE para inactivar la actividad
de la enzima. Posteriormente, se tomó 1 µL de la mezcla (101 419 cpm/ µL.) y
adicionaron a 0.5 mL de líquido de centelleo para cuantificar el marcaje total, el
resto (199 µL) se pasaron a través una columna de Sephadex G25 (1 mL) para
eliminar el nucleótido radioactivo no incorporado y se centrifugó a 400 x g por 5
min para eliminar la marca no incorporada. La sonda quedó a una concentración
de (35940 cpm/ µL). El porcentaje de eficiencia de incorporación fue de 35.5%.
Hibridación
Para realizar la hibridación se agregó la sonda marcada desnaturalizada
(2028380 cpm/mL) al buffer de hibridación (fosfato de sodio 0.5 M, pH 7.5; SDS
al 7%) y se incubó a 55 °C durante la noche en un horno de hibridación rotatorio
marca mrc. Transcurrido el periodo de hibridación, la membrana se lavó a 55 ºC
en SSC 2X conteniendo SDS al 0.1%.

Visualización
La membrana húmeda se cubrió con plástico de envoltura de alimentos
(Plastipack) y se expuso a una película Imaging Screen K de Bio Rad por 24 h
y se leyó en un sistema Personal Molecular Imager FX (Bio-Rad) (E. Sykorova
2003). La imagen se exportó como jpg.
6.3 Análisis estadístico
Se realizó un ANOVA de una vía, para conocer si existen diferencias estadísticas
significativas a través del programa IBM SPSS Statistics, con prueba de Duncan
con α de 0.05. Para la elaboración de las figuras se empleó el programa Excel
de Microsoft Office. En las gráficas y tablas que se muestra el promedio y el error
estándar de cada replica.
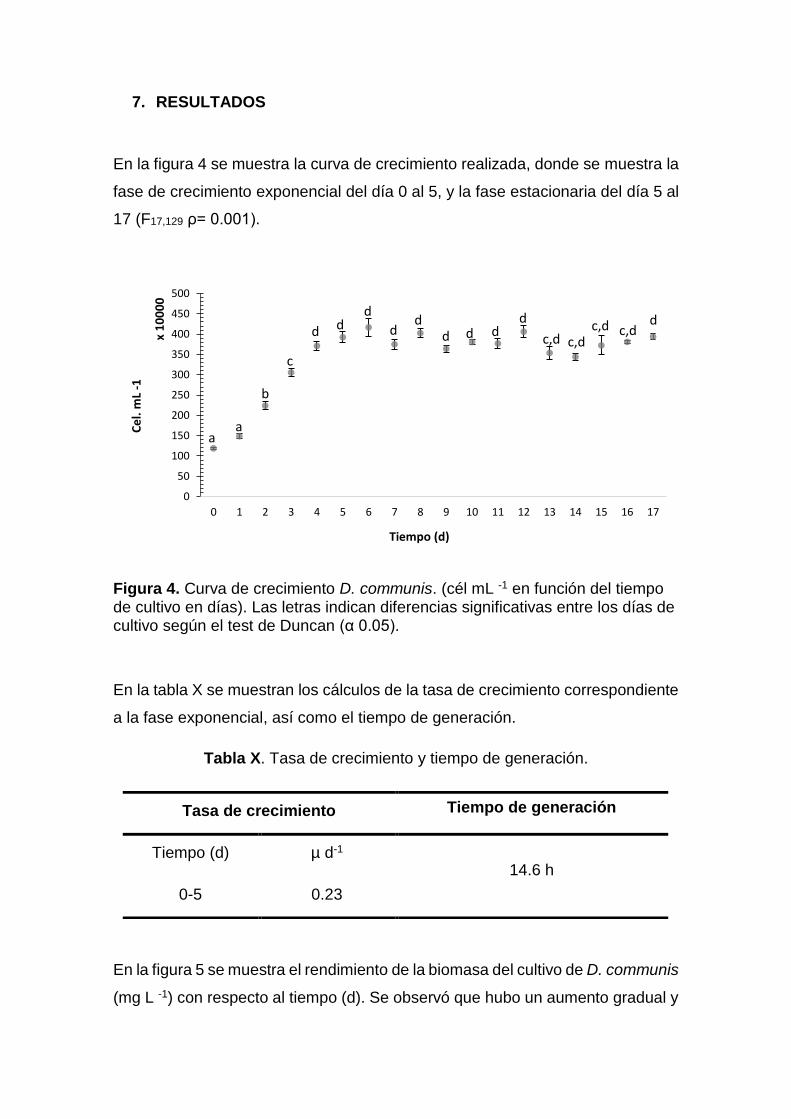
7. RESULTADOS
En la figura 4 se muestra la curva de crecimiento realizada, donde se muestra la
fase de crecimiento exponencial del día 0 al 5, y la fase estacionaria del día 5 al
17 (F17,129 ρ= 0.001).
Figura 4. Curva de crecimiento D. communis. (cél mL -1 en función del tiempo de cultivo en días). Las letras indican diferencias significativas entre los días de cultivo según el test de Duncan (α 0.05).
En la tabla X se muestran los cálculos de la tasa de crecimiento correspondiente
a la fase exponencial, así como el tiempo de generación.
Tabla X. Tasa de crecimiento y tiempo de generación.
En la figura 5 se muestra el rendimiento de la biomasa del cultivo de D. communis
(mg L -1) con respecto al tiempo (d). Se observó que hubo un aumento gradual y
0
50
100
150
200
250
300
350
400
450
500
0 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17
Ce
l. m
L -1
x 1
00
00
Tiempo (d)
aa
b
c
d dd
dd
d d dd
c,d
d
c,dc,d c,d
Tasa de crecimiento Tiempo de generación
Tiempo (d) µ d-1 14.6 h
0-5 0.23

significativo (F4,10 ρ= 0.001) en los distintos días de cultivo; iniciando en el día 0
con 148.4 mg L-1 y para el día 17 se obtuvo 552.73 mg L-1.
Figura 5. Rendimiento de la biomasa del cultivo de D. communis mg L-1, en función del tiempo de cultivo (d). Las letras indican diferencias significativas entre los días de cultivo según el test de Duncan (α 0.05).
La figura 6 muestra la productividad de la biomasa (mg L-1 d-1), observando que
el valor mayor se obtuvo en el día 5 (53±2.3 mg L-1 d-1), disminuyendo
significativamente (F4,10 ρ= 0.001) hasta 32.53±0.79 mg L-1 d-1 en el día 17.
Figura 6. Productividad de la biomasa del cultivo de D. communis (mg L-1 d-1) en función del tiempo de cultivo (d). Las letras indican diferencias significativas entre los días de cosecha según el test de Duncan (α 0.05).
0
100
200
300
400
500
600
700
0 5 10 12 17
mg
L-1
Tiempo (d)
a
b
c
de
0
10
20
30
40
50
60
5 10 12 17
mg
L-1
d-1
Tiempo (d)
b
cc
d

Determinación de peso seco, orgánico e inorgánico
En la figura 7 se muestra que de un porcentaje del 100% que representa al peso
seco, en el tiempo 0 está conformado de un 80.43% ± 0.001 de peso orgánico
y un 19.57% ± .001 de peso inorgánico. Tanto el peso orgánico como el peso
inorgánico variaron significativamente (F4,10 ρ= 0.001) en el día 5, disminuyendo
a 75.86% ± 1.46 y aumentado a 24.14% ± 1.46, respectivamente. En siguientes
días hubo un aumento significativo en el peso orgánico, con 87.44% ± 1.11 en el
día 10, 89.47% ± 0.77 en el día 12 y 90.15% ± 1.04 en el día 17, y una
disminución significativa en el peso inorgánico con 12.56% ± 1.11 en el día 10,
10.53% ± 0.77 en el día 12 y 9.85% ± 1.04 en el día 17, en comparación con los
días 0 y 5, tanto el peso orgánico, como el inorgánico se mantuvieron sin
modificaciones significativas entre los días 10, 12 y 17.
Figura 7. Porcentaje de peso orgánico y peso inorgánico de la biomasa del cultivo de D. communis con respecto al tiempo (d). Las letras indican diferencias significativas entre los días de cosecha según el test de Duncan (α 0.05).
Determinación de nitratos
En la tabla XI se muestran los resultados de la cuantificación de nitratos del
sobrenadante del cultivo de D. communis obtenido al inicio del experimento (día
0
10
20
30
40
50
60
70
80
90
100
0 5 10 12 17
Po
rce
nta
je (
%)
Tiempo (d)
Orgánico
Inorgánico
C C C
AB
ab
c c c

0) y en los días de muestreo (5, 10, 12 y 17). La concentración inicial de nitratos
en el día 0 fue de 445 ± 9.9 µM. Se observó que para el día 5 hubo una
disminución drástica y significativa (F4,10 ρ= 0.001) donde se obtuvo 3.2 ± 0.8
µM, mientras que para los días 10 y 12, no se obtuvieron diferencias
significativas. Para el día 17 no fue posible detectar nitratos debido a que su
concentración en el medio fue muy baja o nula.
Tabla XI. Concentración de nitratos (µM) en medio de cultivo de D .communis, en los diferentes tiempos de muestreo (d). Las letras indican diferencias significativas entre los días de cosecha según el test de Duncan (α 0.05).
Tiempo (d) µM
0 445± 9.9a
5 3.2± 0.8b
10 4.3 ± 1.2c
12 1.5 ± 0.3b
17 ND
Determinación de la relación carbono nitrógeno intracelular
A continuación se muestra (Tabla XII) la relación carbono: nitrógeno intracelular
realizado con la biomasa obtenida en los días de cultivo 0, 5, 10, 12 y 17, en
donde podemos observar que la relación carbono: nitrógeno presentó aumento
significativo (F4,5 ρ= 0.001) entre los días 0 , 5 y 10 habiendo en el día 0, 8.25 ±
0.05 átomos de carbono por cada átomo de nitrógeno, y en el día 5, 12.4 ± 0.80
átomos de carbono por cada átomo de nitrógeno y en el día 10, 24.15 ± 0.05
átomos de carbono por cada átomo de nitrógeno. Entre los días 12 y 17 no se
presentó una disminución significativa en la relación C: N, habiendo 28.25 ±
0.45, y 29.70 ± 0.45 átomos de carbono por cada átomo de nitrógeno,
respectivamente.

Tabla XII. Relación carbono: nitrógeno intracelular de la biomasa del cultivo de D. communis. Las letras indican diferencias significativas entre los días de cosecha según el test de Duncan (α 0.05).
Tiempo (d) C:N
0 8.25± 0.05a
5 12.4± 0.80b
10 24.15± 0.05c
12 28.25± 0.45d
17 29.70± 0.30d
En la tabla XIII se muestra la relación intracelular del isotopo estable N15 y N14
realizada con biomasa obtenida en los días de cultivo 0, 5, 10, 12 y 17. En donde
se observa aumento significativo (F4,5 ρ= 0.001) entre el día 0 con 4.185 ± 0.025,
y el resto de los días de cultivo.
Tabla XIII. Relación 15N: 14N de la biomasa del cultivo de D. communis. Las letras indican diferencias significativas entre los días de cosecha según el test de Duncan (α 0.05).
Tiempo (d) 15 N:14N
0 4.185 ± 0.025a
5 5.975 ± 0.85b
10 5.625 ± 0.215b c
12 5.525 ± 0.075c
17 5.585 ± 0.045b c
En la tabla XIV se muestra la relación intracelular del isotopo estable C13 y C12
realizada con biomasa obtenida en los días de cultivo 0, 5, 10, 12 y 17. En donde
se observa disminución significativa (F4,5 ρ= 0.004) entre el día 0 con -10.73 ±
0.026, y el resto de los días de cultivo, día 5 con 9,76 ± 0.04, día 10 con -8,94 ±
0.03, día 12 con -9,1 ± 0.25 y día 17 con -9,72 ± 0.07.

Tabla XIV. Relación 13C: 12C de la biomasa del cultivo de D. communis. Las letras indican diferencias significativas entre los días de cosecha según el test de Duncan (α 0.05).
Tiempo (d) 13 C: 12C
0 -10.73 ± 0.26a
5 -9,76 ± 0.04b
10 -8,94 ± 0.03c
12 -9,1 ± 0.25c
17 -9,72 ± 0.07b
Determinación de proteínas
En la figura 8 se presenta el porcentaje de proteínas con respecto al peso seco
de la biomasa de D. communis. Al inicio del cultivo (día 0) fue de 10.71 % ± 1.13.
Para el día 5 fue de 8.95 % ± 0.35. Se observó una disminución significativa (F4,5
ρ= 0.002) en los días 10,12 y 17 con respecto a los días 0 y 5, pero se mantuvo
sin cambios significativos entre esos días.
Figura 8. Porcentaje de proteínas con respecto al peso seco de D. communis con respecto al tiempo de muestreo (0, 5, 10, 12 y 17 días). Las letras indican diferencias significativas entre los días de muestreo según el test de Duncan (α 0.05).
0
2
4
6
8
10
12
0 5 10 12 17
Po
rce
nta
je (
%)
Tiempo (d)
bb b
a
a

Determinación de carbohidratos
En cuanto al porcentaje de carbohidratos se observó un aumento significativo
(F4,5 ρ= 0.001). Al inicio del cultivo (día 0) fue de 9.83% ± 0.62. En el día 5 se
obtuvo 23.52% ± 3.35, mientras que el valor máximo fue de 34.86% ± 0.97 en
el día 10, manteniéndose sin cambios significativos entre los días 10,12 y 17
(Fig. 9).
Figura 9. Porcentaje de carbohidratos de D. communis con respecto al peso seco en los diferentes tiempos de muestreo (0, 5, 10, 12 y 17 días). Las letras indican diferencias significativas entre los días de muestreo según el test de Duncan (α 0.05).
Determinación de pigmentos, lípidos totales y ácidos grasos
En la figura 10 se muestra el perfil cuantitativo de pigmentos de D. communis. Al
inicio del cultivo (día 0) se encontraron clorofila a, clorofila b, luteína, feofitina b,
violaxantina, cis- neoxantina y feoforbides (feoforbide 1 y feoforbide 2). Siendo
la clorofila b el pigmento más abundante seguido de la luteína y la clorofila a. En
el día 5 se presentó una drástica disminución en la clorofila a, clorofila b,
feofitina b, la cual se agotó en una totalidad. En cuanto a la luteína y la
violaxantina, éstas disminuyeron significativamente (F4,5 ρ= 0.002). La cis-
neoxantina no presentó diferencias significativas, sin embargo los feoforbides 1
y 2 aumentaron significativamente. En el día 10, se observa una disminución
significativa en la concentración de los pigmentos feoforbides 1 y 2. En cuanto
al pigmento cis-neoxantina éste disminuyó significativamente con respecto al día
0. Para los días 12 y 17, estos no presentaron diferencias significativas con
respecto a los pigmentos del día 10.
0
10
20
30
40
50
0 5 10 12 17
Po
rcen
taje
(%
)
Tiempo (d)
a
b
cc c

Pigmentos
Figura 10 . Perfil cuantitativo de pigmentos de D. communis (µg mg-1) en los diferentes tiempos de muestreo del cultivo (0, 5, 10,
12 y 17 días). Las letras indican diferencias significativas según el test de Duncan (α 0.05).
0
0,5
1
1,5
2
2,5
3
3,5
Feo
forb
ide
1
Feo
forb
ide
2
cis-
ne
oxa
nti
na
vio
laxa
nti
na
Lute
ina
Clo
rofi
la b
Clo
rofi
la a
Feo
fiti
na
b
µg/
mg
0 5 10 12 17
aa
b
aa
ab
b
aa a b
ab
aaa a bb b b
a
b b b bb b bb bbbbbb
ba
a
a
b
50

51
Lípidos totales
En la figura 11 se muestra el porcentaje de lípidos de D. communis con respecto
al peso seco en los diferentes tiempos de muestreo (0, 5, 10, 12 y 17 días). El
porcentaje aumentó significativamente (F4,5 ρ= 0.05). Al inicio (día 0) con 19.55%
± 0.35. En el día 5 se obtuvo 33% ± 5.3, sin embargo disminuyó para el día 10
con 18.1% ± 2.9, pero manteniéndose sin diferencia significativa con respeto al
día 0. En los días 12 y 17 no se presentaron diferencias significativas con respeto
al día 10.
Figura 11. Porcentaje de lípidos de D. communis con respecto al peso seco en los diferentes tiempos de muestreo (0, 5, 10, 12 y 17 días). Las letras indican diferencias significativas entre los días de cosecha según el test Duncan (α 0.05).
Ácidos grasos
En la tabla XV se muestra una comparativa de los porcentajes de los ácidos
grasos saturados, monoinsaturados y poliinsaturados con respecto al total de
ácidos grasos en función del tiempo de cultivo (días) con cambios significativos
(F4,5 ρ= 0.001). Al inicio (día 0) podemos observar a los ácidos grasos saturados
con un 34.2% ± 0.04 y este fue aumentando encontrado su máximo el día 17
con 49.2% ± 0.10. En cuanto a los monoinsaturados estos aumentaron del inicio
(día 0) con 26.1 ± 1.00 a 32% ± 0.01 en el día 17. En cuanto a los poliinsaturados
0
5
10
15
20
25
30
35
40
45
0 5 10 12 17
Po
rcen
taje
(%
)
Tiempo (d)
a
b
aa,b a

52
estos disminuyeron del inicio (día 0) con 41.5% ± 0.21 al 18% ± 0.45 en el día
17.
Tabla XV. Porcentaje de ácidos grasos saturados, monoinsaturados y poliinsaturados con respecto a los ácidos grasos totales en función del tiempo de cultivo (días). Los valores son el promedio ± error estándar. Las letras indican diferencias significativas entre los días de cosecha según el test Duncan (α 0.05).
T (d) Saturados Monoinsaturados Poliinsaturados
0 34.2 ± 0.04a 26.1 ± 1.00a 41.5 ± 0.21a
5 29.6 ±0.40b 32.0 ± 0.30b 39.7 ± 1.44b
10 27.6 ± 0.10c 36.1 ± 1.56c 38.0 ± 0.47c
12 30.9 ± 0.38d 35.1 ± 1.53d 35.9 ± 2.13d
17 49.2 ± 0.10e 32.9 ± 0.01e 18.0 ± 0.45e
Los resultados de la cuantificación de ácidos grasos presentes en la microalga
Desmodesmus communis en los días de cosecha 0, 5, 10, 12 y 17 se muestran
en la figura 12, en donde podemos observar que el 14:0 (ácido miristico)
disminuyo de manera significativa del día 0 a los días 5, 10, 12 y 17
permaneciendo sin cambios significativos en estos últimos días. En cuanto a los
ácidos grasos 15:0, 17:0 y 17:1, la suma de estos no presentó variaciones
significativas entre ninguno de los días de cosecha. El 16:0 (ácido palmítico), no
presentó cambios significativos entre los días 0, 5,10 y 12, sin embargo en el día
17 se presentó un aumento significativo. En cuanto al 16:4 ω3, este fue
disminuyendo poco a poco habiendo diferencias significativas entre el día 0 y el
día 17. En cuanto al 18:0 (ácido esteárico), este se modificó aumentando
ligeramente pero de manera significativa en los días 10 y 12, disminuyendo para
el día 17 a una proporción similar a la que se presentó en el día 0. En cuanto al
18:1 ω9 (ácido oleico), este fue aumentando de manera significativa de los días
0 al 5, 5 al 10, entre en 10 y 12 no se observaron diferencias significativas, y en
el día 17 disminuyó de manera significativa con respecto a los días 10 y 12,
alcanzando un proporción similar a la que se presentó el día 5. En cuanto al 18:2
ω6 c (ácido linoleico), aumentó significativamente del día 0 al 10, y disminuyó
significativamente del día 10 al 17 encontrándose en este último día en su menor

53
proporción. En cuanto al 18:3 ω3 (ácido linolenico), este disminuyó
gradualmente entre los días, con diferencias significativas entre los días 0 y 5, y
entre los días 5 y 17, siendo este último día el de más baja proporción.
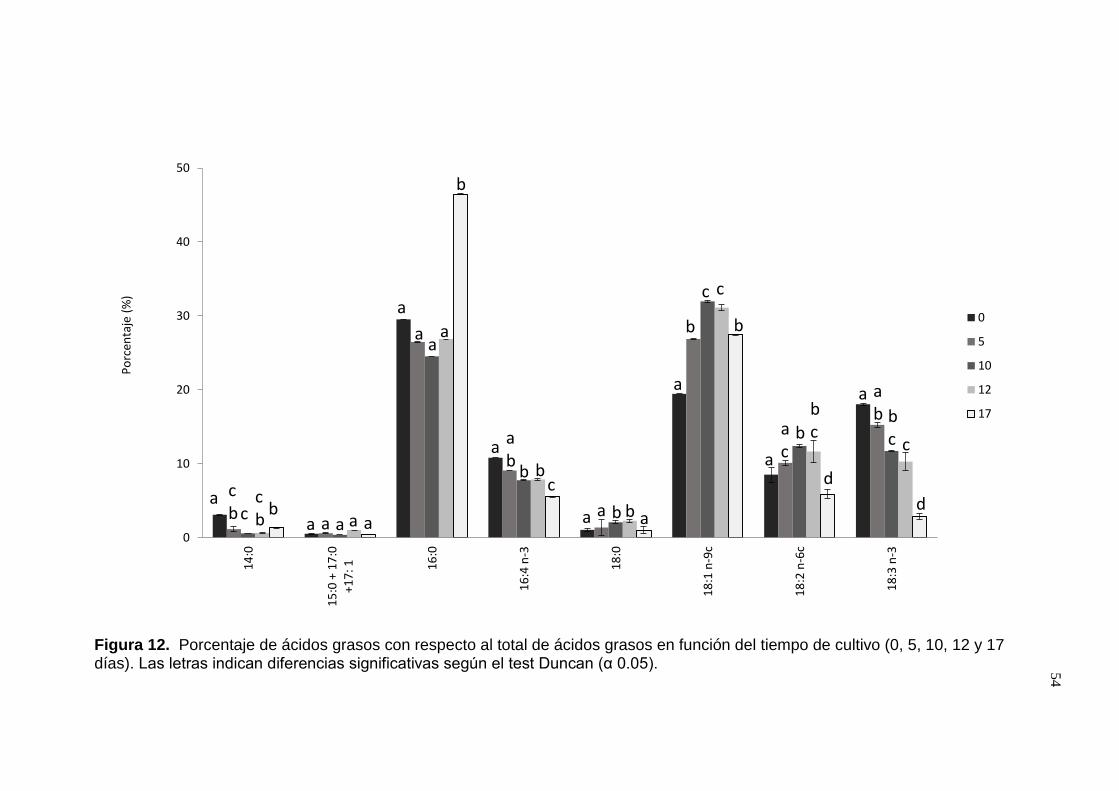
Figura 12. Porcentaje de ácidos grasos con respecto al total de ácidos grasos en función del tiempo de cultivo (0, 5, 10, 12 y 17 días). Las letras indican diferencias significativas según el test Duncan (α 0.05).
0
10
20
30
40
50
14
:0
15
:0 +
17
:0+
17
: 1 16
:0
16
:4 n
-3
18
:0
18
:1 n
-9c
18
:2 n
-6c
18
:3 n
-3
Po
rcen
taje
(%
)
0
5
10
12
17
a
ab
a a a ab
cbc
cb
a
aa
a
a
b
bb
a
c
a a ab b
b b
cc
a
d
c
a ab b
c
d
bcba
ca
54

55
Determinación de bacterias presentes
A continuación (Tabla XVI) se muestras las bacterias presentes en el cultivo
microalgal en todos los días de cosecha, capaces de crecer en agar nutritivo, las
cuales fueron enviadas a Macrogen para una secuenciación parcial del gen
ribosomal 16S cuya secuencia fue comparada mediante Blast con la secuencias
reportadas, arrojando una similitud de entre el 98 -100% con las bacterias
mostradas. De las cuales se destaca Rhizobium rosettiformans puesto que tiene
la capacidad de fijar nitrógeno, y Ancylobacter aquaticus por su capacidad para
degradar el 1,2-dicloroetano y producir haloalcano deshalogenasa.
Tabla XVI. Bacterias cultivables presentes en el cultivo de D. communis.
N.Cepa Bacteria %Similitud Descripción
Cepa 1 Blastomonas natatoria
100 α-proteobacteria fotoheterotrofica y
aerobia
Cepa 2 Rhizobium rosettiforma
ns
100 α-proteobacteria fotótrofo
Se reporta como fijadora de N
Cepa 3 Ancylobacter aquaticus
100 α-proteobacteria, degradar el 1,2-dicloroetano y
produce haloalcano deshalogenasa
Cepa 4 Bosea vaviloviae
100 α-proteobacteria fotótrofo
Cepa 5 Larkinella bovis
98 Flavobacteria
Aislada en fermento bovino en 2011

56
SSCP
En la figura 13 se muestra el gel de acrilamida obtenido posterior a la
electroforesis y tinción con plata realizada, mostrando la comunidad microbiana
presente. Podemos observar que el patrón de la comunidad bacteriana
permaneció muy similar en todos los días de cosecha.
Figura 13. Electroforesis en gel de acrilamida técnica SSCP. T0: día 0, T1: día 5, T2: día 10, T3: día 12 y T4: día 17. M: marcador bacteriano. En las numeraciones del 1-10 se señalan las bandas que se cortaron y se enviaron a secuenciar.
En la tabla XVII se muestran los resultados de las secuencias parciales a partir
de las bandas seleccionadas obtenidas del gel de acrilamida con la técnica
SSCP. Tres de las baterías, Rhizobium rosettiformans, Mesorhizobium
waimense y Microvirga lotononidis son bacterias con la capacidad de fijar
nitrógeno.
M

57
Tabla XVII. Bacterias identificadas mediante la técnica SSCP.
N. Banda
Bacteria % Similitud
Descripción
Banda 1
Rhizobium rosettiformans 100
α-proteobacteria fotótrofo
Se reporta como fijadora de N
Banda 2
Paracoccus fistulariae 93
α-proteobacteria fotótrofo
Banda 3
Bacillus aryabhattai 100
Producción de fitohormonas
Banda 4
Rhodobaca barguzinensis
99 α-proteobacteria
fotosintetica
Banda 5
Herbaspirillum chlorophenolicum 99
β-proteobacteria
degradadora de 4-clorofenol
Banda 6
Roseomonas tokyonensis strain
99 α-proteobacteria fotótrofo
Banda 7
Mesorhizobium waimense
98
Propuesta en 2015 nueva especie
α-proteobacteria
Se reporta como fijadora de N
Banda 8
Blastomonas natatoria 100
α-proteobacteria fotoheterotrofica y aerobia
Banda 9
Microvirga lotononidis
83
α-proteobacteria
Se reporta como fijadora de N
Banda 10
Solitalea canadensis 90
Bacteroidete Heterotrófico Reduce nitratos

58
Determinación de la longitud de los telómeros
De la determinación de los telómeros se observó que la longitud de estos se
mantuvo constante en todos los días de cosecha, es decir no hubo acortamiento
en los telómeros comparando entre el día 0 y el resto de los días de cosecha
(Fig. 14b). En la figura 14a se puede observar el producto de la digestión del
ADN total con las endonucleasas y en la figura 14c se puede observar que la
trasferencia de ADN a la membrana de nylon se realizó de la manera adecuada,
porque no quedó ADN en el gel de agarosa.
Figura 14. Técnica TRF. En la figura 14a: electroforesis en agarosa de las digestiones enzimáticas, donde se muestra en el carril M el marcador de peso molecular, de los carriles 0-17 representando respectivamente los días de cosecha 0, 5, 10, 12 y 17. 14b: autor radiografía de la membrana de nylon hibridada con la sonda telomérica donde se muestra en el carril M el marcador de peso molecular, de los carriles 0-17 representando, respectivamente, los días de cosecha 0, 5, 10, 12 y 17. 14c gel de agarosa después de la trasferencia del ADN a la membrana de nylon.
500
1000 1500 2000 3000
pb M 0 5 10 12 17
A
M 0 5 10 12
C
M 0 5 10 12 17
500
1000 1500 2000 3000
B
pb

59
8. DISCUSIÓN
En la curva de crecimiento se presentó la fase de crecimiento exponencial y la
fase estacionaria. Coincidiendo el día de término de la fase exponencial con una
disminución casi en la totalidad de la concentración de nitrógeno en el medio, sin
embargo se comprobó que aún cuando el nitrógeno del medio de cultivo se
consumió, las células de la microalga almacenaron conservando una relación
C:N de 12.40 ± 0.8 que se presenta cuando no hay limitación de nitrógeno en el
fitoplancton (Bittar et al., 2013). Sin embrago, conforme transcurrieron los días
el nitrógeno almacenado se redujo significativamente hasta encontrarse
deficiente también a nivel intracelular, llegando a una relación de C:N de
29.70±0.30, dicha deficiencia se ha demostrado también en diatomeas con una
relación C:N de 17±1 cuando la concentración de nitrógeno es 4 μM (Bittar et al.,
2013; Grobbelaar, 2010).
La condición de estrés ocasionada por el agotamiento del nitrógeno en el medio
de cultivo provocó una disminución en el porcentaje de proteínas con respecto a
la biomasa seca al final de la fase exponencial, ya que el nitrógeno limitó la
formación de aminoácidos que son las unidades básicas de las proteínas, las
que están involucradas en muchos aspectos del crecimiento, incluyendo
procesos estructurales y metabólicos (Doucette y Harrison, 1991; Li et al., 2012;
Simionato et al., 2013).
En cuanto a los carbohidratos, aumentaron en respuesta a la reducción de
nitrógeno en el medio; dichos resultados coinciden con los hallazgos de Coruzzi
y Zhou (2001) quienes demostraron que la asimilación del nitrógeno es un
proceso íntimamente conectado al metabolismo del carbono y dado que en los
organismos fotosintéticos, el carbono asimilado se reparte entre la formación de
carbohidratos y la síntesis de aminoácidos, al disminuir la síntesis de proteínas
y aminoácidos, la vía del metabolismo del carbono se redirigió a la síntesis de
carbohidratos (Coruzzi y Zhou, 2001).
La capacidad de las microalgas para sobrevivir en diversas condiciones
extremas se refleja en la enorme diversidad y a veces en su patrón inusual de
los lípidos obtenidos (Sato et al., 2000). Asimismo, las microalgas incrementan

60
su contenido de lípidos al ser sometidas a condiciones de estrés, en particular
bajo restricciones de nutrientes como el nitrógeno (Gonzales y Magallanes,
2012).
Los resultados obtenidos en la presente investigación indican que mientras la
cantidad de lípidos totales se incrementó significativamente como respuesta a la
limitación de nitrógeno en el día 5, la tasa de crecimiento o producción de
biomasa disminuyó, lo cual no es rentable cuando se quiere tener una alta
productividad de aceites usando microalgas oleaginosas. Estos resultados
concuerdan con los obtenidos por Huntley y Redalje (2007).
En cuanto a la proporción de ácidos grasos se observó que la disminución de la
concentración de nitrógeno provocó una disminución en la proporción de ácidos
grasos polinsaturados y un aumento en los ácidos grasos saturados y
monoinsaturados. Este comportamiento es de gran interés ya que los ácidos
grasos saturados y monoinsaturados pueden ser utilizados en la industria de los
biocombustibles, dado que los triglicéridos ricos en ácidos grasos poliinsaturados
son menos deseables para la producción de biodiesel, pues disminuyen el
número de cetano y la estabilidad a la oxidación del combustible, mientras que
triglicéridos ricos en ácidos grasos saturados (C16 y C20) y monoinsaturados
(C18:1, ácido oleico), resultan ideales para la producción de biodiesel
(Imahara et al., 2006; Meng et al., 2009, Ramos et al., 2009)
En cuanto a la acumulación de carotenoides secundarios, en las microalgas
cuando son cultivadas en medios con limitación del nitrógeno, se genera también
una disminución del contenido de clorofila de las células (Richmond y Hu, 2013).
En este trabajo, los pigmentos presentes al inicio del cultivo (luteína, clorofila a y
b, neoxantina entre otras) disminuyeron con respecto al tiempo hasta agotarse,
observando en esa etapa, la presencia de feoforbides, que son las formas
degradadas de la clorofila. Al degradarse la clorofila (a partir del día 5), no hubo
fotosíntesis, por lo que la microalga desvió su metabolismo. De manera
fotoautotrófica, las microalgas utilizan la luz como fuente de energía y el
CO2 como fuente de carbono para obtener energía química a través de la
fotosíntesis (Li et al., 2009); mientras que heterotróficamente , las microalgas en
ausencia de luz utilizan solo compuestos orgánicos como fuente de carbono y

61
energía, (Chojnacka y Marquez-Rocha, 2004) mixotróficamente, utilizan carbono
orgánico e inorgánico como fuente de carbono y captan la energía de la luz y del
carbono orgánico (Chen et al., 2017). La principal diferencia entre cultivos
mixotróficos y fotoheterotróficos es que estos últimos requieren de luz para la
incorporación del carbono orgánico. Por lo tanto, este tipo de cultivo necesita de
luz y azúcares al mismo tiempo, la utilización de este tipo de cultivo para la
producción de lípidos en microalgas es poco común. El cultivo mixotrófico
también ha mostrado ser una buena estrategia para aumentar la productividad
de lípidos (Cheirsilp y Torpee, 2012; Liang et al., 2009; Yeh y Chang, 2012)
Probablemente Desmodesmus communis bajo las condiciones experimentales
de este trabajo, modifico su nutrición hacia la mixotrofía, después de terminada
la fase exponencial. Por otro lado, la interacción con las bacterias presentes en
el cultivo, sugieren un apoyo para permanecer en fase estacionaria durante el
resto del experimento, ya que las bacterias podrían estar proporcionando
nutrientes (Watanabe et al., 2005). Se presentó un patrón muy similar de
comunidad bacteriana durante el metabolismo fotosintético y mixotrófico, lo que
puede indicar que los nutrientes proporcionados son necesarios en ambos
metabolismos. En cuanto a Rhizobium rosettiformans, Mesorhizobium waimense
y Microvirga lotononidis que se reporta son capaces de fijar nitrógeno, pudieran
ayudar a mitigar el estrés por consumo de nitrógeno. A pesar de que la actividad
de la nitrogenasa se vea inactivada en ambientes aerobios, los microorganismos
que fijan nitrógeno en ambientes aeróbicos han desarrollado diferentes
adaptaciones metabólicas y estructurales para poder llevar a cabo la fijación de
nitrógeno en un ambiente oxidante (Atlas y Bartha, 1992; Brock et al, 1994). El
crecimiento de la microalga también puede verse estimulado por la bacteria
Bacillus aryabhattai ya que en trabajos anteriores se comprobó la producción de
fitohormonas como ácido abscísico y ácido indólico, por lo que se considera un
recurso valioso para la incorporación en biofertilizantes (Park et al., 2017) . Las
bacterias presentes en el cultivo también pueden jugar un papel distinto al de
proporcionar elementos nutricionales y jugar un rol protector, lo que podría ser el
caso de Ancylobacter aquaticus. Esta bacteria es productora de haloalcano
deshidrogenasa una enzima con potencial uso biotecnológico, que en conjunto
con las microalgas, puede potencializar su eficacia en biorremediación, ya que

62
varios compuestos halogenados son subproductos industriales ambientalmente
tóxicos, y se ha sugerido que las deshalogenasas de haloalcano pueden ser
catalizadores útiles para su biodegradación, con posibles aplicaciones en la
biorremediación (Hesseler et al., 2011), como ocurre con Herbaspirillum
chlorophenolicum, una bacteria capaz de degradar 4-clorofenol, comúnmente
usado en herbicidas y pesticidas (Im et al., 2004). Del total de 13 bacterias
identificadas, 9 pertenecen al grupo de α-proteobacterias, un bacteroide y una β-
proteobacteria, siendo las primeras dos una clase de bacterias que se ha
reportado en estudios previos que es abundante en la relación entre bacterias y
diatomeas, y la última presente en menor medida. Las proteobacterias y los
bacteroides son los principales filos bacterianos heterotróficos asociados en
microalgas (Amin et al., 2012).
La propuesta de la fijación de nitrógeno por las bacterias se respalda en que la
proporción cuantificada intracelularmente de la relación isotópica 15N:14N en el
tiempo 0 fue 4.185 ±0.025 aproximado a los valores reportados con anterioridad
para fitoplancton marino de 5.1 ± 0.6 (Carreón-Palau et al., 2013) , y en el tiempo
5 fue de 5.975 ± 0.85, mostrando un aumento significativo
Se ha demostrado que la composición isotópica de un organismo está
determinada por el valor isotópico del alimento que consume (Deniro y Epstein,
1981), indicando con esto que la fuente de nitrógeno utilizada por la microalga
cambió, y dado que los organismos consumidores se enriquecen de los isótopos
como el 15N (Martínez del Rio et al., 2009), al enriquecerse sugiere que utiliza
una fuente de nitrógeno proveniente de otro organismo. La relación isotópica de
13C también se vio modificada, siendo menos negativa el día 0 con -10.73 ±
0.026, y valor menos negativo el día 10 con -8,94 ± 0.03, indicando que la fuente
de carbono cambió (Martínez del Rio et al., 2009).
Al agotarse los pigmentos con acción antioxidante y al aumentar el estrés por el
consumo de nitrógeno esperábamos que las especies reactivas de oxigeno
causaran daños, observando una disminución en la longitud de los telómeros, ya
que la enzima encargada de elongarlos y conservar su longitud puede verse
afectada por la disminución de nitrógeno al ser una ribonucleoproteína. Sin
embargo esto no sucedió, y la longitud de los telómeros permaneció igual

63
durante todas las etapas de cosecha, esto pudo deberse a mecanismos
antioxidantes no relacionados con los pigmentos, el sistema de defensa
antioxidante enzimático, dentro del que se encuentran las siguientes enzimas:
superóxido dismutasa (SOD). Esta enzima posee como función catalizar la
dismutación del RL O2 •- a H2O2, el cual es menos reactivo y puede llegar a ser
degradado por otras enzimas como la catalasa o la glutatión peroxidasa
(Bandyopadhyay et al., 1998; Constanza et al., 2012) La catalasa es una
hemoproteína que se concentra principalmente en los peroxisomas y en las
mitocondrias, se encarga de catalizar la descomposición del peróxido de
hidrógeno en agua y oxígeno (Yu, 1994). Glutatión peroxidasa (GPx), es una
selenoproteína que en presencia de GSH como agente reductor, cataliza la
reducción del peróxido de hidrógeno y otros hidroperóxidos orgánicos en agua y
alcohol, respectivamente (Xu et al., 2010). Las enzimas anteriores no fueron
medidas en este trabajo, sin embargo las mencionamos ya que estas
representan una herramienta celular importante para mitigar el estrés oxidativo
(Valavanidis et al., 2006).

64
9. CONCLUSIONES
1. Se evaluó el consumo de nitrógeno en el medio de cultivo encontrado que
este se agotó al finalizar la fase exponencial, provocando en los siguientes
días de cultivo deficiencia de nitrógeno a nivel intracelular en la microalga
Desmodesmus communis.
2. Se evaluó el efecto por consumo de nitrógeno en el cultivo batch de la
microalga D. communis encontrando que al disminuir el nitrógeno del
medio de cultivo con respeto al tiempo, la tasa de crecimiento disminuyó,
entrando en la fase estacionaria. Este efecto se reflejó en la composición
bioquímica: las proteínas disminuyeron significativamente, mientras que
los carbohidratos aumentaron y lípidos totales aumentaron.
3. Se encontró que al disminuir el nitrógeno el perfil de ácidos graso se
modificó, incrementando los ácidos grasas saturados y disminuyendo los
poliinsaturados. Los más abundantes fueron 18:1 ω9cis y 16:0, los cuales
tienen potencial biotecnológico en el área de los biocombustibles. En
cuanto a los pigmentos se agotó de manera súbita la clorofila a y b,
encontrándose un aumento en los feoforbides.
4. Se determinó la presencia de bacterias asociadas al crecimiento de la
microalga D. communis en diferentes fases de crecimiento encontrándose
que las mismas permanecieron presentes de manera constante en todas
las fases, siendo las α-proteobacterias las más abundantes.
5. Se evaluó el efecto por consumo de nitrógeno en el medio de cultivo de
la microalga D. communis sobre la longitud de sus telómeros, encontrando
que a pesar de la disminución de nitrógeno y con esto el aumento del
estrés, la longitud no se vio modificada, sugiriendo que intervinieron otros
mecanismos antioxidantes para mantener la integridad de los telómeros.
A partir de los estudios realizados en esta tesis las perspectivas para este
trabajo a futuro se pueden orientar en 2 direcciones: complementar el análisis
de bacterias para conocer de qué manera interactúan con la microalga D.
communis, y medir las especies reactivas de oxigeno así como la actividad
antioxidante enzimática para complementar el estudio.

65
10. LITERATURA CITADA
Amin, S.A., M.S. Parker, E.V. Armbrust. 2012. Interactions between diatoms and
bacteria. MMBR. 76(3):667-84.
Arias-Peñaranda, M.T., E. Cristiani-Urbina, C. Montes-Horcasitas, F. Esparza-
Garcıa, G. Torzillo, R.O. Cañizares-Villanueva. 2013. Scenedesmus
incrassatulus CLHE-Si01: A potential source of renewable lipid for high
quality biodiesel production. BIOREOURCE TECHNOL. 140:158-164.
Arias Peñaranda, M.T., R.O. Cañizares Villanueva, A. de J. Martínez Roldán.
2013. Procucción de biodisel a partir de microalgas: parámetros del cultivo
que afectan la procucción de lípidos. ACTA BIO COLOMB. 18(1):43-68
Arredondo-Vega, B.O, D. Voltolina. 2017. Concentracion, Recuento Celular y
Tasa de Crecimiento En: Arredondo-Vega B.O. Métodos y Herramientas
Analíticas en la Evaluación de la Biomasa Microalgal pp44-50.
Ashen, J.B. L.J. Goff. 2000. Molecular and ecological evidence for species
specificity and coevolution in a group of marine algal-bacterial symbioses.
APPL ENVIRON MICROB. 66(7):3024-30.
Atlas, R.M, R. Bartha. 1992. Hydrocarbon Biodegradation and Oil Spill
Bioremediation. En: Marshall K.C. (eds) Advances in Microbial Ecology. ADV
MICROB ECOL. 12:287-338.
Azam, F. 1998. OCEANOGRAPHY: Microbial Control of Oceanic Carbon Flux:
The Plot Thickens. SCIENCE. 280(5364):694-696.
Bandyopadhyay, U., D. Das, R. K Banerjee. 1998. Reactive oxygen species:
Oxidative damage and pathogenesis. CURR SCI INDIA. 77:658-666
Beer, L.L., E.S. Boyd, J.W. Peters, M.C. Posewitz. 2009. Engineering algae for
biohydrogen and biofuel production. CURR OPIN BIOTECH. 20(3):264-271.
Ben-Amotz, A., A. Katz, M. Avron. 1982. Accumulation of ß carotene in
halotolerant algae : purification and characterization of carotene-rich
globules from Dunaliela bardawil (CHLOROPHYCEAE). J PHICOL.

66
18(4):529-537.
Benov, L., A.F. Beema. 2003. Superoxide-dependence of the short chain sugars-
induced mutagenesis. FREE RADIC BIOL MED. 34(4):429-33
Bhosale, P. 2004. Environmental and cultural stimulants in the production of
carotenoids from microorganisms. APPL MICROBIOL BIOTECHNOL.
63(4):351-361.
Bitog, J.P., I.-B. Lee, C.-G. Lee, K.-S. Kim, H.-S. Hwang, S.-W. Hong, I.-H. Seo,
K.-S. Kwon, E. Mostafa. 2011. Application of computational fluid dynamics
for modeling and designing photobioreactors for microalgae production: A
review. COMPUT ELECTRON AGR. 76(2):131-147.
Bittar, T.B., Y. Lin, L.R. Sassano, B.J. Wheeler, S.L. Brown, W.P. Cochlan, Z.I.
Johnson. 2013. Carbon allocation under light and nitrogen resource
gradients in two model marine phytoplankton. J PHICOL. 49(3):523-535.
Blackburn, E.H. 1991. Structure and function of telomeres. NATURE.
350(6319):569-573.
Borowitzka, M.A. 2016. Algal Physiology and Large-Scale Outdoor Cultures of
Microalgae. En: Borowitzka M., Beardall J., Raven J. (eds) The Physiology
of Microalgae. Springer International Publishing Cham. J APPL PHYCOL.
6:601-652.
Borowitzka, M.A., L.J. Borowitzka. 1988. Micro-algal biotechnology. Cambridge
University Press, Cambridge, 1988. pp. 477
Breen, A.P., J.A. Murphy. 1995. Reactions of oxyl radicals with DNA. FREE
RADIC BIOL MED. 18(6):1033-77.
Bumbak, F., S. Cook, V. Zachleder, S. Hauser, K. Kovar. 2011. Best practices in
heterotrophic high-cell-density microalgal processes: achievements,
potential and possible limitations. APPL MICROBIOL BIOTECHNOL.
91(1):31-46.
Bunker, V.W. 1992. Free radicals, antioxidants and ageing. MLS. 49(4):299-312.

67
Camargo, J.A., Á. Alonso. 2006. Ecological and toxicological effects of inorganic
nitrogen pollution in aquatic ecosystems: A global assessment. ENVIRON
INT. 32(6):831-849.
Caradonna, F., M. Mauro. 2016. Role of the antioxidant defence system and
telomerase in arsenic-induced genomic instability. MUTAGENESIS.
31(6):661-667.
Carlucci, A.F., S.B. Silbernagel, P.M. McNally. 1969. Influnce of temperature and
solar radiation on persistence of vitamin B 12 , thiamine , and biotin in
seawater. J PHICOL. 5(4):302-305.
Carocho, M., I.C.F.R. Ferreira. 2013. A review on antioxidants, prooxidants and
related controversy: Natural and synthetic compounds, screening and
analysis methodologies and future perspectives. FOOD CHEM TOXICOL.
51:15-25
Carreón-Palau, L., C.C. Parrish, J.A. del Angel-Rodríguez, H. Pérez-España, S.
Aguiñiga-García. 2013. Revealing organic carbon sources fueling a coral
reef food web in the Gulf of Mexico using stable isotopes and fatty acids.
LIMNOL. OCEANOGR.. 58(2):593-612.
Cheirsilp, B., S. Torpee. 2012. Enhanced growth and lipid production of
microalgae under mixotrophic culture condition: Effect of light intensity,
glucose concentration and fed-batch cultivation. BIOREOURCE TECHNOL.
110:510-516.
Chen, J., H. Cui, Y. Hou, Y. Cui, Y. Wang, Y. Xu, Q. Wang, S. Qin. 2017.
Molecular cloning, expression and characterisation of glucokinase gene from
the mixotrophic green alga Chlorella kessleri. PHYCOLOGIA. 56(5):477-
486.
Chojnacka, K., F.-J. Marquez-Rocha. 2004. Kinetic and Stoichiometric
Relationships of the Energy and Carbon Metabolism in the Culture of
Microalgae. BIOTECHNOL J. 3(1):21-34.
Constanza, L., C. Msc, M. María, M. Ariza. 2012. Estrés oxidativo: origen,
evolución y consecuencias de la toxicidad del oxígeno. NOVA 10:(18)135-

68
250.
Coruzzi, G.M., L. Zhou. 2001. Carbon and nitrogen sensing and signaling in
plants: emerging ‘matrix effects’. CURR OPIN PLANT BIOL. 4(3):247-253.
Croft, M.T., A.D. Lawrence, E. Raux-Deery, M.J. Warren, A.G. Smith. 2005.
Algae acquire vitamin B12 through a symbiotic relationship with bacteria.
NATURE. 438(7064):90-93.
D’Souza, Y., T.W. Chu, C. Autexier. 2013. A translocation-defective telomerase
with low levels of activity and processivity stabilizes short telomeres and
confers immortalization. MOL. BIOL. CELL. 24(9):1469-1479.
Dae-Hyun, C., R. Rishiram, H. Jina, L. Jimin, K. Byung-Hyuk, O. Hee-Mock, y K.
Hee-Sik 2014. Enhancing microalgal biomass productivity by engineering a
microalgal–bacterial community. BIORESOUR TECHNOL. 175 (2015):578–
585.
de-Bashan, L.E., Y. Bashan, M. Moreno, V.K. Lebsky, J.J. Bustillos. 2002.
Increased pigment and lipid content, lipid variety, and cell and population
size of the microalgae Chlorella spp. when co-immobilized in alginate beads
with the microalgae-growth-promoting bacterium Azospirillum brasilense.
CAN J MICROBIOL. 48(6):514-521.
Deniro, M.J., S. Epstein. 1981. Influence of diet on the distribution of nitrogen
isotopes in animals. GEOCHIM COSMOCHM AC. 45(3):341-351.
Dizdaroglu, M., P. Jaruga, M. Birincioglu, H. Rodriguez. 2002. Free radical-
induced damage to DNA: mechanisms and measurement. FREE RADIC
BIOL MED. 32(11):1102-15
Dizdaroglu, M., P. Jaruga. 2012. Mechanisms of free radical-induced damage to
DNA. FREE RADIC RES. 46(4):382-419.
Doucette, G.J., P.J. Harrison. 1991. Aspects of iron and nitrogen NUTRITION in
the red tide dinoflagellate Gymnodinium sanguineum. MAR BIOL.
110(2):165-173.
Droop, M.R. 1973. Some thoughts on nutrient limitation in algae. J PHICOL.

69
9(3):264-272.
Droop, M.R. 2007. Vitamins, phytoplankton and bacteria: symbiosis or
scavenging?. J PLANKTON RES. 29(2):107-113.
DuBois, M., K.A. Gilles, J.K. Hamilton, P.A. Rebers, F. Smith. 1956. Colorimetric
Method for Determination of Sugars and Related Substances. ANAL CHEM.
28(3):350-356.
Ethier, S., K. Woisard, D. Vaughan, Z. Wen. 2011. Continuous culture of the
microalgae Schizochytrium limacinum on biodiesel-derived crude glycerol for
producing docosahexaenoic acid. BIOREOURCE TECHNOL. 102(1):88-93.
Fischer, C.R., D. Klein-Marcuschamer, G. Stephanopoulos. 2008. Selection and
optimization of microbial hosts for biofuels production. METAB ENG.
10(6):295-304.
Foyer, C.H., G. Noctor. 2003. Redox sensing and signalling associated with
reactive oxygen in chloroplasts, peroxisomes and mitochondria. PHYSIOL
PLANTARUM. 119(3):355-364.
Fujita, M., Y. Fujita, Y. Noutoshi, F. Takahashi, Y. Narusaka, K. Yamaguchi-
Shinozaki, K. Shinozaki. 2006. Crosstalk between abiotic and biotic stress
responses: a current view from the points of convergence in the stress
signaling networks. CURR OPIN PLANT BIOL. 9(4):436-442.
Fulnečková, J., T. Hasíková, J. Fajkus, A. Lukešová, M. Eliáš, E. Sýkorová. 2012.
Dynamic Evolution of Telomeric Sequences in the Green Algal Order
Chlamydomonadales. GENOME BIOL EVOL. 4(3):248-264.
Gavia García, G., z H. González Martínez. 2015. Oxidative damage and
antioxidant defense in thymus of malnourished lactating rats. NUTRITION.
31(11-12):1408-1415.
Grobbelaar, J.U. 2010. Microalgal biomass production: challenges and realities.
PHOTOSYNTH RES. 106(1-2):135-144.
Guiry, M.D., G.M. Guiry. 2018. About AlgaeBase :: Algaebase. World-wide
electronic publication, National University of Ireland, Galway,

70
http://www.algaebase.org/about/p.
Guschina, I.A., J.L. Harwood. 2006. Lipids and lipid metabolism in eukaryotic
algae. PROG LIPID RES. 45(2):160-186.
Haines, K.C., R.R.L. Guillard. 1974. Growth of vitamin B 12- requiring marine
diatoms in mixed laboratory culture with vitamin B 12- producing marine
bacteria. J PHICOL. 10(3):245-252.
Halliwell, B. 1991. Reactive oxygen species in living systems: source,
biochemistry, and role in human disease. AM J MED. 91(3C):14S-22S.
Halliwell, B., S. Chirico. 1993. Lipid peroxidation: its mechanism, measurement,
and significance. AM J CLIN NUTR. 57(5 Suppl):715S-724S; discussion
724S-725S
Harun, R., M. Singh, G.M. Forde, M.K. Danquah. 2010. Bioprocess engineering
of microalgae to produce a variety of consumer products. RENEW SUST
ENERGREV. 14(3):1037-1047.
Hayflick, L. 1985. Theories of biological aging. EXP GERONTOL. 20(3-4):145-
159.
Hesseler, M., X. Bogdanović, A. Hidalgo, J. Berenguer, G.J. Palm, W. Hinrichs,
U.T. Bornscheuer. 2011. Cloning, functional expression, biochemical
characterization, and structural analysis of a haloalkane dehalogenase from
Plesiocystis pacifica SIR-1. APPL MICROBIOL BIOTECHNOL. 91(4):1049-
1060.
Ho, S.-H., W.-M. Chen, J.-S. Chang. 2010. Scenedesmus obliquus CNW-N as a
potential candidate for CO2 mitigation and biodiesel production.
BIOREOURCE TECHNOL. 101(22):8725-8730.
Holzinger, A., M.C. Allen, D.D. Deheyn. 2016. Hyperspectral imaging of snow
algae and green algae from aeroterrestrial habitats. J PHOTOCHEM
PHOTOBIOL. 162:412-420.
Hu, Q., M. Sommerfeld, E. Jarvis, M. Ghirardi, M. Posewitz, M. Seibert, A.
Darzins. 2008. Microalgal triacylglycerols as feedstocks for biofuel

71
production: perspectives and advances. PLANT J. 54(4):621-639.
Im, W.-T., H.-S. Bae, A. Yokota, S.T. Lee. 2004. Herbaspirillum chlorophenolicum
sp. nov., a 4-chlorophenol-degrading bacterium. J SYST EVOL
MICROBIOL. 54(3):851-855.
Imahara, H., E. Minami, S. Hari, S. Saka. 2007. Thermal stability of biodiesel in
supercritical methanol. FUEL .87(1):1-6
Imlay, J.A., S. Linn. 1988. DNA damage and oxygen radical toxicity. SCIENCE.
240(4857):1302-9.
Jasso, A., P. Piña, M. Valdez, J.F. Soto, Arzola, M. González, Guerrero, Ibarra.
2012. La importancia de las microalgas. CONABIO. Biodiversitas. 103:1-5.
Jeyapalan, J.C., J.M. Sedivy. 2008. Cellular senescence and organismal aging.
MECH AGEING DEV. 129(7-8):467-474.
Jialal, I., E.P. Norkus, L. Cristol, S.M. Grundy. 1991. β-Carotene inhibits the
oxidative modification of low-density lipoprotein. BBA - LIPID LIPID METAB.
1086(1):134-138.
Krivtsov, V., D.C. Sigee. 2005. Importance of Biological and Abiotic Factors for
Geochemical Cycling in a Freshwater Eutrophic Lake. BIOGEOCHEMISTR.
74: 205-230.
Lamers, P.P., C.C.W. van de Laak, P.S. Kaasenbrood, J. Lorier, M. Janssen,
R.C.H. De Vos, R.J. Bino, R.H. Wijffels. 2010. Carotenoid and fatty acid
metabolism in light-stressed Dunaliella salina. BIOTECHNOL BIOENG.
106(4):638-648.
Lampert, W., U. Sommer. 2007. Limnoecology. Oxford University Press Inc, New
York, United States. 336p.
Lang, I., L. Hodac, T. Friedl, I. Feussner. 2011. Fatty acid profiles and their
distribution patterns in microalgae: a comprehensive analysis of more than
2000 strains from the SAG culture collection. BMC Plant Biology. 11(1):124.
Lavín, P.L., S.O. Lourenço. 2005. An evaluation of the accumulation of

72
intracellular inorganic nitrogen pools by marine microalgae in batch cultures.
BRAZ J OCEANOGR. 53(1-2):55-68.
Lazar, T. 2003. Taiz, L. and Zeiger, E. Plant physiology. 3rd edn. Annals of
Botany. 91(6):750-751.
Lee, R.E. 2008. Phycology. Cambridge University Press. 4th edition. Colorado
State University. 560p
Lenaz, G. 1998. Role of mitochondria in oxidative stress and ageing. BIOCHIM
BIOPHYS ACTA. 1366:53-67.
Li, Y., X. Fei, X. Deng. 2012. Novel molecular insights into nitrogen starvation-
induced triacylglycerols accumulation revealed by differential gene
expression analysis in green algae Micractinium pusillum. BIOMASS
BIOENERGY. 42:199-211.
Li, Y., J. Huang, G. Sandmann, F. Chen. 2009. High-Light and Sodium Chloride
Stress Differentially Regulate the Bioynthesis of Astaxanthin in Chlorella
zofigiensis (Chlorophyceae). J PHICOL. 45(3):635-641.
Liang, Y., N. Sarkany, Y. Cui. 2009. Biomass and lipid productivities of Chlorella
vulgaris under autotrophic, heterotrophic and mixotrophic growth conditions.
BIOTECHNOL LETT. 31(7):1043-1049.
Lin, Q., J. Lin. 2011. Effects of nitrogen source and concentration on biomass
and oil production of a Scenedesmus rubescens like microalga.
BIOREOURCE TECHNOL. 102(2):1615-1621.
Lobo, V., A. Patil, A. Phatak, N. Chandra. 2010. Free radicals, antioxidants and
functional foods: Impact on human health. PHARMACOGN REV. 4(8):118
Lourenço, S.O., E. Barbarino, P.L. Lavín, U.M. Lanfer Marquez, E. Aidar. 2004.
Distribution of intracellular nitrogen in marine microalgae: Calculation of new
nitrogen to protein conversion factors. European J PHICOL. 39(1):17-32.
Lowry, O.H., N.J. Rosebrough, A.L. Farr, R.J. Randall. 1951. Protein
measurement with the Folin phenol reagent. J BIOL CHEM. 193(1):265-75.

73
Lu, W., Y. Zhang, D. Liu, Z. Songyang, M. Wan. 2013. Telomeres—structure,
function, and regulation. EXP CELL RES. 319(2):133-141.
Maldonado. J.M. 2008. Asimilación del nitrógeno y del azufre. En: Azcón-Biento
J. M. Talón (eds). Fundamento de fisiología vegetaly editorial McGraw- Hill.
Interamericana de España. Madrid. 287-304
Marsh, J.B., D.B. Weinstein. 1966. Simple charring method for determination of
lipids. J LIPID RES. 7(4):574-6.
Martínez del Rio, C., N. Wolf, S.A. Carleton, L.Z. Gannes. 2009. Isotopic ecology
ten years after a call for more laboratory experiments. BIOL REV. 84(1):91-
111.
Medvedev, Z.A. 1990. An attempt at a rational classification of theories of ageing.
BIOL REV. 65(3):375-398.
Meléndez-Martínez, A.J., I.M. Vicario, F.J. Heredia. 2007. Pigmentos
carotenoides: Consideraciones estructurales y fisicoquímicas. ALAN.
57(2):109-117.
Metting, F.B. 1996. Biodiversity and application of microalgae. J IND MICROBIOL
BIOTECHNOL. 17(5-6):477-489.
Mondello, C., A.I. Scovassi. 2004. Telomeres, telomerase, and apoptosis.
BIOCHEM CELL BIOL. 82(4):498-507.
Moo-Puc, R., D. Robledo, Y. Freile-Pelegrín. 2009. In vitro cytotoxic and
antiproliferative activities of marine macroalgae from Yucatán, México.
Ciencias marinas. 35(4):345-358.
Mouget, J., A. Dakhama, M.C. Lavoie, J. de la Noüe. 1995. Algal growth
enhancement by bacteria: Is consumption of photosynthetic oxygen
involved?. FEMS MICROBIOL ECOL. 18(1):35-43.
Norton, T.A., M. Melkonian, R.A. Andersen. 1996. Algal biodiversity.
PHYCOLOGIA. 35(4):308-326.
O’Brien, J.A., A. Vega, E. Bouguyon, G. Krouk, A. Gojon, G. Coruzzi, R.A.

74
Gutiérrez. 2016. Nitrate Transport, Sensing, and Responses in Plants. MOL
PLANT. 9(6):837-856.
Ohara, S., K. Fukami, Y. Ishida. 1993. Algal Effects on the Bacterial Assemblage
in Lake Biwa. JPn. J LIMNOL. 54(4):261-268.
Ohwada, K. 1973. Seasonal Cycles of Vitamin B12, Thiamine and Biotin in Lake
Sagami. Patterns of Their Distribution and Ecological Significance. INT REV
GESAMTEN HYDROBIOL. 58(6):851-871.
Ortiz-Marquez, J.C.F., M. Do Nascimento, M. de L.A. Dublan, L. Curatti. 2012.
Association with an ammonium-excreting bacterium allows diazotrophic
culture of oil-rich eukaryotic microalgae. APPL ENVIRON MICROB.
78(7):2345-52.
Park, Y.-G., B.-G. Mun, S.-M. Kang, A. Hussain, R. Shahzad, C.-W. Seo, A.-Y.
Kim, S.-U. Lee, K.Y. Oh, D.Y. Lee, I.-J. Lee, B.-W. Yun. 2017. Bacillus
aryabhattai SRB02 tolerates oxidative and nitrosative stress and promotes
the growth of soybean by modulating the production of phytohormones. PloS
one. 12(3):e0173203.
Peña-Castro, J.M., F. Martınez-Jerónimo, F. Esparza-Garcıa, R.O. Cañizares-
Villanueva. 2004. Heavy metals removal by the microalga Scenedesmus
incrassatulus in continuous cultures. BIOREOURCE TECHNOL. 94(2):219-
222.
Perales-Vela, H.V., J.M. Peña-Castro, R.O. Cañizares-Villanueva. 2006. Heavy
metal detoxification in eukaryotic microalgae. CHEMOSPHERE. 64(1):1-10.
Peterson, B. J., B. Fry. 1987. Stable isotopes in Ecosystem Studies. ANNU REV
ECOL SYST. 18: 293-320
Pietta, P.G. 2000. Flavonoids as antioxidants. J NAT PROD. 63(7):1035-42.
Pirastru, L., M. Darwish, F.L. Chu, F. Perreault, L. Sirois, L. Sleno, R. Popovic.
2012. Carotenoid production and change of photosynthetic functions in
Scenedesmus sp. exposed to nitrogen limitation and acetate treatment. J
APPL PHYCOL. 24(1):117-124.

75
Prison, A., H. Lorenzen. 1966. Synchronized Dividing Algae. ANNU REV PLANT
PHYSIOL. 17(1):439-458.
Qin, S., G.-X. Liu, Z.-Y. Hu. 2008. The accumulation and metabolism of
astaxanthin in Scenedesmus obliquus (Chlorophyceae). PROCESS
BIOCHEM. 795–802.
Ramanan, R., B.-H. Kim, D.-H. Cho, H.-M. Oh, H.-S. Kim. 2016. Algae–bacteria
interactions: Evolution, ecology and emerging applications. BIOTECHNOL
ADV. 34(1):14-29.
Ratnam, D.V., D.D. Ankola, V. Bhardwaj, D.K. Sahana, M.N.V.R. Kumar. 2006.
Role of antioxidants in prophylaxis and therapy: A pharmaceutical
perspective. J CONTROL RELEASE. 113(3):189-207.
Ríos, M.D.C. 2003. El estrés oxidativo y el destino celular. Quimica viva. 2(1):1
Rouzic, B. Le, G. Bertru. 1997. Phytoplankton community growth in enrichment
bioassays: Possible role of the nutrient intracellular pools. ACTA OECOL.
18(2):121-133.
Ryther, J.H., R.R.L. Guillard. 1962. Studies of marine planktonic diatoms:II. use
of cyclotella nana hustedt for assays of vitamin B 12 in sea water. CAN J
MICROBIOL. 8(4):437-445.
Sahnoun, Z., K. Jamoussi, K.M. Zeghal. 1997. Free radicals and antioxidants:
human physiology, pathology and therapeutic aspects. THERAPIE.
52(4):251-70.
Sánchez, J.F., J.M. Fernández-Sevilla, F.G. Acién, M.C. Cerón, J. Pérez-Parra,
E. Molina-Grima. 2008. Biomass and lutein productivity of Scenedesmus
almeriensis: influence of irradiance, dilution rate and temperature. APPL
MICROBIOL BIOTECHNOL. 79(5):719-729.
Sapp, M., A.S. Schwaderer, K.H. Wiltshire, H.-G. Hoppe, G. Gerdts, A. Wichels.
2007. Species-Specific Bacterial Communities in the Phycosphere of
Microalgae?. MICROB ECOL. 53(4):683-699.
Sato, N., N. Murata. 1988. Membrane lipids. METHODS ENZYMOL. 167:251-

76
259.
Schumpert, C., J. Nelson, E. Kim, J.L. Dudycha, R.C. Patel. 2015. Telomerase
Activity and Telomere Length in Daphnia. PLOS ONE. 10(5):e0127196.
Schwieger, F., C.C. Tebbe. 1998. A new approach to utilize PCR-single-strand-
conformation polymorphism for 16S rRNA gene-based microbial community
analysis. APPL ENVIRON MICROB. 64(12):4870-6.
Scott, S.A., M.P. Davey, J.S. Dennis, I. Horst, C.J. Howe, D.J. Lea-Smith, A.G.
Smith. 2010. Biodiesel from algae: challenges and prospects. CURR OPIN
BIOTECH. 21(3):277-286.
Ševčíková, T., K. Bišová, M. Fojtová, A. Lukešová, K. Hrčková, E. Sýkorová.
2013. Completion of cell division is associated with maximum telomerase
activity in naturally synchronized cultures of the green alga Desmodesmus
quadricauda. FEBS Letters. 587(6):743-748.
Shen, Y., Z. Pei, W. Yuan, E. Mao. 2009. Effect of nitrogen and extraction method
on algae lipid yield. IJABE. 2(1):51-57.
Sieburth, J.M. 1971. An instance of bacterial inhibition in oceanic surface water.
MAR BIOL. 11(1):98-100.
Sieburth, J.Mc. 1959. Antibacterial Activity of Antarctic Marine Phytoplankton1.
LIMNOL. OCEANOGR.. 4(4):419-424.
Simionato, D., M.A. Block, N. La Rocca, J. Jouhet, E. Maréchal, G. Finazzi, T.
Morosinotto. 2013. The response of Nannochloropsis gaditana to nitrogen
starvation includes de novo biosynthesis of triacylglycerols, a decrease of
chloroplast galactolipids, and reorganization of the photosynthetic
apparatus. EUKARYOTIC CELL. 12(5):665-76.
Sorokin, Y.I. 1973. On the Feeding of Some Scleractinian Corals with Bacteria
and Dissolved Organic Matter. LIMNOL. OCEANOGR., 18, 380-385p.
Strickland, J., T. Parsons. 1972. A Practical Handbook of Seawater Analysis. J
FISH RES BOARD CAN. 157:310.

77
Tamiya, H. 1966. Synchronous Cultures Of Algae. PLANT PHYSIOL. 17:1-27.
Tan, Y., J. Lin. 2011. Biomass production and fatty acid profile of a Scenedesmus
rubescens-like microalga. BIOREOURCE TECHNOL. 102(21):10131-
10135.
Thompson, G.A. 1996. Lipids and membrane function in green algae. BIOCHIM
BIOPHYS ACTA. 1302(1):17-45.
Valavanidis, A., T. Vlahogianni, M. Dassenakis, M. Scoullos. 2006. Molecular
biomarkers of oxidative stress in aquatic organisms in relation to toxic
environmental pollutants. ECOTOXICOL ENVIRON SAF. 64(2):178-189.
Valko, M., M. Izakovic, M. Mazur, C.J. Rhodes, J. Telser. 2004. Role of oxygen
radicals in DNA damage and cancer incidence. MOL CELL BIOCHEM.
266(1-2):37-56
Vaulot, D., W. Eikrem, M. Viprey, H. Moreau. 2008. The diversity of small
eukaryotic phytoplankton (≤3 μm) in marine ecosystems. FEMS MICROBIOL
REV. 32(5):795-820.
Vilar-Rojas, C., A.M. Guzman-Grenfell, J.J. Hicks. 1996. Participation of oxygen-
free radicals in the oxido-reduction of proteins. ARCH MED RES. 27(1):1-6.
Vitòria, L., N. Neus Otero, A. Soler, À. Canals. 2004. Fertilizer Characterization:
Isotopic Data (N, S, O, C, and Sr). ENVIRON SCI TECHNOL. 38(12):3254-
3262.
Watanabe, K., N. Takihana, H. Aoyagi, S. Hanada, Y. Watanabe, N. Ohmura, H.
Saiki, H. Tanaka. 2005. Symbiotic association in Chlorella culture. FEMS
MICROBIOL ECOL. 51(2):187-196.
Wehr, J.D. 2007. Algae: Anatomy, Biochemistry, and Biotechnology by Barsanti;
Gualtieri, P. J PHICOL. 43(2):412-414.
Xin, L., H. Hong-ying, G. Ke, S. Ying-xue. 2010. Effects of different nitrogen and
phosphorus concentrations on the growth, nutrient uptake, and lipid
accumulation of a freshwater microalga Scenedesmus sp. BIOREOURCE
TECHNOL. 101(14):5494-5500.

78
Xu, J., Y.-S. Tian, R.-H. Peng, A.-S. Xiong, B. Zhu, X.-L. Hou, Q.-H. Yao. 2010.
Cyanobacteria MT gene SmtA enhance zinc tolerance in Arabidopsis. MOL
BIOL REP. 37(2):1105-1110.
Yeh, K.-L. J.-S. Chang. 2012. Effects of cultivation conditions and media
composition on cell growth and lipid productivity of indigenous microalga
Chlorella vulgaris ESP-31. BIOREOURCE TECHNOL. 105:120-127.
Yu, B.P. 1994. Cellular defenses against damage from reactive oxygen species.
PHYSIOL REV. 74(1):139-162.
Zakian, V.A. 1995. Telomeres: beginning to understand the end. SCIENCE (New
York, N.Y.). 270(5242):1601-7.
Zeiher, A.M., S. Dimmeler, J. Haendeler, J. Hoffmann, J.F. Diehl, M. Vasa, I.
Spyridopoulos. 2004. Replicative Senescence of Endothelial Cells
Antioxidants Inhibit Nuclear Export of Telomerase Reverse Transcriptase
and Delay Antioxidants Inhibit Nuclear Export of Telomerase Reverse
Transcriptase and Delay Replicative Senescence of Endothelial Cells. CIRC
RES. 94:768-775.
Von Zglinicki, T. 2002. Oxidative stress shortens telomeres. TRENDS BIOCHEM
SCI. 27(7):339-344.