
Efecto de la suplementación con harina de arroz y semilla ... · Efecto de la suplementación con...
120
Efecto de la suplementación con harina de arroz y semilla de algodón sobre las concentraciones de los ácidos transvaccénico y ruménico en la grasa láctea de vacas en pastoreo Gastón Adolfo Castaño Jiménez Universidad Nacional de Colombia Facultad de Medicina Veterinaria y de Zootecnia, Departamento de Producción Animal Bogotá D.C., Colombia 2013
Transcript of Efecto de la suplementación con harina de arroz y semilla ... · Efecto de la suplementación con...

Efecto de la suplementación con harina de arroz y semilla de algodón
sobre las concentraciones de los ácidos transvaccénico y ruménico
en la grasa láctea de vacas en pastoreo
Gastón Adolfo Castaño Jiménez
Universidad Nacional de Colombia
Facultad de Medicina Veterinaria y de Zootecnia, Departamento de Producción Animal
Bogotá D.C., Colombia
2013


Efecto de la suplementación con harina de arroz y semilla de algodón
sobre las concentraciones de los ácidos transvaccénico y ruménico
en la grasa láctea de vacas en pastoreo
Gastón Adolfo Castaño Jiménez
Tesis presentada como requisito parcial para optar al título de:
Magíster en Producción Animal
Línea de Profundización Nutrición de Rumiantes
Director:
Ph.D. Juan Evangelista Carulla Fornaguera
Codirectora:
Ph.D. Martha Lucía Pabón Restrepo
Línea de Investigación: Nutrición Animal
Grupo de Investigación: Nutrición Animal
Universidad Nacional de Colombia
Facultad de Medicina Veterinaria y de Zootecnia, Departamento de Producción Animal
Bogotá D.C., Colombia
2013


Dedicatoria y Lema
V
A mis padres, a mi esposa y a mi hermano.
“Nunca consideres el estudio como una
obligación, sino como una oportunidad para
penetrar en el bello y maravilloso mundo del
saber”.
Albert Einstein

VI
Efecto de la suplementación con harina de arroz y semilla de algodón sobre la concentración de los ácidos transvaccénico y ruménico
en la grasa láctea de vacas en pastoreo

Agradecimientos
VII
Agradecimientos
A los Doctores Juan Carulla y Martha Pabón directores de la investigación, por sus
valiosas orientaciones, apoyo y confianza.
Al Ministerio de Agricultura y Desarrollo Rural por el apoyo económico para la
elaboración de este trabajo.
Al Laboratorio de Nutrición Animal de la Facultad de Medicina Veterinaria y de Zootecnia
Unal, Sede Bogotá por el apoyo para realizar los análisis químicos.
Al Centro de Investigación Agropecuario Marengo Unal, Sede Bogotá por facilitar los
animales y el área necesaria para la fase experimental del trabajo.
Al Grupo de Investigación en Nutrición Animal de la Facultad de Medicina Veterinaria y
de Zootecnia Unal, Sede Bogotá por el apoyo durante la fase experimental y trabajo en
el laboratorio.

VIII
Efecto de la suplementación con harina de arroz y semilla de algodón sobre la concentración de los ácidos transvaccénico y ruménico
en la grasa láctea de vacas en pastoreo

Resumen
IX
Resumen
Se realizaron dos experimentos con el fin de determinar el efecto de suplementar con
harina de arroz (HA) o semilla de algodón (SA; experimento 1) y de suministrar dos
niveles de ácido linoleico (Li) proveniente de la mezcla de HA y SA (experimento 2)
sobre el contenido de los ácidos grasos transvaccénico (TVa; C18:1t11) y ruménico (Ru;
C18:2c9,t11) en la grasa láctea de vacas lecheras que pastan P. clandestinum. En el
primer experimento se evaluaron tres suplementos: uno control (C) con baja
concentración de Li y otros con HA o SA como la principal fuente de Li (1.59, 2.91 y 3.69
g de Li / 100 g de MS para los tratamientos C, HA y SA respectivamente); se utilizaron
seis vacas Holstein que fueron distribuidas a una réplica de un diseño experimental
Cuadrado Latino 3 x 3 doble (3 vacas x 3 periodos x 3 tratamientos x 2 cuadrados). En el
segundo experimento se evaluaron dos suplementos que contenían una mezcla de HA y
SA (relación 4.8 : 1) como la principal fuente de Li: uno con baja concentración de Li y
otro con alta (2.64 y 4.08 g de Li / 100 g de MS respectivamente); se utilizaron seis vacas
Holstein distribuidas según un diseño Crossover 6 x 2 x 2 x 2 (6 vacas x 2 periodos x 2
tratamientos x 2 ordenamientos). En cada experimento se emplearon periodos de 28 d
(20 d de acostumbramiento a las dietas experimentales y 8 d para la recolección de
muestras), los animales fueron ordeñados dos veces al día (05:00 y 14:00 h),
permanecieron con agua a voluntad, pastorearon en P. clandestinum y recibieron durante
cada ordeño 60 g de sal mineralizada, 3 kg de suplemento según el tratamiento y 5 g de
Cr2O3. Con la HA se produjo grasa láctea con mayor concentración de TVa (3.11 %;
p<0.01), Ru (1.41 %; p<0.05) y de ácidos grasos insaturados (34.9 %; p<0.01), se
presentó un mayor índice calculado de actividad de la Δ9desaturasa (0.37; p<0.01) y
menores índices de aterogenicidad (1.85; p<0.05) y trombogenicidad (3.09; p<0.01).
Cuando las vacas recibieron SA no se afectaron las concentraciones de TVa y Ru en la
grasa láctea, ni los índices de aterogenicidad, de trombogenicidad y de actividad de la
Δ9desaturasa, pero se obtuvieron menores índices Ru / TVa (0.39; p<0.01) y C14:1 /
C14:0 (0.08; p<0.05). En el experimento 2, al incrementar el nivel de Li no se afectaron
las concentraciones de TVa y Ru en la grasa láctea, pero aumentaron los ácidos grasos

X
Efecto de la suplementación con harina de arroz y semilla de algodón sobre la concentración de ácidos transvaccénico y ruménico
en la grasa láctea de vacas en pastoreo
insaturados (35.1 %; p<0.05) y se presentó un mayor índice calculado de actividad de la
Δ9desaturasa y menores índices de aterogenicidad y trombogenicidad (0.37, 1.79 y 3.22
respectivamente; p<0.05). Suplementar vacas lecheras que pastan P. clandestinum con
HA es una buena alternativa para producir leche con beneficios para la salud humana a
diferencia de la SA; por otro lado, al incrementar la concentración de Li utilizando una
mezcla de HA y SA como principal fuente lipídica de Li no logró incrementar el contenido
de TVa y Ru en la grasa láctea.
Palabras Clave: ácido linoleico, ácido linoleico conjugado, ácidos grasos lácteos, vaca
lechera
Abstract
Two experiments were conducted to determine the effect of supplementing the diet with
rice bran (RB) or cottonseed (CS; experiment 1), and supplementing the diet with two
levels of linoleic acid (Li) from the mixture of RB and CS (experiment 2) on transvaccenic
(TVa; C18:1t11) and rumenic (Ru; C18:2c9,t11) fatty acids content in milk fat from dairy
cows grazing on P. clandestinum. Experiment 1, three supplements were evaluated: a
control (C) with low concentration of Li and others with RB or CS as the main source of Li
(1.59, 2.91 and 3.69 g of Li / 100 g DM for treatments C, RB and CS respectively); we
used six Holstein cows distributed to one replica of a double Latin Square design 3 x 3.
Experiment 2, tested two supplements containing a mixture of RB and CS (4.8 : 1 ratio)
as the main source of Li: one with low concentration of Li and other with high level (2.64
and 4.08 g of Li/100 g DM respectively); we used six Holstein cows distributed in a
Crossover design 6 x 2 x 2. Periods of 28 d were used in each experiment (20 d of
adaptation to the experimental diets and 8 d for collection samples), animals were milked
twice a day (05:00 and 14:00 h), water ad libitum, grazed on P. clandestinum and
received 60 g of mineralized salt, 3 kg of supplement according to the treatment and 5 g
of Cr2O3 during each milking. With the RB produced more milk fat concentrations of TVa
(3.11 %, p<0.01), Ru (1.41 %, p<0.05) and unsaturated fatty acids (34.9 %, p<0.01), had
higher Δ9desaturase activity index (0.37, p<0.01), and lower atherogenicity (1.85,
p<0.05) and thrombogenicity (3.09, p<0.01) indexes. When cows received CS the milk fat
concentrations of TVa and ARu were not affected, likewise for atherogenicity,
thrombogenicity and activity of Δ9desaturase indexes, but lower Ru / TVa (0.39, p<0.01)
and C14:1 / C14:0 (0.08, p<0.05) rates were obtained. In experiment 2, increase level of

Contenido XI
Li not affect milk fat concentrations of TVa and Ru, but increased the unsaturated fatty
acids (35.1 %; p<0.05) and had higher Δ9desaturase activity index and lower of
atherogenicity and thrombogenicity indixes (0.37, 1.79 and 3.22 respectively, p <0.05).
Supplementing dairy cows grazing on P. clandestinum with RB is a good alternative to
produce milk with human health benefits in contrast to CS, on the other hand, by
increasing the concentration of Li using a mixture of RB and CS as the main source of Li
failed to increase the content TVa and Ru in milk fat.
Key Words: linoleic acid, conjugated linoleic acid, milk fatty acid, dairy cow.

XII
Efecto de la suplementación con harina de arroz y semilla de algodón sobre la concentración de ácidos transvaccénico y ruménico
en la grasa láctea de vacas en pastoreo

Contenido
XIII
Contenido
Pág.
Resumen ......................................................................................................................... IX
Lista de tablas ............................................................................................................ XVII
Lista de anexos ........................................................................................................... XIX
Lista de abreviaturas ................................................................................................... XXI
Introducción................................................................................................................... 23
1. Capítulo 1: Efectos de la suplementación con semilla de algodón y harina de arroz sobre las concentraciones de los ácidos transvaccénico (C18:1t11) y ruménico (C18:2c9,t11) en la grasa láctea de vacas en pastoreo Revisión ..................................................................................................................... 25
Resumen ................................................................................................................. 25 Abstract .................................................................................................................. 26 Abreviaturas. .......................................................................................................... 26 1.1 Introducción .................................................................................................... 26 1.2 Los ácidos trans-vaccénico y ruménico y su relación con la salud
humana ............................................................................................................ 27 1.3 Metabolismo de los lípidos en el rumen y su relación con los ácidos
trans-vaccénico y ruménico en la leche de vaca .......................................... 28
1.3.1 Lipólisis ............................................................................................... 29 1.3.2 Biohidrogenación ................................................................................ 29
1.4 La enzima Δ9-desaturasa y la síntesis endógena de ácido ruménico ......... 33
1.4.1 Actividad de la enzima Δ9-desaturasa ................................................. 33 1.4.2 Intermediarios de la biohidrogenación en el rumen que inhiben la
actividad de la enzima Δ9-desaturasa y la síntesis de grasa láctea ..... 33 1.4.3 Inhibición de la enzima Δ9-desaturasa a través de ácidos
ciclopropenos ...................................................................................... 35
1.5 Factores que afectan la concentración de los ácidos trans-vaccénico y ruménico en la grasa láctea ........................................................................ 35
1.5.1 Factores relacionados con la dieta ...................................................... 36 1.5.2 Factores relacionados con el animal ................................................... 40 1.5.3 Factores relacionados con los procesos post-ordeño .......................... 40

XIV
Efecto de la suplementación con harina de arroz y semilla de algodón sobre la concentración de ácidos transvaccénico y ruménico
en la grasa láctea de vacas en pastoreo
1.6 La harina de arroz y su potencial para incrementar la concentración de los ácidos trans-vaccénico y ruménico en la leche de vaca ...................40
1.7 La semilla de algodón y su potencial para incrementar la concentración de los ácidos trans-vaccénico y ruménico en la leche de vaca .............................................................................................................41
1.8 Conclusiones ...................................................................................................42 1.9 Referencias ......................................................................................................43
2. Capítulo 2: Efecto de la suplementación con harina de arroz o semilla de algodón sobre el contenido de los ácidos transvaccénico (C18:1t11) y ruménico (C18:2c9,t11) en la leche de vacas en pastoreo .......................................53
Resumen .................................................................................................................53 Abstract ...................................................................................................................54 Abreviaturas. ...........................................................................................................55 2.1 Introducción .....................................................................................................55 2.2 Materiales y Métodos ......................................................................................56
2.2.1 Localización .........................................................................................56 2.2.2 Periodo experimental y tratamientos ....................................................57 2.2.3 Animales y manejo...............................................................................58 2.2.4 Mediciones, toma de muestras y análisis de laboratorio ......................59 2.2.5 Cálculos ...............................................................................................62 2.2.6 Análisis estadístico ..............................................................................64
2.3 Resultados .......................................................................................................65
2.3.1 Consumo de alimento ..........................................................................65 2.3.2 Consumo de precursores de los ácidos trans-vaccénico y
ruménico ..............................................................................................66 2.3.3 Producción y calidad de la leche ..........................................................68 2.3.4 Perfil de ácidos grasos de la grasa láctea ............................................69 2.3.5 Peso vivo, condición corporal y fluido ruminal ......................................71
2.4 Discusión .........................................................................................................71
2.4.1 Consumo de alimento ..........................................................................71 2.4.2 Producción y calidad de la leche ..........................................................72 2.4.3 Perfil de ácidos grasos de la grasa láctea ............................................73 2.4.4 Actividad de la Δ9desaturasa ..............................................................76
2.5 Conclusiones ...................................................................................................77 2.6 Agradecimientos .............................................................................................78 2.7 Referencias ......................................................................................................78
3. Capítulo 3: Efecto de la utilización de dos niveles de inclusión de la mezcla harina de arroz y semilla de algodón sobre la concentración de los ácidos transvaccénico (C18:1t11) y ruménico (C18:2c9,t11) en la leche de vacas en pastoreo .....................................................................................................................87
Resumen .................................................................................................................87 Abstract ...................................................................................................................88 Abreviaturas. ...........................................................................................................89 3.1 Introducción .....................................................................................................89

Contenido XV
3.2 Materiales y métodos ..................................................................................... 90
3.2.1 Localización ........................................................................................ 90 3.2.2 Periodo experimental y tratamientos ................................................... 91 3.2.3 Animales y manejo .............................................................................. 92 3.2.4 Mediciones, toma de muestras y análisis de laboratorio ...................... 93 3.2.5 Cálculos .............................................................................................. 95 3.2.6 Análisis estadístico .............................................................................. 97
3.3 Resultados ...................................................................................................... 97
3.3.1 Consumo de alimento ......................................................................... 97 3.3.2 Consumo de precursores de los ácidos trans-vaccénico y
ruménico ............................................................................................. 99 3.3.3 Producción y calidad de la leche ....................................................... 100 3.3.4 Perfil de ácidos grasos de la grasa láctea ......................................... 101 3.3.5 Peso vivo y fluido ruminal .................................................................. 104
3.4 Discusión ...................................................................................................... 104
3.4.1 Consumo de alimento ....................................................................... 105 3.4.1 Producción y calidad de la leche ....................................................... 106 3.4.2 Perfil de ácidos grasos de la grasa láctea ......................................... 107
3.5 Conclusiones ................................................................................................ 109 3.6 Agradecimientos ........................................................................................... 109 3.7 Referencias ................................................................................................... 110
4. Conclusiones y recomendaciones ........................................................................ 117
4.1 Conclusiones ................................................................................................ 117 4.2 Recomendaciones ........................................................................................ 118
5. Anexos..................................................................................................................... 119

XVI
Efecto de la suplementación con harina de arroz y semilla de algodón sobre la concentración de ácidos transvaccénico y ruménico
en la grasa láctea de vacas en pastoreo

Lista de tablas
XVII
Lista de tablas
Pág.
Tabla 2-1: Composición de ingredientes y nutrientes de los suplementos alimenticios utilizados en el experimento 58
Tabla 2-2: Perfil de ácidos grasos (AG) de los suplementos alimenticios y del
forraje utilizado en el experimento 58
Tabla 2-3: Efecto del uso de suplementos con harina de arroz (HA) o semilla de algodón (SA) como las principales fuentes de ácido linoleico sobre el consumo de alimento de vacas lactantes que pastaban P. clandestinum 66
Tabla 2-4: Efecto del uso de suplementos con harina de arroz (HA) o semilla de
algodón (SA) como las principales fuentes de ácido linoleico sobre el consumo de los ácidos linoleico y linolénico, precursores de los ácidos trans-vaccénico y ruménico en la leche de vacas que pastaban P. clandestinum 67
Tabla 2-5: Efecto del uso de suplementos con harina de arroz (HA) o semilla de
algodón (SA) como las principales fuentes de ácido linoleico sobre la producción y calidad de la leche de vacas que pastaban P. clandestinum 68
Tabla 2-6: Efecto del uso de suplementos con harina de arroz (HA) o semilla de algodón (SA) como las principales fuentes de ácido linoleico sobre el perfil de ácidos grasos (AG) de la grasa láctea de vacas que pastaban P. clandestinum 70
Tabla 2-7: Efecto del uso de suplementos con harina de arroz (HA) o semilla de algodón (SA) como las principales fuentes de ácido linoleico sobre el peso, la condición corporal y el fluido ruminal de de vacas lactantes que pastan P. clandestinum 71
Tabla 3-1: Composición de ingredientes y nutrientes de los suplementos
alimenticios utilizados en el experimento 92
Tabla 3-2: Perfil de ácidos grasos (AG) de los suplementos alimenticios y del forraje utilizado en el experimento 92
Tabla 3-3: Efecto del uso de suplementos con dos niveles de ácido linoleico (bajo
y alto) y que contienen como principal fuente lipídica una mezcla de

XVIII
Efecto de la suplementación con harina de arroz y semilla de algodón sobre la concentración de ácidos transvaccénico y ruménico
en la grasa láctea de vacas en pastoreo
harina de arroz y semilla de algodón (relación 4.8 : 1) sobre el consumo de alimento de vacas lactantes que pastaban P. clandestinum 99
Tabla 3-4: Efecto del uso de suplementos con dos niveles de ácido linoleico (bajo
y alto) y que contienen como principal fuente lipídica una mezcla de harina de arroz y semilla de algodón (relación 4.8 : 1) sobre el consumo del ácido linoleico, ácido linolénico y precursores de los ácidos transvaccénico y ruménico en la leche de vacas que pastaban P. clandestinum 100
Tabla 3-5: Efecto del uso de suplementos con dos niveles de ácido linoleico (bajo
y alto) y que contienen como principal fuente lipídica una mezcla de harina de arroz y semilla de algodón (relación 4.8 : 1) sobre la producción y calidad de la leche de vacas que pastaban P. clandestinum 101
Tabla 3-6: Efecto del uso de suplementos con dos niveles de ácido linoleico (bajo
y alto) y que contienen como principal fuente lipídica una mezcla de harina de arroz y semilla de algodón (relación 4.8 : 1) sobre el perfil de ácidos grasos (AG) de la grasa láctea de vacas que pastaban P. clandestinum 103
Tabla 3-7: Efecto del uso de suplementos con dos niveles de ácido linoleico (bajo
y alto) y que contienen como principal fuente lipídica una mezcla de harina de arroz y semilla de algodón (relación 4.8 : 1) sobre el peso vivo y el fluido ruminal de vacas lactantes que pastan P. clandestinum 104

Lista de anexos
XIX
Lista de anexos
Pág.
Anexo 1. Características de los animales al iniciar el experimento y distribución de los animales a los tratamientos en el primer experimento 119
Anexo 2. Características de los animales al iniciar el experimento y distribución de
los animales a los tratamientos en el segundo experimento 120

XX
Efecto de la suplementación con harina de arroz y semilla de algodón sobre la concentración de ácidos transvaccénico y ruménico
en la grasa láctea de vacas en pastoreo

Lista de abreviaturas
XXI
Lista de abreviaturas
Abreviatura Término
A Suplemento con alto nivel de ácido linoleico (Experimento 2) Es Ácido esteárico AG Ácido graso ALC Ácido linoleico conjugado. Li Ácidolinoleico. Ln Ácido linolénico. Ol Ácido oleico. Ru Ácido ruménico. TVa Ácidotransvaccénico. B Suplemento con bajo nivel de ácido linoleico (Experimento 2) BH Biohidrogenación C Tratamiento control (Experimento 1) CNFn Carbohidratos no fibrosos corregidos por N. DHA Ácido docosahexaenoico. ENL Energía neta de lactancia. EPA Ácido eicosapentaenoico. FDAi Fibra detergente ácido indigerible. FDAn Fibra detergente ácida corregida pornitrógenoN. HA Harina de arroz. MS Materia seca. SA Semilla de algodón.

XXII
Efecto de la suplementación con harina de arroz y semilla de algodón sobre la concentración de ácidos transvaccénico y ruménico
en la grasa láctea de vacas en pastoreo

Introducción
23
Introducción
La grasa láctea posee una elevada concentración de ácidos grasos (AG) saturados y
contiene AG trans, lo cual ha conducido a cuestionar el consumo de productos lácteos
debido al efecto negativo de este tipo de AG sobre salud humana. Sin embargo, en la
leche se encuentran los AG transvaccénico (TVa; C18:1t-11) y ruménico (Ru; C18:2c-9, t-11)
que están relacionados con efectos benéficos para la salud del hombre, especialmente
por sus propiedades anticarcinogénicas. El TVa y el Ru son producidos como
intermediarios de la biohidrogenación de los AG insaturados en el rumen y el Ru puede
ser sintetizado endógenamente gracias a la actividad de Δ9-desaturasa sobre el TVa. Los
alimentos derivados de los rumiantes son la principal fuente de TVa y Ru.
La dieta es el mayor determinante de la concentración láctea de TVa y Ru. Los animales
bajo el sistema pastoril y que reciben suplementos ricos en AG insaturados pueden
producir leche con elevada concentración de estos AG. El efecto del forraje está
relacionado con el ácido linolénico, mientras que el de la suplementación con el ácido
linoleico (Li).
La producción de leche en el trópico alto colombiano se fundamenta en sistemas
pastoriles, en donde el kikuyo (Pennisetum clandestinum) es la gramínea predominante y
posee una concentración elevada de ácido linolénico. El pastoreo en P. clandestinum,
sumado al aporte de lípidos del suplemento (cerca del 67%), son características de la
ganadería de leche especializada en la sabana de Bogotá que indican un alto potencial
para producir leches ricas en TVa y Ru.
La harina de arroz y la semilla de algodón se utilizan frecuentemente en la elaboración de
suplementos alimenticios para vacas lecheras en Colombia y son fuente de AG
insaturados con elevada concentración de Li, lo cual sugiere que estos dos recursos
podrían ser empleados para producir leche con altos niveles de TVa y Ru.

24
Efecto de la suplementación con harina de arroz o semilla de algodón sobre el contenido de los ácidos transvaccénico y ruménico
en la leche de vacas en pastoreo
Se hipotetiza que la acumulación dentro de los contenidos ruminales Li induce la
activación de la vía metabólica que produce Ru y TVa como intermediarios de la
biohidrogenación a esteárico. Lo cual favorece la concentración de Ru en la leche, ya
sea por la incorporación directa de Ru proveniente del rumen o por la síntesis endógena
en la glándula mamaria a través de la acción de la enzima Δ9-desaturasa sobre el TVa.
Consecuentemente, se espera que al aumentar el aporte Li al suplementar con harina de
arroz o semilla de algodón se produzca un aumento Ru y TVa en la grasa láctea de
vacas que pastan P. clandestinum.
Se realizaron dos experimentos con el fin de determinar el efecto de suplementar con
harina de arroz o semilla de algodón (experimento 1) y de suministrar dos niveles de Li
proveniente de la mezcla de estos dos recursos (experimento 2) sobre el contenido de
TVa y Ru en la grasa láctea de vacas lecheras que pastan P. clandestinum.

Capítulo 1
25
1. Capítulo 1: Efectos de la suplementación con semilla de algodón y harina de arroz sobre las concentraciones de los ácidos transvaccénico (C18:1t11) y ruménico (C18:2c9,t11) en la grasa láctea de vacas en pastoreo Revisión
Gastón A. Castaño J.1,2, Martha L. Pabón R.1,3 y Juan E. Carulla F.1,4
1 Grupo de Investigación en Nutrición Animal. Universidad Nacional de Colombia, Sede Bogotá.
2 Candidato Magíster en Producción Animal. Universidad Nacional de Colombia, Sede Bogotá. E-mail: [email protected]
3 Profesora Titular. Departamento de Química. Facultad de Ciencias. Universidad Nacional de Colombia, Sede Bogotá. E-mail: [email protected]
4 Profesor Titular. Departamento de Producción Animal. Facultad de Medicina Veterinaria
y de Zootecnia. Universidad Nacional de Colombia, Sede Bogotá. E-mail:
Resumen
El creciente interés sobre el ácido ruménico (Ru) se atribuye a sus potenciales beneficios
sobre la salud, por lo que se buscan estrategias para incrementar los niveles de este
ácido graso en la carne y leche proveniente de los rumiantes. El Ru es producido como
intermediario en la biohidrogenación del ácido linoleico y también se sintetiza en la
glándula mamaria por la actividad de la enzima Δ9-desaturasa sobre el ácido trans-
vaccénico (TVa). En esta revisión se describen los ácidos TVa y Ru, así como su
importancia en la salud humana, el metabolismo de los lípidos en el rumen y su relación
con los ácidos TVa y Ru en la leche de vaca, y la síntesis de Ru en la glándula mamaria.
También se tratan los factores que aumentan la concentración de estos ácidos grasos y
las características de la harina de arroz y la semilla de algodón que potencialmente
podrían aumentar la concentración de TVa y Ru en la grasa láctea.

26
Efecto de la suplementación con harina de arroz o semilla de algodón sobre el contenido de los ácidos transvaccénico y ruménico
en la leche de vacas en pastoreo
Palabras clave: ácido linoleico, ácido linoleico conjugado, ácidos grasos lácteos, vaca
lechera.
Abstract
The growing interest in the rumenic fatty acid (Ru) is attributed to its potential health
benefits, so there is an interest in looking for strategies to increase the levels of this fatty
acid in meat and milk from ruminants. The Ru is produced as an intermediate in
biohydrogenation of linoleic acid and is also synthesized in the mammary gland by the
activity of Δ9-desaturase enzyme on trans-vaccenic fatty acid (TVa). This review
describes TVa and Ru fatty acids and their importance in human health, the rumen lipid
metabolism and its relationship with TVa and Ru acids in cow’s milk, and the synthesis of
Ru in mammary gland. It also discusses factors that increase the concentration of these
fatty acids and characteristics of rice bran and cottonseed that could potentially increase
the concentration of TVa and Ru in milk fat.
Key words: linoleic acid, conjugated linoleic acid, milk fatty acid, dairy cow.
Abreviaturas.
BH: Biohidrogenación. Es: ácido esteárico. AG: ácido graso. ALC: ácido linoleico
conjugado. Li: ácido linoleico. Ln: ácido linolénico. Ol: ácido oleico. Ru: ácido ruménico.
TVa: ácido trans-vaccénico. DHA: ácido docosahexaenoico. EPA: ácido
eicosapentaenoico.
1.1 Introducción
El ácido graso (AG) ruménico (Ru; C18:2c-9,t-11) forma parte del grupo de isómeros
conocidos como ácido linoleico conjugado (ALC; Ip et al., 1999) y los alimentos derivados
de los rumiantes son la principal fuente para el hombre (Khanal y Olson, 2004). El Ru es
producido como intermediario en la biohidrogenación (BH) del ácido linoleico (Li;
AbuGhazaleh, 2008) y también se sintetiza endógenamente gracias a la actividad de la
enzima Δ9-desaturasa sobre el ácido trans-vaccéncio (TVa; C18:1t-11), esta vía representa
entre el 75 al 90 % del total de Ru lácteo (Bauman et al., 2008). El creciente interés sobre
el Ru se debe a sus beneficios potenciales sobre la salud humana (Khanal y Olson,
2004) y por eso se buscan estrategias para incrementar los niveles de este AG en la

Capítulo 1
27
carne y leche de los rumiantes.
Existen diferentes factores que afectan la concentración de TVa y Ru en la grasa láctea,
su conocimiento es definitivo para establecer estrategias conducentes a incrementar
estos AG. En la presente revisión se describen los TVa y Ru, así como su importancia en
la salud humana, el metabolismo de los lípidos en el rumen y su relación con los ácidos
TVa y Ru en la leche de vaca y la síntesis de Ru en la glándula mamaria. También se
tratan los factores que afectan la concentración de estos AG y las características de la
harina de arroz y la semilla de algodón que potencialmente podrían aumentar la
concentración de TVa y Ru en la grasa láctea.
1.2 Los ácidos trans-vaccénico y ruménico y su relación con la salud humana
El Ru forma parte del grupo de isómeros conocidos como ALC, este último término se
refiere a la colección de isómeros posicionales y geométricos del AG octadecadienoico
(C18:2) con enlaces conjugados (Ip et al., 1999), que pueden adoptar la forma cis o trans
(Bauman et al., 2004) en diferentes posiciones de la cadena (Khanal y Olson, 2004). El
Ru es la estructura de ALC más común que existe en la naturaleza (Sanhueza et al.,
2002) y los alimentos derivados de los rumiantes son la principal fuente para el hombre
(Khanal y Olson, 2004).
En la leche existen más de 24 isómeros de ALC (Bauman et al., 2008), dentro de los
cuales el Ru y el C18:2t-7,c-9 constituyen la mayor proporción del ALC (Bauman et al.,
2004). El Ru representa del 80 al 90 % del ALC, el C18:2t-7,c-9 del 3 al 16 % y el C18:2t-
10,c-12 cantidades menores al 5 % (Khanal y Olson, 2004), mientras otras formas
representan menos del 1 % (Bauman et al., 2004).
El Ru es producido como intermediario en la BH del Li por las bacterias del rumen
(AbuGhazaleh, 2008) y es sintetizado en la glándula mamaria por la enzima Δ9-
desaturasa que actúa sobre el TVa (Bauman et al., 2008). El TVa representa alrededor
del 53 % del total de los isómeros trans C18:1 de la leche (Paradis et al., 2008) y el 90 %
de los dos isómeros C18:1t-10 y C18:1t-11 (Collomb et al., 2002). El TVa es producido como
intermediario de la BH del Li y Ln en el rumen, además se convierte en Ru en la
glándula mamaria. La relación de TVa : Ru en la grasa de la leche generalmente es 3 : 1

28
Efecto de la suplementación con harina de arroz o semilla de algodón sobre el contenido de los ácidos transvaccénico y ruménico
en la leche de vacas en pastoreo
(Bauman et al., 2008) y la proporción Ru / (TVa + Ru) tiende a permanecer estable
(Paradis et al., 2008).
El creciente interés por el Ru se atribuye a sus potenciales beneficios sobre la salud
humana, especialmente por tener propiedades anticarcinogénicas (Ip et al., 1999; Park et
al., 2001), antiaterogénicas, antidiabetogénicas (Houseknecht et al., 1998; Munday et al.,
1999), antiobesidad (Khanal y Olson, 2004), regulación del sistema inmune (Moya-
Camarena et al., 1999) y mejoramiento de la mineralización del hueso (Jensen, 2002).
Pariza et al. (1979) reportaron por primera vez el efecto anticancerígeno del Ru y la
Academia Nacional de Ciencias de Estados Unidos (NRC, 1996) concluye que "el ALC
es el único AG que ha mostrado sin dejar ninguna duda un efecto anticancerígeno en
animales experimentales". El consumo de mantequilla enriquecida con isómeros ALC
(incluyendo el ARu) es efectivo para reducir la incidencia de tumores cancerígenos en
ratas (Ip et al., 1999) y se ha encontrado un efecto del Ru en la inhibición de la formación
de papilomas de piel, cáncer de estómago, cáncer de colon y tumores mamarios en
modelos animales (Ip et al., 1996; 1999; Park et al., 2001).
El Ru disminuye el riesgo de ateroesclerosis y diabetes (Jensen, 2002). Estudios
realizados en animales han demostrado que el Ru disminuye los niveles de colesterol-
LDL y aumenta la relación colesterol-HDL / colesterol total (Munday et al., 1999). El Ru
reduce los niveles de leptina (Belury, 2002) y disminuye la producción de citoquinas (Yu
et al., 2002) lo cual podría explicar los efectos benéficos de este compuesto frente a la
diabetes y resistencia a la insulina.
El Ru estimula el sistema inmune (Jensen, 2002), aumentando la producción de
anticuerpos y modulando vías intracelulares (Moya-Camarena et al., 1999). Además,
tiene efectos benéficos sobre la formación del hueso (Jensen, 2002).
1.3 Metabolismo de los lípidos en el rumen y su relación con los ácidos trans-vaccénico y ruménico en la leche de vaca
La utilización de las grasas por parte de los rumiantes se caracteriza por eventos que
ocurren en el rumen antes de que éstos sean absorbidos en el intestino (Doreau y
Chilliard, 1997). La BH de los AG insaturados en el rumen produce marcadas diferencias

Capítulo 1
29
entre los perfiles de AG de la leche y el alimento (Enjalbert et al., 1998). Durante su
permanencia en el rumen, las grasas son transformadas tanto en cantidad como en
composición (Doreau y Chilliard, 1997). Los microorganismos ruminales transforman los
lípidos que llegan al rumen a través de dos procesos principales la lipólisis y la BH
(Jenkins et al., 2008). La presencia de isómeros de AG trans y de ALC es consecuencia
de la BH de los AG poliinsaturados por parte de las bacterias del rumen.
1.3.1 Lipólisis
Los ésteres de AG de la dieta (galactolípidos, fosfolípidos y triglicéridos) inicialmente son
hidrolizados en el rumen (Jensen, 2002), sólo los AG no esterificados de cadenas largas
pueden ser biohidrogenados en el rumen. Por lo tanto, los AG que se encuentran en tri-,
di- o monoglicéridos y monogaloctosil glicerol deben ser liberados a través de la lipólisis
antes de su BH en el rumen (Moate et al., 2008). Las enzimas producidas por bacterias
Anaerovibrio lipolytica y por protozoarios provocan la lipólisis, dichas enzimas que liberan
AG son lipasas, galactosidasas y fosfolipasas (Doreau y Chilliard, 1997), el Butyrivibrio
fibrisolvens tiene actividad estearasa. Los microorganismos del rumen son los principales
productores de lipasas; sin embargo, las lipasas de los forrajes también son importantes
(Jenkins et al., 2008). La lipólisis de los triglicéridos se produce por pasos, el triglicérido
es hidrolizado para producir un diglicérido y un AG no esterificado, posteriormente el
diglicérido es hidrolizado para producir un monoglicérido y un AG no esterificado y por
último el monoglicérido es hidrolizado hasta producir glicerol y un AG no esterificado
(Moate et al., 2008).
Las tasas de la lipólisis y de la BH dependen de la grasa liberada en el rumen y del pH
ruminal (Jenkins et al., 2008), los antibióticos y la disminución del pH reducen la lipólisis
(Doreau y Chilliard, 1997) y la BH (Dewhurst et al., 2006).
1.3.2 Biohidrogenación
Los AG insaturados pueden ser hidrogenados una vez que son liberados (Moate et al.,
2008), produciendo un descenso en los AG insaturados, un incremento en los AG
saturados (Emanuele et al., 1991) y de los isómeros trans y cis, principalmente de C18:1
(Jensen, 2002). La completa BH de los ácidos Li y Ln produce el ácido esteárico (Es) y
en dicho proceso se produce una isomerización inicial que da como resultado varios

30
Efecto de la suplementación con harina de arroz o semilla de algodón sobre el contenido de los ácidos transvaccénico y ruménico
en la leche de vacas en pastoreo
isómeros cis y trans (Doreau y Chilliard, 1997). La reacción de isomerización ocurre lejos
de cualquier grupo funcional y no requiere de cofactor (Jenkins et al., 2008).
Las bacterias del rumen son simbióticas ya que intercambian intermediarios de la BH
entre las poblaciones (Mosley et al., 2002) y no se reconocen especies simples de
bacterias por tener la capacidad de llevar a cabo todos los pasos de la BH (Mosley et al.,
2002). Las bacterias ruminales están agrupadas de acuerdo al sustrato que utilizan
durante la BH (Mosley et al., 2002), el grupo A que hidrogena los AG poliinsaturados al
AG trans-11 (Vlaeminck et al., 2008) o bacterias que producen BH sólo hasta C18:1
(Jenkins et al., 2008) y el grupo B que hidrogena el TVa a Es y el C18:2t-11,c-15 a C18:1t-11
ó C18:1c-15 (Vlaeminck et al., 2008), estas últimas son identificadas como Fusocillus spp
(Jenkins et al., 2008) y son bacilos Gram (-) (Mosley et al., 2002). En el proceso de BH
del Ln intervienen los dos grupos, mientras que para la del Li intervienen principalmente
las bacterias del grupo A y algunas del B pero de forma transitoria. Las bacterias más
representativas del grupo A en la isomerización del Li y Ln son la B. fibrisolvens y
Eubacterium, aunque otras bacterias hacen esta isomerización de una manera transitoria
como es el caso del Ruminococcus albus (Rojas et al., 2005). Adicional a esta vía
aparecen otras (Vlaeminck et al., 2008).
La BH de las bacterias Gram (-) producen Es a partir de Li y ácido oleico (Ol), pero no a
partir de Ln. La B. fibrisolvens y el Clostridium proteoclasticum producen TVa, pero sólo
el C. proteoclasticum también produce Es. Todas las bacterias que producen
sustanciales cantidades de Ru y TVa, son productoras de butirato, aunque no todas las
productoras de butirato producen Ru y TVa (p.e Eubacterium y Clostridium spp.). Pocas
bacterias forman Ru y TVa también forman Es (Durmic et al., 2008).
El primer paso de la BH del Ln es la isomerización del enlace cis-12 a la posición del C11
ó C13 (Jenkins et al., 2008) produciendo C18:3c-9,t-11,c-15 (Rojas et al., 2005) ó C18:3c-9,t-13,c-
15, luego uno de los dobles enlaces es hidrogenado para producir un C18:2 (Jenkins et
al., 2008), el C18:2t-11,c-15 (Rojas et al., 2005) o el C18:2t-13,c-15, posteriormente se produce
la hidrogenación de otro enlace produciendo un C18:1 (Jenkins et al., 2008), el trans-15,
el cis-11 o el trans-11 (Rojas et al., 2005) que es hidrogenado para producir el Es como
producto final (Jenkins et al., 2008). La BH del Ln produce TVa pero no Ru.

Capítulo 1
31
La principal vía de BH del Li consiste en una isomerización inicial para producir Ru, el
cual es biohidrogenado a TVa, que luego es biohidrogenado para producir Es (Moate et
al., 2008). El TVa es un intermediario común de la BH incompleta del Li y el Ln (Bauman
et al., 2004) y es el más importante de los isómeros C18:1 (Doreau y Chilliard, 1997). En
sistemas in vitro representa entre el 88 al 95 % del total de trans C18:1 (AbuGhazaleh y
Jenkins, 2004a). Cuando existe mucho Li disponible la BH se detiene, impidiendo que los
diferentes isómeros se saturen (Doreau y Chilliard, 1997) y permitiendo que una porción
de TVa y Ru no se sature en el rumen y pase al abomaso (Baumgard et al., 2001), de tal
manera que la elevada concentración de TVa inhibe su propia BH (Moate et al., 2008).
El B. fibrisolvens produce la BH del Li y Ln (Kepler y Tove, 1967) y no puede modificar el
Ol de ninguna manera (Mosley et al., 2002). En el caso del Li la BH se produce en dos
pasos, primero se produce la BH del Li a Ru y luego ocurre la hidrogenación de este
ALC para formar TVa (Hughes et al., 1982). El Es no se forma por acción del B.
fibrisolovens. La reductasa y la isomerasa no forman parte de un complejo enzimático en
el B. fibrisolvens pero forman parte de la membrana (Jenkins et al., 2008). El pH del
rumen tiene un papel importante en el mantenimiento de un ambiente ruminal para B.
fibrisolvens. Un pH superior a 6.0 tiene un efecto positivo en la producción de Ru
(Tsiplakou et al., 2008).
Los ácidos Li y Ln no son los únicos precursores de ALC, el Ol puede ser precursor de
diversos AG trans (AbuGhazaleh et al., 2003) entre el carbono 6 y 16 (Mosley et al.,
2002) incluyendo el TVa (AbuGhazaleh et al., 2003). Cuando el pH es bajo se produce un
doble enlace en un C<10 (Jenkins et al., 2008) y las Pseudomnas isomerizan el Ol a
trans-10 (Mosley et al., 2002) y bacterias de los géneros Propionibacterium,
Streptococcus y lactobacilus convierten el Li en C18:2t-10,c-12 (Jenkins et al., 2008).
El incremento en docosahexaenoico (DHA) y eicosapentaenoico (EPA) aumenta la BH in
vitro de Li, pero reduce la del Ol. El DHA y EPA reducen la actividad de la enzima
reductasa de los microorganismos responsables de la hidrogenación terminal de Ol a Es
y promueve la conversión hacia trans C18:1, especialmente hacia TVa; en este sentido
tiene una mayor capacidad el DHA frente al EPA (AbuGhazaleh y Jenkins, 2004a). La
adición in vitro de DHA incrementa la concentración de TVa y la hidrogenación in vitro del
trans-11 sólo ocurre en ausencia de DHA. El DHA es un potente inhibidor de bacterias,

32
Efecto de la suplementación con harina de arroz o semilla de algodón sobre el contenido de los ácidos transvaccénico y ruménico
en la leche de vacas en pastoreo
enzimas o ambos involucrados en la BH de isómeros C18:2 no conjugados
intermediarios de la BH como el C18:2t-11,t-15, el C18:2c-9,t-13 y C18:2t-11,c-15 (Vlaeminck et
al., 2008).
Las bacterias, hongos y protozoarios pueden sintetizar o incorporar AG, dentro de
aquellos que son sintetizados los C15 y los C17 son los más característicos (Doreau y
Chilliard, 1997). Los AG de las bacterias raramente son poliinsaturados (Voet y Voet,
1990) y los AG trans monoinsaturados pueden resultar de la desaturación de los AG
saturados. Una vez sintetizados o incorporados los AG son esterificados como
fosfolípidos y esterol ésteres, para constituir lípidos estructurales (Doreau y Chilliard,
1997). Los Ru y TVa en las bacterias pueden constituir entre el 0.51 al 1.37 % y el 3.7 al
12.7 % respectivamente (Váradyová et al., 2008).
Más de la mitad de la masa microbial puede ser protozoal y aproximadamente ¾ partes
de los AG presentes en el rumen pueden estar presentes en los protozoarios. Estos
poseen proporcionalmente mayor cantidad de AG insaturados que las bacterias. Los
protozoarios no sintetizan Ru ni TVa a partir del Es, a través de la ingesta de bacterias
incorporan Ru y TVa formado (Jenkins et al., 2008). La fracción de protozoarios en el
rumen tiene un mayor contenido (2 a 3 veces) de Ru y TVa que las bacterias. Los ácidos
Ru y TVa en los protozoarios pueden constituir entre el 1.5 al 5.9 % y el 7.4 al 17.4 %
respectivamente (Váradyová et al., 2008).
Los hongos anaeróbicos contienen elevados niveles de C18:1, se ha demostrado que el
Piromyces communis convierte el Es en Ol. La incubación de P. communis y
Neocallimastix frontalis produce Ru (Jenkins et al., 2008).
El suministro de AG saturados, palmítico y Es, tiene poco efecto en la fermentación
ruminal de los AG insaturados, y son absorbidos a través del intestino; por lo tanto,
aportan energía que no impacta negativamente en el metabolismo de los
microorganismos del rumen (Warntjes et al., 2008).

Capítulo 1
33
1.4 La enzima Δ9-desaturasa y la síntesis endógena de ácido ruménico
1.4.1 Actividad de la enzima Δ9-desaturasa
La acil-CoA desaturasa (Δ9-desaturasa) cataliza la reacción oxidativa que introduce un
doble enlace en el C9 de la cadena del AG (Voet y Voet, 1990; Jacobs et al., 2011). La
glándula mamaria es el principal sitio de acción de la Δ9-desaturasa en vacas lactantes
(Corl et al., 2001) y la producción de AG insaturados de la leche, particularmente algunos
moninsaturados y casi todo el ALC es regulado por la actividad de esta enzima (Soyeurt
et al., 2008). En la glándula mamaria el Es es convertido a Ol y el TVa a Ru a través de
la Δ9-desaturasa (Corl et al., 2001).
La variación del Ru está relacionada con diferencias en la producción ruminal de TVa y a
diferencias en la acción de la Δ9-desaturasa (Corl et al., 2001). Entre el 75 al 90 % del Ru
en la leche se deriva de la Δ9-desaturación del TVa (Bauman et al., 2008). El incremento
en la cantidad de TVa que llega a la glándula mamaria tiene un efecto positivo sobre la
cantidad Ru en la leche (Bauman et al., 2008), de tal manera al incrementar la
producción de TVa en el rumen mejora la cantidad de Ru lácteo (AbuGhazaleh y Jenkins,
2004b).
La glándula mamaria sólo puede secretar grasa láctea con una adecuada fluidez (Gama
et al., 2008), para ello existe la acción de la Δ9-desaturasa a nivel mamario (Griinari et al.,
2000) que juega un papel importante al generar plasticidad de los triglicéridos para que
puedan ser secretados (Corl et al., 2001). La depresión de grasa láctea puede ser un
mecanismo de adaptación para prevenir la secreción de leche con elevado punto de
fusión (Gama et al., 2008).
1.4.2 Intermediarios de la biohidrogenación en el rumen que inhiben la actividad de la enzima Δ9-desaturasa y la síntesis de grasa láctea
Dietas con y elevado contenido de DHA incrementan C18:2t-10,c-12 (Jenkins et al., 2008).
La primera isomeración del Li en enlace doble cis-12 produce trans-11 (Baumgard et al.,
2001); sin embargo, cambios en el ambiente ruminal producen una isomerización del

34
Efecto de la suplementación con harina de arroz o semilla de algodón sobre el contenido de los ácidos transvaccénico y ruménico
en la leche de vacas en pastoreo
enlace doble cis-9 del Li produciendo trans-10 que luego se convierte en C18:2t-10,c-12, a
diferencia de la típica isomerización del enlace doble cis-12 que produce Ru (Baumgard
et al., 2001), situación que normalmente se presenta en dietas con concentrado (Jenkins
et al., 2008). El DHA disminuye la BH de los C18:1 e incrementa la concentración de
C18:1t-10 y C18:2t-10,c-12 (Or-Rashid et al., 2008).
El C18:2t-10,c-12 inhibe la actividad de la Δ9-desaturasa (Eder et al., 2002; Lock et al., 2007)
disminuyendo la proporción de Ru (Griinari et al., 2000) y la producción de grasa láctea
(Lock et al., 2006). La infusión abomasal de C18:2t-10,c-12 disminuye los AG de cadena
corta, aumenta los AG de cadena larga e incrementa las relaciones entre los AG mirístico
: miristoleico, palmítico : palmitoleico, Es : Ol y TVa : Ru (Baumgard et al., 2000; Bauman
et al., 2000); las relaciones entre los sustratos y los productos de esta enzima en la grasa
láctea pueden ser un indicador de su actividad (Jacobs et al., 2011) o de una mayor
síntesis en la glándula mamaria (Feng et al., 2007). Existe una relación inversa entre el
porcentaje de grasa en leche y el contenido lácteo de C18:2 t-10,c-12 (Baumgard et al.,
2001; Bauman et al., 2008).
Ciertas dietas causan una marcada depresión de grasa láctea reduciendo la producción
de grasa por encima del 50 %, debido a una menor síntesis AG y utilización de AG
preformados (Bauman et al., 2008). Estas dietas están relacionadas con incrementos en
el contenido de C18:1t-10 y C18:2t-10,c-12 en la leche (Griinari et al., 2000). El C18:2t-10,c-12
baja la producción grasa láctea (Lock et al., 2006) pues reduce la cantidad del RNAm de
los genes asociados con la síntesis de grasa láctea (Peterson et al., 2003); sin alterar la
producción de leche y de proteína (Baumgard et al., 2000), aunque se ha encontrado un
incremento en la producción láctea y reducción en la concentración de proteína a causa
del suministro de este AG (Hutchinson et al., 2012). Inicialmente se estableció que la
depresión de grasa láctea estaba asociada específicamente a un incremento en el
C18:1t-10 en la leche (AbuGhazaleh et al., 2003), pero se ha demostrado que el C18:1t-10
no reduce la grasa láctea, mientras el C18:2 t-10,c-12 sí; por tal motivo, la relación el C18:1t-
10 con la depresión de grasa láctea se debe a que el C18:2 t-10,c-12 forma parte de la vía de
BH que produce el C18:1t-10 (Baumgard et al., 2001). Es probable que una producción de
C18:2t-10,c-12 por las bacterias del rumen pueda ser la responsable de muchos, quizás la
mayoría, de las situaciones alimenticias que producen depresión de grasa láctea
(Baumgard et al., 2000).

Capítulo 1
35
No se requiere de un incremento en C18:2t-10,c-12 para producir depresión de grasa láctea
y se sugiere que existen otros componentes que también producen este efecto (Piperova
et al., 2004). En ciertas situaciones las dietas que inducen depresión de grasa láctea, el
contenido C18:2t-10,c-12 y la magnitud de la reducción en la producción de grasa láctea no
concuerdan con la curva generada cuando se realiza infusión abomasal con diferentes
cantidades de C18:2t-10,c-12, lo cual sugiere que en estas situaciones este sólo AG no
explica completamente la magnitud de la reducción en la grasa láctea (Bauman et al.,
2008) y se sugiere que otros isómeros ALC pueden estar involucrados (Piperova et al.,
2000).
El suministro C18:2t-10,c-12 se ha empleado durante la lactancia temprana para mejorar el
desempeño reproductivo posparto, ya que al limitar la síntesis de grasa láctea se
disminuye la cantidad de energía liberada en la leche y se atenúa el balance negativo
posparto (de Veth et al., 2009; Hutchinson et al., 2011). En algunos casos no se mejora
el desempeño reproductivo o en el status energético del animal debido a que la energía
ahorrada en la síntesis de grasa se destina hacia una mayor producción láctea
(Hutchinson et al., 2012).
1.4.3 Inhibición de la enzima Δ9-desaturasa a través de ácidos ciclopropenos
Los AG ciclopropenos estercúlico (C19:1) y malválico (C18:1) inhiben la Δ9-desaturasa,
evitando que TVa sea transformado a Ru (Corl et al., 2001). El suministro de
ciclopropeno en forma “by pass” inactiva la Δ9-desaturasa (Cook et al., 1976; Ashes et
al., 1997), aunque si este compuesto no está protegido se satura en el rumen (Ashes et
al., 1997) produciendo el ácido ciclopropano que no inhibe la Δ9-desaturasa (Cook et al.,
1976).
1.5 Factores que afectan la concentración de los ácidos trans-vaccénico y ruménico en la grasa láctea
Los factores que afectan el contenido de Ru en la leche se pueden dividir en tres grupos,
los relacionados con la dieta, con el animal y con el manejo post-ordeño (Khanal y Olson,
2004).

36
Efecto de la suplementación con harina de arroz o semilla de algodón sobre el contenido de los ácidos transvaccénico y ruménico
en la leche de vacas en pastoreo
1.5.1 Factores relacionados con la dieta
La dieta es el mayor determinante de la concentración láctea de Ru (Khanal y Olson,
2004; Rico et al., 2007). El pH en el rumen, el tipo de lípidos ingeridos y el tipo de
bacterias presentes en el rumen afectan la BH o limitan la conversión de TVa a Es
(Chilliard et al., 2000; Tsiplakou et al., 2008). La suplementación con fuentes lipídicas de
AG insaturados, las características de los forrajes, la relación concentrado : forraje, el
sistema de alimentación (pastoreo – dietas totalmente mezcladas) y el suministro de
antibióticos son factores relacionados con la dieta que afectan la concentración de TVa y
Ru en la leche.
1.5.1.1 Suplementación con fuentes lipídicas de ácidos grasos insaturados
El suministro de lípidos con elevado contenido de AG insaturados tiene un efecto positivo
sobre el contenido de Ru y TVa en la leche. Según Dhiman et al. (2000) tanto las
semillas como los aceites aumentan el contenido de Ru y TVa, siempre y cuando el
aceite esté disponible a los microorganismos del rumen para la BH.
Existen diferencias en el tipo de fuente y la manera como han sido procesadas dichas
fuentes. Bu et al. (2007) demuestran que el suministro de fuentes con niveles elevados
de Li son más eficientes en producir Ru y TVa que aquellas ricas en Ln. Khanal y Olson
(2004) indican que suplementar con semillas de oleaginosas calentadas o extruidas
incrementa el el contenido de Ru debido a que dichos procesos facilitan el acceso de los
lípidos por parte de los microorganismos; lo cual es soportado por los resultados de
Paradis et al. (2008).
A continuación se citan algunas fuentes lipídicas que tienen efecto sobre la producción
de Ru o TVa en la leche.
Soya. El aceite de soya posee una gran proporción de Li (51%) y es efectivo para
incrementar el contenido de Ru (Huang et al., 2008) y TVa (Bu et al., 2007) en la leche
de bovinos, cabras (Bouattour et al., 2008) y ovejas (Gómez-Cortés et al., 2008).
Linaza. El aceite de linaza tiene una alta concentración de Ln (50 %; Ueda et al.,
2003; Bu et al., 2007) e incrementa el contenido de Ru y TVa en bovinos (Loor et al.,
2005) y cabras (Vasta et al., 2008) pero en menor intensidad que la soya (Bu et al.,

Capítulo 1
37
2007).
Girasol. El aceite de girasol contiene elevada proporción de Li (60 %) e incrementa
las concentraciones de Ru y TVa en la leche (AbuGhazaleh et al., 2002; Gagliostro et
al., 2006; AbuGhazaleh y Holmes, 2007).
Maíz. El principal AG del aceite de maíz es el Li (53 %) e incrementa las
concentraciones de Ru y TVa en la leche (Bharathan et al., 2008).
Aceite de pescado. La adición de aceites marinos inhiben la reducción de trans
C18:1 a Es (Vlaeminck et al., 2008). El aceite de pescado contiene aproximadamente
10 % de DHA (AbuGhazaleh et al., 2004) y este AG y/o sus derivados promueven la
acumulación de TVa cuando se incuban AG insaturados provistos por otras fuentes
(AbuGhazaleh y Jenkins, 2004a), aumentando la concentración de Ru en la leche
(AbuGhazaleh et al., 2003).
Algas. Las algas inhiben la reducción de trans C18:1 a Es (Vlaeminck et al., 2008) y
tienen un efecto cuadrático sobre el TVa y Ru, lo cual está relacionado con su elevado
contenido de DHA (5.2 %; AlZahal et al., 2008).
Mezclas de fuentes lipídicas. Al mezclar fuentes de Li y Ln con aceite de pescado
que contiene DHA y EPA se incrementa el contenido de Ru y TVa en comparación al
suministro de estas fuentes de manera separada (AbuGhazaleh et al., 2002;
AbuGhazaleh et al., 2003; Gagliostro et al., 2006). Este efecto es mayor si el aceite de
pescado se suministra en combinación con fuentes ricas en Li que cuando se
suministra con fuentes ricas en Ol y Ln (AbuGhazaleh et al., 2003). La
suplementación con semillas de linaza y el aceite de girasol incrementa Ru y TVa en
la leche (Hervás et al., 2008).
Grasas sobrepasantes. Suministrar Ru como sales de calcio produce mayor
concentración de Ru en la grasa láctea que cuando se suministra Ru como AG libre
(Huang et al., 2008). Algunas fuentes de grasas sobrepasantes pueden ser
biohidrogenadas en el rumen y aumentar la cantidad de precursores, de tal manera
que aumenta los ácidos TVa y Ru en leche (Schroeder et al., 2003; Lundy et al., 2004;
Juchem et al., 2008).
1.5.1.2 Características de los forrajes
La variación en el contenido de Ru es directamente proporcional al contenido de Ln
presente en los forrajes (Mel´uchová et al., 2008); la época del año, la conservación, la

38
Efecto de la suplementación con harina de arroz o semilla de algodón sobre el contenido de los ácidos transvaccénico y ruménico
en la leche de vacas en pastoreo
madurez y el manejo del forraje son factores que afectan la concentración de este AG. La
presencia de metabolitos secundarios también afecta la concentración de Ru en leche.
Época del año. Existen diferencias en la concentración de Ru y TVa a través del año
(Collomb et al., 2008) y estas diferencias van acompañadas de un mayor consumo de
Ln (Vibart et al., 2008; Mel´uchová et al., 2008). Los factores ambientales afectan el
contenido de AG de la serie omega 3 (Dewhurst et al., 2006) y parece que las
pasturas con la vía del carbono C3 incrementan el Ru con relación las de la vía C4
debido a su mayor concentración de Ln (Khanal y Olson, 2004).
Conservación de forrajes. La leche proveniente de animales en pastoreo contiene
mayores niveles de Ru que aquella proveniente de animales que consumen ensilaje
(Elgersma et al., 2006). El proceso de ensilaje produce pérdida de lípidos, de Li y de
Ln (Dewhurst y King, 1998; Dewhurst et al., 2006; Rego et al., 2008) lo cual afecta el
contenido de Ru y TVa en la leche. El ensilaje posee mayor cantidad de AG
insaturados no esterificados que pueden ser fácilmente hidrogenados a Es (Dewhurst
et al., 2006). El efecto de los ensilajes sobre la BH varía según la composición
botánica de los forrajes utilizados para elaborar el ensilaje (Lourenço et al., 2007).
Madurez del forraje. La madurez está asociada con la disminución en la proporción
de Ln y aumento concomitante de Ol y Li (Khanal y Olson, 2004; Vanhatalo et al.,
2007), lo cual tiene un efecto negativo sobre la concentración Ru lácteo.
Manejo. La fertilización con N incrementa el Ln, mientras que al incrementar los
periodos de rebrote se disminuye (Elgersma et al., 2006; Dewhurst et al., 2006).
Metabolitos secundarios. Una gran variedad de extractos de plantas tienen efecto
inhibitorio selectivo sobre C. proteoclasticum que forma Es a partir de Li sin afectar el
B. fibrisolvens (Durmic et al., 2008), lo cual incrementa el flujo de TVa. El contenido de
taninos afecta la composición de AG en la carne y la leche (Vasta et al., 2008) ya que
reducen la BH de la planta (Dewhurst et al., 2006). Así mismo, la polifenol oxidasa
presente en algunos forrajes como el trébol rojo (Trifolium pratense L.) disminuye la
lipólisis (Lourenço et al., 2007).
1.5.1.3 Relación concentrado : forraje
Se presentan mayores concentraciones de Ru y TVa cuando las vacas están pastando,
en relación con aquellas que se encuentran estabuladas (Kelly et al., 1998; Dhiman et al.,
1999a; Schroeder et al., 2003; Rego et al., 2004; Couvreur et al., 2007; Craninx et al.,

Capítulo 1
39
2008; Juchem et al., 2008; Khanal et al., 2008; AlZahal et al., 2008) y lo mismo ocurre en
ovejas; esto se encuentra relacionado con el perfil de AG de la dieta (Tsiplakou et al.,
2008) y con la mayor concentración de Ln en el forraje (Rego et al., 2008; Biondi et al.,
2008). Al incrementar la proporción de forraje consumido en pastoreo se incrementa
linealmente la concentración de Ru en la leche (Vibart et al., 2008).
Existen diferentes factores que pueden afectar el ambiente ruminal y la población
microbiana que podría diferenciar a los animales en pastoreo como son: la tasa de
pasaje y la dilución de nutrientes debido al elevado consumo de agua asociado con el
pastoreo (Schroeder et al., 2004). El tamaño del alimento, la frecuencia de consumo, el
tamaño del bocado y el tiempo dedicado a la rumia pueden diferenciar a las vacas en
pastoreo (Kelly et al., 1998). En el sistema de pastoreo se producen mayores niveles de
AG poliinsaturados y mayores contenidos de Ru en la grasa láctea comparado con el
suministro de forraje conservado (Noni y Battelli, 2008).
La pastura disminuye el tamaño del glóbulo graso, y por lo tanto incrementa los AG
monoinsaturados y poliinsaturados (Couvreur et al., 2007). El tamaño del glóbulo de
grasa disminuye al incrementar la proporción de forraje fresco en la dieta (Couvreur et al.,
2006).
1.5.1.4 Suministro de antibióticos
Los ionóforos inhiben la BH ruminal (AlZahal et al., 2008) e incrementan el total de
isómeros C18:1, específicamente C18:1t-10 y aumentan el Ru (Odongo et al., 2007; Aldai
et al., 2008) especialmente cuando interactúa con el aceite de soya (AlZahal et al., 2008)
y de pescado (Dhiman et al., 1999a; Wang et al., 2005), ya que este efecto es mayor en
dietas con elevada concentración de AG insaturados (Duffield et al., 2008).
La monensina es un antibiótico ionóforo que inhibe la lipólisis (AlZahal et al., 2008), el
crecimiento de las bacterias Gram (+) (Wang et al., 2005) que realizan la BH del Li a Ol,
mientras la reducción del Ol a Es es realizada por bacterias Gram (-), de tal manera que
la acumulación de C18:1 no puede ser atribuida al efecto de la monensina sobre las
Gram (+) (AlZahal et al., 2008).
La monensina que en asociación con suplementación lipídica puede incrementar el

40
Efecto de la suplementación con harina de arroz o semilla de algodón sobre el contenido de los ácidos transvaccénico y ruménico
en la leche de vacas en pastoreo
contenido de Ru (AlZahal et al., 2008) reduce la presencia de AG de cadena corta y de la
grasa láctea (Duffield et al., 2008).
1.5.2 Factores relacionados con el animal
Existe un cambio progresivo en la concentración de los AG de la leche (Craninx et al.,
2008) y un pequeño incremento en los índices de actividad de la Δ9-desaturasa
relacionados con el estado de la lactancia (Soyeurt et al., 2008). También existe una
relación positiva entre el número de partos y el contenido de Ru (Khanal y Olson, 2004).
Se presentan variaciones entre razas, las vacas Holstein producen mayor cantidad de Ru
que las vacas Jersey o Normando, aunque se ha encontrado que si la dieta es la misma
las variaciones son mínimas (Khanal y Olson, 2004). Los animales que producen
glóbulos grasos pequeños producen mayor cantidad de Ru y TVa que aquellas con
glóbulos grasos grandes (Couvreur et al., 2007). Si las vacas producen menos cantidad
de grasa poseen glóbulos grasos más pequeños y producen mayor proporción de Ru
(Weisbjerg et al., 2008). Se ha encontrado una gran variación en los TVa y Ru (Rico et
al., 2007) de 0.3 a 4.1 y de 0.7 a 7.0 respectivamente, lo cual ha sido atribuido a
diferencias individuales de la enzima Δ9-desaturasa (Rego et al., 2008).
1.5.3 Factores relacionados con los procesos post-ordeño
El efecto de los procesos post-ordeño sobre el contenido de Ru es controversial. Se
puede argumentar que el contenido de los productos depende del contenido original de la
leche. Sin embargo, es probable que los cultivos microbianos utilizados para elaborar
productos lácteos pueden producir enzimas que isomerizan el Li en Ru (Rico et al.,
2007).
1.6 La harina de arroz y su potencial para incrementar la concentración de los ácidos trans-vaccénico y ruménico en la leche de vaca
La harina de arroz es el nombre que se le da en Colombia al cilindro de arroz o salvado
de arroz. Es un subproducto industrial de la molienda del arroz (Gadberry et al., 2005; de
Campos et al., 2007), corresponde a la capa color café presente en el arroz
(AbuGhazaleh et al., 2009) e incluye embrión, parte del endospermo almidonado,

Capítulo 1
41
endospermo y un poco de cáscara (Warren y Farrell, 1990). Su composición química
varía ampliamente probablemente por adulteración con la casca del arroz (Warren y
Farrell, 1990) pero la concentración de nutrientes (almidón del 39.2 %, Wilks et al., 1991;
FDN del 54.9 %, Dung et al., 2002; proteína entre el 11 al 15 % y extracto etéreo entre el
15 al 20 %, de Campos et al., 2007) y su bajo costo (de Campos et al., 2007) permiten su
fácil inclusión en el alimento para animales.
La principal característica del salvado de arroz es el aceite, pues es uno de los más
nutritivos debido a su composición favorable de AG insaturados (Lin et al., 2009) y si se
extrae pronto después de la molienda puede ser utilizado como un alimento útil para el
hombre (Warren y Farrell, 1990). La concentración de extracto etéreo puede variar (29 %,
Dung et al., 2002; entre el 15 al 20 %, de Campos et al., 2007) pero se caracteriza por
una elevada concentración de Ol (43 %, Warren y Farrell, 1990; 39 %, Bravi et al., 2006;
del 34 al 37 %, de Campos et al., 2007) y de Li (39 %, Warren y Farrell, 1990; 29 %,
Bravi et al., 2006; del 36 al 42 %, de Campos et al., 2007), además presenta elementos
naturales con actividad antioxidante (Lin et al., 2009).
Esta materia prima es frecuentemente utilizada en la formulación de alimentos
balanceados para vacas lecheras en Colombia y debido a la elevada concentración de Li
puede tener potencial para incrementar las concentraciones lácteas de los ácidos Ru y
TVa de vacas en pastoreo.
1.7 La semilla de algodón y su potencial para incrementar la concentración de los ácidos trans-vaccénico y ruménico en la leche de vaca
La semilla de algodón es un alimento derivado de la industria de la fibra de algodón, que
frecuentemente se utiliza en las raciones de rumiantes. La semilla de algodón, posee
gosipol (C30H30O8), un compuesto polifenólico amarillo que se encuentra de manera libre
y ligada. El gosipol libre es tóxico para animales monogástricos, pero los rumiantes lo
ligan a una proteína del rumen, evitando su absorción. La forma libre también se puede
convertir a ligada debido al calor, humedad y presión durante la extrusión (Zhang et al.,
2007).
La semilla de algodón tiene el 19 % de grasa que contiene el 23 % de palmítico, el 17 %

42
Efecto de la suplementación con harina de arroz o semilla de algodón sobre el contenido de los ácidos transvaccénico y ruménico
en la leche de vacas en pastoreo
de Ol y el 57 % de Li (Dhiman et al., 1999a; Dhiman et al., 1999b; Khanal y Olson, 2004).
Esto convierte a este recurso alimenticio en un buen candidato para incrementar las
concentraciones de Ru en la leche debido a sus elevados niveles de Li. Sin embargo,
Rico et al. (2007) encontraron una correlación negativa entre la presencia de semilla de
algodón en la dieta y la concentración de Ru en la leche (-0.7). Estos resultados no
significan una relación causa efecto y deben ser revisados con precaución pues los
mismos autores afirman que aunque existe una asociación negativa entre el consumo de
semilla de algodón y la concentración de Ru, el contenido de AG saturados e insaturados
no parece ser suficiente para explicar dicho efecto.
El aceite de algodón posee de ácidos ciclopropenos (Corl et al., 2001) como malválico y
estercúlico (alrededor del 1 % del aceite; Coppock et al., 1987) que inhiben la actividad
de la enzima Δ9-desaturasa (Cook et al., 1976). La BH de estos ácidos en el rumen
produce ciclopropano que no es capaz de inhibir la Δ9-desaturasa (Cook et al., 1976); por
tal motivo, los rumiantes no experimentan efectos negativos con el suministro de semilla
de algodón sobre la Δ9-desaturasa (Archibeque et al., 2005; Liu et al., 2008; Vasta et al.,
2008) debido a que los ciclopropenos representan una menor proporción del total de la
dieta y la BH ruminal los inactiva (Corl et al., 2001).
La suplementación con semilla de algodón no genera efectos negativos sobre las
concentraciones de Ru y TVa en leche y en tejido adiposo (Archibeque et al., 2005), no
produce variación en el perfil de AG en la leche (Vasta et al., 2008), pero animales que
consumieron 2.7 kg de semilla de algodón por día presentaron un aumento en la
concentración de Ru en la leche (Liu et al., 2007).
Por lo tanto, no existen evidencias suficientes para asegurar que el consumo de semilla
de algodón pueda producir disminución en la síntesis del Ru en la glándula mamaria. La
concentración de AG insaturados sugiere que esta fuente lipídica puede utilizarse para
incrementar la concentración de Ru y TVa en la leche, teniendo en cuenta que esta
materia prima se usa en la fabricación de alimentos balanceados para vacas lecheras del
trópico alto colombiano
1.8 Conclusiones
La alimentación es el principal factor para incrementar la concentración de los ácidos

Capítulo 1
43
trans-vaccénico y ruménico en la leche. Aumentar el consumo de forraje fresco,
incrementar el consumo de ácido linoleico utilizando diferentes suplementos lipídicos y el
consumo de ácido linolénico proveniente del forraje permite incrementar la concentración
láctea de los ácidos trans-vaccénico y ruménico. El control de las condiciones ruminales
con ionóforos y con ácido docosahexaenoico potencializa el suministro de ácidos grasos
insaturados. La harina de arroz y la semilla de algodón son dos fuentes lipídicas que
aparentemente pueden incrementar la concentración láctea de los ácidos trans-vaccénico
y ruménico.
1.9 Referencias
ABUGHAZALEH, A. A. 2008. Effect of fish oil and sunflower oil supplementation on milk conjugated linoleic acid content for grazing dairy cows. Anim. Feed Sci. and Technol. 141(3-4):220-232.
ABUGHAZALEH, A. A. y HOLMES, L. D. 2007. Supplementation with fish oil and sunflower oil to increase conjugated linoleic acid levels in milk fat of partially grazing dairy cows. J. Dairy Sci. 90(6):2897-2904.
ABUGHAZALEH, A. A. y JENKINS, T. C. 2004a. Disappearance of docosahexaenoic and eicosapentaenoic acids from cultures of mixed ruminal microorganisms. J. Dairy Sci. 87(3):645-651.
ABUGHAZALEH, A. A. y JENKINS, T. C. 2004b. Short communication: Docosahexaenoic acid promotes vaccenic acid accumulation in mixed ruminal cultures when incubated with linoleic acid. J. Dairy Sci. 87(4):1047-1050.
ABUGHAZALEH, A. A.; POTU, R. B. y IBRAHIM, S. 2009. Short communication: The effect of substituting fish oil in dairy cow diets with docosahexaenoic acid-micro algae on milk composition and fatty acids profile. J. Dairy Sci. 92(12):6156-6159.
ABUGHAZALEH, A. A.; SCHINGOETHE, D. J.; HIPPEN, A. R. y KALSCHEUR, K. F. 2003. Milk conjugated linoleic acid response to fish oil supplementation of diets differing in fatty acid profiles. J. Dairy Sci. 86(3):944-953.
ABUGHAZALEH, A. A.; SCHINGOETHE, D. J.; HIPPEN, A. R. y KALSCHEUR, K. F. 2004. Conjugated linoleic acid increases in milk when cows fed fish meal and extruded soybeans for an extended period of time. J. Dairy Sci. 87(6):1758-1766.
ABUGHAZALEH, A. A.; SCHINGOETHE, D. J.; HIPPEN, A. R.; KALSCHEUR, K. F. y WHITLOCK, L. A. 2002. Fatty acid profiles of milk and rumen digesta from cows fed fish oil, extruded soybeans or their blend. J. Dairy Sci. 85(9):2266-2276.
ALDAI, N.; DUGAN, M. E. R.; KRAMER, J. K. G.; MIR, P. S. y MCALLISTER, T. A. 2008. Non-ionophore antibiotics do not affect the trans-18:1 and CLA composition in beef adipose tissue. J. Anim Sci. 86(12):3522-3532.
ALZAHAL, O.; ODONGO, N. E.; MUTSVANGWA, T.; OR-RASHID, M. M.; DUFFIELD, T. F.; BAGG, R.; DICK, P.; VESSIE, G. y MCBRIDE, B. W. 2008. Effects of monensin and dietary soybean oil on milk fat percentage and milk fatty acid profile in lactating dairy cows. J. Dairy Sci. 91(3):1166-1174.

44
Efecto de la suplementación con harina de arroz o semilla de algodón sobre el contenido de los ácidos transvaccénico y ruménico
en la leche de vacas en pastoreo
ARCHIBEQUE, S. L.; LUNT, D. K.; GILBERT, C. D.; TUME, R. K. y SMITH, S. B. 2005. Fatty acid indices of stearoyl-CoA desaturase do not reflect actual stearoyl-CoA desaturase enzyme activities in adipose tissues of beef steers finished with corn-, flaxseed-, or sorghum-based diets. J. Anim Sci. 83(5):1153-1166.
ASHES, J. R.; GULATI, S. K. y SCOTT, T. W. 1997. Potential to alter the content and composition of milk fat through nutrition. J. Dairy Sci. 80(9):2204-2212.
BAUMAN, D. E.; BAUMGARD, L. H.; CORL, B. A. y GRIINARI, J. M. 2000. Biosynthesis of conjugated linoleic acid in ruminants. J. Anim Sci. 77(E-Suppl):1-ae.
BAUMAN, D. E.; PERFIELD, J. W., II; HARVATINE, K. J. y BAUMGARD, L. H. 2008. Regulation of fat synthesis by conjugated linoleic acid: lactation and the ruminant model. J. Nutr. 138(2):403-409.
BAUMAN, D. E.; PERFIELD, J. W., II y LOCK, A. L. 2004. Effect of trans fatty acids on milk fat and their impact on human health. Pages 42-54 in Cornell University, New York.
BAUMGARD, L. H.; CORL, B. A.; DWYER, D. A.; SAEBO, A. y BAUMAN, D. E. 2000. Identification of the conjugated linoleic acid isomer that inhibits milk fat synthesis. Am. J. Physiol. Regul. Integr. Comp. Physiol. 278(1):R179-R184.
BAUMGARD, L. H.; SANGSTER, J. K. y BAUMAN, D. E. 2001. Milk fat synthesis in dairy cows is progressively reduced by increasing supplemental amounts of trans-10, cis-12 conjugated linoleic acid (CLA). J. Nutr. 131(6):1764-1769.
BELURY, M. A. 2002. Dietary conjugated linoleic acd and health: Physiological effects and mechanisms of action. Ann. Rev. Nutr. 22(1):505-531.
BHARATHAN, M.; SCHINGOETHE, D. J.; HIPPEN, A. R.; KALSCHEUR, K. F.; GIBSON, M. L. y KARGES, K. 2008. Conjugated linoleic acid increases in milk from cows fed condensed corn distillers solubles and fish oil. J. Dairy Sci. 91(7):2796-2807.
BIONDI, L.; VALVO, M. A.; DI GLORIA, M.; TENGHI, E. S.; GALOFARO, V. y PRIOLO, A. 2008. Changes in ewe milk fatty acids following turning out to pasture. Small Ruminant Res. 75(1):17-23.
BOUATTOUR, M. A.; CASALS, R.; ALBANELL, E.; SUCH, X. y CAJA, G. 2008. Feeding soybean oil to dairy goats increases conjugated linoleic acid in milk. J. Dairy Sci.
91(6):2399-2407.
BRAVI, E.; PERRETTI, G. y MONTANARI, L. 2006. Fatty acids by high-performance liquid chromatography and evaporative light-scattering detector. J. Chromatogr. A.
1134(1-2):210-214.
BU, D. P.; WANG, J. Q.; DHIMAN, T. R. y LIU, S. J. 2007. Effectiveness of oils rich in linoleic and linolenic acids to enhance conjugated linoleic acid in milk from dairy cows. J. Dairy Sci. 90(2):998-1007.
CHILLIARD, Y.; FERLAY, A.; MANSBRIDGE, R. M. y DOREAU, M. 2000. Ruminant milk fat plasticity: nutritional control of saturated, polyunsaturated, trans and conjugated fatty acids. Ann. Zootech. 49(3):181-205.
COLLOMB, M.; BISIG, W.; BÜTIKOFER, U.; SIEBER, R.; BREGY, M. y ETTER, L. 2008. Fatty acid composition of mountain milk from Switzerland: Comparison of organic and integrated farming systems. Int. Dairy J. 18(10-11):976-982.
COLLOMB, M.; BÜTIKOFER, U.; SIEBER, R.; JEANGROS, B. y BOSSET, J. O. 2002.

Capítulo 1
45
Composition of fatty acids in cow's milk fat produced in the lowlands, mountains and highlands of Switzerland using high-resolution gas chromatography. Int. Dairy J. 12(8):649-659.
COOK, L.; SCOTT, T.; MILLS, S.; FOGERTY, A. y JOHNSON, A. 1976. Effects of protected cyclopropene fatty acids on the composition of ruminant milk fat. Lipids.
11(9):705-711.
COPPOCK, C. E.; LANHAM, J. K. y HORNER, J. L. 1987. A review of the nutritive value and utilization of whole cottonseed, cottonseed meal and associated by-products by dairy cattle. Anim. Feed Sci. and Technol. 18(2):89-129.
CORL, B. A.; BAUMGARD, L. H.; DWYER, D. A.; GRIINARI, J. M.; PHILLIPS, B. S. y
BAUMAN, D. E. 2001. The role of -desaturase in the production of cis-9, trans-11 CLA. J. Nutr. Biochem. 12(11):622-630.
COUVREUR, S.; HURTAUD, C.; LOPEZ, C.; DELABY, L. y PEYRAUD, J. L. 2006. The linear relationship between the proportion of fresh grass in the cow diet, milk fatty acid composition, and butter properties. J. Dairy Sci. 89(6):1956-1969.
COUVREUR, S.; HURTAUD, C.; MARNET, P. G.; FAVERDIN, P. y PEYRAUD, J. L. 2007. Composition of milk fat from cows selected for milk fat globule size and offered either fresh pasture or a corn silage-based diet. J. Dairy Sci. 90(1):392-
403.
CRANINX, M.; STEEN, A.; VAN LAAR, H.; VAN NESPEN, T.; MARTIN-TERESO, J.; DE BAETS, B. y FIEVEZ, V. 2008. Effect of lactation stage on the odd- and branched-chain milk fatty acids of dairy cattle under grazing and indoor conditions. J. Dairy Sci. 91(7):2662-2677.
DE CAMPOS, R. M. L.; HIERRO, E.; ORDÓÑEZ, J. A.; BERTOL, T. M.; TERRA, N. N. y DE LA HOZ, L. 2007. Fatty acid and volatile compounds from salami manufactured with yerba mate (Ilex paraguariensis) extract and pork back fat and meat from pigs fed on diets with partial replacement of maize with rice bran. Food Chem. 103(4):1159-1167.
DE VETH, M. J.; BAUMAN, D. E.; KOCH, W.; MANN, G. E.; PFEIFFER, A. M. y BUTLER, W. R. 2009. Efficacy of conjugated linoleic acid for improving reproduction: A multi-study analysis in early-lactation dairy cows. J. Dairy Sci. 92(6):2662-2669.
DEWHURST, R. J. y KING, P. J. 1998. Effects of extended wilting, shading and chemical additives on fatty acids in laboratory grass silages. Grass Forage Sci. 53:219-225.
DEWHURST, R. J.; SHINGFIELD, K. J.; LEE, M. R. F. y SCOLLAN, N. D. 2006. Increasing the concentrations of beneficial polyunsaturated fatty acids in milk produced by dairy cows in high-forage systems. Anim. Feed Sci. and Technol.
131(3-4):168-206.
DHIMAN, T. R.; ANAND, G. R.; SATTER, L. D. y PARIZA, M. W. 1999a. Conjugated linoleic acid content of milk from cows fed different diets. J. Dairy Sci. 82(10):2146-2156.
DHIMAN, T. R.; HELMINK, E. D.; MCMAHON, D. J.; FIFE, R. L. y PARIZA, M. W. 1999b. Conjugated linoleic acid content of milk and cheese from cows fed extruded oilseeds. J. Dairy Sci. 82(2):412-419.
DHIMAN, T. R.; SATTER, L. D.; PARIZA, M. W.; GALLI, M. P.; ALBRIGHT, K. y TOLOSA, M. X. 2000. Conjugated linoleic acid (CLA) content of milk from cows

46
Efecto de la suplementación con harina de arroz o semilla de algodón sobre el contenido de los ácidos transvaccénico y ruménico
en la leche de vacas en pastoreo
offered diets rich in linoleic and linolenic acid. J. Dairy Sci. 83(5):1016-1027.
DOREAU, M. y CHILLIARD, Y. 1997. Digestion and metabolism of dietary fat in farm animals. Br. J. Nutr. 78(01):S15-S35.
DUFFIELD, T. F.; RABIEE, A. R. y LEAN, I. J. 2008. A Meta-Analysis of the impact of monensin in lactating dairy cattle. Part 2. Production effects. J. Dairy Sci.
91(4):1347-1360.
DUNG, N. N. X.; MANH, L. H. y UDÉN, P. 2002. Tropical fibre sources for pigs-digestibility, digesta retention and estimation of fibre digestibility in vitro. Anim. Feed Sci. and Technol. 102(1-4):109-124.
DURMIC, Z.; MCSWEENEY, C. S.; KEMP, G. W.; HUTTON, P.; WALLACE, R. J. y VERCOE, P. E. 2008. Australian plants with potential to inhibit bacteria and processes involved in ruminal biohydrogenation of fatty acids. Anim. Feed Sci. and Technol. 145(1-4):271-284.
EDER, K.; SLOMMA, N. y BECKER, K. 2002. trans-10,cis-12 Conjugated linoleic acid
suppresses the desaturation of linoleic and -linolenic acids in HepG2 cells. J. Nutr. 132(6):1115-1121.
ELGERSMA, A.; TAMMINGA, S. y ELLEN, G. 2006. Modifying milk composition through forage. Anim. Feed Sci. and Technol. 131(3-4):207-225.
EMANUELE, S. M.; STAPLES, C. R. y WILCOX, C. J. 1991. Extent and site of mineral release from six forage species incubated in mobile dacron bags. J. Anim Sci.
69(2):801-810.
ENJALBERT, F.; NICOT, M. C.; BAYOURTHE, C. y MONCOULON, R. 1998. Duodenal infusions of palmitic, stearic or oleic acids differently affect mammary gland metabolism of fatty acids in lactating dairy cows. J. Nutr. 128(9):1525-1532.
FENG, S.; SALTER, A. M.; PARR, T. y GARNSWORTHY, P. C. 2007. Extraction and quantitative analysis of stearoyl-Coenzyme A desaturase mRNA from dairy cow milk somatic sells. J. Dairy Sci. 90(9):4128-4136.
GADBERRY, M. S.; BECK, P. A.; KELLOGG, D. W. y GUNTER, S. A. 2005. Digestion characteristics and growth of steers fed a corn-grain based supplement compared to a de-oiled rice bran plus cottonseed supplement with or without extrusion processing. Anim. Feed Sci. and Technol. 118(3-4):267-277.
GAGLIOSTRO, G. A.; RODRÍGUEZ, A.; PELLEGRINI, P.; GATTI, P.; MUSSET, G.; CASTAÑEDA, R. A.; COLOMBO, D. y CHILLIARD, Y. 2006. Efectos del suministro de aceite de pescado sólo o en combinación con aceite de girasol sobre las concentraciones de ácido linoleico conjugado (CLA) y omega 3 (n-3) en leche de cabra. Rev. Argent. Prod. Anim. 26:71-87.
GAMA, M. A. S.; GARNSWORTHY, P. C.; GRIINARI, J. M.; LEME, P. R.; RODRIGUES, P. H. M.; SOUZA, L. W. O. y LANNA, D. P. D. 2008. Diet-induced milk fat depression: Association with changes in milk fatty acid composition and fluidity of milk fat. Livest. Sci. 115(2-3):319-331.
GÓMEZ-CORTÉS, P.; FRUTOS, P.; MANTECON, A. R.; JUÁREZ, M.; DE LA FUENTE, M. A. y HERVÁS, G. 2008. Milk production, conjugated linoleic acid content, and in vitro ruminal fermentation in response to high levels of soybean oil in dairy ewe diet. J. Dairy Sci. 91(4):1560-1569.

Capítulo 1
47
GRIINARI, J. M.; CORL, B. A.; LACY, S. H.; CHOUINARD, P. Y.; NURMELA, K. V. V. y BAUMAN, D. E. 2000. Conjugated linoleic acid is synthesized endogenously in
lactating dairy cows by 9-Desaturase. J. Nutr. 130(9):2285-2291.
HERVÁS, G.; LUNA, P.; ÁNGEL, R.; ARES, N.; DE LA FUENTE, M. A.; REZ, M. y FRUTOS, P. 2008. Effect of diet supplementation with sunflower oil on milk production, fatty acid profile and ruminal fermentation in lactating dairy ewes. J. Dairy Res. 75(4):399-405.
HOUSEKNECHT, K. L.; HEUVEL, J. P. V.; MOYA-CAMARENA, S. Y.; PORTOCARRERO, C. P.; PECK, L. W.; NICKEL, K. P. y BELURY, M. A. 1998. Dietary conjugated linoleic acid normalizes impaired glucose tolerance in the zucker diabetic fatty fa/fa rat. Biochem. Biophysi. Res. Comm. 244(3):678-682.
HUANG, Y.; SCHOONMAKER, J. P.; BRADFORD, B. J. y BEITZ, D. C. 2008. Response of milk fatty acid composition to dietary supplementation of soy oil, conjugated linoleic acid, or both. J. Dairy Sci. 91(1):260-270.
HUGHES, P. E.; HUNTER, W. J. y TOVE, S. B. 1982. Biohydrogenation of unsaturated fatty acids. Purification and properties of cis-9,trans-11-octadecadienoate reductase. J. Biol. Chem. 257(7):3643-3649.
HUTCHINSON, I. A.; HENNESSY, A. A.; DEWHURST, R. J.; EVANS, A. C. O.; LONERGAN, P. y BUTLER, S. T. 2012. The effect of strategic supplementation with trans-10,cis-12 conjugated linoleic acid on the milk production, estrous cycle characteristics, and reproductive performance of lactating dairy cattle. J. Dairy Sci.
95(5):2442-2451.
HUTCHINSON, I.; DE VETH, M. J.; STANTON, C.; DEWHURST, R. J.; LONERGAN, P.; EVANS, A. C. O. y BUTLER, S. T. 2011. Effects of lipid-encapsulated conjugated linoleic acid supplementation on milk production, bioenergetic status and indicators of reproductive performance in lactating dairy cows. J. Dairy Res.
78(03):308-317.
IP, C.; BANNI, S.; ANGIONI, E.; CARTA, G.; MCGINLEY, J.; THOMPSON, H. J.; BARBANO, D. y BAUMAN, D. 1999. Conjugated linoleic acid-enriched butter fat alters mammary gland morphogenesis and reduces cancer risk in rats. J. Nutr.
129(12):2135-2142.
IP, C.; BRIGGS, S. P.; HAEGELE, A. D.; THOMPSON, H. J.; STORKSON, J. y SCIMECA, J. A. 1996. The efficacy of conjugated linoleic acid in mammary cancer prevention is independent of the level or type of fat in the diet. Carcinogenesis.
17(5):1045-1050.
JACOBS, A. A. A.; VAN BAAL, J.; SMITS, M. A.; TAWEEL, H. Z. H.; HENDRIKS, W. H.; VAN VUUREN, A. M. y DIJKSTRA, J. 2011. Effects of feeding rapeseed oil, soybean oil, or linseed oil on stearoyl-CoA desaturase expression in the mammary gland of dairy cows. J. Dairy Sci. 94(2):874-887.
JENKINS, T. C.; WALLACE, R. J.; MOATE, P. J. y MOSLEY, E. E. 2008. Board-Invited review: Recent advances in biohydrogenation of unsaturated fatty acids within the rumen microbial ecosystem. J. Anim Sci. 86(2):397-412.
JENSEN, R. G. 2002. The composition of bovine milk lipids: january 1995 to december 2000. J. Dairy Sci. 85(2):295-350.
JUCHEM, S. O.; SANTOS, J. E. P.; CERRI, R. L. A.; CHEBEL, R. C.; GALVO, K. N.;

48
Efecto de la suplementación con harina de arroz o semilla de algodón sobre el contenido de los ácidos transvaccénico y ruménico
en la leche de vacas en pastoreo
BRUNO, R.; DEPETERS, E. J.; SCOTT, T.; THATCHER, W. W. y LUCHINI, D. 2008. Effect of calcium salts of fish and palm oils on lactational performance of Holstein cows. Anim. Feed Sci. and Technol. 140(1-2):18-38.
KELLY, M. L.; KOLVER, E. S.; BAUMAN, D. E.; VAN AMBURGH, M. E. y MULLER, L. D. 1998. Effect of intake of pasture on concentrations of conjugated linoleic acid in milk of lactating cows. J. Dairy Sci. 81(6):1630-1636.
KEPLER, C. R. y TOVE, S. B. 1967. Biohydrogenation of Unsaturated Fatty Acids. III.
Purification and propierties of linoleate 12-cis,
11-trans-isomerasa from butyrivibrio fibrisolvens. J. Biol. Chem. 242(24):5686-5692.
KHANAL, R. C.; DHIMAN, T. R. y BOMAN, R. L. 2008. Changes in fatty acid composition of milk from lactating dairy cows during transition to and from pasture. Livest. Sci.
114(2-3):164-175.
KHANAL, R. C. y OLSON, K. C. 2004. Factors affecting conjugated linoleic acid (CLA) content in milk, meat and egg- A review. Pak. J. Nutr. 3(2):82-98.
LIN, L.; YING, D.; CHAITEP, S. y VITTAYAPADUNG, S. 2009. Biodiesel production from crude rice bran oil and properties as fuel. App. Energy. 86(5):681-688.
LIU, J.; STIPANOVIC, R. D.; BELL, A. A.; PUCKHABER, L. S. y MAGILL, C. W. 2008. Stereoselective coupling of hemigossypol to form (+)-gossypol in moco cotton is mediated by a dirigent protein. Phytochemistry. 69(18):3038-3042.
LIU, S. J.; WANG, J. Q.; BU, D. P.; WEI, H. Y.; ZHOU, L. Y. y LUO, Q. J. 2007. The effect of dietary vegetable oilseeds supplement on fatty acid profiles in milk fat from lactating dairy cows. Agri. Sci. China. 6(8):1002-1008.
LOCK, A. L.; TELES, B. M.; PERFIELD, J. W., II; BAUMAN, D. E. y SINCLAIR, L. A. 2006. A conjugated linoleic acid supplement containing trans-10, cis-12 reduces milk fat synthesis in lactating sheep. J. Dairy Sci. 89(5):1525-1532.
LOCK, A. L.; TYBURCZY, C.; DWYER, D. A.; HARVATINE, K. J.; DESTAILLATS, F.; MOULOUNGUI, Z.; CANDY, L. y BAUMAN, D. E. 2007. trans-10 octadecenoic acid does not reduce milk fat synthesis in dairy cows. J. Nutr. 137(1):71-76.
LOOR, J. J.; FERLAY, A.; OLLIER, A.; DOREAU, M. y CHILLIARD, Y. 2005. Relationship among trans and conjugated fatty acids and bovine milk fat yield due to dietary concentrate and linseed oil. J. Dairy Sci. 88(2):726-740.
LOURENÇO, M.; DE SMET, S.; RAES, K. y FIEVEZ, V. 2007. Effect of botanical composition of silages on rumen fatty acid metabolism and fatty acid composition in longissimus muscle and subcutaneous fat of lambs. Animal. 1(06):911-921.
LUNDY, F. P.; BLOCK, E.; BRIDGES, W. C., Jr.; BERTRAND, J. A. y JENKINS, T. C. 2004. Ruminal biohydrogenation in holstein cows fed soybean fatty acids as amides or calcium salts. J. Dairy Sci. 87(4):1038-1046.
MEL´UCHOVÁ, B.; BLASKO, J.; KUBINEC, R.; GÓROVÁ, R.; DUBRAVSKÁ, J.; MARGETÍN, M. y SOJÁK, L. 2008. Seasonal variations in fatty acid composition of pasture forage plants and CLA content in ewe milk fat. Small Ruminant Res. 78(1-
3):56-65.
MOATE, P. J.; BOSTON, R. C.; JENKINS, T. C. y LEAN, I. J. 2008. Kinetics of ruminal lipolysis of triacylglycerol and biohydrogenation of long-chain fatty acids: New insights from old data. J. Dairy Sci. 91(2):731-742.

Capítulo 1
49
MOSLEY, E. E.; POWELL, G. L.; RILEY, M. B. y JENKINS, T. C. 2002. Microbial biohydrogenation of oleic acid to trans isomers in vitro. J. Lipid Res. 43(2):290-
296.
MOYA-CAMARENA, S. Y.; HEUVEL, J. P. V.; BLANCHARD, S. G.; LEESNITZER, L. A. y BELURY, M. A. 1999. Conjugated linoleic acid is a potent naturally occurring
ligand and activator of PPAR. J. Lipid Res. 40(8):1426-1433.
MUNDAY, J. S.; THOMPSON, K. G. y JAMES, K. A. C. 1999. Dietary conjugated linoleic acids promote fatty streak formation in the C57BL/6 mouse atherosclerosis model. Br. J. Nutr. 81(03):251-255.
NONI, I. D. y BATTELLI, G. 2008. Terpenes and fatty acid profiles of milk fat and "Bitto" cheese as affected by transhumance of cows on different mountain pastures. Food Chem. 109(2):299-309.
NRC. 1996. Carcinogens and anticarcinogens in the human diet. National Academy Press, Washington DC.
ODONGO, N. E.; OR-RASHID, M. M.; BAGG, R.; VESSIE, G.; DICK, P.; KEBREAB, E.; FRANCE, J. y MCBRIDE, B. W. 2007. Long-effects of feeding monensin on milk fatty acid composition in lactating dairy cows. J. Dairy Sci. 90(11):5126-5133.
OR-RASHID, M. M.; KRAMER, J. K. G.; WOOD, M. A. y MCBRIDE, B. W. 2008. Supplemental algal meal alters the ruminal trans-18:1 fatty acid and conjugated linoleic acid composition in cattle. J. Anim Sci. 86(1):187-196.
PARADIS, C.; BERTHIAUME, R.; LAFRENIERE, C.; GERVAIS, R. y CHOUINARD, P. Y. 2008. Conjugated linoleic acid content in adipose tissue of calves suckling beef cows on pasture and supplemented with raw or extruded soybeans. J. Anim Sci.
86(7):1624-1636.
PARIZA, M. W.; ASHOOR, S. H.; CHU, F. S. y LUND, D. B. 1979. Effects of temperature and time on mutagen formation in pan-fried hamburger. Cancer Letters. 7(2-3):63-
69.
PARK, H. S.; RYU, J. H.; HA, Y. L. y PARK, J. H. Y. 2001. Dietary conjugated linoleic acid (CLA) induces apoptosis of colonic mucosa in 1,2-dimethylhydrazine-treated rats: a possible mechanism of the anticarcinogenic effect by CLA. Br. J. Nutr.
86(05):549-555.
PETERSON, D. G.; MATITASHVILI, E. A. y BAUMAN, D. E. 2003. Diet-Induced milk fat depression in dairy cows results in increased trans-10, cis-12 CLA in milk fat and
coordinate suppression of mRNA abundance for mammary enzymes involved in milk fat synthesis. J. Nutr. 133(10):3098-3102.
PIPEROVA, L. S.; MOALLEM, U.; TETER, B. B.; SAMPUGNA, J.; YURAWECZ, M. P.; MOREHOUSE, K. M.; LUCHINI, D. y ERDMAN, R. A. 2004. Changes in milk fat in response to dietary supplementation with calcium salts of trans-18:1 or conjugated linoleic fatty acids in lactating dairy cows. J. Dairy Sci. 87(11):3836-3844.
PIPEROVA, L. S.; TETER, B. B.; BRUCKENTAL, I.; SAMPUGNA, J.; MILLS, S. E.; YURAWECZ, M. P.; FRITSCHE, J.; KU, K. y ERDMAN, R. A. 2000. Mammary lipogenic enzyme activity, trans fatty acids and conjugated linoleic acids are altered in lactating dairy cows fed a milk fat-depressing diet. J. Nutr. 130(10):2568-
2574.
REGO, O. A.; REGALO, S. M. M.; ROSA, H. J. D.; ALVES, S. P.; BORBA, A. E. S.;

50
Efecto de la suplementación con harina de arroz o semilla de algodón sobre el contenido de los ácidos transvaccénico y ruménico
en la leche de vacas en pastoreo
BESSA, R. J. B.; CABRITA, A. R. J. y FONSECA, A. J. M. 2008. Effects of grass silage and soybean meal supplementation on milk production and milk fatty acid profiles of grazing dairy cows. J. Dairy Sci. 91(7):2736-2743.
REGO, O. A.; PORTUGAL, P. V.; SOUSA, M. B.; ROSA, H. J.; VOUZELA, C. M.; BORVA, A. y BESSA, R. J. 2004. Effects of diet on the fatty acid patttern of milk from dairy cows. Anim. Res. 53(3):213-220.
RICO, J. E.; MORENO, B.; PABÓN, M. L. y CARULLA, J. E. 2007. Composición de la grasa láctea en la sabana de Bogotá con énfasis en ácido ruménico -CLA cis-9, trans-11. Rev. Col. Cien. Pec. 20(1):30-39.
ROJAS, C. I.; PABÓN, M. L. y CARULLA, J. E. 2005. Ácido linolénico conjugado (ALC): Factores dietarios que afectan su contenido en la leche. pp. 91-110 En:M. L. PABÓN y J. OSSA LONDOÑO(ed.). (ed.).Bioquímica, nutrición y alimentación de la vaca. Medellín.
SANHUEZA, C.; NIETO, K. y VALENZUELA, B. 2002. Ácido linoleico conjugado: un ácido graso con isometría trans potencialmente beneficioso. Rev. Chil. Nutr.
29:98-105.
SCHROEDER, G. F.; DELAHOY, J. E.; VIDAURRETA, I.; BARGO, F.; GAGLIOSTRO, G. A. y MULLER, L. D. 2003. Milk fatty acid composition of cows fed a total mixed ration or pasture plus concentrates replacing corn with fat. J. Dairy Sci.
86(10):3237-3248.
SCHROEDER, G. F.; GAGLIOSTRO, G. A.; BARGO, F.; DELAHOY, J. E. y MULLER, L. D. 2004. Effects of fat supplementation on milk production and composition by dairy cows on pasture: a review. Livest. Prod. Sci. 86(1-3):1-18.
SOYEURT, H.; DEHARENG, F.; MAYERES, P.; BERTOZZI, C. y GENGLER, N. 2008.
Variation of 9-desaturase activity in dairy cattle. J. Dairy Sci. 91(8):3211-3224.
TSIPLAKOU, E.; KOMINAKIS, A. y ZERVAS, G. 2008. The interaction between breed and diet on CLA and fatty acids content of milk fat of four sheep breeds kept indoors or at grass. Small Ruminant Res. 74(1-3):179-187.
UEDA, K.; FERLAY, A.; CHABROT, J.; LOOR, J. J.; CHILLIARD, Y. y DOREAU, M. 2003. Effect of linseed oil supplementation on ruminal digestion in dairy cows fed diets with different forage:concentrate ratios. J. Dairy Sci. 86(12):3999-4007.
VANHATALO, A.; KUOPPALA, K.; TOIVONEN, V.; SHIOHARA, T. y SHINGFIELD, K. J. 2007. Effects of forage species and stage of maturity on bovine fatty acid composition. Eur. J. Sci. Technol. 109:856-867.
VÁRADYOVÁ, Z.; KIŠIDAYOVÁ, S.; SIROKA, P. y JALC, D. 2008. Comparison of fatty acid composition of bacterial and protozoal fractions in rumen fluid of sheep fed diet supplemented with sunflower, rapeseed and linseed oils. Anim. Feed Sci. and Technol. 144(1-2):44-54.
VASTA, V.; NUDDA, A.; CANNAS, A.; LANZA, M. y PRIOLO, A. 2008. Alternative feed resources and their effects on the quality of meat and milk from small ruminants. Anim. Feed Sci. and Technol. 147(1-3):223-246.
VIBART, R. E.; FELLNER, V.; BURNS, J. C.; HUNTINGTON, G. B. y GREEN, J. T. 2008. Performance of lactating dairy cows fed varying levels of total mixed ration and pasture. J. Dairy Res. 75(4):471-480.

Capítulo 1
51
VLAEMINCK, B.; MENGISTU, G.; FIEVEZ, V.; DE JONGE, L. y DIJKSTRA, J. 2008. Effect of in vitro docosahexaenoic acid supplementation to marine algae-adapted
and unadapted rumen inoculum on the biohydrogenation of unsaturated fatty acids in freeze-dried grass. J. Dairy Sci. 91(3):1122-1132.
VOET, D. y VOET, J. G. 1990. Biochemsitry., New York.
WANG, J. H.; ZHU, B. W.; SONG, M. K. y CHOI, Y. J. 2005. Effect of monensin, fish oil or their combination on in vitro fermentation and conjugated linoleic acid (CLA) production by ruminal bacteria. Anim. Feed Sci. and Technol. 120(3-4):341-349.
WARNTJES, J. L.; ROBINSON, P. H.; GALO, E.; DEPETERS, E. J. y HOWES, D. 2008. Effects of feeding supplemental palmitic acid (C16:0) on performance and milk fatty acid profile of lactating dairy cows under summer heat. Anim. Feed Sci. and Technol. 140(3-4):241-257.
WARREN, B. E. y FARRELL, D. J. 1990. The nutritive value of full-fat and defatted Australian rice bran. I. Chemical composition. Anim. Feed Sci. and Technol. 27(3):219-228.
WEISBJERG, M. R.; WIKING, L.; KRISTENSEN, N. B. y LUND, P. 2008. Effects of supplemental dietary fatty acids on milk yield and fatty acid composition in high and medium yielding cows. J. Dairy Res. 75(02):142-152.
WILKS, D. L.; COPPOCK, C. E.; BROOKS, K. N. y GATES, C. E. 1991. Effects of differences in starch content of diets with whole cottonseed or rice bran on milk casein. J. Dairy Sci. 74(4):1314-1320.
YU, Y.; CORRELL, P. H. y VANDEN HEUVEL, J. P. 2002. Conjugated linoleic acid decreases production of pro-inflammatory products in macrophages: evidence for
a PPARdependent mechanism. Biochim. Biophys. Acta (BBA) - Molecular and Cell Biology of Lipids. 1581(3):89-99.
ZHANG, W. J.; XU, Z. R.; PAN, X. L.; YAN, X. H. y WANG, Y. b. 2007. Advances in gossypol toxicity and processing effects of whole cottonseed in dairy cows feeding. Livest. Sci. 111(1-2):1-9.
.

52
Efecto de la suplementación con harina de arroz o semilla de algodón sobre el contenido de los ácidos transvaccénico y ruménico
en la leche de vacas en pastoreo

Capítulo 2
53
2. Capítulo 2: Efecto de la suplementación con harina de arroz o semilla de algodón sobre el contenido de los ácidos transvaccénico (C18:1t11) y ruménico (C18:2c9,t11) en la leche de vacas en pastoreo
Gastón A. Castaño J.1,2, Martha L. Pabón R.1,3 y Juan E. Carulla F.1,4
1 Grupo de Investigación en Nutrición Animal. Universidad Nacional de Colombia, Sede Bogotá.
2 Candidato Magíster en Producción Animal. Universidad Nacional de Colombia, Sede Bogotá. E-mail: [email protected]
3 Profesora Titular. Departamento de Química. Facultad de Ciencias. Universidad Nacional de Colombia, Sede Bogotá. E-mail: [email protected]
4 Profesor Titular. Departamento de Producción Animal. Facultad de Medicina Veterinaria
y de Zootecnia. Universidad Nacional de Colombia, Sede Bogotá. E-mail:
Resumen
Con el fin de determinar el efecto de suplementar con harina de arroz (HA) o semilla de
algodón (SA) sobre el contenido de los ácidos grasos transvaccénico (TVa, C18:1t11) y
ruménico (Ru, C18:2c9,t11) en la grasa láctea de vacas lecheras que pastan en P.
clandestinum se evaluaron tres suplementos: un control (C) con baja concentración de
ácido linoleico (Li) y otros con HA o SA como la principal fuente de Li (1.59, 2.91 y 3.69 g
de Li/100 g de MS para los tratamientos C, HA y SA respectivamente); se utilizaron seis
vacas Holstein (4.2 ± 1.7 años de edad, 533 ± 51 Kg de peso vivo, 125 ± 29 d de
lactancia y una producción láctea de 21.7 ± 5.8 Kg/d; valor medio ± DE) que fueron
distribuidas a una réplica de un diseño Cuadrado Latino 3 x 3 doble (3 vacas x 3 periodos
x 3 tratamientos x 2 cuadrados). Cada periodo tuvo una duración de 28 d (20 d de
acostumbramiento a las dietas experimentales y 8 d para la recolección de muestras), los

54
Efecto de la suplementación con harina de arroz o semilla de algodón sobre el contenido de los ácidos transvaccénico y ruménico
en la leche de vacas en pastoreo
animales fueron ordeñados dos veces al día (05:00 y 14:00 h), permanecieron con agua
a voluntad, pastorearon en P. clandestinum y recibieron durante cada ordeño 60 g de sal
mineralizada, 3 Kg de suplemento según el tratamiento y 5 g de Cr2O3. Con la HA se
produjo grasa láctea con mayor concentración de TVa (3.11 %; p<0.01), Ru (1.41 %;
p<0.05) y de ácidos grasos insaturados (34.9 %; p<0.01), se presentó un mayor índice
calculado de actividad de la Δ9desaturasa (0.37; p<0.01) y menores índices de
aterogenicidad (1.85; p<0.05) y trombogenicidad (3.09; p<0.01). Cuando las vacas
recibieron SA no se afectaron las concentraciones de TVa y Ru en la grasa láctea, ni los
índices de aterogenicidad, de trombogenicidad y de actividad de la Δ9desaturasa, pero
se obtuvieron menores índices Ru / TVa (0.39; p<0.01) y C14:1 / C14:0 (0.08; p<0.05).
Suplementar vacas lecheras que pastan P. clandestinum con HA es una buena
alternativa para producir leche con beneficios para la salud humana a diferencia de la
SA.
Palabras Clave: ácido linoleico, ácido linoleico conjugado, ácidos grasos lácteos, vaca lechera.
Abstract
To determine the effect of dietary supplementation with rice bran (RB) or cottonseed (CS)
on transvaccenic (TVa, C18:1t11) and rumenic (Ru, C18:2c9,t11) fatty acids content in
milk fat from dairy cows grazing on P. clandestinum three supplements were evaluated: a
control (C) with low concentration of Li and others with RB or CS as the main source of Li
(1.59, 2.91 and 3.69 g of Li / 100 g DM for treatments C, RB and CS respectively); we
used six Holstein (4.2 ± 1.7 years of age, 533 ± 51 Kg of body weight, 125 ± 29 d in milk
and milk production of 21.7 ± 5.8 Kg / d, average ± SD) cows distributed to one replica of
a double Latin Square design 3 x 3 (3 cows x 3 periods x 3 treatments x 2 squares).
Periods of 28 d were used (20 d of adaptation to the experimental diets and 8 d for
collection samples); animals were milked twice a day (05:00 and 14:00 h), water ad
libitum, grazed on P. clandestinum and received during each milking 60 g of mineralized
salt, 3 Kg of concentrate according to the treatment and 5 g of Cr2O3. With the RB
produced more TVa (3.11 %; p<0.01), Ru (1.41 %; p<0.05) and unsaturated fatty acids
(34.9 %; p<0.01) milk fat concentration, higher Δ9desaturase activity index (0.37; p<0.01)
and lower atherogenicity (1.85; p<0.05) and thrombogenicity (3.09; p<0.01) indexes.
When cows received CS the TVa and the Ru milk fat concentration were not affected,

Capítulo 2
55
also for atherogenicity, thrombogenicity and Δ9desaturase activity indexes, but milk fat
had lower rates Ru / TVa (0.39; p<0.01) and C14:1 / C14:0 (0.08; p<0.05). Supplementing
dairy cows grazing P. clandestinum with HA is a good alternative to produce milk with
human health benefits in contrast to SA.
Key words: linoleic acid, conjugated linoleic acid, milk fatty acid, dairy cow.
Abreviaturas.
AG: ácido graso. Li: ácido linoleico. Ln: ácido linolénico. Ru: ácido ruménico. TVa:
ácido trans-vaccénico. C: tratamiento control. CNFn: carbohidratos no fibrosos corregidos
por N. ENL: energía neta de lactancia. FDAi: fibra detergente ácido indigerible. FDAn:
fibra detergente ácida corregida por N. FDNn: fibra detergente neutro corregida por N.
HA: tratamiento harina de arroz. MS: materia seca. SA: tratamiento semilla de algodón.
2.1 Introducción
Los ácidos grasos (AG) transvaccénico (TVa, C18:1t11) y ruménico (Ru, C18:2c9,t11) se
encuentran en la grasa láctea y se consideran como compuestos funcionales por tener
propiedades anticarcinogénicas (Ip et al., 1999; Park et al., 2001), antiaterogénicas,
antidiabetogénicas (Houseknecht et al., 1998; Munday et al., 1999), antiobesidad (Khanal
y Olson, 2004), regulación del sistema inmune (Moya-Camarena et al., 1999; Jensen,
2002) y mejoramiento de la mineralización del hueso (Jensen, 2002).
La dieta es el mayor determinante de la concentración láctea de Ru (Khanal y Olson,
2004). La suplementación con fuentes lipídicas de AG insaturados (Dhiman et al., 2000;
Khanal y Olson, 2004), las características de los forrajes (Mel´uchová et al., 2008; Vasta
et al., 2008), la relación concentrado : forraje (Ueda et al., 2003; Loor et al., 2005b), el
sistema de alimentación ya sea pastoreo o dietas totalmente mezcladas (TMR por sus
siglas en inglés; Kelly et al., 1998; Schroeder et al., 2003; Khanal et al., 2008) y el
suministro de antibióticos (Odongo et al., 2007; AlZahal et al., 2008) son factores
relacionados con la dieta que afectan la concentración de TVa y Ru en leche.
Adicionalmente, el pH ruminal, el tipo de lípidos ingeridos y el tipo de bacterias presentes
en el rumen afectan la biohidrogenación o limitan la conversión de TVa a ácido esteárico
(Chilliard et al., 2000; Tsiplakou et al., 2008).

56
Efecto de la suplementación con harina de arroz o semilla de algodón sobre el contenido de los ácidos transvaccénico y ruménico
en la leche de vacas en pastoreo
La concentración de TVa y Ru se incrementa en la leche cuando se suministran
suplementos ricos en AG insaturados (Dhiman et al., 2000). Se han evaluado diferentes
fuentes lipídicas, entre las cuales se resaltan la soya (Huang et al., 2008; Bouattour et al.,
2008), la linaza (Loor et al., 2005a; Flowers et al., 2008), el girasol (AbuGhazaleh et al.,
2007; Hervás et al., 2008), el maíz (Dewhurst et al., 2006; Bharathan et al., 2008) y el
aceite de pescado (AbuGhazaleh et al., 2003; AbuGhazaleh et al., 2004). El efecto es
diferente según el tipo de fuente lipídica, aquellas con elevado contenido de ácido
linoleico (Li) son más eficientes en producir incremento en la concentración de TVa y Ru
que las ricas en Ln (Bu et al., 2007).
La harina de arroz y la semilla de algodón se utilizan frecuentemente en la elaboración de
suplementos alimenticios para vacas lecheras en Colombia y son fuente de AG
insaturados (Dhiman et al., 1999; Lin et al., 2009). La elevada concentración de Li en la
grasa de estos dos recursos, 36 - 42 % para la harina de arroz (de Campos et al., 2007) y
57.3 % para la semilla de algodón (Dhiman et al., 1999; Khanal y Olson, 2004), son
cualidades que podrían ser aprovechadas para producir leche con altos niveles de TVa y
Ru. En la revisión de literatura no se encontraron estudios donde se hayan evaluado
estos dos recursos como suplemento alimenticio de vacas lecheras en pastoreo con el fin
de mejorar el perfil lipídico de la leche.
La producción de leche en el trópico alto colombiano se fundamenta en sistemas
pastoriles, en donde el kikuyo (Pennisetum clandestinum) es la gramínea predominante
(Correa et al., 2008) con una concentración de Ln en la grasa entre el 53.0 al 59.5 %
(Aguilar et al., 2009). El pastoreo en P. clandestinum, sumado al aporte de lípidos del
suplemento (cerca del 67 %), son características de la ganadería de leche especializada
en la Sabana de Bogotá que sugieren que este sistema tiene alto potencial para producir
leches ricas en TVa y Ru. El objetivo de este experimento fue determinar el efecto de
suplementar harina de arroz o semilla de algodón como principales fuentes de Li sobre el
contenido de TVa y Ru en la leche de vacas que pastan P. clandestinum.
2.2 Materiales y Métodos
2.2.1 Localización
El experimento se realizó en el Centro de Investigación Agropecuario Marengo de la

Capítulo 2
57
Universidad Nacional de Colombia; localizado a 4° 40’ 89’’ de latitud norte y 74° 13’ 13’’
de longitud oeste y a una altitud de 2540 msnm. La temperatura promedio es de 13 °C
(con variaciones de 0 a 20 ºC), la humedad relativa oscila entre el 80 al 85 % y la
precipitación promedio anual es 900 mm / año con dos épocas marcadas de invierno
(abril - mayo y septiembre - noviembre). El experimento fue desarrollado entre mayo y
julio de 2009.
2.2.2 Periodo experimental y tratamientos
El experimento tuvo una duración de 84 d, divididos en tres periodos de 28 d (20 d de
acostumbramiento a las dietas experimentales y 8 d para la recolección de muestras).
Los tratamientos consistieron en tres suplementos suministrados a vacas pastando P.
clandestinum: un control (C) con baja concentración de Li y otros con harina de arroz
(HA) o semilla de algodón (SA) como la principal fuente de Li (1.59, 2.91 y 3.69 g de Li /
100 g de MS para los tratamientos C, HA y SA respectivamente). Se suministraron 6 Kg
de suplemento al día, repartidos durante los dos ordeños (05:00 y 14:00 h). Las dietas
eran isoenergéticas pero contenían diferente concentración de proteína cruda. Los
ingredientes, la composición química y el perfil de AG de los suplementos se pueden
apreciar en las Tablas 2-1 y 2-2.

58
Efecto de la suplementación con harina de arroz o semilla de algodón sobre el contenido de los ácidos transvaccénico y ruménico
en la leche de vacas en pastoreo
Tabla 2-1: Composición de ingredientes y nutrientes de los suplementos alimenticios utilizados
en el experimento
Ítem Suplemento
Control Harina de Arroz Semilla de Algodón
Ingrediente, % Harina de arroz - 47.2 - Semilla de algodón - - 28.0 Aceite de palma 8.0 1.0 4.8 Torta de soya 28.7 23.7 21.3 Harina de yuca 30.0 9.6 30.6 Salvado de trigo 0.2 0.2 0.3 Palmiste (solvente) 5.1 12.9 - Cascarilla de cacao 22.6 - 9.5 Melaza de caña 5.0 5.0 5.0 Premezcla de vitaminas y minerales 0.5 0.5 0.5
Composición química Materia seca (MS), % 89.4 88.1 89.7 Proteína cruda, % MS 15.9 18.4 21.9 Fibra detergente neutro
1, % MS 22.6 24.8 20.8
Fibra detergente ácido1, % MS 20.3 14.9 18.9
Lignina, % MS 9.9 9.1 9.7 Carbohidratos no fibrosos
1, % MS 40.8 38.0 38.1
Extracto etéreo, % MS 11.6 10.3 11.4 Cenizas, % MS 9.1 8.6 7.9 ENL1X
2, Mcal / Kg MS 2.0 1.9 2.1
Ácido linoleico, % MS 1.59 2.91 3.69 1Corregidos por N (NRC, 2001).
2Energía neta de lactancia a 1X el consumo de mantenimiento
(NRC, 2001).
Tabla 2-2: Perfil de ácidos grasos (AG) de los suplementos alimenticios y del forraje utilizado en
el experimento
AG, g/100 g AG Suplemento
Forraje Control Harina de Arroz Semilla de Algodón
C10:0 0.20 - 0.03 - C12:0 1.85 0.85 0.60 2.78 C14:0 1.66 0.71 0.75 0.94 C14:1 0.04 - - - C15:0 0.09 - - - C16:0 32.29 22.59 24.54 24.39 C16:1 0.39 0.16 0.30 1.97 C17:1 0.04 - - - C18:0 7.57 2.83 5.04 6.88 C18:1t9 0.12 - - - C18:1c9 38.24 38.25 31.82 2.91 C18:1c11 0.50 0.55 0.51 - C18:2c9,c12 14.42 30.27 34.28 10.55 C18:3c 9,c12,c15 0.81 1.33 1.00 36.88 Otros 1.78 2.44 1.50 4.25
2.2.3 Animales y manejo
Todos los procesos y procedimientos del experimento fueron aprobados por el Comité de
Bioética de la Facultad de Medicina Veterinaria y de Zootecnia de la Universidad

Capítulo 2
59
Nacional de Colombia sede Bogotá (Acta 001 de 2009). Se utilizaron 6 vacas Holstein
(4.2 ± 1.7 años de edad, 533 ± 51 Kg de peso vivo, 125 ± 29 d de lactancia y una
producción láctea de 21.7 ± 5.8 Kg/d; valor medio ± DE) que fueron asignadas según la
edad, el peso, los días de lactancia y la producción de leche a una réplica de un diseño
Cuadrado Latino doble (3 vacas x 3 periodos x 3 tratamientos x 2 cuadrados). Las vacas
fueron ordeñadas dos veces al día (05:00 y 14:00 h) utilizando un sistema de ordeño
mecánico. Los animales permanecieron con agua a voluntad y recibieron durante cada
ordeño 60 g de sal mineralizada, 3 Kg de suplemento según el tratamiento y 5 g de Cr2O3
para calcular el consumo de forraje.
Antes de iniciar el experimento (50 d) se cortó la pradera y pasados 5 d se realizó
fertilización nitrogenada (46 Kg de N / Ha). Durante el experimento los animales pastaron
en un lote 2.5 Ha de P. clandestinum bajo un sistema de pastoreo en franjas utilizando
cerca eléctrica donde se corrió la cuerda dos veces al día (a.m. y p.m.). Para establecer
la masa forrajera, se determinó la proporción de la pastura con producciones de forraje
baja, media o alta de acuerdo con su altura; luego se cortaron 3 cuadrantes de 0.5 m2
representativos de cada altura y los pesos obtenidos fueron utilizados para determinar el
promedio de cada nivel de producción bajo, medio o alto; dichos promedios se
multiplicaron por la proporción de cada nivel en la pastura con el fin de estimar la
biomasa total disponible. Se utilizó un equipo de geo-posicionamiento GPSMAP® 76CSx
(Garmin Ltda., Kansas, EUA) para la medición de las áreas de pastoreo que se ajustaron
con el fin de garantizar una oferta diaria de 4 Kg de materia seca (MS) por cada 100 Kg
de peso vivo.
2.2.4 Mediciones, toma de muestras y análisis de laboratorio
Mediciones. Se pesaron los animales (día 28 de cada periodo) antes de recibir el
alimento (05:00 h) y se realizó evaluación de la condición corporal según Edmonson et al.
(1989).
Recolección de muestras.
Leche. Se pesó la producción láctea en cada ordeño (a.m. y p.m.) entre los días 21 y
28 de cada periodo. Se colectaron muestras de leche en cada ordeño de los días 21 y
28, que inmediatamente después del ordeño de la tarde se mezclaron de manera

60
Efecto de la suplementación con harina de arroz o semilla de algodón sobre el contenido de los ácidos transvaccénico y ruménico
en la leche de vacas en pastoreo
proporcional a la producción del día hasta completar 200 mL de leche, que se
dividieron en dos alícuotas de 100 mL. A una alícuota se le adicionó 1 mL de solución
de dicromato de potasio al 6 % y se conservó a - 20°C. La otra alícuota fue remitida
inmediatamente al laboratorio para la evaluación de la grasa.
Forraje. Los días 20 y 27 de cada periodo y previo al pastoreo se recolectaron
muestras de forraje (aprox. 500 g) utilizando el sistema “hand plucking” (Cook, 1964;
Muir, 2002). Las muestras se empacaron al vacío en bolsas plásticas y se
conservaron a - 20°C, posteriormente fueron secadas en un liofilizador (Martin Christ®
Alpha 14LD Plus), molidas con un molino Romer® con criba de 1 mm.
Adicionalmente, se tomaron muestras los días 13, 16, 19, 22, 25 y 28, se secaron a 60
°C por 48 h, se molieron utilizando un molino Romer® con criba de 1 mm y se
mezclaron homogéneamente para formar una muestra compuesta que fue utilizada
para determinar el contenido de fibra detergente ácido indigerible (FDAi).
Suplementos. El día 24 de cada periodo se colectó una muestra de cada suplemento
(aprox. 500 g), se molió utilizando un molino Romer® con criba de 1 mm. Se recolectó
el desperdicio de suplemento entre el día 21 y 28, se empacó en bolsas plásticas y se
conservó a 8 °C hasta ser pesado y enviado al laboratorio para determinar la
humedad.
Fluido ruminal. Se colectó fluido ruminal (aprox. 350 mL) a las 16:30 h del día 25 de
cada periodo utilizando una sonda ororuminal (Haumptner®); los primeros 250 mL
fueron eliminados y los 100 mL restantes fueron filtrados a través de dos capas de
gasa. Una alícuota de 70 mL fue utilizada para medir inmediatamente el pH con un
potenciómetro (Beckman®) y a una segunda alícuota de 30 mL se le adicionaron 3 mL
de ácido clorhídrico 6 N (Huang et al., 2009), se congeló a – 20 °C y posteriormente
fue utilizada para determinar amonio según adaptación al método Kjeldahl (Thiex
et al., 2002).
Heces. Se recolectaron dos muestras diarias (05:00 y 13:00 h) de heces entre los días
22 y 27. Las muestras se empacaron en bolsas plásticas y se conservaron a – 20 °C,
posteriormente fueron secadas a 60 °C por 48 h, molidas utilizando un molino Romer®
con criba de 1 mm y mezcladas homogéneamente para formar una muestra
compuesta de heces por animal.
Extracción de grasa y metilación

Capítulo 2
61
Leche. La extracción se realizó a través de modificaciones a las técnicas de
separación mecánica descritas por Hurley et al. (1987) y Díaz-González et al. (2002).
Se centrifugaron 100 mL de leche (15 min a 3000 rpm) y se retiró la fracción acuosa;
el sobrenadante cremoso se mezcló con 15 mL de solución detergente (50 g de
hexametafosfato de sodio y 24 mL de Tritón X100 disueltos en 1 L de agua), se agitó,
se colocó en baño de María (10 min a 90 °C), nuevamente se agitó y se puso en baño
de María (10 min a 90 °C); la grasa de la capa superficial se retiró cuidadosamente
con una micropipeta y se almacenó a – 20 °C. La metilación se hizo adaptando lo
propuesto por McCreary et al. (1978). Se recolectaron 100 µL de grasa y se
solubilizaron en 1000 µL de clorformo : mentanol (1 : 1 v/v). Una alícuota de 20 µL de
la solución de lípidos fue introducida en un vial con insertos cónicos, derivatizada con
20 µL del reactivo de metil esterificiación MethPrep II® (0.2 M
mtrifluorometilfeniltrimetilamonio hidróxido en metanol, Alltech Associates Inc.,
Deerfield, IL, USA) y 160 µL de cloroformo metanol 1 : 1 (v/v).
Suplemento. La extracción se realizó según Folch et al. (1957) y para la metilación de
los AG se utilizó el mismo proceso descrito para la grasa láctea.
Forraje. La extracción y metilación de los AG se realizó adaptando las técnicas
descritas por Garces y Mancha (1993) y por Yamasaki et al. (1999). Al forraje (50 mg)
se le adicionaron 2150 µL de metanol absoluto, 990 µL de tolueno, 66 µL de ácido
sulfúrico al 99.9 %, 1000 µL de N,Ndimetilformamida y 2 mL de nhexano; la mezcla
se colocó en un baño de María (2 h a 80°C), se dejó en reposo (5 - 10 min), se agitó y
se recuperó el sobrenadante. El nhexano del sobrenadante se evaporó mediante
corriente de N. Al residuo se le adicionaron 300 μL de diclorometano.
Cuantificación de AG (leche, forraje y suplementos). Los ésteres de AG metilados
(FAMES por sus iniciales en inglés) de la grasa de la leche, del forraje y de los
suplementos fueron cuantificados por cromatografía de gases, utilizando un cromatógrafo
de gases marca Shimadzu® modelo GC2014 con auto inyector AOC20i y auto
muestreador AOC20C. Los FAMES fueron separados en columna capilar (Rt 2560; 100
m x 0.25 mm di x 0.2 µm espesor de la capa; Restek®). Las temperaturas del inyector y
del FID fueron 260 °C y 270 °C respectivamente. El programa de temperatura fue: 140 °C
por 5 min, se incrementó 4 °C / min hasta 190 °C y se mantuvo por 32.5 min. El Split ratio
fue de 1 : 100 y el He fue el gas de arrastre con una presión de 40.4 psi. Los tiempos de
retención fueron comparados con estándares conocidos (Food Industry FAMEX Mix cat

62
Efecto de la suplementación con harina de arroz o semilla de algodón sobre el contenido de los ácidos transvaccénico y ruménico
en la leche de vacas en pastoreo
35077).
Análisis químicos
Leche. Se determinó el contenido de sólidos totales y de cenizas por gravimetría
(métodos AOAC925.23 y AOAC945.46 respectivamente; AOAC, 2006), de proteína
cruda por el método Kjeldahl (Thiex et al., 2002) y de grasa láctea por el método
Gerber (método AOAC-2000.18; AOAC, 2006). La lactosa se calculó por diferencia
(lactosa = 100 - grasa - proteína cruda - cenizas) según Lynch et al. (2007).
Forraje, suplementos y desperdicio. Se determinó humedad y cenizas por
gravimetría (métodos AOAC925.23 y AOAC945.46 respectivamente; AOAC, 2006),
proteína cruda por el método Kjeldahl (Thiex et al., 2002), extracto etéreo (método
AOAC-930.39; AOAC, 2006), fibra detergente neutro corregida por N (FDNn), fibra
detergente ácido corregida por N (FDAn; Van Soest et al., 1991; NRC, 2001), lignina
(Van Soest et al., 1991), proteína cruda insoluble en detergente neutro, proteína cruda
insoluble en detergente ácido (Licitra et al., 1996) y FDAi (Sunvold y Cochran, 1991).
Los carbohidratos no fibrosos corregidos por N (CNFn) y la energía neta de lactancia
(ENL) se estimaron según el NRC (2001). Al desperdicio sólo se le determinó la
humedad.
Fluido ruminal. Muestras frescas se usaron para medir el pH con un potenciómetro
(Beckman®) y las muestras congeladas para determinar amonio según adaptación
al método Kjeldahl (Thiex et al., 2002).
Heces. Se determinó la concentración de FDAi (Sunvold y Cochran, 1991) y de Cr por
absorción atómica (Williams et al., 1962) con un espectrofotómetro de absorción
atómica Shimadzu® modelo AA7000.
2.2.5 Cálculos
Consumo de forraje. Para determinar el consumo de forraje se utilizó Cr2O3 como
marcador indigerible externo (Holden et al., 1994) y FDAi como marcador indigerible
interno (Sunvold y Cochran, 1991). La producción de heces (Kg / d) se determinó usando
la ecuación (2-1) ajustando lo propuesto por Holden et al. (1994) teniendo en cuenta la
tasa de recuperación de Cr reportada por Correa et al. (2009).
PH = DME x [MEH]-1
x TR x 1000-1
(2-1)

Capítulo 2
63
Donde: PH es la producción de heces (Kg de MS / d); DME es la dosis del marcador
externo (g de Cr / d); [MEH] es la concentración del marcador externo en las heces (g de
Cr / g de MS de heces); TR es la tasa de recuperación del Cr, que según Correa et al.
(2009) es de 0.794. La concentración de Cr en el Cr2O3 fue 50.9 % en base húmeda.
El consumo de forraje se determinó a través de la ecuación (2-2) según lo descrito por
Aguilar et al. (2009).
CF = (PH x [FDAi]H – CS x [FDAi]S) x [FDAi]F-1
(2-2)
Donde: CF es el consumo de forraje (Kg de MS / d); PH es la producción de heces (Kg de
MS / d); [FDAi]H, [FDAi]S y [FDAi]F son las concentraciones de FDAi en heces,
suplemento y forraje respectivamente (g de FDAi / g de MS de heces, suplemento o
forraje); CS es el consumo de suplemento (Kg de MS / d).
Consumo de AG. Para determinar el consumo de AG se utilizó la ecuación (2-3).
CAG = (CF x [TAG]F x [AG]F) + (CS x [TAG]S x [AG]S) (2-3)
Donde: CAG es el consumo total de cada AG (g de AG / d); CF y CS son los consumos de
forraje y de suplemento respectivamente (g de MS de forraje o suplemento / d); [TAG]F y
[TAG]S son las concentraciones del total de AG en el forraje y en el suplemento
respectivamente (g del total de AG / g de MS de forraje o suplemento); [AG]F y [AG]S son
las concentraciones de cada AG en el total de AG de forraje y del suplemento
respectivamente (g de cada AG / g del total de AG del forraje o del suplemento). El total
de AG se estimó según el contenido de extracto etéreo con la ecuación (2-4) propuesta
por Allen (2000).
TAG = -0.98 + 1.03 x EE (2-4)
Donde: TAG es el porcentaje del total de AG en base seca (g de AG / 100 g de MS) y EE
es el porcentaje de extracto etéreo en base seca (g de EE / 100 g de MS).
Índice de actividad Δ9desaturasa. Se calculó con la ecuación (2-5) según Gagliostro et
al. (2006).

64
Efecto de la suplementación con harina de arroz o semilla de algodón sobre el contenido de los ácidos transvaccénico y ruménico
en la leche de vacas en pastoreo
IΔ9 = PΔ
9 x (PΔ
9 + SΔ
9)
-1 (2-5)
Donde: IΔ9 es el índice de actividad de la enzima Δ9desaturasa; PΔ9 es la sumatoria de
productos de la Δ9desaturasa (C14:1c9 + C16:1c9 + C18:1c9 + C18:2c9,t11); SΔ9 es la
sumatoria de sustratos de la Δ9desaturasa (C14:0 + C16:0 + C18:0 + C18:1t11).
Índice de aterogenicidad. Se calculó con la ecuación (2-6) según Ulbricht y Southgate
(1991).
IA = (SC12 + 4SC14 + SC16) x AGI-1 (2-6)
Donde: IA es el índice de aterogenicidad; SC12, SC14 y SC16 AG saturados de 12, 14 y 16 C
respectivamente; AGI es la sumatoria de los AG insaturados.
Índice de trombogenicidad. Se calculó con la ecuación (2-7) según Ulbricht y Southgate
(1991).
IT = (SC14 + SC16 + SC18) x [0.5MI + 0.5ω6 + 3ω3 + (ω3/ω6)]-1 (2-7)
Donde: IT es el índice de trombogenicidad; SC14, SC16 y SC18 AG saturados de 14, 16 y
18 C respectivamente; MI sumatoria AG monoinsaturados; ω3 y ω6 AG de la serie
omega 3 y 6 respectivamente.
2.2.6 Análisis estadístico
Los datos fueron analizados estadísticamente como un diseño Cuadrado Latino 3 x 3
doble (Kaps y Lamberson, 2004) de acuerdo con el modelo presentado en la ecuación (2-
8).
yij(k)m = µ + ϒm + η(ϒ)im + λ(ϒ)jm + τ(k) + εij(k)m (2-8)
Donde:
yij(k)m = Valor de la observación del i-ésimo periodo (i = 1, 2, 3), j-ésimo animal (j = 1, 2,
…, 6), k-ésimo tipo de suplemento (k = C, HA, SA) y m-ésimo cuadrado (m = 1,

Capítulo 2
65
2)
µ = Promedio de la población
ϒm = Efecto del m-ésimo cuadrado (m = 1, 2)
η(ϒ)im = Efecto del i-ésimo periodo (i = 1, 2, 3) dentro del m-ésimo cuadrado (m = 1, 2)
λ(ϒ)jm = Efecto del j-ésimo animal (j = 1, 2, 3) dentro del m-ésimo cuadrado (m = 1, 2)
τ(k) = Efecto del k-ésimo tipo de suplemento (k = C, HA, SA)
εij(k)m = Error experimental
Los supuestos fueron probados con la metodología indicada para cada caso. Cuando el
modelo resultó estadísticamente significativo se procedió con un análisis de comparación
de medias usando la prueba Tukey. Para el análisis de los resultados se utilizó el
programa SAS versión 9.0 (Copyright© 2002 by SAS Institute Inc., Cary, NC, USA).
2.3 Resultados
2.3.1 Consumo de alimento
Los consumos de MS, de proteína cruda, FDNn, FDAn, Lignina, CNFn, extracto etéreo y
ENL no se vieron afectados por el tipo de suplemento (Tabla 2-3).

66
Efecto de la suplementación con harina de arroz o semilla de algodón sobre el contenido de los ácidos transvaccénico y ruménico
en la leche de vacas en pastoreo
Tabla 2-3: Efecto del uso de suplementos con harina de arroz (HA) o semilla de algodón (SA)
como las principales fuentes de ácido linoleico sobre el consumo de alimento de vacas lactantes que pastaban P. clandestinum
Fracción del alimento Suplemento
ESM1 RC
2 p
3
Control HA SA
Materia seca Suplemento, Kg/d 5.1 5.1 4.8 0.70 0.51 NS Forraje, Kg/d 11.7 11.6 12.6 1.85 0.83 NS Total
Kg/d 16.8 16.6 17.4 1.94 0.77 NS Kg/100 Kg de PV
4 3.0 3.0 3.1 0.32 0.68 NS
Proteína cruda Suplemento, Kg/d 0.8 0.9 1.0 0.15 0.69 NS Forraje, Kg/d 2.2 2.2 2.4 0.35 0.84 NS Total, Kg/d 3.0 3.1 3.4 0.37 0.80 NS
ENLP5
Suplemento, Mcal/d 9.4 9.3 9.4 1.32 0.52 NS Forraje, Mcal/d 17.3 17.1 18.5 2.62 0.84 NS Total, Mcal/d 26.7 26.4 27.9 2.75 0.77 NS
Fibra detergente neutro6
Suplemento, Kg/d 1.1 1.3 1.0 0.15 0.71 NS Forraje, Kg/d 5.6 5.5 6.0 0.87 0.82 NS Total
Kg/d 6.7 6.8 7.0 0.89 0.79 NS Kg/100 Kg de PV
4 1.2 1.2 1.2 0.15 0.69 NS
Fibra detergente ácido6
Suplemento, Kg/d 1.0 0.8 0.9 0.13 0.76 NS Forraje, Kg/d 3.2 3.1 3.4 0.50 0.80 NS Total, Kg/d 4.2 3.9 4.3 0.51 0.77 NS
Lignina Suplemento, g/d 501 463 463 68.1 0.53 NS Forraje, g/d 286 282 306 44.9 0.82 NS Total, g/d 787 745 769 80.1 0.41 NS
Carbohidratos no fibrosos6
Suplemento, Kg/d 2.1 1.9 1.8 0.27 0.58 NS Forraje, Kg/d 2.2 2.1 2.3 0.34 0.83 NS Total, Kg/d 4.2 4.1 4.1 0.42 0.64 NS
Extracto etéreo Suplemento, g/d 590 522 545 79.9 0.57 NS Forraje, g/d 430 417 459 68.6 0.90 NS Total, g/d 1021 939 1003 97.2 0.74 NS
1Error estándar de la media de los tratamientos.
2R cuadrado.
3Significancia; NS: no significativa;
letras diferentes en la misma fila indican diferencias estadísticas *: p<0.05, **: p<0.01 y ***: p<0.001.
4Peso vivo.
5Energía neta de lactancia corregida al nivel de consumo (NRC, 2001).
6Corregido por N (NRC, 2001).
2.3.2 Consumo de precursores de los ácidos trans-vaccénico y
ruménico
En el experimento, el consumo promedio de lípidos provenientes del suplemento y del
forraje fue 552 y 435 g / d respectivamente; de tal manera que el 55.9 % de los lípidos de
la dieta provenían del suplemento, aunque éste sólo aportó el 29.6 % de la MS. Los

Capítulo 2
67
animales que recibieron el suplemento HA o SA aumentaron casi dos veces el consumo
de Li comparativamente con aquellos que consumieron el suplemento C (113, 179 y 211
g / d para los tratamientos C, HA y SA respectivamente; p<0.01). Este incremento se
debió principalmente a la suplementación ya que los consumos de Li y Ln provenientes
del forraje fueron similares entre tratamientos. Como consecuencia del mayor consumo
de Li en los suplementos HA y SA (148 y 176 g / d respectivamente; p<0.01) se
incrementó el consumo total de precursores de TVa y Ru, en comparación al obtenido
con el suplemento C (235, 300 y 343 g/d para los tratamientos C, HA y SA
respectivamente; p<0.01; Tabla 2-4).
Tabla 2-4: Efecto del uso de suplementos con harina de arroz (HA) o semilla de algodón (SA)
como las principales fuentes de ácido linoleico sobre el consumo de los ácidos linoleico y linolénico, precursores de los ácidos trans-vaccénico y ruménico en la leche de vacas que pastaban P. clandestinum
Ítem Suplemento
ESM1
RC2 p
3
Control HA SA
Consumo, g/d Linoleico
Suplemento 80b 148
a 176
a 24.6 0.90 **
Forraje 32 32 35 5.3 0.92 NS
Total 113b 179
a 211
a 23.6 0.90 **
Linolénico Suplemento 5 6 5 0.76 0.82 NS
Forraje 118 115 126 19.3 0.92 NS
Total 123 121 132 19.2 0.92 NS
Total precursores4
Suplemento 85b 154
a 181
a 25.3 0.90 **
Forraje 150 146 161 24.6 0.92 NS
Total 235b 300
a 343
a 30.6 0.91 **
Aporte relativo5, %
Linoleico Suplemento 72.2
b 82.5
a 82.6
a 3.74 0.94 **
Forraje 27.8a 17.5
b 17.4
b 3.74 0.94 **
Linolénico Suplemento 4.1
b 5.5
a 4.5
ab 0.73 0.92 *
Forraje 95.9a 94.5
b 95.5
ab 0.73 0.92 *
Total precursores4
Suplemento 32.6b 46.3
a 48.6
a 5.29 0.92 **
Forraje 67.4a 53.7
b 51.4
b 5.29 0.92 **
1Error estándar de la media de los tratamientos.
2R cuadrado.
3Significancia; NS: no significativa;
letras diferentes en la misma fila indican diferencias estadísticas *: p<0.05, **: p<0.01 y ***: p<0.001.
4Sumatoria de los ácidos grasos linoleico y linolénico.
5g de ácido graso proveniente del
suplemento o del forraje / 100 g de precursores consumidos al día

68
Efecto de la suplementación con harina de arroz o semilla de algodón sobre el contenido de los ácidos transvaccénico y ruménico
en la leche de vacas en pastoreo
2.3.3 Producción y calidad de la leche
La producción diaria de leche disminuyó cuando las vacas recibieron SA (20.2 Kg / d;
p<0.01) frente a la obtenida con los otros tratamientos. La concentración de sólidos
totales, proteína cruda, cenizas y lactosa en la leche fue similar entre tratamientos. Sin
embargo, las vacas que consumieron SA presentaron mayores concentraciones de grasa
en la leche comparadas con HA (3.6 y 3.2 % respectivamente; p<0.05; Tabla 2-5).
La producción de leche expresada en sólidos totales fue superior para HA en
comparación con SA (2.58 y 2.40 Kg/d respectivamente; p<0.05). La mayor producción
en estos tratamientos se explicó por un aumento en la producción de lactosa y cenizas ya
que las producciones de grasa y proteína fueron similares entre los tratamientos. Las
vacas que consumieron HA produjeron leche con mayores concentraciones de TVa (0.10
%; p<0.01) y Ru (0.05%; p<0.05) lo que se reflejó en una mayor producción diaria de
estos AG (21.8 y 9.8 g/d respectivamente; p<0.01; Tabla 2-5).
Tabla 2-5: Efecto del uso de suplementos con harina de arroz (HA) o semilla de algodón (SA)
como las principales fuentes de ácido linoleico sobre la producción y calidad de la leche de vacas que pastaban P. clandestinum
Ítem Suplemento
ESM1 RC
2 p
3
Control HA SA Leche
Producción, Kg/d 21.2a 22.0
a 20.2
b 0.62 0.99 **
Sólidos totales Concentración, % 12.1 11.8 11.9 0.18 0.97 NS Producción, Kg/d 2.56
ab 2.58
a 2.40
b 0.103 0.96 *
Proteína cruda Concentración, % 2.9 2.8 2.9 0.16 0.95 NS Producción, Kg/d 0.61 0.60 0.58 0.029 0.89 NS
Grasa Concentración , % 3.4
ab 3.2
b 3.6
a 0.15 0.96 *
Producción, Kg/d 0.71 0.70 0.73 0.045 0.94 NS Cenizas
Concentración , % 0.8 0.8 0.7 0.03 0.77 NS Producción, Kg/d 0.16
a 0.17
a 0.15
b 0.008 0.97 *
Lactosa Concentración, % 5.0 5.0 4.7 0.21 0.79 NS Producción, Kg/d 1.07
ab 1.10
a 0.95
b 0.072 0.94 *
Ácido trans-vaccénico Concentración, % 0.08
b 0.10
a 0.09
b 0.007 0.95 **
Producción, g/d 16.9b 21.8
a 18.4
b 1.51 0.95 **
Ácido ruménico Concentración, % 0.04
b 0.05
a 0.04
b 0.004 0.92 *
Producción, g/d 8.0b 9.8
a 7.2
b 0.65 0.94 **
1Error estándar de la media de los tratamientos.
2R cuadrado.
3Significancia; NS: no significativa;
letras diferentes en la misma fila indican diferencias estadísticas *: p<0.05 y **: p<0.01.

Capítulo 2
69
2.3.4 Perfil de ácidos grasos de la grasa láctea
La concentración de los AG C4:0, C6:0, C8:0, C10:0, C11:0, C13:0, C15:0, C17:0,
C18:2c-9,c-12 y C18:3c-9,c-12,c-15 en la grasa láctea fue similar entre tratamientos. La grasa
láctea de las vacas que consumieron HA presentó una mayor concentración de los AG
C12:0 (2.27 %; p<0.01), C14:0 (8.85 %; p<0.01), C18:1c-9, (26.44 %; p<0.01), C18:1t-11
(TVa; 3.11 %; p<0.01;) y C18:2c-9,t-11 (Ru; 1.41 %; p<0.001), mientras la concentración de
C16:0 fue menor (26.35 %; p<0.001). Con HA se incrementó la concentración de C14:1c-9
(0.79 %; p<0.05), C18:1c-11 (0.28 %; p<0.05) y C18:2c-9,t-12 (0.22 %; p<0.01) en
comparación a SA, pero no frente al C. La concentración de C16:1c-9 fue mayor con el
tratamiento C (1.41 %; p<0.01). La concentración de C18:0 fue diferente al comparar los
tres tratamientos, siendo menor para C y mayor para HA (14.05 y 16.31 %
respectivamente; p<0.001; Tabla 2-6).
Cuando las vacas recibieron HA produjeron grasa láctea con mayor cantidad de AG
insaturados y menor de saturados (34.9 y 60.8% respectivamente; p<0.01). Los
tratamientos no afectaron la concentración de AG de cadena corta (<C12), pero sí el
porcentaje de los AG de cadena media (C12-16) y larga (>C16). El porcentaje de AG de
cadena media fue menor para HA y mayor para el C (40.7 y 44.6 % respectivamente;
p<0.001), este último tratamiento presentó un menor porcentaje de AG de cadena larga
(44.9%; p<0.001; Tabla 2-6).
La grasa láctea presentó una menor concentración de AG preformados (>C17) cuando las
vacas fueron alimentadas con el C (44.5%; p<0.001) y mayor concentración de AG de
novo (<C16) cuando recibieron HA (19.8%; p<0.01; Tabla 2-6).
Los índices calculados Ru / TVa (0.39; p<0.01), C14:1 / C14:0 (0.08; p<0.05) y C16:1 /
C16:0 (0.042; p<0.05) fueron menores para la leche de animales que recibieron SA. Con
el tratamiento HA se encontró un mayor índice de actividad de la Δ9-desaturasa (0.37;
p<0.01), una mayor concentración de los productos de esta enzima (31.49%; p<0.01) y
menor de los sustratos (53.31; p<0.001; Tabla 2-6).
El índice de aterogenicidad fue mayor para la grasa láctea de las vacas que
recibieron C (2.03; p<0.05) frente al HA pero no frente a SA. El índice de
trombogenicidad fue menor para HA (3.09; p<0.01; Tabla 2-6).

70
Efecto de la suplementación con harina de arroz o semilla de algodón sobre el contenido de los ácidos transvaccénico y ruménico
en la leche de vacas en pastoreo
Tabla 2-6: Efecto del uso de suplementos con harina de arroz (HA) o semilla de algodón (SA) como las principales fuentes de ácido linoleico sobre el perfil de ácidos grasos (AG) de la grasa láctea de vacas que pastaban P. clandestinum
Ítem Suplemento
ESM1 RC
2 p
3
Control HA SA
AG, g/100 g de AG C4:0 2.48 2.46 2.55 0.154 0.91 NS C6:0 1.50 1.53 1.55 0.090 0.94 NS C8:0 0.81 0.85 0.83 0.055 0.95 NS C10:0 1.59 1.71 1.60 0.123 0.95 NS C11:0 0.20 0.20 0.19 0.014 0.97 NS C12:0 1.99
b 2.27
a 1.87
b 0.139 0.94 **
C13:0 0.10 0.10 0.09 0.013 0.87 NS C14:0 8.07
b 8.85
a 7.99
b 0.298 0.94 **
C14:1c-9 0.72ab
0.79a 0.63
b 0.074 0.94 *
C15:0 1.06 1.08 1.03 0.079 0.81 NS C16:0 31.30
a 26.35
b 29.36
a 0.512 0.99 ***
C16:1c-9 1.41a 1.22
b 1.22
b 0.060 0.95 **
C17:0 0.43 0.44 0.47 0.035 0.73 NS C18:0 14.05
c 15.00
b 16.31
a 0.531 0.96 ***
C18:1c-9 25.09b 26.44
a 25.07
b 0.515 0.98 **
C18:1c-11 0.27ab
0.28a 0.25
b 0.013 0.93 *
C18:1t-11 (trans-vaccénico) 2.38b 3.11
a 2.52
b 0.209 0.90 **
C18:2c-9,c-12 0.96 1.02 1.08 0.085 0.92 NS C18:2c-9,t-11 (ruménico) 1.13
b 1.41
a 0.98
b 0.106 0.93 ***
C18:2c-9,t-12 0.19ab
0.22a 0.17
b 0.018 0.93 **
C18:3c-9,c-12,c-15 0.40 0.37 0.37 0.051 0.85 NS Otros 3.88 4.29 3.89 0.251 0.85 NS
AG según su saturación, g/100 g de AG
AG insaturados 32.5b 34.9
a 32.3
b 0.9 0.96 **
AG saturados 63.6a 60.8
b 63.8
a 0.9 0.96 **
AG según su longitud, g/100 g de AG AG de cadena corta (<C12) 6.6 6.7 6.7 0.4 0.95 NS AG de cadena media (C12-16) 44.6
a 40.7
c 42.2
b 0.7 0.97 ***
AG de cadena larga (>C16) 44.9b 48.3
a 47.2
a 0.7 0.97 ***
AG según su origen, g/100 g de AG De novo (<C16) 18.5
b 19.8
a 18.3
b 0.6 0.96 **
Preformados (>C17) 44.5b 47.9
a 46.7
a 0.7 0.98 ***
Indicadores actividad Δ9-Desaturasa
Índice Ru / ATVa 0.48a 0.46
a 0.39
b 0.03 0.92 **
Índice C14:1 / C14:0 0.09a 0.09
a 0.08
b 0.01 0.97 *
Índice C16:1 / C16:0 0.045a 0.047
a 0.042
b 0.003 0.92 *
Productos de la Δ9-Desaturasa
4 29.90
b 31.49
a 29.52
b 0.74 0.97 **
Sustratos de la Δ9-Desaturasa
5 55.80
a 53.31
b 56.17
a 0.65 0.97 ***
Índice Δ9-Desaturasa
6 0.35
b 0.37
a 0.34
b 0.008 0.97 **
Índice Aterotogénicidad7 2.03
a 1.85
b 1.97
ab 0.08 0.96 *
Índice Trombogenicidad8 3.43
a 3.09
b 3.50
a 0.11 0.97 **
1Error estándar de la media de los tratamientos.
2R cuadrado.
3Significancia; NS: no significativa;
letras diferentes en la misma fila indican diferencias estadísticas *: p<0.05, **: p<0.01 y ***: p<0.001.
4C14:1c-9 + C16:1c-9 + C18:1c-9 + C18:2c-9,t-11.
5C14:0 + C16:0 + C18:0 + C18:1t-11.
6productos de la Δ
9-desaturasa/(sustratos de la Δ
9-desaturasa + productos de la Δ
9desaturasa)
-1;
Gagliostro et al. (2006). 7(C12:0 + 4C14:0 + C16:0) / AG insaturados; Ulbricht y Southgate (1991).
8 (C14:0 + C16:0 + C18:0) x [0.5 monoinsaturados + 0.5ω-6 + 3ω-3 + (ω-3/ω-6)]
-1; Ulbricht y
Southgate (1991).

Capítulo 2
71
2.3.5 Peso vivo, condición corporal y fluido ruminal
El peso vivo, la condición corporal y el fluido ruminal no se vieron afectados por el tipo de
suplemento (Tabla 2-7).
Tabla 2-7: Efecto del uso de suplementos con harina de arroz (HA) o semilla de algodón (SA) como las principales fuentes de ácido linoleico sobre el peso, la condición corporal y el fluido ruminal de de vacas lactantes que pastan P. clandestinum
Ítem Suplemento
ESM1
RC2 p
3
Control HA SA
Peso vivo, Kg 559 561 562 10.7 0.97 NS Condición corporal 3.5 3.5 3.6 0.17 0.96 NS Fluido ruminal
pH 6.6 6.6 6.9 0.25 0.91 NS N-NH3, mg/dL 13.7 14.5 13.8 1.59 0.88 NS
1Error estándar de la media de los tratamientos.
2R cuadrado.
3Significancia; NS: no significativa;
letras diferentes en la misma fila indican diferencias estadísticas *: p<0.05, **: p<0.01 y ***: p<0.001.
2.4 Discusión
Este trabajo buscaba comparar dos fuentes tradicionalmente usadas en la
suplementación de vacas lecheras (SA y HA) que contienen cantidades apreciables de Li
sobre el perfil de AG de la leche. La literatura es abundante en señalar que el incremento
del Li en la ración aumenta las concentraciones de Ru y TVa en la leche (Dhiman et al.,
2000; Bu et al., 2007). Se elaboraron dos suplementos con SA o HA, con similares
cantidades de Li y semejante composición nutricional. Se incluyó un control con
concentración similar de lípidos y con un aporte menor de Li. De esta manera se
esperaba documentar el efecto de la fuente del Li (SA o HA) sobre el perfil de AG de la
leche.
2.4.1 Consumo de alimento
No se presentaron diferencias en el consumo de MS asociadas al origen de los lípidos en
el suplemento (HA o SA) o a las concentraciones de Li. Algunos autores han sugerido
que al incrementar la grasa en la dieta se puede disminuir el consumo de MS (Schauff y
Clark, 1992) y que este efecto aumenta con el incremento en la proporción de AG
insaturados en el suplemento (Firkins y Eastridge, 1994; Bremmer et al., 1998) lo cual no
fue observado en nuestro experimento. Concentraciones mayores al 8 ó 9 % de grasa en
la dieta disminuyen el consumo de MS (Palmquist y Jenkins, 1980), mientras que en las

72
Efecto de la suplementación con harina de arroz o semilla de algodón sobre el contenido de los ácidos transvaccénico y ruménico
en la leche de vacas en pastoreo
revisiones de experimentos con vacas en pastoreo Schroeder et al. (2004) y Bargo et al.
(2003) no encontraron diferencias en el consumo MS al suplementar con grasa. En
nuestro trabajo, la grasa representó el 5.8 % del total de la MS consumida y la grasa
proveniente de los suplementos fue cercano al 3 %. El NRC (2001) recomienda que no
se debe sobrepasar entre 3 a 4 % de grasa en la dieta proveniente del suplemento.
La ausencia de diferencias en los consumos de MS y de nutrientes entre tratamientos
sugiere que los principales efectos que se presentaron en este estudio estarían
principalmente asociados al perfil de AG y al origen de los mismos. Se puede señalar
que el consumo de Li fue cerca del doble en los tratamientos de HA o SA al compararlos
con los del C mientras que los consumos de C16:0 fueron mayores para del C (p < 0.01;
datos no incluidos). Los consumos de HA y SA fueron similares en todos los nutrientes
incluyendo el Li aunque su concentración fue superior para SA. El suplemento aportó 46
y 49 % de los precursores para los tratamientos HA o SA respectivamente, pero sólo el
33 % para el C. La composición de los forrajes puede afectar la concentración de Ru en
la leche (Lourenço et al., 2007) ya que el forraje es una fuente rica en Ln (Mohammed et
al., 2009) y su concentración en los forrajes es directamente proporcional al contenido de
Ru en ésta (Mel´uchová et al., 2008). El consumo de Ln proveniente del forraje no se vio
afectado por la adición de HA o SA por lo tanto las variaciones en TVa y Ru no estarían
relacionadas con el consumo de Ln.
2.4.2 Producción y calidad de la leche
La producción de leche y sólidos totales fue menor para SA. Las diferencias en el aporte
de Li entre los tratamientos no explicaron las diferencias en la producción de leche y los
sólidos totales entre tratamientos ya que el consumo de este AG en SA y HA fue similar
y diferente al control. Las diferencias en la producción de leche han sido asociadas con el
consumo de energía (van Knegsel et al., 2007). En este trabajo, los consumos de energía
digestible estimada con el uso de los marcadores o energía neta por fórmulas (NRC,
2001) no fueron diferentes entre los tratamientos. Sin embargo, la menor producción de
leche en el tratamiento con SA implicaría de alguna manera una menor eficiencia en el
uso de la energía digestible por parte de los animales en este tratamiento.
La semilla de algodón posee factores tóxicos como el gosipol (Zhang et al., 2007), pero
las vacas lecheras son menos susceptibles de manifestar síntomas de intoxicación que

Capítulo 2
73
los no rumiantes (Coppock et al., 1987; Blauwiekel et al., 1997), pues el gosipol se
detoxifica en el rumen (Reiser y Fu, 1962). Sin embargo, se ha evidenciado que parte del
gosipol escapa de la detoxificación (Hawkins et al., 1985; Mena et al., 2004) y puede
afectar la salud o el desempeño animal (Risco et al., 2002). Algunos autores sugieren
que en vacas lecheras se puede usar semilla de algodón hasta niveles del 15 % de la
ración o entre 3 a 4 Kg / vaca / d sin que se presenten efectos negativos (Coppock et al.,
1987; Arieli, 1998; Zhang et al., 2007) como fragilidad osmótica del eritrocito (Risco et al.,
2002), bajas concentraciones de hemoglobina y hematocrito (Velasquez-Pereira et al.,
1999), limitación del sistema antioxidante de la célula (Risco et al., 2002), disminución en
el desempeño reproductivo (Santos et al., 2003; Villaseñor et al., 2008), reducción de la
producción láctea, dificultad respiratoria y hasta la muerte (Lindsey et al., 1980). Aunque
en este trabajo no se determinó el gosipol es probable que los efectos tóxicos de este
compuesto modifiquen la eficiencia del uso de la energía en el animal. El consumo de
semilla de algodón fue de 1.5 Kg / vaca / d (8.6 % de la MS consumida) que fue inferior al
máximo recomendado en la literatura (Coppock et al., 1987; Arieli, 1998; Zhang et al.,
2007). Sin embargo, las concentraciones de gosipol son variables y están asociadas a
las variedades de algodón (Zhang et al., 2007).
Las vacas suplementadas con SA produjeron leche con mayor porcentaje de grasa, pero
debido a la menor producción de leche en este tratamiento, la producción diaria de grasa
no fue diferente a los demás tratamientos. El efecto de la suplementación con semilla de
algodón sobre la concentración y producción de grasa no es consistente (Arieli, 1998).
Algunos trabajos no encuentran efectos sobre la concentración y producción de grasa
láctea (Bitman et al., 1996), mientras que otros señalan un incremento (Belibasakis y
Tsirgogianni, 1995; Harrison et al., 1995) o una disminución (Wilks et al., 1991; Smith et
al., 1993).
2.4.3 Perfil de ácidos grasos de la grasa láctea
El perfil de AG de la grasa láctea se modificó por el mayor consumo de Li en HA pero no
en SA que fue similar al perfil de AG del tratamiento C para la mayoría de AG. Los AG
en la leche que se modificaron debido al consumo de HA fueron el C12:0, C14:0,
C18:1c9, TVa, Ru y C18:2c9,t12 que se encontraron en mayor concentración y el C16:0 en
menor concentración que en los demás tratamientos. Varios trabajos han señalado que la
adición de AG insaturados en forma de semillas enteras tiene un menor impacto sobre la

74
Efecto de la suplementación con harina de arroz o semilla de algodón sobre el contenido de los ácidos transvaccénico y ruménico
en la leche de vacas en pastoreo
presencia de AG en la leche como el TVa y el Ru que cuando el AG se suministra en una
semilla molida o en forma de aceite no protegido (Dhiman et al., 2000). Esto podría
explicar parcialmente porque se presentaron diferentes concentraciones de AG con HA y
no con SA donde la semilla se proporcionó entera.
Algunos autores han sugerido que la menor concentración de TVa y Ru en la grasa
láctea de vacas consumiendo semillas enteras comparadas con aquellas consumiendo
semillas procesadas se debe a una menor disponibilidad ruminal de los AG insaturados
en las semillas enteras que están protegidos de la biohidrogenación ruminal (Dhiman et
al., 2000; Khanal et al., 2005; Paradis et al., 2008). Esta protección de los AG
insaturados se refleja en una mayor concentración de estos AG en la leche. Sin
embargo, las concentraciones de Li en la grasa láctea fueron iguales para el control y el
SA. Por lo tanto, la menor concentración de TVa, Ru, C18:1c-9, C18:1c-11 y C18:2c-9,t-12 en
la leche para el tratamiento SA en comparación con HA no se podría atribuir a una menor
disponibilidad de los AG en el rumen.
Otra posible explicación es que la biohidrogenación ruminal para el Li en la SA fue mayor
que en HA. Algunos autores han reportado una mayor biohidrogenación para la semilla
de algodón. Pires et al. (1997) encontraron una mayor biohidrogenación del Li a C18:0
cuando se suministra semilla de algodón sin procesar y una menor concentración de
C18:2 en la leche. Harvatine et al. (2002) encontraron un incremento lineal en la
biohidrogenación y flujo de C18:0 hacia el intestino al incrementar la semilla de algodón
de 0 a 15 % en la dieta. Pires et al. (1997) indican que posiblemente se incrementa la
saturación del Li debido a que la semilla de algodón permanece mayor tiempo en el
rumen (tasas de paso más lentas). En nuestro estudio, la mayor concentración de C18:0
en la grasa láctea de vacas que consumieron el suplemento con semillas de algodón
reafirmaría esta teoría. De otra parte, sugeriría que la harina de arroz sería una mejor
alternativa para modificar el perfil de los ácidos grasos hacia TVa y Ru que la semilla de
algodón cuando aportan cantidades similares de Li debido a tasas de paso diferentes
entre estos dos recursos.
Las explicaciones para otras modificaciones en el perfil de la grasa láctea en nuestro
experimento son menos claras. Las mayores concentraciones de C12:0 y C14:0 y
menores de C16:0 en la grasa láctea de animales suplementados con HA al compararlos

Capítulo 2
75
con el C o SA implicarían cambios en la fermentación o en el metabolismo de los lípidos
en el animal para este tratamiento comparativamente con los otros dos. Normalmente los
alimentos y los microorganismos tienen concentraciones muy bajas de C12:0 y C14:0
(AbuGhazaleh et al., 2002), de tal manera que estos AG provienen principalmente de la
síntesis de novo en la glándula mamaria (Garnsworthy et al., 2006), siendo el C14:0 el
principal producto de la misma (Barber et al., 1997). La síntesis de novo es mayor
cuando los animales se encuentran en un mejor balance energético y dependen en
menor grado de las reservas corporales (Palmquist et al., 1993). Bajo estas condiciones,
se esperaría un menor aporte de AG preformados (Garnsworthy et al., 2006) que no fue
el caso de este estudio ya que el suplemento con HA presentó mayores concentraciones
de AG preformados. El aporte similar de energía y producciones de leche entre HA y C
tampoco podría justificar una mayor movilización de tejido en uno o en el otro. El
consumo de C12:0 y C14:0 fue mayor para C en comparación con los otros tratamientos
(p<0.001; datos no incluidos), de tal manera que diferencias en el consumo de C12:0 y
C14:0 no explican la mayor concentración de estos AG con HA. Una variación en la
población microbiana en el rumen podría estar relacionada con la mayor concentración
de C12:0 y C14:0 para HA, pues los lípidos de los microorganismos pueden ser
generados por la síntesis de novo (Schmidely et al., 2008; Váradyová et al., 2008) y
teniendo en cuenta que las bacterias tienen más concentración de estos AG con relación
a los protozoarios (Or-Rashid et al., 2007) es probable que variaciones se debieron a una
disminución en la población de protozoarios.
El C16:0 en la grasa láctea de vacas alimentadas con suplemento que contenía HA fue
menor que SA y C. El C16:0 puede provenir de la dieta, la síntesis de novo, de la
movilización de tejido adiposo o de los microorganismos ruminales (Linzell y Peaker,
1971; Barber et al., 1997). La diferencia con C podría explicarse por un menor consumo
de C16:0 en HA. Sin embargo, la diferencia entre HA y SA no se podría relacionar con
variaciones en el consumo de C16:0, ya que los dos tratamientos presentaron niveles
similares de C16:0 en la dieta. Adicionalmente, C y SA tuvieron concentraciones
similares de C16:0 en la leche. Difícilmente, la menor concentración de C16:0 en HA
podría explicarse por una menor síntesis de novo ya que las concentraciones de C12 y
C14 provenientes de esta ruta metabólica fueron mayores en este tratamiento. Las
menores concentraciones de C16:0 tampoco podrían explicarse por una menor
movilización de lípidos en los tejidos ya que los AG preformados en la grasa láctea

76
Efecto de la suplementación con harina de arroz o semilla de algodón sobre el contenido de los ácidos transvaccénico y ruménico
en la leche de vacas en pastoreo
también fueron mayores para HA. Una menor concentración de C16:0 en los
microorganismos del rumen podría explicar la disminución de este AG para HA pues el
C16:0 es uno de los AG predominantes en los microorganismos ruminales (Bas et al.,
2003; Or-Rashid et al., 2007; Váradyová et al., 2008) y puede ser producido a través de
la síntesis de novo en el rumen (Williams, 1986). La suplementación con diferentes
aceites produce cambios en la composición de AG del contenido ruminal asociados a
modificaciones en las poblaciones microbianas (Toral et al., 2012). Aproximadamente del
7.5 al 15 % de los lípidos presentes en la digesta ruminal (Or-Rashid et al., 2007) y tres
cuartas partes de los AG presentes en los microorganismos del rumen (Jenkins et al.,
2008) provienen de los protozoarios. Teniendo en cuenta que los protozoarios
representan una fuente importante de lípidos para el animal hospedero (Or-Rashid et al.,
2007) y que poseen mayor concentración de C16:0 que la bacterias (Or-Rashid et al.,
2007; Váradyová et al., 2008) es probable que la suplementación con HA produjera una
disminución de los protozoarios en el rumen, lo cual podría explicar la baja concentración
de C16:0 en la grasa láctea para este tratamiento. En apoyo de esta hipótesis se ha
observado que el suministro de grasa disminuyó la población de protozoarios en el rumen
dependiendo del tipo y disponibilidad de AG (Reveneau et al., 2012) y la adición de
aceite de girasol sólo o en combinación con algas marinas disminuyeron la concentración
de C16:0 en el fluido ruminal de ovejas (Toral et al., 2009).
La HA presentó el menor índice de aterogenicidad y trombogenicidad en la grasa láctea,
las diferencias fueron significativas frente al tratamiento C y se presentó una tendencia
frente a la SA (Tabla 2-6). Los menores índices de aterogenicidad y trombogenicidad del
HA pueden ser explicados por la menor concentración de C16:0 y la mayor
concentración de AG insaturados. Un aumento en la proporción de AG insaturados
puede ser benéfico para la salud humana (Jacobs et al., 2011). El uso de leches con
mejores índices de aterogenicidad y trombosidad serían una opción para mercados
saludables ya que las enfermedades coronarias en la mayoría de los casos se deben a la
obstrucción de los vasos por aterosclerosis o trombosis, de manera aislada o en
combinación (Ulbricht y Southgate, 1991).
2.4.4 Actividad de la Δ9desaturasa
En este trabajo no se midió la actividad de la Δ9desaturasa. Sin embargo, algunos
autores sugieren que las relaciones entre los sustratos y los productos de esta enzima en

Capítulo 2
77
la grasa láctea pueden ser un indicador de su actividad (Jacobs et al., 2011) o de una
mayor concentración en la glándula mamaria (Feng et al., 2004). Las relaciones entre los
principales sustratos y productos de esta enzima fueron menores para la SA lo que
sugeriría una menor actividad de esta enzima en la glándula mamaria de las vacas que
consumieron el suplemento con SA y mayor para C y HA. Algunos autores han
demostrado que la presencia de los AG ciclopropenos malválico y estercúlico, presentes
en semilla de algodón (Coppock et al., 1987), inhiben la enzima Δ9desaturasa (Allen et
al., 1967). Su efecto depende de la biohidrogenación de estos ácidos en el rumen ya que
ésta produce ciclopropano que no es capaz de inhibir la Δ9desaturasa (Cook et al.,
1976). Aunque no se conocen las concentraciones de los ácidos malválico y estercúlico,
nuestros resultados podrían sugerir que éstos AG inactivaron la acción de la enzima
Δ9desaturasa similar a lo reportado por Hawkins et al. (1985), debido a que algunos AG
ciclopropenos escapan de la hidrogenación en el rumen (Yang et al., 1999). La diferencia
entre los índices de la Δ9desaturasa de la HA y SA no es suficiente para argumentar una
inactivación de esta enzima con la SA pues no existe diferencia entre SA y C para este
índice. Sin embargo, la relación entre C14:1 / C14:0 fue menor para la SA frente a los
otros dos tratamientos, apoyando la teoría de que los AG ciclopropenos de la SA
inactivaron la acción de la Δ9desaturasa ya que el C14:0 es sintetizado principalmente
de novo por la glándula mamaria y el C14:1 se origina de la desaturación del C14:0 por la
Δ9desaturasa (Corl et al., 2001) de tal manera que la reducción en el índice C14:1 /
C14:0 provee un estimativo de la inhibición de la Δ9desaturasa. La menor actividad de la
Δ9desaturasa contribuiría a que la concentración de C18:0 fuera mayor y la de Ru menor
en la grasa láctea de vacas que consumieron SA como fue el caso en nuestro ensayo.
Otros autores sugieren que en los rumiantes no hay efectos negativos sobre la
Δ9desaturasa debido al suministro de semilla de algodón (Archibeque et al., 2005; Liu et
al., 2008; Vasta et al., 2008) debido a que los ácidos ciclopropenoicos representan una
menor proporción del total de la dieta y la biohidrogenación ruminal los convierte en
compuestos inactivos (Corl et al., 2001) que aparentemente no fue el caso en este
experimento.
2.5 Conclusiones
El tratamiento con harina de arroz incrementó la concentración de los ácidos
transvaccénico (3.11 %; p<0.01) y ruménico (1.41 %; p<0.001) en la grasa láctea;
además, disminuyó los índices de aterogenicidad (1.85; p<0.05) y trombogenicidad (3.09;

78
Efecto de la suplementación con harina de arroz o semilla de algodón sobre el contenido de los ácidos transvaccénico y ruménico
en la leche de vacas en pastoreo
p<0.01) en la leche. La semilla de algodón no logró aumentar la proporción de estos
ácidos grasos y de ácidos grasos insaturados, ni mejorar los índices de aterogenicidad y
trombogenicidad a pesar de que el consumo de ácido linoleico y linolénico fue similar a
los del tratamiento harina de arroz. Por el contrario, la semilla de algodón presentó
mayores proporciones de C18:0 (16.31 %; p<0.001) y menores relaciones ruménico /
transvaccénico (0.39; p<0.01) y C14:1 / C14:0 (0.08; p<0.05) indicando mayor
biohidrogenación y menor actividad de la enzima Δ9desaturasa. A diferencia de la
semilla de algodón, la harina de arroz es una buena alternativa de suplementación para
las vacas lecheras en pastoreo pues induce una mayor concentración de transvaccénico
y ruménico en la leche, y mayor concentración de ácidos grasos insaturados (34.9;
p<0.01) que se sugiere son mejores para la salud del consumidor.
2.6 Agradecimientos
Al Ministerio de Agricultura y Desarrollo Rural por el apoyo económico para la
elaboración de este trabajo. Al Laboratorio de Nutrición Animal de la Facultad de
Medicina Veterinaria y de Zootecnia -Unal, Sede Bogotá-, al Centro de Investigación
Agropecuario Marengo -Unal, Sede Bogotá- y al Grupo de Investigación en Nutrición
Animal de la Facultad de Medicina Veterinaria de y de Zootecnia -Unal, Sede Bogotá- por
el apoyo en la fase experimental y el trabajo en el laboratorio.
2.7 Referencias
ABUGHAZALEH, A. A.; FELTON, D. O. y IBRAHIM, S. A. 2007. Milk conjugated linoleic acid response to fish oil and sunflower oil supplementation to dairy cows managed under two feeding systems. J. Dairy Sci. 90(10):4763-4769.
ABUGHAZALEH, A. A.; SCHINGOETHE, D. J.; HIPPEN, A. R. y KALSCHEUR, K. F. 2003. Milk conjugated linoleic acid response to fish oil supplementation of diets differing in fatty acid profiles. J. Dairy Sci. 86(3):944-953.
ABUGHAZALEH, A. A.; SCHINGOETHE, D. J.; HIPPEN, A. R. y KALSCHEUR, K. F. 2004. Conjugated linoleic acid increases in milk when cows fed fish meal and extruded soybeans for an extended period of time. J. Dairy Sci. 87(6):1758-1766.
ABUGHAZALEH, A. A.; SCHINGOETHE, D. J.; HIPPEN, A. R.; KALSCHEUR, K. F. y WHITLOCK, L. A. 2002. Fatty acid profiles of milk and rumen digesta from cows fed fish oil, extruded soybeans or their blend. J. Dairy Sci. 85(9):2266-2276.
AGUILAR, O. X.; MORENO, B. M.; PABÓN, M. L. y CARULLA, J. E. 2009. Efecto del consumo de kikuyo (Pennisetum clandestinum) o raigrás (Lolium hibridum) sobre
la concentración de ácido linoléico conjugado y el perfil de ácidos grasos de la grasa láctea. Livest. Res. Rural Dev. 21(49).

Capítulo 2
79
ALLEN, E.; JOHNSON, A.; FOGERTY, A.; PEARSON, J. y SHENSTONE, F. 1967. Inhibition by cyclopropene fatty acids of the desaturation of stearic acid in hen liver. Lipids. 2(5):419-423.
ALLEN, M. S. 2000. Effects of diet on short-term regulation of feed intake by lactating dairy cattle. J. Dairy Sci. 83(7):1598-1624.
ALZAHAL, O.; ODONGO, N. E.; MUTSVANGWA, T.; OR-RASHID, M. M.; DUFFIELD, T. F.; BAGG, R.; DICK, P.; VESSIE, G. y MCBRIDE, B. W. 2008. Effects of monensin and dietary soybean oil on milk fat percentage and milk fatty acid profile in lactating dairy cows. J. Dairy Sci. 91(3):1166-1174.
AOAC. 2006. Official methods of analysis of AOAC International. 18a ed. Association Official to Analytical Chemistry Intenational, Maryland.
ARCHIBEQUE, S. L.; LUNT, D. K.; GILBERT, C. D.; TUME, R. K. y SMITH, S. B. 2005. Fatty acid indices of stearoyl-CoA desaturase do not reflect actual stearoyl-CoA desaturase enzyme activities in adipose tissues of beef steers finished with corn-, flaxseed-, or sorghum-based diets. J. Anim Sci. 83(5):1153-1166.
ARIELI, A. 1998. Whole cottonseed in dairy cattle feeding: a review. Anim. Feed Sci. and Technol. 72(1-2):97-110.
BARBER, M. C.; CLEGG, R. A.; TRAVERS, M. T. y VERNON, R. G. 1997. Lipid metabolism in the lactating mammary gland. Biochim. Biophys. Acta (BBA) - Lipids and Lipid Metabolism. 1347(2-3):101-126.
BARGO, F.; MULLER, L. D.; KOLVER, E. S. y DELAHOY, J. E. 2003. Invited review: Production and digestion of supplemented dairy cows on pasture. J. Dairy Sci.
86(1):1-42.
BAS, P.; ARCHIMÈDE, H.; ROUZEAU, A. y SAUVANT, D. 2003. Fatty acid composition of mixed-rumen bacteria: effect of concentration and type of forage. J. Dairy Sci.
86(9):2940-2948.
BELIBASAKIS, N. G. y TSIRGOGIANNI, D. 1995. Effects of whole cottonseeds on milk yield, milk composition, and blood components of dairy cows in hot weather. Anim.
Feed Sci. and Technol. 52(34):227-235.
BHARATHAN, M.; SCHINGOETHE, D. J.; HIPPEN, A. R.; KALSCHEUR, K. F.; GIBSON, M. L. y KARGES, K. 2008. Conjugated linoleic acid increases in milk from cows fed condensed corn distillers solubles and fish oil. J. Dairy Sci. 91(7):2796-2807.
BITMAN, J.; WOOD, D. L.; MILLER, R. H.; TYRRELL, H. F.; REYNOLDS, C. K. y BAXTER, H. D. 1996. Comparison of milk and blood lipids in jersey and holstein cows fed total mixed rations with or without whole cottonseed. J. Dairy Sci. 79(9):1596-1602.
BLAUWIEKEL, R.; XU, S.; HARRISON, J. H.; LONEY, K. A.; RILEY, R. E. y CALHOUN, M. C. 1997. Effect of whole cottonseed, gossypol, and ruminally protected lysine supplementation on milk yield and composition. J. Dairy Sci. 80(7):1358-1365.
BOUATTOUR, M. A.; CASALS, R.; ALBANELL, E.; SUCH, X. y CAJA, G. 2008. Feeding soybean oil to dairy goats increases conjugated linoleic acid in milk. J. Dairy Sci.
91(6):2399-2407.
BREMMER, D. R.; RUPPERT, L. D.; CLARK, J. H. y DRACKLEY, J. K. 1998. Effects of chain length and unsaturation of fatty acid mixtures infused into the abomasum of

80
Efecto de la suplementación con harina de arroz o semilla de algodón sobre el contenido de los ácidos transvaccénico y ruménico
en la leche de vacas en pastoreo
lactating dairy cows. J. Dairy Sci. 81(1):176-188.
BU, D. P.; WANG, J. Q.; DHIMAN, T. R. y LIU, S. J. 2007. Effectiveness of oils rich in linoleic and linolenic acids to enhance conjugated linoleic acid in milk from dairy cows. J. Dairy Sci. 90(2):998-1007.
CHILLIARD, Y.; FERLAY, A.; MANSBRIDGE, R. M. y DOREAU, M. 2000. Ruminant milk fat plasticity: nutritional control of saturated, polyunsaturated, trans and conjugated fatty acids. Ann. Zootech. 49(3):181-205.
COOK, C. W. 1964. Symposium on nutrition of forages and pastures: collecting forage samples representative of ingested material of grazing animals for nutritional studies. J. Anim Sci. 23(1):265-270.
COOK, L.; SCOTT, T.; MILLS, S.; FOGERTY, A. y JOHNSON, A. 1976. Effects of protected cyclopropene fatty acids on the composition of ruminant milk fat. Lipids.
11(9):705-711.
COPPOCK, C. E.; LANHAM, J. K. y HORNER, J. L. 1987. A review of the nutritive value and utilization of whole cottonseed, cottonseed meal and associated by-products by dairy cattle. Anim. Feed Sci. and Technol. 18(2):89-129.
CORL, B. A.; BAUMGARD, L. H.; DWYER, D. A.; GRIINARI, J. M.; PHILLIPS, B. S. y
BAUMAN, D. E. 2001. The role of -desaturase in the production of cis-9, trans-11 CLA. J. Nutr. Biochem. 12(11):622-630.
CORREA, H. J.; PABÓN, M. L. y CARULLA, J. E. 2008. Valor nutricional del pasto kikuyo (Pennisetum clandestinum Hoechst Ex Chiov.) para la producción de leche en
Colombia (Una revisión): I - Composición química y digestibilidad ruminal y posruminal. Livest. Res. Rural Dev. 20(4).
CORREA, H. J.; PABÓN, M. L. y CARULLA, J. E. 2009. Estimación del consumo de materia seca en vacas Holstein bajo pastoreo en el trópico alto de Antioquia. Livest. Res. Rural Dev. 21(4).
DE CAMPOS, R. M. L.; HIERRO, E.; ORDÓÑEZ, J. A.; BERTOL, T. M.; TERRA, N. N. y DE LA HOZ, L. 2007. Fatty acid and volatile compounds from salami manufactured with yerba mate (Ilex paraguariensis) extract and pork back fat and meat from pigs fed on diets with partial replacement of maize with rice bran. Food Chem. 103(4):1159-1167.
DEWHURST, R. J.; SHINGFIELD, K. J.; LEE, M. R. F. y SCOLLAN, N. D. 2006. Increasing the concentrations of beneficial polyunsaturated fatty acids in milk produced by dairy cows in high-forage systems. Anim. Feed Sci. and Technol.
131(3-4):168-206.
DHIMAN, T. R.; HELMINK, E. D.; MCMAHON, D. J.; FIFE, R. L. y PARIZA, M. W. 1999. Conjugated linoleic acid content of milk and cheese from cows fed extruded oilseeds. J. Dairy Sci. 82(2):412-419.
DHIMAN, T. R.; SATTER, L. D.; PARIZA, M. W.; GALLI, M. P.; ALBRIGHT, K. y TOLOSA, M. X. 2000. Conjugated linoleic acid (CLA) content of milk from cows offered diets rich in linoleic and linolenic acid. J. Dairy Sci. 83(5):1016-1027.
DÍAZ-GONZÁLEZ, G.; GUTIÉRREZ, R.; PÉREZ, N.; VEGA, S.; LEÓN, S.; GONZÁLEZ, M.; PRADO, G.; URBÁN, G.; RAMÍREZ, A. y PINTO, M. 2002. Detección de adulteraciones en la grasa de leche pasteurizada mexicana. Rev. Salud Anim.
24(1):54-59.

Capítulo 2
81
EDMONSON, A. J.; LEAN, I. J.; WEAVER, L. D.; FARVER, T. y WEBSTER, G. 1989. A body condition scoring chart for Holstein dairy cows. J. Dairy Sci. 72(1):68-78.
FENG, S.; LOCK, A. L. y GARNSWORTHY, P. C. 2004. Technical note: a rapid lipid separation method for determining fatty acid composition of milk. J. Dairy Sci. 87(11):3785-3788.
FIRKINS, J. L. y EASTRIDGE, M. L. 1994. Assessment of the effects of iodine value on fatty acid digestibility, feed intake, and milk production. J. Dairy Sci. 77(8):2357-2366.
FLOWERS, G.; IBRAHIM, S. A. y ABUGHAZALEH, A. A. 2008. Milk fatty acid composition of grazing dairy cows when supplemented with linseed oil. J. Dairy Sci. 91(2):722-730.
FOLCH, J.; LEES, M. y STANLEY, G. H. S. 1957. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 226(1):497-509.
GAGLIOSTRO, G. A.; RODRÍGUEZ, A.; PELLEGRINI, P.; GATTI, P.; MUSSET, G.; CASTAÑEDA, R. A.; COLOMBO, D. y CHILLIARD, Y. 2006. Efectos del suministro de aceite de pescado sólo o en combinación con aceite de girasol sobre las concentraciones de ácido linoleico conjugado (CLA) y omega 3 (n-3) en leche de cabra. Rev. Argent. Prod Anim. 26:71-87.
GARCES, R. y MANCHA, M. 1993. One-step lipid extraction and fatty acid methyl esters preparation from fresh plant tissues. Anal. Biochem. 211(1):139-143.
GARNSWORTHY, P. C.; MASSON, L. L.; LOCK, A. L. y MOTTRAM, T. T. 2006. Variation of milk citrate with stage of lactation and de novo fatty acid synthesis in dairy cows. J. Dairy Sci. 89(5):1604-1612.
HARRISON, J. H.; KINCAID, R. L.; MCNAMARA, J. P.; WALTNER, S.; LONEY, K. A.; RILEY, R. E. y CRONRATH, J. D. 1995. Effect of whole cottonseeds and calcium salts of long-chain fatty acids on performance of lactating dairy cows. J. Dairy Sci.
78(1):181-193.
HARVATINE, D. I.; FIRKINS, J. L. y EASTRIDGE, M. L. 2002. Whole linted cottonseed as a forage substitute fed with ground or steam-flaked corn: digestibility and performance. J. Dairy Sci. 85(8):1976-1987.
HAWKINS, G. E.; CUMMINS, K. A.; SILVERIO, M. y JILEK, J. J. 1985. Physiological effects of whole cottonseed in the diet of lactating dairy cows. J. Dairy Sci.
68(10):2608-2614.
HERVÁS, G.; LUNA, P.; ÁNGEL, R.; ARES, N.; DE LA FUENTE, M. A.; REZ, M. y FRUTOS, P. 2008. Effect of diet supplementation with sunflower oil on milk production, fatty acid profile and ruminal fermentation in lactating dairy ewes. J. Dairy Res. 75(4):399-405.
HOLDEN, L. A.; MULLER, L. D. y FALES, S. L. 1994. Estimation of intake in high producing Holstein cows grazing grass pasture. J. Dairy Sci. 77(8):2332-2340.
HOUSEKNECHT, K. L.; HEUVEL, J. P. V.; MOYA-CAMARENA, S. Y.; PORTOCARRERO, C. P.; PECK, L. W.; NICKEL, K. P. y BELURY, M. A. 1998. Dietary conjugated linoleic acid normalizes impaired glucose tolerance in the zucker diabetic fatty fa/fa rat. Biochem. Biophysi. Res. Comm. 244(3):678-682.
HUANG, Y.; SCHOONMAKER, J. P.; BRADFORD, B. J. y BEITZ, D. C. 2008. Response

82
Efecto de la suplementación con harina de arroz o semilla de algodón sobre el contenido de los ácidos transvaccénico y ruménico
en la leche de vacas en pastoreo
of milk fatty acid composition to dietary supplementation of soy oil, conjugated linoleic acid, or both. J. Dairy Sci. 91(1):260-270.
HUANG, Y.; SCHOONMAKER, J. P.; OREN, S. L.; TRENKLE, A. y BEITZ, D. C. 2009. Calcium salts of CLA improve availability of dietary CLA. Livest. Sci. 122(1):1-7.
HURLEY, W. L.; WARNER, G. J. y GRUMMER, R. R. 1987. Changes in triglyceride fatty acid composition of mammary secretions during involution. J. Dairy Sci.
70(11):2406-2410.
IP, C.; BANNI, S.; ANGIONI, E.; CARTA, G.; MCGINLEY, J.; THOMPSON, H. J.; BARBANO, D. y BAUMAN, D. 1999. Conjugated linoleic acid-enriched butter fat alters mammary gland morphogenesis and reduces cancer risk in rats. J. Nutr. 129(12):2135-2142.
JACOBS, A. A. A.; VAN BAAL, J.; SMITS, M. A.; TAWEEL, H. Z. H.; HENDRIKS, W. H.; VAN VUUREN, A. M. y DIJKSTRA, J. 2011. Effects of feeding rapeseed oil, soybean oil, or linseed oil on stearoyl-CoA desaturase expression in the mammary gland of dairy cows. J. Dairy Sci. 94(2):874-887.
JENKINS, T. C.; WALLACE, R. J.; MOATE, P. J. y MOSLEY, E. E. 2008. Board-Invited review: Recent advances in biohydrogenation of unsaturated fatty acids within the rumen microbial ecosystem. J. Anim Sci. 86(2):397-412.
JENSEN, R. G. 2002. The composition of bovine milk lipids: january 1995 to december 2000. J. Dairy Sci. 85(2):295-350.
KAPS, M. y LAMBERSON, W. R. 2004. Biostatistics for animal science., London.
KELLY, M. L.; KOLVER, E. S.; BAUMAN, D. E.; VAN AMBURGH, M. E. y MULLER, L. D. 1998. Effect of intake of pasture on concentrations of conjugated linoleic acid in milk of lactating cows. J. Dairy Sci. 81(6):1630-1636.
KHANAL, R. C.; DHIMAN, T. R. y BOMAN, R. L. 2008. Changes in fatty acid composition of milk from lactating dairy cows during transition to and from pasture. Livest. Sci.
114(2-3):164-175.
KHANAL, R. C.; DHIMAN, T. R.; URE, A. L.; BRENNAND, C. P.; BOMAN, R. L. y MCMAHON, D. J. 2005. Consumer acceptability of conjugated linoleic acid-enriched milk and cheddar cheese from cows grazing on pasture. J. Dairy Sci.
88(5):1837-1847.
KHANAL, R. C. y OLSON, K. C. 2004. Factors affecting conjugated linoleic acid (CLA) content in milk, meat and egg- A review. Pak. J. Nutr. 3(2):82-98.
LICITRA, G.; HERNANDEZ, T. M. y VAN SOEST, P. J. 1996. Standardization of procedures for nitrogen fractionation of ruminant feeds. Anim. Feed Sci. and Technol. 57(4):347-358.
LIN, L.; YING, D.; CHAITEP, S. y VITTAYAPADUNG, S. 2009. Biodiesel production from crude rice bran oil and properties as fuel. App. Energy. 86(5):681-688.
LINDSEY, T. O.; HAWKINS, G. E. y GUTHRIE, L. D. 1980. Physiological responses of lactating cows to gossypol from cottonseed meal rations. J. Dairy Sci. 63(4):562-
573.
LINZELL, J. L. y PEAKER, M. 1971. Mechanism of milk secretion. Physiol. Rev.
51(3):564-597.

Capítulo 2
83
LIU, Z. L.; CHEN, P.; LI, J. M.; LIN, S. B.; WANG, D. M.; ZHU, L. P. y YANG, D. P. 2008. Conjugated linoleic acids (CLA) moderate negative responses of heat-stressed cows. Livest. Sci. 118(3):255-261.
LOOR, J. J.; FERLAY, A.; OLLIER, A.; DOREAU, M. y CHILLIARD, Y. 2005a. Relationship among trans and conjugated fatty acids and bovine milk fat yield due to dietary concentrate and linseed oil. J. Dairy Sci. 88(2):726-740.
LOOR, J. J.; FERLAY, A.; OLLIER, A.; UEDA, K.; DOREAU, M. y CHILLIARD, Y. 2005b. High-concentrate diets and polyunsaturated oils alter trans and conjugated isomers in bovine rumen, blood, and milk. J. Dairy Sci. 88(11):3986-3999.
LOURENÇO, M.; VAN RANST, G.; DE SMET, S.; RAES, K. y FIEVEZ, V. 2007. Effect of grazing pastures with different botanical composition by lambs on rumen fatty acid metabolism and fatty acid pattern of longissimus muscle and subcutaneous fat. Animal. 1(04):537-545.
LYNCH, J. M.; BARBANO, D. M. y FLEMING, J. R. 2007. Determination of the lactose content of fluid milk by spectrophotometric enzymatic analysis using weight additions and path length Adjustment: Collaborative Study. Journal of AOAC International. 90(1):196-215.
MCCREARY, D. K.; KOSSA, W. C.; RAMACHANDRAN, S. y KURTZ, R. R. 1978. A novel and rapid method for the preparation of methyl rsters for gas chromatography: application to the determination of the fatty acids of edible fats and oils. J. Chromat. Sci. 16(8):329-331.
MEL´UCHOVÁ, B.; BLASKO, J.; KUBINEC, R.; GÓROVÁ, R.; DUBRAVSKÁ, J.; MARGETÍN, M. y SOJÁK, L. 2008. Seasonal variations in fatty acid composition of pasture forage plants and CLA content in ewe milk fat. Small Ruminant Res. 78(1-3):56-65.
MENA, H.; SANTOS, J. E. P.; HUBER, J. T.; TARAZON, M. y CALHOUN, M. C. 2004. The effects of varying gossypol intake from whole cottonseed and cottonseed meal on lactation and blood parameters in lactating dairy cows. J. Dairy Sci.
87(8):2506-2518.
MOHAMMED, R.; STANTON, C. S.; KENNELLY, J. J.; KRAMER, J. K. G.; MEE, J. F.; GLIMM, D. R.; O'DONOVAN, M. y MURPHY, J. J. 2009. Grazing cows are more efficient than zero-grazed and grass silage-fed cows in milk rumenic acid production. J. Dairy Sci. 92(8):3874-3893.
MOYA-CAMARENA, S. Y.; HEUVEL, J. P. V.; BLANCHARD, S. G.; LEESNITZER, L. A. y BELURY, M. A. 1999. Conjugated linoleic acid is a potent naturally occurring
ligand and activator of PPAR. J. Lipid Res. 40(8):1426-1433.
MUIR, J. P. 2002. Hand-Plucked forage yield and quality and seed production from annual and short-lived perennial warm-season legumes fertilized with composted manure. Crop Sci. 42(3):897-904.
MUNDAY, J. S.; THOMPSON, K. G. y JAMES, K. A. C. 1999. Dietary conjugated linoleic acids promote fatty streak formation in the C57BL/6 mouse atherosclerosis model. Br. J. Nutr. 81(03):251-255.
NRC. 2001. Nutrient requerimients of dairy cattle. 7a ed. National Academy Press, Washington DC.
ODONGO, N. E.; OR-RASHID, M. M.; BAGG, R.; VESSIE, G.; DICK, P.; KEBREAB, E.;

84
Efecto de la suplementación con harina de arroz o semilla de algodón sobre el contenido de los ácidos transvaccénico y ruménico
en la leche de vacas en pastoreo
FRANCE, J. y MCBRIDE, B. W. 2007. Long-effects of feeding monensin on milk fatty acid composition in lactating dairy cows. J. Dairy Sci. 90(11):5126-5133.
OR-RASHID, M. M.; ODONGO, N. E. y MCBRIDE, B. W. 2007. Fatty acid composition of ruminal bacteria and protozoa, with emphasis on conjugated linoleic acid, vaccenic acid, and odd-chain and branched-chain fatty acids. J. Anim Sci.
85(5):1228-1234.
PALMQUIST, D. L.; BEAULIEU, A. D. y BARBANO, D. M. 1993. Feed and animal factors influencing milk fat composition. J. Dairy Sci. 76(6):1753-1771.
PALMQUIST, D. L. y JENKINS, T. C. 1980. Fat in lactation rations: Review. J. Dairy Sci.
63(1):1-14.
PARADIS, C.; BERTHIAUME, R.; LAFRENIERE, C.; GERVAIS, R. y CHOUINARD, P. Y. 2008. Conjugated linoleic acid content in adipose tissue of calves suckling beef cows on pasture and supplemented with raw or extruded soybeans. J. Anim Sci.
86(7):1624-1636.
PARK, H. S.; RYU, J. H.; HA, Y. L. y PARK, J. H. Y. 2001. Dietary conjugated linoleic acid (CLA) induces apoptosis of colonic mucosa in 1,2-dimethylhydrazine-treated rats: a possible mechanism of the anticarcinogenic effect by CLA. Br. J. Nutr. 86(05):549-555.
PIRES, A. V.; EASTRIDGE, M. L.; FIRKINS, J. L. y LIN, Y. C. 1997. Effects of heat treatment and physical processing of cottonseed on nutrient digestibility and production performance by lactating cows. J. Dairy Sci. 80(8):1685-1694.
REISER, R. y FU, H. C. 1962. The mechanism of gossypol detoxification by ruminant animals. J. Nutr. 76(2):215-218.
REVENEAU, C.; RIBEIRO, C. V. D. M.; EASTRIDGE, M. L. y FIRKINS, J. L. 2012. Interaction of unsaturated fat or coconut oil with monensin in lactating dairy cows fed 12 times daily. II. Fatty acid flow to the omasum and milk fatty acid profile. J. Dairy Sci. 95(4):2061-2069.
RISCO, C. A.; ADAMS, A. L.; SEEBOHM, S.; THATCHER, M. J.; STAPLES, C. R.; VAN HORN, H. H.; MCDOWELL, L. R.; CALHOUN, M. C. y THATCHER, W. W. 2002.
Effects of gossypol from cottonseed on hematological responses and plasma -tocopherol concentration of dairy cows. J. Dairy Sci. 85(12):3395-3402.
SANTOS, J. E. P.; VILLASENOR, M.; ROBINSON, P. H.; DEPETERS, E. J. y HOLMBERG, C. A. 2003. Type of cottonseed and level of gossypol in diets of lactating dairy cows: plasma gossypol, health, and reproductive performance. J. Dairy Sci. 86(3):892-905.
SCHAUFF, D. J. y CLARK, J. H. 1992. Effects of feeding diets containing calcium salts of long-chain fatty acids to lactating dairy cows. J. Dairy Sci. 75(11):2990-3002.
SCHMIDELY, P.; GLASSER, F.; DOREAU, M. y SAUVANT, D. 2008. Digestion of fatty acids in ruminants: a meta-analysis of flows and variation factors. 1. Total fatty acids. Animal. 2(05):677-690.
SCHROEDER, G. F.; DELAHOY, J. E.; VIDAURRETA, I.; BARGO, F.; GAGLIOSTRO, G. A. y MULLER, L. D. 2003. Milk fatty acid composition of cows fed a total mixed ration or pasture plus concentrates replacing corn with fat. J. Dairy Sci.
86(10):3237-3248.

Capítulo 2
85
SCHROEDER, G. F.; GAGLIOSTRO, G. A.; BARGO, F.; DELAHOY, J. E. y MULLER, L. D. 2004. Effects of fat supplementation on milk production and composition by dairy cows on pasture: a review. Livest. Prod. Sci. 86(1-3):1-18.
SMITH, W. A.; HARRIS, B.; VAN HORN, H. H. y WILCOX, C. J. 1993. Effects of forage type on production of dairy cows supplemented with whole cottonseed, tallow, and yeast. J. Dairy Sci. 76(1):205-215.
SUNVOLD, G. D. y COCHRAN, R. C. 1991. Technical note: evaluation of acid detergent lignin, alkaline peroxide lignin, acid insoluble ash, and indigestible acid detergent fiber as internal markers for prediction of alfalfa, bromegrass, and prairie hay digestibility by beef steers. J. Anim Sci. 69(12):4951-4955.
THIEX, N. J.; MANSON, H.; ANDERSON, S. y PETACCHI, F. 2002. Determination of crude protein in animal feed, forage, grain, and oilseeds by using block digestion with a copper catalyst and steam distillation into boric acid: collaborative study. J. AOAC Inter. 85(2):309-317.
TORAL, P. G.; BELENGUER, A.; FRUTOS, P. y HERVÁS, G. 2009. Effect of the supplementation of a high-concentrate diet with sunflower and fish oils on ruminal fermentation in sheep. Small Ruminant Res. 81(2-3):119-125.
TORAL, P. G.; BELENGUER, A.; SHINGFIELD, K. J.; HERVÁS, G.; TOIVONEN, V. y FRUTOS, P. 2012. Fatty acid composition and bacterial community changes in the rumen fluid of lactating sheep fed sunflower oil plus incremental levels of marine algae. J. Dairy Sci. 95(2):794-806.
TSIPLAKOU, E.; KOMINAKIS, A. y ZERVAS, G. 2008. The interaction between breed and diet on CLA and fatty acids content of milk fat of four sheep breeds kept indoors or at grass. Small Ruminant Res. 74(1-3):179-187.
UEDA, K.; FERLAY, A.; CHABROT, J.; LOOR, J. J.; CHILLIARD, Y. y DOREAU, M. 2003. Effect of linseed oil supplementation on ruminal digestion in dairy cows fed diets with different forage:concentrate ratios. J. Dairy Sci. 86(12):3999-4007.
ULBRICHT, T. L. V. y SOUTHGATE, D. A. T. 1991. Coronary heart disease: seven dietary factors. The Lancet. 338(8773):985-992.
VAN KNEGSEL, A. T. M.; VAN DEN BRAND, H.; DIJKSTRA, J.; VAN STRAALEN, W. M.; HEETKAMP, M. J. W.; TAMMINGA, S. y KEMP, B. 2007. Dietary energy source in dairy cows in early lactation: energy partitioning and milk composition. J. Dairy Sci.
90(3):1467-1476.
VAN SOEST, P. J.; ROBERTSON, J. B. y LEWIS, B. A. 1991. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J. Dairy Sci. 74(10):3583-3597.
VÁRADYOVÁ, Z.; KIŠIDAYOVÁ, S.; SIROKA, P. y JALC, D. 2008. Comparison of fatty acid composition of bacterial and protozoal fractions in rumen fluid of sheep fed diet supplemented with sunflower, rapeseed and linseed oils. Anim. Feed Sci. and Technol. 144(1-2):44-54.
VASTA, V.; NUDDA, A.; CANNAS, A.; LANZA, M. y PRIOLO, A. 2008. Alternative feed resources and their effects on the quality of meat and milk from small ruminants. Anim. Feed Sci. and Technol. 147(1-3):223-246.
VELASQUEZ-PEREIRA, J.; RISCO, C. A.; MCDOWELL, L. R.; STAPLES, C. R.; PRICHARD, D.; CHENOWETH, P. J.; MARTIN, F. G.; WILLIAMS, S. N.; ROJAS,

86
Efecto de la suplementación con harina de arroz o semilla de algodón sobre el contenido de los ácidos transvaccénico y ruménico
en la leche de vacas en pastoreo
L. X.; CALHOUN, M. C. y WILKINSON, N. S. 1999. Long-term effects of feeding gossypol and vitamin E to dairy calves. J. Dairy Sci. 82(6):1240-1251.
VILLASEÑOR, M.; COSCIONI, A. C.; GALVÃO, K. N.; CHEBEL, R. C. y SANTOS, J. E. P. 2008. Gossypol Disrupts Embryo Development in Heifers. J. Dairy Sci. 91(8):3015-3024.
WILKS, D. L.; COPPOCK, C. E.; BROOKS, K. N. y GATES, C. E. 1991. Effects of differences in starch content of diets with whole cottonseed or rice bran on milk casein. J. Dairy Sci. 74(4):1314-1320.
WILLIAMS, A. G. 1986. Rumen holotrich ciliate protozoa. Microbio. Rev. 50(1):25-49.
WILLIAMS, C. H.; DAVID, D. J. y IISMAA, O. 1962. The determination of chromic oxide in faeces samples by atomic absorption spectrophotometry. J. Agr. Sci. 59(03):381-
385.
YAMASAKI, M.; KISHIHARA, K.; IKEDA, I.; SUGANO, M. y YAMADA, K. 1999. A recommended esterification method for gas chromatographic measurement of conjugated linoleic acid. J. American Oil Chemists' Society. 76(8):933-938.
YANG, A.; LARSEN, T.; SMITH, S. y TUME, R. 1999. Ä9 desaturase activity in bovine
subcutaneous adipose tissue of different fatty acid compositiondesaturase activity in bovine subcutaneous adipose tissue of different fatty acid composition. Lipids. 34(9):971-978.
ZHANG, W. J.; XU, Z. R.; PAN, X. L.; YAN, X. H. y WANG, Y. b. 2007. Advances in gossypol toxicity and processing effects of whole cottonseed in dairy cows feeding. Livest. Sci. 111(1-2):1-9.
.

Capítulo 3
87
3. Capítulo 3: Efecto de la utilización de dos niveles de inclusión de la mezcla harina de arroz y semilla de algodón sobre la concentración de los ácidos transvaccénico (C18:1t11) y ruménico (C18:2c9,t11) en la leche de vacas en pastoreo
Gastón A. Castaño J.1,2, Martha L. Pabón R.1,3 y Juan E. Carulla F.1,4
1 Grupo de Investigación en Nutrición Animal. Universidad Nacional de Colombia, Sede Bogotá.
2 Candidato Magíster en Producción Animal. Universidad Nacional de Colombia, Sede Bogotá. E-mail: [email protected]
3 Profesora Titular. Departamento de Química. Facultad de Ciencias. Universidad Nacional de Colombia, Sede Bogotá. E-mail: [email protected]
4 Profesor Titular. Departamento de Producción Animal. Facultad de Medicina Veterinaria y de Zootecnia. Universidad Nacional de Colombia, Sede Bogotá. E-mail: [email protected]
Resumen
Con el fin de determinar el efecto de suplementar con dos niveles de ácido linoleico (Li)
proveniente de la mezcla de harina de arroz (HA) y semilla de algodón (SA) sobre el
contenido de los ácidos grasos transvaccénico (TVa; C18:1t11) y ruménico (Ru
C18:2c9,t11) en la grasa láctea de vacas lecheras que pastan P. clandestinum se
evaluaron dos suplementos que contenían una mezcla de HA y SA (relación 4.8 : 1)
como la principal fuente lipídica de Li: uno con baja concentración de Li y otro con alta
(2.64 y 4.08 g de Li / 100 g de MS respectivamente). Se utilizaron seis vacas Holstein
(4.2 ± 1.7 años de edad, 544 ± 35 Kg de peso vivo, 168 ± 40 d de lactancia y una
producción láctea de 19.2 ± 0.9 Kg / d; valor medio ± DE) que fueron distribuidas según
un diseño Crossover 6 x 2 x 2 x 2 (6 vacas x 2 periodos x 2 tratamientos x 2

88
Efecto de la suplementación con harina de arroz o semilla de algodón sobre el contenido de los ácidos transvaccénico y ruménico
en la leche de vacas en pastoreo
ordenamientos). Cada periodo tuvo una duración de 28 d (20 d de acostumbramiento a
las dietas experimentales y 8 d para la recolección de muestras), los animales fueron
ordeñados dos veces al día (05:00 y 14:00 h), permanecieron con agua a voluntad,
pastorearon en P. clandestinum y recibieron durante cada ordeño 60 g de sal
mineralizada, 3 Kg de suplemento según el tratamiento y 5 g de Cr2O3. Al incrementar el
nivel de Li no se afectaron las concentraciones de TVa y Ru en la grasa láctea, pero
aumentaron los ácidos grasos insaturados (35.1 %; p<0.05) y se presentó un mayor
índice calculado de actividad de la Δ9desaturasa y menores índices de aterogenicidad y
trombogenicidad (0.37, 1.79 y 3.22 respectivamente; p<0.05). Incrementar la
concentración de Li en la dieta de vacas lecheras que pastan P. clandestinum utilizando
un suplemento alimenticio que contiene una mezcla de HA y SA como la principal fuente
lipídica de Li no logró incrementar el contenido de TVa y Ru en la grasa láctea.
Palabras claves: ácido linoleico, ácido linoleico conjugado, ácidos grasos lácteos, vaca lechera.
Abstract
In order to determine the effect of supplementing the diet with two levels of linoleic acid
(Li) from the mixture of rice bran (RB) and cottonseed (CS) on transvaccenic (TVa;
C18:1t11) and rumenic (Ru; C18:2c9,t11) fatty acids content in milk fat from dairy cows
grazing on P. clandestinum two supplements containing a mixture of RB and CS (4.8 : 1
ratio) as the main source of Li was tested: one with low concentration of Li and other with
high level (2.64 and 4.08 g Li / 100 g DM respectively). We used six Holstein (4.2 ± 1.7
years old, 544 ± 35 Kg live weight, 168 ± 40 d of lactation and 19.2 ± 0.9 Kg / d of milk
production; average ± SD) distributed in a Crossover design 6 x 2 x 2 x 2 (6 cows x 2
periods x 2 treatments x 2 orders). Periods of 28 d were used (20 d of adaptation to the
experimental diets and 8 d for collection samples), animals were milked twice a day
(05:00 and 14:00 h), water ad libitum, grazed on P. clandestinum and received during
each milking 60 g of mineralized salt, 3 Kg of supplement according to the treatment and
5 g of Cr2O3. Increasing the level of Li not affect concentrations of TVa and Ru in milk fat,
but increased the unsaturated fatty acids (35.1 %; p<0.05) and had higher Δ9desaturase
activity index and lower of atherogenicity and thrombogenicity indixes (0.37, 1.79 and
3.22 respectively; p <0.05). Increasing the concentration of Li using a mixture of HA and
SA as the main source of Li failed to increase the content TVa and Ru in milk fat.

Capítulo 3
89
Key words: linoleic acid, conjugated linoleic acid, milk fatty acid, dairy cow.
Abreviaturas.
A: tratamiento nivel alto. AG: ácido graso. Li: ácido linoleico. Ln: ácido linolénico. Ru:
ácido ruménico. TVa: ácido transvaccénico. B: tratamiento nivel bajo. CNFn:
carbohidratos no fibrosos corregidos por N. ENL: energía neta de lactancia. FDAi: fibra
detergente ácido indigerible. FDAn: fibra detergente ácido corregida por N. FDNn: fibra
detergente neutro corregida por N. HA: harina de arroz. SA: semilla de algodón.
3.1 Introducción
Los ácidos grasos (AG) transvaccénico (TVa, C18:1t11) y ruménico (Ru, C18:2c9,t11) se
encuentran en la grasa láctea y se consideran como compuestos funcionales por tener
propiedades anticarcinogénicas (Ip et al., 1999; Park et al., 2001), antiaterogénicas,
antidiabetogénicas (Houseknecht et al., 1998; Munday et al., 1999), antiobesidad (Khanal
y Olson, 2004), regulación del sistema inmune (Moya-Camarena et al., 1999; Jensen,
2002) y mejoramiento de la mineralización del hueso (Jensen, 2002). La dieta es el
mayor determinante de la concentración láctea de Ru (Khanal y Olson, 2004). La
suplementación con fuentes lipídicas AG insaturados (Dhiman et al., 2000; Khanal y
Olson, 2004), las características de los forrajes (Mel´uchová et al., 2008; Vasta et al.,
2008), la relación concentrado : forraje (Ueda et al., 2003; Loor et al., 2005b), el sistema
de alimentación ya sea pastoreo o dietas totalmente mezcladas (TMR por sus siglas en
inglés; Kelly et al., 1998; Schroeder et al., 2003; Khanal et al., 2008) y el suministro de
antibióticos (Odongo et al., 2007; AlZahal et al., 2008) son factores relacionados con la
dieta que afectan la concentración de TVa y Ru en la leche. Adicionalmente, el pH
ruminal, el tipo de lípidos ingeridos y el tipo de bacterias presentes en el rumen afectan la
biohidrogenación o limitan la conversión de TVa a ácido esteárico (Chilliard et al., 2000;
Tsiplakou et al., 2008).
La concentración de TVa y Ru se incrementa en la leche cuando se suministran
suplementos ricos en AG insaturados (Dhiman et al., 2000). Se han evaluado diferentes
fuentes lipídicas con AG insaturados, entre las cuales se resaltan la soya (Huang et al.,
2008; Bouattour et al., 2008), la linaza (Loor et al., 2005a; Flowers et al., 2008), el girasol
(AbuGhazaleh et al., 2007; Hervás et al., 2008), el maíz (Dewhurst et al., 2006;
Bharathan et al., 2008) y el aceite de pescado (AbuGhazaleh et al., 2003; AbuGhazaleh

90
Efecto de la suplementación con harina de arroz o semilla de algodón sobre el contenido de los ácidos transvaccénico y ruménico
en la leche de vacas en pastoreo
et al., 2004). El efecto es diferente según el tipo de fuente lipídica, aquellas con elevadas
concentraciones de ácido linoleico (ALi) son más eficientes en producir incremento en la
concentración de TVa y Ru que las ricas en Ln (Bu et al., 2007).
La producción lechera en el trópico alto colombiano tiene alto potencial para producir
leches ricas en TVa y Ru debido a las características del forraje y al aporte de lípidos a
través del suplemento (Castaño et al., 2013). La harina de arroz (HA) y la semilla de
algodón (SA) se utilizan frecuentemente en la elaboración de suplementos alimenticios
para vacas lecheras en Colombia y son fuente de AG insaturados (Dhiman et al., 1999;
Lin et al., 2009). La elevada concentración de Li en la grasa de estos dos recursos, del
36 al 42% para la HA (de Campos et al., 2007) y del 57.3% para la SA (Dhiman et al.,
1999; Khanal y Olson, 2004), son cualidades que podrían ser aprovechadas para
producir leche con altos niveles de TVa y Ru. Castaño et al. (2013) evaluaron el efecto
de suplementar con HA y SA sobre el contenido de TVa y Ru en la leche de vacas en
pastoreo y encontraron que la HA logró incrementar los niveles de estos AG, mientras la
SA no. Los autores sugieren que estos resultados se podrían haber presentado por una
intensa biohidrogenación ruminal de los AG provenientes de la SA y por la presencia de
AG ciclopropenos en esta semilla, ya que éstos inactivan la actividad de la Δ9desaturasa
(Cook et al., 1976). Otros autores indican que no es posible disminuir la actividad de la
Δ9desaturasa con la adición de SA (Archibeque et al., 2005) ya que los ciclopropenos
representan una menor proporción de la dieta y la biohidrogenación ruminal los inactiva
(Corl et al., 2001). El mezclar SA con HA permite aumentar la cantidad Li en el
suplemento y si se maneja una mayor proporción de HA con respecto a la SA se podría
evitar el efecto negativo de la SA sobre la concentración TVa y Ru encontrado por
Castaño et al. (2013), lo cual podría ser una alternativa para producir leche benéfica para
la salud humana. El objetivo de este experimento fue determinar el efecto de suplementar
con dos niveles de Li a través de suplementos que contienen como principal fuente
lipídica una mezcla de HA y SA (relación 4.8 : 1) sobre el contenido de TVa y Ru en la
leche de vacas que pastan P. clandestinum.
3.2 Materiales y métodos
3.2.1 Localización
El experimento se realizó en el Centro de Investigación Agropecuario Marengo de la

Capítulo 3
91
Universidad Nacional de Colombia; localizado a 4° 40’ 89’’ de latitud norte, 74° 13’ 13’’ de
longitud oeste y a una altitud de 2540 msnm. La temperatura promedio es de 13 °C (con
variaciones de 0 a 20 ºC), la humedad relativa oscila entre 80 a 85 % y la precipitación
promedio anual es 900 mm / año con dos épocas marcadas de invierno (abril - mayo y
septiembre - noviembre). El experimento fue desarrollado entre septiembre y noviembre
de 2009.
3.2.2 Periodo experimental y tratamientos
El experimento tuvo una duración de 55 d, divididos en tres periodos de 28 d (20 d de
acostumbramiento a las dietas experimentales y 8 d para la recolección de muestras).
Los tratamientos consistieron en dos diferentes suplementos suministrados a vacas
pastando P. clandestinum, que contenían como principal fuente lipídica de Li una mezcla
de HA y SA (relación 4.8 : 1): uno con baja concentración de Li (B) y otro con alta (A;
2.64 y 4.08 g de Li / 100 g de MS respectivamente). Se suministraron 6 Kg de
suplemento al día, repartidos durante los dos ordeños (05:00 y 14:00 h). Los
suplementos eran isoenergéticos e isoproteicos, pero con diferente concentración de
grasa. Los ingredientes, la composición química y el perfil de AG de los suplementos se
pueden apreciar las Tablas 3-1 y 3-2.

92
Efecto de la suplementación con harina de arroz o semilla de algodón sobre el contenido de los ácidos transvaccénico y ruménico
en la leche de vacas en pastoreo
Tabla 3-1: Composición de ingredientes y nutrientes de los suplementos alimenticios utilizados
en el experimento
Ítem Suplemento
Bajo Alto
Ingrediente, %
Harina de arroz 39.3 65.5 Semilla de algodón 8.1 13.5 Aceite de palma 2.1 0.2 Torta de soya 17.1 11.8 Harina de yuca 11.0 - Palmiste (solvente) 16.3 3.0 Melaza de caña 5.0 5.0 Premezcla de vitaminas y minerales 0.5 0.5
Composición química Materia seca (MS), % 89.6 90.3 Proteína cruda, % MS 20.2 19.4 Fibra detergente neutro
1, % MS 28.9 28.9
Fibra detergente ácido1, % MS 19.8 17.8
Lignina, % MS 4.4 6.2 Carbohidratos no fibrosos
1, % MS 28.5 26.5
Extracto etéreo, % MS 9.6 12.6 Cenizas, % MS 7.3 6.9 ENL1X
2, Mcal/Kg MS 2.2 2.2
Ácido linoleico, % MS 2.64 4.08 1Corregidos por N (NRC, 2001).
2Energía neta de lactancia a 1X el consumo de mantenimiento
(NRC, 2001).
Tabla 3-2: Perfil de ácidos grasos (AG) de los suplementos alimenticios y del forraje utilizado en
el experimento
AG, g / 100 g AG Suplemento
Forraje Bajo Alto
C12:0 1.81 0.95 3.34 C14:0 1.33 0.91 0.73 C16:0 24.07 22.57 21.65 C16:1 0.11 0.19 1.94 C18:0 3.45 2.81 3.98 C18:1c9 36.94 35.27 3.23 C18:2c9,c12 29.74 33.83 13.44 C18:3c9,c12,c15 1.00 1.26 43.75 Otros 1.6 2.2 7.94
3.2.3 Animales y manejo
Todos los procesos y procedimientos del experimento fueron aprobados por el Comité de
Bioética de la Facultad de Medicina Veterinaria y de Zootecnia de la Universidad
Nacional de Colombia sede Bogotá (Acta 001 de 2009). Se utilizaron 6 vacas Holstein
(4.2 ± 1.7 años de edad, 544 ± 35 Kg de peso vivo, 168 ± 40 d de lactancia y una
producción láctea de 19.2 ± 0.9 Kg/d; valor medio ± DE) que fueron asignadas
aleatoriamente a dos grupos de 3 animales, cada grupo corresponde a un ordenamiento
en un diseño Cross-over 6 x 2 x 2 x 2 (6 animales x 2 periodos x 2 tratamientos x 2

Capítulo 3
93
ordenamientos). Las vacas fueron ordeñadas dos veces al día (05:00 y 14:00 h)
utilizando un sistema de ordeño mecánico. Los animales permanecieron con agua a
voluntad y recibieron durante cada ordeño 60 g de sal mineralizada, 3 Kg de suplemento
según el tratamiento y 5 g de Cr2O3 para calcular el consumo de forraje.
Durante el experimento los animales pastaron en un lote 2.5 Ha de P. clandestinum bajo
un sistema de pastoreo en franjas donde se corrió la cuerda dos veces al día (a.m. y
p.m.). Para establecer la masa forrajera se tuvo en cuenta la proporción de la pastura con
producciones de forraje baja, media o alta según lo descrito por Castaño et al. (2013). Se
utilizó un equipo de geo-posicionamiento GPSMAP® 76CSx (Garmin Ltda., Kansas,
EUA) para la medición de las áreas de pastoreo que se ajustaron con el fin de garantizar
una oferta diaria de 4 Kg de materia seca (MS) por cada 100 Kg de peso vivo.
3.2.4 Mediciones, toma de muestras y análisis de laboratorio
Mediciones. Se pesaron los animales (día 28 de cada periodo) antes de recibir el
alimento (05:00 h).
Toma de muestras.
Leche. Se pesó la producción láctea en cada ordeño (a.m. y p.m.) entre los días 21 y
28 de cada periodo. Se colectaron muestras de leche en cada ordeño de los días 21 y
28, que inmediatamente después del ordeño de la tarde se mezclaron de manera
proporcional a la producción del día hasta completar 200 mL de leche, que se
dividieron en dos alícuotas de 100 mL. A una alícuota se le adicionó 1 mL de solución
de dicromato de potasio al 6 % y se conservó a - 20 °C. La otra alícuota fue remitida
inmediatamente al laboratorio para la evaluación de la grasa.
Forraje. Los días 20 y 27 de cada periodo y previo al pastoreo se recolectaron
muestras de forraje (aprox. 500 g) utilizando el sistema “hand plucking” (Cook, 1964;
Muir, 2002). Las muestras se empacaron al vacío en bolsas plásticas y se
conservaron a - 20 °C, posteriormente fueron secadas en un liofilizador (Martin
Christ® Alpha 14LD Plus), molidas con un molino Romer® con criba de 1 mm.
Adicionalmente, se tomaron muestras los días 13, 16, 19, 22, 25 y 28, se secaron a 60
°C por 48 h, se molieron utilizando un molino Romer® con criba de 1 mm y se
mezclaron homogéneamente para formar una muestra compuesta que fue utilizada

94
Efecto de la suplementación con harina de arroz o semilla de algodón sobre el contenido de los ácidos transvaccénico y ruménico
en la leche de vacas en pastoreo
para determinar el contenido de fibra detergente ácido indigerible (FDAi).
Suplementos. El día 24 de cada periodo se colectó una muestra de cada suplemento
(aprox. 500 g), se molió utilizando un molino Romer® con criba de 1 mm. Se recolectó
el desperdicio de suplemento entre el día 21 y 28, se empacó en bolsas plásticas y se
conservó a 8 °C hasta ser pesado y enviado al laboratorio para determinar la
humedad.
Fluido ruminal. Se colectó fluido ruminal (aprox. 350 mL) a las 14:30 h del día 25 de
cada periodo utilizando una sonda ororuminal (Haumptner®); los primeros 250 mL
fueron eliminados y los 100 mL restantes fueron filtrados a través de dos capas de
gasa. Una alícuota de 70 mL fue utilizada para medir inmediatamente el pH con un
potenciómetro (Beckman®) y a una segunda alícuota de 30 mL se le adicionaron 3 mL
de ácido clorhídrico 6 N (Huang et al., 2009), se congeló a - 20 °C y posteriormente
fue utilizada para determinar amonio según adaptación al método Kjeldahl (Thiex
et al., 2002).
Heces. Se recolectaron dos muestras diarias (05:00 y 13:00 h) de heces entre los días
22 y 27. Las muestras se empacaron en bolsas plásticas y se conservaron a – 20 °C,
posteriormente fueron secadas a 60 °C por 48 h, molidas utilizando un molino Romer®
con criba de 1 mm y mezcladas homogéneamente para formar una muestra
compuesta de heces por animal.
Extracción de grasa y metilación (leche, forraje y suplementos). Se realizó según lo
descrito por Castaño et al. (2013).
Cuantificación de AG (leche, forraje y suplementos). Los ésteres de AG metilados
(FAMES por sus iniciales en inglés) de la grasa de la leche, del forraje y de los
suplementos fueron cuantificados por cromatografía de gases, utilizando un cromatógrafo
de gases marca Shimadzu® modelo GC2014 con auto inyector AOC20i y auto
muestreador AOC20C. Los FAMES fueron separados en columna capilar (Rt 2560; 100
m x 0.25 mm di x 0.2 µm espesor de la capa; Restek®). Las temperaturas del inyector y
del FID fueron 260 °C y 270 °C respectivamente. El programa de temperatura fue: 140 °C
por 5 min, se incrementó 4 °C/min hasta 190 °C y se mantuvo por 32.5 min. El Split ratio
fue de 1 : 100 y el He fue el gas de arrastre con una presión de 40.4 psi. Los tiempos de
retención fueron comparados con estándares conocidos (Food Industry FAMEX Mix cat
35077).

Capítulo 3
95
Análisis químicos
Leche. Se determinó el contenido de sólidos totales y de cenizas por gravimetría
(métodos AOAC925.23 y AOAC945.46 respectivamente; AOAC, 2006), de proteína
cruda por el método Kjeldahl (Thiex et al., 2002) y de grasa láctea por el método
Gerber (método AOAC-2000.18; AOAC, 2006). La lactosa se calculó por diferencia
(lactosa = 100 - grasa - proteína cruda - cenizas) según Lynch et al. (2007).
Forraje, suplementos y desperdicio. Se determinó humedad y cenizas por
gravimetría (métodos AOAC925.23 y AOAC945.46 respectivamente; AOAC, 2006),
proteína cruda por el método Kjeldahl (Thiex et al., 2002), extracto etéreo (método
AOAC-930.39; AOAC, 2006), fibra detergente neutro corregida por N (FDNn), fibra
detergente ácido corregida por N (FDAn; Van Soest et al., 1991; NRC, 2001), lignina
(Van Soest et al., 1991), proteína cruda insoluble en detergente neutro, proteína cruda
insoluble en detergente ácido (Licitra et al., 1996) y FDAi (Sunvold y Cochran, 1991).
Los carbohidratos no fibrosos corregidos por N (CNFn) y la energía neta de lactancia
(ENL) se estimaron según el NRC (2001). Al desperdicio sólo se le determinó la
humedad.
Fluido ruminal. Muestras frescas se usaron para medir el pH con un potenciómetro
(Beckman®) y las muestras congeladas para determinar amonio según adaptación
al método Kjeldahl (Thiex et al., 2002).
Heces. Se determinó la concentración de FDAi (Sunvold y Cochran, 1991) y de Cr por
absorción atómica (Williams et al., 1962) con un espectrofotómetro de absorción
atómica Shimadzu® modelo AA7000.
3.2.5 Cálculos
Consumo de forraje. Para determinar el consumo de forraje se utilizó Cr2O3 como
marcador indigerible externo (Holden et al., 1994) y FDAi como marcador indigerible
interno (Sunvold y Cochran, 1991). Los cálculos se realizaron según lo descrito por
Castaño et al. (2013).
Consumo de AG. Para determinar el consumo de AG se utilizó la ecuación (3-1).
CAG = (CF x [TAG]F x [AG]F) + (CS x [TAG]S x [AG]S) (3-1)

96
Efecto de la suplementación con harina de arroz o semilla de algodón sobre el contenido de los ácidos transvaccénico y ruménico
en la leche de vacas en pastoreo
Donde: CAG es el consumo total de cada AG (g AG / d); CF y CS son los consumos de
forraje y de suplemento respectivamente (g de MS de forraje o suplemento/d); [TAG]F y
[TAG]S son las concentraciones del total de AG en el forraje y en el suplemento
respectivamente (g del total de AG / g de MS de forraje o suplemento); [AG]F y [AG]S son
las concentraciones de cada AG en el total de AG del forraje y del suplemento
respectivamente (g de cada AG / g del total de AG del forraje o del suplemento). El total
de AG se estimó según el contenido de extracto etéreo con la ecuación (3-2) propuesta
por Allen (2000).
TAG = -0.98 + 1.03 x EE (3-2)
Donde: TAG es el porcentaje del total de AG en base seca (g de AG / 100 g de MS) y EE
es el porcentaje de extracto etéreo en base seca (g de EE / 100 g de MS).
Índice de actividad Δ9desaturasa. Se calculó con la ecuación (3-3) según Gagliostro et
al. (2006).
IΔ9 = PΔ
9 x (PΔ
9 + SΔ
9)
-1 (3-3)
Donde: IΔ9 es el índice de actividad de la enzima Δ9desaturasa; PΔ9 es la sumatoria de
productos de la Δ9desaturasa (C14:1c9 + C16:1c9 + C18:1c9 + C18:2c9,t11); SΔ9 es la
sumatoria de sustratos de la Δ9desaturasa (C14:0 + C16:0 + C18:0 + C18:1t11).
Índice de aterogenicidad. Se calculó con la ecuación (3-4) según Ulbricht y Southgate
(1991).
IA = (SC12 + 4SC14 + SC16) x AGI-1 (3-4)
Donde: IA es el índice de aterogenicidad; SC12, SC14 y SC16 AG saturados de 12, 14 y 16 C
respectivamente; AGI es la sumatoria de los AG insaturados.
Índice de trombogenicidad. Se calculó con la ecuación (3-5) según Ulbricht y Southgate
(1991).

Capítulo 3
97
IT = (SC14 + SC16 + SC18) x [0.5MI + 0.5ω6 + 3ω3 + (ω3/ω6)]-1 (3-5)
Donde: IT es el índice de trombogenicidad; SC14, SC16 y SC18 AG saturados de 14, 16 y
18 C respectivamente; MI sumatoria AG monoinsaturados; ω3 y ω6 AG de la serie
omega 3 y 6 respectivamente.
3.2.6 Análisis estadístico
Los datos fueron analizados estadísticamente como un diseño diseño Cross-over 6 x 2 x
2 x 2 (animales x periodos x tratamientos x ordenamientos; Kaps y Lamberson, 2004) de
acuerdo con el modelo presentado en la ecuación (2-8).
yijkl = µ + τi + ϒk + λ(ϒ)jk +ωl + εijkl (3-6)
Donde:
yijkl = Valor de la observación del i-ésimo nivel (i = A, B), j-ésimo animal (j = 1, 2, …,
6), k-ésimo ordenamiento (k = 1, 2) y l-ésimo periodo (l = 1, 2)
µ = Promedio de la población
τi = Efecto del i-ésimo nivel (m = A, B)
ϒk = Efecto del k-ésimo ordenamiento (k = 1, 2)
λ(ϒ)jk = Efecto del j-ésimo animal (j = 1, 2, …, 6) dentro del k-ésimo ordenamiento (k = 1,
2)
ωl = Efecto del l-ésimo periodo (l = 1, 2)
εijkl = Error experimental
Los supuestos fueron probados con la metodología indicada para cada caso. Para el
análisis de los resultados se utilizó el programa SAS versión 9.0 (Copyright© 2002 by
SAS Institute Inc., Cary, NC, USA).
3.3 Resultados
3.3.1 Consumo de alimento
Cuando las vacas recibieron el suplemento A consumieron mayor cantidad de lípidos

98
Efecto de la suplementación con harina de arroz o semilla de algodón sobre el contenido de los ácidos transvaccénico y ruménico
en la leche de vacas en pastoreo
(940 g / d; p<0.05) que se explica por un incremento en la ingestión de extracto etéreo
proveniente del suplemento (618 g / d; p<0.01), ya que los tratamientos no afectaron el
consumo de esta fracción proveniente del forraje. El consumo promedio de lípidos
provenientes del suplemento para el tratamiento B fue 495 g / d y para el A 618 g / d, que
corresponden al 59.6 % y 65.7 % de los lípidos ingeridos por los tratamientos B y A
respectivamente, aunque los suplementos sólo aportaron el 33 % de la MS consumida.
Los consumos de FDA y CNF provenientes del suplemento fueron mayores para el
tratamiento B (1.0 y 1.5 Kg / d respectivamente; p<0.05), mientras que el de lignina fue
menor (457 g / d; p<0.05); sin embargo, el consumo total para cada una de estas
fracciones no se vio afectado. Los consumos de MS, PC, ENL y FDN no se vieron
afectados por el tipo de suplemento (Tabla 3-3).

Capítulo 3
99
Tabla 3-3: Efecto del uso de suplementos con dos niveles de ácido linoleico (bajo y alto) y que
contienen como principal fuente lipídica una mezcla de harina de arroz y semilla de algodón (relación 4.8 : 1) sobre el consumo de alimento de vacas lactantes que pastaban P. clandestinum
Fracción del alimento Nivel
1
ESM2 p
3
Bajo Alto
Materia seca Suplemento, Kg/d 5.2 4.9 0.1 NS Forraje, Kg/d 10.3 9.9 1.5 NS Total
Kg/d 15.5 14.7 1.4 NS Kg/100 Kg de PV
4 2.9 2.8 0.3 NS
Proteína cruda Suplemento, Kg/d 1.0 0.9 0.1 NS Forraje, Kg/d 1.9 1.8 0.3 NS Total, Kg/d 2.9 2.7 0.2 NS
ENLP5
Suplemento, Mcal/d 10.1 10.2 0.7 NS Forraje, Mcal/d 14.8 14.2 2.1 NS Total, Mcal/d 25.0 24.4 1.8 NS
Fibra detergente neutro6
Suplemento, Kg/d 1.5 1.4 0.1 NS Forraje, Kg/d 5.2 5.0 0.7 NS Total
Kg/d 6.7 6.4 0.7 NS Kg/100 Kg de PV
4 1.2 1.2 0.2 NS
Fibra detergente ácido6
Suplemento, Kg/d 1.0 0.9 0.1 * Forraje, Kg/d 2.9 2.8 0.4 NS Total, Kg/d 4.0 3.7 0.4 NS
Lignina Suplemento, g/d 457 610 79 * Forraje, g/d 284 272 40 NS Total, g/d 741 882 120 NS
Carbohidratos no fibrosos6
Suplemento, Kg/d 1.5 1.3 0.1 * Forraje, Kg/d 1.8 1.8 0.3 NS Total, Kg/d 3.3 3.1 0.2 NS
Extracto etéreo Suplemento, g/d 495 618 45 ** Forraje, g/d 335 322 49 NS Total, g/d 830 940 42 *
12.64 g y 4.08 g de ácido linoleico / 100 g MS para los niveles Bajo y Alto respectivamente.
2 Error
estándar de la media de los tratamientos. 3Significancia; NS: no significativa, *: p<0.05, **: p<0.01
y ***: p<0.001. 4Peso vivo.
5Energía neta de lactancia corregida al nivel de consumo (NRC, 2001).
6Corregido por N (NRC, 2001).
3.3.2 Consumo de precursores de los ácidos trans-vaccénico
y ruménico
Cuando los animales recibieron el suplemento A se incrementó el consumo de Li (253 g /
d; p<0.01) y de precursores de los TVa y Ru (341 g / d; p<0.01). Estos incrementos se

100
Efecto de la suplementación con harina de arroz o semilla de algodón sobre el contenido de los ácidos transvaccénico y ruménico
en la leche de vacas en pastoreo
debieron al suplemento, pues el consumo de Li proveniente del forraje fue similar entre
los tratamientos. Aunque se presentó un mayor consumo de Ln proveniente del
suplemento A (7 g / d; p<0.001), el consumo total de este AG no se vio afectado por el
tratamiento. Como consecuencia del mayor consumo de Li del suplemento, se
incrementó el consumo total de precursores de TVa y Ru en comparación a B (Tabla 3-
4).
Tabla 3-4: Efecto del uso de suplementos con dos niveles de ácido linoleico (bajo y alto) y que contienen como principal fuente lipídica una mezcla de harina de arroz y semilla de algodón (relación 4.8 : 1) sobre el consumo del ácido linoleico, ácido linolénico y precursores de los ácidos transvaccénico y ruménico en la leche de vacas que pastaban P. clandestinum
Ítem Nivel
1
ESM2 p
3
Bajo Alto
Consumo, g/d Linoleico
Suplemento 137 199 14 ** Forraje 33 32 5 NS Total 170 231 13 **
Linolénico Suplemento 5 7 0.5 *** Forraje 107 103 16 NS Total 111 110 16 NS
Total precursores4
Suplemento 141 206 15 ** Forraje 140 134 21 NS Total 281 341 17
**
Aporte relativo5, %
Linoleico Suplemento 80.7 86.2 2.9 * Forraje 19.3 13.8 2.5 *
Linolénico Suplemento 4.2 6.9 1.2 * Forraje 95.8 93.1 1.2 *
Total precursores4
Suplemento 50.6 60.5 4.8 *
Forraje 49.4 39.5 4.8 * 12.64 g y 4.08 g de ácido linoleico / 100 g MS para los niveles Bajo y Alto respectivamente.
2Error
estándar de la media de los tratamientos. 3Significancia; NS: no significativa,*: p<0.05, **: p<0.01 y
***: p<0.001. 4Sumatoria de los ácidos grasos linoleico y linolénico.
5g de ácido graso proveniente
del suplemento o del forraje/100 g de precursores consumidos al día
3.3.3 Producción y calidad de la leche
La producción de leche y su composición química no se vieron afectadas por los
tratamientos (Tabla 3-5).

Capítulo 3
101
Tabla 3-5: Efecto del uso de suplementos con dos niveles de ácido linoleico (bajo y alto) y que
contienen como principal fuente lipídica una mezcla de harina de arroz y semilla de algodón (relación 4.8 : 1) sobre la producción y calidad de la leche de vacas que pastaban P. clandestinum
Ítem Nivel
1
ESM2 p
3
Bajo Alto
Leche, Kg/d 17.8 17.4 1.5 NS Sólidos totales
Concentración, % 12.1 12.2 0.7 NS Producción, Kg/d 2.16 2.13 0.23 NS
Proteína cruda Concentración, % 3.2 3.1 0.2 NS Producción, Kg/d 0.56 0.54 0.03 NS
Grasa Concentración , % 3.9 3.7 0.6 NS Producción, Kg/d 0.69 0.63 0.11 NS
Cenizas Concentración , % 0.8 0.8 0.03 NS Producción, Kg/d 0.14 0.14 0.01 NS
Lactosa Concentración, % 4.3 4.6 0.3 NS Producción, Kg/d 0.77 0.82 0.11 NS
Ácido transvaccénico Concentración, % 0.09 0.09 0.01 NS Producción, g/d 16.26 15.13 2.98 NS
Ácido ruménico Concentración, % 0.04 0.04 0.002 NS Producción, g/d 7.00 7.18 0.92 NS
12.64 g y 4.08 g de ácido linoleico / 100 g MS para los niveles Bajo y Alto respectivamente.
2Error
estándar de la media de los tratamientos. 3Significancia; NS: no significativa, *: p<0.05, **: p<0.01
y ***: p<0.001.
3.3.4 Perfil de ácidos grasos de la grasa láctea
Los tratamientos no afectaron la concentración de los AG C11:0, C13:0, C14:0, C14:1c-9,
C15:0, C16:1c9, C17:0, C18:0, C18:1c11, C18:1t11 (TVa), C18:2c9,c12, C18:2c9,t11 (Ru) y
C18:3c9,c12,c15. Con el tratamiento A se presentó una tendencia a incrementar la
concentración en la grasa láctea de C18:1c9 (27.71 %; p=0.08) y a disminuir C8:0 (0.67
%; p=0.08) y C10:0 (1.35 %; p=0.06; Tabla 3-6).
El tratamiento A incrementó los AG insaturados y disminuyó los saturados (35.1 y
60.0 % respectivamente; p<0.05), presentó una tendencia a disminuir los AG de
cadena corta (3.4 %; p=0.06) y media (41.4 %; p=0.06) e incrementar los AG de
cadena larga (50.1 %; p=0.09). Los AG de novo no se vieron afectados por los
tratamientos, pero se presentó una tendencia a incrementar los AG preformados
con el tratamiento A (49.5 %; p=0.08).

102
Efecto de la suplementación con harina de arroz o semilla de algodón sobre el contenido de los ácidos transvaccénico y ruménico
en la leche de vacas en pastoreo
Los índices Ru / TVa, C14:1 / C14:0 y C16:1 / C16:0 no fueron alterados por los
tratamientos, pero el índice calculado de actividad de la Δ9desaturasa fue mayor
para el tratamiento A (0.37 %; p<0.05) debido a que presentó mayor
concentración de productos de la enzima y menor de sustratos (32.5 y 55.7 %
respectivamente; p<0.05). Los índices de aterogenicidad y trombogenicidad
fueron mayores para el tratamiento B (1.79 y 3.22 respectivamente; p<0.05; Tabla 3-
6).
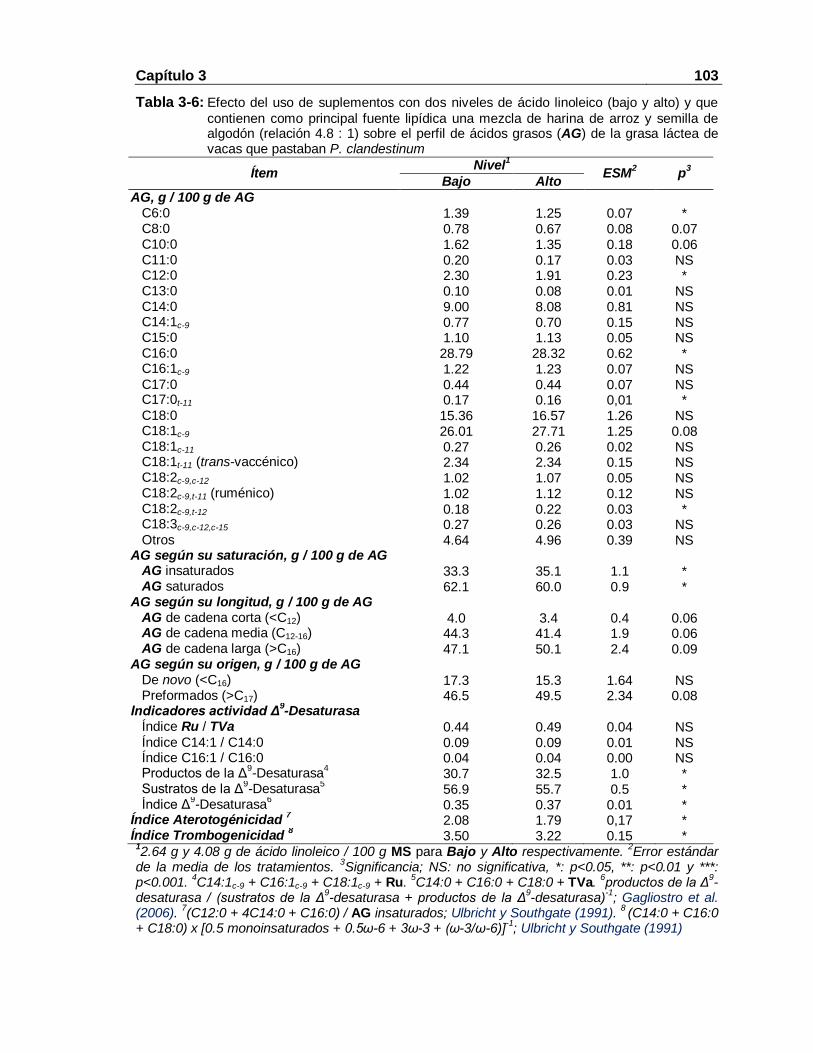
Capítulo 3
103
Tabla 3-6: Efecto del uso de suplementos con dos niveles de ácido linoleico (bajo y alto) y que
contienen como principal fuente lipídica una mezcla de harina de arroz y semilla de algodón (relación 4.8 : 1) sobre el perfil de ácidos grasos (AG) de la grasa láctea de vacas que pastaban P. clandestinum
Ítem Nivel
1
ESM2 p
3
Bajo Alto
AG, g / 100 g de AG C6:0 1.39 1.25 0.07 * C8:0 0.78 0.67 0.08 0.07 C10:0 1.62 1.35 0.18 0.06 C11:0 0.20 0.17 0.03 NS C12:0 2.30 1.91 0.23 * C13:0 0.10 0.08 0.01 NS C14:0 9.00 8.08 0.81 NS C14:1c9 0.77 0.70 0.15 NS C15:0 1.10 1.13 0.05 NS C16:0 28.79 28.32 0.62 * C16:1c9 1.22 1.23 0.07 NS C17:0 0.44 0.44 0.07 NS C17:0t11 0.17 0.16 0,01 * C18:0 15.36 16.57 1.26 NS C18:1c9 26.01 27.71 1.25 0.08 C18:1c11 0.27 0.26 0.02 NS C18:1t11 (trans-vaccénico) 2.34 2.34 0.15 NS C18:2c9,c12 1.02 1.07 0.05 NS C18:2c9,t11 (ruménico) 1.02 1.12 0.12 NS C18:2c9,t12 0.18 0.22 0.03 * C18:3c9,c12,c15 0.27 0.26 0.03 NS Otros 4.64 4.96 0.39 NS
AG según su saturación, g / 100 g de AG
AG insaturados 33.3 35.1 1.1 * AG saturados 62.1 60.0 0.9 *
AG según su longitud, g / 100 g de AG AG de cadena corta (<C12) 4.0 3.4 0.4 0.06 AG de cadena media (C12-16) 44.3 41.4 1.9 0.06 AG de cadena larga (>C16) 47.1 50.1 2.4 0.09
AG según su origen, g / 100 g de AG De novo (<C16) 17.3 15.3 1.64 NS Preformados (>C17) 46.5 49.5 2.34 0.08
Indicadores actividad Δ9Desaturasa
Índice Ru / TVa 0.44 0.49 0.04 NS Índice C14:1 / C14:0 0.09 0.09 0.01 NS Índice C16:1 / C16:0 0.04 0.04 0.00 NS Productos de la Δ
9Desaturasa
4 30.7 32.5 1.0 *
Sustratos de la Δ9Desaturasa
5 56.9 55.7 0.5 *
Índice Δ9Desaturasa
6 0.35 0.37 0.01 *
Índice Aterotogénicidad 7 2.08 1.79 0,17 *
Índice Trombogenicidad 8 3.50 3.22 0.15 *
12.64 g y 4.08 g de ácido linoleico / 100 g MS para Bajo y Alto respectivamente.
2Error estándar
de la media de los tratamientos. 3Significancia; NS: no significativa, *: p<0.05, **: p<0.01 y ***:
p<0.001. 4C14:1c-9 + C16:1c-9 + C18:1c-9 + Ru.
5C14:0 + C16:0 + C18:0 + TVa.
6productos de la Δ
9-
desaturasa / (sustratos de la Δ9-desaturasa + productos de la Δ
9desaturasa)
-1; Gagliostro et al.
(2006). 7(C12:0 + 4C14:0 + C16:0) / AG insaturados; Ulbricht y Southgate (1991).
8 (C14:0 + C16:0
+ C18:0) x [0.5 monoinsaturados + 0.5ω-6 + 3ω-3 + (ω-3/ω-6)]-1; Ulbricht y Southgate (1991)

104
Efecto de la suplementación con harina de arroz o semilla de algodón sobre el contenido de los ácidos transvaccénico y ruménico
en la leche de vacas en pastoreo
3.3.5 Peso vivo y fluido ruminal
El peso vivo y el fluido ruminal no se vieron afectados por el tipo de suplemento (Tabla 3-
7).
Tabla 3-7: Efecto del uso de suplementos con dos niveles de ácido linoleico (bajo y alto) y que contienen como principal fuente lipídica una mezcla de harina de arroz y semilla de algodón (relación 4.8 : 1) sobre el peso vivo y el fluido ruminal de vacas lactantes que pastan P. clandestinum
Ítem Nivel
ESM2 p
3
Bajo Alto
Peso vivo, Kg 536 538 4 NS Fluido ruminal
pH 6.7 6.8 0.2 NS NNH3, mg/dl 14.2 13.1 3.7 NS
12.64 g y 4.08 g de ácido linoleico / 100 g MS para los niveles Bajo y Alto respectivamente.
2Error
estándar de la media de los tratamientos. 3Significancia; NS: no significativa, *: p<0.05, **: p<0.01
y ***: p<0.001.
3.4 Discusión
Este trabajo buscaba comparar la suplementación con dos niveles de Li utilizando como
principal fuente lipídica una mezcla de dos recursos tradicionalmente usados en la
suplementación de vacas lecheras (HA y SA) sobre el perfil de AG de la leche, en
especial sobre la concentración de Ru y TVa. La literatura es abundante en señalar que
el incremento del Li en el suplemento aumenta las concentraciones de Ru y TVa en la
leche (AbuGhazaleh et al., 2003; Bu et al., 2007; Huang et al., 2008); sin embargo, los
efectos son variables según la fuente de Li (Khanal y Olson, 2004; Paradis et al., 2008).
Castaño et al. (2013) evaluaron la SA y la HA como principales fuentes lipídicas de Li y
encontraron que a diferencia de la SA, la HA es una buena alternativa para incrementar
los niveles de Ru y TVa.
Se preparó un suplemento con un contenido de Li similar al utilizado por Castaño et al.
(2013), con el cual dichos autores evidenciaron un incremento en la concentración de Ru
y TVa en la grasa láctea al utilizar HA como principal fuente lipídica y otro cuya
composición nutricional fuera similar al anterior pero con mayor concentración de Li y de
lípidos. En los dos suplementos se utilizó como principal fuente de Li la mezcla de HA y
SA (relación 4.8 : 1) presumiendo que la mayor proporción de HA con respecto a la SA
podría evitar el efecto negativo de la SA sobre la concentración TVa y Ru encontrado por
Castaño et al. (2013). De tal manera se esperaba documentar el efecto de aumentar la

Capítulo 3
105
concentración de Li utilizando una mezcla de las dos fuentes (HA y SA) sobre el perfil de
AG lácteos.
3.4.1 Consumo de alimento
No se presentaron diferencias en el consumo de MS asociadas a las concentraciones de
Li o de lípidos que fueron mayores en el tratamiento A. Algunos autores han sugerido
que al incrementar la grasa en la dieta se puede disminuir el consumo de MS (Schauff y
Clark, 1992) y que este efecto aumenta con el incremento en la proporción de AG
insaturados en el suplemento (Firkins y Eastridge, 1994; Bremmer et al., 1998) lo cual no
fue observado en nuestro experimento. Concentraciones mayores del 8 ó 9 % de grasa
en la dieta disminuyen el consumo de MS (Palmquist y Jenkins, 1980), mientras que en
las revisiones de experimentos con vacas en pastoreo Schroeder et al. (2004) y Bargo et
al. (2003) no encontraron diferencias en el consumo MS al suplementar con grasa. En
nuestro trabajo, la grasa representó el 5.4 y 6.4 % del total de la MS consumida para los
tratamientos B y A respectivamente, mientras que la grasa proveniente de los
suplementos fue 3.2 y 4.2 % para los tratamientos B y A respectivamente. El NRC (2001)
recomienda que no se debe sobrepasar el 3 al 4 % de grasa en la dieta proveniente del
suplemento.
El consumo de FDAn y de CNFn proveniente del suplemento fue menor para el
tratamiento A, mientras que el consumo de lignina fue mayor; sin embargo, el consumo
total de estas fracciones del alimento fue similar entre los tratamientos. La ausencia de
diferencias en los consumos de MS y nutrientes sugiere que los principales efectos que
se presentaron en este estudio en otras variables estarían principalmente asociados al
perfil de AG. En este sentido se puede señalar que el consumo de Li en el tratamiento A
fue un 31 % superior al B.
El incremento en el consumo de grasa se debió al suplemento, ya que los consumos de
extracto etéreo provenientes del forraje fueron similares entre los dos tratamientos. El
suplemento B aportó el 59.0 % de la grasa en la dieta y el A el 66 %; aunque estos
suplementos sólo aportaron en promedio el 33.1 % de la MS consumida. El mayor
consumo de grasa del tratamiento A sugiere un mayor consumo de precursores.
La biosíntesis de Ru en la leche requiere una fuente de AG poliinsaturados como el Li y

106
Efecto de la suplementación con harina de arroz o semilla de algodón sobre el contenido de los ácidos transvaccénico y ruménico
en la leche de vacas en pastoreo
Ln en el alimento, los cuales son biohidrogenados en el rumen supliendo una
combinación del precursor TVa y pequeñas cantidades del producto Ru en la glándula
mamaria (Mohammed et al., 2009). Un mayor consumo Li y Ln está relacionado con
elevados niveles TVa y Ru en la leche, aunque el consumo de precursores no es el único
factor que afecta la producción de Ru (Mohammed et al., 2009). El consumo total de
precursores fue mayor para el tratamiento A pero las concentraciones de TVa y Ru en la
leche no fueron mayores. Castaño et al. (2013).encontraron incrementos en TVa y Ru
cuando las vacas consumieron 300 g / d de precursores y 179 g / d de Li, cantidades
similares a las del tratamiento B (281 g / d y 193 g / d de precursores y de Li
respectivamente). El incremento en el consumo de Li y de precursores TVa y Ru del
tratamiento A con respecto al tratamiento B fue del 31.1 % y 21.4 % respectivamente, lo
cual no fue suficiente para generar un incremento en la concentración de TVa y Ru
lácteos.
La composición de los forrajes puede afectar la concentración de Ru en la leche
(Lourenço et al., 2007). El forraje fresco es una fuente rica en Ln (Mohammed et al.,
2009) y su concentración en los forrajes es directamente proporcional al contenido de Ru
en leche (Mel´uchová et al., 2008). El forraje fue la principal fuente de Ln y el consumo
de Ln proveniente del forraje no se vio afectado por los tratamientos, de tal manera que
no es posible asociar el consumo de Ln proveniente del forraje con variaciones en Ru de
la leche.
3.4.1 Producción y calidad de la leche
La producción y composición de la leche fue similar para los dos tratamientos. La SA
contiene factores tóxicos como el gosipol (Zhang et al., 2007b) que puede afectar la
producción láctea (Lindsey et al., 1980) si se suministra por encima de los niveles
recomendados (Coppock et al., 1987; Arieli, 1998; Zhang et al., 2007a). Aunque en este
trabajo no se determinó la concentración de gosipol, es probable que los niveles
consumidos fueran lo suficientemente bajos como para modificar la eficiencia en el uso
de la energía por parte del animal. En este trabajo el consumo de SA fue de 0.47 y 0.73
Kg / d para los tratamiento B y A respectivamente, que corresponden al 3 y al 5 % de la
MS consumida en los tratamientos B y A respectivamente; estos valores son inferiores a
lo utilizado por Castaño et al. (2013) quienes con 1.5 Kg de SA / d (8.6 % de la MS
consumida) encontraron disminución en la producción láctea y también son inferiores al

Capítulo 3
107
máximo recomendado en la literatura (15 % de la ración ó 3 a 4 Kg/vaca/d; Coppock et
al., 1987; Arieli, 1998; Zhang et al., 2007a).
3.4.2 Perfil de ácidos grasos de la grasa láctea
La composición de AG puede indicar sobre la fuente de precursores (Barber et al., 1997).
Los AG hasta C12, la mayoría de C14:0 y cerca del 50 % de C16:0 son sintetizados en la
glándula mamaria a partir del acetato y butirato producidos por la degradación ruminal de
los carbohidratos de la dieta, mientras que una pequeña cantidad de C14:0, cerca del 50
% de C16:0 y todos los AG de C18 son extraídos de la sangre arterial (Enjalbert et al.,
1998). El tratamiento A presentó menor concentración de los AG C6:0, C12:0 y C16:0,
además de una tendencia hacia menor concentración de C8:0 y C10:0 que como se
mencionó anteriormente son sintetizados en la glándula mamaria. Aunque no se encontró
una diferencia significativa entre los tratamientos para la concentración de los AG de
novo, hay evidencia para suponer que el tratamiento A presentó menor síntesis de AG,
debido a la menor concentración C6:0, C12:0 y C16:0, una tendencia a una menor
concentración de C8:0, C10:0 y de AG de cadena corta (<C12), además a una tendencia
a mayor concentración de AG de preformados.
La biosíntesis de Ru en la leche depende de la cantidad de sustrato consumido, del
grado de biohidrogenación en el rumen y de la actividad de la enzima Δ9desaturasa en
el tejido mamario (Mohammed et al., 2009). El suministro de lípidos con elevado
contenido de AG insaturados tiene un efecto positivo sobre el contenido de Ru y TVa en
la leche (Dhiman et al., 2000) especialmente si las fuentes lipídicas contienen niveles
elevados de Li (Bu et al., 2007). En nuestro experimento, las concentraciones y las
producciones diarias de TVa y Ru no se afectaron al incrementar el nivel de Li en el
suplemento, de tal manera que es probable que otros aspectos relacionados con la
biohidrogenación en el rumen y con la actividad de la Δ9desaturasa a nivel de la
glándula mamaria se asocien con la ausencia de un incremento en la concentración de
Ru al suministrar el tratamiento A.
La variación del Ru está relacionada con diferencias en la acción de la Δ9desaturasa
(Corl et al., 2001) pues entre el 75 al 90% del Ru en la leche se deriva de la
Δ9desaturación del TVa (Bauman et al., 2008) y al inhibir la actividad de la
Δ9desaturasa se disminuye la proporción de Ru en la leche (Griinari et al., 2000). Dos

108
Efecto de la suplementación con harina de arroz o semilla de algodón sobre el contenido de los ácidos transvaccénico y ruménico
en la leche de vacas en pastoreo
AG ciclopropenos, el malválico y el estercúlico, se presentan en la SA (alrededor de 1%
del aceite; Coppock et al., 1987) e inhiben la enzima Δ9desaturasa (Cook et al., 1976). El
mayor consumo de SA con el tratamiento A podría explicar la ausencia de un incremento
de Ru en la leche al suministrar el suplemento A. Aunque no se midió la actividad de la
Δ9desaturasa, los resultados sugieren que esta enzima fue más activa con el tratamiento
A ya que se encontró menor concentración de los sustratos y mayor de los productos de
la enzima, sumado a un mayor índice calculado de actividad de la Δ9desaturasa. La
biohidrogenación en el rumen produce ciclopropano que no es capaz de inhibir la
Δ9desaturasa (Cook et al., 1976) y nuestros resultados indican que aparentemente los
AG ciclopropenos se inactivaron en el rumen o que se consumieron en una cantidad
insuficiente como para inactivar la actividad de la Δ9desaturasa.
El Ru y el TVa son producidos como intermediarios de la biohidrogenación de AG
insaturados en el rumen (AbuGhazaleh, 2008; Bauman et al., 2008), de tal manera que el
suministro de lípidos con elevado contenido de AG insaturados tiene un efecto positivo
sobre el contenido de Ru y TVa en la leche, siempre y cuando el aceite esté disponible a
los microorganismos del rumen para la biohidrogenación (Dhiman et al., 2000; Khanal y
Olson, 2004; Paradis et al., 2008). Sin embargo, la completa biohidrogenación de los Li y
Ln produce el ácido esteárico (Doreau y Chilliard, 1997), por lo que se han buscado
estrategias para reducir la biohidrogenación ruminal hacia ácido esteárico (Vlaeminck et
al., 2008) y permitir un mayor flujo de intermediarios hacia el intestino delgado
(especialmente Ru y el TVa; Odongo et al., 2007; Aldai et al., 2008), dentro de dichas
estrategias resalta el suministro de ionóforos (Odongo et al., 2007; Aldai et al., 2008),
aceites marinos (AbuGhazaleh et al., 2003; Vlaeminck et al., 2008) y de algas (Vlaeminck
et al., 2008; AlZahal et al., 2008). Una mayor biohidrogenación en el rumen podría
explicar la ausencia de un incremento en la concentración de TVa y Ru al suministrar el
tratamiento A debido al mayor consumo de SA, pues Pires et al. (1997) y Harvatine et al.
(2002) indican que existe una extensa biohidrogenación de los AG de la SA; además,
Castaño et al. (2013) encontraron que la SA no logró incrementar la concentración de Ru
en la leche y los autores sugieren que probablemente esto se debió en parte a una mayor
biohidrogenación de los AG de la SA.
Otros cambios en el perfil de AG son aún más difíciles de explicar. La concentración de
C17:1t9 en la grasa láctea fue menor para el tratamiento A, lo cual indica posibles

Capítulo 3
109
diferencias en los procesos de biohidrogenación. Se presentó una tendencia a
incrementar la concentración de C18:1c9 con el tratamiento A y este AG puede provenir
de la sangre (Enjalbert et al., 1998) o a gracias a la acción de la Δ9desaturasa en la
glándula mamaria sobre ácido esteárico (Corl et al., 2001). La información obtenida no
permite explicar el motivo de estos cambios.
La relación entre el tipo de grasa en la dieta y varias enfermedades crónicas en humanos
se ha investigado ampliamente, incluyendo las enfermedades cardiovaculares (Jacobs et
al., 2011). Las enfermedades coronarias en la mayoría de los casos se deben a la
obstrucción de los vasos por aterosclerosis o trombosis, de manera aislada o en
combinación (Ulbricht y Southgate, 1991). El A presentó el menor índice de
aterogenicidad que concuerda con la menor concentración de C12:0 y C16:0, y mayor
concentración de AG insaturados para este tratamiento. El índice de trombogenicidad fue
menor para el tratamiento A lo cual también puede ser explicado por la menor
concentración de C16:0 y mayor concentración de AG insaturados para este tratamiento.
Una disminución en la proporción de AG saturados puede ser benéfico para la salud
humana (Jacobs et al., 2011). Un aspecto importante en el perfil de AG y su relación con
la salud humana es la concentración de AG saturados de cadena corta, como el C12:0 y
C14:0, ya que según Liu et al. (2007) al reducir estos AG se observan efectos positivos
sobre la salud humana.
3.5 Conclusiones
El tratamiento alto no logró incrementar la concentración de los ácidos trans-
vaccénico y ruménico; además, presentó un mayor índice calculado de actividad
de la Δ9desaturasa (0.37; p<0.05), lo cual sugiere una extensa biohidrogenación
en el rumen. El tratamiento A incrementó los AG insaturados, disminuyó los
saturados (35.1 y 60.0 % respectivamente; p<0.05) y presentó menores índices
de aterogenicidad y trombogenicidad (1.79 y 3.22 respectivamente; p<0.05).
3.6 Agradecimientos
Al Ministerio de Agricultura y Desarrollo Rural por el apoyo económico para la
elaboración de este trabajo. Al Laboratorio de Nutrición Animal de la Facultad de
Medicina Veterinaria y de Zootecnia -Unal, Sede Bogotá-, al Centro de Investigación

110
Efecto de la suplementación con harina de arroz o semilla de algodón sobre el contenido de los ácidos transvaccénico y ruménico
en la leche de vacas en pastoreo
Agropecuario Marengo -Unal, Sede Bogotá- y al Grupo de Investigación en Nutrición
Animal de la Facultad de Medicina Veterinaria de y de Zootecnia -Unal, Sede Bogotá- por
el apoyo en la fase experimental y trabajo en el laboratorio.
3.7 Referencias
ABUGHAZALEH, A. A. 2008. Effect of fish oil and sunflower oil supplementation on milk conjugated linoleic acid content for grazing dairy cows. Anim. Feed Sci. and Technol. 141(3-4):220-232.
ABUGHAZALEH, A. A.; FELTON, D. O. y IBRAHIM, S. A. 2007. Milk conjugated linoleic acid response to fish oil and sunflower oil supplementation to dairy cows managed under two feeding systems. J. Dairy Sci. 90(10):4763-4769.
ABUGHAZALEH, A. A.; SCHINGOETHE, D. J.; HIPPEN, A. R. y KALSCHEUR, K. F. 2003. Milk conjugated linoleic acid response to fish oil supplementation of diets differing in fatty acid profiles. J. Dairy Sci. 86(3):944-953.
ABUGHAZALEH, A. A.; SCHINGOETHE, D. J.; HIPPEN, A. R. y KALSCHEUR, K. F. 2004. Conjugated linoleic acid increases in milk when cows fed fish meal and extruded soybeans for an extended period of time. J. Dairy Sci. 87(6):1758-1766.
ALDAI, N.; DUGAN, M. E. R.; KRAMER, J. K. G.; MIR, P. S. y MCALLISTER, T. A. 2008. Non-ionophore antibiotics do not affect the trans-18:1 and CLA composition in beef adipose tissue. J. Anim Sci. 86(12):3522-3532.
ALLEN, M. S. 2000. Effects of diet on short-term regulation of feed intake by lactating dairy cattle. J. Dairy Sci. 83(7):1598-1624.
ALZAHAL, O.; ODONGO, N. E.; MUTSVANGWA, T.; OR-RASHID, M. M.; DUFFIELD, T. F.; BAGG, R.; DICK, P.; VESSIE, G. y MCBRIDE, B. W. 2008. Effects of monensin and dietary soybean oil on milk fat percentage and milk fatty acid profile in lactating dairy cows. J. Dairy Sci. 91(3):1166-1174.
AOAC. 2006. Official methods of analysis of AOAC International. 18a ed. Association Official to Analytical Chemistry Intenational, Maryland.
ARCHIBEQUE, S. L.; LUNT, D. K.; GILBERT, C. D.; TUME, R. K. y SMITH, S. B. 2005. Fatty acid indices of stearoyl-CoA desaturase do not reflect actual stearoyl-CoA desaturase enzyme activities in adipose tissues of beef steers finished with corn-, flaxseed-, or sorghum-based diets. J. Anim Sci. 83(5):1153-1166.
ARIELI, A. 1998. Whole cottonseed in dairy cattle feeding: a review. Anim. Feed Sci. and Technol. 72(1-2):97-110.
BARBER, M. C.; CLEGG, R. A.; TRAVERS, M. T. y VERNON, R. G. 1997. Lipid metabolism in the lactating mammary gland. Biochim. Biophys. Acta (BBA) - Lipids and Lipid Metabolism. 1347(2-3):101-126.
BARGO, F.; MULLER, L. D.; KOLVER, E. S. y DELAHOY, J. E. 2003. Invited review: Production and digestion of supplemented dairy cows on pasture. J. Dairy Sci.
86(1):1-42.
BAUMAN, D. E.; PERFIELD, J. W., II; HARVATINE, K. J. y BAUMGARD, L. H. 2008. Regulation of fat synthesis by conjugated linoleic acid: lactation and the ruminant

Capítulo 3
111
model. J. Nutr. 138(2):403-409.
BHARATHAN, M.; SCHINGOETHE, D. J.; HIPPEN, A. R.; KALSCHEUR, K. F.; GIBSON, M. L. y KARGES, K. 2008. Conjugated linoleic acid increases in milk from cows fed condensed corn distillers solubles and fish oil. J. Dairy Sci. 91(7):2796-2807.
BOUATTOUR, M. A.; CASALS, R.; ALBANELL, E.; SUCH, X. y CAJA, G. 2008. Feeding soybean oil to dairy goats increases conjugated linoleic acid in milk. J. Dairy Sci.
91(6):2399-2407.
BREMMER, D. R.; RUPPERT, L. D.; CLARK, J. H. y DRACKLEY, J. K. 1998. Effects of chain length and unsaturation of fatty acid mixtures infused into the abomasum of lactating dairy cows. J. Dairy Sci. 81(1):176-188.
BU, D. P.; WANG, J. Q.; DHIMAN, T. R. y LIU, S. J. 2007. Effectiveness of oils rich in linoleic and linolenic acids to enhance conjugated linoleic acid in milk from dairy cows. J. Dairy Sci. 90(2):998-1007.
CASTAÑO, G. A.; PABÓN R., M. L. y CARULLA F., J. E. 2013. Efecto de la suplementación con harina de arroz y semilla de algodón sobre el contenido de los ácidos trans-vaccénico (C18:1trans-11) y ruménico (C18:2cis-9, trans-11) en la leche de vacas pastando Pennisetum clandestinum. Master Science Producción Animal, Universidad Nacional de Colombia, Colombia.
CHILLIARD, Y.; FERLAY, A.; MANSBRIDGE, R. M. y DOREAU, M. 2000. Ruminant milk fat plasticity: nutritional control of saturated, polyunsaturated, trans and conjugated fatty acids. Ann. Zootech. 49(3):181-205.
COOK, C. W. 1964. Symposium on nutrition of forages and pastures: collecting forage samples representative of ingested material of grazing animals for nutritional studies. J. Anim Sci. 23(1):265-270.
COOK, L.; SCOTT, T.; MILLS, S.; FOGERTY, A. y JOHNSON, A. 1976. Effects of protected cyclopropene fatty acids on the composition of ruminant milk fat. Lipids.
11(9):705-711.
COPPOCK, C. E.; LANHAM, J. K. y HORNER, J. L. 1987. A review of the nutritive value and utilization of whole cottonseed, cottonseed meal and associated by-products by dairy cattle. Anim. Feed Sci. and Technol. 18(2):89-129.
CORL, B. A.; BAUMGARD, L. H.; DWYER, D. A.; GRIINARI, J. M.; PHILLIPS, B. S. y
BAUMAN, D. E. 2001. The role of -desaturase in the production of cis-9, trans-11 CLA. J. Nutr. Biochem. 12(11):622-630.
DE CAMPOS, R. M. L.; HIERRO, E.; ORDÓÑEZ, J. A.; BERTOL, T. M.; TERRA, N. N. y DE LA HOZ, L. 2007. Fatty acid and volatile compounds from salami manufactured with yerba mate (Ilex paraguariensis) extract and pork back fat and meat from pigs fed on diets with partial replacement of maize with rice bran. Food Chem. 103(4):1159-1167.
DEWHURST, R. J.; SHINGFIELD, K. J.; LEE, M. R. F. y SCOLLAN, N. D. 2006. Increasing the concentrations of beneficial polyunsaturated fatty acids in milk produced by dairy cows in high-forage systems. Anim. Feed Sci. and Technol.
131(3-4):168-206.
DHIMAN, T. R.; HELMINK, E. D.; MCMAHON, D. J.; FIFE, R. L. y PARIZA, M. W. 1999. Conjugated linoleic acid content of milk and cheese from cows fed extruded oilseeds. J. Dairy Sci. 82(2):412-419.

112
Efecto de la suplementación con harina de arroz o semilla de algodón sobre el contenido de los ácidos transvaccénico y ruménico
en la leche de vacas en pastoreo
DHIMAN, T. R.; SATTER, L. D.; PARIZA, M. W.; GALLI, M. P.; ALBRIGHT, K. y TOLOSA, M. X. 2000. Conjugated linoleic acid (CLA) content of milk from cows offered diets rich in linoleic and linolenic acid. J. Dairy Sci. 83(5):1016-1027.
DOREAU, M. y CHILLIARD, Y. 1997. Digestion and metabolism of dietary fat in farm animals. Br. J. Nutr. 78(01):S15-S35.
ENJALBERT, F.; NICOT, M. C.; BAYOURTHE, C. y MONCOULON, R. 1998. Duodenal infusions of palmitic, stearic or oleic acids differently affect mammary gland metabolism of fatty acids in lactating dairy cows. J. Nutr. 128(9):1525-1532.
FIRKINS, J. L. y EASTRIDGE, M. L. 1994. Assessment of the effects of iodine value on fatty acid digestibility, feed intake, and milk production. J. Dairy Sci. 77(8):2357-2366.
FLOWERS, G.; IBRAHIM, S. A. y ABUGHAZALEH, A. A. 2008. Milk fatty acid composition of grazing dairy cows when supplemented with linseed oil. J. Dairy Sci. 91(2):722-730.
GAGLIOSTRO, G. A.; RODRÍGUEZ, A.; PELLEGRINI, P.; GATTI, P.; MUSSET, G.; CASTAÑEDA, R. A.; COLOMBO, D. y CHILLIARD, Y. 2006. Efectos del suministro de aceite de pescado sólo o en combinación con aceite de girasol sobre las concentraciones de ácido linoleico conjugado (CLA) y omega 3 (n-3) en leche de cabra. Rev. Argent. Prod. Anim. 26:71-87.
GRIINARI, J. M.; CORL, B. A.; LACY, S. H.; CHOUINARD, P. Y.; NURMELA, K. V. V. y BAUMAN, D. E. 2000. Conjugated linoleic acid is synthesized endogenously in
lactating dairy cows by 9-Desaturase. J. Nutr. 130(9):2285-2291.
HARVATINE, D. I.; FIRKINS, J. L. y EASTRIDGE, M. L. 2002. Whole linted cottonseed as a forage substitute fed with ground or steam-flaked corn: digestibility and performance. J. Dairy Sci. 85(8):1976-1987.
HERVÁS, G.; LUNA, P.; ÁNGEL, R.; ARES, N.; DE LA FUENTE, M. A.; REZ, M. y FRUTOS, P. 2008. Effect of diet supplementation with sunflower oil on milk production, fatty acid profile and ruminal fermentation in lactating dairy ewes. J. Dairy Res. 75(4):399-405.
HOLDEN, L. A.; MULLER, L. D. y FALES, S. L. 1994. Estimation of intake in high producing Holstein cows grazing grass pasture. J. Dairy Sci. 77(8):2332-2340.
HOUSEKNECHT, K. L.; HEUVEL, J. P. V.; MOYA-CAMARENA, S. Y.; PORTOCARRERO, C. P.; PECK, L. W.; NICKEL, K. P. y BELURY, M. A. 1998. Dietary conjugated linoleic acid normalizes impaired glucose tolerance in the zucker diabetic fatty fa/fa rat. Biochem. Biophysi. Res. Comm. 244(3):678-682.
HUANG, Y.; SCHOONMAKER, J. P.; BRADFORD, B. J. y BEITZ, D. C. 2008. Response of milk fatty acid composition to dietary supplementation of soy oil, conjugated linoleic acid, or both. J. Dairy Sci. 91(1):260-270.
HUANG, Y.; SCHOONMAKER, J. P.; OREN, S. L.; TRENKLE, A. y BEITZ, D. C. 2009. Calcium salts of CLA improve availability of dietary CLA. Livest. Sci. 122(1):1-7.
IP, C.; BANNI, S.; ANGIONI, E.; CARTA, G.; MCGINLEY, J.; THOMPSON, H. J.; BARBANO, D. y BAUMAN, D. 1999. Conjugated linoleic acid-enriched butter fat alters mammary gland morphogenesis and reduces cancer risk in rats. J. Nutr.
129(12):2135-2142.

Capítulo 3
113
JACOBS, A. A. A.; VAN BAAL, J.; SMITS, M. A.; TAWEEL, H. Z. H.; HENDRIKS, W. H.; VAN VUUREN, A. M. y DIJKSTRA, J. 2011. Effects of feeding rapeseed oil, soybean oil, or linseed oil on stearoyl-CoA desaturase expression in the mammary gland of dairy cows. J. Dairy Sci. 94(2):874-887.
JENSEN, R. G. 2002. The composition of bovine milk lipids: january 1995 to december 2000. J. Dairy Sci. 85(2):295-350.
KAPS, M. y LAMBERSON, W. R. 2004. Biostatistics for animal science., London.
KELLY, M. L.; KOLVER, E. S.; BAUMAN, D. E.; VAN AMBURGH, M. E. y MULLER, L. D. 1998. Effect of intake of pasture on concentrations of conjugated linoleic acid in milk of lactating cows. J. Dairy Sci. 81(6):1630-1636.
KHANAL, R. C.; DHIMAN, T. R. y BOMAN, R. L. 2008. Changes in fatty acid composition of milk from lactating dairy cows during transition to and from pasture. Livest. Sci.
114(2-3):164-175.
KHANAL, R. C. y OLSON, K. C. 2004. Factors affecting conjugated linoleic acid (CLA) content in milk, meat and egg- A review. Pak. J. Nutr. 3(2):82-98.
LICITRA, G.; HERNANDEZ, T. M. y VAN SOEST, P. J. 1996. Standardization of procedures for nitrogen fractionation of ruminant feeds. Anim. Feed Sci. and Technol. 57(4):347-358.
LIN, L.; YING, D.; CHAITEP, S. y VITTAYAPADUNG, S. 2009. Biodiesel production from crude rice bran oil and properties as fuel. App. Energy. 86(5):681-688.
LINDSEY, T. O.; HAWKINS, G. E. y GUTHRIE, L. D. 1980. Physiological responses of lactating cows to gossypol from cottonseed meal rations. J. Dairy Sci. 63(4):562-
573.
LOOR, J. J.; FERLAY, A.; OLLIER, A.; DOREAU, M. y CHILLIARD, Y. 2005a. Relationship among trans and conjugated fatty acids and bovine milk fat yield due to dietary concentrate and linseed oil. J. Dairy Sci. 88(2):726-740.
LOOR, J. J.; FERLAY, A.; OLLIER, A.; UEDA, K.; DOREAU, M. y CHILLIARD, Y. 2005b. High-concentrate diets and polyunsaturated oils alter trans and conjugated isomers in bovine rumen, blood, and milk. J. Dairy Sci. 88(11):3986-3999.
LOURENÇO, M.; VAN RANST, G.; DE SMET, S.; RAES, K. y FIEVEZ, V. 2007. Effect of grazing pastures with different botanical composition by lambs on rumen fatty acid metabolism and fatty acid pattern of longissimus muscle and subcutaneous fat. Animal. 1(04):537-545.
LYNCH, J. M.; BARBANO, D. M. y FLEMING, J. R. 2007. Determination of the Lactose Content of Fluid Milk by Spectrophotometric Enzymatic Analysis Using Weight Additions and Path Length Adjustment: Collaborative Study. J. AOAC Inter.
90(1):196-215.
MEL´UCHOVÁ, B.; BLASKO, J.; KUBINEC, R.; GÓROVÁ, R.; DUBRAVSKÁ, J.; MARGETÍN, M. y SOJÁK, L. 2008. Seasonal variations in fatty acid composition of pasture forage plants and CLA content in ewe milk fat. Small Ruminant Res. 78(1-
3):56-65.
MOHAMMED, R.; STANTON, C. S.; KENNELLY, J. J.; KRAMER, J. K. G.; MEE, J. F.; GLIMM, D. R.; O'DONOVAN, M. y MURPHY, J. J. 2009. Grazing cows are more efficient than zero-grazed and grass silage-fed cows in milk rumenic acid

114
Efecto de la suplementación con harina de arroz o semilla de algodón sobre el contenido de los ácidos transvaccénico y ruménico
en la leche de vacas en pastoreo
production. J. Dairy Sci. 92(8):3874-3893.
MOYA-CAMARENA, S. Y.; HEUVEL, J. P. V.; BLANCHARD, S. G.; LEESNITZER, L. A. y BELURY, M. A. 1999. Conjugated linoleic acid is a potent naturally occurring
ligand and activator of PPAR. J. Lipid Res. 40(8):1426-1433.
MUIR, J. P. 2002. Hand-Plucked forage yield and quality and seed production from annual and short-lived perennial warm-season legumes fertilized with composted manure. Crop Sci. 42(3):897-904.
MUNDAY, J. S.; THOMPSON, K. G. y JAMES, K. A. C. 1999. Dietary conjugated linoleic acids promote fatty streak formation in the C57BL/6 mouse atherosclerosis model. Br. J. Nutr. 81(03):251-255.
NRC. 2001. Nutrient requerimients of dairy cattle. 7a ed. National Academy Press, Washington DC.
ODONGO, N. E.; OR-RASHID, M. M.; BAGG, R.; VESSIE, G.; DICK, P.; KEBREAB, E.; FRANCE, J. y MCBRIDE, B. W. 2007. Long-effects of feeding monensin on milk fatty acid composition in lactating dairy cows. J. Dairy Sci. 90(11):5126-5133.
PALMQUIST, D. L. y JENKINS, T. C. 1980. Fat in lactation rations: Review. J. Dairy Sci.
63(1):1-14.
PARADIS, C.; BERTHIAUME, R.; LAFRENIERE, C.; GERVAIS, R. y CHOUINARD, P. Y. 2008. Conjugated linoleic acid content in adipose tissue of calves suckling beef cows on pasture and supplemented with raw or extruded soybeans. J. Anim Sci.
86(7):1624-1636.
PARK, H. S.; RYU, J. H.; HA, Y. L. y PARK, J. H. Y. 2001. Dietary conjugated linoleic acid (CLA) induces apoptosis of colonic mucosa in 1,2-dimethylhydrazine-treated rats: a possible mechanism of the anticarcinogenic effect by CLA. Br. J. Nutr.
86(05):549-555.
PIRES, A. V.; EASTRIDGE, M. L.; FIRKINS, J. L. y LIN, Y. C. 1997. Effects of heat treatment and physical processing of cottonseed on nutrient digestibility and production performance by lactating cows. J. Dairy Sci. 80(8):1685-1694.
SCHAUFF, D. J. y CLARK, J. H. 1992. Effects of feeding diets containing calcium salts of long-chain fatty acids to lactating dairy cows. J. Dairy Sci. 75(11):2990-3002.
SCHROEDER, G. F.; DELAHOY, J. E.; VIDAURRETA, I.; BARGO, F.; GAGLIOSTRO, G. A. y MULLER, L. D. 2003. Milk fatty acid composition of cows fed a total mixed ration or pasture plus concentrates replacing corn with fat. J. Dairy Sci.
86(10):3237-3248.
SCHROEDER, G. F.; GAGLIOSTRO, G. A.; BARGO, F.; DELAHOY, J. E. y MULLER, L. D. 2004. Effects of fat supplementation on milk production and composition by dairy cows on pasture: a review. Livest. Prod. Sci. 86(1-3):1-18.
SUNVOLD, G. D. y COCHRAN, R. C. 1991. Technical note: evaluation of acid detergent lignin, alkaline peroxide lignin, acid insoluble ash, and indigestible acid detergent fiber as internal markers for prediction of alfalfa, bromegrass, and prairie hay digestibility by beef steers. J. Anim Sci. 69(12):4951-4955.
THIEX, N. J.; MANSON, H.; ANDERSON, S. y PETACCHI, F. 2002. Determination of crude protein in animal feed, forage, grain, and oilseeds by using block digestion with a copper catalyst and steam distillation into boric acid: collaborative study. J.

Capítulo 3
115
AOAC Inter. 85(2):309-317.
TSIPLAKOU, E.; KOMINAKIS, A. y ZERVAS, G. 2008. The interaction between breed and diet on CLA and fatty acids content of milk fat of four sheep breeds kept indoors or at grass. Small Ruminant Res. 74(1-3):179-187.
UEDA, K.; FERLAY, A.; CHABROT, J.; LOOR, J. J.; CHILLIARD, Y. y DOREAU, M. 2003. Effect of linseed oil supplementation on ruminal digestion in dairy cows fed diets with different forage:concentrate ratios. J. Dairy Sci. 86(12):3999-4007.
ULBRICHT, T. L. V. y SOUTHGATE, D. A. T. 1991. Coronary heart disease: seven dietary factors. The Lancet. 338(8773):985-992.
VAN SOEST, P. J.; ROBERTSON, J. B. y LEWIS, B. A. 1991. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J. Dairy Sci. 74(10):3583-3597.
VASTA, V.; NUDDA, A.; CANNAS, A.; LANZA, M. y PRIOLO, A. 2008. Alternative feed resources and their effects on the quality of meat and milk from small ruminants. Anim. Feed Sci. and Technol. 147(1-3):223-246.
VLAEMINCK, B.; MENGISTU, G.; FIEVEZ, V.; DE JONGE, L. y DIJKSTRA, J. 2008. Effect of in vitro docosahexaenoic acid supplementation to marine algae-adapted and unadapted rumen inoculum on the biohydrogenation of unsaturated fatty acids in freeze-dried grass. J. Dairy Sci. 91(3):1122-1132.
WILLIAMS, C. H.; DAVID, D. J. y IISMAA, O. 1962. The determination of chromic oxide in faeces samples by atomic absorption spectrophotometry. J. Agri. Sci. 59(03):381-
385.
ZHANG, W. J.; XU, Z. R.; PAN, X. L.; YAN, X. H. y WANG, Y. b. 2007a. Advances in gossypol toxicity and processing effects of whole cottonseed in dairy cows feeding. Livest. Sci. 111(1-2):1-9.
ZHANG, W. J.; XU, Z. R.; ZHAO, S. H.; SUN, J. Y. y YANG, X. 2007b. Development of a microbial fermentation process for detoxification of gossypol in cottonseed meal. Anim. Feed Sci. and Technol. 135(1-2):176-186.
.

116
Efecto de la suplementación con harina de arroz o semilla de algodón sobre el contenido de los ácidos transvaccénico y ruménico
en la leche de vacas en pastoreo

Conclusiones y recomendaciones
117
4. Conclusiones y recomendaciones
4.1 Conclusiones
La alimentación es el principal factor para incrementar la concentración delos ácidos
transvaccénico y ruménico en leche. Aumentar el consumo de forraje fresco,
incrementar el consumo de ácido linoleico utilizando diferentes fuentes lipídicas y de
ácido linolénico del forraje, permite incrementar el contenido lácteo de los ácidos
transvaccénico y ruménico. El control de las condiciones ruminales con ionóforos y ácido
docosahexaenoico potencializa el suministro de ácidos grasos insaturados. La harina de
arroz y la semilla de algodón son dos fuentes lipídicas que aparentemente pueden
incrementar la concentración de estos ácidos grasos.
El suministro de harina de arroz incrementó la concentración de los ácidos
transvaccénico (3.11 %; p<0.01) y ruménico (1.41 %; p<0.001) en la grasa láctea;
además, disminuyó los índices de aterogenicidad (1.85; p<0.05) y trombogenicidad (3.09;
p<0.01). La semilla de algodón no logró aumentarla proporción de estos ácidos grasos y
de ácidos grasos insaturados, ni mejorar los índices de aterogenicidad y trombogenicidad
a pesar de que el consumo de ácido linoleico y linolénico fue similar a los del tratamiento
harina de arroz. Por el contrario, la semilla de algodón presentó mayores proporciones de
C18:0 (16.31 %; p<0.001) y menores relaciones ruménico / transvaccénico (0.39;
p<0.01) y C14:1 / C14:0 (0.08; p<0.05) indicando mayor biohidrogenación y menor
actividad de la enzimaΔ9desaturasa. A diferencia de la semilla de algodón, la harina de
arroz es una buena alternativa de suplementación para las vacas lecheras en pastoreo
pues induce una mayor concentración de transvaccénico y ruménico en la leche, y
mayor concentración de ácidos grasos insaturados (34.9; p<0.01) que se sugiere son
mejores para la salud del consumidor.
Cuando se suministró un concentrado con elevada concentración de ácido linoleico
(4.28% de la materia seca) que contenían como principal fuente lipídica de ácido linoleico

118
Efecto de la suplementación con harina de arroz o semilla de algodón sobre el contenido de los ácidos transvaccénico y ruménico
en la leche de vacas en pastoreo
una mezcla de harina de arroz y semilla de algodón (relación 4.8 : 1) no se logró
incrementar la concentración de los ácidos transvaccénico y ruménico y se presentó un
mayor índice calculado de actividad de la enzima Δ9desaturasa (0.37; p<0.05), lo cual
sugiere una extensa biohidrogenación en el rumen. También incrementó los ácidos
grasos insaturados, disminuyó los saturados (35.1 y 60.0% respectivamente; p<0.05) y
presentó menores índices de aterogenicidad y trombogenicidad (1.79 y 3.22
respectivamente; p<0.05).
4.2 Recomendaciones
La harina de arroz incrementa la concentración de los transvaccénico y ruménico en la
grasa láctea, pero sería importante determinar si su combinación con aceite de pescado
o ionóforos permite maximizar su efecto sobre la concentración de estos ácidos grasos.
La semilla de algodón no presentó el efecto esperado sobre la concentración de
transvaccénico y ruménico en la grasa láctea, probablemente por una mayor
biohidrogenación o por el efecto de los ácidos ciclopropeno. Por lo tanto, evaluar el
ambiente ruminal y la actividad de la enzima Δ9desaturasa de manera que se pueda
determinar el motivo por el cual en condiciones pastoriles se presentó menor
concentración de estos ácidos grasos.
El suministro de La semilla de algodón ya sea como principal fuente lipídica o como parte
de una mezcla de harina de arroz y semilla de algodón (relación 4.8 : 1) no incrementó la
concentración de transvaccénico y ruménico en la grasa láctea, probablemente debido a
una extensa biohidrogenación. Sería importante evaluar si la adición de aceites vegetales
podría disminuir la biohidrogenación e incrementar el flujo de transvaccénico y ruménico
hacia la glándula mamaria.

Anexos
119
5. Anexos
Anexo 1. Características de los animales al iniciar el experimento y distribución de los animales a
los tratamientos en el primer experimento
Ítem
Edad(años)
Días de lactancia
Peso vivo(kg)
Producción láctea(kg/d)
Distribución de los tratamiento según el
periodo1
1 2 3
Cuadrado 1
Vaca
1 6,06 95 593,00 20,7 C HA SA 2 6,01 98 539,00 30,7 SA C HA 3 4,95 103 591,00 26,6 HA SA C
Promedio 5,7 98,7 574,3 26,0 DE 0,6 4,0 30,6 5,0
Cuadrado 2 Vaca
1 2,58 149 472,50 19,5 HA SA C
2 2,92 152 498,00 15,8 SA C HA 3 2,60 153 501,50 17,1 C HA SA
Promedio 2,7 151,3 490,7 17,5 DE 0,2 2,1 15,8 1,9 General Promedio 4,2 125,0 532,5 21,7 DE 1,7 29,0 50,7 5,8
1C: suplemento con bajo nivel de ácido linoléico; HA: suplemento con harina de arroz como la
principal fuente de ácido linoléico; SA: suplemento con semilla de algodón como la principal fuente de ácido linoléico.

120
Efecto de la suplementación con harina de arroz o semilla de algodón sobre el contenido de los ácidos transvaccénico y ruménico
en la leche de vacas en pastoreo
Anexo 2. Características de los animales al iniciar el experimento y distribución de los animales a
los tratamientos en el segundo experimento
Ítem Grupo Edad(años
) Peso
vivo(kg)
Días de lactanci
a
Producción láctea(kg/d
)
Distribución de los tratamientos alPeriodo
1
1 2
Vaca
1 1 6,1 499 151 18,2 A B 2 2 6,0 570 174 19,2 B A 3 1 4,9 497 147 19,2 A B 4 2 2,6 588 232 18,3 B A 5 1 2,9 566 107 19,5 A B 6 2 2,6 542 197 21,0 B A Promedio
4,2 544 168 19,2
DE 1,7 35 40 0,9
1B:suministro de un suplemento con bajo nivel de ácido linoleico (2.64 g / 100 g MS) y A:
suministro de un suplemento con alto nivel bajo de ácido linoleico (4.08 g / 100 gMS).


















