
Cubierta Pirineos 166:Layout 2 - alvorada-pt.comalvorada-pt.com/Filipe/Ungulados/5_Pirineos 166 -...
178
www.publicaciones.csic.es CONSEJO SUPERIOR DE INVESTIGACIONES CIENTÍFICAS CSIC INSTITUTO PIRENAICO DE ECOLOGÍA Volumen 166 | 2011 | Jaca Sumario Presentación ACEVEDO, P., REAL, R. & GORTÁZAR, C.: Favorabilidad ecogeográfica para el corzo: distribución y abundancia. ACEVEDO, P., SANTOS, J. P., REAL, R. & VICENTE, J.: Evaluación del estado de la población de cabra montés de los montes de Toledo: relaciones con el ciervo. BUENO, C. G., BARRIO, I. C., GARCÍA-GONZÁLEZ, R., ALADOS, C. L. & GÓMEZ-GARCÍA, D.: Assessment of wild boar rooting on ecological and pastoral values of alpine pyrenean grasslands. FERRERES, J., ESCUDERO, M. A., FERRER, E. & MARCO, J.: Gestión del corzo en Zaragoza: de conservación a control poblacional. GARCÍA-GONZÁLEZ, R.: Elementos para una filogeografía de la cabra montés ibérica (capra pyrenaica schinz, 1838). HERRERO, J., GARCÍA-SERRANO, A., PRADA, C. & FERNÁNDEZ-ARBERAS, O.: Using block counts and distance sampling to estimate populations of chamois. MARCO, J., HERRERO, J., ESCUDERO, M. A., FERNÁNDEZ-ARBERAS, O., FERRERES, J., GARCÍA-SERRANO, A., GIMÉNEZ-ANAYA, A., LABARTA, J. L., MONRABAL, L. & PRADA C.: Veinte años de seguimiento poblacional de ungulados silvestres de Aragón. ALARCÓN, J. L., FERNÁNDEZ DE LUCO, D., GARCÍA-GONZÁLEZ, R., GARCÍA-SERRANO, A. & HERRERO, J., organizadorres: Primera Reunión sobre el rebeco cantábrico y el sarrio pirenaico. Benasque (Huesca, España), 20-22 de mayo de 2011. PIERRE BARRÈRE, en el recuerdo (1921-2011). Revista de ecología de montaña A journal on mountain ecology Revue d´ecologie de montagne Jaca (España) ISSN: 0373-2568 enero-diciembre 2011 Volumen 166 Revista de ecología de montaña A journal on mountain ecology Revue d´ecologie de montagne 192 págs. ISSN: 0373-2568 enero-diciembre 2011 Volumen 166 Pirineos Pirineos Pirineos CSIC CSIC http://pirineos.revistas.csic.es
Transcript of Cubierta Pirineos 166:Layout 2 - alvorada-pt.comalvorada-pt.com/Filipe/Ungulados/5_Pirineos 166 -...

www.publicaciones.csic.es CONSEJO SUPERIOR DE INVESTIGACIONES CIENTÍFICAS
CSIC
INSTITUTO PIRENAICO DE ECOLOGÍA
Volu
men
166
| 2
011
| J
aca
SumarioPresentaciónACEVEDO, P., REAL, R. & GORTÁZAR, C.: Favorabilidad ecogeográfica parael corzo: distribución y abundancia.ACEVEDO, P., SANTOS, J. P., REAL, R. & VICENTE, J.: Evaluación del estadode la población de cabra montés de los montes de Toledo: relaciones con elciervo.BUENO, C. G., BARRIO, I. C., GARCÍA-GONZÁLEZ, R., ALADOS, C. L. &GÓMEZ-GARCÍA, D.: Assessment of wild boar rooting on ecological and pastoralvalues of alpine pyrenean grasslands.FERRERES, J., ESCUDERO, M. A., FERRER, E. & MARCO, J.: Gestión delcorzo en Zaragoza: de conservación a control poblacional.GARCÍA-GONZÁLEZ, R.: Elementos para una filogeografía de la cabra montésibérica (capra pyrenaica schinz, 1838).HERRERO, J., GARCÍA-SERRANO, A., PRADA, C. & FERNÁNDEZ-ARBERAS,O.: Using block counts and distance sampling to estimate populations ofchamois.MARCO, J., HERRERO, J., ESCUDERO, M. A., FERNÁNDEZ-ARBERAS, O.,FERRERES, J., GARCÍA-SERRANO, A., GIMÉNEZ-ANAYA, A., LABARTA, J.L., MONRABAL, L. & PRADA C.: Veinte años de seguimiento poblacional deungulados silvestres de Aragón.ALARCÓN, J. L., FERNÁNDEZ DE LUCO, D., GARCÍA-GONZÁLEZ, R.,GARCÍA-SERRANO, A. & HERRERO, J., organizadorres: Primera Reuniónsobre el rebeco cantábrico y el sarrio pirenaico. Benasque (Huesca, España),20-22 de mayo de 2011.PIERRE BARRÈRE, en el recuerdo (1921-2011).
R e v i s t a d e e co l og í a d e mon t a ñaA j o u r n a l o n moun t a i n e co l og yR e v u e d ´ e co l og i e d e mon t agne
Jaca (España) ISSN: 0373-2568enero-diciembre 2011Volumen 166
R e v i s t a d e e co l og í a d e mon t a ñaA j o u r n a l o n moun t a i n e co l og yR e v u e d ´ e co l og i e d e mon t agne
192 págs. ISSN: 0373-2568enero-diciembre 2011Volumen 166
P i r i n e o s P i r i n e o s
Pirineos
CSICCSIC
http://pirineos.revistas.csic.es

P i r i n e o sVolumen 166 enero-diciembre 2011 Jaca (España) ISSN: 0373-2568
R e v i s t a d e e co l og í a d e mon t a ñaA j o u r n a l o n moun t a i n e co l og yR e v u e d ´ e co l og i e d e mon t agne
CONSEJO SUPERIOR DE INVESTIGACIONES CIENTÍFICAS

Volumen 166 Enero-diciembre 2011 Jaca (España) ISSN: 0373-2568
PIRINEOSRevista de Ecología de Montaña/A Journal on Mountain Ecology/Revue d’Écologie deMontagne
Pirineos, Revista de Ecología de Montaña aparece anualmente. Tiene por objeto la publicación detrabajos originales relacionados con la dinámica de ecosistemas de montaña y trata de aportarinformación sobre el funcionamiento y la organización específica de los recursos en regionesmontañosas.La revista establecerá intercambio con otras publicaciones de características similares.
Director: Teodoro Lasanta Martínez (I.P.E.)Secretario: Sergio M. Vicente Serrano (I.P.E.)
Consejo de Redacción
Francisco Lloret (U.A.B.) Juan Ignacio López Moreno (I.P.E.)Jesús Julio Camarero Martínez (I.P.E.) Enrique Navarro Rodríguez (I.P.E.)José María Cuadrat Prats (U.Z.) Yolanda Pueyo Estaún (I.P.E.)Adrián Escudero (U.R. J. C.) Carlos Sancho Marcén (U.Z.)José Daniel Gómez García (I.P.E.) Juan Manuel Vilaplana Fernández (U.B.)Penélope González Sampériz (I.P.E.)
Consejo Asesor (Advisory Board)
Dr. P. MONTSERRAT, InstitutoPirenaico de Ecología (CSIC), Jaca,Spain.Dr. B. MESSERLI, Universität Bern,Switzerland.Dr. M. MONASTERIO, Universidadde Los Andes, Mérida, Venezuela.Dr. L. KERGOAT, Centre d’EtudesSpatiales de la Biosphere (CNES),Toulousse, France.Dr. U. MOLAU, Notanical Institute,Göteborg Universitat, Sweden.Dr. F. GALLART, Instituto de Cienciasde la Tierra “Jaume Almera” (CSIC),Barcelona, España.Dr. B. DEVERAUX, University of Cambridge, Cambridge, UnitedKingdom.Dr. J. MARTÍN-VIDE, Universidadde Barcelona, Barcelona, España.Dr. F. SARMIENTO, University ofGeorgia, United States.
Dr. F. DÍAZ PINEDA, Univ. Complutense de Madrid, España.Dr. S. WHITE, Cranfield University,United Kingdom.Dr. H. ROMERO ARAVENA, Universidadde Chile, Chile. Dr. J.O. RABASSA, Centro Australde Investigaciones Científicas, Ushuaia,Argentina.Dr. J. TERRADAS, Universidad Autónomade Barcelona, España.Dr. M.A. LENZI, University of Padova,Padova, Italia.Dr. A. CÉRNUSCA. Institut für Botanik,Universitat Innsbruck, Austria.Dr. K. GROSSENBACHER, Naturhistoris-ches Museum, Bern, Switzerland.Dr. M. BENISTON, University of Geneva,Geneva, Switzerland.Dr. M. PRICE, Centre for MountainStudies, Perth College, Perth, UnitedKingdom.

SUBSCRIPCIONES E INTERCAMBIO DE PUBLICACIONES(Subscriptions and exchange of publications)Instituto Pirenaico de EcologíaCampus de Aula DeiApartado de Correos 1303450080 Zaragoza (España)
SERVICIOS DE INFORMACIÓNPirineos. A Journal of Mountain Ecology is indexed or abstracted by Geo-Abstracts, Scopus,Thomson ISI-Journal Search Soils and Fertilizers, Bibliographie GeographiqueInternationale.
© CSIC, 2011
Las opiniones y hechos consignados en cada artículo son de la exclusiva responsabilidad desus autores. El Consejo Superior de Investigaciones Científicas no se hace responsable enningún caso de la credibilidad y autenticidad de los trabajos.
Los originales de la revista Pirineos, publicados en papel y en la versión electrónica, sonpropiedad del Consejo Superior de Investigaciones Científicas, siendo necesario citar laprocedencia en cualquier reproducción total o parcial.
ISSN: 0373-2568e ISSN: 1988-4281NIPO (en Papel): 472-11-058-9NIPO (en línea): 472-11-057-3Depósito legal: M. 828-1958Impreso en EspañaSdad. Coop. de Artes Gráficas Librería General
Pedro Cerbuna, 2350009 Zaragoza

166
ÍNDICE/CONTENTS
7-8
9-27
29-49
51-67
69-86
87-122
123-133
Volumen 166 Enero-diciembre 2011 Jaca (España) ISSN: 0373-2568
PresentaciónACEVEDO, P., REAL, R. & GORTÁZAR, C.: Favorabilidad ecogeográfica para el
corzo: distribución y abundancia.Ecogeographical favourability for roe deer: population abundance and speciesdistribution.
ACEVEDO, P., SANTOS, J. P., REAL, R. & VICENTE, J.: Evaluación del estadode la población de cabra montés de los montes de Toledo: relaciones con elciervo.Assessing the status of Iberian wild goat population in “Montes de Toledo” and itsrelationships with red deer.
BUENO, C. G., BARRIO, I. C., GARCÍA-GONZÁLEZ, R., ALADOS, C. L. &GÓMEZ-GARCÍA, D.: Assessment of wild boar rooting on ecological andpastoral values of alpine pyrenean grasslands.Efecto de las hozaduras de jabalí sobre el valor ecológico y pastoral de pastossupraforestales pirenaicos.
FERRERES, J., ESCUDERO, M. A., FERRER, E. & MARCO, J.: Gestión del corzoen Zaragoza: de conservación a control poblacional.Monagement of roe deer in Zaragoza: from conservation to population control.
GARCÍA-GONZÁLEZ, R.: Elementos para una filogeografía de la cabra montésibérica (capra pyrenaica schinz, 1838).Elements for a phylogeography of Iberian wild goat (Capra pyrenaica Schinz, 1838).
HERRERO, J., GARCÍA-SERRANO, A., PRADA, C. & FERNÁNDEZ-ARBERAS,O.: Using block counts and distance sampling to estimate populations ofchamois.Uso del conteo simultáneo supraforestal y del muestreo de distancias para estimarpoblaciones de sarrio.
P i r i n e o sR e v i s t a d e e co l og í a d e mon t a ñaA j o u r n a l o n moun t a i n e co l og yR e v u e d ´ e co l og i e d e mon t agne

135-153
155-177
179-180
MARCO, J., HERRERO, J., ESCUDERO, M. A., FERNÁNDEZ-ARBERAS, O.,FERRERES, J., GARCÍA-SERRANO, A., GIMÉNEZ-ANAYA, A., LABARTA, J.L., MONRABAL, L. & PRADA C.: Veinte años de seguimiento poblacional deungulados silvestres de Aragón.Twenty years of wild ungulate’s population monitoring in Aragon.
ALARCÓN, J.L., FERNÁNDEZ DE LUCO, D., GARCÍA-GONZÁLEZ, R.,GARCÍA-SERRANO, A. & HERRERO, J., organizadores: Primera Reuniónsobre el rebeco cantábrico y el sarrio pirenaico. Benasque (Huesca, España),20-22 de mayo de 2011.First Meeting on Cantabrian and Pyrenean chamois. Benasque (Huesca, Spain), 20-22 May 2011.
PIERRE BARRÈRE, en el recuerdo (1921-2011)Pierre Barrère, in the Memory (1921-2011)

7
Presentación
El presente volumen es un compendio de algunas de las comunicacionespresentadas en la Primera Reunión sobre Ungulados Silvestres Ibéricos (RUSI1) celebrada en la Facultad de Veterinaria de la Universidad de Zaragoza el10 de septiembre 2010. Ya en 2002, Pirineos. A Journal of Mountain Ecology sehizo eco de las comunicaciones de la “Third World Conference on MountainUngulates” celebrada en Zaragoza. En aquella ocasión se pudieron escuchary debatir los progresos sobre el estudio de los ungulados, a escala mundial, yconocer el estado de conocimientos de este grupo animal tan relevante porsus impactos económicos y ecológicos.
En la Península Ibérica, al igual que en muchos territorios, la fragmenta-ción intra e interadministrativa ha producido una cierta descoordinación yfalta de información sobre las problemáticas medioambientales que generandicho grupo de animales. La RUSI 1 surgió por la necesidad de mantener uncontacto más estrecho entre los diferentes especialistas en ungulados ibéricos,con el fin de poder compartir experiencias, informar sobre recientes avancesen la materia, debatir problemáticas metodológicas comunes, establecerpuentes entre investigación y gestión, y en definitiva, para crear un foro dedebate presencial y de intercambio informativo. RUSI 1 iba dirigido a las dife-rentes personas o grupos de investigación dedicados al estudio de los ungu-lados ibéricos silvestres y asilvestrados, autóctonos o introducidos de laPenínsula Ibérica y sus islas (jabalí, ciervo, corzo, sarrio, rebeco, cabra mon-tesa, cabra asilvestrada, besta y garrano, muflón y arrui), desde la perspecti-va de la ecología, veterinaria o cualquier otra disciplina científica.
La Reunión puso de manifiesto, entre otros aspectos, cómo los unguladossilvestres en Iberia, se muestran cada vez más dependientes del hombre, locual conduce a su vez, a una progresiva artificialización de sus poblaciones.Esta artificialización adopta diferentes aspectos según el ámbito geográfico yestá relacionada con los usos y propiedad de la tierra. A grandes rasgos, pue-den distinguirse dos grandes tipologías: en el Centro-Sur de la Península, laexistencia de grandes fincas privadas dedicadas a la actividad cinegética, pro-voca altas densidades de ungulados, alteraciones de la conducta y presionesselectivas probablemente muy diferentes a las “naturales”, variando segúnlas diferentes modalidades de gestión. En la mitad Norte de la península

8
Ibérica, la mayor proporción de montes comunales, la baja incidencia dedepredadores, el mayor acceso de los herbívoros a los cultivos (descenso dela población humana en las últimas décadas), han disparado los censos de laspoblaciones. La sobre-abundancia está generando problemas en la actividadagraria y un aumento de accidentes de tráfico. La invasión o proximidad deungulados en zonas periurbanas, crea nuevos conflictos y nuevos desafíos entérminos de gestión.
Ante esta situación, muchos comentarios e intervenciones apuntaron lanecesidad de mejorar el conocimiento del funcionamiento de las poblacionesy de como operan en ellas los factores de selección. Es la información cientí-fica y técnica la que debe servir de base para la gestión y en muchas ocasio-nes las decisiones trascendentales se siguen tomando sin este tipo de infor-mación. La necesidad de realizar controles drásticos de la densidad en deter-minados territorios, o la simple planificación de la actividad cinegética,requiere un buen conocimiento de la influencia de la mortalidad en la estruc-tura y dinámica poblacional. En este sentido, el mantenimiento y estudio depoblaciones en condiciones no extractivas (poblaciones testigo como las man-tenidas en algunos parques nacionales), puede ser de gran ayuda en el cono-cimiento del funcionamiento de las poblaciones y en la mejora de la gestiónde las poblaciones cazadas.
Por otra parte y dada la gran autonomía gestora en caza y vida silvestrede las diferentes Comunidades Autónomas, en el transcurso de la Reunión, seha puesto de manifiesto la conveniencia de crear foros de intercambio deinformación y propuestas de soluciones a problemas comunes, entre losdiversos agentes interesados en los ungulados de la Península ibérica. Lafutura realización de la II Reunión sobre Ungulados Ibéricos en Aveiro(Portugal) durante el verano de 2011, augura un buen futuro para este tipo dereuniones. Los artículos que presentamos en este volumen son una pequeñamuestra del interés científico y aplicado que presenta el estudio de los ungu-lados ibéricos.
Ricardo García-GonzálezJuan Herrero Cortés
Co-editores para el presente número de Pirineos. A Journal of Mountain Ecology.

9
FAVORABILIDAD ECOGEOGRÁFICAPARA EL CORZO: DISTRIBUCIÓN Y ABUNDANCIA
P. ACEVEDO1,2*, R. REAL1 & C. GORTÁZAR2
1 Departamento de Biología Animal, Facultad de Ciencias, Universidad de Málaga, 29071 Málaga*[email protected]; [email protected]
2 Instituto de Investigación en Recursos Cinegéticos (CSIC-UCLM-JCCM), 13071 Ciudad Real
ABSTRACT.– Species distribution models (SDM) are widely used in ecology,biogeography and conservation. Here, we modelled the distribution of roe deer(Capreolus capreolus) in mainland Spain in order to i) determine its macro-eco-logical requirements, ii) assess the favourability for this species and iii) explore themodel usefulness as an index of the species relative abundance. Our results showedthat roe deer distribution can be explained by predictors related with topography, cli-mate, lithology, geography and human activity. Climate was the more relevant fac-tor, followed by human activity, topography, spatial location and lithology. Themodel accurately predicted the species probability of occurrence and also, but weak-ly, the species’ relative abundance.
Keywords: Abundance, roe deer, distribution, favourability function,Capreolus capreolus.
RESUMEN.– Los modelos de distribución de especies (SDM) están siendo cadavez más usados para abordar cuestiones relacionadas con la ecología, biogeografía yconservación de las especies. En este estudio nos proponemos realizar un modelo dedistribución para el corzo (Capreolus capreolus) en la España peninsular paraestudiar los determinantes macroambientales de su rango de distribución, evaluar lafavorabilidad para la especie, y la utilidad del modelo como indicador de su abun-dancia relativa. El modelo indica que su distribución se puede explicar en términostopográficos, climáticos, litológicos, geográficos y de presión humana. El clima fue elfactor que en su efecto puro explicó un mayor porcentaje de variación, y le siguen lapresión humana, la topografía, la localización espacial y la litología. El modelo obte-nido predijo con elevada precisión la probabilidad de presencia de la especie y concierto detalle su abundancia relativa.
Palabras clave: Abundancia, corzo, distribución, función de favorabilidad,Capreolus capreolus.
Pirineos.Revista de Ecología de MontañaVol. 166, 9-27
Jaca, Enero-Diciembre, 2011ISSN: 0373-2568
eISSN: 1988-4281doi: 10.3989/Pirineos.2011.166001

10
P. ACEVEDO, R. REAL & C. GORTÁZAR
1. Introducción
Las relaciones entre los gradientes ambientales y la supervivencia de laspoblaciones de una especie pueden ser usadas para modelizar la respuestapotencial de la especie a tales gradientes (Austin et al., 1990). De este modo sepueden establecer mapas predictivos de potencialidad ambiental para laespecie, así como describir las características macroambientales que las defi-nen (Acevedo et al., 2007a). Esto es lo que algunos autores denominan distri-bución potencial –o incluso nicho ecológico– de la especie (Soberón &Peterson, 2005), y a este tipo de herramientas analíticas se las conoce comomodelos de distribución de especies o modelos de distribución de hábitat(Guisan & Zimmermann, 2000).
Los modelos de distribución de especies (en adelante SDM) están siendocada vez más usados para abordar cuestiones relacionadas con la ecología,biogeografía y conservación de las especies. Sus aplicaciones son muy varia-das y –refiriéndonos por ejemplo a estudios realizados sobre ungulados ibé-ricos– abarcan desde el estudio de los requerimientos macroambientales deuna especie o de un grupo de especies (Acevedo et al., 2007a), la evaluaciónde la potencialidad del territorio para especies invasoras (Cassinello et al.,2006), o el estudio de las relaciones biogeográficas entre diferentes taxones(Acevedo et al., 2007b; Delibes-Mateos et al., 2009; Acevedo & Real, 2011),hasta la evaluación del efecto del cambio en el clima (Real et al., 2010) o de losusos del suelo (Acevedo et al., 2011) sobre su área de distribución.
La información necesaria para realizar un SDM generalmente proviene deatlas de distribución (Palomo et al., 2007), sobre todo cuando la escala espa-cial del análisis es regional o nacional, y de las coberturas digitales de infor-mación ambiental, disponibles muchas de ellas de manera gratuita en servi-dores de Internet (http://edit.csic.es/GISdownloads.html). Desde esta infor-mación, la modelización permite obtener una medida continua del grado deadecuación del área de estudio para la especie. Pero la interpretación de loque realmente representa dicha adecuación no puede realizarse de maneradirecta ya que precisa de ciertas consideraciones relacionadas tanto con elprocedimiento de modelización (Jiménez-Valverde et al., 2008) como con laecología de la especie que se está estudiando (Múñoz & Real, 2006).
Algunos de los SDM implementados en programas amigables, como porejemplo ENFA (Hirzel et al., 2002) o Maxent (Phillips et al., 2006), producenvalores de adecuación de difícil interpretación debido a que en estos procedi-mientos se realiza un reescalamiento –se asigna a la localidad de máximaprobabilidad el valor máximo que puede tomar el índice– de los valores deadecuación. Por ello, estos valores de adecuación sólo pueden interpretarsecomo valores relativos cuya utilidad consiste en el establecimiento de com-
(Pirineos, 2011, Vol. 166, 9-27, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166001)

paraciones entre localidades (Acevedo et al., 2007a). Para poder ser interpre-tados en términos absolutos –en relación a alguna característica biológica dela especie–, es preciso realizar una validación previa de los valores predichospor el modelo con datos independientes. Esta validación consistirá en el aná-lisis de la relación existente entre los valores de adecuación y los de un pará-metro de la población (probabilidad de presencia, abundancia poblacional,calidad de la dieta,…) que estamos interesados en predecir. Sin embargo, losvalores predichos mediante regresiones logísticas o modelos generalizadosaditivos tienen en si mismos una interpretación directa ya que indican la pro-babilidad de presencia de la especie en cada una de las localidades (Acevedo& Cassinello, 2009). En estos casos, se debe evaluar la calibración (Pearce &Ferrier, 2000), es decir, testar el grado en el que los valores observados (pre-valencia de la especie) se ajustan a los valores predichos por el modelo, porejemplo ver si la especie está presente en el 40% de las localidades en las queel modelo predice una probabilidad de 0.4.
Una interpretación similar –aunque no idéntica– a la probabilidad de pre-sencia puede obtenerse usando la función de favorabilidad (Real et al., 2006),en la que los valores predichos son indicativos del grado en el que la proba-bilidad de presencia de una especie en una determinada localidad difiere dela probabilidad de encontrarla por azar en el área de estudio (prevalencia dela especie). Así, valores de favorabilidad superiores a 0.5 indican una proba-bilidad de presencia superior a la prevalencia (localidades con característicasambientales que favorecen la presencia de la especie), y por debajo de dichovalor muestran una probabilidad inferior a la prevalencia (localidades concaracterísticas ambientales desfavorables).
Pero los SDM no se quedan aquí, ya que en ocasiones se da un paso másy se evalúa su capacidad para predecir otros parámetros de la dinámica de laespecie como por ejemplo la abundancia relativa de las poblaciones (Nielsenet al., 2005). Para ello podría ser usado cualquiera de los modelos anterior-mente mencionados, ya que en este caso el interés no radica en el valor abso-luto de las predicciones, tal y como ocurría con la probabilidad y la favorabi-lidad, sino en su valor relativo respecto al resto del área de estudio. En estepunto, trabajos recientes han demostrado la utilidad de los modelos de hábi-tat para predecir la abundancia local de plantas y animales (Boyce et al., 2001;Pearce & Ferrier 2001; Real et al., 2009). Sin embargo, los resultados mostra-dos en otros estudios muestran que la aplicabilidad de este tipo de modelosdepende de numerosos factores, muchos de ellos intrínsecos de la especie amodelizar, por lo que a la hora de afrontar un modelo de abundancia el dise-ño del modelo debe contemplar, en mayor medida que los modelos dirigidosa determinar la distribución, aspectos de la biología de la especie (Pearce &Ferrier, 2001; Nielsen et al., 2005), y las predicciones deben ser validadas con
FAVORABILIDAD ECOGEOGRÁFICA PARA EL CORZO: DISTRIBUCIÓN Y ABUNDANCIA
11(Pirineos, 2011, Vol. 166, 9-27, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166001)

datos de abundancia, preferiblemente provenientes de muestreos indepen-dientes.
En este contexto nos proponemos realizar un modelo de distribución parael corzo (Capreolus capreolus) en la España peninsular con un triple objetivo: i)conocer los determinantes macroambientales de su actual rango de distribu-ción, ii) evaluar la favorabilidad ambiental del territorio para la especie, y iii)determinar la utilidad del modelo como indicador de la abundancia relativade la especie para así poder interpretar correctamente los patrones obtenidosdesde el punto de vista de la gestión global de la especie.
2. Métodos
2.1. Datos de distribución y caracterización ambiental
El área de estudio sobre la que se ha realizado este análisis es la Españapeninsular, y las cuadrículas UTM de 100 km2 han sido las unidades territo-riales consideradas (n=5273). Los datos sobre la distribución de la especie hansido tomados del Atlas y Libro Rojo de los Mamíferos Terrestres de España(Palomo et al., 2007), y en Cuenca y Guadalajara esta información se ha actua-lizado con datos provenientes de muestreos de campo (Acevedo et al., 2010a;ver Figura 1).
Para la modelación de la distribución de corzo se han reunido 24 variablesrelacionadas con la localización espacial, la topografía, el clima, la litología yla actividad humana (Tabla 1). Las variables se eligieron en función de su dis-ponibilidad a esta escala y de su potencial poder predictivo (ver Acevedo &Real, 2011), asumiéndose que están al menos correlacionadas con factorescausales de la distribución de las especies. Como Robertson et al. (2003) hansugerido, los modelos que dependen de relaciones indirectas entre variablesambientales y la presencia de las especies pueden predecir las distribucionesincluso mejor que los modelos mecanísticos –que utilizan variables más direc-tamente relacionadas con la fisiología de las especies–, razón por la que, eneste estudio se han considerado variables ambientales. La explicación sobre lametodología seguida para la obtención de las variables puede encontrarse enBarbosa et al. (2003).
2.2. El modelo estadístico
La modelización se ha realizado mediante regresión logística (Hosmer &Lemeshow, 1989), y se ha aplicado la función propuesta por Real et al. (2006)
P. ACEVEDO, R. REAL & C. GORTÁZAR
12 (Pirineos, 2011, Vol. 166, 9-27, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166001)

para obtener la favorabilidad del territorio para la especie. El modelo fueconstruido en dos pasos. En el primero de ellos se ha evaluado la relación decada variable de manera independiente con la variable dependiente (presen-cia [n=2043] o ausencia [n=3230] de corzo en cada UTM de 100 km2). Tras esteprimer paso se ha considerado la tasa de descubrimientos falsos (FDR, deFalse Discovery Rate; Benjamini & Hochberg, 1995), con el fin de minimizar laprobabilidad de cometer un error de tipo I (ver otras aplicaciones por ejem-plo en Acevedo & Real 2011). Las variables seleccionadas tras el FDR han sidoincluidas en el modelo final que se ha construido utilizando un procedimien-to de selección por pasos hacia adelante-hacia atrás, con un umbral de signi-ficación de 0,05 para inclusión de una variable en el modelo y de 0,10 para su
FAVORABILIDAD ECOGEOGRÁFICA PARA EL CORZO: DISTRIBUCIÓN Y ABUNDANCIA
13(Pirineos, 2011, Vol. 166, 9-27, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166001)
Figura 1. Las cuadrículas sombreadas en gris representan el área de distribución del corzo en laEspaña peninsular (adaptado de Palomo et al., 2007). En Cuenca y Guadalajara estos datos fue-ron actualizados con los provenientes de Acevedo et al. (2010a). El tamaño de los círculos indicala abundancia relativa de las poblaciones de corzo consideradas en el estudio de Acevedo
et al. (2010a) que serán las usadas para validar el modelo.Figure 1. Roe deer distribution in mainland Spain (adapted from Palomo et al., 2007). These data wereupdated in Cuenca and Guadalajara according to Acevedo et al. (2010a). Circles size shows the roe deer
relative abundance (Acevedo et al., 2010a) in those localities used to validate the model.

eliminación. En una fase final de la modelización se ha evaluado la existenciade autocorrelación espacial sobre los residuos del modelo usando el estadís-tico I de Moran y siguiendo los procedimientos y razonamientos descritos enDiniz-Filho et al. (2003).
Los valores de probabilidad obtenidos con la regresión logística no depen-den exclusivamente de las variables independientes, sino también de la pro-babilidad al azar debida a la proporción de presencias de la especie en el áreade estudio (Hosmer & Lemeshow, 1989). Cuando el número de presencias yel de ausencias en el área de estudio es diferente, los valores de probabilidad(P) están sesgados hacia la categoría que presenta un mayor número de casos.Para obtener un valor predicho que fuera independiente de la prevalencia dela especie se ha descrito la función de favorabilidad (Real et al., 2006):
P. ACEVEDO, R. REAL & C. GORTÁZAR
14 (Pirineos, 2011, Vol. 166, 9-27, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166001)
Tabla 1. Variables utilizadas para modelar la distribución de corzo en cuadrículas UTM de 10x10km en la España peninsular.
Table 1. Variables used to model roe deer distribution in UTM 10x10 km squares in mainland Spain.
Factor Variable CódigoTopografía Altitud media (m) Alti
Pendiente media (grados) PendClima Número medio anual de días de helada DiHe
Número medio anual de días de granizo DGraNúmero medio anual de días de niebla DNieVariación anual de temperatura [=T Julio – T Enero] VtemInsolación media anual (horas/año) InsoRadiación solar global media anual (Kwh/m2/día) RadSEvapotranspiración potencial media anual (mm) ETPEvapotranspiración real media anual (mm) [=min. entre Prec y ETP] ETRPrecipitación media anual (mm) PrecPrecipitación máxima en 24 horas (mm) PM24Precipitación máxima relativa (mm) [=PM24 / PMed] PMRNúmero medio anual de días de precipitación >0,1 mm DiPrIrregularidad pluviométrica interanual IrrPÍndice de humedad relativa IHumHumedad relativa media en enero a las 07:00 horas (%) HuEnHumedad relativa media en julio a las 07:00 horas (%) HuJu
Litología Permeabilidad del suelo PermPresión Distancia mínima a una autopista o autovía (km) DAuthumana Distancia a un centro urbano de más de 100.000 habitantes (km) U100
Distancia a un centro urbano de más de 500.000 habitantes (km) U500Situación Latitud media (grados N) Latiespacial Longitud media (grados E) Long

Cuando el número de presencias (n1) es igual al número de ausencias (n0),F=P, pero si el número de presencias es menor que el de ausencias F>P, einversamente, cuando el número de presencias es mayor que el de ausenciasF<P. El umbral ambiental (F=0,5) corresponde al valor esperado bajo igualfavorabilidad en todas las localidades. Esto sería equivalente a asignar elvalor F=0.5 a las condiciones ambientales por las cuales la P es igual a la pro-porción de presencias de la especie en el área de estudio. Como anteriormen-te se ha comentado, los valores obtenidos utilizando la función de favorabili-dad no reflejan probabilidad de presencia, sino favorabilidad del territorio,que es lo que normalmente se busca con la modelación de las distribuciones,principalmente en aquellas ocasiones en las que se precisa catalogar comofavorable un área no ocupada por la especie o incluso como desfavorable unazona donde ésta sí ocurre. El modelo de favorabilidad es más adecuado quela probabilidad para dilucidar tendencias biogeográficas, para determinar elvalor de conservación de distintos territorios, o para seleccionar las zonasmás apropiadas para la reintroducción de especies.
El modelo desarrollado en este estudio tiene una componente predictiva yotra explicativa. El mejor modelo predictivo se obtiene considerando todoslos predictores disponibles independientemente de la colinealidad existenteentre ellos (Graham, 2003), tal y como hemos realizado en este estudio. Sinembargo, la colinealidad entre predictores afecta a la componente explicativadel modelo e imposibilita la descripción de relaciones simples entre cada pre-dictor y la variable dependiente. En este caso, y para potenciar la capacidadexplicativa del modelo, se ha realizado una partición de la variación con laque poder explorar la naturaleza funcional de las colinearidades (Borcard etal., 1992). Este procedimiento permite cuantificar la variación de una variabledependiente –en este caso los valores predichos por el modelo final– que esexplicada por el efecto puro de cada factor, esto es, independientemente alresto de factores, así como la variación que se explica por el efecto combina-do de dos o más factores. En este trabajo se ha seguido el procedimiento desustracción descrito en Real et al. (2003), pero no se ha cuantificado la varia-ción explicada por el efecto combinado entre dos o más factores debido a queel elevado número de factores considerado en este estudio (5; Tabla 1) difi-culta mucho su interpretación (Acevedo & Real, 2011).
Por otro lado, los valores predichos por el modelo han sido validadoscomo indicadores de la probabilidad de presencia de la especie y tambiéncomo indicadores de su abundancia relativa (Real et al., 2009). Lo primero se
FAVORABILIDAD ECOGEOGRÁFICA PARA EL CORZO: DISTRIBUCIÓN Y ABUNDANCIA
15(Pirineos, 2011, Vol. 166, 9-27, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166001)

ha realizado mediante la evaluación del nivel de calibración del modelo parapredecir la presencia de la especie (Pearce & Ferrier, 2000). Para ello las loca-lidades se han clasificado en 10 grupos de igual amplitud (0,1) en función dela probabilidad predicha por el modelo en cada una de ellas. Sobre las locali-dades pertenecientes a cada una de estos grupos se ha calculado la prevalen-cia observada de la especie, medida como el porcentaje de las localidades per-tenecientes al grupo en las que la especie está presente. Enfrentando la pro-babilidad predicha por el modelo con la prevalencia observada en cada unode los grupos obtenemos el gráfico de calibración, en el que el ajuste perfecto(perfecta calibración) se obtiene cuando todos los puntos se sitúan en la líneade igualdad entre ambos ejes (diagonal). Hay descritos numerosos índicesque permiten cuantificar el grado de ajuste a la diagonal, como por ejemploel de Miller –mejores ajustes cuando toma valores más próximos a cero–(Miller et al., 1991), que es el que ha sido usado en este estudio.
Para validar los valores predichos por el modelo como indicadores de laabundancia relativa de corzo se han usado datos provenientes de muestreosde campo, basados en el recuento de indicios de presencia de la especie, rea-lizados en Cuenca y Guadalajara y que han sido publicados en Acevedo et al.(2010a). De este trabajo se pueden extraer datos para 61 localidades en las quela abundancia de corzo es altamente variable (rango 0,01 - 20,74 corzos/100ha [media±ET: 5,56±0,75]). Dicho muestreo resulta muy adecuado ya que estábasado en la misma unidad territorial que la usada en este estudio, es decir,las cuadrículas UTM de 100 km2. Por tanto la validación ha consistido en elanálisis mediante una correlación de Spearman de la relación existente entrelos valores de probabilidad/favorabilidad y la abundancia medida para lascuadrículas muestreadas en el trabajo anteriormente mencionado.
3. Resultados
Tras la exploración del FDR se excluyeron 3 variables (HuEn, PM24 yVtem) del análisis multivariante. El modelo final se ha construido en 19 pasosque han incluido 17 variables en el modelo final para explicar la distribuciónde corzo en la España peninsular (Tabla 2). Los 5 factores considerados, estoes, situación espacial, topografía, clima, litología y actividad humana, hanquedado representados en el modelo final. Los valores de probabilidad y defavorabilidad predichos para la especie se muestran en la Figura 2. En estecaso, debido a que la prevalencia de la especie no difiere mucho de 0,5 losvalores de probabilidad y favorabilidad son similares. Los resultados delestadístico I de Moran sólo evidenciaron la existencia de una autocorrelaciónespacial significativa a distancias cortas (menos de 20 km) y con unos índices
P. ACEVEDO, R. REAL & C. GORTÁZAR
16 (Pirineos, 2011, Vol. 166, 9-27, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166001)

de Moran muy bajos (I media=-0,0002, rango -0,026 0,037). Por tanto, el mode-lo presentado en este trabajo, dada la reducida magnitud de las I de Moran yla reducida distancia a la que ha sido detectada la autocorrelación espacial enrelación al tamaño de las unidades territoriales, la autocorrelación espacial noes considerado un problema (Diniz-Filho et al., 2003).
FAVORABILIDAD ECOGEOGRÁFICA PARA EL CORZO: DISTRIBUCIÓN Y ABUNDANCIA
17(Pirineos, 2011, Vol. 166, 9-27, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166001)
Tabla 2. Variables incluidas en el modelo final y sus parámetros estadísticos. Las abreviaturas delas variables como en la Tabla 1.
Table 2. Variables retained in the final model and their statistical parameters. Variables coded as in Table 1.
Variable Estimador Wald P-valorPend 0.159 74.533 <0.001DGra -0.107 26.595 <0.001DiHe 0.027 104.990 <0.001DNie -1.75E-09 5.608 0.018RadS 0.016 55.922 <0.001ETP -0.003 7.022 0.008ETR 0.004 98.894 <0.001DiPr -0.011 15.848 <0.001IrrP -0.234 141.724 <0.001
IHum 5.87E-08 14.886 <0.001HuJu 0.035 32.500 <0.001Perm -0.262 22.690 <0.001DAut -0.008 17.924 <0.001U100 0.014 86.563 <0.001U500 0.002 5.667 0.017Long -1.65E-06 38.288 <0.001Lati 3.32E-06 52.327 <0.001
Constante -18.644 44.170 <0.001
La partición de la variación indica que de forma independiente al resto defactores, esto es su efecto puro, el factor climático es el que más variaciónexplica del modelo final: 25,7%. La contribución del resto de factores en suforma pura es más reducida, siendo 3,4% para el factor presión humana, 2,7%para el factor topográfico, el 2,6% lo explica la situación espacial y el 0,8% lalitología. La reducida cantidad de variación explicada por el efecto puro delos factores considerados indica que la mayor parte de la variación se debe alefecto combinado de 2 o más factores.
El modelo obtenido para el corzo se muestra como perfectamente calibra-do en lo que a probabilidad de presencia se refiere. El valor del índice deMiller es de 1E-06 y el gráfico de evaluación de la calibración se muestra enla Figura 3 y en él se aprecia un buen ajuste a lo largo de todo el rango de pro-babilidad predicha por el modelo.
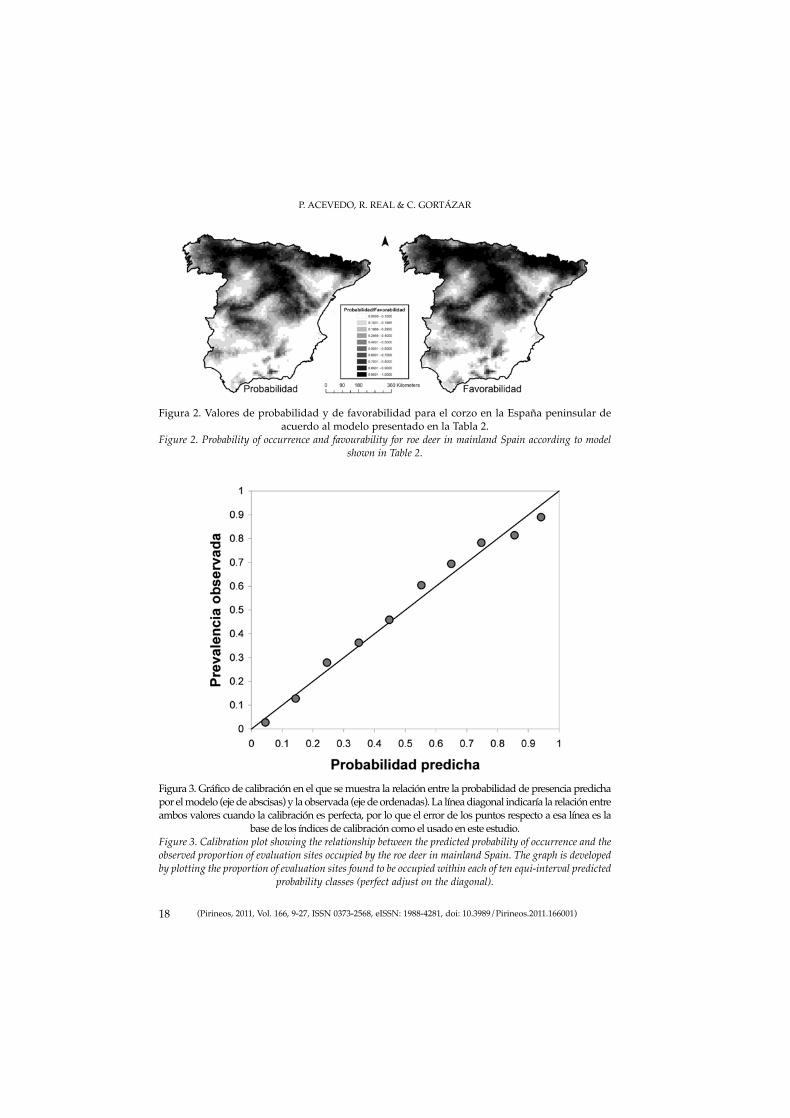
P. ACEVEDO, R. REAL & C. GORTÁZAR
18 (Pirineos, 2011, Vol. 166, 9-27, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166001)
Figura 2. Valores de probabilidad y de favorabilidad para el corzo en la España peninsular deacuerdo al modelo presentado en la Tabla 2.
Figure 2. Probability of occurrence and favourability for roe deer in mainland Spain according to modelshown in Table 2.
Figura 3. Gráfico de calibración en el que se muestra la relación entre la probabilidad de presencia predichapor el modelo (eje de abscisas) y la observada (eje de ordenadas). La línea diagonal indicaría la relación entreambos valores cuando la calibración es perfecta, por lo que el error de los puntos respecto a esa línea es la
base de los índices de calibración como el usado en este estudio.Figure 3. Calibration plot showing the relationship between the predicted probability of occurrence and theobserved proportion of evaluation sites occupied by the roe deer in mainland Spain. The graph is developedby plotting the proportion of evaluation sites found to be occupied within each of ten equi-interval predicted
probability classes (perfect adjust on the diagonal).

Respecto a la abundancia, los resultados indican que los valores predichospor el modelo se pueden consideran como un índice indirecto de la abun-dancia relativa de las poblaciones de corzo al estar correlacionadas significa-tivamente con los valores de abundancia independientes (rs=0,323; p=0,011;n=61). Por tanto, los valores más altos de probabilidad/favorabilidad estánseñalando las localidades con mayor abundancia relativa de la especie.
4. Discusión
4.1. Determinantes macroambientales y favorabilidad para el corzo
El modelo obtenido indica que la distribución del corzo, y por tanto lafavorabilidad ecogeográfica del territorio para esta especie, se puede explicaren términos topográficos, climáticos, litológicos, geográficos y de presiónhumana, aunque la importancia relativa del efecto puro de cada uno de ellosresultó ser bien distinta. Así el clima fue el factor que en su efecto puro expli-có un mayor porcentaje de variación, y en orden de importancia le siguen lapresión humana, la topografía, la situación espacial y la litología.
Es bien conocido el efecto del clima como determinante de la dinámica delas poblaciones de corzo, en particular, la precipitación diaria y la coberturade nieve, y la temperatura y el viento han sido variables descritas previa-mente como condicionantes para esta especie (Aragón et al., 1995; Latham etal., 1997; Acevedo et al., 2005, 2010b). Estos trabajos evidencian el elevadopoder explicativo que presentan las variables climáticas a la hora de estudiarlos patrones de ocupación de las especies (Múñoz et al., 2005; Acevedo et al.,2010a). Los resultados aquí obtenidos indican que el agua útil es un requeri-miento importante para el corzo ya que además de la utilidad directa para laespecie, de manera indirecta se relaciona por ejemplo con la composición flo-rística de las áreas ocupadas por corzo. La evapotranspiración real es la can-tidad de agua que se evapora para la atmósfera, y es una medida de la dis-ponibilidad simultánea de agua –relacionada a su vez con la permeabilidaddel suelo y con la pendiente del terreno– y de energía biológicamente útiles(Márquez, 1996). Por su parte la irregularidad pluviométrica es el coeficientede variación interanual de la precipitación y es una medida de la predecibili-dad de las precipitaciones (Romero & Real, 1996). La relación negativa detec-tada entre la irregularidad pluviométrica y la favorabilidad ecogeográficapara el corzo indica que las áreas por las que el corzo se distribuye presentanun régimen pluviométrico internanual más constante, es decir zonas en lasque la influencia atlántica es más notoria. Análogamente, una relación nega-
FAVORABILIDAD ECOGEOGRÁFICA PARA EL CORZO: DISTRIBUCIÓN Y ABUNDANCIA
19(Pirineos, 2011, Vol. 166, 9-27, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166001)

tiva entre la mediterraneidad y la presencia de corzo fue detectada estudian-do la distribución del corzo en Francia (Gaudin et al., 1997).
El elevado peso relativo que presenta el factor climático explicando la dis-tribución de esta especie sugiere una elevada sensibilidad de ésta a cambiosen el clima (Real et al., 2010). Sería esperable, por tanto, que el corzo en laPenínsula Ibérica viese alterada en una elevada magnitud su área de distri-bución en el futuro. Este resultado es coherente en el marco de la expansiónque está experimentando la especie en la Península y que está modulada, enbuena medida, por factores climáticos (Acevedo et al., 2005). Estudios reali-zados sobre otros ungulados ibéricos, por ejemplo la cabra montés Capra pyre-naica (Real et al., 2010), indican que su distribución se verá aumentada en unfuturo próximo de acuerdo a los escenarios de cambio climático descritos porel Panel Intergubernamental de Cambio Climático (Nakicenovic et al., 2000).Además del clima, un aumento del área de distribución del corzo en un futu-ro reciente fue también predicho en relación a simulaciones de cambios en losusos del suelo (Acevedo et al., 2011).
De acuerdo con trabajos previos, la presión humana ha sido otro factordeterminante de la distribución y abundancia del corzo (Hewison et al., 2001;pero ver Acevedo et al., 2005). La favorabilidad ecogeográfica de la Penínsulapara el corzo aumenta conforme lo hace la distancia a los núcleos urbanos.Los núcleos urbanos, además de ser una fuente de molestias para la fauna sil-vestre, son las áreas en las que más fuertemente se dejan notar las alteracio-nes del medio debido a actividades humanas. Por tanto son áreas poco ade-cuadas para esta especie que tradicionalmente ha estado vinculada a mediosboscosos y escasamente alterados por el hombre (Aragón et al., 1995;Cargnelutti et al., 2002), si bien es cierto que la reciente expansión que estáexperimentando la especie hace que esta percepción del corzo como animalvinculado al bosque deba ser reconsiderada.
El hecho de que o bien variables geográficas puras o la combinación de lasmismas en un polinomio (Legendre, 1993) queden retenidas en los modelossuele ser considerado como un reflejo indirecto de factores históricos que con-dicionaron fuertemente la distribución de la especie en el pasado (Real et al.,2008). En este caso concreto del corzo, esta especie se hallaba relegada en elsiglo XIX en áreas muy concretas debido a un pastoreo intensivo, quemas ytalas del monte, etc. Sin embargo dicha situación fue más crítica en la décadade los 60 ya que, y presumiblemente debido a una elevada presión cinegéti-ca, se produjo una nueva retracción del área de distribución de la especie(Gortázar et al., 2000). Por ello, el corzo quedó restringido a determinadosenclaves desde los que, y principalmente en las últimas décadas, comenzó unrápido proceso de recolonización (Acevedo et al., 2005) estando ahora amplia-mente distribuido por la Península Ibérica, especialmente en su mitad norte
P. ACEVEDO, R. REAL & C. GORTÁZAR
20 (Pirineos, 2011, Vol. 166, 9-27, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166001)

(Figura 1). La distancia a estos núcleos de expansión ha sido descrita como elfactor más relevante a la hora de explicar la expansión de la especie (Acevedoet al., 2005), estando esto en concordancia con los resultados del modelo pre-dictivo aquí presentado.
En lo que a la interpretabilidad de los modelos se refiere, los resultadosindican que, con elevada fiabilidad, los valores predichos por el modelo pue-den ser considerados como indicadores de la probabilidad de presencia de laespecie (Figura 3). Estos valores de probabilidad de presencia son de utilidadpara completar datos de distribución provenientes de muestreos incompletos,y establecer una información de base en la que basar el diseño de posterioresmuestreos (Guisan et al., 2006). En otras ocasiones el interés radica en conocerel grado en el que las condiciones ambientales de una determinada localidadpotencian la presencia de la especie, y eso se obtiene aplicando la función defavorabilidad (Real et al., 2006). Este concepto de favorabilidad es muy rele-vante desde el punto de vista biogeográfico y su utilidad aumenta aun máscuando el interés del estudio consiste en realizar comparaciones entre espe-cies/modelos (Acevedo et al., 2010b). La posibilidad de realizar comparacio-nes viene dada por la independencia que presenta la favorabilidad de la pre-valencia de la especie lo que hace que los valores predichos de diferentesmodelos sean directamente comparables, y que puedan ser usadas las reglasde la lógica difusa para poder relacionarlos (Real et al., 2009; Acevedo et al.,2010b, 2011). Conviene tener claros por tanto ambos conceptos (probabilidady favorabilidad) con el fin de poder aplicar una función y otra –o las dos comoen este caso– según las necesidades de la investigación. En este estudio, y debi-do a que la prevalencia de la especie es próxima a 0,5 (Real et al., 2006), elpatrón de probabilidad y el de favorabilidad y así como sus valores son muysimilares (Figura 2). Sin embargo, creímos necesario incluir ambas funcionesen este manuscrito para clarificar ambos conceptos en el ámbito de los SDM.
4.2. Abundancia relativa
Los resultados indican que las predicciones del modelo realizado para elcorzo pueden ser interpretadas como indicadores indirectos de la abundanciarelativa de las poblaciones de corzo, al menos en las provincias de Cuenca yGuadalajara que es donde el modelo ha sido validado. Disponer de una esti-mación de la abundancia relativa de una especie para un territorio amplio essumamente costoso en términos económicos y de esfuerzo por lo que, habi-tualmente, para ello se suele recurrir a la colaboración de voluntarios pararealizar los muestreos de campo (Daniels, 2006). Por otro lado, trabajando conespecies cinegéticas se pueden usar los datos de las capturas en las activida-
FAVORABILIDAD ECOGEOGRÁFICA PARA EL CORZO: DISTRIBUCIÓN Y ABUNDANCIA
21(Pirineos, 2011, Vol. 166, 9-27, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166001)

des cinegéticas ya que éstos suponen una información de utilidad para tal fin(Mysterud et al., 2007), si bien el éxito de las actividades cinegéticas no depen-de exclusivamente de la abundancia de la especie (Acevedo et al., 2009) por loque dicha información debe ser manejada con cautela sobretodo cuando seconsideran territorios sometidos a diferentes regímenes de explotación cine-gética. En este contexto, la determinación de un indicador de abundanciapoblacional a lo largo del territorio que pueda ser obtenido desde datos dedistribución de la especie –como el aquí evaluado– se muestra como unaalternativa rentable y eficaz (Real et al., 2009) que, dada su relevancia, debe-ría ser objeto de más profundas investigaciones.
La necesidad de disponer de información sobre la abundancia de unaespecie a lo largo del territorio se articula a muy diversos niveles. Desde elpunto de vista de la fauna cinegética, resulta necesario en muchas ocasionesdisponer de una estimación de la abundancia de la especie con el fin de poderadecuar los planes de gestión a las particularidades de cada localidad, perosin perder de vista el contexto global que presenta la especie en todo su áreade distribución (Farfán et al., 2008; Real et al., 2009). Por otro lado, la tasa detransmisión de patógenos en una población está íntimamente relacionada consu abundancia relativa (ver ejemplos en Gortázar et al., 2006). Así, la estima-ción de la abundancia de una especie en todo su área de distribución es unproceso clave para poder comprender el ciclo epidemiológico de los patóge-nos en los que dicha especie juega un papel relevante bien como vector u hos-pedador. Por ejemplo, la abundancia de ciervo (Cervus elaphus), estimadamediante SDM, ha sido recientemente utilizada para estudiar el papel queésta juega en la abundancia del principal vector del virus de la lengua azul(Culicoides imicola) en la Península Ibérica (Acevedo et al., 2010c). Desde elpunto de vista de la sanidad animal es necesario también conocer la abun-dancia de las poblaciones para poder establecer planes de contingencia fren-te a la entrada de un patógeno en el territorio como los que actualmente estánemergiendo en la Península Ibérica (Ruiz-Fons et al., 2008).
Agradecimientos
Este trabajo ha sido promovido y financiado por la Consejería de MedioAmbiente de la Junta de Comunidades de Castilla – La Mancha. No podemosdejar de agradecer la ayuda proporcionada a todos los propietarios y gesto-res de fincas privadas que nos han facilitado el trabajo de campo realizado. P.Acevedo disfruta de un contrato dentro del programa Juan de la Cierva –Fondo Social Europeo y sus investigaciones están parcialmente financiadaspor el proyecto del Plan Nacional CGL2009-11316 - FEDER.
P. ACEVEDO, R. REAL & C. GORTÁZAR
22 (Pirineos, 2011, Vol. 166, 9-27, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166001)

Referencias
Acevedo, P. & Cassinello, J., 2009. Human-induced range expansion of wildungulates causes niche overlap between previously allopatric species: reddeer and Iberian ibex in mountainous regions of southern Spain. AnnalesZoologici Fennici, 46: 39-50.
Acevedo, P. & Real, R., 2011. Biogeographical differences between the twoCapra pyrenaica subspecies, C. p. victoriae and C. p. hispanica, inhabiting inthe Iberian Peninsula: conservation implications. Ecological Modelling, 222:814-823.
Acevedo, P., Delibes-Mateos, M., Escudero, M.A., Vicente, J., Marco, J. &Gortázar, C., 2005. Environmental constraints in the colonization sequenceof roe deer (Capreolus capreolus Linnaeus, 1758) across the IberianMountains, Spain. Journal of Biogeography, 32: 1671-1680.
Acevedo, P., Cassinello, J. & Gortázar, C., 2007a. The Iberian ibex is under anexpansion trend but displaced to suboptimal habitats by the presence ofextensive goat livestock in central Spain. Biodiversity and Conservation, 16:3361-3376.
Acevedo, P., Cassinello, J., Hortal, J. & Gortázar, C., 2007b. Invasive exoticaoudad (Ammotragus lervia) as a major threat to native Iberian ibex (Caprapyrenaica): a habitat suitability model approach. Diversity and Distributions,13: 587-597.
Acevedo, P., Vicente, J., Alzaga, V. & Gortázar, C., 2009. Wild boar abundanceand hunting effectiveness in Atlantic Spain: Environmental constraints.Galemys, 21: 13-29.
Acevedo, P., Ferreres, J., Jaroso, R., Durán, M., Escudero, M.A., Marco, J. &Gortázar, C., 2010a. Estimating roe deer abundance from pellet groupcounts in Spain: An assessment of methods suitable for Mediterraneanwoodlands. Ecological Indicators, 10: 1226-1230.
Acevedo, P., Ward, A.I., Real, R. & Smith, G.C., 2010b. Assessing biogeo-graphical relationships of ecologically related species using favourabilityfunctions: a case study on British deer. Diversity and Distributions, 16: 515-528.
Acevedo, P., Farfán, M.A., Márquez, A.L., Delibes-Mateos, M., Real, R. &VargasS, J.M., 2011. Past, present and future of wild ungulates in rela-tion to changes in land use. Landscape Ecology, DOI: 10.1007/s10980-010-9538-2.
Acevedo, P., Ruiz-Fons, F., Estrada, R., Márquez, A.L., Miranda, M.A.,Gortázar, C. & Lucientes, J., 2010c. A broad assessment of factors deter-mining Culicoides imicola abundance: modelling the present and forecast-
FAVORABILIDAD ECOGEOGRÁFICA PARA EL CORZO: DISTRIBUCIÓN Y ABUNDANCIA
23(Pirineos, 2011, Vol. 166, 9-27, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166001)

ing its future in climate change scenarios. PLoS ONE 5 (12): e14236.doi:10.1371/journal.pone.0014236
Aragon, S., Braza, F. & San José, C., 1995. Socioeconomic, physiognomic, andclimate factors determining the distribution pattern of roe deer Capreoluscapreolus in Spain. Acta Theriologica, 40: 37-43.
Austin, M.P., Nicholls, A.O. & Margules, C.R., 1990. Measurement of the real-ized qualitative niche: environmental niches of five Eucalyptus species.Ecological Monographs, 60: 161-177.
Barbosa, A.M., Real, R., Olivero, J. & Vargas, J.M., 2003. Otter (Lutra lutra) dis-tribution modeling at two resolution scales suited to conservation plan-ning in the Iberian Peninsula. Biological Conservation, 114: 377-387.
Benjamini, Y. & Hochberg, Y., 1995. Controlling the false discovery rate: Apractical and powerful approach to multiple testing. Journal of the RoyalStatistical Society, Series B, 57: 289-300.
Borcard, D., Legendre, P. & Drapeau, P., 1992. Partialling out the spatial com-ponent of ecological variation. Ecology, 73: 1045-1055.
Boyce, M.S., Mackenzie, D.I., Manly, B.F.J., Haroldson, M.A. & Moody, D.,2001. Negative binomial models for abundance estimation of multipleclosed populations. Journal of Wildlife Management, 65: 498-509.
Cargnelutti, B., Reby, D., Desneux, L. & Angibault, J.M., 2002. Space use byroe deer in a fragmented landscape, some preliminary results. RevueEcologie-Terre et Vie, 57: 29-37.
Cassinello, J., Acevedo, P. & Hortal, J., 2006. Prospects for population expan-sion of the exotic aoudad (Ammotragus lervia; Bovidae) in the IberianPeninsula: clues from habitat suitability modelling. Diversity andDistributions, 12: 666-678.
Daniels, M.J, 2006. Estimating red deer Cervus elaphus populations: an analy-sis of variation and cost-effectiveness of counting methods. MammalReview, 36: 235-247.
Delibes-Mateos, M., Farfán, M.A., Olivero, J., Márquez, A.L. & Vargas, J.M.,2009. Long-term changes in game species over a long period of transfor-mation in the Iberian Mediterranean landscape. Environmental Mana-gement, 43: 1256-1268.
Diniz-Filho, J.A.F., Bini, L.M. & Hawkins, B.A., 2003. Spatial autocorrelationand red herrings in geographical ecology. Global Ecology and Biogeography,12: 53-64.
Farfán, M.A., Vargas, J.M., Guerrero, J.C., Barbosa, A.M., Duarte, J. & Real, R.,2008. Distribution modelling of wild rabbit hunting yields in its originalarea (S Iberian Peninsula). Italian Journal of Zoology, 75: 161-172.
P. ACEVEDO, R. REAL & C. GORTÁZAR
24 (Pirineos, 2011, Vol. 166, 9-27, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166001)

Gaudin, J.C., Maillard, D. & Gaillard, J.M., 1997. Colonisation par le chevreuil(Capreolus capreolus L.) de la zone méditerranéenne Française. Revued’Ecologie – la Terre et la Vie, 52: 113-122.
Gortázar, C., Herrero, J., Villafuerte, R. & Marco, J., 2000. Historical examina-tion of the distribution of large mammals in Aragón, Northeastern Spain.Mammalia, 61: 411-422.
Gortázar, C., Acevedo, P., Ruiz-Fons, F. & Vicente, J., 2006. Disease risks andoverabundance of game species. European Journal of Wildlife Research, 52:81-87.
Graham, M. H. (2003). Confronting multicollinearity in ecological multipleregression. Ecology, 84(11): 2809-2815.
Guisan, A. & Zimmermann, N.E., 2000. Predictive habitat distribution mod-els in ecology. Ecological Modelling, 135: 147-186.
Guisan, A., Broennimann, O., Engler, R., Vust, M., Yoccoz, N.G., Lehmann, A.& Zimmermann, N.E., 2006. Using niche-based models to improve thesampling of rare species. Conservation Biology, 20: 501-511.
Hewison, A.J.M., Vincent, J.P., Joachim, J., Angibault, J.M., Cargnelutti, B. &Cibien, C., 2001. The effects of woodland fragmentation and human activ-ity on roe deer distribution in agricultural landscapes. Canadian Journal ofZoology - Revue Cannadiene de Zoologie, 79: 679-689.
Hirzel, A.H., Hausser, J., Chessel, D. & Perrin, N., 2002. Ecological-niche fac-tor analysis: How to compute habitat suitability maps without absencedata? Ecology, 83: 2027-2036.
Hosmer, D.W. & Lemeshow, S., 1989. Applied logistic regression. Wiley, NewYork.
Jiménez-Valverde, A., Lobo, J.M. & Hortal, J., 2008. Not as good as they seem:the importance of concepts in species distribution modelling. Diversity andDistributions, 14:885-890.
Latham, J., Staines, B.W. & Gorman, M.L., 1997. Correlations of red (Cervuselaphus) and roe (Capreolus capreolus) deer densities in Scottish forests withenvironmental variables. Journal of Zoology, 242: 681-704.
Legendre, P., 1993. Spatial autocorrelation: trouble or new paradigm? Ecology,74: 1659-1673
Márquez, A.L., 1996. Análisis biogeográfico de la pteridoflora ibérica. TesisDoctoral, Universidad de Málaga.
Miller, M.E., Hui, S.L. & Tierney, W.M., 1991. Validation techniques for logis-tic regression models. Statistics in Medicine, 10: 1213-1226.
Muñoz, A.R., Real, R., Barbosa, A.M., & Vargas, J.M., 2005. Modelling the dis-tribution of Bonelli’s eagle in Spain: implications for conservation plan-ning. Diversity and Distributions, 11: 477-486.
FAVORABILIDAD ECOGEOGRÁFICA PARA EL CORZO: DISTRIBUCIÓN Y ABUNDANCIA
25(Pirineos, 2011, Vol. 166, 9-27, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166001)

Muñoz, A.R. & Real, R., 2006. Assessing the potential range expansion of theexotic monk parakeet in Spain. Diversity and Distributions, 12: 656-665.
Mysterud, A., Meisingset, E.L., Veiberg, V., Langvatn, R., Solberg, E.J., Loe,E.L. & Stenseth, N.C., 2007. Monitoring population size of red deer Cervuselaphus: an evaluation of two types of census data from Norway. WildlifeBiology, 13: 285-298.
Nakicenovic, N., Alcamo, J., Davis, G., De Vries, B., Fenhann, J., Gaffin, S.,Gregory, K., Grübler, A., Jung, T.Y., Kram, T., La Rover, E.L., Michaelis, L.,Mori, S., Morita, T., Pepper, W., Pitcher, H., Price, L., Raihi, K., Roehrl, A.,Rogner, H.H., Sankovski, A., Schlesinger, M., Shukla, P., Smith, S., Swart,R., Van Rooijen, S., Victor, N. & Dadi, Z., 2000. IPCC special report on emis-sions scenarios. Cambridge University Press, Cambridge.
Nielsen, S.E., Johnson, C.J., Heard, D.C. & Boyce, M.S., 2005. Can models ofpresence-absence be used to scale abundance? Two cases studies consid-ering extremes in life history. Ecography, 28: 1-12.
Palomo, L.J., Blanco, J.C. & Gisbert, J., 2007. Atlas y libro rojo de los mamíferosterretres de España. Dirección General de Conservación de la Naturaleza-SECEM-SECEMU, Madrid.
Pearce, J. & Ferrier, S., 2000. Evaluating the predictive performance of habitatmodels developed using logistic regression. Ecological Modelling, 133: 225-245.
Pearce, J. & Ferrier, S., 2001. The practical value of modelling relative abun-dance of species for regional conservation planning: a case study. BiologicalConservation, 98: 33-43.
Phillips, S.J., Anderson, R.P. & Schapire, R.E., 2006. Maximum entropy mod-eling of species geographic distributions. Ecological Modelling, 190: 231-259.
Real, R., Barbosa, A.M., Porras, D., Kin, M.S., Márquez, A.L., Guerrero, J.C.,Palomo, L.J., Justo, E.R. & Vargas, J.M., 2003. Relative importance of envi-ronment, human activity and spatial situation in determining the distri-bution of terrestrial mammal diversity in Argentina. Journal ofBiogeography, 30: 939-947.
Real, R., Barbosa, A.M. & Vargas, J.M., 2006. Obtaining environmentalfavourability functions from logistic regression. Enviromental and EcologicalStatistics, 13: 237-245.
Real, R., Márquez, A.L., Estrada, A., Román Múñoz, A. & Vargas, J.M., 2008.Modelling chorotypes of invasive vertebrates in mainland Spain. Diversityand Distributions, 14: 364-373.
Real, R., Barbosa, A.M., Rodríguez, A., García, F.J., Vargas, J.M., Palomo, L.J.& Delibes, M., 2009. Conservation biogeography of ecologically interact-
P. ACEVEDO, R. REAL & C. GORTÁZAR
26 (Pirineos, 2011, Vol. 166, 9-27, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166001)

ing species: the case of the Iberian lynx and the European Rabbit. Diversityand Distributions, 5: 390-400.
Real, R., Márquez, A.L., Olivero, J. & Estrada, A., 2010. Are species distribu-tion models in climate warming scenarios useful for informing emissionpolicy planning? An uncertainty assessment using fuzzy logic. Ecography,33: 304-314.
Robertson, M.P., Peter, C.I., Villet, M.H. & Ripley, B.S., 2003. Comparing mod-els for predicting species’ potential distributions: a case study using cor-relative and mechanistic predictive modelling techniques. EcologicalModelling, 164: 153-167
Romero, J. & Real, R. 1996. Macroenvironmental factors as ultimate determi-nants of the distribution of common toad and netterjack toad in the southof Spain. Ecography, 19: 305-312
Ruiz-Fons, F., Reyes-García, A.R., Alcaide, V. & Gortázar, C., 2008. Spatial andtemporal evolution of bluetongue virus in wild ruminants, Spain.Emerging Infectious Diseases, 14: 951-953.
Soberón, J. & Peterson, A.T., 2005. Interpretation of models of fundamentalecological niches and species’ distribution areas. Biodiversity Informatics, 2:1-10.
FAVORABILIDAD ECOGEOGRÁFICA PARA EL CORZO: DISTRIBUCIÓN Y ABUNDANCIA
27(Pirineos, 2011, Vol. 166, 9-27, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166001)

29
EVALUACIÓN DEL ESTADO DE LA POBLACIÓNDE CABRA MONTÉS DE LOS MONTES DE TOLEDO:
RELACIONES CON EL CIERVO
P. ACEVEDO1,2*, J. P. SANTOS2,3, R. REAL1 & J. VICENTE2
1 Departamento de Biología Animal, Facultad de Ciencias, Universidad de Málaga, 29071 Málaga*[email protected]; [email protected]
2 Instituto de Investigación en Recursos Cinegéticos (CSIC-UCLM-JCCM), 13071 Ciudad Real3 Departamento de Biologia/CESAM, Universidade de Aveiro, 3810-193 Aveiro, Portugal
ABSTRACT.– Iberian wild goat subsp. Capra pyrenaica victoriae inhabitsfewer and smaller areas than subs. C. p. hispanica reason why the former is consid-ered as Vulnerable by IUCN. Therefore studies aimed to assess the status of C. p. vic-toriae are very valuables. We studied the habitat use, parasite excretion rates and dietquality of the Montes de Toledo’s C. p. victoriae population. The red deer is a poten-tial competitor of the Iberian wild goat and subsequently was included in this study.Our results showed that both species had a good population status (good corporalcondition, high quality diet and suitable population density). According to the indi-cators considered in this study we do not expect that red deer suppose a relevant riskfor the viability of the Iberian wild goat population. The implications of the resultsto Iberian wild goat management are deeper discussed.
Keywords: Iberian wild goat; diet quality; red deer; bronchopulmonarynematodes; habitat use.
RESUMEN.– La Capra pyrenaica victoriae presenta pocas poblaciones y éstasocupan áreas reducidas, siendo considerada por ello por la IUCN como vulnerable.Son necesarios, por tanto, estudios locales encaminados a evaluar el estado de dichaspoblaciones. En este trabajo estudiamos la población presente en los Montes deToledo a través del análisis del uso que hace del hábitat, del nivel de parasitación yde la calidad de su dieta. El ciervo fue también incluido en este esquema de monito-rización por ser un potencial competidor para la cabra montés. Los resultados indi-can que ambas especies presentan un adecuado estado poblacional (buena condicióncorporal, dieta de elevada calidad y valores de densidad razonables). Según los indi-cadores considerados no sería esperable que el ciervo supusiera un riesgo relevante
Pirineos.Revista de Ecología de MontañaVol. 166, 29-49
Jaca, Enero-Diciembre, 2011ISSN: 0373-2568
eISSN: 1988-4281doi: 10.3989/Pirineos.2011.166002

30
P. ACEVEDO, J. P. SANTOS, R. REAL & J. VICENTE
para la viabilidad de la población de cabra montés. Se discuten las implicaciones delos resultados para la gestión cinegética de la cabra montés.
Palabras clave: Cabra; calidad de dieta; ciervo; parásitos; uso del hábitat.
1. Introducción
Las poblaciones de caprinos silvestres son particularmente vulnerables ala extinción debido a su aislamiento genético, sus requerimientos de hábitat ysu reducida tasa reproductiva (Shackleton, 1997). Además de éstos, la cabramontés (Capra pyrenaica) se ve afectada por otros factores de riesgo adiciona-les como son el incremento del ganado doméstico y ungulados exóticos, quepodrían transmitir enfermedades a la fauna nativa (ver Gortázar et al., 2006)y competir por recursos (Acevedo et al., 2007a, 2007b; Acevedo & Cassinello,2009a), y la presión turística (R.C. Soriguer, comunicación personal), que ten-drán efectos más marcados cuanto peor sea el estatus de las poblaciones.Lamentablemente este hecho ha quedado recientemente corroborado con laextinción, a finales de los 90, de la subespecie C. p. pyrenaica que habitaba elPirineo (García-González & Herrero, 1999).
La cabra montés estaba representada en la Península Ibérica por 4 subes-pecies de las que sólo 2 permanecen en la actualidad, la C. p. victoriae, vincu-lada al noroeste, y la C. p. hispanica que se distribuye por el arco montañosodel este peninsular (Figura 1). La reducida área de distribución es una de lasrazones por las que la IUCN considera Vulnerable (VUd2) a la subespecie delnoroeste, que “presenta pocas poblaciones y éstas ocupan áreas reducidas”(IUCN, 2007). Sin embargo, la situación de la otra subespecie es bien distinta,con distribución amplia y un elevado número de núcleos poblacionales, razo-nes por las que la IUCN la considera como de Bajo Riesgo, aunque próxima aconsiderarse Vulnerable (LR/nt; IUCN, 2007). Recientemente la IUCN hacambiado el estatus de la especie considerándola en el grupo dePreocupación Menor al detectar un incremento de sus poblaciones (IUCN,2009). Pese a ello, la situación de ambas subespecies es claramente distinta,siendo la C. p. victoriae la que muestra un estado más crítico debido a la vul-nerabilidad de sus poblaciones (Escós et al., 2008; Acevedo & Cassinello,2009b; Acevedo & Real, en prensa). Así, la subespecie C. p. victoriae se distri-buye en sólo 7 de los ya más de 50 núcleos poblacionales presentes en laPenínsula Ibérica (Pérez et al., 2002).
Por otro lado, estudios recientes han evidenciado que, a escala peninsular,las poblaciones de la subespecie C. p. victoriae se encuentran restringidas a
(Pirineos, 2011, Vol. 166, 29-49, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166002)

determinados enclaves dentro de áreas ambientalmente adecuadas de mayoramplitud (Acevedo & Real, en prensa). Esto podría estar mostrando la exis-tencia de algunos factores, bióticos u abióticos, que podrían estar limitando laaccesibilidad de las monteses a otras áreas favorables que permanecen des-habitadas. Así, estudios realizados a escala local, y encaminados a determinarel estado de las poblaciones, aportarán información de utilidad para com-prender los mecanismos que regulan la dinámica de esta subespecie.
Aunque no se han realizado estudios genéticos que lo corroboren, la esca-sa información disponible indica que la población de cabra montés delParque Nacional de Cabañeros pertenece a la subespecie C. p. victoriae. Así, setiene constancia de introducciones realizadas antes de 1995 (Pérez et al., 2002)con animales procedentes supuestamente de Gredos. En este mismo sentido,
EVALUACIÓN DEL ESTADO DE LA POBLACIÓN DE CABRA MONTÉS DE LOS MONTES DE TOLEDO:...
31(Pirineos, 2011, Vol. 166, 29-49, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166002)
Figura 1. Distribución actual de la cabra montés en la Península Ibérica. La línea discontinua divi-de la distribución de ambas subespecies, al noroeste la C. p. victoriae, y en el este y el sur la C. p.hispanica. Las flechas muestran translocaciones. Adaptada de Acevedo & Cassinello
(2009b).Figure 1. Current distribution of Iberian wild goat in the Iberian Peninsula. Grey lines delimit boundariesof the provinces. The distribution of the two subspecies is represented by the discontinuous line, C. p. vic-toriae in the north-west, and C. p. hispanica in the south and east. Arrows indicate translocations from
labelled populations. Figure adapted from Acevedo & Cassinello (2009b).

análisis biogeográficos realizados recientemente son congruentes con dichaprocedencia ya que muestran que las cabras monteses de los Montes deToledo ocupan áreas ambientalmente más próximas a las que ocupan las C. p.victoriae de Gredos o Riaño que a las zonas ocupadas por C. p. hispanica enSierra Morena o en el Alto Tajo (Acevedo & Real, en prensa).
Pérez et al. (2002), basándose en información proveniente de conversacio-nes personales con trabajadores del Parque, describen que las cabras en esteterritorio se distribuyen por las sierras de Las Parrillas, Muelas y Ciguiñuelas.Estos mismos autores estimaron una población de 15-20 ejemplares para estenúcleo en 1999. Posteriormente, un muestreo realizado en 2003 ha confirma-do la presencia de la especie en las inmediaciones del Pico Rocigalgo (verAcevedo et al., 2005). Dicho muestreo mostró una reducida abundancia rela-tiva en este núcleo en comparación con otras poblaciones de cabra montés deCastilla-La Mancha (CLM).
Además de estimar la abundancia relativa de las poblaciones en el mues-treo realizado en 2003 se realizó un estudio parasitario de las poblaciones decabra de CLM. Las prevalencias obtenidas para la población de Montes deToledo fueron más elevadas de lo que cabía esperar dada la abundancia rela-tiva de cabra montés en este núcleo. Así el 72% de las muestras analizadaspresentaron formas de excreción de la Familia Protostrongylidae y el 33% deDictyocaulus spp. (Acevedo et al., 2005). Esto, considerando la clara denso-dependencia que muestran estos grupos de parásitos en este hospedador,podría indicar que las abundancias estimadas entonces pudieran estar infra-valorando la abundancia real de la población.
En este contexto, el objetivo principal de este estudio es conocer el estadode la población de cabra montés presente en el Parque Nacional deCabañeros, y estudiar el uso que ésta hace del hábitat. Esto permitirá laimplantación de un sistema de seguimiento de dicha población a lo largo deltiempo. Además, se cree necesario incorporar al ciervo en este sistema deseguimiento para así poder detectar las posibles relaciones interespecíficasque medien la simpatría de ambas especies.
2. Metodología
2.1. Encuestas de distribución
Son numerosos los trabajos en los que se han empleado datos provenien-tes de encuestas para conocer la distribución y tendencias poblacionales deungulados cinegéticos (Gortázar et al., 2000). Así, en este estudio se han dis-tribuido encuestas con el fin de recopilar información sobre la población de
P. ACEVEDO, J. P. SANTOS, R. REAL & J. VICENTE
32 (Pirineos, 2011, Vol. 166, 29-49, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166002)

cabra montés presente en el Parque Nacional de Cabañeros y sus inmedia-ciones (área de estudio). En las encuestas se ha solicitado información cuali-tativa sobre la procedencia y características del núcleo fundador de la pobla-ción, y también sobre la percepción personal de la tendencia que ésta ha expe-rimentado en los últimos años. Además de los formularios, las encuestas hanido acompañadas de mapas de la zona donde se les pedía a los colaborado-res que señalasen los parajes en los que suelen avistar cabra montés.
2.2. Uso del hábitat y densidad poblacional
Los muestreos de excrementos en parcelas pueden ser usados para estu-diar los patrones de uso y preferencias de hábitat en ungulados (Edge &Marcum, 1989; Hemami et al., 2005). Actualmente están disponibles nuevasherramientas, basadas en sistemas de posicionamiento global (GPS), que per-miten realizar estudios más precisos (Moen et al., 1997), pero lamentablemen-te éstas no están todavía al alcance de todos los proyectos de investigación.Por otro lado, el recuento de excrementos en parcelas es un método usadopara estimar la abundancia de las poblaciones de ungulados (Acevedo et al.,2008, 2010), siendo posible incluso estimar valores de densidad siempre quela tasa de deposición de excrementos y el tiempo de permanencia de losexcrementos en el medio sean parámetros conocidos (Smart et al., 2004). Eneste trabajo, mediante el recuento de excrementos en parcelas se han estudia-do los patrones de uso del hábitat de cabras monteses y ciervos dentro delárea de distribución de la cabra montés en el Parque a lo largo de un cicloanual (desde julio de 2009 hasta junio de 2010).
Una vez determinada el área de distribución de la cabra montés en elParque, a lo largo de 4 recorridos se fijaron 32 parcelas sobre las que realizarel recuento de excrementos (Figura 2), aplicando un esfuerzo de muestreoaproximado de 4 parcelas por cada 500 ha de territorio. De estos 4 recorridos,2 de ellos discurrían por zonas con cierto manejo cinegético que se manifes-taba en el aporte de alimentación suplementaria. De acuerdo a experienciasprevias con esta metodología en poblaciones de ciervo (Acevedo et al., 2008)y corzo (Acevedo et al., 2010), y dada la reducida densidad esperada de cabramontés en este núcleo poblacional, las parcelas han sido de 5 m x 5 m, es decirde 25 m2 (se han muestreado 800 m2).
Todas las parcelas han sido revisadas con una periodicidad que ha varia-do entre los 42 y los 55 días, con la excepción de los meses de invierno en losque las condiciones climáticas adversas no permitieron hacer una de las revi-siones. En cada revisión se ha registrado el número de grupos fecales de cadauna de las especies consideradas –cabra montés y ciervo– presentes en cada
EVALUACIÓN DEL ESTADO DE LA POBLACIÓN DE CABRA MONTÉS DE LOS MONTES DE TOLEDO:...
33(Pirineos, 2011, Vol. 166, 29-49, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166002)

parcela. Tras el recuento, los excrementos eran eliminados de las parcelas, porlo que se ha usado el Faecal Accumulation Rate (Smart et al., 2004) como índicede abundancia. A partir de este método se puede inferir un valor de densidadpoblacional siempre que se conozca la tasa de defecación de estas especies.Para el caso del ciervo, en este tipo de ambientes mediterráneos, puede con-siderarse de 11.8 grupos/día (Acevedo et al., 2008), y para la cabra, aunquedesconocida, podría situarse entre la del ciervo y la del corzo, y por tantotomar valores próximos a 15.0 grupos/día (son 20 para el caso del corzo;Acevedo et al., 2010). Los recuentos de excrementos se han aprovechado paracoger muestras de heces frescas con las que realizar estudios parasitarios y decalidad de dieta (ver abajo).
P. ACEVEDO, J. P. SANTOS, R. REAL & J. VICENTE
34 (Pirineos, 2011, Vol. 166, 29-49, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166002)
Figura 2. Localización del área de estudio y de las parcelas de recuento de excrementos a lo largode los recorridos diseñados, dos de ellos por zonas sin manejo cinegético (líneas discontinuas) ydos por zonas manejadas (líneas continuas). Las curvas de nivel (cada 200 m) son incluidas para
mostrar la orografía del área de estudio.Figure 2. The location of sampling plots in which Iberian wild goat population was monitored from faeces.Unmanaged (discontinuous lines) and intensively managed (continuous lines) areas are differentiated. The
orography of the study area is shown with the elevation model.

2.3. Parásitos y calidad de la dieta
Con las muestras de excrementos tomadas en el campo y los análisis rea-lizados en el laboratorio (n=115) se ha podido caracterizar la variación anualde excreción de las principales formas parásitas presentes en ambas especies(Vicente et al., 2005). La extracción de larvas L1 de nematodos de las heces fuerealizada en menos de 24-48 horas tras su recolección. A partir de 8-10 gramosde heces, se empleó el método Baermann de extracción larvaria modificadopor Forrester & Lankester (1997). Este método, consiste en inducir la migra-ción de las larvas en agua en un vaso de precipitados de 250 ml durante 24horas. Las heces pesadas se colocan envueltas por una tela de vinilo de 2 mmde retícula cuadrada a modo de sobre grapado. Las larvas fueron cuantifica-das en cámara de Favatti y se expresan como larvas por gramo de heces. Laslarvas fueron identificadas hasta el nivel de género según su morfología(Melhorn et al., 1992; Cordero del Campillo & Rojo, 1999; Anderson, 2000).
Por otra parte, la monitorización de la calidad de la dieta puede ofrecer undiagnóstico adicional del estado de la población. Los altos requerimientos deproteína de los animales en relación a su disponibilidad en las plantas, con-vierten al nitrógeno ingerido en un buen indicador de la calidad proteica dela dieta (Mattson, 1980; Putman, 1984), y por tanto al nitrógeno fecal en unparámetro de gran utilidad como herramienta para la gestión de unguladossilvestres (Massey et al., 1994; Hamel et al., 2009). En este estudio se ha usadoel nitrógeno fecal como indicador de la calidad de la dieta. Los análisis paracuantificar el nitrógeno se han realizado sobre los grupos fecales recogidosdurante los muestreos de campo siempre que la cantidad de muestra era sufi-ciente para la realización de ambos análisis. Se ha cuantificado el nitrógenode 163 grupos fecales, entre ciervo y cabra montés. Para ello se ha usado elanalizador elemental LECO CHNS-932 que determina de forma cuantitativael carbono, hidrógeno, nitrógeno y azufre de las muestras. La técnica estábasada en la completa e instantánea oxidación térmica de 1 a 3 mg de lamuestra mediante una combustión en una atmósfera controlada en presión yenriquecida con oxígeno puro a una temperatura de 1100 ºC, generando pro-ductos gaseosos, CO2, H2O, SO2 y NO2 que, una vez separados, se cuantificanmediante métodos selectivos de absorción de infrarrojos y de conductividadtérmica. Una vez registrados los datos, se procesan junto al peso de la mues-tra y a los datos proporcionados por una muestra patrón, tras lo cual obten-dremos el contenido porcentual de cada elemento de la muestra –en este casonitrógeno– con respecto al peso analizado.
EVALUACIÓN DEL ESTADO DE LA POBLACIÓN DE CABRA MONTÉS DE LOS MONTES DE TOLEDO:...
35(Pirineos, 2011, Vol. 166, 29-49, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166002)

3. Resultados
3.1. Encuestas de distribución
Se han cumplimetado 10 encuestas de distribución realizadas a personalde diversos sectores profesionales, todos ellos con una estrecha vinculacióncon el medio natural y con el área de estudio. En términos generales, se puededecir que entre los encuestados existe conocimiento de la existencia de lapoblación de cabra montés, pero ésta no es considerada como especialmenterelevante por ninguno de los sectores encuestados, entre los que se incluyenlugareños, personal técnico y guardas de caza de fincas próximas.
Todos los encuestados que aportaron información sobre el origen de lapoblación señalaron que la procedencia de los animales fue Gredos. El 66% delos encuestados considera que la población de cabra montés objeto de estudiopresenta una tendencia al aumento de sus efectivos, siendo esta percepciónmás evidente en los últimos años. El porcentaje restante perciben estabilidady ninguno de ellos disminución. El tamaño máximo de grupo visto por loscolaboradores fue de 8 ejemplares y el 100% de los encuestados avistaronhembras con crías.
3.2. Uso del hábitat y densidad poblacional
Con las 7 revisiones realizadas de las 32 parcelas de recuento de excre-mentos se han podido detectar 775 grupos fecales de ciervo y 108 grupos feca-les de cabra montés. El número medio de grupos detectados por revisión–excluyendo los datos de la primera en la que se limpiaron las parcelas– fuede 78±19 grupos fecales de ciervo y 8±2 de cabra montés, no apreciándosediferencias estadísticamente significativas entre revisiones, ni en el caso delciervo (Kruskal-Wallis: Chi2=4.39, 5 gl, p=0.495), ni en el de la cabra montés(Kruskal-Wallis: Chi2=4.91, 5 gl, p=0.427).
En relación a los diferentes recorridos considerados en el estudio, se hanencontrado diferencias significativas en lo que respecta al índice de abun-dancia, medido como el número medio de grupos fecales detectados por par-cela y mes, para el caso del ciervo (Kruskal-Wallis: Chi2=26.72, 3 gl, p<0.001),sin embargo las diferencias no fueron significativas para el caso de la cabramontés, mostrándose que esta especie presenta una abundancia similar entodos ellos (Kruskal-Wallis: Chi2=3.81, 3 gl, p=0.283).
El uso que cada una de las especies hace de las parcelas ha sido modelado(modelos lineales generalizados con distribución de Poisson y función de vín-
P. ACEVEDO, J. P. SANTOS, R. REAL & J. VICENTE
36 (Pirineos, 2011, Vol. 166, 29-49, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166002)

culo logarítmica) con variables relacionadas con la orografía y los usos delsuelo (Tabla 1). El modelo indica que la pendiente del terreno es determinan-te del uso que los ciervos hacen del territorio, registrándose un mayor uso de
EVALUACIÓN DEL ESTADO DE LA POBLACIÓN DE CABRA MONTÉS DE LOS MONTES DE TOLEDO:...
37(Pirineos, 2011, Vol. 166, 29-49, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166002)
Tabla 1. Resultados de los modelos lineales generalizados realizados para analizar el efecto devariables ambientales sobre el uso, a nivel de parcela, que cada especie hace del territorio.
Table 1. Variables retained in the generalized lineal models developed to explain the red deer and Iberianibex habitat use at plot level.
Variable Wald gl p(Intersección) 240.328 1 <0.001Recorrido 57.373 3 <0.001Orientación 56.427 6 <0.001Vegetación 45.287 4 <0.001Pendiente 13.485 1 0.001
Ciervo(Intersección) 0.430 1 0.512Recorrido 1.302 2 0.522Orientación 4.899 5 0.428Vegetación 17.424 4 0.002Pendiente 0.604 1 0.437
Cabra
las parcelas con menor pendiente. Además, los resultados indican que losciervos son más frecuentes en aquellas parcelas situadas en orientación suro-este y menos en las zonas de bosque claro (ver Figura 3). Además se handetectado diferencias significativas entre recorridos. Por su parte, el modelorealizado para la cabra montés muestra que no existen diferencias entre fin-cas, ni entre tipos de orientación, ni en relación a la pendiente. Sólo se obser-varon diferencias significativas en el uso en relación al tipo de vegetación,siendo las zonas de vegetación rala y matorral arbolado las más frecuentadaspor las monteses, y los bosques claros los menos usados (ver Figura 3).
En base al recuento de excremento se ha podido estimar una densidadmedia de ciervo para todo el área de estudio de 14.62±2.27 ind/km2, y de1.39±0.17 ind/km2 para la cabra montés. Por tanto, considerando que el áreade estudio son aproximadamente 4250 ha, el tamaño de la población de cabramontés objeto de estudio se podría situar entre los 52 y 66 individuos.
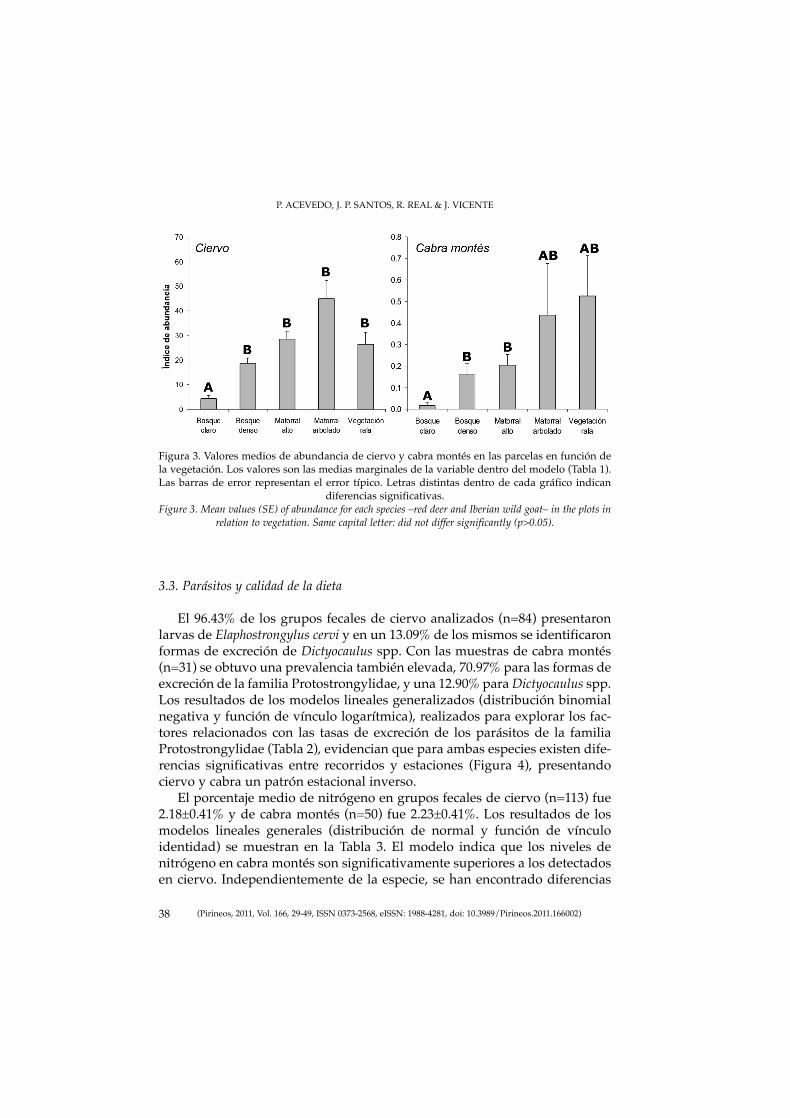
3.3. Parásitos y calidad de la dieta
El 96.43% de los grupos fecales de ciervo analizados (n=84) presentaronlarvas de Elaphostrongylus cervi y en un 13.09% de los mismos se identificaronformas de excreción de Dictyocaulus spp. Con las muestras de cabra montés(n=31) se obtuvo una prevalencia también elevada, 70.97% para las formas deexcreción de la familia Protostrongylidae, y una 12.90% para Dictyocaulus spp.Los resultados de los modelos lineales generalizados (distribución binomialnegativa y función de vínculo logarítmica), realizados para explorar los fac-tores relacionados con las tasas de excreción de los parásitos de la familiaProtostrongylidae (Tabla 2), evidencian que para ambas especies existen dife-rencias significativas entre recorridos y estaciones (Figura 4), presentandociervo y cabra un patrón estacional inverso.
El porcentaje medio de nitrógeno en grupos fecales de ciervo (n=113) fue2.18±0.41% y de cabra montés (n=50) fue 2.23±0.41%. Los resultados de losmodelos lineales generales (distribución de normal y función de vínculoidentidad) se muestran en la Tabla 3. El modelo indica que los niveles denitrógeno en cabra montés son significativamente superiores a los detectadosen ciervo. Independientemente de la especie, se han encontrado diferencias
P. ACEVEDO, J. P. SANTOS, R. REAL & J. VICENTE
38 (Pirineos, 2011, Vol. 166, 29-49, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166002)
Figura 3. Valores medios de abundancia de ciervo y cabra montés en las parcelas en función dela vegetación. Los valores son las medias marginales de la variable dentro del modelo (Tabla 1).Las barras de error representan el error típico. Letras distintas dentro de cada gráfico indican
diferencias significativas.Figure 3. Mean values (SE) of abundance for each species –red deer and Iberian wild goat– in the plots in
relation to vegetation. Same capital letter: did not differ significantly (p>0.05).

EVALUACIÓN DEL ESTADO DE LA POBLACIÓN DE CABRA MONTÉS DE LOS MONTES DE TOLEDO:...
39(Pirineos, 2011, Vol. 166, 29-49, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166002)
Tabla 2. Resultados de los modelos lineales generalizados realizados para analizar el efecto definca y estación sobre la abundancia de protostongílidos.
Table 2. Variables retained in the generalized lineal models developed to explain the excretionrates of Elaphostrongylus cervi and Protostrongylidae.
Variable Wald gl p(Intersección) 766.286 1 <0.001Recorrido 7.964 3 0.047Estación 32.359 2 <0.001
Ciervo – E. cervi(Intersección) 4.687 1 0.030Recorrido 11.000 3 0.012Estación 8.258 2 0.016
Cabra – Protostrongylidae
Figura 4. Diferencias entre estaciones y especies en las tasas de excreción de parásitos protostron-gílidos. Los valores son las medias marginales de las variables dentro del modelo (Tabla 2). Lasbarras de error representan el error típico. Letras distintas, mayúsculas (ciervo) y minúsculas
(cabra), indican diferencias significativas.Figure 4. Seasonal patterns of Protostrongylidae’s excretion rates in both red deer and Iberian wild goat.
Same capital letter or lower case letter: did not differ significantly (p>0.05).
significativas entre estaciones y entre recorridos con y sin manejo cinegético.Así, los mayores niveles se detectaron en primavera y en aquellos recorridosen los que el manejo cinegético es inexistente (Figura 5).
P. ACEVEDO, J. P. SANTOS, R. REAL & J. VICENTE
40 (Pirineos, 2011, Vol. 166, 29-49, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166002)
Figura 5. Diferencias entre especies y recorridos (en relación al manejo cinegético) en los nivelesde nitrógeno fecal. Los valores son las medias marginales de la interacción entre estas variables
dentro del modelo (Tabla 3). Las barras de error representan el error típico.Figure 5. Mean values (SE) of faecal nitrogen levels in relation to game management and monitored species
–red deer and Iberian wild goat–.
Tabla 3. Resultados de los modelos lineales generalizados realizados para analizar el efecto deespecie, fincas (manejadas o no manejadas) y estación sobre los niveles de nitrógeno fecal.
Table 3. Variables retained in the generalized lineal models developed to explain the individual levels offaecal nitrogen.
Variable F gl p(Intersección) 4879.621 1 <0.001Especie 2.815 1 0.045Estación 34.022 3 <0.001Recorrido 4.256 1 0.041Especie*Estación 1.399 3 0.245Especie*Recorrido 3.878 1 0.045

4. Discusión
4.1. Sobre la población en general
Los estudios realizados han permitido realizar una primera caracteriza-ción, en función de ciertos indicadores biológicos, del estado de la poblaciónde cabra montés presente en el Parque Nacional de Cabañeros. Mediante lasencuestas de distribución se ha podido corroborar que dicha población pare-ce tener su origen en introducciones realizadas con animales procedentes deGredos (Pérez et al., 2002). En general, los encuestados perciben que estapoblación en la actualidad se encuentra en aumento, si bien es cierto que noes un crecimiento especialmente acusado a tenor de las densidades observa-das. Esta tendencia poblacional es congruente con la descrita para otraspoblaciones de cabra montés de CLM (Acevedo et al., 2007a) y otras pobla-ciones peninsulares de esta subespecie (Escós et al., 2008).
En el caso de la población de Cabañeros, su crecimiento podría estar ralen-tizado por la presencia de elevadas densidades de ciervo, pero son muchaslas causas que pueden estar restringiendo la expansión numérica de estenúcleo poblacional y permanecen desconocidas. Escós et al. (2008) analizandola evolución temporal de varias poblaciones de cabra montés, mostraron quetodas las poblaciones de C. p. hispanica están en notoria expansión numérica,pero la población de C. p. victoriae monitorizada, la de Gredos, se mantieneestable en los últimos años. Esto indica que podrían existir razones ecológicasdiferentes a la competencia que explicaran que las poblaciones de esta subes-pecie no tuvieran la misma potencialidad que la C. p. hispanica para expan-dirse. Los estudios parasitarios y de calidad de dieta realizados aquí, aunqueno son conclusivos, aportan un poco de luz sobre los efectos de esta relacióninterespecífica (ver abajo).
4.2. Uso del hábitat y densidad poblacional
Los modelos realizados para determinar el uso que los ciervos y las cabrashacen del territorio han mostrado diferencias claras entre ambas especies. Así,el ciervo parece que hace un uso acorde con sus requerimientos naturalesseleccionando zonas calientes (orientación SW), de matorral arbolado y conpoca pendiente (ver Acevedo & Cassinello, 2009a), mientras que el uso quehace la cabra montés indica que esta especie es más frecuente en zonas devegetación escasa (Acevedo et al., 2007a). Aunque con este tipo de estudios esdifícil inferir competencia entre especies, la segregación observada podría
EVALUACIÓN DEL ESTADO DE LA POBLACIÓN DE CABRA MONTÉS DE LOS MONTES DE TOLEDO:...
41(Pirineos, 2011, Vol. 166, 29-49, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166002)

estar indicando que i) ambas especies presentan diferentes requerimientosambientales –poco probable dadas las similitudes ecológicas previamentecomentadas–, pero también que ii) las cabras se han quedado relegadas aaquellos ambientes poco frecuentados por los ciervos. Los datos disponiblesno permiten inferir cual de estas causas puede ser la que esté mediando el usoque las monteses hacen del territorio, siendo necesarios para ello estudiosbasados en el radioseguimiento de los animales de ambas especies junto conestudios de dieta realizados para ambas especies en este territorio.
La densidad media de ciervo en el área de estudio puede considerarsemedia, o incluso media-alta, en relación a otras fincas de caza de CLM(Acevedo et al., 2008). Como anteriormente se ha comentado, elevadas densi-dades de ciervo podrían determinar la tendencia de las poblaciones de cabramontés. Esta misma interpretación podría también extraerse del artículo dePérez et al. (2002) en los que muestran que las poblaciones de cabra montés engeneral están en aumento, con la salvedad de la población de Sierra Morenaque es precisamente la que convive con densidades elevadas –en ocasionespor encima de la capacidad de carga del medio– de ciervo. La densidad decabra montés en el área de estudio se puede considerar baja en relación a lasestimadas para otras poblaciones manchegas de las cuencas de los ríos Seguray Mundo en la provincia de Albacete. Trabajos realizados en 2006 en dichaspoblaciones han mostrado densidades que oscilaban entre 6 y 15 ind/km2
(Acevedo et al., 2007c), siendo estos valores notoriamente más elevados quelos de la población de Cabañeros.
4.3. Parásitos y calidad de la dieta
Los resultados indican que la prevalencia media de E. cervi detectada enlas poblaciones de estudio es elevada (media 96.43%) y congruente con laobtenida en estudios previos realizados sobre estas mismas poblaciones (verVicente et al., 2006), y también sobre otras poblaciones de similares caracte-rísticas en cuanto a manejo y área geográfica (Tayce et al., 2008). En el marcode estos estudios se puede decir que las poblaciones estudiadas se sitúanentre el 25% de las poblaciones con prevalencias más elevadas de una selec-ción de 18 de este mismo entorno (Vicente et al., 2006). La clara vinculaciónexistente entre la prevalencia de este nemátodo y la condición corporal de losciervos (Vicente et al., 2007) indica que estas poblaciones presentan una buenacondición corporal individual, lo que podría estar relacionado con dos facto-res principales: i) las poblaciones presentan una densidad adecuada a lapotencialidad del medio, y ii) en el caso de presentar una densidad más ele-vada, las poblaciones son manejadas con el fin de mitigar los efectos negati-
P. ACEVEDO, J. P. SANTOS, R. REAL & J. VICENTE
42 (Pirineos, 2011, Vol. 166, 29-49, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166002)

vos que la sobreabundancia poblacional tiene sobre la condición corporal delos individuos.
En lo que respecta al ciclo de excreción anual (Figura 4), los resultados deeste estudio destacan la presencia de un pico de excreción en el invierno(Vicente et al., 2005). Los picos estacionales pueden ser interpretados comouna respuesta del parásito, más o menos rápida, a las condiciones meteoroló-gicas manteniéndose el máximo anual durante el periodo de lluvias. Losresultados aquí mostrados son coherentes con investigaciones anteriores yponen de manifiesto que el periodo crítico para estos animales en estas lati-tudes no se centra en el invierno tal y como ocurre en otras poblaciones delnorte de Europa, por ejemplo Polonia (Demiaszkiewicz, 1987), aquí es el vera-no (principios de otoño) el periodo de menor disponibilidad de alimento.
Por otro lado, la prevalencia de Protostrongylidae obtenida en las mues-tras de cabra montés fue también elevada (71%) y muy similar a las obtenidasen el muestreo realizado en 2003 (70%; Acevedo et al., 2005). Al igual queanteriormente se ha comentado para el caso de los ciervos, las tasas de excre-ción detectadas en las muestras de cabra son elevadas si se contextualizan enel marco de otras poblaciones de CLM. Podemos decir que la población deestudio está entre el 30% de las poblaciones con prevalencias más elevadas deuna selección de 14 fincas de este mismo entorno (Acevedo et al., 2005). Otrosautores han estudiado las tasas de infestación y excreción de protostongílidosen poblaciones de cabra montés de Andalucía y han obtenido prevalenciasincluso más elevadas que las presentadas aquí (Alasaad et al., 2009).
En la Figura 4 se muestra la estacionalidad de la excreción de larvas encabra montés. En ella se muestra una situación claramente distinta, casiopuesta, a la comentada anteriormente para el ciervo. En este caso, el pico deexcreción se detecta en otoño y no en invierno. No existen estudios previossobre la estacionalidad de estos nematodos en cabra montés con lo que no sepueden establecer comparaciones y resulta, por tanto, más complicado reali-zar una interpretación certera del patrón observado. El patrón podría deber-se a que es a finales de otoño cuando ocurre el celo en esta especie por lo queeste es el periodo de máxima agregación de la poblaciones, máxime en el áreade estudio donde dado el reducido tamaño poblacional es posible que todaslas cabras del núcleo se agrupen en 2-3 rebaños en esta época. Sería necesariorealizar un estudio continuado en el tiempo –idealmente usando collaresGPS– para corroborar estos resultados ya que, al carecer de réplica, por elmomento deben ser tomados con cierta cautela.
La cantidad de nitrógeno fecal nos informa sobre la calidad proteica de ladieta de los individuos. Las poblaciones de ciervo estudiadas presentan valo-res elevados, con un 2.19% de nitrógeno de media, se sitúan entre el 25% delas fincas con mayores niveles de nitrógeno (Toledano, 2008). Las de cabra,
EVALUACIÓN DEL ESTADO DE LA POBLACIÓN DE CABRA MONTÉS DE LOS MONTES DE TOLEDO:...
43(Pirineos, 2011, Vol. 166, 29-49, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166002)

con un 2.32% de media, presentan niveles ligeramente superiores a todas laspoblaciones de ciervo de dicho estudio en el que el máximo valor obtenidofue 2.30%. Por tanto, los resultados parecen estar indicando que las poblacio-nes estudiadas muestran indicios de disponer de una dieta de buena calidadproteica. Este resultado es congruente con la discusión previamente plantea-da sobre las tasas de excreción de Protostrongylidae como indicadores de unabuena condición corporal de los animales. Al igual que antes, los altos nive-les detectados aquí indicarían que la densidad poblacional se ajusta a la capa-cidad de carga del medio y que, en el caso de sobrepasarla, la alimentaciónsuplementaria estaría mitigando el efecto también a este nivel de estudio.
Tal y como previamente ya ha sido sugerido, la cantidad de nitrógenoexcretado en heces de cabra fue significativamente más elevada que el excre-tado en heces de ciervo, siendo la diferencia entre especies más acentuada enlos recorridos sin manejo cinegético (ver Figura 5). Aunque las diferenciasentre especies pudieran ser esperables dada la determinación de estrategiasalimentarias diferentes en función de su tamaño corporal (Gordon, 1989),ambas especies presentan un elevado solapamiento de dieta (Martínez et al.,1992) por lo que podrían ser igualmente esperables niveles de nitrógeno simi-lares entre ambas especies. En este caso concreto parece más razonable pen-sar que las diferencias observadas entre especies pudieran estar más relacio-nadas con el uso del hábitat que con diferencias fisiológicas. Así, los mayoresniveles de nitrógeno se han detectado en los recorridos que discurren porterrenos sin manejo cinegético, siendo esto especialmente notorio en el casode la cabra montés. Este resultado contrasta con estudios previos en los quese pusieron de manifiesto mayores niveles de nitrógeno en fincas con suple-mentación (Toledano, 2008). Las diferencias entre los resultados aquí presen-tados y los estudios previos pueden ser debidas a: i) considerar además delciervo a la cabra montés, ii) el alimento aportado a los animales no es de sufi-ciente calidad proteica, y iii) el alimento no es suficiente para mitigar los efec-tos de la mayor densidad sobre la calidad de dieta. A la vista de la Figura 5parece que las diferencias entre manejos se deben principalmente a la inclu-sión de la cabra, ya que los niveles de nitrógeno en las muestras de ciervo sonbastante similares en ambos sistemas de manejo y elevadas en relación a otrasfincas de similares características, por lo que parece que el efecto negativo dela mayor densidad de ciervos sobre la calidad de la dieta cuando los recursosson limitantes (ver Masahiko & Keiji, 1999) queda mitigado con la suplemen-tación.
En este mismo sentido, los resultados aquí presentados parecen indicarque la alimentación suplementaria no tiene efectos directos sobre la calidadde la dieta de las cabras monteses. De no ser así, y dada la ausencia de dife-rencias en densidad entre fincas, las cabras excretarían mayores niveles de
P. ACEVEDO, J. P. SANTOS, R. REAL & J. VICENTE
44 (Pirineos, 2011, Vol. 166, 29-49, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166002)

nitrógeno en las fincas con alimentación suplementaria. Posiblemente la ali-mentación no esté disponible para las cabras debido a una elevada presiónpor parte de los ciervos, y el hecho de que los valores sean significativamen-te más elevados en las fincas sin manejo pudiera estar relacionado con ciertacompetencia entre especies dado el solapamiento de dieta existente. Portanto, en los recorridos sin manejo, donde las densidades de ciervos son másreducidas, las cabras podrían ser más selectivas a la hora de seleccionar losrecursos y por ello ingerir una dieta de mayor calidad proteica que los cier-vos. Otras interpretaciones a estos resultados podrían buscarse en relación ala fisiología ruminal de ambas especies.
Respecto al ciclo estacional de los niveles de excreción de nitrógeno, losvalores obtenidos en este estudio muestran un patrón lógico y coincidentecon la productividad de estos ambientes. La primavera es el periodo dondemayores valores de nitrógeno se obtienen y el otoño y el invierno las que pre-sentan los menores valores de productividad, y por tanto, de calidad de dieta.Los resultados obtenidos indican que para monitorizar los niveles de nitró-geno de una población en años sucesivos o de varias poblaciones, todas lasmuestras deben ser tomadas en un corto periodo de tiempo para evitar lavariación estacional. La primavera al aportar el máximo valor del año semuestra el periodo más adecuado para ese propósito. Quizás muestrear enotoño-invierno pueda resultar también recomendable ya que indicarán lacalidad de dieta mínima del año y será ésta la que pueda indicar la necesidadde establecer estrategias de gestión encaminadas a potenciarla.
5. Implicaciones para la gestión
Los estudios realizados sugieren que, para los indicadores biológicosempleados, las poblaciones de ciervo y cabra montés objeto de estudio mues-tran un adecuado estado poblacional. En este sentido sería esperable que lapoblación de cabra montés continuara en ligero aumento en los próximosaños a tenor de los indicadores obtenidos y de la percepción social sobre latendencia de esta población.
El uso que las cabras monteses hacen del territorio podría estar determi-nado, en cierta medida, por la abundancia de ciervo y ésta, indirectamente,podría también estar determinando la calidad de la dieta de las montesesaunque por el momento no de forma alarmante. Por tanto, no sería esperableque el ciervo supusiera un riesgo relevante para la viabilidad de esta pobla-ción de cabra montés, al menos si la gestión de las poblaciones de ciervos semantiene como hasta el momento y las poblaciones no se disparan demográ-ficamente.
EVALUACIÓN DEL ESTADO DE LA POBLACIÓN DE CABRA MONTÉS DE LOS MONTES DE TOLEDO:...
45(Pirineos, 2011, Vol. 166, 29-49, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166002)

Finalmente, dado que las estrategias de gestión aplicadas en las fincas deestudio, centradas principalmente en potenciar las poblaciones de ciervo,parecen no tener efectos sobre la cabra montés, por lo que si fuese necesarioaplicar alguna medida a la población de cabra, ésta tendría que hacerse demanera específica para esta especie, ya que de no ser así difícilmente estaríaaccesible para las cabras dada la elevada presión de los ciervos en estos terri-torios.
Agradecimientos
Este trabajo ha sido promovido y financiado por el Parque Nacional deCabañeros (Organismo Autónomo de Parques Nacionales) en el marco delconvenio de colaboración 8.06/5.46.3324. La realización de los trabajos decampo no hubiera sido posible sin la colaboración de Ángel Gómez y guíasdel Parque Nacional, Adolfo y Jesús de Parrillas y Francisco y Jorge deMuelas, muchas gracias por el apoyo logístico a pie de campo. Igualmentedebemos agradecer la colaboración de “voluntarios” del IREC en los trabajosde campo, Luca, Ricardo, Alba, Virginia y Mónica… y también a Javi Urra dela UMA. Ya en el laboratorio el mérito es de Paqui Talavera en el preprocesa-do de las muestras para los análisis parasitológicos. P. Acevedo disfruta de uncontrato dentro del programa Juan de la Cierva – Fondo Social Europeo.
Referencias
Acevedo, P. & Cassinello, J., 2009a. Human-induced range expansion of wildungulates causes niche overlap between previously allopatric species: reddeer and Iberian ibex in mountainous regions of southern Spain. AnnalesZoologici Fennici, 46: 39-50.
Acevedo, P. & Cassinello, J., 2009b. Biology, ecology and status of Iberian ibexCapra pyrenaica: a critical review and research prospectus. Mammal Review,39: 17-32.
Acevedo, P., Cassinello, J. & Gortázar, C., 2007a. The Iberian ibex is under anexpansion trend but displaced to suboptimal habitats by the presence ofextensive goat livestock in central Spain. Biodiversity and Conservation, 16:3361-3376.
Acevedo, P., Cassinello, J., Hortal, J. & Gortázar, C., 2007b. Invasive exoticaoudad (Ammotragus lervia) as a major threat to native Iberian ibex (Caprapyrenaica): a habitat suitability model approach. Diversity and Distributions,13: 587-597.
P. ACEVEDO, J. P. SANTOS, R. REAL & J. VICENTE
46 (Pirineos, 2011, Vol. 166, 29-49, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166002)

Acevedo, P., Ferreres, J., Jaroso, R., Durán, M., Escudero, M.A., Marco, J. &Gortázar, C., 2010. Estimating roe deer abundance from pellet groupcounts in Spain: An assessment of methods suitable for Mediterraneanwoodlands. Ecological Indicators, 10: 1226-1230.
Acevedo, P. & Real, R. en prensa. Biogeographycal differences between thetwo Capra pyrenaica subspecies, C. p. victoriae and C. p. hispanica, inhabitingin the Iberian Peninsula: conservation implications. Ecological Modelling.
Acevedo, P., Ruiz-Fons, F., Reyes-García, A.R. & Gortázar, C., 2007c. Estudiocomparativo de métodos para la estima de la abundancia de ungulados de monta-ña: la cabra montés. VIII Jornadas de la Sociedad Española para laConservación y Estudio de los Mamíferos. Huelva.
Acevedo, P., Ruiz-Fons, F., Vicente, J., Reyes-García, A.R., Alzaga, V. &Gortázar, C., 2008. Estimating red deer abundance in a wide range ofmanagement situations in Mediterranean habitats. Journal of Zoology, 276:37-47.
Acevedo, P., Vicente, J., Alzaga, V. & Gortázar, C., 2005. Relationship betweenbronchopulmonary nematode larvae and relative abundances of Spanishibex (Capra pyrenaica hispanica) from Castilla-La Mancha, Spain. Journal ofHelminthology, 79: 113-118.
Alasaad, S., Morrondo, P., Dacal-Rivas, V., Soriguer, R.C., Granados, J.E.,Serrano, E., Zhu, X.Q., Rossi, L. & Pérez, J.M., 2009. Brochopulmonarynematode infection of Capra pyrenaica in the Sierra Nevada massif, Spain.Veterinary Parasitology, 164: 340-343.
Anderson, R.C., 2000. Nematode parasites of vertebrates: their development andtransmission. 672 pp. CABI Publishing Wallingford, Oxon.
Cordero del Campillo, M. & Rojo, F.A., 1999. Parasitología veterinaria. Spain,McGraw Hill, Interamericana.
Demiaszkiewicz, A., 1987. Epizootiolgy of Elaphostrongylus cervi infection ofred deer in the Bialowieza forest (in polish). Med Weter, 43: 208-211.
Edge, W.D. & Marcum, C.L., 1989. Determining elk distribution with pellet-group and telemetry techniques. Journal of Wildlife Management, 53: 621-624.
Escós, J.M., Alados, C.L., Pulido, A., Romera, J., González-Sánchez, N. &Martínez, F., 2008. Estimating population trends using population viabili-ty analyses for the conservation of Capra pyrenaica. Acta Theriologica, 53:275-286.
Forrester, S.G. & Lankester, M.W., 1997. Extracting Protostrongylid nematodefrom ungulate feces. Journal of Wildlife Diseases, 33: 511–516.
García-González, R. & Herrero, J., 1999. El bucardo de los Pirineos: historia deuna extinción. Galemys, 11: 17–26.
EVALUACIÓN DEL ESTADO DE LA POBLACIÓN DE CABRA MONTÉS DE LOS MONTES DE TOLEDO:...
47(Pirineos, 2011, Vol. 166, 29-49, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166002)

Gordon, I.J., 1989. Vegetation community selection by ungulates on the IsleRhum. II. Vegetation community selection. Journal of Applied Ecology, 26:53-64.
Gortázar, C., Acevedo, P., Ruiz-Fons, F. & Vicente, J., 2006. Disease risks andoverabundance of game species. European Journal of Wildlife Research, 52:81-87.
Gortázar, C., Herrero, J., Villafuerte, R. & Marco, J., 2000. Historical examina-tion of the distribution of large mammals in Aragón, Northeastern Spain.Mammalia 61: 411-422.
Hamel, S., Garel, M., Festa-Bianchet, M., Gaillard, J.M. & Côté, S.D., 2009.Spring Normalized Difference Vegetation Index (NDVI) predicts annualvariation in timing of peak faecal crude protein in mountain ungulates.Journal of Applied Ecology, 46: 582-589.
Hemami, M.R., Watkinson, A.R. & Dolman, P.M., 2005. Population densitiesand habitat associations of introduced muntjac Muntiacus reevesi andnative roe deer Capreolus capreolus in a lowland pine forest. Forest Ecologyand Management 215: 224-238.
Martínez, T., Soriguer, R.C. & Fandos, P., 1992. Trophics relationships betweenwild goat (Capra pyrenaica) and the red deer (Cervus elaphus) in the south-east Spain (S. Cazorla). En: Proceedings of the International Congress of thegenus Capra in Europe: 225-228. Ronda, España.
Masahico, A. & Keiji, O., 1999. Nitrogen content in feces and diet of sika deeron the Boso Peninsula, central Japan. Ecological Research, 14: 249-253.
Massey, B.N., Weckerly, F.W., Vaughn, C.E. & Mccullough, D.R., 1994.Correlations between fecal nitrogen and diet composition in free-rangingblack-tailed deer. The Southwestern Naturalist, 39: 165-170.
Mattson, W.J.J., 1980. Herbivory in relation to plant nitrogen content. AnnualReview on Ecology and Systematics, 11: 119-161.
Melhorn, H., Düwel, D. & Raether, W., 1992. Atlas de parasitologia veterinaria.Barcelona, Grass.
Moen, R., Pastor, J. & Cohen, Y., 1997. Accuracy of GPS telemetry collar loca-tions with differential correction. Journal of Wildlife Management, 61: 530-539.
Pérez, J.M., Granados, J.E., Soriguer, R.C., Fandos, P., Márquez, F.J. & Crampe,J.P., 2002. Distribution, status and conservation problems of the SpanishIbex, Capra pyrenaica (Mammalia: Artiodactyla). Mammal Review, 32 : 26-39.
Putman, R.J., 1984. Facts from faeces. Mammal Review, 14: 79-97.Shackleton, D.M., 1997. Wild Sheep and Goats and their Relatives: Status Survey
and Conservation Action Plan for Caprinae. IUCN, Gland, Switzerland.Smart, J.C.R., Ward, A.I. & White, P.C.L., 2004. Monitoring woodland deer
populations in the UK: an imprecise science. Mammal Review, 34: 99-114.
P. ACEVEDO, J. P. SANTOS, R. REAL & J. VICENTE
48 (Pirineos, 2011, Vol. 166, 29-49, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166002)

Tayce, J., Acevedo, P., Vicente, J. & Gortázar, C., 2008. Minimum samplingeffort for reliable non-invasive estimations of excretion abundance ofElaphostrongylus cervi L1 in red deer (Cervus elaphus) populations. Journal ofHelminthology, 82: 255–261.
Toledano, P., 2008. Relaciones entre la cantidad de nitrógeno fecal y característicasbiométricas, de condición corporal, demográficas y de hábitat en poblaciones deciervo (Cervus elaphus) del centro sur de España. Trabajo fin de Carrera,EUITA, Universidad de Castilla-La Mancha.
Vicente, J., Fernández-de-Mera, I.G. & Gortázar, C., 2006. Epidemiology andrisk factors analysis of elaphostrongylosis in red deer (Cervus elaphus) fromSpain. Parasitology Research, 98: 77-85.
VIcente, J., Fierro, Y. & Gortázar, C., 2005. Seasonal dynamics of the faecalexcretion of Elaphostrongylus cervi (Nematoda, Metastrongyloidea) firststage larvae in Iberian red deer from South Spain. Parasitology Research, 95:60-64.
Vicente, J., Höfle, U., Fernández-de-Mera, I.G. & Gortázar, C., 2007. Theimportance of parasite life history and host density in predicting theimpact of infections in red deer. Oecologia, 152: 655-664.
EVALUACIÓN DEL ESTADO DE LA POBLACIÓN DE CABRA MONTÉS DE LOS MONTES DE TOLEDO:...
49(Pirineos, 2011, Vol. 166, 29-49, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166002)

51
ASSESSMENT OF WILD BOAR ROOTINGON ECOLOGICAL AND PASTORAL VALUES
OF ALPINE PYRENEAN GRASSLANDS
C. G. BUENO1*, I. C. BARRIO1,2, R. GARCÍA-GONZÁLEZ1,C. L. ALADOS3 & D. GÓMEZ-GARCÍA1
1 Pyrenean Institute of Ecology, Spanish National Research Council (CSIC),Avda. Regmto. Galicia, s/n P. O. Box 64, Jaca 22700, Huesca, Spain
2 Departament of Zoology, University of Córdoba, Campus de Rabanales, 14071, Córdoba, Spain3 Pyrenean Institute of Ecology, Spanish National Research Council (CSIC),
Avda. Montañana 1005, P. O. Box 13034, 50192 Zaragoza, Spain*Corresponding author: E-mail [email protected]
ABSTRACT.– Wild boar rooting is nowadays one of the main disturbances inPyrenean alpine grasslands. Its consequences for the ecosystem are not perfectlyunderstood yet despite alpine grasslands in the Pyrenees have an important eco-nomic role and a priority conservation interest. The ecosystem services of this habi-tat lay mainly on pastoral and ecological values that wild boar rooting seems toaffect. In this study, we measured those ecological and pastoral values at differentscales to improve our understanding of the reach of these disturbances in this sensi-tive ecosystem. At landscape and community scales we compare disturbed andundisturbed areas in pastoral, ecological and community maps of the study area bymeans of a geographic information system. At a local scale we compare ecologicaland pastoral values of different plant groups (based on species abundance), withinand outside wild boar rootings. A preference for areas of high pastoral and interme-diate ecological values was found for wild boar rooting at the landscape level.However at the community level, disturbances notably reduced pastoral and ecolog-ical values in all communities. At the local level, the ecological value of bulbs and thepastoral value of annual dicots increased within disturbances, suggesting that dis-turbances may favour functional group diversity. In sum, wild boar rooting affectsPyrenean alpine grasslands moderately, with higher affection to pastoral than eco-logical values at all levels, what should be considered for the management andpreservation of these habitats since these disturbances are likely to increase.
Keywords: Sus scrofa; disturbance assessment; multi-scale evaluation; Ordesaand Monte Perdido National Park.
Pirineos.Revista de Ecología de MontañaVol. 166, 51-67
Jaca, Enero-Diciembre, 2011ISSN: 0373-2568
eISSN: 1988-4281doi: 10.3989/Pirineos.2011.166003

52
C. G. BUENO, I. C. BARRIO, R. GARCÍA-GONZÁLEZ, C. L. ALADOS & D. GÓMEZ-GARCÍA
RESUMEN.– Las hozaduras de jabalí son una de las mayores perturbacionesactuales de los pastos supraforestales pirenaicos. Sus consecuencias para el ecosiste-ma no están todavía perfectamente descritas, a pesar de ser uno de los hábitats demayor interés de conservación y que juegan un importante papel en las economíaslocales. Los bienes y servicios que provee dicho hábitat están claramente relacionadostanto con su valor ecológico como pastoral, los cuales parecen estar afectados por lascitadas perturbaciones. En este trabajo se midieron dichos valores dentro y fuera deperturbaciones a diferentes escalas espaciales para incrementar el conocimiento de larepercusión de las hozaduras de jabalí en el ecosistema de los pastos alpinos, parti-cularmente sensibles a la remoción del suelo. A escala de paisaje y de comunidad,comparamos las áreas perturbadas y no perturbadas por el jabalí, en mapas del valorpastoral, valor ecológico y de comunidades pascícolas de la zona de estudio, pormedio de un sistema de información geográfica. A escala local comparamos los valo-res ecológico y pastoral de los grupos de plantas (sobre la base de la abundancia deespecies), dentro y fuera de las hozaduras de jabalí. A escala de paisaje se encontróuna preferencia por zonas de alto valor pastoral y valores ecológicos intermedios. Sinembargo, a escala de comunidad las perturbaciones redujeron notablemente el valorpastoral y ecológico en todas las comunidades. A escala local, se encontró un aumen-to del valor ecológico de los bulbos y del valor pastoral de las dicotiledóneas anualesdentro de las perturbaciones, lo que sugiere que éstas pueden favorecer la diversidadde grupos funcionales. Las hozaduras de jabalí afectan moderadamente a los pastosalpinos pirenaicos, con mayor afección sobre los valores pastorales que sobre los eco-lógicos en todas las escalas estudiadas, lo que debiera tenerse en cuenta para el mane-jo y preservación de estos hábitats, ya que dichas perturbaciones se están, muy pro-bablemente, incrementando.
Palabras clave: Hozaduras de jabalí; evaluación de las perturbaciones; valorecológico; valor pastoral; evaluación multi-escala; Parque Nacional de Ordesay Monte Perdido.
1. Introduction
A crucial step to value an ecosystem is to accurately report how it maycontribute to human wellbeing, through measuring what have been called‘ecosystem services’. This term includes both the naturally-occurring ecolog-ical processes and the benefits to humans that can be derived from them(Balmford et al., 2008). However, we still lack the theoretical basis that linksecological diversity to ecosystem services underlying human wellbeing(Carpenter et al., 2006). The need to develop indicators that synthesize andsimplify the ecological complexity has been recently claimed, to aid monitor-ing biological, physical and social changes related to the ecosystem services(Carpenter et al., 2006).
(Pirineos, 2011, Vol. 166, 51-67, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166003)

The main ecosystem services provided by mountainous areas are relatedto the provision of high quality water and food, control of climate and ero-sion, biodiversity and carbon storage, as well as recreation, aesthetic and spir-itual values (Millennium Ecosystem Assessment Board, 2005). The greatesthistorical environmental change in these areas has been the spread of grass-lands through scrubland and forest reduction from the Middle Ages on (Fillatet al., 2008). This progressive change due to livestock activities has developeda seminatural system with high ecological and pastoral values in a dynamictrade-off based on plant-herbivore interactions (Olff & Ritchie, 1998;Austrheim & Eriksson, 2001; Fillat et al., 2008). More recently, livestock graz-ers have changed in European mountain systems from almost exclusivesheep herds to comparatively higher numbers of cattle heads (Macdonald etal., 2000; Lasanta-Martínez et al., 2005). Alongside with this trend, a sharpdecline of extensive husbandry has occurred in the last decades in favour ofmore productive intensive farming systems (Luick, 1998; Körner, 1999). Inparticular, this last trend has been related with drastic changes in the ecosys-tem structure, such as shrub encroachment and forest recolonization(Lasanta-Martínez et al., 2005; Lasanta-Martínez et al., 2006), what in turn mayincrease fire events (Carcaillet et al., 2009), reduce biodiversity (Helm et al.,2006), and boost some forest animal populations beyond an equilibriumthreshold (Laiolo et al., 2004).
Wild boar is a native forest species whose populations have been increas-ing from the sixties in their European distribution range (Sáez-Royuela &Tellería, 1986) being nowadays considered a pest in several areas within andoutside its native distribution range (Oliver & Leus, 2008). The main effect ofits presence is an extensive disturbance that consists in turning over the soilwhile searching for underground feeding resources. These disturbances(hereafter ‘rooting’) affect directly some ecosystem elements, such as soils(Lacki & Lancia, 1983; Bueno et al., unpublished-b) and vegetation (Bratton,1975; Bueno et al., unpublished-a), and may indirectly alter some ecosystemprocesses such as nutrient cycling and species turnover (Kotanen, 1995;Bueno et al., unpublished-b). Rooting can be worrisome in some places, as itmay affect habitats particularly sensitive to soil disturbances such asPyrenean alpine grasslands (García-González, 2008) with indirect conse-quences to traditional grazing activities that preserve the system and holdlocal economies (Bueno et al., 2010). Therefore, evaluating the direct and mainindirect implications of these disturbances is basic to guide adequate man-agement actions and understand future consequences (Carpenter, 2002).Despite its relevance for conservation and management, little attention hasbeen given to the evaluation of alpine grasslands ecologically and in terms offorage quality (Gartzia et al., 2005; García-González et al., 2007) and even less
ASSESSMENT OF WILD BOAR ROOTING ON ECOLOGICAL AND PASTORAL VALUES OF ALPINE...
53(Pirineos, 2011, Vol. 166, 51-67, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166003)

the approaches that take into account the main disturbances that affect suchsensitive areas (García-González et al., 2003).
The main objective of the present study is to determine the effect of wildboar rooting in terms of pastoral and ecological values of Pyrenean alpinegrasslands. We are especially interested in measuring its effect on ecologicaland pastoral attributes of grasslands at the landscape (1), the plant commu-nity (2) and plant functional group (3) levels within and outside wild boardisturbances. The integration of these three levels may provide insights intothe global assessment of this animal impact to a sensitive ecosystem within aprotected area such as Ordesa and Monte Perdido National Park (OMPNP).
2. Methods
2.1. Study area
The study was carried out in the alpine grasslands (above the present tree-line; sensu Körner 1999) within Ordesa and Monte Perdido National Park(OMPNP; 42º 36’N, 0º 00’), located in the Spanish Central Pyrenees. Theextent of the study area is 3.865 ha (25% of the entire National Park), rangingfrom 1500 to 2800 m a.s.l. The climate in this area is a proper high-mountainclimate with 5 ºC and 1720 mm of annual average temperature and precipita-tion respectively (García-González et al., 2007). Lithology comprises mainlycalcareous substrates such as limestone, sandstone and flysch (an overlayingcomplex of marlstone and sandstones). Pyrenean alpine communities withinOMPNP can be structurally separated into sparse and dense grasslandsdepending on plant cover and soil depth (Bueno et al., 2009). For communitycomparisons, we selected the five most disturbed plant communities, with atotal extent of 2309.4 ha, close to 2/3 of the total surface, but receiving morethan 96 % of wild boar disturbances in the study area (Bueno et al., 2009).Plant communities were classified at alliance levels following phytosociolog-ical procedures (Braun-Blanquet, 1979; Benito, 2006). The selected communi-ties are all dense grasslands located at sites with smooth topography and ele-vations lower than 2400 m a.s.l. Communities mainly differ in their diversity-dominance composition of species and their livestock use. Three of them aredominated by one grass species: Festucion eskiae (FE), Festucion paniculatae(FP) and Nardion strictae (NS) by Festuca eskia, F. paniculata spadicea and Nardusstricta, respectively. The other communities have tree or more co-dominantspecies: Bromion erecti (BE; Festuca rubra nigrescens, Agrostis capillaris, Plantagomedia, etc) and Rumicion pseudoalpini (RP; Chenopodium bonus-henricus, Rumex
C. G. BUENO, I. C. BARRIO, R. GARCÍA-GONZÁLEZ, C. L. ALADOS & D. GÓMEZ-GARCÍA
54 (Pirineos, 2011, Vol. 166, 51-67, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166003)

alpinus, Poa supina, etc) (Fillat et al., 2008). Among these plant communities agrazing gradient can be set, from the most to the least used by livestock: RP> BE > FP > NS > FE (Fillat et al., 2008). Grazing use and plant diversity areintrinsically linked especially in RP and BE. The occurrence of RP is totallydependent on the high presence of livestock and the extent of BE dependspartly on moderate intensities of grazing (Fillat et al., 2008). Extensive hus-bandry is the main land use within OMPNP, mainly by cattle and sheep withlittle contribution of horses. Wild boar is relatively abundant in the sur-roundings of OMPNP with densities from 3.3 to 3.8 boars/km2 (Herrero et al.,2005; Giménez-Anaya et al., 2010). It lives in the forest but visits alpine grass-lands whenever forest feeding resources are scarce and soils are wet enoughto be easily grubbed (Welander, 2000; HERRERO et al., 2005). Therefore wildboar disturbances occur mainly in late spring, with the snow-melting effecton soil, and early Autumn, with higher rainfall events (García-González et al.,2003). Also it has been suggested that protected areas where hunting activityis forbidden can act as refuges for wild boars (Acevedo et al., 2006; Herrero etal., 2006) so higher wild boar disturbances within OMPNP can be expected(Bueno et al., 2009).
2.2. Wild boar rooting, ecological and pastoral maps
Wild boar rooting in the study area was mapped during summer 2005(June-August; for more details see Bueno et al., 2009). The whole study areawas explored, and each disturbance patch over one squared meter was drawnover an aerial photograph (scale 1:3500). Most disturbance patches were alsogeoreferenced by GPS (more than 1600 GPS data; Figure 1). Then the map waselaborated crossing both sources of information (drawings and GPS data) ina Geographic Information System (Bueno et al., 2009; Figure 1). The accuracyof the map was also evaluated taking 120 random points clearly falling with-in disturbance patches and 120 outside those patches (i.e. clearly outside dis-turbances) and calculating the overall accuracy of the map through a confu-sion matrix (Congalton & Green, 2009). The overall accuracy of the map was90.6 % of correct classification (Bueno et al., 2010).
For the ecological and pastoral maps we used a previous vegetation mapof the area of a similar scale (1:5000; García-González et al., 2007). The extentof each plant community was replaced by the ecological and pastoral valuesof each community previously calculated for the Spanish Central Pyrenees(García-González et al., 2007). Those values were categorized into three levels;low, medium and high, taking three equal intervals of the ecological and pas-toral range values for the study area (Figure 1).
ASSESSMENT OF WILD BOAR ROOTING ON ECOLOGICAL AND PASTORAL VALUES OF ALPINE...
55(Pirineos, 2011, Vol. 166, 51-67, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166003)

2.3. Sampling design and data collection
To determine the composition of species, and their relative abundance,comparatively within and outside disturbances a paired stratified samplingdesign was used. 40 sampling units were randomly located within distur-bance patches in each community. For each sampling unit within distur-bances another sampling unit outside disturbances was set the closest possi-ble to the previous (400 sampling units in total). Therefore the design wasstratified by plant communities and paired within/outside disturbances. Thesampling unit was composed by two square frames of 25x25 cm, 5 cm grid.The size of the square frame was selected as a proxy of the minimum wildboar disturbance size (unit of disturbances) found in this habitat (CGB,pers.obs). In each square, 25 point-intercept contacts were set in the bottom-left corner of each grid (50 point-intercept contacts per sampling unit, 20,000in total). To quantify the abundance of species, all individuals contacted by avertical nail at each point-intercept were recorded (Goodall, 1952). Among thespecies found, we distinguished four types of functional groups: annual andperennial dicots, graminoids (most of them perennial in the study area), andbulbs.
C. G. BUENO, I. C. BARRIO, R. GARCÍA-GONZÁLEZ, C. L. ALADOS & D. GÓMEZ-GARCÍA
56 (Pirineos, 2011, Vol. 166, 51-67, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166003)
Figure 1. Wild boar rooting, ecological (a) and pastoral (b) values in the study area within OMPNP.Ecological and pastoral values were reclassified into 3 categories from three equal intervals of the
ecological and pastoral range values of the study area.Figura 1. Hozaduras de jabalí y valores ecológicos y pastorales en la zona de estudio dentro del PNOMP.Los valores ecológicos y pastorales fueron reclasificados en 3 categorías correspondientes a 3 intervalos
equitativos del rango de valores ecológicos y pastorales de la zona de estudio.
a) Ecological Values b) Pastoral Values

2.4. Ecological and pastoral values for community and plant functional groups
Community pastoral and ecological values were calculated as the averagevalues obtained multiplying the frequency of the species for each samplingunit in each community within and outside disturbances (previously sam-pled, see data collection section) by the species ecological (EV) and pastoralvalues (PV; see below) previously calculated in this area (García-González etal., 2007). We refer as pastoral value to what García-González (2007) referredas potential pastoral value, without considering the selection of the herbivore.
Ecological and pastoral values of plant functional groups were obtainedfrom calculating the total abundance of each species within and outside dis-turbances, separately. These abundance values were then multiplied by theircorresponding values of EV and PV previously calculated. For each function-al group, its percentage was calculated by adding up the values (EV and PVseparately) for each species belonging to each functional group and dividedby the total value of all species.
EV was estimated as the average of three ordinal components (valuesranging from 0 to 5) related to plant species distribution in Europe, theIberian Peninsula and the Spanish Pyrenees (Gómez-García et al., 2002).
PV was calculated by dividing the quality of each species by a proper con-version factor to get comparable values to ecological values (in our study theconversion factor was equal to ten). Quality of each species is obtained bysumming up the percentages of total plant nitrogen (N) and total plant phos-phorous (P), multiplied by its digestibility (Dg), expressed as the percentageof dry matter; Q=(N+P) x Dg (Gómez-García et al., 2002; García-González etal., 2007; Fillat et al., 2008). Pastoral values for community level in this study,differed from Gómez-García et al., (2002) in that they were calculated takinginto account the quality values of the most abundant species and their fre-quencies within communities, instead of analyzing the quality and produc-tion from community samples per se.
2.5. Statistical analyses
To determine whether the occurrence wild boar rooting on the OMPNPgrasslands was related to areas comprising low, medium or high ecologicaland pastoral values, we used resource selection analysis (RSA) (Manly et al.,1993). RSA compares the used area (disturbed by wild boar) to the available,where the null hypothesis is that wild boars root up the soil independently ofits ecological and pastoral values, i.e. in proportion to their availability in thestudy area. RSA is based first on a Chi-squared Test, to test the null hypothe-
ASSESSMENT OF WILD BOAR ROOTING ON ECOLOGICAL AND PASTORAL VALUES OF ALPINE...
57(Pirineos, 2011, Vol. 166, 51-67, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166003)

sis and after rejection, Bonferroni confidence intervals are used to detectwhich level (low, medium or high) of ecological and pastoral values are (moreor less) selected relative to their availability (Manly et al., 1993; Alldredge &Griswold, 2006).
To assess the differences between disturbed and undisturbed plant com-munities in ecological and pastoral values, we used paired t-tests owing tothe pair-matched sampling design (disturbed-undisturbed). Ecological val-ues were square root transformed to achieve normality. All statistical analy-ses were performed with R 2.10.1 (R Development Core Team, 2010).
3. Results
At the landscape level, wild boar significantly selected areas with inter-mediate ecological values, avoiding those with high ecological values andused those with low ecological values in proportion to their availability(Figure 2a). On the contrary, areas with high pastoral values were activelyselected by wild boar to root while low and intermediate values were signif-icantly avoided (Figure 2b).
C. G. BUENO, I. C. BARRIO, R. GARCÍA-GONZÁLEZ, C. L. ALADOS & D. GÓMEZ-GARCÍA
58 (Pirineos, 2011, Vol. 166, 51-67, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166003)
Figure 2. Percentage of disturbed areas by wild boar rooting in comparison to percentage ofavailable grasslands at low, medium and high ecological (a) and pastoral (b) values. Error barsrepresent Bonferroni confident intervals (BCI). Percentage of disturbed areas higher or lowerthan BCI, means selection or avoidance of wild boar to root in each level respectively. All pairsof comparisons were significant at p<0.001 except low ecological values that were not significant.Figura 2. Porcentaje de las zonas perturbadas por las hozaduras de jabalí en comparación con el porcenta-je de la superficie de pastos disponibles clasificados según su valor ecológico bajo, medio y alto (a) y segúnsu valor pastoral bajo, medio y alto (b). Las barras de error representan intervalos de confianza deBonferroni (BCI). Porcentajes de las zonas alteradas superiores o inferiores al BCI, significan preferencia orechazo del jabalí para hozar. Todos los pares de comparaciones fueron significativos (p < 0,001), excepto
para los valores ecológicos bajos, que no fueron significativos.
a) Ecological Value b) Pastoral Value
At the community scale, ecological values were higher in undisturbedareas than in disturbed ones when considering all plant communities togeth-er (t=-16.02; p=0.000). This trend was consistent within each plant communi-ty (Figure 3a). In relation to the pastoral value, a similar trend was detectedtowards a decrease in pastoral value within disturbances for all communitiestaken together (t=-20.04; p=0.000) and separately (Figure 3b).
ASSESSMENT OF WILD BOAR ROOTING ON ECOLOGICAL AND PASTORAL VALUES OF ALPINE...
59(Pirineos, 2011, Vol. 166, 51-67, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166003)
a) Ecological value
b) Pastoral value
Figure 3. Ecological (a) and pastoral (b) values within and outside wild boar disturbances in thefive grassland communities most affected by rooting. Mean values (±SE) are shown.Significance of comparisons between disturbed and undisturbed areas within each plant com-munity was assessed using paired t-tests and is indicated by the asterisks. ** p<0.01, *** p<0.001.
For community abbreviations see Methods.Figura 3. Valores ecológicos (a) y pastorales (b) dentro y fuera de las perturbaciones de jabalí en las cincocomunidades de pastos más afectadas por las hozaduras. Se muestran los valores medios ± el error están-dar. La significación de las comparaciones entre las zonas perturbadas y sin perturbar dentro de cada comu-nidad de pastos se evaluó mediante pruebas de la t-pareadas y se indica mediante asteriscos. ** p< 0,01, ***
p< 0,001. Para las abreviaturas de la comunidad véase Métodos.

For plant functional groups, both ecological and pastoral values outsidedisturbances were mainly due to graminoids and perennial dicots. Withindisturbances the relative contribution changed, being perennial dicots themain group in determining both ecological and pastoral values. Interestingly,the ecological value of bulbs and the pastoral value of annual dicots increasedwithin disturbances (Figure 4).
C. G. BUENO, I. C. BARRIO, R. GARCÍA-GONZÁLEZ, C. L. ALADOS & D. GÓMEZ-GARCÍA
60 (Pirineos, 2011, Vol. 166, 51-67, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166003)
a) Ecological value
b) Pastoral value
Figure 4. Distribution of ecological (a) and pastoral (b) values within and outside wild boar dis-turbances for plant functional groups. PD= perennial dicots, G= graminoids, AD= annual dicots,
B= bulbs.Figura 4. Distribución de los valores ecológicos (a) y pastorales totales (b) dentro y fuera de las hozadu-ras de jabalí por grupos funcionales de plantas. PD= dicotiledóneas perennes, G= graminoides, AD=
dicotiledóneas anuales, B= bulbos.

4. Discussion
The multi-scale approach used in the present study has let us assess theeffects of wild boar rooting on ecological and pastoral values of Pyreneanalpine grasslands. At a landscape level, rooting affects mainly areas of highpastoral value, but it is least concerning from an ecological point of view.While searching for underground feeding resources rooting reduces plantcover at the community level, with the subsequent reduction in ecologicaland pastoral values. However, when detailed analyses are performed at theplant functional group level, responses differ within disturbances, evenincreasing the values for annual dicots and bulbs despite the reduction inplant cover.
At the landscape level, we found that wild boars select those areas thathave a great pastoral value, which directly affect traditional grazing activities.Areas with higher pastoral values are those that are more used by livestock(García-González et al., 2007), hold the most nutrient-rich plants, and mightbe preferred by wild boars simply due to physiological constraints. Wildboars are monogastric animals, so that they cannot absorb nutrients fromplant digestion as efficiently as do ruminants, which have a much longer andcomplex digestive system (Hofmann, 1989). Therefore, wild boars may needto look actively for high-nutrient content foods, which they would more eas-ily find in these highly-valuable pastoral areas, as plants in highly grazedareas are suggested to have higher nutrient content, both above and below-ground (Bryant et al., 1983; Gibson, 2009). However, this feeding selection fornutrient-rich food items may in part not fit previous results showing a pref-erence of wild boars to root in livestock grazing areas that withstand a mod-erate stocking pressure instead of areas with high stocking pressure (Bueno etal., 2010). Assuming the most grazed areas i.e. areas with high stocking rate,have plants with higher nutrient content (Milchunas & Laurenroth, 1993;Gibson, 2009), why would wild boars not prefer intensively-stocked insteadof intermediate grazed areas? There are at least two plausible explanations forthis. The first one relates to the physical properties of soils under high live-stock pressure, since heavily grazed areas are more soil-compacted due tolivestock trampling (Van Haveren, 1983; Albon et al., 2007; Bueno et al.,unpublished-b), what would make it harder for wild boars to dig in them(Bueno et al., 2009). A second explanation would be that intensively-grazedplant communities are dominated by herbivore-tolerant species (Ritchie et al.,1998; Olofsson & Oksanen, 2002). These plant species compensate the loss ofbiomass caused by herbivory through faster nutrient uptake rates that allowfaster re-growth rates and the production of nutrient-rich tissues (Tilman,1988; Holland et al., 1992). A key point here is where nutrient resources are
ASSESSMENT OF WILD BOAR ROOTING ON ECOLOGICAL AND PASTORAL VALUES OF ALPINE...
61(Pirineos, 2011, Vol. 166, 51-67, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166003)

allocated. In herbivore-tolerant species nutrient resources are mainly allocatedaboveground, compensating for herbivory loss, while belowground biomassreceives fewer nutrient resources (Holland & Detling, 1990; Ritchie et al., 1998).This may explain why these areas dominated by herbivore-tolerant species areof less interest to rooting wild boars that instead display a grazing behaviourin these sites (C.G. Bueno pers. obs.). On the contrary, for intermediate grazedareas with moderate-to-low herbivore densities a decelerating effect on nutri-ent cycling can be expected (Tilman, 1988; Ritchie et al., 1998). In these casesherbivores may be more selective and feed on nutrient-rich plant species, sub-sequently increasing the dominance of nutrient-poor species or plants withphysical or chemical defences against herbivory (Pastor & Neiman, 1992;Ritchie et al., 1998; Hanley et al., 2007). These species may in turn have moreresources belowground as they invest more in rooting competence (Tilman,1988), being potentially more attractive to rooting wild boars.
On the other hand, wild boar rooting mainly affected areas with an inter-mediate ecological value. This might be related to the origin and dynamics ofthese grasslands in the Middle Ages, when lands were gained to the forest forgrazing activities (Fillat et al., 2008; Bueno et al., 2010). The floristic composi-tion of those communities might have changed from stress-tolerant species inthe early stages, to grazing-tolerant species spread or even brought alongfrom lowland areas by livestock (Austrheim et al., 1999). These species intro-duced by livestock have a very wide distribution being some of them sub-cos-mopolitan, leading thus to plant communities with lower ecological values.Currently, species and communities with higher ecological value would bemore restricted to rocky or high elevation habitats, where livestock does notgraze (García-González et al., 2007; García & Gómez-García, 2007) and wildboar does not root (Bueno et al., 2009).
Regarding the effect of rooting in the ecological and pastoral value of plantcommunities, a decreasing trend was detected for all plant communities.Lower ecological and pastoral values within disturbed areas can be simplyexplained by a sharp decrease in plant cover, which would reduce the poten-tial pastoral biomass together with the occurrence of ruderal or fugitivespecies within disturbances (Kotanen, 1995; Bueno et al., unpublished-a).When studying plant functional groups in more detail, the trend is similar;graminoids, the main component of undisturbed grasslands, decrease, whileperennial dicots and to a lesser extent bulbs and annual dicots, more relatedwith the regenerative niches, notably increase their pastoral and ecologicalvalues within disturbances.
In conclusion, wild boar mainly affects to pastoral and, to a lesser extent,to ecological values of Pyrenean grasslands at the landscape and communitylevel. For ecological values, despite finding a clear trend of ecological degra-
C. G. BUENO, I. C. BARRIO, R. GARCÍA-GONZÁLEZ, C. L. ALADOS & D. GÓMEZ-GARCÍA
62 (Pirineos, 2011, Vol. 166, 51-67, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166003)

dation due to disturbances, areas of high ecological value remained unaffect-ed. Therefore, notwithstanding the vast extent of wild boar rooting (321.5 hain the study area; Bueno et al., 2009) and the evident affection to grazing inthis protected area (16.2 % of livestock grazing area affected by wild boarrooting; Bueno et al., 2010), the impact of wild boar may be considered asmoderate in terms of current conservation value. Nevertheless, the potential-ly increasing trend of this disturbance (Bueno et al., 2010) may lead to someconcerns regarding the preservation of these habitats in the near future.
Acknowledgements
We would like to thank María Bueno, Ricardo Pineda and Dorleta Jiménezfor field assistance, Maite Gartzia for digitalizing and ground-truthing vege-tation maps and the staff of Ordesa and Monte Perdido National Park for thesupport and data provided. CGB was funded by a FPU fellowship from theSpanish Ministry of Education. Additional comments provided by twoanonymous reviewers greatly improved earlier versions of this manuscript.We gratefully acknowledge the support of the Spanish Science andInnovation Ministry (PN-MCI) (CGL2008-00655/BOS), the SpanishEnvironmental Ministry, and the National Parks Network (MMAMRM002/2007).
References
Acevedo, P., Escudero, M.A., Múñoz, R. & Gortázar, C., 2006. Factors affect-ing wild boar abundance across an environmental gradient in Spain. ActaTheriologica, 51: 327-336.
Albon, S.D., Brewer, M.J., O’brien, S., Nolan, A.J. & Cope, D., 2007.Quantifying the grazing impacts associated with different herbivores onrangelands. Journal of Applied Ecology, 44: 1176-1187.
Alldredge, J.R. & Griswold, J., 2006. Design and analysis of resource selectionstudies for categorical resource variables. The Journal of WildlifeManagement, 70: 337-345.
Austrheim, G. & Eriksson, O., 2001. Plant species diversity and grazing in theScandinavian mountains-patterns and processes at different spatial scales.Ecography, 24: 683–695.
Austrheim, G., Gunilla, E., Olsson, A. & Grøntvedt, E., 1999. Land-use impacton plant communities in semi-natural sub-alpine grasslands of Budalen,central Norway. Biological Conservation, 87: 369-379.
ASSESSMENT OF WILD BOAR ROOTING ON ECOLOGICAL AND PASTORAL VALUES OF ALPINE...
63(Pirineos, 2011, Vol. 166, 51-67, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166003)

Balmford, A., Rodrigues, A., Walpole, M., Ten Brink, P., Kettunen, M., Braat,L. & Groot, R., 2008. Review on the economics of biodiversity loss: scoping thescience. European Commission, Cambridge.
Benito, J.L., 2006. Vegetación del Parque Nacional de Ordesa y Monte Perdido(Sobrarbe, Pirineo central aragonés). Publicaciones del Consejo de Protecciónde la Naturaleza de Aragón, 421 pp., Zaragoza.
Bratton, S.P., 1975. The effect of the European wild boar, Sus scrofa, on GrayBeech Forest in the Great Smoky Mountains. Ecology, 56: 1356-1366.
Braun-Blanquet, J., 1979. Fitosociología, bases para el estudio de las comunidadesvegetales. Blume Ediciones, 820 pp. Madrid.
Bryant, J.P., Chapin, F.S. & Klein, D.R., 1983. Carbon/nutrient balance of bore-al plants in relation to vertebrate herbivory. Oikos, 40: 357-368.
Bueno, C.G., Alados, C.L., Gómez-García, D. & Barrio, I.C., unpublished-a.Effects of wild boar rooting on plant diversity, composition and spatialvariation in Pyrenean alpine grasslands.
Bueno, C.G.; Alados, C.L., Gómez-García, D., Barrio, I.C. & García-González,R., 2009. Understanding the main factors in the extent and distribution ofwild boar rooting on alpine grasslands. Journal of Zoology, 279: 195-202.
Bueno, C.G., Azorín, J., Gómez-García, D., Alados, C.L. & Badía, D., unpu-blished-b. Soil responses to large wild boar disturbances in Pyreneanalpine grasslands.
Bueno, C.G., Barrio, I.C., García-González, R., Alados, C.L. & Gómez-García,D., 2010. Is wild boar rooting affecting livestock grazing in alpine grass-lands? European Journal of Wildlife Management, 56: 765-770.
Carcaillet, C., Ali, A.A., Blarquez, O., Genries, A.L., Mourier, B. & Bremond,L., 2009. Spatial Variability of Fire History in Subalpine Forests: FromNatural to Cultural Regimes. Ecoscience, 16: 1-12.
Carpenter, S.R., 2002. Ecological futures: building an ecology on the long now.Ecology, 83: 2069-2083.
Carpenter, S.R., Defries, R., Dietz, T., Mooney, H.A., Polasky, S. & Reid, W.V.& Scholes, R.J., 2006. Millennium Ecosystem Assessment: Research Needs.Science, 314: 257-258.
Congalton, R.G. & Green, K., 2009. Assesing the accuracy of remotely sensed data:principles and practices. CRC Press, BocaRaton.
Fillat, F., García-González, R., Gómez, D. & Reiné, R., 2008. Pastos del Pirineo.CSIC - Diputación de Huesca, 319 pp., Madrid.
García-González, R., 2008. Management of Natura 2000 habitats. 6170 Alpine andsubalpine calcareous grasslands. In: European Commission (Ed.), TechnicalReport 2008 11/24, 23 pp. http://ec.europa.eu/environment/ nature/natura2000/ management/habitats/models_en.htm.
C. G. BUENO, I. C. BARRIO, R. GARCÍA-GONZÁLEZ, C. L. ALADOS & D. GÓMEZ-GARCÍA
64 (Pirineos, 2011, Vol. 166, 51-67, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166003)

García-González, R., Alados, C.L., Bueno, G., Fillat, F., Gartzia, M., Gómez,D., Komac, B., Marinas, A. & Saint-Jean, N., 2007. Valoración ecológica yproductiva de los pastos supraforestales en el Parque Nacional de Ordesay Monte Perdido In: Ramirez, L. & Asensio, B. (Eds.), Proyectos de investi-gación en Parques Nacionales: 2003-2006. pp. 105-128, OAPN-MMA,Madrid.
García González, R., Marinas Murillo, A. & Sanz, G., 2003. Valoración delimpacto de las hozaduras del jabalí en los pastos supraforestales del Parc Nationaldes Pyrénées. Unpublished report, pp. 86. Parc National des Pyrénées -Instituto Pirenaico de Ecología, Jaca.
García, M.B. & Gómez-García, D., 2007. Flora del Pirineo aragonés, patronesespaciales de biodiversidad y su relevancia para la conservación. Pirineos,162: 71-88.
Gartzia, M., Marinas, A., Campo, A. & Gómez-García, D., 2005. Valoracióneco-pastoral de los pastos supraforestales de Aísa (Pirineo Occidental). In:De la Roza, B. et al. (eds.), Producciones agroganaderas: Gestión eficiente y con-servación del medio natural. SERIDA, pp. 817-824, Gijón
Gibson, D.J., 2009. Grasses and grassland ecology. Oxford University Press, 305pp., Oxford.
Giménez-Anaya, A., Herrero, J., García-Serrano, A., Prada, C. & García-González, R., 2010. Long-term wild boar monitoring in the Pyreneesreveals a positive trend. 8th International Symposium on wild boar and othersuids, York, UK, p. 83.
Gómez-García, D.; GARCÍA-GONZÁLEZ, R.; MARINAS, A. & ALDEZA-BAL, A., 2002. An eco-pastoral index for evaluating Pyrenean mountaingrasslands. In: Durand, J.L., Emile, J.C., Huyghe, C., Lemaire, G. (Eds.),Multi-Function Grasslands. Quality Forages, Animal Products and Landscape.Association Française Production Fourragère, pp. 922-923, Poitiers,France.
Goodall, D.W., 1952. Some considerations in the use of point quadrats for theanalysis of vegetation. Aust. J. Sci. Res. Bull., 5: 1–41.
Hanley, M.E., Lamont, B.B., Fairbanks, M.M. & Rafferty, C.M., 2007. Plantstructural traits and their role in anti-herbivore defence. Perspectives inPlant Ecology, Evolution and Systematics, 8: 157-178.
Helm, A., Hanski, I. & Pärtel, M., 2006. Slow response of plant species rich-ness to habitat loss and fragmentation. Ecology Letters, 9: 72-77.
Herrero, J., Irizar, I.; Laskurain, N.A., García-Serrano, A. & García González,R., 2005. Fruits and roots: wild boar foods during the cold season in thesouthwestern Pyrenees. Italian Journal of Zoology, 72: 49-52.
ASSESSMENT OF WILD BOAR ROOTING ON ECOLOGICAL AND PASTORAL VALUES OF ALPINE...
65(Pirineos, 2011, Vol. 166, 51-67, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166003)

Herrero, J., Prada, C., García-Serrano, A. & Fernández Arberas, O., 2006.Mamíferos de Guara. Publicaciones del Consejo de Protección de laNaturaleza de Aragón, 165 pp., Zaragoza.
Hofmann, R.R., 1989. Evolutionary steps of ecophysiological adaptation anddiversification of ruminants: a comparative view of their digestive system.Oecologia, 78: 443-457.
Holland, E.A. & Detling, J.K., 1990. Plant Response to Herbivory andBelowground Nitrogen Cycling. Ecology, 71: 1040-1049.
Holland, E.A., Parton, W.J., Detling, J.K. & Coppock, D.L., 1992. Physiologicalresponses of plant populations to herbivory and their consequences forecosystem nutrient flow. The American Naturalist, 140: 685-706.
Körner, C., 1999. Alpine plant life. Functional plant ecology of high mountainecosystems. Springer-Verlag: 338 pp., Berlin.
Kotanen, P.M., 1995. Responses of vegetation to changing regime of distur-bance: effects of feral pigs in a Californian coastal prairie. Ecography, 18:190-199.
Lacki, M.J., Lancia, R.A., 1983. Changes in soil properties of forests rooted bywild boar. Proc. Annu. Conf. Southeasth. Assoc. Fish and Wildl. Agencies 37,228-236.
Laiolo, P., Dondero, F., Ciliento, E. & Rolando, A., 2004. Consequences of pas-toral abandonment for the structure and diversity of the alpine avifauna.Journal of Applied Ecology, 41: 294-304.
Lasanta-Martínez, T., González-Hidalgo, J.C., Vicente-Serrano, S.M. & Sferi,E., 2006. Using landscape ecology to evaluate an alternative managementscenario in abandoned Mediterranean areas. Landscape ad Urban Planning,78: 101-114.
Lasanta-Martínez, T., Vicente-Serrano, S.M. & Cuadrat-Prats, J.M., 2005.Mountain Mediterranean landscape evolution caused by the abandon-ment of traditional primary activities: a study of the Spanish CentralPyrenees. Applied Geography, 25: 47-65.
Luick, R., 1998. Ecological and socio-economic implications of livestock-keep-ing systems on extensive grasslands in south-western Germany. Journal ofApplied Ecology, 35: 979.
Macdonald, D., Crabtree, J.R., Wiesinger, G., Dax, T., Stamou, N., Fleury, P.,Gutiérrez-Lazpita, J. & Gibon, A., 2000. Agricultural abandonment inmountain areas of Europe: Environmental consequences and policyresponse. Journal of Environmental Management, 59: 47-69.
Manly, B.F.J., Mcdonald, L.L., Thomas, D.L., Mcdonald, T.L. & Erikson, W.P.,1993. Resource selection by animals: statistical design and analysis for field stud-ies. Kluwer Academic Publishers: 236 pp., New York.
C. G. BUENO, I. C. BARRIO, R. GARCÍA-GONZÁLEZ, C. L. ALADOS & D. GÓMEZ-GARCÍA
66 (Pirineos, 2011, Vol. 166, 51-67, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166003)

Milchunas, D.G. & Laurenroth, W.K., 1993. Quantitive effects of grazing onvegetation and soils over a global range of environments. EcologicalMonographs, 63: 327-266.
Millennium Ecosystem Assessment Board, 2005. Living Beyond Our Means:Natural Assets and Human Well-Being, Statements from the board. www.mil-lenniumassessment.org/proxy/document.429.aspx. Accessed on 2ndFebruary 2006. United Nation Fundation.
Moore, D.S., Mccabe, G.P., Craig, B., 2008. Introduction to the practice of statis-tics. W.H Freeman. 100 pp., New York.
Olff, H. & Ritchie, M.E., 1998. Effects of herbivores on grassland plant diver-sity. TREE, 13: 261-265.
Oliver, W. & Leus, K., 2008. Sus scrofa. IUCN 2010. IUCN Red List ofThreatened Species. Version 2010.3. <www.iucnredlist.org>
Olofsson, J. & Oksanen, L., 2002. Role of litter decomposition for the increasedprimary production in areas heavily grazed by reindeer: a litterbag exper-iment. Oikos, 96: 507-515.
Pastor, J. & Neiman, R.J., 1992. Selective foraging and ecosystem processes inboreal forest. The American Naturalist, 139: 690-705.
R Development Core Team, 2010. R: A language and environment for statisticalcomputing. R Foundation for Statistical ComputingVienna, Austria. ISBN3-900051-07-0, URL http://www.R-project.org.
Ritchie, M.E., Tilman, D. & Knops, J.M.H., 1998. Herbivore effects on plantnitrogen dynamics in Oak Savanna. Ecology, 79: 165-177.
Sáez-Royuela, C. & Tellería, J.L., 1986. The increased population of Wild Boar(Sus scrofa) in Europe. Mammal Review, 16: 97-101.
Tilman, D., 1988. Plant strategies and the dynamics and structure of plant com-munties. Princeton University Press, Princeton, New Jersey, USA.
Van Haveren, B.P., 1983. Soil Bulk Density as Influenced by Grazing Intensityand Soil Type on a Shortgrass Prairie Site. Journal of Range Management, 36:586-588.
Welander, J., 2000. Spatial and temporal dynamics of a disturbance regime: wildboar (Sus scrofa) rooting and its effects on plant species diversity. PhD Thesis,Swedish University of Agricultural SciencesUtgivningsort, Sweden.
ASSESSMENT OF WILD BOAR ROOTING ON ECOLOGICAL AND PASTORAL VALUES OF ALPINE...
67(Pirineos, 2011, Vol. 166, 51-67, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166003)

69
GESTIÓN DEL CORZO EN ZARAGOZA:DE CONSERVACIÓN A CONTROL POBLACIONAL
J. FERRERES1, M. A. ESCUDERO1, E. FERRER2 & J. MARCO1
1 Ebronatura S.L. Calle del Arpa, 6, 50003 Zaragoza. [email protected] SODEMASA, Avda. César Augusto, 3, 50004 Zaragoza. [email protected]
ABSTRACT.– As it has happened with other wild ungulates in Europe, the roedeer (Capreolus capreolus) population in Aragon has suffered a considerableexpansion during the last two decades. In the Iberian Mountains of Zaragoza (SIZ)this process come along with an increasing sporting interest in the species and theraising of conflicts due to crop damage, mainly on fruit trees and vineyards.
The Government of Aragon is responsible of managing big game populationsand fixes the hunting quotas. During the elaboration of hunting plans it has to con-sider different and even opposite interests and points of view, so availability of reli-able population trends it is essential for the management of these populations. It hasbeen developed a scheme for monitoring roe deer population in Aragon based on striptransect counts of pellet groups. This information is complemented by the sightingsof roe deer during a fox and lagomorph monitoring scheme based on spotlight countsfrom car. In the Iberian Mountain System of Zaragoza, between 2005 and 2009, thesurvey results point that the roe deer abundance has doubled, and the mean densityreaches 5.4 ± 0.33 roe deer/km2.
Initially, management plans followed a conservative criterion in order to keepthe population growing, with a moderate extraction rate, still hunt and a balancedsex-specific quota. Due to the positive trend of the species and the necessity of reduc-ing damage caused to agriculture, mainly fruit trees, the management goals changeto population control and it was decided to increase hunting pressure: hunting sea-son was extended, hunting drives with dogs are authorized and the extraction rateis raised. Although the hunting quota fulfillment is close to 80%, and the number ofshot deer has increased from 193 in 2005 to 1.084 in 2009, until now there is noreduction in the population growth.
Keywords: Roe deer; Capreolus capreolus; Management; Hunting quota;Population monitoring; Fecal abundance; Spotlight counts; Aragon.
Pirineos.Revista de Ecología de MontañaVol. 166, 69-86
Jaca, Enero-Diciembre, 2011ISSN: 0373-2568
eISSN: 1988-4281doi: 10.3989/Pirineos.2011.166004

70
J. FERRERES, M. A. ESCUDERO, E. FERRER & J. MARCO
RESUMEN.– Al igual que ha ocurrido con otros ungulados en el resto deEuropa, durante las dos últimas décadas la población de corzo (Capreolus capreo-lus) ha experimentado una considerable expansión en Aragón. En el Sistema Ibéricozaragozano (SIZ) este proceso se ha visto acompañado por un incremento del interéscinegético hacia la especie y por una creciente conflictividad asociada a los daños encultivos de leñosas.
En Aragón es la Administración la responsable de planificar el aprovechamien-to cinegético de la caza, asignando los cupos de captura a los cotos de caza. Durantela elaboración de los planes de caza se debe atender a posiciones e intereses muchasveces contrapuestos, por lo que para gestionar estas poblaciones es imprescindiblecontar con datos objetivos sobre su estado. Para ello se ha desarrollado un plan deseguimiento del corzo en Aragón basado en la realización periódica de transectos derecuento de grupos fecales sobre recorridos fijos. Esta información se complementacon las observaciones de corzo registradas en una red de fareos nocturnos diseñadaoriginalmente para el seguimiento de lagomorfos y zorro. Los datos obtenidos en elSIZ indican que entre 2005 y 2009 la abundancia media de corzo se ha duplicado yque su densidad media alcanza 5,4 ± 0,33 corzos/km2.
Inicialmente los planes de caza seguían criterios conservadores que permitiesenmantener la población en crecimiento, con una tasa de extracción moderada, un cupoequilibrado entre machos y hembras y una caza selectiva a rececho. Ante la evoluciónpositiva de la especie y la necesidad de atenuar las protestas por daños en cultivos,fundamentalmente frutales, se ha cambiado a una estrategia de control poblacionalcon un notable incremento de la presión cinegética: se amplía el periodo hábil de caza,se autoriza la caza en batidas y se aumenta la tasa de extracción, aunque se ha man-tenido la sex-ratio equilibrada en los cupos. A pesar de que el cumplimiento del cupose sitúa en torno al 80%, y se ha pasado de 193 capturas en 2005 a 1.084 en 2009,hasta ahora no se ha conseguido frenar el crecimiento poblacional.
Palabras clave: Corzo; Capreolus capreolus; gestión; seguimiento poblacional;cupos de caza; abundancia fecal; recorridos nocturnos con foco; Aragón.
1. Introducción
El incremento de las poblaciones de ungulados silvestres es un fenómenogeneralizado en todo el continente europeo y que afecta prácticamente atodas las especies de cérvidos (Apolonio et al., 2010). El corzo es un caso para-digmático, pues durante las últimas décadas además de recuperar zonas enlas que había desaparecido debido a la presión humana, se ha extendido detal manera que probablemente nunca ha tenido una distribución tan ampliacomo la actual (Linell et al., 1998). Está ocupando hábitats que hasta hace pocono se consideraban adecuados para la especie, como agrosistemas de llanurasin cobertura vegetal, zonas alpinas y ambientes semidesérticos mediterráne-os (Linell et al., 1998).
(Pirineos, 2011, Vol. 166, 69-86, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166004)

En Aragón la evolución de la especie ha sido similar a la observada en elresto del continente. A mediados del siglo XIX, la distribución del corzo selimitaba a algunas áreas montañosas de los Pirineos y los sectores más occi-dentales del Sistema Ibérico. Un siglo más tarde, hacia 1960 su área de distri-bución se había reducido hasta ocupar un tercio de la estimada en 1850.Desde entonces se produce una rápida expansión y para 1998 se estima quela superficie ocupada por el corzo en Aragón se multiplica por 13 respecto ala de 1960 y cuadruplica la estimada en el siglo XIX hasta alcanzar el 43% delterritorio aragonés (Gortázar et al., 2000). En 2005, Osuna et al., (2006) com-prueban que este porcentaje ha subido al 60%. Este proceso sigue activo y enlos últimos años se van sucediendo las citas y observaciones en nuevas loca-lidades (Ferreres et al., 2010).
En el Sistema Ibérico zaragozano (SIZ) la rapidez del proceso ha sido nota-ble. Las primeras observaciones de la especie no aparecen antes de 1985 e ini-cialmente se limitan a un número reducido de localizaciones en el límite occi-dental de la provincia (Moncayo y Alto Jalón). Para el año 1999 ocupa prácti-camente la totalidad del SIZ de forma que en menos de 15 años el corzo hacolonizado 5.800 km2. Aunque en las fases iniciales la especie selecciona lossectores con características ecológicas más favorables, zonas con precipitacio-nes más altas y mayor diversidad paisajística, conforme avanza el proceso elcorzo ha ido ocupando toda el área (Acevedo et al., 2005).
En esta situación tan dinámica, en la que en pocos años se ha pasado de laausencia de la especie a una presencia generalizada, la administración arago-nesa se ve en la necesidad de desarrollar un programa de seguimiento delcorzo que permita obtener unos datos fiables sobre la abundancia y la ten-dencia poblacional en las que basar su gestión de la especie. Inicialmentedesde el Gobierno de Aragón se promovieron diferentes estudios centradosen conocer la distribución y la secuencia de colonización de la especie a nivellocal (Marco y Jordán, 1990; Gortázar y Marco, 1998). Más tarde ante el incre-mento de las poblaciones de corzo se hace necesario un verdadero plan deseguimiento que permita obtener datos fiables de tendencia poblacional yque además, aporte información sobre la densidad absoluta que sirva de basepara fijar los cupos de caza. En el diseño de este plan de seguimiento se hanconsiderado condicionantes como la gran extensión de la zona de estudio, lasbajas densidades de las poblaciones de corzo, los hábitos forestales de la espe-cie que unidos a su carácter esquivo hacen que la especie sea difícilmentedetectable y la disponibilidad de recursos humanos y materiales que, como eshabitual, es reducida. Se plantea un seguimiento a gran escala que abarca latotalidad de su área de distribución en Aragón (más de 30.000 km2).Atendiendo a características bio-geográficas y extensión se divide el territo-rio de Aragón en una serie de grandes unidades (Sistema Ibérico Zaragozano,
GESTIÓN DEL CORZO EN ZARAGOZA: DE CONSERVACIÓN A CONTROL POBLACIONAL
71(Pirineos, 2011, Vol. 166, 69-86, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166004)

Pirineo Oriental etc.) en los que los muestreos se van alternando y repitiendocon periodicidad plurianual (dependiendo de la unidad entre 2 y 4 años). Deesta forma se concentra el esfuerzo de muestreo en unos sectores concretos, ala vez que se dejan transcurrir varios años entre dos revisiones del mismo sec-tor. Se consigue así mejorar la precisión de las estimas de abundancia y la fia-bilidad de las tendencias aumenta (Smart et al., 2004; Centenera, 2002).
Un plan de gestión de cualquier especie debería contemplar los siguien-tes aspectos: cuál es el estado de la población, cuales son los objetivos delplan, qué medidas se van a tomar para alcanzar esos objetivos y finalmente,debería establecer un programa de monitorización para determinar en quémedida se han alcanzado esos objetivos (Strickland et al., 1994; Putman, 2003).La decisión de los objetivos de este plan no responde únicamente a argu-mentos técnicos, y en el fondo es un juicio de valor en el que confluyen aspec-tos sociales, económicos, biológicos y políticos (Sinclair et al., 2006). En el casodel SIZ, con una considerable ocupación humana del territorio, los objetivosde la gestión de una especie cinegética estarían condicionados por la capaci-dad de carga social, definida como la densidad máxima que la sociedad con-sidera aceptable en un área determinada (Strickland et al., 1994). Se trataría deun valor subjetivo y variable en el tiempo, que refleja un equilibrio fluidoentre los distintos grupos de interés (cazadores, agricultores, etc.).
En este artículo se presentan los resultados obtenidos en el programa deseguimiento del corzo en el Sistema Ibérico Zaragozano, se describe cómohan evolucionado los objetivos de los planes de gestión de la especie, se ana-liza la efectividad de las medidas adoptadas y finalmente se discuten las pers-pectivas futuras de su gestión.
2. Material y métodos
2.1. Área de estudio
El área de estudio se localiza en la provincia de Zaragoza, en el noreste deEspaña (42o 00’ 25” - 40o 56’ 00” N, 2º 10’ 28” – 0º 56’ 56” W) sobre las estriba-ciones del Sistema Ibérico. En total son cerca de 5.800 km2 repartidos en unamplio rango altitudinal que abarca desde los 300 a los 2300 m.s.n.m en lospisos climáticos mesomediterráneo y supramediterráneo (Sampietro, 2000).Las precipitaciones son escasas, en la mayor parte del territorio: la precipita-ción anual se sitúa en torno a los 400 mm y sólo en las sierras más elevadassupera los 700 (Ascaso & Cuadrat, 1981). El paisaje está dominado por agro-sistemas abiertos, entre los que se intercalan áreas de matorral (Rosmarinus
J. FERRERES, M. A. ESCUDERO, E. FERRER & J. MARCO
72 (Pirineos, 2011, Vol. 166, 69-86, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166004)

officinalis, Quercus coccifera, Juniperus spp, Erica spp) y zonas arboladas, funda-mentalmente Quercus ilex y repoblaciones de Pinus spp. Estas manchas bosco-sas y de matorral generalmente están asociadas a zonas de mayor relieve ycubren 1.299 km2, algo menos del 25 % de la superficie total.
Los datos de abundancia se han obtenido a partir del programa de segui-miento basado en conteos de grupos fecales, complementado con datos deobservaciones de corzo recopilados durante la realización de recorridos noc-turnos.
2.2. Estima de densidad por conteo de grupos fecales
Se han tomado como unidades de muestreo las cuadriculas UTM de 100km2 (n = 64). A partir del Mapa Forestal de Aragón (Escudero y Franchés,2005) se ha caracterizado cada una de las cuadrículas desde el punto de vistade la oferta de refugio y la disponibilidad de alimento. En ambientes medite-rráneos los hábitat forestados se consideran más adecuados para la especiepor ofrecer simultáneamente alimentación y refugio (Acevedo et al., 2005).Así, las teselas que presentan árboles o matorral de más de 1,5 m de altura sehan considerado favorables para el corzo y el resto desfavorables.
Para cada cuadrícula se seleccionó sobre el Mapa Forestal de Aragón unpunto representativo de la disponibilidad de hábitat favorable para el corzo.En una primera fase del estudio en 1999, en cada punto de muestreo se reali-zó un recorrido de 2000 m para localizar indicios de presencia de la especie:grupos fecales, escodaduras, huellas, encames etc. Con estos datos, ademásde determinar la presencia o ausencia de corzo, se obtuvo un índice kilomé-trico de abundancia (IKA) a partir del número de grupos fecales de corzodetectados por cada kilómetro de transecto realizado (Mayle et al., 2000). Seconsideró grupo de heces cuando seis o más excrementos parecían productode la misma defecación. Este índice es una estima de abundancia relativa que,aunque permite estimaciones locales de la tendencia poblacional, al no consi-derar la influencia del medio sobre la detectabilidad de los grupos fecales, nosería útil para hacer comparaciones entre poblaciones (Cederlund et al., 1998).El IKA es un índice lineal que no permite estimar la densidad absoluta de gru-pos fecales y por tanto, tampoco la densidad poblacional absoluta (Mayle etal., 1999).
Posteriormente se diseñó una metodología que permitiera valorar lasuperficie revisada a lo largo del transecto, para poder estimar la densidad degrupos fecales y calcular la densidad de corzo. La metodología empleada,transectos de anchura de banda variable, es una adaptación del recuento deexcrementos en transectos en banda, en el que se contabilizan los grupos feca-
GESTIÓN DEL CORZO EN ZARAGOZA: DE CONSERVACIÓN A CONTROL POBLACIONAL
73(Pirineos, 2011, Vol. 166, 69-86, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166004)

les que se detectan en una banda de anchura fija a cada lado de la línea deprogresión (Mayle et al., 1999). En esta adaptación, no se pretende hacer unarevisión exhaustiva de la totalidad de la banda en la que se hace el recuentode grupos fecales sino que, para ganar rapidez, únicamente se cuentan losgrupos fecales dentro del ancho de banda que se asume se puede revisar sinralentizar el proceso. En función de la cobertura vegetal se va asignando acada tramo del transecto (10 m) un ancho de banda estandarizado en clasesde 0, 0,5, 1 ó 2 m, basándose en experiencias previas sobre detectabilidad delos grupos fecales. Se trataría de un transecto en banda en el que el ancho debanda es variable en cada tramo de forma que la superficie revisada es esti-mada subjetivamente y presenta una relación coste-resultados superior aotros métodos similares. Una descripción más completa, junto a una compa-rativa frente a otros métodos basados en recuentos de grupos fecales, sepuede encontrar en Acevedo et al. (2010).
La superficie total revisada en un transecto sería:
S = ∑bi x 10
donde “b” es el ancho de banda de cada tramo de 10 m.A partir de este dato y conociendo la tasa de persistencia de los grupos
fecales y la tasa de defecación se calcula la densidad de corzo (Mayle et al.,1999):
D = gf / S x td x pp
Donde S es la superficie revisada, gf es el número de grupos fecales detec-tados, td es la tasa de defecación (20 g.f./día, Mayle et al.,1999), y pp el perio-do de persistencia, aplicado en nuestro caso, 135 días. Este último valor seobtuvo a partir de una experiencia previa realizada en el Parque Natural delMoncayo (SIZ) en la que se estimó una tasa de desaparición conjunta para unamplio abanico de hábitats, desde coscojales a hayedos (Escudero et al., 2007).Es un valor promedio que se sitúa dentro de los márgenes estimados enambientes mediterráneos en Grecia, entre 115 y 164 días según tipo de hábi-tat (Tsaparis et al., 2009), y es inferior a los calculados en el centro de Portugalen un clima con mayor influencia atlántica, entre 151 y 297 días (Carvalho,2007). Sin embargo, según la estación, se han citado valores entre 26 y 118 enPolonia (Aulak y Babinska, 1990), y dentro del contexto del Reino Unido,entre 64 y 165 días en áreas con menores precipitaciones, y entre 98 y 537 enáreas más lluviosas (Mayle et al., 1999).
Sobre los puntos seleccionados en 1999 se han realizado muestreos basa-dos en transectos de anchura de banda variable que se repiten con periodici-
J. FERRERES, M. A. ESCUDERO, E. FERRER & J. MARCO
74 (Pirineos, 2011, Vol. 166, 69-86, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166004)

dad bienal a partir de 2005 (hasta ahora tres campañas: 2005, 2007 y 2009). En2005 se rediseñaron los transectos originales de forma que atravesasen áreasrepresentativas del hábitat de la zona, evitando itinerarios paralelos a arro-yos, sendas, linderos y otras zonas que puedan influir sobre la abundancia decorzos (Mayle et al., 1999). En cada punto de muestreo se fijaron dos transec-tos de 700 m y se georreferenciaron para facilitar su repetición en el tiempo yevitar la variabilidad de los resultados asociada a efectos topográficos. Losmuestreos se realizaron en dos periodos: mayo-junio y octubre-noviembre.Para reducir un posible efecto estacional sobre cobertura vegetal, la tasa depersistencia de los grupos fecales o la tasa de defecación, las revisiones decada punto se realizaron siempre en la misma estación. Con el fin de minimi-zar el efecto observador, que podría ser importante ya que existe una compo-nente subjetiva en la valoración del ancho de banda, todas las revisiones sehan realizado por un solo equipo formado por los mismos tres observadores(M. A. Escudero, E .Ferrer y J. Ferreres), coautores de este trabajo.
2.3. Estima de la abundancia a partir de avistamientos nocturnos
La información obtenida a partir de los recorridos de abundancia fecal, secomplementa con los datos de avistamientos ocasionales de corzo durante larealización de recorridos, en el marco del Plan de Monitorización deMamíferos de Aragón (PMMA) (Gortázar et al., 1994). Este plan está diseña-do para monitorizar a gran escala las poblaciones de mesomamíferos, funda-mentalmente lagomorfos (conejo Oryctolagus cuniculus, liebre europea Lepuseuropaeus, liebre ibérica Lepus granatensis y zorro Vulpes vulpes) y consiste enuna red de 60 localidades repartidas por todo el territorio, en las que se reali-zan conteos nocturnos desde vehículo equipado con faro adicional. Los itine-rarios son recorridos fijos que discurren por pistas agrícolas en agrosistemasabiertos, tienen una longitud aproximada de 30 km y se repiten dos veces almes entre enero y abril (8 repeticiones anuales con un total de 240 km) por elmismo equipo, formado por dos Agentes de Protección de la Naturaleza delGobierno de Aragón. El faro de 100 W es manejado por el copiloto que barrede manera sistemática con el haz de luz un arco de 270º en torno al vehículo,y cuando es necesario la identificación de los animales se confirma medianteel uso de prismáticos 8x42. Los recorridos se realizan con periodicidad anual,lo que posibilita el cálculo eficaz de los índices de abundancia relativa y lastendencias demográficas de cada especie (Williams et al., 2007, Gortázar et al.,2007). Durante los recorridos, además de las especies diana se anotan todaslas observaciones de otras especies de interés, entre ellas el corzo.
GESTIÓN DEL CORZO EN ZARAGOZA: DE CONSERVACIÓN A CONTROL POBLACIONAL
75(Pirineos, 2011, Vol. 166, 69-86, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166004)

Se han recopilado las observaciones de corzo en los 12 puntos de monito-rización localizados en el área de estudio. A partir del número de corzos avis-tados y la longitud total recorrida en los transectos se ha calculado un índicekilométrico de abundancia (IKA) expresado en corzos/100 km (Acevedo etal., 2008). Los datos se han agrupado por bienios.
2.4. Estadísticas cinegéticas
Las estadísticas cinegéticas se han elaborado a partir de los datos sumi-nistrados por el Instituto Aragonés de Gestión Ambiental del Gobierno deAragón. Los cupos son los asignados por la Dirección General de DesarrolloSostenible y Biodiversidad del Gobierno de Aragón, mientras que los resulta-dos se obtienen a partir de las declaraciones de resultados que los titulares delos cotos de caza comunican a la administración al finalizar la temporada decaza. Los objetivos de gestión y tasas de extracción se han obtenido de infor-mes internos del Gobierno de Aragón sobre la gestión del corzo (Ferreres etal., 2010).
2.5. Análisis de los datos
Se han utilizado tests no paramétricos (prueba de Kruskal-Wallis) parahacer comparaciones entre periodos y regresiones lineales para estudiar latendencia poblacional. Con el fin de unificar periodos (datos disponibles decupos, capturas, datos de abundancia) las tendencias poblacionales se hancalculado con los datos disponibles a partir de 1999.
3. Resultados
3.1. Transectos de abundancia fecal
Se han realizado transectos en 64 puntos de muestreo. Las cuatro campa-ñas de trabajo de campo entre 1999 y 2009, muestran el incremento poblacio-nal de la especie (Tabla 1). En un periodo de 6 años, entre 1999 y 2005, el corzopasa de ser detectado en la mitad de los puntos de muestreo a más del 90%de puntos positivos, y dos años más tarde se alcanza el 100% de puntos posi-tivos. Los índices de abundancia confirman un continuo crecimiento pobla-cional, tanto inicialmente, entre 1999 y 2005, como en los últimos años, cuan-do los muestreos se realizan con periodicidad bienal (Figura 1).
J. FERRERES, M. A. ESCUDERO, E. FERRER & J. MARCO
76 (Pirineos, 2011, Vol. 166, 69-86, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166004)

GESTIÓN DEL CORZO EN ZARAGOZA: DE CONSERVACIÓN A CONTROL POBLACIONAL
77(Pirineos, 2011, Vol. 166, 69-86, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166004)
Tabla 1. Evolución de los cupos, capturas y abundancia de corzo estimada a partir de transectosde abundancia fecal. Se indica el factor de crecimiento observado entre los distintos periodos
(Nfinal/ Ninicial).Table 1. Evolution of roe deer hunting quota, hunting bag and abundance estimated from fecal strip counts.
The growth factor is also indicated.
Increment Increment Increment1999 2005 2007 2009 o 1999-2009 o 1999-2005 o 2005-2009% puntos muestreo
positivos 54,6 93,9 100 100 x 1,8 x 1,7 x 1,1
IKA fecal (g.f./km) 0,8 12,3 17,5 21,5 x 26,9 x 15,4 x 1,7Densidad
(corzos/km2) – 2,6 3,9 5,4 – – x 2,1
Cupo total (corzos) 46 258 700 1.272 x 27,5 x 11,2 x 4,9Capturas (corzos) 23 193 533 1.084 x 47,1 x 8,4 x 5,6
Figura 1. Abundancia de corzo en el Sistema Ibérico Zaragozano (grupos fecales/ 1000 m de tran-secto), (media, media ± EE, media ± 1,96*EE). La abundancia se ha estimado a partir de transectos
a pie realizados en 64 puntos de muestreo fijos.Figure 1. Roe deer abundance in the SIZ (pellet group/1000m), (median, median ± SE, median ± 1,96*SE).
Abundance is estimated from fecal standing crop transects in 64 fixed points.

Este incremento de la abundancia se refleja en un incremento de las den-sidades estimadas que pasan de 2,6 ± 0,29 corzos/km2 en 2005 a 3,9 ± 0,35 en2007 y alcanza 5,4 ± 0,33 en 2009 (no hay estimas de densidad en 1999), condiferencias interanuales significativas (KW, H = 27,95; p = 0,000). La tenden-cia poblacional es positiva, con una tasa anual de crecimiento del 15,8 %, (R2
= 0,997; p = 0,002; β = 2,07).
3.2 Avistamientos nocturnos
Los datos obtenidos a partir de observaciones de corzo durante los reco-rridos nocturnos del PMMA reflejan la expansión de la población de corzo enel SIZ, apreciable tanto por el aumento del área de distribución como en laabundancia. Hasta 1999 sólo se habían observado corzos en dos de las 12 loca-lidades donde se realizan muestreos, desde entonces se va detectando laespecie en un número creciente de localidades, 5 en 2001, 8 en 2004 y en 2007ya se cita la especie en todas las localidades prospectadas. La evolución de laabundancia sigue una tendencia similar (Figura 2), valores de IKA muy redu-
J. FERRERES, M. A. ESCUDERO, E. FERRER & J. MARCO
78 (Pirineos, 2011, Vol. 166, 69-86, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166004)
Figura 2. Abundancia de corzo en el SIZ (corzos/100 km), (media, media ± EE, media ± 1,96* EE),estimada a partir de recorridos nocturnos en vehículo en 12 localidades (180 km por localidad yaño). Figure 2. Roe deer abundance in the SIZ (deer/100 km), (median, median ± SE, median ± 1,96* SE).
Abundance is estimated from spotlight counts in 12 localities (180 km per locality and year).

cidos, por debajo de 0,5 corzos/100 km en los años 90, y a partir de entoncesse dispara, alcanzando los 9 corzos/100km en 2009 (1999: 0,3 ± 0,14 cor-zos/100 km; 2005: 3,5 ± 0,67; 2007: 6,1 ± 0,97; 2009: 9,4 ± 1,21). Diferenciasentre periodos significativas (KW, H = 215,98, p = 0,000). La tendencia pobla-cional es positiva, con una tasa anual de crecimiento entre 2005 y 2009 del 21,9%, (R2 = 0,896, p = 0,004; β = 0,896).
3.3 Estadísticas cinegéticas
Los cupos y las capturas de corzo en el SIZ han seguido un patrón similaral que muestra la evolución de la abundancia de la especie (Figura 3).
GESTIÓN DEL CORZO EN ZARAGOZA: DE CONSERVACIÓN A CONTROL POBLACIONAL
79(Pirineos, 2011, Vol. 166, 69-86, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166004)
Figura 3. Cupos de corzo en el Sistema Ibérico Zaragozano (SIZ). Se indica en gris el cupo auto-rizado por el Gobierno de Aragón y en negro las capturas declaradas por los cazadores (no hay
datos disponibles entre 2001 y 2004).Figure 3. Hunting quotas in the SIZ. In grey the hunting quotas set by the Aragon Government, in black
the hunting bags reported by hunters (no data available in the 2001-2004 period).

Entre 1999 y 2005, los cupos asignados y las capturas son reducidos. Enesta fase inicial del aprovechamiento cinegético, por precaución y debido a lafalta de datos objetivos sobre los que calcular los cupos, éstos se asignansiguiendo criterios conservadores, las capturas no llegan a 200 ejemplares portemporada. A partir de 2005 se obtienen las primeras estimaciones de densi-dad, con valores más altos que los supuestos hasta entonces, lo que permiteincrementar considerablemente los cupos. Los sucesivos estudios van mos-trando un crecimiento poblacional sostenido entre 2005 y 2009, lo que unidoa una creciente conflictividad motivada por daños agrícolas, hace que se deci-da incrementar la presión cinegética sobre la especie: se amplía el periodohábil de caza hasta 11 meses, se favorece la caza en batidas, y especialmenteen las dos últimas temporadas de caza se incrementan los cupos de caza sus-tancialmente, con una tasa de extracción que supone el 30% de la poblacióntotal estimada y que en algunos sectores donde la presencia de cultivos defrutal es mayor, se incrementa hasta superar el 40% de la población (Ferrereset al., 2010). Mientras que entre 2005 y 2009 la densidad se multiplica por dos,en este mismo periodo los cupos autorizados se multiplican por 5. En el casode las capturas declaradas el incremento es aún mayor, y en 2009 se superanlos 1.000 ejemplares cazados. El grado de cumplimiento de los cupos deextracción ha ido creciendo y se situaría en torno al 80% (Tabla 1). Según lasdeclaraciones de resultados de caza el 52% de los 3.189 animales abatidos sonmachos.
4. Discusión
Los muestreos de abundancia fecal basados en transectos de anchuravariable están basados en apreciaciones subjetivas del ancho de banda y pre-sentan como gran inconveniente su susceptibilidad a errores debidos al efec-to observador (Acevedo et al., 2010). Por otro lado, posibles variaciones de latasa de defecación ligadas a factores como la alimentación, densidad, estruc-tura poblacional y variaciones estacionales de la calidad de la dieta podríanafectar a la validez de los cálculos de la densidad de la población a partir dela densidad de grupos fecales, por lo que, en el mejor de los casos, los conte-os de grupos fecales deberían ser usados como un indicador de cambios en ladensidad a nivel local (Cederlund et al., 1998). Sin embargo, el diseño delseguimiento del corzo en Aragón, en el que se repiten los mismos transectosen las mismas fechas y por los mismos observadores, reduce al mínimo lavariabilidad asociada a efectos locales, estacionales o del observador. Paracompletar los datos de abundancia obtenidos a partir del seguimiento decorzo mediante grupos fecales, se ha estimado la evolución de la población de
J. FERRERES, M. A. ESCUDERO, E. FERRER & J. MARCO
80 (Pirineos, 2011, Vol. 166, 69-86, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166004)

corzo a partir de un índice kilométrico de avistamientos (IKA) durante reco-rridos nocturnos en automóvil. Aunque diversos autores desaconsejan estemétodo para estimar tendencias poblacionales de cérvidos debido a su bajafiabilidad (Mayle et al., 1999; Cederlund et al., 1998; Van Laere et al., 1999),otros autores defienden su utilidad para el seguimiento de las poblaciones deciervo en ambientes mediterráneos (Acevedo et al., 2008). El hecho de que dosmétodos completamente distintos, uno indirecto basado en indicios de pre-sencia y otro directo, muestren resultados coincidentes, confirmaría la validezde la tendencia observada. Por otro lado, el esfuerzo de muestreo empleadoen el programa de seguimiento, ha sido suficiente para detectar un incre-mento significativo en la abundancia de corzo.
Los valores de densidad obtenidos deben interpretarse como una aproxi-mación general a la densidad media, ya que en el cálculo se ha empleado unúnico valor para el periodo de persistencia en todos los puntos de muestreo,y no se han empleado tasas de desaparición específicas para los principalestipos de vegetación presentes en el área de estudio, lo que habría mejorado laexactitud de las estimas (Mayle et al., 1999; Putman, 2003; Campbell et al.,2004). Como ya se ha comentado, se trata de un valor promedio (135 días)estimado a partir de una experiencia previa en una parte del área de estudio,y se sitúa dentro de los márgenes estimados o es inferior a los calculados pordiversos autores en ambientes mediterráneos (Tsaparis et al., 2009; Carvalho,2007). Con estos datos sería esperable que los valores de densidad estimadosfuesen cercanos a los reales o se produjese una ligera sobrestima. La densidadestimada en 2009, en torno a 5,5 corzos/km2 es similar a las de otras pobla-ciones del interior de la Península Ibérica, 6,5 en Guadalajara (Burón et al.,2005) y 5,6 en Castilla - La Mancha (Acevedo et al., 2010). Es la más alta obser-vada en Aragón: entre 0,5 y 1,7 en Teruel, y entre 1,8 y 2,3 en Huesca (Ferrereset al., 2010) y supera ampliamente la estimada en sectores con mayor influen-cia eurosiberiana como las Reservas de Caza del Pirineo aragonés, 2,4 cor-zos/km2 (Prada et al., 2010). Este hecho contrasta con los valores observadosen otros puntos de la Península Ibérica y Francia, en donde se describenabundancias más bajas en los sectores con mayor carácter mediterráneo(Aragón et al., 1995, Virgós y Tellería 1998, Gaudin et al., 1997, Acevedo et al.,2005).
En esta situación de expansión poblacional y densidades moderadas, seplanteó la gestión del corzo en el SIZ. Existen cuatro posibles opciones al ges-tionar una especie: fomentar el incremento poblacional (conservación), perse-guir una reducción en su densidad (control), realizar una extracción sosteni-ble (explotación) y no tomar ninguna medida pero monitorizar su evolución(vigilancia) (Sinclair et al., 2006). La gestión del corzo en el SIZ ha pasado porestas cuatro opciones. En la fase inicial del proceso simplemente se vigilaba la
GESTIÓN DEL CORZO EN ZARAGOZA: DE CONSERVACIÓN A CONTROL POBLACIONAL
81(Pirineos, 2011, Vol. 166, 69-86, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166004)

evolución de la especie y a partir de 1999 se decidió comenzar el aprovecha-miento cinegético en unas condiciones con las que se perseguía favorecer laexpansión de las poblaciones. Posteriormente se fue comprobando que laabundancia del corzo aumentaba y surgió una considerable problemáticaasociada a daños en los cultivos de frutales (Ferreres et al., 2010). En este con-texto el objetivo de la gestión pasa a ser el control del crecimiento poblacionalmediante un incremento de la presión cinegética. Aunque el aumento en loscupos ha sido notable durante los últimos años, muy superior al de la abun-dancia, hasta ahora no ha sido suficiente para contener el crecimiento pobla-cional.
El éxito de un plan de gestión se puede valorar como el grado de conse-cución de los objetivos marcados (Sinclair et al., 2006). Bajo este punto devista, los planes de gestión del corzo en el SIZ fueron adecuados mientras sepersiguió favorecer la expansión de la especie y el fomento de su aprovecha-miento cinegético. Sin embargo, cuando los objetivos cambiaron y se intentóestabilizar el tamaño poblacional para reducir el nivel de daños, los resulta-dos no están siendo los deseados. Al igual que está ocurriendo en muchaszonas de Europa, los planes de gestión de los ungulados están encontrandomuchos problemas a la hora de controlar sus poblaciones (Apollonio et al.,2010). En una población en expansión la tasa anual de crecimiento teórica sesituaría en torno al 35 % (Sáez de Buruaga, 1997; Boisaubert y Muron, 1997),por lo que la tasa de crecimiento observada en el SIZ, el 20%, sería mayor quela esperable tras aplicar una tasa de extracción cercana al 30%. Diversos fac-tores pueden estar influyendo en esta situación. En primer lugar, la extracciónpodría ser inferior a la prevista en el plan de caza, ya que según declaran loscazadores, el grado de cumplimiento del cupo está en el 80%. Por otra parte,parece existir cierta incidencia de caza furtiva, principalmente sobre machosy posiblemente se están declarando capturas que en realidad no se han pro-ducido, especialmente en el caso de hembras, lo que provocaría un desequi-librio en la sex-ratio, y que la población aumentara con mayor rapidez (Saézde Buruaga, 1997; Putman, 2003). Finalmente no es descartable que se estésubestimando el tamaño poblacional, pues se desconoce la exactitud de lametodología que se está empleando para estimar la densidad (Acevedo et al.,2010) y tampoco se ha cuantificado el número de corzos fuera de las áreasfavorables, ya que aunque la densidad pueda ser baja, estos hábitats suponenel 75% de la superficie total.
Para controlar la población de corzo probablemente sea necesario incre-mentar aún más la presión cinegética sobre la especie. Si actualmente el gradode cumplimiento del cupo se sitúa en torno al 80%, y viendo la evolución deotras especies de ungulados en Aragón (ver Marco et al., 2011 en este mismonúmero), parece problemático conseguir frenar la tendencia positiva de la
J. FERRERES, M. A. ESCUDERO, E. FERRER & J. MARCO
82 (Pirineos, 2011, Vol. 166, 69-86, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166004)

población sin la adopción de medidas drásticas como podría ser la autoriza-ción de la caza sin cupos. Estas medidas circunscritas a los sectores más con-flictivos deberían ser acompañadas por un seguimiento riguroso, tanto deevolución de las poblaciones como de los daños agrícolas. Aunque hastaahora se han marcado los objetivos de gestión influidos por la conflictividadasociada a los daños y no por la valoración objetiva de los daños, sería con-veniente desarrollar un sistema de recogida de datos que permita cuantificarsu importancia real y la efectividad de las medidas de prevención adoptadas.En este sistema se debería incluir también los datos de accidentes de tráficocon corzos.
Además de mantener el programa actual de seguimiento del corzo basa-do en indicios de presencia, parece necesario completar esta información condatos fiables sobre la estructura poblacional, basada en un número suficientede observaciones, que sea representativo del conjunto del área de estudio yespecialmente en los sectores donde se autorice la caza sin cupo, ya que estetipo de caza podría desequilibrar fuertemente la sex-ratio hacia las hembras,reduciendo la eficacia de las medidas de control adoptadas.
Referencias
Aragón, S., Braza, F. & San José, C., 1995. Socioeconomic, physiognomic andclimatic factors determining the distribution pattern of roe deer Capreoluscapreolus in Spain, Acta Theriologica, 40: 37-43.
Acevedo, P., Delibes-Mateos, M., Escudero, M. A., Vicente, J., Marco, J. &Gortázar, C., 2005. Environmental constrains in the colonization sequenceof roe deer (Capreolus capreolus Linnaeus, 1785) across the IberianMountains, Spain. Journal of Biogeography, 32: 1671-1680.
Acevedo P., Ruiz-Fons, F., Vicente, J., Reyes-García, A.R., Alzaga, V. &Gortázar, C., 2008. Estimating red deer abundance in a wide range ofmanagement situations in Mediterranean habitats. Journal of Zoology, 276:37-47.
Acevedo P., Ferreres, J., Jaroso, R., Durán, M., Escudero, M.A., Marco, J. &Gortázar, C., 2010. Estimating roe deer abundance for pellet groups countsin Spain: an assessment of methods suitable for Mediterranean wood-lands. Ecological Indicators, 10: 1226-1230.
Apollonio M., Anderson, R. & Putman, R., 2010. European Ungulates and theirmanagement in the 21st century. Cambridge University Press, 618 pp.Cambridge.
Ascaso, A. & Cuadrat, J.M., 1981. Geografía de Aragón. T.I. Guara Ed. 275 pp.Zaragoza.
GESTIÓN DEL CORZO EN ZARAGOZA: DE CONSERVACIÓN A CONTROL POBLACIONAL
83(Pirineos, 2011, Vol. 166, 69-86, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166004)

Aulak, W. & Babinska-Werka, J., 1990. Estimation of roe deer density based onthe abundance and rate of disapperance of their faeces from de forest.Actha Theriologica, 35 (1-2): 111-120.
Boisaubert, B. & Mouron, B., 1997. La situation du chevreuil en France.Bulletin Mensuel de l´Office National de la Chasse, 218 : 22-25.
Burón, D., Fernández, L., Martínez, A. L., López, J. C. & Fernández-Mellado,R., 2005. Status of roe deer population in Northen Guadalajara (Spain).Acta of the 7th European Roe Deer Meeting. Jerez (España).
Campbell, D., Swanson, J. & Sales, J., 2004. Comparing the precission andcost-effectiveness of faecal pellet group count methods. Journal of AppliedEcology, 41: 1185-1196.
Carvalho, P., 2007. Ecologia de uma população reintroduzida de corços(Capreolus capreolus L.). Tese de Mestrado em Ecologia. Universidade de Coimbra. Coimbra. Portugal.
Cederlund, G., Bergovist, J., Kjellander, P, Gill, R., Gaillard, J. M., Boisuabert,B., Ballon, P. & Duncan, P., 1998. Managing roe deer and their impact onthe enviroment: maximising the net benefits to society. The european roedeer: the biology of success. (ed. by R. Andersen, P. Duncan and J.D.C.Linnell), 338-372. Scandinavian University Press, Oslo (Noruega).
Centenera, R., 2002. Consideraciones en torno a la caza en la conservación delcorzo.
II Simposio sobre el corzo en España. Asociación del Corzo Español.195-204.Luarca. España.
Escudero, M. A., Ferreres, J. & Marco, J., 2007. Situación de la población de corzoen el Parque Natural del Moncayo. Informe interno del Gobierno de Aragón.
Escudero, O. & Franchés, M. J., 2005. Memoria de síntesis del Mapa Forestal deAragón. Serie Difusión nº 11, Consejo de Protección de la Naturaleza 119pp, Zaragoza.
Ferreres, J., Marco, J., Escudero M. A., Labarta, J. L. & Monrabal, L., 2010.Propuesta de plan de aprovechamiento cinegético del corzo para las temporadas2005-2011. Informes internos del Gobierno de Aragón.
Gaudin, J.C., Maillard, J.M., & Gailiard, G., 1997. Colonisation par leChevreuil (Capreolus capreolus) de la zone méditerrannéenne française. RevEcol. (Terre Vie), 52: 113-122.
Gortázar, C., Marco, J. & Martín, M., 1994. Seguimiento del Plan deMonitorización de Abundancias de Mamíferos. Factores ecológicos limitantes ydinámica de las poblaciones vulpinas. Informe interno del Gobierno deAragón.
Gortázar, C. & Marco, J., 1998. Distribución de grandes mamíferos en laComunidad Autónoma Aragonesa. Informe inédito del Gobierno de Aragón.
J. FERRERES, M. A. ESCUDERO, E. FERRER & J. MARCO
84 (Pirineos, 2011, Vol. 166, 69-86, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166004)

Gortázar C., Herrero, J., Villafuerte, R. & Marco, J., 2000. Historical examina-tion of the status of large mammals in Aragon, Spain. Mammalia, 64: 411-422.
Gortázar, C., Millán, J., Acevedo, P., Escudero, M. A., Marco, J. & Fernándezde Lucio, D., 2007. A large-scale survey of brown hare Lepus europaeus andIberian hare Lepus granatensis population at the limit of their ranges.Wildlife Biology, 13 (3): 244-250.
Linell, J. D. C., Duncan, P. & Andersen, R., 1998. The European roe deer: Aportait of a successful species. The european roe deer: the biology of success. (R.Andersen, P. Duncan and J.D.C. Linnell editores): 11-22. ScandinavianUniversity Press, Oslo (Noruega).
Marco, J., Herrero, J., Escudero, M. A., Fernández-Arberas, O., Ferreres, J.,García-Serrano, A., Giménez-Anaya, A., Labarta, J. L., Monrabal, L. & Prada,C., 2011. Veinte años de seguimiento poblacional de ungulados silvestresen Aragón. Pirineos, 166: 135-157.
Marco, J. & Jordán, G., 1990. Estudio de la población de corzo en el Macizo delMoncayo y área de influencia. Informe interno del Gobierno de Aragón.
Mayle, B. A., Putman, R. J. & Wyllie, I., 2000. The use of trackway counts toestablish an index of deer presence. Mammal Review, 30: 233-237.
Mayle, B.A., Peace, J.A. & Gill, R.M., 1999. How many deer? A field guide to esti-mating deer population size. UK: Forestry Commission. Field Book, 18: 93pp. Edimburgo (Reino Unido).
Osuna, D., Prada, C., Herrero, J. & Marco, J., 2006. Distribución de los ungu-lados silvestres en Aragón (2001-2005) determinada a partir de encuestas.Lucas Mallada, 13: 191-212.
Prada, C., García-Serrano, A., Fernández-Arberas, O. & Herrero, J., 2010.Seguimiento del corzo Capreolus capreolus en el Pirineo aragonés (2002-2010).Informe interno del Gobierno de Aragón.
Putman, R., 2003. The deer manager´s companion. Swan Hill Press, 180 pp. Bath.Reino Unido.
Sáez de Buruaga, M., 1997. La ordenación cinegética en caza mayor. I Curso deGestión de Cotos. Fundación para el estudio y defensa de la naturaleza y dela caza mayor, 37-56. Madrid. España.
Sampietro, F. J., 2000. Descripción fisiográfica de la Comunidad Autónoma deAragón. En: Aves de Aragón. Atlas de especies nidificantes. DiputaciónGeneral de Aragón, segunda edición: 21-38. Zaragoza.
Sinclair, A.R., Fryxell, J. & Caughley, G., 2006. Wilflife Ecology, Conservation,and Management. Blackwell Publishing: 450 pp. Oxford (Reino Unido).
Smart, J. C. R., Ward, A. I. & White, P. C. L., 2004. Monitoring woodland deerpopulations in the UK: an imprecise science. Mammal Review, 34: 99-114.
GESTIÓN DEL CORZO EN ZARAGOZA: DE CONSERVACIÓN A CONTROL POBLACIONAL
85(Pirineos, 2011, Vol. 166, 69-86, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166004)

Strickland, M. D., Harju, H., McCaffert, K., Miller, H., Smith, L. & Stoll, R.,1994. Harvest management. Research and management techniques for wildlifeand habitats. (ed. T.A. Bookhout) ,445-473. The Wildlife Society, Bethesa,Md. Estados Unidos.
Tsaparis, D., Katsanevakis, S., Ntolka, E. & Legakis, A., 2009. Estimating dungdecay rates of roe deer (Capreolus capreolus) in different habitat types of aMediterranean ecosystem: an information theory approach. EuropeanJournal of Wildlife Research, 55: 167-172
Van Laere, G., Maillard, D., Boutin, J. M. & Delorne, D., 1999. Le suivi despopulations de chevreuils: des methodes traditionnelles d´estimation auxindicateurs population-enviroment. Bulletin Mensuel de l´Office National dela Chasse, 244 : 46-53.
Virgós, E. & Tellería, J. L., 1998. Roe deer habitat selection in Spain: constraintson the distribution of a species. Canadian Journal of Zoology, 76: 1294-1299.
Williams, D., Acevedo, P., Gortázar, C., Escudero, M. A., Labarta, J. L., Marco,J. & Villafuerte, R., 2007. Hunting for answers: rabbit (Oryctolagus cinicu-lus) population trends in northeastern Spain. Journal of Wildlife Research, 53:19-28.
J. FERRERES, M. A. ESCUDERO, E. FERRER & J. MARCO
86 (Pirineos, 2011, Vol. 166, 69-86, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166004)

87
ELEMENTOS PARA UNA FILOGEOGRAFÍADE LA CABRA MONTÉS IBÉRICA
(CAPRA PYRENAICA SCHINZ, 1838)
RICARDO GARCÍA-GONZÁLEZ
Instituto Pirenaico de Ecología (IPE-CSIC). Apdo. 64. E-22700 [email protected]
ABSTRACT.– In order to provide some issues for a future study of Capra pyre-naica phylogeography I review classic and recent works considering data of geneticstudies known to date, as well as recent findings related to paleontology and bio-metrics. C. pyrenaica has experienced several population bottlenecks throughout itshistory that have led to a loss of genetic diversity. In addition, numerous recenttranslocations and population reinforcements can complicate the clarification ofintraspecific genetic differentiation. I review current theories about the evolution ofthe species. One of them establishes a recent evolution (past 18,000 years) fromancestors related to C. caucasica. I analyze in some detail the morphological char-acters (mainly teeth and horns) on which this theory is based, showing their highvariability. Present genetic studies show a clear relationship between C. pyrenaicaand Alpine ibex (C. i. ibex). They also show a north-south genetic differentiationbetween Iberian goats, occupying the Pyrenean goats (C. p. pyrenaica) an inter-mediate position between Alpine ibex and southern Iberian goats. Recent paleonto-logical findings evidence an ancient presence (Lower Pleistocene) of Capra in theIberian Peninsula. I propose the hypothesis that the descendants of these ancientgoats, may have come into contact with C. ibex in the Pyrenees and southernFrance, one or more times during the Upper Pleistocene. Intermediate characteris-tics, both genetic and biometric, of the extinct Pyrenean goats, would agree with thishypothesis. New paleontological records and more genetic studies (including fossiland recent materials) are necessary, to elucidate both the phylogeny of C. pyrenaicaand its intraspecific diversification.
Keywords: Caprinae, Iberian wild goat, distribution, phylogeny, horn biom-etry, palaeontology, genetics.
RESUMEN.– En este trabajo se realiza una revisión sintética de los trabajosclásicos y recientes que conciernen a la filogeografía de Capra pyrenaica, aportan-
Pirineos.Revista de Ecología de MontañaVol. 166, 87-122
Jaca, Enero-Diciembre, 2011ISSN: 0373-2568
eISSN: 1988-4281doi: 10.3989/Pirineos.2011.166005

88
RICARDO GARCÍA-GONZÁLEZ
do datos de los estudios genéticos conocidos hasta la fecha, así como de nuevos hallaz-gos relativos a su paleontología y biometría. C. pyrenaica ha experimentado varioscuellos de botella poblacionales a lo largo de su historia que han producido pérdida dediversidad genética. Al mismo tiempo, numerosas traslocaciones y reforzamientospoblacionales pueden complicar la clarificación de su diferenciación genética a nivelintraespecífico. Se revisan algunas teorías actuales sobre la evolución de la especie,que contemplan su evolución reciente (últimos 18.000 años) a partir de ancestros afi-nes a C. caucasica. Se examinan con cierto detalle los caracteres morfológicos en losque se basa dicha teoría (dientes y cuernos principalmente) y se evidencia la altavariabilidad de los mismos. Por otra parte, los estudios genéticos muestran una clararelación entre C. pyrenaica y el íbice de los Alpes (C. i. ibex). También muestran unadiferenciación genética norte-sur entre las cabras ibéricas, ocupando el bucardo pire-naico (C. p. pyrenaica) una posición intermedia. Recientes hallazgos paleontológicosponen de manifiesto una presencia muy antigua de Capra en la península Ibérica(Pleistoceno Inferior). Se propone la hipótesis de que los descendientes de estas cabrasantiguas, podrían haber entrado en contacto con cabras afines a C. ibex en losPirineos y sur de Francia, una o varias veces a lo largo del Pleistoceno Superior. Lascaracterísticas intermedias, tanto genéticas como biométricas, de las extintas cabrasdel Pirineo, coincidirían con esta hipótesis. Se requieren más estudios paleontológicosy genéticos (incluyendo materiales fósiles y actuales), para poder dilucidar, tanto lafilogenia de C. pyrenaica como su diversificación y relaciones intraespecíficas.
Palabras clave: caprinos, distribución, filogenia, biometría, paleontología,genética, bucardo.
1. Introducción
La filogeografía es la ciencia que estudia los principios y procesos quegobiernan las distribuciones geográficas de las líneas genealógicas dentro delas especies y entre especies relacionadas (Domínguez et al., 2009). Este tipode estudios utiliza el análisis geográfico y la información genealógica de lostaxones en cuestión, para dilucidar los procesos históricos que han condicio-nado su distribución actual y su variación genética. Recientes avances en elcampo de la genética de poblaciones y nuevos hallazgos paleontológicos, hanimpulsado el desarrollo de esta ciencia que ha dado lugar a una nueva visiónde la distribución y filogenia para varias especies de ungulados europeos,incluidas algunas considerablemente manipuladas por el hombre como elcorzo Capreolus capreolus (Randi et al., 2004), el ciervo Cervus elaphus (Skog etal., 2009) o el rebeco Rupicapra sp. (Rodríguez et al., 2010).
En el caso de la cabra montes ibérica (Capra pyrenaica Schinz, 1838), tantosu distribución pasada (Cabrera, 1911) como actual (Acevedo & Cassinello,2009), así como su diversificación genética (Pérez et al., 2002), han sido objetode numerosos estudios, siendo todavía un tema controvertido, empezando
(Pirineos, 2011, Vol. 166, 87-122, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166005)

por la misma validez de las subespecies establecidas por Cabrera (1914). Estacontroversia se debe, parcial y recientemente, a las numerosas traslocacionesy refuerzos poblacionales que ha experimentado esta especie, como conse-cuencia de su gran interés cinegético (Crampe, 1991; Acevedo & Cassinello,2009), y que han permitido la hibridación entre diferentes poblaciones yvariedades geográficamente alejadas. Por otra parte, la baja variabilidadgenética, fruto de su reciente diversificación y de los numerosos cuellos debotella poblacionales que ha soportado a lo largo de su historia, podría estardificultando la identificación y diferenciación de taxones intraespecíficos(Fandos et al., 1992). La cabra montés siempre ha sido un objeto de caza codi-ciado ya desde la prehistoria, como lo atestiguan importantes cazaderos delPleistoceno superior (Pailhaugue, 1995; Castaños, 1993). Además, histórica-mente, ha sufrido el acoso y la presión del ganado doméstico que ha reduci-do su hábitat (Acevedo et al., 2007). La consecuencia de todo ello son las extin-ciones, cuellos de botella y pérdida de diversidad genética general experi-mentada por la especie (Amills et al., 2004).
Por lo que se refiere al parentesco de C. pyrenaica con el resto de especiesdel género, existe también un profundo debate entre las teorías de algunospaleontólogos que atribuyen su origen a cabras afines a C. caucasica (Crégut-Bonneure, 1992, 2009; Rivals, 2002) y estudios genéticos que establecen unmayor parentesco con el íbice de los Alpes (C. i. ibex) (Hartl et al., 1992;Manceau et al., 1999a; Pidancier et al., 2006). En la compresión de este proce-so de especiación y diferenciación genética de las cabras del sudoeste euro-peo, podría haber tenido un papel relevante la cabra ibérica de los Pirineos(C. p. pyrenaica). Tanto la situación Sur de la península Ibérica (P.I. en adelan-te), alejada del Norte y Centro de Europa, como el efecto de barrera orográfi-ca de los Pirineos, permitieron a la Península constituirse como refugio gené-tico durante las glaciaciones cuaternarias para varias especies animales yvegetales (Taberlet et al., 1998; Hewitt, 2004; Centeno-Cuadros et al., 2009). Porotra parte, algunos taxones orófilos propios de los Pirineos, se han encontra-do en una posición muy favorable para establecer complejos hibridógenosentre los taxones mediterráneos de la P.I. y los propiamente centroeuropeos(Rodríguez et al., 2010). Desafortunadamente el bucardo (C. p. pyrenaica), seextinguió en el año 2000 sin apenas haber podido revelarnos casi nada de suscaracterísticas genéticas, morfológicas o ecológicas (García-González &Herrero, 1999; García-González, 2009).
El presente trabajo pretende ser una revisión y puesta al día de la infor-mación existente sobre la distribución, características genéticas e historia evo-lutiva de la cabra montés ibérica, con objeto de aportar materiales y sugerirhipótesis que puedan ser contrastadas en futuros estudios específicos sobresu filogeografía.
ELEMENTOS PARA UNA FILOGEOGRAFÍA DE LA CABRA MONTÉS IBÉRICA (CAPRA PYRENAICA...
89(Pirineos, 2011, Vol. 166, 87-122, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166005)

2. Las subespecies de Capra pyrenaica
Las subespecies admitidas actualmente por el grupo de caprinos de laUICN son las originalmente descritas por Cabrera (1911, 1914) (Herrero &Pérez, 2008). Aunque la UICN reconoce su problemática taxonomía, tambiénaconseja que su diversificación genética debiera formar parte de las futurasestrategias de conservación (Shackleton, 1997). Como es conocido, de las cua-tro subespecies definidas originalmente por Cabrera (1914), solo dos sobrevi-ven en la actualidad: C. p. victoriae y C. p. hispanica. La primera se distribuyepor el centro y noroeste de la P.I. y la segunda por el sur y el este de laPenínsula (Acevedo & Cassinello, 2009). C. p. lusitanica que habitaba enGalicia y norte de Portugal, se extinguió a principios del siglo pasado (França1917) y C. p. pyrenaica, propia de los Pirineos y que sirvió para definir el tipode la especie, se extinguió en el año 2000 (García-González, 2009).
La definición de las subespecies realizada por Cabrera (1914), se funda-mentó en un número relativamente pequeño de individuos y se basó en ladescripción de caracteres morfológicamente muy variables, tales como los deldiseño del pelaje y la forma de los cuernos en los machos. Esta circunstanciaha hecho que dicha clasificación haya sido puesta en duda reiteradamente alo largo del último siglo. Algunos autores (De la Cerda & De la Peña, 1971)sugieren que es mejor no hablar de subespecies de la cabra montés ibérica,sino de sus tipos y razas geográficas, llegando a proponer una nueva clasifi-cación basada también en la forma de los cuernos. Otros autores, apoyándo-se en la facilidad de hibridación entre prácticamente todas las especies delgénero (y por tanto su unidad específica), defendida por Couturier, (1962),invalidan las subespecies ibéricas y han reclamado reiteradamente la repo-blación en los Pirineos a partir de cualquier taxón ibérico (Clouet & Pfeffer,1979; Crampe, 1991). Las sociedades de cazadores francesas apoyaban tam-bién firmemente esta reivindicación.
Particularmente interesante por desconocidas y originales fueron las hipó-tesis de Camerano (1917), contemporáneo de Cabrera y en parte valedor de supropuesta. Según este autor, “las dos estirpes más antiguas en la P.I. serían lapirenaica y la hispánica, a las cuales habría que conceder categoría de espe-cies: C. pyrenaica y C. hispanica. El resto de las formas (victoriae, lusitanica ycabrerae) procederían de cruces entre las primeras y habría que conferirlescategoría de subespecies, asignándolas a la especie (pyrenaica o hispanica) quemás hubiera intervenido (o que hubiera dejado más caracteres) en su forma-ción”. En su trabajo expone una serie de caracteres craneales diferenciadoresentre estirpes.
Sería demasiado prolijo, extenderme aquí en las sucesivas revisiones, cam-bios de nombre y propuestas taxonómicas (Wyrwoll, 1999) realizadas duran-
RICARDO GARCÍA-GONZÁLEZ
90 (Pirineos, 2011, Vol. 166, 87-122, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166005)

te el pasado siglo sobre la catalogación intraespecífica de la cabra montés ibé-rica. El lector interesado puede encontrar una excelente revisión en Granadoset al., (2001). En este texto he preferido adoptar la clasificación seguida por laUICN (Shackleton, 1997; Herrero & Pérez, 2008), por ser ampliamente admi-tida, por coincidir con las propuestas clásicas y por su concordancia con losdatos genéticos, biométricos y paleontológicos que se van conociendo por elmomento. Lo cual no excluye que dicha clasificación pueda ser revocada enel futuro.
3. Distribución y abundancia de Capra pyrenaica
En cuanto a su distribución y abundancia, las poblaciones de C. pyrenaicahan pasado por muchas vicisitudes y aquí solo podemos hacer un breve men-ción a ellas (ver por ejemplo: Fandos, 1991; Alados & Escós, 1996; Pérez et al.,2002). Uno de los primeros en describir su distribución en la P.I. fue Cabrera(1911), quien dice que, en el pasado (refiriéndose probablemente al siglo XIX),prácticamente se extendía por todas las localidades con hábitat favorable,como lo prueban la gran cantidad de topónimos relacionados con el términocabra repartidos por toda la Península. En el mapa de distribución que adjun-ta (basado parcialmente en información del libro de Chapman & Buck, 1910),su distribución pasada en la P.I. era casi continua (fig. 1).
Aunque muy especulativa, puede ser interesante reproducir la descrip-ción que Cabrera (1911) realiza de las posibles vías de dispersión y coloniza-ción de la P.I. Desde los Pirineos la cabra montés alcanzó Galicia siguiendo lacornisa cantábrica. Desde allí descendió hacia el Sur por las sierras portugue-sas (Serra da Estrela) y Montes de Toledo hasta llegar a las sierras andaluzas.Esta ruta colonizadora le parece más probable que la de Burgos (Silos) yGuadarrama. Desde Andalucía y ante la barrera del estrecho de Gibraltarascendió por el Levante español hasta el río Ebro, que también constituyó unabarrera geográfica importante. Según Cabrera las cabras del Levante ibéricoson muy diferentes a las pirenaicas (a pesar de su proximidad) y muy simila-res a las andaluzas (op. cit. p. 965). Estas vías de dispersión se han represen-tado con flechas de trazo grueso en la figura 1. Aunque completamente espe-culativa, el interés de la hipótesis dispersiva de Cabrera, es que ofrece unabase teórica para la diversificación genética en subespecies de la cabra mon-tés ibérica. Así, la C. p. hispanica de Beceite sería la más alejada genéticamen-te de C. p. pyrenaica, y C. p. victoriae presentaría algunos caracteres morfológi-cos intermedios de las anteriores (pelaje), ocupando también una posicióngeográfica intermedia. Cabe mencionar que algunos resultados bioquímicosy genéticos mas recientes, coinciden parcialmente con esta hipótesis (Hartl et
ELEMENTOS PARA UNA FILOGEOGRAFÍA DE LA CABRA MONTÉS IBÉRICA (CAPRA PYRENAICA...
91(Pirineos, 2011, Vol. 166, 87-122, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166005)

al., 1992; Manceau et al., 1999b), aunque otros señalan mayor similitud gené-tica entre el bucardo pirenaico y la población de Tortosa (Jiménez et al., 1999).
La fuerte presión cinegética durante el periodo 1940-1970, mermó consi-derablemente las poblaciones de la cabra montés ibérica. En 1970, De la Cerda& De la Peña (1971) estiman el censo en 11.660 individuos repartidos en variaspoblaciones que representan en un detallado mapa. Preconizan y fomentan lacreación de Reservas de Caza y paulatinamente los efectivos fueron aumen-tando. Paralelamente numerosas reintroducciones, a veces incontroladas,tuvieron lugar hasta época reciente, algunas de las cuales quedaron reflejadasen el mapa de Crampe (1991) y más recientemente en el de Acevedo &Cassinello (2009). Posiblemente se produjeron también otras traslocacionesno constatadas oficialmente y muy probablemente ha habido intercambio deindividuos entre poblaciones e hibridaciones de estirpes de diferentes oríge-
RICARDO GARCÍA-GONZÁLEZ
92 (Pirineos, 2011, Vol. 166, 87-122, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166005)
Figura 1. Distribución de Capra pyrenaica en la península Ibérica a principios del siglo XX (líneasverticales) y anteriormente (punteado) según Cabrera (1911). Se han redibujado las probables
vías de dispersión (flechas) según interpretación del texto de Cabrera.Figure 1. Distribution of Capra pyrenaica in the Iberian Peninsula in the early XX century (verticallines) and previously (dotted) according to Cabrera (1911). Possible pathways (arrows) have been redrawn
from Cabrera’s text.

nes geográficos. A principio de 2000, sus efectivos se cifraban en torno a los50.000 individuos, según Pérez et al., (2002), quienes proporcionan además undetallado mapa de los núcleos poblacionales existentes entonces. En la actua-lidad, el censo podría sobrepasar ampliamente esa cifra, con más cantidad denúcleos poblacionales (J. Herrero, com. personal). El dictamen más recientede la UICN cataloga a la especie como Least concern (Herrero & Pérez, 2008).
C. pyrenaica ha demostrado una gran capacidad colonizadora cuando lascondiciones le han sido favorables (control de la caza, ausencia de depreda-dores, ausencia de cabras domésticas). Algunas de sus poblaciones han expe-rimentado una extraordinaria expansión natural (Gortazar et al., 2000;Granados et al., 2001; Pérez et al., 2002; Acevedo et al., 2007; Marco et al., 2011),lo cual puede hacer verosímil una amplia distribución, casi continua, por laP.I. como proponía Cabrera (1911). Ello hace pensar en un flujo genético rela-tivamente alto y en cierta dificultad para la diferenciación filogenética, excep-to cuando se produjeron reducciones de efectivos, aislamiento geográfico ycuellos de botella poblacionales, como así ha sucedido a lo largo de su histo-ria (Pérez et al., 2002; Amills et al., 2004; Sánchez, 2006). Sin embargo, lospocos estudios genéticos realizados hasta el momento sobre la especie, apo-yan las subespecies de Cabrera, como se expondrá más adelante (Manceau etal., 1999b; Jiménez et al., 1999; Pérez et al., 2002). Es posible que estas subes-pecies o variedades posean además requerimientos ecológicos diferenciados(Acevedo & Real, 2011). Algunas subespecies, como el bucardo o la cabra lusi-tánica, que presentaban rasgos morfológicos bien diferenciados (Cabrera,1914; França, 1917), apenas han podido ser estudiados debido a la escasez demateriales. Posiblemente el avance de las técnicas genéticas y los estudiosconsiguientes, permitan establecer en el futuro un panorama más claro sobrela diversificación filogenética de las cabras ibéricas.
4. Teorías sobre los orígenes de C. pyrenaica
Según el esquema de Cabrera (1911) expuesto anteriormente, C. pyrenaicahabría llegado por el Norte hasta los Pirineos y de ahí se habría expandido alresto de la península Ibérica. Esta visión sería coincidente con la hipótesis deCrégut-Bonneure (1992), según la cual, los ancestros de Capra ibex (íbice de losAlpes) habrían llegado a Francia en una primera oleada migratoria proce-dente de Oriente Próximo durante la glaciación de Riss, junto con Rupicaprarupicapra (R. pyrenaica había llegado ya en la glaciación de Mindel). A princi-pios del Würm, en una segunda oleada, llegó otra cabra del tipo Capra cauca-sica, la cual evolucionó, dando lugar a C. pyrenaica al final del Würm. La lle-gada de C. caucasica se hizo primero por el Macizo Central y después alcanzó
ELEMENTOS PARA UNA FILOGEOGRAFÍA DE LA CABRA MONTÉS IBÉRICA (CAPRA PYRENAICA...
93(Pirineos, 2011, Vol. 166, 87-122, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166005)

los Pirineos por el corredor del Ródano, siendo la causa de la extinción deHemitragus. Según Rivals (2002, p. 48, 109), la presencia de C. ibex en los Alpeshabría impedido, a la segunda migración de C. caucasica, que ésta se estable-ciese allí, por lo que continuó hasta el Macizo Central y luego al Pirineo orien-tal, en donde evolucionó a C. pyrenaica al final del Würm.
En la fig. 2 se expone un esquema espacio-temporal de esta doble migra-ción. La que dio lugar al íbice de los Alpes (C. i. ibex) habría tenido lugar haceunos 300.000 años (300 ka) y habría dejado restos fósiles en Turingia(Alemania) y Petralona (Grecia), bajo los nombre de C. camburgenis y C. mace-donica respectivamente, con las que estaría emparentadas según Crégut-Bonneure (2006). Una segunda oleada de un taxón afín a C. caucasica, habríatenido lugar durante el interglaciar Riss-Würm hace unos 120 ka. A estetaxón, establecido en el Macizo Central francés y el corredor occidental del
RICARDO GARCÍA-GONZÁLEZ
94 (Pirineos, 2011, Vol. 166, 87-122, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166005)
Figura 2. Esquema de la hipótesis de Crégut-Bonneure (1992) sobre el origen de las cabras salva-jes del sudoeste europeo durante el Pleistoceno Superior (ver texto). Distribución de C. ibex (pun-teado), C. pyrenaica (rayado) y extensión de los hielos. Localización de restos de Capra del
Pleistoceno Inferior (estrellas). Mapa de base según Griggo (1991).Figure 2. Diagram of the Crégut-Bonneure (1992) hypothesis about the origin of the SW European wildgoats during the Upper Pleistocene (see text). Distribution of C. ibex (dotted), C. pyrenaica (dashed) andice extent. Location of remains of Capra in the Early Pleistocene (stars). Base map from Griggo (1991).

Ródano, se le ha dado el nombre de C. caucasica-praepyrenaica y durante elúltimo periodo glaciar evolucionó gradualmente hasta originar C. pyrenaica,la cual estaría completamente diferenciada en los Pirineos y estribaciones, aprincipios del Magdaleniense, hace unos 18 ka (Rivals, 2002; Crégut-Bonneure, 2006). Supuestamente (aunque los autores mencionados no lodicen expresamente), desde los Pirineos C. pyrenaica habría colonizado elresto de la península Ibérica.
Esta hipótesis se basa en el estudio de los abundantes restos fósiles deja-dos por Capra en sur de Francia durante el último periodo glaciar, y tendríasu precedente en el artículo de Pales (1976-77) describiendo los cráneos deSoulabé. Sin embargo, no todos los paleontólogos la suscriben. Véase porejemplo Griggo (1991), Chaix & Desse (1994) o Arribas & Garrido (2008).
En contraposición a la teoría de Crégut-Bonneure (1992), otros autoresafirman que C. pyrenaica estaría ya asentada desde muy antiguo en la P.I. Asípor ejemplo, Couturier (1962), citando varios trabajos de principios del sigloXX, expone que el género Capra aparece por primera vez durante la glaciaciónde Riss bajo la forma de C. camburgensis, la cual pobló los Alpes antes de laglaciación de Würm. “Esto hace pensar que C. pyrenaica había adquirido suscaracteres en una época precedente, en la cual su hábitat no comunicaba conel de la forma alpina, como fue el caso durante el periodo del Würm” (op. cit.p. 537).
También para otros autores (Koby, 1958; Kurtén, 1968; Geist, 1987a; Chaix& Desse, 1994) C. camburgensis sería la cabra más antigua de Europa (230 kaaños), la cual poseería caracteres de C. pyrenaica y C. ibex, y podría ser el ori-gen común de ambas. Como veremos más adelante, los estudios genéticosapoyarían esta hipótesis (Manceau et al., 1999a). C. camburgensis y posterioreshallazgos de cabras fósiles del Riss o contemporáneos de ella (Petralona,Grecia ó Kozarnika, Bulgaria), han sido considerados exclusivamente ances-tros de C. ibex por Crégut-Bonneure (2006, 2009).
En los epígrafes siguientes, expondré varios argumentos que entrarían encontradicción con la teoría de Crégut-Bonneure, relacionados con la variabi-lidad morfológica en dientes y forma de los cuernos, la presencia antigua delgenero Capra en la P.I. y los resultados de los análisis genéticos.
5. Argumentos de la teoría de la doble migración de Crégut-Bonneure
5.1. Morfología dental de Capra ibex, C. caucasica y C. pyrenaica
Según Crégut-Bonneure (1992) es posible diferenciar la línea de C. caucási-ca-praepirenaica y C. ibex por determinados caracteres dentales y más en con-
ELEMENTOS PARA UNA FILOGEOGRAFÍA DE LA CABRA MONTÉS IBÉRICA (CAPRA PYRENAICA...
95(Pirineos, 2011, Vol. 166, 87-122, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166005)

creto, por la forma del molar M3 (“el diente más útil en ausencia del cráneo”).En la línea caucásico-pirenaica el ala metastilar es poco aparente y las super-ficies interestilares son de la misma anchura. Además los estilos son reduci-dos y estrechos (Rivals & Testu 2006, p. 713). En la línea ibex el ala metastilares muy saliente y la superficie interestilar del lado distal es mucho más anchaque la del lado mesial. También, el premolar P3 tiene un metacónido en formade columnita en el grupo caucásico-pirenaico y ninguno en el grupo alpino.“Con el tiempo parece que esta situación se invierte” (?) (Crégut-Bonneure,2009, p. 498). Por último, P2 es triradicular en la línea caucásica y biradicularen la alpina. C. caucasica-praepyrenaica y C. pyrenaica sensu stricto no presenta-rían diferencias dentales según Crégut-Bonneure (2009, p. 504).
En contraposición, Arribas & Garrido (2008) señalan el carácter “deriva-do” del ala metastilar de M3 y por tanto, hay algunas Capra que lo tienen yotras que no, por ejemplo C. baetica o C. alba (Moyà-Solà, 1987), a la cual con-sideran su descendiente. Por el contrario en C. pyrenaica y C. ibex sí estaríapresente, lo cual contradeciría las afirmaciones de Crégut-Bonneure. TambiénCarlos Calero et al., (2006) señalan que algunos ejemplares de C. alba deQuibas (Murcia) la presentan y otros no. En los ejemplares de C. caucasica-praepirenaica de la grotte Tournal (Languedoc) examinados por Magniez(2009), este carácter, al igual que los demás caracteres dentales señalados porCrégut-Bonneure, presenta una elevada variabilidad y en algún caso, como elde la desigualdad de las superficies interestilares en la línea caucásico-praepi-renaica, las diferencias son significativas.
El material de la cueva del Moro de Gabasa (Huesca) examinado por elautor, datado de alrededor de 40 ka (Blasco & Montes, 1997) y asignado a C.pyrenaica (Blasco, 1995), presenta en casi todos los casos una gran ala metasti-lar (fig. 3). En los cráneos de ejemplares actuales de C. p. pyrenaica este carác-ter también es muy variable (observación personal).
5.2. Morfología córnea de Capra ibex, C. caucasica y C. pyrenaica
Las diferencias en la morfología de los cuernos, entre las tres especies deCapra implicadas, son bien conocidas y se refieren especialmente a la diver-gencia y forma de los estuches córneos. Los machos de C. ibex presentan cuer-nos en forma de arco, relativamente juntos y con marcadas nudosidades en lacara anterior. Los cuernos de C. pyrenaica presentan la característica triple cur-vatura hacia afuera, hacia atrás y adentro, y C. caucasica tiene los cuernos muyabiertos desde la base para luego curvarse hacia atrás y hacia dentro(Couturier, 1962; Schaller, 1977). En el caso de los restos fósiles, normalmentelos estuches no están disponibles y las características se refieren a los núcleos
RICARDO GARCÍA-GONZÁLEZ
96 (Pirineos, 2011, Vol. 166, 87-122, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166005)

óseos o clavijas córneas. Los caracteres más apreciables en estos, se refieren ala forma y dimensiones de la sección transversal y al ángulo de divergenciaentre los núcleos óseos. En raras ocasiones en donde los núcleos se conservancasi enteros, pueden apreciarse pequeños efectos de torsión relacionables conlas curvaturas de los estuches (Griggo, 1991).
La divergencia lateral es más abierta en C. caucasica, débil y regular en C.ibex, débil en la base y luego acusada en C. pyrenaica (Crégut-Bonneure, 2009).Rivals (2002, p. 127) señala también las diferencias de C. caucasica-praepyre-naica (Capra aff. caucasica) respecto a C. pyrenaica: pivotes óseos muy diver-gentes; sección de los núcleos subtriangular; la cara anterior se inserta obli-cuamente en relación al plano frontal; y la cara antero-interna de los pivoteses cóncava y la postero-interna y externa son convexas. Todos estos caracte-res estarían presentes también en C. caucasica (la original del Caucaso).
ELEMENTOS PARA UNA FILOGEOGRAFÍA DE LA CABRA MONTÉS IBÉRICA (CAPRA PYRENAICA...
97(Pirineos, 2011, Vol. 166, 87-122, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166005)
Figura 3. Varias muestras de M3 de Capra pyrenaica de la Cueva del Moro de Gabasa (Norte deHuesca, 40 ka), presentando un ala metastilar muy desarrollada (Museo Provincial de Huesca).
Foto: R. García-González.Figure 3. Several samples of the M3 of Capra pyrenaica from Gabasa Cave (North Huesca province, 40ka), showing a well developed metastilar wing (Huesca Provincial Museum). Photo by R. García-
González.

Efectivamente los cráneos de Capra del sur de Francia comprendidos entre80 y 30 ka (OIS 3 y 4) presentan un gran ángulo de divergencia entre los cuer-nos: 65º en el cráneo de Portel (Rivals, 2002), 56º el cráneo de Arche (Rivals &Testu, 2006), 63º el de Tournal (Magniez, 2009), 66º y 76º los cráneos deSoulabé (Pales, 1976-77), ó 65º del más reciente (OIS 3) cráneo de Malarnaud(Griggo, 1991). Este último, asignado por Crampe & Crégut-Bonnoure (1994)a C. caucasica-pyrenaica es considerado por Griggo (1991) como C. pyrenaicasensu stricto. Expone dos argumentos: 1) aunque el núcleo óseo está roto, enla parte más distal se aprecia una ligera torsión típica de C. pyrenaica: “C. cau-casica nunca la tiene”, y 2) el ángulo de divergencia es un carácter muy varia-ble, tal como señalan también otros autores (Couturier 1962). A este respectoSchaller (1977, p. 23) expone: “Horns have been the main taxonomic charac-ter used to classify goats. New species and subspecies have sometimes beendesignated on the basis of a minor variation in the shape of one set of horns,clearly an undesirable tactic with a structure that is highly malleable in evo-lution”. Esta plasticidad morfológica queda también patente en los ejemplosexpuestos por Crampe (1991, p. 25 y sigs). Muchos ejemplares del Sur deEspaña tienen los cuernos divergentes, incluso algunos expertos en caza, pro-pusieron la creación de un tipo geográfico especial de trofeo para este carác-ter (tipo Lujar y Almijara de De la Cerda & De la Peña, 1971 o tipo “Rondeño”(Salas et al., 2008).
6. La sección de la base de los cuernos en Capra
Uno de los principales criterios para la diferenciación taxonómica de lascabras del Pleistoceno se basa en la forma de la sección de la base de los núcle-os óseos. Como veremos, la subjetividad en la apreciación de este carácter,junto con la falta de estudios estadísticos de su variabilidad, conceden por elmomento un valor muy limitado al mismo. Las diferencias entre C. caucasica-praepyrenaica y C. pyrenaica, en cuanto a la sección de los cuernos, consistiríanen un “redondeamiento” de los núcleos según Crégut-Bonneure (1992) o biendel paso de una sección sub-triangular a redondeada (Rivals, 2002). Sinembargo, años más tarde, Crégut-Bonneure (2009) asigna una sección sub-cuadrangular a C. ibex y oval para C. pirenaica y C. caucasica. Magniez (2009)compara la sección del cuerno del cráneo de Tournal con la de Portel y la deArche (asignados a C. caucasica-prapyrenaica, Rivals & Testu, 2006). Todas sonsubtriangulares y de forma parecida. Todas corresponderían a la línea caucá-sica antigua de Portel (OIS 4). En cambio, en el cráneo de Saulabé, que es másmoderno, la sección córnea es menos triangular, y todavía menos lo esMalarnaud (Griggo, 1991) que es “claramente oval”.
RICARDO GARCÍA-GONZÁLEZ
98 (Pirineos, 2011, Vol. 166, 87-122, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166005)

Según Pailhaugue (1995) la sección de los cuernos de C. ibex fósil (Mónaco)sería triangular, mientras que los de C. pyrenaica de la grotte de la Vache(Magdaleniense) son sub-cuadrangulares. En esa misma línea, Clot (1988)asegura que la sección del cuerno es oval en C. pyrenaica (comprimida late-ralmente) e “irregularmente triangular y ligeramente aplastada en sentidotransversal” en C. ibex, “donde se pueden distinguir tres caras y tres bordes”.Sin embargo, en la figura 3c de su trabajo reproduce una sección del cuernode C. ibex tomada de Couturier (1962) que es completamente ovalada (?). ParaCouturier (1962, p. 210) la sección del núcleo óseo en C. ibex es irregularmen-te triangular con tres caras y tres bordes.
Según parece, la forma de la sección de los cuernos, argüida por muchosautores como carácter diagnóstico entre especies y líneas evolutivas, adolecede bastante subjetividad. Lo deseable sería poder establecer comparacionescuantitativas del perímetro completo de las secciones córneas, tal como se hautilizado para las secciones de los dientes (Michaux et al., 2007; Caumul et al.,2009). Sin embargo, escasean las comparaciones con sentido estadístico enfósiles de Capra. A falta de trabajos en ese sentido, la única cuantificaciónensayada hasta el momento, sería la comparación de los diámetros longitudi-nal (DAP) y transversal (DLM) de la base de los núcleos óseos (como se rea-liza en la sección siguiente), lo cual nos proporciona un índice de compresióno achatamiento (DAP/DLM) de los mismos. Otra aproximación son las rectasde regresión entre estas variables, expuestas en algunos trabajos recientes(Rivals, 2002; Made et al., 2008). Según Rivals (2002) y Magniez (2009) dichasrectas serían claramente divergentes para la línea íbice y la línea caucásico-pirenaica. Cabe recordar que muchos autores (Koby, 1958; Couturier, 1962;Schaller, 1977) no atribuyen ningún carácter diagnóstico taxonómico a laforma de la sección transversal de los cuernos.
6.1. Comparaciones del índice de compresión de la sección de los cuernos
En la figura 4 se han representado los valores del índice de compresiónlateral (DAP/DLM > 1) o achatamiento (DAP/DLM < 1), para varios taxonesy complejos poblacionales (fósiles y actuales), tomados de la bibliografía, asícomo de datos propios. Los datos de C. pyrenaica del Holoceno correspondena la media de dos ejemplares de la Gouffre des Bouquetins (Clot, 1988), dosfósiles de Barbat y Annie (vertiente norte de los Pirineos, datos propios) y dosde la vertiente Sur (Larra y Millaris, García-González, in prep.). Los delMagdaleniense del Sur de Francia (Grotte de la Vache) corresponden aPailhaugue (1995). La media y la varianza del índice de estos últimos, se cal-cularon a partir de Mood et al., (1977, p.181), al igual que para los datos deChaix & Desse (1983). Los datos de C. caucasica-praepyrenaica corresponden a
ELEMENTOS PARA UNA FILOGEOGRAFÍA DE LA CABRA MONTÉS IBÉRICA (CAPRA PYRENAICA...
99(Pirineos, 2011, Vol. 166, 87-122, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166005)

una media de los cráneos de Malarnoud, Tournal, Arche-98 (3) y Portel (2)según Griggo (1991), Magniez (2009) y Rivals (2002).
Como puede apreciarse (fig. 4), los núcleos más comprimidos correspon-den a Capra alba, una cabra antigua de hace 1,7 Ma del sudeste de la P.I. des-crita por primera vez por Moyà-Solà (1987). Los datos pertenecen a dos indi-viduos de Venta Micena (Granada) y de Quibas (Murcia) (Made et al., 2008).Aunque estos autores no dudan en asignarlos al género Capra, Crégut-Bonneure (1999, 2006) asegura que se trata de Hemitragus cuya presencia hasido señalada recientemente en el Levante ibérico (Rivals & Blasco, 2008). Porlo que se refiere a la compresión de sus núcleos óseos, estarían más cercanasa los Hemitragus actuales que a Capra (fig. 4).
RICARDO GARCÍA-GONZÁLEZ
100 (Pirineos, 2011, Vol. 166, 87-122, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166005)
Figura 4. Índice de compresión (DAP/DLM) de la sección de los núcleos óseos de ejemplaresmachos para varios taxones de Capra fósil y actual. DAP: diámetro longitudinal, DLM diámetrotransversal. Valores medios (línea vertical), límites de confianza al 95% (barra gris) y rango devariación (barra blanca). La línea a trazos indicaría la forma circular (DAP = DLM). Entre parén-tesis nº de datos. (*) Por razones de espacio las referencias bibliográficas y elaboración de los
datos se indican en el texto.Figure 4. Flattening index (DAP/DLM) of horn core section of males for several fossil and current Caprataxa. DAP: longitudinal diameter, DLM: transverse diameter. Mean values (vertical line), 95% confidencelimits (gray bar) and variation range (white bar). The dashed line indicates the circular form (DAP =DLM). Number of data in brackets. (*) For space reasons, references and data processing are given in the
text.

C. baetica, también del sudeste ibérico y datada de unos 2,0 Ma, ha sidodescrita como la cabra más antigua encontrada hasta el momento en la P.I. yposiblemente en Europa (Arribas & Garrido, 2008). Los autores no tienenduda de que se trata de Capra y no de Hemitragus como afirma Crégut-Bonneure (2009). Sus núcleos óseos tienen un achatamiento considerable,curiosamente coincidente con el de las poblaciones actuales de C. pyrenaicadel sudeste ibérico (C. p. hispanica de la Sierra de Cazorla). Las cabras actua-les de Gredos (C. p. victoriae) poseen valores intermedios entre la subespeciepyrenaica y la hispanica, y, en cuanto a ese carácter, coincidirían con la hipóte-sis de Camerano (1917), que considera a victoriae como el resultado de unahibridación entre pyrenaica e hispanica (ver más arriba).
Los valores del índice de compresión de los ejemplares actuales de Pirineos(C. p. pyrenaica) coinciden mejor con los rangos de variación de las poblacionesfósiles del Holoceno y Pleistoceno Superior de la línea C. pyrenaica, incluidaslas consideradas como C. caucasica-praepyrenaica, si bien es verdad que estasúltimas también son similares a los valores de las cabras caucásicas actuales.Aunque estas últimas tienden más a la redondez, sus rangos de variación sola-pan todos entre sí, e incluso son coincidentes con los de otra cabra antigua,datada de 1,7 Ma, hallada también en la región del Caucaso (C. dalii deDmanisi, Georgia, Bukhsianidze & Vekua, 2006 en Made et al., 2008).
De la fig. 4 se desprende una cierta homogeneidad para los valores delíndice de compresión entre determinados taxones o formas geográficas. Asípor ejemplo, no hay diferencias significativas entre las formas bastante com-primidas de los íbices fósiles y actuales. Tampoco entre las formas afines a C.pyrenaica de los Pirineos y su entorno (Sur de Francia y Norte de Huesca) enlos últimos 80 ka, con un índice ligeramente comprimido, entre 1,0 y 1,1 apro-ximadamente. Precisamente el taxón que muestra más divergencia para estecarácter es C. pyrenaica (sensu lato), con formas muy achatadas (C. p. hispanica)y muy comprimidas (C. p. pyrenaica), sugiriendo una notable divergenciamorfológica entre las subespecies ibéricas reconocidas actualmente (García-González et al., in prep).
El escenario expuesto, podría coincidir con la hipótesis que propongo másadelante, según la cual, unas formas antiguas de C. pyrenaica, que habríanocupado el Sur y Centro de la P.I. (la cual habría actuado como refugio gené-tico durante las sucesivas glaciaciones del Cuaternario), habrían entrado encontacto con formas afines a C. ibex en los Pirineos y su entorno (Sur deFrancia), una o varias veces, en momentos no precisados del Pleistoceno,dando lugar a las cabras pirenaicas sensu stricto (C. p. pyrenaica). Sin embargo,los datos expuestos (fig. 4), tampoco permitirían descartar una hipotéticainfluencia de taxones relacionados con cabras de origen caucásico de secciónun poco más redondeada y cuernos más divergentes.
ELEMENTOS PARA UNA FILOGEOGRAFÍA DE LA CABRA MONTÉS IBÉRICA (CAPRA PYRENAICA...
101(Pirineos, 2011, Vol. 166, 87-122, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166005)

Las diferencias del índice de compresión de la sección de los cuernos(núcleos óseos) no parecen relacionadas con el tamaño del animal (lo cual lasharía clino- o denso-dependientes). Los ejemplares pirenaicos (C. p. pyrenaica)son por lo general de menor tamaño que los íbices (C. ibex) y su índice decompresión es menor, pero el tamaño de C. caucasica es mayor a ambos(Couturier, 1962) y sin embargo el índice de compresión de sus cuernos esmenor a C. ibex y similar a C. p. pyrenaica (fig. 4). Fandos (1995) asegura quela forma de la sección de los cuernos puede variar con la edad del individuo.En realidad, la variación debida a la edad es aplicable solo a los machos, loscuales muestran una tendencia al achatamiento con la edad, pero no para lashembras que no experimentan variaciones significativas en la forma de labase de los núcleos óseos en la edad adulta (García-González et al., in prep).Por otra parte, los machos están sujetos a una fuerte presión selectiva, que lesconduciría a aumentar el tamaño de los cuernos en condiciones de abundan-cia de recursos para aumentar su éxito reproductivo, mientras que las hem-bras no están sujetas a esta presión selectiva y maximizan el nivel de proteí-na en la leche (Geist, 1987b).
Por las razones anteriores, cabría pensar que el estudio de la variación dela forma de los núcleos óseos en hembras, podría ser más revelador desde elpunto de vista filogenético. Los datos de hembras fósiles de Capra son más
RICARDO GARCÍA-GONZÁLEZ
102 (Pirineos, 2011, Vol. 166, 87-122, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166005)
Figura 5. Índice de compresión (DAP/DLM) de la sección de los núcleos óseos de ejemplareshembras para varios taxones de Capra fósil y actual. Símbolos como en fig. 4. Capra ibex antiguas
(Petralona-Camburg) según Made et al., (2008).Figure 5. Flattening index (DAP/DLM) of horn core section of females for several fossil and current Caprataxa. Symbols as in figure 4. Ancient Capra ibex (Petralona-Camburg) according to Made et al., (2008).

escasos que los de los machos, pero en la figura 5 se exponen algunos valoresdel índice de compresión de los núcleos óseos, para varios taxones y pobla-ciones, tomados de la bibliografía y de datos propios. Los datos de C. ibexantiguas (Petralona, C. camburgensis) se han extraído a partir de Made et al.,(2008). Los de C. pyrenaica del Pleistoceno superior son una mezcla de seisindividuos (alguno también actual), pero parece que en su mayoría proce-dentes del Levante ibérico (Made et al., 2008) y su índice de achatamiento esmuy parecido al de las cabras de Gedros actuales (C. p. victoriae). El dato ais-lado de C. pyrenaica del Holoceno corresponde a un cráneo de 7.000 añoshallado en Ordesa (Pirineo Occidental) (García-González, in prep.) y es muysimilar a las hembras pirenaicas actuales (datos propios) y a las magdale-nienses de la Grotte de la Vache (Pailhaugue, 1995). Como puede observarselas tendencias generales son muy parecidas a las encontradas para losmachos, con cuernos más comprimidos en la línea alpina (C. ibex y afines) ycuernos más redondeados en C. pyrenaica, tanto más cuanto más al Sur de laP.I. nos trasladamos. Desafortunadamente, no poseemos por el momentodatos de hembras de C. baetica, pero si fueran similares a los achatados núcle-os óseos de los machos (fig. 4), tendríamos un gradiente de achatamientoNorte-Sur en cuyos extremos estarían las estirpes más antiguas: C. cambur-gensis y afines (quizás precursoras de C. ibex) y C. baetica (quizás precursorade las cabras ibéricas mediterráneas). Dicho gradiente es compatible con laidea de unas poblaciones pirenaicas con caracteres intermedios entre las alpi-nas y las del Sur de la P.I. y quizás resultado de la hibridación entre ambas.
7. La presencia de Capra en la península Ibérica durante el Pleistoceno
Uno de los principales problemas para aceptar la hipótesis de la doblemigración de Crégut-Bonneure (1992), radica en el hecho de que la presenciadel género Capra en la P.I., parece mucho más antigua de lo que se deduce desu teoría. Si C. pyrenaica se diferenció durante el Magdaleniense a partir de unantecesor afín a C. caucasica en el sur de Francia, ¿cual es el significado de lapresencia de Capra en la P.I. antes de ese periodo? Sin embargo, parece que lasevidencias de esa presencia, incluso en períodos tan antiguos como elPleistoceno inferior, son palpables y están sustentadas en numerosos hallaz-gos paleontológicos.
En la tabla 1 se exponen algunos de los yacimientos más destacables en laP.I. Como se ha señalado anteriormente, dos cabras antiguas C. baetica y C.alba han sido descritas y estudiadas recientemente en el sudeste ibérico yespecialmente en la cuenca de Guadix-Baza (si bien C. alba ya fue descrita porMoyà-Solà en 1987). Aunque Crégut-Bonneure (1999, 2006) asigna estos res-
ELEMENTOS PARA UNA FILOGEOGRAFÍA DE LA CABRA MONTÉS IBÉRICA (CAPRA PYRENAICA...
103(Pirineos, 2011, Vol. 166, 87-122, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166005)
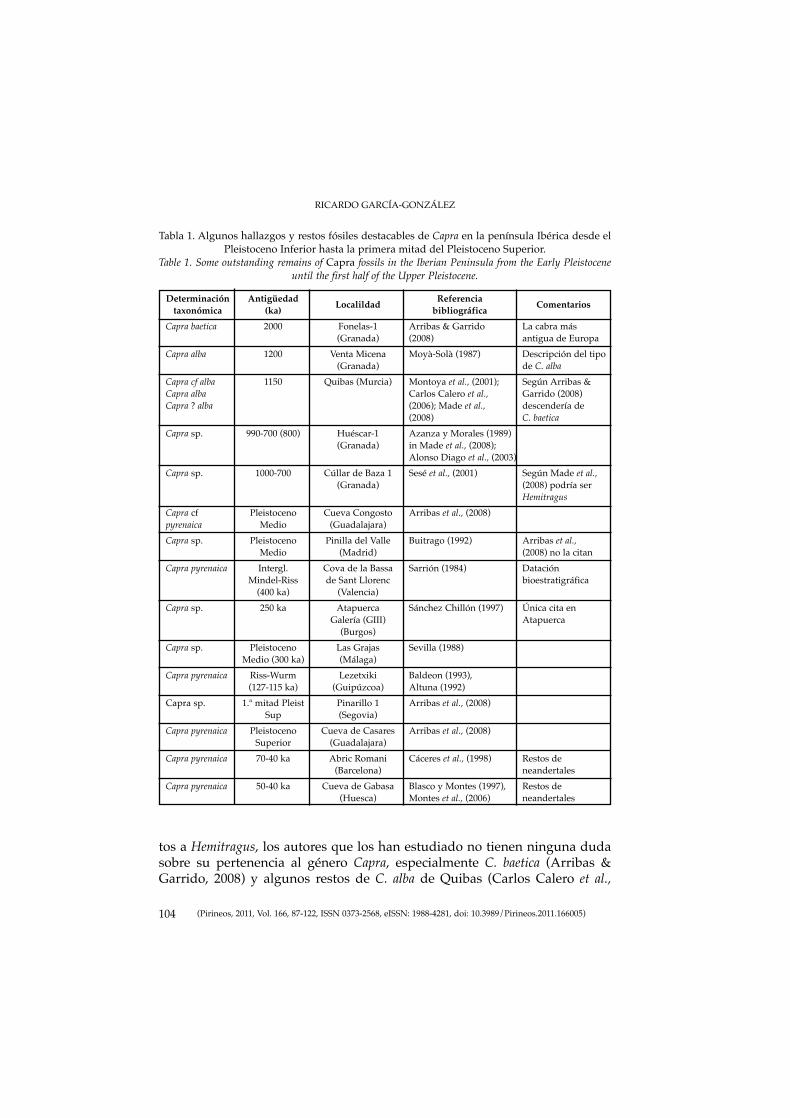
tos a Hemitragus, los autores que los han estudiado no tienen ninguna dudasobre su pertenencia al género Capra, especialmente C. baetica (Arribas &Garrido, 2008) y algunos restos de C. alba de Quibas (Carlos Calero et al.,
RICARDO GARCÍA-GONZÁLEZ
104 (Pirineos, 2011, Vol. 166, 87-122, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166005)
Tabla 1. Algunos hallazgos y restos fósiles destacables de Capra en la península Ibérica desde elPleistoceno Inferior hasta la primera mitad del Pleistoceno Superior.
Table 1. Some outstanding remains of Capra fossils in the Iberian Peninsula from the Early Pleistoceneuntil the first half of the Upper Pleistocene.
Determinación Antigüedad Referenciataxonómica (ka) Localildad bibliográfica Comentarios
Capra baetica 2000 Fonelas-1 Arribas & Garrido La cabra más(Granada) (2008) antigua de Europa
Capra alba 1200 Venta Micena Moyà-Solà (1987) Descripción del tipo(Granada) de C. alba
Capra cf alba 1150 Quibas (Murcia) Montoya et al., (2001); Según Arribas &Capra alba Carlos Calero et al., Garrido (2008)Capra ? alba (2006); Made et al., descendería de
(2008) C. baeticaCapra sp. 990-700 (800) Huéscar-1 Azanza y Morales (1989)
(Granada) in Made et al., (2008);Alonso Diago et al., (2003)
Capra sp. 1000-700 Cúllar de Baza 1 Sesé et al., (2001) Según Made et al.,(Granada) (2008) podría ser
HemitragusCapra cf Pleistoceno Cueva Congosto Arribas et al., (2008)pyrenaica Medio (Guadalajara)Capra sp. Pleistoceno Pinilla del Valle Buitrago (1992) Arribas et al.,
Medio (Madrid) (2008) no la citanCapra pyrenaica Intergl. Cova de la Bassa Sarrión (1984) Datación
Mindel-Riss de Sant Llorenc bioestratigráfica(400 ka) (Valencia)
Capra sp. 250 ka Atapuerca Sánchez Chillón (1997) Única cita enGalería (GIII) Atapuerca
(Burgos)Capra sp. Pleistoceno Las Grajas Sevilla (1988)
Medio (300 ka) (Málaga)Capra pyrenaica Riss-Wurm Lezetxiki Baldeon (1993),
(127-115 ka) (Guipúzcoa) Altuna (1992)Capra sp. 1.ª mitad Pleist Pinarillo 1 Arribas et al., (2008)
Sup (Segovia)Capra pyrenaica Pleistoceno Cueva de Casares Arribas et al., (2008)
Superior (Guadalajara)Capra pyrenaica 70-40 ka Abric Romani Cáceres et al., (1998) Restos de
(Barcelona) neandertalesCapra pyrenaica 50-40 ka Cueva de Gabasa Blasco y Montes (1997), Restos de
(Huesca) Montes et al., (2006) neandertales

2006). Por otra parte, en el artículo de Rivals & Blasco (2008) en el que descri-be un Hemitragus en Valencia por primera vez en la P.I., no se hace referenciaa esta asignación taxonómica de Crégut-Bonneure.
Durante el Pleistoceno Medio (1000-130 ka) los restos de Capra en la P.I.son escasos, tal vez debido a que el nivel de prospección no ha sido tan inten-so como en otras regiones (p.e. Sur de Francia). También, porque al tratarsede una especie vinculada a roquedos y medios abruptos, sus restos son másdifíciles de fosilizar (Garrido, 2008), excepto cuando se trata de yacimientoscorrespondientes a cazaderos humanos o de carnívoros salvajes (Pailhaugue,1995; Blasco, 1995). Aun así, existen unos cuantos yacimientos de ese periodoen los que se cita el género Capra (tabla 1).
Durante el Pleistoceno Superior (130 - 10 ka), los hallazgos de restos fósi-les de Capra son más abundantes. En la tabla 1 se han señalado algunos de losmás conocidos en la primera mitad de este periodo, pero se han descrito otrosyacimientos repartidos por toda la geografía ibérica: Amalda en Guipúzcoa(Altuna, 1990); Pinarillo-1 en Segovia (Arribas et al., 2008); Valdeboga enBurgos (Quam et al., 2001). También Yravedra (2004-5) señala varios yaci-mientos del Musteriense (90-45 ka) con Capra en Andalucía y Levante, porseñalar solo algunos anteriores al Magdaleniense (18-9 ka), cuando supuesta-mente se inició la diferenciación de C. pyrenaica.
¿Los restos de Capra encontrados en la P.I. durante el Pleistoceno son deCapra pyrenaica? o, ¿cual es su relación con las cabras ibéricas actuales?.Muchos de los restos hallados durante el Pleistoceno superior son asignadosa Capra pyrenaica, a veces de forma un poco automática. Lo más probable esque todavía no tengamos la información suficiente, como para conocer endetalle la diferenciación de estas cabras fósiles hasta su transformación en lascabras ibéricas actuales. Necesitamos aumentar los datos procedentes delregistro fósil y acompañarlos, si cabe, de análisis genéticos de esos mismosmateriales. Por el momento, solo algunas hipótesis pueden ser esbozadas.
8. ¿Como llegó el género Capra a la península Ibérica durante elPleistoceno?
Los estudios sobre las vías de colonización de la fauna del sudoeste euro-peo durante el Pleistoceno, suelen referirse a la dispersión del hombre y desus faunas asociadas (Bailey et al., 2008). Algunos de esos trabajos sugierenque la expansión de Homo estuvo influida por el seguimiento de los rebañosde grandes herbívoros a los que cazaba (p.e. bisonte) o bien de las carroñasdejadas por los grandes depredadores como el tigre dientes de sable (Arribas& Palmqvist, 1999). Las hipótesis más convencionales sugieren que esa vía de
ELEMENTOS PARA UNA FILOGEOGRAFÍA DE LA CABRA MONTÉS IBÉRICA (CAPRA PYRENAICA...
105(Pirineos, 2011, Vol. 166, 87-122, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166005)

colonización discurre por la ribera norte del Mediterráneo, procedentes deÁfrica, pero pasando por Oriente Próximo (“Levant corridor”) (Carbonell etal., 2006; Van der Made & Mateos, 2009). Los restos de Homo en Dmanisi(Georgia) datado de 1,7 Ma, en donde también se encontró una cabra (C.dalii), sería uno de los apoyos a esta hipótesis (Gabunia & Vekua, 1995).
Sin embargo, otros autores sugieren que esa colonización del sudeste euro-peo por Homo y faunas asociadas, pudo hacerse por el Estrecho de Gibraltar(Martínez-Navarro & Palmqvist, 1995; Cosson et al., 2005). Los recienteshallazgos de las mismas especies de fauna en el sur de la P.I. y en el norte deÁfrica, así como de las fluctuaciones del nivel del mar durante el Cuaternario,son los principales argumentos de los partidarios de la vía africana (Gibert etal., 2008). Junto a los restos de Homo en el SE ibérico del Pleistoceno inferior(considerados entre los más antiguos de Europa), se han encontrado restos defauna africana, tales como: Theropithecus cf. oswaldi (un papión), Megantereonwaitei (tigre dientes de sable), Hippopotamus antiquus, Equus granatensis yPachicrucuta brevirrostris. Otros datos interesantes son la aparición de Ursusetruscus (típico taxón euroasiático) en depósitos del Plioceno tardío deMarruecos (Casablanca) (Gibert et al., 2003).
En el máximo del último periodo glaciar, el nivel del mar era 135 m menorque en la actualidad y la distancia entre Marruecos y España de tan solo 7 km.En el Estrecho de Gibraltar había varias islas emergidas con vegetación rela-tivamente grandes (fig. 6), las cuales pudieron hacer de puente para la dis-persión de varias especies desde el Norte de África a Europa (Collina-Girard,2001; Cosson et al., 2005), durante el último periodo glaciar o anteriormentedurante el Pleistoceno. Aunque el Estrecho de Gibraltar constituye una barre-ra natural notable, la dispersión de mamíferos de talla grande a través delocéano (“oceanic dispersal theory”) parece que ha tenido lugar en gruposcomo los carnívoros de Madagascar (Yoder et al., 2003) o los monos platirri-nos del Nuevo Mundo (Schrago & Russo, 2005).
Por lo que se refiere al género Capra, los recientes hallazgos del SE ibérico(Made et al., 2008; Arribas & Garrido, 2008), así como de algunos restos encon-trados en el sur de Túnez (C. primaeva in Moyà-Solà, 1987) y en Marruecos(Aulagnier, 1992), permiten especular con una posible entrada de Capra por lavía del Estrecho de Gibraltar. Estás cabras antiguas, de cuernos muy redon-deados o incluso aplastados (fig. 4), podrían haber permanecido en la P.I. (quehabría actuado como refugio glaciar durante el Pleistoceno), y haber entradoen contacto ocasionalmente con cabras del tipo ibex en el norte de la P.I. y surde Francia. La gran proximidad genética entre C. pyrenaica y C. ibex mostradapor los últimos estudios genéticos sobre la filogenia del género (Pidancier etal., 2006), podrían apoyar una entrada de C. pyrenaica por el Norte, pero esodepende de la heterogeneidad genética de las cabras ibéricas y de en que
RICARDO GARCÍA-GONZÁLEZ
106 (Pirineos, 2011, Vol. 166, 87-122, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166005)

estirpes de C. pyrenaica se hayan obtenido las muestras genéticas para esosanálisis.
9. Resultados de los estudios genéticos
Los trabajos clásicos sobre la genética de Capra, basados en análisis bio-químicos de ejemplares actuales, se realizaron normalmente en el nivel taxo-nómico de género o familia (Hartl, 1990; Randi et al., 1991), pero desafortuna-damente no incluyen muestras de C. pyrenaica. En trabajos más recientes(Manceau et al., 1999a; Pidancier et al., 2006; Kazanskaya et al., 2007) se tratade establecer relaciones filogéneticas del género Capra, pero incluyendo ya aC. pyrenaica. En estos últimos, las conclusiones suelen ser coincidentes: a) C.ibex y C. pyrenaica tienen un origen monofilético, por lo que están fuertemen-te emparentadas. Las dos especies forman un clado muy sólido que excluyea otras especies como C. caucasica (Hartl, 1992; Manceau et al., 1999a). b) son
ELEMENTOS PARA UNA FILOGEOGRAFÍA DE LA CABRA MONTÉS IBÉRICA (CAPRA PYRENAICA...
107(Pirineos, 2011, Vol. 166, 87-122, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166005)
Figura 6. Oscilaciones del nivel del agua en el Estrecho de Gibraltar durante el último máximoglaciar hace 20 ka. Zonas emergidas en sombreado; en trazo grueso perfil de la costa actual (a
partir de Collina-Girard, 2001).Figure 6. Water level oscillations in the Strait of Gibraltar during the last glacial maximum (20 ka).
Shaded: emerged areas; thick outline: present coast (redrawn from Collina-Girard, 2001).

partidarios de la teoría de “una sola onda de migración” del genero Caprahacia el oeste de Europa y no de dos como sostiene Crégut-Bonneure (1992).Sus resultados serían más coincidentes con el escenario paleontológico de unancestro común del tipo C. camburgensis, que daría origen a las dos especiesactuales, planteados por otros autores (ver más arriba). Manceau et al.,(1999b) estiman que la inmigración de la especie única antecesora de C. ibex yC. pyrenaica) a Europa, sucedió entre 0,5 y 2,5 Ma y que la separación entre C.p. pyrenaica y el resto de las ibéricas fue contemporánea o se produjo muypoco después. A esta separación siguió un cierto aislamiento de C. p. pyrenai-ca en relación a las demás ibéricas.
La clara y fuerte relación genética entre C. ibex y C. pyrenaica podría ir endetrimento de la hipótesis, anteriormente expuesta, de una posible hibrida-ción entre ancestros de C. pyrenaica residentes en la P.I. y ancestros de C. ibexdurante periodos indeterminados del Pleistoceno. Sin embargo, es necesarioanalizar con más detalle las relaciones genéticas entre las diferentes poblacio-nes ibéricas y especialmente las poblaciones muestreadas. Actualmente, solounos pocos estudios se encuentran disponibles sobre la genética de las pobla-ciones ibéricas actuales y el íbice de los Alpes (C. i. ibex). Dos de ellos se basanen secuencias del ADN mitocondrial (Villalta et al., 1997; Manceau, 1999b) yotros dos en marcadores nucleares (microsatélites) (Jiménez et al., 1999;Amills et al., 2004). Otro estudio analiza los haplotipos de las poblaciones delsur de la P.I. (Márquez et al., 2002). Los dos primeros proporcionaron resulta-dos similares: la población pirenaica se muestra claramente diferenciada delresto de las ibéricas, con una distancia genética equidistante entre estas y elíbice de los Alpes. La distancia genética entre pirenaicas e ibéricas es de 4,9%,similar a la distancia entre bucardo e íbice (5,3%).
En los estudios de ADN nuclear (Jiménez et al., 1999; Amills et al., 2004),realizados con menos poblaciones que los de ADN mitocondrial, las diferen-cias entre las ibéricas se mantienen, pero la distancia genética con el íbice delos Alpes es mucho mayor y la población de Sierra Nevada queda netamentediferenciada de entre las ibéricas. Los cuatro estudios reconocen, además dela estirpe pirenaica, dos unidades genéticas de conservación (ESU) que agru-parían a las poblaciones del Norte peninsular (Gredos, Beceite, Muela deCortes) frente a las del Sur (Sra. Nevada, Cazorla, Ronda).
El estudio de Márquez et al., (2002) se basa en el análisis del gen del cito-cromo b en el ADN mitocondrial de 276 individuos de varias poblacionesandaluzas. Deducen que en el sur de la P.I. existen diez haplotipos, de los cua-les siete se encuentran en Sierra Nevada (esta población parece que no hapasado por cuellos de botella, al contrario que las restantes andaluzas anali-zadas). La mayor diversidad genética encontrada en la población de SierraNevada estaría en coincidencia con los resultados de Manceau et al., (1999b).
RICARDO GARCÍA-GONZÁLEZ
108 (Pirineos, 2011, Vol. 166, 87-122, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166005)

Basándose en el estudio de Márquez et al. (2002), Pérez et al. (2002) establecentambién tres ESU: Pirineos, Gredos y alrededores, y cabras del sudeste ibéri-co, lo cual estaría de acuerdo con las subespecies de Cabrera (1914).
Los resultados anteriores serían compatibles con la teoría de una pobla-ción de origen sur-mediterráneo antigua, que habría inmigrado durante elPleistoceno inferior, la cual habría quedado refugiada en la P.I. (Taberlet et al.,1998), y que durante alguno o varios periodos glaciares (especialmente el últi-mo), habría entrado en contacto con C. ibex, originando las poblaciones delNorte de España y sur de Francia (C. p. pyrenaica), con caracteres genéticos ymorfológicos intermedios. La hibridación interespecífica como factor evoluti-vo en el genero Capra, ha sido admitido en otros estudios (Pidancier et al.,2006, Ropiquet & Hassanin, 2006). También en otras especies próximas(Rupicapra), parece que ha podido suceder un proceso similar (Rodríguez etal., 2010). Los análisis genéticos sugieren la expansión y concentración de laspoblaciones durante el Cuaternario al ritmo de los periodos glaciares e inter-glaciares, así como la hibridación entre los clados del Oeste de Europa y losAlpes.
10. La península Ibérica como refugio de faunas pleistocenas y los Pirineoscomo área de hibridación mediterráneo-centroeuropea
Cada vez existe un mayor cuerpo de evidencias acerca del papel comorefugio genético jugado por las penínsulas del sur de Europa durante elPleistoceno. Taberlet et al., (1998) a partir de un análisis filogeográfico en elque incluyen 10 taxones genéticamente muy diferentes (mamíferos, anfibios,artrópodos y árboles), concluyen que la P.I., Italia y los Balcanes habían actua-do como reservorios de biodiversidad durante la última glaciación y que lasregiones del Norte de Europa habían sido colonizadas desde la P.I. y losBalcanes (la península Itálica tuvo menos influencia debido a la barrera de losAlpes). Los Pirineos podrían haber actuado como una zona de “sutura” endonde confluyen dos o más rutas migratorias y podrían haber sido un área dehibridación.
Hewitt (2004) sugiere también que el oso pardo (Ursus arctos) salió deIberia y colonizó todo el oeste de Europa hasta que se hibridó con los quevenían del Este. Sommer et al., (2008) en un exhaustivo estudio sobre la filo-geografía del ciervo (Cervus elaphus) en Europa, llegan a conclusiones muysimilares: el ciervo se expandió rápidamente a principios del Holoceno haciael centro y norte de Europa a partir de sus refugios glaciares (Iberia, Italia,Balcanes), en donde habían permanecido por lo menos durante la segundamitad del último periodo glaciar. Lo mismo ha sido propuesto para especies
ELEMENTOS PARA UNA FILOGEOGRAFÍA DE LA CABRA MONTÉS IBÉRICA (CAPRA PYRENAICA...
109(Pirineos, 2011, Vol. 166, 87-122, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166005)
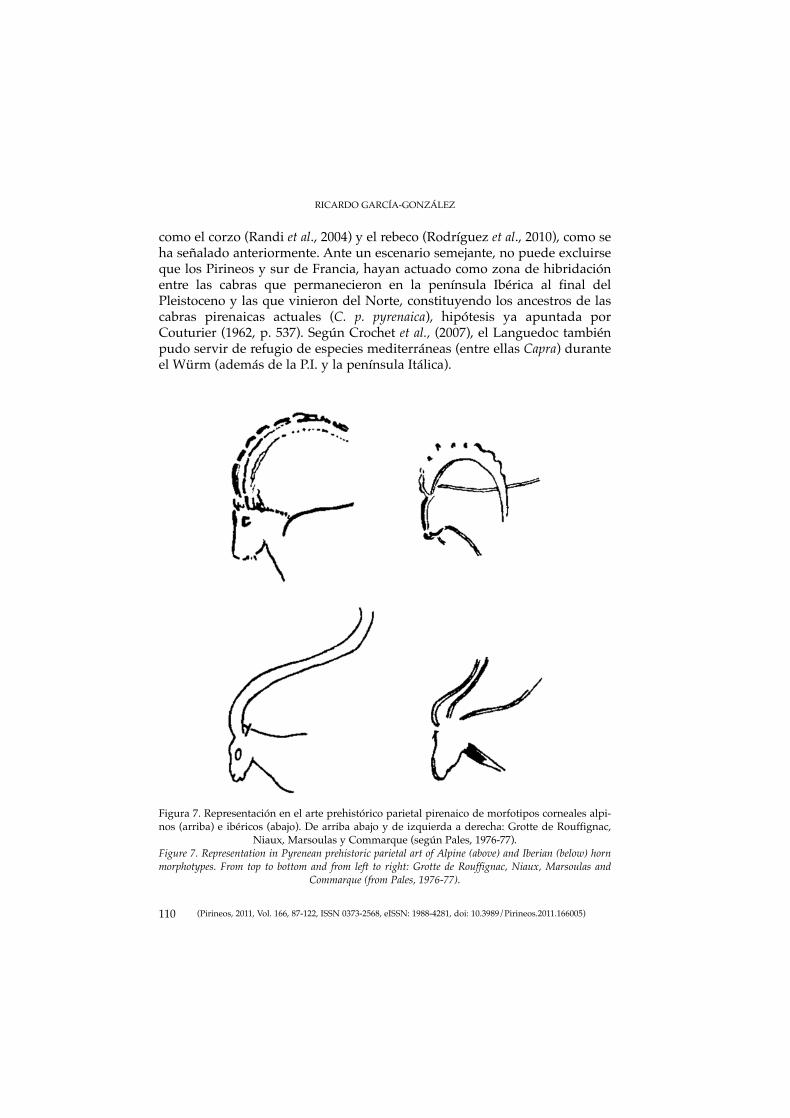
como el corzo (Randi et al., 2004) y el rebeco (Rodríguez et al., 2010), como seha señalado anteriormente. Ante un escenario semejante, no puede excluirseque los Pirineos y sur de Francia, hayan actuado como zona de hibridaciónentre las cabras que permanecieron en la península Ibérica al final delPleistoceno y las que vinieron del Norte, constituyendo los ancestros de lascabras pirenaicas actuales (C. p. pyrenaica), hipótesis ya apuntada porCouturier (1962, p. 537). Según Crochet et al., (2007), el Languedoc tambiénpudo servir de refugio de especies mediterráneas (entre ellas Capra) duranteel Würm (además de la P.I. y la península Itálica).
RICARDO GARCÍA-GONZÁLEZ
110 (Pirineos, 2011, Vol. 166, 87-122, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166005)
Figura 7. Representación en el arte prehistórico parietal pirenaico de morfotipos corneales alpi-nos (arriba) e ibéricos (abajo). De arriba abajo y de izquierda a derecha: Grotte de Rouffignac,
Niaux, Marsoulas y Commarque (según Pales, 1976-77).Figure 7. Representation in Pyrenean prehistoric parietal art of Alpine (above) and Iberian (below) hornmorphotypes. From top to bottom and from left to right: Grotte de Rouffignac, Niaux, Marsoulas and
Commarque (from Pales, 1976-77).

La posible presencia de C. ibex en los Pirineos había sido señalada desdehace tiempo en la literatura especializada: Koby (1958), Couturier (1962),Pales (1976-77), Clot & Duranthon (1990). Dichas afirmaciones habían sidonormalmente invalidadas, por estar basadas en su mayoría en pinturasrupestres dejadas por el hombre prehistórico en grutas del periodoMagdaleniense. La subjetividad del pintor y la variabilidad córnea de C. pyre-naica pudo ser la causa de que la misma especie presentara cuernos en lira yen arco, como sucede en la actualidad. Sin embargo, hay un carácter muy típi-co de los íbices (C. ibex), que es la presencia de nudosidades o protuberanciasen la superficie anterior de los estuches córneos, que no pasó desapercibidapara el hombre prehistórico. Como tampoco lo hizo, la característica dobleinflexión de las cabras ibéricas. Pales (1976-77) y Couturier (1962) muestranuna amplia colección de dibujos en los que se representan ambos caracteres(fig. 7). Curiosamente, en la extensa colección de dibujos de Capra del arteparietal andaluz, mostrada por Carrasco et al., (2004), ningún ejemplar pre-senta nudosidades en los cuernos. Tampoco aparecen en las cabras del arteparietal levantino (J. Herrero, com. pers.).
Quizás más convincente que la figuración parietal, puede ser la apariciónde algún ejemplar actual con nudos en los cuernos entre el escaso material de
ELEMENTOS PARA UNA FILOGEOGRAFÍA DE LA CABRA MONTÉS IBÉRICA (CAPRA PYRENAICA...
111(Pirineos, 2011, Vol. 166, 87-122, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166005)
Figura 8. Izq.) Morfotipo actual de cuernos alpinos (Capra i. ibex). Der.) Morfotipo actual de cuer-nos alpinos en un cráneo de Capra p. pyrenaica (PNOMP-03)(fotos: D. Gómez y R. García-
González).Figure 8. Left: Current morphotype of Alpine horns (Capra i. ibex). Right: Alpine morphotype horn inCapra p. pyrenaica (PNOMP-03, IPE-CSIC collections) (Photo by D. Gómez & R. García-González).

C. p. pyrenaica que se conserva en la actualidad, como puede apreciarse en lafigura 8. Más que la presencia física de íbices (tal como los conocemos hoy) enlos Pirineos al final de la última glaciación, una interpretación mas lógica con-sistiría en que, al tratarse de una zona hibridógena entre ambas formas, losPirineos podrían haber albergado individuos con alelos de la forma de cuer-nos alpina, aunque la frecuencia génica hiciera dominante las formas en lira,típicas de la forma pirenaica. Estos alelos raros en C. p. pyrenaica podríanhaberse manifestado esporádicamente a lo largo de su historia. La clarifica-ción de esta y otras hipótesis expuestas tendrá que esperar a la realización deestudios genéticos, que incluyan también material fósil.
11. Conclusiones y propuestas de investigación
Capra pyrenaica es considerada una especie diferenciada dentro del géneroy endémica de la península Ibérica. Los análisis genéticos realizados hasta elmomento la sitúan muy emparentada con el íbice de los Alpes (C. i. ibex), perotambién manifiestan una notable variabilidad genética intraespecífica conuna clara diferenciación norte-sur. La cabra montés ibérica siempre ha sidoun codiciado objetivo de caza, tanto para el hombre prehistórico como para elactual. Este hecho, unido a la presión ejercida por las cabras domésticas, haconducido a algunas de sus poblaciones a drásticas reducciones de número,ocasionando cuellos de botella poblacionales con pérdida de diversidad gené-tica. La presión humana (caza, pastoreo), ha conducido a algunas de sussubespecies o variedades a la desaparición. Más recientemente, algunaspoblaciones han sido reintroducidas o traslocadas para reforzar a otras, pro-duciendo mezcla del material genético. Las vicisitudes de su historia recientedificultan la interpretación, tanto de su distribución actual como de las rela-ciones de parentesco entre poblaciones y variedades, aunque por el momen-to se admiten dos grandes clados, ocupando aproximadamente la mitadnord-occidental y sud-oriental de la península Ibérica, respectivamente, y quecoincidirían con las subespecies victoriae e hispanica descritas por Cabrera(1914).
Algunas teorías basadas en estudios paleontológicos sitúan el origen de C.pyrenaica en una época muy cercana (18 ka) y derivando de un antepasadoafín a C. caucasica. Ésta habría emigrado del Cercano Oriente hacia el oeste deEuropa durante el interglaciar Riss-Würm, alcanzando el sur de Francia y losPirineos, en donde habría evolucionado hacia C. pyrenaica sin haber manteni-do contacto con el íbice de los Alpes, instalados allí desde una migraciónanterior hace aproximadamente 300 ka. Tanto los estudios genéticos, como la
RICARDO GARCÍA-GONZÁLEZ
112 (Pirineos, 2011, Vol. 166, 87-122, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166005)

variabilidad de los caracteres morfológicos en los que se sustenta, confierenpor el momento poca consistencia a esta teoría. Por otra parte, en el registrofósil de C. pyrenaica en la península Ibérica, se aprecia una notable continui-dad durante todo el Pleistoceno superior, y recientemente se han encontradorestos mucho más antiguos de Capra datados del Pleistoceno Inferior. Losmateriales del Pleistoceno Medio son escasos, pero ello puede deberse a lainsuficiente prospección y a la dificultad de dejar restos fósiles en mediosrocosos. Sin embargo, la presencia de Capra en la P.I. a lo largo del todo elCuaternario parece sumamente factible.
La relación de estas cabras antiguas con Capra pyrenaica es difícil de preci-sar con la información disponible hasta el momento, pero es probable que suscaracteres estuvieran ya bien definidos durante el Pleistoceno superior. Su lle-gada a la P.I. podría haberse realizado por el Norte, aunque tampoco es des-cartable la vía del Estrecho de Gibraltar. En cualquier caso, esta cabra “ibéro-mediterránea” se encontraría desde hace varios cientos de miles de años en lapenínsula Ibérica, que habría actuado como refugio europeo durante las gla-ciaciones. En periodos climáticamente favorables del Pleistoceno, podríanhaber entrado en contacto con cabras de centro-Europa, una o varias veces,hibridándose en el área de “sutura” de los Pirineos y sur de Francia (en expre-sión de Taberlet). Este contacto podría haberse producido con los antecesoresde C. ibex del tipo C. camburgensis, hace 200 o 300 ka, o más recientemente, alfinal del último periodo glaciar, con cabras más similares a los íbices alpinosactuales. Como resultado de esta hibridación o hibridaciones, las cabras pire-naicas (el extinto bucardo C. p. pyrenaica) presentarían características genéti-cas y morfológicas intermedias entre las poblaciones más meridionales de laP.I. (quizás más ligadas a las cabras “ibéro-mediterráneas” antiguas) y el íbicede los Alpes. Este escenario es concordante con los datos conocidos hasta elmomento, tanto de la genética como de la morfometría de los cuernos.
Numerosas lagunas existen todavía para conocer con cierto detalle el ori-gen, filogenia y diversificación de las cabras monteses ibéricas y de las rela-ciones con sus vecinas europeas o africanas. Es necesario profundizar muchomás en el registro fósil y realizar nuevos hallazgos paleontológicos quecubran los enormes huecos temporales existentes en la actualidad. Los análi-sis morfométricos del material fósil deberían realizarse con aproximaciónestadística, siempre que su abundancia y estado de conservación lo permitan.Al mismo tiempo, sería necesario realizar más estudios genéticos que inclu-yeran tanto material actual como fósil. Posiblemente el avance y simplifica-ción de las técnicas genéticas y los estudios consiguientes, permitan estable-cer en el futuro un panorama más clarificador sobre la diversificación filoge-nética de las cabras ibéricas.
ELEMENTOS PARA UNA FILOGEOGRAFÍA DE LA CABRA MONTÉS IBÉRICA (CAPRA PYRENAICA...
113(Pirineos, 2011, Vol. 166, 87-122, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166005)

Agradecimientos
Deseo expresar mi más sincero agradecimiento a las siguientes institucionesy organizaciones, que amablemente me han facilitado el acceso al material oste-ológico en el que se basa el presente estudio: Museo Provincial de Huesca(Gobierno de Aragón), Parque Nacional de Ordesa y Monte Perdido (Gobiernode Aragón), Parc National des Pyrénées (Ministère de l’Environnement,Francia), Grup Espeleològic Pedraforca (Barcelona) y Grupo Espeleológico deEstella (Navarra). En especial quisiera agradecer a Jean Paul Crampe (PNP) yElena Villagrasa (PNOMP) las facilidades prestadas para el estudio de dichomaterial. Lourdes Montes (Universidad de Zaragoza) me orientó y ayudó parael examen de los restos de la Cueva del Moro de Gabasa (Huesca) y MontserratEsteban (Institut Català de Paleoecologia Humana i Evolució Social) me prestóapoyo bibliográfico. Agradezco también la revisión y comentarios a la primeraversión del manuscrito, realizados por Juan Herrero (Universidad deZaragoza), Pelayo Acevedo (Universidad de Málaga), Jean Paul Crampe (PNP)y José Enrique Granados (Junta de Andalucía), que han contribuido notable-mente a la mejora del artículo. Una parte de este estudio se ha realizado confondos provistos por el Plan de Recuperación del bucardo (Gobierno deAragón) durante la década de 1990 a 2000.
Referencias
Acevedo, P. & Cassinello, J., 2009. Biology, ecology and status of Iberian ibexCapra pyrenaica: a critical review and research prospectus. Mammal Rev., 39:17–32.
Acevedo, P., Cassinello, J. & Gortazar, C., 2007. The Iberian ibex is under anexpansion trend but displaced to suboptimal habitats by the presence ofextensive goat livestock in central Spain. Biodiversity and Conservation, 16:3361–3376.
Acevedo, P. & Real, R., 2011. Biogeographical differences between the twoCapra pyrenaica subspecies, C. p. victoriae and C. p. hispanica, inhabiting inthe Iberian Peninsula: conservation implications. Ecological Modelling, 222:814-823.
Alados, C.L. & Escós, J., 1996. Ecología y comportamiento de la cabra montés: con-sideraciones para su gestión. Museo Nacional de Ciencias Naturales.Monografía nº 11, 329 pp., Madrid.
Alonso Diago, M.A., Hoyos, M. & Alberdi, M.T., 2003. Tafonomía del yaci-miento de vertebrados pleistoceno de Huéscar-l (Granada, España).Estudios Geológicos, 59: 213-227.
RICARDO GARCÍA-GONZÁLEZ
114 (Pirineos, 2011, Vol. 166, 87-122, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166005)

Altuna, J., 1990. La caza de los herbívoros durante el Paleolítico y Mesolíticodel País Vasco. Munibe, 42: 229-240.
Altuna, J., 1992. El medio ambiente durante el Pleistoceno Superior en laregión cantábrica con referencia especial a sus faunas de mamíferos.Munibe, 44: 13-29.
Amills, M., Jiménez, N., Jordana, J., Riccardi, A., Fernández-Arias, A., Guiral,J., Bouzat, J.L., Folch, J. & Sánchez, A., 2004. Low diversity in the major his-tocompatibility complex class II DRB1 gene of the Spanish ibex, Caprapyrenaica. Heredity, 93: 266-272.
Arribas, A. & Garrido, G., 2008. Los representantes más antiguos del géneroCapra (Bovidae, Artiodactyla, Mammalia) en el registro euroasiático(Fonelas P-1, Cuenca de Guadix, España). En: Arribas, A. (ed.): Vertebradosdel Plioceno superior terminal en el suroeste de Europa: Fonelas P-1 y el ProyectoFonelas, pp. 461-473. Instituto Geológico y Minero de España, Madrid.
Arribas, A. & Palmqvist, P., 1999. On the Ecological Connection BetweenSabre-tooths and Hominids: Faunal Dispersal Events for the First HumanArrival in Europe. Journal of Archaelogical Science, 26: 571-585.
Arribas, A., Solano, J.G., Garrido, G. & Carroza, J.A., 2008. El registro dife-rencial de carnívoros y homínidos en los karst del Pleistoceno del SistemaCentral español: condicionantes geológicos y espaciales. En: Diez, J. C.(ed.): Zooarqueología hoy. Encuentros Hispano-Argentinos, pp. 39-52.Universidad de Burgos, Burgos.
Aulagnier, S., 1992. Zoogeography and status of the moroccan wildUngulates. En: Spitz, F., Janeau, G., González, G. & Aulagnier, S. (eds.):Ongulés/Ungulates 91, pp. 365-369. S.F.E.P.M. - I.R.G.M., Paris - Toulouse.
Bailey, G., Carrión, J.S., Fa, D. A., Finlayson, C., Finlayson, G. & Rodríguez-Vidal, J., 2008. The coastal shelf of the Mediterranean and beyond:Corridor and refugium for human populations in the Pleistocene.Quaternary Science Reviews, 27: 2095-2099.
Baldeon, A., 1993. El yacimiento de Lezetxiki (Guipuzcoa, País Vasco). Losniveles musterienses. Munibe, 45: 3-97.
Blasco, F., 1995. Hombres, fieras, y presas. Estudio arqueozoologico y tafono-mico del yacimiento del Paleolitico Medio de la Cueva de Gabasa 1(Huesca). Monografias Arqueologicas, 38: 1-205.
Blasco, F. & Montes, L., 1997. Los hiénidos del yacimiento musteriense deGabasa 1 (Huesca, España). Bolskan, 14: 9-27.
Buitrago, A.M., 1992. Estudio de los artiodáctilos del yacimiento del PleistocenoMedio de Pinilla del Valle (Madrid). Tesis doctoral, UniversidadComplutense. Madrid.
Cabrera, A., 1911. The subespecies of the Spanish ibex. Proceed. Zool. Soc.London, 66: 963-977.
ELEMENTOS PARA UNA FILOGEOGRAFÍA DE LA CABRA MONTÉS IBÉRICA (CAPRA PYRENAICA...
115(Pirineos, 2011, Vol. 166, 87-122, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166005)

Cabrera, A., 1914. Fauna ibérica: Mamíferos. Museo Nacional de CienciasNaturales, 441 pp, Madrid.
Cáceres, I., Rosell, J. & Huguet, R., 1998. Séquence d’utilisation de la biomas-se animale dans le gisement de l’Abric Romani (Barcelone, Espagne).Quaternaire, 9: 379-383.
Camerano, L., 1917. Contributo allo studio degli Stembecchi Iberici. Bollettinodei Musei di Zoologia ed Anatomia comparata della R. Università di Torino, 32:1-30.
Carbonell, E. & Rodríguez, X.P., 2006. The first human settlementof Mediterranean Europe. C. R. Palevol., 5: 291-298.
Carlos Calero, J.A., Made, J.V. D., Mancheño, M.A., Montoya, P. & Romero,G., 2006. Capra alba Moyà-Solà, 1987 del Pleistoceno Inferior de la Sierra deQuibas (Albanilla, Murcia, España). Estudios Geológicos, 62: 571-578.
Carrasco, J., Riquelme, J.A., Sanchidrián, J.L., Pachón, J.A. & Soledad, M.S.,2004. La cabra montés (Capra pyrenaica, Schinz 1838) en el registro delPleistoceno Superior y Holoceno de Andalucía y su incidencia en el ArtePrehistórico. Antiquitas, 16: 27-66.
Castaños, P.M., 1993. Estudios de los macromamíferos de los niveles paleolí-ticos de “Chaves”. Bolskan, 10: 9-30.
Caumul, R., Polly, P.D. & Janis, C., 2009. Phylogenetic and environmentalcomponents of morphological variation: skull, mandible, and molar shapein marmots (Marmota, Rodentia). Evolution, 59: 2460-2472.
Centeno-Cuadros, A.; Delibes, M. & Godoy, J.A., 2009. Phylogeography ofSouthern Water Vole (Arvicola sapidus): evidence for refugia within theIberian glacial refugium? Molecular Ecology, 18: 3652-3667.
Chaix, L. & Desse, J., 1983. Les bouquetins del’Observatoire (Monaco) et desBaoussé-Roussé (Grimaldi, Italie). Première partie: cranium, atlas, epistro-pheus. Bulletin du Musée d’Anthropologie Préhistorique de Monaco, 26: 41-74.
Chaix, L. & Desse, J., 1994. Les Bouquetins fossiles du Sud-Est de la France.Travaux Scientifiques du Parc National de la Vanoise, 13: 17-30.
Chapman, A. & Buck, W. J., 1910. Unexplored Spain, General Books, London.Clot, A., 1988. Bouquetins quaternaires dans les Pyrénées. 1.- Restes osseux
du gouffre des Bouquetins (Accous, Pyrénées). Revue de Comminges, 101:5-13.
Clot, A. & Duranthon, F., 1990. Les mammifères fossiles du Quaternaire dans lesPyrénées. Museum d’Histoire Naturelle de Toulouse, 159 pp., Toulouse.
Clouet, M. & Pfeffer, P.P., 1979. Le bouquetin d’Espagne: systématique et pro-jet de réintroduction. Bull. Mensuel Office Nat. Chasse, 25: 12-17.
Collina-Girard, J., 2001. Atlantis off the Gibraltar Strait? Myth and geology.Comptes Rendus de l’Académie des Sciences Serie, II, 333: 233–240.
RICARDO GARCÍA-GONZÁLEZ
116 (Pirineos, 2011, Vol. 166, 87-122, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166005)

Cosson, J.F., Hutterer, R.; Libois, R.; Sarà, M.; Taberlet, P. & Vogel, P., 2005.Phylogeographical footprints of the Strait of Gibraltar and Quaternary cli-matic fluctuations in the western Mediterranean: a case study with thegreater white-toothed shrew, Crocidura russula (Mammalia: Soricidae).Molecular Ecology, 14: 1151-1162.
Couturier, A.J., 1962. Le bouquetin des Alpes (Capra aegagrus ibex ibex L.). Ed.par l’ auteur., 1564 pp., Grenoble.
Crampe, J.P., 1991. Le bouquetin iberique. Elements pour une réintroductionau versant Nord des Pyrénées Occidentales. Documents Scientifiques duParc National des Pyrénées, 26: 1-187.
Crampe, J.P. & Crégut-Bonnoure, E., 1994. Le massif des Pyrénées, habitatnaturel du bouquetin ibérique (Capra pyrenaïca, Schinz, 1838). Evolutiontemporo-spatiale de l’espèce de la préhistoire a nos jours. Ibex SpecialeG.S.E., 2: 39-48.
Crégut-Bonnoure, E., 1992. Intérêt biostratigraphique de la morphologie den-taire de Capra (Mammalia, Bovidae). Ann. Zool. Fennici, 28: 273-290.
Crégut-Bonnoure, E., 1999. Les petits Bovidae de Venta Micena (Andalousie)et de Cueva Victoria (Murcia). En: Gibert, J., Sánchez, F., Gibert, L. & Ribot,F. (eds), The Hominids and their environment during the lower and middlePleistocene of Eurasia, pp. 191-228. Museo Prehistoria y Paleontología “J.Gibert”, Orce (Granad).
Crégut-Bonnoure, E., 2006. Nouvelles donnés paléogéographiques et chrono-logiques sur les Caprinae (Mammalia, Bovidae) du Pléistocène moyen etsupérieur d’Europe. Munibe, 57: 205-219.
Crégut-Bonnoure, E., 2009. Biochronologie et grands mammifères auPléistocène moyen et supérieur en Europe occidentale: l’apport desCaprinae de la tribu des Caprini. Quaternaire, 20: 481-508.
Crochet, J.Y., Gence, J., Boulbes, N., Boutié, P., Cretin, C., Crégut-Bonnoure, É.,Duzer, D., Jolly, D., Laudet, F., Lefévre, D., Mourer-Chauviré, C.,Rousselières, F. & Thouand, E., 2007. Nouvelles données paléoenvironne-mentales dans le Sud de la France vers 30000 ans 14C BP: le cas de la grot-te Marie (Hérault). C. R. Palevol., 6: 241-251.
De la Cerda, J.M. & De la Peña, J., 1971. La cabra montés española. Montes,159: 219-227.
Domínguez, A., Rodríguez, F., Albornoz, J. & Pérez, T., 2009. El rebeco cantá-brico, filogeografía e historia evolutiva. En: Pérez-Barbería, F.J. & Palacios,B. (eds.): El rebeco cantábrico Rupicapra pyrenaica parva. Conservación y ges-tión, pp. 70-102. Organismo Autónomo de Parques Nacionales MMA,Madrid.
Fandos, P., 1991. La Cabra Montés (Capra pyrenaica) en el Parque Natural de lasSierras de Cazorla, Segura y Las Villas. ICONA - C.S.I.C., 176 pp., Madrid.
ELEMENTOS PARA UNA FILOGEOGRAFÍA DE LA CABRA MONTÉS IBÉRICA (CAPRA PYRENAICA...
117(Pirineos, 2011, Vol. 166, 87-122, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166005)

Fandos, P., 1995. Factors affecting horn growth in male Spanish ibex (Caprapyrenaica). Mammalia, 59: 229-235.
Fandos, P., Soriguer, R.C. & Martínez, T., 1992. Implicaciones Genéticas yDemográficas en la Gestión de las Poblaciones de Cabra Montés. En:International Congress on the Genus Capra in Europe, pp. 159-162. AMA -Junta de Andalucía, Ronda (Málaga).
França, C., 1917. Le Bouquetin du Gerez. (Capra Lusitana). Notes sur une espè-ce éteinte. Arquivo da Universidade de Lisboa, 6: 19-54 + 10 planches.
Gabunia, L. & Vekua, A., 1995. A Plio-Pleistocene hominid from Dmanisi, EastGeorgia, Caucasus. Nature, 373: 509-512.
García-González, R., 2009. La extinción del bucardo, cabra montés de losPirineos. En: de Camps, J. R. (ed.): Conversaciones sobre el macho montés, pp.22-49. Cabrame 98 S.L., Barcelona.
García-González, R., in prep. New Holocene Capra pyrenaica skulls fromSouthern Pyrenees: its implication for C. pyrenaica filogeny.
García-González, R. & Herrero, J., 1999. El Bucardo de los Pirineos: historia deuna extinción. Galemys, 11: 17-26.
García-González, R., Vigal, C. R. & Fandos, P., in prep. Skull characteristics ofthe extinct Capra pyrenaica pyrenaica Cabrera, 1914, and its relationshipswith other neibouring Capra taxa.
Garrido, G., 2008. Generalidades sobre los artiodáctilos del Villafran-quiensesuperior en relación con el registro fósil de Fonelas P-1. Cuadernos delMuseo Geominero, 10: 279-335.
Geist, V., 1987a. On the evolution of the Caprinae. En: Soma, H. (ed.): TheBiology and Management of Capricornis and related mountain antelopes. pp. 3-40, Croom Helm, London.
Geist, V., 1987b. On speciation in Ice Age mammals, with special reference tocervids and caprids. Canadian Journal of Zoology, 65: 1067-1084.
Gibert, J., Gibert, L. & Iglesias, A., 2003. The Gibraltar Strait: A Pleistocenedoor of Europe? Human Evolution, 18: 147-160.
Gibert, J., Sánchez, F., Ribot, F., Gibert, L., Iglesias, A. & Hamouti, N.E., 2008.Dispersion du Genre Homo au sud d’Ibérie et au Maghreb. Nouvelles don-nées à propos des fossiles VM-1960 et BL-0. L’Anthropologie, 112: 48-73.
Gortázar, C., Herrero, J., Villafuerte, R. & Marco, J., 2000. Historical examina-tion of the status of large mammals in Aragon, Es. Mammalia, 64: 411-422.
Granados, J.E., Pérez, J. M., Márquez, F.J., Serrano, E., Soriguer, R.C. &Fandos, P., 2001. La cabra montés (Capra pyrenaica, Schinz 1838). Galemys,13: 3-38.
GRIGGO, C., 1991. Le bouquetin de Malarnaud (Ariège); implications paléo-biogéographiques. Quaternaire, 2: 76-82.
Hartl, G.B., Burger, H.; Willing, R. & Suchentrunk, F., 1990. On the biochemi-
RICARDO GARCÍA-GONZÁLEZ
118 (Pirineos, 2011, Vol. 166, 87-122, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166005)

cal systematics of the Caprini and the Rupicaprini. Biochemical Systematicand Ecology, 18: 175-182.
Hartl, G.B., Meneguz, P.G., Apollonio, M., Marco-Sánchez, I.; Nadlinger, K. &Suchentrunk, F., 1992. Molecular Systematics of Ibex in Western Europe.En: International Congress on the Genus Capra in Europe, pp. 21-26. Junta deAndalucia, Agencia de Medio Ambiente, Ronda (Málaga).
Herrero, J. & Pérez, J.M., 2008. Capra pyrenaica. In: IUCN 2010. IUCN Red Listof Threatened Species. Version 2010.4. www.iucnredlist.org. Downloadedon 13 january 2011.
Hewitt, G.M., 2004. Genetic Consequences of Climatic Oscillations in theQuaternary. Philosophical Transactions: Biological Sciences, 359: 183-195.
Jiménez, N., Folch, J., Fernández-Arias, A., Guiral, J. & Sánchez, A., 1999.Estudio genético mediante marcadores microsatélites de las poblacionesde cabra montés. ITEA, Vol. Extra, 20: 300-302.
Kazanskaya, E., Kuznetsova, M. & Danilkin, A., 2007. Phylogenetic recon-structions in the genus Capra (Bovidae, Artiodactyla) based on the mito-chondrial DNA analysis. Russian Journal of Genetics, 43: 181-189.
Koby, F. E., 1958. Ostéologie de la Chevre fossile des Pyrénées (Capra pyrenai-ca Schinz). Eclogae Geolog. Helveticae, 51: 475-480.
Kurtén, B., 1968. Pleistocene mammals of Europe. Weidenfeld & Nicolson, 317pp., London.
Made, J.V.D., Carlos Calero, J.A. & Mancheño, M.A., 2008. New material ofthe goat Capra ? alba from the lower Pleistocene of Quibas (Spain): noteson sexual dimorphism, stratigraphic distribution and systematics.Bollettino della Società Paleontologica Italiana, 47: 13-23.
Magniez, P., 2009. Nouvelles données sur le genre Capra Linné, 1758(Mammalia, Bovidae) du Pléistocène supérieur de la grotte Tournal (Bize-Minervois, France): Implications biochronologiques et évolutives.Quaternaire, 20: 509 -525.
Manceau, V., Crampe, J.P., Boursot, P. & Taberlet, P., 1999b. Identification ofevolutionary significant units in the Spanish wild goat, Capra pyrenaica(Mammalia, Artiodactyla). Animal Conservation, 2: 33-39.
Manceau, V., Despres, L., Bouvet, J. & Taberlet, P., 1999a. Systematics of theGenus Capra Inferred from Mitochondrial DNA Sequence Data. MolecularPhylogenetics and Evolution, 13: 504-510.
Marco, J., Escudero, E., Herrero, J., Escudero, M.A., Fernández, O., Ferreres,J., García-Serrano, A., Giménez-Anaya, A., Labarta, J.L., Monrabal, L. &Prada, C., 2011. Veinte años de seguimiento poblacional de ungulados sil-vestres de Aragón. Pirineos, 166 (este volumen).
Márquez, F.J., Pérez, J.M., Granados, J.E., Soriguer, R. & Fandos, P., 2002.Estudios genéticos de las poblaciones de cabra montés. En: Pérez, J. M.
ELEMENTOS PARA UNA FILOGEOGRAFÍA DE LA CABRA MONTÉS IBÉRICA (CAPRA PYRENAICA...
119(Pirineos, 2011, Vol. 166, 87-122, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166005)

(coord.): Distribución, genética y estatus sanitario de las poblaciones andaluzasde cabra montés, pp. 257-274. Universidad de Jaén. Junta de Andalucía,Consejería de Medio Ambiente. Sevilla, Jaén.
Martínez-Navarro, B. & Palmqvist, P., 1995. Presence of the Africanmachairodont Megantereon whitei (Broom, 1937) (Felidae, Carnivora,Mammalia) in the lower Pleistocene site of Venta Micena (Orce, Granada,Spain), with some considerations on the origin, evolution and dispersal ofthe genus. Journal of Archaeological Science, 22: 569-582.
Michaux, J., Cucchi, T., Renaud, S., García-Talavera, F. & Hutterer, R., 2007.Evolution of an invasive rodent on an archipelago as revealed by molarshape analysis: the house mouse in the Canary Islands. Journal ofBiogeography, 34: 1412-1425.
Montes, L., Utrilla, P. & Martínez-Bea, M., 2006. Trabajos recientes en yaci-mientos musterienses de Aragón: Una revisión de la transición PaleolíticoMedio/Superior en el Valle del Ebro. En: Maillo, J. M. & Baquedano, E.(eds.): Miscelánea en homenaje a Victoria Cabrera. pp. 214-233.
Montoya, P., Alberdi, M.T., Barbadillo, L.J., Van der Made, J., Morales, J.,Murelaga, X., Penalver, E., Robles, F., Ruiz Bustos, A. & Sánchez, A., 2001.Une faune très diversifiée du Pleistocene Inferieur de la Sierra de Quibas(province de Murcia, Espagne). Comptes Rendus de l’Academie des Sciences -Series IIA - Earth and Planetary Science, 332: 387-393.
Mood, A.M., Graybil, F.A. & Boes, D.C., 1974. Introduction to the theory of sta-tistics. McGraw-Hillpp., New York.
Moyà-Solà, S., 1987. Los bóvidos (Artiodactyla, Mammalia) del yacimientodel Pleistoceno inferior de Venta Micena (Orce, Granada, España). Paleont.i Evol., Mem. Esp. 1: 181-236.
Pailhaugue, N., 1995. La faune de la Salle Monique, Grotte de La Vache (Alliat,Ariège). Bulletin de la Societé Préhistorique Ariège-Pyrénées, 50: 225-290.
PALES, L., 1976-77. Les ovicaprides prehistoriques franco-iberiques au natu-rel et figures. Sautuola, 1: 67-105.
Pérez, J.M., Granados, J.E., Soriguer, R.C., Fandos, P., Márquez, F.J. & Crampe,J.P., 2002. Distribution, status and conservation problems of the Spanishibex, Capra pyrenaica (Mammalia: Artiodactyla). Mammal Rev. 32: 26-39.
Pidancier, N., Jordan, S., Luikart, G. & Taberlet, P., 2006. Evolutionary historyof the genus Capra (Mammalia, Artiodactyla): Discordance between mito-chondrial DNA and Y-chromosome phylogenies. Molecular Phylogeneticsand Evolution, 40: 739-749.
Quam, R.M., Arsuaga, J.L., Bermúdez de Castro, J.M., Díez, C.J., Lorenzo, C.,Carretero, M., García, N. & Ortega, A.I., 2001. Human remains fromValdegoba Cave (Huérmeces, Burgos, Spain). Journal of Human Evolution,41: 385-435.
RICARDO GARCÍA-GONZÁLEZ
120 (Pirineos, 2011, Vol. 166, 87-122, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166005)

Randi, E., Fusco, G., Lorenzini, R., Toso, S. & Tosi, G., 1991. Allozyme diver-gence and phylogenetic relationships among Capra, Ovis and Rupicapra(Artyodactyla, Bovidae). Heredity, 67: 281-286.
Randi, E., Alves, P.C., Carranza, J., Milosevic-Zlatanovic, S., Sfougaris, A. &Mucci, N., 2004. Phylogeography of roe deer (Capreolus capreolus) popula-tions: the effects of historical genetic subdivisions and recent nonequilib-rium dynamics. Molecular Ecology, 13: 3071-3083.
Rivals, F., 2002. Les petits bovidés pléistocènes dans le bassin méditerranéen et leCaucase. Etude paléontologique, biostratigraphique, archéozoologique et paléoéco-logique. Thèse doctorale, Université de Perpignan. Perpignan (France).
Rivals, F. & Blasco, R., 2008. Presence of Hemitragus aff. cedrensis (Mammalia,Bovidae) in the Iberian Peninsula: biochronological and biogeographicalimplications of its discovery at Bolomor Cave (Valencia, Spain). C. R.Palevol., 7: 391-399.
Rivals, F. & Testu, A., 2006. Un nouveau gisement paléontologique à Capracaucasica praepyrenaica: la grotte de l’Arche à Bugarach (Aude, France). C.R. Palevol., 5: 711–719.
Rodríguez, T., Pérez, T., Hammer, S.E., Albornoz, J. & Domínguez, A., 2010.Integrating phylogeographic patterns of microsatellite and mtDNA diver-gence to infer the evolutionary history of chamois (genus Rupicapra). BMCEvolutionary Biology, 10.
Ropiquet, A. & Hassanin, A., 2006. Hybrid origin of the Pliocene ancestor ofwild goats. Molecular Phylogenetics and Evolution, 41: 395-404.
Salas, R., Arenas, A. & Zamora, R., 2008. Comparativa gráfica de los alzadosfrontales y laterales de los cuernos de machos monteses. En: Granados,J.E., Cano-Manuel, J., Fandos, P. & Llano, R.C. (eds): Tendencias actuales enel Estudio y Conservación de los Caprinos Europeos (Suplemento). II Cong Int delgen Capra en Europa, pp. 59-76. Junta de Andalucía, Consejería de MedioAmbiente, Granada.
Sánchez Chillón, B., 1997. Primera cita del género Capra en el PleistocenoMedio de la Sierra de Atapuerca (Burgos, España). Geogaceta, 22: 210-212.
Sánchez, L., 2006. Las hijas del Almanzor. La herencia de la Sierra de Gredos.Linde y Ribera, 37 (Julio): 70-75.
Sarrión, I., 1984. Nota preliminar sobre los yacimientos paleontológicos pleis-tocénicos en la Ribera Baixa, Valencia. Cuadernos de Geografía, 35: 163-174.
Schaller, G.B., 1977. Mountain monarchs. Wild sheep and goats of the Himalaya.University Chicago Press, 425 pp., Chicago & London.
Schrago, C.G. & Russo, C.A.M., 2003. Timing the Origin of New WorldMonkeys. Mol Biol Evol, 20: 1620-1625.
ELEMENTOS PARA UNA FILOGEOGRAFÍA DE LA CABRA MONTÉS IBÉRICA (CAPRA PYRENAICA...
121(Pirineos, 2011, Vol. 166, 87-122, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166005)

Sesé, C., Alberdi, M.T., Mazo, A. & Morales, J., 2001. Mamíferos del Mioceno,Plioceno y Pleistoceno de la Cuenca de Guadix-Baza (Granada, España):revisión de las asociaciones faunísticas más características. Paleontologia iEvolució, 32-33: 31-36.
Sevilla, P., 1988. Estudio paleontológico de los Quirópteros del Cuaternarioespañol. Paleontologia i Evolució, 22: 113-233.
Shackleton, D.M., 1997. Conservation Priorities and Options. En: Shackleton,D.M. (ed.): Wild Sheep and Goats and their relatives. Status survey andConservation Action Plan for Caprinae, pp. 318-330. IUCN/SSC CaprinaeSpecialist Group, Gland & Cambridge.
Skog, A., Zachos, F.E., Rueness, E.K., Feulner, P.G.D., Mysterud, A., Langvatn,R., Lorenzini, R., Hmwe, S.S., Lehoczky, I., Hartl, G.B., Stenseth, N.C. &JAKOBSEN, K. S., 2009. Phylogeography of red deer (Cervus elaphus) inEurope. Journal of Biogeography, 36: 66-77.
Sommer, R.S., Zachos, F.E., Street, M., Jöris, O., Skog, A. & Benecke, N., 2008.Late Quaternary distribution dynamics and phylogeography of the reddeer (Cervus elaphus) in Europe. Quaternary Science Reviews, 27: 714-733.
Taberlet, P., Fumagalli, L., Wust-Saucy, A.G. & Cosson, J.F., 1998. Comparativephylogeography and postglacial colonization routes in Europe. MolecularEcology, 7: 453-464.
Van der Made, J. & Mateos, A., 2009. Longstanding biogeographic patternsand the dispersal of early Homo out of Africa and into Europe. QuaternaryInternational, 223-224: 195-200.
Villalta, M., Folch, J. & Alabart, J.L., 1997. Estudio genético molecular de las pobla-ciones de cabra montés de la Península ibérica. Informe inédito. Plan deRecuperación del Bucardo LIFE. Servicio Investigación Agraria (SIA-DGA).
Wyrwoll, T. W., 1999. Eine Neubeschreibung des sudspanischen Steinbocks.Saugetierkundliche Mitteilungen, 44: 93-98.
Yoder, A.D., Burns, M.M., Zehr, S., Delefosse, T., Veron, G., Goodman, S.M. &Flynn, J.J., 2003. Single origin of Malagasy Carnivora from an Africanancestor. Nature, 421: 734-737.
Yravedra Sáinz de los Terreros, J., 2004-2005. Estado de la cuestión sobre lasubsistencia del Musteriense de la vertiente mediterránea de la PenínsulaIbérica. Lvcentvm, 23-24: 7-17.
RICARDO GARCÍA-GONZÁLEZ
122 (Pirineos, 2011, Vol. 166, 87-122, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166005)

123
USING BLOCK COUNTS AND DISTANCE SAMPLINGTO ESTIMATE POPULATIONS OF CHAMOIS
J. HERRERO1, A. GARCÍA-SERRANO2, C. PRADA2 & O. FERNÁNDEZ-ARBERAS2
1 Area of Ecology. Technical School of Huesca. University of Zaragoza. E-22071 [email protected]
2 Ega Wildlife Consultants. Sierra de Vicort, 31, 1.º A. E-50003 [email protected]
ABSTRACT.– This study compared the efficacies of total block counts (BC) anddistance sampling (DS) procedures to estimate the abundance of chamois popula-tions in two mountain massifs, Posets and Maladeta, Spain. In 2002, 2006, 2007,and 2008, chamois populations were surveyed along itineraries above the timberline,twice per year: in July, after the parturition period, and in November, during the rut.The latter survey was used to estimate the sex ratio and to correct the proportion ofmales present in July. In 2008, poor weather prevented surveys in November. In theDS procedure, we used the data collected using BC and calculated the distances ofthe mapped groups to the itinerary, using a Geographical Information System. InPosets, estimates of population densities derived using BC ranged from 5.5 to 9.1chamois km-2, while those derived using DS ranged from 7.5 to 9.7 chamois km-2. InMaladeta, the estimates ranged from 3.4 to 5.4 chamois km-2 (BC) and from 4.6 to8.5 chamois km-2 (DS). Coefficients of variation of DS ranged between 14% and26%. In five of eight cases the counts of population size derived from BC were with-in the 95% confidence interval of the estimate derived from DS. In two of the otherthree cases, weather conditions created poor visibility during the rut, and few cham-ois were seen and, consequently, the rut sex ratio could not be estimated. BC pro-vided objective, high-quality counts of chamois populations and it is easy to obtain,even if its efficacy can be constrained by the need of simultaneous itineraries and anunderestimation of unknown magnitude. DS does not require sampling throughoutthe entire area above the timberline, and generates an estimate and a confidenceinterval; however, calculations require some skill and sample size must be high(n>300 groups observed to produce a CV <15%), which represents a sampling effortat least as large as the one derived from BC. BC represents a valuable tool and, itshould continue to be used to estimate minimum population size and the derivedhunting quotas.
Keywords: Rupicapra p. pyrenaica, block count, distance sampling, above thetimberline, mountain environments, mountain ungulates, Pyrenees.
Pirineos.Revista de Ecología de MontañaVol. 166, 123-133
Jaca, Enero-Diciembre, 2011ISSN: 0373-2568
eISSN: 1988-4281doi: 10.3989/Pirineos.2011.166006

124
J. HERRERO, A. GARCÍA-SERRANO, C. PRADA & O. FERNÁNDEZ-ARBERAS
RESUMEN.– Se comparan los resultados de estimar la abundancia de sarrioaplicando el conteo simultáneo supraforestal (BC) y el muestreo de distancias (DS)a lo largo de itinerarios, durante cuatro años (2002, 2006, 2007 y 2008) en dos maci-zos pirenaicos, Posets y Maladeta. Se realizaron dos operaciones anuales, una enjulio (tras la paridera) y otra en noviembre (en el celo), esta última para conocer laproporción de sexos que se utiliza para corregir el número de sarrios adultos de julio.En 2008 la operación de noviembre no se pudo realizar por inclemencias meteoroló-gicas. Para el DS se utilizan los datos del BC, calculando las distancias perpendicu-lares al recorrido de los grupos observados y cartografiados, con la ayuda de unSistema de Información Geográfica. Las densidades de julio de Posets oscilaron entre5,5 y 9,1 sarrios km-2 (BC) y entre 7,5 y 9,7 sarrios km-2 (DS) y en Maladeta entre3,4 y 5,4 sarrios km-2 (BC) y entre 4,6 y 8,5 sarrios km-2 (DS), con coeficientes devariación (CV) del DS del 14 al 26%. El número de animales mínimo estimado conel BC está dentro del intervalo de confianza del 95% de la densidad calculada por elDS en 5 de los 8 casos. En los otros tres casos, las condiciones meteorológicas adver-sas durante el celo no hicieron posible el cálculo de la sex ratio. El BC aporta unnúmero objetivo de indudable calidad y su cálculo es sencillo aunque cuenta con elinconveniente de la necesaria simultaneidad en la realización de los recorridos, lacoincidencia para ello de buenas condiciones de visibilidad y la existencia de unasubestima de magnitud desconocida. El DS, sin embargo, no requiere batir todo elterritorio supraforestal y produce una estima de la densidad con su intervalo de con-fianza. No obstante, el cálculo numérico requiere cierta destreza y dedicación y eltamaño muestral debe ser elevado (n>300 grupos avistados para un CV <15%) quesupone un esfuerzo de muestreo al menos igual al que se realiza en el BC. El BCconstituye una herramienta valiosa y su uso debería continuar para estimar el tama-ño mínimo poblacional y los cupos de caza derivados.
Palabras clave: Rupicapra p. pyrenaica, conteo simultáneo, muestreo de dis-tancias, ambientes supraforestales, montañas, ungulados de montaña,Pirineos.
1. Introduction
Historically, populations of mountain ungulates such as chamoisRupicapra sp. (Berducou, 1982; García-González et al., 1992,) and Iberian wildgoat Capra pyrenaica (Pérez et al., 2002) have been monitored and quantifiedusing coordinated block counts (BC), a type of total counts in which severalobserver teams beat the complete study area simultaneously, which are per-formed when the weather is favorable (e.g., good visibility, lack of strongwind, rain, or snow). In areas that have dense tree or shrub vegetation, theability to detect animals can be restricted and BC can significantly underesti-mate population size (Housin et al., 1994; Cano et al., 2009). The effects of lowvisibility are overcome by performing observations from vantage points in
(Pirineos, 2011, Vol. 166, 123-133, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166006)

rocky areas (Escós & Alados, 1988) using a Distance Sampling (DS) procedure(Buckland et al., 2001) along itineraries in open areas (Moço et al., 2006; Pérezet al., 2002) or mainly forested areas (García-González et at., 1992; Garín &Herrero, 1997).
Recently new experiences with mountain ungulates have been undertak-en using itineraries (Cano et al., 2009; Afonso et at., 2008; Monteiro et al., 2010).DS might have advantages over BC because, in the former, not all of the indi-viduals have to be counted to generate a reliable estimate of a population and,theoretically, the time and cost of fieldwork will be less than with the BC pro-cedure. In addition, DS generates an estimate of population density that hasa given precision (Borchers et al., 2002).
The vast majority of chamois populations are hunted (Aulagnier et al.,2008; Herrero et álli, 2008). Even non-huntable populations within protectedareas are hunted when they venture outside these areas to make altitudinalmigrations (Pañella et al., 2010). The prudent or responsible use of this impor-tant wildlife resource relies on accurate estimates of population sizes, and theefficacy of BC above the timberline has not been evaluated thoroughly. BCproduces estimates of the minimum number of animals that occupy the openareas above the timberline, but the extent to which this procedure underesti-mates population size is unknown. Although BC is the main procedure forthe long-term monitoring and management of populations, uncertainty overthe magnitude of the underestimation raises questions about the quality ofthe data. For that reason, this study evaluated the efficacy of BC and DS inestimating chamois populations in open areas above the timberline, as part ofthe long-term monitoring of Pyrenean chamois organized in Aragon andNavarre by their Regional Governments.
2. Study area
Chamois populations were surveyed in two mountain massifs in theSpanish Pyrenees, Posets and Maladeta (Fig. 1). Mountain massifs are the nat-ural management units in the region. In Posets and Maladeta, the amounts ofarea above 1,600 m a.s.l. are 21,199 ha and 28,964 ha, respectively, and mostlies within Posets - Maladeta Nature Park, where the highest elevation is>3,000 m. In our study in the Pyrenees, all of the area above 1,600 m was con-sidered suitable habitat for chamois, which facilitated comparisons betweenmassifs, rather than between administrative units (Herrero et al., 2002). In thevillages in the valley between the massifs, average annual precipitation is1,215 mm. The vegetation is mainly forests of mountain pine Pinus uncinata,with Rhododendron ferrugineum shrubs. Above the timberline, over 2,000 m,
USING BLOCK COUNTS AND DISTANCE SAMPLING TO ESTIMATE POPULATIONS OF CHAMOIS
125(Pirineos, 2011, Vol. 166, 123-133, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166006)

the habitat is pasture and rocky areas. Prominent fauna in the region includebearded vulture Gypaetus barbatus, golden eagle Aquila chrysaetos, rockptarmigan Lagopus mutus, and other wild ungulates such as wild boar Susscrofa and roe deer Capreolus capreolus. Traditional economic activities are live-stock farming and forest exploitation although, today, the main economicactivity is tourism. Game hunters mainly target ungulates.
3. Materials and methods
BC (Berducou et al., 1982) involve counting animals simultaneously inopen habitats, with natural features such as massifs as the sampling unit. Inour study, typically, pairs of rangers with experience with chamois and thearea performed the counts. Each pair of observers had binoculars, spottingscopes, specific cartography, and experience with chamois and the area. In theanalysis, duplicate counts were removed based on the groups mapped.
J. HERRERO, A. GARCÍA-SERRANO, C. PRADA & O. FERNÁNDEZ-ARBERAS
126 (Pirineos, 2011, Vol. 166, 123-133, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166006)
Figure 1. Study area.Figura 1. Área de estudio.

Estimates of minimum population size were derived from two surveys: onein July, after parturition, which estimated the number of females, yearlings,and kids, and another in November, during the rut, which estimated thenumber of adult males. The rut sex-ratio is applied to the females seen in Julyto estimate the number of males. Using this estimation we calculate the den-sity of chamois. During four years (2002, 2006, 2007 and 2008), the chamoispopulations in the Posets and Maladeta mountain massifs, Spanish Pyreneeswere surveyed using the BC procedure. In 2008, poor weather prevented asuitable survey in November.
DS comprises a set of methods for estimating the density of biologicalpopulations (Buckland et al., 2001; Thomas et al.,, 2002, 2010). Some surveysof chamois populations have used the line-transect method, in which theobservers perform a standardized visual survey along a series of itineraries,searching for objects of interest (in this case, individual or groups of chamois)and recording on a map their location. The objective in the DS procedure is tofit a detection function, g(y), to the observed distances, which reflects theprobability that a cluster is detected, given that it is at distance y from thetransect line. The fitted function is used to estimate the proportion of objectsthat were missed by the survey, which contributes to more accurate estimatesof population size and structure. The surveys were analyzed using the soft-ware Distance (http://www.ruwpa.st-and.ac.uk/distance). The DS proce-dure has three main assumptions: (1) all of the individuals on the route aredetected with certainty, (2) the distance between an animal or groups of ani-mals and the transect is measured accurately, and (3) animals do not move inresponse to the presence of the observer. Another assumption is that the loca-tions of animals are independent of the locations of the survey routes, whichwill be met if there is a sufficient number of transects and they are randomlydistributed (Thomas et al.,, 2010). In this study, there were two factors to con-sider: randomly distributed line transects cannot be used in rugged mountainenvironments, and itineraries are used instead, and observers have to haveexperience with the terrain, chamois, and its habits, locally.
The DS procedure was used retrospectively in calculating the distancesbetween groups of chamois and the itinerary using a Geographic InformationSystem. We used ArcView software to calculate the perpendicular distancebetween the itineraries and the chamois or the center of detected groups dur-ing the counts using the BC method. The data were analyzed using Distance 5(Thomas et al., 2006). The detection function was selected from among a set ofsuite of parametric distribution models: uniform, half-normal, and hazard-rate, which included adjustment terms to improve the fit (cosine expansion).We chose the model that fit best Akaike’s Criteria (AIC); viz., low coefficient ofvariation (CV), small number of parameters, and statistical non-significance.
USING BLOCK COUNTS AND DISTANCE SAMPLING TO ESTIMATE POPULATIONS OF CHAMOIS
127(Pirineos, 2011, Vol. 166, 123-133, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166006)

4. Results
In the Posets and Maladeta mountain massifs, 14 samplings were under-taken (Table 1). In calculating the estimates of population density with DSprocedure, the 5% of the groups that were farthest from the itineraries wereexcluded. The detection functions were half-normal (n=6) and hazard rate(n=8) with a cosine expansion. CVs were 14-23%, except for Posets in July2002 (CV=26%), when poor weather limited to 15 the number of itineraries.To reduce it to 10%, the distance covered by all of the itineraries should be 320km, which reflects a 33% greater sampling effort.
In the three years in which both annual surveys occurred in both of themassifs, the CI 95% of the estimate of population density derived from DS inJuly and November overlapped and the first included the estimates derivedfrom BC in five of the six site-years (Table 1). In Maladeta 2002 the estimatefrom BC were very near the DS lower confidence interval limit. In 2008, dur-ing the rut, weather conditions did not permitted doing the itineraries cor-rectly and the adult sex ratio could not be estimated. In this case the BC esti-mates were very near the DS lower confidence interval limit (Figure 2).
5. Discussion
Aspects of the retrospective analysis of BC data using DS are relevant tothe monitoring and management of Rupicapra populations. The calculated CVcan be considered small (Franzetti & Foccardi, 2006), and more precise popu-lation estimate require a much larger sampling effort, which is impractical inmany cases by the existing ranger staff. Coordinated monitoring of chamoispopulations in both regions involves 16 mountain massifs (Herrero et al.,2004), of which only five have enough chamois at a sufficiently high densityto be able to have a DS analysis given the current sampling effort. Other prob-lems to overcome when DS is used are: (i) the need for technical experiencein data analysis, (ii) the deskwork derived from fieldwork, which increasesthe cost of the monitoring program, and (iii) the need for a number of long-range telemeters, which are expensive. The estimates derived from BC werenot appreciably different from the estimates derived using DS, which indi-cates that the underestimation bias of BC is small and this has a consequencefor management: hunting quotas are based on population sizes that probablywere not significantly underestimated. Thus, the data from BC can remain theprimary source of information when annual hunting quotas are set.
We conclude that, with the sampling effort used in our study, the DS pro-cedure can be applied in few massifs where chamois are numerous and at a
J. HERRERO, A. GARCÍA-SERRANO, C. PRADA & O. FERNÁNDEZ-ARBERAS
128 (Pirineos, 2011, Vol. 166, 123-133, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166006)

129(Pirineos, 2011, Vol. 166, 123-133, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166006)
BCD
S
Mas
sifSu
rface
(ha)
Year
Seas
onIti
nera
ries
Effo
rtO
bser
ved
Obs
erve
dN
.ºCh
amoi
sCh
amoi
sCI
95
%CV
(km
)gr
oups
cham
ois
estim
ated
km-2
km-2
cham
ois k
m-2
Part
uriti
on23
199,
219
716
2619
319,
118,
55,
7-12
,520
%20
02Ru
t15
118,
710
962
26,
63,
9-11
,126
%Pa
rtur
ition
2621
4,1
270
1647
1783
8,41
9,7
7,1-
13,3
16 %
Pose
ts21
.199
2006
Rut
2621
4,2
293
1362
9,3
6,6-
13,3
18 %
Part
uriti
on26
214,
129
817
8417
848,
427,
75,
9-10
,214
%20
07Ru
t23
179,
126
214
085,
74,
0-8,
118
%20
08Pa
rtur
ition
2722
3,6
280
1156
1156
5,45
7,5
5,6-
10,1
15 %
Part
uriti
on22
170,
318
313
3915
695,
428,
55,
8-12
,620
%20
02Ru
t23
179,
119
592
85,
64,
0-7,
917
%Pa
rtur
ition
2317
9,1
167
935
993
3,43
4,6
3,1-
7,0
21 %
2006
Mal
adet
a28
.964
Rut
2217
1,2
184
764
4,2
2,6-
6,7
23 %
Part
uriti
on23
179,
118
711
5712
604,
355,
64,
0-7,
917
%20
07Ru
t22
172,
418
884
96,
34,
0-9,
923
%20
08Pa
rtur
ition
2418
6,8
188
1280
1280
4,42
7,1
4,7-
10,6
21 %
Tabl
e 1.
Res
ults
of
cham
ois
estim
atio
ns u
sing
tot
al b
lock
cou
nts
(BC
) an
d di
stan
ce s
ampl
ing
(DS)
proc
edur
es i
n Po
sets
and
Mal
adet
a m
assi
fs(P
yren
ees)
. CI:
conf
iden
ce in
terv
al; C
V: v
aria
tion
coef
ficie
nt.
Tabl
a 1.
Res
ulta
dos
de lo
s co
nteo
s y
estim
a de
la d
ensid
ad d
e sa
rrio
s us
ando
el m
étod
o de
l con
teo
tota
l sim
ultá
neo
(BC)
y el
mue
stre
o de
dist
ancia
s (D
S)en
los
mac
izos
de P
oset
s y M
alad
eta
(Piri
neos
). CI
: int
erva
lo d
e con
fianz
a; CV
: coe
ficien
te d
e var
iació
n.
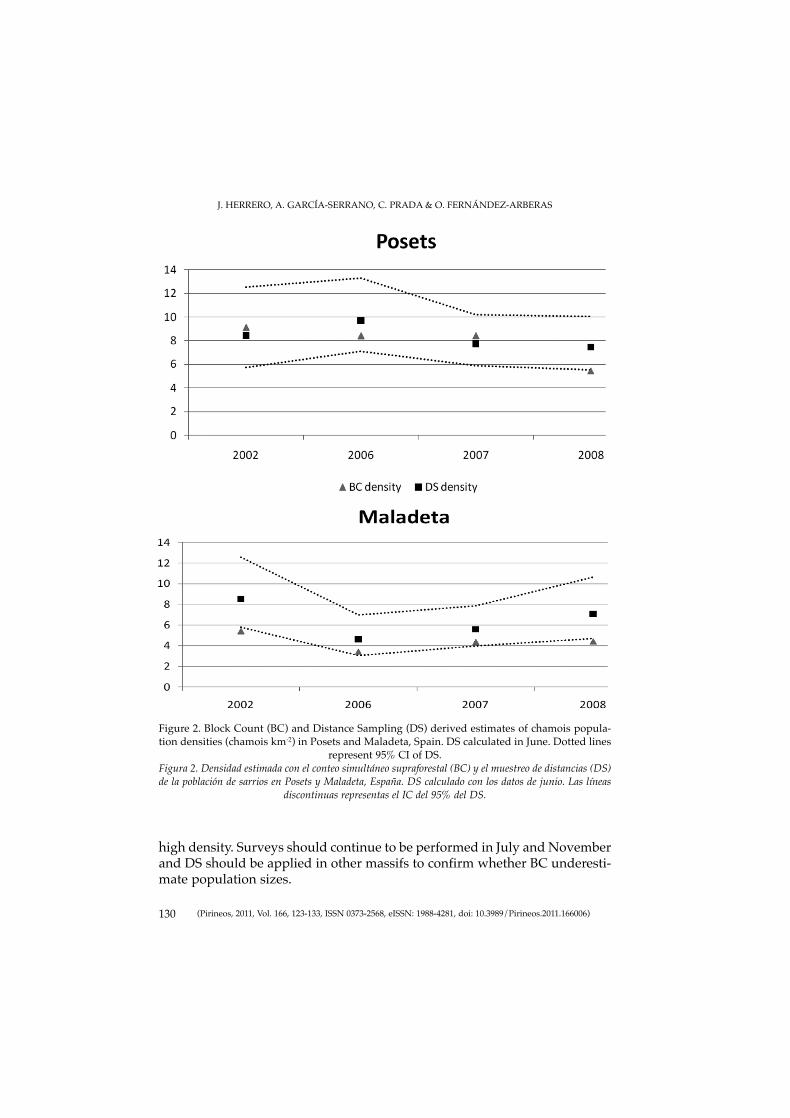
high density. Surveys should continue to be performed in July and Novemberand DS should be applied in other massifs to confirm whether BC underesti-mate population sizes.
J. HERRERO, A. GARCÍA-SERRANO, C. PRADA & O. FERNÁNDEZ-ARBERAS
130 (Pirineos, 2011, Vol. 166, 123-133, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166006)
Figure 2. Block Count (BC) and Distance Sampling (DS) derived estimates of chamois popula-tion densities (chamois km-2) in Posets and Maladeta, Spain. DS calculated in June. Dotted lines
represent 95% CI of DS.Figura 2. Densidad estimada con el conteo simultáneo supraforestal (BC) y el muestreo de distancias (DS)de la población de sarrios en Posets y Maladeta, España. DS calculado con los datos de junio. Las líneas
discontinuas representas el IC del 95% del DS.

Acknowledgments
This research was the product of the ongoing, long-term monitoring ofchamois populations in Aragon and Navarre (Pyrenees, Spain. We greatlyappreciate the invaluable contribution made by the rangers in Posets andMaladeta.
References
Afonso, A., Alpizar-Jara, M., Moço, G., Granados, J.E., & Pérez, J.M., 2008.Dimensión de la muestra y estimación de la variancia en muestreo por dis-tancias: una aplicación en la estima de abundancia de poblaciones deungulados de montaña. In: J.E. Granados, J. Cano-Manuel León, P. FandosParís, R. Cadenas de Llano Aguilar (Eds.). Tendencias actuales en el Estudioy Conservación de los Caprinos Europeos. Junta de Andalucía, Consejería deMedio Ambiente. Pp. 15-30.
Aulagnier, S., Giannatos, G. & Herrero, J., 2008. Rupicapra rupicapra. In: IUCNRed List of Threatened Species. Version 2010.4. <www.iucnredlist.org>.Downloaded on 19 November 2010
Berducou, C., Besson, C., & Guardas monitores del PNP, 1982. Dynamiquedes populations d’isards du Parc National des Pyrénées. Acta BiologicaMontana 1: 153-175.
Borchers, D. L., Buckland, S.T. & Zucchini, W., 2002. Estimating animal abun-dance. Closed populations. Springer-Verlag, London, U.K.
Buckland, S.T., Anderson, D.R., Burnham, K.P., Laake, J.L., Borchers, D.L. &Thomas, L., 2001. Introduction to distance sampling. Oxford UniversityPress, Oxford, U.K.
Cano, M., García-Rovés, P., González-Quirós, P. & Nores, C., 2009.Experiencias de estimación de poblaciones de rebeco en la CordilleraCantábrica. En: F.J. Pérez-Barbería y B. Palacios (Eds.). El rebeco cantábricoRupicapra pyrenaica parva. Conservación y Gestión de sus poblaciones.Naturaleza y Parques Nacionales. Serie Técnica.
Escós, J. & Alados, C.L., 1988. Estimating mountain ungulate density inSierras de Cazorla y Segura. Mammalia, 52(3): 425-428.
Franzetti, B. & Focardi, S., 2006. La stima di popolazione degli ungulati medianteDistance sampling e termocamera a infrarossi. Min. Politiche Agricole,Alimentari e Forestali – Ist. Naz. Fauna Selvatica, Documenti Tecnici, 26:1-88.
García-González, R., Hidalgo, R., Herrero, J. & Ameztoy, J.M., 1992. Census,population structure and habitat use of a chamois population living in
USING BLOCK COUNTS AND DISTANCE SAMPLING TO ESTIMATE POPULATIONS OF CHAMOIS
131(Pirineos, 2011, Vol. 166, 123-133, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166006)

sympatry with Pyrenean wild goat. In: Ongulés/Ungulates. 91. Pp. 321-325.F. Spitz, G. Janeau, G. Gonzalez and S. Aulaguier (Eds). SFEPM-IRGM.Paris, Toulouse.
Garin, I. & Herrero, J., 1997. Distribution, abundance and demographicparameters of the Pyrenean chamois (Rupicapra p. pyrenaica) in Navarre,Western Pyrenees. Mammalia, 61(1): 55-63.
Herrero, J., Escudero, E., García, J.M., García-Serrano, A., Prada, C. & Couto,S., 2004. Gestión y seguimiento demográfico del sarrio en el Pirineo ara-gonés. In: J. Herrero., E. Escudero, D. Fernández de Luco, R. García-González (Eds.). El sarrio: biología, patología y gestión. Consejo de Protecciónde la Naturaleza de Aragón, Gobierno de Aragón, Zaragoza. Pp. 69-82.
Herrero, J., Lovari, S. & Berducou, C., 2008. Rupicapra pyrenaica. In: IUCN RedList of Threatened Species. Version 2010.4. <www.iucnredlist.org>.Downloaded on 19 November 2010.
Houssin, H., Loison, A., Jullien, J.M. & Gaillard, J.M. 1994. Validité de la méth-ode du pointage-flash pour l’estimation des effectifs de chamois(Rupicapra rupicapra). Gibier Faune Sauvage, 11 (4): 287-298.
Moço, G., Guerreiro, M., Ferreira, A.F., Rebelo, A., Loureiro, A., Petrucci-Fonseca, F. & Pérez, J.M., 2006. The ibex Capra pyrenica returns to its for-mer Portuguese range. Oryx, 40 (3): 351-354.
Monteiro, J.F., Alpizar-Jara, R., Serrano, E., Crampe, J.P. & Pérez, J.M., 2008.Estimación Bayesiana de go usando el muestreo por distancias y su apli-cación en las estimas de densidad de ungulados de montaña. In: J.Granados, J. Cano-Manuel León, P. Fandos París, R. Cadenas de LlanoAguilar (Eds.). Tendencias actuales en el Estudio y Conservación de los CaprinosEuropeos. Junta de Andalucía, Consejería de Medio Ambiente. Pp 207-218.
Pañella, P., Herrero, J., Canut, J. & García-Serrano, A., 2011. Long-term moni-toring of Pyrenean chamois in a protected area reveals a fluctuating pop-ulation. Hystrix (in press).
Pérez, J.M., Granados, J.E., Soriguer, R.C., Fandos, P., Márquez, F.J. &Crampe, J.-P., 2002. Distribution, status and conservation problems of theSpanish Ibex, Capra pyrenaica (Mammalia: Artiodactyla) Mammal Review,32(1): 26-39.
Thomas, L., Buckland, S.T., Burnham, K.P., Anderson, D.R., Laake, J.L.,Borchers, D.L. & Strindberg, S., 2002. Distance sampling. In: El-ShaarawiA.H, and W.W. Piegorsch (Eds). Encyclopedia of Environmetrics. Volume 1,pp 544-552.
Thomas, L., Laake, J.L., Strindberg, S., Marques, F.C., Buckland, S.T.,Borchers, D.L., Anderson, D.R., Burnham, K.P., Hedley, S.L., Pollard, J.H.,Bishop, J.R.B. & Marques, T.A., 2006. Distance 5.0. Release 2. Research Unit
J. HERRERO, A. GARCÍA-SERRANO, C. PRADA & O. FERNÁNDEZ-ARBERAS
132 (Pirineos, 2011, Vol. 166, 123-133, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166006)

for Wildlife Population Assessment, University of St. Andrews, UK.http://www.ruwpa.st-and.ac.uk/distance/.
Thomas, L., Buckland, S., Rexstad, E.A., Laake, J.L., Strindberg, S., Hedley,S.L., Bishop, J.R.B., Marques, T.A. & Burnham, F.P., 2010. DistanceSoftware: design and analysis of distance sampling surveys for estimatingpopulation size. Journal of Applied Ecology, 47: 5-14.
USING BLOCK COUNTS AND DISTANCE SAMPLING TO ESTIMATE POPULATIONS OF CHAMOIS
133(Pirineos, 2011, Vol. 166, 123-133, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166006)

135
VEINTE AÑOS DE SEGUIMIENTO POBLACIONALDE UNGULADOS SILVESTRES DE ARAGÓN
J. MARCO1, J. HERRERO2, M. A. ESCUDERO1, O. FERNÁNDEZ-ARBERAS3,J. FERRERES1, A. GARCÍA-SERRANO3, A. GIMÉNEZ-ANAYA3, J. L. LABARTA1,
L. MONRABAL1 & C. PRADA3
1 Ebronatura S.L. C/del Arpa, 6 (local). E-50003 [email protected]
2 Área de Ecología. Departamento de Agricultura y Economía Agraria.Escuela Politécnica Superior de Huesca. Universidad de Zaragoza. E-22071 Huesca
[email protected] Ega Consultores. Sierra de Vicort, 31, 1.º A. E-50003 Zaragoza. [email protected]
ABSTRACT.– The remarkable increase of wild ungulates populations inAragon during the last 50 years, both in numbers and distribution, has made popu-lation monitoring a need for effective management of these species. In parallel totheir raising numbers, problems related to crop damage and collisions with vehiclesare growing. In this situation there has been a shift in the management goals fromconservation to population control. Different methods have been used in the surveys:postal enquiries for distribution, battues, distance sampling, night spotlight surveysand indirect ones based on transect and plot counts of pellet groups. Surveys start-ed in 1985 for red deer, 1990 for wild boar and Iberian wild goat, 1995 for chamoisand 2002 for roe deer, gradually becoming a population monitoring program. Thesurveys are carried out by rangers and wildlife consultants under the coordinationand supervision of the Fish and Game Service of the Aragon Government. The ungu-late populations are still growing (wild boar, wild goat, red and roe deer), are rela-tively stabilised (fallow deer, feral goat), fluctuate due to diseases (chamois) or showsmall populations limited to fenced states (Barbary sheep, mouflon). Hunting bagswent from 1,100 to 36,000 during an eleven-year period (1990-2009): 303 chamois,1,173 wild goats, 2,141 roe deer, 2,424 red deer and 29,595 wild boars, showing atrend which is still increasing. The main tool is therefore hunting quota and its ver-ification through hunting bags declarations, damages to crops and car crashes, andpopulation trend and abundance.
Keywords: Capra pyrenaica; Capra hircus; Capreolus capreolus; Cervus elaphus;Dama dama; Ovis aries; Sus scrofa; Ammotragus laervia; hunting bags.
Pirineos.Revista de Ecología de MontañaVol. 166, 135-153
Jaca, Enero-Diciembre, 2011ISSN: 0373-2568
eISSN: 1988-4281doi: 10.3989/Pirineos.2011.166007

136
J. MARCO et al.
RESUMEN.– La importante expansión de las poblaciones de ungulados silves-tres en Aragón desde los años 50 del siglo XX hasta la actualidad, tanto en superfi-cie ocupada como en abundancia de ejemplares, ha hecho necesario el seguimiento delas mismas para su adecuada gestión. Su incremento en las dos últimas décadas,junto con el importante aumento de daños agrícolas y colisiones con vehículos, hamotivado un importante cambio en los criterios gestores; pasando de la conservacióny fomento al control e incluso reducción de efectivos. Los métodos utilizados para elseguimiento de las poblaciones fueron: encuestas quincenales de distribución, bati-das, muestreos de distancia, captura – recaptura, fareos nocturnos y muestreos indi-rectos basados en el conteo de grupos de excrementos en transectos y parcelas fijas.Los seguimientos se comenzaron a realizar en 1985 en el caso del ciervo, 1990 en eljabalí y cabra montesa, 1995 en el sarrio y 2002 en el corzo, habiéndose convertidopaulatinamente en una verdadera monitorización poblacional. Los seguimientos sonrealizados por Agentes de Protección de la Naturaleza del Gobierno de Aragón yasistencias técnicas externas, bajo la supervisión y coordinación de los responsablesadministrativos. Los resultados obtenidos muestran que, algunas poblaciones conti-núan su expansión e incremento (jabalí, ciervo, corzo, cabra montesa), otras seencuentran relativamente estabilizadas (gamo, cabra doméstica asilvestrada), y unaspocas fluctúan debido a enfermedades (sarrio) o muestran presencias testimonialesprácticamente reducidas a cercados (arruí, muflón). Las capturas declaradas hanpasado de 1.100 a 36.000 en 11 años (1999-2009): 303 sarrios, 1.173 cabras monte-sas, 2.141 corzos, 2.424 ciervos y 29.595 jabalíes, con una tendencia al incrementoque aún no ha finalizado. El instrumento fundamental por lo tanto son los cupos decaza y su verificación se realiza a partir de las declaraciones de capturas, los daños ala agricultura y accidentes automovilísticos y la tendencia y abundancia poblacional.
Palabras clave: Capra pyrenaica; Capra hircus; Capreolus capreolus; Cervus ela-phus; Dama dama; Ovis aries; Sus scrofa; Ammotragus laervia; capturas cinegéticas.
1. Introducción
Los ungulados silvestres han sufrido una importante expansión e incre-mento demográfico en toda Europa durante las últimas décadas, pasando engran medida de áreas marginales a ocupar gran parte del territorio(Apollonio et al., 2010). Hoy juegan un papel importante como consumidoresprimarios, especies cinegéticas y origen de daños agrícolas y a los vehículos.En Aragón sus poblaciones son objeto de seguimiento demográfico y sanita-rio y la información obtenida marca la gestión desarrollada, particularmentesobre los cupos de extracción. Las especies y poblaciones presentes son: intro-ducidas (gamo Dama dama, arrui Ammotragus laervia); asilvestradas (muflónOvis aries y cabra doméstica asilvestrada Capra hircus) y autóctonas (jabalí Susscrofa, sarrio Rupicapra p.pyrenaica, cabra montesa Capra pyrenaica, ciervoCervus elaphus y corzo Capreolus capreolus).
(Pirineos, 2011, Vol. 166, 135-153, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166007)

Los daños a los cultivos agrícolas son pagados por la administración autó-noma en Reservas de Caza y Áreas Naturales Protegidas y ascienden a180.000 € en 2008 y 90.000 en 2009, constituyendo el principal problema degestión, sobre todo para el caso de corzos y ciervos y en menor medida, decabra montesa. Los atropellos también van en aumento, suponiendo para elerario público un coste aproximado de 500.000 € en 2009 y una cantidad tam-bién importante, pero no conocida en la actualidad, pagada por los cotos.
Los resultados de estos seguimientos permiten establecer anualmentecupos de capturas adaptados a las posibilidades poblacionales, sobre la basede datos actualizados, sin por ello perder de vista otros condicionantes denaturaleza antrópica que puedan influir, en zonas y momentos concretos, loscriterios de asignación de dichos cupos.
En este trabajo pretendemos mostrar por primera vez en su conjunto losresultados básicos de más de 20 años de seguimientos demográficos de ungu-lados silvestres en Aragón, sintetizando la información disponible sobre dis-tribución, abundancia y capturas de las diferentes especies y poblaciones,base de las medidas de gestión adoptadas.
2. Área de estudio
El área de estudio la conforma la Comunidad Autónoma de Aragón con47.669 km2 y 1.269.027 habitantes en 2006. El 70% de la población vive en laprovincia de Zaragoza y aproximadamente la mitad del total en su capital,Zaragoza, existiendo grandes superficies con densidades inferiores a los 10habitantes km-2. Los municipios rurales presentan un perfil demográficodeterminado por la emigración y el envejecimiento. El territorio puede divi-dirse en tres grandes unidades de relieve: el Valle Medio del Ebro; el Pirineoy Prepirineo y el Sistema Ibérico (Sampietro, 2000; De la Riva, 1997).
En el Pirineo las precipitaciones anuales se sitúan por debajo de los 800mm sólo en las zonas bajas de los grandes ríos. La temperatura media anuales inferior a 12º. El relieve impone un gran número de variaciones locales,entre las que destacan la alternancia entre solana y umbría, las inversionestérmicas y el encajonamiento de los vientos. La cubierta vegetal, mayoritaria-mente incluida en el dominio eurosiberiano, denota la transición hacia elmediterráneo. La sucesión altitudinal es clara: escasos pastizales alpinos, pisosubalpino muy transformado en matorrales y pastos alpinizados, piso mon-tano (hayedos, abetos, fresnos y extensos pinares de Pinus sylvestris y pisosubmediterráneo (quejigo Quercus faginea y encina Quercus ilex, con grandeforestación para uso agrícola y ganadero). De carácter mediterráneo conti-nental, el clima del Prepirineo es más suave y húmedo que el del centro del
VEINTE AÑOS DE SEGUIMIENTO POBLACIONAL DE UNGULADOS SILVESTRES DE ARAGÓN
137(Pirineos, 2011, Vol. 166, 135-153, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166007)

Valle del Ebro. El encinar, coscojar y la sabina albar (Juniperus thurifera) con-forman una vegetación potencial intensamente desplazada por los cultivos odegradada hacia matorrales de jara (Cistus ladanifer), retama (Retama sphaero-carpa) o romero (Rosmarinus officinalis).
El Valle Medio del Ebro se caracteriza por su marcada aridez, amplificadapor la acción de vientos desecantes, fríos en invierno y frescos en verano. Aeste ambiente mediterráneo continental muy seco con tendencia a subdesér-tico, se adapta la sabina albar y el coscojar. En las áreas endorreicas de lacuenca del Ebro en ambientes hidrófilos, la salinidad es determinante para lacaracterización de los hábitats y las especies dominantes.
El Sistema Ibérico, montaña mediterránea con fuerte matiz continental,presenta precipitaciones menores que el Pirineo. Aquí se registran las tempe-raturas más bajas de toda la Península. En cuanto a la vegetación, el quejigoha desplazado a la encina en un ambiente cultivado o muy degradado haciamatorrales. La sabina albar responde al frío intenso, pero predomina laexpansión del pinar de silvestres, rodenos (Pinus pinaster) y laricios (Pinusnigra). La vegetación del somontano del Ibérico se encuentra muy influidapor la altitud, que en las zonas elevadas le proporciona una pluviometríamayor. Las temperaturas son ligeramente más bajas y las precipitaciones infe-riores a las del somontano pirenaico (Sampietro, 2000). Excepto la cabradoméstica asilvestrada, el resto de ungulados silvestres son especies cinegéti-cas en Aragón. El territorio se halla acotado para la práctica cinegética en un91% de su superficie.
3. Métodos
En 2006 llevamos a cabo una encuesta a los Agentes de Protección de laNaturaleza (APN) del Gobierno de Aragón (n=242 encuestas), en la que seconsignaron la presencia o ausencia de ungulados silvestres en cuadrículasUTM de 10 x 10 km en Aragón durante el quinquenio anterior (2001-2005)(Osuna et al., 2006-08). Esta encuesta se repetirá cada cinco años.
Los fareos nocturnos, para establecer la abundancia de ciervos, comenza-ron en 1985 en la Reserva de Caza de Montes Universales (RCMU) y a media-dos de los 90 se incorporaron los tres grandes núcleos restantes: Reserva deCaza de la Garcipollera (RCG) y alrededores, Montes de Agüero; cotos delBajo Aragón y la zona adyacente a la RCMU (Figura 1). Los cuatro núcleosmonitorizados son objeto de seguimientos mediante muestreos directos noc-turnos realizados desde vehículos todo terreno provistos de foco manual, conun mínimo de dos observadores por vehículo, utilizando para el cálculo dedensidades el estimador de Kelker (Tellería, 1986) y con una velocidad de
J. MARCO et al.
138 (Pirineos, 2011, Vol. 166, 135-153, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166007)
VEINTE AÑOS DE SEGUIMIENTO POBLACIONAL DE UNGULADOS SILVESTRES DE ARAGÓN
139(Pirineos, 2011, Vol. 166, 135-153, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166007)
Figura 1. Aragón y las diferentes áreas de seguimiento de poblaciones de ungulados. Corzos y cabras montesas son objeto de seguimiento en toda su área de distribución (ver fig. 2).
Figure 1. Aragon and its different ungulate monitoring areas. Roe deer and Iberian wild goats are monitoredall over their distribution area (see fig. 2).
Límites provinciales.Cotos Sociales.Reservas de Caza.Áreas Naturales Protegidas.Poblaciones de ciervomonitorizadas en Aragón.

unos 20 km h-1. Cada núcleo se muestreó dos veces en abril y dos en sep-tiembre, época de berrea, por equipos de entre cuatro y doce personas, depen-diendo de las dimensiones del territorio.
En la RCMU, RCG y Montes de Agüero se realizan estimas de abundanciaa partir de muestreos indirectos basados en el seguimiento de parcelas fijas deacumulación fecal (n=139) cada 3-4 meses según zonas (Marco et al., 2005). Elmétodo consiste en contar el número de grupos de excrementos que se hanacumulado durante un tiempo controlado y en una superficie conocida yconstante (que se limpia tras cada revisión), conocida la tasa de defecación dela especie “diana” en el medio concreto que interesa al estudio (Garin, 2000).
El conteo directo en sarrio y cabra montesa se lleva a cabo en toda el áreade distribución de las dos especies, a partir de recorridos a pie (mayoritaria-mente), puntos fijos de observación, recorridos en vehículo y, anecdóticamen-te, en barca. Los conteos se realizan dos veces al año tras la paridera y en elcelo (sarrio); cuatro veces (antes de la paridera, tras ella, antes del celo y en elcelo) en la cabra montesa del Parque Natural de los Cañones y Sierra de Guara(PNG) y Zaragoza (Herrero et al., 2004; Herrero et al., 2007; García-Serrano etal., 2010a), dos veces (celo y primavera) en el Alto Matarraña (Teruel) (Marcoet al., 2004-2010) y una vez (celo) en el resto de la mitad oriental de la provin-cia de Teruel (Servicio Provincial de Medio Ambiente de Teruel, 1994-2009). Elseguimiento afecta a la totalidad de la población de sarrio; de cabra en Huescay Zaragoza, y la mayor parte de la población de cabra en Teruel.
Durante la década de los noventa, los estudios sobre el corzo en Aragónestuvieron reducidos a verificar la presencia de la especie en parajes cada vezmás numerosos. A partir de 2002 se inició un seguimiento de la abundancia dela especie en toda la comunidad autónoma, abarcando 8 grandes unidadesterritoriales de entre 150.000 y 450.000 ha, a partir de transectos de recuento degrupos fecales, para realizar los muestreos a razón de una o dosunidades/año, según sus dimensiones. Como índice de abundancia se ha cal-culado la densidad de grupos fecales, estimada a partir de transectos deanchura de banda variable (Acevedo et al., 2010). Estos datos se han transfor-mado en densidad de corzo a partir de una tasa de defecación de 20 gruposfecales día-1 (Mayle et al., 1999) y un periodo de persistencia medio de 135 días(Escudero et al., 2007) y dentro del rango de entre 116 y 164 días estimado enGrecia por Tsaparis et al., (2009). Hasta 2009 se han establecido un total de332 puntos de muestreo con dos transectos de 700 m cada uno, estando pre-visto llegar casi a los 500 puntos cuando se completen las ocho unidades defi-nidas para toda la comunidad autónoma. En los espacios donde existen par-celas fijas de conteo de excrementos orientadas a ciervo, se obtienen resulta-dos de corzo que enriquecen la precisión de las estimas de abundancia de estaespecie a nivel más local.
J. MARCO et al.
140 (Pirineos, 2011, Vol. 166, 135-153, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166007)

El método de las batidas cinegéticas de jabalí consiste en calcular la super-ficie batida durante su caza así como el número de jabalíes y otros animalesque son vistos durante la misma (Herrero, 2003). Ha sido utilizada en lasReservas de Caza del Pirineo aragonés (Herrero et al., 2008) y en el ParqueNatural del Moncayo (PNM) (Giménez-Anaya et al., 2009).
El muestro de distancias o distance sampling, está basado en el cálculo de ladistancia a la que es avistado un ejemplar con respecto al observador, en pun-tos o en transectos de muestreo (Buckland et al., 2001). Esta segunda aproxi-mación se ha llevado a cabo para el sarrio durante la estima poblacional debucardo Capra p. pyrenaica (García-González et al., 1992) en el Parque Nacionalde Ordesa y Monte Perdido, así como en el mismo macizo, como parte delseguimiento anual del sarrio desde 1998 y en el corzo en las Reservas de Cazapirenaicas desde 2002 y en el PNM desde 2009. La cabra doméstica asilves-trada también ha sido estimada a partir del muestreo de distancias en el PNGen 1995, a partir de transectos (Gortázar et al., 1997) y puntos fijos de obser-vación (Herrero et al., 2009a) y lo será de esta forma quinquenalmente.
Las capturas cinegéticas (1999-2009, 11 años) se calculan a partir del núme-ro de ejemplares que los titulares de los terrenos cinegéticos (sociedades decazadores, ayuntamientos, propietarios privados o administración autonómi-ca) declaran haber capturado durante la temporada cinegética. Generalmenteen estas declaraciones no se incluyen las capturas obtenidas fuera de tempo-rada en controles excepcionales por daños agrícolas u otras causas. Todos losungulados cinegéticos menos el jabalí y el gamo son objeto de caza a partir decupos determinados por la Administración para cada territorio cinegético.Cada ejemplar cazado es precintado a excepción, nuevamente, del jabalí.
4. Resultados
El jabalí ocupa la totalidad del territorio aragonés (Figura 2), con densida-des no muy dispares (Tabla 1) y sus capturas se han triplicado, siendo la espe-cie con mayor distribución y capturas (83% en 2009) (Tabla 5).
El ciervo, ocupa el 30,1% del territorio (Figura 2) y sus capturas se han tri-plicado también (Tabla 5) (Marco et al., 2010). Las mayores poblaciones seencuentran en la RCMU y sus cotos colindantes (Cotos SW prov. Teruel),seguidas de los Cotos del Bajo Aragón, la RCG y Agüero, en orden decre-ciente. La mayor densidad aparece en la RCG. Considerando los cuatro núcle-os existentes y la titularidad de los terrenos, los que son gestionados por laAdministración autonómica presentan mayores densidades (Tabla 2). Loscupos de capturas han podido reducir el tamaño poblacional en la Reserva deCaza de La Garcipollera, tras una intensa presión cinegética en un espacio de
VEINTE AÑOS DE SEGUIMIENTO POBLACIONAL DE UNGULADOS SILVESTRES DE ARAGÓN
141(Pirineos, 2011, Vol. 166, 135-153, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166007)

J. MARCO et al.
142 (Pirineos, 2011, Vol. 166, 135-153, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166007)
Figu
ra 2
. Dis
trib
ució
n de
los u
ngul
ados
silv
estr
es e
n A
ragó
n (O
suna
et á
lii, 2
008-
10).
Figu
re 2
. Wild
ung
ulat
e dist
ribut
ion
in A
rago
n (O
suna
et á
lii, 2
008-
2010
).
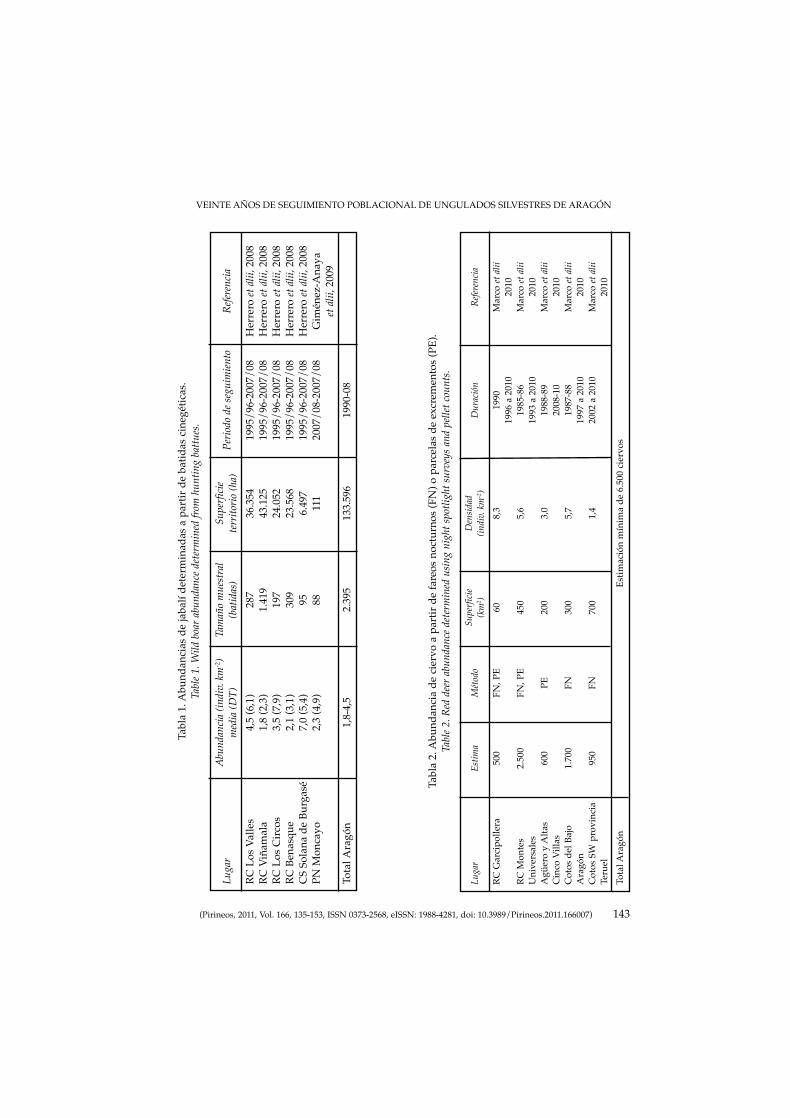
VEINTE AÑOS DE SEGUIMIENTO POBLACIONAL DE UNGULADOS SILVESTRES DE ARAGÓN
143(Pirineos, 2011, Vol. 166, 135-153, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166007)
Abu
ndan
cia (i
ndiv
. km
-2)
Tam
año
mue
stra
lSu
perfi
cieLu
gar
med
ia (D
T)(b
atid
as)
terr
itorio
(ha)
Perio
do d
e seg
uim
iento
Refer
encia
RC L
os V
alle
s4,
5 (6
,1)
287
36.3
5419
95/9
6-20
07/0
8H
erre
ro et
álii
,200
8RC
Viñ
amal
a1,
8 (2
,3)
1.41
943
.125
1995
/96-
2007
/08
Her
rero
et á
lii, 2
008
RC L
os C
ircos
3,5
(7,9
)19
724
.052
1995
/96-
2007
/08
Her
rero
et á
lii, 2
008
RC B
enas
que
2,1
(3,1
)30
923
.568
1995
/96-
2007
/08
Her
rero
et á
lii, 2
008
CS
Sola
na d
e Bu
rgas
é7,
0 (5
,4)
956.
497
1995
/96-
2007
/08
Her
rero
et á
lii, 2
008
PN M
onca
yo2,
3 (4
,9)
8811
120
07/0
8-20
07/0
8G
imén
ez-A
naya
et á
lii, 2
009
Tota
l Ara
gón
1,8-
4,5
2.39
513
3.59
619
90-0
8
Tabl
a 1.
Abu
ndan
cias
de
jaba
lí de
term
inad
as a
par
tir d
e ba
tidas
cin
egét
icas
.Ta
ble 1
. Wild
boa
r abu
ndan
ce d
eter
min
ed fr
om h
untin
g ba
ttues
.
Supe
rficie
Den
sidad
Luga
rEs
tima
Mét
odo
(km
2 )(in
div.
km-2)
Dur
ació
nRe
feren
cia
RC G
arci
polle
ra50
0FN
, PE
608,
319
90M
arco
et á
lii19
96 a
201
020
10RC
Mon
tes
2.50
0FN
, PE
450
5,6
1985
-86
Mar
co et
álii
Uni
vers
ales
1993
a 2
010
2010
Agü
ero
y A
ltas
600
PE20
03,
019
88-8
9M
arco
et á
liiC
inco
Vill
as20
08-1
020
10C
otos
del
Baj
o1.
700
FN30
05,
719
87-8
8M
arco
et á
liiA
ragó
n19
97 a
201
020
10C
otos
SW
pro
vinc
ia95
0FN
700
1,4
2002
a 2
010
Mar
co et
álii
Teru
el20
10To
tal A
ragó
nEs
timac
ión
mín
ima
de 6
.500
cie
rvos
Tabl
a 2.
Abu
ndan
cia
de c
ierv
o a
part
ir de
fare
os n
octu
rnos
(FN
) o p
arce
las d
e ex
crem
ento
s (PE
).Ta
ble 2
. Red
dee
r abu
ndan
ce d
eter
min
ed u
sing
nigh
t spo
tligh
t sur
veys
and
pell
et co
unts
.
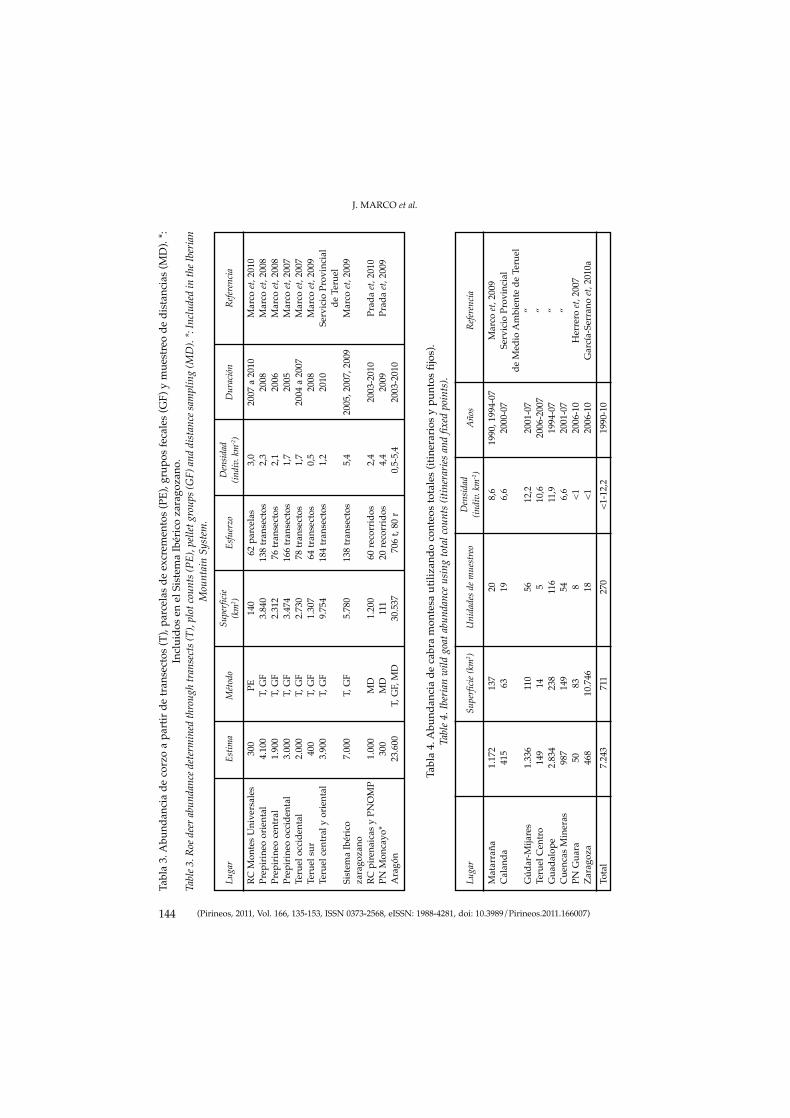
J. MARCO et al.
144 (Pirineos, 2011, Vol. 166, 135-153, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166007)
Luga
rEs
tima
Mét
odo
Supe
rficie
Esfu
erzo
Den
sidad
Dur
ació
nRe
feren
cia(k
m2 )
(indi
v. km
-2)
RC M
onte
s Uni
vers
ales
300
PE14
062
par
cela
s3,
020
07 a
201
0M
arco
et, 2
010
Prep
irine
o or
ient
al4.
100
T, G
F3.
840
138
tran
sect
os2,
320
08M
arco
et, 2
008
Prep
irine
o ce
ntra
l1.
900
T, G
F2.
312
76 tr
anse
ctos
2,1
2006
Mar
co et
, 200
8Pr
epiri
neo
occi
dent
al3.
000
T, G
F3.
474
166
tran
sect
os1,
720
05M
arco
et, 2
007
Teru
el o
ccid
enta
l2.
000
T, G
F2.
730
78 tr
anse
ctos
1,7
2004
a 2
007
Mar
co et
, 200
7Te
ruel
sur
400
T, G
F1.
307
64 tr
anse
ctos
0,5
2008
Mar
co et
, 200
9Te
ruel
cen
tral
y o
rient
al3.
900
T, G
F9.
754
184
tran
sect
os1,
220
10Se
rvic
io P
rovi
ncia
lde
Ter
uel
Sist
ema
Ibér
ico
7.00
0T,
GF
5.78
013
8 tr
anse
ctos
5,4
2005
, 200
7, 2
009
Mar
co et
, 200
9za
rago
zano
RC p
irena
icas
y P
NO
MP
1.00
0M
D1.
200
60 re
corr
idos
2,4
2003
-201
0Pr
ada
et, 2
010
PN M
onca
yo*
300
MD
111
20 re
corr
idos
4,4
2009
Prad
a et
, 200
9A
ragó
n23
.600
T, G
F, M
D30
.537
706
t, 80
r0,
5-5,
420
03-2
010
Tabl
a 3.
Abu
ndan
cia
de c
orzo
a p
artir
de
tran
sect
os (T
), pa
rcel
as d
e ex
crem
ento
s (PE
), gr
upos
feca
les (
GF)
y m
uest
reo
de d
ista
ncia
s (M
D).
*:In
clui
dos e
n el
Sis
tem
a Ib
éric
o za
rago
zano
.Ta
ble 3
. Roe
dee
r abu
ndan
ce d
eter
min
ed th
roug
h tr
anse
cts (
T), p
lot c
ount
s (PE
), pe
lletg
roup
s(G
F) an
d di
stan
ce sa
mpl
ing
(MD
). *:
Inclu
ded
in th
e Ibe
rian
Mou
ntai
n Sy
stem
.
Luga
rSu
perfi
cie (k
m2 )
Uni
dade
s de m
uest
reo
Den
sidad
Año
sRe
feren
cia(in
div.
km-2)
Mat
arra
ña1.
172
137
208,
619
90, 1
994-
07M
arco
et, 2
009
Cal
anda
415
6319
6,6
2000
-07
Serv
icio
Pro
vinc
ial
de M
edio
Am
bien
te d
e Te
ruel
Gúd
ar-M
ijare
s1.
336
110
5612
,220
01-0
7“
Teru
el C
entr
o14
914
510
,620
06-2
007
“G
uada
lope
2.83
423
811
611
,919
94-0
7“
Cue
ncas
Min
eras
987
149
546,
620
01-0
7“
PN G
uara
5083
8<1
2006
-10
Her
rero
et, 2
007
Zara
goza
468
10.7
4618
<120
06-1
0G
arcí
a-Se
rran
o et
, 201
0aTo
tal
7.24
371
127
0<1
-12,
219
90-1
0
Tabl
a 4.
Abu
ndan
cia
de c
abra
mon
tesa
util
izan
do c
onte
os to
tale
s (iti
nera
rios y
pun
tos f
ijos)
.Ta
ble 4
. Ibe
rian
wild
goa
t abu
ndan
ce u
sing
tota
l cou
nts (
itine
rarie
s and
fixe
d po
ints
).

VEINTE AÑOS DE SEGUIMIENTO POBLACIONAL DE UNGULADOS SILVESTRES DE ARAGÓN
145(Pirineos, 2011, Vol. 166, 135-153, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166007)
Año
Jaba
líCo
rzo
Cier
voSa
rrio
Cabr
a m
onte
saTo
tal
1999
10.2
8412
982
9–
6311
.305
2000
11.4
0630
493
672
311
113
.480
2001
14.5
4936
694
671
417
816
.753
2002
17.9
0943
71.
304
646
196
20.4
9220
0319
.328
530
1.31
364
625
822
.075
2004
20.4
4160
81.
333
653
264
23.2
9920
0522
.283
758
1.62
765
952
325
.850
2006
25.2
5286
91.
677
613
727
29.1
3820
0726
.597
1.04
01.
996
518
860
31.0
1120
0826
.424
1.49
62.
463
312
984
31.6
7920
0929
.595
2.14
12.
424
303
1.17
335
.636
Tasa
anu
alcr
ecim
ient
o10
%25
,6 %
11,8
%-7
,5 %
30,2
%10
,6 %
Tabl
a 5.
Cap
tura
s de
ungu
lado
s silv
estr
es d
uran
te la
tem
pora
da c
ineg
étic
a (1
999-
2009
).Ta
ble 5
. Wild
ung
ulat
es h
untin
g ba
gs h
unte
d du
ring
hunt
ing
seas
ons(
1999
-200
9).

discreta extensión (5.580 ha). La expansión territorial y demográfica pareceasimismo contenida en las poblaciones prepirenaicas occidentales.
El corzo está presente en el 59,3 % del territorio (Figura 2) y presenta abun-dancias dispares (Tabla 3). Sus capturas se han multiplicado por 16,6 (1999-2009), si bien se parte de unos cupos asignados con poca información en 1999(129 ejemplares). A partir de 2007, la información disponible es más precisa yafecta a gran parte de la comunidad autónoma, pasando los cupos de 1.040 a2.141 ejemplares en 2009, esto es duplicando el cupo (Tabla 3) (Ferreres et al.,2010).
La cabra montesa ocupa el 23,2 % del territorio (Figura 2). En Huesca lasubespecie autóctona Capra p. pyrenaica se extinguió en 2000 (García-González& Herrero, 2000). Desde mediados de los 90 un escape de cabras pertenecien-tes a la subespecie C. p. hispanica procedentes de Cazorla, comenzaron a repo-blar el PNG. En 2006 comenzó su seguimiento y en 2010 su población se esti-ma en no menos de 50 ejemplares en pleno proceso de expansión (Herrero etal., 2007). En Zaragoza la presencia de la cabra montés comienza hacia prin-cipios de los 90 como resultado de la expansión natural de la especie desdeTeruel. En 2006 comienza su seguimiento y en 2010 se estiman un mínimo de300 cabras para toda la provincia, también en pleno proceso de expansión(García-Serrano et al.,, 2010a). En Teruel y a partir de 1994, se inicia una ruti-na de estimas poblacionales, en años alternos, en época de celo, por medio derecorridos y puntos de observación, en unidades previamente diseñadas eidentificadas cartográficamente (Informes inéditos, Gobierno de Aragón). Amedida que la especie ha colonizado nuevos territorios se ha ido ampliandola superficie de estudio hasta alcanzar las casi cien mil ha actuales. Se esti-maron unas 7.000 cabras en 2007, aunque no todas las subpoblaciones sonobjeto de seguimiento. A partir de 2004 se estandarizan y generalizan losseguimientos en gran parte de Teruel, de forma que se llega a proponer unaextracción de 1.173 ejemplares en 2009; esto es, en cinco años el cupo se hamultiplicado por 4,5 (Tabla 4).
El sarrio ocupa la práctica totalidad de su área potencial (Figura 2). Supoblación se ha estimado a partir de conteos totales de forma puntual desde1983. Desde entonces diversos trabajos han ido aproximándose al tamaño desus poblaciones. En 1995-96 comienza un seguimiento anual propiamentedicho de forma coordinada entre Aragón y Navarra (Herrero et al., 2010) y en2000 todo el conjunto de subpoblaciones están integradas en él. Desde enton-ces pueden distinguirse tres periodos en cuanto a su tendencia: incrementomoderado (2000-2007); decremento por una epidemia de queratoconjuntivitisinfecciosa (2007-2009) y comienzo de la recuperación (2010) (Figura 3). Estoha llevado parejo la adecuación del cupo, que en ausencia de brotes epidémi-
J. MARCO et al.
146 (Pirineos, 2011, Vol. 166, 135-153, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166007)

cos con mortalidad relevante rondan el 5%. (Tabla 5). En 2010 el número míni-mo de ejemplares se estima en unos 13.000 (García-Serrano et al., 2010b).
La cabra doméstica asilvestrada ocupa tan solo el 3,6 % del territorio ara-gonés (Figura 2) y está presente en el Pirineo y Prepirineo. El conjunto de supoblación no ha sido estimado, aunque en el PNG su número asciende a 941animales y su densidad a 2,1 cabras km-2 (Herrero et al., 2009), lo que coinci-de con lo estimado en 1995 por Gortázar et al., (1997).
La presencia de arruí se circunscribe en Aragón al coto cercado deBastarás, en el PNG (Figura 2), en el que en 2009 había alrededor de unos 30ejemplares (Herrero et al., 2009b). El muflón está también circunscrito a estecercado y algún otro, no manteniendo poblaciones viables fuera de los mis-mos. El gamo también tiene una presencia marginal, circunscrita a la RCMUy alrededores y procedente de algunos escapes (Figura 2).
5. Discusión
Todos los ungulados silvestres presentes en Aragón tienen una distribu-ción conocida y los de abundancia relevante son objeto de seguimiento demo-gráfico en la práctica totalidad de su área de distribución. La extraordinariarecuperación de la comunidad de ungulados aragonesa desde mediados delsiglo XIX hasta finales del XX (Madoz, 1848; Gortázar et al., 2000) probable-mente continuará en el futuro, lo cual plantea un escenario muy dinámico enel que la monitorización de las poblaciones es crucial para su gestión. El papelesencial lo juegan afortunadamente las especies autóctonas, siendo las espe-cies y poblaciones introducidas bastante marginales, aunque el papel de lacabra doméstica asilvestrada posee cierta importancia (Aldezabal & Garin,2000). Salvo en el caso del sarrio, con poblaciones bastante estabilizadas ofluctuantes y un área potencial ya ocupada, el incremento en las capturas delas cuatro especies principales (jabalí, ciervo, corzo, cabra montesa) muestra(Figura 3), por un lado, su recuperación, y por otro, la necesidad de controlarsu abundancia y expansión, en plena fase inicial de incremento; algo eviden-temente difícil y que requiere un notable esfuerzo.
Esta expansión, unida a los cambios de cultivo de secano a regadío (cere-ales y frutales) en amplias zonas del territorio, está planteando un nivel deincompatibilidad entre la agricultura y la recuperación de los ungulados sil-vestres que muestra un nuevo escenario: la necesidad de zonificar el territo-rio en función de la capacidad de carga social de estas especies, influida deforma determinante por la relevancia de los daños agrícolas y los atropellosde vehículos. En pocos años, el modelo ha pasado del fomento de la conser-
VEINTE AÑOS DE SEGUIMIENTO POBLACIONAL DE UNGULADOS SILVESTRES DE ARAGÓN
147(Pirineos, 2011, Vol. 166, 135-153, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166007)

vación, a la promoción del control de muchas especies, tal y como ocurre enel resto de Europa (Apollonio et ál., 2010; Ferreres et ál., este mismo número).
Las poblaciones de ungulados silvestres han crecido y colonizado nuevosterritorios, no sólo por el seguimiento de criterios de extracción conservado-res y orientados a la promoción poblacional, sino por la creciente disponibili-dad de superficies con buenas condiciones de refugio y tranquilidad, conse-cuencia del abandono de usos tradicionales y de la presencia humana enzonas de montaña y terrenos con rendimientos marginales; lugares en los quela caza mayor se ha convertido en el aprovechamiento rústico mejor pagado,muy por encima del pastoril o del forestal. Sin embargo, en las zonas en lasque se ha modificado recientemente la vocación de algunos cultivos, graciasa la creación de nuevos regadíos, las especies de caza mayor son rechazadaspor el agricultor, el cual no asume la preexistencia de éstas y los condicio-nantes que conlleva su presencia en lo que concierne a medidas de protecciónde cultivos, pretendiendo que sea el sector cinegético o la Administraciónquienes satisfagan sus pérdidas o costeen dichas medidas de protección,incluyendo la eliminación de la especie problema.
Los cupos de caza han sido claramente conservadores con carácter gene-ral en toda la Comunidad Autónoma, si bien, debido a la conflictividad conlos productores de cultivos leñosos (principalmente cerezos y viñas) en elSistema Ibérico zaragozano, desde 2008 se vienen aplicando tasas de extrac-ción mucho más severas en estas zonas.
El periodo de caza del jabalí no ha hecho más que ampliarse y no existenapenas condicionantes para autorizar batidas y esperas por daños fuera detemporada. Podría afirmarse, con independencia de las variaciones locales dela abundancia, que el número de jabalíes cazados depende de la capacidad ydedicación de los cazadores ya que éstos no tienen limitadas las capturas,pese a lo cual éstas no han hecho más que crecer en los últimos once años(Figura 3).
La población de ciervo del Bajo Aragón ha visto duplicados sus efectivosen diez años pese a la presión cinegética ejercida, lo cual, junto a la crecienteimplantación de nuevos regadíos en la zona, ha hecho necesario casi duplicarla tasa de extracción en las dos últimas temporadas de caza, en pos de unadrástica reducción del tamaño poblacional a corto plazo. En la Reserva deCaza de Montes Universales la importante presión de caza que se mantienedesde hace cinco años, tan sólo consigue contener la explosión demográficade la especie, sin evitar un discreto pero constante crecimiento, habida cuen-ta del importante flujo de animales que se produce con las vecinas Cuenca yGuadalajara, en las que los criterios de gestión poblacional no tienen por quécoincidir con los seguidos en dicha Reserva.
J. MARCO et al.
148 (Pirineos, 2011, Vol. 166, 135-153, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166007)
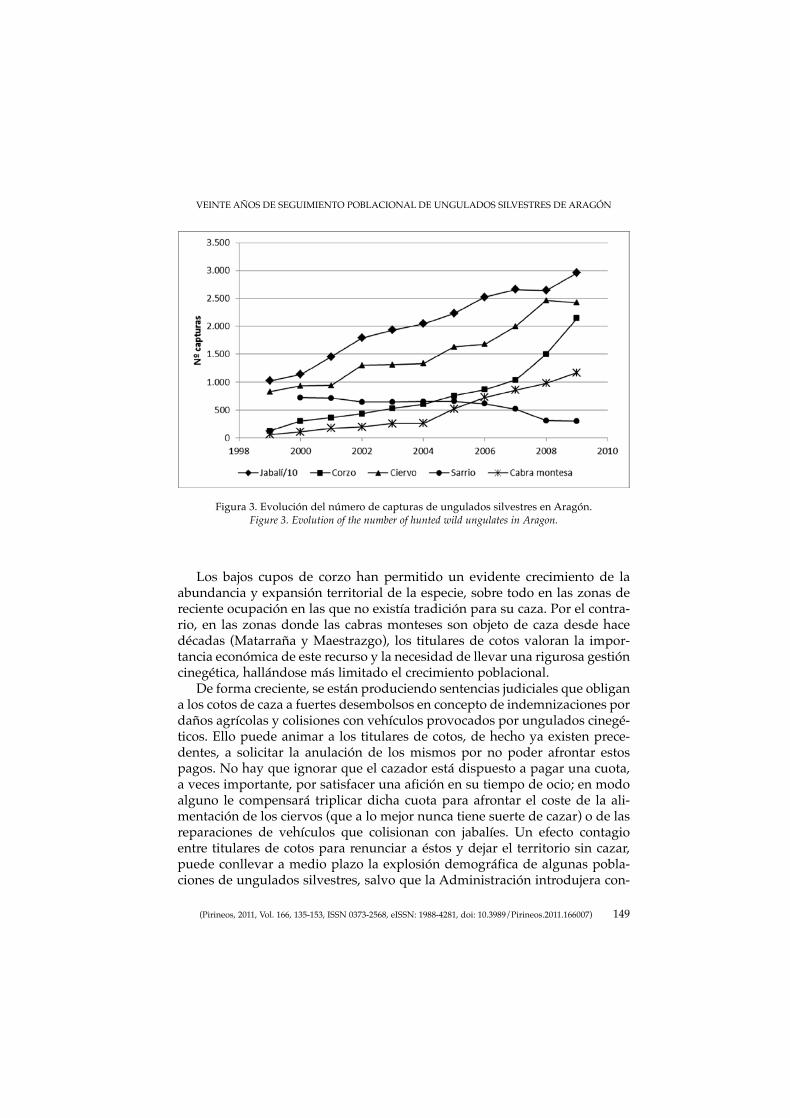
Los bajos cupos de corzo han permitido un evidente crecimiento de laabundancia y expansión territorial de la especie, sobre todo en las zonas dereciente ocupación en las que no existía tradición para su caza. Por el contra-rio, en las zonas donde las cabras monteses son objeto de caza desde hacedécadas (Matarraña y Maestrazgo), los titulares de cotos valoran la impor-tancia económica de este recurso y la necesidad de llevar una rigurosa gestióncinegética, hallándose más limitado el crecimiento poblacional.
De forma creciente, se están produciendo sentencias judiciales que obligana los cotos de caza a fuertes desembolsos en concepto de indemnizaciones pordaños agrícolas y colisiones con vehículos provocados por ungulados cinegé-ticos. Ello puede animar a los titulares de cotos, de hecho ya existen prece-dentes, a solicitar la anulación de los mismos por no poder afrontar estospagos. No hay que ignorar que el cazador está dispuesto a pagar una cuota,a veces importante, por satisfacer una afición en su tiempo de ocio; en modoalguno le compensará triplicar dicha cuota para afrontar el coste de la ali-mentación de los ciervos (que a lo mejor nunca tiene suerte de cazar) o de lasreparaciones de vehículos que colisionan con jabalíes. Un efecto contagioentre titulares de cotos para renunciar a éstos y dejar el territorio sin cazar,puede conllevar a medio plazo la explosión demográfica de algunas pobla-ciones de ungulados silvestres, salvo que la Administración introdujera con-
VEINTE AÑOS DE SEGUIMIENTO POBLACIONAL DE UNGULADOS SILVESTRES DE ARAGÓN
149(Pirineos, 2011, Vol. 166, 135-153, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166007)
Figura 3. Evolución del número de capturas de ungulados silvestres en Aragón.Figure 3. Evolution of the number of hunted wild ungulates in Aragon.

troles con medios propios o externos, con el coste que ello podría suponer.Cierto es que llegados a situaciones de sobreabundancia, las poblacionespodrían sufrir la acción de mecanismos de regulación dependientes de ladensidad, pero mientras eso ocurre ¿Soportarían los particulares o el erariopúblico un crecimiento geométrico de los daños?
Sea como fuere, parece evidente que el ejercicio de la caza mayor, sin per-der su componente de actividad lúdica, con cierta repercusión económicapara los propietarios de terrenos cinegéticos y otros sectores, se ha converti-do en una herramienta de control de ciertas poblaciones de ungulados sil-vestres (al uso de lo que ocurre con el conejo de monte allí donde es muyabundante). La adecuada utilización de esta herramienta, va a ser de gran uti-lidad para mantener dichas poblaciones en niveles compatibles con su propiaconservación, manteniendo unas afecciones sobre intereses antrópicos queresulten soportables o asumibles, por quienes las padecen y por quienesdeban satisfacer su coste.
Agradecimiento
Los trabajos que han servido de base a este artículo son mayoritariamenteresultado de asistencias técnicas financiadas por el Gobierno de Aragón.Agradecemos el esfuerzo llevado a cabo por los Agentes de Protección de laNaturaleza, vigilantes contratados y técnicos del Gobierno de Aragón. Losseguimientos poblacionales de ungulados comenzaron a mediados de losaños ochenta en Aragón, gracias al empeño de personas como Julio Guiral yMatilde Cabrera, técnicos ambos del Gobierno de Aragón. Posteriormenteotros responsables administrativos en materia cinegética desde las reservasde caza, unidades de caza y pesca de las tres provincias y sucesivas direccio-nes generales competentes en la materia, han continuado con la labor de pro-mover y coordinar estos seguimientos, por encima de no pocas adversidadespresupuestarias y de la frecuente incomprensión de propios y extraños antela utilidad de tanto esfuerzo. Vaya para todos ellos nuestro más sincero reco-nocimiento.
Referencias
Acevedo, P., Ferreres, J., Jaroso, R., Durán, M., Escudero, M.A., Marco, J. &Gortázar, C., 2010. Estimating roe deer abundance for pellet groups countsin Spain: an assessment of methods suitable for Mediterranean wood-lands. Ecological indicators, 10: 1226-1230.
J. MARCO et al.
150 (Pirineos, 2011, Vol. 166, 135-153, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166007)

Aldezabal, A. & Garin, I., 2000. Browsing preferences of feral goats (Capra hir-cus L.) in a Mediterranean mountain scrubland. Journal of AridEnvironments, 44 (1): 133-142.
Apollonio, M., Anderson, R. & Putman, R., 2010. European Ungulates and theirManagement in the 21st Century. Cambridge University Press, Cambridge(Reino Unido).
Buckland S.T., Anderson, D.R., Burnham, K.P., Laake, J.L., Borchers, D.L. &Thomas, L., 2001. Introduction to Distance Sampling. Oxford UniversityPress, Oxford, Reino Unido.
De la Riva, J., 1997. Atlas Geográfico de Aragón en Internet. Disponible en http:// 155.210.60.15/geoatlas/atlas.htm (consulta: julio de 2006).
Escudero, M.A., Ferreres, J. & Marco, J., 2007. Situación de la población de corzoen el Parque Natural del Moncayo. Informe interno del Gobierno de Aragón.Zaragoza.
Ferreres, J., Escudero, M.A., Marco, J. & Escudero, E., 2011. Gestión del corzoen Zaragoza: de conservación a control poblacional. Pirineos, 166: 69-86.
García-González, R. & Herrero, J., 1999. El bucardo de los Pirineos: historia deuna extinción. Galemys, 11: 17 -26.
García-González, R., Hidalgo, R., Ameztoy, J.M. & Herrero, J., 1992. Census,population structure and habitat use of a chamois population in OrdesaN.P. living in sympatry with Pyrenean wild goat. Proceedings of theSymposium Ongulés/Ungulates 91. F. Spitz, G. Janeau, G. Gonzalez, S.Aulagnier, Eds.: 321-25.
García-Serrano A., Prada, C., Fernández-Arberas, O. & Herrero, J., 2010.Seguimiento de la cabra montesa en Zaragoza durante 2010. Informe internodel Gobierno de Aragón. Zaragoza.
García-Serrano, A., Prada C., Fernández-Arberas, O. & Herrero, J., 2010b.Seguimiento de los ungulados en el Pirineo aragonés. Informe interno delGobierno de Aragón. Zaragoza.
Garín, I., 2000. El ciervo de la Reserva de Caza de la Garcipollera. Publicacionesdel Consejo de Protección de la Naturaleza de Aragón, serie Investigaciónnº 22.
Giménez-Anaya, A., Revilla, M., Prada, C., Arnal, M.C., García-Serrano, A.,Martínez, D., Fernández de Luco, D. & Herrero, J., 2009. El jabalí Sus scro-fa y su caza en el Parque Natural del Moncayo. Un ejemplo de seguimien-to poblacional en un espacio natural protegido. Naturaleza Aragonesa, 22:20-25.
Gortázar, C., Ferreres, J., Marco, J., Fernández de Luco, D., Garin, I. &Aldezabal, A., 1997. The feral goat in Guara (Aragón, Spain): conservationor erradication. BIPAS, 17: 73-79.
VEINTE AÑOS DE SEGUIMIENTO POBLACIONAL DE UNGULADOS SILVESTRES DE ARAGÓN
151(Pirineos, 2011, Vol. 166, 135-153, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166007)

Gortázar, C., Herrero, J., Villafuerte, R. & Marco, J., 2000. Historical examina-tion of the status of large mammals in Aragon, Spain. Mammalia, 64: 411-422.
Herrero, J., 2003. Adaptación funcional del jabalí a un ecosistema forestal y a un sis-tema agrario intensivo en Aragón. Publicaciones del Consejo de Protecciónde la Naturaleza de Aragón. Serie Investigación, número 41. 159 páginas.
Herrero, J., Escudero, E., García, J.M., García-Serrano, A., Prada, C. & Couto,S., 2004. Gestión y seguimiento demográfico del sarrio en el Pirineo ara-gonés. En: J. Herrero, E. Escudero, D. Fernández de Luco, R. García-González (Eds.). El sarrio pirenaico Rupicapra p. pyrenaica: biología, patolo-gía y gestión. Actas de las Primeras Jornadas sobre el Sarrio Pirenaico.Publicaciones del Consejo de Protección de la Naturaleza de Aragón. SerieInvestigación, número 46, 263 páginas. Páginas: 69-82.
Herrero, J., Garin, I., Prada, C. & García-Serrano, A., 2010. Interagency coor-dination fosters the recovery of the Pyrenean chamois Rupicapra p. pyre-naica at its western limit. Oryx, 44 (4): 529-532.
Herrero, J., Giménez-Anaya, A., García-Serrano, A., Prada, C. & Fernández-Arberas, O., 2008. Seguimiento del jabalí Sus scrofa L. en las Reservas de Cazade Los Valles, Viñamala, Los Circos y Benasque y el Coto Social de la Solana deBurgasé. Informe interno del Gobierno de Aragón. Zaragoza.
Herrero, J., Prada, C., Fernández-Arberas, O. & García-Serrano, A., 2009a.Estimación poblacional de la cabra doméstica asilvestrada Capra hircus en elParque Natural de la Sierra y Cañones de Guara durante 2009. Informe internodel Gobierno de Aragón. Zaragoza.
Herrero J., Prada, C., Fernández-Arberas, O. & García-Serrano, A., 2009b. Elarruí Ammotragus laervia en el Parque Natural de la Sierra y Cañones deGuara. Informe interno del Gobierno de Aragón. Zaragoza.
Herrero, J., Prada, C., Fernández-Arberas, O. & García-Serrano, A., 2007. Thebeginning of the recovery of the Iberian wild goat Capra pyrenaica in thePyrenees. Newsletter of the IUCN/SSC Caprinae Specialist Group, Marzo2007: 6-7.
Madoz, P., 1845-1850. Diccionario geográfico-estadístico-histórico de España y susposesiones de ultramar. La Ilustración, Madrid. [Edición facsímil a cargo deSánchez Zurro, D. (ed.). Diccionario geográfico-estadístico-histórico:Aragón. 3 volúmenes. Ámbito Ediciones S. A., Valladolid. 307pp. Año 1985.
Marco, J., Escudero, M.A., Ferreres, J., Labarta, J.L. & Monrabal, L., 1996-2010.Censo de las poblaciones de ciervo en Aragón: Propuesta de Plan de Caza para lastemporadas de caza 1996-2010. Informes internos del Gobierno de Aragón.Zaragoza.
Marco, J., Escudero, M.A., Labarta, J.L. & Monrabal, L., 2005. Determinación deltamaño de la población de ciervo (Cervus elaphus l.) en la Reserva de Caza de la
J. MARCO et al.
152 (Pirineos, 2011, Vol. 166, 135-153, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166007)

Garcipollera (Huesca) mediante métodos de censo indirectos. Informe internodel Gobierno de Aragón. Zaragoza.
Marco, J. & Jordán, G., 1990. Estudio de la población de corzo en el Macizo delMoncayo y área de influencia. Informe interno del Gobierno de Aragón.
MArco, J., Ferreres, J., Escudero, M.A., Labarta, J.L. & Monrabal, L., 2004-2010. Estimación de abundancias de corzo en Aragón según sectores entre 2004y 2010. Informes internos del Gobierno de Aragón. Zaragoza.
Marco, J., Escudero, M.A., Ferreres, F., Labarta, J.L. & Monrabal, L., 2009-2010.Seguimiento de la población de cabra montés en el núcleo poblacional del AltoMatarraña: Propuesta de capturas temporada 2009-2010. Informe interno delGobierno de Aragón. Zaragoza.
Mayle, B.A., Peace, J.A. & Gill, R.M., 1999. How many deer? A field guide to esti-mating deer population size. UK: Forestry Commision, Edingburgh. FieldBook 18.
Osuna, D., Prada, C., Herrero, J. & Marco, J., 2006-2008. Distribución de losungulados silvestres en Aragón (2001-2005) determinada a partir deencuestas. Lucas Mallada, 13: 191-212.
Prada, C., García-Serrano, A., Fernández-Arberas, O. & Herrero, J., 2010.Seguimiento del corzo Capreolus capreolus en el Pirineo aragonés. Informeinterno del Gobierno de Aragón. Zaragoza.
Prada, C., Herrero, J., Fernández-Arberas, O. & García-Serrano, A., 2009.Protocolos para el Seguimiento ecológico del Parque Natural del Moncayo.Informe interno del Gobierno de Aragón. Zaragoza.
Sampietro Latorre, F.J., Pelayo Zueco, E., Hernández Fernández, F., CabreraMillet, M. & Guiral Pelegrín, J., 2000. Descripción fisiográfica de laComunidad Autónoma de Aragón. En: Aves de Aragón. Atlas de especiesnidificantes. Diputación General de Aragón, segunda edición. 566 páginas.
Tellería, J.L., 1986. Manual para el censo de vertebrados terrestres. Ed. Raíces.Madrid
Tsaparis, D., Katsanevakis, S., Ntolka, E. & Legakis, A., 2009. Estimating dungdecay rates of roe deer (Capreolus capreolus) in different habitat types of aMediterranean ecosystem: an information theory aproach. Eur. J. WildlifeRes., 55, 167-172.
VEINTE AÑOS DE SEGUIMIENTO POBLACIONAL DE UNGULADOS SILVESTRES DE ARAGÓN
153(Pirineos, 2011, Vol. 166, 135-153, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166007)

155
PRIMERA REUNIÓN SOBRE EL REBECO CANTÁBRICOY EL SARRIO PIRENAICO.
BENASQUE (HUESCA, ESPAÑA), 20-22 DE MAYO 2011
J.L. ALARCÓN1, D. FERNÁNDEZ DE LUCO 2, R. GARCÍA-GONZÁLEZ3,A. GARCÍA-SERRANO4 & J. HERRERO5 (COMITÉ ORGANIZADOR)
1. Gobierno de Aragón 2. Departamento de Patología. Universidad de Zaragoza. Miguel Servet 177. E-50013 Zaragoza.
[email protected]. Instituto Pirenaico de Ecología, CSIC. Apdo. Correos 64, 22700 Jaca (Spain), [email protected]
4. Ega Consultores en Vida Silvestre. Sierra de Vicort 31. E-50003 Zaragoza, Spain. [email protected]. Área de Ecología. Escuela Politécnica Superior de Huesca. Universidad de Zaragoza. E-22071
Zaragoza. [email protected]
Antecedentes y objetivos
En 2002 se celebraron las primeras jornadas sobre el sarrio pirenaico en Jaca(Huesca). Constituyó de encuentro importante para todos los gestores, cientí-ficos y cazadores interesados en la especie, que permitió compartir experien-cias y establecer relaciones entre especialistas de ambas vertientes de la cade-na pirenaica. Las jornadas fueron posteriormente publicadas en un volumenmonográfico que aunaba el conocimiento básico sobre la biología del sarriopirenaico (Rupicapra pyrenaica pyrenaica) con aspectos de la patología y gestiónde sus poblaciones (Herrero et al. 2004). Pocos años más tarde surgió una ini-ciativa similar sobre el rebeco cantábrico (R. p. parva). Tras unas jornadas cele-bradas en Asturias en 2006, se publicó también un libro con las principalescontribuciones junto con otros trabajos posteriores (Pérez-Barbería & Palacios,2009). Tras casi una década desde la primera iniciativa y con el fin de actuali-zar el estado del conocimiento de las poblaciones de rupicaprinos del sudoes-te europeo, se celebraron unas nuevas jornadas en Benasque (Huesca) enmayo de 2011, que reunieron a diferentes sectores interesados en este ungula-do autóctono de alto valor ecológico y económico. A continuación se transcri-ben los resúmenes de las comunicaciones presentadas.
Pirineos.Revista de Ecología de MontañaVol. 166, 155-177
Jaca, Enero-Diciembre, 2011ISSN: 0373-2568
eISSN: 1988-4281doi: 10.3989/Pirineos.2011.166008

LOS NOMBRES POPULARES DEL GÉNERO RUPICAPRA:REBECO Y SARRIO
R. VIDALLER TRICAS
Caldearenas 18. E-22600 Samianigo
Existe lo que tiene nombre, y el nombre encuadra a lo que existe en taxo-nomías que permiten organizar el saber, manejarlo, dentro de marcos cultu-rales concretos. El análisis de estas culturas nos remite a formas diversas dever y entender el entorno. Los nombres comunes del género Rupicapra, a par-tir del aragonés “sarrio”, nos permiten descubrir formas diferentes de rela-cionarnos con estas especies y de replantearnos la clasificación científica, queal fin y al cabo es una forma cultural más de organizar la biodiversidad.
Así, tanto la descripción de “sarrio” como “crapa chicota” (cabra pequeña),como los nombres secundarios (crapa... para las hembras, crapitos... para lascrías), nos llevan a buscar entre los nombres de las diferentes cabras y cabraspequeñas (= Capreolus) indoeuropeos, donde encontramos denominacionescomo sarne, serne, sergue, surna o srne que comparten sarrios y corzos en laEuropa central y oriental, o los derivados de ‘cabra’ en las zonas sudorienta-les, desde los Alpes al Caucaso (koza, keçi, ayts...). A su vez, nombres cercanostaxonómicamente al sarrio como “bucardo” y “erco” (Capra pyrenaica), nosremiten a otros europeos de la misma cabra (bucardo), o del ciervo y el alce(erco), mientras que el gamo nos recuerda a la gamuza centroeuropea.
Al fin, se dibujaría un panorama en el que el corzo se separa del ciervopara ir junto a las cabras, mientras el ciervo y el alce forman grupo aparte conreminiscencias comunes. Una visión popular previa y diferente de la científi-ca que nos obliga a replantear, por ejemplo, lo oportuno de asignar los nom-bres comunes a la taxonomía linneana (un ‘sarrio’ lo es tanto en Cantabriacomo en el Caucaso), que se organiza desde otros principios, por cierto, amenudo replanteados. Por otro lado, la existencia o no de determinados nom-bres populares puede correlacionar con la presencia histórica de algunasespecies, como en el caso del gamo.
Palabras clave: sarrio, rebeco, Rupicapra, nombre común.
JORNADAS
156 (Pirineos, 2011, Vol. 166, 155-177, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166008)

REINTRODUCCIÓN DEL REBECO CANTÁBRICOEN LA MONTAÑA ORIENTAL, CANTABRIA
C. SÁNCHEZ MARTÍNEZ, L. GARCÍA FUENTE, B. SERRANO GARCÍA& M. RODRÍGUEZ
Biogestión-consultora ambiental. Fundación Naturaleza y [email protected]. www.fnyh.org
Fundación Naturaleza y Hombre (FNYH), ONG ambiental, estudió y rea-lizó la reintroducción de rebeco cantábrico Rupicapra pyrenaica parva en laMontaña Oriental de Cantabria, de donde se extinguió hace más de 150 años.Este proyecto se enmarcó en el LIFE-Naturaleza Recuperación y conservaciónde la biodiversidad en la cuenca del río Asón (2002-2005), promovido porFNYH con la financiación de la Comisión Europea. La Montaña Oriental deCantabria se ubica en el sector más oriental de la Cordillera Cantábrica, inclu-ye las cuencas fluviales de los ríos Pas, Miera y Asón y Castro Valnera (1718m) es la cumbre más elevada. El proyecto se concentró en los términos muni-cipales de San Roque de Riomiera, Soba, Ruesga y Arredondo, donde se libe-raron los rebecos y donde posteriormente se han asentado los grupos depoblación.
La desaparición del rebeco del área del Alto Asón comenzó en el sigloXVIII debido a la devastación de su hábitat, provocada por la tala masiva deárboles destinados a la industria naval y a la presión cinegética. La especie noconsiguió adaptarse, lo que provocó su acantonamiento en el área de Picos deEuropa. Los últimos rebecos de los que se tiene testimonio en la MontañaOriental databan de 1845 en Lanchares y San Pedro del Romeral. En la actua-lidad, las presiones al ecosistema que ocupaba el rebeco han disminuido y hasido posible realizar su reintroducción. Entre 2002 y 2003 se liberaron un totalde 26 rebecos procedentes de la zona cántabra del Macizo Central o de losUrrieles, en los Puertos de Salvorón, el Monte Quebres y los Puertos de Áliva.Cada ejemplar se liberaba con un collar radiotransmisor que permitió elseguimiento del proceso de adaptación de los rebecos a su nuevo hábitat. Enla actualidad, la población asciende a 95 rebecos, con 19 nuevas crías en 2010.
Palabras clave: reintroducción, rebeco cantábrico, Montaña Oriental,Cantabria, ONG.
JORNADAS
157(Pirineos, 2011, Vol. 166, 155-177, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166008)

ASPECTOS ECONÓMICOS DE LA GESTIÓNDE LAS RESERVAS DE CAZA DE SARRIO
M. J. DOMÍNGUEZ & A. PARÍS
Aneto Promoción y Comunicación SL, Zaragoza. www.aneto.biz, [email protected]
Las Reservas de Caza son espacios para la promoción, fomento y protec-ción de determinadas especies cinegéticas. En el Pirineo aragonés existen cua-tro reservas para el sarrio, creadas en 1966 por sus “excepcionales posibilida-des cinegéticas” con el objeto de “garantizar la difícil pervivencia de espe-cies”. En conjunto abarcan 131.804 has y la titularidad cinegética correspondepor ley al Gobierno de Aragón. El departamento competente en medioambiente gestiona estos terrenos cinegéticos de forma directa velando por laconservación de las especies y “subordinando a esta finalidad el aprovecha-miento cinegético”.
Los costes de la gestión se estiman en promedio en 1.042.369,68 €/año (7,9€/ha) para el periodo 2004-2010, desglosados en los siguientes conceptos:38% personal, 33% funcionamiento, 23% subvenciones a los ayuntamientos y6% asistencias técnicas. La evolución del coste anual estimado es relativa-mente estable. En los municipios, la repercusión socioeconómica directaalcanzaría tres cuartas partes de los costes de gestión, correspondientes alpersonal, las subvenciones y al menos la mitad de las partidas de funciona-miento. A estas aportaciones directas hay que sumar los ingresos obtenidosde las subastas de los permisos asignados a los propietarios y la cuota com-plementaria por pieza cazada. Además, el turismo asociado a la caza originaotras aportaciones económicas directas e indirectas. La caza en las Reservasde Caza origina unos ingresos a la Administración gestora por permisos con-cedidos. Según el tipo de cazador se distribuyen de la siguiente manera: 62%de cazadores nacionales, 20% de los permisos de los propietarios y 18% decazadores locales. El ratio ingresos/costes sólo alcanza un promedio del 7%.La gestión prioriza la conservación de recursos.
Palabras clave: economía, inversión, costes, ingresos, tarifas, rendimiento.
JORNADAS
158 (Pirineos, 2011, Vol. 166, 155-177, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166008)

TRAJECTOMETRIE DES CHAMOIS: COMMENTBOUGERAIS-JE SI J’ETAIS UN CHAMOIS?
Z. TABLADO1, A. LOISON1, BOURGOING2, G. GARELL 3 & DUBRAY, D.4
1 CNRS UMR 5553 – Laboratoire d’Ecologie Alpine –Le Bourget du Lac 733762 VetAgro Sup – Campus vétérinaire de Lyon – CNRS UMR 5558 Marcy l’Etoile 69280
3 Office National de la Chasse et de la Faune Sauvage- CNERA FM – 147 route de Lodève - Juvignac34990
4 Office National de la Chasse et de la Faune Sauvage, 95 rue Pierre Flourens - Montpellier 34098
Les chamois utilisent leurs capacités de déplacement pour interagir avecleur environnent. Ces mouvements dépendent à la fois des caractéristiques etmotivations propres de l’individu, comme l’âge, le patron d’activité ou le sta-tut de reproduction, et des caractéristiques environnementales, telles que ladisponibilité de ressources alimentaires ou de refuge, et le climat. En identi-fiant quels sont les facteurs qui affectent les déplacements des chamois, onpourra comprendre comment ils répondront à des altérations futures de leurhabitat et du climat.
Nous avons ainsi étudié les trajectoires de chamois à partir de localisationsde 30 femelles équipes avec des colliers GPS dans la Reserve National deChasse et Faune Sauvage des Bauges (France). Nous avons examiné le tauxde déplacements des individus à différentes échelles temporelles en regar-dant les associations entre mouvement, activité et caractéristiques intrin-sèques et externes. Nous nous sommes intéressés plus spécifiquement auxeffets du climat (température, hauteur de neige) et du dérangement par letourisme et nous discutons aussi les possibles implications que le futur régi-me climatique pourra avoir sur les patrons de mouvement et d’activité decette espèce.
Mots clés: chamois, Rupicapra rupicapra, déplacements, localisations GPS, cli-mat, activité.
JORNADAS
159(Pirineos, 2011, Vol. 166, 155-177, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166008)

LES INDICATEURS DE CHANGEMENT ECOLOGIQUEPOUR LA GESTION DES POPULATIONS D’ISARDS
D. MAILLARD1, M. GAREL1, A. LOISON3, D. DUBRAY1, J. APPOLINAIRE, 1
& G. BOURGOIN2
1 Office National de la Chasse et de la Faune Sauvage - CNERA FM - 147 route de Lodève - Juvignac34990, France
2 École Nationale Vétérinaire de Lyon – CNRS UMR 5558 Marcy l’Etoile 69280, France3 CNRS UMR 5553 – Laboratoire d’Écologie Alpine – Le Bourget du Lac 73376, France
Grâce aux marquages d’un grand nombre de chamois et d’isards réalisésdepuis plus de 20 ans sur nos territoires d’étude, nous avons montré que lescomptages traditionnels sous-estimaient, parfois fortement, l’effectif réel despopulations suivies. Ces résultats remettent en cause l’utilisation d’estima-tions d’effectifs basées uniquement sur des dénombrements pour définir desplans de gestion. Nous proposons plutôt aujourd’hui de baser ces plans degestion sur le suivi à long terme d’un faisceau d’indicateurs caractérisant larelation population-environnement: indicateurs d’abondance, indicateurs decondition et indicateurs de consommation.
Dans ce but, nous avons mis en évidence chez le chamois et l’isard quel’observation répétée d’animaux le long d’itinéraire pédestre pré-déterminépouvait être utilisée comme un indicateur des variations d’abondance de lapopulation suivie (Loison et al., 2006, Dubray et Groupe ICE, 2008). Le poidsdes jeunes de l’année est quant à lui un indicateur sensible aux conditionsenvironnementale et à la densité d’animaux (Garel et al., sous presse) et peutdonc à ce titre servir d’indicateur de performance pour le suivi des popula-tions. Enfin l’indice de reproduction (nombre de jeunes observés par femelleadulte) pourrait jouer le même rôle que le suivi du poids des jeunes dans lamesure où chez les ongulés la performance reproductive est étroitement liéeaux conditions environnementales et aux variations de densité. La pertinen-ce de cet indicateur comme mesure de la performance reproductive réelle dela population est à l’heure actuelle en test dans un de nos territoires de réfé-rence pour le suivi de l’isard.
Cette approche basée sur le suivi simultanée de plusieurs indicateurs estdépendante de l’application rigoureuse des protocoles si l’on veut pouvoirréaliser une gestion efficace et durable des populations d’isards et de cha-mois.
Mots clés: isard, Rupicapra pyrenaica, ICE, IPS, poids, gestion des populations.
JORNADAS
160 (Pirineos, 2011, Vol. 166, 155-177, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166008)

NITRÓGENO FECAL, PARÁSITOS Y ESTATUSNUTRICIONAL EN EL REBECO
D. GASSÓ1, J. BARTOLOMÉ2, D. FERRER3, G. MENTABERRE1, G. GÁLVEZ-CERÓNA2,N. NAVARRO-GONZÁLEZ1, L. FERNÁNDEZ-SIRERA1, J.R. LÓPEZ-OLVERA1,
X. FERNÁNDEZ-AGUILAR1, L. ROSSI4, S. LAVÍN1, I. MARCO1 & E. SERRANO1
1 Servei d’ Ecopatología de Fauna Salvatge (SEFaS), Departament de Medicina i Cirurgia Animals,Universitat Autònoma de Barcelona (UAB), E-08193, Bellaterra, Barcelona, Spain. * emmanuel.serra-
[email protected] Departament de Ciència Animal i dels Aliments, Universitat Autònoma de Barcelona (UAB), E-08193,
Bellaterra Barcelona, España.3 Laboratori de Parasitologia Veterinaria, Departament de Sanitat i d’Anatomia Animals, Bellaterra,
Barcelona, España.4 Dipartimento de Produzioni Animali, Epidemiologia ed Ecologia, Universtità di Torino, Italia.
De forma general, los nematodos gastrointestinales (NGI) influyen sobrela condición nutricional de los rumiantes, reduciendo el apetito (anorexia) oalterando la capacidad de absorción de nutrientes. Este efecto negativo de losNGI, ha motivado el desarrollo de varias investigaciones para averiguar quéestrategias alimentarias maximizan la cantidad de energía ingerida con unmínimo riesgo de depredación o parasitismo (“Foraging theory”). Los rebe-cos viven en ecosistemas donde la disponibilidad de alimento sufre fuertesvariaciones estacionales. Según esta teoría, en verano los rebecos deberíandecidir si alimentarse en los pastos más nutritivos, pero quizás muy utiliza-dos y probablemente con muchos NGI, o comer en zonas de peor calidadpero más seguras. La determinación del nitrógeno fecal (Nf) es la técnica máshabitual para evaluar la calidad nutricional de las poblaciones de unguladossalvajes, ya que existe una relación directa entre la calidad del forraje y el por-centaje de nitrógeno en las heces. Pero los NGI interfieren en la absorción denutrientes, y una mayor intensidad de parasitación puede también significaruna mayor pérdida de proteínas y un aumento de Nf. Durante mayo de 2009a septiembre de 2010 estudiamos los patrones estacionales de cargas de NGIy porcentaje de Nf en heces de rebeco (Rupicapra pyrenaica) y de oveja en laRNC de Freser-Setcases (Pirineo Catalán). En contradicción con la “Foragingtheroy”, las mayores cargas parasitarias de rebecos y ovejas coincidieron conlas mejores calidades nutricionales (mayor Nf). Posiblemente los animalesprefirieron comer mejor aunque con más parásitos, o el Nf aumentó debido auna peor absorción del alimento. En esta comunicación, discutiremos lospatrones estacionales de NGI y Nf de rebecos y ovejas y presentaremos undiseño experimental para aclarar los usos y limitaciones del Nf para monito-rizar el efecto de los parásitos en poblaciones de ungulados salvajes.
Palabras clave: carga parasitaria, “Foraging theory”, indicadores fecales.
JORNADAS
161(Pirineos, 2011, Vol. 166, 155-177, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166008)

AZAPERONA Y MORTALIDAD DE REBECOS DURANTECAPTURAS CON RED VERTICAL
G. MENTABERRE, E. SERRANO, J.R. LÓPEZ-OLVERA, E. CASAS-DÍAZ, R. VELARDE,I. MARCO & S. LAVÍN
Servei d’Ecopatología de la Fauna Salvatge (SEFaS), Facultat de Veterinària,Universitat Autònoma de Barcelona, 08193, Bellaterra, Barcelona
La captura de animales salvajes puede tener distintas motivaciones.Independientemente de éstas, es necesario tener en cuenta el bienestar animaly minimizar el estrés de la captura. El uso de tranquilizantes puede ayudar aeste fin, si bien se deben realizar estudios para determinar los principios acti-vos y dosis más adecuados para cada especie y método de captura.
En esta comunicación, presentamos el análisis de las causas que determi-naron varios episodios de mortalidad durante operaciones de captura físicacon red vertical de rebecos pirenaicos (Rupicapra pyrenaica pyrenaica) destina-das a evaluar la acción de distintos tranquilizantes. Los parámetros clínicosde los rebecos que murieron se compararon con los del resto de animalesmediante análisis de la varianza. Además, utilizamos una selección de mode-los estadísticos mediante el criterio de información de Akaike para evaluar lainfluencia del tipo de tranquilizante (acepromazina, haloperidol y azapero-na), la dosis utilizada, el número de operarios por animal y la zona.
La concentración sérica de lactato -indicador de fatiga- fue significativa-mente superior en los rebecos que murieron y los dos modelos seleccionadosindican que las dosis altas de azaperona y, en menor medida, de haloperidolaumentan el riesgo de muerte. Estos resultados, junto a las alteraciones obser-vadas en las necropsias, sugieren que los animales que llegaron más fatigadosa la red de captura fueron más sensibles a los tranquilizantes y experimenta-ron efectos secundarios adversos que incrementaron su probabilidad demorir. Esto nos lleva a pensar que las butirofenonas, especialmente la azape-rona, tienen un margen de seguridad estrecho en el rebeco, contrariamente alo descrito para otras especies de ungulados salvajes. Dada la ausencia debibliografía, alertamos de que esta sensibilidad a las butirofenonas tambiénpodría darse en otras especies de caprinos.
Palabras clave: acepromazina, azaperona, captura física, haloperidol, lactato,mortalidad, rebeco, Rupicapra pyrenaica.
JORNADAS
162 (Pirineos, 2011, Vol. 166, 155-177, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166008)

IDENTIFICACIÓN DE INDICADORES DE CAMBIOCLIMÁTICO EN UNA POBLACIÓN DE ISARD(RUPICAPRA P. PYRENAICA) EN ANDORRA
L. RIBA MANDICÓ
Àrea d’Hàbitats i Fauna, Departament d’Agricultura i Patrimoni Natural, Governd’Andorra. [email protected]
La población de isard de la reserva de caza de Enclar (Andorra) ha sidogestionada desde su creación apoyando las decisiones de gestión en los resul-tados de los censos simultáneos. Para poder trabajar con más parámetros ymejorar el nivel de conocimiento de la dinámica de la población, se planteódurante el año 2006 analizar los datos biométricos, parasitológicos y sanita-rios, obtenidos en todos los animales cazados desde 2001. El objetivo fueidentificar indicadores que permitieran “observar” los cambios en la pobla-ción, inducidos por diferentes parámetros ambientales. En este trabajo se pre-sentan los resultados preliminares del análisis realizado a partir de los ani-males cazados entre el 2001 y 2010, así como, las tendencias observadas enalgunas de las variables analizadas en relación con parámetros de tipoambiental o demográfico. Estos resultados preliminares permiten identificarqué parámetros podrían ser los mejores indicadores para la población estu-diada, destacando como posibles variables válidas, el peso de la canal y lasmedidas asociadas con los cuernos (longitud, altura y perímetro de la base),siendo a la vez las menos sujetas a un “efecto manipulador” en relación aotras variables morfológicas. La tendencia mostrada por estas variables en losanimales que se encuentran en crecimiento (en teoría, más sensibles a loscambios ambientales), indica una disminución progresiva de los valores en eltiempo, mostrando además una correlación negativa con la densidad estima-da de la población, durante los años anteriores al muestreo, así como conalgunas variables climatológicas analizadas. Otros datos de tipo clínico ofisiológico (valores de excreción parasitaria, morfología de las glándulasadrenales) se encuentran actualmente en estudio.
Palabras claves: isard, Andorra, Enclar, indicadores.
JORNADAS
163(Pirineos, 2011, Vol. 166, 155-177, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166008)

VARIACIONES ESTACIONALES DE NITRÓGENO FECALEN EL REBECO MEDIANTE NIRS
A. GÁLVEZ-CERÓN1, E. ALBANELL1, E. SERRANO2, G. MENTABERRE2, X.FERNÁNDEZ-AGUILAR2, L. FERNÁNDEZ2, N. NAVARRO-GONZÁLEZ2, J.R. LÓPEZ-
OLVERA2, S. LAVÍN4, I. MARCO 4 & J. BARTOLOMÉ
1 Departament de Ciència Animal i dels Aliments, Universitat Autònoma de Barcelona (UAB), E-08193,Bellaterra, Barcelona, Spain. [email protected]
2 Servei d’ Ecopatología de Fauna Salvatge (SEFaS), Departament de Medicina i Cirurgia Animals,Universitat Autònoma de Barcelona (UAB), E-08193, Bellaterra, Barcelona, Spain.
Para maximizar su aptitud biológica, los rebecos (Rupicapra pyrenaica)incrementan su condición física cuando la productividad primaria es máxi-ma. El nitrógeno fecal (Nf) es un excelente indicador de la calidad de la dietade los herbívoros. Monitorizar las variaciones estacionales de Nf, puede serútil para comprender la ecología de esta especie y establecer planes de ges-tión adaptados a las condiciones locales. En esta comunicación, presentamoslos patrones anuales de Nf de rebeco pirenaico a partir de 192 muestras deheces recolectadas desde mayo de 2009 hasta septiembre de 2010 en la RNCde Freser-Setcases (Pirineo Catalán). Las muestras se analizaron por el méto-do tradicional DUMAS/LECO y se calibraron posteriormente medianteespectroscopía en el infrarrojo cercano (NIRS) por regresión MPLS (mínimoscuadrados parciales modificados). El NIRS ofrece ciertas ventajas adicionalesrespecto al DUMAS/LECO, ya que se puede reutilizar la muestra y no pro-duce residuos. La dieta de los rebecos mostró un marcado patrón estacional,siendo el Nf máximo entre abril-agosto y mínimo entre septiembre-marzo.Tras observar este patrón, decidimos hacer dos calibraciones con el NIRS: [1]incluyendo muestras de todo el año y [2] dividendo el año en dos periodos[abril-agosto (Nf alto) y septiembre-marzo (Nf bajo)]. Aunque los resultadospara la calibración general [coeficiente determinación (R2) 0,97; error estándarde calibración (SEC) 0,076; coeficiente determinación de validación (r2) 0,95 yerror estándar de predicción (SEP) 0,109], garantizan una predicción adecua-da del Nf, las calibraciones parciales de los dos períodos del año (Nf alto y Nfbajo) mejoraron algunos parámetros estadísticos, especialmente el SEC y SEP.Los resultados obtenidos indican que la tecnología NIRS puede ser de granutilidad para evaluar con suficiente precisión el nitrógeno fecal en rebecos.
Palabras clave: calidad nutricional, heces, infrarrojo cercano.
JORNADAS
164 (Pirineos, 2011, Vol. 166, 155-177, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166008)

REBECOS Y PARÁSITOS, UN ESBOZO MEDIANTESISTEMAS DE ECUACIONES ESTRUCTURALES
E. SERRANO1, D. FERRER2, G. MENTABERRE1, N. NAVARRO-GONZÁLEZ1, L.FERNÁNDEZ-SIRERA1, J.R. LÓPEZ-OLVERA1, X. FERNÁNDEZ-AGUILAR1, A. GÁLVEZ-
CERON3, J. BARTOLOMÉ3, L. ROSSI4, J. PÉREZ5, S. LAVÍN1 & I. MARCO1
1 Servei d’ Ecopatologia de Fauna Salvatge (SEFaS), Departament de Medicina i Cirurgia Animals,Universitat Autònoma de Barcelona (UAB), E-08193, Bellaterra, Barcelona, Spain. *emmanuel.serra-
[email protected] Laboratori de Parasitologia Veterinaria, Departament de Sanitat i d’Anatomia Animals, Bellaterra,
Barcelona, Spain.3 Departament de Ciència Animal i dels Aliments, Universitat Autònoma de Barcelona (UAB), E-08193,
Bellaterra, Barcelona, Spain.4 Dipartimento de Produzioni Animali, Epidemiologia ed Ecologia, Universtità di Torino, Italy.
5 Departamento de Biología Animal, Biología Vegetal y Ecología, Universidad de Jaén, E- 23071, Jaén,Spain.
En la fauna salvaje los patrones de infección no son homogéneos, ni en eltiempo ni en el espacio. Este hecho se debe principalmente a tres mecanismos:[1] hay una gran diversidad de factores de riesgo ligados a la infección (p. e.,ambientales, poblacionales y de tipo humano), [2] todos los individuos de lapoblación no tienen la misma probabilidad de entrar en contacto con el agen-te patógeno o, [3] no manifiestan la misma susceptibilidad a ser infectados. Laimportancia relativa de estos mecanismos no está esclarecida para la mayoríade la enfermedades de la fauna salvaje. En esta comunicación presentamoslos path-pls, una herramienta estadística emergente para explorar relacionescausa-efecto entre factores espaciales (altitud y localidad), de la fenologíalocal (mes del año, calidad nutricional) y poblacionales (tamaño de grupo ydensidad), sobre las cargas de coccidios y nematodos gastrointestinales, endos poblaciones de rebeco pirenaico (Rupicapra pyrenaica) de la ReservaNacional de Caza de Freser-Setcases (Pirineo catalán). Entre otras ventajas,respecto a las herramientas estadísticas convencionales, los path-pls ponen demanifiesto relaciones indirectas entre variables que estén fuertemente rela-cionadas entre sí (colinealidad).
Palabras clave: modelos path-pls, relación parasito-hospedador, Rupicaprapyrenaica.
JORNADAS
165(Pirineos, 2011, Vol. 166, 155-177, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166008)

VARIATIONS DEMOGRAPHIQUES CHEZ L’ISARD(RUPICAPRA P. PYRENAICA) D’APRES UN SUIVI PAR
RECENSEMENTS HIVERNAUX STANDARDISES
J. P. CRAMPE
Parc National des Pyrénées, 2 rue du IV septembre 65000 Tarbes, [email protected]
Un suivi hivernal par recensements répétés a été appliqué à la populationd’isard protégée de Cauterets, dans le Parc National des Pyrénées. De 1992 à2010, 290 recensements standardisés ont été réalisés de décembre à avril surune zone d’hivernage appelée « Clot-Cayan ». La stabilité spatiale des isardssur cette zone d’hivernage a été démontrée par un suivi par marquage surplusieurs années. Les données basées sur le comptage des effectifs des diffé-rentes classes d’âge et sexe ont permis de mettre en évidence des variationsrelativement importantes de certains paramètres démographiques. Parmi lesfacteurs invoqués, certains paraissent relever de la densité, tandis qued’autres sont de nature stochastiques et dépendent plutôt du climat hivernalou d’évènements passagers. Ces facteurs de nature différente paraissent agiren synergie. Le passage d’une épizootie de kérato-conjonctivite de 2007 à2008 a permis de constater un impact relativement important de cette mala-die sur la taille de la population.
JORNADAS
166 (Pirineos, 2011, Vol. 166, 155-177, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166008)

CALENTAMIENTO GLOBAL Y HELMINTOS DELREBECO: UNA APROXIMACIÓN EXPERIMENTAL
A. GÁLVEZ-CERÓN1, J. BARTOLOMÉ1, D. FERRER2, L. ROSSI3, G. MENTABERRE4, S.LAVÍN4, I. MARCO4 & E. SERRANO4
1 Departament de Ciència Animal i dels Aliments, Universitat Autònoma de Barcelona (UAB), E-08193,Bellaterra, Barcelona, Spain. *[email protected]
2 Laboratori de Parasitologia Veterinaria, Departament de Sanitat i d’Anatomia Animals, Bellaterra,Barcelona, Spain.
3 Dipartimento de Produzioni Animali, Epidemiologia ed Ecologia, Universtità di Torino, Italy.4 Servei d’ Ecopatologia de Fauna Salvatge (SEFaS), Departament de Medicina i Cirurgia Animals,
Universitat Autònoma de Barcelona (UAB), E-08193, Bellaterra, Barcelona, Spain.
El calentamiento global está alterando la ecología de muchas especies deparásitos de ungulados domésticos y salvajes de todo el mundo. En las últi-mas décadas, leves incrementos en la temperatura media anual han sido sufi-cientes para explicar el aumento de helmintiasis en el Reino Unido o la apa-rición de nuevas especies de nematodos en ungulados salvajes del Ártico. Dehecho, incrementos de 1,5 ºC (tº media diaria) son suficientes para acelerar eldesarrollo de parásitos de ciclo directo o favorecer la dispersión de hospeda-dores intermediarios. No obstante, los efectos del calentamiento global sobrela dinámica parásito-hospedador no parecen sencillos. Un aumento de la tem-peratura también puede desecar las fases infectivas de los parásitos, o mejo-rar el estado nutricional de los hospedadores (p.ej., aumentando la producti-vidad primaria). Esta compleja interacción entre el calentamiento global y ladinámica parásito-hospedador es difícil de abordar mediante la observaciónde patrones naturales, pero se puede estudiar con la ayuda de una manipu-lación en condiciones naturales. En esta comunicación, revisaremos los efec-tos del calentamiento global sobre la dinámica y distribución de los parásitosde varias especies de ungulados domésticos y salvajes, y presentaremos eldiseño de una simulación de calentamiento global en la Reserva Nacional deCaza de Freser-Setcases (Pirineo Catalán). Con este experimento pretende-mos estudiar la influencia de un aumento de temperatura sobre: [1] la estruc-tura y composición de los pastos y [2] el desarrollo y supervivencia de losnematodos gastrointestinales del rebeco pirenaico (Rupicapra p. pyrenaica) yde otros ungulados domésticos (ovejas, vacas y caballos).
Palabras clave: nematodos gastrointestinales, ganadería de montaña,Rupicapra pyrenaica.
JORNADAS
167(Pirineos, 2011, Vol. 166, 155-177, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166008)

FACTORES DE RIESGO ASOCIADOS A LA PESTIVIROSISDEL REBECO PIRENAICO
I. MARCO1, O. CABEZÓN1,2, L. FERNÁNDEZ-SIRERA1,2, J.R. LÓPEZ-OLVERA1,G. MENTABERRE1, E. CASAS1, E. SERRANO1, R. VELARDE1, R. ROSELL2,3 & S. LAVÍN1
1 Servei d’Ecopatologia de Fauna Salvatge, Facultat de Veterinària, Universitat Autònoma de Barcelona,E-08193, Bellaterra, Barcelona, Spain. * [email protected]
2 Centre de Recerca en Sanitat Animal (CRESA), Universitat Autònoma de Barcelona, E-08193,Bellaterra, Barcelona, Spain
3 Departament d’Agricultura, Ramaderia, Pesca, Alimentació i Medi Natural, Generalitat de Catalunya,Gran Via de les Corts Catalanes 612-614, E-08007, Barcelona, Spain
En 2001 un Virus de la Enfermedad de la Frontera (Género Pestivirus,Familia Flaviviridae) se asoció con una nueva enfermedad en el rebeco pire-naico (Rupicapra pyrenaca pyrenaica). Desde entonces se han sucedido nume-rosos brotes de enfermedad en una gran parte del Pirineo central y orientalde España, Francia y Andorra, con graves consecuencias demográficas. Losfactores observados, que afectan a la aparición y gravedad de la enfermedad,son los relacionados con el propio virus, la aparición de infecciones secunda-rias, la inmunidad poblacional, la ecología y el comportamiento del rebeco yla variabilidad genética. Otros factores que también pueden tener influenciason las cepas del virus y los individuos persistentemente infectados. En con-clusión, la Enfermedad de la Frontera del rebeco pirenaico es una enfermedademergente que representa la principal amenaza para las poblaciones de estaespecie. A pesar de que es una enfermedad difícilmente controlable, el análi-sis de los factores de riesgo puede ayudar a predecir la aparición de nuevosbrotes de la enfermedad.
Palabras clave: Ecologia de las enfermedades, Epidemiologia, Pestivirus,Rupicapra pyrenaica.
JORNADAS
168 (Pirineos, 2011, Vol. 166, 155-177, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166008)

EFECTO DE LAS ENFERMEDADES SOBRE LA DINÁMICAPOBLACIONAL DEL REBECO
J.R. LÓPEZ-OLVERA1, I. MARCO1, O. CABEZÓN1,2, N. NAVARRO-GONZÁLEZ1,X. FERNÁNDEZ-AGUILAR1, L. FERNÁNDEZ-SIRERA1,2, G. MENTABERRE1, E. CASAS1,
R. VELARDE1, S. LAVÍN1 & E. SERRANO1
1 Servei d’ Ecopatología de Fauna Salvatge, Departament de Medicina i Cirurgia Animals, UniversitatAutònoma de Barcelona, E-08193, Bellaterra, Barcelona, Spain. *[email protected]
2 Centre de Recerca en Sanitat Animal (CRESA), Universitat Autònoma de Barcelona, E-08193,Bellaterra, Barcelona, Spain
La evolución del tamaño y la estructura de las poblaciones constituyenuno de los aspectos centrales de la ecología. En los ungulados se observa quelos factores estocásticos y dependientes de la densidad afectan principalmen-te a la supervivencia de los juveniles (reclutamiento), ya que la supervivenciade los adultos es prácticamente constante y la productividad de las hembrases poco variable entre años. Aunque se han descrito numerosas enfermeda-des en los ungulados salvajes, su efecto sobre las poblaciones es bastante des-conocido, ya que no se ha valorado su efecto sobre cada una de las fases delciclo vital: nacimiento, mortalidad pre-destete (supervivencia estival), morta-lidad post-destete (supervivencia invernal), supervivencia entre uno y dosaños, supervivencia adulta y supervivencia en la senescencia, así como la pro-ductividad de las hembras en cada una de las cuatro fases de su vida repro-ductora (primala, joven, adulta y senescente). En la presente comunicaciónabordamos una revisión de enfermedades descritas (o que podrían afectar) ala eficacia biológica y la dinámica poblacional de las especies del géneroRupicapra.
Palabras clave: Ecología de las enfermedades, dinámica poblacional.
JORNADAS
169(Pirineos, 2011, Vol. 166, 155-177, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166008)

INFLUENCE DU PESTIVIRUS SUR LA DYNAMIQUE DESPOPULATIONS D’ISARDS PYRENEENS
M. PIOZ1, E. GILOT-FROMONT2, A. LOISON3, D. DUBRAY4 & V. GROSBOIS5
1 Institut National pour la Recherche Agronomique- Unité d’Epidémiologie Animale – Centre deClermont-Ferrand Theix - St Genès Champanelle 63122 [email protected]
2 Ecole Nationale Vétérinaire de Lyon – CNRS UMR 5558 Marcy l’Etoile 69280 [email protected] CNRS UMR 5553 – Laboratoire d’Ecologie Alpine –Le Bourget du Lac 73376 anne.loison@univ-
savoie.fr4 Office National de la Chasse et de la Faune Sauvage- CNERA FM – 147 route de Lodève - Juvignac
34990 [email protected] CIRAD – UR AGIRs – Campus International de Baillarguet - Montpellier cedex 5 34398
Depuis l’année 2001, des épisodes de mortalité massive associés à ladécouverte d’isards (Rupicapra pyrenaica) malades ont été rapportés dans lesPyrénées en Espagne, en Andorre et en France. Les investigations réaliséesont permis l’isolement chez une partie des individus malades d’un Pestivirusidentifié comme un Border Disease Virus (BDV). Ce virus, soupçonné d’êtreresponsable des symptômes et mortalités observés, fut classé dans le sous-groupe Border Disease Virus-4 (BDV-4).Des études sérologiques rétrospectives ont montré que le virus circulaitdepuis au moins 1995 dans la population d’isards de la Réserve Nationale deChasse et de Faune Sauvage d’Orlu alors qu’aucune mortalité anormalen’avait été rapportée. Depuis, 90% des animaux de plus de 2 ans ont eu uncontact avec le virus et la présence, chaque année, d’animaux virémiques sug-gère que le virus est devenu endémique dans cette population. Nous avonscherché à comprendre comment ce virus influençait la dynamique de lapopulation d’isards en étudiant l’évolution de la survie et de la fécondité decette population.
Motsclés: Rupicapra pyrenaica, Border Disease Virus, infection, survie, fécon-dité.
JORNADAS
170 (Pirineos, 2011, Vol. 166, 155-177, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166008)

DIEZ AÑOS DE VIGILANCIA SANITARIA, ACTIVAY PASIVA, EN EL SARRIO DEL PIRINEO ARAGONÉS*
M.C. ARNAL1, M. REVILLA1, D. MARTÍNEZ1, M.J. DE MIGUEL2, P.M. MUÑOZ2
& D. FERNÁNDEZ DE LUCO1
1 Centro de Investigación en encefalopatías y enfermedades trasmisibles emergentes. Departamento dePatología Animal. Facultad de Veterinaria. Universidad de Zaragoza. C/ Miguel Servet 177, E-50013
Zaragoza, España2 Centro de Investigación y Tecnología Agroalimentaria (CITA). Gobierno de Aragón.
*Estudio financiado por el Dpto. de Medio Ambiente y Dpto. de Agricultura y Alimentación. Gobiernode Aragón.
La red de vigilancia sanitaria del sarrio en el Pirineo aragonés permiteconocer la situación de procesos epidémicos, como pestivirosis, sarna y que-ratoconjuntivitis infecciosa (QCI); zoonosis como brucelosis, tuberculosis ehidatidosis; y otros procesos patológicos en la especie. En un periodo de 10años (2000-09) el número de sarrios enfermos-muertos analizados ha sido de187, además de 117 animales cazados y 1.852 muestras de sarrios cazados(n=2.156). El seguimiento sobre pestivirosis se ha realizado en 1.459 sarrios,no detectándose casos clínicos ni sarrios infectados. El estudio serológicofrente a pestivirus muestra una mayor proporción de animales seropositivosen las RC de Benasque (18,5%) que en el resto de reservas más occidentales(2,3%). Un brote de QCI afectó a sarrios de Aragón y Navarra desde la pri-mavera de 2006 hasta finales de 2008. Se recogieron 109 animales afectados,siendo M. conjunctivae el principal agente causal identificado, (89,4%, n=104),aunque Chlamydophila spp. también fue detectado en un menor porcentaje(12,5%, n=104). Un 4,2% (8/190) de los sarrios cazados y sin lesiones oculares(2006-09) portaban M. conjunctivae, al igual que un 25,8% (159/617) del gana-do ovino-caprino muestreado (17 rebaños, 2008-09). Los ciervos y corzoscazados de las zonas afectadas durante el 2008 fueron negativos a la presen-cia de M. conjunctivae. Un 0,57% (11/1.924) de los sarrios fue positivo frente abrucelosis siendo negativo el cultivo bacteriológico. La necropsia de 187sarrios enfermos-muertos mostró un 58,82% (110) con QCI, un 10,2% (19) conprocesos bronco-neumónicos no parasitarios, un 9,1% (17) con politraumatis-mo, un 2,1% (4) con procesos digestivos, un 1,6% (3) con otras patologías, yun 18,2% (34) con procesos indeterminados en animales incompletos. Otraslesiones observadas esporádicamente son dermatofilosis, feocromocitoma,
JORNADAS
171(Pirineos, 2011, Vol. 166, 155-177, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166008)

hidatidosis y pseudotuberculosis. Los animales cazados (n=117) presentabanneumonía verminosa, cisticercosis visceral, coccidios y nematodos (Ostertagiaspp.) en abomaso. La presencia de áreas con neumonía catarral crónica enlóbulos apicales y Haemonchus spp. en abomaso era frecuente. En los sarriosestudiados no se observaron lesiones sospechosas de tuberculosis ni sarna(n=219). Ningún sarrio de los analizados (n=128) en 2008 y 2009 fue seropo-sitivo frente a lengua azul.
JORNADAS
172 (Pirineos, 2011, Vol. 166, 155-177, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166008)

DINÁMICA DE UN BROTE DE QUERATOCONJUNTIVITISINFECCIOSA POR MYCOPLASMA CONJUNCTIVAE
EN EL SARRIORUPICAPRA P. PYRENAICA
J. HERRERO1, C. PRADA2, O. FERNÁNDEZ-ARBERAS31 & A. GARCÍA-SERRANO2
1. Área de Ecología. Escuela Politécnica Superior de Huesca. Universidad de Zaragoza. E-22071Zaragoza. [email protected]
2. Ega Consultores en Vida Silvestre. Sierra de Vicort 31. E-50003 Zaragoza, Spain. [email protected]. Instituto Pirenaico de Ecología. Ap. 64. E-22700 Jaca. [email protected]
Entre 2006 y 2008 tuvo lugar un brote de queratoconjuntivitis infecciosa(QCI) en el sarrio en Aragón y Navarra, así como en el Pirineo francés. El áreade estudio fue el Pirineo aragonés y navarro, constituido por 15 macizos mon-tañosos, con un seguimiento poblacional previo, con presencia estable de laespecie y en los que 10 de ellos tuvieron casos de QCI. En cinco de ellos huboademás un brote importante acompañado de fuerte mortalidad. Los dosmacizos más afectados tuvieron además un impacto en la proporción desexos en la edad adulta, que favorecía a los machos. El conjunto de la ten-dencia poblacional entre 2000 y 2007 fue de λ = 1.026, disminuyendo entre2007 y 2008 a λ = 0.8, 0.85 en 2008-2009 y comenzando su recuperación entre2009 y 2010 con λ = 1.1. El número de sarrios encontrados muertos (2006-10)fue de 470: 31 en 2006, un año sin casi mortalidad debida a la QCI; 220 (2007)y 169 (2008), dos años con fuerte mortalidad por QCI; 34 en 2009 y 16 en 2010,dos años sin mortalidad debida a la enfermedad. Un total de 105 animalesenfermos o muertos fueron recogidos y Mycoplasma conjunctivae fue identifi-cado en 93 (88.5%). El brote se extendió en mancha de aceite, hacia el E y elW desde el punto donde fue detectado por primera vez. Se detectaron dospicos: uno en 2007, que afectó fundamentalmente a un macizo y otro en 2008que afectó a tres. Los picos fueron estacionales en tres macizos y continuarondesde la primavera hasta el otoño del siguiente año en uno. No encontramosrelación entre la densidad, el número y la tendencia con el decrecimiento o elnúmero de animales encontrados muertos, sugiriendo que otros factores jue-gan un papel en la difusión e impacto de la enfermedad. La caza quedó prác-ticamente cerrada en los macizos afectados por importante mortalidad, concupos inferiores al 1%. Se abrió de nuevo al finalizar el brote y realizar nue-vas estimaciones poblacionales. Aunque algunos animales recuperados o node la enfermedad se encuentran todos los años, este es el primer brote de QCIdesde finales de los 70 y principios de los 80 del siglo pasado, hace 30 años,cuando estimas de la guardería indicaron una pérdida del 50% de la pobla-ción de las áreas afectadas.
Palabras clave: gestión cinegética, seguimiento poblacional, mortalidad.
JORNADAS
173(Pirineos, 2011, Vol. 166, 155-177, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166008)

DISTANCE SAMPLING EN LA ESTIMADE ABUNDANCIA DE REBECO
J.M. PÉREZ1, J.M. LÓPEZ MARTÍN2, J. XIFRA COROMINAS3, R. ALPIZAR JARA4,I. MARCO5, G. MENTABERRE5, S. LAVÍN5 & E. SERRANO5
1Departamento de Biología Animal, Biología Vegetal y Ecología, Universidad de Jaén, Campus LasLagunillas, s.n., 23071, Jaén, Spain. [email protected].
2Direcció General de Medi Natural i Biodiversidad, Departament d’ Agricultura, Ramadería, PescaAlimentació i Medi Natura, E- 08017, Barcelona, España.
3Reserva Nacional de Caça Freser-Setcases, Girona, España.4Centro de Investigação em Matemática e Aplicações, Departamento de Matemática, Universidade de
Évora, Rua Romão Ramalho, 59, P-7000-671, Évora, Portugal.5Servei d’Ecopatología de la Fauna Salvatge (SEFaS), Facultat de Veterinària, Universitat Autònoma de
Barcelona, 08193, Bellaterra, Barcelona, España.
La estima de abundancia de una población animal es básica para detectarcambios y tendencias demográficas a diferentes escalas de tiempo. Esta labores especialmente importante en especies que se someten a aprovechamientocinegético, ya que los cupos de caza se basan en una estima previa del tama-ño de la población. En Europa, las estimas poblacionales de rebeco y de sarrio(Rupicapra spp.) se suelen basar en recuentos totales (censos), que tienden ainfraestimar el tamaño total de la población. Además, rara vez se cumplen lossupuestos de esta técnica: (i) es imposible muestrear el 100% del área de estu-dio, (ii) la probabilidad de observar a todos y cada uno de los individuos noes igual a 1 y (iii) no se puede garantizar que ningún individuo se observa yregistra más de una vez.
Durante las últimas décadas, el muestreo por distancias (Distance Sampling- DS) presenta una alternativa a los censos. Esta metodología requiere de laobservación de una fracción de la población (muestra) y del registro de la dis-tancia (radial, si se observa desde un punto; o perpendicular a la línea de tran-secto) al animal o al grupo. A los datos de las distancias observadas son ajus-tados diferentes modelos probabilísticos que permiten obtener una estima dela densidad y la precisión asociada. A pesar de que esta técnica requiere pocoesfuerzo de muestreo, durante la etapa de análisis de los datos hay que tomarvarias decisiones que por lo general no están estandarizadas. En este trabajoproponemos un protocolo de obtención de datos y de análisis bajo la metodo-logía DS. Además discutiremos los supuestos del método, la influencia deltamaño de muestra, la importancia del tamaño del área de estudio y la posibi-lidad de combinar DS con la metodología de captura-marcaje-recaptura, entreotros aspectos.Palabras clave: censos, Distance Sampling, estima de abundancia, Rupicapraspp.
JORNADAS
174 (Pirineos, 2011, Vol. 166, 155-177, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166008)

Conclusiones
Cuarenta y ocho participantes del Pirineo y la Cordillera Cantábrica, caza-dores, investigadores, guardas, técnicos de las administraciones y consulto-res, pertenecientes a estos territorios, intercambiaron experiencias y actuali-zaron su conocimiento multidisciplinar sobre el rebeco cantábrico y el sarriopirenaico. Algunos de los temas tratados a lo largo de las cuatro sesiones dela jornada han aportado información sobre los siguientes aspectos:
Consideración del sarrio y del rebeco como elementos relevantes de la cul-tura, historia, paisaje y naturaleza de las montañas ibéricas, valorada y apre-ciada por sus habitantes y visitantes.
Aportación económica del sarrio a las economías locales de los municipiosafectados por las Reservas de Caza.
Incorporación de nuevos actores, tales como grupos conservacionistas, ala conservación de las poblaciones, con experiencias de reintroducción, segui-miento y apoyo a la gestión.
Relevancia de los trabajos de seguimiento a largo plazo y del marcajepoblacional, indispensables para la interpretación de los cambios ocurridosen los diversos parámetros objeto de seguimiento (abundancia, tendencia,tamaño, crecimiento, estado sanitario, etc.).
Existencia de nuevas herramientas de análisis de datos (estadísticas, com-putacionales, Sistemas de Información Territorial, imágenes satelitales, etc.) yun creciente enfoque multidisciplinar de los fenómenos naturales (ecología,botánica, veterinaria, alimentación, etc.), que deben ser incorporadas a losseguimientos e investigación del sarrio y el rebeco.
Importancia del establecimiento de un seguimiento sanitario adecuadodel ganado doméstico estivante en los pastos de montaña, especialmente elovino, por su relevancia en el mantenimiento y transmisión de enfermedadesa la fauna silvestre, particularmente a los ungulados salvajes.
Necesidad de instaurar múltiples indicadores que permitan interpretar losfenómenos naturales que afectan a la evolución de las poblaciones, como losefectos de las cohortes, el cambio global, etc.
Comprobación de cambios de gran magnitud en los ecosistemas de mon-taña, como son: la matorralización, la disminución de la cabaña ganadera,particularmente de ovino, la composición de la comunidad de ungulados ode la caza menor y la posible modificación de la calidad del pasto.
Destacar la trascendencia de la divulgación del trabajo realizado, tanto alas comunidades locales como a los medios de comunicación, aportandoinformación de calidad e incluyendo el estado de las poblaciones de rebeco ysarrio.
JORNADAS
175(Pirineos, 2011, Vol. 166, 155-177, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166008)

Relevancia de la existencia de poblaciones no cazadas y cazadas, y de laoportunidad que aportan para la investigación y gestión.
Incomprensión de los cazadores por parte de la sociedad como conse-cuencia de los cambios socioeconómicos y necesidad de comunicar correcta-mente los intereses del colectivo y su papel.
Trascendencia de la comprensión del papel que juegan las enfermedadesen la dinámica poblacional y urgencia en establecer una red de seguimientosanitario coordinada.
Considerando las conclusiones y recomendaciones de las PrimerasJornadas pirenaicas sobre el sarrio Rupicapra p. pyrenaica celebradas en Jacaentre el 12 y el 14 de marzo de 2003, se han llevado a cabo cuatro talleres,cuyas principales conclusiones han sido:
Taller sobre el muestreo de distancias (distance sampling). Esta herra-mienta de estima poblacional ofrece interesantes alternativas a los conteossupraforestales simultáneos tradicionales y su aplicabilidad está siendo estu-diada. Un taller en la Reunión sobre Ungulados Silvestres Ibéricos (RUSI)parece una vía adecuada para compartir experiencias sobre su utilización.
Taller sanitario. La normativa europea establece la obligatoriedad de unavigilancia sanitaria (activa y pasiva) de la fauna silvestre. Es necesario refle-xionar sobre las enfermedades que deben ser objeto de seguimiento y esta-blecer una formación adecuada de los agentes de campo en lo relativo a latoma de muestras.
Taller sobre cráneos de sarrio. Es necesario mejorar la recogida de datosrelativos a la biometría corporal y medición de los cuernos de los animalescapturados y cazados, para que aumente el interés de su tratamiento. Estoimplica formación y homogeneización en la toma de datos.
Taller sobre gestión poblacional. Algunos temas sobre los que convienetrabajar en los próximos años son: el concepto y desarrollo de la caza selecti-va; la mejora de la comercialización de la carne de caza; la actualización delestado de las poblaciones (demográfico y sanitario) unido al intercambio deesta información; la homogeneización de los criterios de gestión entre territo-rios que comparten poblaciones y la coordinación de los seguimientos pobla-cionales.
JORNADAS
176 (Pirineos, 2011, Vol. 166, 155-177, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166008)

Referencias
Herrero, J., Escudero, E., Fernández De Luco, D. & García-González, R. (eds.),2004. El sarrio pirenaico Rupicapra p. pyrenaica: Biología, Patología y Gestión.Consejo de Protección de la Naturaleza de Aragón, vol. 46, 263 pp.,Zaragoza.
Pérez-Barbería, F. J. & Palacios, B. (eds.), 2009. El rebeco cantábrico Rupicaprapyrenaica parva. Conservación y gestión. Organismo Autónomo de ParquesNacionales MMA, 501 pp., Madrid.
JORNADAS
177(Pirineos, 2011, Vol. 166, 155-177, ISSN 0373-2568, eISSN: 1988-4281, doi: 10.3989/Pirineos.2011.166008)

179
PIERRE BARRÈRE, EN EL RECUERDO (1921-2011)
Recientemente nos llegó la triste noticia del fallecimiento del Profesor PierreBarrère, uno de los geógrafos franceses más importantes de la segunda mitad del sigloXX y un referente para la Geografía española, especialmente en el campo de laGeomorfología. Para el Instituto Pirenaico de Ecología ha representado además unapérdida muy sensible, por cuanto el Profesor Pierre Barrère mantuvo relaciones muyestrechas con nuestra institución hasta comienzos de los años noventa del pasadosiglo.
Pierre Barrère nació el 12 de junio de 1921 y falleció el 27 de febrero de 2011. Suvida profesional estuvo vinculada a la Geografía, primero como Profesor Agregado en1948 y como Ayudante entre 1950 y 1953 en la Facultad de Letras de la Universidad deToulouse. Desde allí pasó definitivamente a la Universidad Michel de MontaigneBordeaux 3, donde fue primero Maître de Conferences y luego Professeur. En esaUniversidad fue Director del Institut de Géographie Louis-Papy (1969-1977) yDirector del Laboratorio de Geografía Física Aplicada (1975-1989). Además fueDirector de la Maison des Sciences de l’Homme d’Aquitaine y Presidente del Comitépour l’Environnement de la Gironde. Fue también codirector de la Revue Géographiquedes Pyrénées et du Sud-Ouest, más tarde denominada Sud-Ouest Européen. Maestro entrelos grandes, quienes le conocimos personalmente sabemos de su rigor, sencillez ypasión por la naturaleza y los hombres. Era sobre todo un geógrafo de campo clásico,un ejemplo para todos nosotros por su extraordinaria capacidad de observación.
Inicialmente se sintió atraído por la Geografía Urbana, realizando su TesisDoctoral sobre “La banlieue maraichère de Bordeaux” (1949). Su interés por laGeografía Urbana se mantuvo a lo largo de su vida, dirigiendo varias Tesis Doctorales.Mostró una especial preocupación por su ciudad, Burdeos, y su entorno próximo;fruto de ello es la creación del CESURB (Centre d’Études sur les Espaces Urbains) ysus trabajos “Nouveau visage de Bordeaux” (1972) y “Evolution récente du secteursauvagardé de Bordeaux” (1980), entre otros. Su capacidad didáctica quedó plasmadaen el libro “Les villes françaises”, Edit. Masson (1980), escrito junto a MichelineCassou-Mounat, que fue utilizado como manual por varias generaciones deestudiantes de Geografía.
Pero su verdadera pasión, donde demostró su grandes capacidades científicas, fueen el campo de la geomorfología y particularmente los Pirineos. Su Tesis de Doctoradode Estado versó sobre “Le relief des Pyrénées Centrales franco-espagnoles”, quepuede considerarse uno de los trabajos de mayor valor para entender los grandesrasgos del relieve en el sector central de la vertiente meridional de los Pirineos. Fuepublicada en 1971 por el Institut de Géographie de l’Université de Bordeaux. Lacartografía geomorfológica que acompaña a la Tesis (12 mapas en color, a escala1:50.000) es verdaderamente excepcional, con una gran capacidad para transmitir alobservador las formas de relieve. Con ella creó escuela, curiosamente más en Españaque en Francia. Hoy sigue siendo un trabajo de consulta imprescindible para losgeomorfólogos españoles, como también otros estudios de enorme transcendencia,sobre todo en el campo del glaciarismo. Así, cabe destacar los siguientes trabajos:

180
Barrère, P. (1952): Evolution mécanique et nivation sur les versants calcaires de lahaute montagne pyrénéenne. Pirineos, 24: 201-213.
Barrère, P. (1953): Équilibre glaciaire actuel et quaternaire Dans l’Ouest desPyrénées centrales. Revue Géographique des Pyrénées et du Sud-Ouest, 116-134.
Barrère P. (1963): La période glaciaire dans l’Ouest des Pyrénées Centrales franco-espagnoles. Bulletin de la Société Géologique de France, 7 (5): 516-526.
Barrère, P. (1964): Le relief karstique dans l’ouest des Pyrénées Centrales. RevueBelge de Géographie, 88 (1-2): 9-62.
Barrère, P. (1966): La morphologie quaternaire de la région de Biescas et deSabiñánigo (Haut Aragon). Bulletin de l’Association Française pour l’Étude duQuaternaire, 2: 83-93.
Barrère, P. (1975): Terrasses et glacis d’érosion en roches tendres dans lesmontagnes du Haut Aragon. Études Géographiques Mélanges Georges Viers, Universitéde Toulouse, pp. 29-43, Toulouse.
Todos ellos siguen siendo referentes importantes para la geomorfología pirenaica,aunque algunas de sus tesis más originales hayan perdido recientemente vigencia.Pero aún así deslumbra su capacidad para entender los estadios glaciares y susrelaciones con terrazas y glacis en un tiempo en que había pocos recursos analíticos ymenos aún para obtener dataciones que hoy son imprescindibles.
Su vinculación con el Pirineo y España no terminó ahí. Fue secretario de la UniónInternacional de Estudios Pirenaicos, institución hispano-francesa encargada deorganizar cada dos años los congresos internacionales de estudios pirenaicos, en losque participó muy activamente. No pocos cursos traía a sus estudiantes a España,explicándoles el relieve, y también los nuevos regadíos, los pueblos de colonización,el impacto del turismo, el éxodo rural y el crecimiento urbano. Compartióexperiencias con los profesores Alfredo Floristán, con el que coincidió en Burdeossiendo ambos recién licenciados, Salvador Ménsua, Eusebio García Manrique, ManuelFerrer y Antonio Higueras. A finales de los años ochenta, uno de nosotros disfrutó deuna beca postdoctoral en el Institut de Géographie, bajo su dirección, demostrandouna vez más su abierta disposición a ayudar a los más jóvenes, con la generosidad quele caracterizaba. El profesor Barrère estaba muy próximo a la jubilación, peromantenía un gran entusiasmo por el trabajo y la difusión de la Geografía a la sociedad,organizando cada lunes una conferencia (Les Lundis de la Géographie), que formabaparte de la vida cultural de Burdeos.
Fue un gran viajero, sobre todo por el norte de África y América Latina. El litoralde Aquitania y la dinámica de sus dunas fue su espacial obsesión en los últimos añosde su actividad científica, incorporando técnicas modernas de investigación. Su atlassobre los tipos de dunas del litoral de Aquitania (en colaboración con F. Pouget),publicado en 1990, es otro referente que ayuda a transmitir su imagen de geógrafocompleto, alguien excepcional que dejó no sólo una obra científica imperecedera, sinotambién su humanidad y su presencia como hombre de bien, respetado por todos losque le tratamos personalmente.
José M. García-RuizTeodoro Lasanta
Instituto Pirenaico de Ecología, CSIC, Zaragoza, Mayo de 2011.

181
PIRINEOS. NORMAS DE PUBLICACIÓN
La revista Pirineos publica trabajos originales relacionados con la dinámica de los ecosiste-mas de montaña, es decir, con los procesos y las relaciones que se establecen entre los seresvivos y entre estos últimos y el medio ambiente, en un espacio tan peculiar como el de las regio-nes más elevadas del planeta. Trata, por lo tanto, de contribuir a explicar el funcionamiento glo-bal de las regiones de montaña y la organización espacial de recursos y procesos, con una pers-pectiva integradora en la que el hombre ejerce también un papel destacado. La revista aceptatrabajos orginales con datos científicos nuevos e importantes para el conjunto de la Ecología deMontaña, especialmente aquellos que aportan información sobre los flujos existentes entre sub-sistemas, la influencia del hombre en la transformación del medio ambiente montano, las rela-ciones tróficas entre los seres vivos y el funcionamiento de los diferentes aspectos que consti-tuyen la base de los ecosistemas de montaña.
Para ser aceptados, los trabajos recibidos serán enviados al menos a dos evaluadoresexternos, aplicando el sistema de evaluación “por pares y doble ciego”.
Artículos. Los trabajos serán entregados en soporte impreso y electrónico (de preferencia enWord), deben enviarse en español, inglés o francés y no podrán tener una extensión superior a25 páginas (de 30 líneas y 70 espacios). No obstante, a la vista de la importancia de un trabajoconcreto, la Dirección de Pirineos podrá aceptar excepcionalmente artículos de dimensión supe-rior a 25 páginas. En todo caso irán precedidos de un resumen (máximo 12 líneas) en españoly en inglés. Si el autor tiene problemas para elaborar el resumen en español, la Dirección dePirineos se encargará de la traducción.
Debajo del título y autor(es) se consignará la dirección completa del autor o autores.Los gráficos se enviarán en soporte electrónico, en formato tiff o jpg. La leyenda o pie de las
figuras debe colocarse en página aparte, numeradas y con texto en el idioma original del artícu-lo y en inglés. Si el artículo está escrito en inglés, el Consejo de Redacción elaborará la leyen-da en español. Con las fotografías deben seguirse las mismas normas. En los mapas ha de indi-carse siempre la escala gráfica y el Norte. Figuras y fotografías en color, asi como cambios queeleven el coste de la edición, sólo podrán admitirse si van a cargo del autor.
En las referencias bibliográficas en el texto, se citará al autor en minúsculas y a continua-ción, el año de referencia del trabajo citado y, en su caso, la página; o bien el autor, año y pági-na, todo entre paréntesis. Al final del texto se incluirá la bibliografía en minúsculas –sólo la cita-da en el artículo– por orden alfabético de autores, seguidos del año de publicación. A continua-ción se señalará el título completo del trabajo, el nombre de la revista, el número del volumen yla página inicial y final. Por ejemplo:
Ackroyd, P. 1986. Debris transport by avalanche. Torlesse Range, New Zealand, Z.Geomorph, 30(1): 1-14.
Si la cita corresponde a un libro, se subrayará el título del mismo y se incluirá a continuaciónel nombre de la editorial, el número total de páginas y la ciudad de edición. Por ejemplo:
Creus, J. 1983. El clima del Alto Aragón Occidental. Instituto de Estudios Pirenaicos, 233 pp.,Jaca.
Si en las referencias hay dos o más libros o artículos del mismo autor y del mismo año, seañadirán minúsculas a, b, etc. al año de publicación, tanto en la lista bibliográfica final como enlas referencias en el texto.
Notas breves. Pirineos contará con una sección de notas breves que en lo referente al idio-ma y a las citas bibliográficas se ajustarán a lo señalado para los artículos. Su extensión no reba-sará las 8 páginas; no incluirán resumen y deberán reducir en lo posible las referencias. Iránacompañadas de un máximo de 2 figuras.
Reseñas. Se admitirán reseñas bibliográficas sobre obras o conjunto de obras de gran inte-rés. Se aconseja que cada reseña no supere la extensión de 3 páginas. El autor no recibiráseparatas aunque sí un ejemplar de la revista Pirineos en la que se incluya su reseña.
Los autores aceptan la cesión a la Revista de los derechos de reproducción y explotaciónde sus artículos. Las opiniones y hechos consignados en cada artículo son de exclusiva res-ponsabilidad de sus autores. El Consejo Superior de Investigaciones Científicas no se hace res-ponsable en ningún caso de la credibilidad y autenticidad de los trabajos.

182
PIRINEOS. INSTRUCTIONS AUX AUTEURS
La revue Pirineos publie des travaux concernant la dynamique des écosystèmes de mon-tagne, c’est à-dire, les processus et les rapports établis entre les êtres vivants, et entre ces der-niers et l’environnement, sur un territoire aussi spécial que celui des hautes altitudes.
C’est pour cela que la revue essaye d’expliquer le fonctionnement global des regions demontagne et l’organization spatiale des ressources et des processus, avec une perspectivesynthétique dans laquelle l’homme joue un rôle important. Pirineos accepte des travaux originauxapportant des donneés scientifiques nouvelles et importantes pour l’ensemble de l’Ecologie deMontagne, spécialement des informations sur le flux entre sous-systèmes, sur l’influence del’homme dans la transformation de l’environnement montagnard, sur les relations trophiquesentre les êtres vivants et sur le fonctionnement des différentes caractéristiques qui constituent labase des écosystèmes de montagne.
Avant d’être acceptés, les travaux reçus seront révisés par, du moins, deux chercheurs exter-nes, en train de suivre le système d’évaluation “en double avengle”.
Articles. Les travaux doivent être rédigés en espagnol, en anglais ou en français, et ne pou-rront dépasser 25 pages. Pourtant, la Direction de Pirineos pourra accepter, exceptionnellement,des travaux de plus grande extension, en fonction de l’importance de l’article envoyé. Dans tousles cas, ceux-ci seron précédés d’un resumé (pas plus de 12 lignes) en espagnol et en anglais.Si l’auteur a des problèmes pour rédiger le resumé en espagnol, la traduction en sera faite parles soins de l’éditeur.
Au bas du titre et auteur(s) doit être signalée l’adresse complete de l’auteur(s).Les illustrations (designées comme figures) seront présentées dans des fichiers aux formats
tiff ou gif. La légende, ou pied des figures, doit apparaître en page à part, dans la langue origi-nelle de l’article et en anglais. Si l’article est écrit en anglais; les éditeurs élaboreront aussi untexte des figures en espagnol. Pour les photographies il est necessaire de procéder de la mêmefaçon. Sur les cartes, on indiquera toujours l’échelle graphique et le Nord. La revue ne peutaccepter des figures ni des photographies en couleur.
Dans les renvois dans le texte à la bibliographie suivront le système: auteur en minusculessuivi, de l’année d’édition du travail cité; ou bien auteur et année, le tout entre parentheses. Labibliographie (uniquement celle citée dans le texte) doit figurer à la fin du travail, et être présen-tée par ordre alphabétique des noms d’auteurs, suivis inmédiatement de l’année de parution.Ensuite, le titre complet du travail cité, le nom souligné de la revue, le volume et les numéros despages du début et de la fin de l’article. Exemple:
Ackroyd, P. 1986. Debris transport by avalanche. Torlesse Range, New Zeland. Z.Geomorph., 30 (1): 1-14.
Si la citation appartient à un livre, après le nom de l’auteur et l’anneé de parution, on doit sou-ligner le titre, accompagné du nom de la maison d’édition, du nombre total de pages et de la villed’édition. Exemple:
Creus, J. 1983. El clima del Alto Aragón Occidental. Instituto de Estudios Pirenaicos, 223 pp.,Jaca.
Quand un auteur a plus d’une référence de la même année, des caractères minuscules sui-vront l’année d’édition, tant dans le texte que dans la bibliographie finale 1986 a, 1986 b, ...
Notes breves. Pirineos comptera sur une section de notes breves qui doivent s’ajuster auxnormes signalées pour les articles en ce qui concerne la langue et les références bibliographi-ques. Leur extension ne doit pas dépasser 8 pages. Elles ne doivent pas comporter de resuméet le nombre de figures ne peut être supérieur à deux.
Comptes-rendus. Des comptes-rendus bibliographiques sur des travaux très intéressantsseront bien reçus. Il est conseillé que chacun d’eux ne depasse pas 3 pages d’extension. Lesauteurs n’en recevront aucun tiré-à-part, mais par contre il leur sera envoyé un exemplaire dePirineos dans lequel le compte-rendu est publié. Les auteurs acceptent de céder à la Revue tousles droits de reproduction et exploitation de leurs articles. Les opinions et faits inclus à chaquearticle sont responsabilité, uniquement des auteurs. Le Conseil Supérieur de RecherchesScientifiques n’est jamais responsable, ni de la crédibilité ni de l’authenticité des travaux.

183
PIRINEOS. GUIDE FOR AUTHORS
The journal Pirineos publishes original works dealing with the dynamics of mountain ecos-ystems, that is, with processes and relationships established between living beings and theirenvironment, in the highest regions on Earth. Therefore it tries to explain the global functioningof mountainous areas and the spatial organization of processes and resources, with an integra-ted perspective in which man performs an important role. The journal accepts original works withnew and important scientific data on Mountain Ecology, especially those containing informationabout the extant fluxes between subsystems, the influence of man in the evolution and dynamicsof mountain environment, the trophic relationships between living beings and the functioning ofdifferent features that constitute the basis of mountain ecosystems.
Manucripts submitted for publication are peer-revieweb by at least two referees.Articles. Manuscripts may be submitted in Spanish, English or French and their length should
not exceed 25 pages. Nevertheless, when a longer paper proves to be of extraordinary impor-tance, the Direction of Pirineos may exceptionally accept it. All articles should include, after thetitle and the name(s) of the author(s), a summary (12 lines maximum) in Spanish and in English.Should the author find any difficulties in the writing of the summary in Spanish, the Direction ofPirineos can undertake translation.
The complete address of the authors sould be clearly indicated below the list of authors.Suitable formats for figures are tiff or jpg. Each illustration should have one arabic number.
The text of the legend at the foot of each drawing should be written on a separate sheet in theoriginal language of the paper and in English. If the article is written in English, the editors shouldalso include the text acompanying the figures in Spanish. Photographs should observe the samerules. Maps should always indicate the scale and North. Authors will be charged when they sub-mit colour drawings or colour photographs.
References to publications in the text should be made by name of author, the year of publi-cation and, when necessary, the page numbers, the last two should be in parentheses.
The list of references, alphabetically ordered should appear at the end of the paper. In thislist the name of the author should be followed by the year of publication, the complete title of thework, the name of the journal in italics, and both volume and page numbers. For example:
Ackroyd, P. 1986. Debris transport by avalanche. Torlesse Range, New Zealand. Z.Geomorph., 30 (1): 1-14.
For books, titles should be in italics and should be followed by the name of the publisher, thenumber of pages and the place of publication. For example:
Creus, J. 1983. El clima del Alto Aragón Occidental. Instituto de Estudios Pirenaicos, 223 pp.,Jaca.
If, in the references, there are two or more books or papers with the same author and thesame year of publication, a lower case a, b, etc, should be added to the year of publication bothin the reference list and in the references in the body of the text.
Notes. Pirineos also includes a section of Notes that will be subjected to the same rules aslonger papers. They should not exceed 8 pages and should not be accompanied by a summary.Only a maximum of two drawings willbe accepted. Authors will receive 50 free copies.
Reviews. Bibliographical reviews on very interesting topics will be accepted. Their lengthshould not exceed 3 pages. Authors will not receive any reprint, but will receive a copy of the jour-nal Pirineos in which his review is published. Opinions expressed in the articles are those of theAuthors and the CSIC does not endorse them.

PIRINEOS
SUSCRIPCIONES Y PEDIDOSINFORMACIÓN GENERAL
Periodicidad: ANUALPrecio Suscripciones:
España: 24 eurosExtranjero: 29 euros
A estos precios se les añadirá el 4% de IVA
Números sueltos anteriores: Especificar en pedido adjunto volumen y número y se servirán en fun-ción del stock. A los precios establecidos se le añadirán gastos de envío y los impuestos que procedan.
Forma de Pago:
Transferencia bancaria a la Cta. Número: C/c 0049 2777 27 2214052221
SWIFT/BIC CODE: BSCHESMMIBAN NUMBER: ES98 0049 2777 27 2214052221
Tarjeta de Crédito, VISA / Master Card / Eurocard / 4B
Número: _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ Fecha de caducidad: _ _ / _ _
Redacción, subscripción e intercambio:Instituto Pirenaico de EcologíaDirección: Campus de Aula Dei, Apdo. 13034, 50080, Zaragoza (España)Correo electrónico: bib_jaca @ipe.csic.es
DATOS DEL PETICIONARIO
Nombre y Apellidos:...........................................................................................................................................................................................................................Razón Social:...............................................................................................................................................................................................................................................NIF / CIF:.......................................................................................................................................................................................................................................................Dirección: CP:.............................................................................................................................................................................................................................................Localidad:..................................................................................... Provincia:........................................................ País/Estado:..................................Teléfono:.......................................................................................... Fax:.............................................................................. e-mail:..................................................Fecha de la solicitud:...........................................................................................................................................................................................................................Firma:.................................................................................................................................................................................................................................................................


















