
Control del crecimiento y desarrollo de las plantas por la ...Control del crecimiento y desarrollo...
115
Dirección: Dirección: Biblioteca Central Dr. Luis F. Leloir, Facultad de Ciencias Exactas y Naturales, Universidad de Buenos Aires. Intendente Güiraldes 2160 - C1428EGA - Tel. (++54 +11) 4789-9293 Contacto: Contacto: [email protected] Tesis de Posgrado Control del crecimiento y Control del crecimiento y desarrollo de las plantas por la luz: desarrollo de las plantas por la luz: Funciones fotoperceptivas y Funciones fotoperceptivas y mecanismos de acción del mecanismos de acción del fitocromo A fitocromo A Yanovsky, Marcelo Javier 1999 Tesis presentada para obtener el grado de Doctor en Ciencias Químicas de la Universidad de Buenos Aires Este documento forma parte de la colección de tesis doctorales y de maestría de la Biblioteca Central Dr. Luis Federico Leloir, disponible en digital.bl.fcen.uba.ar. Su utilización debe ser acompañada por la cita bibliográfica con reconocimiento de la fuente. This document is part of the doctoral theses collection of the Central Library Dr. Luis Federico Leloir, available in digital.bl.fcen.uba.ar. It should be used accompanied by the corresponding citation acknowledging the source. Cita tipo APA: Yanovsky, Marcelo Javier. (1999). Control del crecimiento y desarrollo de las plantas por la luz: Funciones fotoperceptivas y mecanismos de acción del fitocromo A. Facultad de Ciencias Exactas y Naturales. Universidad de Buenos Aires. http://digital.bl.fcen.uba.ar/Download/Tesis/Tesis_3154_Yanovsky.pdf Cita tipo Chicago: Yanovsky, Marcelo Javier. "Control del crecimiento y desarrollo de las plantas por la luz: Funciones fotoperceptivas y mecanismos de acción del fitocromo A". Tesis de Doctor. Facultad de Ciencias Exactas y Naturales. Universidad de Buenos Aires. 1999. http://digital.bl.fcen.uba.ar/Download/Tesis/Tesis_3154_Yanovsky.pdf
Transcript of Control del crecimiento y desarrollo de las plantas por la ...Control del crecimiento y desarrollo...

Di r ecci ó n:Di r ecci ó n: Biblioteca Central Dr. Luis F. Leloir, Facultad de Ciencias Exactas y Naturales, Universidad de Buenos Aires. Intendente Güiraldes 2160 - C1428EGA - Tel. (++54 +11) 4789-9293
Co nta cto :Co nta cto : [email protected]
Tesis de Posgrado
Control del crecimiento yControl del crecimiento ydesarrollo de las plantas por la luz:desarrollo de las plantas por la luz:
Funciones fotoperceptivas yFunciones fotoperceptivas ymecanismos de acción delmecanismos de acción del
fitocromo Afitocromo A
Yanovsky, Marcelo Javier
1999
Tesis presentada para obtener el grado de Doctor en CienciasQuímicas de la Universidad de Buenos Aires
Este documento forma parte de la colección de tesis doctorales y de maestría de la BibliotecaCentral Dr. Luis Federico Leloir, disponible en digital.bl.fcen.uba.ar. Su utilización debe seracompañada por la cita bibliográfica con reconocimiento de la fuente.
This document is part of the doctoral theses collection of the Central Library Dr. Luis FedericoLeloir, available in digital.bl.fcen.uba.ar. It should be used accompanied by the correspondingcitation acknowledging the source.
Cita tipo APA:Yanovsky, Marcelo Javier. (1999). Control del crecimiento y desarrollo de las plantas por la luz:Funciones fotoperceptivas y mecanismos de acción del fitocromo A. Facultad de CienciasExactas y Naturales. Universidad de Buenos Aires.http://digital.bl.fcen.uba.ar/Download/Tesis/Tesis_3154_Yanovsky.pdf
Cita tipo Chicago:Yanovsky, Marcelo Javier. "Control del crecimiento y desarrollo de las plantas por la luz:Funciones fotoperceptivas y mecanismos de acción del fitocromo A". Tesis de Doctor. Facultadde Ciencias Exactas y Naturales. Universidad de Buenos Aires. 1999.http://digital.bl.fcen.uba.ar/Download/Tesis/Tesis_3154_Yanovsky.pdf

.7....__...-._...._..—..—.._.. . - 'O-“o‘ —su
"Hull asumirá:Universidad de Buenos Aires
Facultad de Ciencias Exactas y Naturales
4--....” .- .‘
lí .
Control del Crecimiento y Desarrollo de las
Plantas por la Luz:
Funciones fotoperceptivas y mecanismos de acción del
fltocromo A
Tesis presentada para optar al grado de Doctor de 1aUniversidad de Buenos Aires
Licenciado Marcelo Javier Yanovsky
Director: Dr. Jorge José Casal
Wi
Instituto de Investigaciones Fisiológicas y Ecológicas Vinculadas a la Agricultura
(IFEVA)
Departamento de Ecología
Facultad de AgronomíaUniversidad de Buenos Aires1999

Dam,m1amor slempre
mis padres, por su constante apo

ÍNDICE
i. RESUMEN
ii. SUMMARY
iii. ABREVIATURAS
l. INTRODUCCIÓN GENERAL
1.1.1. Señales lumínicas
1.1.1. Señales lumínicas asociadas a la emergencia del suelo
1.1.2. Señales lumínicas asociadas a la presencia de plantas vecinas1.1.3. Señales lumínicas asociadas a las estaciones climáticas
1.2. Mecanismos de percepción
1.2.1 Fotorreceptores de UV-B
1.2.2. Fotorreceptores de luz azul
1.2.3. Fotorreceptores de R y RL, los fitocromos
1.2.3.1. Propiedades estructurales y fotobiológicas del fitocromo1.2.3.2. Efecto de la luz sobre el fitocromo
1.2.3.3. Modos de acción del fitocromo
1.2.3.4. Los fitocromos, una familia de proteínas1.2.3.5. Funciones sensoriales de los distintos fitocromos
1.3. Mecanimos de acción y vías de transducción de señales de losfitocromos
2. OBJETIVOS DE LA TESIS
3. FUNCIONES SENSORIALES (I)3. 1 Introducción
3.2. Resultados
3.2. Discusión
3.3. Materiales y métodos
3. FUNCIONES SENSORIALES (II)3.1. Introducción
27
28
28
29
39
48
48

3.2.
3.3.
3.4.
Resultados
Discusión
Materiales y métodos
4. MECANISMOS DE ACCIÓN (I)4.1.
4.2.
4.3.
4.4.
Introducción
Resultados
Discusión
Materiales y métodos
5. MECANISMOS DE ACCIÓN (II)5.1.
5.2.
5.3.
5.4.
Introducción
Resultados
Discusión
Materiales y métodos
6. MECANISMOS DE ACCIÓN (III)6.1.
6.2.
6.3.
6.4.
Introducción
Resultados
Discusión
Materiales y métodos
7. DISCUSIÓN GENERAL
8. BIBLIOGRAFÍA
52
58
60
63
63
64
71
73
76
76
77
82
84
86
86
87
92
95
98
102

RESUMEN
Iítulg; Control del crecimiento y desarrollo de las plantas por la luz: Funciones fotoperceptivas ymecanismos de acción del fitocromo A.
Autor; Marcelo J. Yanovsky
M luz,fitocromoA, desetiolación,percepciónde vecinas,relojcircadíano,transduccióndeseñales, Arabidopsis thalíana, papa.
La luz es un factor ambiental crítico para el crecimiento y desarrollo de las plantas. Provee no solo la
energía lumínica necesaria para el proceso fotosíntético, sino también señales informativas que las plantas
utilizan para coordinar su crecimiento y desarrollo con el ambiente circundante. La percepción de las
señales lumínicas esta mediada por fotorreceptores específicos como los fitocromos. Estos son una
pequeña familia de proteínas codificada por cinco genes en Arabidopsis thaliana, FITOCROMO A-E
(PH YA-E).
El objetivo de esta tesis es dilucidar las funciones sensoriales y los mecanismos de acción de uno de los
fitocromos más abundantes, el fitocromo A (phyA).
Las funciones fotosensorias del phyA en condiciones naturales de radiación fueron investigadas
comparando el comportamiento de plantas con niveles nonnales de phyA y de plantas mutantes y/o
transgénicas con niveles incrementados o reducidos de este fotorreceptor. De los experimentos se puede
concluir que el phyA actúa como un sensor de cambios en la irradiancia. En plantas etioladas el phyA es
el único fotorreceptor que permite a las plantas distinguir la transición entre la oscuridad reinante bajo la
superficie del suelo y la escasa luz presente bajo canopeos muy densos. Los cambios morfológicos y
fisiológicos desencadenados por el phyA en respuesta a la percepción de dicho cambio en el ambiente
lumínico incrementan notablemente la supervivencia de las plantas emergidas en tales condiciones. El
phyA, además, está involucrado en la percepción de los cambios en el ambiente lumínico producidos por
la presencia de plantas vecinas: percibe directamente la disminución de la cantidad de luz roja (R) y rojo
lejana (RL) que se produce por el sombreado y modula, indirectamente, la sensibilidad a reducciones de la
relación R/RL que se producen por la relexión selectiva de RL en las hojas de plantas vecinas. Estas
acciones del phyA conducen finalmente a un incremento en la tasa de alargamiento del tallo que
disminuyen el riesgo de sombreado. En plantas verdes el phyA también está involucrado en la percepción
de las transiciones cotidianas oscuridad/luz que se producen al amanacer. En respuesta a la percepción de
dicha señal el phyA sincroniza al reloj endógeno que controla el movimiento de las hojas y probablemente
la actividad de numerosos procesos vitales en las plantas.
El phyA actúa a través de dos modos de acción, el VLFR (por Very Low Fluence Responses) que se
satura con muy bajos flujos de luz, y el HIR (por High Irradiance Responses) que se satura con altas
irradiancias de RL. Aquí presento evidencias genéticas que indican que 1aacción del phyA en el VLFR y
e] HIR involucra dos mecanismos moleculares diferentes. Por un lado se identificaron dos loci, vlfl (en el
cromosoma 2) y vle (en el cromosoma 5), polimórficos entre los ecotipos Landsberg erecta y Columbia

de Arabidopsis thaliana, que afectan las respuestas VLFR pero no las HIR. También se caracterizaron las
respuestas VLFR y HIR en dos mutantes de transduccíón de señales del phyA previamente identificados y
se observó que uno de ellos, elfhyI, afectaba de manera similar las respuestas VLFR y HIR mientras que
el otro, el jhy3, afectaba mucho más las respuestas HIR que las VLFR. En conjunto, estos resultados
sugieren que el phyA desencadena las respuestas VLFR y HIR a través de dos vías de transduccíón de
señales diferentes que poseen algunos elementos en común. Por otro lado, utilizando una estrategia
novedosa de búsqueda, se identificaron dos mutantes débiles de phyA, denominadas phyA-301 y phyA
302, que afectaban mucho más las respuestas HIR que las VLFR. Tanto la mutante phyA-30] como la
mutante phyA-302 presentaron niveles reducidos de phyA en ensayos de inmunodetección. La
secuenciación del gen PH YAen el mutante phyA-301, reveló una mutación que da lugar a una proteína
truncada que carece de 142 aa del extremo C-terminal. La secuenciación del gen PHYA en el mutante
phyA-302 reveló una mutación puntual que convierte el glutamato 778 en lisina. Esta mutación se
encuentra en una región previamente descripta como importante para la actividad regulatoria de los
fitocromos. Estos resultados indican que las respuestas a muy bajos flujos, es decir las VLFR, y las
respuestas a altas irradiancias, es decir las HIR, se diferenciar-lan en los niveles de phyA y/o en los
dominios de ésta molécula necesarios para inducir cada una de las respuestas.
En resúmen, hemos demostrado que el phyA está involucrado en la percepción de diferentes cambios
en el ambiente lumínico durante distintas etapas del ciclo de vida de las plantas y hemos encontrado
evidencias genéticas que indican que el phyA actuaría a través de dos vías de transduccíón de señales
distintas, una inducida en respuesta a muy bajos flujos de luz y otra inducida en respuesta a altasirradiancias.

SllMMARX
11119;Light control of plant grth and development: photosensory functions and mechanisms of action
of phytochrome A.
Ainhoa Marcelo J. Yanovsky
Key words; light, phytochrome A, de-etiolation, neighbor perception, circadian rhythms, signal
transduction, Arabidopsis thalíana, potato.
Light is a critical enviromental factor for plants. It provides not only the radiant energy for
photosynthesis, but also the infonnational signals that plants use to adapt and optimize their grth and
development in response to the ambient conditions. Perception of light signals is mediated by specific
photoreceptors such as phytochromes. Phytochromes are a small family of proteins encoded by five genes
in Arabidopsis thaliana (PH YA-PHYE).
The aim of this thesis is to investigate the photosensory functions of phytochrome A (phyA) and to
improve our understanding of the mechanisms by which this photoreceptor transduces different light
signals.
The photoperceptive functions of phyA were analyzed by comparing the behavior of mutant and
transgenic plants with either reduced or increased phyA levels to that of wild type (WT) plants that have
normal levels of this photoreceptor. It can be concluded, from the experimental results, that phyA acts as a
sensor of irradiance. In etiolated plants phyA is the only photoreceptor capable of perceiving the transition
from the dark envirrnonment present below the earth surface to the low light levels present beneath very
dense canopies. In response to the perception of that signal phyA induces several changes in the
developmental pattem that are crucial for seedling survival. In light-grown plants phyA is directly
involved in the perception of changes in the amount of red light (R) and far-red light (FR) produced as a
consequence of vegetation shading, and modulates the sensitivity to changes in the R/FR ratio produced
by the selective reflection of FR in the leaves of nearby plants. These actions of phyA lead to an increment
in stem elongation that helps preventing future shading. In light-grown plants phyA is also involved in the
perception of the dark/light transitions that occure daily at dawn. In response to such signal phyA
sinchronizes the endogenous clock that controls the rhythm of leaf movement, and probably the activity of
other vital processes.
PhyA acts through two different action modes. The first, VLFR (for Very Low Fluence Responses) is
saturated with very low fluences of either R or FR light. The second, HIR (for High Irradiance Reactions)
is only saturated with high irradiances of FR. Here I present genetic evidence indicating that the action of
phyA in the VLFR and HIR involve two different molecular mechanisms. First, I have identified two loci
polyrnorphic between ecotypes Landsberg erecta and Columbia of Arabídopsis thalíana, vlfl (on
chromosome 2) and vaZ (on chromosome 5), that affect VLFR but not HIR. Second, I have characterized
two previously identified phyA signal transduction mutants,fhy1 and fhy3, and found that fhy3 is more
affected in the HIR than in the VLFR, while fhy! is similarly affected in both phyA action modes. These

resulsts suggest that phyA elicits VLFR and HIR through two different signal transduction pathways that
have some common component(s). Finally, using a novel screening strategy, I have identified two mutants
lacking HIR but retaining VLFR. The genetic complementation analysis indicated that they were weak
alleles of phyA and therefore they were named phyA-301 and phyA-302. Western blott analysis revealed
that both mutants had reduced phyA levels. The sequence analysis of the phyA-301 mutant indicated that
the PH YAgene beared a mutation leading to a truncated phyA protein, lackingl42 aa from the C-terminal
domain. The sequence analysis of the phyA-302 mutant revealed a mutation that converted glutamic acid
778 into lysine. These results indicate that the responses to very low fluences of light and the responses to
high irradiances of FR are different either in the levels of phyA or in the domains of this molecule that are
necessary to induce each type of response.
In summary, we have shown that phyA is involved in the perception of different changes in the light
environment during the whole life cycle of plants and we have found genetic evidence supporting the
hypothesis that phyA activates two different signal transduction pathways, one in response to very low
fluences of light and the other in response to high irradiances.

BREV
CR Y
cry
cry
fllylfhy3HIR
LFR
pefPH Y(A-E)
Phy (A-E)
phyA-I
phyA-ZÜI
Phy (A-E)
Pfr
Pr
P
Pfr/P
PFD
VLFR
WT
gen que codifica para el fotorreceptor de luz azul denominado criptocromo
mutante de criptocromo
critpocromo (proteína + cromóforos)mutante de la cadena de transducción de señales del fitocromo A
mutante de la cadena de transducción de señales del fitocromo A
Respuestas a altas irradiancias de rojo lejano continuo
Respuestas a flujos moderados de luz rojamutantes de la cadena de transducción de señales de los fitocromos
genes que codifican para los fitocromos (A-E)
mutantes de fitocromo (A-E)
mutante de fitocromo A, alelo 1
mutante de fitocromo A, alelo 201
fitocromos (A-E)
forma activa del fitocromo; ¡máx= 730 nm
forma inactiva del fitocromo; ¡»máx= 660 nm
fitocromo total = Pr + Pfr
Proporción del fitocromo total que se encuentra en la forma activa
pulso de luz dado al final del día
luz roja
luz rojo-lejana
respuestas a muy bajos flujos de luz
genotipo salvaje

Los resultados de esta tesis han sido total o parcialmente incluidos en los siguientestrabajos:
1) M.J. Yanovsky, J.J. Casal and G.C. Whitelam (1995). Phytochrome A,Phytochrome B and HY4 are involved in hypocotyl-growth responses to naturalradiation in Arabidopsis. Weak de- etiolation of the phyA mutant under densecanopies. Plant, Cell & Environment 18, 788-794.
2) MJ. Yanovsky, J.J. Casal, J.P. Luppi (1997) The VLF loci, polyrnorphicbetween ecotypes Landsberg erecta and Columbia dissect two branches ofphytochrome A signalling pathways that correspond to the very-low fluenceand high-irradiance responses of phytochrome. Plant Journal, 12, 659-667.
3) J.J. Casal, R.A. Sanchez, M.J. Yanovsky (1997) The function ofphytochrome A. Plant, Cell & Environment, 20, 813-819.
4) M.J. Yanovsky, T.M. Alconada-Magliano, M.A. Mazzella, C. Gatz, B.Thomasand J.J. Casal (1998) Phytochrome A is required for normal growth,anthocyanin synthesis, sucrose-phosphate- synthase activity and neighbourdetection in sunlight-grown potato. Planta 205, 235-241.
S) P.D. Cerdán, M.J. Yanovsky, C. Reymundo, A. Nagatani, R. Staneloni, G.C. Whitelam,and J.J. Casal (1999) Regulation of phytochrome B signaling by phytochrome A andFHYl. Plant Journal, 18 (499-508).

l. INTRODUCCIÓN GENERAL
Las plantas son organismos sésiles y por lo tanto no pueden ir en busca de un hábitat favorable
si las condiciones ambientales se tornan adversas. En lugar de moverse, las plantas responden a
los cambios de su entorno con modificaciones en el desarrollo que parecen mejorar su adaptación
al ambiente. La coordinación del desarrollo con los cambios ambientales es un proceso
sumamente complejo que involucra:
1. señales inequívocamente asociadas a los cambios ambientales,
2. sofisticados mecanismos de percepción,
3. sistemas de transducción que conviertan las señales percibidas en modificaciones del
crecimiento y desarrollo.
1.1 SEÑALES LUMINICAS
La luz es uno de los estímulos ambientales que mayor efecto tiene sobre las plantas. No sólo
representa una fuente de energía para su crecimiento, sino también una de las principales fuentes
de información. A los fenómenos por medio de los cuales las plantas sensan y responden a la
información provista por el ambiente lumínico se los denomina colectivamente
FOTOMORFOGÉNESIS, que literalmente significa influencia de la luz (foto) en el desarrollo
(génesis) de la forma (morfo). Los aspectos del ambiente luminoso que pueden proveer
información son: la irradiancia (cantidad de luz por unidad de tiempo y superficie), la
composición espectral (cantidad de luz en cada zona del espectro), el fotoperíodo (duración diaria
del período lumínico) y la dirección de incidencia. Los cambios en estos parámetros del ambiente
luminoso constituyen señales que permiten a las plantas detectar (muchas veces en forma
anticipada) situaciones ecológicas tan variadas como la proximidad a la superficie del suelo
durante el proceso de emergencia, la proximidad de plantas vecinas que representan un riesgo de
sombreado futuro, y la llegada de la estación climática más favorable para el crecimiento
(Kendrick & Kronenberg, 1994).
1.1.1. Señales lumínicas asociadas al proceso de emergencia del suelo
Durante el proceso de emergencia del suelo las plántulas sufren cambios dramáticos en el
ambiente al que están expuestas y, por lo tanto, no resulta sorprendente que el patrón de10

desarrollo cambie radicalmente en ese momento. En general, el crecimiento de las plántulas
debajo de la superficie del suelo se caracteriza por una pronunciada tasa de alargamiento de los
tallos y un escaso desarrollo de los cotiledones y de la maquinaria fotosíntética en general. Este
patrón de desarrollo parece favorecer el rápido crecimiento de las plantas a través de la tierra,
permitiendo que éstas alcancen la superficie terrestre antes que se agoten las reservas disponibles
en los cotiledones. Al aproximarse las plántulas a la superficie se inhibe el alargamiento del tallo
y se promueve la expansión de los cotiledones y el desarrollo de toda la maquinaria necesaria
para el proceso fotosíntético. Una de las señales ambientales inequívocamente asociada a la
proximidad de la superficie del suelo es la transición desde la oscuridad completa a la luz provista
por el sol. Cuando las plántulas se acercan a la superficie, la detección de los primeros fotones
gatilla los cambios morfológicos, fisiológicos y moleculares anteriormente mencionados que
parecen preparar a las plántulas para crecer al aire libre (Mohr, 1972).
1.1.2. Señales lumínicas asociadas a la presencia de plantas vecinas
Una vez que la plántula ha emergido y se ha establecido, su capacidad de crecer y reproducirse
dependerá de su habilidad para captar luz y convertirla en energía química por medio del proceso
fotosíntético. Dado que la luz disponible disminuye con el incremento en la densidad de plantas,
éstas deben competir con sus vecinas para obtener dicho recurso. En general, las plantas
características de hábitats abiertos responden al incremento en la densidad de plantas vecinas con
aumentos dramáticos en la tasa de extensión de los tallos y una marcada reducción en la
dominancia apical. Estos cambios están frecuentemente acompañados de modificaciones en el
patrón de distribución de fotoasimilados, aumentando la cantidad destinada al crecimiento de los
tallos y disminuyendo la destinada al crecimiento de las hojas. A este patrón de desarrollo se lo
conoce como Síndrome de Escape del Sombreado (en inglés, Shade Avoidance Responses) y
constituye una estrategia exitosa en ambientes naturales altamente competitivos ya que facilita la
proyección de las hojas de las plantas sombreadas hacia un ambiente sin limitaciones lumínicas
(Smith, 1994).
Una de las señales ambientales que permite a las plantas percibir la presencia de vecinas son
los cambios que éstas provocan en la composición espectral del ambiente luminoso circundante.
Cuando la luz solar es interceptada por las hojas de las plantas, ésta es absorbida, reflejada y
transmitida en forma diferencial debido a que los pigmentos fotosíntéticos absorben
principalmente luz roja (R) y azul, mientras que reflejan y transmiten luz rojo lejana (RL) y verde
ll

(Smith, 1982). En etapas tempranas del desarrollo de un canopeo la relación R/RL de la luz
propagada en forma horizontal se reduce como consecuencia del RL que reflejan las hojas de las
vecinas, aún con anterioridad a que las plantas se sombreen entre sí (Ballaré et al., 1987, 1989).
Cuando el canopeo se hace más denso las plantas comienzan a sombrearse entre sí y la absorción
de la luz en las hojas de las vecinas reduce tanto la relación R/RL como la cantidad de azul, R y
RL que incide sobre los tallos y posteriormente, también la cantidad de luz que incide sobre las
hojas (Ballaré et al., 1991).
La importancia que tiene la relación R/RL como señal desencadenante del síndrome de evitado
del sombreado ha sido puesta de manifiesto de varias maneras. Por un lado, se han podido inducir
respuestas de evitado del sombreado en plantas cultivadas bajo altas irradiancias de luz blanca,
adicionando artificialmente RL al ambiente luminoso de manera de reducir la relación R/RL para
que se asemeje a la presente en cultivos densos (Smith, 1982; Casal & Smith, 1989). Por otro
lado, se han podido bloquear las respuestas desencadenadas por la presencia de plantas vecinas
filtrando selectivamente el RL reflejado por éstas de modo de mantener una alta relación R/RL
(Ballaré et al., 1990).
1.1.3. Señales lumínicas asociadas a las estaciones climáticas
Numerosos factores vitales como la luz, el agua y la temperatura fluctúan estacionalmente. Por
lo tanto, coordinar el ciclo de vida con las estaciones climáticas favorables es crucial para la
supervivencia de las plantas. Una señal ambiental que permite a las plantas anticipar
confiablemente los cambios en las estaciones es la duración del período lumínico ó fotoperíodo.
Si bien en las regiones ecuatoriales la salida y la puesta del sol se produce a la misma hora todos
los días del año, a medida que uno se desplaza desde el ecuador hacia los polos, los días de
verano se alargan y los de invierno se acortan. Procesos claves del desarrollo de las plantas tales
como la floración, la senescencia, la dormición de yemas, etc., están influenciados por la duración
de los días de manera que sólo se manifiestan, o se aceleran, cuando la longitud del día es mayor
o menor (dependiendo del proceso y de la especie) que cierto valor crítico (Thomas & Vince
Prue, 1997). A las respuestas de las plantas a la longitud del día se las conoce como respuestas
fotoperiódicas y constituyen uno de los mecanismos que permite a las plantas coordinar su
crecimiento y desarrollo con las estaciones climáticas favorables.
1.2. MECANISMOS DE PERCEPCIÓN

Las señales lumínicas sólo pueden proveer información a las plantas si éstas poseen receptores
adecuados para las mismas. Hasta el momento se han identificado en las plantas tres grupos de
fotorreceptores: los fitocromos, que perciben en el rango del R y el RL, los fotorreceptores
del azul y el UV-A, y el (los) fotorreceptor(es) de ultravioleta-B (UV-B) (Mohr, 1994).
1.2.1 Fotorreceptores de UV-B
El UV-B (280 —320 nm) tiene influencia en numerosos aspectos del crecimiento y desarrollo
de las plantas. Si bien a altas intensidades produce daño a las macromoléculas como el ADN y las
proteínas (Jordan, 1995), en niveles moderados tiene efectos específicos sobre el desarrollo
(Tevini and Terarnura, 1989; Ballaré et al., 1995; Ballaré et al., 1996). Entre otras cosas, el UV-B
promueve la síntesis y la acumulación de pigmentos que absorben el UV-B, y por ende protegen a
las plantas de sus consecuencias dañinas (Jenkins, 1997). El UV-B es percibido por
fotorreceptores específicos, distintos de los que perciben el UV-A y el azul. Existe, sin embargo,
poca información sobre la naturaleza molecular de estos fotorreceptores y sus mecanismos deacción.
1.2.2. Fotorreceptores de luz azul
Numerosos procesos son desencadenados por la luz azul (400-500 nm) en las plantas (Short &
Briggs, 1994). Entre estos se encuentran las respuestas fototrópicas, la inhibición del
alargamiento del tallo, la apertura de los estomas, la expresión de genes, etc. Tres fotorreceptores
de luz azul han sido identificados hasta ahora a nivel molecular: cryl, cry2 (por criptocromo l y
2) y nphl (por non-photoptropic hypocotyl l). Cryl y cry2 son flavoproteínas con homología a
las fotoliasas bacterianas, pero carecen de actividad reparadora del ADN (Ahmad and Cashmore,
1996). A1 igual que las fotoliasas, los criptocromos poseen dos cromóforos, uno de ellos una
flavina y el otro una pterina (Ahmad, 1999). La caracterización de mutantes deficientes en estas
proteínas, así como de plantas transgénicas que las sobreexpresan, indican que el cryl y el cry2
están involucradas en el control del alargamiento del tallo, la expresión de genes y la floración en
respuesta a la luz azul (Cashmore, 1999). El gen NPHI, por otro lado, parece codificar al
fotorreceptor de luz azul involucrado en las respuestas fototrópicas. El nphl es una flavoproteína
con poca homología a los criptocromos, capaz de autofosfon'larse en respuesta a la luz azul
(Christie et al., 1998). Si bien en estos últimos años se ha avanzado notablemente en la
identificación a nivel molecular de los fotorreceptores de la luz azul, aún se conoce muy poco
13

sobre los mecanismos por medio de los cuales 1a excitación de estos fotorreceptores por la luz
desencadena finalmente cambios en el desarrollo de las plantas.
1.2.3. Fotorreceptores de R y RL, los fitocromos
El R (600-700 nm) y el RL (700-800 nm) afectan marcadamente diversos aspectos del
desarrollo de las plantas. Espectros de acción llevados a cabo hace aproximadamente 50 años
revelaron que la luz de 660 nm era la más efectiva para controlar la floración en plantas de día
corto y la desetiolación de plántulas de arveja (Parker et al., 1946; 1949). La similitud de
espectros de acción para procesos tan distintos sugería que un mismo fotorreceptor controlaba
ambas respuestas. La identificación y caracterización de este fotorreceptor surgió finalmente de
estudios sobre el fotocontrol de la germinación de las semillas de lechuga. En 1952, Borthwick y
col. encontraron que la promoción de 1a germinación por pulsos de R podía ser prevenida por
pulsos de RL dados en forma inmediata luego del R. Lo más llamativo de esta respuesta dual a la
luz era que si se daba en forma altemada una serie de pulsos de R y RL, la respuesta fisiológica
(promoción o inhibición de 1a germinación) dependía de las características del último pulso de
luz. Si la secuencia terminaba con R, la mayoría de las semillas germinaban durante la incubación
posterior en oscuridad; en cambio, si la serie terminaba con RL, la mayoría de las semillas
permanecían dormidas.
OSCURIDAD R R —)RL
Figura 1: Germinación de las semillas de lechuga en oscuridad, después de un pulso de R y después de
un pulso de R seguido de RL, Borthwick y col., 1952.
14
Los autores concluyeron brillantemente que la respuesta debía estar mediada por un único
fotorreceptor, luego denominado fitocromo, que podía existir en dos estados llamados Pr y Pfr,
interconvertibles por R y RL. En 1959 Butler y col. fueron capaces de purificar parcialmente el
pigmento fotorreversible y demostraron que el mismo era una proteína.
1.2.3.1. Propiedades estructurales y fotobiológicas del fitocromo
Los fitocromos son proteínas solubles que existen como homodímeros de un polipéptido de
aproximadamente 1100 aminoácidos (120-124 KDa). La apoproteína de cada monómero se
encuentra unida covalentemente a un cromóforo, un tetrapirrol de cadena abierta denominado
fitocromobilina (Quail, 1991). En cada monómero se pueden distinguir dos dominios
estructurales y funcionales: el dominio N-terminal (aproximadamente 70 KDa) contiene los sitios
necesarios para la unión del cromóforo a la apoproteína, y el dominio C-terminal
(aproximadamente 55 KDa) contiene los sitios necesarios para la dimerización (Quail, 1997). Los
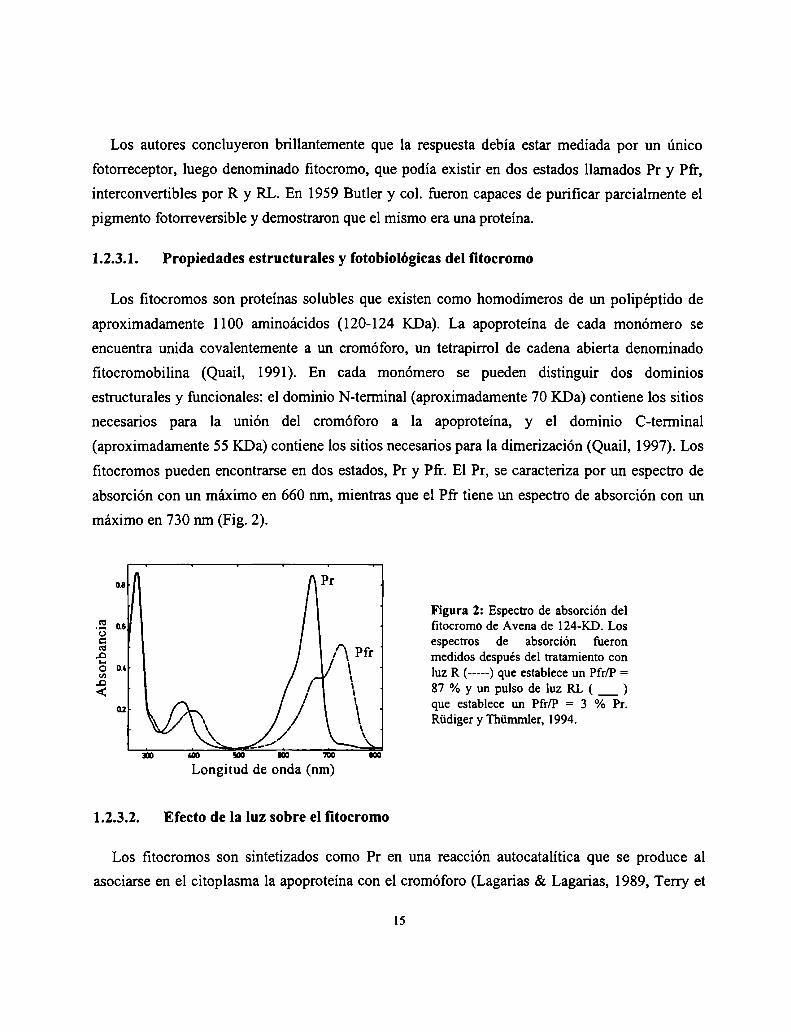
fitocromos pueden encontrarse en dos estados, Pr y Pfr. El Pr, se caracteriza por un espectro de
absorción con un máximo en 660 nm, mientras que el Pfr tiene un espectro de absorción con un
máximo en 730 nm (Fig. 2).
Figura 2: Espectro de absorción delfitocromo de Avena de 124-KD. Losespectros de absorción fueronmedidos después del tratamiento conluz R (-----) que establece un Pfr/P =87%yunpulsodelquL(_)que establece un Pfr/P = 3 % Pr.Rüdiger y Thümmler, 1994.
Absorbancia
¿no son son no
Longitud de onda (nm)
1.2.3.2. Efecto de la luz sobre el fítocromo
Los fitocromos son sintetizados como Pr en una reacción autocatalítica que se produce al
asociarse en el citoplasma la apoproteína con el cromóforo (Lagan'as & Lagarias, 1989, Terry et
15

al., 1993). En las plántulas etioladas el 100 % de los fitocromos se encuentra en la forma Pr. Al
exponer plántulas etioladas a la luz, el Pr se fotoconvierte en Pfr. La fototransformación involucra
la isomerización cís-trans de uno de los doble enlaces del cromóforo. Esta isomerización provoca
a su vez múltiples rearreglos conformacionales de la proteína que conducen, luego de pasar por
varios estados intermediarios, a la formación del Pfr (Furuya & Song, 1994). La
fototransformación es fotorreversible y, por lo tanto, al absorber luz el Pfr se convierte
nuevamente en Pr (Figura 3).
Amáx. 660 nm
TranscripciónTraducción —>PHY —> —> PHY Pr Pfr' \Amáx.730 nm \
cromóforo \destrucción
reversión
Figura 3. La expresión del gen PHY da lugar a la apoproteína PHY que al acoplarseautocatalíticamente con el cromóforo origina el Pr. La luz fotoconvíerte el Pr a Pfr yviceversa. El Pfr también puede destruirse o convertirse en Pr en una reacciónindependiente de la luz conocida como reversión.
Dado que los espectros de absorción de las dos formas están parcialmente solapadas (Fig. 2),
al irradiar una planta se establece un fotoequilibn'o dinámico entre el Pr y el Pfr. La proporción de
Pfr respecto al fitocromo total (Pfr/P) establecida por cada tratamiento lumínico dependerá de la
composición espectral del mismo (Mancinelli, 1994). Luz monocromática de 660 nm convierte
un 87 % del fitocromo a la forma Pfr, mientras que luz monocromática de 730 nm establece un 3
% de Pfr/P. Cuando se exponen las plántulas a radiación policromática, el Pfr/P estará
determinado principalmente por la proporción de R y RL de la luz incidente (Mancinelli, 1994).
Cuanto mayor sea la relación R/RL de la luz incidente, mayor será el Pfr/P.
Los niveles de Pfr también son afectados por dos reacciones que no dependen de la luz: la
destrucción y la reversión a Pr (Fig. 3). La destrucción del Pfr ocurre a través de la vía
16

proteolítica mediada por las ubiquitinas (Clough & Vierstra, 1997). Esta reacción, sin embargo,
no afecta en forma homogénea a todas las moléculas de fitocromo, siendo muy importante para el
fitocromo predominante en plántulas etioladas y muy poco importante para el fitocromo presente
en plantas des-etioladas. Por último, la reversión del Pfr a Pr es una reacción muy lenta
comparada con la fotoconversión Pr (—>Pfr y, por lo tanto, sólo es cuantitativamente importante
en tratamientos que incluyen períodos prolongados de incubación en oscuridad ó exposición a luz
continua de muy bajos flujos (Vierstra, 1994).
1.2.3.3. Modos de acción de los fitocromos
Cuando las plantas etioladas son expuestas a la luz, la fotoconversión del Pr a Pfr desencadena
numerosos cambios morfológicos, fisiológicos, y moleculares. La inducción de dichos cambios
podn’aser consecuencia de la formación del Pfr, de la desaparición del Pr o de ambos eventos a la
vez. Sin embargo, el fenotipo de plantas transgénicas sobreexpresantes de fitocromo y de plantas
mutantes, con niveles reducidos o nulos de fitocromo, no se diferencia del fenotipo de las plantas
salvajes cuando todas son cultivadas en oscuridad. Por el contrario, las plantas transgénicas
sobreexpresantes de fitocromo presentan una respuesta exagerada a la luz, mientras que las
plantas mutantes con niveles reducidos o nulos de fitocromo presentan una respuesta disminuida
a la luz. El hecho que los niveles alterados de fitocromo no afecten el fenotipo en oscuridad,
cuando el fitocromo se encuentra en un 100% como Pr, y si lo hagan cuando las plantas son
expuestas a la luz, cuando parte del fitocromo se convierte de Pr a Pfr, indica que la formación
del Pfr, y no la desaparición del Pr, desencadena las respuestas (Fankhauser & Chory, 1997).
No obstante, la forma de actuar de los fitocromos es compleja y las respuestas inducidas por
este fotorreceptor no son una función sencilla de los niveles de Pfr establecidos por los
tratamientos lumínicos. Por ejemplo, cuando se exponen semillas o plántulas etioladas a pulsos
breves de luz que establecen niveles crecientes de Pfr/P, la curva que relaciona la magnitud de las
respuestas desencadenadas con los niveles de Pfr/P presenta dos fases (Fig. 4). La primera fase se
induce con niveles de Pfr/P inferiores al 0.01 % y alcanza la saturación entre el 0.1 y el 10% de
Pfr/P. La segunda fase comienza a observarse con niveles de Pfr/P superiores al 20 % y se saturacon niveles cercanos al 60 %. La observación de una relación bifásica entre los niveles de
Pfr/P y la magnitud de las respuestas revela la existencia de dos modos de acción del fitocromo
(Casal et al., 1998).
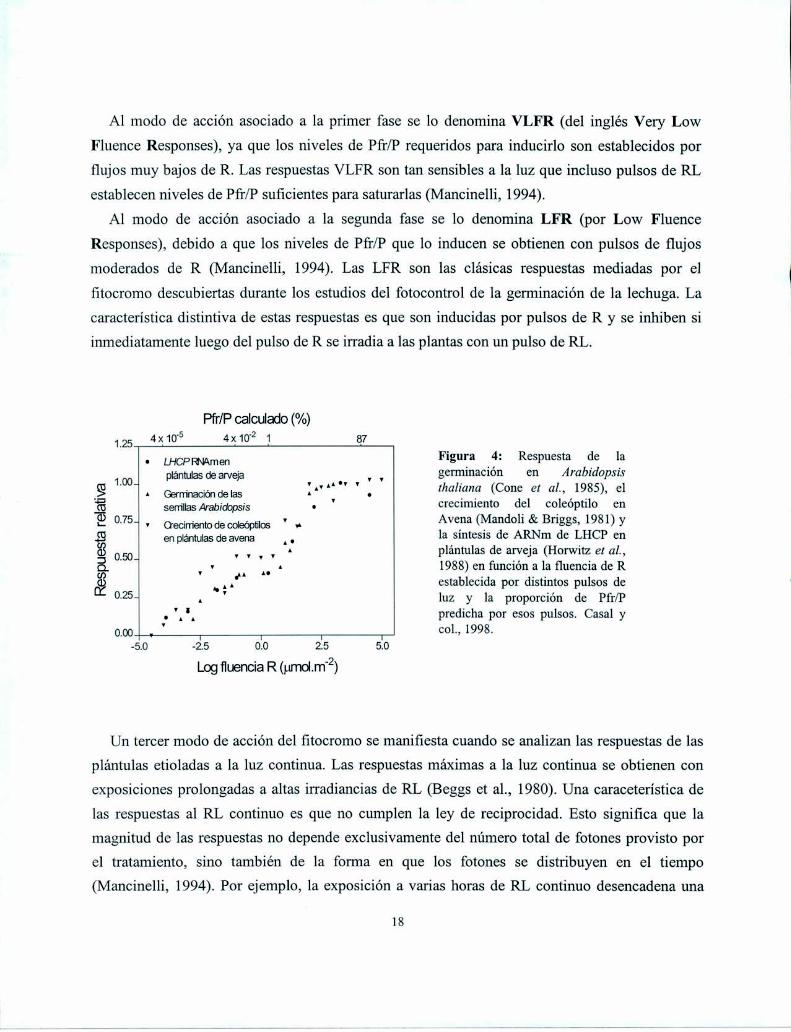
Al modo de acción asociado a la primer fase se lo denomina VLFR (del inglés Very Low
Fluence Responses), ya que los niveles de Pfr/P requeridos para inducirlo son establecidos por
flujos muy bajos de R. Las respuestas VLFR son tan sensibles a la luz que incluso pulsos de RL
establecen niveles de Pfr/P suficientes para saturarlas (Mancinelli, 1994).
A1 modo de acción asociado a la segunda fase se lo denomina LFR (por Low Fluence
Responses), debido a que los niveles de Pfr/P que lo inducen se obtienen con pulsos de flujos
moderados de R (Mancinelli, 1994). Las LFR son las clásicas respuestas mediadas por el
fitocromo descubiertas durante los estudios del fotocontrol de la germinación de la lechuga. La
característica distintiva de estas respuestas es que son inducidas por pulsos de R y se inhiben si
inmediatamente luego del pulso de R se irradia a las plantas con un pulso de RL.
Pfr/P calculado (%)
1,; 4x10'5 4x10'2 1 87
_ LHCPFNAmen Figura 4: Respuesta de laplánuflasdeaweja v v germinación en Arabidopsis
g 1.00a A Germnadóndelas In A‘" ' . thaliana (Cone et aL, 1985), el'ÏU sem-"asArab¡dops¡s . ' crecimiento del coleóptilo en
(¿D0.75- V Oecimnto de compu“ v _ Avena (Mandoli & Briggs, 1981) y
É enplánuasdeavena A. la síntesis de ARNm de LHCP en3 0.3L ' v V v plántulas de arveja (Horwitz et aL,Q v y A n A 1988) en función a la fluencia de R
É h AA" establecida por distintos pulsos de0-25- A ' luz y la proporción de Pfr/P
, 'A'A predicha por esos pulsos. Casal y
nm v 1 I l T COL,-E.O -2.5 0.0 2.5 5.0
Logfluencia R (mmm-2)
Un tercer modo de acción del fitocromo se manifiesta cuando se analizan las respuestas de las
plántulas etioladas a la luz continua. Las respuestas máximas a la luz continua se obtienen con
exposiciones prolongadas a altas irradiancias de RL (Beggs et al., 1980). Una caraceterística de
las respuestas al RL continuo es que no cumplen la ley de reciprocidad. Esto significa que la
magnitud de las respuestas no depende exclusivamente del número total de fotones provisto por
el tratamiento, sino también de la forma en que los fotones se distribuyen en el tiempo
(Mancinelli, 1994). Por ejemplo, la exposición a varias horas de RL continuo desencadena una
18

respuesta significativamente mayor que la exposición a pulsos breves de RL repetidos en forma
horan'a, aún cuando ambos tratamientos provean la misma cantidad total de fotones. La falla a la
ley de reciprocidad no se observa en las respuestas al R, donde los pulsos de luz y la luz continua
tienen el mismo efecto (Mazzella et al., 1997). Este hecho sugiere que las respuestas al RL
involucran dos modos de acción distintos, el VLFR que se activa con breves pulsos de luz, y otro
modo de acción diferente que se activa sólo con luz continua. Debido a que las respuestas al RL
continuo dependen fuertemente de la irradiancia, a este modo de acción se lo denomina HIR (por
High Irradiance Responses).
No obstante, dado que el fitocromo de las plántulas etioladas es inestable en la forma Pfr
porque se destruye o revierte a Pr, el RL continuo podría ser más efectivo que los pulsos de RL
simplemente porque la luz continua mantendría constantes los niveles de Pfr. Sin embargo, el RL
continuo es mucho más efectivo incluso que muy bajos flujos de R continuo que (debido al
incremento en la proporción Pfr que revierte a Pr a muy bajas irradiancias) establecen niveles
similares y constantes de Pfr en el tiempo (Heim & Scháfer, 1984). Esto revela que las respuestas
HIR, a diferencia de las VLFR y las LFR, no pueden ser explicadas simplemente por los niveles
de Pfr. Si bien elegantes experimentos fotobiológicos de la década del 60 demostraron que las
respuestas al RL continuo estaban medíadas por el fitocromo, no está claro aún que parámetro del
fitocromo las desencadena (Hartmann, 1966). La dependencia de las respuestas HIR de la
irradiancias y de luz continua ha llevado a algunos autores a proponer que las mismas serían
desencadenadas por un intermediario inestable producido durante la fotoconversión entre las
formas Pr y Pfr (Johnson & Tasker, 1979).
1.2.3.4. Los fitocromos, una familia de proteínas
La caracterización inmunológica del fitocromo llevada a cabo a comienzos de los 80' reveló
que, incluso en una misma especie, existía más de un tipo de fitocromo. Una vez que los genes de
los fitocromos comenzaron a ser clonados y caracterizados quedó en evidencia que los fitocromos
constituían una famila de genes relacionados. En la planta superior Arabidopsís thaliana, una
dicotiledónea, existen y se expresan cinco genes de fitocromo distintos denominados PHYA a
PH YE (Clack et al., 1994; Shar-rockand Quail, 1989). En otras dicotiledóneas, como el tomate,
también sen’an cinco los genes que codifican fitocromos (homólogos a los genes PHYA-E de
Arabidopsis), mientras que en las monocotiledoneas sólo existirían tres genes de fitocromos,
homólogos al PHYA, PHYB y PHYC (Pratt et al., 1997; Mathews & Shanok, 1997). La
19

comparación de secuencias entre los distintos fitocromos indicó que los fitocromos de distintas
especies (incluso si se compara mocotiledóneas con dicotiledóneas) son más parecidos entre sí
que los distintos fitocromos de una misma especie. Estos análisis sugieren que los distintos
fitocromos se originaron a partir de un único ancestro común, a través de sucesivas duplicaciones
(Mathews & Sharrok, 1997).
El phyA y el phyB son los fitocromos más abundantes y por lo tanto son los que más han
caracterizados a nivel bioquímico, biofisico y molecular. El phyA y el phyB tienen espectros
similares de absorción y se supone que ambos compartirían el mismo cromóforo (Braslavsky et
al., 1997; Terry et al, 1997). En cuanto a su distribución espacial, el gen PHYA se expresa
intensamente en todo tipo de tejidos (Somers & Quail, 1995). La proteína se acumula en altas
cantidades en plántulas etioladas, pero sus niveles disminuyen dramáticamente luego de la
exposición a la luz (Somers et al., 1991). La caída en los niveles de phyA a la luz se debe a que el
Pfr del phyA es rápidamente degradado por la vía de las ubiquitina y el proteasoma 268 (Clough
& Vierstra, 1997). La disminución de los niveles de phyA a la luz también es consecuencia de la
represión mediada por el Pfr de la expresión del gen PHYA (Colbert et al., 1983). El patrón
espacial de expresión del phyB parece ser similar al del PHYA, pero el phyB se diferenciar-ía del
phyA en cuanto a su abundancia relativa (Somers et al, 1995). Los niveles del phyB son 100
veces más bajos que los del phyA en plantas etioladas, pero similares o superiores a éste en
plantas verdes. Esto se debe a que el phyB es mucho más estables que el phyA en la forma Pfr
(Somers et al., 1991) y, si bien sus niveles de expresión son más bajos, los niveles de su proteína
se mantienen constantes en el tiempo.
La localización celular de los fitocromos ha sido objeto de debate por mucho tiempo. A pesar
de la existencia de algunos trabajos que sugerían una asociación a las membranas, la mayoría de
los estudios indican que los fitocromos son proteínas solubles con localización citoplasmática
(Quail et al., 1995). Recientemente, sin embargo, se han obtenido evidencias que tanto el phyA
como el phyB tienen localización citosólica sólo en oscuridad, y migran al núcleo al ser
expuestos a la luz (Yamaguchi et al., 1999; Kircher et al., 1999).
1.2.3.5. Funciones sensoriales de los distintos fítocromos
La existencia de más de un tipo de fitocromo llevó rápidamente a proponer que la variedad de
señales lumínicas percibidas por este fotorreceptor reflejaría la acción de distintos fitocromos
(Smith & Whitelam, 1990). La obtención de plantas mutantes y/o transgénicas con niveles
20

incrementados o reducidos de phyA, phyB, phyC, phyD ó phyE, ha permitido investigar esta
hipótesis en detalle (Koomneef et al., 1980; Adamse et al., 1988; Boylan & Quail, 1989; Kay et
al., 1989; Keller et al., 1989; Reed et al., 1993; Parks & Quail, 1993; Nagatani et al., 1993;
Whitelam et al., 1993; van Tuinen et al, 1995 a; van Tuinen et al., 1995 b; Halliday et al., 1997;
Aukerman et al., 1997; Quin et al., 1997; Devlin et al., 1998).
La caracterización de plantas con niveles alterados de phyB indica que el phyB afecta el
desarrollo de las plantas durante todo su ciclo de vida. Los mutantes de phyB presentan
deficiencias en la germinación y en el proceso de desetiolación (desarrollo de los cotiledones,
inhibición del alargamiento de los hipocotilos) en respuesta al R en el rango del LFR, pero
responden normalmente al RL continuo (HIR) y a los pulsos de RL (VLFR) (Reed et al., 1993;
Botto et al., 1995; Shinomura et al., 1996). En el estadío adulto, las plantas mutantes de phyB
presentan atenuadas las respuestas a bajas relaciones R/RL y a pulsos de RL dados al final del
día, dos tratamientos que simulan los cambios en el ambiente luminoso asociados a la presencia
de plantas vecinas, sugiriendo que el phyB sería uno de los principales fotorreceptores de estas
señales (Ballaré et al., 1991; Whitelam & Smith, 1991; Robson et al., 1993).
La caracterización de plantas mutantes y transgénicas con niveles alterados de los fitocromos
C, D y E indica que estos fotorreceptores tienen fimciones sensoriales similares al phyB, es decir
que están involucrados en la percepción del R en el rango del LFR pero, en algunos casos, se
diferenciarían del phyB en el grupo de procesos fisiológicos que controlan (p. ej., el phyC estaría
específicamente involucrado en el control del crecimiento de las hojas y no en el crecimiento de
los hipocotilos) (Halliday et al., 1997; Quin et al., 1997; Aukerrnan et al., 1997; Devlin et al.,
1998)
Las funciones fotoperceptivas del phyA parecen ser diferentes de las del resto de los
fitocromos. Estudios realizados con plantas mutantes y transgénicas con niveles alterados de
phyA revelaron que este fotorreceptor controla la germinación y la desetiolación de las plántulas
en respuesta a los pulsos de RL (VLFR) y al RL continuo (HIR) (Dehesh et al., 1993; Heyer et
al., 1995; Nagatani et al., 1993; van Tuinen et al., 1995; Whitelam et al., 1993). La ausencia de
phyA, sin embargo, no afecta la desetiolación bajo R en el rango del LFR, ni bajo luz blanca
provista por tubos fluorescentes; tampoco afecta el fenotipo de plantas adultas ni reduce las
respuestas a pulsos de RL dados al final del día ó a bajas relaciones R/RL establecidas
artificialmente durante el fotoperíodo (Nagatani et al., 1993; Whitelam et al., 1993; van Tuinen et
al., 1995; Heyer et al., 1995). Estas observaciones, sumadas al hecho que los niveles de phyA se
21

reducen notablemente en plantas expuestas a la luz, llevaron a proponer que en condiciones
naturales el phyA tendría un rol secundario en la desetiolación y no partíciparía del control del
desarrollo en las plantas adultas.
No obstante, si bien la caracterización de plantas mutantes y transgénicas con fuentes
artificiales de luz es útil como primera aproximación para investigar el rol sensorial de los
fotorreceptores, debe tenerse mucha cautela al extrapolar las conclusiones derivadas de estos
experimentos hacia interpretaciones sobre el significado ecológico de la acción de los
fotorreceptores. Por ejemplo, asumir que los fenotipos de plantas crecidas bajo luz blanca
provista por tubos fluorescentes serán iguales a los fenotipos de las mismas cuando son cultivadas
bajo radiación natural es apresurado. Las diferencias entre la luz solar y la luz provista por tubos
fluorescentes no son triviales ya que esta última tiene una alta proporción de luz azul y roja, pero,
a diferencia de la luz solar, carece casi por completo de RL. Siendo el RL la longitud de onda más
eficiente para activar el phyA, es posible suponer que en condiciones naturales de radiación la
ausencia del phyA tenga un impacto importante sobre el crecimiento y el desarrollo de las
plantas.
En las plántulas etioladas, el phyA es el único fotorreceptor capaz de percibir el RL. Por lo
tanto, es posible pensar que la acción de este fotorreceptor podría ser muy importante para la
desetiolación de plántulas que emergen bajo canopeos muy densos. Por otro lado, las plantas
adultas responden no Sólo a los cambios en la calidad, sino también a los cambios en la cantidad
de luz que provocan las plantas vecinas. Dado que las respuestas mediadas por el phyA dependen
de la irradiancia, este fotorreceptor podría estar involucrado en percibir el sombreado provocado
por la presencia de plantas vecinas. Por último, debido a su inestabilidad, los niveles de Pfr del
phyA se reducen al comenzar la noche (se destruye el Pfr presente y no hay luz para
fototransformar el nuevo Pr que se sintetiza) y se incrementan al amanecer. Esto hace del phyA
un gran candidato a participar del proceso de medición de la longitud de los días.
1.3. MECANISMOS DE ACCIÓN Y VÍAS DE TRANSDUCCIÓN DE SEÑALES DE
LOS FITOCROMOS
Si bien el fitocromo fire descubierto hace más de 40 años, los mecanismos por medio de los
cuales regula el desarrollo de las plantas son todavía mayoritariamente desconocidos. Aún no
sabemos con certeza cuál es el mecanismo bioquímico que se pone en marcha cuando el
fitocromo es excitado por la luz, ni quién(es) es(son) el blanco incial de acción del mismo. Por
22

otro lado, dado que los fitocromos son una familia de proteínas, otra pregunta que surge es si el
mecanismo de acción es el mismo para los distintos fitocromos o no.
La posibilidad de que el fitocromo fuera una enzima cuya actividad estuviera regulada por la
luz fue planteada hace casi 40 años (Butler et al., 1959). Aunque esta hipótesis fue abandonada
tempranamente, nuevas evidencias sugieren que el fitocromo podn'a actuar como una kinasa de
proteinas. Recientemente se descubrió que la cianobactena Synechocístis sp. PCC6803 posee un
gen con un 30 % de homología a los fitocromos en el dominio N-terminal, mientras que eldominio C-terminal tiene una alta similitud con las histidin-kinasas de los sistemas sensorios
regulatorios bacterianos (Yeh et al., 1997). Esta proteína de Synechocístis es capaz de unirse
autocataliticamente al cromóforo de los fitocromos y su espectro de absorción es modificado
fotorreversiblemente por el R y RL, indicando que funcionalmente sería un fitocromo. El
fitocromo de Synechocistis es capaz de autofosforilarse y de transferir el fosfato a una proteína
codificada en el mismo operón (Yeh et al., 1997). Estos hallazgos llevaron a postular que los
fitocromos de plantas superiores actuarían también como kinasas de proteínas (Elich & Chory,
1997). Existen en los fitocromos de plantas superiores dos dominios característicos de kinasas de
proteinas, uno de ellos similar a las kinasas de histidina y el otro con homología a las kinasas de
ser-¡nay treonina. Estos dominios habrían evolucianado a partir de la duplicación de un antecesor
común, similar al dominio con actividad de kinasa de histidinas de Synechocístis.
PRD HKRDFitocromodeplantas superiores : 14-15 : 13.17%.»_Mmm de lSynechocisüs
HKD
Figura 5. Modelo de evolución molecular de fitocromos eucan'óticos, por medio de laduplicación del modulo con homología a kinasas de histidinas presente en el fitocromo deSynechocistis.
Recientemente se ha demostrado que el fitocromo A de avena tiene actividad serin-treonin
kinasa (Yeh & Lagarias, 1998). El fitocromo A de avena es capaz de autofosforilarse y de
fosforilar otras proteínas, siendo esta fosfon'lación dependiente de la luz. Sin embargo aún no hay
evidencias de que la actividad kinasa esté vinculada a la actividad biológica de este fotorreceptor
y, por lo tanto, el mecanismo de acción de los fitocromos es aún objeto de debate.
23

Si bien no conocemos con certeza cual es el mecanismo bioquímico de acción de los
fitocromos, existen evidencias indirectas que sugieren que el phyA y el phyB actuarían de manera
similar. Esta idea proviene de experimentos donde se analizó el efecto de la sobreexpresión de
phyA, phyB y phy quiméricos que tenían el dominio N-terminal del phyA y el C-terminal del
phyB y viceversa. En estos experimentos se observó que los dominios N-terminal eran
responsables de la especificidad fotosensoria de los fitocromos. Los dominios C-terminales, en
cambio, son necesarios para la actividad regulatoria y no presentan especificidad alguna. El hecho
que los dominios C-terminal pudieran ser intercambiados sin problema alguno sugiere que la
actividad regulatoria sen’ala misma para ambos fitocromos (Wagner et al., 1996).
Otra evidencia de la existencia de un mecanismo de acción común para el phyA y el phyB
proviene de una aproximación en la que se buscó identificar mutantes de phyA y phyB que
tuvieran afectada la capacidad de desencadenar respuestas biológicas pero que conservaran los
niveles y las propiedades fotobiológicas normales de estos fotorreceptores. Sorprendentemente, la
mayoría de las mutaciones con estas caracteristicas estaban concentradas en 160 aminoácidos del
extremo C-terminal de la proteína, sugiriendo que esa región estan'a involucrada en la transmisión
de la señal. El hecho que las mutaciones se concentraron en esa región tanto del phyA como del
phyB, indicaria que ambos fitocromos compartirían el mismo mecanismo de acción (Quail et al.,
1995)
"73‘" A710Voso
mas G7 crias
phyA ym” cmv cazanusw ¡me
P100L "TS ym
PhYBGTÜE enla
I l l I I l l l I I l | I
100 200 30 400 500 600 700 800 900 1000 1100 1200
Figura 6. Mapa de las sustituciones aminoacídicas detectadas en mutantes regulatorios de
phyA y phyB (Quail, 1997).24

Uno de los debates fotomorfogénicos más antiguos es si los fitocromos poseen una ó
múltiples acciones primarias. La hipótesis que sostiene que los fitocromos son capaces de activar
múltiples vías de transducción de señales fue planteada originalmente para explicar la diversidad
de modos de acción de estos fotorreceptores. Según esta hipótesis los cambios morfológicos,
fisiológicos y moleculares producidos en respuesta a los muy bajos flujos de R (VLFR), a los
flujos moderados de R (LFR) y a las altas irradiancia de RL (HIR) serían el reflejo de distintas
vías de transducción de señales desencadenadas por los fitocromos.
Hoy en día sabemos que el phyA desencadena el VLFR y el HIR mientras que el phyB es el
responsable del LFR. Por lo tanto, de acuerdo a la hipótesis recién mencionada, el Pfr del phyB
formado en respuesta a flujos moderados de R desencadenaría una vía de transducción de señales
distinta a la del phyA. A su vez, el phyA tendría dos vías de acción diferentes. La primera, el
VLFR, sería desencadenada por el Pfr del phyA. Esta Vía seria muy sensible e incluso
tratamientos lumínicos que establecen muy bajos niveles de Pfr serian suficientes para saturarla.
La otra vía, el HIR, sería desencadenada por altas irradiancias de RL continuo. Este tratamiento
permitiría la acumulación de un intermediario excitado e inestable resultante del ciclado entre el
Pr y el Pfr, el cuál pondría en marcha una cadena de transducción de señales distinta a la
desencadenada por el Pfr del phyA (Fig. 7).Elementos del modelo
PliyA PhyBPr Pfr cfcctorno Pr Pfr VLFR LFR HIR0o Ü Ó DEE
Pulsos de RL ó muy bajos flujos de R
00000000o) o)I 3 3
Ü Ü Ü D D phyAyelphyBactivadospor diferentes
Blanco de acción del
Figura 7. Modelo de lasVLFR bases molecularesde los
modos de acción del
Pulsos de R ó flujos moderados de R continuo . , .tratamlentos luminlcos.
O . . . VLFR LamagnitudrelativadeE ïl E las respuestas VLFR,
Ü LFRyHIRestádadaporel grosor de las flechas.
Altos Flujos de RL continuo
oooooooo.3 .3;
Ü Ü D D
VL FR
HIR

En los últimos tiempos diferentes aproximaciones han permitido identificar proteínas que
constituirían los blancos de la acción inicial de los fitocromos, asi como también moléculas que
actuarían como componenetes de las vías de transducción de señales de estos fotorreceptores.
La utilización del sistema de dos híbridos de levaduras ha permitido encontrar proteínas que
interactúan específicamente con los fitocromos (Ni et al., 1998; Fankhauser et al., 1999). Algunas
de estas proteínas han sidos denominadas PIF, por Phytochrome Interacting Factors, y se especula
que podn'an ser los blancos iniciales de acción de los fitocromos. In vitro PIF3 interactúa tanto
con el phyB como con el phyA y las plantas transgénicas con el gen PIF3 en antisentido se
caracterizan por una sensibilidad reducida al R y al RL. Por el contrario, PIF2 interactúa
específicamente con el phyB y no con el phyA. Estos resultados sugieren que el phyA y el phyB
tendrían algunos blancos de acción en comúm, mientras que otros serían específicos.
Conclusiones similares se han obtenido utilizando aproximaciones genéticas. Tres tipos de
mutantes de las vías de transducción de señales lumínicas han sido aislados hasta ahora. El primer
grupo está integrado por mutantes que tienen una sensibilidad disminuida a la luz, el segundo
grupo esta constituido por mutantes que tienen una respuesta exagerada a la luz, y el tercer grupo
por mutantes que se comportan a la luz como si estuvieran en la oscuridad, es decir presentan
respuestas constitutivas a la luz (Quail, 1998). Dentro del primer grupo, los mutantes red], pe]? y
pef3 se caracterizan por presentar respuestas reducidas al R en el rango del LFR (Ahmad &
Cashrnore., 1996; Wagner et al., 1997), mientras que los mutantes fhyI y fhy3 sólo presentan
reducidas las respuestas HIR, es decir las respuestas a altas irradiancias de RL (Whitelam et al.,
1993). Las respuestas de estos mutantes en el rango del VLFR no han sido analizadas aún. La
existencia de mutantes que afectan las respuestas LFR y no las HIR y viceversa sugiere
fuertemente que el phyA y el phyB activan diferentes vías de transducción de señales. Por otro
lado, la mutante pef] tiene respuestas reducidas tanto al R como al RL indicando nuevamente que
algunos de los componentes las vías de transducción de señales del phyA y del phyB son
comunes y otros son específicos (Ahmad & Cashmore, 1996).
Posibles componentes bioquímicos de las vías de transducción de señales de los fitocromos
fueron identificados recientemente analizando el efecto de la microinyección de potenciales
moléculas mensajeras (conocidas por su acción en otros sistemas) sobre las respuestas mediadas
por este fotorreceptor. Este enfoque reveló la existencia de tres vías de transducción de señales
diferentes que operarian en forma posterior a la acción de las proteínas G (Bowler & Chua, 1994).
La primera requiere GMPc como intermediario y conduce a la activacion del gen CHS y a la
26

síntesis de antocianas. El calcio y la calmodulina forman parte de la segunda cascada, que
conduce a la activación del gen CAB y al desarrollo parcial de los cloroplastos. Por último, tanto
el calcio como el GMPc son necesarios en forma conjunta para la activación del gen FNR y la
maduración completa de los cloroplastos. La posibilidad que estas diferentes cascadas detransduccíón de señales estén asociadas a los distintos modos de acción identificados
fotobiológicamente ha sido recientemente planteada, pero no ha sido investigada aún (Mustilli &
Bowler., 1997).
En resúmen, las evidencias genéticas, moleculares y bioquímicas disponibles son consistentes
con la hipótesis que sostiene que las respuestas LFR desencadenadas por el phyB y las respuestas
desencadenadas por el phyA están mediadas por vías de transduccíón de señales diferentes,
aunque con algunos elementos en común.
Hasta ahora, sin embargo, no se ha explorado debidamente por medio de aproximaciones
genéticas, bioquímicas o moleculares la posibilidad de que el phyA actúe a través de dos vías
distintas de transduccíón de señales, que se corresponder-lancon las respuestas VLFR y HIR.
1.4. OBJETIVOS DE LA TESIS
Esta tesis tiene como objetivo central incrementar el conocimiento disponible sobre las
funciones sensoriales y los mecanismos de acción del phyA.
En la primer parte de la tesis se investigará el rol que cumple el phyA durante la desetiolación
y el posterior crecimiento y desarrollo de las plantas en condiciones naturales de radiación. La
hipótesis que se pone a prueba es que el phyA participa del ajuste del desarrollo de las plantas en
condiciones naturales en respuesta a cambios en la cantidad y en la duración de la luz incidente.
En la segunda parte de la tesis se intentará dilucidar los mecanismos a través de los cuales el
phyA transduce las señales lumínicas percibidas en cambios en el patrón de crecimiento y
desarrollo. La hipótesis es que el phyA actúa a través de dos vías de acción. Una, el VLFR, es
desencadenada por el Pfr del phyA y se satura con muy bajos flujos de luz. La otra, el HIR, es
desencadenada por un estado aún no identificado del fitocromo que se acumula en respuesta aaltas irradiancias de RL.

FUNCIONES SENSORIALES DEL FITOCROMO A (l)
ROL DEL FITOCROMO A EN LA DES-ETIOLACIÓN Y EN LA PERCEPCIÓN DE
PLANTAS VECINAS BAJO CONDICIONES NATURALES DE RADIACION
INTRODUCCION
Los fitocromos constituyen una familia de fotorreceptores, encontrándose en Arabidopsis
thalíana y en tomate, representada por 5 genes distintos (Clack et al., 1994; Sharrock & Quail,
1989, Pratt et al., 1997). Los más caracterizados de estos son el phyA y el phyB. La utilización de
mutantes con niveles reducidos (o nulos) de phyA o phyB y de plantas transgénicas que
sobreexpresan o subexpresan estos fotorreceptores, en combinación con tratamientos lumínicos
clásicos de laboratorio, ha permitido revelar los posibles roles fotoperceptivos de estos
fitocromos. Se sabe que la fuerte inhibición del alargamiento del tallo causada por RL continuo
en plántulas etioladas está mediada por el phyA, mientras que el phyB parece ser totalmente
innecesario para esta respuesta (Parks & Quail., 1993; Dehesh et al., 1993; McCormac et al.,
1993; Nagatani et al., 1993; Parks & Quail, 1993; Whitelam et al., 1993). En contraste, las
repuestas al R continuo en plantas etioladas, y las respuestas a pulsos de RL dados al final del día
en plantas des-etioladas parecen estar mayoritariamente mediadas por el phyB (López Juez et al.,
1992; McCormac et al., 1993; Nagatani et al., 1991; Parks & Quail, 1993).
Si bien la información obtenida con experimentos de laboratorio bajo condiciones controlados
constituye la base para nuestra comprensión de la función de los fotorreceptores, las diferencias
cualitativas y cuantitativas entre la iluminación artificial y la radiación natural no son triviales y,
en consecuencia, el significado ecológico de los resultados obtenidos en condiciones artificiales
de radiación es incierto. Por ejemplo, los mutantes de phyA presentan un fenotipo etiolado bajo
RL, pero tienen un fenotipo des-etiolado indistiguible de las plantas salvajes (WT) cuando se las
cultiva bajo luz blanca continua provista por tubos fluorescente (van Tuinen et al., 1995;
Whitelam et al., 1993). Este resultado ha llevado a varios investigadores a sugerir
apresuradamente que el phyA no cumplir-ía un rol importante durante la des-etiolación y el
posterior desarrollo de las plantas en condiciones naturales. Sin embargo, asumir que los
fenotipos de las plantas crecidas bajo luz blanca provista por tubos fluoresentes serán iguales a
los fenotipos de las mismas plantas cultivadas bajo radiación natural no es aconsejable. La luz
provista por los tubos fluorescentes tiene una alta proporción de luz azul y roja, pero, a diferencia
28

de la luz solar, carece casi por completo de RL. Siendo el RL la longitud de onda más eficiente
para activar el phyA, es posible suponer que en condiciones naturales de radiación (que poseen
una proporción de RL similar a la de R) la ausencia de phyA tenga un impacto importante en el
proceso de des-etiolación y en el posterior desarrollo de las plantas.
En esta parte de la tesis se investigará el rol del phyA en el control del desarrollo en plantas
cultivadas en condiciones naturales de radicación. En particular se analizará el rol del phyA en la
percepción de la transición oscuridad/luz que ocurre durante la emergencia de las plántulas así
como también el rol que cumple el phyA en la percepción de los cambios en el ambiente
luminoso producidos por la presencia de plantas vecinas. Para dilucidar el rol del phyA en dichos
procesos se compararán las respuestas a distintos estímulos lumínicos de plantas normales y
plantas mutantes de Arabidopsís thalíana y/o de plantas de papa (Solanum tuberosum L.)
transgénicas con niveles reducidos o incrementados de phyA.
RESULTADOS
Rol del phyA durante la emergencia de plantas en suelos cubiertos de vegetación
Uno de los cambios más marcados en el desarrollo de una planta que germina bajo la
superficie del suelo es la inhibición del crecimiento longitudinal del hipocotilo o del tallo que se
produce cuando la planta emerge y queda expuesta a la luz. Si bien las semillas de numerosas
especies requieren luz directa para gerrninar, un porcentaje variable del banco de semillas del
suelo germina aún en la oscuridad reinante varios centímetros por debajo de la superficie. Con
frecuencia, las plántulas emergerán en una superficie ya cubierta por otras plantas. La luz que
llega a la superficie de un suelo con abundante vegetación es muy escasa y, por lo tanto, las
plántulas deben poseer un mecanismo especial que les permita distinguir entre la oscuridad que
encuentran bajo tierra y la lluz escasa que encuentran bajo canopeos muy densos. Los tejidos
fotosíntéticos absorben principalmente la luz roja y azul, pero poco la RL, de modo que el
ambiente lumínico bajo canopeos muy densos está altamente enriquecido en ésta última longitud
de onda. El phyA es, en condiciones de laboratorio, el único fotorreceptor capaz de distinguir la
diferencia entre oscuridad y RL, y suponemos que este fotorreceptor sería el responsable de
inducir la des-etiolación de plántulas bajo canopeos muy densos.
Para investigar el proceso de des-etiolación en plantas que emergían en suelos ya cubiertos con
otras plantas analizamos el crecimiento del hipocotilo de plántulas de Arabidopsis cultivadas bajo
29
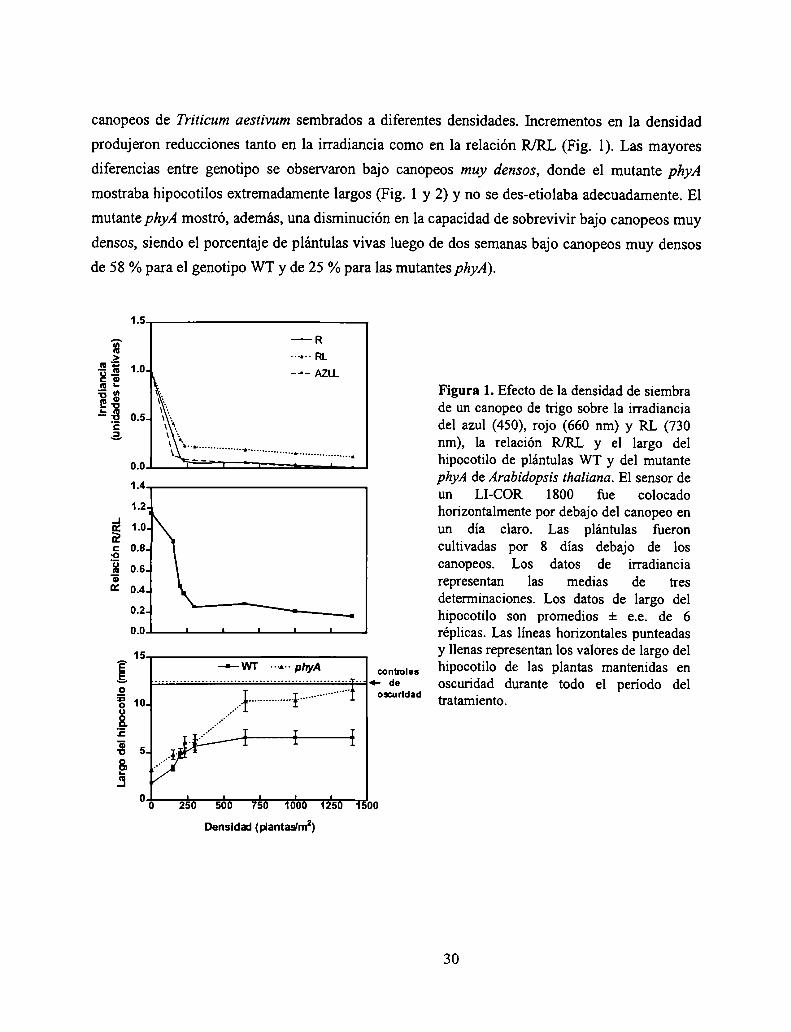
canopeos de Triticum aestivum sembrados a diferentes densidades. Incrementos en la densidad
produjeron reducciones tanto en la irradiancia como en la relación R/RL (Fig. 1). Las mayores
diferencias entre genotipo se observaron bajo canopeos muy densos, donde el mutante phyA
mostraba hipocotilos extremadamente largos (Fig. 1 y 2) y no se des-etiolaba adecuadamente. El
mutante phyA mostró, además, una disminución en la capacidad de sobrevivir bajo canopeos muy
densos, siendo el porcentaje de plántulas vivas luego de dos semanas bajo canopeos muy densos
de 58 % para el genotipo WT y de 25 % para las mutantes phyA).
1 n:
í.2
Lg a
g E Figura 1. Efecto de la densidad de siembra
E É de un canopeo de trigo sobre la irradiancia"E del azul (450), rojo (660 nm) y RL (7303 nm), la relación R/RL y el largo del
hipoootilo de plántulas WT y del mutante1 A phyA de Arabidopsis thalíana. El sensor de
un LI-COR 1800 fue colocado1'2 horizontalmente por debajo del canopeo en
.1 l ,
¡É 1.0- un dia claro. Las plantulas fueronc o,3_ cultivadas por 8 días debajo de losÉ 0_6_ canopeos. Los datos de irradiancia
g 04 representan las medias de tres' ' determinaciones. Los datos de largo del
0-2“ hipocotilo son promedios :l: e.e. de 60 n ' ' ' ' ' réplicas. Las líneas horizontales punteadasu y llenas representan los valores de largo del
É confio.“ hipocotilo de las plantas mantenidas enZ : de oscuridad durante todo el período delT3 “md” tratamiento.0.3SÉ
É!5
oo
Densidad (plantadmz)
30

Figura 2. Plántulas de Arabidopsis thalíana WT (derecha) y mutante phyA-1 (izquierda)emergiendo debajo de un canopeo muy denso de trigo.
Rol del phyA en el control del crecimiento de plántulas que emergen en suelos descubiertos
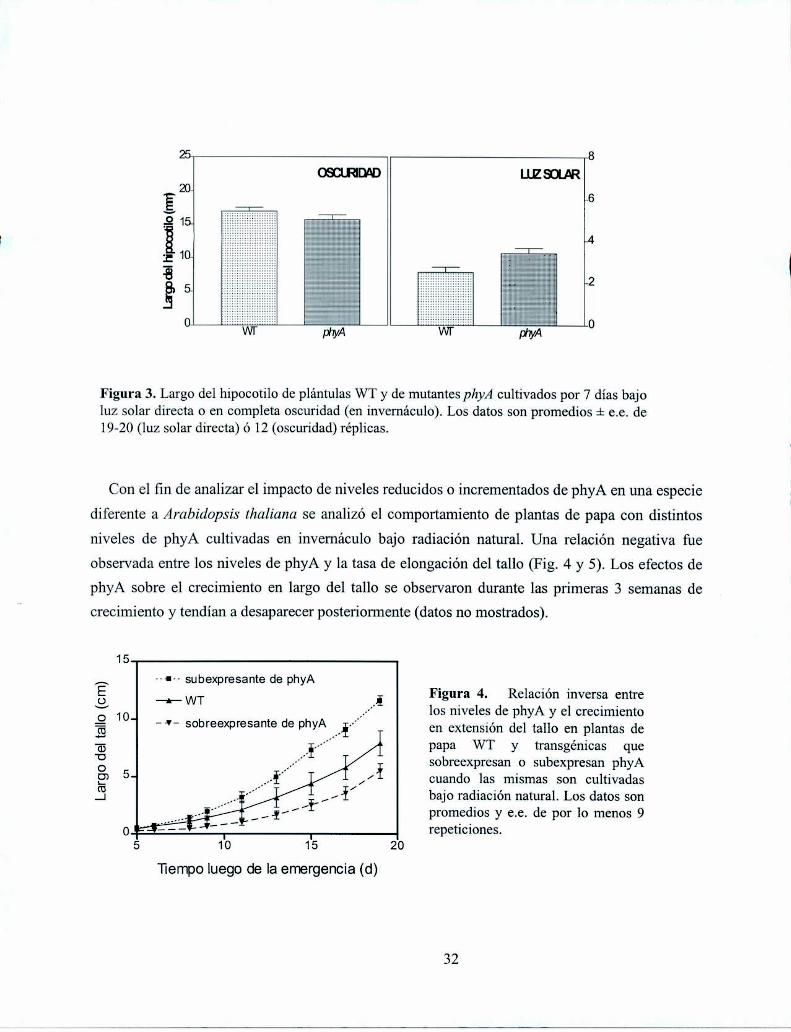
La luz solar plena tiene un componente importante de RL y, por lo tanto, es posible que
también el desarrollo de las plantas con niveles reducidos o incrementados de phyA se vea
alterado cuando éstas emergen en suelos descubiertos. En plantas de Arabídopsís thaliana el
alargamiento del hipocotilo fue significativamente reducido por la exposición directa a
fotoperíodos naturales, tanto en las plantas del genotipo salvaje como en la mutante deficiente en
phyA (Fig. 3). Sin embargo, si bien la inhibición del alargamiento del hipocotilo por la luz solar
fue muy importante en el mutante de phyA, la inhibición del alargamiento por la luz fue mayor en
el WT, observándose esto en el hecho que el largo del hipocotilo de las plantas WT es
significativamente mayor que el de las mutantes de phyA cuando ambas son cultivadas bajo luz
solar directa. Este resultado indica que el phyA participa en el control del crecimiento en plantas
que emergen en suelos descubiertos y sugiere que otros fotorreceptores (otros fitocromos y/o
criptocromos) también participan del control de este proceso.
31
B
a
pu
La‘gocblripooofilo(mr)
a
p
Figura 3. Largo del hipocotilo de plántulas WT y de mutantes phyA cultivados por 7 días bajoluz solar directa o en completa oscuridad (en invemáculo). Los datos son promedios i e.e. de19-20 (luz solar directa) ó 12 (oscuridad) réplicas.
Con el fin de analizar el impacto de niveles reducidos o incrementados de phyA en una especie
diferente a Arabidopsz's thaliana se analizó el comportamiento de plantas de papa con distintos
niveles de phyA cultivadas en invemáculo bajo radiación natural. Una relación negativa fue
observada entre los niveles de phyA y la tasa de elongación del tallo (Fig. 4 y 5). Los efectos de
phyA sobre el crecimiento en largo del tallo se observaron durante las primeras 3 semanas de
crecimiento y tendían a desaparecer posteriormente (datos no mostrados).
1::
--I-- subexpresante de phyA .WT j Figura 4. Relac1on inversa entre
10 los niveles de phyA y el crecimiento' -J'- sobreexpresantede phyA en extensión del tallo en plantas de
papa WT y transgénicas quesobreexpresan o subexpresan phyAcuando las mismas son cultivadasbajo radiación natural. Los datos sonpromedios y e.e. de por lo menos 9repeticiones.
Largodeltallo(cm)
Tiempo luego de la emergencia (d)
32

Figura 5. Plantas de papa WT y transgénicas con niveles incrementados (+phyA) o reducidos(-phyA) de phyA cultivadas durante dos semanas bajo luz solar directa.
El crecimiento radial del tallo, por otro lado, fue mayor en las plantas con niveles
incrementados de phyA y menor en las plantas con niveles disminuidos de phyA, comparadas
con las WT, en plantas cultivadas 1 mes bajo radiación natural (Fig. 6).
7K
5.0..
2.5
Diámetrodeltallo(mm)
subexpresante WT sooreexpresantede phyA de phyA
Figura 6. Relación directa entre los niveles de phyA y el crecimiento radial del tallo enplantas de papa WT y transgénicas que sobreexpresan o subexpresan phyA cuando lasmismas son cultivadas por un mes bajo radiación natural. Los datos son promedios y e.e. de8 repeticiones.
33
Rol del phyA en la percepción de las plantas vecinas
Percepción de las señales tempranas
La disminución de la relación R/RL de la luz propagada en forma horizontal que se produce
como consecuencia de la reflexión selectiva de RL por las hojas de las plantas (Ballaré et al.,
1987) constituye una señal temprana de la presencia de plantas vecinas, observable aún con
anterioridad a que las plantas se sombreen entre sí. Uno de los cambios más importantes que se
produce en las plantas en respuesta a la reducción de la relación R/RL provocada por la presencia
de plantas vecinas es la promoción del alargamiento del tallo. Con el objetivo de investigar si el
phyA cumple algún rol en esta respuesta se cultivaron plántulas de Arabídopsis WT y mutantes
phyA, ya sea en forma aislada de la vegetación circundante o hacia el norte de un canopeo de
Chenopodium quinoa (en la figura 6 se puede ver un esquema de un diseño experimental similar
al utilizado en este caso). Las plantas de Arabidopsis ubicadas hacia el norte del canopeo recibían
RL reflejado por este pero no eran sombreadas por el mismo. La relación R/RL de la luz
propagada en forma horizontal se redujo de 0.93 a 0.86, mientras que la irradiancia en el azul y en
el rojo no fue afectada. La presencia de vecinos promovió el alargamiento del hipocotilo en el
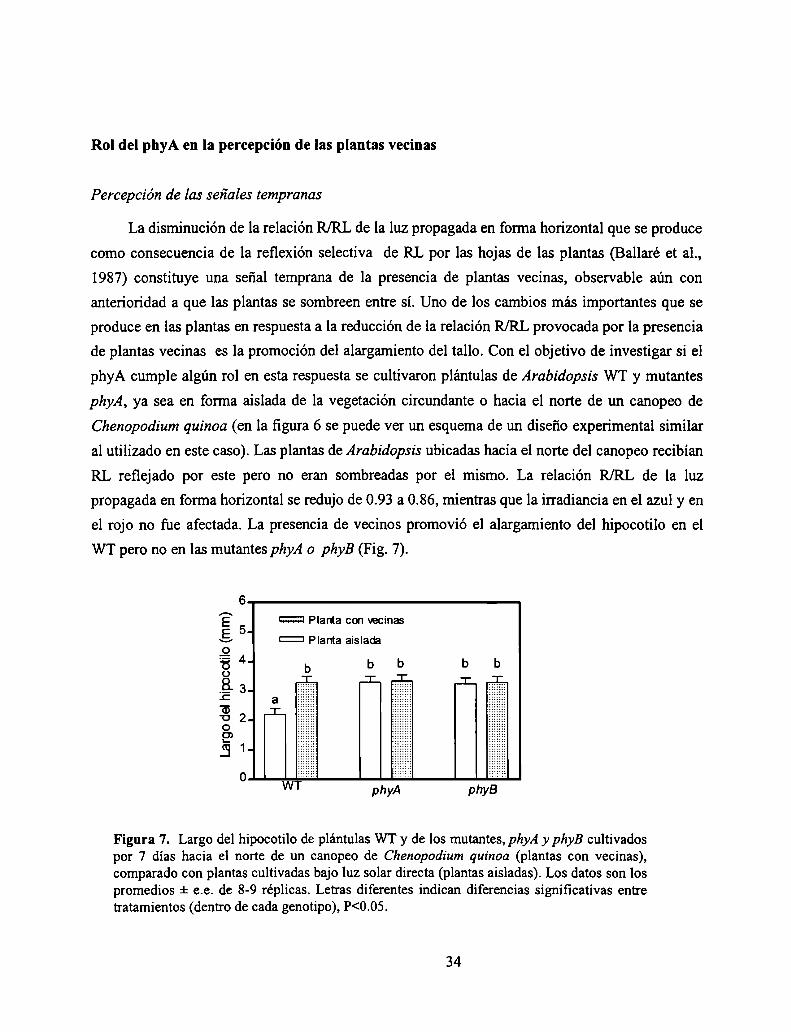
WT pero no en las mutantes phyA o phyB (Fig. 7).
m Plantaconvecinas=| Plantaaislada
Largodelhipocotilo(mm)
ph yA phyB
Figura 7. Largo del hipocotilo de plántulas WT y de los mutantes, phyAy phyB cultivadospor 7 días hacia el norte de un canopeo de Chenopodium quinoa (plantas con vecinas),comparado con plantas cultivadas bajo luz solar directa (plantas aisladas). Los datos son lospromedios :t e.e. de 8-9 réplicas. Letras diferentes indican diferencias significativas entretratamientos (dentro de cada genotipo), P<0.05.
34

Adicionalmente, se analizó la respuesta de plántulas de Arabídopsis a reducciones artificiales
de la relación R/RL de la luz propagada horizontalmente. La adición lateral de RL (140 umoles
m‘2s") durante el fotoperíodo redujo el R/RL de la luz propagada horizontalmente de 0.93 a 0.83.
Esta reducción de la relación R/RL, a1igual que la provocada por el RL reflejado por vecinos no
sombreantes (Fig. 7), promovió el alargamiento en las plántulas WT pero no en el mutante phyB
(Fig. 8). Por el contrario, el mutante phyA que no había mostrado respuesta alguna a los vecinos
que reflejaban RL sin sombrear respondió normalmente a la reducción simulada del R/RL
provocada por la fiiente artificial de RL (Fig. 8).
PLANTA AISLADA
L Figura 8. Promocióndel alargamientodelPLANTACONRLADICIONAL hipocotilo de plantas de Arabidopsis thaliana
provocada por bajas relaciones R/RL establecidasdurante todo el día por medio de una fuenteartificial de RL. Plantas WT, mutantes de phyA ymutantes de phyB fueron cultivadas por 7 días bajoluz solar directa o bajo luz solar más RL provistoen forma horizontal por una fuente artificial de luz.La promoción fue calculada para cada genotipocomo la diferencia entre el largo del hipocotilo deplantas cultivadas con RL adicional menos el largodel hipocotilo de plantas cultivadas sin RLadicional. *=promoción significativa (P<0.05), ns=no significativa. Los datos son promedios i e.e. de16 réplicas.
Promocióndel
alarganientoporelRL
adicional(¡mi
La discrepancia observada entre los resultados obtenidos con plantas vecinas y con reducciones
artificiales de la relación R/RL podría deberse a que si bien el valor de la reducción en la relación
R/RL fue similar en ambos casos cuando se midió durante el mediodía de un día soleado, la
disminución de la relación R/RL con el RL adicionado artificialmente file constante, mientras
que aquella provocada por la presencia de plantas vecinas seguramente varía con la hora del día y
las condiciones climáticas. El hecho que el mutante phyB no responda en ninguno de los dos
casos a la reducción de la relación R/RL sugiere que este fotorreceptor participa directamente de
35
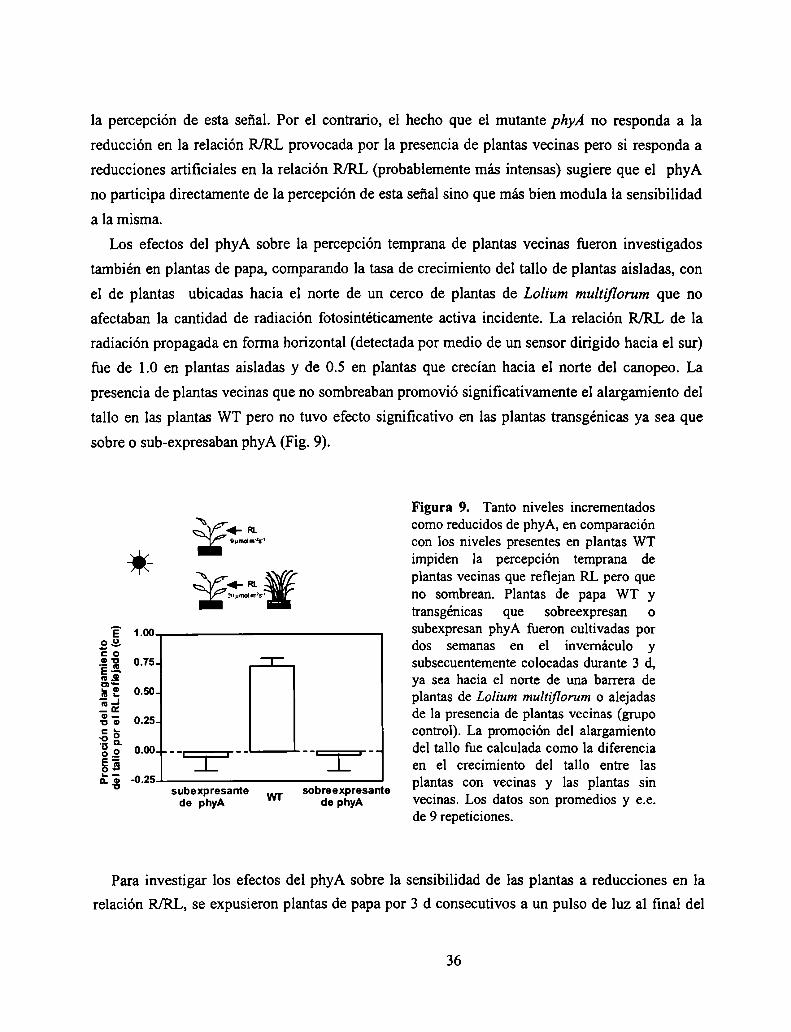
la percepción de esta señal. Por el contrario, el hecho que el mutante phyA no responda a la
reducción en la relación R/RL provocada por la presencia de plantas vecinas pero si responda a
reducciones artificiales en la relación R/RL (probablemente más intensas) sugiere que el phyA
no participa directamente de la percepción de esta señal sino que más bien modula la sensibilidad
a la misma.
Los efectos del phyA sobre la percepción temprana de plantas vecinas fueron investigados
también en plantas de papa, comparando la tasa de crecimiento del tallo de plantas aisladas, con
el de plantas ubicadas hacia el norte de un cerco de plantas de Lolium multíflorum que noafectaban la cantidad de radiación fotosíntéticamente activa incidente. La relación R/RL de la
radiación propagada en forma horizontal (detectada por medio de un sensor dirigido hacia el sur)
fue de 1.0 en plantas aisladas y de 0.5 en plantas que crecían hacia el norte del canopeo. La
presencia de plantas vecinas que no sombreaban promovió significativamente el alargamiento del
tallo en las plantas WT pero no tuvo efecto significativo en las plantas transgénicas ya sea que
sobre o sub-expresaban phyA (Fig. 9).
Figura 9. Tanto niveles incrementados\ <_ RL como reducidos de phyA, en comparación
ÏDuMm‘" con los niveles presentes en plantas WT_*_ impiden la percepción temprana de
\ <_ RLy plantasvecinasque reflejan RL pero quemmm' no sombrean. Plantas de papa WT ytransgénicas que sobreexpresan o
'g 1 nn subexpresan phyA fueron cultivadas por.37‘: dos semanas en el invemáculo y
0.75- subsecuentementecolocadas durante 3 d,38 ya sea hacia el norte de una barrera de¡Las 0'50“ plantas de Lolium multiflorum o alejadas3'; 0.2L de la presencia de plantas vecinas (grupo3 5 control). La promoción del alargamiento'g 3 n nn 7 del tallo fue calculada como la diferenciaEE I en el crecimiento del tallo entre lasE é .n al: plantas con vecinas y las plantas sin
subexpresante WT sobreexpresante _ .de phyA de phyA vecmas. Los datos son promedios y e.e.de 9 repeticiones.
Para investigar los efectos del phyA sobre la sensibilidad de las plantas a reducciones en la
relación R/RL, se expusieron plantas de papa por 3 d consecutivos a un pulso de luz al final del
36
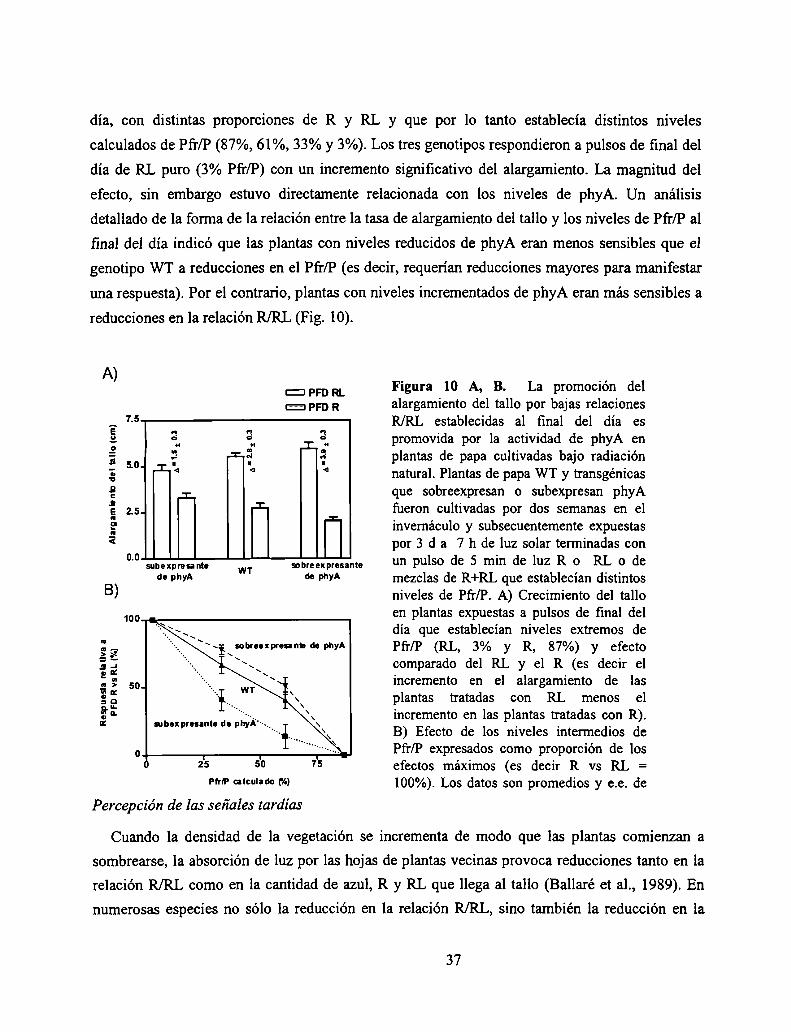
día, con distintas proporciones de R y RL y que por lo tanto establecía distintos niveles
calculados de Pfr/P (87%, 61%, 33% y 3%). Los tres genotipos respondieron a pulsos de final del
día de RL puro (3% Pfr/P) con un incremento significativo del alargamiento. La magnitud del
efecto, sin embargo estuvo directamente relacionada con los niveles de phyA. Un análisis
detallado de la forma de la relación entre la tasa de alargamiento del tallo y los niveles de Pfr/P al
final del día indicó que las plantas con niveles reducidos de phyA eran menos sensibles que el
genotipo WT a reducciones en el Pfr/P (es decir, requerían reducciones mayores para manifestar
una reSpuesta). Por el contrario, plantas con niveles incrementados de phyA eran más sensibles a
reducciones en la relación R/RL (Fig. 10).
A) ¡:¡PFDRL Figura 10 A, B. La promoción delpFD R alargamientodel tallo por bajas relaciones
E ""‘ R/RL establecidas al final del día esa 3“ promovida por la actividad de phyA en
É Mi ¿"3 ¿É E plantas de papa cultivadas bajo radiacióne ‘1 " “ natural. Plantas de papa WT y transgémcasg que sobreexpresan o subexpresan phyAÉ ¿5- fueron cultivadas por dos semanas en elÉ invemáculo y subsecuentemente expuestas7‘ por 3 d a 7 h de luz solar terminadas con
n subexpresante WT sobreexpresante un pUIso de 5 mín de luz R o dede phyA 6° WA mezclas de R+RL que establecían distintos
B) niveles de Pfr/P. A) Crecimiento del tallomn _ en plantas expuestas a pulsos de final del
" día que establecían niveles extremos deEG ‘xífobroexprennh dePhyA Pfr/P (RL, 3% y R, 87%) y efectoÉ; \ comparado del RL y el R (es decir elE g, 50- incremento en el alargamiento de las
É: WT plantas tratadas con RL menos elSÉ albums.” “¿han incrementoen las plantas tratadas con R).
B) Efecto de los niveles intermediosden ...............__ Pfr/P expresados como proporción de los
2'5 5'0 7.5 efectos máximos (es decir R vs RL =PMP“Mi” W 100%). Los datos son promedios y e.e. de
Percepción de las señales tardías
Cuando la densidad de la vegetación se incrementa de modo que las plantas comienzan a
sombrearse, la absorción de luz por las hojas de plantas vecinas provoca reducciones tanto en la
relación R/RL como en la cantidad de azul, R y RL que llega al tallo (Ballaré et al., 1989). En
numerosas especies no sólo la reducción en la relación R/RL, sino también la reducción en la
37
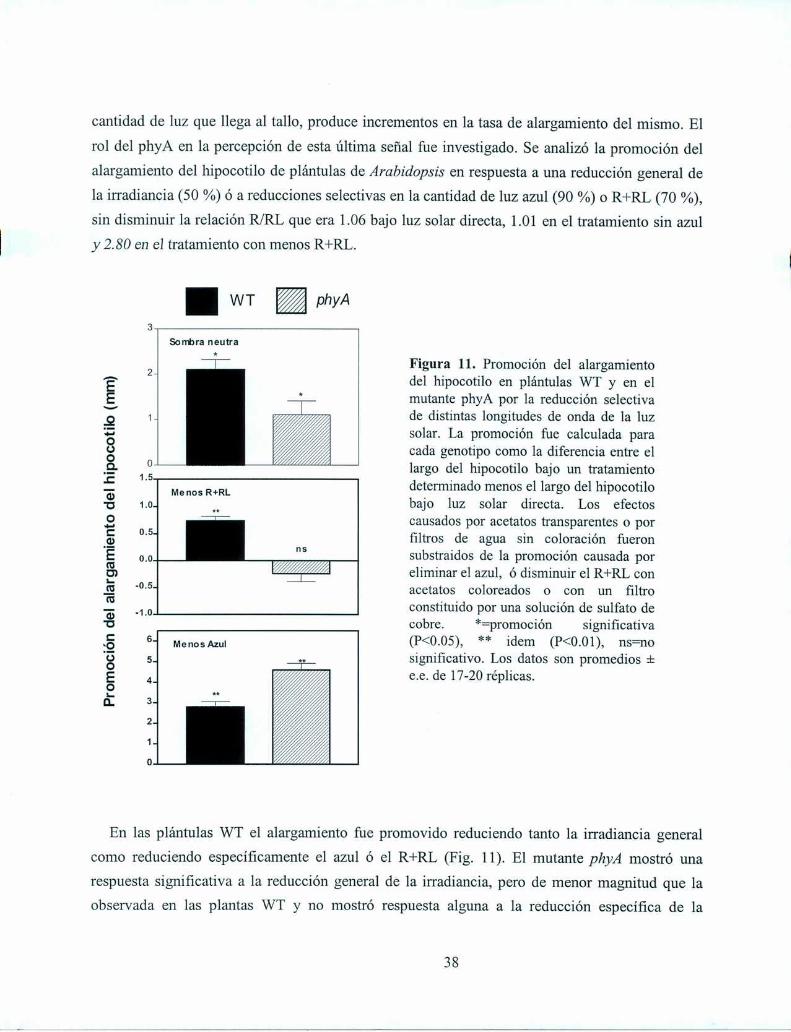
cantidad de luz que llega al tallo, produce incrementos en la tasa de alargamiento del mismo. El
rol del phyA en la percepción de esta última señal fue investigado. Se analizó la promoción del
alargamiento del hipocotilo de plántulas de Arabidopsís en respuesta a una reducción general de
la irradiancia (50 %) ó a reducciones selectivas en la cantidad de luz azul (90 %) o R+RL (70 %),
sin disminuir la relación R/RL que era 1.06 bajo luz solar directa, 1.01 en el tratamiento sin azul
y 2.80 en el tratamiento con menos R+RL.
- WT phyA
Sorrbra neutra'k
Figura ll. Promoción del alargamientodel hipocotilo en plántulas WT y en elmutante phyA por la reducción selectivade distintas longitudes de onda de la luzsolar. La promoción fue calculada paracada genotipo como la diferencia entre ellargo del hipocotilo bajo un tratamiento
MenosR+RL determinado menos el largo del hipocotilo1.o_ fl bajo luz solar directa. Los efectos
causados por acetatos transparentes o porfiltros de agua sin coloración fueronsubstraidos de la promoción causada poreliminar el azul, ó disminuir el R+RL con
0-5- acetatos coloreados o con un filtro_1n constituido por una solución de sulfato de
cobre. *=promoción significativa6- MenosAZu. (P<0.05), ** idem (P<0.01), ns=no
significativo. Los datos son promedios d:e.e. de 17-20 réplicas.
Promocióndelalargamientodelhipocotilo(mm)
En las plántulas WT el alargamiento fue promovido reduciendo tanto la irradiancia general
como reduciendo específicamente el azul ó el R+RL (Fig. ll). El mutante phyA mostró una
respuesta significativa a la reducción general de la irradiancia, pero de menor magnitud que la
observada en las plantas WT y no mostró respuesta alguna a la reducción específica de la
38
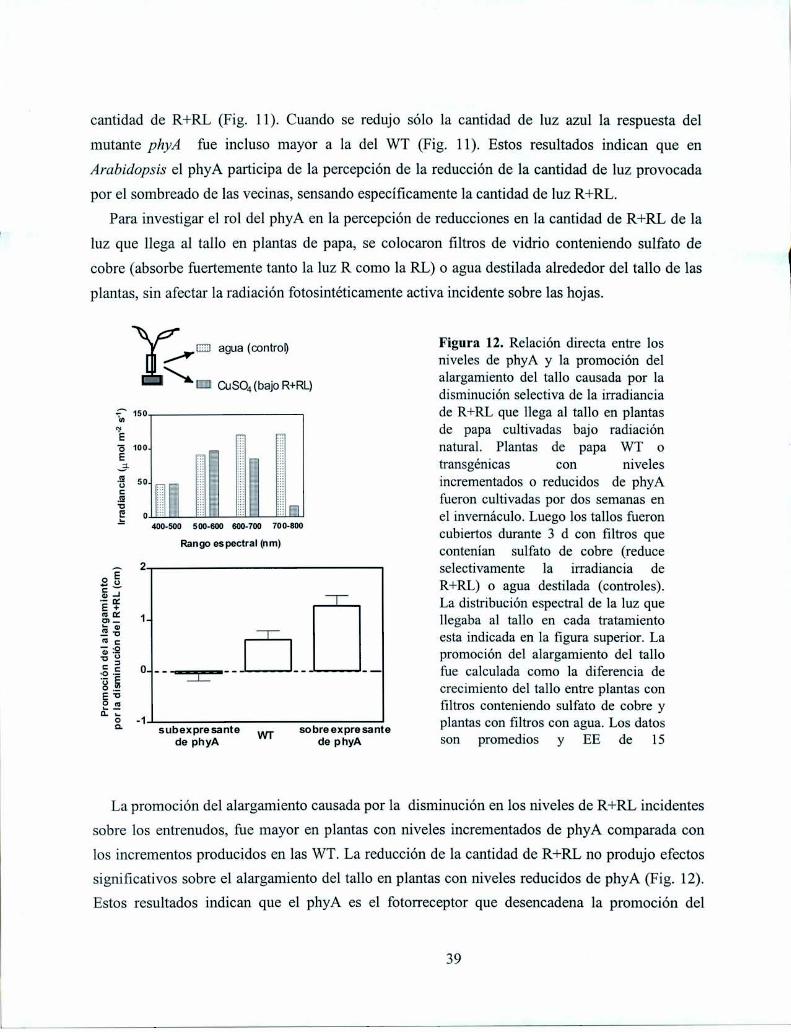
cantidad de R+RL (Fig. ll). Cuando se redujo sólo la cantidad de luz azul la respuesta del
mutante phyA fue incluso mayor a la del WT (Fig. ll). Estos resultados indican que en
Arabidopsís el phyA participa de la percepción de la reducción de la cantidad de luz provocada
por el sombreado de las vecinas, sensando específicamente la cantidad de luz R+RL.
Para investigar el rol del phyA en la percepción de reducciones en la cantidad de R+RL de la
luz que llega al tallo en plantas de papa, se colocaron filtros de vidrio conteniendo sulfato de
cobre (absorbe fuertemente tanto 1aluz R como la RL) o agua destilada alrededor del tallo de las
plantas, sin afectar la radiación fotosíntéticamente activa incidente sobre las hojas.
agua (control) Figura 12. Relación directa entre los/ nivelesde phyA y la promocióndel\
alargamiento del tallo causada por ladisminución selectiva de la irradianciade R+RL que llega al tallo en plantasde papa cultivadas bajo radiaciónnatural. Plantas de papa WT otransgénicas con nivelesincrementados o reducidos de phyAfueron cultivadas por dos semanas enel invemáculo. Luego los tallos fueroncubiertos durante 3 d con filtros quecontenían sulfato de cobre (reduceselectivamente la irradiancia de
R+RL) o agua destilada (controles).T La distribuciónespectral de la luz que
llegaba al tallo en cada tratamiento
IL] esta indicadaen la figurasuperior.Lapromoción del alargamiento del tallofue calculada como la diferencia de
CuSO4(bajo R+RL)
Irradiancia(umolm'2s")o sz :.. ÉÉÉ
400-500 500-600 600-700 100-800
Rango es pectral (nm)
0
..\ I
0I l 0 l lPromocióndelalargamiento
porladisminucióndelR+RL(cm)
_l_crecimiento del tallo entre plantas confiltros conteniendo sulfato de cobre y
-‘ plantas con filtros con agua. Los datossubexpresante wr sobreexpresante .
de phyA de phyA son promedlos y EE de 15
La promoción del alargamiento causada por la disminución en los niveles de R+RL incidentes
sobre los entrenudos, fue mayor en plantas con niveles incrementados de phyA comparada con
los incrementos producidos en las WT. La reducción de la cantidad de R+RL no produjo efectos
significativos sobre el alargamiento del tallo en plantas con niveles reducidos de phyA (Fig. 12).
Estos resultados indican que el phyA es el fotorreceptor que desencadena la promoción del
39

alargamiento del tallo de las plantas en respuesta a la reducción de la cantidad de luz R y RL que
se produce por el sombreado entre las plantas.
DISCUSIÓN
El análisis del desarrollo de plantas de Arabidopsis thaliana y de plantas de papa con distintos
niveles de phyA reveló que el phyA contribuye a controlar el crecimiento en condiciones
naturales de radiación durante distintas etapas del ciclo de vida de las plantas y en respuesta a
diferentes señales ambientales.
1. El phyA sería el principal fotorreceptor responsable de desencadenar la des-etiolacíón de
plantas que emergen bajo canopeos muydensos.
Mientras que en las plantas de Arabidopsis thalíana del genotipo salvaje (Fig. 1), y en los
mutantes phyB y cry] (datos no mostrados), la escasa luz presente bajo canopeos muy densos fue
capaz de inhibir el crecimiento del hipocotilo comparado con plantas cultivadas en oscuridad, el
crecimiento en extensión del hipocotilo fue virtualmente inalterado por la luz presente bajo
canopeos densos en el mutante phyA (Fig. 1). La idea que el phyA podria ser importante para la
des-etiolación en ambientes naturalmente enriquecidos en RL, como el presente bajo canopeos
muy densos, había sido propuesta con anterioridad por otros investigadores basándose en el
hecho que en condiciones controladas este fotorreceptor mediaba las respuestas al RL continuo.
Sin embargo, esta idea era considerada con cautela debido a la falta de evidencia experimental
(Parks & Quail, 1993; Smith & Whitelam, 1990). En este trabajo no sólo mostramos que el phyA
cumple un rol fundamental en el control de la des-etiolación bajo canopeos densos, sino que
observamos que la incapacidad del mutante phyA de des-etiolarse normalmente en esas
condiciones redujo la tasa de supervivencia de ese genotipo, comparado al salvaje. Este resultado
indica que la acción del phyA constituye efectivamente una ventaja adaptativa para aquellas
plántulas que emergen bajo canopeos verdes muy densos.
2. El phyA también afecta el crecimiento de plantas que emergen bajo luz solar directa.
Esto se deduce de observar que en Arabidopsis thalíana, el mutante phyA fué más alto que el
WT incluso en plántulas expuestas a radiación solar directa (aunque la diferencia fué
sustancialmente menor que bajo canopeos muy densos) (Fig. 3). Resultados similares se
obtuvieron comparando el desarrollo de plantas de papa salvajes y plantas transgénicas con
niveles elevados (sobreexpresantes) o reducidos (subexpresantes) de phyA (Fig. 4, S y 6). Cuando
40

las plantas de papa fueron cultivadas bajo radiación natural el largo del tallo se correlacionó
negativamente, y el radio positivamente, con los niveles de phyA de cada genotipo.
Los resultados obtenidos con plantas transgénicas, no obstante, deben ser analizados con
cautela. Los mismos podrían ser resultado de variación somaclonal ocurrida durante el proceso de
transformación y no específicamente por los efectos de alterar los niveles de phyA. Sin embargo,
el hecho que en la mayoría de las respuestas analizadas la sobreexpresión y la subexpresíón de
phyA tuvieron efectos opuestos y el WT mostró respuestas intermedias sugiere que los efectos
estuvieron específicamente relacionados con los niveles de phyA, siendo improbable que este
patrón de respuestas haya resultado de variación somaclonal. Por otro lado, los resultados podrían
deberse a efectos no deseados de la transgenesis de PH YAsobre los niveles de otros fitocromos.
Sin embargo, los niveles de phyB no están afectados en las plantas transgénicas que
sobreexpresan phyA ni en las transgénicas con el gen de phyA en antisentido (Heyer et al., 1995).
Aunque la posibilidad de que las plantas transgénicas con el gen de phyA en antisentido tengan
afectados los niveles de miembros aún no identificados de la familia de los fitocromos distintos
de phyB (probablemente presentes en la papa), el grado de homología entre miembros conocidos
de la familia de los fitocromos en otras especies hace que esto sea improbable. En resumen, la
correlación entre los niveles de phyA y la magnitud de las respuestas de los distintos genotipos
favorece la interpretación que los fenotipos se debieron a efectos específicos de la transgenesis
sobre los niveles de phyA y no a efectos indirectos sobre los niveles de otros fitocromos.
El hecho que el phyA no afecte el crecimiento de plantas cultivadas bajo luz blanca continua
provista por tubos fluorescentes pero si afecte el crecimiento de plantas cultivadas bajo luz solar
directa puede tener dos orígenes. Por un lado, en condiciones naturales las plantas están expuestas
a períodos altemados de luz y oscuridad. Debido a que el phyA es inestable en la forma Pfr, los
niveles de phyA son constantemente bajos en plantas cultivadas en luz continua. En cambio, en
plantas cultivadas con fotoperíodos el phyA puede acumularse nuevamente durante la noche,
haciendo que en estas plantas los niveles de phyA sean más altos que en las primeras. Por otro
lado, la luz natural posee una proporción importante de RL mientras que los tubos fluorescentes
carecen casi por completo de este componente de la luz. Siendo el RL la luz que más
eficientemente activa al phyA, es posible que las diferencias entre los experimentos realizados
con tubos fluorescentes y los experimentos realizados bajo luz natural se deban a las diferencias
en contenido de RL de ambas fuentes lumínicas. Al parecer, ambas posibilidades parecen ser
ciertas ya que se ha observado que el mutante phyA tiene hipocotilos más largos que el WT
cuando es cultivado en condiciones controladas bajo ciclos de luz-oscuridad (utilizando tubos
41

fluorescentes) y también se ha observado que el mutante phyA de Arabidospsis y las plantas de
papa transgénicas con niveles reducidos de phyA son más altas que sus respectivos genotipos
salvajes cuando son cultivadas bajo luz blanca continua provista por tubos fluorescente
suplementada con RL (Johnson et al., 1994).
3. ElphyA participa en la detección temprana de vecinos.
Con anterioridad a que ocurra sombreado mutuo entre las plantas de un canopeo en desarrollo,
se produce una reducción de la relación R/RL de la luz que llega al tallo como consecuencia de la
reflexión selectiva del RL en los tejidos verdes de las plantas vecinas (Ballaré et al., 1987). La
presencia de plantas vecinas que reflejan RL sin sombrear promovió el crecimiento del hipocotilo
en plantas del genotipo salvaje de Arabidopsís thaliana, pero no en las mutantes phyA o phyB
(Fig. 7). La falta de respuesta en el mutante phyB al RL reflejado está de acuerdo con
observaciones previas que reportan respuestas nulas (Ballaré et al., 1991) o reducidas (Smith et
al., 1992; Whitelam & Smith, 1991) a la reducción en la relación R/RL en mutantes de pepino y
Arabidopsis deficientes en phyB. Aunque las plantas mutantes de phyA no respondan a las
reducción de la relación R/RL provocada por la presencia de plantas vecinas, es poco probable
que el phyA tenga un rol directo en la percepción de esta señal. Esto se debe a que bajo radiación
continua la concentración del phyA en su forma activa (Pfr) no se correlaciona bien con la
relación R/RL (Smith, 1981). En lugar de participar directamente de la percepción de la relación
R/RL, los resultados presentados aquí sugieren que el phyA actuar-íamodulando la sensibilidad
de las plantas a las reducciones en la relación R/RL percibidas por el phyB. Esta idea se basa en
que en este trabajo se observó que el mutante phyA, pero no el phyB, sí respondió a reducciones
obtenidas en forma artificial de la relación R/RL que proveían señales más intensas (ya sea en
téminos geométricos o en magnitud) que los cambios sutiles causados por la presencia de plantas
vecinas que reflejan RL sin sombrear (Fig. 8). Esta idea además es consistente con trabajos
previos que muestran que la disminución de la radiación absorbida por el fitocromo
(posiblemente percibida por el phyA) reduce el grado de respuesta a la reducción de la relación
R/RL (Casal & Kendrik, 1993), y que las plantas des-etioladas del mutante aurea de tomate, que
tienen phyB espectralmente activo pero carecen de phyA espectralmente activo (Shar-ma et al.,
1993), también muestra una sensibilidad menor hacia la reducción de la relación R/RL (Casal &
Kendrik, 1993).
Resultados similares a los obtenidos en Arabidopsis fueron observados en plantas de papa.
Comparadas con las plantas WT, las plantas trangénicas que sobre o subexpresan phyA no
42

mostraron incrementos en el alargamiento del tallo en respuesta a la presencia de plantas vecinas
que no producen sombra (Fig. 9). La falta de respuesta a la presencia de plantas vecinas que
reflejan RL tuvo, probablemente, un orígen distintos en las plantas que sobre y subexpresaban
phyA. Tanto las plantas de papa (Fig. 9), como las plantas de tabaco que sobreexpresan phyA
(Casal & Sánchez, 1994) parecen interpretar los incrementos en RL debido a la reflexión en los
tejidos verdes de las plantas vecinas como más luz, produciendo una respuesta HIR que
contrabalancea el efecto de la reducción en la relación R/RL (Casal et al., 1997). La falta de
respuesta al RL reflejado por las plantas vecinas no-sombreantes en las plantas de papa con el gen
PH YAantisentido (Fig. 9) puede ser explicada como consecuencia de una menor sensibilidad a
las reducciones en la relación R/RL percibida por los fitocromos estables. A favor de esta
interpretación se haya el hecho de haber observado que la magnitud de la respuesta al RL de final
del día (Fig. 10A) y la sensibilidad en las respuestas a reducciones en los niveles de Pfr/P (Fig.
lOB) estaban directamente relacionadas con los niveles de phyA en las plantas de papa. La
sensibilidad a reducciones en la relación R/RL al final del día también es alta en plantas de tabaco
y plantas de tomate que sobreexpresan phyA (Casal et al., 1995) y menor en mutantes de phyA de
Arabidopsis (Casal, 1996), comparadas con las respuestas de sus líneas isogénicas WT. En
resumen, los resultados obtenidos aquí más las obsevaciones de otros trabajos son consistentes
con un efecto del phyA sobre la sensibilidad frente a los cambios en la relación R/RL más que un
rol directo en la percepción de la relación R/RL.
4. El phyA es importante en la detección de cambios en Ia irradíancia provocados por el
sombreado de plantas vecinas.
A medida que el desarrollo de un canopeo progresa, el ambiente luminoso se ve afectado no
sólo por la reflexión de la luz sino también por la absorción de la misma en las hojas de las
plantas vecinas, es decir por el sombreado. La sombra de los canopeos reduce tanto la cantidad de
luz azul como la cantidad de radiación R+RL. Existen evidencias que indican que las plantas no
sólo perciben y responden a la presencia de plantas vecinas a través de los cambios en la relación
R/RL de la luz sino también a través de los cambios en la cantidad de la misma. En plantas de
Sínapis alba (Ballaré et al., 1991), Datura ferox (Ballaré et al., 1991), tomate (Casal, 1994) y
tabaco (Casal & Sánchez, 1994), reducciones selectivas en la cantidad azul y de R+RL que llega
al tallo de plantas cultivadas bajo radiación natural, promueven significativamente el crecimiento
del mismo. En este trabajo observamos que en Arabídopsis, la reducción de la cantidad de luz sin
afectar la relación R/RL promovió el alargamiento del hipocotilo de las plántulas. La respuesta a
43

reducciones en la cantidad de luz fue significativamente mayor en plantas WT que en el mutante
phyA indicando que el phyA es uno de los fotorreceptores involucrados en la percepción de esta
señal (Fig. 11). El phyA es capaz de percibir tanto la cantidad de R+RL como la cantidad de azul.
Dado que la sombra provocada por las plantas vecinas reduce simultáneamente la luz azul y el
R+RL, analizamos el efecto de estas señales por separado. La reducción del azul promovió
significativamente el alargamiento en las plantas WT y en el mutante phyA (Fig. ll) pero no en el
mutante cry] (datos no mostrados). La disminución del R+RL promovió el alargamiento en las
plantas WT pero no tuvo un efecto significativo en el mutante phyA. Estas observaciones son
consistentes con roles específicos para el cryl y el phyA en la percepción de los cambios en azul
y en R+RL que se producen bajo la sombra de un canopeo.
El rol del phyA en la percepción de reducciones en la cantidad de R+RL que llega al tallo
también fue estudiado en plantas de papa. La reducción de la cantidad de R+RL promovió
significativamente el crecimiento del tallo en las plantas WT. Comparado con las plantas WT, la
promoción se ve incrementada en plantas que sobreexpresan phyA y está ausente en plantas
transgénicas de papa con niveles reducidos de phyA (Fig. 12). Esto sugiere fuertemente que el
phyA participa de la percepción de la reducción de la cantidad de luz que llega a los tallos como
consecuencia de la presencia de plantas vecinas, aún antes que ocurra sombreado mutuo entre las
hojas de las plantas.
MATERIALES Y MÉTODOS
Material Vegetal
Las semillas de Arabidopsis thaliana (L.) Heynh, cultivar Landsberg erecta, y la de los mutantes
phyA-I y phyB-I se obtuvieron haciendo crecer a las plantas bajo luz blanca fluorescente. Las
muestras originales de las semillas WT yphyB-I fueron provistas por los profesores Koomneef y
Kendrick (Universidad de Wageningen, Holanda) y la muestra original de phyA-1 por el Doctor
Whitelam (Universidad de Leicester, Inglaterra).
Las plantas de papa (Solanum tuberosum L.) utilizadas en estos experimentos, tanto las salvajes
(WT) de la variedad Desiree, como las transgénícas de la misma variedad con niveles elevados
(sobreexpresantes) o reducidos (subexpresantes) de phyA fueron previamente descriptas por
Heyer et al. 1995.

Condiciones de cultivo
Todas las plantas fueron cultivadas en un invemáculo con temperatura semicontrolada,
perteneciente a la cátedra de Fisiología Vegetal de la Facultad de Agronomía, UBA. Para los
experimentos realizados con Arabidopsis thaliana, se sembraron 3 semillas en cada maceta
plástica (2.5 cm de diámetro y 3.5 cm de altura) llena con tierra humedecida a capacidad de
campo. Los potes fueron incubados en oscuridad a 7°C por 3 días, recibiendo luego 20 minutos
de rojo seguidos de un día en oscuridad a 25°C antes de ser transferidos a los distintos
tratamientos. Plántulas de l día fueron transferidas al invemáculo. El fotoperíodo fue de 10-12
horas. La radiación fotosintéticamente activa al mediodía de días claros era de 750 umol. m'2.s".
Los controles en oscuridad fueron crecidos en una caja negra (48 dm’) cubierta con papel
aluminio. Para los experimentos realizados con las papas, tubérculos de aproximadamente 2 cm
de diámetros fueron sembrados en macetas plásticas de 650 cm3 que contenían una mezcla de
arena y tierra. La tierra fue llevada diariamente a capacidad de campo. Plantas de 1 mes de edad
fueron utilizadas para las mediciones del diámetro del tallo y los niveles de antocianas y clorofila.
Para los experimentos donde el ambiente lumínico fue modificado, las plantas fireron cultivadas
por dos semanas en el invemáculo y luego fueron expuestas a los distintos tratamientos por 3 d.
En mediodías dCSpejados, la cantidad de radiación fotosintéticamente activa era superior a los
1000 umoles de fotones rn'2s". Los fotoperíodos variaron desde lO a 13 h d".
Mediciones de crecimiento
En Arabz'dopsis thaliana, la altura del hipocotilo sobre el nivel del suelo fue medida diariamente
con regla. Para los controles en oscuridad sólo se tomaron medidas al final del experimento. Cada
experimento fiJé realizado un mínimo de 3 veces.
En las papas, el largo del tallo fue medido con una regla desde el suelo (o una marca hecha con
tinta sobre el tallo cerca del suelo) hasta la inserción del pecíolo de la hoja más joven. Los
incrementos en el alargamiento del tallo fueron estimados como la diferencia entre el largo del
tallo en un momento determinado y el largo del tallo al comienzo del tratamiento. Las
determinaciones del diámetro del tallo fueron realizadas con un calibre, 10 cm por debajo de la
inserción de la hoja más joven.
45

Tratamientos lumínicos
Canopeos naturales de distintas densidades:
Se obtuvieron canopeos naturales sembrando Triticum aestívum a distintas densidades en
recipientes cerámicos de 800 cm2 y 10 cm de altura. Las macetas conteniendo semillas de
Arabidopsís fueron almacenadas 3 d a 7 °C, expuestas a 5 min de R e incubadas en oscuridad
para inducir la germinación. Al día siguiente las macetas fueron llevadas al invemáculo y
colocadas debajo de los distintos canopeos.
Plantas vecinas que no sombrean:
Para investigar el efecto que tenían plantas vecinas que no sombrean sobre el crecimiento del
hipocotilo de plantas de Arabidopsís thaliana se utilizaron dos hileras de plantas de
Chenopodium quínoa (de 40 cm de altura) cultivadas en recipientes plásticos de 400 cm’. La
mitad de las plantas de Arabidopsis thaliana fue colocada 5 cm al norte de la barrera de plantas
de quinoa y la otra mitad permaneció alejada de la vegetación circundante. El canopeo de quinoa
no interfería con la intercepción de radiación solar directa que llegaba a las plantas de
Arabidopsís. Para estudiar el efecto que tenían plantas vecinas que no sombrean sobre el
crecimiento del tallo de las plantas de papa se utilizó un diseño experimental similar al utilizado
para las plantas de Arabidopsis thaliana, pero en lugar de utilizar plantas de Chenopodium
quinoa como vecinas se utilizaron plantas de Lolium multzflorum (de 45 cm de altura). En ambos
experimentos la relación R/RL y la cantidad de RL (entre 725 y 735 nrn) propagada en forma
horizontal fue medida con un medidor Skye (SKR 100, sensor remoto SKR 110; Skye
instruments, Llandrindod Wells, Powys, UK), cuyo sensor fue colocado en forma vertical
apuntando hacia el sur.
Reducción de la relación R/RL adicionando RL:
Para analizar el efecto que tenía una reducción constante de la relación R/RL (sin alterar la
radiación fotosintéticamente activa que llegaba a las hojas) sobre el alargamiento del hipocotilo
en plantas de Arabiodpsís thaliana se suplementó la luz solar directa con RL provisto
horizontalmente. La suplementación de RL fue obtenida por medio de un reflector incandescente
de 150 W combinado con un filtro de agua de 15 cm, un acrílico azul y filtros de acetato rojo.
46

Pulsos de luz dados alfinal del día para establecer distintos nivelesde Pfr/P:
Para investigar la sensibilidad de plantas de papa con distintos niveles de phyA a la reducción de
los niveles de Pfr al final del fotoperíodo, se expusieron plantas de aproximadamente 2 semanas
de edad a fotoperíodos de 7 h bajo radiación natural, terminados con pulsos lO minutos que
establecían distintos niveles de Pfr/P, utilizando las fuentes de luz descriptas por Casal et al.
(1991). El alargamiento del tallo producido entre el primer y el tercer día del tratamiento fue
medido con regla.
Reducciónselectiva dela irradiancia:
Para reducir la irradiancia de la luz incidente sobre las plantas de Arabídopsis thalíana se
colocaron distintos filtros en forma horizontal, por encima de las plantas. Una red de plástico gris
se utilizó como filtro neutro. Un acetato naranja sumado a un acetato amarillo (La Casa de
Acetato) se usó para reducir la luz azul, un acetato celeste se utilizó para reducir la luz roja, y con
una solución de sulfato de cobre 2.5% de 1.2 crn de altura se redujo el R+RL. Las mediciones
lumínicas fiieron obtenidas con un espectrorradiómetro (LI-COR inc. Lincoln, Nebraska).
En las plantas de papa se redujo selectivamente la cantidad de luz R+RL propagada en forma
horizontal que llegaba al tallo, sin afectar la luz percibida por las hojas, colocando el tallo de cada
planta entre dos filtros que contenían una solución de sulfato de cobre al 3 %. Los filtros (de 7.5
cm de largo) fueron obtenidos agrupando tubos de vidrio de 1 cm de diámetro en forma
horizontal, uno arriba del otro, de modo de cubrir el tallo. Como controles se utilizaron filtros
similares conteniendo agua destilada en lugar de sulfato de cobre.
Análisis estadístico
Cada experimento fue repetido por lo menos en tres ocasiones. Los datos correspondientes a
un mismo tratamiento fueron promediados y las comparaciones entre los distintos tratamientos
fueron realizadas por medio de la técnica de ANOVA, previa verificación de los supuestos,
utilizando el programa GRAPHPAD/PRISM.
47

FUNCIONES SENSORIALES DEL FITOCROMO A (II)
EL FITOCROMO A PERCIBE LAS TRANSICIONES OSCURIDAD/LUZ
QUE SINCRONIZAN EL RELOJ CIRCADIANO EN LAS PLANTAS
INTRODUCCIÓN
Diversos aspectos del desarrollo de las plantas (la floración, la tuberización, etc.) se
manifiestan sólo cuando la longitud del día es mayor (en las plantas de día largo) o menor (en las
plantas de día corto) que cierto valor crítico. Las plantas miden la duración de los días por medio
de relojes endógenos. Estos relojes son del tipo pendular u oscilatorio y actúan estableciendo un
ritmo diario de sensibilidad a la luz (baja durante la mayor parte del día y alta durante parte de la
noche). Si el fotoperíodo sólo coincide con la fase de baja sensibilidad a la luz, las plantas
interpretan esto como día corto. Por el contrario, si el fotoperíodo abarca la fase de baja
sensibilidad y una parte de la fase de alta sensibilidad a la luz, las plantas interpretan esto como
día largo (Thomas & Vince-Prue, 1997). A este modelo se lo conoce como de coincidencia
externa porque implica la coincidencia de una fase de un ritmo interno (de sensibilidad a la luz)
con la fase de un ritmo externo (de altemancia de luz y oscuridad). En este modelo la luz juega un
doble rol. Por un lado el fotoperíodo tiene un efecto directo, de inducción (en plantas de día
largo) o de represión (en plantas de día corto) de las respuestas fotoperiódicas, si la luz está
presente en el momento de mayor sensibilidad. Por otro lado el fotoperíodo determina el
momento del día en el que la luz es más efectiva a través de un efecto sobre el reloj que establece
el ritmo de sensibilidad (Pittendrigh & Minis, 1964).
Se ha observado que las plantas con niveles reducidos de phyA tienen alteradas las respuestas
a la longitud del día (Johnson et al., 1994; Weller et al., 1997). En principio, el phyA podría
afectar las respuestas fotoperiódicas ya sea porque está involucrado en la inducción fotoperiódica
propiamente dicha, o porque participa controlando al reloj que establece el ritmo de sensibilidad a
la luz. En esta parte de la tesis evaluaremos esta última posibilidad, es decir si el phyA participa
en la regulación del reloj de las plantas.
Los relojes asociados a la medición del fotoperíodo también son responsables de controlar
43

diversos procesos como la fotosíntesis, la apertura estomática, etc., de modo que éstos procesos
se manifiesten con mayor intensidad en momentos específicos del día (Kreps & Kay, 1997). Este
tipo de relojes no es exclusivo de las plantas. En la mayoría de los seres vivos, tanto procan'otas
como eucariotas, numerosos aspectos comportamentales, fisiológicos, metabólicos y/o
moleculares oscilan diariamente. Estas oscilaciones persisten bajo condiciones constantes de luz
y temperatura revelando que las mismas son generadas por osciladores endógenos, denominados
relojes círcadianos (Marques et al., 1997).
En los últimos años ha crecido notablemente el conocimiento sobre los mecanismos
moleculares asociados al funcionamiento de los relojes círcadianos en diversos sistemas. Las
oscilaciones son generadas por genes que regulan negativamente su propia expresión, como PER
y TIM en Drosophila melanogaster y FRQ en Neurospora crassa. Las proteínas codificadas por
estos genes controlan a su vez la expresión de otros numerosos genes, generándose así los ritmos
en distintos procesos (Dunlap et al., 1999).
Debido a que en condiciones constantes de luz y temperatura los relojes círcadianos tienen
períodos cercanos pero no iguales a 24h, los mismos deben ser diariamente sincronizados por los
ciclos naturales de luüoscuridad para mantener una sincronía estricta con el ambiente. La diaria
sincronización permite, a su vez, que los diferentes organismos sean siempre capaces de anticipar
el amanecer y el atardecer a pesar de los cambios que ocurren durante el año en la duración de los
días (Pittendrigh, 1981).
En comparación con los conocimientos disponibles sobre los mecanismos moleculares
asociados al funcionamiento de los relojes círcadianos, sabemos muy poco sobre los mecanismos
por medio de los cuales las transiciones luz/oscuridad soncronizan diariamente los mismos. El
primer paso en esta dirección es determinar cuáles son los fotorreceptores que controlan la
sincronía de los relojes.
Los espectros de acción realizados para analizar la regulación por la luz del ritmo circadiano
de emergencia de las pupas en Drosophila pseudoobscura, de conidiación en Neurospora y de
locomoción en hamsters, indicaron que las flavinas podrían estar involucradas en los primeros
dos sistemas y la rodopsina en el último (Sargent & Briggs, 1967; Klemm & Ninnemann, 1976;
Takahashi et al., 1984).
49

Recientemente se han identificado en los mamíferos genes con homología a los criptocromos
originalmente descubiertos en las plantas. Estos genes parecerían expresarse específicamente en
tejidos que carecen de opsinas pero que presentan una elevada sensibilidad a tratamientos
lumínicos que sincronizan al reloj. Este hallazgo proporciona evidencia correlativa para los
criptocromos, más que para la rodopsina, como los pigmentos involucrados en sincronizar los
relojes circadianos en los mamíferos (Miyamoto & Sancar, 1998). Sin embargo, experimentos
con ratones knock-out para el CRY2 revelaron que la sensibilidad a pulsos de luz que retrasan al
reloj fue incluso mayor en los ratones sin cry2 que en los ratones WT, es decir al revés de lo
esperado (Thresher et al., 1998). Esto sugiere que el cry2 no participa directamente en la
percepción de señales que sincronizan los relojes y el cryl u otro(s) fotorreceptores no
identificado(s) estaría(n) involucrado(s).
Un gen homólogo al criptocromo de plantas también ha sido recientemente identificado en
Drosophila melanogaster y se observó que moscas transgénicas que sobreexpresan este
fotorreceptor tienen incrementada la sensibilidad a pulsos de luz que sincronizan el reloj,
mientras que las moscas mutantes, carentes del fotorreceptor, tienen reducida la sensibilidad
(Emery et al., 1998; Stanewsky et al., 1998). Estos resultados indican que este criptocromo sí es
uno de los fotorreceptores que controlan al reloj circadiano en las moscas.
Mutantes de fotorreceptores no han sido inequívocamente identificados en Neurospora. Hasta
el momento se han identificado sólo dos loci que corresponden a mutantes insensibles a todo tipo
de señales lumínicas, WC-I y WC-2, y se ha sugerido que estos loci podrían codificar para genes
de fotorreceptores (Ballario et al., 1996; Linden & Macino, 1997).
En las plantas se conocen hasta ahora tres grupos de fotorreceptores: fitocromos, criptocromos,
y fotorreceptor(es) de UV-B (Mohr, 1994). En Samanea saman, pulsos de R resetean el ritmo del
movimiento de las hojas y este efecto es cancelado cuando el R es seguido inmediatamente por
un pulso de RL, indicando la participación de los fitocromos (Simon et al., 1976). En algunos
casos, incluso pulsos de RL son capaces de re-sincronizar el ritmo, sugiriendo la participación las
respuestas a muy bajos flujos de luz mediadas por el fitocromo (Hillman, 1971; Nagy et al., 1993;
Satter et al., 1981). Recientemente se ha observado que mutantes afectados en el cromóforo de los
fitocromos, y por lo tanto deficientes en todos los miembros de esta familia de fotorreceptores,
presentan un período alterado del ritmo circadiano de la expresión del gen CABZ, cuando son
incubadas bajo luz R (Millar et al., 1995). Estos resultados en conjunto proveen evidencia tanto
50

fotobiológica como genética indicando que la familia de los fitocromos controlan la
sincronización de los ritmos circadianos. Sin embargo no se sabe con precisión que miembros dela familia de los fitocromos están involucrados.
Las funciones fotoperceptivas del phyA parecen diferir en parte de las del resto de los
fitocromos (Casal et al., 1997). Al igual que los demás fitocromos, el phyA es sintetizado como
Pr, pero una vez convertido a la forma Pfr el phyA es rápidamente degradado (Quail et aL, 1973).
Esto hace que los niveles de Pfr (la forma fisiológicamente activa del fitocromo) del phyA sean
bajos durante la noche (cuando sólo hay destrucción del Pfr) pero aumenten durante el día (ya que
aunque la destrucción del Pfr continúa, nuevo Pfr es producido por la fototransforrnación del Pr
sintetizado). Los cambios en la actividad del phyA con las transiciones luz/oscuridad hacen de
este fotorreceptor un buen candidato para mediar la sincronización de los ritmos circadianos por
señales como el atardecer o el amanecer en las plantas.
A continuación se describe la utilización de plantas de papa transformadas con el gen de phyA
homólogo en sentido y en antisentido (Heyer et al., 1995) para evaluar el rol de este fotorreceptor
en la re-sincronización del reloj circadiano en las plantas.

RESULTADOS
Uno de los ritmos circadianos más caracterizados es el ritmo observado en el movimiento de
las hojas. En las plantas de papa, al igual que en muchas especies, la lámina asume una posición
horizontal durante el día y vertical durante la noche. Esta oscilación persiste en condiciones
constantes de luz y temperatura indicando que la misma es generada por un oscilador endógeno.
Para investigar el rol del phyA en el control de la fase del ritmo del movimiento de las hojas se
cultivaron plantas de papa bajo fotoperíodos y termoperíodos naturales. Un grupo de plantas (el
control) fue transferido directamente a condiciones constantes de luz (luz blanca continua) y
temperatura (25 °C) al amanecer. Los grupos restantes fueron expuestos a 5 h de diferentes
tratamientos lumínicos antes de ser transferidos junto al grupo control a condiciones constantes
de luz y temperatura (Fig. 1). Protocolos similares han sido utilizados exitosamente para
investigar el efecto de diferentes calidades de luz en la sincronización del reloj circadiano en
Chlamydomonas (Kondo et al., 1991) y Gonyaulax (Roenneberg & Deng, 1997). En estos
sistemas, al igual que en muchos otros, se ha observado que pulsos de luz dados al final de la
noche (como los utilizados en este protocolo) producen avances en la fase de los ritmos, mientras
que pulsos de luz dados al comienzo de la noche producen retrasos en la fase de los ritmos
(Pittendrigh, 1981).
control (oscuridad)RL
RL+RRL+R+AZUL
Figura 1. Protocolo experimental. Plantas de papa WT y transgénicas transformadascon el gen PHYA en sentido o antisentído fueron cultivadas en el invernáculo poraproximadamente 2 semanas y luego transferidas (durante el amanecer del día quecomenzaba el experimento) a una cámara de luz blanca contínua y temperatura constante de25 °C, con o sin (control) una exposición previa a 5 h de RL, RL+R ó RL+R+azul.
52
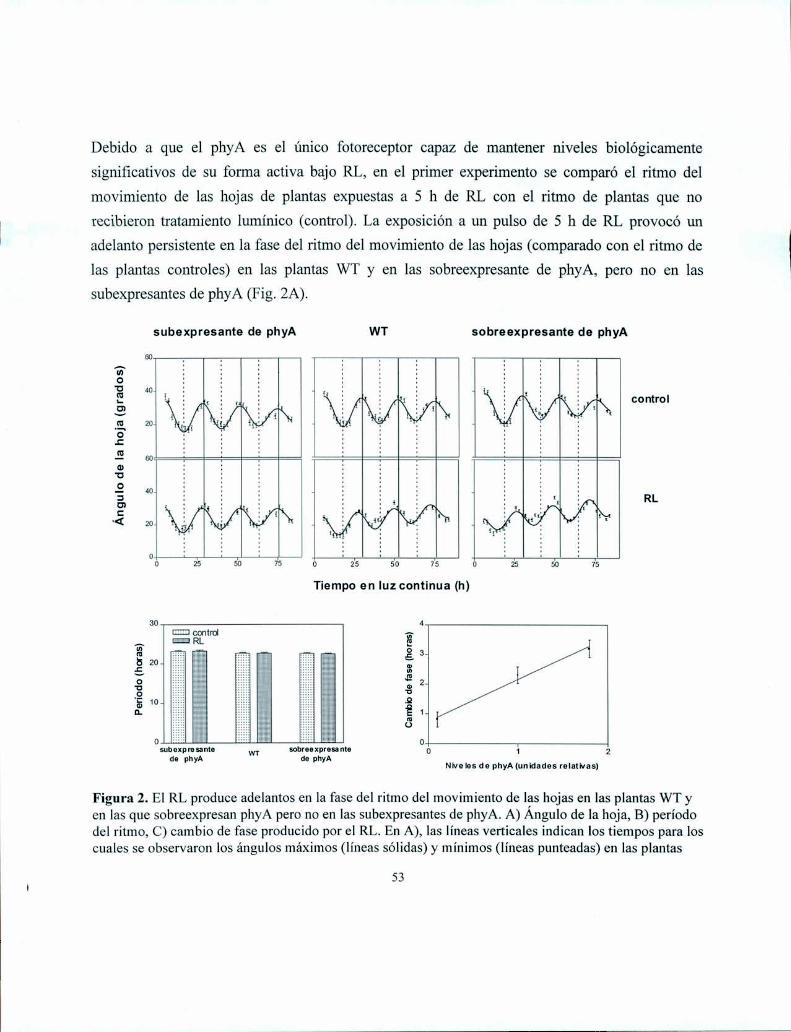
Debido a que el phyA es el único fotoreceptor capaz de mantener niveles biológicamente
significativos de su forma activa bajo RL, en el primer experimento se comparó el ritmo del
movimiento de las hojas de plantas expuestas a 5 h de RL con el ritmo de plantas que no
recibieron tratamiento lumínico (control). La exposición a un pulso de 5 h de RL provocó un
adelanto persistente en la fase del ritmo del movimiento de las hojas (comparado con el ritmo de
las plantas controles) en las plantas WT y en las sobreexpresante de phyA, pero no en las
subexpresantes de phyA (Fig. 2A).
subexpresante de phyA WT sobreexpresante de phyA
M
:Vva N/VVX:VVVx
Ángulodelahoja(grados) X/kfXfiN xfa/“VN _wfiwfiuk
Tiempo en luz continua (h)
JO D r.
Período(horas)
{o(¡o
_¿l
Carrindefase(horas)
D0subexpresanto WT sobranxpresa me i
de phyANiveles de phyA(unidades relativas)
Figura 2. El RL produce adelantos en la fase del ritmo del movimiento de las hojas en las plantas WT yen las que sobreexpresan phyA pero no en las subexpresantes de phyA. A) Ángulo de la hoja, B) períododel ritmo, C) cambio de fase producido por el RL. En A), las líneas verticales indican los tiempos para loscuales se observaron los ángulos máximos (líneas sólidas) y mínimos (líneas punteadas) en las plantas
53

control. Los datos son promedios i e.e. de por lo menos 15 réplicas.
El período del ritmo bajo luz blanca continua no fue afectado significativamente por el
pretratamiento de RL ni por el genotipo (Fig. 2B) pero una relación lineal positiva fue observada
entre los niveles de phyA y la magnitud del adelanto de fase producida por el tratamiento de RL
(Fig. 2C). El cambio de fase producido por el RL fue significativamente mayor en las plantas
sobreexpresantes de phyA que en las WT (P < 0.05) y menor en las subexpresantes de phyA que
en las WT (P < 0.05).
Un único pulso de RL de 5 min dado 5 h antes de transferir las plantas a la luz continua no
tuvo ningún efecto sobre la fase del ritmo del movimiento de las hojas (datos no mostrados). Esto
indica que el adelanto de fase producido por la exposición a 5 h de RL en forma continua no fuela consecuencia de la reducción de los niveles de Pfr de los fitocromos estables.
De esto modo, tanto evidencias genéticas (ausencia de efecto de la luz sobre el ritmo de las
plantas transgénicas con niveles reducidos de phyA) como fisiológicas (efecto del RL continuo
pero no de un pulso corto de RL sobre la fase del ritmo de movimiento de las hojas) indican que
el phyA es uno de los fotorreceptores involucrados en la sincronización del reloj circadiano en
plantas adultas de papa.
Cabe mencionar, sin embargo, que la exposición al RL continuo es una herramienta
fotobiológicamente útil para investigar la participación del phyA en el control de un proceso
determinado, pero sólo plantas que crecen bajo canopeos muy densos están expuestas a ambientes
lumínicos semejantes. Para evaluar el rol del phyA en la sincronización del ritmo circadiano del
movimiento de las hojas en condiciones lumínicas similares a las que experimentan plantas que
crecen en lugares abiertos, se analizó el efecto de exponer a las plantas a una mezcla de RL+R
que proveía una relación R/RL similar a la presente bajo luz solar directa.
El tratamiento con RL+R provocó un adelanto persistente en el ritmo del movimiento de las
hojas de las plantas WT y de las sobreexpresantes de phyA comparado con el ritmo del
movimiento de las hojas de plantas que no fueron expuestas a la luz (control). De manera similar
a lo ocurrido con el RL sólo, el RL+R no produjo un cambio de fase estadísticamente
significativo en las plantas transgénicas con el gen PH YAen antisentido (Fig 3).
54
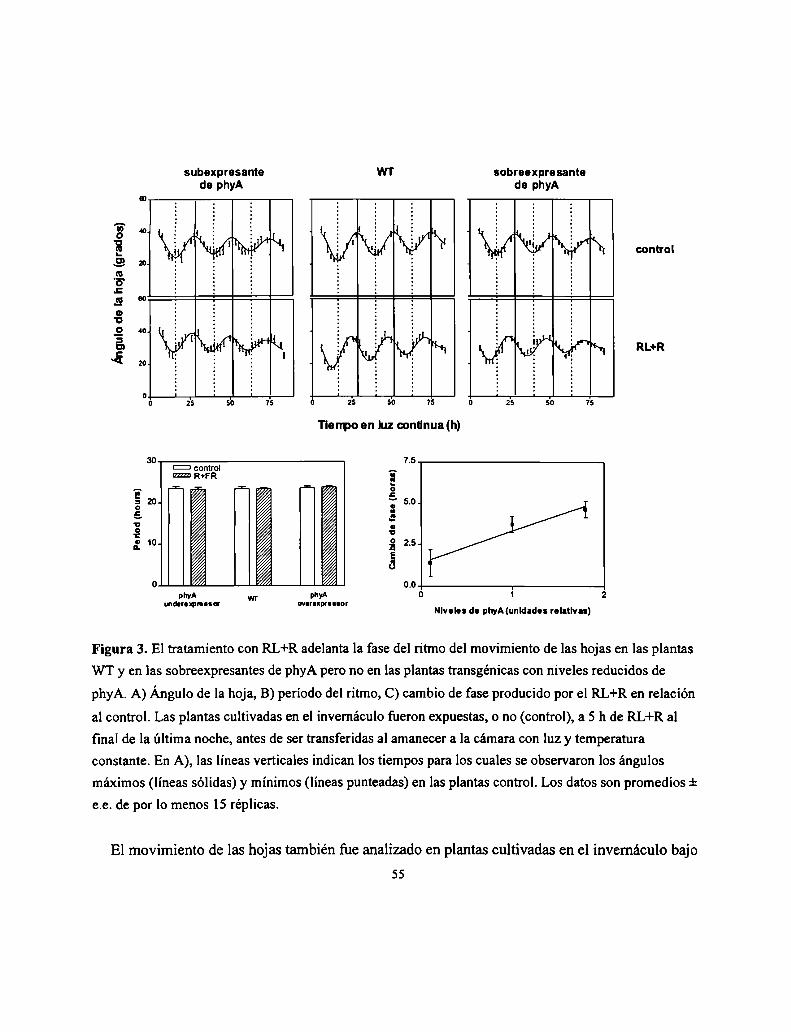
subexpresante WT sobreexprosantedo phyA de phyA
"'WWW TV¿flA2,71:i“:nf :“Xfx, control
Ángqudelahoja(grados) .o- M/“kuflïwlhh RL+R2'5 s' 15 2'5 si 75 z's 5'o 15
Tlenpo en luz continua (h)
1 q
1:t!A 0
g 5 5.og 8; e1°: 8
¡1’ g 2.5.E¿l
fl fl I
phyA w, phyA t 1 :maromnuu MIOIWOIIOF
vaoln de phyA(unld¡dosrolatlvao)
Figura 3. El tratamiento con RL+R adelanta la fase del ritmo del movimiento de las hojas en las plantas
WT y en las sobreexpresantes de phyA pero no en las plantas transgénicas con niveles reducidos de
phyA. A) Ángulo de la hoja, B) período del ritmo, C) cambio de fase producido por el RL+R en relación
al control. Las plantas cultivadas en el invemáculo fueron expuestas, o no (control), a 5 h de RL+R al
final de la última noche, antes de ser transferidas al amanecer a la cámara con luz y temperatura
constante. En A), las líneas verticales indican los tiempos para los cuales se observaron los ángulos
máximos (líneas sólidas) y mínimos (líneas punteadas) en las plantas control. Los datos son promedios :l:
e.e. de por lo menos 15 réplicas.
El movimiento de las hojas también fue analizado en plantas cultivadas en el invemáculo bajo55
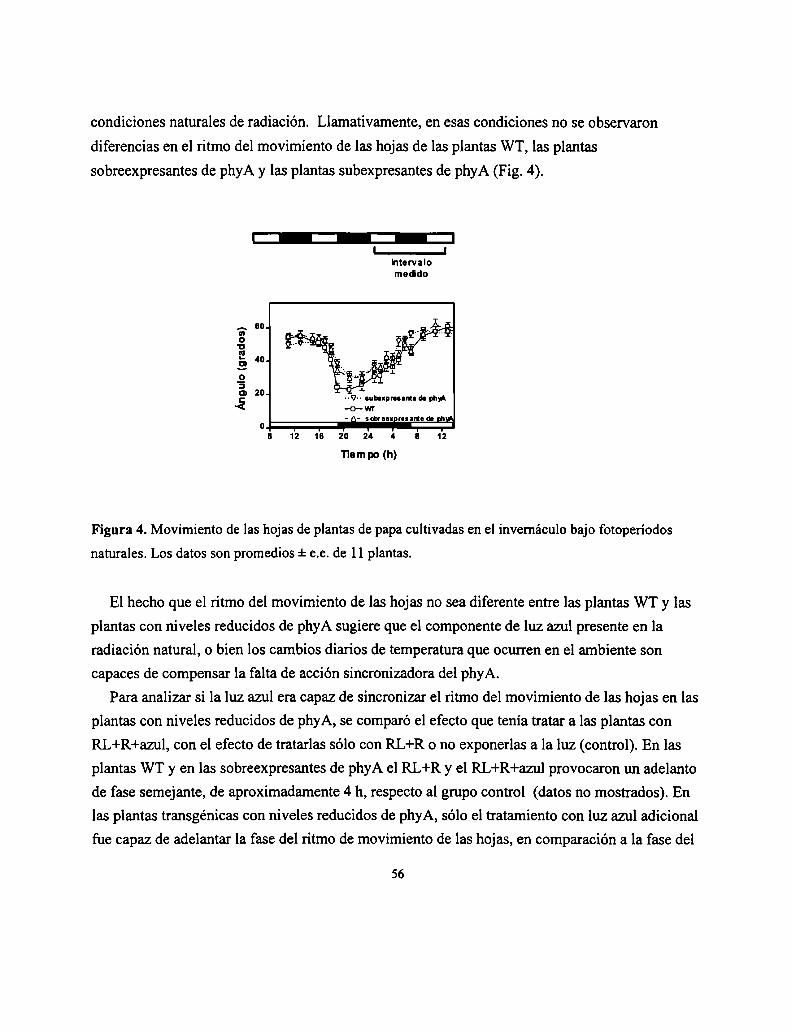
condiciones naturales de radiación. Llarnativamente, en esas condiciones no se observaron
diferencias en el ritmo del movimiento de las hojas de las plantas WT, las plantas
sobreexpresantes de phyA y las plantas subexpresantes de phyA (Fig. 4).1-2-2l—lhtervalomeddo
Ángulo(grados)
Tiempo (h)
Figura 4. Movimiento de las hojas de plantas de papa cultivadas en el invemáculo bajo fotoperiodos
naturales. Los datos son promedios i e.e. de ll plantas.
El hecho que el ritmo del movimiento de las hojas no sea diferente entre las plantas WT y las
plantas con niveles reducidos de phyA sugiere que el componente de luz azul presente en la
radiación natural, o bien los cambios diarios de temperatura que ocurren en el ambiente son
capaces de compensar la falta de acción sincronizadora del phyA.
Para analizar si la luz azul era capaz de sincronizar el ritmo del movimiento de las hojas en las
plantas con niveles reducidos de phyA, se comparó el efecto que tenía tratar alas plantas con
RL+R+azul, con el efecto de tratarlas sólo con RL+R o no exponerlas a la luz (control). En las
plantas WT y en las sobreexpresantes de phyA el RL+R y el RL+R+azul provocaron un adelanto
de fase semejante, de aproximadamente 4 h, respecto al grupo control (datos no mostrados). En
las plantas transgénicas con niveles reducidos de phyA, sólo el tratamiento con luz azul adicional
fue capaz de adelantar la fase del ritmo de movimiento de las hojas, en comparación a la fase del
56
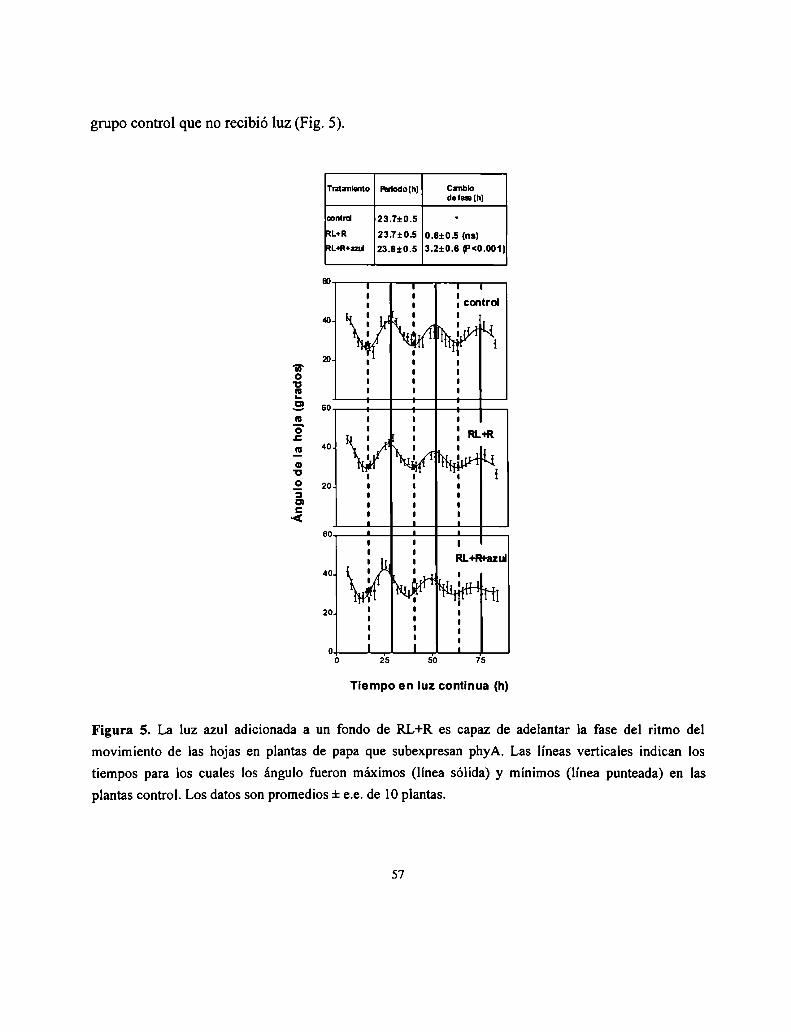
grupo control que no recibió luz (Fig. 5).
Tnta'nlemo Modo(h) Cantilodelas (h)
contra 23110.5
UR 23.7205 0.810.5(ns)¡iman! 23.0205 3210.0 No.001)
m l
control
‘°' ai
zo
¿Ti
n-n-q
Ángulodelahoja(grados)
en
40.
wm> 2'5 5'0 5
Tiempo en luz contlnua (h)
Figura 5. La luz azul adicionada a un fondo de RL+R es capaz de adelantar la fase del ritmo del
movimiento de las hojas en plantas de papa que subexpresan phyA. Las líneas verticales indican los
tiempos para los cuales los ángulo fueron máximos (linea sólida) y mínimos (línea punteada) en las
plantas control. Los datos son promedios :i:e.e. de 10 plantas.

DISCUSIÓN
Los resultados presentados aquí proporcionan una fuerte evidencia genética de que el phyA es
uno de los fotorreceptores que controlan el reloj circadiano en plantas de papa. En este trabajo
demostramos que la exposición de plantas de papa a 5 h de RL al final de la noche
(inmediatamente antes del amanecer) adelantó la fase del ritmo circadiano del movimiento de las
hojas en plantas con niveles normales de phyA y en aquellas que sobreexpresan phyA pero no en
las transgénicas con niveles reducidos de phyA. La correlación entre los niveles de phyA y la
magnitud del cambio de fase entre los genotipos salvajes, transgénicos con PHYA en antisentido y
transgénicos con PHYA en sentido, favorecen la interpretación que los fenotipos se deben
directamente a los niveles alterados de phyA y no a una posible variación somaclonal ocurrida
durante el proceso de obtención de las plantas transgénicas ó a un posible efecto del gen PHYA en
antisenstido sobre los niveles de expresión de otros miembros de la familia de fitocromos. Se
sabe, además, que los niveles de phyB no están afectados en las plantas con PHYA en antisentido
(Heyer et al., 1995).
Dado que el phyA es el único fotorreceptor capaz de ser activado por RL, el hecho que la
exposición a 5 h de RL provoque un adelanto en la fase del ritmo del movimiento de las hojas por
sí mismo proporciona evidencia fotofisiológica de que el phyA es uno de los fotorreceptores que
sincroniza al reloj circadiano. No obstante, sería posible que el RL afecte la fase del ritmo del
movimiento de las hojas no por su efecto sobre el phyA, sino porque reduce los niveles de Pfr de
los fitocromos estables. Sin embargo, un único pulso de RL dado en el momento en el que
normalmente comenzaría el período de 5 h de tratamiento lumínico no fue efectivo en inducir un
cambio de fase en las plantas WT sugiriendo que el RL contínuo actuó específicamente activando
al phyA y no reduciendo los niveles de Pfr de los fitocromos estables.
El hecho que la mezcla de RL+R con una relación R/RL similar a la provista por la luz solar
fuera capaz de adelantar la fase del ritmo del movimiento de las hojas en plantas WT y
transgénicas que sobreexpresan phyA pero no en plantas con niveles reducidos de phyA sugiere
que el phyA es el principal fitocromo involucrado en la sincronización del reloj en las plantas de
papa. No obstante, a pesar de que el RL+R (que se supone activaría a todos los miembros de la
familia de los fitocromos), ó un pulso breve de RL (que reduciría los niveles de Pfr de los
fitocromos estables) no fueron capaces de resetear el ritmo del movimiento de las hojas en plantas
58

transgénicas con niveles reducidos de phyA, sería prematuro concluir que otros fitocromos no
juegan un rol en el control del reloj. Experimentos recientes han demostrado que el phyB es
importante en el control fotoperiódico de la tuberización en la papa (Jackson et al., 1996).
También se ha observado que el phyB regula la amplitud de la oscilación de la expresión del gen
CAB (Childs et aL, 1995) y de la producción de etileno (Finlayson et al., 1998) en Sorghum
bicolor. Además, Somers et al. (1998) han analizado recientemente el efecto de la luz sobre el
período del ritmo círcadiano de la expresión del gen CABZen plantas de Arabidopsis thaliana
WT y en los mutantes phyA y phyB. Ellos observaron que incrementos en el flujo de R producían
una reducción en el período del ritmo en todos los genotipos. No obstante el período era mayor en
el mutante phyA que en el WT a bajos flujos de R, y mayor en el mutante phyB que en el WT a
altos flujos de R. Si bien estos resultados permiten concluir que tanto el phyA como el phyB
controlan la marcha del reloj, no se puede saber de que forma lo hacen. Esto se debe a que los
efectos de la luz continua sobre el período de los ritmos circadianos es el resultado de la
integración de los efectos de la luz en distintos momentos del día. En la noche temprana la luz
atrasa el reloj y en la noche tardía la luz adelanta el reloj. En los organismos que tienen mayor
actividad durante el día que durante la noche, como es el caso de las plantas, los adelantos son
mayores que los atrasos y, en consecuencia, el período de los ritmos se acorta con la exposición a
la luz continua. Un período mayor en los ritmos de las mutantes comparadas con las plantas WT
puede originarse tanto por una reducción en la magnitud de los adelantos como por un incremento
en los retrasos de los ritmos. Por lo tanto, no es posible descartar la posibilidad de que en
Arabídopsis thaliana el phyA y el phyB no participen directamente de la percepción de señales
que adelantan el reloj sino que regulen negativamente la acción de otro fotorreceptor que retrasa
al reloj.
Con el sistema experimental utilizado en este caso se puede concluir efectivamente que el
phyA participa directamente de la percepción de las señales lumínicas que producen adelantos del
reloj. En el futuro podrian realizarse experimentos con plantas con niveles alterados de los demás
fitocromos (con protocolos similares a los descriptos aquí) para conocer el rol de otros miembros
de esta familia de fotorreceptores en la sincronización de los ritmos circadianos.
Por último, en este trabajo se demostró que la luz azul era capaz de adelantar el reloj en
plantas con niveles reducidos de phyA indicando por un lado que estas plantas no muestran un
problema generalizado en la sincronización de los ritmos y, por otro lado, que fotorreceptres de
S9

luz azul también estarían involucrados en el control del reloj circadiano.
Si bien el efecto sincronizador de la luz azul sobre el ritmo del movimiento de las hojas había
sido observado con anterioridad (Halaban 1969), no era posible determinar si el fotorreceptor
involucrado era un fotorreceptor específico de la luz o un fitocromo. El motivo de tal
incertidumbre estaba dado por que los fitocromos también absorben en rango del azul y esta luz
por si misma es capaz de modificar distintos parámetros de los fitocromos cuando se la da sola.
Este no fue el caso de la aproximación experimental utilizada aquí, donde se dió luz azul sobre un
fondo de RL+R que satura la absorción de luz por parte de los fitocromos (Thomas & Dickinson,
1979). En estos experimentos la proporción de Pfr/P establecida por el tratamiento de RL+R+azul
fue similar al establecido por el RL+R sólo (69%), y la luz azul adicionada al fondo de RL+R
solo incrementó la tasa de fotoconversión entre el Pr y el Pfr en un 15%. Además, incrementar la
fotoconversión entre Pr y Pfr por adición de más RL+R al fondo de luz no pudo imitar el efecto
de la luz azul en plantas con niveles reducidos de phyA (datos no mostrados). Es decir, el efecto
fue específico para la luz azul y por lo tanto es evidencia inequívoca de la participación de uno ó
más fotorreceptores de luz azul en el control del reloj circadiano. Somers y col. (1998) también
observaron que, a bajos flujos de luz azul, el período del ritmo de expresión de CAB2 era mayor
en el mutante cry] que en el genotipo WT de Arabidopsis thaliana, pero por los motivos
anteriormente descriptos no se puede determinar si el cryl afecta directa o indirectamente al reloj.
El hecho que los niveles incrementados o reducidos de phyA no afectaron la fase ni la
amplitud del ritmo bajo luz blanca continua (Fig. l) o bajo fotoperíodos naturales (Fig. 4 A)
conteniendo luz azul indican que fotorreceptores específicos de luz azul pueden compensar la
ausencia de phyA (por lo menos bajo estas condiciones experimentales) y por lo tanto las
acciones de estos dos fotorreceptores serían redundantes. Esta redundancia permitiría mantener la
homeostasis del sistema, ya sea cuando alguno de los fotorreceptores pierda su capacidad de
actuar debido a una mutación o en condiciones ambientales diferentes, donde las señales
lumínicas preponderantes favorezcan la actividad de uno u otro fotorreceptor.
60

MATERIALES Y METODOS
Material vegetal y condiciones de cultivos
Las plantas de papa (Solanum tuberosum) tanto del genotipo WT (cultivar Désirée) como las
transformadas con el gen PHYA en sentido (línea PS4) o antísentido (línea APl l), son iguales a
las descriptas por Heyer et al. (1995). Las plantas fueron propagadas a partir de tubérculos
pequeños y cultivadas en un invemáculo de temperatura semi-controlada, en macetas plásticas de
650 cm3 conteniendo una mezcla de tierra y arena. Se utilizaron para los experimentos plantas de
aproximadamente 3 semanas de edad.
Mediciones del ángulo de las hojas
La tercer hoja más joven fue marcada en cada planta. El ángulo formado ente la inserción del
pecíolo en la lámina y una línea normal a la superficie del suelo fue medido cada 2 h durante los
experimentos. Cada experimento fue repetido al menos en tres oportunidades y los datos fueron
promediados para el análisis estadístico. Los datos se ajustaron, de acuerdo a Millar et al. (1995),
a la ecuación: L(t) = (ca+ clt) + (ao-an) sin [Zn/T (t-ó)]; donde L es el ángulo de inserción de la
hoja, co es el valor estimado del ángulo a t = 0, c, es la estimación de la tasa lineal de cambio en
el ángulo de la hoja, a0 es el valor estimado de la amplitud a t = 0, a] es la tasa de cambio
estimada de la amplitud de oscilación, t es el tiempo, T es el período estimado, y d)es la fase
estimada a t = 0. El componente lineal de la ecuación, C¡t,sólo fue utilizado para ajustar los datos
de la figura l.
Tratamientos lumínicos
La posición de las hojas fue medido tanto en el invemáculo como en una cámara de cultivo a 25
°C bajo luz continua (50 pmol.m'2.s") provista por lámparas de sodio de alta presión en
combinación con filtros de agua destilada. Estas lámparas proveen luz en el visible y en el
infrarrojo cercano, con picos en el naranja y el azul (ver Casal & Sánchez 1994, por detalle del
espectro). Los tratamientos de RL, R+RL y R+RL+AZUL fueron provistos en el invemáculo. En61

los experimentos donde las mediciones se llevaron a cabo bajo condiciones constantes de luz y
temperatura, las plantas fueron transferidas desde el invemáculo hacia la cámara de crecimiento
al amanecer. Un banco de luces incandescente, en combinación con filtros de agua, dos acetatos
rojos (La Casa del Acetato, Buenos Aires, Argentina) y un filtro de sulfato de cobre (1.5 % de
sulfato de cobre en solución acuosa de 5 mm de espesor) fue usado para proveer RL+R (30
pmol.m'2.s'l) con una relación WRL similar a la del día bajo radiación natural (1.1). Para proveer
RL solo (100 umol.m'2.s'l), la luz de las lámparas incandescentes fue filtrada a través de filtros
Roscolux (Rosco Laboratories, Port Chester, NY), No. 83 (Medium Blue), No. 12 (Straw), y No.
42 (Deep Salmon). Para obtener RL+R+AZUL, se utilizó, además de la fuente de RL+R, una
fuente de luz azul (22 umol.m'2.s"), consistente en un banco de tubos fluorescentes (Philips, TLF
40W/54) combinados con un filtro de acetato azul.
62

LOS LOCI VLFI Y VLF2 AFECTAN DIFERENCIALMENTE LAS RESPUESTAS
VLFR Y HIR DESENCADENADASPOR EL FITOCROMO A
INTRODUCCIÓN
Dos modos de acción del phyA han sido descriptos en estudios fotomorfogénicos clásicos.
Por un lado el phyA media las respuestas a muy bajos flujos de luz, conocidas como VLFR
(Casal et al., 1994; Botto et al., 1996; Shinomura et al., 1996). Estas respuestas son tan
sensibles a la luz que se saturan con pulsos de RL que fototransforrnan menos de un 10 % del
phyA de sus forma inactiva (Pr) a su forma activa (Pfr). Por otro lado, el phyA también media
las respuestas HIR (Dehesh et al., 1993; Heyer et al., 1995; Nagatani et al., 1993; van Tuinen
et al., 1995; Whitelam et al., 1993). Estas respuestas sólo se saturan con exposiciones
prolongadas a altas irradiancias de RL continuo (Mancinelli, 1994). Una característica
frecuente de la acción del phyA es que las respuestas al RL continuo son mucho mayores que
aquellas producidas por pulsos (de 3 min) horarios de RL, aún cuando ambos tratamientos
provean igual cantidad de fotones. Este hecho, conocido como falla a la ley de reciprocidad
sugiere la existencia de dos mecanismos distintos involucrados en las respuestas VLFR y HIR
(Casal et al., 1998).
No obstante, dado que el phyA en su forma Pfr puede destruirse o revertir a la forma Pr
(Mancinelli, 1994), el RL continuo podría ser más efectivo que los pulsos horarios de RL
porque permitiría mantener niveles constantes de Pfr en exposiciones prolongadas a la luz
(Heim and Sch'áfer, 1984; Heim and Scháfer, 1982). Sin embargo, se ha observado que altas
irradiancias de RL continuo son significativamente más efectivas que muy bajos fljuos de R
que establecen niveles similares y constantes de Pfr (Heim and Scháfer, 1984). Esto sugiere
que el RL continuo hace algo más que mantener niveles bajos pero constantes de Pfr en el
tiempo. Debido a que el HIR depende fuertemente de la irradiancia, se ha propuesto que ese
algo más que hace el RL continuo es aumentar la tasa de ciclado entre las formas Pr y Pfr.
La hipótesis de este trabajo es que el VLFR y el HIR involucran dos mecanismos
moleculares distintos puestos en marcha por el phyA. El Pfr sería la forma activa del phyA en
las respuestas inductivas ó VLFR y un efector distinto al Pfr, posiblemente un intermediario
inestable formado durante el ciclado del phyA entre las formas Pr y Pfr, sería el que
desencadena las respuestas al RL contínuo o HIR (Heim and Schafer, 1984; Johnson and
Tasker, 1979). A su vez, las vias de transducción de señales activadas por el Pfr del phyA en
el VLFR y por la forma aún no identificada del phyA en el HIR podrían ser diferentes. Si esta
hipótesis fuera correcta debería ser posible identificar variantes génicas que afectendiferencialmente a estos modos de acción.
La búsqueda de variantes génicas que afecten diferencialmente al VLFR y al HIR fue
63

llevada a cabo en Arabidopsis thaliana. Ésta es una planta herbácea muy pequeña
perteneciente a la familia de las crucíferas que en los últimos años ha sido intensamente
utilizada como sistema modelo para estudios de genética vegetal por su tamaño reducido, su
corto ciclo de vida y su pequeño genoma Las bases genéticas de diversos procesos
fisiológicos han comenzado a ser dilucidadas gracias a la identificación de mutantes
específicos. Hasta ahora, la mayor parte de los mutantes han sido aislados a partir de
poblaciones donde la variabilidad genética había sido inducida artificialmente por agentes
químicos o fisicos. Recientemente, sin embargo, se ha comenzado a explorar la enorme fuente
de variabilidad genética presente en poblaciones naturales de Arabidopsis thaliana. Las
poblaciones de esta especie están distribuidas en un amplio rango geográfico que abarca desde
el norte de Escandinavia hasta el desierto de Áfi'ica y el medio oriente. Su adaptación a
ambientes tan distintos ha generado por lo tanto una gran variabilidad genética en la
población. Esta variabilidad ha permitido identificar loci asociados al control de la floración
(Blanco et al., 1998), a la defensa de patógenos (Parker et al., 1993), etc. Dado que distintas
poblaciones pueden haber estado expuestas a ambientes luminosos muy variados es posible
que la magnitud de las respuestas a distintas señales luminosas, y por lo tanto los loci
involucrados en estas respuestas, podría variar entre ecotipos (poblaciones de una misma
especie que habitan distintos ambientes).
A continuación se describe la caracterización de las respuestas fotomorfogénicas de
Landsberg erceta y Columbia, dos de los ecotipos más utilizados en los laboratorios. El
análisis detallado de las respuestas a la luz de estos ecotipos reveló la existencia de un
comportamiento diferencial en las respuestas a las señales lumínicas que actúan en el rango
del VLFR, pero no en las respuestas LFR y HIR. Utilizando lineas endocríadas
recombinanates obtenidas anteriormente para estos ecotipos (Lister & Dean, 1993) pudimos
identificar y mapear los loci que afectan diferencialmente a estos modos de acción delfitocromo.
RESULTADOS
La primer parte de la caracterización fotomorfogénica de los ecotipos consistió en analizar
las respuestas a pulsos de luz que establecían distintos niveles de Pfr/P, es decir el VLFR y el
LFR. Se analizaron procesos como la germinación, la apertura de los cotiledones, la
inhibición del alargamiento del hipocotilo y la potenciación de la síntesis de clorofila en
plántulas etioladas.
Germínación
Para analizar la promoción de la germinación por la luz se expusieron semillas de64
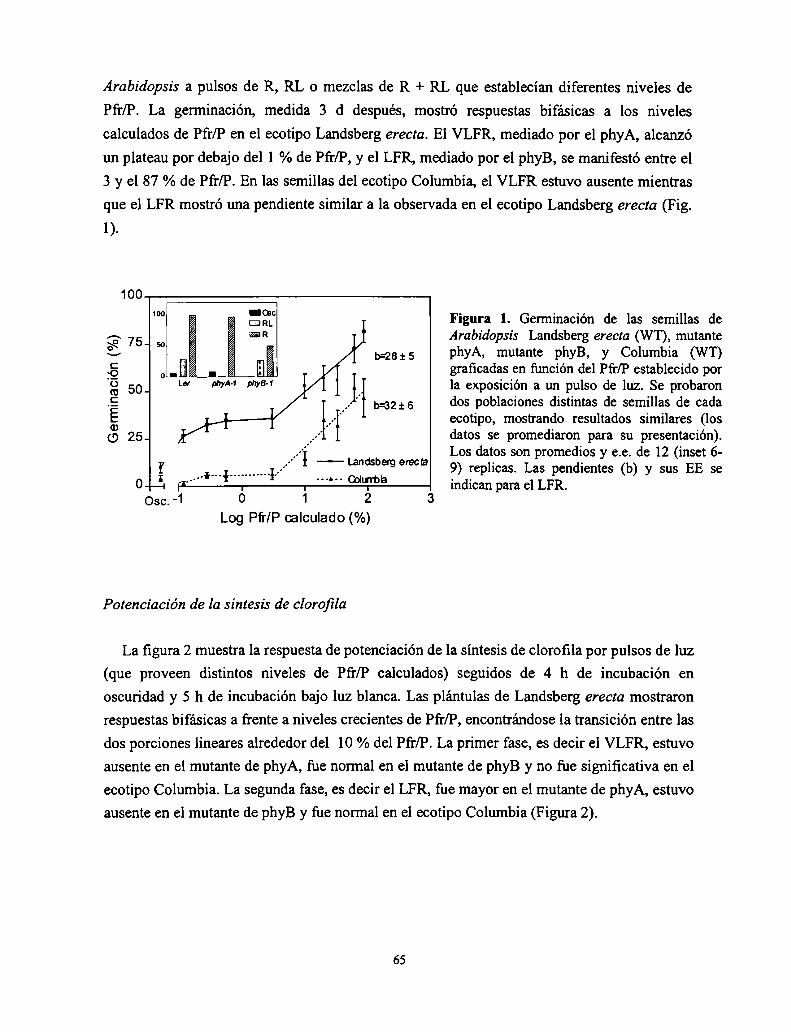
Arabidopsís a pulsos de R, RL o mezclas de R + RL que establecían diferentes niveles de
Pfr/P. La germinación, medida 3 d después, mostró respuestas bifásicas a los niveles
calculados de Pfr/P en el ecotipo Landsberg erecta. El VLFR, mediado por el phyA, alcanzó
un plateau por debajo del l % de Pfr/P, y el LFR, mediado por el phyB, se manifestó entre el
3 y el 87 % de Pfr/P. En las semillas del ecotipo Columbia, el VLFR estuvo ausente mientras
que el LFR mostró una pendiente similar a la observada en el ecotipo Landsberg erecta (Fig.
b=28i5
Germinación(%)
1- mi,lFigura l. Germinación de las semillas deArabidopsís Landsberg erecta (WT), mutantephyA, mutante phyB, y Columbia (WT)graficadas en función del Pfr/P establecido porla exposición a un pulso de luz. Se probarondos poblaciones distintas de semillas de cadaecotipo, mostrando resultados similares (losdatos se promediaron para su presentación).
Los datos son promedios y e.e. de 12 (inset 6Í ¿mi _________ LandSbÉrgemm 9) replicas. Las pendientes (b) y sus EE se
0-—¡i ¡"‘"I . I (“me indicanparael LFR.Osc.-1 0 1 2 3
Log Pfr/P lculado (%)
Potenciación dela síntesis de clorofila
La figura 2 muestra la respuesta de potenciación de la síntesis de clorofila por pulsos de luz
(que proveen distintos niveles de Pfr/P calculados) seguidos de 4 h de incubación en
oscuridad y 5 h de incubación bajo luz blanca. Las plántulas de Landsberg erecta mostraron
respuestas bifásicas a frente a niveles crecientes de Pfr/P, encontrándose la transición entre las
dos porciones lineares alrededor del 10 % del Pfr/P. La primer fase, es decir el VLFR, estuvo
ausente en el mutante de phyA, fue normal en el mutante de phyB y no fue significativa en el
ecotipo Columbia. La segunda fase, es decir el LFR, fue mayor en el mutante de phyA, estuvo
ausente en el mutante de phyB y fue normal en el ecotipo Columbia (Figura 2).
65
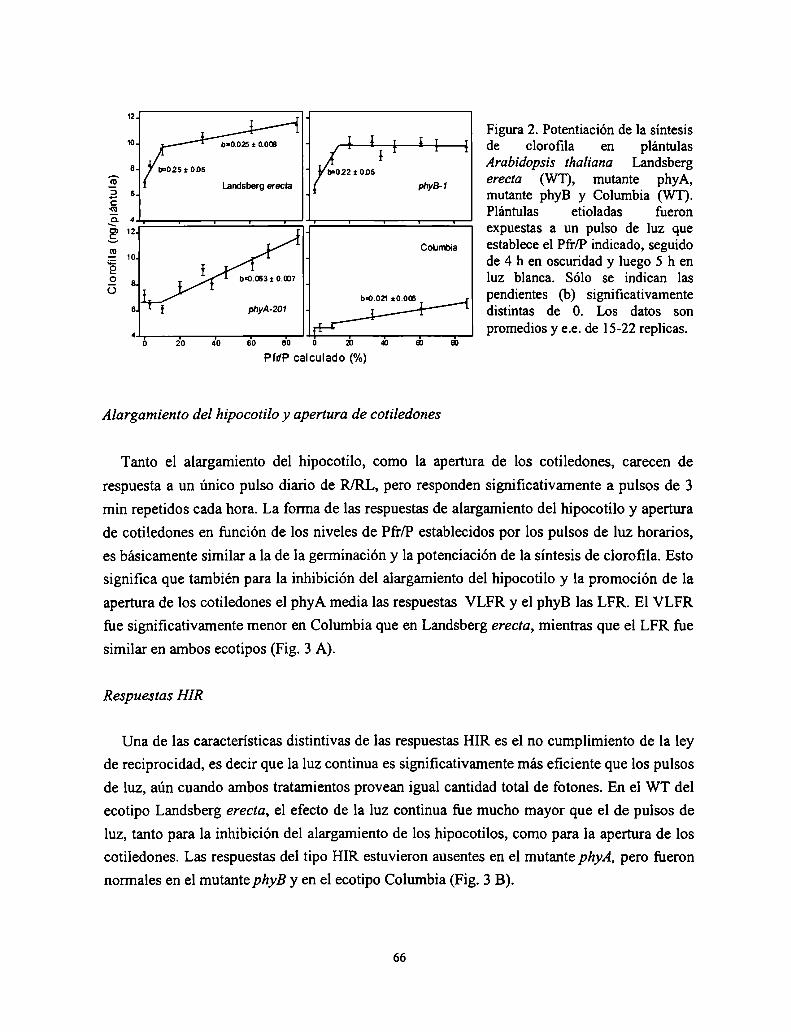
Figura 2. Potentiación de la síntesisde clorofila en plántulas
mmm“ _ “10221095 Arabzdopsís thaliana LandsbergLandsbergmeta WM erecta (WT), mutante phyA,
6- - mutante phyB y Columbia (WT).Plántulas etioladas fueronexpuestas a un pulso de luz que
Com” establece el Pfr/P indicado, seguidol de 4 h en oscuridad y luego S h en
luz blanca. Sólo se indican lasmomo“ pendientes (b) significativamente
6- Í phyA-201-H/ distintasde 0. Los datossonA promedios y e.e. de 15-22 replicas.ó 2'0 4'0 e'o a'o ó zb F a': ÏI'J
PfrfP calculado (%)
¡”0.05 2 0.0(6l
Clorofila(ng/plántula)
bI0.(B3 t o.m7 .
Alargamiento del hipocotiloy apertura de cotiledones
Tanto el alargamiento del hipocotilo, como la apertura de los cotiledones, carecen de
respuesta a un único pulso diario de R/RL, pero responden significativamente a pulsos de 3
min repetidos cada hora. La forma de las respuestas de alargamiento del hipocotilo y apertura
de cotiledones en función de los niveles de Pfr/P establecidos por los pulsos de luz horarios,
es básicamente similar a la de la germinación y la potenciación de la síntesis de clorofila. Esto
significa que también para la inhibición del alargamiento del hipocotilo y la promoción de la
apertura de los cotiledones el phyA media las respuestas VLFR y el phyB las LFR. El VLFR
fue significativamente menor en Columbia que en Landsberg erecta, mientras que el LFR fue
similar en ambos ecotipos (Fig. 3 A).
Respuestas HIR
Una de las características distintivas de las respuestas HIR es el no cumplimiento de la ley
de reciprocidad, es decir que la luz continua es significativamente más eficiente que los pulsos
de luz, aún cuando ambos tratamientos provean igual cantidad total de fotones. En el WT del
ecotipo Landsberg erecta, el efecto de la luz continua fue mucho mayor que el de pulsos de
luz, tanto para la inhibición del alargamiento de los hipocotilos, como para la apertura de los
cotiledones. Las respuestas del tipo HIR estuvieron ausentes en el mutante phyA, pero fueron
normales en el mutante phyB y en el ecotipo Columbia (Fig. 3 B).
66
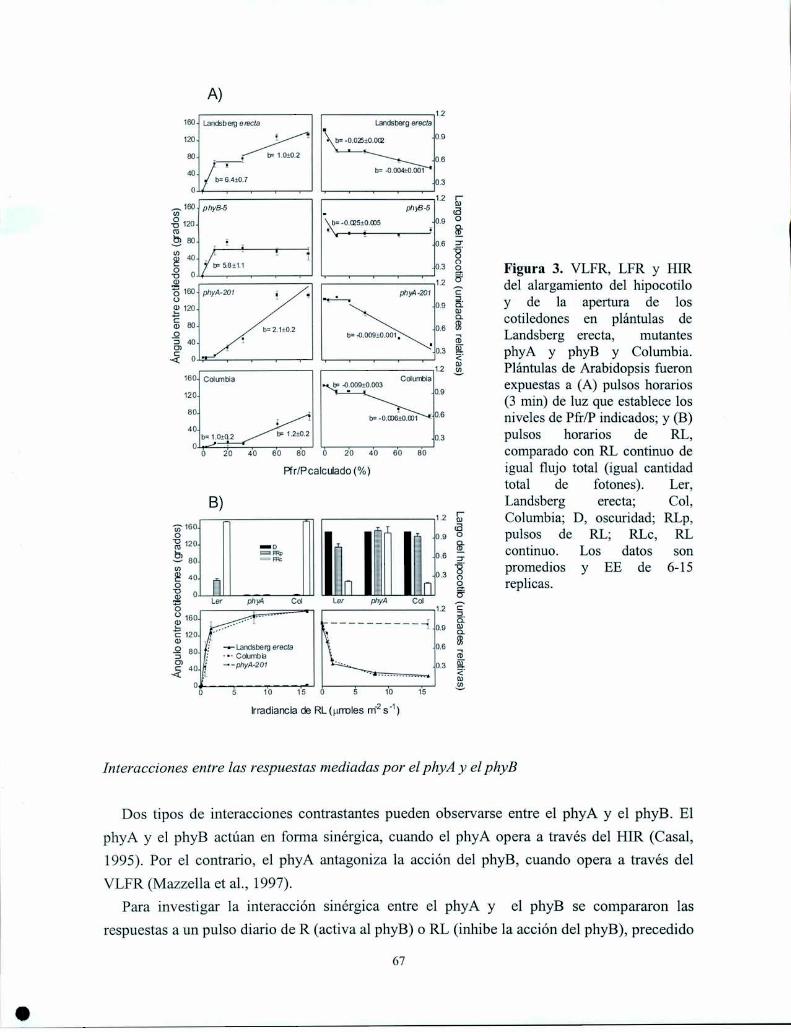
A)
160- Lanchbetg erecta ' "'" a ‘I
120- / Xb=-olozsioiooz -°-930- 1 "F"0m \ o640_ Í b: 4004:0001
0: 6,410] .0_3n v l l v I I I I I I
1 7
7;; ‘50 phyBó phyfió
8 120 \l b=-o.azsto.oo5 ‘09
Figura 3. VLFR, LFR y HIRdel alargamiento del hípocotíloy de la apertura de loscotiledones en plántulas deLandsberg erecta, mutantesphyA y phyB y Columbia.Plántulas de Arabidopsis fueron
ph yA.201
To” b= 2.1i0.2b=0009:0001.
,3
(SEADBIE’Jsepamun)qnooodmlap05121
150 Columbia Colurmia5000010.003 expuestas a (A) pulsos horarios
120_ ' .0.9 _(3 min) de luz que establece los
80- . . .“0000010.001-°-6 n1veles de Pfr/P indicados; y (B)
0- .4 0:10: 2 IF1210-2 .03 pulsos horarios de RL,./' .n 0 2'0 4'0 0'0 0'0 ó 2'0 4'0 0'0 a'o comparado con RL continuo de
Pfr/Pca|cu|ado(%) igual flujo total (igual cantidadtotal de fotones). Ler,
B) Landsberg erecta; Col,
A160 a 3': Columbia; D, oscuridad; RLp,5120 o pulsos de RL; RLc, RLg %%p É continuo. Los datos sonv 80 :FRC 2'. .
É 13 promedios y EE de 6-1540 O .
o É g replicas.8 0 5,-_, Ler phyA Col A8 5e 160-1 '
É 120. 5Q 80 +Landsberg erecta _‘
a) ' 4; - Coumb‘a %-‘- ¡4-201
¿(z 40.: Phy “2’.n '_- U)
5 1o 15 5 10 15 V
Irradiancia de RL(pm.')les m'zs'l)
Interacciones entre las respuestas mediadas por el phyAy elphyB
Dos tipos de interacciones contrastantes pueden observarse entre el phyA y el phyB. El
phyA y el phyB actúan en forma sinérgica, cuando el phyA opera a través del HIR (Casal,
1995). Por el contrario, el phyA antagoníza la acción del phyB, cuando opera a través del
VLFR (Mazzella et aL, 1997).
Para investigar la interacción sinérgica entre el phyA y el phyB se compararon las
respuestas a un pulso diario de R (activa al phyB) o RL (inhibe la acción del phyB), precedido
67
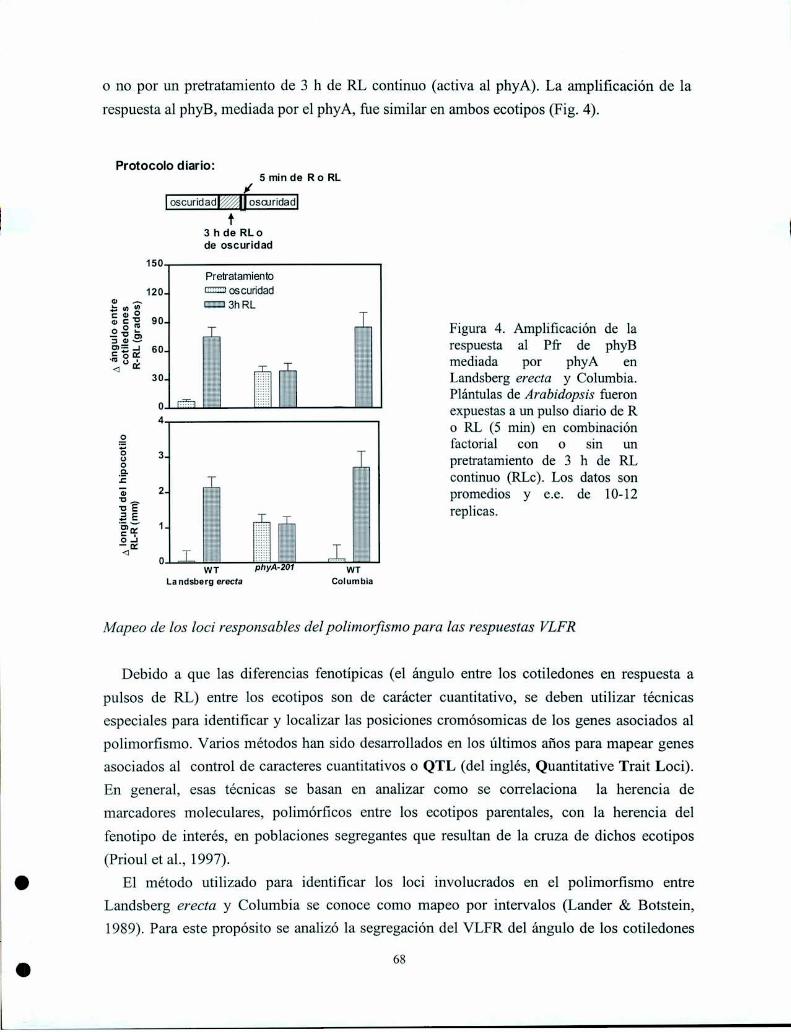
o no por un pretratamiento de 3 h de RL continuo (activa al phyA). La amplificación de la
respuesta a1phyB, mediada por el phyA, fue similar en ambos ecotipos (Fig. 4).
Protocolo diario:5 min de R o RLWE
f3 h de RLode oscuridad
1:0Pretratamientooscuridad3h RL
_¡ N o I
Figura 4. Amplificación de larespuesta al Pfr de phyBmediada por phyA enLandsberg erecta y Columbia.Plántulas de Arabidopsís fueronexpuestas a un pulso diario de Ro RL (5 min) en combinación
Aanguloentre
cotiledones
R-RL(grados)
O . .
É factorial con o sm ung 3' pretratamíento de 3 h de RLQ .E contlnuo (RLc). Los datos sonEA 2- promedios y e.e. de 10-123 E replicas.EN; 1'O _l_ n:4 o
- WT phyA-201 WTLandsberg erecta Columbia
Mapeo de los locí responsables delpolimorfismo para las respuestas VLFR
Debido a que las diferencias fenotípicas (el ángulo entre los cotiledones en respuesta a
pulsos de RL) entre los ecotipos son de carácter cuantitativo, se deben utilizar técnicas
especiales para identificar y localizar las posiciones cromósomicas de los genes asociados al
polimorfismo. Varios métodos han sido desarrollados en los últimos años para mapear genes
asociados al control de caracteres cuantitativos o QTL (del inglés, Quantitative Trait Loci).
En general, esas técnicas se basan en analizar como se correlaciona la herencia de
marcadores moleculares, polimórficos entre los ecotipos parentales, con 1a herencia del
fenotipo de interés, en poblaciones segregantes que resultan de la cruza de dichos ecotipos
(Prioul et al., 1997).
El método utilizado para identificar los loci involucrados en el polimorfismo entre
Landsberg erecta y Columbia se conoce como mapeo por intervalos (Lander & Botstein,
1989). Para este propósito se analizó 1a segregación del VLFR del ángulo de los cotiledones
68

en líneas endocriadas recombinantes o RILs (por Recombinanat Inbred Lines) generadas a
partir de la cruza de estos dos ecotipos, en relación a la segregación de un gran número de
marcadores moleculares que ya habían sido mapeados en estas líneas (Lister & Dean, 1993).
Para realizar el análisis de mapeo por intervalos se utilizó el programa MAPMAKER/QTL
(Lincoln et al., 1992; Patterson et al., 1988), junto con datos del VLFR de la apertura de los
cotiledones de 98 RILs (calculado como el ángulo promedio de los cotiledones de plantas de
cada línea recombinante expuestas a pulsos de RL menos el ángulo promedio de plantas de
dicha línea en oscuridad), y datos de la segregación de 76 marcadores en dichas líneas. Este
programa analiza en forma simultánea los efectos de diferentes grupos de marcadores ligados
sobre el carácter y calcula un índice, denominado LOD score (por likelihood odd ratio) para
cada posición del genoma analizada. El LOD score esencialmente indica el logaritmo de la
probabilidad de que efectivamente exista un QTL en ese segmento genómico en relación a la
probabilidad de que no exista. En este caso, declaramos la existencia de un QTL cuando el
LOD score excedió un valor predeterminado de 2.4 (calculado de acuerdo a (Lander and
Botstein, 1989)).
Figura 5. Mapeo de los loci VLF en"' Arabidopsis. Mapas de probabilidadesCmmt Cramsnn'az . _a - para locr asocrados a caractereso. . cuantitativos, producidos por el
programa MAPMAKER/QTL (en laabcisa solo se muestran graficados2. .
_'_/_'/\ algunosdelosmarcadoresusadosparagg; g g i; g g ¿Las 3 el mapeo). La línea punteada indica el
m ' umbral estadístico (2.4 LOD score)& emana l Cmm4 utilizado para definir la existencia de
g de un QTL (Lander and Botstein,3 L ' 1989). Las barras y las líneas8 " ' extendidas a partir de las mismas-' 2- indican los intervalos de confianza (l
n . ¿ á 1.g. ¿ _-¿- o 2 LOD)para la localizaciónde losm 3 r e i r a i ie i 5 1 = QTLs (Lander and Botstein, 1989). La
o VLF: carisma!) longitud de la abscisa corresponde a150; 60; 80; 100 y 125 cM para los
°'/\ cromosomasl a 5.2 \/__/\
vs(se;mi ¡su
¡nm
mu ¡Im uns
Los resultados indican que dos loci mayoritarios están involucrados en el control del
VLFR de la apertura de los cotiledones, uno en el cromosoma 2, que denominamos VLF], y
otro en el cromosoma 5, que denominamos VLFZ(Fig. 5). No se encontraron loci adicionales
asociados al control en oscuridad o por el VLFR, LFR o HIR de la apertura de los
69

cotiledones, inhibición del alargamiento del hipocotilo o la germinación (datos no mostrados).
Se analizaron los efectos de los alelos VLF1 y VLF2 de los dos ecotipos sobre tres
caracteres diferentes, utilizando la técnica de análisis de varianza (para el caso de la apertura
de los cotiledones o el alargamiento de los hipocotilos) o el test de chi cuadrado (en el caso de
la germinación). El alelo VLF1 de Landsberg erecta promovió el VLFR de la apertura de los
cotiledones y de la germinación de las semillas (Fig. 6). El alelo VLF2 de Landsberg erecta
promovió el VLFR de la apertura de los cotiledones y el crecimiento de los hipocotilos (Fig.
6). Ninguno de estos QTLs promovió el LFR o el HIR. No se observaron interacciones obvias
entre VLF1 y VLF2. Además, el alelo VLF1 de Landsberg erecta redujo la germinación en
oscuridad (un efecto también observado en el LFR de la germinación) y el crecimiento del
hipocotilo en oscuridad (Fig. 6).
VLF1 VLF2
p<0.01 Figura 6. Efectos alélicos de VLF1 yVLF2. Efectos alélicos principales delos loci VLFl y VLF2 sobre elVLFR y el LFR de la germinación
o. _ _ _ . (calculado sobre la base de lassemillas germinadas en oscuridad oluego de un pulso de RL,respectivamente) y el VLFR, LFR yHIR de la apertura de los cotiledonese inhibición del alargamiento delhipocotilo. Los niveles basales en loscontroles de oscuridad estánindicados. ns, no significtivo; Ler,Landsberg erecta; Col, Columbia.
Lmtsha'gerecraram Oohnüa
p<0.(D1
Genninación(lla)
Ánguloentrecdiledores
(grados)
Largoddhipomtilo(mn)
Dada la proximidad de la localización más probable del locus VLF1al marcador erecta, se
evaluó la posibilidad que los loci VLF1 y erecta fueran el mismo locus. Sin embargo, no se
encontraron diferencias en la apertura de cotiledones bajo pulsos de RL entre Columbia
ERECTA y Columbia erecta or entre Landsberg ERECTA y Landsberg erecta (datos no
mostrados). En cada caso, la magnitud de la respuesta estuvo dada por el background
Columbia o Landsberg, es decir VLF1 no es ERECTA.
Se realizaron siete cruzas utilizando Landsberg erecta como dador de polen y Columbia
como aceptor y seis cruzas en el orden recíproco. La generación F1 mostó muy poca apertura
de cotiledones bajo pulsos de RL, al igual que el parental Columbia (Figura 7).
En la generación siguiente o F2 approximadamente un 60 % de las plántulas mostró
70
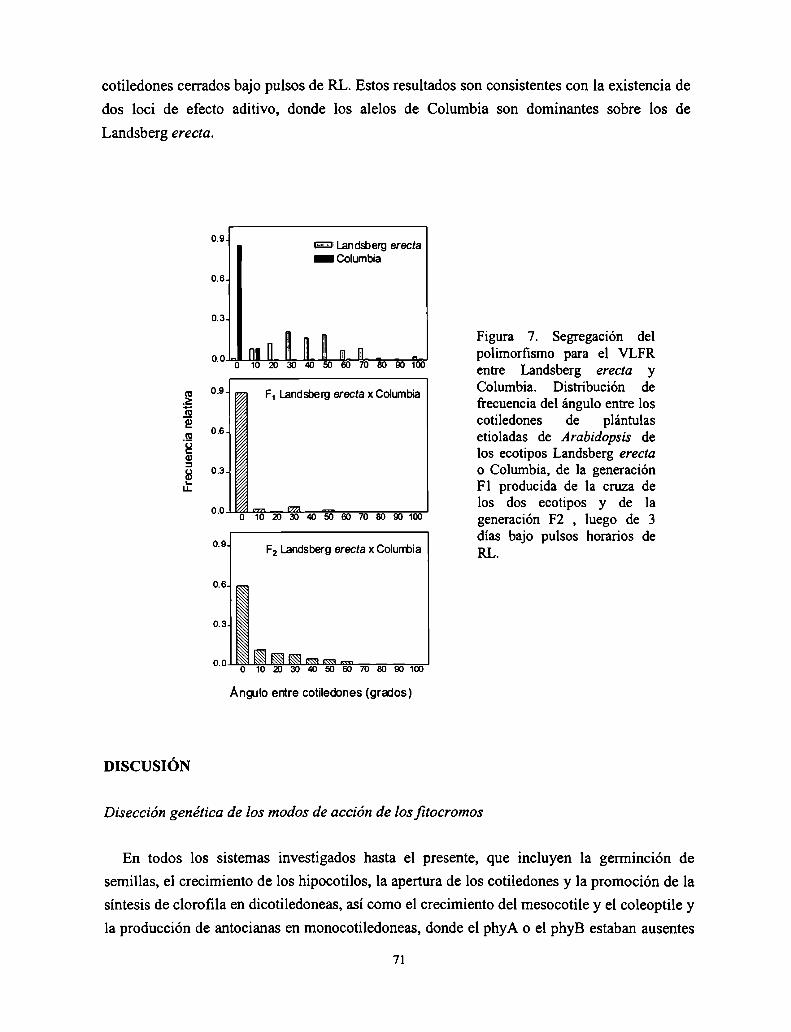
cotiledones cerrados bajo pulsos de RL. Estos resultados son consistentes con la existencia de
dos loci de efecto aditivo, donde los alelos de Columbia son dominantes sobre los de
Landsberg erecta.
asi Landsberg erecta- Columbia
Figura 7. Segregacíón delpolimorfismo para el VLFRentre Landsberg erecta yColumbia. Distribución defrecuencia del ángulo entre loscotiledones de plántulasetioladas de Arabidopsis delos ecotipos Landsberg erectao Columbia, de la generaciónFl producida de la cruza delos dos ecotipos y de lageneración F2 , luego de 3días bajo pulsos horarios de
F2 Landsberg erecta x Colun'bia RL.
Frecuenciarelativa
0.9
Angulo entre cotiledones (grados)
DISCUSIÓN
Disección genética de los modos de acción de losfitocromos
En todos los sistemas investigados hasta el presente, que incluyen la germinción de
semillas, el crecimiento de los hipocotilos, la apertura de los cotiledones y la promoción de la
síntesis de clorofila en dicotiledoneas, así como el crecimiento del mesocotile y el coleoptile y
la producción de antocianas en monocotiledoneas, donde el phyA o el phyB estaban ausentes
7l

o sobreexpresados, los resultados son consistentes con la participación del phyA en el VLFR
(Casal et al., 1994; Botto et al., 1996; Clough et al., 1995; Casal et al., 1996; Shinomura et al.,
1996; Mazzella et al., 1997) y el HIR (Dehesh et al., 1993; Nagatani et al., 1993; Parks and
Quail, 1993; Whitelam et al., 1993; van Tuinen et al., 1995; Casal et al., 1996), y del phyB en
el LFR (McCormac et al., 1993; Botto et al., 1995; Mazzella et al., 1997). Los loci VLF están
selectivamente involucrados en el VLFR, y no afectan (por lo menos en la misma dirección)
el HIR o el LFR. De esta forma, usando los mutantes phyA y phyB, asi como los loci VLF
polimórficos entre Landsberg erecta y Columbia, los tres modos de acción del fitocromo han
sido separados genéticamente en este trabajo por primera vez.
Posición jerarquica de los loci VLF]y VLF2en la cadena de transducción de señales de los
fitocromos
Los loci VLF no afectan al phyA en si mismo, más bien operan río abajo de este
fotorreceptor. Esta afirmación se basa en que: 1) VLF] y VLF2 mapean en los cromosomas 2
y 5 respectivamente, mientras que el gen que codifica para la apoproteína del phyA está en el
cromosoma 1; 2) los ecotipos Landsberg erecta y Columbia no muestran diferencias
significativas en los niveles inmudetectables o espectrales de phyA ((Whitelarn et al., 1993) y
datos no mostrados). Los efectos selectivos de los loci VLF en el VLFR sugieren que lacadena de transducción de señales asociada al VLFR involucra elementos distintos de la usada
por el LFR y el HIR o es iniciciada en distintos tipos celulares. Los productos de los loci VLF
podn’an ser componentes específicos de la vía de transducción de señales asociada al VLFR o
modificar la actividad de alguno de sus componentes. Loci como los VLF, donde los alelos
dominantes tienen efectos negativos sobre las respuestas, podrían estar involucrados en la
cadena de transducción de señales si, por ejemplo, codifican para subunidades de una proteína
multimérica. El complejo proteico se vería inactivado en este caso por la presencia del alelo
del ecotipo Columbia. Altemativamente, los loci VLFpodrían estar directamente involucrados
en la represión de las respuestas VLFR. En ese caso, el alelo de Landsberg erecta carecería de
efecto en la respuesta negativa.
Debido a su efecto específico sobre el VLFR los productos de los loci VLF debieran ser
clasificados como componentes relativamente tempranos de la cadena de transducción de
señales del phyA. Sin embargo, el hecho que los loci VLF no afecten simultáneamente todas
las respuestas VLFR (el VLF] afecta, por lo menos la apertura de los cotiledones y el
crecimiento de los hipocotilos pero no afecta la germinación y el VLFZafecta por lo menos, la
apertura de cotiledones y la germinación de semillas pero no el crecimiento de los
hipocotilos) sugiere que no estarían implicados en los primeros eventos de la cadena de
transducción de señales del phyA. Los loci VLF tampoco parecen afectar la interacción
72

negativa entre el VLFR y el LFR. Esta interacción negativa se evidencia en que las pendientes
de las regresiones en el rango del LFR son generalmente más empinadas cuando el VLFR está
ausente, es decir son mayores en el mutante phyA que en el WT del ecotipo Landsberg erecta
(Botto et al., 1996; Mazzella et al., 1997 y Figuras 2 y 3). El hecho que no se observen
diferencias en las pendientes del LFR entre Landsberg erecta y Columbia, sugire que los loci
VLF] y VLF2 operan río abajo de los elementos del VLFR que interactúan negativamente conel LFR.
Luz continua versus pulsos de luz
La idea de que diferentes mecanismos podrían estar involucrados en las respuestas
mediadas por el fitocromo a pulsos de luz y a luz continua (en particular RL) data de varias
décadas (de Lint, 1957; Hendricks et al., 1968; Negbi and Koller, 1964). La mayoría de los
modelos considera que el Pfr es la forma activa del fitocromo involucrada en la percepción de
los pulsos de luz, mientras que otro componente(s) (el ciclado entre Pfr y Pr, algún estado
exitado del Pfr, etc.) podría estar involucrado en la respuesta a la luz continua (Heim and
Scháfer, 1984; Johnson and Tasker, 1979). Los loci VLFI y VLF2 de Arabidopsís afectan la
acción del phyA en respuesta a pulsos de RL (VLFR), pero no en respuesta a luz RL continua
(HIR) y constituyen la evidencia más contundente para sostener la hipótesis que existen
diferencias en los mecanismos moleculares de las respuestas a pulsos de luz o a luz continua.
Si se lograra clonar los loci VLF,aspectos importantes de la cadena de transducción de
señales del phyA podrían ser revelados, proporcionando un ejemplo de como los conceptos
fisiológicos tradicionales (los modos de acción del fitocromo) ofrecen una base sólida para la
utilización de las nuevas herramientas genéticas y moleculares.
MATERIALES Y MÉTODOS
Material Vegetal
Las semillas de Arabidopsís thalíana (L.) Heynh de los ecotipos Landsberg erecta y
Columbia fueron proporcionadas por los Drs R. E. Kendrick and M. Koomneef (Wageningen
University, The Netherlands) y por el Arabidopsís Biological Research Center de la
Universidad de Ohio (ABRC), respectively. Batches adicionales de ambos ecotipos fueron
comprados a la compañía Lehle Seeds (Texas). Las semillas de los mutantes phyA-201 y
phyA-I fueron proporcionadas por los Drs J. Chory (The Salk Institute, La Jolla, California) y
G. Whitelam (Department of Botany, University of Leicester, UK) respectivamente, mientras
73

que las semillas de los mutantes phyB-I y phyB-5 fueron proporcionadas por el ABRC. Las
semillas de las 98 líneas endocriadas recombínantes fueron obtenidas por los Dres Drs C.
Lister and C. Dean (Lister and Dean, 1993) usando Landsberg erecta y Columbia como
ecotipos parentales y fueron proporcionadas por el ABRC. Aproximadamente 25 (en los
experimentos de germinación), 80 (en los de clorofila) o 15 (en los experimentos de
crecimiento del hipocotilo y apertura de los cotiledoness) semillas de cada ecotipo fueron
sembradas en cajas plásticas transparentes (40 x 33 mm2 x 15 mm de altura) sobre dos capas
de papel de filtro saturado con agua destilada. Las líneas endocriadas recombínantes fueron
sembradas sobre agar en placas de elisa y colocadas dentro de cajas con cubiertas
transparentes. Las cajas fueron icubadas por 3 d a 7 °C.
Procedimientos experimentalesy observaciones
Para los experimentos de germinación, las semillas previamente incubadas 3 d a 7 °C
fueron llevadas a 35 °C por 4, 8 o 24 hr en oscuridad, expuestas a pulsos de R/RL que
establecían distintos niveles de Pfr/P, y transferidas a oscuridad por 4 d a 25 °C, antes de
contar las semillas germinadas (Botto et al., 1995, 1996). En los experimentos de
crecimientos del hipocotilo y apertura de los cotiledones, las semillas fireron expuestas a un
pulso de R (para inducir la germinación), incubadas en oscuridad por 24 hr a 25 °C y
transferidas a los tratamientos lumínicos o a oscuridad por 3 d. El largo del hipocotilo de las
10 plántulas más altas de cada caja (siendo cada caja una repetición) fire medido con regla de
presición de 0.5 mm y fueron promediados (Casal, 1995). El ángulo entre los cotiledones fue
medido con un transportador en las mismas plantas utilizadas para medir el largo de los
cotiledones y los diez datos obtenidos por caja también fueron promediados (constituyendo
una repetición) previamente a realizar el análisis estadístico. En los experimentos de
clorofila, las semillas fueron expuestas a un pulso de R (para inducir la germinación),
incubadas en oscuridad por 3 d a 25 °C, expuestas a pulsos de 5 min de R/RL que establecían
diferentes Pfr/P, incubadas en oscuridad por 4 h a 25 °C, y finalmente transferidas por 5 h a
una cámara de luz blanca fluorescente (100 umol m'2 s") . La clorofila fue determinada luego
de la extracción de la misma con N N’ dimetilforrnamida (Moran, 1982). El número exacto de
plántulas fue luego contado para expresar los datos de clorofila sobre la base de una plántula.
Fuentes de luz
Las fuentes de luz utilizadas para dar los pulsos saturantes (5-10 min, por lo menos 2.5
mmol m'z) o subsaturantes (RG9 RL), pulsos horarios (3 min/h, de por lo menos ll pmol rn'2
s") y luz R o RL continua fueron iguales a las descriptas previamente por Casal y col. (Botto
74

et al., 1995; Casal, 1995; Casal et al., 1991). Estas fuentes de R, RL o mezclas de R y RL
proporcionan distintos niveles calculados de Pfr/P (Casal et al., 1991). Para la germinación de
semillas, los espectros de las fuentes de luz fueron multiplicados por un factor que corregía
por la atenuación de la transmitancia causada por la cubierta de las semillas (Botto et al.,
1995).
Mapeo
El ángulo entre los cotiledones y el largo del hipocotilo fue medido en plántulas de las 98
líneas endocriadas recombinantes cultivadas ya sea en oscuridad, bajo pulsos horarios de 3
min de RL o R+RL (Pfr/P calculado= lO y 61%, respectivamente) o bajo luz RL continua (l
umol m'2 s"). La tasa de germinación fue estimada en semillas de las líneas endocriadas
recombinantes incubadas por 3 d a 7 °C y por 4 h a 35 °C, y expuestas a R, RL o a ningún
tratamiento lumínico (control de oscuridad) seguidos de 4 d en oscuridad a 25 °C. El VLFR
(la diferencia entre el pulso de RL y la oscuridad), el LFR (la diferencia entre pulsos de R+RL
y pulsos de RL) y el HIR (la diferencia entre RL continuo y pulsos de RL) fue calculada para
el crecimiento del hipocotilo y la apertura de los cotiledones. El valor promedio de cada línea
para cada carácter fue usado para el análisis genético por medio del programa
MAPMAKEPJQTL (Lincoln et al., 1992; Patterson et al., 1988) en combinación con 76
marcadores, cuya segregación en las líneas fue obtenida a partir del Arabidopsis thaliana
Genome Center (University of Pennsylvania). Se utilizó la técnica de análisis de varianza para
testear los efectos de cada alelo de los loci identificados sobre el crecimiento del hipocotilo y
la apertura de los cotiledones en oscuridad o en el VLFR, LFR y HIR. Se utilizó el test de
chi-cuadrado para testear el efecto de los alelos de cada loci sobre la germinación de las
semillas en oscuridad, en respuesta al VLFR (la diferencia entre la germinación bajo pulsos
de RL y oscuridad) y al LFR (la diferencia entre el pulso de R y RL). Un grupo de ligamiento
de tres marcadores fue usado para analizar los efectos alélicos, incluyendo sólo las líneas que
tenían los grupos alélicos de un mismo ectotipo para estos marcadores.

MODOS DE ACCIÓN DEL FITOCROMO A (II)
ANÁLISIS DE LOS MODOS DE ACCION DE LOS FITOCROMOS
EN LAS MUTANTESfhy] yfhy3
INTRODUCCION
El aislamiento y la caracterización de mutantes que presentan respuestas fotomorfogénicas
alteradas sin afectar a los fotorreceptores, ha constituido una poderosa herramienta para
identificar componentes de las vías de transducción de señales lumínicas. Existen tres tipos de
mutantes con estas características. El primero tiene respuestas fotomorfogénicas constitutivas, es
decir que se comporta en oscuridad como si estuviera a la luz. El segundo tiene reducida la
sensibilidad a la luz y se comporta a la luz como si estuviera en oscuridad. El último grupo
presenta una respuesta exagerada a la luz.
De los mutantes con una sensibilidad lumínica reducida, algunos como el hy5 afectan
simultáneamente las respuestas a varias longitudes de onda. Este tipo de mutantes probablemente
afecte componentes tardíos de la cadena de transducción de señales lumínicas comunes a varios
fotorreceptores. Otros mutantes, sin embargo, sólo afectan las respuestas a longitudes de onda
específicas. Por ejemplo, los mutantes red], pef2 y pef3 presentan disminuidas las respuestas al
rojo en el rango del LFR, pero no las respuestas al RL. Los mutantes fhyl y jhy3, en cambio,
presentan respuestas disminuidas al RL y no al R en el rango del LFR. Dado que el phyB media
las respuestas al R en el rango del LFR y el phyA las respuestas al RL, la existencia de mutantes
que afectan diferencialmente estas respuestas sugiere que estos fotorreceptores actúan a través de
vías de transducción de señales diferentes. La posibilidad de disectar genéticamente el LFR
mediado por el phyB de las respuestas medíadas por el phyA, es consistente con la hipótesis que
sostiene que los modos de acción de los fitocromos son el reflejo de la existencia de vías distintasde transducción de señales.
El phyA actúa a través de dos modos de acción, el VLFR y el HIR. Muy poco se ha hecho
hasta ahora para analizar si existen loci que afecten específicamente a las respuestas VLFR y no a
las HIR y viceversa. Las primeras evidencias en tal dirección fueron presentadas en el capítulo
previo. Las mutantesjhy] yfhy3 afectan las respuestas mediadas por el phyA. Estas mutantes han
sido muy poco caracterizadas fotobiológicarnente y desconocemos si afectan los dos modos de
76
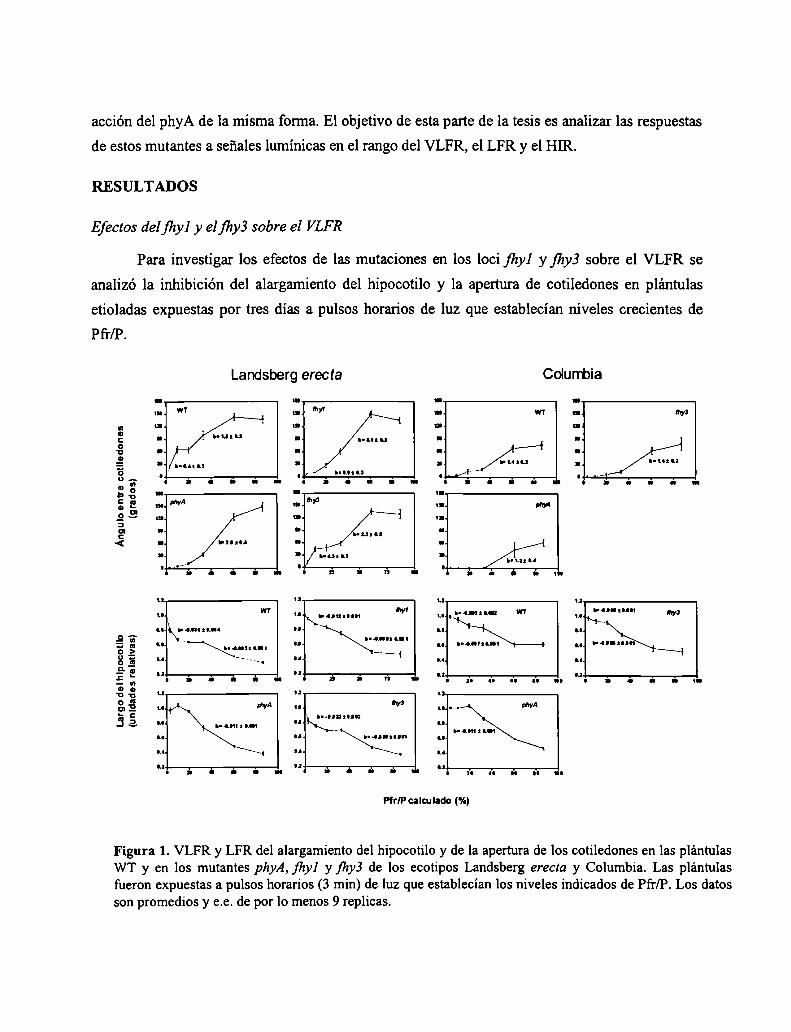
acción del phyA de la misma forma. El objetivo de esta parte de la tesis es analizar las respuestas
de estos mutantes a señales lumínicas en el rango del VLFR, el LFR y el HlR.
RESULTADOS
Efectos delfliyl y elflty3 sobre el VLFR
Para investigar los efectos de las mutaciones en los loci jhyI y jhy3 sobre el VLFR se
analizó la inhibición del alargamiento del hipocotilo y la apertura de cotiledones en plántulas
etioladas expuestas por tres días a pulsos horarios de luz que establecían niveles crecientes de
Pfr/P.
Landsberg erecta Colurrbla
un. v" a. n. WT a My:
n un. m. ¡a DJg . “tuu . . ..1°: I o o ",4 n.g ’ b-¡uu ’ a ¡uuu ._ ¡uuuo A ’ . ¡no.nu _ l" ‘ _ ¡g ï . Ó . - I. I . . . Í. . . - . Í. . . - . Í.o ¡a u '- ...5 1! m "7‘ a ma 1a. W2 3 u, m Ki ¡a= ng ' u-mu .
.4 g han“ I a {
A .zn n a n u l a i Y! II n o o ó 1p
u vn u \_“‘“"“ M u .:.,..-g¡...¡ w-r u ¿“una my,
u Ku-¿Clint-l u k u N\1\1—‘ u.9 Á ':‘6E M ‘N_“u‘| H . :Ï'Ï:m u sum-unn u hasta“. i8 fi u \r-»t,. H 1 u u2-3 ., ..f 5 I . - . I. á . n '. ¡O CI I. I. Í. I . . I l.o o u.u v ..8, 5 u - W‘ 1! m u A-o WAu c ‘ ¡u-nnauu3 er n _ ¡adjust-1 u u “im ‘m
t. CI. II.I.-QII" t. R
0.4 \\1 U \»\. 0.4a a n n u n 5 I I I II n n u u u n
PfrIP calculado ('l.)
Figura 1. VLFR y LFR del alargamiento del hipocotilo y de la apertura de los cotiledones en las plántulasWT y en los mutantes phyA, flzyl y fliy3 de los ecotipos Landsberg erecta y Columbia. Las plántulasfueron expuestas a pulsos horarios (3 mín) de luz que establecían los niveles indicados de Pfr/P. Los datosson promedios y e.e. de por lo menos 9 replicas.
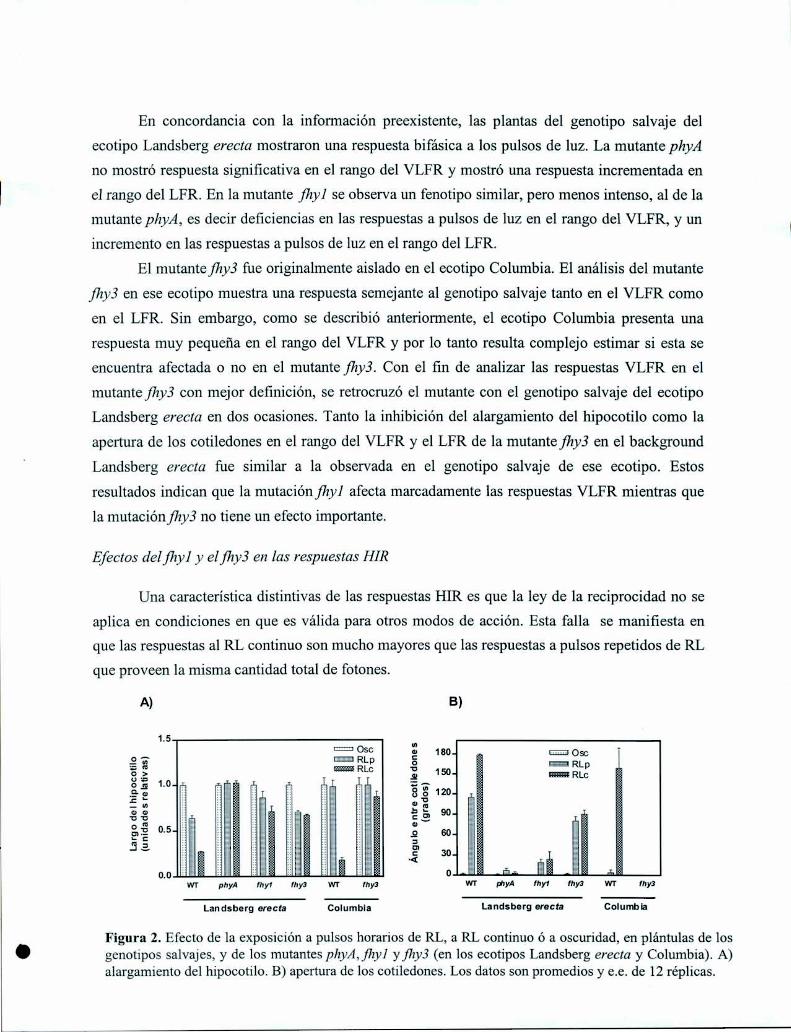
En concordancia con la información preexistente, las plantas del genotipo salvaje del
ecotipo Landsberg erecta mostraron una respuesta bifásica a los pulsos de luz. La mutante phyA
no mostró respuesta significativa en el rango del VLFR y mostró una respuesta incrementada en
el rango del LFR. En la mutante fhy] se observa un fenotipo similar, pero menos intenso, al de la
mutante phyA, es decir deficiencias en las respuestas a pulsos de luz en el rango del VLFR, y un
incremento en las respuestas a pulsos de luz en el rango del LFR.
El mutante fhy3 fue originalmente aislado en el ecotipo Columbia. El análisis del mutante
flzy3 en ese ecotipo muestra una respuesta semejante al genotipo salvaje tanto en el VLFR como
en el LFR. Sin embargo, como se describió anteriormente, el ecotipo Columbia presenta una
respuesta muy pequeña en el rango del VLFR y por lo tanto resulta complejo estimar si esta se
encuentra afectada o no en el mutante jhy3. Con el fin de analizar las respuestas VLFR en el
mutante flzy3 con mejor definición, se retrocruzó el mutante con el genotipo salvaje del ecotipo
Landsberg erecta en dos ocasiones. Tanto la inhibición del alargamiento del hipocotilo como la
apertura de los cotiledones en el rango del VLFR y el LFR de la mutante fhy3 en el background
Landsberg erecta fue similar a la observada en el genotipo salvaje de ese ecotipo. Estos
resultados indican que la mutación fhy] afecta marcadamente las respuestas VLFR mientras que
la mutación fhy3 no tiene un efecto importante.
Efectos delflzy] y elflzy3 en las respuestas HIR
Una caracteristica distintivas de las respuestas HIR es que la ley de la reciprocidad no se
aplica en condiciones en que es válida para otros modos de acción. Esta falla se manifiesta en
que las respuestas al RL continuo son mucho mayores que las respuestas a pulsos repetidos de RL
que proveen la misma cantidad total de fotones.
A) B)
g 180
“ 150¿A.9- 9 3 3 120Í (la q, gI8 8 3gg 90
m 0g) 33 ,9 60|- ¡: a3 3' 8’ 30
"< 333;
o a H 5555 -:-: ‘
phyA fhy‘! fhy3 wr nm WT phyA nun "un wr fhyJ
Landsberg erecta Columbia LaHÓSbergafecta COlumbia
Figura 2. Efecto de la exposición a pulsos horarios de RL, a RL continuo ó a oscuridad, en plántulas de los. genotipos salvajes, y de los mutantes phyA,jhy] yfhy3 (en los ecotipos Landsberg erecta y Columbia). A)
alargamiento del hipocotilo. B) apertura de los cotiledones. Los datos son promedios y e.e. de 12 réplicas.
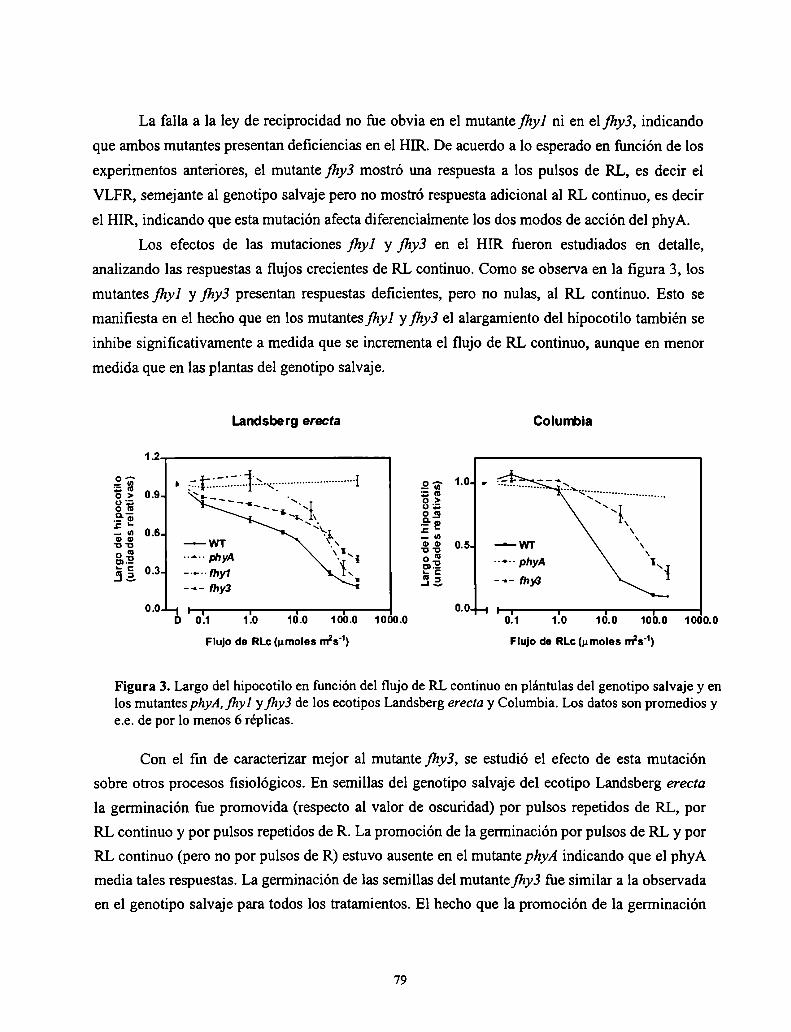
La falla a la ley de reciprocidad no fue obvia en el mutante flin ni en elflzy3, indicando
que ambos mutantes presentan deficiencias en el HIR. De acuerdo alo esperado en función de los
experimentos anteriores, el mutante flzy3 mostró una respuesta a los pulsos de RL, es decir el
VLFR, semejante al genotipo salvaje pero no mostró respuesta adicional al RL continuo, es decir
el HIR, indicando que esta mutación afecta diferencialmente los dos modos de acción del phyA.
Los efectos de las mutaciones fliyl y fliy3 en el HIR fueron estudiados en detalle,
analizando las respuestas a flujos crecientes de RL continuo. Como se observa en la figura 3, los
mutantes fhy] y fhy3 presentan respuestas deficientes, pero no nulas, al RL continuo. Esto se
manifiesta en el hecho que en los mutantes jhy] yfhy3 el alargamiento del hípocotilo también se
inhibe significativamente a medida que se incrementa el flujo de RL continuo, aunque en menor
medida que en las plantas del genotipo salvaje.
Landsberg erecta Colurrbia
1 9
gg a ...... .........................“I g? 1.o.Ío > 0.9 .. mu ':. o 3o g g 7€Ee si3 g o s f cn1: 1:: -—WT g .3 0.5.3.5 - o s¡- C 9...S a 0.3 _....my1 5 g
--A- fhy3 V L0.0 - 0.0- ;D 0.1 1Ïo 16.0 100.0 1000.0 0Ï1 1Ïo 16.0 100.0 1oco.o
Flujo do RLe(pmoles mzs") Flujo de RLe(pmoles mas")
Figura 3. Largo del hípocotilo en función del flujo de RL continuo en plántulas del genotipo salvaje y enlos mutantes phyA,fhyl yjhy3 de los ecotipos Landsberg erecta y Columbia. Los datos son promedios ye.e. de por lo menos 6 réplicas.
Con el fin de caracterizar mejor al mutante fhy3, se estudió el efecto de esta mutación
sobre otros procesos fisiológicos. En semillas del genotipo salvaje del ecotipo Landsberg erecta
la germinación fiie promovida (respecto al valor de oscuridad) por pulsos repetidos de RL, por
RL continuo y por pulsos repetidos de R. La promoción de la germinación por pulsos de RL y por
RL continuo (pero no por pulsos de R) estuvo ausente en el mutante phyA indicando que el phyA
media tales respuestas. La germinación de las semillas del mutantefhy3 fue similar a la observada
en el genotipo salvaje para todos los tratamientos. El hecho que la promoción de la germinación
79
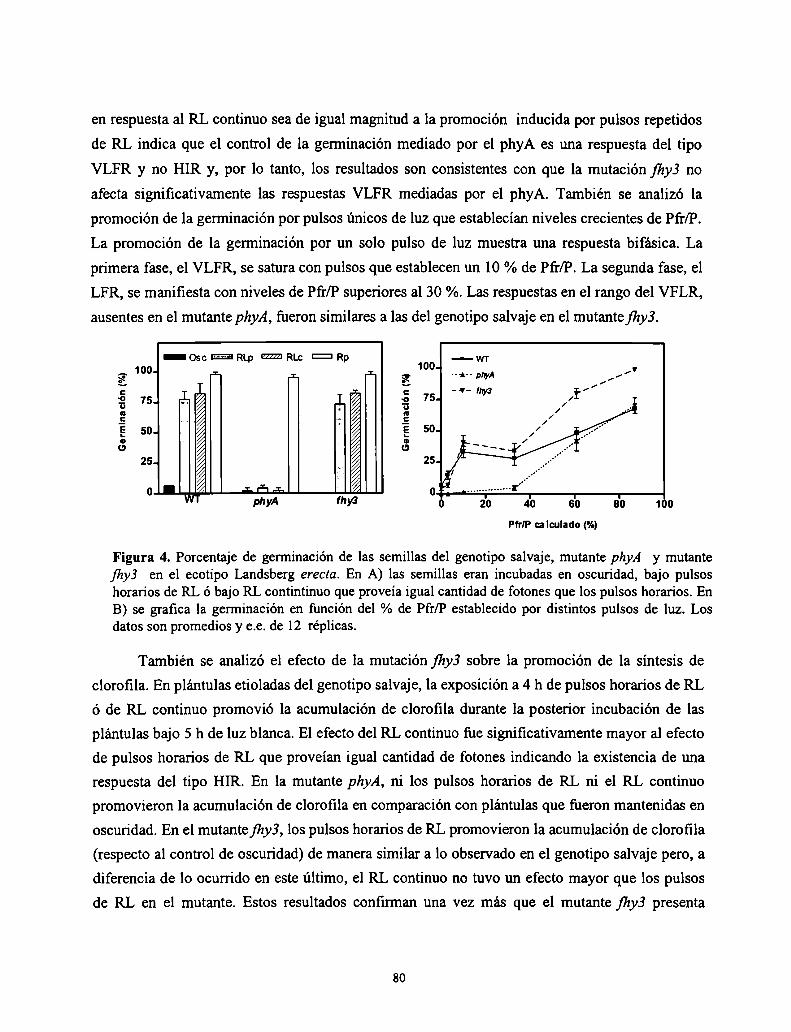
en respuesta al RL continuo sea de igual magnitud a la promoción inducida por pulsos repetidos
de RL indica que el control de la germinación mediado por el phyA es una respuesta del tipo
VLFR y no HIR y, por lo tanto, los resultados son consistentes con que la mutación fliy3 no
afecta significativamente las respuestas VLFR mediadas por el phyA. También se analizó la
promoción de la germinación por pulsos únicos de luz que establecían niveles crecientes de Pfr/P.
La promoción de la germinación por un solo pulso de luz muestra una respuesta bifásica. La
primera fase, el VLFR, se satura con pulsos que establecen un 10 % de Pfr/P. La segunda fase, el
LFR, se manifiesta con niveles de Pfr/P superiores al 30 %. Las respuestas en el rango del VFLR,
ausentes en el mutante phyA, fiJeron similares a las del genotipo salvaje en el mutante jhy3.
.n
NOUIO
UIO
Germlnaclón(%) Germlnaclón(7)
N U'I
PfrlP a Ieulado (7.)
Figura 4. Porcentaje de germinación de las semillas del genotipo salvaje, mutante phyA y mutantefliy3 en el ecotipo Landsberg erecta. En A) las Semillas eran incubadas en oscuridad, bajo pulsoshorarios de RL ó bajo RL contintinuo que proveía igual cantidad de fotones que los pulsos horarios. EnB) se grafica la germinación en función del % de Pfr/P establecido por distintos pulsos de luz. Losdatos son promedios y e.e. de 12 réplicas.
También se analizó el efecto de la mutación jhy3 sobre la promoción de la síntesis de
clorofila. En plántulas etioladas del genotipo salvaje, la exposición a 4 h de pulsos horarios de RL
ó de RL continuo promovió la acumulación de clorofila durante la posterior incubación de las
plántulas bajo 5 h de luz blanca. El efecto del RL continuo fue significativamente mayor al efecto
de pulsos horarios de RL que proveían igual cantidad de fotones indicando la existencia de una
respuesta del tipo HIR. En la mutante phyA, ni los pulsos horarios de RL ni el RL continuo
promovieron la acumulación de cloroñla en comparación con plántulas que fueron mantenidas en
oscuridad. En el mutantejhy3, los pulsos horarios de RL promovieron la acumulación de clorofila
(respecto al control de oscuridad) de manera similar a lo observado en el genotipo salvaje pero, a
diferencia de lo ocurrido en este último, el RL continuo no tuvo un efecto mayor que los pulsos
de RL en el mutante. Estos resultados confirman una vez más que el mutante jhy3 presenta
80
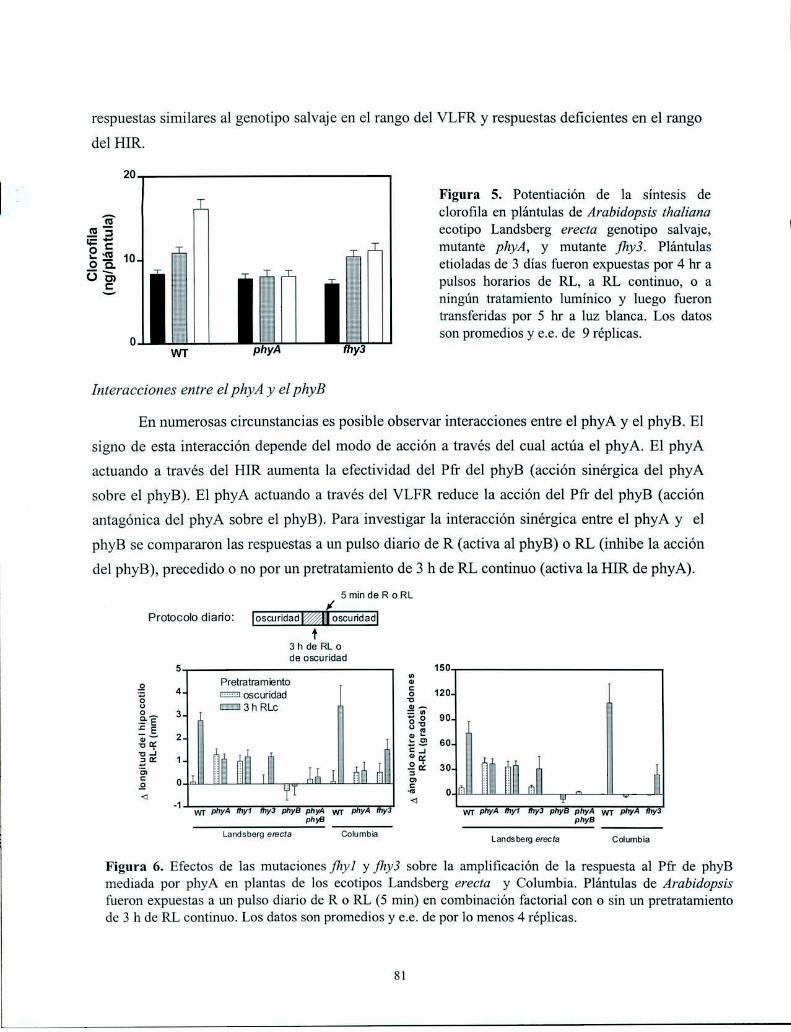
respuestas similares al genotipo salvaje en el rango del VLFR y respuestas deficientes en el rango
del HIR.
90
Cloroflla
(ng/plántula)
:5
Interacciones entre elphyAy elphyB
WT phyA fl1y3
Figura 5. Potentiación de la síntesis declorofila en plántulas de Arabídopsis thalianaecotipo Landsberg erecta genotipo salvaje,mutante phyA, y mutante fhy3. Plántulasetioladas de 3 días fueron expuestas por 4 hr apulsos horarios de RL, a RL continuo, o aningún tratamiento lumínico y luego fuerontransferidas por 5 hr a luz blanca. Los datosson promedios y e.e. de 9 réplicas.
En numerosas circunstancias es posible observar interacciones entre el phyA y el phyB. El
signo de esta interacción depende del modo de acción a través del cual actúa el phyA. El phyA
actuando a través del HIR aumenta la efectividad del Pfr del phyB (acción sinérgica del phyA
sobre el phyB). El phyA actuando a través del VLFR reduce la acción del Pfr del phyB (acción
antagónica del phyA sobre el phyB). Para investigar la interacción sinérgica entre el phyA y el
phyB se compararon las respuestas a un pulso diario de R (activa al phyB) o RL (inhibe la acción
del phyB), precedido o no por un pretratamiento de 3 h de RL continuo (activa la HIR de phyA).
Protocolo diario:
Alongituddelhipocotilo
RL-R(mm)
5mindeRoRLWBf
3 h de RL ode oscuridad
Pretratramientooscuridad3 h RLc
wr phyA M1 M3 phyB pag wr phyA ¡hy-1p
Landsberg electa Columbia
Aánguloentrecotiledones
R-RL(grados)
p phyA wr " yphyB
Landsberg erecta Columbia
Figura 6. Efectos de las mutaciones fhy] y fhy3 sobre la amplificación de la respuesta al Pfr de phyBmediada por phyA en plantas de los ecotipos Landsberg erecta y Columbia. Plántulas de Arabidopsisfueron expuestas a un pulso diario de R o RL (5 min) en combinación factorial con o sin un pretratamientode 3 h de RL continuo. Los datos son promedios y e.e. de por lo menos 4 réplicas.
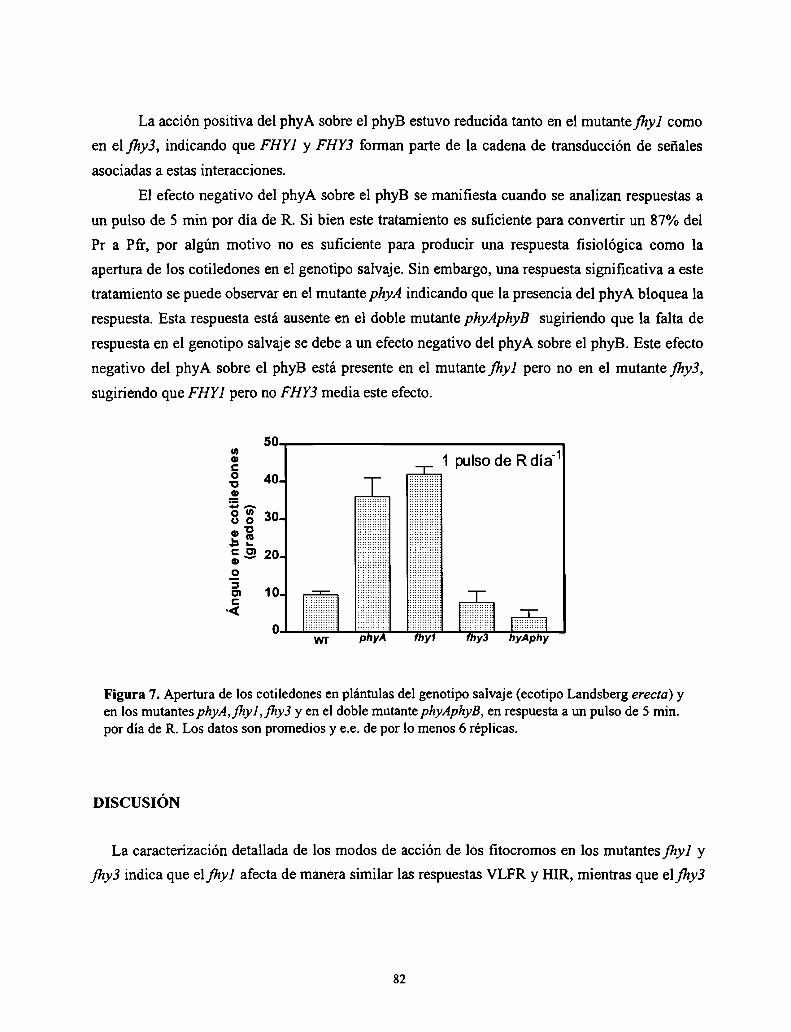
La acción positiva del phyA sobre el phyB estuvo reducida tanto en el mutante fhyl como
en el fhy3, indicando que FHYI y FH Y3 forman parte de la cadena de transducción de señales
asociadas a estas interacciones.
El efecto negativo del phyA sobre el phyB se manifiesta cuando se analizan respuestas a
un pulso de 5 min por día de R. Si bien este tratamiento es suficiente para convertir un 87% del
Pr a Pfr, por algún motivo no es suficiente para producir una respuesta fisiológica como la
apertura de los cotiledones en el genotipo salvaje. Sin embargo, una respuesta significativa a este
tratamiento se puede observar en el mutante phyA indicando que la presencia del phyA bloquea la
respuesta. Esta respuesta está ausente en el doble mutante phyAphyB sugiriendo que la falta de
respuesta en el genotipo salvaje se debe a un efecto negativo del phyA sobre el phyB. Este efecto
negativo del phyA sobre el phyB está presente en el mutante fhyl pero no en el mutante fliy3,
sugiriendo que FH Y] pero no FHY3 media este efecto.
nn
1 pulso de R día’1
Ánguloentrecotiledones
(grados)
Figura 7. Apertura de los cotiledones en plántulas de] genotipo salvaje (ecotipo Landsberg erecta) yen los mutantes phyA,/hy1,fhy3 y en el doble mutante phyAphyB, en respuesta a un pulso de 5 min.por día de R. Los datos son promedios y e.e. de por lo menos 6 réplicas.
DISCUSIÓN
La caracterización detallada de los modos de acción de los fitocromos en los mutantes fhy] y
jhy3 indica que elflzy] afecta de manera similar las respuestas VLFR y HIR, mientras que eljhy3
82

afecta las respuestas HIR y no afecta, ó afecta muy poco (dependiendo de la respuesta analizada),
las respuestas VLFR.
El hecho que la mutación fhy3 afecte significativamente a las respuestas HIR y no afecte
sustancialmente las respuestas VLFR sugiere que estos dos modos de acción del phyA serian el
reflejo de dos vías de transducción de señales distintas activadas por el phyA y el FHY3
participar-íaprincipalmente en la correspondiente al HIR. La observación que el fhyI afecta los
dos modos de acción desencadenados por el phyA no argumenta en contra de la posibilidad de
que los mismos representen distintas vias de transducción de señales. En Drosophila
melanogaster y en Caenorbditis elegans, diferentes miembros de la familia de receptores del
factor transformante del crecimiento B, activan miembros especificos de la familia de
transductores de señales Smad. Estos Smads específicos (Smads l, 2, 3 o 5) luego se asocian con
un Smad común (Smad4) y se translocan al núcleo donde regulan la transcripción. FHYl podría
ser un componente regulatorio común al VLFR y al HIR, de manera homóloga a la acción de
Smad4 en los sistemas recién descriptos.
Algunos autores han descripto efectos diferenciales de la mutación jhy] sobre varias
respuestas fisiológicas y moleculares. Por ejemplo, el mutante fhy] tiene afectado en mayor
medida la inhibición del alargamiento del hipocotilo que la promoción de la germinación en
respuesta al RL continuo (Johnson et al., 1994). Por otro lado, la mutación fhyI afecta
significativamente la promoción por el RL continuo de la expresión de los genes CHS y CHI pero
afecta muy poco la promoción de la expresión de los genes CABy NR (Barnes et al., 1996). Estos
resultados llevaron a proponer que la cadena de transducción de señales del phyA estaría
ramificada y que el FHYl sólo formaría parte de una de estas ramas (Barnes et al., 1996). Existe,
sin embargo, una forma alternativa de interpretar dichos resultados. En este trabajo observamos
que el mutante fhy] tiene una sensibilidad disminuida, pero no nula, al RL continuo. Por lo tanto,
el efecto diferencial delflzyI sobre diferentes procesos podria ser simplemente la consecuencia de
un umbral diferente de activación por el RL para cada respuesta. Esta posibilidad podría ser
facilmente investigada realizando curvas de flujo de RL en todos los procesos.
Por último, las mutaciones flryI yfhy3 también afectan los efectos del phyA sobre el phyB. El
phyA tiene un efecto positivo sobre el phyB cuando actúa a través del HIR y tiene un efecto
negativo cuando actúa a través del VLFR. En este trabajo observamos que tanto el jhyI como el
jhy3 tienen reducida la acción sinérgica del phyA sobre el phyB, pero sólo elflryl tiene afectada
la acción antagónica del phyA sobre el phyB. Estos resultados son consistentes con una
participación del fhy] en el VLFR y en el HIR y una participación diferencial del jhy3 en estos
83

modos de acción. A su vez, estos resultados demuestran que tanto la coacción como el
antagonismo entre el phyA y el phyB no son la consecuencia de una interacción directa entre
estas moléculas y sí la resultante de una interacción entre las cadenas de transducción de señales
de ambos fotorreceptores.
La existencia de “cross-talk” entre los elementos de la cadena de transducción de señales
de los fitocromos no debería sorprendernos. Las interacciones entre distintas cadenas de
transducción de señales permite integrar la información proporcionada por múltiples sistemas
sensorias. Esto constituye una ventaja ya que permite obtener una información del ambiente
distinta y adicional a la que proporciona cada señal por separado, lo que permite reducir
significativamente la ambigüedad de la información.
MATERIALES Y MÉTODOS
Material Vegetal
Las semillas de Arabidopsis thaliana (L.) Heynh de los ecotipos Landsberg erecta y Columbia
fueron proporcionadas por los Drs R. E. Kendrick and M. Koomneef (Wageningen University,
The Netherlands) y por el Arabidopsis Biological Research Centre de la Universidad de Ohio
(ABRC), respectivamente. Las semillas de los mutantes phyA del ecotipo Landsberg erecta y de
los mutantesfliy] (ecotipo Landsberg erecta) yflry3 (ecotipo Columbia) fueron proporcionadas el
Dr. Garry Whitelam (Department of Botany, University of Leicester, UK). Las semillas del
mutante phyA en el ecotipo Columbia y las semillas de los mutantes phyB y phyAphyB (ecotipo
Landsberg erecto) fueron proporcionadas por el ABRC.
Aproximadamente 15 (en los experimentos de crecimiento del hipocotilo, apertura de
cotiledones y germinación) u 80 (en los de clorofila) semillas de cada ecotipo fueron sembradas
en cajas plásticas transparentes (40 x 33 mrn2 x 15 mm de altura) sobre papel de filtro saturado
con agua destilada. Las cajas fueron icubadas por 3 d a 7 °C.
Procedimientos Experimentales y Observaciones
En los experimentos donde se midió el largo del hipocotilo y la apertura de los
cotiledones, 15 semillas de cada genotipo fueron sembradas en cajas de plástico transparente (40
x 33 mm2 x 15 mm height) sobre dos capas de papel de filtro saturado en agua destilada. Las
cajas fueron incubadas en oscuridad a 7 °C por 3 d, recibieron luego un pulso de 5 min de R y
84

fueron icubadas en oscuridad a 25 °C por 24 h y luego transferidas por 3 d a la oscuridad (control)
o a los distintos tratamientos lumínicos (Casal, 1995). EL largo del hipocotilo fue medido con
una regla de 0.5 mm de precisión y los datos de las 10 plántulas más altas de cada caja fueron
promediados constituyendo una repetición para el análisis estadísitico. El ángulo entre los
cotiledones fue medido con un transportador en ls mismas plántulas utilizadas para la medición
del largo del hipocotilo y los 10 valores obtenidos por caja fileron promediados, constituyendo
una repetición a ser utilizada para el análisis estadístico.
Para los experimentos de germinación las semillas previamente incubadas por 3 d a 7°C
fueron expuestas a un pulso breve de luz que establecían distintos niveles de Pfr/P, y transferidas
a oscuridad, o fueron expuestas a RL continuo, pulsos de RL u oscuridad por 4 d a 25 °C antes de
contar las semillas germinadas (Botto et al., 1995, 1996.
En los experimentos de clorofila, las semillas fueron expuestas a un pulso de R (para
inducir la germinación), incubadas en oscuridad por 3 d a 25 °C, expuestas a pulsos de 5 min de
R/RL que establecían diferentes Pfr/P, incubadas en oscuridad por 4 hr a 25 °C, y finalmente
transferidas por 5 h a una cámara de luz blanca fluorescente (100 mol m'2 s") . La clorofila fue
determinada luego de la extracción de la misma con N N’ dimetilforrnamida (Moran, 1982).
Fuentes de luz
Las fuentes de luz utilizadas fueron iguales a las descriptas en el capítulo anterior.

MECANISMOS DE ACCIÓN DEL PHYA (III)
AISLAMIENTO Y CARACTERIZACIÓN DE NUEVOS MUTANTES QUE AFECTANDIFERENCIALMENTE LAS RESPUESTAS HIR Y VLFR
INTRODUCCIÓN
En forma adicional a la caracterización de los modos de acción del phyA en mutantes de
transducción de señales previamente identificados, como eljhyl y elfliy3, en esta parte de la tesis
se buscó identificar y caracterizar nuevos mutantes de Arabídopsis thaliana afectados
diferencialmente en las respuestas HIR y VLFR. La estrategia para obtener dichos mutantes
consistió en incubar semillas mutagenizadas de Arabz'dopsís thaliana bajo RL continuo (Fig. l ).
La germinación bajo RL es una VLFR mediada por el phyA ya que los pulsos de RL y el RL
continuo tienen el mismo efecto (Figura 4, capítulo 5). La inhibición del hipocotilo bajo RL
continuo es predominantemente un HIR porque el RL continuo es significativamente mas
efectivo que los pulsos (Figura .., capítulo 4, y figura... capítulo 5). Por lo tanto, aquellas
plántulas que germinen bajo RL, pero que sean altas en esta condición, deberían retener el VLFR
y no el HIR.
Semillas M2expuestas a RLe
Semillas no
germmadas Pláqtm'as PlántulasphyA, mutantes de la baJas- mas
biosíntesis del WT ‘cromóforo. Figura l. Dlseño del
l protocolo utilizado paraPlántulas altas: .
mmamesde ¡abiosímesisdel semillas M3 identificar mutantes afectadoscromóforo,mutantesde‘_ expuestasa RP más fuertemente en el HIRtransducciónde seí’ales IVLFRd l h A
comunes al phyAy al phyB. l que en e e p y '
Plántulasbajas:
mutantesbuscados
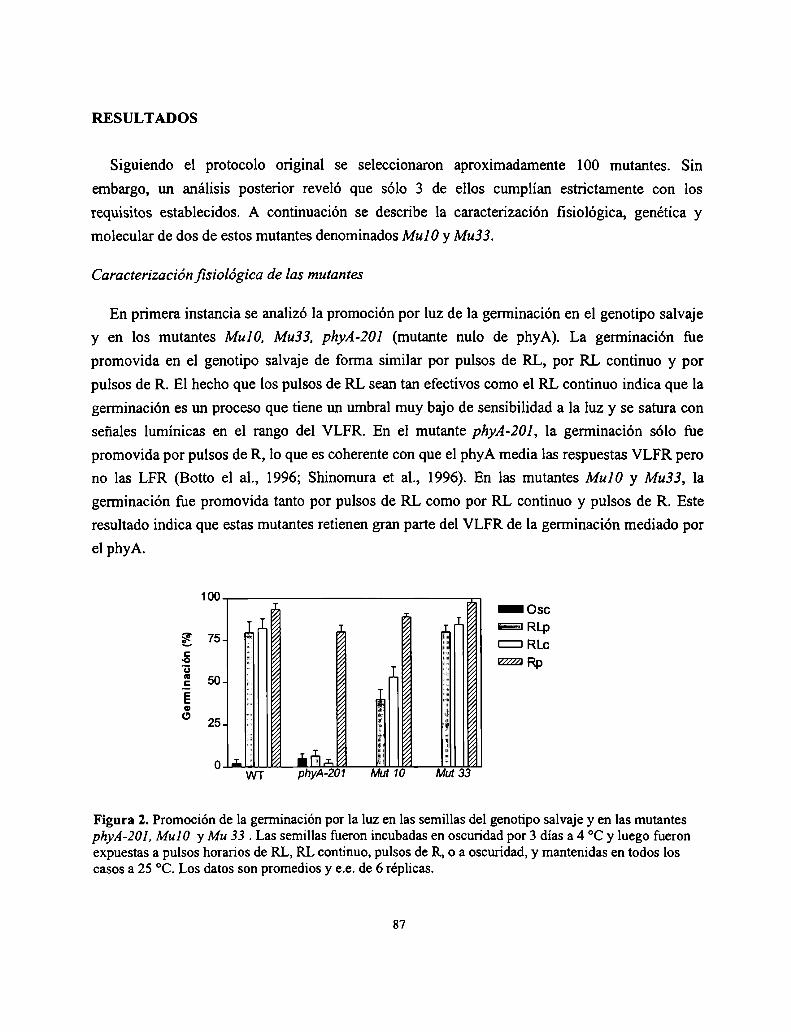
RESULTADOS
Siguiendo el protocolo original se seleccionaron aproximadamente 100 mutantes. Sin
embargo, un análisis posterior reveló que sólo 3 de ellos cumplían estrictamente con los
requisitos establecidos. A continuación se describe la caracterización fisiológica, genética y
molecular de dos de estos mutantes denominados Mu] 0 y Mu33.
Caracterización fisiológica de las mutantes
En primera instancia se analizó la promoción por luz de la germinación en el genotipo salvaje
y en los mutantes Mu10, Mu33, phyA-201 (mutante nulo de phyA). La germinación fue
promovida en el genotipo salvaje de forma similar por pulsos de RL, por RL continuo y por
pulsos de R. El hecho que los pulsos de RL sean tan efectivos como el RL continuo indica que la
germinación es un proceso que tiene un umbral muy bajo de sensibilidad a la luz y se satura con
señales lumínicas en el rango del VLFR. En el mutante phyA-201, la germinación sólo fue
promovida por pulsos de R, lo que es coherente con que el phyA media las respuestas VLFR pero
no las LFR (Botto el al., 1996; Shinomura et al., 1996). En las mutantes Mu10 y Mu33, la
germinación fue promovida tanto por pulsos de RL como por RL continuo y pulsos de R. Este
resultado indica que estas mutantes retienen gran parte del VLFR de la germinación mediado por
el phyA.
-OscRLpl: RLcRp
Germlnación(°/.)
Figura 2. Promoción de la germinación por la luz en las semillas del genotipo salvaje y en las mutantesphyA-201, Mu10 y Mu 33 . Las semillas fueron incubadas en oscuridad por 3 días a 4 °C y luego fueronexpuestas a pulsos horarios de RL, RL continuo, pulsos de R, o a oscuridad, y mantenidas en todos loscasos a 25 °C. Los datos son promedios y e.e. de 6 réplicas.
87
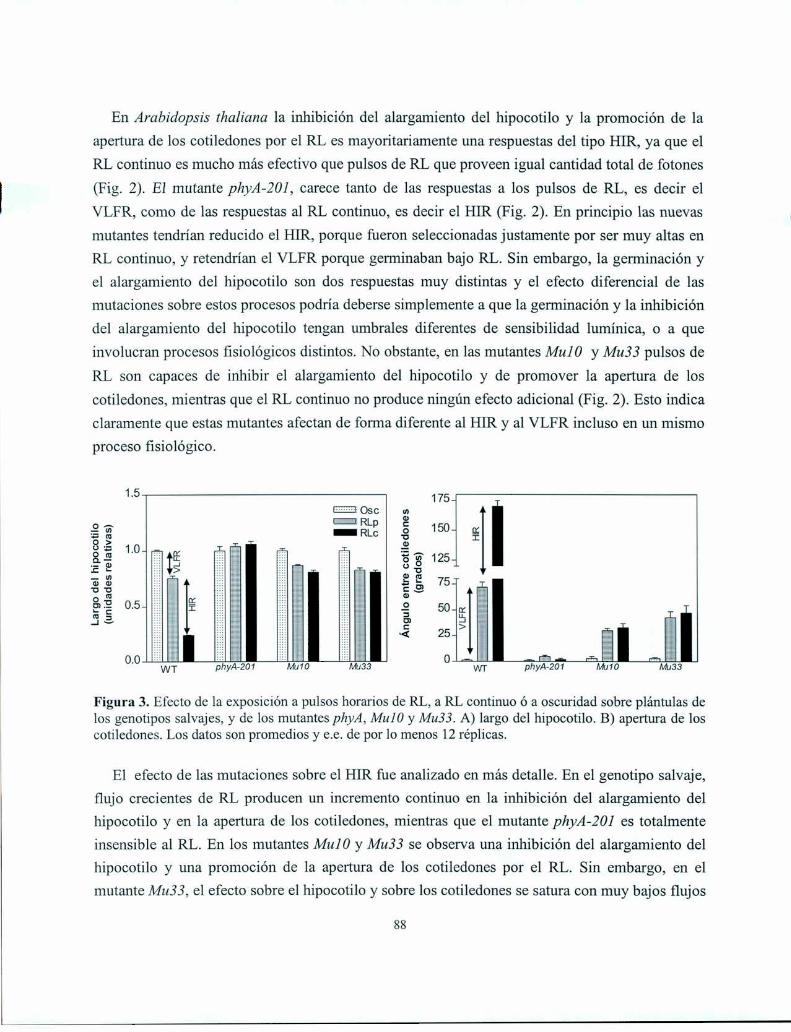
En Arabia’opsis thaliana la inhibición del alargamiento del hipocotilo y la promoción de la
apertura de los cotiledones por el RL es mayoritariamente una respuestas del tipo HIR, ya que el
RL continuo es mucho más efectivo que pulsos de RL que proveen igual cantidad total de fotones
(Fig. 2). El mutante phyA-ZO], carece tanto de las respuestas a los pulsos de RL, es decir el
VLFR, como de las respuestas a1RL continuo, es decir el HIR (Fig. 2). En principio las nuevas
mutantes tendrían reducido el HIR, porque fueron seleccionadas justamente por ser muy altas en
RL continuo, y retendrían el VLFR porque germinaban bajo RL. Sin embargo, la germinación y
el alargamiento del hipocotilo son dos respuestas muy distintas y el efecto diferencial de las
mutaciones sobre estos procesos podría deberse simplemente a que la germinación y la inhibición
del alargamiento del hipocotilo tengan umbrales diferentes de sensibilidad lumínica, o a que
involucran procesos fisiológicos distintos. No obstante, en las mutantes M1410 y Mu33 pulsos de
RL son capaces de inhibir el alargamiento del hipocotilo y de promover la apertura de los
cotiledones, mientras que el RL continuo no produce ningún efecto adicional (Fig. 2). Esto indica
claramente que estas mutantes afectan de forma diferente al HIR y al VLFR incluso en un mismo
proceso fisiológico.
i;1. 175
Osc gRLp r:OAEQ -RLc .8 150“ %3.2 °3‘“? E_ O
LE E o_ 0oa "ug sEL? 2N 5 a,_|v C
n<
Figura 3. Efecto de la exposición a pulsos horarios de RL, a RL continuo ó a oscuridad sobre plántulas delos genotipos salvajes, y de los mutantes phyA, Mu] 0 y Mu33. A) largo del hipocotilo. B) apertura de loscotiledones. Los datos son promedios y e.e. de por lo menos 12 réplicas.
E1 efecto de las mutaciones sobre el HIR fue analizado en más detalle. En el genotipo salvaje,
flujo crecientes de RL producen un incremento continuo en la inhibición del alargamiento del
hipocotilo y en la apertura de los cotiledones, mientras que el mutante phyA-201 es totalmente
insensible al RL. En los mutantes Mu] 0 y Mu33 se observa una inhibición del alargamiento del
hipocotilo y una promoción de la apertura de los cotiledones por el RL. Sin embargo, en el
mutante Mu33, el efecto sobre el hipocotilo y sobre los cotiledones se satura con muy bajos flujos
88
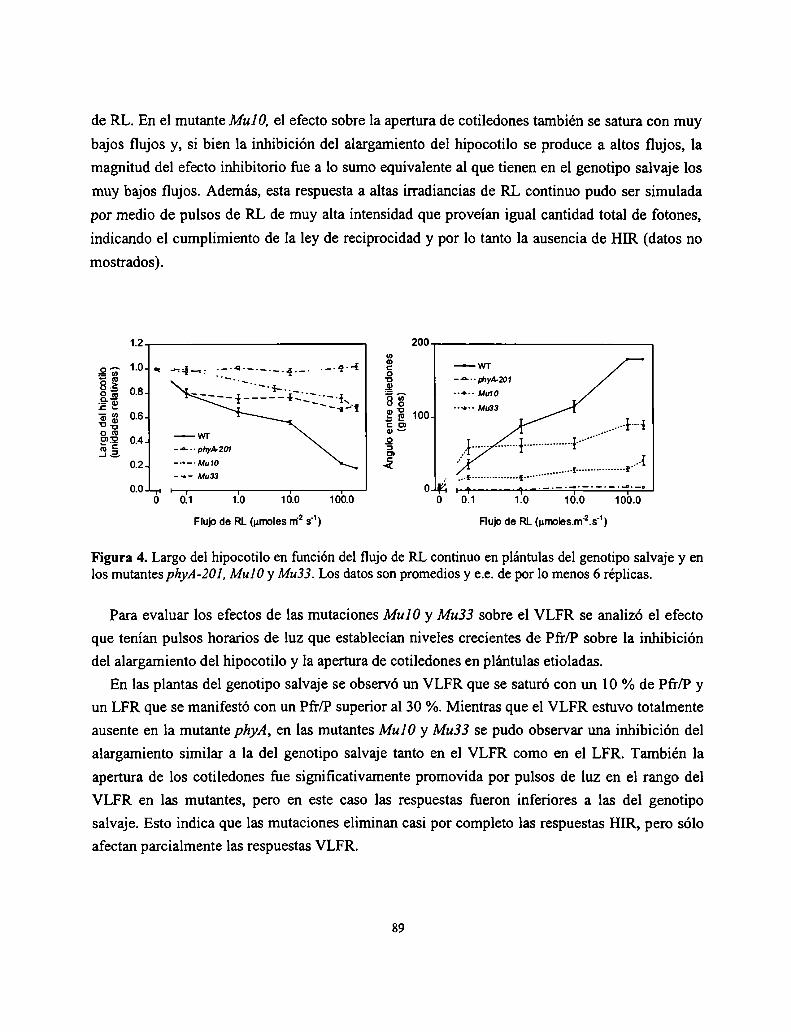
de RL. En el mutante Mu10, el efecto sobre la apertura de cotiledones también se satura con muy
bajos flujos y, si bien la inhibición del alargamiento del hipocotilo se produce a altos flujos, la
magnitud del efecto inhibitorio fue a lo sumo equivalente al que tienen en el genotipo salvaje los
muy bajos flujos. Además, esta respuesta a altas irradiancias de RL continuo pudo ser simulada
por medio de pulsos de RL de muy alta intensidad que proveían igual cantidad total de fotones,
indicando el cumplimiento de la ley de reciprocidad y por lo tanto la ausencia de HIR (datos no
mostrados).
1 9 onn
Largodelhipocotilo(unidadesrelativas)
Anguloentrewtiledones
(grados)
80
o ' oÏ1 1.o 16.0 1ob.o o 'oÏ1 1Ïo 16.0 1ob.o
Flujo de RL (urnoles rn"2s") Flujo de RL (umolesm'25'1)
Figura 4. Largo del hipocotilo en función del flujo de RL continuo en plántulas del genotipo salvaje y enlos mutantes phyA-201, Mu10 y Mu33. Los datos son promedios y e.e. de por lo menos 6 réplicas.
Para evaluar los efectos de las mutaciones Mu10 y Mu33 sobre el VLFR se analizó el efecto
que tenían pulsos horarios de luz que establecían niveles crecientes de Pfr/P sobre la inhibición
del alargamiento del hipocotilo y la apertura de cotiledones en plántulas etioladas.
En las plantas del genotipo salvaje se observó un VLFR que se saturó con un 10 % de Pfr/P y
un LFR que se manifestó con un Pfr/P superior al 30 %. Mientras que el VLFR estuvo totalmente
ausente en la mutante phyA, en las mutantes M1410y Mu33 se pudo observar una inhibición del
alargamiento similar a la del genotipo salvaje tanto en el VLFR como en el LFR. También la
apertura de los cotiledones fue significativamente promovida por pulsos de luz en el rango del
VLFR en las mutantes, pero en este caso las respuestas fueron inferiores a las del genotipo
salvaje. Esto indica que las mutaciones eliminan casi por completo las respuestas HIR, pero sólo
afectan parcialmente las respuestas VLFR.
89
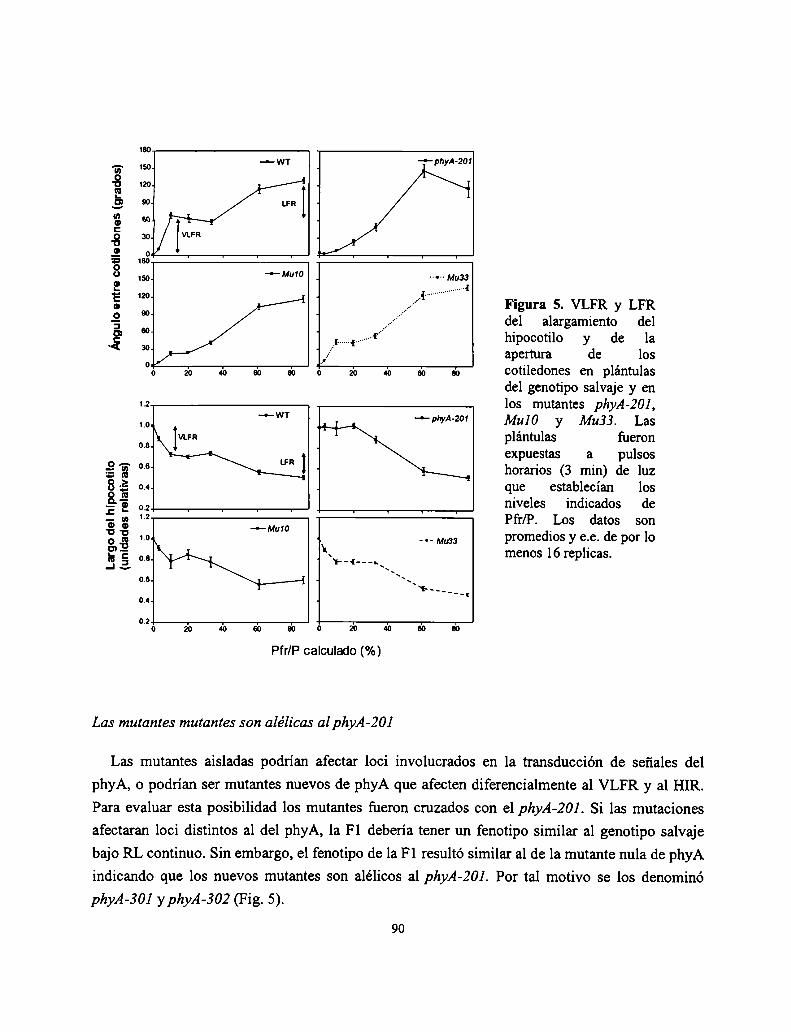
A 15° _._WT - -°-phyA-201m .
g .20.
g 901 LFR‘g uC
30 VLFR
2 o l .g 1M
g 15°- --Mu10 _ ..... “"33l. ..................--{
É, ‘2°' l Figura 5. VLFR y LFR° 9°“ d l l ' d l3 e a argamiento eg °°' hípocotiloy de la‘ 3°' ' apertura de los
“ ¿o ¡o ¿o ¿o ' ¿o ¡o ¿o ¿o cotíledones en plántulasdel genotipo salvaje y en
19 los mutantes phyA-201,m. +WT 4 --phyA-201 Mu10 y Mu33. Las
Ivan plántulas fueron°'° LFR ' expuestas a pulsos
g fi °-° I horarios (3 min) de luz8.5 o... _ que establecían losgg M I l I I niveles indicados deÉ m 1° ' Pfr/P. Los datos son'03 +MU1° romedios ee de or lo0.3 ‘-°' ---Muc3 P y :- Pg’; oa _\ menos 16replicas.J a P-{---o.\
0.6 “x‘h‘ ‘ ' ‘ *-I
0.4
"° 2'o ¡o e'o a'o 2'0 ¡o e'o ao
Pfr/P calculado (%)
Las mutantes mutantes son alélicas al phyA-201
Las mutantes aisladas podrían afectar loci involucrados en la transducción de señales del
phyA, o podn'an ser mutantes nuevos de phyA que afecten diferencialmente al VLFR y al HIR.
Para evaluar esta posibilidad los mutantes fueron cruzados con el phyA-201. Si las mutaciones
afectaran loci distintos al del phyA, la Fl debería tener un fenotipo similar al genotipo salvaje
bajo RL continuo. Sin embargo, el fenotipo de la Fl resultó similar al de la mutante nula de phyA
indicando que los nuevos mutantes son alélicos al phyA-201. Por tal motivo se los denominó
phyA-301 y phyA-302 (Fig. 5).
90
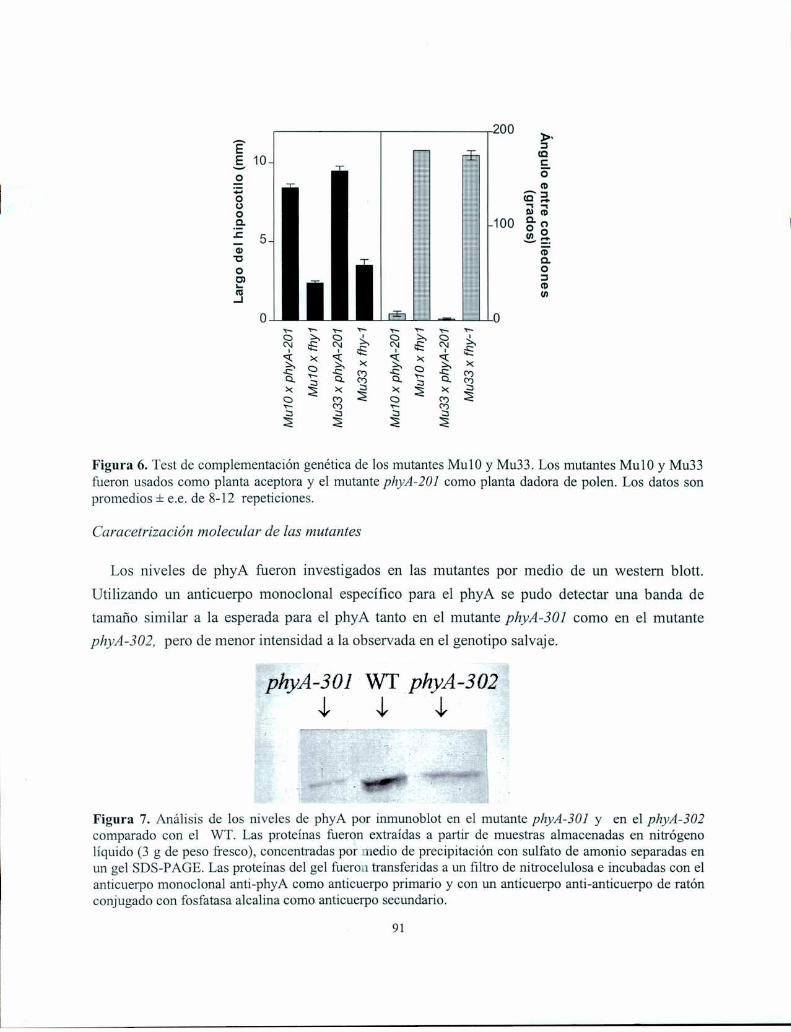
0D 3>.A
E .3E10- cV _o oE A0s «25o 30D- o._ oof 5- 3:.o _"o 3
o8, :É CD
_¡ cn
0_ ma 2 s T s 2 s YN 5 N É N E N É¿Z >< ¿t "" ¿t >< ¿Z kA o >s. >< >». Q >s ><.2 F c cv) c F —: mo. 3 O. m Q. 3 Q. m>< 3 >< É >< 3 >< ÉÉ ï É ïE E E E
Figura 6. Test de complementación genética de los mutantes MulO y Mu33. Los mutantes MulO y Mu33fueron usados como planta aceptora y el mutante phyA-20] como planta dadora de polen. Los datos sonpromedios i e.e. de 8-12 repeticiones.
Caracetrización molecular de las mutantes
Los niveles de phyA fueron investigados en las mutantes por medio de un western blott.
Utilizando un anticuerpo monoclonal específico para el phyA se pudo detectar una banda de
tamaño similar a la esperada para el phyA tanto en el mutante phyA-301 como en el mutante
phyA-302, pero de menor intensidad a la observada en el genotipo salvaje.
phyA-301 WT phyA-3 02J, i J,
I ..w wa? - .
Figura 7. Análisis de los niveles de phyA por inmunoblot en el mutante phyA-301 y en el phyA-302comparado con el WT. Las proteínas fueron extraídas a partir de muestras almacenadas en nitrógenolíquido (3 g de peso fresco), concentradas por medio de precipitación con sulfato de amonio separadas enun gel SDS-PAGE. Las proteínas del gel fueron transferidas a un filtro de nitrocelulosa e incubadas con elanticuerpo monoclonal anti-phyA como anticuerpo primario y con un anticuerpo anti-anticuerpo de ratónconjugado con fosfatasa alcalina como anticuerpo secundario.
91

Para identificar las mutaciones phyA-301 y phyA-302 a nivel molecular, se clonó por PCR y se
secuenció completamente (intrones + exones) las dos hebras del gen PHYA de las mutantes (en
material proveniente de por lo menos dos extracciones independientes) y del genotipo salvaje. La
comparación de las secuencias reveló la existencia en la mutante phyA-301 de un cambio C X T
que condujo a la aparición de un codón de terminación prematuro en lugar de la glutarnina 980,
dando lugar a una proteína truncada con 142 aa menos en el extremo C-terminal (Fig. 6 ).
C T K W N P
phyA-302 tgc aca aag tgg aat ccaWT tgc aca gagtgg aatcca775 C T E W N P/
PHYA——I—ñ IL H;H:l—
978 T S Q V M MWT aca agtcaagtaatgatg
phyA-301 aca agt taa gta atg agT S STOP
Figura 8. Secuencias del tercer y cuarto exón del gen PH YA WT y de los mutantes phyA-301 y phyA-302.
Los exones están representados como cuadrados y los intrones como líneas llenas. Las regiones 5’y 3’no
codificantes están presentadas como recuadros negros. Las líneas punteadas indican la posición de lassecuencias mostradas.
En la mutante phyA-302 se detectó una mutación puntual que derivó en un cambio en el
arninoacido 978 de lisina por ácido glutámico.
DISCUSIÓN
El phyA media tanto las respuestas a pulsos de RL ó VLFR, como las respuestas al RL
continuo ó HIR. El hecho que el RL continuo sea mucho más efectivo que pulsos de RL que
proveen igual cantidad de fotones sugiere que el VLFR y el HIR son el resultado de la activaciónde dos mecanismos moleculares distintos.
Aquí se describe la identificación y caracterización de dos nuevos mutantes el phyA-301 y el
phyA-302. Mientras que en las plántulas WT el RL continuo es mucho más efectivo que los
92

pulsos de RL en inhibir el alargamiento del hipocotilo, y en promover la apertura de los
cotiledones, en las mutantes phyA-301 y phyA-302, 3-min por hora de RL tuvieron el mismo
efecto que el RL continuo. Si el RL continuo fuera más efectivo que los pulsos de RL
simplemente porque mantiene niveles constantes de Pfr de phyA en el tiempo, los mutantes
phyA-301 y phyA-302 que incluso tienen reducidos los niveles totales de phyA, deberían afectar
en forma similar las respuestas a los pulsos de RL y al RL continuo. EL hecho de haber podido
identificar mutantes que retengan la respuesta a los pulsos de RL y pierdan la respuesta al RL
continuo indicar-ía que el HIR y el VLFR son el resultado de la activación de dos mecanismosdiferenciables.
Las mutantes phyA-301 y phyA-302, no obstante, tienen niveles reducidos de phyA
comparadas al WT. En consecuencia, estas mutantes podrían carecer de HIR pero retener una
parte importante del VFR en caso que el HIR requiera mayores niveles de phyA que el VLFR
para activarse. A favor de esta idea, en las plantas transgénicas de arroz que sobreexpresan phyA
de avena, las respuestas del crecimiento del coleoptile y el contenido de antocianas en respuesta a
pulsos de RL parece saturarse con los niveles endógenos de phyA, mientras que las respuestas al
RL continua son significativamente promovidas por la sobreexpresión del phyA de avena (Casal
et al., 1996). Sin embargo, no cualquier reducción en los niveles o en la actividad de phyA afecta
más al HIR que al VLFR. Tanto en las mutantes de cromóforo de Arabidopsis thaliana, como en
un alelo débil del phyA, el phyA-205, las respuestas VLFR y HIR se ven afectadas de forma
similar (Casal & Yanovsky, datos no publicados).
La hipótesis que sostiene que las respuestas VLFR y HIR involucran distintas acciones de la
molécula de phyA puede proponerse en términos moleculares sugiriendo que las respuestas a los
pulsos de RL serian desencadenadas por el Pfr del phyA y las respuestas al RL continuo serían
desencadenadas por un efector desconocido, posiblemente un intermediario inestable formado
durante el ciclado entre el Pr y el Pfr. Si esta hipótesis fuera cierta, las mutaciones phyA-301 y
phyA-302 afectarían poco la acción del Pfr del phyA pero reducirían notablemente la eficacia del
efector del HIR, el hipotético intermediario del ciclado entre el Pr y el Pfr.
La mutación phyA-302 involucra un cambio de un ácido glutámico por una lisina en el
aminoácido 778. Este aminoácido se encuentra en el extremo derecho de la Quail Box (QB), una
región que Quail y colaboladores revelaron como importante para la acción regulatoria del
fitocromo (Quail et al., 1995; Fig. 6, Capítulo 1). Estos resultados indican que la QB sería más
importante para las respuestas HIR que para las respuestas VLFR. El rol regulatorio de la QB es
desconocido. En principio esta región podría afectar directamente la actividad bioquímica del
fitocromo (por ejemplo su actividad kinasa), o podría afectar la capacidad del fitocromo de
93

interactuar con su blanco de acción inicial. Sin embargo, no parecen existir en la QB regiones con
secuencias homologas a los sitios activos de las protein kinasas. Por el contrario, se ha
demostrado que mutaciones en la QB disminuyen la capacidad de los fitocromos de interactuar
en el sistema de dos híbridos de levaduras con PIF3. Este dato sugiere que la QB estaría
vinculada a la interacción fisica con el blanco de acción inicial más que con su modificación
bioquímica. El hecho que la mutación phyA-302 situada en el borde derecho de la QB afecte
diferencialmente al HIR en mayor medida que al VLFR sugiere que el(los) factor(es) con el(los)
que interactúa el phyA en el modo HIR sen’a(n)distinto(s) a el(los) factor(es) del VLFR.
La mutación phyA-301 convierte el codón que da lugar a la glutamina 980 en un codón de
terminación. Esto da lugar a un phyA con 142 aminoácidos menos en el extremo C-terminal.
Llamativamente, la mutación identificada en este mutante es igual a la presente en el mutante
nulo de phyA, el phyA-201. Este hallazgo es sorprendente ya que el phyA-201 es un mutante que
carece totalmente de fitocromo detectable y no muestra respuesta alguna en el VLFR ni en el
HIR. Sin embargo, a pesar de compartir la misma mutación, el phyA-301 presenta niveles
reducidos pero detectables de phyA y presenta respuestas significativas en el VLFR. La ausencia
de niveles detectables de phyA en el mutante phyA-201 es consistente con la información previa
que indica que el extremo C-terminal es importante para la estabilidad del fitocromo. La hipótesis
que propongo para explicar la diferencia en el fenotipo fisiológico (presencia de VLFR) y
molecular (presencia de niveles detectables de proteína) entre el phyA-201 y el phyA-301 es que
este último mutante posee una segunda mutación que estabilice la proteína truncada. En la
actualidad se están haciendo experimentos para investigar esta posibilidad en detalle.
Independientemente de cuál sea la explicación para la controversia, el hecho que la mutante
phyA-301 tenga un phyA que carece de 142 aa del extremo C-terminal pero que retiene una parte
importante de la respuesta a pulsos de RL mientras que pierde la respuesta al RL continuo indica
que esta región es más importante para las respuestas HIR que para las VLFR.
Recientemente se han reportado resultados con implicancias similares a las derivadas del
mutante phyA-301. El grupo de Minami Matsui en Riken, Japón, ha estado investigando el efecto
fisiológico de sobreexpresar, en Arabidopsis, phyA con deleciones en el extremo C-terminal.
Ellos han observado que la sobreexpresión del phyA completo incrementa las respuestas al R y
al RL continuo, mientras que la sobreexpresión de phyA con deleciones en el C-terminal no sólo
no incrementa, sino que reduce las respuestas al RL continuo, pero mantiene normales las
respuestas al R. Se sabe que el incremento en la sensibilidad al R en las plantas que
sobreexpresan phyA es una respuesta de tipo VLFR, mientras que el incremento en la
sensibilidad al RL continuo es una respuesta HIR. Esos resultados son consistentes con la idea
94

que el extremo C-terminal es crucial para las respuestas HIR pero no es tan importante para las
respuestas VLFR.
Dado que la zona C-terminal el phyA involucrada específicamente en el HIR contiene el
dominio con homología a histidin-kínasas pero no el homólogo a serin-treonin-kinasas, es posible
especular que el HIR pero no el VLFR requiera de la actividad histidin-kinasa. Por otro lado,
parte de la región delecionada en el mutante phyA-301 también ha sido involucrada en la
dimerización del fitocromo. Si bien no lo sabemos con certeza, es probable que el phyA presente
en este mutante se encuentre como monomero y, por lo tanto, cabe la posibilidad que la acción
del phyA en el HIR involucre el dímero de phyA mientras que el monómero sería suficiente parael VLFR.
Resumiendo, hemos identificado mutaciones en el phyA que afectan diferencialmente al
VLFR del HIR. Este resultado es consistente con la hipótesis que sostiene que estos modos de
acción serían desencadenados por dos formas distintas del phyA, y la activación por parte del
phyA de las respuestas involucraría distintas regiones de esta molécula.
MATERIALES Y MÉTODOS
Material Vegetal
Las semillas de Arabidopsis thaliana (L.) Heynh del ecotipo Landsberg erecta fueron
proporcionadas por los Drs R. E. Kendrick and M. Koomneef (Wageningen University, The
Netherlands). Las semillas del mutante phyA-201 fueron proporcionadas por la Dra J. Chory (The
Salk Institute, La Jolla, California) y las semillas del mutante phyA-205 fueron proporcionadas
por el Arabidopsis Biological Research Centre de la Universidad de Ohio (ABRC). Las semillas
mutagenizadas (ecotipo Landsberg erecta) fueron adquiridas a la firma Lehle Seeds (Texas).
Procedimientos Experimentales
En los experimentos donde se midió el largo del hipocotilo y la apertura de los cotiledones, 15
semillas de cada genotipo fueron sembradas en cajas de plástico transparente (40 x 33 mm2 x 15
mm height) sobre dos capas de papel de filtro saturado en agua destilada. Las cajas fueron
incubadas en oscuridad a 7 °C por 3 d, recibieron luego un pulso de 5 min de R y fireron icubadas
en oscuridad a 25 °C por 24 h y luego transferidas por 3 d a la oscuridad (control) o a los distintos
tratamientos lumínicos (Casal, 1995). EL largo del hipocotilo fire medido con una regla de 0.5
mm de precisión y los datos de las 10 plántulas más altas de cada caja fueron promediados95

constituyendo una repetición para el análisis estadísitico. El ángulo entre los cotiledones fire
medido con un transportador en ls mismas plántulas utilizadas para la medición del largo del
hípocotilo y los 10 valores obtenidos por caja fueron promediados, constituyendo una repetición
a ser utilizada para el análisis estadístico.
Las fuentes de luz utilizadas fueron iguales a las descriptas en los capítulos anteriores.
Extracción de proteínas e Inmunobloting.
Se prepararon extractos proteicos enriquecidos en fitocromo a partir de muestra almacenadas en
nitrógeno líquido de acuerdo a los procedimientos seguidos por Somers y col. (Somers et al.,
1991). Los extractos protéicos fueron corridos en un gel SDS-PAGE (de 1,5-mm- de espesor,
7.5%) (Mini Proteam II, Bio-Rad, Richmond, CA). Las proteínas fiieron transferidas por
electroblotting desde el gel hacia filtros de nitrocelulosa (con un tamaño de poro de 0.45u m,
Sigma, ST Louis, Mo.) por medio del sistema Milliblot-SDE (Millipore, Bedford, Mass.)
siguiendo las instrucciones del fabricante. El filtro fue bloqueado incubándolo con leche
descremada (1%), 50 mM Tris-Cl, 200 mM NaCl, pH 7.4 por 30 min a 37 °C. El filtro fue luego
incubado a 7 °C con 10 ml del anticuerpo primario anti-phyA a una dilución de 111000. El
anticuerpo monoclonal realizado en ratón contra phyA de Arabidopsis fue proporcionado por el
Dr Peter H. Quail (University of Berkeley, USA). Luego de lavada, la membrana fue incubada
con una dilución 12500 de anticuerpo anti anti-IgG de ratón conjugado con fofatasa alcalina
(Sigma). Las bandas fueron visualizadas incubando las membranas en 0.1 M Tris (pH 9.5), 100
mM NaCl, 5 mM MgClz conteniendo 0.165 mg/ ml de BCIP (5-bromo-4chloro-3-indoyl
phosphate, p-toluidine salt) y 0.33 mg/ml de NBT (nitro blue tetrazolium), ambos productos de
Sigma.
Clonado y secuenciación del gen PH YA
Se aisló ADN genómico de plantas WT y de las mutantes phyA-301 y phyA-302 siguiendo el
protocolo descripto por Neff y col. (1998). El gen PH YAcompleto (incluyendo exones e intrones)
fue amplificado por PCR utilizando dos primers diseñados a partir de la secuencia completa del
gen ya publicada (Dehesh et al., 1993). Las condiciones de la PCR fueron las siguientes: Tris
HCl 20 mM pH 8.4, KCl 50 mM, MgClZ 2mM, 200 uM de cada dNTP, 25 pmoles de cada
primer, 2 ul de la preparación de ADN genómico y 2.5. unidades de Taq Polimersa (GIBCO
BRL) en un volumen final de 25 ul. La mezcla de reacción se calentó a 94 °C por 4 min y se
sometió a 35 ciclos de 30 s a 94 °C, 30 s a 55 °C y 3 min 30 s a 72 °C. Los productos de reacción
96

fueron analizados en geles de agarosa al 0.8 %. Los mismos tenían un tamaño de
aproximadamente 4.8 Kb, similar al esperado en función de la secuencia conocida del gen. Los
fragmentos correspondientes fiJeron extraídos del gel y sub-clonados, siguiendo protocolos de
rutina, en el plásmido pUC19 (Sambrock et al., 1989). Con los plásmidos recombinantes como
moldes se secuenciaron las dos hebras de ADN de cada uno de los genes PHYA clonados
utilizando un secuenciador automático y 10 pares de primers internos (diseñados utilizando la
secuencia publicada por Dehesh et al., 1993), superpuestos de modo de cubrir las 4.8 Kb del gen.

DISCUSIÓN GENERAL
Las plantas utilizan la información provista por el ambiente lumínico (cantidad, calidad,
duración y/o dirección de la luz) para adaptarse a los cambios que ocurren en su entorno. El
ambiente luminoso es sumamente complejo y las plantas posean múltiples sistemas fotosensorios
para percibirlo. Los fitocromos, cromoproteínas que absorben en el rango del R y el RL, son los
fotorreceptores más caracterizados. En Arabidopsis y en tomate (las dicotiledóneas más
estudiadas en términos fotomorfogénicos) existen cinco genes distintos que codifican fitocromos,
PHYA, PHYB, PHYC, PHYD y PHYE. La existencia de diferentes fitocromos en una misma
especie llevó a proponer que estos fotorreceptores cumplírían funciones sensoriales y/o
regulatorias distintas.
En esta tesis se investigaron en detalle las fiinciones fotoperceptivas y los mecanismos de
acción del phyA. La caracterización fotofisiológica de plantas mutantes y transgénicas con
niveles reducidos o incrementados de phyA ha permitido identificar diferentes circunstancias
donde la acción del phyA es importante:
l) el phyA está involucrado en promover la des-etiolación de las plántulas. La señal que
induce al phyA a promover esta respuesta es la transición oscuridad/luz. La acción del
phyA se manifiesta tanto en plántulas que emergen en suelos descubiertos, es decir bajo
luz solar directa, como en plántulas que emergen bajo canopeos muy densos, es decir en
ambientes con muy poca luz, enriquecidos en RL.
2V el phyA, tanto en forma independiente, como mediante su interacción con otros
fotorreceptores como el phyB, participa del control de las respuestas de las plantas a la
presencia de plantas vecinas. Por un lado, el phyA está involucrado directamente en la
percepción de reducciones en la irradiancia (en particular la cantidad de R+RL) que se
producen por la absorción de luz en las hojas de las plantas vecinas. Por otro lado, el phyA
modula la capacidad del phyB de percibir reducciones pequeñas en la relación R/RL
producidas por el RL reflejado en las hojas de las plantas vecinas.
3V el phyA sincroniza el reloj endógeno que controla la fase del ritmo del movimiento de las
hojas en las plantas de papa. En las plantas, los relojes edógenos también controlan
procesos como la fotosíntesis, la expresión de genes, e incluso están involucrados en el
mecanismo de percepción de la longitud del día que permite a las plantas coordinar su
98

ciclo de vida con las estaciones climáticas favorables para el crecimiento. Una de las
señales que estimula al phyA a sincronizar el reloj circadiano que controla el ritmo del
movimiento de las hojas es la transición oscuridad/luz que ocurre diariamente al amanecer.
Los resultados presentados aquí indican que el phyA participa del control de diversos aspectos
del desarrollo de las plantas durante etapas muy distintas de su ciclo de vida y, además, que las
funciones fotoperceptivas del phyA son diferentes a las del resto de los fitocromos. Mientras que
el phyB, phyC, phyD y phyE participan de la percepción de los cambios en la relación R/RL, el
phyA está involucrado en percibir principalmente cambios en la irradiancia como los que ocurren
durante la emergencia de las plántulas o en la transición entre la noche y el día.
Es interesante observar que el phyA participa tanto en la detección de cambios muy pequeños
en la irradiancia (como los que perciben las semillas de malezas cuando son expuestas por
milisegundos a la luz solar durante la arada), como en la detección de cambios intensos (por
ejemplo, los que se producen como consecuencia del sombreado por plantas vecinas). Estas
acciones del phyA ocurren a través de dos mecanismos, el VLFR que se satura con muy poca luz,
y el HIR que se manifiesta frente a exposiciones prolongadas a altas irradiancias de RL. Los
resultados obtenidos en la segunda parte de la tesis son consistentes con que las respuestas VLFR
y HIR involucra dos acciones distintas de este fotorreceptor, que se diferenciarían tanto en las
regiones de la molécula de phyA involucradas, como en las vías de transducción de señalesactivadas.
En esta tesis se ha demostrado que es posible separar genéticamente el VLFR del HIR. Por un
lado, los loci VLF] y VLF2polimórficos entre los ecotipos Landsberg erecta y Columbia, afectan
al VLFR pero no al HIR (Yanovsky et al., 1997) y son la prueba más contundente para sostener
que estos dos modos de acción involucran mecanismos moleculares diferentes. Estos loci podrían
codificar proteínas que actúen normalmente como represores del VLFR. Se ha demostrado que
deleciones o sustituciones de serinas que se encuentran en el extremo N-terminal de la molécula
de phyA aumentan la sensibilidad de este fotorreceptor. Esas serinas serían fosforiladas luego de
la exposición del phyA a la luz y de alguna manera la fosforilación reduciría la efectividad del
phyA en las respuestas a muy bajos flujos. Por lo tanto, los loci VLF presentes en el ecotipo
Columbia podrían codificar proteín kinasas que fosforilen al phyA en el extremo N-terminal,
reduciendo su efectividad regulatoria.
Además, hemos identificado los mutantes phyA-301 y phyA-302 que tienen un efecto mucho
más marcado sobre el HIR que sobre el VLFR. Esto sugiere que la Quail Box, posiblemente
alterada en el mutante phyA-302, como el dominio con homología a kinasas de histidina, ausente
99

en el mutante phyA-301, serían más importantes para la acción del phyA en el HIR que en el
VLFR. Estos mutantes también tienen niveles reducidos de phyA y por lo tanto no es posible
descartar que el efecto diferencial de estas mutaciones sobre las respuestas VLFR y HIR se deba
a diferentes requerimientos de phyA para inducir ambas respuestas. Por otro lado, hemos
caracterizado las respuestas VLFR y HIR en dos loci previamente identificados, los cuales se
hayan afectados específicamente en la transducción de señales del phyA. Nuestros resultados
indican que uno de estos loci, eljhyI afecta similaimente al VLFR y al HIR, mientras que elfliy3
afecta mucho más al HIR que al VLFR.
Todos estos resultados son compatibles con un modelo (Fig. 1) donde los pulsos de RL
actuando a través del Pfr del phyA desencadenan las respuestas VLFR y el RL continuo,
actuando a través del hipotético intermediario formado durante el ciclado entre el Pr y el Pfr
desencadena las respuestas HIR. Los dominios de la molécula de phyA afectados en las mutantes
phyA-301 y phyA-302 serían más importantes para la acción del efector del HIR que para la del
efector del VLFR. A su vez, los efectores del VLFR y del HIR iniciarían dos cadenas de
transducción de señales que tendrían algunos elementos en común (FHYl) y otros diferentes
(FHY3, VLFl y VLF2).
phyA-30
RLcontimn -> PhyA
HIR phyA_30¡ Inhibición delalargamiento
. . > del h' cofilo,Pulsos de RL —> phyA | | V apertiïa de
VLFR vlfl' vlfl los cotibdones,etc.
R continm0 -> h B
Pulsos de R p y
[FR
Figura l. Modelo que resume los resultados presentados en la segunda parte de la tesis, es decir la
disección genética de las respuestas VLFR y HIR. Las líneas terminadas como flechas representan
acciones positivas. Las líneas terminadas en T representan acciones negativas.
La posibilidad del phyA de desencadenar el VLFR y el HIR a través de dos vías de
transducción de señales diferentes permitiría una regulación más fina de cada uno de los procesos
por separado. Esto es importante ya que si bien en procesos como la inhibición del alargamiento
100

del hipocotilo y la apertura de cotiledones las respuestas VLFR y HIR van en la misma dirección,
en otros procesos esto no es así. Por ejemplo, el phyA tiene un efecto positivo sobre la acción del
phyB cuando actúa a través del HIR, pero tiene un efecto negativo cuando actúa a través del
VLFR. En algunas especies como Datura ferox (pero no Arabidopsis) el VLFR promueve la
germinación mientras que el HIR la inhibe. Esto es interesante ya que bajos niveles de Pfr pueden
ser establecidos en las semillas por circunstancias tan diversas como: (a) pulsos muy breves de
luz solar (con una alta relación R/RL) experimentada por parte del banco de semillas como
consecuencia de las labranzas (Scopel et al., 1991); (b) exposiciones prolongadas a la luz
presente bajo canopeos muy densos (con una relación R/RL muy baja). El hecho que la
germinación sea promovida por pulsos de luz pero inhibida por exposiciones prolongadas al RL
constituye un mecanismo que permite a las semillas distinguir entre un canopeo recién cultivado
donde toda la vegetación ha sido removida y un canopeo muy denso. La existencia de diferentes
vías de transducción de señales para las respuestas a pulsos de luz o a luz continua ofrecer-íapor
lo tanto una manera de regular diferencialmente la sensibilidad a diferentes señales naturales.
En resúmen, las plantas poseen múltiples fotorreceptores que cumplen funciones sensoriales y
regulatorias diferentes. A su vez, estos fotorreceptores desencadenan una o más vías de
transducción de señales diferentes que, en ciertas circunstancias, interactúan entre sí. Esta
multiplicidad de fotorreceptores y de vías de transducción de señales independientes y a la vez
interactuantes permite a las plantas interpretar de manera más completa y eficiente toda la
información provista por el ambiente luminoso.

BIBLIOGRAFÍA
Adamse, P., Jaspers, P. A. M., Bakker, J. A., Kendrick, R. E., and Koomneef, M. (1988).Photophysiology and phytochrome content of long-hypocotyl mutant and wild-type cucumberseedlings. Plant Physiology 87, 264-268.
Ahmad, M. (1999). Seeing the world in red and blue: insight into plant vision and photoreceptors.Current Opinion in Plant Biology 2, 230-235.
Ahmad, M., and Cashmore, A. R. (1996). The pef mutants of Arabidopsis thaliana define lesionsearly in the phytochrome signaling pathway. Plant Journal 10, l 103-1l 10.
Ahmad, M., and Cashmore, A. R. (1996). Seeing blue: the discovery of cryptochrome. PlantMolecular Biology 30, 851-861.
Aukerman, M. J., Hirschfeld, M., Wester, L., Weaver, M., Clack, T., Amasino, R. M., andShar-rock,R. A. (1997). A deletion in the PH YDgene of the Arabidopsis Wassilewskij a ecotypedefines a role for phytochrome D in red/far-red light sensing. Plant Cell 9, 1317-1326.
Ballaré, C. L., Sánchez, R. A., Scopel, A. L., Casal, J. J., and Ghersa, C. M. (1987). Earlydetection of neighbour plants by phytochrome perception of spectral changes in reflectedsunlight. Plant, Cell and Environment 10, 551-557.
Ballaré, C. L., Scopel, A. L., and Sánchez, R. A. (1989). Photomodulation of axis extension insparse canopies. Plant Physiology 89, 1324-1330.
Ballaré, C. L., Scopel, A. L., and Sánchez, R. A. (1990). Far-red radiation reflected from adjacentleaves: an early signal of competition in plant canopies. Science 247, 329-332.
Ballaré, C. L., Casal, J. J., and Kendrick, R. E. (1991). Responses of light-grown wild-type andlong-hypocotyl mutant cucumber seedlings to natural and simulated shade light. Photochemistryand Photobiology 54, 819-826.
Ballaré, C. L., Scopel, A. L., and Sánchez, R. A. (1991). Photocontrol of stem elongation inplant neighbourhoods: effects of photon fluence rate under natural conditions of radiation. Plant,Cell and Environment 14, 57-65.
Ballaré, C. L., Barnes, P. W., and Flínt, S. D. (1995). Inhibition of hypocotyl elongation byultraviolet-B radiation in de-etiolating tomato seedlings .l. The photoreceptor. Physiol Plant 93,584-592.
Ballaré, C. L., Scopel, A. L., Stapleton, A. E., and Yanovsky, M. J. (1996). Solar ultraviolet-Bradiation affects seedling emergence, DNA integrity, plant morphology, growth rate, andattractiveness to herbivore insects in Daturaferox. Plant Physiology 112, 161-170.
102

Ballario, P., Vittorioso, P., Magrelli, A., Talora, C., Cabibbo, A., and Macino, G. (1996). Whitecollar-l , a central regulator of blue light responses in Neurospora, is a zinc finger protein. EMBOJournal 15, 1650-1657.
Borthwick, H. A., Hendricks, S. B., Parker, M. W., Toole, E. H., and Toole, V. K. (1952). Areversible photoreaction controllíng seed germination. Proceedings of the National Academy ofScience, USA 38, 662-666.
Botto, J. F., Sánchez, R. A., and Casal, J. J. (1995).Role of phytochrome B in the inductíon ofseed germination by light in Arabidopsis thaliana. Journal of Plant Physiology 146,307-312.
Botto, J. F., Sánchez, R. A., Whitelam, G. C., and Casal, J. J. (1996).Phytochrome A mediatesthe promotion of seed germination by very low fluences of light and canopy shade light inArabidopsis. Plant Physiology 110, 439-444.
Bowler, C., and Chua, N. H. (1994). Emerging themes of plant signal transduction. Plant Cell 6,1529-1541.
Boylan, M. T., and Quail, P. H. (1989). Oat phytochrome is biologically active in transgenictomatoes. The Plant Cell 1, 765-773.
Braslavsky, S. E., Gartner, W., and Schaffner, K. (1997). Phytochrome photoconversion. Plant,Cell and Environment 20, 700-706.
Butler, W., Norris, K., Siegelman, H., and Hendricks, S. (1959). Detection, assay, andpreliminary purification of the pigment controllíng photoresponsive development of plants.Biochemistry 45, 1703-1708.
Casal, J. J. (1994). Stern extension-growth responses to blue-light require Pfr in tomato seedlingsbut are not reduced by the low phytochrome levels of the aurea mutant. Physiologia Plantar-um91, 263-267.
Casal, J. J. (1995). Coupling of phytochrome B to the control of hypocotyl growth inArabidopsis. Planta 196, 23-29.
Casal, J. J. (1996). Phytochrome A enhances the promotion of hypocotyl growth caused byreductions of phytochrome B Pfr levels in light-grown Arabidopsis thaliana. Plant Physiology112, 965-973.
Casal, J. J., and Smith, H. (1989). The function, action and adaptive significance of phytochromein light-grown plants. Plant, Cell and Environment 12, 855-862.
103

Casal, J. J., Sánchez, R. A., Benedetto, D., and De Miguel, L. C. (1991). Light promotion of seedgerrnination in Datura ferox is mediated by a highly stable pool of phytochrome. Photochemistryand Photobiology 53, 249-254.
Casal, J. J., and Kendrik, R. E. (1993). Impaired phytocrome-meditaed shade avoidanceresponses in the aurea mutant of tomato. Plant, Cell and Environment 16, 703-710.
Casal, J. J., Sánchez, R. A., and Vierstra, R. D. (1994). Avena phytochrome A overexpressed intransgenic tobacco seedlings differentially affects red/far-red reversible and very-low-fluenceresponses (cotyledon unfolding) during de-etiolation. Planta 192, 306-309.
Casal, J. J., and Sánchez, R. A. (1994). Impaired stem-growth responses to blue light irradiancein light-grown transgenic tobacco seedlings overexpressing Avena phytochrome A. PhysiologiaPlantarum 91, 268-272.
Casal, J. J., and Sánchez, R. A. (1994). Overexpression of oat phytochrome A gene differentiallyaffects stem grth responses to red/far-redratio signals characteristicof sparse or densecanopies. Plant, Cell and Environment 17, 409-417.
Casal, J. J., and Boccalandro, H. (1995). Co-action between phytochrome B and HY4 inArabidopsis thaliana. Planta 197, 213-218.
Casal, J. J., Sánchez, R. A., Boylan, M., Vierstra, R. D., and Quail, P. I-I.(1995). Is the far-redabsorbing form of Avena phytochrome A that is present at the end of the day able to sustainstem-growth inhibition during the night in transgenic tobacco and tomato seedlings? Planta 197,225-232.
Casal, J. J., Clough, R. C., and Vierstra, R. D. (1996). High-irradiance responses induced by farred light in grass seedlings of the wild type or overexpressing phytochrome A. Planta 200, 132137.
Casal, J. J., Sánchez, R. A., and Yanovsky, M. J. (1997). The fimction of phytochrome A. Plant,Cell and Environment 20, 813-819.
Casal, J. J., Sánchez, R. A., and Botto, J.F. (1998). Modes of action of phytochromes. Journal ofExperimental Botany 49, 127-138.
Cashmore, A. R., Jarrillo, J. A., Wu, Y.-J., and Liu, D. (1999). Cryptochromes: blue lightreceptors for plants and animals. Science 284, 760-765.
Childs, K. L., Lu, J. L., Mullet, J. E., and Morgan, P. W. (1995). Genetic regulation ofdevelopment in Sorghum bicolor X. Greatly attenuated photoperiod sensitivity in a phytochromedeficient sorghum possessing a biological clock but lacking a red light-high irradiance response.Plant Physiology 108, 345-351.
104

Christie, J. M., Reymond, P., Powell, G. K., Bemasconi, P., Raibekas, A. A., Liscum, E., andBriggs, W. R. (1998). Arabídopsis NPHl: a flavoprotein with the properties of a photoreceptorfor phototropísm. Science 282, 1698-1701.
Clack, T., Mathews, S., and Sharrock, R. A. (1994). The phytochrome apoprotein family inArabidopsis is encoded by five genes: the sequences and expression of PHYD and PHYE. PlantMolecular Biology 25, 413-427.
Clough, R. C., Casal, J. J., Jordan, E. T., Christou, P., and Vierstra, R. D. (1995). Expression offunctional phytochrome A in transgenic rice. Plant Physiology 109, 1039-1043.
Clough, R. C., and Viestra, R. D. (1997). Phytochrome degradation. Plant, Cell and Enviroment20, 713-721.
Colbert, J. T., Hershey, H. P., and Quail, P. H. (1983). Autoregulatory control of translatablephytochrome mRNA levels. Plant Physiology 80, 2248-2252.
de Lint, P. J. A. L. (1957). Double action of near infrared in length growth of the Avenacoleoptile. . Mendedelingen van de Landbouwhogeschool te Wageningen/Nederland 57, 1-9.
Dehesh, K., Franci, C., Parks, B. M., Seeley, K. A., Short, T. W., Tepperrnan, J. M., and Quail, P.H. (1993). Arabidopsis hy8 locus encodes phytochrome A. Plant Cell 5, 1081-1088.
Devlin, P. F., Patel, S. R., and Whitelam, G. C. (1998). Phytochrome E influences intemodeelongation and flowering time in Arabídopsis. Plant Cell 10, 1479-1487.
Dunlap, J. C. (1999). Molecular bases for circadian clocks. Cell 96, 271-290.
Elich, T. D., and Chory, J. (1997). Phytochrome: if it looks and smells like a histidine kinase, is ita histidine kinase? Cell 91, 713-716.
Emery, P., So, W. V., Kaneko, M., Hall, J. C., and Rosbasch, M. (1998). Cry, a Drosophila clockand light regulated cryptochrome, is a major contributor to circadian rhythm resetting andphotosensitivity. Cell 95, 669-679.
Fankhauser, C., and Chory, J. (1997). Light control of plant development. Annual Review of Celland Developmental Biology 13, 203-229.
Finlayson, S. A., Lee, I. J., and Morgan, P. W. (1998). Phytochrome B and the regulation ofcircadian ethylene production in Sorghum. Plant Physiology 116, 17-25.
Furuya, M., and Song, P.-S. (1994). Assembly and properties of holophytochrome. InPhotomorphogenesis in plants, R. E. Kendrick and G. H. M. Kronenberg, eds. (Dordrecht, TheNetherlands: Kluwer Academic Publishers), pp. 105-140.
105

Halaban, R. (1969). Effects of light quality on the circadian rhythm of leaf movement of a shortday plant. Plant Physiology 44, 973-977.
Halliday, K. J., Thomas, B., and Whitelam, G. C. (1997). Expression of heterlogousphytochromes A, B, or C in transgenic tobacco plants alters vegetative development andflowering time. Plant Journal 12, 1079-1090.
Hartmann, K. M. (1966). A general hypothesis to interpret ‘high energy phenomena’ ofphotomorphogenesis on the basis of phytocrome. Photochem. Photobiol. 5, 349-366.
Heim, B., and Scháfer, E. (1982). Light-controlled inhibition of hypocotyl growth in Sinapis albaL. seedlings. Fluence rate dependency of hourly light pulses and continuous irradiation. Planta154,150-155.
Heim, B., and Scháfer, E. (1984). The effect of red and far-red light in the high irradiancereaction of phytochrome (hypocotyl growth in dark-grown Sinapis alba L.). Plant, Cell andEnvironment 7, 39-44.
Hendricks, S. B., Toole, V. K., and Borthwick, H. A. (1968). Opposíng actions of light in seedgennination of Poa pratensis and Amaranthus arenicola. Plant Physiol. 43, 2023-2028.
Heyer, A. G., Mozley, D., Landschütze, V., Thomas, B., and Gatz, C. (1995). Function ofphytochrome A in Solanum tuberosum as revealed through the study of transgenic plants. PlantPhysiology 109, 53-61.
Hillman, W. S. (1971). Entrainment of Lemna C02 Output through phytochrome. PlantPhysiology 48, 770-774.
Jackson, S. D., Heyer, A., Dietze, J., and Prat, S. (1996). Phy'tochrome B mediates thephotoperiodic control of tuber formation in potato. Plant Journal 9, 159-166.
Jenkins, G. I. (1997). UV and blue light signal transduction in Arabidopsis. Plant, Cell andEnvironment 20, 773-778.
Johnson, C. B., and Tasker, R. (1979). A scheme to account quantitatively for the action ofphytochrome in etiolated and light- grown plants. Plant, Cell Environ. 2, 259-265.
Johnson, E., Bradley, M., Harberd, P., and Whitelam, G. C. (1994). Photoresponses of lightgrown phyA mutants of Arabidopsis. Phytochrome A is required for the perception of daylengthextensions. Plant Physiology 105, 141-149.
Jordan, B. R. (1995). The effects of ultraviolet-B radiation on plants: a molecular perspective.Advan. Bot. Res. 22, 97-162.
106

Kay, S. A., Nagatani, A., Keith, B., Furuya, M., and Chua, N. H. (1989). Rice phytochrome isbiologically active in transgenic tobacco. The Plant Cell 1, 775-782.
Keller, J. M., Shaklin, J., Vierstra, R., and Hershey, H. P. (1989). Expression of a functionalmonocotyledonous phytochrome in transgenic tobacco. . EMBO Journal 8, 1005-1012.
Kendrick, R. E., and Kronemberg, G. H. M. (1994). Photomorfogenesis in plants, 2 Edition(Dordrecht, The Netherlands: Kluwer Academic Publishers).
Kircher, S., Kozma-Bognar, L., Kim, L., Adam, E., Harter, K., Schafer, E., and Nagy, F. (1999).Light quality dependent nuclear import of the plant photoreceptors phytochrome A and B. PlantCell (inpress).
Klemm, E., and Ninnemann, H. (1976). Detailed action sectrum for the delay shift in pupaeemergence of Drosophila pseudoobscura. Photochemistry and Photobiology 24, 369-317.
Kondo, T., Johnson, C. H., and Hastings, J. W. (1991). Action spectrum for resetting thecircadian phototaxis rhythm in the CW15 strain of Chlamydomonas. I. Cells in darkness. PlantPhysiology 95, 197-205.
Koomneef, M., Rolf, E., and Spruit, C. J. P. (1980). Genetic control of light-inhibited hypocotylelongation in Arabidopsis thaliana (L.) Heynh. Zeitschrift fur Pflanzenphysiologie 100, 147-160.
Kreps, J. A., and Simon, A. E. (1997). Environmental and genetic effects on circadian clockregulated gene expression in Arabidopsis. Plant Cell 9, 297-304.
Lander, E. S., and Botstein, D. (1989). Mapping mendelian factors underlying quantitative traitsusing RFLP linkage maps. Genetics 121, 185-199.
Lincoln, S., Daly, M., and Lander, E. (1992). Mapping genes controlling quantitative traits withMAPMAKER/QTL. Whitehead Institute Technical Report, 2nd Edn.
Linden, H., and Macino, G. (1997). White collar 2, a partner in blue-light signal transduction,controlling expression of light-regulated genes in Neurospora crassa. EMBO Journal 16, 98-109.
Lister, C., and Dean, C. (1993). Recombinant inbred lines for mapping RFLP and phenotypicmarkers in Arabidopsis thaliana. Plant J. 4, 745-750.
López Juez, E., Nagatani, A., Tomizawa, K.-I., Deak, M., Kem, R., Kendrick, R. E., and Furuya,M. (1992). The cucumber long hypocotyl mutant lacks a light-stable PHYB-like phytochrome.Plant Cell 4, 241-251.
Mancinelli, A. L. (1994). The physiology of phytochrome action. In Photomorphogenesis inplants, R. E. Kendrick and G. H. M. Kronenberg, eds. (Dordrecht, The Netherlands: KluwerAcademic Publishers), pp. 211-269.
107

Marques, N., Mena-Barreto, L., and Golombek, D. A. (1997). Cronobiología (Buenos Aires Argentina: Eudeba).
Mathews, S., and Shar-rock,R. A. (1997). Phytochrome gene diversity. Plant, Cell andEnvironment 20, 666-671.
Mazzella, M. A., Alconada Magliano, T. M., and Casal, J. J. (1997). Dual effect of phytochromeA on hypocotyl growth under continuous red light. Plant, Cell and Environment 20, 261-267.
McCormac, A. C., Wagner, D., Boylan, M. T., Quail, P. H., Smith, H., and Whitelam, G. C.(1993). Photoresponses of transgenic Arabidopsis seedlings expressíng introduced phytochromeB- encoding cDNAs: evidence that phytochrome A and phytochrome B have distinctphotoregulatory fimctions. Plant Journal 4, 19-27.
Millar, A. J., Straume, M., Chory, J., Chua, N. H., and Kay, S. A. (1995). The regulation ofcircadian period by phototransduction pathways in Arabidopsis. Science 267, 1163-1166.
Miyamoto, Y., and Sancar, A. (1998). Vitamin B2-based blue-light photoreceptors in theretinohypothalamic tract as the photoactive pigments for setting the circadian clock in marnmals.Proceedings of the National Academy of Science, USA 95, 6097-6102.
Mohr, H. (1972). Lectures on Photomorphogenesis (New York, USA: Springer-Verlag).
Mohr, H. (1994). Coactíon between pigment systems. In Photomorphogenesis in Plants, R. E.Kendrick and G. H. M. Kronenberg, eds. (Dordrecht, The Netherlands: Kluwer AcademicPublishers), pp. 353-373.
Moran, R. (1982). Formulae for determination of chlorophyllous pigments extracted with N,Ndimethylformamide. Plant Physiol. 69, 1376-1381.
Mustilli, A. C., and Bowler, C. (1997). Tuning in to the signals controlling photoregulated geneexpression in plants. EMBO J. 16, 5801-5806.
Nagatani, A., Chory, J., and Furuya, M. (1991). Phytochrome B is not detectable in the hy3mutant of Arabidopsis, which is deficient in responding to end-of-day far- red light treatments.Plant Cell Physiol. 32, 1119-1122.
Nagatani, A., Reed, J. W., and Chory, J. (1993). Isolation and initial characterization ofArabidopsis mutants that are deficient in phytochrome A. Plant Physiology 102, 269-277.
Nagy, F., Fejes, E., Wehmeyer, B., Dallman, G., and Scháfer, E. (1993). The circadian oscillatoris regulated by a very low fluence response of phytochrome in wheat. Proceedings of theNational Academy of Sciences USA 90, 6290-6294.
108

Negbi, M., and Koller, D. (1964). Dual action of white light in the photocontrol of germinationof Oryzopsis miliacea. . Plant Physiology 39, 247-253.
Ni, M., Tepperrnan, J. M., and Quail, P. H. (1998). PIF3, a phytochrome-interacting factornecessary for normal photoínduced signal transduction, is a novel basic helix-loop-helix protein.Cell 95, 657-667.
Parker, M. W., Hendricks, S. B., Borthwick, H. A., and Scully, N. J. (1946). Action spectrum forthe photoperiodic control of floral initiation of short-day plants. Botanical Gazzette 108, 1-26.
Parker, M. W., Hendricks, S. B., Borthwick, H. A., and Went, F. W. (1949). Spectral sensitivityfor leaf and stem growth of etiolated pea seedlings and their similarity to action spectra forphotoperiodism. American Journal of Botany 39, 194-204.
Parker, J. E., Szabo, V., Staskawicz, B. J., Lister, C., Dean, C., Daniels, M. J., and Jones, J. D. G.(1993). Phenotypic characterization and molecular mapping of the Arabidopsis thaliana locusRPP5, ditermining disease resistance to Peronospora parasitíca. The Plant Journal 4, 821-831.
Parks, B. M., and Quail, P. H. (1993). hy8, a new class of Arabidopsis long hypocotyl mutantsdeficient in functional phytochrome A. The Plant Cell 5, 39-48.
Patterson, A. H., Lander, E. S., Hewitt, J. D., Peterson, S., Lincoln, S. E., and Tansksley, S. D.(1988). Resolution of quantitative traits into mendelian factors by using a complete linkage mapof restriction fragment length polymorphisms. Nature 335, 721-726. .
Pittendrigh, C. S., and Minis, D. H. (1964). The entrainment of circadian oscillations by light andtheir role as photoperiodic clocks. The American Naturalist 98, 261-294.
Pittendrigh, C. S. (1981). Circadian systems: entrainment. In Handbook of BehavioralNeurobiology, J. Aschoff, ed. (New York: Plenum Press), pp. 95-124.
Pratt, L. H., Cordonier-Pratt, M.-M., Kelmenson, P. M., Lazarova, G. I., Kubota, T., and Alba, R.M. (1997). The phytochrome gene family in tomato (Solanum lycopersicum L.). Plant, Cell andEnvironment 20, 672-677.
Prioul, J. L., Quarrie, S., Causse, M., and de Vienne, D. (1997). Dissecting complexphysiological functions through the use of molecular quantitative genetics. Journal ofExperimental Botany 48, 1151-1163.
Quail, P. H., Scháfer, E., and Marmé, D. (1973). Tumover of phytochrome in pumpkincotyledons. Plant Physiology 52, 128-134.
Quail, P. H. (1991). Phytochrome: a light-activated molecular switch that regulates plant geneexpression. Annu. Rev. Genet. 25, 389-409.
109

Quail, P. H., Boylan, M. T., Parks, B. M., Short, T. W., Xu, Y., and Wagner, D. (1995).Phytochromes: Photosensory perception and signal transduction. Science 268, 675-680.
Quail, P. H. (1997). An emerging molecular map of the phytochromes. Plant, Cell andEnvironment 20, 657-665.
Quail, P. H. (1998). The phytochrome family: dissection of functional roles and signallingpathways among family members. Phil. Trans. R. Soc. London. B. 353, 1399-1403.
Quin, M., Kuhn, R., Moran, S., and Quail, P. H. (1997). Overexpressed phytochrome C hassimilar photosensory specificity to phytochrome B but a distinct capacity to enhance primary leafexpansion. Plant Journal 12, 1163-1172.
Reed, J. W., Nagpal, P., Poole, D. S., Furuya, M., and Chory, J. (1993). Mutations in the gene forthe Red/Far-Red light receptor phytochrome B alter cell elongation and physiological responsesthroughout Arabidopsis development. Plant Cell 5, 147-157.
Robson, P. R. H., Whitelam, G. C., and Smith, H. (1993). Selected components of the shadeavoidance syndrome are displayed in a normal manner in mutants of Arabidopsis thaliana andBrassica rapa deficient in phytochrome B. . Plant Physiology 102, 1179-1184.
Roenneberg, T., and Deng, T.-S. (1997). Photobiology of the Gonyaulax circadian system. I.Different phase response curves for red and blue light. Planta 202, 494-501.
Rüdiger, W., and Thümmler, F. (1994). The phytochrome chromophore. In Photomorphogenesisin plants, R. E. Kendrick and G. H. M. Kronenberg, eds. (Dordrecht: Kluwer AcademicPublishers), pp. 51-69.
Sargent, M. L., and Briggs, W. R. (1967). The effects of light on a circadian rhythm ofconidiation in Neurospora. Plant Physiology 42, 1504-1510.
Satter, R. L., Guggino, S. E., Lonergan, T. A., and Galston, A. W. (1981). The effects of Blueand Far-red light on rhythmic leaflet movements in Samanea and Albizzia. Plant Physiology 67,965-958.
Sharrna, R., Lepez Juez, E., Nagatani, A., and Furuya, M. (1993). Identification of photo-inactívephytochrome A in etiolated seedlings and photo-active phytochrome B in green leaves of theaurea mutant of tomato. Plant Journal 4, 1035-1042.
Sharrock, R. A., and Quail, P. H. (1989). Novel phytochrome sequences in Arabidopsis thaliana:structure, evolution and differential expression of a plant regulatory photoreceptor family. Genesand Development 3, 1745-1757.
Shinomura, T., Nagatani, A., Hanzawa, H., Kubota, M., Watanabe, M., and Furuya, M. (1996).Action spectra for phytochrome A- and phytochrome B-specific photoinduction of seed
110

gennination in Arabidopsis thaliana. Proceedings of the National Academy of Sciences USA 93,8129-8133.
Short, T. W., and Bn'ggs, W. L. (1994). The transductíon of blue light signals in higher plants.Annu. Rev. Plant Physiol. Plant Molec. Biol. 45, 143-171.
Simon, E., Satter, R. L., and Galston, A. W. (1976). Circadian rhythmicity in excised Samaneapulvini. Plant Phisiology 58, 421-425.
Smith, H. (1982). Light quality, photoperception and plant strategy. . Annual Review of PlantPhysiology 33, 481-518.
Smith, H., and Whitelam, G. C. (1990). Phytochrome, a family of photoreceptors with multiplephysiological roles. Plant, Cell and Environment 13, 695-707.
Smith, H., Tumball, M., and Kendrick, R. E. (1992). Light-grown plants of cucumber longhypocotyl mutant exhibit both long term and rapid elongationgrth responses to irradiationwith supplementary far-red light. Photochemistry and Photobiology 569, 607-610.
Smith, H. (1994). Sensing the light environment: the functions of the phytochrome family. InPhotomorphogenesis in Plants, R. E. Kendrick and G. H. M. Kronenberg, eds. (Dordrecth, TheNetherlands: Kluwer Acadarnic Publishers), pp. 377-416.
Somers, D. E., Sharrock, R. A., Tepperrnan, J. M., and Quail, P. H. (1991). The hy3 longhypocotyl mutant of Arabidopsís is deficient in phytochrome B. Plant Cell 3, 1263-1274.
Somers, D. E., and Quail, P. H. (1995). Temporal and spatial expression patterns of PHYA andPHYB genes in Arabidopsis. Plant Journal 7, 413-427.
Somers, D. E., Devlin, P. F., and Kay, S. A. (1998). Phytochromes and cryptochromes in theentrainment of the Arabidopsís circadian clock. Science 282, 1488-1490.
Stanewsky, R., Kaneko, M., Emery, P., Beretta, B., Wager-Smith, K., Kay, S. A., Rosbasch, M.,and Hall, J. C. (1998). The cryb mutation identifies cryptochrome as a circadian photoreceptor inDrosophila. Cell 95, 681-692.
Takahashi, J. S., DeCoursey, P. J., Bauman, L., and Menaker, M. (1984). Spectral sensitivity of anovel photoperceptive system mediating entrainment of mammalian circadian rhythms. Nature308, 186-188.
Terry, M. J. (1997). Phytochrome chromophore-deficient mutants. Plant, Cell and Environment20, 740-745.
Tevini, M., and Teramura, A. H. (1989). UV-B effects on terrestrial plants. Photochemistry andphotobiology 50, 479-487.
lll

Thomas, B., and Dickinson, H. G. (1979). Evidence for two photoreceptors controlling growth inde-etiolated seedlings. Planta 146, 545-550.
Thomas, B., and Vince-Prue, D. (1997). Photoperiodism in Plants, 2 Edition (San Diego, USA:Academic Press).
Thresher, R. J., Vitatema, M. H., Miyamoto, Y., Kazantsev, A., Hsu, D. S., Petit, C., Selby, C. P.,Dawut, L., Smithies, 0., Takahashi, J. S., and Sancar, A. (1998). Role of mouse cryptochromeblue-light photoreceptor in circadian photoresponses. Science 282, 1490-1494.
van Tuinen, A., Kerckhoffs, L. H. J., Nagatani, A., Kendrick, R. E., and Koomneef, M. (1995).Far-red light insensitive, phytochrome A deficient mutants of tomato. Molecular and GeneralGenetics 246, 133-141.
van Tuinen, A., Kerckhoffs, L. H. J., Nagatani, A., Kendn'ck, R. E., and Koomneef, M. (1995). Atemporarin red light-insensitive mutant of tomato lacks a light-stable, B-Like phytochrome.Plant Physiol 108, 939-947.
Vierstra, R. D. (1994). Phytochrome degradation. In Photomorphogenesis in Plants, R. E.Kendrick and G. H. M. Kronenberg, eds. (Dordrecht, The Netherlands: Kluwer AcademicPublishers), pp. 141-162.
Wagner, D., Fairchild, C. D., Kuhn, R. T., and Quail, P. I-I.(1996). Chromophore-bearíng NH2terminal domains of phytochromes A and B determines their photosensory specificity anddifferential light lability. Proceedings of the National Academy of Science USA 93, 4011-4015.
Wagner, D., Hoecker, E., and Quail, P. H. (1997). RED] is necessary for phytochrome Bmediated red light specific signal transduction in Arabidopsis. Plant Cell 9, 731-743.
Weller, J. D., Murfet, I. C., and Reid, J. B. (1997). Pea mutants with reduced sensitivity to far-redlight define an important role for phytochrome A in day-length detection. Plant Physiology 114,1225-1236.
Whitelam, G. C., and Smith, H. (1991). Retention of phytochrome-mediated shade avoidanceresponses in phytochrome-deficient mutants of Arabidopsis, cucumber and tomato. . Joumal ofPlant Physiology 139, 119-125.
Whitelam, G. C., Johnson, E., Peng, J., Carol, P., Anderson, M. L., Cowl, J. S., and Harberd, N.P. (1993). Phytochrome A null mutants of Arabidopsis display a wild-type phenotype in whitelight. Plant Cell 5, 757-768.
Yamaguchi, R., Nakamura, M., Mochizuki, N., Kay, S. A., and Nagatani, A. (1999). Lightdependent translocation of a phytochrome B-GFP fusion protein to the nucleus in transgenicArabidopsis. The Joumal of Cell Biology 145, 437-445.
112

Yeh, K.-C., Wu, S.-H., Murphy, J. T., and Lagarias, J. C. (1997). A cyanobacterial phytochrometwo-component light sensory system. Science 277, 1505-1508.
Yeh, K.-C., and Lagarias, C. J. (1998). Eukaryotic phytochromes: light-regulated serine/threoninprotein kinases with histidine kinase ancestry. Proceedings of the National Academy of Science,USA 95, 13976-1398].


















