
CONTROL DE CALIDAD DEL PARASITOIDE DE … · durante mi estancia como estudiante y asesoramiento de...
76
CONTROL DE CALIDAD DEL PARASITOIDE DE Anastrepha spp.: Utetes anastrephae Viereck (HYMENOPTERA: BRACONIDAE) EN METAPA, CHIAPAS. TESIS Como requisito parcial para obtener el titulo de INGENIERO AGRÓNOMO ESPECIALISTA EN PARASITOLOGÍA AGRÍCOLA Presenta: MARÍA DOLORES GARCÍA CANCINO Chapingo, Estado de México, Diciembre de 2012
Transcript of CONTROL DE CALIDAD DEL PARASITOIDE DE … · durante mi estancia como estudiante y asesoramiento de...

CONTROL DE CALIDAD DEL PARASITOIDE DE Anastrepha spp.: Utetes anastrephae Viereck
(HYMENOPTERA: BRACONIDAE) EN METAPA, CHIAPAS.
TESIS
Como requisito parcial para obtener el titulo de
INGENIERO AGRÓNOMO ESPECIALISTA EN
PARASITOLOGÍA AGRÍCOLA
Presenta:
MARÍA DOLORES GARCÍA CANCINO
Chapingo, Estado de México, Diciembre de 2012



AGRADECIMIENTOS
A la Universidad Autónoma Chapingo por darme la inmensa dicha de formarme
profesionalmente en sus aulas.
Al M.C. José Manuel Gutiérrez Ruelas miembro de la Dirección de la Campaña contra
Moscas de la fruta y al Programa Moscamed por la oportunidad de realizar mis
estudios prácticos pre-profesionales en sus instalaciones y la convivencia con el
personal que en ahí laboran.
Al Dr. Jorge Luis Cancino Díaz por el inmenso apoyo, sus sugerencias, aportaciones y
consejos en éste trabajo. A la M.C. Patricia López, M.C. Lía Ruíz, Ing. Florida López,
los técnicos Patricia Rosario, Elvia Padilla, Belisario y Floriberto por su gran apoyo
durante mi estancia en el laboratorio de Control Biológico.
A la M.C. Amalia Pérez Valdez por sus valiosas recomendaciones y apreciable apoyo
durante mi estancia como estudiante y asesoramiento de esta tesis.
Al comité revisor por el grato apoyo y correcciones realizadas para la culminación del
presente trabajo de tesis.
A todos y cada una de las personas que participaron en mi formación como
profesionista; a mis profesores de Preparatoria y de Parasitología Agrícola.
Gracias por sus buenas enseñanzas.
A mis Padres Evelio García Avalos y Amalia Cancino Villarreal por darme la vida, por
sus buenos consejos y su inmenso apoyo en mis decisiones.
A mis hermanos Ignacio, Rodrigo Rosendo, Evelio y José Nahúm por ser tan cariñosos,
por esta familia tan sólida y amorosa; a mi Tía Lilia que me acompañó en las buenas y
en las malas durante mi estancia en Chapingo.
A mis muy estimados amigos Jorge A. Arcos Rangel, Luz María Hermoso Santamaría,
Ramés Salcedo Baca, María López López y Carlos Diego León Arellano por todas las
experiencias vividas.
A mis colegas y grandiosos amigos Carmen Herrera, Ángeles Cruz, Luisa Cristóbal,
Monserrat Tuxtla, Marilyn Hernández, Israel Cueto y Pablo Galván, gracias por las
aventuras y por el fabuloso apoyo durante los días de estudiantes.

Dedicado a…
José Darío
Por ser la inspiración que conduce mi vida.

i
INDICE GENERAL INDICE GENERAL ........................................................................................................................ i
INDICE DE FIGURAS ................................................................................................................ iii
INDICE DE CUADROS ............................................................................................................... iv
INDICE DE APENDICE ............................................................................................................... iv
RESUMEN .................................................................................................................................... v
1. INTRODUCCIÓN ................................................................................................................. 1
2. OBJETIVOS ......................................................................................................................... 3
2.1. GENERAL ..................................................................................................................... 3
2.2. PARTÍCULARES ........................................................................................................... 3
3. HIPOTESIS .......................................................................................................................... 3
4. REVISION DE LITERATURA .............................................................................................. 4
4.1. Moscas de la fruta ......................................................................................................... 4
4.1.1. Importancia económica ......................................................................................... 4
4.1.2. Distribución y hospederos ......................................................................................... 4
4.1.3. Morfología de larvas de Anastrepha spp. ............................................................. 5
4.1.4. Taxonomía ............................................................................................................. 6
4.1.5. Ciclo biológico y hábitos de Anastrepha ................................................................... 8
4.1.6. Manejo integrado de Anastrepha spp. ...................................................................... 9
4.1.7. Control biológico de moscas de la fruta .................................................................... 9
Antecedentes ..................................................................................................................... 10
4.2. Cría masiva de parasitoides........................................................................................ 11
4.2.1. Unidades de oviposición:..................................................................................... 12
4.2.2. Tiempo de exposición .......................................................................................... 12
4.2.3. Jaulas de producción: .......................................................................................... 13
4.2.4. Condiciones de desarrollo: .................................................................................. 13
4.2.5. Mantenimiento de la colonia de adultos .............................................................. 14
4.2.6. Alimentación de adultos ...................................................................................... 14
4.2.7. Implicaciones de la Cría Masiva .......................................................................... 15
4.3. Relación Huésped – Parasitoide ................................................................................. 15
4.3.1. Atributos ............................................................................................................... 16
4.3.2. Capacidad de búsqueda...................................................................................... 16
4.3.3. Especificidad. ....................................................................................................... 17
4.3.4. Capacidad reproductiva ....................................................................................... 17
4.3.5. Adaptabilidad ....................................................................................................... 18
4.3.6. Calidad del hospedero ......................................................................................... 18
4.3.7. Oviposición, emergencia y proporción de sexos. ............................................... 19

ii
4.3.8. Comportamiento .................................................................................................. 20
4.3.9. Efecto de la irradiación ........................................................................................ 21
4.3.10. Muerte larvaria ................................................................................................. 22
4.3.11. Uso de huéspedes alternantes (no nativos) .................................................... 23
4.4. Demografía .................................................................................................................. 24
4.4.1. Tabla de vida de adultos ..................................................................................... 24
4.5. Uso de Parasitoides .................................................................................................... 26
4.5.1. Parasitoides de Anastrepha ................................................................................ 27
4.5.2. Familia Braconidae .............................................................................................. 27
4.6. Utetes anastrephae ..................................................................................................... 28
4.6.1. Origen e Importancia ........................................................................................... 28
4.6.2. Distribución y Hospederos................................................................................... 28
4.6.3. Morfología ............................................................................................................ 29
4.6.4. Taxonomía ........................................................................................................... 33
4.6.5. Ciclo biológico y hábitos de Utetes. .................................................................... 34
5. MATERIALES Y MÉTODOS ............................................................................................ 36
5.1. Localización ................................................................................................................. 36
5.2. Evaluación del hospedero ........................................................................................... 37
5.3. Efecto de la Irradiación ............................................................................................... 38
5.4. Tiempo de exposición ................................................................................................. 38
5.5. Tiempos y frecuencia de exposición ........................................................................... 39
5.6. Tabla de vida de adultos ............................................................................................. 39
5.7. Análisis estadístico: ..................................................................................................... 40
6. RESULTADOS Y DISCUSIÓN .......................................................................................... 41
6.1. Selección de hospederos. ........................................................................................... 41
6.2. Efecto de la Irradiación. .............................................................................................. 43
6.3. Tiempos de exposición ............................................................................................... 44
6.4. Tiempo y Frecuencia de oviposición........................................................................... 46
6.5. Tabla de vida y fertilidad de Utetes anastrephae ....................................................... 48
7. CONCLUSIÓNES .............................................................................................................. 54
8. BIBLIOGRAFÍA ................................................................................................................. 55
9. APENDICE ......................................................................................................................... 60
9.1. Clave taxonómica para determinar géneros de Anastrepha de importancia económica. ............................................................................................................................. 60
9.2. Morfología comparativa de Anastrepha ludens y Anastrepha obliqua. ...................... 63
9.3. Clave para la determinación de las especies de parasitoides (Hymenoptera: Braconidae) de Anastrepha spp. ....................................................................................... 65

iii
INDICE DE FIGURAS
Fig. 1.- Regiones del país con registro de la plaga………………………………..5
Fig. 2.- Larvas de tercer instar de A. ludens y A. obliqua…………………………6
Fig. 3.- Ciclo biológico de Anastrepha spp………………………………………….8
Fig. 4.- Alas de Utetes anastrephae………………………………………………..30
Fig. 5.- Margen de la caryna hypostomal………………………………………….31
Fig. 6.- Tórax de U. anastrephae…………………………………………………...31
Fig. 7.- Ovipositor con válvulas o fundas………………………………………….31
Fig. 8.- Huevo de U. anastrephae………………………………………………….32
Fig. 9.- Adulto hembra de U. anastrephae………………………………………...32
Fig. 10.- Adulto macho de U. anastrephae………………………………………..33
Fig. 11.- Ciclo biológico de los parasitoides de larvas…………………………...34
Fig. 12.- Ubicación de las instalaciones Moscafrut………………………………36
Fig. 13.- Trabajo de laboratorio y manejo de material biológico………………..40
Fig. 14.- Gráfica comparativa de selección de hospederos……………………..43
Fig. 15.- Gráfica comparativa de los efectos de irradiación……………………..44
Fig. 16.- Gráfica comparativa de Tiempos de exposición……………………….45
Fig. 17.- Promedio de tiempo de oviposición de U. anastrephae……………….47
Fig. 18.- Comportamiento de U. anastrephae durante la búsqueda de
hospedero………………………………………………………………………………...47
Fig. 19.- Tasa de supervivencia de U. anastrephae……………………………..48
Fig. 20.- Esperanza de vida de hembras de U. anastrephae…………………...49
Fig. 21.- Tasa de fertilidad de U. anastrephae en los huéspedes
comparados……………………..………………………………………….…………….50
Fig. 22.- Adultos de A. ludens y A. obliqua………………………………………..63
Fig. 23.- Tórax característico de A. ludens y A. obliqua…………………………64
Fig. 24.- Alas características de A. ludens y A. obliqua…………………………64
Fig. 25.- Morfología del ovipositor y extremo apical de A. ludens y A. obliqua..64
Fig. 26.- Espiráculos anteriores característicos de las larvas de A. ludens y A.
obliqua……………………………………………………………………………………65

iv
INDICE DE CUADROS
Cuadro 1.- Análisis comparativo de los caracteres de larvas maduras (tercer
instar) (Montoya, et al. 2010)…………………………………………………………….5
Cuadro 2.- Clasificación Taxonómica de los hospederos utilizados en este estudio
(Aluja, 1994)…………………………………………………………………………….....6
Cuadro 3.- Clasificación Taxonomía de la especie parasitoide……………………34
Cuadro 4.- U. anastrephae en pose de parasitación…..…………………………....47
Cuadro 5.- Tabla de vida de cohorte de U. anastrephae en A. obliqua…….……..52
Cuadro 6.- Tabla de vida de cohorte de U. anastrephae en A. ludens….………...53
INDICE DE APENDICE Apéndice 1.- Clave taxonómica de parasitoides nativos de moscas de la fruta………………………………………………………………………………………60 Apéndice 2.- Morfología de los adultos de Anastrepha…………………………..63 Apéndice 2.- Clave taxonómica de géneros de Anastrepha……………………...65

v
CONTROL DE CALIDAD DEL PARASITOIDE DE Anastrepha spp.: Utetes
anastrephae Viereck (HYMENOPTERA: BRACONIDAE) EN METAPA, CHIAPAS.
RESUMEN
La presente investigación tuvo la finalidad de determinar los parámetros en la calidad del parasitoide Utetes anastrephae en su reproducción masiva en laboratorio, en Anastrepha obliqua y Anastrepha ludens, a través de: el porcentaje de parasitismo, proporción de sexos, longevidad, fecundidad, esperanza de vida, tiempo de exposición, tiempo de oviposición, efecto de la irradiación, capacidad de búsqueda, selección de huéspedes y mortalidad larvaria. Cada una con procedimiento y técnicas distintas. Los resultados fueron: porcentaje de parasitismo de 40 a 60%, proporción sexual promedio de 2 hembras por macho, longevidad máxima de adulto de 22 días, fecundidad máxima en A. obliqua de 22 hijos por hembra y en A. ludens de 36, esperanza de vida de 11 días, tiempo de exposición de 2 horas, tiempo de oviposición de 1.8 minutos, la irradiación no mostró efecto negativo o positivo en la evaluación, capacidad de búsqueda de oviposición máxima fue de 1 hora y la promedio fue de 2 minutos, mortalidad larvaria entre 5 a 15%; los requerimientos de cría masiva de U. anastrephae tales como: las adecuadas unidades de oviposición, sitios de las jaulas de producción, alimentación del adulto, condiciones de desarrollo y mantenimiento de la colonia lo ubican como una alternativa más del control biológico de moscas de la fruta. Palabras clave: cría masiva, parasitismo, mortalidad larvaria, A. obliqua y A. ludens.
ABSTRACT The present investigation was intended to determine the quality parameters of the parasitoid Utetes anastrephae in mass reproduction laboratory, Anastrepha obliqua and Anastrepha ludens, across the percentage of parasitism, sex ratio, longevity, fertility, hope life, exposure time, oviposition time, effect of irradiation, search capability, selection of hosts and larval mortality. Each with different procedures and techniques. The results were: percentage of parasitism of 40-60%, average sex ratio of 2 females per male, adult longevity maximum of 22 days, maximum fertility in A. obliqua of 22 offspring per female and A. ludens of 36, life expectancy of 11 days exposure time of 2 hours, oviposition time of 1.8 minutes irradiation showed no negative or positive effect on the evaluation, search capability maximum oviposition was 1 hour and the average was 2 minutes, larval mortality between 5 to 15% mass rearing requirements of U. anastrephae such as the units suitable oviposition sites of production cages, adult feeding, development and maintenance conditions of the colony place it as an alternative biological control of fruit flies. Keywords: mass rearing, parasitism, larval mortality, A. obliqua and A. ludens.

1
1. INTRODUCCIÓN
La fruticultura es muy importante dentro de la economía de México y las moscas
de la fruta (Díptera: Tephritiidae) son una plaga que devasta sorprendentemente la
calidad del producto en campo (Montoya, 2004). En México, este problema es mas
que evidente ya que se tiene la presencia de los nativos del género Anastrepha,
que es una fuerte amenaza en los cultivos frutícolas del país y da lugar a la actual
Campaña Nacional Contra Moscas de la Fruta con sustento jurídico y técnico en la
NOM-023-FITO-1995 y NOM-075-FITO-1997(Gutiérrez, et al 1995; Cancino &
Montoya, 2004).
El control biológico de moscas de la fruta representa una de las técnicas más
importantes dentro del manejo integrado de esta plaga. Su aplicación ha sido
principalmente por medio de las liberaciones aumentativas de parasitoides
(Montoya et al. 2000), para lo cual es importante contar con una cría masiva que
provea constantemente de los parasitoides.
Las liberaciones de parasitoides para el control de moscas del genero Anastrepha
spp. se iniciaron con el fin de establecer especies de parasitoides exóticos
(Jiménez et al., 1990). De esta actividad se obtuvo que los parasitoides
Diachasmimorpha longicaudata (Ashmead) y Aceratoneuromyia indica (Silvestri)
(Martínez, et al. 2003) fueron los que tuvieron un establecimiento notable.
Partiendo de este antecedente, D. longicaudata ha sido la especie más empleada
en el control de poblaciones de Anastrepha (Montoya et al., 2007). Sin embargo
existe un amplio gremio de parasitoides nativos del genero Anastrepha que
pueden tener importancia en el control de poblaciones, principalmente en áreas
silvestres de moscas de Anastrepha. Como resultado de un proceso evolutivo,
estas especies se encuentran muy asociadas a especies de Anastrepha en
diferentes hospederos nativos (Aluja et al., 2003, Sivinski et al., 1999). El
parasitoide de larvas Utetes anastrephae (Viereck) es un enemigo nativo de
moscas del genero Anastrepha que se distribuye ampliamente en toda la región
Neotropical (Ovruski, et al., 2000). Su importancia radica a que está asociado con

2
el parasitismo de larvas de Anastrepha obliqua (Mcquart), la cual se reporta como
una de las cuatro especies más importantes económicamente en nuestro país y la
principal plaga en la producción de mango en áreas tropicales (Teodoro, 2011).
Actualmente en la Planta Moscafrut se tiene una cría de U. anastrephae para
realizar estudios de su importancia. Esta cría se mantiene, empleando como
hospedero alternativo a larvas de moscas de Anastrepha ludens (Loew), la cual
funciona como un hospedero ficticio. Diferentes problemas se han encontrado que
pueden ser atribuidos a esta relación forzosa. Por ejemplo es notable la mortalidad
de larvas hospederas después de la exposición, lo cual puede deberse a un factor
fisiológico para el desarrollo de U. anastrephae. De acuerdo a lo anterior se
llevaron a cabo una serie de evaluaciones comparativas empleando larvas de A.
ludens y larvas de A. obliqua (el hospedero nativo de U. anastrephae) para poder
analizar si el empleo de un hospedero ficticio puede ser la razón de los problemas
en la cría masiva de U. anastrephae, y determinando cuantitativamente los
diferentes factores y parámetros que estiman el control de calidad de la
producción masiva del parasitoide.

3
2. OBJETIVOS
2.1. GENERAL Conocer las atribuciones de Utetes anastrephae (Viereck) desarrollado en su
hospedero nativo Anastrepha obliqua y en un hospedero ficticio Anastrepha
ludens, determinando los parámetros que éste requiere para su reproducción en
laboratorio.
2.2. PARTÍCULARES Obtener indicativos que permitan conocer cual es la causa de la mortalidad de
larvas hospederas expuestas a U. anastrephae en base a evaluaciones de, la
relación hospedero-parasitoide, comportamiento, efecto de la irradiación y tiempos
de exposición requeridos.
Conocer y determinar parámetros demográficos a través de una tabla de vida de
adultos del parasitoide empleando A. ludens y A. obliqua como hospederos.
3. HIPOTESIS
a) La especie Anastrepha obliqua es más apta que Anastrepha ludens como
hospedero en la reproducción del parasitoide Utetes anastrephae en
condiciones de cría masiva.
a‟) La especie Anastrepha obliqua es igual o menos apta que A. ludens como
hospedero en la reproducción del parasitoide Utetes anastrephae en
condiciones de cría masiva.
b) La mortalidad larvaria se debe a la calidad y resistencia por parte del
huésped.
b’) La mortalidad larvaria no se debe a la calidad y resistencia por parte del
huésped, sino a otros factores.

4
4. REVISION DE LITERATURA
4.1. Moscas de la fruta
Es una plaga de cultivos frutícolas, en México principalmente ataca a frutos
comerciales como cítricos y mango; causan daño a través de las larvas que al
alimentarse de los frutos, los destruyen completamente; además tienen un gran
número de hospederos, lo que unido a su capacidad de dispersión y alta
reproducción permiten su permanencia en el campo en altas poblaciones y
constituyendo una seria limitante para la comercialización de productos agrícolas
(Montoya, et al. 2010).
Por lo que se requiere la aplicación armónica de actividades encaminadas a lograr
el establecimiento de zonas libres y de baja prevalencia de la plaga.
4.1.1. Importancia económica
Las moscas de Anastrepha ocasionan grandes pérdidas económicas a la
fruticultura que ascienden a 100 millones de dólares anuales (Peña, et al. 2002);
reducen el rendimiento productivo de los cultivos, pérdida de cosechas, reduce el
valor comercial, incrementan gastos por tratamiento, etc. (Montoya, et al. 2010).
Esta plaga es considerada de importancia cuarentenaria, afectando el ingreso de
sus dependientes económicos, reduce el volumen y obstaculiza la
comercialización de los productos hacia el exterior, sobre todo cuando esos países
están sujetos a medidas cuarentenarias impuestas en los países importadores.
(Salcedo, 2010).
4.1.2. Distribución y hospederos
Las especies de Anastrepha spp. más abundantes en México afectan
especialmente cítricos y mango. Su distribución natural comprende del sur de
Estados Unidos hasta Sudamérica, donde las poblaciones alcanzan índices altos
de la plaga, si las medidas de control no se practican (Figura 1).

5
Fig. 1.- Zonas del país con registro de la plaga.
Estados libres: Baja California Norte y Sur, Sonora, Chihuahua y Coahuila
(Montoya, et al. 2010).
Hospederos:
A. obliqua; Mango, Jobo, ciruela (Spondias spp.), marañón, Almendro americano,
Cerezo, Eugenia (pomarrosa, guayaba, caimito), carambola, naranja dulce.
A. ludens; Cítricos, mango, caimito, guayaba, carambola, durazno silvestre,
(Rocha, et al. 2009).
4.1.3. Morfología de larvas de Anastrepha spp.
Las larvas son color blanco, alargadas y cilíndricas, por lo general recurvadas
ventralmente y con ganchos bucales, aplanadas en el extremo caudal, ocho áreas
fusiformes ventrales (indistinto entre el tórax y el abdomen, con 11 segmentos en
el cuerpo). Con anchura de hipostoma casi igual en todas las especies. (Vea
Figura 2, Cuadro 1.)
Presencia de moscas de la fruta en México.

6
Fig. 2.- A) Larvas de tercer instar de A. ludens y B), A. obliqua. (Fotografiado por
García C. M.D. 2012)
Cuadro 1.- Análisis comparativo de los caracteres de larvas maduras (tercer instar) (Montoya, et al. 2010).
Especie
hospedera
Longitud
de larva
Carinas
orales
Espiráculos
anteriores
Espiráculos
posteriores
Procesos
espiraculares
Lóbulos
anales
A. ludens 9-11mm 12-16 15-21 dígitos 3-3.5 veces
más largo
que ancho.
Dorsal y ventral:
10-12 ramas
cortas.
Mediales: 6-11
ramas cortas.
Bífidos
A. obliqua 8-10mm 7-10 12-15 dígitos 3.5-4 veces
más largo
que ancho.
Dorsal y ventral:
10-12 ramas
largas.
Mediales: 6-11
ramas largas.
Enteros
4.1.4. Taxonomía
Tephritidae: Perteneciente al orden Díptera; la más diversa dentro de la
superfamilia Tephritoidea, está constituida aproximadamente por 4223 especies
(Norrbom, et al. 1998). Según López, et al. (2010) en el Continente Americano
existen cerca de 977 especies de las cuales el 73% han sido registradas para la
región Neotropical (incluyendo desde México hasta Argentina).
A B

7
Subfamilia Tephritinae: En el Continente Americano existe una tercera parte de
la diversidad global del grupo (619 especies). En virtud del alto grado de
especialización biológica hacia sus huéspedes, se considera como la subfamilia
mas evolucionada de Tephritidae (Headrick & Goeden 1998, Montoya, et al. 2010).
Género Anastrepha Schiner: incluye cuatro especies de importancia económica
en los trópicos americanos, Anastrepha ludens (mosca Mexicana de la fruta), A.
obliqua (mosca de las Indias Occidentales), A. serpentina (mosca de las
sapotaceas), A. striata (mosca de la guayaba) (Hernández- Ortiz 1992; Aluja
1993). (Cuadro 2).
Cuadro 2.- Clasificación Taxonómica de los hospederos utilizados en este estudio (Aluja, 1994). Especie Huésped 1:
Clase: Insecta.
Orden: Díptera.
Familia: Tephritidae
Subfamilia: Tephritinae
Tribu: Toxotrypanini
Género: Anastrepha
Especie: Anastrepha ludens low.
Especie Huésped 2:
Clase: Insecta.
Orden: Díptera.
Familia: Tephritidae
Subfamilia: Tephritinae
Tribu: Toxotrypanini
Género: Anastrepha
Especie: Anastrepha obliqua Maquart.

8
4.1.5. Ciclo biológico y hábitos de Anastrepha
Las moscas de la fruta son insectos holometábolos. Los adultos de Anastrepha
son medianos (8-10 mm) a grandes (12-14 mm) de color marrón amarillentos a
negro. Al poco tiempo de emerger, se activan y vuelan en busca de agua y
alimento, el cual lo pueden encontrar en frutos maduros o fermentados,
secreciones de troncos u hojas, excrementos de los pájaros silvestres y
secreciones de áfidos u otros insectos chupadores. Una vez que alcanza la
madurez sexual (5 a 20 días), se inicia la cópula, que en general consiste en que
el macho secreta una feromona sexual para atraer a la hembra y al quedar
cargada deposita sus huevos en un fruto próximo a madurar, depositando una
feromona de marcaje que impide que el fruto pueda ser utilizado por otra hembra,
aunque esto no se cumple cuando las poblaciones son muy elevadas. Su alta
capacidad de vuelo y adaptación a diferentes medios es una característica de los
adultos que pueden movilizarse por más de 200 Km ayudados por el viento. Los
huevos son pequeños (0,5 mm), alargados y blanquecinos (3 a 6 días). Larvas
muscidiformes, cremosas, miden de 1 a 1,5 cm. (15 a 30 días). Presentan tres
instares larvales y al completar su desarrollo salen del fruto cuando éste cae al
suelo, se entierran a unos 2-5 cm para formar la pupa que presenta un pupario
como una cápsula (barrilito) de 3-5 mm, amarillo oscuro hasta castaño oscuro (15
a 25 días). Los adultos llegan para reproducir una generación nuevamente (Fig. 3),
viven alrededor de 60-90 días.
Fig. 3.- Ciclo biológico de Anastrepha spp.

9
4.1.6. Manejo integrado de Anastrepha spp.
Desde1993 se reproduce en las instalaciones de la Planta Moscafrut la especie
Anastrepha ludens con niveles de producción de 100 millones de moscas por
semana. Estas moscas se liberan para control autocida como una de las
actividades de la Campaña Nacional contra Moscas de la Fruta. Además se tiene
la producción de 50 millones de pupas del parasitoide D. longicaudata, para el
impulso del control biológico en México.
Con el fin de fortalecer las medidas de mitigación de riesgo de zonas libres y de
baja prevalencia de moscas de la fruta, el 23 de abril de 1998 se publicó la NOM-
075-FITO-1997, por la que se establecen los requisitos y especificaciones para la
movilización de frutos hospedantes de moscas de la fruta (Gutiérrez, 2010). En
ésta se incluyen diversas técnicas que aportan alternativas de control en la región
sureste del país para regular y controlar la plaga que se encuentra presente como
una amenaza a la agricultura.
En plantaciones de frutos comerciales hospederos de esta plaga, la Junta Local de
Sanidad Vegetal y el programa Moscafrut instalan trampas con proteína
hidrolizada, cebos alimenticios, aspersión de formulados con insecticida (malation
o spinosad), se realiza monitoreo continuo y se estimula a los productores con
actividades de control cultural para que las apliquen a nivel regional.
4.1.7. Control biológico de moscas de la fruta
Control biológico: Es una forma de manejar poblaciones de animales o plantas.
Consiste en el uso de uno o más organismos para reducir la densidad de una
planta o animal que causa daño al hombre (DeBach, 1964). Así, el control
biológico puede definirse como el uso de organismos benéficos (enemigos
naturales) contra aquellos que causan daño (plagas).
DeBach (1964), mencionó que “la acción de los parasitoides, depredadores o
patógenos para mantener la densidad de la población de un organismo plaga a un
promedio más bajo del que ocurriría en su ausencia”; describe un fenómeno
natural que al ser utilizado por el hombre en el manejo de plagas, malezas y

10
enfermedades en los agroecosistemas, constituye la base de la ciencia del control
biológico.
En moscas de la fruta se aplican dos conceptos fundamentales:
Control Biológico Clásico, que se define como la “introducción y establecimiento
permanente de un enemigo natural para el control o supresión, a largo término, de
la población de una plaga” e involucra la búsqueda de enemigos naturales en su
lugar de origen o fuera de él (Van Driesche, 2007).
El segundo concepto es el Control Biológico por Aumento ya que en algunas
ocasiones los enemigos naturales nativos o exóticos se establecen, pero sus
números son insuficientes para reducir la plaga a un grado que no cause daño.
Por esto, los enemigos naturales pueden incrementarse mediante la cría masiva
en laboratorio que resulte de bajo costo, para su posterior liberación ya sea
ocasional o repetida (Van Driesche, 2007). Este concepto se considera la manera
mas adecuada de aplicar el control biológico en Moscas de la fruta.
Otros conceptos relacionados son: el Control Biológico Fortuito, que trata de las
especies de enemigos naturales nativos o exóticos que aparecen
esporádicamente y, por tanto, ejercen un factor de mortalidad que provee un
control a veces fuerte o en otras ocasiones muy débil sobre la plaga. Y el
Control Biológico por Conservación, que consiste en la adopción de prácticas
culturales que fomentan la aparición y abundancia de enemigos naturales ya
establecidos (Nicholls, 2008).
Antecedentes
El primer programa de control biológico de moscas de la fruta se efectuó en
Australia en 1902 para combatir a Ceratitis capitata (Wiedemann), pero los
resultados del programa no fueron satisfactorios (Wharton, 1989, Leyva 1999). En
México se formó la comisión de Parasitología Agrícola donde De la Barrera,
encuentra el parasitoide Incrastopilas rudibunda (Ashmead) ahora conocido como
el parasitoide nativo Doryctobracon crawfordi (Viereck) atacando larvas de
Anastrepha spp. en Cuernavaca Morelos (Herrera, 1905, citado por Aluja, 2008).

11
En 1913, ante la cuarentena que promulga E.U.A. contra la exportación de frutas
de México, el Gobierno Federal y la Compañía de Ferrocarril Nacional apoyaron la
asociación cooperativo de productores de fruta para que buscaran solución al
problema de la cuarentena por la plaga, en este intento se realizan estudios por el
Dr. David L. Crawford que reporta encontrar enemigos naturales como hormiga
grande de rapiña que devora larvas de fruta caída, algunos pájaros, gallinas,
guajolotes que consumen pupas de mosca ejerciendo cierto control (Crawford,
1918; Aluja, et al. 2008). Fue hasta la década de los 40´s cuando se inicia el
interés en nuestro país por combatir biológicamente a las moscas de la fruta
mediante la importación de enemigos naturales. En 1929, se reporta a Coptera sp.
(Hymenoptera: Diapriidae), Anthrax scylla (Díptera: Bombyliidae) y Eucoila sp.,
obtenidos de pupas colectadas en campo. En 1954 se introdujeron cuando menos
10 especies de parasitoides, la mayoría de ellos originarios de la Región Indo
Asiática y Australiana (Wharton, 1987).
Se consignan mas de 80 especies parasíticas que atacan moscas de la fruta en el
mundo y al menos 10 especies fueron introducidas al país, tres se consideran
establecidas, el Bracónido Diachasmimorpha longicaudata, Aceratoneuromya
indica (Eulophidae) y Pachycrepoideus vindemiae (Pteromalido) (Leyva, 1999), en
conjunto con las especies nativas en las que se encuentran Utetes anastrephae,
U. divergens, U. vierecki, Doryctobracon areolatus, Opius hirtus (Hymenoptera:
Braconidae), Coptera haywardi, Aganaspis pelleranoi, Odontosema anastrephae y
Tribliographa sp. (Hymenoptera: Eucoilidae) que redundan en efectuar un control
eficiente como especies totalmente adaptadas a las regiones frutícolas del país.
4.2. Cría masiva de parasitoides
La cría masiva consiste en un proceso donde una colonia de parasitoides es
establecida de manera artificial dándose las condiciones adecuadas para el
desarrollo e incremento de la población, utilizando hospederos apropiados y
nutridos para el desarrollo y crecimi8eto poblacional del endoparasitoide.

12
El desarrollo tecnológico en la cría de D. longicaudata ha permitido establecer la
producción más grande del mundo de esta especie en México. Gracias a esta
producción se lleva a cabo el control biológico por medio de liberaciones
aumentativas con resultados favorables en el control de las poblaciones plaga de
moscas de la fruta (Montoya et al. 2007).
Algunas de las técnicas básicas de cría para la producción masiva de parasitoides
de moscas de la fruta son:
4.2.1. Unidades de oviposición:
En el caso de parasitoides de larvas, se realiza con unidades planas cubiertas con
telas porosas (por uno o por ambos lados) y aprisionadas con cintas elásticas
(Wong et al. 1991). En la parte media de las unidades se colocan larvas con o sin
dieta. Estas unidades tienen un grosor aproximado de 0.1 a 0.5 cm, para que el
hospedero pueda ser alcanzado por el ovipositor de la hembra. Para esta técnica
el parasitoide generalmente encuentra al hospedero por medio de las vibraciones
producidas por el movimiento y la actividad de alimentación (Lawrence 1981).
4.2.2. Tiempo de exposición
Dentro de la cría masiva las hembras requieren de tiempos específicos para
localizar, aceptar y ovipositar a su hospedero (Wong et al. 1990) lo cual tiene
suma importancia en el proceso, ya que existe un fenómeno común en la cría de
parasitoides de moscas de la fruta y es la presencia de superparasitismo
(Lawrence, 1988; Montoya et al. 2000; Cancino, 2010).
La alta incidencia del superparasitismo incrementa la mortalidad por alta
competencia (González, et al. 2007), por lo que el tiempo de exposición esta
directamente relacionado con el superparasitismo. Un decremento en la
emergencia también puede ser resultado de un periodo de exposición corto que no
permite un parasitismo suficiente de los hospederos expuestos (Cancino, 2010).

13
4.2.3. Jaulas de producción:
Los modelos básicos consisten en jaulas de forma cubica, con dimensiones
variables, con longitudes que van de los 25 a los 40 cm. La luz de la malla en cada
caso también es variable pero en todos los casos debe impedir la fuga de adultos
y permitir la aireación suficiente de acuerdo a la densidad de parasitoides en el
interior, quedan en juego la ubicación estratégica de los materiales como
bebederos, alimento, y sobre todo las unidades de oviposición (Wong et al. 1992).
4.2.4. Condiciones de desarrollo:
Los parasitoides de larvas de tercer estadio (próximas a pupar), requieren de
mantener visibles y al alcanze de su ovipositor a las larvas huéspedes por lo que
solo requieren ser colocadas en contenedores con un sustrato de pupación (por
ejemplo, vermiculita o suelo). En este periodo las condiciones ambientales tienen
un papel preponderante.
En primer lugar está la temperatura, sugiriéndose emplear un intervalo de 24 a 28
°C. El incremento en la temperatura implica un aceleramiento en el desarrollo, lo
que provoca en muchas ocasiones aumento de mortalidad. Un descenso en la
temperatura resulta en la prolongación del tiempo de desarrollo. En cría masiva, la
temperatura debe estar dentro de un intervalo muy corto, de lo contrario se retrasa
o se acelera el desarrollo, lo que puede originar problemas de emergencia
temprana de adultos. Cuando se tiene un programa estructurado de liberaciones,
el atraso o adelanto de la emergencia causa serios problemas (Hurtrel, et al.
2001).
La humedad relativa es otro factor importante. En la mayoría de los parasitoides la
humedad se debe mantener en un intervalo de 60-80%. En lugares con alta
humedad ambiental se pueden presentar problemas de contaminación por la
proliferación de microorganismos (hongos, ácaros, etc.) (Ashley et al. 1976), lo
que puede provocar descenso en la emergencia de adultos. La disminución
notable de la humedad también puede incrementar la mortalidad por
deshidratación (Cancino, 2010).

14
4.2.5. Mantenimiento de la colonia de adultos
La temperatura en que debe oscilar la sala es de 24-26 °C (Cancino, 2010);
cuando se ha obtenido un porcentaje de emergencia superior al 50%, es
recomendable en algunas especies como D. longicaudata y D. tryoni, cambiar los
adultos a otra sala con temperatura de 20 °C, 60-80% de humedad relativa, y un
fotoperiodo de 12:12 h luz: oscuridad. Estas condiciones disminuyen la mortalidad
durante el periodo de oviposicion de los parasitoides y generan una adecuada
producción de descendientes (Ramadan, et al. 1989, Cancino 2002). En la cría de
F. arisanus, mantener una adecuada proporción sexual es un problema
constante. Se considera que la feromona emitida por los machos disminuye su
efectividad en un medio saturado y se requiere de aireación forzosa en las jaulas.
En el caso de D. longicaudata el ruido puede ser un factor negativo (Sivinski y
Webb 1991) porque se pierden los sonidos producidos por las vibraciones de las
alas de los machos que son una fuente de estimulo para la hembra.
4.2.6. Alimentación de adultos
La mayoría de los parasitoides de moscas de la fruta que se conocen
(posiblemente con la excepción de Aceratoneuromyia indica (Silvestri)) son
clasificados como sinovigénicos, o sea, parasitoides que en estado adulto
requieren de alimentación para efectuar posteriormente su actividad de
oviposicion.
Al emerger los adultos es importante proporcionar alimento ya que de ello
dependerán su desarrollo y atributos. En la elaboración hay que cuidar dos
aspectos: 1) Que sea práctico para su manejo, y 2) que permita a los adultos
caminar y comer sin dificultad (Cancino, 2010).
En la mayoría de los casos la miel de abeja natural o enriquecida con proteínas o
vitaminas ofrece buenos resultados.

15
4.2.7. Implicaciones de la Cría Masiva
La cría masiva de parasitoides presenta diversos problemas y primeramente es
disponer de un lugar adecuado y acondicionado. Los insectos criados deben
mantener todas sus características para que una vez liberados, interactúen
competentemente con los insectos de la población que se quiere controlar
(Martínez, 1992).
La habilidad de los insectos para localizar su alimento y sitios de oviposición se
reduce, se restringe la dispersión, la disponibilidad de alimento puede no ser
suficiente o adecuado (Mackauer, 1976), nutrición inadecuada, enfermedades y
mala manipulación del material biológico.
La revisión continua de la colonia en crecimiento debe incluir componentes de la
biología y comportamiento de la especie que serán importantes para su
supervivencia, como: ciclo de vida, dispersión, supervivencia, localización de sitios
de cortejo, apareamiento y oviposición (Martínez 1992).
Chambers, 1977 considera que las causas que contribuyen al desplome genético
de los insectos de laboratorio son: la endogamia, la deriva genética y la selección.
Que al paso de las generaciones repercuten en la poca adaptabilidad en el
ambiente distinto al del laboratorio, mutaciones deletéreas y perdidas de la
variabilidad genética.
La mayoría de especies criadas en laboratorio presentan tasas reproductivas altas
y vida corta comparada con las cepas silvestres. Estas diferencias seguramente
se deben a la selección que ocurre durante la colonización, en donde se favorece
la alta fecundidad y el menor tiempo de desarrollo.
4.3. Relación Huésped – Parasitoide
Algunos datos importantes de los parasitoides en cuanto a sus huéspedes Van
Driesche, (2007) son: alta especificidad en cuanto a su huésped. Generalmente
mas pequeños que su huésped. Únicamente la hembra busca al huésped.

16
Varias especies diferentes de parasitoides pueden atacar las diferentes etapas del
ciclo de vida del huésped. Los huevos o larvas (estados inmaduros) se
desarrollan dentro del huésped y se alimenta de él, exterminándolo.
4.3.1. Atributos
Los atributos de un enemigo natural nos permiten conocer particularidades
propias, ayudándonos a conocer que tan efectivo puede ser regulando una
población de insectos a bajas densidades, actuando de forma densodependiente
(Martínez, 1992).
Un enemigo natural efectivo es altamente densodependiente aun cuando la
densidad del huésped sea baja y responde rápidamente a cualquier tendencia de
éste a sobrepasar las densidades, esto es: mientras la población plaga sea más
densa la respuesta numérica del enemigo natural ascenderá de manera
simultánea. Sus atributos claves deben ser alta capacidad de búsqueda,
especificidad, poder de incremento y adaptabilidad (Doutt y De Bach, 1964, Rosen
y Huffaker, 1982, Martínez 1992).
4.3.2. Capacidad de búsqueda.
El comportamiento de búsqueda de huésped es un buen punto de unión en la
biología de estos organismos para localizar sus huéspedes aun cuando sean
escasos, siendo el atributo más importante para un enemigo natural efectivo
(Doutt y DeBach, 1964). Los parasitoides utilizan principalmente semioquímicos
que son emitidos desde la fruta infestada o vibraciones. Estas sustancias
generalmente son volátiles aromáticos que se dispersan fácilmente en el
ambiente. Posteriormente al arribo de los parasitoides a la fruta infestada, éstos
son guiados por pistas más específicas. La capacidad de búsqueda depende en
gran medida de la movilidad que tenga el himenóptero, habilidad para dispersarse
rápidamente, longevidad durante la etapa de búsqueda de huéspedes habilidad
para utilizar su ovipositor, la deposición de huevecillos y el rápido desarrollo en
comparación con su huésped (Vinson, 1976; Romani, et al. 2002; Cancino, 2010).

17
Se considera que las hembras más aptas para reproducción son las que
desarrollan más rápidamente la capacidad de búsqueda.
En la cría masiva de D. longicaudata se observó que las hembras mas grandes y
mejor desarrolladas responden de manera mas directa y rápida hacia la búsqueda
del hospedero (Cancino, et al. 2006).
4.3.3. Especificidad.
Un alto grado de especificidad indica que el parasitoide tiene buena adaptación
fisiológica hacia el huésped y una dependencia directa sobre los cambios en el
huésped (Doutt y DeBach, 1964). Un parasitoide debe ser muy estricto en su
ataque, no puede mostrar una débil propensión al hacerlo, ni atacar a ningún otro
organismo benéfico. Aunque ser un parasitoide generalista también tiene sus
ventajas ya que al escasearse su huésped natural puede sobrevivir utilizando otra
especie como huésped alternativo. Este segundo tipo de parasitoide puede ser de
suficiente utilidad en el caso de control biológico de un complejo de plagas (Rosen
y Huffaker, 1982).
La capacidad para reconocer sus huéspedes es un atributo deseable en los
enemigos naturales. En los parasitoides incluye reconocer si el huésped ya esta
parasitado o aun no y con ello evitar el superparasitismo (Rosen y Huffaker, 1982).
4.3.4. Capacidad reproductiva
En ambientes inestables o alterados es deseable que un enemigo natural posea
una tasa alta de incremento, en comparación con la de la plaga, cuando se
requiere de una efectiva explotación de la población huésped (Rosen y Huffaker,
1982; Martínez, 1992).
En ambientes inestables o en campo el poder de incremento se ve afectado por la
fecundidad, la velocidad de desarrollo, capacidad de búsqueda, y la adaptación a
las condiciones de un hábitat en particular, requiriéndose en muchos casos que

18
las especies parasitas se vuelvan oportunistas y buenos competidores (Rosen y
Huffaker, 1982).
4.3.5. Adaptabilidad
Esta habilidad también es de suma importancia pues seria un excelente
parasitoide al adaptarse a todos los nichos donde habite su huésped y al mismo
tiempo tener alta supervivencia, es decir, que sea capaz de tolerar condiciones
climáticas extremas (incluso donde intervenga el hombre) y que se sincronice
adecuadamente con la biología y fenología de su huésped, siguiéndolo hacia
donde se disperse la plaga. Los enemigos naturales también deben evolucionar
junto con su huésped, para que puedan sobreponer cualquier resistencia (Doutt y
DeBach, 1964; Martínez, 1992). Todos estos atributos logran la posibilidad de que
el enemigo natural pueda criarse en laboratorio, lo cual facilita la distribución y
colonización y hace posible un control oportuno de la plaga (Doutt y DeBach,
1964).
4.3.6. Calidad del hospedero
Tamaño del cuerpo
En el caso de los huéspedes de parasitoides el tamaño si influye, pues hay un
incremento consistente en la producción de huevos y mayor sobrevivencia
conforme el tamaño de cuerpo se incrementa.
Las diferencias en el tamaño del cuerpo pueden ser interpretadas como el efecto
de competencia intraespecífica, a mayor densidad, mayor competencia y al no
disponer de suficientes recursos, menor tamaño. Siendo así, el tamaño del
hospedero en la cría de parasitoides repercute principalmente en los porcentajes
de emergencia y la proporción sexual, de manera general se considera que los
hospederos pequeños son utilizados para la producción de machos y los mas
grandes para emergencia de hembras (Godfray, 1994). Para ello es importante

19
tomar en cuenta el peso del hospedero en términos de producción masiva
(Cancino, et al. 2006).
Selección de hospederos
En la cría masiva la calidad del huésped es un requisito indispensable para
asegurar buenos resultados en la eficiencia de la producción de los parasitoides
(Godfray, 1994). En algunos estudios se ha mostrado que existe una relación
directa entre factores como la edad, el tamaño y el peso del hospedero con la
proporción sexual y la emergencia del parasitoide (Wong y Ramadan, 1992;
Cancino, et al. 2002). Es importante conocer los estándares de calidad que tendrá
la larva huésped a emplear en la producción masiva y asegurar una excelente
calidad de producción (Cancino, et al. 2006). Se deben cuidar los parámetros de
edad de la larva, preferentemente en 3er instar larval, el peso de la larva
dependiendo de la especie, porcentaje de pupación y mortalidad a las 72 horas.
El peso del huésped y el número de larvas en 10 ml, es un indicador del desarrollo
larvario durante la cría de moscas. Factores como la calidad de la dieta, su
composición y las condiciones ambientales donde se mantuvo el lote de larvas
son indicadores básicos que se relacionan con la calidad de la larva y los
parámetros inmediatos de los parasitoides adultos. Comparando con los
estándares de calidad se debe decidir la aceptación o rechazo de un lote de larvas
huésped, pudiendo evitar pérdidas económicas considerables, ante la obtención
de un lote de parasitoides de mala calidad (Cancino, et al. 2006).
4.3.7. Oviposición, emergencia y proporción de sexos.
La emergencia de adultos se considera el parámetro más indicativo de la calidad
del proceso de cría. La emergencia de adultos es también un complemento en la
evaluación de parámetros como proporción sexual y habilidad de vuelo. Estos dos
primeros factores se evalúan simultáneamente en la mayoría de los casos; en la
producción de Diachasmimorpha longicaudata es importante mantener una
proporción sexual inclinada a las hembras, sin embargo, la proporción debe ser

20
adecuada para evitar un desequilibrio que propicie una inclinación a machos en la
progenie de las siguientes generaciones (Cancino, et al. 2002).
Martínez (1992), menciona que la proporción de sexos puede ser influida por
diversos factores como: Densidad de sitios de oviposición (número de huéspedes),
periodo previo al apareamiento después de la emergencia, cópulas excesivas,
presencia de sitios preferenciales para la oviposición, tasa de oviposición, cantidad
de huevecillos depositados por cada inserción del ovipositor y cantidad de
huevecillos en los ovarios listos para su deposición. Albajes (1984), menciona al
analizar Opius concolor que de acuerdo a los días en que se realice la oviposición
será la proporción de machos y hembras el primer día dará origen solo a machos y
en días subsecuentes disminuirá gradualmente.
Como se menciona en páginas anteriores la capacidad de búsqueda juega un
papel importante también en la oviposición, ya que al realizar una oviposición
eficaz en tiempo y forma se logrará un buen desarrollo del parasitoide. En
ocasiones la disminución de la rapidez para desarrollar capacidad de oviposición
es relacionada con malformaciones presentes en parasitoides adultos. Sin
embargo, el promedio general es definido por las características previas de
desarrollo como baja mortalidad, alta pupación a las 72 h, tamaño y peso del
hospedero, para lograr liberar en campo un enemigo natural fuerte con suficientes
atributos que permitan un efecto eficiente en la supresión de la plaga (Cancino, et
al. 2006).
4.3.8. Comportamiento
El comportamiento de cortejo y cópula es un aspecto de la reproducción sexual
encaminado a la descendencia (Van Den Assem, 1986). Se requiere de la
fertilización para la producción de progenie viable; ya que en el caso de los
himenópteros los machos son resultado de huevecillos no fertilizados. Al tener
contacto sexual los espermatozoides se almacenan en la espermateca hasta su
salida para la fertilización de los huevecillos, las hembras potenciales para

21
inseminación no admiten la copula directa ya que los machos previamente deben
exhibirse con un cortejo previo (Van Den Assem, 1986).
En Utetes anastrephae emergen primero los machos y tres días después las
hembras. Las cuales requieren de 7 días para que alcancen su madurez sexual;
hasta los diez días de edad inician los apareamientos y comportamiento de cortejo
en los especímenes.
El acto de cortejo es similar entre los Bracónidos, y como sucede en D.
longicaudata, incluye: atracción, reconocimiento, orientación, aleteo, vibración de
alas, monta, movimiento de las antenas, copula y finalmente acicalamiento
postcopulatorio.
La copula en D. longicaudata se lleva a cabo en un lapso realmente corto de
menos de un minuto (Matthews, 1976). Donde el macho monta a la hembra
dorsalmente.
En el caso de U. anastrephae, esta posición puede permanecer desde 10
segundos hasta 2 horas, la hembra con una postura rígida del cuerpo, antenas
abiertas en un ángulo de 45 ° hacia el frente, el mancho montado dorsalmente
aleteando constantemente.
Es posible que U. anastrephae esté incluido en el grupo de parasitoides que
menciona (Hagen 1953), que producen sustancias volátiles o feromonas de
atracción que sirve de estimulo para el reconocimiento de su especie y de copula
entre machos. Puesto que el macho de Utetes anastrephae recorre distancias
extremas entre la colonia y se dirige lentamente donde esta la hembra que
permanece recta, al llegar el macho se detiene frente a la hembra se mueve
lentamente e inicia los movimientos del cortejo. En otras ocasiones cuando el
macho esta cerca, gira alrededor de la hembra y se posiciona en un lugar para
mostrar el movimiento continuo de alas, así una hembra puede tener hasta tres
machos, realizando esta danza.
4.3.9. Efecto de la irradiación

22
Durante la exposición no todos los huéspedes llegan a ser parasitados lo cual
representa un alto riesgo para los programas de control pues implicaría la
liberación de moscas adultas. Una propuesta viable para evitarlo es la aplicación
de irradiación a las larvas hospederas previas a la exposición. Sivinski y Smittle
(1990), observaron la supresión total del desarrollo de Anastrepha suspensa
(Loew) cuando irradiaron al hospedero. Resultados similares reporta Cancino et al.
2002 con Anastrepha ludens (Loew). En ambos trabajos las larvas fueron
empleadas como hospederos de D. longicaudata.
La irradiación de larvas previa a la exposición a parasitoides afecta solamente el
desarrollo posterior del hospedero sin que se presenten resultados desfavorables
en la emergencia, la fecundidad y supervivencia de los parasitoides emergidos.
Desde los 20 Gy aplicadas en larvas se puede suprimir la emergencia de moscas,
sin embargo, en condiciones de cría masiva se requiere incrementar esta dosis a
40 Gy. El incremento es recomendado debido a los mayores volúmenes con lo
cual se dispersa mas la intensidad de la radiación (Cancino, et al. 2002). Las dosis
dependen de la especie de huésped.
Las ventajas del uso de irradiación en la cría masiva son amplias, ya que da
muchas facilidades para el manejo de la colonia y el procedimiento general de
cría. El uso de irradiación en la cría masiva de parasitoides de moscas de la fruta
se ha ampliado a huevecillos y pupas. En cada caso se han determinado dosis
para la efectiva reducción de la viabilidad de los hospederos no parasitados y el
desarrollo adecuado de los parasitoides (J. Cancino, datos no publicados).
4.3.10. Muerte larvaria
La sobrevivencia de la larva y del pupario en moscas de la fruta dependen
fundamentalmente de la calidad de la larva, influidos por el manejo durante y
después de la exposición a parasitación. La mortalidad larvaria a las 72 horas de
exposición se puede dar por condiciones inadecuadas en el medio de pupación
como contaminación, altas densidades de larva, humedad, etc. El incremento de la
mortalidad es un grave problema en el cuarto de desarrollo de inmaduros, permite

23
el incremento dinámico de contaminantes como ácaros, hongos, bacterias y otros
microrganismos, siendo este parámetro la principal razón del decremento en la
producción de parasitoides (Cancino, et al. 2002).
Alta mortalidad larvaria o bajo porcentaje de pupación pueden ser resultado de
una mala selección de hospederos, problemas durante el proceso de exposición
larvaria o durante la pupación e inclusive de una presencia excesiva de
superparasitismo. Cuando los niveles de mortalidad son altos se pueden tomar
decisiones acerca del retiro del lote completo de producción para evitar el riesgo
de contaminar el resto de la producción en la sala de inmaduros. Los datos
óptimos o esperados en un lote de producción masiva, son bajos niveles de
mortalidad larvaria y altos porcentajes de pupación efectiva. Un porcentaje de
larva muerta arriba del 20% en cría masiva se relaciona con presencia de
contaminantes por microrganismos en el lote de producción (Cancino, et al. 2006).
4.3.11. Uso de huéspedes alternantes (no nativos)
Los factores bióticos son los principales agentes reguladores de una población
plaga. Regular significa que las poblaciones del insecto, al ser diezmadas por un
parasitoide, por ejemplo, disminuyen a tal punto que, por escasez de alimento, la
población del parasitoide también declina o busca un hospedero similar para
mantener su población con vida (Nicholls, 2008).
Estudiando la conducta de los parasitoides Van Driesche, (2007) menciona que,
los parasitoides aprenden y usan lo que aprenden para encontrar hospederos.
La experimentación previa del parasitoide, también puede influenciar la
preferencia hacia un huésped sobre otro. Muchos parasitoides adultos contactan
las kairomonas del huésped durante la emergencia y si las preferencias de un
parasitoide están débilmente determinadas genéticamente, el contacto con la
especie huésped de donde emergió o con sus productos puede reforzar la
preferencia por esa especie. Consecuentemente, los parasitoides criados en
hospederos alternantes pudieran funcionar menos contra la plaga a controlar. Para

24
los parasitoides especializados cuyas preferencias de hospedero están
fuertemente determinadas genéticamente, el acondicionamiento puede tener poco
efecto (Van Bergeijk et al., 1989; Van Driesche, 2007).
Establecer una nueva especie se facilita cuando son expuestos primero a la plaga
hospedera a la cual se asocian. Similarmente, la exposición de enemigos
naturales criados en masa a la plaga por controlar antes de la liberación, puede
corregir cualquier pérdida de eficacia al ser criados en un hospedero alternante
(Matadha et al., 2005).
4.4. Demografía
La demografía es definida como el estudio de las poblaciones y los procesos que
determinan sus atributos (Carey, 1993).
Carey (1982) inició los estudios sobre la demografía y dinámica de poblaciones de
la mosca del Mediterráneo, después de él, se dio continuidad a la temática en
materia de plaga de moscas de la fruta.
El manejo moderno de plagas es esencialmente el desarrollo y aplicación de
tecnologías como está que influyen en la fecundidad, desarrollo y mortalidad de la
población (Montoya, et al. 2010.)
La estructura de una población esta determinada por los nacimientos y muertes,
incluyendo aquellos parámetros como fecundidad, fertilidad y crecimiento
poblacional.
4.4.1. Tabla de vida de adultos
La tabla de vida fue presentada a los ecólogos por Pearl desde 1921, la cual se
define como un catálogo, sumario o inventario que describe la supervivencia y las
tasas de mortalidad de los individuos de una población según la edad de éstos.
Las tablas fueron desarrolladas primeramente por demógrafos para describir
poblaciones humanas con el fin de conocer la esperanza media de vida.
Posteriormente esta técnica se ha venido aplicando a muchas poblaciones de
animales y plantas (Montoya, 2006).

25
Una tabla de vida de cohorte, es aquella que toma un cierto número (n) de
individuos recién nacidos y se observa la supervivencia durante cada unidad de
tiempo pre-establecida por el investigador (horas, días, semanas, meses, etc.)
hasta la muerte del último individuo; una vez realizado esto, se definen los
estadísticos (Vera, et al. 2002):
x: Intervalo de edad en unidades de tiempo
nx: Número de individuos vivos al inicio del intervalo x a x+1
dx: Número de individuos muertos durante el intervalo x a x+1
qx: Tasa de mortalidad durante el intervalo x a x+1
Lx: Número promedio de individuos vivos durante el intervalo x a x+1
Tx: Suma acumulativa de L
x para obtener valores expresados en número de
individuos por unidades de tiempo
ex: Esperanza media de vida de los individuos de la población al inicio del intervalo
x; o bien, unidades de tiempo que le quedan por vivir, en promedio, a
cualquier individuo que haya cumplido cierta edad x
lx: Tasa (o probabilidad) de supervivencia al inicio del intervalo x.
Dentro del conocimiento del ciclo de vida del insecto están implicados dos
parámetros de suma importancia como la supervivencia y la fecundidad.
La supervivencia es un parámetro básico, posibilita conocer el tiempo de vida del
insecto, el cual debe considerarse largo de manera que permita llegar a la
madurez sexual, localizar microclimas adecuados para su reproducción, buscar
alimento, aparearse y reproducirse (Cancino, et al. 2002). También permite al
parasitoide mayores oportunidades para llevar a cabo la búsqueda de hospederos
y un mayor numero de oviposiciones (Cancino, et al. 2006).
La fecundidad va encaminada a la producción de huevecillos por hembra por día.
No existen registros anteriores de considerarlo un parámetro de control de calidad.
Sin embargo Cancino, et al. (2006) lo considera como un parámetro importante
para el control de calidad en la cría masiva de parasitoides de moscas de la fruta,
ya que el resultado de la fecundidad que se tenga en laboratorio servirá de

26
indicativo para inferir sobre el parasitismo que se obtenga en campo, mostrando
una perspectiva inmediata de la efectividad de un lote de parasitoides.
La esperanza de vida, es la media de la cantidad de tiempo (horas, días, meses,
años, etc.), que vive una cierta población en un cierto período (Vera, et al. 2002).
4.5. Uso de Parasitoides
Los parasitoides son a menudo los enemigos naturales más eficientes de los
insectos plaga (Van Driesche, 2007), su aprovechamiento es la primera opción a
ser considerada en programas de Manejo Integrado de Plagas (MIP) y han sido la
más explorada a nivel mundial (Clausen, 1978).
Esta modalidad se aplica en México y en moscas de la fruta al liberar
inundativamente enemigos naturales exóticos con el fin de controlar de manera
permanente a la especie plaga nativa.
De las 26 familias de parasitoides, los géneros usados más frecuentemente en
control biológico son Braconidae, Ichneumonidae, Eulophidae, Pteromalidae,
Encyrtidae y Aphelinidae (Hymenoptera) y Tachinidae (Diptera) (Greathead, 1986;
Van Driesche, 2007).
Cerca de 82 especies de parasitoides pertenecientes a diferentes familias han
sido obtenidos de moscas de las frutas, pero la mayoría de ellos y los más
importantes pertenecen a la familia Braconidae, dentro de la cual una gran parte
pertenecen a la subfamilia Opiinae (Wharton, 1989).
La familia Braconidae es una de las más importantes del orden Hymenoptera, con
cerca de 15000 especies conocidas, la mayoría de ellas parasitoides de insectos
(Wharton, 1993).

27
4.5.1. Parasitoides de Anastrepha
Las familias que contienen un mayor número de especies que atacan moscas de
la fruta son: Braconidae, Figitidae, Diapriidae, Eulophidae, Pteromalidae y
Chalcididae. Aproximadamente el 59% de las 46 especies de parasitoides de
Anastrepha pertenecen a la familia Braconidae, el 19,5% a la Figitidae, el 10,8% a
la Diapriidae, un 8,6% a la Pteromalidae y 2,1 a la Eulophidae (Ovruski, et al,
2000).
Entre los principales parasitoides de moscas de la fruta del genero Anastrepha se
encuentran Diachasmimorpha longicaudata (Ashmead), D. tryoni (Borgmeier)
Aceratoneuromyia indica (Silvestri), Opius hirtus (Fisher), Doryctobracon crawfordy
(Szépligeti), Utetes anastrephae (Viereck) que parasitan larvas , Coptera haywardi,
C. lopezi (Masner), A. pelleranoi (Brethes), Odontosema anastrephae,
Pachycrepoideus vindemiae (Rondan) que parasitan pupas y Fopius arisanus que
parasita huevecillos (Sonan) (Ovruski, 2000; Cancino, 2006); siendo los
parasitoides de pupa los primeros enemigos naturales de moscas de la fruta en
ser descritos (Sivinski, et al. 1997).
4.5.2. Familia Braconidae
Los Bracónidos pertencen a la familia de himenópteros apócritos de la super-
familia Ichneumonoidea. Tiene distribución mundial y es diversa en todas las áreas
(Wikipedia, 2012).
La mayoría de los Bracónidos son endoparasitoides, aunque hay un número
importante de ectoparásitoides. Los endoparásitos no paralizan a su huésped (o
sólo lo hacen momentáneamente) y dejan sus huevos dentro del cuerpo del
huésped.
En países como México y Centroamérica se han realizado estudios para
determinar las tasas de parasitismo en áreas naturales y cultivadas (Aluja et al.,
1990; Steck et al., 1986) dentro de la familia Braconidae se encuentran los

28
porcentajes de parasitismo altos, como 56% y bajos de 5%; resultados similares
se presentaron en los análisis realizados en Ecuador (Núñez, et al. 2004).
En estudios en Ecuador, Arias, et al., (2003); encontró a nuestra especie
estudiada Utetes anastrephae (Viereck) como la especie Bracónida más común,
atacando larvas de Anastrepha obliqua (Macquart) en Spondias spp. y mango.
En 2004 se realizó un estudio de distribución de parasitoides en la misma zona
encontrando que D. crawfordi es la especie predominante en los pisos altos,
seguida por Anagaspis pelleranoi, mientras que, en tierras bajas predomina
Utetes anastrephae, lo que se considera también aplica en las costas del pacifico
sur Mexicano (Arias, et al., 2003).
4.6. Utetes anastrephae
4.6.1. Origen e Importancia
Es un endoparasitoide nativo de las regiones tropicales de América capaz de
controlar diversas especies del complejo de moscas de la fruta.
Parasita larvas de Anastrepha spp. Prefiere Anastrepha obliqua (Macquart), A.
ludens y A. alveata (Aluja, 1998). Hasta la fecha solamente emergidos de „Jobos‟,
„Ciruelas‟ (Spondias mombin and S. purpurea, Anacardiaceae) y Mango
(Manguifera indica L). Con un porcentaje de parasitismo de 6 a 92% de (Colmart,
2006).
Huéspedes: A. manihoti; A. obliqua, A. suspensa, A. sororcula, A. ludens,
Rhagoletotrypeta sp., A. fraterculus y A. striata (Insuasty B. et al. 2007).
4.6.2. Distribución y Hospederos
Los primeros reportes de Utetes anastrephae (Viereck) fueron en México y
Republica Dominicana (Colmart, 2006). Mas tarde fue registrado en Estados
Unidos de Norteamérica, Costa Rica, Guatemala, El Salvador, Puerto Rico.
Introducido a Hawái en 1978 pero no se estableció (Wharton y Gilstrap, 1983).

29
Actualmente establecido en América Central, Norteamérica, Colombia, Brasil y
Argentina. (Wharton & Marsh, 1978; Colmart, 2006). Abarcando toda la región
neotropical.
4.6.3. Morfología
Wharton (1988) caracterizó al género Utetes por el margen de la carina
hypostomal bien desarrollada (Fig. 5); surco malar ausente; pronoto con una
depresión, margen anterior del clípeo cóncavo, presentando espacio entre éste y
las mandíbulas cerradas; estigma en forma de cuña con la inserción de la R1
levemente desplazada del medio a la base; con segunda célula cubital
ensanchada; ausencia de posnervelo. Este autor también dividió el género en dos
subgéneros, Bracanastrepha y Utetes y definió el primero por la total ausencia
de carina occipital.
Utetes anastrephae (Viereck, 1913) marca una coloración general predominante
rojo-amarillenta (fig. 9 y 10). Antenas, ojos, ocelos, extremidad distal de las
mandíbulas (Fig. 5.a y 5.b), tarsoapicales del primer y segundo par de patas y
funda del ovipositor, negros (Fig,7). Alas hialinas con venas negras; segunda
vena radial mas larga que la primera intercubital; vena recurrente alcanzando la
segunda célula cubital; ausencia de posnervelo (fig. 4); Propodeo areolado
anteriormente (Fig. 6).
Longitud corporal: 2.80 mm – 3.75mm.
Longitud de alas: 4.5 mm
Longitud de ovipositor: 1.6 mm (Sivinski et al. 2001).
Longitud de antenas: 4.5 – 5.5 mm
Coloración: cabeza y mitad anterior del cuerpo de color anaranjado; mitad
posterior del cuerpo y patas naranja-amarillentas; antenas, ápice de las
mandíbulas y los tarsos, triangulo ocelar, tegulas y vainas del ovipositor
oscurecidos, alas anteriores y posteriores pubescentes y hialinas, con venas y
estigma castaño oscuro.

30
Fig. 4.- Alas de Utetes anastrephae. (Fotografías tomadas por Valdez C. J. y
García C., M.D. 2012. Colegio de Postgraduados, Montecillo, México).
Fig. 5.- Margen de la carina hypostomal bien desarrollada.

31
Fig. 6.- Tórax.
Machos con manchas negras sobre el pronoto (Ovruski, 1996).
Fig. 7.- Ovipositor con válvulas. (Fotografiado por Valdez C., J. y García C., M. D.
2012. Colegio de Postgraduados, Montecillo. México).

32
.
Fig. 8.- Huevo de Utetes anastrephae (Viereck). Fotografiado por Charles Stuhl, USDA-ARS-CMAVE Gainesville, Florida.
Fig. 9.- Adulto hembra de U. anastrephae (Viereck). Fotografiado por Valdez C., J.
y García C., M.D.,2012.

33
Fig. 10.- Adulto macho de U. anastrephae (Viereck). Fotografiado por Valdez C., J.
y García C., M.D. 2012.
4.6.4. Taxonomía
Esta especie fue descrita originalmente como Opius anastrephae por Viereck
(1913). Se transfirió posteriormente al género Bracanastrepha por Fischer (1977),
pasando por el nombre de Opius mombinpraeoptantis Fischer (1966) y finalmente
a Utetes por Wharton (1988).
Cuadro 3.- Clasificación Taxonómica de la especie parasitoide.
Clase: Insecta.
Orden: Hymenoptera.
Familia: Braconidae
Subfamilia: Opiinae
Genero: Utetes
Especie: Utetes anastrephae (Viereck, 1913)

34
4.6.5. Ciclo biológico y hábitos de Utetes.
Parasitoide nativo de México ataca larvas maduras de 3er instar de mosca de la
fruta de algunas especies del género Anastrepha; A. ludens, A. obliqua y A. striata.
Su supervivencia es alrededor de 3 semanas como adulto (fig. 9 y 10), se trata de
un parasitoide sinovigénicos, el cual requiere de un tiempo en el estado adulto
para el desarrollo de su carga de huevecillos (Godfray 1994, Cancino 2010). Así
mismo es una especie de himenóptero con reproducción partenogenética haploide
con la variante arrenotoquia, primero produce los huevecillos del cual se originan
únicamente machos y posteriormente al ser fertilizados se da origen a las
hembras (Doutt, 1959; Martínez, 1992). En especies arrenotocas la proporción de
sexos puede llegar a ser un problema fuerte, lo que ocasiona conflictos en los
proyectos de control biológico, crías de insectarios y colonización de parasitoides.
Las hembras de Utetes anastrephae, son atraídas por los semioquímicos volátiles
de la fruta en la búsqueda de alimento y con ello de las larvas huéspedes que
atraves de la vibraciones buscan y precisan el ataque a través de sus antenas
sondeando con el ovipositor para detectar compuestos únicos, ovipositan un solo
huevo en el interior del cuerpo de las larvas de mosca que se encuentran dentro
del fruto (Stuhl et al. 2011).
Al romper el corion aguado del huevecillo, la larva parasitoide se mantiene en su
primera etapa de desarrollo hasta que el huésped comienza a pupar. El tiempo de
desarrollo de huevo a adulto del parasitoide depende de la temperatura, pero por
lo general toma alrededor de dos semanas y en estado adultos tres semanas. Los
zumos de frutas y otras sustancias producidas en las plantas como néctares
florales son la fuente de alimento de este parasitoide (Ovruski, et al. 2000).
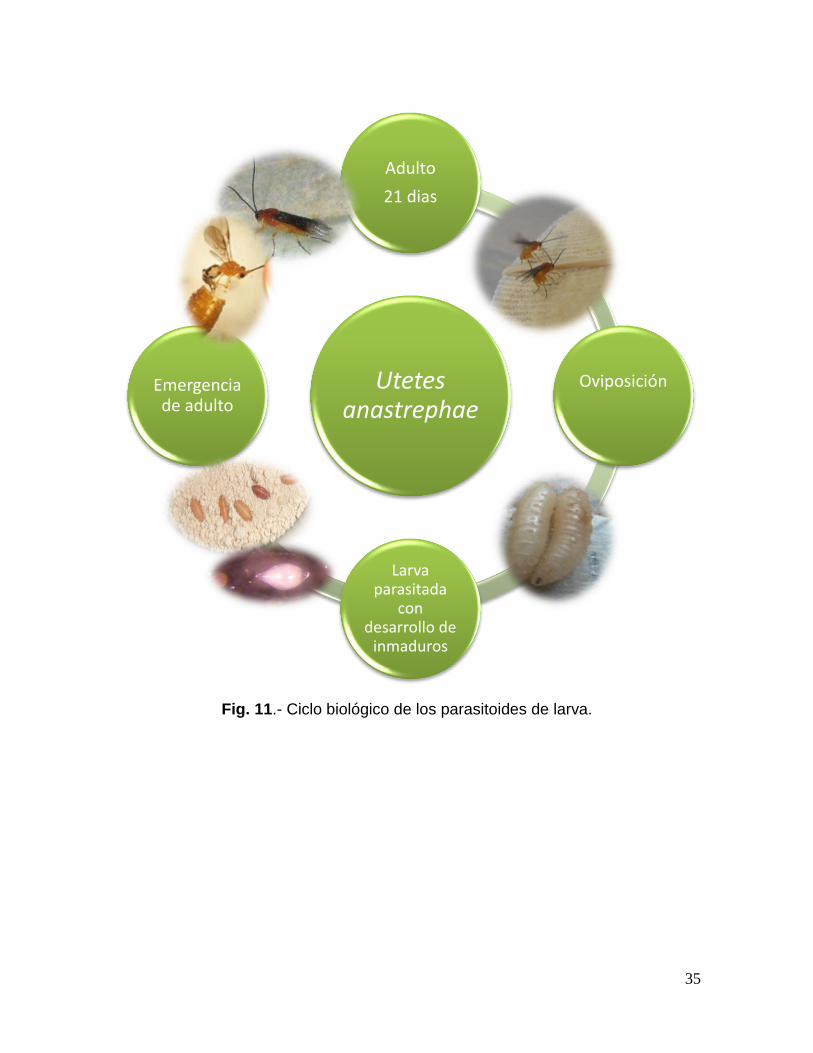
35
Utetes anastrephae
Adulto
21 dias
Oviposición
Larva parasitada
con desarrollo de
inmaduros
Emergencia de adulto
Fig. 11.- Ciclo biológico de los parasitoides de larva.

36
5. MATERIALES Y MÉTODOS
5.1. Localización Las evaluaciones se llevaron a cabo en el Laboratorio de Control Biológico de la
Subdirección de Desarrollo de Métodos del Programa Moscafrut (SAGARPA-IICA),
ubicados en el municipio de Metapa de Domínguez, Chiapas.
Fig. 12.- Ubicación de las instalaciones.
Las condiciones ambientales que tuvo este trabajo fue de 24±2 °C y 60-80% de H:
R. y un fotoperiodo de 12:12 h de relación luz: obscuridad.
Se utilizaron especímenes de la colonia de U. anastrephae mantenidos en
condiciones de cría por alrededor de 80 generaciones.
Se emplearon parasitoides adultos de cinco días de edad, copulados y
alimentados con miel de abeja. Como hospederos se emplearon larvas de 7 a 8
días de edad de Anastrepha obliqua con peso promedio de 15 mg y A. ludens con
peso promedio de 25 mg. obtenidos de la cría masiva de la Planta Moscafrut.
Algunas de estas larvas se sometieron a irradiación a una dosis de 4 Krad para
evitar el desarrollo de moscas de larvas no parasitadas.

37
Las jaulas utilizadas durante el experimento consistían en un diseño denominado
tipo Hawái (Fig. 13a); son cubicas de 27 cm por lado, cubiertas con malla
mosquitera en cuatro de sus caras, piso de madera y frente con vidrio corredizo.
En el piso de la jaula se colocó una porción de alimento a base de miel de abeja y
proteínas (Fig.13b), sobre una caja Petri de 5 cm de diámetro. Un bebedero de
vidrio con tapa de plástico y un trozo de papel filtro que permite el ascenso del
agua por medio de capilaridad, así como la unidad de exposición que consistió en
un disco de plástico de 10 cm de diámetro, cubierto por tela organdí que contenía
un numero determinado de larvas (100 o 200) en su interior con un poco de dieta
larvaria (realizada en la planta moscafrut) (Fig. 13e) sujetado firmemente por una
liga plástica. El tiempo de exposición varió según el experimento desde 1 hora
hasta 8 horas, terminando el tiempo de exposición las larvas se vaciaron en un
contenedor plástico de 30 y 50 ml de capacidad con tapa plástica que cuenta con
aireación a través de una ventana de tela organdí. En el que permanecían 3 días
antes de convertirse totalmente en pupas (Fig. 13d). Una vez convertidas en
pupas, se lavan y se cambia el medio de reposo de ser dieta por una cantidad
mínima de vermiculita que por su consistencia permite el secado rápido. En estos
recipientes se esperó la emergencia de los adultos de la progenie. Se calculó el
porcentaje de parasitismo con base en la emergencia de adultos, la proporción
sexual y algunos atributos de los parasitoides.
5.2. Evaluación del hospedero
Se aplicaron dos tratamientos uno con selección a hospedero y otro sin selección.
En cada caso se colocó una proporción de 30 hembras y 15 machos de U.
anastrephae en una jaula tipo Hawái. Los tratamientos aplicados fueron:
Unidad de exposición con 200 larvas de Anastrepha obliqua.
Unidad de exposición con 200 larvas de Anastrepha ludens.
Dos unidades de exposición con 100 larvas de A. ludens y otra con 100 de A.
obliqua.

38
Cada muestra consistió en la colocación de 200 larvas sobre una base de caja
Petri de plástico de 10 cm de diámetro cubierto con tela tricopt y atado con una
liga para asegurar su consistencia. La muestra se colocó dentro de una jaula, con
2 horas de exposición diarias, después de la oviposición fueron retiradas y
colocadas en contenedores de plástico con capacidad de 50 ml con dieta especial
elaborada para cada una de las especies de mosca. Dos días después la larva es
separada por lavado y tamizada para cambiarla a vermiculita como medio de
pupación, dentro del mismo contenedor se contabilizó la larva muerta hasta ese
momento. Esta evaluación se mantuvo con el mismo lote de hembras en la edad
de 5 a 10 días.
5.3. Efecto de la Irradiación
Se utilizaron cuatro jaulas (Fig. 13f) con una proporción de 30 hembras y 15
machos de parasitoides en cada uno de los tratamientos. Dos jaulas sometidas al
hospedero de A. ludens, una jaula con 200 larvas irradiadas y otra con 200 larvas
no irradiadas, los mismos tratamientos se aplicaron, con A. obliqua. En todos los
casos se aplicó un tiempo de exposición de 2 horas.
5.4. Tiempo de exposición
Se aplicaron diferentes tiempos de exposición en las larvas de Anastrepha ludens
y A. obliqua, se tomaron cuatro muestras con 200 larvas de mosca cada una, que
fueron introducidas por separado en una jaula tipo Hawái con 30 hembras y 15
machos de parasitoides U. anastrephae. Los tiempos de exposición fueron
diferentes para cada muestra, se emplearon tiempos de exposición de 2, 4, 6 y 8
horas. Después de la exposición las larvas se mantuvieron de manera similar al
experimento anterior hasta la emergencia.

39
5.5. Tiempos y frecuencia de exposición
En esta prueba se evaluó la frecuencia de oviposición contando el número de
veces que un parasitoide introdujo su ovipositor en las larvas, Se emplearon
larvas de A. ludens y A. obliqua. Una muestra de 10 larvas fueron colocadas en
una caja Petri de 5 cm de diámetro con dieta larvaria. En una jaula de 10 cm por
lado se colocaron 10 parasitoides por muestra, relación 1:1. Se observó y se
midió el tiempo que tardó una hembra en posición de parasitación sobre la
muestra y la observación del comportamiento de esa hembra al encontrar larvas
activas. Otras actividades observadas fueron: cuánto tarda por picadura y cuantas
oviposiciones tiene una larva expuesta. Ésta prueba se desarrolló con
observaciones realizadas en un período de una hora.
5.6. Tabla de vida de adultos
Se colocaron una hembra y un macho recién emergidos, dentro de una pequeña
jaula de vidrio 10 cm3 con una pared de tela organdí, realizando 30 repeticiones de
esta observación a las cuales, se les suministró agua por medio de un algodón y
alimento suficiente en pequeños contenedores de plástico. Cada día se observó el
número de hembras muertas, las cuales fueron retiradas del experimento.
Se expuso un total de 30 larvas colocadas en la unidad de exposición por día. A
15 jaulas se les colocó larvas de Anastrepha ludens, y al resto se les expuso
larvas de A. obliqua. La exposición se llevó a cabo en una tapa de caja Petri de 5
cm de diámetro atadas con una tela suave y cascara de guayaba como atrayente
que simula la corteza suave de un fruto. La exposición fue por un período de 2
horas. De esta manera se obtuvieron datos de supervivencia de adultos y con la
emergencia el número de hijos por hembra.
En nuestros tratamientos comparativos las muestras (cohortes) a comparar fueron
representativas, aleatorias e independientes entre sí. La escala de medición del
tiempo fue continua, medido en días; el total de unidades (individuos) fue
homogéneo, aunque no todos sobrevivieron al término de la prueba.

40
5.7. Análisis estadístico:
Obtenidos los resultados se procedió al ingreso de los datos en tablas de Microsoft
Excel, posteriormente se corrió en el programa SAS, con un diseño de
experimentos completamente al azar y en los casos donde hubo diferencia
stadística por varianzas y medias se corroboró a través de la prueba Tukey,
logrando realizar conclusiones concisas del comportamiento de los experimentos,
Así mismo se realizó obtención de promedios en tiempos de exposición y números
de oviposiciones.
Fig. 13.- Trabajo de laboratorio y manejo de muestras. A) Muestra jaula tipo
Hawái. B) acondicionamiento de jaula. C) parasitismo en selección de huéspedes.
D) larvas de A. ludens irradiadas. E) parasitismo en jaulas de conservación de la
colonia. F) implementación del experimento en el laboratorio.
A B
C D
C F

41
6. RESULTADOS Y DISCUSIÓN
Utetes anastrephae es un parasitoide nativo que no tiene registros de haber sido
evaluado en México por tanto los datos aquí proporcionados considero sirvan de
base para posteriores estudios.
Se utilizaron cuatro tratamientos para las pruebas 1 y 2 los cuales se definen
como:
Tratamiento Uno: Hospedero A. ludens sin selección.
Tratamiento Dos: A. ludens con selección.
Tratamiento Tres: A. obliqua con selección.
Tratamiento Cuatro: A. obliqua sin selección.
Para la prueba 3, los tratamientos utilizados se agrupan en:
Especie 1(A. ludens): tratamientos de 2horas, 4 horas, 6 horas y 8 horas.
Especie 2(A. obliqua): tratamientos de 2 horas, 4 horas, 6 horas y 8 horas.
6.1. Selección de hospederos.
En este experimento se realizó un análisis estadístico por medio del programa
SAS (Stadistical Analysis System) usando el cuadro estadístico Completamente al
Azar, con la prueba de Tukey se evaluaron tres variables dependientes en 26
repeticiones; la muerte larvaria de hospederos, el porcentaje de parasitismo y la
proporción sexual, utilizando una confiabilidad del 95%, lo cual arrojó los
siguientes resultados.
Para la variable muerte larvaria en la prueba „Selección de hospederos‟ no hay
diferencias entre un tratamiento y el otro, es decir, significa lo mismo para los
propósitos de producción utilizar de larva hospedera a la especie A. ludens sola,
A. obliqua sola, a las dos especies en conjunto ó a las dos especies en diferentes
unidades de oviposición dentro de la misma jaula, los porcentajes de muerte
larvaria varían del 5 al 10% en el conteo a las 72 horas de reposo. Sin embargo,

42
después de 15 días o al término del reposo, el porcentaje de larva o pupa que
murió radica del 40 al 50% del total de especímenes por unidad de exposición.
En la variable proporción sexual se utilizó el mismo método de análisis y los
resultados de la regla de decisión marcan que no existe diferencia estadística
entre los tratamientos, mostrando como modelo que en la emergencia se obtienen
2 hembras por cada macho emergido, aunque en el uso de A. ludens sin
selección, se observó que la emergencia del adulto hembra fue un poco mas
vigorosa y mas grande comparado con el caso de A. obliqua sin selección como lo
menciona Godfray (1994), que las hembras fértiles tendrán cierta preferencia a
los hospederos mas grandes para emergencia de hembras y confirma Cancino,
(2002), que el peso o tamaño del huésped ocasionalmente repercuten en la
producción y pueden influir en la proporción sexual. Aunque para los tratamientos
que tienen selección de hospederos la tendencia de proporción de sexos fue
menos notoria ya que la emergencia fue similar en ambas especies, mientras se
encontraran expuestas a U. anastrephae.
El porcentaje de parasitismo estuvo marcado por la emergencia total de los
adultos que lograron salir del pupario completamente y tuvieron la capacidad de
volar, de manera particular el parasitismo de U. anastrephae en el huésped del
tratamiento uno, fue de 50.7%, para el tratamiento dos de 58.2%, para el
tratamiento tres de 44% y para el tratamiento cuatro de 48%; lo cual en estadística
no se marca una diferencia intrínseca, en resumen y de manera general se obtuvo
un porcentaje de parasitismo de 50%.
Podemos observar en la Fig. 14, que la gráfica a simple vista nos muestra que la
especie A. ludens, puede representar ser un mejor huésped por sus tendencias de
mayor parasitismo; sin embargo estadísticamente no es relevante ya que la
especie A. obliqua también se encuentra por arriba del rango de confiabilidad,
interpretándose estadísticamente igual para ambos casos.
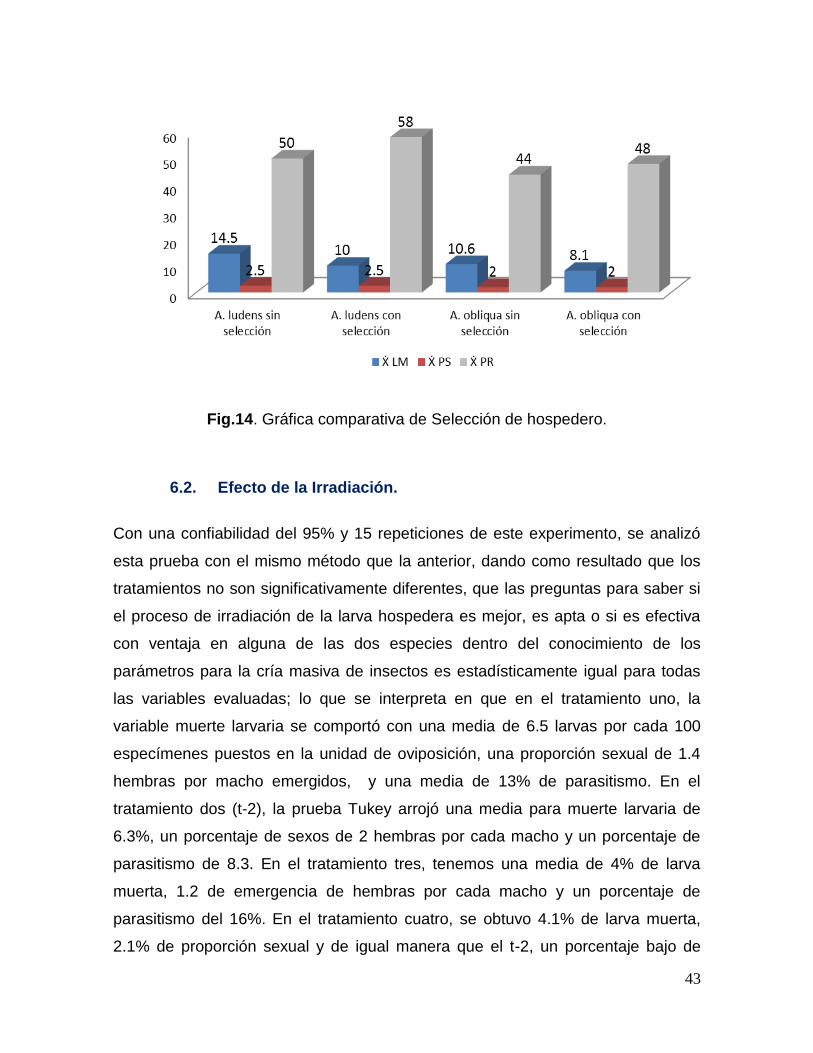
43
Fig.14. Gráfica comparativa de Selección de hospedero.
6.2. Efecto de la Irradiación.
Con una confiabilidad del 95% y 15 repeticiones de este experimento, se analizó
esta prueba con el mismo método que la anterior, dando como resultado que los
tratamientos no son significativamente diferentes, que las preguntas para saber si
el proceso de irradiación de la larva hospedera es mejor, es apta o si es efectiva
con ventaja en alguna de las dos especies dentro del conocimiento de los
parámetros para la cría masiva de insectos es estadísticamente igual para todas
las variables evaluadas; lo que se interpreta en que en el tratamiento uno, la
variable muerte larvaria se comportó con una media de 6.5 larvas por cada 100
especímenes puestos en la unidad de oviposición, una proporción sexual de 1.4
hembras por macho emergidos, y una media de 13% de parasitismo. En el
tratamiento dos (t-2), la prueba Tukey arrojó una media para muerte larvaria de
6.3%, un porcentaje de sexos de 2 hembras por cada macho y un porcentaje de
parasitismo de 8.3. En el tratamiento tres, tenemos una media de 4% de larva
muerta, 1.2 de emergencia de hembras por cada macho y un porcentaje de
parasitismo del 16%. En el tratamiento cuatro, se obtuvo 4.1% de larva muerta,
2.1% de proporción sexual y de igual manera que el t-2, un porcentaje bajo de

44
parasitismo del 6.2% (Fig. 15). Como menciona Cancino, et al. (2002). No hubo
una relación directa entre el efecto negativo de la irradiación, al contrario, el efecto
que tuvimos fue apropiado que facilitó el conteo de emergencia de adultos fértiles
del parasitoide y, las larvas y pupas muertas al termino de las evaluaciones a
diferencia de donde hubo emergencia de adultos de moscas; en otro aspecto la
dosis que se empleo en el experimento fue de 4 Gy. A diferencia del parasitismo
que se muestra en otras pruebas, en ésta se considera bajo, la emergencia de
parasitoides no fue superior del 20%, lo cual fue mas bajo que los obtenidos en
experimentos anteriores.
Fig. 15. Gráfica comparativa de „Efectos de la Irradiación‟.
6.3. Tiempos de exposición
En este experimento se aplicó un diseño completo al azar, analizado con prueba
de Tukey, en 12 repeticiones, se realizó una comparación de los tiempos de
exposición medido en horas y se determinó la diferencia de parasitación por
especie dependiendo del tiempo que las larvas duraron expuestas a la oviposición
de U. anastrephae.
En la agrupación uno, la evaluación del tiempo de 2 horas se determinó que la
muerte larvaria del hospedero tuvo una media de 6.5%, en exposiciones de 4
0
2
4
6
8
10
12
14
16
A. ludens sinirradiación
A. ludens conirradiación
A. obliqua sinirradiación
A. obliqua conirradiación
6.5 6.3
4 4.1
1.4 2 1.2
2.1
13
8.3
16
6.2
Ẋ LM Ẋ PS Ẋ PR

45
horas, ascendió a 10.5%, a las 6 horas 8.5%, y en la evaluación de 8 horas
10.66% de mortandad.
En la especie dos, este mismo parámetro evaluado a las 72 horas de exposición
mostró que si existe diferencia estadística entre los tratamientos.
Tenemos una mortandad larvaria de 4.6% al ser expuestas durante un tiempo de 2
horas, 9.4% a las 4 horas, 11.5% a las 6 horas y 12% a las 8 horas de exposición.
Para el parámetro de proporción de sexos, en la especie uno y dos, no hubo
diferencia estadística, el indicativo general en todas las evaluaciones del tiempo es
de 2 hembras por macho.
En la evaluación del parasitismo para la especie uno, las medias son:
A las 2 horas de exposición un porcentaje del 49%, a las 4 horas 57%, a las 6
horas 53% y a las 8 hora 73.5%.
En la especie dos si se marca una diferencia estadística en el análisis de
varianzas al 95% de confiabilidad ya que mientras mas tiempo transcurrió hubo
mayor parasitismo al igual que un pequeño incremento en la muerte larvaria.
A las 2 horas de exposición el porcentaje de parasitismo fue de 26%, a las 4 horas
de 48%, a las 6 horas de 67.3% y a las 8 horas de 70%. (Figura 16).
Fig. 16. Gráfica comparativa de „Tiempos de exposición‟.
0
10
20
30
40
50
60
70
80
A. ludens2H
A. ludens4H
A. ludens6H
A. ludens8H
A. obliqua2H
A. obliqua4H
A. obliqua6H
A. obliqua8H
6.5 10.5 8.5 10.6
4.6 9.4 11.5 12
2 2 2 2 2 2 2 2
49 57
53
73.5
26
48
67.3 70
LM PS PR

46
6.4. Tiempo y Frecuencia de oviposición
En esta prueba no se presentaron diferencias estadísticas notables. No hubo un
patrón de tiempo de oviposición de estricta diferencia entre la larva de A. obliqua y
la de A. ludens. En términos generales el tiempo de exploración fue muy
prolongado ya que hubieron parasitoides que permanecieron en la muestra hasta
por una hora y el tiempo promedio de oviposición se prolongo a 1.8 minutos, con
un mínimo de 5 segundos, que se muestra como conocimiento previo o ensayos y
un tiempo máximo de 7 minutos. (Figura 17)
Estos tiempos son muy importantes para definir la acción de Utetes anastrephae
en su proceso de búsqueda de huésped.
Van Driesche, (2007) menciona que: la experimentación previa del parasitoide,
también puede influenciar la preferencia de un hospedero sobre otro. Muchos
parasitoides adultos contactan las kairomonas del huésped durante la emergencia
y si las preferencias de un parasitoide están débilmente determinadas
genéticamente, el contacto con la especie huésped de donde emergió puede
reforzar la preferencia por esa especie. Consecuentemente, los parasitoides
criados en huéspedes alternantes pudieran funcionar menos contra la plaga a
controlar.
Los tiempos de búsqueda de U. anastrephae son largos y el tiempo de oviposición
corto, pero garantiza un parasitismo máximo hasta del 90%. (Figura 18)
Esto se puede comparar con otros parasitoides de moscas de la fruta que tiene un
tiempo promedio de oviposición que no rebasa el minuto (Cuadro 4). Por ejemplo
en D. longicaudata se reporta un tiempo promedio de 1 minuto (Matthews, 1976).
En medidas de laboratorio puede convertirse en una desventaja para U.
anastrephae al invertir en él, un tiempo muy prolongado, con una evolución muy
lenta del proceso de reproducción masiva.
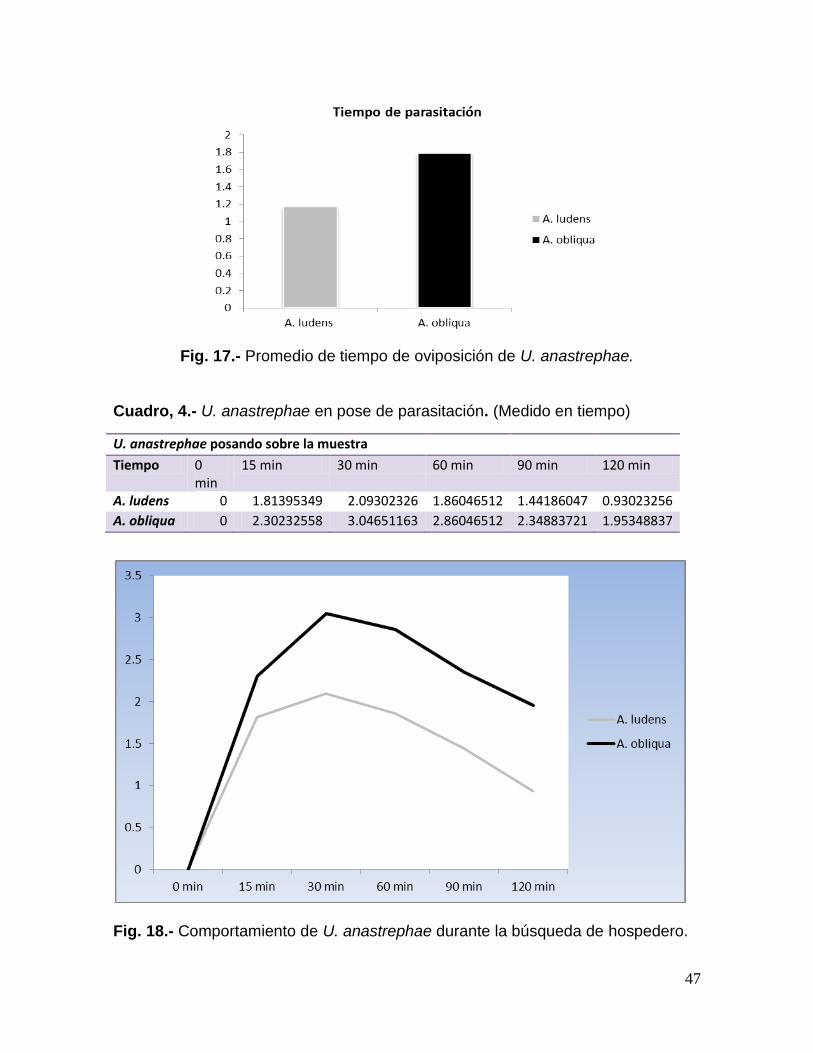
47
Fig. 17.- Promedio de tiempo de oviposición de U. anastrephae.
Cuadro, 4.- U. anastrephae en pose de parasitación. (Medido en tiempo)
U. anastrephae posando sobre la muestra Tiempo 0
min 15 min 30 min 60 min 90 min 120 min
A. ludens 0 1.81395349 2.09302326 1.86046512 1.44186047 0.93023256 A. obliqua 0 2.30232558 3.04651163 2.86046512 2.34883721 1.95348837
Fig. 18.- Comportamiento de U. anastrephae durante la búsqueda de hospedero.

48
Utetes anastrephae al tener un rango de hospederos amplio puede considerarse
generalista no especifico de A. obliqua y, ser un parasito generalista, lo cual
también tiene sus ventajas ya que al escasearse su hospedero natural puede
sobrevivir utilizando otra especie como hospedero alternativo. Este segundo tipo
de parasitoide puede ser de suficiente utilidad en el caso de control biológico de
un complejo de plagas (Rosen y Huffaker, 1982).
6.5. Tabla de vida y fertilidad de Utetes anastrephae
Durante la observación del comportamiento de los parasitoides en un medio
controlado y con una densidad conocida se obtuvo una tabla de vida que nos
permitió obtener algunos parámetros importantes en el ciclo de vida de los
parasitoides, como:
La curva de sobrevivencia (Fig.19); que es la representación gráfica de una tabla
de vida en un sistema de coordenadas, en donde la nx corresponde a las
ordenadas y x (intervalo de edad) a las abscisas. Observemos la siguiente figura:
Fig. 19.- Tasa de supervivencia comparativa de Utetes anastrephae en dos huéspedes.
Según Krebs, (1978) a través de Vera, (2002). El comportamiento que tenemos
anteriormente de las tasa de supervivencia es característico de algunos insectos
0
2
4
6
8
10
12
14
16
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23
supervivencia A. obliqua
supervivencia A. ludens

49
que atacan frutos y granos almacenados; ejemplos de este tipo se pueden ver en
Pérez et al., 1989, para moscas de la fruta.
Así mismo conocemos la esperanza de vida de U. anastrephae (Figura. 20), que tiene como máximo 13 días.
Fig. 20.- Esperanza de vida de hembras de U. anastrephae.
La fertilidad también es un parámetro dentro de una tabla de vida y el valor
estadístico que representa la fertilidad se denomina mx, se define como: el
número de hijas producidas por unidad de tiempo por hembra madre de edad x.
(Figura 21) la ecuación que utilizamos es:
hx: número de hijas producidas durante el intervalo x a x + 1.
nx: número de hembras de edad x.
La tasa de fertilidad estuvo definida por la siguiente ecuación:
Al combinar las tablas de vida y fertilidad se obtuvo la tasa neta de reproducción y
se representó como:
0
2
4
6
8
10
12
14
16
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23
ludens
obliqua
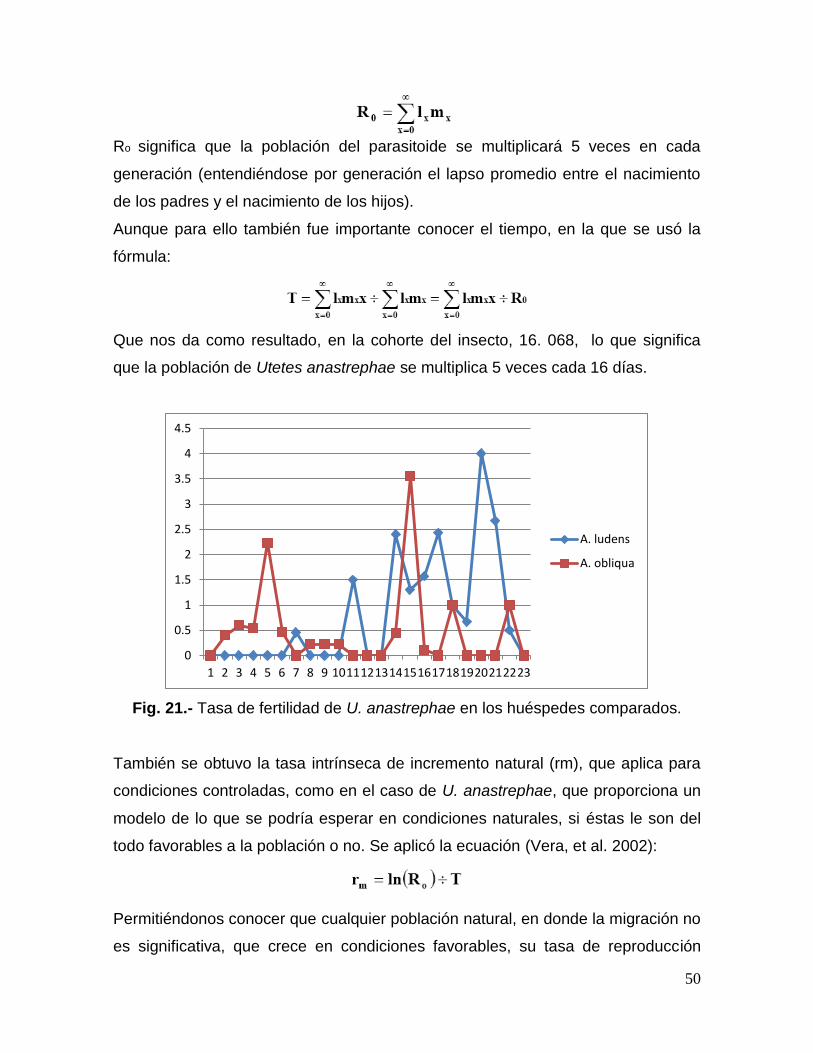
50
Ro significa que la población del parasitoide se multiplicará 5 veces en cada
generación (entendiéndose por generación el lapso promedio entre el nacimiento
de los padres y el nacimiento de los hijos).
Aunque para ello también fue importante conocer el tiempo, en la que se usó la
fórmula:
Que nos da como resultado, en la cohorte del insecto, 16. 068, lo que significa
que la población de Utetes anastrephae se multiplica 5 veces cada 16 días.
Fig. 21.- Tasa de fertilidad de U. anastrephae en los huéspedes comparados.
También se obtuvo la tasa intrínseca de incremento natural (rm), que aplica para
condiciones controladas, como en el caso de U. anastrephae, que proporciona un
modelo de lo que se podría esperar en condiciones naturales, si éstas le son del
todo favorables a la población o no. Se aplicó la ecuación (Vera, et al. 2002):
Permitiéndonos conocer que cualquier población natural, en donde la migración no
es significativa, que crece en condiciones favorables, su tasa de reproducción
0
0.5
1
1.5
2
2.5
3
3.5
4
4.5
1 2 3 4 5 6 7 8 9 1011121314151617181920212223
A. ludens
A. obliqua

51
permanecerá constante y su densidad se incrementará sin límite (teóricamente) y
de forma geométrica.
Para U. anastrephae se obtuvo en las hembras emergidas de A. ludens una tasa
de 0.16 y en A. obliqua de 0.10; esto quiere decir que en cualquier instante el
número de insectos de la población se está incrementando a una tasa tal, que se
espera un crecimiento del 16 % (aproximadamente) en A. ludens y de 10% en A.
obliqua de una unidad de tiempo a la siguiente. Las últimas evaluaciones fueron
las tasas finitas que nos indican por cuanto se multiplica una población por unidad
de tiempo; en los parasitoide emergidos de A. ludens se registró 1.17 y para A.
obliqua 1.10; lo que significa que por cada parasitoide presente en un momento
dado (t), habrá casi 1.17 en la siguiente unidad de tiempo (t+1).
Estos valores obtenidos mediante la aplicación de las fórmulas de cálculo se
encuentran señalados de manera específica en los cuadros 5 y 6.
La tabla de vida sirve a los entomólogos como punto de referencia de los
parámetros del ciclo de vida del insecto para comparar periódicamente la calidad
de la población en crianza. La tabla de cohorte se usa habitualmente en el análisis
de sobrevivencia, que se realiza sobre muestras de población pequeñas y durante
un tiempo corto (Cano, V., 1992).
De acuerdo a las características de Utetes que se evaluaron y que se observaron
durante el periodo de los experimentos decimos que es importante la utilización de
unidades de exposición planas en la forma geométrica que se desee pero que
permita al parasitoide introducir su pequeño ovipositor de 1.6 mm de largo, para
que alcanze a las larvas huéspedes. En la especificidad no tiene problemas, pues
se adapta fácilmente a cualquiera de las especies del complejo Anastrepha (A.
alveata, A. bahiensis, A. cebra, A. fraterculus, A. ludens, A. obliqua, A. serpentina,
A. striata, A. suspensa y Rhagoletis spp. U. anastrephae se adapta fácilmente en
toda América ya que radica en un margen amplio de altitud y latitud.
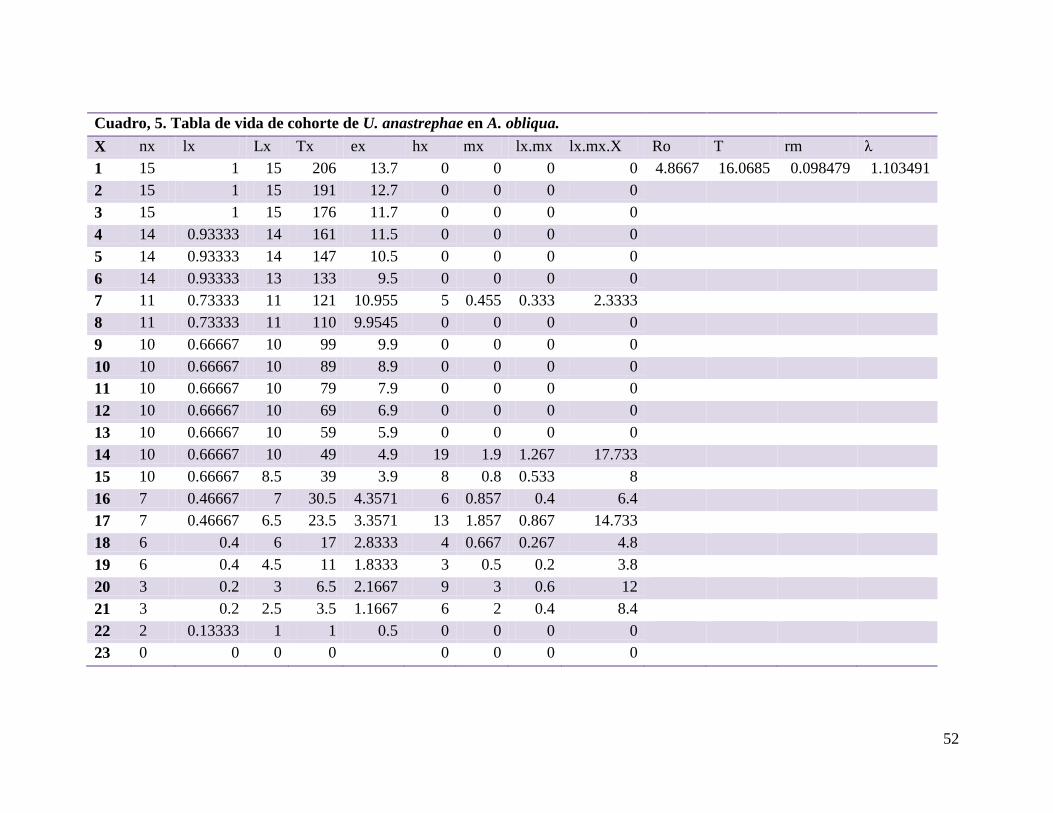
52
Cuadro, 5. Tabla de vida de cohorte de U. anastrephae en A. obliqua.
X nx lx Lx Tx ex hx mx lx.mx lx.mx.X Ro T rm λ
1 15 1 15 206 13.7 0 0 0 0 4.8667 16.0685 0.098479 1.103491
2 15 1 15 191 12.7 0 0 0 0
3 15 1 15 176 11.7 0 0 0 0
4 14 0.93333 14 161 11.5 0 0 0 0
5 14 0.93333 14 147 10.5 0 0 0 0
6 14 0.93333 13 133 9.5 0 0 0 0
7 11 0.73333 11 121 10.955 5 0.455 0.333 2.3333
8 11 0.73333 11 110 9.9545 0 0 0 0
9 10 0.66667 10 99 9.9 0 0 0 0
10 10 0.66667 10 89 8.9 0 0 0 0
11 10 0.66667 10 79 7.9 0 0 0 0
12 10 0.66667 10 69 6.9 0 0 0 0
13 10 0.66667 10 59 5.9 0 0 0 0
14 10 0.66667 10 49 4.9 19 1.9 1.267 17.733
15 10 0.66667 8.5 39 3.9 8 0.8 0.533 8
16 7 0.46667 7 30.5 4.3571 6 0.857 0.4 6.4
17 7 0.46667 6.5 23.5 3.3571 13 1.857 0.867 14.733
18 6 0.4 6 17 2.8333 4 0.667 0.267 4.8
19 6 0.4 4.5 11 1.8333 3 0.5 0.2 3.8
20 3 0.2 3 6.5 2.1667 9 3 0.6 12
21 3 0.2 2.5 3.5 1.1667 6 2 0.4 8.4
22 2 0.13333 1 1 0.5 0 0 0 0
23 0 0 0 0 0 0 0 0
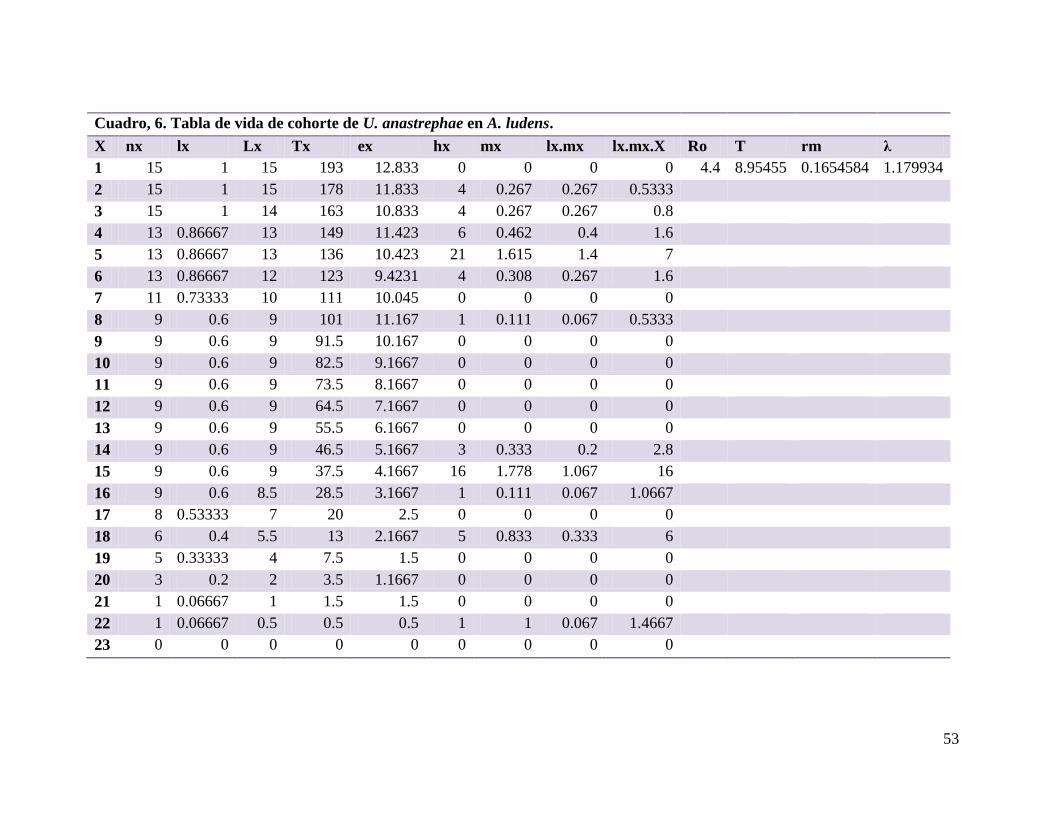
53
Cuadro, 6. Tabla de vida de cohorte de U. anastrephae en A. ludens.
X nx lx Lx Tx ex hx mx lx.mx lx.mx.X Ro T rm λ
1 15 1 15 193 12.833 0 0 0 0 4.4 8.95455 0.1654584 1.179934
2 15 1 15 178 11.833 4 0.267 0.267 0.5333
3 15 1 14 163 10.833 4 0.267 0.267 0.8
4 13 0.86667 13 149 11.423 6 0.462 0.4 1.6
5 13 0.86667 13 136 10.423 21 1.615 1.4 7
6 13 0.86667 12 123 9.4231 4 0.308 0.267 1.6
7 11 0.73333 10 111 10.045 0 0 0 0
8 9 0.6 9 101 11.167 1 0.111 0.067 0.5333
9 9 0.6 9 91.5 10.167 0 0 0 0
10 9 0.6 9 82.5 9.1667 0 0 0 0
11 9 0.6 9 73.5 8.1667 0 0 0 0
12 9 0.6 9 64.5 7.1667 0 0 0 0
13 9 0.6 9 55.5 6.1667 0 0 0 0
14 9 0.6 9 46.5 5.1667 3 0.333 0.2 2.8
15 9 0.6 9 37.5 4.1667 16 1.778 1.067 16
16 9 0.6 8.5 28.5 3.1667 1 0.111 0.067 1.0667
17 8 0.53333 7 20 2.5 0 0 0 0
18 6 0.4 5.5 13 2.1667 5 0.833 0.333 6
19 5 0.33333 4 7.5 1.5 0 0 0 0
20 3 0.2 2 3.5 1.1667 0 0 0 0
21 1 0.06667 1 1.5 1.5 0 0 0 0
22 1 0.06667 0.5 0.5 0.5 1 1 0.067 1.4667
23 0 0 0 0 0 0 0 0 0

54
7. CONCLUSIÓNES
1.- Utetes anastrephae Viereck, se reproduce de manera similar en ambas especies
Anastrepa obliqua y A. ludens, cumpliéndose la hipótesis a´ y b´.
2.- U. anastrephae mostró un porcentaje de parasitismo entre el 40 a 60%, y una muerte
larvaria de 5 a 15% en las especies A. obliqua y A. ludens.
3.-En el efecto de la irradiación a 4 krad fue similar en todos los tratamientos, con o sin
irradiación de las especies A. ludens y A. obliqua.
4.- U. anastrephae mostró un tiempo de exposición de 2 horas. Con A. obliqua, tuvo
relación directa, mientras más tiempo se exponen, mayor es el porcentaje de parasitismo,
mayor la muerte larvaria e interés de las hembras fértiles por la búsqueda de huéspedes,
con una proporción sexual de 2 hembras por macho.
5.- En comportamiento no hay diferencia por ovipositar en larvas de A. obliqua y A.
ludens. En términos generales el tiempo de exploración fue muy prolongado con hembras
que permanecieron explorando hasta una hora y en tiempo promedio de oviposición a 1.8
minutos, con un mínimo de 5 segundos, que se mostró como conocimiento previo o
ensayos y un tiempo máximo de 7 minutos.
6.- Utetes anastrephae Viereck, es un endoparasitoides alternante, capaz de desarrollarse
en varios huéspedes, sin problemas por copulación efectiva o fecundidad, con una
longevidad de vida suficiente para la madurez y capacidad de búsqueda de hospederos,
una esperanza de vida promedio de 11 días en condiciones controladas de laboratorio.
7.-Apto para reproducción masiva, y liberación en campo; por sus porcentajes de
parasitismo altos del 73% y bajos del 10%. Su tiempo promedio de oviposición de 1.8
minutos y proporción sexual de 2 hembras por macho. Adaptable a condiciones tropicales,
cálido - húmedas de regiones costeras, medias templadas, entre otras, donde se
encuentra su huésped nativa A. obliqua y el complejo de Anastrepha spp.

55
8. BIBLIOGRAFÍA
Aluja, M. M. López & J. Sivinski. 1998. Ecological evidence for diapause in four
native and one exotic species of larval-pupal fruit fly (Diptera: Tephritidae)
parasitoids in tropical environments. Annals of the Entomological Society of
America 91: 821-833.
Aluja, et al. 2003. Fruit flies of the genus Anastrepha (Díptera: Tephritidae) and
associated native parasitoids in the tropical rain forest biosphere reserve of Montes
Azules, Chiapas. México. Rev. Enviromental Entomlogical 32: 1377-1385.
Ashley, T.R., P.D. Greany & D.L. Chambers. 1976. Adult emergence of Biosteres
(Opius) longicaudatus and Anastrepha suspensa in relation to the temperature and
moisture concentration of the pupation medium. Florida Entomologist 59: 391-395
Cancino, D. J., et al. 2002. Quality control parameters of wild and mass reared
Diachasmimorpha longicaudata (Ashmead), a fruit fly parasitoid. In Leppla N. C.,
K. A. Bloem, R. F. Luck.(Eds.) Quality Control for Mass-reared
Arthropods.Proceeding of the Eighth and Ninth Workshops of the international
Organization for Biological Control Working Group on Quality Control of Mass-
reared Arthropods.Pp. 84-94.
Cancino, J. and Montoya, P. 2004. Desirable attributes of mass reared parasitoids
for fruit fly control: a comment. Review Vedalia, International Journal of Biological
Control. 11: 53- 58.
Cancino, et al. 2006. Control de calidad en la cría masiva de Diachasmimorpha
longicaudata (Hymenoptera: Braconidae). Fundamentos y procedimientos.
SAGARPA. México. 54 págs.

56
Cancino, J. 2010. Biología y comportamiento de parasitoides. En Montoya et al,
2010. Moscas de la fruta: Fundamentos y procedimientos para su manejo. . S y G.
Editores. México, D. F. Pp. 113-129.
Cano, V. 1992. Tabla de vida en laboratorio y liberación en el campo de
Trichogramma pretiosum riley (Hymenoptera: Trichogrammatidae) en Nicaragua.
Rev. Nica. Ent., (1992), 21:43-56.
Carey, J.R. 1993. Applied Demography for Biologists with Special Emphasis on
Insects. Oxford Univ. Press. Oxford, UK. 296 p.
Doutt R., L. y P. De Bach. 1964. Algunos conceptos y preguntas sobre control
biológico. En control biológico de las plagas de insectos y malas hierbas. CECSA.
Mexico. Pp. 175.
Godfray, H. C. J. 1994. Parasitoids.Behavioral and evolutionary ecology.Princeton
University Pres. New Jersey.473 Págs.
Gutiérrez S., J., et al. 1995. Plan Nacional contra Moscas de la Fruta. Memorias de
IX Curso Internacional contra Moscas de la Fruta. Programa Moscamed. DGSV-
SAGARPA . México. Pp. 17-18.
Hernández O., V. 1992. El género Anastrepha Schiner en México (Diptera:
Tephritidae). Taxonomía, Distribución y sus plantas huéspedes. Instituto de
Ecología. Publicación. 33. 162 p.
Insuasty B. et al. 2007. Manejo Integrado de Moscas de la Fruta de la Guayaba
(Anastrepha spp.) CORPOICA. Colombia. 28 pp.

57
Krebs, J.C. 1978. Ecology .The experimental analysis of distribution and
abundance.Second Edition.Harper & Row, Publishers. New York, Evanston, San
Francisco, London.
Lawrence, P.O. 1981. Host vibration: a cue to host location by the parasite
Biosteres longicaudatus. Oecologia 48: 249-251.
Leyva V., J. 1999. Control biológico de moscas de la fruta: uso de parasitoides.
Vedalia 6: 15-21.
Mackahuer, M. 1976. Genetic problems in the production of biological control
agents.Annu. Rev. Entomol. 21: 369-385.
Martinez M., L. 1992. Parasitismo, Supervivencia y Reproducción de
Diachasmimorpha longicaudata (Aschmead) (Hymenoptera: Braconidae),
parasitoide de moscas de la fruta (díptera: Tephritidae). Tesis de maestría en
ciencias. Colegio de posgraduados. México. 73 Págs.
Montoya, P. et al. 2000. Biological control of Anastrepha spp. (Diptera: Tephritidae)
in mango Orchards through augmentative releases of Diachasmimorpha
longicaudata (Ashmead) (Hymenoptera: Braconidae). Biol. Control 18: 216-224.
Montoya, P. & J. Cancino. 2004. Control Biológico por aumento en moscas de la
fruta (Diptera: Tephritidae). Folia Entomológica Mexicana 43: 257-270.
Montoya, P., J. Cancino, M. Zenil, G. Santiago and J.M. Gutierrez. 2007. The
augmentative biological control component in the Mexican National Campaign
against Anastrepha spp. fruit flies, pp. 661-670. En Vreysen, M.J.B, A.S. Robinson
& J. Hendrichs (edits), Area-Wide Control of Insect Pests. From Research to Field
Implementation. Springer, The Netherlands.

58
Montoya, et al. 2010. Moscas de la fruta: fundamentos y procedimientos para su
manejo. S y G. Editores. México, D. F. Pp. 394.
Nicholls E., C. 2008. Control biológico de insectos: un enfoque agroecológico.
Medellín: Editorial Universidad de Antioquia, Colombia. 282 págs.
Ovruski, S., M. Aluja, J. Sivinski and R. Wharton, 2000. Hymenopteran parasitoids
on fruit-infesting tephritidae in Latin America and Southern united states: Diversity,
distribution, taxonomic, and their uses in fruit fly biological control. Integrated pests
management. Review 5:81-107.
Pérez R., A.; J. Vera G. y H. González. 1989. Tasas de supervivencia y
reproducción de Anastrepha ludens (Loew) en diferentes hospedantes.
Agrociencia No.76: 175-181.
Rocha L. M., et al. 2009. Diversidade de moscas-das-frutas, suas plantas
hospedeiras e seus parasitoides nas regiões Norte e Noroeste do Estado do Rio
de Janeiro, Brasil. Revista Ciência Rural, Santa Maria, v.39 n.3, Brazil. Pp. 627-
634.
Sivinski, J. 1993. The influence of host fruit morphology on parasitization rates in
the Caribbean fruit fly Anastrepha suspensa. Entomophaga 36: 447-454.
Sivinski, J., M. Aluja & M. Lopez. 1997. The spatial and temporal distributions of
parasitoids of Mexican Anastrepha species (Diptera: Tephritidae) within the
canopies of fruit trees. Annals of the Entomological Society of America 90:05.
Stuhl C, Sivinski J, Teal P, Paranhos B, Aluja M. 2011. A compound produced by
frugivorous Tephritidae (Diptera) larvae promotes oviposition behavior by the
biological control agent Diachasmimorpha longicaudata (Hymenoptera:
Braconidae). Environmental Entomology 40: 727-736.

59
Teodoro C., V. 2011. Evolución genética de cuatro especies de Anastrepha spp.
(Diptera: Tephritidae). Tesis profesional. Colegio de posgraduados. México. 89 pp.
Van de Assem, J. 1986. Mating. Behavior in parasitic wasp. In: Insect parasitoids.
J. Waage y D. Greathead (Eds). Academic Press. Great Britain. Pp 137-167.
Van Driesche, R. G., 2007. Control de plagas y malezas por enemigos
naturales.Forest Health Technology Enterprise Team. USDA. 757 pp.
Vera G., J. 2002. Ecología de poblaciones de insectos. Colegio de posgraduados.
México. 137 pp.
Viereck H. L. 1913. Descriptions of ten new genera and twenty-three new species
of Ichneumon-flies. Proceedings of the United States National Museum 44: 555-
568.
Wharton, R. A. & Gilstrap, F. E., 1983. Key to and status of opiine braconid
(Hymenoptera) parasitoids used in biological control of Ceratitis and Dacus s. l.
(Diptera: Tephritidae) Annals of the Entomological Society of America 76: 721-742
Wong, T.Y.Y., & J. Herr. 1991. Augmentative releases of Diachasmimorpha tryoni
(Hymenoptera: Braconidae) to suppress a Mediterranean fruit fly (Diptera:
Tephritidae) population in Kula, Maui, Hawaii. Biological Control 1: 2-7.
Wong, T. T. Y. & M.M. Ramadan. 1992. Mass rearing of larval parasitoids
(Hymenoptera: Braconidae: Opinae) of tephritid flies (Diptera: Tephritidae) in
Hawaii, 425-426 pp. En Anderson, T.E. & N. Leppla (edits.), Advances in Insects
Rearing for Research and Pest Management. Westview Press, Oxford, England.
Wikipedia:http://entomology.ifas.ufl.edu/creatures/fruit/tropical/mexicanfruitfly.htm,
Consultada el 7 de agosto de 2012.

60
9. APENDICE
9.1. Clave taxonómica para determinar géneros de Anastrepha de importancia económica.
Fotos de caracterización morfotaxonómica : Figuras 22-26.
1. Ala con la banda Costal (C) interrumpida justo al final de la vena R1 por una
marca hialina bien delimitada en la celda r1; brazo distal de la banda V
generalmente presente al menos de manera parcial, pero si esta ausente,
entonces el patrón alar es de color pardo oscuro a negruzco; seda orbital anterior
y posterior presentes............................................................................... (2)
1‟ Ala con la banda Costal (C) completa a lo largo de la vena costal, desde la base
hasta el ápice del ala, algunas veces difusa; brazo distal de la banda V ausente.
Todos los siguientes caracteres presentes: Mitad basal de la banda S (sobre la
celda discal) completa desde el ápice de la celda bcua través de la vena cruzada
r-m, conectándose anteriormente con la banda C; celda r2+3 completamente
pigmentada; celda br hialina entre las venas BM-Cu y R-M; seda orbital posterior
generalmente ausente.
Terguitos abdominales amarillos, escudo con franjas dorsocentrales pardo
oscuras; aculeus largo de 5.3 - 6.2 mm de longitud, usualmente > 0.10 mm de
ancho; márgenes laterales de la punta del aculeus no aserrados. Distribución.
Argentina, Bolivia,Brasil, Colombia, Ecuador, Panamá, Paraguay, Perú y
Venezuela (ref: Norrbom 1991).
“Mosca del Melon”.................................................Anastrepha grandis (Macquart)
2. Terguitos abdominales con amplias franjas pardo oscuras o negruzcas, pero si
es amarillo, entonces el escudo presenta un patron de franjas negras bien
diferenciado......................................................................................................... (3)
2‟ Terguitos abdominales de un solo color usualmente amarillos o anaranjados,
escudo del mismo color y sin franjas negras dorsales, excepto en ocasiones con
una mancha central sobre la sutura escudo-escutelar………………………….. (4)

61
3 Patrón alar de color pardo oscuro o negruzco; brazo distal de la banda V
completamente ausente; terguitos abdominales con franjas obscuras; manchas
negras en la pleura que contrastan con otras manchas amarillas. Aculeus de 2.8-
3.7 mm longitud, márgenes laterales de la punta del aculeus finamente aserrados.
Distribución: Posee algunos registros históricos en EUA; México, América Central
y Sudamérica, Trinidad y Tobago y Curazao (refs: Footeet al. 1993, Norrbom
2002).
“Mosca de los Zapotes”..............................Anastrepha serpentina (Wiedemann).
3‟ Patrón alar con franjas primordialmente amarillas; brazo distal de la banda V
presente de manera parcial o apenas evidente basalmente; terguitos abdominales
y pleuras amarillos; escudo con dos franjas amplias dorsocentrales conectadas en
el margen posterior en forma de “U”. Aculeus de 2.05-2.30 mm de longitud.
Distribución: Posee registros esporádicos en EUA; México al sur hasta Bolivia y
Brasil (refs: Footeet al. 1993, Norrbom 2002).
“Mosca de la Guayaba”………..................................Anastrepha striata Schiner.
4. Banda apical anterior (= sección distal de la banda S) normalmente
desarrollada, nunca alcanza el ápice de la vena M; banda V usualmente
desconectada de la banda S y con los brazos separados en el vértice superior,
pero si están unidos, entonces los brazos siempre dejan un área hialina amplia en
su parte inferior sobre la celda m.
Mancha negra central en la sutura escudo-escutelar ausente, pero si esta
presente, nunca es redondeada; medioterguito generalmente con manchas
laterales; aculeusvariable.................................................................................... (5).
4‟ Banda apical anterior (= sección distal de la banda S) extremadamente ancha y
alcanzando el ápice de la vena M; banda V ancha, completa y siempre conectada
a la banda S, con los brazos ampliamente conectados en el vértice superior y

62
ocupando prácticamente toda la celda m. Sutura escudo-escutelar usualmente
presenta un gran punto negro redondeado en su parte media; manchas laterales
del medio terguito presentes o ausentes; aculeus de 1.4 - 1.6 m longitud.
Distribución. Florida (EUA), Antillas mayores, Bahamas (ref: Foote et al. 1993).
“Mosca del Caribe”.................................................. Anastrepha suspensa (Loew).
5. Longitud total del aculeus menor o igual a 2.0 mm (generalmente entre 1.4-1.9
mm); otros caracteres variables...........................................................................(6).
5‟ Longitud total del aculeus mayor de 2.5 mm (generalmente entre 3.1-4.9 mm);
punta del aculeus alargada con 0.34-0.38 mm de longitud y con una constricción
moderada cerca de su parte media, con dientes en sus márgenes laterales mas
omenos simétricos y apicalmente redondeados; subescutelo siempre con
manchas negras laterales que en ocasiones se extienden en el medio terguito con
una reducción gradual inferiormente. Distribución: Texas (EUA), México y América
central hasta Costa Rica (refs: Hernández-Ortiz 1992, Foote et al. 1993).
“Mosca Mexicana de la Fruta”.................................... Anastrepha ludens (Loew).
6 Subescutelo completamente amarillo, solo el medio terguito presenta manchas
negras laterales; mancha central en la sutura escudo-escutelar ausente; punta del
aculeus con una longitud de 0.16-0.20 mm, provista de dientes marginales que
ocupan2/3 o 4/5 de toda su longitud. Patrón alar como en la Distribución: Posee
registros desde EUA (cayos de Florida, sur de Texas, California); México, América
central, Sudamérica hasta Brasil, Antillas Mayores y Menores (refs: Foote et al.
1993, Hernández-Ortiz & Aluja 1993).
“Mosca de las Indias Occidentales”.....................Anastrepha obliqua (Macquart).
6‟ Subescutelo y medio terguito, ambos con manchas negras laterales; mancha
central de la sutura escudo-escutelar usualmente presente; especímenes con
todos los siguientes caracteres presentes: aculeus de 1.4-1.9 mm de longitud;
punta del aculeus de 0.20-0.28 mm de largo, con márgenes laterales provistos de
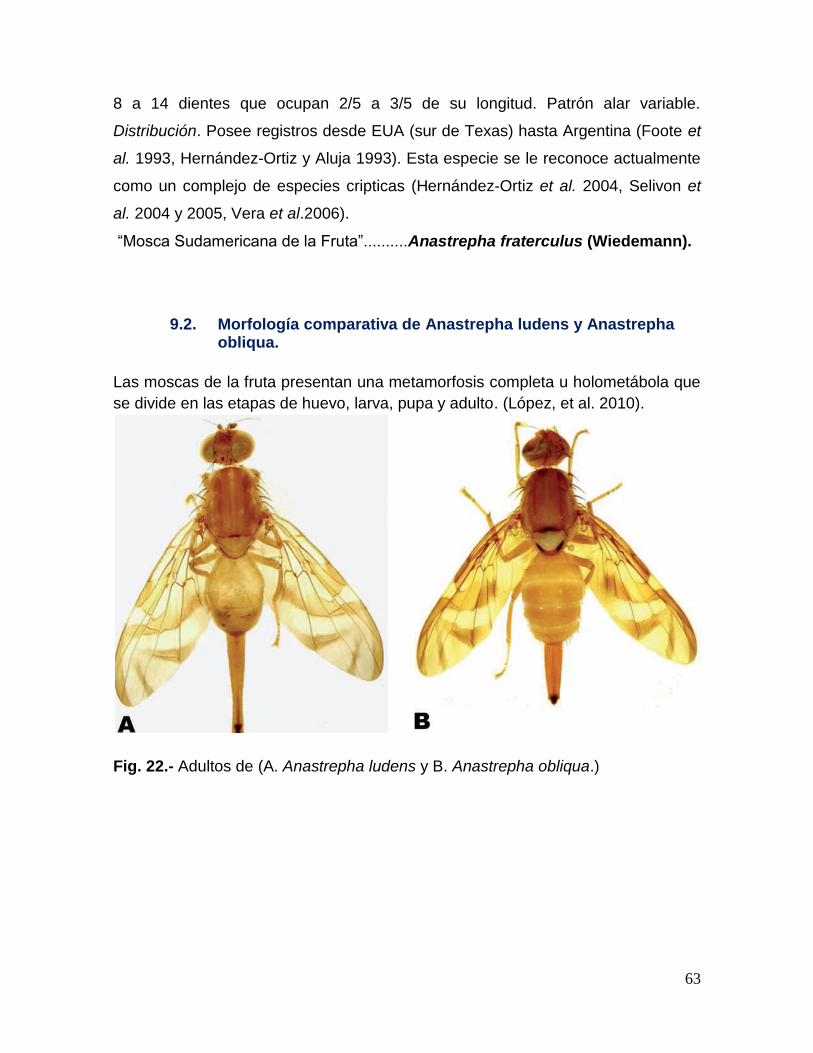
63
8 a 14 dientes que ocupan 2/5 a 3/5 de su longitud. Patrón alar variable.
Distribución. Posee registros desde EUA (sur de Texas) hasta Argentina (Foote et
al. 1993, Hernández-Ortiz y Aluja 1993). Esta especie se le reconoce actualmente
como un complejo de especies cripticas (Hernández-Ortiz et al. 2004, Selivon et
al. 2004 y 2005, Vera et al.2006).
“Mosca Sudamericana de la Fruta”..........Anastrepha fraterculus (Wiedemann).
9.2. Morfología comparativa de Anastrepha ludens y Anastrepha obliqua.
Las moscas de la fruta presentan una metamorfosis completa u holometábola que
se divide en las etapas de huevo, larva, pupa y adulto. (López, et al. 2010).
Fig. 22.- Adultos de (A. Anastrepha ludens y B. Anastrepha obliqua.)

64
Fig. 23.- A) Muestra el tórax característico de A. ludens y B el torax de A. obliqua.
Fig. 24.- A) Muestra el tipo de ala característico de A. ludens y B la de A. obliqua.
Fig. 25.- A y b. Morfología del ovipositor y su extremo apical en Anastrepha ludens
y obliqua. C y D Membrana eversible mostrando los ganchos esclerosados para
las dos especies hospederas. (Montoya, et al., 2010).
A B
A B
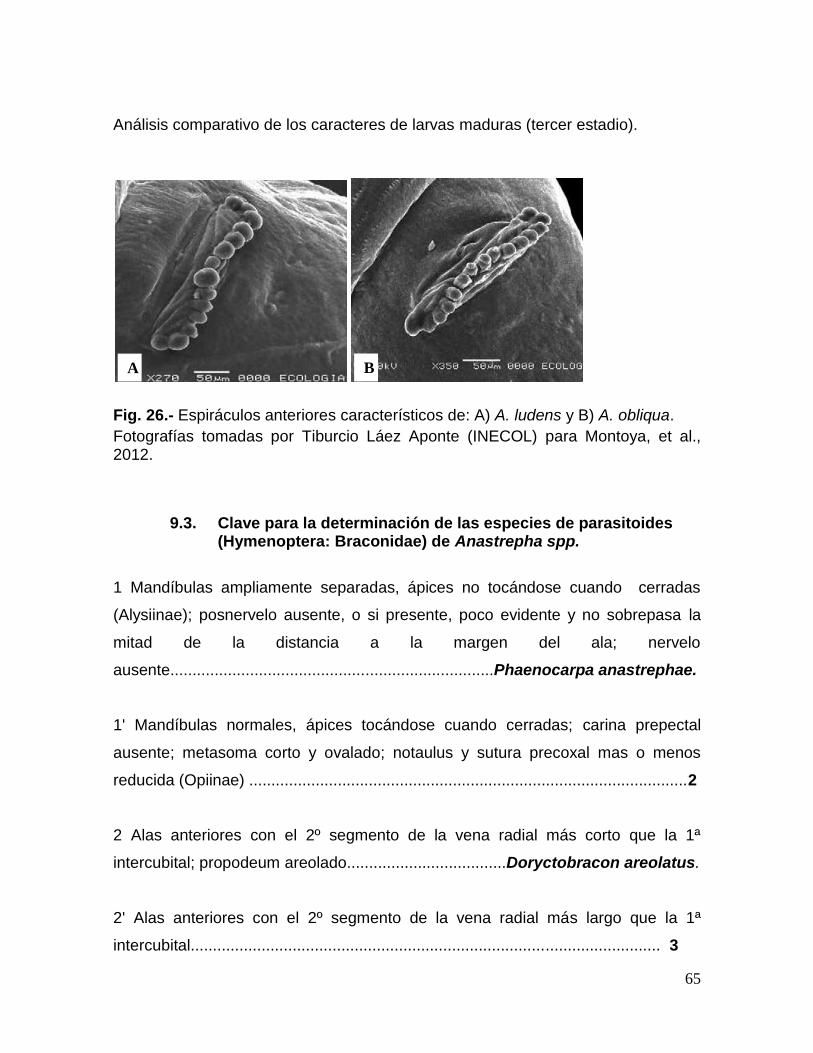
65
Análisis comparativo de los caracteres de larvas maduras (tercer estadio).
Fig. 26.- Espiráculos anteriores característicos de: A) A. ludens y B) A. obliqua.
Fotografías tomadas por Tiburcio Láez Aponte (INECOL) para Montoya, et al., 2012.
9.3. Clave para la determinación de las especies de parasitoides (Hymenoptera: Braconidae) de Anastrepha spp.
1 Mandíbulas ampliamente separadas, ápices no tocándose cuando cerradas
(Alysiinae); posnervelo ausente, o si presente, poco evidente y no sobrepasa la
mitad de la distancia a la margen del ala; nervelo
ausente.........................................................................Phaenocarpa anastrephae.
1' Mandíbulas normales, ápices tocándose cuando cerradas; carina prepectal
ausente; metasoma corto y ovalado; notaulus y sutura precoxal mas o menos
reducida (Opiinae) ...................................................................................................2
2 Alas anteriores con el 2º segmento de la vena radial más corto que la 1ª
intercubital; propodeum areolado....................................Doryctobracon areolatus.
2' Alas anteriores con el 2º segmento de la vena radial más largo que la 1ª
intercubital.......................................................................................................... 3
B A

66
3 Alas anteriores con la vena recurrente alcanzando la 2ª célula cubital;
propodeum areolado anteriormente.........................................Utetes anastrephae.
3' Alas anteriores con la vena recurrente alcanzando la 1ª célula cubital;
propodeum con carina medio-longitudinal………….................................................4
4 Tibia posterior negra en la base y en el
ápice.....................................................................................................Opius bellus.
4' Tibia posterior rojo-amarillenta…............................................................Opius sp.


















