
Clm 2do Avance
28
PONTIFICIA UNIVERSIDAD CATÓLICA DE VALPARAISO FACULTAD DE INGENIERÍA ESCUELA DE INGENIERÍA BIOQUÍMICA PRODUCCION DE METABOLITOS SEGUNDO AVANCE “PRODUCCIÓN DE CLORANFENICOL” Alumnos: POLETTE AGUILAR M. NICOLE ARRAÑO U. JUAN IGNACIO RAMÍREZ. Profesores: CLAUDIA ALTAMIRANO JULIO BERRIOS
-
Upload
jn-ignacio -
Category
Documents
-
view
53 -
download
2
Transcript of Clm 2do Avance

PONTIFICIA UNIVERSIDAD CATÓLICA DE VALPARAISO
FACULTAD DE INGENIERÍA
ESCUELA DE INGENIERÍA BIOQUÍMICA
PRODUCCION DE METABOLITOS
SEGUNDO AVANCE“PRODUCCIÓN DE CLORANFENICOL”
Alumnos:
POLETTE AGUILAR M.
NICOLE ARRAÑO U.
JUAN IGNACIO RAMÍREZ.
Profesores:
CLAUDIA ALTAMIRANO
JULIO BERRIOS
ABRIL 2023

i
INDICE
1. INTRODUCCIÓN.........................................................................................................................................1
2. MÉTODOS DE PRODUCCIÓN DE CLORANFENICOL....................................................................................1
2.1. PRODUCCIÓN MEDIANTE SÍNTESIS QUÍMICA...................................................................................12.2. PRODUCCIÓN MEDIANTE BIOSÍNTESIS.............................................................................................2
3. RESPUESTA BACTERIANA ANTE LA ACCIÓN DEL CLORANFENICOL............................................................3
4. TÉCNICAS PARA LA PRODUCCIÓN DE CLORANFENICOL............................................................................4
4.1. MANIPULACIÓN DE LAS CONDICIONES AMBIENTALES.....................................................................44.2. FORMULACIÓN DE UN MEDIO DE CULTIVO PARA UNA FERMENTACIÓN OPTIMA...........................5
5. POTENCIALES ALTERNATIVAS PARA PRODUCCIÓN DE CLORANFENICOL..................................................5
5.1. MANIPULACIONES GENÉTICAS: EL PLÁSMIDO DE STREPTOMYCES VENEZUELAE......................................55.2. MANIPULACIONES GENÉTICAS EN EL GÉNERO STREPTOMYCES...........................................................6
6. DISCUSIÓN BIBLIOGRÁFICA.......................................................................................................................6
7. CONCLUSIÓN.............................................................................................................................................8
8. REFERENCIAS.............................................................................................................................................9
9. ANEXOS...................................................................................................................................................11

ii
INDICE DE FIGURAS
Figura 9.1.1: Estructura del cloranfenicol y sus derivados..................................................................................................12Figura 9.1.2: Mecanismo de acción del cloranfenicol a nivel de ribosomas........................................................................12Figura 9.1.3: Etapas de la síntesis química de cloranfenicol...............................................................................................13Figura 9.1.4: Etapas de la biosíntesis en la producción de cloranfenicol............................................................................14Figura 9.1.5: Agrupación de genes que participan en la producción de cloranfenicol........................................................15Figura 9.1.6: Comparación entre el crecimiento celular y la producción de cloranfenicol en dos medios de cultivo distintos .............................................................................................................................................................................15Figura 9.1.7: Producción de cloranfenicol y pH en cultivo por lotes en S. venezuelae.......................................................16Figura 9.1.8: Producción de cloranfenicol y pH en cultivo continuo en S.venezuelae ........................................................16Figura 9.1.9: Mapa de restricción de pUC3 en Streptomyces venezuelae..........................................................................17

1
1. INTRODUCCIÓN
El cloranfenicol es un antibiótico de amplio espectro producido por diversas especies del género bacteriano Streptomyces (Morales, Herrera, & Muñoz, 2007), y fue aislado por primera vez en el año 1947 del microrganismo Streptomyces venezuelae desde una muestra de suelo traída de Caracas, Venezuela por Paul Burkholder, la que posteriormente fue llevada al Laboratorio Parke Davis, Estados Unidos (Korolkovas & Burckhalter, 2006).
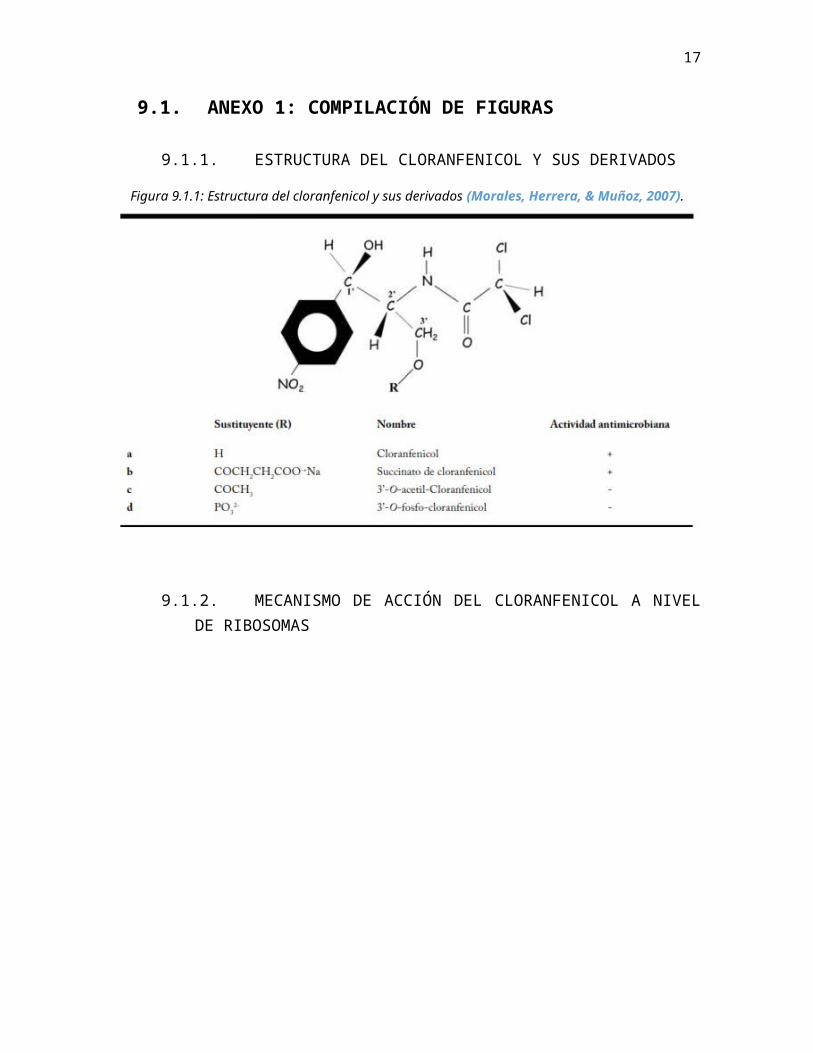
La molécula de cloranfenicol es pequeña y con características poco polares (ver anexo 9.1.1), lo que le permite difundir con facilidad a través de las membranas de células eucariotas y paredes de células procariotas (Morales, Herrera, & Muñoz, 2007).La estereoquímica de este antibiótico juega un rol preponderante en su acción, ya que posee dos carbonos asimétricos que hace posible que hayan cuatro isómeros; no obstante, sólo el isómero natural posee una marcada actividad antibacteriana. Se han hecho múltiples modificaciones moleculares, mas no se ha mejorado el producto (Korolkovas & Burckhalter, 2006). De hecho, la solubilidad lipídica de este antibiótico es la que permite que penetre bien dentro del cerebro, razón por las que sus potencialidades se aumentan para tratar infecciones en estos sitios donde otros antibióticos no podrían llegar a producir los efectos deseados (Morales, Herrera, & Muñoz, 2007). Por ello se ha reservado su uso para pacientes con infecciones graves como meningitis, tifus y fiebre tifoidea.
El mecanismo de acción principal del cloranfenicol sobre diversas bacterias (ver anexo 9.1.2), involucra la inhibición de la síntesis proteínas en las cepas sensibles. El antibiótico se une a la subunidad 50S bloqueando las funciones principales de los ribosomas, como la reacción en la que participa la peptidiltransferasa (TPasa), la unión y el movimiento de los sustratos a través del centro de esta enzima, la terminación de la traducción en cualquier momento. En conclusión, el cloranfenicol impide el ensamblaje de los aminoacil-tRNA que transportan los aminoácidos al ribosoma, inhibiendo la formación del enlace peptídico, lo que permite deducir que este compuesto tiene un efecto bacteriostático sobre las células (Morales, Herrera, & Muñoz, 2007).
2. MÉTODOS DE PRODUCCIÓN DE CLORANFENICOL
2.1. PRODUCCIÓN MEDIANTE SÍNTESIS QUÍMICA
La síntesis descrita a continuación permite obtener (-)-cloranfenicol puro, y corresponde a uno de los procesos más rápidos y prácticos informados hasta la fecha, ya que sólo consta de 4 etapas (ver anexo 9.1.3). El proceso trata principalmente de la aziridinación asimétrica de iminas, lo que permite obtener aziridina; a éste se le inserta un grupo clave que es la dicloroacetamida, y después de ciertos procedimientos se llega finalmente al compuesto del cloranfenicol.
La reacción de azidirinación es llevada a cabo por catalizadores quirales, obtenidos principalmente del compuestotri-fenilborato y el ligando (R)-VAPOL; también pueden ser generados a partir de los compuestos VANOL, BANOL, BINOL, 6,6-difenil-VAPOL. Estos catalizadores se obtienen a partir del

2
calentamiento de VAPOL y tri-fenilborato a 55°C durante una hora, y posterior eliminación de todos los compuestos volátiles a esta misma temperatura bajo vacío (Loncaric & Wulff, 2001).
Comienza transformando el p-nitrobenzaldehído a su imina correspondiente, a través de una cristalización a partir de etanol, con un 80% de rendimiento. La reacción de aziridinación se lleva a cabo con 10% en moles del catalizador, el cual es preparado a partir de VAPOL en tolueno a 0°C, para finalmente obtener aziridina con un 80% de rendimiento. El producto de esta reacción consta de un 96% enantiómeros de aziridina, con especies cis y trans que se encuentran en una relación de 30:1 respectivamente (Loncaric & Wulff, 2001).
La siguiente etapa consiste en la apertura de la aziridina, que permite la introducción del grupo dicloro acetamida. Ésta se realiza a través de un tratamiento simple que hace uso de ácido dicloroacético (Loncaric & Wulff, 2001). Este paso se lleva a cabo con 10 equivalentes de ácido dicloroacético y 1,2-dicloroetano en reflujo durante 1 hora, obteniendo hidroxi-acetamida como un único diastereómero, con un porcentaje de rendimiento del 80%.Finalmente se reduce el éster de etilo con el compuesto borohidruro de sodio, generándose (-)-cloranfenicol con 74% de rendimiento, con más de un 99% de exceso enantiomérico.
2.2. PRODUCCIÓN MEDIANTE BIOSÍNTESIS
El cloranfenicol es producido por diversas especies del género bacteriano Streptomyces, y a continuación se presentará la ruta biosintética del microorganismo Streptomyces venezuelae, que ha sido una de las más estudiadas a través del tiempo. El origen de este antibiótico se encuentra en la vía del shikimato, la que tiene como precursor al ácido shikímico, que se somete a una serie de reacciones enzimáticas, propias de la ruta del corismato, llegando a ser ácido corísmico. A partir de este compuesto, distintas rutas han evolucionado por separado para proveer a las células bacterianas del cofactor del ácido fólico denominado PABA (ácido p-aminobenzoico) y de aminoácidos aromáticos tales como: fenilalanina, tirosina, y triptófano (Vining & Westlake, 1984).
La reacción inicial de la síntesis de cloranfenicol está catalizada por un complejo enzimático formado de la asociación de los productos obtenidos de la expresión de los genes pabA y pabB (ver anexo 9.1.5). Este complejo llamado amino-desoxicorismatosintasa, convierte el ácido corísmico en ácido 4-amino-4-deoxicorísmico (ADC) (Brown, Aidoo, & Vining, 1996), el cual constituye un punto metabólico de bifurcación, ya que se generan dos productos: PABA a través de una reacción catalizada por un enzima de tipo liasa, y ácido 4-amino-4-desoxiprefénico a través de una reacción catalizada por una enzima del tipo mutasa.
Para la obtención del cloranfenicol, se debe seguir la ruta en la que se convierte el ácido 4-amino-4-desoxiprefénico en ácido p-aminofenilpirúvico mediante la acción de la prefanato deshidrogenasa. El ácido obtenido es transformado en p-aminofenilalanina (PAPA) gracias a la acción de la enzima arilamina sintetasa; de esta forma PAPA servirá como precursor del antibiótico. Una vez que se dispone de PAPA, este compuesto comienza a sufrir una serie de modificaciones tales como, la adición de AMP, y una posterior alteración e hidroxilación de sus grupos funcionales (Morales, Herrera, & Muñoz, 2007).

3
Al intermediario generado de las reacciones que se mencionaron anteriormente, se le transfiere el grupo acetilo del compuesto dicloroacetilCoA gracias a la acción de la N-Acetiltransferasa, y posteriormente se reduce mediante la oxidación del equivalente reductor NADH, para obtenerse N-dicloroacetil-p-aminofenilserinol. El dicloroacetilCoA proviene de una ruta que ha sido recientemente dilucidada en lo que se refiere a la producción de cloranfenicol. El operón de la biosíntesis de cloranfenicol en Streptomyces venezuelae codifica la cloranfenicol halogenasa(Podzelinska, Latimer, Bhattacharya, Vining, Zechel, & Jia, 2010), y se ha demostrado que esta enzima es esencial para la formación del dicloroacetilCoA a partir intermediarios derivados de la glucosa (Chang, Sun, He, & L., 2001).
Finalmente, el intemediario N-dicloroacetil-p-aminofenilserinal es reducido mediante una reacción enzimática a N-dicloroacetil-p-aminofenilseriaol, y sobre este último actúa una oxidasa que lo lleva a ser cloranfenicol (ver anexo 9.1.4).
3. RESPUESTA BACTERIANA ANTE LA ACCIÓN DEL CLORANFENICOL
Hoy en día el cloranfenicol no es preferido como un antibiótico de primera selección debido a su toxicidad y especificidad en el tratamiento de ciertas enfermedades, razón por la cual sus límites de producción son bastante acotados (Morales, Herrera, & Muñoz, 2007). Por otra parte, se sabe que la alternativa escogida para su producción es la síntesis química, ya que de la síntesis biológica, se obtiene cloranfenicol y derivados de éste. Esto se basa en el hecho de que la célula responde a la actividad antimicrobiana utilizando mecanismos de resistencia al cloranfenicol, y por ello éste es sometido a una serie de modificaciones, de las que se obtienen sus derivados.
Los microorganismos han desarrollado distintas estrategias para competir por los nutrientes. Diversas son las sustancias antagónicas que éstos mismos producen para dominar en su hábitat, por ejemplo, los antibióticos de amplio espectro, como lo es el cloranfenicol (Morales, Herrera, &Muñoz, 2007).
En este caso, el antibiótico es capaz de unirse a los ribosomas de S. venezuelae e inhibir la biosíntesis de proteínas (Morales, Herrera, & Muñoz, 2007). Sin embargo, este microorganismo ha desarrollado métodos de resistencia para evadir el efecto del cloranfenicol, y se presentan a continuación:
a) Resistencia por expulsión inespecífica y específica del cloranfenicol al ambiente extracelular: El flujo activo debido a transporadores multidroga es una de las formas más usadas por diversas bacterias para evadir el efecto de los compuestos tóxicos tales como los antibióticos, por ejemplo, el cloranfenicol. La inducción del flujo activo por trasportadores multidroga parece involucrar proteínas “sensoras de droga” que se unen a una gran variedad de agentes antimicrobianos que no tienen una relación estructural. En el caso de S. venezuelae, se ha reportado un flujo específico de cloranfenicol al exterior de la célula que representa una forma activa de resistencia al antibiótico, el cual involucra al gen cmlF (ver anexo 9.1.5) (Morales, Herrera, & Muñoz, 2007).

4
b) Resistencia mediante fosforilación: En 1995 mediante técnicas de ingeniería genética, se le insertó a una cepa hipersensible a cloranfenicol un vector plasmídico responsable de conferir la resistencia al antibiótico (Moshe, Camp, Yang, Brown, & Shaw, 1995). Los cultivos de esta cepa metabolizaron rápidamente el cloranfenicol a un producto fosforilado, llamado 3’-O-fosfo-cloranfenicol, el cual no presentó actividad antimicrobiana. Se descubrió que la enzima encargada de esta fosforilación corresponde a cloranfenicol 3’-O-fosfotransferasa (Moshe, Camp, Yang, Brown, & Shaw, 1995), y ciertos estudios han comprobado que el flujo activo y específico del cloranfenicol puro, es menor que el del 3’-O-fosfo-cloranfenicol (ver anexo 9.1.1). Este es el principal mecanismo de resistencia utilizado por S.venezuelae (Morales, Herrera, & Muñoz, 2007).
En términos de producción, estos mecanismos no son ventajosos para la obtención del producto deseado, ya que el cloranfenicol sufre una serie de modificaciones. Además, otra desventaja es que a medida que el microorganismo crece y produce clorafenicol, aumenta simultáneamente su resistencia a la acción del antibiótico (Malik & Vining, 1971).
Por otra parte, durante el crecimiento de S. venezuelae bajo condiciones donde el cloranfenicol no es producido por la bacteria, ésta es relativamente sensible al antibiótico. No obstante, se inducen niveles elevados de resistencia por la exposición a cloranfenicol. En los cultivos que crecen bajo condiciones de producción de cloranfenicol, la resistencia aumenta simultáneamente con la síntesis del antibiótico (Malik & Vining, 1971).
4. TÉCNICAS PARA LA PRODUCCIÓN DE CLORANFENICOL
4.1. MANIPULACIÓN DE LAS CONDICIONES AMBIENTALES
En el año 1958 la viabilidad de producción de antibióticos por fermentación continua de una sola etapa fue explorada por medio de una planta piloto especialmente diseñada, y el cloranfenicol parecía estar bien adaptado a esta modalidad de cultivo. Paralelamente, a esta modalidad de cultivo continuo, se planteó producir el antibiótico de forma tradicional utilizando un cultivo por lotes, para finalmente poder comparar ambas experiencias (Bartlett & Gerhardt, 1959).
En el caso del caso del cultivo por lote se alcanzó una concentración máxima de 400 (µg/ml) en 4 días (ver anexo 9.1.7), lo cual fue satisfactorio para la cepa que se estaba utilizando, a su vez se pudo demostrar que a la aparición temprana de contaminantes a veces no tiene ningún efecto perjudicial sobre la formación de clorafenicol. No obstante, los cambios de pH anormales pudieron haber sido ocasionados por los agentes contaminantes. Dentro de las principales desventajas de esta modalidad de cultivo, se tiene que las hifas de S. venezuelae productoras de cloranfenicol tienden a fragmentarse y lisar durante la última etapa del ciclo de crecimiento por lote, y el microorganismo presenta una alta frecuencia de reversión a mutantes con baja capacidad de síntesis. En base a esta última información se procedió a realizar un cultivo continuo; para ello se preparó el inóculo dentro del reactor durante 24 horas, para luego comenzar la alimentación en forma continua de este mismo medio. A medida que se desarrollaba la fermentación, se probaron

5
distintas tasas de dilución, y con valores bajos de este parámetro, se obtuvo que la concentración de producto alcanzada fue más baja que en el cultivo por lote. Finalmente se determinó que con una tasa de 1 (d-1), se tenían las concentraciones más altas de producto, en un rango entre 80-110 (µg/ml); y por otra parte, esta modalidad de cultivo propició que el pH no sufriera grandes alteraciones (ver anexo 9.1.8). Este cultivo, comenzó a presentar agentes contaminantes alrededor del día 18, y por ello se finalizó al día 20 (Bartlett & Gerhardt, 1959).
Durante la tercera semana se obtuvieron bajas concentraciones de cloranfenicol, y se pensó que esto podía fundamentarse en el hecho de que las cepas habían sufrido mutación; para comprobarlo se hicieron 2 cultivos durante 10 días, en el primero se utilizaron las cepas originales (día 1), y en el segundo, se utilizaron cepas recuperadas del cultivo continuo (día 17). Éstos se analizaron día tras día, para determinar si es que existía la presencia de alguna mutante que hiciera que disminuyera la concentración del antibiótico, y concluyeron que no había evidencia para atribuirle esta disminución a la acción de una cepa mutada (Bartlett & Gerhardt, 1959).
4.2. FORMULACIÓN DE UN MEDIO DE CULTIVO PARA UNA FERMENTACIÓN ÓPTIMA
La optimización de un cultivo utilizando Actinomicetes productores de antibióticos, la formulación del medio y el ajuste de las condiciones ambientales, son clave para el proceso de fermentación; permiten obtener la máxima producción del antibiótico, ya que el principal objetivo es apoyar el crecimiento eficiente de los microrganismos (Nanjwade, Chandrashekhara, Goudanavar,Shamarez, & Manvi, 2010).
Con el fin de lograr una máxima producción, se deben tener en cuenta los siguientes parámetros: concentración de fuente de carbono, nitrógeno, oxigeno disuelto y micronutrientes, además de temperatura y pH. Una investigación reveló que la fuente de carbono, no apoyó la producción de antibióticos, ya que la síntesis no depende totalmente del crecimiento del microrganismo, sino que depende de las condiciones adversas impuestas por el ambiente o sus alrededores. Algunos parámetros para la producción eficiente de antibióticos formulados por Actinomicetes recopilados en este estudio son: 28°C de temperatura, pH de 7 y una concentración de oxigeno disuelto de un 60% (Nanjwade, Chandrashekhara, Goudanavar, Shamarez, & Manvi, 2010).
5. POTENCIALES ALTERNATIVAS PARA PRODUCCIÓN DE CLORANFENICOL
5.1. MANIPULACIONES GENÉTICAS: EL PLÁSMIDO DE Streptomyces venezuelae
En el año 1983 se estudió la resistencia al cloranfenicol por parte del microrganismo S. venezuelae, y se planteó que la inactivación del antibiótico era mediante una hidrólisis; sin embargo, esta hipótesis no fue concluyente, ya que S. venezuelae seguía siendo sensible a su presencia en el medio cuando ésta no lo estaba produciendo. En tanto, la resistencia y la producción del antibiótico se inducirían simultáneamente (Malik V. , 1983).
Se sugirió que el cambio de las cepas del estado sensible a resistente, se originaba en la pérdida de algún elemento genético (Malik V. , 1983). S. venezuelae contiene un plásmido (pUC3)(ver

6
anexo 9.1.9) cuyo mapa de restricción fue realizado con varias enzimas, para luego determinar si existen elementos genéticos involucrados tanto en el proceso de producción como en el de inducción a la resistencia. La obtención de fragmentos con las enzimas de restricción EcoR1 y BamH1, fue la más satisfactoria. La metodología consistió en tomar los fragmentos obtenidos desde pUC3 e incorporarlos en el sitio de restricción de pBR322 en E. coli, donde no fue posible establecer si pUC3 contiene los genes implicados en la biosíntesis de cloranfenicol (Malik V. ,1983).
Contribuyendo a la búsqueda de genes involucrados en la producción en el plásmido pUC3, también se realizó una comparación del elemento extracromosomal entre una cepa normal y una mutante. Los resultados sugieren que la producción de antibióticos no tiene que estar codificada en un plásmido, y los genes codificadores para la biosíntesis de cloranfenicol y su regulación se encuentran en el cromosoma (Ahmed & Vining, 1983).
5.2. MANIPULACIONES GENÉTICAS EN EL GÉNERO Streptomyces
La clonación de genes y la recombinación en el género de Streptomyces ofrece una buena posibilidad de incrementar la producción de metabolitos secundarios y la creación de nuevos antibióticos. Este tipo de manipulación genética se facilita gracias a la construcción de diferentes vectores de clonación como plásmidos, fagos transductantes y elementos transposables, entre otros. El aumento en la producción industrial de muchos antibióticos se puede lograr mediante la introducción de las múltiples copias de genes reguladores positivos. El género de Streptomyces es bastante versátil, e incluye un amplio número de especies que han sido clasificadas por sus características fisiológicas, morfológicas y genéticas. Dentro de estas manipulaciones se encuentra el uso de vectores plasmídicos; por ejemplo, una cepa de Streptomyces lividans pudo ser transformada con la inserción del plásmido pIJ702 proveniente de la librería genómica de S. venezuelae. Se demostró que las células transformantes habían incorporado el vector recombinante que contenía 2,5 kb del fragmento de DNA de S. venezuelae. El fago es uno de los usos más ampliamente utilizado en el caso de la clonación de genes de Streptomyces, y tanto S. avermilitis como S. verticillus han demostrado ser huéspedes competentes para fagos transductantes. En el caso de los tranposones recombinantes, éstos son construídos para la clonación de genes en Streptomyces, y son insertados en el sitio neutral del cromosoma huésped, lo que no tiene un efecto negativo sobre la producción de metabolitos secundarios. Un ejemplo en el que la transposición ocurre es S. lividans, quien presenta una frecuencia de transposición de un 3%, y la inserción ocurre en el genoma de la célula diana azarosamente (Bhattacharyya & Sen,2004).
6. DISCUSIÓN BIBLIOGRÁFICA
Esta investigación en términos de producción por biosíntesis está basada en la ruta metabólica presente en Streptomyces venezuelae, ya que fue el primer microorganismo en el que se descubrió la producción del antibiótico, y con el que se siguió trabajando posteriormente para conocer todos los aspectos del cloranfenicol.

7
En primer lugar, hoy en día se prefiere una síntesis química ya que presentaría un mejor rendimiento global que una biológica (Loncaric & Wulff, 2001). Esto se debe a que en la biológica existen mecanismos de resistencia propios de las cepas de Streptomyces ante la actividad antimicrobiana del antibiótico. La respuesta de la célula para evitar la acción del cloranfenicol sobre los ribosomas, consiste en una serie de modificaciones que se le hacen al antibiótico; y éstas implican la obtención de una serie de derivados del producto, los cuales a su vez no son útiles para el tratamiento de enfermedades, ya que pierden la característica de ser agentes antimicrobianos. A diferencia de la síntesis química, en la que hay certeza de que el producto final va a tener una mayor cantidad de cloranfenicol puro, con un mínimo de interferencias.
Por otra parte, para la síntesis química las materias primas utilizadas son de fácil obtención en el comercio, y el proceso que debe llevarse a cabo es práctico y convencional. Mientras que en la biosíntesis el proceso tendría que ser mucho más minucioso, ya que se trata de agentes biológicos con mecanismos de resistencia, y por ende habría que plantear una última etapa de recuperación de los derivados de cloranfenicol obtenidos, revirtiendo las modificaciones que se les hicieron para que vuelvan a ser el antibiótico puro, y a su vez vuelvan a tener actividad antimicrobiana.
En términos de productividad, la biosíntesis sería más demorosa que la síntesis química, y este hecho no podría compensarse con la posibilidad de que el cloranfenicol producido biosintéticamente se comercialice a un precio elevado, ya que hoy en día éste no se usa masivamente en tratamientos debido a su toxicidad; y por ende su demanda no sería alta.
También se determinó que no se puede tener una sobreproducción del antibiótico mediante la biosíntesis ya que a medida que se va incrementando la producción de cloranfenicol en un cultivo, las células activan cada vez más sus mecanismos de resistencia contra este agente antimicrobiano, y es por esto que la concentración del antibiótico se estanca, quedando en un valor fijo (ver anexo 9.1.6), que no puede considerarse como una sobreproducción (Malik & Vining, 1971). En el caso de S. venezuelae, el principal mecanismo de resistencia utilizado es la fosforilación, disminuyendo la concentración de cloranfenicol puro ya que éste se va convirtiendo en un derivado del antibiótico, hasta que finalmente se llega a un valor constante (ver anexo 9.1.6).
Otro de los motivos que podría explicar por qué hoy en día no se privilegia la producción biosintética radica en el hecho de que hace 50 años atrás se probó una modalidad de cultivo continuo para producir cloranfenicol utilizando S. venezuelae (Bartlett & Gerhardt, 1959); sin embargo, ésta demostró no ser muy efectiva, ya que las concentraciones de producto alcanzadas no eran tan altas como se esperaría que fuesen. Por otra parte, se desconocía qué consecuencias podía traer el cultivo de cepas que sufrieran algún tipo de mutación. Además, los agentes contaminantes se incrementaban a medida que pasaba el tiempo del cultivo, y esto a su vez generaba que la concentración de cloranfenicol fuera disminuyendo. De todas maneras, esta modalidad de producción presentó algunas ventajas, tales como: cambios de pH moderados durante toda la etapa de cultivo, una suspensión de células de fluido, producto estable, entre otras. Estas ventajas llevan a pensar que se podría volver a retomar esta investigación, ya que hoy se dispone de tecnología y estrategias de cultivo más avanzadas para poder controlar ciertos

8
parámetros, como el pH, la concentración de contaminantes, y que las cepas se mantengan estables, es decir, sin el riesgo de sufrir mutación. Las condiciones planteadas en estudios recientes concuerdan con las investigaciones que se realizaron hace 50 años tanto en los valores de pH y temperatura.
Al descartarse la presencia de los genes involucrados en la producción de cloranfenicol en el plásmido pUC3, se dificulta el avance en cuanto a la manipulación de los genes para producir el antibiótico, ya que no se lograron atenuar los mecanismos de resistencia y la sensibilidad al cloranfenicol, para poder producir el antibiótico sin que sea modificado.
Como se puede apreciar en cuanto a la mutagénesis en el género de Streptomyces, ésta tiene diversas alternativas tales como el uso de tranposones, inserción de plásmidos en el cromosoma, fagos transductores, entre otras; y en la mayoría de los casos se ha demostrado que son efectivas en distintas especies de Streptomyces, como por ejemplo S. lividans, que fue transformada con genes provenientes de S. venezuelae. Por lo tanto, se deduce que al conocer el mapa genético de este microorganismo, se podría aplicar alguna de las manipulaciones que se hicieron a otras cepas del mismo género (que también producen otros antibióticos) de tal manera que produzca cloranfenicol, sin que éste genere ningún tipo de inhibición, ni efecto negativo sobre la célula.
7. CONCLUSIÓN
Farmacológicamente el cloranfenicol es un antibiótico efectivo, sin embargo, no se prefiere como uno de primera elección, a causa de su toxicidad sobre las células sanguíneas y médula ósea. Se ha reservado su uso para pacientes con infecciones graves como meningitis, tifus y fiebre tifoidea
Respecto a su producción se evidenció como principal ruta la síntesis química por sobre la biológica, tal cual como ha sucedido por años con varios productos de interés comercial. Su estructura química le confiere propiedades antimicrobianas, y cualquier cambio en ella hace que pierda total efecto. Esta es una de las razones por las cuales se produce su síntesis de forma química antes que la biosíntesis.
Los mecanismos de defensa de los microorganismos son diversos, y S. venezuelae no se excluye de ellos. Este punto es relevante en la producción biológica de cloranfenicol, ya que el perfil de producción del antibiótico llega a un punto de saturación, y esto se fundamenta en el hecho de que a medida que el microorganismo va produciendo cloranfenicol también activa sus mecanismos de resistencia, modificándolo para obtener un producto que no tiene actividad antimicrobiana.
A pesar que se han evidenciado distintas modificaciones genéticas en el género Streptomyces, aún no se tiene una alternativa para mejorar la producción de cloranfenicol aplicando alguna de estas modificaciones. Además, se probaron modalidades de cultivo las cuales tampoco fueron satisfactorias en cuanto al nivel de producción del antibiótico. No obstante hoy en día con la información recopilada a través de los años, se podría plantear una estrategia que permita la obtención del metabolito deseado.

9
8. REFERENCIAS
Ahmed, Z., & Vining, L. (1983). Evidence for a chromosomal location of the genes coding for chloramphenicol production in Streptomyces venezuelae. Journal of Bacteriology., 239-244.
Bartlett, M., & Gerhardt, P. (1959). Continuous Antibiotic Fermentation-Design of a 20 litre, Single-stage Pilot Plant and Trials with Two Contrasting Processes. Journal of Biochemical and Microbiological Technology and Engineering, 359-377.
Bhattacharyya, B., & Sen, S. (2004). Gene manipulation in Streptomyces. Indian Journal of Biotecnology, 22-28.
Brown, M., Aidoo, K., & Vining, L. (1996). A role for pabAB, a p-aminobenzoate synthase gene of Streptomyces venezuelae ISP5230 in chloramphenicol biosynthesis. Microbiology, 142:1345-1355.
Chang, Z., Sun, Y., He, J., & L., V. (2001). p-Aminobenzoic acid and chloramphenicol biosynthesis in Streptomyces venezuelae: gene sets for a key enzyme, 4-amino-4-deoxychorismate synthase. Microbiology, 147(8):2113-2126.
Korolkovas, A., & Burckhalter, J. H. (2006). Compendio Esencial de Quimica Farmaceutica. In A. Korolkovas, & J. H. Burckhalter, Compendio Esencial de Quimica Farmaceutica (pp. 643-644). España: Reverté S.A.
Loncaric, C., & Wulff, W. (2001). An Efficient Synthesis of (−)-Chloramphenicol via Asymmetric Catalytic Aziridination: A Comparison of Catalysts Prepared from Triphenylborate and Various Linear and Vaulted Biaryls. ORGANIC LETTERS, 3675-3678.
Malik, V. (1983). Biochemistry and genetics of chloramphenicoi production. Enzyme and Microbial Technology, 332-336.
Malik, V., & Vining, L. (1971). Effect of chloramphenicol on its biosynthesis by Streptomyces species 3022a. Canadian Journal of Microbiology, 137-143.
Morales, Y., Herrera, M. C., & Muñoz, J. (2007). Cloranfenicol, un antibiótico clásico como una alternativa del presente. Revista Mexicana de Ciencias Farmacéuticas, 58-69.
Moshe, R., Camp, D., Yang, K., Brown, M. P., & Shaw, W. (1995). Inactivation of Chloramphenicol by O-Phosphorylation. The Journal of Biological Chemistry, 27000–27006.
Nanjwade, B., Chandrashekhara, S., Goudanavar, P., Shamarez, A., & Manvi, F. (2010). Production of Antibiotics from Soil-Isolated Actinomycetes and Evaluation of their Antimicrobial Activities. Tropical Journal of Pharmaceutical Research, 373-377.

10
Podzelinska, K., Latimer, R., Bhattacharya, A., Vining, L., Zechel, D., & Jia, Z. (2010). Chloramphenicol Biosynthesis: The Structure of CmlS, a Flavin-Dependent Halogenase Showing a Covalent Flavin–Aspartate Bond. Journal of molecular biology, 316–331.
Vining, L., & Westlake, D. (1984). Chloramphenicol: properties, biosynthesis and fermentation. In W. D. Vining L., Chloramphenicol: properties, biosynthesis and fermentation. (pp. 387-411). New York.

11
9. ANEXOS
ÍNDICE DE ANEXOS
9. ANEXOS...................................................................................................................................................11
9.1. ANEXO 1: COMPILACIÓN DE FIGURAS............................................................................................129.1.1. ESTRUCTURA DEL CLORANFENICOL Y SUS DERIVADOS..........................................................129.1.2. MECANISMO DE ACCIÓN DEL CLORANFENICOL A NIVEL DE RIBOSOMAS..............................129.1.3. ETAPAS DE LA SÍNTESIS QUÍMICA DE CLORANFENICOL..........................................................139.1.4. ETAPAS DE LA BIOSÍNTESIS EN LA PRODUCCIÓN DE CLORANFENICOL...................................149.1.5. AGRUPACIÓN DE GENES QUE PARTICIPAN EN LA PRODUCCION DE CLORANFENICOL...........159.1.6. COMPARACIÓN ENTRE EL CRECIMIENTO CELULAR Y LA PRODUCCIÓN DE CLORANFENICOL.159.1.7. PRODUCCIÓN DE CLORANFENICOL EN CULTIVO POR LOTES..................................................169.1.8. PRODUCCIÓN DE CLORANFENICOL EN CULTIVO CONTINUO.................................................169.1.9. MAPA DE RESTRICCIÓN DE pUC3 EN S. venezuelae...............................................................17

12
9.1. ANEXO 1: COMPILACIÓN DE FIGURAS
9.1.1. ESTRUCTURA DEL CLORANFENICOL Y SUS DERIVADOS
Figura 9.1.1: Estructura del cloranfenicol y sus derivados (Morales, Herrera, &Muñoz, 2007).
9.1.2. MECANISMO DE ACCIÓN DEL CLORANFENICOL A NIVEL DE RIBOSOMAS
Figura 9.1.2: Mecanismo de acción del cloranfenicol a nivel de ribosomas (Morales,Herrera, & Muñoz, 2007).

13

14
9.1.3. ETAPAS DE LA SÍNTESIS QUÍMICA DE CLORANFENICOL
Figura 9.1.3: Etapas de la síntesis química de cloranfenicol (Loncaric & Wulff, 2001).

15
9.1.4. ETAPAS DE LA BIOSÍNTESIS EN LA PRODUCCIÓN DE CLORANFENICOL
Figura 9.1.4: Etapas de la biosíntesis en la producción de cloranfenicol (Morales,Herrera, & Muñoz, 2007).

16
9.1.5. AGRUPACIÓN DE GENES QUE PARTICIPAN EN LA PRODUCCION DE CLORANFENICOL.
Figura 9.1.5: Agrupación de genes que participan en la producción de cloranfenicol(Morales, Herrera, & Muñoz, 2007).
La función sugerida para cada gen se enuncia a continuación: cmlF, Expulsión del Cm; cmlE, desoxi-arabino-heptulosonato-7-fosfato sintasa; cmlD, corismatomutasa; cmlC, prefenatodeshidrogenasa; pabAB, amino-desoxicorismatosintasa; cmlA, no identificada; cmlP, péptido con actividad sintetasa no ribosómica; cmlH, no identificada; cmlI, transporte de membrana tipo ABC; cmlJ, cetoreductasa; cmlK, AMP-ligasa; cmlS, halogenasa.
9.1.6. COMPARACIÓN ENTRE EL CRECIMIENTO CELULAR Y LA PRODUCCIÓN DE CLORANFENICOL.
Figura 9.1.6: Comparación entre el crecimiento celular y la producción de cloranfenicol en dos medios de cultivo distintos (Malik & Vining, 1971).

17
9.1.7. PRODUCCIÓN DE CLORANFENICOL EN CULTIVO POR LOTES
Figura 9.1.7: Producción de cloranfenicol y pH en cultivo por lotes en S. venezuelae(Bartlett & Gerhardt, 1959)
9.1.8. PRODUCCIÓN DE CLORANFENICOL EN CULTIVO CONTINUO
Figura 9.1.8: Producción de cloranfenicol y pH en cultivo continuo en S.venezuelae (Bartlett & Gerhardt, 1959)
18
9.1.9. MAPA DE RESTRICCIÓN DE pUC3 EN S. venezuelae
Figura 9.1.9: Mapa de restricción de pUC3 en Streptomyces venezuelae (Malik V. ,1983)


















