
Clase 4 2da parte
66
Bioquímica y Biología Molecular Maestría en Bioinformática Laboratorio de Interacciones Moleculares Facultad de Ciencias, UdelaR Iguá 4225,11400 Montevideo, Uruguay tel: + 598 2 525 86 18-22 ext 237 Bioenergética, Enzimas, Metabolismo….
Transcript of Clase 4 2da parte

Bioquímica y Biología Molecular
Maestría en Bioinformática
Laboratorio de Interacciones Moleculares
Facultad de Ciencias, UdelaR
Iguá 4225,11400 Montevideo, Uruguay
tel: + 598 2 525 86 18-22 ext 237
Bioenergética, Enzimas, Metabolismo….
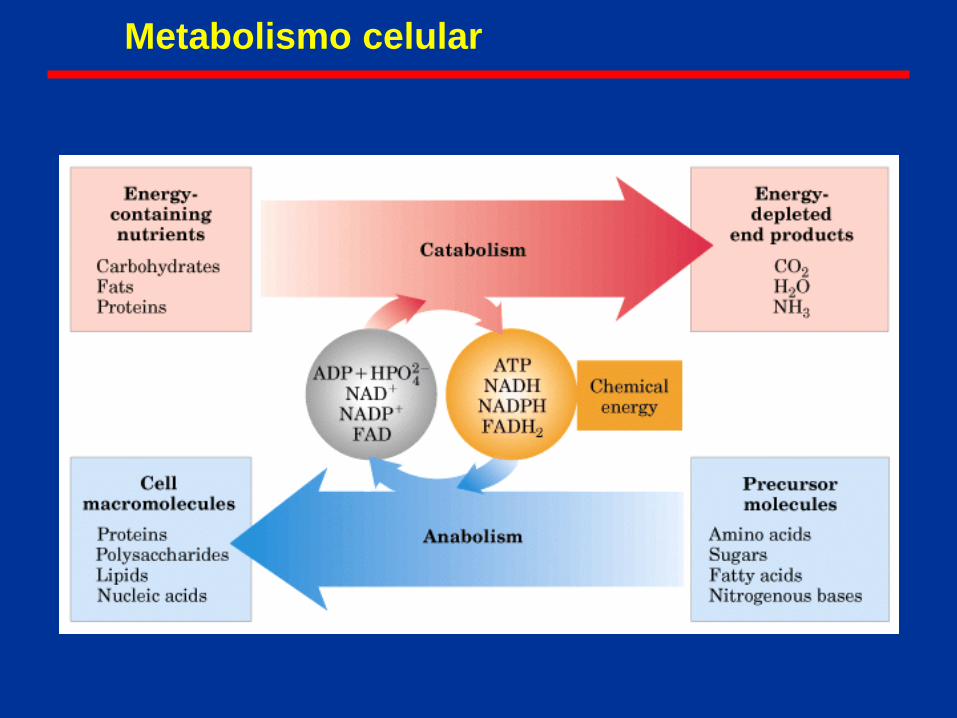
Metabolismo celular
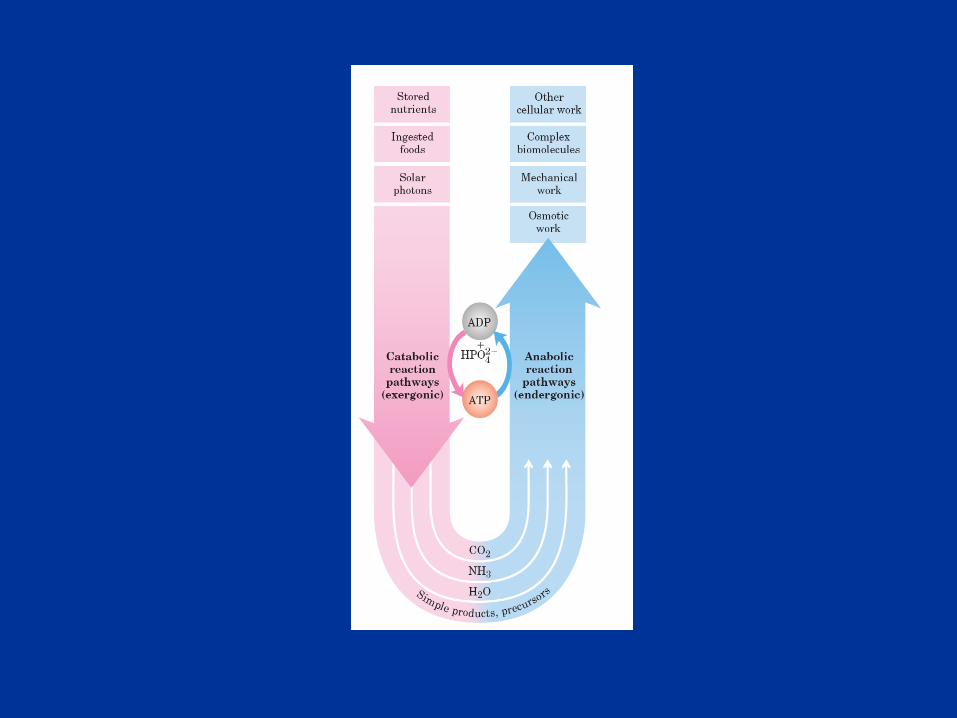

Bioenergética: es el estudio cuantitativo de las transferencias energéticas
que se producen en las células así como la naturaleza y función de los procesos
químicos implicados en estas transferencia de energía.
Leyes de la Termodinámica:
Primera ley o principio de la conservación de la energía: en cualquier cambio físico o
químico, la cantidad total de energía del universo permanece constante.
Segunda ley: en todo los procesos la entropía del universo se incrementa o la
entropía de un sistema aislado tenderá a aumentar hacia un valor máximo.

SISTEMA: Es la porción de universo que tomamos como objeto de estudio.
Existen tres tipos de sistemas:
SISTEMAS AISLADOS (no intercambia materia ni energía)
SISTEMAS CERRADOS (no intercambia materia si energía)
SISTEMAS ABIERTOS (intercambia materia y energía)
ESTADO DE UN SISTEMA: es el conjunto de
propiedades que permiten definirlo (ej.: P, V, T)
SISTEMA + ENTORNO= UNIVERSO
¿Qué tipo de sistema es una célula?

Algunas definiciones:
Entalpía
H o entalpía, expresa el contenido de calor en una reacción a presión constante,
se mide como la diferencia entre: H(productos) – H(reactivos) = H
Cuando se libera calor se dice que es una reacción exotérmica y H es negativo
ya que el contenido de calor de los productos es menor que los reactivos; si la
reacción absorbe calor del medio se habla de una reacción endotérmica y
H es positivo. H es equivalente a E cuando no hay cambios de volumen.
Energía Libre
G o energía libre de Gibbs, expresa la cantidad de energía capaz de realizar
trabajo, se mide como la diferencia de energía entre
G(productos) – G(reactivos) = G,
si G es negativo si dice que es una reacción exergónica, si G es positivo la
reacción es endergónica.
Entropía
S o entropía, es una magnitud del desorden en un sistema, cuando los productos
son menos complejos y más desordenados que los reactivos la entropía aumenta,
S(productos) – S(reactivos) = S

Estas magnitudes (bajo condiciones de temperatura y presión constantes)
están relacionadas entre si de acuerdo con la siguiente ecuación:
G = H - T S
energía libre entalpía entropía
donde T es la temperatura absoluta (en grados K).
Todo proceso esta termodinámicamente favorecido cuando G es negativo
o es exergónico, cuando G = 0 el proceso esta en equilibrio.
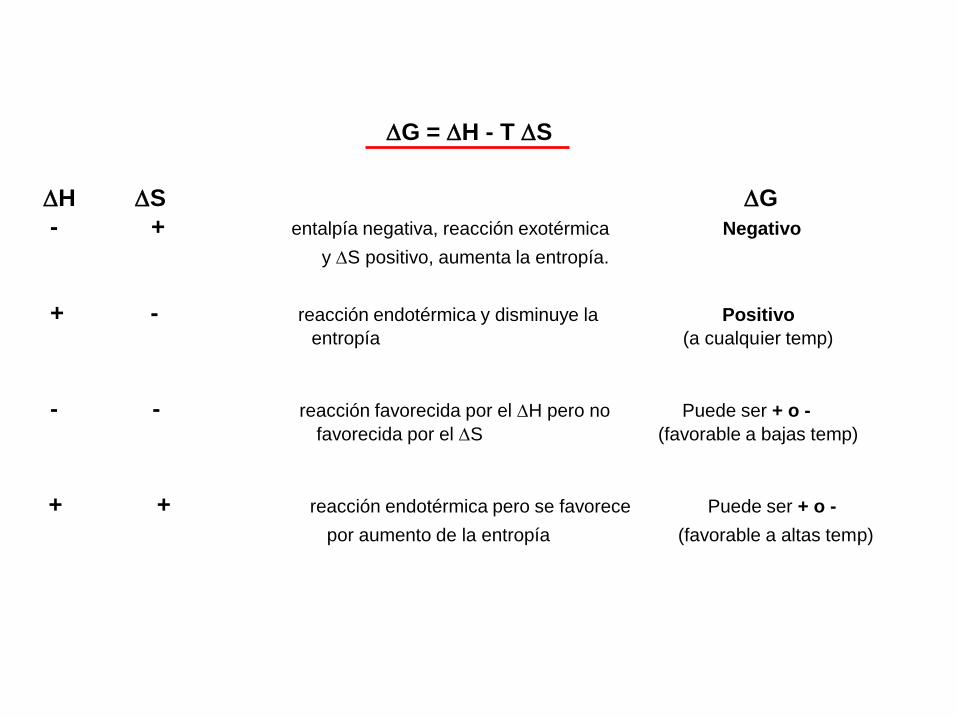
G = H - T S
H S G
- + entalpía negativa, reacción exotérmica Negativo
y S positivo, aumenta la entropía.
+ - reacción endotérmica y disminuye la Positivo
entropía (a cualquier temp)
- - reacción favorecida por el H pero no Puede ser + o -
favorecida por el S (favorable a bajas temp)
+ + reacción endotérmica pero se favorece Puede ser + o -
por aumento de la entropía (favorable a altas temp)
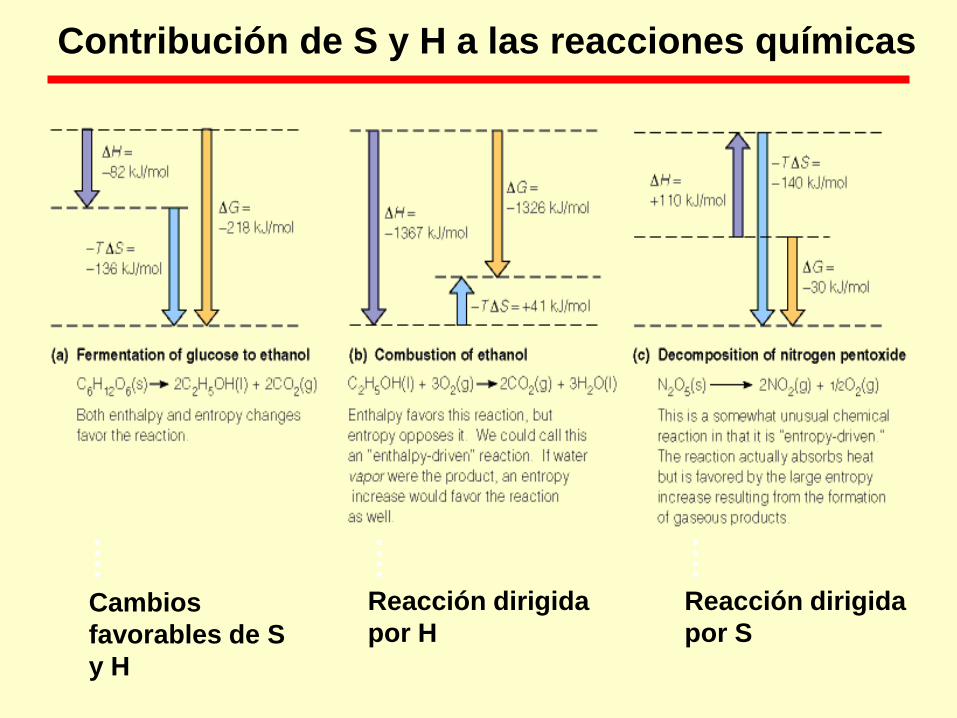
Cambios
favorables de S
y H
Reacción dirigida
por H
Reacción dirigida
por S
Contribución de S y H a las reacciones químicas

Dependencia de G con la concentración de reactivos y productos
A + B C + D
CH3COOH CH3COO- + H+
[C][D] [CH3COO-][H+]
Keq = Ka =
[A][B] [CH3COOH]
Ka = 1.74 x 10-5
Cuando se alcanza el equilibrio, el valor de G = O

El potencial químico de una sustancia esta determinado por:
GA = GoA + RT ln[A] donde Go
A es el potencial químico en condiciones estándar
A + B C + D
G = G productos - G reactivos
(GC + GD) (GA + GB)
[C][D] [Productos]
G = GO + RT ln = GO + RT ln
[A][B] [Reactivos]
Go es la energía libre en condiciones
estándar, 1 M de reactivos y
productos, 25oC
R = 8.314 J/K.mol
T = temperatura absoluta en oK
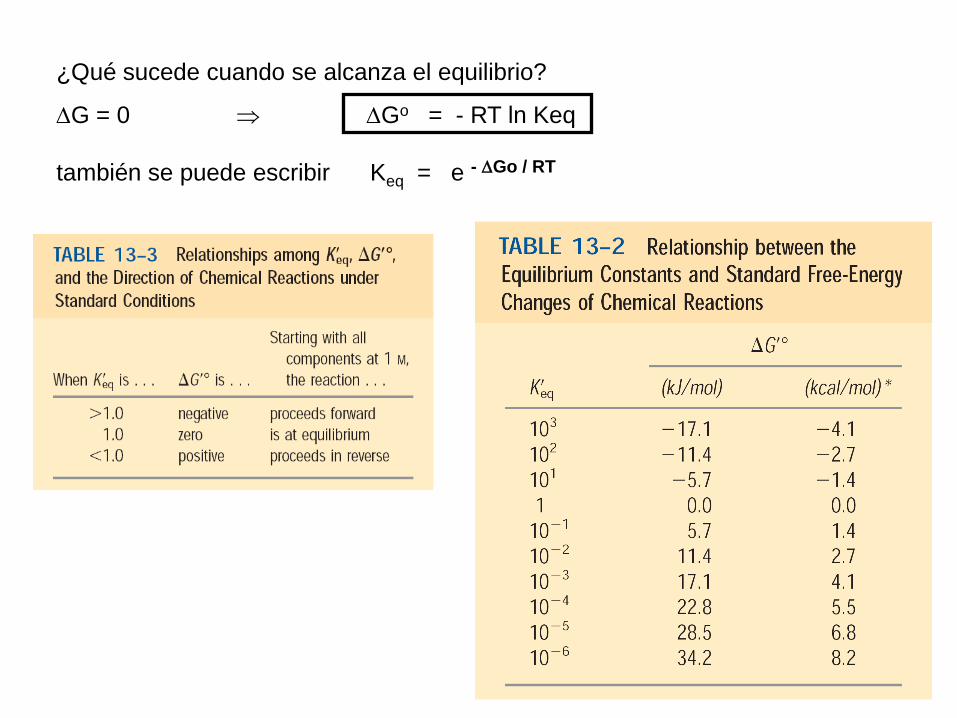
G = 0 Go = - RT ln Keq
también se puede escribir Keq = e - Go / RT
¿Qué sucede cuando se alcanza el equilibrio?

Reacciones acopladas

Una cantidad termodinámica (ej: G, H o
S) nos indica si una reacción es permitida,
A B está “permitida”;
B A no es espontánea, a menos que se le
acople otra reacción favorecida (ej: ATP
ADP)
Sin embargo, para que la reacción se
produzca, la energía neta debe descender
(i.e., G total debe ser negativa.)
Reacciones acopladas
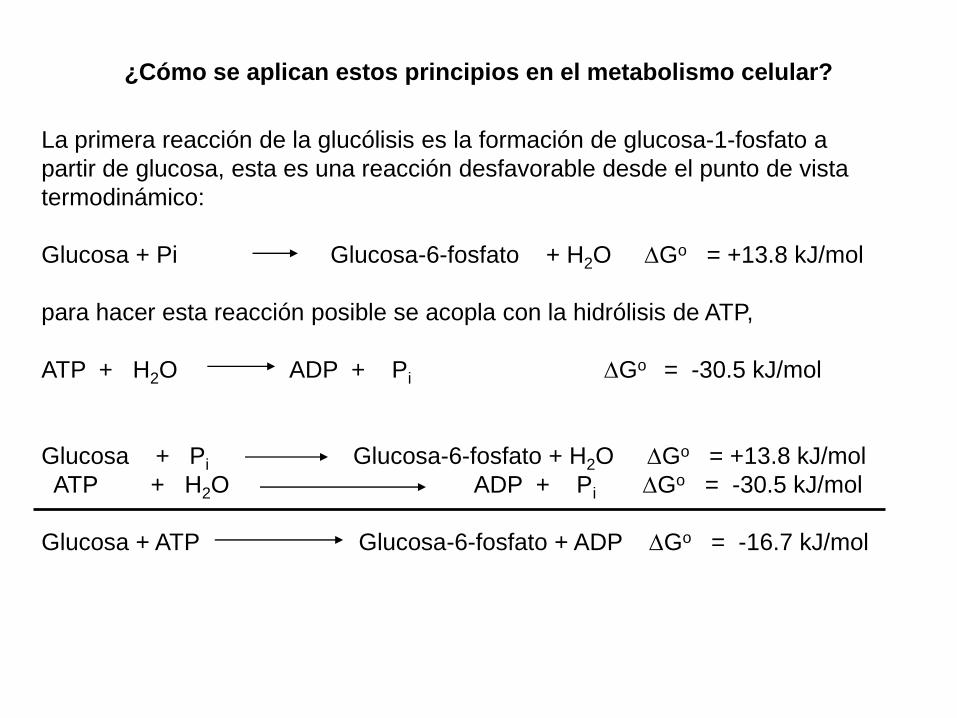
¿Cómo se aplican estos principios en el metabolismo celular?
La primera reacción de la glucólisis es la formación de glucosa-1-fosfato a
partir de glucosa, esta es una reacción desfavorable desde el punto de vista
termodinámico:
Glucosa + Pi Glucosa-6-fosfato + H2O Go = +13.8 kJ/mol
para hacer esta reacción posible se acopla con la hidrólisis de ATP,
ATP + H2O ADP + Pi Go = -30.5 kJ/mol
Glucosa + Pi Glucosa-6-fosfato + H2O Go = +13.8 kJ/mol
ATP + H2O ADP + Pi Go = -30.5 kJ/mol
Glucosa + ATP Glucosa-6-fosfato + ADP Go = -16.7 kJ/mol
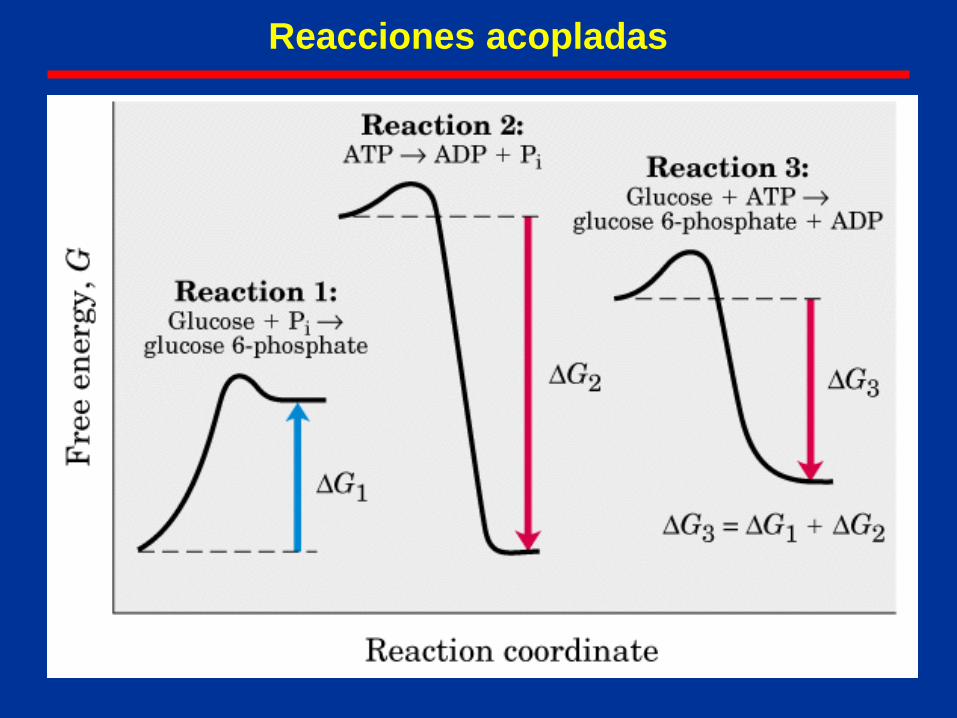
Reacciones acopladas

Glucosa + 6O2 6CO2 + 6H20
si Ho = -2816 kJ/mol y So = +0.181 kJ/mol
¿Cuál es el valor de Go a 37oC?
¿Si se aumenta la temperatura, puede TSo igualar a Ho y hacer Go
cero ?
¿Si el Go de hidrólisis de ATP es -31 kJ/mol, cuál es el máximo de
moles de ATP que se podrían generar si se acopla a la oxidación
completa de glucosa la síntesis de ATP a partir de ADP y Pi?
1) Go = Ho - TSo = - 2816 – (310º x 0.181 ) = - 2872 kJ/mol
2) No, es imposible porque So es + 0.181 y - T So siempre será un
valor negativo
3) 2872 kJ/mol / 31 kJ/mol = 92.6 ATP !!
¿Cuál es el rendimiento a nivel biológico?
38 ATP o sea 41 %

Gº’ = -RT ln Keq Keq = e - Gº’ / RT = 0.52
Quiere decir que el equilibrio hay [F6P]/[G6P] = 0.52
o sea hay 34% de F6P y 66% de G6P
¿Qué sucede en la célula?
Glucosa Glucosa-6-P Fructosa-6-P Piruvato
Glucosa-6-P Fructosa-6-P Go = + 1.7 kJ/mol
¿Cuál es la Keq de esta reacción?
83 µM 14 µM concentraciones intracelulares
[14x10-3M]
G = +1.7 kJ/mol + 0.0083 kJ//K.mol x 310oK ln = -2.9 kJ/mol
[83x10-3M]

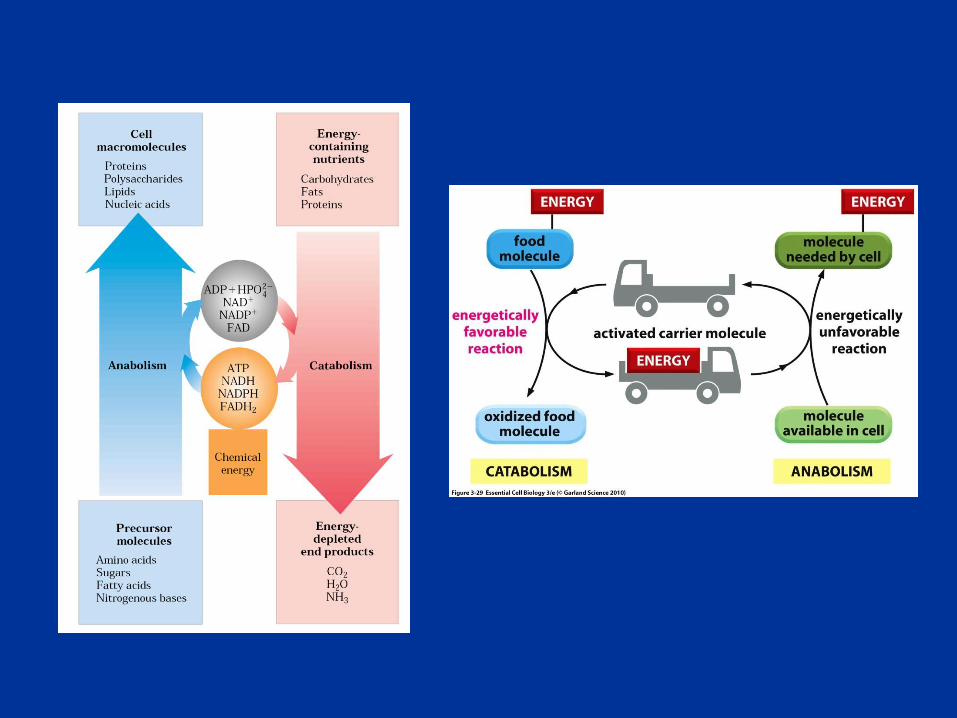
Actividad celular muy coordinada y dirigida en la que
muchos sistemas multi-enzimáticos cooperan para
cumplir 4 funciones:
• obtener energía química a partir de nutrientes ricos en
energía
• convertir moléculas nutrientes en moléculas características
de la propia célula
• polimerizar precursores monoméricos a proteínas, ácidos
nucleicos, lípidos, polisacáridos y otros
• Sintetizar y degradar biomoléculas requeridas en funciones
celulares especializadas
Metabolismo

Características del Metabolismo
1. Las vías metabólicas son irreversibles
2. Las vías anabólicas y catabólicas deben ser
diferentes
3. Cada vía metabólica tiene un primer paso limitante
4. Todas las vías metabólicas están reguladas
finamente
5. En los eucariotas las vías metabólicas transcurren
en localizaciones celulares específicas
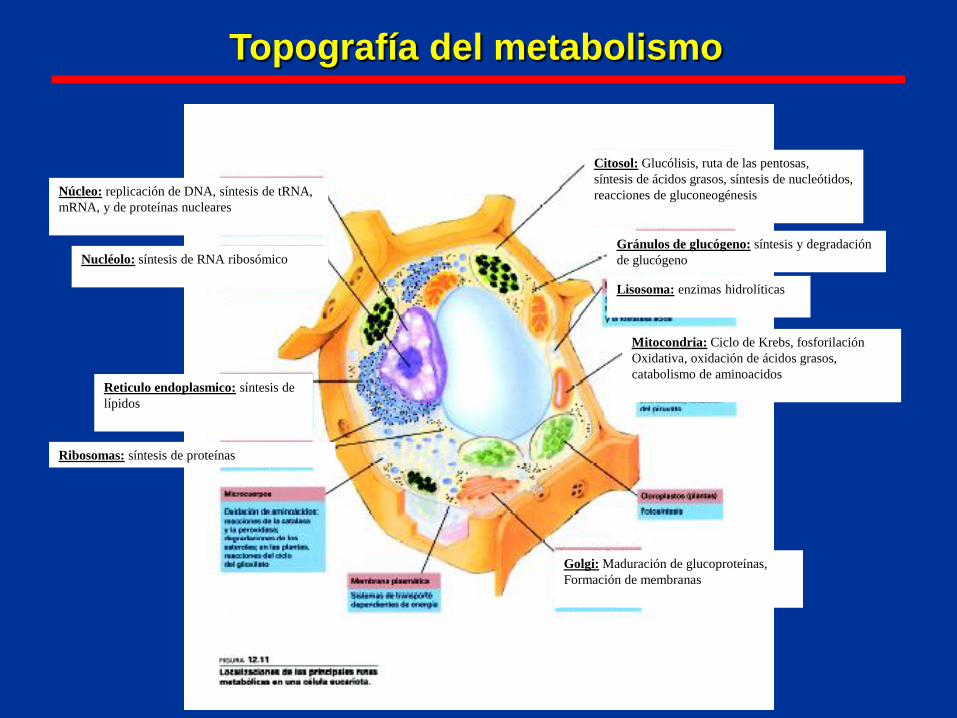
Topografía del metabolismo
Citosol: Glucólisis, ruta de las pentosas,
síntesis de ácidos grasos, síntesis de nucleótidos,
reacciones de gluconeogénesis
Gránulos de glucógeno: síntesis y degradación
de glucógeno
Lisosoma: enzimas hidrolíticas
Mitocondria: Ciclo de Krebs, fosforilación
Oxidativa, oxidación de ácidos grasos,
catabolismo de aminoacidos
Golgi: Maduración de glucoproteínas,
Formación de membranas
Reticulo endoplasmico: síntesis de
lípidos
Ribosomas: síntesis de proteínas
Nucléolo: síntesis de RNA ribosómico
Núcleo: replicación de DNA, síntesis de tRNA,
mRNA, y de proteínas nucleares
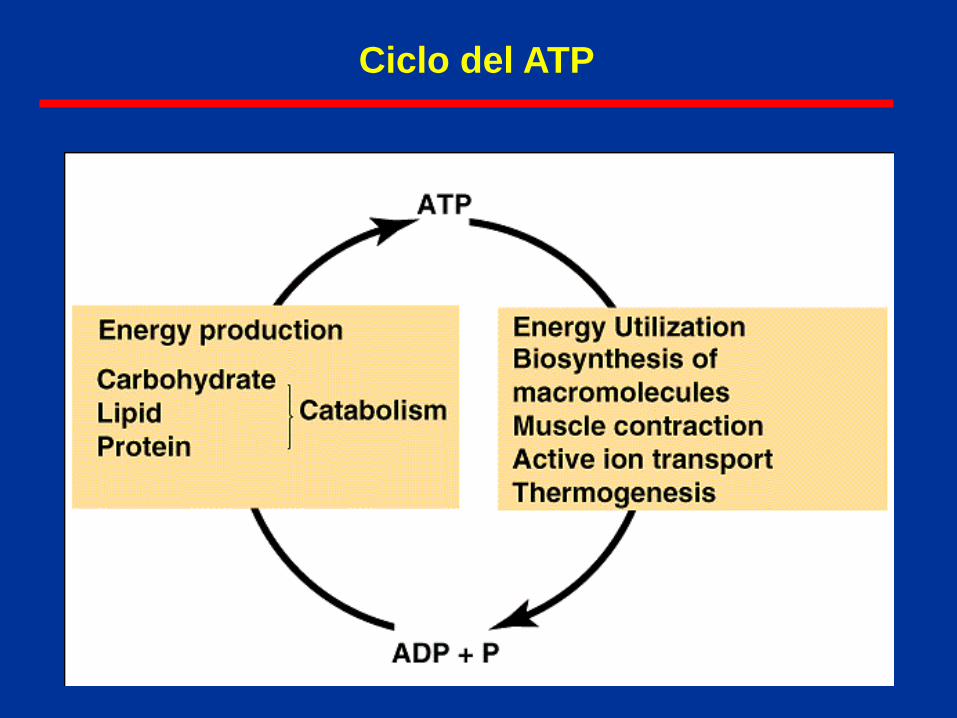
Ciclo del ATP

1. Repulsión electrostática
2. Estabilización por
resonancia del Pi saliente
3. Ionización del ADP
4. Mayor solvatación de
ADP + Pi que ATP
Factores que influyen en el G de hidrólisis del ATP
G = Go’ + RT ln [ADP][Pi]
[ATP]
[ADP] = 0,25 x 10-3 M
[Pi] = 1,65 x 10-3 M
[ATP] = 2,25 x 10-3 M
G = -51,8 KJ/mol
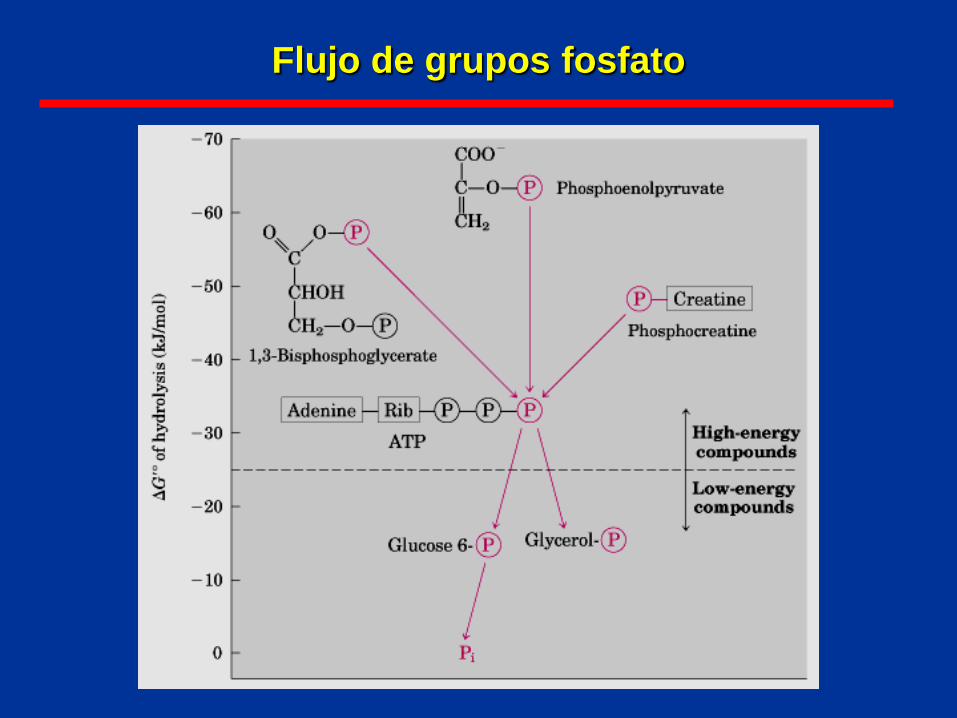
Flujo de grupos fosfato
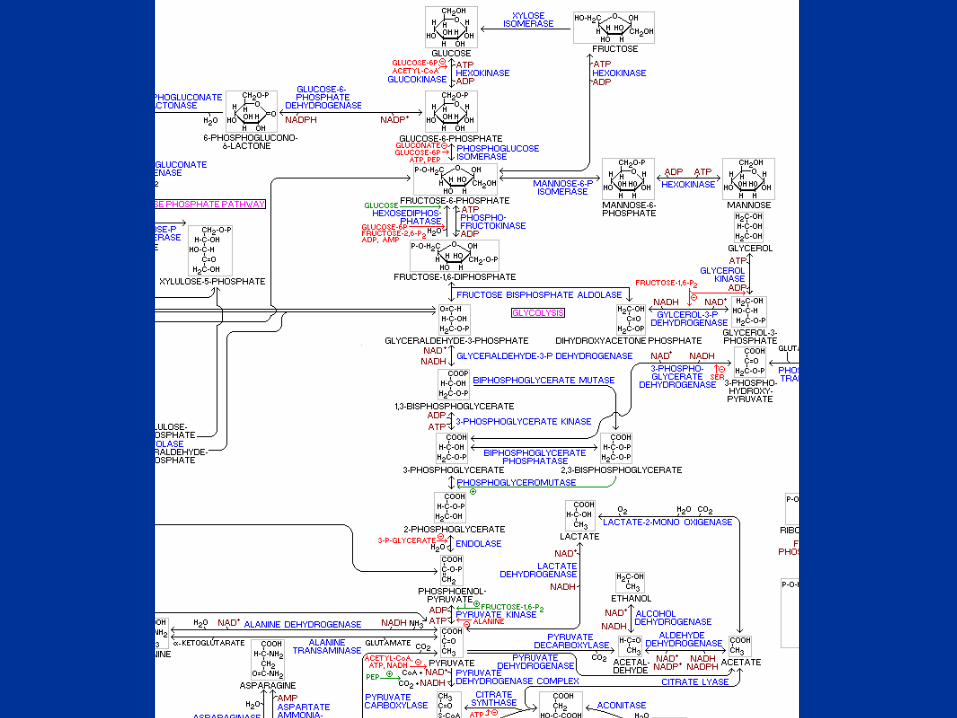
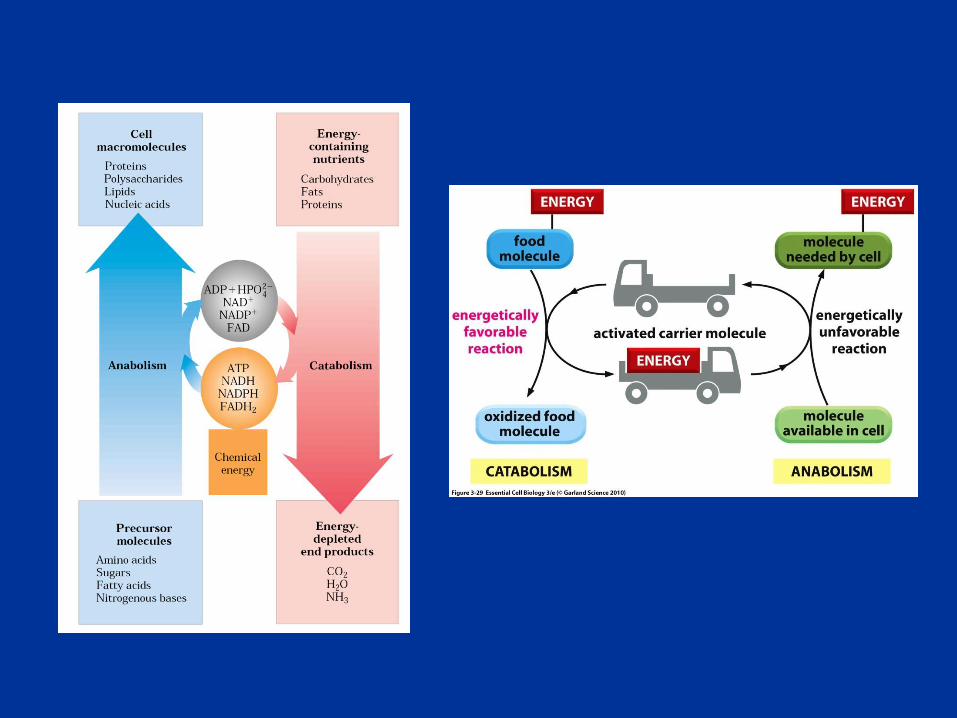

Oxidaciones y Generación de Energía Celular
Durante el metabolismo celular se producen oxidaciones de los sustratos
metabólicos (con la concomitante reducción de intermediarios) y estas
reacciones se utilizan para obtener energía.
Un compuesto que se oxida cede electrones (reductor)
Un compuesto que se reduce recibe electrones (oxidante)
Ejemplo: Fe2+ + Cu2+ Fe3+ + Cu1+
Hay dos semi reacciones:
Fe2+ Fe3+ + 1e- oxidación
Cu2+ + 1e- Cu1+ reducción
Al igual que los ácidos y las bases, siempre que hay una oxidación (perdida de
electrones) debe haber una reducción (ganancia de electrones).
¿Quién se lleva los electrones?
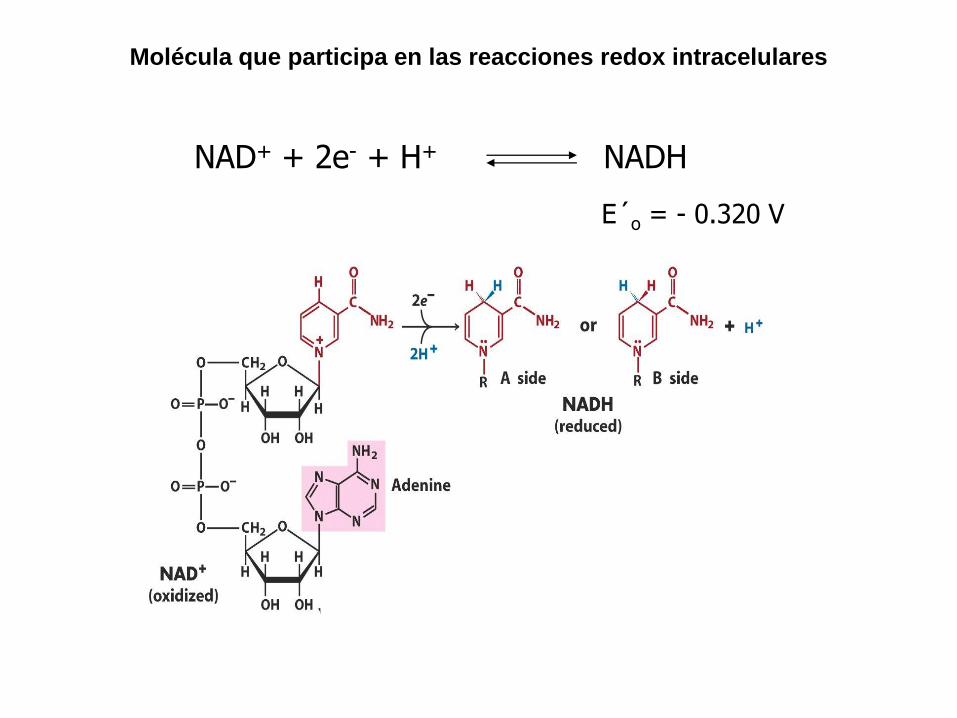
NAD+ + 2e- + H+ NADH
E´o = - 0.320 V
Molécula que participa en las reacciones redox intracelulares
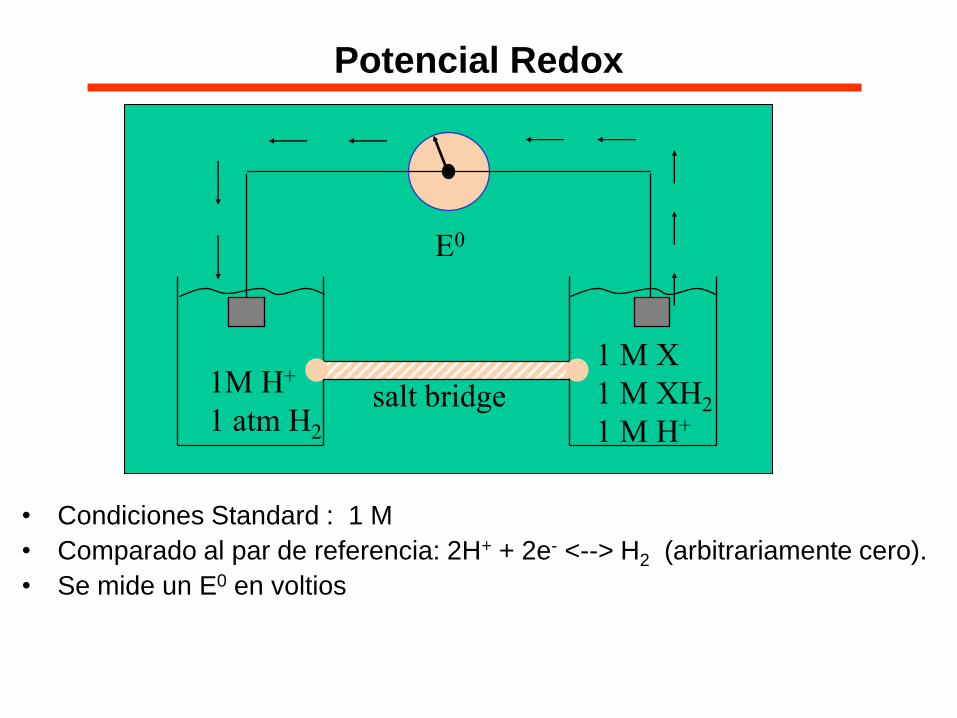
• Condiciones Standard : 1 M
• Comparado al par de referencia: 2H+ + 2e- <--> H2 (arbitrariamente cero).
• Se mide un E0 en voltios
1M H+
1 atm H2
1 M X
1 M XH2
1 M H+ salt bridge
E0
Potencial Redox

¿Quién se lleva los electrones?
A diferencia del G,
cuanto más grande y positivo
el potencial redox (Eo), mayor la
tendencia a aceptar electrones
(actuar como agente oxidante).
Hay una relación directa entre el
potencial redox y la energía libre:
G0 = -n F E0
Donde
E0 = Eo(aceptor)- Eo(donador)
n = número de electrones
F constante de Faraday
96.5 kJ/mol.V
Un valor de E positivo generará
valores de G negativos
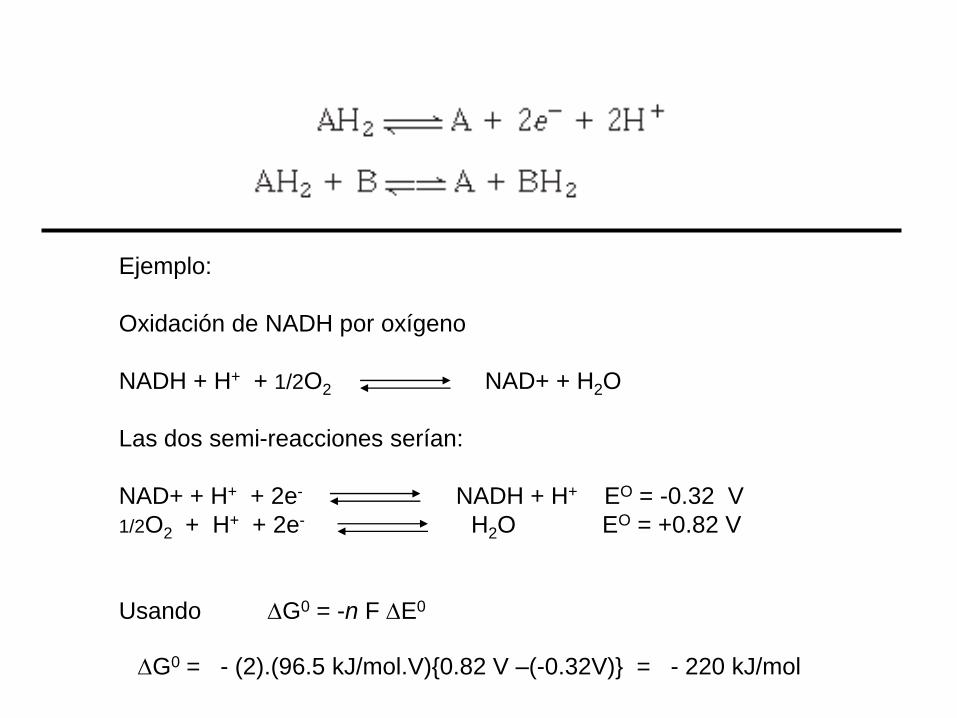
Ejemplo:
Oxidación de NADH por oxígeno
NADH + H+ + 1/2O2 NAD+ + H2O
Las dos semi-reacciones serían:
NAD+ + H+ + 2e- NADH + H+ EO = -0.32 V
1/2O2 + H+ + 2e- H2O EO = +0.82 V
Usando G0 = -n F E0
G0 = - (2).(96.5 kJ/mol.V){0.82 V –(-0.32V)} = - 220 kJ/mol

A. Propiedades generales de las enzimas
B. Principios fundamentales de su acción catalítica
C. Introducción a la cinética enzimática
D. Enzimas reguladores
Enzimas

Propiedades generales de las enzimas
1. Son los catalizadores de las reacciones químicas en
los sistemas biológicos
2. Aceleran muchísimo la velocidad de las reacciones
(106 – 1014 veces).
3. Poseen un elevado grado de especificidad de
sustrato
4. La actividad catalítica depende de la integridad de la
estructura nativa así como del pH y temperatura.

La mayoría de las enzimas son proteínas:
• La función depende de la integridad de la
conformación proteica nativa
• Existen enzimas que son proteínas simples y otras
que requieren componentes químicos adicionales:
- Cofactores: - iones inorgánicos (Fe2+, Mg2+, Mn2+ o Zn2+)
- complejos orgánicos o metaloorgánicos
(coenzimas)
Los cofactores unidos covalentemente: grupos prostéticos
• Holoenzima / Apoenzima
Propiedades generales de las enzimas

Propiedades generales de las enzimas
Las enzimas se clasifican según la reacción
catalizada: • Nomenclatura:
- Número clasificatorio de 4 dígitos (E.C.)
- Nombre sistemático
- Nombre trivial
ATP + D-Glucosa ADP + D-Glucosa-fosfato
Número clasificatorio: E.C. 2.7.1.1. (E.C: Enzyme Commission of the IUB:
International Union of Biiochemistry)
2. Clase: Transferasa
7. Subclase: Fosfotransferasa
1. Fosfotransferasas con OH como aceptor
1. D-glucosa como aceptor del fosfato
Nombre sistemático: ATP:glucosa fosfotransferasa
Nombre trivial: hexoquinasa
Nomenclatura: se adiciona sufijo “asa” al nombre del sustrato o de la reacción que cataliza


A. Propiedades generales de las enzimas
B. Principios fundamentales de su acción catalítica
C. Introducción a la cinética enzimática
E. Enzimas reguladores
Enzimas
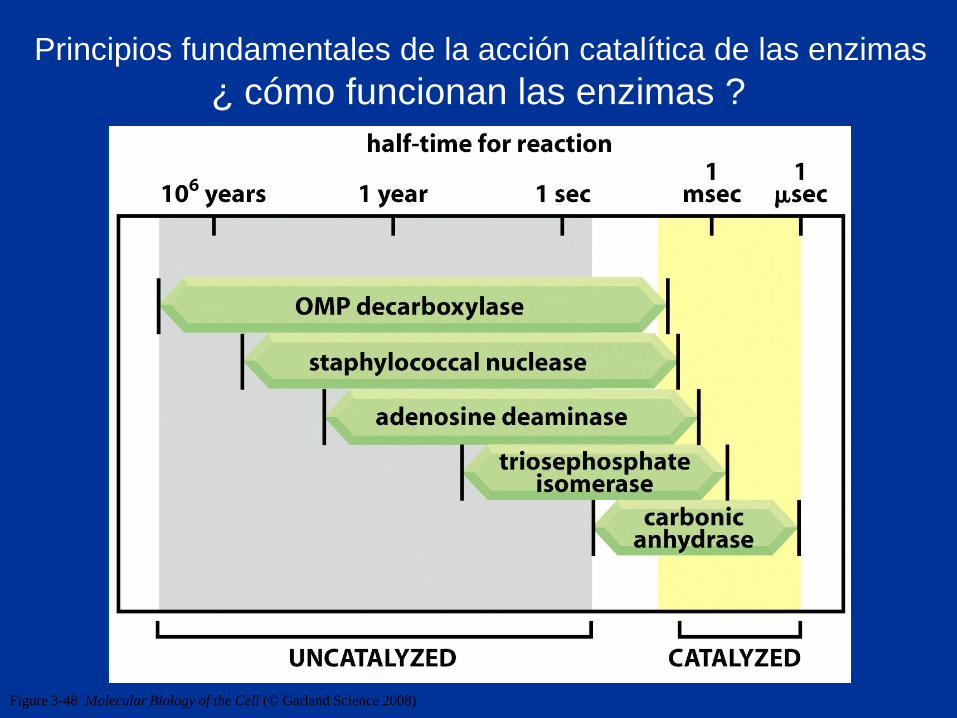
Figure 3-48 Molecular Biology of the Cell (© Garland Science 2008)
¿ cómo funcionan las enzimas ?
Principios fundamentales de la acción catalítica de las enzimas
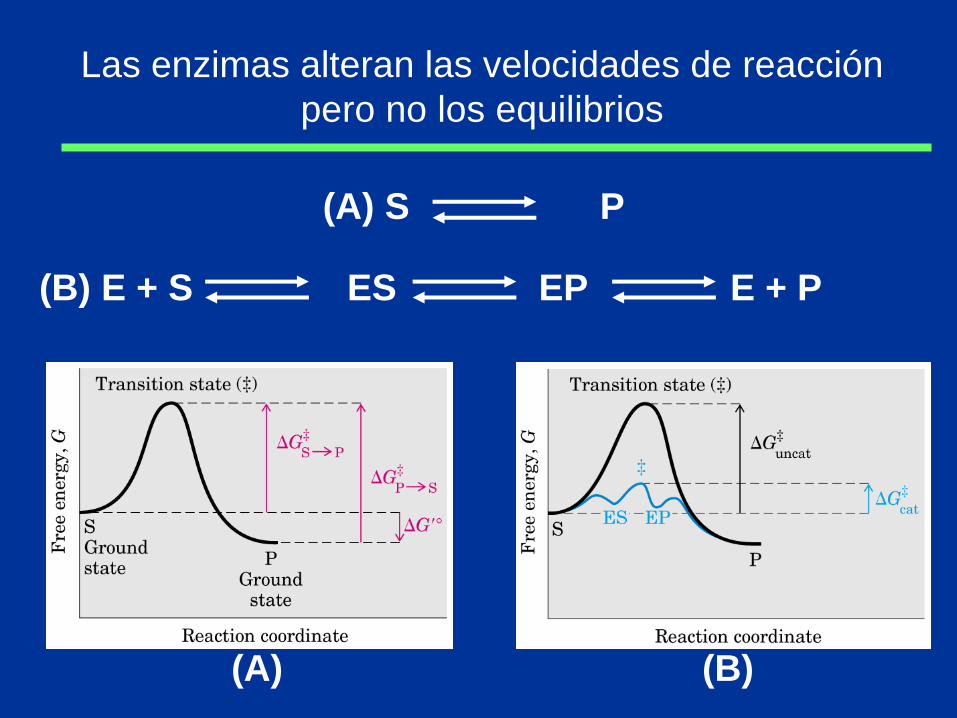
Las enzimas alteran las velocidades de reacción
pero no los equilibrios
(B) E + S ES EP E + P
(A) S P
(A) (B)

Las velocidades de reacción y los equilibrios
tienen definiciones termodinámicas precisas
• Equilibrio de reacción depende ΔG’o
• Velocidad de reacción depende de ΔG‡
S P Keq’ = [P]
[S] G’o = - RT ln Keq’
V = k [S] k = h
e - G‡ / RT KT
K = constante de Boltzmann
h = constante de Planck
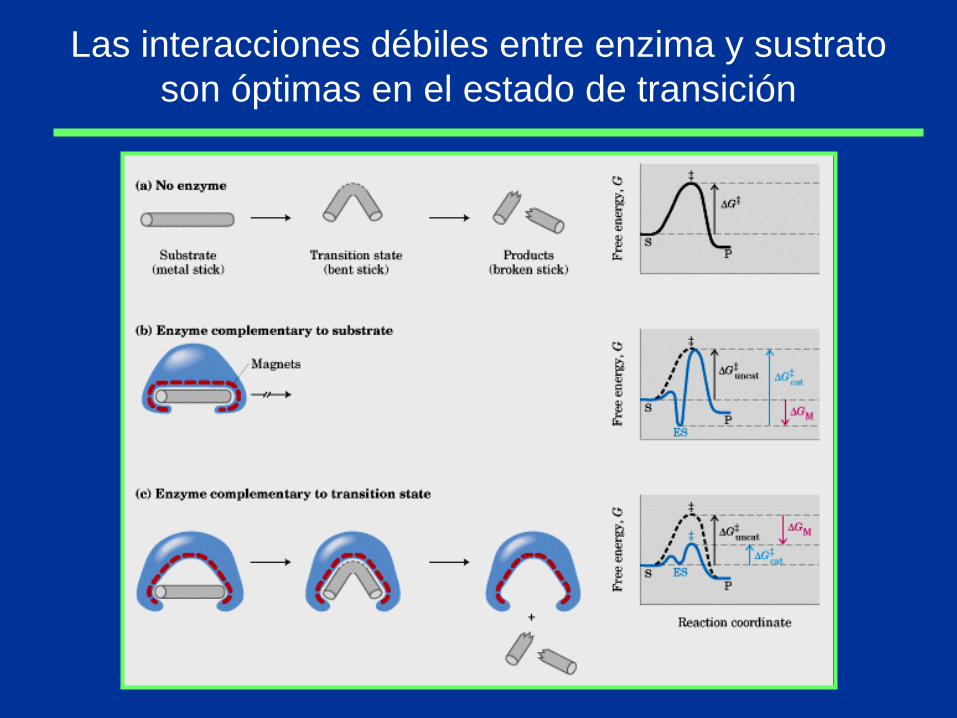
Las interacciones débiles entre enzima y sustrato
son óptimas en el estado de transición

A. Propiedades generales de las enzimas
B. Principios fundamentales de su acción catalítica
C. Introducción a la cinética enzimática
D. Enzimas reguladores
Enzimas

El modelo de Michaelis-Menten (1913)
Leonor Michaelis Maud Menten
Postularon que la enzima se combina en primer lugar
con el sustrato, de forma reversible
El complejo se descompone en una reacción más
lenta, dando lugar al producto y enzima libre

Cinética del estado estacionario
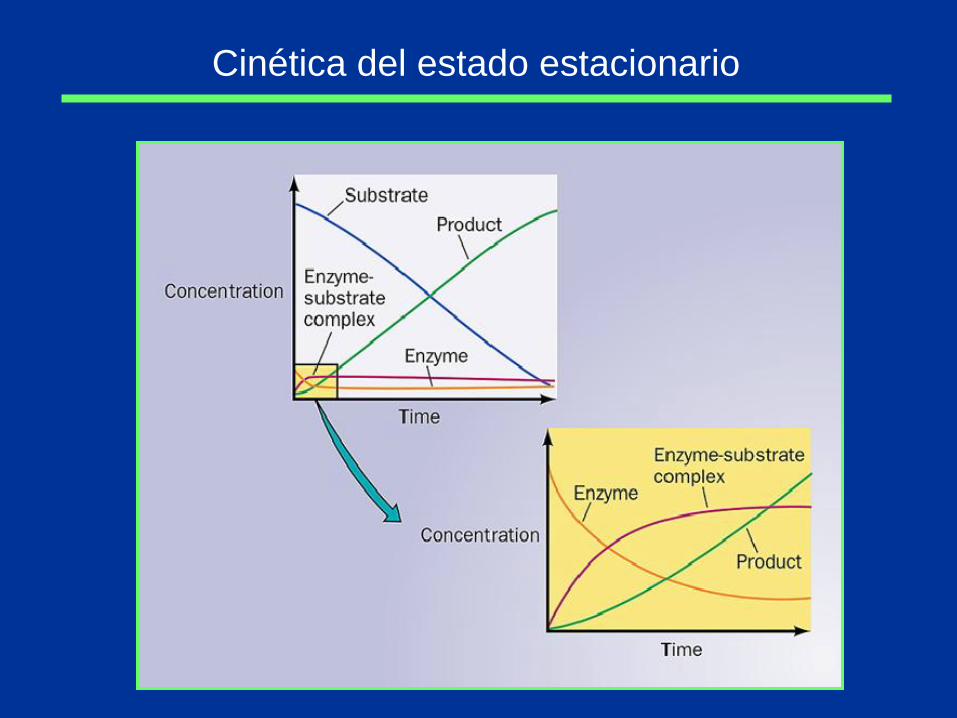
Cinética del estado estacionario

Cinética del estado estacionario
d[ES] = k1 [E][S] dt
- d[ES] = k-1 [ES] + k2[ES]
dt
E + S ↔ ES → E + P k1
k-1
k2

ET = EL + ES
EL = ET - ES
E + S ↔ ES → E + P
Formación de ES: k1[EL][S]
Descomposición de ES: k-1 [ES] + k2 [ES]
k1[EL][S] = k-1 [ES] + k2 [ES]
k-1+ k2 = [EL][S]
k1 [ES]

k-1+ k2 = [ET - ES][S]
k1 [ES] KM =
Constante de Michaelis:
Constante de disociación del complejo ES:
k-1 = [EL][S]
k1 [ES] Ks =

Relación entre concentración de sustrato y
velocidad de reacción enzimática
La velocidad inicial de la reacción siempre
corresponde a la ecuación:
vo = k2 [ES]
Cuando toda la enzima se encuentra formando
complejo ES:
vmax = k2 [ET]

Relación entre concentración de sustrato y
velocidad de reacción enzimática
k1[EL][S] = k-1 [ES] + k2 [ES]
k1[ET - ES][S] = k-1 [ES] + k2 [ES]
k1[ET][S] – k1[ES][S] = k-1 [ES] + k2 [ES]
k1[ET] [S] = k-1 [ES] + k2 [ES] + k1[ES][S]
k1[ET] [S] = [ES] (k-1 + k2 + k1[S])
Dividimos por k1: [ET] [S] = [ES] ((k-1 + k2 ) + [S])
k1
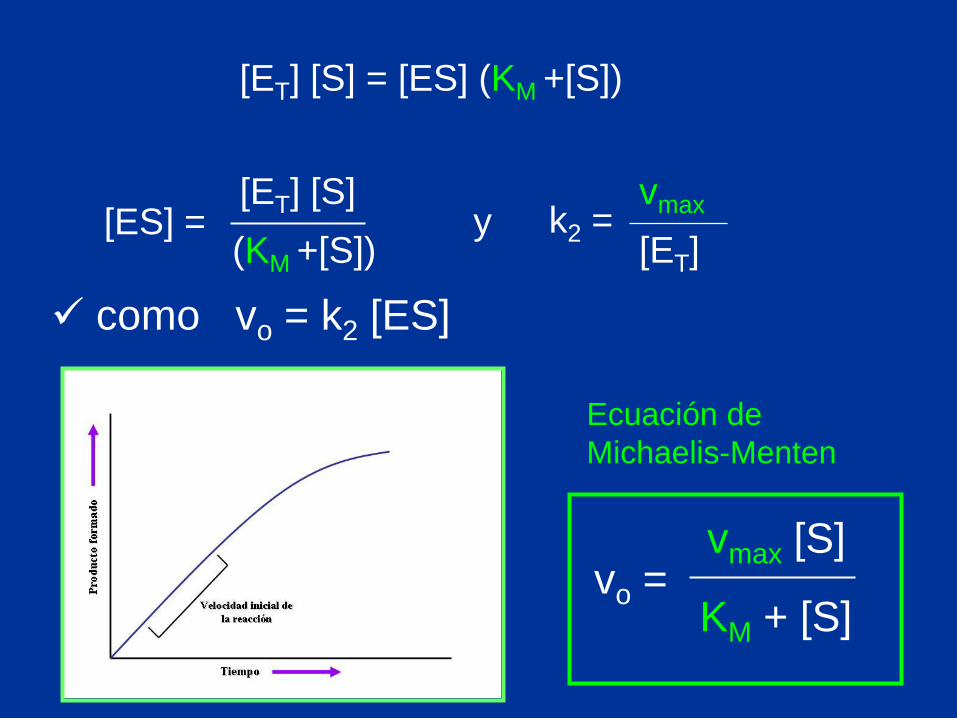
[ET] [S] = [ES] (KM +[S])
(KM +[S])
[ET] [S] [ES] = y
[ET]
vmax k2 =
como vo = k2 [ES]
vo = vmax [S]
KM + [S]
Ecuación de
Michaelis-Menten
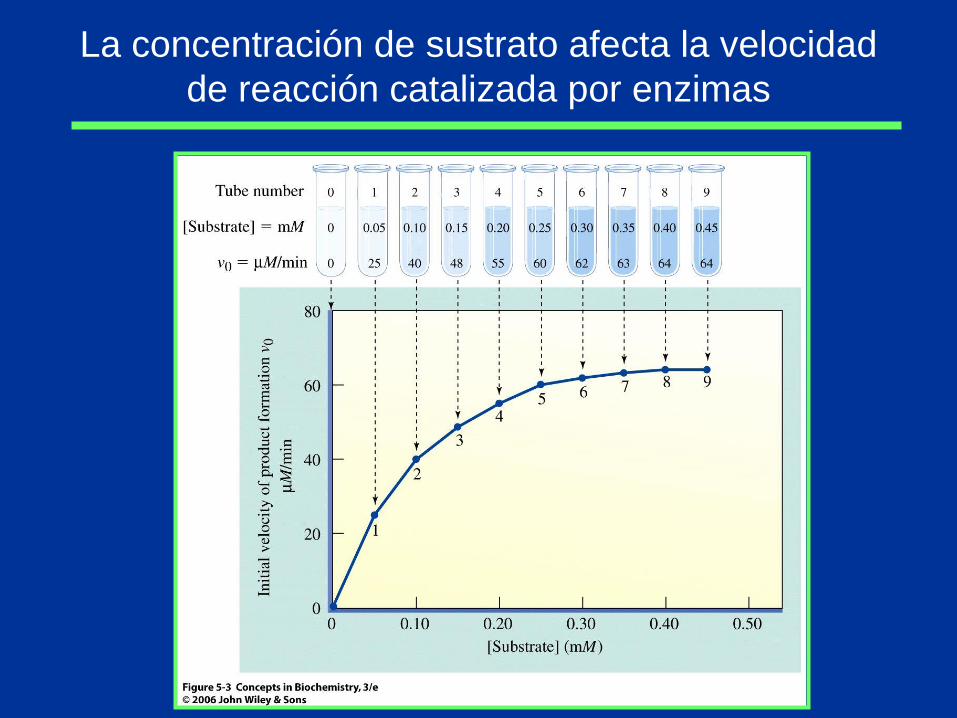
La concentración de sustrato afecta la velocidad
de reacción catalizada por enzimas
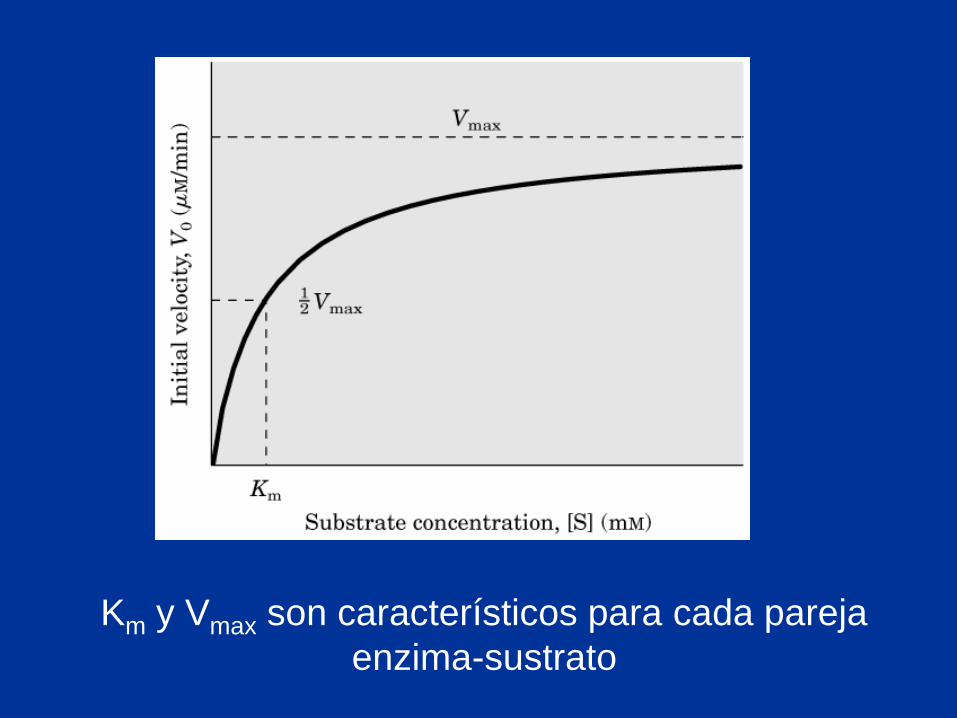
Km y Vmax son característicos para cada pareja
enzima-sustrato
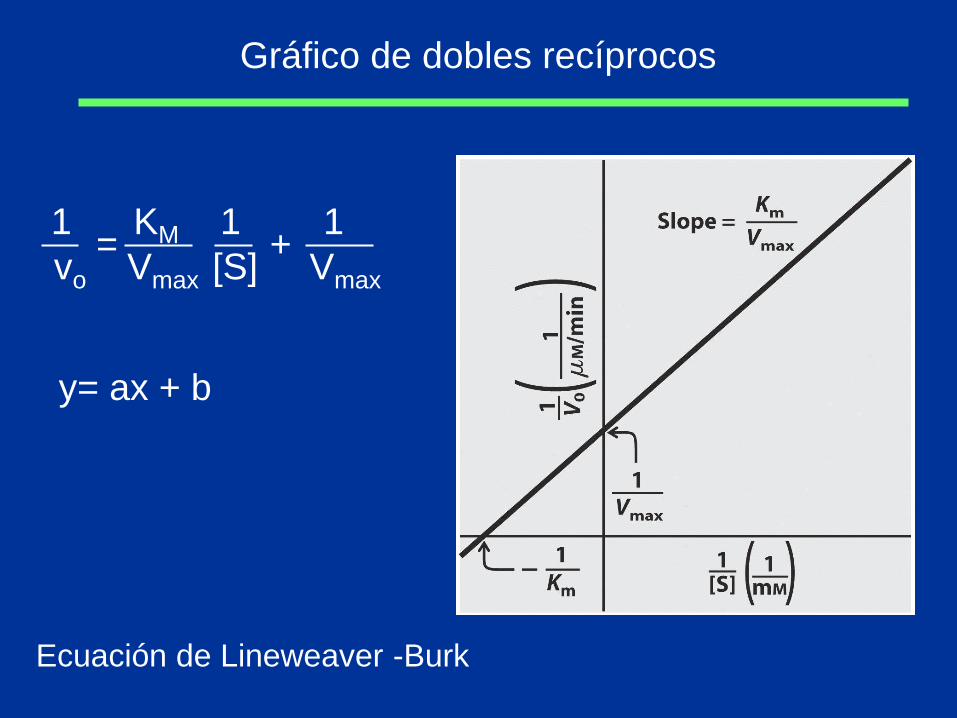
Gráfico de dobles recíprocos
1 KM 1 1
vo Vmax [S] Vmax
= +
Ecuación de Lineweaver -Burk
y= ax + b

El Nº de recambio es el número de moléculas
de sustrato convertidas en producto por una
molécula de enzima, cuando la enzima está
saturada.
Si k2 es la constante del paso limitante de
reacción entonces:
k2 = kcat o Número de recambio
1/kcat es el tiempo que dura un ciclo catalítico.

Muchas enzimas catalizan reacciones con dos
o más sustratos
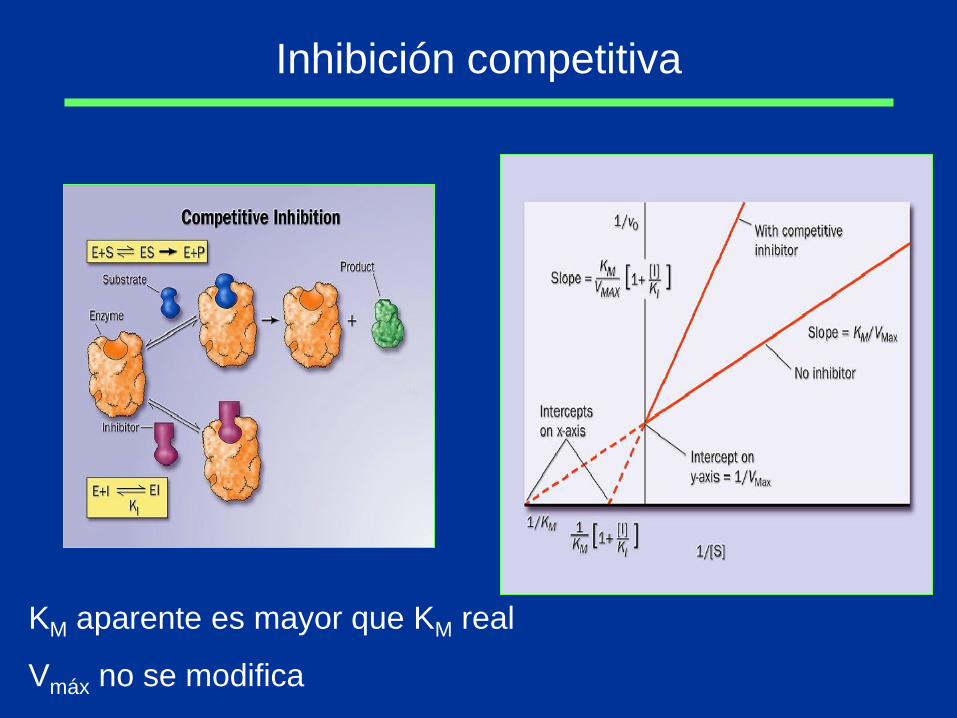
Inhibición competitiva
KM aparente es mayor que KM real
Vmáx no se modifica
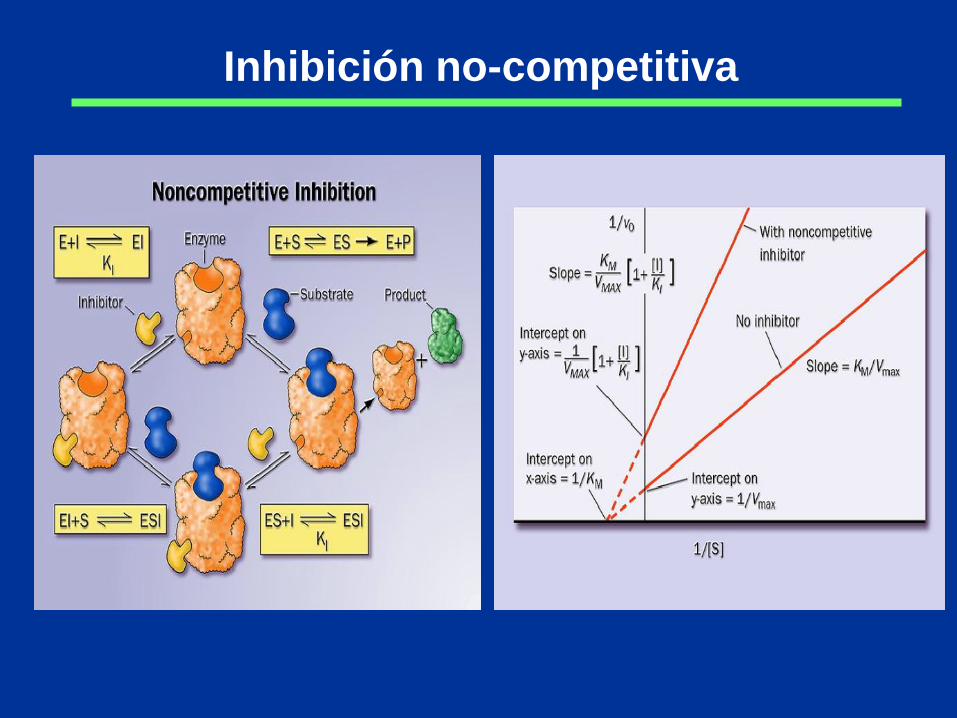
Inhibición no-competitiva
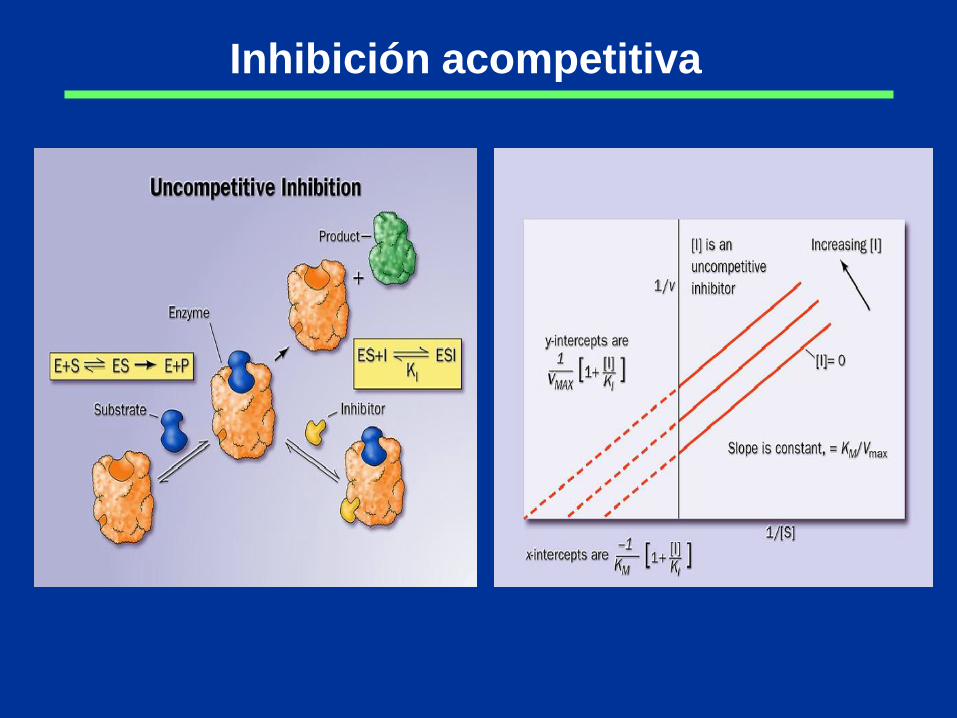
Inhibición acompetitiva
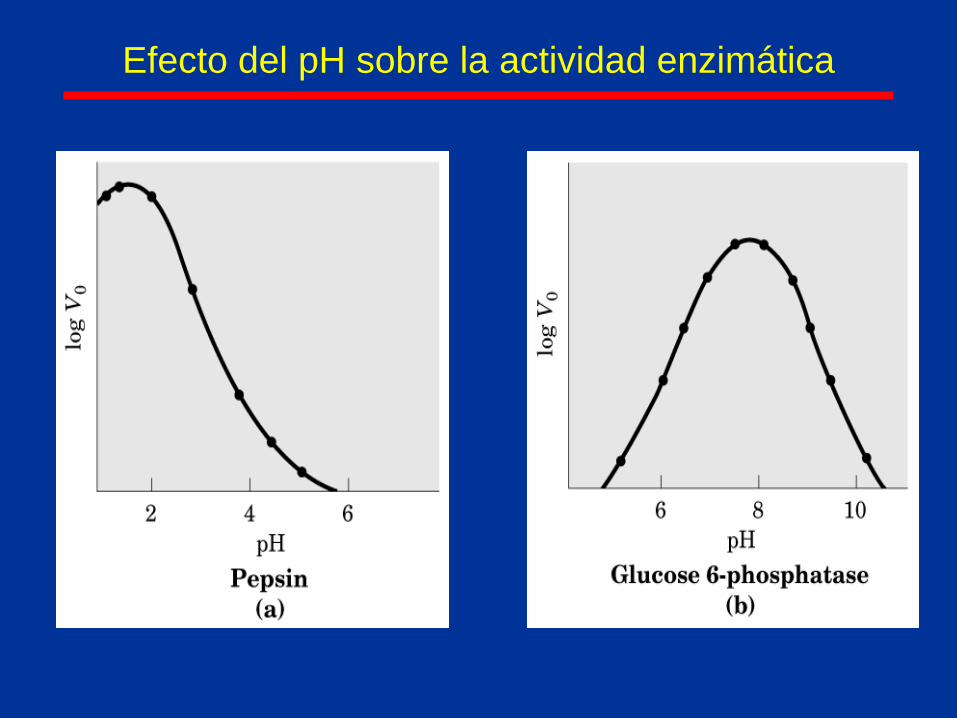
Efecto del pH sobre la actividad enzimática

A. Propiedades generales de las enzimas
B. Principios fundamentales de su acción catalítica
C. Introducción a la cinética enzimática
D. Enzimas reguladoras
Enzimas
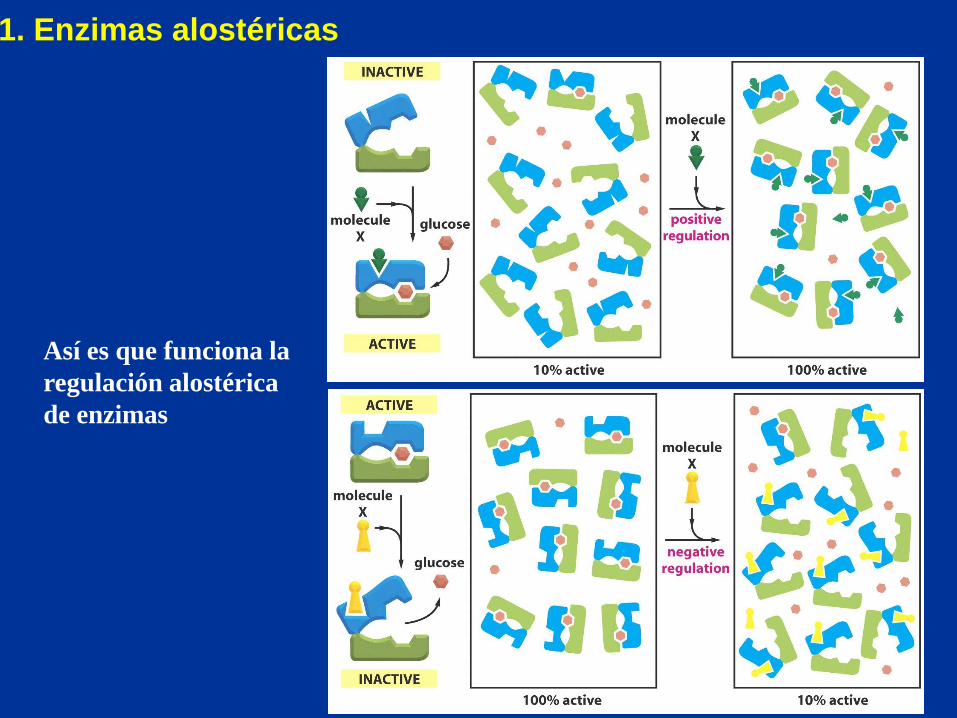
Así es que funciona la
regulación alostérica
de enzimas
1. Enzimas alostéricas

Enzimas reguladoras
2. Modulación covalente reversible






![2da clase[1]](https://static.fdocuments.mx/doc/165x107/559f31a31a28aba6168b47ca/2da-clase1.jpg)









![2da clase[1]2](https://static.fdocuments.mx/doc/165x107/5497af77b479596e058b49c7/2da-clase12.jpg)

