Celulas exitables clase 3
49
1 Células Excitables Clase 3 Ernesto Cristina
-
Upload
biologia-intermedio-1c-ipa -
Category
Education
-
view
465 -
download
1
Transcript of Celulas exitables clase 3

1
Células Excitables
Clase 3Ernesto Cristina

2

3
4

5
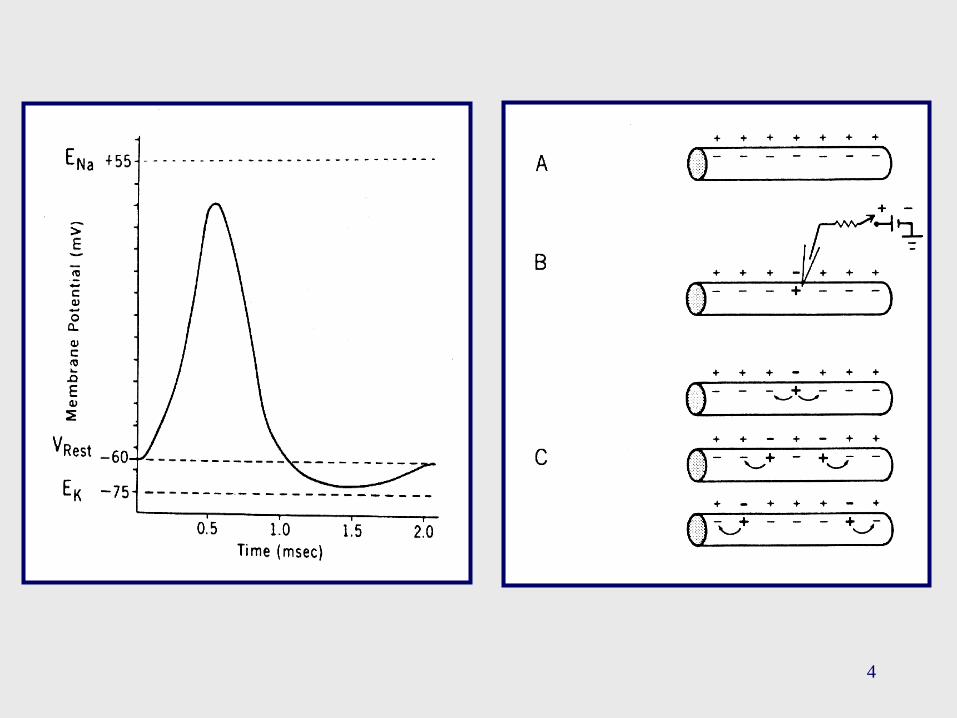
Una observación importante acerca del PA es que el
cambio en el PM atraviesa una región que se halla limitada por
ENa en uno de sus extremos, y por EK en el otro extremo.

6
• Problema: cuando se alcanza el umbral durante una
despolarización, y se dispara el PA, se generan
corrientes a través de la membrana. Como consecuencia,
resulta muy difícil poder medir el cambio en la corriente
resultante, debido a que todo esto ocurre muy
rápidamente.
• Este fue el principal obstáculo que enfrentaron los
investigadores de la época para lograr el posterior
análisis de los mecanismos iónicos que gobiernan la
generación y propagación del PA.

7
• Hodgkin, Huxley, y sus colegas, perfeccionaron un
dispositivo que les permitió estabilizar el PM en diferentes
valores de voltaje, por prolongados períodos de tiempo.
Emplearon un dispositivo electrónico de retroalimentación
que denominaron “voltage clamp amplifier” (controlador o
regulador de voltaje).
• Este dispositivo toma la diferencia entre el registro actual del
PM y el valor deseado (valor al cual se quiere llevar el PM),
y genera una corriente hiperpolarizante o despolarizante,
según sea el caso, para minimizar la diferencia. La cantidad
de corriente necesaria para mantener el PM en el valor
deseado representa la corriente que atraviesa a la membrana,
para ese “step” (paso) específico de voltaje.

8
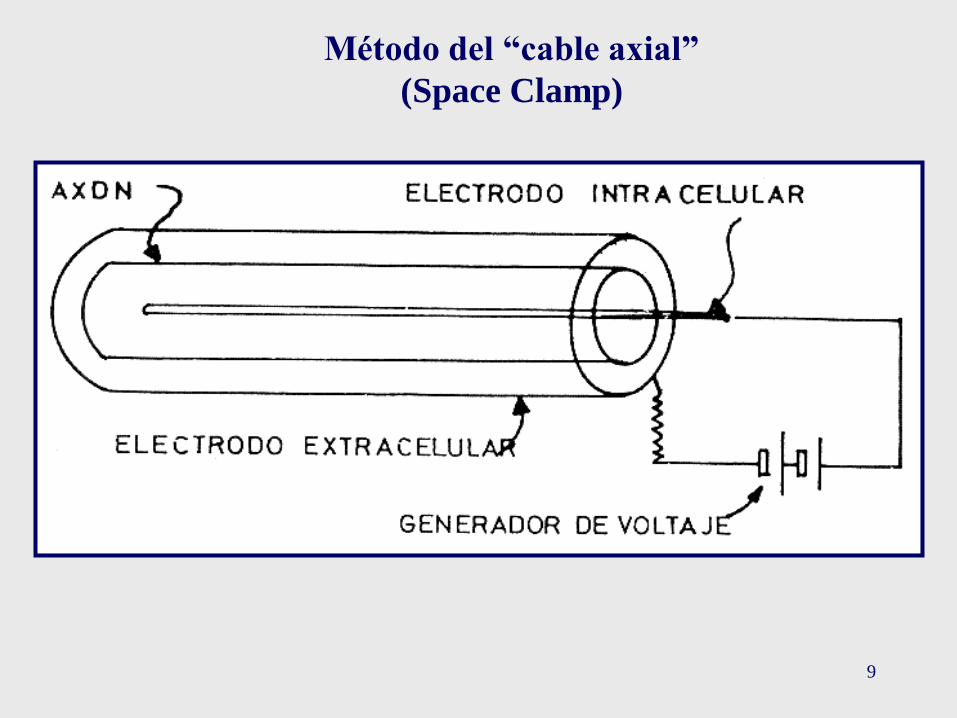
9
Método del “cable axial”
(Space Clamp)

10
Registros de la corriente de membrana medidos en el AGC
(PM controlado mediante la técnica de “voltage clamp”)
(T = 3.8 ºC)

11
• La técnica de “voltage clamp” ofreció por primera vez la
posibilidad de medir las corrientes iónicas que atraviesan las
membranas de las células excitables, ante los diferentes valores
de potencial eléctrico impuestos por el experimentador.
• Hodgkin y Huxley (HH) (1952) desarrollaron una serie de
experimentos para poder determinar cuáles eran los iones que
transportaban estas corrientes, y de qué forma los cambios en la
permeabilidad de la membrana se encontraban relacionados con
las mismas.
• Como se trataba de un área de investigación totalmente
novedosa, debieron formular nuevas estrategias para dilucidar
los mecanismos involucrados en la generación del PA.

12
Hipótesis formuladas por HH
Hipótesis 1. Asumieron que los iones involucrados en lacorriente total que atraviesa a la membrana del axón duranteun PA se desplazaban pasivamente a favor de su gradienteelectroquímico. En consecuencia, basándose en argumentostermodinámicos se podría predecir si el movimiento de un iónespecífico ocurrirá hacia el interior o hacia el exterior de lacélula.
Por ejemplo: corrientes transportadas por iones Na+ ocurriríanhacia el interior celular a PM menores a ENa, y hacia elexterior celular a PM mayores a ENa. A un PM igual a ENa, nohabría ninguna contribución de este ión a la corriente total queatravesaría la membrana.
Argumentos similares se aplicarían a otros iones (K+, Cl-,Ca2+, etc.), teniendo en consideración sus respectivospotenciales de equilibrio.

13
Corrientes registradas a partir de la técnica de “voltage clamp” para
distintos “steps” de voltaje
(T = 6.6 ºC)

14
Hipótesis 2. Iones pueden ser removidos de la soluciónexterna en la que se halla el axón, por lo tanto, la remoción deiones específicos en esta solución podría ocasionar cambiosen la corriente total que atraviesa a la membrana, para un“step” de voltaje establecido mediante la técnica de “voltageclamp”.
Por ejemplo: considerando una situación extrema, si un iónpermeable se sustituye completamente por uno no permeable,alguno de los componentes de la corriente total deberíadesaparecer, si este ión transporta parte de esa corriente.
La identificación de INa fue confirmada mediante elreemplazo del cloruro de sodio del medio externo porcloruro de colina, en experimentos de “voltage clamp” comolos descritos anteriormente.
Nota: 10 años después (1961- 1962) de los experimentos deHH, se desarrollaron métodos para modificar lasconcentraciones de los iones a nivel intracelular.
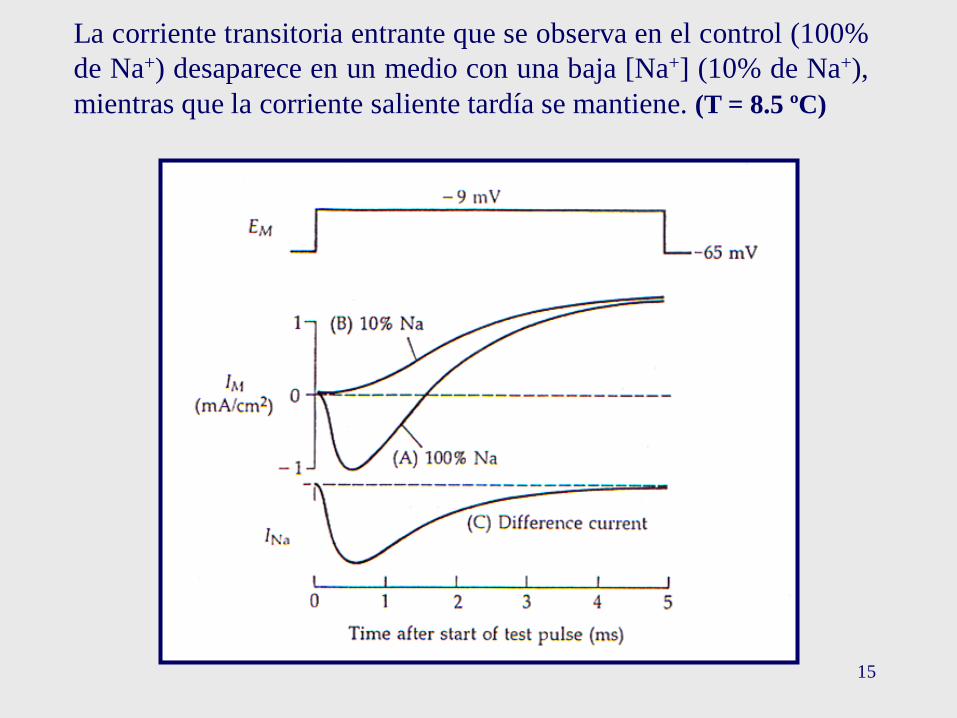
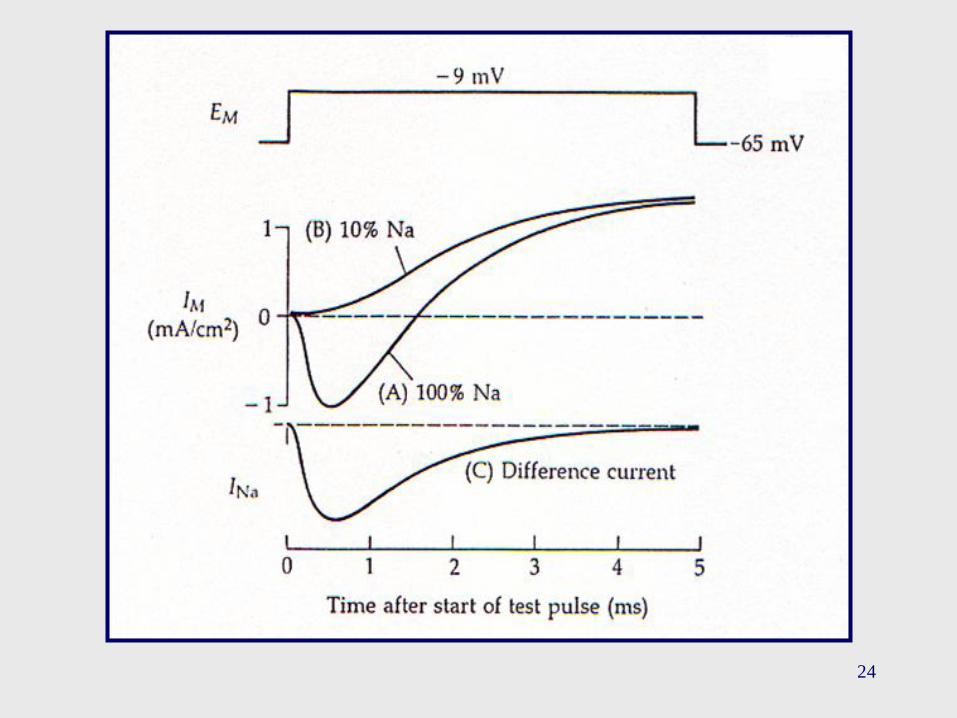
15
La corriente transitoria entrante que se observa en el control (100%
de Na+) desaparece en un medio con una baja [Na+] (10% de Na+),
mientras que la corriente saliente tardía se mantiene. (T = 8.5 ºC)

16
• Aunque HH no intentaron alterar las concentraciones externa o
interna de K+, investigaciones posteriores permitieron identificar
la corriente saliente tardía con el ión K+.
• Por lo tanto, el registro de corriente obtenido en un medio con
una baja [Na+] corresponde casi completamente a IK.

17
Componente Entrante de la corriente total:
Corriente de Na+.
Componente Saliente de la corriente total:
Corriente de K+.

18
• Hodgkin y sus colegas sugirieron, en base a los resultados
obtenidos, que durante la fase inicial de un PA la membrana se
vuelve selectivamente permeable al Na+. Es decir, la membrana
pasa de un estado de alta permeabilidad al K+ (condiciones de
“reposo”), hacia un estado de alta permeabilidad al Na+.
• Pregunta: si la membrana se vuelve altamente permeable al Na+,
¿qué valor de Vm se podría predecir?
• Respuesta: el valor de Vm se aproximará al valor de ENa
(aproximadamente +55 mV para el AGC).
ggEgEg
VKNa
KKNaNa
m

19
Pregunta: ¿Cómo puede ser que la
membrana sea altamente permeable al K+ en
un instante de tiempo y poco tiempo después,
como consecuencia de un estímulo
despolarizante, se vuelva altamente
permeable al Na+?
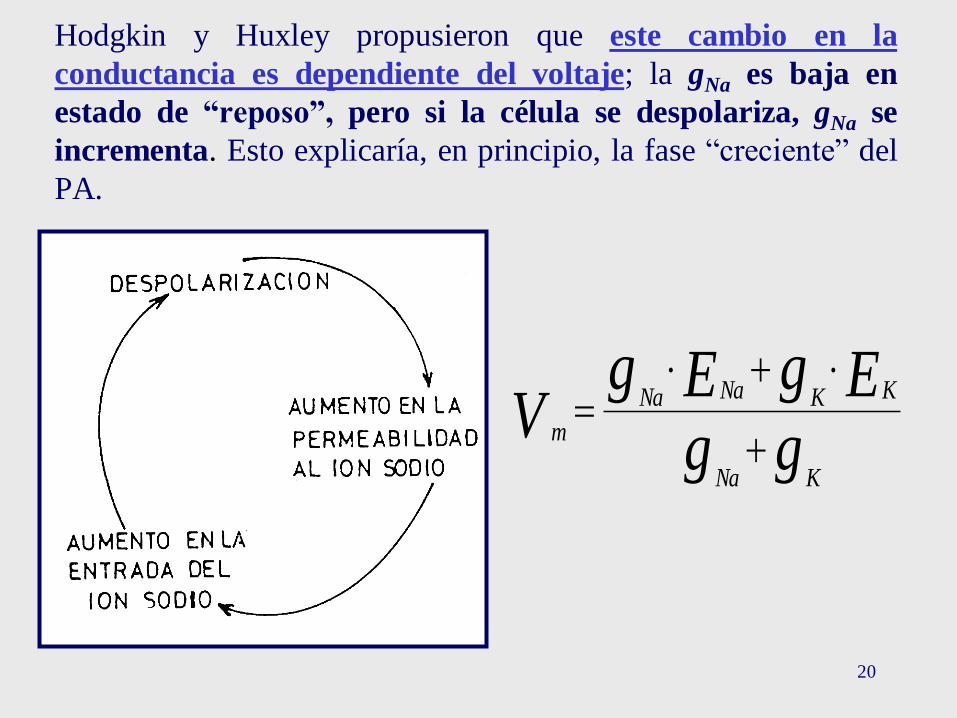
20
Hodgkin y Huxley propusieron que este cambio en la
conductancia es dependiente del voltaje; la gNa es baja en
estado de “reposo”, pero si la célula se despolariza, gNa se
incrementa. Esto explicaría, en principio, la fase “creciente” del
PA.
ggEgEg
VKNa
KKNaNa
m

21
• Hodgkin y sus colegas sugirieron, además, que durante un PA
también se produce un aumento en la permeabilidad de la
membrana al K+, fundamentalmente durante la fase de
repolarización.
• Pregunta: si la membrana se vuelve más permeable al K+, ¿qué
valor de Vm se podría predecir?
• Respuesta: el valor de Vm se aproximará al valor de EK
(aproximadamente -75 mV para el AGC).
ggEgEg
VKNa
KKNaNa
m

22
Pregunta: ¿Cómo se podría verificar la
dependencia de la conductancia de Na+, y la
de K+, con respecto al voltaje?

23
• Midiendo la corriente iónica en función del tiempo, I(t);
conociendo Vm (el cual se establece por “voltage clamp”), y
efectuando, por ejemplo, los experimentos de sustitución de Na+
extracelular, la conductancias de Na+ y de K+ en función del
tiempo, gNa (t) y gK (t), se pueden calcular simplemente aplicando
la Ley de Ohm (para corrientes iónicas):
• Cambiando Vm mediante el dispositivo de control de voltaje, se
pueden determinar las conductancias de Na+ y de K+
correspondientes a una gran variedad de potenciales de
membrana impuestos por el experimentador, y obtener de esta
manera, mediante el empleo de las ecuaciones anteriores, el
cambio en la conductancia de la membrana a estos iones ante
diferentes valores de potencial.
EV Nam
Na
Na
tt Ig
)()(
EV Km
K
K
tt Ig
)()(
24
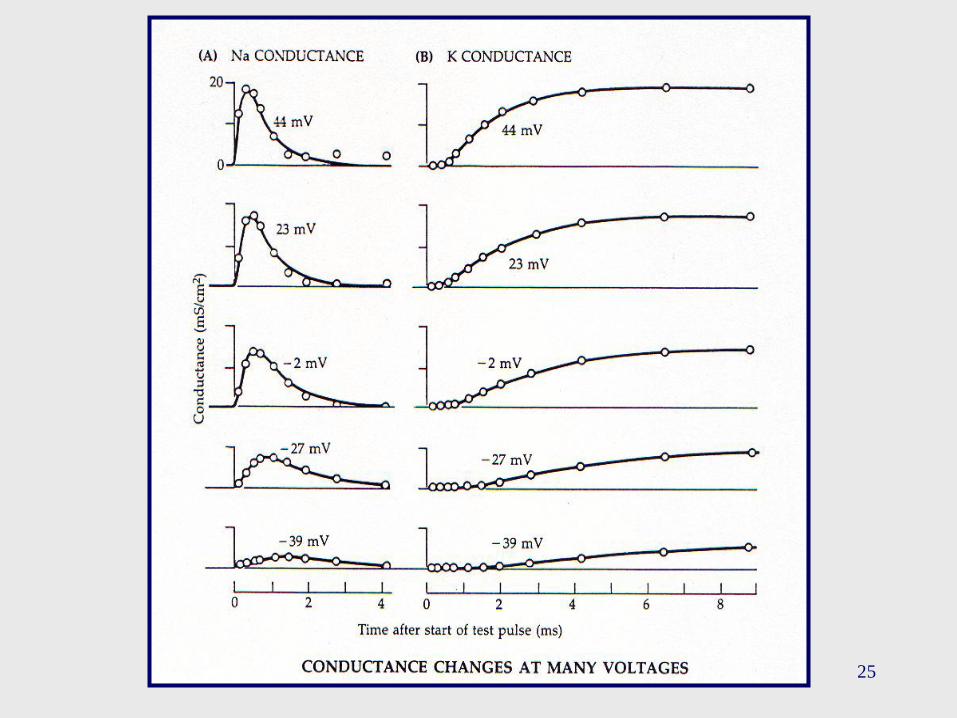
25
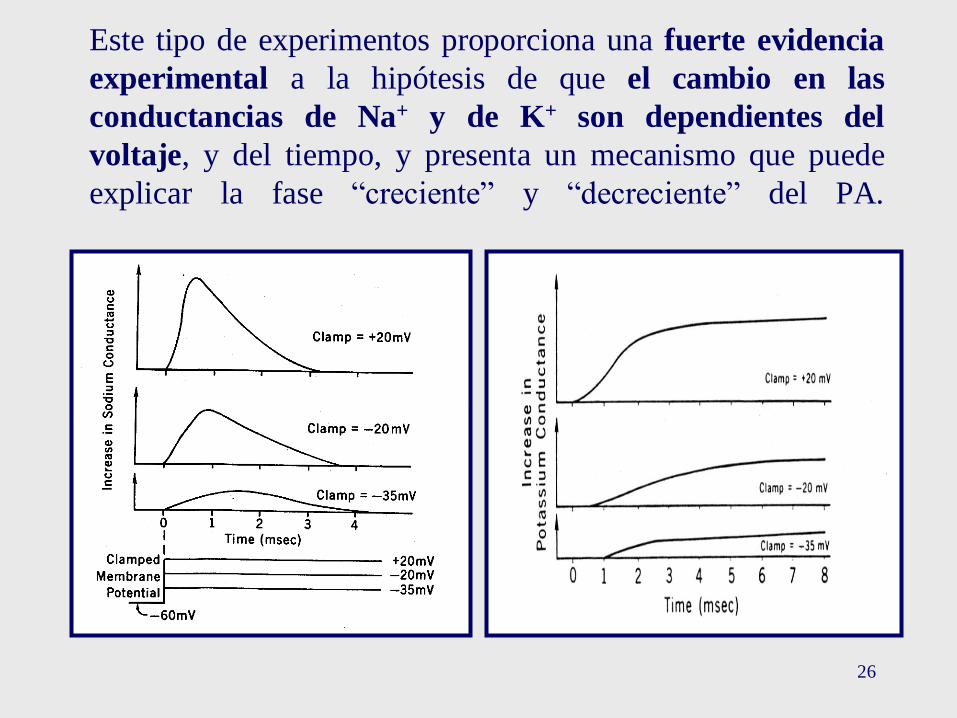
26
Este tipo de experimentos proporciona una fuerte evidencia
experimental a la hipótesis de que el cambio en las
conductancias de Na+ y de K+ son dependientes del
voltaje, y del tiempo, y presenta un mecanismo que puede
explicar la fase “creciente” y “decreciente” del PA.
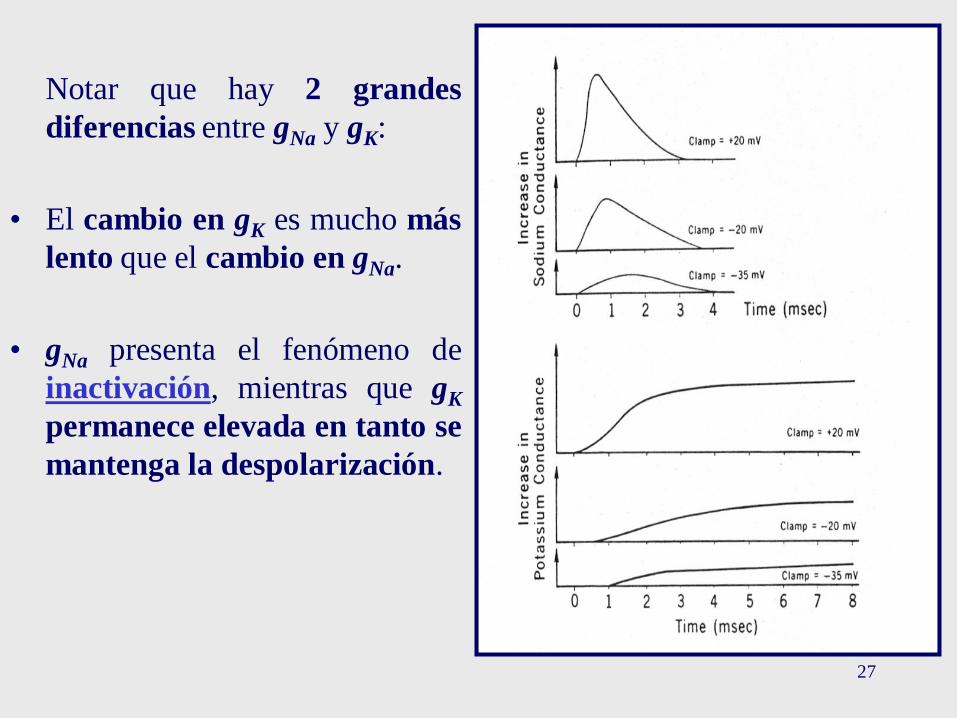
27
Notar que hay 2 grandes
diferencias entre gNa y gK:
• El cambio en gK es mucho más
lento que el cambio en gNa.
• gNa presenta el fenómeno de
inactivación, mientras que gK
permanece elevada en tanto se
mantenga la despolarización.

28
• Inactivación de gNa: aunque el PM permanece despolarizado durantetodo el tiempo que dura el estímulo, la gNa decae a su valor de reposo.El incremento en la gNa es sólo transitorio. A esto se lo conocecomo Inactivación.
• Actualmente resulta evidente que este flujo de Na+ hacia el interiorcelular, así como el flujo de K+ hacia el exterior celular, se verifican através de canales de Na+ y de K+ que presentan la particularidad deser sensibles al voltaje.
• En la molécula proteica existen estructuras cargadas capaces demovilizarse, según el campo eléctrico de la membrana. Los primerosbiofísicos que estudiaron este tipo de proteínas llamaron a estasestructuras “compuertas” (gates).
• En el caso concreto del canal de Na+ se puede pensar que posee dostipos de compuertas: de “activación” y de “inactivación”. En el casodel canal de K+ podemos asumir que presenta exclusivamentecompuertas de “activación”.

29

30
• Si la gNa decae, la despolarización se reducirá, y esta reducción
promoverá un descenso en el ingreso de Na+ a la célula. Como
resultado, un nuevo ciclo se inicia, tendiente a repolarizar a la
membrana.
• Aunque debemos notar lo siguiente:
1. La duración del PA es de aproximadamente 1 mseg., mientras
que el cambio en la gNa dura alrededor de 4 mseg.. Según esto,
el PA debería durar un período de tiempo mayor a 1 mseg..
2. Resulta difícil explicar el efecto de hiperpolarización que se
verifica en el PA, teniendo en cuenta exclusivamente a la
inactivación de Na+.
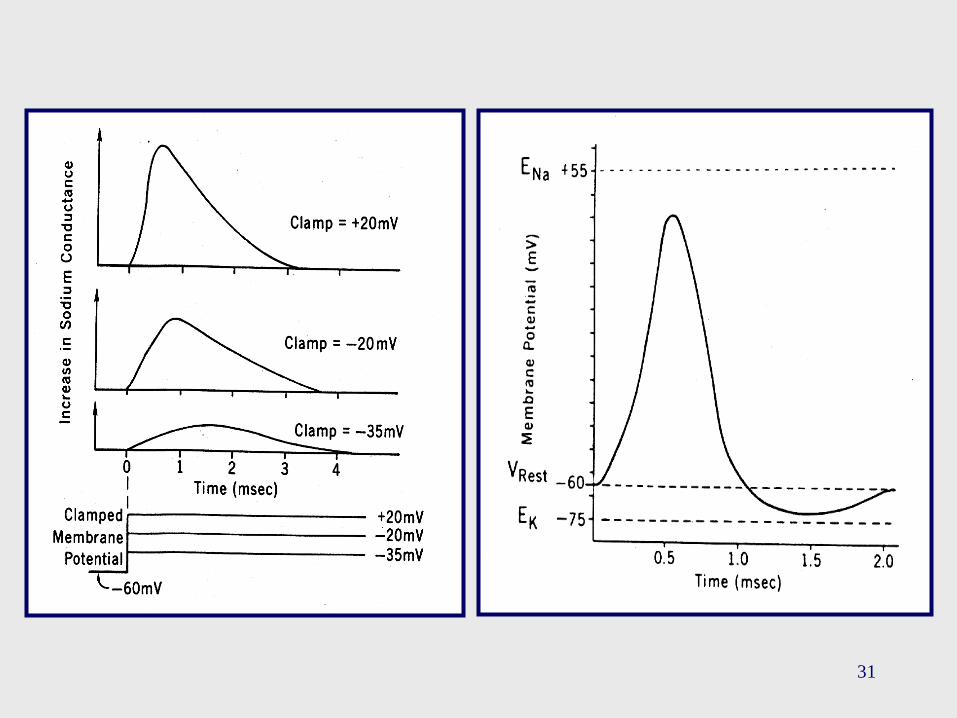
31
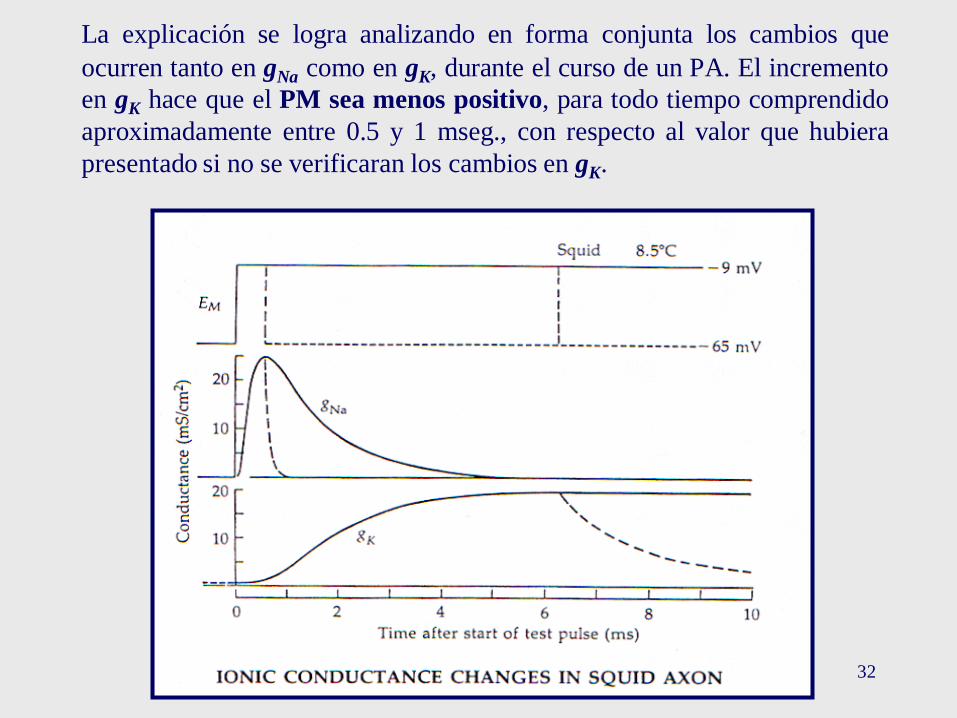
32
La explicación se logra analizando en forma conjunta los cambios que
ocurren tanto en gNa como en gK, durante el curso de un PA. El incremento
en gK hace que el PM sea menos positivo, para todo tiempo comprendido
aproximadamente entre 0.5 y 1 mseg., con respecto al valor que hubiera
presentado si no se verificaran los cambios en gK.

33
Los cambios en gK permiten que el PM se repolarice más rápidamente, ya
que ahora existen dos factores que contribuyen a la repolarización de la
membrana: 1) la inactivación de los canales de sodio, y 2) el incremento en
gK. Estos dos fenómenos explican la corta duración del PA.
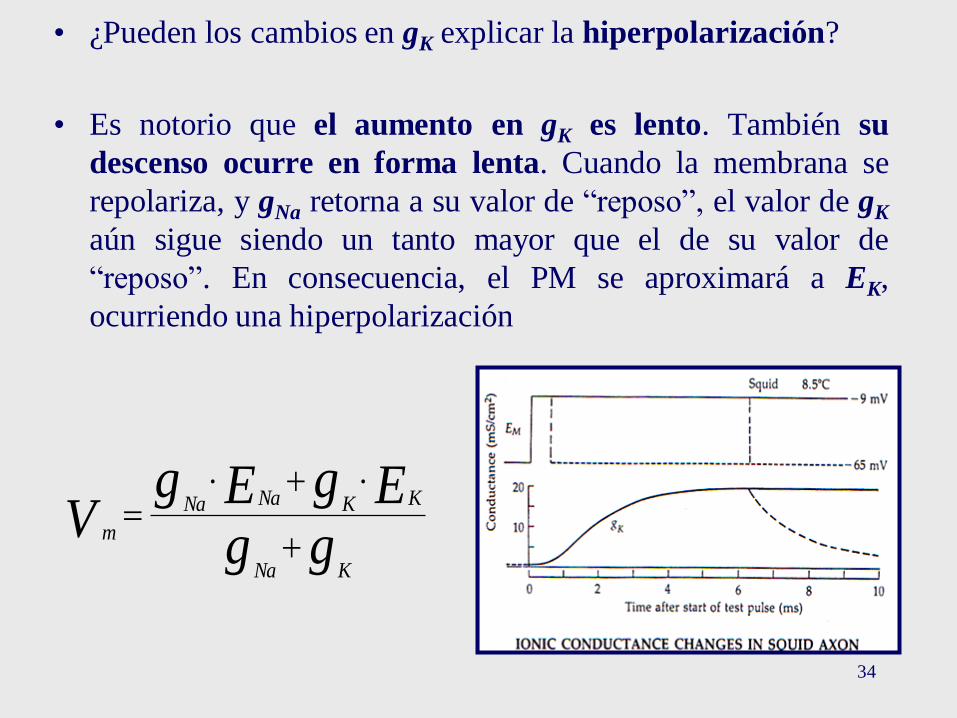
34
• ¿Pueden los cambios en gK explicar la hiperpolarización?
• Es notorio que el aumento en gK es lento. También su
descenso ocurre en forma lenta. Cuando la membrana se
repolariza, y gNa retorna a su valor de “reposo”, el valor de gK
aún sigue siendo un tanto mayor que el de su valor de
“reposo”. En consecuencia, el PM se aproximará a EK,
ocurriendo una hiperpolarización
ggEgEg
VKNa
KKNaNa
m

35
• Hodgkin y Huxley buscaron “testear” en forma más rigurosalos resultados obtenidos a partir de la técnica de “voltageclamp”. Para ello, desarrollaron un modelo matemático del PAbasado en las medidas experimentales de los cambios en lasconductancias a los iones Na+ y K+.
• Se preguntaron si un modelo matemático que considerara,exclusivamente, los cambios en las conductancias de los dosiones mencionados, sería capaz de “generar” un PA.
• En la época en la que HH desarrollaron sus investigaciones sedesconocía la existencia de los canales iónicos. Sin embargo,su trabajo teórico (modelo matemático) suponía la presenciade cierta clase de “partículas cargadas”, vinculadas a lamembrana, que debían encontrarse en una posición específica,en un tiempo dado, para que se produjera el cambio en laconductancia de la membrana.
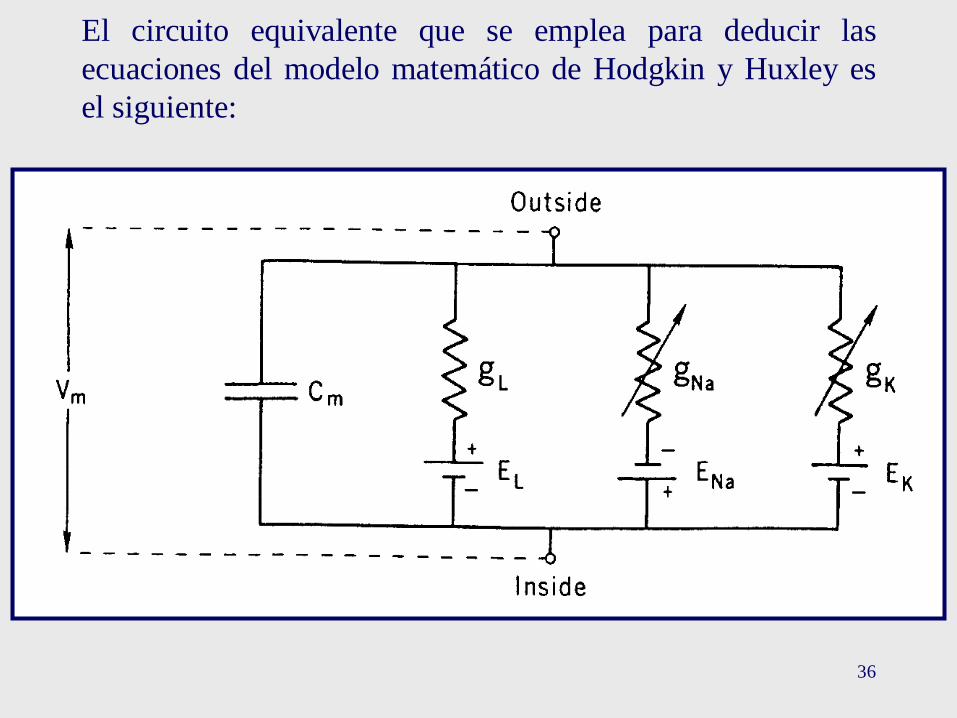
36
El circuito equivalente que se emplea para deducir las
ecuaciones del modelo matemático de Hodgkin y Huxley es
el siguiente:

37
Algunos de los resultados de la simulación de su modelo
se muestran en el siguiente gráfico:

38
• Como resultado de un PA el cambio en la [Na+]i por cm2
de área de membrana es de aproximadamente 1 pM, y ese
cambio se halla restringido a la superficie interna de la
membrana. Dicho cambio es despreciable en comparación
con la [Na+]i que se halla en el orden de mM. Algo similar
ocurre con el K+.
• Si la bomba de Na+ y K+ fuera “bloqueada” en el AGC,
sería posible iniciar más de 500.000 PA antes de que se
pudieran evidenciar cambios en el PM o en la amplitud del
PA (condiciones de laboratorio).
• En general, el rol de la bomba de Na+ y K+ se basa en
mantener los gradientes iónicos necesarios para que se
produzca esta señal electroquímica, pero no participa
directamente en el mecanismo que da lugar al PA.

39
• Umbral: se alcanza cuando el flujo entrante de Na+ supera alflujo saliente de K+, durante la despolarización.
• Período Refractario Absoluto: hace referencia al período detiempo (entre 0.5 - 1 mseg), luego de iniciado un PA, durante elcual no es posible generar un nuevo PA, independientementede la intensidad del estímulo despolarizante. Esto se debe a quese requiere un cierto número de canales de sodio que haya“salido” de su inactivación para poder iniciar un nuevo PA.
• Período Refractario Relativo: hace referencia al período detiempo, luego de iniciado un PA, durante el cual es posibleiniciar un nuevo PA, pero únicamente con un estímulo demayor intensidad con respecto al empleado para lainiciación del primer PA. En parte se explica teniendo encuenta que la membrana se encuentra hiperpolarizada.
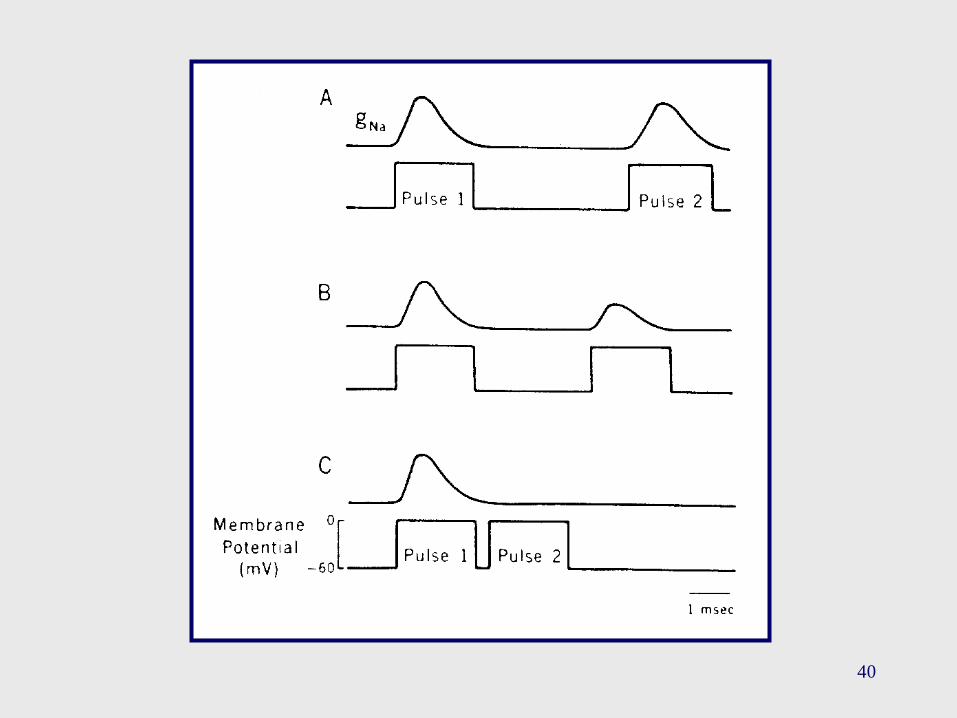
40

41
• Acomodación: se define como el cambio en el umbral de la
membrana de una célula excitable cuando una lenta
despolarización es aplicada. De hecho, si la despolarización es
lo suficientemente lenta ningún PA se generará. El proceso de
inactivación de Na+ contribuye a este fenómeno.

42
• Básicamente, un proceso de lenta despolarización provee de
suficiente tiempo para que los canales de Na+ pasen al estado
“abierto inactivado” antes de que se alcance el número necesario
de canales de Na+ en el estado abierto, para que la corriente
entrante de Na+ supere a la corriente saliente de K+. Es decir,
habría un número insuficiente de canales de Na+ con sus
compuertas de inactivación abiertas, ya que la mayoría de estos
canales se encontraría en el estado abierto pero “inactivado”.

43
Electrotono (respuesta “pasiva”) y PA
• Electrotono: 1) es graduado; si el pulso es de menor
amplitud, la respuesta es de menor tamaño; 2) decae con la
distancia.
• Potencial de Acción: 1) es de tipo “todo o nada”, y sus
características básicas, una vez generado, son independientes
de la amplitud del estímulo; 2) se propaga sin sufrir
modificación (sin decremento) a lo largo de grandes
distancias en las fibras axónicas.

44
Propagación del PA en axones con mielina

45
Toxinas
46

47
• Name: Alan Lloyd Hodgkin.
• Birth Date: February 5, 1914.
• Death Date: December 20, 1998.
• Nationality: English.
• Occupations: Physiologist and
Biophysicist.
• Name: Andrew Fielding Huxley.
• Birth Date: 1917.
• Nationality: English.
• Occupations: Physiologist and
Biophysicist.

48
Algunos datos relativos al AGC
• [Na+]i = 50 mM; [Na+]e = 440 mM.
• [K+]i = 400 mM; [K+]e = 20 mM.
• EK = -75 mV; ENa = +55 mV.
• Vrest = -60 mV.

49
Bibliografía consultada (de la cual se extrajeron las figuras de las tres clases)
• A quantitative description of membrane current and its application to conduction and excitation innerve. A. L. Hodgkin and A. F. Huxley. J. Physiol. (1952) 117, 500-544.
• An Introduction to Membrane Transport and Bioelectricity. Second Edition. John H. Byrne,Stanley G. Schultz. Raven Press. 1988.
• Biophysics of Computation. Information Processing in Single Neurons. Christof Koch. OxfordUniversity Press. 1998.
• Cellular Biophysics – Electrical Properties. Thomas Fischer Weiss. The MIT Press. Cambridge,Massachusetts. London, England. 1996.
• Foundations of Cellular Neurophysiology. Daniel Johnston and Miao-Sin Wu. MIT. 1994.
• Ionic Channels of Excitable Membranes. Second Edition. Bertil Hille. SINAUER ASSOCIATESINC. Publishers Sunderland, Massachusetts. 1992.
• Medical Physiology, 14th Edition. Vernon B. Mountcastle. The C. V. Mosby Company. 1979.
• Methods in Neuronal Modeling. Edited by Christof Koch and Idan Segev. MIT. 1999.
• Transporte y Excitabilidad. Eduardo Ríos. División Publicaciones y Ediciones, Facultad deOdontología, UdelaR. 1983.