
BIÓLOGO MARINO - biblio.uabcs.mxbiblio.uabcs.mx/tesis/te2840.pdf · Cromatografía en columna:...
59
UNIVERSIDAD AUTÓNOMA DE BAJA CALIFORNIA SUR ÁREA DE CONOCIMIENTO DE CIENCIAS DEL MAR DEPARTAMENTO ACADEMICO DE BIOLOGÍA MARINA TESIS Efecto in vitro del extracto etanólico de Macrocystis pyrifera sobre el desarrollo de microepífitos. QUE COMO REQUISITO PARA OBTENER EL TÍTULO DE BIÓLOGO MARINO PRESENTA: Lucero Gómez Alcalá DIRECTOR: Dr. Iván Murillo Álvarez LA PAZ, BAJA CALIFORNIA SUR, DICIEMBRE DEL 2012
Transcript of BIÓLOGO MARINO - biblio.uabcs.mxbiblio.uabcs.mx/tesis/te2840.pdf · Cromatografía en columna:...

UNIVERSIDAD AUTÓNOMA DE BAJA CALIFORNIA SUR
ÁREA DE CONOCIMIENTO DE CIENCIAS DEL MAR
DEPARTAMENTO ACADEMICO DE BIOLOGÍA MARINA
TESIS
Efecto in vitro del extracto etanólico de Macrocystis pyrifera sobre el
desarrollo de microepífitos.
QUE COMO REQUISITO PARA OBTENER EL TÍTULO DE
BIÓLOGO MARINO
PRESENTA:
Lucero Gómez Alcalá
DIRECTOR:
Dr. Iván Murillo Álvarez
LA PAZ, BAJA CALIFORNIA SUR, DICIEMBRE DEL 2012


Agradecimientos
A mi comité revisor, Iván Murillo, Mauricio Muñoz, Francisco Nieto, Juan Manuel López y
Rafael Riosmena agradezco la oportunidad que me dieron de trabajar con ustedes, el tiempo que
dedicaron, el basto conocimiento que me compartieron, la cordialidad y su amistad. Aprovecho
también para agradecer por los trabajos de Lorena Durán y Gaël Quesseveur, los cuales fueron la
base primordial para elaborar este trabajo. Del mismo modo, agradezco a todo aquel que
compartió su conocimiento, experiencia, apoyo y amistad para terminar este proyecto que es el
inicio de algo mayor.
GRACIAS

Índice
Lista de Figuras…………………………………………………………………………………………… II
Lista de Tablas………………………………………………………………………………………........ II
Glosario…………………………………………………………………………………………………….. III
Resumen…………………………………………………………………………………………………… VII
1. Introducción…..………………………………………………………………………………………… 1
2. Antecedentes…..………………………………………………………………………………………. 5
3. Distribución y descripción de Macrocystis pyrifera…………..…………………………………...... 10
4. Justificación…..………………………………………………………………………………………… 14
5. Objetivo……..…………………………………………………………………………………………... 15
6. Metodología...………………………………………………………………………………………….. 15
6.1 Zona de estudio y recolecta de Macrocystis pyrifera…………………………………………….. 15
6.2 Obtención del extracto etanólico……………………………………………………………………. 16
6.3 Fraccionamiento inicial del extracto etanólico…………………………………………………….. 17
6.4 Evaluación de la actividad antibacteriana y anti-incrustante..…………………………………… 18
6.4.1 Ensayo de difusión en agar con discos………………………………………………………….. 18
6.4.2 Ensayo de microdilución en placa……………………………………………………………….. 20
6.5 Fraccionamiento de 06-019 41CC2F3……………………………………………………………... 22
6.6 Identificación estructural del compuesto activo………………………………………………….... 23
7. Resultados…..………………………………………………………………………………………..... 25
7.1 Actividad biológica………………………………………………………………………………….... 25
7.2 Caracterización estructural………………………………………………………………………….. 30
8. Discusión…………..…………………………………………………………………………………… 31
9. Conclusiones…………………………………………………………………………………………… 37
10. Literatura citada……………………………………………………………………………………… 38
11. Anexos………………………………………………………………………………………………… 49

II
LISTA DE FIGURAS
Figura 1. Macrocystis pyrifera. Al norte, se distribuye sobre las costas del Pacífico de
Norteamérica, México, EU y Alaska. Al sur tiene una distribución circunsubantártica
desde Perú hasta Cabo de Hornos en América, Sudáfrica, Australia y Nueva Zelanda……
11
Figura 2. Punta Eugenia, BCS. Sitio de colecta de M. pyrifera (mapa de Durán-Riveron,
2009).……..…………………………………………………………………………………………..
16
Figura 3. Fraccionamiento guiado por bioensayo de la fracción 06-019 41CC2 F3
derivada de la fase orgánica del extracto de M. pyrifera……………………………………….
24
Figura 4. Espectro infrarrojo de la fracción 06-019 41CC3F2. Se observan cuatro señales
principales en la zona general del espectro y pequeñas señales en la zona de la huella
dactilar, entre 600m-1
y 1400m-1
…………………...................................................................
31
LISTA DE TABLAS
Tabla I. Actividad anti-incrustante de M. pyrifera, método de dilución en
microplacas…………………………………………………………………………………
27
Tabla II. Efecto de M. pyrifera sobre el crecimiento microalgal, método de
microdilución en placa…………………………………………………………………….
28
Tabla III. Actividad antibacteriana de M. pyrifera, método de difusión en agar……. 29

III
GLOSARIO
Ácidos grasos: son biomoléculas de naturaleza lipídica que consisten en un grupo
carboxilo (polar) unido a una cadena hidrocarbonada lineal con número par de átomos
de carbono. La mayoría de los ácidos grasos encontrados en la naturaleza contienen
entre 12-24 átomos de carbono. La principal función de los ácidos grasos es como
reserva de energía.
Agar: es un polisacárido compuesto de agarosa (70%) y agaropectina, que se extrae
de algunas algas rojas. Los geles de agar son transparentes, duros y quebradizos,
sirven como coloides y espesantes, y son un valioso medio de cultivo utilizado en
bacteriología.
Agar Mueller Hinton: es un medio utilizado para realizar las pruebas de
susceptibilidad antimicrobiana en microorganismos aeróbicos. El Agar Mueller Hinton
cumple con los requerimientos de la Organización Mundial de la Salud y está
especificado en la FDA (Food and Drug Administration). Este medio es el
seleccionado por la NCCLS (National Commitee for Clinical Laboratory Standards)
para realizar las pruebas de susceptibilidad, por su alta reproducibilidad, su bajo
contenido de substancias inhibidoras y el crecimiento satisfactorio que presentan la
mayoría de los patógenos.
Anti-incrustante: se refiere a las sustancias que tienen actividad contra organismos
involucrados en el epifitismo.
Biocidas: definidos por la legislación Europea actual, como: “las sustancias activas y
preparados que contengan una o más sustancias activas, presentados en la forma en
que son suministrados al usuario, destinados a destruir, contrarrestar, neutralizar,

IV
impedir la acción o ejercer un control de otro tipo sobre cualquier organismo nocivo
por medios químicos o biológicos.”
Biofouling: deterioro o degradación ocasionada por organismos vivos (bacterias,
algas, protozoos y crustáceos) sobre superficies tales como tuberías, tanques y
cascos de buques que se encuentran sumergidas en el agua, lo que resulta en
corrosión, obstrucción, contaminación o disminución en la eficiencia de piezas
móviles.
Bosques de kelps: son ecosistemas marinos, uno de los hábitat más productivos del
mar que ofrecen protección y alimento a una gran cantidad de organismos. Destaca la
alta tasa de crecimiento del género Macrocystis y la especie Nereocystis luetkeana.
Cromatografía: es el método físico de separación de los componentes de una
muestra. Es un conjunto de técnicas basadas en el principio de retención selectiva,
donde los componentes se distribuyen entre dos fases, una estacionaria y otra móvil.
Los componentes de la mezcla interaccionan en distinta forma con la fase
estacionaria, lo que hace que atraviesen esta fase a distintas velocidades y así se
separan los componentes. La fase estacionaria puede ser un sólido o un líquido y
puede estar empaquetada en una columna, extendida en una capa o distribuida como
una película.
Cromatografía en columna: donde la fase estacionaria se sitúa dentro de una
columna, y según el fluido empleado, puede ser cromatografía de líquidos o gases.
Cromatografía plana: donde la fase estacionaria se sitúa sobre una placa plana o
sobre papel (en papel o capa fina).

V
Epifitismo: forma de vida en donde un organismo vegetal, el epífito, utiliza como
sustrato para vivir a otro organismo vegetal. Este término se deriva de epi (sobre) y fito
(vegetal).
Epibiosis: es la relación, permanente o no, que se establece entre dos especies
diferentes en la que una sirve de sustrato de fijación para la otra, el epibionte.
También se aplica el concepto como ensuciamiento biológico, o bien, asentamiento de
algún organismo en cualquier estructura sumergida en el agua.
Espectrometría de infrarrojo: La espectrometría de infrarrojos (espectroscopia IV) es
un tipo de espectrometría de absorción que utiliza la región infrarroja del espectro
electromagnético.
Grupo funcional: Conjunto de átomos, enlazados de una determinada forma, que
presentan una estructura y propiedades físico-químicas determinadas que
caracterizan a los compuestos orgánicos que los contienen.
Metabolismo: es el conjunto de reacciones químicas que ocurren en el interior de la
célula.
Metabolismo primario: se refiere al conjunto de procesos metabólicos que
desempeñan una función esencial en el vegetal, tal como la fotosíntesis, respiración y
el transporte de solutos, los productos de este metabolismo (metabolitos primarios)
tienen una distribución universal en todas las plantas, como aminoácidos, nucleótidos,
lípidos, etc.
Metabolismo secundario: no es fundamental para el crecimiento, desarrollo o
reproducción del organismo, y sus productos, metabolitos secundarios, no poseen
una distribución universal, sin embargo se cree que la función principal es de
incrementar la probabilidad de la sobrevivencia de los organismos.

VI
Método de difusión en agar con disco: método generalmente utilizado y aprobado
por el NCCLS para efectuar las pruebas de sensibilidad.
Principios activos: es la sustancia responsable de la actividad biológica, ya sean
drogas, antibacterianos, etc. y regularmente derivan de recursos naturales para
emplearse en los fármacos, sin embargo existen principios activos sintéticos.
Productos naturales: compuestos producidos por los organismos vivos, son los
metabolitos secundarios, y se ha observado que tienen un valor terapéutico causando
algún efecto sobre los organismos vivos, actuando como antibióticos, antitumorales,
antivirales, insecticidas, sustancias citotóxicas y neurotóxicas, además de anti
incrustantes.
Protocolo de aislamiento guiado por bioensayo: metodología que consiste en el
fraccionamiento de una mezcla para aislar los principios activos, a partir de ensayos
de actividad biológica previos a cada fraccionamiento; de este modo se eligen
únicamente fracciones activas y/o las más activas.
Sensidiscos: son discos de papel filtro que se emplean para los ensayos de actividad
biológica.
Sílica: el gel de sílice es una forma granular y porosa de dióxido de silicio fabricado
sintéticamente a partir de silicato sódico. En química se usa en la cromatografía como
la fase estacionaria.

VII
Resumen
La epibiosis, o bien, el asentamiento de los organismos ya sea sobre otros organismos
o sobre estructuras creadas por el hombre, suelen tener efectos negativos, de modo
que en la acuacultura y en la transportación naval principalmente, se han valido del
conocimiento de los productos naturales que se obtienen a partir del metabolismo
secundario de algunas especies, principalmente marinos, para elaborar revestimientos
que eviten o disminuyan el encrustamiento de organismos epibiontes. Con el objetivo
de identificar metabolitos con actividad anti-incrustante en el alga parda Macrocystis
pyrifera se eligió la fracción de baja polaridad 06-019 41CC2F3 resultado del
fraccionamiento que realizó Durán-Riverol, la cual presentó actividad contra varias
cepas bacterianas. Para continuar con el fraccionamiento guiado por bioensayo, se
realizó la técnica de cromatografía en placas TLC y se estableció el sistema de
fraccionamiento para realizar la técnica de cromatografía en columna sobre sílica. En
total se realizaron dos fraccionamientos, se obtuvieron ocho fracciones y se determinó
que M. pyrifera puede inhibir el crecimiento de Staphylococcus aureus y Vibrio
parahaemolyticus, bacterias que están relacionadas en el microepifitismo. Mediante
espectroscopia de infrarrojo se analizó la fracción más activa y se determinó que está
compuesta por ácidos grasos.
Palabras clave: Ácidos grasos, Macrocystis pyrifera, actividad anti-incrustante,
productos naturales, cromatografía.

1
1. Introducción
El epifitismo se define como una forma de vida que ocurre cuando un
organismo, el epífito, utiliza como sustrato para vivir a otro organismo vegetal
(Borowitzka & Lethbridge, 1989). Se sabe que el epífito busca recursos tales como
espacio, refugio y alimento en los hospederos; la mayoría de las publicaciones
sugieren que estos organismos pueden tener una marcada y negativa influencia
en el alga hospedera al interferir en su susceptibilidad, en la esporulación o en la
captación de luz o nutrientes (Dixon et al., 1981; Scheibling et al., 1999). Esta
información ha hecho suponer que los epífitos tienen un efecto negativo en el
crecimiento, sobrevivencia y fecundidad del alga colonizada, cuando en realidad
no existen tales evidencias (Hepburn & Hurd, 2005). Por el contrario, se ha
demostrado una posible relación de mutuo beneficio, donde las algas proveen
protección y alimento (De Burg & Frankboner, 1978); al mismo tiempo que
aprovechan el amonio y dióxido de carbono que producen sus epífitos cuando la
concentración de estas moléculas en el ambiente se encuentra limitada (Hurd et
al., 1994; Taylor & Ress, 1988).
El fenómeno de epifitismo, sin embargo, no se restringe a los vegetales
marinos; cualquier otro organismo o superficie sólida sumergida en el mar es
susceptible a la epibiosis, o bien, al asentamiento de otras especies. Para que
cualquier comunidad biótica de bacterias, algas o invertebrados se asiente sobre
una superficie, existen varias etapas que implican en su fase inicial, un micro-
epifitismo, es decir una primera colonización por microorganismos, ya sean

2
bacterias o microalgas. La primera colonización comprende cuatro pasos: primero
los biopolímeros que están en el agua se adsorben a la superficie, luego, si hay
atracción química con alguna bacteria se da una adsorción reversible donde
únicamente hay un asentamiento en la superficie, casi inmediatamente la
adsorción es irreversible cuando la bacteria se une a la superficie con estructuras
celulares y finaliza con la formación de colonias bacterianas. Con el crecimiento
bacteriano se produce un mucílago o sustancia exopolimérica (EPS) que les
permite a los nuevos epibiontes establecerse sobre un sustrato más adecuado, y
ocurre entonces una sucesión ecológica (Bakus et al., 1986; Henschell & Cook,
1990).
Hay ciertas especies de bacterias, cuyo asentamiento tiene efectos negativos
sobre el organismo hospedero, de modo que muchos organismos en el ambiente
marino han evolucionado a mecanismos de defensa contra el asentamiento y
crecimiento de estas especies, lo que en farmacognosia se conoce como
capacidad anti-incrustante (Scardino & de Nys, 2010). La gran mayoría de
organismos sésiles, tales como esponjas, cnidarios y algas, que no presentan
formas motoras de defensa, se ven obligadas a desarrollar adaptaciones
metabólicas que les permitan sintetizar naturalmente compuestos de defensa que
les proporcionan ventajas en el ambiente marino. A la elaboración de estos
compuestos se le denomina metabolismo secundario, y a lo que resulta de él, se
conoce como metabolitos secundarios o bien productos naturales. Se define como
metabolismo secundario porque a diferencia del metabolismo primario, no es un

3
proceso fundamental para el crecimiento o reproducción del organismo, sin
embargo, los compuestos resultantes tienen un papel ecológico importante en
contra de la herbivoría, ataque de patógenos o depredadores, competencia y/o en
la protección en relación con el estrés biótico, como cambios térmicos o lumínicos
y deficiencias nutricionales (Darías-Jerez, 1998; Márquez et al., 2003; Maschek &
Baker, 2008). Por eso la ocurrencia y concentración de los metabolitos
secundarios depende de condiciones externas como el modo de vida y el hábitat;
por lo que los organismos que habiten ambientes menos estables y altamente
competitivos tienden a producir una mayor cantidad de defensas químicas (Hay &
Fenical, 1996). Por ejemplo, Bakus & Kawaguachi (1984) demostraron que los
organismos marinos tropicales tienen un potencial mayor como anti-incrustantes
que el de especies de otras latitudes. Por otra parte, Nakatsu et al. (1983), al igual
que McCaffrey & Endean (1985) observaron que organismos con superficies libres
de epibiontes contienen una mayor cantidad de compuestos derivados del
metabolismo secundario que organismos epifitados, lo que comprueba el papel
ecológico de los metabolitos secundarios.
Es por la importancia ecológica de estos organismos que se obtienen
productos naturales que no solamente son anti-epífitos o anti-incrustantes, sino
que también son valiosos para la elaboración de antibióticos, antitumorales,
antivirales, insecticidas, sustancias citotóxicas y neurotóxicas (Donia & Hamann,
2003). En los últimos cuarenta años, el auge en las investigaciones sobre la
química y actividad biológica se ha enfocado principalmente en los organismos

4
marinos, pues se ha determinado que estos contienen una gran cantidad y
variedad de metabolitos secundarios con estructuras químicas diferentes a las
encontradas en organismos terrestres y además con diferentes actividades
biológicas (Blunt et al., 2009; Faulkner, 2000); lo que probablemente se debe a las
condiciones físicas y químicas del mar.
Un recurso marino de gran relevancia son las algas, quienes han tenido una
participación muy importante en la química de productos naturales. Hoy en día se
conocen casi 3,000 compuestos derivados del metabolismo secundario de algas
marinas, que representa un 20% de los productos naturales marinos (Amsler,
2009), en donde se incluye una variedad de polisacáridos, fenoles bromados,
esteroles, polifenoles y péptidos que tienen aplicaciones alimenticias e
industriales, como alginatos, agar y carragenanos; o son de interés farmacológico
por su potencial de aplicación como moduladores en la coagulación sanguínea,
como el fucoidan; o presentan actividad anti-bacteriana, citotóxica y anti-
cancerígena (Maschek & Baker, 2008; Muñoz-Ochoa, 2010). De estos
compuestos, más de 1,100 se han extraído de las algas pardas, clase
Phaeophyceae, incluyendo terpenos, florotaninos y pequeñas acetogeninas (Blunt
et al., 2009). La mayor parte de los compuestos aislados de estas algas han sido
diterpenos lineales como el crinitol; y principalmente fenoles y meroterpenos
fenólicos derivados del floroglucinol, que son los metabolitos secundarios más
abundantes en las algas pardas, se han reportado en casi todos los órdenes y
pueden llegar a constituir hasta el 20% del peso seco del alga, además se ha

5
observado que actúan como defensa química contra la fijación de larvas de
animales marinos y bacterias (Maschek & Baker, 2008).
Dentro de las algas pardas, Macrocystis pyrifera se considera como la base
del ecosistema de los bosques de kelps cuya productividad y valor ecológico son
muy altos, ya que en estos hábitat pueden soportar hasta 270 diferentes especies.
Se sabe que tiene actividad anti-microepífita porque también experimenta una
gran variedad de condiciones ambientales que la hacen propensa al epifitismo a lo
largo de sus extensas frondas que llegan a medir hasta 50m (Zimmerman &
Kremer, 1986).
2. Antecedentes
M. pyrifera ha sido un recurso marino sumamente explotado, pues tiene
muchas aplicaciones industriales: se ha utilizado principalmente como fuente
importante para la obtención de ficocoloides, especialmente de alginatos que son
usados como agentes espesantes, gelificantes y estabilizantes; tiene una amplia
gama de aplicaciones en diversas industrias como la alimenticia, cosmética, textil,
papelera, cervecera y en medicina se emplea para obtener emulsiones,
encapsulados, pastillas e impresiones dentales (Casas-Valdéz, 2001).
Estados Unidos, México, Chile y Perú fueron los primeros en emplearla
para la obtención de alginatos; en Argentina la utilizan como alimento; y en
Australia la aprovechan en la agricultura. Mientras, otros países como Japón,

6
China, Francia o Reino Unido la importan para procesarla de distintas maneras
(Zemke-White & Ohno, 1999).
En México se empezó a cosechar esta alga desde 1958, y aunque tiene
una distribución restringida solo a la parte norte de la Península de Baja California,
que va de Isla Coronado a Isla San Martín, BC, México llegó a ser de los
principales exportadores del recurso como materia prima (Zemke-White & Ohno,
1999).
En la década de 1950’s, se determinó el primer caso de sobreexplotación
en California, EU: con una cosecha anual de aproximadamente 100 mil toneladas
en peso húmedo de M. pyrifera, se había mermado casi un 70% del recurso en
solo 50 años (North, 1987). Al dañar todo un ecosistema altamente productivo y
que además es hábitat para una amplia variedad de mamíferos, aves, peces,
invertebrados y otras algas, surge una inquietud por mantener el recurso, ya que
empezaron a haber pérdidas en otras actividades como la pesca, haciéndose
evidente que era mayor el valor económico de los organismos que crecen en los
bosques de M. pyrifera, al valor del alga cosechada.
A partir de la década de 1960´s se iniciaron varios programas de
investigación sobre M. pyrifera en California, más tarde en British Columbia, Chile
y México. En México se establece hasta 1982 el programa “Evaluación y
Aprovechamiento de M. pyrifera” por parte del Centro Interdisciplinario de Ciencias
Marinas (CICIMAR), donde se determinó la composición química de proteína, fibra
cruda, extracto etéreo, cenizas, carbohidratos e inclusive de humedad de acuerdo

7
a las condiciones ambientales, distribución y cambio estacional (Rodríguez-
Montesinos & Hernández-Carmona, 1991; Serviere-Zaragoza et al., 2002).
También se estudió la abundancia del alga en relación a la variabilidad climática,
según la temperatura superficial del agua, nivel medio del mar e índice de
surgencias. Del mismo modo, se le dio mucha importancia a los estudios
relacionados con la sobrevivencia y reclutamiento de M. pyrifera (Hernández-
Carmona et al., 2006) y el efecto que tiene sobre este proceso la disponibilidad de
nutrientes (Hernández-Carmona et al., 2001). Con el fenómeno “El Niño 1997-
1998” se evaluaron los efectos en la sobrevivencia de estadios microscópicos, en
la sobrevivencia del kelp en aguas profundas (Ladah & Zertuche, 2004), en el
reclutamiento (Ladah et al., 1999), y se abordaron técnicas de restauración del
recurso en México (Hernández-Carmona et al., 2000). Además se realizaron
investigaciones sobre el método de extracción de alginatos (Hernández-Carmona
et al., 2002), y el uso de ellos en materiales de impresión dental (Reyes-Tisnado et
al., 2004). En otros países, se enfocaron más a aspectos ecológicos (Graham et
al., 2007), se estudiaron los procesos físicos, químicos y biológicos que regulaban
la abundancia y distribución de M. pyrifera (Moreno & Sutherland, 1982; Plana et
al., 2007); mecanismos de reclutamiento del kelp (Kinlan et al., 2003) y sus
limitantes (Reed et al., 2004). Asimismo la influencia del medio ambiente (Barrales
& Lobban, 1975) y del tipo de vida multifacética (Schiel & Foster, 2006) sobre la
biología poblacional del alga así como en la estructura y organización de las
comunidades de bosques de kelp. Y en menor medida, se ha puesto interés en

8
establecer la relación de M. pyrifera con otros géneros de laminariales desde un
punto de vista evolutivo (Coyer et al, 2001).
Pero poco se había estudiado sobre el potencial biomédico de sus
metabolitos secundarios. Hasta mediados de la década de 1980´s se publicó por
primera vez la actividad biológica de un extracto de M. pyrifera (Mayer & Panick,
1984; Mayer et al., 1986), donde el extracto crudo mostró tener actividad
antitumoral, citotóxica, genotóxica y antiviral, y como se había observado actividad
en el fucoidan de otras especies de algas pardas, las investigaciones
subsecuentes se enfocaron en purificar el fucoidan, y observar si era el
responsable de la actividad antitumoral o citotóxica (Mayer et al., 1987a),
genotóxica (Larripa et al., 1987) y antiviral (Mayer et al., 1987b); además probaron
la actividad biológica del alginato de sodio y laminaran. Debido a sus resultados
negativos, concluyeron que la actividad biológica en M. pyrifera se debe a una
compleja interacción de metales y polisacáridos, por lo que no se continuó con
estudios más finos, y mucho menos con intenciones de aislar el compuesto activo,
de modo que la explotación de este recurso continuó basándose en productos
para la industria agroalimentaria, y el único interés biomédico fue en la impresión
dental y en sus propiedades espesantes.
Pero en la investigación de productos naturales es menester la constante
producción de los mismos, ya sea evaluando nuevos organismos o empleando
distintas metodologías. Por lo que M. pyrifera no deja de ser candidata para
evaluar su potencial biológico. Por el contrario, algunas evidencias de otras algas

9
con actividad antibacteriana (Culioli et al., 2008; Etahiri et al., 2001; Ganti et al.,
2006; Jennings & Steinberg, 1997; Plouguerné et al., 2008; Plouguerné et al.,
2010) sostienen que es posible que M. pyrifera posea compuestos que inhiban las
primeras etapas del micro-epifitismo, ya que se trata de especies con ecología
similar. También han surgido nuevas metodologías en el estudio de la actividad
anti-epífita, que son distintas a las de Mayer et al. (1987a) y Larripa et al. (1987),
que podrían emplearse para reanudar nuevas investigaciones con M. pyrifera
(Dhanasekaran et al., 2009; Feng et al., 2007; Hsu, 1982; Silkina et al., 2009).
Durán-Riverol (2009) evaluando el protocolo de aislamiento guiado por
bioensayo, observó entre otras algas, que M. pyrifera tiene actividad frente a
bacterias patógenas; dicho estudio dejó asentada la actividad del alga contra
Vibrio alginolyticus, V. campbellii, V. harveyi, V. parahaemolyticus, Staphylococcus
aureus, Bacillus subtilis y Exiguobacterium sp. La autora obtuvo 10 fracciones
cromatográficas de M. pyrifera, todas ellas con actividad biológica al menos contra
una cepa bacteriana.
Mientras, en la Universidad de Portsmouth en Reino Unido, en colaboración
con el CICIMAR, Quesseveur (2009) evalúo el efecto de 79 extractos provenientes
de 42 especies marinas colectadas en aguas del Pacífico mexicano sobre el
crecimiento de bacterias marinas y terrestres comúnmente implicadas en las
primeras etapas del proceso de colonización o asentamiento; entre las sustancias
probadas, se encontró que el extracto de M. pyrifera y las diez fracciones

10
cromatográficas obtenidas del fraccionamiento realizado por Durán-Riverol (2009)
presentaron actividad anti-epífita.
Aunque el trabajo de Quesseveur (2009) es de los primeros en considerar a
M. pyrifera como anti-incrustante, existen una gran cantidad de estudios que
demuestran la actividad en otras algas, dentro de las algas pardas que presentan
actividad contra algún tipo de microorganismo involucrado en el epifitismo, se
encuentra el trabajo de Jennings & Steinberg (1997), quienes observan que la
cantidad de florotaninos influye en la abundancia de los epífitos sobre el kelp
Ecklonia radiata. Ganti y colaboradores (2006) aislaron y caracterizaron agentes
anti-incrustantes en el alga Sargassum confusum. En el 2008, Culioli y
colaboradores encontraron en Halidrys siliquosa productos derivados de los
terpenos con actividad anti-incrustante; en el mismo año, Plouguerné et al. (2008)
extraen sustancias activas contra el micro-epifitismo de las algas invasivas
Grateloupia turuturu y Sargassum muticum. Plouguerné et al. (2010) encuentran
ácido palmítico, linoleico y palmitoleico en Sargassum muticum, además de
galactoglicerolípidos e hidrocarbonos que inhibieron el crecimiento de microalgas,
bacterias y hongos implicados en el micro-epifitismo.
3. Distribución y descripción de Macrocystis pyrifera
Macrocystis pyrifera (Figura 1) es un alga parda, pertenece al orden de las
laminariales, se caracteriza por formar densos bosques submarinos que son el
hogar de muchas especies marinas que dependen de las algas directamente para

11
la alimentación y la vivienda o indirectamente como un lugar de caza (Grant-
Burges et al., 2003).
Figura 1. Macrocystis pyrifera. Al norte, se distribuye sobre las costas del Pacífico de
Norteamérica, México, EU y Alaska. Al sur tiene una distribución circunsubantártica desde
Perú hasta Cabo de Hornos en América, Sudáfrica, Australia y Nueva Zelanda.

12
Debido a su fisiología muy variable e historia de vida, M. pyrifera ocupa una
amplia variedad de ambientes. Habita en ambos hemisferios: se localiza
principalmente a lo largo de las costas del Pacífico de Norteamérica, México, EU y
Alaska. Al sur tiene una distribución circunsubantártica que va de Perú hasta Cabo
de Hornos en América, Sudáfrica, Australia y Nueva Zelanda. En México, los
mantos de M. pyrifera se localizan en la Península de Baja California, desde Islas
Coronado (23’ 25’ N; 117’’ 15’0) hasta Bahía Asunción (27’ 08’ N; 114’’ 18’0).
Aunque esta especie prospera en aguas frías, donde la temperatura del
agua se mantiene por debajo de los 21°C, su distribución en altas latitudes parece
estar limitada por el oleaje y la baja radiación solar; mientras que los límites que
determinan su distribución en latitudes bajas parecen ser la poca disponibilidad de
nutrientes en aguas cálidas donde no ocurren surgencias y/o la competencia con
especies que son más termo-tolerantes (Grant-Burges et al., 2003).
Dentro de estos límites, su distribución lateral corresponde a cambios
abruptos en la batimetría y/o en la composición del sustrato; y su distribución
superficial se ve influida por la desecación y radiación ultravioleta principalmente,
aunque el oleaje, el pastoreo y la competencia con otras algas en las zonas más
someras también puede ser importante. Así, se encuentra desde la zona
intermareal hasta 30 m de profundidad (Grant-Burges et al., 2003).
M. pyrifera puede llegar a medir hasta más de 50 m de longitud,
dependiendo el ambiente donde se desarrolle. Su estructura (Figura 2) consiste en
un rizoide perene mediante el cual se sujeta a un sustrato generalmente rocoso;

13
de este rizoide se desprenden varias ramificaciones llamadas estipes, a lo largo de
estos se encuentran los neumatocistos que son pequeñas vesículas llenas de aire
que le sirven como flotadores; a partir de estas estructuras crecen las láminas o
filoides, los cuales miden alrededor de medio metro de largo, y todo esto compone
una fronda. Las frondas son efímeras con una vida media de aproximadamente 6
meses. Las frondas jóvenes crecen hasta la superficie, con nuevas láminas
formadas a intervalos regulares; una vez que la fronda alcanza la superficie, el
crecimiento continúa por un corto tiempo formando doseles en la superficie del
mar, luego cesa la formación de nuevas láminas y finalmente llega a la
senescencia.
Esta alga tiene una gran importancia ecológica en los lugares donde habita:
por su gran tamaño y densidad altera de manera significativa la disponibilidad de
la luz, el flujo de las corrientes oceánicas y la química de las aguas del océano en
la zona donde crecen. Su presencia es crucial para mantener la organización y
diversidad de comunidades ecológicas, por ello es considerada una especie clave
que puede albergar y dar protección a una gran diversidad de organismos,
incluyendo algas, peces, moluscos, entre otros. Además se ha determinado que
M. pyrifera posee un elevado contenido de aminoácidos esenciales y ácidos
grasos, y la calidad de sus proteínas y lípidos lo hace comparable con las de otras
fuentes vegetales (Grant-Burges et al., 2003).

14
4. Justificación
Existe desde hace mucho tiempo un esfuerzo por evitar el biofouling o
epibiosis, es decir, el asentamiento de organismos sobre superficies y estructuras
creadas por el hombre; en un inicio se comenzaron a añadir compuestos tóxicos
como mercurio, plomo, arsénico y cloro en los revestimientos de barcos y
estanques que fueron sustituidos en su mayoría por el tributilo de estaño (TBT) a
principios de 1950´s, porque tenía un menor impacto ecológico mientras
retrasaban el desarrollo de organismos adheridos a sus superficies; así las
pinturas adicionadas con TBT permitieron el ahorro en combustible y
mantenimiento, ya que permitieron intervalos entre operaciones de repintado más
largos (Barreiro et al., 2004). No obstante, seguían causando daños a la biota
marina, desde alteraciones bioquímicas hasta extinciones a escala local; se
acumulaban en los sedimentos durante años y finalmente su uso se restringió en
el 2008; el sustituto del TBT en las pinturas fue el cobre, que siguió ocasionando
problemas ambientales (Fent, 1996); por lo que se optó por el empleo de otras
tecnologías como limpiadores de agua a presión, antifouling por ultrasonido o
vibraciones, y la limpieza criogénica que consiste en pequeños cristales de hielo
seco (Fondear, 2011), sin embargo, estas tecnologías terminan teniendo costos
elevados y aplicados en la acuacultura resultaría poco práctico porque se requiere
un continuo mantenimiento. De modo que una alternativa que ofrece eficacia y
respeto con el medio ambiente además de bajo costo es la implementación de
anti-incrustantes naturales en las pinturas, tal como extractos de algas marinas.

15
Ya varios autores han incorporado estos productos en pinturas y otros
revestimientos; aunque hace falta definir la química de los extractos utilizados
(Grant-Burges et al., 2003). Por consiguiente es necesario continuar con el
aislamiento e identificación de los principios activos, que si bien, representan un
desafío académico, también representan el más exitoso acercamiento al
descubrimiento de nuevas moléculas para el beneficio de la humanidad (Lock,
2011).
5. Objetivo
Identificar metabolitos presentes en el extracto etanólico de Macrocystis
pyrifera que inhiban el crecimiento de organismos involucrados en el
microepifitismo sobre estructuras sumergidas en cuerpos de agua dulce y/o
marina.
6. Metodología
6.1 Zona de estudio y colecta de Macrocystis pyrifera.
El material algal fue recolectado en aguas superficiales en Punta Eugenia,
Municipio de Mulegé, Baja California Sur (Figura 2) en agosto de 2006. El material
algal recolectado fue identificado in situ como M. pyrifera por el Dr. Rafael
Riosmena Rodríguez del Laboratorio de Botánica Marina de la Universidad
Autónoma de Baja California Sur.

16
Figura 2. Punta Eugenia, BCS. Sitio de colecta de M. pyrifera (mapa de Durán-Riveron,
2009).
6.2 Obtención del extracto etanólico.
Después de lavar y eliminar los epífitos conspicuos y ser secado al sol, se
realizó el extracto crudo con etanol destilado (EtOHd) en las instalaciones del
CICIMAR. Se obtuvo el extracto etanólico a partir de la maceración de 5.6 kg de
M. pyrifera con 8 L de EtOHd en una primera extracción, y con 7 L del disolvente

17
recuperado en una segunda extracción. Después de filtrar el sobrenadante y
concentrar a sequedad con rotavapor a presiones bajas y temperaturas de 40°C,
resultaron 306.7 g de extracto etanólico.
6.3 Fraccionamiento inicial del extracto etanólico
Del extracto obtenido, mediante filtración simple sobre papel, se obtuvo 29
g de una porción de sales insolubles en EtOHd y 277 g de extracto orgánico libre
de sales; este último sirvió para obtener tres fracciones: fase orgánica, emulsión y
fase acuosa. Por la actividad que presentó contra Vibrio campbellii y V.
parahaemolyticus, en el trabajo descrito por Durán-Riverol (2009), la fase orgánica
se sometió a extracción en fase sólida utilizando diclorometano (CH2Cl2) y EtOHd
como disolventes en mezclas con creciente polaridad, obteniendo 5 fracciones
(06-019 41 EFS2 Fn); de las cuales la fracción de baja polaridad 06-019 41 EFS2
F1, se seleccionó para ser fraccionada en búsqueda de principios activos. A partir
de un tercer fraccionamiento, se obtuvieron 10 fracciones (06-019 41CC2 Fn)
según la polaridad.
Una parte de estas fracciones se conservaron en el CICIMAR, y se evaluó
su actividad antibacteriana. Otra parte de las fracciones se enviaron a la
Universidad de Portsmouth para su evaluación anti-incrustante contra bacterias y
microalgas.

18
6.4 Evaluación de la actividad antibacteriana y anti-incrustante
Se determinó la actividad antibacteriana por el método de difusión en agar
con disco en un medio sólido, descrito y realizado por Durán-Riverol (2009). La
actividad anti-incrustante se evaluó mediante la técnica microdilución en placas,
realizada por Quesseveur (2009).
6.4.1 Ensayo de difusión en agar con discos
Los microorganismos probados mediante este ensayo fueron Vibrio
alginolyticus (X56576), V. campbellii (ATCC 25920), V. harveyi (ATCC 14126), V.
parahaemolyticus (ATCC 17802), Staphylococcus aureus (ATCC BAA-42),
Bacillus subtilis (ATCC 6051), Escherichia coli (ATCC BAA-196), obtenidas de la
American Type Culture Collection: http://www.atcc.org/. Exiguobacterium sp. fue
aislada de Artemia sp. por Carmona-Pérez (2006). Todas las cepas fueron
preservadas a -70°C con 50% de glicerol hasta el momento de reactivarse.
Para reactivar las cepas, fueron sembradas en agar marino [1 g de extracto
de levadura (DIFCO), 5 g de peptona de carne (DIFCO), 15 g de Bacto agar
(DIFCO) en 1 L de agua de mar artificial (Coral Reef)] y sembradas en agar
infusión cerebro-corazón (BHI) (DIFCO) y agar tripticaseina-soya (TSA) (DIFCO)
respectivamente las bacterias marinas y no marinas. Posteriormente se incubaron
durante 24 h. Una vez transcurrido el período de incubación, se sembraron en

19
tubos de agar inclinado (dos tubos de trabajo y uno de reserva) en agar marino.
Se incubaron nuevamente durante 24 h y posteriormente fueron almacenados a
11°C.
Para la preparación de inóculos, se sembraron cultivos masivos en agar; se
utilizó agar marino para las bacterias marinas y agar infusión cerebro-corazón
(BHI) (DIFCO) y agar tripticaseina-soya (TSA) (DIFCO) para las bacterias no
marinas, y se incubaron durante 24 h. Una vez obtenida suficiente biomasa
bacteriana de cada especie, se tomó una parte de ella con un hisopo estéril y se
suspendió en solución salina estéril (2.5% NaCl para bacterias marinas, y 0.85%
para bacterias no marinas), hasta obtener una densidad óptica de
aproximadamente 1.00 UA, a una longitud de onda 585 nm.
Con cada especie bacteriana se inocularon masivamente placas de agar
Muller-Hinton (Merck ®) (2.5% NaCl), sobre las cuales se colocaron los
sensidiscos con los extractos de prueba y los controles positivos y negativos.
Estas placas se dejaron a temperatura ambiente (24ºC aproximadamente) por 1.5
h para permitir la difusión del compuesto en el agar y posteriormente se incubaron
a 30°C por 24 h. El efecto sobre las bacterias fue registrado como negativo
cuando no se observó inhibición del crecimiento y en los casos positivos, las
zonas de inhibición alrededor de cada disco fueron medidas en mm.
Para los sensidiscos, se emplearon discos de papel filtro Whatman No. 4,
de 8 mm, esterilizados en autoclave, se impregnaron con aproximadamente 2 mg
de los extracto/fracción/compuesto a probar, diluidos en los disolventes

20
apropiados. La preparación se llevó a cabo en campana de flujo laminar
desinfectada con benzal, sobre papel aluminio lavado con etanol, para asegurar la
esterilidad. En esas condiciones los discos se dejaron evaporar y se almacenaron,
a -18°C por un período no mayor a 48 h. De igual manera se prepararon controles
negativos con los disolventes puros utilizados y controles positivos con el
antibiótico comercial Terramicina ® (oxitetraciclina), cápuslas de 500mg Pfizer.
6.4.2 Ensayo de microdilución en placa
En este ensayo, fueron evaluadas tres bacterias marinas: Polaribacter
irgensii (ATCC 700398), Pseudoalteromonas elyakovii (ATCC 700519) y Vibrio
aestuarianus (ATCC 35048); tres bacterias terrestres: Escherichia coli (ATCC
23176), Enterobacter aerogenes (ATCC 13048) y Staphylococcus aureus (ATCC
25923); y tres microalgas de agua dulce: Fragilaria crotonensis, Cosmarium sp. y
Scenedesmus armatus, todas ellas obtenidas de la colección de la Universidad de
Portsmout.
La actividad de cada extracto se probó a cinco diferentes concentraciones
(0.1, 1, 10, 25 y 50 g/mL) con el fin de evaluar la Concentración Inhibitoria
Mínima (CIM). Las concentraciones se obtuvieron siguiendo un protocolo de
dilución en serie: la primera concentración de 1 mg/mL; consecuentes fueron de
100 y 10 g/mL; de 100 g/mL se obtuvieron concentraciones de 25 y 50 g/mL y
de 10 g/mL se obtuvieron las concentraciones de 0.1, 1 y 10 g/mL. Así, se

21
evaluó la CIM por el método de dilución utilizando placas para microdilución de 96
celdas.
Para los ensayos de actividad anti-bacteriana, se utilizó el medio de cultivo
para bacterias marinas que se preparó con 5 g de peptona diluida en 1 L de agua
de mar filtrada y estéril (10 m, Whatman); y para bacterias terrestres, el medio
que se utilizó estaba compuesto de un caldo nutritivo CM 0067 No. 2 de 25 g/mL
(Oxoid). Las bacterias se incubaron durante 5 días a 30°C para determinar la
densidad bacteriana por espectrometría con el método de Amsterdam (1996);
todos los medios de cultivo se inocularon con la misma cantidad de bacterias de
cada cepa y se dejaron incubar a temperatura constante de 30°C.
Para los ensayos de actividad anti-microalgal, se elaboró un medio de
cultivo compuesto de macronutrientes WC (calcio, sodio, magnesio, potasio,
carbonato y fosfato), micronutrientes y vitaminas disueltas en agua destilada
estéril. Por su parte se dejaron incubar las microalgas en una cámara de
incubación a 25°C, con iluminación constante durante 3 semanas; para inocular
los medios de cultivo con una misma concentración, se midió la concentración de
clorofila a mediante métodos espectrofotométricos y la ecuación de Strickland y
Parson (1968); se dejaron incubar durante 2 semanas a 30°C.
Para probar cada extracto se realizaron 6 réplicas de cada una de las
concentraciones obtenidas además de los dos controles, uno sin extracto y otro
con metanol para probar el efecto del disolvente; de modo que por cepa hubo 42
ensayos (7 x 6). Así se comparó la turbidez de los tubos control con los del

22
extracto para evaluar la actividad anti-bacteriana y anti-microalgal, y dependiendo
el número de réplicas activas, el criterio para calificar la actividad fue de la
siguiente manera:
0 a 2 de las 6 réplicas = sin actividad
3 ó 4 de las 6 réplicas = - (inhibidora) ó (activadora) +
5 ó 6 de las 6 réplicas = - - (inhibidora) ó (activadora) + +
6.5 Fraccionamiento de 06-019 41CC2F3
La fracción de baja polaridad 06-019 41CC2F3 se obtuvo en el tercer
fraccionamiento de M. pyrifera, a partir de 06-019 41 EFS2 F1 derivada de la fase
orgánica (Anexo I); y se eligió para continuar su fraccionamiento por la actividad
que mostró frente a Vibrio alginolyticus, V. campbellii, V. parahaemolyticus y
Staphylococcus aureus; por el rendimiento (0.767 mg) y por su apariencia
aceitosa.
Primero se determinó la solubilidad de la fracción en CH2Cl2 utilizando
aproximadamente 1 mg de la muestra; luego se estableció el patrón de
fraccionamiento de la muestra en cromatografía en placas (TLC) y se fraccionó por
cromatografía en columna sobre sílica (60ª230-400 mesh ASTM) con una relación
muestra adsorbente de 1:100 (76.7 mg de muestra); la elución fue realizada con
hexano:diclorometano (Hex:CH2Cl2) con una relación (5:5); y continuó con un
gradiente de polaridad de diclorometano:metanol (CH2Cl2:MeOH) iniciando
(CH2Cl2 100%), (98:2), (95:5), (9:1) (8:2) (7:3) y se lavó con MeOH (100%).

23
Simultáneamente se realizó una cromatografía en capa fina, y se determinaron 8
fracciones que fueron sometidas a la prueba de actividad contra la bacteria
Staphylococcus aureus (como se describe en la sección 6.4.1). Se decidió hacer la
prueba únicamente para las fracciones de mediana polaridad: 06-019 41CC3 F3,
CC3 F4, CC3 F6 y CC3 F7, tomando como criterio el perfil cromatográfico, la
apariencia y el rendimiento de la fracción.
Se continuó con el fraccionamiento de 06-019 41CC3F4 por ser la fracción
más activa, esta vez la relación muestra-adsorbente fue de 1:200 (con 9.4 mg de
muestra). La elusión fue realizada con CH2Cl2 100% y continuó con un gradiente
de polaridad CH2Cl2:MeOH (98:2), (95:5), (9:1) y (8:2). Mediante cromatografía en
capa fina se determinaron 4 fracciones: 06-019 41CC4 F1, CC4 F2, CC4 F3 y CC4
F4. Se evaluó la actividad de 06-019 CC4F2 y 06-019 41CC4F3 contra Vibrio
parahaemolyticus como se describe en la sección 6.4.1, las cuales resultaron ser
muy parecidas en cromatografía de capa fina. El proceso de fraccionamiento a
partir de 06-19 41CC2F3 se desglosa en la Figura 3.
6.6 Identificación estructural del compuesto activo
Este análisis se realizó por espectroscopia de infrarrojo (IR); las fracciones
analizadas fueron 06-019 41CC4 F2 y CC4 F3. El espectro (IR) se obtuvo en las
instalaciones del laboratorio de Farmacognosia Marina en la Universidad
Autónoma de Baja California Sur con un espectrómetro Paragon 500 FT-IR en
forma de película sobre pastilla de Bromuro de potasio con dominio del tiempo por

24
transformada de fourier. Y la estructura molecular se determinó comparando el IR
contra tablas de frecuencias de absorción para determinados grupos funcionales.
Figura 3. Fraccionamiento guiado por bioensayo de la fracción de baja polaridad
06-019 41CC2 F3 derivada de la fase orgánica del extracto de M. pyrifera.
CC4 Si-gel fase normal 60 230-400 Relación muestra:sílica 1:200 CH2Cl2 100%; CH2Cl2:MeOH (98:2, 95:5, 9:1, 8:2)
06-019 41 CC2 F3 76.7 mg
CC3 Si-gel fase normal 60 230-400 Relación muestra:sílica 1:100 CH2Cl2 100%; CH2Cl2:MeOH (98:2, 95:5, 9:1, 8:2, 7:3)
F1 0.96 mg
F2 2.33 mg
F8 2.36 mg
F4 9.4 mg
S. aureus
F3 19.2 mg S. aureus
F7 22.79 mg S. aureus
F5 3.89 mg
F6 11.5 mg S. aureus
F2 29.5 mg
V. parahaemolyticus *11.6 mm
F1 3.2 mg
F3 24.2 mg
V. parahaemolyticus *11.5 mm
F4 3.5 mg
IR: Ácidos grasos
*8.3 mm *13.1 mm *12.6 mm *9.1 mm

25
7 Resultados
7.1 Actividad biológica
Se determinó que M. pyrifera es un alga capaz de sintetizar varios
compuestos activos contra organismos marinos, terrestres y dulceacuícolas, tanto
bacterias Gram-positivas como Gram-negativas. Si se analizan las Tablas, se
observa que en general las fracciones fueron más activas contra organismos
marinos, y no se observó que exista una especificidad de acción de las sustancias
de acuerdo a la naturaleza de la cepa bacteriana por ser Gram-positiva o Gram-
negativa. Aunque sí se observó que diferentes fracciones tuvieron diferente
actividad sobre cada microorganismo.
La fase orgánica del extracto etanólico de M. pyrifera inhibió el crecimiento
de las bacterias Vibrio campbelili, V. parahaemolyticus, Poliaribacter irgensii, V.
aestuarianus y Pseudoalteromonas elyakovii; y promovió el crecimiento de las
microalgas Fragilaria crotonensis y Scenedesmus armatus.
El extracto etanólico (06-019) no fue activo contra ninguna bacteria
terrestre, sin embargo, varias fracciones del extracto etanólico de M. pyrifera
inhibieron a Enterobacter aerogenes y Staphylococcus aureus. En la Tabla I
vemos que las fracciones 06-019 41CC2F6 y CC2F9 fueron activas contra E.
aerogenes, y en la Tabla III se observan las fracciones 06-019 41CC2F1, CC2F5,
CC2F7, CC2F8, CC2F9 y CC2F10 que inhibieron a S. aureus. Por su parte, las
fracciones 06-019 41CC2F3, CC2F9 y CC2F10 promovieron el crecimiento de

26
Cosmarium sp. a pesar de que no se determinó actividad en esta cepa por parte
del extracto etanólico (Tabla II). Escherichia coli fue la única bacteria que mostró
resistencia tanto al extracto como a las fracciones de M. pyrifera (Tabla III).
En la Tabla III se puede observar que el extracto etanólico de M. pyrifera
promovió el crecimiento algal de Scenedesmus armatus a distintas
concentraciones, y las fracciones 06-019 41CC2F2, CC2F3 y CC2F10,
contrariamente, inhibieron su crecimiento a concentraciones de 10 y 25 g/mL.
Con respecto a la fracción 06-019 41CC2F3, en la Tabla I se muestra la
actividad contra V. aestuarianus, en la Tabla III se muestra la actividad contra las
bacterias Vibrio parahaemolyticus, V. campbellii, V. alginolyticus, Exiguobacterium
y Staphylococcus aureus mediante el ensayo de difusión en agar por disco. Sin
embargo, en la Tabla I, con el método de dilución en microplacas, no presenta
actividad contra S. aureus. En la Tabla II se muestra que inhibe el crecimiento de
la microalga Scenedesmus armatus, pero tiene un efecto positivo sobre el
crecimiento de Fragilaria crotonensis y Cosmarium sp.
A partir del fraccionamiento de 06-019 41 CC2F3, se obtuvieron 8
fracciones (06-019 41 CC3Fn1-8). Y solamente se evaluó la actividad contra
Staphylococcus aureus de cuatro de estas fracciones, las cuales presentaron
distinta intensidad inhibitoria, lo que se muestra en la Tabla III. Donde también se
observa cómo en 06-019 41CC3F4 y 06-019 41CC3F6 la actividad se incrementó
en comparación con la fracción 06-019 41 CC2F3, y sin embargo, disminuye con
la fracción 06-019 CC3F3.

27
Se obtuvieron cuatro fracciones a partir de 06-019 41 CC3F4 (fracción más
activa contra S. aureus). Dos de estas fracciones (06-019 41CC4F2 y CC4F3)
inhibieron el crecimiento de V. parahaemolyticus, con un halo de inhibición de 11.5
mm. No se evaluó la actividad de las fracciones 06-019 41 CC4 F1 y 06-019 41
CC4 F4 porque en el caso de la primera fracción, la TLC no mostró que hubiera un
compuesto evidente; y la segunda fracción mostró ser una mezcla muy compleja,
y con poco rendimiento.
Tabla I. Actividad anti-incrustante de M. pyrifera, método de microdilución en placa.
Bacterias terrestres Fracción de M. pyrifera
Concentración mínima
inhibitoria g/mL
E. coli
E. aerogenes 06-019 41CC2F6 0.1
06-019 41CC2F9 10
S. aureus
06-019 41CC2F1 0.1
06-019 41CC2F5 25
06-019 41CC2F7 10
06-019 41CC2F8 50
06-019 41CC2F9 1
06-019 41CC2F10 0.1
Bacterias marinas
P. irgensii 06-019 0.1
P. elyakovii 06-019 0.1
V. aestuarianus
06-019 0.1
06-019 41CC2F1 1
06-019 41CC2F2 10
06-019 41CC2F3 0.1
06-019 41CC2F4 1
06-019 41CC2F5 0.1
06-019 41CC2F6 10
06-019 41CC2F7 0.1
06-019 41CC2F8 0.1
06-019 41CC2F9 0.1
06-019 41CC2F10 0.1

28
Tabla II. Efecto de M. pyrifera sobre el crecimiento microalgal, método de microdilución en
placa.
Nota:Un signo (+) significa que promueve el crecimiento, un signo (-) significa que lo inhibe; y el
número de signos refiere al número de réplicas que fueron promotoras o inhibidoras; un signo
equivale a 3 ó 4 réplicas activas y dos signos equivale a 5 ó 6 réplicas.
Microalgas Fracción de M. pyrifera Concentración activa
g/ mL
Efecto sobre el crecimiento
F. crotonensis
06-019 0.1 +
06-019 41CC2F1 10 +
06-019 41CC2F2
0.1 +
1 +
10 ++
25 ++
50 ++
06-019 41CC2F3
01 ++
1 ++
10 ++
25 ++
50 ++
06-019 41CC2F4
0.1 +
1 +
10 +
25 +
50 +
06-019 41CC2F5
1 ++
10 +
25 +
50 +
06-019 41CC2F6
01 +
1 +
10 +
25 +
50 +
06-019 41CC2F7
0.1 +
1 +
10 +
25 +
50 +
06-019 41CC2F10 25 ++
50 ++
Cosmarium sp.
06-019 41CC2F3 50 ++
06-019 41CC2F4 50 ++
06-019 41CC2F10
0.1 +
1 +
10 +
25 ++
50 ++
S. armatus 06-019 0.1 +

29
1 +
10 +
25 +
50 +
06-019 41CC2F2 10 - -
25 - -
06-019 41CC2F3 50 - -
06-019 41CC2F10 25 - -
50 - -
Tabla III. Actividad antibacteriana de M. pyrifera, método de difusión en agar.
Nota: El número es el diámetro promedio del halo de inhibición expresado en mm. NA significa que
no se observó actividad; el signo (-) significa que no se evaluó la actividad de las fracciones para la
cepa bacteriana.
Fracción S. aureus V. para V. algino Exiguobact E. coli V. campb
06-019 41CC2F1 9 11 8.3 11 NA 7.8
06-019 41CC2F2 10.5 11.5 9.5 17 NA 10.3
06-019 41CC2F3 11 11 9.5 14 NA 9
06-019 41CC2F4 14.5 8.5 7.6 11.5 NA 7.5
06-019 41CC2F5 10 8 NA 10.5 NA 7.5
06-019 41CC2F6 10.5 NA NA 12 NA NA
06-019 41CC2F7 14 9.5 8 20.5 NA 9
06-019 41CC2F8 13 9.5 8.5 20.5 NA 9
06-019 41CC2F9 9.5 8 7.5 16.5 NA 8
06-019 41CC2F10 7.5 NA NA 13 NA NA
06-019 41CC3F3 8 - - - - -
06-019 41CC3F4 12.5 - - - - -
06-019 41CC3F6 12 - - - - -
06-019 41CC3F7 9 - - - - -
06-019 41CC4F2 - 11.5 - - - -
06-019 41CC4F3 - 11.5 - - - -

30
Después de las pruebas bacterianas se obtuvo el espectro en infrarrojo (IR)
de 06-019 41 CC4 F2 y 06-019 41 CC4 F3, las cuales presentaron mucha similitud
en la TLC, y aunque ambas eran aún mezclas, la cantidad de muestra no permitía
continuar con el fraccionamiento y los ensayos de actividad biológica.
7.2 Caracterización estructural
De las señales observadas en el espectro de infrarrojo (IR) se distinguen
cuatro bandas de absorción principales en la zona general del espectro: la primera
se encuentra a 3432 m-1, que corresponde a la tensión de grupos OH. Dos
señales más se observan a 2930 m-1 y 2850 m-1: la primera señal, 2930 m-1, indica
el estiramiento asimétrico de enlaces C-H en los alcanos (CH3), y la señal 2850 m-
1 indica estiramientos simétricos en los enlaces del grupo metilo (Silverstein et al.,
2005). También se identificó una absorción en 1713 m-1, la cual puede atribuirse a
carbonilos en general, a esteres o ácidos carboxílicos (Silverstein et al., 2005).
En la zona denominada “huella dactilar”, aparecen, las señales 1244 m-1 y
1176 m-1, las cuales indican la presencia ya sea de alcanos, alcoholes, esteres,
eteres y/o ácidos carboxílicos (Silverstein et al., 2005).

31
Figura 4. Espectro infrarrojo de la fracción 06-019 41CC3F2. Se observan cuatro
señales principales en la zona general del espectro y pequeñas señales en la zona
de la huella dactilar, entre 600m-1 y 1400m-1.
Estas evidencias aunadas a la apariencia y baja polaridad de la fracción,
sugieren que se trata de algún derivado de ácido graso.
8 Discusión
Aunque ya se había reportado en el alga parda M. pyrifera actividad
citotóxica, antiviral, antitumoral (Mayer & Panick, 1984), antibacteriana (Durán-
Riverol, 2009) y anti-incrustante (Quesseveur, 2009), este estudio es de los
primeros en tratar de aislar y averiguar el tipo de compuesto específico que inhibe

32
el crecimiento de dos bacterias: la bacteria terrestre (Gram-positiva)
Staphylococcus aureus y la bacteria marina (Gram-negativa) Vibrio
parahaemolyticus, ambas involucradas en el micro-epifitismo y en infecciones
graves en humanos. En el trabajo precedente, Durán-Riverol (2009) dejó
comprobada la existencia de compuestos en M. pyrifera que inhiben el crecimiento
de varias cepas bacterianas, tanto Gram-positivas como Gram-negativas: S.
aureus, V. parahaemolyticus, V. alginolyticus, Exiguobacterium y V. campbellii. Los
halos de inhibición fueron relativamente iguales para las bacterias, por lo tanto no
existe una especificidad de acción evidente en relación a la naturaleza de la cepa
bacteriana Gram-positiva y Gram-negativa.
Por su parte, Quesseveur (2009), indica que M. pyrifera posee compuestos
que inhiben el desarrollo de bacterias marinas y terrestres y puede ser promotora
y/o inhibidora del crecimiento microalgal. En dicho estudio, las bacterias marinas
mostraron más sensibilidad a más fracciones de M. pyrifera y a concentraciones
más bajas que las bacterias terrestres, lo cual parece razonable porque
pertenecen al mismo ecosistema.
La actividad biológica que presenta el extracto etanólico y las fracciones de
M. pyrifera es diferencial sobre cada microorganismo; así se observan fracciones
que inhiben el crecimiento de más de una cepa y microorganismos que son
sensibles a varias fracciones diferentes. Esto se debe a que por el proceso de
purificación de compuestos activos mediante el protocolo de aislamiento guiado
por bioensayo, las sustancias activas se van separando y concentrando en ciertas

33
fracciones, y es por eso que algunas fracciones inhibieron el crecimiento de
microorganismos que el extracto crudo de M. pyrifera no inhibió, ya que en 2
g/mL de extracto crudo existe el principio activo en menor porcentaje que en las
fracciones; por eso también el diámetro del halo de inhibición contra S. aureus es
mayor en ciertas fracciones que en el resto, incluyendo la fracción de procedencia.
Esto podría representar un problema porque empleando un método no muy
sensible, y utilizando bajas concentraciones del principio activo, puede pasar por
desapercibida la actividad en el extracto.
Otro efecto que se observa por la concentración de la sustancia activa es
con las microalgas, específicamente con Scenedesmus armatus, ya que su
crecimiento se vio promovido por el extracto crudo y sin embargo, inhibido por tres
de sus fracciones. Lo que estimula el crecimiento de la microalga es la alta
concentración de auxina y citocinina en M. pyrifera (Stirk et al., 2004), que son
hormonas reguladoras del crecimiento vegetal, y al momento de fraccionar, los
compuestos con actividad inhibitoria se concentran (Bhakuni & Rawat, 2005).
Así como puede surgir una fracción activa contra una cepa que no haya
sido sensible al extracto crudo por cuestiones de concentración, puede que en el
fraccionamiento ninguna de las fracciones obtenidas presente la actividad del
extracto sobre alguna bacteria, como Poliaribacter irgensii y Pseudoalteromonas
elyakovii; entonces se puede decir que se pierde la actividad porque los principios
activos tienen grupos funcionales reactivos que al reaccionar originan
subproductos inactivos, o porque la actividad biológica se debe precisamente a la

34
sinergia entre los componentes de la mezcla, entonces al separarla, los
compuestos individuales no son activos (Bhakuni & Rawat, 2005).
Se obtuvo el espectro infrarrojo de 06-019 41CC4F2 y CC4F3 aunque no
eran compuestos puros, sin embargo el bajo rendimiento de la muestra no
permitía continuar con los fraccionamientos, y el espectro mostró varias señales
con las que se infiere que es un ácido graso.
Se sabe que los ácidos grasos en los seres vivos vienen en pares de
carbonos y los más comunes van de los 14-24 carbonos, y mientras más larga la
cadena es menor la solubilidad y más sólido el compuesto. En el trabajo de Rosell
& Srivastava (1987), donde queda establecida la actividad antibacteriana de
ácidos grasos de algas pardas, se identificaron en su mayoría ácidos grasos mono
y poli-insaturados de 18 y 20 carbonos, que además de ser los más abundantes
eran los que tenían mayor actividad. Entonces, basándonos en las señales del IR,
concluimos que se trata de ácidos grasos, y tomando en cuenta las
observaciones de Rosel & Srivastava (1987), determinamos que probablemente
se trata de un compuesto de cadena hidrocarbonada entre 14-24 carbonos,
porque las fracciones tenían una apariencia aceitosa y eran de olor agradable.
Determinando así que a partir de una serie de fraccionamientos de 06-019 CC2F3
(fracción poco polar derivada de la fase orgánica de M. pyrifera) se obtienen
ácidos grasos que presentan actividad contra Vibrio parahaemolyticus y
Staphylococcus aureus.

35
La mayoría de los estudios que reportan actividad anti-incrustante en algas
pardas se enfocan en la bioactividad de los florotaninos, sin embargo, se han
aislado otros compuestos anti-incrustantes de Phaeophyceae (Plouguerné et al.,
2010), como terpenos de algunas especies de Dictyota (Schmitt et al., 1995;
Barbosa et al., 2007). En este trabajo se encontraron ácidos grasos con actividad
antibacteriana, lo cual no es nuevo en algas pardas; Rosell & Srivastava (1987)
demostraron la importancia de los ácidos grasos como anti-incrustantes con
ácidos grasos insaturados derivados de Desmaresteia ligulata, al igual, Katsuoka
& colaboradores (1990) aislaron galactosil y sulfoquinovosil-diacilgliceroles de las
algas Costaria costata y Undaria pinnatifida, que exhibieron actividad anti-
macroepífita contra el bivalvo Mytilus edulis. Y en investigaciones más recientes,
Ganti & colaboradores (2006), demostraron la actividad anti-incrustante de los
derivados de ácidos grasos y ácido ftálico en el alga parda Sargassum confusum.
Plouguerné et al. (2010), aislaron compuestos identificados como ácidos grasos y
galactoglicerolípidos que exhibieron actividad anti-incrustante en Sargassum
muticum, así como hidrocarburos saturados y poli-insaturados con interesante
actividad antibacteriana, antifúngica y anti-microalgal.
Dichos estudios recalcan el rol que tienen los ácidos grasos en la ecología
de las algas; porque en realidad se consideran productos del metabolismo
primario, y que tienen funciones esenciales en los seres vivos: su función principal
en las algas es la reserva de energías y brindar soporte a las paredes celulares
formando parte de la estructura de fosfolípidos y gucolípidos en las membranas

36
biológicas; aunque también hay derivados de ácidos grasos que actúan como
hormonas o mensajeros intracelulares, y algunos ácidos grasos dirigen a las
proteínas hacia posiciones en las membranas (Stryer, 1995). Sin embargo, ahora
destaca su papel en la ecología del alga, ya que se ha demostrado que es mayor
la actividad biológica de los ácidos grasos que la de los mismos florotaninos (Deal
et al., 2003).
La función ecológica de los ácidos grasos en M. pyrifera puede explicarse
considerando precisamente la ecología del alga. M. pyrifera es parte de un
ecosistema muy complejo, que son los bosques de kelps; debido al gran tamaño
de las algas y a su densidad poblacional, las condiciones fisicoquímicas de las
aguas en la zona donde crecen estas algas, se alteran de manera significativa,
como la disponibilidad de luz, la concentración de nutrientes y el flujo de la
corriente. Al disminuir considerablemente el flujo de la corriente, M. pyrifera es
más propensa al epifitismo porque es aprovechada como sustrato y refugio para
organismos bentónicos y como alimento para toda una comunidad (Zimmerman &
Kremer, 1986). También la competencia por espacio y por recursos obliga a M.
pyrifera a biosintetizar metabolitos que funcionen como defensa ante la
colonización de organismos incrustantes y/o para conferir una ventaja competitiva
en poblaciones muy densas (Grant-Burges et al., 2003). Estas sustancias o
metabolitos con capacidad anti-incrustante pueden aislarse para generar
alternativas a los biocidas tóxicos que se utilizan como anti-incrustantes en las
pinturas y revestimientos de estructuras navales y de acuacultura para el control y

37
prevención del biofouling con tecnologías que respeten al medio ambiente (Grant-
Burges et al., 2003). Así ya se ha reportado un gran número de compuestos anti-
incrustantes que provienen de organismos marinos, desde corales, moluscos,
equinodermos, piel de ballena o tiburón y algas, cuya aplicación en pinturas y
revestimientos han tenido resultados positivos, sin embargo hace falta determinar
la estructura de los compuestos activos, pues es el siguiente paso a la síntesis de
moléculas activas en laboratorios (Bhosale et al., 2002; Scardino & de Nys, 2010).
9. Conclusiones
El alga parda Macrocystis pyrifera posee actividad antibacteriana y anti-
incrustante; también puede inhibir y/o promover el crecimiento microalgal.
Macrocystis pyrifera es capaz de sintetizar varios compuestos activos
contra organismos marinos, terrestres y dulceacuícolas; e inhibe por igual a
bacterias Gram-positivas y Gram-negativas.
Macrocystis pyrifera posee ácidos grasos en fracciones de baja polaridad,
que inhiben el crecimiento de Vibrio parahaemolyticus, bacteria implicada en el
microepifitismo.

38
10. Literatura citada
Amsler, ChD (2009) Algal Chemical Ecology. Springer. USA. 311pp.
Bakus, GJ & M Kawaguachi (1984) Toxins from marine organisms: Studies on
antifouling, EN: Bakus, GJ, NM Targett & B Schulte (1986) Chemical
ecology of marine organisms: an overview. Journal of Chemical Ecology.
12(5):951-987.
Bakus, GJ, NM Targett & B Schulte (1986) Chemical ecology of marine organisms:
an overview. Journal of Chemical Ecology. 12(5):951-987.
Barbosa JP, BG Fleury, BA Da Gama, VL Teixeira & RC Pereira (2007) Natural
products as antifoulants in the Brazilian brown alga Dictyota pfaffii
(Phaeophyta, Dictyotales). Biochemical Systematics and Ecology. 35:549-
553.
Barrales, HL & ChS Lobban (1975) The comparative ecology of Macrocystis
pyrifera, with emphasis on the forest of Chubut, Argentina. The Journal of
Ecology. 62(2):657-677.
Barreiro, R, M Quintela & JM Ruiz (2004) TBT e imposex en Galicia: Los efectos
de un disruptor endocrino en poblaciones de gasterópodos marinos.
Ecosistemas. 13(003):1-17.
Bhakuni, DS & DS Rawat (2005) Bioactive Marine Natural Products. Springer. New
Delhi, India. 400pp.

39
Bhosale, SH, VL Nagle & TG Jagtap (2002) Antifouling Potential of Some Marine
Organisms from India Against Species of Bacillus and Pseudomonas.
Marine biotechnology. 4:111-118.
Blunt, JW, BR Copp, WP Hu, MHG Munro, PT Northcote & MR Prinsep (2009)
Marine natural products. Natural Product Reports. (26):170-244.
Borowitzka, MA & RC, Lethbridge (1989) Seagrasses epiphytes, Chp. 14. En
Ortuño-Aguirre, CR, Riosmena-Rodríguez (2007) Dinámica del epifitismo en
Padina concrescens (Dictyotales: Phaeophyta) en el suroeste de la
Península de Baja California, México. Ciencias Marinas. 33(003):311-317.
Casas-Valdéz, MM (2001) Efecto de la variabilidad climática sobre la abundancia
de Macrocystis pyrifera y Gelidium robustum en México. Tesis doctoral.
Universidad de Colima, Colima. 148 pp.
Coyer, JA, GJ Smith & RA Andersen (2001) Evolution of Macrocystis sp.
(Phaeophyceae) as determined by ITS1 and ITS2 sequences. Journal of
Phycology. 37:574-585.
Culioli G, A Ortalo-Magné, C Hellio, AS Clare, L Piovetti (2008) Antifouling activity
of meroditerpenoids from he marine brown alga Halidrys siliquosa. Journal
of Natural Products. 71:1121-1126.
Darías-Jerez, J (1998) La biodiversidad de las algas marinas como fuente de
interés farmacológico. Medio Ambiente Canarias. 9:15-19.
De Burg, ME & Fankboner, PV (1978) A nutritional association between the bull
kelp Nereocystis luetkeana and its epizoic bryozoan Membranipora

40
membranacea. EN: Hepburn, ChD & CL Hurd (2005) Conditional mutualism
between the giant kelp Macrocystis pyrifera and colonial epifauna. Marine
Ecology Progress Series. 302:37-48.
Deal, MS, ME Hay, D Wilson & W Fenical (2003) Galactolipids rather than
phlorotannins as herbivore deterrents in the brown seaweed Fucus
vesiculosus. Oecologia. 136:107-114.
Dhanasekaran, D, N Thajuddin, M Rashmi, TL Deepika & M Gunasekaran (2009)
Screening of biofouling activity in marine bacterial isolate from ship hull.
International Journal of Environmental Science and Technology, 6(2):197-
202.
Dixon, J, SC Schroeter & J Kastendick (1981) Effects of the encrusting bryozoan,
Membranipora membranacea on the loss of blades and fronds by the giant
kelp Macrocystis pyrifera (Laminariales). Journal of Phycology. 17:341-345.
Donia, M & MT Hamann (2003) Review Marine natural products and their potential
applications as anti-infective agents. The Lancet Infectious Diseases.
3(6):338-348.
Durán-Riverol, LM (2009) Estudio de la actividad frente a bacterias patógenas de
cinco especies algales: pros y contras del protocolo de aislamiento guiado
por bioensayo. Tesis de Maestría. CICIMAR. 126pp.
Etahiri, SV, Bultel-Ponce, C Caux, M Guyot (2001) New bromoditerpenes from the
red alga Sphaerococcus coronopifolius. Journal of Natural Products.
64:1024-1027.

41
Faulkner, DJ (2000) Marine natural products. Natural Product Reports. (17):7-55.
Feng, D, C Ke, S Li & S Zhou (2007). Study on antifouling activity of extracts from
zingiber officinale roscoe. Journal of Xiamen University.Natural
science/Xiamen Daxue Xuebao, 46(1), 135-140.
Fent, K (1996) Ecotoxicology of organotin compounds. En: Barreiro, R, M Quintela
& JM Ruiz (2004) TBT e imposex en Galicia: Los efectos de un disruptor
endocrino en poblaciones de gasterópodos marinos. Ecosistemas.
13(003):1-17.
Fondear (2011) e-magazine Portal Náutico. www.fondear.com.
Ganti, VS, KH Kim, HD Bhattarai & HW Shin (2006) Isolation and characterisation
of some antifouling agents from the brown alga Sargassum confusum.
Journal of Asian Natural Products Research. 8:309-315.
Graham, MH, J Vásquez & AH Bushmann (2007) Global ecology of the giant kelp
Macrocystis: From ecotypes to ecosystems. Oceanography and Marine
Biology. 45:39-38.
Grant-Burges, JG, KG Boyd, E Armstrong, Z Jiang, L Yan, M Berggen, U May, T
Pisacane, K Granmo & DR Adams (2003) The Development of a Marine
Natural Product-based Antifouling Paint. Biofouling. 19:197-205.
Hay, ME & W Fenical (1996) Chemical ecology and marine biodiversity: insights
and products from the sea. Oceanography. 9(1):10–20.
Henschell, JR & PA Cook (1990) The development of a marine fouling community
in relation to the primary film of microorganisms. Biofouling. 2(1):1-11.

42
Hepburn, ChD & CL Hurd (2005) Conditional mutualism between the giant kelp
Macrocystis pyrifera and colonial epifauna. Marine Ecology Progress Series.
302:37-48.
Hernández-Carmona, G, O García, D Robledo & MS Foster (2000) Restoration
techniques for Macrocystis pyrifera (Phaeophyceae) populations at the
southern limit of their distribution in México. Botanica Marina. 43:273–284.
Hernández-Carmona, G, B Hughes & MH Graham (2006) Reproductive longevity
of drifting kelp Macrocystis pyrifera (Phaeophyceae) in Monterey Bay,
U.S.A. Journal of Phycology. 42:1199–1207.
Hernández-Carmona, G, DJ McHugh, DL Arvizu-Higuera & YE Rodríguez-
Montesinos (2002) Pilot plant scale extraction of alginates from Macrocystis
pyrifera 4. Conversion of alginic acid to sodium alginate, drying and milling.
Journal of Applied Phycology. 14:445-451.
Hernández-Carmona, G, D Robledo & E Serviere-Zaragoza (2001) Effect of
Nutrient Availability on Macrocystis pyrifera recruitment and survival near its
southern limit off Baja California. Botánica Marina. 44:221-229.
Hsu, K (1982) New marine antifouling coatings containing organotin and
chlorophenol compounds as antifouling agents.
Hurd, CI, KM Durante, FS Chia & PJ Harrison (1994) Effect of bryozoan
colonization on inorganic nitrogen acquisition by the kelps Agarum
fimbriatum and Macrocystis integrifolia. Marine Biology. 121:167-173.

43
Jennings, JG & PD Steinberg (1997) Phlorotannins vs. others factors affecting
epiphyte abundance on kelp Ecklonia radiata. Oecología. 109:461-473.
Katsuoka, M, C Ogura, H Etoh & K Sakata (1990) Galactosyl and sulfoquinovosyl-
diacylglycerols isolated from the brown algae Undaria pinnatifida and
Costaria costata as repellents of blue mussel Mytilus edulis. Agricultural
Biology and Chemistry. 54:3043-3044.
Kinlan, BP, MH Graham, E Sala & PK Dayton (2003) Arrested development of
giant kelp (Macrocystis pyrifera, Phaeophyceae) embryonic sporophytes: a
mechanism for delayed recruitment in perennial kelps?. Journal of
Phycology. 39:47-57.
Ladah, LB & JA Zertuche-González (2004) Giant kelp (Macrocystis pyrifera)
survival in deep water (25–40 m) during El Niño of 1997–1998 in Baja
California, México. Botanica Marina. 47:367-372.
Ladah, LB, JA Zertuche-González & G Hernández-Carmona (1999) Giant kelp
(Macrocystis pyrifera, Phaeophyceae) recruitment near its southern limit in
Baja California after mass disappearance during ENSO 1997-1998. Journal
of Phycology. 35:1106-1112.
Larripa, IB, MM de Pargament, ML de Vinuesa & AMS Mayer (1987) Biological
activity in Macrocystis pyrifera from Argentina, sodium alginate, fucoidan
and laminaran. II. Genotoxicity. Hidrobiologia. 151/152:491-496.
Lock, O (2011) Investigación Fitoquímica: Métodos en el Estudio de Productos
Naturales. Agenda Virtual; revisado en 2011.

44
Maschek JA & BJ Baker (2008) The chemistry of algal secondary metabolism. En:
Amsler, ChD (2009) Algal Chemical Ecology. Springer. USA. 311pp.
Márquez, DM, E Galeano & A Martínez (2003) Natural products with antimicrobial
activity. Part I. VITAE. 10(2):61-71.
Mayer, AMS & B Panick (1984) Antitumor evaluation of marine algae in Argentina.
Proceedings International Seaweed Symposium. 11:529- 533.
Mayer, AMS, B Panick, RD Bonfil, MA Espeche, ER Fraile, A Diaz, A Pesce, M
Criscuolo, JF Groisman & RM de Lederkremer (1986) Screening of
antitumor, cytotoxic, immunologic, antimicrobial and antiviral activity in
Macrocystis pyrifera, a Patagonian marine alga of economic importance.
Actas II Congr. Algas Marinas Chilenas: 177-183.
Mayer, AMS, L Krotz, RD Bonfil, OD Bustuobad, JF Groisman, RM de
Lederkremer & DB Stierle (1987a) Biological activity in Macrocystis pyrifera
from Argentina: sodium alginate, fucoidan and laminarian. I. Antitumor,
cytotoxicity and humoral immune response. Hidrobiologia. 151/152:483-489.
Mayer AMS, A Díaz, A Pesce, M Criscuolo, JI Groisman & RM de Lederkreimer
(1987b). Biological activity in Macrocystis pyrifera from Argentina: sodium
alginate, fucoidan and laminarian. III. Antiviral activity. Hidrobiología.
151/152:497-500.
McCaffrey, E. J. & R. Endean. 1985. Antimicrobial activity of tropical and
subtropical sponges. Marine Biology. 89:1-8.

45
Moreno, CA & JP Sutherland (1982) Physical and Biological Processes in a
Macrocystis pyrifera Community Near Valdivia, Chile. Oecologia. 55:1-6.
Muñoz Ochoa, M (2010) Potencial farmacológico de algas marinas de Baja
California Sur, México. Tesis de Doctorado. La Paz, Baja California Sur,
México, Centro Interdisciplinario de Ciencias Marinas. 73 pp.
Nakatsu, T, RP Walker, JE Thompson & DJ Faulkner (1983) Biologically active
sterol sulfates from marine sponge Toxadocia zumi. Experientia. 39:759-
761.
North, WJ (1987) Biology of the Macrocystis resource in North America. In Case
Studies of Seven Commercial Seaweed Resources. EN: Graham, MH, J
Vásquez & AH Bushmann (2007) Global ecology of the giant kelp
Macrocystis: From ecotypes to ecosystems. Oceanography and Marine
Biology. 45:39-38.
Plana, J, A Mansilla, M Palacios & NP Navarro (2007) Population study of
Macrocystis pyrifera (L.) C. Agardh (Laminariales: Phaeophyta) in wave
protected and exposed environments in Tierra del Fuego. Gayana 71(1):66-
75.
Plouguerné, E, C Hellio, E Deslandes, B Veron, V Stiger-Pouvreau (2008) Anti-
microfouling activities of extracts of two invasive algae: Grateloupia turuturu
and Sargassum mutcum. Botanica Marina. 51:202-208.
Plouguerné, E, E Ioannou, P Georgantea, C Vagias, V Roussis, C Hellio, E Kraffe
& V Stiger-Pouvreau (2010) Anti-microfouling Activity of Lipidic Metabolites

46
from the Invasive Brown Alga Sargassum muticum (Yendo) Fensholt.
Marine biotechnology. 12:52-61.
Quesseveur, G (2009) Criblage d’activité antimicrobienne à partir d’extraits
d’algues et d’éponges du littoral mexicain. Tesis de maestría. Universidad
de Portsmouth, UK. 59pp.
Reed, DC, SC Schroeter & PT Raimondi (2004) Spore supply and habitat
availability as sources of recruitment, limitation in the giant kelp Macrocystis
pyrifera (Phaeophyceae). Journal of Phycology. 40:275-284.
Reyes-Tisnado, R, G Hernández-Carmona, F López-Gutiérrez, EJ Vernon-Carter
& P Castro-Moroyoqui (2004) Sodium and potassium alginates extracted
from Macrocystis pyrifera algae for use in dental impression materials.
Ciencias Marinas. 30(01B):189-199.
Rodríguez-Montesinos, E & G Hernández-Carmona (1991) Seasonal and
geographic variations on Macrocystis pyrifera chemical composition at the
western coast of Baja California. Ciencias Marinas. 17(3):91-107.
Rosell, KG & LM Srivastava (1987) Fatty acids as antimicrobial substances in
brown algae. Hidrobiologia. 151/152:471-475.
Scardino, AJ & R.de Nys (2010) Mini review: Biomimetic models and bioinspired
surfaces for fouling control. Biofouling. 27(1):73-86.
Scheibling, RE, AW Hennigar & T Balch (1999) Destructive grazing, epiphytism,
and disease: the dynamics of sea urchin-kelp interactions in Nova Scotia.
Canadian Journal of Fischeries and Aquatic Sciences. 56:2300-2314.

47
Schiel, DR & MS Foster (2006) The population biology of large brown algae:
ecological consequences of multiphase life histories in dynamic coastal
environments. Annual Review of Ecology Evolution and Systematics.
37:343–372.
Schmitt TM, ME Hay & N Lindquist (1995) Constraints on chemically mediated
coevolution: multiple functions for seaweed secondary metabolites. Ecology.
76:107-123.
Serviere-Zaragoza, E, D Gómez-López & G Ponce-Díaz (2002) Gross chemical
composition of three common macroalgae and a sea grass on the pacific
coast of Baja California, México. Hidrobiológica. 12(002):113-117.
Silkina, A, A Bazes, F Vouve, V Le Tilly, P Douzenel, JL Mouget & N Bourgougnon
(2009) Antifouling activity of macroalgal extracts on fragilaria pinnata
(bacillariophyceae): A comparison with diuron. Aquatic Toxicology, 94(4),
245-254.
Silverstein, RM, FX Webster & D Kiemle (2005) Spectrometric identification of
organic compounds. 7a edición. Wiley. USA. 512 pp.
Stirk, WA, GD Arthur, AF Lourens, O Novak, M Strnad, J Van Staden (2004)
Changes in cytokinin and auxin concentrations in seaweed concentrates
when stored at elevated temperature. Journal of Applied Phycology. 16:31-
39.
Stryer, R (1995) Bioquímica. Cuarta Edición, Tomo II. Editorial Reverté. Barcelona,
España. 1010pp.

48
Taylor, RB & TA Rees (1998) Excretory products of mobile epifauna as a nitrogen
source for seaweeds. Limnology & Oceanography. 43:600-606.
Zemke-White, W & M Ohno (1999) World seaweed utilisation: An end-of-century
summary. Journal of Applied Phycology. 11:369-376.
Zimmerman, RC & JN Kremer (1986) In situ growth and chemical composition of
the giant kelp, Macrocystis pyrifera: response to temporal changes in
ambient nutrient availability. Marine Ecology Progress Series. 27:277-285.
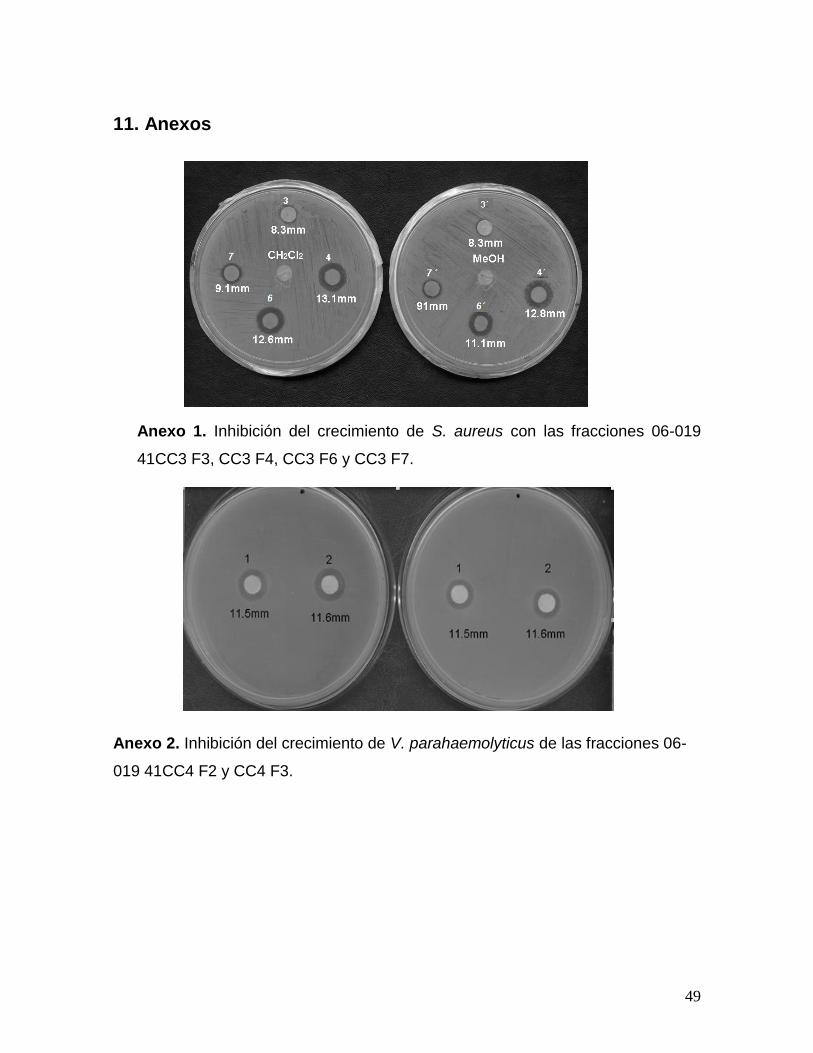
49
11. Anexos
Anexo 1. Inhibición del crecimiento de S. aureus con las fracciones 06-019
41CC3 F3, CC3 F4, CC3 F6 y CC3 F7.
Anexo 2. Inhibición del crecimiento de V. parahaemolyticus de las fracciones 06-
019 41CC4 F2 y CC4 F3.


















