
APUNTES DE BIOLOGÍA LA CÉLULA EUCARIÓTICA · total de la envoltura nuclear y se piensa que son...
22
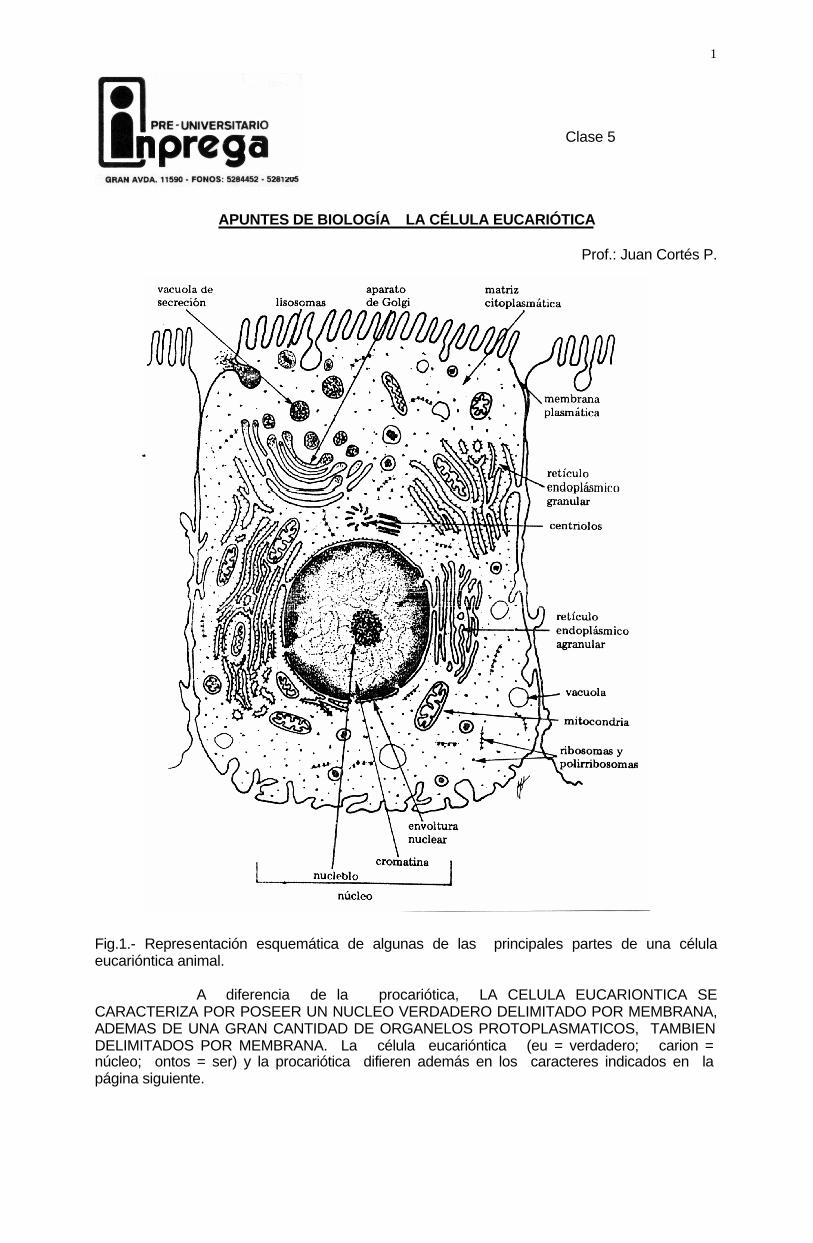
1 Clase 5 APUNTES DE BIOLOGÍA LA CÉLULA EUCARIÓTICA Prof.: Juan Cortés P. Fig.1.- Representación esquemática de algunas de las principales partes de una célula eucarióntica animal. A diferencia de la procariótica, LA CELULA EUCARIONTICA SE CARACTERIZA POR POSEER UN NUCLEO VERDADERO DELIMITADO POR MEMBRANA, ADEMAS DE UNA GRAN CANTIDAD DE ORGANELOS PROTOPLASMATICOS, TAMBIEN DELIMITADOS POR MEMBRANA. La célula eucarióntica (eu = verdadero; carion = núcleo; ontos = ser) y la procariótica difieren además en los caracteres indicados en la página siguiente.
Transcript of APUNTES DE BIOLOGÍA LA CÉLULA EUCARIÓTICA · total de la envoltura nuclear y se piensa que son...
1 Clase 5
APUNTES DE BIOLOGÍA LA CÉLULA EUCARIÓTICA
Prof.: Juan Cortés P. Fig.1.- Representación esquemática de algunas de las principales partes de una célula eucarióntica animal. A diferencia de la procariótica, LA CELULA EUCARIONTICA SE CARACTERIZA POR POSEER UN NUCLEO VERDADERO DELIMITADO POR MEMBRANA, ADEMAS DE UNA GRAN CANTIDAD DE ORGANELOS PROTOPLASMATICOS, TAMBIEN DELIMITADOS POR MEMBRANA. La célula eucarióntica (eu = verdadero; carion = núcleo; ontos = ser) y la procariótica difieren además en los caracteres indicados en la página siguiente.

2 CARACTER CELULA PROCARIONTICA CELULA EUCARIONTICA -Grupo en donde se Moneras Protistas, Hongos, Ve- encuentran getales y Animales. -Metabolismo Aero o anaeróbico Aeróbico -Envoltura nuclear Ausente Presente -ADN Desnudo, circular y Combinado a proteínas básicas
cerrado (histonas). También se observa ADN en mitocondrias y cloro-
plastos. -Núcleo Ausente. Sólo un cuerpo Verdadero núcleo delimitado nuclear o nucleoide por membrana. -Retículo endo- Ausente Presente plásmico -Cromosomas Unicos Múltiples (siempre más de uno) -Nucleólo Ausente Presente -División celular Amitosis Mitosis o meiosis -Ribosomas 70 S (200 A.) 80 S (de unos 250 A.) -Endomembranas Ausentes Presentes -Mitocondrias Ausentes. Respiración y Presentes fotosíntesis en plasmalema -Cloroplastos Ausentes Presentes en vegetales y algunos protistas autótrofos. -Pared celular No celulósica, excepto en Celulósica. Sólo en hongos, algas verdeazules algas y vegetales. -Exo y endocitosis No se observa. Sí se observa -Locomoción Pili y flagelos formados por Cilios y flagelos verdaderos proteínas simples. Formados por microtúbulos -Mesosomas Presentes Ausentes -Seudópodos Ausentes Presentes en algunas como amebas -Vacuolas Ausentes Presentes -Recombinación Conjugación Meiosis y fecundación sexual -Centríolos Ausentes Presentes en animales y vegetales inferiores -Cápsula o vaina Presente Ausente -Golgi, lisosomas, Ausentes Presentes peroxisomas, etc.

3 Fig. 2. - Concepción artística de una célula eucariónte animal. Fig. 3. - Se observa al lado una célula eucarió- tica animal completa. Se describe a continuación, la estructura típica de una célula eucarióntica, incluyendo sus principales organoides (organelos, organitos u organillos celulares). Los ORGANELOS son estructuras celulares membranosas, de presencia constante (durante toda o la mayor parte de la vida de la célula), de morfología y composición química definidas y que llevan a cabo funciones características y particulares. Estos órganos membranosos de la célula (organelos) se presentan como compartimentos donde se realizan diferentes funciones. Una célula vegetal típica posee un núcleo, unos 20 plastidios, 700 mitocondrias, 400 dictiosomas, 500 mil ribosomas y muchos otros organelos, los que obedecen al coordinador central: el NUCLEO.
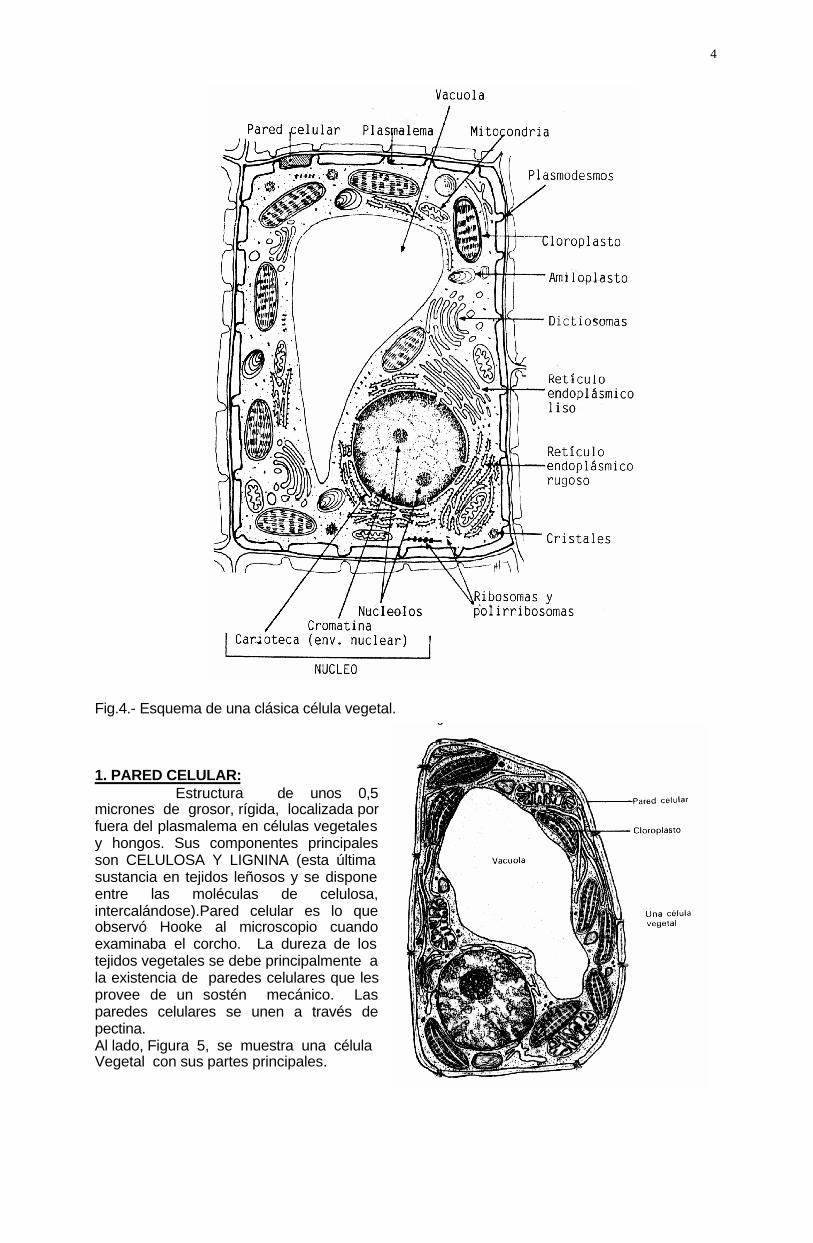
4 Fig.4.- Esquema de una clásica célula vegetal. 1. PARED CELULAR: Estructura de unos 0,5 micrones de grosor, rígida, localizada por fuera del plasmalema en células vegetales y hongos. Sus componentes principales son CELULOSA Y LIGNINA (esta última sustancia en tejidos leñosos y se dispone entre las moléculas de celulosa, intercalándose).Pared celular es lo que observó Hooke al microscopio cuando examinaba el corcho. La dureza de los tejidos vegetales se debe principalmente a la existencia de paredes celulares que les provee de un sostén mecánico. Las paredes celulares se unen a través de pectina. Al lado, Figura 5, se muestra una célula Vegetal con sus partes principales.

5 Es frecuente que una célula produzca varias paredes. La más externa (pared primaria) es menos rígida y crece junto con la célula. La pared secundaria, mucho más rígida, se estructura una vez que la célula ha alcanzado su tamaño definitivo. Fig.6.- Esquema de la pared celular de vegetales. La pared celular de células epidérmicas también puede contener cutinas y ceras, de carácter impermeabilizante. Muchos hongos y levaduras tienen una pared formada por quitina (polímero de glucosamina). La pared celular es un producto de la actividad citoplasmática y tiene por función proteger a la célula de cambios osmóticos, soporte y resistencia. También determina la forma celular. Su permeabilidad no es selectiva debido a su gran porosidad. Entre paredes de células contiguas es posible observar una LAMINILLA MEDIA de función cementante, formada por derivados de Ca (pectatos de Calcio) y Mg (Magnesio), permitiendo adhesión entre células. Una célula sin pared celular forma un PROTOPLASTO, TOTIPOTENCIAL de forma esférica, y puede crecer, mantenerse, dividirse e incluso llegar a formar un individuo adulto completo. Si la remoción de la pared es incompleta se forma un ESFEROPLASTO. En realidad, una célula puede vivir sin cápsula, pared celular cilios y flagelos. Aún sin núcleo podría mantenerse un tiempo, pero sin poder crecer ni dividirse, para finalmente morir. La pared celular se estructura a partir de la placa celular que se empieza a observar cuando la célula va a dividirse. 2. PLASMALEMA o membrana citoplasmática o celular: Estructura lipoproteica, formada principalmente por proteínas globulares y lípidos (fosfolípidos y esteroides), de apariencia trilaminar al microscopio electrónico, de unos 75 a 100 A. de grosor. En general todas las estructuras membranosas tienen una apariencia similar, sea cual sea su ubicación. Sin embargo, las proyecciones con forma de hongo, que a veces se visualizan en la membrana de bacterias, nunca se encuentran en eucarióntes. La membrana citoplasmática tiene por función MANTENER LA CONSTANCIA DEL MEDIO CELULAR INTERNO (homeostasis celular), controlando todo lo que entra y sale de la célula, y la velocidad a que lo hacen. Las membranas también ayudan a determinar la forma celular . Las membranas serán estudiadas con detalle en un capítulo especial.
Esquema de la Membrana celular

6 Fig.7.- Estructura de la membrana plasmática según el modelo de Singer (Modelo del Mosaico Fluido). 3. NUCLEO: El elemento interno más prominente, y de variadas formas, en la mayor parte de las células eucariónticas es el núcleo, descubierto por Robert Brown en 1831. El núcleo, que constituye el depósito de casi toda la información genética de la célula, está delimitado por una envoltura nuclear. Esta envoltura está compuesta por dos membranas similares, cada una de las cuales tiene alrededor de 100 A. de espesor, separadas por un espacio de unos 150 A. Individualmente, cada una de las membranas es similar a la membrana plasmática; no obstante, las dos membranas de la envoltura nuclear están unidas regularmente, fusionadas, formando aberturas circulares llamadas ANNULLI o POROS o complejos de poro. El tamaño y espaciado de los poros varía, pero es muy común el diámetro de 0,1 micrón (1.000 A.). Frecuentemente, los annulli ocupan una superficie equivalente a 1/3 del total de la envoltura nuclear y se piensa que son los responsables del pasaje selectivo hacia dentro y fuera del núcleo. Es decir, a través de estos poros se comunica directamente el carioplasma con el citoplasma, permitiendo el paso de macromoléculas como el ARNm. El espacio comprendido entre ambas membranas nucleares, es el espacio perinuclear. La cara nuclear de la membrana interna está cubierta por una capa proteica, que se conoce con el nombre de lámina, y a ella se encuentra unida cromatina condensada. La prominencia e importancia del núcleo se pone de manifiesto en la nomenclatura usada para describir el contenido celular: Todo lo encerrado dentro de la membrana citoplasmática, colectivamente se llama PROTOPLASMA, pero sólo aquella parte del protoplasma que se encuentra fuera del núcleo se llama CITOPLASMA. El contenido nuclear es el nucleoplasma, carioplasma o jugo nuclear. Asociados a la porción más externa de la carioteca, se encuentran abundantes ribosomas adoptando una gran similitud con el Retículo endoplásmico rugoso. Fig.8.- En la página siguiente, se observa parte del núcleo, que muestra las dos capas de la envoltura nuclear y, cerca, los dos centríolos, más partes de otros organelos. Al lado, se observa la ultraestructura de la envoltura nuclear. La membrana externa es similar a la del REG. Incluso posee ribosomas.

7 Fig.9.- Al lado, ultraestructura del núcleo celular mostrando las conexiones a que se hace alusión en párrafos anteriores. A veces, la forma del núcleo está relacionada con la forma de la célula, pero puede ser completamente irregular. El tamaño del núcleo, aunque variable, en general guarda relación con el volumen citoplasmático, de acuerdo al valor de índice o relación nucleoplasmática (NP): Vn NP = Vc-Vn Vn = volumen nuclear, Vc = volumen celular Al expresar NP un equilibrio entre los volúmenes nuclear y citoplasmático, cuando aumenta uno, el otro también debe aumentar. EL DESEQUILIBRIO DE ESTA RELACION SERIA UN ESTIMULO PARA LA DIVISION CELULAR. Existen también células binucleadas (como células hepáticas y cartilaginosas) y polinucleadas (hasta un centenar en células de la médula ósea (osteocitos)).En los plasmodios, que son grandes masas protoplasmáticas no divididas en territorios celulares, los núcleos son aún más numerosos. Tal es el caso de la fibra muscular estriada y el de ciertas algas. Las células animales multinucleadas se llaman sinciciales y las multinucleadas de vegetales y hongos se llaman cenocitos. La posición del núcleo es variable, pero en general es característica y constante para cada tipo celular. En las células embrionarias, casi siempre ocupa el centro geométrico, pero por lo común se desplaza cuando se produce la diferenciación, o por la formación de sustancias específicas o de reserva. En las células secretoras, se sitúa en la parte basal, y los gránulos de secreciones ocupan la zona apical (extremo superior). En vegetales gran parte del volumen del centro de la célula es ocupado por una gran vacuola central por lo que el núcleo es relegado a la periferia. Cualquiera sea la ubicación del núcleo, siempre está rodeado de citoplasma. Cuando se extirpa el núcleo a una célula por microdisección, ésta sigue con vida algún tiempo, pero no puede crecer ni dividirse, y finalmente muere. El núcleo (en realidad el ADN) se tiñe con verde de metilo.

8 4. Cromatina y cromosomas: Casi todos los genes de la célula se hallan en el núcleo asociados a los filamentos de CROMATINA. La cromatina está compuesta principalmente por DNA y proteínas básicas (histonas) y poca cantidad de ARN (Acido Ribonucleico o RNA). El ADN y las histonas se agregan entre sí en proporciones regulares, para conformar cuerpos semiesféricos conocidos como NUCLEOSOMAS. Cada nucleosoma está constituido por dos Tetrámeros de histonas. Por entre y sobre las proteínas histónicas se ubica el ADN. Estas proteínas básicas, con gran afinidad con los ácidos nucleicos, y que conforman el nucleosoma, se denominan H2A, H2B, H3 y H4. La cromatina se puede encontrar dispersa en el núcleo o agrupada en cuerpos discretos y compactos llamados CROMOSOMAS (cromos = color; soma = cuerpo), es decir cuerpos que se tiñen fácilmente. Los genes de una célula humana típica por ejemplo se hallan distribuidos en los 46 cromosomas. La cromatina se encuentra dispersa en el núcleo interfásico, en tanto que durante la división celular (mitosis o meiosis), se compacta formando los cromosomas. Los filamentos individuales de cromatina miden entre 100 y 400 A. de grosor (visibles sólo al microscopio electrónico). Fig.10.- Representación esquemática de cromatina y cromosomas. En a, está el modelo de la disposición del ADN y las proteínas en la cromatina. En b, aspecto de la cromatina al microscopio electrónico. En c, aspecto de un cromosoma. Evidencias recientes demuestran que la cromatina en parte se encuentra dispersa y se tiñe débilmente. A esta se le denomina EUCROMATINA y es la que sé está transcribiendo (formando ARNm), es decir la eucromatina tiene información genética que sé está utilizando, es la cromatina genéticamente activa. Otros filamentos de cromatina se condensan y tiñen con mayor facilidad constituyendo la HETEROCROMATINA (contiene información genética que no se está transcribiendo, no se está utilizando). Algunos filamentos de heterocromatina se pueden encontrar delimitando al nucléolo o asociada a la envoltura interna de la carioteca. A esta heterocromatina se le denomina HETEROCROMATINA CONSTITUTIVA. Otra porción de la heterocromatina puede convertirse en eucromatina y se le denomina HETEROCROMATINA FACULTATIVA. En relación a las histonas se debe señalar que forman unidades globulares, alrededor de las cuales se enrolla una fibra de ADN dando dos vueltas. Estos agrupamientos o abultamientos se llaman NUCLEOSOMAS y son los que le dan a la cromatina una apariencia similar a la de un collar de cuentas (de bolitas). Por enrollamientos sucesivos la cromatina se condensa y compacta. Se debe destacar que los filamentos de cromatina se condensan para formar los cromosomas sólo cuando la célula se va a dividir y como el material genético ya se ha duplicado en la interfase, cada cromosoma consta de 2 filamentos idénticos llamados cromátidas hermanas y que se encuentran unidas por el centrómero.

9 5. NUCLEOLO: Es la estructura más claramente definida en el interior del núcleo, el que en ocasiones puede observarse unido a la envoltura nuclear. La composición del nucléolo es muy similar a la de la cromatina, excepto por la presencia de numerosos gránulos ricos en ARN. Estos gránulos son PRECURSORES DE LOS RIBOSOMAS. La función del nucleólo es ARMAR las subunidades de los ribosomas, por lo que el tamaño del nucleólo y a veces el número de nucleólos por célula, varía con los requerimientos de síntesis de ribosomas. En el nucleólo, entonces, se forman, procesan y maduran los ARNr. (ARN ribosomal) El nucleólo se forma a partir de las regiones organizadoras nucleolares (NOR) localizadas en algunos cromosomas. El nucleólo no posee membrana propia, y no sería visible al microscopio óptico si no fuera por el empaquetamiento denso de sus fibras y gránulos. Durante la replicación celular, en tanto que la cromatina se condensa para formar los cuerpos discretos llamados cromosomas, el material del nucleólo normalmente se dispersa, volviendo a condensarse en las células hijas. Los nucleólos desaparecen al comienzo de la división celular y reaparecen durante la telofase. En el nucleólo, al microscopio electrónico, se observan 2 grandes zonas: a) Zona fibrilar: corresponde a la región central del nucleólo, en la que se encuentra el ADN que permite formar los ARNr. b) Zona granular: Es la región periférica del nucleólo, delimitada por heterocromatina. En esta zona se observan los precursores ribosómicos en diferentes etapas de producción. 6. MATRIZ CITOPLASMATICA: Llamada también HIALOPLASMA o citosol, es una estructura de naturaleza semilíquida, formada por un citoesqueleto, en el que se encuentran sostenidas las estructuras intracelulares. Está formada principalmente por agua, iones y pequeñas moléculas en solución verdadera, macromoléculas y enzimas solubles en solución coloidal, además de los componentes del citoesqueleto (microfilamentos, filamentos y microtúbulos) y filamentos microtrabeculares a los cuales se encuentran anclados los organelos, tal como se muestra en el siguiente esquema. El estado físicoquímico del protoplasma es considerado un coloide. De esto son responsables principalmente proteínas macromoléculas en general de un tamaño entre 0.0001 a 0,0000001 mm. Proteínas filamentosas se unen en ciertos puntos por enlaces débiles constituyendo una Red Tridimensional, dando en gran parte la consistencia COLOIDAL. En esta matriz ocurre gran parte de las reacciones metabólicas (glucólisis, fermentaciones y activación de aminoácidos por ej.). La parte más externa puede adquirir un estado más solidificado llamado GEL y constituye el ECTOPLASMA. La zona más interna de la matriz constituye el ENDOPLASMA, mucho más líquida llamada SOL. El paso GEL - SOL es reversible por reacciones tixotrópicas.
Fig.11.- Sistema microtrabecular y elementos del citoesqueleto al cual se encuentran anclados algunos organelos.

10 La estructura físico-química de una célula corresponde a la de un COLOIDE, con características intermedias entre GEL y SOL; es decir sin formar una solución verdadera, pero tampoco una solución grosera (precipitado). Las moléculas que forman un coloide miden entre 0.1 y 0.0001 micrón de diámetro (también se denominan micelas). Son soluciones coloidales la manteca, mayonesa, el jabón, la crema, la gelatina, la niebla y por supuesto la "cola" (pegamento), término del cual deriva coloide. Específicamente el citosol es el espacio comprendido entre los organelos membranosos. 7. COMPLEJO O APARATO DE GOLGI (Dictiosoma en vegetales): Casi todas las células eucarióntes poseen un complejo de vesículas y membranas llamado aparato o complejo de Golgi, en honor a Camilo Golgi quien lo describió por primera vez en 1898, denominándolo aparato reticular interno.
Fig.12.- Ultraestructura de complejo de Golgi visto al microscopio electrónico. El complejo de Golgi se presenta como un APILAMIENTO DE SACOS APLANADOS, separados entre sí por 200 - 300 A., con vesículas internas y vacuolas cerca de sus bordes laterales generalmente abultados. (Una vesícula es una pequeña esfera rodeada por una membrana. Una vacuola es una vesícula grande. Una cisterna es una cavidad). La función principal del Complejo de Golgi es la de SECRETAR . Por ejemplo, la secreción de material de la pared celular en plantas y de enzimas digestivas en páncreas, frecuentemente implica la liberación de vesículas desde los apilamientos del Golgi, y la unión de estas vesículas con otras desprendidas del retículo endoplásmico, para formar en conjunto una vacuola o gránulo de secreción. La vacuola de secreción, descarga su contenido fusionándose con la membrana plasmática, y abriéndose al exterior en un proceso llamado EXOCITOSIS (hacia afuera). El complejo de Golgi en células secretoras se encuentra polarizado entre el núcleo y el polo apical.
Fig.13.- A la izquierda se observa el complejo de Golgi en una ameba, con vesículas desprendiéndose de los bordes de las cisternas. A la derecha un dictiosoma de células apicales de la raíz de maíz, dedicado activamente a la formación de vesículas secretoras.

11 En general, se acepta que la función general del Complejo de Golgi es ALMACENAR, CONCENTRAR sustancias que le llegan principalmente por el retículo endoplásmico y EMPAQUETARLAS para su posterior SECRECION o para mantenerlas aisladas en una vesícula, como ocurre con los lisosomas, peroxisomas, etc. En realidad todas las sustancias que le llegan al Golgi, son rodeadas finalmente por membranas para ser secretadas o almacenadas. Se ha demostrado que el complejo de Golgi en células secretoras está polarizado estructural y funcionalmente, distinguiéndose una CARA DE FORMACION (zona cis o de entrada) próxima al RER y una CARA DE MADURACION (zona trans o de salida) orientada hacia el polo apical de la célula. Las moléculas desde el RE (retículo endoplásmico), llegan al Golgi por la cara de formación y salen por la cara de MADURACION. Otras funciones del Complejo de Golgi: - Forma el acrosoma de los espermatozoides - Formación del fragmoplasto (estructura que forma la placa celular de división en células vegetales). - Concentra enzimas hidrolíticas (proteolíticas, lipolíticas, etc.) en lisosomas (lisosomas primarios). - Forma carbohidratos que luego une a proteínas para formar glucoproteínas (glicoproteínas) de secreción. Esto se denomina Glicosilación . - Forma el glicocálix (glucocálix) celular. (Organelo Nro. 24). - Síntesis de carbohidratos del tipo de los polisacáridos como celulosa. - Transporte intracelular. - Aumento del área de la superficie celular. - Reciclaje y rehuso de componentes de membrana. Las células animales, normalmente presentan uno o unos pocos complejo de Golgi. Otros textos indican que las células animales poseen 10 a 20 Complejos de Golgi. Las células vegetales, normalmente presentan más de uno. 8. RETICULO ENDOPLASMATICO:

12 Fig. 15.- En el esquema de página anterior, se representa el RETICULO ENDOPLASMICO junto a otros organelos celulares. El RETICULO ENDOPLASMICO es una compleja red de membranas citoplasmáticas que en ciertos puntos se conecta con el plasmalema y la carioteca, formando una verdadera red (retículo) en el citoplasma. Es endoplásmico pues en ésa área celular se le encuentra principalmente. Cuando se rompe una célula y se separan sus componentes, se puede identificar una fracción microsomal compuesta principalmente por retículo endoplásmico.
El retículo puede adoptar forma tubular, cisternas, surcos, vesículas y sacos aplanados, es decir, áreas cerradas y semicerradas. Estos canales y cámaras actúan en el almacenamiento y principalmente TRANSPORTE INTRACELULAR de sustancias que no se liberan al citoplasma. Muchos ribosomas se encuentran unidos a parte del retículo y por la apariencia que adopta, se le denomina retículo endoplásmico (endoplasmático) RUGOSO o GRANULOSO, para diferenciarlo del retículo endoplásmico LISO O AGRANULAR, que no posee ribosomas adheridos. Fig.16.- Ultraestructura del retículo endoplásmico rugoso o granular. A la derecha, la fig.17 muestra ribosomas del retículo endoplásmico rugoso en plena síntesis proteica. Los materiales destinados a ser secretados se transportan en vesículas por el citoplasma, y que se han desprendido del retículo. Estas vesículas pueden fusionarse con otras del Golgi como ya se indicó para formar gránulos de secreción. El RETICULO ENDOPLASMICO puede considerarse como la FABRICA DE MEMBRANAS celulares. Los ribosomas se unen al retículo endoplásmico a través de su subunidad mayor y TODAS LAS PROTEINAS SINTETIZADAS POS ESTOS RIBOSOMAS UNIDOS AL RETICULO, CAEN AL RETICULO PARA SER FINALMENTE SECRETADAS O ALMACENADAS RODEADAS DE UNA MEMBRANA, COMO SE OBSERVA EN LISOSOMAS POR EJEMPLO, CON AYUDA DEL COMPLEJO DE GOLGI. Fig.18.- Ultraestructura del retículo endoplásmico liso o agranular
Recuerda que los MICROSOMAS son vesículas derivadas del RER, de mayor densidad que los microsomas lisos provenientes del REL. Estos debido a la presencia de los ribosomas.
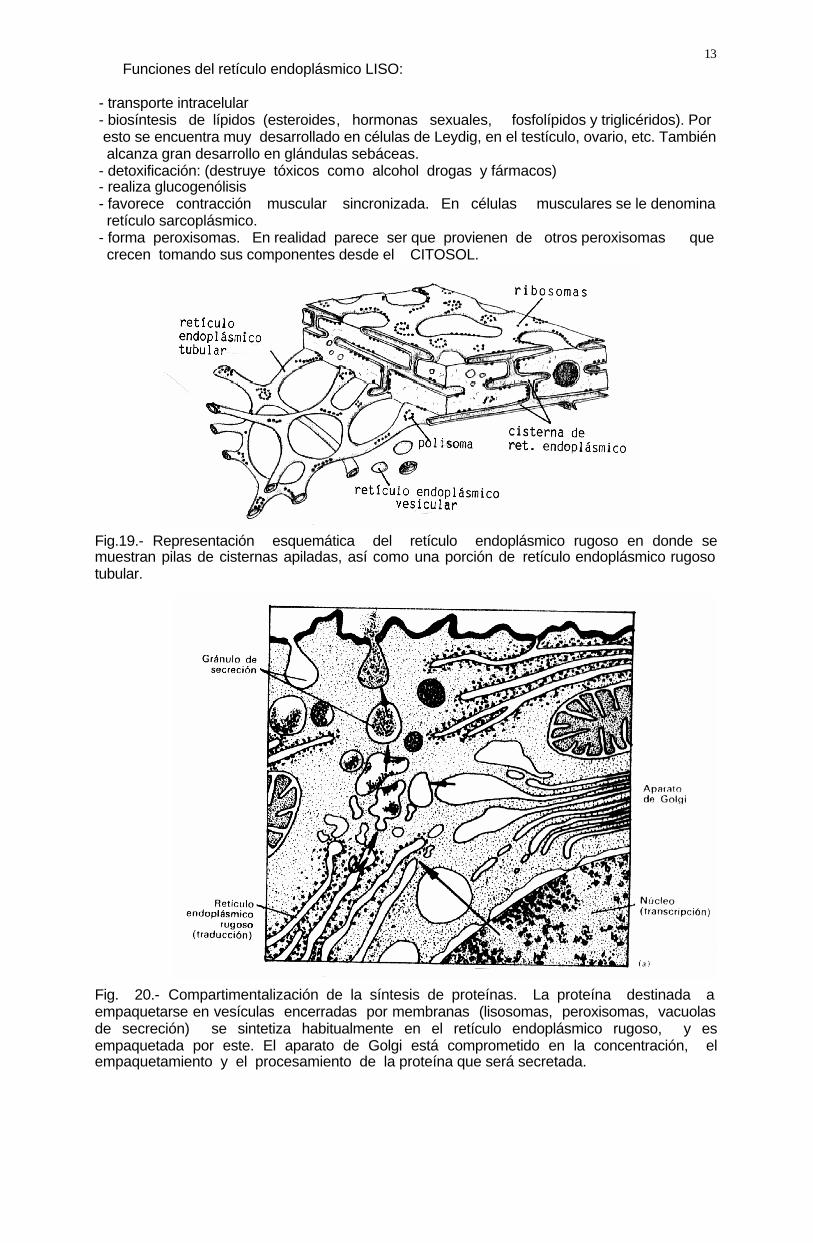
13 Funciones del retículo endoplásmico LISO: - transporte intracelular - biosíntesis de lípidos (esteroides, hormonas sexuales, fosfolípidos y triglicéridos). Por esto se encuentra muy desarrollado en células de Leydig, en el testículo, ovario, etc. También alcanza gran desarrollo en glándulas sebáceas. - detoxificación: (destruye tóxicos como alcohol drogas y fármacos) - realiza glucogenólisis - favorece contracción muscular sincronizada. En células musculares se le denomina retículo sarcoplásmico. - forma peroxisomas. En realidad parece ser que provienen de otros peroxisomas que crecen tomando sus componentes desde el CITOSOL. Fig.19.- Representación esquemática del retículo endoplásmico rugoso en donde se muestran pilas de cisternas apiladas, así como una porción de retículo endoplásmico rugoso tubular. Fig. 20.- Compartimentalización de la síntesis de proteínas. La proteína destinada a empaquetarse en vesículas encerradas por membranas (lisosomas, peroxisomas, vacuolas de secreción) se sintetiza habitualmente en el retículo endoplásmico rugoso, y es empaquetada por este. El aparato de Golgi está comprometido en la concentración, el empaquetamiento y el procesamiento de la proteína que será secretada.

14 Funciones del R. E. RUGOSO: - Transporte intracelular - Síntesis de proteínas de exportación o que van a ser delimitadas, encerradas por membranas. - Síntesis de hormonas proteicas. El retículo endoplásmico también se conoce como ERGASTOPLASMA, sustancia basófila o tigroide o sustancia de Nissl. Estos dos últimos sinónimos son usados especialmente en neuronas. El Retículo endoplásmico, Complejo de Golgi y Carioteca conforman el SISTEMA VACUOLAR O ENDOMEMBRANOSO y permiten el transporte de sustancias sin que éstas entren en contacto directo con el hialoplasma. En este sistema vacuolar es posible observar un FLUJO DE MEMBRANA, en donde un trozo de membrana de la carioteca puede luego encontrarse formando parte del retículo y luego del complejo de Golgi. El Ret. End. y la membrana externa de la envoltura nuclear presentan una continuidad física. Por otra parte, R. E. , Complejo de Golgi y Plasmalema se comunican entre sí a través de vesículas citoplasmáticas. 9. RIBOSOMAS : Partículas de unos 200 - 250 A. de diámetro, visibles sólo al microscopio electrónico, y compuestos principalmente por ARNr (ARN ribosomal) y proteínas. Fig.21.- Al lado, modelo actual de la estructura de un ribosoma. La subunidad menor está señalada con gris, y la subunidad mayor (por supuesto más grande) está señalada de color más blanco. Abajo se muestra el esquema clásico de un ribosoma. Los RIBOSOMAS constituyen el lugar físico para la SINTESIS DE PROTEINAS. Este es el proceso por medio del cual los aminoácidos (unidades constitutivas de las proteínas) son unidos en secuencias definidas para producir las cadenas polipeptídicas (proteínas). Los ribosomas son un constituyente esencial de todos los seres vivos, y están formados por dos subunidades, cada una formada por ARNr + proteínas. Estas subunidades se "arman" en el NUCLEOLO gracias a la existencia de 3 factores, y luego como subunidades difunden al citoplasma en donde se unen para formar al ribosoma, siempre y cuando exista ARN mensajero (ARNm). Los ribosomas pueden encontrarse en el citoplasma en forma independiente o unidos al retículo endoplásmico rugoso. Todas las proteínas que se sintetizan en los ribosomas del retículo son "exportadas" (secretadas) o almacenadas en vesículas como las de los lisosomas. Las proteínas fabricadas en los ribosomas libres, quedarán en la matriz celular o formando parte de otra estructura protoplasmática. Los principales componentes de los ribosomas son proteínas y ARNr que conforman las RIBONUCLEOPROTEINAS. Proteínas y ARNr se encuentran en proporciones equivalentes. Las proteínas guardan semejanza con las histonas (proteínas asociadas al ADN). Los ribosomas eucariónticos son 80 S y resultan de la unión de las subunidades 60 S y 40 S. Los procarióntes tienen ribosomas 70 S, y resultan de la unión de subunidades 50S y 30S. "S" indica coeficiente de sedimentación o de Sbedberg. Las subunidades se unen cuando existe ARNm, es decir, cuando existen necesidades proteicas. Para la unión de estas subunidades se requiere magnesio.

15 Varios ribosomas unidos constituyen un POLISOMA o polirribosoma, y en ellos tiene lugar la síntesis proteica. MICROCUERPOS: Incluyen a Peroxisomas y Glioxisomas: 10. PEROXISOMAS: Delimitados por membrana, miden entre unos 0,5 y 1,5 micrones de diámetro, y contienen la mayor parte de las enzimas capaces de producir y degradar peróxidos. De su contenido sobresale la catalasa, cuya función es degradar peróxidos de oxígeno (H2O2) a oxígeno y agua. En otros seres vivos también es posible encontrar peroxisomas conteniendo enzimas de importantes vías metabólicas. Los peroxisomas también permiten la oxidación de una gran variedad de sustancias que usan al oxígeno como agente oxidante. Una de las enzimas que permite esto es la uratooxidasa. La mitad del etanol consumido por un individuo es detoxificado en peroxisomas de células hepáticas. También degrada lípidos. (mineduc.cl) 11. GLIOXISOMAS : (Peroxisomas especializados). Vesículas semiesféricas de unos 0,5 a 1,5 micrones de diámetro y contienen las enzimas necesarias para la conversión de grasas y aceites en hidratos de carbono (ciclo del glioxilato). Estos organelos, análogos a peroxisomas son típicos de microorganismos y vegetales superiores. Por esto también, suele decirse que los peroxisomas OXIDAN ACIDOS GRASOS. 12. LISOSOMAS (BOLSAS SUICIDAS O VACUOLAS AUTOFAGICAS): Fueron descubiertos por Christian de Duve en 1952, y poseen un diámetro de unos 0,03 a 0,8 micrones. Se originan en el complejo de Golgi, y su función es MANTENER AISLADAS ENZIMAS DIGESTIVAS (Proteolíticas, lipolíticas, etc. Hidrolíticas en definitiva), capaces de degradar una gran variedad de sustancias. Es decir, los lisosomas mantienen aisladas aquellas enzimas que signifiquen riesgo para su metabolismo, o para el órgano al cual pertenece. El pH lisosómico es 5 aproximadamente (ácido). Estas enzimas hidrolíticas de los lisosomas incluyen fosfatasas, lipasas (hidrolizan lípidos), glucosidasas (hidrolizan polisacáridos), proteasas o proteolíticas como la catepsina (hidrolizan proteínas) y nucleasas como la ADNasa y ARNasa que hidrolizan ácidos nucleicos.
Fig.22.- Diagrama de la estructura del ribosoma y la influencia del magnesio (Mg). Se muestra también un polirribosoma formado por cinco ribosomas. Se considera como ARN mensajero (ARNm) al filamento que une los ribosomas. Se indican también las constantes de sedimentación (S) de las diversas partículas. Recordemos que las subunidades ribosomales se arman en la zona fibrilar (central) del nucleólo. Algunas células como los reticulocitos pueden presentar hasta 100.000 ribosomas.

16 El lisosoma con sus enzimas inactivas es un LISOSOMA PRIMARIO. En períodos de ayuno prolongado, la proteína de nuestras células musculares se destruye fácilmente con ayuda de los lisosomas, y es reutilizada por la célula. Las sustancias a digerir también pueden ser introducidas a la célula por el proceso de la ENDOCITOSIS, encerrándolas en una vacuola (fagosoma o pinosoma. ¿Recuerdas los esquemas hechos en clase?. De todas formas en estas páginas tienes otros esquemas). El fagosoma o pinosoma, se funde con un lisosoma para constituir una vacuola digestiva (llamada también LISOSOMA SECUNDARIO O LISOSOMA ACTIVADO). Como ya vimos en clase, la endocitosis de líquido se llama PINOCITOSIS y la de material sólido FAGOCITOSIS. La vacuola que se forma se llama PINOSOMA y FAGOSOMA respectivamente. Lo no digerible ni utilizable de la vacuola digestiva constituye una vacuola de desecho o vacuola residual, la que luego es eliminada por exocitosis (¿excreción?). La exocitosis de material útil se llama secreción. En la exocitosis, la vacuola se funde con el plasmalema, vertiendo su contenido al medio externo. Se observa prácticamente en todo tipo de células eucariótas.
En ocasiones, los lisosomas (también llamados bolsas suicidas y vacuolas autofágicas), al romperse en el interior de la célula, liberan las enzimas que pueden producir la muerte y destrucción celular (AUTOLISIS). Esto permite la reabsorción de la cola del renacuajo durante la metamorfosis de los sapos por ejemplo. Los glóbulos blancos pueden ingerir hasta más de un centenar de gérmenes, los cuales son fusionados con lisosomas para su posterior destrucción. Cuando el fagosoma contiene sustancias provenientes del exterior, se llaman HETEROFAGOSOMAS y cuando contienen sustancias provenientes de la propia célula, se llaman AUTOFAGOSOMAS .
Fig. 23.- Endocitosis. En a, endocitosis de material semisólido (FAGOCITOSIS). En b, endocitosis de material líquido (PINOCITOSIS).
Fig. 24. Exocitosis: salida de sustancias desde la célula, vía vesículas. Puede tratarse de secreción o excreción. Se debe destacar que las sustancias a secretar provienen del ANABOLISMO, en tanto que las sustancias a excretar, provienen del CATABOLISMO.

17 13. VACUOLAS : Además de los ya descritos, se encuentran muchos otros sacos o vacuolas rodeados por membranas en diferentes tipos de células, especialmente en vegetales. En realidad se acepta que las vacuolas son típicas de células vegetales. En células animales se observan sólo vesículas (menor tamaño). (Recuerda que las vesículas son vacuolas chicas). Las vacuolas están delimitadas por una membrana transparente y elástica llamada TONOPLASTO, y la función de vacuolas es la de actuar como SITIOS DE ALMACENAMIENTO. En realidad, la mayoría de las células vegetales tiene una vacuola única que ocupa el 80 - 90% del total del volumen celular. Su contenido se denomina jugo celular e incluye principalmente agua, a la cual almacena, compuestos inorgánicos, azúcares, etc. También colaboran en la mantención de la turgencia celular. En vegetales, la vacuola henchida de agua ejerce una presión sobre la pared, que es determinante para la firmeza y sostén de los tejidos (FUNCION ESTRUCTURAL), ya que al henchirse de agua, aumenta la presión (presión de turgor = TURGENCIA) sobre la pared de la célula. Siempre en vegetales, la vacuola puede actuar almacenando desperdicios, cristales de calcio y sodio entre otras. En las VACUOLAS de vegetales, también existe una DIGESTION INTRACELULAR , es decir en esto se parecen a lisosomas. GUTACION: Transpiración en que el agua abandona la hoja en forma gaseosa. 14. CLOROPLASTOS (PLASTIDIOS): Organelos típicos de vegetales, aunque también es posible observarlos en algunos protistas como la euglena. Los plastidios poseen una doble membrana; ADN circular, cerrado y desnudo; ARN; ribosomas (70 S) y variadas inclusiones. Los plástidos se originan de un precursor más pequeño, llamado PROPLASTIDO, o bien por desdiferenciación de otro plástido ya maduro. Algunos plástidos poseen color, y se llaman CROMOPLASTOS. Los incoloros se llaman LEUCOPLASTOS. De los cromoplastos, el más notable es el CLOROPLASTO, frecuentemente de unos 3 a l0 micrones de largo. Estos plástidos, y más específicamente la clorofila que contienen es la responsable del color verde de los vegetales. Pero los cloroplastos también tienen pigmentos de otros colores, opacados por el verde de la clorofila, pero cuando una hoja muere en otoño, o madura un tomate, se destruye primero la clorofila, descubriendo los otros pigmentos. Los cloroplastos son el sitio en donde tiene lugar la FOTOSINTESIS, proceso que usa la energía lumínica para fijar CO2 y junto al agua producir azúcares como la glucosa y otros compuestos orgánicos, liberándose oxígeno en el proceso. Como los cloroplastos poseen ADN, son organelos celulares semiautónomos al igual que las mitocondrias. Serán estudiados en detalle en la unidad de fotosíntesis.
Fig. 25. Acción de un lisosoma primario, formado en el aparato de Golgi, sobre una vacuola fagocítica. En a, formación de una vacuola fagocítica. En b, vacuola fagocítica (fagosoma). En c, lisosoma primario. En d, fusión de un lisosoma primario con el fagosoma, originando una vacuola digestiva o lisosoma secundaria. En f, vacuola o cuerpo residual. En g, exocitosis de desechos: excreción o "defecación".
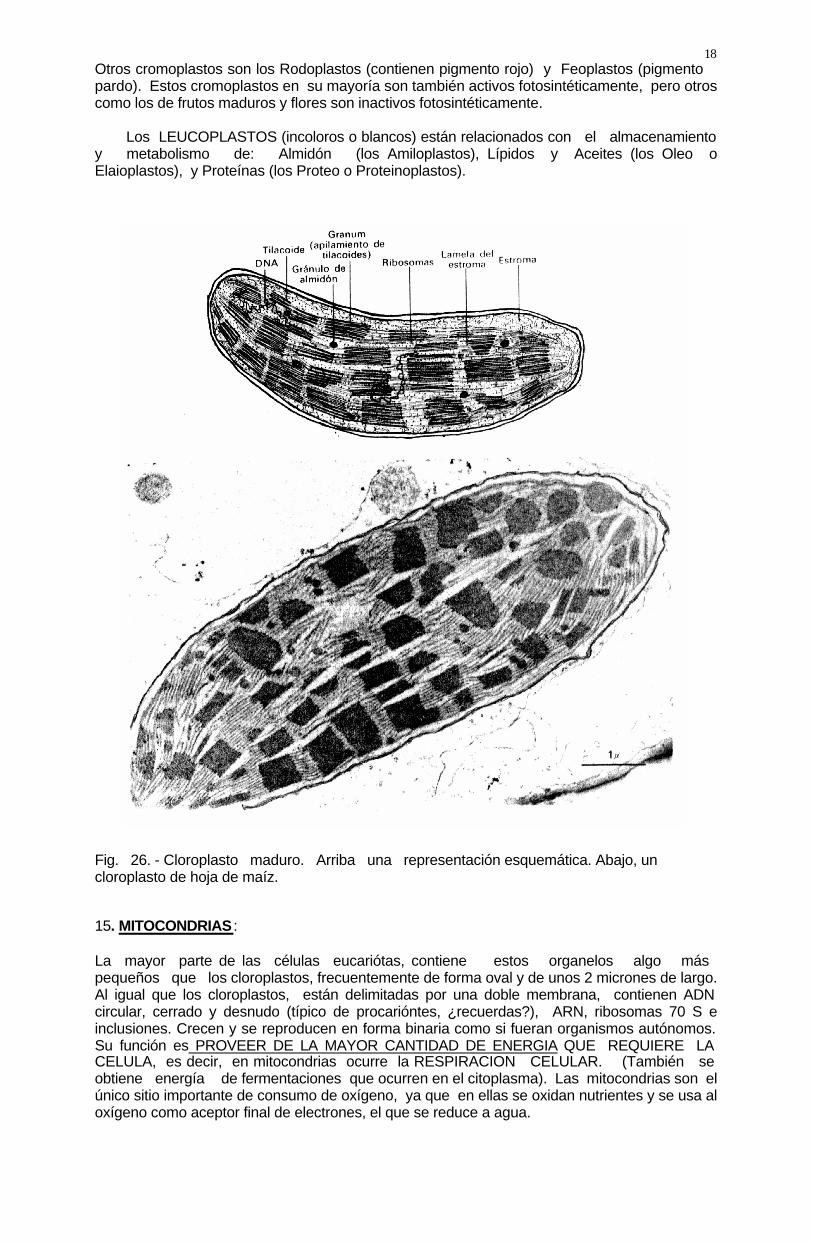
18 Otros cromoplastos son los Rodoplastos (contienen pigmento rojo) y Feoplastos (pigmento pardo). Estos cromoplastos en su mayoría son también activos fotosintéticamente, pero otros como los de frutos maduros y flores son inactivos fotosintéticamente. Los LEUCOPLASTOS (incoloros o blancos) están relacionados con el almacenamiento y metabolismo de: Almidón (los Amiloplastos), Lípidos y Aceites (los Oleo o Elaioplastos), y Proteínas (los Proteo o Proteinoplastos). Fig. 26. - Cloroplasto maduro. Arriba una representación esquemática. Abajo, un cloroplasto de hoja de maíz. 15. MITOCONDRIAS : La mayor parte de las células eucariótas, contiene estos organelos algo más pequeños que los cloroplastos, frecuentemente de forma oval y de unos 2 micrones de largo. Al igual que los cloroplastos, están delimitadas por una doble membrana, contienen ADN circular, cerrado y desnudo (típico de procarióntes, ¿recuerdas?), ARN, ribosomas 70 S e inclusiones. Crecen y se reproducen en forma binaria como si fueran organismos autónomos. Su función es PROVEER DE LA MAYOR CANTIDAD DE ENERGIA QUE REQUIERE LA CELULA, es decir, en mitocondrias ocurre la RESPIRACION CELULAR. (También se obtiene energía de fermentaciones que ocurren en el citoplasma). Las mitocondrias son el único sitio importante de consumo de oxígeno, ya que en ellas se oxidan nutrientes y se usa al oxígeno como aceptor final de electrones, el que se reduce a agua.

19 Fig. 27.- Representación diagramática de la estructura de una mitocondria. Veremos más detalles al estudiarlas en metabolismo de la glucosa. Fig. 28. - Comparación entre mitocondrias, cloroplastos y células procarióticas. Tal como ya lo explicamos en clases, parece ser que en algún momento mitocondrias y cloroplastos fueron organismos procariónticos de vida independiente, hasta ser "fagocitados", pero no digeridos por otra célula, de la cual finalmente se hicieron mutuamente dependientes. Las células que no contienen mitocondrias (como los glóbulos rojos), no pueden usar oxígeno. Las mitocondrias, también llamados condriosomas o sarcosomas en músculos, se reconocen fácilmente pues poseen en su interior membranas que conforman las crestas mitocondriales, que dividen su interior en compartimentos casi aislados. Se estudiarán con más detalle en la unidad de respiración celular. El DNA que existe en mitocondrias, posee el material genético que permite formar el 10% del total de proteínas mitocondriales. 16. FILAMENTOS CITOPLASMATICOS: Además de los microtúbulos, estos filamentos forman parte del CITOESQUELETO. Este citoesqueleto que se muestra en figuras anteriores, tiene por función SOSTENER los organelos y permitir el movimiento del protoplasma.
Como ya estudiaremos, parece ser que las mitocondrias y cloroplastos fueron alguna vez organismos moneras, tal como se pone en evidencia con el siguiente esquema.

20 Sigamos ahora con los filamentos. Estos pueden ser de 2 tipos: a) Microfilamentos (ACTINA PRINCIPALMENTE): miden entre 40 y 60 A. de diámetro, y presentan un aspecto de simples hebras. Entre sus funciones se debe destacar: - Forman haces en el ECTOPLASMA, haciéndolo más sólido, mas GEL. El ectoplasma es la región inmediatamente por debajo celular o segmentación. - Forman parte de la emisión de seudópodos (responsables del movimiento ameboídeo). - Estabilizan la estructura de las microvellosidades celulares. - Se relacionan con movimientos citoplasmáticos de células vegetales como por ej. ciclosis. Estos microfilamentos están formados por actina, miosina y otras proteínas contráctiles, y de ahí su capacidad para participar en contracciones y movimiento. (Recuerda que actina y miosina son proteínas contráctiles típicas de células musculares). b) Filamentos o filamentos intermedios: Miden unos 80 A 100 A. de diámetro y son de forma tubular. Han sido estudiados principalmente en neuronas en donde se cree, participan en TRANSPORTE DE SUSTANCIAS, y estabilizando la forma y estructura celular. En células nerviosas se les conoce como NEUROFILAMENTOS. En células epiteliales constituyen los tonofilamentos o DESMOSOMAS y las UNIONES INTERMEDIAS, tipos de unión entre células que estudiaremos más adelante. La QUERATINA también es un filamento intermedio. También existen filamentos gruesos de miosina en la mayoría de las células eucariónticas. Los tres elementos del citoesqueleto se conectan con el sistema microtrabecular. 17. MICROTUBULOS: Estructuras tubulares de unos 250 A., con un núcleo hueco de unos 150 A.. Sus paredes están formadas por 13 protofilamentos individuales, cada uno de los cuales tiene unos 40 A. de diámetro. Entre sus funciones podemos destacar: La tubulina es una proteína globular, y es la subunidad constitutiva de los microtúbulos - Son los componentes básicos de centríolos, cuerpos basales, cilios, flagelos y huso mitótico. - Son los responsables de la forma definida que presentan algunas células, como los hematíes por ej. (planos bicóncavos) - Transporte de sustancias a lo largo del axón en neuronas. - Con sus proteínas contráctiles permite el movimiento de cilios y flagelos. 18. SISTEMA MICROTRABECULAR : (Ver FIG. 11): Formado por fibras de 30 a 60 A. de diámetro y de naturaleza proteica. Este sistema permite anclarse a las otras partes del citoesqueleto (microtúbulos y ambos tipos de filamentos) formando una red en donde se encuentran los organelos orientados de acuerdo a la actividad metabólica celular. El sistema microtrabecular es como la musculatura celular dinámica (se mueve). 19. CENTRIOLOS Y CUERPOS BASALES: Las células animales (pero no las células vegetales, excepto algunas algas primitivas) contienen habitualmente cerca del núcleo dos cuerpos cilíndricos huecos, de unos 0,4 micrones de largo por 0,15 de diámetro, generalmente ubicados en forma perpendicular entre sí, alrededor del núcleo. La ubicación de estos centríolos define el CENTROSOMA o CENTROSFERA, zona clara y melificada que aparece alrededor de los centríolos durante la división celular. La zona clara alrededor del áster se llama ASTROSFERA.
Fig.29.- Esquema en que se muestra la ultraestructura de centríolos.

21 Las paredes de centríolos y cuerpos basales están formadas por 9 conjuntos de microtúbulos (cada conjunto formado por 3 microtúbulos), estructura conocida como "9 + 0". Se cree que microtúbulos, así como toda estructura tubular derivan siempre de los centríolos o cuerpos basales. Estos 2 organelos parecen ser autoduplicables. De entre las funciones de los centríolos, podemos destacar: - organiza huso acromático durante la división celular. - origina cuerpos basales de estructura similar y a partir de los cuales surgen cilios y flagelos. 20. CILIOS Y FLAGELOS: Los cilios miden generalmente entre 3 y l0 micrones de longitud por 0,5 de diámetro, y normalmente se encuentran en gran número en aquellas células que los poseen. Los flagelos son de un diámetro similar, pudiendo medir entre 100 y 200 micrones de longitud. En general, las células no presentan más de uno o dos flagelos. Cilios y flagelos son típicos de protistas y células animales. No se observan en células vegetales ni en hongos, (tampoco presentan centríolos ni cuerpos basales). La estructura de cilios y flagelos está dada principalmente por microtúbulos dispuestos en 9 pares periféricos, más dos microtúbulos simples centrales, estructura conocida como "9 + 2". Cilios y flagelos son apéndices locomotores, que deben su movimiento a la flexión del axonema formado por los microtúbulos y proteínas asociadas. Los cilios cumplen también una función protectora, como se observa en cilios de las células de vías respiratorias (10 mil millones por cm2), en donde son capaces de captar bacterias y partículas nocivas para que sean fagocitadas por glóbulos blancos o sean "barridas" hacia el medio ambiente externo. En oviducto también existen cilios. Normalmente alrededor del flagelo existe gran cantidad de mitocondrias que le están aportando la energía para el movimiento. En la Fig.30 siguiente, se representa la estructura de un cuerpo basal con su raicilla correspondiente. También se observa un corte transversal por cuerpo basal y por cilio con las estructuras clásicas "9 + 0" y "9 +2" respectivamente. También se alcanzan a observar otros organelos que identificas fácilmente, ¿verdad?.

22 21. VACUOLA PULSATIL O CONTRACTIL: Organelo típico de protistas y que tiene por función eliminar el exceso de agua celular. Fig.31.- Esquema de un paramecio, protista, en el que se muestran, entre otros organelos una vacuola pulsátil. Al lado se representa una vacuola pulsátil "llena". 22. INCLUSIONES: Estructuras protoplasmáticas no regulares (por ello no se consideran organelos) pues constituyen reservas de sustancias nutritivas. 23. DEUTEROPLASMA: Inclusiones caracterizadas por ser de gran tamaño, como por ej. vitelo de un huevo. 24. GLUCOCALIX O GLICOCALIX: Envoltura por fuera del plasmalema, típica de células animales y análoga a pared celular de vegetales. Está formada principalmente por proteínas (glucoproteinas) y polisacáridos. Participa en la adhesión entre células, le OTORGA INDIVIDUALIDAD A LA CELULA Y CONTIENE ANTIGENOS. 25. HIDROGENOSOMAS : Encargado de regular la concentración de H+ en el medio, almacenando más o menos. Participa así en la regulación de pH. TAMAÑOS CELULARES: Diámetros promedio Tipos celulares 0.1 a 1 micrón micoplasmas, rickettsias y clamidias (bacterias más pequeñas) 0.5 a 5 " bacterias y algas verdeazules. 5 " levaduras (hongos unicelulares). 5 a 25 " células humanas. 10 a 60 " células animales en general. 10 a 100 " células vegetales en general. 10 a 200 " protistas en general (protozoos y algas unicelulares). Otros tamaños: 5 " Escherichia coli 7 " eritrocitos (glóbulos rojos) 54 " espermatozoide humano. 75 " huevo de erizo. 100 " óvulo humano. 100 " grosor de un cabello. 75 mm óvulo de avestruz. 80 a 100 mm Acetabularia (alga unicelular)


















