
APRENDIZAJE SOCIAL Y ESTRATEGIAS DE FORRAJEO - … · 111 aprendizaje social y estrategias de...
12
Revista Mexicana de Psicología ISSN: 0185-6073 [email protected] Sociedad Mexicana de Psicología A.C. México Cabrera, Rosalva; Durán, Ángel; Nieto, Javier Aprendizaje social y estrategias de forrajeo en parvadas de palomas: Efectos de la cantidad de alimento Revista Mexicana de Psicología, vol. 23, núm. 1, junio, 2006, pp. 111-121 Sociedad Mexicana de Psicología A.C. Distrito Federal, México Disponible en: http://www.redalyc.org/articulo.oa?id=243020646013 Cómo citar el artículo Número completo Más información del artículo Página de la revista en redalyc.org Sistema de Información Científica Red de Revistas Científicas de América Latina, el Caribe, España y Portugal Proyecto académico sin fines de lucro, desarrollado bajo la iniciativa de acceso abierto
Transcript of APRENDIZAJE SOCIAL Y ESTRATEGIAS DE FORRAJEO - … · 111 aprendizaje social y estrategias de...

Revista Mexicana de Psicología
ISSN: 0185-6073
Sociedad Mexicana de Psicología A.C.
México
Cabrera, Rosalva; Durán, Ángel; Nieto, Javier
Aprendizaje social y estrategias de forrajeo en parvadas de palomas: Efectos de la cantidad de
alimento
Revista Mexicana de Psicología, vol. 23, núm. 1, junio, 2006, pp. 111-121
Sociedad Mexicana de Psicología A.C.
Distrito Federal, México
Disponible en: http://www.redalyc.org/articulo.oa?id=243020646013
Cómo citar el artículo
Número completo
Más información del artículo
Página de la revista en redalyc.org
Sistema de Información Científica
Red de Revistas Científicas de América Latina, el Caribe, España y Portugal
Proyecto académico sin fines de lucro, desarrollado bajo la iniciativa de acceso abierto

111
APRENDIZAJE SOCIAL Y ESTRATEGIAS DE FORRAJEOEN PARVADAS DE PALOMAS:
EFECTOS DE LA CANTIDAD DE ALIMENTO
SOCIAL LEARNING AND FORAGING STRATEGIES IN FLOCKSOF PIGEONS: EFFECTS OF FOOD AMOUNT
Rosalva Cabrera,1 Ángel Durán y Javier NietoUniversidad Nacional Autónoma de México
Revista Mexicana de Psicología, Junio 2006Volumen 23, Número 1, 111-121
Resumen: Se evaluó el efecto de variar la cantidad de granocolocada en diferentes depósitos sobre la adquisición de unarespuesta para obtener alimento y sobre las estrategias de forrajeode grupos de palomas expuestas a un coespecífico experto. Par-vadas de palomas ingenuas fueron expuestas a una paloma en-trenada en abrir los sellos de depósitos de alimento de diferentecolor, cada color se correlacionó con una cantidad de granodiferente. Posteriormente, la paloma entrenada fue retirada,manteniéndose la correlación color-cantidad de alimento. Losresultados mostraron que las palomas adquirieron la respuestapara obtener alimento y que la ejecutaron discriminadamente,respondiendo en mayor proporción a los colores que señalaronmayor cantidad de alimento. También jugaron tanto papeles deproductores como de parásitos, los cuales variaron en funciónde la composición del grupo. Los datos son congruentes conhallazgos previos sobre aprendizaje social y con lo propuestopor la teoría de forrajeo óptimo.Palabras clave: aprendizaje, cantidad de alimento, discrimina-ción, forrajeo, palomas
Abstract: This experiment evaluated the effect of varying theamount of grain placed into food containers on the acquisitionof a response to obtain food, and on the foraging strategies offlocks of pigeons given the opportunity to observe foraging inan expert conspecific. Flocks of naïve pigeons were first ex-posed to an experienced pigeon trained to open food cachesidentified by different colored seals, where colors identifieddifferent amounts of grain. Later, the experienced pigeon wasremoved from the experimental setting, and the naïve pigeonswere tested with the correlation between colors and amounts offood maintained. The results showed that the naïve pigeonsacquired the food-obtaining response, and that they performedit discriminately by responding in a greater proportion to thestimulus associated with a larger food cache. With respect toforaging strategies, the pigeons adopted the roles of producerand scrounger; varying as a function of group composition. Thesefindings are consistent with previous studies on social learningand with the theory of optimal foraging.Key words: learning, food amount, discrimination, foraging,social learning, pigeons
La teoría de forrajeo óptimo da cuenta de cómo los orga-nismos utilizan los recursos disponibles en su entorno(Krebs & Davies, 1984). Shettleworth (1984) señala queel punto de inicio de la teoría de forrajeo óptimo es unaformalización de las cosas que un organismo necesitaaprender para maximizar su tasa neta de consumo de ener-gía mientras forrajea. Así, un animal que está buscandoalimento tiene que elegir qué parcela deberá visitar, cuán-to tiempo permanecerá en esa parcela consumiendo el
alimento y en qué momento deberá abandonarla parabuscar otra alternativa más rica.
La teoría de la distribución libre ideal de Fretwell yLucas (1970), derivada de la teoría de forrajeo óptimo,propone que los animales van a distribuirse entre las al-ternativas existentes en relación con la calidad de esasalternativas, es decir, cada animal va a elegir la alternati-va que le proporcione el máximo beneficio. Por lo tanto,un animal va a desarrollar la estrategia de forrajeo que le
1 Este trabajo fue financiado por DGAPA-UNAM a través del proyecto IN309402. Los autores agradecen a Abel Javier Zamora, Elizabeth Vázquezy Martha Elisa López su asistencia durante los experimentos y registros. También agradecen a dos revisores anónimos por sus sugerencias paraenriquecer el manuscrito. Los primeros dos autores están adscritos a la Facultad de Estudios Superiores Iztacala, UNAM y el tercero a la Facultadde Psicología, UNAM. Dirigir correspondencia a: Dra. Rosalva Cabrera, Facultad de Estudios Superiores Iztacala, Av. de los Reyes s/n, Tlalnepantla,Edo. de Méx. Dirección electrónica: [email protected]

112 Cabrera et al.
VOL. 23, NÚM 1, JUNIO 2006
permita obtener el máximo beneficio al menor costo, esdecir, va a optimizar los recursos disponibles en su am-biente, eligiendo siempre la mejor de las opciones dispo-nibles. La elección del organismo va a depender de variosfactores, entre ellos: 1) la cantidad de alimento disponi-ble en la parcela; 2) la distancia existente entre parcelas,ya que si el tiempo de viaje incrementa el tiempo óptimopara estar en una parcela también incrementa y 3) elnúmero de predadores presentes al mismo tiempo (Krebs& McCleery, 1984; Parker, 1984; Pulliam & Caraco,1984).
Por otra parte, Valone (1989) y Avery (1994) planteanque los organismos que forrajean en grupo pueden dis-frutar tasas de alimentación superiores debido a que ellosaprenden la localización de fuentes de alimento a partirde observar a otros sujetos del grupo. Los forrajeadoressociales se benefician al estimar mejor y/o más rápido lacalidad de las fuentes de alimentación en relación conlos forrajeadores solitarios (Clark & Mangel, 1984). Así,cada miembro del grupo puede usar tres tipos de infor-mación para evaluar la calidad de una fuente de alimen-to: 1) la distribución de recursos en el ambiente; 2) elmuestreo directo de la fuente y 3) la observación de otrosindividuos. Los primeros dos tipos de información son“individuales” y el último “social”.
En situaciones experimentales de búsqueda de alimen-to, Giraldeau y Lefebvre (1986) describieron que palo-mas expuestas en grupo a una condición en la que tienenque aprender por observación una técnica particular paraobtener alimento, desarrollan un sistema de productoresy parásitos. Este sistema consiste en que ciertos indivi-duos (productores) invierten tiempo y energía en obte-ner el alimento, mientras que otros individuos (parásitos)consumen el alimento producido por otros. Asimismo,los autores reportaron que los roles de productor-parási-to no son fijos en un mismo grupo, es decir, los sujetosparásitos pueden funcionar como productores cuando losproductores originales son retirados del grupo, o bien,cuando cambia la técnica requerida para la obtención dealimento. En otro estudio, Giraldeau y Lefebvre (1987)reportaron que en situaciones de aprendizaje por obser-vación, los observadores no aprenden una respuestanovedosa cuando juegan la estrategia de parasitar el ali-mento producido por el modelo. Por el contrario, otroestudio demostró que aun cuando los grupos de observa-dores parasitaron el alimento producido por el modelo,fueron capaces de ejecutar la respuesta, sobre todo cuan-
do el modelo fue retirado de la situación experimental(Cabrera, Nieto, & Zamora, 2002).
Por otra parte, Delestrade (1999) mencionó que sonpocos los experimentos que han evaluado los efectos com-binados de cantidad de alimento y de distribución espa-cial del mismo y que ello es fundamental, ya que la teoríade la distribución libre ideal expresa que los individuostienden a optimizar el consumo de energía por ir a unaparcela, estacionarse en ella o buscar una nueva si elloslogran la ganancia neta más alta, la cual, si dos o másparcelas ofrecen el mismo tipo de presa, elegirán aquellaen la que la tasa de captura sea más alta (véase tambiénBaum & Kraft, 1998).
En un experimento, Delestrade (1999) evaluó las es-trategias de tres parvadas de chovas pinariegas alpinasviviendo en libertad. La autora proporcionó cantidadesvariables de manzanas (2, 4, o 20 kg), distribuidas en unnúmero variable de parcelas (1, 6, o 12). Sus hallazgosprincipales mostraron que el número máximo de avesque visitaron el sitio experimental incrementó con la can-tidad de alimento, pero no fue afectado por la distribu-ción de alimento. La proporción de aves que nointeractuaron con otras no fue afectada por la variaciónen la cantidad de alimento, el número de parcelas o eltamaño de la parvada. La proporción de aves que notuvo acceso a la comida fue afectada por la cantidad dealimento y por el tamaño de la parvada, pero no por elnúmero de parcelas. Tanto las hembras como los machostrataron de ganar acceso al alimento, independientementede que en estas aves las hembras son dominadas por losmachos.
Dado que generalmente los animales forrajean en gru-po y que la conducta de forrajeo requiere del aprendiza-je y de la discriminación que los animales realicen de lacalidad de las diferentes alternativas disponibles, resultainteresante diseñar una situación experimental que per-mita evaluar las estrategias de comportamiento de gru-pos de animales expuestos a un procedimiento deaprendizaje social, en el cual sólo uno de los integrantesdel grupo ejecuta una respuesta discriminada para acce-der a diferentes fuentes de alimento. Este diseño permi-tirá conocer si a través de la ejecución de un congénere,los observadores son capaces de identificar no sólo cuáles la respuesta a realizar para obtener alimento, sino tam-bién cuál de las opciones disponibles es la que debe serelegida para obtener mayor cantidad de alimento, lo cualimplica aprender por observación a realizar respuestas

Aprendizaje social y estrategias de forrajeo 113
VOL. 23, NÚM 1, JUNIO 2006
discriminadas. Este efecto sólo ha sido reportado conobservadores expuestos individualmente a demostradoresentrenados (Campbell, Heyes, & Goldsmith, 1999; Heyes& Dawson, 1990; Heyes, Dawson, & Nokes, 1992; Nicol& Pope, 1992, 1993; Nieto & Cabrera 1993; Simmons &Lejeune, 1997). Si se obtiene evidencia de aprendizajesocial discriminado en observadores expuestos en grupoa un congénere entrenado, tendrá mayor fortaleza empí-rica el argumento de que el aprendizaje por observaciónpuede ser considerado un tipo de aprendizaje instrumen-tal (Zentall, 1996). Adicionalmente, los datos de este es-tudio permitirán identificar la dinámica de las estrategiasproductor-parásito, pues éstas serán analizadas para cadaobservador tanto en presencia como en ausencia delmodelo. Por lo tanto, este experimento tuvo tres objeti-vos: primero, conocer si palomas expuestas en grupo aun demostrador entrenado aprenden una respuesta paraacceder a diferentes fuentes de alimento. Segundo, iden-tificar si la conducta alimenticia de las palomas observa-doras es discriminada en función de diferentes estímulosque señalan fuentes ricas o pobres de alimento. Tercero,evaluar el desarrollo de estrategias de forrajeo en gruposde palomas bajo situaciones controladas de laboratorio.
MÉTODO
Sujetos
Los observadores fueron 24 palomas criollas, machos,de aproximadamente dos años de edad y experimental-mente ingenuas. Los modeladores fueron dos palomascriollas, machos, de aproximadamente dos años de edady experimentalmente ingenuas.
Aparatos
Como aparato experimental se utilizó una tarima demadera de 120 cm de ancho por 180 cm de largo, con 12perforaciones de un diámetro de 4 cm. La separaciónmínima entre perforaciones fue de 30 cm y la máximafue de 150 cm. Debajo de cada perforación se pegó unrecipiente de plástico de 4.5 cm de profundidad dondese podía depositar grano mixto (compuesto de maíz que-brado, trigo, linaza y mijo rojo en proporciones iguales).Seis de los depósitos estuvieron sellados con dos capas
de papel de china amarillo y los otros seis con dos capas depapel de china azul. La distribución de los colores en losdiferentes depósitos se determinó aleatoriamente a travésde las sesiones experimentales. La Figura 1 muestra unafotografía del aparato usado. Se utilizó una videocámaramarca Panasonic para filmar cada una de las sesionesexperimentales y una reproductora de video marca Sonypara ver las filmaciones. Los datos se registraron me-diante una computadora personal VAIO Sony.
Procedimiento
El experimento se realizó en un aviario techado de 3 m x3 m x 3 m. Tres paredes del aviario fueron de concreto yla otra pared de malla de acero. El aviario estuvo ilumi-nado con luz artificial y se mantuvo a una temperaturaambiente. En el piso del aviario fue colocada la tarimade madera, tal como puede apreciarse en la Figura 1. Lavideocámara fue colocada en un tripié por fuera delaviario, frente a la pared de malla de acero.
Inicialmente los 24 observadores fueron aleato-riamente asignados a seis diferentes grupos de cuatropalomas cada uno. Posteriormente se conformaron tresgrupos integrados cada uno por dos de los grupos origi-nales o por dos parvadas de cuatro palomas. La exposiciónde los grupos a las diferentes condiciones experimentales
Figura 1. Fotografía del aparato empleado. Pueden verse losdepósitos sellados con papel de china, así como la parvada depalomas.

114 Cabrera et al.
VOL. 23, NÚM 1, JUNIO 2006
se llevó a cabo de lunes a domingo en un horario entrelas 9:00 y las 11:00 am. El experimento constó de unpreentrenamiento a los demostradores y de tres fasesexperimentales: habituación, modelamiento y prueba. Acontinuación se describe cada una de estas fases.
Preentrenamiento. Durante esta fase, se entrenó poraproximaciones sucesivas a los demostradores en la res-puesta de perforar los sellos de los recipientes de alimen-to. El entrenamiento inició colocando sellos en losdepósitos de comida con una capa de papel de china blan-co y colocando grano sobre ellos, de tal manera que elsujeto al picotear el grano rompiera el papel y tuvieraacceso al alimento contenido en el depósito. Una vez queesto ocurrió, se aumentó a dos el número de hojas queselló cada depósito sin colocar alimento sobre el mismo.Cuando el sujeto abrió el 100% de los 12 depósitos du-rante dos sesiones consecutivas, se procedió al entrena-miento discriminado. Este entrenamiento consistió enque la mitad de los depósitos estuvo sellada con papel dechina azul y la otra mitad con papel de china amarillo.Cada color estuvo correlacionado con una cantidad dealimento diferente. Este entrenamiento se llevó a cabohasta que durante dos sesiones consecutivas los demostra-dores tuvieron un índice de discriminación superior a.80. Cada vez que de acuerdo a las condiciones demodelamiento se varió la correlación color-cantidad dealimento se reentrenó a cada uno de los modelos segúnfue necesario.
Fase de habituación. Esta fase consistió en que du-rante 14 días consecutivos, los observadores fueron so-metidos a un régimen de privación de alimento de 23:20hrs por 40 min de acceso a grano mixto. Cada día, to-dos los sujetos de un mismo grupo fueron introducidosen una jaula habitación para ser trasladados conjunta-mente a la situación experimental. Se introdujo al grupode palomas a la caja experimental, en donde perma-necieron 20 minutos. Cada grupo (n = 4) tuvo acceso a40 g de grano mixto, el cual se presentó en una charolade aluminio de 20 x 30 cm que estuvo colocada en elpiso del aviario. Cada vez que un grupo salía del aviariose barría el piso y se lavaba con agua y detergente lacharola.
Fase de modelamiento. Durante esta fase, los gru-pos de observadores fueron introducidos a la situaciónexperimental al mismo tiempo que un modelo entrena-do en la respuesta de perforar los sellos de papel de losdepósitos de alimento. Los depósitos estuvieron sella-
dos con papel de diferente color y dependiendo del colorpodían tener acceso a una cantidad de alimento mayoro menor. El Grupo 1 estuvo conformado por dosparvadas diferentes de cuatro palomas cada una y fueexpuesto al demostrador M21. Para estos observadoresen los seis depósitos ricos en alimento hubo 10 g degrano mixto, mientras que en los seis depósitos pobresno hubo alimento. Con la finalidad de balancear la re-lación color-cantidad de alimento entre las parvadas,para una primera parvada (n = 4) el color de los depósi-tos ricos fue amarillo y el color de los depósitos pobresfue azul. Para la segunda parvada de palomas que con-formó el Grupo 1 (n = 4), el estímulo azul selló los de-pósitos ricos y el color amarillo los depósitos pobres. ElGrupo 2 también estuvo integrado por dos diferentesparvadas de cuatro palomas cada una y ambos gruposde parvadas fueron expuestas al demostrador M30. Parael Grupo 2, los depósitos ricos contuvieron 7 g de gra-no mixto y los depósitos pobres contuvieron 3 g de gra-no. Para la primera parvada (n = 4) el color amarilloselló los depósitos ricos y el color azul los depósitospobres. Para la segunda parvada (n = 4) el azul fue elcolor para los depósitos ricos y el amarillo el color delos depósitos pobres. Finalmente, los observadores delGrupo 3, compuesto al igual que los otros dos grupospor dos diferentes parvadas, también fueron expuestosal demostrador M30. Para este grupo, el color de lossellos no señaló diferentes cantidades de alimento enlos depósitos, ya que para ambas parvadas todos losdepósitos de alimento (amarillos y azules) contuvieron5 g de grano mixto. La fase de modelamiento tuvo unaduración de cinco sesiones diarias de 20 minutos cadauna, en las cuales tanto el modelo como los observado-res podían abrir los depósitos y consumir el alimentocontenido en ellos. Las primeras parvadas de cada gru-po fueron expuestas a las diferentes condiciones expe-rimentales en los mismos días y se varió aleatoriamenteel orden en que cada grupo fue sometido a la situaciónexperimental en cada una de las cinco sesiones. Unasemana después, se trabajó con las segundas parvadasde cada uno de los tres grupos.
Fase de prueba. Durante esta fase se introdujo alaviaro a todos los grupos de observadores, en ausenciadel modelo, durante cuatro sesiones diarias de 20 minu-tos cada una. Las proporciones de alimento correla-cionadas con cada color, azul o amarillo, fueron lasmismas que durante la fase de modelamiento. Igualmen-

Aprendizaje social y estrategias de forrajeo 115
VOL. 23, NÚM 1, JUNIO 2006
te, primero se trabajó con las primeras parvadas de cadagrupo y una semana después con las segundas parvadasde cada grupo.
Todas las sesiones fueron videograbadas, posterior-mente las videograbaciones fueron revisadas por dosobservadores independientes y sólo se consideraron parasu análisis los registros que tuvieron una concordanciaentre observadores superior a 85%. El índice de concor-dancia se calculó dividiendo el número de acuerdos so-bre la suma del número de acuerdos más el número dedesacuerdos en cada uno de los registros.
Los observadores al realizar el registro hicieron usode un Sistema de Registro Computarizado (Torres, López& Zarabozo, 1991) para almacenar los datos y despuésrecuperarlos. Este sistema permitió cuantificar la frecuen-cia y la duración absolutas de hasta 10 categoríasconductuales por sesión y su formato permitió exportarlos datos a paquetes estadísticos como el SYSTAT®. Conbase en estos registros, se cuantificaron los depósitos abier-tos por sesión, se identificaron los observadores que abrie-ron depósitos, el orden en el cual fueron abiertos y laposición de cada sujeto en el dispositivo experimentalcuando un depósito fue abierto. Al sujeto que abrió un
depósito se le consideró un productor y al sujeto que sealimentó de un depósito abierto por otro sujeto se le con-sideró como parásito.
RESULTADOS
La Figura 2 muestra el porcentaje de observadores decada grupo que abrió los sellos de los depósitos de comi-da. Se muestra este porcentaje para la primera y la últi-ma sesión de las fases de modelamiento (1Mod y 5Mod)y de prueba (1Pru y 5 Pru). Como muestra esta figura, enlos tres grupos el porcentaje de observadores que ejecu-tó la respuesta de abrir los sellos incrementó de la prime-ra a la quinta sesión de la fase de modelamiento. Para laquinta sesión, el 100% de los observadores de los Grupos1 y 2 y el 75% de los observadores del Grupo 3 emitióesta respuesta. Por tanto, los observadores de todos losgrupos adquirieron la respuesta de perforar los sellosdesde la fase de modelamiento. Un análisis de varianzasimple por bloques que contrastó el número promediode observadores por grupo que ejecutó la respuesta deabrir los sellos de los depósitos de comida mostró que
Figura 2. Porcentaje de observadores que abrieron los sellos de los depósitos de comida en cada uno de los grupos. Se muestranlos porcentajes durante las sesiones inicial y final de la fase de modelamiento (1Mod y 5Mod, respectivamente) e inicial y final dela fase de prueba (1 Pru y 4Pru).

116 Cabrera et al.
VOL. 23, NÚM 1, JUNIO 2006
hubo diferencias significativas F(4, 8) = 13.27, p < 0.01.Una comparación de los promedios de la última sesiónde modelamiento, de acuerdo a una prueba de Tukey,mostró que no hubo diferencias significativas entre losgrupos (p > 0.05) en esta sesión.
Como muestra también la Figura 2, en la fase de prue-ba en los tres grupos el porcentaje de observadores emi-tiendo la respuesta de perforar los sellos tambiénincrementó de la primera a la última sesión de esta fase.En la Sesión 4 de prueba el 100% de observadores delGrupo 2, el 87.5% de observadores del Grupo 1 y el 75%de observadores en el Grupo 3 ejecutó la respuesta deperforar los sellos. Un análisis de varianza simple porbloques que contrastó el número promedio de observa-dores por grupo que ejecutó la respuesta de abrir los se-llos de los depósitos de comida mostró que no hubodiferencias significativas entre los tres diferentes gruposF(2, 8) = 2.85, p = 0.13.
Con respecto a las ejecuciones discriminadas de los ob-servadores, en la Figura 3 se presenta el índice de discrimi-nación promedio para los observadores de los diferentesgrupos en las sesiones iniciales y finales de las fases demodelamiento (1Mod y 5Mod) y de prueba (1Pru y 5Pru).
Como muestra esta figura, los observadores del Grupo 1en la sesión inicial de la fase de modelamiento respon-dieron con un nivel de oportunidad ante los estímulospresentados (.50). Este valor incrementó en la última se-sión a .65. En la fase de prueba, tanto en la sesión inicialcomo en la final, el índice de discriminación fue 1.0 paralos observadores del Grupo 1. Por su parte, los observa-dores del Grupo 2 en la sesión inicial de la fase de modela-miento respondieron de manera completamentediscriminada, ya que su índice fue de 1.0. Este valordecrementó en las sesiones subsiguientes, registrando enla sesión final de esta fase un valor de .8. Durante la fasede prueba el índice de discriminación para este grupo fuemayor a .8 tanto en la sesión inicial como en la final. Losobservadores del Grupo 3 tuvieron un índice de discrimi-nación de .5 en cada una de las sesiones de las dos fases.
Un análisis de varianza simple por bloques que con-trastó los índices de discriminación promedio indicó quehubo diferencias significativas entre los índices de discri-minación de los diferentes grupos tanto en la fase demodelamiento F(4, 8) = 11.69, p = 0.004, como en la fasede prueba F(2, 8) = 70.60, p < 0.001. Pruebas post hoc deTukey mostraron que en ambas fases los valores prome-
Figura 3. Índice de discriminación promedio para los observadores de los diferentes grupos en la primera y última sesión demodelamiento (1Mod y 5Mod) y en la primera y última sesión de prueba (1Pru y 4Pru).
Aprendizaje social y estrategias de forrajeo 117
VOL. 23, NÚM 1, JUNIO 2006
dio del Grupo 3 fueron significativamente menores (p <0.05) que los valores promedio de los Grupos 1 y 2.
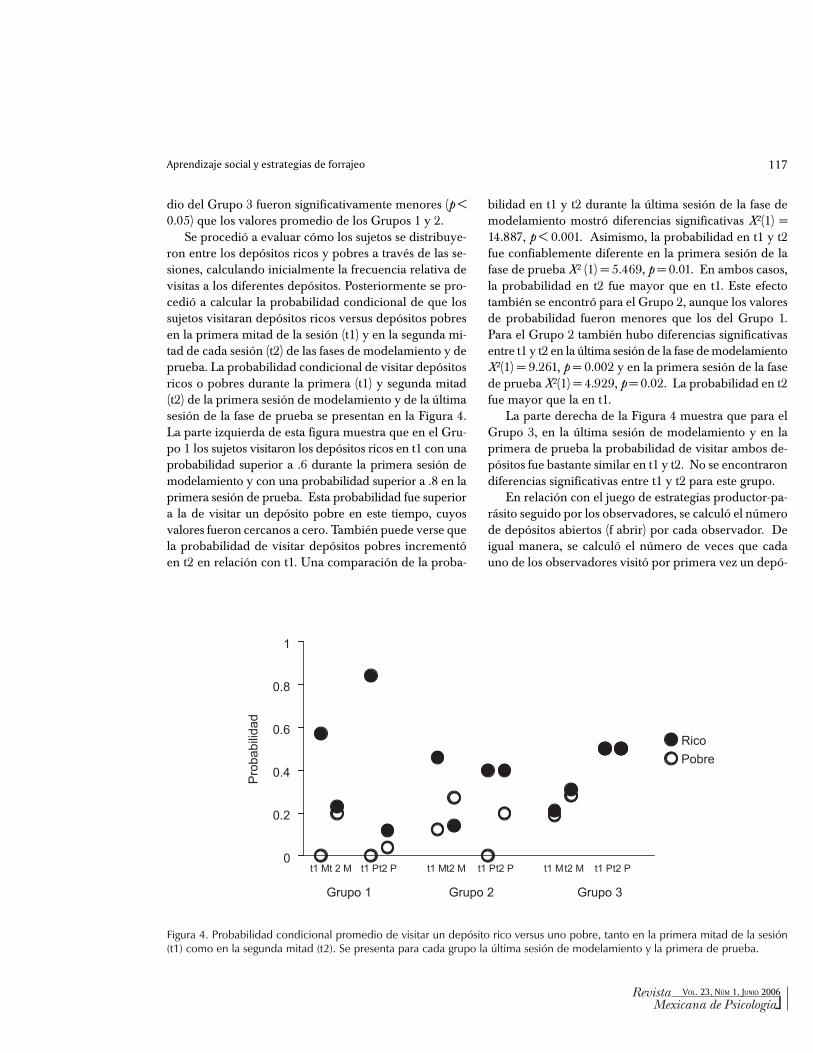
Se procedió a evaluar cómo los sujetos se distribuye-ron entre los depósitos ricos y pobres a través de las se-siones, calculando inicialmente la frecuencia relativa devisitas a los diferentes depósitos. Posteriormente se pro-cedió a calcular la probabilidad condicional de que lossujetos visitaran depósitos ricos versus depósitos pobresen la primera mitad de la sesión (t1) y en la segunda mi-tad de cada sesión (t2) de las fases de modelamiento y deprueba. La probabilidad condicional de visitar depósitosricos o pobres durante la primera (t1) y segunda mitad(t2) de la primera sesión de modelamiento y de la últimasesión de la fase de prueba se presentan en la Figura 4.La parte izquierda de esta figura muestra que en el Gru-po 1 los sujetos visitaron los depósitos ricos en t1 con unaprobabilidad superior a .6 durante la primera sesión demodelamiento y con una probabilidad superior a .8 en laprimera sesión de prueba. Esta probabilidad fue superiora la de visitar un depósito pobre en este tiempo, cuyosvalores fueron cercanos a cero. También puede verse quela probabilidad de visitar depósitos pobres incrementóen t2 en relación con t1. Una comparación de la proba-
bilidad en t1 y t2 durante la última sesión de la fase demodelamiento mostró diferencias significativas X2(1) =14.887, p < 0.001. Asimismo, la probabilidad en t1 y t2fue confiablemente diferente en la primera sesión de lafase de prueba X2 (1) = 5.469, p = 0.01. En ambos casos,la probabilidad en t2 fue mayor que en t1. Este efectotambién se encontró para el Grupo 2, aunque los valoresde probabilidad fueron menores que los del Grupo 1.Para el Grupo 2 también hubo diferencias significativasentre t1 y t2 en la última sesión de la fase de modelamientoX2(1) = 9.261, p = 0.002 y en la primera sesión de la fasede prueba X2(1) = 4.929, p = 0.02. La probabilidad en t2fue mayor que la en t1.
La parte derecha de la Figura 4 muestra que para elGrupo 3, en la última sesión de modelamiento y en laprimera de prueba la probabilidad de visitar ambos de-pósitos fue bastante similar en t1 y t2. No se encontrarondiferencias significativas entre t1 y t2 para este grupo.
En relación con el juego de estrategias productor-pa-rásito seguido por los observadores, se calculó el númerode depósitos abiertos (f abrir) por cada observador. Deigual manera, se calculó el número de veces que cadauno de los observadores visitó por primera vez un depó-
Figura 4. Probabilidad condicional promedio de visitar un depósito rico versus uno pobre, tanto en la primera mitad de la sesión(t1) como en la segunda mitad (t2). Se presenta para cada grupo la última sesión de modelamiento y la primera de prueba.

118 Cabrera et al.
VOL. 23, NÚM 1, JUNIO 2006
sito recién abierto por otra paloma (f parasitar). Con es-tos datos se realizó un cálculo de probabilidad para cadaobservador, en cada una de las fases del experimento, deacuerdo a la siguiente fórmula:
p abrir = f abrir/f abrir + f parasitar (1)
La Figura 5 muestra la probabilidad de abrir de losobservadores de los tres grupos. En la parte izquierdapuede verse que en la fase de modelamiento sólo dos delos observadores del Grupo 1 mostraron una probabili-dad superior a .6, lo cual permitió considerarlos comolos mejores productores de este grupo. La probabilidadde abrir para el resto de los observadores fue inferior a.5, lo cual permitió identificarlos como parásitos. Ahorabien, en la fase de prueba hubo una tendencia en la mayo-ría de los observadores a incrementar la probabilidad deabrir en relación con la de parasitar, aun cuando sólo dosobservadores tuvieron valores superiores a .7. Medianteuna prueba de chi cuadrada se encontró que la probabi-lidad de abrir fue diferente entre la fase de modelamientoy la de prueba X2(1) = 4.073, p = 0.04, siendo mayor du-rante la fase de prueba.
En la parte central de la Figura 5 se muestra que du-rante el modelamiento en el Grupo 2 las probabilidadesde todos los observadores fueron inferiores a .4, es decir,estos sujetos siguieron una estrategia de parásitos. En lafase de prueba todos los valores de probabilidad incremen-taron, lo cual indicó que los observadores modificaron suestrategia al ser retirado el modelo. Adicionalmente, dossujetos tuvieron valores cercanos a 1.0, lo cual indicó quefueron los mejores productores. Las diferencias en los va-lores de probabilidad registrados por cada observador entrelas fases de modelamiento y prueba fueron estadísticamentesignificativas X2(1) = 15.865, p < 0.001, siendo mayoresdurante la fase de prueba.
En la parte derecha de la Figura 5 se muestra quepara el Grupo 3, los valores de probabilidad en la fase demodelamiento fueron inferiores a .5, lo cual indicó quelos observadores siguieron una estrategia de parásitos.En la fase de prueba dos sujetos cambiaron esta estrate-gia, pues el valor de probabilidad condicional fue de 1.0,lo cual indicó que abrieron depósitos y que nuncaparasitaron un depósito. El resto de los observadores semantuvo siguiendo una estrategia de parásito, con valo-res inferiores a .4. Las diferencias entre los valores de
Figura 5. Probabilidad condicional de abrir un depósito versus parasitario de cada uno de los observadores de los diferentes grupos.Se muestran los valores promedio de cada observador tanto en la fase de modelamiento como en la de prueba.

Aprendizaje social y estrategias de forrajeo 119
VOL. 23, NÚM 1, JUNIO 2006
probabilidad de cada observador en las dos fases fueronsignificativas, X2(1) = 15.699, p < 0.001. Nuevamente, laprobabilidad de abrir fue mayor durante la fase de prue-ba que durante la fase de modelamiento.
DISCUSIÓN
Los datos sobre el porcentaje de observadores que ejecu-tó la respuesta de abrir los depósitos de alimento en lasfases de modelamiento y de prueba demuestran que gru-pos de observadores expuestos a un coespecífico expertoen la ejecución de una respuesta para obtener alimentoaprendieron a ejecutar esa respuesta, lo cual permite su-gerir la presencia de un proceso de aprendizaje porobservación (Galef, 1988; Heyes, 1994, 1996). De igualmanera, estos resultados son consistentes con lo propuestopor autores que proponen que cuando los animales es-tán hambrientos y buscan alimento usan la informaciónque les proporcionan sus coespecíficos —informaciónsocial— para tener acceso al alimento (Clark & Mangel,1984; Delestrade, 1999; Galef & Giraldeau, 2001;Valone, 1989).
Los observadores de los grupos en los cuales el colordel sello se correlacionó con diferente cantidad de ali-mento mostraron una ejecución discriminada, respon-diendo preferentemente al estímulo asignado al depósitocon mayor cantidad de alimento (rico). Esto demuestraque es posible el aprendizaje por observación de ejecu-ciones discriminadas cuando grupos de observadores sonexpuestos a un modelo entrenado. La ejecución de losobservadores fue diferencial ante dos propiedades deestímulo, de manera similar a como ocurre cuando unsólo observador es expuesto a un modelo (Campbell etal., 1999; Heyes & Dawson, 1990; Heyes et al., 1992; Nicol& Pope, 1992, 1993; Nieto & Cabrera 1993; Simmons &Lejeune, 1997). Los resultados encontrados en este tra-bajo sobre ejecuciones discriminadas también se ajustana lo propuesto por la teoría de la distribución libre idealde Fretwell y Lucas (1970) en el sentido de que los indivi-duos tenderán a ir y/o a permanecer en la parcela en la queellos logren la ganancia más alta.
También puede decirse que los resultados de este ex-perimento guardan una relación entre el número de de-pósitos visitados y la cantidad de alimento ofrecida porestos depósitos, similar a la observada en situaciones semi-naturales por Delestrade (1999) y en experimentos ope-
rantes de elección (Herrnstein, 1970). Esta semejanza esen el sentido de que proporcionalmente respondieronmás a la opción que tenía disponible una proporciónmayor de alimento. Esta analogía entre el modelo deforrajeo de la distribución libre ideal y la ley de iguala-ción de Herrnstein (1970) ha sido señalada previamente(Krebs & Davies, 1984; Shettleworth, 1984, 1998).
Los observadores en este experimento jugaron tantoestrategias de productores como de parásitos (Parker, 1984).Al respecto, hay cuatro cuestiones importantes que comen-tar. Primero, a pesar de que los observadores aprendierona emitir la respuesta modelada, lo cual sugiere la presen-cia de un proceso de difusión de respuestas novedosas engrupos animales (Laland, Richerson, & Boyd, 1993;Lefebvre, 1986), no todos los observadores la ejecutaronal mismo nivel a lo largo de las sesiones. Es más, huboobservadores que sólo ejecutaron la respuesta en una delas fases. Esto es, sólo uno o dos integrantes de cada gru-po fueron los que abrieron un mayor número de depósi-tos de alimento —mejores productores— y los demás sujetosno ejecutaron la respuesta, o abrieron pocos depósitos(véase Giraldeau & Lefebvre 1986, 1987). Segundo, elhecho de que los observadores que fueron mejores pro-ductores en la fase de modelamiento no necesariamentelo fueron en la fase de prueba, puede indicar que la diná-mica de comportamiento de todo el grupo cambia con lapresencia o ausencia de uno de sus integrantes, en estecaso el modelo. Los roles de productor y de parásito tam-bién son intercambiables en función de la composicióndel grupo (Giraldeau & Lefebvre, 1987; Parker, 1984;Pulliam & Caraco, 1984). Tercero, aquellos observadoresque realizaron el mayor número de visitas a depósitosabiertos por un congénere fueron malos productores dela respuesta modelada, lo cual indica que niveles altos deparasitismo por parte de los observadores interfieren conla ejecución de la respuesta modelada (Giraldeau &Lefebvre, 1987), mas no necesariamente con la adquisi-ción de la misma, pues en algunos casos hubo observa-dores parásitos que sí ejecutaron la respuesta, aunque aniveles muy bajos. Cuarto, los observadores que ejecuta-ron la respuesta modelada a niveles altos —mejores pro-ductores— fueron sujetos que previamente habían visitadodepósitos abiertos por otro sujeto, pero lo hicieron a unnivel medio o bajo. Lo anterior puede ser un indicadorde que es necesario que los observadores visiten depósi-tos con alimento, pues ello les posibilita “identificar” larelación respuesta-reforzador, la cual se ha demostrado

120 Cabrera et al.
VOL. 23, NÚM 1, JUNIO 2006
que es necesaria para que el aprendizaje por observa-ción tenga lugar (Nieto & Cabrera, 1994). Adicional-mente, los datos aquí analizados demuestran que lasestrategias productor/parásito pueden coexistir en el mis-mo sujeto en periodos de tiempo tan cortos como lo fuela sesión experimental. Al respecto, Parker (1984) men-cionó que en ambos, productor y parásito, los pagos sonuna función de la frecuencia de las dos estrategias y quecoexistirán a un equilibrio estable y la aptitud media decada estrategia puede ser similar si el tamaño del grupoafecta las habilidades competitivas en maneras opuestasen productor y parásito.
REFERENCIAS
Avery, M. I. (1994). Finding good food and avoiding bad food:Does it help to associate with experienced flockmates? Ani-mal Behaviour, 48, 1371-1378.
Baum, W. M., & Kraft, J. R. (1998). Group choice: Competi-tion, travel, and the ideal free distribution. Journal of theExperimental Analysis of Behavior, 69, 227-245.
Cabrera, R., Nieto, J., & Zamora, A. (2002). Aprendizaje porobservación en grupos de palomas: Un análisis de diferentespatrones de respuesta. En J. C. P. Arriaga, R. Hernández,& F. López (Eds.), Perspectivas de la psicología experimental enMéxico. II. (pp. 25-44). México: UNAM.
Campbell, F. M., Heyes, C. M., & Goldsmith, A. R. (1999).Stimulus learning and response learning by observation inthe European starling, in a two-object/two-action test. Ani-mal Behaviour, 58, 151-158.
Clark, C. W., & Mangel, M. (1984). Foraging and flocking strat-egies: Information in an uncertain environment. AmericanNaturalist, 123, 626-641.
Delestrade, A. (1999). Foraging strategy in a social bird, thealpine chough: Effect of variation in quantity and distribu-tion of food. Animal Behaviour, 57, 299-305.
Fretwell, S. D., & Lucas, H. L. (1970). On territorial behaviourand other factors influencing habitat distribution in birds.Acta Biotheoretica, 19,16-36.
Galef, B. G. Jr. (1988). Communication of information con-cerning distant diets in a social, central-place foraging spe-cies: Rattus norvegicus. En T. R. Zentall, & B. G. Galef Jr.(Eds.), Social Learning: Psychological and Biological Perspectives.(pp. 3-28). New Jersey, Estados Unidos: Lawrence EribaumAssociates Publishers.
Galef, B. G. Jr., & Giraldeau, L. A. (2001). Social influences onforaging in vertebrates: Causal mechanisms and adaptivefunctions. Animal Behaviour, 61, 3-15.
Giraldeau, L. A., & Lefebvre, L. (1986). Exchangeable pro-ducer and scrounger roles in captive flock of feral pigeons:A case for the skill pool effect. Animal Behaviour, 34, 797-803.
Giraldeau, L. A., & Lefebvre, L. (1987). Scrounging preventscultural transmission of a food-finding behavior in pigeons.Animal Behaviour, 35, 387-394.
Herrnstein, J. R. (1970). On the law of effect. Journal of the Ex-perimental Analysis of Behavior. 13, 243-266.
Heyes, C. M. (1994). Social learning in animals: Categoriesand mechanisms. Biology Review, 69, 207-231.
Heyes, C. M. (1996). Genuine imitation? En C. M. Heyes, & B.G. Jr. Galef (Eds.), Social learning in animals: The roots of cul-ture. (pp. 371-389). USA: Academic Press.
Heyes, C. M., & Dawson, G. R. (1990). A demonstration ofobservational learning in rats using a bidirectional con-trol. Quarterly Journal of Experimental Psychology, 42 b, 59-71.
Heyes, C. M., Dawson, G. R., & Nokes, T. (1992). Imitation inrats: Initial responding and transfer evidence. Quarterly Jour-nal of Experimental Psychology, 45 b, 229-240.
Krebs, J. R., & Davies, N. B. (1984). Behavioural ecology: An evo-lutionary approach. (2a. Ed.). Oxford: Blackwell ScientificPublications.
Krebs, J. R., & McCleery, R. H. (1984). Optimization in be-havioral ecology. En J. R. Krebs, & N. B. Davies (Eds.),Behavioural ecology: An evolutionary approach. (2a. Ed. pp.91-121). Oxford: Blackwell. Scientific Publications.
Laland, K. M., Richerson, P. J., & Boyd, R. (1993). Animalsocial learning: Toward a new theoretical approach. Per-spective in Ethology, 10, 249-277.
Lefebvre, L. (1986). Cultural diffusion of a novel food findingbehavior in urban pigeons: An experimental field test. Ethol-ogy, 71, 295-303.
Nicol, C. J., & Pope, S. J. (1992). Effects of social learning onthe acquisition of discriminatory keypecking in hens. Bul-letin of Psychonomic Society, 30, 293-296.
Nicol, C. J., & Pope, S. J. (1993). Food deprivation during ob-servation reduces social learning in hens. Animal Behaviour,45, 193-196.
Nieto, J., & Cabrera, R. (1993). Adquisición de unadiscriminación operante mediante observación en palomas.Revista Latinoamericana de Psicología, 25, 467-478.
Nieto, J., & Cabrera, R. (1994). La evolución cultural enanimales. En J. L. Díaz (Ed.), La mente y el comportamientoanimal: Ensayos de etología cognitiva (pp. 91-109). México:Fondo de Cultura Económica.
Parker, G. A. (1984). Evolutionary stable strategies. En J. R.Krebs, & N. B. Davies (Eds.), Behavioural ecology: An evolu-tionary approach. (2a. Ed., pp. 30-61). Oxford: BlackwellScientific Publications.
Pulliam, H. R., & Caraco, T. (1984). Living in groups: Is therean optimal group size? En J. R. Krebs, & N. B. Davies (Eds.),Behavioural ecology: An evolutionary approach. (2a. Ed., pp. 122-147). Oxford: Blackwell Scientific Publications.
Shettlewoth, S. J. (1984). Learning and behavioural ecology.En J. R. Krebs, & N. B. Davies (Eds.), Behavioural ecology:An evolutionary approach. (2a. Ed., pp. 179-194). Oxford:Blackwell Scientific Publications.

Aprendizaje social y estrategias de forrajeo 121
VOL. 23, NÚM 1, JUNIO 2006
Shettleworth, S. J. (1998). Cognition, evolution and behavior. Ox-ford: Oxford University Press.
Simmons, F., & Lejeune, H. (1997). Observational learning ofduration discrimination in rats (Rattus norvegicus). BehaviouralProcesses, 41, 97-105.
Torres, A., López, F., & Zarabozo, D. (1991). Registroobservacional a través de computadora. Revista Mexicanade Análisis de la Conducta, 17, 147-161.
Valone, T. J. (1989). Group foraging, public information, andpatch estimation. Oikos, 56, 357-363.
Zentall, T. R. (1996). An analysis of imitative learning in ani-mals. En C. M. Heyes, & B. G. Jr. Galef (Eds.), Social learn-ing in animals: The roots of culture (pp.221-243). USA:Academic Press.
Recibido 14, 07, 2005Aceptación final 31, 03, 2006


















