
APORTES DE LA ANATOMÍA COMPARADA A LA RESOLUCIÓN DE ...
52
UNIVERSIDAD NACIONAL DE CÓRDOBA CIENCIAS BIOLÓGICAS APORTES DE LA ANATOMÍA COMPARADA A LA RESOLUCIÓN DE PROBLEMAS TAXONÓMICOS A DOS REPRESENTANTES DE TRICHOCEREUS (CACTACEAE) DE FAMATINA, LA RIOJA. Tesinista: FERNÁNDEZ, GIMENA NIEVES Firma:_________ Directora: DELBÓN, NATALIA Firma:_________ Co-directora: STIEFKENS, LAURA Firma: _________ CÁTEDRA DE MORFOLOGÍA VEGETAL ESCUELA DE BIOLOGÍA INSTITUTO MULTIDISCIPLINARIO DE BIOLOGÍA VEGETAL FACULTAD DE CIENCIAS EXACTAS FÍSICAS Y NATURALES CÓRDOBA, ARGENTINA, AÑO 2017.
Transcript of APORTES DE LA ANATOMÍA COMPARADA A LA RESOLUCIÓN DE ...

UNIVERSIDAD NACIONAL DE CÓRDOBA
CIENCIAS BIOLÓGICAS
APORTES DE LA ANATOMÍA COMPARADA
A LA RESOLUCIÓN DE PROBLEMAS TAXONÓMICOS
A DOS REPRESENTANTES DE TRICHOCEREUS (CACTACEAE)
DE FAMATINA, LA RIOJA.
Tesinista: FERNÁNDEZ, GIMENA NIEVES Firma:_________
Directora: DELBÓN, NATALIA Firma:_________
Co-directora: STIEFKENS, LAURA Firma: _________
CÁTEDRA DE MORFOLOGÍA VEGETAL
ESCUELA DE BIOLOGÍA
INSTITUTO MULTIDISCIPLINARIO DE BIOLOGÍA VEGETAL
FACULTAD DE CIENCIAS EXACTAS FÍSICAS Y NATURALES
CÓRDOBA, ARGENTINA,
AÑO 2017.

APORTES DE LA ANATOMÍA COMPARADA A LA RESOLUCIÓN DE
PROBLEMAS TAXONÓMICOS A DOS REPRESENTANTES DE TRICHOCEREUS
(CACTACEAE) DE FAMATINA, LA RIOJA.
Tribunal Examinador:
Nombre:………………………………. Firma: ……………………….
Nombre:………………………………. Firma: ……………………….
Nombre:………………………………. Firma: ……………………….
Calificación: ……………………………………...
Fecha: ………………………………………………
2

A la memoria de mi Abuelo
3

ÍNDICE
ÍNDICE 4
RESUMEN 5
Palabras clave 6
INTRODUCCIÓN 7
Descripciones generales de los taxones 13
Trichocereus candicans (Gillies) Britton et Rose 13
Trichocereus pseudocandicans (Back.) ex Kiesling 14
OBJETIVOS 16
Objetivos generales 16
Objetivos específicos 16
MATERIALES Y MÉTODOS 17
RESULTADOS 20
Análisis Cuantitativos 28
FIGURAS 32
DISCUSIÓN 39
CONCLUSIONES 44
Consideraciones finales 45
BIBLIOGRAFÍA 46
AGRADECIMIENTOS 52
4

RESUMEN
La Rioja posee varios sistemas de sierras y montañas, entre las que se
destacan las Sierras de Famatina por su pico de más de 6000 msnm. En este
ambiente árido la familia Cactaceae se encuentra bien representada y en particular
el género Trichocereus; aquí T. candicans y T. pseudocandicans conviven, siendo el
último un endemismo restringido de esta zona. Históricamente la identidad de
estos taxones se encuentra en discusión, dependiendo de los autores se las
considera dos especies diferentes o dos subespecies de T. candicans (subsp.
candicans y subsp. pseudocandicans). Se ha planteado también que T.
pseudocandicans podría ser un híbrido de reciente formación entre T. candicans y
quizás T. vatteri o T. strigosus. Frente a este panorama complejo, se planteó el
estudio interdisciplinario de estas entidades con el objetivo de dilucidar su
identidad taxonómica.
En el presente trabajo se realiza una comparación morfoanatómica de sus
órganos reproductivos con el fin de aportar datos confiables que ayuden a su
delimitación. Para ello se realizaron preparados permanentes y temporarios de las
flores y los frutos en diferentes estadios de desarrollo, mediante el uso de técnicas
histológicas clásicas.
Los resultados de nuestro estudio indican que T. candicans y T.
pseudocandicans son anatómica y morfológicamente similares. Se observaron
abundantes conductos secretores esquizógenos en escamas florales, tépalos, ovario
y estilo. Además, se encontraron numerosos cristales, de los tipos piramidales,
conglomerados y drusas en la gran mayoría de los órganos analizados. Los taxones
se diferencian mayormente en las características medidas para el estilo,
pudiéndose deber a que las flores de T. candicans son un tanto mayores que las de
T. pseudocandicans.
Por último, 10 de las 43 variables analizadas fueron significativamente
diferentes entre las especies; es decir, no obtuvimos pruebas suficientes para
considerar que sean dos entidades completamente diferenciadas.5

Palabras clave: Trichocereus candicans – Trichocereus pseudocandicans –
Cactaceae – Trichocereus – flor – fruto – morfoanatomía – taxonomía.
6

INTRODUCCIÓN
La Provincia de La Rioja posee varios sistemas de sierras y montañas entre
las que se destacan las Sierras de Famatina, cuyo pico más elevado es el Cerro Gral.
Belgrano, más conocido por el “Nevado de Famatina” con 6097 msnm. Este cordón
montañoso se encuentra incluido en los dominios fitogeográficos más áridos y con
mayor amplitud térmica de Argentina: el Dominio Chaqueño y el Dominio Andino-
Patagónico, representados aquí por las Provincias del Monte en valles y bolsones,
la Provincia de la Prepuna en las sierras de poca altitud y las provincias Puneña y
Altoandina en las zonas más elevadas (Aceñolaza, 1996; Biurrun et al., 2012;
Barboza et al., 2016).
Famatina presenta un clima seco y cálido, con gran amplitud térmica, tanto
diaria como anual, alta radiación solar y suelos empobrecidos (Cabrera, 1971;
Lopez de Casenave, 2001). La vegetación asociada a este tipo de ambientes
desérticos o semidesérticos es denominada xerófita. Las especies que viven en
estos ambientes, sobreviven a las condiciones adversas gracias a una serie de
características estructurales, anatómicas y fisiológicas muy especializadas (Fahn &
Cutler, 1992; Dickison, 2000).
Cactaceae se encuentra bien representada en estos ambientes, ya que
soporta favorablemente las inclemencias climáticas y los hábitats extremos. Esto es
posible gracias a que desarrollaron múltiples adaptaciones morfofisiológicas, como
la reducción de las hojas y el desarrollo de espinas, la reducción de la superficie
transpiratoria, la presencia de tallos suculentos y acanalados para la contracción o
expansión según el agua disponible, el engrosamiento de la cutícula, la presencia
de ceras, la disminución y disposición hundida de los estomas, las raíces extensas y
extremadamente ramificadas con alta capacidad de absorción, el metabolismo
ácido de las crasuláceas el cual minimiza la pérdida de agua por parte de la planta,
entre otras (Gibson & Nobel, 1986; Anderson, 2001; Mazzola et al., 2013).
7

La familia es nativa del continente americano y nuclea a ca. 1600 especies
distribuidas desde Canadá hasta la Patagonia, donde México es el mayor centro de
diversificación (Bravo-Hollis & Scheinvar, 1999; Becerra, 2000; Mandujano et al.,
2002; Hunt et al., 2006). Sus representantes suelen ser plantas longevas y se
presentan en una variedad de formas desde simples a ramificadas, de globosas a
columnares y algunas son exclusivamente subterráneas (Anderson, 2001). Sus
flores son grandes, con numerosas piezas y se encuentran solitarias en los tallos,
por lo general son muy llamativas gracias a sus pigmentos iridiscentes y
usualmente son hermafroditas (Kiesling, 1978; Anderson, 2001). La polinización es
realizada en general por abejas, aunque también por otros insectos y murciélagos.
Poseen frutos simples, ya que derivan de una sola flor con gineceo gamocarpelar y
complejos, ya que las flores son de ovario ínfero, por lo que la pared de dichos
frutos está formada por, además del ovario, por el tejido de origen axial caulinar al
cual llamaremos pericarpelo o hipanto donde está embebido el ovario (Cerén-
López et al., 2015). Los frutos suelen ser globosos, carnosos y pueden estar
recubiertos por espinas o pelos. Las semillas están protegidas por dos tegumentos,
son de color castaño claro o amarillento hasta oscuro o negro (Kiesling, 1978;
Kiesling & Ferrari, 2009). Por su parte, tanto los órganos vegetativos como
reproductivos se caracterizan por la presencia de betalaínas y de un gran número
de alcaloides, la presencia de células idioblásticas, estructuras secretoras, drusas y
agregados de cristales de oxalato de calcio, entre otras (Anderson, 2001; Esquivel,
2004).
Dentro de Cactaceae, Trichocereus (Berg.) Riccob. (Tribu Trichocereeae -
Subtribu Trichocereinae) cobra importancia por ser exclusivamente Sudamericano.
Incluye aproximadamente 45 especies distribuidas en ambientes xéricos de la
cordillera de Ecuador, Perú, Bolivia, Chile y Argentina (Brako & Zarucchi, 1993;
Navarro, 1996; Hoffmann & Walter, 2004, Kiesling et al., 2008); en nuestro país
habita en el noroeste y centro, hasta La Pampa y el sudoeste de Buenos Aires
(Kiesling et al., 2008).
Según Kiesling (1978), el género representaría el taxón más basal dentro de
su tribu, debido a caracteres tales como flores grandes, actinomorfas, acampanadas
8

y con muchas piezas, el receptáculo del ovario grueso, las axilas de las escamas
florales con cerdas o lana, los frutos pilosos, tallos postrados o prismáticos,
presencia de numerosas costillas y numerosas espinas de disposición indefinida. El
sistema de cruzamiento más común observado es la xenogamia y se asocia la
polinización con animales tales como polillas, abejas, avispas, aves y mamíferos
(Badano & Schlumpberger, 2001). En cuanto al uso de distintas especies de
Trichocereus, se destaca la utilización de la madera para carpintería y ebanistería.
Muchas especies también son usadas como forraje y los frutos son comestibles, ya
que suelen ser dulces (Kiesling, 1978).
Actualmente, el género comprende 58 taxones (38 especies, 10 subespecies,
8 variedades y 2 formas) (Albesiano, 2015). Pero su delimitación, así como también
la de sus grupos hermanos, ha estado históricamente en discusión y ha sido
modificada numerosas veces (Albesiano & Terrazas, 2012). Algunos autores
incluyen a Trichocereus, junto a otros grupos, dentro del género Echinopsis Zucc.
(ej. Friedrich, 1974; Hunt & Taylor, 1990; Anderson, 2001; Hunt et al., 2006;
Schlumpberger & Renner, 2012); mientras que, por el contrario, otros lo reconocen
como un género diferente (ej. Kiesling, 1978; Gibson & Nobel, 1986; Albesiano &
Terrazas, 2012; Hunt, 2013, Albesiano, 2015), criterio adoptado en el presente
trabajo.
Dentro de este conflictivo género, existen dos representantes de los cuales
se discute su identidad taxonómica, Trichocereus candicans (Gillies) Britton et Rose
y T. pseudocandicans (Back.) ex Kiesling. Kiesling (1978) las considera como
especies diferentes y propone que T. pseudocandicans podría ser un híbrido de
reciente formación, derivado de T. candicans, no estabilizado genéticamente.
Contradiciendo lo antes dicho, Hunt y colaboradores (2006) proponen que T.
pseudocandicans es sinónimo de T. candicans; en tanto que Schlumpberger y
Renner (2012) en su filogenia molecular incluyen solamente ejemplares de T.
candicans (Echinopsis candicans (Gillies ex Salm-Dyck) D.R.Hunt), sin referirse al
otro taxón. Recientemente, Albesiano (2015) en su tesis doctoral concluye que, si
bien las entidades son similares, las diferencias exomorfológicas entre ellas son
9

suficientes para considerar a T. pseudocandicans como una subespecie de T.
candicans.
Ambos taxones comparten numerosas características exomorfológicas,
entre las que se destacan la forma de crecimiento en matas de hasta 3 metros de
diámetro, tallos cilíndricos, ascendentes, ramificados en la base y areolas con
abundante tomento blanco. Flores grandes de ovario ínfero, escamas externas
carnosas con pelos largos y abundantes en sus axilas, tépalos en tres ciclos
diferentes, estilo con estigma lobulado y anteras en dos ciclos. Frutos globosos,
carnosos, dehiscentes por una rajadura longitudinal, con numerosas semillas
negras (Kiesling, 1978).
Por su parte, las entidades presentan ciertas características que no
comparten entre sí, las cuales hacen posible su diferenciación. A continuación se
mencionan las más relevantes:
- forma de las espinas: subuladas en T. candicans y aciculares en T.
pseudocandicans (Kiesling, 1978);
- cantidad de costillas: entre 9 y 11 en T. candicans y entre 11 y 13 en T.
pseudocandicans. Cabe destacar que según Kiesling el número de costillas puede
variar, ya que aumenta la cantidad de costillas en suelos ricos y disminuye en
suelos pobres (Albesiano, 2015);
- color de las piezas interiores de la corola: blanco níveo en T. candicans y de
amarillento a anaranjado hasta rojizo en T. pseudocandicans (Kiesling, 1978);
- tiempo de apertura floral: las flores de T. candicans duran sólo una noche, en
cambio las de T. pseudocandicans permanecen abiertas dos o tres días (Albesiano,
2015);
- tamaño floral: las flores de T. candicans pueden medir 18-23 cm de longitud y
11-19 cm de diámetro; mientras que las de T. pseudocandicans son menores,
alcanzan entre 15-18 cm de longitud y 11-14 cm de diámetro (Kiesling, 1978).
10

- otros caracteres: T. candicans posee flores acampanadas con aroma similar a
jazmín o rosas y su fruto mantiene la flor seca adherida, en cambio, las flores de T.
pseudocandicans son infundibuliformes, inodoras y su fruto pierde el perianto al
madurar (Kiesling, 1978; Albesiano, 2015).
- distribución: T. candicans presenta una distribución amplia, en Mendoza, San
Juan, Córdoba, La Rioja, La Pampa, Buenos Aires y San Luis; desde los cerros bajos
de la precordillera y de las Sierras Pampeanas, hasta zonas llanas pedregosas o
salinas. Por el contrario, T. pseudocandicans constituye un endemismo restringido a
La Rioja, habita solamente desde el norte de Famatina hasta la cuesta de Miranda,
entre el matorral bajo con grandes espacios de suelo desnudo (Kiesling, 1978;
Kiesling et al., 2008).
Frente a este panorama complejo, donde el problema taxonómico de
muchos géneros de la familia, sobre todo de la tribu Trichocereeae representa un
gran desafío para los especialistas en la actualidad (Albesiano, 2015), donde la
anatomía sistemática no ha sido muy desarrollada para Cactaceae (Gibson & Horak,
1978), y donde según la bibliografía consultada, no hay registros de estudios
anatómicos de órganos reproductivos para ninguna especie de este género, es la
razón por la que en el presente trabajo nos propusimos realizar una comparación
cualitativa y cuantitativa a nivel morfoanatómico y morfométrico de las flores y los
frutos de T. candicans y T. pseudocandicans con el objetivo de aportar caracteres
confiables que ayuden a su delimitación taxonómica.
Más allá de la contribución al conocimiento básico de las entidades,
pretendemos aportar información útil para, además de seguir dilucidando
problemas taxonómicos, contribuir a la conservación de la familia Cactaceae, con
énfasis en especies endémicas como las estudiadas aquí, como así también de las
zonas áridas donde habitan y de esta manera revalorizar la biodiversidad de las
mismas.
11

Descripciones generales de los taxones:
Trichocereus candicans (Gillies) Britton et Rose
Son plantas ascendentes, con tallos de 25-60 cm de alto y 12-16 cm de
diámetro, cilíndricos, de color verde-pálido. Costillas del ápice 9, obtusas, 2-3 cm
de ancho, 1 cm de alto; con areolas redondas, 5 mm de alto y 1 cm de ancho, con
tomento blanco, gris-claro y oscuro; espinas centrales 4, dispuestas en cruz, 2,5-6,0
cm de largo, 1-2 mm de diámetro, subuladas, rectas o ligeramente arqueadas,
amarillas con base marrón y luego gris; espinas radiales 13, de 0,7-4,5 cm de largo ,
1 mm de diámetro, amarillas con base marrón. Costillas de la base 8-9, obtusas, 2,8
cm de ancho y 1,8 cm de alto, con areolas circulares, 2 mm de alto y 1 cm de ancho,
con tomento gris-oscuro, 3-5 cm de distancia entre ellas; con 4 espinas centrales, 4
cm de largo y 1 mm de diámetro, subuladas, marrones; espinas radiales 11, de 1,7-
3 cm de largo, subuladas, 1 mm de diámetro, amarillas.
Flores generalmente subapicales, de 18-23 cm de largo, 8-19 cm de
diámetro, apenas aromáticas, con abundante pilosidad marrón-claro en el ovario y
tubo floral, cada tricoma o pelo de 1 a 2 cm de longitud; ovario y tubo floral de 2,5-
2,6 cm de ancho; escamas florales de 0,7 a 2 cm de largo, 4-8 mm de ancho, de
color verde o verde-amarillento con ápice marrón; tépalos externos linear-
lanceolados, 5-8 cm de longitud, 1 cm de ancho en la base, verdes con la base rojiza,
tépalos intermedios rosados, internos blancos, espatulados, 7 cm de largo, 3 cm de
ancho, mucronados; estambres superiores con filamentos de 3 cm de longitud y 1
mm de diámetro, blancos; anteras de 2,5 mm de largo, 1,5 mm de ancho, amarillas;
estambres inferiores con filamentos blanco-verdosos, 8 cm de longitud, 1 mm de
diámetro; cámara nectarial de 1 mm de alto, blanco-verdosa; estilo de 17 cm de
longitud, 0,6 cm de diámetro, blanco-amarillento; estigma con 17-20 lóbulos, de
1,2-1,7 cm de largo y 1,5-2,0 mm de diámetro, amarillos.
Fruto pseudobaya mayormente lateral, de dehiscencia longitudinal, 7 cm de
largo, 5-7 cm de diámetro; parte externa amarilla tornándose rosada, 6 mm de
12

espesor, cavidad ovárica rellena de funículos carnosos, blancos, de 2 cm de
diámetro. Semillas de 1,5 mm de largo, 1 mm de alto y 0,7 mm de ancho; testa con
células hexagonales, verrugosas, con estrías transversales (Kiesling, 1978).
Floración nocturna, entre octubre y febrero. Sus frutos maduran entre
diciembre y marzo. Tanto la floración como la fructificación dependen de la
disponibilidad de agua, ya que luego de épocas secas la floración se demora o
incluso pueden no florecen (Albesiano, 2015).
Trichocereus pseudocandicans (Back.) ex Kiesling
Son plantas ascendentes, con tallos de 60-100 cm de alto, 8-10 cm de
diámetro, cilíndricos, ascendentes, de color verde-oscuro, ramificados en la base.
Costillas del ápice obtusas, de un centímetro de alto; con areolas de 8 mm de alto;
una espina central, acicular de 5,3 cm de largo, 1 mm de diámetro, amarilla, a veces
amarillento-amarronada; espinas radiales 8, aciculares, de 1 mm de diámetro, 4 cm
de largo, de igual color que la espina central. Costillas de la base 11-13, obtusas, 2-
3 cm de ancho; con areolas circulares, de 1 cm de ancho, con abundante pilosidad
blanca y separadas entre sí por 2 cm; espinas aciculares y subuladas, radiales 12,
de 2 cm de largo y 0,5 mm de ancho, centrales 4, de 4-8 cm de longitud y 1 mm de
diámetro, amarillas, rígidas y en algunos casos no se diferencian las radiales de las
centrales.
Flores de 15-18 cm de largo, inodoras, corola de 11-14 cm de diámetro,
ovario de 2 cm de ancho, tubo floral de 3-5 cm de diámetro, con escamas carnosas
de 5 mm de largo, de cuyas axilas emergen pelos blancos o castaños. Tépalos
exteriores de 3-5 cm de longitud, mayormente rojizos, tépalos intermedios e
interiores de 6-8 cm de largo, de color variable: desde blanco-amarillentos,
amarillo-claro hasta oscuro, rosados y hasta rojos; estambres superiores con
filamentos de 2 cm de longitud, amarillo-verdosos; estambres inferiores de 7 cm de
longitud, 1 mm de diámetro; estilo de 11-12 cm de longitud, 0,5 cm de diámetro;
estigma con 15 lóbulos, de 1,3-1,8 cm de largo y 5 mm de diámetro, amarillos.
13

Pseudobaya con dehiscencia lateral, 5-6 cm de diámetro; parte externa verde-
rosada a amarilla y cavidad ovárica con funículos blancos. Semillas negras, 1,3 mm
de largo y 1 mm de ancho, testa finamente punteada y brillante (Kiesling, 1978).
Floración diurna, entre octubre y febrero. Los frutos maduran entre enero y
marzo (Albesiano, 2015).
14

OBJETIVOS
Objetivos generales:
El objetivo general de la presente tesina fue indagar sobre la morfoanatomía
de los órganos reproductivos de Trichocereus candicans y Trichocereus
pseudocandicans que habitan en las sierras de Famatina, La Rioja, con el fin de
aportar datos confiables que ayuden a resolver su identidad taxonómica.
Además, se pretende ampliar lo que se conoce de la historia natural de
ambas especies, lo que podrá aportar en estudios futuros relacionados a la biología
floral y polinización, al hábitat y su conservación, entre otros.
Objetivos específicos:
Analizar y comparar las características exomorfológicas y anatómicas de las
flores y los frutos jóvenes de diferentes poblaciones de T. candicans y T.
pseudocandicans.
Realizar un estudio morfométrico analizando diferentes variables para
comparar estadísticamente a las entidades.
15

MATERIALES Y MÉTODOS
Se estudiaron dos taxones, Trichocereus candicans (Gillies) Britton et Rose y
Trichocereus pseudocandicans (Back.) ex Kiesling, provenientes de tres poblaciones
ubicadas en las Sierras de Famatina, del Departamento Famatina, Provincia de La
Rioja. Fueron recolectados entre el 27 y 29 de noviembre de 2015 y los respectivos
ejemplares de herbario se encuentran depositados en el Herbario de Córdoba
(CORD). Se recolectaron los órganos reproductivos (flores y frutos) de 9 individuos,
los cuales fueron fijados inmediatamente en FAA (formol, ácido acético y alcohol).
Los datos de recolección y la ubicación se sintetizan en la tabla 1.
Tabla 1. Datos de colección y ubicación geográfica de los individuos recolectados.
Se realizó un análisis exomorfológico y anatómico exhaustivo de las flores y
frutos colectados. Bajo lupa, se contabilizó el número de lóbulos estigmáticos de
cada estilo y los tricomas de las escamas florales.
16
Especie
Población
Coordenadas
GPS (° ' ")
Altura (msnm)
Dato de
recolección
T. pseudocandicans 1 28 51 58 8 / 67 35 44 3 1889 Las Peñas M.L. et al 593, 27/XI/2015
T. pseudocandicans 1 28 50 06 4 / 67 38 56 3 2150 Las Peñas M.L. et al
605, 27/XI/2015
T. pseudocandicans 1 28 51 00 4 / 67 36 54 4 1963 Las Peñas M.L. et al 606, 27/XI/2015
T. candicans 1 28 52 00 3 / 67 35 45 4 1894 Las Peñas M.L. et al594, 27/XI/2015
T. candicans 1 28 51 54 7 / 67 35 37 7 1884 Las Peñas M.L. et al
603, 27/XI/2015
T. candicans 1 28 51 41 1 / 67 35 18 1 1884 Las Peñas M.L. et al 604, 27/XI/2015
T. candicans 2 28 53 15 2 / 67 32 45 8 1708 Las Peñas M.L. et al616, 28/XI/2015
T. candicans 3 29 20 31 0 /67 46 05 7 2053 Las Peñas M.L. et al 628, 29/XI/2015
T. candicans 3 30 20 31 0 /67 46 05 7 2053 Las Peñas M.L. et al
631, 29/XI/2015

Luego, se realizaron cortes transversales a mano alzada del tubo floral,
escamas, tépalos externos e internos, estilo, ovario y frutos. Los cortes fueron
teñidos con los colorantes azul astral y fucsina básica y luego montados con
glicerina diluida al 50% (D’ Ambrogio de Argüeso, 1986). Las observaciones se
realizaron con microscopio óptico Carl Zeiss® (Primo Star) y Arcano®, lupa Carl
Zeiss® (West Germany) y las fotografías digitales se tomaron con las cámaras
Nikon® Coolpix S630 y Nikon® Coolpix 5200.
Por último, se confeccionaron preparados permanentes de cortes
transversales de ovario, estilo, estigma y fruto joven. Las muestras se
deshidrataron en una serie de alcohol etílico en graduación creciente (50º, 60º,
70º, 80º, 90º, 95º y 100º) durante una hora cada paso. Se dejó toda una noche en
alcohol butílico. Posteriormente las muestras se clarificaron con dos pasos de xilol
puro de una hora cada uno, se incluyeron en Paramat de la siguiente forma:
primero 3:1 de xilol-Paramat (5 h), segundo 3:1 Paramat-xilol toda la noche,
tercero Paramat puro I 24 h, Paramat puro II 24 h. A los ovarios y frutos jóvenes se
los sometió a una serie de inclusiones en Paramat por una semana. (Zarlavsky,
2014).
Con el material incluido se realizaron cortes transversales seriados de 12 a
15 micras de espesor con micrótomo de rotación tipo Minhot, los mismos fueron
adheridos a portaobjetos con agua destilada y se estiraron en una platina
termostatizada a 37º. Luego se dejaron secar y se desparafinaron con xilol, se
hidrataron con una serie decreciente de alcohol etílico y se tiñeron con azul astral y
fucsina básica y se montaron con bálsamo de Canadá artificial (Kraus et. al., 1998;
Zarlavsky, 2014).
Las fotomicrografías de estos cortes se tomaron en un microscopio Olympus
BX61 software Cytovision® (Leica Biosystems); y en una lupa Olympus SZX16 con
una cámara Olympus DP71.
Finalmente, mediante el uso de los preparados temporarios, se llevaron a
cabo análisis cuantitativos de todos los órganos florales de los 9 individuos, con 5
17

mediciones (réplicas) de cada órgano/corte. En ellos se realizaron conteos y
mediciones con el empleo de un ocular métrico. Los órganos observados y las
variables medidas fueron:
Escama y Tépalo externo: ancho y espesor de cada órgano, espesor de la
epidermis e hipodermis de las caras adaxial y abaxial, número de conductos
secretores y de hacecillos. En las escamas florales, las cuales fueron tomadas a la
altura media del tubo floral, se contó el número de tricomas de las axilas.
Estilo y Estigma: el largo total del estilo más el estigma y cantidad de
lóbulos del estigma. El estilo se dividió en tres zonas, alta (inmediatamente debajo
del estigma), media (porción intermedia) y baja (justo sobre el ovario) y en cada
una se midió el diámetro total, espesor de la pared estilar, espesor de epidermis e
hipodermis, número de conductos y de haces vasculares.
Ovario y fruto: el espesor de la epidermis e hipodermis y el espesor de las
distintas zonas, hipanto, pared del ovario propiamente dicha, clamidocarpo y
pericarpo.
Los datos obtenidos de las mediciones fueron procesados con el programa
Infostat (Di Rienzo et al., 2017) y se trataron como un diseño con estructura
anidada, para ser sometidos a un análisis de la varianza (ANOVA) y a un análisis de
componentes principales (PCA).
Previamente se verificaron los supuestos de homogeneidad de medias y
normalidad. En los casos en que algún supuesto o ambos no se cumplieron, se
transformaron los datos con log 10. Como así mismo, en algunas variables los
supuestos siguieron sin cumplirse, se realizó un análisis de la varianza no
paramétrico mediante la prueba de Kruskal-Wallis.
18

RESULTADOS
T. candicans y T. pseudocandicans presentaron una exomorfología y anatomía
similar, con gran cantidad de tricomas, conductos secretores esquizógenos,
cristales y drusas en la mayoría de los órganos analizados.
Se describen a continuación las características compartidas y las diferencias
encontradas entre ambos taxones para cada uno de los órganos estudiados.
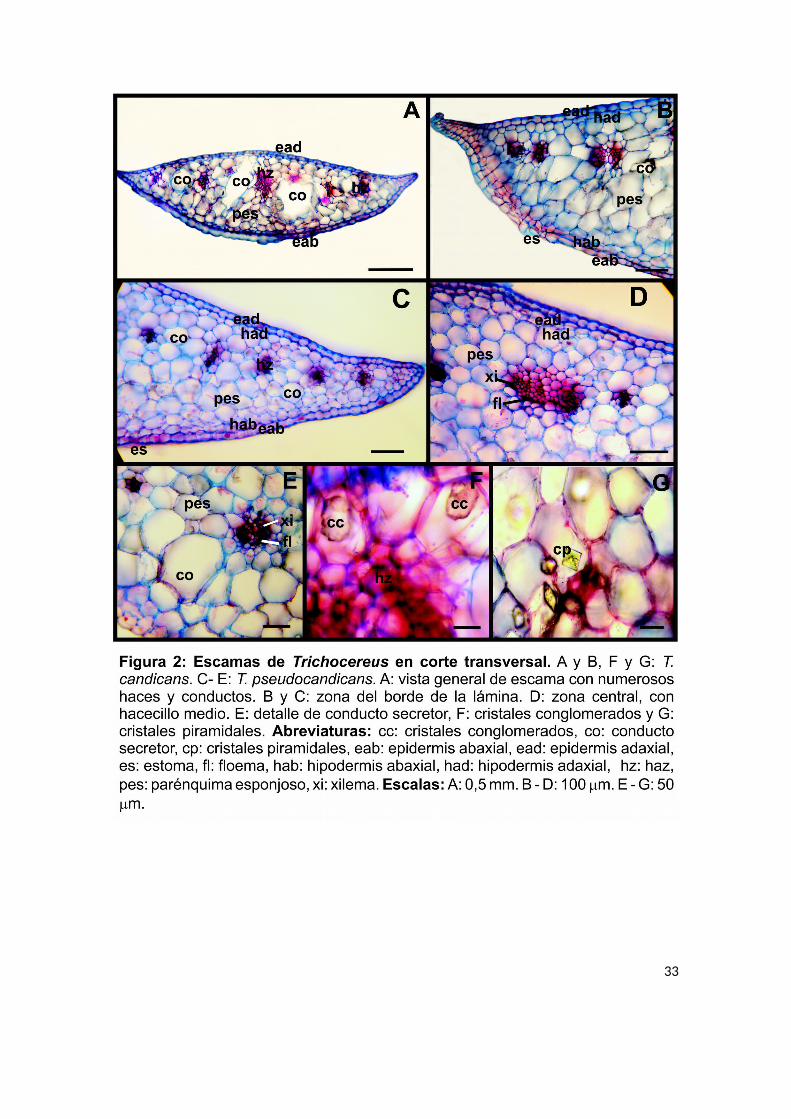
Escamas:
En T. candicans y T. pseudocandicans las escamas fueron numerosas y se
observaron alrededor de la cara externa del ovario y la zona baja del tubo floral
(Fig. 1 B y E); resultaron de color verdoso-amarillento, de forma subuladas y de
espesor considerable.
En las axilas de cada escama (Fig. 1 G) se observaron grupos de tricomas
eglandulares, pluricelulares, uniseriados, de 1,5 cm. de largo en promedio, de color
castaño claro. Algunos se agruparon en paquetes de hasta 5 tricomas, muy
adheridos entre sí. Fueron muy abundantes, con un promedio de 115,47 – 98,47
tricomas por escamas para T. candicans y T. pseudacandicans respectivamente
(Tabla 2).
En ambas especies, las escamas en corte transversal presentaron igual
anatomía y organización que una hoja (Fig. 2 A). La epidermis fue uniestratificada y
colenquimatosa con cutícula fina, formada por células pequeñas, con las paredes
periclinales más largas que las anticlinales. Inmediatamente por debajo se observó
una hipodermis también uniestratificada y colenquimatosa, de células un tanto
más grandes que la epidermis, pero de paredes más delgadas (Fig. 2 B y C).
La estructura foliar fue homogénea y compacta, las células de parénquima
esponjoso fueron grandes e isodiamétricas (Fig. 2 C). Los hacecillos se ubicaron
hacia la cara adaxial, fueron numerosos, aproximadamente 10 haces por escama
(Tabla 2), pequeños, del tipo colateral cerrado y rodeados de una vaina de células
19

parenquimáticas poco diferenciada (Fig. 2 C); el haz central se destacó por su
mayor tamaño (Fig. 2 D).
Relacionados a los haces, se observaron conductos secretores de origen
esquizógeno, con un promedio de 6,7 y 8,4 para cada especie (Tabla 2, Fig. 2 E),
además de gran cantidad de cristales, en su mayoría del tipo conglomerado (Fig. 2
F) y piramidales (Fig. 2 G).
Con respecto a los análisis cuantitativos, ninguna de las variables analizadas
en este órgano mostró diferencias significativas entre las especies (Tabla 2), lo cual
concuerda con su gran similitud anatómica. Sin embargo, T. candicans presentó
mayores valores en la mayoría de las mediciones y conteos realizados (Tabla 2).
Tépalos:
En ambas especies los tépalos fueron muy numerosos, se encontraron
fusionados en la base de la flor, formando un tubo floral, y libres en el ápice (Fig. 1
B, C, E y F).
Fueron de tres tipos: externos, intermedios e internos; diferenciados por su
tamaño y espesor (Tabla 2, Fig. 1 H). Los externos fueron iguales en ambas
especies, de color anaranjado-rojizo, pequeños, pero con gran espesor, similares a
las escamas. Los internos resultaron de mayores dimensiones, pero de menor
espesor y aspecto petaloide, resultaron diferentes entre las especies, de color
blancos en T. candicans y desde amarillentos a rojizos en T. pseudocandicans (Fig. 1
C y F). Los tépalos intermedios presentaron características medias entre los
externos e internos (Fig. 1 H).
Anatómicamente, los tépalos fueron similares entre sí, así como también
similares a las escamas. La epidermis e hipodermis fueron uniestratificadas
colenquimatosas, de células pequeñas en los tépalos externos (Fig. 3 B y E) y algo
mayores en los tépalos internos (Fig. 3 F). Las células del parénquima esponjoso
fueron irregulares en los tépalos externos (Fig. 3 B y C); en tanto que en los
internos fueron más redondeadas, grandes y distribuidas en pocos estratos (Fig. 3
F). Los hacecillos fueron numerosos y pequeños, ubicados hacia la cara adaxial en
20

los tépalos externos (Fig. 3 A y C) y en la línea central en los internos (Fig. 3 F).
Fueron colaterales cerrados con vaina (Fig. 3 G), acompañados de numerosos
conductos secretores (Fig. 3 A y F) y algunos cristales (Fig. 3 D).
Los análisis cuantitativos en los tépalos externos mostraron diferencias
significativas solamente en una variable analizada, el espesor de la epidermis
abaxial resultó mayor en T. candicans. Si bien el resto de las variables no fueron
estadísticamente diferentes, se observó que la mayoría de ellas presentaron
valores mayores en T. candicans (Tabla 2).
Por su parte, todas las mediciones realizadas en ambas especies resultaron
mayores en los tépalos que en las escamas (Tabla 2).
Estilo y estigma:
En ambas especies, los estilos y estigmas fueron de gran tamaño,
sobrepasaron la altura de los tépalos con una longitud total promedio de 10,8 - 11
cm para T. candicans y T. pseudacandicans respectivamente (Tabla 2).
Los estilos se dividieron en su extremo terminal, donde forman numerosos
lóbulos estigmáticos (Fig. 1 I). La cantidad de ellos resultó estadísticamente
diferente entre las especies estudiadas, siendo mayor en T. pseudocandicans (20 cm
± 1,46 cm) que en T. candicans (18,33 cm ± 1,4 cm) (Tabla 2).
Anatómicamente, cada lóbulo estuvo rodeado de un tejido epidérmico
papiloso (Fig. 4 A y B). Inmerso en el tejido parenquimático esponjoso se observó
un solo haz vascular de escasas dimensiones, acompañado por dos a cinco
conductos secretores esquizógenos (Fig. 4 C).
El estilo se observó circular en sección transversal. Presentó una epidermis
unistratificada con cutícula y paredes algo engrosadas, con un espesor promedio
de entre 35,0 y 49,19 µm (Tabla 2). Por debajo se observó una hipodermis, también
unistratificada pero con células isodiamétricas, redondeadas, con paredes
primarias delgadas, con un espesor promedio algo mayor, de entre 37,54 a 50,54
µm T. candicans y T. pseudacandicans respectivamente (Tabla 2).
21

La pared del estilo estuvo formada por parénquima esponjoso con
conductos secretores, variando en sus características según la zona. Inmerso en él
se encontraron pequeños haces vasculares concéntricos anficribales o colaterales
cerrados, los que generalmente presentaron forma de herradura (Fig. 5 D). Fueron
numerosos, con un promedio de 19,67 haces para T. pseudocandicans y 18,67 para
T. candicans, siendo la diferencia entre ambas estadísticamente significativa (Tabla
2).
El centro del estilo presentó tejido de transmisión con células filamentosas,
largas y delgadas y un canal estilar revestido por un tejido secretor papiloso (Fig. 4
E).
A lo largo del estilo se pudieron diferenciar tres zonas (alta, media y baja),
cada una con características particulares, las cuales se detallan a continuación.
Estilo alto o estigmático:
Fue seccionado justo por debajo del estigma, al cortarlo se notó mayor
rigidez del tejido que en las porciones inferiores. En esta zona, la pared estilar fue
muy angosta (0,81 – 0,75 mm para cada especie, Tabla 2), pero con una gran
superficie de tejido transmisor (Fig. 4 D).
En la pared se encontraron gran cantidad de conductos (48,83 – 26,87
conductos para cada especie, Tabla 2) y de notable tamaño, ubicados mayormente
en la zona cercana al tejido de transmisión. Los hacecillos no se ubicaron en el
centro de la pared, sino más cercanos a la epidermis interna. Esta última presentó
ondulaciones bien marcadas, en forma de crestas (Fig. 4 E y F). El tejido de
transmisión se observó muy laxo, cuyas características histológicas resultaron
difíciles de observar (Fig. 4 D y E).
En esta porción del estilo, se observaron diferencias significativas entre
ambas especies en todas las variables indagadas, menos en el espesor de la
hipodermis. Todas resultaron con mayores valores en T. candicans (Tabla 2).
22

Estilo medio:
Esta zona resultó con un diámetro total levemente menor que las restantes
(3,34 – 3,01 mm para T. candicans y T. pseudocandicans). En contraste, la pared
estilar fue de mayor grosor que en el estilo alto (0,92 – 0,85 mm para cada especie),
y en consecuencia el tejido transmisor fue menor (Tabla 2, Fig. 5 A).
Anatómicamente, la pared estilar presentó mayor cantidad de estratos
celulares, el parénquima fue compacto y los hacecillos se ubicaron en el centro de
la pared (Fig. 5 B y C). Los conductos secretores fueron menos numerosos (38,33 –
25,4 en cada especie) y se observaron en mayor proporción en la zona interna de la
pared, entre los hacecillos y la epidermis interna (Fig. 5 C y D).
Esta última presentó pequeñas ondulaciones, producidas por la presencia
de los conductos (Fig. 5 E). El tejido transmisor fue más compacto y el tejido
secretor se observó bien definido, con gran cantidad de papilas secretoras
recubriendo el canal estilar (Fig. 5 E y F).
Los análisis estadísticos en esta porción del estilo mostraron diferencias
significativas solo para dos variables, de las cinco observadas, en el espesor de la
pared estilar y en la cantidad de conductos que ella presentó. Asimismo, todas las
variables resultaron con mayores valores en T. candicans (Tabla 2).
Estilo bajo u ovárico:
Su diámetro resultó similar al diámetro del estilo alto (3,61 – 3,24 mm para
cada especie), pero el ancho de la pared estilar resultó con mayores valores
respecto a las zonas restantes (1,11 – 1,01 mm para T. candicans y T.
pseudocandicans) (Tabla 2, Fig. 5 G y H). En contraste, el tejido de transmisión, el
secretor y el canal estilar fueron muy reducidos (Fig. 5 H).
La pared estilar en esta porción presentó gran cantidad de estratos
parenquimáticos, muy compactos y formados por pequeñas células. Estas se
observaron de menor tamaño hacia la epidermis interna, la cual fue poco
diferenciada, pero sin ningún tipo de ondulación (Fig. 5 H e I). Los conductos
23

fueron menos numerosos (29,67 – 38,87 en cada especie) y más pequeños que en
las zonas restantes (Fig. 5 I).
Cuantitativamente, los estilos bajos de ambas especies resultaron similares,
ya que no se encontraron diferencias significativas en ninguna variable analizada.
Asimismo, todas presentaron valores mayores en T. candicans, a excepción del
número de conductos secretores que fue mayor en T. pseudocandicans (Tabla 2).
Ovario:
En ambos taxones, el ovario estuvo rodeado externamente de numerosas
escamas y tricomas (Fig. 1 B, E, y J). En corte longitudinal, se observó claramente
un ovario ínfero, con un disco nectarial por encima de él, justo en la base del estilo
(Fig. 1 J). En corte transversal resultó multicarpelar y de placentación parietal, su
pared presentó dos tejidos bien diferenciados, pero íntimamente soldados, el
hipanto y la pared del ovario propiamente dicha (Fig. 6 A). Los óvulos fueron
numerosos, unidos a las placentas mediante los funículos (Fig. 1 K, 6 A y D).
Anatómicamente, el contorno fue circular, pero con ondulaciones más o
menos marcadas, provocadas por la presencia de las escamas. La epidermis e
hipodermis estuvieron presentes, con iguales características que en las descriptas
para escamas, aunque con un mayor espesor (Fig. 6 B, Tabla 2).
El tejido parenquimático del hipanto estuvo formado por células grandes,
alargadas, algunas con los extremos en forma de huso, dispuestas perpendicular al
eje. Los hacecillos fueron escasos y muy reducidos, dispersos sin un patrón
definido. Por su parte, se observaron en esta zona gran cantidad de conductos y de
gran tamaño, además de cristales tipo drusas, prismáticos y conglomerados (Fig. 6
B).
La pared del ovario propiamente dicha estuvo formada por parénquima
compacto, con células notablemente más pequeñas e isodiamétricas. No presentó
conductos, ni cristales (Fig. 6 C y D). Entre el hipanto y la pared del ovario se
observó una zona de transición, formada por numerosos pero pequeños hacecillos
(Fig. 6 C). Cerca de la zona de placentación, las células parenquimáticas fueron aún
24

más pequeñas y se encontraron gran cantidad de haces que inervan los funículos
(Fig. 6 D).
Los óvulos fueron del tipo circinótropo, siendo este una leve variación del
tipo anátropo, aquí el óvulo sufre una rotación de hasta 360° gracias a que el
funículo se curva aún más. Se observó claramente el tegumento, la nucela y el saco
embrionario (Fig. 6 E). Los óvulos se presentaron agrupados en ramilletes de entre
8 a 12 óvulos, unidos por sus largos y respectivos funículos (Fig. 6 F).
Los análisis estadísticos no mostraron diferencias significativas entre las
especies en las mediciones tomadas, sin embargo T. pseudocandicans presentó
mayores valores en el espesor de la epidermis e hipodermis, y menores en el
espesor del hipanto y pared del ovario (Tabla 2).
Fruto:
Para ambos taxones, luego de la fecundación, la pared del fruto comenzó a
aumentar su espesor; el hipanto dio lugar al clamidocarpo, mientras que la pared
del ovario formó el pericarpo (Fig. 6 G). Cuantitativamente, todas las variables
medidas presentaron mayores valores en este estadio de desarrollo en
comparación con el anterior (Tabla 2).
En general, el fruto joven presentó características anatómicas similares a las
descriptas para el ovario. Se destaca la epidermis con paredes celulares más
engrosadas, el clamidocarpo con un parénquima más compacto, con mayor
cantidad de conductos secretores y cristales (Fig. 6 G y H); en tanto que el
pericarpo también se observó más compacto y con haces de mayores dimensiones
(Fig. 6 G e I).
Las semillas se diferenciaron de los óvulos por su mayor tamaño. Si bien el
embrión se encontró en estado de incipiente desarrollo, el episperma o tegumento
seminal se destacó por sus grandes células con paredes lignificadas (Fig. 6 I).
Los análisis estadísticos en esta etapa del fruto mostraron diferencias
significativas entre las especies solamente en el espesor de la hipodermis.
25

Asimismo, la mayoría de las variables mostraron valores mayores en T.
pseudocandicans (Tabla 2).
26

Análisis Cuantitativos:
De las 43 variables analizadas, 28 (65,12%) resultaron con valores mayores
en T. candicans, en tanto que solo 15 (34,88%) fueron mayores en T.
pseudocandicans.
Por su parte, se encontraron diferencias estadísticamente significativas solo
en 10 variables (23%). De las 10 variables significativamente diferentes, 7
resultaron con valores mayores en T. candicans, en tanto que solo 3 variables
fueron mayores en T. pseudocandicans.
Particularmente se observó en la parte alta del estilo, la mayor cantidad de
variables con diferencias estadísticamente significativas, siendo todas mayores
para T. candicans.
Los resultados confirman que las flores en T. candicans presentan mayores
dimensiones que en T. pseudocandicans, no así los frutos jóvenes, los cuales
presentaron mayores dimensiones en T. pseudocandicans aunque la mayoría de las
diferencias fueron pequeñas, y estadísticamente insuficientes.
A continuación, se presentan de las mediciones realizadas a los órganos deambas especies, las medias con su respectivo desvío estándar.
27

Tabla 2. Características cuantitativas en T. candicans y T.pseudocandicans. Las variables de cada órgano analizado se indican comopromedios ± desvío estándar. * indican diferencia significativa entre las especies,para un Análisis de la Varianza y Kruskal Wallis con p ≤ 0,05.
28

Por último, a través del análisis de componentes principales (PCA) se
observó algo similar a lo que se analizó anteriormente, es decir, que a través de las
variables analizadas no se puede explicar una diferenciación absoluta de
Trichocereus candicans respecto Trichocereus pseudocandicans, ya que en este
análisis los individuos de ambos taxones se encontraron sumamente dispersos a
través del gráfico, inmersos entre las variables, también distribuidas de manera
homogénea. Cabe destacar que Trichocereus pseudocandicans, (los puntos de color
fucsia) estuvieron mejor representados por el eje x (CP1), en cambio los individuos
de Trichocereus candicans (los puntos de color azul) estuvieron representados de
manera equitativa por ambos ejes x e y (CP1 y CP2), (Gráfico 1).
A continuación se presenta el gráfico Biplot de componentes principales
(CP1 y CP2) con su respectiva tabla de autovectores (Tabla 3 y 4).
Gráfico 1. Biplot de componentes principales 1 (CP1) y 2 (CP2). Los cuadrados(■) fucsia numerados del 1-3 representan a T. pseudocandicans, los cuadrados (■)azules numerados del 4-9 representan a T. candicans y los triángulos (▲) amarillosrepresentan a cada una de las variables medidas para ambos taxones.
29

Tabla 3. Autovectores.
Tabla 4. Referencias de las variables medidas.
30

FIGURAS
31

32
33

34

35

36

37

DISCUSIÓN
Las flores y los frutos de T. candicans y T. pseudocandicans comparten la
mayoría de las características exomorfologicas y anatómicas, las cuales se detallan
a continuación:
Las flores de las especies estudiadas fueron de gran tamaño, muy vistosas y
con gran cantidad de piezas periánticas, como es común en la familia (Kiesling,
1978; Anderson, 2001; Kiesling & Ferrari, 2009). Presentaron gran cantidad de
escamas y tricomas eglandulares en la superficie externa del ovario y la zona baja
del tubo floral, los cuales se mantienen en el fruto luego de la fecundación. Estas
características son comunes en el género Trichocereus, pero infrecuentes en el
resto de la familia, lo cual es considerado un carácter taxonómico importante
(Kiesling, 1978, Kiesling & Ferrari, 2009). Los tricomas se encontraron agrupados
en un conjunto bien empaquetado y tendrían la función de proteger a la planta del
sobrecalentamiento por la radiación solar, siendo una adaptación a las condiciones
áridas (Dickison, 2000; Kalashnyk et al., 2016) y su presencia, particularmente en
la flor, estaría relacionada con la atracción y sujeción de los polinizadores
(Bernacki et al., 2015). El tubo floral estuvo compuesto por numerosos tépalos
fusionados en su base y libres hacia el ápice, reconociéndose tres tipos diferentes
(Kiesling, 1978; Anderson, 2001). En la familia, los tépalos son considerados
brácteas (escamas) derivadas y su principal función es proteger las partes
reproductivas de la flor en desarrollo (Anderson, 2001; Fuentes-Pérez et al., 2009).
El gineceo resultó de grandes dimensiones, compuesto por un ovario ínfero,
un estilo muy desarrollado y un estigma formado por numerosos lóbulos (Dickison,
2000; Albesiano, 2015). El estilo de las especies estudiadas se caracterizó por la
presencia de tejido de transmisión y canal estilar. El tejido de transmisión presentó
células filamentosas, largas y delgadas, a diferencia de lo encontrado en Opuntia
Mill., donde las células fueron bien redondeadas, pequeñas y compactas (Fuentes-38

Pérez et al., 2009). Este tejido es fundamental para el desarrollo del tubo polínico,
es rico en pectinas y su consistencia es mucilaginosa (González & Cristóbal, 2006).
El canal estilar se observó recubierto internamente por una epidermis papilosa
(Buxbaum, 1953), lo que también se reportó en Opuntia, Pereskia Mill. y en varios
miembros de Cactoidea (Boke, 1963, 1966, 1968; Fuentes-Pérez, 2004; Fuentes-
Pérez et al., 2009); con la diferencia que las especies aquí estudiadas carecen del
tejido parenquimatoso entre el tejido de transmisión y la epidermis papilosa, que sí
se presentó en Opuntia (Fuentes-Pérez et al., 2009).
La pared del ovario se encontró recubierta por el hipanto o pericarpelo, el
tejido caulinar que rodea al ovario ínfero de las Cactáceas (Buxbaum, 1953;
Barthlott & Hunt, 1993; Anderson, 2001). En las especies analizadas se observó a
estos tejidos íntimamente fusionados, incluso luego de la fecundación y formación
del fruto, a diferencia de Pereskia donde éstos aparentan estar soldados, pero
realmente lo están sólo en la base del ovario (Da Rosa & De Souza ., 2003). Entre el
hipanto y la pared del ovario propiamente dicho se puede observar un anillo de
haces colaterales como en Opuntia y en Epiphyllum phyllanthus (L.) Haw. (Fuentes-
Pérez et al., 2009; García de Almeida et al., 2010).
La placentación fue parietal, con numerosas placentas y sin distinción entre
los carpelos, característica común en el género y en la familia (Dickison, 2000;
Anderson, 2001; Albesiano, 2015). El número de carpelos puede estimarse con un
promedio de 19, tomando como referencia el número de hacecillos del estilo y el
número de los lóbulos estigmáticos (Boke, 1964; Roth, 1977; García de Almeida et
al., 2010). Los óvulos fueron del tipo circinótropos, lo que es una pequeña
variación del tipo anátropo, el más común en la familia. Aquí el funículo continúa
curvándose todavía más (Johri et al, 1992), lo mismo ocurre en Epiphillum
phyllanthus (García de Almeida et al., 2010). Por ser mínima la variación entre los
dos tipo de óvulo, algunos autores como Kiesling (1978), menciona que los óvulos
para Trichocereus son del tipo anátropos.
39

En cuanto al fruto, se diferencian a simple vista ya que el de T. candicans
mantiene la flor seca adherida y es de color amarillo-dorado a rosado, en cambio el
fruto de T. pseudocandicans pierde la flor al madurar y es verde-rosado a
amarillento. Llamativamente observamos que el fruto de T. pseudocandicans es un
tanto mayor que el de T. candicans donde coincidimos con Kiesling (1978) a pesar
que sus flores son más pequeñas. Los frutos de todas las especies del género son
comestibles, de forma globosa u ovoide, son carnosos, dehiscentes por un lado, las
escamas se hacen más pequeñas que en la flor y se mantienen los tricomas
(Kiesling, 1978). Según Kiesling (1978) debería llamarse “pseudofruto” o
“pseudobaya” debido al tejido pedicelar que rodea al ovario: el pericarpelo o
hipanto.
Anatómicamente, algunas características encontradas merecen especial
mención, por haber sido observadas en todos los órganos analizados, de ambas
especies, y por ser consideradas caracteres xerófitos de importancia taxonómica.
Los mismos se describen a continuación:
Epidermis: Se observó una epidermis uniestratificada colenquimatosa con
cutícula delgada, al igual que en Opuntia brasiliensis (Azevedo et al., 2013), en
especies de Lepismium Pfeiff. (Torres-Boeger et al., 2010) y en Backebergia
militaris (Audot) Bravo ex Sánchez-Mej. (Vázquez-Sánchez et al., 2015). Las células
presentaron pared periclinal externa recta también en todos los órganos
analizados.
Tanto epidermis uniestratificada como células con pared periclinal externa
recta son características predominantes en la familia Cactaceae (Gibson & Nobel,
1986; Mauseth, 2006; Gonçalves da Silva et al., 2013). Por otra parte, el grosor de la
cutícula es una característica muy variable dentro de Cactoideae, la cual junto con
epidermis e hipodermis cumplen una importante función de protección y de
retención de agua (Mauseth, 2006; Gonçalves da Silva et al., 2013).
40

Hipodermis: Se observó hipodermis uniestratificada colenquimatosa, lo
cual es una característica común en Cactaceae (Vázquez-Sánchez et al., 2015).
Algunas especies de Lepismium (Torres-Boeger et al. 2010), de Rhipsalis Gaertn., de
Cereus Mill. y varias especies de la tribu Pachycereeae (Guerra & Medri, 2015;
Kalashnyk et al., 2016) también tienen este tipo de hipodermis, a diferencia de por
ejemplo de Backebergia militaris donde se observaron dos estratos (Vázquez-
Sánchez et al., 2015) y en plántulas de Epiphyllum donde fue ausente (Kalashnyk et
al., 2016). La presencia de hipodermis puede afectar la penetración de la radiación
solar al clorénquima subyacente, regulando así la pérdida de agua en la planta
(Kalashnyk et al., 2016). Tanto el número de capas como el grosor de la hipodermis
son muy variables dentro de Cactaceae (Gonçalves da Silva et al., 2013) por lo que
estas variables son sumamente importantes para la delimitación de taxones (Monje
& Baran, 2000; Terrazas-Salgado et al., 2005).
Conductos: Se observaron abundantes conductos secretores esquizógenos,
presumiblemente con mucílagos. Estos son altamente hidrófilos por lo que se los
relaciona con el almacenamiento de agua y afectan las relaciones hídricas en la
planta, lo que constituye una importante adaptación a los ambientes xerofíticos y
son característicos de muchos grupos de cactus (Gibson & Nobel, 1986; Anderson,
2001; Soffiatti & Angyalossy, 2007; Kalashnyk et al., 2016) Como por ejemplo en
especies de Ariocarpus Scheidw., Opuntia y Lepismium (Anderson, 2001; Guerra &
Medri, 2015). Se piensa que los mucílagos han evolucionado por separado en cada
tribu de Cactaceae (Soffiatti & Angyalossy, 2007).
Cristales: En ambos taxones se observaron gran cantidad de cristales de los
tipos conglomerado, prismático y drusas; donde estas últimas son comunes en la
subfamilia Cactoideae (Gibson, 1973; Kalashnyk et al., 2016). Los cristales tienen la
función de ser soporte mecánico, defensa ante la herbivoría y además podrían
evitar el colapso cuando el tejido pierde la suculencia (Cenizo et al., 2013). A su vez
controlan el equilibrio iónico en la planta, regulan la presión osmótica, la absorción
y la reflexión de la luz (Soffiatti & Angyalossy, 2007; Arruda, 2010; Azevedo et al.,
41

2013; Kalashnyk et al., 2016). Respecto a la cuantificación de los cristales
utilizados como una variable de importancia taxonómica, se los utilizó para dividir
a Pilosocereus (Byles & Rowley) en dos subgéneros (Silva & Alves, 1999) y también
a Pachycereeae en dos subtribus (Soffiatti & Angyalossy, 2007). Sin embargo, otros
autores afirman que no suelen ser importantes para la filogenética y la taxonomía
(Ogburn & Edwards, 2009; Guerra & Medri, 2015; Kalashnyk et al., 2016).
42

CONCLUSIONES
Los resultados obtenidos en el presente trabajo muestran una gran similitud
morfoanatómica en las flores y frutos jóvenes de Trichocereus candicans y
Trichocereus pseudocandicans, es decir, no se encontraron pruebas suficientes para
considerar a los taxones como dos especies completamente distintas.
Sin embargo, apoyamos la teoría que Trichocereus pseudocandicans podría
ser una subespecie de reciente formación, e inferimos que la posterior
diferenciación podría ser causada por diferentes presiones selectivas ejercidas por
sus polinizadores, como se evidencia en sus respectivos biotipos florales: las flores
de T. candicans son de gran tamaño, blancas, de antesis nocturna y levemente
perfumadas, mientras que las de T. pseudocandicans son más pequeñas, de color
amarillento a rojizo, de antesis diurna y escasa o imperceptible fragancia.
A su vez, proponemos que esta diferenciación podría estar acrecentada por
cuestiones biogeográficas, ya que T. pseudocandicans es endémica de las Sierras de
Famatina, donde se observa un microclima propio de sierras de altura.
43

Consideraciones finales:
Consideramos que para lograr dilucidar la identidad taxonómica real de las
entidades estudiadas se deberían llevar a cabo análisis complementarios sobre la
morfoanatomía de los órganos vegetativos, así como de los frutos maduros y
semillas. Además, sería de suma importancia el estudio de la biología reproductiva,
para conocer aspectos relacionados a los visitantes florales, sistema de
compatibilidad del polen, diseminación y germinación de semillas, estudios
citogenéticos, etc.
Finalmente, sería interesante estudiar la estructura anatómica y
morfológica de distintas especies del género, afines a las aquí estudiadas, por
ejemplo Trichocereus vatteri, T. strigosus, T. lamprochlorus, T. pectiniferus, T.
spinibarbis y T. desertícola; para seguir estableciendo semejanzas y diferencias, y a
su vez relacionarlas con su filogenia y evolución.
44

BIBLIOGRAFÍA
ACEÑOLAZA, P. G. 1996. Vegetación. In: ACEÑOLAZA, F. G., H. MILLER & A. TOSELLI
(eds.), Geología del sistema de Famatina, pp.19-22. Münchner Geologische Hefte
19 (Reihe A). Múnich.
ALBESIANO, S. & T. TERRAZAS. 2012. Cladistic analysis of Trichocereus (Cactaceae:
Cactoideae: Trichocereeae) based on morphological data and chloroplast Dna
sequences. Haseltonia 17: 3-23.
ALBESIANO, A. S. 2015. Taxonomía y filogenia de Trichocereus (Berg.) Riccob.
(Trichocereeae-Cactaceae). (Tesis doctoral). Universidad Nacional de La Plata,
La Plata, Argentina.
ANDERSON, E. 2001. The Cactus Family. Timber Press. Portland, Oregon. (Pp. 777).
ARRUDA, E. 2010. Histogênese de segmentos caulinares de espécies de Opuntioideae
(Cactaceae). (Tesis doctoral). Universidad Nacional de San Pablo. San Pablo,
Brasil.
AZEVEDO, C. F., K. R. GOMES DA SILVA, R. L. ALCANTARA-BRUNO, Z. G. & MACIEL-
QUIRINO. 2013. Morfoanatomía vegetativa de Opuntia brasiliensis (Willd) Haw.
Ambiência 9: 73-82.
BADANO, E. I., & B. O. S SCHLUMPBERGER. 2001. Sistema de cruzamiento y
estimaciones en la eficiencia de polinización sobre Trichocereus pasacana
(Cactaceae) en dos poblaciones del noroeste argentino. Gayana Bot. 58: 115-
122.
BARBOZA, G., F. CHIARINI & L. ARIZA ESPINAR (eds.). 2016. Plantas del Famatina.
Series Naturales. Editorial de la Universidad Nacional de Córdoba. Córdoba,
Argentina.
BARTHLOTT, W. & D. R. HUNT. 1993. Cactaceae. In: KUBITZKI K., J. G. ROHWER &
V. BITTRICH (eds.), Flowering Plants. Dicotyledons. The Families and Genera
of Vascular Plants, vol 2, pp. 161-197. Springer Verlag, Berlin.
BECERRA, R. 2000. Las Cactáceas, plantas amenazadas por su belleza. Biodiversitas
32: 1-5.
45

BERNACKI, F. G., P. L. ALBORNOZ, M. VALOY & M. ORDANO. 2015. Anatomía de flor y
fruto de Vassobia breviflora (Solanaceae) en el sur de las Yungas australes
(Argentina). Phyton 84: 478-487.
BIURRUN, F. N., W. D. AGÜERO & D. F. TERUEL. 2012. Consideraciones fitogeográficas
sobre la vegetación de los llanos de La Rioja . Ediciones INTA, Serie: estudios
sobre el ambiente y el territorio. Catamarca, Argentina.
BRAKO, L. & J. ZARUCCHI. 1993. Catalogue of the flowering plants and gymnosperms
of Peru. Monogr. Syst. Bot. Missouri Bot. Gard 45: 1-1286.
BRAVO HOLLIS, H. & L. SCHEINVAR. 1999. El interesante mundo de las Cactáceas. 2da
ed. Fondo de Cultura Económica, México. (Pp. 233).
BOKE, N. H. 1963. Anatomy and development of the flower and fruit of Pereskia
pititache. Amer. J. Bot. 50: 843-858.
BOKE, N. H. 1964. The Cactus Gynoecium: A New Interpretation. Amer. J. Bot. 51: 598-
610.
BOKE, N. H. 1966. Ontogeny and structure of the flower and fruit of Pereskia aculeate.
Amer. J. Bot. 53: 534-542.
BOKE, N. H. 1968. Structure and Development of the Flower and Fruit of Pereskia
Diaz-Romeroana. Amer. J. Bot. 55: 1254-1260.
BUXBAUM, F. 1953. Morphology of Cacti. Abbey Garden Press, Pasadena, California.
(Pp. 223).
CABRERA, L. A. 1971. Fitogeografía de la República Argentina. Bol. Soc. Argent. Bot.
14: 23-33.
CENIZO, V. J., M. B. MAZZOLA, B. M. CORRO-MOLAS & A. G. KIN. 2013.
Características morfológicas y anatómicas de las plántulas de Trichocereus
candicans (Cactaceae). Bol. Soc. Argent. Bot. 48: 443-451.
CERÉN-LÓPEZ, J. G., J. E. MENJÍVAR-CRUZ & S. ARIAS. 2015. Diversidad de cactus del
Salvador. Colección de Historia Natural Vol. 3, 1era ed. Dirección Nacional de
Patrimonio Cultural y Natural, El Salvador. (Pp. 148).
D’ AMBROGIO DE ARGÜESO, A. (ed.). 1986. Manual de técnicas de Histología Vegetal.
Hemisferio Sur, Buenos Aires. (Pp. 83).
46

DA ROSA, S. M. & L. A. DE SOUZA. 2003. Morfo-anatomia do fruto (hipanto, pericarpo
e semente) em desenvolvimento de Pereskia aculeata Miller (Cactaceae). Acta
Sci. Biol. Sci. 25: 415-428.
DICKISON, W. C. (ed.). 2000. Integrative Plant Anatomy. Academic Press, Orlando,
Florida.
DI RIENZO, J., F. CASANOVES, M. BALZARINI, L. GONZALEZ, M. TABLADA & C.
ROBLEDO. 2017. Infostat, versión 2017, Grupo Infostat, F. C. A. (UNC), Córdoba,
Argentina.
ESQUIVEL, P. 2004. Los frutos de las cactáceas y su potencial como materia prima.
Agron. Mesoam. 15: 215-219.
FAHN, A. & D. F. CUTLER (eds.). 1992. Xerophytes. Gebruder Borntraeger, Berlín. (Pp.
176).
FRIEDRICH, H. 1974. Zur taxonomie und phylogenie der Echinopsidinae
(Trichocereinae). I. O. S. Bull. 3: 79-93.
FUENTES-PÉREZ, M. 2004. Anatomía floral de algunas especies de la tribu
Pachycereeae (Cactaceae). (Tesis de licenciatura). Facultad de Estudios
Superiores Iztacala, Universidad Nacional Autónoma de México, México.
FUENTES-PÉREZ, M., T. TERRAZA & S. ARIAS. 2009. Anatomía floral de cinco
especies de Opuntia (Opuntioideae, Cactaceae) de México. Polibotanica 27: 89-
102.
GARCÍA DE ALMEIDA, O. J.; A. A. SARTORI-PAOLI & L. A. DA SOUZA. 2010. Morfo-
anatomía de la flor de Epiphyllum phyllanthus (Cactaceae). Revista Mex.
Biodivers. 81: 65-80.
GIBSON, A. C. 1973. Comparative Anatomy of Secondary Xylem in Cactoideae
(Cactaceae). Biotropica 5: 29-65.
GIBSON, A. C. & K. E. HORAK. 1978. Systematic anatomy and phylogeny of Mexican
columnar Cacti. Ann. Missouri Bot. Gard. 64: 999-1057.
GIBSON, A. & P. NOBEL (ed.). 1986. The cactus primer. Harvard University Press.
Cambridge. (Pp. 286)
47

GONÇALVES DA SILVA, C.H., A. CALVENTE, A. SOLLER, L. T. MARANHO, M.R.
TORRES-BOEGER & P. SOFFIATTI. 2013. Anatomia de Rhipsalis subgênero
Erythrorhipsalis A. Berger (Rhipsalideae, Cactaceae). Iheringia, Bot. 68: 249-259.
GONZALEZ, A. M. & C. L. CRISTOBAL. 2006. Ontogenia del óvulo y semilla de Sterculia
striata (Sterculiaceae). Bonplandia 15: 63-77.
GUERRA, A. & C. MEDRI. 2015. Diferenciação morfoanatômica de Lepismium
cruciforme (Vell.) Miq. (Cactaceae) do dosel e do sub-bosque. Rev. Saúde e Biol.
10: 43-51.
HOFFMANN, A. & H. WALTER. 2004. Cactáceas en la flora silvestre de Chile. 2da ed.
Ediciones Fundación Claudio Gay. Santiago de Chile. (Pp. 307).
HUNT, D. R. & N. TAYLOR. 1990. The genera of Cactaceae: progress towards
consensus. Bradleya 8: 85-107.
HUNT, D. R., N. TAYLOR & G. CHARLES (eds.). 2006. The New Cactus Lexicon. DH
Books, Milborne Port. (Pp. 527).
HUNT, D. R. 2013. The New Cactus Lexicon Illustrations. DH Books, Milborne Port.
JOHRI, B. M., AMBEGAOKAR K. B. & SRIVASTAVA P. S. 1992. Comparative
Embryology of Angiosperms. Vol. 1/2. Springer Verlag, Berlin. (Pp. 1221).
KALASHNYK, H., N. NUZHYNA & M. GAIDARZHY. 2016. Anatomical and
morphological features of seedlings of some Cactoidea Eaton (Cactaceae Juss)
species. Acta Agrobot. 69: 1697.
KIESLING, R. 1978. El género Trichocereus (Cactaceae). Apéndice I: Las especies de la
República Argentina. Darwiniana 21: 263-330.
KIESLING, R, J. LAROCCA, L. FAÚNDEZ, D. METZING & S. ALBESIANO. 2008.
Cactaceae. In: ZUOLAGA F. O., O. MORRONE & M. J. BELGRANO (eds.) Catálogo de
las Plantas Vasculares del Cono Sur. Vol. 2, pp. 1715-1830. Missouri Botanical
Garden Press, Saint Louis.
KIESLING, R. & FERRARI O. E. (eds.). 2009. 100 cactus argentinos. Albatros, Buenos
Aires. (Pp. 128).
KRAUS, J., H. DE SOUSA, M. REZENDE, N. CASTRO, C. VECCHI & R. LUQUE. 1998.
Astra Blue and Basic Fuchsin double staining of plant materials. Biotechnic &
Histochemistry 73: 235-243.
48

LOPEZ DE CASENAVE, J. N. 2001. Estructura gremial y organización de un ensamble de
aves del Desierto del Monte. (Tesis doctoral). Facultad de Ciencias Exactas y
Naturales, Universidad de Buenos Aires, Argentina.
MANDUJANO, M. C., A. FLORES-MARTÍNEZ, J. GOLUBOV & E. ESCURRA. 2002.
Spatial distribution of three globose cacti in relation to different nurse-plant
canopies and bare areas. S. W. Naturalist 47: 162-168.
MAUSETH, J. D. 2006. Structure–function relationships in highly modified shoots of
Cactaceae. Ann. Bot. 98: 901–926.
MAZZOLA, M. B., V. J. CENIZO & A. G. KIN. 2013. Factores que afectan la germinación
de Trichocereus candicans (Cactaceae). Bol. Soc. Argent. Bot. 48: 515-523.
MONJE, P. V. & E. J. BARAN. 2000. First evidences of the bioaccumulation of α-quartz
in Cactaceae. J. Pl. Physiol. 157: 457-460.
NAVARRO, G. 1996. Catálogo ecológico preliminar de las Cactáceas de Bolivia. Lazaroa
11: 33-84.
ROTH, I. 1977. Fruits of angiosperms. In: K. LINSBAUER (ed.) Encyclopedia of plant
anatomy, (pp. 675). Gebrüder Borntrager, Berlin.
SCHLUMPBERGER, B. O. & S. S. RENNER. 2012. Molecular phylogenetics of Echinopsis
(Cactaceae): Polyphyly at all levels and converget evolution of pollination modes
and growth forms. Amer. J. Bot. 99: 1335–1349.
SILVA, D. & J. L. ALVES. 1999. Anatomia dos órgãos vegetativos de seis espécies de
Pilosocereus Byles & Rowley (Cactaceae). Bol. Bot. Univ. São Paulo 18: 53-60.
SOFFIATTI, P. & V. ANGYALOSSY. 2007. Anatomy of Brazilian Cereeae (subfamily
Cactoideae, Cactaceae): Arrojadoa Britton & Rose, Stephanocereus A. Berger and
Brasilicereus Backeberg. Acta Bot. Bras. 21: 813-822.
TERRAZAS-SALGADO, T., S. LOZA-CORNEJO & H. J. ARREOLA-NAVA. 2005. Stem
anatomy of the species of the genus Stenocereus (Cactaceae). Acta Bot. Venez. 28:
321-336.
TORRES-BOEGER, M. R., P. SOFFIATTI, M. A. GOMES-SOUTO, M. BUDCHEN, K. P.
BAGATINI & M. DAL FORNO. 2010. Morfología funcional de dos especies de
Lepismium (Rhipsalideae, Cactaceae). Revista Mex. Biodivers. 81: 393-400.
49

OGBURN, R. M. & E. J. EDWARDS (eds.). 2009. Anatomical variation in Cactaceae and
relatives: Trait lability and evolutionary innovation. Amer. J. Bot. 96: 391-408.
VÁZQUEZ-SÁNCHEZ, M., T. TERRAZAS & S. ARIAS. 2015. Comparative morphology
and anatomy of Backebergia militaris (Echinocereeae–Cactaceae) cephalium.
Plant. Syst. Evol. 302: 245-256.
ZARLAVSKY, G. E. (ed). 2014. Histología Vegetal: técnicas simples y complejas, 1era ed.
Sociedad Argentina de Botánica. Gráfica Arte, Buenos Aires.
50

AGRADECIMIENTOS
A mi Directora Natalia Delbón y Co-directora Laura Stiefkens, por guiarme,
por tenerme paciencia y ayudarme en este proceso. A Teresa Cosa por
permitirme hacer mi tesina en la cátedra. A Laura Las Peñas, Alejandra y a
Adriana por enseñarme las técnicas y recibirme siempre con una sonrisa. A
mis compañeras de cátedra. Al laboratorio de Morfología Vegetal por
proveerme las instalaciones y materiales.
A mi querida UNC, por la educación pública.
A mis padres Silvia y Walter, por la vida, la educación, el apoyo, por guiarme,
por respetar siempre mis elecciones y acompañarme con amor. A mi
hermano, mis tíos y primos. A mi madre por su amor absoluto.
A mi querida Nona Elida, a mis abuelos Mari y Dardo. A los tres, gracias por
alimentarme rico antes de clase tantas veces, por darme cobijo, por el amor
de abuelos. A mi viejito Don Luis que desde donde sea que esté me siguen
llegando sus bendiciones y protección. Abuelo Dardo, este trabajo te lo
dedico a vos, sé cuanto deseabas verme recibida.
A mis amigos, las chicas de la facu, los amigos de la vida, los del trabajo, a
todos, siempre presentes en cada etapa, gracias por los abrazos, por la
diversión, por el aguante, y a muchos de ellos gracias por colaborar de
alguna manera con este trabajo.
A mis jefes de todos los trabajos durante estos años, sobre todo los últimos,
por la paciencia y la confianza.
Gracias a todos los seres que me acompañaron con tanto amor y paciencia
durante estos años.
A la Naturaleza, por inspirarme y enamorarme cada día.
51

52


















