1. DESCRIPCION DE LAS ACTIVIDADES …sappi.ipn.mx/cgpi/archivos_anexo/20060587_3419.pdf ·...
12
1. DESCRIPCION DE LAS ACTIVIDADES REALIZADAS. El trabajo se planteo desde su inicio para TRES años • Se tiene publicado el articulo Ramón Arteaga Garibay , Ma Guadalupe Aguilera Arreola, Armando Navarro Ocaña, Silvia Giono Cerezo, Miroslava Sánchez, José M olina, Carlos Eslava, Alejandro Cravioto, Graciela Castro Escarpulli * . 2006. Serogroups, K1 antigen, and antimicrobial resistance patterns of Aeromonas spp. strains isolated from different sources in Mexico . Memorias do Instituto Oswaldo Cruz : 101(2):157-161 PMID 16830708 ISSN 0074-0276 • Se envió al comité editorial para su consideración el artículo: Subashkumar, R., Tha. Thayumanavan , G. Vivekanandhan, D. J. Colquhoun, G. Castro-Escarpulli and P. Lakshmanaperumalsamy . 2006. Persistence of genetic diversity in Aeromonas hydrophila from children diarrhoea by Random Amplified Polymorphic DNA PCR and Enterobacterial Repetitive Intergenic Consensus Sequence PCR. FEMS Microbiology Letters (SOMETIDO) • Se tiene en preparación en articulo Ma. Guadalupe Aguilera-Arreola, César Hernández-Rodríguez, Gerardo Zúñiga, María José Figueras , Rafael Garduño Graciela Castro-Escarpulli . Virulence potential and genetic diversity of A. hydrophila, A. caviae and A. veronii clinical isolates from Mexico and Spain: A comp arative study • Se tiene en proceso cinco tesis de licenciatura MARIA ISABEL PORTILLA MUÑOZ Evaluación de un medio cromogénico para el aislamiento e identificación presuntiva de Aeromonas.10 de Octubre del 2006 ERIKA BEATRIZ ANGELES MORALES Determinación de factores extrínsecos relacionados con la formación de biopelícula de Aeromonas caviae Sch3 . 7 de Agosto del 2006 MARÍA ALEJANDRA ALVARADO HERNÁNDEZ Búsqueda del gen cag A en Aeromonas asiladas de medio ambiente. 15 de Febrero 2006 CARLOS SANCHEZ GARIBAY Demostración de β-lactamasas de espectro expendido en cepas de Aeromonas spp .15 de Febrero 2006 NIRYA RAMIREZ ZETINA Expresión fenotípica del Sistema Flagelar de Aeromonas spp y su relación con la formación biofilm. • Se tiene en proceso dos Tesinas de Licenciatura ROSALBA GUERRERO GONZALEZ El flagelo bacteriano. 21 de Agosro del 2006 ROSALIA BAEZ ARIAS Sistemas de Secreción bacterianos. 21 de Agosro del 2006 • Se tiene en proceso una tesis de doctorado RAMÓN IGNACIO ARTEAGA GARIBAY El biofilm de Aeromonas y su relación con el sistema flagelar de cómo mecanismo de patogenicidad y resistencia en Aeromonas. 23 de Noviembre 2006 • Se terminó una tesis de doctorado. Ma GUADALUPE AGUILERA ARREOLA Doctoral Diversidad genética intraespecífica de Aeromonas hydrophila 6 Septiembre 2006
Transcript of 1. DESCRIPCION DE LAS ACTIVIDADES …sappi.ipn.mx/cgpi/archivos_anexo/20060587_3419.pdf ·...

1. DESCRIPCION DE LAS ACTIVIDADES REALIZADAS.
El trabajo se planteo desde su inicio para TRES años
• Se tiene publicado el articulo Ramón Arteaga Garibay , Ma Guadalupe Aguilera Arreola, Armando Navarro Ocaña, Silvia Giono Cerezo, Miroslava Sánchez, José M olina, Carlos Eslava, Alejandro Cravioto, Graciela Castro Escarpulli *. 2006. Serogroups, K1 antigen, and antimicrobial resistance patterns of Aeromonas spp. strains isolated from different sources in Mexico. Memorias do Instituto Oswaldo Cruz : 101(2):157-161 PMID 16830708 ISSN 0074-0276
• Se envió al comité editorial para su consideración el artículo: Subashkumar, R., Tha. Thayumanavan , G. Vivekanandhan, D. J. Colquhoun, G. Castro-Escarpulli and P. Lakshmanaperumalsamy . 2006. Persistence of genetic diversity in Aeromonas hydrophila from children diarrhoea by Random Amplified Polymorphic DNA PCR and Enterobacterial Repetitive Intergenic Consensus Sequence PCR. FEMS Microbiology Letters (SOMETIDO)
• Se tiene en preparación en articulo
Ma. Guadalupe Aguilera-Arreola, César Hernández-Rodríguez, Gerardo Zúñiga, María José Figueras , Rafael Garduño Graciela Castro-Escarpulli . Virulence potential and genetic diversity of A. hydrophila, A. caviae and A. veronii clinical isolates from Mexico and Spain: A comp arative study
• Se tiene en proceso cinco tesis de licenciatura MARIA ISABEL PORTILLA MUÑOZ Evaluación de un medio cromogénico para el aislamiento e identificación presuntiva de Aeromonas.10 de Octubre del 2006 ERIKA BEATRIZ ANGELES MORALES Determinación de factores extrínsecos relacionados con la formación de biopelícula de Aeromonas caviae Sch3 . 7 de Agosto del 2006 MARÍA ALEJANDRA ALVARADO HERNÁNDEZ Búsqueda del gen cag A en Aeromonas asiladas de medio ambiente.15 de Febrero 2006 CARLOS SANCHEZ GARIBAY Demostración de β−lactamasas de espectro expendido en cepas de Aeromonas spp .15 de Febrero 2006 NIRYA RAMIREZ ZETINA Expresión fenotípica del Sistema Flagelar de Aeromonas spp y su relación con
la formación biofilm.
• Se tiene en proceso dos Tesinas de Licenciatura
ROSALBA GUERRERO GONZALEZ El flagelo bacteriano.21 de Agosro del 2006 ROSALIA BAEZ ARIAS Sistemas de Secreción bacterianos. 21 de Agosro del 2006
• Se tiene en proceso una tesis de doctorado RAMÓN IGNACIO ARTEAGA GARIBAY
El biofilm de Aeromonas y su relación con el sistema flagelar de cómo mecanismo de patogenicidad y resistencia en Aeromonas. 23 de Noviembre 2006 • Se terminó una tesis de doctorado.
Ma GUADALUPE AGUILERA ARREOLA Doctoral Diversidad genética intraespecífica de Aeromonas hydrophila
6 Septiembre 2006

2
• Se dictaron las siguientes conferencias: Toxinas bacterianas y Sistemas de Secreción Centro de Investigación y Estudios avanzados del IPN Curso Microbiología Programa de Posgrado del depto Genética y Biología Molecular 7 Septiembre 2006 Aeromonas spp y Plesiomonas shigelloides Centro de Estudios Científicos y Tecnológicos Diodoro Antunez Echegaray Instituto Politécnico Nacional 20 Junio 2006 Aeromonas 3er semana de Actualización en Microbiología Instituto Mexicano del seguro social Hospital general de Zona 1-A Venados 31 de Julio al 4 de Agosto 2006 Aeromonas un patógeno importante en México IV Congreso Internacional de Ingeniería Bioquímica XV Congreso Nacional de Ingeniería Bioquímica 4-7 Abril 2006 Importancia y Mecanismos de patogenicidad de Aeromonas Conferencia Magistral Programa de aseguramiento de la Calidad (PACAL) 16 Marzo 2006 El género Aeromonas y su mecanismo de patogenicidad” Escuela Nacional de Medicina y Homeopatía Sección de Posgrado
14 de Febrero 2006
• Representaron los siguientes trabajos en en seminario academico
MARIA ISABEL PORTILLA MUÑOZ Evaluación de un medio cromogénico para el aislamiento e identificación presuntiva de Aeromonas. 13 de Diciembre 2006 ERIKA BEATRIZ ANGELES MORALES Determinación de factores extrínsecos relacionados con la formación de biopelícula de Aeromonas caviae Sch3.13 de Diciembre 2006 MARÍA ALEJANDRA ALVARADO HERNÁNDEZ Búsqueda del gen cag A en Aeromonas asiladas de medio ambiente. 22 de Septiembre 2006 CARLOS SANCHEZ GARIBAY Demostración de β−lactamasas de espectro expendido en cepas de Aeromonas spp 22 de Septiembre 2006
• Representaron los siguientes trabajos en congresos nacionales e internacionales:
R.I. Arteaga Garibay. M.G.Aguilera Arreola , S.Giono cerezo, A. Navarro Ocaña y G.Castro-Escarpulli. 2006. El Flagelo lateral en la adherencia y la formación de Biofilm de Aeromonas spp. XXXV Congreso Nacional de Microbiología.
Arteaga -Garibay RI, Méndez Tenorio A, Maldonado Rodríguez R, Cervantes Alarcón M , Castro Escarpulli G. 2006. Predicción de estructuras secundaria y terciaria de la flagelina lateral de Aeromonas hydrophila. IV Congreso Internacional de Ingeniería Bioquímica y XV Congreso Nacional de Ingeniería Bioquímica. Michoacán México Cervantes-Alarcón, Martha; Méndez-Tenorio, Alfonso; Maldonado-Rodríguez Rogelio; Aguilera-Arreola, Ma Guadalupe; Castro-Escarpulli Graciela. 2006. Búsqueda de secuencias de proteínas homólogas del sistema de secreción tipo III (SSTIII) DE Aeromonas hydrophila en el genoma completo de Yersinia pestis. IV Congreso Internacional de Ingeniería Bioquímica y XV Congreso Nacional de Ingeniería Bioquímica. Michoacán México
3
• En la parte experimental planteada :
Se tiene una sonda para el gen cag A
Se tiene algunas cepas Beta lactamasas de espectro extendido positivas
Se secuenciaron los espacios intergénico
METODOLOGIA
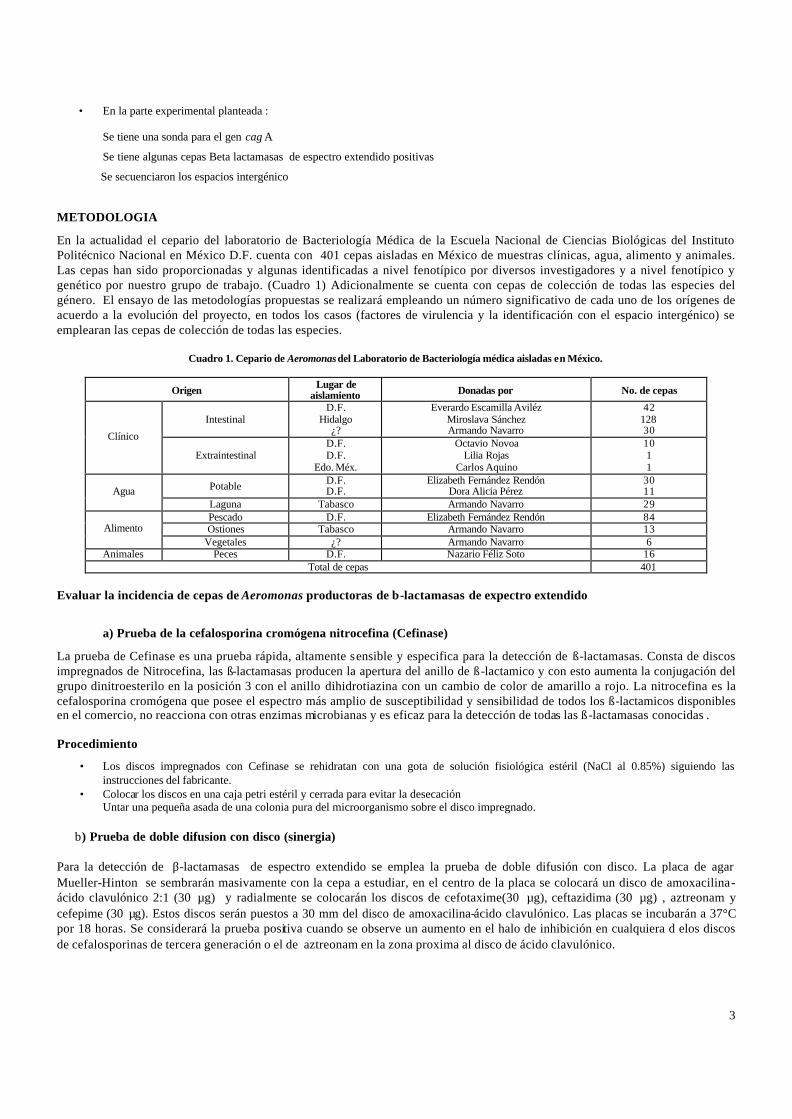
En la actualidad el cepario del laboratorio de Bacteriología Médica de la Escuela Nacional de Ciencias Biológicas del Instituto Politécnico Nacional en México D.F. cuenta con 401 cepas aisladas en México de muestras clínicas, agua, alimento y animales. Las cepas han sido proporcionadas y algunas identificadas a nivel fenotípico por diversos investigadores y a nivel fenotípico y genético por nuestro grupo de trabajo. (Cuadro 1) Adicionalmente se cuenta con cepas de colección de todas las especies del género. El ensayo de las metodologías propuestas se realizará empleando un número significativo de cada uno de los orígenes de acuerdo a la evolución del proyecto, en todos los casos (factores de virulencia y la identificación con el espacio intergénico) se emplearan las cepas de colección de todas las especies.
Cuadro 1. Cepario de Aeromonas del Laboratorio de Bacteriología médica aisladas en México.
Origen Lugar de aislamiento Donadas por No. de cepas
Intestinal D.F.
Hidalgo ¿?
Everardo Escamilla Aviléz Miroslava Sánchez Armando Navarro
42 128 30 Clínico
Extraintestinal D.F. D.F.
Edo. Méx.
Octavio Novoa Lilia Rojas
Carlos Aquino
10 1 1
Potable D.F. D.F.
Elizabeth Fernández Rendón Dora Alicia Pérez
30 11 Agua
Laguna Tabasco Armando Navarro 29 Pescado D.F. Elizabeth Fernández Rendón 84 Ostiones Tabasco Armando Navarro 13 Alimento
Vegetales ¿? Armando Navarro 6 Animales Peces D.F. Nazario Féliz Soto 16
Total de cepas 401 Evaluar la incidencia de cepas de Aeromonas productoras de β-lactamasas de expectro extendido
a) Prueba de la cefalosporina cromógena nitrocefina (Cefinase)
La prueba de Cefinase es una prueba rápida, altamente sensible y especifica para la detección de ß-lactamasas. Consta de discos impregnados de Nitrocefina, las ß-lactamasas producen la apertura del anillo de ß-lactamico y con esto aumenta la conjugación del grupo dinitroesterilo en la posición 3 con el anillo dihidrotiazina con un cambio de color de amarillo a rojo. La nitrocefina es la cefalosporina cromógena que posee el espectro más amplio de susceptibilidad y sensibilidad de todos los ß-lactamicos disponibles en el comercio, no reacciona con otras enzimas microbianas y es eficaz para la detección de todas las ß-lactamasas conocidas . Procedimiento
• Los discos impregnados con Cefinase se rehidratan con una gota de solución fisiológica estéril (NaCl al 0.85%) siguiendo las instrucciones del fabricante.
• Colocar los discos en una caja petri estéril y cerrada para evitar la desecación Untar una pequeña asada de una colonia pura del microorganismo sobre el disco impregnado.
b) Prueba de doble difusion con disco (sinergia) Para la detección de β-lactamasas de espectro extendido se emplea la prueba de doble difusión con disco. La placa de agar Mueller-Hinton se sembrarán masivamente con la cepa a estudiar, en el centro de la placa se colocará un disco de amoxacilina-ácido clavulónico 2:1 (30 µg) y radialmente se colocarán los discos de cefotaxime(30 µg), ceftazidima (30 µg) , aztreonam y cefepime (30 µg). Estos discos serán puestos a 30 mm del disco de amoxacilina-ácido clavulónico. Las placas se incubarán a 37°C por 18 horas. Se considerará la prueba positiva cuando se observe un aumento en el halo de inhibición en cualquiera d elos discos de cefalosporinas de tercera generación o el de aztreonam en la zona proxima al disco de ácido clavulónico.

4
Determinar la distribución del gen cagA en cepas de origen ambiental. El gen cag A genes que se buscarán mediante la técnica de la reacción en cadena de la polimerasa y Dot Blot. Con los iniciadores siguientes : 5´GAATCAGTATTTTTCAGAC) y Cr2 ( 5´GGGTTGTATGATATTTTCC) Los productos de amplificación serán analizados por electroforesis en geles de agarosa al 1% (agarosa D-1 Baja EEO, ) disuelta en una solución reguladora TAE 1x (trisma base-ácido acético-EDTA) se calentará hasta su disolución, se adicionará 5µl de bromuro de etidio (0.5 µg/ml), después de su gelificación las muestras se colocarán dentro de los pozos, mezclando en papel parafilm 2 µl de densificante con azul de bromofenol al 0.25% con 5 µl de la muestra amplificada con ayuda de una micropipeta, estas se correrán a 55 V/cm durante treinta minutos. A término del tiempo se visualizarán en un documentador de imágenes con luz ultravioleta DISEÑO DE SONDA
La marcación de la sonda, con Dioxigenin 11 dUTP(Dig-dUTP) a partir de la amplificación del gen cag A que se obtuvo por PCR: La mezcla de reacción que se utilizará para esto se muestra en el cuadro 2.
Cuadro 2 Mezcla y condicones de Reacción para el marcaje d ela Sonda DOT BLOT Se Cuantificación del DNA con Genequant. Preparar una solución de 10µl con 10 µg de DNA genómico de cada cepa a ensayar Depositar 10µl de DNA genómico de cada cepa a ensayar sobre la membrana de nylon. Esta membrana previamente se habrá autoclavado y dividido en secciones con el uso de un lápiz, en cada una de las cuales habrá DNA de cada una de las cepas. Situar en 1 de las secciones 10 µl del producto de PCR del gen a analizar (control positivo).la membrana se colocará sobre un papel filtro empapado con una solución de SDS (sodio dodecil sulfato) al 10% durante 3 min. Seguidamente la membrana se colocará sobre un papel de filtro empapado con una solución desnaturalizante (0.5 N NaOH, 1.5M NaCl), durante 5 min a temperatura ambiente. Al término del tiempo, se neutralizará la membrana sobre un papel filtro impregnado con una solución neutralizante (1.5 M NaCl. 0.5 M tris-Cl pH 7.4) durante 5 min. A continuación se paso la membrana a un papel empapado en 10 X SSC durante 5 min. Finalmente se fijará el DNA a la membrana de forma permanente, mediante la exposición de la misma a luz ultravioleta durante 3 min., se enjuagaron con agua las membranas y se dejaron secar a temperatura ambiente. La membrana se prehibridará con 20 ml/cm2 de Dig Easy Hyb (Boehringer Mannhein) a 68ºC en un horno de hibridación durante 30 min. La sonda marcada con digoxigenina se someterá a ebullición durante 5 min, y se colocará en hielo inmediatamente. Se eliminará la solución de prehibridación y se adicionará 5 ml de Dig Easy Hyb con 125 ng de la sonda marcada y se incubará durante toda la noche a 68ºC. Al día siguiente, se lavará a temperatura ambiente la membrana sumergiéndola dos veces durante 15 min, en agitación con la solución (2 X SSC, 0.1% SDS) seguida de un segundo lavado 2 X 15 min con una solución (0.5 X SSC, 0.1% SDS) y se realizará un ultimó lavado con una solución (0.1 X SSC, 0.1% SDS) 2 X 15 min. Para la detección inmunológica, se incubará la membrana durante 5 min en agitación suave, en una solución de 0.1 M ácido maléico, 0.15 M NaCl a pH 7.5 a 20ºC y 0.3% (v/v) Tween 20. A continuación se incubará 1 h, en 150 ml de solución de bloqueo (Boehringer Mannhein) agitando suavemente y a temperatura ambiente. Seguidamente, se añadirá a la membrana el anticuerpo anti-digoxigenina (dilución 1:5000) acoplado a fosfatasa alcalina en 20 ml de solución de bloqueo y se incub ará 1 h a temperatura ambiente y en agitación suave. Finalizando este paso, se lavará la membrana dos veces con ácido maléico 0.1 M-NaCl 0.15 M, pH 7.5 a 20°C durante 30 min y se equilibrará con 25 ml de solución de Tris -HCl 0.1 M, NaCl 0.1 M pH 9.5 durante 5 min. La detección de la señal se hizo en la oscuridad con 25 ml de una solución recién preparada de 20 µl NBT (nitro blue tetrazolium chloride). A los 5 min se parará la reacción de color con 50 ml de agua destilada. La membrana se dejará secar a temp eratura ambiente.
• Secuenciación de los espacios intérgenicos
a) Reacción en cadena de la polimerasa del ITS 16/23S rRNA.
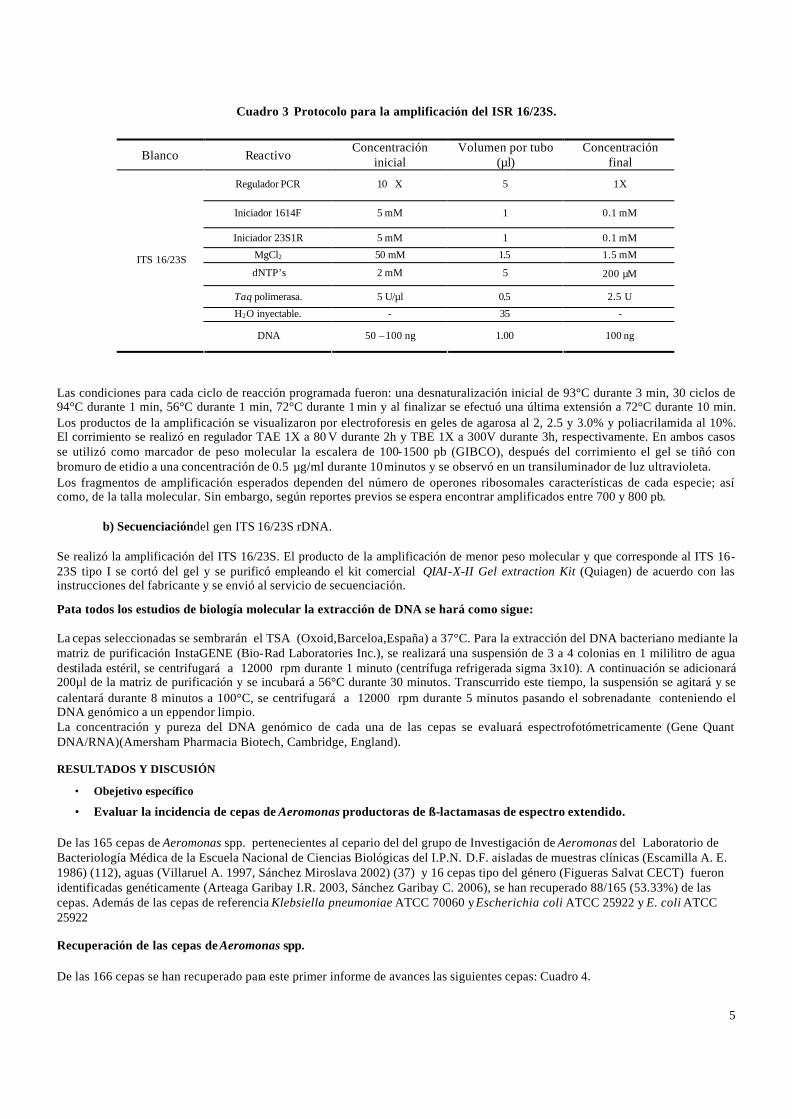
La amplificación del espacio intergénico 16/23S rDNA se realizó empleando los iniciadores 1614F-5’-CTT GTA CAC ACC GCC CGT C-3’ (1389-1407 pb) y 23S1R- 5’-GGG TTT CCC CAT TCG GAA ATC-3’ (124 -103 pb). Las concentraciones de cada uno de los reactivos y las condiciones para la amplificación se detallan en el cuadro 3.
PCR buffer (10x) 5 µL MgCl2 (50mM) 1.8µL DNTPs (2mM) 250M Primer(0.1 µM) 1µl Primer (0.1 µM) 1µL Taq polimerasa (5U/ µl) 0.2µL Dig-11-UTP(0.5 nmol) 0.5µL H2O 20µL
94ºC 3minutos 94ºC 1 minuto 55 ºC 1 minuto 30 ciclos 72ºC 1 minuto 72ºC 5 minutos
5
Cuadro 3 Protocolo para la amplificación del ISR 16/23S. Las condiciones para cada ciclo de reacción programada fueron: una desnaturalización inicial de 93°C durante 3 min, 30 ciclos de 94°C durante 1 min, 56°C durante 1 min, 72°C durante 1 min y al finalizar se efectuó una última extensión a 72°C durante 10 min. Los productos de la amplificación se visualizaron por electroforesis en geles de agarosa al 2, 2.5 y 3.0% y poliacrilamida al 10%. El corrimiento se realizó en regulador TAE 1X a 80 V durante 2h y TBE 1X a 300V durante 3h, respectivamente. En ambos casos se utilizó como marcador de peso molecular la escalera de 100-1500 pb (GIBCO), después del corrimiento el gel se tiñó con bromuro de etidio a una concentración de 0.5 µg/ml durante 10 minutos y se observó en un transiluminador de luz ultravioleta. Los fragmentos de amplificación esperados dependen del número de operones ribosomales características de cada especie; así como, de la talla molecular. Sin embargo, según reportes previos se espera encontrar amplificados entre 700 y 800 pb.
b) Secuenciacióndel gen ITS 16/23S rDNA.
Se realizó la amplificación del ITS 16/23S. El producto de la amplificación de menor peso molecular y que corresponde al ITS 16-23S tipo I se cortó del gel y se purificó empleando el kit comercial QIAI-X-II Gel extraction Kit (Quiagen) de acuerdo con las instrucciones del fabricante y se envió al servicio de secuenciación.
Pata todos los estudios de biología molecular la extracción de DNA se hará como sigue: La cepas seleccionadas se sembrarán el TSA (Oxoid,Barceloa,España) a 37°C. Para la extracción del DNA bacteriano mediante la matriz de purificación InstaGENE (Bio-Rad Laboratories Inc.), se realizará una suspensión de 3 a 4 colonias en 1 mililitro de agua destilada estéril, se centrifugará a 12000 rpm durante 1 minuto (centrífuga refrigerada sigma 3x10). A continuación se adicionará 200µl de la matriz de purificación y se incubará a 56°C durante 30 minutos. Transcurrido este tiempo, la suspensión se agitará y se calentará durante 8 minutos a 100°C, se centrifugará a 12000 rpm durante 5 minutos pasando el sobrenadante conteniendo el DNA genómico a un eppendor limpio. La concentración y pureza del DNA genómico de cada una de las cepas se evaluará espectrofotómetricamente (Gene Quant DNA/RNA)(Amersham Pharmacia Biotech, Cambridge, England). RESULTADOS Y DISCUSIÓN
• Obejetivo específico
• Evaluar la incidencia de cepas de Aeromonas productoras de ß-lactamasas de espectro extendido. De las 165 cepas de Aeromonas spp. pertenecientes al cepario del del grupo de Investigación de Aeromonas del Laboratorio de Bacteriología Médica de la Escuela Nacional de Ciencias Biológicas del I.P.N. D.F. aisladas de muestras clínicas (Escamilla A. E. 1986) (112), aguas (Villaruel A. 1997, Sánchez Miroslava 2002) (37) y 16 cepas tipo del género (Figueras Salvat CECT) fueron identificadas genéticamente (Arteaga Garibay I.R. 2003, Sánchez Garibay C. 2006), se han recuperado 88/165 (53.33%) de las cepas. Además de las cepas de referencia Klebsiella pneumoniae ATCC 70060 y Escherichia coli ATCC 25922 y E. coli ATCC 25922 Recuperación de las cepas de Aeromonas spp. De las 166 cepas se han recuperado para este primer informe de avances las siguientes cepas: Cuadro 4.
Blanco Reactivo Concentración
inicial Volumen por tubo
(µl) Concentración
final
Regulador PCR 10 X 5 1X
Iniciador 1614F 5 mM 1 0.1 mM
Iniciador 23S1R 5 mM 1 0.1 mM
MgCl2 50 mM 1.5 1.5 mM
dNTP’s 2 mM 5 200 µM
Taq polimerasa. 5 U/µl 0.5 2.5 U
H2O inyectable. - 35 -
ITS 16/23S
DNA 50 – 100 ng 1.00 100 ng

6
Estas cepas se encontraban conservadas por congelación a -80 ºC en caldo Todd– Hewitt con glicerol al 40 % como crioprotector y fueron resembradas en agar Mueller-Hinton. Después de la incubación se comprobó su pureza por tinción de Gram e identidad con la prueba de oxidasa, ambas pruebas presentan resultado negativo. Estas cepas fueron conservadas a corto plazo para su uso.
Cuadro 4. Genoespecies de Aeromonas spp. Recuperadas
Especies Cepas clínicas
n = 53 n (%)
Cepas de agua n = 19 n (%)
Cepas tipo n=16
A. hydrophila 21 (39.62%) 6 (31.58%) A. bestiarum ——— 8 (42.11%)
A. cavie 22 (41.51%) 4 (21.05%) A. trota 1 (1.89%) ———
A. veronii biovar. sobria 9 (16.98%) 1 (5.26%)
TOTAL 53 19 16 88 Detección de cepas productoras de ß-lactamasas por la prueba de la cefalosporina cromógena nitrocefina (Cefinase©). La prueba de cefinase© es una prueba rápida, altamente sensible y especifica para la detección de ß-lactamasas. Consta de discos impregnados de Nitrocefina, las ß-lactamasas producen la apertura del anillo de ß-lactámico y con esto aumenta la conjugación del grupo dinitroesterilo en la posición 3 con el anillo dihidrotiazina con un cambio de color de amarillo a rojo De las cepas de Aeromonas spp. probadas 87/88 (98.86%) (Figura 5) fueron productoras de ß-lactamasas. Cabe mencionar que encontramos una cepa identificada genéticamente como Aeromonas trota la cual es reportada como sensible a la penicilina, en el presente trabajo se detectó con el fenotipo cefinase© positiva. Además una Aeromonas hydrophila cefinase negativa.
Figura No.1 Fotografía de las cepas productoras de ß-lactamasas.a- b: Cepas dereferencia c Cepas utilizadas en este trabajo
b) Detección de cepas productoras de BLEE´s por la prueba de difusión con doble disco (sinergia).
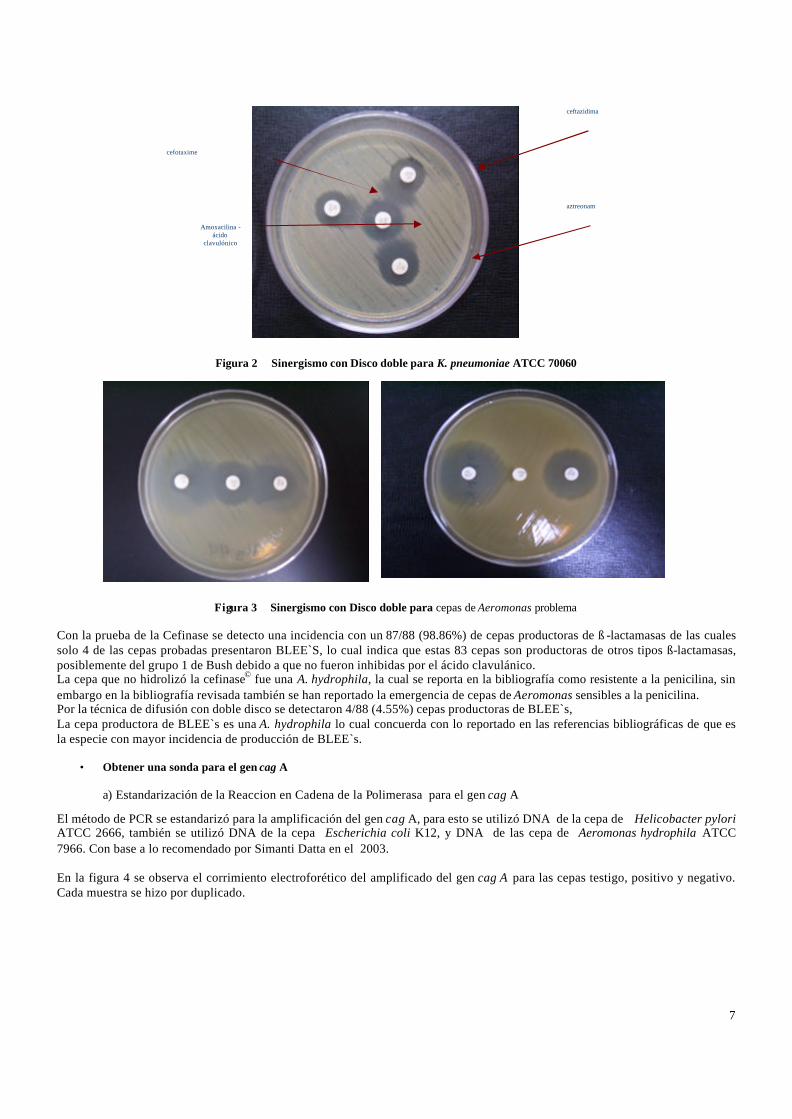
Este método inicialmente fue realizado de acuerdo a los criterios descritos por Roxana Alinga (2001), sin embargo no presentó los resultados esperados ya que la distancia de 30 mm entre los discos de las cefalosporinas y amoxicilina - ac. clavulánico 2:1 30µg (AMC) fue muy grande y no se observo el sinergismo de los antibióticos en las cepas de referencia, por lo que decidimos seguir las recomendaciones de la CLSI (Cuadro 5). Se colocó en el centro de la placa el disco de AMC y alrededor de este a 20 mm los discos con Aztreonam 30µg (ATM), Cefotaxime 30 µg (CTX) y Ceftazidima 30µg (CAZ) y a esta distancia se pudo observar el sinergismo . Se probó la cepa de referencia K. pneumoniae ATCC 70060 donde se presentó el sinergismo entre el disco de AMC y las cefalosporinas debido a la producción de ß-lactamasas de espectro extendido (BLEE´s). Y en las cepas de E. coli ATCC 25922 en donde no se presentó el sinergismo ya que no son cepas productoras de BLEE´s ( Figura 2). . A partir de las 88 cepas que fueron estudiadas por este método solo 4 cepas fueron productoras de BLEE´s, obteniendo así 2 A. caviae (2.27%) y 2 A. hydrophila (2.27%) (Figura 3).
E.coli ATCC 25922 K. pneumoniae ATCC 70060 Aeromonas spp
7
Figura 2 Sinergismo con Disco doble para K. pneumoniae ATCC 70060
Figura 3 Sinergismo con Disco doble para cepas de Aeromonas problema Con la prueba de la Cefinase se detecto una incidencia con un 87/88 (98.86%) de cepas productoras de ß -lactamasas de las cuales solo 4 de las cepas probadas presentaron BLEE`S, lo cual indica que estas 83 cepas son productoras de otros tipos ß-lactamasas, posiblemente del grupo 1 de Bush debido a que no fueron inhibidas por el ácido clavulánico. La cepa que no hidrolizó la cefinase© fue una A. hydrophila, la cual se reporta en la bibliografía como resistente a la penicilina, sin embargo en la bibliografía revisada también se han reportado la emergencia de cepas de Aeromonas sensibles a la penicilina. Por la técnica de difusión con doble disco se detectaron 4/88 (4.55%) cepas productoras de BLEE`s, La cepa productora de BLEE`s es una A. hydrophila lo cual concuerda con lo reportado en las referencias bibliográficas de que es la especie con mayor incidencia de producción de BLEE`s.
• Obtener una sonda para el gen cag A
a) Estandarización de la Reaccion en Cadena de la Polimerasa para el gen cag A
El método de PCR se estandarizó para la amplificación del gen cag A, para esto se utilizó DNA de la cepa de Helicobacter pylori ATCC 2666, también se utilizó DNA de la cepa Escherichia coli K12, y DNA de las cepa de Aeromonas hydrophila ATCC 7966. Con base a lo recomendado por Simanti Datta en el 2003. En la figura 4 se observa el corrimiento electroforético del amplificado del gen cag A para las cepas testigo, positivo y negativo. Cada muestra se hizo por duplicado.
aztreonam
Amoxacilina - ácido
clavulónico
cefotaxime
ceftazidima

8
1 2 3 4 5 6 7 8
Fig. 3. Electroforesis del gel que muestra los productos de PCR para el amplificado de 208 pb del gen cag A.En 1 Marcador de Peso molecular, 2 y 5 Producto de PCR de Helicobacter pylori , 3 y 7 Producto de PCR de Escherichia coli, 4 y 8 Producto de PCR de Aeromonas ATCC 7966
Por lo que a la luz de los resultados se cambiaron las condiciones de reacción para volver a repetir la PCR; de esta manera logramos obtener la estandarización del método para el gen cag A Una vez cambiadas las condiciones se llego a obtener la estandarización del método y esto se muestra en la figura 5, en donde es posible observar una banda de aproximadamente 208 pb en el pozo 1, que se refiere a la muestra de Helicobcter pylori, y que se esta utilizando como control positivo, es decir las condiciones de reacción son las optimas para obtener el amplificado esperado
Fig 5 Electroferograma de los productos de la segunda PCR para el amplificado de 208 pb del gen cag A, (1) muestra marcador de Peso Molecular de 100, (2) Producto de PCR de Helicobacter pylori, (3) Producto de PCR de Aeromonas ATCC 7966, (4) Producto de PCR de Escherichia coli . Para la elaboración de la sonda, se usaron las mismas condiciones de reacción ya estandarizadas para el método de PCR est, la sonda se marco con Dioxigenina 11 UTP (dig-UTP ). El tamaño de la sonda es un poco más pesado que el amplificado, por lo que se ve una banda mas gruesa, según se aprecia en la figura 6; mientras que el amplificado sin marcar es menos pesado que el marcado, por lo que la banda que se observa es menos marcada.
Amplificado 208 pb

9
1 2 3 4 5 6 7 8
208 pb
Fig. 6 Electroforesis en gel de agarosa de la sonda marcada para cag A. carril 8 marcador de peso Molecular, carril 2, 3 y 4 amplificado de cag A sin marcar, carriles 5, 6 y 7 sonda marcada. Análisis del espaciador intergénico 16/23S rRNA y secuenciación del ITS 16/23S rRNA tipo I; tRNAIle, Ala
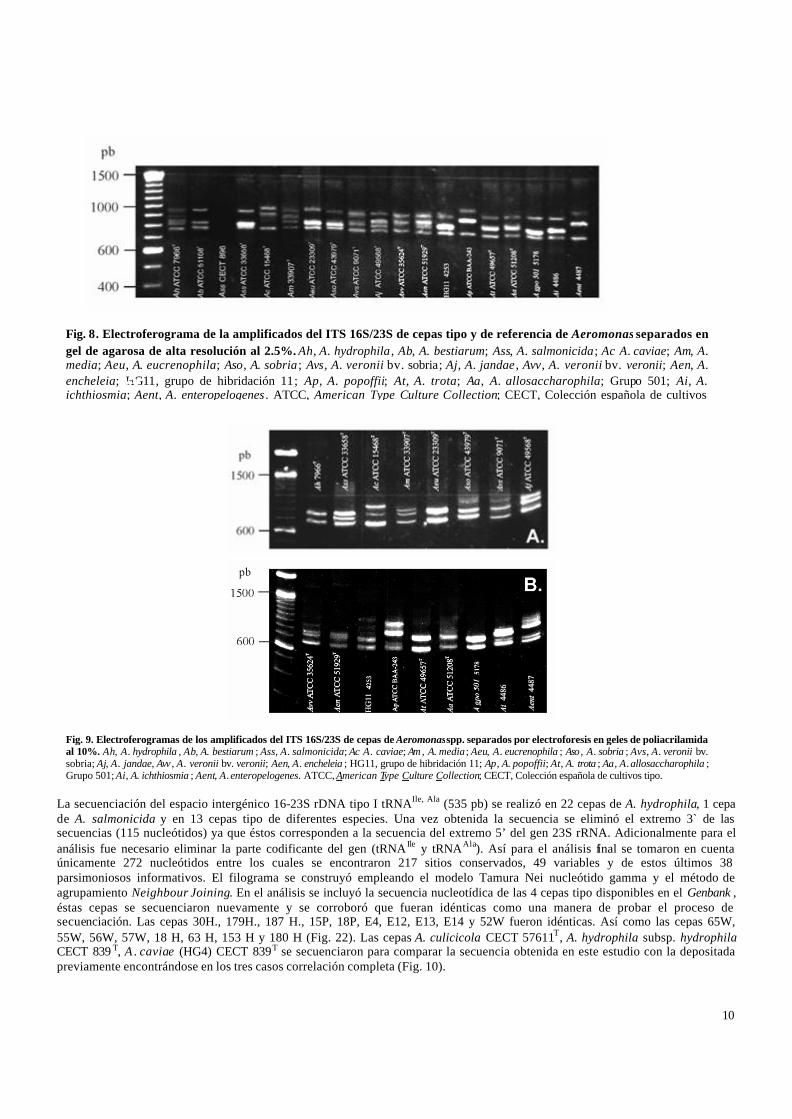
La amplificación del espacio intergénico ITS 16-23S rDNA como se esperaba produjo en la mayoría de las cepas tres bandas correspondientes a los tres diferentes tipos de ITS descritos únicamente en 4 especies. La separación de los productos de amplificación se realizó inicialmente en geles de agarosa al 2 %. Sin embargo, en este soporte no fue posible la separación debido a que la talla molecular de los productos es muy similar y se observó una sola banda muy gruesa. La separación se intentó entonces en geles de agarosa de alta resolución al 2.5% (Fig. 7 y 8). Asimismo, se corrieron geles de poliacrilamida al 10% con el mismo propósito (Fig. 9). En éstos últimos se observó que las bandas con diferencia de PM de más de 100 pb se separaban mejor, sin embargo las dos bandas que sólo se diferencian en un menor número de bases (aproximadamente 20-30 pb) tampoco se separaron idealmente bajo estas condiciones en algunos de los aislamientos.Si bien se observó que algunas de las cepas tipo de las diferentes especies del género producen patrones diferentes entre ellas, en otras tantas el perfil de amplificación es muy similar.
208 pb
10
La secuenciación del espacio intergénico 16-23S rDNA tipo I tRNAIle, Ala (535 pb) se realizó en 22 cepas de A. hydrophila, 1 cepa de A. salmonicida y en 13 cepas tipo de diferentes especies. Una vez obtenida la secuencia se eliminó el extremo 3` de las secuencias (115 nucleótidos) ya que éstos corresponden a la secuencia del extremo 5’ del gen 23S rRNA. Adicionalmente para el análisis fue necesario eliminar la parte codificante del gen (tRNA Ile y tRNAAla). Así para el análisis final se tomaron en cuenta únicamente 272 nucleótidos entre los cuales se encontraron 217 sitios conservados, 49 variables y de estos últimos 38 parsimoniosos informativos. El filograma se construyó empleando el modelo Tamura Nei nucleótido gamma y el método de agrupamiento Neighbour Joining. En el análisis se incluyó la secuencia nucleotídica de las 4 cepas tipo disponibles en el Genbank , éstas cepas se secuenciaron nuevamente y se corroboró que fueran idénticas como una manera de probar el proceso de secuenciación. Las cepas 30H., 179H., 187 H., 15P, 18P, E4, E12, E13, E14 y 52W fueron idénticas. Así como las cepas 65W, 55W, 56W, 57W, 18 H, 63 H, 153 H y 180 H (Fig. 22). Las cepas A. culicicola CECT 57611T, A. hydrophila subsp. hydrophila CECT 839 T, A . caviae (HG4) CECT 839T se secuenciaron para comparar la secuencia obtenida en este estudio con la depositada previamente encontrándose en los tres casos correlación completa (Fig. 10).
Fig. 8. Electroferograma de la amplificados del ITS 16S/23S de cepas tipo y de referencia de Aeromonas separados en gel de agarosa de alta resolución al 2.5%. Ah, A. hydrophila , Ab, A. bestiarum; Ass, A. salmonicida; Ac A. caviae; Am, A. media; Aeu, A. eucrenophila; Aso, A. sobria ; Avs, A. veronii bv. sobria; Aj, A. jandae , Avv, A. veronii bv. veronii; Aen, A. encheleia; HG11, grupo de hibridación 11; Ap, A. popoffii; At, A. trota; Aa, A. allosaccharophila; Grupo 501; Ai, A. ichthiosmia; Aent, A. enteropelogenes . ATCC, American Type Culture Collection; CECT, Colección española de cultivos
A
Fig. 9. Electroferogramas de los amplificados del ITS 16S/23S de cepas de Aeromonas spp. separados por electroforesis en geles de poliacrilamida al 10%. Ah, A. hydrophila , Ab, A. bestiarum ; Ass, A. salmonicida; Ac A. caviae; Am , A. media ; Aeu, A. eucrenophila ; Aso , A. sobria ; Avs, A. veronii bv. sobria; Aj, A. jandae, Avv , A. veronii bv. veronii; Aen, A. encheleia ; HG11, grupo de hibridación 11; Ap, A. popoffii; At, A. trota ; Aa, A. allosaccharophila ; Grupo 501; Ai, A. ichthiosmia ; Aent, A. enteropelogenes. ATCC, American Type Culture Collection; CECT, Colección española de cultivos tipo.

11
Fig. 23. Filograma basado en el análisis de la secuencia del espacio intergénico 16/23S rDNA tipo I tRNAIle, Ala con el modelo Tamura Nei y Neighbour Joining. P, cepas aisladas de pescado congelado, W, cepas aisladas de agua; H, cepas aisladas de heces diarreicas; E, cepas aisladas de cuadros clínicos extraintestinales. ATCC, American type culture collection . CECT, Colección española de cepas tipo. El cuadro enmarca las cepas de A. hydrophila estudiadas. Las flechas apuntan las cepas tipo cuyas secuencias fueron obtenidas en el GenBank y que posteriormente se compararon con la secuencia obtenida en el laboratorio.
El espacio intergénico 16/23S rRNA se amplificó en cepas tipo y de referencia descritas en el género Aeromonas. El número de productos de la amplificación del ITS y el peso molecular de las mismas fue como se esperaba variable en cada especie probada. Esto debido a que el número de bandas obtenidas depende del número de operones ribosómicos que posea cada especie o cepa analizada y en función de que el ITS en cada uno de ellos sea de diferente peso molecular. No obstante, según algunas secuencias reportadas en el GenBank y dos publicaciones previas en el género (East y Collins, 1993; Pidiyar y col., 2003) a este respecto, se esperaba encontrar amplicones entre 700 y 800 pb. Con esta información y basándonos en que en otras bacterias como Vibrio spp. se han descrito diferentes tipos de ITS se deduce que en Aeromonas existen al menos tres tipos de ITS. La región espaciadora entre los genes ribosomales 16S y 23S varían ampliamente en tamaño y secuencia nucleotídica y han sido útiles para tipificar e identificar diversas especies bacterianas (Kiebre-Toe y col., 2005; Laganowska y Kaznowski, 2004; Pérez-Luz y col., 2004; Gütler y Stanisich, 1996; Jensen y col., 1993). Sin embargo, el análisis del ITS de Aeromonas spp. realizado en el presente trabajo mostró que este no es un buen blanco molecular en la identificación de todas las especies descritas hasta el momento, ya que sólo se encontró un patrón especie específico para 8 de las especies lo cual concuerda con los resultados reportados por Kong y col., (1999) quienes lograron describir iniciadores específicos para la amplificación de 7 especies. Adicionalmente, el análisis de los patrones producidos por las diferentes cepas de A. hydrophila mostró que existe una cierta diversidad de este blanco molecular dentro de la especie por lo cual no sería útil su aplicación como un método de identificación. La secuenciación de uno de los tipos del ITS (tipo I tRNA Ile, Ala) se realizó tanto en cepas tipo del género como en cepas de A. hydrophila. Ya que en las bases de datos no se cuenta con secuencias depositadas de las cepas tipo (a excepción de A. caviae, A. hydrophila, A. veronii bv. sobria y A. culicicola) o de A. hydrophila. El análisis de las secuencia de A. hydrophila demostró que existe heterogeneidad nucleotídica en este marcador entre algunas de las cepas analizadas. Lo anterior coincide con algunos estudios realizados en otras bacterias que señalan que el cambio de la secuencia del ITS no es uniforme ni a través de la regió n misma ni en especies estrechamente relacionadas y por lo tanto el nivel de divergencia observada es apropiado para detectar diferencias entre individuos o tipificación (Vogler y DeSalle, 1994). Sin embargo, 18 de las 21 cepas analizadas de A. hydrophila mostraron secuencia idéntica entre sí. Dado que la evolución concertada ocurre de manera significativa en casi todas las familias de multigenes, como es el caso los genes ribosomales, genera la homogenización o pérdida de los cambios en los diferentes operones este tipo de genes generalmente presentan reducida variación intraespecífica (Ganley y scout, 1998). Sin embargo, recientemente Morandi y col., 2005 reportaron que en algunas especies de Aeromonas existe heterogeneidad intragenómica del gen 16S rRNA, datos que coinciden de manera indirecta con lo observado en la secuenciación del gene 16S rRNA e ITS tipo I tRNA Ile,Ala de este estudio y contrasta con la poca heterogeneidad intragenómica detectada por Coenye y Vandamme (2003) en los diferentes

12
operones ribosomales de los genomas de bacterias que se habían secuenciado completamente hasta ese año. Finalmente los valores de bootstrap obtenidos en el análisis de las secuencias del ITS tipo I son en la mayoría de los casos muy bajos y no aseguran que no hubiera distorsión de los datos durante el análisis, razón por la cual no es posible hacer mayores conclusiones acerca de los resultados obtenidos en las cepas. No obstante se observan todas las cepas de A. hydrophila agrupadas y separadas del resto de las especies. CONCLUSIONES DE LOS AVANCES HASTA LA FECHA
• El criterio del sinergismo para detectar BLEE`s en Gramnegativos es rápido y sencillo pero se debe hacer cuidadosamente pues se debe tomar en cuenta la distancia a la que se colocaran los discos de las cefalosporinas del AMC y determinar que el sinergismo se deba a la acción de el ácido clavulánico y de la cefalosporina.
• Se pueden encontrar Aeromonas productora de BLEE`s en casos clínicos de cuadros gastrointestinales.
• Se estandarizó la PCR para amplificar al gen cag A • Se tiene una sonda marcada no radiactiva para el gen cag A.
• • El espacio intergénico ITS 16S/23S no es una herramienta útil en la identificación de todas las especies del género.
BIBLIOGRAFÍA
1. Aguilera Arreola G. 2000. Caracterización de cepas de Aeromonas aisladas de pescado congelado. Tesis (Licenciatura) México, D.F. I.P.N. E.N.C.B.
2. Aguilera Arreola G. 2002. Diversidad genética de Aeromonas hydrophila aisladas de diferentes orígenes. Tesis (Maestría) México, D.F. I.P.N. E.N.C.B.
3. Castro Escarpulli G. 1998. Determinación de algunos factores de virulencia de cepas de Aeromonas aisladas de muestras clínicas y de agua. Tesis. (Maestría) Microbiología. México D.F. I.P.N. E.N.C.B
4. Castro Escarpulli G. 2002. Aplicación de técnicas molculares en cepas de Aeromonas aisladas en México.. Tesis. (Doctoral) Microbiología. México D.F. I.P.N. E.N.C.B
5. Castro-Escarpulli G. Peña del Barrio D, Castañeda N, García A, Morier-Dias L, Aguilera-Arreola G, Bravo F L. 2002. Virulence factors of A. caviae strains isolated from acute diarrheic disease in Cuba. Rev. Lat. Microbiol. 44 : 11-13.
6. Castro-Escarpulli G , Aguilera-Arreola Ma. G, Giono Cerezo S, Hernández-Rodríguez CH, Chacón Rodríguez M, Soler Falgás L, Aparicio Ozores G, Figueras S MJ . 2002. El género Aeromonas. ¿Un patógeno importante en México?. Rev. Enf. Infe. y Microbiol.44(4)
7. Castro-Escarpulli G, Figueras MJ, Aguilera-Arreola G, Soler L, Fernández-Rendón E, Aparicio OG, Guarro J, Chacón RM. 2003. Characterisation of Aeromonas spp. isolated from frozen fish intended for human consumption in Mexico. Int. J. Food. Microbiol. 84:41-49
8. Castro-Escarpulli G, Aparicio Ozores G. 2003. Aeromonas y Plesiomonas. En José Tomás Hernández Méndez (eds). Bacteriología Médica diagnóstica. 2ª Edición. ISBN 968-7022-34-5
9. Chacón RM, Castro-Escarpulli G, Soler L, Guarro J, Figueras MJ. 2002. A DNA probe specific for Aeromonas colonies. Diag. Microbio. and Infec. Dis.. 44 (3): 221- 225
10. Chacón RM, Figueras MJ, Castro-Escarpulli G, Soler L, Guarro J. 2003. Distribution of virulence genes in clinical and environmental isolates of Aeromonas spp . Antonie van Leeuwenhoek, International Journal of General and Molecular Microbiology
11. Vilches S., Urgell C.,Merino S., Chacón R.M., Soler L., Castro-Escarpulli G., Figueras M.J., Tomás J.M. (2004) The complete type III secretion system of a mesophilic Aeromonas hydrophila. Appl. Environ. Microbiol. 70:6914-9